Область техники
Настоящее изобретение относится к области онкологии или иммунотерапии рака. В частности, настоящее изобретение обеспечивает рекомбинантное гуманизированное антитело против рецептора-1 запрограммированной гибели клеток (PD-1), которое можно использовать в иммунотерапии опухолей или рака. Настоящее изобретение также обеспечивает последовательности нуклеиновых кислот, кодирующих указанное антитело, векторы, содержащие указанные последовательности нуклеиновых кислот, фармацевтические композиции и наборы.
Технологии предшествующего уровня техники
PD-1 представляет собой трансмембранный гликопротеин типа I, состоящий из 288 аминокислот с молекулярным весом примерно 50 кДа и является членом семейства CD28, также известным как CD279. Он функционирует как регулятор запрограммированной гибели клеток и экспрессируется в основном на поверхности зрелых CD4+ и CD8+ Т-клеток, но также и на природных Т-клетках-киллерах, В-клетках, моноцитах и некоторых дендритных клетках.
PD-1 имеет два лиганда, PD-L1 (то есть В7-Н1, также известный как CD274) и PD-L2 (то есть, B7-DC, также известный как CD273), оба из которых являются трансмембранными белковыми молекулами семейства В7. По количеству и действию PD-L1 является основным лигандом PD-1. PD-L1 широко экспрессируется на поверхности различных типов клеток, включая гематопоэтические клетки, такие как дендритные клетки, В-клетки и Т-клетки, а также негематопоэтические клетки, такие как эпителиальные и эндотелиальные клетки. Экспрессия PD-L1 на поверхности опухолевых клеток является высокой, и она может быть усилена воспалительными цитокинами, такими как интерферон-γ и ФНО-α. Лиганд PD-L2 имеет высокое сходство с PD-L1, но его распределение относительно ограничено, в основном экспрессируется на поверхности иммунных клеток, таких как макрофаги, дендритные клетки и мастоциты, и его экспрессия низкая. PD-L2 связывает PD-1 с более высокой аффинностью, примерно в три раза большей, чем PD-L1.
PD-1 играет свою биологическую роль, связываясь со своими лигандами, ингибируя Т-клеточную пролиферацию Т-клеток, активацию и секрецию цитокинов, тем самым подавляя начальную и эффекторную фазы иммунного ответа, поддерживая иммунную стабильность и предотвращая развитие аутоиммунных заболеваний. Когда PD-1 связывается со своим лигандом, остатки тирозина внутри иммунорецепторного тирозинового переключающего мотива (ITSM), одного домена цитоплазматической области PD-1, фосфорилируются, затем фосфорилированный ITSM рекрутирует фосфатазу SHP-2, что приводит к ингибированию важных нижестоящих путей посредством дефосфорилирования, такому как блокирование активации фосфоинозитид-3-киназы (PI3K) и ее нижестоящей протеинкиназы В (РКВ или Akt), ингибирование метаболизма глюкозы, выработка цитокина интерлейкина-2 (ИЛ-2) и экспрессия антиапоптотического белка Bc1-x1; таким образом он подавляет пролиферацию и активацию Т- и В-клеток и секрецию иммуноглобулинов, в результате подавляется аутоиммунный ответ. Опухолевые клетки используют этот иммуносупрессивный механизм для ускользания от иммунного ответа за счет связывания PD-L1, в высокой степени экспрессируемого на молекулах PD-1 на поверхности лимфоцитов, они уклоняются от иммунного распознавания и удаления организмом.
Моноклональные антитела, нацеленные на PD-1, в настоящее время являются актуальной темой в исследованиях иммунотерапии опухолей или рака. Блокируя связывание PD-1 с его лигандом, эти моноклональные антитела могут увеличивать секрецию Т-клеток, а также интерферон-γ и ИЛ-2 на участках опухоли, уменьшать долю клеток-супрессоров миелоидного происхождения (MDSC), изменять микроокружение опухоли, восстанавливать и усиливать иммунную убивающую функцию Т-клеток и, таким образом, подавляют рост опухоли. В настоящее время управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) одобрило регистрационное удостоверение антител к PD-1, в том числе Opdivo от Bristol-Myers Squibb (МНН ниволумаб) и Keytruda от Merck & Co's (МНН пембролизумаб). Opdivo одобрен для лечения меланомы, немелкоклеточного рака легкого, плоскоклеточного рака головы и шеи, классической лимфомы Ходжкина, уротелиальной карциномы, карциномы с высокой микросателлитной нестабильностью, прогрессирующей почечно-клеточной карциномы и гепатоцеллюлярной карциномы; Keytruda одобрена для лечения меланомы, немелкоклеточного рака легкого, плоскоклеточного рака головы и шеи, классической лимфомы Ходжкина, уротелиальной карциномы, высокой микросателлитной нестабильности и рака желудка. Кроме того, отечественные и международные антитела против PD-1 в клинических исследованиях включают REGN2810 от Regeneron/Sanofi, JS001 от Top Alliance, SHR-1210 от Hengrui, BGB-A317 от Beigene, IBI308 от Cinda и GLS-010 от Gloria/WuXiPharmaTech и многие другие антитела против PD-1 и против PD-L1, проходящие в доклинические исследования.
В заключение, исследования моноклональных антител против PD-1 достигли определенного прогресса. Однако все еще существует потребность в улучшении их эффективности, безопасности и * создании конкурентоспособных моноклональных биопрепаратов в онкологии или иммунотерапии рака.
Описание настоящего изобретения
Технические термины, использованные в настоящем изобретении, и их соответствующие сокращения приведены в Таблице 1.
Первый аспект настоящего изобретения обеспечивает выделенное антитело против PD-1 или его антиген-связывающий фрагмент, содержащий вариабельный участок легкой цепи или его часть и/или вариабельный участок тяжелой цепи или его часть;
при этом указанный вариабельный участок легкой цепи или его часть содержит CDR1 легкой цепи, имеющий последовательность аминокислот SEQ ID NO: 10, CDR2 легкой цепи, имеющий последовательность аминокислот SEQ ID NO: 11, и CDR3 легкой цепи, имеющий последовательность аминокислот SEQ ID NO: 12; и
указанный вариабельный участок тяжелой цепи или его часть содержит CDR1 тяжелой цепи, имеющий последовательность аминокислот SEQ ID NO: 13, CDR2 тяжелой цепи, имеющий последовательность аминокислот SEQ ID NO: 14, и CDR3 тяжелой цепи, имеющий последовательность аминокислот SEQ ID NO: 15.
В конкретном варианте осуществления указанное антитело или его антиген-связывающий фрагмент содержит, состоит из или по существу состоит из: последовательности аминокислот, имеющей по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью вариабельного участка легкой цепи SEQ ID NO: 23 указанного антитела против PD-1, и последовательности аминокислот, имеющей по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью вариабельного участка тяжелой цепи SEQ ID NO: 22 указанного антитела против PD-1.
В определенном варианте осуществления указанное антитело дополнительно содержит константный участок легкой цепи и константный участок тяжелой цепи; при этом указанное антитело дополнительно содержит константный участок легкой цепи и константный участок тяжелой цепи, предпочтительно указанный константный участок легкой цепи имеет последовательность аминокислот, имеющую по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью константного участка каппа легкой цепи с последовательностью аминокислот SEQ ID NO: 25, и/или указанный константный участок тяжелой цепи имеет последовательность аминокислот, имеющую по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью константного участка тяжелой цепи IgG4 с последовательностью аминокислот SEQ ID NO: 24. В конкретном варианте осуществления указанным антителом является антитело IgG. В еще одном конкретном варианте осуществления указанным антителом является антитело IgG4.
В еще одном конкретном варианте осуществления указанным антителом или его антиген-связывающим фрагментом является моноклональное антитело или его антиген-связывающий фрагмент.
В конкретном варианте осуществления указанное антитело или его антиген-связывающий фрагмент связывается с указанным рекомбинантным белком PD-1 человека со средней аффинностью KD примерно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200 пМ или выше, или в любых диапазонах с указанными выше значениями в качестве конечных точек, например примерно 20-200 пМ, или примерно 60-70 пМ и т.д., или с любыми их значениями, такими как примерно 64,8 пМ или примерно 108 пМ и т.д. Метод определения аффинности связывания KD описан в примерах данной заявки.
В еще одном конкретном варианте осуществления указанное антитело или его антиген-связывающий фрагмент специфически связывается с молекулой белка PD-1, содержащей последовательность аминокислот SEQ ID NO: 1 или с молекулой белка с последовательностью аминокислот, имеющей по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью SEQ ID NO: 1.
Второй аспект настоящего изобретения обеспечивает выделенное антитело против PD-1 или его антиген-связывающий фрагмент, который специфически связывается с последовательностью пептида, выбранного из по меньшей мере одного из внеклеточных участков белка PD-1 60SESFV64, 78KLAAFPEDRSQP89, 128LAPKAQI134.
В конкретном варианте осуществления обеспечивается выделенное антитело или его антиген-связывающий фрагмент, который специфически связывается с остатком аминокислоты, выбранной из по меньшей мере одного из остатка внеклеточного участка Е61, K78, D85, Р130 указанного белка PD-1.
В еще одном конкретном варианте осуществления указанный антиген-связывающий фрагмент присутствует в форме Fv, Fab, Fab', Fab'-SH, F(ab')2, Fd фрагмента, Fd' фрагмента, молекулы одноцепочечного антитела или антитела с единичным доменом; в еще одном конкретном варианте осуществления, указанной молекулой одноцепочечного антитела является scFv, di-scFv, tri-scFv, диатело или scFab.
В дополнительном конкретном варианте осуществления указанное антитело или его антиген-связывающий фрагмент из указанных выше вариантов осуществления образует ковалентный или нековалентный конъюгат или рекомбинантное многоцелевое слитое с другой молекулой лекарственное средство, образующее таким образом модифицированную молекулу лекарственного средства, где указанная другая молекула выбрана из низкомолекулярного соединения и/или биомакромолекулы.
В третьем аспекте настоящее изобретение обеспечивает выделенную нуклеиновую кислоту, чья последовательность нуклеотидов кодирует указанное антитело и/или антиген-связывающий фрагмент из первого и второго аспектов.
Четвертый аспект настоящего изобретения обеспечивает вектор, содержащий указанную нуклеиновую кислоту из третьего аспекта.
Пятый аспект настоящего изобретения обеспечивает выделенную клетку, экспрессирующую указанное антитело и/или антиген-связывающий фрагмент из первого и второго аспектов, и/или содержащую указанную нуклеиновую кислоту из третьего аспекта или указанный вектор из четвертого аспекта.
В еще одном конкретном варианте осуществления указанными клетками являются прокариотические или эукариотические клетки.
Шестой аспект настоящего изобретения обеспечивает способ получения указанного антитела и/или антиген-связывающего фрагмента из первого и второго аспектов, при этом указанный способ включает выращивание клеток из пятого аспекта и выделение указанных антител.
Седьмой аспект настоящего изобретения обеспечивает применение указанного антитела и/или антиген-связывающего фрагмента и/или модифицированной молекулы лекарственного средства из первого и второго аспектов для приготовления лекарственного средства для лечения опухоли или рака.
В еще одном конкретном варианте осуществления указанной опухолью или раком является рак толстой кишки.
Восьмой аспект настоящего изобретения обеспечивает применение указанного антитела и/или антиген-связывающего фрагмента и/или модифицированной молекулы лекарственного средства из первого аспекта для лечения опухоли или рака.
В еще одном конкретном варианте осуществления указанной опухолью или раком является рак толстой кишки.
Девятый аспект настоящего изобретения обеспечивает фармацевтическую композицию, содержащую указанное антитело и/или антиген-связывающий фрагмент и/или модифицированную молекулу лекарственного средства из первого и второго аспектов.
Десятый аспект настоящего изобретения обеспечивает фармацевтическую комбинацию, содержащую указанную фармацевтическую композицию из девятого аспекта и одно или более терапевтически активное соединение.
Одиннадцатый аспект настоящего изобретения обеспечивает набор, содержащий указанное антитело и/или антиген-связывающий фрагмент и/или модифицированную молекулу лекарственного средства из первого и второго аспектов, или указанную фармацевтическую композицию из девятого аспекта или указанную фармацевтическую комбинацию из десятого аспекта, предпочтительно дополнительно содержащий устройство для введения.
Двенадцатый аспект настоящего изобретения обеспечивает способ лечения опухоли или рака, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества указанного выделенного антитела против PD-1 или его антиген-связывающего фрагмента или модифицированной молекулы лекарственного средства из первого и второго аспектов, или фармацевтической композиции из девятого аспекта, или фармацевтической комбинации из десятого аспекта, или набора из одиннадцатого аспекта для лечения указанной опухоли или рака, при этом указанная опухоль или рак предпочтительно представляет собой рак толстой кишки.
Краткое описание чертежей
Фиг. 1: Блокирование антителом PD1-M944 мыши связывания PD-L1 (А) и PD-L2 (В) с белком PD-1.
Фиг. 2: Определение с помощью иммуноферментного анализа (ELISA) связывания гуманизированного антитела PD1-H944 с рекомбинантным белком PD-1 человека (А, В, С, n=3).
Фиг. 3: Определение с помощью сортировки клеток с активированной флуоресценцией (FACS) связывания PD1-H944 с клетками с рекомбинантным Jurkat/PD-1.
Фиг. 4: Октетный анализ по определению аффинности PD1-H944 (A1, B1, С1, n=3) и ниволумаба (А2, В2, С2, n=3) с рекомбинантным белком PD-1 человека.
Фиг. 5: Анализ ELISA по связыванию PD1-H944 (А), ниволумаба (В) и отрицательного контроля (С) с рекомбинантным белком PD-1 человека, обезьяны и мыши.
Фиг. 6: Связывание PD1-H944 и ниволумаба при различных концентрациях с белком PD-1 обезьяны (А) и белком PD-1 мыши (В).
Фиг. 7: Определение с помощью иммуноферментного анализа (ELISA) блокирования связывания PD-L1 человека (А) и PD-L2 человека (В) с рекомбинантным белком PD-1 человека с помощью PD1-H944.
Фиг. 8: Анализ FACS по определению блокирования связывания PD-L1 человека с клетками Jurkat/PD-1 человека с помощью PD1-H944.
Фиг. 9: Определение с помощью ELISA связывания PD1-H944 с рекомбинантным белком CD 16а.
Фиг. 10: Определение с помощью ELISA связывания PD1-H944 с рекомбинантным белком C1q.
Фиг. 11: Определение с помощью ELISA связывания PD1-H944 с белком FcRn человека.
Фиг. 12: Секреция ИЛ-2 (сверху) и интерферона-γ (снизу) в анализе мешанной лимфы (А, В, С представляют собой CD4+ Т-клетки от 3 доноров), стимулированная антителом против PD-1.
Фиг. 13: Активирующая функция PD1-H944 в клеточной системе с репортерным геном рекомбинантного PD-1 человека.
Фиг. 14: Функция ADCC PD1-H944, определенная в клеточной системе с репортерным геном рекомбинантного CD 16а (А и В являются результатами различных партий анализа; С и D представляют собой значения биолюминесцентной интенсивности при наибольшей концентрации антитела; ** означает Р<0.01).
Фиг.15: Эффект CDC PD1-H944 на клетках CHO-PD-1.
Фиг. 16: Кривая изменений веса тела животных после введения.
Фиг. 17: Кривая роста объема опухоли после введения.
Фиг. 18: График веса опухоли в конце анализа (*Р<0.05).
Фиг. 19: График изменений веса тела животных после введения.
Фиг. 20: График роста объема опухоли после введения лекарства.
Фиг. 21: График веса опухоли в конце анализа.
Фиг. 22: Моделирование гомологии PD1-H944, связанного с кристаллической структурой PD1.
Фиг. 23: Модель белка PD1, демонстрирующая участок с мутацией.
Фиг. 24: Связывание мутантных белков PD-1 с PD-L1, ниволумабом и PD1-Н944.
Фиг. 25А-В: Участки аминокислот, в которым PD1-H944, PD-L1 и ниволумаб специфически связываются с PD-1.
Подробное описание настоящего изобретения
Определения
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют значение, обычно понимаемое специалистом в данной области техники, к которой принадлежит настоящее изобретение. Для целей настоящего изобретения определены следующие термины.
При использовании в настоящем описании и в прилагаемой формуле изобретения формы единственного числа «один», «другой» и «указанный» включают в себя множественное обозначение объекта, если контекст явно не указывает иное.
Термин «антитело» относится к молекуле иммуноглобулина и относится к любой форме антитела, которая проявляет желаемую биологическую активность. К ним относятся, помимо прочего, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела и мультиспецифические антитела (например, биспецифические антитела) и даже фрагменты антител. Обычно полноразмерные структуры антитела предпочтительно включают четыре полипептидных цепи, две тяжелые (Н) цепи и две легкие (L) цепи, обычно соединенные дисульфидными связями. Каждая тяжелая цепь содержит вариабельный участок тяжелой цепи и константный участок тяжелой цепи. Каждая легкая цепь содержит вариабельный участок легкой цепи и константный участок легкой цепи. Помимо этой типичной полноразмерной структуры антитела, указанная структура также включает другие производные формы.
Термин «вариабельный участок» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные области тяжелой и легкой цепей природных антител (VH и VL, соответственно) обычно имеют аналогичную структуру и могут быть подразделены на сильно вариабельные участки (называемые участками, определяющими комплементарность (CDR)), перемежающимися с более консервативными областями (называемыми каркасными участками (FR)).
Термин «участок, определяющий комплементарность» (CDR, например, CDR1, CDR2 и CDR3) относится к таким остаткам аминокислот в вариабельном участке антитела, присутствие которых необходимо для связывания антигена. Каждый вариабельный участок обычно имеет три участка CDR, обозначенных как CDR1, CDR2 и CDR3. Каждый участок, определяющий комплементарность, может содержать остатки аминокислот из «участка, определяющего комплементарность», определенного Кабат (Kabat и др., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. 1991) и/или остатки аминокислот из «высоковариабельной петли» (Chothia и Lesk; J Mol Biol 196: 901-917 (1987)).
Термин остатки аминокислот «каркасного» или «FR» участка относится к остаткам в вариабельном участке, отличным от остатков в CDR, как это определено в настоящем документе.
Каждый вариабельный участок тяжелой цепи и вариабельный участок легкой цепи обычно содержит 3 CDR и до 4 FR, причем указанные CDR и FR расположены от аминоконца до карбоксильного конца в следующем порядке, например: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
Участок, определяющий комплементарность, (CDR) и каркасная область (FR) конкретного антитела могут быть идентифицированы с использованием системы Kabat. (Kabat и др.: Sequences of Proteins of Immunological Interest, 5th edition, US Department of Health and Human Services, PHS, NIH, NIH Publication No. 91- 3242, 1991).
Термин «константный участок» относится к таким последовательностям аминокислот в легкой и тяжелой цепях антитела, которые непосредственно не участвуют в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как антитело-зависимая цитотоксичность.
В соответствии с последовательностью аминокислот константного участка их тяжелых цепей интактные антитела можно разделить на пять классов антител: IgA, IgD, IgE, IgG и IgM, из которых IgG и IgA можно условно разделить на подклассы (изоформы), такие как IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Соответственно, тяжелые цепи пяти классов антител подразделяются на цепи α, δ, ε, γ и μ, соответственно. Легкие цепи антител классифицируются на κ и λ, в соответствии с последовательностью аминокислот их константного участка легкой цепи.
«Антиген-связывающий фрагмент антитела» содержит часть молекулы интактного антитела, которая сохраняет по меньшей мере некоторую специфичность связывания родительского антитела, и обычно включает в меньшей мере часть антигенсвязывающего участка или вариабельного участка (например, один или более CDR) родительского антитела. Примеры антиген-связывающих фрагментов включают Fv, Fab, Fab', Fab'-SH, F(ab')2, фрагменты Fd, фрагменты Fd', молекулы одноцепочечных антител (например, scFv, di-scFv или tri-scFv, диатело или scFab), однодоменные антитела, но не ограничиваются ими.
«Фрагмент антитела» представляет собой молекулу неинтактного антитела, которая по меньшей мере сохраняет некоторые биологические свойства исходного антитела, включая, помимо прочего, фрагмент Fc, в дополнение фрагментам, которые описаны выше в качестве «антиген-связывающих фрагментов».
Термин «модифицированная молекула лекарственного средства» означает конъюгат или рекомбинантное слитое лекарственное средство для множества мишеней, образованное ковалентным или нековалентным соединением антитела или его фрагмента, такого как антиген-связывающий фрагмент, с другой молекулой, которая представляет собой низкомолекулярное соединение или биомакромолекулу.
Термин «химерные» антитела относится к антителам, в которых часть тяжелой и/или легкой цепи получена из определенного источника или вида, а остальная часть получена из другого источника или вида. «Гуманизированные антитела» - это подмножество «химерных антител».
Термин «гуманизированное антитело» или «гуманизированный антиген-связывающий фрагмент» в настоящем описании означает антитело или фрагмент антитела, который: (i) получен из источника, не являющегося человеком, (например, трансгенная мышь, имеющая гетерологичную иммунную систему) и основано на последовательности зародышевой линии человека; или (ii) химерное антитело, где вариабельная область получена из источника, не являющегося человеком, а константная область получена от человека; или (iii) трансплантат CDR, где CDR вариабельной области получена из источника, не являющегося человеком, а одна или более каркасные области вариабельной области получена от человека, и константная область, если таковая имеется, получена от человека. Целью «гуманизации» является устранение иммуногенности антител, полученных из источника, не являющегося человеком, в организме человека при сохранении максимально возможной аффинности. В качестве матрицы для гуманизации выгодно выбрать последовательность каркасной области человека, которая наиболее похожа на последовательность каркасной области не относящегося к человеку исходного антитела. В некоторых случаях может быть необходимо заменить одну или более аминокислот в последовательности каркасной области человека соответствующими остатками в конструкции, полученной из источника, не являющегося человеком, чтобы избежать потери аффинности.
