I. УРОВЕНЬ ТЕХНИКИ
1. В последние годы получила развитие иммунотерапия рака. Генно модифицированные Т-клетки химерного антигенного рецептора (CAR) являются отличным примером сконструированных иммунных клеток, успешно применяемых в иммунотерапии рака. Эти клетки были недавно одобрены FDA (Комиссия по контролю за лекарствами и питательными веществами, США) для лечения злокачественных новообразований CD19 + В-клеток, но до сих пор эффективность ограничивалась заболеваниями, несущими несколько целевых антигенов, и нацеливание на такой ограниченный антигенный репертуар подвержено неудачам из-за ускользания от иммунного ответа. Дополнительно, CAR Т-клетки были ориентированы на использование аутологичных Т-клеток из-за риска болезни «трансплантат против хозяина», вызванной аллогенными Т-клетками. Напротив, NK-клетки способны убивать опухолевые мишени антиген-независимым образом и не вызывают GvHD, что позволяет рассматривать их как потенциально эффективные для иммунотерапии рака.
2. Технология CRISPR/Cas9 недавно использовалась для конструирования иммунных клеток, но генетическое перепрограммирование NK клеток с помощью плазмид всегда было сложной задачей. Это происходит из-за трудностей доставки трансгена ДНК-зависимым образом, такого как лентивирусная и ретровирусная трансдукция, вызывающая существенный связанный с процедурой апоптоз NK-клеток и ограниченное производство геннетически сконструированных NK-клеток. Необходимы новые способы получения NK-клеток способами генной инженерии.
II. СУЩНОСТЬ ИЗОБРЕТЕНИЯ
3. Раскрыты способы и композиции, относящиеся к генетически модифицированным NK-клеткам.
4. В одном аспекте в данной заявке раскрыты способы генной модификации NK-клетки (такой как, например, первичная или размножаемая NK-клетка), включающие: а) получение направляющей РНК (гРНК), специфичной для целевой последовательности ДНК в NK-клетке (такой как, например, трансформирующий фактор роста-β рецептор 2 (TGFBR2) или гипоксантинфосфорибозилтрансфераза 1 (HPRT1); и б) введение посредством электропорации в целевую NK-клетку комплекса рибонуклеопротеина (RNP), содержащего эндонуклеазу CRISPR/Cas (Cas9) класса 2, связанную в комплекс с соответствующей направляющей РНК CRISPR/Cas, который гибридизуется с целевой последовательностью в геномной ДНК NK-клетки.
5. В данной заявке также раскрыты способы в соответствии с любым предыдущим аспектом, в которых геном NK-клетки модифицирован путем вставки или делеции одной или более пар оснований, вставки фрагмента гетерологичной ДНК (например, донорного полинуклеотида), путем делеции фрагмента эндогенной ДНК, путем инверсии или транслокации фрагмента эндогенной ДНК, или их комбинациями.
6. В одном аспекте в данной заявке раскрыты способы генной модификации NK-клетки в соответствии с любым предыдущим аспектом, где NK-клетки (например, первичные или размножаемые NK-клетки) инкубируют в присутствии IL-2 и/или облученных питающих клеток в течение 4, 5, 6 или 7 дней до трансдукции (например, электропорации).
7. В данной заявке также раскрыты способы генной модификации NK-клетки в соответствии с любым из предшествующих аспектов, дополнительно включающие размножение модифицированных NK-клеток с помощью облученных мембраносвязанных питающих клеток, экспрессирующих интерлейкин-21 (mbIL-21), после проведения электропорации.
8. В одном аспекте в данной заявке раскрыты модифицированные NK-клетки, полученные способом в соответствии с любым из предшествующих аспектов. В одном аспекте модифицированная NK-клетка может содержать нокаут гена, кодирующего трансформирующий фактор роста-β рецептор 2 (TGFBR2) или гипоксантинфосфорибозилтрансферазу 1 (HPRT1).
9. В данной заявке также раскрыты способы лечения рака, включающие введение субъекту, страдающему от рака, модифицированной NK-клетки в соответствии с любым из предшествующих аспектов.
10. В одном аспекте в данной заявке раскрыты способы адоптивной передачи сконструированных NK-клеток субъекту, нуждающемуся в этом, где указанный способ включает: а) получение целевой NK-клетки (такой как первичная NK-клетка или размноженная NK-клетка) для модификации; б) получение гРНК, специфичной для целевой последовательности ДНК; в) введение посредством электропорации в целевую NK-клетку комплекса RNP, содержащего эндонуклеазу CRISPR/Cas (Cas9) класса 2, связанную в комплекс с соответствующей гРНК CRISPR/Cas, который гибридизуется с целевой последовательностью в геномной ДНК целевой NK-клетки, создавая сконструированную NK-клетку; и г) перенос сконструированной NK-клетки в субъект.
11. В данной заявке также раскрыты способы адоптивного переноса сконструированных NK-клеток нуждающемуся в этом субъекту, где NK-клетка является первичной NK-клеткой (такой как, например, аутологичная NK-клетка или NK-клетка из аллогенного донорского источника), которые были модифицированы ex vivo, и после модификации перенесены в субъект.
12. В одном аспекте в данной заявке раскрыты способы адоптивного переноса сконструированных NK-клеток субъекту, нуждающемуся в этом, в соответствии с любым из предшествующих аспектов, при этом NK-клетка размножается с помощью облученных питающих клеток, экспрессирующих mbIL-21, или путем введения IL-21 до, одновременно с, или после введения модифицированных NK-клеток субъекту.
13. В одном аспекте в данной заявке раскрыты способы адоптивного переноса сконструированных NK-клеток нуждающемуся в этом субъекту, при этом субъект, получающий адоптивно перенесенные модифицированные NK-клетки, болен раком.
III. КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
14. Сопроводительные чертежи, которые включены в данное описание и составляют его часть, иллюстрируют несколько вариантов воплощения изобретения и вместе с описанием иллюстрируют раскрытые композиции и способы.
15. Фигура 1 демонстрирует эффективность электропорации миРНК и плазмидной ДНК, экспрессирующих GFP, в NK-клетках с использованием программы EN-138. Как видно в данной заявке, жизнеспособность NK-клеток составляет 77,5% и 35% живых клеток были GFP-позитивными.
16. Фигура 2 демонстрирует жизнеспособность и эффективность еще одной из 16 программ (DN-100), протестированных для оптимизации электропорации.
17. Фигура 3 демонстрирует опосредованный Cas9/RNP нокаут TGFBR2 в размножающихся (а) первичных NK-клетках (б), измеренный с помощью анализа мутации Т7Е1. Фермент Т7Е1 распознает и расщепляет несовпадающую ДНК. Каждая небольшая полоса (синие стрелки) представляет собой расщепленные фрагменты ДНК, содержащие вставку-делецию.
18. Фигура 4 демонстрирует опосредованное Cas9/RNP разрушение HPRT в размножающихся NK-клетках, измеренное с помощью анализа мутации Т7Е1.
19. Фигура 5 демонстрирует уровень экспрессии мРНК эктодомена TGFBR2 в NK-клетках, модифицированных CRISPR, введенных с помощью Cas9/RNP (гРНК1 + гРНК2) с использованием ОТ-ПЦР. GAPDH использовали как эндогенный контрольный ген. Снижение уровней РНК указывает на разрушение гена TGFBR2.
20. Фигура 6А демонстрирует анализ цитотоксичности модифицированных Cas9/RNP (гРНК1 + гРНК2, гРНК2 и гРНК3) клеток, показывающий, что инкубация клеток с TGFB в течение ночи существенно не снижает их способность к лизису клеток DAOY.
21. Фигура 6В демонстрирует, что по сравнению с немодифицированными NK-клетками, клетки, модифицированные Cas9/RNP (гРНК2 и гРНК3), менее чувствительны к TGFB.
22. Фигура 7 демонстрирует экзон 2 гена SOCS3 и гРНК, используемые для нацеливания на экзон 2 гена SOCS3.
23. Фигура 8 демонстрирует относительный нормализованный уровень экспрессии Socs3 в нокаутированных NK-клетках по сравнению с NK-клетками дикого типа.
24. Фигуры 9А, 9В, 9С и 9D демонстрируют повышенные размножение и цитотоксичность NK-клеток SOCS3-KO. Фигура 9А демонстрирует результаты Incucyte NK-клеток SOCS3-KO в отношении AML. Фигуры 9В и 9С демонстрируют результаты цитотоксичности 3 доноров в отношении клеток DAOY (9В) и линии клеток нейробластомы NB1643 (9С). Фигура 9D демонстрирует фактическое количество мертвых клеток для каждой клеточной линии и условия лечения на Фигурах 9В и 9С.
25. Фигура 10 демонстрирует анализ пролиферации, показывающий влияние КО SOCS3 на размножение NK-клеток.
26. Фигура 11 демонстрирует экспрессию CD38 на NK-клетках дикого типа и NK-клетках с нокаутом CD38.
27. Фигура 12 демонстрирует устойчивость к фратрациду, опосредованному даратумумабом.
28. Фигура 13 демонстрирует, что платформа Cas9/RNP успешно нацеливается на локус AAVS1 в NK-клетках.
29. Фигура 14 демонстрирует, как интеграцию репортерного гена mCherry в локус AAVS1 первичных человеческих NK-клеток оценивали с помощью ПЦР.
30. Фигура 15 демонстрирует стабильную экспрессию гена mCherry после размножения и сортировки, изученных с помощью проточной цитометрии и флуоресцентной микроскопии. *Результаты представляют 2 из 12 разработанных конструктов AAV.
31. Фигура 16 демонстрирует стабильную экспрессию гена mCherry после размножения, причем сортировку первичных NK-клеток человека с использованием различных условий культивирования оценивали с помощью проточной цитометрии. Первичные NK-клетки подвергали электропорации с помощью CAS9/RNP и трансдуцировали 300K MOI AAV6 SS800-mCherry и культивировали в среде RPMI + фетальная бычья сыворотка (FBS) или в бессывороточной среде AIMV.
IV. ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
32. Перед тем как настоящие соединения, композиции, изделия, устройства и/или способы будут раскрыты и описаны, следует понимать, что они не ограничиваются конкретными синтетическими способами или конкретными способами рекомбинантной биотехнологии, если не указано иное, или конкретными реагентами, если не указано иное, поскольку таковые, конечно, могут варьироваться. Также следует понимать, что используемая в данной заявке терминология предназначена только для описания конкретных вариантов воплощения изобретения и не предназначена для его ограничения.
А. Определения
33. Используемые в описании и прилагаемой формуле изобретения, формы единственного числа включают ссылки на множественное число, если контекст явно не требует иного. Таким образом, например, ссылка на «фармацевтический носитель» включает смеси двух или более таких носителей и тому подобное.
34. В данной заявке диапазоны могут быть выражены как от «приблизительно» одного конкретного значения и/или до «приблизительно» другого конкретного значения. Когда такой диапазон явно выражен, другой вариант воплощения включает от одного конкретного значения и/или до другого конкретного значения. Точно так же, когда значения выражаются как приблизительные величины с использованием определения «приблизительно», следует понимать, что конкретное значение образует другой вариант воплощения изобретения. Далее следует понимать, что конечные точки каждого из диапазонов значимы как по отношению к другой конечной точке, так и независимо от другой конечной точки. Также понятно, что, при наличии ряда определенных значений, представленных в данной заявке, каждое значение также предусматривается в данной заявке как «приблизительное» от этого конкретного значения в дополнение к самому значению. Например, если представлено значение «10», то также предусматривается «приблизительно 10». Также понятно, что, когда представлено определенное значение, также подразумеваются значения «менее чем или равные» указанному значению, «более чем или равные значению» и возможные диапазоны между значениями, как надлежащим образом понимает специалист в данной области техники. Например, если представлено значение «10», также подразмевается «менее или равный 10», а также «более или равный 10». Также понятно, что во всей данной заявке данные предоставляются в нескольких различных форматах, и что эти данные представляют конечные и начальные точки, а также диапазоны для любой комбинации точек замера. Например, если представлена конкретная точка замера «10» и конкретная точка замера 15, подразумевается, что раскрыты значения превышающие, превышающие или равные, меньшие чем, меньшие чем или равные, и равные 10 и 15, а также значения между 10 и 15. Также понятно, что также предусмотрена каждая величина между двумя конкретными значениями. Например, если представлены значения 10 и 15, то также предусмотрены значения 11, 12, 13 и 14.
35. В данном описании и в приведенной ниже формуле изобретения будет выполнена ссылка на ряд терминов, которые должны иметь следующие значения:
36. «Необязательный» или «необязательно» означает, что описанное ниже событие или обстоятельство может или не может произойти, и что описание включает случаи, когда указанное событие или обстоятельство происходит, и случаи, когда не происходит.
37. «Праймеры» представляют собой подгруппу зондов, которые способны поддерживать некоторый тип ферментативной манипуляции и которые могут гибридизоваться с целевой нуклеиновой кислотой, так что может происходить ферментативная манипуляция. Праймер может быть изготовлен из любой комбинации нуклеотидов или производных нуклеотидов или аналогов, доступных в данной области техники, которые не мешают ферментативным манипуляциям.
38. Зонды» представляют собой молекулы, способные взаимодействовать с целевой нуклеиновой кислотой, обычно специфическим для последовательности образом, например, посредством гибридизации. Гибридизация нуклеиновых кислот хорошо известна в данной области техники и обсуждается в данной заявке. Обычно зонд может быть изготовлен из любой комбинации нуклеотидов или производных нуклеотидов или аналогов, доступных в данной области техники.
39. Последовательность ДНК, которая «кодирует» конкретную РНК, представляет собой последовательность нуклеиновой кислоты ДНК, которая транскрибируется в РНК. Полинуклеотид ДНК может кодировать РНК (мРНК), которая транслируется в белок (и, следовательно, ДНК и мРНК оба кодируют белок), или полинуклеотид ДНК может кодировать РНК, которая не транслируется в белок (например, тРНК, рРНК, микроРНК (миРНК), «некодирующая» РНК (нкРНК), направляющая РНК и т.д.).
40. «Последовательность, кодирующая белок» или последовательность, которая кодирует конкретный белок или полипептид, представляет собой последовательность нуклеиновой кислоты, которая транскрибируется в мРНК (в случае ДНК) и транслируется (в случае мРНК) в полипептид in vitro или in vivo при помещении под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяются стартовым кодоном на 5'-конце (N-конец) и бессмысленным кодоном остановки трансляции на 3'-конце (С-конец). Кодирующая последовательность может включать, без ограничения, кДНК из прокариотической или эукариотической мРНК, последовательности геномной ДНК из прокариотической или эукариотической ДНК и синтетические нуклеиновые кислоты. Последовательность терминации транскрипции обычно располагается в положении 3' от кодирующей последовательности.
41. В контексте данного документа, термины «встречающийся в природе», «немодифицированный» или «дикого типа» в применении к нуклеиновой кислоте, полипептиду, клетке или организму, относится к нуклеиновой кислоте, полипептиду, клетке или организму, найденным в природе. Например, полипептидная или полинуклеотидная последовательность, присутствующая в организме (включая вирусы), которая может быть выделена из природного источника и которая не была намеренно модифицирована человеком в лаборатории, относится к дикому типу (и встречается в природе).
42. «Введение» субъекту включает любой маршрут введения или доставки агента субъекту. Введение может осуществляться любым подходящим маршрутом, включая пероральный, местный, внутривенный, подкожный, чрескожный, трансдермальный, внутримышечный, внутрисуставной, парентеральный, внутриартериольный, внутрикожный, внутрижелудочковый, внутричерепной, внутрибрюшинный, внутриочаговый, интраназальный, ректальный, вагинальный, путем ингаляции, через имплантированный резервуар, парентеральный (например, подкожные, внутривенные, внутримышечные, внутрисуставные, внутрисиновиальные, внутригрудинные, интратекальные, внутрибрюшинные, внутрипеченочные, внутриочаговые и внутричерепные инъекциии или способы инфузии) и т.п. «Параллельное введение», «введение в комбинации», «одновременное введение» или «вводимые одновременно» в контексте настоящего описания означают, что соединения вводят в один и тот же момент времени или по существу сразу после друг друга. В последнем случае два соединения вводят в моменты времени, достаточно близкие, чтобы наблюдаемые результаты были неотличимы от результатов, достигнутых при введении соединений в один и тот же момент времени. «Системное введение» относится к введению или доставке агента субъекту посредством маршрута, который вводит или доставляет агент в обширные участки тела субъекта (например, более чем 50% тела), например, через вход в кровеносную или лимфатическую системы. Напротив, «местное введение» относится к введению или доставке агента субъекту таким маршрутом, который вводит или доставляет агент в участок введения или участок, непосредственно примыкающий к месту введения, и не вводит агент системно в терапевтически значимом количестве. Например, агенты, вводимые местно, легко обнаруживаются в непосредственной близости от места введения, но не обнаруживаются или обнаруживаются в незначительных количествах в дистальных частях тела субъекта. Введение включает самостоятельное введение и введение другим лицом.
43. «Эффективное количество» агента относится к количеству агента, достаточному для обеспечения желаемого эффекта. Количество агента, которое является «эффективным», будет варьироваться в зависимости от субъекта и в зависимости от многих факторов, таких как возраст и общее состояние субъекта, конкретный агент или агенты и тому подобное. Таким образом, не всегда возможно указать количественное «эффективное количество». Однако подходящее «эффективное количество» в любом случае для субъекта может быть определено специалистом в данной области техники с использованием рутинных экспериментов. Также, как используют в данной заявке, если специально не указано иное, «эффективное количество» агента может также относиться к количеству, охватывающему как терапевтически эффективные количества, так и профилактически эффективные количества. «Эффективное количество» агента, необходимое для достижения терапевтического эффекта, может варьироваться в зависимости от таких факторов, как возраст, пол и вес субъекта. Режимы дозирования можно регулировать для обеспечения оптимального терапевтического ответа. Например, можно вводить несколько разделенных доз ежедневно, или доза может быть пропорционально уменьшена в зависимости от необходимости терапевтической ситуации.
