ОБЛАСТЬ ТЕХНИКИ
[001] Предложены способы направления рекомбинации ДНК между гомологичными хромосомами в соматических клетках растений, при этом указанные клетки растений могут представлять собой выделенные клетки, часть выделенной ткани растения, часть целого растения или целое растение, при этом последовательность-мишень может соответствовать эухроматину или гетерохроматину.
УРОВЕНЬ ТЕХНИКИ
[002] Двухнитевые разрывы ДНК (ДНР, DSB) являются одной из мощных сил, которая влияет на состав геномов растений. Эти DSB могут происходить в течение всего жизненного цикла растения в соматических или мейотических клетках, спонтанно во время движения репликативных вилок или под контролем процессов развития, как на ранних стадиях первого мейоза. Они также могут быть вызваны ионизирующим излучением, генотоксическими лекарственными средствами или активацией эндонуклеаз. Нерепарированный ДНР ДНК может вызывать критические виды повреждений, включая потерю хромосом, что приводит к стерильности гамет или гибели клеток. Репарация ДНР также может быть связана с инсерционно-делеционными (индел) мутациями. Следовательно, механизмы репарации ДНР являются ключевым элементом поддержания целостности генома. Понимание этих механизмов имеет решающее значение для обеспечения возможности точного конструирования геномов, например, для направленного мутагенеза, таргетинга генов или для других типов направленной «перетасовки» хромосом.
[003] Механизмы репарации ДНР ДНК широко изучены во многих организмах, включая растения. Исследования у растений позволили охарактеризовать гены, участвующие в репарации ДНР посредством негомологичного соединения концов (non-homologous end joining, NHEJ) или гомологичной рекомбинации (homologous recombination, HR), и исследовать результаты репарации ДНР как в соматических, так и в мейотических тканях. NHEJ было охарактеризовано в большом числе видов и тканей (в основном соматических) с использованием множества агентов, индуцирующих ДНР, включая сайт-специфичные мегануклеазы, вырезание транспозонов и специально сконструированные нуклеазы, такие как нуклеазы с цинковыми пальцами (ZFN), эффекторные нуклеазы, подобные активаторам транскрипции (TALEN), и белок, ассоциированный со сгруппированными короткими палиндромными повторами Cas9 (CRISPR-Cas). На основании картины, вырисовывающейся из этих работ, предполагается, что NHEJ является значимым путем репарации в соматических клетках растений. Этот подверженный ошибкам механизм включает индел-мутации в диапазоне от нескольких пар оснований (п.о.) до нескольких тысяч п. о. в сайте ДНР и часто связан с микрогомологиями. Кроме того, системы на основе CRISPR-Cas доказали свою высокую эффективность у широкого спектра видов растений, включая томат.
[004] Частота встречающейся в природе гомологичной рекомбинации в соматических клетках растений является очень низкой и почти равна нулю, если рассматривать соматическую рекомбинацию в конкретном локусе. Считается, что эта низкая частота гомологичной рекомбинации важна для поддержания стабильности больших геномов растений, содержащих повторяющиеся элементы. Несколько исследований, посвященных механизму репарации ДНР посредством HR в соматических тканях, были проведены на Arabidopsis с использованием трансгенных анализов, в которых были протестированы такие механизмы репарации, как внутрихромосомная рекомбинация и неравный кроссинговер. Во всех случаях индукция ДНР повышала частоту репарации посредством HR. Частота рекомбинации в анализе неравного кроссинговера была намного ниже, чем для внутрихромосомной рекомбинации. Соматическую репарацию ДНР с помощью гомологичной хромосомы с использованием аллельной последовательности также изучали на трансгенных растениях табака с использованием разрывов, индуцируемых перемещаемым генетическим элементом: репарация посредством HR происходила после вырезания транспозона, но не была обнаружена посредством неподвижного генетического элемента. Индукция ДНР может также вызывать HR-опосредованную репарацию с использованием матрицы эктопической геномной последовательности, хотя и с очень низкой частотой.
[005] Индукция HR между эндогенными (нетрансгенными) партнерами по рекомбинации посредством ДНР была показана на кукурузе при вырезании элементов Activator (Ac) или Mutator. В обоих случаях рекомбинация происходила в цис-положении между повторами, фланкирующими транспозон в соматических тканях. Напротив, активность Ac зародышевой линии не стимулировала частоту мейотической рекомбинации между гомологичными хромосомами в локусе bronze кукурузы. Этот результат может быть связан с отсутствием координации между вырезанием Ac и мейотической рекомбинацией, предпочтением Spo11-индуцированных разрывов мейотической HR или другой неизвестной причиной. Способность индуцировать HR между гомологичными хромосомами в конкретной локализации в геноме предоставит генетикам и селекционерам мощный инструмент для направленной индукции кроссинговера или генной конверсии.
[006] Таким образом, существует потребность в способах, которые обеспечат направленную HR между гомологичными хромосомами для точной селекции культурных растений. Одним из применений направленной HR является направленная генная конверсия, а именно, перенос гена из одной хромосомы в ее гомолог. Такие способы должны также учитывать размер популяции растений, которая обычно может быть использована для достижения этой цели. Кроме того, процесс повторного обратного скрещивания для достижения изогенных линий может также приводить к перетягиванию больших сегментов нежелательной ДНК, фланкирующих желаемый ген, в растение-потомок. В настоящей заявке раскрыты способы направленной рекомбинации между гомологичными хромосомами, которые могут быть осуществлены с относительно небольшими популяциями растений и без необходимости извлечения больших сегментов нежелательной ДНК.
[007] Хромосомы растений обладают как высококонденсированным гетерохроматином, выраженным в перицентромерных областях и соответствующим холодным точкам мейотической рекомбинации, так и в значительной степени деконденсированными эухроматическими областями, часто соответствующими дистальным, субтеломерным областям и горячим точкам мейотической рекомбинации. Хотя гетерохроматин часто ассоциируется с отсутствием транскрипционной активности и подавленной генетической рекомбинацией, он все же содержит транскрипционно активные гены. Направленная индуцированная рекомбинация между гомологичными хромосомами в областях гетерохроматина была бы преимуществом в селекции растений, так как в отсутствие такой рекомбинации нежелательные гены не могут быть отделены от полезных генов. В настоящей заявке раскрыты способы направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения, в которых, как было показано, направленная ДНР-индуцированная рекомбинация происходит как в эухроматических, так и гетерохроматических сайтах-мишенях.
[008] Еще одним потенциальным применением ДНР-индуцированной соматической HR является «направленный кроссинговер», то есть взаимный обмен большими сегментами хромосом в точно определенном сайте. Современные методы селекции основаны на случайном кроссинговере и поиске редких событий рекомбинации, для получения которых в случае связанных генов могут потребоваться десятки тысяч растений, причем процент гомологичной рекомбинации в естественной ненаправленной HR в любом конкретном сайте в геноме близится к 0% (встречается менее 1 раза на каждые 105-106 событий встречающейся в природе HR).
[009] В настоящей заявке предложены способы, которые могут применяться в направленной соматической HR для объединения желательных признаков от родителей и для отделения от нежелательных генетических связей с использованием небольших популяций растений.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0010] В одном из аспектов в настоящей заявке предложен способ направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения, включающий следующие стадии:
(а) осуществление экспрессии системы нуклеаз в указанной клетке растения, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей;
(b) осуществление анализа потомства указанной клетки растения, или ткани растения, выращенной из указанной клетки растения, или растения, выращенного из указанной клетки, или потомства указанного растения на гомологичную рекомбинацию между гомологичными хромосомами, где указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером); и
(c) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых произошла направленная гомологичная рекомбинация.
[0011] В одном из аспектов способы, раскрытые в настоящей заявке, обеспечивают получение растения, обладающего комбинацией полезных признаков или качеств, где способ включает направленную рекомбинацию ДНК между гомологичными хромосомами в гибридной соматической клетке растения, где указанный способ включает следующие стадии:
(а) осуществление экспрессии системы нуклеаз в указанной клетке растения, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей;
(b) осуществление анализа потомства указанной клетки растения, или ткани растения, выращенной из указанной клетки растения, или растения, выращенного из указанной клетки, или потомства указанного растения на гомологичную рекомбинацию между гомологичными хромосомами, где указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером);
(c) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых произошла направленная гомологичная рекомбинация;
(d) выращивание указанной клетки растения, или полученной из нее ткани растения, или полученного из нее растения, или потомства указанного растения с получением растения, содержащего указанную направленную гомологичную рекомбинацию, где указанное растение обладает комбинацией полезных качеств или признаков, не присутствующей ни у одного из родительских растений, из которых происходила гибридная соматическая клетка.
[0012] В одном из аспектов в настоящей заявке раскрыт способ получения растения-потомка, обладающего комбинацией полезных признаков или качеств, не присутствующей ни у одного из родительских растений, где указанный способ включает:
выбор родительских растений, где каждый из указанных родителей обладает по меньшей мере одним полезным признаком, где указанные полезные признаки не идентичны, и где указанные родители полиморфны по по меньшей мере одному указанному полезному признаку;
скрещивание указанных родительских растений для создания гибридного растения;
сбор соматических клеток из гибридного растения;
экспрессию системы нуклеаз в указанных соматических клетках, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей, где гомологичный кроссинговер или генная конверсия (не сопровождающаяся кроссинговером) в указанном целевом предварительно выбранном эндогенном сайте-мишени приводит к обмену ДНК, экспрессирующей или регулирующей экспрессию по меньшей мере одного из указанных полезных признаков или качеств;
анализ потомства указанных клеток растения, или ткани растения, выращенной из указанных клеток растения, или растения, выращенного из указанных клеток, или потомства указанного растения, на указанное событие кроссинговера или генной конверсии (не сопровождающейся кроссинговером), в которых экспрессируется указанная комбинация признаков;
выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых экспрессируется комбинация признаков; и
выращивание указанной клетки растения, полученной из нее ткани растения, полученного из нее растения, с получением растения-потомка, обладающего указанной комбинацией полезных признаков или качеств.
[0013] В связанном аспекте система нуклеаз содержит систему нуклеаз с цинковыми пальцами (ZFN), систему эффекторных нуклеаз, подобных активаторам транскрипции (TALEN), или систему сгруппированных коротких палиндромных повторов (CRISPR)/белков, ассоциированных с CRISPR (Cas).
[0014] В еще одном аспекте система нуклеаз содержит нуклеазу с цинковыми пальцами (ZFN), содержащую ДНК-связывающий домен цинкового пальца и ДНК-расщепляющий домен нуклеазы, где указанный ДНК-связывающий домен цинкового пальца связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым ДНК-расщепляющий домен нуклеазы на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени. В еще одном аспекте система нуклеаз содержит систему нуклеаз, подобных активаторам транскрипции (TALEN), содержащую ДНК-связывающий домен TAL-эффектора и ДНК-расщепляющий домен, где указанный ДНК-связывающий домен TAL-эффектора связывается с указанным предварительно выбранным эндогенным сайтом-мишенью, направляя тем самым ДНК-расщепляющий домен на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени. В еще одном аспекте система нуклеаз содержит систему нуклеаз CRISPR/Cas, содержащую CRISPR-ассоциированную эндонуклеазу и молекулу гРНК (гидовой РНК), где указанная молекула гРНК связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную CRISPR-ассоциированную эндонуклеазу на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени. В еще одном аспекте CRISPR-ассоциированная эндонуклеаза (нуклеаза Cas) выбрана из группы, содержащей Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, Cpf1, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, C2c1, CasX, NgAgo, Csf1, Csf2, Csf3 и Csf4, их гомологи или их модифицированные версии.
[0015] В связанном аспекте соматическая клетка растения происходит из существующей гибридной или гетерозиготной клетки растения, имеющей полиморфные аллели в указанном предварительно выбранном сайте. В еще одном аспекте существующая гибридная или гетерозиготная клетка растения происходит из растения дикого типа.
[0016] В связанном аспекте способ, раскрытый в настоящей заявке, обеспечивает получение соматической клетки растения, содержащей направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную соматическую клетку растения, или растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или плода, полученного от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или семян, полученных от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или любой их комбинации, обладающих комбинацией родительских признаков, не присутствующей ни у одного из родителей. В еще одном аспекте родительский признак включает повышенную засухоустойчивость, повышенную устойчивость к сельскохозяйственным вредителям, повышенную устойчивость к патогенам, улучшенное содержание питательных веществ или улучшенные параметры роста, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени.
[0017] В связанном аспекте соматическая клетка растения происходит от клетки из потомства, полученного при скрещивании двух растений, где каждая из указанных родительских клеток растения содержит полиморфный аллель по сравнению с указанным партнером в указанном предварительно выбранном сайте.
[0018] В связанном аспекте способ, раскрытый в настоящей заявке, обеспечивает получение соматической клетки растения, содержащей направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную соматическую клетку растения, или растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или плода, полученного от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или семян, полученных от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или любой их комбинации, обладающих полученной в результате комбинацией родительских признаков, не присутствующей ни у одного из родителей. В еще одном аспекте родительские признаки, рекомбинированные посредством указанной направленной гомологичной рекомбинации, включают повышенную засухоустойчивость, повышенную устойчивость к сельскохозяйственным вредителям, повышенную устойчивость к патогенам, улучшенное содержание питательных веществ или улучшенные параметры роста, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени.
[0019] В связанном аспекте одна из указанных родительских соматических клеток растения содержит указанную систему нуклеаз, где ДНК-расщепляющая активность указанной системы нуклеаз направлена на полиморфный аллель, присутствующий в другой родительской клетке растения, не содержащей указанную систему нуклеаз.
[0020] В еще одном связанном аспекте одна из указанных родительских соматических клеток растения содержит нуклеазу Cas, а другая из указанных родительских соматических клеток растения содержит молекулу гРНК, связывающуюся в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени.
[0021] В еще одном связанном аспекте соматическая клетка растения включает клетку из потомства растения, полученного при скрещивании двух полиморфных родительских линий, приводящем к созданию гибридного растения, где каждая из указанных родительских линий растений содержит полиморфный аллель в указанном предварительно выбранном эндогенном сайте-мишени, и где только одна из родительских линий содержит указанную систему нуклеаз.
[0022] В еще одном связанном аспекте способ, раскрытый в настоящей заявке, обеспечивает получение соматической клетки растения, содержащей направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную соматическую клетку растения, или растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или плода, полученного от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или семян, полученных от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, или любой их комбинации, обладающих комбинацией родительских признаков, не присутствующей ни у одного из родителей. В еще одном аспекте родительские признаки включают повышенную засухоустойчивость, повышенную устойчивость к сельскохозяйственным вредителям, повышенную устойчивость к патогенам, улучшенное содержание питательных веществ или улучшенные параметры роста, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени.
[0023] В еще одном связанном аспекте система нуклеаз содержит нуклеазу Cas и молекулу гРНК, связывающуюся в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени, где ДНК-расщепляющая активность указанной системы нуклеаз проявляется только на гетерологичном аллеле, присутствующем в родительской клетке растения дикого типа.
[0024] В еще одном связанном аспекте соматическая клетка растения содержится в ткани растения или в целом растении. В еще одном связанном аспекте соматическая клетка растения включает протопласт. В еще одном связанном аспекте соматическая клетка растения включает клетку культурного растения.
[0025] В еще одном связанном аспекте предварительно выбранный эндогенный сайт-мишень содержит ДНК, содержащую ген, часть гена или регуляторную расположенную в 3'-5' направлении или расположенную в 5'-3' направлении последовательность гена, или любую их комбинацию, и где экспрессия указанного гена или ее отсутствие влияет на рост, засухоустойчивость, устойчивость к сельскохозяйственным вредителям, устойчивость к патогенам или содержание питательных веществ, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени, или любую их комбинацию. В еще одном связанном аспекте предварительно выбранный эндогенный сайт-мишень содержит область эухроматина или гетерохроматина.
[0026] В еще одном связанном аспекте экспрессия включает конститутивную индукцию экспрессии, индуцибельную индукцию экспрессии, тканеспецифичную индукцию экспрессии или специфичную для определенных условий индукцию экспрессии, или любую их комбинацию.
[0027] В связанном аспекте анализ указанного растения включает анализ части указанного растения или его потомства, включая лист, стебель, почку, плод, семя.
[0028] В связанном аспекте стадия выбора потомства включает поколения F1, F2 или F3, или любое последующее поколение, или обратные скрещивания на 1-3 поколения, или любое последующее поколение обратного скрещивания.
[0029] В связанном аспекте способ, раскрытый в настоящей заявке, обеспечивает получение соматической клетки растения, содержащей указанную направленную гомологичную рекомбинацию в указанном предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или растения, содержащего указанную направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или плода, полученного от растения, содержащего направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или семян, полученных от растения, содержащего указанную направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или любой их комбинации, где указанные клетка, ткань, растение или его потомство, плод или семя содержат гены для повышенной засухоустойчивости, повышенной устойчивости к сельскохозяйственным вредителям, повышенной устойчивости к патогенам, улучшенного содержания питательных веществ, параметров роста или любого другого признака, полезного для клетки растения, ткани растения, растения или его потомства, плода или семени, или любой их комбинации, по сравнению с контрольными клеткой растения, растением или его потомством, плодом или семенем. В связанном аспекте предварительно выбранный эндогенный сайт-мишень содержит область эухроматина или гетерохроматина.
[0030] В еще одном аспекте в настоящей заявке раскрыто растение, обладающее комбинацией полезных признаков или качеств, полученное способом, включающим направленную рекомбинацию ДНК между гомологичными хромосомами в гибридной соматической клетке растения, где указанный способ включает следующие стадии: (а) осуществление экспрессии системы нуклеаз в указанной клетке растения, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей;
(b) осуществление анализа потомства указанной клетки растения, или ткани растения, выращенной из указанной клетки растения, или растения, выращенного из указанной клетки, или потомства указанного растения на гомологичную рекомбинацию между гомологичными хромосомами, где указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером);
(c) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых произошла направленная гомологичная рекомбинация;
(d) выращивание указанной клетки растения, или полученной из нее ткани растения, или полученного из нее растения, или потомства указанного растения с получением растения, содержащего указанную направленную гомологичную рекомбинацию, где указанное растение обладает комбинацией полезных качеств или признаков, не присутствующей ни у одного из родительских растений, из которых происходила гибридная соматическая клетка. В связанном аспекте предварительно выбранный эндогенный сайт-мишень содержит область эухроматина или гетерохроматина.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0031] Следующие графические материалы являются частью настоящего описания и включены для дополнительной демонстрации отдельных вариантов реализации настоящего изобретения, способы, описанные в настоящей заявке, могут быть более понятными со ссылкой на один или более из этих графических материалов в сочетании с подробным описанием конкретных вариантов реализации, представленным в настоящей заявке.
[0032] На Фиг. 1 представлена схема репарации двухцепочечного разрыва (ДНР), которая может происходить посредством негомологичного соединения концов (NHEJ) или гомологичной рекомбинации (HR).
[0033] На Фиг. 2 представлена схема варианта реализации репарации направленного ДНР с помощью гомологичной рекомбинации (HR).
[0034] На Фиг. 3 представлена блок-схема, включающая некоторые варианты реализации индукции рекомбинации между гомологичными хромосомами. Индукция двухнитевых разрывов ДНК показана желтой молнией.
[0035] На Фиг. 4А-Е представлены результаты анализа окраски плодов томата и молекулярного анализа результатов событий репарации двухнитевых разрывов (ДНР) ДНК. Фиг. 4А: ожидается, что скрещивание желтая мякотьe3756 35S:Cas9 и двухцветныйcc383 u6-26:Ps#1-огРНК даст растения F1 с фенотипом бледных двухцветных плодов. Были отобраны растения F1, экспрессирующие как Cas9, так и гРНК. гРНК была разработана для направленной индукции ДНР (показана черной молнией) в обоих аллелях между мутациями желтая мякотьe3756 и двухцветныйcc383(*). В случае репарации аллеля двухцветныйcc383 посредством негомологичного соединения концов (NHEJ) ожидалось, что окраска плода будет желтой после подверженной ошибкам репарации, оставляющей следы от индел (инсерция/делеция)-мутаций (синяя линия). В случаях отсутствия кроссинговера или кроссинговера ожидалось, что окраска плода будет красной или, в случае последнего события, двухцветной с красными пятнами. Фиг. 4В: распределение фенотипа плодов в растениях F1 и контрольных растениях: двухцветные плоды показаны оранжевыми прямоугольниками; желтые плоды показаны желтыми прямоугольниками; плоды с красными участками (предположительно, с соматической HR) показаны прямоугольниками с красной штриховкой. Каждый столбец представляет собой популяцию плодов, полученных от растений F1, происходящих от скрещивания между независимыми трансгенными линиями Cas9 и данной линией u6-26:Ps#1-огРНК. Количество плодов, проанализированных для каждого скрещивания, показано на столбцах черным цветом. Фиг. 4C: последовательности следов репарации ДНР посредством NHEJ показаны справа, а их относительная частота показана на круговой диаграмме. Последовательность-мишень для CRISPR-Cas из PSY1 показана вверху. Местоположение ДНР показано черной молнией; инициирующий кодон PSY1 показан красным, а PAM - мотив, примыкающий к протоспейсеру, -показан синим. Верхняя круговая диаграмма представляет собой среднее прочтений, полученных с помощью illumina Hiseq, для 22 различных растений F1, происходящих от скрещивания желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК. В этом скрещивании 88% последовательностей отклоняются от последовательности WT (дикого типа). Нижняя круговая диаграмма представляет собой среднее прочтений, полученных с помощью illumina Hiseq, для 2 растений из контрольной популяции F1 (растения желтая мякоть e3756 x двухцветныйcc383 F1 без компонентов CRISPR-Cas). Оранжевая делеция CTTG является преимущественным следом NHEJ. Фиг. 4D: схема ПЦР (полимеразной цепной реакции) с обратной транскрипцией для идентификации фрагментов рекомбинантной ДНК. (1) ДНК из отдельных образцов листьев сначала расщепляли по ApaI(A) и HindIII(H), а затем затупляли ее концы. (2) Каждый образец самолигировали. (3) Каждый образец амплифицировали двумя различными наборами праймеров (показаны зеленым и желтым). Синий - аллель двухцветный; красный - аллель желтая мякоть; пунктирная синяя линия - делеция двухцветный, * - мутация желтая мякоть, молния - сайт ДНР. Фиг. 4E. Соотношение родительского (P) и рекомбинантного (R) типов (полученных в секции C) у отдельных растений. Растения 1-15 - растения F1, происходящие от скрещивания желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК; растение 16 - контроль синтетическим кроссинговером; растения 17-18 - растения F1, происходящие от скрещивания желтая мякоть x двухцветный (Cas9-).
[0036] На Фиг. 5 представлена репарация посредством NHEJ в соматических клетках. Распределение следов NHEJ у отдельных растений F1 и у контрольных растений (желтая мякоть e3756 x двухцветныйcc383), полученных путем секвенирования продуктов ПЦР, амплифицированных вокруг вызванного CRISPR-Cas9 ДНР (молния) с праймерами, показанными красными стрелками. Каждая круговая диаграмма представляет собой общее количество прочтений, полученных с помощью illumina Hiseq, для одного растения (250 000-850 000 прочтений на растение).
[0037] На Фиг. 6A-6B представлен анализ ОНП (однонуклеотидных полиморфизмов) томатов на события репарации двухнитевых разрывов (ДНР) зародышевой ДНК. Фиг. 6А. Гомозиготный мутант M82 CRISPR (+A,+A), экспрессирующий 35S:Cas9 и u6-26:Ps#2-огРНК, скрещивали с S. pimpinellifoliumLA1578. Ожидается, что F1 даст красные плоды без ДНР ДНК и желтые плоды в случае, если разрыв был репарирован посредством NHEJ, не сопровождающегося кроссинговером или сопровождающегося им. Паттерн ОНП позволяет различать механизмы репарации. Треугольники обозначают ОНП; молния обозначает сайт ДНР; синяя линия обозначает индел-мутации, вызванные NHEJ. Фиг. 6B. Анализ маркеров, фланкирующих ДНР ДНК у растений F2 и F3. Красный - гомозигота по ОНП S.pimpinellifoliumLA1578; желтый - гомозигота по ОНП M82 (включая мутант +A CRISPR-Cas9); оранжевый - гетерозигота; пустые ячейки означают отсутствующие данные; молния - сайт ДНР.
[0038] На Фиг. 7 представлен анализ ОНП томатов на аллель-специфичную репарацию ДНР ДНК. ДНК выделяли из 4 листьев M82 35S:Cas9 u6-26:Ps#2-огРНК psy1+A/ psy1+A, S. pimpinellifoliumLA1578 и 5 растений их инбредной популяции F1. Было проведено секвенирование с помощью illumina, и каждая круговая диаграмма представляет собой сводные данные для 600 000-900 000 прочтений на растение.
[0039] На Фиг. 8 представлена схематическая карта фенотипов окраски плодов на протяжении развития и секвенирования следов репарации ДНР ДНК из тканей околоплодника плода с использованием секвенирования с помощью illumina. Пример приведен для растения № 1, которое представляет собой растение F1 M82 35S:Cas9 u6-26:Ps#2-огРНК psy1+A/ psy1+A x S. pimpinellifoliumLA1578. Фенотип окраски плода менялся от красного до красного с маленькими или большими желтыми участками до желтого. Каждая круговая диаграмма была составлена из 15 000-50 000 прочтений секвенирования с помощью illumina на плод.
[0040] На Фиг. 9A и 9B представлена количественная оценка аллель-зависимой репарации. Фиг. 9А. Выращивали две популяции растений, обе на фоне генотипа M82: одна была гомозиготной по PSY1/PSY, а другая - гетерозиготной по генотипу PSY1/psy+A. Потомство этих растений может давать +A ОНП в сайте разрыва (молния) или любую другую мутацию (*). ДНК выделяли из листьев 4-недельных растений из обеих популяций и секвенировали с помощью illumina. В растениях PSY1/PSY1 мишенью могут быть оба аллеля, тогда как в растениях PSY1/psy+A мишенью является только аллель WT PSY1. Фиг. 9B. Процент мутации +A на аллель WT в растениях PSY1/PSY1 служил в качестве ожидаемого значения для аллель-независимой мутации +A. Его рассчитывали по следующим уравнениям: Ожидаемый=(%(прочтений +A)T=4 недели (wt,wt))/2. Для оценки наблюдаемой встречаемости мутации +A, когда второй аллель имеет мутацию +A (в гетерозиготных растениях M82-WT PSY1/ M82 psy1+A), как показано на Фиг. 9A, использовали следующее уравнение: Наблюдаемый=%(прочтений +A)T=4 недели, (wt,+A)-50%. Столбцы соответствуют стандартной ошибке для 22 растений PSY1/PSY1 и 14 растений PSY1/psy+A. Разница между средними значениями была значимой (р-значение (критерий суммы рангов Уилкоксона)=0,009).
[0041] На Фиг. 10 показано событие репарации ДНР ДНК, за которым следуют фенотип плода и специфичное секвенирование околоплодника с помощью illumina - растение № 2. Все подробности аналогичны Фиг. 8. Это растение показало высокий уровень psy1+A. Продукты конверсии на Фиг. 6B представляют собой потомство этого растения.
[0042] На Фиг. 11 представлена таблица 10, в которую занесены мишени ДНР CRISPR на 3 хромосоме Arabidopsis.
[0043] На Фиг. 12A-12C показана система Arabidopsis для индукции ДНР соматической ДНК в горячих и холодных точках рекомбинации. (Фиг. 12A) Двенадцать мишеней мейотической рекомбинации в областях, считающихся горячими или холодными точками между маркерами семян GFP (зеленый флуоресцентный белок) и RFP (красный флуоресцентный белок). Горячие (показаны красным) и холодные (показаны синим) мишени имели признаки эухроматина или гетерохроматина, характерные для горячих точек или холодных точек рекомбинации, соответственно. Показаны координаты мишеней и их распределение на 3 хромосоме между маркерами GFP и RFP. На Фиг. 12B показана программа проведения эксперимента: двенадцать гомозиготных тестерных линий Columbia, экспрессирующих кассету 35Sx2: гигромицин, u6-26:гРНК, каждая из которых кодирует гРНК, направленную на конкретную последовательность горячей/холодной точки, скрещивали с линиями Columbia WT, экспрессирующими nos:nptII:nos Ubi:spCas9. Частоту рекомбинации рассчитывали на основе семян самоопыленных растений F2, которые использовали для расчета частоты кроссинговера между маркерами GFP и RFP - слева (результаты показаны на Фиг. 12C). Кроме того, растения F1 скрещивали с растениями Ландсберга дикого типа и выделяли ДНК из соматических тканей для определения частоты соматичности и механизма репарации ДНР ДНК вокруг ДНР посредством секвенирования PacBio (результаты показаны на Фиг. 13A-13Q). (Фиг. 12C) Частота кроссинговера в сантиморганах (ось Y) между маркерами GFP и RFP после индукции ДНР CRISPR-Cas9 в мишенях, показанных на оси X, с номером координаты для горячих (красные) или холодных (синие) сайтов. Контроли в отсутствие индукции ДНР показаны черным. Большой красный ромб обозначает среднюю степень кроссинговера для каждой популяции.
[0044] На Фиг. 13A-13Q представлен молекулярный анализ репарации ДНР в горячих мишенях -chr3:1854159 с использованием секвенирования Pacbio. ДНК выделяли из молодых почек (на стадии премейоза), стеблей и ткани верхних листьев каждого растения из популяций обратного скрещивания тестера Columbia x Landsberg. Фрагменты размером 5 тысяч п.о., фланкирующие сайт ДНР ДНК, амплифицировали с помощью ПЦР и секвенировали с использованием PacBio. Прочтения строк группировали в консенсусные последовательности с использованием анализа длинных ампликонов PacBio, а затем выравнивали с геномом Arabidopsis с использованием программного обеспечения для выравнивания последовательностей Burrows-Wheeler Aligner (BWA) и наносили на график. Красные полосы представляют собой однонуклеотидные полиморфизмы (ОНП) Columbia (Col), а синие полосы представляют собой ОНП Landsberg (Ler). Сайт ДНР в мишени № 1854179 на 3 хромосоме показан пунктирной линией. Желтая линия указывает на следы NHEJ в сайте ДНР. Зеленые линии представляют собой последовательности, не принадлежащие ни одному из родителей. Для каждого растения (Фиг. 13A-13Q, где каждый прямоугольник представляет собой отдельное растение, а Фиг. 13O-13Q представляют собой контрольные растения) выделенную ДНК штрихкодировали (отдельные прямоугольники с указанным штрих-кодом), сотни или тысячи отдельных молекул секвенировали и группировали в соответствии с последовательностью (включая паттерны ОНП). Этот способ позволяет различать родительское происхождение каждой молекулы. В некоторых растениях (например, штрихкод 89 в верхнем левом квадрате) не было обнаружено каких-либо изменений, и родительские аллели присутствовали более или менее в равной пропорции. В других растениях (например, штрих-код 90 - второе растение сверху, начиная с левой стороны) имелись свидетельства кроссинговера, фланкирующего разрыв, на что указывает переход от красных ОНП к синим в 10-12% молекул. Три контрольных растения F1 Ler x тестер Col также секвенировали и анализировали таким же образом, и у них не было обнаружено никаких признаков события кроссинговера или генной конверсии.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0045] Для обеспечения полного понимания способов, представленных в настоящей заявке, в нижеследующем подробном описании поясняется множество отдельно взятых подробностей. Однако специалистам в данной области техники должно быть понятно, что эти способы направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения, или полученной из нее ткани, или полученном из нее растении могут применяться на практике без этих отдельно взятых подробностей. В других случаях хорошо известные способы, процедуры и компоненты не были описаны подробно, чтобы не затруднить понимания способов и полученных в результате клеток растения и полученных из них растений, содержащих ДНК, содержащую направленную гомологичную рекомбинацию, как раскрыто в настоящей заявке.
[0046] В одном из вариантов реализации в настоящей заявке раскрыт способ направления рекомбинации ДНК между гомологичными хромосомами в соматических клетках растения, включающий следующие стадии: (а) осуществление экспрессии системы нуклеаз в указанной клетке растения, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей; (b) осуществление анализа потомства указанной клетки растения, или ткани растения, выращенной из указанной клетки растения, или растения, выращенного из указанной клетки, или потомства указанного растения на гомологичную рекомбинацию между гомологичными хромосомами, где указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером); и (c) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых произошла гомологичная рекомбинация.
[0047] В одном из вариантов реализации способы, раскрытые в настоящей заявке, обеспечивают получение растения, обладающего комбинацией полезных признаков или качеств, где указанный способ включает направленную рекомбинацию ДНК между гомологичными хромосомами в гибридной соматической клетке растения, содержащей полиморфные аллели на указанных гомологичных хромосомах, где указанный способ включает следующие стадии: (a) осуществление экспрессии системы нуклеаз в указанной клетке растения, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей; (b) осуществление анализа потомства указанной клетки растения, или ткани растения, выращенной из указанной клетки растения, или растения, выращенного из указанной клетки, или потомства указанного растения на гомологичную рекомбинацию между гомологичными хромосомами, где указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером); (c) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых произошла направленная гомологичная рекомбинация; (d) выращивание указанной клетки растения, или полученной из нее ткани растения, или полученного из нее растения, или потомства указанного растения с получением растения, содержащего указанную направленную гомологичную рекомбинацию, где указанное растение обладает комбинацией полезных качеств или признаков, не присутствующих ни у одного из родительских растений, из которых происходила гибридная соматическая клетка.
[0048] В одном из вариантов реализации способ, раскрытый в настоящей заявке, включает получение растения-потомка, обладающего комбинацией полезных признаков или качеств, не присутствующей ни у одного из родительских растений, из которых происходит потомство, где указанный способ включает: (a) выбор родительских растений, где каждый из указанных родителей обладает по меньшей мере одним полезным признаком, где указанный по меньшей мере один полезный признак не идентичен, и где указанные родители полиморфны по одному из указанного по меньшей мере одного полезного признака; (b) скрещивание указанных родительских растений для создания гибридного растения; (c) сбор соматических клеток из гибридного растения; (d) экспрессию системы нуклеаз в указанных соматических клетках, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей, где гомологичный кроссинговер или генная конверсия (не сопровождающаяся кроссинговером) в указанном целевом предварительно выбранном эндогенном сайте-мишени приводит к обмену ДНК, экспрессирующей или регулирующей экспрессию по меньшей мере одного из указанных полезных признаков или качеств; (e) анализ потомства указанных клеток растения, или ткани растения, выращенной из указанных клеток растения, или растения, выращенного из указанных клеток, или потомства указанного растения на указанное событие кросингсовера или генной конверсии (не сопровождающейся кроссинговером), при которых экспрессируется указанная комбинация признаков; (f) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, в которых экспрессируется комбинация признаков; и (g) осуществление размножения указанной клетки растения, полученной из нее ткани растения, полученного из нее растения с получением растения-потомка, обладающего указанной комбинацией полезных признаков или качеств.
[0049] В одном из вариантов реализации клетка растения представляет собой выделенную клетку растения. В еще одном варианте реализации клетка растения содержится в ткани растения. В еще одном варианте реализации клетка растения содержится в целом растении. Специалисту в данной области техники должно быть понятно, что использование термина «клетка растения» на всем протяжении настоящего документа включает в различных вариантах реализации выделенную клетку растения, клетку растения, содержащуюся в ткани растения, или клетку растения, содержащуюся в целом растении, или их комбинацию.
[0050] В некоторых вариантах реализации клетка растения, описанная в настоящей заявке, происходит из растения дикого типа. В некоторых вариантах реализации клетка растения происходит из культивируемого растения, которое было отобрано в отношении желательных характеристик, которые можно поддерживать путем размножения. Культивируемые растения также могут быть известны как культурные сорта, хотя некоторые культурные сорта возникли в дикой природе.
[0051] Специалисту в данной области техники должно быть понятно, что способы направления рекомбинации ДНК между гомологичными хромосомами в соматических клетках растения, как описано в настоящей заявке, могут охватывать применения для точной селекции культурных растений.
[0052] В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к делеции определенного аллеля или его части. В некоторых вариантах реализации аллель кодирует полипептид, экспрессия которого обеспечивает признак или качество, полезное для растения или продукта растения, например, плода или цветка. В некоторых вариантах реализации аллель кодирует полипептид, экспрессия которого усиливает полезный признак или качество в растении. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к добавлению определенного аллеля или его части. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к введению в аллель мутации ДНК. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к замене одного аллеля другим аллелем. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к делеции регуляторной расположенной в 3'-5' направлении последовательности гена аллеля. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к делеции расположенной в 5'-3' направлении последовательности гена аллеля. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к добавлению регуляторной расположенной в 3'-5' направлении последовательности гена аллеля. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к расположенной в 5'-3' направлении последовательности гена аллеля. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к мутации регуляторной расположенной в 3'-5' направлении последовательности гена. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к расположенной в 5'-3' направлении последовательности гена. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к делеции определенного аллеля или его части, или добавлению определенного аллеля или его части, или введению в аллель мутации ДНК, или замене одного аллеля другим аллелем, или делеции регуляторной расположенной в 3'-5' направлении последовательности гена аллеля, или делеции расположенной в 5'-3' направлении последовательности гена аллеля, или добавлению регуляторной расположенной в 3'-5' направлении последовательности гена аллеля или регуляторной расположенной в 5'-3' направлении последовательности гена аллеля, или мутации регуляторной расположенной в 3'-5' направлении последовательности гена или регуляторной расположенной в 5'-3' направлении последовательности гена, или любой их комбинации в аллеле.
