Область настоящего изобретения
Настоящее изобретение в основном относится к антигенсвязывающим рецепторам, способным специфически связываться с мутированными Fc доменами, которые характеризуются сниженным связыванием с Fc рецептором, и Т-клеткам, экспрессирующим эти антигенсвязывающие рецепторы. Более конкретно, в настоящем изобретении предлагаются Т-клетки, трансфицированные/трандуцированные антигенсвязывающим рецептором, которые накапливаются при специфическом связывании/взаимодействии с мутированным Fc доменом терапевтических антител. Более того, изобретение относится к набору, включающему Т-клетки по изобретению и/или молекулы нуклеиновой кислоты, векторы, экспрессирующие антигенсвязывающие рецепторы по настоящему изобретению, и направленные к опухоли антитело/антитела, включающие мутированный Fc домен. В изобретении также предлагается продуцирование и применение Т-клеток в способе лечения конкретных заболеваний в сочетании с опухолеспецифичными антителами, а также фармацевтические композиции/лекарственные средства, включающие Т-клетки и/или терапевтические антитела, где Т-клетки предназначены для введения в комбинации с терапевтическим/направленным к опухоли антителом (антителами), включающим мутированный Fc домен со сниженным связыванием с Fc рецептором.
Предпосылки создания настоящего изобретения
Адаптивная Т-клеточная терапия (АКТ) является перспективным подходом к лечению с использованием опухолеспецифичных Т-клеток (Rosenberg и Restifo, Science 348(6230), 62-68 (2015)). При лечении методом АКТ можно использовать природные опухолеспецифические Т-клетки или Т-клетки, которым придают специфичность методом генной инженерии с использованием химерных рецепторов антигена (Rosenberg и Restifo, Science 348(6230) (2015), 62-68). Метод АКТ можно успешно использовать для лечения и индукции ремиссии у пациентов, страдающих даже от прогрессирующих и иных резистентных к лечению заболеваний, таких как острый лифобластный лейкоз, неходжкинские лимфомы или меланомы (Dudley и др., J Clin Oncol 26(32), 5233-5239 (2008), Grupp и др., N Engl J Med 368 (16) (2013), 1509-1518; Kochenderfer и др., J Clin Oncol. 33(6):540-549 (2015), doi: 10.1200/JCO.2014.56.2025. публ. 2014 август 25).
Однако, несмотря на впечатляющую клиническую эффективность, АКТ ограничена связанными с лечением токсичными действиями. Специфичность и наблюдаемые в результате целевые и побочные действия рекомбинантных Т-клеток в ходе АКТ обычно осуществляются с помощью направленного к опухоли антигенсвязывающего фрагмента, включенного в химерный антигенсвязывающий рецептор (CAR). Неполная экспрессия опухолевого антигена или временное различие уровней экспрессии может привести к тяжелым побочным действиям или даже к прерыванию АКТ в связи с непереносимой токсичностью лечения.
Кроме того, доступность опухолеспецифичных Т-клеток для эффективного лизиса опухолевых клеток зависит от способности рекомбинантных Т-клеток к долгосрочному выживанию и пролиферации in vivo. С другой стороны, долгосрочное выживание и пролиферация Т-клеток in vivo может привести к нежелательным долгосрочным эффектам за счет устойчивости неконтролируемого CAR-T-ответа (Grupp и др. N Engl J Med 368(16):1509-1518 (2013), Maude и др. N Engl J Med 371 (16): 1507-1517 (2014)).
Один из подходов для снижения тяжелых связанных с лечением токсичностей и для улучшения безопасности АКТ заключается в ограничении активации и пролиферации CAR-T клеток с помощью введения молекул-адапторов в иммунологический синапс. Такие молекулы-адапторы включают бимодулярное переключение низкомолекулярных соединений, например, как недавно описанное переключение фолат-FITC (Kim и др. J Am Chem Soc; 137:2832-2835 (2015)). Другой подход включает искусственно модифицированные антитела, включающие метку для контроля и направления специфичности CAR-T клеток в отношении опухолевых клеток-мишеней (Ма и др. PNAS; 113(4):Е450-458 (2016), Сао и др. Angew Chem; 128:1-6 (2016), Rogers и др. PNAS; 113(4):Е459-468 (2016), Tamada и др. Clin Cancer Res; 18(23):6436-6445 (2012)).
Однако существующие подходы ограничены по ряду причин. Для иммунологических синапсов, основанных на молекулярном переключении, требуется введение дополнительных элементов, которые могут вызвать иммунный ответ или привести к неспецифическому побочному эффекту. Более того, сложность таких многокомпонентных систем может ограничить эффективность и переносимость лечения. С другой стороны, введение структуры метки в существующие терапевтические моноклональные антитела могут влиять на эффективность и профиль безопасности таких конструктов.
Соответственно, существует необходимость в улучшении направленной против опухоли терапии, прежде всего адаптивной Т-клеточной терапии, чтобы удовлетворить потребности онкологических пациентов. Таким образом, все еще существует необходимость в разработке улучшенных средств, способных улучшить безопасность и эффективность АКТ и исключить указанные выше недостатки.
Краткое описание настоящего изобретения
Настоящее изобретение в основном относится к антигенсвязывающим рецепторам, способным специфически связываться с мутированными Fc доменами, которые характеризуются сниженным связыванием с Fc рецептором, и Т-клеткам, экспрессирующим эти антигенсвязывающие рецепторы.
В одном объекте настоящего изобретения предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен, и внеклеточный домен, включающий антигенсвязывающий фрагмент, где антигенсвязывающий фрагмент способен специфически связываться с мутированным фрагментом кристаллизующегося (Fc) домена, но не способный связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает по меньшей мере одну аминокислотную замену по сравнению с немутированным исходным Fc доменом.
В одном варианте связывание Fc рецептора с мутированным Fc доменом снижено по сравнению со связыванием Fc рецептора с немутированным исходным Fc доменом, прежде всего, где Fc рецептор является Feγ рецептором или неонатальным Fc рецептором (FcRn). В одном варианте связывание Fc рецептора измеряют методом поверхностного плазмонного резонанса (ППР) при 25°С.
В одном варианте антигенсвязывающий фрагмент представляет собой scFv, Fab, crossFab или scFab фрагмент. В предпочтительном варианте антигенсвязывающий фрагмент представляет собой scFv. В другом предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab или crossFab.
В одном варианте осуществления настоящего изобретения заякоренный трансмембранный домен представляет собой трансмембранный домен, выбранный из группы, состоящей из CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 трансмембранного домена или их фрагментов.
В одном варианте заякоренный трансмембранный домен представляет собой трансмембранный домен CD8, прежде всего, где заякоренный трансмембранный домен включает аминокислотную последовательность SEQ ID NO: 11.
В одном варианте осуществления настоящего изобретения антигенсвязывающий рецептор дополнительно включает по меньшей мере один стимулирующий сигнальный домен и/или по меньшей мере костимулирующий сигнальный домен. В одном варианте осуществления настоящего изобретения по меньшей мере один симулирующий сигнальный домен индивидуально выбирают из группы, состоящий из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов. В одном варианте осуществления настоящего изобретения по меньшей мере один стимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z, прежде всего, где по меньшей мере один стимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 13. В одном варианте осуществления настоящего изобретения по меньшей мере один костимулирующий сигнальный домен индивидуально выбирают из группы, состоящий из внутриклеточного домена CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12 или их фрагментов. В одном варианте осуществления настоящего изобретения по меньшей мере один костимулирующий сигнальный домен представляет собой внутриклеточный домен CD28. В одном варианте осуществления настоящего изобретения антигенсвязывающий рецептор включает один стимулирующий сигнальный домен, включающий внутриклеточный домен CD3z или его фрагмент, и где антигенсвязывающий рецептор включает костимулирующий сигнальный домен, включающий внутриклеточный домен CD28 или его фрагмент. В одном варианте осуществления настоящего изобретения стимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 13 и костимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 12.
В одном варианте осуществления настоящего изобретения внеклеточный домен присоединен к заякоренному трансмембранному домену, необязательно через пептидный линкер. В одном варианте осуществления настоящего изобретения пептидный линкер включает аминокислотную последовательность GGGGS (SEQ ID NO: 17). В одном варианте осуществления настоящего изобретения заякоренный трансмембранный домен присоединен к косигнальному домену или к сигнальному домену, необязательно через пептидный линкер. В одном варианте осуществления настоящего изобретения сигнальный и/или косигнальный домены соединены, необязательно по меньшей мере через один пептидный линкер.
В одном варианте осуществления настоящего изобретения антигенсвязывающий фрагмент представляет собой scFv фрагмент, присоединенный через С-концевой остаток к N-концевому остатку заякоренный трансмембранного домена, необязательно через пептидный линкер.
В одном варианте осуществления настоящего изобретения антигенсвязывающий фрагмент представляет собой Fab фрагмент или crossFab фрагмент, где Fab или crossFab фрагмент присоединен через С-концевой остаток тяжелой цепи к N-концевому остатку заякоренный трансмембранного домена, необязательно через пептидный линкер.
В одном варианте осуществления настоящего изобретения антигенсвязывающий рецептор включает один косигнальный домен, где косигнальный домен присоединен через N-концевой остаток к С-концевому остатку заякоренного трансмембранного домена. В одном варианте осуществления настоящего изобретения антигенсвязывающий рецептор включает один стимулирующий сигнальный домен, где стимулирующий сигнальный домен присоединен через N-концевой остаток к С-концевому остатку костимулирующего сигнального домена.
В одном варианте немутированный Fc домен представляет собой IgG1 или IgG4 Fc домен, прежде всего IgG1 Fc домен человека. В одном варианте мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235, I253, Н310, Р331, Р329 и/или Н435 согласно EU нумерации, прежде всего, где аминокислотной мутацией является L234A, L235A, I253A, N297A, Н310А, P329G и/или Н435А.
В одном варианте мутированный Fc домен включает по меньшей мере одну мутацию в положении, выбранном из группы, состоящей из L234, L235 и Р329 согласно EU нумерации, прежде всего аминокислотные мутации L234A, L235A и P329G ("PGLALA").
В одном варианте мутированный Fc домен включает аминокислотную мутацию P329G согласно EU нумерации, где связывание Feγ рецептора с мутированным Fc доменом снижается по сравнению со связыванием Feγ рецептора с немутированным исходным Fc доменом, прежде всего, где Feγ рецептор представляет собой FcγRIIIa и/или FcγRIIa человека.
В одном варианте мутированный Fc домен включает по меньшей мере одну, аминокислотную мутацию в положении, выбранном из группы, состоящей из I253, Н310 и Н435 согласно EU нумерации, прежде всего аминокислотные мутации I253A, Н310А и Н435А ("AAA"), где связывание FcRn с мутированным Fc доменом снижается по сравнению со связыванием FcRn с немутированным исходным Fc доменом.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным Fc доменом, где антигенсвязывающий фрагмент включает:
(i) вариабельную область (VH) тяжелой цепи, включающую
(а) определяющую комплементарность область тяжелой цепи 1 (CDR Н) с аминокислотной последовательностью RYWMN (SEQ ID NO: 1),
(б) CDR Н2 с аминокислотной последовательностью EITPDSSTINYTPSLKD (SEQ ID NO: 2) и
(в) CDR Н3 с аминокислотной последовательностью PYDYGAWFAS (SEQ ID NO: 3), и
(ii) вариабельную область легкой цепи (VL), включающую
(г) определяющую комплементарность область легкой цепи 1 (CDR L) с аминокислотной последовательностью RSSTGAVTTSNYAN (SEQ ID NO: 4),
(д) CDR L2 с аминокислотной последовательностью GTNKRAP (SEQ ID NO: 5) и
(е) CDR L3 с аминокислотной последовательностью ALWYSNHWV (SEQ ID NO: 6).
В одном варианте по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным Fc доменом, где антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи (VH), включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 8 и SEQ ID NO: 32, и вариабельную область легкой цепи (VL), включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 9 и SEQ ID NO: 33.
В одном варианте осуществления настоящего изобретения антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи (VH) SEQ ID NO: 8 и вариабельную область легкой цепи (VL) SEQ ID NO: 9.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv, который способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 7 и SEQ ID NO: 31. В одном варианте антигенсвязывающий рецептор включает аминокислотную последовательность SEQ ID NO: 7.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент представляет собой Fab фрагмент, который способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает
а) гибридный полипептид тяжелой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 39 и SEQ ID NO: 48, и
б) полипептид легкой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 41 и SEQ ID NO: 50.
В одном варианте антигенсвязывающий рецептор включает
а) гибридный полипептид тяжелой цепи SEQ ID NO: 39 и
б) полипептид легкой цепи SEQ ID NO: 41.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим аминокислотные мутации I253A, Н310А и Н435А ("AAA"), но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий фрагмент включает:
(i) вариабельную область (VH) тяжелой цепи, включающую
(а) определяющую комплементарность область тяжелой цепи 1 (CDR Н) с аминокислотной последовательностью SYGMS (SEQ ID NO: 53),
(б) CDR Н2 с аминокислотной последовательностью SSGGSY (SEQ ID NO: 54) и
(в) CDR Н3 с аминокислотной последовательностью LGMITTGYAMDY (SEQ ID NO: 55), и
(ii) вариабельную область легкой цепи (VL), включающую
(г) определяющую комплементарность область легкой цепи 1 (CDR L) с аминокислотной последовательностью RSSQTIVHSTGHTYLE (SEQ ID NO: 56),
(д) CDR L2 с аминокислотной последовательностью KVSNRFS (SEQ ID NO: 57) и
(е) CDR L3 с аминокислотной последовательностью FQGSHVPYT (SEQ ID NO: 58).
В одном варианте по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим аминокислотные мутации I253A, Н310А и Н435А ("AAA"), но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи (VH), включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 61, и вариабельную область легкой цепи (VL), включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 62.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент включает
а) вариабельную область (VH) тяжелой цепи SEQ ID NO: 61 и
б) вариабельную область (VL) легкой цепи SEQ ID NO: 62.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv, который способен специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А ("AAA"), но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 59. В одном варианте антигенсвязывающий рецептор включает аминокислотную последовательность SEQ ID NO: 59.
В одном варианте по меньшей мере один антигенсвязывающий фрагмент представляет собой Fab фрагмент, который способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает
а) гибридный полипептид тяжелой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности SEQ ID NO: 39, и
б) полипептид легкой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности SEQ ID NO: 41.
В одном варианте антигенсвязывающий рецептор включает
а) гибридный полипептид тяжелой цепи SEQ ID NO: 39 и
б) полипептид легкой цепи SEQ ID NO: 41.
В одном варианте предлагается выделенный полинуклеотид, кодирующий антигенсвязывающий рецептор, как описано в данном контексте. В одном варианте предлагается выделенный полинуклеотид, кодирующий гибридный полипептид тяжелой цепи или полипептид легкой цепи антигенсвязывающего рецептора, как описано в данном контексте. В одном варианте предлагается композиция, кодирующая антигенсвязывающий рецептор, как описано в данном контексте, включающая первый выделенный полинуклеотид, кодирующий гибридный полипептид тяжелой цепи, и второй выделенный полинуклеотид, кодирующий полипептид легкой цепи.
В одном варианте предлагается полипептид, кодированный полинуклеотидом, как описано в данном контексте, или композицией, как описано в данном контексте.
В одном варианте предлагается вектор, прежде всего экспрессионный вектор, включающий полинуклеотид (полинуклеотиды), как описано в данном контексте.
В одном варианте предлагается трансдуцированная Т-клетка, включающая полинуклеотид (полинуклеотиды), как описано в данном контексте, или вектор, как описано в данном контексте. В одном варианте предлагается трансдуцированная Т-клетка, способная экспрессировать антигенсвязывающий рецептор, как описано в данном контексте. В одном варианте предлагается трансдуцированная Т-клетка, как описано в данном контексте, при этом трансдуцированную Т-клетку ко-трансдуцируют с Т-клеточным рецептором (TCR), способным специфически связываться с антигеном-мишенью.
В одном варианте предлагается набор, включающий
(А) трансдуцированную Т-клетку, способную экспрессировать антигенсвязывающий рецептор, как описано в данном контексте, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В одном варианте предлагается набор, включающий
(А) выделенный полинуклеотид, кодирующий антигенсвязывающий рецептор, как писано в данном контексте, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В одном варианте предлагается набор, включающий
(А) композицию или вектор, как описано в данном контексте, кодирующий антигенсвязывающий рецептор, как писано в данном контексте, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В одном варианте немутированный исходный Fc домен представляет собой IgG1 или IgG4 Fc домен, прежде всего IgG1 Fc домен человека. В одном варианте предлагается мутированный Fc домен, включающий по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235, I253, Н310, Р331, Р329 и/или Н435 согласно EU нумерации, прежде всего, где аминокислотной мутацией является L234A, L235A, I253A, N297A, Н310А, P329G и/или Н435А. В одном варианте мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235 и Р329 согласно EU нумерации, прежде всего аминокислотные мутации L234A, L235A и P329G ("PGLALA"). В одном варианте мутированный Fc домен включает аминокислотную мутацию P329G согласно EU нумерации. В одном варианте мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из I253, Н310 и Н435 согласно EU нумерации, прежде всего аминокислотные мутации I253A, Н310А и Н435А ("AAA").
В одном варианте антитело, включающее мутированный Fc домен, способный специфически связываться с антигеном на поверхности опухолевой клетки, прежде всего, где антиген выбирают из группы, включающей FAP, СЕА, р95, ВСМА, EpCAM, MSLN, MCSP, HER-1, HER-2, HER-3, CD19, CD20, CD22, CD33, CD38, CD52Flt3, FOLR1, Trop-2, CA-12-5, HLA-DR, MUC-1 (муцин), A33-антигена, PSMA, PSCA, рецептора трансферрина, TNC (тенасцин), CA-IX и/или с пептидом, присоединенным к молекуле главного комплекса гистосовместимости человека (МНС). В одном варианте осуществления настоящего изобретения антитело, включающее мутированный Fc домен, способно специфически связываться с антигеном, выбранным из группы, состоящей из белка активации фибробластов (FAP), карциноэмбрионального антигена (СЕА), мезотелина (MSLN), CD20, рецептора 1 фолиевой кислоты (FOLR1) и тенасцина (TNC).
В одном варианте предлагается набор, как описано в данном контексте, для применения в качестве лекарственного средства.
В одном варианте предлагается антигенсвязывающий рецептор или трансдуцированная Т-клетка, как описано в данном контексте, для применения в качестве лекарственного средства, где трансдуцированную Т-клетку, экспрессирующую антигенсвязывающий рецептор, вводят до, одновременно с ним или после введения антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В одном варианте предлагается набор, как описано в данном контексте, для применения при лечении злокачественного заболевания. В одном варианте предлагается антигенсвязывающий рецептор или трансдуцированная Т-клетка, как описано в данном контексте, для применения при лечении злокачественного заболевания, где лечение включает введение трансдуцированной Т-клетки, экспрессирующей антигенсвязывающий рецептор, до, одновременно с ним или после введения антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В одном варианте указанное злокачественное заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и злокачественного заболевания крови.
В одном варианте трансдуцированную Т-клетку получают из клетки, выделенной из организма субъекта, которого предполагается лечить. В одном варианте трансдуцированную Т-клетку не получают из клетки, выделенной из организма субъекта, которого предполагается лечить.
В одном варианте предлагается способ лечения заболевания у субъекта, включающий введение субъекту трансдуцированной Т-клетки, способной экспрессировать антигенсвязывающий рецептор, как описано в данном контексте, и введение до, одновременно с ним или после введения трансдуцированной Т-клетки, терапевтически эффективного количества антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом. В одном варианте Т-клетку дополнительно выделяют из организма субъекта и трансдуцированную Т-клетку получают с помощью трансдукции выделенной Т-клетки с использованием полинуклеотида, композиции или вектора, как описано в данном контексте. В одном варианте Т-клетку трансдуцируют конструктом ретровирусного или лентивирусного вектора или конструктом невирусного вектора. В одном варианте конструктом невирусного вектора является миникольцевой вектор «спящая красавица».
В одном варианте трансдуцированную Т-клетку вводят субъекту внутривенным вливанием. В одном варианте до введения субъекту трансдуцированная Т-клетка контактирует с антителами анти-CD3 и/или анти-CD28. В одном варианте до введения субъекту трансдуцированная Т-клетка контактирует по меньшей мере с одним цитокином, предпочтительно с интерлейкином-2 (IL-2), интерлейкином-7 (IL-7), интерлейкином-15 (IL-15) и/или с интерлейкином-21 (IL-21 или их вариантами.
В одном варианте заболеванием является злокачественное заболевание. В одном варианте злокачественное заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и злокачественного заболевания крови.
В одном варианте предлагается способ индукции лизиса клетки-мишени, включающий контактирование клетки-мишени с трансдуцированной Т-клеткой, способной экспрессировать антигенсвязывающий рецептор, как описано в данном контексте, в присутствии антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В одном варианте клеткой-мишенью является опухолевая клетка. В одном варианте клетка-мишень экспрессирует антиген, выбранный из группы, состоящей из FAP, СЕА, р95, BCMA, EpCAM, MSLN, MCSP, HER-1, HER-2, HER-3, CD19, CD20, CD22, CD33, CD38, CD52Flt3, FOLR1, Trop-2, CA-12-5, HLA-DR, MUC-1 (муцин), А33-антигена, PSMA, PSCA, рецептора трансферрина, TNC (тенасцин) и CA-IX. В одном варианте клетка-мишень экспрессирует антиген, выбранный из группы, состоящей из карциноэмбрионального антигена (СЕА), мезотелина (MSLN), CD20, рецептора 1 фолиевой кислоты (FOLR1) и тенасцина (TNC).
В одном варианте полинуклеотиды или трансдуцированную Т-клетку, как описано в данном контексте, используют для получения лекарственного средства. В одном варианте лекарственное средство предназначено для лечения злокачественного заболевания.
Краткое описание фигур
На фиг. 1 показана структура типичных антигенсвязывающих рецепторов по настоящему изобретению. На фиг. 1А показана структура формата анти-P329G-scFv-CD28ATD-CD28CSD-CD3zSSD и формата анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD. Указан внеклеточный домен, включающий антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G. Антигенсвязывающий фрагмент состоит из вариабельной тяжелой цепи и вариабельной легкой цепи. Обе цепи соединены через линкер(Gly4Ser)4. Линкер (Gly4Ser)4, присоединенный к вариабельной легкой цепи, соединяет распознающий антиген домен с CD28 трансмембранным доменом (ТМ), который присоединен к внутриклеточному костимулирующему сигнальному домену (CDS) в составе CD28, который в свою очередь присоединен к стимулирующему сигнальному домену (SSD) в составе CD3z. На фиг. 1Б показана структура форматов анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD и анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD. Указан внеклеточный домен, включающий антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G. Антигенсвязывающий фрагмент состоит из тяжелой цепи Ig и легкой цепи Ig. Присоединенный к тяжелой цепи линкер(Gly4Ser)4 соединяет распознающий антиген домен с CD28 трансмембранным доменом, который присоединен к внутриклеточному костимулирующему сигнальному домену в составе CD28, который в свою очередь присоединен к стимулирующему сигнальному домену в составе CD3z.
На фиг. 2 показано схематическое изображение, иллюстрирующее модульную композицию типичных экспрессионных конструктов, кодирующих антигенсвязывающие рецепторы по настоящему изобретению. На фиг. 2А показан направленный к P392G формат scFab. На фиг. 2Б показан направленный к P392G формат Fab.
На фиг. 3 показана типичная молекула IgG1, несущая мутацию P329G в Fc домене, которая распознается анти-P329G антигенсвязывающим рецептором по настоящему изобретению.
На фиг. 4 показано схематическое изображение опухолеассоциированного антигена (ТАА), присоединенного к IgG, несущему мутацию P329G. Это антитело в свою очередь распознается Т-клеткой, экспрессирующей анти-P329G антигенсвязывающий рецептор, при этом Т-клетка активируется.
На фиг. 5 показано схематическое изображение метода анализа по репортерному гену Т-клеток Jurkat NFAT. ТАА, присоединенный к IgG, несущему мутацию P329G, можно распознавать с использованием анти-P329G антигенсвязывающего рецептора, экспрессируемого Т-клеткой Jurkat NFAT. Такое распознавание приводит к активации клетки, которую можно детектировать по интенсивности люминесценции (имп./с).
На фиг. 6 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием опухолевых клеток SUDHDL4, экспрессирующих CD20, в качестве клеток-мишеней. Использовали антитело anti-CD20 IgG (GA101), несущее мутацию P329G, которое с одной стороны распознает опухолеассоциированный антиген, а с другой стороны распознается антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. На фиг. 6А отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток. На фиг. 6Б отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 7 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием CD20 опухолевых клеток в качестве клеток-мишеней. Использовали антитело anti-CD20 IgG (GA101), несущее мутацию P329G, которое распознает опухолеассоциированный антиген и распознается антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. На фиг. 7А единый клон 5 Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки WSUDLCL2 были использованы в качестве опухолевых клеток. На фиг. 7Б единый клон 2 Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки WSUDLCL2 были использованы в качестве опухолевых клеток. На фиг. 7В единый клон 5 Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки SUDHL4 были использованы в качестве опухолевых клеток. На фиг. 7Г единый клон 2 Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 8 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием адгезивных опухолевых клеток NIH/3T3-huFAP cl 19, экспрессирующих FAP, в качестве клеток-мишеней. Был использован клон 4В9 антитела anti-FAP IgG, несущего мутацию P329G, которое распознает опухолеассоциированный антиген и распознается антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. Для контроля изотипов был включен IgG DP47/vk3, несущий мутацию P329G. На фиг. 8А отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток. На фиг. 8Б отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток. На фиг. 8В отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток. На фиг. 8Г отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 9 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием адгезивных опухолевых клеток MKN45, экспрессирующих СЕА, в качестве клеток-мишеней. Был использован либо клон А5В7 анти-СЕА IgG либо клон Т84 LCHA анти-СЕА IgG, которые оба несут мутацию P329G, и которые распознают опухолеассоциированный антиген и распознаются антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. Далее для контроля изотипов был включен IgG DP47/vk3, несущий мутацию P329G. На фиг. 9А и фиг. 9Б отсортированный пул клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSS, был использован в качестве эффекторных клеток. На фиг. 9В и фиг. 9Г отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 10 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием адгезивных опухолевых клеток MKN45, экспрессирующих СЕА, в качестве клеток-мишеней. Был использован либо клон anti-CEA СН1А1А 98 99 либо клон hMN14 IgG анти-СЕА IgG, которые оба несут мутацию P329G, и которые распознают опухолеассоциированный антиген и распознаются антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. Далее для контроля изотипов был включен IgG DP47/vk3, несущий мутацию P329G. На фиг. 10А и фиг. 10Б отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток. На фиг. 10В и фиг. 10Г отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 11 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием адгезивных опухолевых клеток CT26TNC cl 19, экспрессирующих TNC, в качестве клеток-мишеней. Клон анти-TNC IgG А2В10, несущего мутацию P329G, был использован в качестве антитела IgG, которое распознает опухолеассоциированный антиген и распознается антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. Далее для контроля изотипов был включен IgG DP47/vk3, несущий мутацию P329G. На фиг. 11А и фиг. 11Б отсортированный пул клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток. На фиг. 11В и фиг. 11Г отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 12А и фиг. 12Б показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием адгезивных опухолевых клеток CT26TNC cl 19, экспрессирующих TNC, в качестве клеток-мишеней. Клон anti-TNC IgG А2В10, несущего мутацию P329G, был использован в качестве антитела IgG, которое распознает опухолеассоциированный антиген и распознается антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. Далее для контроля изотипов был включен IgG DP47/vk3, несущий мутацию P329G. Отсортированный пул клеток Jurkat NFAT, экспрессирующих anti-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток.
На фиг. 13 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием CD20 опухолевых клеток в качестве клеток-мишеней. Использовали либо антитело anti-CD20 IgG (GA101), несущее мутацию P329G и мутацию LALA, мутацию P329G и D265A, только мутацию LALA, или вообще не несущее мутаций, которые можно использовать для детекции опухолеассоциированного антигена, и которые распознаются антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. На фиг. 13А пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки SUDHL4 были использованы в качестве опухолевых клеток. На фиг. 13Б пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки SUDHL4 были использованы в качестве опухолевых клеток.
На фиг. 14 показан метод анализа Т-клеток Jurkat NFAT по репортерному гену с использованием CD20 опухолевых клеток в качестве клеток-мишеней. Использовали либо антитело анти-CD20 IgG (GA101), несущее мутацию P329G и мутацию LALA, только мутацию P329G, только мутацию LALA или вообще не несущее мутаций, которые можно использовать для детектирования опухолеассоциированного антигена, и которые распознаются антигенсвязывающими рецепторами по изобретению, экспрессируемыми Т-клетками Jurkat NFAT. На фиг. 14А пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки SUDHL4 были использованы в качестве опухолевых клеток. На фиг. 14Б пул Т-клеток Jurkat NFAT, экспрессирующих anti-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, был использован в качестве эффекторных клеток, а клетки SUDHL4 были использованы в качестве опухолевых клеток.
Подробное описание настоящего изобретение
Определения
Использованные в данном контексте термины в основном представляют собой стандартные термины, используемые в уровне техники, если иначе не определено в следующем разделе описания настоящего изобретения.
Термин «активирующий Fc рецептор» означает Fc рецептор, который после взаимодействия с Fc доменом антитела испускает сигналы, которые стимулируют несущую рецептор клетку для выполнения эффекторных функций. Активирующие Fc рецепторы человека включают FcγRIIIa (CD16a), FcγRI (CD64), FcγRIIa (CD32) и FcαRI (CD89).
Термин «антителозависимая опосредованная клетками цитотоксичность» (ADCC) означает иммунный механизм, приводящий к лизису покрытых антителом клеток-мишеней под действием иммунных эффекторных клеток. Клетки-мишени представляют собой клетки, с которыми специфически связываются антитела или их производные, содержащие Fc область, в основном через N-концевой остаток белковой части, в направлении к Fc области. Использованный в данном контексте термин «сниженная ADCC» определен или как снижение числа клеток-мишеней, которые лизируют в определенный момент времени при определенной концентрации антитела в среде, окружающей клетки-мишени, по механизму ADCC, как определено выше, и/или повышение концентрации антитела в среде, окружающей клетки-мишени, которая требуется для осуществления лизиса определенного числа клеток-мишеней в определенный момент времени по механизму ADCC. Снижение ADCC определяют относительно ADCC, опосредованной тем же самым антителом, которое продуцируется тем же самым типом клеток хозяина, с использованием стандартных методов продуцирования, очистки, переработки и хранения (которые известны специалистам в данной области техники), но которое не подвергалось мутации. Например, снижение ADCC, опосредованное антителом, включающим в своем Fc домене аминокислотную мутацию, которая снижает ADCC, определяют относительно ADCC, опосредованной тем же самым антителом, не содержащим указанную аминокислотную мутацию в Fc домене. Пригодные методы определения ADCC известны в данной области техники (см., например, заявку РСТ WO 2006/082515 или заявку РСТ WO 2012/130831).
Термин «эффективное количество» агента (например, антитело) означает количество, которое требуется для осуществления физиологического изменения в клетке или ткани, в которые его вводят.
Термин «аффинность» означает силу суммарных нековалентных взаимодействий между одним участком связывания молекулы (например, рецептор) и его связывающим партнером (например, лиганд). Если не указано иное, использованный в данном контексте термин «связывающая аффинность» означает характерную связывающую аффинность, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антигенсвязывающий фрагмент и антиген, или рецептор и его лиганд). Аффинность молекулы X в отношении ее партнера У в основном можно представить в виде константы диссоциации (KD), которая равна соотношению константы скорости диссоциации и константы скорости ассоциации (koff и kon, соответственно). Таким образом, значения эквивалентных аффинностей могут включать различные константы скоростей, при условии, что соотношение констант скоростей остается одинаковым. Аффинность можно определить хорошо известными методами, описанными в данной области техники, включая методы, описанные в данном контексте. Предпочтительным методом определения аффинности является поверхностный плазмонный резонанс (ППР), а предпочтительная температура измерения составляет 25°.
Термин «аминокислота» относится к природным или синтетическим аминокислотам, а также к аналогам аминокислот и миметикам аминокислот, которые проявляют функцию способом, аналогичным природным аминокислотам. Природными аминокислотами являются аминокислоты, кодируемые генетическим кодом, и такие аминокислоты, которые были позже модифицированы, например, гидроксипролин, γ-карбоксиглютамат и О-фосфосерин. Аналогами аминокислот являются соединения, обладающие основной химической структурой, одинаковой с природными аминокислотами, то есть у которых присутствует атом α углерода, связанный с водородом, карбоксильной группой и R группой, например, гомосерин, норлейцин, сульфоксид метионина, метилсульфоний метионина. Такие аналоги содержат модифицированные R группы (например, норлейцин) или модифицированную пептидною цепь, но сохраняют основную химическую структуру, что и природная аминокислота. Аминокислотными миметиками являются химические соединения, обладающие структурой, которая отличается от основной химической структуры аминокислот, но проявляют функцию, аналогичную природным аминокислотам. Аминокислоты, указанные в данном контексте, можно обозначать либо с помощью широко известного трехбуквенного кода, либо с помощью однобуквенного кода, рекомендованного Комиссией по биохимической номенклатуре IUPAC-IUB.
Термин «аминокислотная мутация», использованный в данном контексте, включает замены, делеции, вставки и модификации аминокислот. Можно выполнить любую комбинацию замены, делеции, вставки и модификации аминокислоты, чтобы получить конечный конструкт, который обеспечивает проявление конечным конструктом требуемых характеристик, например, сниженное связывание с Fc рецептором. Делеции и вставки аминокислотной последовательности включают делеции и вставки амино- и/или карбокси-концевых аминокислот. Конкретными аминокислотными мутациями являются аминокислотные замены. С целью изменения, например, связывающих характеристик Fc области, прежде всего предпочтительными являются замены неконсервативных аминокислот, то есть замена одной аминокислоты на другую аминокислоту с различными структурными и/или химическими свойствами. Аминокислотные замены включают замену на неприродные аминокислоты или на производные одной из двадцати стандартных природных аминокислот (например, 4-гидроксипролин, 3-метилгистидин, орнитин, гомосерин, 5-гидроксилизин). Аминокислотные мутации можно получить с использованием генетических или химических методов, известных в данной области техники. Генетические методы могут включать сайт-направленный мутагенез, ПЦР, синтез генов и т.п. Полагают, что также могут быть использованы методы изменения боковой группы аминокислотной цепи с использованием отличающихся от генной инженерии методик, таких как химическая модификация. Для обозначения аналогичной мутации можно использовать различные обозначения. Например, замещение пролина в положении 329 в Fc домене на глицин можно обозначить как 329G, G329, G329, P329G, или Pro329Gly.
Термин «антитело» в данном контексте используется в самом широком смысле и включает различные структуры антител, включая, но, не ограничиваясь только ими, моноклональные антитела, поликлональные антитела и фрагменты антител, при условии, что они проявляют требуемую антигенсвязывающую активность. Соответственно, в контексте настоящего изобретения термин антитело относится к полноразмерным молекулам иммуноглобулина, а также частям таких молекул иммуноглобулина. Более того, термин относится, как описано в данном контексте, к модифицированным и/или измененным молекулам антитела, прежде всего к мутированным молекулам антитела. Термин также относится к полученным рекомбинантным или синтетическим методом/синтезированным антителам. В контексте настоящего изобретения термин антитело используется взаимозаменяемо с термином иммуноглобулин.
Термин «фрагмент антитела» означает молекулу, отличающуюся от интактного антитела, включающую часть интактного антитела, которая связывается с антигеном, с которым связывается интактное антитело. Примеры фрагментов антитела включают, но, не ограничиваясь только ими, Fv, Fab, Fab', Fab'-SH, F(ab')2, диатела, линейные антитела, одноцепочечные антитела (например, scFv), и содержащие один домен антитела. См. обзор, в котором описаны определенные фрагменты антител (Hudson и др., Nat. Med., 9, сс. 129-134 (2003)). Обзор scFv фрагментов приведен, например, в статье  , в книге The Pharmacology of Monoclonal Antibodies, т. 113, Rosenburg and Moore изд., Springer-Verlag, New York, cc. 269-315 (1994), см. также заявку WO 93/16185 и патенты US №5571894 и US №5587458. Диатела представляют собой фрагменты антител с двумя антигенсвязывающими участками, которые могут быть бивалентными или биспецифическими. См., например, патент ЕР 404097, заявку WO 1993/01161, статьи Hudson и др., Nat. Med., 9, сс. 129-134 (2003) и Hollinger и др., Proc. Natl. Acad. Sci. USA, 90, сс. 6444-6448 (1993). Тритела и тетратела описаны в статье Hudson и др., Nat. Med., 9, сс. 129-134 (2003). Антитела с одним доменом представляют собой фрагменты антител, включающие весь вариабельный домен тяжелой цепи или часть вариабельного домена тяжелой цепи или весь вариабельный домен легкой цепи или часть вариабельного домена легкой цепи антитела (Domantis, Inc., Waltham, MA, см., например, патент US 6248516 В1). Фрагменты антител можно получить различными методами, включая, но, не ограничиваясь только ими, расщепление протеолитическими ферментами интактного антитела, а также продуцирование рекомбинантными клетками-хозяина (например, Е. coli или фаг), как описано в данном контексте.
, в книге The Pharmacology of Monoclonal Antibodies, т. 113, Rosenburg and Moore изд., Springer-Verlag, New York, cc. 269-315 (1994), см. также заявку WO 93/16185 и патенты US №5571894 и US №5587458. Диатела представляют собой фрагменты антител с двумя антигенсвязывающими участками, которые могут быть бивалентными или биспецифическими. См., например, патент ЕР 404097, заявку WO 1993/01161, статьи Hudson и др., Nat. Med., 9, сс. 129-134 (2003) и Hollinger и др., Proc. Natl. Acad. Sci. USA, 90, сс. 6444-6448 (1993). Тритела и тетратела описаны в статье Hudson и др., Nat. Med., 9, сс. 129-134 (2003). Антитела с одним доменом представляют собой фрагменты антител, включающие весь вариабельный домен тяжелой цепи или часть вариабельного домена тяжелой цепи или весь вариабельный домен легкой цепи или часть вариабельного домена легкой цепи антитела (Domantis, Inc., Waltham, MA, см., например, патент US 6248516 В1). Фрагменты антител можно получить различными методами, включая, но, не ограничиваясь только ими, расщепление протеолитическими ферментами интактного антитела, а также продуцирование рекомбинантными клетками-хозяина (например, Е. coli или фаг), как описано в данном контексте.
Термин "антигенсвязывающая молекула", использованный в данном контексте в самом широком смысле, означает молекулу, которая специфически связывается с антигенной детерминантой. Примеры антигенсвязывающих молекул включают иммуноглобулины и их производные, например, их фрагменты, а также антигенсвязывающие рецепторы и их производные.
Как использовано в данном контексте, термин «антигенсвязывающий фрагмент» означает полипептидную молекулу, которая специфически связывается с антигенной детерминантой. В одном варианте антигенсвязывающий фрагмент способен направлять всю структурную единицу, к которой он присоединен (например, иммуноглобулин или антигенсвязывающий рецептор), к участку-мишени, например, к специфическому типу опухолевой клетки или к опухолевой строме, несущей антигенную детерминанту, или к иммуноглобулину, связывающемуся с антигенной детерминантой на опухолевой клетке. В другом варианте антигенсвязывающий фрагмент способен активировать передачу сигнала через его антиген-мишень, например передача сигнала, активируется после связывания антигенной детерминанты с антигенсвязывающим рецептором на Т-клетке. В контексте настоящего изобретения антигенсвязывающие фрагменты могут быть включены в состав антител и их фрагментов, а также в состав антигенсвязывающих рецепторов и их фрагментов, как будет дополнительно определено в данном контексте. Антигенсвязывающие фрагменты включают антигенсвязывающий домен, включающий вариабельную область тяжелой цепи иммуноглобулина и вариабельную область легкой цепи иммуноглобулина. В некоторых вариантах антигенсвязывающие фрагменты могут включать консервативные области иммуноглобулина, как будет дополнительно определено в данном контексте и известно в уровне техники. Пригодные консервативные области тяжелой цепи включают любой из пяти изотипов:: α, δ, ε, γ или μ. Пригодные консервативные области легкой цепи включают любой из двух изотипов: κ и λ.
В контексте настоящего изобретения термин «антигенсвязывающий рецептор» относится к антигенсвязывающей молекуле, включающей заякоренный трансмембранный домен и внеклеточный домен, включающий по меньшей мере один антигенсвязывающий фрагмент. Антигенсвязывающий рецептор можно получить из полипептидных частей из различных источников. Соответственно, следует также понимать, что этот термин можно определить, как «гибридный белок» и/или «химерный белок». Обычно, гибридные белки являются белками, которые конструируют при соединении двух или более генов (или предпочтительно кДНК), которые исходно кодируют отдельные белки. Трансляция этого гибридного гена (или гибридной кДНК) приводит к образованию одного полипептида, предпочтительно с функциональными свойствами, характерными для каждого из этих исходных белков. Рекомбинантные гибридные белки конструируют искусственно с использованием технологии рекомбинантных ДНК для применения в биологических исследованиях или при получении медицинских препаратов. Более подробное описание антигенсвязывающих рецепторов по настоящему изобретению представлено ниже в данном контексте. В контексте настоящего изобретения CAR (химерный антигенный рецептор) является антигенсвязывающим рецептором, включающим внеклеточную часть, которая включает антигенсвязывающий фрагмент, присоединенный через спейсерную последовательность к заякоренному трансмембранному домену, который сам по себе присоединен к внутриклеточным сигнальным доменам CD3z и CD28.
Термин «антигенсвязывающий сайт» относится к сайту, то есть к одному или более аминокислотных остатков в составе антигенсвязывающей молекулы, которые обеспечивают взаимодействие с антигеном. Например, антигенсвязывающий сайт антитела или антигенсвязывающего рецептора включает аминокислотные остатки из определяющих комплементарность областей (CDR). Молекула природного иммуноглобулина обычно содержит два антигенсвязывающих сайта, а молекула Fab или scFv обычно содержит один антигенсвязывающий сайт.
Термин «антигенсвязывающий домен» означает часть антитела или антигенсвязывающего рецептора, включающую область, которая специфически связывается с частью или интактным антигеном и является комплементарной части антигена или интактному антигену. Антигенсвязывающий домен может содержать, например, один или более вариабельных доменов иммуноглобулина (также называемые вариабельные области). Более подробно, антигенсвязывающий домен включает вариабельную область легкой цепи (VL) иммуноглобулина и вариабельный домен тяжелой цепи (VH) иммуноглобулина.
Термин "вариабельная область" или "вариабельный домен" означает домен тяжелой или легкой цепи иммуноглобулина, которые принимают участие в связывании с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) природного антитела обычно характеризуются одинаковыми структурами, при этом каждый домен включает 4 консервативные каркасные области (FR) и 3 гипервариабельные области (HVR). См., например, Kindt и др., Kuby Immunology, 6е изд., Freeman W.H. and Co., с. 91 (2007). Для придания антигенсвязывающей специфичности может быть достаточно одного VH или VL домена.
Термин "ATD", использованный в данном контексте, относится к «заякоренному трансмембранному домену», который имеет полипептидный «тяж», способный интегрироваться в клеточную мембрану (мембраны). ATM можно присоединять к дополнительным внеклеточным и/или внутриклеточным полипептидным доменам, причем эти внеклеточные и/или внутриклеточные полипептидные домены равным образом будут включены в клеточную мембрану. В контексте антигенсвязывающих рецепторов по настоящему изобретению ATM обеспечивает присоединение к мембране и удерживание в мембране антигенсвязывающего рецептора по настоящему изобретению. Антигенсвязывающие рецепторы по настоящему изобретению включают по меньшей мере один ATM и внеклеточный домен, включающий антигенсвязывающий фрагмент. Кроме того, ATM можно присоединять к дополнительным внутриклеточным сигнальным доменам.
Термин «связывание», использованный в контексте антигенсвязывающих рецепторов по настоящему изобретению, определяет связывание (взаимодействие) «взаимодействующего с антигеном сайта» и антигена друг с другом. Термин «взаимодействующий с антигеном сайт» определяет, в отношении антигенсвязывающих рецепторов по настоящему изобретению, полипептидный мотив, который характеризуется способностью специфически взаимодействовать с определенным антигеном или специфической группой антигенов (например, мутированные Fc домены). Соответственно, следует понимать, что указанное связывание/взаимодействие можно определить, как «специфическое распознавание». Термин «специфическое распознавание», в соответствии с настоящим изобретением, означает, что антигенсвязывающий рецептор способен специфически взаимодействовать и/или связываться с модифицированной молекулой, как определено в данном контексте, но не распознает немодифицированную молекулу. Антигенсвязывающий фрагмент в составе антигенсвязывающего рецептора может распознавать различные эпитопы, взаимодействовать и/или связываться с ними на одной и той же молекуле. Этот термин относится к специфичности антигенсвязывающего рецептора, то есть к его способности различать определенные области модифицированной молекулы, то есть в мутированном Fc домене, как определено в данном контексте. Специфическое взаимодействие взаимодействующего с антигеном сайта с его специфическим антигеном может привести к инициации сигнала, например, за счет индукции изменения конформации полипептида, включающего антиген, олигомеризации полипептида, включающего антиген, олигомеризации антигенсвязывающего рецептора и т.п. Таким образом специфический мотив в аминокислотной последовательности взаимодействующего с антигеном сайта и антиген связываются друг с другом за счет их первичной, вторичной или третичной структуры, а также за счет вторичных модификаций указанной структуры. Соответственно термин «связывание» относится не только к линейному эпитопу, но также может относиться к конформационному эпитопу, структурному эпитопу или прерывистому эпитопу, состоящему из двух областей молекул-мишеней или их частей. В контексте настоящего изобретения конформационный эпитоп определяется двумя или более отдельными аминокислотными последовательностями, разделенными в первичной последовательности, которые сближаются на поверхности молекулы, когда полипептид сворачивается в природный белок (Sela, Science 166 1365 (1969) и Laver, Cell 61, 553-536 (1990)). Более того, термин «связывание» используется в данном контексте взаимозаменяемо с термином «взаимодействие». Способность антигенсвязывающего фрагмента (например, Fab или scFv домена) в составе антигенсвязывающего рецептора или антитела связываться со специфической антигенной детерминантой-мишенью, можно оценивать методом твердофазного иммуноферментного анализа (ИФА) или другими методами, известными специалисту в данной области техники, например, метод поверхностного плазмонного резонанса (ППР) (анализ проводят в системе BIAcore) (Liljeblad и др., Glyco J 17, 323-329 (2000)), и традиционных методов анализа связывания (Heeley, Endocr Res 28, 217-229 (2002)). В одном варианте степень связывания антигенсвязывающего фрагмента с неродственным белком составляет менее приблизительно 10% связывания антигенсвязывающего фрагмента с антигеном мишенью по результатам измерения, прежде всего методом ППР. В некоторых вариантах антигенсвязывающий фрагмент, который связывается с антигеном-мишенью, характеризуется константой диссоциации (KD) ≤ 1 мкМ, ≤ 100 нМ, ≤ 10 нМ, ≤ 1 нМ, ≤ 0,1 нМ, ≤ 0,01 нМ или ≤ 0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М). Термин «специфическое связывание», использованный в соответствии с настоящим изобретении, означает, что молекулы по изобретению не проявляют или в основном не проявляют перекрестную реактивность с (поли)пептидами с аналогичными структурами, то есть с немутированным исходным Fc доменом, где антигенсвязывающий рецептор по изобретению способен специфически связываться с мутированным Fc доменом. Соответственно антигенсвязывающий рецептор по изобретению специфически связывается/взаимодействует с мутированным Fc доменом. Перекрестную реактивность панели исследуемых конструктов можно определять, например, при оценке связывания панели антигенсвязывающих фрагментов в стандартных условиях (см. например, Harlow и Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, (1988) and Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, (1999)) связывания с исследуемым мутированным Fc доменом, а также с исходным немутированным Fc доменом. Только такие конструкты (то есть Fab фрагменты, scFv и т.п.), которые связываются с исследуемым мутированным Fc доменом, но не связываются или в основном не связываются с немутированным исходным Fc доменом, рассматриваются как специфические в отношении исходного мутированного Fc домена, и такие конструкты выбирают для дальнейших исследований в соответствии со способом, описанным в данном контексте. Эти методы могут включать, наряду с прочими, исследования связывания, блокирования или конкурентного связывания с использованием структурно и/или функционально близких Fc доменов. Исследование связывания также включает анализ сортировки клеток с активированной флуоресценцией (FACS), ППР (например, с использованием биосенсоров BIAcore®), аналитическое ультрацентрифугирование, изотермическую титрационную калориметрию, флуоресцентную анизотропию, флуоресцентную спектроскопию или использование лиганда с радиоактивной меткой.
Термин «CDR», использованный в данном контексте, относится к «определяющей комплементарность области», которая хорошо известна в уровне техники. CDR означают части иммуноглобулинов или антигенсвязывающих рецепторов, которые определяют специфичность указанных молекул и осуществляют контактирование со специфическим лигандом. CDR представляют собой наиболее вариабельную часть молекулы и вносят вклад в антигенсвязывающее разнообразие этих молекул. Существуют три области CDR: CDR1, CDR2 и CDR3 в каждом V домене. CDR-H представляет собой CDR область вариабельной тяжелой цепи, a CDR-L представляет собой CDR область вариабельной области легкой цепи. VH означает вариабельную тяжелую цепь, а VL означает вариабельную легкую цепь. Области CDR в области Ig можно определить, как описано у «Кабата» (Sequences of Proteins of Immunological Interest", 5th edit. NIH Publication no. 91-3242 U.S. Department of Health and Human Services (1991), Chothia J. Mol. Biol. 196, 901-917 (1987)) или "Chothia" (Nature 342, 877-883 (1989)).
Термин "CD3z" относится к зета-цепи CD3 гликопротеина Т-клеточной поверхности, также известный как "зета-цепь Т-клеточного рецептора Т3" и "CD247".
Термин «химерный рецептор антигена» или «химерный рецептор» или «CAR» относится к антигенсвязывающему рецептору, представляющему собой внеклеточную часть антигенсвязывающего фрагмента (например, одноцепочечный домен антитела), соединенную через последовательность спейсера с внутриклеточными сигнальными доменами в составе CD3z и CD28. В изобретении дополнительно предлагаются антигенсвязывающие рецепторы, причем антигенсвязывающим фрагментом является Fab или crossFab фрагмент. Термин «CAR» следует понимать в самом широком смысле, то есть этот термин включает антигенсвязывающие рецепторы, представляющие собой внеклеточную часть, включающую антигенсвязывающий фрагмент, соединенный с CD3z и его фрагментом и с CD28 и его фрагментом необязательно через один или несколько пептидных линкеров.
Термин «класс» антитела или иммуноглобулина означает тип консервативного домена или консервативной области, характерный для тяжелой цепи антитела. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно подразделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Консервативные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
Термин «кроссоверная Fab молекула» (также называемая «crossFab» или кроссоверный Fab фрагмент») означает Fab молекулу, в которой либо вариабельные области, либо консервативные области тяжелой и легкой цепей Fab заменены друг на друга, то есть crossFab фрагмент включает пептидную цепь, состоящую из вариабельной области легкой цепи и консервативной области тяжелой цепи, и пептидную цепь, состоящую из вариабельной области тяжелой цепи и консервативной области легкой цепи. Для ясности, в crossFab фрагменте, где вариабельные области легкой цепи Fab и тяжелой цепи Fab заменены друг на друга, причем пептидная цепь, включающая консервативную область тяжелой цепи, в данном контексте называется тяжелой цепью кроссоверной Fab молекулы. И наоборот, в crossFab фрагменте, где консервативные области легкой цепи Fab и тяжелой цепи Fab заменены друг на друга, причем пептидная цепь, включающая вариабельную область тяжелой цепи, в данном контексте называется тяжелой цепью кроссоверной Fab молекулы. Соответственно crossFab фрагмент включает тяжелую или легкую цепь, состоящую из вариабельной области тяжелой цепи и консервативной области легкой цепи (VH-CL), и тяжелую или легкую цепь, состоящую из вариабельной области легкой цепи и консервативной области тяжелой цепи (VL-СН1). И наоборот, термин «стандартная Fab» молекула означает Fab молекулу в ее природном формате, то есть включающую тяжелую цепь, состоящую из вариабельной области тяжелой цепи и консервативной области тяжелой цепи (VH-CH1), и легкую цепь, состоящую из вариабельной области легкой цепи и консервативной области легкой цепи (VL-CL).
Термин «CSD», использованный в данном контексте, означает костимулирующий сигнальный домен.
Термин «эффекторные функции» означает биологические функции, которые связаны с Fc областью антитела, и которые изменяются в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и зависимую от комплемента цитотоксичность (CDC), связывание Fc рецептора, антителозависимую опосредованную клетками цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP), секрецию цитокинов, опосредованный иммунным комплексом захват антигена антиген-презентирующими клетками, регуляцию с понижением активности рецепторов клеточной поверхности (например, рецептор В-клеток) и активацию В-клеток.
Использованные в данном контексте термины «генно-инженерный, созданный методом генной инженерии, сконструированный» включают любые действия на пептидную цепь или пост-трансляционные модификации природного или рекомбинантного полипептида или его фрагмента. «Конструирование» включает модификации аминокислотной последовательности, профиля гликозилирования или боковой группы индивидуальных аминокислот, а также комбинации таких подходов.
Термин «экспрессионная кассета» относится к полинуклеотиду, полученному рекомбинантным или синтетическим методом, с наборами специфических элементов нуклеиновых кислот, которые позволяют осуществлять транскрипцию конкретной нуклеиновой кислоты в клетке-мишени. Рекомбинантную экспрессионную кассету можно включить в плазмиду, хромосому, митохондриальную ДНК, пластидную ДНК, вирус или фрагмент нуклеиновой кислоты. Обычно часть рекомбинантной экспрессионной кассеты экспрессионного вектора включает, наряду с другими последовательностями, последовательность нуклеиновой кислоты, предназначенной для транскрипции, и промотор. В некоторых вариантах экспрессионная кассета по настоящему изобретению включает полинуклеотидные последовательности, которые кодируют антигенсвязывающие молекулы по настоящему изобретению или их фрагменты.
Термин «Fab молекула» означает белок, состоящий из VH и СН1 домена тяжелой цепи ("Fab тяжелая цепь") и VL и CL домена легкой цепи («Fab легкая цепь») антигенсвязывающей молекулы.
Термин "Fc домен" или "Fc область", использованный в данном контексте, означает С-концевую область тяжелой цепи иммуноглобулина, содержащую по меньшей мере часть консервативной области. Термин включает природные последовательности Fc областей и варианты последовательностей Fc областей. Хотя границы Fc области тяжелой цепи IgG могут в незначительной степени изменяться, обычно Fc область тяжелой цепи IgG человека расположена от Cys226, или от Pro230, до С-концевого остатка тяжелой цепи. Однако С-концевой лизин (Lys447) Fc области может как присутствовать, так и отсутствовать. Если в данном контексте не указано иное, нумерация аминокислотных остатков в Fc области или в консервативной области представлена согласно системе EU нумерации, называемой также EU индексом, как описано в статье Kabat и др., Sequences of Proteins of Immunological Interest, 5oe изд., Public Health Service, National Institutes of Health, Bethesda, MD (1991). Термин «субъединица Fc домена», использованный в данном контексте, означает один из двух полипептидов, формирующих димерный Fc домен, то есть полипептид, включающий С-концевые консервативные области тяжелой цепи иммуноглобулина, способные к стабильной самоассоциации. Например, субъединица Fc домена IgG включает консервативные домены IgG СН2 и IgG СН3.
Термин «каркасный участок» или «FR» означает остатки вариабельного домена, отличающиеся от остатков гипервариабельной области (HVR). Участок FR вариабельного домена в основном состоит из четырех FR доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в основном присутствуют в следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термин «полноразмерное антитело» означает антитело, состоящее из двух «тяжелых цепей полноразмерного антитела» и двух «легких цепей полноразмерного антитела». «Тяжелая цепь полноразмерного антитела» представляет собой полипептид, включающий, в направлении от N-концевого остатка до С-концевого остатка, вариабельный домен тяжелой цепи антитела (VH), консервативный домен 1 тяжелой цепи антитела (СН1), шарнирную область антитела (HR), консервативный домен 2 тяжелой цепи антитела (СН2) и консервативный домен 3 тяжелой цепи антитела (СН3), сокращенно VH-CH1-HR-CH2-CH3, и не обязательно консервативный домен 4 (СН4) тяжелой цепи антитела в случае антитела подкласса IgE. Предпочтительно «тяжелая цепь полноразмерного антитела» представляет собой полипептид, включающий, в направлении от N-концевого до С-концевого остатка, VH, CH1, HR, СН2 и СН3. «Легкая цепь полноразмерного антитела» представляет собой полипептид, включающий, в направлении от N-концевого до С-концевого остатка, вариабельный домен легкой цепи антитела (VL) и консервативный домен легкой цепи антитела (CL), сокращенно VL-CL. Консервативный домен легкой цепи антитела (CL) может представлять собой изотип к (каппа) или X (лямбда). Две цепи полноразмерного антитела связаны друг с другом через межпептидные дисульфидные связи между CL доменом и СН1 доменом и между шарнирными областями тяжелых цепей полноразмерного антитела. Примеры типичных полноразмерных антител представляют собой природные антитела, такие как IgG (например, IgG1 и IgG2), IgM, IgA, IgD и IgE). Полноразмерные антитела, использованные согласно изобретению, могут быть получены из одного вида, например, человека, или они могут быть получены в виде химерных или гуманизированных антител. В некоторых вариантах полноразмерные антитела, использованные согласно изобретению, то есть терапевтическое антитело, включающее мутированный Fc домен, включают два антигенсвязывающих сайта, каждый из которых сформирован из пары VH и VL, и которые оба специфически связываются с одним и тем же антигеном. В другом варианте полноразмерные антитела, использованные согласно изобретению, включают два антигенсвязывающих сайта, каждый из которых сформирован из пары VH и VL, где два антигенсвязывающих сайта связываются с различными антигенами, например, где антитела являются биспецифические. С-концевой остаток тяжелой или легкой цепи указанного полноразмерного антитела означает последний аминокислотный остаток в С-концевом фрагменте указанной тяжелой или легкой цепи.
Термин «соединенный» означает, что компоненты (например, Fab и трансмембранный домен) соединены пептидными связями либо напрямую, либо через один или более пептидных линкеров.
Термины «клетка хозяина», «линия клетки хозяина» и «культура клетки хозяина» используют взаимозаменяемо и они означают клетки, в которые включена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки хозяина включают «трансформанты» и «трансформированные клетки», которые включают первичные трансформированные клетки и их потомство без учета числа пересевов. Потомство может быть не полностью идентичным исходной клетке по содержанию нуклеиновых кислот и может содержать мутации. В объем изобретения включено мутантное потомство, которое проявляет ту же самую функцию или биологическую активность по данным скрининга или селекции для исходной трансформированной клетки. Клетка хозяина представляет собой любой тип клеточной системы, которую можно использовать для получения антитела, используемого согласно настоящему изобретению. Клетки хозяина включают, перечисляя только некоторые из них, культивируемые клетки, например, культивируемые клетки млекопитающих, такие как клетки СНО, клетки BHK, клетки NS0, клетки SP2/0, миеломные клетки YO, миеломные клетки мыши Р3Х63, клетки PER, клетки PER.C6 или гибридомные клетки, дрожжевые клетки, клетки насекомых и клетки растений, но также и клетки, включенные в трансгенное животное, трансгенное растение или культивированное растение или ткань животного.
Термин «гипервариабельная область» или «HVR», использованный в данном контексте, означает каждую из областей вариабельного домена антитела, которая характеризуется гипервариабельностью последовательности и/или формирует структурно определенные петли («гипервариабельные петли»). В основном природные четырехцепочечные антитела включают шесть HVR, три в VH домене (H1, Н2, Н3) и три в VL домене (L1, L2, L3). Области HVR в основном включают аминокислотные остатки из гипервариабельных петель и/или из определяющих комплементарность областей (CDR), причем последние характеризуются наибольшей вариабельностью последовательностей и/или принимают участие в распознавании антигена. За исключением CDR1 в домене VH, области CDR в основном включают аминокислотные остатки, которые формируют гипервариабельные петли. Области HVR также называют «определяющими комплементарность областями» (CDR), и эти термины используются в данном контексте взаимозаменяемо со ссылкой на части вариабельной области, которые формируют антигенсвязывающие области. Эта конкретная область описана в статьях Kabat и др., U.S. Dept. of Health and Human Services, Sequences of Proteins of Immunological Interest (1983) и Chothia и др., J Mol Biol 196:901-917 (1987), где определения включают перекрывание или подклассы аминокислотных остатков при сравнении друг с другом. Тем не менее, применение каждого определения в отношении CDR антитела и/или антигенсвязывающего рецептора или их вариантов находится в пределах объема термина, определенного и использованного в данном контексте. Соответствующие аминокислотные остатки, которые включены в последовательности CDR, как определено в каждом из цитированных выше документов, перечислены ниже в таблице 1 для сравнения. Точные положения каждого из остатков, которые определяют конкретную область CDR, изменяются в зависимости от последовательности и размера CDR. Специалисты в данной области техники могут легко установить, какие остатки включают конкретную область CDR в данной аминокислотной последовательности вариабельной области антитела.
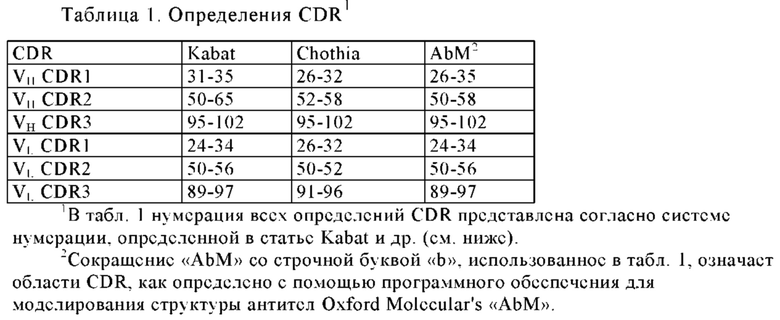
В статье Kabat и др. также определена система нумерации для последовательностей вариабельной области, которую можно применять для любого антитела. Специалист в данной области техники может однозначно использовать эту систему «нумерации Кабата» к любой последовательности вариабельной области без любых экспериментальных данных, не относящихся к самой последовательности. Термин «нумерация Кабата», использованный в данном контексте, означает систему нумерации, установленную в статье Kabat и др., U.S. Dept. of Health and Human Services, Sequences of Proteins of Immunological Interest (1983). Если не указано иное, все номера положений определенных аминокислотных остатков в вариабельной области антитела представлены согласно системе нумерации Кабата. Полипептидные последовательности в перечне последовательностей не пронумерованы согласно системе нумерации Кабата. Однако специалисту в данной области техники представляется очевидным преобразовать нумерацию последовательностей в перечне последовательностей согласно системе нумерации Кабата.
Термины «индивидуум» или «субъект» означают млекопитающее. Млекопитающие включают, но, не ограничиваясь только ими, домашних животных (например, крупный рогатый скот, овцы, собаки, кошки и лошади), приматов (например, человек и приматы, не относящиеся к человеку, такие как нечеловекообразные обезьяны), кроликов и грызунов (например, мыши и крысы). Прежде всего, индивидуумом или субъектом является человек.