Термин «моноклональное антитело» относится к антителу, полученному из практически гомогенной популяции антител, т.е. каждое отдельное антитело, входящее в популяцию, идентично, за исключением возможных мутаций (например, естественных мутаций), которые могут присутствовать в очень небольших количествах. Таким образом, термин «моноклональное» указывает на природу рассматриваемого антитела, то есть не на смесь неродственных антител. В отличие от препаратов поликлональных антител, которые обычно содержат разные антитела против разных эпитопов, каждое моноклональное антитело в препарате моноклональное антитело направлено против одного эпитопа в антигене. Помимо специфичности, препараты моноклональных антител обладают тем преимуществом, что они обычно не содержат других антител. Термин «моноклональное» не следует понимать как требующее получения указанного антитела каким-либо конкретным методом. Термин моноклональное антитело специфически включает химерные антитела, гуманизированные антитела и антитела человека.
Антитело «специфически связывается» с антигеном-мишенью, таким как опухоль-ассоциированная пептидная антигенная мишень (в данном случае PD-1), т.е. связывает указанный антиген с достаточной аффинностью, чтобы указанное антитело можно было использовать в качестве терапевтического агента, нацеленного на клетку или ткань, экспрессирующую указанный антиген, и не вступает в значительную перекрестную реакцию с другими белками, или не вступает в значительную перекрестную реакцию с белками, отличными от гомологов и вариантов целевых белков, упомянутых выше (например, мутантные формы, варианты сплайсинга или укороченные в результате гидролиза формы белка).
Термин «аффинность связывания» относится к силе суммы нековалентных взаимодействий между отдельными сайтами связывания молекулы и ее партнерами по связыванию. Если не указано иное, «аффинность связывания» в контексте настоящего описания относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между членами пары связывания (например, антитело и антиген). «KD», «константа скорости связывания Kon» и «константа скорости диссоциации Koff» обычно используются для описания сродства между молекулой (например, антителом) и ее партнером по связыванию (например, антигеном). Аффинность - это степень силы, в которой лиганд связывает определенный белок. На аффинность связывания оказывают влияние нековалентные межмолекулярные взаимодействия, такие как водородная связь, электростатические взаимодействия, гидрофобные силы и силы Ван-дер-Ваальса между двумя молекулами. Кроме того, на аффинность связывания между лигандом и его молекулой-мишенью может влиять присутствие других молекул. Аффинность можно анализировать обычными методами, известными в данной области, включая метод ELISA, описанный в настоящем документе.
Термин «эпитоп» включает любой детерминантный кластер белка, который специфически связывается с антителом или Т-клеточным рецептором. Детерминантные кластеры эпитопа обычно состоят из химически активных поверхностных групп молекулы (например, боковых цепей аминокислот или Сахаров, или их комбинации) и часто имеют специфические трехмерные структурные особенности, а также специфические характеристики заряда.
«Выделенное» антитело представляет собой идентифицированное антитело и антитело, выделенное из клетки, которая естественным образом экспрессирует указанное антитело. Выделенное антитело включает антитела in situ в рекомбинантных клетках и антитела, которые обычно получают по меньшей мере за одну стадию очистки.
«Идентичность последовательностей» между двумя полипептидами или последовательностями нуклеиновых кислот указывает количество остатков, которые идентичны между указанными последовательностями, в процентах от общего количества остатков. При вычислении процентной идентичности выравниваемые последовательности сопоставляются таким образом, чтобы обеспечить максимальное совпадение между последовательностями, при этом пробелы в совпадении (если они есть) устраняются с помощью определенного алгоритма. Предпочтительные компьютерные программные методы для определения идентичности двух последовательностей включают программные пакеты GCG, включая GAP, BLASTP, BLASTN и FASTA (Altschul и др., 1990, J. Mol. Biol. 215: 403-410), но не ограничиваются ими. Вышеуказанные процедуры общедоступны в Национальном центре биотехнологической информации (NCBI) и из других источников. Известный алгоритм Смита Ватермана также можно использовать для определения идентичности.
Термин «рецептор Fc» или «FcR» относится к рецептору, который связывается с областью Fc антитела. Предпочтительны FcR человека с природной последовательностью, и предпочтительно рецепторы, связывающиеся с антителами IgG (гамма-рецепторы), которые включают изоформы FcγRI, FcγRII и FcγRIII, а также варианты этих рецепторов. Все другие FcR включены в термин «FcR». Указанный термин также включает неонатальный рецептор (FcRn), который отвечает за транспорт материнского IgG к плоду (Guyer и др., Journal of Immunology 117: 587 (1976) и Kim и др., Journal of Immunology 24: 249 (1994)).
Термин «неонатальный рецептор Fc», сокращенно «FcRn», связывается с Fc-областью антител IgG. Указанный неонатальный рецептор Fc (FcRn) играет важную роль в метаболической судьбе IgG-подобных антител in vivo. FcRn предохраняет IgG от пути лизосомной деградации, тем самым уменьшая его клиренс в сыворотке и удлиняя его период полужизни. Следовательно, свойства/характеристики связывания FcRn in vitro с IgG указывают на его фармакокинетические свойства in vivo в кровотоке.
Термин «эффекторная функция» относится к тем биологическим активностям, приписываемым Fc-области антитела, которые варьируют от изотипа к изотипу.
Примеры эффекторных функций антитела включают связывание C1q и комплемент-зависимую цитотоксичность (CDC), связывание с рецептором Fc, антитело-зависимую клеточно-опосредованную цитотоксичность (ADCC), антитело-зависимый клеточный фагоцитоз (ADCP), секрецию цитокинов, опосредованное иммунным комплексом поглощение антигена антигенпрезентирующими клетками, подавление рецепторов поверхности клеток (например, рецепторов В-клеток) и активация В-клеток.
Термин «эффекторная клетка» относится к лейкоциту, который экспрессирует один или более FcR и выполняет эффекторные функции. В одном аспекте указанные эффекторные клетки в меньшей степени экспрессируют FcγRIII и выполняют эффекторные функции ADCC. Примеры указанных лейкоцитов человека являются лейкоциты, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (РВМС), естественные киллерные (NK) клетки, моноциты, цитотоксические Т-клетки и нейтрофилы. Эффекторные клетки могут быть выделены из природных источников, например, крови. Эффекторные клетки обычно представляют собой лимфоциты, связанные с эффекторной фазой, и функционируют для выработки цитокинов (хелперные Т-клетки), уничтожению клеток, инфицированных патогенами (цитотоксические Т-клетки), или секреции антител (дифференцированные В-клетки).
«Иммунные клетки» включают клетки, которые имеют гемопоэтическое происхождение и принимают участие в иммунном ответе. Иммунные клетки включают: лимфоциты, такие как В-клетки и Т-клетки; естественные клетки-киллеры; и миелоидные клетки, такие как моноциты, макрофаги, эозинофилы, мастоциты, базофилы и гранулоциты.
«Антитело-зависимая клеточно-опосредованная цитотоксичность» или «ADCC» относится к форме цитотоксичности, при которой секретируемый Ig связывается с рецепторами Fcγ, представленными на определенных цитотоксических клетках (например, NK-клетках, нейтрофилах и макрофагах), что позволяет этим цитотоксическим эффекторным клеткам специфически связываться с клетками-мишенями, несущими антигены, и впоследствии убивать указанные клетки-мишени, используя, например, цитотоксин. Для оценки активности ADCC целевого антитела могут быть выполнены анализы ADCC in vitro, такие как анализы ADCC in vitro, описанные в патенте США №5,500,362 или 5,821,337 или в патенте США №6,737,056 (Presta), методы, задокументированные в вариантах осуществления настоящей заявки. Подходящие эффекторные клетки для использования в таких анализах включают РВМС и NK-клетки.
«Комплемент-зависимая цитотоксичность» или «CDC» относится к лизису клеток-мишеней в присутствии комплемента. Активация классического пути комплемента инициируется связыванием первого компонента системы комплемента (C1q) с антителом (соответствующего подкласса), в котором указанное антитело связывается с его соответствующим антигеном. Для оценки активации комплемента может быть проведен анализ CDC, например анализ CDC, описанный в Gazzano-Santoro и др., J. Immunol Methods 202: 163 (1996), методы, задокументированные в вариантах осуществления настоящей заявки, например, методы, описанные в патенте США №6,194,551 В1 и WO 1999/51642, где описаны варианты полипептидов, имеющие измененные последовательность аминокислот области Fc (полипептиды, имеющие вариант области Fc), и варианты полипептидов, демонстрирующие повышенное или пониженное связывание C1q.
Последовательность аминокислот и последовательность нуклеотидов указанного антитела согласно настоящему изобретению
В настоящем изобретении использовали рекомбинантный белок PD-1 человека для иммунизации мыши, а затем получали клон с антителом М944 scFv, которое специфически связывается с рекомбинантным PD-1 человека с высокой аффинностью, посредством скрининга фаговой библиотеки антител. Последовательности нуклеотидов, кодирующих вариабельные участки тяжелые цепи и легкой цепи указанного антитела М944 scFv, собирали посредством ПЦР с последовательностями, кодирующими константный участок тяжелой цепи IgG1 мыши и константный участок легкой цепи каппа мыши, соответственно, и полученную собранную последовательность вставляли в вектор для транзиентной экспрессии в НЕК-293, культивируемых для экспрессии. Мышиное антитело PD1-M944 высокой чистоты очищали с использованием колонки для очистки белка А. Метод ELISA показал, что антитело PD1-M944 мыши способно блокировать связывание PD-1 с его лигандом.
Затем, используя классический метод трансплантации гуманизированных CDR, в качестве матрицы были выбраны легкая цепь антитела человека или вариабельный участок тяжелой цепи, ближайшие к легкой цепи или вариабельному участку тяжелой цепи антитела мыши. В одном варианте осуществления IGKV3-11*01 выбран в качестве гуманизированного участка вариабельного участка легкой цепи, a IGHV3-21*02 - в качестве вариабельного участка тяжелой цепи. Последовательности гуманизированного вариабельного участка легкой цепи (VL) и вариабельного участка тяжелой цепи (VH) получали путем вставки каждой из трех CDR легкой цепи/тяжелой цепи антитела мыши в соответствующие положения вышеуказанных матриц антител человека. Поскольку ключевой сайт каркасного участка антитела мыши необходим для поддержания активности CDR, ключевой сайт подвергали обратной мутации в последовательность антитела мыши. Указанные последовательность аминокислот и последовательность нуклеотидов гуманизированного антитела PD1-H944 получали последовательным сплайсингом последовательности легкой цепи/сигнального пептида тяжелой цепи, последовательности вариабельного участка легкой цепи/тяжелой цепи гуманизированного антитела с обратной мутацией и последовательности константного участка тяжелой цепи антитела человека/константного участка легкой цепи каппа человека, соответственно.
Нуклеиновые кислоты согласно настоящему изобретению
Настоящее изобретение также относится к молекуле нуклеиновой кислоты, кодирующей антитела или их части согласно настоящему изобретению. Указанные последовательности этих молекул нуклеиновой кислоты включают SEQ ID NO: 3-7 и 26-33, но не ограничиваются ими.
Указанные молекулы нуклеиновой кислоты согласно настоящему изобретению не ограничиваются последовательностями, указанными в настоящем описании, но также включают их варианты. Варианты настоящего изобретения могут быть описаны со ссылкой на их физические свойства при гибридизации. Специалистам в данной области будет понятно, что с использованием методов гибридизации нуклеиновых кислот, указанную нуклеиновую кислоту можно использовать для идентификации нуклеиновых кислот, комплементарных им, а также их эквивалентов или гомологов. Также следует понимать, что гибридизация может происходить при комплементарности менее 100%. Однако при соответствующем выборе условий методы гибридизации могут быть использованы для различения указанных последовательностей ДНК на основе структурной релевантности последовательности ДНК для конкретного зонда. Для получения информации о таких условиях см. Sambrook и др., Molecular Cloning: А Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, Cold Spring Harbor, N. Y., 1989 и Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Sedman, J.G., Smith, J.A., & Struhl, K. eds. (1995)б Current Protocols in Molecular Biology. New York: John Wiley и Sons.
Рекомбинантное векторы и векторы экспрессии
Настоящее изобретение также обеспечивает рекомбинантные конструкции, содержащие одну или более последовательностей нуклеотидов согласно настоящему изобретению. Указанные рекомбинантные конструкции согласно настоящему изобретению могут быть использованы с векторами, при этом указанными векторами могут быть, например, плазмиды, фагмиды, фаги или вирусные векторы, а указанные молекулы нуклеиновой кислоты, кодирующие антитела согласно настоящему изобретению вставлены в указанные векторы.
Представленные в настоящем описании антитела могут быть получены путем рекомбинантной экспрессии последовательностей нуклеотидов, кодирующих легкие и тяжелые цепи или их части в клетке-хозяине. Чтобы рекомбинантно экспрессировать это антитело, клетка-хозяин может быть трансфицирована одним или более вектором рекомбинантной экспрессией, несущим последовательности нуклеотидов, кодирующих легкие и/или тяжелые цепи или их части, так что указанные легкие и тяжелые цепи экспрессируются в указанной клетке-хозяине. Для подготовки и/или получения нуклеиновых кислот, кодирующих тяжелые и легкие цепи, для включения этих нуклеиновых кислот в векторы рекомбинантной экспрессии и для введения указанных векторов в клетки-хозяева, используют стандартные рекомбинантные методики ДНК, описанные, например, в Sambrook, Fritsch и Maniatis (eds.), Molecular Cloning; A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989), Ausubel, F.M. и др. (eds.) Current Protocols in Molecular Biology, Greene Publishing Associates, (1989) и методики, задокументированные в патенте США №4,816,397 (Boss) и др.
Дополнительно, последовательности нуклеотидов, кодирующих вариабельный участок указанной тяжелой и/или легкой цепи, могут быть преобразованы в последовательности нуклеотидов, кодирующих, например, полноразмерную цепь антитела, фрагмент Fab или ScFv: например, фрагмент ДНК, кодирующий вариабельную часть легкой цепи или вариабельную часть тяжелой цепи может быть функционально лигирован (так что последовательность аминокислот, кодируемая обоими фрагментами ДНК, находится в одной рамке считывания) с другим фрагментом ДНК, кодирующим, например, константный участок антитела или гибкий линкер. Указанные последовательности константных участков тяжелой и легкой цепи человека известны в данной области техники (смотри, например, Kabat, Е.А., и др. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), а фрагменты ДНК, включающие эти участки, могут быть получены с помощью стандартной ПЦР-амплификации.
Для экспрессии антител могут быть использованы стандартные рекомбинантные методы экспрессии ДНК (смотри, например, Goeddel; Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, Calif. (1990)). Например, последовательность нуклеотидов, кодирующая желаемое антитело, может быть вставлена в вектор экспрессии, который впоследствии трансфицируют в подходящую клетку-хозяин. Подходящими клетками-хозяевами являются прокариотические и эукариотические клетки. Примерами прокариотических клеток-хозяев являются бактерии, а примерами эукариотических клеток-хозяев являются клетки дрожжей, насекомых или млекопитающих. Следует понимать, что дизайн вектора экспрессии, включая выбор регуляторной последовательности, определяется рядом факторов, таких как выбор клетки-хозяина, уровень экспрессии желаемого белка и то, является ли экспрессия конститутивной или индуцируемой.
Антитела согласно настоящему изобретению могут быть выделены и очищены из рекомбинантных культур клеток хорошо известными методами, при этом указанные хорошо известные методы включают осаждение сульфатом аммония или этанолом, кислотную экстракцию, аффинную хроматографию с протеином А, аффинную хроматографию с протеином G, анионную или катионообменную хроматографию, хроматографию на фосфате целлюлозы, хроматографию гидрофобного взаимодействия, аффинную хроматографию, гидроксиапатитную хроматографию и пектиновую хроматографию, но не ограничиваются ими. Для очистки также может использоваться высокоэффективная жидкостная хроматография (ВЭЖХ). Смотри, например, Colligan, Current Protocols in Immunology, или Current Protocols in Protein Science, John Wiley & Sons, NY, N.Y., (1997-2001), например, главы 1, 4, 6, 8, 9, 10, каждая из которых целиком включена в настоящий документ посредством ссылки.
Антитела согласно настоящему изобретению включают естественно очищенные продукты, продукты методов химического синтеза и продукты, полученные рекомбинантными методами из прокариотических и эукариотических хозяев, при этом указанные эукариотические хозяева включают, например, дрожжи, клетки высших растений, насекомых и млекопитающих. Антитела согласно настоящему изобретению могут быть гликозилированными или негликозилированными. Такие методы задокументированы во многих стандартных лабораторных руководствах, например, Sambrook, указанный выше, разделы 17.37-17.42; Ausubel, указанный выше, главы 10, 12, 13, 16, 18 и 20.
Таким образом, варианты осуществления согласно настоящему изобретению также могут быть клетками-хозяевами, содержащими указанный вектор, или молекулами нуклеиновой кислоты, при этом указанными клетками-хозяевами могут быть клетки-хозяева высших эукариот, такие как клетки млекопитающих, клетки-хозяева низших эукариот, такие как клетки дрожжей, и могут быть прокариотическими клетками, такими как как бактериальные клетки.
Свойства и функции антител согласно настоящему изобретению
Был проведен анализ свойств и функций указанного антитела PD1-H944. Анализ показал, что указанное антитело согласно настоящему изобретению имеет следующие преимущества: (1) оно способно связывать PD-1 человека с высокой аффинностью и специфичностью и имеет низкую скорость диссоциации, что обеспечивает хорошую противоопухолевую эффективность; (2) оно способно блокировать связывание PD-L1 с PD-1 более сильно, чем ниволумаб; (3) по сравнению с ниволумабом связывает рекомбинантное PD-1 человека с большей чувствительностью и специфичностью; перекрестно связывается с рекомбинантным PD-1 обезьяны, при этом сравнительно не связывается с PD-1 мыши; (4) оно способен эффективно блокировать связывание рекомбинантного PD-1 человека с его лигандами Pd-L1 и Pd-L2, (5) оно способно эффективно повторно активировать иммунодепрессивные Т-клетки и активировать рекомбинантную репортерную PD-1 систему клеток человека; (6) демонстрирует превосходный противоопухолевый эффект на модели на мышах с гуманизированным PD-1, несущей опухоль рака толстой кишки МС38, намного лучший, чем ниволумаб (Sino Biological, Inc.) и сравнимый с эффектом пембролизумаба (Sino Biological, Inc.); (7) оно проявляет как низкую активность ADCC, так и низкую активность CDC, при этом его активность ADCC ниже, чем у ниволумаба.
Применения
Антитела согласно настоящему изобретению могут быть использованы для лечения множества опухолей или рака или для приготовления лекарств для лечения множества опухолей или рака, специфически нацеленных на аберрантно экспрессируемые рецепторы PD-1. Неограничивающие примеры указанных опухолей или рака включают рак толстой кишки. Фармацевтические композиции
Антитела согласно настоящему изобретению могут быть приготовлены с, по меньшей мере, одним другим агентом (например, стабилизирующим соединением) с образованием фармацевтических композиций, содержащих антитела согласно настоящему изобретению и один или более фармацевтически приемлемых носителей, разбавителей или наполнителей.
Наборы
Настоящее изобретение также относится к фармацевтическим упаковкам и наборам, содержащим один или более контейнеров, содержащих вышеизложенные фармацевтические композиции согласно настоящему изобретению. С такими контейнерами могут быть связаны спецификации в форме, предписанной правительственным агентством, регулирующим производство, использование или распространение лекарственного средства или биологического продукта, которые выдают одобрение для его применения для людей в стране, в которой указанный продукт производится, используется или распространяется.
Изготовление и хранение
Фармацевтические композиции согласно настоящему изобретению могут быть приготовлены способами, известными в данной области, например методами обычного смешивания, растворения, гранулирования, приготовления пастилок, измельчения, эмульгирования, инкапсулирования, заливки или лиофилизации.
Уже приготовленные фармацевтические композиции, содержащие соединения согласно настоящему изобретению, изготовленные в виде лекарственной формы с приемлемым носителем, могут быть помещены в соответствующие контейнеры и помечены для лечения указанного состояния. Такая маркировка будет включать информацию о количестве, частоте и пути введения лекарственного средства.
Комбинации
Фармацевтические композиции, содержащие антитела согласно настоящему изобретению, описанные выше, также комбинируют с одним или несколькими другими терапевтическими агентами, такими как противоопухолевые агенты, при этом полученная комбинация не вызывает неприемлемых побочных эффектов.
Следующие ниже примеры приведены для иллюстрации настоящего изобретения и не предназначены для ограничения настоящего изобретения каким-либо образом.
Примеры
Пример 1: Скрининг антител, полученных из мыши и блокирующих связывание PD-1 с PD-L1/PD-L2, с использованием библиотеки фаговых антител
1.1. Иммунизация мыши
Рекомбинантный белок PD-1 человека (Sino Biological, Inc, кат. №10377-Н08Н) использовали для иммунизации мыши. Последовательность аминокислот внеклеточного участка этого белка PD-1 человека (UniProtKB Q15116) - Metl-Glnl67 (SEQ ID NO: 1).