[0053] В некоторых вариантах реализации способы направленной рекомбинации ДНК между гомологичными хромосомами приводят к замене аллеля. В одном из вариантов реализации замена аллеля включает замену гена дикого типа мутантным аллелем в эндогенном локусе. В еще одном варианте реализации замена аллеля включает замену мутантного аллеля аллелем дикого типа в эндогенном локусе. В еще одном варианте реализации замена аллеля включает замену мутантного аллеля другим мутантным аллелем в эндогенном локусе. В некоторых вариантах реализации замена аллеля приводит к экспрессии признака или качества, полезного для клетки растения, полученной из нее ткани, полученного из нее растения или его потомства. Преимущество способов замены аллелей, раскрытых в настоящей заявке, состоит в том, что нет необходимости разрабатывать экзогенную последовательность нуклеиновой кислоты, содержащую заменяющий аллель, например, векторы, содержащие заменяющие аллели. Обмен аллельным материалом происходит между гомологичными хромосомами в клетке, где хромосомы содержат полиморфные аллели.
[0054] В некоторых вариантах реализации способы направленной рекомбинации ДНК между гомологичными хромосомами приводят к замене однонуклеотидным полиморфизмом (ОНП). В одном из вариантов реализации замена ОНП включает создание миссенс-мутации в гене. В еще одном варианте реализации замена ОНП включает размещение миссенс-мутации с нуклеотидом дикого типа. В еще одном варианте реализации замена ОНП включает создание миссенс-мутации в гене, усиливающей функцию кодируемого полипептида. В еще одном варианте реализации замена ОНП включает создание миссенс-мутации в гене, снижающей функцию кодируемого полипептида. В еще одном варианте реализации замена ОНП включает создание миссенс-мутации в гене, усиливающей экспрессию кодируемого полипептида. В еще одном варианте реализации замена ОНП включает создание миссенс-мутации в гене, снижающей экспрессию кодируемого полипептида. В некоторых вариантах реализации замена ОНП приводит к экспрессии признака или качества, полезного для клетки растения, полученной из нее ткани, полученного из нее растения или его потомства. Преимущество способов замены ОНП, раскрытых в настоящей заявке, состоит в том, что нет необходимости разрабатывать экзогенную последовательность нуклеиновой кислоты, содержащую заменяющий ОНП, например, векторы, содержащие заменяющий ОНП. Обмен последовательностями нуклеиновой кислоты, содержащей ОНП, происходит между гомологичными хромосомами в клетке, где хромосомы содержат полиморфные аллели.
[0055] В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к переносу одного локуса из одной хромосомы в ее гомолог посредством гомологичной рекомбинации (HR), причем в клетке растения-потомка создается новая желаемая комбинация признаков. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к переносу одного локуса из одной хромосомы в ее гомолог посредством гомологичной рекомбинации (HR), причем в ткани растения-потомка создается новая желаемая комбинация признаков. В некоторых вариантах реализации способы направления рекомбинации ДНК между гомологичными хромосомами приводят к переносу одного локуса из одной хромосомы в ее гомолог посредством гомологичной рекомбинации (HR), причем в растении-потомке создается новая желаемая комбинация признаков. В некоторых вариантах реализации перенос одного локуса включает перестановку хромосомных фрагментов из одной хромосомы в ее гомолог посредством гомологичной рекомбинации (HR), причем в клетке растения-потомка создается новая желаемая комбинация признаков. В некоторых вариантах реализации перенос одного локуса включает перестановку хромосомных фрагментов из одной хромосомы в ее гомолог посредством гомологичной рекомбинации (HR), причем в ткани растения-потомка создается новая желаемая комбинация признаков. В некоторых вариантах реализации перенос одного локуса включает перестановку хромосомных фрагментов из одной хромосомы в ее гомолог посредством гомологичной рекомбинации (HR), причем в растении-потомке создается новая желаемая комбинация признаков. В некоторых вариантах реализации комбинация признаков не присутствует ни у одного из родителей.
[0056] В некоторых вариантах реализации локус содержит аллель. В некоторых вариантах реализации локус содержит часть аллеля. В некоторых вариантах реализации локус содержит расположенную в 3'-5' направлении последовательность аллеля. В некоторых вариантах реализации локус содержит расположенную в 5'-3' направлении последовательность аллеля. В некоторых вариантах реализации локус содержит один ОНП в аллеле. В некоторых вариантах реализации локус содержит несколько ОНП в аллеле. В некоторых вариантах реализации локус содержит последовательность смежных нуклеиновых кислот, содержащую аллель, расположенную в 3'-5' направлении последовательность аллеля, расположенную в 5'-3' направлении последовательность аллеля, регуляторную последовательность аллеля или ОНП в аллеле, или любую их комбинацию.
[0057] Новый желаемый признак или комбинацию признаков трудно получить посредством естественной рекомбинации, например, во время культивирования растений, при которой рекомбинация не направлена на конкретный локус, и при которой рекомбинация в конкретном локусе происходит менее чем 105-106 раз на событие естественной рекомбинации.
[0058] В одном из вариантов реализации один локус содержит ген. В одном из вариантов реализации один локус содержит аллель. В одном из вариантов реализации один локус содержит часть гена. В одном из вариантов реализации один локус содержит часть аллеля. В одном из вариантов реализации один локус содержит промотор гена. В одном из вариантов реализации один локус содержит экзон гена. В одном из вариантов реализации один локус содержит по меньшей мере один экзон гена. В одном из вариантов реализации один локус содержит по меньшей мере два экзона гена. В одном из вариантов реализации один локус содержит по меньшей мере три экзона гена. В одном из вариантов реализации один локус содержит интрон гена. В одном из вариантов реализации один локус содержит по меньшей мере один интрон гена. В одном из вариантов реализации один локус содержит по меньшей мере два интрона гена. В одном из вариантов реализации один локус содержит по меньшей мере три интрона гена. В одном из вариантов реализации один локус содержит по меньшей мере один экзон и один интрон гена. В одном из вариантов реализации один локус содержит любую комбинацию экзона(ов) и интрона(ов) гена. В одном из вариантов реализации один локус содержит последовательность ДНК, кодирующую малую РНК. В одном из вариантов реализации один локус содержит последовательность ДНК, кодирующую микроРНК. В одном из вариантов реализации один локус содержит последовательность ДНК, кодирующую тРНК (транспортную РНК). В одном из вариантов реализации один локус содержит последовательность ДНК, кодирующую регуляторную последовательность или регуляторные последовательности гена.
[0059] В одном из вариантов реализации способы направленной рекомбинации между гомологичными хромосомами приводят к делеции определенного аллеля или его части. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к добавлению определенного аллеля или его части. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к введению в аллель мутации ДНК. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к замене одного аллеля другим аллелем. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к делеции регуляторной расположенной в 3'-5' направлении или расположенной в 5'-3' направлении последовательности гена аллеля. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к добавлению регуляторной расположенной в 3'-5' направлении или расположенной в 5'-3' направлении последовательности гена аллеля. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к мутации регуляторной расположенной в 3'-5' направлении или расположенной в 5'-3' направлении последовательности гена.
[0060] В еще одном варианте реализации мутация включает точечную мутацию, делеционную мутацию, мутацию по типу замены или инсерционную мутацию, или любую их комбинацию. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к точечной мутации. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к делеционной мутации. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к мутации по типу замены. В еще одном варианте реализации способы направленной рекомбинации между гомологичными хромосомами приводят к инсерционной мутации.
[0061] В некоторых вариантах реализации способы, раскрытые в настоящей заявке, приводят к «нокауту» гена, причем специалисту в данной области техники должно быть понятно, что «нокаут» гена заключается в том, чтобы сделать неработоспособным ген в геноме растения. В некоторых вариантах реализации нокаут гена приводит к экспрессии полезного качества или признака в растении. В некоторых вариантах реализации нокаут гена приводит к повышенной экспрессии полезного качества или признака в растении. В некоторых вариантах реализации нокаут гена приводит к понижению экспрессии нежелательного качества или признака в растении. В некоторых вариантах реализации нокаут гена приводит к отсутствию экспрессии не являющегося полезным качества или признака в растении. В некоторых вариантах реализации нокаут состоит в обмене полиморфными аллелями гена.
[0062] В некоторых вариантах реализации способы, раскрытые в настоящей заявке, приводят к «включению» гена, причем специалисту в данной области техники должно быть понятно, что «включение» гена заключается в том, чтобы сделать работоспособным ген в геноме растения, который в нем ранее не экспрессировался. В некоторых вариантах реализации включение гена приводит к экспрессии полезного качества или признака в растении. В некоторых вариантах реализации включение гена приводит к повышенной экспрессии полезного качества или признака в растении. В некоторых вариантах реализации включение гена приводит к понижению экспрессии нежелательного качества или признака в растении. В некоторых вариантах реализации включение гена приводит к отсутствию экспрессии не являющегося полезным качества или признака в растении. В некоторых вариантах реализации включение состоит в обмене полиморфными аллелями гена.
[0063] Специалисту в данной области техники должно быть понятно, что «гомологичная рекомбинация» включает механизм генетической рекомбинации, в котором две цепи ДНК, содержащие схожие нуклеотидные последовательности, обмениваются генетическим материалом. Клетки используют гомологичную рекомбинацию во время мейоза, где она служит для перестройки ДНК с целью создания совершенно уникального набора гаплоидных хромосом. Соматические клетки могут использовать гомологичную рекомбинацию для репарации поврежденной ДНК, в частности для репарации двухнитевых разрывов (ДНР). В одном из вариантов реализации, как описано в настоящей заявке, гомологичную рекомбинацию индуцируют между гомологичными хромосомами, содержащими полиморфные аллели в соматической клетке. Событие гомологичной рекомбинации может быть использовано для изменения эндогенного гена любым количеством способов. В некоторых вариантах реализации гомологичная рекомбинация может приводить к генной конверсии (не сопровождающейся кроссинговером). В некоторых вариантах реализации гомологичная рекомбинация может приводить к инактивации эндогенного гена. В некоторых вариантах гомологичная рекомбинация может приводить к получению рекомбинантного локуса, например, аллеля, полученного из двух родственных генов. Новый рекомбинантный аллель может в одном из вариантов реализации иметь новую активность по сравнению с любым из генов, из которых он был получен. Изменения в паттернах метилирования ДНК могут приводить к изменениям в экспрессии гена или генов. В некоторых случаях это может быть полезным, в то время как в других случаях было показано, что изменения в паттернах метилирования связаны с такими патологическими состояниями, как рак. В некоторых вариантах реализации способы направленной гомологичной рекомбинации, раскрытые в настоящей заявке, могут приводить к изменениям метилирования на эпигенетическом уровне, то есть изменению паттерна метилирования. В других вариантах реализации способы направленной гомологичной рекомбинации не приводят к изменениям метилирования на эпигенетическом уровне, то есть нет изменений в паттерне метилирования.
[0064] В некоторых вариантах реализации направленная рекомбинация ДНК между гомологичными хромосомами в соматической клетке, где сайт-мишень для рекомбинации содержит полиморфные аллели, является наследуемой, причем событие рекомбинации передается потомству. Таким образом, после того, как клетка растения, ткань растения, полученная размножением клетки, или растение, полученное размножением клетки, проанализированы и отобраны как содержащие событие направленной гомологичной рекомбинации, может быть получено потомство, содержащее это событие направленной рекомбинации. В одном из вариантов реализации событие рекомбинации наследуется через семена посредством зародышевой линии растения, полученного выращиванием из клетки или ткани, содержащей направленную рекомбинацию ДНК, как раскрыто в настоящей заявке. В еще одном варианте реализации событие рекомбинации наследуется посредством регенерации вегетативной ткани, содержащей событие рекомбинации. В еще одном варианте реализации событие рекомбинации наследуется посредством размножения вегетативных тканей, содержащих наследуемое событие. Неограничивающие примеры размножения вегетативной ткани, содержащей события рекомбинации, раскрытые в настоящей заявке, включают применение ветви, содержащей рекомбинантное событие, для среза дерева или для прививки на дерево, и применение каллуса, содержащего событие рекомбинации, для регенерации растения банана.
[0065] ДНР ДНК может служить мощным инструментом для изменения и контроля геномов растений. У растений большинство двухнитевых разрывов ДНК будет репарироваться с помощью механизма NHEJ, который обычно оставляет небольшие индел-мутации в месте разрыва. (Фиг. 1) Разрыв также может быть репарирован с помощью гомологичной рекомбинации (HR). (Фиг. 1, Фиг. 2 и Фиг. 3 справа). В одном из вариантов реализации, когда HR осуществляется путем синтез-зависимого отжига цепей, результатом является генная конверсия (перенос локуса из одной хромосомы в другую хромосому; также известная как событие, не сопровождающееся кроссинговером). В еще одном варианте реализации, когда HR осуществляется путем образования структур Холлидея, результатом является событие генной конверсии или событие кроссинговера, в зависимости от того, как была разрешена структура Холлидея. Специалисту в данной области техники должно быть понятно, что событие «кроссинговера» гомологичной рекомбинации между гомологичными хромосомами включает обмен цепями между последовательностями ДНК. В одном из вариантов реализации событие кроссинговера включает обмен между последовательностями ДНК, содержащими по существу схожий состав нуклеотидов. В еще одном варианте реализации событие кроссинговера включает обмен между последовательностями ДНК гомологичных хромосом, содержащих полиморфные аллели, где событие кроссинговера включает обмен цепями между последовательностями ДНК, содержащими полиморфный аллель. Другими словами, гомологичная рекомбинация путем кроссинговера гомологичных хромосом, содержащих полиморфный аллель, может привести к расширенному обмену последовательностью ДНК, где последовательность включает последовательность, содержащую отличающийся состав нуклеотидов. Кроме того, события кроссинговера гомологичной рекомбинации, в еще одном варианте реализации, обеспечивают обмен последовательностью ДНК, фланкирующей ДНР.
[0066] В некоторых вариантах реализации раскрытые в настоящей заявке способы направленной гомологичной рекомбинации в эндогенном сайте-мишени включают обмен непрерывной последовательностью ДНК, где указанная непрерывная последовательность ДНК содержит приблизительно 0,01-20 тыс. п.о. ДНК. В некоторых вариантах реализации раскрытые в настоящей заявке способы направленной гомологичной рекомбинации в эндогенном сайте-мишени включают обмен непрерывной последовательности ДНК, где указанная непрерывная последовательность ДНК содержит приблизительно 0,1-20 тыс. п.о. ДНК. В некоторых вариантах реализации раскрытые в настоящей заявке способы направленной гомологичной рекомбинации в эндогенном сайте-мишени включают обмен непрерывной последовательностью ДНК, где указанная непрерывная последовательность ДНК содержит приблизительно 1-20 тыс. п.о. ДНК.
[0067] В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен приблизительно 1-5 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен приблизительно 5-10 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен приблизительно 10-15 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен приблизительно 15-20 тыс. п.о.
[0068] В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 1 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 2 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 3 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 4 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 5 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 6 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 7 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 8 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 9 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 10 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 11 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 12 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 13 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 14 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 15 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 16 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 17 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 18 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 19 тыс. п.о. В некоторых вариантах реализации способы направленной гомологичной рекомбинации включают обмен по меньшей мере приблизительно 20 тыс. п.о.
[0069] На Фиг. 2 схематично показано, как направленная репарация разрывов может быть использована в качестве инструмента точной селекции. Продукты репарации этих разрывов могут быть очень полезны для селекции. В одном из вариантов реализации гомологичную рекомбинацию применяют для доставки признаков из сорта растений дикого типа в известный культурный сорт. В еще одном варианте реализации гомологичную рекомбинацию применяют для доставки признаков из одного известного культурного сорта, имеющего определенный признак, во второй известный культурный сорт, не имеющий определенного признака. В некоторых вариантах реализации репарация ДНР посредством гомологичной рекомбинации «разрушает» очень тесную генетическую связь 2 генов, вовлеченных в важные признаки. Например: когда ген, участвующий в устойчивости к заболеванию (Фиг. 2, «R»), находится рядом с геном, участвующим в обеспечении высокого урожая (Фиг. 2, «Y»), естественный мейотический кроссинговер может быть очень низким. Индукция направленного ДНР между этими двумя генами, между Yr на синей хромосоме и yR на красной хромосоме, с последующей репарацией гомологичным кроссинговером будет разделяться между двумя связанными признаками, позволяя создавать новую рекомбинантную комбинацию в потомстве с высоким выходом и устойчивостью к заболеванию. Это также позволяет минимизировать длину хромосомных сегментов от сортов дикого типа, которые могут содержать нежелательные гены.
[0070] В одном из вариантов реализации способы направленной рекомбинации между гомологичными хромосомами, как раскрыто в данном настоящей заявке, отличаются от способов генного таргетинга путем гомологичной рекомбинации, включающих обмен генетической информацией между молекулами геномной и экзогенной дезоксирибонуклеиновой кислоты (ДНК) посредством гомологичной рекомбинации. В еще одном варианте реализации способ направленной рекомбинации между гомологичными хромосомами, как раскрыто в настоящей заявке, не включает, или не требует, или не использует экзогенный гомологичный фрагмент ДНК в качестве матрицы для гомологичной рекомбинации.
[0071] Способы, описанные в настоящей заявке, обладают преимуществами по сравнению со способами генного таргетинга, известными в данной области техники, для которых необходимы экзогенные фрагменты ДНК в качестве матриц для обмена генетической информацией между молекулами геномной и экзогенной дезоксирибонуклеиновой кислоты (ДНК) посредством гомологичной рекомбинации. Еще одно преимущество состоит в том, что получающаяся в результате клетка растения, из которой может быть получена ткань растения-потомка и целые растения, не является трансгенной. В одном из вариантов реализации клетка-потомок растения, ткань растения или целое растение, полученные с использованием способов, описанных в настоящей заявке, не содержат чужеродной ДНК из системы нуклеаз, которая может быть удалена, например, путем генетической сегрегации, или которая может быть обеспечена временной экспрессией. Описанная в настоящей заявке направленная гомологичная рекомбинация имитирует естественное явление гомологичной рекомбинации, но поскольку ДНР является мишенью, событие рекомбинации ДНК направлено на обмен, например, предпочтительными признаками. Еще одно преимущество применения способов направленной рекомбинации, описанных в настоящей заявке, заключается в том, что получение растения, содержащего желаемое событие, повлечет за собой скрининг десятков тысяч растений для идентификации растения, содержащего определенный обмен признаками, при естественной (неиндуцированной/ненаправленной) гомологичной рекомбинации. Еще дополнительное преимущество перед способами, использующими экзогенную ДНК, заключается в том, что было показано, что экзогенная ДНК может изменять паттерн метилирования ДНК в месте инсерции. В одном из вариантов реализации способы направленной рекомбинации, раскрытые в настоящей заявке, изменяют паттерн метилирования ДНК в месте генной конверсии. В еще одном варианте реализации способы направленной рекомбинации, раскрытые в настоящей заявке, изменяют паттерн метилирования ДНК в сайте кроссинговера. В одном из вариантов реализации способы направленной рекомбинации, раскрытые в настоящей заявке, не изменяют паттерн метилирования ДНК в месте генной конверсии. В еще одном варианте реализации способы направленной рекомбинации, раскрытые в настоящей заявке, не изменяют паттерн метилирования ДНК в сайте кроссинговера.
[0072] В одном из вариантов реализации способы, раскрытые в настоящей заявке, применяют с соматическими клетками растения. Специалисту в данной области техники должно быть понятно, что соматическая клетка растения включает любую клетку растения, кроме зародышевых клеток. В еще одном варианте реализации соматические клетки растения выбраны из группы, включающей клетки корня, клетки ризоида, клетки луковицы, стволовые клетки, клетки листа, клетки почки, клетки семенной коробочки или клетки плода. В некоторых вариантах реализации соматическая клетка растения, полученная способами, раскрытыми в настоящей заявке, может быть выращена в надлежащих условиях, известных в данной области техники, для создания ткани растения, содержащей ДНК, содержащую событие направленной HR, например, событие генной конверсии или кроссинговера. В одном из вариантов реализации ткань растения включает ткань корня, ткань ризоида, ткань луковицы, ткань ствола, ткань листа, ткань почки, ткань клубня, срез дерева, каллус растения, семя или семенную коробочку, или ткань плода, или любую их комбинацию. В еще одном варианте реализации ткань растения, выращенная из клетки растения, полученной способами, раскрытыми в настоящей заявке, может быть использована для получения растения-потомка, например, срез может быть использован для получения дерева или части дерева в случае прививки.
[0073] В некоторых вариантах реализации соматическая клетка растения, содержащая направленную рекомбинацию ДНК с использованием способов, раскрытых в настоящей заявке, может быть выращена в надлежащих условиях, известных в данной области техники, для получения целого растения, содержащего полученную в результате направленную рекомбинацию ДНК.
[0074] В еще одном варианте реализации целое растение, содержащее полученную в результате направленную рекомбинацию ДНК, содержит рекомбинантную ДНК в тканях всего растения. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в тканях только в части растения. Например, в еще одном варианте реализации целое растение содержит рекомбинантную ДНК в плодах. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в семенах. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в семенных коробочках. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в пыльце. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в листьях. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в ткани корня. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в ткани ризоида. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в ткани луковицы. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в стволах. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в почках. В еще одном варианте реализации целое растение содержит рекомбинантную ДНК в плодах, семенах, семенных коробочках, листьях, ткани корня, ткани ризоида, ткани луковицы, стеблях или почках, или любой их комбинации.
[0075] В некоторых вариантах реализации соматическая клетка растения включает протопласт. Специалисту в данной области техники должно быть понятно, что протопласт включает клетку растения, защитная клеточная стенка которой была частично или полностью удалена, например, путем ферментативной обработки, обеспечивающей получение неповрежденной биохимически компетентной единицы живого растения, которая может регенерировать клеточную стенку и в дальнейшем в надлежащих условиях выращивания вырасти в целое растение. Клеточная стенка растения также может быть частично или полностью удалена с использованием механических обработок, где интактная биохимическая компетентная единица живого растения представляет собой продукт, который может регенерировать клеточную стенку и в дальнейшем в надлежащих условиях выращивания вырасти в целое растение.
[0076] В некоторых вариантах реализации раскрытые в настоящей заявке способы получения соматической клетки растения, содержащей ДНК, содержащую событие направленной гомологичной рекомбинации, как раскрыто в настоящей заявке, где указанная клетка растения включает протопласт, могут применяться для получения ткани растения путем выращивания протопласта в надлежащих условиях выращивания, известных в данной области техники, для регенерации клеточной стенки и затем выращивания ткани растения. В некоторых вариантах реализации раскрытый в настоящей заявке способ получения соматической клетки растения, содержащей ДНК, содержащую событие направленной гомологичной рекомбинации, как раскрыто в настоящей заявке, где указанная клетка растения включает протопласт, могут применяться для получения целого растения путем выращивания протопласта в надлежащих условиях выращивания, известных в данной области техники, для регенерации клеточной стенки и затем выращивания целого растения.
[0077] В некоторых вариантах реализации в описанных в настоящей заявке способах используют направленную рекомбинацию между гомологичными хромосомами. Специалисту в данной области техники должно быть понятно, что термин «гомологичные хромосомы» охватывает хромосомы, содержащие информацию об одних и тех же биологических признаках и содержащие одни и те же гены в тех же локусах, но, возможно, разные аллели этих генов. В некоторых вариантах реализации гомологичные хромосомы включают хромосомы, содержащие информацию об одних и тех же биологических признаках и содержащие одни и те же гены в тех же локусах, но имеющие разные паттерны метилирования для этих генов, что может влиять на уровни экспрессии генов.
[0078] Специалисту в данной области техники должно быть понятно, что термин «аллель(и)» может охватывать любую из одной или более альтернативных форм гена в конкретном локусе. В диплоидной (или амфидиплоидной) клетке растения аллели данного гена расположены в определенном месте или локусе (локусах во множественном числе) на хромосоме. Один аллель присутствует на каждой хромосоме пары гомологичных хромосом. В одном из вариантов реализации полиморфные аллели включают аллели, которые отличаются в соответствующих хромосомных локусах. Термин «полиморфные аллели» может использоваться взаимозаменяемо с «гетерологичными аллелями» или «гетерозиготными аллелями», имеющими все те же значения и качества.
[0079] Кроме того, специалисту в данной области техники должно быть понятно, что термин «локус» (локусы во множественном числе) охватывает конкретное место или места, или сайт на хромосоме, где находится, например, ген или генетический маркер. В некоторых вариантах реализации локус содержится в области эухроматической ДНК. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит область гетерохроматической ДНК. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит область эухроматической ДНК или гетерохроматической ДНК.
[0080] В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит локус на хромосоме, где находится ген или генетический маркер. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит экзон гена. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит интрон гена. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит множество экзонов и интронов гена. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит область, включающую границу между по меньшей мере одним экзоном и одним интроном. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит регуляторные расположенные в 3'-5' направлении последовательности. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит регуляторные расположенные в 5'-3' направлении последовательности. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит регуляторные последовательности, расположенные в локусе гена. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит расположенные в 3'-5' направлении последовательности. В еще одном варианте реализации предварительно выбранный эндогенный сайт-мишень содержит расположенные в 5'-3' направлении последовательности.
[0081] В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит область эухроматической ДНК. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит область гетерохроматической ДНК. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит область эухроматической ДНК или гетерохроматической ДНК.
[0082] Специалисту в данной области техники должно быть понятно, что хромосома растения обладает как высококонденсированным перицентромерным гетерохроматическим плечом, так и в значительной степени деконденсированным эухроматическим плечом. Гетерохроматин часто ассоциируется с отсутствием транскрипционной активности и подавленной генетической рекомбинацией. Тем не менее, хотя гетерохроматин может быть беден генами по сравнению с эухроматином, он все же содержит транскрипционно активные гены. У растений, помимо гетерохроматина, расположенного в центромерной и перицентромерной областях, гетерохроматин находится в ядрышковом организаторе, на вздутиях и вдоль B-хромосом кукурузы (Zea mays). В геномах растений местоположение потенциально активных генов было идентифицировано в гетерохроматине, например, в структурах вздутий и в перицентромерной области. Тем не менее, хотя гетерохроматин может быть беден генами по сравнению с эухроматином, он все же содержит транскрипционно активные гены. Неожиданные результаты, представленные ниже в примере 5, демонстрируют, что способы, раскрытые в настоящей заявке для сайт-специфичного направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения, работают как для эухроматина, так и, неожиданно, для гетерохроматина, где рекомбинация обычно подавляется.
[0083] В еще одном варианте реализации нуклеазу, раскрытую в настоящей заявке, направляют в область в предварительно выбранном эндогенном сайте-мишени, где длина указанной области-мишени составляет приблизительно 20 п.о. В еще одном варианте реализации длина области-мишени составляет приблизительно 30 п.о. В еще одном варианте реализации длина области-мишени составляет менее 20 п.о. В еще одном варианте реализации длина области-мишени для ДНР составляет более 20 п.о. В некоторых вариантах реализации нуклеазу, раскрытую в настоящей заявке, направляют в область-мишень для создания ДНР.
[0084] В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень содержит полиморфный аллель. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень является смежным с полиморфным аллелем. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень расположен выше полиморфного аллеля. В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень расположен ниже полиморфного аллеля.
[0085] В некоторых вариантах реализации направленная гомологичная рекомбинация между гомологичными хромосомами включает обмены ДНК, управляемые гомологичными последовательностями, присутствующими на гомологичных хромосомах, присутствующих в геноме клеток растения, и на нее воздействует ферментативный механизм клетки (Фиг. 3; Фиг. 4A; Фиг. 6A). В одном из вариантов реализации обмен ДНК включает ДНК в предварительно выбранном эндогенном сайте-мишени. В еще одном варианте реализации обмен ДНК включает, не ограничиваясь перечисленным, ДНК в предварительно выбранном эндогенном сайте-мишени. В еще одном варианте реализации обмен ДНК включает ДНК в предварительно выбранном эндогенном сайте-мишени и ДНК, смежную с предварительно выбранным эндогенным сайтом-мишенью. В еще одном варианте реализации обмен ДНК включает ДНК, содержащую весь предварительно выбранный эндогенный сайт-мишень. В еще одном варианте реализации обмен ДНК включает ДНК, содержащую весь предварительно выбранный эндогенный сайт-мишень, и ДНК, смежную с предварительно выбранным эндогенным сайтом-мишенью. В еще одном варианте реализации обмен ДНК включает ДНК, содержащую только часть предварительно выбранного эндогенного сайта-мишени. В еще одном варианте реализации обмен ДНК включает ДНК, содержащую только часть предварительно выбранного эндогенного сайта-мишени, и ДНК, смежную с предварительно выбранным эндогенным сайтом-мишенью. В еще одном варианте реализации обмен ДНК включает 3'-ДНК в ДНР. В еще одном варианте реализации обмен ДНК включает 5'-ДНК в ДНР.
[0086] В некоторых вариантах реализации клетка растения, используемая в описанных в настоящей заявке способах, имеет мутации в ее генах и/или их регуляторных элементах, которые необходимы для репарации гомологичной ДНК по пути негомологичного соединения концов (NHEJ) после ДНР. Например, в некоторых вариантах реализации клетка растения может иметь мутацию в гене ku (например, ku70 и/или ku80). В некоторых вариантах реализации клетка растения может иметь мутацию в lig4. В некоторых вариантах реализации клетка растения может иметь мутацию в любом гене или регуляторном элементе, где мутация приводит к снижению репарации посредством NHEJ после ДНР.
[0087] На Фиг. 3 схематически представлены некоторые варианты реализации способа индукции гомологичной рекомбинации между гомологичными хромосомами, например, гомологичными хромосомами в клетке растения, ткани растения или целом растении. Способ направленной рекомбинации между гомологичными хромосомами в геноме соматической клетки, например, клетки растения, включает три стадии: (1) экспрессию системы нуклеаз в клетке растения; (2) индукцию двухцепочечного разрыва ДНК в одном или обоих аллелях предварительно выбранного сайта; и (3) репарацию ДНК путем рекомбинации между гомологичными хромосомами. В одном из вариантов реализации в настоящей заявке раскрыт способ направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения, включающий следующие стадии: (а) осуществление экспрессии системы нуклеаз в указанной клетке растения, где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, где при экспрессии системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, где указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв (ДНР) в ДНК по меньшей мере одного из полиморфных аллелей; (b) осуществление анализа потомства указанной клетки растения, или ткани растения, выращенной из указанной клетки растения, или растения, выращенного из указанной клетки, или потомства указанного растения на гомологичную рекомбинацию между гомологичными хромосомами, где гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером); и (c) выбор клетки растения, полученной из нее ткани растения, полученного из нее растения или потомства указанного растения, содержащих ДНК, содержащую указанное событие направленной гомологичной рекомбинации.
[0088] Фиг. 3 - Стадия 1: экспрессия системы нуклеаз. Система нуклеаз, подлежащая экспрессии, может включать любую систему нуклеаз, способную направлять активность двухцепочечного расщепления на предварительно выбранный сайт в ДНК по меньшей мере одного аллеля гомологичных хромосом.
[0089] Например, в некоторых вариантах реализации система нуклеаз, используемая в способе, раскрытом в настоящей заявке, включает систему нуклеаз с цинковыми пальцами (ZFN), систему эффекторных нуклеаз, подобных активаторам транскрипции (TALEN), или систему сгруппированных коротких палиндромных повторов (CRISPR)/белков, ассоциированных с CRISPR (Cas). В других вариантах реализации система нуклеаз, используемая в способе, раскрытом в настоящей заявке, включает любую систему нуклеаз, способную нацеливать нуклеазу, способную к двухцепочечному расщеплению ДНК, на предварительно выбранный сайт в ДНК. В еще одном варианте реализации система нуклеаз содержит бактериальный Argonaute и ДНК-гид. В еще одном варианте реализации двухцепочечная нуклеаза расщепляет ДНК с образованием тупых концов. В еще одном варианте реализации двухцепочечная нуклеаза расщепляет ДНК с образованием зубчатых концов.
[0090] В еще одном варианте реализации двухцепочечная нуклеаза расщепляет ДНК в полиморфном аллеле. В еще одном варианте реализации двухцепочечная нуклеаза расщепляет ДНК выше полиморфного аллеля. В еще одном варианте реализации двухцепочечная нуклеаза расщепляет ДНК ниже полиморфного аллеля. В еще одном варианте реализации система нуклеаз содержит нуклеазу с цинковыми пальцами (ZFN), где ZFN может быть известной в данной области техники или вновь созданной для расщепления предварительно выбранного сайта. В еще одном варианте реализации система нуклеаз включает эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), где TALEN может быть известной в данной области техники или вновь созданной для расщепления предварительно выбранного сайта. В еще одном варианте реализации система нуклеаз содержит систему сгруппированных коротких палиндромных повторов (CRISPR)/белков, ассоциированных с CRISPR (Cas) (CRISPR/Cas), где огРНК и/или Cas могут быть известными в данной области техники или вновь созданными для расщепления в предварительно выбранном сайте.
[0091] Специалисту в данной области техники должно быть понятно, что термины «одиночная гидовая РНК», «огРНК» и «гРНК» являются взаимозаменяемыми и имеют одни и те же качества и значения, где сгРНК может включать молекулу химерной РНК, состоящую из CRISPR-РНК (crRNA) и транскодируемой CRISPR-РНК (tracrRNA). В некоторых вариантах реализации crRNA комплементарна области ДНК в предварительно выбранном эндогенном сайте-мишени на по меньшей мере одной из гомологичных хромосом, где crRNA «нацеливает» белок нуклеазы, ассоциированный с CRISPR (Cas), на предварительно выбранный эндогенный сайт-мишень.
[0092] В одном из вариантов реализации длина комплементарной последовательности crRNA составляет 19-22 нуклеотидов, например, 19-22 последовательных нуклеотидов, комплементарных мишени. В еще одном варианте длина последовательности crRNA, комплементарной области ДНК, составляет приблизительно 15-30 нуклеотидов. В еще одном варианте реализации длина последовательности crRNA, комплементарной области ДНК, составляет приблизительно 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидов. В еще одном варианте длина последовательности crRNA, комплементарной области ДНК, составляет 20 нуклеотидов. В одном из вариантов реализации crRNA расположена на 5'-конце молекулы огРНК. В еще одном варианте реализации crRNA содержит 100% комплементацию в предварительно выбранной последовательности-мишени. В еще одном варианте реализации crRNA содержит по меньшей мере 80% комплементации в предварительно выбранной последовательности-мишени. В еще одном варианте реализации crRNA содержит по меньшей мере 85% комплементацию в предварительно выбранной последовательности-мишени. В еще одном варианте реализации crRNA содержит по меньшей мере 90% комплементацию в предварительно выбранной последовательности-мишени. В еще одном варианте реализации crRNA содержит по меньшей мере 95% комплементацию в предварительно выбранной последовательности-мишени. В еще одном варианте реализации crRNA содержит по меньшей мере 97% комплементацию предварительно выбранной последовательности-мишени. В еще одном варианте реализации crRNA содержит по меньшей мере 99% комплементацию в предварительно выбранной последовательности-мишени.
[0093] В еще одном варианте реализации tracrRNA имеет длину 100-300 рибонуклеотидов и обеспечивает сайт связывания для нуклеазы Cas, например, белка Cas9, образующего комплекс CRISPR/Cas9.
[0094] В одном из вариантов реализации система нуклеаз содержит нуклеазу с цинковыми пальцами (ZFN), содержащую ДНК-связывающий домен цинкового пальца и ДНК-расщепляющий домен нуклеазы, где указанный ДНК-связывающий домен цинкового пальца связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым ДНК-расщепляющий домен нуклеазы на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени.
[0095] Специалисту в данной области техники должно быть понятно, что термины «нуклеаза с цинковыми пальцами» или «ZFN» являются взаимозаменяемыми и имеют одни и те же значения и качества, причем ZFN включает молекулу химерного белка, содержащую по меньшей мере один ДНК-связывающий домен цинкового пальца, функционально связанный с по меньшей мере одной нуклеазой, способной к двухцепочечному расщеплению ДНК. В одном из вариантов реализации нуклеаза с цинковыми пальцами создает двухнитевой разрыв в предварительно выбранном эндогенном сайте-мишени. В еще одном варианте реализации нуклеаза с цинковыми пальцами содержит ДНК-связывающий домен и ДНК-расщепляющий домен, где ДНК-связывающий домен состоит по меньшей мере из одного цинкового пальца и функционально связан с ДНК-расщепляющим доменом. В еще одном варианте реализации ДНК-связывающий домен цинкового пальца находится на N-конце молекулы химерного белка, а ДНК-расщепляющий домен расположен на С-конце молекулы. В еще одном варианте реализации ДНК-связывающий домен цинкового пальца находится на C-конце молекулы химерного белка, а ДНК-расщепляющий домен расположен на N-конце молекулы. В еще одном варианте реализации связывающий домен цинкового пальца охватывает область в нуклеазе с цинковыми пальцами, способную связываться с локусом-мишенью, например, предварительно выбранным эндогенным сайтом-мишенью, как раскрыто в настоящей заявке. В еще одном варианте реализации ДНК-связывающий домен цинкового пальца содержит белковый домен, связывающийся с предварительно выбранным эндогенным сайтом-мишенью на по меньшей мере одной гомологичной хромосоме. В еще одном варианте реализации ДНК-связывающий домен цинкового пальца содержит белковый домен, связывающийся с полиморфным аллелем на по меньшей мере одной гомологичной хромосоме. В еще одном варианте реализации ДНК-связывающий домен цинкового пальца содержит белковый домен, связывающийся с предварительно выбранным эндогенным сайтом-мишенью на обеих гомологичных хромосомах. В еще одном варианте реализации ДНК-связывающий домен цинкового пальца содержит белковый домен, связывающийся с полиморфными аллелями на обеих гомологичных хромосомах.