Термин «выделенная молекула нуклеиновой кислоты» или «выделенный» полинуклеотид означает молекулу нуклеиновой кислоты, ДНК или РНК, которые были удалены из своей природной окружающей среды. Например, рекомбинантный кодирующий полипептид полинуклеотид, содержащийся в векторе, означает выделенный для целей настоящего изобретения. Другие примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, поддерживаемые в гетерологичных клетках хозяина или очищенные (частично или в существенной степени очищенные) полинуклеотиды в растворе. Выделенный полинуклеотид включает молекулу полинуклеотида, содержащуюся в клетках, которые обычно содержат молекулу полинуклеотида, но молекула полинуклеотида присутствует вне хромосомы или расположена в хромосоме, но в положении, которое отличается от природного расположения в хромосоме. Выделенные молекулы РНК включают in vivo или in vitro транскрипты РНК по настоящему изобретению, а также положительные и отрицательные формы цепей, и двухцепочечные формы. Выделенные полинуклеотиды или нуклеиновые кислоты по настоящему изобретению дополнительно включают указанные молекулы, полученные синтетическим методом. Кроме того, полинуклеотид или нуклеиновая кислота могут представлять собой или могут включать регуляторный элемент, такой как промотор, участок связывания рибосомы или терминатор транскрипции.
Нуклеиновая кислота или полинуклеотид с нуклеотидной последовательностью, которая по сравнению с эталонной полинуклеотидной последовательностью по настоящему изобретению «идентична», например, по меньшей мере на 95%, означает, что нуклеотидная последовательность идентична эталонной последовательности за исключением того, что полинуклеотидная последовательность может включать вплоть до пяти точечных мутаций на каждые 100 нуклеотидов эталонной нуклеотидной последовательности. Другими словами, чтобы получить полинуклеотид с нуклеотидной последовательностью, которая по сравнению с эталонной нуклеотидной последовательностью идентична по меньшей мере на 95%, и в которой можно удалить или заменить на другой нуклеотид вплоть до 5% нуклеотидов в эталонной последовательности, или вплоть до 5% нуклеотидов от общего числа нуклеотидов в эталонной последовательности можно вставить в эталонную последовательность. Эти изменения эталонной последовательности можно проводить в 5'- или 3'-концевых положениях эталонной нуклеотидной последовательности или в любом положении между этими концевыми положениями, распределенные либо индивидуально среди остатков в эталонной последовательности, либо в одной или более соседних групп в эталонной последовательности. На практике идентичность любой конкретной нуклеотидной последовательности по сравнению с нуклеотидной последовательностью по настоящему изобретению, которая составляет по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99%, можно определять стандартным методом с использованием известных компьютерных программ, таких как программы, описанные выше для полипептидов (например, ALIGN-2).
Термин «выделенный полипептид» или его вариант или производное означает, что он находится не в своей природной среде. Не требуется конкретный уровень очистки. Например, выделенный полипептид можно удалить из его нативной или природной окружающей среды. Считается, что полученные рекомбинантным методом полипептиды и белки, экспрессированные в клетках хозяина, выделены для целей настоящего изобретения в виде природных или рекомбинантных полипептидов, которые были разделены, фракционированы или частично очищены любым пригодным методом.
«Идентичность аминокислотной последовательности в процентах (%)» в отношении эталонной полипептидной последовательности определяют как процентное содержание аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в эталонной полипептидной последовательности после выравнивания последовательностей и при необходимости включении гэпов для обеспечения максимальной идентичности последовательностей в процентах и без учета любых консервативных замен в качестве части идентичности последовательностей. Выравнивание с целью определения идентичности аминокислотных последовательностей в % можно осуществлять различными способами, известными в данной области техники, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, которые требуются для достижения максимального выравнивания вдоль всей длины сравниваемых последовательностей. Однако согласно настоящему изобретению значения идентичности аминокислотных последовательностей в % определяют с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 создана фирмой Genentech Inc., и пользователь может найти исходный код в документации пользователей U.S. Copyright Office, Washington D.C., 20559, где он зарегистрирован под номером U.S. Copyright Registration № TXU510087. Программа ALIGN-2 является общедоступной на фирме Genentech, Inc., South San Francisco, Калифорния, или ее можно скомпилировать по исходному коду. Программу ALIGN-2 можно также скомпилировать для использования в операционной системе UNIX, включая управляющую программу UNIX V4.0D. Все параметры сравнения последовательностей установлены в программе ALIGN-2 и не изменяются. В случаях, когда для сравнения аминокислотных последовательностей используют программу ALIGN-2, идентичность данной аминокислотной последовательности А в % с данной аминокислотной последовательностью В (которую в другом варианте можно назвать как данная аминокислотная последовательность А, которая имеет или включает определенную идентичность аминокислотной последовательности в % по сравнению с данной аминокислотной последовательностью В) рассчитывают по формуле:
100 × частное X/Y
где X означает число аминокислотных остатков, рассчитанных как идентичные совпадения при сравнении последовательностей А и B в программе ALIGN-2, и где Y означает общее число аминокислотных остатков в последовательности В. Следует понимать, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то идентичность в % при сравнении А с В не равна идентичности аминокислотной последовательности в % при сравнении В с А. Если специально не указано иное, все значения идентичности аминокислотных последовательностей в %, использованные в данном контексте, получены, как описано в предыдущем абзаце с использованием компьютерной программы ALIGN-2.
Термин «молекула нуклеиновой кислоты» означает последовательность оснований, включающую пуриновые и пиримидиновые основания, которые составляют последовательность полинуклеотида, причем указанные основания представляют первичную структуру молекулы нуклеиновой кислоты. В данном контексте термин молекула нуклеиновой кислоты включает ДНК, кДНК, геномную ДНК, РНК, синтетические формы ДНК и смешанные полимеры, включающие две или более этих молекул. Кроме того, термин молекула нуклеиновой кислоты включает как смысловые, так и антисмысловые цепи. Более того описанная в данном контексте молекула нуклеиновой кислоты может содержать неприродные нуклеотидные основания или их производные, как представляется очевидным специалистам в данной области техники.
Термин «вкладыш в упаковке» означает инструкции, обычно содержащиеся в коммерческих упаковках терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждения в отношении применения таких терапевтических продуктов.
Термин «фармацевтическая композиция» означает препарат, который в указанной форме обеспечивает биологическую активность активного ингредиента, содержащегося в этом препарате и проявляющего эффективность, и который не содержит никаких дополнительных компонентов, оказывающих неприемлемую токсичность на организм субъекта, которому вводят этот препарат. Фармацевтическая композиция обычно содержит один или более фармацевтически приемлемых носителей.
«Фармацевтически приемлемый носитель» означает ингредиент в фармацевтической композиции, который не является активным ингредиентом и который не оказывает токсическое действие на субъекта. Фармацевтически приемлемый носитель включает, но, не ограничиваясь только ими, буферное вещество, эксципиент, стабилизатор или консервант.
Использованный в данном контексте термин «полипептид» означает молекулу, состоящую из мономеров (аминокислот), соединенных в линейную цепь амидными связями (известными также как пептидные связи). Термин «полипептид» означает любую цепь из двух или более аминокислот и не означает конкретную длину продукта. Таким образом, в определение «полипептид» включены пептиды, дипептиды, олигопептиды, белок, аминокислотная цепь или любой другой термин, использованный для обозначения цепи из двух или более аминокислот, и термин «полипептид» можно использовать вместо любого из указанных терминов или взаимозаменяемо с ними. Термин «полипептид» можно также использовать для обозначения продуктов пост-экспрессионных модификаций полипептида, включая, но, не ограничиваясь только ими, гликозилирование, ацетилирование, фосфорилирование, амидирование, модификацию с использованием известных защитных/блокирующих групп, протеолитическое расщепление или модификацию с использованием неприродных аминокислот. Полипептид можно получить из природного биологического источника или получить рекомбинантной технологией, но не обязательно транслировать из указанной последовательности нуклеиновой кислоты. Полипептид можно получить любым образом, включая химический синтез. Размер полипептида по настоящему изобретению может составлять от приблизительно 3 или более, 5 или более, 10 или более, 20 или более, 25 или более, 50 или более, 75 или более, 100 или более, 200 или более, 500 или более, 1000 или более, 2000 или более аминокислот. Полипептиды могут характеризоваться определенной трехмерной структурой, хотя необязательно они могут характеризоваться такой структурой. Полипептиды с определенной трехмерной структурой обозначают как свернутые, а полипептиды, которые не характеризуются определенной трехмерной структурой, но могут присутствовать во множестве различных конформаций, обозначают как развернутые.
Термин «полинуклеотид» означает выделенную молекулу нуклеиновой кислоты или конструкта, например, матричную РНК (мРНК), РНК вирусного происхождения или плазмидную ДНК (пДНК). Полинуклеотид может включать стандартную фосфодиэфирную связь или нестандартную связь (например, амидную связь, такую как обнаруженная в пептид-нуклеиновых кислотах (ПНК)). Термин «молекула нуклеиновой кислоты» означает любой один или более сегментов нуклеиновой кислоты, например, фрагменты ДНК или РНК, присутствующие в полинуклеотиде.
«Сниженное связывание», например, сниженное связывание с Fc рецептором, означает уменьшение аффинности в отношении соответствующего взаимодействия по данным, например, ППР. Для ясности, указанный термин включает также снижение аффинности до нуля (или ниже предела обнаружения аналитического метода), то есть до полного отсутствия взаимодействия. Наоборот, «повышенное связывание» означает увеличение связывающей аффинности в отношении соответствующего взаимодействия.
Термин «регуляторная последовательность» означает последовательности ДНК, которые необходимы для влияния на экспрессию кодирующих последовательностей, с которыми они лигированы. Природа таких контролирующих последовательностей различается в зависимости от организма хозяина. У прокариотов, контрольные последовательности в основном включают промотор, сайт связывания рибосомы и терминаторы. У эукариотов контрольные последовательности в основном включают промоторы, терминаторы и в некоторых случаях энхансеры, трансактиваторы или факторы транскрипции. Термин «контрольная последовательность» включает как минимум все компоненты, присутствие которых необходимо для экспрессии, и может включать дополнительные полезные компоненты.
Использованный в данном контексте термин «одноцепочечный» относится к молекуле, содержащей аминокислотные мономеры, связанные в линейную цепь пептидными связями. В определенных вариантах один из антигенсвязывающих остатков является scFv фрагментом, то есть VH доменом и VL доменом, соединенными через пептидный линкер. В определенных вариантах один из антигенсвязывающих фрагментов является одноцепочечной Fab молекулой, то есть Fab молекулой, где легкая цепь Fab и тяжелая цепь Fab соединены пептидным линкером, при этом образуется одна пептидная цепь. В таком конкретном варианте в одноцепочечной Fab молекуле С-концевой аминокислотный остаток легкой цепи Fab присоединен к N-концевому аминокислотному остатку тяжелой цепи Fab.
Термин «SSD», использованный в данном контексте, относится к стимулирующему сигнальному домену.
Использованный в данном контексте термин «лечение» (и его грамматические варианты, такие как «лечебный» или «терапия») означают клиническое вмешательство с целью изменить природный ход заболевания у индивидуума, нуждающегося в лечении, которое можно осуществлять либо для профилактики, либо в ходе клинической патологии. Требуемые действия лечения включают, но, не ограничиваясь только ими, профилактику возникновения или рецидива заболевания, снижение интенсивности симптомов, снижение любых прямых или косвенных патологических последствий заболевания, профилактику метастазирования, снижение скорости прогрессирования заболевания, смягчение или облегчение патологического состояния, а также ремиссию или улучшенный прогноз. В некоторых вариантах экспрессируемые клеткой антигенсвязывающие рецепторы по настоящему изобретению используют совместно с терапевтическими антителами, включающими мутированный Fc домен, для замедления развития заболевания или для замедления прогрессирования заболевания.
Термин «антигенная детерминанта-мишень» является синонимом терминов «антиген-мишень», «эпитоп-мишень» и «клеточный антиген-мишень», и относится к участку (например, непрерывная (линейная) последовательность аминокислотных остатков или конформационная конфигурация, образованная из различных областей не-соседних (нелинейных) аминокислотных остатков) в полипептидной макромолекуле, с которой связывается антитело, при этом формируется комплекс антигенсвязывающего фрагмента и антигена. Пригодные антигенные детерминанты можно найти, например, на поверхности опухолевых клеток, на поверхности инфицированных вирусом клеток, на поверхности других пораженных заболеванием клеток, на поверхности иммунных клеток, в виде свободных структур в сыворотке крови и/или во внеклеточном матриксе (ЕСМ). Белки, определенные в данном контексте как антигены (например, CD20, CEA, FAP, TNC), могут находиться в любой природной форме белков из любого источника позвоночных, включая млекопитающих, таких как приматы (например, человек) и грызуны (например, мыши и крысы), если не указано иное. В конкретном варианте антигеном-мишенью является белок человека. Если в данном контексте упоминается определенный белок-мишень, то термин включает «полноразмерный» непроцессированный белок-мишень, а также любую форму белка-мишени, которая образуется в результате процессинга в клетке-мишени. Термин также включает природные варианты белка-мишени, например варианты, после сплайсинга или аллельные варианты. Типичные белки-мишени человека, использованные в качестве антигенов, включают, но, не ограничиваясь только ими, CD20, CEA, FAP, TNC, MSLN, FolR1, HER1 and HER2. Способность антигенсвязывающего рецептора связываться со специфической антигенной детерминантой-мишенью можно измерить методом твердофазного иммуноферментного анализа (ИФА) иди другими методами, известными специалисту в данной области техники, например, метод поверхностного плазмонного резонанса (ППР) (анализ проводят в системе BIAcore) (Liljeblad и др., Glyco J 17, 323-329 (2000)), и традиционные методы анализа связывания (Heeley, Endocr Res 28, 217-229 (2002)). В одном варианте степень связывания антитела с неродственным белком составляет менее приблизительно 10% связывания антитела с антигеном-мишенью по результатам измерения, например, методом ППР. В некоторых вариантах антитело, которое связывается с антигеном-мишенью, характеризуется аффинной константой диссоциации (KD) ≤1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ, ≤0,1 нМ, ≤0,01 нМ или ≤0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 М, например, от 10-9 М до 10-13 М).
«Антитела, содержащие мутированный Fc домен» по настоящему изобретению, то есть терапевтические антитела, могут содержать один, два, три или более связывающих доменов и могут являться моноспецифичными, диспецифичными или мультиспецифичными. Антитела могут быть получены в виде полноразмерных антител из одного вида, например, человека, или они могут быть получены в виде химерных или гуманизированных антител. В случае антител, содержащих более двух антигенсвязывающих доменов, некоторые связывающие домены могут быть идентичными и/или характеризоваться одинаковой специфичностью.
Использованный в данном контексте термин «активация Т-клеток» относится к одной или более ответных клеточных реакций Т лимфоцита, прежде всего цитотоксического Т лимфоцита, выбранных из пролиферации, дифференциации, секреции цитокинов, высвобождения цитотоксических эффекторных соединений, цитотоксической активности и экспрессии маркеров активации. Антигенсвязывающие рецепторы по настоящему изобретению способны индуцировать активацию Т-клеток. Пригодные методы анализа активации Т-клеток известны в данной области техники и описаны в данном контексте.
Согласно настоящему изобретению, термин «Т-клеточный рецептор» или "TCR" известны в уровне техники. Прежде всего использованный в данном контексте термин «Т-клеточный рецептор» относится к любому Т-клеточному рецептору, который соответствует трем следующим критериям: (i) опухолевая специфичность, (ii) распознавание (большинства) опухолевых клеток, что означает, что антиген или мишень должны экспрессироваться в (большинстве) опухолевых клетках и (iii) TCR соответствует HLA-типу субъектов, нуждающихся в таком лечении. В данном контексте пригодные Т-клеточные рецепторы, которые соответствуют указанным выше трем критериям, известны в уровне техники, такие как рецепторы, распознающие NY-ESO-1 (см. информацию о последовательности, например, в PCT/GB 2005/001924) и/или HER2neu (см. информацию о последовательности в WO-A1 2011/0280894).
Термин «терапевтически эффективное количество» агента, например, фармацевтической композиции, означает количество, проявляющее эффективность в дозировках и в течение периодов времени, когда достигается требуемый лечебный или профилактический результат. Терапевтически эффективное количество агента, например, устраняет, снижает, замедляет развитие, сводит к минимуму или предотвращает отрицательные действия заболевания.
Термин «вектор» или «экспрессионный вектор» является синонимом термину «экспрессионный конструкт» и означает молекулу ДНК, которую используют для включения и прямой экспрессии специфического гена, с которым она функционально связана в клетке-мишени. Термин включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки хозяина, в которую он включен. Экспрессионный вектор по настоящему изобретению включает кассету экспрессии. Экспрессионные векторы позволяют осуществлять транскрипцию больших количеств стабильной мРНК. При попадании экспрессионного вектора внутрь клетки-мишени рибонуклеиновая кислота или белок, которые кодируются геном, продуцируются в ходе клеточной транскрипции и/или с использованием трансляционного комплекса. В одном варианте экспрессионный вектор по настоящему изобретению включает кассету экспрессии, которая включает полинуклеотидные последовательности, кодирующие антигенсвязывающие рецепторы по настоящему изобретению или их фрагменты.
Антигенсвязывающие рецепторы, способные специфически связываться с мутированным Fc доменом (доменами)
Настоящее изобретение относится к антигенсвязывающим рецепторам, способным специфически связываться с мутированным Fc доменом антитела, то есть терапевтическим антителом, которое направленно связывается с опухолевой клеткой. Прежде всего, настоящее изобретение относится к антигенсвязывающим рецепторам, которые включают внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с исходным немутированным Fc доменом. В предпочтительных вариантах мутированный Fc домен включает по меньшей мере одну аминокислотную замену по сравнению с немутированным исходным Fc доменом, где связывание Fc рецептора с мутированным Fc доменом снижено по сравнению со связыванием Fc рецептора с немутированным Fc доменом. В конкретных вариантах настоящее изобретение относится к антигенсвязывающим рецепторам, которые включают внеклеточный домен, включающий по меньшей мере один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, где по меньшей мере один антигенсвязывающий фрагмент не способен специфически связываться с исходным немутированным Fc доменом, где мутированный Fc домен включает по меньшей мере одну аминокислотную замену, которую выбирают из группы, состоящей из L234, L235, I253, Н310, Р331, Р329 и Н435, прежде всего, где аминокислотной мутацией является L234A, L235A, I253A, N297A, Н310А, P329G и/или Н435А, по сравнению с немутированным исходным Fc доменом, где связывание Fc рецептора с мутированным Fc доменом снижено по сравнению со связыванием Fc рецептора с немутированным Fc доменом. В одном предпочтительном варианте аминокислотной мутацией является P329G, где связывание с Fcγ рецептором снижено по данным ППР при 25°С. В другом предпочтительном варианте аминокислотными мутациями являются I253A, Н310А и Н435А, где связывание с неонатальным Fc рецептором (FcRn) снижено по данным ППР при 25°С.
Настоящее изобретение также относится к трансдукции Т-клеток, таких как CD8 + Т-клетки, CD4 + Т-клетки, CD3 + Т-клетки, γδ Т-клетки или природные Т-клетки-киллеры (NK), предпочтительно CD8 + Т-клетки, с использованием антигенсвязывающего рецептора, как описано в данном контексте, и их целевому накапливанию, например, в опухоли с помощью антитела, например, терапевтического антитела, включающего мутированный Fc домен. В одном варианте антитело способно специфически связываться с опухолеспецифическим антигеном, который присутствует на поверхности природной опухолевой клетки.
В прилагаемых примерах в качестве подтверждения концепции изобретения сконструирован антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен по настоящему изобретению pETR17096 (SEQ ID NO: 7, кодируемая последовательностью ДНК SEQ ID NO: 19), способный специфически связываться с терапевтическим антителом (представляет собой анти-CD20 антитело, включающее тяжелую цепь с последовательностью SEQ ID NO ID: 112, и легкую цепь с последовательностью SEQ ID NO: 113), которые включают мутацию P329G. Трансдуцированные Т-клетки (Т-клетки Jurkat NFAT), экспрессирующие белок анти-P329G-scFv-CD28ATD-CD28CSD-CD3zSSD (SEQ ID NO: 7, кодируемая последовательностью ДНК SEQ ID NO: 19), могут существенно активироваться при совместной инкубации с анти-CD20 антителом, включающим мутацию P329G в составе Fc домена, и CD20 положительными опухолевыми клетками. Для подтверждения концепции изобретения авторы изобретения также предлагают множество форматов антигенсвязывающего рецептора, способного специфически связываться с мутированным Fc доменом, но не способного специфически связываться с немутированным исходным Fc доменом.
Обработка опухолевых клеток комбинацией антитела, направленного против опухолевого антигена, где антитело, включает мутацию P329G, и трансдуцированными Т-клетками, экспрессирующими белок анти-P329G-Fab-ds-CD28ATD-CD28CSD-CD3zSSD (SEQ ID NO: 44 (ДНК) и 39, 41 (белок)), неожиданно приводит к существенной активации трансдуцированной Т-клетки по сравнению с трансдуцированными Т-клетками, экспрессирующими анти-P329G-scFv-CD28ATD-CD28CSD-CD3zSSD (SEQ ID NO: 19 (ДНК) и 7 (белок)) гибридный белок (см., например, фиг. 6 и 8-11).
Соответственно, неожиданно и непредсказуемо было обнаружено, что Т-клетки, предпочтительно CD8 + Т-клетки, которые трансдуцированы антигенсвязывающим рецептором по настоящему изобретению, можно специфически стимулировать с использованием опухолеспецифического антитела, включающего мутированный Fc домен, и накапливать с использованием опухолеспецифического антитела, которое выступает в качестве элемента связывания с опухолевой клеткой. Таким образом, в настоящем изобретении неожиданно и непредсказуемо было установлено, что объединение опухолеспецифического антитела, то есть терапевтического антитела, включающего мутированный Fc домен, с Т-клетками, трансдуцированными антигенсвязывающим рецептором, который включает/состоит из внеклеточного домена, включающего антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, приводит к специфической активации Т-клеток и последующему лизису опухолевой клетки. Указанный подход характеризуется значительными преимуществами в плане безопасности по сравнению с традиционными подходами, основанными на применении Т-клеток, поскольку Т-клетка будет инертной в отсутствии антитела, содержащего мутированный Fc домен, и их наличие можно контролировать, выбирая формат молекулы антитела (т.е. использовать низкомолекулярные молекулы с более коротким периодом полураспада и наоборот). Соответственно, в изобретении предлагается многофункциональная терапевтическая платформа, где антитела типа IgG используются для маркирования или введения метки в опухолевые клетки с целью формирования сигнала для целевой активации Т-клетки, и где действие трансдуцированных Т-клеток специфически направлено на опухолевые клетки за счет обеспечения специфичности мутированного Fc домена антитела типа IgG. После связывания с мутированным Fc доменом антитела на поверхности опухолевой клетки, трансдуцированная Т-клетка, как описано в данном контексте, активируется и затем происходит лизис. Платформа является универсальной и специфичной, что позволяет использовать разнообразные (существующие или вновь разработанные) антитела-мишени или совместно применять несколько антител с различной специфичностью к антигену, но содержащих одну и ту же мутацию в Fc домене. Степень активации Т-клеток можно дополнительно отрегулировать за счет корректировки дозы совместно применяемого терапевтического антитела или за счет использования антител с различной специфичностью или различных форматов антител. Трансдуцированные Т-клетки по изобретению являются инертными в случае применения без антитела-мишени, содержащего мутированный Fc домен, поскольку мутации в Fc домене, как описано в данном контексте, не встречаются в природных или немутантных иммуноглобулинах. Соответственно, в одном варианте мутированный Fc домен не обнаруживается в природных иммуноглобулинах дикого типа.
Соответственно, настоящее изобретение относится к антигенсвязывающему рецептору, который включает внеклеточный домен, включающий по меньшей мере один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, где по меньшей мере один антигенсвязывающий фрагмент не способен специфически связываться с исходным немутированным Fc доменом, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию по сравнению с немутированным исходным Fc доменом, причем связывание Fc рецептора с мутированным Fc доменом и/или эффекторная функция, индуцированные мутированным Fc доменом, снижены по сравнению со связыванием Fc рецептора и/или эффекторной функцией, индуцированными немутированным Fc доменом. При лечении рака, прежде всего, желательно использовать терапевтические антитела с пониженной эффекторной функцией, поскольку эффекторная функция может приводить к серьезным побочным эффектам в ходе противоопухолевого лечения на основе антител, как дополнительно описано в данном контексте.
В настоящем изобретении антигенсвязывающий рецептор включает внеклеточный домен, который не обнаруживается в природных Т-клетках или на их поверхности. Таким образом, антигенсвязывающий рецептор способен обеспечить заданную специфичность связывания с клетками, экспрессирующими антигенсвязывающий рецептор по настоящему изобретению. Клетки, например, Т-клетки, трансдуцированные антигенсвязывающим рецептором (рецепторами) по изобретению способны специфически связываться с мутированным Fc доменом, но не с исходным немутированным Fc доменом. Специфичность обеспечивается антигенсвязывающим фрагментом внеклеточного домена антигенсвязывающего рецептора, при этом указанные антигенсвязывающие фрагменты считаются специфичными в отношении мутированного Fc домена, как описано в данном контексте. В настоящем изобретении и, как разъясняется в настоящем описании, антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, связывается/взаимодействует с мутированным Fc доменом, но не с исходным немутированным Fc доменом.
Антигенсвязывающие фрагменты
В иллюстративном варианте по настоящему изобретению в качестве доказательства концепции, предлагаются антигенсвязывающие рецепторы, включающие заякоренный трансмембранный домен и внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент, где по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает по меньшей мере одну аминокислотную замену по сравнению с немутированным исходным Fc доменом.
В определенном варианте, по меньшей мере, один из антигенсвязывающих фрагментов представляет собой стандартный Fab фрагмент, то есть Fab молекулу, состоящую из легкой цепи Fab и тяжелой цепи Fab. В определенном варианте, по меньшей мере, один из антигенсвязывающих фрагментов представляет собой crossFab фрагмент, то есть Fab молекулу, состоящую из легкой цепи Fab и тяжелой цепи Fab, где либо вариабельные области, либо консервативные области тяжелой и легкой цепей Fab заменены друг на друга. В некоторых вариантах по меньшей мере один из антигенсвязывающих фрагментов представляет собой scFv фрагмент. В таком конкретном варианте в scFv фрагменте С-концевой аминокислотный остаток вариабельной области (VH) тяжелой цепи присоединен к N-концевому аминокислотному остатку вариабельной области легкой цепи (VL) в молекуле scFv, необязательно через пептидный линкер. В некоторых вариантах по меньшей мере один из антигенсвязывающих фрагментов представляет собой одноцепочечную Fab молекулу, то есть Fab молекулу, где легкая цепь Fab и тяжелая цепь Fab соединены пептидным линкером, при этом образуется одна пептидная цепь. В таком конкретном варианте в одноцепочечной Fab молекуле С-концевой аминокислотный остаток легкой цепи Fab присоединен к N-концевому аминокислотному остатку тяжелой цепи Fab, необязательно через пептидный линкер.
Соответственно, в настоящем изобретении антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает по меньшей мере одну аминокислотную замену по сравнению с немутированным исходным Fc доменом.
Антигенсвязывающие фрагменты, способные специфически связываться с мутированным Fc доменом, можно получить методом иммунизации, например, методами иммунизации млекопитающих. Указанные методы известны в данной области техники и описаны, например, в статье Burns, Methods in Molecular Biology, 295:1-12 (2005). В другом варианте антигенсвязывающие фрагменты по изобретению могут быть выделены путем скрининга комбинаторных библиотек для антител с требуемой активностью или активностями. Способы скрининга комбинаторных библиотек описаны в литературе, например, в статье Lerner и др., Nature Reviews, 16: 498-508 (2016). Например, в данной области техники известно множество способов создания библиотек фагового дисплея и скрининга таких библиотек для антигенсвязывающих фрагментов, обладающих требуемыми характеристиками связывания. Указанные способы описаны, например, в статьях Frenzel и др., mAbs, 8: 1177-1194 (2016), Bazan и др., Human Vaccines and Immunotherapeutics, 8: 1817-1828 (2012) и Zhao и др., Critical Reviews in Biotechnology 36: 276-289 (2016), а также в статье Hoogenboom и др., Methods in Molecular Biology, 178: 1-37 (O'Brien и др., изд. Human Press, Totowa, NJ, 2001), и дополнительно описаны, например, в статьях McCafferty и др., Nature, 348: 552-554, Clackson и др., Nature, 352: 624-628 (1991), Marks и др., J. Mol. Biol., 222: 581-597 (1992) и Marks и Bradbury, Methods in Molecular Biology, 248: 161-175 (Lo, изд. Human Press, Totowa, NJ, 2003), Sidhu и др., J. Mol. Biol., 338(2): 299-310 (2004), Lee и др., J. Mol. Biol., 340(5): 1073-1093 (2004), Fellouse, Proc. Natl. Acad. Sci. USA, 101(34): 12467-12472 (2004) и Lee и др., J. Immunol. Methods, 284(1-2): 119-132 (2004). В некоторых способах фагового дисплея репертуары генов VH и VL раздельно клонируют при помощи полимеразной цепной реакции (ПЦР) и произвольным образом рекомбинируют в фаговых библиотеках, и затем проводят их скрининг для антигенсвязывающего фага, согласно описанию Winter и др., Annual Review of Immunology, 12: 433-455 (1994). Как правило, фаги воспроизводят фрагменты антител либо в виде одноцепочечных Fv (scFv) фрагментов, либо в виде Fab фрагментов. Библиотеки из иммунизированных источников предоставляют высокоаффинные антигенсвязывающие фрагменты в отношении иммуногена без необходимости конструирования гибридом. В другом варианте можно клонировать наивный репертуар (например, человека), чтобы обеспечить единый источник антигенсвязывающих фрагментов к широкому диапазону "чужеродных", а также "собственных" антигенов без какой-либо иммунизации, как описано в статье Griffiths и др., EMBO Journal, 12: 725-734 (1993). Наконец, наивные библиотеки также можно получить синтетическим методом с использованием клонирования сегментов неперегруппированного V-гена из стволовых клеток, а также с использованием ПЦР-праймеров, содержащих случайную последовательность, с тем, чтобы кодировать высоковариабельные CDR3 области и осуществить перегруппировку in vitro, как описано в статье Hoogenboom и Winterin, Journal of Molecular Biology, 227: 381-388 (1992). Фаговые библиотеки антител человека описаны, например, в патентах US 5750373, US 7985840, US 7785903 и US 8679490, а также патентах US 2005/0079574, US 2007/0117126, US 2007/0237764, US 2007/0292936 и US 2009/0002360. Дополнительные примеры способов скрининга комбинаторных библиотек для антител с требуемой активностью или активностями, известные в данной области техники, включают рибосомальные и мРНК дисплеи, а также способы селекции антител на бактериальных дисплеях, дисплеях клеток млекопитающих, клеток насекомых или дрожжевых клеток. Методы дисплея на поверхности дрожжевых клеток описаны, например, в статьях Scholler и др., Methods in Molecular Biology, 503: 135-156 (2012) и Cherf и др., Methods in Molecular biology, 1319: 155-175 (2015), а также в статье Zhao и др., Methods in Molecular Biology, 889: 73-84 (2012). Методы рибосомального дисплея описаны, например, в статьях Не и др., Nucleic Acids Research, 25: 5132-5134 (1997) и Hanes и др., PNAS, 94: 4937-4942 (1997).
В настоящем изобретении предлагаются антигенсвязывающие рецепторы, включающие, по меньшей мере, один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc домен. Соответственно, трансдуцированные клетки, то есть Т-клетки, экспрессирующие антигенсвязывающий рецептор по настоящему изобретению, способны специфически связываться с мутированным Fc доменом антитела, то есть терапевтическим антителом. Fc домен придает антителам, то есть терапевтическим антителам благоприятные фармакокинетические свойства, включая длительный период полураспада в сыворотке крови, что способствует хорошему накоплению в ткани-мишени и благоприятному коэффициенту распределения в ткани и крови. В то же время, однако, он может приводить к нежелательному направленному взаимодействию терапевтических антител с клетками, экспрессирующими рецепторы Fc фрагмента, а не к связыванию с предпочтительными антигенпрезентирующими клетками. Более того, коактивация сигнальных путей Fc рецептора может приводить к высвобождению цитокинов, что может приводить к избыточной активации рецепторов цитокинов и серьезным побочным эффектам при системном введении терапевтических антител. Активация презентирующих Fc рецептор иммунных клеток, отличных от Т-клеток, может приводить даже к снижению эффективности терапевтических антител в результате возможного разрушения иммунных клеток. Соответственно, терапевтические антитела, известные в данной области техники, сконструированы или мутированы таким образом, чтобы проявлять пониженную связывающую аффинность в отношении Fc рецептора и/или пониженную эффекторную функцию по сравнению, например, с нативным Fc доменом IgG1. Антигенсвязывающие рецепторы по настоящему изобретению можно использовать для направленного действия эффекторных клеток, например, Т-клеток, экспрессирующих антигенсвязывающие рецепторы по настоящему изобретению in vitro и/или in vivo на клетки-мишени, то есть опухолевые клетки, которые маркированы антителом, способным специфически связываться с клетками-мишенями, где антитело включает сконструированный и/или мутированный Fc домен, как описано в данном контексте.
В иллюстративном варианте по настоящему изобретению в качестве подтверждения концепции предлагаются антигенсвязывающие рецепторы, способные специфически связываться с мутированным Fc доменом, включающим аминокислотную мутацию P329G, а также эффекторные клетки, экспрессирующие указанные антигенсвязывающие рецепторы. Мутация P329G снижает уровень связывания с Fcγ рецепторами и ассоциированную с ними эффекторную функцию. Соответственно, мутированный Fc домен, включающий мутацию P329G, характеризуется пониженной связывающей аффинностью в отношении Fcγ рецепторов или ее отсутствием по сравнению с немутированным Fc доменом. В другом иллюстративном варианте по настоящему изобретению в качестве подтверждения концепции предлагаются антигенсвязывающие рецепторы, способные специфически связываться с мутированным Fc доменом, содержащим аминокислотные замены I253A, Н310А и Н435А ("AAA"). Мутации AAA практически полностью подавляют связывание с FcRn.
Однако в данной области техники широко используются антитела, характеризующиеся пониженным, повышенным или уменьшенным уровнем связывания с Fc рецепторами (FcR) и/или эффекторной функции, содержащие мутированный Fc домен. Соответственно, в данном контексте предлагаются антигенсвязывающие рецепторы, способные специфически связываться с антителами, включающими мутированный Fc домен, и указанные антитела в данном контексте также называются антитела-мишени. Соответственно, в одном варианте осуществления настоящего изобретения антигенсвязывающий рецептор по настоящему изобретению способен специфически связываться с антителом-мишенью, включающим мутированный Fc домен, и которое характеризуется уменьшенной связывающей аффинностью с Fc рецептором и/или сниженной эффекторной функцией. Антитела-мишени, характеризующиеся сниженной эффекторной функцией, включают такие Fc домены, которые содержат одну или более мутаций в положении аминокислот 238, 265, 269, 270, 297, 327 и 329 (см. патент US 6737056). Такие мутированные Fc включают мутированные Fc с мутациями в двух или более положениях аминокислот 265, 269, 270, 297 и 327, включая так называемый "DANA" мутированный Fc, в котором аминокислотные остатки в положении 265 и 297 заменены на аланин (см. патент US 7332581). Определенные варианты антител, характеризующиеся повышенным или уменьшенным связыванием с рецепторами FcR, описаны в литературе (см., например, патент US 6737056, заявку WO 2004/056312 и статью Shields и др., J. Biol. Chem., 9(2): 6591-6604 (2001)). В некоторых вариантах предлагается антигенсвязывающий рецептор, способный специфически связываться с вариантом антитела, которое включает Fc домен, содержащий одну или более аминокислотных мутаций, улучшающих ADCC, например, Fc область, которая включает мутации в положениях 298, 333 и/или 334 (EU система нумерации аминокислотных остатков). В некоторых вариантах вариант антитела-мишени включает Fc область, которая содержит одну или более аминокислотных мутаций, которые понижают или уменьшают связывание с FcRn, например, Fc область, которая включает мутации в положениях 253 и/или 310 и/или 435 (EU система нумерации аминокислотных остатков). В некоторых вариантах вариант антитела-мишени включает Fc область, которая содержит аминокислотные мутации в положениях 253, 310 и 435. В одном варианте Fc область, полученная из Fc области IgG1 человека, включает мутации I253A, Н310А и Н435А, см., например, статью Grevys А. и др., J. Immunol., 194, 5497-5508 (2015).
В некоторых вариантах, предлагается антигенсвязывающий рецептор, способный специфически связываться с вариантом антитела, включающим Fc область, которая содержит одну или более аминокислотных мутаций, которые понижают или уменьшают связывание с FcRn, например, Fc область, которая включает одну мутацию в одном из положений 310 и/или 433 и/или 436 (EU система нумерации аминокислотных остатков). В некоторых вариантах вариант антитела-мишени включает Fc область, которая содержит аминокислотные мутации в положениях 310, 433 и 436. В одном варианте Fc область, полученная из Fc области IgG1 человека, включает мутации Н310А, Н433А и Y436A. В некоторых вариантах вариант антитела-мишени включает Fc область, которая содержит одну или более аминокислотных мутаций, которые усиливают связывание с FcRn, например, Fc область, содержащую мутации в положениях 252 и/или 254 и/или 256 (EU система нумерации аминокислотных остатков). В некоторых вариантах вариант антитела-мишени включает Fc область, которая содержит аминокислотные мутации в положениях 252, 254 и 256. В одном варианте Fc область, полученная из Fc области IgG1 человека, включает мутации M252Y, S254T и Т256Е. В некоторых вариантах предлагается антигенсвязывающий рецептор, способный специфически связываться с вариантом антитела, включающим Fc область, содержащую аминокислотные мутации, которые уменьшают связывание с FcγR, например, Fc область, содержащую мутации в положениях 234, 235 и 329 (EU система нумерации аминокислотных остатков). В одном варианте мутации представляют собой мутации L234A и L235A (LALA). В некоторых вариантах Fc область варианта антитела-мишени, полученная из Fc области IgG1 человека, дополнительно включает мутацию D265A и/или P329G. В одном варианте Fc область, полученная из Fc области IgG1 человека, включает мутацию P329G ("PG"). В другом варианте Fc область, полученная из Fc области IgG1 человека, включает мутации I253A, Н310А и Н435А ("AAA").
В одном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, состоящим из первой и второй субъединиц, способных к стабильной ассоциации. В одном варианте Fc домен представляет собой Fc домен IgG, конкретно Fc домен IgG1 или IgG4. В одном варианте Fc домен представляет собой Fc домен человека. В одном варианте мутированный Fc домен проявляет сниженную связывающую аффинность в отношении Fc рецептора и/или сниженную эффекторную функцию, по сравнению с нативным Fc доменом IgG1. В одном варианте осуществления настоящего изобретения Fc домен включает одну или более аминокислотных мутаций, которые снижают связывание с Fc рецептором и/или эффекторную функцию.
В одном предпочтительном варианте одна или более аминокислотных мутаций расположены в одном или более положений, выбранных из группы L234, L235 и Р329 (согласно системе нумерации Кабата). В одном конкретном варианте каждая субъединица Fc домена включает три аминокислотных мутаций, которые снижают связывание с активирующим Fc рецептором и/или эффекторную функцию, где указанные аминокислотные мутации включают L234A, L235A и P329G. В одном конкретном варианте Fc рецептором является Fcγ рецептор. В одном варианте эффекторная функция представляет собой антителозависимую опосредованную клетками цитотоксичность (ADCC).
В конкретном варианте мутированный Fc домен включает мутацию P329G. Соответственно, мутированный Fc домен, включающий мутацию P329G, характеризуется пониженной связывающей аффинностью в отношении Fcγ рецепторов или ее отсутствием по сравнению с немутированным Fc доменом. В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую, по меньшей мере, одну:
(а) определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью RYWMN (SEQ ID NO: 1),
(б) CDR Н2 с аминокислотной последовательностью EITPDSSTINYTPSLKD (SEQ ID NO: 2) и
(в) CDR Н3 с аминокислотной последовательностью PYDYGAWFAS (SEQ ID NO: 3).
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую, по меньшей мере, одну:
(г) легкую цепь 1 (CDR L) с аминокислотной последовательностью RSSTGAVTTSNYAN (SEQ ID NO: 4),
(д) CDR L2 с аминокислотной последовательностью GTNKRAP (SEQ ID NO: 5) и
(е) CDR L3 с аминокислотной последовательностью ALWYSNHWV (SEQ ID NO: 6).
В одном варианте осуществления настоящего изобретения внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает по меньшей мере одну определяющую комплементарность область тяжелой цепи (CDR), включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, и, по меньшей мере, одну CDR легкой цепи, выбранную из группы, включающей последовательность SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6.
В одном варианте осуществления настоящего изобретения внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает определяющие комплементарность области тяжелой цепи (CDR) с последовательностью SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, и CDR легкой цепи с последовательностью SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6.
В одном предпочтительном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую:
(а) определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью RYWMN (SEQ ID NO: 1),
(б) CDR Н2 с аминокислотной последовательностью EITPDSSTINYTPSLKD (SEQ ID NO: 2),
(в) CDR Н3 с аминокислотной последовательностью PYDYGAWFAS (SEQ ID NO: 3),
и вариабельную область легкой цепи, включающую:
(г) легкую цепь 1 (CDR L) с аминокислотной последовательностью RSSTGAVTTSNYAN (SEQ ID NO: 4),
(д) CDR L2 с аминокислотной последовательностью GTNKRAP (SEQ ID NO: 5) и
(e) CDR L3 с аминокислотной последовательностью ALWYSNHWV (SEQ ID NO: 6).
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из SEQ ID NO: 8 и SEQ ID NO: 32, и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из SEQ ID NO: 9 и SEQ ID NO: 33.
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность, выбранную из SEQ ID NO: 8 и SEQ ID NO: 32, и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность, выбранную из SEQ ID NO: 9 и SEQ ID NO: 33.
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 32, и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 33.
В одном предпочтительном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 8, и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 9.
В одном варианте осуществления настоящего изобретения, по меньшей мере, один антигенсвязывающий фрагмент представляет собой scFv, Fab, crossFab или scFab фрагмент. В одном варианте осуществления настоящего изобретения внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент представляет собой Fab фрагмент.
В предпочтительном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где Fab фрагмент включает тяжелую цепь с последовательностью SEQ ID NO: 40 и легкую цепь с последовательностью SEQ ID NO: 41.
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv фрагмент, который является полипептидом, состоящим из вариабельного домена тяжелой цепи (VH), вариабельного домена легкой цепи (VL) и линкера, где указанные вариабельные домены и указанный линкер характеризуются одной из следующих конфигураций в направлении от N-концевого остатка к С-концевому остатку: а) VH-линкер-VL или б) VL-линкер-VH. В предпочтительном варианте scFv фрагмент характеризуется конфигурацией VH-линкер-VL.
В предпочтительном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где scFv фрагмент включает аминокислотную последовательность SEQ ID NO: 10.
В альтернативном конкретном варианте осуществления настоящего изобретения мутированный Fc домен включает мутации I253A, Н310А и Н435А ("AAA"). Мутации AAA снижают уровень связывания с неонатальным Fc рецептором (FcRn). Соответственно, мутированный Fc домен, включающий мутации AAA, характеризуется пониженной связывающей аффинностью в отношении FcRn или ее отсутствием по сравнению с немутированным Fc доменом.
Соответственно, в одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую, по меньшей мере, одну:
(а) определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью SYGMS (SEQ ID NO: 53),
(б) CDR Н2 с аминокислотной последовательностью SSGGSY (SEQ ID NO: 54) и
(в) CDR Н3 с аминокислотной последовательностью LGMITTGYAMDY (SEQ ID NO: 55).
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает вариабельную область легкой цепи, включающую, по меньшей мере, одну:
(г) легкую цепь 1 (CDR L) с аминокислотной последовательностью RSSQTIVHSTGHTYLE (SEQ ID NO: 56),
(д) CDR L2 с аминокислотной последовательностью KVSNRFS (SEQ ID NO: 57) и
(е) CDR L3 с аминокислотной последовательностью FQGSHVPYT (SEQ ID NO: 58).
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает по меньшей мере одну определяющую комплементарность область тяжелой цепи (CDR), включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 53, SEQ ID NO: 54 и SEQ ID NO: 55, и, по меньшей мере, одну CDR легкой цепи, выбранную из группы, включающей последовательность SEQ ID NO: 56, SEQ ID NO: 57 и SEQ ID NO: 58.
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает определяющие комплементарность области тяжелой цепи (CDR) с последовательностью SEQ ID NO: 53, SEQ ID NO: 54 и SEQ ID NO: 55, и CDR легкой цепи с последовательностью SEQ ID NO: 56, SEQ ID NO: 57 и SEQ ID NO: 58.
В предпочтительном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи, включающую:
(а) определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью SYGMS (SEQ ID NO: 53),
(б) CDR Н2 с аминокислотной последовательностью SSGGSY (SEQ ID NO: 54),
(в) CDR Н3 с аминокислотной последовательностью LGMITTGYAMDY (SEQ ID NO: 55), и вариабельную область легкой цепи, включающую:
(г) легкую цепь 1 (CDR L) с аминокислотной последовательностью RSSQTIVHSTGHTYLE (SEQ ID NO: 56),
(д) CDR L2 с аминокислотной последовательностью KVSNRFS (SEQ ID NO: 57) и
(е) CDR L3 с аминокислотной последовательностью FQGSHVPYT (SEQ ID NO: 58).
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 61 и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из SEQ ID NO: 62.
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 61, и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 62.
В одном варианте осуществления настоящего изобретения, по меньшей мере, один антигенсвязывающий фрагмент представляет собой scFv, Fab, crossFab или scFab фрагмент. В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где по меньшей мере антигенсвязывающий фрагмент представляет собой Fab фрагмент. В конкретном варианте осуществления настоящего изобретения внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где Fab фрагмент включает тяжелую цепь с последовательностью SEQ ID NO: 64 и легкую цепь с последовательностью SEQ ID NO: 65.
В одном варианте внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv фрагмент. В конкретном варианте осуществления настоящего изобретения внеклеточный домен антигенсвязывающего рецептора включает антигенсвязывающий фрагмент, способный специфически связываться с Fc доменом, включающим мутации 1253А, Н310А и Н435А, где scFv фрагмент включает аминокислотную последовательность SEQ ID NO: 60.
В других вариантах осуществления настоящего изобретения антигенсвязывающий фрагмент в составе внеклеточного домена представляет собой одноцепочечный Fab фрагмент или scFab фрагмент.
Fab и scFab фрагменты стабилизируются природной дисульфидной связью между CL доменом и СН1 доменом. Антигенсвязывающие фрагменты, включающие вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), такие как Fab, crossFab, scFv и scFab фрагменты, как описано в данном контексте, можно дополнительно стабилизировать за счет введения межцепочечных дисульфидных мостиков между VH и VL доменами. Соответственно, в одном варианте Fab фрагмент (фрагменты), crossFab фрагмент (фрагменты), scFv фрагмент (фрагменты) и/или scFab фрагмент (фрагменты) в составе антигенсвязывающих рецепторов по настоящему изобретению можно дополнительно стабилизировать за счет формирования межцепочечных дисульфидных связей посредством введения остатков цистеина (например, в положение 44 вариабельной области тяжелой цепи и в положение 100 вариабельной области легкой цепи согласно системе нумерации Кабата). В прилагаемых примерах и на фигурах такие стабилизированные антигенсвязывающие фрагменты обозначаются термином "ds".
Заякоренный трансмембранный домен
В настоящем изобретении заякоренный трансмембранный домен антигенсвязывающих рецепторов по настоящему изобретению может характеризоваться отсутствием сайта расщепления протеазами млекопитающих. В настоящем изобретении термин "протеазы" обозначает протеолитические ферменты, которые способны гидролизовать аминокислотную последовательность трансмембранного домена, включающую сайт расщепления протеазой. Термин "протеазы" включает как эндопептидазы, так и экзопептидазы. В настоящем изобретении любой заякоренный трансмембранный домен трансмембранного белка, определенный, среди прочих, в CD-номенклатуре, можно использовать для получения антигенсвязывающих рецепторов по изобретению, которые активируют Т-клетки, предпочтительно CD8 + Т-клетки, после связывания с мутированным Fc доменом, как описано в данном контексте.
Соответственно, в настоящем изобретении заякоренный трансмембранный домен может включать часть трансмембранного домена крысы/мыши или предпочтительно человека. Примером такого заякоренного трансмембранного домена является трансмембранный домен CD28, например, характеризующийся аминокислотной последовательностью, как описано в данном контексте последовательностью SEQ ID NO: 11 (кодируемой последовательностью ДНК SEQ ID NO: 24). В настоящем изобретении трансмембранный домен антигенсвязывающего рецептора по настоящему изобретению может включать/состоит из аминокислотной последовательности SEQ ID NO: 11 (кодируемой последовательностью ДНК SEQ ID NO: 24).
В иллюстративном варианте по настоящему изобретению в качестве подтверждения концепции предлагается антигенсвязывающий рецептор, включающий антигенсвязывающий фрагмент, содержащий аминокислотную последовательность SEQ ID NO: 10 (кодируемую последовательностью ДНК SEQ ID NO: 22), и фрагмент/часть полипептида CD28 (номер доступа в базе данных Uniprot для CD28 человека Р10747 (номер версии 173, версия последовательности 1)), как описано в данном контексте последовательностью SEQ ID NO: 71 (кодируемой последовательностью ДНК SEQ ID NO: 70). В другом варианте любой белок, содержащий трансмембранный домен, определенный, среди прочих, CD-номенклатуре, можно использовать в качестве заякоренного трансмембранного домена белкового антигенсвязывающего рецептора по изобретению. Как описано выше, представленный в данном контексте антигенсвязывающий рецептор может включать заякоренный трансмембранный домен CD28, который расположен в положениях аминокислотных остатков 153-179, 154-179, 155-179, 156-179, 157-179, 158-179, 159-179, 160-179, 161-179, 162-179, 163-179, 164-179, 165-179, 166-179, 167-179, 168-179, 169-179, 170-179, 171-179, 172-179, 173-179, 174-179, 175-179, 176-179, 177-179 или 178-179 полноразмерного белка CD28 человека с последовательностью SEQ ID NO: 71 (кодируемой кДНК с последовательностью ID NO: 70). Соответственно, в настоящем изобретении заякоренный трансмембранный домен может включать или состоит из аминокислотной последовательности SEQ ID NO: 11 (кодируемой последовательностью ДНК SEQ ID NO: 24).
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий Fab фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий рецептор включает:
(а) тяжелую цепь, включающую аминокислотную последовательность SEQ ID NO: 64, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17, и
(б) легкую цепь, включающую аминокислотную последовательность SEQ ID NO: 65.
В одном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий Fab фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает:
(а) тяжелую цепь, включающую аминокислотную последовательность, выбранную из SEQ ID NO: 40 и SEQ ID NO: 49, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17, и
(б) легкую цепь, включающую аминокислотную последовательность, выбранную из SEQ ID NO: 41 и SEQ ID NO: 50.
В одном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий Fab фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает:
(а) тяжелую цепь, включающую аминокислотную последовательность SEQ ID NO: 40, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17, и
(б) легкую цепь, включающую аминокислотную последовательность SEQ ID NO: 41.
В одном предпочтительном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий Fab фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает:
(а) тяжелую цепь, включающую аминокислотную последовательность SEQ ID NO: 49, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17, и
(б) легкую цепь, включающую аминокислотную последовательность SEQ ID NO: 50.
В одном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий scFv фрагмент, способный специфически связываться с Fc доменом, включающим мутации I253A, Н310А и Н435А, но не способный специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность SEQ ID NO: 60, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17.
В одном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий scFv фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, но не способный специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность, выбранную из SEQ ID NO: 10 и SEQ ID NO: 34, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17.
В одном предпочтительном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий scFv фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, но не способный специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность SEQ ID NO: 10, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17.
В одном варианте предлагается антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий scFab фрагмент, способный специфически связываться с Fc доменом, включающим мутацию P329G, но не способный специфически связываться с немутированным исходным Fc доменом, где scFv фрагмент включает аминокислотную последовательность SEQ ID NO: 34, присоединенную через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена с последовательностью SEQ ID NO: 11, необязательно через пептидный линкер с последовательностью SEQ ID NO: 17.
Стимулирующий сигнальный домен (SSD) и костимулирующий сигнальный домен (CSD)
Предпочтительно антигенсвязывающий рецептор по настоящему изобретению включает по меньшей мере один стимулирующий сигнальный домен и/или, по меньшей мере, один костимулирующий сигнальный домен. Соответственно, представленный в данном контексте антигенсвязывающий рецептор предпочтительно включает стимулирующий сигнальный домен, который обеспечивает активацию Т-клетки. Представленный в данном контексте антигенсвязывающий рецептор может включать стимулирующий сигнальный домен, который представляет собой фрагмент/часть полипептида CD3z крысы/мыши или человека (номер доступа в базе данных Uniprot для CD3z человека Р20963 (номер версии 177, номер последовательности 2, номер доступа в базе данных Uniprot для CD3z крысы/мыши Р24161 (первичный цитируемый номер доступа) или Q9D3G3 (вторичный цитируемый номер доступа), номер версии 143, номер последовательности 1)), FCGR3A (номер доступа в базе данных Uniprot для FCGR3A человека Р08637 (номер версии 178, номер последовательности 2)) или NKG2D (номер доступа в базе данных Uniprot для NKG2D человека Р26718 (номер версии 151, номер последовательности 1), номер доступа в базе данных Uniprot для NKG2D крысы/мыши 054709 (номер версии 132, номер последовательности 2)).
Таким образом, стимулирующий сигнальный домен, который содержится в представленном в данном контексте антигенсвязывающем рецепторе, может представлять собой фрагмент/часть полипептида полноразмерного CD3z, FCGR3A или NKG2D. Аминокислотные последовательности полноразмерного CD3z или NKG2D крысы/мыши, как описано в данном контексте, являются последовательностями SEQ ID NO: 96 (CD3z), SEQ ID NO: 100 (FCGR3A) или SEQ ID NO: 104 (NKG2D) (последовательности крысы/мыши кодируются последовательностями ДНК SEQ ID NO: 97 (CD3z), SEQ ID NO: 101 (FCGR3A) или SEQ ID NO: 105 (NKG2D). Аминокислотные последовательности полноразмерных CD3z, FCGR3A или NKG2D человека, как описано в данном контексте, являются последовательностями SEQ ID NO: 94 (CD3z), SEQ ID NO: 98 (FCGR3A) или SEQ ID NO: 102 (NKG2D) (последовательности человека кодируются последовательностями ДНК SEQ ID NO: 95 (CD3z), SEQ ID NO: 99 (FCGR3A) или SEQ ID NO: 103 (NKG2D)). Антигенсвязывающий рецептор по настоящему изобретению может включать фрагменты CD3z, FCGR3A или NKG2D в качестве стимулирующего домена, при условии, что они содержат, по меньшей мере, один сигнальный домен. Предпочтительно любая часть/фрагмент CD3z, FCGR3A или NKG2D пригодны для использования в качестве стимулирующего домена при условии, что они содержат, по меньшей мере, один сигнальный мотив. Однако более предпочтительно антигенсвязывающий рецептор по настоящему изобретению включает полипептиды человека. Таким образом, более предпочтительно представленный в данном контексте антигенсвязывающий рецептор включает аминокислотные последовательности, как описано в данном контексте, с последовательностями SEQ ID NO: 94 (CD3z), SEQ ID NO: 98 (FCGR3A) или SEQ ID NO: 102 (NKG2D) (последовательности человека кодируются последовательностями ДНК SEQ ID NO: 95 (CD3z), SEQ ID NO: 99 (FCGR3A) или SEQ ID NO: 103 (NKG2D)). Например, фрагмент/часть полипептида CD3z человека, которая может содержаться в антигенсвязывающем рецепторе по настоящему изобретению, может включать или состоит из аминокислотной последовательности SEQ ID NO: 13 (кодируемой последовательностью ДНК SEQ ID NO: 26). Соответственно, в одном варианте антигенсвязывающий рецептор включает последовательность SEQ ID NO: 13, или последовательность, которая содержит от 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 23, 24, 25, 26, 27, 28, 29 или 30 замен, делеций или вставок по сравнению с последовательностью SEQ ID NO: 13, и который характеризуется стимулирующей сигнальной активностью. Конкретные конфигурации антигенсвязывающих рецепторов, включающих стимулирующий сигнальный домен (SSD), представлены ниже в данном контексте, а также в примерах и на фигурах. Стимулирующую сигнальную активность можно определить, например, по повышению уровня высвобождения цитокинов, оцененного методом твердофазного иммуноферментного анализа (ИФА) (IL-2, IFNγ, TNFα), повышенной пролиферативной активности (определяют по увеличению числа клеток) или повышенной литической активности, определяемой методами анализа по высвобождению LDH.
Кроме того, представленный в данном контексте антигенсвязывающий рецептор предпочтительно включает по меньшей мере один костимулирующий сигнальный домен, который обеспечивает дополнительную активацию Т-клетки. Представленный в данном контексте антигенсвязывающий рецептор может включать костимулирующий сигнальный домен, который представляет собой фрагмент/часть полипептида CD28 крысы/мыши или человека (номер доступа в базе данных Uniprot для CD28 человека Р10747 (номер версии 173, номер последовательности 1), номер доступа в базе данных Uniprot для CD28 крысы/мыши Р31041 (номер версии 134, номер последовательности 2)), CD137 (номер доступа в базе данных Uniprot для CD137 человека Q07011 (номер версии 145, номер последовательности 1), номер доступа в базе данных Uniprot для CD137 крысы/мыши Р20334 (номер версии 139, номер последовательности 1)), ОХ40 (номер доступа в базе данных Uniprot для ОХ40 человека Р23510 (номер версии 138, номер последовательности 1), номер доступа в базе данных Uniprot для ОХ40 крысы/мыши Р43488 (номер версии 119, номер последовательности 1)), ICOS (номер доступа в базе данных Uniprot для ICOS человека Q9Y6W8 (номер версии 126, номер последовательности 1)), номер доступа в базе данных Uniprot для ICOS крысы/мыши Q9WV40 (первичный цитируемый номер доступа) или Q9JL17 (вторичный цитируемый номер доступа), номер версии 102, версия последовательности 2)), CD27 (номер доступа в базе данных Uniprot для CD27 человека Р26842 (номер версии 160, номер последовательности 2), номер доступа в базе данных Uniprot для CD27 крысы/мыши Р41272 (номер версии 137, версия последовательности 1)), 4-1-ВВ (номер доступа в базе данных Uniprot для 4-1-ВВ крысы/мыши Р20334 (номер версии 140, версия последовательности 1), номер доступа в базе данных Uniprot для 4-1-ВВ человека Q07011 (номер версии 146, версия последовательности)), DAP10 (номер доступа в базе данных Uniprot для DAP10 человека Q9UBJ5 (номер версии 25, номер последовательности 1), номер доступа в базе данных Uniprot для DAP10 крысы/мыши Q9QUJ0 (первичный цитируемый номер доступа) или Q9R1E7 (вторичный цитируемый номер доступа), номер версии 101, номер последовательности 1)) или DAP12 (номер доступа в базе данных Uniprot для DAP12 человека O43914 (номер версии 146, номер последовательности 1), номер доступа в базе данных Uniprot для DAP12 крысы/мыши O054885 (первичный цитируемый номер доступа) или Q9R1E7 (вторичный цитируемый номер доступа), номер версии 123, номер последовательности 1). В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий рецептор по настоящему изобретению может включать один или более, то есть 1, 2, 3, 4, 5, 6 или 7 описанных в данном контексте костимулирующих сигнальных доменов. Соответственно, в настоящем изобретении антигенсвязывающий рецептор по настоящему изобретению может включать, в качестве первого костимулирующего сигнального домена, фрагмент/часть полипептида CD28 крысы/мыши или предпочтительно человека, а второй костимулирующий сигнальный домен выбирают из группы, состоящей из CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12 крысы/мыши или предпочтительно человека, или их фрагментов. Предпочтительно антигенсвязывающий рецептор по настоящему изобретению включает костимулирующий сигнальный домен, полученный из организма человека. Таким образом, более предпочтительно костимулирующий сигнальный домен (домены), который включен в антигенсвязывающий рецептор по настоящему изобретению, включает или состоит из аминокислотной последовательности SEQ ID NO: 12 (кодируемой последовательностью ДНК SEQ ID NO: 25).
Таким образом, костимулирующий сигнальный домен, который необязательно включен в представленный в данном контексте антигенсвязывающий рецептор, представляет собой фрагмент/часть полноразмерного полипептида CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12. Аминокислотные последовательности полноразмерных CD27, CD28, CD137, ОХ40, ICOS, CD27, DAP10 или DAP12 крысы/мыши представлены в последовательностях SEQ ID NO: 69 (CD27), SEQ ID NO: 73 (CD28), SEQ ID NO: 77 (CD137), SEQ ID NO: 81 (OX40), SEQ ID NO: 85 (ICOS), SEQ ID NO: 89 (DAP10) или SEQ ID NO: 93 (DAP12) (полипептиды крысы/мыши кодируются последовательностями ДНК, представленными в последовательностях SEQ ID NO: 68 (CD27), SEQ ID NO: 72 (CD28), SEQ ID NO: 76 (CD137), SEQ ID NO: 80 (OX40), SEQ ID NO: 84 (ICOS), SEQ ID NO: 88 (DAP10) или SEQ ID NO: 92 (DAP12)). Однако, так как в настоящем изобретении последовательности полипептидов человека являются более предпочтительными, костимулирующий сигнальный домен, который необязательно включен в представленный в данном контексте антигенсвязывающий белковый рецептор, представляет собой фрагмент/часть полноразмерного CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 человека. Аминокислотные последовательности полноразмерного CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 человека в данном контексте представлены в последовательностях SEQ ID NO: 67 (CD27), SEQ ID NO: 71 (CD28), SEQ ID NO: 75 (CD137), SEQ ID NO: 79 (OX40), SEQ ID NO: 83 (ICOS), SEQ ID NO: 87 (DAP10) или SEQ ID NO: 91 (DAP12) (полипептиды человека кодируются последовательностями ДНК, представленными в последовательностях SEQ ID NO: 66 (CD27), SEQ ID NO: 70 (CD28), SEQ ID NO: 74 (CD137), SEQ ID NO: 78 (OX40), SEQ ID NO: 82 (ICOS), SEQ ID NO: 86 (DAP10) или SEQ ID NO: 90 (DAP12)).
В одном предпочтительном варианте, антигенсвязывающий рецептор включает CD28 или его фрагмент в качестве костимулирующего сигнального домена. Представленный в данном контексте антигенсвязывающий рецептор может включать фрагмент CD28 в качестве костимулирующего сигнального домена, при условии, что он включает по меньшей мере один сигнальный домен CD28. Предпочтительно любая часть/фрагмент CD28 пригодны для использования в качестве части антигенсвязывающего рецептора по изобретению при условии, что они содержат, по меньшей мере, один из сигнальных мотивов CD28. Например, полипептид CD28, который включен в антигенсвязывающий белковый рецептор по настоящему изобретению, может включать или состоит из аминокислотной последовательности SEQ ID NO: 12 (кодируемой последовательностью ДНК SEQ ID NO: 25). Согласно настоящему изобретению, внутриклеточный домен CD28, который проявляет активность костимулирующего сигнального домена, может включать последовательность (последовательности) из внутриклеточного домена полипептида CD28, YMNM (SEQ ID NO: 106) и/или PYAP (SEQ ID NO: 107). Предпочтительно антигенсвязывающий рецептор по настоящему изобретению включает полипептиды, которые получают из организма человека. Например, фрагмент/часть полипептида CD28 человека, который может быть включен в антигенсвязывающий рецептор по настоящему изобретению, может включать или состоит из аминокислотной последовательности SEQ ID NO: 12 (кодируемой последовательностью ДНК SEQ ID NO: 25). Соответственно, в настоящем изобретении антигенсвязывающий рецептор включает последовательность SEQ ID NO: 12, или последовательность, которая содержит от 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 замен, делеций или вставок по сравнению с последовательностью SEQ ID NO: 12, и который характеризуется костимулирующей сигнальной активностью. Конкретные конфигурации антигенсвязывающих рецепторов, включающих костимулирующий сигнальный домен (CSD), представлены ниже в данном контексте, а также в примерах и на фигурах. Костимулирующую сигнальную активность можно определить, например, по повышению уровня высвобождения цитокинов, по данным метода твердофазного иммуноферментного анализа (ИФА) (IL-2, IFNγ, TNFα), повышенной пролиферативной активности (по увеличению числа клеток) или повышенной литической активности, по данным метода оценки уровня высвобождения LDH.