44. «Фармацевтически приемлемый» компонент может относиться к компоненту, который не имеет нежелательного эффекта с биологической или иной точки зрения, то есть компонент может быть включен в фармацевтическую композицию в соответствии с данным изобретением и введен субъекту, как описано в данной заявке, не вызывая значительных нежелательных биологических эффектов или не взаимодействуя вредным образом с любым из других компонентов композиции, в которой он содержится. При использовании в отношении введения человеку этот термин обычно подразумевает, что компонент соответствует требуемым стандартам токсикологических и производственных испытаний или что он включен в Руководство по неактивным ингредиентам, подготовленное Управлением по контролю за продуктами и лекарствами США.
45. «Фармацевтически приемлемый носитель» (иногда называемый «носителем») означает носитель или эксципиент, который полезен при приготовлении фармацевтической или терапевтической композиции, которая в целом безопасна и нетоксична, и содержит носитель, приемлемый для ветеринарии и/или фармацевтического или терапевтического использования человеком. Термины «носитель» или «фармацевтически приемлемый носитель» могут включать, но не ограничиваются ими, забуференный фосфатом физиологический раствор, воду, эмульсии (такие как эмульсия типа масло/вода или вода/масло) и/или различные типы смачивающих агентов. Используемый в данной заявке термин «носитель» охватывает, но не ограничивается ими, любой эксципиент, разбавитель, наполнитель, соль, буфер, стабилизатор, солюбилизатор, липид, стабилизатор или другой материал, хорошо известный в данной области техники для использования в фармацевтических композициях и как описано ниже.
46. «Фармакологически активный» (или просто «активный»), как в «фармакологически активном» производном или аналоге, может относиться к производному или аналогу (например, соли, сложному эфиру, амиду, конъюгату, метаболиту, изомеру, фрагменту и т.д.), имеющему тот же тип фармакологической активности, что и исходное соединение, и приблизительно эквивалентному по степени.
47. «Терапевтический агент» относится к любой композиции, обладающей благоприятным биологическим эффектом. Благоприятные биологические эффекты включают как терапевтические эффекты, например, лечение расстройства или другого нежелательного физиологического состояния, так и профилактические эффекты, например, предотвращение расстройства или другого нежелательного физиологического состояния (например, неиммуногенного рака). Термины также охватывают фармацевтически приемлемые, фармакологически активные производные лечебных агентов, конкретно указанных в данной заявке, включая, но не ограничиваясь ими, соли, сложные эфиры, амиды, проагенты, активные метаболиты, изомеры, фрагменты, аналоги и тому подобное. Когда используется термин «терапевтический агент», или если конкретный агент конкретно идентифицирован, следует понимать, что термин включает агент как таковой, а также фармацевтически приемлемые, фармакологически активные соли, сложные эфиры, амиды, проагенты, конъюгаты, активные метаболиты, изомеры, фрагменты, аналоги и др.
48. «Терапевтически эффективное количество» или «терапевтически эффективная доза» композиции (например, композиции, содержащей агент) относится к количеству, которое эффективно для достижения желаемого терапевтического результата. В некоторых вариантах воплощения изобретения, желаемый терапевтический результат представляет собой контроль диабета I типа. В некоторых вариантах воплощения, желаемый терапевтический результат представляет собой контроль ожирения. Терапевтически эффективные количества данного терапевтического агента обычно будут варьироваться в зависимости от таких факторов, как тип и тяжесть расстройства или заболевания, которое лечат, а также возраст, пол и вес субъекта. Термин также может относиться к количеству терапевтического агента или скорости доставки терапевтического агента (например, количество в зависимости от времени), эффективных для облегчения желаемого терапевтического эффекта, такого как облегчение боли. Точный желаемый терапевтический эффект будет варьироваться в зависимости от состояния, которое необходимо лечить, переносимости субъекта, вводимого агента и/или состава агента (например, эффективности терапевтического агента, концентрации агента в композиции и тому подобное) и множество других факторов, которые оценят специалисты в данной области техники. В некоторых случаях желаемый биологический или медицинский ответ достигается после введения нескольких доз композиции субъекту в течение нескольких дней, недель или лет.
49. В данной заявке приведены ссылки на различные публикации. Раскрытие этих публикаций во всей их полноте настоящим включено в качестве ссылки в данную заявку, чтобы более полно описать состояние техники, к которой относится данное изобретение. Раскрытые ссылки также индивидуально и конкретно включены в данную заявку в качестве ссылки на содержащийся в них материал, который обсуждается в предложении, в котором приведена данная ссылка.
В. Способы генной модификации NK-клеток
50. Генетическое перепрограммирование NK-клеток с помощью плазмид всегда было сложной задачей из-за трудностей с доставкой трансгена ДНК-зависимым образом, таким как лентивирусная и ретровирусная трансдукция, вызывающая существенный связанный с процедурой апоптоз NK-клеток и ограниченное производство сконструированных при помощи генной инженерии NK-клеток. В данной заявке описаны способы использования редактирования генома, не содержащего ДНК, первичных и размножаемых NK-клеток человека с использованием эндонуклеазных рибонуклеопротеиновых комплексов (таких как, например, Cas9/RNP) для репрограммирования (т.е. инженерии или модификации) NK-клеток.
51. Эндонуклеазы/RNP (например, Cas9/RNP) состоят из трех компонентов: рекомбинантного эндонуклеазного белка (например, эндонуклеазы Cas9), связанной в комплекс с локусами CRISPR. Эндонуклеаза, связанная в комплекс с локусами CRISPR, может называться направляющей РНК CRISPR/Cas. Локусы CRISPR содержат синтетическую однонаправляющую РНК (гРНК), состоящую из РНК, которая может гибридизоваться с комплементарной повторной РНК (крРНК) в комплексе с целевой последовательностью и транс-комплементарной повторной РНК (tracrРНК). Соответственно, направляющая РНК CRISPR/Cas гибридизуется с целевой последовательностью в геномной ДНК клетки. В некоторых случаях эндонуклеаза CRISPR/Cas класса 2 является эндонуклеазой CRISPR/Cas типа II. В некоторых случаях эндонуклеаза CRISPR/Cas класса 2 представляет собой полипептид Cas9, а соответствующая направляющая РНК CRISPR/Cas представляет собой направляющую РНК Cas9. Эти Cas9/RNP способны расщеплять геномные мишени с более высокой эффективностью по сравнению с чужеродными ДНК-зависимыми подходами благодаря их доставке в виде функциональных комплексов. Дополнительно, быстрый клиренс Cas9/RNP из клеток может снизить нецелевые эффекты, такие как индукция апоптоза. Соответственно, в одном аспекте в данной заявке раскрыты способы генной модификации NK-клетки, включающие: а) получение направляющей РНК (гРНК), специфичной для целевой последовательности ДНК в NK-клетке; и б) трансдукцию (например, введения посредством электропорации) в целевую NK-клетку, комплекса рибонуклеопротеина (RNP), содержащего эндонуклеазу CRISPR/Cas (Cas9) класса 2 в комплексе с соответствующей направляющей РНК CRISPR/Cas, которая гибридизуется с целевой последовательностью в геномной ДНК NK-клетки.
52. Известно и в данной заявке предполагается, что для нацеливания активности нуклеазы Cas9 на целевой сайт-мишень, а также для расщепления донорной плазмиды, чтобы сделать возможным рекомбинацию донорного трансгена в ДНК хозяина, используется crisprРНК (крРНК). В некоторых случаях крРНК комбинируется с tracrРНК с образованием направляющей РНК (гРНК). Раскрытые плазмиды используют интеграцию AAV, интрон 1 протеинфосфатазы 1, гена регуляторной субъединицы 12С (PPP1R12C) на хромосоме 19 человека, которая называется AAVS1, в качестве сайта-мишени для интеграции трансгена. Этот локус является «геном безопасной гавани» и обеспечивает стабильную долгосрочную экспрессию трансгена во многих типах клеток. Поскольку разрушение PPP1R12C не связано с каким-либо известным заболеванием, локус AAVS1 часто считается безопасной гаванью для нацеливания трансгена. Поскольку сайт AAVS1 используется в качестве целевого местоположения, CRSPR РНК (крРНК) должна нацеливаться на указанную ДНК. В данной заявке направляющая РНК, используемая в раскрытых плазмидах, включает GGGGCCACTAGGGACAGGAT (SEQ ID NO: 9) или любой его 10-нуклеотидный смысловой или антисмысловой смежный фрагмент. Хотя AAVS1 используется в данной заявке в иллюстративных целях, известно и подразумевается, что другие «гены-безопасные гавани» могут использоваться с эквивалентными результатами и могут быть заменены на AAVS1, если это более целесообразно, с учетом конкретного типа трансфицируемых клеток или трансгена. Примеры других безопасных генов включают, но не ограничиваются ими, рецептор хемокина С-С типа 5 (CCR5), локус ROSA26 и TRAC.
53. Известно и в данной заявке предполагается, что могут существовать ограничения по размеру донорской трансгенной конструкции, доставленной в целевой геном. Одним из способов увеличения допустимого размера трансгена является создание дополнительного пространства путем обмена обычно используемого Cas9 пиогенного стрептококка (SpCas9) на синтетический Cas9 или Cas9 из другого бактериального источника. Замещение Cas9 также может использоваться для повышения специфичности нацеливания, поэтому необходимо использовать меньшее количество гРНК. Так, например, Cas9 может происходить из золотистого стафилокока (Staphylococcus aureus) (SaCas9), Acidaminococcus sp. (AsCpf1), Lachnospiracase bacterium (LbCpf1), менингококка (Neisseria meningitidis) (NmCas9), термофильного стрептококка (Streptococcus thermophilus) (StCas9), кампилобактер еюни (Campylobacter jejuni) (CjCas9), размножаемых SpCas9 (eSpCas9), SpCas9-HF1, Fokl-Fused dCas9, размножаемых Cas9 (xCas9), и/или каталитически мертвых Cas9 (dCas9).
54. Известно и в данной заявке предполагается, что использование конкретного Cas9 может изменять последовательность РАМ, которую эндонуклеаза Cas9 (или альтернатива) использует для скрининга мишеней. Используемые в данной заявке подходящие последовательности РАМ включают NGG (SpCas9 РАМ) NNGRRT (SaCas9 РАМ) NNNNGATT (NmCAs9 РАМ), NNNNRYAC (CjCas9 РАМ), NNAGAAW (St), TTTV (LbCpf1 РАМ и AsCpf1 РАМ); TYCV (LbCpf1 РАМ вариант и AsCpf1 РАМ вариант); где N может представлять собой любой нуклеотид; V=А, С, или G; Y=С или Т; W=А или Т; и R=А или G.
55. Для получения комплекса RNP, крРНК и tracrРНК могут быть смешаны в соотношении 1:1, 2:1 или 1:2 концентраций от приблизительно 50 мкМ до приблизительно 500 мкМ (например, 50, 60, 70, 80, 90, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475 или 500 мкм), предпочтительно от 100 мкМ до приблизительно 300 мкМ, наиболее предпочтительно приблизительно 200 мкМ при 95°С в течение приблизительно 5 минут для образования комплекса крРНК:tracrРНК (т.е. направляющая РНК). Комплекс крРНК:tracrРНК затем может быть смешан с приблизительно 20 мкМ и приблизительно 50 мкМ (например, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 мкМ) конечным разведением Cas-эндонуклеазы (такой как, например, Cas9).
56. После связывания с целевой последовательностью в целевой клетке локусы CRISPR можно генно модифицировать путем введения в целевую ДНК вставки или делеции одной или более пар оснований, путем вставки фрагмента гетерологичной ДНК (например, донорного полинуклеотида), посредством делеции фрагмента эндогенной ДНК, путем инверсии или транслокации фрагмента эндогенной ДНК или их комбинации. Таким образом, описанные способы можно использовать для создания нокаутов или нокинов при объединении с ДНК для гомологичной рекомбинации. В данной заявке показано, что трансдукция посредством электропорации Cas9/RNP представляет собой простой и относительно эффективный способ, который преодолевает предыдущие ограничения генной модификации в NK-клетках.
57. Известно и в данной заявке предполагается, что раскрытые способы можно использовать с любым типом клеток, включая естественные клетки-киллеры (NK-клетки), Т-клетки, В-клетки, макрофаги, фибробласты, остеобласты, гепатоциты, нейрональные клетки, эпителиальные клетки и/или мышечные клетки. Человеческие NK-клетки представляют собой субпопуляцию лимфоцитов периферической крови, определяемую экспрессией CD56 или CD16 и отсутствием рецептора Т-клеток (CD3). NK-клетки обнаруживают и убивают целевые клетки, в которых отсутствуют молекулы класса I главного комплекса гистосовместимости (МНС). Рецепторы, активирующие NK-клетки, включают, среди прочего, рецепторы естественной цитотоксичности (NKp30, NKp44 и NKp46) и лектин-подобные рецепторы NKG2D и DNAM-1. Их лиганды экспрессируются на стрессированных, трансформированных или инфицированных клетках, но не на нормальных клетках, что делает нормальные клетки устойчивыми к уничтожению NK-клеток. Активация NK-клеток негативно регулируется через ингибирующие рецепторы, такие как киллерные иммуноглобин (Ig)-подобные рецепторы (KIR), NKG2A/CD94, TGFβ, и лейкоцитный Ig-подобный рецептор-1 (LIR-1). В одном аспекте целевые клетки могут быть первичными NK-клетками из донорского источника (такого как, например, аллогенный донорский источник для адоптивной трансферной терапии или аутологичный донорский источник (т.е. конечный реципиент модифицированных NK-клеток), линия NK-клеток (включая, но не ограничиваясь приведенным, NK RPMI8866; HFWT, K562 и EBV-LCL) или из источника размножаемых NK-клеток, полученных из первичного источника NK-клеток или NK-клеточной линии.
58. Перед трансдукцией NK-клеток, NK-клетки можно инкубировать в среде, подходящей для размножения NK-клеток. Известно и в данной заявке предполагается, что условия культивирования могут включать добавление цитокинов, антител и/или питающих клеток. Таким образом, в одном аспекте в данной заявке раскрыты способы генной модификации NK-клетки, дополнительно включающие инкубацию NK-клеток для 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 или 14 дней до трансдукции клеток в среде, которая поддерживает размножение NK-клеток; где среда дополнительно содержит цитокины, антитела и/или питающие клетки. Например, среда может содержать IL-2, IL-12, IL-15, IL-18 и/или IL-21. В одном аспекте среда также может содержать анти-CD3-антитело. В одном аспекте питающие клетки могут быть очищены от питающих клеток, которые стимулируют NK-клетки. Представленные в данной заявке питающие клетки, стимулирующие NK-клетки, при использовании в заявляемом изобретении, могут быть либо облученными аутологичными или аллогенными мононуклеарными клетками периферической крови (РВМС), либо необлученными аутологичными или РВМС; RPMI8866; HFWT, K562; клетками K562, трансфицированными мембраносвязанными IL-15 и 41BBL, или IL-21 или любой их комбинацией; или EBV-LCL. В некоторых аспектах питающие клетки NK-клетки предоставлены в комбинации с раствором IL-21, IL-15 и/или 41BBL. Питающие клетки можно засеять в культуру NK-клеток в соотношении 1:2, 1:1 или 2:1. Известно и в данной заявке предполагается, что период культивирования может составлять от 1 до 14 дней после электропорации (т.е. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, или 14 дней), предпочтительно от 3 до 7 дней, наиболее предпочтительно от 4 до 6 дней.
59. Известно и в данной заявке предполагается, что условия инкубации первичных NK-клеток и размноженных NK-клеток могут быть разными. В одном аспекте культивирование первичных NK-клеток перед электропорацией включает среду и цитокины (такие как, например, IL-2, IL-12, IL-15, IL-18 и/или IL-21) и/или антитела к CD3 менее 5 дней (например, 1, 2, 3 или 4 дня). Для размноженных NK-клеток культивирование может происходить в присутствии питающих NK-клеток (например, в соотношении 1:1) в дополнение или вместо цитокинов (таких как, например, IL-2, IL-12, IL-15, IL-18 и/или IL-21) и/или антитело к CD3. Культивирование увеличившихся NK-клеток может происходить за 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дней до трансдукции. Таким образом, в одном аспекте в настоящем документе раскрыты способы генной модификации NK-клетки, включающие инкубацию первичных NK-клеток в течение 4 дней в присутствии IL-2 перед электропорацией или инкубацию увеличенных NK-клеток в присутствии облученных питающих клеток в течение 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 30, 36, 42, 48, 54, 60 часов, 3, 4, 5, 6 или 7 дней до электропорации.