[0096] Специалисту в данной области техники должно быть понятно, что термин «химерный белок» используется для описания белка, который получен в результате экспрессии молекулы ДНК, которая была создана путем функционального соединения двух или более фрагментов ДНК. Фрагменты ДНК могут происходить от одного и того же вида, или они могут происходить от разных видов. Фрагменты ДНК могут происходить из одного и того же или разных генов.
[0097] Специалисту в данной области техники должно быть понятно, что термин «ДНК-расщепляющий домен» ZFN охватывает область в нуклеазе с цинковыми пальцами, способную разрушать химические связи между нуклеиновыми кислотами в нуклеотидной цепи. Примеры белков, содержащих расщепляющие домены, включают ферменты рестрикции, топоизомеразы, рекомбиназы, интегразы и ДНКазы.
[0098] В одном из вариантов реализации система нуклеаз включает систему эффекторных нуклеаз, подобных активаторам транскрипции (TALEN), содержащую ДНК-связывающий домен TAL-эффектора и ДНК-расщепляющий домен, где указанный ДНК-связывающий домен TAL-эффектора связывается с указанным предварительно выбранным эндогенным сайтом-мишенью, направляя тем самым ДНК-расщепляющий домен на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени.
[0099] Специалисту в данной области техники должно быть понятно, что термины «нуклеазы, подобные активаторам транскрипции», «TALEN» и «эффекторная нуклеаза TAL» могут использоваться взаимозаменяемо, имея одни и те же значения и качества, причем TALEN включает нуклеазу, способную распознавать и расщеплять ее сайт-мишень, например, предварительно выбранный эндогенный сайт-мишень, как описано в настоящей заявке. В еще одном варианте реализации TALEN содержит слитый белок, содержащий домен TALE и нуклеотид-расщепляющий домен. В еще одном варианте реализации домен TALE содержит белковый домен, связывающийся с нуклеотидом специфичным для последовательности образом через один или несколько повторяющихся модулей TALE. В еще одном варианте реализации домен TALE содержит белковый домен, связывающийся с предварительно выбранным эндогенным сайтом-мишенью на по меньшей мере одной гомологичной хромосоме. В еще одном варианте реализации домен TALE содержит белковый домен, связывающийся с полиморфным аллелем на по меньшей мере одной гомологичной хромосоме. В еще одном варианте реализации домен TALE содержит белковый домен, связывающийся с предварительно выбранным эндогенным сайтом-мишенью на обеих гомологичных хромосомах. В еще одном варианте реализации домен TALE содержит белковый домен, связывающийся с полиморфными аллелями на обеих гомологичных хромосомах.
[00100] В одном из вариантов реализации домен TALE содержит по меньшей мере один из повторяющихся модулей TALE. В еще одном варианте реализации домен TALE содержит от одного до тридцати повторяющихся модулей TALE. В еще одном варианте реализации домен TALE содержит более тридцати повторяющихся модулей.
[00101] В еще одном варианте реализации слитый белок TALEN содержит N-концевой домен, один или более повторяющихся модулей TALE, за которыми следуют модуль полуповтора, линкер и нуклеотид-расщепляющий домен.
[00102] В одном из вариантов реализации система нуклеаз содержит систему CRISPR/Cas. В еще одном варианте реализации система CRISPR/Cas содержит нуклеазу Cas и молекулу гРНК, где указанная молекула гРНК связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени.
[00103] В некоторых вариантах реализации система CRISPR/Cas содержит ферментную систему, включающую последовательность гидовой РНК («гРНК» или «огРНК»), содержащую нуклеотидную последовательность, комплементарную или по существу комплементарную области полинуклеотида-мишени, например, предварительно выбранному эндогенному сайту-мишени, и белок с нуклеазной активностью.
[00104] В еще одном варианте реализации система CRISPR/Cas содержит систему CRISPR-Cas I типа, или систему CRISPR-Cas II типа, или систему CRISPR-Cas III типа, или их производные. В еще одном варианте реализации система CRISPR-Cas содержит сконструированные и/или запрограммированные системы нуклеаз, полученные из природных систем CRISPR-Cas. В еще одном варианте реализации система CRISPR-Cas содержит сконструированные и/или мутированные белки Cas. В еще одном варианте реализации система CRISPR-Cas содержит сконструированную и/или запрограммированную гидовую РНК.
[00105] Специалисту в данной области техники должно быть понятно, что термин «гидовая РНК» охватывает РНК, содержащую последовательность, комплементарную или по существу комплементарную области последовательности ДНК-мишени. Гидовая РНК может содержать нуклеотидные последовательности, отличные от области, комплементарной или по существу комплементарной области последовательности ДНК-мишени, например, предварительно выбранному эндогенному сайту-мишени. В еще одном варианте реализации гидовая РНК содержит crRNA или ее производное. В еще одном варианте реализации гидовая РНК содержит химеру crRNA: tracrRNA.
[00106] В еще одном варианте реализации молекула гРНК содержит домен, комплементарный и связывающийся с предварительно выбранным эндогенным сайтом-мишенью на по меньшей мере одной гомологичной хромосоме. В еще одном варианте реализации молекула гРНК содержит домен, комплементарный и связывающийся с полиморфным аллелем на по меньшей мере одной гомологичной хромосоме. В еще одном варианте реализации молекула гРНК содержит домен, комплементарный и связывающийся с предварительно выбранным эндогенным сайтом-мишенью на обеих гомологичных хромосомах. В еще одном варианте реализации молекула гРНК содержит домен, комплементарный и связывающийся с полиморфным аллелем на обеих гомологичных хромосомах.
[00107] Ферменты Cas содержат направляемую РНК ДНК-эндонуклеазу, способную создавать двухнитевые разрывы (ДНР) в ДНК. Термин «фермент Cas» может использоваться взаимозаменяемо с терминами «CRISPR-ассоциированные эндонуклеазы» или «CRISPR-ассоциированные полипептиды», имеющими одни те же качества и значения. В одном из вариантов реализации фермент Cas выбран из группы, содержащей Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, C2cl, CasX, NgAgo, Cpf1, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3 и Csf4, их гомологи или их модифицированные версии. В еще одном варианте реализации фермент Cas включает Cas9. В еще одном варианте реализации фермент Cas включает Cas1. В еще одном варианте реализации фермент Cas включает Cas1B. В еще одном варианте реализации фермент Cas включает Cas2. В еще одном варианте реализации фермент Cas включает Cas3. В еще одном варианте реализации фермент Cas включает Cas4. В еще одном варианте реализации фермент Cas включает Cas5. В еще одном варианте реализации фермент Cas включает Cas6. В еще одном варианте реализации фермент Cas включает Cas7. В еще одном варианте реализации фермент Cas включает Cas8. В еще одном варианте реализации фермент Cas включает Cas10. В еще одном варианте реализации фермент Cas включает Cpf1. В еще одном варианте реализации фермент Cas включает Csy1. В еще одном варианте реализации фермент Cas включает Csy2. В еще одном варианте реализации фермент Cas включает Csy3. В еще одном варианте реализации фермент Cas включает Cse1. В еще одном варианте реализации фермент Cas включает Cse2. В еще одном варианте реализации фермент Cas включает Csc1. В еще одном варианте реализации фермент Cas включает Csc2. В еще одном варианте реализации фермент Cas включает Csa5. В еще одном варианте реализации фермент Cas включает Csn2. В еще одном варианте реализации фермент Cas включает Csm2. В еще одном варианте реализации фермент Cas включает Csm3. В еще одном варианте реализации фермент Cas включает Csm4. В еще одном варианте реализации фермент Cas включает Csm5. В еще одном варианте реализации фермент Cas включает Csm6. В еще одном варианте реализации фермент Cas включает Cmr1. В еще одном варианте реализации фермент Cas включает Cmr3. В еще одном варианте реализации фермент Cas включает Cmr4. В еще одном варианте реализации фермент Cas включает Cmr5. В еще одном варианте реализации фермент Cas включает Cmr6. В еще одном варианте реализации фермент Cas включает Csb1. В еще одном варианте реализации фермент Cas включает Csb2. В еще одном варианте реализации фермент Cas включает Csb3. В еще одном варианте реализации фермент Cas включает Csx17. В еще одном варианте реализации фермент Cas включает Csx14. В еще одном варианте реализации фермент Cas включает Csx10. В еще одном варианте реализации фермент Cas включает Csx16, CsaX. В еще одном варианте реализации фермент Cas включает Csx3. В еще одном варианте реализации фермент Cas включает Csx1, Csx15, Csf1. В еще одном варианте реализации фермент Cas включает Csf2. В еще одном варианте реализации фермент Cas включает Csf3. В еще одном варианте реализации фермент Cas включает Csf4. В еще одном варианте реализации фермент Cas включает Cpf1. В еще одном варианте реализации фермент Cas включает C2cl. В еще одном варианте реализации фермент Cas включает CasX. В еще одном варианте реализации фермент Cas включает NgAgo. В еще одном варианте реализации фермент Cas представляет собой гомолог Cas. В еще одном варианте реализации фермент Cas представляет собой ортолог Cas. В еще одном варианте реализации фермент Cas представляет собой модифицированный фермент Cas. В еще одном варианте реализации фермент Cas представляет собой любые CRISPR-ассоциированные эндонуклеазы, известные в данной области техники.
[00108] В одном из вариантов реализации соматическую клетку растения трансформируют для экспрессии системы нуклеаз или ее компонента. В еще одном варианте реализации по меньшей мере одну родительскую клетку соматической клетки растения трансформируют для экспрессии системы нуклеаз или ее компонента. В еще одном варианте реализации одного из родителей соматической клетки растения трансформируют для экспрессии системы нуклеаз или ее компонента. В еще одном варианте реализации каждого из родителей соматической клетки растения трансформируют для экспрессии компонента системы нуклеаз. В еще одном варианте реализации одного из родителей трансформируют для экспрессии обоих компонентов системы нуклеаз.
[00109] В некоторых вариантах реализации после события гомологичной рекомбинации клетки-потомки, ткани и/или растения больше не содержат мишени для системы нуклеаз. (Например, см. Фиг. 6A, где дополнительные ДНР не возникают, так как предварительно выбранный эндогенный сайт-мишень больше не существует. Секвенирование ДНК показало отсутствие дополнительных событий ДНР.) В некоторых вариантах реализации после гомологичной рекомбинации, индуцированной ДНР в эндогенном сайте-мишени, последовательность эндогенного сайта-мишени была изменена посредством HR. В некоторых вариантах реализации после гомологичной рекомбинации, индуцированной ДНР, система нуклеаз является нефункциональной. В некоторых вариантах реализации после гомологичной рекомбинации, индуцированной ДНР, система нуклеаз не обладает способностью направлять нуклеазную активность на эндогенный сайт-мишень.
[00110] В еще одном варианте реализации соматическая клетка растения содержится в гибридном растении или в гетерозиготном растении, где указанная клетка содержит полиморфные аллели. В еще одном варианте реализации соматическая клетка растения содержит существующую гибридную или гетерозиготную клетку растения, имеющую полиморфные аллели в предварительно выбранном эндогенном сайте-мишени. В еще одном варианте реализации соматическую клетку растения, содержащую полиморфные аллели, трансформируют для экспрессии системы нуклеаз.
[00111] В одном из вариантов реализации соматическую клетку трансформируют ДНК, кодирующей систему нуклеаз или ее компонент. В еще одном варианте реализации выделенную ткань растения трансформируют ДНК, кодирующей систему нуклеаз или ее компонент. В еще одном варианте реализации родительскую клетку трансформируют ДНК, кодирующей систему нуклеаз или ее компонент. В еще одном варианте реализации обе родительские клетки трансформируют ДНК, кодирующей систему нуклеаз или ее компонент.
[00112] В одном из вариантов реализации соматическую клетку трансформируют РНК, кодирующей систему нуклеаз или ее компонент. В еще одном варианте реализации выделенную ткань растения трансформируют РНК, кодирующей систему нуклеаз или ее компонент. В еще одном варианте реализации родительскую клетку трансформируют РНК, кодирующей систему нуклеаз или ее компонент. В еще одном варианте реализации обе родительские клетки трансформируют РНК, кодирующей систему нуклеаз или ее компонент.
[00113] В одном из вариантов реализации соматическую клетку трансформируют полипептидом, содержащим систему нуклеаз или ее компонент. В еще одном варианте реализации выделенную ткань растения трансформируют полипептидом, содержащим систему нуклеаз или ее компонент. В еще одном варианте реализации родительскую клетку трансформируют полипептидом, содержащим систему нуклеаз или ее компонент. В еще одном варианте реализации обе родительские клетки трансформируют полипептидом, содержащим систему нуклеаз или ее компонент.
[00114] В некоторых вариантах реализации трансформацию клетки растения или выделенной ткани растения осуществляют любым способом, известным в данной области техники. В еще одном варианте реализации трансформация приводит к временной экспрессии. В еще одном варианте трансформация приводит к стабильной экспрессии. В еще одном варианте реализации стабильную трансформацию осуществляют способом с использованием Agrobacterium. В еще одном варианте реализации трансформация включает прямую трансформацию. В еще одном варианте реализации прямая трансформация включает применение полиэтиленгликоля (ПЭГ). В еще одном варианте реализации прямая трансформация включает применение электропорации путем бомбардировки.
[00115] В некоторых вариантах реализации ДНК, введенная в клетку растения, например ДНК, кодирующая систему нуклеаз, может быть удалена из генома растения путем генетической сегрегации. В качестве альтернативы, в некоторых вариантах реализации ДНК экспрессируется временно и, таким образом, не остается в клетке растения.
[00116] На Фиг. 3 в блоке (а) показано, что трансформация может происходить для обоих родителей, где, например, каждого из них трансформируют одним компонентом нуклеазы, например, нуклеазы CRISPR/Cas, которая становится активной при гибридизации. Оба компонента нуклеазы могут быть введены в одного из родителей (Фиг. 3, блок (b)). В варианте реализации, показанном на Фиг. 3, блок (b), нуклеаза должна быть «молчащей» и становиться активированной в гибриде (с использованием индуцибельной системы). В еще одном варианте реализации, показанном на Фиг. 3, блок (b), система нуклеаз может быть направлена на аллель второго родителя, в то время как она не расщепляет аллель трансформированной родительской клетки растения, таким образом, она приобретает аллель-специфичную активность в гибриде. В еще одном варианте реализации трансформация может быть осуществлена на существующем гибриде или гетерозиготном растении (Фиг. 3, блок (с)) со всеми компонентами нуклеазы.
[00117] В некоторых вариантах реализации активность или активация системы нуклеаз является индуцибельной. В еще одном варианте реализации в индуцибельной системе нуклеаз могут использоваться индуцибельные промоторы. В еще одном варианте реализации индуцибельный промотор может быть тканеспецифичным. В еще одном варианте реализации индуцибельный промотор может быть индуцирован (включен) в условиях, стрессовых для клетки растения или ткани. В некоторых вариантах реализации активность или активация системы нуклеаз является конститутивной. В еще одном варианте реализации активность или активация могут быть тканеспецифичными. В еще одном варианте реализации экспрессия системы нуклеаз или ее части регулируется. В еще одном варианте реализации конститутивный промотор используют для экспрессии всех компонентов системы нуклеаз, раскрытой в настоящей заявке. В еще одном варианте реализации любой регулируемый растительный промотор, известный в данной области техники, используют для экспрессии всех компонентов системы нуклеаз, раскрытой в настоящей заявке. В еще одном варианте реализации любой регулируемый растительный промотор, известный в данной области техники, используют для экспрессии по меньшей мере одного компонента системы нуклеаз, раскрытой в настоящей заявке. В еще одном варианте реализации любой регулируемый промотор, известный в данной области техники и функционирующий в клетке растения, используют для экспрессии всех компонентов системы нуклеаз, раскрытой в настоящей заявке. В еще одном варианте реализации любой регулируемый промотор, известный в данной области техники и функционирующий в клетке растения, используют для экспрессии по меньшей мере одного компонента системы нуклеаз, раскрытой в настоящей заявке.
[00118] В некоторых вариантах реализации соматическая клетка растения включает клетку из потомства, полученного при скрещивании двух клеток или растений культурного сорта, где каждая из указанных родительских клеток растения содержит полиморфный аллель по сравнению с указанным партнером в указанном предварительно выбранном эндогенном сайте-мишени. В некоторых вариантах реализации соматическая клетка растения включает клетку из потомства растений, полученного при скрещивании двух полиморфных родительских линий, приводящем к созданию гибридного растения, где каждая из указанных родительских линий растений содержит полиморфный аллель в указанном предварительно выбранном эндогенном сайте-мишени, и где только одна из родительских линий содержит указанную систему нуклеаз. В некоторых вариантах реализации соматическая клетка растения включает клетку из потомства растений, полученного при скрещивании двух полиморфных родительских линий, приводящем к созданию гибридного растения, где каждая из указанных родительских линий растений содержит полиморфный аллель в указанном предварительно выбранном эндогенном сайте-мишени, и где каждая из родительских линий содержит компонент системы нуклеаз.
[00119] Специалисту в данной области техники должно быть понятно, что термин «потомство» в контексте настоящей заявки охватывает потомство, полученное при самоопылении или скрещивании, и включает прямое потомство первого поколения (например, F1), а также более поздние поколения (например, F2, F3 и т.д.), а также поколения обратного скрещивания, например, на 1-3 поколения. В одном из вариантов реализации потомство включает любое поколение растений или клеток растения, полученных из растения, в котором произошла индуцированная направленная гомологичная рекомбинация, как раскрыто в настоящей заявке.
[00120] В еще одном варианте реализации потомство включает поколение F1. В еще одном варианте реализации потомство включает поколение F2. В еще одном варианте реализации потомство включает поколение F3. В еще одном варианте реализации потомство включает поколение F4. В еще одном варианте реализации потомство включает несколько поколений, выбранных из поколений F1-F4. В еще одном варианте реализации потомство включает 1-е поколение обратного скрещивания. В еще одном варианте реализации потомство включает 2-е поколение обратного скрещивания. В еще одном варианте реализации потомство включает 3-е поколение обратного скрещивания. В еще одном варианте реализации потомство включает 4-е поколение обратного скрещивания. В еще одном варианте реализации потомство включает несколько поколений обратного скрещивания, выбранных из 1-4 поколений обратного скрещивания.
[00121] В еще одном варианте реализации одна из указанных родительских соматических клеток растения содержит указанную систему нуклеаз, где активность расщепления ДНК указанной системы нуклеаз направлена на полиморфный аллель, присутствующий в другой родительской клетке растения, не содержащей указанную систему нуклеаз. В еще одном варианте реализации одна из указанных родительских соматических клеток растения содержит нуклеазу Cas, а другая из указанных родительских соматических клеток растения содержит молекулу гРНК, связывающуюся в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени. В еще одном варианте реализации одна из указанных родительских соматических клеток растения содержит нуклеазу Cas9, а другая из указанных родительских соматических клеток растения содержит молекулу гРНК, связывающуюся в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas9 на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени.
[00122] В еще одном варианте реализации соматическая клетка растения включает клетку из потомства, полученного в результате скрещивания клетки растения культурного сорта с клеткой растения дикого типа, где каждая из указанных родительских клеток растения содержит полиморфный аллель по сравнению с указанным партнером в указанном предварительно выбранном эндогенном сайте-мишени, и где указанная клетка растения культурного сорта содержит указанную систему нуклеаз. В еще одном варианте реализации, где соматическая клетка растения содержит клетку из потомства, полученного в результате скрещивания клетки растения культурного сорта с клеткой растения дикого типа, активность расщепления ДНК указанной системы нуклеаз проявляется исключительно в отношении полиморфного аллеля, присутствующего в родительской клетке растения дикого типа. В еще одном варианте реализации система нуклеаз содержит ZFN, где активность расщепления ДНК указанной системы нуклеаз проявляется исключительно в отношении полиморфного аллеля, присутствующего в родительской клетке растения дикого типа. В еще одном варианте реализации система нуклеаз содержит TALEN, где активность расщепления ДНК указанной системы нуклеаз проявляется исключительно в отношении полиморфного аллеля, присутствующего в родительской клетке растения дикого типа.
[00123] В еще одном варианте реализации соматическая клетка растения, имеющая полиморфные аллели, создана любым способом, известным в данной области техники. В еще одном варианте реализации соматическая клетка растения, имеющая полиморфные аллели, представляет собой клетку растения, полученную из культурного сорта.
[00124] Фиг. 3 - стадия 2: индукция двухцепочечного разрыва (ДНР) ДНК, которая представлена в виде желтой молнии. В некоторых вариантах реализации ДНР происходит в одном аллеле указанных полиморфных аллелей. В некоторых вариантах реализации ДНР происходит в обоих аллелях указанных полиморфных аллелей в случае диплоида. В некоторых вариантах реализации ДНР происходит только в одном аллеле указанных полиморфных аллелей в случае диплоида или клетки с более высокой плоидностью, например, триплоида. В некоторых вариантах реализации ДНР происходит в двух аллелях указанных полиморфных аллелей в случае диплоида или клетки с более высокой плоидностью, например, триплоида. В некоторых вариантах реализации ДНР происходит в каждом аллеле указанных полиморфных аллелей, например, два ДНР в диплоиде, три ДНР в триплоиде и т.д.
[00125] Хотя в примерах, представленных ниже, и на Фиг. 1 показано ДНР-расщепление с использованием системы CRISPR/Cas, специалисту в данной области техники должно быть понятно, что эти примеры не являются ограничивающими и что можно использовать любую другую сайт-специфичную нуклеазу, например, ZFN или TALEN. В еще одном варианте реализации индукцию ДНР осуществляют с помощью системы ZFN. В еще одном варианте реализации индукцию ДНР осуществляют с помощью системы TALEN. В еще одном варианте реализации индукцию ДНР осуществляют с помощью системы CRISPR/Cas. В еще одном варианте реализации индукцию ДНР осуществляют с помощью любой системы нуклеаз, которая может быть направлена на предварительно выбранный эндогенный сайт-мишень и способна создавать ДНР в ДНК.
[00126] В некоторых вариантах реализации ДНР может быть вызван в любых тканях растения или клетках и на любых стадиях клеточного цикла. Например, в одном из вариантов реализации конститутивный промотор может быть использован для активации нуклеазы, так что индукция ДНР может происходить уже в зиготе гибрида. В еще одном варианте реализации ДНР происходит на ранней стадии развития соматической ткани растения. В еще одном варианте реализации ДНР происходит на поздней стадии развития соматической ткани. В еще одном варианте реализации ДНР происходит между ранней и поздней стадиями развития соматической ткани.
[00127] В еще одном варианте реализации протопласт трансформируют ДНК- или РНК-вектором, или комплексом очищенного белка и гРНК, или очищенным белком в случае однокомпонентной системы нуклеаз, например, ZFN или TALEN.
[00128] В одном из вариантов реализации индукция включает конститутивную индукцию. В еще одном варианте реализации индукция включает неконститутивную индукцию. В еще одном варианте реализации индукция включает тканеспецифичную индукцию. В еще одном варианте реализации индукция включает специфичную для определенных условий индукцию. В еще одном варианте реализации индукция включает зависимую от клеточного цикла индукцию. В еще одном варианте реализации индукция включает конститутивную индукцию, неконститутивную индукцию, тканеспецифичную индукцию или зависимую от клеточного цикла индукцию, или любую их комбинацию.
[00129] Фиг. 3 - стадия 3: внутри клетки индуцированный ДНР может быть репарирован посредством негомологичного соединения концов (Фиг. 1) или посредством гомологичной рекомбинации (HR) между гомологичными хромосомами (матрица для эндогенной репарации, Фиг. 1, Фиг. 2 и Фиг. 3). В некоторых вариантах реализации ДНР репарируется посредством негомологичного соединения концов (NHEJ). В других вариантах реализации ДНР репарируется посредством гомологичной рекомбинации (HR). Неожиданно, как показано в приведенных ниже примерах, применение способа направленной рекомбинации между гомологичными хромосомами, как раскрыто в настоящей заявке, позволяет достичь значительно более высокой частоте репарации посредством HR, чем можно было бы ожидать естественным путем в отсутствие индукции ДНР.
[00130] В одном из вариантов реализации результат репарации ДНР включает генную конверсию (также известную как не сопровождающаяся кроссинговером). В еще одном варианте реализации результат репарации ДНР включает кроссинговер. Продукты репарации ДНР посредством HR могут быть идентифицированы с помощью различных анализов, например, с помощью генетических маркеров, изменения паттерна ОНП, методами секвенирования ДНК, потерей фенотипов гетерозиготности (LOH), или фенотипическим или любым другим маркером, или их комбинацией.
[00131] Чтобы определить и отобрать клетки растения, в которых произошла HR, можно проанализировать потомство указанных клеток. Анализ, в одном из вариантов реализации, может включать анализ клеток-потомков. В еще одном варианте реализации анализ включает анализ клеток растения, полученных из указанной соматической клетки или полученного из нее растения-потомка или ткани растения. В еще одном варианте реализации анализ включает анализ ткани растения, полученной из указанной соматической клетки, или полученного из нее растения-потомка или ткани растения. В еще одном варианте реализации анализ включает анализ ткани растения. В еще одном варианте реализации анализ включает анализ растительного потомства указанной соматической клетки или полученной из нее ткани или клетки. Клетки любого типа могут быть подвергнуты скринингу, в зависимости от растительной системы и желаемой области применения. Например, в растениях, размножающихся половым путем, семена, зерна, плоды или даже пыльцевые зерна могут быть подвергнуты скринингу для выявления аллелей, репарированных посредством HR, которые были унаследованы следующим поколением. У деревьев или растений, размножающихся вегетативным путем, протопласты, каллусы, листья, стебли и т.д. могут быть подвергнуты скринингу и затем регенерированы. В некоторых вариантах реализации анализ указанного растения включает анализ части указанного растения или его потомства, включающей лист, стебель, почку, плод, семя или пыльцу, или любую их комбинацию. В результате способ применим к любым видам растений.
[00132] В некоторых вариантах реализации соматическая клетка растения содержится в ткани растения или растении. Специалисту в данной области техники должно быть понятно, что термин «растение» охватывает любые виды древесных, травянистых, многолетних или однолетних растений. В одном из вариантов реализации соматическая клетка растения, раскрытая в настоящей заявке, происходит от растения, включающего любые виды древесных, травянистых, многолетних или однолетних растений. Термин «растение» может также охватывать множество клеток растения, которые в значительной степени дифференцированы в структуру, которая находится на стадии развития растения, способной производить культурное растение.
[00133] В одном из вариантов реализации соматическая клетка, раскрытая в настоящей заявке, происходит от культурного растения. В некоторых вариантах реализации соматическая клетка растения включает клетку культурного растения. Специалисту в данной области техники должно быть понятно, что термин «культурное растение» охватывает растение, по меньшей мере одна часть которого имеет коммерческую ценность. В одном из вариантов реализации культурное растение включает растения, производящие съедобные плоды (включая овощи), растения, производящие зерно (в качестве пищи, корма и для производства масла), растения, производящие цветы и декоративные растения, бобовые, корнеплоды, клубнеплоды или листовые культуры, и тому подобное.
[00134] В одном из вариантов реализации растение включает люцерну, яблоко, абрикос, Arabidopsis, артишок, рукколу, спаржу, авокадо, банан, ячмень, фасоль, свеклу, ежевику, чернику, брокколи, брюссельскую капусту, капусту, канолу, канталупу, морковь, маниок, клещевину, цветную капусту, сельдерей, вишню, цикорий, кинзу, цитрусовые, Clementines, клевер, кокос, кофе, кукурузу, хлопок, клюкву, огурец, Douglas fir, баклажаны, цикорий, эскарол, эвкалипт, фенхель, инжир, чеснок, тыкву, виноград, грейпфрут, белую мускатную дыню, хикаму, киви, латук, лук-порей, лимон, лайм, Loblolly pine, лен обыкновенный, манго, дыню, гриб, нектарин, орех, овес, масличную пальму, масличный рапс, окру, маслину, лук, апельсин, декоративное растение, пальму, папайю, петрушку, пастернак, горох, персик, арахис, грушу, перец, хурму, сосну, ананас, подорожник, сливу, гранат, тополь, картофель, тыкву, айву, сосну лучистую, радиккьо, редис, рапс, малину, рис, рожь, сорго, Southern pine, сою, шпинат, кабачки, клубнику, сахарную свеклу, сахарный тростник, подсолнечник, сладкий картофель, ликвидамбар, просо, мандарин, чай, табак, томат, тритикале, дерн, турнепс, виноград, арбуз, пшеницу, ямс и цукини.
[00135] В некоторых вариантах реализации способы направленной рекомбинации между гомологичными хромосомами соматической клетки растения включают способы точной селекции культурных растений. В некоторых вариантах реализации способы получения соматической клетки растения, содержащей ДНК, содержащую событие направленной HR (например, событие генной конверсии или кроссинговера), с использованием направленной рекомбинации между гомологичными хромосомами, включают способы точной селекции культурных растений. Способы, раскрытые в настоящей заявке, могут точно вводить качества и/или признаки, которые ранее не присутствовали в соматической клетке растения, полученных из нее тканях, полученных из нее растениях или ее потомстве. Такие качества присутствуют, например, в одной из родительских клеток указанной соматической клетки. Например, с помощью способов, раскрытых в настоящей заявке, фермер или селекционер может создать более выносливое растение или растение, устойчивое к опасным природным явлениям, таким как сельскохозяйственные вредители, патогены, засуха или плохие почвенные условия, или любой их комбинации. В другом варианте реализации способы, раскрытые в настоящей заявке, могут давать урожай, например, плод или овощ, имеющий улучшенные пищевые свойства, или повышенную устойчивость к сельскохозяйственным вредителям или патогенам, или более устойчивый во времени для улучшения качества плодоовощной продукции, перевозимой из одного места в другое. В некоторых вариантах реализации желаемое качество или признак присутствуют в популяции растения дикого типа. В некоторых вариантах реализации желаемое качество или признак присутствуют в культурном сорте растения. В некоторых вариантах реализации желаемое качество или признак присутствуют в видах растения дикого типа, но не в соответствующем культурном сорте. В некоторых вариантах реализации желаемое качество или признак присутствуют в одном культурном сорте вида растения, но не в соответствующем культурном сорте.
[00136] В некоторых вариантах реализации соматическая клетка растения, содержащая ДНК, содержащую указанное событие HR, или ткань растения, содержащая указанную клетку, содержащую ДНК, содержащую указанное событие HR, или растение, содержащее указанную клетку, или его растение-потомок, содержащее ДНК, содержащую указанное событие HR, или плод, полученный от растения, содержащего указанную клетку, или его растения-потомка, содержащего ДНК, содержащую указанное событие HR, или семена, полученные от растения, содержащего указанную клетку, или его растения-потомка, содержащего ДНК, содержащую указанное событие HR, или любая их комбинация обладают повышенной засухоустойчивостью, повышенной устойчивостью к сельскохозяйственным вредителям, повышенной устойчивостью к патогенам, улучшенным содержанием питательных веществ, улучшенными параметрами роста или любой их комбинацией по сравнению с контрольной клеткой растения, растением или его потомством. В одном из вариантов реализации контрольные клетка растения, растение или его потомство представляют собой родительскую клетку, растение или его потомство. В еще одном варианте реализации контрольные клетка растения, растение или его потомство представляют собой соматическую клетку, растение или его потомство, в которых указанный ДНР не происходит или не происходил.
[00137] В некоторых вариантах реализации предварительно выбранный эндогенный сайт-мишень, служащий в качестве мишени в способах, раскрытых в настоящей заявке, содержит ДНК, содержащую локус, часть локуса, ген, часть гена, регуляторную расположенную в 3'-5' направлении последовательность гена, регуляторные расположенные в 5'-3' направлении последовательности гена, расположенную в 5'-3' направлении последовательность гена или любую их комбинацию, где экспрессия указанного гена или ее отсутствие влияет на рост, засухоустойчивость, устойчивость к сельскохозяйственным вредителям, устойчивость к патогенам или содержание питательных веществ, или любую их комбинацию, указанной клетки растения, содержащей ДНК, содержащую событие направленной HR, или ее потомства по сравнению с контрольными клеткой растения или ее потомством, тканью растения, растением или его потомством.
[00138] В некоторых вариантах реализации потомство, отобранное на стадии (е), выбрано из группы, содержащей F1, F2, F3, F4, 1-е поколение обратного скрещивания, 2-е поколение обратного скрещивания, 3-е поколение обратного скрещивания и 4-е поколение обратного скрещивания.
[00139] В некоторых вариантах реализации способ, раскрытый в настоящей заявке, обеспечивает получение соматической клетки растения, содержащей ДНК, содержащую событие направленной HR, или ткани растения, содержащей указанную клетку, содержащую ДНК, содержащую событие направленной HR, или растения, содержащего указанную клетку, содержащую ДНК, содержащую событие направленной HR, или его растения-потомка, содержащего ДНК, содержащую событие направленной HR, или плода, полученного от растения, содержащего указанную клетку, содержащую ДНК, содержащую событие направленной HR, или его растения-потомка, содержащего ДНК, содержащую событие направленной HR, или семян, полученных от растения, содержащего указанную клетку, содержащую ДНК, содержащую событие направленной HR, или их растения-потомка, содержащего ДНК, содержащую событие направленной HR, или любой их комбинации, где указанная клетка, указанная ткань, указанное растение или его потомство обладают повышенной засухоустойчивостью, повышенной устойчивостью к сельскохозяйственным вредителям, повышенной устойчивостью к патогенам, улучшенным содержанием питательных веществ, улучшенными параметрами роста или любой их комбинацией по сравнению с контрольными клеткой растения, тканью растения, растением или его потомством, плодом или семенем.
ПРИМЕРЫ
[00140] Материалы и методы - для примеров 1-4
[00141] Растительный материал
[00142] Все растения томата выращивали в тепличных условиях при температуре от 18 до 25°C. Мутантные линии томатов (S. lycopersicum) с желтая мякоть e3756, двухцветныйcc383, M82 и Solanum pimpinellifoliumLA1578 были любезно предоставлены лабораториями профессора Джозефа Хиршберга (Joseph Hirschberg) и профессора Даниэля Замира (Daniel Zamir) из Еврейского университета в Иерусалиме (Kachanovsky, D. E., Filler, S., Isaacson, T. & Hirschberg, J. Epistasis in tomato color mutations involves regulation of phytoene synthase 1 expression by cis-carotenoids. Proc. Natl. Acad. Sci. U. S. A. 109, 19021-6 (2012)).
[00143] Плазмиды и трансформация растений
[00144] Конструкции 35s:Cas9 b u6-26:огРНК были ранее клонированы Ross A. Johnson (Johnson, R. A., Gurevich, V., Filler, S., Samach, A. & Levy, A. A. Comparative assessments of CRISPR-Cas nucleases' cleavage efficiency in planta. Plant Mol. Biol. 87, 143-56 (2015)). Праймеры, использованные для конструирования мишеней ps#1 огРНК ps#2f, указаны в списке праймеров, приведенном в Johnson et al., (2015) (тот же источник) и представлены в настоящей заявке в таблице 1.
[00145] Таблица 1: Праймеры для мишеней огРНК
[00146] Последовательность ДНК, кодирующая огРНК, использованную в примерах, представлена в таблице 2.
[00147] Таблица 2:
[00148] Все линии томатов были трансформированы с помощью Agrobacterium tumefaciens GV3101 с трансформацией семядолей согласно McCormick (McCormick, S. в Plant Tissue Culture Manual 311-319 (Springer Netherlands, 1991). doi:10.1007/978-94-009-0103-2_17).