Указанный рекомбинантный белок PD-1 человека смешивали с адъювантом Фрейнда, и смесь, которую вводили подкожно с интервалами 2 недели, 3 недели, 2 недели и 3 недели, использовали для иммунизации мышей 5 раз в дозировке 20 мкг каждая. Начиная со второй иммунизации, кровь отбирали через семь дней после каждой иммунизации через медиальное кантальное сплетение глаза. Сывороточный титр анти-PD-1 мыши измеряли с помощью ELISA с использованием покрытия рекомбинантного белка PD-1 человека. Титр сыворотки достиг 8000-кратного разведения после пятой иммунизации, и мышам внутривенно вводили 25 мкг рекомбинантного белка PD-1 человека через 9 недель после пятой иммунизации. Через 4 дня мышей умерщвляли, а ткань селезенки замораживали в жидком азоте.
1.2. Скрининг библиотеки фагового дисплея
РНК экстрагировали из ткани селезенки мыши с использованием TriPure Isolation Reagent (Roche), а кДНК получали путем обратной транскрипции с использованием набора для обратной транскрипции (Invitrogen). Соответствующие последовательности нуклеотидов, кодирующие вариабельные участки легкой и тяжелой цепей антитела мыши, амплифицировали и собирали в последовательность нуклеотидов, кодирующих их scFv, с помощью метода ПЦР с перекрывающимся удлинением, в котором соответствующие последовательности нуклеотидов, кодирующие вариабельные участки легкой и тяжелой цепей связывали посредством линкера 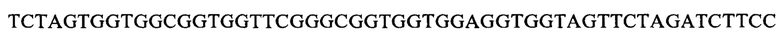 (SEQ ID NO:2) (ссылка: Rapid PCR-cloning of full-length mouse immunoglobulin variable regions; Cloning immunoglobulin variable domains for expression by the polymerase chain reaction T Jones и др., Bio/Technolgy 9(6):579, July 1991), затем ферментативно лигировали с фаговым вектором pComb3x (Sino Biological, Inc.) с использованием эндонуклеазы рестрикции SƒiI (Fermentas), и затем электротрансформировали компетентные клетки Х-Blue для конструирования библиотеки антител scFv фагового дисплея для иммунизированных мышей. Посредством покрытия планшетов для ELISA рекомбинантным белком PD-1 человека получали обогащенные антителами против PD-1 фаговые библиотеки после процесса нанесения фаговых антител, (ссылка: Antibody Phage Display: Methods and Protocols, Philippa M. O'Brien и Robert Aitken (Eds), Humana Press, ISBN: 9780896037113). Моноклональные фаги, выбранные из обогащенной библиотеки, экспрессировали, а затем тестировали их связывание с рекомбинантным белком PD-1 человека с помощью ELISA. Клоны сильносвязывающего антитела М944 scFv, которое специфически связывается с рекомбинантным PD-1 человека, отбирали и заказали компании по секвенированию определить последовательности нуклеотидов scFv антитела М944 (SEQ ID NO:3). 1.3. Получение антитела PD1-M944 мыши
(SEQ ID NO:2) (ссылка: Rapid PCR-cloning of full-length mouse immunoglobulin variable regions; Cloning immunoglobulin variable domains for expression by the polymerase chain reaction T Jones и др., Bio/Technolgy 9(6):579, July 1991), затем ферментативно лигировали с фаговым вектором pComb3x (Sino Biological, Inc.) с использованием эндонуклеазы рестрикции SƒiI (Fermentas), и затем электротрансформировали компетентные клетки Х-Blue для конструирования библиотеки антител scFv фагового дисплея для иммунизированных мышей. Посредством покрытия планшетов для ELISA рекомбинантным белком PD-1 человека получали обогащенные антителами против PD-1 фаговые библиотеки после процесса нанесения фаговых антител, (ссылка: Antibody Phage Display: Methods and Protocols, Philippa M. O'Brien и Robert Aitken (Eds), Humana Press, ISBN: 9780896037113). Моноклональные фаги, выбранные из обогащенной библиотеки, экспрессировали, а затем тестировали их связывание с рекомбинантным белком PD-1 человека с помощью ELISA. Клоны сильносвязывающего антитела М944 scFv, которое специфически связывается с рекомбинантным PD-1 человека, отбирали и заказали компании по секвенированию определить последовательности нуклеотидов scFv антитела М944 (SEQ ID NO:3). 1.3. Получение антитела PD1-M944 мыши
Последовательности нуклеотидов вариабельных участков тяжелой и легкой цепей антитела М944 scFv (SEQ ID NO: 4/5) объединяли с помощью ПЦР с последовательностями нуклеотидов константного участка тяжелой цепи антитела IgG1 мыши (SEQ ID NO: 6) и константного участка легкой цепи каппа мыши (SEQ ID NO: 7), соответственно. Затем полученные последовательности нуклеотидов ферментативно расщепляли с использованием HindIII и XbaI (Fermentas), и встраивали в вектор транзиентной экспрессии pSTEP2 (Sino Biological, Inc), а указанную плазмиду экстрагировали и трансфицировали в клетки HEK-293, которые вьфащивали 7 суток. Супернатант культуры очищали на колонке с белком А с получением антитела высокой чистоты.
1.4 Функциональный анализ антитела PD1-M944 мыши
(1) Блокирование связывания рекомбинантного PD-1 человека с PD-L1: Раствор рекомбинантного белка PD-1 человека концентрации 1 мкг/мл наносили на 96-луночные планшеты в количестве 100 мл на лунку в течение ночи при 4°С. Указанные планшеты отмывали на следующий день и блокировали при комнатной температуре в течение 1 ч. 100 мкл раствора 10 мкг/мл биотинилированного белка PD-L1-Fc-биотин (SinoBiological, Inc.) совместно инкубировали с различными концентрациями (0,4 мкг/мл, 2 мкг/мл и 10 мкг/мл) PD1-M944 или ниволумаба (Sino Biological, Inc). Указанные планшеты отмывали для удаления несвязавшихся антител. Указанные планшеты инкубировали с антибиотиком стрептавидином/пероксидаза хрена (Beijing Zhong Shan - Golden Bridge Biological Technology Co., Ltd) и затем несколько раз промывали, и раствор с хромогенным субстратом добавляли для получения окраски. После остановки реакции определяли OD450. Результаты продемонстрировали, что указанный рекомбинантный белок PD-L1 может эффективно связываться с покрытием белка PD-1, и связывание рекомбинантного белка PD-L1 с PD-1 эффективно ингибировалось путем добавления различных концентраций PD1-M944 или ниволумаба (Фиг. 1А). Эти результаты указывают на то, что указанное антитело PD1-M944 мыши обладает хорошей активностью по блокированию связывания PD-1 с его лигандом PD-L1.
(2) Блокирование связывания рекомбинантного PD-1 человека с PD-L2: Раствор рекомбинантного белка PD-1 человека концентрации 1 мкг/мл наносили на 96-луночный планшет в количестве 100 мл на лунку, в течение ночи при 4°С, на следующий день отмывали и блокировали при комнатной температуре в течение 1 ч. 100 мкл 0,5 мкг/мл биотинилированного белка PD-L2-Fc-биотин (Sino Biological, Inc.) совместно инкубировали с различными концентрациями (0,5 мкг/мл, 2,5 мкг/мл и 12,5 мкг/мл) PD1-М944 или ниволумаба (Sino Biological, Inc.). Указанные планшеты отмывали для удаления несвязавшихся антител, инкубировали с антибиотиком стрептавидином/пероксидаза хрена (Beijing Zhong Shan - Golden Bridge Biological Technology Co., Ltd), несколько раз промывали, и раствор с хромогенным субстратом добавляли для получения окраски, и после остановки реакции определяли OD450. Результаты продемонстрировали, что указанный рекомбинантный белок PD-L2 может эффективно связываются с покрытием белка PD-1, и связывание рекомбинантного белка PD-L2 с PD-1 эффективно ингибировалось путем добавления различных концентраций PD1-M944 или ниволумаба (Фиг. 1В). Эти результаты указывают на то, что указанное антитело PD1-M944 мыши обладает хорошей активностью по блокированию связывания PD-1 с его лигандом PD-L2.
Пример 2: Гуманизация последовательности антитела PD1-M944 мыши для получения последовательности гуманизированного антитела PD1-H944
2.1 Определение участков CDR для каждой из легкой и тяжелой цепей антитела PD1-М944 мыши
Последовательность нуклеотидов антитела М944 scFv определяли в Примере 1.2, в котором определяли последовательности аминокислот вариабельных участков тяжелой и легкой цепей антитела PD1-M944 scFv, смотри SEQ ID NO: 8 и 9.
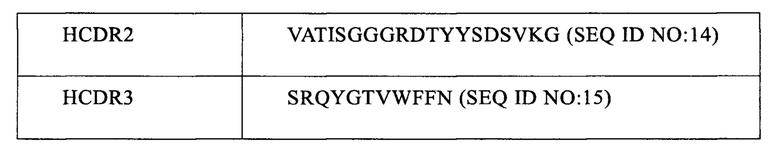
Последовательности аминокислот 3-х участков CDR каждой из легкой и тяжелой цепей антитела PD1-M944 мыши определяли со ссылкой на схему нумерации Кабат, смотри Таблицу 1. Указанные 3 участка CDR каждой из вышеуказанных легкой и тяжелой цепей переносили и вставляли в конечное гуманизированное антитело PD1-H944 последовательными стадиями, смотри Примеры 2.2 и 2.3.

2.2 Гуманизация антитела PD1-M944 мыши трансплантацией CDR
Гуманизацию антитела мыши проводили с использованием классического метода гуманизации трансплантацией CDR. Человеческий антитело Вариабельные участки легкой или тяжелой цепей антитела человека, которые наиболее близки к вариабельным участкам легкой или тяжелой цепи мыши, выбрали в качестве матрицы, и три CDR (Таблица 1) из каждой из легкой или тяжелой цепей антитела мыши вставляли в вариабельный участок антитела человека для получения последовательностей гуманизированные вариабельный участок легкой цепи (VL) и вариабельный участок тяжелой цепи (VH). Указанной матрицей для каждого из вариабельных участков легкой цепи PD1-M944 являлось антитело IGKV3-11*01, которое на 73,48% гомологично легкой цепи PD1-M944, а указанной матрицей для каждого из вариабельных участков тяжелой цепи являлось антитело IGHV3-21*02, которое на 81,94% гомологично тяжелой цепи PD1-М944.
2.3 Обратная мутация каркасной области последовательности гуманизированного вариабельного участка
Поскольку ключевые сайты в каркасной области антитела мыши необходимы для поддержания активности CDR, проводили обратную мутацию соответствующих сайтов на те, которые показаны в последовательности антитела мыши, где положение 89 в легкой цепи было обратно мутировано на М и положение 91 - на F, в тяжелой цепи положение 44 - на R и положение 78 - на N.
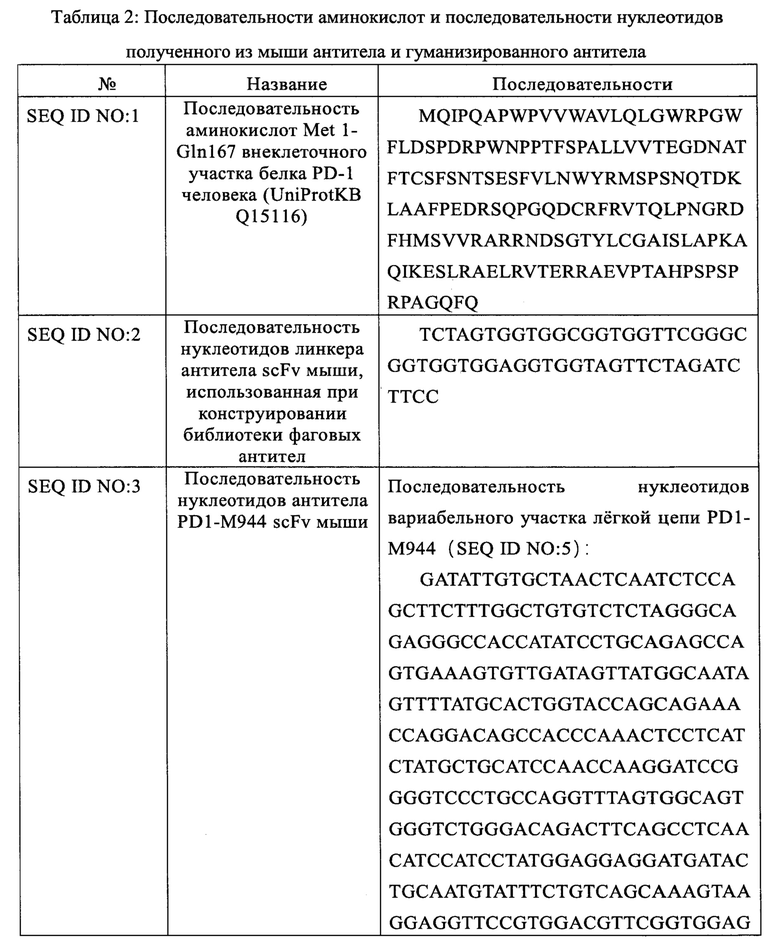
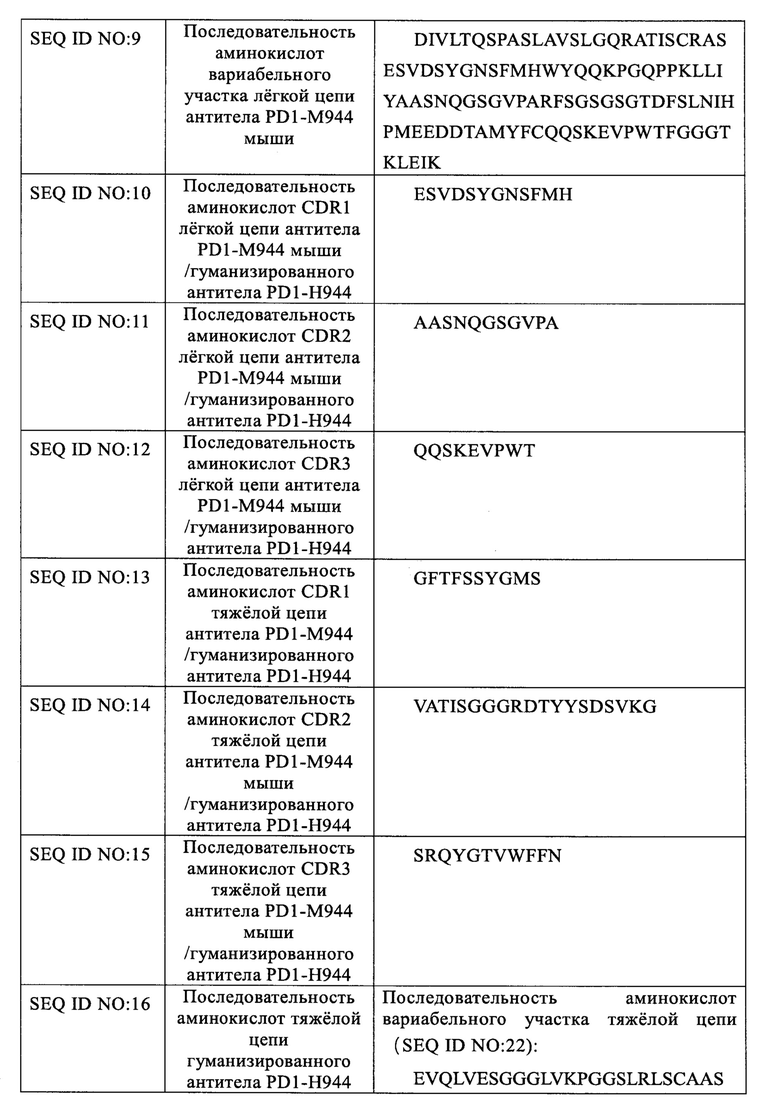
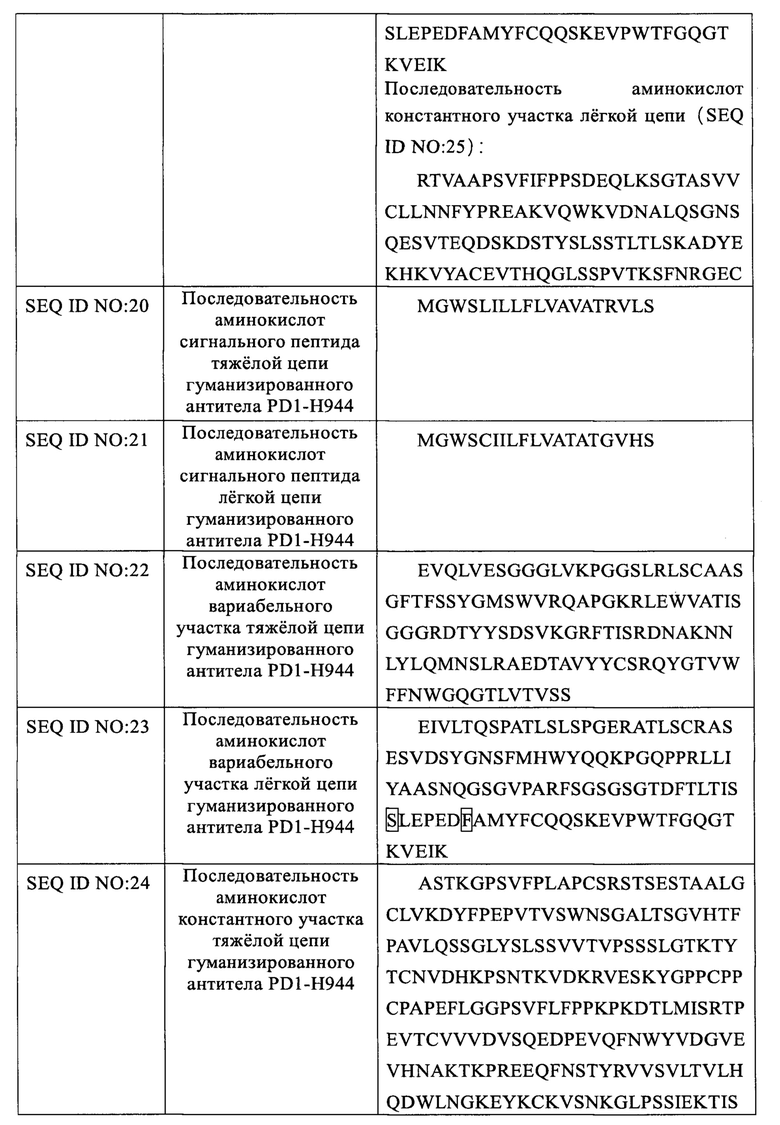
Указанное гуманизированное антитело PD1-H944 получали гуманизированной трансплантацией CDR и обратной мутацией каркасной области, а последовательности аминокислот его тяжелой и легкой цепи показаны в SEQ ID NO: 16 и 17, соответственно; последовательности аминокислот его тяжелой и легкой цепи в форме, содержащей сигнальный пептид, показаны в SEQ ID NO: 18 и 19, соответственно, содержащей последовательно связанные последовательности сигнального пептида тяжелой/легкой цепи (SEQ ID NO: 20 и 21); последовательность вариабельного участка тяжелой цепи/легкой цепи гуманизированного антитела (SEQ ID NO: 22 и 23); и последовательность константного участка гуманизированного антитела, которая представляет собой константный участок тяжелой цепи антитела IgG4 человека/константный участок легкой цепи каппа человека, соответственно (SEQ ID NO: 24 и 25).












Пример 3: Получение гуманизированного антитела PD1-H944
После ПЦР-амплификации, соответствующие указанные выше последовательности нуклеотидов тяжелой и легкой цепей, кодирующие PD1-H944 антитело, содержащее сигнальный пептид, (SEQ ID NO:26 и 27), которые содержат последовательности нуклеотидов, кодирующие последовательно соединенные сигнальный пептид тяжелой/легкой цепи (SEQ ID NO:28/29), вариабельный участок тяжелой/легкой цепи гуманизированного антитела (SEQ ID NO:30/31) и константный участок тяжелой цепи IgG4 человека/ константный участок легкой цепи каппа человека (SEQ ID NO:32/33), соответственно, расщепляли двумя эндонуклеазами рестрикции HindIII viXbaI (Fermentas), затем вставляли в коммерческий вектор pcDNA3 (Invitrogen). После экстракции плазмиды, получали фрагмент размером 1,8 т.о. путем тройного расщепления вектора pCDNA3 с легкой цепью с помощью NruI+NaeI+DraI, затем вставляли в вектор pSSE системы экспрессии CHO/dhfr (Sino Biological, Inc), а затем получали фрагмент размером 2,5 путем тройного расщепления вектора pCDNA3 с тяжелой цепью с помощью NruI+NaeI+DraI (Fermentas). Затем также вводили в вектор pSSE (Sino Biological, Inc) с легкой цепью, полученный на предыдущей стадии для получения полного вектора. Указанный вектор экспрессии является эукариотическим вектором экспрессии, содержащим элемент амплифицированной ДНК гена dhƒr, ген устойчивости NeoR и элементы экспрессии легкой и тяжелой цепей указанного антитела. Указанный вектор экспрессии трансфицировали в клетки dhfr-DG44, Р01-Н944-положительные линии клеток получали посредством скрининга G418, линии клеток с высокой экспрессией PD1-H944 получали посредством скрининга со ступенчатым повышением давления отбора МТХ, а затем одомашнивали в бессывороточной культуре для клонального скрининга. На каждой стадии клоны с высокой экспрессией антитела отбирали на основании результатов анализа ELISA в сочетании со статусом роста клеток и ключевыми атрибутами качества указанного антитела.