В варианте осуществления настоящего изобретения, как описано выше, костимулирующий сигнальный домен антигенсвязывающего рецептора можно получить из гена CD28 человека (номер доступа в базе данных Uniprot Р10747 (номер версии 173, версия последовательности 1)) и проявляет CD28 активность, то есть продуцирование цитокинов, пролиферацию и литическую активность трансдуцированной клетки, как описано в данном контексте, например, трансдуцированной Т-клетки. Активность CD28 можно оценить по уровню высвобождения цитокинов методом ИФА или методом проточной цитометрии, определяя уровень цитокинов, таких как гамма-интерферон (IFN-γ) или интерлейкин 2 (IL-2), по уровню пролиферации Т-клеток, оцененного, например, по уровню маркера пролиферации ki67, числу клеток, по данным метода проточной цитометрии, или уровню литической активности, по данным измерения импеданса клеток-мишеней в режиме реального времени (с использованием анализатора ICELLligence, как описано, например, в статьях Thakur и др., Biosens Bioelectron., 35(1), 503-506 (2012), Krutzik и др., Methods Mol Biol., 699, 179-202 (2011), Ekkens и др., Infect Immun., 75(5), 2291-2296 (2007), Ge и др., Proc Natl Acad Sci USA., 99(5), 2983-2988 (2002),  и др., Cell Death Differ., 21(12), 1825-1837 (2014), Erratum, Cell Death Differ., 21(12), 161 (2014)). Костимулирующие сигнальные домены PYAP (AA 208-211 в последовательности SEQ ID NO: 107 и YMNM (AA 191-194 в последовательности SEQ ID NO: 106) оказывают благотворное действие на функцию полипептида CD28 и функциональные эффекты, описанные выше. Аминокислотная последовательность YMNM домена представлена в последовательности SEQ ID NO: 106, аминокислотная последовательность PYAP домена представлена в последовательности SEQ ID NO: 107. Соответственно, в антигенсвязывающем рецепторе по настоящему изобретению полипептид CD28 предпочтительно включает последовательность из внутриклеточного домена полипептида CD28 с последовательностями YMNM (SEQ ID NO: 106) и/или PYAP (SEQ ID NO: 107). В настоящем изобретении внутриклеточный домен полипептида CD28 с последовательностями YMNM (SEQ ID NO: 106) и/или PYAP (SEQ ID NO: 107), проявляет CD28 активность, то есть продуцирование цитокинов, пролиферацию и литическую активность трансдуцированной клетки, как описано в данном контексте, например, трансдуцированной Т-клетки. Соответственно, в настоящем изобретении костимулирующий сигнальный домен антигенсвязывающих рецепторов по настоящему изобретению, характеризуется аминокислотной последовательностью SEQ ID NO: 12 (человека) (кодируемую последовательностью ДНК SEQ ID NO: 25). Однако в антигенсвязывающем рецепторе по настоящему изобретению один или оба указанных домена могут представлять собой мутированные домены, такие как FMNM (SEQ ID NO: 108) и/или AYAA (SEQ ID NO: 109), соответственно. Любая из указанных мутаций снижает способность трансдуцированной клетки, содержащей антигенсвязывающий рецептор, высвобождать цитокины, не влияя на ее пролиферацию, и преимущество заключается в повышении жизнеспособности и, следовательно, терапевтической эффективности трансдуцированных клеток. Или, другими словами, указанная нефункциональная мутация предпочтительно повышает устойчивость клеток, которые трансдуцированы с помощью представленного в данном контексте антигенсвязывающего рецептора, in vivo. Однако указанные сигнальные мотивы могут присутствовать в любом сайте внутриклеточного домена антигенсвязывающего рецептора, представленного в данном контексте.
и др., Cell Death Differ., 21(12), 1825-1837 (2014), Erratum, Cell Death Differ., 21(12), 161 (2014)). Костимулирующие сигнальные домены PYAP (AA 208-211 в последовательности SEQ ID NO: 107 и YMNM (AA 191-194 в последовательности SEQ ID NO: 106) оказывают благотворное действие на функцию полипептида CD28 и функциональные эффекты, описанные выше. Аминокислотная последовательность YMNM домена представлена в последовательности SEQ ID NO: 106, аминокислотная последовательность PYAP домена представлена в последовательности SEQ ID NO: 107. Соответственно, в антигенсвязывающем рецепторе по настоящему изобретению полипептид CD28 предпочтительно включает последовательность из внутриклеточного домена полипептида CD28 с последовательностями YMNM (SEQ ID NO: 106) и/или PYAP (SEQ ID NO: 107). В настоящем изобретении внутриклеточный домен полипептида CD28 с последовательностями YMNM (SEQ ID NO: 106) и/или PYAP (SEQ ID NO: 107), проявляет CD28 активность, то есть продуцирование цитокинов, пролиферацию и литическую активность трансдуцированной клетки, как описано в данном контексте, например, трансдуцированной Т-клетки. Соответственно, в настоящем изобретении костимулирующий сигнальный домен антигенсвязывающих рецепторов по настоящему изобретению, характеризуется аминокислотной последовательностью SEQ ID NO: 12 (человека) (кодируемую последовательностью ДНК SEQ ID NO: 25). Однако в антигенсвязывающем рецепторе по настоящему изобретению один или оба указанных домена могут представлять собой мутированные домены, такие как FMNM (SEQ ID NO: 108) и/или AYAA (SEQ ID NO: 109), соответственно. Любая из указанных мутаций снижает способность трансдуцированной клетки, содержащей антигенсвязывающий рецептор, высвобождать цитокины, не влияя на ее пролиферацию, и преимущество заключается в повышении жизнеспособности и, следовательно, терапевтической эффективности трансдуцированных клеток. Или, другими словами, указанная нефункциональная мутация предпочтительно повышает устойчивость клеток, которые трансдуцированы с помощью представленного в данном контексте антигенсвязывающего рецептора, in vivo. Однако указанные сигнальные мотивы могут присутствовать в любом сайте внутриклеточного домена антигенсвязывающего рецептора, представленного в данном контексте.
Линкер и сигнальные пептиды
Представленный в данном контексте антигенсвязывающий рецептор, кроме того, может включать по меньшей мере один линкер (или "спейсер"). Линкер обычно представляет собой пептид, включающий до 20 аминокислотных остатков. Соответственно, в настоящем изобретении линкер может включать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислотных остатков. Например, представленный в данном контексте антигенсвязывающий рецептор может включать линкер между внеклеточным доменом, включающим по меньшей мере один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, заякоренным трансмембранным доменом, костимулирующим сигнальным доменом и/или стимулирующим сигнальным доменом. Преимущество указанных линкеров заключается в том, что они повышают вероятность того, что различные полипептиды антигенсвязывающего рецептора (то есть внеклеточный домен, включающий по меньшей мере один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, заякоренный трансмембранный домен, костимулирующий сигнальный домен и/или стимулирующий сигнальный домен) независимо сворачиваются в определенную структуру и характеризуются ожидаемым действием. Таким образом, в настоящем изобретении внеклеточный домен, включающий по меньшей мере один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, заякоренный трансмембранный домен, который не содержит сайт расщепления протеазами млекопитающих, костимулирующий сигнальный домен и стимулирующий сигнальный домен, может быть включен в одноцепочечный полифункциональный полипептид. Одноцепочечный гибридный конструкт, например, может состоять из полипептида (полипептидов), включающего внеклеточный домен (домены), содержащий, по меньшей мере, один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, заякоренный трансмембранный домен (домены), костимулирующий сигнальный домен (домены) и/или стимулирующий сигнальный домен (домены). В других вариантах антигенсвязывающий рецептор включает антигенсвязывающий фрагмент, который не является одноцепочечным гибридным конструктом, то есть антигенсвязывающий фрагмент представляет собой Fab или crossFab фрагмент. В таких вариантах антигенсвязывающий рецептор не является одноцепочечным гибридным конструктом, включающим только одну полипептидную цепь. Предпочтительно такие конструкты будут включать одноцепочечный гибридный полипептид тяжелой цепи, соединенный с легкой цепью иммуноглобулина, как описано в данном контексте, например, гибридный полипептид тяжелой цепи включает тяжелую цепь (цепи) иммуноглобулина, заякоренный трансмембранный домен (домены), костимулирующий сигнальный домен (домены) и/или стимулирующий сигнальный домен (домены) и соединен с легкой цепью (цепями) иммуноглобулина. Соответственно, антигенсвязывающий фрагмент, заякоренный трансмембранный домен, костимулирующий сигнальный домен и стимулирующий сигнальный домен могут быть соединены с использованием одного или более одинаковых или различных пептидных линкеров, как описано в данном контексте. Например, в представленном в данном контексте антигенсвязывающем рецепторе линкер между внеклеточным доменом, включающим по меньшей мере один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, и заякоренным трансмембранным доменом может включать аминокислотную последовательность SEQ ID NO: 17 или состоять из нее. Соответственно, заякоренный трансмембранный домен, костимулирующий сигнального домен и/или стимулирующий домен могут быть соединены друг с другом через пептидные линкеры или в другом варианте домены соединены непосредственно друг с другом.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий фрагмент, содержащийся во внеклеточном домене, представляет собой одноцепочечный вариабельный фрагмент (scFv), который является гибридным белком вариабельных областей тяжелой (VH) и легкой (VL) цепей антитела, соединенный с коротким пептидным линкером, который содержит от 10 до приблизительно 25 аминокислотных остатков. Линкер обычно обогащен остатками глицина, что обеспечивает гибкость, а также серином или треонином для улучшения растворимости, и может соединять N-концевой остаток VH с С-концевым остатком VL или наоборот. Например, в представленном в данном контексте антигенсвязывающем рецепторе линкер может характеризоваться аминокислотной последовательностью SEQ ID NO: 16. scFv антигенсвязывающий фрагмент, как описано в данном контексте, сохраняет специфичность исходного антитела, несмотря на удаление консервативных областей и введение линкера. scFv антитела описаны, например, в статье Houston J.S., Methods in Enzymol., 203, 46-96 (1991).
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий фрагмент, включенный во внеклеточный домен, представляет собой одноцепочечный Fab или scFab фрагмент, который является полипептидом, состоящим из вариабельного домена тяжелой цепи (VH), консервативного домена 1 (СН1) антитела, вариабельного домена легкой цепи (VL) антитела, консервативного домена легкой цепи (CL) антитела и линкера, где указанные домены антитела и указанный линкер расположены в одном из следующих порядков в направлении от N-концевого остатка к С-концевому остатку: а) VH-СН1-линкер-VL-CL, б) VL-CL-линкер-VH-СН1, в) VH-CL-линкер-VL-CH1 или г) VL-СН1-линкер-VH-CL, и, где указанный линкер представляет собой полипептид, включающий по меньшей мере 30 аминокислотных остатков, предпочтительно от 32 до 50 аминокислотных остатков. Указанные одноцепочечные Fab фрагменты стабилизируют с использованием природной дисульфидной связи между CL доменом и СН1 доменом.
В некоторых вариантах осуществления настоящего изобретения антигенсвязывающий фрагмент, содержащийся во внеклеточном домене, представляет собой кроссоверный одноцепочечный Fab фрагмент, который является полипептидом, состоящим из вариабельного домена тяжелой цепи (VH) антитела, консервативного домена 1 (CH1) антитела, вариабельного домена легкой цепи (VL) антитела, консервативного домена легкой цепи (CL) антитела и линкера, где указанные домены антитела и указанный линкер расположены в одном из следующих порядков в направлении от N-концевого остатка к С-концевому остатку: а) VH-CL-линкер-VL-CH1 и б) VL-CH1-линкер-VH-CL, где VH и VL вместе образуют антигенсвязывающий сайт, который специфически связывается с антигеном, и где указанный линкер представляет собой полипептид, содержащий по меньшей мере 30 аминокислотных остатков.
Представленный в данном контексте антигенсвязывающий рецептор или его части могут включать сигнальный пептид. Такой сигнальный пептид обеспечивает доставку белка на поверхность мембраны Т-клетки. Например, в представленном в данном контексте антигенсвязывающем рецепторе сигнальный пептид может характеризоваться аминокислотной последовательностью SEQ ID NO: 110 (кодируемой последовательностью ДНК SEQ ID NO: 111).
Активирующие Т-клетки антигенсвязывающие рецепторы, способные специфически связываться с мутированными Fc доменами
Компоненты антигенсвязывающих рецепторов, как описано в данном контексте, могут быть соединены друг с другом в различных конфигурациях для получения активирующих Т-клетки антигенсвязывающих рецепторов.
В некоторых вариантах антигенсвязывающий рецептор включает внеклеточный домен, состоящий из вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL), соединенных с заякоренный трансмембранным доменом. В некоторых вариантах VH домен присоединен через С-концевой остаток к N-концевому остатку VL домена, необязательно через пептидный линкер. В других вариантах антигенсвязывающий рецептор дополнительно включает стимулирующий сигнальный домен и/или костимулирующий сигнальный домен. В таком конкретном варианте антигенсвязывающий рецептор в основном состоит из VH домена и VL домена, заякоренного трансмембранного домена и необязательно стимулирующего сигнальный домена, соединенных через один или более пептидных линкеров, где VH домен присоединен через С-концевой остаток к N-концевому остатку домена VL, и VL домен присоединен через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена, где заякоренный трансмембранный домен присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена. Необязательно антигенсвязывающий рецептор дополнительно включает костимулирующий сигнальный домен. В одном таком конкретном варианте антигенсвязывающий рецептор в основном состоит из VH домена и VL домена, заякоренного трансмембранного домена, стимулирующего сигнальный домена и костимулирующего сигнального домена, соединенных через один или более пептидных линкеров, где VH домен присоединен через С-концевой остаток к N-концевому остатку VL домена, и VL домен присоединен через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена, где заякоренный трансмембранный домен присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена, где стимулирующий сигнальный домен присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена. В другом варианте костимулирующий сигнальный домен соединен с заякоренным трансмембранным доменом, а не со стимулирующим сигнальным доменом. В предпочтительном варианте антигенсвязывающий рецептор в основном состоит из VH домена и VL домена, заякоренного трансмембранного домена, костимулирующего сигнального домена и стимулирующего сигнального домена, соединенных через один или более пептидных линкеров, где VH домен присоединен через С-концевой остаток к N-концевому остатку VL домена, и VL домен присоединен через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена, где заякоренный трансмембранный домен присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена, где костимулирующий сигнальный домен присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнальный домена.
В предпочтительных вариантах один из связывающих фрагментов представляет собой Fab фрагмент или crossFab фрагмент. В одном предпочтительном варианте антигенсвязывающий фрагмент присоединен через С-концевой остаток тяжелой цепи Fab или crossFab к N-концевому остатку заякоренного трансмембранного домена, необязательно через пептидный линкер. В другом варианте, антигенсвязывающий фрагмент присоединен через С-концевой остаток легкой цепи Fab или crossFab к N-концевому остатку заякоренного трансмембранного домена, необязательно через пептидный линкер. В других вариантах антигенсвязывающий рецептор дополнительно включает стимулирующий сигнальный домен и/или костимулирующий сигнальный домен. В таком конкретном варианте антигенсвязывающий рецептор в основном состоит из Fab или crossFab фрагмента, заякоренного трансмембранного домена и необязательно стимулирующего сигнального домена, соединенных через один или более пептидных линкеров, где Fab или crossFab фрагмент присоединен через С-концевой остаток тяжелой или легкой цепи к N-концевому остатку заякоренного трансмембранного домена, где заякоренный трансмембранный домен присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена. Предпочтительно антигенсвязывающий рецептор дополнительно включает костимулирующий сигнальный домен. В одном из указанных вариантов антигенсвязывающий рецептор в основном состоит из Fab или crossFab фрагмента, заякоренного трансмембранного домена, стимулирующего сигнального домена и костимулирующего сигнального домена, соединенных через один или более пептидных линкеров, где Fab или crossFab фрагмент присоединен через С-концевой остаток тяжелой или легкой цепи к N-концевому остатку заякоренного трансмембранного домена, где стимулирующий сигнальный домен присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена. В предпочтительном варианте костимулирующий сигнальный домен соединен с заякоренным трансмембранным доменом вместо стимулирующего сигнального домена. В более предпочтительном варианте антигенсвязывающий рецептор в основном состоит из Fab или crossFab фрагмента, заякоренного трансмембранного домена, костимулирующего сигнального домена и стимулирующего сигнального домена, где Fab или crossFab фрагмент присоединен через С-концевой остаток тяжелой цепи к N-концевому остатку заякоренного трансмембранного домена через пептидный линкер, где заякоренный трансмембранный домен присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена, где костимулирующий сигнальный домен присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена.
Антигенсвязывающий фрагмент, заякоренный трансмембранный домен и стимулирующий сигнальный и/или костимулирующий сигнальный домены соединены друг с другом напрямую или через один или более пептидных линкеров, включающих один или более аминокислотных остатков, обычно приблизительно 2-20 аминокислотных остатков. Пептидные линкеры известны в данной области техники и описаны в данном контексте. Пригодные неиммуногенные пептидные линкеры включают, например, (G4S)n, (SG4)n, (G4S)n или G4(SG4)n пептидные линкеры, где "n" в основном обозначает число от 1 до 10, обычно от 2 до 4. Предпочтительным пептидным линкером для соединения антигенсвязывающего фрагмента и заякоренного трансмембранного фрагмента является линкер GGGGS (G4S), характеризующийся последовательностью SEQ ID NO 17. Примером пептидного линкера, пригодного для соединения вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи является линкер GGGSGGGSGGGSGGGS (G4S)4, характеризующийся последовательностью SEQ ID NO 16.
Линкеры могут дополнительно содержать шарнирную область (часть) иммуноглобулина. Предпочтительно, если антигенсвязывающий фрагмент присоединен к N-концевому остатку заякоренного трансмембранного домена, он может быть присоединен через шарнирную область иммуноглобулина или ее часть, с использованием дополнительного пептидного линкера или без него.
Как описано в данном контексте, антигенсвязывающие рецепторы по настоящему изобретению включают внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент. Антигенсвязывающий рецептор с одним антигенсвязывающим фрагментом, способным специфически связываться с антигеном клетки-мишени, является пригодным и предпочтительным прежде всего в случаях, когда требуется высокая экспрессия антигенсвязывающего рецептора. В таких случаях присутствие более одного антигенсвязывающего фрагмента, специфичного в отношении антигена клетки-мишени, может ограничить эффективность экспрессии антигенсвязывающего рецептора. Однако, в других случаях, выгодно использовать антигенсвязывающий рецептор, содержащий два или более антигенсвязывающих фрагмента, специфичных в отношении антигена клетки-мишени, например, для оптимизации воздействия, направленного к сайту-мишени, или для обеспечения сшивания антигенов клетки-мишени.
В одном конкретном варианте антигенсвязывающий рецептор включает один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, предпочтительно Fc доменом IgG1, содержащим мутацию P329G. В одном варианте антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с немутированным исходным Fc доменом, представляет собой scFv, Fab или crossFab.
В одном варианте антигенсвязывающий фрагмент присоединен через С-концевой остаток scFv фрагмента или через С-концевой остаток тяжелой цепи Fab или crossFab к N-концевому остатку заякоренного трансмембранного домена, необязательно через пептидный линкер. В одном варианте пептидный линкер включает аминокислотную последовательность GGGGS (SEQ ID NO: 16). В одном варианте заякоренный трансмембранный домен представляет собой трансмембранный домен, который выбирают из группы, состоящей из трансмембранного домена CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 или его фрагмента. В предпочтительном варианте заякоренным трансмембранным доменом является трансмембранный домен CD28 или его фрагмент. В конкретном варианте заякоренный трансмембранный домен включает или состоит из аминокислотной последовательности FWVLVVVGGVLACYSLLVTVAFIIFWV (SEQ ID NO: 11). В одном варианте антигенсвязывающий рецептор дополнительно включает костимулирующий сигнальный домен (CSD). В одном варианте заякоренный трансмембранный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена. В одном варианте костимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12 или их фрагментов, как описано выше в данном контексте. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD28 или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO: 12). В одном варианте антигенсвязывающий рецептор дополнительно включает стимулирующий сигнальный домен. В одном варианте костимулирующий сигнальный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена. В одном варианте, по меньшей мере, один стимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO: 13).
В одном варианте осуществления настоящего изобретения антигенсвязывающий рецептор присоединен к репортерному белку, прежде всего к GFP или его усиленным аналогам. В одном варианте антигенсвязывающий рецептор присоединен через С-концевой остаток к N-концевому остатку eGFP (усиленный зеленый флуоресцентный белок), необязательно через пептидный линкер, как описано в данном контексте. В предпочтительном варианте пептидный линкер представляет собой пептид GEGRGSLLTCGDVEENPGP (Т2А), представленный в последовательности SEQ ID NO: 18.
В конкретном варианте антигенсвязывающий рецептор включает заякоренный трансмембранный домен и внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент, где по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает мутацию P329G. Мутация P329G снижает связывание с рецептором Fcγ. В одном варианте антигенсвязывающий рецептор по изобретению включает заякоренный трансмембранный домен (ATD), костимулирующий сигнальный домен (CSD) и стимулирующий сигнальный домен (SSD). В одном из указанных вариантов антигенсвязывающий рецептор характеризуется конфигурацией scFv-ATD-CSD-SSD. В предпочтительном варианте антигенсвязывающий рецептор характеризуется конфигурацией scFv-G4S-ATD-CSD-SSD, где G4S представляет собой линкер, включающий последовательность GGGGS SEQ ID NO: 17. Репортерный белок необязательно присоединен к С-концевому остатку антигенсвязывающего рецептора, необязательно через пептидный линкер.
В конкретном варианте антигенсвязывающий фрагмент представляет собой scFv фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает по меньшей мере одну определяющую комплементарность область тяжелой цепи (CDR), которую выбирают из группы, состоящей из последовательности SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, и, по меньшей мере, одну CDR легкой цепи, выбранную из группы, включающей последовательность SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой scFv, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью RYWMN (SEQ ID NO: 1), CDR Н2 с аминокислотной последовательностью EITPDSSTINYTPSLKD (SEQ ID NO: 2), CDR Н3 с аминокислотной последовательностью PYDYGAWFAS (SEQ ID NO: 3), определяющую комплементарность область 1 легкой цепи (CDR L) с аминокислотной последовательностью RSSTGAVTTSNYAN (SEQ ID NO: 4), CDR L2 с аминокислотной последовательностью GTNKRAP (SEQ ID NO: 5) и CDR L3 с аминокислотной последовательностью ALWYSNHWV (SEQ ID NO: 6).
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой scFv фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где scFv фрагмент включает вариабельную область (VH) тяжелой цепи, содержащую определяющую комплементарность область 1 тяжелой цепи (CDR) с последовательностью SEQ ID NO: 1, CDR 2 тяжелой цепи с последовательностью SEQ ID NO: 2, CDR 3 тяжелой цепи с последовательностью SEQ ID NO: 3, и вариабельную область (VH) легкой цепи, содержащую CDR 1 легкой цепи с последовательностью SEQ ID NO: 4, CDR 2 легкой цепи с последовательностью SEQ ID NO: 5 и CDR 3 легкой цепи с последовательностью SEQ ID NO: 6,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу scFv, способную специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где scFv включает вариабельный домен тяжелой цепи (VH), выбранный из последовательности SEQ ID NO: 8 и SEQ ID NO: 32, и вариабельный домен легкой цепи (VL), выбранный из последовательности SEQ ID NO: 9 и SEQ ID NO: 33,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В предпочтительном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу scFv, способную специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где scFv включает вариабельный домен тяжелой цепи (VH) с последовательностью SEQ ID NO: 8, и вариабельный домен легкой цепи (VL) с последовательностью SEQ ID NO: 9,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В предпочтительном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу scFv, способную специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где scFv включает аминокислотную последовательность SEQ ID NO: 10 или SEQ ID NO: 34,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В конкретном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 31.
В предпочтительном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает аминокислотную последовательность SEQ ID NO: 31
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент. В одном варианте антигенсвязывающий фрагмент присоединен через С-концевой аминокислотный остаток тяжелой цепи Fab к N-концевому аминокислотному остатку заякоренного трансмембранного домена. В одном варианте заякоренный трансмембранный домен представляет собой трансмембранный домен, который выбирают из группы, состоящей из трансмембранного домена CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 или его фрагмента. В предпочтительном варианте заякоренным трансмембранным доменом является CD28 трансмембранный домен или его фрагмент. В конкретном варианте заякоренным трансмембранным доменом является последовательность FWVLVVVGGVLACYSLLVTVAFIIFWV (SEQ ID NO: 11). В одном варианте антигенсвязывающий рецептор дополнительно включает костимулирующий сигнальный домен (CSD). В одном варианте заякоренный трансмембранный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена. В одном варианте костимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12 или их фрагментов, как описано выше в данном контексте. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD28 или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO: 12). В одном варианте антигенсвязывающий рецептор дополнительно включает стимулирующий сигнальный домен. В одном варианте костимулирующий сигнальный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена. В одном варианте, по меньшей мере, один стимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO: 13).
В одном варианте антигенсвязывающий рецептор присоединен к репортерному белку, прежде всего к GFP или его усиленным аналогам. В одном варианте антигенсвязывающий рецептор присоединен через С-концевой остаток к N-концевому остатку eGFP (усиленный зеленый флуоресцентный белок), необязательно через пептидный линкер, как описано в данном контексте. В предпочтительном варианте пептидный линкер представляет собой последовательность GEGRGSLLTCGDVEENPGP (Т2А) SEQ ID NO: 18.
В конкретном варианте антигенсвязывающий рецептор включает заякоренный трансмембранный домен и внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент, где по меньшей мере один антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает мутацию P329G, где мутация P329G снижает связывание с Fcγ рецептором. В одном варианте антигенсвязывающий рецептор по изобретению включает заякоренный трансмембранный домен (ATD), костимулирующий сигнальный домен (CSD) и стимулирующий сигнальный домен (SSD). В одном из указанных вариантов антигенсвязывающий рецептор характеризуется конфигурацией Fab-ATD-CSD-SSD. В предпочтительном варианте антигенсвязывающий рецептор характеризуется конфигурацией Fab-G4S-ATD-CSD-SSD, где G4S представляет собой линкер, включающий последовательность GGGGS SEQ ID NO: 17. Репортерный белок необязательно присоединен к С-концевому остатку антигенсвязывающего рецептора, необязательно через пептидный линкер.
В конкретном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент представляет собой Fab фрагмент, включающий по меньшей мере одну определяющую комплементарность область тяжелой цепи (CDR), которую выбирают из группы, состоящей из последовательности SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, а также по меньшей мере одну CDR легкой цепи, выбранную из группы, включающей последовательность SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий фрагмент включает определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью RYWMN (SEQ ID NO: 1), CDR Н2 с аминокислотной последовательностью EITPDSSTINYTPSLKD (SEQ ID NO: 2), CDR Н3 с аминокислотной последовательностью PYDYGAWFAS (SEQ ID NO: 3), определяющую комплементарность область 1 легкой цепи (CDR L) с аминокислотной последовательностью RSSTGAVTTSNYAN (SEQ ID NO: 4), CDR L2 с аминокислотной последовательностью GTNKRAP (SEQ ID NO: 5) и CDR L3 с аминокислотной последовательностью ALWYSNHWV (SEQ ID NO: 6).
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу Fab, способную специфически связываться с мутированным Fc доменом, включающим мутацию P329G, содержащий определяющую комплементарность область 1 тяжелой цепи (CDR) с последовательностью SEQ ID NO: 1, CDR 2 тяжелой цепи с последовательностью SEQ ID NO: 2, CDR 3 тяжелой цепи с последовательностью SEQ ID NO: 3, CDR 1 легкой цепи с последовательностью SEQ ID NO: 4, CDR 2 легкой цепи с последовательностью SEQ ID NO: 5 и CDR 3 легкой цепи с последовательностью SEQ ID NO: 6,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий:
а) гибридный полипептид тяжелой цепи, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) тяжелую цепь, которая включает определяющую комплементарность область 1 тяжелой цепи (CDR) с последовательностью SEQ ID NO: 1, CDR 2 тяжелой цепи с последовательностью SEQ ID NO: 2, CDR 3 тяжелой цепи с последовательностью SEQ ID NO: 3,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13, а также
б) легкую цепь, которая включает CDR 1 легкой цепи с последовательностью SEQ ID NO: 4, CDR 2 легкой цепи с последовательностью SEQ ID NO: 5 и CDR 3 легкой цепи с последовательностью SEQ ID NO: 6.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий:
а) гибридный полипептид тяжелой цепи, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) вариабельный домен тяжелой цепи (VH) с последовательностью SEQ ID NO: 8,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13, а также
б) вариабельный домен легкой цепи (VL) с последовательностью SEQ ID NO: 9.
В одном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, содержащий тяжелую цепь, включающую или состоящую из аминокислотной последовательности SEQ ID NO: 40 или SEQ ID NO: 49, и легкую цепь, включающую или состоящую из аминокислотной последовательности SEQ ID NO: 41 или SEQ ID NO: 50. В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, содержащий тяжелую цепь, включающую или состоящую из аминокислотной последовательности SEQ ID NO: 40, а также легкую цепь, включающую или состоящую из аминокислотной последовательности SEQ ID NO: 41.
В конкретном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает: гибридный полипептид тяжелой цепи, включающий аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, которую выбирают из группы, включающей последовательность SEQ ID NO: 39 и SEQ ID NO: 48, а также полипептид легкой цепи, включающий аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, которую выбирают из группы, включающей последовательность SEQ ID NO: 41 и SEQ ID NO: 50.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, где антигенсвязывающий рецептор включает: гибридный полипептид тяжелой цепи, включающий аминокислотную последовательность SEQ ID NO: 39, а также полипептид легкой цепи, включающий аминокислотную последовательность SEQ ID NO: 41.
В другом варианте антигенсвязывающий рецептор включает один антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, предпочтительно Fc доменом IgG1, включающим мутации I253A, Н310А и Н435А ("AAA"), В одном варианте антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с немутированным исходным Fc доменом, представляет собой scFv, Fab или crossFab.
В одном варианте антигенсвязывающий фрагмент присоединен через С-концевой остаток scFv фрагмента или через С-концевой остаток тяжелой цепи Fab или crossFab к N-концевому остатку заякоренного трансмембранного домена, необязательно через пептидный линкер. В одном варианте осуществления настоящего изобретения пептидный линкер включает аминокислотную последовательность GGGGS (SEQ ID NO: 16). В одном варианте заякоренный трансмембранный домен представляет собой трансмембранный домен, который выбирают из группы, состоящей из трансмембранного домена CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 или его фрагмента. В предпочтительном варианте заякоренный трансмембранный домен представляет собой CD28 трансмембранный домен или его фрагмент. В конкретном варианте заякоренный трансмембранный домен включает или состоит из аминокислотной последовательности FWVLVVVGGVLACYSLLVTVAFIIFWV (SEQ ID NO: 11). В одном варианте антигенсвязывающий рецептор дополнительно включает костимулирующий сигнальный домен (CSD). В одном варианте заякоренный трансмембранный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена. В одном варианте костимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12 или их фрагментов, как описано выше в данном контексте. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD28 или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO: 12). В одном варианте антигенсвязывающий рецептор дополнительно включает стимулирующий сигнальный домен. В одном варианте костимулирующий сигнальный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнального домена. В одном варианте, по меньшей мере, один стимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO: 13).
В одном варианте антигенсвязывающий рецептор присоединен к репортерному белку, прежде всего к GFP или его усиленным аналогам. В одном варианте антигенсвязывающий рецептор присоединен через С-концевой остаток к N-концевому остатку eGFP (усиленный зеленый флуоресцентный белок), необязательно через пептидный линкер, как описано в данном контексте. В предпочтительном варианте пептидный линкер представляет собой последовательность GEGRGSLLTCGDVEENPGP (Т2А) SEQ ID NO: 18.
В конкретном варианте антигенсвязывающий рецептор включает заякоренный трансмембранный домен и внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент, где по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает мутации I253A, Н310А и Н435А. Мутации I253A, Н310А и Н435А снижают связывание с FcRn рецептором. В одном варианте антигенсвязывающий рецептор по изобретению включает заякоренный трансмембранный домен (ATD), костимулирующий сигнальный домен (CSD) и стимулирующий сигнальный домен (SSD). В одном из указанных вариантов антигенсвязывающий рецептор характеризуется конфигурацией scFv-ATD-CSD-SSD. В предпочтительном варианте антигенсвязывающий рецептор характеризуется конфигурацией scFv-G4S-ATD-CSD-SSD, где G4S представляет собой линкер, включающий последовательность GGGGS SEQ ID NO: 17. Репортерный белок необязательно присоединен к С-концевому остатку антигенсвязывающего рецептора, необязательно через пептидный линкер.
В конкретном варианте антигенсвязывающий фрагмент представляет собой scFv фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает по меньшей мере одну определяющую комплементарность область тяжелой цепи (CDR), которую выбирают из группы, состоящей из последовательности SEQ ID NO: 53, SEQ ID NO: 54 и SEQ ID NO: 55, а также по меньшей мере одну CDR легкой цепи, выбранную из группы, включающей последовательность SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой scFv, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает определяющую комплементарность область 1 (CDR Н) с аминокислотной последовательностью SYGMS (SEQ ID NO: 53), CDR Н2 с аминокислотной последовательностью SSGGSY (SEQ ID NO: 54), CDR Н3 с аминокислотной последовательностью LGMITTGYAMDY (SEQ ID NO: 55), определяющую комплементарность область 1 легкой цепи (CDR L) с аминокислотной последовательностью RSSQTIVHSTGHTYLE (SEQ ID NO: 56), CDR L2 с аминокислотной последовательностью KVSNRFS (SEQ ID NO: 57) и CDR L3 с аминокислотной последовательностью FQGSHVPYT (SEQ ID NO: 58).
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой scFv фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где scFv фрагмент включает вариабельную область (VH) тяжелой цепи, содержащую определяющую комплементарность область 1 тяжелой цепи (CDR) с последовательностью SEQ ID NO: 53, CDR 2 тяжелой цепи с последовательностью SEQ ID NO: 54, CDR 3 тяжелой цепи с последовательностью SEQ ID NO: 55, а также вариабельную область (VH) легкой цепи, содержащую CDR 1 легкой цепи с последовательностью SEQ ID NO: 56, CDR 2 легкой цепи с последовательностью SEQ ID NO: 57 и CDR 3 легкой цепи с последовательностью SEQ ID NO: 58,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу scFv, способную специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где scFv включает вариабельный домен тяжелой цепи (VH) с последовательностью SEQ ID NO: 61, и вариабельный домен легкой цепи (VL) с последовательностью SEQ ID NO: 62,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу scFv, способную специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где scFv включает аминокислотную последовательность SEQ ID NO: 60,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В конкретном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий рецептор включает аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 59.
В предпочтительном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий рецептор включает аминокислотную последовательность SEQ ID NO: 595.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент. В одном варианте антигенсвязывающий фрагмент присоединен через С-концевой аминокислотный остаток тяжелой цепи Fab к N-концевому аминокислотному остатку заякоренного трансмембранного домена. В одном варианте заякоренный трансмембранный домен представляет собой трансмембранный домен, который выбирают из группы, состоящей из трансмембранного домена CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 или его фрагмента. В предпочтительном варианте заякоренным трансмембранным доменом является CD28 трансмембранный домен или его фрагмент. В конкретном варианте заякоренным трансмембранным доменом является последовательность FWVLVVVGGVLACYSLLVTVAFIIFWV (SEQ ID NO: 11). В одном варианте антигенсвязывающий рецептор дополнительно включает костимулирующий сигнальный домен (CSD). В одном варианте заякоренный трансмембранный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку костимулирующего сигнального домена. В одном варианте костимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12 или их фрагментов, как описано выше в данном контексте. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD28 или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS (SEQ ID NO: 12). В одном варианте антигенсвязывающий рецептор дополнительно включает стимулирующий сигнальный домен. В одном варианте костимулирующий сигнальный домен антигенсвязывающего рецептора присоединен через С-концевой остаток к N-концевому остатку стимулирующего сигнал домена. В одном варианте, по меньшей мере, один стимулирующий сигнального домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов. В предпочтительном варианте костимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z или его фрагмент. В конкретном варианте костимулирующий сигнальный домен включает или состоит из последовательности RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR (SEQ ID NO: 13).
В одном варианте антигенсвязывающий рецептор присоединен к репортерному белку, прежде всего к GFP или его усиленным аналогам. В одном варианте антигенсвязывающий рецептор присоединен через С-концевой остаток к N-концевому остатку eGFP (усиленный зеленый флуоресцентный белок), необязательно через пептидный линкер, как описано в данном контексте. В предпочтительном варианте пептидный линкер представляет собой последовательность GEGRGSLLTCGDVEENPGP (Т2А) SEQ ID NO: 18.
В конкретном варианте антигенсвязывающий рецептор включает заякоренный трансмембранный домен и внеклеточный домен, содержащий, по меньшей мере, один антигенсвязывающий фрагмент, где по меньшей мере один антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, но не способный специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает мутации I253A, Н310А и Н435А, где мутации I253A, Н310А и Н435А снижают связывание с FcRn рецептором. В одном варианте антигенсвязывающий рецептор по изобретению включает заякоренный трансмембранный домен (ATD), костимулирующий сигнальный домен (CSD) и стимулирующий сигнальный домен (SSD). В одном из указанных вариантов антигенсвязывающий рецептор характеризуется конфигурацией Fab-ATD-CSD-SSD. В предпочтительном варианте антигенсвязывающий рецептор характеризуется конфигурацией Fab-G4S-ATD-CSD-SSD, где G4S представляет собой линкер, включающий последовательность GGGGS SEQ ID NO: 17. Репортерный белок необязательно присоединен к С-концевому остатку антигенсвязывающего рецептора, необязательно через пептидный линкер.
В конкретном варианте антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент представляет собой Fab фрагмент, включающий по меньшей мере одну определяющую комплементарность область тяжелой цепи (CDR), которую выбирают из группы, состоящей из последовательности SEQ ID NO: 53, SEQ ID NO: 54 и SEQ ID NO: 55, а также по меньшей мере одну CDR легкой цепи, которую выбирают из группы, включающей последовательность SEQ ID NO: 56, SEQ ID NO: 57, SEQ ID NO: 58.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий фрагмент включает определяющую комплементарность область 1 (CDR Н) с аминокислотной последовательностью SYGMS (SEQ ID NO: 53), CDR Н2 с аминокислотной последовательностью SSGGSY (SEQ ID NO: 54), CDR Н3 с аминокислотной последовательностью LGMITTGYAMDY (SEQ ID NO: 55), определяющую комплементарность область 1 легкой цепи (CDR L) с аминокислотной последовательностью RSSQTIVHSTGHTYLE (SEQ ID NO: 56), CDR L2 с аминокислотной последовательностью KVSNRFS (SEQ ID NO: 57) и CDR L3 с аминокислотной последовательностью FQGSHVPYT (SEQ ID NO: 58).
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) антигенсвязывающий фрагмент, который представляет собой молекулу Fab, способную специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, содержащую определяющую комплементарность область 1 тяжелой цепи (CDR) с последовательностью SEQ ID NO: 53, CDR 2 тяжелой цепи с последовательностью SEQ ID NO: 54, CDR 3 тяжелой цепи с последовательностью SEQ ID NO: 55, CDR 1 легкой цепи с последовательностью SEQ ID NO: 56, CDR 2 легкой цепи с последовательностью SEQ ID NO: 57 и CDR 3 легкой цепи с последовательностью SEQ ID NO: 58,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий:
а) гибридный полипептид тяжелой цепи, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) тяжелую цепь, включающую определяющую комплементарность область 1 тяжелой цепи (CDR) с последовательностью SEQ ID NO: 53, CDR 2 тяжелой цепи с последовательностью SEQ ID NO: 54, CDR 3 тяжелой цепи с последовательностью SEQ ID NO: 55,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13, а также
б) легкую цепь, которая включает CDR 1 легкой цепи с последовательностью SEQ ID NO: 56, CDR 2 легкой цепи с последовательностью SEQ ID NO: 57 и CDR 3 легкой цепи с последовательностью SEQ ID NO: 58.
В одном варианте осуществления настоящего изобретения предлагается антигенсвязывающий рецептор, включающий:
а) гибридный полипептид тяжелой цепи, включающий в направлении от N-концевого остатка к С-концевому остатку:
(i) вариабельный домен тяжелой цепи (VH) с последовательностью SEQ ID NO: 61,
(ii) пептидный линкер, предпочтительно пептидный линкер с последовательностью SEQ ID NO: 17,
(iii) заякоренный трансмембранный домен, предпочтительно заякоренный трансмембранный домен с последовательностью SEQ ID NO: 11,
(iii) костимулирующий сигнальный домен, предпочтительно костимулирующий сигнальный домен с последовательностью SEQ ID NO: 12, и
(iv) стимулирующий сигнальный домен, предпочтительно стимулирующий сигнальный домен с последовательностью SEQ ID NO: 13, а также
б) вариабельный домен легкой цепи (VL) с последовательностью SEQ ID NO: 62.
В одном конкретном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, содержащий тяжелую цепь, которая включает или состоит из аминокислотной последовательности SEQ ID NO: 64, и легкую цепь, которая включает или состоит из аминокислотной последовательности SEQ ID NO: 65.
В конкретном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий рецептор включает гибридный полипептид тяжелой цепи, включающий аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 63, и полипептид легкой цепи, включающий аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 65.
В предпочтительном варианте антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А, где антигенсвязывающий рецептор включает гибридный полипептид тяжелой цепи, включающий аминокислотную последовательность SEQ ID NO: 63, и полипептид легкой цепи, включающий аминокислотную последовательность SEQ ID NO: 65.
В некоторых других вариантах антигенсвязывающий рецептор по изобретению, полипептид Fab легкой цепи и гибридный полипептид Fab тяжелой цепи соединены друг с другом, необязательно через пептидный линкер. Соединение Fab тяжелой и легкой цепей может улучшить объединение Fab тяжелой и легкой цепей, а также снизить число плазмид, необходимых для экспрессии некоторых антигенсвязывающих рецепторов по изобретению. Альтернативным подходом снижения числа плазмид, необходимых для экспрессии антигенсвязывающего рецептора, является использование внутреннего сайта связывания рибосомы, чтобы обеспечить экспрессию конструктов как тяжелой, так и легкой цепи из одной и той же плазмиды, как показано, например, на фиг. 2.
В некоторых вариантах антигенсвязывающий рецептор включает полипептид, где С-концевой остаток вариабельной области легкой цепи Fab антигенсвязывающего фрагмента связан пептидной связью с консервативной областью тяжелой цепи Fab антигенсвязывающего фрагмента (то есть антигенсвязывающий фрагмент включает тяжелую цепь crossFab, где вариабельная область тяжелой цепи заменена вариабельной областью легкой цепи), С-концевой остаток которой в свою очередь связан пептидной связью с заякоренный трансмембранным доменом (VL(1)-CH1(1)-ATD). В некоторых вариантах антигенсвязывающий рецептор дополнительно включает полипептид, где С-концевой остаток вариабельной области тяжелой цепи Fab первого антигенсвязывающего фрагмента связан пептидной связью с консервативной областью легкой цепи Fab первого антигенсвязывающего фрагмента (VH(1)-CL(1)). В некоторых вариантах полипептиды ковалентно связаны, например, с помощью дисульфидной связи. В других вариантах антигенсвязывающий рецептор включает полипептид, где С-концевой остаток вариабельной области тяжелой цепи Fab антигенсвязывающего фрагмента связан пептидной связью с консервативной областью легкой цепи Fab антигенсвязывающего фрагмента (то есть антигенсвязывающий фрагмент включает тяжелую цепь crossFab, где консервативная область тяжелой цепи заменена консервативной областью легкой цепи), С-концевой остаток которой в свою очередь связан пептидной связью с заякоренным трансмембранным доменом (VH(1)-CL(1)-ATD). В некоторых вариантах антигенсвязывающий рецептор дополнительно включает полипептид, где С-концевой остаток вариабельной области легкой цепи Fab антигенсвязывающего фрагмента связан пептидной связью с консервативной областью тяжелой цепи Fab антигенсвязывающего фрагмента (VL(1)-CH1(1)). В некоторых вариантах полипептиды ковалентно связаны, например, с помощью дисульфидной связи.
В соответствии с любым из вышеуказанных вариантов компоненты антигенсвязывающего рецептора (например, VH и VL, антигенсвязывающий фрагмент, заякоренный трансмембранный домен, костимулирующий сигнальный домен, стимулирующий сигнальный домен) могут быть соединены друг с другом напрямую или через различные линкеры, прежде всего пептидные линкеры, содержащие один или более аминокислотных остатков, обычно приблизительно 2-20 аминокислотных остатков, как описано в данном контексте или известно в данной области техники. Пригодные неиммуногенные пептидные линкеры включают, например, (G4S)n, (SG4)n, (G4S)n или G4(SG4)n пептидные линкеры, где n в основном обозначает число от 1 до 10, предпочтительно от 1 до 4.
Типичные активирующие Т-клетки антигенсвязывающие рецепторы
В прилагаемых иллюстративных примерах и на фиг. 1А в качестве подтверждения концепции настоящего изобретения предлагается сконструированный антигенсвязывающий рецептор "анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD pETR17096" (SEQ ID NO: 7), который включает один стабилизированный scFv антигенсвязывающий фрагмент, который связывается/направлен против антитела/взаимодействует с антителом, включающим мутацию P329G в составе Fc домена. Конструкт дополнительно включает CD28 трансмембранный домен, фрагмент CD28 в качестве костимулирующего сигнального домена и а фрагмент CD3z в качестве стимулирующего сигнального домена. Последовательности (аминокислотные и кДНК) антителосвязывающей молекулы, "анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD pETR17096" представлены в таблицах 2 и 3.
Более того, как представлено на фиг. 1Б, в качестве дополнительного подтверждения концепции настоящего изобретения сконструирован антигенсвязывающий рецептор "анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD pETR17100" (SEQ ID NO: 39, SEQ ID NO: 41), который включает один стабилизированный антигенсвязывающий Fab фрагмент, который связывается/направлен против антитела/взаимодействует с антителом, включающим мутации P329G в составе Fc домена. Конструкт дополнительно включает CD28 трансмембранный домен, фрагмент CD28 в качестве костимулирующего сигнального домена и фрагмент CD3z в качестве стимулирующего сигнального домена. Последовательности (аминокислотные и кДНК) антигенсвязывающего рецептора, "анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD pETR17100" представлены в таблицах 4 и 5.
В качестве еще одного подтверждения концепции настоящего изобретения сконструирован антигенсвязывающий рецептор, "анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD pETR17594" (SEQ ID NO: 48, SEQ ID NO: 50), который включает один антигенсвязывающий Fab фрагмент, который связывается/направлен против антитела/взаимодействует с антителом, включающим мутацию P329G в составе Fc домена. Конструкт дополнительно включает CD28 трансмембранный домен, фрагмент CD28 в качестве костимулирующего сигнального домена и фрагмент CD3z в качестве стимулирующего сигнального домена. Последовательности (аминокислотные и кДНК) антигенсвязывающего рецептора "анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD pETR17594" представлены в таблицах 6 и 7.
В качестве дополнительного подтверждения концепции настоящего изобретения сконструирован антигенсвязывающий рецептор, "анти-ААА scFv" (SEQ ID NO: 59), который включает один антигенсвязывающий scFv фрагмент, который связывается/направлен против антитела/взаимодействует с антителом, включающим мутации I253A, Н310А и Н435 в составе Fc домена. Конструкт дополнительно включает CD28 трансмембранный домен, фрагмент CD28 в качестве костимулирующего сигнального домена и фрагмент CD3z в качестве стимулирующего сигнального домена. Последовательности (аминокислотные и кДНК) антителосвязывающей молекулы, "анти-ААА scFv" представлены в таблицах 8 и 9.
В качестве дополнительного подтверждения концепции настоящего изобретения сконструирован антигенсвязывающий рецептор, "анти-ААА Fab" (SEQ ID NO: 63, SEQ ID NO: 65), который включает один антигенсвязывающий Fab фрагмент, который связывается/направлен против антитела/взаимодействует с антителом, включающим мутации I253A, Н310А и Н435А в составе Fc домена. Конструкт дополнительно включает CD28 трансмембранный домен, фрагмент CD28 в качестве костимулирующего сигнального домена и фрагмент CD3z в качестве стимулирующего сигнального домена. Последовательности (аминокислотные и кДНК) антителосвязывающей молекулы, "анти-ААА scFv" представлены в таблицах 10 и 11.
В изобретении также предлагается молекула (молекулы) нуклеиновой кислоты, кодирующей антигенсвязывающие рецепторы по изобретению, как описано в данном контексте. Настоящее изобретение также относится к молекуле (молекулам) нуклеиновой кислоты, кодирующей антигенсвязывающие рецепторы по изобретению, а также наборам, включающим молекулу (молекулы) нуклеиновой кислоты по изобретению, как дополнительно описано в данном контексте.
Наборы
В другом объекте настоящего изобретения предлагаются наборы, включающие или состоящие из нуклеиновой кислоты, кодирующей антигенсвязывающий рецептор по изобретению, и/или клеток, предпочтительно Т-клеток, трансдуцированных антигенсвязывающими рецепторами по изобретению, и необязательно включающие антитело/антитела, включающие мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом.
Соответственно, предлагается набор, включающий:
(А) трансдуцированную Т-клетку, способную экспрессировать антигенсвязывающий рецептор по изобретению, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
Также предлагается набор, включающий:
(А) выделенный полинуклеотид и/или вектор, кодирующий антигенсвязывающий рецептор по изобретению, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
В данном контексте наборы по изобретению могут включать трансдуцированные Т-клетки, выделенные полинуклеотиды и/или векторы и одно или более антител, включающих мутированный Fc домен. В конкретных вариантах антитело представляет собой терапевтическое антитело, например, опухолеспецифическое антитело. Опухолеспецифические антигены известны в данной области техники и описаны в данном контексте. В настоящем изобретении антитело вводят до, одновременно с введением или после введения трансдуцированной Т-клетки, экспрессирующей антигенсвязывающий рецептор по изобретению. Наборы по настоящему изобретению включают трансдуцированные Т-клетки или полинуклеотиды/векторы для получения трансдуцированных Т-клеток. В данном контексте трансдуцированные Т-клетки являются универсальными Т-клетками поскольку они не являются специфичными в отношении определенной опухоли, но могут быть направлены к любой опухоли в зависимости от терапевтического антитела, включающего мутированный Fc домен. В данном контексте предлагаются примеры антител, включающих мутированный Fc домен, однако, в представленные в данном контексте наборы можно включать любое антитело, включающее мутированный Fc домен, как описано в данном контексте. В конкретных вариантах мутированный Fc домен антител проявляет сниженную связывающую аффинностью в отношении Fc рецептора и/или сниженную эффекторную функцию, по сравнению с нативным Fc доменом IgG1. В одном из указанных вариантов мутированный Fc домен (или антитело, включающее указанный мутированный Fc домен) характеризуется связывающей аффинностью в отношении Fc рецептора на уровне менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и еще более предпочтительно менее 5%, по сравнению с нативным Fc доменом IgG1 (или антителом, включающим нативный Fc домен IgG1), и/или эффекторной функцией на уровне менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и еще более предпочтительно менее 5%, по сравнению с нативным Fc доменом IgG1 (или антителом, включающим нативный Fc домен IgG1). В одном варианте мутированный Fc домен (или антитело, включающее указанный мутированный Fc домен) практически не связывается с Fc рецептором и/или не индуцирует эффекторную функцию. В конкретном варианте осуществления настоящего изобретения Fc рецептором является Fcγ рецептор. В одном варианте осуществления настоящего изобретения Fc рецептором является Fc рецептор человека. В одном варианте осуществления настоящего изобретения Fc рецептором является активирующий Fc рецептор. В конкретном варианте Fc рецептором является активирующий Fcγ рецептор человека, более конкретно FcγRIIIa, FcγRI или FcγRIIa человека, еще более конкретно FcγRIIIa человека. В одном варианте осуществления настоящего изобретения эффекторная функция представляет собой одну или более функций, выбранных из группы, включающей CDC, ADCC, ADCP и секрецию цитокинов. В конкретном варианте осуществления настоящего изобретения эффекторной функцией является ADCC. В одном варианте осуществления настоящего изобретения мутированный Fc домен характеризуется существенно измененной связывающей аффинностью в отношении неонатального Fc рецептора (FcRn), по сравнению со связывающей аффинностью нативного Fc домена IgG1. В одном варианте осуществления настоящего изобретения антитело, включающее мутированный Fc домен, характеризуется связывающей аффинностью в отношении Fc рецептора на уровне менее 20%, предпочтительно менее 10%, более предпочтительно менее 5% связывающей аффинности антитела, включающего несконструированный Fc домен. В конкретном варианте осуществления настоящего изобретения Fc рецептором является Fcγ рецептор. В некоторых вариантах Fc рецептором является Fc рецептор человека. В некоторых вариантах Fc рецептором является активирующий Fc рецептор. В конкретном варианте Fc рецептором является активирующий Fcγ рецептор человека, более конкретно FcγRIIIa, FcγRI или FcγRIIa человека, еще более конкретно FcγRIIIa человека. Предпочтительно, связывание с каждым из указанных рецепторов снижается. В некоторых вариантах связывающая аффинность в отношении компонента комплемента, например, связывающая аффинность в отношении C1q также снижается.
В некоторых вариантах в Fc домен антитела вводили мутации для снижения эффекторной функции по сравнению с немутированным Fc доменом. Сниженная эффекторная функция может включать, но, не ограничиваясь только ими, одну или несколько из следующих функций: пониженная комплементзависимая цитотоксичность (CDC), пониженная антителозависимая обусловленная клетками цитотоксичность (ADCC), пониженный антителозависимый клеточный фагоцитоз (ADCP), пониженная секреция цитокинов, понижение опосредованного иммунным комплексом захвата антигена антигенпрезентирующими клетками, пониженное связывание с NK клетками, пониженное связывание с макрофагами, снижение связывания с моноцитами, снижение связывания с полиморфно-ядерными клетками, снижение прямой передачи сигналов, индуцирующих апоптоз, снижение сшивки антител, связанных с мишенью, снижение созревания дендритных клеток или снижение примирования Т-клеток. В одном варианте сниженной эффекторной функцией является одна или более функций, выбранных из группы, включающей сниженную CDC, сниженную ADCC, сниженную ADCP и снижение секреции цитокинов. В конкретном варианте сниженной эффекторной функцией является сниженная ADCC. В одном варианте снижение ADCC составляет менее 20% от уровня ADCC, индуцированной несконструируемым Fc доменом (или антителом, содержащим несконструированный Fc).
В одном варианте осуществления настоящего изобретения аминокислотная мутация, которая снижает связывающую аффинность Fc домена с Fc рецептором и/или эффекторную функцию, представляет собой аминокислотную замену. В одном варианте осуществления настоящего изобретения Fc домен включает аминокислотную замену, расположенную в положении, выбранном из группы Е233, L234, L235, N297, Р331 и Р329. В более конкретном варианте Fc домен включает аминокислотную замену, расположенную в положении, выбранном из группы L234, L235 и Р329. В некоторых вариантах Fc домен включает аминокислотные замены L234A и L235A. В одном из указанных вариантов Fc домен представляет собой Fc домен IgG1, прежде всего Fc домен IgG1 человека. В одном варианте осуществления настоящего изобретения Fc домен включает аминокислотную замену в положении Р329. В более конкретном варианте аминокислотная замена представляет собой замену Р329А или P329G, прежде всего P329G. В одном варианте осуществления настоящего изобретения Fc домен включает аминокислотную замену в положении Р329 и дополнительную аминокислотную замену в положении, выбранном из Е233, L234, L235, N297 и Р331. В более конкретном варианте дополнительная аминокислотная замена представляет собой замену Е233Р, L234A, L235A, L235E, N297A, N297D или P331S. В конкретных вариантах Fc домен включает аминокислотные замены в положениях Р329, L234 и L235. В одном варианте осуществления настоящего изобретения Fc домен включает аминокислотные мутации L234A, L235A и P329G ("P329G LALA"). В одном из указанных вариантов Fc домен представляет собой Fc домен IgG1, прежде всего Fc домен IgG1 человека. Комбинация аминокислотных замен "P329G LALA" практически полностью подавляет связывание Fcγ рецептора (а также комплемента) с Fc доменом IgG1 человека, как описано в заявке РСТ WO 2012/130831, которая включена в полном объеме в настоящее описание в качестве ссылки. В заявке WO 2012/130831 также описаны способы получения указанных мутированных Fc доменов и способы определения их свойств, таких как связывание с Fc рецептором или эффекторные функции.
В конкретном варианте осуществления настоящего изобретения Fc домен, характеризующийся сниженной связывающей аффинностью в отношении Fc рецептора и/или сниженной эффекторной функцией, по сравнению с нативным Fc доменом IgG1, представляет собой Fc домен IgG1 человека, содержащий аминокислотные мутации L234A, L235A и необязательно P329G, или Fc домен IgG4 человека, содержащий аминокислотные замены S228P, L235E и необязательно P329G.
В некоторых вариантах проводили де-N-гликозилирование Fc домена. В одном из указанных вариантов Fc домен включает аминокислотную мутацию в положении N297, прежде всего аминокислотную мутацию, обеспечивающую замену аспарагина на аланин (N297A) или на аспарагиновую кислоту (N297D).
Кроме Fc доменов, описанных выше, а также в заявке WO 2012/130831, Fc домены, характеризующиеся сниженными уровнем связывания с Fc рецептором и/или эффекторной функцией, также включают такие Fc домены, которые включают одну или более мутаций в положении 238, 265, 269, 270, 297, 327 и 329 (см. патент US 6737056). Указанные мутированные Fc включают мутированные Fc с мутациями в двух или более положениях аминокислот 265, 269, 270, 297 и 327, включая так называемый "DANA" мутированный Fc, в котором аминокислотные остатки в положении 265 и 297 заменены на аланин (см. патент US 7332581).
Мутированные Fc домены можно получить за счет делеции, замены, вставки или модификации аминокислот с использованием генно-инженерных или химических способов, известных специалисту в данной области техники. Генно-инженерные способы могут включать сайт-направленный мутагенез кодирующей последовательности ДНК, ПЦР, синтез генов и тому подобное. Правильные нуклеотидные изменения можно подтвердить, например, секвенированием.
Связывание с Fc рецепторами можно легко определить, например, методом твердофазного иммуноферментного анализа (ИФА), или поверхностного плазмонного резонанса (ППР) с использованием стандартного оборудования, такого как установка BIAcore (фирмы GE Healthcare), с использованием Fc рецепторов, которые можно получить методом рекомбинантной экспрессии. В другом варианте, связывающую аффинность Fc доменов или биспецифичных связывающих активирующий клетки антиген соединений, содержащих Fc домен для Fc рецепторов, можно оценить с использованием известных клеточных линий для экспрессии конкретных Fc рецепторов, таких как NK клетки человека, экспрессирующие FcγIIIa рецептор.
Эффекторную функцию Fc домена или антитела, содержащего Fc домен, можно оценить методами, известными в данной области техники. Другие примеры анализов in vitro для оценки активности ADCC исследуемого соединения описаны в патенте US 5500362, статье Hellstrom Proc. Natl. Acad. Sci. USA, 83, 7059-7063 (1986) и статье Hellstrom и др., Proc. Natl. Acad. Sci. USA, 82, 1499-1502 (1985), патенте US 5821337, статье Bruggemann и др., J. Exp. Med., 166, 1351-1361 (1987). В другом варианте можно использовать методы нерадиоактивного анализа (см., например, нерадиоактивный анализ цитотоксичности ACTI™ для проточной цитометрии (фирмы CellTechnology, Inc. Mountain View, CA) и нерадиоактивный анализ цитотоксичности CytoTox 96® (фирмы Promega, Madison, WI)). Пригодные эффекторные клетки для указанных анализов включают мононуклеарные клетки периферической крови (РВМС) и природные клетки-киллеры (NK клетки). В другом варианте или дополнительно ADCC активность исследуемого соединения можно оценить in vivo, например, на модели животных, такой как модель, описанная в статье Clynes и др., Proc. Natl. Acad. Sci. USA, 95, 652-656 (1998).
В некоторых вариантах уменьшается связывание Fc домена с компонентом комплемента, прежде всего с C1q. Соответственно, в некоторых вариантах, где Fc домен сконструирован таким образом, чтобы снизить эффекторную функцию, указанная сниженная эффекторная функция включает снижение CDC. Для оценки способности антитела связываться с C1q и, следовательно, проявлять активность CDC, можно проводить анализ связывания C1q, см., например, ИФА анализ для оценки связывания C1q и С3с, описанный в заявках WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно проводить анализ CDC (см., например, статьи Gazzano-Santoro и др., J. Immunol. Methods, 202, 163 (1996), Cragg и др., Blood, 101, 1045-1052 (2003) и Cragg и Glennie, Blood, 103, 2738-2743 (2004)).
В одном варианте снижается связывающая аффинность с неонатальным Fc рецептором (FcRn). В конкретных вариантах мутированный Fc домен по настоящему изобретению проявляет сниженную связывающую аффинность в отношении FcRn рецептора, по сравнению с нативным Fc доменом IgG1. В одном из указанных вариантов Fc домен (или антитело, включающее указанный Fc домен) характеризуется связывающей аффинностью в отношении неонатального Fc рецептора на уровне менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и еще более предпочтительно менее 5%, по сравнению с нативным Fc доменом IgG1 (или антителом, включающим нативный Fc домен IgG1), и/или эффекторной функцией на уровне менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и еще более предпочтительно менее 5%, по сравнению с нативным Fc доменом IgG1 (или антителом, включающим нативный Fc домен IgG1). В одном варианте мутированный Fc домен (или антитело, включающее указанный мутированный Fc домен) в основном не связывается с неонатальным Fc рецептором. В конкретном варианте Fc рецептором является FcRn рецептор. В одном варианте Fc рецептором является FcRn рецептор человека. В конкретных вариантах Fc домен включает аминокислотные замены в положениях I253, Н310 и Н435. В более конкретном варианте Fc домен включает аминокислотные мутации I253A, Н310А и Н435А ("AAA"). В одном из указанных вариантов Fc домен представляет собой Fc домен IgG1, прежде всего Fc домен IgG1 человека. Комбинация аминокислотных замен "AAA" практически полностью подавляет связывание FcRn рецептора с Fc доменом IgG1 человека.
В конкретном варианте антитело, включающее мутированный Fc домен, способно специфически связываться с CD20 и включает тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. В одном варианте антитело, включающее мутированный Fc домен, способно специфически связываться с FAP и включает тяжелую цепь с последовательностью SEQ ID NO: 114, и легкую цепь с последовательностью SEQ ID NO: 115. В одном варианте антитело, включающее мутированный Fc домен, способно специфически связываться с СЕА и включает тяжелую цепь с последовательностью SEQ ID NO: 116, и легкую цепь с последовательностью SEQ ID NO: 117, тяжелую цепь с последовательностью SEQ ID NO: 118, и легкую цепь с последовательностью SEQ ID NO: 119, тяжелую цепь с последовательностью SEQ ID NO: 120, и легкую цепь с последовательностью SEQ ID NO: 121, или тяжелую цепь с последовательностью SEQ ID NO: 122, и легкую цепь с последовательностью SEQ ID NO: 123. В других вариантах антитело, включающее мутированный Fc домен, способно специфически связываться с тенасцином (TNC) и включает тяжелую цепь с последовательностью SEQ ID NO: 124, и легкую цепь с последовательностью SEQ ID NO: 125.