[00149] ПЦР с обратной транскрипцией для обнаружения гомологичной рекомбинации (HR)
[00150] Образцы ДНК для анализа методом ПЦР с обратной транскрипцией выделяли с использованием набора для очистки ДНК (MACHEREY-NAGEL®). Для каждого растения 300 нг ДНК из образца листьев или контрольной плазмиды обрабатывали по отдельности: сначала их инкубировали в течение ночи с буфером 10xFD, ApaLI (ThermoFisher scientific) и HindIII-HF (New England BioLabs®). Спустя 20 минут инактивации при 80°С 150 нг расщепленных фрагментов затупляли ДНК-полимеразой Т4 (New England BioLabs®) в течение 2 часов при комнатной температуре. ДНК-полимеразу Т4 инактивировали при 75°С в течение 10 минут, и линейную ДНК самолигировали с ДНК-лигазой Quick T4 (New England BioLabs®) в течение 30 минут при комнатной температуре. Контрольные плазмиды разбавляли DDW (водой, обедненной по дейтерию) в соотношении 1:10000 и смешивали для имитации «гетерозиготности». Затем все образцы амплифицировали с помощью 18 циклов ПЦР с высокоточной ДНК-полимеразой Phusion® High-Fidelity (New England BioLabs®) (праймеры см. в таблице 3).
[00151] Таблица 3: Праймеры для обнаружения гомологичной рекомбинации с помощью ПЦР с обратной транскрипцией
[00152] Праймеры для этого анализа были разработаны для аллель-специфичной амплификации. Образцы объединяли и секвенировали с помощью секвенирования с высокой пропускной способностью.
[00153] Для клонирования контрольных плазмид для синтетического кроссинговера амплифицировали два фрагмента ПЦР из образцов ДНК желтая мякоть e3756 и двухцветныйcc383 с использованием высокоточной ДНК-полимеразы Phusion® High-Fidelity (New England BioLabs®) (последовательность праймеров см. в таблице 4) и затем клонировали с использованием система клонирования GoldenBraid (Sarrion-Perdigones, A. et al. GoldenBraid: An Iterative Cloning System for Standardized Assembly of Reusable Genetic Modules. PLoS One 6, e21622 (2011)).
[00154] Таблица 4: Праймеры для контрольных плазмид для синтетического кроссинговера

[00155] Сначала каждый из четырех ампликонов клонировали в плазмиду pUPD. Затем плазмиду pUPD2 с сегментом «ps» из желтая мякоть e3756 клонировали с плазмидой pUPD2 с сегментом «y1» из двухцветныйcc383 в плазмиду pDGB3_alpha1. Параллельно плазмиду pUPD2 с сегментом «ps» из двухцветныйcc383 клонировали с плазмидой pUPD2 с сегментом «y1» из желтая мякоть e3756 в плазмиду pDGB3_alpha1. Эти две плазмиды с «синтетическими аллелями» объединяли и подвергали процессу ПЦР с обратной транскрипцией и секвенированию. Последовательность ДНК этих плазмид с «синтетическими аллелями» представлена в таблице 5.
[00156] Таблица 5: Плазмиды для контроля синтетическим кроссинговером
[00157] Амплификация и секвенирование ДНК
[00158] Образцы ДНК для секвенирования с высокой пропускной способностью амплифицировали, используя высокоточную ДНК-полимеразу Phusion® High-Fidelity (New England BioLabs®) и 18 циклов ПЦР (конкретные праймеры, использованные в каждом эксперименте, см. в таблице 6).
[00159] Таблица 6: Праймеры для секвенирования с высокой пропускной способностью
* n представляет собой A, T, C или G.
[00160] Библиотеки были подготовлены согласно Blecher-Gonen et al. (Blecher-Gonen, R. et al. High-throughput chromatin immunoprecipitation for genome-wide mapping of in vivo protein-DNA interactions and epigenomic states. Nat. Protoc. 8, (2013)). Секвенирование с высокой пропускной способностью проводили в подразделении G-INCPM в Научном институте Вейцмана с помощью платформы Illumina HiSeq 2500 для 2×125 прочтений спаренных концов.
[00161] Образцы ДНК для секвенирования по Сэнгеру амплифицировали с использованием REDTaq® (SIGMA-ALDRICH) с 35 циклами ПЦР (праймеры см. в таблице 7).
[00162] Таблица 7: Праймер для секвенирования по Сэнгеру
[00163] После проведения ПЦР ДНК «очищали» экзонуклеазой I и щелочной фосфатазой креветки (rSAP) (New England BioLabs®). Секвенирование проводили в подразделении биологических услуг Научного института Вейцмана с помощью анализатора ДНК Applied Biosystems 3730.
Пример 1. Анализ окраски плодов томата для анализа репарации двухцепочеченого разрыва ДНК (ДНР).
[00164] Цель: Оценить частоту репарации двухцепочечного разрыва (ДНР) посредством соматического негомологичного соединения концов (NHEJ) по сравнению с гомологичной рекомбинацией (HR) в эндогенном локусе растения.
[00165] Методы: Для оценки частоты репарации ДНР посредством соматического NHEJ по сравнению с HR в эндогенном локусе растения был разработан анализ окраски плодов. Использовали две мутантные линии томата, каждая с отличной мутацией в гене Phytoene synthase 1 (PSY1). Аллель желтая мякоть e3756 является индуцированным ЭМС (этилметансульфонатом) мутантом с преждевременным стоп-кодоном в PSY1, приводящим к фенотипу желтых плодов (Kachanovsky, D. E., Filler, S., Isaacson, T. & Hirschberg, J. Epistasis in tomato color mutations involves regulation of phytoene synthase 1 expression by cis-carotenoids. Proc. Natl. Acad. Sci. U. S. A. 109, 19021-6 (2012)). Аллель двухцветныйcc383 является мутантом с делецией 3,7 тыс. п. о. в промоторе PSY1, приводящей к фенотипу желто-красных линялых плодов (Фиг. 4А).
[00166] Для мониторинга CRISPR-Cas-индуцированных мутаций в процессе развития растений, начиная с оплодотворения, были получены трансгенные линии желтая мякоть e3756, экспрессировавшие 35S:Cas9 (SEQ ID NO: 59), и трансгенные линии двухцветныйcc383, экспрессировавшие одиночную гидовую РНК PSY1 (u6-26:Ps#1-огРНК; последовательность плазмиды - SEQ ID NO: 60; PS#1-огРНК - SEQ ID NO: 68). Эта u6-26:Ps#1-огРНК была разработана для индукции ДНР ДНК между мутациями двухцветныйcc383 и желтая мякоть e3756, на обоих аллелях (Фиг. 4A). Ожидается, что скрещивание между желтая мякоть e3756идвухцветныйcc383 u6-26:Ps#1-огРНК даст растения F1 с доминантным фенотипом плодов двухцветныйcc383. То же самое ожидается для контрольных растений, не экспрессирующих 35S:Cas9 либо u6-26:Ps#1-огРНК. Ожидаются отклонения от этого фенотипа у растений, экспрессирующих как Cas9, так и Ps#1-огРНК, вследствие индукции ДНР на одном или обоих аллелях с последующей подверженной ошибкам репарацией ДНК. Репарация аллеля двухцветныйcc383 посредством NHEJ должна привести к фенотипу желтых плодов (участки или целые плоды). Результатом репарации ДНР с помощью механизмов на основе HR (событий, сопровождающихся или не сопровождающихся кроссинговером) должен быть красный плод в случае события HR, произошедшего на ранней стадии развития, или желтые плоды с красными пятнами или участками в случае поздних событий (Фиг. 4A).
[00167] Результаты: После индукции ДНР популяция из 50 растений F1 желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК дала двухцветные, желтые и желтые с красными пятнами плоды. Распределение фенотипов плодов варьировалось при использовании различных трансгенных линий желтая мякоть e3756 35S:Cas9 (Фиг. 4B). Как и ожидалось, в отсутствие индукции ДНР контрольная популяция 6 растений F1 желтая мякоть e3756 x двухцветныйcc383 продемонстрировала только двухцветные плоды (Фиг. 4B). Одним из преимуществ анализа окраски плодов является его способность прогнозировать наследование продуктов репарации в следующем поколении. Действительно, ожидалось, что семена F2, взятые из полностью желтых плодов, вызовут передаваемую через зародышевую линию мутацию.
[00168] Чтобы подтвердить, что желтые плоды свидетельствуют о событиях NHEJ в зародышевой линии, были выращены растения F2, полученные из желтых плодов. С использованием аллель-специфичной ПЦР-амплификации аллелей желтая мякоть e3756 и двухцветныеcc383 и секвенирования продуктов ПЦР было показано, что во всех исследованных случаях семена из желтых плодов давали семена, несущие передаваемую через зародышевую линию мутацию в сайте ДНР аллеля двухцветныйcc383 (таблица 9). В таблице 8 представлены использованные праймеры для ПЦР.
[00169] Таблица 8: Праймеры для событий NHEJ в зародышевой линии
[00170] Таблица 9: События NHEJ в зародышевой линии
[00171] Аллель-специфичные продукты ПЦР растений F2 из желтых плодов F1 желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК из таблицы 9 секвенировали с помощью секвенирования по Сэнгеру. Все растения имели индел-мутации (инсерции и/или делеции) в аллеле двухцветный. Четыре из 13 также имели индел-мутации в аллеле желтая мякоть.
[00172] Некоторые представители потомства желтых плодов также продемонстрировали мутацию в аллеле желтая мякоть e3756 (таблица 9). Хотя было обнаружено много желтых плодов с небольшими красными участками (Фиг. 4В), среди растений F1 не было обнаружено полностью красных плодов. Кроме того, была выращена популяция из 400 растений F2, полученных из плодов с красными пятнами, что наводит на мысль о соматической гомологичной рекомбинации (HR), но не было обнаружено полностью красных плодов, которые бы указывали на передаваемые через зародышевую линию события HR.
Пример 2: Высокая степень индукции ДНР ДНК посредством CRISPR-Cas9 приводит к репарации посредством как соматического негомологичного соединения концов (NHEJ), так и гомологичной рекомбинации (HR)
[00173] Цель: выявить, охарактеризовать и количественно оценить события соматического NHEJ в F1.
[00174] Методы: чтобы идентифицировать, охарактеризовать и количественно оценить события соматического NHEJ в F1 использовали 22 растения из популяции желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК и 2 растения желтая мякоть e3756 x двухцветныйcc383 в качестве контроля. Были использованы четыре (4) листа из разных ветвей растений. Затем их ДНК выделяли, область, фланкирующую индуцированный ДНР обоих аллелей, амплифицировали с помощью ПЦР, и полученные продукты секвенировали с использованием секвенирования с высокой пропускной способностью на платформе Illumina HiSeq 2500.
[00175] Для измерения репарации HR был разработан метод ПЦР с обратной транскрипцией, который позволил секвенировать две аллель-специфичные мутации, находящиеся на расстоянии 1,7 тыс. п.о. (желтая мякоть e3756 и двухцветныйcc383), позволил отличить родительские молекулы от рекомбинантных (Фиг. 4D) и минимизировал формирование ложноположительных продуктов ПЦР. Те же самые образцы ДНК, которые использовались для секвенирования соматического NHEJ (Фиг. 4С), использовали для ПЦР с обратной транскрипцией. Кроме того, были клонированы два синтетических положительных контроля (подобные рекомбинантным клоны), которые также обрабатывали тем же способом ПЦР с обратной транскрипцией. Продукты ПЦР с обратной транскрипцией каждой реакции (как показано на Фиг. 4D) секвенировали посредством секвенирования парных прочтений Illumina HiSeq 2500.
[00176] Результаты: из 250 000-850 000 прочтений на растение (образец ПЦР) в среднем 88% прочтений растений желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК содержали мутацию в сайте ДНР CRISPR, в то время как только 2% прочтений illumina растений желтая мякоть e3756 x двухцветныйcc383отклонялись от последовательности WT, предположительно из-за ошибок ПЦР и секвенирования (Фиг. 4C). Высокий уровень индукции ДНР CRISPR-Cas в системе приводил к широкому спектру мутаций в результате различных событий репарации посредством NHEJ. Кроме того, было обнаружено, что некоторые признаки NHEJ, такие как делеция CTTG величиной 4 п.о., были предпочтительнее других в этом локусе (Фиг. 4C, Фиг. 5).
[00177] В анализе измерения репарации HR было получено 5 000-50 000 прочтений на растение. Отрицательные контроли желтая мякоть e3756 x двухцветныйcc383 показали только родительские аллели в отсутствие индукции ДНР, в то время как положительный синтетический контроль показал рекомбинантные аллели (Фиг. 4E). Большинство растений F1 из желтая мякоть e3756 35S:Cas9 x двухцветныйcc383 u6-26:Ps#1-огРНК показали только родительские аллели, но у некоторых из них был обнаружен один из рекомбинантных аллелей, что указывает на репарацию на основе соматической HR.
[00178] Пример 3: Аллель-специфичная индукция ДНР и анализ репарации с высоким разрешением
[00179] Цель: создать способ, который позволил бы различать сломанную хромосому и матрицу для репарации.
[00180] Вышеуказанное скрещивание между желтая мякоть e3756 x двухцветныйcc383 не дало достаточного количества ОНП для детального анализа продуктов репарации посредством HR. Более того, оно не позволило выполнить аллель-специфичный разрыв, необходимый для проведения точного эксперимента, в котором можно различить сломанную хромосому и матрицу для репарации.
[00181] Метод: таким образом, был разработан новый анализ репарации ДНР, обеспечивающий несколько ОНП вокруг места разрыва, а также в дистальных областях, с помощью Solanum pimpinellifoliumLA1578, образца дикого томата с маленькими красными плодами, секвенированный геном которого демонстрирует наличие и местоположение множества ОНП по сравнению с Solanum lycopersicum, съедобным томатом. Для обеспечения аллель-специфичного разрыва был получен аллель на фоне генотипа сорта S. lycopersicum M82, который является невосприимчивым к u6-26:Ps#2-огРНК. Для этого красные плоды сорта М82 трансформировали 35S:Cas9 (последовательность плазмиды представляет собой SEQ ID NO: 59; последовательность, кодирующая Cas9, представляет собой SEQ ID NO: 65) и u6-26:Ps#2-огРНК (последовательность плазмиды представляет собой SEQ ID NO: 60; последовательность огРНК кодируется SEQ ID NO: 68). Затем отбирали желтые плоды в T0 и выращивали их семена T1. Из этой популяции T1 выделяли гомозиготное растение с инсерцией аденина (+A) в сайте ДНР CRISPR-Cas9, которое скрещивали с образцом дикого томата.
[00182] Последовательность молекулы гРНК, использованной для аллель-специфичной индукции ДНР и аллель-зависимой репарации, представлена в SEQ ID NO: 61.
[00183] В этом анализе S. pimpinellifoliumLA1578 является единственной мишенью для ДНР ДНК из-за инсерции +A в аллеле M82 psy1, которая нарушает мотив, прилежащий к протоспейсеру (PAM), и предотвращает расщепление посредством Cas9. Мутация +A аллеля M82 является рецессивной, и поэтому ожидается, что растения F1 будут иметь маленькие красные плоды. Репарация ДНР в PSY1 посредством NHEJ или HR (сопровождающейся или не сопровождающейся кроссинговером) приводит к желтым плодам или красным плодами с желтыми участками, в зависимости от стадии развития плода, когда произошла репарация. Ожидается, что события репарации посредством NHEJ оставят небольшие индел-мутации в сайте ДНР, в то время как события, сопровождающиеся и не сопровождающиеся кроссинговером, могут быть идентифицированы по разнице в паттернах ОНП с обеих сторон ДНР ДНК (Фиг. 6A).
[00184] Для анализа соматической репарации ДНР область ДНР ДНК из листьев обоих родителей (M82 35S:Cas9, U6-26:гРНК, +A гомозигота и S.pimpinellifoliumLA1578) и из пяти растений F1 амплифицировали с помощью ПЦР и секвенировали с помощью секвенирования парных прочтений Illumina HiSeq 2500 (Фиг. 7). Выход этого секвенирования составил 600 000-900 000 прочтений на растение.
[00185] В данном анализе показано, что аллель M82 psy1+A был невосприимчив к индукции ДНР и у родителя M82 (M82 35S:Cas9 и u6-26:Ps#2-огРНК, +A гомозигота) практически отсутствовали следы ДНР, что подтверждает полученную специфичность гРНК к аллелю (Фиг. 7). Кроме того, по меньшей мере 50% прочтений дали инсерцию +A, в то время как аллель S.pimpinellifolium был мутирован (красный цвет на круговой диаграмме на Фиг. 7). Было обнаружено, что только 7-18% прочтений растений F1 представляли собой WT, а остальные дали различные паттерны индел-мутаций (Фиг. 7). Чтобы оценить частоту развития событий в зародышевой линии, документировали окраску плодов на разных ветвях, а ткань околоплодника плода секвенировали с помощью иллюмина (Фиг. 8).
[00186] В этом анализе полностью желтые плоды могут содержать семена, которые являются событиями репарации в зародышевой линии посредством NHEJ или HR (Фиг. 6А). Более того, события, сопровождающиеся или не сопровождающиеся кроссинговером, должны давать +A,+A гомозиготное растение, поскольку матрицей для репарации является аллель M82 psy1+A (Фиг. 9A). В одном из растений F1 были обнаружены желтые плоды, которые при секвенировании illumina и секвенировании по Сэнгеру показали высокое содержание +A,+A (Фиг. 10). Потомство F2 этих растений выращивали и секвенировали по методу Сэнгера. Секвенирование выявило растения F2 с паттернами ОНП, соответствующими событиям HR в зародышевой линии (Фиг. 6B). Растения № 2 и № 7 выглядят как явные случаи отсутствия кроссинговера, оба из них имеют треки конверсии размером по меньшей мере 5 тыс. п.о. Растение № 11 похоже на случай кроссинговера (Фиг. 6B), однако, анализ фланкирующих маркеров (индел-мутаций и ОНП), расположенных в растении № 11 на расстоянии более 20 тыс. п.о. с обеих сторон сайта ДНР, не мог быть проведен из-за гибели растения по этой причине, поэтому этот случай был назван предполагаемым кроссинговером.
[00187] Для идентификации продуктов гомозиготной генной конверсии и для лучшей характеристики границ трека конверсии секвенировали растения F3 из потомства растения F2 № 7. Один из представителей потомства F3 растения F2 № 7 (показан внизу на Фиг. 6B) является гомозиготным продуктом репарации посредством генной конверсии с подтвержденным треком конверсии длиной 5-6 тыс. п.о.
Пример 4: Количественная оценка степени аллель-зависимой репарации
[00188] Цель: исследовать аллель-зависимую репарацию. Благодаря разработанной выше в примере 3 аллель-специфичной индукции ДНР, которая является отличительной для HR, удалось исследовать аллель-зависимую репарацию. Индукция ДНР ДНК в аллеле S. pimpinellifoliumLA1578 показала сигнатуру +A, схожую с аллелем M82 psy1+A, в поврежденном сайте во многих секвенированных плодах и листьях (Фиг. 7 и 8). Этот избыток репарации +A может быть обусловлен предпочтительным паттерном репарации посредством NHEJ или аллель-зависимой репарацией, опосредованной HR.
[00189] Методы: чтобы различить эти две возможности, было выращено несколько растений сорта М82, все они были потомками одного и того же 35S:Cas9 u6-26:Ps#2-огРНК. В этой популяции 22 растения первоначально были гомозиготными по аллелю M82 WT PSY1, тогда как 14 растений изначально были гетерозиготными M82-WT PSY1/ M82 psy1+A. Растения выращивали до возраста 4 недель, и с каждого из них собирали по 4 листа. Затем ДНК вокруг ДНР амплифицировали с помощью ПЦР и секвенировали продукты ПЦР с помощью платформы Illumina HiSeq 2500. Для каждого растения был рассчитан процент каждой индел-мутации из общего числа прочтений. Если мутация +A происходит независимо в каждой хромосоме, должно быть в два раза больше прочтений с новыми мутациями +A в WT (который имеет две потенциальные мишени), чем в гетерозиготе, где доступна только одна мишень (Фиг. 9A).
[00190] Для измерения ожидаемого аллель-независимого следа +A NHEJ использовали 22 гомозиготных растения WT и для получения значения встречаемости мутации +A на аллель рассчитывали процент прочтений +A, деленный на 2. Использовали следующее уравнение: Ожидаемый=(%(прочтений +A)T=4 недели (wt,wt))/2. Встречаемость новой мутации +A в аллеле WT, когда второй аллель содержит мутацию +A (в гетерозиготных растениях M82-WT PSY1/ M82 psy1+A), рассчитывали, взяв % прочтений +A в растениях M82-WT PSY1/ M82 psy1+A, и рассчитав 50% (исходный процент прочтений, происходящих из аллеля M82 psy1+A). Следующее уравнение использовали для наблюдаемой частоты мутации +A у гетерозиготных растений M82-WT PSY1/ M82 psy1+A: Наблюдаемый=% (прочтений +A)T=4 недели, (wt,+A) - 50%.
[00191] Результаты: при сравнении ожидаемого и наблюдаемого следа +A было обнаружено, что в гетерозиготной популяции частота новых мутаций +A значительно выше, чем ожидалось (p=0,009). Учитывая, что две популяции являются изогенными, это говорит о том, что репарация в сайте ДНР зависит от последовательности его гомологичного аллеля (Фиг. 9B). Частота аллель-независимого следа +A составляла 4% на аллель в M82-WT, в то время как частота следа +A в аллеле M82-WT PSY1 в гетерозиготе M82-WT PSY1/ M82 psy1+A составила 18% (Фиг. 9B). Это указывает на то, что 18-4=~14% событий репарации ДНР являются аллель-зависимыми (события репарации посредством гомологичной рекомбинации), а остальные происходят посредством NHEJ аллель-независимым образом.
[00192] Подведение итогов для примеров 1-4
[00193] Соматическая репарация ДНР
[00194] Более ранние исследования репарации посредством гомологичной HR, индуцированной ДНР, проводились в основном с помощью трансгенных анализов внутрихромосомной рекомбинации или неравного кроссинговера между хроматидами. Важно отметить, что способы, раскрытые и приведенные в качестве примера в настоящей заявке, были осуществлены в контексте эндогенного генома, где происхождение матрицы для репарации можно было отследить на гомологичной хромосоме. Результаты примеров 1-4 показывают, что направленные ДНР могут быть репарированы посредством соматической гомологичной рекомбинации с использованием гомологичной хромосомы в качестве матрицы. Это значительно отличается от генного таргетинга с использованием экзогенных матриц.
[00195] Кроме того, было продемонстрировано, что некоторые из этих событий репарации могут передаваться через зародышевую линию следующим поколениям. В одном из наборов скрещиваний было показано, что аллель WT может быть восстановлена путем внутригенной рекомбинации между двумя дефектными по psy1 родительскими аллелями (двухцветныйcc383 и желтая мякоть e3756) - событие, проявляющегося как красные пятна (пример 1, Фиг. 4A), и охарактеризованного с помощью анализа последовательности (пример 2, Фиг. 4C). В этом скрещивании не были получены полностью красные плоды, что соответствовало бы ранним событиям в зародышевой линии. Это может быть связано с геномным контекстом большой делеции в двухцветном аллеле или, в качестве альтернативы, «излеченный» рекомбинантный аллель WT подвергся второму раунду NHEJ в ходе развития (сайт-мишень не был разрушен в ходе HR), что привело бы к потере функции (желтый) аллеля посредством NHEJ. Учитывая высокую эффективность NHEJ, этот сценарий вполне правдоподобен. Кроме того, в анализе аллель-специфичной индукции ДНР в гибриде F1 S. pimpinellifolium X S. lycopersicum были обнаружены три случая HR-зависимой репарации, которые были переданы через зародышевую линию поколениям F2 и F3. Два случая соответствовали событиям, не сопровождавшимся кроссинговером, с треками конверсии размером 5-6 тыс. п.о. (пример 3, Фиг. 6B). Третий случай (растение F2 № 11) - это событие HR в зародышевой линии, которое может как сопровождаться, так и не сопровождаться кроссинговером, что не могло быть продемонстрировано из-за гибели растения. Наконец, для попытки количественной оценки частоты HR по сравнению с NHEJ была разработана огРНК для аллель-специфичной индукции ДНР на фоне генотипа S. lycopersicum. Эта экспериментальная схема позволила измерить избыток следов репарации, происходящих от гомологичного аллеля по сравнению с ожидаемым, что позволяет предположить, что из всех обнаруживаемых событий репарации ДНР 14% являются аллель-зависимыми, а остальные негомологичны. Величина четырнадцать процентов аллель-зависимой репарации HR была неожиданной, так как использованный способ неожиданно обеспечивал значительно больше HR, чем ожидалось.
[00196] Соматическая HR по сравнению с мейотической
[00197] Интересно сравнить HR-опосредованную репарацию в соматических и мейотических клетках. В целом мало что известно о рекомбинации между гомологами в соматических тканях, вероятно, из-за низкой частоты таких событий, отсутствия фенотипических маркеров и сложности поиска событий в зародышевой линии. Красные участки не были обнаружены в отсутствие ДНР, а присутствие внутригенных рекомбинантных молекул было нулевым или незначительным. Это согласуется с более ранними исследованиями на табаке, показавшими, что встречаемость соматической HR является очень низкой в отсутствие индукции ДНР для событий как взаимной, так и невзаимной HR. Низкие степени соматической HR между гомологичными хромосомами могут указывать на сдерживающие факторы, такие как отсутствие механизма HR, обнаруженного в мейозе, который контролирует спаривание гомологов, образование синаптонемного комплекса и т.д. Результаты, показывающие неожиданную относительно высокую степень репарации посредством HR, основанные как на тематических исследованиях, так и на количественных оценках, указывают на то, что ДНР являются основным сдерживающим фактором, неожиданно вызывая соматическую HR от 0% (в отсутствие разрывов) до ~14% на аллель (аллель-зависимая репарация, измеренная в примере 4, Фиг. 9B), и что ДНР-индуцированная HR между гомологами может происходить в отсутствие механизма мейотической HR.
[00198] Частота репарации ДНР посредством HR, отраженная в настоящей заявке (составляющая 14% на аллель), по-видимому, выше, чем описанная в ходе мейоза. Действительно, только небольшая доля мейотических разрывов (~3-5%) развивается в кроссинговер, и аналогичная доля подвергается репарации в отсутствие кроссинговера.
[00199] Аналогичным образом, представлены данные о встречаемости ДНР-опосредованной репарации HR, однако в большинстве анализов было невозможно различить механизмы репарации, сопровождающиеся и не сопровождающиеся кроссинговером. Анализ 3 событий в зародышевой линии на полиморфном фоне позволил провести это различие, но выборка (из 2 конверсий и одного предполагаемого кроссинговера) слишком мала, чтобы делать выводы.
[00200] Существенная разница по сравнению с более ранними сообщениями о мейотической рекомбинации заключается в том, что длина конверсионных треков в описанных в настоящей заявке не сопровождающихся кроссинговером соматических событиях составляла ~ 5 тыс. п.о. по сравнению со средним треком, составляющим 552 п.о., описанным для мейотических событий HR. Эти длинные треки конверсии могут отражать разницу между видами (томат по сравнению с арабидопсисом) или между мейотическими и соматическими клетками. Возможно также, что связывание Spo11 с концами ДНР более эффективно, чем у Cas9, для защиты концов от деградации и уменьшения длины трека конверсии. Наконец, это первое сообщение о направленной HR между эндогенными гомологичными хромосомами, в то время как ранее не сообщалось о направленной мейотической рекомбинации.
[00201] Соматический кроссинговер действительно встречается у растений и даже может достигать высоких уровней у некоторых мутантов, что указывает на то, что механизм кроссинговера между гомологами доступен в соматических тканях, и что направленный кроссинговер возможен. Интересно, что даже несмотря на то, что механизм мейотического кроссинговера был оптимизирован в ходе эволюции, направленная индукция данного ДНР во время мейоза должна была бы конкурировать с сотнями других естественных разрывов, служащих основой кроссинговера, и, вразрез с интуитивными представлениями, может оказаться менее эффективным для направленной индукции кроссинговера, чем соматическая HR.
[00202] Использование соматической HR для точной селекции
[00203] Результаты показывают, что специально разработанные нуклеазы, такие как CRISPR-Cas, могут быть использованы для точной перестановки хромосомных сегментов между гомологичными хромосомами в соматических клетках. Например, может быть возможен перенос гена устойчивости к заболеванию от дикого в культурное растение без длительного процесса обратного скрещивания, который не только занимает несколько поколений для достижения изогенных линий, но также перетягивает большие сегменты нежелательной ДНК, фланкирующие желаемый ген. Таким образом, применение способов, раскрытых в настоящей заявке, обеспечивает преимущество при использовании для получения специально разработанной рекомбинации в растениях с событием направленной HR (сопровождающейся кроссинговером или генной конверсией), где качество или признак добавляются или удаляются из полученного растения по сравнению с родительским растением, не подвергавшимся событию направленной HR. Кроме того, растение получают в более короткие сроки и со значительно меньшим размером популяции по сравнению со скринингом событий естественной рекомбинации, и событие рекомбинации в представляющем интерес растении осуществляется более точно, без добавления при этом нежелательной ДНК. В некоторых вариантах реализации событие HR, сопровождающейся кроссинговером или генной конверсией, вводит ген или регуляторный элемент, который не может быть с легкостью введен с помощью естественной HR из-за тесной связи между генами или генными элементами. Другими словами, способы, описанные и приведенные в качестве примеров в настоящей заявке, демонстрируют, что соматическая HR может быть использована для замены аллеля.
Пример 5: Направленный ДНР-индуцированный кроссинговер в соматических тканях в областях эухроматина и гетерохроматина у арабидопсиса
[00204] Цель: результаты вышеприведенных примеров 1-4 на томате показали высокий уровень репарации на основе HR при ДНР соматической ДНК. Эти результаты были ограничены исследованием одного локуса (PSY1), расположенного в субтеломерной области 3 хромосомы, обычно соответствующей областям эухроматина (открытого хроматина). Более того, наблюдаемая зависимая от гомологии репарация могла происходить либо путем кроссинговера, либо путем генной конверсии, поскольку экспериментальная схема не позволяла различать эти два механизма. Наконец, после изучения одного локуса у одного вида не было известно, насколько общим было явление и будет ли HR-индуцирующий эффект ДНР в соматических тканях наблюдаться как в эухроматических, так и в гетерохроматических (с плотно упакованным хроматином) областях. Целью в данном примере было изучение репарации на основе HR при ДНР соматической ДНК у других видов, а также в областях как эухроматина, так и гетерохроматина.
[00205] Известно, что области гетерохроматина подавляются при рекомбинации ДНК. У некоторых видов гетерохроматин составляет 80% генома (например, кукуруза и пшеница). Гетерохроматин преобладает вокруг центромеры и может содержать до 25% всех генов. Отсутствие рекомбинации в этих областях является препятствием для селекции растений, поскольку вредные гены не могут быть отделены от хороших генов.
[00206] В настоящей заявке описаны примеры направленной HR, сопровождающейся как кроссинговером, так и генной конверсией, в нескольких генетических локусах, включая локусы с модификациями хроматина, соответствующими эухроматину (низкое метилирование цитозина, низкая занятость нуклеосом, ди-или триметилирование гистона 3 по лизину 4 (H3K4me2/3)), и локусы с гетерохроматическими признаками (высокое метилирование цитозина, высокая занятость нуклеосом, H3K9me2/3, H3K27me3, как известно в данной области техники). Было показано, что эти области эухроматина и гетерохроматина соответствуют мейотическим горячим точкам или холодным точкам, соответственно) (Shilo et al., 2015 "DNA Crossover Motifs Associated with Epigenetic Modifications Delineate Open Chromatin Regions in Arabidopsis" Plant Cell, Sep;27(9):2427-36).
[00207] Методы: для исследования свойств репарации ДНР ДНК в областях с эухроматическими и гетерохроматическими признаками использовали тестовую кроссинговерную линию (Melamed-Bessudo et al. 2005 "A new seed-based assay for meiotic recombination in Arabidopsis thaliana" Plant J. 43(3):458-66), линию Arabidopsis экотипа Columbia с маркерами GFP и RFP, разделенными расстоянием 5 миллионов п.о. на 3 хромосоме, экспрессируемую под воздействием специфичного для семян промотора Napine и дающей семена, обладающие красной и зеленой флуоресценцией (родительские типы), или только красной или только зеленой флуоресценцией (кроссинговерные рекомбинантные типы).
[00208] На основании генетических мотивов и особенностей эпигенетики было выбрано 12 различных мишеней для индукции ДНР между маркерами GFP и RFP (таблица 10 представлена на Фиг. 11); четыре сайта в «холодных областях» (с гетерохроматическими признаками, типичными для холодных точек рекомбинации) и восемь мишеней в «горячих областях» (эухроматические признаки, типичные для горячих точек рекомбинации) (Фиг. 12A).
[00209] Сначала двенадцать тестерных рекомбинантных линий Columbia, которые включали 12 различных мишеней, трансформировали для экспрессии малых гидовых РНК, соответствующих мишеням ДНР (35Sx2: гигромицин, конструкция u6-26:гРНК). Кроме того, были сконструированы линии Columbia WT для экспрессии Cas9 под действием конститутивного промотора убиквитина, активного в соматических тканях (nos:nptII:nos Ubi:spCas9). Затем двенадцать линий, экспрессирующих гРНК, скрещивали с линиями Columbia WT, экспрессирующими Cas9 и F1, выращивали популяции растений, устойчивых как к гигромицину, так и к канамицину (т.е. содержащих как гРНК, так и Cas9), и собирали вместе с контрольными популяциями F1 без гРНК и F1 из тестерной линии экотипов Columbia и Landsberg (Фиг. 12B). В этом анализе разрыв ДНР индуцируется уже в соматических тканях, а результат измеряется в семенах. Следовательно, будут измеряться ранние события соматического кроссинговера, которые передаются на зародышевую линию. Для каждого растения F1 300-500 его семян F2 подсчитывали на наличие только красных (рекомбинантный тип), только зеленых (рекомбинантный тип), красных и зеленых (родительский тип) и нефлуоресцентных (родительский тип) семян. На основании этих подсчетов была рассчитана частота рекомбинации между маркерами GFP и RFP (в сМ).
[00210] Результаты: неожиданно результаты этого теста (Фиг. 12C) показали, что для всех подсчитанных горячих и холодных мишеней была обнаружена сопоставимая или повышенная степень кроссинговера по сравнению с контрольной популяцией F1 Columbia Ubi:cas9 x контрольная тестерная линия Columbia. В этом анализе оба родителя были на фоне генотипа Columbia, и только маркеры были полиморфными.
[00211] Чтобы охарактеризовать точки прерывания рекомбинации, растения F1 подвергали обратному скрещиванию с полиморфным экотипом WT Landsberg, выращивали популяции обратного скрещивания F2 (растения, устойчивые к гигромицину и канамицину), выделяли ДНК из соматических тканей этих растений и собирали их семена F3 (Фиг. 12B). С помощью PacBio фрагменты величиной 5 тыс. п.о., фланкирующие целевую область этих полученных в результате обратного скрещивания растений тестера Columbia x Landsberg, секвенировали для характеристики событий репарации ДНР ДНК с высоким разрешением (Фиг. 13). В целом, эти результаты подтверждают направленную ДНР-индуцированную рекомбинацию в сайтах репарации как эухроматина, так и гетерохроматина.
[00212] Тогда как в настоящей заявке проиллюстрированы и описаны некоторые признаки, раскрытые в настоящей заявке, специалистам в данной области должно быть понятно, что могут иметь место многие модификации, замещения, изменения и эквиваленты. Следовательно, следует понимать, что прилагаемая формула изобретения распространяется на все такие модификации и изменения, которые находятся в рамках сущности и объема генетически модифицированных растений и способов, раскрытых в настоящей заявке.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> YEDA RESEARCH AND DEVELOPMENT CO. LTD.
LEVY, Avraham A.