Указанную линию клеток СНО, продуцирующую PD1-H944, выращивали в бессывороточной суспензионной культуре для получения антитела PD1-H944 высокой чистоты и качества.
Пример 4: Характеристика гуманизированного антитела PD1-H944
4.1 Анализ аффинности связывания PD1-H944 с антигеном PD-1 человека, мыши и обезьяны
(1) Способность связывания PD1-H944 с рекомбинантным белком PD-1 человека Непрямой анализ ELISA использовали для обнаружения специфического связывания
PD1-H944 с рекомбинантным белком PD-1 человека. Различные концентрации (0,16 нг/мл, 0,49 нг/мл, 1,48 нг/мл, 4,44 нг/мл, 13,33 нг/мл, 40 нг/мл, 120 нг/мл, 360 нг/мл, 1080 нг/мл, 3240 нг/мл и 9720 нг/мл) рекомбинантного белком PD-1 человека наносили на 96-луночные планшеты в течение ночи при 4°С. На следующий день, указанные планшеты отмывали и блокировали при комнатной температуре в течение 1 ч. После инкубации с 100 мкл раствора 2 мкг/мл PD1-Н944, ниволумаба (Bristol-Myers Squibb) и антитела H7N9-R1-IgG4 в качестве отрицательного контроля (Sino Biological, Inc.), соответственно, указанные планшеты отмывали для удаления несвязавшегося антитела, затем инкубировали с антителом козы против IgG F(ab)2 человека/пероксидаза хрена (JACKSON) и последовательно отмывали, а раствор с хромогенным субстратом добавляли для получения окраски и детекции OD450. ЕС50 для связывания PD-1-H944 и ниволумаба с рекомбинантным белком PD-1 человека составлял 31,5 нг/мл, R2=0,998 и 179,0 нг/мл, R2=0,997, соответственно. Это указывает на то, что PD1-H944 связывается с рекомбинантным белком PD-1 человека существенно лучше, чем ниволумаб (Фиг. 2А-В).
(2) Способность связывания PD1-H944 с рекомбинантными клетками Jurkat/PD-1
Способность связывания PD1-H944 с рекомбинантными клетками Jurkat/PD-1 измеряли с помощью клеточной цитометрии с использованием линии клеток Jurkat/PD-1 со стабильной экспрессией рекомбинантного PD-1 человека (SinoCellTech Ltd) в качестве экспериментального материала. Различные концентрации PD1-H944, ниволумаба и антитела H7N9-R1-IgG4 в качестве отрицательного контроля (0,195 мкг/мл, 0,391 мкг/мл, 0,781 мкг/мл, 1,562 мкг/мл, 3,125 мкг/мл, 6,25 мкг/мл, 12,5 мкг/мл, 25 мкг/мл, 50 мкг/мл, 100 мкг/мл и 200 мкг/мл) добавляли к 5×105/пробирку рекомбинантным клеткам Jurkat/PD-1 в логарифмической фазе роста, перемешивали и инкубировали при 4°С, а затем отмывали и центрифугировали для удаления несвязавшегося антитела. Связывание указанного антитела с указанными клетками определяли путем добавления антитела козы против IgG человека с Fc-FITC (Sino Biological, Inc.). Полученные результаты продемонстрировали, что PD1-H944 специфически связывалось с клетками Jurkat/PD-1 с ЕС50 9,63 мкг/мл, R2=1,000, ниволумаб специфически связывалось с клетками Jurkat/PD-1 с ЕС50 10,18 мкг/мл, R2=0,994, а антитело H7N9-R1- IgG4 в качестве отрицательного контроля не связывался с клетками Jurkat/PD-1 (Фиг. 3). Это указывает на то, что PD1-H944 обладает хорошей способностью связываться с PD-1, экспрессированным клетками Jurkat/PD-1, и что его способность связывания немного лучше, чем у ниволумаба.
(3) Аффинность связывания PD1-H944 с рекомбинантным белком PD-1 человека
Аффинность связывания PD1-H944 (0,90 нМ, 1,79 нМ, 3,66 нМ, 7,24 нМ), ниволумаба (0,51 нМ, 1,02 нМ, 2,05 нМ, 4,10 нМ) с PD-1 определяли с использованием с использованием системы Octet Biomolecular Interaction вместе с антителом H7N9-R1-IgG4 (13,30 нМ) в качестве отрицательного контроля. Полученные результаты продемонстрировали, что средняя Ко аффинности связывания PD1-H944 с рекомбинантным белком PD-1 человека составляла 64,8 пМ, среднее значение константы скорости ассоциации kon составляло 3,01Е+05 М-1с-1 и среднее значение константы скорости диссоциации koff составляло 1,95Е-05 с-1; среднее значение аффинности связывания KD ниволумаба с белком PD-1 составляло 74,1 мМ, среднее значение константы скорости ассоциации kon составляло 6.92Е+05 M-1c-1 и среднее значение константы скорости диссоциации koff составляло 5,12Е-05 с-1(Фиг. 4). PD1-H944 имеет более сильное сродство, чем ниволумаб, что примерно в 1,14 раза больше, чем у ниволумаба, и PD1-H944 имеет более низкую скорость диссоциации, так что PD1-H944 обладает более высокой способностью к связыванию с белком PD-1, чем ниволумаб.
(4) Способность связывания PD1-H944 с рекомбинантным белком PD-1 обезьяны и мыши
Непрямой анализ ELISA использовали для обнаружения специфического связывания PD1-H944 с рекомбинантным белком PD-1 обезьяны и мыши. Рекомбинантные белки PD-1 человека, обезьяны и мыши (Sino Biological, Inc.) в различных концентрациях (0,16 нг/мл, 0,49 нг/мл, 1,48 нг/мл, 4,44 нг/мл, 13,33 нг/мл, 40 нг/мл, 120 нг/мл, 360 нг/мл, 1080 нг/мл, 3240 нг/мл и 9720 нг/мл) наносили на 96-луночные планшеты, 100 мкл на лунку, выдерживали в течение ночи при 4°С, отмывали на следующий день и блокировали при комнатной температуре в течение 1 ч. После инкубации с 100 мкл PD1-H944, ниволумаба и антитела H7N9-R1-IgG4 в качестве отрицательного контроля (все в концентрации 2 мкг/мл), указанные планшеты отмывали и вторичное антитело, антитело козы против IgG F(ab)2 человека/пероксидаза хрена, добавляли для получения окраски, определяли OD450, указанный анализ проводили в трех повторах. Полученные результаты продемонстрировали, что PD1-H944 связывалось с рекомбинантным белком PD-1 человека и рекомбинантным белком PD-1 обезьяны с ЕС50 25,8 нг/мл (R2=0,999) и 32,7 нг/мл (R2=0,997), соответственно, но не связывалось с рекомбинантным белком PD-1 мыши (Фиг. 5А); ниволумаб связывался с рекомбинантным белком PD-1 человека и рекомбинантным белком обезьяны PD-1 с ЕС50 113,2 нг/мл (R2=0,997) и 80,2 нг/мл (R2=0,997), соответственно, и не связывался с белком PD-1 мыши (Фиг. 5Б); указанное выше антитело, использованное в виде отрицательного контроля, не связывалось с рекомбинантным белком PD-1 человека, рекомбинантным белком PD-1 обезьяны или рекомбинантным белком PD-1 мыши (Фиг. 5). Хорошее связывание PD1-H944 с белком PD-1 обезьяны обеспечивает применение обезьян для оценки безопасности этого препарата.
(5) Аффинность связывания PD1-H944 с рекомбинантными белками PD-1 обезьяны и мыши
Аффинность PD1-H944 и ниволумаба по отношению к биотинилированным белкам PD-1 обезьяны и мыши (Sino Biological, Inc.) при различных градиентах концентраций (смотри Фиг. 6А-В) измеряли с использованием системы Octet и анализировали для получения значений KD. Полученные результаты продемонстрировали, что указанное значение аффинности KD PD1-H944 с рекомбинантным белком PD-1 обезьяны составляло 108 пМ, а значение аффинности KD ниволумаба с рекомбинантным белком PD-1 обезьяны составляло 131 пМ, оба значения сопоставимы (Фиг. 6А). Как PD1-H944, так и ниволумаб не связывались с рекомбинантным белком PD-1 мыши (Фиг. 6Б).
4.2. PD1-H944 блокирует связывание PD-1 лигандов (PD-L1 и PD-L2) с белком PD-1 человека
Рекомбинантный белок PD-1 человека наносили на 96-луночный планшет в количестве 100 мл на лунку и оставляли на ночь при 4°С. Указанный планшет отмывали на следующий день и блокировали при комнатной температуре в течение 1 часа перед добавлением 1 мкг/мл биотинилированного PD-L1 человека (Sino Biological, Inc.) или 2 мкг/мл биотинилированного PD-L2 человека (Sino Biological, Inc.). Затем PD1-H944, ниволумаб или антитело H7N9-R1-IgG4 в качестве отрицательного контроля добавляли при различных концентрациях (0,003 мкг/мл, 0,008 мкг/мл, 0,025 мкг/мл, 0,074 мкг/мл, 0,222 мкг/мл, 0,667 мкг/мл, 2 мкг/мл, 6 мкг/мл, 18 мкг/мл), инкубировали, добавляли антибиотик стрептавидин/пероксидаза хрена, инкубировали, и определяли OD450. Для каждой группы указанный тест осуществляли в двух повторах. Полученные результаты продемонстрировали, что биотинилированные рекомбинантные белки PD-L1 человека и PD-L2 эффективно связывались с покрытием из белка PD-1 человека, а добавление различных концентраций как PD1-H944, так и ниволумаба эффективно ингибировало связывание рекомбинантного белка PD-L1 человека (Фиг. 7А) и рекомбинантного белка PD-L2 человека (Фиг. 7Б) с PD-1 человека. PD1-H944 и ниволумаб ингибировали рекомбинантный белок PD-L1 человека с IC50 0,116 мкг/мл (R2=0,995) и 0,129 мкг/мл (R2=0,997), соответственно, и рекомбинантный белок PD-L2 человека с IC50 0,446 мкг/мл (R2=0,994) и 0,486 мкг/мл (R2=0,996), соответственно. Способность PD1-H944 ингибировать связывание PD-1 человека с PD-L1 человека или PD-L2 человека существенно не отличалась от такой способности ниволумаба. Для антитела, использованного в качестве отрицательного контроля, существенного ингибирования не наблюдалось.
4.3. PD1-H944 блокирует связывание лиганда PD-1 человека (PD-L1) с клетками, экспрессирующими PD-1 человека
Клетки Jurkat/PD-1, стабильно экспрессирующие рекомбинантный PD-1 человека, в количестве 5 × 105 клеток на пробирку в логарифмической фазе роста добавляли к PD1-Н944, ниволумабу и антителу H7N9-R1-IgG4 в качестве отрицательного контроля в различных концентрациях (0,260 мкг/мл, 0,390 мкг/мл, 0,585 мкг/мл, 0,878 мкг/мл, 1,317 мкг/мл, 1,975 мкг/мл, 2,963 мкг/мл, 4,444 мкг/мл, 6,667 мкг/мл или 10,000 мкг/мл) и инкубировали при 4°С.Добавляли 10 мкл 0,4 мкг/мл B7H1-Fc-6hothh (Sino Biological, Inc.), отмывали фосфатно-солевым раствором и удаляли несвязавшееся антитело центрифугированием. Добавляли конъюгат стрептавидина Alexa Fluor® 488 (Life Technologies), инкубировали при 4°С в течение 20 минут, последовательно отмывали и центрифугировали, и 200 мкл фосфатно-солевого раствора добавлял, чтобы ресуспендировать клетки для детекции в проточном цитометре. Полученные результаты продемонстрировали, что указанный рекомбинантный белок PD-L1 человека может эффективно связываться с клетками Jurkat/PD-1, а связывание рекомбинантного белка PD-L1 человека с клетками Jurkat/PD-1 эффективно ингибировалось при добавлении различных концентраций PD1-H944 и ниволумаба (Фиг. 8). Концентрации ингибирования IC50 составляли 1,78 мкг/мл (R2=0,994) и 2,48 мкг/мл (R2=0,989), соответственно. Антитело PD1-H944 ингибировало связывание PD-1 человека и PD-L1 человека немного лучше, чем ниволумаб. При добавлении антитела, использовавшегося в качестве отрицательного контроля, существенного ингибирования не наблюдалось. 4.4. Связывание PD1-H944 с CD16a
Белок аффинного рецептора CD 16а (FcγRIIIa) является основным рецептором Fc, опосредующим эффекты ADCC, при этом CD 16а, содержащий сайт V158 SNP, обладает относительно высокой аффинностью. Связывание PD1-H944 с указанным рекомбинантным белком CD16a (V158) измеряли с помощью ELISA для оценки потенциального влияния PD1-H944 на эффекты антитело-зависимой клеточно-опосредованной цитотоксичности (ADCC).
Положительным контролем PD1-H944-1-IgG1 (о), использованным в этом анализе является вариабельный участок PD1-H944, связанный с природным IgG1, а отрицательным контролем PD1-H944-1-IgG1 является вариабельный участок PD1-H944, связанный с N297A мутантным IgG1, чей фрагмент Fc лишен способности связываться с CD 16а ввиду делеции N-гликозида. Рекомбинантный белок PD-1 человека (10 мкг/мл, 100 мкл на лунку) наносили в 96-луночные планшеты в течение ночи при 4°С, отмывали на следующий день и блокировали при комнатной температуре в течение 1 ч. PD1-H944, ниволумаб, антитело PD1-H944-1-IgG1(o) (Sino Biological, Inc.), использованное в качестве положительного контроля, и антитело PD1-H944-1-IgG1 (Sino Biological, Inc.), использованное в качестве отрицательного контроля добавляли при различных концентрациях (0,020 мкг/мл, 0,078 мкг/мл, 0,3125 мкг/мл, 1,25 мкг/мл, 5 мкг/мл, 20 мкг/мл и 80 мкг/мл) (смотри, Фиг. 9), и указанные планшеты отмывали для удаления несвязавшегося антитело после инкубирования. 10 мкг/мл рекомбинантного белка CD16a-AVI-His (V158)+BirA (Sino Biological, Inc.) добавляли в луки после инкубации с 1 мкг/мл стрептавидина/пероксидаза хрена, и определяли OD450 цветной реакции после инкубирования.
Полученные результаты продемонстрировали, что связывание антитела, использованного в качестве положительного контроля по отношению к рекомбинантному белку CD16a, повышалось с увеличением концентрации антитела. PD1-H944, ниволумаб и антитело, использованное в качестве отрицательного контроля, не связывались с рекомбинантным белком CD16a при любой из исследованных концентраций (Фиг. 9). Это позволяет предположить, что как PD1-H944, сконструированное как антитело подтипа IgG4, так и ниволумаб обладают очень низкой способностью к связыванию CD16a, и позволяет предположить, что PD1-H944 вызывает очень низкий эффект ADCC или не вызывает его вовсе.
4.5. Связывание PD1-H944 с C1q
Связывание PD1-H944 с рекомбинантным белком C1q измеряли с помощью ELISA, оценивая таким образом потенциал влияния PD1-H944 на эффекты комплемент-зависимой цитотоксичности (CDC).
PD1-H944-1-IgG1(o), полученный присоединением вариабельного участка PD1-Н944 к природному IgG1, использовали в качестве антитела, являющегося положительным контролем (Sino Biological, Inc.), a H7N9-R1-IgG4 (Sino Biological, Inc.) использовали в качестве антитела, являющегося контролем изотипа. Различные концентрации (20 мкг/мл, 10 мкг/мл, 5 мкг/мл, 2,5 мкг/мл, 1,25 мкг/мл, 0,625 мкг/мл, 0,313 мкг/мл, 0,156 мкг/мл и 0,078 мкг/мл) (смотри Фиг. 10) PD1-H944, ниволумаба, антитела, использованного в качестве положительного контроля, и антитела, использованного в качестве контроля изотипа, наносили в 96-луночные планшеты в количестве 100 мл на лунку в течение ночи при 4°С. На следующий день, указанные планшеты отмывали, блокировали при комнатной температуре в течение 1 часа и добавляли 100 мкл раствора 5 мкг/мл рекомбинантного белка C1q (Quidel Corporation), соответственно, и указанные планшеты инкубировали в течение 3 часов. Добавляли 100 мкл раствора вторичного антитела против C1q/пероксидаза хрена (Abeam) при разведении 1:400, инкубировали в течение 1 часа, проводили цветную реакцию и детектировали OD450.
Полученные результаты продемонстрировали, что связывание антитела, использованного в качестве положительного контроля, с рекомбинантным белком C1q повышалось с повышением концентрации антитела. PD1-H944, ниволумаб и антитело, использованное в качестве контроля изотипа, имели сравнимую способность к связыванию с рекомбинантным белком C1q, при значительно сниженной способности к связыванию антитела типа IgG1, использованного в качестве положительного контроля (Фиг. 10).
4.6. Связывание PD1-H944 с FcRn
Связывание PD1-H944 с рекомбинантным белком рецептора Fc (FcRn) измеряли с помощью ELISA и оценивали таким образом фармакокинетику PD1-H944 у человека.
Белок против биотина Avidin (Thermo Fisher Scientific) в концентрации 10 мкг/мл наносили на 96-луночные планшеты в количестве 100 мл на лунку, инкубировали в течение ночи при 4°С и отмывали на следующий день, блокировали при комнатной температуре в течение 1 часа и инкубировали с раствором 10 мкг/мл биотинилированного белка FCGRT-His+B2M (Sino Biological, Inc.) в течение 1 часа, затем PD1-H944, ниволумаб или isotype control антитело H7N9-R1-IgG4 (Sino Biological, Inc.), использованное в качестве контроля изотипа, соответственно, добавляли при различных концентрациях (0,123 мкг/мл, 0,37 мкг/мл, 1,11 мкг/мл, 3,33 мкг/мл, 10 мкг/мл, 30 мкг/мл, 90 мкг/мл, 270 мкг/мл) (смотри Фиг. 11), инкубировали в течение 1 ч, отмывали и добавляли 250 нг/мл антитела козы против IgG Fc человека/пероксидаза хрена (Sino Biological Inc.). Процесс от разбавления антитела до инкубирования со вторичным антителом проводили, поддерживая рН, равный 6,0, и измеряли OD450.
Полученные результаты продемонстрировали, что PD1-H944 и ниволумаб были способны связываться с рекомбинантным белком FcRn человека, и способность связывания повышалась с повышением концентрации; при высоких концентрациях (270 мкг/мл), связывание PD1-H944 с рекомбинантным белком FcRn было примерно в 1,62 выше, чем связывание ниволумаба (Фиг. 11). На соновании этих результатов было высказано предположение, что PD1-H944 обладает хорошей фармакокинетикой в человеке.
Пример 5: Функциональный анализ гуманизированного антитела PD1-H944 5.1. Активация CD4+ Т-клеток в смешанных лифоидных ответах, стимулированных PD1-H944
Нейтрализующую активность антитела против PD-1 человека определяли с помощью анализа смеси CD4+Т-лимфоцитов с дендритными клетками. Мононуклеарные клетки периферической крови человека (РВМС) выделяли центрифугированием в градиенте плотности Ficoll, затем мононуклеарные клетки получали адгезионным методом культивирования в среде для культивирования клеток 1640 (GIBCO) (содержащей 10% ФБС, 100 Ед/мл пенициллина, 100 мкг/мл стрептомицина), содержащей 160 нг/мл rhIL-4 (Sino Biological, Inc.), затем к полученным мононуклеарным клеткам добавляли 20 нг/мл rhGM-CSF (R&D Systems) и инкубировали в инкубаторе с СО2 в течение трех суток, затем меняли половину среды для инкубации. Через 6 суток инкубации собирали суспензионые клетки, известные как дендритные клетки (DC). Плотность DC доводили до 2×106 клеток/мл, добавляли митомицин до конечной концентрации 50 мкг/мл, выдерживали при 37°С в течение 20 минут, отмывали три раза средой 1640.
CD4+ Т-лимфоциты отсортировывали из РВМС, выделенных из периферической крови другого человека с использованием набора для сортировки CD4+ Т-лимфоцитов (MiltenyiBiotec). CD4+ Т-лимфоциты от каждого из трех доноров крови использовали в каждой из партий для этого анализа.
Отсортированные CD4+ Т-лимфоциты перемешивали с DC, предварительно обработанными 50 мкг/мл митомицина в соотношении 10:1, и 105 клеток на лунку равномерно вносили в 96-луночные планшеты с PD1-H944, ниволумабом и антителом H7N9-R1-IgG4, использованным в качестве отрицательного контроля (Sino Biological, Inc.), соответственно, и контрольную группу смешанных лимфоцитов, то есть группу только с CD4+ Т-лимфоцитами и смесью DC без антитела, контрольную группу клеток DC, то есть группу только с клетками DC без антитела и CD4+ Т-лимфоцитов, контрольную группу CD4+ Т-клеток, то есть группу только с CD4+ Т-клетками без антитела и клеток DC, в том же объеме среды 1640 для разведения образцов, конечный градиент концентрации каждого антитела составлял 1 мкг/мл, 0,1 мкг/мл, 0,01 мкг/мл, и инкубировали в инкубаторе с СО2 при 37°С и 5% СО2 в течение 5 суток. Супернатанты культуральной жидкости собирали и измеряли уровни экспрессии ИЛ-2 и интерферона-γ в указанных супернатантах культуральной жидкости с помощью ELISA.