60. Известно и в данной заявке предполагается, что способы трансдукции для модификации NK-клеток в раскрытых способах ограничены. Благодаря своей иммунной функции NK-клетки устойчивы к вирусным и бактериальным векторам, а также к индукции апоптоза NK-клеток указанными векторами. Таким образом, до настоящих способов модификация NK-клеток CRISPR/Cas была неудачной. Чтобы обойти проблемы с вирусными векторами, раскрытые способы трансформируют целевые NK-клетки с помощью электропорации. Электропорация представляет собой способ, при котором к клеткам прикладывают электрическое поле для увеличения проницаемости клеточной мембраны. Приложение электрического поля вызывает зарядный градиент сквозь мембрану, который втягивает заряженные молекулы, такие как нуклеиновая кислота, сквозь клеточную мембрану. Таким образом, в одном аспекте в данной заявке раскрыты способы генной модификации NK-клетки, включающие: а) получение направляющей РНК (гРНК), специфичной для целевой последовательности ДНК в NK-клетке; и б) введение посредством электропорации в целевую NK-клетку, комплекса рибонуклеопротеина (RNP), содержащего CRISPR/Cas эндонуклеазу (Cas9) класса 2, связанную в комплекс с соответствующей CRISPR/Cas направляющей РНК, которая гибридизуется с целевой последовательностью в геномной ДНК NK-клетки.
61. После трансдукции (например, электропорации) NK-клетки, теперь модифицированная NK-клетка может размножаться в среде, содержащей питающие клетки, которые стимулируют модифицированные NK-клетки. Таким образом, модифицированные клетки сохраняют жизнеспособность и пролиферативный потенциал, так как они могут быть размножены после электропорации с использованием облученных питающих клеток. Питающие клетки, стимулирующие NK-клетки, для использования в заявленном изобретении, раскрытом в данной заявке, могут быть либо облученными аутологичными или аллогенными мононуклеарными клетками периферической крови (РВМС), либо необлученными аутологичными или РВМС; RPMI8866; HFWT, K562; клетками K562, трансфицированными мембраносвязанными IL-15 и 41BBL, или IL-21 или любой их комбинацией; или EBV-LCL. В некоторых аспектах питающие клетки NK-клетки предоставлены в комбинации с раствором IL-21, IL-15 и/или 41BBL. Питающие клетки можно засеять в культуру NK-клеток в соотношении 1:2, 1:1 или 2:1. Известно и в данной заявке предполагается, что период культивирования может составлять от 1 до 14 дней после электропорации (т.е. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, или 14 дней), предпочтительно от 3 до 7 дней, наиболее предпочтительно от 4 до 6 дней. В некоторых аспектах среды для культивирования модифицированных NK-клеток могут дополнительно содержать цитокины, такие как, например, IL-2, IL-12, IL-15, IL-18 и/или IL-21.
62. В одном аспекте известно и предполагается, что одна из целей раскрытых способов генной модификации NK-клетки состоит в том, чтобы производить модифицированную NK-клетку. Соответственно, в данной заявке раскрыты модифицированные NK-клетки, полученные описанными способами.
63. Как отмечалось выше, активация NK-клеток негативно регулируется посредством ингибирующих рецепторов, таких как киллерные иммуноглобин(Ig)-подобные рецепторы (KIR), NKG2A /CD94, TGFβ, и лейкоцитарный Ig-подобный рецептор-1 (LIR-1). Для предотвращения лизиса мишени может быть достаточно вовлечения одного ингибирующего рецептора. Следовательно, NK-клетки эффективно нацелены на клетки, которые экспрессируют многие лиганды, индуцированные стрессом, и несколько лигандов МНС класса I. TGFβ является основным иммунодепрессивным цитокином, который подавляет активацию и функции NK-клеток. Таким образом, известно и в данной заявке предполагается, что одна модификация NK-клеток, которая могла бы быть полезной, представляет собой подавление ингибирующих рецепторов, таких как киллерные иммуноглобин(Ig)-подобные рецепторы (KIR), NKG2A /CD94, TGFβ и лейкоцитарный Ig-подобный рецептор-1 (LIR-1), поэтому негативная регуляция NK-клеток будет подавлена. Такие модифицированные клетки были бы очень полезны в иммунотерапии любого заболевания или состояния, которое можно лечить добавлением NK-клеток. Таким образом, в одном аспекте в данной заявке раскрыты генетически модифицированные NK-клетки, содержащие нокаут гена, кодирующего трансформирующий фактор роста-β рецептор 2 (TGFBR2) или гипоксантинфосфорибозилтрансферазу 1 (HPRT1).
64. Как отмечается в настоящем описании, раскрытые модифицированные NK-клетки идеально подходят для использования в иммунотерапии, такой как адоптивный перенос модифицированных (то есть сконструированных) NK-клеток субъекту, нуждающемуся в этом. Таким образом, в одном аспекте в данной заявке раскрыты способы адоптивной передачи сконструированных NK-клеток нуждающемуся в этом субъекту, где указанный способ включает а) получение целевой NK-клетки, которая должна быть модифицирована; б) получение гРНК, специфичной для целевой последовательности ДНК; в) введение посредством электропорации в целевую NK-клетку комплекса RNP, содержащего эндонуклеазу CRISPR/Cas (Cas9) класса 2, связанную в комплекс с соответствующей гРНК CRISPR/Cas, которая гибридизуется с целевой последовательностью в геномной ДНК целевой NK-клетки, создавая сконструированную NK-клетку; и г) перенос сконструированной NK-клетки субъекту.
65. В одном аспекте модифицированные NK-клетки, используемые в раскрытых способах иммунотерапии, могут быть первичными NK-клетками из донорского источника [такого как, например, аллогенный донорский источник для адоптивной трансферной терапии или аутологичный донорский источник (т.е. конечный реципиент) модифицированных NK-клеток], NK-клеточной линией (включая, помимо прочего, NK RPMI8866; HFWT, K562 и EBV-LCL) или из источника размножаемых NK-клеток, полученных из первичного источника NK-клеток или NK-клеточной линии. Известно и в данной заявке подразумевается, что, поскольку могут использоваться первичные NK-клетки, представленные модификации NK-клетки могут быть получены ex vivo или in vitro.
66. После трансдукции NK-клеток модифицированные NK-клетки можно размножать и стимулировать перед введением модифицированных (т.е. сконструированных) NK-клеток субъекту. Например, в данной заявке раскрыты способы адоптивного переноса NK-клеток субъекту, нуждающемуся в этом, где NK-клетка размножается облученными питающими клетками, экспрессирующими mbIL-21, перед введением субъекту. В некоторых аспектах понимается и подразумевается, что стимуляция и размножение модифицированных (то есть сконструированных) NK-клеток может происходить in vivo после или одновременно с введением модифицированных NK-клеток субъекту. В соответствии с этим в данной заявке раскрыты способы иммунотерапии, в которых NK-клетки размножаются у субъекта после переноса NK-клеток субъекту посредством введения IL-21 или облученных питающих клеток, экспрессирующих mbIL-21.
67. Было показано, что нацеливание на маршрут TGFβ может увеличить функции иммунных клеток. Мишенью был участок, кодирующий эктодомен TGBR2, связывающий TGFβ. Репрезентативные результаты показывают значительное снижение уровня экспрессии мРНК этого гена и дополнительно демонстрируют, что модифицированные NK-клетки становятся устойчивыми к TGFβ. Соответственно, в данной заявке раскрыты способы иммунотерапии, в которых комплекс RNP нацелен на ген TGRBR2 или HPRT1.
68. Известно и в данной заявке предполагается, что раскрытые способы модифицированной NK-клетки и адоптивного переноса модифицированных NK-клеток могут быть эффективной иммунотерапией против рака. Раскрытые способы и композиции можно использовать для лечения любого заболевания, при котором происходит неконтролируемая клеточная пролиферация, такая, как рак. Неограничивающий список различных типов рака следующий: лимфомы (Ходжкина и неходжкинские), лейкемии, карциномы, карциномы твердых тканей, плоскоклеточные карциномы, аденокарциномы, саркомы, глиомы, глиомы высокой степени злокачественности, бластомы, нейробластомы, плазмацитомы, гистиоцитомы, меланомы, аденомы, гипоксические опухоли, миеломы, лимфомы или саркомы, связанные со СПИД, метастатические злокачественные опухоли или рак в общем.
69. Репрезентативный, но не ограничивающий список видов рака, для лечения которых могут быть использованы раскрытые композиции, следующий: лимфома, В-клеточная лимфома, Т-клеточная лимфома, грибовидный микоз, болезнь Ходжкина, миелоидный лейкоз, рак мочевого пузыря, рак мозга, рак нервной системы, рак головы и шеи, плоскоклеточный рак головы и шеи, рак легких, такой как мелкоклеточный рак легкого и немелкоклеточный рак легкого, нейробластома/глиобластома, рак яичников, рак кожи, рак печени, меланома, плоскоклеточный рак ротовой полости, горла, гортани и легких, рак шейки матки, карцинома шейки матки, рак груди и рак эпителия, рак почек, рак мочеполовой системы, рак легких, карциному пищевода, рак головы и шеи, рак толстой кишки, рак систем кроветворения; рак яичек; рак толстой кишки, рак прямой кишки, рак простаты или рак поджелудочной железы. Соответственно, в одном аспекте в данной заявке раскрыты способы лечения рака у субъекта, включающие введение субъекту NK-клетки, которая была модифицирована, с тем, чтобы включать нок-аут гена TGFBR2.
70. Термины «лечить», «лечение» и их грамматические вариации, в контексте данного документа, включают введение композиции с намерением или целью частичного или полного предотвращения, замедления (отсрочки), лечения, заживления, облегчения, смягчения, изменения, исцеления, улучшения, регрессии, стабилизации, паллиации и/или снижения интенсивности или частоты одного или более заболеваний или состояний, симптома заболевания или состояния или основной причины заболевания или состояния. Лечебные процедуры по настоящему изобретению можно применять для предотвращения, профилактики, паллиативно или для лечения. Профилактическое лечение назначают субъекту до начала (например, до появления явных признаков рака), во время раннего начала (например, при начальных признаках и симптомах рака) или после установленного развития рака. Профилактическое введение может происходить за день (дни) или за годы до проявления симптомов инфекции.
1. Гибридизация/селективная гибридизация
71. Термин «гибридизация» обычно означает управляемое последовательностью взаимодействие между по меньшей мере двумя молекулами нуклеиновой кислоты, такими как праймер или зонд и ген. Взаимодействие, управляемое последовательностью, означает взаимодействие, которое происходит между двумя нуклеотидами или аналогами нуклеотидов или производными нуклеотидов специфическим для нуклеотидов образом. Например, G, взаимодействующий с С или А, взаимодействующий с Т, являются взаимодействиями, управляемыми последовательностями. Обычно взаимодействия, управляемые последовательностью, происходят на стороне Уотсона-Крика или на стороне Хугстина нуклеотида. На гибридизацию двух нуклеиновых кислот влияет ряд условий и параметров, известных специалистам в данной области техники. Например, концентрация солей, рН и температура реакции влияют на то, будут ли две молекулы нуклеиновой кислоты гибридизоваться.
72. Параметры селективной гибридизации между двумя молекулами нуклеиновой кислоты хорошо известны специалистам в данной области техники. Например, в некоторых вариантах воплощения изобретения, условия селективной гибридизации можно определить как жесткие условия гибридизации. Например, жесткость гибридизации контролируется как температурой, так и концентрацией соли на одном или обоих этапах гибридизации и промывания. Например, условия гибридизации для достижения селективной гибридизации могут включать гибридизацию в растворе с высокой ионной силой (6Х SSC или 6Х SSPE) при температуре приблизительно на 12-25°С ниже Тпл (температура плавления, при которой половина молекул диссоциируют от своих партнеров по гибридизации) с последующим промыванием при комбинации температуры и концентрации соли, выбранной таким образом, чтобы температура промывания была на приблизительно 5-20°С ниже Тпл. Температуру и солевые условия легко определить эмпирически в предварительных экспериментах, в которых образцы эталонной ДНК, иммобилизованные на фильтрах, гибридизируют с меченой рассматриваемой нуклеиновой кислотой, а затем промывают в условиях различной жесткости. Температуры гибридизации обычно выше для гибридизации ДНК-РНК и РНК-РНК. Условия могут использоваться, как описано выше, для достижения жесткости, или как известно в данной области техники. Предпочтительные жесткие условия гибридизации для гибридизации ДНК:ДНК могут составлять приблизительно 68°С (в водном растворе) в 6Х SSC или 6Х SSPE с последующим промыванем при 68°С. Жесткость гибридизации и промывания, если желательно, может быть соответственно уменьшена по мере уменьшения желаемой степени комплементарности и, кроме того, в зависимости от насыщенности G-C или А-Т любого участка, где ищется изменчивость. Аналогичным образом, жесткость гибридизации и промывания, если желательно, может быть соответственно увеличена по мере увеличения желаемой гомологии и, кроме того, в зависимости от G-C или А-Т насыщенности любого участка, где желательна высокая гомология, как известно в данной области техники.
73. Другим способом определения селективной гибридизации является обзор количества (процента) одной из нуклеиновых кислот, связанного с другой нуклеиновой кислотой. Например, в некоторых вариантах воплощения изобретения условия селективной гибридизации будут такими, когда по меньшей мере приблизительно 60, 65, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 процентов лимитирующей нуклеиновой кислоты связано с неограничивающей нуклеиновой кислотой. Обычно неограничивающий праймер имеет, например, 10-, 100-или 1000-кратный избыток. Этот тип анализа можно проводить в условиях, когда как ограничивающий, так и неограничивающий праймеры, например, в 10 или 100 или 1000 раз ниже их kd, или когда только одна из молекул нуклеиновой кислоты в 10 или 100 раз или в 1000 раз, или когда одна или обе молекулы нуклеиновой кислоты, превышают их kd.
74. Другим способом определения селективной гибридизации является обзор процента праймера, который подвергается ферментативной манипуляции в условиях, когда гибридизация требуется для стимулирования желаемой ферментативной манипуляции. Например, в некоторых вариантах воплощения изобретения, условия селективной гибридизации будут такими, когда по меньшей мере приблизительно 60, 65, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 процентов праймера подвергаются ферментативной обработке в условиях, которые способствуют ферментативной манипуляции, например, если ферментативная манипуляция представляет собой удлинение ДНК, тогда условия селективной гибридизации будут такими, когда по меньшей мере приблизительно 60, 65, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 процентов молекул праймера удлиняются. Предпочтительные условия также включают условия, предложенные производителем или указанные в данной области техники, как подходящие для фермента, выполняющего манипуляцию.
75. Как и в случае с гомологией, известно, что существует множество описанных в данной заявке способов определения уровня гибридизации между двумя молекулами нуклеиновой кислоты. Известно, что эти способы и условия могут обеспечивать разные проценты гибридизации между двумя молекулами нуклеиновой кислоты, но, если не указано иное, соответствия параметрам любого из способов будет достаточно. Например, если требуется 80% гибридизация и до тех пор, пока гибридизация происходит в пределах требуемых параметров в любом из этих способов, это считается раскрытым в данной заявке.
76. Специалистам в данной области техники понятно, что если композиция или способ удовлетворяют любому из этих критериев для определения гибридизации в совокупности или по отдельности, то это композиция или способ, которые раскрыты в данной заявке.
2. Нуклеиновые кислоты
77. Существует множество раскрытых в данной заявке молекул, которые основаны на нуклеиновых кислотах, включая, например, нуклеиновые кислоты, которые кодируют, например, TGFβR2, или любую из нуклеиновых кислот, описанных в данной заявке, для создания нокаутов TGFβR2 или их фрагментов в виде различных функциональных нуклеиновых кислот. Раскрытые нуклеиновые кислоты состоят, например, из нуклеотидов, аналогов нуклеотидов или заместителей нуклеотидов. В данной заявке обсуждаются неограничивающие примеры этих и других молекул. Понятно, что, например, когда вектор экспрессируется в клетке, эта экспрессируемая мРНК обычно будет состоять из А, С, G и U. Аналогично, понятно, что если, например, антисмысловая молекула введена в клетку или клеточную среду посредством, например, экзогенной доставки, предпочтительно, чтобы антисмысловая молекула состояла из аналогов нуклеотидов, которые уменьшают распад антисмысловой молекулы в клеточной среде.
а) Нуклеотиды и родственные молекулы
78. Нуклеотид представляет собой молекулу, которая содержит фрагмент основания, сахарный фрагмент и фосфатный фрагмент. Нуклеотиды могут быть связаны друг с другом через их фосфатные фрагменты и сахарные фрагменты, создавая межнуклеозидную связь. Фрагмент основания нуклеотида может представлять собой аденин-9-ил (А), цитозин-1-ил (С), гуанин-9-ил (G), урацил-1-ил (U) и тимин-1-ил (Т). Сахарный фрагмент нуклеотида представляет собой рибозу или дезоксирибозу. Фосфатный фрагмент нуклеотида представляет собой пятивалентный фосфат. Неограничивающим примером нуклеотида может быть 3'-АМФ (3'-аденозинмонофосфат) или 5'-ГМФ (5'-гуанозинмонофосфат). Существует множество разновидностей этих типов молекул, доступных в данной области техники и представленных в данной заявке.
79. Аналог нуклеотида представляет собой нуклеотид, который содержит некоторый тип модификации любого из фрагмента основания, сахарного основания или фосфатного основания. Модификации нуклеотидов хорошо известны в данной области техники и могут включать, например, 5-метилцитозин (5-me-С), 5-гидроксиметилцитозин, ксантин, гипоксантин и 2-аминоаденин, а также модификации сахарных или фосфатных фрагментов. Существует множество разновидностей этих типов молекул, доступных в данной области техники и представленных в данной заявке.
80. Нуклеотидные заместители представляют собой молекулы, имеющие сходные функциональные свойства с нуклеотидами, но не содержащие фосфатный фрагмент, например пептидная нуклеиновая кислота (ПНК). Нуклеотидные заместители представляют собой молекулы, которые распознают нуклеиновые кислоты по методу Уотсона-Крика или Хугстина, но которые связаны друг с другом посредством фрагмента, отличного от фосфатного фрагмента. Нуклеотидные заместители способны соответствовать структуре типа двойной спирали при взаимодействии с соответствующей целевой нуклеиновой кислотой. Существует множество разновидностей этих типов молекул, доступных в данной области техники и представленных в данной заявке.