MELAMED-BESSUDO, Cathy
FILLER-HAYUT, Shdema
<120> НАПРАВЛЕННАЯ РЕКОМБИНАЦИЯ МЕЖДУ ГОМОЛОГИЧНЫМИ ХРОМОСОМАМИ И
ЕЕ ПРИМЕНЕНИЯ
<130> P-80701-PC
<150> 62/444,827
<151> 2017-01-11
<160> 80
<170> PatentIn версия 3.5
<210> 1
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ps#1огРНК F
<400> 1
attggaatgt ctgttgcctt gtta 24
<210> 2
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ps#1 огРНК R
<400> 2
aaactaacaa ggcaacagac att 23
<210> 3
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ps#2 огРНК F
<400> 3
attggagcgt atataatgct gctt 24
<210> 4
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ps#2 огРНК R
<400> 4
aaacaagcag cattatatac gct 23
<210> 5
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> a 3756 bic hr f
<400> 5
tcagctatgc taatgactcc cgag 24
<210> 6
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> a bic hr r
<400> 6
agtccattct ctattccgca tagtga 26
<210> 7
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> a 3756 r
<400> 7
tgacaaccga cctaaatcga tccg 24
<210> 8
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> b bic hr r
<400> 8
actgcattct ctattccgca tagtga 26
<210> 9
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> b 3756 r
<400> 9
gactactgaa ccgacctaaa tcgatccg 28
<210> 10
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pupd_y1_f
<400> 10
gcgccgtctc gctcgtactc gaacgagggt catc 34
<210> 11
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pupd_y1_r
<400> 11
gcgccgtctc gctcgagcgc cataattgga acactcatca a 41
<210> 12
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pupd_ps_f
<400> 12
gcgccgtctc gctcgggagc aaccttattt tgtactt 37
<210> 13
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pupd_ps_r
<400> 13
gcgccgtctc gctcgagtac aacatatcaa aataggtat 39
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp f
<400> 14
ggtttgcctg tctgtggtct 20
<210> 15
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp r1
<400> 15
agtcccatga aacttgtccc atttg 25
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp r2
<400> 16
tcagccatga aacttgtccc atttg 25
<210> 17
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp r3
<400> 17
actgccatga aacttgtccc atttg 25
<210> 18
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp r4
<400> 18
tgacccatga aacttgtccc atttg 25
<210> 19
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp r5
<400> 19
gactccatga aacttgtccc atttg 25
<210> 20
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 t1 htp r6
<400> 20
ctgaccatga aacttgtccc atttg 25
<210> 21
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> nhej_psy1_t2_r
<400> 21
gcctaaatac ggcacttcca 20
<210> 22
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> a_nhej_psy1_t2_f
<400> 22
agtcgtatcg cccctgaatc aaag 24
<210> 23
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> b_nhej_psy1_t2_f
<400> 23
tcaggtatcg cccctgaatc aaag 24
<210> 24
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> c_nhej_psy1_t2_f
<400> 24
tgacgtatcg cccctgaatc aaag 24
<210> 25
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> d_nhej_psy1_t2_f
<400> 25
actggtatcg cccctgaatc aaag 24
<210> 26
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> e_nhej_psy1_t2_f
<400> 26
gactgtatcg cccctgaatc aaag 24
<210> 27
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> f_nhej_psy1_t2_f
<400> 27
ctgagtatcg cccctgaatc aaag 24
<210> 28
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> nnn_a_nhej_psy1_t2_f
<220>
<221> misc_feature
<222> (1)..(6)
<223> n is a, c, g, or t
<400> 28
nnnnnnagtc gtatcgcccc tgaatcaaag 30
<210> 29
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> nnn_b_nhej_psy1_t2_f
<220>
<221> misc_feature
<222> (1)..(6)
<223> n is a, c, g, or t
<400> 29
nnnnnntcag gtatcgcccc tgaatcaaag 30
<210> 30
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> nnn_nhej_psy1_t2_r
<220>
<221> misc_feature
<222> (1)..(6)
<223> n is a, c, g, or t
<400> 30
nnnnnngcct aaatacggca cttcca 26
<210> 31
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1_1_F
<400> 31
tttgcagaag tcaagaaaca gg 22
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1_t4_ident_R
<400> 32
gatgtcatcg tccgttctcc 20
<210> 33
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1 psnps F
<400> 33
acggtatctt cccaccttca 20
<210> 34
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1 psnps R2
<400> 34
atagtgttaa ttgtgtaggc tcctt 25
<210> 35
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1 psnps F2
<400> 35
cgacgaggag taaggtttgc 20
<210> 36
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1 psnps R
<400> 36
tcagtccatt tcgttttcgt 20
<210> 37
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1_t123_f
<400> 37
atgttgcagc cattcagaga 20
<210> 38
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1_t123_r
<400> 38
tgatcatggc tcgtcactgt 20
<210> 39
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 term f
<400> 39
acaagtaccc tgggttggag 20
<210> 40
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1_term_r2
<400> 40
gcagtttttg taggaggcac a 21
<210> 41
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1_term_f2
<400> 41
tgtgcctcct acaaaaactg c 21
<210> 42
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> psy1 term r
<400> 42
tggattgaat cgaatttgga taa 23
<210> 43
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pimpixm82_co_f
<400> 43
ctttgcactt ggttactcag a 21
<210> 44
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pb_psy1_r
<400> 44
agcctacggc ccaaactatt 20
<210> 45
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 14036_f
<400> 45
tgctaatggg gcaggaaata 20
<210> 46
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 14036_r
<400> 46
tcaagtaacg taaaacacgt tgaaa 25
<210> 47
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5kb_up_t2_F
<400> 47
ttcatttgac gagcgatctg 20
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 5kb_up_t2_R
<400> 48
ttggctgctt tgaccttacc 20
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 40kb_down_t2_f
<400> 49
cattatccta agagtgcagt cagc 24
<210> 50
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 40kb_down_t2_r
<400> 50
tggtttctcg attacctctt tca 23
<210> 51
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 20kb_down_t2_f
<400> 51
tgacaccaat ccatccaatc 20
<210> 52
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 20kb_down_t2_r
<400> 52
ctgctacctg cactggctct 20
<210> 53
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 20kb_up_t2_f
<400> 53
tacgtccccg aagaaatcac 20
<210> 54
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 20kb_up_t2_r
<400> 54
cccttaggct ccgaagttgt 20
<210> 55
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 40kb_up_t2_f
<400> 55
cacataagag gacacgttta ttca 24
<210> 56
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 40kb_up_t2_r
<400> 56
gccacggaga aaatagttga 20
<210> 57
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> JF_F
<400> 57
tgcaaagtgc tacgtgtcct 20
<210> 58
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> PSY1_1_R
<400> 58
aatgtgaaca gcaacgcaaa 20
<210> 59
<211> 18063
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Плазмида 35S:Cas9
<400> 59
ggggacaagt ttgtacaaaa aagcaggcta tggatccccg ggatcatcta cttctgaaga 60
ctcagactca gactaagcag gtgacgaacg tcaccaatcc caattcgatc tacatcgata 120
agaagtactc tatcggactc gatatcggaa ctaactctgt gggatgggct gtgatcaccg 180
atgagtacaa ggtgccatct aagaagttca aggttctcgg aaacaccgat aggcactcta 240
tcaagaaaaa ccttatcggt gctctcctct tcgattctgg tgaaactgct gaggctacca 300
gactcaagag aaccgctaga agaaggtaca ccagaagaaa gaacaggatc tgctacctcc 360
aagagatctt ctctaacgag atggctaaag tggatgattc attcttccac aggctcgaag 420
agtcattcct cgtggaagaa gataagaagc acgagaggca ccctatcttc ggaaacatcg 480
ttgatgaggt ggcataccac gagaagtacc ctactatcta ccacctcaga aagaagctcg 540
ttgattctac tgataaggct gatctcaggc tcatctacct cgctctcgct cacatgatca 600
agttcagagg acacttcctc atcgagggtg atctcaaccc tgataactct gatgtggata 660
agttgttcat ccagctcgtg cagacctaca accagctttt cgaagagaac cctatcaacg 720
cttcaggtgt ggatgctaag gctatcctct ctgctaggct ctctaagtca agaaggcttg 780
agaacctcat tgctcagctc cctggtgaga agaagaacgg acttttcgga aacttgatcg 840
ctctctctct cggactcacc cctaacttca agtctaactt cgatctcgct gaggatgcaa 900
agctccagct ctcaaaggat acctacgatg atgatctcga taacctcctc gctcagatcg 960
gagatcagta cgctgatttg ttcctcgctg ctaagaacct ctctgatgct atcctcctca 1020
gtgatatcct cagagtgaac accgagatca ccaaggctcc actctcagct tctatgatca 1080
agagatacga tgagcaccac caggatctca cacttctcaa ggctcttgtt agacagcagc 1140
tcccagagaa gtacaaagag attttcttcg atcagtctaa gaacggatac gctggttaca 1200
tcgatggtgg tgcatctcaa gaagagttct acaagttcat caagcctatc ctcgagaaga 1260
tggatggaac cgaggaactc ctcgtgaagc tcaatagaga ggatcttctc agaaagcaga 1320
ggaccttcga taacggatct atccctcatc agatccacct cggagagttg cacgctatcc 1380
ttagaaggca agaggatttc tacccattcc tcaaggataa cagggaaaag attgagaaga 1440
ttctcacctt cagaatccct tactacgtgg gacctctcgc tagaggaaac tcaagattcg 1500
cttggatgac cagaaagtct gaggaaacca tcaccccttg gaacttcgaa gaggtggtgg 1560
ataagggtgc tagtgctcag tctttcatcg agaggatgac caacttcgat aagaaccttc 1620
caaacgagaa ggtgctccct aagcactctt tgctctacga gtacttcacc gtgtacaacg 1680
agttgaccaa ggttaagtac gtgaccgagg gaatgaggaa gcctgctttt ttgtcaggtg 1740
agcaaaagaa ggctatcgtt gatctcttgt tcaagaccaa cagaaaggtg accgtgaagc 1800
agctcaaaga ggattacttc aagaaaatcg agtgcttcga ttcagttgag atttctggtg 1860
ttgaggatag gttcaacgca tctctcggaa cctaccacga tctcctcaag atcattaagg 1920
ataaggattt cttggataac gaggaaaacg aggatatctt ggaggatatc gttcttaccc 1980
tcaccctctt tgaagataga gagatgattg aagaaaggct caagacctac gctcatctct 2040
tcgatgataa ggtgatgaag cagttgaaga gaagaagata cactggttgg ggaaggctct 2100
caagaaagct cattaacgga atcagggata agcagtctgg aaagacaatc cttgatttcc 2160
tcaagtctga tggattcgct aacagaaact tcatgcagct catccacgat gattctctca 2220
cctttaaaga ggatatccag aaggctcagg tttcaggaca gggtgatagt ctccatgagc 2280
atatcgctaa cctcgctgga tctcctgcaa tcaagaaggg aatcctccag actgtgaagg 2340
ttgtggatga gttggtgaag gtgatgggaa ggcataagcc tgagaacatc gtgatcgaaa 2400
tggctagaga gaaccagacc actcagaagg gacagaagaa ctctagggaa aggatgaaga 2460
ggatcgagga aggtatcaaa gagcttggat ctcagatcct caaagagcac cctgttgaga 2520
acactcagct ccagaatgag aagctctacc tctactacct ccagaacgga agggatatgt 2580
atgtggatca agagttggat atcaacaggc tctctgatta cgatgttgat catatcgtgc 2640
cacagtcatt cttgaaggat gattctatcg ataacaaggt gctcaccagg tctgataaga 2700
acaggggtaa gagtgataac gtgccaagtg aagaggttgt gaagaaaatg aagaactatt 2760
ggaggcagct cctcaacgct aagctcatca ctcagagaaa gttcgataac ttgactaagg 2820
ctgagagggg aggactctct gaattggata aggcaggatt catcaagagg cagcttgtgg 2880
aaaccaggca gatcactaag cacgttgcac agatcctcga ttctaggatg aacaccaagt 2940
acgatgagaa cgataagttg atcagggaag tgaaggttat caccctcaag tcaaagctcg 3000
tgtctgattt cagaaaggat ttccaattct acaaggtgag ggaaatcaac aactaccacc 3060
acgctcacga tgcttacctt aacgctgttg ttggaaccgc tctcatcaag aagtatccta 3120
agctcgagtc agagttcgtg tacggtgatt acaaggtgta cgatgtgagg aagatgatcg 3180
ctaagtctga gcaagagatc ggaaaggcta ccgctaagta tttcttctac tctaacatca 3240
tgaatttctt caagaccgag attaccctcg ctaacggtga gatcagaaag aggccactca 3300
tcgagacaaa cggtgaaaca ggtgagatcg tgtgggataa gggaagggat ttcgctaccg 3360
ttagaaaggt gctctctatg ccacaggtga acatcgttaa gaaaaccgag gtgcagaccg 3420
gtggattctc taaagagtct atcctcccta agaggaactc tgataagctc attgctagga 3480
agaaggattg ggaccctaag aaatacggtg gtttcgattc tcctaccgtg gcttactctg 3540
ttctcgttgt ggctaaggtt gagaagggaa agagtaagaa gctcaagtct gttaaggaac 3600
ttctcggaat cactatcatg gaaaggtcat ctttcgagaa gaacccaatc gatttcctcg 3660
aggctaaggg atacaaagag gttaagaagg atctcatcat caagctccca aagtactcac 3720
tcttcgaact cgagaacggt agaaagagga tgctcgcttc tgctggtgag cttcaaaagg 3780
gaaacgagct tgctctccca tctaagtacg ttaactttct ttacctcgct tctcactacg 3840
agaagttgaa gggatctcca gaagataacg agcagaagca acttttcgtt gagcagcaca 3900
agcactactt ggatgagatc atcgagcaga tctctgagtt ctctaaaagg gtgatcctcg 3960
ctgatgcaaa cctcgataag gtgttgtctg cttacaacaa gcacagagat aagcctatca 4020
gggaacaggc agagaacatc atccatctct tcacccttac caacctcggt gctcctgctg 4080
ctttcaagta cttcgataca accatcgata ggaagagata cacctctacc aaagaagtgc 4140
tcgatgctac cctcatccat cagtctatca ctggactcta cgagactagg atcgatctct 4200
cacagctcgg tggtgattca agggctgatc ctaagaagaa gaggaaggtt tgaacccagc 4260
tttcttgtac aaagtggggg ttcgaaatcg ataagcttgg atcctctaga gtcctgcttt 4320
aatgagatat gcgagacgcc tatgatcgca tgatatttgc tttcaattct gttgtgcacg 4380
ttgtaaaaaa cctgagcatg tgtagctcag atccttaccg ccggtttcgg ttcattctaa 4440
tgaatatatc acccgttact atcgtatttt tatgaataat attctccgtt caatttactg 4500
attgtaccct actacttata tgtacaatat taaaatgaaa acaatatatt gtgctgaata 4560
ggtttatagc gacatctatg atagagcgcc acaataacaa acaattgcgt tttattatta 4620
caaatccaat tttaaaaaaa gcggcagaac cggtcaaacc taaaagactg attacataaa 4680
tcttattcaa atttcaaaag gccccagggg ctagtatcta cgacacaccg agcggcgaac 4740
taataacgtt cactgaaggg aactccggtt ccccgccggc gcgcatgggt gagattcctt 4800
gaagttgagt attggccgtc cgctctaccg aaagttacgg gcaccattca acccggtcca 4860
gcacggcggc cgggtaaccg acttgctgcc ccgagaatta tgcagcattt ttttggtgta 4920
tgtgggcccc aaatgaagtg caggtcaaac cttgacagtg acgacaaatc gttgggcggg 4980
tccagggcga attttgcgac aacatgtcga ggctcagcag gacctgcagg catgcaagct 5040
agcttactag tgatgcatat tctatagtgt cacctaaatc tgcggccgca ctagtgatat 5100
cccgcggcca tggcggccgg gagcatgcga cgtcgggccc aattcgccct atagtgagtc 5160
gtattacaat tcactggccg tcgttttaca acgtcgtgac tgggaaaacc ctggcgttac 5220
ccaacttaat cgccttgcag cacatccccc tttcgccagc tggcgtaata gcgaagaggc 5280
ccgcaccgat cgcccttccc aacagttgcg cagcctgaat ggcgaatgga aattgtaaac 5340
gttaatgggt ttctggagtt taatgagcta agcacatacg tcagaaacca ttattgcgcg 5400
ttcaaaagtc gcctaaggtc actatcagct agcaaatatt tcttgtcaaa aatgctccac 5460
tgacgttcca taaattcccc tcggtatcca attagagtct catattcact ctcaatccaa 5520
ataatctgca atggcaatta ccttatccgc aacttcttta cctatttccg cccggatccg 5580
ggcaggttct ccggccgctt gggtggagag gctattcggc tatgactggg cacaacagac 5640
aatcggctgc tctgatgccg ccgtgttccg gctgtcagcg caggggcgcc cggttctttt 5700
tgtcaagacc gacctgtccg gtgccctgaa tgaactgcag gacgaggcag cgcggctatc 5760
gtggctggcc acgacgggcg ttccttgcgc agctgtgctc gacgttgtca ctgaagcggg 5820
aagggactgg ctgctattgg gcgaagtgcc ggggcaggat ctcctgtcat ctcaccttgc 5880
tcctgccgag aaagtatcca tcatggctga tgcaatgcgg cggctgcata cgcttgatcc 5940
ggctacctgc ccattcgacc accaagcgaa acatcgcatc gagcgagcac gtactcggat 6000
ggaagccggt cttgtcgatc aggatgatct ggacgaagag catcaggggc tcgcgccagc 6060
cgaactgttc gccaggctca aggcgcgcat gcccgacggc gaggatctcg tcgtgaccca 6120
tggcgatgcc tgcttgccga atatcatggt ggaaaatggc cgcttttctg gattcatcga 6180
ctgtggccgg ctgggtgtgg cggaccgcta tcaggacata gcgttggcta cccgtgatat 6240
tgctgaagag cttggcggcg aatgggctga ccgcttcctc gtgctttacg gtatcgccgc 6300
tcccgattcg cagcgcatcg ccttctatcg ccttcttgac gagttcttct gagcgggact 6360
ctggggttcg aaatgaccga ccaagcgacg cccaacctgc catcacgaga tttcgattcc 6420
accgccgcct tctatgaaag gttgggcttc ggaatcgttt tccgggacgc cggctggatg 6480
atcctccagc gcggggatct catgctggag ttcttcgccc accccgatcc aacacttacg 6540
tttgcaacgt ccaagagcaa atagaccacg aacgccggaa ggttgccgca gcgtgtggat 6600
tgcgtctcaa ttctctcttg caggaatgca atgatgaata tgatactgac tatgaaactt 6660
tgagggaata ctgcctagca ccgtcacctc ataacgtgca tcatgcatgc cctgacaaca 6720
tggaacatcg ctatttttct gaagaattat gctcgttgga ggatgtcgcg gcaattgcag 6780
ctattgccaa catcgaacta cccctcacgc atgcattcat caatattatt catgcgggga 6840
aaggcaagat taatccaact ggcaaatcat ccagcgtgat tggtaacttc agttccagcg 6900
acttgattcg ttttggtgct acccacgttt tcaataagga cgagatggtg gagtaaagaa 6960
ggagtgcgtc gaagcagatc gttcaaacat ttggcaataa agtttcttaa gattgaatcc 7020
tgttgccggt cttgcgatga ttatcatata atttctgttg aattacgtta agcatgtaat 7080
aattaacatg taatgcatga cgttatttat gagatgggtt tttatgatta gagtcccgca 7140
attatacatt taatacgcga tagaaaacaa aatatagcgc gcaaactagg ataaattatc 7200
gcgcgcggtg tcatctatgt tactagatcg aattaattca gtacattaaa aacgtccgca 7260
atgtgttatt aagttgtcta agcgtcaatt tgtttacacc acaatatatc ctgccaccag 7320
ccagccaaca gctccccgac cggcagctcg gcacaaaatc accactcgat acaggcagcc 7380
catcagtccg ggacggcgtc agcgggagag ccgttgtaag gcggcagact ttgctcatgt 7440
taccgatgct attcggaaga acggcaacta agctgccggg tttgaaacac ggatgatctc 7500
gcggagggta gcatgttgat tgtaacgatg acagagcgtt gctgcctgtg atcaaatatc 7560
atctccctcg cagagatccg aattatcagc cttcttattc atttctcgct taaccgtgac 7620
aggctgtcga tcttgagaac tatgccgaca taataggaaa tcgctggata aagccgctga 7680
ggaagctgag tggcgctatt tctttagaag tgaacgttga cgatgtcgac ggatcttttc 7740
cgctgcataa ccctgcttcg gggtcattat agcgattttt tcggtatatc catccttttt 7800
cgcacgatat acaggatttt gccaaagggt tcgtgtagac tttccttggt gtatccaacg 7860
gcgtcagccg ggcaggatag gtgaagtagg cccacccgcg agcgggtgtt ccttcttcac 7920
tgtcccttat tcgcacctgg cggtgctcaa cgggaatcct gctctgcgag gctggccggc 7980
taccgccggc gtaacagatg agggcaagcg gatggctgat gaaaccaagc caaccagggg 8040
tgatgctgcc aacttactga tttagtgtat gatggtgttt ttgaggtgct ccagtggctt 8100
ctgtttctat cagctgtccc tcctgttcag ctactgacgg ggtggtgcgt aacggcaaaa 8160
gcaccgccgg acatcagcgc tatctctgct ctcactgccg taaaacatgg caactgcagt 8220
tcacttacac cgcttctcaa cccggtacgc accagaaaat cattgatatg gccatgaatg 8280
gcgttggatg ccgggcaaca gcccgcatta tgggcgttgg cctcaacacg attttacgtc 8340
acttaaaaaa ctcaggccgc agtcggtaac ctcgcgcata cagccgggca gtgacgtcat 8400
cgtctgcgcg gaaatggacg aacagtgggg ctatgtcggg gctaaatcgc gccagcgctg 8460
gctgttttac gcgtatgaca gtctccggaa gacggttgtt gcgcacgtat tcggtgaacg 8520
cactatggcg acgctggggc gtcttatgag cctgctgtca ccctttgacg tggtgatatg 8580
gatgacggat ggctggccgc tgtatgaatc ccgcctgaag ggaaagctgc acgtaatcag 8640
caagcgatat acgcagcgaa ttgagcggca taacctgaat ctgaggcagc acctggcacg 8700
gctgggacgg aagtcgctgt cgttctcaaa atcggtggag ctgcatgaca aagtcatcgg 8760
gcattatctg aacataaaac actatcaata agttggagtc attacccaac caggaagggc 8820
agcccaccta tcaaggtgta ctgccttcca gacgaacgaa gagcgattga ggaaaaggcg 8880
gcggcggccg gcatgagcct gtcggcctac ctgctggccg tcggccaggg ctacaaaatc 8940
acgggcgtcg tggactatga gcacgtccgc gagctggccc gcatcaatgg cgacctgggc 9000
cgcctgggcg gcctgctgaa actctggctc accgacgacc cgcgcacggc gcggttcggt 9060
gatgccacga tcctcgccct gctggcgaag atcgaagaga agcaggacga gcttggcaag 9120
gtcatgatgg gcgtggtccg cccgagggca gagccatgac ttttttagcc gctaaaacgg 9180
ccggggggtg cgcgtgattg ccaagcacgt ccccatgcgc tccatcaaga agagcgactt 9240
cgcggagctg gtattcgtgc agggcaagat tcggaatacc aagtacgaga aggacggcca 9300
gacggtctac gggaccgact tcattgccga taaggtggat tatctggaca ccaaggcacc 9360
aggcgggtca aatcaggaat aagggcacat tgccccggcg tgagtcgggg caatcccgca 9420
aggagggtga atgaatcgga cgtttgaccg gaaggcatac aggcaagaac tgatcgacgc 9480
ggggttttcc gccgaggatg ccgaaaccat cgcaagccgc accgtcatgc gtgcgccccg 9540
cgaaaccttc cagtccgtcg gctcgatggt ccagcaagct acggccaaga tcgagcgcga 9600
cagcgtgcaa ctggctcccc ctgccctgcc cgcgccatcg gccgccgtgg agcgttcgcg 9660
tcgtctcgaa caggaggcgg caggtttggc gaagtcgatg accatcgaca cgcgaggaac 9720
tatgacgacc aagaagcgaa aaaccgccgg cgaggacctg gcaaaacagg tcagcgaggc 9780
caagcaggcc gcgttgctga aacacacgaa gcagcagatc aaggaaatgc agctttcctt 9840
gttcgatatt gcgccgtggc cggacacgat gcgagcgatg ccaaacgaca cggcccgctc 9900
tgccctgttc accacgcgca acaagaaaat cccgcgcgag gcgctgcaaa acaaggtcat 9960
tttccacgtc aacaaggacg tgaagatcac ctacaccggc gtcgagctgc gggccgacga 10020
tgacgaactg gtgtggcagc aggtgttgga gtacgcgaag cgcaccccta tcggcgagcc 10080
gatcaccttc acgttctacg agctttgcca ggacctgggc tggtcgatca atggccggta 10140
ttacacgaag gccgaggaat gcctgtcgcg cctacaggcg acggcgatgg gcttcacgtc 10200
cgaccgcgtt gggcacctgg aatcggtgtc gctgctgcac cgcttccgcg tcctggaccg 10260
tggcaagaaa acgtcccgtt gccaggtcct gatcgacgag gaaatcgtcg tgctgtttgc 10320
tggcgaccac tacacgaaat tcatatggga gaagtaccgc aagctgtcgc cgacggcccg 10380
acggatgttc gactatttca gctcgcaccg ggagccgtac ccgctcaagc tggaaacctt 10440
ccgcctcatg tgcggatcgg attccacccg cgtgaagaag tggcgcgagc aggtcggcga 10500
agcctgcgaa gagttgcgag gcagcggcct ggtggaacac gcctgggtca atgatgacct 10560
ggtgcattgc aaacgctagg gccttgtggg gtcagttccg gctgggggtt cagcagccag 10620
cgctttactg gcatttcagg aacaagcggg cactgctcga cgcacttgct tcgctcagta 10680
tcgctcggga cgcacggcgc gctctacgaa ctgccgataa acagaggatt aaaattgaca 10740
attgtgatta aggctcagat tcgacggctt ggagcggccg acgtgcagga tttccgcgag 10800
atccgattgt cggccctgaa gaaagctcca gagatgttcg ggtccgttta cgagcacgag 10860
gagaaaaagc ccatggaggc gttcgctgaa cggttgcgag atgccgtggc attcggcgcc 10920
tacatcgacg gcgagatcat tgggctgtcg gtcttcaaac aggaggacgg ccccaaggac 10980
gctcacaagg cgcatctgtc cggcgttttc gtggagcccg aacagcgagg ccgaggggtc 11040
gccggtatgc tgctgcgggc gttgccggcg ggtttattgc tcgtgatgat cgtccgacag 11100
attccaacgg gaatctggtg gatgcgcatc ttcatcctcg gcgcacttaa tatttcgcta 11160
ttctggagct tgttgtttat ttcggtctac cgcctgccgg gcggggtcgc ggcgacggta 11220
ggcgctgtgc agccgctgat ggtcgtgttc atctctgccg ctctgctagg tagcccgata 11280
cgattgatgg cggtcctggg ggctatttgc ggaactgcgg gcgtggcgct gttggtgttg 11340
acaccaaacg cagcgctaga tcctgtcggc gtcgcagcgg gcctggcggg ggcggtttcc 11400
atggcgttcg gaaccgtgct gacccgcaag tggcaacctc ccgtgcctct gctcaccttt 11460
accgcctggc aactggcggc cggaggactt ctgctcgttc cagtagcttt agtgtttgat 11520
ccgccaatcc cgatgcctac aggaaccaat gttctcggcc tggcgtggct cggcctgatc 11580
ggagcgggtt taacctactt cctttggttc cgggggatct cgcgactcga acctacagtt 11640
gtttccttac tgggctttct cagccgggat ggcgctaaga agctattgcc gccgatcttc 11700
atatgcggtg tgaaataccg cacagatgcg taaggagaaa ataccgcatc aggcgctctt 11760
ccgcttcctc gctcactgac tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag 11820
ctcactcaaa ggcggtaata cggttatcca cagaatcagg ggataacgca ggaaagaaca 11880
tgtgagcaaa aggccagcaa aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt 11940
tccataggct ccgcccccct gacgagcatc acaaaaatcg acgctcaagt cagaggtggc 12000
gaaacccgac aggactataa agataccagg cgtttccccc tggaagctcc ctcgtgcgct 12060
ctcctgttcc gaccctgccg cttaccggat acctgtccgc ctttctccct tcgggaagcg 12120
tggcgctttc tcaatgctca cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca 12180
agctgggctg tgtgcacgaa ccccccgttc agcccgaccg ctgcgcctta tccggtaact 12240
atcgtcttga gtccaacccg gtaagacacg acttatcgcc actggcagca gccactggta 12300
acaggattag cagagcgagg tatgtaggcg gtgctacaga gttcttgaag tggtggccta 12360
actacggcta cactagaagg acagtatttg gtatctgcgc tctgctgaag ccagttacct 12420
tcggaaaaag agttggtagc tcttgatccg gcaaacaaac caccgctggt agcggtggtt 12480
tttttgtttg caagcagcag attacgcgca gaaaaaaagg atatcaagaa gatcctttga 12540
tcttttctac ggggtctgac gctcagtgga acgaaaactc acgttaaggg attttggtca 12600
tgagattatc aaaaaggatc ttcacctaga tccttttaaa ttaaaaatga agttttaaat 12660
caatctaaag tatatatgag taaacttggt ctgacagtta ccaatgctta atcagtgagg 12720
cacctatctc agcgatctgt ctatttcgtt catccatagt tgcctgactc cccgtcgtgt 12780
agataactac gatacgggag ggcttaccat ctggccccag tgctgcaatg ataccgcgag 12840
acccacgctc accggctcca gatttatcag caataaacca gccagccgga agggccgagc 12900
gcagaagtgg tcctgcaact ttatccgcct ccatccagtc tattaaacaa gtggcagcaa 12960
cggattcgca aacctgtcac gccttttgtg ccaaaagccg cgccaggttt gcgatccgct 13020
gtgccaggcg ttaggcgtca tatgaagatt tcggtgatcc ctgagcaggt ggcggaaaca 13080
ttggatgctg agaaccattt cattgttcgt gaagtgttcg atgtgcacct atccgaccaa 13140
ggctttgaac tatctaccag aagtgtgagc ccctaccgga aggattacat ctcggatgat 13200
gactctgatg aagactctgc ttgctatggc gcattcatcg accaagagct tgtcgggaag 13260
attgaactca actcaacatg gaacgatcta gcctctatcg aacacattgt tgtgtcgcac 13320
acgcaccgag gcaaaggagt cgcgcacagt ctcatcgaat ttgcgaaaaa gtgggcacta 13380
agcagacagc tccttggcat acgattagag acacaaacga acaatgtacc tgcctgcaat 13440
ttgtacgcaa aatgtggctt tactctcggc ggcattgacc tgttcacgta taaaactaga 13500
cctcaagtct cgaacgaaac agcgatgtac tggtactggt tctcgggagc acaggatgac 13560
gcctaacaat tcattcaagc cgacaccgct tcgcggcgcg gcttaattca ggagttaaac 13620
atcatgaggg aagcggtgat cgccgaagta tcgactcaac tatcagaggt agttggcgtc 13680
atcgagcgcc atctcgaacc gacgttgctg gccgtacatt tgtacggctc cgcagtggat 13740
ggcggcctga agccacacag tgatattgat ttgctggtta cggtgaccgt aaggcttgat 13800
gaaacaacgc ggcgagcttt gatcaacgac cttttggaaa cttcggcttc ccctggagag 13860
agcgagattc tccgcgctgt agaagtcacc attgttgtgc acgacgacat cattccgtgg 13920
cgttatccag ctaagcgcga actgcaattt ggagaatggc agcgcaatga cattcttgca 13980
ggtatcttcg agccagccac gatcgacatt gatctggcta tcttgctgac aaaagcaaga 14040
gaacatagcg ttgccttggt aggtccagcg gcggaggaac tctttgatcc ggttcctgaa 14100
caggatctat ttgaggcgct aaatgaaacc ttaacgctat ggaactcgcc gcccgactgg 14160
gctggcgatg agcgaaatgt agtgcttacg ttgtcccgca tttggtacag cgcagtaacc 14220
ggcaaaatcg cgccgaagga tgtcgctgcc gactgggcaa tggagcgcct gccggcccag 14280
tatcagcccg tcatacttga agctaggcag gcttatcttg gacaagaaga tcgcttggcc 14340
tcgcgcgcag atcagttgga agaatttgtt cactacgtga aaggcgagat caccaaggta 14400
gtcggcaaat aatgtctaac aattcgttca agccgacgcc gcttcgcggc gcggcttaac 14460
tcaagcgtta gagagctggg gaagactatg cgcgatctgt tgaaggtggt tctaagcctc 14520
gtacttgcga tggcatcggg gcaggcactt gctgacctgc caattgtttt agtggatgaa 14580
gctcgtcttc cctatgacta ctccccatcc aactacgaca tttctccaag caactacgac 14640
aactccataa gcaattacga caatagtcca tcaaattacg acaactctga gagcaactac 14700
gataatagtt catccaatta cgacaatagt cgcaacggaa atcgtaggct tatatatagc 14760
gcaaatgggt ctcgcacttt cgccggctac tacgtcattg ccaacaatgg gacaacgaac 14820
ttcttttcca catctggcaa aaggatgttc tacaccccaa aaggggggcg cggcgtctat 14880