Результаты продемонстрировали, что секреция интерферона-γ и ИЛ-2 не детектировалась в супернатантах культур CD4+ Т-клеток без примеси DC, в то время как в контрольной группе смеси лимфоцитов, смесь CD4+ Т-клеток и DC существенно повышали секрецию интерферона-γ и ИЛ-2 CD4+ Т-клетками. Когда антитело против PD-1 добавляли к системе со смесью лимфоцитов, активация CD4+ Т-клеток в смеси лимфотического ответа CD4+ Т-клеток и DC существенно усиливалась с высокой секрецией интерферона-γ и ИЛ-2. В противоположность как PD1-H944, так и ниволумаба, которые демонстрировали существенный эффект, антитело, использованное в качестве отрицательного контроля, не демонстрировало этот эффект. Полученные результаты показаны на Фиг. 12, указывающие на то, что PD1-H944 обладает лучшей функциональной активностью, чем ниволумаб, может эффективно реактивировать иммуносупрессированные Т-клетки.
5.2. PD1-H944 стимулирует активацию репортерных генов в сигнальном пути ИЛ2 ниже PD1
В этом анализе эффекторные клетки Jurkat-NFAT-Luc2p-PD-1 (SinoCellTech Ltd) и клетки-мишени CHO-K1-PD-L1-CD3E (SinoCellTech Ltd) использовали в качестве экспериментальных материалов. Взаимодействие PD-1/PD-L1 в результате совместного культивирования этих двух клеток ингибирует передачу сигналов TCR и биолюминесценцию, опосредованную NFAT-RE, а добавление антитела против PD-1 блокировало взаимодействие PD-1/PD-L1 и, таким образом, снимало это ингибирование. Функцию активации PD1-H944 в рекомбинантной системе репортерных клеток PD-1 человека определяли путем измерения интенсивности биолюминесценции (R1U) в эффекторных клетках.
Клетки-мишени CHO-K1-PD-L1-CD3E инокулировали по 2×104 на лунку в 96-луночные планшеты и культивировали в течение ночи в среде DMEM, содержащей 10% ФБС, затем супернатант удаляли. PD1-H944, ниволумаб и антитело H7N9-R1-IgG4 (Sino Biological, Inc.), использованное в качестве отрицательного контроля, в различных концентрациях (0,007 мкг/мл, 0,023 мкг/мл, 0,082 мкг/мл, 0,286 мкг/мл, 1,000 мкг/мл, 3,499 мкг/мл, 12,245 мкг/мл, 42,857 мкг/мл, 150 мкг/мл) добавляли в количестве 40 мкл на лунку (смотри Фиг. 13). 7,5 × 104 эффекторных клеток Jurkat-NFAT-Luc2p-PD-1 последовательно добавляли в количестве 40 мкл на лунку и инкубировали в инкубаторе с СО2 в течение 6 часов. Каждый анализ проводили в трех повторах для клеток-мишеней, эффекторных клеток и отрицательного контроля. Через 6 часов инкубации добавляли 5-кратный буфер для пассивного лизиса (Promega) в количестве 20 мкл на лунку и 96-луночные планшеты затем помещали в морозильник при -80°С для дополнительной детекции. Для этого анализа 96-луночные планшеты помещали на -80°С. Оставляли оттаивать при комнатной температуре, перемешивали и 20 мкл супернатанта из лунок переносили в 96-луночный планшет с белым дном для детекции люминесценции с помощью анализа LB960-Microplate Luminol.
Полученные результаты продемонстрировали, что как PD1-H944, так и контрольное антитело ниволумаб вызывали существенную активацию клеточной системы репортерных генов рекомбинантного PD-1 человека, как показано на Фиг. 13. В диапазоне концентраций 0,007-150 мкг/мл, ЕС50 PD1-H944 составляла 0,49 мкг/мл, а у ниволумаба - 1,08 мкг/мл. PD1-H944 имел значительно более низкую ЕС50, чем ниволумаб, его активность выше в 2,21 раза, чем у ниволумаба. Антитело, использованное в качестве отрицательного контроля, не имело активирующей функции клеточной системы репортерных генов рекомбинантного PD-1 человека.
5.3. PD1-H944 не стимулирует в значительной степени функцию ADCC рецептора Fc пути CD16a
В этом анализе, эффекторные клетки Jurkat-NFAT-Luc2p-CD16A (SinoCellTech Ltd) и клетки-мишени CHO-PD-1 (SinoCellTech Ltd) использовали в качестве материала для анализа для измерения опосредованного PD1-H944 эффекта ADCC с помощью метода с использованием системы рекомбинантного высокоактивного репортерного гена CD16a (V158). Механизм заключается в том, что когда два вида клеток совместно культивируются и PD1-H944 добавляется одновременно, Fab-фрагмент PD1-H944 связывается с PD-1, сверхэкспрессированным клетками-мишенями, таким образом, его Fc-фрагмент связывается с эффекторными клетками, несущими Fcγ-рецептор CD16A, следовательно, активирует эффекторные клетки Jurkat-NFAT-Luc2p-CD16A и способствует хемилюминесценции, опосредованной NFAT-RE.
Клетки-мишени CHO-PD-1 инокулировали в количестве 2×104 на лунку в 96-луночный планшет и культивировали в течение ночи в среде DMEM, содержащей 10% ФБС, и супернатант удаляли. Указанные планшеты отмывали дважды средой RPMI 1640 (без фенола красного), содержащей 0,1% БСА, и антитела PD1-Н944-1-IgG 1 (о), PD1-Н944, ниволумаб и отрицательный контроль H7N9-R1-IgG4 (Sino Biological, Inc.) добавляли в различных концентрациях (см. Фиг. 14А-Б) в количестве 40 мкл на лунку, добавляли 1×106 эффекторных клеток Jurkat-NFAT-Luc2p-CD16A в количестве 40 мкл на лунку, и помещали в инкубатор с СО2 при 37°С и 5% СО2 на 4 часа. Каждый анализ проводили в трех повторах для клеток-мишеней, эффекторных клеток и отрицательного контроля. По истечении 4 часов добавляли 20 мкл на лунку 5-кратного буфера для пассивного лизиса (Promega), а 96-луночные планшеты помещали в морозильник с температурой -80°С для тестирования. Для анализа 96-луночные планшеты оставляли оттаивать до комнатной температуры, встряхивали и 20 мкл супернатанта из лунки переносили в 96-луночный планшет с белым дном для детекции люминесценции с помощью анализа LB960-Microplate Luminal.
Полученные результаты продемонстрировали, что указанное антитело PD1-H944-1-IgG1(о), использованное в качестве положительного контроля, вызывает существенный эффект ADCC, ниволумаб вызывает более слабый эффект ADCC, a PD1-H944 вызывает еще более слабый эффект ADCC, чем ниволумаб в диапазоне концентраций 0,018-300 нг/мл (р<0,01). Фиг. 14А и Фиг. 14Б демонстрируют результаты с различными партиями анализа. На Фиг. 14В и 14Г показана интенсивность биолюминесценции (RLU), соответствующей наивысшей точке концентрации антитела (300 нг/мл) с Фиг. 14А и 15Б, соответственно. В этом чувствительном функциональном анализе ADCC, клеточная активация, вызванная связыванием PD1-H944 с CD 16а, была значительно ниже, чем у ниволумаба, при этом результаты были повторяемыми. Низкая ADCC-активность препарата PD1-H944 является хорошим подтверждением эффективности и безопасности этого антитела в клинике.
5.4. PD1-H944 существенно не активирует функцию CDC пути комплемента C1q
После связывания с опухолевыми клетками, сверхэкспрессирующими PD-1, PD1-Н944 может активировать классический путь комплемента для уничтожения опухолевых клеток, что приводит к их гибели. В этом анализе CDC-эффект антитела PD1-H944 исследовали с использованием метода WST-8.
Клетки CHO-PD-1 ресуспендировали в среде RPMI 1640, содержащей 0,1% БСА (SinoCellTech Ltd), и равномерно инокулировали в 96-ночные планшеты в количестве 5×104 на лунку, с последующим добавлением антител при различных концентрациях (Фиг. 15) в количестве 50 мкл на лунку, а затем добавляли комплемент C1q при разведении 1:4 в количестве 50 мкл на лунку (One lambda). Антитела представляли собой: антитело PD1-H944-1-IgG1 (о) (Sino Biological, Inc.), использованное в качестве положительного контроля, PD1-H944, ниволумаб и антитело PD1-H944-1-IgG1 (Sino Biological, Inc.), использованное в качестве отрицательного контроля, с анализом пустой лунки В (без клеток) и отрицательного контроля М (с клетками, но без антитела). После инкубации в течение 3 часов при 37°С 15 мкл окрашивающего раствора WST-8 добавляли в каждую лунку и измеряли оптическую плотность при 450 нМ и 630 нМ на анализаторе ELISA после стабилизации окраски. Результаты рассчитывали с использованием значений оптической плотности OD450-OD630 с вычитанием значения для пустой лунки. % гибели=(OD отрицательного контроля М - OD образца)/OD отрицательного контроля М × 100%.
Полученные результаты продемонстрировали, что PD1-H944, ниволумаб и антитело, использованное в качестве отрицательного контроля, не вызывали эффект CDC в отношении опухолевых клеток CHO-PD-1, сверхэкспрессирующих PD-1, в то время как антитело, использованное в качестве положительного контроля, вызвало эффект гибели клеток CHO-PD-1 с максимальной степенью гибели 82,1%, результаты показаны на Фиг. 15. Анализ CDC также подтвердил, что PD1-H944 не вызывает эффект CDC в отношении клеток-мишеней, экспрессирующих PD-1, демонстрируя хорошую безопасность антитела.
5.5 PD1-H944 эффективно ингибирует рост опухоли в модели подкожного трансплантата МС38 рака толстой кишки у мышей с гуманизированным PD-1 in vivo
(1) Исследование фармакодинамики I антитела PD1-H944 в модели подкожного трансплантата МС38 рака толстой кишки у мышей с гуманизированным PD-1
Клетки МС38 на стадии логарифмического роста (Sun Ran Shanghai Biotechnology Co., Ltd.) использовали для инокуляции опухоли. Клетки МС38, ресуспендированные в фосфатно-солевом растворе, инокулировали в количестве 5×105 клеток/0,1 мл в мышей с гуманизированным B-hPD-1 (Biocytogen) (полученным путем замены части экзона 2 гена PD-1 мышей C57BL/6 соответствующей частью генома PD-1 человека) подкожно на правой стороне грудной клетки мышей, всего 47 мышей. Когда объем опухоли вырос примерно до 100 мм3, мышей отбирали в соответствии с их индивидуальным объемом опухоли, случайным образом распределяли на 5 групп с использованием программного обеспечения Excel, по 8 животных в каждой группе. Группам 4 и 5 вводили другие антитела против PD-1, не относящиеся к данной заявке, поэтому их данные не представлены в настоящей заявке. Введение дозировки начали в день группировки. Мышам вводили внутрибрюшинную инъекцию (LP.) один раз в 3 дня в течение 6 последовательных доз, затем мышей умерщвляли через 10 дней после введения последней дозы и опухолевую ткань удаляли для взвешивания. Конкретный режим дозирования показан в Таблице 3 ниже. Противоопухолевый эффект препарата оценивали путем расчета степени ингибирования роста опухоли TGI (%). TGI (%)<60% считали неэффективным; если TGI (%) ≥60% и объем опухоли в обработанной группе был статистически значимо ниже, чем у контрольной группы растворителя (Р<0,05), этот уровень считался эффективным, то есть антитело оказывало значительное ингибирующее действие на рост опухоли. TGI (%) рассчитывали следующим образом:
TGI (%)=[1-(Ti-T0)/(Vi-V0)] × 100, где
Ti: среднее значение объема опухоли в группе лечения в день i,
Т0: среднее значение объема опухоли в группе лечения в день 0,
Vi: среднее значение объема опухоли в контрольной группе с растворителем в день i,
V0: среднее значение объема опухоли в контрольной группе с растворителем в день 0.
Животных умерщвляли в конце анализа, взвешивали опухоль и рассчитывали степень ингибирования веса опухоли IRTW% с IRTW%>60% в качестве контрольного показателя эффективности, рассчитанного следующим образом. Степень ингибирования веса опухоли IRTW (%)=(W контрольная группа с растворителем - W группа обработки) / W контрольная группа с растворителем × 100, W - вес опухоли.

Все подопытные животные находились в хорошем общем состоянии с точки зрения активности и кормления в период дозирования, и их вес тела в некоторой степени увеличивался. После введения исследуемого препарата (группа 3) и контрольного препарата (группа 2) достоверной разницы (Р>0,05) в весе тела животных не было. Изменения веса тела всех животных показаны на Фиг. 16 и в Таблице 4.
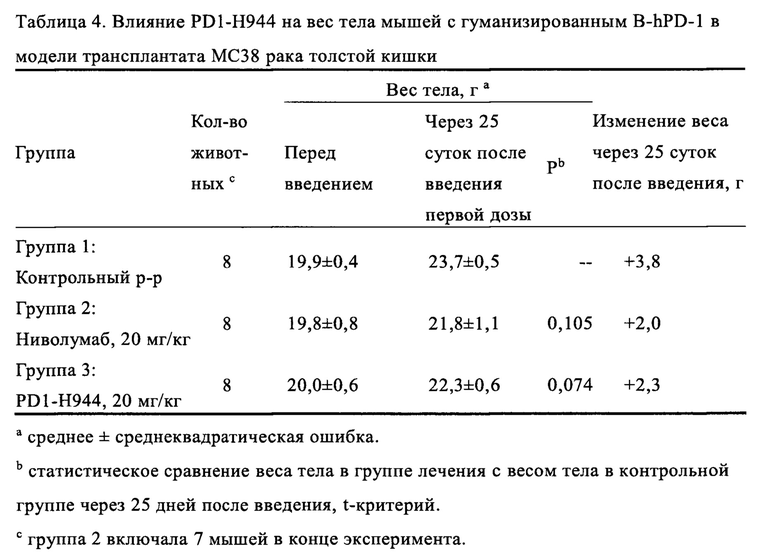
Результаты измерения объема опухоли для каждой группы в испытании показаны в Таблице 5 и на Фиг. 17.
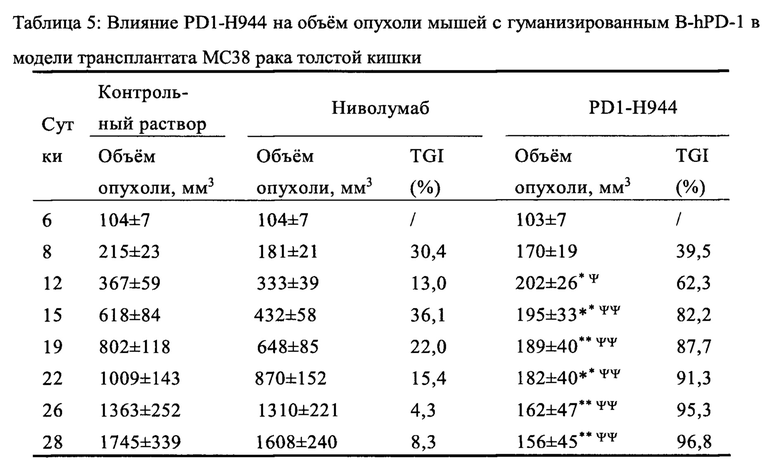

Сутки: количество суток после инокуляции клеток
Через 25 дней после введения первой дозы всех животных умерщвляли, опухоли удаляли, взвешивали и фотографировали. Вес опухолей статистически сравнивали между группами, и результаты суммировали в Таблице 6 и на Фиг. 18.

В соответствии с результатами измерения объема опухоли, ниволумаб не оказывал значительного ингибирования объема опухоли в этой модели, в то время как среднее значение веса опухоли в группе PD1-H944 составляло 0,044±0,017 г. Его степень ингибирования веса опухоли (IRTW) достигала 97,4%. Анализ данных показал, что опухоли в группе PD1-H944 значительно отличались от таковых для контрольной группы с растворителем (Р<0,05), демонстрируя значительную противоопухолевую эффективность PD1-H944.
В конце анализа среднее значение объема опухоли контрольной группы с растворителем составило 2171±430 мм3, масса опухоли 1,719±0,310 г. Среднее значение объема опухоли группы положительного контроля с ниволумабом (20 мг/кг) составило 1802±228 мм3, TGI% - 17,8%, вес опухоли - 1,492±0,274 г. Степень ингибирования веса опухоли IRTW ниволумабом составляла 13,2%, что существенно не отличалось от этого значения опухоли для контрольной группы с растворителем (Р=0,480). Напротив, среднее значение объема опухоли в группе PD1-H944 составляло 171±51 мм3, TGI% составляло 96,7%, вес опухоли составлял 0,044±0,017 г, а степень ингибирования веса опухоли IRTW составляла 97,4%, что значительно отличалось от этого значения опухоли для контрольной группы растворителя (Р<0,05), что указывает на то, что указанное антитело PD1-H944 связывается с PD-1 эпитопом, и демонстрирует значительное ингибирование рака толстой кишки подкожных опухолей трансплантата МС38 при уровне дозировки 20 мг/кг.
(2) Фармакодинамическое исследование II антитела PD1-H944 в модели подкожного трансплантата МС38 рака толстой кишки у мышей с гуманизированным PD-1
Клетки МС38 на стадии логарифмического роста (Sun Ran Shanghai Biotechnology Co., Ltd.) использовали для инокуляции опухоли. Клетки МС38, ресуспендированные в фосфатно-солевом растворе, инокулировали в количестве 5×105 клеток/0,1 мл гуманизированным мышам B-hPD-1 (Biocytogen) (мыши, полученные путем замены части экзона 2 гена PD-1 мышей C57BL/6 соответствующей частью генома PD-1 человека). В общей сложности 80 мышей были инокулированы подкожно в правую часть грудной клетки. Когда объем опухоли вырос примерно до 100 мм3, мышей отбирали в соответствии с их индивидуальным объемом опухоли, случайным образом распределяли по 8 группам с использованием программного обеспечения Excel, по 8 животных в каждой группе. Группам 3,4, 7 и 8 вводили с другие антителами против PD-1, не имеющими отношения к данной заявке, поэтому их данные не представлены. Введение дозы начинали в день группировки. Мышам вводили внутрибрюшинную инъекцию (LP.) один раз в 3 дня в течение 6 последовательных доз, мышей умерщвляли через 10 дней после введения последней дозы, опухолевую ткань удаляли для взвешивания. Конкретный режим дозирования показан в Таблице 7 ниже.


Противоопухолевый эффект препарата оценивали путем расчета степени ингибирования роста опухоли TG (%). TGI (%)<60% считали неэффективным; если TGI (%)>60% и объем опухоли в обработанной группе был статистически значимо ниже, чем у контрольной группы растворителя (Р<0,05), этот уровень считался эффективным, то есть антитело оказывало значительное ингибирующее действие на рост опухоли. TGI (%) рассчитывали следующим образом.
TGI (%)=[1-(Ti-T0)/(Vi-V0)] × 100, где
Ti: среднее значение объема опухоли в группе лечения в день i,
Т0: среднее значение объема опухоли в группе лечения в день 0,
Vi: среднее значение объема опухоли в контрольной группе с растворителем в день i,
V0: среднее значение объема опухоли в контрольной группе с растворителем в день 0.
Животных умерщвляли в конце анализа, взвешивали опухоль и рассчитывали степень ингибирования веса опухоли IRTW% с IRTW%>60% в качестве контрольного показателя эффективности, рассчитанного следующим образом. Степень ингибирования веса опухоли IRTW (%)=(W контрольная группа с растворителем - W группа обработки) / W контрольная группа с растворителем × 100, W - вес опухоли.
Все подопытные животные находились в хорошем общем состоянии с точки зрения активности и кормления во время введения препарата, и в некоторой степени вес их тела увеличился. Достоверной разницы в весе тела животных после введения тестовых и контрольных препаратов не было (Р>0,05) (Таблица 8 и Фиг. 19).

Объемы опухолей для каждой группы в анализе показаны в Таблице 9 и на Фиг. 20.
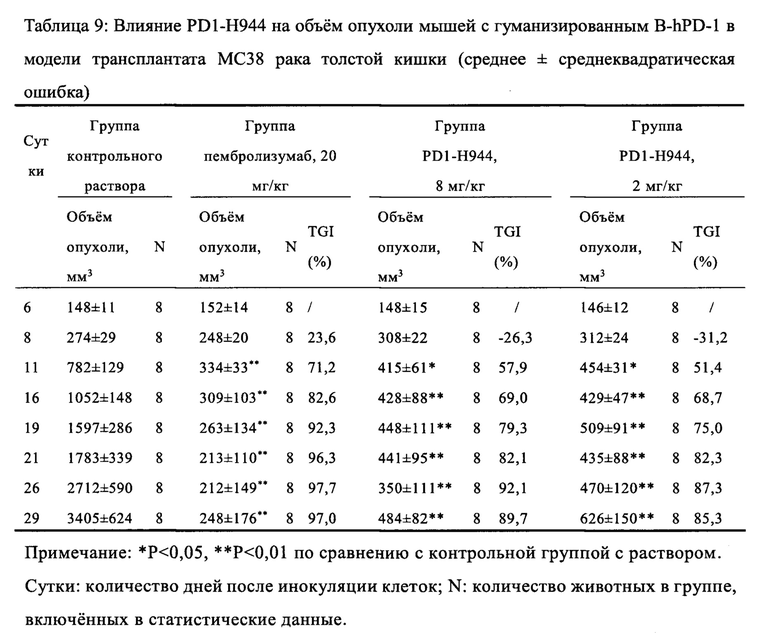
Через 24 дня после введения первой дозы всех животных умерщвляли, опухоли удаляли, взвешивали и фотографировали. Статистически сравнивали веса опухолей между группами и полученные результаты представлены в Таблице 10 и на Фиг. 21.