81. Также возможно связать другие типы молекул (конъюгаты) с нуклеотидами или аналогами нуклеотидов, например, для усиления клеточного поглощения. Конъюгаты могут быть химически связаны с нуклеотидами или нуклеотидными аналогами. Такие конъюгаты включают, но не ограничиваются приведенным, липидные фрагменты, такие как холестериновый фрагмент. (Letsinger et al., Proc. Natl. Acad. Sci. USA, 1989, 86, 6553-6556). Существует множество разновидностей этих типов молекул, доступных в данной области техники и представленных в данной заявке.
82. Взаимодействие Уотсона-Крика представляет собой по меньшей мере одно взаимодействие со стороной Уотсона-Крика нуклеотида, аналога нуклеотида или нуклеотидного заместителя. Сторона Уотсона-Крика нуклеотида, аналога нуклеотида или нуклеотидного заместителя включает положения С2, N1 и С6 нуклеотида на основе пурина, аналога нуклеотида или нуклеотидного заместителя, и положения С2, N3, С4 нуклеотида на основе пиримидина, аналога нуклеотида или нуклеотидного заместителя.
83. Взаимодействие Хугстина представляет собой взаимодействие, которое происходит на стороне Хугстина нуклеотида или аналога нуклеотида, расположенного в большой бороздке двунитевой ДНК. Сторона Хугстина включает положение N7 и реактивные группы (NH2 или О) в положении С6 пуриновых нуклеотидов.
б) Последовательности
84. Существует множество последовательностей, относящихся к белковым молекулам, участвующим в раскрытых в данной заявке сигнальных путях, например TGFβR2, все из которых кодируются нуклеиновыми кислотами или являются нуклеиновыми кислотами. Последовательности человеческих аналогов этих генов, а также других аналогов и аллелей этих генов, а также вариантов сплайсинга и других типов вариантов доступны во множестве баз данных белков и генов, включая Genbank. Специалисты в данной области техники понимают, как разрешить несоответствия и различия последовательностей и скорректировать композиции и способах, относящиеся к конкретной последовательности, к другим родственным последовательностям. Праймеры и/или зонды могут быть сконструированы для любой данной последовательности с учетом информации, раскрытой в данной заявке и известной в данной области техники.
в) Праймеры и зонды
85. Раскрыты композиции, содержащие праймеры и зонды, которые способны взаимодействовать с раскрытыми нуклеиновыми кислотами, такими как TGFβR2 и/или HPRT1, как описано в данной заявке. В некоторых вариантах воплощения изобретения, праймеры используются для поддержки реакций амплификации ДНК. Обычно праймеры могут удлиняться специфическим для последовательности образом. Удлинение праймера специфическим для последовательности образом включает любые способы, в которых последовательность и/или композиция молекулы нуклеиновой кислоты, с которой гибридизируется или иным образом ассоциирован праймер, направляет или влияет на композицию или последовательность продукта, полученного удлинением праймера. Удлинение праймера специфическим для последовательности образом, таким образом, включает, но не ограничивается, ПЦР, секвенирование ДНК, удлинение ДНК, полимеризацию ДНК, транскрипцию РНК или обратную транскрипцию. Предпочтительны способы и условия, которые амплифицируют праймер специфическим для последовательности образом. В некоторых вариантах воплощения, праймеры используются для реакций амплификации ДНК, таких как ПЦР или прямое секвенирование. Понятно, что, в некоторых вариантах воплощения, праймеры также могут быть удлинены с использованием неферментативных способов, при которых, например, нуклеотиды или олигонуклеотиды, используемые для удлинения праймера, модифицированы таким образом, что они будут химически реагировать для удлинения праймера специфическим для последовательности образом. Обычно описанные праймеры гибридизуются с описанными нуклеиновыми кислотами или участком нуклеиновых кислот, или они гибридизуются с комплементом нуклеиновых кислот или комплементом участка нуклеиновых кислот.
86. Размер праймеров или зондов для взаимодействия с нуклеиновыми кислотами, в определенных вариантах воплощения изобретения, может быть любого размера, который поддерживает желаемые ферментативные манипуляции с праймером, такие как амплификация ДНК или простая гибридизация зонда или праймера. Типичный праймер или зонд должен иметь длину по меньшей мере 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1250, 1500, 1750, 2000, 2250, 2500, 2750, 3000, 3500 или 4000 нуклеотидов.
87. В других вариантах воплощения изобретения, праймер или зонд может иметь длину меньшую или равную 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1250, 1500, 1750, 2000, 2250, 2500, 2750, 3000, 3500 или 4000 нуклеотидов.
88. Праймеры для гена TGFβR2 и HPRT1 обычно будут использоваться для получения продукта амплифицированной ДНК, который содержит область гена TGFβR2 и HPRT1 или полный ген. Обычно размер продукта будет таким, чтобы размер можно было точно определить с точностью до 3, 2 или 1 нуклеотида.
89. В определенных вариантах воплощения, этот продукт имеет длину по меьшей мере 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1250, 1500, 1750, 2000, 2250, 2500, 2750, 3000, 3500 или 4000 нуклеотидов.
90. В других вариантах воплощения, продукт имеет длину меньше или равную 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1250, 1500, 1750, 2000, 2250, 2500, 2750, 3000, 3500 или 4000 нуклеотидов.
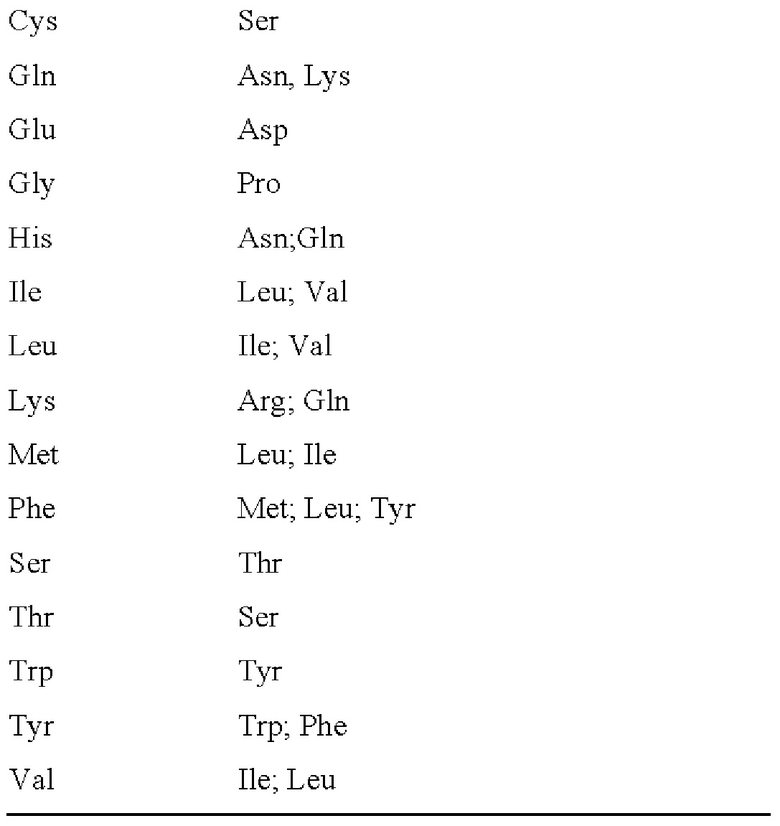
3. Системы экспрессии
91. Нуклеиновые кислоты, которые доставляются в клетки, обычно содержат системы контроля экспрессии. Например, встроенные гены в вирусные и ретровирусные системы обычно содержат промоторы и/или энхансеры, помогающие контролировать экспрессию целевого генного продукта. Промотор, как правило, представляет собой последовательность или последовательности ДНК, которые функционируют в относительно фиксированном положении относительно сайта начала транскрипции. Промотор содержит основные элементы, необходимые для базового взаимодействия РНК-полимеразы и факторов транскрипции, и может содержать вышестоящие элементы и элементы ответа.
а) Вирусные промоторы и энхасеры
92. Предпочтительные промоторы, контролирующие транскрипцию векторов в клетках-хозяевах млекопитающих, могут быть получены из различных источников, например, геномов вирусов, таких как полиома, обезьяний вирус 40 (SV40), аденовирус, ретровирусы, вирус гепатита В и наиболее предпочтительно цитомегаловирус, или из гетерологичных промоторов млекопитающих, например промотора бета-актина. Ранний и поздний промоторы вируса SV40 удобно получать в виде рестрикционного фрагмента SV40, который также содержит вирусный ориджин репликации SV40 (Fiers et al., Nature, 273: 113 (1978)). Непосредственно ранний промотор цитомегаловируса человека удобно получать в виде рестрикционного фрагмента HindIII Е (Greenway, P.J. et al., Gene 18: 355 360 (1982)). Конечно, в данной заявке также могут быть использованы промоторы из клетки-хозяина или родственных видов.
93. Энхансер обычно относится к последовательности ДНК, которая функционирует без фиксированного расстояния от сайта начала транскрипции и может быть либо 5' (Laimins, L. et al., Proc. Natl. Acad. Sci. 78: 993 (1981)), либо 3' (Lusky, M.L., et al., Mol. Cell Bio. 3: 1108 (1983)) к единице транскрипции. Дополнительно, энхансеры могут находиться внутри интрона (Banerji, J.L. et al., Cell 33: 729 (1983)), а также внутри самой кодирующей последовательности (Osborne, T.F., et al., Mol. Cell Bio. 4: 1293 (1984)). Обычно они имеют длину от 10 до 300 п.н. и функционируют в цис. Энхансеры работают, чтобы увеличить транскрипцию с близлежащих промоторов. Энхансеры также часто содержат ответные элементы, которые опосредуют регуляцию транскрипции. Промоторы также могут содержать элементы ответа, которые опосредуют регуляцию транскрипции. Энхансеры часто определяют регуляцию экспрессии гена. Хотя в настоящее время известно много энхансерных последовательностей генов млекопитающих (глобин, эластаза, альбумин, фетопротеин и инсулин), обычно для общей экспрессии используют энхансер вируса эукариотических клеток. Предпочтительными примерами являются энхансер SV40 на поздней стороне источника репликации (bp 100-270), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне источника репликации и энхансеры аденовируса.
94. Промотор и/или энхансер может специфически активироваться светом или определенными химическими событиями, которые запускают их функцию. Системы можно регулировать с помощью таких реагентов, как тетрациклин и дексаметазон. Существуют также способы усиления экспрессии генов вирусных векторов путем воздействия облучения, такого как гамма-облучение, или алкилирования химиотерапевтических препаратов.
95. В некоторых вариантах воплощения изобретения, промоторная и/или энхансерная область может действовать как конститутивный промотор и/или энхансер, чтобы максимизировать экспрессию области транскрибируемой единицы. В определенных конструкциях промоторная и/или энхансерная область активна во всех типах эукариотических клеток, даже если она экспрессируется только в конкретном типе клеток в определенное время. Предпочтительным промотором этого типа является промотор CMV (650 оснований). Другими предпочтительными промоторами являются промоторы SV40, цитоме гало вирус (полноразмерный промотор) и ретровирусный вектор LTR.
96. Было установлено, что все специфические регуляторные элементы могут быть клонированы и использованы для создания векторов экспрессии, которые селективно экспрессируются в определенных типах клеток, таких как клетки меланомы. Промотор глиального фибриллярного кислого белка (GFAP) использовался для селективной экспрессии генов в клетках глиального происхождения.
97. Векторы экспрессии, используемые в эукариотических клетках-хозяевах (дрожжах, грибах, насекомых, растениях, животных, людях или ядросодержащих клетках), также могут содержать последовательности, необходимые для прекращения транскрипции, которые могут влиять на экспрессию мРНК. Эти области транскрибируются как полиаденилированные сегменты в нетранслируемой части мРНК, кодирующей белок тканевого фактора. 3'-нетранслируемые области также включают сайты терминации транскрипции. Предпочтительно, чтобы единица транскрипции также содержала область полиаденилирования. Одним из преимуществ этой области является то, что она увеличивает вероятность того, что транскрибируемая единица будет процессироваться и транспортироваться подобно мРНК. Идентификация и использование сигналов полиаденилирования в экспрессионных конструкциях хорошо изучены. Предпочтительно, чтобы в трансгенных конструкциях использовались гомологичные сигналы полиаденилирования. В некоторых единицах транскрипции область полиаденилирования происходит из сигнала раннего полиаденилирования SV40 и состоит из приблизительно 400 оснований. Также предпочтительно, чтобы транскрибируемые единицы содержали другие стандартные последовательности по отдельности или в комбинации с вышеуказанными последовательностями, улучшали экспрессию или стабильность конструкции.
б) Маркеры
98. Вирусные векторы могут включать последовательность нуклеиновой кислоты, кодирующую маркерный продукт. Этот маркерный продукт используется для определения того, был ли ген доставлен в клетку и после доставки экспрессируется. Предпочтительными маркерными генами являются ген Е. Coli lacZ, который кодирует β-галактозидазу, и зеленый флуоресцентный белок.
99. В некоторых вариантах воплощения изобретения маркер может быть селектируемым маркером. Примерами подходящих селектируемых маркеров для клеток млекопитающих являются дигидрофолатредуктаза (DHFR), тимидинкиназа, неомицин, аналог неомицина G418, гидромицин и пуромицин. Когда такие селектируемые маркеры успешно переносятся в клетку-хозяин млекопитающего, трансформированная клетка-хозяин млекопитающего может выжить, если поместить ее под селективное давление. Существует две широко используемых различных категории селективных режимов. Первая категория основана на метаболизме клетки и использовании мутантной клеточной линии, у которой отсутствует способность расти независимо от среды с добавками. Двумя примерами являются: клетки СНО DHFR и клетки мышиные LTK. Эти клетки не способны расти без добавления таких питательных веществ, как тимидин или гипоксантин. Поскольку в этих клетках отсутствуют определенные гены, необходимые для полного пути синтеза нуклеотидов, они не могут выжить, если недостающие нуклеотиды не будут добавлены в среду с добавками. Альтернативой добавлению среды является введение интактного гена DHFR или ТК в клетки, в которых отсутствуют соответствующие гены, что изменяет их потребности в росте. Отдельные клетки, которые не были трансформированы геном DHFR или ТК, не будут способны выживать в средах без добавок.
100. Вторая категория представляет собой доминантный отбор, который относится к схеме отбора, используемой в любом типе клеток, и не требует использования мутантной клеточной линии. В этих схемах обычно используется лекарство для остановки роста клетки-хозяина. Те клетки, которые имеют новый ген, будут экспрессировать белок, передающий лекарственную устойчивость, и выживут при отборе. Примеры такого доминирующего отбора включают препараты неомицин (Southern P. and Berg, P., J. Molec. Appl. Genet. 1: 327 (1982)), микофеноловую кислоту (Mulligan, R.C. and Berg, P. Science 209: 1422 (1980)) или гигромицин (Sugden, В. et al., Mol. Cell. Biol. 5: 410-413 (1985)). В трех примерах используются бактериальные гены под эукариотическим контролем для передачи устойчивости к соответствующему препарату G418 или неомицину (генетицину), xgpt (микофеноловая кислота) или гигромицину соответственно. Другие включают аналог неомицина G418 и пурамицин.
4. Пептиды
а) Белковые варианты
101. Варианты и производные белков хорошо известны специалистам в данной области техники и могут включать модификации аминокислотной последовательности. Например, модификации аминокислотной последовательности обычно относятся к одному или более из трех классов: варианты с заменами, вставками или делециями. Вставки включают амино- и/или карбоксильные концевые слияния, а также вставки внутри последовательности одного или более аминокислотных остатков. Вставки обычно представляют собой вставки меньшего размера, чем вставки амино- или карбоксильных концевых слияний, например, порядка одного-четырех остатков. Иммуногенные производные слитого белка, такие как описанные в примерах, получают путем слияния полипептида, достаточно большого для придания иммуногенности целевой последовательности, путем поперечного сшивания in vitro или с помощью культуры рекомбинантных клеток, трансформированных ДНК, кодирующей слияние. Делеции характеризуются удалением одного или более аминокислотных остатков из белковой последовательности. Обычно не более чем приблизительно от 2 до 6 остатков удаляется в любом одном сайте в молекуле белка. Эти варианты обычно получают путем сайт-специфического мутагенеза нуклеотидов в ДНК, кодирующей белок, в результате чего получают ДНК, кодирующую вариант, и после этого экспрессируют ДНК в культуре рекомбинантных клеток. Способы создания заменяющих мутаций в заранее определенных участках ДНК, имеющих известную последовательность, хорошо известны, например, мутагенез праймера М13 и мутагенез ПЦР. Аминокислотные замены обычно представляют собой одиночные остатки, но могут происходить одновременно в нескольких различных местах; вставки обычно имеют порядок приблизительно от 1 до 10 аминокислотных остатков; и делеции будут варьироваться от 1 до 30 остатков. Делеции или вставки предпочтительно производятся в соседних парах, т.е. делеция 2 остатков или вставка 2 остатков. Замещения, делеции, вставки или любые их комбинации могут быть объединены для получения конечного конструкта. Мутации не должны выводить последовательность за пределы рамки считывания и предпочтительно не будут создавать комплементарные области, которые могут продуцировать вторичную структуру мРНК. Варианты с замещением представляют собой варианты, в которых по меньшей мере один остаток был удален, а на его место вставлен другой остаток. Такие замены обычно выполняются в соответствии с приведенными ниже Таблицами 1 и 2 и называются консервативными замещениями.