ggcggcaaag atgggagctt ctgcggggca ttggtcgtca taaatggcca attttcgctt 14940
gccctgacag ataacggcct gaagatcatg tatctaagca actagcctgc tctctaataa 15000
aatgttagga gcttggctgc catttttggg gtgaggccgt tcgcggccga ggggcgcagc 15060
ccctgggggg atgggaggcc cgcgttagcg ggccgggagg gttcgagaag ggggggcacc 15120
ccccttcggc gtgcgcggtc acgcgccagg gcgcagccct ggttaaaaac aaggtttata 15180
aatattggtt taaaagcagg ttaaaagaca ggttagcggt ggccgaaaaa cgggcggaaa 15240
cccttgcaaa tgctggattt tctgcctgtg gacagcccct caaatgtcaa taggtgcgcc 15300
cctcatctgt cagcactctg cccctcaagt gtcaaggatc gcgcccctca tctgtcagta 15360
gtcgcgcccc tcaagtgtca ataccgcagg gcacttatcc ccaggcttgt ccacatcatc 15420
tgtgggaaac tcgcgtaaaa tcaggcgttt tcgccgattt gcgaggctgg ccagctccac 15480
gtcgccggcc gaaatcgagc ctgcccctca tctgtcaacg ccgcgccggg tgagtcggcc 15540
cctcaagtgt caacgtccgc ccctcatctg tcagtgaggg ccaagttttc cgcgaggtat 15600
ccacaacgcc ggcggccggc cgcggtgtct cgcacacggc ttcgacggcg tttctggcgc 15660
gtttgcaggg ccatagacgg ccgccagccc agcggcgagg gcaaccagcc cggtgagcgt 15720
cggaaagggt cgacatcttg ctgcgttcgg atattttcgt ggagttcccg ccacagaccc 15780
ggattgaagg cgagatccag caactcgcgc cagatcatcc tgtgacggaa ctttggcgcg 15840
tgatgactgg ccaggacgtc ggccgaaaga gcgacaagca gatcacgatt ttcgacagcg 15900
tcggatttgc gatcgaggat ttttcggcgc tgcgctacgt ccgcgaccgc gttgagggat 15960
caagccacag cagcccactc gaccttctag ccgacccaga cgagccaagg gatctttttg 16020
gaatgctgct ccgtcgtcag gctttccgac gtttgggtgg ttgaacagaa gtcattatcg 16080
tacggaatgc cagcactccc gaggggaacc ctgtggttgg catgcacata caaatggacg 16140
aacggataaa ccttttcacg cccttttaaa tatccgttat tctaataaac gctcttttct 16200
cttaggttta cccgccaata tatcctgtca aacactgata gtttaaactg aaggcgggaa 16260
acgacaatct gatcatgagc ggagaattaa gggagtcacg ttatgacccc cgccgatgac 16320
gcgggacaag ccgttttacg tttggaactg acagaaccgc aacgattgaa ggagccactc 16380
agccccaata cgcaaaccgc ctctccccgc gcgttggccg attcattaat gcagctggca 16440
cgacaggttt cccgactgga aagcgggcag tgagcgcaac gcaattaatg tgagttagct 16500
cactcattag gcaccccagg ctttacactt tatgcttccg gctcgtatgt tgtgtggaat 16560
tgtgagcgga taacaatttc acacaggaaa cagctatgac catgattacg ccaagctatt 16620
taggtgacac tatagaatac tcaagctatg catccaacgc gttgggagct ctcccatatc 16680
gacctgcagg cggccgctcg acgaattaat tccaatccca caaaaatctg agcttaacag 16740
cacagttgct cctctcagag cagaatcggg tattcaacac cctcatatca actactacgt 16800
tgtgtataac ggtccacatg ccggtatata cgatgactgg ggttgtacaa aggcggcaac 16860
aaacggcgtt cccggagttg cacacaagaa atttgccact attacagagg caagagcagc 16920
agctgacgcg tacacaacaa gtcagcaaac agacaggttg aacttcatcc ccaaaggaga 16980
agctcaactc aagcccaaga gctttgctaa ggccctaaca agcccaccaa agcaaaaagc 17040
ccactggctc acgctaggaa ccaaaaggcc cagcagtgat ccagccccaa aagagatctc 17100
ctttgccccg gagattacaa tggacgattt cctctatctt tacgatctag gaaggaagtt 17160
cgaaggtgaa ggtgacgaca ctatgttcac cactgataat gagaaggtta gcctcttcaa 17220
tttcagaaag aatgctgacc cacagatggt tagagaggcc tacgcagcag gtctcatcaa 17280
gacgatctac ccgagtaaca atctccagga gatcaaatac cttcccaaga aggttaaaga 17340
tgcagtcaaa agattcagga ctaattgcat caagaacaca gagaaagaca tatttctcaa 17400
gatcagaagt actattccag tatggacgat tcaaggcttg cttcataaac caaggcaagt 17460
aatagagatt ggagtctcta aaaaggtagt tcctactgaa tctaaggcca tgcatggagt 17520
ctaagattca aatcgaggat ctaacagaac tcgccgtgaa gactggcgaa cagttcatac 17580
agagtctttt acgactcaat gacaagaaga aaatcttcgt caacatggtg gagcacgaca 17640
ctctggtcta ctccaaaaat gtcaaagata cagtctcaga agaccaaagg gctattgaga 17700
cttttcaaca aaggataatt tcgggaaacc tcctcggatt ccattgccca gctatctgtc 17760
acttcatcga aaggacagta gaaaaggaag gtggctccta caaatgccat cattgcgata 17820
aaggaaaggc tatcattcaa gatctctctg ccgacagtgg tcccaaagat ggacccccac 17880
ccacgaggag catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt 17940
gatgtgacat ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc 18000
cttcctctat ataaggaagt tcatttcatt tggagaggac acgctcgagg aattcggtac 18060
ccc 18063
<210> 60
<211> 12150
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ps#1 гРНК промотора U6-26
<400> 60
acctagggag ctctcccata tcgacctgca ggcggccgca ctagtgatat cccgcggcca 60
tggcggccgg gagcatgcga cgtcgggccc aattcgccct atagtgagtc gtattacaat 120
tcactggccg tcgttttaca acgtcgtgac tgggaaaacc ctggcgttac ccaacttaat 180
cgccttgcag cacatccccc tttcgccagc tggcgtaata gcgaagaggc ccgcaccgat 240
cgcccttccc aacagttgcg cagcctgaat ggcgaatgga aattgtaaac gttaatgggt 300
ttctggagtt taatgagcta agcacatacg tcagaaacca ttattgcgcg ttcaaaagtc 360
gcctaaggtc actatcagct agcaaatatt tcttgtcaaa aatgctccac tgacgttcca 420
taaattcccc tcggtatcca attagagtct catattcact ctcaatccaa ataatctgca 480
atggcaatta ccttatccgc aacttcttta cctatttccg cccggatccg ggcaggttct 540
ccggccgctt gggtggagag gctattcggc tatgactggg cacaacagac aatcggctgc 600
tctgatgccg ccgtgttccg gctgtcagcg caggggcgcc cggttctttt tgtcaagacc 660
gacctgtccg gtgccctgaa tgaactgcag gacgaggcag cgcggctatc gtggctggcc 720
acgacgggcg ttccttgcgc agctgtgctc gacgttgtca ctgaagcggg aagggactgg 780
ctgctattgg gcgaagtgcc ggggcaggat ctcctgtcat ctcaccttgc tcctgccgag 840
aaagtatcca tcatggctga tgcaatgcgg cggctgcata cgcttgatcc ggctacctgc 900
ccattcgacc accaagcgaa acatcgcatc gagcgagcac gtactcggat ggaagccggt 960
cttgtcgatc aggatgatct ggacgaagag catcaggggc tcgcgccagc cgaactgttc 1020
gccaggctca aggcgcgcat gcccgacggc gaggatctcg tcgtgaccca tggcgatgcc 1080
tgcttgccga atatcatggt ggaaaatggc cgcttttctg gattcatcga ctgtggccgg 1140
ctgggtgtgg cggaccgcta tcaggacata gcgttggcta cccgtgatat tgctgaagag 1200
cttggcggcg aatgggctga ccgcttcctc gtgctttacg gtatcgccgc tcccgattcg 1260
cagcgcatcg ccttctatcg ccttcttgac gagttcttct gagcgggact ctggggttcg 1320
aaatgaccga ccaagcgacg cccaacctgc catcacgaga tttcgattcc accgccgcct 1380
tctatgaaag gttgggcttc ggaatcgttt tccgggacgc cggctggatg atcctccagc 1440
gcggggatct catgctggag ttcttcgccc accccgatcc aacacttacg tttgcaacgt 1500
ccaagagcaa atagaccacg aacgccggaa ggttgccgca gcgtgtggat tgcgtctcaa 1560
ttctctcttg caggaatgca atgatgaata tgatactgac tatgaaactt tgagggaata 1620
ctgcctagca ccgtcacctc ataacgtgca tcatgcatgc cctgacaaca tggaacatcg 1680
ctatttttct gaagaattat gctcgttgga ggatgtcgcg gcaattgcag ctattgccaa 1740
catcgaacta cccctcacgc atgcattcat caatattatt catgcgggga aaggcaagat 1800
taatccaact ggcaaatcat ccagcgtgat tggtaacttc agttccagcg acttgattcg 1860
ttttggtgct acccacgttt tcaataagga cgagatggtg gagtaaagaa ggagtgcgtc 1920
gaagcagatc gttcaaacat ttggcaataa agtttcttaa gattgaatcc tgttgccggt 1980
cttgcgatga ttatcatata atttctgttg aattacgtta agcatgtaat aattaacatg 2040
taatgcatga cgttatttat gagatgggtt tttatgatta gagtcccgca attatacatt 2100
taatacgcga tagaaaacaa aatatagcgc gcaaactagg ataaattatc gcgcgcggtg 2160
tcatctatgt tactagatcg aattaattca gtacattaaa aacgtccgca atgtgttatt 2220
aagttgtcta agcgtcaatt tgtttacacc acaatatatc ctgccaccag ccagccaaca 2280
gctccccgac cggcagctcg gcacaaaatc accactcgat acaggcagcc catcagtccg 2340
ggacggcgtc agcgggagag ccgttgtaag gcggcagact ttgctcatgt taccgatgct 2400
attcggaaga acggcaacta agctgccggg tttgaaacac ggatgatctc gcggagggta 2460
gcatgttgat tgtaacgatg acagagcgtt gctgcctgtg atcaaatatc atctccctcg 2520
cagagatccg aattatcagc cttcttattc atttctcgct taaccgtgac aggctgtcga 2580
tcttgagaac tatgccgaca taataggaaa tcgctggata aagccgctga ggaagctgag 2640
tggcgctatt tctttagaag tgaacgttga cgatgtcgac ggatcttttc cgctgcataa 2700
ccctgcttcg gggtcattat agcgattttt tcggtatatc catccttttt cgcacgatat 2760
acaggatttt gccaaagggt tcgtgtagac tttccttggt gtatccaacg gcgtcagccg 2820
ggcaggatag gtgaagtagg cccacccgcg agcgggtgtt ccttcttcac tgtcccttat 2880
tcgcacctgg cggtgctcaa cgggaatcct gctctgcgag gctggccggc taccgccggc 2940
gtaacagatg agggcaagcg gatggctgat gaaaccaagc caaccagggg tgatgctgcc 3000
aacttactga tttagtgtat gatggtgttt ttgaggtgct ccagtggctt ctgtttctat 3060
cagctgtccc tcctgttcag ctactgacgg ggtggtgcgt aacggcaaaa gcaccgccgg 3120
acatcagcgc tatctctgct ctcactgccg taaaacatgg caactgcagt tcacttacac 3180
cgcttctcaa cccggtacgc accagaaaat cattgatatg gccatgaatg gcgttggatg 3240
ccgggcaaca gcccgcatta tgggcgttgg cctcaacacg attttacgtc acttaaaaaa 3300
ctcaggccgc agtcggtaac ctcgcgcata cagccgggca gtgacgtcat cgtctgcgcg 3360
gaaatggacg aacagtgggg ctatgtcggg gctaaatcgc gccagcgctg gctgttttac 3420
gcgtatgaca gtctccggaa gacggttgtt gcgcacgtat tcggtgaacg cactatggcg 3480
acgctggggc gtcttatgag cctgctgtca ccctttgacg tggtgatatg gatgacggat 3540
ggctggccgc tgtatgaatc ccgcctgaag ggaaagctgc acgtaatcag caagcgatat 3600
acgcagcgaa ttgagcggca taacctgaat ctgaggcagc acctggcacg gctgggacgg 3660
aagtcgctgt cgttctcaaa atcggtggag ctgcatgaca aagtcatcgg gcattatctg 3720
aacataaaac actatcaata agttggagtc attacccaac caggaagggc agcccaccta 3780
tcaaggtgta ctgccttcca gacgaacgaa gagcgattga ggaaaaggcg gcggcggccg 3840
gcatgagcct gtcggcctac ctgctggccg tcggccaggg ctacaaaatc acgggcgtcg 3900
tggactatga gcacgtccgc gagctggccc gcatcaatgg cgacctgggc cgcctgggcg 3960
gcctgctgaa actctggctc accgacgacc cgcgcacggc gcggttcggt gatgccacga 4020
tcctcgccct gctggcgaag atcgaagaga agcaggacga gcttggcaag gtcatgatgg 4080
gcgtggtccg cccgagggca gagccatgac ttttttagcc gctaaaacgg ccggggggtg 4140
cgcgtgattg ccaagcacgt ccccatgcgc tccatcaaga agagcgactt cgcggagctg 4200
gtattcgtgc agggcaagat tcggaatacc aagtacgaga aggacggcca gacggtctac 4260
gggaccgact tcattgccga taaggtggat tatctggaca ccaaggcacc aggcgggtca 4320
aatcaggaat aagggcacat tgccccggcg tgagtcgggg caatcccgca aggagggtga 4380
atgaatcgga cgtttgaccg gaaggcatac aggcaagaac tgatcgacgc ggggttttcc 4440
gccgaggatg ccgaaaccat cgcaagccgc accgtcatgc gtgcgccccg cgaaaccttc 4500
cagtccgtcg gctcgatggt ccagcaagct acggccaaga tcgagcgcga cagcgtgcaa 4560
ctggctcccc ctgccctgcc cgcgccatcg gccgccgtgg agcgttcgcg tcgtctcgaa 4620
caggaggcgg caggtttggc gaagtcgatg accatcgaca cgcgaggaac tatgacgacc 4680
aagaagcgaa aaaccgccgg cgaggacctg gcaaaacagg tcagcgaggc caagcaggcc 4740
gcgttgctga aacacacgaa gcagcagatc aaggaaatgc agctttcctt gttcgatatt 4800
gcgccgtggc cggacacgat gcgagcgatg ccaaacgaca cggcccgctc tgccctgttc 4860
accacgcgca acaagaaaat cccgcgcgag gcgctgcaaa acaaggtcat tttccacgtc 4920
aacaaggacg tgaagatcac ctacaccggc gtcgagctgc gggccgacga tgacgaactg 4980
gtgtggcagc aggtgttgga gtacgcgaag cgcaccccta tcggcgagcc gatcaccttc 5040
acgttctacg agctttgcca ggacctgggc tggtcgatca atggccggta ttacacgaag 5100
gccgaggaat gcctgtcgcg cctacaggcg acggcgatgg gcttcacgtc cgaccgcgtt 5160
gggcacctgg aatcggtgtc gctgctgcac cgcttccgcg tcctggaccg tggcaagaaa 5220
acgtcccgtt gccaggtcct gatcgacgag gaaatcgtcg tgctgtttgc tggcgaccac 5280
tacacgaaat tcatatggga gaagtaccgc aagctgtcgc cgacggcccg acggatgttc 5340
gactatttca gctcgcaccg ggagccgtac ccgctcaagc tggaaacctt ccgcctcatg 5400
tgcggatcgg attccacccg cgtgaagaag tggcgcgagc aggtcggcga agcctgcgaa 5460
gagttgcgag gcagcggcct ggtggaacac gcctgggtca atgatgacct ggtgcattgc 5520
aaacgctagg gccttgtggg gtcagttccg gctgggggtt cagcagccag cgctttactg 5580
gcatttcagg aacaagcggg cactgctcga cgcacttgct tcgctcagta tcgctcggga 5640
cgcacggcgc gctctacgaa ctgccgataa acagaggatt aaaattgaca attgtgatta 5700
aggctcagat tcgacggctt ggagcggccg acgtgcagga tttccgcgag atccgattgt 5760
cggccctgaa gaaagctcca gagatgttcg ggtccgttta cgagcacgag gagaaaaagc 5820
ccatggaggc gttcgctgaa cggttgcgag atgccgtggc attcggcgcc tacatcgacg 5880
gcgagatcat tgggctgtcg gtcttcaaac aggaggacgg ccccaaggac gctcacaagg 5940
cgcatctgtc cggcgttttc gtggagcccg aacagcgagg ccgaggggtc gccggtatgc 6000
tgctgcgggc gttgccggcg ggtttattgc tcgtgatgat cgtccgacag attccaacgg 6060
gaatctggtg gatgcgcatc ttcatcctcg gcgcacttaa tatttcgcta ttctggagct 6120
tgttgtttat ttcggtctac cgcctgccgg gcggggtcgc ggcgacggta ggcgctgtgc 6180
agccgctgat ggtcgtgttc atctctgccg ctctgctagg tagcccgata cgattgatgg 6240
cggtcctggg ggctatttgc ggaactgcgg gcgtggcgct gttggtgttg acaccaaacg 6300
cagcgctaga tcctgtcggc gtcgcagcgg gcctggcggg ggcggtttcc atggcgttcg 6360
gaaccgtgct gacccgcaag tggcaacctc ccgtgcctct gctcaccttt accgcctggc 6420
aactggcggc cggaggactt ctgctcgttc cagtagcttt agtgtttgat ccgccaatcc 6480
cgatgcctac aggaaccaat gttctcggcc tggcgtggct cggcctgatc ggagcgggtt 6540
taacctactt cctttggttc cgggggatct cgcgactcga acctacagtt gtttccttac 6600
tgggctttct cagccgggat ggcgctaaga agctattgcc gccgatcttc atatgcggtg 6660
tgaaataccg cacagatgcg taaggagaaa ataccgcatc aggcgctctt ccgcttcctc 6720
gctcactgac tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag ctcactcaaa 6780
ggcggtaata cggttatcca cagaatcagg ggataacgca ggaaagaaca tgtgagcaaa 6840
aggccagcaa aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct 6900
ccgcccccct gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac 6960
aggactataa agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc 7020
gaccctgccg cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc 7080
tcaatgctca cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg 7140
tgtgcacgaa ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga 7200
gtccaacccg gtaagacacg acttatcgcc actggcagca gccactggta acaggattag 7260
cagagcgagg tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta 7320
cactagaagg acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag 7380
agttggtagc tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg 7440
caagcagcag attacgcgca gaaaaaaagg atatcaagaa gatcctttga tcttttctac 7500
ggggtctgac gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc 7560
aaaaaggatc ttcacctaga tccttttaaa ttaaaaatga agttttaaat caatctaaag 7620
tatatatgag taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc 7680
agcgatctgt ctatttcgtt catccatagt tgcctgactc cccgtcgtgt agataactac 7740
gatacgggag ggcttaccat ctggccccag tgctgcaatg ataccgcgag acccacgctc 7800
accggctcca gatttatcag caataaacca gccagccgga agggccgagc gcagaagtgg 7860
tcctgcaact ttatccgcct ccatccagtc tattaaacaa gtggcagcaa cggattcgca 7920
aacctgtcac gccttttgtg ccaaaagccg cgccaggttt gcgatccgct gtgccaggcg 7980
ttaggcgtca tatgaagatt tcggtgatcc ctgagcaggt ggcggaaaca ttggatgctg 8040
agaaccattt cattgttcgt gaagtgttcg atgtgcacct atccgaccaa ggctttgaac 8100
tatctaccag aagtgtgagc ccctaccgga aggattacat ctcggatgat gactctgatg 8160
aagactctgc ttgctatggc gcattcatcg accaagagct tgtcgggaag attgaactca 8220
actcaacatg gaacgatcta gcctctatcg aacacattgt tgtgtcgcac acgcaccgag 8280
gcaaaggagt cgcgcacagt ctcatcgaat ttgcgaaaaa gtgggcacta agcagacagc 8340
tccttggcat acgattagag acacaaacga acaatgtacc tgcctgcaat ttgtacgcaa 8400
aatgtggctt tactctcggc ggcattgacc tgttcacgta taaaactaga cctcaagtct 8460
cgaacgaaac agcgatgtac tggtactggt tctcgggagc acaggatgac gcctaacaat 8520
tcattcaagc cgacaccgct tcgcggcgcg gcttaattca ggagttaaac atcatgaggg 8580
aagcggtgat cgccgaagta tcgactcaac tatcagaggt agttggcgtc atcgagcgcc 8640
atctcgaacc gacgttgctg gccgtacatt tgtacggctc cgcagtggat ggcggcctga 8700
agccacacag tgatattgat ttgctggtta cggtgaccgt aaggcttgat gaaacaacgc 8760
ggcgagcttt gatcaacgac cttttggaaa cttcggcttc ccctggagag agcgagattc 8820
tccgcgctgt agaagtcacc attgttgtgc acgacgacat cattccgtgg cgttatccag 8880
ctaagcgcga actgcaattt ggagaatggc agcgcaatga cattcttgca ggtatcttcg 8940
agccagccac gatcgacatt gatctggcta tcttgctgac aaaagcaaga gaacatagcg 9000
ttgccttggt aggtccagcg gcggaggaac tctttgatcc ggttcctgaa caggatctat 9060
ttgaggcgct aaatgaaacc ttaacgctat ggaactcgcc gcccgactgg gctggcgatg 9120
agcgaaatgt agtgcttacg ttgtcccgca tttggtacag cgcagtaacc ggcaaaatcg 9180
cgccgaagga tgtcgctgcc gactgggcaa tggagcgcct gccggcccag tatcagcccg 9240
tcatacttga agctaggcag gcttatcttg gacaagaaga tcgcttggcc tcgcgcgcag 9300
atcagttgga agaatttgtt cactacgtga aaggcgagat caccaaggta gtcggcaaat 9360
aatgtctaac aattcgttca agccgacgcc gcttcgcggc gcggcttaac tcaagcgtta 9420
gagagctggg gaagactatg cgcgatctgt tgaaggtggt tctaagcctc gtacttgcga 9480
tggcatcggg gcaggcactt gctgacctgc caattgtttt agtggatgaa gctcgtcttc 9540
cctatgacta ctccccatcc aactacgaca tttctccaag caactacgac aactccataa 9600
gcaattacga caatagtcca tcaaattacg acaactctga gagcaactac gataatagtt 9660
catccaatta cgacaatagt cgcaacggaa atcgtaggct tatatatagc gcaaatgggt 9720
ctcgcacttt cgccggctac tacgtcattg ccaacaatgg gacaacgaac ttcttttcca 9780
catctggcaa aaggatgttc tacaccccaa aaggggggcg cggcgtctat ggcggcaaag 9840
atgggagctt ctgcggggca ttggtcgtca taaatggcca attttcgctt gccctgacag 9900
ataacggcct gaagatcatg tatctaagca actagcctgc tctctaataa aatgttagga 9960
gcttggctgc catttttggg gtgaggccgt tcgcggccga ggggcgcagc ccctgggggg 10020
atgggaggcc cgcgttagcg ggccgggagg gttcgagaag ggggggcacc ccccttcggc 10080
gtgcgcggtc acgcgccagg gcgcagccct ggttaaaaac aaggtttata aatattggtt 10140
taaaagcagg ttaaaagaca ggttagcggt ggccgaaaaa cgggcggaaa cccttgcaaa 10200
tgctggattt tctgcctgtg gacagcccct caaatgtcaa taggtgcgcc cctcatctgt 10260
cagcactctg cccctcaagt gtcaaggatc gcgcccctca tctgtcagta gtcgcgcccc 10320
tcaagtgtca ataccgcagg gcacttatcc ccaggcttgt ccacatcatc tgtgggaaac 10380
tcgcgtaaaa tcaggcgttt tcgccgattt gcgaggctgg ccagctccac gtcgccggcc 10440
gaaatcgagc ctgcccctca tctgtcaacg ccgcgccggg tgagtcggcc cctcaagtgt 10500
caacgtccgc ccctcatctg tcagtgaggg ccaagttttc cgcgaggtat ccacaacgcc 10560
ggcggccggc cgcggtgtct cgcacacggc ttcgacggcg tttctggcgc gtttgcaggg 10620
ccatagacgg ccgccagccc agcggcgagg gcaaccagcc cggtgagcgt cggaaagggt 10680
cgacatcttg ctgcgttcgg atattttcgt ggagttcccg ccacagaccc ggattgaagg 10740
cgagatccag caactcgcgc cagatcatcc tgtgacggaa ctttggcgcg tgatgactgg 10800
ccaggacgtc ggccgaaaga gcgacaagca gatcacgatt ttcgacagcg tcggatttgc 10860
gatcgaggat ttttcggcgc tgcgctacgt ccgcgaccgc gttgagggat caagccacag 10920
cagcccactc gaccttctag ccgacccaga cgagccaagg gatctttttg gaatgctgct 10980
ccgtcgtcag gctttccgac gtttgggtgg ttgaacagaa gtcattatcg tacggaatgc 11040
cagcactccc gaggggaacc ctgtggttgg catgcacata caaatggacg aacggataaa 11100
ccttttcacg cccttttaaa tatccgttat tctaataaac gctcttttct cttaggttta 11160
cccgccaata tatcctgtca aacactgata gtttaaactg aaggcgggaa acgacaatct 11220
gatcatgagc ggagaattaa gggagtcacg ttatgacccc cgccgatgac gcgggacaag 11280
ccgttttacg tttggaactg acagaaccgc aacgattgaa ggagccactc agccccaata 11340
cgcaaaccgc ctctccccgc gcgttggccg attcattaat gcagctggca cgacaggttt 11400
cccgactgga aagcgggcag tgagcgcaac gcaattaatg tgagttagct cactcattag 11460
gcaccccagg ctttacactt tatgcttccg gctcgtatgt tgtgtggaat tgtgagcgga 11520
taacaatttc acacaggaaa cagctatgac catgattacg ccaagctatt taggtgacac 11580
tatagaatac tcaagctatg catccaacgc gttgggagct ccctaggctt tttttcttct 11640
tcttcgttca tacagttttt ttttgtttat cagcttacat tttcttgaac cgtagctttc 11700
gttttcttct ttttaacttt ccattcggag tttttgtatc ttgtttcata gtttgtccca 11760
ggattagaat gattaggcat cgaaccttca agaatttgat tgaataaaac atcttcattc 11820
ttaagatatg aagataatct tcaaaaggcc cctgggaatc tgaaagaaga gaagcaggcc 11880
catttatatg ggaaagaaca atagtatttc ttatataggc ccatttaagt tgaaaacaat 11940
cttcaaaagt cccacatcgc ttagataaga aaacgaagct gagtttatat acagctagag 12000
tcgaagtagt gattgaatgt ctgttgcctt gttagtttta gagctagaaa tagcaagtta 12060
aaataaggct agtccgttat caacttgaaa aagtggcacc gagtcggtgc tttttttcta 12120
gacccagctt tcttgtacaa agttggcatt 12150
<210> 61
<211> 104
<212> ДНК
<213> Искусственная последовательность
<220>
<223> молекула гРНК для аллель-специфического DSB+аллель-зависимой репарации
<400> 61
ggagcgtata taatgctgct tgttttagag ctagaaatag caagttaaaa taaggctagt 60
ccgttatcaa cttgaaaaag tggcaccgag tcggtgcttt tttt 104
<210> 62
<211> 13688
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Плазмида R1
<400> 62
ggagcaacct tattttgtac tttaaaaaat tcattttttt tattttttcg actttaaagc 60
caaaattatc ctttatttat gaaagtggat gtatttttat ccctttaata taaggttgag 120
ttttctttaa gtttgtaatg ttaagtgggg cactttatat tagtccaatt aacagaattg 180
acttagaaat tgatggcgat tacttgaaaa aatcgctaca aaaacataat aaaataaaat 240
ttcatttgat ttttaaacca aaagaaaagt gacatgacta acgatttgat cccatcttct 300
ttttttttta agaaaaaaat taaaagaaaa atttattaaa ctggatgatt aattttaaca 360
taaaaattaa aaaaataaaa atcaatcgag ctcactattt gatattatta ttttcatctt 420
gatattgtta cttcatatca ctgaattata aaatgcgtta acggcaatta tcatagacgc 480
agactgaaag cataaaatta caactattcc atttttgttt tttttttgcc aacccaaaaa 540
aaactaataa tttatttaca taattataga aaattggaat ttattcatgt ttttggacca 600
ttcagataac cccacaataa aaaaaaagtg cacaaaagtg caaagtgcta cgtgtcctaa 660
ttatagccat actacatttg tgcataaaat tagccaccat tatttttaag attattcttt 720
ttcttgtttt aatttatatg ccacactttc ttattaatcc atcttaaaca gaatgacaca 780
ctttttattt gacaaatatt taatgacatt attaacattt tatcctcgtt agatttcact 840
tatttggtta aaggtggtgt acttttctaa tctaaaatca ttaattttag gtataatttt 900
agaacattgc aaagcctttt catatattta aacaatgtac ataattatgt aattatgtta 960
tgttatataa attgagacga aaaaaatatt atttatcagg tacgagtgtc actttacact 1020
agaggtgttt acggacagag acgaatccaa gatttgaact tcatggtgtc aagtatacac 1080
attgcttaga ttattaaatc aactttaggt ttcaattccg cacaaaatct ataaaacata 1140
acctttttaa aaataatttt actaatatta taaacatata ttattcattt ataatagttc 1200
tcgacgagtt tacatatttt caaaactgtc aattagaaca tcattactta tatttgtaaa 1260
tgtatcactt actacgtaac aaactaaaca aatcatttga atatcactca tcactcttat 1320
ttatatttta tttttttatc tacttcacca aagaaaatga taatttcaca attgcgattg 1380
ctaccagaaa aatatttact taatcaattt taaaaagatt aagtaataga aatatttttt 1440
attttattaa gaaaagagga ttttatcata tctaaatata aattaaagat gaagttacta 1500
tgatattatt taaaaagtaa atgacattat tctatttaaa ttacaaaatg agattaagaa 1560
aagtataact atttccattt ttttagctaa aaggatttca aaaaaaaaag taacaagaaa 1620
gagatttttt aaaaataata attttattag aaatatatat aaaaaattaa taatattcgt 1680
tagaaaaagt gaattttcaa aataataaag taataacacg aatactttta gggtcaaaca 1740
attaatagtg gagaatggag atatttttag tacaaattat ttatacaaaa tatctttatt 1800
caatgtgaca taattttaag ctatttttaa cacttttcca tcaaataaat aattaataac 1860
aaattattaa aaaatcaaag attaataaat taaattaatg aactataatt agttattaaa 1920
accaagccaa ctagaattaa atagaaaaaa ggaaatataa tcttcaaaat atgctttgtt 1980
gaagtataat tcctcgtcta tttaaagctt cggagattta aattggcaag cgaattacac 2040
aaagcaaatg aaactacacc ttttggttct gtgggtgtca tttgcttatt tataaacatt 2100
gtacatatat acacctatat gtatatcacg tttcgatcga agctcgtggg tggcgtggca 2160
cccataaatc gtagtagatt cgcccctgtt tacggatcga tttaggtcgg ttatagatca 2220
aaatcaaatc aatttaatta attttaccaa tcaaaccaaa tcaaatatac tatacattta 2280
ttggtttggt tgttcgaatt tcgatttgat ttgattcaat tattcgatta tcaatcatac 2340
agaaataaaa gaaacaattc attttaaaga ggaacaaaag aggcaagaac tcattcaaaa 2400
tgaaaaatac ttaaaatagc cgaatttata aaacttttta ttactaaatt tactgtgaat 2460
aaaactttta aattcaccat tttagcctag aagtaagaga gaaaaacatt tccaccaaga 2520
attgtcaaga atactcatat acatgaaacc aatatgataa atgttctctt caggacggtt 2580
cataaataaa ttatgataaa catatataag atatttaaaa taaaataaaa tatatatata 2640
taaaatttat cggttcgatt atttcttcaa ctttttaaat aaaatcaaag actatcaatt 2700
cttaagaatc ttagaacaaa ctaaaccaaa taaaattaaa cgatttaatc aacttaattt 2760
aatttttcaa tgtgggtcaa tttttatcca aatactttta cacccacttt tcaccatcac 2820
aaggggtaag aactagcaaa ttattttgcc ttaacaaaaa cttactattg gtgtagagaa 2880
aattggttga ttttcttatc aatatatata atttagtacc caattttccc aaaactaaat 2940
aaaatataga atatccacta aggtggttac acgtgtacac tcaaccaatg acggcacttc 3000
aaattcttga taacggtatc ttcccacctt catatatttc aacattttat tgttaaaata 3060
aaatcgcacg ctctatttta tctaaatttt atttatagaa ttatattaag tatgttatta 3120
ttaattttta cgtataaata tatttcatat taaaatcata taagttgaca tcactaagtt 3180
cgttggattt gcatgtagac ccaattattc agatactcaa catgactcat ttaatattgg 3240
attattgaaa tatttgtaat acattaagaa tataattgtt aactatttta atttttaatt 3300
atacgtaaat atcaaacaaa aatattaata tgatcgctat attagatgat aactataagg 3360
agcctacaca attaacacta tttaactcta ttctttgcat ttataaaaag ttactttagt 3420
cttaggttca caatgtcaaa atctaaacaa ctaaaaacga cgaggagtaa ggtttgcaac 3480
gacgataaca aggattaggc aacaattaga gttgtgaatt gtgagtatta actatacttt 3540
tactatatta ggcagaattt ttgcactcaa tgagtaactt gatttattta ttttttattt 3600
cgccctaaat tattggacaa gtcatatatt tgttttgaaa acattctttt attggctaaa 3660
tcgaaaattg aatcgttaaa gatcaaaaat caataacaaa tatcttattg gtttaacata 3720
tttaaaaata aaaaaccaat aaatctaact aataatattt aatacgaaaa cgaaatggac 3780
tgacacacat tcctaaattt ttggtcaaaa ttttttcata atttccctaa aatctaaaat 3840
attaaatatt tgacggaaac aaaaaattca cttttaataa attatttgaa ggactaaaac 3900
agtggaagaa tatatttaag aagctaattt gaacctagtg ccaaatataa agggaccatt 3960
tttgtcattt ttcaacttga aaatctacgt gtcttaatat aacaccaaag aattaatatt 4020
tactgaaaaa atgtaaaaat gaggatatgg attctgaatc actcaattcc aatcagcaaa 4080
aataaaataa aataaaataa aataaaattt aaaaaataat aataaatgct ataaaatgac 4140
caaaatgtgt ggagcaaaaa gtgcagaaaa aaccaacaaa ttgcattctc cattcttgga 4200
agtggccatt cttgatttct tgaaacaaag gtttgtttcc cttcacttct tgatatgtaa 4260
agttgcaatc tttataactt tctattgctt tgctagtgtt tttgttatat acagggggtg 4320
gagttagagg gtaagttacg catttagtcg taactttagt caaacttcgt aataatttag 4380
taagttaaaa tatattagaa attttcagaa ttcataaact ttaaatttta aattttgact 4440
tcgctttgtg tgactataca attacagaaa ttcagagtgg ccattgttga aagagagggt 4500
ggaatttgtg taagttttgt ttcctttcag ttcttgatat ataaagttgc aatctttaac 4560
attctttgtt cactttctat aggtttgcta ggttcggtta aattcagtag ctttagttta 4620
aaccctatgc ggaatagaga atgtgtaaac tttaaacttc aaattttggc tccgcatacg 4680
actagcgact atataataat aggaattgag cacttggctt ttgtatatag cttctatgtg 4740
taccaaaatt agaaaatcag gcgattatta taatcttgtt gactaaatat agaatgcatc 4800
cattaccccc aaaaagtgtg attccactgt cataggaggt tttttttatt tcattttatt 4860
tgtgctttca ataatgtaga gtagttttac aaagatcctt tctttgtgac acatggtagg 4920
taatattgct gattttgttg tagttttggg gttataaagt ttcaaattat ttatactgga 4980
gggtaggggt gggggttgtc tataatgcag gttatggttt tacgtgaact caataattat 5040
tgtagatact aagaaatcca ctcagtgttc ttgcggtgtc ttgcttttga tttcagcatc 5100