Для контрольной группы с растворителем: объем опухоли продолжал расти после образования группы и среднее значение объема опухоли в конце исследования составило 3405±624 мм3.
Для группы положительного контроля: после введения пембролизумаба в дозировке 20 мг/кг объем опухоли увеличивался медленно по сравнению с контрольной группой с растворителем, со значительной разницей в объеме опухоли с 5-го дня введения (Р<0,05); TGI% в конце испытания составлял 97,0%, а степень ингибирования веса опухоли составляла 93,5%, Р<0,05 по сравнению с контрольной группой с растворителя. Пембролизумаб продемонстрировал явное ингибирование опухоли на этой модели оценки эффективности, что указывает на то, что эта модель может быть использована для оценки эффективности лекарственного препарата антитела Keytruda, связывающего эпитоп.
Для группы PD1-H944, 8 мг/кг: после введения PD1-H944 в дозировке 8 мг/кг объем опухоли медленно увеличивался по сравнению с контрольной группой с растворителем, объем опухоли статистически значимо отличался по сравнению с 5-м днем введения (Р<0,05); TGI% в конце теста составлял 89,7%, а степень ингибирования веса опухоли составляла 87,8%, Р<0,05 по сравнению с контрольной группой с растворителем.
Для группы PD1-H944, 2 мг/кг: после введения PD1-H944 в дозировке 2 мг/кг объем опухоли медленно увеличивался по сравнению с контрольной группой с растворителем, объем опухоли статистически значимо отличался по сравнению с 5-м днем введения (р<0,05); TGI% в конце испытания составлял 85,3%, а степень ингибирования веса опухоли составляла 84,7%, р<0,05 с контрольной группой с растворителем. Это снова свидетельствует о том, что связывание указанного антитела PD1-H944 с эпитопом PD-1 эффективно в этой модели, демонстрируя значительный эффект подавления опухоли подкожного трансплантата рака толстой кишки МС38, демонстрируя эффект, коррелирующий с дозировкой. Противоопухолевые эффективности PD1-H944 и пембролизумаба в этом анализе также были подтверждены с точки зрения веса опухоли.
Результаты этого анализа показали, что внутрибрюшинное введение PD1-H944 в дозировке 8 мг/кг или 2 мг/кг каждые 3 дня в течение 6 последовательных доз значительно подавляло рост трансплантированных опухолей у мышей с гуманизированным В-hPD-l в модели трансплантата МС38 (Р<0,05), а характеристики животных и веса их тела в группе, получавшей лечение, не отличались от таковых у животных из модельной группы. Учитывая предпочтение дизайна модели для различных эпитопов антител, в этом анализе не оценивалась разница в эффективности между PD1-H944 и пембролизумабом.
В исследовании фармакодинамики in vivo изучали противоопухолевый эффект одного PD1-H944 на мышах C57BL/6 с гуманизированным PD-1 в модели трансплантата МС38 рака толстой кишки. Мышь с гуманизированным В-hPD-l представляет собой мышь C57BL/6, у которой был гуманизированный ген PD-1, что делает эту мышь подходящей для оценки in vivo эффективности антитела против PD-1 человека. Модель была селективной в отношении эффективности антител против различных эпитопов PD-1, и пембролизумаб продемонстрировал значительное ингибирование опухоли в этой модели. Результаты продемонстрировали, что PD1-H944 сам по себе (2, 8 и 20 мг/кг, вводимые каждые 3 дня в виде 6 последовательных доз) значительно ингибирует рост трансплантированных опухолей МС38, а сводные данные представлены в Таблицах 11 и 12.
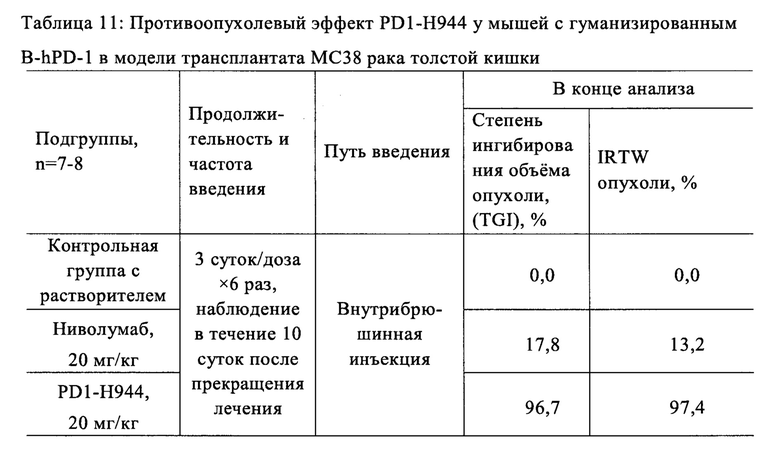
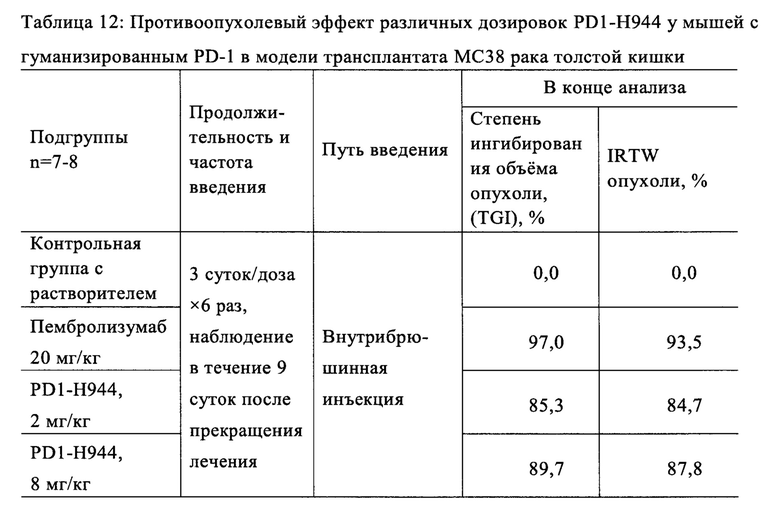
Пример 6. Идентификация сайтов аминокислот, специфически связывающихся с PD1-H944
Данные из Пример 4.2 демонстрируют, что PD1-H944 эффективно конкурирует с PD-L1 (Фиг. 7А) или PD-L2 (Фиг. 7Б) за связывание с белком PD-1, демонстрируя перекрывание между связыванием эпитопа в PD-1, нацеленного на PD1-H944, и эпитопа, который является целью лиганда PD-L1 или PD-L2.
6.1 Молекулярное моделирование, предсказывающее конформационные эпитопы PD1-H944
Чтобы исследовать взаимодействие между PD1-H944 и белком PD-1, в этом примере ZDOCK было осуществлено связывание PD1-H944 и белка PD-1. Основываясь на том факте, что связывающий эпитоп белка PD-1. нацеленный на PD1-H944, перекрывается с эпитопом, на который нацелен лиганд PD-L1. моделирование гомологии для PD1-H944 было выполнено с использованием программы моделирования антител в DS 4.0 (Accelrys Software Inc.), и окончательная обоснованность конструкции модели была проверена с помощью карты Рамачандрана. Трехмерная структура белка PD-1 была загружена из базы данных PDB (PDB TD: 4ZQK) и инициализирована программой Protein Preparation. Модель связывания модельного PD1-H944 и структуры PD-1 осуществляли с помощью программы ZDOCK. Первые по количеству очков десять вариантов были оптимизированы с использованием RDOCK. Лучшая модель была дополнительно проанализирована программой анализа интерфейса белков (Фиг. 22). Модель стыковки взаимодействия поверхностей продемонстрировала, что двумя основными пептидными последовательностями, связывающими PD1-H944 с белком PD-1, были СС и FG складчатые слои (Таблица 13).

6.2 Подтверждение функционального эпитопа связывания антитела PD1-H944, проверенного на мутантах белка PD-1
Для более точного подтверждения функционального эпитопа PD1-H944 на основе последовательностей пептидов, которые, как предсказано в Таблице 13, являются основными сайтами связывания PD1-H944 с белком PD-1, была приготовлена серия мутантов белка PD-1 с мутацией аланина и проанализирована в анализе ELISA. Чтобы проверить точность модели, несколько сайтов вне модели Примера 6.1 также были выбраны для этого исследования (Таблица 14), и эти мутантные сайты были сгруппированы в пять пространственных конформационных эпитопов (Фиг. 23).

Связывание каждого мутанта из Таблицы 14 с PD1-H944 определяли с помощью ELISA в соответствии с методом из Примера 4.2. Мутанты имеют различную степень снижения связывания по сравнению с немутантным белком PD-1. Данные были проанализированы и нанесены на график с использованием Excel 2007 с профилем мутанта белка PD-1 в качестве горизонтальной координаты и скоростью связывания в
Качестве Вертикальной координаты [скорость связывания=OD(мутантный белок PD-1)/OD(белок PD-1)×100%)]. Полученные результаты продемонстрировали, что основными сайтами связывания белка PD-1 с лигандом PD-L1 были Сайт 2, Сайт 3, Сайт 4 и Сайт 5 на складчатых слоях СС и FG белка PD-1, которые соответствовали сайтам из анализа кристаллического комплекса (PDB ID: 4ZQK). Сайты связывания для положительного контроля связывание ниволумаба с белком PD-1 в основном располагалось на Сайте 1 и Сайте 4 его N-петли и складчатого слоя FG, и этот результат также согласуется с сайтами из анализа кристаллического комплекса (PDB ID: 5WT9); основными сайтами связывания PD1-H944 с белком PD-1 были Е61, K78, D85 и Р130 на Сайте 3 и Сайте 4. Полученные результаты также соответствовали результатам модели связывания в приведенном выше Примере (Фиг. 24 и Таблица 15), а точность связывания модели между PD1-H944 и белком PD-1 подтверждена.
Модель связывания между PD1-H944 и белком PD-1 показывает, что связывающий эпитоп на белке PD-1, на который нацелено PD1-H944, перекрывается с эпитопом лиганда PD-L1. Этот факт подтверждает идею о том, что PD1-H944 выступает в роли прямого пространственного стерического препятствия. Между тем, пересечение между эпитопом, на который нацелены как PD1-H944, так и ниволумаб, предполагало, что PD1-H944 имеет более крупную область для блокирования связывания PD-L1 с эпитопами (Фиг. 25А-В). Следовательно, PD1-H944 может иметь превосходную терапевтическую эффективность при лечении онкологии.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> СиноСеллТех Лтд.
<120> Гуманизированное антитело против PD-1 и его применение
<130> FP1180486P
<141> 2018-12-21
<160> 35
<170> SIPOSequenceListing 1.0
<210> 1
<211> 167