102. Существенные изменения в функции или иммунологической идентичности производятся путем выбора замещений, которые менее консервативны, чем в Таблице 2, т.е. выбора остатков, которые более существенно различаются по своему влиянию на поддержание (а) структуры полипептидного каркаса в области замещения, например, в виде листовой или спиральной конформации, (б) заряда или гидрофобности молекулы в целевом сайте или (в) основной части боковой цепи. Замещения, которые в общем, как ожидается, приведут к наибольшим изменениям свойств белка, будут замещениями, в которых (а) гидрофильный остаток, например серил или треонил замещен на (или) гидрофобным остатком, например лейцилом, изолейцилом, фенилаланилом, валилом или аланилом; (б) цистеин или пролин замещен на (или) любым другим остатком; (в) остаток, имеющий электроположительную боковую цепь, например лизил, аргинил или гистидил, замещен на (или) электроотрицательным остатком, например глутамилом или аспартилом; или (г) остаток, имеющий объемную боковую цепь, например, фенилаланин, замещен (или) остатком, не имеющим боковой цепи, например, глицином, в этом случае, (д) путем увеличения количества сайтов для сульфатирования и/или гликозилирования.
103. Например, замещение одного аминокислотного остатка другим, биологически и/или химически подобным, известно специалистам в данной области техники как консервативное замещение. Например, консервативное замещение будет замещать один гидрофобный остаток на другой или один полярный остаток на другой. Замещения включают комбинации, такие как, например, Gly, Ala; Val, Ile, Leu; Asp, Glu; Asn, Gln; Ser, Thr; Lys, Arg; и Phe, Tyr. Такие консервативно замещенные варианты каждой явно раскрытой последовательности включены в мозаичные полипептиды, представленные в данной заявке.
104. Замещающий или делеционный мутагенез может использоваться для вставки сайтов для N-гликозилирования (Asn-X-Thr/Ser) или О-гликозилирования (Ser или Thr). Также может быть желательным удаление цистеина или других лабильных остатков. Делеции или замены потенциальных сайтов протеолиза, например Arg достигаются, например, удалением одного из основных остатков или заменой одного остатками глутаминила или гистидила.
105. Определенные посттрансляционные дериватизации являются результатом действия рекомбинантных клеток-хозяев на экспрессируемый полипептид. Остатки глутаминила и аспарагинила часто дезамидируются посттрансляционно до соответствующих остатков глутамила и аспарила. Альтернативно, эти остатки дезамидируют в умеренно кислых условиях. Другие посттрансляционные модификации включают гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп остатков серила или треонила, метилирование о-аминогрупп боковых цепей лизина, аргинина и гистидина (Т.Е. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco pp 79-86 [1983]), ацетилирование N-концевого амина и, в некоторых случаях, амидирование С-концевого карбоксила.
106. Известно, что одним из способов определения вариантов и производных описанных в данной заявке белков является определение вариантов и производных с точки зрения гомологии/идентичности с конкретными известными последовательностями. В частности, раскрыты варианты этих и других описанных в данной заявке белков, которые имеют по меньшей мере 70, 75, 80, 85, 90 или 95% гомологии с указанной последовательностью. Специалисты в данной области техники легко поймут, как определить гомологию двух белков. Например, гомологию можно рассчитать после выравнивания двух последовательностей, чтобы гомология была на самом высоком уровне.
107. Другой способ вычисления гомологии может быть выполнен с помощью опубликованных алгоритмов. Оптимальное выравнивание последовательностей для сравнения можно провести с помощью алгоритма локальной гомологии Smith and Waterman Adv. Appl. Math. 2: 482 (1981), с помощью алгоритма выравнивания гомологии Needleman and Wunsch, J. MoL Biol. 48: 443 (1970), способом поиска сходства Pearson and Lipman, Proc. Natl. Acad. Sci. U.S.A. 85: 2444 (1988), компьютеризированной реализацией этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI) или путем проверки.
108. Такие же типы гомологии могут быть получены для нуклеиновых кислот, например, с помощью алгоритмов, раскрытых в Zuker, М. Science 244:48-52, 1989, Jaeger et al. Proc. Natl. Acad. Sci. USA 86:7706-7710, 1989, Jaeger et al. Methods Enzymol. 183:281-306, 1989.
109. Известно, что описание консервативных мутаций и гомологии может быть объединено в любую комбинацию, например варианты воплощения изобретения, которые имеют по меньшей мере 70% гомологии с конкретной последовательностью, где варианты представляют собой консервативные мутации.
110. Поскольку в этом описании обсуждаются различные белки и белковые последовательности, понятно, что также раскрыты нуклеиновые кислоты, которые могут кодировать эти белковые последовательности. Это может включать все вырожденные последовательности, относящиеся к конкретной последовательности белка, то есть все нуклеиновые кислоты, имеющие последовательность, которая кодирует одну конкретную последовательность белка, а также все нуклеиновые кислоты, включая вырожденные нуклеиновые кислоты, кодирующие раскрытые варианты и производные последовательностей белка. Таким образом, хотя каждая конкретная последовательность нуклеиновой кислоты не может быть записана в данной заявке, следует понимать, что все и каждая последовательность фактически раскрыта и описана в данной заявке посредством раскрытой последовательности белка. Также понятно, что хотя никакая аминокислотная последовательность не указывает, какая конкретная последовательность ДНК кодирует этот белок в организме, где в данной заявке раскрыты конкретные варианты раскрытого белка, известная последовательность нуклеиновой кислоты, которая кодирует этот белок, также известна и раскрыта и описана в данной заявке.
111. Известно, что существует множество аналогов аминокислот и пептидов, которые могут быть включены в раскрытые композиции. Например, существует множество D-аминокислот или аминокислот, которые имеют другой функциональный заместитель, чем аминокислоты, показанные в Таблице 1 и Таблице 2. Раскрыты противоположные стереоизомеры природных пептидов, а также стереоизомеры пептидных аналогов. Эти аминокислоты можно легко включить в полипептидные цепи, заряжая молекулы тРНК выбранной аминокислотой и создавая генетические конструкции, которые используют, например, янтарные кодоны, для вставки аналоговой аминокислоты в пептидную цепь сайт-специфическим способом.
112. Могут быть получены молекулы, напоминающие пептиды, но не связанные естественной пептидной связью. Например, связи для аминокислот или аналогов аминокислот могут включать CH2NH--, --CH2S--, --СН2--СН2--, --СН=СН-- (цис и транс), --СОСН2--, --СН(ОН)СН2--, и --CHH2SO- (Эти и остальные могут быть найдены у Spatola, A.F. в Chemistry and Biochemistry of Amino Acids, Peptides, and Proteins, B. Weinstein, eds., Marcel Dekker, New York, p. 267 (1983); Spatola, A.F., Vega Data (March 1983), Vol.1, Issue 3, Peptide Backbone Modifications (общий обзор); Morley, Trends Pharm Sci (1980) pp.463-468; Hudson, D. et al., Int J Pept Prot Res 14:177-185 (1979) (--CH2NH--, CH2CH2--); Spatola et al. Life Sci 38:1243-1249 (1986) (--CHH2--S); Hann J. Chem. Soc Perkin Trans. 1307-314(1982) (--CH--CH--, цис и транс); Almquist et al. J. Med. Chem. 23:1392-1398 (1980) (--COCH2--); Jennings-White et al. Tetrahedron Lett 23:2533 (1982) (--COCH2--); Szelke et al. European Appln, EP 45665 CA (1982): 97:39405 (1982) (--CH(OH)CH2--); Holladay et al. Tetrahedron. Lett 24:4401-4404 (1983) (--C(OH)CH2--); и Hruby Life Sci 31:189-199 (1982) (--CH2--S--); каждый из которых включен в данную заявку путем ссылки. Особенно предпочтительной непептидной связью является --CH2NH--. Известно, что аналоги пептидов могут иметь более одного атома между атомами связи, такие как b-аланин, g-аминомасляная кислота и т.п.
113. Аналоги аминокислот и аналоги пептидов часто обладают улучшенными или желательными свойствами, такими как более экономичное производство, более высокая химическая стабильность, улучшенные фармакологические свойства (период полужизни, абсорбция, удельная активность, эффективность и т.д.), измененная специфичность (например, широкий спектр биологической активности), пониженная антигенность и другие.
114. D-аминокислоты можно использовать для получения более стабильных пептидов, поскольку D-аминокислоты не распознаются пептидазами и т.п.Систематическое замещение одной или более аминокислот консенсусной последовательности на D-аминокислоту того же типа (например, D-лизин вместо L-лизина) может быть использовано для получения более стабильных пептидов. Остатки цистеина можно использовать для циклизации или связывания двух или более пептидов вместе. Это может быть полезно для ограничения пептидов в определенных конформациях.
5. Фармацевтические носители/Доставка фармацевтических продуктов
115. Как описано выше, композиции также можно вводить in vivo в фармацевтически приемлемом носителе. Под «фармацевтически приемлемым» подразумевается материал, который не является нежелательным с биологической или иной точки зрения, т.е. материал может быть введен субъекту вместе с нуклеиновой кислотой или вектором, не вызывая каких-либо нежелательных биологических эффектов или не взаимодействуя вредным образом с любыми другими компонентами фармацевтической композиции, в которой он содержится. Носитель, естественно, будет выбран так, чтобы свести к минимуму любой распад активного ингредиента и свести к минимуму любые неблагоприятные побочные эффекты у субъекта, что хорошо известно специалисту в данной области техники.
116. Композиции можно вводить перорально, парентерально (например, внутривенно), внутримышечной инъекцией, внутрибрюшинной инъекцией, трансдермально, экстракорпорально, местно или тому подобное, включая местное интраназальное введение или введение посредством ингаляции. Используемый в данной заявке термин «местное интраназальное введение» означает доставку композиций в нос и носовые ходы через одну или обе ноздри и может включать доставку с помощью механизма распыления или капельного механизма, или путем аэрозолизации нуклеиновой кислоты или вектора. Введение композиций путем ингаляции может осуществляться через нос или рот путем распыления или капельного механизма. Доставка также может осуществляться непосредственно в любую область дыхательной системы (например, в легкие) через интубацию. Точное количество требуемых композиций будет варьироваться в зависимости от субъекта и в зависимости от вида, возраста, веса и общего состояния субъекта, тяжести аллергического расстройства, которое лечат, конкретной используемой нуклеиновой кислоты или вектора, способа его введения и тому подобного. Таким образом, невозможно указать точное количество для каждой композиции. Однако подходящее количество может определить специалист в данной области техники, используя только рутинные эксперименты с учетом изложенных в данной заявке доктрин.
117. Парентеральное введение композиции, если она используется, обычно характеризуется инъекцией. Инъекционные препараты могут быть приготовлены в обычных формах, либо в виде жидких растворов или суспензий, твердых форм, подходящих для растворения суспензии в жидкости перед инъекцией, либо в виде эмульсий. Недавно пересмотренный подход к парентеральному введению включает использование системы медленного или замедленного высвобождения, так что сохраняется постоянная дозировка. См., например, патент США №3610795, который включен в данную заявку посредством ссылки.
118. Материалы могут находиться в растворе, суспензии (например, включенными в микрочастицы, липосомы или клетки). Они могут быть нацелены на определенный тип клеток через антитела, рецепторы или лиганды рецепторов. Следующие ссылки являются примерами использования этой технологии для нацеливания определенных белков на опухолевую ткань (Senter, et al., Bioconjugate Chem., 2:447-451, (1991); Bagshawe, K.D., Br. J. Cancer, 60:275-281, (1989); Bagshawe, et al., Br. J. Cancer, 58:700-703, (1988); Senter, et al., Bioconjugate Chem., 4:3-9, (1993); Battelli, et al., Cancer Immunol. Immunother., 35:421-425, (1992); Pietersz and McKenzie, Immunolog. Reviews, 129:57-80, (1992); and Roffler, et al., Biochem. Pharmacol, 42:2062-2065, (1991)). Основы, такие как «невидимые» и другие липосомы, конъюгированные с антителами (включая липид-опосредованное нацеливание лекарств на карциному толстой кишки), опосредованное рецептором нацеливание ДНК через клеточно-специфические лиганды, нацеливание на лимфоциты опухоли и высокоспецифичное терапевтическое нацеливание ретровирусов на клетки глиомы мышей in vivo. Следующие ссылки являются примерами использования этой технологии для нацеливания определенных белков на опухолевую ткань (Hughes et al., Cancer Research, 49:6214-6220, (1989); and Litzinger and Huang, Biochimica et Biophysica Acta, 1104:179-187, (1992)). Как правило, рецепторы участвуют в маршрутах эндоцитоза, конститутивного, либо лиганд-индуцированного. Эти рецепторы группируются в ямках, окаймленных клатрином, проникают в клетку через покрытые клатрином везикулы, проходят через подкисленную эндосому, в которой сортируются рецепторы, а затем либо возвращаются на поверхность клетки, либо накапливаются внутри клетки, либо разрушаются в лизосомах. Маршруты интернализации выполняют множество функций, таких как поглощение питательных веществ, удаление активированных белков, клиренс макромолекул, условно-патогенное проникновение вирусов и токсинов, диссоциация и распад лиганда и регуляция на уровне рецепторов. Многие рецепторы следуют более чем по одному внутриклеточному пути, в зависимости от типа клетки, концентрации рецептора, типа лиганда, валентности лиганда и концентрации лиганда. Были рассмотрены молекулярные и клеточные механизмы рецептор-опосредованного эндоцитоза (Brown and Greene, DNA and Cell Biology 10:6, 399-409 (1991)).
С. Примеры
119. Приведенные ниже примеры представлены для того, чтобы предоставить специалистам в данной области техники полное раскрытие и описание того, как соединения, композиции, изделия, устройства и/или способы, заявленные в данной заявке, производят и оценивают, и предназначены для чисто иллюстративных целей и не предназначены для ограничения раскрытия. Были предприняты усилия для обеспечения точности чисел (например, количества, температуры и т.д.), но следует учитывать некоторые ошибки и отклонения. Если не указано иное, части являются массовыми частями, температура указана в°С или соответствует температуре окружающей среды, а давление равно атмосферному или близко к нему.
1. Пример 1
а) Способы
(1) Очистка и размножение человеческих NK клеток
120. Лейкоцитарная пленка здоровых доноров была получена в качестве исходного материала от Американского Красного Креста Центрального Огайо (Central Ohio Region American Red Cross). Экспертный совет общенациональной детской больницы (Institutional Review Board of Nationwide Children's Hospital) признал это исследование исключенным.
121. Выделяли РВМС из лейкоцитарной пленки. Вкратце, наносили 35 мл образца лейкоцитарной пленки на 15 мл Ficol-Paque. Центрифугировали при 400 × g в течение 20 минут без перерыва. Промывали восстановленные РВМС три раза PBS. На этом этапе с помощью RosettesSep можно выделить NK-клетки.
122. Размножали NK-клетки путем стимуляции облученными 10×106 питающими клетками, экспрессирующими mbIL21, в 0, 7 и 14 день. Заменяли среду свежей AIMV или RPMI, содержащей 10% FBS, 1% глутаминила, 1% пенициллина, стрептомицина и 100 МЕ/мл IL-2 на весь объем носителя через день.
(2) Разработка и отбор гРНК
123. Выбирали конкретные геномные локусы для нацеливания, используя онлайн-инструменты, например NCBI, Ensemble. В качестве примера использовали эктодомен бета-рецептора 2 трансформирующего фактора роста (TGFBRR2). См. запись: PF08917; См. InterPro: IPR015013; Положение: 49-157 аа. Целевая последовательность: экзон 4 гена TGFBR2 (ENSG00000163513)
124. Для разработки гРНК использовали веб-инструменты разработки CRISPR, такие как http://crispr.mit.edu и Ошибка! Недопустимый объект гиперссылки.. Вводили в последовательность ДНК, выбранную на этапе 2.1. Выбирали геном человека (hg 19) в качестве целевого генома. CRISPR направляющие (20 нуклеотидов, за которыми следует последовательность PAM:NGG) сканировали из последовательности, введенной ранее. Он также показывает возможные несовпадения с мишенями в выбранном геноме. Выбирали три лучших гРНК, которые имеют наивысший балл, исходя из их целевых и нецелевых показателей.
125. В Таблице 1 показаны разработанные РНК CRISPR для нацеливания на экзон 4 гена TGFBR2, указанные с помощью веб-инструментов разработки CRISPR.

126. Упорядочивали РНК CRISPR в виде синтетические крРНК, специфичные для последовательности, и упорядочивали консервативную трансактивирующую РНК (tracrРНК) для взаимодействия через частичную гомологию с крРНК.
(3) Разработка праймеров скрининга делеции
127. Разрабатывали праймеры, охватывающие сайты расщепления гРНК, для анализа мутации Т7Е1. Использовали праймеры на расстоянии по меньшей мере 100 bp от предполагаемого сайта расщепления, для гарантии появления небольших вставок-делеций (инсерций) в целевом сайте sгРНК в 1,5% агарозном геле после анализа мутаций. В Таблице 2 показаны праймеры, которые используются для амплификации эктодомена TGFBR2.