В другом варианте антитело, включающее мутированный Fc домен, представляет собой биспецифическое антитело, например, активирующее Т-клетку биспецифическое антитело. В одном из указанных вариантов биспецифическое антитело включает первый связывающий фрагмент, способный специфически связываться с мишенью, активирующей Т-клетку, предпочтительно CD3, и второй связывающий фрагмент, способный специфически связываться с опухолевым антигеном, как описано в данном контексте.
В одном варианте антитело, включающее мутированный Fc домен, является биспецифическим и способно специфически связываться с Her2, где биспецифическое антитело включает первую тяжелую цепь с последовательностью SEQ ID NO: 126, первую легкую цепь с последовательностью SEQ ID NO: 127, вторую тяжелую цепь с последовательностью SEQ ID NO: 128, и вторую легкую цепь с последовательностью SEQ ID NO: 129.
В иллюстративном варианте осуществления настоящего изобретения в качестве подтверждения концепции изобретения предлагается набор, включающий аминокислотную последовательность SEQ ID NO: 7 ("анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD" (кодируемую последовательностью ДНК SEQ ID NO: 19)), в комбинации с антителом, включающим тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. В другом варианте набор может включать аминокислотную последовательность SEQ ID NO: 31 ("анти-P329G-scFv-CD28ATD-CD28CSD-CD3zSSD" (кодируемую последовательностью ДНК SEQ ID NO: 35)), в комбинации с антителом, включающим тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. Кроме того, в настоящем изобретении набор может включать аминокислотную последовательность SEQ ID NO: 39 ("анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD" (кодируемую последовательностью ДНК SEQ ID NO: 44)) в комбинации с антителом, включающим тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. В другом варианте набор может включать аминокислотную последовательность SEQ ID NO: 48 ("анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD" (кодируемую последовательностью ДНК SEQ ID NO: 51)), в комбинации с антителом, включающим тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. В другом варианте набор может включать аминокислотную последовательность SEQ ID NO: 59 ("анти-ААА-scFv-CD28ATD-CD28CSD-CD3zSSD"), в комбинации с антителом, включающим тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. Кроме того, в настоящем изобретении набор может включать аминокислотную последовательность SEQ ID NO: 63 ("анти-AAA-Fab-CD28ATD-CD28CSD-CD3zSSD") в комбинации с антителом, включающим тяжелую цепь с последовательностью SEQ ID NO: 112, и легкую цепь с последовательностью SEQ ID NO: 113. Кроме того, в настоящем изобретении набор может включать по меньшей мере одно антитело, включающее тяжелую цепь и легкую цепь, которые выбирают из группы, состоящей из последовательности SEQ ID NO: 112 и SEQ ID NO: 113, SEQ ID NO: 114 и SEQ ID NO: 115, SEQ ID NO: 116 и SEQ ID NO: 117, SEQ ID NO: 118 и SEQ ID NO: 119, SEQ ID NO: 120 и SEQ ID NO: 121, SEQ ID NO: 122 и SEQ ID NO: 123, а также SEQ ID NO: 124 и SEQ ID NO: 125. Кроме того, в настоящем изобретении набор может включать биспецифическое антитело, предпочтительно биспецифическое антитело, включающее первую тяжелую цепь с последовательностью SEQ ID NO: 128, первую легкую цепь с последовательностью SEQ ID NO: 129, и вторую тяжелую цепь с последовательностью SEQ ID NO: 130, и вторую легкую цепь с последовательностью SEQ ID NO: 131.
Более того, части набора по изобретению могут быть упакованы индивидуально во флаконы или бутылочки или в виде комбинации в контейнеры или много контейнерную упаковку. В другом варианте набор по настоящему изобретению может включать инкубационную систему для инкубации клеток в (закрытом) контейнере, где клетки пациента, предпочтительно Т-клетки, как описано в данном контексте, можно трансдуцировать антигенсвязывающим рецептором (рецепторами) по изобретению и инкубировать в условиях, согласно правилам GMP (правила производства и контроля качества лекарственных средств, как описано в руководствах о правилах производства и контроля качества лекарственных средств, опубликованных Европейской комиссией на интернет-ресурсе http://ec.europa.eu/health/documents/eudralex/index_en.htm). Более того, набор по настоящему изобретению включает систему для инкубации клеток в (закрытых) контейнерах, где выделенные/полученные Т-клетки пациентов можно трансдуцировать антигенсвязывающим рецептором (рецепторами) по изобретению и инкубировать в условиях, рекомендованных в GMP. Более того, в настоящем изобретении набор может также включать вектор, кодирующий антигенсвязывающий рецептор (рецепторы), как описано в данном контексте. Набор по настоящему изобретению можно эффективно использовать, среди прочего, для осуществления способа по изобретению и можно использовать в различных областях, как описано в данном контексте, например, в качестве научно-исследовательских средств или медицинских средств. Наборы предпочтительно получают по стандартным методикам, которые известны специалисту в данной области техники.
В данном контексте клетки, полученные от пациента, предпочтительно Т-клетки, можно трансдуцировать антигенсвязывающим рецептором по изобретению, способным специфически связываться с мутированным Fc доменом, как описано в данном контексте, с использованием набора, как описано выше. Внеклеточный домен, включающий антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом, отсутствует в Т-клетках или на их поверхности. Соответственно, клетки, полученные от пациента, трансдуцированные с использованием наборов по изобретению, приобретают способность специфически связываться с мутированным Fc доменом антитела, например, терапевтического антитела, и способность индуцировать подавление/лизис клеток-мишеней в результате взаимодействия с терапевтическим антителом, включающим мутированный Fc домен, где терапевтическое антитело способно связываться с природным опухолеспецифическим антигеном (который эндогенно экспрессируется) на поверхности опухолевых клеток. Связывание внеклеточного домена антигенсвязывающего рецептора, как описано в данном контексте, активирует указанную Т-клетку и обеспечивает ее физическое контактирование с опухолевой клеткой через терапевтическое антитело, включающее мутированный Fc домен. Нетрансдуцированные или эндогенные Т-клетки (например, CD8 + Т-клетки) не способны связываться с мутированным Fc доменом терапевтического антитела, включающим мутированный Fc домен. Трансдуцированные Т-клетки, экспрессирующие антигенсвязывающий рецептор, включающий внеклеточный домен, способный специфически связываться с мутированным Fc доменом, не взаимодействуют с терапевтическим антителом, которое не содержит мутации в составе Fc домена, как описано в данном контексте. Соответственно, Т-клетки, экспрессирующие антигенсвязывающий рецептор по изобретению, способны лизировать клетки-мишени in vivo и/или in vitro в присутствии антитела, содержащего мутации в составе Fc домена, как описано в данном контексте. Соответствующие клетки-мишени включают клетки, экспрессирующие поверхностную молекулу, то есть природный опухолеспецифический антиген, расположенный на поверхности природных опухолевых клеток, который распознается, по меньшей мере, одним, предпочтительно двумя, доменами связывания терапевтического антитела, как описано в данном контексте. Указанные поверхностные молекулы описаны ниже в данном контексте.
Лизис клетки-мишени можно детектировать методами, известными в данной области техники. Соответственно, указанные методы включают, среди прочего, анализы физиологических параметров in vitro. Такие анализы физиологических параметров позволяют контролировать гибель клеток, например, по оценке целостности клеточной мембраны (например, анализ на основе FACS с использованием иодида пропидия, анализ проницаемости мембраны с использованием трипанового синего, фотометрический анализ высвобождения меченого фермента (LDH), радиометрический анализ высвобождения 51Cr, флуорометрический анализ высвобождения европия и анализ высвобождения CalceinAM). Дополнительные анализы включают мониторинг жизнеспособности клеток, например, с помощью фотометрических анализов МТТ, ХТТ, WST-1 и alamarBlue, радиометрического анализа включения 3Н-Thd, клоногенного анализа активности деления клеток и флуорометрического анализа митохондриального трансмембранного градиента с использованием родамина-123. Дополнительно можно контролировать уровень апоптоза, например, с помощью анализа на основе FACS воздействия фосфатидилсерина, TUNEL-теста на основе ИФА, анализа активности каспазы (фотометрический, флуориметрический или на основе ИФА) или анализа изменения морфологии клеток (сжатие клетки, пузырение мембраны).
Трансдуцированные Т-клетки, способные экспрессировать антигенсвязывающие рецепторы по изобретению
В другом объекте настоящего изобретения предлагаются трансдуцированные Т-клетки, способные экспрессировать антигенсвязывающий рецептор по настоящему изобретению. Антигенсвязывающие рецепторы, как описано в данном контексте, представляют собой молекулы, которые в природных условиях не содержатся в Т-клетках и/или на их поверхности, и которые не экспрессируются (эндогенно) в нормальных (нетрансдуцированных) Т-клетках или на их поверхности. Таким образом, антигенсвязывающий рецептор по изобретению искусственно вводится в Т-клетки и/или на их поверхность. В настоящем изобретении указанные Т-клетки, предпочтительно CD8 + Т-клетки, можно выделить/получить от субъекта, нуждающегося в лечении, как описано в данном контексте. Соответственно, антигенсвязывающие рецепторы, как описано в данном контексте, которые искусственно вводятся и затем презентируются в указанных Т-клетках и/или на их поверхности, содержат домены, включающие один или более антигенсвязывающих фрагментов, которые являются доступными (in vitro или in vivo) для иммуноглобулинов (lg-производные), предпочтительно для антител, предпочтительно для Fc домена антител. В настоящем изобретении указанные искусственно введенные молекулы презентируются в указанных Т-клетках и/или на их поверхности после (ретровирусной или лентивирусной) трансдукции, как описано ниже в данном контексте. Соответственно, после трансдукции Т-клетки по настоящему изобретению можно активировать иммуноглобулинами, предпочтительно (терапевтическими) антителами, включающими специфические мутации в составе Fc домена, как описано в данном контексте.
В настоящем изобретении также предлагаются трансдуцированные Т-клетки, экспрессирующие антигенсвязывающий рецептор, кодируемый молекулой (молекулами) нуклеиновой кислоты, кодирующей антигенсвязывающий рецептор по настоящему изобретению. Соответственно, в настоящем изобретении трансдуцированная клетка может включать молекулу нуклеиновой кислоты, кодирующую антигенсвязывающий рецептор по настоящему изобретению или вектор по настоящему изобретению, который экспрессирует антигенсвязывающий рецептор по настоящему изобретению.
В настоящем изобретении термин "трансдуцированная Т-клетка" включает генетически модифицированную Т-клетку (то есть Т-клетку, в которую преднамеренно введена молекула нуклеиновой кислоты). Представленная в данном контексте трансдуцированная Т-клетка может включать вектор по настоящему изобретению. Предпочтительно, представленная в данном контексте трансдуцированная Т-клетка включает молекулу нуклеиновой кислоты, кодирующую антигенсвязывающий рецептор по настоящему изобретению, и/или вектор по настоящему изобретению. Трансдуцированная Т-клетка по изобретению может представлять собой Т-клетку, которая временно или стабильно экспрессирует чужеродную ДНК (то есть молекулу нуклеиновой кислоты, которая была введена в Т-клетку). Прежде всего, молекулу нуклеиновой кислоты, кодирующую антигенсвязывающий рецептор по настоящему изобретению, можно стабильно интегрировать в геном Т-клетки в ходе ретровирусной или лентивирусной трансдукции. При использовании трансфекции мРНК молекулу нуклеиновой кислоты, кодирующую антигенсвязывающий рецептор по настоящему изобретению, можно экспрессировать временно. Предпочтительно, представленная в данном контексте трансдуцированная Т-клетка генетически модифицирована за счет введения молекулы нуклеиновой кислоты в Т-клетку с использованием вирусного вектора (например, ретровирусного вектора или лентивирусного вектора). Соответственно, экспрессия антигенсвязывающих рецепторов может являться конститутивной, и внеклеточный домен антигенсвязывающего рецептора можно обнаружить на поверхности клетки. Указанный внеклеточный домен антигенсвязывающего рецептора может включать полноразмерный внеклеточный домен антигенсвязывающего рецептора, как описано в данном контексте, но также и его часть. Требуемым минимальным размером является антигенсвязывающий сайт антигенсвязывающего фрагмента в антигенсвязывающем рецепторе.
Экспрессия также может быть условной или индуцибельной в случае, когда антигенсвязывающий рецептор введен в Т-клетки под контролем индуцибельного или репрессируемого промотора. Примерами указанных индуцибельных или репрессируемых промоторов может являться транскрипционная система, содержащая промотор гена алкогольдегидрогеназы I (alcA) и трансактиваторный белок AlcR. Различные сельскохозяйственные составы на основе спирта используются для контроля экспрессии исследуемого гена, связанного с alcA промотором. Более того, чувствительные к тетрациклину промоторные системы могут проявлять функцию либо для активации, либо для подавления экспрессии генов в присутствии тетрациклина. Некоторые элементы системы включают белок-репрессор тетрациклина (TetR), последовательность оператора тетрациклина (tetO) и гибридный белок-трансактиватор тетрациклина (tTA), который представляет собой гибрид TetR и активирующей последовательности белка 16 (VP16) вируса простого герпеса. Кроме того, можно использовать стероид-чувствительные промоторы, промоторы, регулируемые металлом, или промоторы, которые связаны с белками, связанными с патогенезом (PR).
В зависимости от используемой системы экспрессия может являться конститутивной или конституциональной. Антигенсвязывающие рецепторы по настоящему изобретению могут экспрессироваться на поверхности трансдуцированных Т-клеток, представленных в данном контексте. Внеклеточная часть антигенсвязывающего рецептора (то есть внеклеточный домен антигенсвязывающего рецептора) можно детектировать на поверхности клетки, в то время как внутриклеточная часть (то есть костимулирующий сигнальный домен (домены) и стимулирующий сигнальный домен) не детектируются на поверхности клетки. Обнаружение внеклеточного домена антигенсвязывающего рецептора можно осуществлять с использованием антитела, которое специфически связывается с указанным внеклеточным доменом, или с использованием мутированного Fc-домена, который способен связываться с внеклеточным доменом. Внеклеточный домен можно обнаруживать с использованием указанных антител или Fc-доменов методом проточной цитометрии или микроскопии.
Трансдуцированные клетки по настоящему изобретению могут представлять собой любые иммунные клетки. Указанные клетки включают, но, не ограничиваясь только ими, В-клетки, Т-клетки, природные клетки-киллеры (NK клетки), природные Т-клетки-киллеры (NK), γδ Т-клетки, лимфоидные клетки врожденного иммунитета, макрофаги, моноциты, дендритные клетки или нейтрофилы. Предпочтительно, указанная иммунная клетка представляет собой лимфоцит, предпочтительно NK или Т-клетки. Указанные Т-клетки включают CD4 Т-клетки и CD8 Т-клетки. Введение антигенсвязывающего рецептора по настоящему изобретению на поверхность лейкоцита обеспечивает развитие клеточной цитотоксичности в отношении клетки-мишени в сочетании с терапевтическим антителом, включающим мутированный Fc домен, независимо от исходной линии клетки. Цитотоксичность будет наблюдаться независимо от того, какой стимулирующий сигнальный домен или костимулирующий сигнальный домен выбран для антигенсвязывающего рецептора, и не зависит от экзогенного источника дополнительных цитокинов. Соответственно, трансдуцированная клетка по настоящему изобретению может представлять собой, например, CD4 + Т-клетку, CD8 + -Т-клетку, γδ Т-клетку, природную Т-клетку-киллер (NK), природную клетку-киллер (NK), лейкоцит, опухоль-инфильтрующий лимфоцит (TIL), миелоидную клетку или мезенхимальную стволовую клетку. Предпочтительно, представленная в данном контексте трансдуцированная клетка представляет собой Т-клетку (например, аутологичную Т-клетку), более предпочтительно трансдуцированная клетка представляет собой CD8 + Т-клетку. Соответственно, в контексте настоящего изобретения, трансдуцированная клетка представляет собой CD8 + Т-клетку. Кроме того, в контексте настоящего изобретения, трансдуцированная клетка представляет собой аутологичную Т-клетку. Соответственно, в контексте настоящего изобретения трансдуцированная клетка предпочтительно представляет собой аутологичную CD8 + Т-клетку. Помимо использования аутологичных клеток (например, Т-клеток), выделенных из организма субъекта, в настоящем изобретении также предлагается использование аллогенных клеток. Соответственно, в настоящем изобретении трансдуцированная клетка может также представлять собой аллогенную клетку, такую как аллогенная CD8 + Т-клетка. Использование аллогенных клеток основано на том факте, что клетки, предпочтительно Т-клетки, могут распознавать специфический эпитоп антигена, презентируемый чужеродными антиген-презентирующими клетками (АРС), при условии, что АРС экспрессируют молекулу МНС класса I или класса II, к которой клеточная популяция проявляет специфическую ответную реакцию, то есть популяция Т-клеток ограничена, наряду с эпитопом антигена, распознаваемым Т-клетками. Таким образом, термин "аллогенная" обозначает клетки неродственного происхождения, полученные из неродственного индивидуума/субъекта-донора, которая представляет собой лейкоцитарный антиген человека (HLA), совместимый с индивидуумом/субъектом, нуждающимся в таком лечении, например, антигенсвязывающий рецептор, экспрессирующийся трансдуцированной клеткой, как описано в данном контексте. Аутологичные клетки представляют собой клетки, которые выделяют/получают, как описано выше в данном контексте, из организма субъекта, нуждающегося в лечении трансдуцированной клеткой, как описано в данном контексте.
Трансдуцированную клетку по изобретению можно котрансдуцировать другими молекулами нуклеиновой кислоты, например, молекулой нуклеиновой кислоты, кодирующей рецептор Т-клетки.
В настоящем изобретении предлагается также способ получения трансдуцированной Т-клетки, экспрессирующей антигенсвязывающий рецептор по изобретению, включающий стадии трансдукции Т-клетки вектором по настоящему изобретению, культивирование трансдуцированной Т-клетки в условиях, обеспечивающих экспрессию антигенсвязывающего рецептора в указанной трансдуцированной клетке или на ее поверхности, и выделение указанной трансдуцированной Т-клетки.
В контексте настоящего изобретения, трансдуцированную клетку по настоящему изобретению предпочтительно получают следующим способом: клетки (например, Т-клетки, предпочтительно CD8 + Т-клетки) выделяют/получают от субъекта (предпочтительно пациенты - люди). Методы выделения/получения клеток (например, Т-клеток, предпочтительно CD8 + Т-клеток) от пациентов или от доноров хорошо известны в данной области техники и в контексте настоящего изобретения клетки (например, Т-клетки, предпочтительно CD8 + Т-клетки) от пациентов или от доноров можно выделять при заборе крови или образца костного мозга. После выделения/получения клеток в составе образца ткани пациента, клетки (например, Т-клетки) отделяют от других ингредиентов образца. Известно несколько методов выделения клеток (например, Т-клеток) из образца, которые включают, но, не ограничиваясь только ими, например, лейкаферез для получения клеток из образца периферической крови пациента или донора, выделение/получение клеток с использованием системы FACSort, отделение жизнеспособных клеток от мертвых клеток в свежих биопсийных образцах, содержащих жизнеспособные клетки, вручную или с использованием микроманипулятора (см., например, статьи Dudley, Immunother., 26, 332-342 (2003), Robbins, Clin. Oncol., 29, 917-924 (2011) или Leisegang, J. Mol. Med., 86, 573-658 (2008)). Выделенные/полученные Т-клетки, предпочтительно CD8 + Т-клетки, впоследствии культивируют и размножают с использованием анти-CD3 антитела, с использованием анти-CD3 и анти-CD28 моноклональных антител и/или с использованием анти-CD3 антитела, анти-CD28 антитела и интерлейкина-2 (IL-2) (см., например, статьи Dudley, Immunother., 26, 332-342 (2003) или Dudley, Clin. Oncol., 26, 5233-5239 (2008)).
На следующей стадии клетки (например, Т-клетки) искусственным способом/генетически модифицируют/трансдуцируют методами, известными в данной области техники (см., например, статью Lemoine, J. Gene Med., 6, 374-386 (2004)). Методы трансдукции клеток (например, Т-клеток) известны в данной области техники, и они включают, но, не ограничиваясь только ими, в случае если осуществляют трансдукцию нуклеиновой кислотой или рекомбинантной нуклеиновой кислотой, например, электропорацию, кальций-фосфатный метод, метод с использованием катионных липидов или липосомный метод. Трансдукцию нуклеиновой кислотой можно осуществить стандартным и высокоэффективным методом с использованием коммерческих реагентов для трансфекции, например, липофектамина (выпускается фирмой Invitrogene, номер по каталогу 11668027). В случае, когда используется вектор, трансдукцию вектором можно осуществить аналогичным методом, как описано выше для нуклеиновой кислоты, при условии, что вектор представляет собой плазмидный вектор (то есть вектор, который не является вирусным вектором). В контексте настоящего изобретения способы трансдукции клеток (например, Т-клеток) включают ретровирусную или лентивирусную трансдукцию Т-клеток, трансфекцию невирусными векторами (например, миникольцевой вектор "спящая красавица"), а также трансфекцию мРНК. "Трансфекция мРНК" представляет собой метод, хорошо известный специалистам в данной области, для временной экспрессии исследуемого белка, в данном контексте антигенсвязывающего рецептора по настоящему изобретению, в клетке, подлежащей трансдукции. Краткое описание метода: клетки можно обработать методом электропорации с использованием мРНК, которая кодирует антигенсвязывающий рецептор по настоящему изобретению, с использованием системы электропорации (такой как, например, система Gene Pulser, фирмы Bio-Rad), затем клетки культивируют в соответствии со стандартным протоколом культивирования клеток (например, Т-клеток), как описано выше (см. статью Zhao и др., Mol. Ther., 13(1) 151-159 (2006)). Трансдуцированная клетка по изобретению представляет собой Т-клетку, наиболее предпочтительно CD8 + Т-клетку, и ее получают лентивирусной или наиболее предпочтительно ретровирусной трансдукцией Т-клетки.
В данном контексте ретровирусными векторами, пригодными для трансдукции Т-клеток, являются векторы, известные в данной области техники, такие как SAMEN CMV/SRa (Clay и др., J. Immunol., 163, 507-513 (1999)), LZRS-id3-IHRES (Heemskerk и др., J. Exp. Med., 186, 1597-1602 (1997)), FeLV (Neil и др., Nature, 308, 814-820 (1984)), SAX (Kantoff и др., Proc. Natl. Acad. Sci. USA, 83, 6563-6567 (1986)), pDOL (Desiderio, J. Exp. Med., 167, 372-388 (1988)), N2 (Kasid и др., Proc. Natl. Acad. Sci. USA, 87, 473-477 (1990)), LNL6 (Tiberghien и др., Blood, 84, 1333-1341 (1994)), pZipNEO (Chen и др., J. Immunol., 153, 3630-3638 (1994)), LASN (Mullen и др., Hum. Gene Ther., 7, 1123-1129 (1996)), pG1XsNa (Taylor и др., J. Exp. Med., 184, 2031-2036 (1996)), LCNX (Sun и др., Hum. Gene Ther., 8, 1041-1048 (1997)), SFG (Gallardo и др., Blood, 90 (1997) и LXSN (Sun и др., Hum. Gene Ther., 8, 1041-1048 (1997)), SFG (Gallardo и др., Blood 90, 952-957 (1997)), HMB-Hb-Hu (Vieillard и др., Proc. Natl. Acad. Sci. USA, 94, 11595-11600 (1997)), pMV7 (Cochlovius и др., Cancer Immunol. Immunother., 46, 61-66 (1998)), pSTITCH (Weitjens и др., Gene Ther, 5, 1195-1203 (1998)), pLZR (Yang и др., Hum. Gene Ther., 10, 123-132 (1999)), pBAG (Wu и др., Hum. Gene Ther., 10, 977-982 (1999)), rKat.43.267bn (Gilham и др., J. Immunother., 25, 139-151 (2002)), pLGSN (Engels и др., Hum. Gene Ther., 14, 1155-1168 (2003)), pMP71 (Engels и др., Hum. Gene Ther., 14, 1155-1168 (2003)), pGCSAM (Morgan и др., J. Immunol., 171, 3287-3295 (2003)), pMSGV (Zhao и др., J. Immunol., 174, 4415-4423 (2005)) или pMX (de Witte и др., J. Immunol., 181, 5128-5136 (2008)). В настоящем изобретении лентивирусным вектором, пригодным для трансдукции клеток (например, Т-клеток) являются, например, лентивирусный вектор PL-SIN (Hotta и др., Nat. Methods., 6(5), 370-376 (2009)), p156RRL-sinPPT-CMV-GFP-PRE/NheI (Campeau и др., PLoS One, 4(8), e6529 (2009)), pCMVR8.74 (Addgene, номер по каталогу No.: 22036), FUGW (Lois и др., Science, 295, (5556), 868-872 (2002)), pLVX-EF1 (Addgene, номер по каталогу No.: 64368), pLVE (Brunger и др., Proc. Natl. Acad. Sci. USA, 111(9), E798-806 (2014)), pCDH1-MCS1-EF1 (Hu и др., Mol Cancer Res., 7(11), 1756-1770 (2009)), pSLIK (Wang и др., Nat. Cell. Biol., 16(4), 345-356 (2014)), pLJM1 (Solomon и др., Nat. Genet., 45(12), 1428-1430 (2013)), pLX302 (Kang и др., Sci. Signal., 6(287), rs13 (2013)), pHR-IG (Xie и др., J. Cereb. Blood Flow Metab., 33(12), 1875-1885 (2013)), pRRLSIN (номер no каталогу No.: 62053), pLS (Miyoshi и др., J. Virol., 72(10), 8150-8157 (1998)), pLL3.7 (Lazebnik и др., J. Biol. Chem., 283(7), 11078-11082 (2008)), FRIG (Raissi и др., Mol. Cell Neurosci., 57, 23-32 (2013)), pWPT (Ritz-Laser и др., Diabetologia., 46(6), 810-821 (2003)), pBOB (Marr и др., J. Mol. Neurosci., 22(1-2), 5-11 (2004)) или pLEX (Addgene, номер по каталогу No.: 27976).
Трансдуцированные Т-клетка/Т-клетки по настоящему изобретению предпочтительно выращивают в контролируемых условиях вне естественной среды. Прежде всего, термин "культивирование" обозначает, что клетки (например, трансдуцированная клетка (клетки) по изобретению), которые получены из многоклеточных эукариот (предпочтительно от пациентов - людей), выращивают in vitro. Культивирование клеток - это лабораторный метод выращивания жизнеспособных клеток, которые отделяются от исходного источника ткани. В данном контексте трансдуцированные клетки по настоящему изобретению культивируют в условиях, обеспечивающих экспрессию антигенсвязывающего рецептора по настоящему изобретению в указанных трансдуцированных клетках или на их поверхности. Условия, обеспечивающие экспрессию или трансген (то есть антигенсвязывающий рецептор по настоящему изобретению) являются общеизвестными в данной области техники и включают, например, использование агонистических анти-CD3- и анти-CD28 антител и добавление цитокинов, таких как интерлейкин 2 (IL-2), интерлейкин 7 (IL-7), интерлейкин 12 (IL-12) и/или интерлейкин 15 (IL-15). После экспрессии антигенсвязывающего рецептора по настоящему изобретению в культивируемой трансдуцированной клетке (например, CD8 + Т-клетке) трансдуцированную клетку выделяют (то есть реэкстрагируют) из культуры (то есть из культуральной среды).
Соответственно, в изобретении также предлагается трансдуцированная клетка, предпочтительно Т-клетка, прежде всего CD8 + Т-клетка, экспрессирующая антигенсвязывающий рецептор, кодируемый молекулой нуклеиновой кислоты по настоящему изобретению, полученной способом по настоящему изобретению.
Молекулы нуклеиновой кислоты
В другом объекте настоящего изобретения предлагаются нуклеиновые кислоты и векторы, кодирующие один или несколько антигенсвязывающих рецепторов по настоящему изобретению. Примеры молекул нуклеиновых кислот, кодирующих антигенсвязывающие рецепторы по настоящему изобретению, включают молекулы с последовательностями SEQ ID NO: 19, 30, 35, 38, 44, 47, 51 и 52. Молекулы нуклеиновых кислот по изобретению могут находиться под контролем регуляторных последовательностей. Например, можно использовать промоторы, энхансеры транскрипции и/или последовательности, которые обеспечивают индукцию экспрессии антигенсвязывающего рецептора по изобретению. В настоящем изобретении молекулы нуклеиновых кислот экспрессируются под контролем конститутивного или индуцибельного промотора. Пригодными промоторами являются, например, промотор CMV (Qin и др., PLoS One, 5(5), e10611 (2010)), промотор UBC (Qin и др., PLoS One, 5(5), e10611 (2010)), PGK (Qin и др., PLoS One, 5(5), e10611 (2010)), промотор EF1A (Qin и др., PLoS One, 5(5), e10611 (2010)), промотор CAGG (Qin и др., PLoS One, 5(5), e10611 (2010)), промотор SV40 (Qin и др., PLoS One, 5(5), e10611 (2010)), промотор COPIA (Qin и др., PLoS One, 5(5), е10611 (2010)), промотор АСТ5С (Qin и др., PLoS One, 5(5), е10611 (2010)), промотор TRE (Qin и др., PLoS One, 5(5), e10611 (2010)), промотор Oct3/4 (Chang и др., Molecular Therapy, 9, S367-S367 (2004) (doi: 10.1016/j.ymthe.2004.06.904)) или промотор Nanog (Wu и др., Cell Res., 15(5), 317-324 (2005)). Следовательно, настоящее изобретение также относится к вектору (векторам), включающим нуклеиновую кислоту (нуклеиновые кислоты), описанные в настоящем изобретении. Термин вектор, использованный в данном контексте, включает кольцевую или линейную молекулу нуклеиновой кислоты, которая может автономно реплицироваться в клетке-хозяина (то есть в трансдуцированной клетке), в которую она была введена. Специалистам в области молекулярной биологии известно множество пригодных векторов, выбор которых зависит от требуемой функции, и включает плазмиды, космиды, вирусы, бактериофаги и другие стандартные векторы, используемые в генной инженерии. Методы, которые хорошо известны специалистам в данной области, можно использовать для конструирования различных плазмид и векторов, см., например, методы, описанные в работе Sambrook и др. (loc cit.) и Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989), (1994). В другом варианте полинуклеотиды и векторы по изобретению можно преобразовывать в липосомы для доставки в клетки-мишени. Ниже более подробно обсуждается использование вектора для клонирования для выделения индивидуальных последовательностей ДНК. Соответствующие последовательности можно переносить в векторы экспрессии, где требуется экспрессия конкретного полипептида. Типичные векторы для клонирования включают pBluescript SK, pGEM, pUC9, pBR322, pGA18 и pGBT9. Типичные векторы экспрессии включают pTRE, pCAL-n-EK, pESP-1, pOP13CAT.
В изобретении также предлагается вектор (векторы), включающий молекулу (молекулы) нуклеиновой кислоты, которая представляет собой регуляторную последовательность, функционально связанную с указанной молекулой (молекулами) нуклеиновой кислоты, кодирующей антигенсвязывающий рецептор, как описано в данном контексте. В настоящем изобретении вектор может быть полицистронным. Такие регуляторные последовательности (элементы контроля) известны специалисту в данной области техники и могут включать промотор, сплайс-кассету, кодон инициации трансляции и сайт трансляции и интеграции для встраивания вставки в вектор (векторы). В контексте настоящего изобретения указанная молекула (молекулы) нуклеиновой кислоты функционально связана с указанными последовательностями, контролирующими экспрессию, что обеспечивает экспрессию в эукариотических или прокариотических клетках. Предусматривается, что указанный вектор (векторы) представляет собой вектор (векторы) экспрессии, который включает молекулу (молекулы) нуклеиновой кислоты, кодирующую антигенсвязывающий рецептор, как определено в данном контексте. Термин "функционально связан" означает смежное расположение, при котором взаимодействие описанных компонентов позволяет им функционировать надлежащим образом. Контролирующая последовательность, функционально связанная с кодирующей последовательностью, лигирована таким образом, что экспрессия кодирующей последовательности достигается в условиях, сопоставимых с контролирующей последовательностью. В случае если контролирующая последовательность является промотором, для специалиста, очевидно, что предпочтительно использовать двуцепочечную нуклеиновую кислоту.
В настоящем изобретении указанный вектор (векторы) является вектором (векторами) экспрессии. Вектор экспрессии представляет собой конструкт, который можно использовать для трансформации выбранной клетки и обеспечивать экспрессию кодирующей последовательности в выбранной клетке. Вектор (векторы) экспрессии может являться, например, клонирующим вектором (векторами), бинарным вектором (векторами) или интегрирующим вектором (векторами). Экспрессия включает транскрипцию молекулы нуклеиновой кислоты, предпочтительно транслируемой в мРНК. Регуляторные элементы, обеспечивающие экспрессию в прокариотических и/или эукариотических клетках, хорошо известны специалистам в данной области техники. В случае эукариотических клеток они обычно содержат промоторы, обеспечивающие инициацию транскрипции, и необязательно сигналы поли-А, обеспечивающие терминацию транскрипции и стабилизацию транскрипта. Возможные регуляторные элементы, обеспечивающие экспрессию в прокариотических клетках-хозяина, включают, например, PL, lac, trp или tac промотор для Е. coli, а примерами регуляторных элементов, обеспечивающих экспрессию в эукариотических клетках-хозяина, являются промоторы АОХ1 или GAL1 для дрожжей или промоторы CMV, SV40, RSV (вирус саркомы Рауса), CMV-энхансер, SV40-энхансер или интрон глобина для клеток млекопитающих и клеток других животных.
Помимо элементов, которые отвечают за инициацию транскрипции, такие регуляторные элементы могут также включать сигналы терминации - транскрипции, такие как сайт SV40-поли-А или сайт tk-поли-А, расположенные за последовательностью полинуклеотида. Более того, в зависимости от используемой системы экспрессии, к кодирующей последовательности указанной последовательности нуклеиновой кислоты могут быть добавлены лидерные последовательности, кодирующие сигнальные пептиды, хорошо известные в области техники, способные направлять полипептид в клеточное пространство или его секрецию в среду, см., также, например, прилагаемые примеры.
Лидерная последовательность (последовательности) образует (образуют) соответствующую фазу с транслируемыми, инициирующими и терминирующими последовательностями и предпочтительно, с лидерной последовательностью, способной направлять секрецию транслированного белка или его части в периплазматическое пространство или внеклеточную среду. Необязательно, гетерологичная последовательность может кодировать антигенсвязывающий рецептор с N-концевым сигнальным пептидом, придающим желаемые характеристики, например, для стабилизации или упрощения очистки экспрессируемого рекомбинантного продукта, см. выше. В данном контексте в данной области техники известны пригодные экспрессионные векторы, такие как экспрессионный вектор кДНК Okayama-Berg pcDV1 (фирмы Pharmacia), pCDM8, pRc/CMV, pcDNA1, pcDNA3 (фирмы In-vitrogene), pEF-DHFR, pEF-ADA или pEF-neo (см. статью Raum и др., Cancer Immunol Immunother, 50, 141-150 (2001)) или pSPORT1 (фирмы GIBCO BRL).
В контексте настоящего изобретения последовательности, контролирующие экспрессию, представляют собой системы эукариотических промоторов в векторах, способных трансформировать или трансфектировать эукариотические клетки, однако можно применять и контролирующие последовательности для прокариотических клеток. После введения вектора в соответствующую клетку, ее поддерживают в условиях, пригодных для высокого уровня экспрессии нуклеотидных последовательностей. Дополнительные регуляторные элементы могут включать энхансеры транскрипции и трансляции. Предпочтительно, описанные выше векторы по изобретению включают селективный маркер и/или маркер для подсчета трансформантов. Селективные маркерные гены, придающие устойчивость к антиметаболитам, как основу для селекции, и подходящие для отбора трансформированных клеток и, например, тканей растений и растений, хорошо известны специалистам в данной области и включают, например, ген dhfr, который придает устойчивость к метотрексату (см. статью Reiss, Plant Physiol., (Life Sci. Adv.) 13, 143-149 (1994)), npt, который придает устойчивость к аминогликозидам, таким как неомицин, канамицин и паромицин (см. статью Herrera-Estrella, EMBO J., 2, 987-995 (1983)) и hygro, который придает устойчивость к гигромицину (см. статью Marsh, Gene, 32, 481-485 (1984)). Описаны дополнительные селективные гены, а именно trpB, который позволяет клеткам использовать индол вместо триптофана, hisD, который позволяет клеткам использовать гистинол вместо гистидина (см. статью Hartman, Proc. Natl. Acad. Sci. USA, 85, 8047 (1988)), маннозо-6-фосфат-изомераза, которая позволяет клеткам использовать маннозу (см. заявку WO 94/20627) и ODC (орнитиндекарбоксилаза), которая придает резистентность к ингибитору орнитиндекарбоксилазы, 2-(дифторметил)-DL-орнитину, DFMO (см. работу McConlogue, Current Communications in Molecular Biology, издательство Cold Spring Harbor Laboratory (1987)) или деаминаза из Aspergillus terreus, которая придает резистентность к бластицидину S (см. статью Tamura, Biosci. Biotechnol. Biochem., 59, 2336-2338 (1995)).
Пригодные маркеры для подсчета трансформантов также известны специалистам в данной области техники, и они являются коммерческими продуктами. Преимущественно указанный маркер представляет собой ген, кодирующий люциферазу (см. статьи Giacomin, Pl. Sci., 116, 59-72 (1996), Scikantha, J. Bact., 178, 121 (1996)), зеленый флуоресцентный белок (см. статью Gerdes, FEBS Lett., 389, 44-47 (1996)) или β-глюкуронидазу (см. статью Jefferson, EMBO J., 6, 3901-3907 (1987)). Указанный вариант особенно полезен для простого и быстрого скрининга клеток, тканей и организмов, содержащих указанный вектор.
Как описано выше, указанную молекулу (молекулы) нуклеиновой кислоты можно использовать в отдельности или, как часть вектора (векторов) для экспрессии антигенсвязывающих рецепторов по изобретению в клетках, например, для адаптивной Т-клеточной терапии, но также для целей генной терапии. Нуклеиновые кислоты или вектор (векторы), содержащие последовательность (последовательности) ДНК, колирующую любой один из описанных в данном контексте антигенсвязывающих рецепторов, вводят в клетки, которые в свою очередь продуцируют исследуемый полипептид. Генная терапия, основанная на введении терапевтических генов в клетки методами ех-vivo или in-vivo, представляет собой одно из наиболее важных применений переноса генов. Пригодные векторы, методы или системы доставки генов для методов или систем доставки генов для генной терапии in-vitro или in-vivo описаны в литературе и известны специалистам в данной области техники, см., например, статьи Giordano, Nature Medicine, 2, 534-539 (1996), Schaper, Circ. Res., 79, 911-919 (1996), Anderson, Science, 256, 808-813 (1992), Verma, Nature, 389, 239 (1994), Isner, Lancet, 348, 370-374 (1996), Muhlhauser, Circ. Res., 77, 1077-1086 (1995), Onodera, Blood, 91, 30-36 (1998), Verma, Gene Ther., 5, 692-699 (1998), Nabel, Ann. N.Y. Acad. Sci., 811, 289-292 (1997), Verzeletti, Hum. Gene Ther., 9, 2243-2251 (1998), Wang, Nature Medicine, 2, 714-716 (1996), заявки WO 94/29469, WO 97/00957, патенты US 5580859, US 5589466 или статью Schaper, Current Opinion in Biotechnology, 7, 635-640 (1996). Указанную молекулу (молекулы) нуклеиновой кислоты и вектор (векторы) можно сконструировать для прямого введения в клетку или введения в клетку при помощи липосом или вирусных векторов (например, аденовирусных, ретровирусных). В контексте настоящего изобретения, указанная клетка представляет собой Т-клетки, такие как CD8 + Т-клетки, CD4 + Т-клетки, CD3 + Т-клетки, γδ Т-клетки или природные Т-клетки-киллеры (NK), предпочтительно CD8 + Т-клетки.
В соответствии с вышеизложенным, настоящее изобретение относится к способам получения векторов, прежде всего, плазмид, космид и бактериофагов, обычно используемых в генной инженерии, которые включают молекулу нуклеиновой кислоты, кодирующую последовательность полипептида антигенсвязывающего рецептора, как описано в данном контексте. В контексте настоящего изобретения, указанный вектор представляет собой вектор экспрессии и/или переноса гена или направленный вектор. Для доставки указанных полинуклеотидов или векторов в популяции клеток-мишеней можно использовать векторы экспрессии, полученные на основе вирусов, таких как ретровирусы, вирус осповакцины, аденоассоциированный вирус, вирус герпеса или вирус папилломы крупного рогатого скота.
Методы, хорошо известные специалистам в данной области, можно использовать для конструирования рекомбинантного вектора (векторов), см., например, методики, описанные в работах Sambrook и др. (loc cit.), Ausubel (1989, loc cit.) или в других стандартных руководствах. В другом варианте, указанные молекулы нуклеиновой кислоты и векторы можно встраивать в липосомы для доставки в клетки-мишени. Векторы, содержащие молекулы нуклеиновой кислоты по изобретению, можно переносить в клетки-хозяина с помощью хорошо известных способов, которые изменяются в зависимости от типа клетки-хозяина. Например, трансфекцию с помощью хлорида кальция часто используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно использовать для других клеток-хозяина, см. Sambrook, выше. Указанным вектором, среди прочих, может являться pEF-DHFR, pEF-ADA или pEF-neo. Векторы pEF-DHFR, pEF-ADA и pEF-neo описаны в литературе, например, в статьях Mack и др., Proc. Natl. Acad. Sci. USA, 92, 7021-7025 (1995) и Raum и др., Cancer Immunol. Immunother, 50, 141-150 (2001).
В изобретении предлагается также Т-клетка, трансформированная или трансфектированная вектором, как описано в данном контексте. Указанную Т-клетку можно получить при введении в Т-клетку или ее клетку-предшественник по меньшей мере одного из описанных выше векторов или по меньшей мере одной из описанных выше молекул нуклеиновой кислоты. Присутствие по меньшей мере одного указанного вектора или по меньшей мере одной молекулы нуклеиновой кислоты в Т-клетке может опосредовать экспрессию гена, кодирующего описанный выше антигенсвязывающий рецептор, включающий внеклеточный домен, включающий антигенсвязывающий фрагмент, способный специфически связываться с мутированным Fc доменом. Вектор по настоящему изобретению может являться полицистронным.
Описанную молекулу (молекулы) нуклеиновой кислоты или вектор (векторы), которые вводят в Т-клетку или ее клетку-предшественник, можно либо интегрировать в геном клетки, либо поддерживать его вне хромосомы.
Опухолеспецифические антигены
Как описано выше (Ig-производный) домен (домены) описанного в данном контексте антитела, включающего мутированный Fc домен, может включать сайт взаимодействия с антигеном, специфичный в отношении молекулы клеточной поверхности, то есть в отношении опухолеспецифического антигена, который расположен на поверхности природной опухолевой клетки. В контексте настоящего изобретения, указанные антитела обеспечивают физическое контактирование трансдуцированных Т-клеток, как описано в данном контексте, включающих антигенсвязывающий рецептор по изобретению, и опухолевой клетки, при этом трансдуцированная Т-клетка активируется. Активация трансдуцированных Т-клеток по настоящему изобретению может привести к лизису опухолевой клетки, как описано в данном контексте.
Примеры онкомаркеров, расположенных на поверхности природных опухолевых клеток, приводятся ниже и включают, но, не ограничиваясь только ими, FAP (белок активации фибробластов), СЕА (карциноэмбриональный антиген), р95 (p95HER2), ВСМА (В-клеточный антиген созревания), ЕрСАМ (молекула адгезии эпителиальных клеток), MSLN (мезотелин), MCSP (ассоциированный с меланомой хондроитинсульфат-протеогликан), HER-1 (эпидермальный фактор роста 1 человека), HER-2 (эпидермальный фактор роста 2 человека), HER-3 (эпидермальный фактор роста 3 человека), CD19, CD20, CD22, CD33, CD38, CD52Flt3, рецептор 1 фолиевой кислоты (FOLR1), поверхностный антиген-2 трофобластов человека (Trop-2), опухолевый антиген 12-5 (СА-12-5), лейкоцитарный антиген человека - связанный с D-антигеном (HLA-DR), MUC-1 (муцин-1), А33-антиген, PSMA (простатический специфический мембранный антиген), FMS-подобную тирозинкиназу 3 (FLT-3), PSMA (простат-специфический мембранный антиген), PSCA (антиген стволовых клеток предстательной железы), рецептор трансферрина, TNC (тенасцин), карбоангидразу IX (CA-IX), и/или пептиды, связанные с молекулой главного комплекса гистосовместимости человека (МНС).
Соответственно, в настоящем изобретении антигенсвязывающий рецептор, как описано в данном контексте, связывается с мутированным Fc доменом антитела, то есть терапевтическим антителом, способным специфически связываться с антигеном/маркером, расположенным на поверхности природных опухолевых клеток, который выбирают из группы, состоящей из FAP (белок активации фибробластов), СЕА (карциноэмбриональный антиген), р95 (p95HER2), ВСМА (В-клеточный антиген созревания), ЕрСАМ (молекула адгезии эпителиальных клеток), MSLN (мезотелин), MCSP (ассоциированный с меланомой хондроитинсульфат-протеогликан), HER-1 (эпидермальный фактор роста 1 человека), HER-2 (эпидермальный фактор роста 2 человека), HER-3 (эпидермальный фактор роста 3 человека), CD19, CD20, CD22, CD33, CD38, CD52Flt3, рецептора 1 фолиевой кислоты (FOLR1), поверхностного антигена-2 трофобластов человека (Trop-2), опухолевого антигена 12-5 (СА-12-5), лейкоцитарного антигена человека - связанного с D-антигеном (HLA-DR), MUC-1 (муцин-1), А33-антигена, PSMA (простат-специфический мембранный антиген), FMS-подобной тирозинкиназы 3 (FLT-3), PSMA (простат-специфический мембранный антиген), PSCA (антиген стволовых клеток предстательной железы), рецептора трансферрина, TNC (тенасцин), карбоангидразы IX (CA-IX), и/или пептидов, связанных с молекулой главного комплекса гистосовместимости человека (МНС).
Последовательность (последовательности) элементов антигенов (человека), таких как А33-антиген, ВСМА (В-клеточный антиген созревания), опухолевый антиген 12-5 (СА-12-5), карбоангидраза IX (CA-IX), CD19, CD20, CD22, CD33, CD38, СЕА (карциноэмбриональный антиген), ЕрСАМ (молекула адгезии эпителиальных клеток), FAP (белок активации фибробластов), FMS-подобная тирозинкиназа 3 (FLT-3), рецептор 1 фолиевой кислоты (FOLR1), HER-1 (эпидермальный фактор роста 1 человека), HER-2 (эпидермальный фактор роста 2 человека), HER-3 (эпидермальный фактор роста 3 человека), лейкоцитарный антиген человека - связанный с D-антигеном DR (HLA-DR), MSLN (мезотелин), MCSP (ассоциированный с меланомой хондроитинсульфат-протеогликан), MUC-1 (муцин-1), PSMA (простат-специфический мембранный антиген), PSMA (простат-специфический мембранный антиген), PSCA (антиген стволовых клеток предстательной железы), р95 (p95HER2), рецептор трансферрина, TNC (тенасцин), поверхностный антиген-2 трофобластов человека (Trop-2), доступны в базе данных UniProtKB/Swiss-Prot и могут быть получены на интернет-ресурсе http://www.uniprot.org/uniprot/?query=reviewed%3Ayes. Указанные (белковые) последовательности также могут относиться к аннотированным модифицированным последовательностям. В настоящем изобретении также предлагаются методики и методы, где используются гомологичные последовательности, а также генетические аллельные варианты и т.п. точных последовательностей, описанных в данном контексте. Предпочтительно в данном контексте используются указанные варианты и подобные точные последовательности. Предпочтительно указанными вариантами являются генетические варианты. Специалист в данной области может легко определить соответствующую кодирующую область указанных (белковых) последовательностей в указанных записях баз данных, которые также могут включать запись, относящуюся к геномной ДНК, а также к мРНК/кДНК. Последовательность (последовательности) FAP (белок активации фибробластов) (человека) можно получить из записи Q12884 (версия записи 168, версия последовательности 5) базы данных Swiss-Prot, последовательность (последовательности) СЕА (карциноэмбриональный антиген) (человека) можно получить из записи Р06731 (версия записи 171, версия последовательности 3) базы данных Swiss-Prot, последовательность (последовательности) ЕрСАМ (молекула адгезии эпителиальных клеток) (человека) можно получить из записи Р16422 (версия записи 117, версия последовательности 2) базы данных Swiss-Prot, последовательность (последовательности) MSLN (мезотелин) (человека) можно получить из записи Q13421 (номер версии 132, версия последовательности 2) базы данных UniProt, последовательность (последовательности) FMS-подобной тирозинкиназы 3 (FLT-3) (человека) можно получить из записи Р36888 (первичный цитируемый номер доступа) базы данных Swiss-Prot или Q13414 (вторичный цитируемый номер доступа), версия 165, версия последовательности 2, последовательность MCSP (ассоциированный с меланомой хондроитинсульфат-протеогликан) (человека) можно получить из записи Q6UVK1 (номер версии 118, версия последовательности 2) базы данных UniProt, последовательность (последовательности) рецептора 1 фолиевой кислоты (FOLR1) (человека) можно получить из номера записи Р15328 (первичный цитируемый номер доступа) базы данных UniProt или Q53EW2 (вторичный цитируемый номер доступа), версия 153, версия последовательности 3, последовательность (последовательности) поверхностного антигена-2 трофобластов (Trop-2) (человека) можно получить из номера записи Р09758 (первичный цитируемый номер доступа) базы данных UniProt или Q15658 (вторичный цитируемый номер доступа), версия 172, версия последовательности 3, последовательность (последовательности) PSCA (антиген стволовых клеток предстательной железы) (человека) можно получить из записи 043653 (первичный цитируемый номер доступа) базы данных UniProt или Q6UW92 (вторичный цитируемый номер доступа), версия 134, версия последовательности 1, последовательность (последовательности) HER-1 (эпидермальный фактор роста) (человека) можно получить из записи Р00533 (версия записи 177, версия последовательности 2) базы данных Swiss-Prot, последовательность (последовательности) HER-2 (рецептор тирозинпротеинкиназы erbB-2) (человека) можно получить из записи Р04626 (версия записи 161, версия последовательности 1) базы данных Swiss-Prot, последовательность (последовательности) HER-3 (рецептор тирозинпротеинкиназы erbB-3) (человека) можно получить из записи Р21860 (версия записи 140, версия последовательности 1) базы данных Swiss-Prot, последовательность (последовательности) CD20 (В-лимфоцитарный антиген CD20) (человека) можно получить из записи Р11836 (версия записи 117, версия последовательности 1) базы данных Swiss-Prot, последовательность (последовательности) CD22 (В-лимфоцитарный антиген CD22) (человека) можно получить из записи Р20273 (версия записи 135, версия последовательности 2) базы данных Swiss-Prot, последовательность (последовательности) CD33 (В-лимфоцитарный антиген CD33) (человека) можно получить из записи Р20138 (версия записи 129, версия последовательности 2) базы данных Swiss-Prot, последовательность (последовательности) СА-12-5 (муцин 16) (человека) можно получить из записи Q8WXI7 (версия записи 66, версия последовательности 2) базы данных Swiss-Prot, последовательность (последовательности) HLA-DR (человека) можно получить из записи Q29900 (версия записи 59, версия последовательности 1) базы данных Swiss-Prot, последовательность (последовательности) MUC-1 (муцин-1) (человека) можно получить из записи Р15941 (версия записи 135, версия последовательности 3) базы данных Swiss-Prot, последовательность (последовательности) А33 (антиген А33 клеточной поверхности) (человека) можно получить из записи Q99795 (версия записи 104, версия последовательности 1) базы данных Swiss-Prot, последовательность (последовательности) PSMA (глутаматкарбоксипептидаза 2) (человека) можно получить из записи Q04609 (версия записи 133, версия последовательности 1) базы данных Swiss-Prot, последовательность (последовательности) рецептора трансферрина (человека) можно получить из записи Q9UP52 (версия записи 99, версия последовательности 1) и Р02786 (версия записи 152, версия последовательности 2) базы данных Swiss-Prot, последовательность TNC (тенасцин) (человека) можно получить из записи Р24821 (версия записи 141, версия последовательности 3) базы данных Swiss-Prot или последовательность (последовательности) CA-IX (карбоангидраза IX) (человека) можно получить из записи Q16790 (версия записи 115, версия последовательности 2) базы данных Swiss-Prot.
Терапевтическое применение и способы лечения
Молекулы или конструкты (то есть антигенсвязывающие рецепторы, трансдуцированные Т-клетки и наборы), предложенные в данном контексте, прежде всего пригодны для применения в медицинских учреждениях, предпочтительно для лечения злокачественного заболевания. Например, опухоль можно лечить с использованием трансдуцированной Т-клетки, экспрессирующей антигенсвязывающий рецептор по настоящему изобретению, в сочетании с терапевтическим антителом, специфичным к опухолевой клетке и содержащим мутированный Fc домен. Соответственно, в некоторых вариантах антигенсвязывающий рецептор, трансдуцированная Т-клетка или набор используются для лечения злокачественного заболевания, прежде всего, где злокачественное заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
Опухолеспецифичность лечения обеспечивается терапевтическим антителом, содержащим мутированный Fc домен, где антитело вводят до, одновременно с введением или после введения трансдуцированной Т-клетки, экспрессирующей антигенсвязывающий рецептор по изобретению. В данном контексте, трансдуцированные Т-клетки являются универсальными Т-клетки, поскольку они не являются специфичными в отношении определенной опухоли, но их можно направлять к любой опухоли в зависимости от терапевтического антитела, включающего мутированный Fc домен по настоящему изобретению.
В данном контексте злокачественное заболевание может представлять собой рак/карциному эпителиального, эндотелиального или мезотелиального происхождения или рак крови. В настоящем изобретении рак/карциному выбирают из группы, состоящей из рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака ротовой полости, рака желудка, рака шейки матки, В- и Т-клеточной лимфомы, миелоидного лейкоза, рака яичников, лейкоза, лимфолейкоза, назофарингеальной карциномы, рака ободочной кишки, рака предстательной железы, почечно-клеточного рака, рака головы и шеи, рака кожи (меланома), рака мочеполовой системы, например, рака яичек, рака яичников, эндотелиального рака, рака шейки матки и рака почек, рака желчного протока, рака пищевода, рака слюнных желез и рака щитовидной железы или других опухолевидных заболеваний, таких как гематологические опухоли, глиомы, саркомы или остеосаркомы.
Например, опухолевидные заболевания и/или лимфомы можно лечить с помощью конкретного конструкта, направленного против указанного медицинского показания (показаний). Показания для трансдуцированной Т-клетки по настоящему изобретению в сочетании с терапевтическим антителом, содержащим мутированный Fc домен, определяются специфичностью терапевтического антитела к опухолевому антигену. Например, рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи и/или рак ротовой полости можно лечить с использованием антитела, которое включает мутированный Fc домен, где антитело направлено против ЕрСАМ (человека) (в качестве опухолеспецифического антигена, который расположен на поверхности природной опухолевой клетки).
Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против HER1, предпочтительно HER1 человека. Более того, рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи, глиобластому и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против MCSP, предпочтительно MCSP человека. Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи, глиобластому и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против FOLR1, предпочтительно FOLR1 человека. Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи, глиобластому и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против Trop-2, предпочтительно Trop-2 человека. Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи, глиобластому и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против PSCA, предпочтительно PSCA человека. Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи, глиобластому и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против EGFRvIII, предпочтительно EGFRvIII человека. Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи, глиобластому и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против MSLN, предпочтительно MSLN человека. Рак желудка, рак молочной железы и/или рак шейки матки можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против HER2, предпочтительно HER2 человека. Рак желудка и/или рак легких можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против HER3, предпочтительно HER3 человека. В-клеточную лимфому и/или Т-клеточную лимфому можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против CD20, предпочтительно CD20 человека. В-клеточную лимфому и/или Т-клеточную лимфому можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против CD22, предпочтительно CD22 человека. Миелоидный лейкоз можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против CD33, предпочтительно CD33 человека. Рак яичников, рак легких, рак молочной железы и/или рак желудочно-кишечного тракта можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против СА12-5, предпочтительно СА12-5 человека. Рак желудочно-кишечного тракта, лейкоз и/или назофарингеальную карциному можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против HLA-DR, предпочтительно HLA-DR человека. Рак ободочной кишки, рак молочной железы, рак яичников, рак легких и/или рак поджелудочной железы можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против MUC-1, предпочтительно MUC-1 человека. Рак ободочной кишки можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против А33, предпочтительно А33 человека. Рак предстательной железы можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против PSMA, предпочтительно PSMA человека. Рак желудочно-кишечного тракта, рак поджелудочной железы, холангиоцеллюлярный рак, рак легких, рак молочной железы, рак яичников, рак кожи и/или рак ротовой полости можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против рецептора трансферрина, предпочтительно рецептора трансферрина человека. Рак поджелудочной железы, рак легких и/или рак молочной железы можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против рецептора трансферрина, предпочтительно рецептора трансферрина человека. Рак почек можно лечить с использованием трансдуцированной Т-клетки по настоящему изобретению, вводимой до, одновременно с введением или после введения терапевтического антитела, включающего мутированный Fc домен, где антитело направлено против CA-IX, предпочтительно CA-IX человека.
Соответственно, в изобретении также предлагается способ лечения заболевания, злокачественного заболевания, такого как рак эпителиального, эндотелиального или мезотелиального происхождения и/или рак крови. В контексте настоящего изобретения, указанным субъектом является человек.
В настоящем изобретении конкретный способ лечения заболевания включает стадии:
(а) выделение Т-клеток, предпочтительно CD8 + Т-клеток, из организма субъекта,
(б) трансдукция указанных выделенных Т-клеток, предпочтительно CD8 + Т-клеток, антигенсвязывающим рецептором, как описано в данном контексте, и
(в) введение трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток, указанному субъекту.
В контексте настоящего изобретения указанные трансдуцированные Т-клетки, предпочтительно CD8 + Т-клетки, и/или терапевтическое антитело/антитела вводят указанному субъекту совместно с использованием внутривенного вливания.
Кроме того, в настоящем изобретении предлагается способ лечения заболевания, включающий стадии:
(а) выделение Т-клеток, предпочтительно CD8 + Т-клеток, из организма субъекта,
(б) трансдукция указанных выделенных Т-клеток, предпочтительно CD8 + Т-клеток, антигенсвязывающим рецептором, как описано в данном контексте,
(в) необязательно котрансдукция указанных выделенных Т-клеток, предпочтительно CD8 + Т-клеток, рецептором Т-клетки,
(г) размножение Т-клеток, предпочтительно CD8+Т-клеток, в присутствии анти-CD3 и анти-CD28 антител и
(д) введение трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток, указанному субъекту.
Указанную выше стадию (г) (относится к стадии размножения Т-клеток, таких как TIL, в присутствии анти-CD3 и/или анти-CD28 антител) можно также проводить в присутствии (стимулирующих) цитокинов, таких как интерлейкин-2 и/или интерлейкин-15 (IL-15). В контексте настоящего изобретения указанную стадию (г) (относится к стадии размножения Т-клеток, таких как TIL, в присутствии анти-CD3 и/или анти-CD28 антител) можно также проводить в присутствии интерлейкина-12 (IL-12), интерлейкина-7 (IL-7) и/или интерлейкина-21 (IL-21).
Способ лечения дополнительно включает введение антитела, используемого в соответствии с настоящим изобретением. Указанное антитело можно вводить до, одновременно с введением или после введения трансдуцированных Т-клеток. В настоящем изобретении введение трансдуцированных Т-клеток можно осуществлять с использованием внутривенного вливания. В настоящем изобретении трансдуцированные Т-клетки выделены/получены из организма субъекта, нуждающегося в лечении.
Композиции
Более того, в настоящем изобретении предлагаются композиции (лекарственные средства), включающие молекулу (молекулы) антитела с мутированным Fc доменом (доменами), трансдуцированную Т-клетку (клетки), включающую антигенсвязывающий рецептор по изобретению, молекулу (молекулы) нуклеиновой кислоты и вектор (векторы), кодирующие антигенсвязывающие рецепторы по настоящему изобретению, и/или наборы, включающие одну или более указанных композиций. В контексте настоящего изобретения композиция представляет собой фармацевтическую композицию, которая дополнительно включает необязательно пригодные составы носителя, стабилизаторов и/или эксципиентов. Соответственно, в настоящем изобретении предлагается фармацевтическая композиция (лекарственное средство), которая включает молекулу антитела, включающую мутированный Fc домен, как описано в данном контексте, которую вводят в комбинации с трансдуцированной Т-клеткой, включающей антигенсвязывающий рецептор, как описано в данном контексте, и/или композицию, включающую указанную трансдуцированную Т-клетку, где указанную молекулу антитела вводят до, одновременно с введением или после введения трансдуцированных Т-клеток, включающих антигенсвязывающий рецептор по изобретению.
В соответствии с настоящим изобретением термин "фармацевтическая композиция" обозначает композицию для введения пациенту, предпочтительно человеку. Более того, в настоящем изобретении указанный пациент страдает от заболевания, где указанное заболевание представляет собой злокачественное заболевание, прежде всего онкозаболевания/карциномы эпителиального, эндотелиального или мезотелиального происхождения или рак крови. В настоящем изобретении онкозаболевания/карциномы выбирают из группы, состоящей из рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака ротовой полости, рака желудка, рака шейки матки, В- и Т-клеточной лимфомы, миелоидного лейкоз, рака яичников, лейкоза, лимфолейкоза, назофарингеальной карциномы, рака ободочной кишки, рака предстательной железы, почечно-клеточного рака, рака головы и шеи, рака кожи (меланома), опухоли мочеполовой системы, например, рак яичек, эндотелиальный рак, рак шейки матки и рак почек, рак желчного протока, рак пищевода, рак слюнных желез и рак щитовидной железы, или из других опухолевидных заболеваний, таких как гематологические опухоли, глиомы, саркомы или остеосаркомы.
В предпочтительном варианте фармацевтическая композиция/лекарственное средство включает антитело и/или трансдуцированную Т-клетку, как описано в данном контексте, для парентерального, чрезкожного, внутрипросветного, внутриартериального, внутривенного, внутриоболочечного введения или для инъекции непосредственно в ткань или опухоль. В настоящем изобретении композиция/лекарственное средство включает антитело, включающее мутированный Fc домен, как описано в данном контексте, которое вводят до, одновременно с введением или после введения трансдуцированных Т-клеток, включающих антигенсвязывающий рецептор, как описано в данном контексте. В настоящем изобретении фармацевтическую композицию/лекарственное средство, включающие антитело, как описано в данном контексте, вводят в комбинации с композицией/лекарственным средством, включающими трансдуцированные Т-клетки, включающие антигенсвязывающий рецептор, как описано в данном контексте, где указанные Т-клетки получены из организма субъекта, нуждающегося в таком лечении.