<212> PRT
<213> Человек (Homo sapiens)
<400> 1
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln
165
<210> 2
<211> 54
<212> ДНК
<213> Мышь (Mus musculus)
<400> 2
tctagtggtg gcggtggttc gggcggtggt ggaggtggta gttctagatc ttcc 54
<210> 3
<211> 741
<212> ДНК
<213> Мышь (Mus musculus)
<400> 3
gatattgtgc taactcaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atatcctgca gagccagtga aagtgttgat agttatggca atagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatg ctgcatccaa ccaaggatcc 180
ggggtccctg ccaggtttag tggcagtggg tctgggacag acttcagcct caacatccat 240
cctatggagg aggatgatac tgcaatgtat ttctgtcagc aaagtaagga ggttccgtgg 300
acgttcggtg gaggcaccaa gctggaaatc aaatctagtg gtggcggtgg ttcgggcggt 360
ggtggaggtg gtagttctag atcttccgag gtgcaactgg tggaatctgg gggaggctta 420
gtgaagcctg gagggtccct gaaactctcc tgtgcagcct ctggattcac tttcagttcc 480
tatggcatgt cttgggttcg tcagactccg gagaagaggc tggagtgggt cgcgaccatt 540
agtggtggtg gtcgtgacac ctactattca gacagtgtga aggggcggtt caccgtctcc 600
agagacaatg ccaagaacaa cctgttcctg caaatgagca gtctgaggtc tgaagacacg 660
gccttgtatt attgttcacg tcaatatggt acggtctggt tttttaactg gggccagggg 720
actctggtca ctgtctctgc a 741
<210> 4
<211> 354
<212> ДНК
<213> Мышь (Mus musculus)
<400> 4
gaggtgcaac tggtggaatc tgggggaggc ttagtgaagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt tcctatggca tgtcttgggt tcgtcagact 120
ccggagaaga ggctggagtg ggtcgcgacc attagtggtg gtggtcgtga cacctactat 180
tcagacagtg tgaaggggcg gttcaccgtc tccagagaca atgccaagaa caacctgttc 240
ctgcaaatga gcagtctgag gtctgaagac acggccttgt attattgttc acgtcaatat 300
ggtacggtct ggttttttaa ctggggccag gggactctgg tcactgtctc tgca 354
<210> 5
<211> 333
<212> ДНК
<213> Мышь (Mus musculus)
<400> 5
gatattgtgc taactcaatc tccagcttct ttggctgtgt ctctagggca gagggccacc 60
atatcctgca gagccagtga aagtgttgat agttatggca atagttttat gcactggtac 120
cagcagaaac caggacagcc acccaaactc ctcatctatg ctgcatccaa ccaaggatcc 180
ggggtccctg ccaggtttag tggcagtggg tctgggacag acttcagcct caacatccat 240
cctatggagg aggatgatac tgcaatgtat ttctgtcagc aaagtaagga ggttccgtgg 300
acgttcggtg gaggcaccaa gctggaaatc aaa 333
<210> 6
<211> 972
<212> ДНК
<213> Мышь (Mus musculus)
<400> 6
gccaaaacga cacccccatc tgtctatcca ctggcccctg gatctgctgc ccaaactaac 60
tccatggtga ccctgggatg cctggtcaag ggctatttcc ctgagccagt gacagtgacc 120
tggaactctg gatccctgtc cagcggtgtg cacaccttcc cagctgtcct gcagtctgac 180
ctctacactc tgagcagctc agtgactgtc ccctccagca cctggcccag cgagaccgtc 240
acctgcaacg ttgcccaccc ggccagcagc accaaggtgg acaagaaaat tgtgcccagg 300
gattgtggtt gtaagccttg catatgtaca gtcccagaag tatcatctgt cttcatcttc 360
cccccaaagc ccaaggatgt gctcaccatt actctgactc ctaaggtcac gtgtgttgtg 420
gtagacatca gcaaggatga tcccgaggtc cagttcagct ggtttgtaga tgatgtggag 480
gtgcacacag ctcagacgca accccgggag gagcagttca acagcacttt ccgctcagtc 540
agtgaacttc ccatcatgca ccaggactgg ctcaatggca aggagttcaa atgcagggtc 600
aacagtgcag ctttccctgc ccccatcgag aaaaccatct ccaaaaccaa aggcagaccg 660
aaggctccac aggtgtacac cattccacct cccaaggagc agatggccaa ggataaagtc 720
agtctgacct gcatgataac agacttcttc cctgaagaca ttactgtgga gtggcagtgg 780
aatgggcagc cagcggagaa ctacaagaac actcagccca tcatggacac agatggctct 840
tacttcgtct acagcaagct caatgtgcag aagagcaact gggaggcagg aaatactttc 900
acctgctctg tgttacatga gggcctgcac aaccaccata ctgagaagag cctctcccac 960
tctcctggta aa 972
<210> 7
<211> 321
<212> ДНК
<213> Мышь (Mus musculus)
<400> 7
cgggctgatg ctgcaccaac tgtatccatc ttcccaccat ccagtgagca gttaacatct 60
ggaggtgcct cagtcgtgtg cttcttgaac aacttctacc ccaaagacat caatgtcaag 120
tggaagattg atggcagtga acgacaaaat ggcgtcctga acagttggac tgatcaggac 180
agcaaagaca gcacctacag catgagcagc accctcacgt tgaccaagga cgagtatgaa 240
cgacataaca gctatacctg tgaggccact cacaagacat caacttcacc cattgtcaag 300
agcttcaaca ggaatgagtg t 321
<210> 8
<211> 118
<212> PRT
<213> Мышь (Mus musculus)
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Arg Asp Thr Tyr Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Lys Asn Asn Leu Phe
65 70 75 80
Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ser Arg Gln Tyr Gly Thr Val Trp Phe Phe Asn Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala
115
<210> 9
<211> 111
<212> PRT
<213> Мышь (Mus musculus)
<400> 9
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 10
<211> 12
<212> PRT
<213> Мышь (Mus musculus)
<400> 10
Glu Ser Val Asp Ser Tyr Gly Asn Ser Phe Met His
1 5 10
<210> 11
<211> 11
<212> PRT
<213> Мышь (Mus musculus)
<400> 11
Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
1 5 10
<210> 12
<211> 9
<212> PRT
<213> Мышь (Mus musculus)
<400> 12
Gln Gln Ser Lys Glu Val Pro Trp Thr
1 5
<210> 13
<211> 10
<212> PRT
<213> Мышь (Mus musculus)
<400> 13
Gly Phe Thr Phe Ser Ser Tyr Gly Met Ser
1 5 10
<210> 14
<211> 19
<212> PRT
<213> Мышь (Mus musculus)
<400> 14
Val Ala Thr Ile Ser Gly Gly Gly Arg Asp Thr Tyr Tyr Ser Asp Ser
1 5 10 15
Val Lys Gly
<210> 15
<211> 11
<212> PRT
<213> Мышь (Mus musculus)
<400> 15
Ser Arg Gln Tyr Gly Thr Val Trp Phe Phe Asn
1 5 10
<210> 16
<211> 445
<212> PRT
<213> Человек (Homo sapiens)
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Arg Asp Thr Tyr Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Gln Tyr Gly Thr Val Trp Phe Phe Asn Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
<210> 17
<211> 218
<212> PRT
<213> Человек (Homo sapiens)
<400> 17
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 18
<211> 464
<212> PRT
<213> Человек (Homo sapiens)
<400> 18
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Ser Tyr Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Arg Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Gly Gly Gly Arg Asp Thr Tyr Tyr Ser
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn
85 90 95
Asn Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ser Arg Gln Tyr Gly Thr Val Trp Phe Phe Asn Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
225 230 235 240
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
275 280 285
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
340 345 350
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
355 360 365
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 19
<211> 237
<212> PRT
<213> Человек (Homo sapiens)
<400> 19
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu
20 25 30
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val
35 40 45
Asp Ser Tyr Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly
50 55 60
Gln Pro Pro Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly
65 70 75 80
Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
85 90 95
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Met Tyr Phe Cys Gln
100 105 110
Gln Ser Lys Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu
115 120 125
Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
130 135 140
Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
145 150 155 160
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
165 170 175
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
180 185 190
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
195 200 205
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
210 215 220
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 20
<211> 19
<212> PRT
<213> Человек (Homo sapiens)
<400> 20
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser
<210> 21
<211> 19
<212> PRT
<213> Человек (Homo sapiens)
<400> 21
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 22
<211> 118
<212> PRT
<213> Человек (Homo sapiens)
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Gly Gly Gly Arg Asp Thr Tyr Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Gln Tyr Gly Thr Val Trp Phe Phe Asn Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 23
<211> 111
<212> PRT
<213> Человек (Homo sapiens)
<400> 23
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 24
<211> 327
<212> PRT
<213> Человек (Homo sapiens)
<400> 24
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 25