(4) Трансдукция человеческих первичных и размножаемых NK-клеток
128. Трансдукция элементов Cas9/RNP в NK осуществляется электропорацией с использованием системы 4D-Nucleofector следующим образом:
(5) Приготовление клеток
129. Для первичных NK-клеток инкубировали свежевыделенные NK-клетки в среде RPMI или AIMV в присутствии 100 МЕ/мл IL-2 в течение 4 дней и проводили электропорацию на 5-й день (заменяйте среду через день, как описано ранее, и в день перед трансдукцией). Для размножаемых NK-клеток возможны модификации. Для размножаемых NK-клеток стимулировали клетки в день 0 с помощью облученных питающих клеток в соотношении 1:1 и проводили электропорацию в день 5, 6 или 7. (Заменяли среду через день, как описано ранее, и в день перед трансдукцией). В день электропорации подготавливали колбу Т25, наполненную 8 мл свежего RPMI, содержащего 100 МЕ/мл IL-2 для клеток, подвергающихся электропорации, и предварительно инкубировали колбы в увлажненном инкубаторе 37°С/5% CO2. Размороженные клетки или клетки, подвергшиеся 2ой или 3ей стимуляции, можно подвергнуть электропорации в любое время после их восстановления, как описано. Брали 3-4×106 клеток в условиях для 26 мкл трансдукционной смеси, поскольку очень высокая концентрация NK-клеток в растворе Nucleofector увеличивает скорость трансдукции. Клетки 3 раза промывали PBS, для удаления FBS, который обычно содержит активность РНКазы. Каждый раз центрифугировали их при 300 г в течение 8 минут. Рассмотрим 7 условий электропорации для Cas/RNP как одну гРНК (гРНК1, гРНК2, гРНК3) и комбинацию двух гРНК (гРНК1 + гРНК2, гРНК1 + гРНК3, гРНК2 + гРНК3) и один контроль без Cas9/RNP.
(6) Образование крРНК:trасеrРНК/комплекса
130. Повторно суспендировали крРНК (гРНК1, гРНК2 и гРНК3) и ТrасеrРНК в растворе 1X ТЕ до конечных концентраций 200 мкМ. Смешивали 2,2 мкл каждой 200 мкМ гРНК с 200 мкМ ТrасеrРНК, как показано в Таблице 3. Нагревали образцы при 95°С в течение 5 минут и дайте остыть на столе до комнатной температуры (15-25°С). Хранили ресуспендированные РНК и крРНК:trасеrРНК/комплекс при - 20°С для дальнейшего использования.


(7) Образование комплекса RNP
131. Чтобы сэкономить время, на этапе промывки формировали комплекс RNP. Для одинарной реакции дуплекса крРНК:trасrРНК, разбавляли эндонуклеазу Cas9 до 36 мкМ, как указано в Таблице 4.

132. Для комбинированной трансдукции дуплексов крРНК:tracrРНК разводили эндонуклеазу Cas9 до 36 мкМ, как показано в Таблице 5.

133. Медленно добавляли эндонуклеазу Cas9 к крРНК:tracrРНК дуплексам, вращая кончик пипетки, от 30 секунд до 1 минуты. Инкубировали смесь при комнатной температуре 15-20 мин. Если смесь не используется непосредственно после инкубации, смесь сохраняется на льду до использования.
(8) Электропорация
134. Добавляли всю добавку к раствору Nucleofector Solution Р3 и выдерживали его при комнатной температуре. Повторно суспендировали осадок клеток (3-4×106 клеток) в 20 мкл раствора первичного 4D-нуклеофектора Р3. Следует избегать образования пузырьков воздуха при отборе пипеткой. Клетки не следует надолго оставлять в растворе Р3. Немедленно добавляли 5 мкл комплекса RNP к суспензии клеток. Добавляли 1 мкл 100 мкМ усилителя электропорации Cas9 в Cas9/RNP/клеточную смесь. Переносили Cas9/RNP/клеточную смесь в полоски Nucleocuvette по 20 мкл. Осторожно постукивали по полоскам Nucleocuvette, чтобы убедиться, что образец покрывает нижнюю часть полосок. Запустите 4D-Nucleofector System и выбирали программу EN-138.
(9) Пост-трансдукция
135. Клетки оставляли на 3 минуты на полосках. Добавляли 80 мкл предварительно уравновешенной питательной среды в кювету и осторожно переносили образец в колбы. Через 48 часов после трансдукции извлекали геномную ДНК из 5×105 клеток для скрининга делеций генов. Амплифицировали интересующий ген с помощью праймеров, разработанных на этапе 3.2 с наборами ДНК-полимеразы Taq. Формировали гетеродуплексы ампликона ПЦР для переваривания T7EI и инкубировали продукт в течение 30-60 минут с ферментом T7EI при 37°С. Анализ T7EI является предпочтительным для скрининга, поскольку он быстрый, простой и обеспечивает чистые результаты электрофореза по сравнению с использованием анализа Surveyor. Однако этот способ не может обнаруживать вставки и делеции 2 и менее оснований, которые генерируются активностью негомологичного соединения концов (NHEJ) в экспериментах с Cas9 RNP.
136. Запускали дигестированную ДНК на 1,5% агарозном геле при 110 B в течение 30-45 минут, каждые 15 минут визуализировали Ваш гель. Стимулировали остальные клетки с mbIL21-экспрессирующими питающими клетками в соотношении 1:1. Через пять дней после стимуляции экстрагировали РНК для определения уровня экспрессии гена с помощью кПЦР.
137. Проводили анализы на кальцеин, как сообщалось ранее. Вкратце, нагружали целевые клетки кальцеином AM (в показанном примере использовали 3 мкг/мл/1000000 клеток DAOY). Подготавливали NK-клетки для анализа цитотоксичности, оставив на ночь в IL2 (100 МЕ/мл) плюс или минус 10 нг/мл растворимого TGFβ. Проводили анализы кальцеина в тех же цитокинах, что и NK-клетки в течение ночи.
б) РЕЗУЛЬТАТЫ:
(1) Эффективность электропорации:
138. Для оптимизации 4D-нуклеофекции Cas9/RNP электропорации были протестированы 16 различных программ с трансдукцией нецелевой сиРНК и ДНК-плазмиды GFP в NK-клетки. Анализ проточной цитометрии показал, что EN-138 имел самый высокий процент жизнеспособности клеток и эффективности трансдукции (35% живых GFP-положительных клеток) для обеих частиц. (Фигура 1 и Фигура 2). Интересно, что эффективность использования этой программы для электропорации Cas9/RNP была выше, поскольку наблюдалось 60% снижение уровня экспрессии мРНК TGFBR2 (Фигура 5).
(2) Анализ мутации
139. Cas9/RNP, содержащие гРНК2, гРНК1 + гРНК2 и гРНК3, имели успешный нокаут гена эктодомена TGFBR2, но гРНК1 сама по себе не приводила к детектируемым вставкам/делециям Т7Е1 (Фигура 3). Дополнительно, Фигура 4 показывает успешный нокаут HPRT1 человека (гипоксантинфосфорибозилтрансфераза 1) в размножаемых NK-клетках человека с использованием коммерчески поставляемых гРНК.
(3) Анализ уровня генной экспрессии
140. В качестве репрезентативного результата, на Фигуре 5 показано влияние Cas9/RNP (гРНК1 + гРНК2) на уровень продукции мРНК эктодомена TGFBR2, проанализированное с помощью ОТ-ПЦР. Как видно на графике, уровень экспрессии мРНК целевого гена значительно снизился.
(4) Цитотоксичность
141. Как видно на Фигуре 6, после инкубации клеток, модифицированных гРНК1 + гРНК2, гРНК2 и гРНК3 Cas9/RNP с TGFB, совместно культивированных с клетками DAOY, модифицированные клетки не показали какого-либо значительного снижения уровня их цитотоксичности по сравнению с контрольной группой, которая содержала в среде IL-2 всю ночь. Этот результат демонстрирует, что модифицированные клетки Cas9/RNP сохраняют свою функцию цитотоксичности в присутствии TGFB, и показывает, что модифицированные клетки стали устойчивыми к TGFB.
(5) Анализ РНКseq
142. Анализ РНКseq в наивных и размножаемых NK-клетках выявил активный механизм репарации и репликации ДНК в IL-21 размножаемых NK-клетках. Это указывает на то, что размножаемые NK-клетки могут быть более открытыми для генетических манипуляций с использованием системы Cas9/RNP.
в) ОБСУЖДЕНИЕ:
143. Cas9-опосредованная геномная инженерия произвела революцию в экспериментальной и клинической медицине. Использование этого способа в Т-клетках оказалось успешным, но ДНК-зависимая модификация NK-клеток была сложной задачей. В системе CRISPR/Cas9 ДНК-вектор, который несет кодирующую последовательность для sгРНК, находится под контролем промоторов U6 или H1, поскольку возникающий в результате процесс транскрипции не необходим и не желателен. Доставка ДНК-зависимых трансгенов, таких как лентивирусная и ретровирусная трансфекция, является недостаточной из-за значительного апоптоза NK-клеток, связанного с процедурой, который ограничивает эффективное производство генно-инженерных NK-клеток.
144. Поэтому синтетически предварительно сформированный комплекс рибонуклеопротеина (RNP) и белок Cas9 были введены в виде очищенного белка в первичные и размножаемые NK-клетки.
145. Этот способ позволил устранить кэппинг, присоединение хвостовой части и другие процессы транскрипции и трансляции, запускаемые РНК-полимеразой II, которые могут вызывать существенный, связанный с процедурой апоптоз NK-клеток, который, как считается, имеет место в способах ДНК-зависимой трансдукции. Дополнительно, описанный в данной заявке способ использует очищенный белок Cas9, увеличивая воздействие на цель и уменьшая воздействие вне цели, поскольку Cas9/RNP активны сразу после электропорации и также быстро разлагаются, обеспечивая улучшение существующих протоколов.
146. Таким образом, Cas9/RNP можно использовать для генной модификации первичных и размножаемых NK-клеток человека для иммунотерапии рака с использованием описанного выше способа. Результаты также продемонстрировали, что успешный нокаут гена эктодомена TGFBR2 приводит к тому, что эти модифицированные NK-клетки становятся устойчивыми к TGFB.
147. Комбинирование доставки RNP с источником матричной ДНК (например, донорскими векторами природного рекомбиногенного аденоассоциированного вируса (AAV)) может обеспечить сайт-специфичную вставку гена путем гомологичной рекомбинации.
2. Пример 2: супрессор передачи сигналов цитокинов 3 (SOCS3)
148. Генетическая модификация NK-клеток для усиления иммунотерапии рака применяется для лечения широкого спектра видов рака. Недавно была разработана новая стратегия, в которой элементы CRISPR/Cas9 вводятся в NK-клетки в виде рибонуклеопротеинов (RNP) посредством электропорации с последующим размножением на питающие клетки, экспрессирующие 4-1BBL и связанный с мембраной IL-21, для образования большого количества генетически модифицированных NK-клеток. Этот способ был использован для генной модификации нескольких генов в первичных и размножаемых NK-клетках, включая супрессор передачи сигналов цитокинов 3 (SOCS3). SOCS3 негативно регулирует передачу сигналов цитокинов через путь JAK/STAT. Было высказано предположение, что нарушение SOCS3 в первичных NK-клетках с использованием Cas9/RNP может поддерживать уровни передачи сигналов STAT3 и впоследствии увеличивать их пролиферацию и цитотоксическую функцию.
149. гРНК были сконструированы для нацеливания на экзон 2 (Фигура 7) гена SOCS3 и их электропорировали вместе с белком Cas9 в виде Cas9/RNP в первичные NK-клетки с использованием электропоратора Lonza 4D. Шесть различных состояний гРНК тестировали по отдельности или в комбинации. NK-клетки в контрольной группе подвергали электропорации без Cas9/RNP. После электропорации клетки помещали в культуральную среду с добавлением 100 ME человеческого IL-2 в течение 48 часов, а затем размножали с использованием облученных питающих клеток. На 7 день такое же количество клеток повторно стимулировали облученными питающими клетками, чтобы проверить влияние SOCS3 KO на пролиферацию. Вестерн-блоттинг использовали для оценки нокаутной эффективности на уровне белка. Анализ на кальцеин и IncuCyte Zoom (Essen) были выполнены для измерения цитотоксичности в отношении двух линий раковых клеток, K562 и Daoy.
150. Результаты показали значительное снижение уровней белка SOCS3 в 3 условиях (гРНК1, гРНК3 и гРНК1 + гРНК3) по сравнению с контрольной группой. Относительная нормализованная экспрессия SOCS3 показана на Фигуре 8. Анализ кальцеина и увеличение в IncuCyte показали, что модифицированные NK-клетки SOCS3 KO могут убивать опухоли более эффективно по сравнению с контролем в линиях раковых клеток AML и Daoy (Фигуры 9А, 9В, 9С и 9D). гРНК 1 (G1) показала двукратное уничтожение негативных контролей (NC). В частности, NC показывает гибель 80% при 10:1, тогда как G1 показывает гибель 80% при 5:1 (Фигуры 9В и 9D). Клетки нейробластомы не убивались быстрее при нокауте SOCS3 (Фигуры 9С и 9D). Данные о пролиферации показали, что клетки SOCS3 KO могут расти быстрее, чем контрольная группа (Фигура 10).
151. В заключение, данные демонстрируют роль марушрутов SOCS3 и JAK/STAT в функции NK-клеток и указывают на то, что SOCS3 является хорошей мишенью для генной модификации с целью улучшения иммунотерапии рака с использованием NK-клеток.
3. Пример 3: Генерирование CD38-KO NK-клеток для преодоления фратрицида и повышения ADCC
152. Природные клетки-киллеры играют важную роль в нацеливании на множественную миелому (ММ), экспрессирующую CD38, моноклональным антителом к CD38, даратумамбом (DARA). Чтобы преодолеть фратрицид NK-клеток в терапии DARA, были созданы нокаутированные NK-клетки с использованием Cas9/RNP. Комбинированная терапия ex vivo аутологичных нокаутированных NK-клеток с DARA показала значительное улучшение индуцированного DARA уничтожения опухолевых клеток (Фигуры 11 и 12).
4. Пример 4: AAVS1
153. Эта методология была использована для нацеливания на ген AAVS1 в качестве безопасной гавани для использования в качестве сайта интеграции для любых рассматриваемых генов, включая CAR и репортерные гены, в геном первичных NK-клеток.
154. ICE (Interference of CRISPR Edits) показал высокую эффективность нацеливания на интересующий ген с использованием Cas9/RNP. Нацеливание на AAVS1 не изменяет цитотоксический эффект первичных NK-клеток (Фигура 13).
Использованная гРНК: GGGGCCACTAGGGACAGGAT (SEQ ID NO: 9)
5. Пример 5: Генерирование mCherry позитивных первичных NK-клеток в качестве доказательства концепции для производства CAR-NK с использованием Cas9/RNP доноров
155. С использованием данного подхода были созданы первичные NK-клетки человека, экспрессирующие mCherry. Дополнительно, эти модифицированные первичные NK-клетки были размножены путем стимуляции облученными питающими клетками, экспрессирующими mbIL21, и продемонстрировали стабильную экспрессию репортерного гена (Фигуры 14, 15 и 16). Это подтверждает генерацию первичных и увеличенных клеток CAR-NK.
Использованная гРНК: GGGGCCACTAGGGACAGGAT (SEQ ID NO: 9)
6. Пример 6: Тестирование ненаправленного действия
156. Чтобы определить ненаправленное действие после использования Cas9/RNP в NK-клетках, было выполнено полногеномное секвенирование нормальных и модифицированных NK-клеток (CD38-KO). На основании алгоритма, используемого для прогнозирования генов-кандидатов, WGS показал отсутствие или очень низкое (2 гена) ненаправленное действие.
157. Список подходщих ненаправленных генов, созданный Benching.com, был исследован на предмет гРНК, используемой для нацеливания на CD-38. [5'-CTGAACTCGCAGTTGGCCAT-3' (SEQ ID NO: 11)] и не выявил никакого ненаправленного действия. (Список ненаправленных подходящих генов показан в Таблице 6)
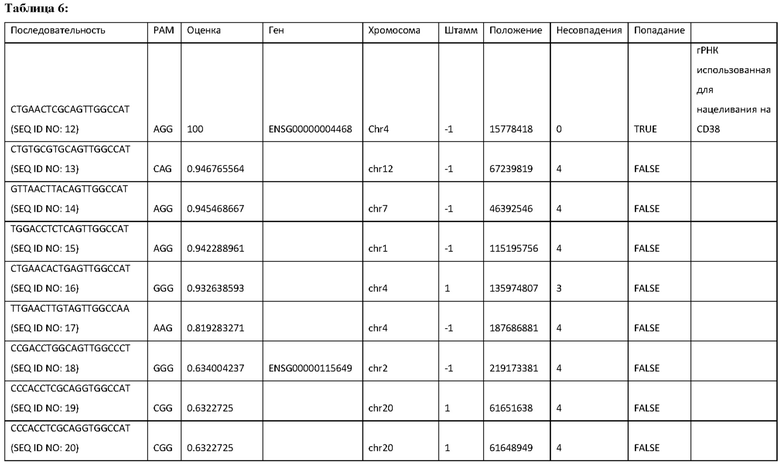
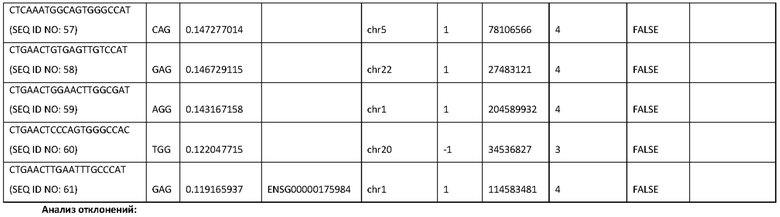
D. Ссылки
Alici, E., Sutlu, Т. & Sirac Dilber, M. Retroviral gene transfer into primary human natural killer cells. Methods Mol Biol. 506 127-137, doi:10.1007/978-1-59745-409-4_10, (2009).