Использование термина "в комбинации" не ограничивает порядок, в котором субъекту следует вводить компоненты, предусмотренные курсом лечения. Соответственно, фармацевтическая композиция/лекарственное средство, описанное в данном контексте, относится к введению антитела, как описано выше в данном контексте, которое вводят до, одновременно с введением или после введения трансдуцированных Т-клеток, включающих антигенсвязывающий рецептор по настоящему изобретению. Термин "в комбинации", использованный в данном контексте, также не ограничивает время между введением антитела, как описано выше в данном контексте, и трансдуцированных Т-клеток, содержащих антигенсвязывающий рецептор, как описано в данном контексте. Таким образом, в случае, когда два компонента не вводят одновременно/в комбинации, интервал между введениями может составлять 1 мин, 5 мин, 15 мин, 30 мин, 45 мин, 1 ч, 2 ч, 4 ч, 6 ч, 12 ч, 24 ч, 48 ч или 72 ч или любой пригодный временной интервал, легко определяемый специалистом в данной области техники, и/или как описано в данном контексте.
В настоящем изобретении термин "в комбинации" также включает случай, когда антитело, как описано в данном контексте, и трансдуцированные Т-клетки, включающие антигенсвязывающий рецептор по настоящему изобретению, предварительно инкубируют в виде смеси перед введением субъекту. Таким образом, два компонента можно предварительно инкубировать перед введением, например, в течение 1 мин, 5 мин, 10 мин, 15 мин, 30 мин, 45 мин или 1 ч или в течение любого пригодного интервала времени, который легко может определить специалист в данной области техники. В другом предпочтительном варианте осуществления настоящего изобретения предлагается курс лечения, когда антитело, как описано в данном контексте, и трансдуцированные Т-клетки, включающие антигенсвязывающий рецептор, как описано в данном контексте, вводят одновременно/в комбинации. В контексте настоящего изобретения, антитело, как описано в данном контексте, можно вводить после введения трансдуцированных Т-клеток, включающих антигенсвязывающий рецептор.
Кроме того, термин "в комбинации", используемый в данном документе, не ограничивает заявленные курсы лечения только введением антитела, как описано в данном контексте, и трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток, включающих антигенсвязывающий рецептор по изобретению, непосредственно друг за другом (то есть сначала вводят один из двух компонентов, а затем (через некоторый промежуток времени) вводят другой компонент, но при этом не вводят другое лекарственное средство и/или не проводят любой другой курс лечения в эти промежутки времени. Таким образом, данные курсы лечения также включают раздельное введение молекулы антитела, как описано в данном контексте, и трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток, включающих антигенсвязывающий рецептор по настоящему изобретению, где введения разделены одним или несколькими курсами лечения, необходимыми и/или пригодными для лечения или профилактики заболевания или его симптома. Примеры таких промежуточных курсов лечения включают, но, не ограничиваясь только ими, введение обезболивающих препаратов, введение химиотерапевтических препаратов, хирургическое вмешательство для лечения заболевания или его симптома. Соответственно, курсы лечения, как описано в данном контексте, включают введение антитела, как описано в данном контексте, и трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток, включающих антигенсвязывающий рецептор, как описано в данном контексте, в сочетании ни с одним, с одним или более курсами лечения, пригодными для лечения или профилактики заболевания или его симптома, как описано в данном контексте или известно из данной области техники.
В частности, предусматривается, что указанную фармацевтическую композицию (композиции)/лекарственное средство (средства) следует вводить пациенту с использованием вливания или инъекции. В настоящем изобретении трансдуцированные Т-клетки, включающие антигенсвязывающий рецептор, как описано в данном контексте, следует вводить с использованием вливания или инъекции. Введение пригодных композиций/лекарственных средств можно осуществлять различными способами, то есть с использованием внутривенного, внутрибрюшинного, подкожного, внутримышечного, местного или внутрикожного введения.
Фармацевтическая композиция/лекарственное средство по настоящему изобретению может также содержать фармацевтически приемлемый носитель. Примеры пригодных фармацевтических носителей хорошо известны в данной области техники и включают фосфатно-солевые буферные растворы, воду, эмульсии, такие как эмульсии типа масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Композиции, содержащие такие носители, можно получать с использованием хорошо известных стандартных способов. Указанные фармацевтические композиции можно вводить субъекту в пригодной дозе. Режим дозировки определяют лечащие врачи в зависимости от клинических факторов. Как известно в медицине, дозировки для любого пациента зависят от множества факторов, включая массу пациента, площадь поверхности тела, возраст, конкретное соединение, предназначенное для введения, пол, время и путь введения, общее состояние здоровья, а также необходимость одновременного введения других лекарственных средств. Как правило, курс лечения может представлять собой обычное введение фармацевтической композиции в диапазоне от 1 мкг до 5 г единиц в сутки. Однако, более предпочтительная дозировка при непрерывном вливании может находиться в диапазоне от 0,01 мкг до 2 мг, предпочтительно от 0,01 мкг до 1 мг, более предпочтительно от 0,01 мкг до 100 мкг, еще более предпочтительно от 0,01 мкг до 50 мкг и наиболее предпочтительно от 0,01 мкг до 10 мкг единиц на килограмм массы тела в час. Наиболее предпочтительные дозировки указаны ниже в данном контексте. Положительную динамику можно контролировать при периодической оценке. Дозировка может изменяться, однако предпочтительная дозировка для внутривенного введения ДНК составляет приблизительно от 106 до 1012 копий молекулы ДНК.
Композиции по изобретению можно вводить местным или системным методом. Введение как правило осуществляют парентерально, например, внутривенно, трансдуцированные Т-клетки также можно вводить непосредственно в участок-мишень, например, катетером через артерию в область участка. Составы для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая солевые и буферные растворы. Растворители для парентерального введения включают раствор хлорида натрия, раствор декстрозы в растворе Рингера, раствор декстрозы в растворе натрия хлорида, раствор Рингер-лактат или нелетучие масла. Растворители для внутривенного введения включают растворы для восполнения дефицита жидкости и питательных веществ, растворы для восполнения дефицита электролитов (например, на основе раствора декстрозы в растворе Рингера) и т.п. Могут также присутствовать консерванты или другие добавки, например, такие как противомикробные агенты, антиоксиданты, хелатирующие агенты, инертные газы и т.п. Кроме того, фармацевтическая композиция по настоящему изобретению может содержать белковые носители, например, сывороточный альбумин или иммуноглобулин, предпочтительно человеческого происхождения. Предусматривается, что кроме белковых конструктов антитела или молекул нуклеиновой кислоты или векторов (описанных в настоящем изобретении), кодирующих их, и/или клеток фармацевтическая композиция по изобретению может содержать другие биологически активные агенты, в зависимости от предполагаемого применения фармацевтической композиции. Такие агенты могут представлять собой лекарственные средства, действующие на желудочно-кишечный тракт, лекарственные средства, действующие как цитостатики, лекарственные средства, предупреждающие гиперурикемию, лекарственные средства, ингибирующие иммунные реакции (например, кортикостероиды), лекарственные средства, действующие на систему кровообращения, и/или агенты, такие как костимулирующие Т-клеточные молекулы или цитокины, известные в данной области техники.
Возможными показаниями к применению композиции (композиций)/лекарственного средства (средств) по изобретению являются злокачественные заболевания, такие как рак эпителиального, эндотелиального или мезотелиального происхождения и рак крови, прежде всего злокачественные заболевания эпителиального происхождения/карциномы, такие как рак молочной железы, рак ободочной кишки, рак предстательной железы, рак головы и шеи, рак кожи (меланома), опухоли мочеполовой системы, например, рак яичников, рак яичек, эндотелиальный рак, рак шейки матки и рак почек, рак легких, рак желудка, рак желчного протока, рак пищевода, рак слюнных желез и рак щитовидной железы или другие опухолевидные заболевания, такие как гематологические опухоли, глиомы, саркомы или остеосаркомы.
В изобретении также предлагаются протоколы совместного введения с другими соединениями, например, молекулами, способными обеспечить активирующий сигнал для эффекторных клеток иммунной системы, клеточной пролиферации или стимуляции клеток. Указанные молекулы могут, например, обеспечивать дополнительный первичный активирующий сигнал для Т-клеток (например, представлять собой дополнительные костимулирующие молекулы: молекулы семейства В7, Ox40L, 4.1 BBL, CD40L, анти-CTLA-4, анти-PD-1) или представлять собой дополнительный интерлейкин из суперсемейства цитокинов (например, IL-2).
Композиция по изобретению, описанная выше, может также представлять собой диагностическую композицию, которая необязательно дополнительно включает средства и способы детекции.
Соответственно, в предпочтительных вариантах, предлагаются набор, антигенсвязывающие рецепторы или трансдуцированная Т-клетка, как описано в данном контексте, для использования в качестве лекарственного средства. В настоящем изобретении предлагается антигенсвязывающий рецептор по настоящему изобретению для применения в качестве лекарственного средства, где одно или более антител, включающих мутированный Fc домен, как описано в данном контексте, вводят до, одновременно с введением или после введения трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток, включающих и/или экспрессирующих антигенсвязывающий рецептор, как описано в данном контексте, и где указанные Т-клетки, предпочтительно CD8 + Т-клетки, получены от субъекта, нуждающемуся в лечении. Указанное лекарственное средство можно использовать в способе лечения злокачественных заболеваний, прежде всего рака/карцином эпителиального, эндотелиального или мезотелиального происхождения или рака крови. В настоящем изобретении рак/карциному выбирают из группы, состоящей из рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака ротовой полости, рака желудка, рака шейки матки, В- и Т-клеточной лимфомы, миелоидного лейкоза, рака яичников, лейкоза, лимфолейкоза, назофарингеальной карциномы, рака ободочной кишки, рака предстательной железы, почечно-клеточного рака, рака головы и шеи, рака кожи (меланомы), опухолей мочеполовой системы, например, рака яичек, рака яичников, эндотелиального рака, рака шейки матки и рака почек, рака желчного протока, рака пищевода, рака слюнных желез и рака щитовидной железы или других опухолевидных заболеваний, таких как гематологические опухоли, глиомы, саркомы или остеосаркомы.
Более того, в контексте настоящего изобретения антитело, как описано в данном контексте, включающее мутированный Fc домен, связывается с опухолеспецифическим антигеном, расположенным на поверхности природной опухолевой клетки, где указанную молекулу антитела вводят до, одновременно с введением или после введения трансдуцированных Т-клеток, предпочтительно CD8 + Т-клеток из указанного организма субъекта, включающих антигенсвязывающий рецептор, как описано в данном контексте. В настоящем изобретении рак/карциному выбирают из группы, состоящей из рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, рака ротовой полости, рака желудка, рака шейки матки, В- и Т-клеточной лимфомы, миелоидного лейкоза, рака яичников, лейкоза, лимфолейкоза, назофарингеальной карциномы, рака ободочной кишки, рака предстательной железы, почечно-клеточного рака, рака головы и шеи, рака кожи (меланомы), опухолей мочеполовой системы, например, рака яичек, рака яичников, эндотелиального рака, рака шейки матки и рака почек, рака желчного протока, рака пищевода, рака слюнных желез и рака щитовидной железы или других опухолевидных заболеваний, таких как гематологические опухоли, глиомы, саркомы или остеосаркомы.
Более того, в настоящем изобретении молекула или конструкт (то есть молекула антитела, описанная в данном контексте), включающие один или два связывающих домена направленных к опухолевому антигену/связывающихся/взаимодействующих с ним, предпочтительно с опухолевым антигеном человека (в качестве опухолеспецифического антигена, расположенным на поверхности природной опухолевой клетки), и включающие мутированный Fc домен, где описанные в данном контексте внеклеточные домены антигенсвязывающего рецептора по настоящему изобретению направлены к мутированному Fc домену/связываются/взаимодействуют с ним, предлагаются для лечения рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака ротовой полости. Таким образом, в контексте настоящего изобретения молекула антитела, включающая два связывающих домена, которые направлены к опухолевому антигену/связываются/взаимодействуют с ним, предпочтительно с опухолевым антигеном человека, и включающая мутированный Fc домен, где описанные в данном контексте внеклеточные домены антигенсвязывающего рецептора направлены к мутированному Fc домену /связываются/взаимодействуют с ним, предлагается для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее два домена связывания, которые направлены к опухолевому антигену/связываются/взаимодействуют с ним, предпочтительно с опухолевым антигеном человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака ротовой полости.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против HER1, предпочтительно HER1 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи и/или рака ротовой полости.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против HER2, предпочтительно HER2 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака желудка, рака молочной железы и/или рака шейки матки.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против HER3, предпочтительно HER3 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака желудка и/или рака легких.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против СЕА, предпочтительно СЕА человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против р95, предпочтительно р95 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против ВСМА, предпочтительно ВСМА человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против MSLN, предпочтительно MSLN человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против MCSP, предпочтительно MCSP человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против CD19, предпочтительно CD19 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против CD20, предпочтительно CD20 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении В-клеточной лимфомы и/или Т-клеточной лимфомы.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против CD22, предпочтительно CD22 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении В-клеточной лимфомы и/или Т-клеточной лимфомы.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против CD38, предпочтительно CD38 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против CD52Flt3, предпочтительно CD52Flt3 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против FolR1, предпочтительно FolR1 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против Trop-2, предпочтительно Trop-2 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака желудочно-кишечного тракта, рака поджелудочной железы, холангиоцеллюлярного рака, рака легких, рака молочной железы, рака яичников, рака кожи, глиобластомы и/или рака ротовой полости.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против СА-12-5, предпочтительно СА-12-5 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака яичников, рака легких, рака молочной железы и/или рака желудочно-кишечного тракта.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против HLA-DR, предпочтительно HLA-DR человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака желудочно-кишечного тракта, лейкоза и/или назофарингеальной карциномы.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против MUC-1, предпочтительно MUC-1 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака ободочной кишки, рака молочной железы, рака яичников, рака легких и/или рака поджелудочной железы.
В одном варианте предлагается: (i) молекула антитела, включающая один или два связывающих домена против А33, предпочтительно А33 человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака ободочной кишки.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против PSMA, предпочтительно PSMA человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака предстательной железы.
В одном варианте предлагается: (i) молекула антитела, включающая один или два связывающих домена против PSCA, предпочтительно PSCA человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) молекула антитела, включающая один или два связывающих домена против рецептора трансферрина, предпочтительно рецептора трансферрина человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) антитело, включающее один или два связывающих домена против тенасцина, предпочтительно тенасцина человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
В одном варианте предлагается: (i) молекула антитела, включающая один или два связывающих домена против CA-IX, предпочтительно XA-IX человека, и мутированный Fc домен, и (ii) антигенсвязывающий рецептор по настоящему изобретению, который направлен к мутированному Fc домену/связывается/взаимодействует с ним, для применения при лечении рака почек.
Типичные варианты осуществления настоящего изобретения
1. Антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий антигенсвязывающий фрагмент, где антигенсвязывающий фрагмент способен специфически связываться с кристаллизующимся (Fc) доменом мутированного фрагмента, но не способен специфически связываться с немутированным исходным Fc доменом, где мутированный Fc домен включает по меньшей мере одну аминокислотную замену по сравнению с немутированным исходным Fc доменом.
2. Антигенсвязывающий рецептор по варианту 1, где связывание Fc рецептора с мутированным Fc доменом снижено по сравнению со связыванием Fc рецептора с немутированным исходным Fc доменом, прежде всего, где Fc рецептором является Fcγ рецептор или неонатальный Fc рецептор (FcRn).
3. Антигенсвязывающий рецептор по любому из вариантов 1 или 2, где связывание с Fc рецептором оценивают методом ППР при 25°С.
4. Антигенсвязывающий рецептор по любому из вариантов 1-3, где антигенсвязывающий фрагмент представляет собой scFv, Fab, crossFab или scFab.
5. Антигенсвязывающий рецептор по любому из вариантов 1-4, где заякоренный трансмембранный домен представляет собой трансмембранный домен, который выбирают из группы, состоящей из трансмембранного домена CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, ОХ40, ICOS, DAP10 или DAP12 или его фрагмента.
6. Антигенсвязывающий рецептор по любому из вариантов 1-5, где заякоренным трансмембранным доменом является CD28 трансмембранный домен, прежде всего, где заякоренный трансмембранный домен включает аминокислотную последовательность SEQ ID NO: 11.
7. Антигенсвязывающий рецептор по любому из вариантов 1-6, который дополнительно включает по меньшей мере один стимулирующий сигнальный домен и/или по меньшей мере один костимулирующий сигнальный домен.
8. Антигенсвязывающий рецептор по любому из вариантов 1-7, где по меньшей мере один стимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов.
9. Антигенсвязывающий рецептор по любому из вариантов 1-8, где по меньшей мере один стимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z или его фрагмент, прежде всего, где по меньшей мере один стимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 13.
10. Антигенсвязывающий рецептор по любому из вариантов 1-9, где по меньшей мере один костимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD27, CD28, CD137, ОХ40, ICOS, DAP10 и DAP12, или их фрагментов.
11. Антигенсвязывающий рецептор по любому из вариантов 1-10, где по меньшей мере один костимулирующий сигнальный домен представляет собой CD28 внутриклеточный домен или его фрагмент, прежде всего, где по меньшей мере один костимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 12.
12. Антигенсвязывающий рецептор по любому из вариантов 1-11, где антигенсвязывающий рецептор включает один стимулирующий сигнальный домен, включающий внутриклеточный домен CD3z, или его фрагмент, и где антигенсвязывающий рецептор включает один костимулирующий сигнал домен, включающий внутриклеточный домен CD28, или его фрагмент.
13. Антигенсвязывающий рецептор по варианту 12, где стимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 13, а костимулирующий сигнальный домен включает аминокислотную последовательность SEQ ID NO: 12.
14. Антигенсвязывающий рецептор по любому из вариантов 1-13, где внеклеточный домен соединен с заякоренным трансмембранным доменом, необязательно через пептидный линкер.
15. Антигенсвязывающий рецептор по варианту 14, где пептидный линкер включает аминокислотную последовательность GGGGS (SEQ ID NO: 17).
16. Антигенсвязывающий рецептор по любому из вариантов 1-15, где заякоренный трансмембранный домен соединен с косигнальным доменом или сигнальным доменом, необязательно через пептидный линкер.
17. Антигенсвязывающий рецептор по любому из вариантов 1-16, где сигнальный и/или косигнальный домены соединены, необязательно через по меньшей мере один пептидный линкер.
18. Антигенсвязывающий рецептор по любому из вариантов 1-17, где антигенсвязывающий фрагмент представляет собой scFv фрагмент, где scFv фрагмент присоединен через С-концевой остаток к N-концевому остатку заякоренного трансмембранного домена, необязательно через пептидный линкер.
19. Антигенсвязывающий рецептор по любому из вариантов 1-17, где антигенсвязывающий фрагмент представляет собой Fab фрагмент или crossFab фрагмент, где Fab или crossFab фрагмент присоединен через С-концевой остаток тяжелой цепи к N-концевому остатку заякоренного трансмембранного домена, необязательно через пептидный линкер.
20. Антигенсвязывающий рецептор по любому из вариантов 7-19, где антигенсвязывающий рецептор включает один косигнальный домен, где косигнальный домен присоединен через N-концевой остаток к С-концевому остатку заякоренного трансмембранного домена.
21. Антигенсвязывающий рецептор по варианту 20, где антигенсвязывающий рецептор дополнительно включает один стимулирующий сигнальный домен, где стимулирующий сигнальный домен присоединен через N-концевой остаток к С-концевому остатку костимулирующего сигнального домена.
22. Антигенсвязывающий рецептор по любому из вариантов 1-21, где немутированный исходный Fc домен представляет собой Fc домен IgG1 или IgG4, прежде всего Fc домен IgG1 человека.
23. Антигенсвязывающий рецептор по любому из вариантов 1-22, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235, I253, Н310, Р331, Р329 и Н435 в соответствии с EU системой нумерации, прежде всего, где аминокислотной мутацией является L234A, L235A, I253A, N297A, Н310А, P329G и/или Н435А.
24. Антигенсвязывающий рецептор по любому из вариантов 1-23, где мутированный Fc домен включает аминокислотную замену в положении, выбранном из группы, состоящей из остатка 117, 118, 136, 180, 193, 212, 214 и 318 Fc IgG1 человека (SEQ ID NO: 130), прежде всего, где аминокислотной мутацией является L117A, L118A, I136A, N180A, Н193А, P212G, P214G и/или Н318А.
25. Антигенсвязывающий рецептор по любому из вариантов 1-24, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235 и Р329 в соответствии с EU системой нумерации, предпочтительно аминокислотные мутации L234A, L235A и P329G ("PGLALA").
26. Антигенсвязывающий рецептор по любому из вариантов 1-25, где мутированный Fc домен включает аминокислотную мутацию P329G в соответствии с EU системой нумерации, где связывание Fcγ рецептора с мутированным Fc доменом снижено по сравнению со связыванием Fcγ рецептора с немутированным исходным Fc доменом, прежде всего, где Fcγ рецептор представляет собой FcγRIIIa и/или FcγRIIa человека.
27. Антигенсвязывающий рецептор по любому из вариантов 1-26, где мутированный Fc домен включает аминокислотную замену в положении 212 Fc IgG1 человека (SEQ ID NO: 130), прежде всего, где аминокислотной мутацией является P212G.
28. Антигенсвязывающий рецептор по любому из вариантов 1-24, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из I253, Н310 и Н435 в соответствии с EU системой нумерации, предпочтительно аминокислотные мутации I253A, Н310А и Н435А ("AAA"), где связывание FcRn с мутированным Fc домен снижено по сравнению со связыванием FcRn с немутированным исходным Fc доменом.
29. Антигенсвязывающий рецептор по любому из вариантов 1-24 или 28, где мутированный Fc домен включает аминокислотную замену в положениях 136, 193 и 318 Fc IgG1 человека (SEQ ID NO: 130), прежде всего, где аминокислотной мутацией является I136A, Н193А и Н318А ("AAA").
30. Антигенсвязывающий рецептор по любому из вариантов 1-27, где по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий фрагмент включает:
(i) вариабельную область (VH) тяжелой цепи, включающую:
(а) определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью RYWMN (SEQ ID NO: 1),
(б) CDR Н2 с аминокислотной последовательностью EITPDSSTINYTPSLKD (SEQ ID NO: 2) и
(в) CDR Н3 с аминокислотной последовательностью PYDYGAWFAS (SEQ ID NO: 3), а также
(ii) вариабельную область (VL) легкой цепи, включающую:
(г) определяющую комплементарность область 1 легкой цепи (CDR L) с аминокислотной последовательностью RSSTGAVTTSNYAN (SEQ ID NO: 4),
(д) CDR L2 с аминокислотной последовательностью GTNKRAP (SEQ ID NO: 5) и
(е) CDR L3 с аминокислотной последовательностью ALWYSNHWV (SEQ ID NO: 6).
31. Антигенсвязывающий рецептор по любому из вариантов 1-27 или 30, где по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 8 и SEQ ID NO: 32, и вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 9 и SEQ ID NO: 33.
32. Антигенсвязывающий рецептор по варианту 1-27, 30 или 31, где по меньшей мере один антигенсвязывающий фрагмент включает вариабельную область тяжелой цепи (VH) с последовательностью SEQ ID NO: 8 и вариабельную область легкой цепи (VL) с последовательностью SEQ ID NO: 9.
33. Антигенсвязывающий рецептор по любому из вариантов 1-27 или 30-32, где по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способный специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 7 и SEQ ID NO: 31.
34. Антигенсвязывающий рецептор по варианту 33, включающий аминокислотную последовательность SEQ ID NO: 7.
35. Антигенсвязывающий рецептор по любому из вариантов 1-27 или 30-32, где по меньшей мере один антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает:
а) гибридный полипептид тяжелой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 39 и SEQ ID NO: 48, и
б) полипептид легкой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности, выбранной из группы, состоящей из последовательности SEQ ID NO: 41 и SEQ ID NO: 50.
36. Антигенсвязывающий рецептор по варианту 35, включающий:
а) гибридный полипептид тяжелой цепи с последовательностью SEQ ID NO: 39 и
б) полипептид легкой цепи с последовательностью SEQ ID NO: 41.
37. Антигенсвязывающий рецептор по любому из вариантов 1-24 или 28-29, где по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А ("AAA"), но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий фрагмент включает:
(i) вариабельную область (VH) тяжелой цепи, включающую:
(а) определяющую комплементарность область 1 тяжелой цепи (CDR Н) с аминокислотной последовательностью SYGMS (SEQ ID NO: 53),
(б) CDR Н2 с аминокислотной последовательностью SSGGSY (SEQ ID NO: 54) и
(в) CDR Н3 с аминокислотной последовательностью LGMITTGYAMDY (SEQ ID NO: 55) и
(ii) вариабельную область (VL) легкой цепи, включающую:
(г) определяющую комплементарность область 1 легкой цепи (CDR L) с аминокислотной последовательностью RSSQTIVHSTGHTYLE (SEQ ID NO: 56),
(д) CDR L2 с аминокислотной последовательностью KVSNRFS (SEQ ID NO: 57) и
(е) CDR L3 с аминокислотной последовательностью FQGSHVPYT (SEQ ID NO: 58).
38. Антигенсвязывающий рецептор по любому из вариантов 1-24, 28, 29 или 37, где по меньшей мере один антигенсвязывающий фрагмент способен специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А ("AAA"), но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий фрагмент включает вариабельную область (VH) тяжелой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 61, а также вариабельную область (VL) легкой цепи, включающую аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 62.
39. Антигенсвязывающий рецептор по варианту 1-24, 28, 29 или 37-38, где по меньшей мере один антигенсвязывающий фрагмент включает:
а) вариабельную область тяжелой цепи (VH) с последовательностью SEQ ID NO: 61 и
б) вариабельную область легкой цепи (VL) с последовательностью SEQ ID NO: 62.
40. Антигенсвязывающий рецептор по любому из вариантов 1-24, 28, 29 или 37-39, где по меньшей мере один антигенсвязывающий фрагмент представляет собой scFv, способный специфически связываться с мутированным Fc доменом, включающим мутации I253A, Н310А и Н435А ("AAA"), но не способен специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает аминокислотную последовательность, которая по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентична аминокислотной последовательности SEQ ID NO: 59.
41. Антигенсвязывающий рецептор по варианту 40, включающий аминокислотную последовательность SEQ ID NO: 59.
42. Антигенсвязывающий рецептор по любому из вариантов 1-27 или 30-32, где по меньшей мере один антигенсвязывающий фрагмент представляет собой Fab фрагмент, способный специфически связываться с мутированным Fc доменом, включающим мутацию P329G, но не способный специфически связываться с немутированным исходным Fc доменом, где антигенсвязывающий рецептор включает:
а) гибридный полипептид тяжелой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности SEQ ID NO: 39, и
б) полипептид легкой цепи, который по меньшей мере приблизительно на 95%, 96%, 97%, 98%, 99% или на 100% идентичен аминокислотной последовательности SEQ ID NO: 41.
43. Антигенсвязывающий рецептор по варианту 42, включающий:
а) гибридный полипептид тяжелой цепи с последовательностью SEQ ID NO: 39 и
б) полипептид легкой цепи с последовательностью SEQ ID NO: 41.
44. Выделенный полинуклеотид, кодирующий антигенсвязывающий рецептор по любому из вариантов 1-43.
45. Выделенный полинуклеотид, кодирующий гибридный полипептид тяжелой цепи или полипептид легкой цепи антигенсвязывающего рецептора по любому из вариантов 1-32, 35-39 и 42-43.
46. Композиция, кодирующая антигенсвязывающий рецептор по любому из вариантов 1-32, 35-39 и 42-43, включающая первый выделенный полинуклеотид, кодирующий гибридный полипептид тяжелой цепи, а также второй выделенный полинуклеотид, кодирующий полипептид легкой цепи.
47. Полипептид, кодируемый полинуклеотидом по любому из вариантов 44 или 45 или композицией по варианту 46.
48. Вектор, прежде всего вектор экспрессии, включающий полинуклеотид по варианту 44 или полинуклеотиды по варианту 45.
49. Трансдуцированная Т-клетка, включающая полинуклеотид по варианту 44, композиция по варианту 46 или вектор по варианту 48.
50. Трансдуцированная Т-клетка, способная экспрессировать антигенсвязывающий рецептор по любому из вариантов 1-43.
51. Трансдуцированная Т-клетка по любому из вариантов 49 или 50, где трансдуцированная Т-клетка является котрансдуцированной с Т-клеточным рецептором (TCR), способным специфически связываться с антигеном-мишенью.
52. Набор, включающий:
(А) трансдуцированную Т-клетку, способную экспрессировать антигенсвязывающий рецептор по любому из вариантов 1-43, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
53. Набор, включающий:
(А) выделенный полинуклеотид, кодирующий антигенсвязывающий рецептор по любому из вариантов 1-43, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
54. Набор, включающий:
(А) композицию по варианту 46 или вектор по варианту 48, кодирующий антигенсвязывающий рецептор по любому из вариантов 1-43, и
(Б) антитело, включающее мутированный Fc домен,
где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
55. Набор по любому из вариантов 52-54, где немутированный исходный Fc домен представляет собой Fc домен IgG1 или IgG4, прежде всего Fc домен IgG1 человека.
56. Набор по любому из вариантов 52-55, где связывание Fc рецептора мутированного Fc домена снижено по сравнению со связыванием Fc рецептора немутированного исходного Fc домена, прежде всего, где Fc рецептором является Fcγ рецептор или неонатальный Fc рецептор (FcRn).
57. Набор по варианту 56, где связывание Fc рецептора определяют методом поверхностного плазмонного резонанса (ППР) при 25°С.
58. Набор по любому из вариантов 52-57, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235, I253, Н310, Р331, Р329 и Н435 в соответствии с EU системой нумерации, прежде всего, где аминокислотной мутацией является L234A, L235A, I253A, N297A, Н310А, P329G и/или Н435А.
59. Набор по любому из вариантов 52-58, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из L234, L235 и Р329 в соответствии с EU системой нумерации, предпочтительно аминокислотные мутации L234A, L235A и P329G ("PGLALA").
60. Набор по любому из вариантов 52-59, где мутированный Fc домен включает аминокислотную мутацию P329G в соответствии с EU системой нумерации.
61. Набор по любому из вариантов 52-60, где мутированный Fc домен включает по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из I253, Н310 и Н435 в соответствии с EU системой нумерации, предпочтительно аминокислотные мутации I253A, Н310А иН435А ("AAA").
62. Набор по любому из вариантов 52-61, где антитело, включающее мутированный Fc домен, способно специфически связываться с антигеном на поверхности опухолевой клетки, прежде всего, где антиген выбирают из группы, состоящей из FAP, СЕА, р95, ВСМА, ЕрСАМ, MSLN, MCSP, HER-1, HER-2, HER-3, CD19, CD20, CD22, CD33, CD38, CD52Flt3, FOLR1, Trop-2, CA-12-5, HLA-DR, MUC-1 (муцин), А33-антигена, PSMA, PSCA, рецептора трансферрина, TNC (тенасцин) и CA-IX, и/или с пептидом, связанным с молекулой главного комплекса гистосовместимости человека (МНС).
63. Набор по любому из вариантов 52-62, где антитело, включающее мутированный Fc домен, способно специфически связываться с антигеном, который выбирают из группы, состоящей из белка активации фибробластов (FAP), карциноэмбрионального антигена (СЕА), мезотелина (MSLN), CD20, рецептора 1 фолиевой кислоты (FOLR1) и тенасцина (TNC).
64. Набор по любому из вариантов 52-63 для применения в качестве лекарственного средства.
65. Антигенсвязывающий рецептор по любому из вариантов 1-43 или трансдуцированная Т-клетка по любому из вариантов 49-51 для применения в качестве лекарственного средства, где трансдуцированную Т-клетку, экспрессирующую антигенсвязывающий рецептор вводят до, одновременно с введением или после введения антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
66. Набор по любому из вариантов 52-63 для применения при лечении заболевания, прежде всего для применения при лечении злокачественного заболевания.
67. Антигенсвязывающий рецептор по любому из вариантов 1-43 или трансдуцированная Т-клетка по любому из вариантов 49-51 для применения при лечении злокачественного заболевания, где лечение включает введение трансдуцированной Т-клетки, экспрессирующей антигенсвязывающий рецептор, до, одновременно с введением или после введения антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
68. Антигенсвязывающий рецептор, трансдуцированная Т-клетка или набор для применения по вариантам 66 или 67, где указанные злокачественное заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
69. Антигенсвязывающий рецептор, трансдуцированная Т-клетка или набор для применения по вариантам 66-68, где антитело, включающее мутированный Fc домен, способно специфически связываться с антигеном на поверхности опухолевых клеток, прежде всего, где антиген выбирают из группы, состоящей из FAP, СЕА, р95, ВСМА, ЕрСАМ, MSLN, MCSP, HER-1, HER-2, HER-3, CD19, CD20, CD22, CD33, CD38, CD52Flt3, FOLR1, Trop-2, СА-12-5, HLA-DR, MUC-1 (муцин), А33-антигена, PSMA, PSCA, рецептора трансферрина, TNC (тенасцин) и CA-IX, и/или с пептидом, связанным с молекулой главного комплекса гистосовместимости человека (МНС).
70. Антигенсвязывающий рецептор, трансдуцированная Т-клетка или набор для применения по вариантам 66-69, где антитело, включающее мутированный Fc домен, способно специфически связываться с антигеном, который выбирают из группы, состоящей из белка активации фибробластов (FAP), карциноэмбрионального антигена (СЕА), мезотелина (MSLN), CD20, рецептора 1 фолиевой кислоты (FOLR1) и тенасцина (TNC).
71. Антигенсвязывающий рецептор, трансдуцированная Т-клетка или набор для применения по любому из вариантов 66-70, где трансдуцированная Т-клетка получена из клетки, выделенной из организма субъекта, нуждающегося в лечении.
72. Антигенсвязывающий рецептор, трансдуцированная Т-клетка или набор для применения по любому из вариантов 66-70, где трансдуцированная Т-клетка не получена из клетки, выделенной из организма субъекта, нуждающегося в лечении.
73. Способ лечения заболевания у пациента, включающий введение субъекту трансдуцированной Т-клетки, способной экспрессировать антигенсвязывающий рецептор по любому из вариантов 1-43, и введение до, одновременно с введением или после введения трансдуцированной Т-клетки, терапевтически эффективного количества антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
74. Способ по варианту 73, дополнительно включающий выделение Т-клетки из организма субъекта и получение трансдуцированной Т-клетки при трансдуцировании выделенной Т-клетки полинуклеотидом по варианту 44, композицией по варианту или вектором по варианту 48.
75. Способ по варианту 74, где Т-клетку трансдуцируют ретровирусным или лентивирусным векторным конструктом или невирусным векторным конструктом.
76. Способ по варианту 75, где невирусный векторный конструкт представляет собой миникольцевой вектор "спящая красавица".
77. Способ по любому из вариантов 73-76, где трансдуцированную Т-клетку вводят субъекту внутривенным вливанием.
78. Способ по любому из вариантов 73-77, где трансдуцированная Т-клетка контактирует с анти-CD3 и/или анти-CD28 антителами до введения субъекту.
79. Способ по любому из вариантов 73-78, где трансдуцированная Т-клетка контактирует по меньшей мере с одним цитокином до введения субъекту, предпочтительно с интерлейкином-2 (IL-2), интерлейкином-7 (IL-7), интерлейкином-15 (IL-15) и/или интерлейкином-21, или их вариантами.
80. Способ по любому из вариантов 73-79, где заболевание представляет собой злокачественное заболевание.
81. Способ по любому из вариантов 73-79, где заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
82. Способ индуцирования лизиса клетки-мишени, включающий контактирование клетки-мишени с трансдуцированной Т-клеткой, способной экспрессировать антигенсвязывающий рецептор по любому из вариантов 1-43 в присутствии антитела, включающего мутированный Fc домен, где антигенсвязывающий рецептор способен специфически связываться с мутированным Fc доменом, но не способен специфически связываться с немутированным исходным Fc доменом.
83. Способ по варианту 82, где клетка-мишень представляет собой опухолевую клетку.
84. Способ по любому из вариантов 82 или 83, где клетка-мишень экспрессирует антиген, который выбирают из группы, состоящей из FAP, СЕА, р95, ВСМА, ЕрСАМ, MSLN, MCSP, HER-1, HER-2, HER-3, CD19, CD20, CD22, CD33, CD38, CD52Flt3, FOLR1, Trop-2, CA-12-5, HLA-DR, MUC-1 (муцин), A33-антигена, PSMA, PSCA, рецептора трансферрина, TNC (тенасцин) и CA-IX.
85. Способ по любому из вариантов 82-84, где клетка-мишень экспрессирует антиген, который выбирают из группы, состоящей из белка активации фибробластов (FAP), карциноэмбрионального антигена (СЕА), мезотелина (MSLN), CD20, рецептора 1 фолиевой кислоты (FOLR1) и тенасцина (TNC).
86. Применение антигенсвязывающего рецептора по любому из вариантов 1-43, полинуклеотидов по любому из вариантов 44 и 45 или трансдуцированной Т-клетки по любому из вариантов 49-51 для получения лекарственного средства.
87. Применение по варианту 86, где лекарственное средство предназначено для лечения злокачественного заболевания.
88. Применение по варианту 86, где лекарственное средство предназначено для лечения заболевания.
89. Применение по варианту 87, отличающееся тем, что указанное злокачественное заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
90. Применение по варианту 88, отличающееся тем, что указанное заболевание выбирают из рака эпителиального, эндотелиального или мезотелиального происхождения и рака крови.
Указанные и другие варианты раскрыты и содержатся в примерах настоящего изобретения. Дополнительную литературу, касающуюся любого из антител, способов, применений и соединений, используемых в соответствии с настоящим изобретением, можно найти в публичных библиотеках и базах данных, например, с помощью электронных устройств. Например, можно использовать открытую базу данных "Medline", доступ к которой можно получить по интернет-ссылке http://www.ncbi.nlm.nih.gov/PubMed/medline.html. Специалистам в данной области техники известны другие базы данных и адреса, например, такие как http://www.ncbi.nlm.nih.gov, http://www.infobiogen.fr/, http://www.fmi.ch/biology/research_tools.html, http://www.tigr.org/, и доступ к ним можно получить, например, по интернет-ссылке http://www.lycos.com.
Примеры
Ниже приведены примеры способов и композиций по настоящему изобретению. Следует понимать, что с учетом общего описания, приведенного выше, на практике можно выполнять и другие варианты осуществления настоящего изобретения.
Технология рекомбинантных ДНК
Для работы с ДНК использовали стандартные методы, описанные в руководстве Sambrook и др., Molecular cloning: A laboratory manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии использовали согласно инструкциям фирм производителей. Общая информация о нуклеотидных последовательностях легких и тяжелых цепей иммуноглобулинов человека приведена в книге: Kabat, Е.А. и др., Sequences of Proteins of Immunological Interest, 5-е изд., NIH Publication No 91-3242 (1991).
Секвенирование ДНК
Последовательности ДНК определяли при секвенировании двуцепочечной матрицы.
Синтез генов
Необходимые генные сегменты получали с помощью ПЦР с использованием соответствующих матриц, либо были получены с использованием набора Geneart AG (Регенсбург, Германия) из синтетических олигонуклеотидов и продуктов ПЦР с использованием системы для автоматизированного синтеза генов. Генные сегменты, фланкированные уникальными сайтами расщепления с использованием эндонуклеазы рестрикции, клонировали в стандартных клонирующих/секвенирующих векторах. Плазмидную ДНК выделяли из трансформированных бактерий и определяли концентрацию УФ спектроскопическим методом. Последовательности ДНК субклонированных генных фрагментов подтверждали (верифицировали) секвенированием ДНК. Генные сегменты конструировали с использованием пригодных сайтов рестрикции, чтобы обеспечить субклонирование в соответствующих экспрессионных векторах. Все конструкты имели 5'-концевую последовательность ДНК, кодирующую лидерный пептид, обеспечивающий секрецию белков в эукариотических клетках.
Очистка белков
После стадии фильтрования белки очищали из отфильтрованных супернатантов клеточных культур стандартными способами. Краткое описание метода: антитела наносили на колонку Protein A-Sepharose (фирмы GE Healthcare) и промывали ФСБ. Антитела элюировали при рН 2,8, затем образцы немедленно нейтрализовали. Белок в виде агрегатов отделяли от мономерных антител методом эксклюзионной хроматографии (колонка Superdex 200, фирмы GE Healthcare), используя ФСБ или 20 мМ гистидин, 150 мМ NaCl, рН 6,0. Фракции, содержащие мономерные антитела, объединяли, концентрировали (при необходимости), например, с использованием центрифужного концентратора Amicon Ultra (30 MWCO, фирмы MILLIPORE), замораживали и хранили при -20 или -80°С. Часть образцов затем была использована для проведения анализа белкового препарата и получения аналитических характеристик, например, методом ДСН-ПААГ электрофореза и эксклюзионной хроматографии (SEC).
ДСН-ПААГ электрофорез
Гелевую систему NuPAGE® Pre-Cast (фирмы Invitrogen) использовали в соответствии с инструкцией фирмы-производителя. Прежде всего, использовали 10% или 4-12% гели NuPAGE® Novex® Bis-TRIS Pre-Cast (pH 6,4) и рабочие буферные растворы NuPAGE® MES (гели в восстанавливающих условиях с антиоксидантной буферной добавкой NuPAGE® или MOPS (гели в невосстанавливающихся условиях).
Аналитическая эксклюзионная хроматография
Эксклюзионную хроматографию (SEC) для определения агрегации и олигомерного состояния антител проводили в системе для высокоэффективной жидкостной хроматографии (ВЭЖХ). Краткое описание метода: антитела, очищенные на колонке с иммобилизованным белком А, наносили на колонку Tosoh TSKgel G3000SW в 300 мМ NaCl, 50 мМ KH2PO4/K2HPO4, рН 7,5, в системе Agilent 1100 для ВЭЖХ, или на колонку Superdex 200 (фирмы GE Healthcare) в 2× ФСБ в системе ВЭЖХ Dionex. Содержание элюированного белка определяли количественно по данным УФ-поглощения и интегрированию площадей пиков. В качестве стандарта использовали набор Gel Filtration Standard, фирмы BioRad, артикул 151-1901.
Получение антител
Мутации Pro329Gly, Leu234Ala и Leu235Ala вводили в консервативную область для устранения связывания с Fc-гамма-рецепторами способом, описанным в заявке WO 2012/130831 A1. Соответственно, мутации I253A, Н310А и Н435А ("AAA") вводили в консервативную область для устранения связывания с FcRn. Соответствующие антитела получали в результате котрансфекции клеток НЕK293-EBNA векторами экспрессии млекопитающих с использованием полиэтиленимина. Клетки трансфектировали соответствующими векторами экспрессии тяжелых и легких цепей в соотношении 1:1.
Лентивирусная трансдукция Т-клеток Jurkat NFAT
Для получения лентивирусных векторов соответствующие последовательности ДНК для правильной сборки антигенсвязывающего рецептора клонировали в лентивирусном полинуклеотидном векторе под контролем конститутивно активного немедленно-раннего промотора цитомегаловируса человека (CMV). Ретровирусный вектор содержал посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE), элемент центрального полипуринового тракта (сРРТ), участок начала репликации pUC и ген, кодирующий устойчивость к антибиотикам, способствующую размножению и селекции в бактериях.
Для получения функциональных вирусных частиц осуществляли трансфекцию на основе Lipofectamine LTX™ с использованием клеток Hek293T (АТСС CRL3216), (с плотностью посадки) характеризующиеся 60-70% уровнем конфлюентности, и CAR-содержащих векторов, а также векторов переноса pCMV-VSV-G: pRSV-REV: pCgpV в соотношении 3:1:1:1. Через 48 ч супернатант собирали, центрифугировали в течение 5 мин при 250g для удаления клеточного дебриса и фильтровали через фильтр из полиэфирсульфона с размером пор 0,45 или 0,22 мкм. Концентрированные вирусные частицы (Lenti-x-Concentrator, Takara) использовали для трансдукции клеток Jurkat NFAT (Signosis). Положительно-трансдуцированные клетки сортировали для получения пула или отдельных клонов с использованием сортера FACSARIA (фирмы BD Bioscience). После размножения клеток до соответствующей плотности Т-клетки Jurkat NFAT использовали в экспериментах.
Пример 1
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием опухолевых клеток SUDHDL4, экспрессирующих CD20, в качестве клеток-мишеней, и отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 6А), или пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 6Б), в качестве клеток-мишеней. В качестве IgG использовали IgG GA101 с мутацией P329G LALA, которое, с одной стороны, распознает опухолевый антиген, а с другой стороны, распознается трансдуцированными Т-клетками Jurkat NFAT. Для положительного контроля лунки 96-луночного планшета (Cellstar Greiner-bio-one, САТ-№655185) обрабатывали CD3-антителами (фирмы Biolegend®) в концентрации 10 мкг/мл в фосфатно-солевом буферном растворе (ФСБ) либо при 4°С в течение ночи, либо в течение по меньшей мере 1 ч при 37°С. Лунки, покрытые CD3, дважды промывали ФСБ, после стадии заключительной промывки ФСБ полностью удаляли. Эффекторные клетки или клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток). Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл среды для культивирования клеток. Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 или 1:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one). На следующей стадии готовили серийные разведения GA101 с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2. После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
При совместном культивировании клеток-мишеней и эффекторных клеток в соотношении 5:1 (точки) или 1:1 (квадраты) в течение 20 ч согласно данным, представленным на графиках, наблюдается дозозависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, а также Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, когда в качестве антитела использовали IgG GA101 с мутацией P329G LALA (см. фиг. 6А и Б, данные, выделенные черным цветом). Если использовали IgG GA101 без мутации P329G LALA (фигуры 6А и Б, данные, выделенные серым цветом), активация трансдуцированных Т-клеток Jurkat NFAT не наблюдалась. Каждая точка обозначает среднее значение по двум повторам биологического образца, каждый из которых технически измеряли три раза. Все значения отображены в виде значений, скорректированных по базовой линии. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 2
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием опухолевых клеток SUDHDL4 (фиг. 7В и 7Г) или WSUDLCL2 (фиг. 7А и 7Б), экспрессирующих CD20, в качестве клеток-мишеней, и единый клон Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, в качестве клеток-мишеней. В качестве IgG использовали IgG GA101 с мутацией P329G LALA, которое, с одной стороны, распознает опухолевый антиген, а с другой стороны, распознается трансдуцированными Т-клетками Jurkat NFAT. Эффекторные клетки или клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток). Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл среды для культивирования клеток. Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 10:1, 5:1 или 1:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one). На следующей стадии готовили серийные разведения GA101 с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2. После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
При совместном культивировании клеток-мишеней и эффекторных клеток в соотношении 10:1 (точки), 5:1 (квадраты) или 1:1 (треугольники) в течение 20 ч согласно данным, представленным на графиках, наблюдается дозозависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, когда использовали IgG GA101 с мутацией P329G LALA (см. фиг. 7А-Г, данные, выделенные черным цветом). Если использовали IgG GA101 без мутации P329G LALA (фиг. 7А-Г, данные, выделенные серым цветом), то обнаружена лишь незначительная активация трансдуцированных Т-клеток Jurkat NFAT при самой высокой концентрации антител 1 мкг/мл. Каждая точка обозначает среднее значение по двум техническим измерениям каждого образца. Все значения отображены в виде значений, скорректированных по базовой линии. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 3
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием адгезивных опухолевых клеток NIH/3T3-huFAP cl 19, экспрессирующих FAP, в качестве клеток-мишеней. В качестве эффекторных клеток использовали отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 8А), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 8В). В качестве IgG использовали IgG FAP 4В9 с мутацией P329G LALA, которое, с одной стороны, распознает опухолевый антиген, а с другой стороны, распознается трансдуцированными Т-клетками Jurkat NFAT. IgG DP47/vk3, содержащие мутацию P329G LALA, использовали в качестве контроля изотипов. Для положительного контроля лунки 96-луночного планшета (Greiner-bio-one, САТ-№655185) обрабатывали CD3-антителами (фирмы Biolegend®) в концентрации 10 мкг/мл в фосфатно-солевом буферном растворе (ФСБ) в течение по меньшей мере 1 ч при 37°С. Лунки, покрытые CD3, дважды промывали ФСБ, после стадии заключительной промывки ФСБ полностью удаляли. Прикрепленные NIH/3T3-huFAP cl 19 клетки-мишени один раз промывали ФСБ и отделяли, используя трипсин. Отделенные клетки ресуспендировали в DMEM + 4,5 г LD-глюкозы + L-глютамина +25 мМ HEPES + 10% ФТС и 1% Glutamax. Эффекторные клетки или Т-клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток). Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл среды для культивирования клеток. Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one). На следующей стадии готовили серийные разведения антител с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2. После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
На фиг. 8Б и 8Г представлены данные совместного культивирования Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 8Г), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 8Б), с клетками-мишенями и антителами FAP 4В9 в концентрации 1 мкг/мл по сравнению с различными контрольными условиями.
Люминесцентный сигнал не обнаружен при инкубации Т-клеток Jurkat NFAT в присутствии 1 мкг/мл FAP 4В9 P329G LALA (фиг. 8Б и 8Г, черный треугольник), а также только клеток-мишеней (фиг. 8Б и 8Г, перевернутый черный треугольник).
Люминесцентный сигнал также не обнаружен при совместном культивировании Т-клеток Jurkat NFAT с клетками-мишенями и в присутствии 1 мкг/мл антител FAP 4В9 (фиг. 8Б и фиг. 8Г, черный ромб). При этом CD3-зависимая активация клеток Jurkat NFAT, совместно культивируемых с клетками-мишенями в присутствии 1 мкг/мл антител FAP 4В9, подтверждает их функциональность по данным детекции люминесцентного сигнала (белые точки).
СD3-зависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 8Б, белые квадраты), и активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 8Г, белые квадраты), совместно культивируемых с клетками-мишенями в присутствии 1 мкг/мл антител FAP 4В9, наиболее выражена, о чем свидетельствуют самые высокие из всех люминесцентные сигналы, поскольку они объединяют CAR-опосредованную активацию с СD3-опосредованной активацией. CD3-опосредованный люминесцентный сигнал также обнаружен, когда CAR инкубируют с клетками-мишенями в присутствии 1 мкг/мл антител DP47/vk3 (фиг. 8Б и фиг. 8Г, перевернутые белые треугольники). Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Все значения отображены в виде значений, скорректированных по базовой линии. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 4
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием адгезивных опухолевых клеток MKN45, экспрессирующих СЕА, в качестве клеток-мишеней. В качестве эффекторных клеток использовали отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 9А) или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 9В). Использовали СЕА А5В7 IgG или СЕА Т84 LCHA IgG, оба из которых включают мутацию P329G LALA. Затем IgG DP47/vk3, содержащие мутацию P329G LALA, использовали в качестве контроля изотипов.
Для положительного контроля лунки 96-луночного планшета (Greiner-bio-one, САТ-№655185) обрабатывали CD3-антителами (фирмы Biolegend®) в концентрации 10 мкг/мл в фосфатно-солевом буферном растворе (ФСБ) в течение 1 ч при 37°С. Лунки, покрытые CD3, дважды промывали ФСБ, после стадии заключительной промывки ФСБ полностью удаляли.
Прикрепленные MKN45 клетки-мишени один раз промывали ФСБ и отделяли, используя трипсин. Отделенные клетки ресуспендировали в DMEM + 4,5 г LD-глюкозы + L-глютамина + 25 мМ HEPES + 10% ФТС и 1% Glutamax.
Эффекторные клетки или клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток).
Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл в RPMI-160 + 10% ФТС + 1% Glutamax.
Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one).
На следующей стадии готовили серийные разведения антител с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2.
После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
При совместном культивировании клеток-мишеней и эффекторных клеток в соотношении 5:1 (фиг. 9А и фиг. 9В, точки) в течение 20 ч согласно данным, представленным на графиках, наблюдается дозозависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, а также Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, когда в качестве антитела использовали СЕА А5В7 с мутацией P329G LALA (см. фиг. 9А и фиг. 9В, серые точки). При использовании СЕА Т84 LCHA с мутацией P329G LALA дозозависимая активация наблюдалась только для Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 9А, черные точки). При этом если использовали антитела с мутацией P329G LALA, то активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, обнаружена только при самой высокой концентрации антител 1 мкг/мл.
В случае использования контрольного антитела DP47/vk3 IgG с мутацией P329G LALA (фиг. 9А и фиг. 9В, черные треугольники) активации Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, не обнаружено. Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
На фиг. 9Б и 9Г представлены данные совместного культивирования Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 9Б), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 9Г), с клетками-мишенями и антителами СЕА Т8 LCHA P329G LALA или СЕА А5В7 P329G LALA в концентрации 1 мкг/мл по сравнению с различными контрольными условиями.
Люминесцентный сигнал не обнаружен при инкубации только CAR Т-клеток Jurkat NFAT в присутствии 1 мкг/мл СЕА Т8 LCHA P329G LALA (фиг. 9Б и 9Г, черный ромб), а также только клеток-мишеней (фиг. 9Б и 9Г, белые круги).
Люминесцентный сигнал также не обнаружен при совместном культивировании Т-клеток Jurkat NFAT с клетками-мишенями в присутствии 1 мкг/мл IgG (фиг. 9Б и фиг. 9Г, белый квадрат и белый ромб). При этом CD3-зависимая активация Т-клеток Jurkat NFAT, совместно культивируемых с клетками-мишенями в присутствии 1 мкг/мл IgG, подтверждает их функциональность по данным детекции люминесцентного сигнала (фиг. 9Б и фиг. 9Г, серый крест).
CD3-зависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 9Б, черная звездочка и серая звездочка), и активация Т-клеток NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 9Г, черная звездочка и серая звездочка), совместно культивируемых с клетками-мишенями в присутствии 1 мкг/мл IgG, наиболее выражена, о чем свидетельствуют самые высокие из всех люминесцентные сигналы, поскольку они объединяют CAR-опосредованную активацию с CD3-опосредованной активацией. CD3-опосредованный люминесцентный сигнал также обнаружен, когда CAR инкубируют с клетками-мишенями в присутствии 1 мкг/мл антител DP47/vk3 (фиг. 9Б и фиг. 9Г, серый плюс). Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 5
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием адгезивных опухолевых клеток MKN4, экспрессирующих СЕА в качестве клеток-мишеней. В качестве эффекторных клеток использовали отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 10В), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 10 А). Использовали СН1А1А 98 99 или СЕА hMN14 IgG, оба с мутацией P329G LALA. Кроме того, IgG DP47/vk3, несущее мутацию P329G LALA, использовали в качестве контроля изотипов.
Для положительного контроля лунки 96-луночного планшета (Greiner-bio-one, САТ-№655185) обрабатывали CD3-антителами (фирмы Biolegend®) в концентрации 10 мкг/мл в фосфатно-солевом буферном растворе (ФСБ) в течение 1 ч при 37°С. Лунки, покрытые CD3, дважды промывали ФСБ, после стадии заключительной промывки ФСБ полностью удаляли.
Прикрепленные MKN45 клетки-мишени один раз промывали ФСБ и отделяли, используя трипсин. Отделенные клетки ресуспендировали в DMEM + 4,5 г LD-глюкозы + L-глютамина + 25 мМ HEPES + 10% ФТС и 1% Glutamax.
Эффекторные клетки или клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток).
Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл в RPMI-160 + 10% ФТС + 1% Glutamax.
Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one).
На следующей стадии готовили серийные разведения антител с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2.
После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
После совместного культивирования в течение 20 ч клеток-мишеней и Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, в соотношении 5:1 (фиг. 10А черные и серые точки) никакой активации не обнаружено, в случае использования антитела СЕА hMN14 или антитела СН1А1А 98 99 (фиг. 9А и фиг. 9Б, серые точки). Т-клетки Jurkat NFAT, экспрессирующие анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, характеризуются небольшой активацией в присутствии антитела СЕА hMN14 или антитела СН1А1А 98 99 в концентрации 0,1 и 1 мкг/мл (фиг. 10В, черные и серые точки).
В случае использования контрольного антитела DP47/vk3 IgG с мутацией P329G LALA (фиг. 10А и фиг. 10В, черные треугольники) активации ни Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, ни Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, не обнаружено. Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Все значения отображены в виде значений, скорректированных по базовой линии. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
На фиг. 10Б и 10Г представлены данные совместного культивирования Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. Г), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 9Г), с клетками-мишенями и антителами СЕА hMN14 или антителами СН1А1А 98 99 в концентрации 1 мкг/мл по сравнению с различными контрольными условиями.
Во всех проведенных контрольных экспериментах не обнаружено какого-либо люминесцентного сигнала, за исключением случаев, когда в качестве активирующего агента использовали CD3. Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 6
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием адгезивных опухолевых клеток CT26TNC cl 19, экспрессирующих TNC, в качестве клеток-мишеней. В качестве эффекторных клеток использовали отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 11В), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 11А). В качестве IgG использовали TNCA2B10 с мутацией P329G LALA. Затем IgG DP47/vk3, содержащие мутацию P329G LALA, использовали в качестве контроля изотипов.
Для положительного контроля лунки 96-луночного планшета (фирмы Greiner-bio-one, САТ-№655185) обрабатывали CD3-антителами (фирмы Biolegend®) в концентрации 10 мкг/мл в фосфатно-солевом буферном растворе (ФСБ) в течение 1 ч при 37°С. Лунки, покрытые CD3, дважды промывали ФСБ, после стадии заключительной промывки ФСБ полностью удаляли.
Прикрепленные CT26TNC cl 19 клетки-мишени один раз промывали ФСБ и отделяли, используя трипсин. Отделенные клетки ресуспендировали в RPMI-1630 + 10% ФТС и 1% Glutamax + 15 мкг/мл пуромицина.
Эффекторные клетки или Т-клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток).
Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл в RPMI-160 + 10% ФТС + 1% Glutamax.
Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one).
На следующей стадии готовили серийные разведения антител с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2.
После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
При совместном культивировании клеток-мишеней и эффекторных клеток в соотношении 5:1 (фиг. 11А и фиг. 11В, черные точки) в течение 20 ч согласно данным, представленным на графиках, наблюдается дозозависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, а также Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, когда в качестве антитела использовали TNC А2В10 с мутацией P329G LALA. Если использовали контрольное антитело DP47/vk3 IgG с мутацией P329G LALA (фиг. 11А и 11В, черные точки), то активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD, или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD, не наблюдалась. Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Все значения отображены в виде значений, скорректированных по базовой линии. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
На фиг. 11Б и 11Г представлены данные совместного культивирования Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 11Г), или Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 11Б), с клетками-мишенями и TNC А2В10 в концентрации 1 мкг/мл по сравнению с различными контрольными условиями.
Люминесцентный сигнал не обнаружен при совместной инкубации Т-клеток Jurkat NFAT и клеток-мишеней в присутствии IgG в концентрации 1 мкг/мл (фиг. 11Б и 11Г, белый треугольник). При этом CD3-зависимая активация клеток Jurkat NFAT, совместно культивируемых с клетками-мишенями в присутствии 1 мкг/мл IgG, подтверждает их функциональность по данным детекции люминесцентного сигнала (фиг. 11Б и фиг. 11Г, белый квадрат).
CD3-зависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 11Б, белый круг), и активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 11Г, белый круг), совместно культивируемых с клетками-мишенями в присутствии 1 мкг/мл IgG, наиболее выражена, о чем свидетельствуют самые высокие из всех люминесцентные сигналы, поскольку они объединяют CAR-опосредованную активацию с CD3-опосредованной активацией. CD3-опосредованный люминесцентный сигнал также обнаружен, когда CAR инкубируют с клетками-мишенями в присутствии 1 мкг/мл антител DP47/vk3 (фиг. 11Б и фиг. 11Г, черный ромб). Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Стандартное отклонение указано Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 7
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием адгезивных опухолевых клеток CT26TNC cl 19, экспрессирующих TNC, в качестве клеток-мишеней. В качестве эффекторных клеток использовали отсортированный пул Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 12А). В качестве IgG использовали TNCA2B10 с мутацией P329G LALA. Затем IgG DP47/vk3, содержащие мутацию P329G LALA, использовали в качестве контроля изотипов.
Для положительного контроля лунки 96-луночного планшета (Greiner-bio-one, САТ-№655185) обрабатывали CD3-антителами (фирмы Biolegend®) в концентрации 10 мкг/мл в фосфатно-солевом буферном растворе (ФСБ) в течение 1 ч при 37°С. Лунки, покрытые CD3, дважды промывали ФСБ, после стадии заключительной промывки ФСБ полностью удаляли.
Прикрепленные CT26TNC cl 19 клетки-мишени один раз промывали ФСБ и отделяли, используя трипсин. Отделенные клетки ресуспендировали в RPMI-1630 + 10% ФТС и 1% Glutamax + 15 мкг/мл пуромицина.
Эффекторные клетки или клетки Jurkat NFAT дикого типа подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax (среда для культивирования клеток).
Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл в RPMI-160 + 10% ФТС + 1% Glutamax.
Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one).
На следующей стадии готовили серийные разведения антител с мутацией P329G LALA, направленной к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 0,0001 мкг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2.
После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке.
При совместном культивировании клеток-мишеней и эффекторных клеток в соотношении 5:1 (фиг. 12А, черные точки) в течение 20 ч согласно данным, представленным на графиках, наблюдается дозозависимая активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD, когда в качестве антитела использовали TNC А2В10 с мутацией P329G LALA, начиная с концентрации 0,01 мкг/мл. Если использовали контрольное антитело DP47/vk3 IgG с мутацией P329G LALA (фиг. 12А и 12В, серые точки), то активация Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD, не наблюдалась. Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Все значения отображены в виде значений, скорректированных по базовой линии. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
На фиг. 12Б представлены данные совместного культивирования Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD, с клетками-мишенями и антителами TNC А2В10 в концентрации 1 мкг/мл по сравнению с различными контрольными условиями.
Люминесцентный сигнал не обнаружен при совместной инкубации Т-клеток Jurkat NFAT, экспрессирующих анти-P329G-Fab-CD28ATD-CD28CSD-CD3zSSD, с клетками-мишенями в отсутствии антител (фиг. 12Б, черный квадрат), а также при инкубировании клеток Jurkat NFAT с клетками-мишенями в присутствии антитела TNC А2В10 в концентрации 1 мкг/мл (фиг. 12Б, белые точки). При этом клетки Jurkat NFAT, культивированные совместно с клетками-мишенями в присутствии TNC А2В10 в концентрации 1 мкг/мл, помещенные в лунки, покрытые CD3, характеризуются четким люминесцентным сигналом.
Кроме того, Т-клетки Jurkat NFAT, экспрессирующие анти-P329G--CD28ATD-CD28CSD-CD3zSSD Fab, культивированные совместно с клетками-мишенями в присутствии либо TNC А2В10 в концентрации 1 мкг/мл либо антитела DP47/vk3 в концентрации 1 мкг/мл, помещенные в лунки, покрытые CD3, характеризуются высоким люминесцентным сигналом. Каждая точка обозначает среднее значение по трем техническим измерениям каждого образца. Значения представлены с учетом стандартного отклонения, указанного в виде линий с маркерами.
Пример 8
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием опухолевых клеток SUDHDL4, экспрессирующих CD20, в качестве клеток-мишеней, и пул клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 13А) или анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 13Б) в качестве эффекторных клеток. В качестве IgG использовали GA101 IgG с мутацией P329G LALA, D265A P29G, только с мутацией LALA или без мутаций, которое, с одной стороны, распознает опухолевый антиген, а с другой стороны, распознается Т-клетками Jurkat NFAT. Эффекторные клетки подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax. Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл среды для культивирования клеток. Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 96-луночный планшет для суспензионных культур (фирмы Greiner-bio one). На следующей стадии готовили серийные разведения различных антител, направленных к исследуемому антигену, в среде для культивирования клеток, используя планшет с глубокими лунками объемом 2 мл (фирмы Axygen®). Для получения конечных концентраций в диапазоне от 1 мкг/мл до 10 пг/мл в конечном образце объемом 200 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 50 мкл в различных разведениях. 96-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2. После инкубации в течение 20 ч содержимое каждой лунки перемешивали, закачивая и выпуская содержимое лунки 10 раз с использованием многоканальной пипетки. 100 мкл клеточной суспензии переносили в новый 96-луночный планшет белого цвета с плоским прозрачным дном (фирмы Greiner-bio-one) и добавляли 100 мкл раствора ONE-Glo™ для проведения люциферазного анализа (фирмы Promega). После инкубации в течение 15 мин в темноте на роторном шейкере при 300 об/мин и КТ измеряли уровень люминесценции с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке. Согласно данным, представленным на графиках, дозозависимая активация клеток-мишеней наблюдается только в случаях, когда используются антитела, включающие мутацию P329G или мутации P329G и LALA, но не в случае, когда присутствует только мутация LALA. Кроме того, активация эффекторных клеток не наблюдалась, если использовали антитело GA101 дикого типа.