acttgtagtt gattgtgttt agattatcac attattctgt ggctgtaact gtatccttgt 5160
tagttgcttt gtttctacac tgttgttttc cctcttttat acctattttg atatgttgta 5220
ctcgaacgag ggtcatcggg gaacaacctc tttacctccg tgaggtagag ctatggtctg 5280
tgtccactct accctcccca gatccctctt gtaggatttc actatattgt aatattaact 5340
tgaggtcact ataggagctc aaaaacttct aattttgaat caatgtctgg ttatactttt 5400
tttgtcataa ctgtatctca aatgtggtgt ttggtttatc tcattttgca gaagtcaaga 5460
aacaggttac tcctgtttga gtgaggaaaa gttggtttgc ctgtctgtgg tctttttata 5520
atctttttct acagaagaga aagtgggtaa ttttgtttga gagtggaaat attctctagt 5580
gggaatctac taggagtaat ttattttcta taaactaagt aaagtttgga aggtgacaaa 5640
aagaaagaca aaaatcttgg aattgtttta gacaaccaag gttttcttgc tcagaatgtc 5700
tgttgccttg ttatgggttg tttctccttg tgacgtctca aatgggacaa gtttcatgga 5760
atcagtccgg gagggaaacc gtttttttga ttcatcgagg cataggaatt tggtgtccaa 5820
tgagagaatc aatagaggtg gtggaaagca aactaataat ggacggaaat tttctgtacg 5880
gtctgctatt ttggctactc catctggaga acggacgatg acatcggaac agatggtcta 5940
tgatgtggtt ttgaggcagg cagccttggt gaagaggcaa ctgagatcta ccaatgagtt 6000
agaagtgaag ccggatatac ctattccggg gaatttgggc ttgttgagtg aagcatatga 6060
taggtgtggt gaagtatgtg cagagtatgc aaagacgttt aacttaggtt agcttcttca 6120
atctattcat tcgtttacca aatattattt ggtaagcact aattatgaat atatatatgt 6180
tcatgttatt gatgaagaca aaatttgatc tttgtttgtt tattcaggaa ctatgctaat 6240
gactcccgag agaagaaggg ctatctgggc aatatatggt gaggtttcta gccatttaat 6300
aacagttacg cgcacaaaca catatgatta atcggggacg agaaaaaaag aaatgaagtt 6360
tgagttttga gggtcatatg taataggtaa atccgagctt gactagcttg agatgtttat 6420
tgtcatatca tgctcaatac taaccaaaac actgaaaaag aacttgatta tatttacata 6480
ctaatatttt catttgcgtt gctgttcaca tttttaccta tggaactggt ttttgtgatt 6540
tgttatactt catattcgat gttaataaaa tatatcattc ctcccttttt ctccacttca 6600
agctttactg tagtgttgaa aggggaaact ccttttaatg attgcatata taaacgaact 6660
tcttgagttg aatagtttct cattatgatc tgtttaaaca gtatggtgca gaagaacaga 6720
tgaacttgtt gatggcccaa acgcatcata tattaccccg gcagccttag ataggtggga 6780
aaataggcta gaagatgttt tcaatgggcg gccatttgac atgctcgatg gtgctttgtc 6840
cgatacagtt tctaactttc cagttgatat tcaggttagt ctaccaattc tatggtcttt 6900
atatttgttc aatttgcgtt tgatgtcact tttgctgagg gcttttctaa tagcttactt 6960
cagcctagcg gaaatgtttg tagttgaatc tctagttctg tctcctatat ctgtttctct 7020
cgtcctagat actacacata cttcatttct gttttaacat tttattcgtc ttttggtgtt 7080
gttttgtatg tgaatcatat atttggaaca gaatcattat tagttcacat gatttcattt 7140
gctttcttca atagcgtaat tgtctaacct tccaatatat gttgcagcca ttcagagata 7200
tgattgaagg aatgcgtatg gacttgagaa aatcgagata caaaaacttc gacgaactat 7260
acctttattg ttattatgtt gctggtacgg ttgggttgat gagtgttcca attatggcgc 7320
tgtcatgaga cgaattctga caggatatat tggcgggtaa acctaagaga aaagagcgtt 7380
tattagaata atcggatatt taaaagggcg tgaaaaggtt tatccgttcg tccatttgta 7440
tgtgcatgcc aaccacaggg ttcccctcgg gatcaaagta ctttgatcca acccctccgc 7500
tgctatagtg cagtcggctt ctgacgttca gtgcagccgt catctgaaaa cgacatgtcg 7560
cacaagtcct aagttacgcg acaggctgcc gccctgccct tttcctggcg ttttcttgtc 7620
gcgtgtttta gtcgcataaa gtagaatact tgcgactaga accggagaca ttacgccatg 7680
aacaagagcg ccgccgctgg cctgctgggc tatgcccgcg tcagcaccga cgaccaggac 7740
ttgaccaacc aacgggccga actgcacgcg gccggctgca ccaagctgtt ttccgagaag 7800
atcaccggca ccaggcgcga ccgcccggag ctggccagga tgcttgacca cctacgccct 7860
ggcgacgttg tgacagtgac caggctagac cgcctggccc gcagcacccg cgacctactg 7920
gacattgccg agcgcatcca ggaggccggc gcgggcctgc gtagcctggc agagccgtgg 7980
gccgacacca ccacgccggc cggccgcatg gtgttgaccg tgttcgccgg cattgccgag 8040
ttcgagcgtt ccctaatcat cgaccgcacc cggagcgggc gcgaggccgc caaggcccga 8100
ggcgtgaagt ttggcccccg ccctaccctc accccggcac agatcgcgca cgcccgcgag 8160
ctgatcgacc aggaaggccg caccgtgaaa gaggcggctg cactgcttgg cgtgcatcgc 8220
tcgaccctgt accgcgcact tgagcgcagc gaggaagtga cgcccaccga ggccaggcgg 8280
cgcggtgcct tccgtgagga cgcattgacc gaggccgacg ccctggcggc cgccgagaat 8340
gaacgccaag aggaacaagc atgaaaccgc accaggacgg ccaggacgaa ccgtttttca 8400
ttaccgaaga gatcgaggcg gagatgatcg cggccgggta cgtgttcgag ccgcccgcgc 8460
acctctcaac cgtgcggctg catgaaatcc tggccggttt gtctgatgcc aagctggcgg 8520
cctggccggc cagcttggcc gctgaagaaa ccgagcgccg ccgtctaaaa aggtgatgtg 8580
tatttgagta aaacagcttg cgtcatgcgg tcgctgcgta tatgatccga tgagtaaata 8640
aacaaatacg caaggggaac gcatgaaggt tatcgctgta cttaaccaga aaggcgggtc 8700
aggcaagacg accatcggaa cccatctagc ccgcgccctg caactcgccg gggccgatgt 8760
tctgttagtc gattccgatc cccagggcag tgcccgcgat tgggcggccg tgcgggaaga 8820
tcaaccgcta accgttgtcg gcatcgaccg cccgacgatt gaccgcgacg tgaaggccat 8880
cggccggcgc gacttcgtag tgatcgacgg agcgccccag gcggcggact tggctgtgtc 8940
cgcgatcaag gcagccgact tcgtgctgat tccggtgcag ccaagccctt acgacatatg 9000
ggccaccgcc gacctggtgg agctggttaa gcagcgcatt gaggtcacgg atggaaggct 9060
acaagcggcc tttgtcgtgt cgcgggcgat caaaggcacg cgcatcggcg gtgaggttgc 9120
cgaggcgctg gccgggtacg agctgcccat tcttgagtcc cgtatcacgc agcgcgtgag 9180
ctacccaggc actgccgccg ccggcacaac cgttcttgaa tcagaacccg agggcgacgc 9240
tgcccgcgag gtccaggcgc tggccgctga aattaaatca aaactcattt gagttaatga 9300
ggtaaagaga aaatgagcaa aagcacaaac acgctaagtg ccggccgtcc gagcgcacgc 9360
agcagcaagg ctgcaacgtt ggccagcctg gcagacacgc cagccatgaa gcgggtcaac 9420
tttcagttgc cggcggagga tcacaccaag ctgaagatgt acgcggtacg ccaaggcaag 9480
accattaccg agctgctatc tgaatagatc gcgcagctac cagagtaaat gagcaaatga 9540
ataaatgagt agatgaattt tagcggctaa aggaggcggc atggaaaatc aagaacaacc 9600
aggcaccgac gccgtggaat gccccatgtg tggaggaacg ggcggttggc caggcgtaag 9660
cggctgggtt gtctgccggc cctgcaatgg cactggaacc cccaagcccg aggaatcggc 9720
gtgacggtcg caaaccatcc ggcccggtac aaatcggcgc ggcgctgggt gatgacctgg 9780
tggagaagtt gaaggccgcg caggccgccc agcggcaacg catcgaggca gaagcacgcc 9840
ccggtgaatc gtggcaagcg gccgctgatc gaatccgcaa agaatcccgg caaccgccgg 9900
cagccggtgc gccgtcgatt aggaagccgc ccaagggcga cgagcaacca gattttttcg 9960
ttccgatgct ctatgacgtg ggcacccgcg atagtcgcag catcatggac gtggccgttt 10020
tccgtctgtc gaagcgtgac cgacgagctg gcgaggtgat ccgctacgag cttccagacg 10080
ggcacgtaga ggtttccgca gggccggccg gcatggccag tgtgtgggat tacgacctgg 10140
tactgatggc ggtttcccat ctaaccgaat ccatgaaccg ataccgggaa gggaagggag 10200
acaagcccgg ccgcgtgttc cgtccacacg ttgcggacgt actcaagttc tgccggcgag 10260
ccgatggcgg aaagcagaaa gacgacctgg tagaaacctg cattcggtta aacaccacgc 10320
acgttgccat gcagcgtacg aagaaggcca agaacggccg cctggtgacg gtatccgagg 10380
gtgaagcctt gattagccgc tacaagatcg taaagagcga aaccgggcgg ccggagtaca 10440
tcgagatcga gctagctgat tggatgtacc gcgagatcac agaaggcaag aacccggacg 10500
tgctgacggt tcaccccgat tactttttga tcgatcccgg catcggccgt tttctctacc 10560
gcctggcacg ccgcgccgca ggcaaggcag aagccagatg gttgttcaag acgatctacg 10620
aacgcagtgg cagcgccgga gagttcaaga agttctgttt caccgtgcgc aagctgatcg 10680
ggtcaaatga cctgccggag tacgatttga aggaggaggc ggggcaggct ggcccgatcc 10740
tagtcatgcg ctaccgcaac ctgatcgagg gcgaagcatc cgccggttcc taatgtacgg 10800
agcagatgct agggcaaatt gccctagcag gggaaaaagg tcgaaaagga ctctttcctg 10860
tggatagcac gtacattggg aacccaaagc cgtacattgg gaaccggaac ccgtacattg 10920
ggaacccaaa gccgtacatt gggaaccggt cacacatgta agtgactgat ataaaagaga 10980
aaaaaggcga tttttccgcc taaaactctt taaaacttat taaaactctt aaaacccgcc 11040
tggcctgtgc ataactgtct ggccagcgca cagccgaaga gctgcaaaaa gcgcctaccc 11100
ttcggtcgct gcgctcccta cgccccgccg cttcgcgtcg gcctatcgcg gccgctggcc 11160
gctcaaaaat ggctggccta cggccaggca atctaccagg gcgcggacaa gccgcgccgt 11220
cgccactcga ccgccggcgc ccacatcaag gcaccctgcc tcgcgcgttt cggtgatgac 11280
ggtgaaaacc tctgacacat gcagctcccg gtgacggtca cagcttgtct gtaagcggat 11340
gccgggagca gacaagcccg tcagggcgcg tcagcgggtg ttggcgggtg tcggggcgca 11400
gccatgaccc agtcacgtag cgatagcgga gtgtatactg gcttaactat gcggcatcag 11460
agcagattgt actgagagtg caccatatgc ggtgtgaaat accgcacaga tgcgtaagga 11520
gaaaataccg catcaggcgc tcttccgctt cctcgctcac tgactcgctg cgctcggtcg 11580
ttcggctgcg gcgagcggta tcagctcact caaaggcggt aatacggtta tccacagaat 11640
caggggataa cgcaggaaag aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta 11700
aaaaggccgc gttgctggcg tttttccata ggctccgccc ccctgacgag catcacaaaa 11760
atcgacgctc aagtcagagg tggcgaaacc cgacaggact ataaagatac caggcgtttc 11820
cccctggaag ctccctcgtg cgctctcctg ttccgaccct gccgcttacc ggatacctgt 11880
ccgcctttct cccttcggga agcgtggcgc tttctcatag ctcacgctgt aggtatctca 11940
gttcggtgta ggtcgttcgc tccaagctgg gctgtgtgca cgaacccccc gttcagcccg 12000
accgctgcgc cttatccggt aactatcgtc ttgagtccaa cccggtaaga cacgacttat 12060
cgccactggc agcagccact ggtaacagga ttagcagagc gaggtatgta ggcggtgcta 12120
cagagttctt gaagtggtgg cctaactacg gctacactag aaggacagta tttggtatct 12180
gcgctctgct gaagccagtt accttcggaa aaagagttgg tagctcttga tccggcaaac 12240
aaaccaccgc tggtagcggt ggtttttttg tttgcaagca gcagattacg cgcagaaaaa 12300
aaggatctca agaagatcct ttgatctttt ctacggggtc tgacgctcag tggaacgaaa 12360
actcacgtta agggattttg gtcatgcatt ctaggtgatt agaaaaactc atcgagcatc 12420
aaatgaaact gcaatttatt catatcagga ttatcaatac catatttttg aaaaagccgt 12480
ttctgtaatg aaggagaaaa ctcaccgagg cagttccata ggatggcaag atcctggtat 12540
cggtctgcga ttccgactcg tccaacatca atacaaccta ttaatttccc ctcgtcaaaa 12600
ataaggttat caagtgagaa atcaccatga gtgacgactg aatccggtga gaatggcaaa 12660
agtttatgca tttctttcca gacttgttca acaggccagc cattacgctc gtcatcaaaa 12720
tcactcgcat caaccaaacc gttattcatt cgtgattgcg cctgagcgag tcgaaatacg 12780
cgatcgctgt taaaaggaca attacaaaca ggaatcgaat gcaaccggcg caggaacact 12840
gccagcgcat caacaatatt ttcacctgaa tcaggatatt cttctaatac ctggaatgct 12900
gttttccctg ggatcgcagt ggtgagtaac catgcatcat caggagtacg gataaaatgc 12960
ttgatggtcg gaagaggcat aaattccgtc agccagttta gtctgaccat ctcatctgta 13020
acatcattgg caacgctacc tttgccatgt ttcagaaaca actctggcgc atcgggcttc 13080
ccatacaatc ggtagattgt cgcacctgat tgcccgacat tatcgcgagc ccatttatac 13140
ccatataaat cagcatccat gttggaattt aatcgcggcc ttgagcaaga cgtttcccgt 13200
tgaatatggc tcataacaga acttattatt tccttcctct tttctacagt atttaaagat 13260
accccaagaa gctaattata acaagacgaa ctccaattca ctgttccttg cattctaaaa 13320
ccttaaatac cagaaaacag ctttttcaaa gttgttttca aagttggcgt ataacatagt 13380
atcgacggag ccgattttga aaccgcggtg atcacaggca gcaacgctct gtcatcgtta 13440
caatcaacat gctaccctcc gcgagatcat ccgtgtttca aacccggcag cttagttgcc 13500
gttcttccga atagcatcgg taacatgagc aaagtctgcc gccttacaac ggctctcccg 13560
ctgacgccgt cccggactga tgggctgcct gtatcgagtg gtgattttgt gccgagctgc 13620
cggtcgggga gctgttggct ggctggtggc aggatatatt gtggtgtaaa cataacgaat 13680
tcgtctca 13688
<210> 63
<211> 9902
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Плазмида R2
<400> 63
ggagcaacct tattttgtac tttaaaaaat tcattttttt tattttttcg actttaaagc 60
caaaattatc ctttatttat gaaagtggat gtatttttat ccctttaata taaggttgag 120
ttttctttaa gtttgtaatg ttaagtgggg cactttatat tagtccaatt aacagaattg 180
acttagaaat tgatggcgat tacttgaaaa aatcgctaca aaaacataat aaaataaaat 240
ttcatttgat ttttaaacca aaagaaaagt gacatgacta acgatttgat cccatcttct 300
ttttttttta agaaaaaaat taaaagaaaa atttattaaa ctggatgatt aattttaaca 360
taaaaattaa aaaaataaaa atcaatcgag ctcactattt gatattatta ttttcatctt 420
gatattgtta cttcatatca ctgaattata aaatgcgtta acggcaatta tcatagacgc 480
agactgaaag cataaaatta caactattcc atttttgttt tttttttgcc aacccaaaaa 540
aaactaataa tttatttaca taattataga aaattggaat ttattcatgt ttttggacca 600
ttcagataac cccacaataa aaaaaaagtg cacaaaagtg caaagtgcta cgtgtcctaa 660
ttatagccat actacatttg tgcataaaat tagccaccat tatttttaag attattcttt 720
ttcttgtttt aatttatatg ccacactttc ttattaatcc atcttaaaca gaatgacaca 780
ctttttattt gacaaatatt taatgacatt attaacattt tatcctcgtt agatttcact 840
atgcggaata gagaatgtgt aaactttaaa cttcaaattt tggctccgca tacgactagc 900
gactatataa taataggaat tgagcacttg gcttttgtat atagcttcta tgtgtaccaa 960
aattagaaaa tcaggcgatt attataatct tgttgactaa atatagaatg catccattac 1020
ccccaaaaag tgtgattcca ctgtcatagg aggttttttt tatttcattt tatttgtgct 1080
ttcaataatg tagagtagtt ttacaaagat cctttctttg tgacacatgg taggtaatat 1140
tgctgatttt gttgtagttt tggggttata aagtttcaaa ttatttatac tggagggtag 1200
gggtgggggt tgtctataat gcaggttatg gttttacgtg aactcaataa ttattgtaga 1260
tactaagaaa tccactcagt gttcttgcgg tgtcttgctt ttgatttcag catcacttgt 1320
agttgattgt gtttagatta tcacattatt ctgtggctgt aactgtatcc ttgttagttg 1380
ctttgtttct acactgttgt tttccctctt ttatacctat tttgatatgt tgtactcgaa 1440
cgagggtcat cggggaacaa cctctttacc tccgtgaggt agagctatgg tctgtgtcca 1500
ctctaccctc cccagatccc tcttgtagga tttcactata ttgtaatatt aacttgaggt 1560
cactatagga gctcaaaaac ttctaatttt gaatcaatgt ctggttatac tttttttgtc 1620
ataactgtat ctcaaatgtg gtgtttggtt tatctcattt tgcagaagtc aagaaacagg 1680
ttactcctgt ttgagtgagg aaaagttggt ttgcctgtct gtggtctttt tataatcttt 1740
ttctacagaa gagaaagtgg gtaattttgt ttgagagtgg aaatattctc tagtgggaat 1800
ctactaggag taatttattt tctataaact aagtaaagtt tggaaggtga caaaaagaaa 1860
gacaaaaatc ttggaattgt tttagacaac caaggttttc ttgctcagaa tgtctgttgc 1920
cttgttatgg gttgtttctc cttgtgacgt ctcaaatggg acaagtttca tggaatcagt 1980
ccgggaggga aaccgttttt ttgattcatc gaggcatagg aatttggtgt ccaatgagag 2040
aatcaataga ggtggtggaa agcaaactaa taatggacgg aaattttctg tacggtctgc 2100
tattttggct actccatctg gagaacggac gatgacatcg gaacagatgg tctatgatgt 2160
ggttttgagg caggcagcct tggtgaagag gcaactgaga tctaccaatg agttagaagt 2220
gaagccggat atacctattc cggggaattt gggcttgttg agtgaagcat atgataggtg 2280
tggtgaagta tgtgcagagt atgcaaagac gtttaactta ggttagcttc ttcaatctat 2340
tcattcgttt accaaatatt atttggtaag cactaattat gaatatatat atgttcatgt 2400
tattgatgaa gacaaaattt gatctttgtt tgtttattca ggaactatgc taatgactcc 2460
cgagagaaga agggctatct aggcaatata tggtgaggtt tctagccatt taataacagt 2520
tacgcgcaca aacacatatg attaatcggg gacgagaaaa aaagaaatga agtttgagtt 2580
ttgagggtca tatgtaatag gtaaatccga gcttgactag cttgagatgt ttattgtcat 2640
atcatgctca atactaacca aaacactgaa aaagaacttg attatattta catactaata 2700
ttttcatttg cgttgctgtt cacattttta cctatggaac tggtttttgt gatttgttat 2760
acttcatatt cgatgttaat aaaatatatc attcctccct ttttctccac ttcaagcttt 2820
actgtagtgt tgaaagggga aactcctttt aatgattgca tatataaacg aacttcttga 2880
gttgaatagt ttctcattat gatctgttta aacagtatgg tgcagaagaa cagatgaact 2940
tgttgatggc ccaaacgcat catatattac cccggcagcc ttagataggt gggaaaatag 3000
gctagaagat gttttcaatg ggcggccatt tgacatgctc gatggtgctt tgtccgatac 3060
agtttctaac tttccagttg atattcaggt tagtctacca attctatggt ctttatattt 3120
gttcaatttg cgtttgatgt cacttttgct gagggctttt ctaatagctt acttcagcct 3180
agcggaaatg tttgtagttg aatctctagt tctgtctcct atatctgttt ctctcgtcct 3240
agatactaca catacttcat ttctgtttta acattttatt cgtcttttgg tgttgttttg 3300
tatgtgaatc atatatttgg aacagaatca ttattagttc acatgatttc atttgctttc 3360
ttcaatagcg taattgtcta accttccaat atatgttgca gccattcaga gatatgattg 3420
aaggaatgcg tatggacttg agaaaatcga gatacaaaaa cttcgacgaa ctataccttt 3480
attgttatta tgttgctggt acggttgggt tgatgagtgt tccaattatg gcgctgtcat 3540
gagacgaatt ctgacaggat atattggcgg gtaaacctaa gagaaaagag cgtttattag 3600
aataatcgga tatttaaaag ggcgtgaaaa ggtttatccg ttcgtccatt tgtatgtgca 3660
tgccaaccac agggttcccc tcgggatcaa agtactttga tccaacccct ccgctgctat 3720
agtgcagtcg gcttctgacg ttcagtgcag ccgtcatctg aaaacgacat gtcgcacaag 3780
tcctaagtta cgcgacaggc tgccgccctg cccttttcct ggcgttttct tgtcgcgtgt 3840
tttagtcgca taaagtagaa tacttgcgac tagaaccgga gacattacgc catgaacaag 3900
agcgccgccg ctggcctgct gggctatgcc cgcgtcagca ccgacgacca ggacttgacc 3960
aaccaacggg ccgaactgca cgcggccggc tgcaccaagc tgttttccga gaagatcacc 4020
ggcaccaggc gcgaccgccc ggagctggcc aggatgcttg accacctacg ccctggcgac 4080
gttgtgacag tgaccaggct agaccgcctg gcccgcagca cccgcgacct actggacatt 4140
gccgagcgca tccaggaggc cggcgcgggc ctgcgtagcc tggcagagcc gtgggccgac 4200
accaccacgc cggccggccg catggtgttg accgtgttcg ccggcattgc cgagttcgag 4260
cgttccctaa tcatcgaccg cacccggagc gggcgcgagg ccgccaaggc ccgaggcgtg 4320
aagtttggcc cccgccctac cctcaccccg gcacagatcg cgcacgcccg cgagctgatc 4380
gaccaggaag gccgcaccgt gaaagaggcg gctgcactgc ttggcgtgca tcgctcgacc 4440
ctgtaccgcg cacttgagcg cagcgaggaa gtgacgccca ccgaggccag gcggcgcggt 4500
gccttccgtg aggacgcatt gaccgaggcc gacgccctgg cggccgccga gaatgaacgc 4560
caagaggaac aagcatgaaa ccgcaccagg acggccagga cgaaccgttt ttcattaccg 4620
aagagatcga ggcggagatg atcgcggccg ggtacgtgtt cgagccgccc gcgcacctct 4680
caaccgtgcg gctgcatgaa atcctggccg gtttgtctga tgccaagctg gcggcctggc 4740
cggccagctt ggccgctgaa gaaaccgagc gccgccgtct aaaaaggtga tgtgtatttg 4800
agtaaaacag cttgcgtcat gcggtcgctg cgtatatgat ccgatgagta aataaacaaa 4860
tacgcaaggg gaacgcatga aggttatcgc tgtacttaac cagaaaggcg ggtcaggcaa 4920
gacgaccatc ggaacccatc tagcccgcgc cctgcaactc gccggggccg atgttctgtt 4980
agtcgattcc gatccccagg gcagtgcccg cgattgggcg gccgtgcggg aagatcaacc 5040
gctaaccgtt gtcggcatcg accgcccgac gattgaccgc gacgtgaagg ccatcggccg 5100
gcgcgacttc gtagtgatcg acggagcgcc ccaggcggcg gacttggctg tgtccgcgat 5160
caaggcagcc gacttcgtgc tgattccggt gcagccaagc ccttacgaca tatgggccac 5220
cgccgacctg gtggagctgg ttaagcagcg cattgaggtc acggatggaa ggctacaagc 5280
ggcctttgtc gtgtcgcggg cgatcaaagg cacgcgcatc ggcggtgagg ttgccgaggc 5340
gctggccggg tacgagctgc ccattcttga gtcccgtatc acgcagcgcg tgagctaccc 5400
aggcactgcc gccgccggca caaccgttct tgaatcagaa cccgagggcg acgctgcccg 5460
cgaggtccag gcgctggccg ctgaaattaa atcaaaactc atttgagtta atgaggtaaa 5520
gagaaaatga gcaaaagcac aaacacgcta agtgccggcc gtccgagcgc acgcagcagc 5580
aaggctgcaa cgttggccag cctggcagac acgccagcca tgaagcgggt caactttcag 5640
ttgccggcgg aggatcacac caagctgaag atgtacgcgg tacgccaagg caagaccatt 5700
accgagctgc tatctgaata gatcgcgcag ctaccagagt aaatgagcaa atgaataaat 5760
gagtagatga attttagcgg ctaaaggagg cggcatggaa aatcaagaac aaccaggcac 5820
cgacgccgtg gaatgcccca tgtgtggagg aacgggcggt tggccaggcg taagcggctg 5880
ggttgtctgc cggccctgca atggcactgg aacccccaag cccgaggaat cggcgtgacg 5940
gtcgcaaacc atccggcccg gtacaaatcg gcgcggcgct gggtgatgac ctggtggaga 6000
agttgaaggc cgcgcaggcc gcccagcggc aacgcatcga ggcagaagca cgccccggtg 6060
aatcgtggca agcggccgct gatcgaatcc gcaaagaatc ccggcaaccg ccggcagccg 6120
gtgcgccgtc gattaggaag ccgcccaagg gcgacgagca accagatttt ttcgttccga 6180
tgctctatga cgtgggcacc cgcgatagtc gcagcatcat ggacgtggcc gttttccgtc 6240
tgtcgaagcg tgaccgacga gctggcgagg tgatccgcta cgagcttcca gacgggcacg 6300
tagaggtttc cgcagggccg gccggcatgg ccagtgtgtg ggattacgac ctggtactga 6360
tggcggtttc ccatctaacc gaatccatga accgataccg ggaagggaag ggagacaagc 6420
ccggccgcgt gttccgtcca cacgttgcgg acgtactcaa gttctgccgg cgagccgatg 6480
gcggaaagca gaaagacgac ctggtagaaa cctgcattcg gttaaacacc acgcacgttg 6540
ccatgcagcg tacgaagaag gccaagaacg gccgcctggt gacggtatcc gagggtgaag 6600
ccttgattag ccgctacaag atcgtaaaga gcgaaaccgg gcggccggag tacatcgaga 6660
tcgagctagc tgattggatg taccgcgaga tcacagaagg caagaacccg gacgtgctga 6720
cggttcaccc cgattacttt ttgatcgatc ccggcatcgg ccgttttctc taccgcctgg 6780
cacgccgcgc cgcaggcaag gcagaagcca gatggttgtt caagacgatc tacgaacgca 6840
gtggcagcgc cggagagttc aagaagttct gtttcaccgt gcgcaagctg atcgggtcaa 6900
atgacctgcc ggagtacgat ttgaaggagg aggcggggca ggctggcccg atcctagtca 6960
tgcgctaccg caacctgatc gagggcgaag catccgccgg ttcctaatgt acggagcaga 7020
tgctagggca aattgcccta gcaggggaaa aaggtcgaaa aggactcttt cctgtggata 7080
gcacgtacat tgggaaccca aagccgtaca ttgggaaccg gaacccgtac attgggaacc 7140
caaagccgta cattgggaac cggtcacaca tgtaagtgac tgatataaaa gagaaaaaag 7200
gcgatttttc cgcctaaaac tctttaaaac ttattaaaac tcttaaaacc cgcctggcct 7260
gtgcataact gtctggccag cgcacagccg aagagctgca aaaagcgcct acccttcggt 7320
cgctgcgctc cctacgcccc gccgcttcgc gtcggcctat cgcggccgct ggccgctcaa 7380
aaatggctgg cctacggcca ggcaatctac cagggcgcgg acaagccgcg ccgtcgccac 7440
tcgaccgccg gcgcccacat caaggcaccc tgcctcgcgc gtttcggtga tgacggtgaa 7500
aacctctgac acatgcagct cccggtgacg gtcacagctt gtctgtaagc ggatgccggg 7560
agcagacaag cccgtcaggg cgcgtcagcg ggtgttggcg ggtgtcgggg cgcagccatg 7620
acccagtcac gtagcgatag cggagtgtat actggcttaa ctatgcggca tcagagcaga 7680
ttgtactgag agtgcaccat atgcggtgtg aaataccgca cagatgcgta aggagaaaat 7740
accgcatcag gcgctcttcc gcttcctcgc tcactgactc gctgcgctcg gtcgttcggc 7800
tgcggcgagc ggtatcagct cactcaaagg cggtaatacg gttatccaca gaatcagggg 7860
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccaggaac cgtaaaaagg 7920
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 7980
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 8040
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 8100
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 8160
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 8220
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 8280
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 8340
tcttgaagtg gtggcctaac tacggctaca ctagaaggac agtatttggt atctgcgctc 8400
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 8460
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 8520
ctcaagaaga tcctttgatc ttttctacgg ggtctgacgc tcagtggaac gaaaactcac 8580
gttaagggat tttggtcatg cattctaggt gattagaaaa actcatcgag catcaaatga 8640
aactgcaatt tattcatatc aggattatca ataccatatt tttgaaaaag ccgtttctgt 8700
aatgaaggag aaaactcacc gaggcagttc cataggatgg caagatcctg gtatcggtct 8760
gcgattccga ctcgtccaac atcaatacaa cctattaatt tcccctcgtc aaaaataagg 8820
ttatcaagtg agaaatcacc atgagtgacg actgaatccg gtgagaatgg caaaagttta 8880
tgcatttctt tccagacttg ttcaacaggc cagccattac gctcgtcatc aaaatcactc 8940
gcatcaacca aaccgttatt cattcgtgat tgcgcctgag cgagtcgaaa tacgcgatcg 9000
ctgttaaaag gacaattaca aacaggaatc gaatgcaacc ggcgcaggaa cactgccagc 9060
gcatcaacaa tattttcacc tgaatcagga tattcttcta atacctggaa tgctgttttc 9120
cctgggatcg cagtggtgag taaccatgca tcatcaggag tacggataaa atgcttgatg 9180
gtcggaagag gcataaattc cgtcagccag tttagtctga ccatctcatc tgtaacatca 9240
ttggcaacgc tacctttgcc atgtttcaga aacaactctg gcgcatcggg cttcccatac 9300
aatcggtaga ttgtcgcacc tgattgcccg acattatcgc gagcccattt atacccatat 9360
aaatcagcat ccatgttgga atttaatcgc ggccttgagc aagacgtttc ccgttgaata 9420
tggctcataa cagaacttat tatttccttc ctcttttcta cagtatttaa agatacccca 9480
agaagctaat tataacaaga cgaactccaa ttcactgttc cttgcattct aaaaccttaa 9540
ataccagaaa acagcttttt caaagttgtt ttcaaagttg gcgtataaca tagtatcgac 9600
ggagccgatt ttgaaaccgc ggtgatcaca ggcagcaacg ctctgtcatc gttacaatca 9660
acatgctacc ctccgcgaga tcatccgtgt ttcaaacccg gcagcttagt tgccgttctt 9720
ccgaatagca tcggtaacat gagcaaagtc tgccgcctta caacggctct cccgctgacg 9780
ccgtcccgga ctgatgggct gcctgtatcg agtggtgatt ttgtgccgag ctgccggtcg 9840
gggagctgtt ggctggctgg tggcaggata tattgtggtg taaacataac gaattcgtct 9900
ca 9902
<210> 64
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CaMV35S
<400> 64
ctcgacgaat taattccaat cccacaaaaa tctgagctta acagcacagt tgctcctctc 60
agagcagaat cgggtattca acaccctcat atcaactact acgttgtgta taacggtcca 120
catgccggta tatacgatga ctggggttgt acaaaggcgg caacaaacgg cgttcccgga 180
gttgcacaca agaaatttgc cactattaca gaggcaagag cagcagctga cgcgtacaca 240
acaagtcagc aaacagacag gttgaacttc atccccaaag gagaagctca actcaagccc 300
aagagctttg ctaaggccct aacaagccca ccaaagcaaa aagcccactg gctcacgcta 360
ggaaccaaaa ggcccagcag tgatccagcc ccaaaagaga tctcctttgc cccggagatt 420
acaatggacg atttcctcta tctttacgat ctaggaagga agttcgaagg tgaaggtgac 480
gacactatgt tcaccactga taatgagaag gttagcctct tcaatttcag aaagaatgct 540
gacccacaga tggttagaga ggcctacgca gcaggtctca tcaagacgat ctacccgagt 600
aacaatctcc aggagatcaa ataccttccc aagaaggtta aagatgcagt caaaagattc 660
aggactaatt gcatcaagaa cacagagaaa gacatatttc tcaagatcag aagtactatt 720
ccagtatgga cgattcaagg cttgcttcat aaaccaaggc aagtaataga gattggagtc 780
tctaaaaagg tagttcctac tgaatctaag gccatgcatg gagtctaaga ttcaaatcga 840
ggatctaaca gaactcgccg tgaagactgg cgaacagttc atacagagtc ttttacgact 900
caatgacaag aagaaaatct tcgtcaacat ggtggagcac gacactctgg tctactccaa 960
aaatgtcaaa gatacagtct cagaagacca aagggctatt gagacttttc aacaaaggat 1020
aatttcggga aacctcctcg gattccattg cccagctatc tgtcacttca tcgaaaggac 1080
agtagaaaag gaaggtggct cctacaaatg ccatcattgc gataaaggaa aggctatcat 1140
tcaagatctc tctgccgaca gtggtcccaa agatggaccc ccacccacga ggagcatcgt 1200
ggaaaaagaa gacgttccaa ccacgtcttc aaagcaagtg gattgatgtg acatctccac 1260
tgacgtaagg gatgacgcac aatcccacta tccttcgcaa gacccttcct ctatataagg 1320
aagttcattt catttggaga ggacacg 1347
<210> 65
<211> 4224
<212> ДНК
<213> Искусственная последовательность
<220>
<223> atCas9
<400> 65
atggatcccc gggatcatct acttctgaag actcagactc agactaagca ggtgacgaac 60
gtcaccaatc ccaattcgat ctacatcgat aagaagtact ctatcggact cgatatcgga 120
actaactctg tgggatgggc tgtgatcacc gatgagtaca aggtgccatc taagaagttc 180
aaggttctcg gaaacaccga taggcactct atcaagaaaa accttatcgg tgctctcctc 240
ttcgattctg gtgaaactgc tgaggctacc agactcaaga gaaccgctag aagaaggtac 300
accagaagaa agaacaggat ctgctacctc caagagatct tctctaacga gatggctaaa 360
gtggatgatt cattcttcca caggctcgaa gagtcattcc tcgtggaaga agataagaag 420
cacgagaggc accctatctt cggaaacatc gttgatgagg tggcatacca cgagaagtac 480
cctactatct accacctcag aaagaagctc gttgattcta ctgataaggc tgatctcagg 540
ctcatctacc tcgctctcgc tcacatgatc aagttcagag gacacttcct catcgagggt 600
gatctcaacc ctgataactc tgatgtggat aagttgttca tccagctcgt gcagacctac 660
aaccagcttt tcgaagagaa ccctatcaac gcttcaggtg tggatgctaa ggctatcctc 720
tctgctaggc tctctaagtc aagaaggctt gagaacctca ttgctcagct ccctggtgag 780
aagaagaacg gacttttcgg aaacttgatc gctctctctc tcggactcac ccctaacttc 840
aagtctaact tcgatctcgc tgaggatgca aagctccagc tctcaaagga tacctacgat 900