Carlsten, M. & Childs, R.W. Genetic Manipulation of NK Cells for Cancer Immunotherapy: Techniques and Clinical Implications. Front Immunol. 6 266, doi: 10.3389/fimmu.2015.00266, (2015).
Chmielecki, J. et al. Genomic Profiling of a Large Set of Diverse Pediatric Cancers Identifies Known and Novel Mutations across Tumor Spectra. Cancer Res. 77 (2), 509-519, doi: 10.115 8/0008-5 472. can-16-1106, (2017).
DeWitt, M.A., Corn, J.E. & Carroll, D. Genome editing via delivery of Cas9 ribonucleoprotein. Methods. 121-122 9-15, (2017).
Eyquem, J. et al. Targeting a CAR to the TRAC locus with CRISPR/Cas9 enhances tumour rejection. Nature. 543 (7643), 113-117, doi: 10.1038/nature21405, (2017).
Guven, H. et al. Efficient gene transfer into primary human natural killer cells by retroviral transduction. Exp Hematol. 33 (11), 1320-1328, doi:10.1016/j.exphem.2005.07.006, (2005).
Kim, S., Kim, D., Cho, S. W., Kim, J. & Kim, J. S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 24 (6), 1012-1019, doi: 10.1101/gr.171322.113, (2014).
Lee, D.A., Verneris, M.R. & Campana, D. Acquisition, preparation, and functional assessment of human NK cells for adoptive immunotherapy. Methods Mol Biol. 651 61-77, doi: 10.1007/978-1-60761-786-0_4, (2010).
Liang, X. et al. Rapid and highly efficient mammalian cell engineering via Cas9 protein transfection. J Biotechnol. 208 44-53, doi:10.1016/j.jbiotec.2015.04.024, (2015).
Liang, Z. et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat Commun. 8 14261, doi:10.1038/ncommsl4261, (2017).
Mehta, R. S. & Rezvani, K. Chimeric Antigen Receptor Expressing Natural Killer Cells for the Immunotherapy of Cancer. Front Immunol. 9 283, doi:10.3389/fimmu.2018.00283, (2018).
Rezvani, K., Rouce, R., Liu, E. & Shpall, E. Engineering Natural Killer Cells for Cancer Immunotherapy. Mol Ther. 25 (8), 1769-1781, doi: 10.1016/j.ymthe.2017.06.012, (2017).
Schultz, L. M., Majzner, R., Davis, K. L. & Mackall, C. New developments in immunotherapy for pediatric solid tumors. Curr Opin Pediatr. 30 (1), 30-39, doi:10.1097/mop.0000000000000564, (2018).
Somanchi, S. S., Senyukov, V. V., Denman, C.J. & Lee, D.A. Expansion, purification, and functional assessment of human peripheral blood NK cells. J Vis Exp.(48), doi: 10.3791/2540, (2011).
Sutlu, T. et al. Inhibition of intracellular antiviral defense mechanisms augments lentiviral transduction of human natural killer cells: implications for gene therapy. Hum Gene Ther. 23 (10), 1090-1100, doi: 10.1089/hum.2012.080, (2012).
Viel, S. et al. TGF-beta inhibits the activation and functions of NK cells by repressing the mTOR pathway. Sci Signal. 9 (415), ral9, doi:10.1126/scisignal.aad1884, (2016).
Vouillot, L., Thelie, A. & Pollet, N. Comparison of T7E1 and surveyor mismatch cleavage assays to detect mutations triggered by engineered nucleases. G3 (Bethesda). 5 (3), 407-415, doi:10.1534/g3.114.015834, (2015).
Wang, M. et al. Efficient delivery of genome-editing proteins using bioreducible lipid nanoparticles. Proc Natl Acad Sci USA. 113 (11), 2868-2873, doi: 10.1073/pnas.1520244113, (2016).
Zuris, J. A. et al. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nat Biotechnol. 33 (1), 73-80, doi: 10.1038/nbt.3081, (2015).
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ | 2019 |
|
RU2835579C2 |
| ИСКУССТВЕННО СОЗДАННАЯ СИСТЕМА УПРАВЛЕНИЯ ФУНКЦИЕЙ ШК | 2017 |
|
RU2768043C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МОДИФИКАЦИИ ЦЕЛЕВОГО ЛОКУСА | 2015 |
|
RU2704283C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ НАЦЕЛЕННОЙ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ С ИСПОЛЬЗОВАНИЕМ ПАРНЫХ ГИДОВЫХ РНК | 2015 |
|
RU2734770C2 |
| НАЦЕЛИВАНИЕ НА SRC-3 В ИММУННЫХ КЛЕТКАХ В КАЧЕСТВЕ ИММУНОМОДУЛИРУЮЩЕГО ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ РАКА | 2021 |
|
RU2828447C1 |
| ИСКУССТВЕННАЯ МОДИФИКАЦИЯ ГЕНОМА ДЛЯ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНА | 2018 |
|
RU2767201C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2829155C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2745563C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2018 |
|
RU2834231C2 |
| ОПОСРЕДОВАННАЯ НУКЛЕАЗОЙ СБОРКА ДНК | 2015 |
|
RU2707911C2 |
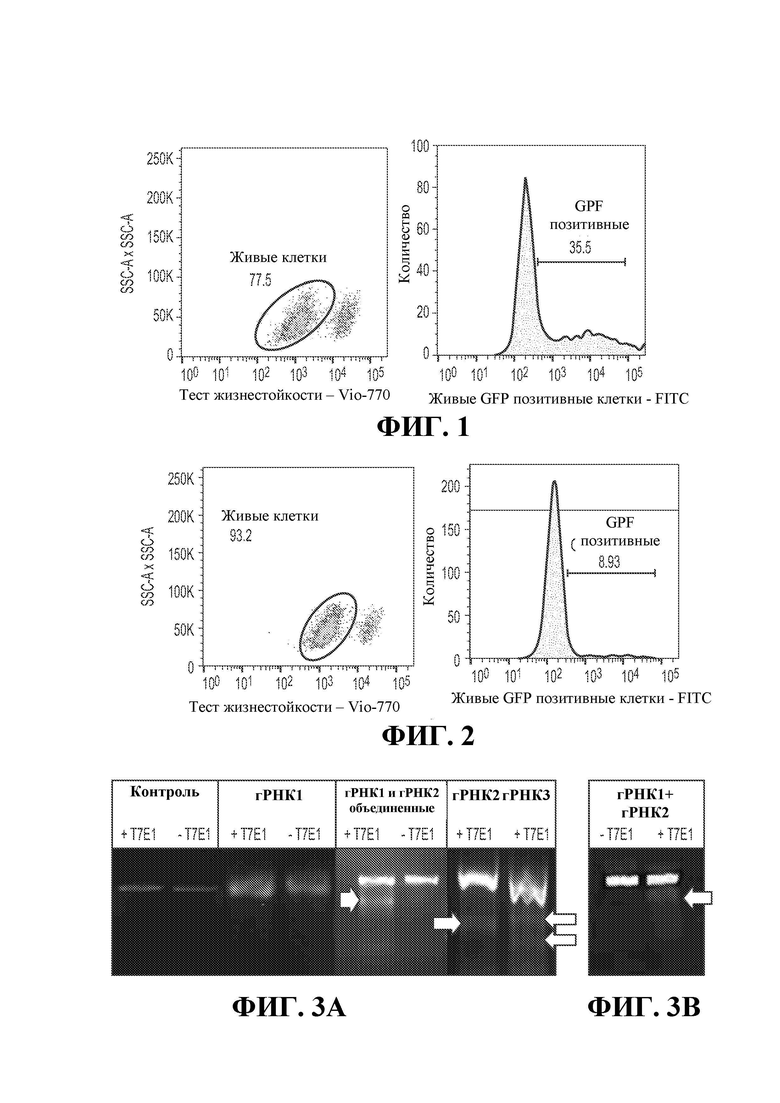
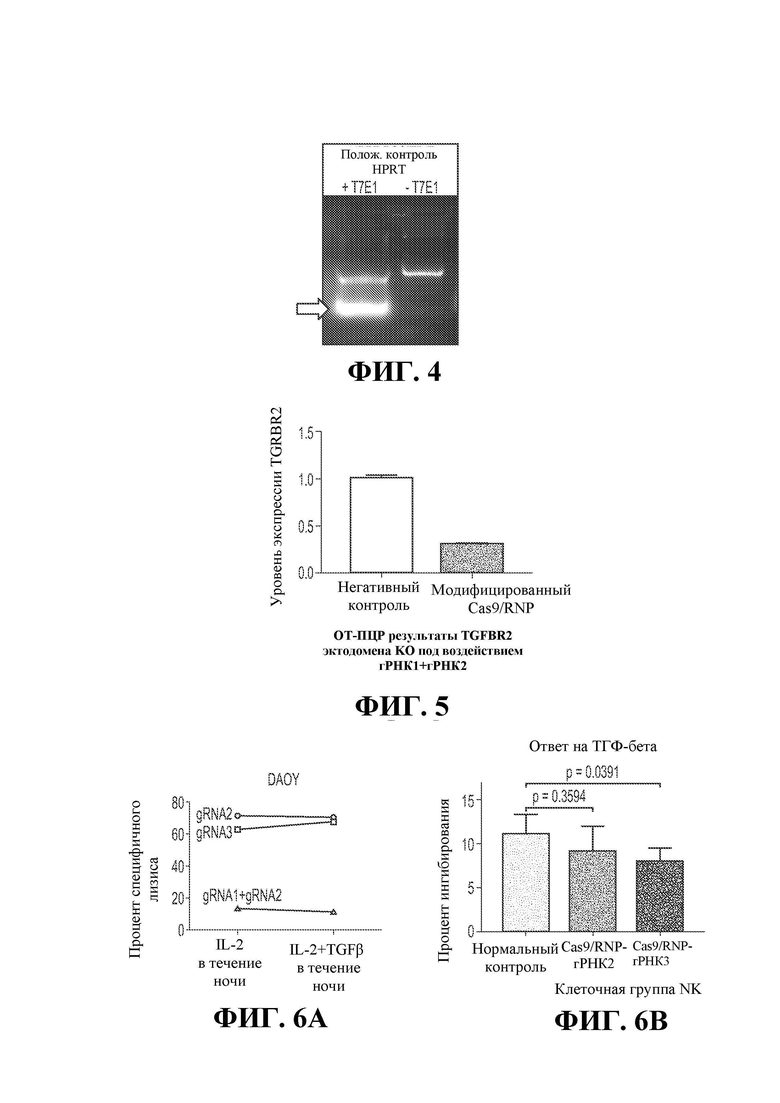

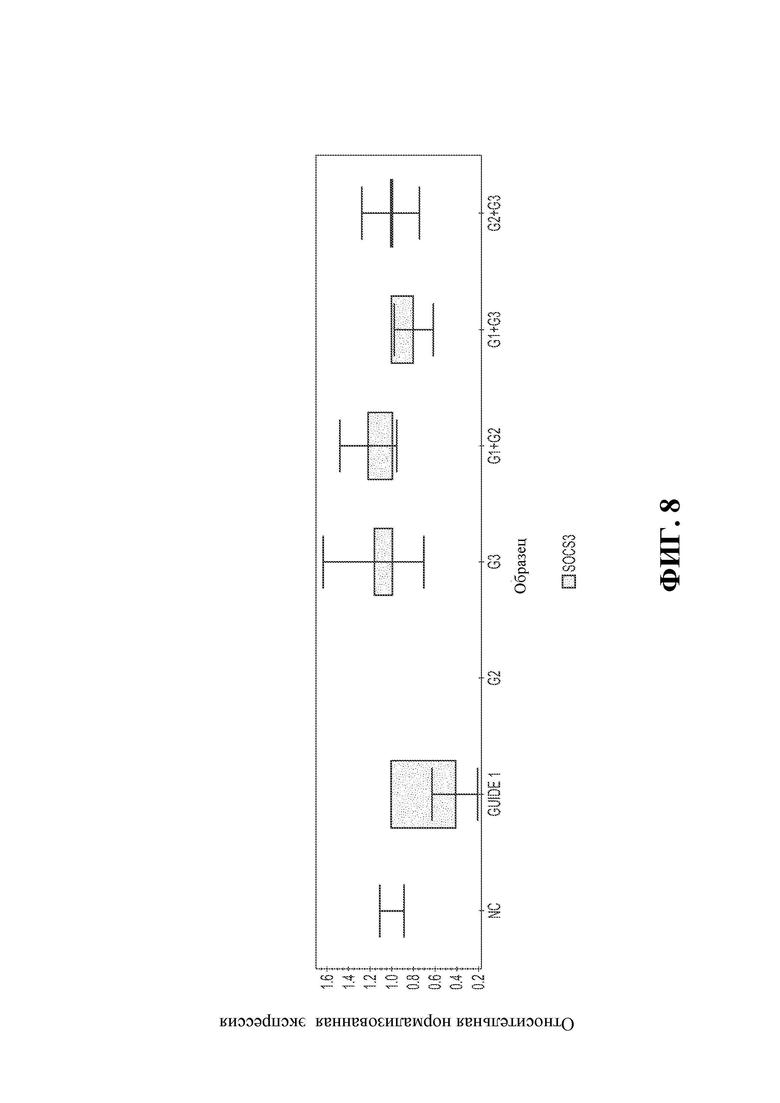
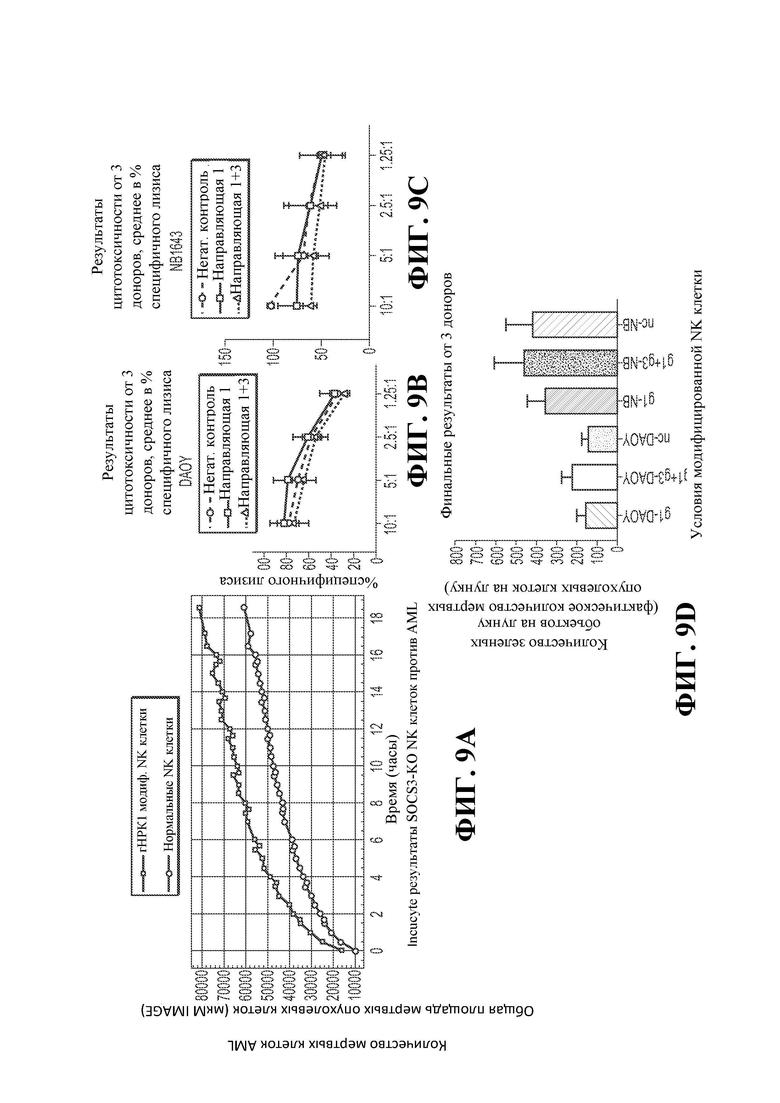
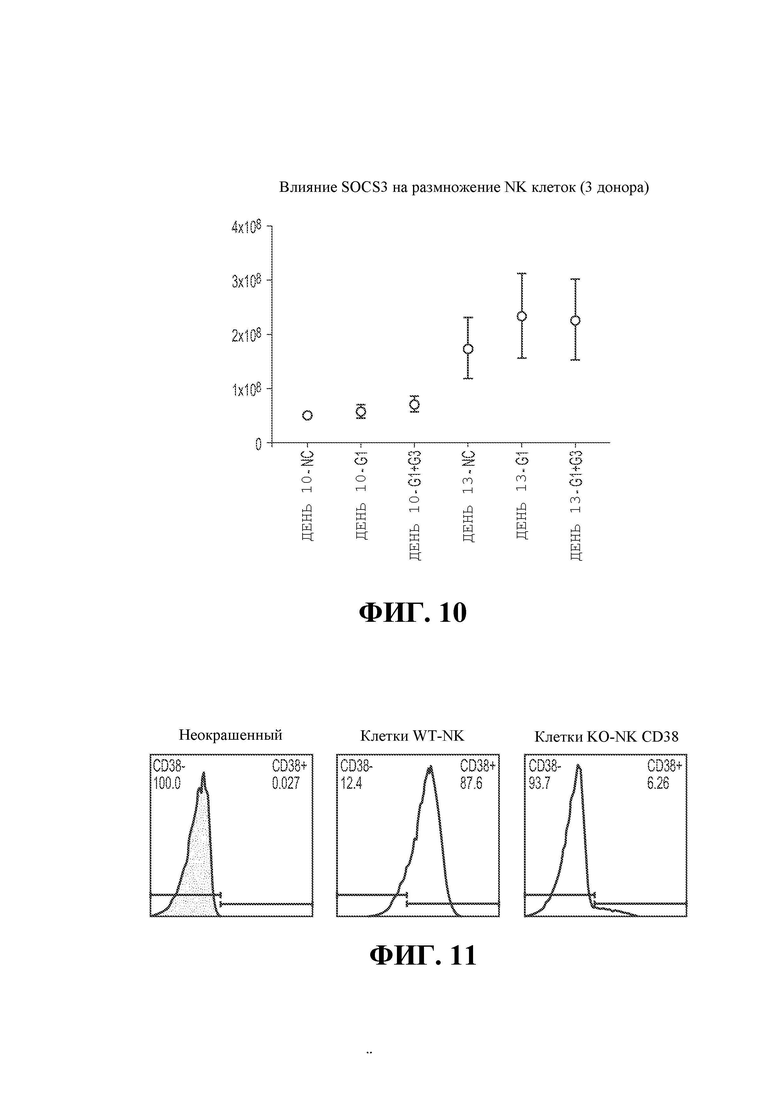
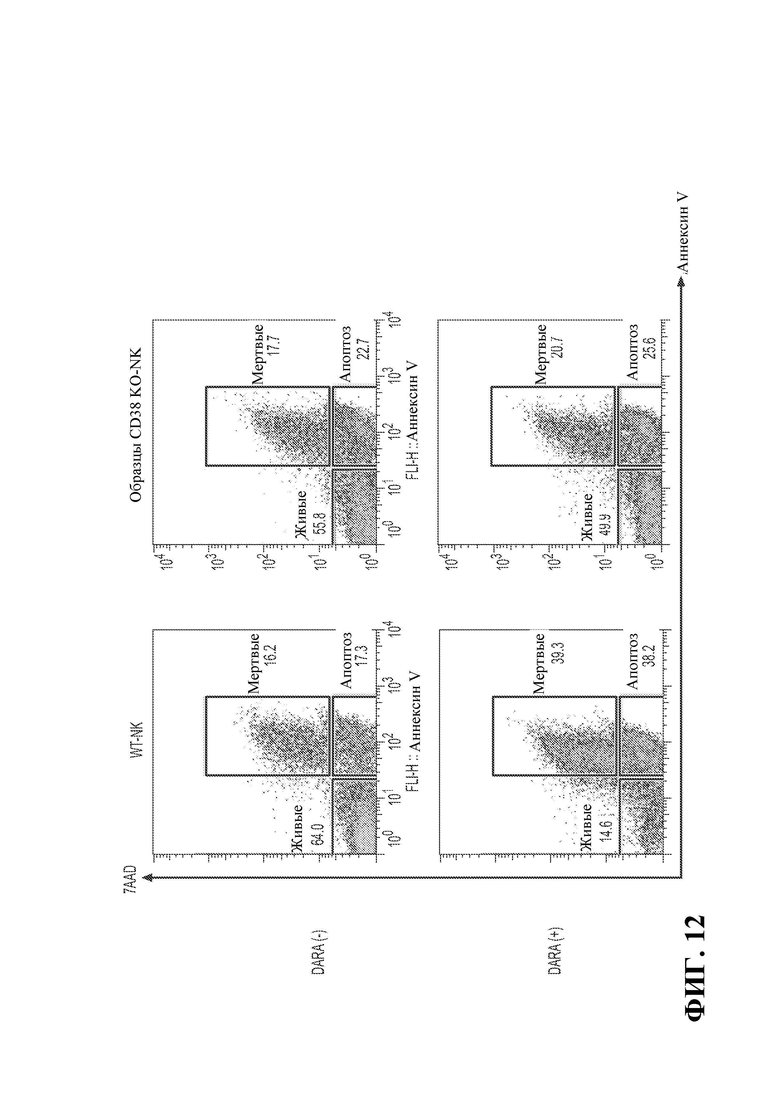
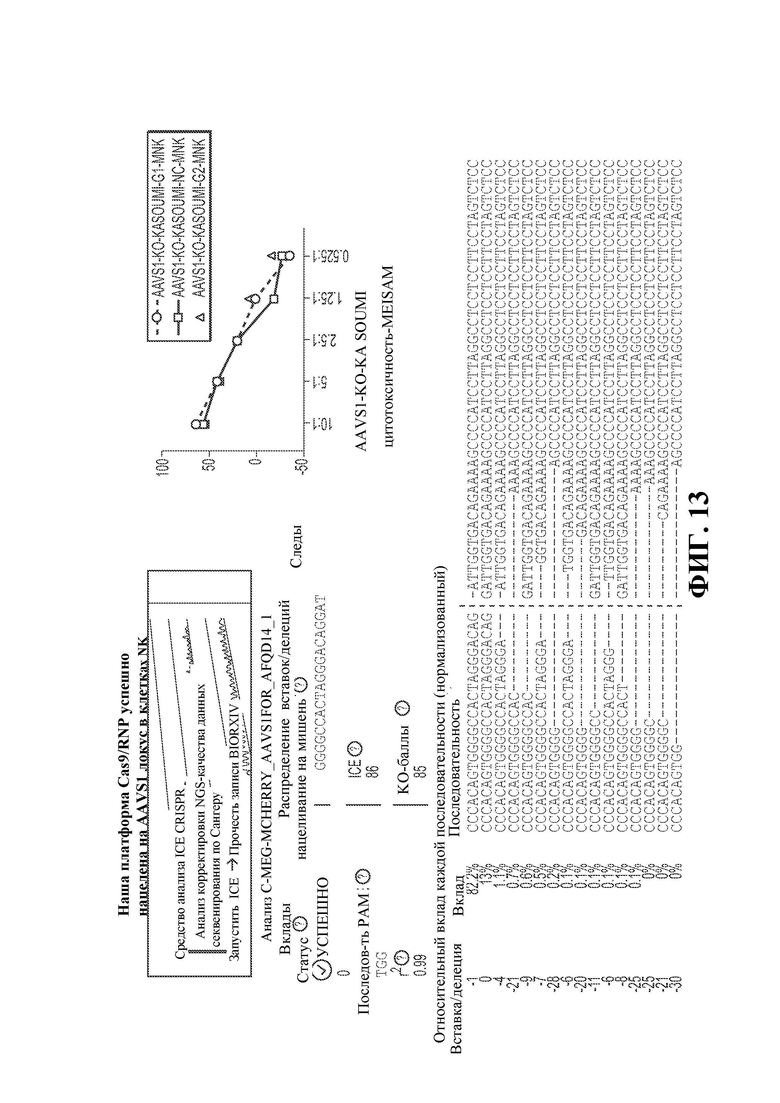

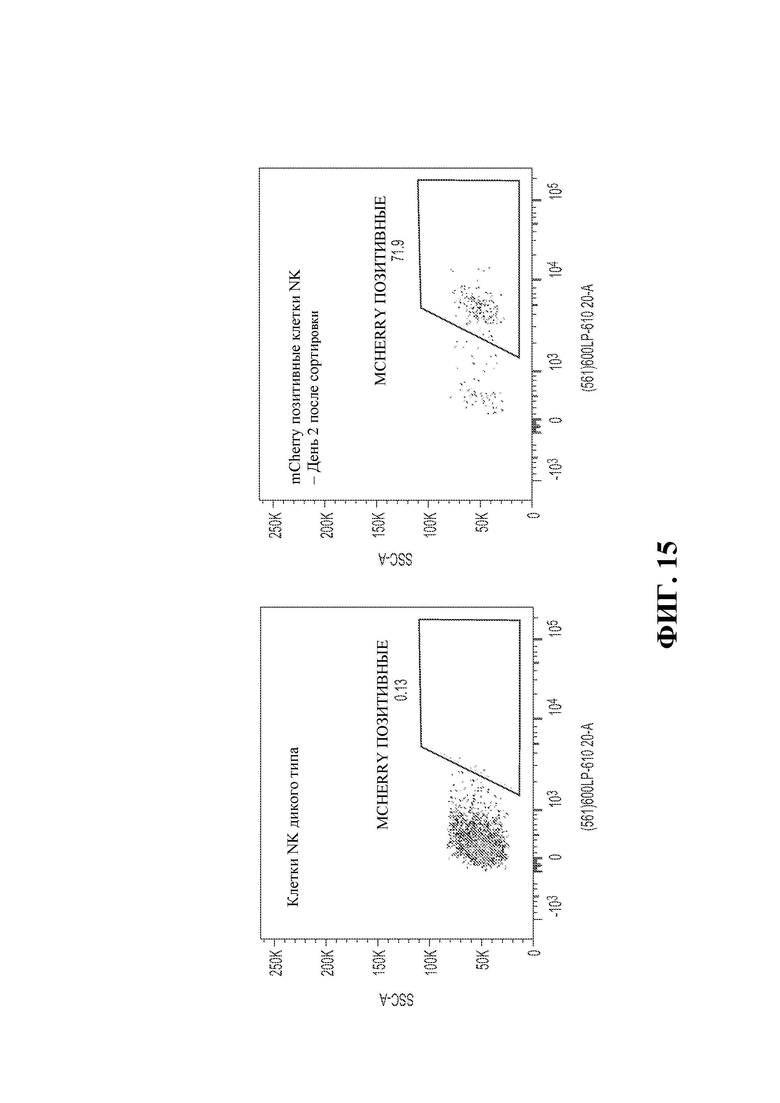
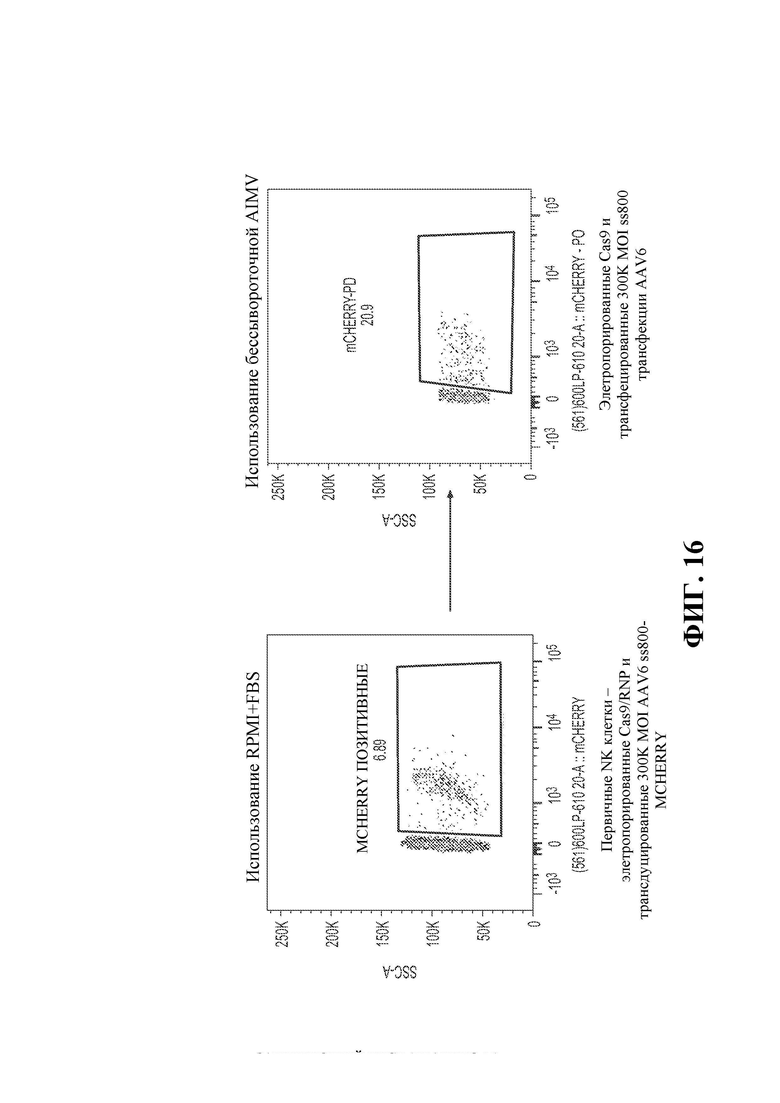