Пример 9
В данном примере описан анализ по репортерному гену Т-клеток Jurkat NFAT с использованием опухолевых клеток SUDHDL4, экспрессирующих CD20, в качестве клеток-мишеней и пул клеток Jurkat NFAT, экспрессирующих анти-P329G-ds-scFv-CD28ATD-CD28CSD-CD3zSSD (фиг. 14А) или анти-P329G-ds-Fab-CD28ATD-CD28CSD-CD3zSSD (фиг. 14Б) в качестве эффекторных клеток. В качестве IgG использовали GA101 IgG с мутацией P329G LALA, только с мутацией P329G, только с мутацией LALA или без мутаций, которое, с одной стороны, распознает опухолевый антиген, а с другой стороны, распознается Т-клетками Jurkat NFAT. Эффекторные клетки подсчитывали и оценивали их жизнеспособность с использованием Cedex HiRes. Число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл. Для этого соответствующую аликвоту суспензии клеток центрифугировали при 210g в течение 5 мин при комнатной температуре (КТ) и осажденные клетки ресуспендировали в свежей среде RPMI-160 + 10% ФТС + 1% Glutamax. Клетки-мишени, экспрессирующие исследуемый антиген, также подсчитывали и проверяли их жизнеспособность. Число клеток подбирали, аналогично тому, как описано для эффекторных клеток, при этом число клеток подбирали до содержания 1×106 жизнеспособных клеток/мл среды для культивирования клеток. Клетки-мишени (М) и эффекторные клетки (Э) высеивали в соотношении Э:М 5:1 (110000 клеток в лунке) в трех повторах в 384-луночный планшет. На следующей стадии готовили серийные разведения различных антител, направленных к исследуемому антигену, в среде для культивирования клеток, используя 96-луночный планшет. Для получения конечных концентраций в диапазоне от 1 мкг/мл до 10 пг/мл в конечном образце объемом 30 мкл в лунке, в соответствующие лунки пипеткой вносили аликвоту объемом 10 мкл в различных разведениях. 384-луночный планшет центрифугировали в течение 2 мин при 190g и КТ. Планшет, закрытый пленкой Parafilm®, инкубировали при 37°С во влажной атмосфере, содержащей 5% СО2. После инкубации в течение 20 ч добавляли 6 мкл раствора ONE-Glo™ для проведения люциферазного анализа и считывание проводили немедленно с использованием планшет-ридера Tecan® Spark10M, время обнаружения составляло 1 с в лунке. Согласно данным, представленным на графиках, дозозависимая активация клеток-мишеней наблюдается только в случаях, когда использовали антитела, включающие мутации P329G или мутацию P329G и LALA, но не в случае, когда присутствует только мутация LALA. Кроме того, активация эффекторных клеток не наблюдалась, если использовали антитело GA101 дикого типа.
Примеры последовательностей

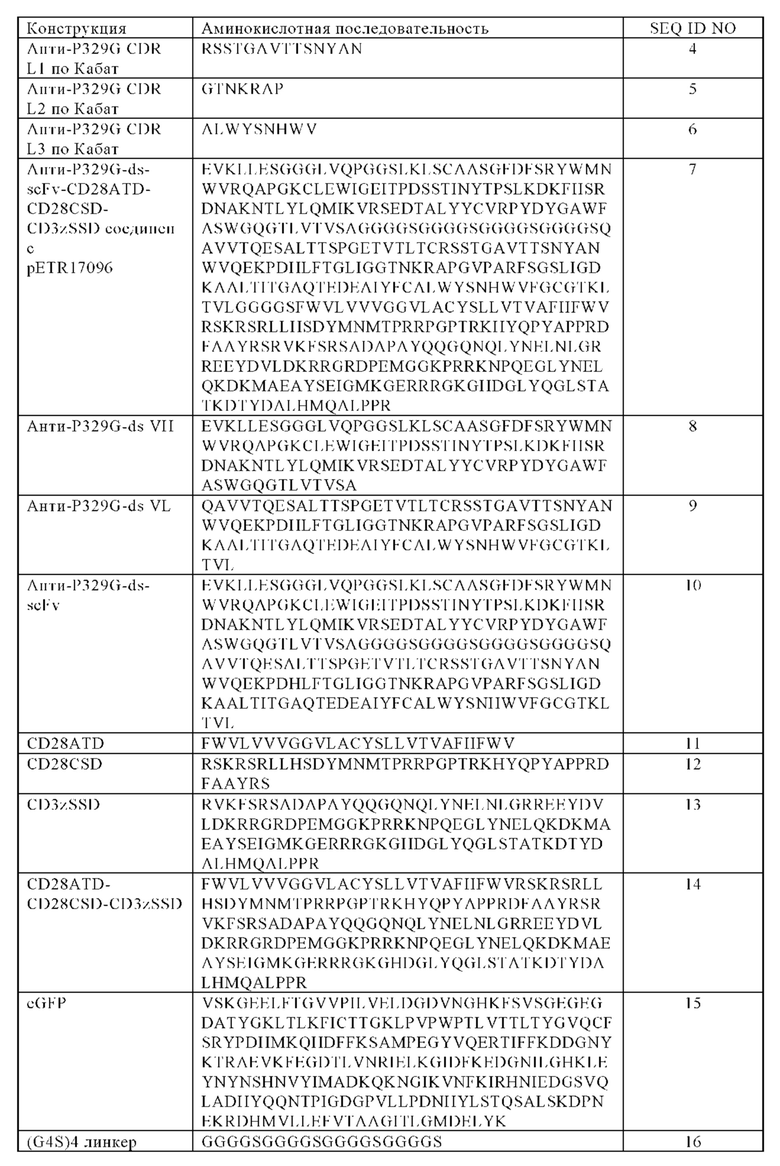





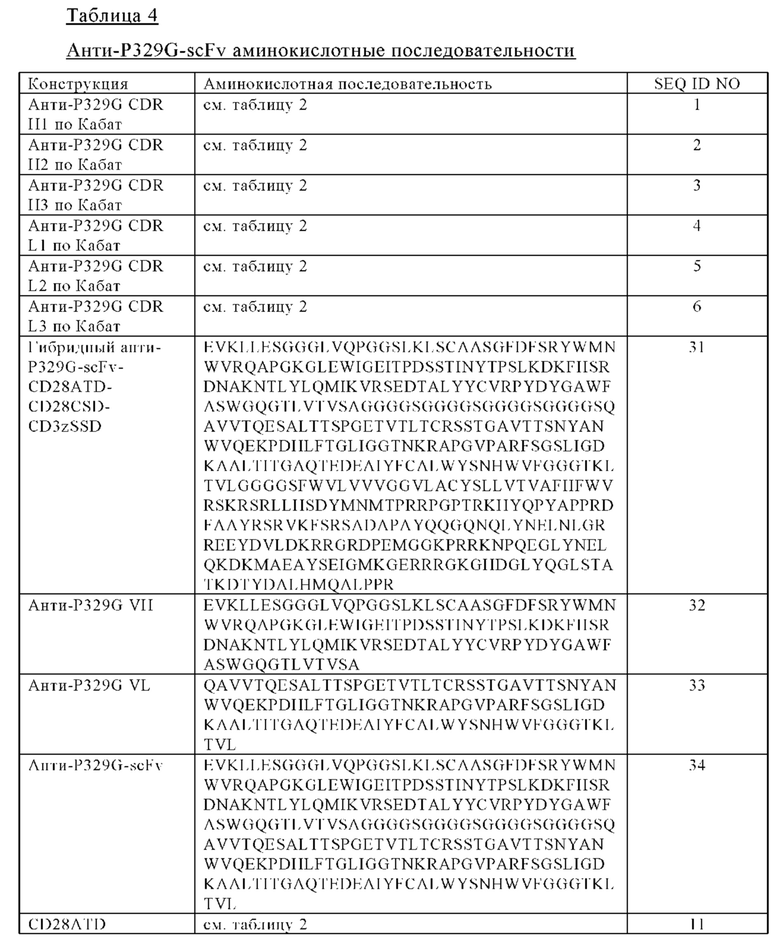


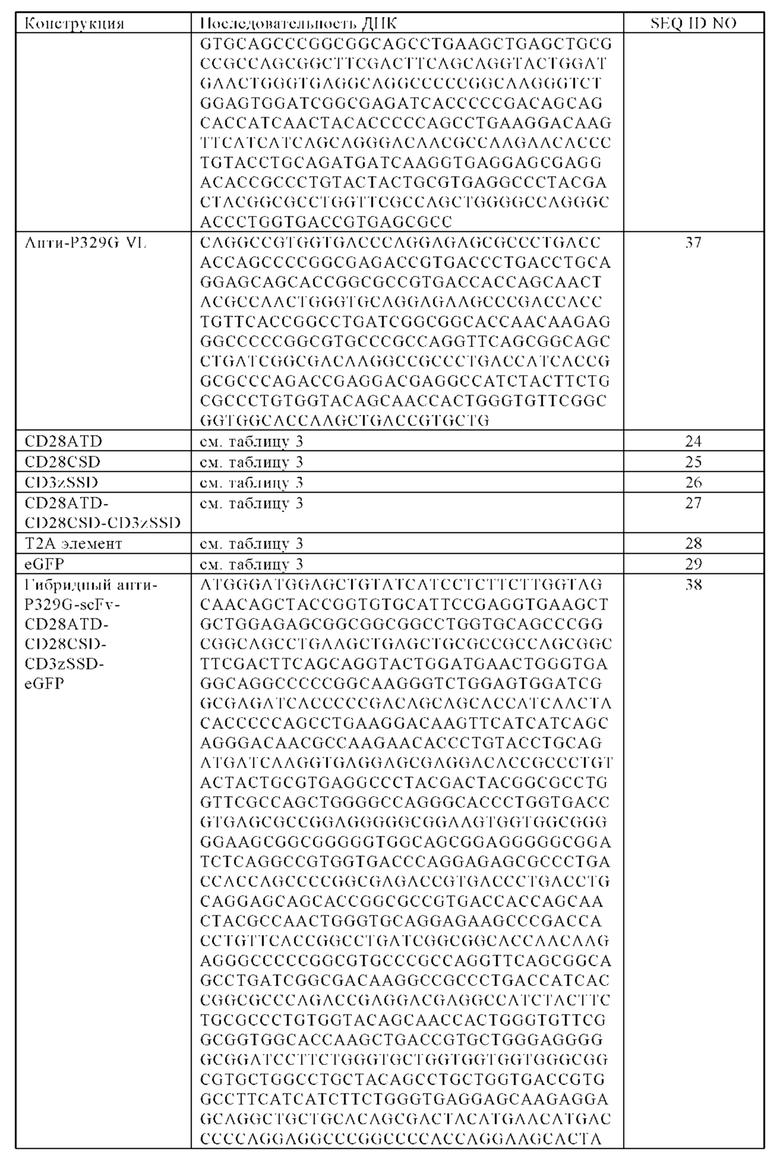
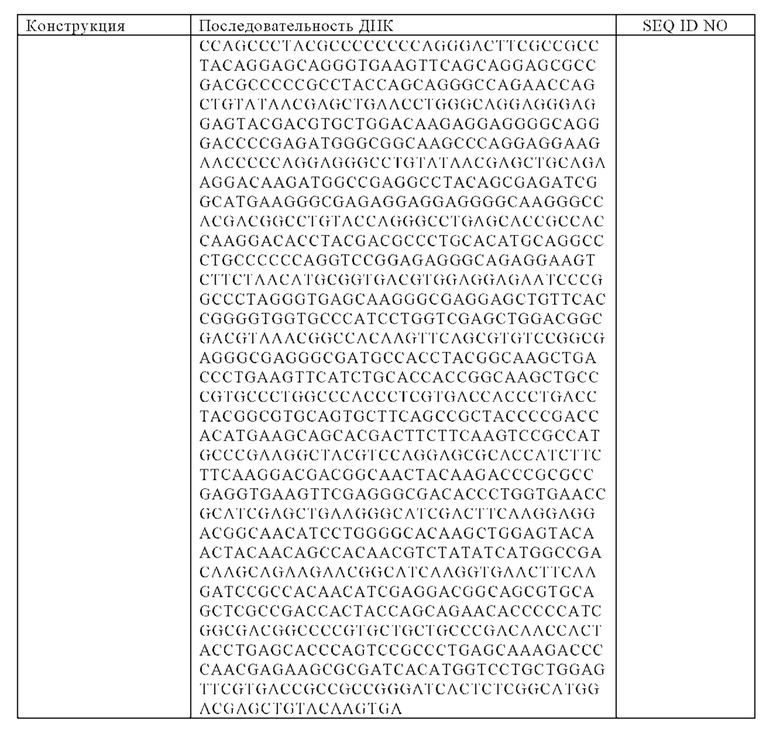


















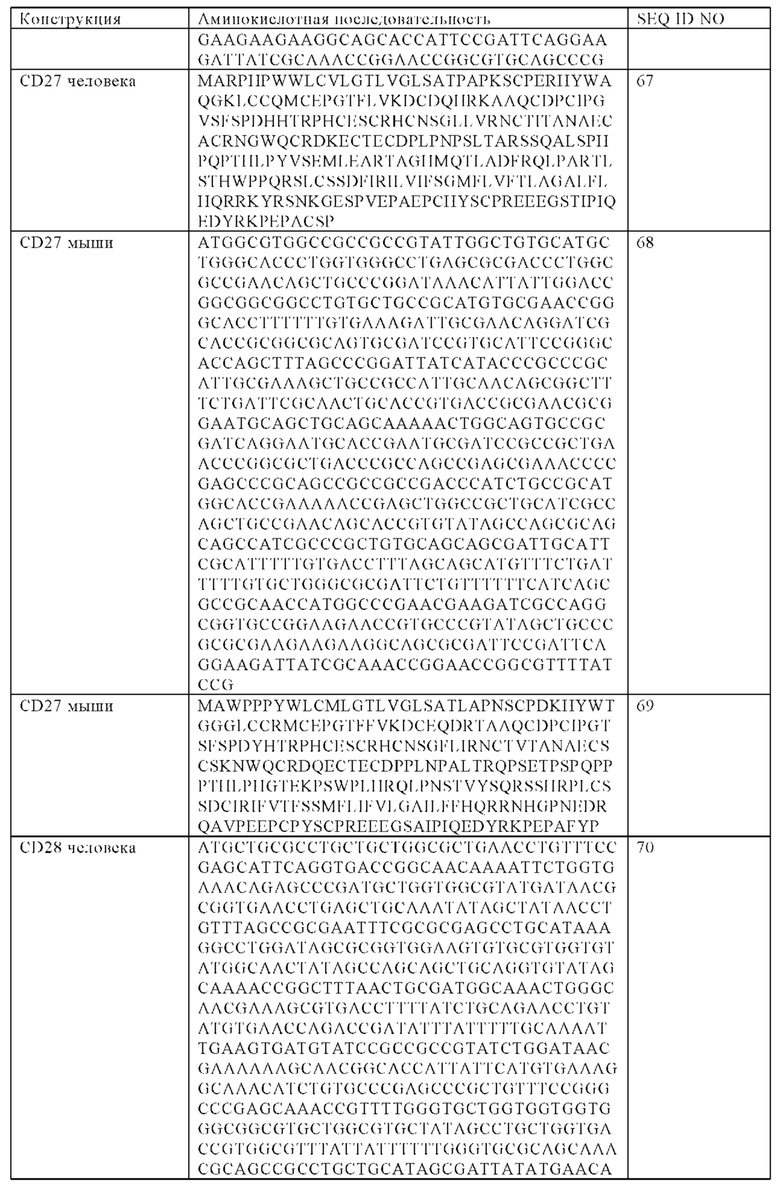

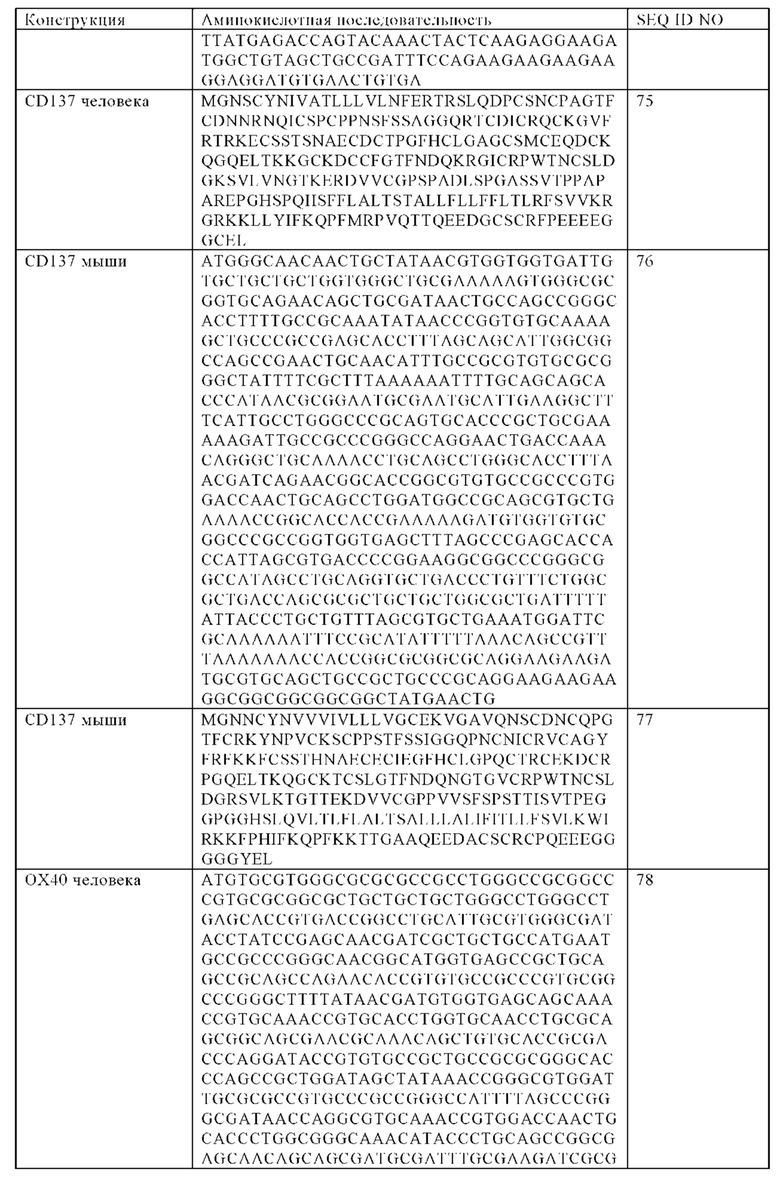



--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф. ХОФФМАНН-ЛЯ РОШ АГ
<120> УЛУЧШЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ РЕЦЕПТОРЫ
<130> P34178
<160> 130
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G CDR H1 Kabat
<400> 1
Arg Tyr Trp Met Asn
1 5
<210> 2
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G CDR H2 Kabat
<400> 2
Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu Lys
1 5 10 15
Asp
<210> 3
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G CDR H3 Kabat
<400> 3
Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser
1 5 10
<210> 4
<211> 14
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G CDR L1 Kabat
<400> 4
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G CDR L2 Kabat
<400> 5
Gly Thr Asn Lys Arg Ala Pro
1 5
<210> 6
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G CDR L3 Kabat
<400> 6
Ala Leu Trp Tyr Ser Asn His Trp Val
1 5
<210> 7
<211> 433
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-scFv-CD28ATM-CD28CSD-CD3zSSD fusion pETR17096
<400> 7
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr
130 135 140
Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu Thr Val Thr Leu Thr
145 150 155 160
Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp
165 170 175
Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly Leu Ile Gly Gly Thr
180 185 190
Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe Ser Gly Ser Leu Ile
195 200 205
Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Thr Glu Asp Glu
210 215 220
Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn His Trp Val Phe Gly
225 230 235 240
Cys Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Phe Trp Val
245 250 255
Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr
260 265 270
Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu
275 280 285
His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg
290 295 300
Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg
305 310 315 320
Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln
325 330 335
Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu
340 345 350
Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly
355 360 365
Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
370 375 380
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
385 390 395 400
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
405 410 415
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
420 425 430
Arg
<210> 8
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-ds VH
<400> 8
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala
115
<210> 9
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-ds VL
<400> 9
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105
<210> 10
<211> 248
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-scFv
<400> 10
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr
130 135 140
Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu Thr Val Thr Leu Thr
145 150 155 160
Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp
165 170 175
Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly Leu Ile Gly Gly Thr
180 185 190
Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe Ser Gly Ser Leu Ile
195 200 205
Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Thr Glu Asp Glu
210 215 220
Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn His Trp Val Phe Gly
225 230 235 240
Cys Gly Thr Lys Leu Thr Val Leu
245
<210> 11
<211> 27
<212> PRT
<213> Artificial sequence
<220>
<223> CD28ATM
<400> 11
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
20 25
<210> 12
<211> 41
<212> PRT
<213> Artificial sequence
<220>
<223> CD28CSD
<400> 12
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
1 5 10 15
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
20 25 30
Pro Arg Asp Phe Ala Ala Tyr Arg Ser
35 40
<210> 13
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> CD3zSSD
<400> 13
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 14
<211> 180
<212> PRT
<213> Artificial sequence
<220>
<223> CD28ATM-CD28-CD3z
<400> 14
Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu
1 5 10 15
Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser
20 25 30
Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly
35 40 45
Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala
50 55 60
Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala
65 70 75 80
Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
85 90 95
Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu
100 105 110
Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
115 120 125
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
130 135 140
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
145 150 155 160
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
165 170 175
Leu Pro Pro Arg
180
<210> 15
<211> 238
<212> PRT
<213> Artificial sequence
<220>
<223> eGFP
<400> 15
Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu Val
1 5 10 15
Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly Glu
20 25 30
Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys
35 40 45
Thr Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu
50 55 60
Thr Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln
65 70 75 80
His Asp Phe Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg
85 90 95
Thr Ile Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val
100 105 110
Lys Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile
115 120 125
Asp Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr Asn
130 135 140
Tyr Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly
145 150 155 160
Ile Lys Val Asn Phe Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val
165 170 175
Gln Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro
180 185 190
Val Leu Leu Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser
195 200 205
Lys Asp Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe Val
210 215 220
Thr Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys
225 230 235
<210> 16
<211> 20
<212> PRT
<213> Artificial sequence
<220>
<223> (G4S)4 linker
<400> 16
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 17
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> G4S linker
<400> 17
Gly Gly Gly Gly Ser
1 5
<210> 18
<211> 19
<212> PRT
<213> Artificial sequence
<220>
<223> T2A linker
<400> 18
Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn
1 5 10 15
Pro Gly Pro
<210> 19
<211> 1356
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-scFv-CD28ATM-CD28CSD-CD3zSSD fusion pETR17096
<400> 19
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctaccggtgt gcattccgag 60
gtgaagctgc tggagagcgg cggcggcctg gtgcagcccg gcggcagcct gaagctgagc 120
tgcgccgcca gcggcttcga cttcagcagg tactggatga actgggtgag gcaggccccc 180
ggcaagtgtc tggagtggat cggcgagatc acccccgaca gcagcaccat caactacacc 240
cccagcctga aggacaagtt catcatcagc agggacaacg ccaagaacac cctgtacctg 300
cagatgatca aggtgaggag cgaggacacc gccctgtact actgcgtgag gccctacgac 360
tacggcgcct ggttcgccag ctggggccag ggcaccctgg tgaccgtgag cgccggaggg 420
ggcggaagtg gtggcggggg aagcggcggg ggtggcagcg gagggggcgg atctcaggcc 480
gtggtgaccc aggagagcgc cctgaccacc agccccggcg agaccgtgac cctgacctgc 540
aggagcagca ccggcgccgt gaccaccagc aactacgcca actgggtgca ggagaagccc 600
gaccacctgt tcaccggcct gatcggcggc accaacaaga gggcccccgg cgtgcccgcc 660
aggttcagcg gcagcctgat cggcgacaag gccgccctga ccatcaccgg cgcccagacc 720
gaggacgagg ccatctactt ctgcgccctg tggtacagca accactgggt gttcggctgt 780
ggcaccaagc tgaccgtgct gggagggggc ggatccttct gggtgctggt ggtggtgggc 840
ggcgtgctgg cctgctacag cctgctggtg accgtggcct tcatcatctt ctgggtgagg 900
agcaagagga gcaggctgct gcacagcgac tacatgaaca tgacccccag gaggcccggc 960
cccaccagga agcactacca gccctacgcc ccccccaggg acttcgccgc ctacaggagc 1020
agggtgaagt tcagcaggag cgccgacgcc cccgcctacc agcagggcca gaaccagctg 1080
tataacgagc tgaacctggg caggagggag gagtacgacg tgctggacaa gaggaggggc 1140
agggaccccg agatgggcgg caagcccagg aggaagaacc cccaggaggg cctgtataac 1200
gagctgcaga aggacaagat ggccgaggcc tacagcgaga tcggcatgaa gggcgagagg 1260
aggaggggca agggccacga cggcctgtac cagggcctga gcaccgccac caaggacacc 1320
tacgacgccc tgcacatgca ggccctgccc cccagg 1356
<210> 20
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds VH
<400> 20
gaggtgaagc tgctggagag cggcggcggc ctggtgcagc ccggcggcag cctgaagctg 60
agctgcgccg ccagcggctt cgacttcagc aggtactgga tgaactgggt gaggcaggcc 120
cccggcaagt gtctggagtg gatcggcgag atcacccccg acagcagcac catcaactac 180
acccccagcc tgaaggacaa gttcatcatc agcagggaca acgccaagaa caccctgtac 240
ctgcagatga tcaaggtgag gagcgaggac accgccctgt actactgcgt gaggccctac 300
gactacggcg cctggttcgc cagctggggc cagggcaccc tggtgaccgt gagcgcc 357
<210> 21
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds VL
<400> 21
caggccgtgg tgacccagga gagcgccctg accaccagcc ccggcgagac cgtgaccctg 60
acctgcagga gcagcaccgg cgccgtgacc accagcaact acgccaactg ggtgcaggag 120
aagcccgacc acctgttcac cggcctgatc ggcggcacca acaagagggc ccccggcgtg 180
cccgccaggt tcagcggcag cctgatcggc gacaaggccg ccctgaccat caccggcgcc 240
cagaccgagg acgaggccat ctacttctgc gccctgtggt acagcaacca ctgggtgttc 300
ggctgtggca ccaagctgac cgtgctg 327
<210> 22
<211> 799
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-scFv
<400> 22
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctaccggtgt gcattccgag 60
gtgaagctgc tggagagcgg cggcggcctg gtgcagcccg gcggcagcct gaagctgagc 120
tgcgccgcca gcggcttcga cttcagcagg tactggatga actgggtgag gcaggccccc 180
ggcaagtgtc tggagtggat cggcgagatc acccccgaca gcagcaccat caactacacc 240
cccagcctga aggacaagtt catcatcagc agggacaacg ccaagaacac cctgtacctg 300
cagatgatca aggtgaggag cgaggacacc gccctgtact actgcgtgag gccctacgac 360
tacggcgcct ggttcgccag ctggggccag ggcaccctgg tgaccgtgag cgccggaggg 420
ggcggaagtg gtggcggggg aagcggcggg ggtggcagcg gagggggcgg atctcaggcc 480
gtggtgaccc aggagagcgc cctgaccacc agccccggcg agaccgtgac cctgacctgc 540
aggagcagca ccggcgccgt gaccaccagc aactacgcca actgggtgca ggagaagccc 600
gaccacctgt tcaccggcct gatcggcggc accaacaaga gggcccccgg cgtgcccgcc 660
aggttcagcg gcagcctgat cggcgacaag gccgccctga ccatcaccgg cgcccagacc 720
gaggacgagg ccatctactt ctgcgccctg tggtacagca accactgggt gttcggctgt 780
ggcaccaagc tgaccgtgc 799
<210> 23
<211> 647
<212> DNA
<213> Artificial sequence
<220>
<223> IRES EV71, internal ribosomal entry side
<400> 23
cccgaagtaa cttagaagct gtaaatcaac gatcaatagc aggtgtggca caccagtcat 60
accttgatca agcacttctg tttccccgga ctgagtatca ataggctgct cgcgcggctg 120
aaggagaaaa cgttcgttac ccgaccaact acttcgagaa gcttagtacc accatgaacg 180
aggcagggtg tttcgctcag cacaacccca gtgtagatca ggctgatgag tcactgcaac 240
ccccatgggc gaccatggca gtggctgcgt tggcggcctg cccatggaga aatccatggg 300
acgctctaat tctgacatgg tgtgaagtgc ctattgagct aactggtagt cctccggccc 360
ctgattgcgg ctaatcctaa ctgcggagca catgctcaca aaccagtggg tggtgtgtcg 420
taacgggcaa ctctgcagcg gaaccgacta ctttgggtgt ccgtgtttcc ttttattcct 480
atattggctg cttatggtga caatcaaaaa gttgttacca tatagctatt ggattggcca 540
tccggtgtgc aacagggcaa ctgtttacct atttattggt tttgtaccat tatcactgaa 600
gtctgtgatc actctcaaat tcattttgac cctcaacaca atcaaac 647
<210> 24
<211> 81
<212> DNA
<213> Artificial sequence
<220>
<223> CD28ATM
<400> 24
ttttgggtgc tggtggtggt tggtggagtc ctggcttgct atagcttgct agtaacagtg 60
gcctttatta ttttctgggt g 81
<210> 25
<211> 123
<212> DNA
<213> Artificial sequence
<220>
<223> CD28CSD
<400> 25
aggagtaaga ggagcaggct cctgcacagt gactacatga acatgactcc ccgccgcccc 60
gggcccaccc gcaagcatta ccagccctat gccccaccac gcgacttcgc agcctatcgc 120
tcc 123
<210> 26
<211> 336
<212> DNA
<213> Artificial sequence
<220>
<223> CD3z SSD
<400> 26
agagtgaagt tcagcaggag cgcagacgcc cccgcgtacc agcagggcca gaaccagctc 60
tataacgagc tcaatctagg acgaagagag gagtacgatg ttttggacaa gagacgtggc 120
cgggaccctg agatgggggg aaagccgaga aggaagaacc ctcaggaagg cctgtacaat 180
gaactgcaga aagataagat ggcggaggcc tacagtgaga ttgggatgaa aggcgagcgc 240
cggaggggca aggggcacga tggcctttac cagggtctca gtacagccac caaggacacc 300
tacgacgccc ttcacatgca ggccctgccc cctcgc 336
<210> 27
<211> 540
<212> DNA
<213> Artificial sequence
<220>
<223> CD28ATM-CD28-CD3z
<400> 27
ttctgggtgc tggtggtggt gggcggcgtg ctggcctgct acagcctgct ggtgaccgtg 60
gccttcatca tcttctgggt gaggagcaag aggagcaggc tgctgcacag cgactacatg 120
aacatgaccc ccaggaggcc cggccccacc aggaagcact accagcccta cgcccccccc 180
agggacttcg ccgcctacag gagcagggtg aagttcagca ggagcgccga cgcccccgcc 240
taccagcagg gccagaacca gctgtataac gagctgaacc tgggcaggag ggaggagtac 300
gacgtgctgg acaagaggag gggcagggac cccgagatgg gcggcaagcc caggaggaag 360
aacccccagg agggcctgta taacgagctg cagaaggaca agatggccga ggcctacagc 420
gagatcggca tgaagggcga gaggaggagg ggcaagggcc acgacggcct gtaccagggc 480
ctgagcaccg ccaccaagga cacctacgac gccctgcaca tgcaggccct gccccccagg 540
<210> 28
<211> 63
<212> DNA
<213> Artificial sequence
<220>
<223> T2A element
<400> 28
tccggagagg gcagaggaag tcttctaaca tgcggtgacg tggaggagaa tcccggccct 60
agg 63
<210> 29
<211> 717
<212> DNA
<213> Artificial sequence
<220>
<223> eGFP
<400> 29
gtgagcaagg gcgaggagct gttcaccggg gtggtgccca tcctggtcga gctggacggc 60
gacgtaaacg gccacaagtt cagcgtgtcc ggcgagggcg agggcgatgc cacctacggc 120
aagctgaccc tgaagttcat ctgcaccacc ggcaagctgc ccgtgccctg gcccaccctc 180
gtgaccaccc tgacctacgg cgtgcagtgc ttcagccgct accccgacca catgaagcag 240
cacgacttct tcaagtccgc catgcccgaa ggctacgtcc aggagcgcac catcttcttc 300
aaggacgacg gcaactacaa gacccgcgcc gaggtgaagt tcgagggcga caccctggtg 360
aaccgcatcg agctgaaggg catcgacttc aaggaggacg gcaacatcct ggggcacaag 420
ctggagtaca actacaacag ccacaacgtc tatatcatgg ccgacaagca gaagaacggc 480
atcaaggtga acttcaagat ccgccacaac atcgaggacg gcagcgtgca gctcgccgac 540
cactaccagc agaacacccc catcggcgac ggccccgtgc tgctgcccga caaccactac 600
ctgagcaccc agtccgccct gagcaaagac cccaacgaga agcgcgatca catggtcctg 660
ctggagttcg tgaccgccgc cgggatcact ctcggcatgg acgagctgta caagtga 717
<210> 30
<211> 2136
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-scFv-CD28ATM-CD28CSD-CD3zSSD-eGFP fusion pETR17096
<400> 30
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctaccggtgt gcattccgag 60
gtgaagctgc tggagagcgg cggcggcctg gtgcagcccg gcggcagcct gaagctgagc 120
tgcgccgcca gcggcttcga cttcagcagg tactggatga actgggtgag gcaggccccc 180
ggcaagtgtc tggagtggat cggcgagatc acccccgaca gcagcaccat caactacacc 240
cccagcctga aggacaagtt catcatcagc agggacaacg ccaagaacac cctgtacctg 300
cagatgatca aggtgaggag cgaggacacc gccctgtact actgcgtgag gccctacgac 360
tacggcgcct ggttcgccag ctggggccag ggcaccctgg tgaccgtgag cgccggaggg 420
ggcggaagtg gtggcggggg aagcggcggg ggtggcagcg gagggggcgg atctcaggcc 480
gtggtgaccc aggagagcgc cctgaccacc agccccggcg agaccgtgac cctgacctgc 540
aggagcagca ccggcgccgt gaccaccagc aactacgcca actgggtgca ggagaagccc 600
gaccacctgt tcaccggcct gatcggcggc accaacaaga gggcccccgg cgtgcccgcc 660
aggttcagcg gcagcctgat cggcgacaag gccgccctga ccatcaccgg cgcccagacc 720
gaggacgagg ccatctactt ctgcgccctg tggtacagca accactgggt gttcggctgt 780
ggcaccaagc tgaccgtgct gggagggggc ggatccttct gggtgctggt ggtggtgggc 840
ggcgtgctgg cctgctacag cctgctggtg accgtggcct tcatcatctt ctgggtgagg 900
agcaagagga gcaggctgct gcacagcgac tacatgaaca tgacccccag gaggcccggc 960
cccaccagga agcactacca gccctacgcc ccccccaggg acttcgccgc ctacaggagc 1020
agggtgaagt tcagcaggag cgccgacgcc cccgcctacc agcagggcca gaaccagctg 1080
tataacgagc tgaacctggg caggagggag gagtacgacg tgctggacaa gaggaggggc 1140
agggaccccg agatgggcgg caagcccagg aggaagaacc cccaggaggg cctgtataac 1200
gagctgcaga aggacaagat ggccgaggcc tacagcgaga tcggcatgaa gggcgagagg 1260
aggaggggca agggccacga cggcctgtac cagggcctga gcaccgccac caaggacacc 1320
tacgacgccc tgcacatgca ggccctgccc cccaggtccg gagagggcag aggaagtctt 1380
ctaacatgcg gtgacgtgga ggagaatccc ggccctaggg tgagcaaggg cgaggagctg 1440
ttcaccgggg tggtgcccat cctggtcgag ctggacggcg acgtaaacgg ccacaagttc 1500
agcgtgtccg gcgagggcga gggcgatgcc acctacggca agctgaccct gaagttcatc 1560
tgcaccaccg gcaagctgcc cgtgccctgg cccaccctcg tgaccaccct gacctacggc 1620
gtgcagtgct tcagccgcta ccccgaccac atgaagcagc acgacttctt caagtccgcc 1680
atgcccgaag gctacgtcca ggagcgcacc atcttcttca aggacgacgg caactacaag 1740
acccgcgccg aggtgaagtt cgagggcgac accctggtga accgcatcga gctgaagggc 1800
atcgacttca aggaggacgg caacatcctg gggcacaagc tggagtacaa ctacaacagc 1860
cacaacgtct atatcatggc cgacaagcag aagaacggca tcaaggtgaa cttcaagatc 1920
cgccacaaca tcgaggacgg cagcgtgcag ctcgccgacc actaccagca gaacaccccc 1980
atcggcgacg gccccgtgct gctgcccgac aaccactacc tgagcaccca gtccgccctg 2040
agcaaagacc ccaacgagaa gcgcgatcac atggtcctgc tggagttcgt gaccgccgcc 2100
gggatcactc tcggcatgga cgagctgtac aagtga 2136
<210> 31
<211> 433
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-scFv- CD28ATM-CD28CSD-CD3zSSD fusion
<400> 31
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr
130 135 140
Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu Thr Val Thr Leu Thr
145 150 155 160
Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp
165 170 175
Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly Leu Ile Gly Gly Thr
180 185 190
Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe Ser Gly Ser Leu Ile
195 200 205
Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Thr Glu Asp Glu
210 215 220
Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn His Trp Val Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Phe Trp Val
245 250 255
Leu Val Val Val Gly Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr
260 265 270
Val Ala Phe Ile Ile Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu
275 280 285
His Ser Asp Tyr Met Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg
290 295 300
Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg
305 310 315 320
Ser Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln
325 330 335
Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu
340 345 350
Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly
355 360 365
Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
370 375 380
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
385 390 395 400
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
405 410 415
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
420 425 430
Arg
<210> 32
<211> 119
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G VH
<400> 32
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala
115
<210> 33
<211> 109
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G VL
<400> 33
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 34
<211> 248
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-scFv
<400> 34
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr
130 135 140
Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu Thr Val Thr Leu Thr
145 150 155 160
Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp
165 170 175
Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly Leu Ile Gly Gly Thr
180 185 190
Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe Ser Gly Ser Leu Ile
195 200 205
Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Thr Glu Asp Glu
210 215 220
Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn His Trp Val Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 35
<211> 1356
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-scFv-CD28ATM-CD28CSD-CD3zSSD fusion
<400> 35
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctaccggtgt gcattccgag 60
gtgaagctgc tggagagcgg cggcggcctg gtgcagcccg gcggcagcct gaagctgagc 120
tgcgccgcca gcggcttcga cttcagcagg tactggatga actgggtgag gcaggccccc 180
ggcaagggtc tggagtggat cggcgagatc acccccgaca gcagcaccat caactacacc 240
cccagcctga aggacaagtt catcatcagc agggacaacg ccaagaacac cctgtacctg 300
cagatgatca aggtgaggag cgaggacacc gccctgtact actgcgtgag gccctacgac 360
tacggcgcct ggttcgccag ctggggccag ggcaccctgg tgaccgtgag cgccggaggg 420
ggcggaagtg gtggcggggg aagcggcggg ggtggcagcg gagggggcgg atctcaggcc 480
gtggtgaccc aggagagcgc cctgaccacc agccccggcg agaccgtgac cctgacctgc 540
aggagcagca ccggcgccgt gaccaccagc aactacgcca actgggtgca ggagaagccc 600
gaccacctgt tcaccggcct gatcggcggc accaacaaga gggcccccgg cgtgcccgcc 660
aggttcagcg gcagcctgat cggcgacaag gccgccctga ccatcaccgg cgcccagacc 720
gaggacgagg ccatctactt ctgcgccctg tggtacagca accactgggt gttcggcggt 780
ggcaccaagc tgaccgtgct gggagggggc ggatccttct gggtgctggt ggtggtgggc 840
ggcgtgctgg cctgctacag cctgctggtg accgtggcct tcatcatctt ctgggtgagg 900
agcaagagga gcaggctgct gcacagcgac tacatgaaca tgacccccag gaggcccggc 960
cccaccagga agcactacca gccctacgcc ccccccaggg acttcgccgc ctacaggagc 1020
agggtgaagt tcagcaggag cgccgacgcc cccgcctacc agcagggcca gaaccagctg 1080
tataacgagc tgaacctggg caggagggag gagtacgacg tgctggacaa gaggaggggc 1140
agggaccccg agatgggcgg caagcccagg aggaagaacc cccaggaggg cctgtataac 1200
gagctgcaga aggacaagat ggccgaggcc tacagcgaga tcggcatgaa gggcgagagg 1260
aggaggggca agggccacga cggcctgtac cagggcctga gcaccgccac caaggacacc 1320
tacgacgccc tgcacatgca ggccctgccc cccagg 1356
<210> 36
<211> 357
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G VH
<400> 36
gaggtgaagc tgctggagag cggcggcggc ctggtgcagc ccggcggcag cctgaagctg 60
agctgcgccg ccagcggctt cgacttcagc aggtactgga tgaactgggt gaggcaggcc 120
cccggcaagg gtctggagtg gatcggcgag atcacccccg acagcagcac catcaactac 180
acccccagcc tgaaggacaa gttcatcatc agcagggaca acgccaagaa caccctgtac 240
ctgcagatga tcaaggtgag gagcgaggac accgccctgt actactgcgt gaggccctac 300
gactacggcg cctggttcgc cagctggggc cagggcaccc tggtgaccgt gagcgcc 357
<210> 37
<211> 327
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G VL
<400> 37
caggccgtgg tgacccagga gagcgccctg accaccagcc ccggcgagac cgtgaccctg 60
acctgcagga gcagcaccgg cgccgtgacc accagcaact acgccaactg ggtgcaggag 120
aagcccgacc acctgttcac cggcctgatc ggcggcacca acaagagggc ccccggcgtg 180
cccgccaggt tcagcggcag cctgatcggc gacaaggccg ccctgaccat caccggcgcc 240
cagaccgagg acgaggccat ctacttctgc gccctgtggt acagcaacca ctgggtgttc 300
ggcggtggca ccaagctgac cgtgctg 327
<210> 38
<211> 2136
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-scFv-CD28ATM-CD28CSD-CD3zSSD-eGFP fusion
<400> 38
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctaccggtgt gcattccgag 60
gtgaagctgc tggagagcgg cggcggcctg gtgcagcccg gcggcagcct gaagctgagc 120
tgcgccgcca gcggcttcga cttcagcagg tactggatga actgggtgag gcaggccccc 180
ggcaagggtc tggagtggat cggcgagatc acccccgaca gcagcaccat caactacacc 240
cccagcctga aggacaagtt catcatcagc agggacaacg ccaagaacac cctgtacctg 300
cagatgatca aggtgaggag cgaggacacc gccctgtact actgcgtgag gccctacgac 360
tacggcgcct ggttcgccag ctggggccag ggcaccctgg tgaccgtgag cgccggaggg 420
ggcggaagtg gtggcggggg aagcggcggg ggtggcagcg gagggggcgg atctcaggcc 480
gtggtgaccc aggagagcgc cctgaccacc agccccggcg agaccgtgac cctgacctgc 540
aggagcagca ccggcgccgt gaccaccagc aactacgcca actgggtgca ggagaagccc 600
gaccacctgt tcaccggcct gatcggcggc accaacaaga gggcccccgg cgtgcccgcc 660
aggttcagcg gcagcctgat cggcgacaag gccgccctga ccatcaccgg cgcccagacc 720
gaggacgagg ccatctactt ctgcgccctg tggtacagca accactgggt gttcggcggt 780
ggcaccaagc tgaccgtgct gggagggggc ggatccttct gggtgctggt ggtggtgggc 840
ggcgtgctgg cctgctacag cctgctggtg accgtggcct tcatcatctt ctgggtgagg 900
agcaagagga gcaggctgct gcacagcgac tacatgaaca tgacccccag gaggcccggc 960
cccaccagga agcactacca gccctacgcc ccccccaggg acttcgccgc ctacaggagc 1020
agggtgaagt tcagcaggag cgccgacgcc cccgcctacc agcagggcca gaaccagctg 1080
tataacgagc tgaacctggg caggagggag gagtacgacg tgctggacaa gaggaggggc 1140
agggaccccg agatgggcgg caagcccagg aggaagaacc cccaggaggg cctgtataac 1200
gagctgcaga aggacaagat ggccgaggcc tacagcgaga tcggcatgaa gggcgagagg 1260
aggaggggca agggccacga cggcctgtac cagggcctga gcaccgccac caaggacacc 1320
tacgacgccc tgcacatgca ggccctgccc cccaggtccg gagagggcag aggaagtctt 1380
ctaacatgcg gtgacgtgga ggagaatccc ggccctaggg tgagcaaggg cgaggagctg 1440
ttcaccgggg tggtgcccat cctggtcgag ctggacggcg acgtaaacgg ccacaagttc 1500
agcgtgtccg gcgagggcga gggcgatgcc acctacggca agctgaccct gaagttcatc 1560
tgcaccaccg gcaagctgcc cgtgccctgg cccaccctcg tgaccaccct gacctacggc 1620
gtgcagtgct tcagccgcta ccccgaccac atgaagcagc acgacttctt caagtccgcc 1680
atgcccgaag gctacgtcca ggagcgcacc atcttcttca aggacgacgg caactacaag 1740
acccgcgccg aggtgaagtt cgagggcgac accctggtga accgcatcga gctgaagggc 1800
atcgacttca aggaggacgg caacatcctg gggcacaagc tggagtacaa ctacaacagc 1860
cacaacgtct atatcatggc cgacaagcag aagaacggca tcaaggtgaa cttcaagatc 1920
cgccacaaca tcgaggacgg cagcgtgcag ctcgccgacc actaccagca gaacaccccc 1980
atcggcgacg gccccgtgct gctgcccgac aaccactacc tgagcaccca gtccgccctg 2040
agcaaagacc ccaacgagaa gcgcgatcac atggtcctgc tggagttcgt gaccgccgcc 2100
gggatcactc tcggcatgga cgagctgtac aagtga 2136
<210> 39
<211> 407
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-Fab- heavy chain-CD28ATM-CD28CSD-CD3zSSD fusion
pETR17100
<400> 39
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys
225 230 235 240
Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser
245 250 255
Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg
260 265 270
Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg
275 280 285
Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp
290 295 300
Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
305 310 315 320
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
325 330 335
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
340 345 350
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
355 360 365
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
370 375 380
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
385 390 395 400
Met Gln Ala Leu Pro Pro Arg
405
<210> 40
<211> 222
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-Fab heavy chain
<400> 40
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 41
<211> 216
<212> PRT
<213> Artificial sequence
<220>
<223> Anti P329G-ds-Fab light chain
<400> 41
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 42
<211> 107
<212> PRT
<213> Artificial sequence
<220>
<223> CL
<400> 42
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 43
<211> 103
<212> PRT
<213> Artificial sequence
<220>
<223> CH1
<400> 43
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys
100
<210> 44
<211> 2645
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-Fab-heavy chain-CD28ATM-CD28CSD-CD3zSSD fusion
pETR17100
<400> 44
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacgggtgt gcattcccag 60
gccgtggtga cccaggagag cgccctgacc accagccccg gcgagaccgt gaccctgacc 120
tgcaggagca gcaccggcgc cgtgaccacc agcaactacg ccaactgggt gcaggagaag 180
cccgaccacc tgttcaccgg cctgatcggc ggcaccaaca agagggcccc cggcgtgccc 240
gccaggttca gcggcagcct gatcggcgac aaggccgccc tgaccatcac cggcgcccag 300
accgaggacg aggccatcta cttctgcgcc ctgtggtaca gcaaccactg ggtgttcggc 360
tgtggcacca agctgaccgt gctgcgtacg gtggctgcac catctgtctt catcttcccg 420
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 480
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 540
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 600
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 660
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttagga attccccgaa 720
gtaacttaga agctgtaaat caacgatcaa tagcaggtgt ggcacaccag tcataccttg 780
atcaagcact tctgtttccc cggactgagt atcaataggc tgctcgcgcg gctgaaggag 840
aaaacgttcg ttacccgacc aactacttcg agaagcttag taccaccatg aacgaggcag 900
ggtgtttcgc tcagcacaac cccagtgtag atcaggctga tgagtcactg caacccccat 960
gggcgaccat ggcagtggct gcgttggcgg cctgcccatg gagaaatcca tgggacgctc 1020
taattctgac atggtgtgaa gtgcctattg agctaactgg tagtcctccg gcccctgatt 1080
gcggctaatc ctaactgcgg agcacatgct cacaaaccag tgggtggtgt gtcgtaacgg 1140
gcaactctgc agcggaaccg actactttgg gtgtccgtgt ttccttttat tcctatattg 1200
gctgcttatg gtgacaatca aaaagttgtt accatatagc tattggattg gccatccggt 1260
gtgcaacagg gcaactgttt acctatttat tggttttgta ccattatcac tgaagtctgt 1320
gatcactctc aaattcattt tgaccctcaa cacaatcaaa cgccaccatg ggatggagct 1380
gtatcatcct cttcttggta gcaacagcta ccggtgtgca ctccgaggtg aagctgctgg 1440
agagcggcgg cggcctggtg cagcccggcg gcagcctgaa gctgagctgc gccgccagcg 1500
gcttcgactt cagcaggtac tggatgaact gggtgaggca ggcccccggc aagtgtctgg 1560
agtggatcgg cgagatcacc cccgacagca gcaccatcaa ctacaccccc agcctgaagg 1620
acaagttcat catcagcagg gacaacgcca agaacaccct gtacctgcag atgatcaagg 1680
tgaggagcga ggacaccgcc ctgtactact gcgtgaggcc ctacgactac ggcgcctggt 1740
tcgccagctg gggccagggc accctggtga ccgtgagcgc cgctagcacc aagggcccct 1800
ccgtgttccc cctggccccc agcagcaaga gcaccagcgg cggcacagcc gctctgggct 1860
gcctggtcaa ggactacttc cccgagcccg tgaccgtgtc ctggaacagc ggagccctga 1920
cctccggcgt gcacaccttc cccgccgtgc tgcagagttc tggcctgtat agcctgagca 1980
gcgtggtcac cgtgccttct agcagcctgg gcacccagac ctacatctgc aacgtgaacc 2040
acaagcccag caacaccaag gtggacaaga aggtggagcc caagagctgc ggagggggcg 2100
gatccttctg ggtgctggtg gtggtgggcg gcgtgctggc ctgctacagc ctgctggtga 2160
ccgtggcctt catcatcttc tgggtgagga gcaagaggag caggctgctg cacagcgact 2220
acatgaacat gacccccagg aggcccggcc ccaccaggaa gcactaccag ccctacgccc 2280
cccccaggga cttcgccgcc tacaggagca gggtgaagtt cagcaggagc gccgacgccc 2340
ccgcctacca gcagggccag aaccagctgt ataacgagct gaacctgggc aggagggagg 2400
agtacgacgt gctggacaag aggaggggca gggaccccga gatgggcggc aagcccagga 2460
ggaagaaccc ccaggagggc ctgtataacg agctgcagaa ggacaagatg gccgaggcct 2520
acagcgagat cggcatgaag ggcgagagga ggaggggcaa gggccacgac ggcctgtacc 2580
agggcctgag caccgccacc aaggacacct acgacgccct gcacatgcag gccctgcccc 2640
ccagg 2645
<210> 45
<211> 324
<212> DNA
<213> Artificial sequence
<220>
<223> CL
<400> 45
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttag 324
<210> 46
<211> 309
<212> DNA
<213> Artificial sequence
<220>
<223> CH1
<400> 46
gctagcacca agggcccctc cgtgttcccc ctggccccca gcagcaagag caccagcggc 60
ggcacagccg ctctgggctg cctggtcaag gactacttcc ccgagcccgt gaccgtgtcc 120
tggaacagcg gagccctgac ctccggcgtg cacaccttcc ccgccgtgct gcagagttct 180
ggcctgtata gcctgagcag cgtggtcacc gtgccttcta gcagcctggg cacccagacc 240
tacatctgca acgtgaacca caagcccagc aacaccaagg tggacaagaa ggtggagccc 300
aagagctgc 309
<210> 47
<211> 3425
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-ds-Fab-heavy chain-CD28TM-CD28CSD-CD3ZSSD-eGFP fusion
pETR17100
<400> 47
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacgggtgt gcattcccag 60
gccgtggtga cccaggagag cgccctgacc accagccccg gcgagaccgt gaccctgacc 120
tgcaggagca gcaccggcgc cgtgaccacc agcaactacg ccaactgggt gcaggagaag 180
cccgaccacc tgttcaccgg cctgatcggc ggcaccaaca agagggcccc cggcgtgccc 240
gccaggttca gcggcagcct gatcggcgac aaggccgccc tgaccatcac cggcgcccag 300
accgaggacg aggccatcta cttctgcgcc ctgtggtaca gcaaccactg ggtgttcggc 360
tgtggcacca agctgaccgt gctgcgtacg gtggctgcac catctgtctt catcttcccg 420
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 480
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 540
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 600
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 660
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttagga attccccgaa 720
gtaacttaga agctgtaaat caacgatcaa tagcaggtgt ggcacaccag tcataccttg 780
atcaagcact tctgtttccc cggactgagt atcaataggc tgctcgcgcg gctgaaggag 840
aaaacgttcg ttacccgacc aactacttcg agaagcttag taccaccatg aacgaggcag 900
ggtgtttcgc tcagcacaac cccagtgtag atcaggctga tgagtcactg caacccccat 960
gggcgaccat ggcagtggct gcgttggcgg cctgcccatg gagaaatcca tgggacgctc 1020
taattctgac atggtgtgaa gtgcctattg agctaactgg tagtcctccg gcccctgatt 1080
gcggctaatc ctaactgcgg agcacatgct cacaaaccag tgggtggtgt gtcgtaacgg 1140
gcaactctgc agcggaaccg actactttgg gtgtccgtgt ttccttttat tcctatattg 1200
gctgcttatg gtgacaatca aaaagttgtt accatatagc tattggattg gccatccggt 1260
gtgcaacagg gcaactgttt acctatttat tggttttgta ccattatcac tgaagtctgt 1320
gatcactctc aaattcattt tgaccctcaa cacaatcaaa cgccaccatg ggatggagct 1380
gtatcatcct cttcttggta gcaacagcta ccggtgtgca ctccgaggtg aagctgctgg 1440
agagcggcgg cggcctggtg cagcccggcg gcagcctgaa gctgagctgc gccgccagcg 1500
gcttcgactt cagcaggtac tggatgaact gggtgaggca ggcccccggc aagtgtctgg 1560
agtggatcgg cgagatcacc cccgacagca gcaccatcaa ctacaccccc agcctgaagg 1620
acaagttcat catcagcagg gacaacgcca agaacaccct gtacctgcag atgatcaagg 1680
tgaggagcga ggacaccgcc ctgtactact gcgtgaggcc ctacgactac ggcgcctggt 1740
tcgccagctg gggccagggc accctggtga ccgtgagcgc cgctagcacc aagggcccct 1800
ccgtgttccc cctggccccc agcagcaaga gcaccagcgg cggcacagcc gctctgggct 1860
gcctggtcaa ggactacttc cccgagcccg tgaccgtgtc ctggaacagc ggagccctga 1920
cctccggcgt gcacaccttc cccgccgtgc tgcagagttc tggcctgtat agcctgagca 1980
gcgtggtcac cgtgccttct agcagcctgg gcacccagac ctacatctgc aacgtgaacc 2040