gatgatctcg ataacctcct cgctcagatc ggagatcagt acgctgattt gttcctcgct 960
gctaagaacc tctctgatgc tatcctcctc agtgatatcc tcagagtgaa caccgagatc 1020
accaaggctc cactctcagc ttctatgatc aagagatacg atgagcacca ccaggatctc 1080
acacttctca aggctcttgt tagacagcag ctcccagaga agtacaaaga gattttcttc 1140
gatcagtcta agaacggata cgctggttac atcgatggtg gtgcatctca agaagagttc 1200
tacaagttca tcaagcctat cctcgagaag atggatggaa ccgaggaact cctcgtgaag 1260
ctcaatagag aggatcttct cagaaagcag aggaccttcg ataacggatc tatccctcat 1320
cagatccacc tcggagagtt gcacgctatc cttagaaggc aagaggattt ctacccattc 1380
ctcaaggata acagggaaaa gattgagaag attctcacct tcagaatccc ttactacgtg 1440
ggacctctcg ctagaggaaa ctcaagattc gcttggatga ccagaaagtc tgaggaaacc 1500
atcacccctt ggaacttcga agaggtggtg gataagggtg ctagtgctca gtctttcatc 1560
gagaggatga ccaacttcga taagaacctt ccaaacgaga aggtgctccc taagcactct 1620
ttgctctacg agtacttcac cgtgtacaac gagttgacca aggttaagta cgtgaccgag 1680
ggaatgagga agcctgcttt tttgtcaggt gagcaaaaga aggctatcgt tgatctcttg 1740
ttcaagacca acagaaaggt gaccgtgaag cagctcaaag aggattactt caagaaaatc 1800
gagtgcttcg attcagttga gatttctggt gttgaggata ggttcaacgc atctctcgga 1860
acctaccacg atctcctcaa gatcattaag gataaggatt tcttggataa cgaggaaaac 1920
gaggatatct tggaggatat cgttcttacc ctcaccctct ttgaagatag agagatgatt 1980
gaagaaaggc tcaagaccta cgctcatctc ttcgatgata aggtgatgaa gcagttgaag 2040
agaagaagat acactggttg gggaaggctc tcaagaaagc tcattaacgg aatcagggat 2100
aagcagtctg gaaagacaat ccttgatttc ctcaagtctg atggattcgc taacagaaac 2160
ttcatgcagc tcatccacga tgattctctc acctttaaag aggatatcca gaaggctcag 2220
gtttcaggac agggtgatag tctccatgag catatcgcta acctcgctgg atctcctgca 2280
atcaagaagg gaatcctcca gactgtgaag gttgtggatg agttggtgaa ggtgatggga 2340
aggcataagc ctgagaacat cgtgatcgaa atggctagag agaaccagac cactcagaag 2400
ggacagaaga actctaggga aaggatgaag aggatcgagg aaggtatcaa agagcttgga 2460
tctcagatcc tcaaagagca ccctgttgag aacactcagc tccagaatga gaagctctac 2520
ctctactacc tccagaacgg aagggatatg tatgtggatc aagagttgga tatcaacagg 2580
ctctctgatt acgatgttga tcatatcgtg ccacagtcat tcttgaagga tgattctatc 2640
gataacaagg tgctcaccag gtctgataag aacaggggta agagtgataa cgtgccaagt 2700
gaagaggttg tgaagaaaat gaagaactat tggaggcagc tcctcaacgc taagctcatc 2760
actcagagaa agttcgataa cttgactaag gctgagaggg gaggactctc tgaattggat 2820
aaggcaggat tcatcaagag gcagcttgtg gaaaccaggc agatcactaa gcacgttgca 2880
cagatcctcg attctaggat gaacaccaag tacgatgaga acgataagtt gatcagggaa 2940
gtgaaggtta tcaccctcaa gtcaaagctc gtgtctgatt tcagaaagga tttccaattc 3000
tacaaggtga gggaaatcaa caactaccac cacgctcacg atgcttacct taacgctgtt 3060
gttggaaccg ctctcatcaa gaagtatcct aagctcgagt cagagttcgt gtacggtgat 3120
tacaaggtgt acgatgtgag gaagatgatc gctaagtctg agcaagagat cggaaaggct 3180
accgctaagt atttcttcta ctctaacatc atgaatttct tcaagaccga gattaccctc 3240
gctaacggtg agatcagaaa gaggccactc atcgagacaa acggtgaaac aggtgagatc 3300
gtgtgggata agggaaggga tttcgctacc gttagaaagg tgctctctat gccacaggtg 3360
aacatcgtta agaaaaccga ggtgcagacc ggtggattct ctaaagagtc tatcctccct 3420
aagaggaact ctgataagct cattgctagg aagaaggatt gggaccctaa gaaatacggt 3480
ggtttcgatt ctcctaccgt ggcttactct gttctcgttg tggctaaggt tgagaaggga 3540
aagagtaaga agctcaagtc tgttaaggaa cttctcggaa tcactatcat ggaaaggtca 3600
tctttcgaga agaacccaat cgatttcctc gaggctaagg gatacaaaga ggttaagaag 3660
gatctcatca tcaagctccc aaagtactca ctcttcgaac tcgagaacgg tagaaagagg 3720
atgctcgctt ctgctggtga gcttcaaaag ggaaacgagc ttgctctccc atctaagtac 3780
gttaactttc tttacctcgc ttctcactac gagaagttga agggatctcc agaagataac 3840
gagcagaagc aacttttcgt tgagcagcac aagcactact tggatgagat catcgagcag 3900
atctctgagt tctctaaaag ggtgatcctc gctgatgcaa acctcgataa ggtgttgtct 3960
gcttacaaca agcacagaga taagcctatc agggaacagg cagagaacat catccatctc 4020
ttcaccctta ccaacctcgg tgctcctgct gctttcaagt acttcgatac aaccatcgat 4080
aggaagagat acacctctac caaagaagtg ctcgatgcta ccctcatcca tcagtctatc 4140
actggactct acgagactag gatcgatctc tcacagctcg gtggtgattc aagggctgat 4200
cctaagaaga agaggaaggt ttga 4224
<210> 66
<211> 771
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Терминатор OCS
<400> 66
gtcctgcttt aatgagatat gcgagacgcc tatgatcgca tgatatttgc tttcaattct 60
gttgtgcacg ttgtaaaaaa cctgagcatg tgtagctcag atccttaccg ccggtttcgg 120
ttcattctaa tgaatatatc acccgttact atcgtatttt tatgaataat attctccgtt 180
caatttactg attgtaccct actacttata tgtacaatat taaaatgaaa acaatatatt 240
gtgctgaata ggtttatagc gacatctatg atagagcgcc acaataacaa acaattgcgt 300
tttattatta caaatccaat tttaaaaaaa gcggcagaac cggtcaaacc taaaagactg 360
attacataaa tcttattcaa atttcaaaag gccccagggg ctagtatcta cgacacaccg 420
agcggcgaac taataacgtt cactgaaggg aactccggtt ccccgccggc gcgcatgggt 480
gagattcctt gaagttgagt attggccgtc cgctctaccg aaagttacgg gcaccattca 540
acccggtcca gcacggcggc cgggtaaccg acttgctgcc ccgagaatta tgcagcattt 600
ttttggtgta tgtgggcccc aaatgaagtg caggtcaaac cttgacagtg acgacaaatc 660
gttgggcggg tccagggcga attttgcgac aacatgtcga ggctcagcag gacctgcagg 720
catgcaagct agcttactag tgatgcatat tctatagtgt cacctaaatc t 771
<210> 67
<211> 387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Промотор U6-26
<400> 67
ctttttttct tcttcttcgt tcatacagtt tttttttgtt tatcagctta cattttcttg 60
aaccgtagct ttcgttttct tctttttaac tttccattcg gagtttttgt atcttgtttc 120
atagtttgtc ccaggattag aatgattagg catcgaacct tcaagaattt gattgaataa 180
aacatcttca ttcttaagat atgaagataa tcttcaaaag gcccctggga atctgaaaga 240
agagaagcag gcccatttat atgggaaaga acaatagtat ttcttatata ggcccattta 300
agttgaaaac aatcttcaaa agtcccacat cgcttagata agaaaacgaa gctgagttta 360
tatacagcta gagtcgaagt agtgatt 387
<210> 68
<211> 104
<212> ДНК
<213> Искусственная последовательность
<220>
<223> огРНК Ps#1
<400> 68
ggaatgtctg ttgccttgtt agttttagag ctagaaatag caagttaaaa taaggctagt 60
ccgttatcaa cttgaaaaag tggcaccgag tcggtgcttt tttt 104
<210> 69
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1228483
<400> 69
aactgctttg aatgtccata tgg 23
<210> 70
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1222177
<400> 70
gctggagaac cgccgtttaa cgg 23
<210> 71
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1261146
<400> 71
cgcttgaatg atgaccactg cgg 23
<210> 72
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1352616
<400> 72
atattgtttt tcatattttt tgg 23
<210> 73
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1352124
<400> 73
ccaaaaaaaa aaaatacagt cgt 23
<210> 74
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1854159
<400> 74
gtttccgcca ccaccgcctc cgg 23
<210> 75
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1843852
<400> 75
tctacaaagt cattgaaggt tgg 23
<210> 76
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1858597
<400> 76
agagttgatc tgtggctgtg gcgg 24
<210> 77
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 4684724
<400> 77
tgactgcagg tgagcttaca cgg 23
<210> 78
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1565357
<400> 78
ccttggaaat tttctcttcc caa 23
<210> 79
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 1559196
<400> 79
ccccgacatt taatgatgtt ttt 23
<210> 80
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 3 хромосома Arabidopsis - 4639826
<400> 80
agaagttcag aaagtcgccc agg 23
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2745563C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2829155C2 |
| РНК-НАПРАВЛЯЕМАЯ ИНЖЕНЕРИЯ ГЕНОМА ЧЕЛОВЕКА | 2013 |
|
RU2699523C2 |
| УЛУЧШЕННЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ ДЛЯ ПОЛУЧЕНИЯ БЕЛКА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2016 |
|
RU2752529C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| ГЕТЕРОЗИГОТНЫЕ ПО CENH3 ОДНОДОЛЬНЫЕ РАСТЕНИЯ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ИНДУКЦИИ ГАПЛОИДОВ И ОДНОВРЕМЕННОГО РЕДАКТИРОВАНИЯ ГЕНОМА | 2021 |
|
RU2832578C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ С ИСПОЛЬЗОВАНИЕМ ПАРНЫХ ГИДОВЫХ РНК | 2015 |
|
RU2734770C2 |
| РНК-НАПРАВЛЯЕМАЯ ИНЖЕНЕРИЯ ГЕНОМА ЧЕЛОВЕКА | 2013 |
|
RU2766685C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ ИЛИ ПОДДЕРЖАНИЯ ПЛЮРИПОТЕНТНЫХ КЛЕТОК | 2015 |
|
RU2706965C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННЫХ ГЕНЕТИЧЕСКИХ МОДИФИКАЦИЙ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2711740C2 |


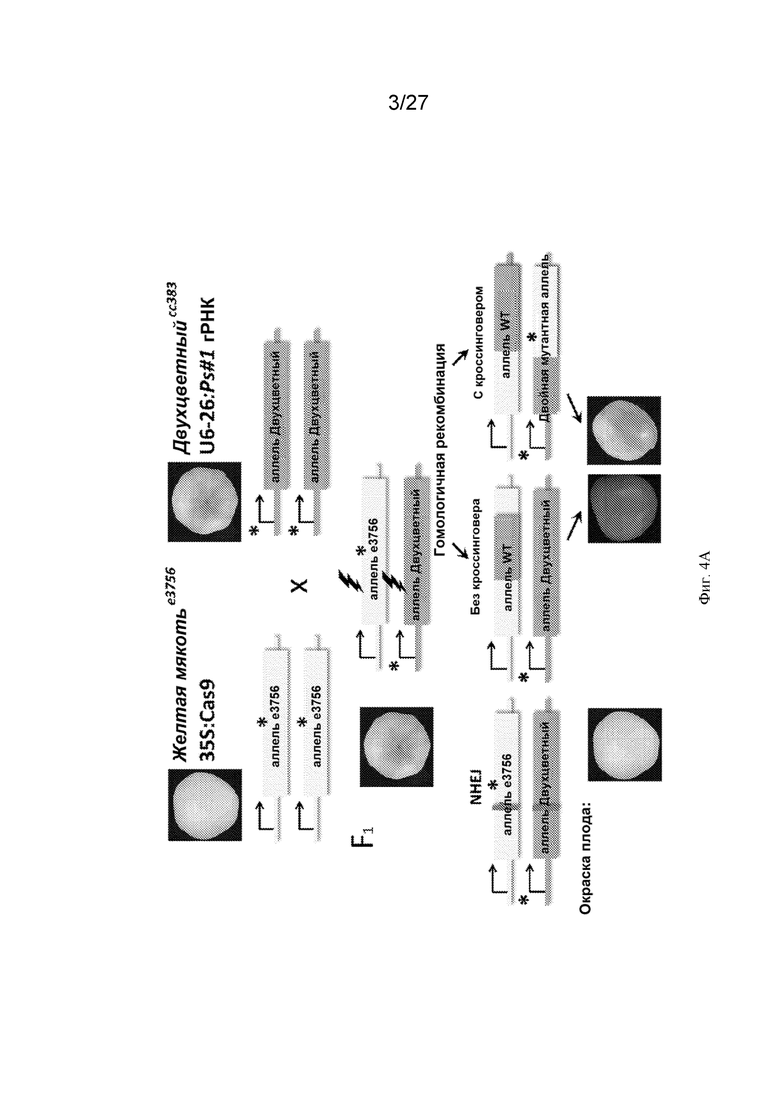

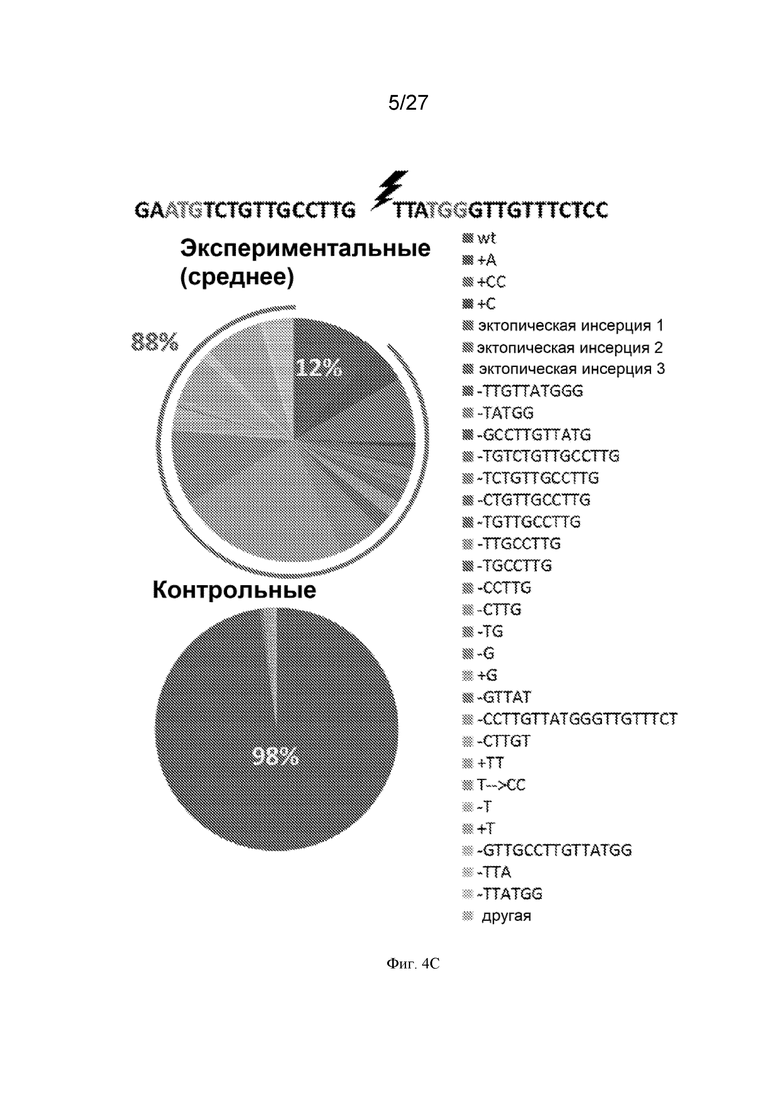


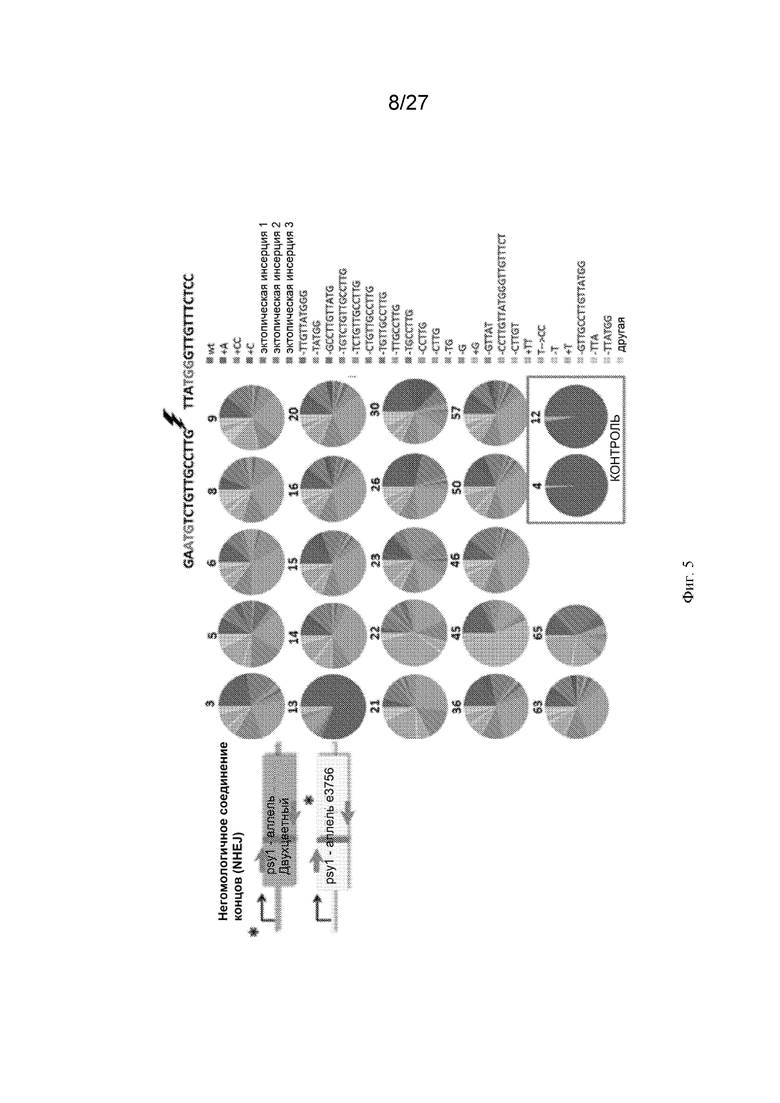







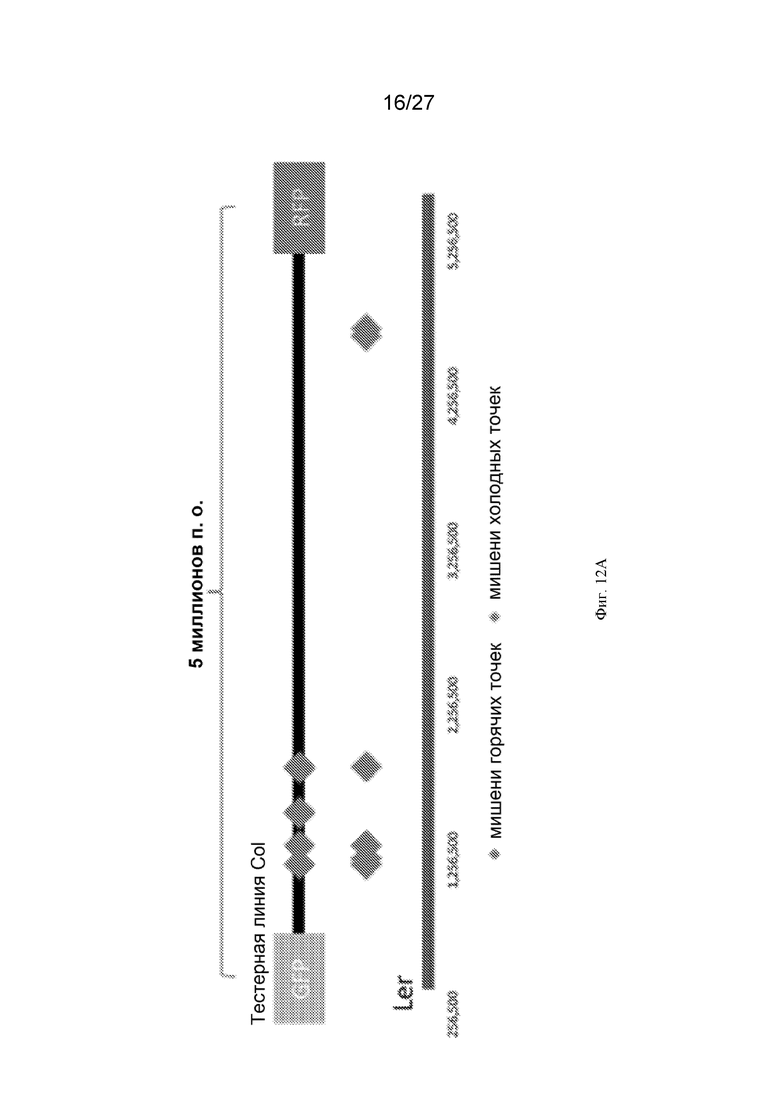



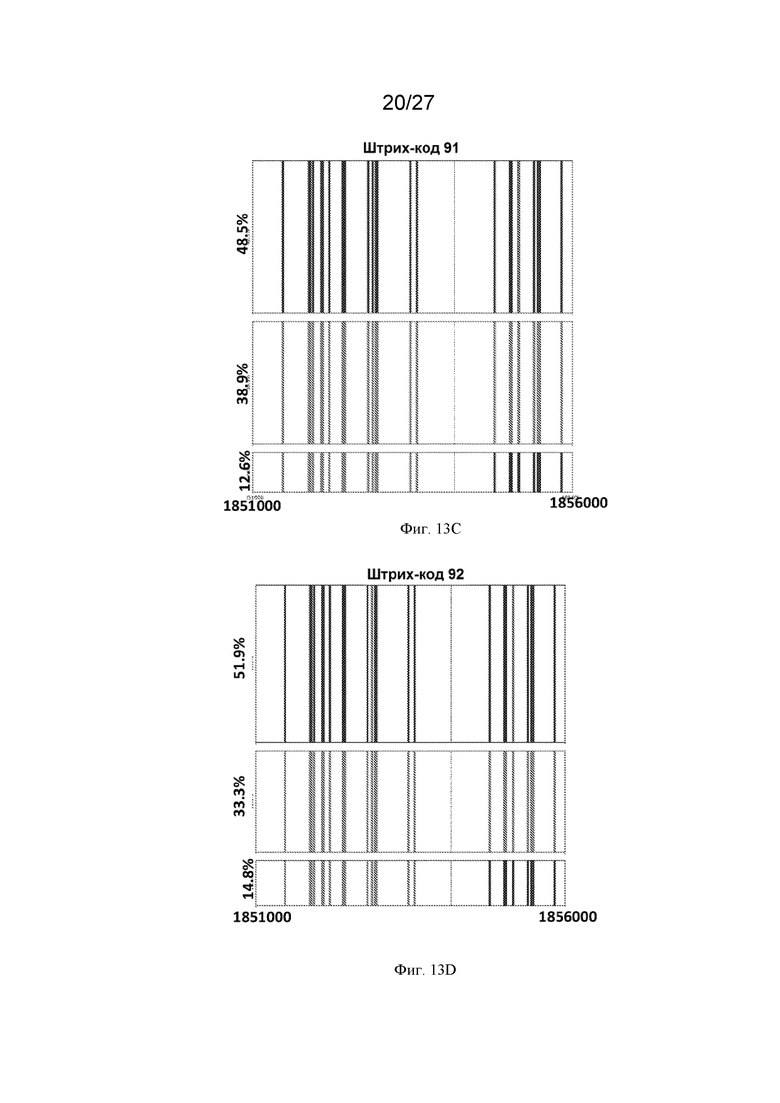



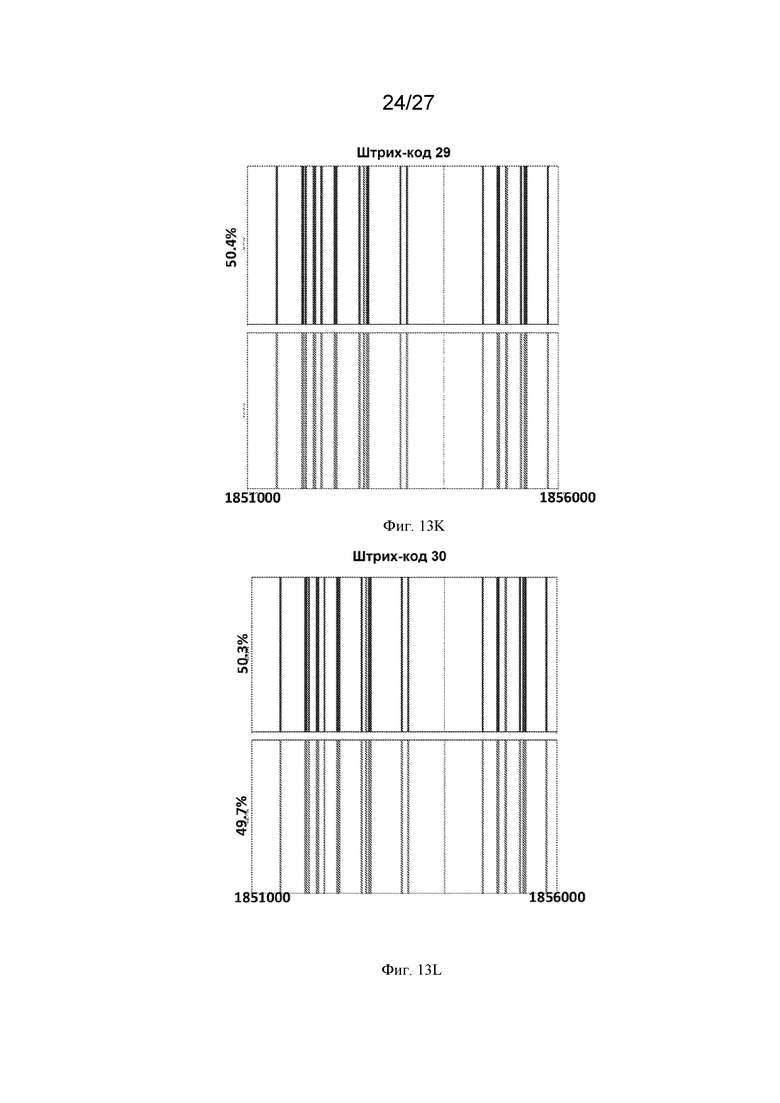

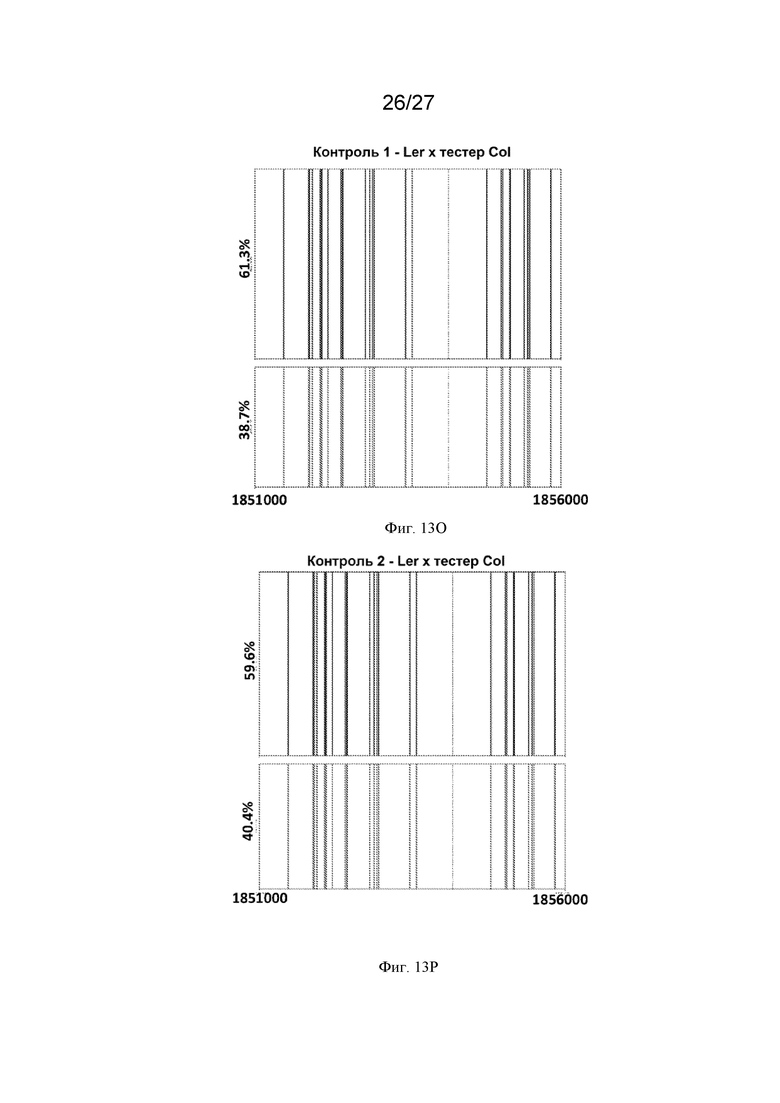

Изобретение относится к области биотехнологии. Описана группа изобретений, включающая способ направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения и генетически сконструированное растение томата, обладающее полезным признаком или качеством, полученное способом, включающим направленную рекомбинацию ДНК между гомологичными хромосомами в гибридной соматической клетке растения. В одном из вариантов реализации способ включает стадии осуществление экспрессии системы нуклеаз в соматической клетке растения, осуществление анализа потомства указанной соматической клетки растения или ткани растения, выращенной из указанной соматической клетки растения, или растения, выращенного из указанной соматической клетки, или потомства указанного растения на гомологичную рекомбинацию между указанными гомологичными хромосомами и выбор указанного потомства указанной соматической клетки растения или растительной ткани, выращенной из указанной соматической клетки растения, или растения, выращенного из указанной соматической клетки, или потомства указанного растения, в которых произошла направленная гомологичная рекомбинация. Изобретение расширяет арсенал средств направленной рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения. 2 н. и 25 з.п. ф-лы, 37 ил., 9 табл., 5 пр.
1. Способ направления рекомбинации ДНК между гомологичными хромосомами в соматической клетке растения, включающий следующие стадии:
(а) осуществление экспрессии системы нуклеаз в указанной соматической клетке растения,
где указанная система нуклеаз содержит систему II типа коротких палиндромных повторов, регулярно расположенных группами (CRISPR)/белка Cas9, ассоциированного с CRISPR (Cas9),
где указанная CRISPR/Cas9 система нуклеаз содержит Cas9 CRISPR-ассоциированную эндонуклеазу или ее гомолог и молекулу гРНК, где указанная молекула гРНК связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную Cas9 CRISPR-ассоциированную эндонуклеазу или ее гомолог на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени,
где указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на указанных гомологичных хромосомах, при этом при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, при этом указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей;
(b) осуществление анализа потомства указанной соматической клетки растения или ткани растения, выращенной из указанной соматической клетки растения, или растения, выращенного из указанной соматической клетки, или потомства указанного растения на гомологичную рекомбинацию между указанными гомологичными хромосомами, где указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером); и
(c) выбор указанного потомства указанной соматической клетки растения или растительной ткани, выращенной из указанной соматической клетки растения, или растения, выращенного из указанной соматической клетки, или потомства указанного растения, в которых произошла направленная гомологичная рекомбинация.
2. Способ по п. 1, отличающийся тем, что указанная соматическая клетка растения происходит из существующей гибридной или гетерозиготной клетки растения, имеющей полиморфные аллели в указанном предварительно выбранном сайте.
3. Способ по п. 2, отличающийся тем, что указанная существующая гибридная или гетерозиготная клетка растения происходит из растения дикого типа.
4. Способ по п. 2, отличающийся тем, что указанный способ обеспечивает получение соматической клетки растения, содержащей направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную соматическую клетку растения, где ткань растения содержит направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или растения, содержащего указанную соматическую клетку растения, или его растения-потомка, где растение содержит направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или плода, полученного от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, где плод содержит направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или семян, полученных от растения, содержащего указанную соматическую клетку растения, или его растения-потомка, где семена содержат направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или любой их комбинации.
5. Способ по п. 4, где соматическая клетка растения, содержащая направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткань растения, содержащая направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или растение, содержащее направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или плод, содержащий направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или семена, содержащие направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или любая их комбинация обладает комбинацией родительских признаков, не присутствующей ни у одного из родителей.
6. Способ по п. 5, отличающийся тем, что указанные родительские признаки включают повышенную засухоустойчивость, повышенную устойчивость к сельскохозяйственным вредителям, повышенную устойчивость к патогенам, улучшенное содержание питательных веществ или улучшенные параметры роста, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени.
7. Способ по п. 1, отличающийся тем, что указанная соматическая клетка растения происходит от клетки из потомства, полученного при скрещивании двух растений, при этом каждая из указанных родительских клеток растения содержит полиморфный аллель по сравнению с указанным партнером в указанном предварительно выбранном сайте.
8. Способ по п. 7, отличающийся тем, что указанный способ обеспечивает получение соматической клетки растения, содержащей направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или растения, содержащего указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или плода, полученного от растения, содержащего указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или семян, полученных от растения, содержащего указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или любой их комбинации, обладающих полученной в результате комбинацией родительских признаков, не присутствующей ни у одного из родителей.
9. Способ по п. 8, отличающийся тем, что указанные родительские признаки, рекомбинированные посредством указанной направленной гомологичной рекомбинации, включают повышенную засухоустойчивость, повышенную устойчивость к сельскохозяйственным вредителям, повышенную устойчивость к патогенам, улучшенное содержание питательных веществ или улучшенные параметры роста, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени.
10. Способ по п. 7, отличающийся тем, что одна из указанных родительских соматических клеток растения содержит указанную систему нуклеаз, и при этом активность расщепления ДНК указанной системы нуклеаз направлена на полиморфный аллель, присутствующий в другой родительской клетке растения, не содержащей указанную систему нуклеаз.
11. Способ по п. 7, отличающийся тем, что одна из указанных родительских соматических клеток растения содержит нуклеазу Cas9, и другая из указанных родительских соматических клеток растения содержит молекулу гРНК, при этом указанная молекула гРНК связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas9 на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени.
12. Способ по п. 1, отличающийся тем, что указанная соматическая клетка растения включает клетку из потомства растений, полученного при скрещивании двух полиморфных родительских линий, приводящем к созданию гибридного растения, при этом каждая из указанных родительских линий растений содержит полиморфный аллель в указанном предварительно выбранном эндогенном сайте-мишени, и при этом только одна из указанных родительских линий содержит указанную систему нуклеаз.
13. Способ по п. 12, отличающийся тем, что указанный способ обеспечивает получение соматической клетки растения, содержащей направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или растения, содержащего указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или плода, полученного от растения, содержащего указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или семян, полученных от растения, содержащего указанную соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или любой их комбинации, обладающих комбинацией родительских признаков, не присутствующей ни у одного из родителей.
14. Способ по п. 13, отличающийся тем, что указанные родительские признаки включают повышенную засухоустойчивость, повышенную устойчивость к сельскохозяйственным вредителям, повышенную устойчивость к патогенам, улучшенное содержание питательных веществ или улучшенные параметры роста, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени.
15. Способ по п. 12, отличающийся тем, что указанная система нуклеаз содержит нуклеазу Cas9 и молекулу гРНК, при этом указанная молекула гРНК связывается в указанном предварительно выбранном эндогенном сайте-мишени, направляя тем самым указанную нуклеазу Cas9 на расщепление ДНК в указанном предварительно выбранном эндогенном сайте-мишени, и при этом ДНК-расщепляющая активность указанной системы нуклеаз проявляется только в отношении гетерологичного аллеля, присутствующего в родительской клетке растения дикого типа.
16. Способ по п. 1, отличающийся тем, что указанная соматическая клетка растения содержится в ткани растения или в целом растении.
17. Способ по п. 1, отличающийся тем, что указанный предварительно выбранный эндогенный сайт-мишень содержит ДНК, содержащую ген, часть гена или регуляторную расположенную в 3'-5' направлении или расположенную в 5'-3' направлении последовательности гена, или любую их комбинацию, и при этом экспрессия указанного гена или ее отсутствие влияет на рост, засухоустойчивость, устойчивость к сельскохозяйственным вредителям, устойчивость к патогенам или содержание питательных веществ, или любой другой признак, полезный для клетки растения, ткани растения, растения, плода или семени, или любую их комбинацию.
18. Способ по п. 1, отличающийся тем, что указанный предварительно выбранный эндогенный сайт-мишень содержит область эухроматина или гетерохроматина.
19. Способ по п. 1, отличающийся тем, что указанное осуществление экспрессии включает конститутивную индукцию экспрессии, индуцибельную индукцию экспрессии, тканеспецифичную индукцию экспрессии или специфичную для определенных условий индукцию экспрессии, или любую их комбинацию.
20. Способ по п. 1, отличающийся тем, что указанная соматическая клетка растения включает протопласт.
21. Способ по п. 1, отличающийся тем, что указанная соматическая клетка растения включает клетку культурного растения.
22. Способ по п. 1, отличающийся тем, что анализ указанного растения включает анализ части указанного растения или его потомства, включая лист, стебель, почку, плод, семя.
23. Способ по п. 1, отличающийся тем, что указанное отобранное потомство согласно стадии (с), включает поколения F1, F2 или F3, или любое последующее поколение, или обратные скрещивания на 1-3 поколения, или любое последующее поколение, полученное при обратном скрещивании.
24. Способ по п. 1, отличающийся тем, что указанный способ обеспечивает получение соматической клетки растения, содержащей указанную направленную гомологичную рекомбинацию в указанном предварительно выбранном эндогенном сайте-мишени, или ткани растения, содержащей соматическую клетку растения, содержащую указанную направленную гомологичную рекомбинацию в указанном предварительно выбранном эндогенном сайте-мишени, или растения, содержащего соматическую клетку растения, содержащую указанную направленную гомологичную рекомбинацию в указанном предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или плода, полученного от растения, содержащего соматическую клетку растения, содержащую направленную гомологичную рекомбинацию в указанном предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или семян, полученных от растения, содержащего соматическую клетку растения, содержащую указанную направленную гомологичную рекомбинацию в предварительно выбранном эндогенном сайте-мишени, или его растения-потомка, или любой их комбинации, при этом указанные клетка, ткань, растение или его потомство, плод или семя имеют комбинацию родительских признаков, где указанная комбинация не присутствует ни у одного из родителей.
25. Способ по п. 24, где указанная клетка, ткань, растение или его потомство, плод или семя содержат гены для повышенной засухоустойчивости, повышенной устойчивости к сельскохозяйственным вредителям, повышенной устойчивости к патогенам, улучшенного содержания питательных веществ, улучшенных параметров роста или любого другого признака, полезного для клетки растения, ткани растения, растения или его потомства, плода или семени, или любой их комбинации, по сравнению с контрольными клеткой растения, растением или его потомством, плодом или семенем.
26. Генетически сконструированное растение томата, обладающее полезным признаком или качеством, полученное способом, включающим направленную рекомбинацию ДНК между гомологичными хромосомами в гибридной соматической клетке растения, при этом указанный способ включает следующие стадии:
(а) осуществление экспрессии CRISPR/Cas9 системы нуклеаз II типа или ее гомолога в указанной гибридной соматической клетке растения, при этом указанная экспрессированная система нуклеаз направлена на предварительно выбранный эндогенный сайт-мишень, содержащий полиморфные аллели на гомологичных хромосомах, при этом при экспрессии указанной системы нуклеаз ДНК по меньшей мере одного из указанных полиморфных аллелей расщепляется в указанном предварительно выбранном эндогенном сайте-мишени, при этом указанная нуклеаза расщепляет ДНК, создавая двухнитевой разрыв в ДНК по меньшей мере одного из указанных полиморфных аллелей;
(b) осуществление анализа потомства указанной соматической клетки растения, или ткани растения, выращенной из указанной соматической клетки растения, или растения, выращенного из указанной соматической клетки растения, или потомства указанного растения на гомологичную рекомбинацию между указанными гомологичными хромосомами, при этом указанная гомологичная рекомбинация включает кроссинговер или генную конверсию (не сопровождающуюся кроссинговером);
(c) выбор клетки растения, включающей потомство указанной соматической клетки растения, ткани растения, включающей потомство указанной соматической клетки растения, полученного из нее растения, включающего потомство указанной соматической клетки растения, или потомства указанного растения, содержащего потомство указанной соматической клетки растения, в которых произошла направленная гомологичная рекомбинация, наблюдаемая при указанном анализе на стадии (b);
(d) выращивание указанной клетки растения, включающей потомство указанной соматической клетки растения, или полученной из нее ткани растения, включающей потомство указанной соматической клетки растения, или полученного из нее растения, включающего потомство указанной соматической клетки растения, или потомства указанного растения, включающего потомство указанной соматической клетки растения, с получением генетически сконструированного растения томата, содержащего указанную направленную гомологичную рекомбинацию,
при этом указанное генетически сконструированное растение томата представляет собой генетически сконструированное растение Solanum lycopersicum, клетки которого экспрессируют гетерологичную CRISPR/Cas9 систему нуклеаз, и обладает полезным качеством или признаком, не присутствующим ни у одного из родительских растений, из которых происходила гибридная соматическая клетка,
где указанные полезное качество или признак представляет собой улучшенное содержание питательных веществ, и указанное генетически сконструированное растение томата пригодно для получения томатов с улучшенным содержанием питательных веществ.
27. Растение по п. 26, отличающееся тем, что указанный предварительно выбранный эндогенный сайт-мишень содержит область эухроматина или гетерохроматина.
| K | |||
| BELHAJ et al | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Способ образования коричневых окрасок на волокне из кашу кубической и подобных производных кашевого ряда | 1922 |
|
SU32A1 |
| Аппарат, предназначенный для летания | 0 |
|
SU76A1 |
| K.S | |||
| MAKAROVA et al | |||
| "Annotation and Classification of CRISPR-Cas Systems" Methods Mol | |||
| Biol., 2015, V | |||
| Устройство для выемки отдельных труб в экономайзерах | 1925 |
|
SU1311A1 |
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| WO 2014013056 A1 23.01.2014 | |||
| WO 2014104878 | |||