Группа изобретений относится к биотехнологии. Раскрыт способ генетической модификации первичной NK-клетки, включающий: получение направляющей РНК (гРНК), специфичной для целевой последовательности ДНК, и введение путем электропорации в целевую NK-клетку комплекса рибонуклеопротеина (RNP), содержащего эндонуклеазу CRISPR/Cas класса 2, связанную в комплекс с соответствующей CRISPR/Cas направляющей РНК, гибридизующей целевую последовательность внутри геномной ДНК NK-клетки. Также представлен способ адоптивного переноса сконструированных NK-клеток субъекту, нуждающемуся в этом, включающий: получение целевой NK-клетки, подлежащей модификации, получение гРНК, специфичной для целевой последовательности ДНК, введение путем электропорации в целевую NK-клетку комплекса RNP и перенос сконструированной NK-клетки в организм субъекта. Изобретения можно эффективно использовать в иммунотерапии рака. 2 н. и 11 з.п. ф-лы, 16 ил., 6 табл., 6 пр.
1. In vitro способ генетической модификации первичной NK-клетки, включающий:
а) получение направляющей РНК (гРНК), специфичной для целевой последовательности ДНК; и
б) введение путем электропорации в целевую NK-клетку комплекса рибонуклеопротеина (RNP), содержащего эндонуклеазу CRISPR/Cas класса 2, связанную в комплекс с соответствующей CRISPR/Cas направляющей РНК, гибридизующей целевую последовательность внутри геномной ДНК NK-клетки, с получением генетически модифицированной NK-клетки,
где первичную NK-клетку инкубируют в течение 2, 3 или 4 дней в присутствии IL-2 до проведения электропорации; или где первичную NK-клетку размножают в течение 4 дней в присутствии облученных питающих клеток до проведения электропорации, и
где указанная модифицированная NK-клетка содержит нокаут гена, кодирующего супрессор передачи сигналов цитокинов 3 (SOCS3).
2. Способ по п. 1, отличающийся тем, что геном NK-клетки модифицируют путем вставки или делеции одной или более пар оснований, путем вставки фрагмента гетерологичной ДНК, путем делеции фрагмента эндогенной ДНК, путем инверсии или транслокации фрагмента эндогенной ДНК, или их комбинации.
3. Способ по п. 1, отличающийся тем, что первичные NK-клетки размножают в течение 4 дней в присутствии облученных питающих клеток до проведения электропорации и где облученные питающие клетки содержат мембраносвязанные IL-15, 41BBL, или IL-21 или любую их комбинацию.
4. Способ по п. 3, отличающийся тем, что облученные питающие клетки содержат мембраносвязанные IL-15, 41BBL и IL-21.
5. Способ по п. 1, дополнительно включающий размножение генетически модифицированных NK-клеток с облученными питающими клетками, экспрессирующими mbIL-21, после проведения электропорации.
6. Способ по п. 1, отличающийся тем, что указанный способ дополнительно включает образование комплекса RNP путем разбавления 36 мкМ Cas9 в растворе 200 мкМ крРНК и TracrРНК.
7. Способ по п. 1, отличающийся тем, что эндонуклеаза CRISPR/Cas класса 2 представляет собой эндонуклеазу Cas9.
8. Способ адоптивного переноса сконструированных NK-клеток субъекту, нуждающемуся в этом, включающий:
а) получение целевой NK-клетки, подлежащей модификации;
б) получение гРНК, специфичной для целевой последовательности ДНК;
в) введение путем электропорации в целевую NK-клетку комплекса RNP,
содержащего эндонуклеазу CRISPR/Cas класса 2, связанную в комплекс с соответствующей CRISPR/Cas направляющей РНК, гибридизующейся с целевой последовательностью внутри геномной ДНК целевой NK-клетки, с образованием сконструированной NK-клетки; и
г) перенос сконструированной NK-клетки в организм субъекта, где
сконструированную NK-клетку размножают в течение 4 дней при помощи облученных питающих клеток, экспрессирующих mbIL-21, до введения субъекту; или где сконструированную NK-клетку размножают в организме субъекта после переноса сконструированной NK-клетки субъекту посредством введения IL-21 или облученных питающих клеток, экспрессирующих mbIL-21, и где указанная модифицированная NK-клетка содержит нокаут гена, кодирующего супрессор передачи сигналов цитокинов 3 (SOCS3).
9. Способ по п. 8, отличающийся тем, что субъект страдает от рака.
10. Способ по п. 8, отличающийся тем, что NK-клетка представляет собой первичную NK-клетку, модифицированную ex vivo, и перенесенную субъекту после модификации.
11. Способ по п. 8, отличающийся тем, что NK-клетка представляет собой аутологичную NK-клетку.
12. Способ по п. 8, отличающийся тем, что NK-клетку получают из аллогенного донорского источника.
13. Способ по п. 8, отличающийся тем, что эндонуклеаза CRISPR/Cas класса 2 представляет собой эндонуклеазу Cas9.
| WO 2017001572 A1, 05.01.2017 | |||
| VIEL S | |||
| et al., TGF-β inhibits the activation and functions of NK cells by repressing the mTOR pathway, Sci Signal, 2016, 16, 9 (415) | |||
| SCHUMANN K | |||
| et al., Generation of knock-in primary human T cells using Cas9 ribonucleoproteins, Proc Natl Acad Sci, 2015, 112 (33) | |||
| ЧЕМЕРИС Д.А | |||
| и др | |||
| Биоинформатические ресурсы для |