acaagcccag caacaccaag gtggacaaga aggtggagcc caagagctgc ggagggggcg 2100
gatccttctg ggtgctggtg gtggtgggcg gcgtgctggc ctgctacagc ctgctggtga 2160
ccgtggcctt catcatcttc tgggtgagga gcaagaggag caggctgctg cacagcgact 2220
acatgaacat gacccccagg aggcccggcc ccaccaggaa gcactaccag ccctacgccc 2280
cccccaggga cttcgccgcc tacaggagca gggtgaagtt cagcaggagc gccgacgccc 2340
ccgcctacca gcagggccag aaccagctgt ataacgagct gaacctgggc aggagggagg 2400
agtacgacgt gctggacaag aggaggggca gggaccccga gatgggcggc aagcccagga 2460
ggaagaaccc ccaggagggc ctgtataacg agctgcagaa ggacaagatg gccgaggcct 2520
acagcgagat cggcatgaag ggcgagagga ggaggggcaa gggccacgac ggcctgtacc 2580
agggcctgag caccgccacc aaggacacct acgacgccct gcacatgcag gccctgcccc 2640
ccaggtccgg agagggcaga ggaagtcttc taacatgcgg tgacgtggag gagaatcccg 2700
gccctagggt gagcaagggc gaggagctgt tcaccggggt ggtgcccatc ctggtcgagc 2760
tggacggcga cgtaaacggc cacaagttca gcgtgtccgg cgagggcgag ggcgatgcca 2820
cctacggcaa gctgaccctg aagttcatct gcaccaccgg caagctgccc gtgccctggc 2880
ccaccctcgt gaccaccctg acctacggcg tgcagtgctt cagccgctac cccgaccaca 2940
tgaagcagca cgacttcttc aagtccgcca tgcccgaagg ctacgtccag gagcgcacca 3000
tcttcttcaa ggacgacggc aactacaaga cccgcgccga ggtgaagttc gagggcgaca 3060
ccctggtgaa ccgcatcgag ctgaagggca tcgacttcaa ggaggacggc aacatcctgg 3120
ggcacaagct ggagtacaac tacaacagcc acaacgtcta tatcatggcc gacaagcaga 3180
agaacggcat caaggtgaac ttcaagatcc gccacaacat cgaggacggc agcgtgcagc 3240
tcgccgacca ctaccagcag aacaccccca tcggcgacgg ccccgtgctg ctgcccgaca 3300
accactacct gagcacccag tccgccctga gcaaagaccc caacgagaag cgcgatcaca 3360
tggtcctgct ggagttcgtg accgccgccg ggatcactct cggcatggac gagctgtaca 3420
agtga 3425
<210> 48
<211> 407
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-Fab-heavy chain-CD28ATM-CD28CSD-CD3zSSD fusion
pETR17594
<400> 48
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Gly Gly
210 215 220
Gly Gly Ser Phe Trp Val Leu Val Val Val Gly Gly Val Leu Ala Cys
225 230 235 240
Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val Arg Ser
245 250 255
Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg
260 265 270
Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg
275 280 285
Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp
290 295 300
Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
305 310 315 320
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
325 330 335
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
340 345 350
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
355 360 365
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
370 375 380
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
385 390 395 400
Met Gln Ala Leu Pro Pro Arg
405
<210> 49
<211> 222
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-Fab heavy chain
<400> 49
Glu Val Lys Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Thr Pro Asp Ser Ser Thr Ile Asn Tyr Thr Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ile Lys Val Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Arg Pro Tyr Asp Tyr Gly Ala Trp Phe Ala Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 50
<211> 216
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-P329G-Fab light chain
<400> 50
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Glu
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Thr Glu Asp Glu Ala Ile Tyr Phe Cys Ala Leu Trp Tyr Ser Asn
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 51
<211> 2645
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-Fab-heavy chain-CD28ATM-CD28CSD-CD3zSSD fusion
pETR17594
<400> 51
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacgggtgt gcattcccag 60
gccgtggtga cccaggagag cgccctgacc accagccccg gcgagaccgt gaccctgacc 120
tgcaggagca gcaccggcgc cgtgaccacc agcaactacg ccaactgggt gcaggagaag 180
cccgaccacc tgttcaccgg cctgatcggc ggcaccaaca agagggcccc cggcgtgccc 240
gccaggttca gcggcagcct gatcggcgac aaggccgccc tgaccatcac cggcgcccag 300
accgaggacg aggccatcta cttctgcgcc ctgtggtaca gcaaccactg ggtgttcggc 360
ggtggcacca agctgaccgt gctgcgtacg gtggctgcac catctgtctt catcttcccg 420
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 480
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 540
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 600
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 660
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttagga attccccgaa 720
gtaacttaga agctgtaaat caacgatcaa tagcaggtgt ggcacaccag tcataccttg 780
atcaagcact tctgtttccc cggactgagt atcaataggc tgctcgcgcg gctgaaggag 840
aaaacgttcg ttacccgacc aactacttcg agaagcttag taccaccatg aacgaggcag 900
ggtgtttcgc tcagcacaac cccagtgtag atcaggctga tgagtcactg caacccccat 960
gggcgaccat ggcagtggct gcgttggcgg cctgcccatg gagaaatcca tgggacgctc 1020
taattctgac atggtgtgaa gtgcctattg agctaactgg tagtcctccg gcccctgatt 1080
gcggctaatc ctaactgcgg agcacatgct cacaaaccag tgggtggtgt gtcgtaacgg 1140
gcaactctgc agcggaaccg actactttgg gtgtccgtgt ttccttttat tcctatattg 1200
gctgcttatg gtgacaatca aaaagttgtt accatatagc tattggattg gccatccggt 1260
gtgcaacagg gcaactgttt acctatttat tggttttgta ccattatcac tgaagtctgt 1320
gatcactctc aaattcattt tgaccctcaa cacaatcaaa cgccaccatg ggatggagct 1380
gtatcatcct cttcttggta gcaacagcta ccggtgtgca ctccgaggtg aagctgctgg 1440
agagcggcgg cggcctggtg cagcccggcg gcagcctgaa gctgagctgc gccgccagcg 1500
gcttcgactt cagcaggtac tggatgaact gggtgaggca ggcccccggc aagggtctgg 1560
agtggatcgg cgagatcacc cccgacagca gcaccatcaa ctacaccccc agcctgaagg 1620
acaagttcat catcagcagg gacaacgcca agaacaccct gtacctgcag atgatcaagg 1680
tgaggagcga ggacaccgcc ctgtactact gcgtgaggcc ctacgactac ggcgcctggt 1740
tcgccagctg gggccagggc accctggtga ccgtgagcgc cgctagcacc aagggcccct 1800
ccgtgttccc cctggccccc agcagcaaga gcaccagcgg cggcacagcc gctctgggct 1860
gcctggtcaa ggactacttc cccgagcccg tgaccgtgtc ctggaacagc ggagccctga 1920
cctccggcgt gcacaccttc cccgccgtgc tgcagagttc tggcctgtat agcctgagca 1980
gcgtggtcac cgtgccttct agcagcctgg gcacccagac ctacatctgc aacgtgaacc 2040
acaagcccag caacaccaag gtggacaaga aggtggagcc caagagctgc ggagggggcg 2100
gatccttctg ggtgctggtg gtggtgggcg gcgtgctggc ctgctacagc ctgctggtga 2160
ccgtggcctt catcatcttc tgggtgagga gcaagaggag caggctgctg cacagcgact 2220
acatgaacat gacccccagg aggcccggcc ccaccaggaa gcactaccag ccctacgccc 2280
cccccaggga cttcgccgcc tacaggagca gggtgaagtt cagcaggagc gccgacgccc 2340
ccgcctacca gcagggccag aaccagctgt ataacgagct gaacctgggc aggagggagg 2400
agtacgacgt gctggacaag aggaggggca gggaccccga gatgggcggc aagcccagga 2460
ggaagaaccc ccaggagggc ctgtataacg agctgcagaa ggacaagatg gccgaggcct 2520
acagcgagat cggcatgaag ggcgagagga ggaggggcaa gggccacgac ggcctgtacc 2580
agggcctgag caccgccacc aaggacacct acgacgccct gcacatgcag gccctgcccc 2640
ccagg 2645
<210> 52
<211> 3425
<212> DNA
<213> Artificial sequence
<220>
<223> Anti-P329G-Fab-heavy chain-CD28ATM-CD28CSD-CD3zSSD-eGFP fusion
pETR17594
<400> 52
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacgggtgt gcattcccag 60
gccgtggtga cccaggagag cgccctgacc accagccccg gcgagaccgt gaccctgacc 120
tgcaggagca gcaccggcgc cgtgaccacc agcaactacg ccaactgggt gcaggagaag 180
cccgaccacc tgttcaccgg cctgatcggc ggcaccaaca agagggcccc cggcgtgccc 240
gccaggttca gcggcagcct gatcggcgac aaggccgccc tgaccatcac cggcgcccag 300
accgaggacg aggccatcta cttctgcgcc ctgtggtaca gcaaccactg ggtgttcggc 360
ggtggcacca agctgaccgt gctgcgtacg gtggctgcac catctgtctt catcttcccg 420
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 480
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 540
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 600
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 660
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttagga attccccgaa 720
gtaacttaga agctgtaaat caacgatcaa tagcaggtgt ggcacaccag tcataccttg 780
atcaagcact tctgtttccc cggactgagt atcaataggc tgctcgcgcg gctgaaggag 840
aaaacgttcg ttacccgacc aactacttcg agaagcttag taccaccatg aacgaggcag 900
ggtgtttcgc tcagcacaac cccagtgtag atcaggctga tgagtcactg caacccccat 960
gggcgaccat ggcagtggct gcgttggcgg cctgcccatg gagaaatcca tgggacgctc 1020
taattctgac atggtgtgaa gtgcctattg agctaactgg tagtcctccg gcccctgatt 1080
gcggctaatc ctaactgcgg agcacatgct cacaaaccag tgggtggtgt gtcgtaacgg 1140
gcaactctgc agcggaaccg actactttgg gtgtccgtgt ttccttttat tcctatattg 1200
gctgcttatg gtgacaatca aaaagttgtt accatatagc tattggattg gccatccggt 1260
gtgcaacagg gcaactgttt acctatttat tggttttgta ccattatcac tgaagtctgt 1320
gatcactctc aaattcattt tgaccctcaa cacaatcaaa cgccaccatg ggatggagct 1380
gtatcatcct cttcttggta gcaacagcta ccggtgtgca ctccgaggtg aagctgctgg 1440
agagcggcgg cggcctggtg cagcccggcg gcagcctgaa gctgagctgc gccgccagcg 1500
gcttcgactt cagcaggtac tggatgaact gggtgaggca ggcccccggc aagggtctgg 1560
agtggatcgg cgagatcacc cccgacagca gcaccatcaa ctacaccccc agcctgaagg 1620
acaagttcat catcagcagg gacaacgcca agaacaccct gtacctgcag atgatcaagg 1680
tgaggagcga ggacaccgcc ctgtactact gcgtgaggcc ctacgactac ggcgcctggt 1740
tcgccagctg gggccagggc accctggtga ccgtgagcgc cgctagcacc aagggcccct 1800
ccgtgttccc cctggccccc agcagcaaga gcaccagcgg cggcacagcc gctctgggct 1860
gcctggtcaa ggactacttc cccgagcccg tgaccgtgtc ctggaacagc ggagccctga 1920
cctccggcgt gcacaccttc cccgccgtgc tgcagagttc tggcctgtat agcctgagca 1980
gcgtggtcac cgtgccttct agcagcctgg gcacccagac ctacatctgc aacgtgaacc 2040
acaagcccag caacaccaag gtggacaaga aggtggagcc caagagctgc ggagggggcg 2100
gatccttctg ggtgctggtg gtggtgggcg gcgtgctggc ctgctacagc ctgctggtga 2160
ccgtggcctt catcatcttc tgggtgagga gcaagaggag caggctgctg cacagcgact 2220
acatgaacat gacccccagg aggcccggcc ccaccaggaa gcactaccag ccctacgccc 2280
cccccaggga cttcgccgcc tacaggagca gggtgaagtt cagcaggagc gccgacgccc 2340
ccgcctacca gcagggccag aaccagctgt ataacgagct gaacctgggc aggagggagg 2400
agtacgacgt gctggacaag aggaggggca gggaccccga gatgggcggc aagcccagga 2460
ggaagaaccc ccaggagggc ctgtataacg agctgcagaa ggacaagatg gccgaggcct 2520
acagcgagat cggcatgaag ggcgagagga ggaggggcaa gggccacgac ggcctgtacc 2580
agggcctgag caccgccacc aaggacacct acgacgccct gcacatgcag gccctgcccc 2640
ccaggtccgg agagggcaga ggaagtcttc taacatgcgg tgacgtggag gagaatcccg 2700
gccctagggt gagcaagggc gaggagctgt tcaccggggt ggtgcccatc ctggtcgagc 2760
tggacggcga cgtaaacggc cacaagttca gcgtgtccgg cgagggcgag ggcgatgcca 2820
cctacggcaa gctgaccctg aagttcatct gcaccaccgg caagctgccc gtgccctggc 2880
ccaccctcgt gaccaccctg acctacggcg tgcagtgctt cagccgctac cccgaccaca 2940
tgaagcagca cgacttcttc aagtccgcca tgcccgaagg ctacgtccag gagcgcacca 3000
tcttcttcaa ggacgacggc aactacaaga cccgcgccga ggtgaagttc gagggcgaca 3060
ccctggtgaa ccgcatcgag ctgaagggca tcgacttcaa ggaggacggc aacatcctgg 3120
ggcacaagct ggagtacaac tacaacagcc acaacgtcta tatcatggcc gacaagcaga 3180
agaacggcat caaggtgaac ttcaagatcc gccacaacat cgaggacggc agcgtgcagc 3240
tcgccgacca ctaccagcag aacaccccca tcggcgacgg ccccgtgctg ctgcccgaca 3300
accactacct gagcacccag tccgccctga gcaaagaccc caacgagaag cgcgatcaca 3360
tggtcctgct ggagttcgtg accgccgccg ggatcactct cggcatggac gagctgtaca 3420
agtga 3425
<210> 53
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA CDR H1 Kabat
<400> 53
Ser Tyr Gly Met Ser
1 5
<210> 54
<211> 6
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA CDR H2 Kabat
<400> 54
Ser Ser Gly Gly Ser Tyr
1 5
<210> 55
<211> 12
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA CDR H3 Kabat
<400> 55
Leu Gly Met Ile Thr Thr Gly Tyr Ala Met Asp Tyr
1 5 10
<210> 56
<211> 16
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA CDR L1 Kabat
<400> 56
Arg Ser Ser Gln Thr Ile Val His Ser Thr Gly His Thr Tyr Leu Glu
1 5 10 15
<210> 57
<211> 7
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA CDR L2 Kabat
<400> 57
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 58
<211> 9
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA CDR L3 Kabat
<400> 58
Phe Gln Gly Ser His Val Pro Tyr Thr
1 5
<210> 59
<211> 457
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA-scFv-CD28ATM-CD28CSD-CD3zSSD fusion
<400> 59
Met Asn Phe Gly Leu Ser Leu Val Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Tyr Ile Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Leu Gly Met Ile Thr Thr Gly Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
165 170 175
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Thr Ile Val His Ser
180 185 190
Thr Gly His Thr Tyr Leu Glu Trp Phe Leu Gln Lys Pro Gly Gln Ser
195 200 205
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
210 215 220
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
225 230 235 240
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
245 250 255
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
260 265 270
Gly Gly Gly Gly Ser Phe Trp Val Leu Val Val Val Gly Gly Val Leu
275 280 285
Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile Phe Trp Val
290 295 300
Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr
305 310 315 320
Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro
325 330 335
Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser
340 345 350
Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu
355 360 365
Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg
370 375 380
Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln
385 390 395 400
Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr
405 410 415
Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp
420 425 430
Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala
435 440 445
Leu His Met Gln Ala Leu Pro Pro Arg
450 455
<210> 60
<211> 272
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA-scFv
<400> 60
Met Asn Phe Gly Leu Ser Leu Val Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Tyr Ile Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Leu Gly Met Ile Thr Thr Gly Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
165 170 175
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Thr Ile Val His Ser
180 185 190
Thr Gly His Thr Tyr Leu Glu Trp Phe Leu Gln Lys Pro Gly Gln Ser
195 200 205
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
210 215 220
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
225 230 235 240
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
245 250 255
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
260 265 270
<210> 61
<211> 140
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA VH
<400> 61
Met Asn Phe Gly Leu Ser Leu Val Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Tyr Ile Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Leu Gly Met Ile Thr Thr Gly Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
130 135 140
<210> 62
<211> 112
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA VL
<400> 62
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Thr Ile Val His Ser
20 25 30
Thr Gly His Thr Tyr Leu Glu Trp Phe Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 63
<211> 428
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA-Fab-heavy chain-CD28ATM-CD28CSD-CD3zSSD fusion
<400> 63
Met Asn Phe Gly Leu Ser Leu Val Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Tyr Ile Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Leu Gly Met Ile Thr Thr Gly Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys Gly Gly Gly Gly Ser Phe Trp Val Leu Val Val Val Gly
245 250 255
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
260 265 270
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
275 280 285
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
290 295 300
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe
305 310 315 320
Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu
325 330 335
Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp
340 345 350
Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys
355 360 365
Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala
370 375 380
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys
385 390 395 400
Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr
405 410 415
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
420 425
<210> 64
<211> 243
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA-Fab heavy chain
<400> 64
Met Asn Phe Gly Leu Ser Leu Val Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Ser Gly Gly Ser Tyr Ile Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met
100 105 110
Tyr Tyr Cys Ala Arg Leu Gly Met Ile Thr Thr Gly Tyr Ala Met Asp
115 120 125
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
225 230 235 240
Lys Ser Cys
<210> 65
<211> 219
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-AAA-Fab light chain
<400> 65
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Thr Ile Val His Ser
20 25 30
Thr Gly His Thr Tyr Leu Glu Trp Phe Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 66
<211> 780
<212> DNA
<213> Homo sapiens
<400> 66
atggcgcgcc cgcatccgtg gtggctgtgc gtgctgggca ccctggtggg cctgagcgcg 60
accccggcgc cgaaaagctg cccggaacgc cattattggg cgcagggcaa actgtgctgc 120
cagatgtgcg aaccgggcac ctttctggtg aaagattgcg atcagcatcg caaagcggcg 180
cagtgcgatc cgtgcattcc gggcgtgagc tttagcccgg atcatcatac ccgcccgcat 240
tgcgaaagct gccgccattg caacagcggc ctgctggtgc gcaactgcac cattaccgcg 300
aacgcggaat gcgcgtgccg caacggctgg cagtgccgcg ataaagaatg caccgaatgc 360
gatccgctgc cgaacccgag cctgaccgcg cgcagcagcc aggcgctgag cccgcatccg 420
cagccgaccc atctgccgta tgtgagcgaa atgctggaag cgcgcaccgc gggccatatg 480
cagaccctgg cggattttcg ccagctgccg gcgcgcaccc tgagcaccca ttggccgccg 540
cagcgcagcc tgtgcagcag cgattttatt cgcattctgg tgatttttag cggcatgttt 600
ctggtgttta ccctggcggg cgcgctgttt ctgcatcagc gccgcaaata tcgcagcaac 660
aaaggcgaaa gcccggtgga accggcggaa ccgtgccatt atagctgccc gcgcgaagaa 720
gaaggcagca ccattccgat tcaggaagat tatcgcaaac cggaaccggc gtgcagcccg 780
<210> 67
<211> 260
<212> PRT
<213> Homo sapiens
<400> 67
Met Ala Arg Pro His Pro Trp Trp Leu Cys Val Leu Gly Thr Leu Val
1 5 10 15
Gly Leu Ser Ala Thr Pro Ala Pro Lys Ser Cys Pro Glu Arg His Tyr
20 25 30
Trp Ala Gln Gly Lys Leu Cys Cys Gln Met Cys Glu Pro Gly Thr Phe
35 40 45
Leu Val Lys Asp Cys Asp Gln His Arg Lys Ala Ala Gln Cys Asp Pro
50 55 60
Cys Ile Pro Gly Val Ser Phe Ser Pro Asp His His Thr Arg Pro His
65 70 75 80
Cys Glu Ser Cys Arg His Cys Asn Ser Gly Leu Leu Val Arg Asn Cys
85 90 95
Thr Ile Thr Ala Asn Ala Glu Cys Ala Cys Arg Asn Gly Trp Gln Cys
100 105 110
Arg Asp Lys Glu Cys Thr Glu Cys Asp Pro Leu Pro Asn Pro Ser Leu
115 120 125
Thr Ala Arg Ser Ser Gln Ala Leu Ser Pro His Pro Gln Pro Thr His
130 135 140
Leu Pro Tyr Val Ser Glu Met Leu Glu Ala Arg Thr Ala Gly His Met
145 150 155 160
Gln Thr Leu Ala Asp Phe Arg Gln Leu Pro Ala Arg Thr Leu Ser Thr
165 170 175
His Trp Pro Pro Gln Arg Ser Leu Cys Ser Ser Asp Phe Ile Arg Ile
180 185 190
Leu Val Ile Phe Ser Gly Met Phe Leu Val Phe Thr Leu Ala Gly Ala
195 200 205
Leu Phe Leu His Gln Arg Arg Lys Tyr Arg Ser Asn Lys Gly Glu Ser
210 215 220
Pro Val Glu Pro Ala Glu Pro Cys His Tyr Ser Cys Pro Arg Glu Glu
225 230 235 240
Glu Gly Ser Thr Ile Pro Ile Gln Glu Asp Tyr Arg Lys Pro Glu Pro
245 250 255
Ala Cys Ser Pro
260
<210> 68
<211> 750
<212> DNA
<213> Mus musculus
<400> 68
atggcgtggc cgccgccgta ttggctgtgc atgctgggca ccctggtggg cctgagcgcg 60
accctggcgc cgaacagctg cccggataaa cattattgga ccggcggcgg cctgtgctgc 120
cgcatgtgcg aaccgggcac cttttttgtg aaagattgcg aacaggatcg caccgcggcg 180
cagtgcgatc cgtgcattcc gggcaccagc tttagcccgg attatcatac ccgcccgcat 240
tgcgaaagct gccgccattg caacagcggc tttctgattc gcaactgcac cgtgaccgcg 300
aacgcggaat gcagctgcag caaaaactgg cagtgccgcg atcaggaatg caccgaatgc 360
gatccgccgc tgaacccggc gctgacccgc cagccgagcg aaaccccgag cccgcagccg 420
ccgccgaccc atctgccgca tggcaccgaa aaaccgagct ggccgctgca tcgccagctg 480
ccgaacagca ccgtgtatag ccagcgcagc agccatcgcc cgctgtgcag cagcgattgc 540
attcgcattt ttgtgacctt tagcagcatg tttctgattt ttgtgctggg cgcgattctg 600
ttttttcatc agcgccgcaa ccatggcccg aacgaagatc gccaggcggt gccggaagaa 660
ccgtgcccgt atagctgccc gcgcgaagaa gaaggcagcg cgattccgat tcaggaagat 720
tatcgcaaac cggaaccggc gttttatccg 750
<210> 69
<211> 250
<212> PRT
<213> Mus musculus
<400> 69
Met Ala Trp Pro Pro Pro Tyr Trp Leu Cys Met Leu Gly Thr Leu Val
1 5 10 15
Gly Leu Ser Ala Thr Leu Ala Pro Asn Ser Cys Pro Asp Lys His Tyr
20 25 30
Trp Thr Gly Gly Gly Leu Cys Cys Arg Met Cys Glu Pro Gly Thr Phe
35 40 45
Phe Val Lys Asp Cys Glu Gln Asp Arg Thr Ala Ala Gln Cys Asp Pro
50 55 60
Cys Ile Pro Gly Thr Ser Phe Ser Pro Asp Tyr His Thr Arg Pro His
65 70 75 80
Cys Glu Ser Cys Arg His Cys Asn Ser Gly Phe Leu Ile Arg Asn Cys
85 90 95
Thr Val Thr Ala Asn Ala Glu Cys Ser Cys Ser Lys Asn Trp Gln Cys
100 105 110
Arg Asp Gln Glu Cys Thr Glu Cys Asp Pro Pro Leu Asn Pro Ala Leu
115 120 125
Thr Arg Gln Pro Ser Glu Thr Pro Ser Pro Gln Pro Pro Pro Thr His
130 135 140
Leu Pro His Gly Thr Glu Lys Pro Ser Trp Pro Leu His Arg Gln Leu
145 150 155 160
Pro Asn Ser Thr Val Tyr Ser Gln Arg Ser Ser His Arg Pro Leu Cys
165 170 175
Ser Ser Asp Cys Ile Arg Ile Phe Val Thr Phe Ser Ser Met Phe Leu
180 185 190
Ile Phe Val Leu Gly Ala Ile Leu Phe Phe His Gln Arg Arg Asn His
195 200 205
Gly Pro Asn Glu Asp Arg Gln Ala Val Pro Glu Glu Pro Cys Pro Tyr
210 215 220
Ser Cys Pro Arg Glu Glu Glu Gly Ser Ala Ile Pro Ile Gln Glu Asp
225 230 235 240
Tyr Arg Lys Pro Glu Pro Ala Phe Tyr Pro
245 250
<210> 70
<211> 660
<212> DNA
<213> Homo sapiens
<400> 70
atgctgcgcc tgctgctggc gctgaacctg tttccgagca ttcaggtgac cggcaacaaa 60
attctggtga aacagagccc gatgctggtg gcgtatgata acgcggtgaa cctgagctgc 120
aaatatagct ataacctgtt tagccgcgaa tttcgcgcga gcctgcataa aggcctggat 180
agcgcggtgg aagtgtgcgt ggtgtatggc aactatagcc agcagctgca ggtgtatagc 240
aaaaccggct ttaactgcga tggcaaactg ggcaacgaaa gcgtgacctt ttatctgcag 300
aacctgtatg tgaaccagac cgatatttat ttttgcaaaa ttgaagtgat gtatccgccg 360
ccgtatctgg ataacgaaaa aagcaacggc accattattc atgtgaaagg caaacatctg 420
tgcccgagcc cgctgtttcc gggcccgagc aaaccgtttt gggtgctggt ggtggtgggc 480
ggcgtgctgg cgtgctatag cctgctggtg accgtggcgt ttattatttt ttgggtgcgc 540
agcaaacgca gccgcctgct gcatagcgat tatatgaaca tgaccccgcg ccgcccgggc 600
ccgacccgca aacattatca gccgtatgcg ccgccgcgcg attttgcggc gtatcgcagc 660
<210> 71
<211> 220
<212> PRT
<213> Homo sapiens
<400> 71
Met Leu Arg Leu Leu Leu Ala Leu Asn Leu Phe Pro Ser Ile Gln Val
1 5 10 15
Thr Gly Asn Lys Ile Leu Val Lys Gln Ser Pro Met Leu Val Ala Tyr
20 25 30
Asp Asn Ala Val Asn Leu Ser Cys Lys Tyr Ser Tyr Asn Leu Phe Ser
35 40 45
Arg Glu Phe Arg Ala Ser Leu His Lys Gly Leu Asp Ser Ala Val Glu
50 55 60
Val Cys Val Val Tyr Gly Asn Tyr Ser Gln Gln Leu Gln Val Tyr Ser
65 70 75 80
Lys Thr Gly Phe Asn Cys Asp Gly Lys Leu Gly Asn Glu Ser Val Thr
85 90 95
Phe Tyr Leu Gln Asn Leu Tyr Val Asn Gln Thr Asp Ile Tyr Phe Cys
100 105 110
Lys Ile Glu Val Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Lys Ser
115 120 125
Asn Gly Thr Ile Ile His Val Lys Gly Lys His Leu Cys Pro Ser Pro
130 135 140
Leu Phe Pro Gly Pro Ser Lys Pro Phe Trp Val Leu Val Val Val Gly
145 150 155 160
Gly Val Leu Ala Cys Tyr Ser Leu Leu Val Thr Val Ala Phe Ile Ile
165 170 175
Phe Trp Val Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met
180 185 190
Asn Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro
195 200 205
Tyr Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser
210 215 220
<210> 72
<211> 654
<212> DNA
<213> Mus musculus
<400> 72
atgaccctgc gcctgctgtt tctggcgctg aactttttta gcgtgcaggt gaccgaaaac 60
aaaattctgg tgaaacagag cccgctgctg gtggtggata gcaacgaagt gagcctgagc 120
tgccgctata gctataacct gctggcgaaa gaatttcgcg cgagcctgta taaaggcgtg 180
aacagcgatg tggaagtgtg cgtgggcaac ggcaacttta cctatcagcc gcagtttcgc 240
agcaacgcgg aatttaactg cgatggcgat tttgataacg aaaccgtgac ctttcgcctg 300
tggaacctgc atgtgaacca taccgatatt tatttttgca aaattgaatt tatgtatccg 360
ccgccgtatc tggataacga acgcagcaac ggcaccatta ttcatattaa agaaaaacat 420
ctgtgccata cccagagcag cccgaaactg ttttgggcgc tggtggtggt ggcgggcgtg 480
ctgttttgct atggcctgct ggtgaccgtg gcgctgtgcg tgatttggac caacagccgc 540
cgcaaccgcc tgctgcagag cgattatatg aacatgaccc cgcgccgccc gggcctgacc 600
cgcaaaccgt atcagccgta tgcgccggcg cgcgattttg cggcgtatcg cccg 654
<210> 73
<211> 218
<212> PRT
<213> Mus musculus
<400> 73
Met Thr Leu Arg Leu Leu Phe Leu Ala Leu Asn Phe Phe Ser Val Gln
1 5 10 15
Val Thr Glu Asn Lys Ile Leu Val Lys Gln Ser Pro Leu Leu Val Val
20 25 30
Asp Ser Asn Glu Val Ser Leu Ser Cys Arg Tyr Ser Tyr Asn Leu Leu
35 40 45
Ala Lys Glu Phe Arg Ala Ser Leu Tyr Lys Gly Val Asn Ser Asp Val
50 55 60
Glu Val Cys Val Gly Asn Gly Asn Phe Thr Tyr Gln Pro Gln Phe Arg
65 70 75 80
Ser Asn Ala Glu Phe Asn Cys Asp Gly Asp Phe Asp Asn Glu Thr Val
85 90 95
Thr Phe Arg Leu Trp Asn Leu His Val Asn His Thr Asp Ile Tyr Phe
100 105 110
Cys Lys Ile Glu Phe Met Tyr Pro Pro Pro Tyr Leu Asp Asn Glu Arg
115 120 125
Ser Asn Gly Thr Ile Ile His Ile Lys Glu Lys His Leu Cys His Thr
130 135 140
Gln Ser Ser Pro Lys Leu Phe Trp Ala Leu Val Val Val Ala Gly Val
145 150 155 160
Leu Phe Cys Tyr Gly Leu Leu Val Thr Val Ala Leu Cys Val Ile Trp
165 170 175
Thr Asn Ser Arg Arg Asn Arg Leu Leu Gln Ser Asp Tyr Met Asn Met
180 185 190
Thr Pro Arg Arg Pro Gly Leu Thr Arg Lys Pro Tyr Gln Pro Tyr Ala
195 200 205
Pro Ala Arg Asp Phe Ala Ala Tyr Arg Pro
210 215
<210> 74
<211> 768
<212> DNA
<213> Homo sapiens
<400> 74
atgggaaaca gctgttacaa catagtagcc actctgttgc tggtcctcaa ctttgagagg 60
acaagatcat tgcaggatcc ttgtagtaac tgcccagctg gtacattctg tgataataac 120
aggaatcaga tttgcagtcc ctgtcctcca aatagtttct ccagcgcagg tggacaaagg 180
acctgtgaca tatgcaggca gtgtaaaggt gttttcagga ccaggaagga gtgttcctcc 240
accagcaatg cagagtgtga ctgcactcca gggtttcact gcctgggggc aggatgcagc 300
atgtgtgaac aggattgtaa acaaggtcaa gaactgacaa aaaaaggttg taaagactgt 360
tgctttggga catttaacga tcagaaacgt ggcatctgtc gaccctggac aaactgttct 420
ttggatggaa agtctgtgct tgtgaatggg acgaaggaga gggacgtggt ctgtggacca 480
tctccagccg acctctctcc gggagcatcc tctgtgaccc cgcctgcccc tgcgagagag 540
ccaggacact ctccgcagat catctccttc tttcttgcgc tgacgtcgac tgcgttgctc 600
ttcctgctgt tcttcctcac gctccgtttc tctgttgtta aacggggcag aaagaaactc 660
ctgtatatat tcaaacaacc atttatgaga ccagtacaaa ctactcaaga ggaagatggc 720
tgtagctgcc gatttccaga agaagaagaa ggaggatgtg aactgtga 768
<210> 75
<211> 255
<212> PRT
<213> Homo sapiens
<400> 75
Met Gly Asn Ser Cys Tyr Asn Ile Val Ala Thr Leu Leu Leu Val Leu
1 5 10 15
Asn Phe Glu Arg Thr Arg Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro
20 25 30
Ala Gly Thr Phe Cys Asp Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys
35 40 45
Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile
50 55 60
Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu Cys Ser Ser
65 70 75 80
Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His Cys Leu Gly
85 90 95
Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu
100 105 110
Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln
115 120 125
Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys
130 135 140
Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro
145 150 155 160
Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala
165 170 175
Pro Ala Arg Glu Pro Gly His Ser Pro Gln Ile Ile Ser Phe Phe Leu
180 185 190
Ala Leu Thr Ser Thr Ala Leu Leu Phe Leu Leu Phe Phe Leu Thr Leu
195 200 205
Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe
210 215 220
Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly
225 230 235 240
Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
245 250 255
<210> 76
<211> 768
<212> DNA
<213> Mus musculus
<400> 76
atgggcaaca actgctataa cgtggtggtg attgtgctgc tgctggtggg ctgcgaaaaa 60
gtgggcgcgg tgcagaacag ctgcgataac tgccagccgg gcaccttttg ccgcaaatat 120
aacccggtgt gcaaaagctg cccgccgagc acctttagca gcattggcgg ccagccgaac 180
tgcaacattt gccgcgtgtg cgcgggctat tttcgcttta aaaaattttg cagcagcacc 240
cataacgcgg aatgcgaatg cattgaaggc tttcattgcc tgggcccgca gtgcacccgc 300
tgcgaaaaag attgccgccc gggccaggaa ctgaccaaac agggctgcaa aacctgcagc 360
ctgggcacct ttaacgatca gaacggcacc ggcgtgtgcc gcccgtggac caactgcagc 420
ctggatggcc gcagcgtgct gaaaaccggc accaccgaaa aagatgtggt gtgcggcccg 480
ccggtggtga gctttagccc gagcaccacc attagcgtga ccccggaagg cggcccgggc 540
ggccatagcc tgcaggtgct gaccctgttt ctggcgctga ccagcgcgct gctgctggcg 600
ctgattttta ttaccctgct gtttagcgtg ctgaaatgga ttcgcaaaaa atttccgcat 660
atttttaaac agccgtttaa aaaaaccacc ggcgcggcgc aggaagaaga tgcgtgcagc 720
tgccgctgcc cgcaggaaga agaaggcggc ggcggcggct atgaactg 768
<210> 77
<211> 256
<212> PRT
<213> Mus musculus
<400> 77
Met Gly Asn Asn Cys Tyr Asn Val Val Val Ile Val Leu Leu Leu Val
1 5 10 15
Gly Cys Glu Lys Val Gly Ala Val Gln Asn Ser Cys Asp Asn Cys Gln
20 25 30
Pro Gly Thr Phe Cys Arg Lys Tyr Asn Pro Val Cys Lys Ser Cys Pro
35 40 45
Pro Ser Thr Phe Ser Ser Ile Gly Gly Gln Pro Asn Cys Asn Ile Cys
50 55 60
Arg Val Cys Ala Gly Tyr Phe Arg Phe Lys Lys Phe Cys Ser Ser Thr
65 70 75 80
His Asn Ala Glu Cys Glu Cys Ile Glu Gly Phe His Cys Leu Gly Pro
85 90 95
Gln Cys Thr Arg Cys Glu Lys Asp Cys Arg Pro Gly Gln Glu Leu Thr
100 105 110
Lys Gln Gly Cys Lys Thr Cys Ser Leu Gly Thr Phe Asn Asp Gln Asn
115 120 125
Gly Thr Gly Val Cys Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Arg
130 135 140
Ser Val Leu Lys Thr Gly Thr Thr Glu Lys Asp Val Val Cys Gly Pro
145 150 155 160
Pro Val Val Ser Phe Ser Pro Ser Thr Thr Ile Ser Val Thr Pro Glu
165 170 175
Gly Gly Pro Gly Gly His Ser Leu Gln Val Leu Thr Leu Phe Leu Ala
180 185 190
Leu Thr Ser Ala Leu Leu Leu Ala Leu Ile Phe Ile Thr Leu Leu Phe
195 200 205
Ser Val Leu Lys Trp Ile Arg Lys Lys Phe Pro His Ile Phe Lys Gln
210 215 220
Pro Phe Lys Lys Thr Thr Gly Ala Ala Gln Glu Glu Asp Ala Cys Ser
225 230 235 240
Cys Arg Cys Pro Gln Glu Glu Glu Gly Gly Gly Gly Gly Tyr Glu Leu
245 250 255
<210> 78
<211> 831
<212> DNA
<213> Homo sapiens
<400> 78
atgtgcgtgg gcgcgcgccg cctgggccgc ggcccgtgcg cggcgctgct gctgctgggc 60
ctgggcctga gcaccgtgac cggcctgcat tgcgtgggcg atacctatcc gagcaacgat 120
cgctgctgcc atgaatgccg cccgggcaac ggcatggtga gccgctgcag ccgcagccag 180
aacaccgtgt gccgcccgtg cggcccgggc ttttataacg atgtggtgag cagcaaaccg 240
tgcaaaccgt gcacctggtg caacctgcgc agcggcagcg aacgcaaaca gctgtgcacc 300
gcgacccagg ataccgtgtg ccgctgccgc gcgggcaccc agccgctgga tagctataaa 360
ccgggcgtgg attgcgcgcc gtgcccgccg ggccatttta gcccgggcga taaccaggcg 420
tgcaaaccgt ggaccaactg caccctggcg ggcaaacata ccctgcagcc ggcgagcaac 480
agcagcgatg cgatttgcga agatcgcgat ccgccggcga cccagccgca ggaaacccag 540
ggcccgccgg cgcgcccgat taccgtgcag ccgaccgaag cgtggccgcg caccagccag 600
ggcccgagca cccgcccggt ggaagtgccg ggcggccgcg cggtggcggc gattctgggc 660
ctgggcctgg tgctgggcct gctgggcccg ctggcgattc tgctggcgct gtatctgctg 720
cgccgcgatc agcgcctgcc gccggatgcg cataaaccgc cgggcggcgg cagctttcgc 780
accccgattc aggaagaaca ggcggatgcg catagcaccc tggcgaaaat t 831
<210> 79
<211> 277
<212> PRT
<213> Homo sapiens
<400> 79
Met Cys Val Gly Ala Arg Arg Leu Gly Arg Gly Pro Cys Ala Ala Leu
1 5 10 15
Leu Leu Leu Gly Leu Gly Leu Ser Thr Val Thr Gly Leu His Cys Val
20 25 30
Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys His Glu Cys Arg Pro
35 40 45
Gly Asn Gly Met Val Ser Arg Cys Ser Arg Ser Gln Asn Thr Val Cys
50 55 60
Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val Ser Ser Lys Pro
65 70 75 80
Cys Lys Pro Cys Thr Trp Cys Asn Leu Arg Ser Gly Ser Glu Arg Lys
85 90 95
Gln Leu Cys Thr Ala Thr Gln Asp Thr Val Cys Arg Cys Arg Ala Gly
100 105 110
Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp Cys Ala Pro Cys
115 120 125
Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala Cys Lys Pro Trp
130 135 140
Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln Pro Ala Ser Asn
145 150 155 160
Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro Ala Thr Gln Pro
165 170 175
Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Ile Thr Val Gln Pro Thr
180 185 190
Glu Ala Trp Pro Arg Thr Ser Gln Gly Pro Ser Thr Arg Pro Val Glu
195 200 205
Val Pro Gly Gly Arg Ala Val Ala Ala Ile Leu Gly Leu Gly Leu Val
210 215 220
Leu Gly Leu Leu Gly Pro Leu Ala Ile Leu Leu Ala Leu Tyr Leu Leu
225 230 235 240
Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly Gly
245 250 255
Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His Ser
260 265 270
Thr Leu Ala Lys Ile
275
<210> 80
<211> 816
<212> DNA
<213> Mus musculus
<400> 80
atgtatgtgt gggtgcagca gccgaccgcg ctgctgctgc tggcgctgac cctgggcgtg 60
accgcgcgcc gcctgaactg cgtgaaacat acctatccga gcggccataa atgctgccgc 120
gaatgccagc cgggccatgg catggtgagc cgctgcgatc atacccgcga taccctgtgc 180
catccgtgcg aaaccggctt ttataacgaa gcggtgaact atgatacctg caaacagtgc 240
acccagtgca accatcgcag cggcagcgaa ctgaaacaga actgcacccc gacccaggat 300
accgtgtgcc gctgccgccc gggcacccag ccgcgccagg atagcggcta taaactgggc 360
gtggattgcg tgccgtgccc gccgggccat tttagcccgg gcaacaacca ggcgtgcaaa 420
ccgtggacca actgcaccct gagcggcaaa cagacccgcc atccggcgag cgatagcctg 480
gatgcggtgt gcgaagatcg cagcctgctg gcgaccctgc tgtgggaaac ccagcgcccg 540
acctttcgcc cgaccaccgt gcagagcacc accgtgtggc cgcgcaccag cgaactgccg 600
agcccgccga ccctggtgac cccggaaggc ccggcgtttg cggtgctgct gggcctgggc 660
ctgggcctgc tggcgccgct gaccgtgctg ctggcgctgt atctgctgcg caaagcgtgg 720
cgcctgccga acaccccgaa accgtgctgg ggcaacagct ttcgcacccc gattcaggaa 780
gaacataccg atgcgcattt taccctggcg aaaatt 816
<210> 81
<211> 272
<212> PRT
<213> Mus musculus
<400> 81
Met Tyr Val Trp Val Gln Gln Pro Thr Ala Leu Leu Leu Leu Ala Leu
1 5 10 15
Thr Leu Gly Val Thr Ala Arg Arg Leu Asn Cys Val Lys His Thr Tyr
20 25 30
Pro Ser Gly His Lys Cys Cys Arg Glu Cys Gln Pro Gly His Gly Met
35 40 45
Val Ser Arg Cys Asp His Thr Arg Asp Thr Leu Cys His Pro Cys Glu
50 55 60
Thr Gly Phe Tyr Asn Glu Ala Val Asn Tyr Asp Thr Cys Lys Gln Cys
65 70 75 80
Thr Gln Cys Asn His Arg Ser Gly Ser Glu Leu Lys Gln Asn Cys Thr
85 90 95
Pro Thr Gln Asp Thr Val Cys Arg Cys Arg Pro Gly Thr Gln Pro Arg
100 105 110
Gln Asp Ser Gly Tyr Lys Leu Gly Val Asp Cys Val Pro Cys Pro Pro
115 120 125
Gly His Phe Ser Pro Gly Asn Asn Gln Ala Cys Lys Pro Trp Thr Asn
130 135 140
Cys Thr Leu Ser Gly Lys Gln Thr Arg His Pro Ala Ser Asp Ser Leu
145 150 155 160
Asp Ala Val Cys Glu Asp Arg Ser Leu Leu Ala Thr Leu Leu Trp Glu
165 170 175
Thr Gln Arg Pro Thr Phe Arg Pro Thr Thr Val Gln Ser Thr Thr Val
180 185 190
Trp Pro Arg Thr Ser Glu Leu Pro Ser Pro Pro Thr Leu Val Thr Pro
195 200 205
Glu Gly Pro Ala Phe Ala Val Leu Leu Gly Leu Gly Leu Gly Leu Leu
210 215 220
Ala Pro Leu Thr Val Leu Leu Ala Leu Tyr Leu Leu Arg Lys Ala Trp
225 230 235 240
Arg Leu Pro Asn Thr Pro Lys Pro Cys Trp Gly Asn Ser Phe Arg Thr
245 250 255
Pro Ile Gln Glu Glu His Thr Asp Ala His Phe Thr Leu Ala Lys Ile
260 265 270
<210> 82
<211> 597
<212> DNA
<213> Homo sapiens
<400> 82
atgaaaagcg gcctgtggta tttttttctg ttttgcctgc gcattaaagt gctgaccggc 60
gaaattaacg gcagcgcgaa ctatgaaatg tttatttttc ataacggcgg cgtgcagatt 120
ctgtgcaaat atccggatat tgtgcagcag tttaaaatgc agctgctgaa aggcggccag 180
attctgtgcg atctgaccaa aaccaaaggc agcggcaaca ccgtgagcat taaaagcctg 240
aaattttgcc atagccagct gagcaacaac agcgtgagct tttttctgta taacctggat 300
catagccatg cgaactatta tttttgcaac ctgagcattt ttgatccgcc gccgtttaaa 360
gtgaccctga ccggcggcta tctgcatatt tatgaaagcc agctgtgctg ccagctgaaa 420
ttttggctgc cgattggctg cgcggcgttt gtggtggtgt gcattctggg ctgcattctg 480
atttgctggc tgaccaaaaa aaaatatagc agcagcgtgc atgatccgaa cggcgaatat 540
atgtttatgc gcgcggtgaa caccgcgaaa aaaagccgcc tgaccgatgt gaccctg 597
<210> 83
<211> 199
<212> PRT
<213> Homo sapiens
<400> 83
Met Lys Ser Gly Leu Trp Tyr Phe Phe Leu Phe Cys Leu Arg Ile Lys
1 5 10 15
Val Leu Thr Gly Glu Ile Asn Gly Ser Ala Asn Tyr Glu Met Phe Ile
20 25 30
Phe His Asn Gly Gly Val Gln Ile Leu Cys Lys Tyr Pro Asp Ile Val
35 40 45
Gln Gln Phe Lys Met Gln Leu Leu Lys Gly Gly Gln Ile Leu Cys Asp
50 55 60
Leu Thr Lys Thr Lys Gly Ser Gly Asn Thr Val Ser Ile Lys Ser Leu
65 70 75 80
Lys Phe Cys His Ser Gln Leu Ser Asn Asn Ser Val Ser Phe Phe Leu
85 90 95
Tyr Asn Leu Asp His Ser His Ala Asn Tyr Tyr Phe Cys Asn Leu Ser
100 105 110
Ile Phe Asp Pro Pro Pro Phe Lys Val Thr Leu Thr Gly Gly Tyr Leu
115 120 125
His Ile Tyr Glu Ser Gln Leu Cys Cys Gln Leu Lys Phe Trp Leu Pro
130 135 140
Ile Gly Cys Ala Ala Phe Val Val Val Cys Ile Leu Gly Cys Ile Leu
145 150 155 160
Ile Cys Trp Leu Thr Lys Lys Lys Tyr Ser Ser Ser Val His Asp Pro
165 170 175
Asn Gly Glu Tyr Met Phe Met Arg Ala Val Asn Thr Ala Lys Lys Ser
180 185 190
Arg Leu Thr Asp Val Thr Leu
195
<210> 84
<211> 600
<212> DNA
<213> Mus musculus
<400> 84
atgaaaccgt atttttgccg cgtgtttgtg ttttgctttc tgattcgcct gctgaccggc 60
gaaattaacg gcagcgcgga tcatcgcatg tttagctttc ataacggcgg cgtgcagatt 120
agctgcaaat atccggaaac cgtgcagcag ctgaaaatgc gcctgtttcg cgaacgcgaa 180
gtgctgtgcg aactgaccaa aaccaaaggc agcggcaacg cggtgagcat taaaaacccg 240
atgctgtgcc tgtatcatct gagcaacaac agcgtgagct tttttctgaa caacccggat 300
agcagccagg gcagctatta tttttgcagc ctgagcattt ttgatccgcc gccgtttcag 360
gaacgcaacc tgagcggcgg ctatctgcat atttatgaaa gccagctgtg ctgccagctg 420
aaactgtggc tgccggtggg ctgcgcggcg tttgtggtgg tgctgctgtt tggctgcatt 480
ctgattattt ggtttagcaa aaaaaaatat ggcagcagcg tgcatgatcc gaacagcgaa 540
tatatgttta tggcggcggt gaacaccaac aaaaaaagcc gcctggcggg cgtgaccagc 600
<210> 85
<211> 200
<212> PRT
<213> Mus musculus
<400> 85
Met Lys Pro Tyr Phe Cys Arg Val Phe Val Phe Cys Phe Leu Ile Arg
1 5 10 15
Leu Leu Thr Gly Glu Ile Asn Gly Ser Ala Asp His Arg Met Phe Ser
20 25 30
Phe His Asn Gly Gly Val Gln Ile Ser Cys Lys Tyr Pro Glu Thr Val
35 40 45
Gln Gln Leu Lys Met Arg Leu Phe Arg Glu Arg Glu Val Leu Cys Glu
50 55 60
Leu Thr Lys Thr Lys Gly Ser Gly Asn Ala Val Ser Ile Lys Asn Pro
65 70 75 80
Met Leu Cys Leu Tyr His Leu Ser Asn Asn Ser Val Ser Phe Phe Leu
85 90 95
Asn Asn Pro Asp Ser Ser Gln Gly Ser Tyr Tyr Phe Cys Ser Leu Ser
100 105 110
Ile Phe Asp Pro Pro Pro Phe Gln Glu Arg Asn Leu Ser Gly Gly Tyr
115 120 125
Leu His Ile Tyr Glu Ser Gln Leu Cys Cys Gln Leu Lys Leu Trp Leu
130 135 140
Pro Val Gly Cys Ala Ala Phe Val Val Val Leu Leu Phe Gly Cys Ile
145 150 155 160
Leu Ile Ile Trp Phe Ser Lys Lys Lys Tyr Gly Ser Ser Val His Asp
165 170 175
Pro Asn Ser Glu Tyr Met Phe Met Ala Ala Val Asn Thr Asn Lys Lys
180 185 190
Ser Arg Leu Ala Gly Val Thr Ser
195 200
<210> 86
<211> 279
<212> DNA
<213> Homo sapiens
<400> 86
atgattcatc tgggccatat tctgtttctg ctgctgctgc cggtggcggc ggcgcagacc 60
accccgggcg aacgcagcag cctgccggcg ttttatccgg gcaccagcgg cagctgcagc 120
ggctgcggca gcctgagcct gccgctgctg gcgggcctgg tggcggcgga tgcggtggcg 180
agcctgctga ttgtgggcgc ggtgtttctg tgcgcgcgcc cgcgccgcag cccggcgcag 240
gaagatggca aagtgtatat taacatgccg ggccgcggc 279
<210> 87
<211> 93
<212> PRT
<213> Homo sapiens
<400> 87
Met Ile His Leu Gly His Ile Leu Phe Leu Leu Leu Leu Pro Val Ala
1 5 10 15
Ala Ala Gln Thr Thr Pro Gly Glu Arg Ser Ser Leu Pro Ala Phe Tyr
20 25 30
Pro Gly Thr Ser Gly Ser Cys Ser Gly Cys Gly Ser Leu Ser Leu Pro
35 40 45
Leu Leu Ala Gly Leu Val Ala Ala Asp Ala Val Ala Ser Leu Leu Ile
50 55 60
Val Gly Ala Val Phe Leu Cys Ala Arg Pro Arg Arg Ser Pro Ala Gln
65 70 75 80
Glu Asp Gly Lys Val Tyr Ile Asn Met Pro Gly Arg Gly
85 90
<210> 88
<211> 237
<212> DNA
<213> Mus musculus
<400> 88
atggatccgc cgggctatct gctgtttctg ctgctgctgc cggtggcggc gagccagacc 60
agcgcgggca gctgcagcgg ctgcggcacc ctgagcctgc cgctgctggc gggcctggtg 120
gcggcggatg cggtgatgag cctgctgatt gtgggcgtgg tgtttgtgtg catgcgcccg 180
catggccgcc cggcgcagga agatggccgc gtgtatatta acatgccggg ccgcggc 237
<210> 89
<211> 79
<212> PRT
<213> Mus musculus
<400> 89
Met Asp Pro Pro Gly Tyr Leu Leu Phe Leu Leu Leu Leu Pro Val Ala
1 5 10 15
Ala Ser Gln Thr Ser Ala Gly Ser Cys Ser Gly Cys Gly Thr Leu Ser
20 25 30
Leu Pro Leu Leu Ala Gly Leu Val Ala Ala Asp Ala Val Met Ser Leu
35 40 45
Leu Ile Val Gly Val Val Phe Val Cys Met Arg Pro His Gly Arg Pro
50 55 60
Ala Gln Glu Asp Gly Arg Val Tyr Ile Asn Met Pro Gly Arg Gly
65 70 75
<210> 90
<211> 342
<212> DNA
<213> Homo sapiens
<400> 90
atggggggac ttgaaccctg cagcaggctc ctgctcctgc ctctcctgct ggctgtaagt 60
ggtctccgtc ctgtccaggc ccaggcccag agcgattgca gttgctctac ggtgagcccg 120
ggcgtgctgg cagggatcgt gatgggagac ctggtgctga cagtgctcat tgccctggcc 180
gtgtacttcc tgggccggct ggtccctcgg gggcgagggg ctgcggaggc agcgacccgg 240
aaacagcgta tcactgagac cgagtcgcct tatcaggagc tccagggtca gaggtcggat 300
gtctacagcg acctcaacac acagaggccg tattacaaat ga 342
<210> 91
<211> 113
<212> PRT
<213> Homo sapiens
<400> 91
Met Gly Gly Leu Glu Pro Cys Ser Arg Leu Leu Leu Leu Pro Leu Leu
1 5 10 15
Leu Ala Val Ser Gly Leu Arg Pro Val Gln Ala Gln Ala Gln Ser Asp
20 25 30
Cys Ser Cys Ser Thr Val Ser Pro Gly Val Leu Ala Gly Ile Val Met
35 40 45
Gly Asp Leu Val Leu Thr Val Leu Ile Ala Leu Ala Val Tyr Phe Leu
50 55 60
Gly Arg Leu Val Pro Arg Gly Arg Gly Ala Ala Glu Ala Ala Thr Arg
65 70 75 80
Lys Gln Arg Ile Thr Glu Thr Glu Ser Pro Tyr Gln Glu Leu Gln Gly
85 90 95
Gln Arg Ser Asp Val Tyr Ser Asp Leu Asn Thr Gln Arg Pro Tyr Tyr
100 105 110
Lys
<210> 92
<211> 345
<212> DNA
<213> Mus musculus
<400> 92
atgggggctc tggagccctc ctggtgcctt ctgttccttc ctgtcctcct gactgtggga 60
ggattaagtc ccgtacaggc ccagagtgac actttcccaa gatgcgactg ttcttccgtg 120
agccctggtg tactggctgg gattgttctg ggtgacttgg tgttgactct gctgattgcc 180
ctggctgtgt actctctggg ccgcctggtc tcccgaggtc aagggacagc ggaagggacc 240
cggaaacaac acattgctga gactgagtcg ccttatcagg agcttcaggg tcagagacca 300
gaagtataca gtgacctcaa cacacagagg caatattaca gatga 345
<210> 93
<211> 114
<212> PRT
<213> Mus musculus
<400> 93
Met Gly Ala Leu Glu Pro Ser Trp Cys Leu Leu Phe Leu Pro Val Leu
1 5 10 15
Leu Thr Val Gly Gly Leu Ser Pro Val Gln Ala Gln Ser Asp Thr Phe
20 25 30
Pro Arg Cys Asp Cys Ser Ser Val Ser Pro Gly Val Leu Ala Gly Ile
35 40 45
Val Leu Gly Asp Leu Val Leu Thr Leu Leu Ile Ala Leu Ala Val Tyr
50 55 60
Ser Leu Gly Arg Leu Val Ser Arg Gly Gln Gly Thr Ala Glu Gly Thr
65 70 75 80
Arg Lys Gln His Ile Ala Glu Thr Glu Ser Pro Tyr Gln Glu Leu Gln
85 90 95
Gly Gln Arg Pro Glu Val Tyr Ser Asp Leu Asn Thr Gln Arg Gln Tyr
100 105 110
Tyr Arg
<210> 94
<211> 164
<212> PRT
<213> Homo sapiens
<400> 94
Met Lys Trp Lys Ala Leu Phe Thr Ala Ala Ile Leu Gln Ala Gln Leu
1 5 10 15
Pro Ile Thr Glu Ala Gln Ser Phe Gly Leu Leu Asp Pro Lys Leu Cys
20 25 30
Tyr Leu Leu Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Leu Thr Ala
35 40 45
Leu Phe Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr
50 55 60
Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
65 70 75 80
Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
85 90 95
Gly Gly Lys Pro Gln Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn
100 105 110
Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met
115 120 125
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
130 135 140
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala
145 150 155 160
Leu Pro Pro Arg
<210> 95
<211> 492
<212> DNA
<213> Homo sapiens
<400> 95
atgaagtgga aggcgctttt caccgcggcc atcctgcagg cacagttgcc gattacagag 60
gcacagagct ttggcctgct ggatcccaaa ctctgctacc tgctggatgg aatcctcttc 120
atctatggtg tcattctcac tgccttgttc ctgagagtga agttcagcag gagcgcagag 180
ccccccgcgt accagcaggg ccagaaccag ctctataacg agctcaatct aggacgaaga 240
gaggagtacg atgttttgga caagagacgt ggccgggacc ctgagatggg gggaaagccg 300
agaaggaaga accctcagga aggcctgtac aatgaactgc agaaagataa gatggcggag 360
gcctacagtg agattgggat gaaaggcgag cgccggaggg gcaaggggca cgatggcctt 420
taccagggtc tcagtacagc caccaaggac acctacgacg cccttcacat gcaggccctg 480
ccccctcgct aa 492
<210> 96
<211> 164
<212> PRT
<213> Mus musculus
<400> 96
Met Lys Trp Lys Val Ser Val Leu Ala Cys Ile Leu His Val Arg Phe
1 5 10 15
Pro Gly Ala Glu Ala Gln Ser Phe Gly Leu Leu Asp Pro Lys Leu Cys
20 25 30
Tyr Leu Leu Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Ile Thr Ala
35 40 45
Leu Tyr Leu Arg Ala Lys Phe Ser Arg Ser Ala Glu Thr Ala Ala Asn
50 55 60
Leu Gln Asp Pro Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg
65 70 75 80
Glu Glu Tyr Asp Val Leu Glu Lys Lys Arg Ala Arg Asp Pro Glu Met
85 90 95
Gly Gly Lys Gln Gln Arg Arg Arg Asn Pro Gln Glu Gly Val Tyr Asn
100 105 110
Ala Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Thr
115 120 125
Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly
130 135 140
Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Thr
145 150 155 160
Leu Ala Pro Arg
<210> 97
<211> 495
<212> DNA
<213> Mus musculus
<400> 97
atgaagtgga aagtgtctgt tctcgcctgc atcctccacg tgcggttccc aggagcagag 60
gcacagagct ttggtctgct ggatcccaaa ctctgctact tgctagatgg aatcctcttc 120
atctacggag tcatcatcac agccctgtac ctgagagcaa aattcagcag gagtgcagag 180
actgctgcca acctgcagga ccccaaccag ctctacaatg agctcaatct agggcgaaga 240
gaggaatatg acgtcttgga gaagaagcgg gctcgggatc cagagatggg aggcaaacag 300
cagaggagga ggaaccccca ggaaggcgta tacaatgcac tgcagaaaga caagatggca 360
gaagcctaca gtgagatcgg cacaaaaggc gagaggcgga gaggcaaggg gcacgatggc 420
ctttaccagg gtctcagcac tgccaccaag gacacctatg atgccctgca tatgcagacc 480
ctggcccctc gctaa 495
<210> 98
<211> 254
<212> PRT
<213> Homo sapiens
<400> 98
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Phe
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 99
<211> 762
<212> DNA
<213> Homo sapiens
<400> 99
atgtggcagc tgctgctgcc gaccgcgctg ctgctgctgg tgagcgcggg catgcgcacc 60
gaagatctgc cgaaagcggt ggtgtttctg gaaccgcagt ggtatcgcgt gctggaaaaa 120
gatagcgtga ccctgaaatg ccagggcgcg tatagcccgg aagataacag cacccagtgg 180
tttcataacg aaagcctgat tagcagccag gcgagcagct attttattga tgcggcgacc 240
gtggatgata gcggcgaata tcgctgccag accaacctga gcaccctgag cgatccggtg 300
cagctggaag tgcatattgg ctggctgctg ctgcaggcgc cgcgctgggt gtttaaagaa 360
gaagatccga ttcatctgcg ctgccatagc tggaaaaaca ccgcgctgca taaagtgacc 420
tatctgcaga acggcaaagg ccgcaaatat tttcatcata acagcgattt ttatattccg 480
aaagcgaccc tgaaagatag cggcagctat ttttgccgcg gcctgtttgg cagcaaaaac 540
gtgagcagcg aaaccgtgaa cattaccatt acccagggcc tggcggtgag caccattagc 600
agcttttttc cgccgggcta tcaggtgagc ttttgcctgg tgatggtgct gctgtttgcg 660
gtggataccg gcctgtattt tagcgtgaaa accaacattc gcagcagcac ccgcgattgg 720
aaagatcata aatttaaatg gcgcaaagat ccgcaggata aa 762
<210> 100
<211> 261
<212> PRT
<213> Mus musculus
<400> 100
Met Phe Gln Asn Ala His Ser Gly Ser Gln Trp Leu Leu Pro Pro Leu
1 5 10 15
Thr Ile Leu Leu Leu Phe Ala Phe Ala Asp Arg Gln Ser Ala Ala Leu
20 25 30
Pro Lys Ala Val Val Lys Leu Asp Pro Pro Trp Ile Gln Val Leu Lys
35 40 45
Glu Asp Met Val Thr Leu Met Cys Glu Gly Thr His Asn Pro Gly Asn
50 55 60
Ser Ser Thr Gln Trp Phe His Asn Gly Arg Ser Ile Arg Ser Gln Val
65 70 75 80
Gln Ala Ser Tyr Thr Phe Lys Ala Thr Val Asn Asp Ser Gly Glu Tyr
85 90 95
Arg Cys Gln Met Glu Gln Thr Arg Leu Ser Asp Pro Val Asp Leu Gly
100 105 110
Val Ile Ser Asp Trp Leu Leu Leu Gln Thr Pro Gln Arg Val Phe Leu
115 120 125
Glu Gly Glu Thr Ile Thr Leu Arg Cys His Ser Trp Arg Asn Lys Leu
130 135 140
Leu Asn Arg Ile Ser Phe Phe His Asn Glu Lys Ser Val Arg Tyr His
145 150 155 160
His Tyr Lys Ser Asn Phe Ser Ile Pro Lys Ala Asn His Ser His Ser
165 170 175
Gly Asp Tyr Tyr Cys Lys Gly Ser Leu Gly Ser Thr Gln His Gln Ser
180 185 190
Lys Pro Val Thr Ile Thr Val Gln Asp Pro Ala Thr Thr Ser Ser Ile
195 200 205
Ser Leu Val Trp Tyr His Thr Ala Phe Ser Leu Val Met Cys Leu Leu
210 215 220
Phe Ala Val Asp Thr Gly Leu Tyr Phe Tyr Val Arg Arg Asn Leu Gln
225 230 235 240
Thr Pro Arg Glu Tyr Trp Arg Lys Ser Leu Ser Ile Arg Lys His Gln
245 250 255
Ala Pro Gln Asp Lys
260
<210> 101
<211> 786
<212> DNA
<213> Mus musculus
<400> 101
atgtttcaga atgcacactc tggaagccaa tggctacttc caccactgac aattctgctg 60
ctgtttgctt ttgcagacag gcagagtgca gctcttccga aggctgtggt gaaactggac 120
cccccatgga tccaggtgct caaggaagac atggtgacac tgatgtgcga agggacccac 180
aaccctggga actcttctac ccagtggttc cacaacggga ggtccatccg gagccaggtc 240
caagccagtt acacgtttaa ggccacagtc aatgacagtg gagaatatcg gtgtcaaatg 300
gagcagaccc gcctcagcga ccctgtagat ctgggagtga tttctgactg gctgctgctc 360
cagacccctc agcgggtgtt tctggaaggg gaaaccatca cgctaaggtg ccatagctgg 420
aggaacaaac tactgaacag gatctcattc ttccataatg aaaaatccgt gaggtatcat 480
cactacaaaa gtaatttctc tatcccaaaa gccaaccaca gtcacagtgg ggactactac 540
tgcaaaggaa gtctaggaag tacacagcac cagtccaagc ctgtcaccat cactgtccaa 600
gatccagcaa ctacatcctc catctctcta gtctggtacc acactgcttt ctccctagtg 660
atgtgcctcc tgtttgcagt ggacacgggc ctttatttct acgtacggag aaatcttcaa 720
accccgaggg agtactggag gaagtccctg tcaatcagaa agcaccaggc tcctcaagac 780
aagtga 786
<210> 102
<211> 216
<212> PRT
<213> Homo sapiens
<400> 102
Met Gly Trp Ile Arg Gly Arg Arg Ser Arg His Ser Trp Glu Met Ser
1 5 10 15
Glu Phe His Asn Tyr Asn Leu Asp Leu Lys Lys Ser Asp Phe Ser Thr
20 25 30
Arg Trp Gln Lys Gln Arg Cys Pro Val Val Lys Ser Lys Cys Arg Glu
35 40 45
Asn Ala Ser Pro Phe Phe Phe Cys Cys Phe Ile Ala Val Ala Met Gly
50 55 60
Ile Arg Phe Ile Ile Met Val Ala Ile Trp Ser Ala Val Phe Leu Asn
65 70 75 80
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
85 90 95
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
100 105 110
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
115 120 125
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
130 135 140
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
145 150 155 160
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
165 170 175
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
180 185 190
Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr Pro Asn Thr
195 200 205
Tyr Ile Cys Met Gln Arg Thr Val
210 215
<210> 103
<211> 648
<212> DNA
<213> Homo sapiens
<400> 103
atgggctgga ttcgcggccg ccgcagccgc catagctggg aaatgagcga atttcataac 60
tataacctgg atctgaaaaa aagcgatttt agcacccgct ggcagaaaca gcgctgcccg 120
gtggtgaaaa gcaaatgccg cgaaaacgcg agcccgtttt ttttttgctg ctttattgcg 180
gtggcgatgg gcattcgctt tattattatg gtggcgattt ggagcgcggt gtttctgaac 240
agcctgttta accaggaagt gcagattccg ctgaccgaaa gctattgcgg cccgtgcccg 300
aaaaactgga tttgctataa aaacaactgc tatcagtttt ttgatgaaag caaaaactgg 360
tatgaaagcc aggcgagctg catgagccag aacgcgagcc tgctgaaagt gtatagcaaa 420
gaagatcagg atctgctgaa actggtgaaa agctatcatt ggatgggcct ggtgcatatt 480
ccgaccaacg gcagctggca gtgggaagat ggcagcattc tgagcccgaa cctgctgacc 540
attattgaaa tgcagaaagg cgattgcgcg ctgtatgcga gcagctttaa aggctatatt 600
gaaaactgca gcaccccgaa cacctatatt tgcatgcagc gcaccgtg 648
<210> 104
<211> 232
<212> PRT
<213> Mus musculus
<400> 104
Met Ala Leu Ile Arg Asp Arg Lys Ser His His Ser Glu Met Ser Lys
1 5 10 15
Cys His Asn Tyr Asp Leu Lys Pro Ala Lys Trp Asp Thr Ser Gln Glu
20 25 30
Gln Gln Lys Gln Arg Leu Ala Leu Thr Thr Ser Gln Pro Gly Glu Asn
35 40 45
Gly Ile Ile Arg Gly Arg Tyr Pro Ile Glu Lys Leu Lys Ile Ser Pro
50 55 60
Met Phe Val Val Arg Val Leu Ala Ile Ala Leu Ala Ile Arg Phe Thr
65 70 75 80
Leu Asn Thr Leu Met Trp Leu Ala Ile Phe Lys Glu Thr Phe Gln Pro
85 90 95
Val Leu Cys Asn Lys Glu Val Pro Val Ser Ser Arg Glu Gly Tyr Cys
100 105 110
Gly Pro Cys Pro Asn Asn Trp Ile Cys His Arg Asn Asn Cys Tyr Gln
115 120 125
Phe Phe Asn Glu Glu Lys Thr Trp Asn Gln Ser Gln Ala Ser Cys Leu
130 135 140
Ser Gln Asn Ser Ser Leu Leu Lys Ile Tyr Ser Lys Glu Glu Gln Asp
145 150 155 160
Phe Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val Gln Ile
165 170 175
Pro Ala Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ser Leu Ser Tyr
180 185 190
Asn Gln Leu Thr Leu Val Glu Ile Pro Lys Gly Ser Cys Ala Val Tyr
195 200 205
Gly Ser Ser Phe Lys Ala Tyr Thr Glu Asp Cys Ala Asn Leu Asn Thr
210 215 220
Tyr Ile Cys Met Lys Arg Ala Val
225 230
<210> 105
<211> 696
<212> DNA
<213> Mus musculus
<400> 105
atggcgctga ttcgcgatcg caaaagccat catagcgaaa tgagcaaatg ccataactat 60
gatctgaaac cggcgaaatg ggataccagc caggaacagc agaaacagcg cctggcgctg 120
accaccagcc agccgggcga aaacggcatt attcgcggcc gctatccgat tgaaaaactg 180
aaaattagcc cgatgtttgt ggtgcgcgtg ctggcgattg cgctggcgat tcgctttacc 240
ctgaacaccc tgatgtggct ggcgattttt aaagaaacct ttcagccggt gctgtgcaac 300
aaagaagtgc cggtgagcag ccgcgaaggc tattgcggcc cgtgcccgaa caactggatt 360
tgccatcgca acaactgcta tcagtttttt aacgaagaaa aaacctggaa ccagagccag 420
gcgagctgcc tgagccagaa cagcagcctg ctgaaaattt atagcaaaga agaacaggat 480
tttctgaaac tggtgaaaag ctatcattgg atgggcctgg tgcagattcc ggcgaacggc 540
agctggcagt gggaagatgg cagcagcctg agctataacc agctgaccct ggtggaaatt 600
ccgaaaggca gctgcgcggt gtatggcagc agctttaaag cgtataccga agattgcgcg 660
aacctgaaca cctatatttg catgaaacgc gcggtg 696
<210> 106
<211> 4
<212> PRT
<213> Artificial sequence
<220>
<223> CD28 YMNM
<400> 106
Tyr Met Asn Met
1
<210> 107
<211> 4
<212> PRT
<213> Artificial sequence
<220>
<223> CD28 PYAP
<400> 107
Pro Tyr Ala Pro
1
<210> 108
<211> 4
<212> PRT
<213> Artificial sequence
<220>
<223> CD28 FMNM
<400> 108
Phe Met Asn Met
1
<210> 109
<211> 4
<212> PRT
<213> Artificial sequence
<220>
<223> CD28 AYAA
<400> 109
Ala Tyr Ala Ala
1
<210> 110
<211> 21
<212> PRT
<213> Artificial sequence
<220>
<223> Signal peptide
<400> 110
Ala Thr Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala
1 5 10 15
Thr Gly Val His Ser
20
<210> 111
<211> 57
<212> DNA
<213> Artificial sequence
<220>
<223> Signal peptide DNA sequence
<400> 111
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctaccggtgt gcactcc 57
<210> 112
<211> 449
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CD20 (GA101) heavy chain
<400> 112
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Tyr Ser
20 25 30
Trp Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Phe Pro Gly Asp Gly Asp Thr Asp Tyr Asn Gly Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Val Phe Asp Gly Tyr Trp Leu Val Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 113
<211> 219
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CD20 (GA101) light chain
<400> 113
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Val Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 114
<211> 447
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-FAP(4B9) PGLALA heavy chain
<400> 114
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 115
<211> 215
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-FAP(4B9) light chain
<400> 115
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 116
<211> 451
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (A5B7) PGLALA heavy chain
<400> 116
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Gly Thr Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Arg Gly Leu Arg Phe Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 117
<211> 223
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (A5B7) light chain
<400> 117
Gln Ala Val Leu Thr Gln Pro Ala Ser Leu Ser Ala Ser Pro Gly Ala
1 5 10 15
Ser Ala Ser Leu Thr Cys Thr Leu Arg Arg Gly Ile Asn Val Gly Ala
20 25 30
Tyr Ser Ile Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Pro Pro Gln Tyr
35 40 45
Leu Leu Arg Tyr Lys Ser Asp Ser Asp Lys Gln Gln Gly Ser Gly Val
50 55 60
Ser Ser Arg Phe Ser Ala Ser Lys Asp Ala Ser Ala Asn Ala Gly Ile
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Met Ile Trp His Ser Gly Ala Ser Ala Val Phe Gly Gly Gly Thr Lys
100 105 110
Leu Thr Val Leu Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro
115 120 125
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
130 135 140
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
145 150 155 160
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
165 170 175
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
180 185 190
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
195 200 205
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 118
<211> 451
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (T84.66LCHA) PGLALA heavy chain
<400> 118
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 119
<211> 218
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (T84.66LCHA) light chain
<400> 119
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Gly Glu Ser Val Asp Ile Phe
20 25 30
Gly Val Gly Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Arg Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Thr Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 120
<211> 451
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (CH1A1A98/992F1) PGLALA heavy chain
<400> 120
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 121
<211> 215
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (CH1A1A98/992F1) light chain
<400> 121
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 122
<211> 449
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (hMN14) PGLALA heavy chain
<400> 122
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Thr Thr Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile His Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Phe Gly Phe Pro Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 123
<211> 213
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-CEA (hMN14) light chain
<400> 123
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Gly Thr Ser
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Thr Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Leu Tyr Arg Ser
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 124
<211> 451
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-TNC (2B10) PGLALA heavy chain
<400> 124
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Tyr Gly Tyr Ala Tyr Tyr Gly Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 125
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-TNC (2B10) light chain
<400> 125
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asn Gly Leu Gln Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 126
<211> 449
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-HER2 (PER) PG LALA heavy chain 1
<400> 126
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 127
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-HER2 (PER) light chain 1
<400> 127
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 128
<211> 449
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-HER2 (PER) PG LALA heavy chain 2
<400> 128
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 129
<211> 214
<212> PRT
<213> Artificial sequence
<220>
<223> Anti-HER2 (PER) light chain 2
<400> 129
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 130
<211> 330
<212> PRT
<213> Homo sapiens
<400> 130
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |


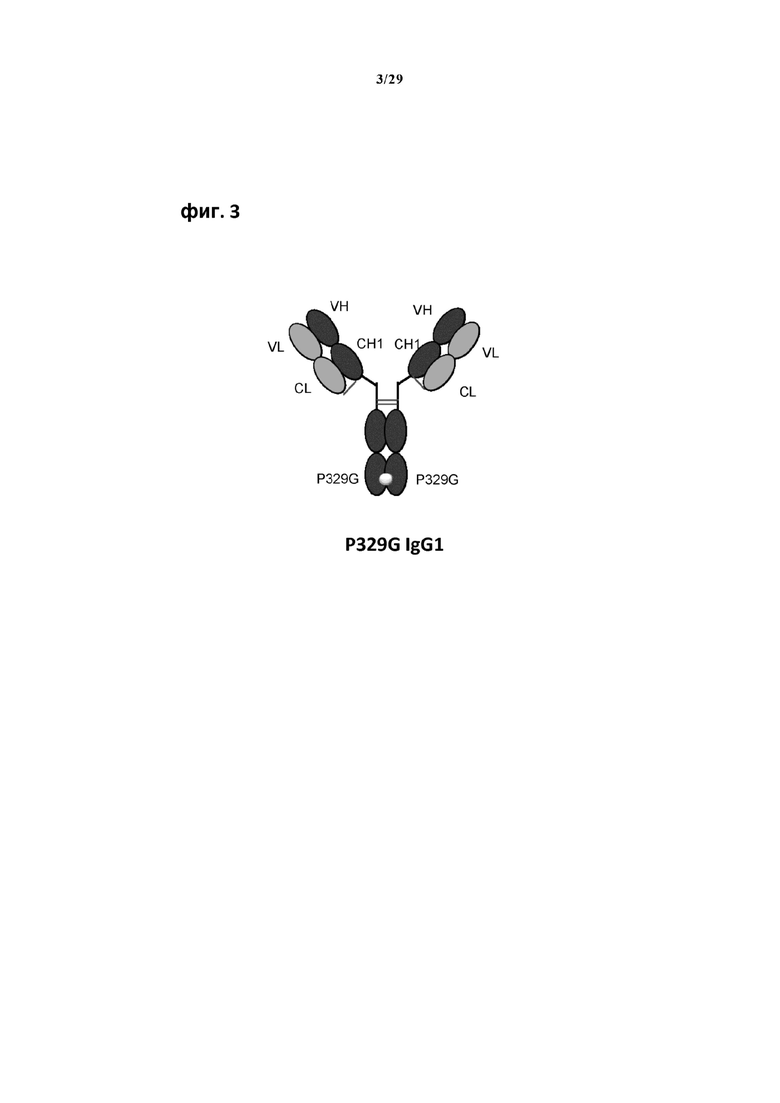

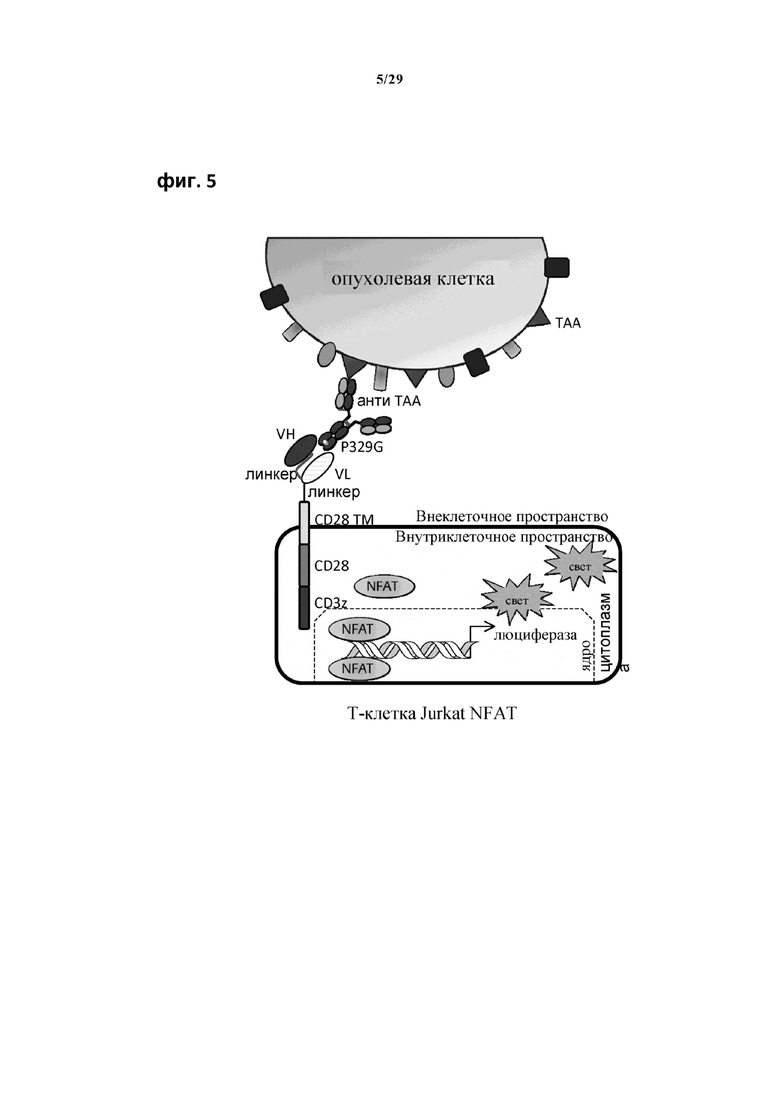
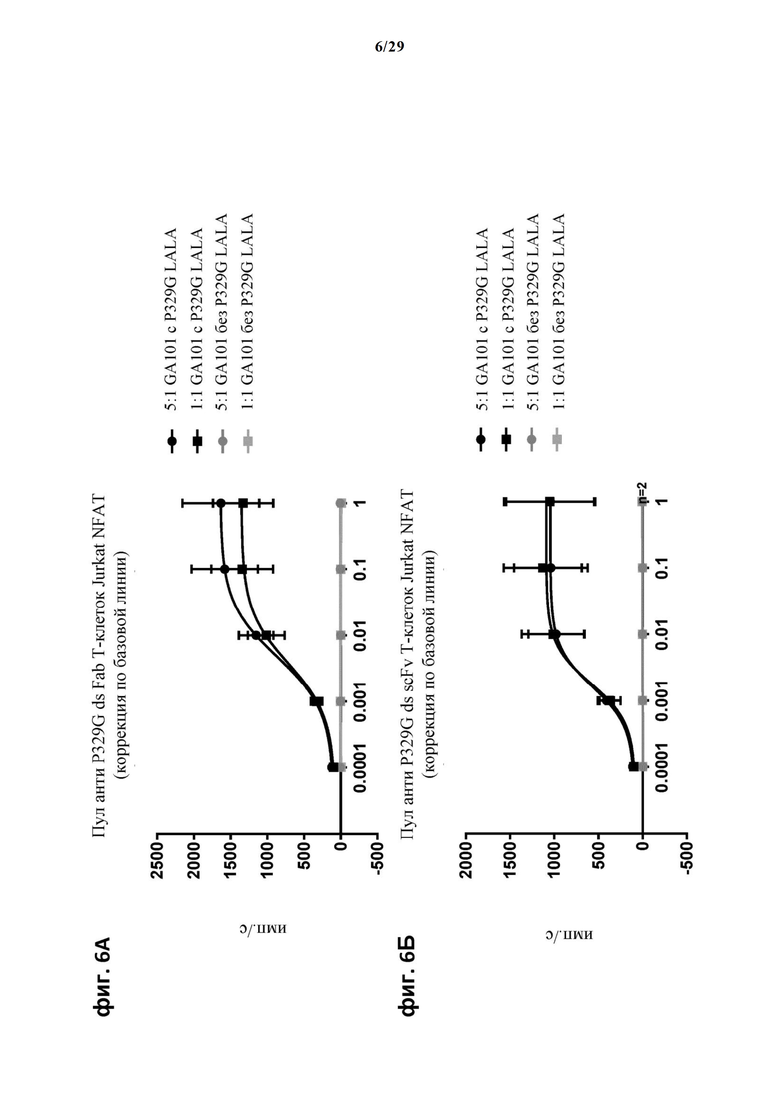
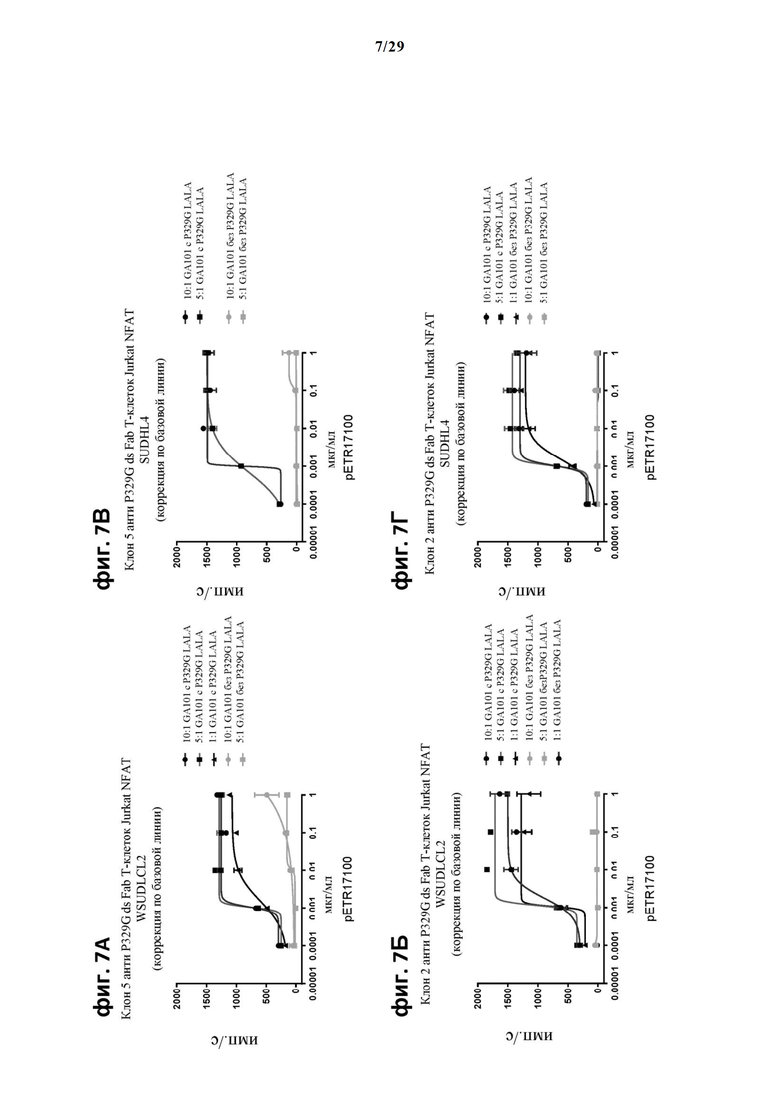
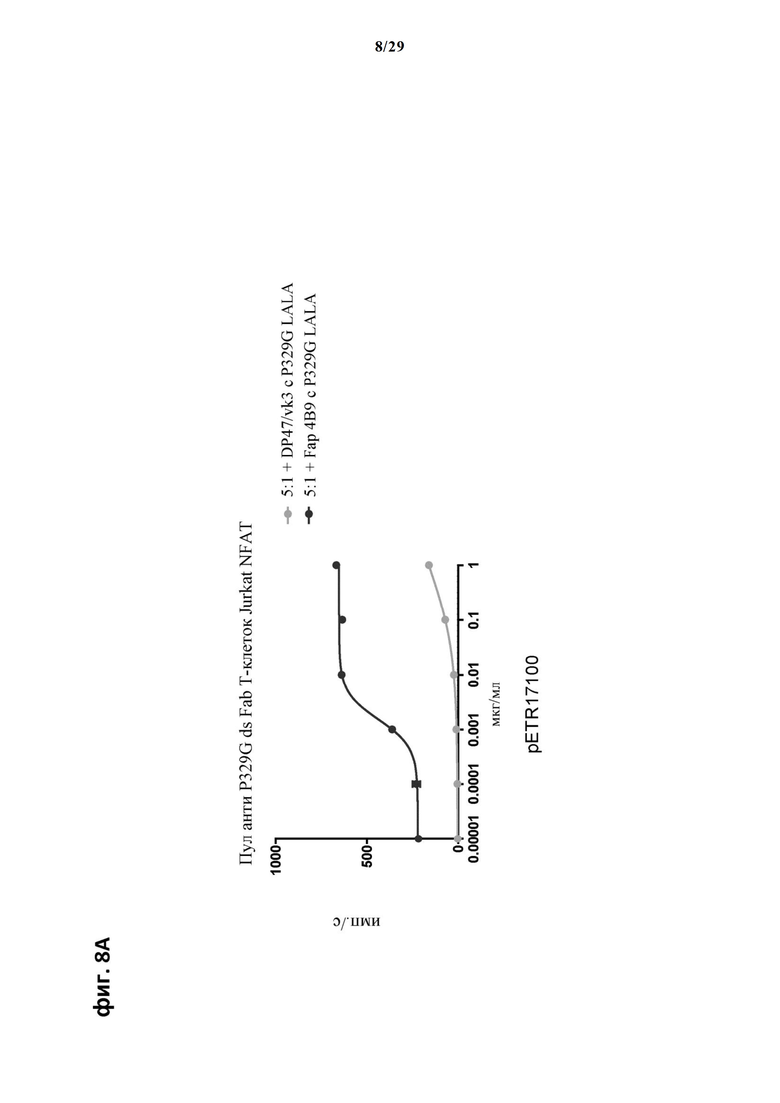
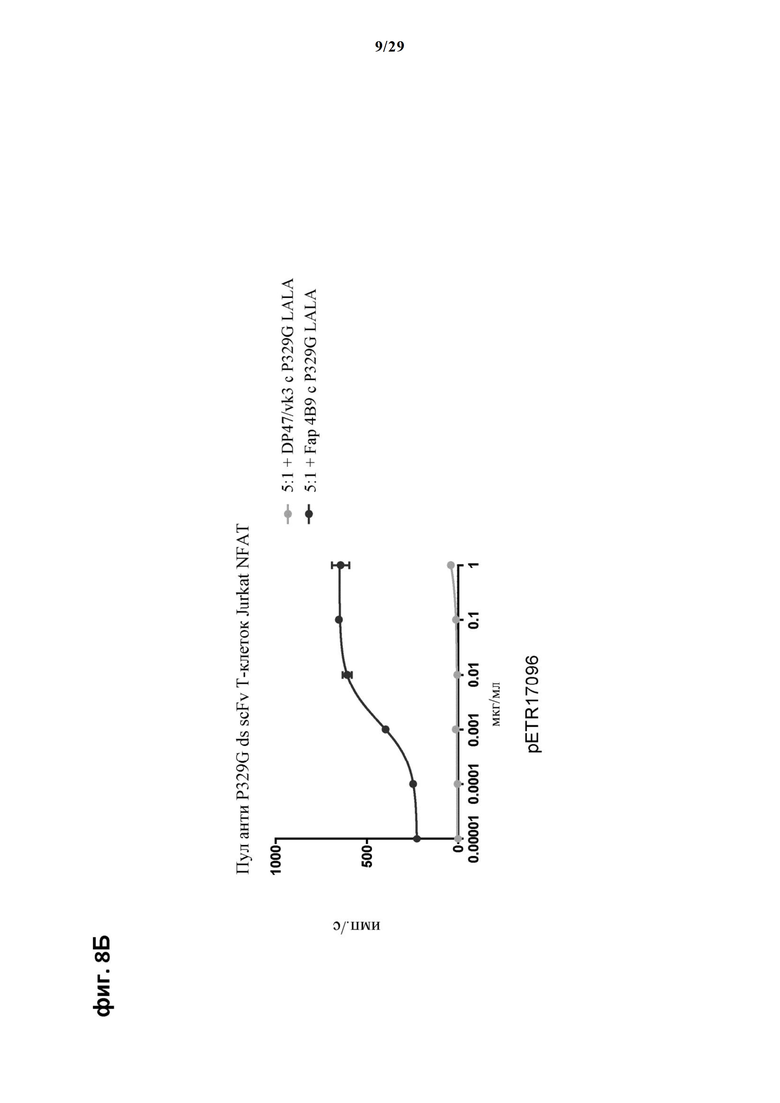
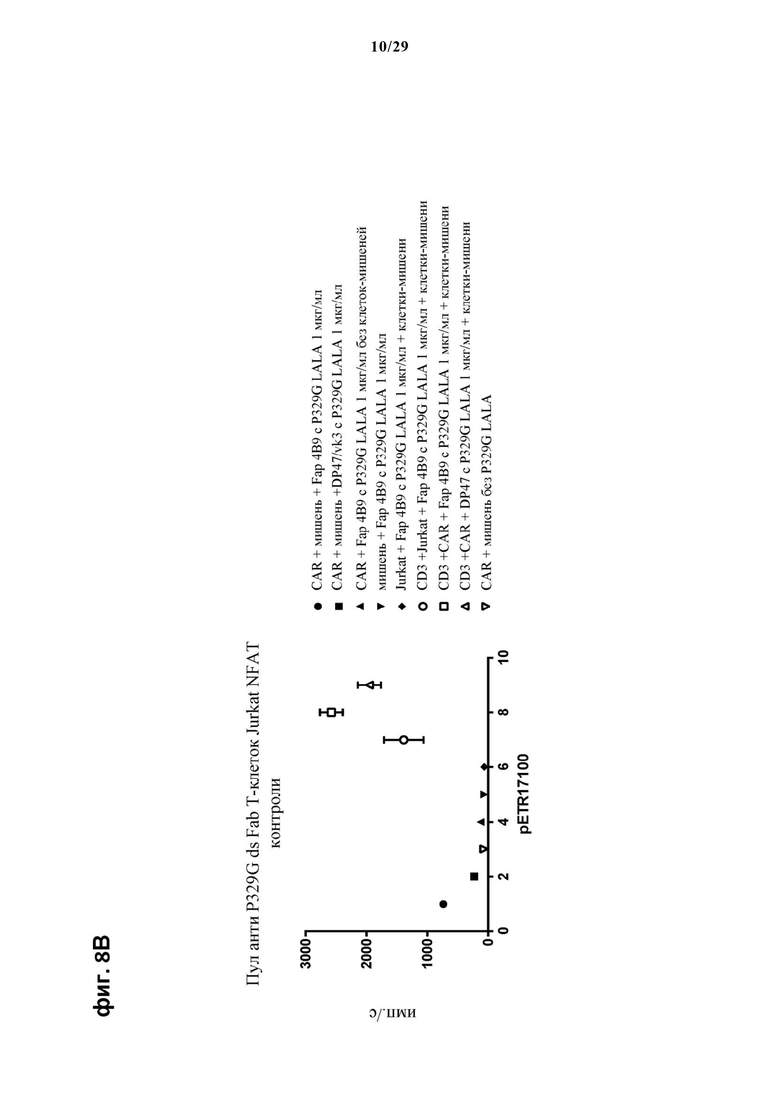



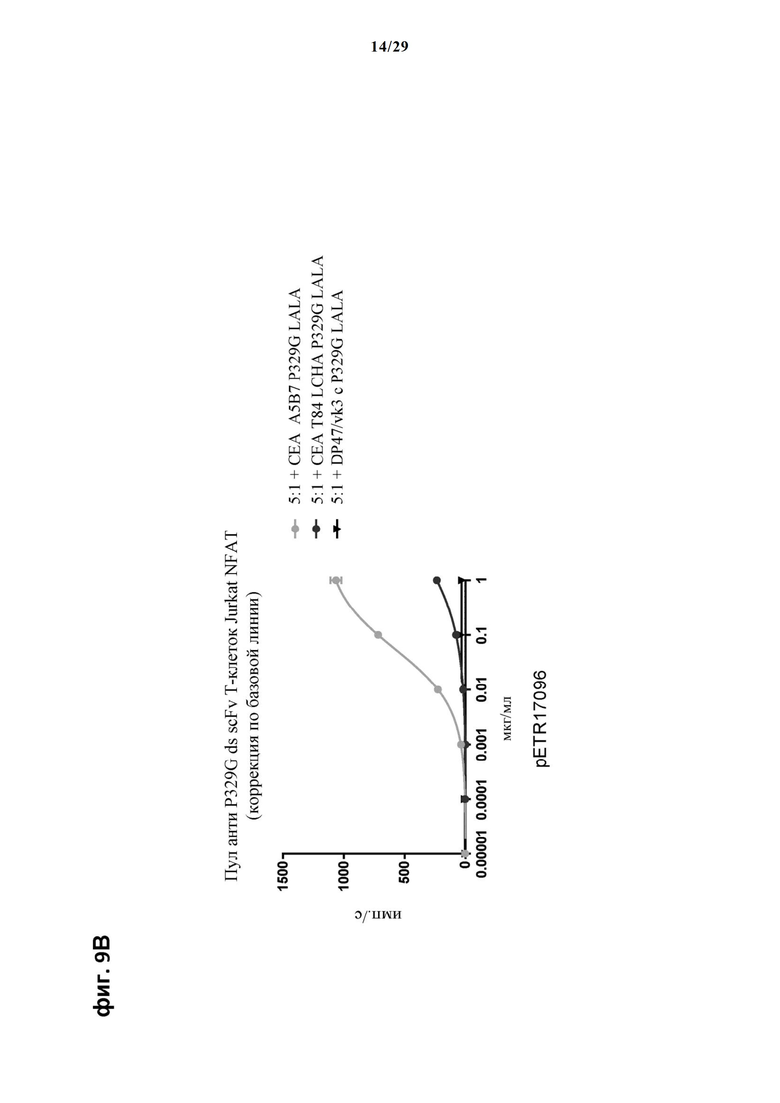
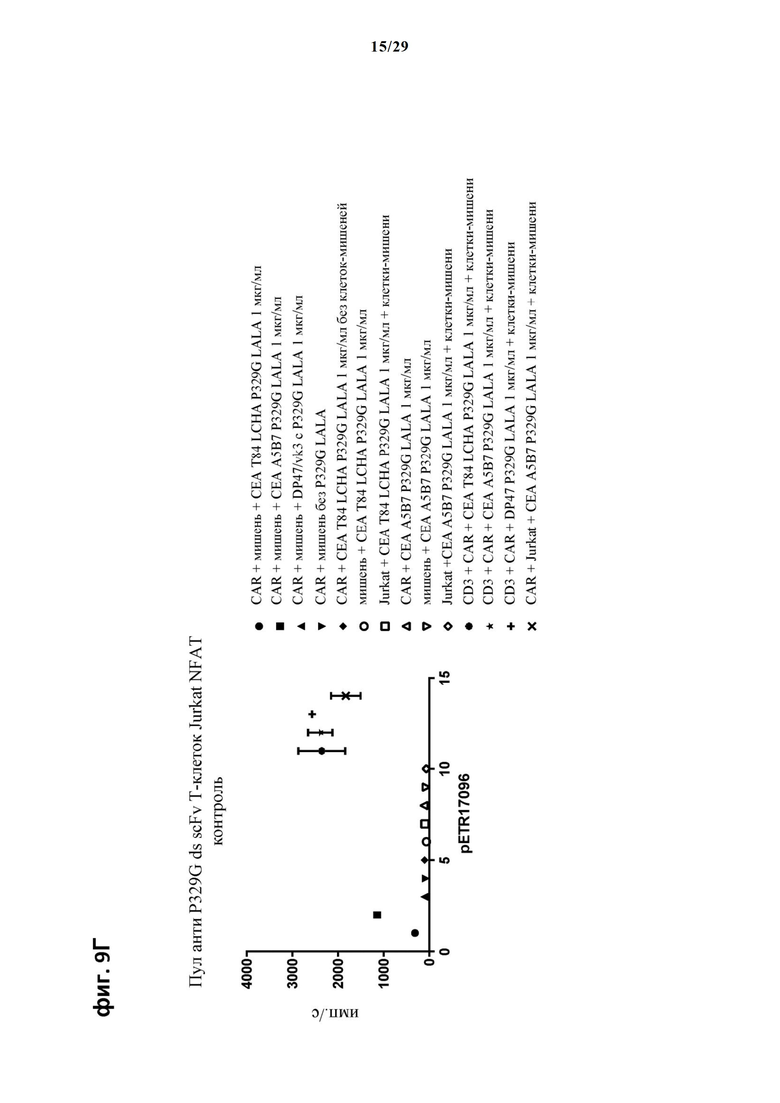
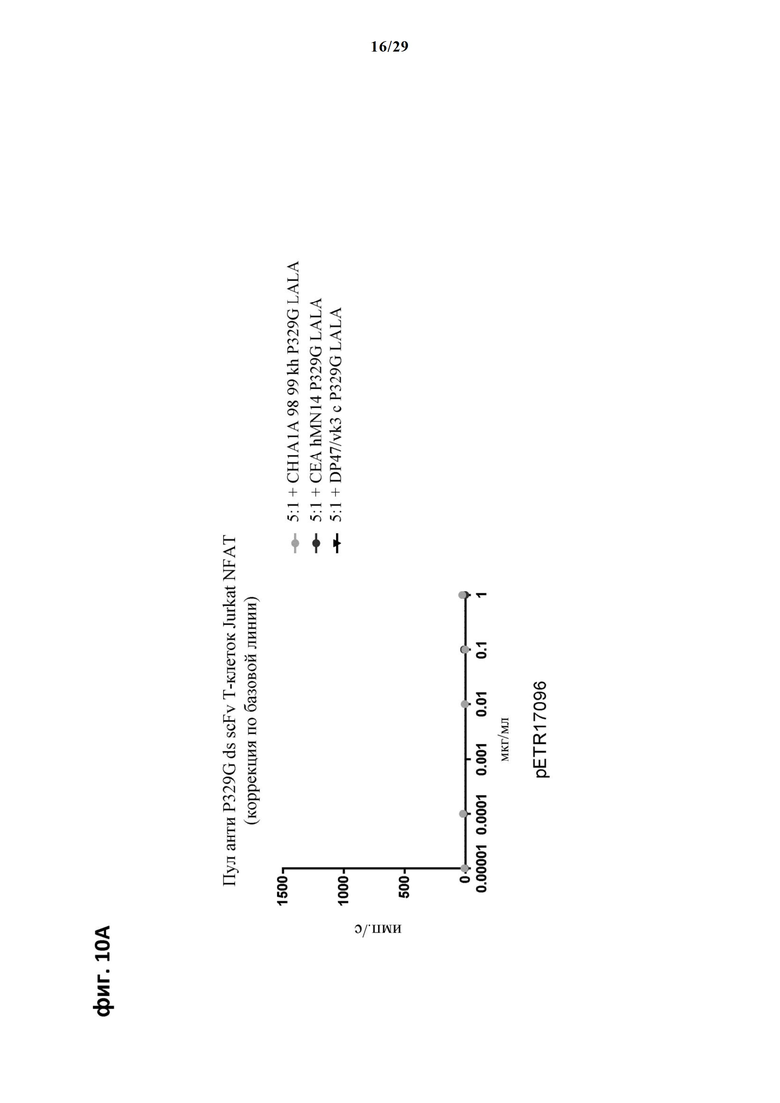


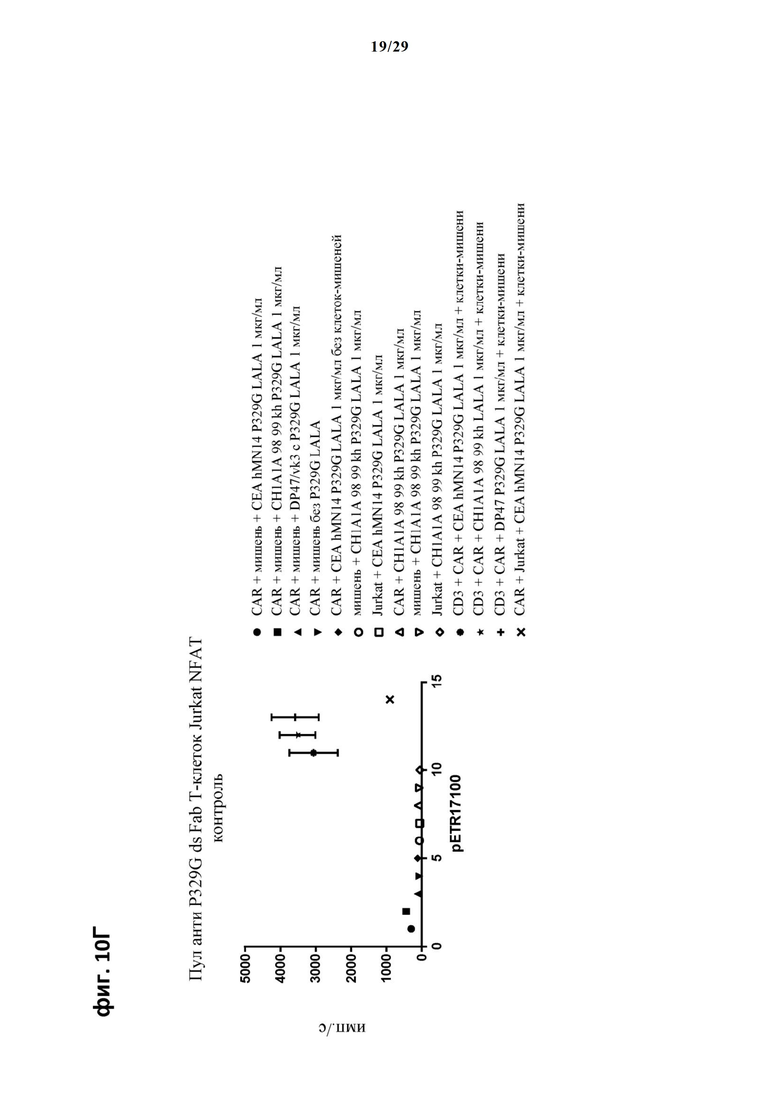
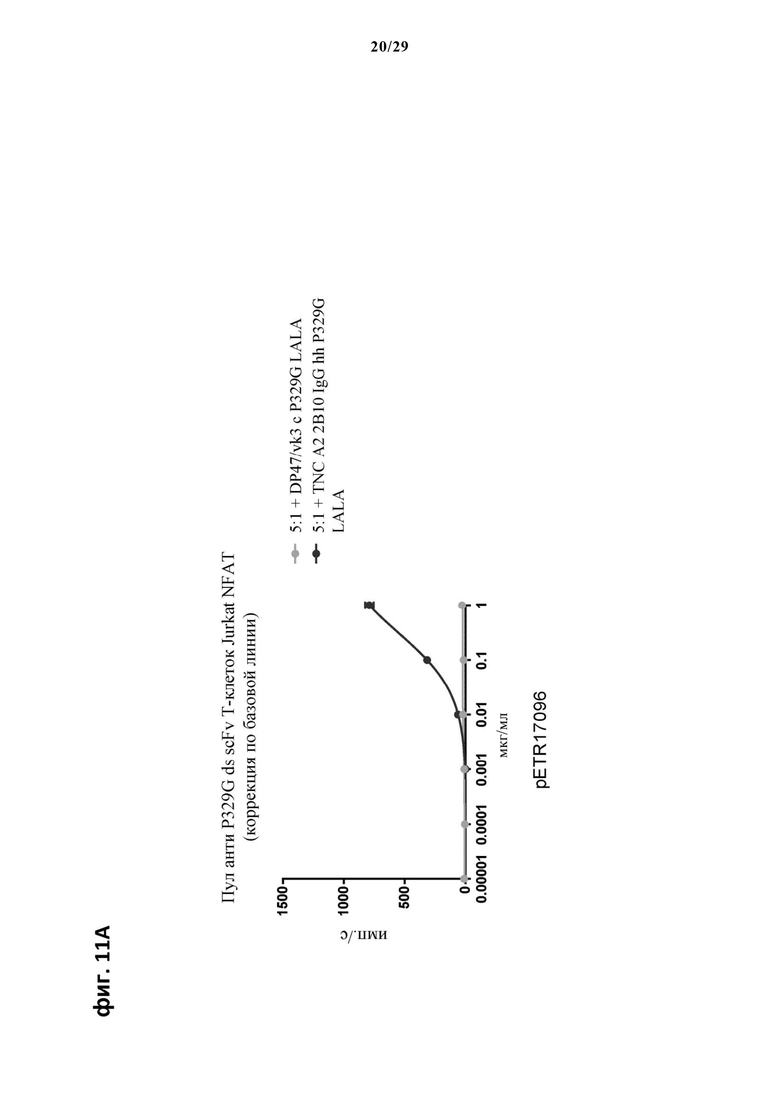
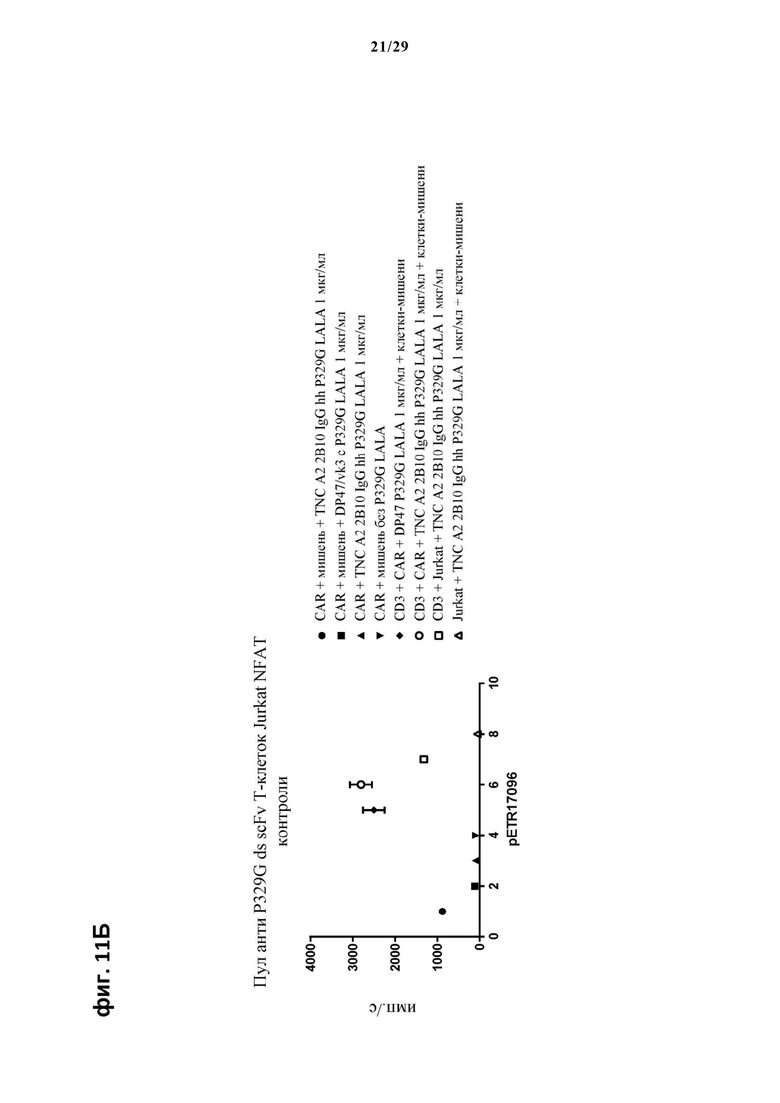
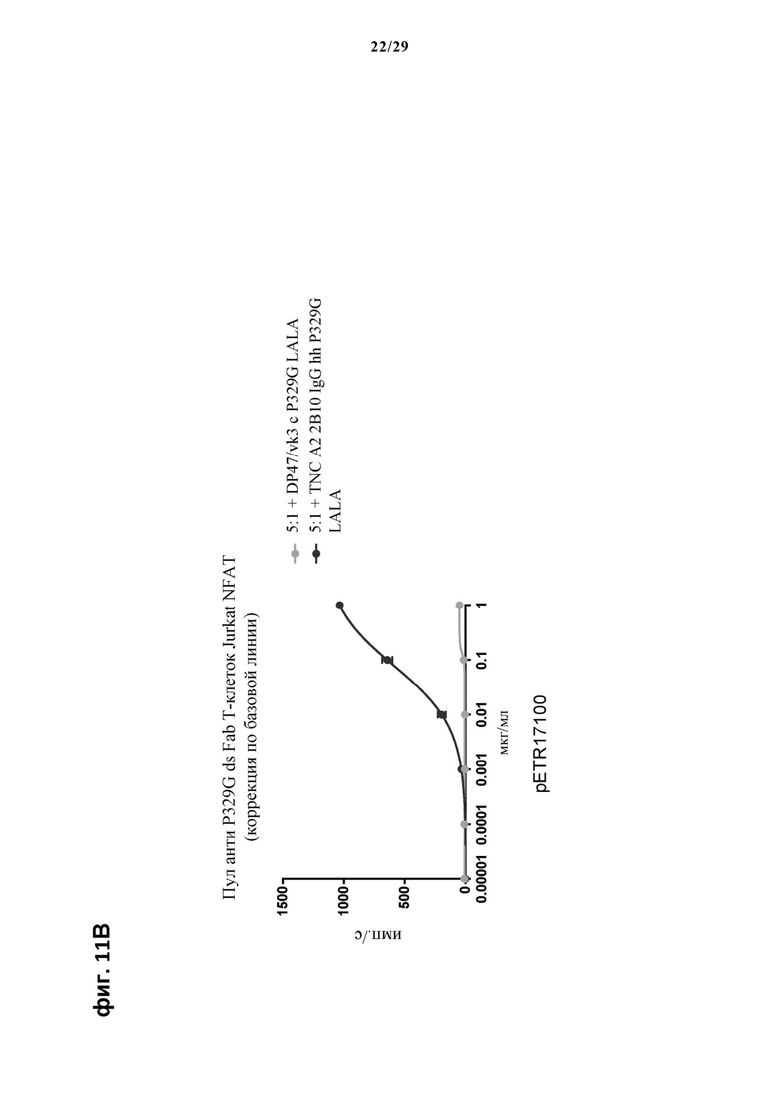

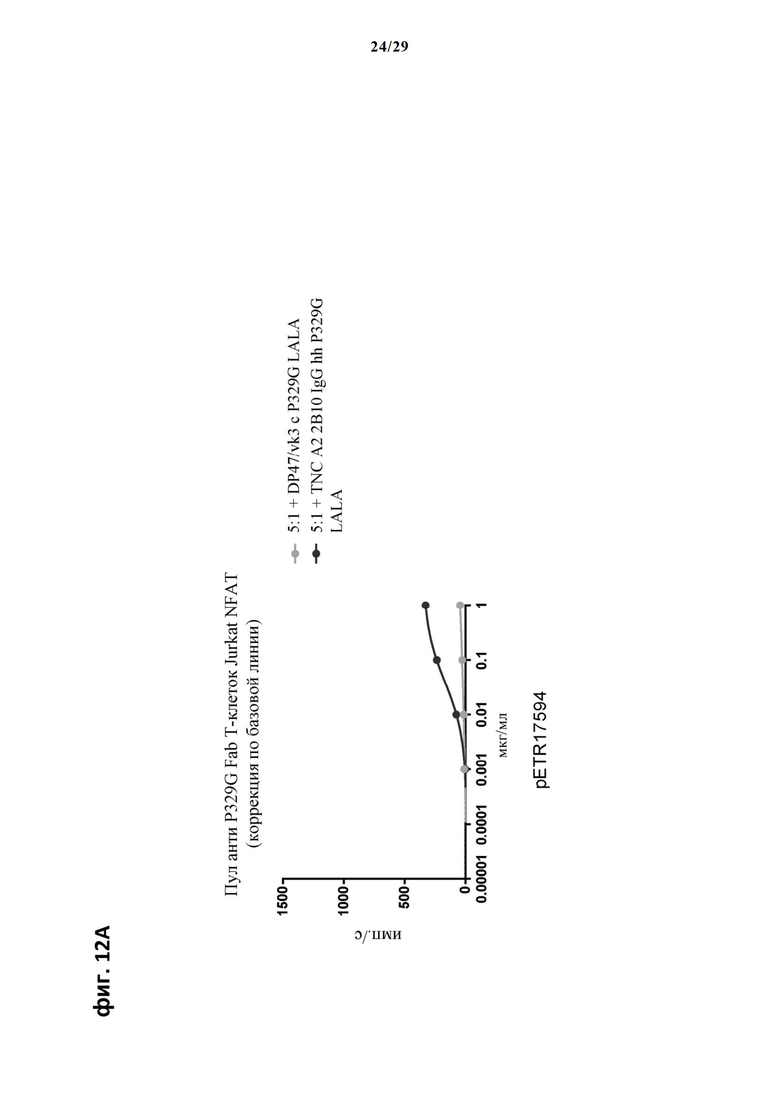
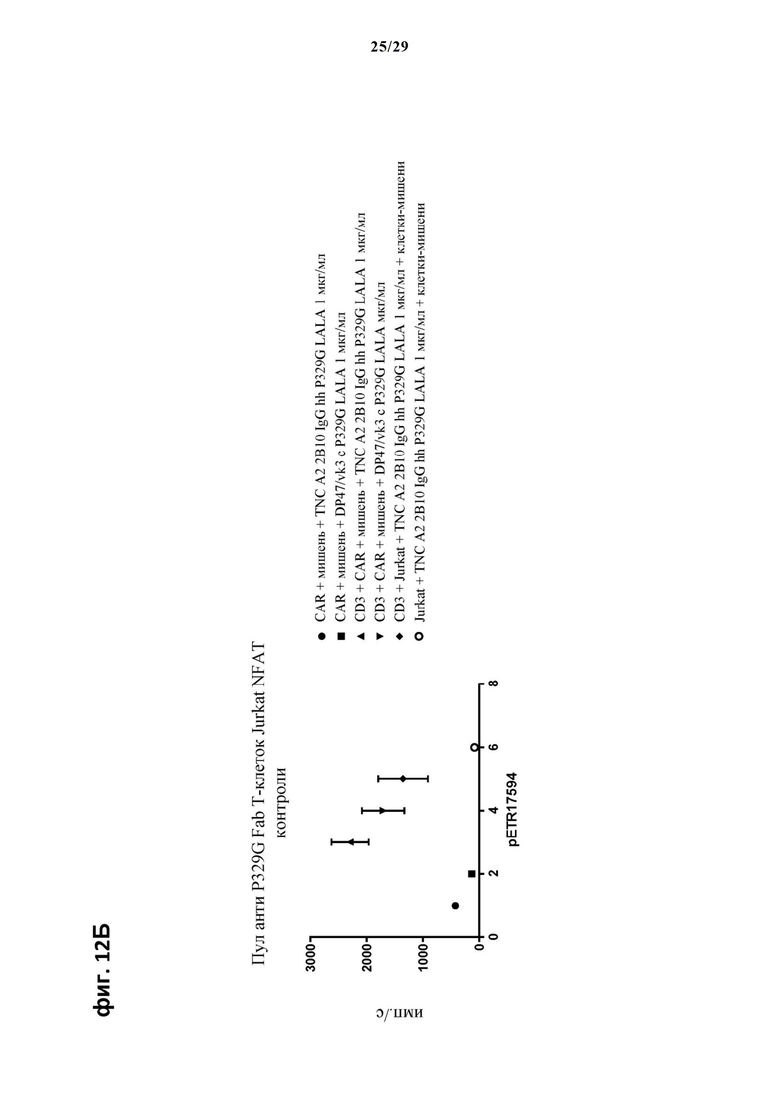
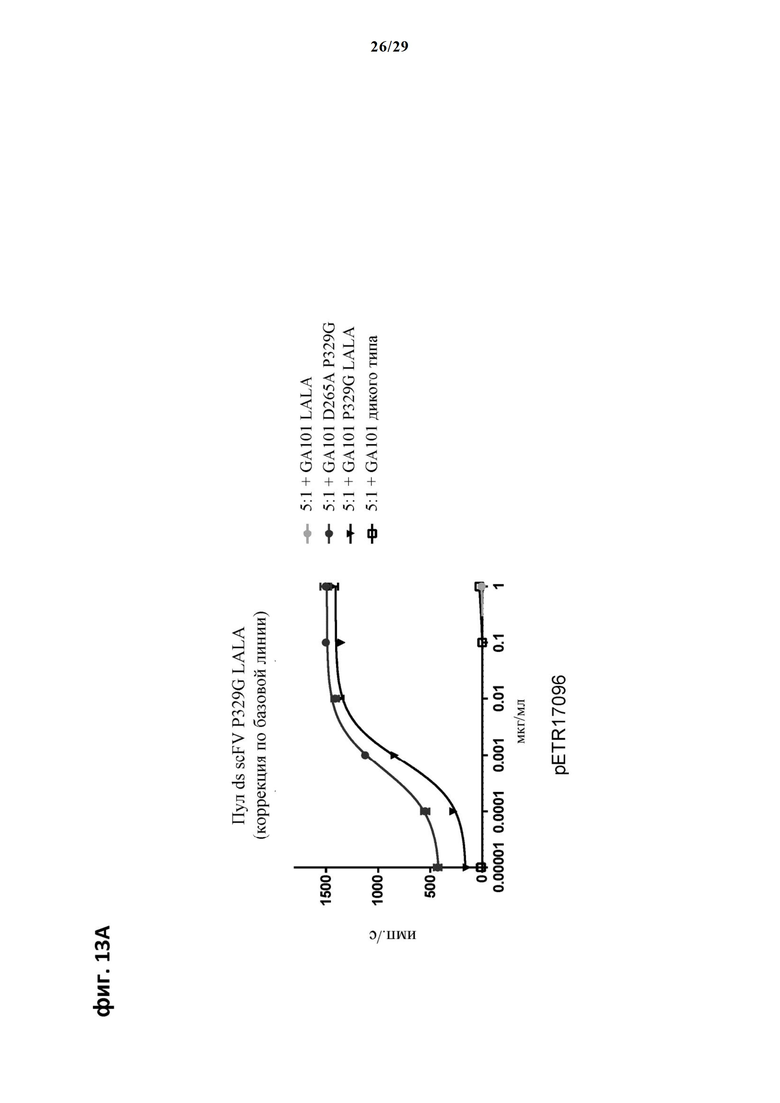


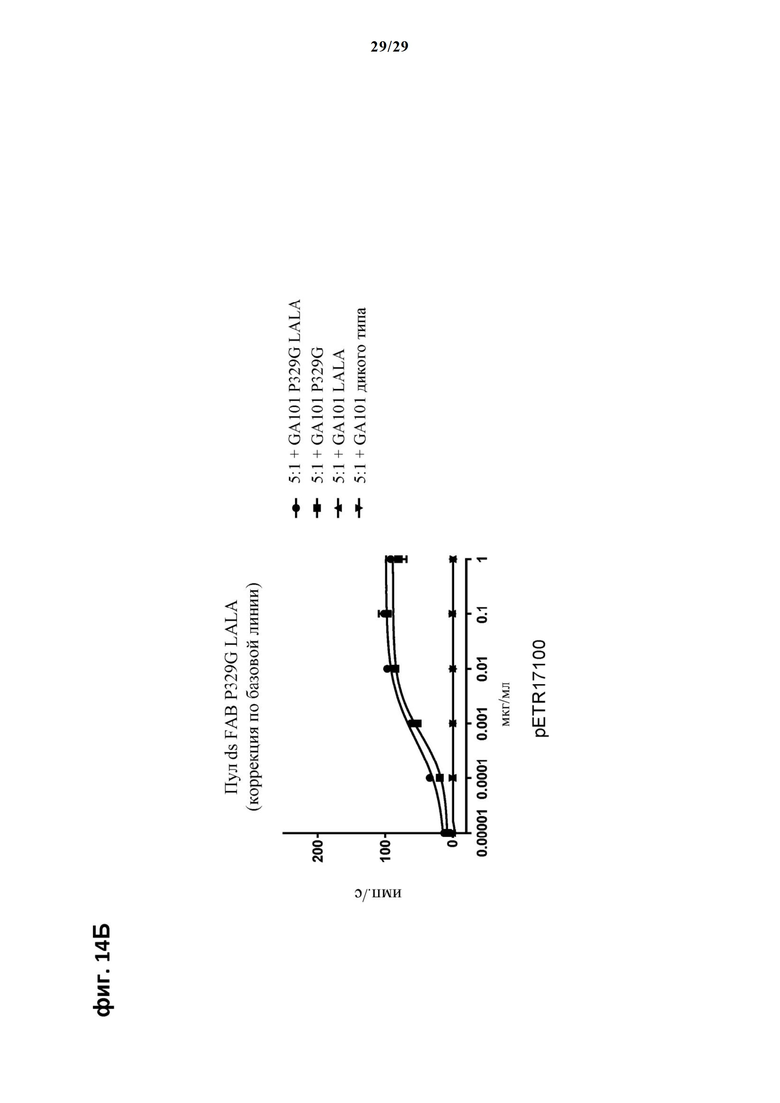
Изобретение относится к области биотехнологии. Предложены антигенсвязывающие рецепторы, содержащие заякоренный трансмембранный домен и внеклеточный домен, включающий антигенсвязывающий фрагмент. Антигенсвязывающие рецепторы специфически связываются с мутированными Fc-доменами, но не связываются с немутированными исходными Fc-доменами. Изобретение обеспечивает специфическую активацию трансдуцированных Т-клеток, экспрессирующих указанные антигенсвязывающие рецепторы, в присутствии антитела IgG с мутированным Fc-доменом для эффективного лизиса опухолевых клеток-мишеней. 4 н. и 27 з.п. ф-лы, 35 ил., 12 табл., 9 пр.
1. Антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий антигенсвязывающий фрагмент, причем антигенсвязывающий рецептор специфически связывается с доменом мутированного кристаллизующегося фрагмента (Fc), но не способен специфически связываться с немутированным исходным Fc-доменом, при этом мутированный Fc-домен включает по меньшей мере одну аминокислотную мутацию P329G согласно нумерации ЕU, причем антигенсвязывающий фрагмент способен специфически связываться с мутированным доменом Fc, содержащим мутацию P329G, но не способен специфически связываться с немутированным исходным доменом Fc, и при этом антигенсвязывающий фрагмент содержит:
(i) вариабельную область тяжелой цепи (VH), содержащую
(a) аминокислотную последовательность RYWMN области тяжелой цепи, определяющей комплементарность (CDR H) 1 (SEQ ID NO: 1);
(b) аминокислотную последовательность CDR H2 EITPDSSTINYTPSLKD (SEQ ID NO: 2) и
(c) аминокислотную последовательность CDR H3 PYDYGAWFAS (SEQ ID NO: 3); и
(ii) вариабельную область легкой цепи (VL), содержащую
(d) аминокислотную последовательность RSSTGAVTTSNYAN области легкой цепи, определяющей комплементарность (CDR L) 1 (SEQ ID NO: 4);
(e) аминокислотную последовательность CDR L2 GTNKRAP (SEQ ID NO: 5) и
(f) аминокислотную последовательность CDR L3 ALWYSNHWV (SEQ ID NO: 6).
2. Антигенсвязывающий рецептор по п. 1, где связывание Fc-рецептора с мутированным Fc-доменом снижено по сравнению со связыванием Fc-рецептора с немутированным исходным Fc-доменом.
3. Антигенсвязывающий рецептор по п. 1, где антигенсвязывающий фрагмент представляет собой scFv, Fab, CrossFab или scFab.
4. Антигенсвязывающий рецептор п. 1, где заякоренный трансмембранный домен представляет собой трансмембранный домен, который выбирают из группы, состоящей из трансмембранного домена CD8, CD3z, FCGR3A, NKG2D, CD27, CD28, CD137, OX40, ICOS, DAP10 или DAP12 или его фрагмента.
5. Антигенсвязывающий рецептор по п. 1, дополнительно включающий по меньшей мере один стимулирующий и/или костимулирующий сигнальный домен.
6. Антигенсвязывающий рецептор по п. 5, где по меньшей мере один стимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD3z, FCGR3A и NKG2D или их фрагментов.
7. Антигенсвязывающий рецептор по п. 5, где по меньшей мере один костимулирующий сигнальный домен индивидуально выбирают из группы, состоящей из внутриклеточного домена CD27, CD28, CD137, OX40, ICOS, DAP10 и DAP12 или их фрагментов.
8. Антигенсвязывающий рецептор по п. 5, где антигенсвязывающий рецептор содержит один стимулирующий сигнальный домен, содержащий внутриклеточный домен CD3z или его фрагмент, и один костимулирующий сигнальный домен, содержащий внутриклеточный домен CD28 или его фрагмент.
9. Антигенсвязывающий рецептор по п. 1, где антигенсвязывающий фрагмент представляет собой scFv-фрагмент, где scFv-фрагмент присоединен через C-концевой остаток к N-концевому остатку заякоренного трансмембранного домена.
10. Антигенсвязывающий рецептор по п. 1, где антигенсвязывающий фрагмент представляет собой Fab или crossFab-фрагмент, где Fab или crossFab-фрагмент присоединен через C-концевой остаток тяжелой цепи к N-концевому остатку заякоренного трансмембранного домена.
11. Антигенсвязывающий рецептор по п. 1, где аминокислотной мутацией является L234A, L235A, Р331 и N297 в соответствии с системой нумерации EU.
12. Антигенсвязывающий рецептор, содержащий заякоренный трансмембранный домен и внеклеточный домен, содержащий антигенсвязывающий фрагмент, причем антигенсвязывающий рецептор специфически связывается с мутированным доменом Fc, но не способен специфически связываться с немутированным исходным доменом Fc, при этом мутированный домен Fc содержит по меньшей мере одну аминокислотную замену по сравнению с немутированным исходным доменом Fc, где мутированный домен Fc содержит аминокислотные мутации I253A, H310A и H435A в соответствии с нумерацией ЕU, причем антигенсвязывающий фрагмент способен специфически связываться с мутированным доменом Fc, содержащим мутации I253A, H310A и H435A, но не способен специфически связываться с немутированным исходным доменом Fc, и при этом антигенсвязывающий фрагмент содержит:
(i) вариабельную область тяжелой цепи (VH), содержащую
(a) аминокислотную последовательность SYGMS области тяжелой цепи, определяющей комплементарность (CDR H) 1 (SEQ ID NO: 53);
(b) аминокислотную последовательность CDR H2 SSGGSY (SEQ ID NO: 54) и
(c) аминокислотную последовательность CDR H3 LGMITTGYAMDY (SEQ ID NO: 55);
и
(ii) вариабельную область легкой цепи (VL), содержащую
(d) аминокислотную последовательность RSSQTIVHSTGHTYLE области легкой цепи, определяющей комплементарность (CDR L) 1 (SEQ ID NO: 56);
(e) аминокислотную последовательность CDR L2 KVSNRFS (SEQ ID NO: 57) и
(f) аминокислотную последовательность CDR L3 FQGSHVPYT (SEQ ID NO: 58).
13. Антигенсвязывающий рецептор по п. 1, в котором VH содержит аминокислотную последовательность SEQ ID NO: 8, а VL содержит аминокислотную последовательность SEQ ID NO: 9.
14. Антигенсвязывающий рецептор по п. 1, в котором антигенсвязывающий фрагмент является гуманизированным.
15. Антигенсвязывающий рецептор по п. 2, где Fc-рецептором является Fcγ-рецептор.
16. Антигенсвязывающий рецептор по п. 3, где антигенсвязывающий фрагмент представляет собой scFv.
17. Антигенсвязывающий рецептор по п. 16, где scFv является гуманизированным.
18. Антигенсвязывающий рецептор по п. 16, где scFv включает аминокислотную последовательность SEQ ID NO: 10.
19. Антигенсвязывающий рецептор по п. 3, где антигенсвязывающий фрагмент представляет собой Fab-фрагмент.
20. Антигенсвязывающий рецептор по п. 19, где Fab-фрагмент является гуманизированным.
21. Антигенсвязывающий рецептор п. 4, где заякоренный трансмембранный домен представляет собой трансмембранный домен CD28 или его фрагмент.
22. Антигенсвязывающий рецептор по п. 6, где по меньшей мере один стимулирующий сигнальный домен представляет собой внутриклеточный домен CD3z или его фрагмент.
23. Антигенсвязывающий рецептор по п. 7, где по меньшей мере один костимулирующий сигнальный домен представляет собой внутриклеточный домен CD28 или его фрагмент.
24. Антигенсвязывающий рецептор по п. 9, где scFv-фрагмент присоединен к заякоренному трансмембранному домену через пептидный линкер.
25. Антигенсвязывающий рецептор по п. 10, где Fab или crossFab-фрагмент присоединен к заякоренному трансмембранному домену через пептидный линкер.
26. Антигенсвязывающий рецептор по п. 11, где аминокислотной мутацией является L234A, L235A, Р331S и/или N297А.
27. Антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий антигенсвязывающий фрагмент, причем антигенсвязывающий рецептор специфически связывается с мутированным доменом Fc, но не способен специфически связываться с немутированным исходным доменом Fc, при этом мутированный Fc-домен включает по меньшей мере одну аминокислотную замену по сравнению с исходным Fc-доменом, где мутированный домен Fc включает по меньшей мере одну аминокислотную мутацию P329G согласно нумерации ЕU, причем антигенсвязывающий фрагмент способен специфически связываться с мутированным доменом Fc, содержащим мутацию P329G, но не способен специфически связываться с немутированным исходным доменом Fc, где антигенсвязывающий фрагмент представляет собой scFv, причем scFv-фрагмент присоединен через C-концевой остаток к N-концевому остатку заякоренного трансмембранного домена через пептидный линкер, при этом антигенсвязывающий фрагмент содержит:
(i) VH, содержащую
(a) аминокислотную последовательность CDR H1 RYWMN (SEQ ID NO: 1);
(b) аминокислотную последовательность CDR H2 EITPDSSTINYTPSLKD (SEQ ID NO: 2) и
(c) аминокислотную последовательность CDR H3 PYDYGAWFAS (SEQ ID NO: 3); и
(ii) VL, содержащую
(d) аминокислотную последовательность CDR L 1 RSSTGAVTTSNYAN (SEQ ID NO: 4);
(e) аминокислотную последовательность CDR L2 GTNKRAP (SEQ ID NO: 5) и
(f) аминокислотную последовательность CDR L3 ALWYSNHWV (SEQ ID NO: 6),
где заякоренный трансмембранный домен представляет собой трансмембранный домен CD28 или его фрагмент и
где антигенсвязывающий рецептор дополнительно включает
по меньшей мере один костимулирующий сигнальный домен, содержащий внутриклеточный домен CD28 или его фрагмент, и
по меньшей мере один стимулирующий сигнальный домен, включающий внутриклеточный домен CD3z или его фрагмент.
28. Антигенсвязывающий рецептор по п. 27, где антигенсвязывающий рецептор содержит аминокислотную последовательность SEQ ID NO: 7.
29. Антигенсвязывающий рецептор, включающий заякоренный трансмембранный домен и внеклеточный домен, включающий антигенсвязывающий фрагмент, причем антигенсвязывающий рецептор специфически связывается с мутированным доменом Fc, но не способен специфически связываться с немутированным исходным доменом Fc, где мутированный Fc-домен включает по меньшей мере одну аминокислотную замену по сравнению с исходным Fc-доменом, причем мутированный домен Fc включает по меньшей мере одну аминокислотную мутацию P329G согласно нумерации ЕU, где антигенсвязывающий фрагмент способен специфически связываться с мутированным доменом Fc, содержащим мутацию P329G, но не способен специфически связываться с немутированным исходным доменом Fc, где антигенсвязывающий фрагмент представляет собой Fab-фрагмент, причем Fab-фрагмент присоединен через C-концевой остаток тяжелой цепи к N-концевому остатку заякоренного трансмембранного домена,
и при этом антигенсвязывающий фрагмент содержит:
(i) VH, содержащую
(a) аминокислотную последовательность CDR H1 RYWMN (SEQ ID NO: 1);
(b) аминокислотную последовательность CDR H2 EITPDSSTINYTPSLKD (SEQ ID NO: 2) и
(c) аминокислотную последовательность CDR H3 PYDYGAWFAS (SEQ ID NO: 3); и
(ii) VL, содержащую
(d) аминокислотную последовательность CDR L1 RSSTGAVTTSNYAN (SEQ ID NO: 4);
(e) аминокислотную последовательность CDR L2 GTNKRAP (SEQ ID NO: 5) и
(f) аминокислотную последовательность CDR L3 ALWYSNHWV (SEQ ID NO: 6),
где заякоренный трансмембранный домен представляет собой трансмембранный домен CD28 или его фрагмент и
где антигенсвязывающий рецептор дополнительно включает
по меньшей мере один костимулирующий сигнальный домен, содержащий внутриклеточный домен CD28 или его фрагмент, и
по меньшей мере один стимулирующий сигнальный домен, включающий внутриклеточный домен CD3z или его фрагмент.
30. Антигенсвязывающий рецептор по п. 29, где Fab-фрагмент включает полипептид тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 40, и полипептид легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 41.
31. Антигенсвязывающий рецептор по п. 29, где антигенсвязывающий рецептор включает полипептид тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 39, и полипептид легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 41.
| WO 2015179833 A1, 26.11.2015 | |||
| SCHLOTHAUER T | |||
| et al., Novel human IgG1 and IgG4 Fc-engineered antibodies with completely abolished immune effector functions, Protein Engineering, Design & Selection, 2016, vol | |||
| Солесос | 1922 |
|
SU29A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| KLEIN C | |||
| et al., The use of CrossMAb technology for the generation of bi- and multispecific antibodies, MABS, 2016, | |||