<211> 107
<212> PRT
<213> Человек (Homo sapiens)
<400> 25
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 26
<211> 1395
<212> ДНК
<213> Человек (Homo sapiens)
<400> 26
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgtctgag 60
gtccaacttg tggagtctgg aggaggactg gtgaagcctg gaggctccct gagactgtcc 120
tgtgctgcct ctggcttcac cttctcctcc tatgggatga gttgggtgag acaggctcct 180
gggaagagat tggagtgggt ggctaccatc tctggaggag gcagggacac ctactactct 240
gactctgtga agggcaggtt cacaatcagc agggacaatg ccaagaacaa cctgtacctc 300
caaatgaact ccctgagggc tgaggacaca gcagtctact actgtagcag acaatatggc 360
acagtgtggt tcttcaactg gggacaaggc accctggtga cagtgtcctc tgctagcacc 420
aagggcccat cggtcttccc gctggcgccc tgctccagga gcacctccga gagcacagcc 480
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca 540
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac 600
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacgaagac ctacacctgc 660
aacgtagatc acaagcccag caacaccaag gtggacaaga gagttgagtc caaatatggt 720
cccccatgcc caccctgccc agcacctgag ttcctggggg gaccatcagt cttcctgttc 780
cccccaaaac ccaaggacac tctcatgatc tcccggaccc ctgaggtcac gtgcgtggtg 840
gtggacgtga gccaggaaga ccccgaggtc cagttcaact ggtacgtgga tggcgtggag 900
gtgcataatg ccaagacaaa gccgcgggag gagcagttca acagcacgta ccgtgtggtc 960
agcgtcctca ccgtcctgca ccaggactgg ctgaacggca aggagtacaa gtgcaaggtc 1020
tccaacaaag gcctcccgtc ctccatcgag aaaaccatct ccaaagccaa agggcagccc 1080
cgagagccac aggtgtacac cctgccccca tcccaggagg agatgaccaa gaaccaggtc 1140
agcctgacct gcctggtcaa aggcttctac cccagcgaca tcgccgtgga gtgggaaagc 1200
aatgggcagc cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc 1260
ttcttcctct acagcaggct aaccgtggac aagagcaggt ggcaggaggg gaatgtcttc 1320
tcatgctccg tgatgcatga ggctctgcac aaccactaca cacagaagag cctctccctg 1380
tctctgggta aataa 1395
<210> 27
<211> 714
<212> ДНК
<213> Человек (Homo sapiens)
<400> 27
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcattctgag 60
attgtgctga cccagagccc tgccaccctg tccctgagcc ctggagagag ggctaccctg 120
tcctgtaggg catctgagtc tgtggactcc tatggcaact cctttatgca ctggtatcaa 180
cagaagcctg gacaaccacc aagactgctg atttatgctg ccagcaacca gggctctgga 240
gtgcctgcca ggttctctgg ctctggctct ggcacagact tcaccctgac catctcctcc 300
ttggaacctg aggactttgc tatgtacttc tgtcaacaga gcaaggaggt gccatggacc 360
tttggacaag gcaccaaggt ggagattaag cgtacggtgg ctgcaccatc tgtcttcatc 420
ttcccgccat ctgatgagca gttgaaatct ggaactgcct ctgttgtgtg cctgctgaat 480
aacttctatc ccagagaggc caaagtacag tggaaggtgg ataacgccct ccaatcgggt 540
aactcccagg agagtgtcac agagcaggac agcaaggaca gcacctacag cctcagcagc 600
accctgacgc tgagcaaagc agactacgag aaacacaaag tctacgcctg cgaagtcacc 660
catcagggcc tgagctcgcc cgtcacaaag agcttcaaca ggggagagtg ttag 714
<210> 28
<211> 57
<212> ДНК
<213> Человек (Homo sapiens)
<400> 28
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgtct 57
<210> 29
<211> 57
<212> ДНК
<213> Человек (Homo sapiens)
<400> 29
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcattct 57
<210> 30
<211> 354
<212> ДНК
<213> Человек (Homo sapiens)
<400> 30
gaggtccaac ttgtggagtc tggaggagga ctggtgaagc ctggaggctc cctgagactg 60
tcctgtgctg cctctggctt caccttctcc tcctatggga tgagttgggt gagacaggct 120
cctgggaaga gattggagtg ggtggctacc atctctggag gaggcaggga cacctactac 180
tctgactctg tgaagggcag gttcacaatc agcagggaca atgccaagaa caacctgtac 240
ctccaaatga actccctgag ggctgaggac acagcagtct actactgtag cagacaatat 300
ggcacagtgt ggttcttcaa ctggggacaa ggcaccctgg tgacagtgtc ctct 354
<210> 31
<211> 333
<212> ДНК
<213> Человек (Homo sapiens)
<400> 31
gagattgtgc tgacccagag ccctgccacc ctgtccctga gccctggaga gagggctacc 60
ctgtcctgta gggcatctga gtctgtggac tcctatggca actcctttat gcactggtat 120
caacagaagc ctggacaacc accaagactg ctgatttatg ctgccagcaa ccagggctct 180
ggagtgcctg ccaggttctc tggctctggc tctggcacag acttcaccct gaccatctcc 240
tccttggaac ctgaggactt tgctatgtac ttctgtcaac agagcaagga ggtgccatgg 300
acctttggac aaggcaccaa ggtggagatt aag 333
<210> 32
<211> 984
<212> ДНК
<213> Человек (Homo sapiens)
<400> 32
gctagcacca agggcccatc ggtcttcccg ctggcgccct gctccaggag cacctccgag 60
agcacagccg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacgaagacc 240
tacacctgca acgtagatca caagcccagc aacaccaagg tggacaagag agttgagtcc 300
aaatatggtc ccccatgccc accctgccca gcacctgagt tcctgggggg accatcagtc 360
ttcctgttcc ccccaaaacc caaggacact ctcatgatct cccggacccc tgaggtcacg 420
tgcgtggtgg tggacgtgag ccaggaagac cccgaggtcc agttcaactg gtacgtggat 480
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagttcaa cagcacgtac 540
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaacggcaa ggagtacaag 600
tgcaaggtct ccaacaaagg cctcccgtcc tccatcgaga aaaccatctc caaagccaaa 660
gggcagcccc gagagccaca ggtgtacacc ctgcccccat cccaggagga gatgaccaag 720
aaccaggtca gcctgacctg cctggtcaaa ggcttctacc ccagcgacat cgccgtggag 780
tgggaaagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc 840
gacggctcct tcttcctcta cagcaggcta accgtggaca agagcaggtg gcaggagggg 900
aatgtcttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc 960
ctctccctgt ctctgggtaa ataa 984
<210> 33
<211> 324
<212> ДНК
<213> Человек (Homo sapiens)
<400> 33
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttag 324
<210> 34
<211> 247
<212> PRT
<213> Мышь (Mus musculus)
<400> 34
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Ser Tyr
20 25 30
Gly Asn Ser Phe Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Gln Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys Gln Gln Ser Lys
85 90 95
Glu Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Ser
100 105 110
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg Ser
115 120 125
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly
130 135 140
Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser
145 150 155 160
Tyr Gly Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp
165 170 175
Val Ala Thr Ile Ser Gly Gly Gly Arg Asp Thr Tyr Tyr Ser Asp Ser
180 185 190
Val Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Lys Asn Asn Leu
195 200 205
Phe Leu Gln Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Leu Tyr Tyr
210 215 220
Cys Ser Arg Gln Tyr Gly Thr Val Trp Phe Phe Asn Trp Gly Gln Gly
225 230 235 240
Thr Leu Val Thr Val Ser Ala
245
<210> 35
<211> 18
<212> PRT
<213> Мышь (Mus musculus)
<400> 35
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg
1 5 10 15
Ser Ser
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕЙТРАЛИЗАЦИЯ ИНГИБИТОРНЫХ ПУТЕЙ В ЛИМФОЦИТАХ | 2015 |
|
RU2734771C2 |
| НЕЙТРАЛИЗАЦИЯ ИНГИБИТОРНЫХ ПУТЕЙ В ЛИМФОЦИТАХ | 2017 |
|
RU2769569C2 |
| СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2812280C1 |
| СВЯЗЫВАЮЩИЕСЯ С PD-1 АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2773758C2 |
| АНТИТЕЛА К PD1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2746409C1 |
| ГУМАНИЗОВАННОЕ АНТИТЕЛО ПРОТИВ PD-1 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ПРИМЕНЕНИЕ | 2022 |
|
RU2829508C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| СТАБИЛЬНАЯ КОМПОЗИЦИЯ РЕКОМБИНАНТНОГО МОНОКЛОНАЛЬНОГО АНТИТЕЛА ПРОТИВ PD-1 | 2021 |
|
RU2827610C1 |
| Антитело против PD-1 и его применение | 2017 |
|
RU2739610C1 |
| ИММУНОТЕРАПИЯ С ПРИМЕНЕНИЕМ АНТИТЕЛ, СВЯЗЫВАЮЩИХ БЕЛОК 1 ПРОГРАММИРУЕМОЙ СМЕРТИ КЛЕТОК (PD-1) | 2017 |
|
RU2768404C2 |
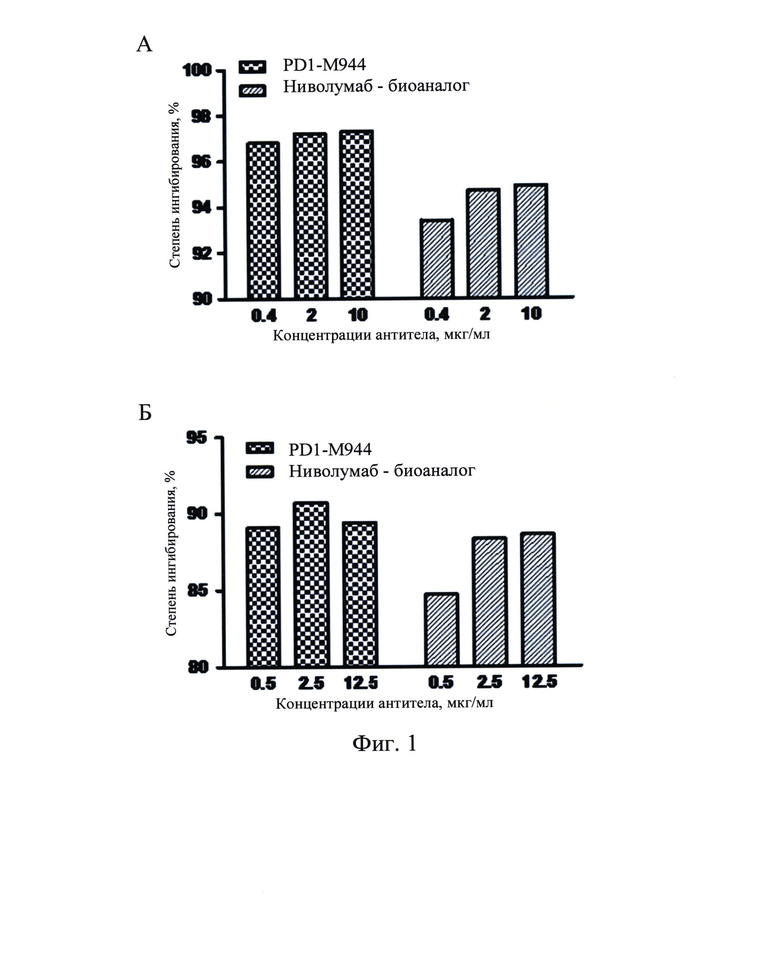
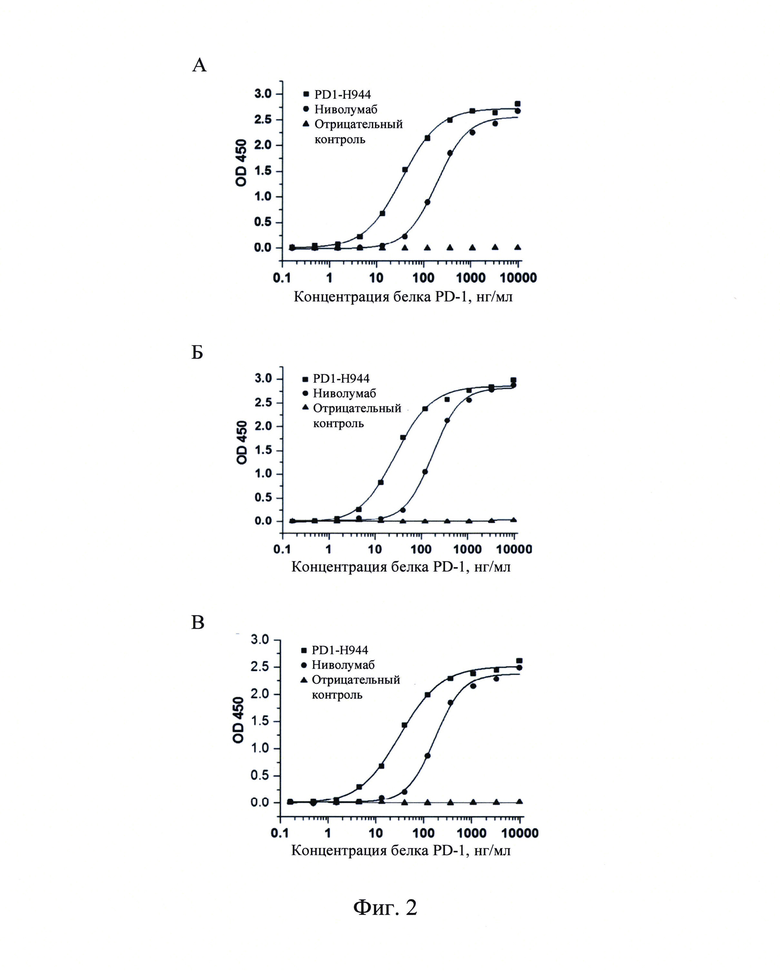
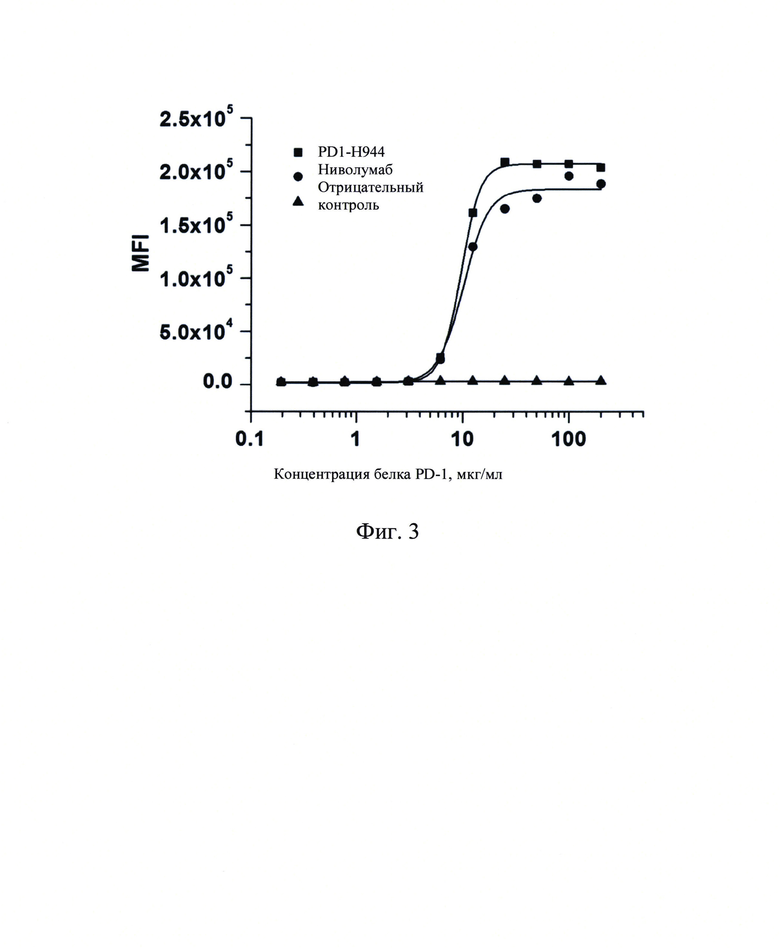
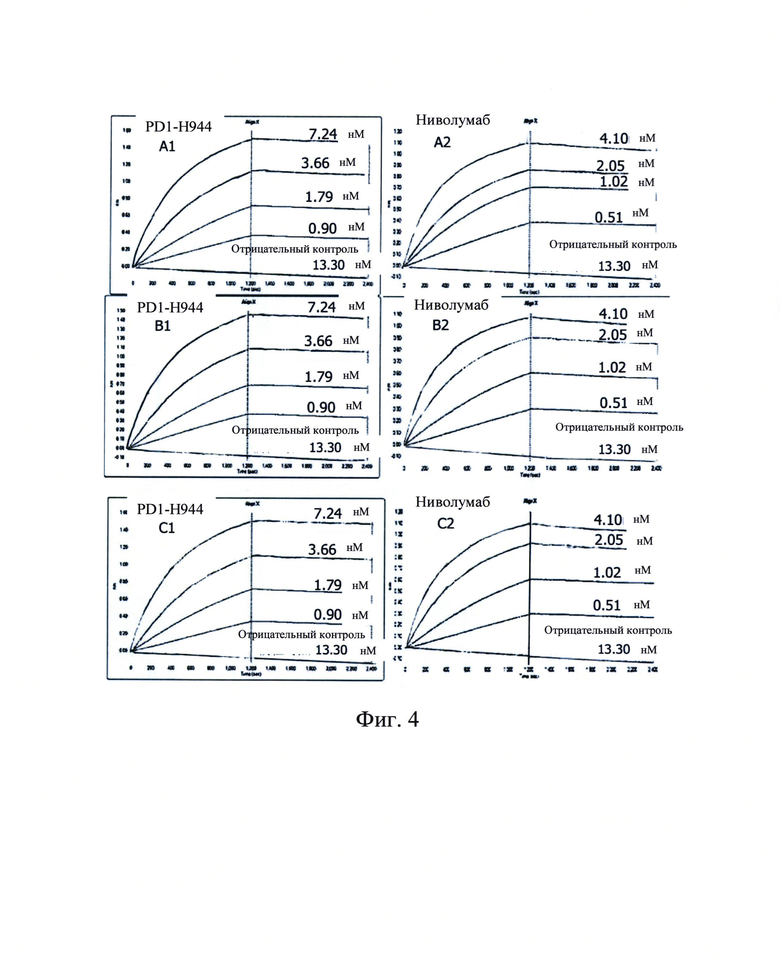
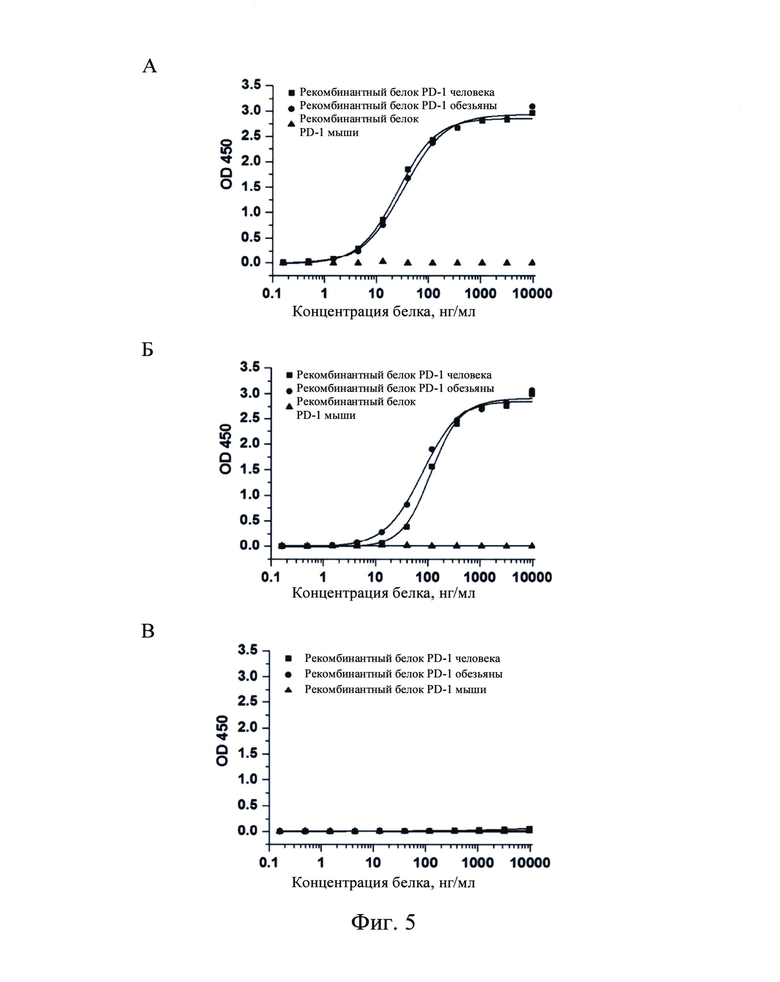
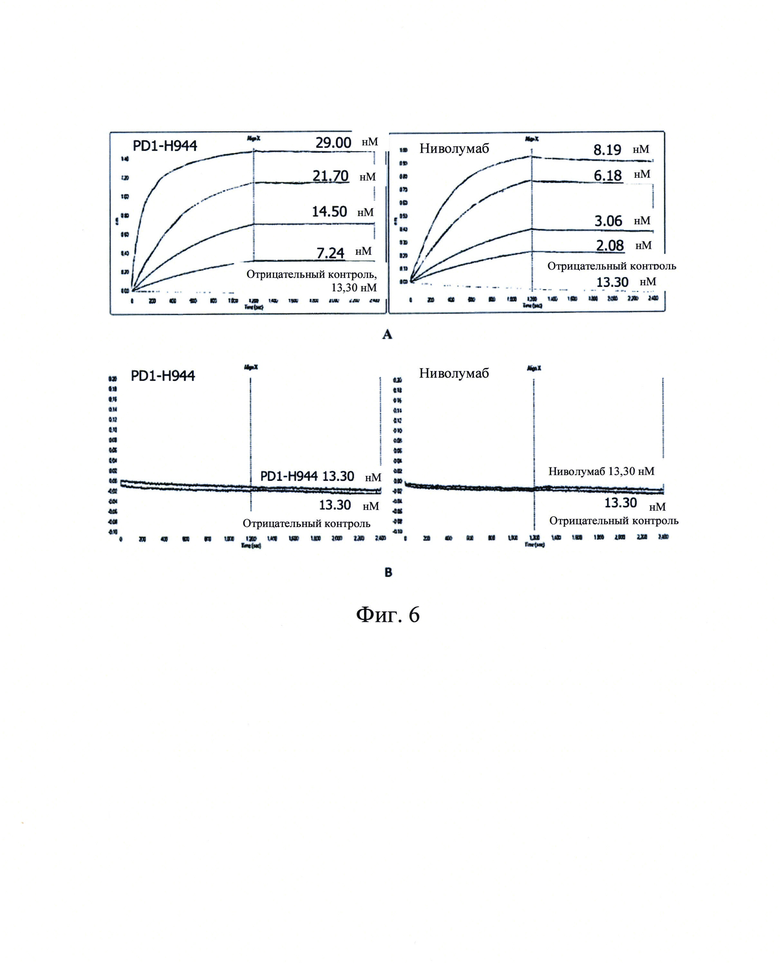
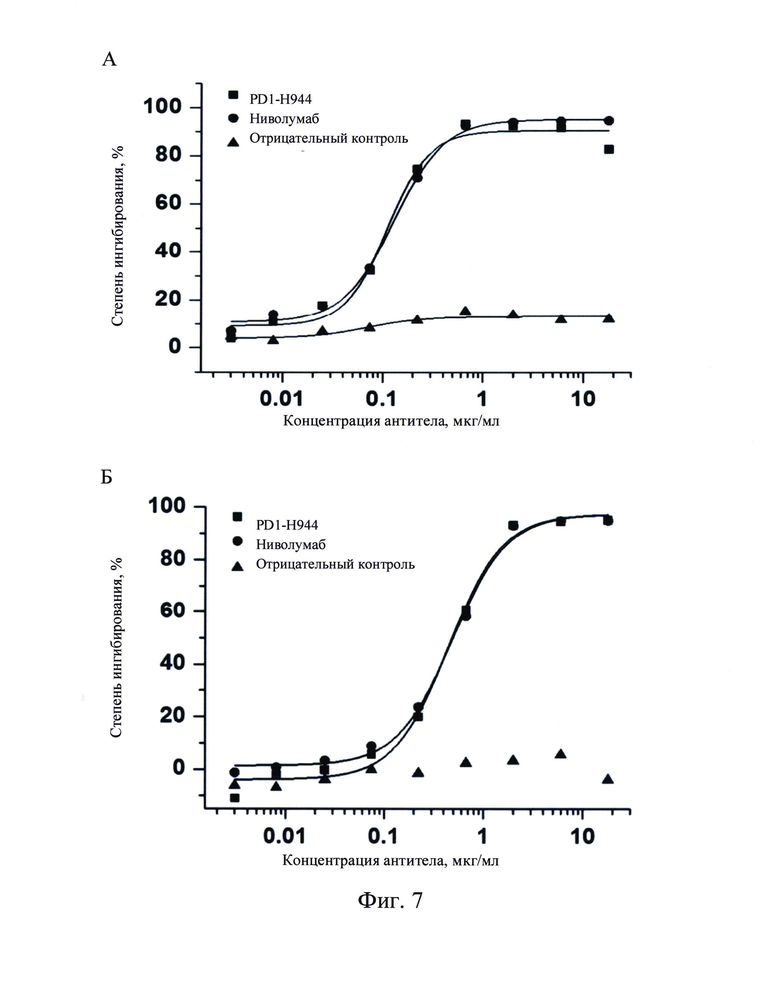
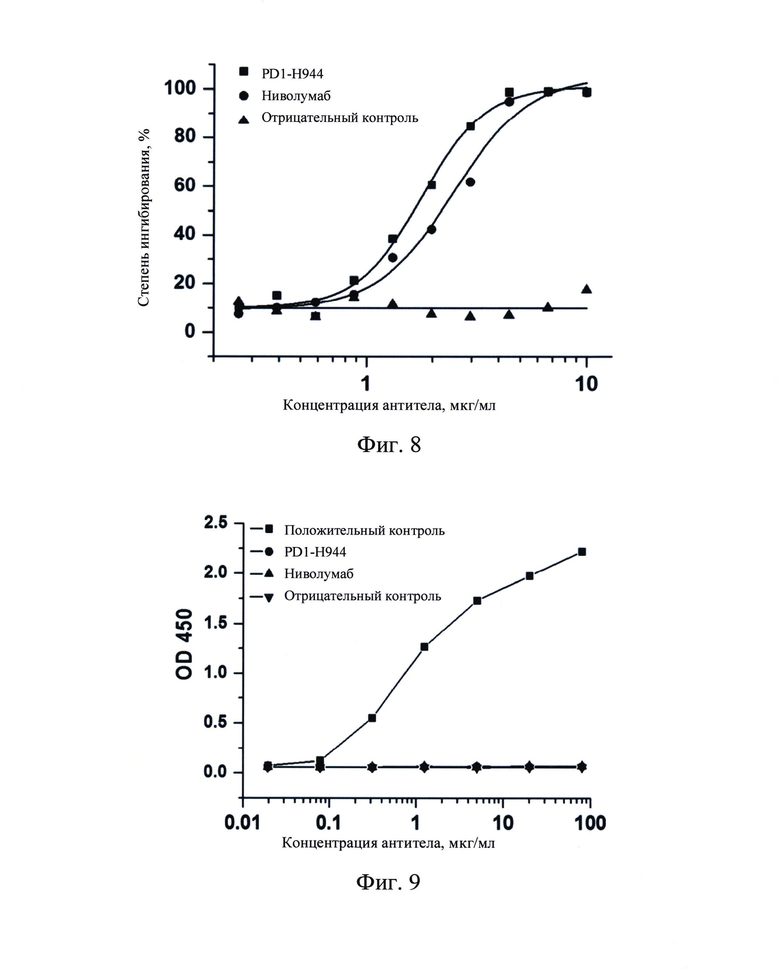
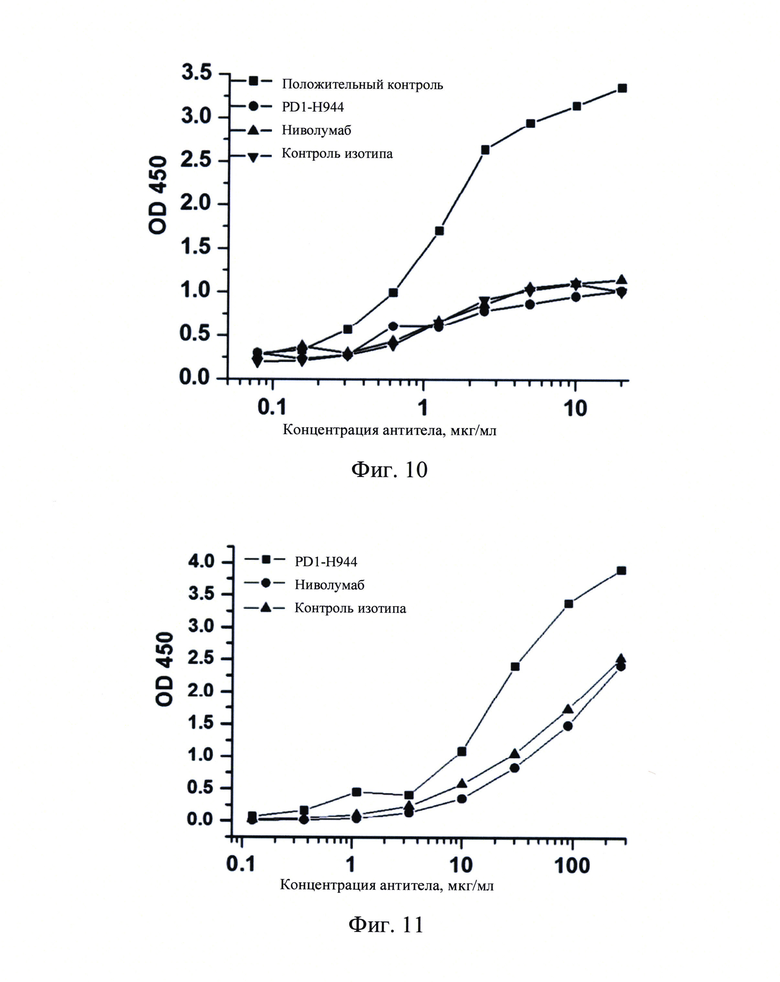
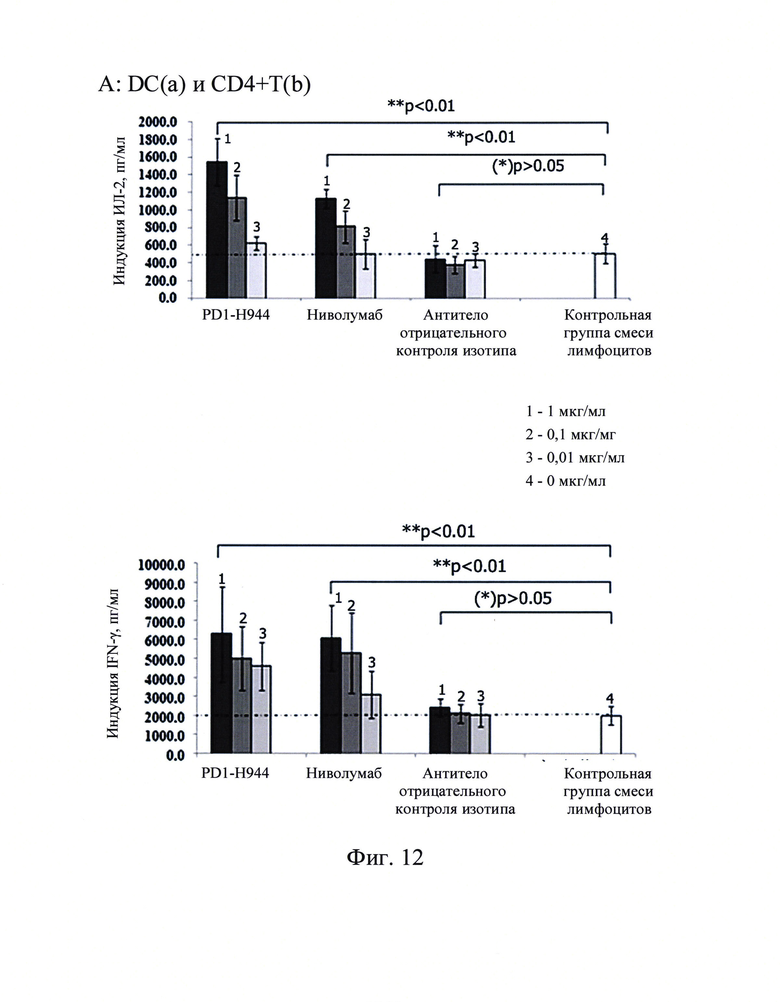
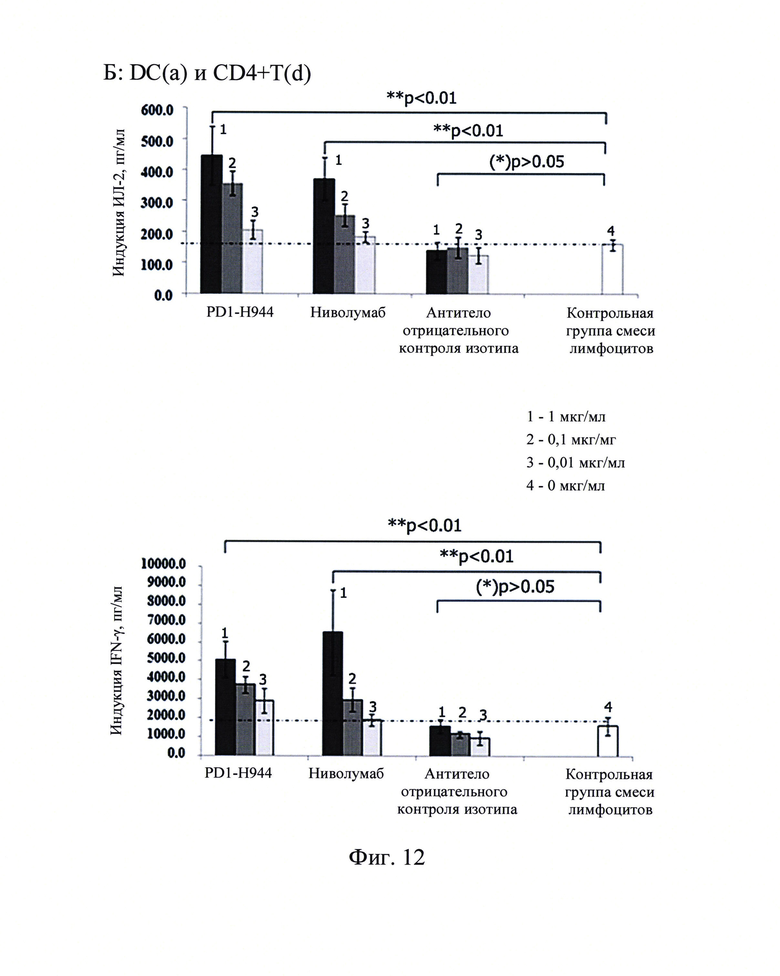
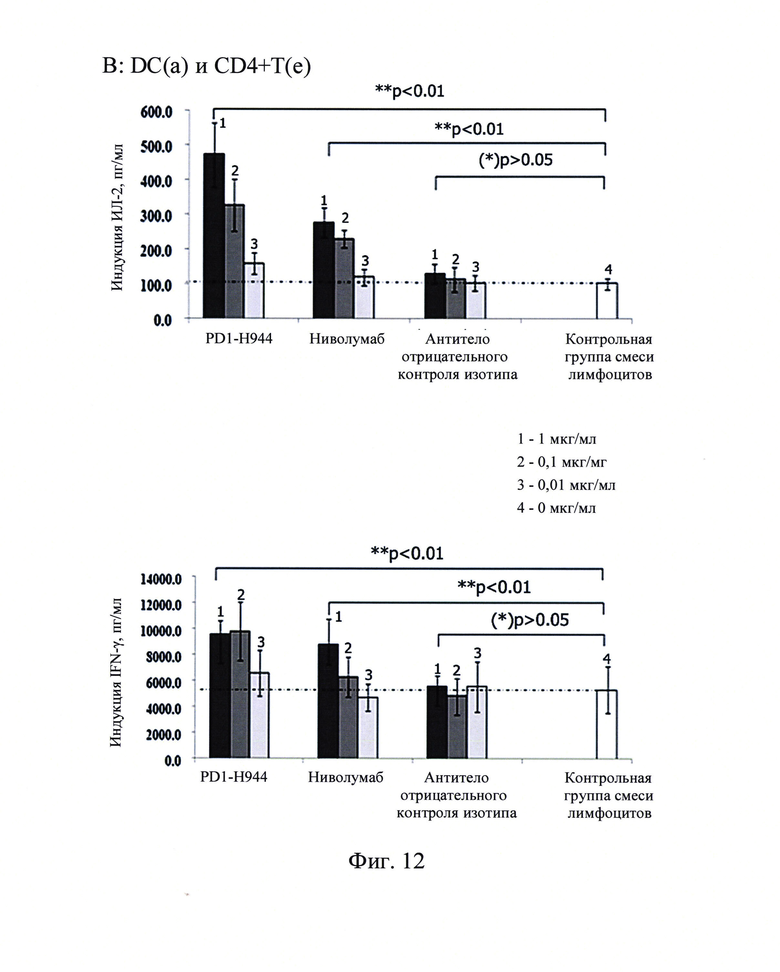
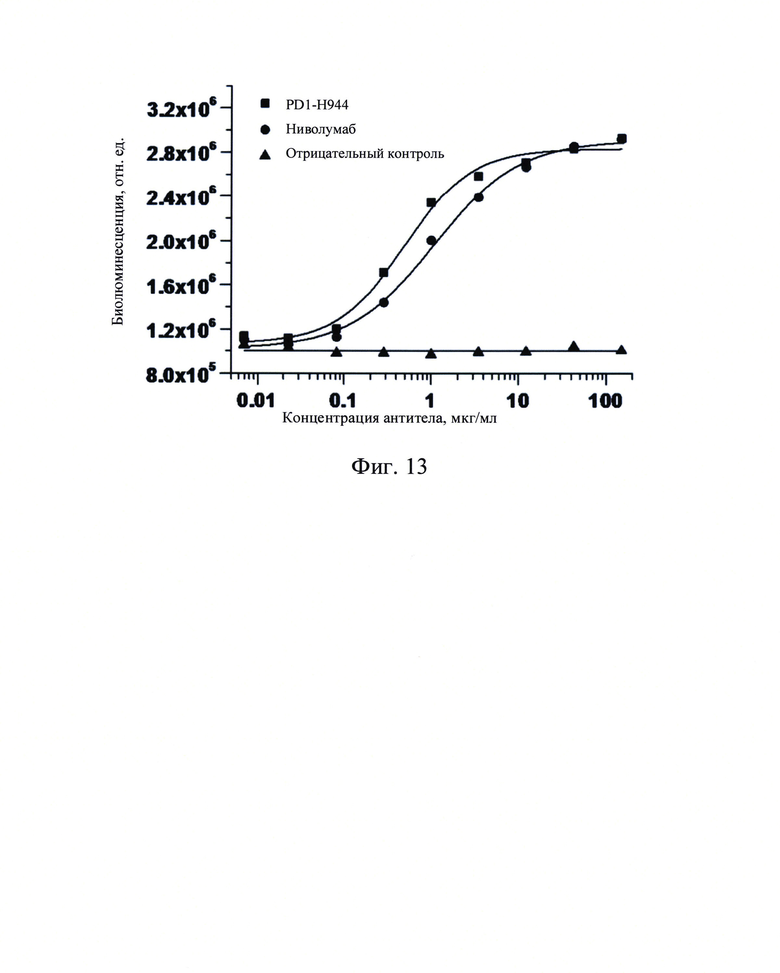
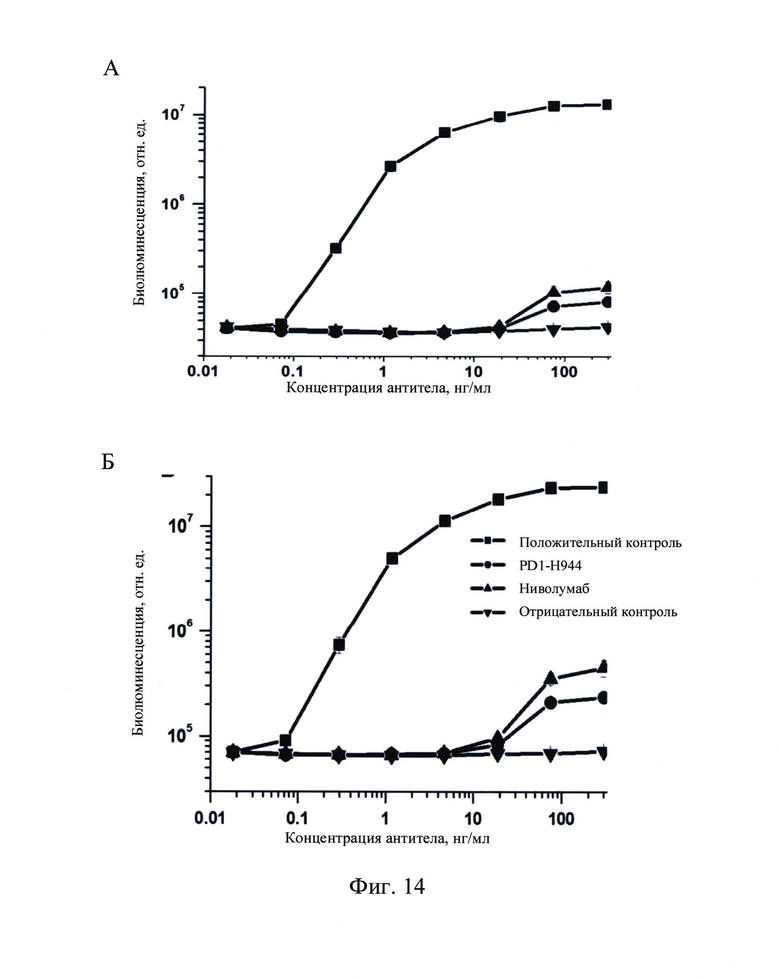
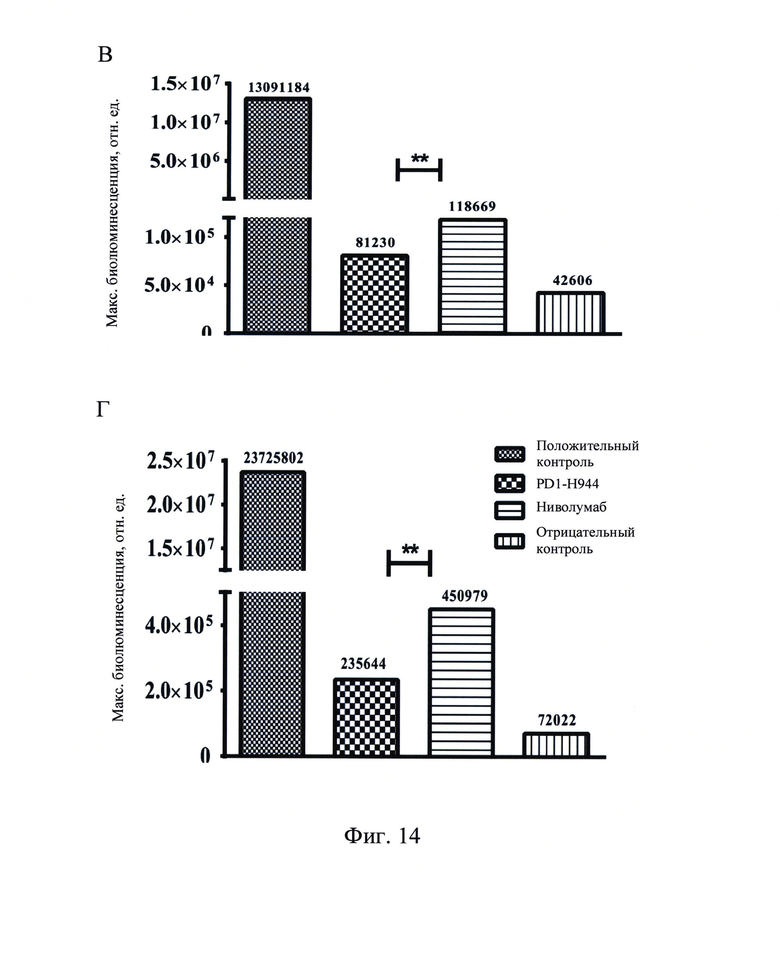
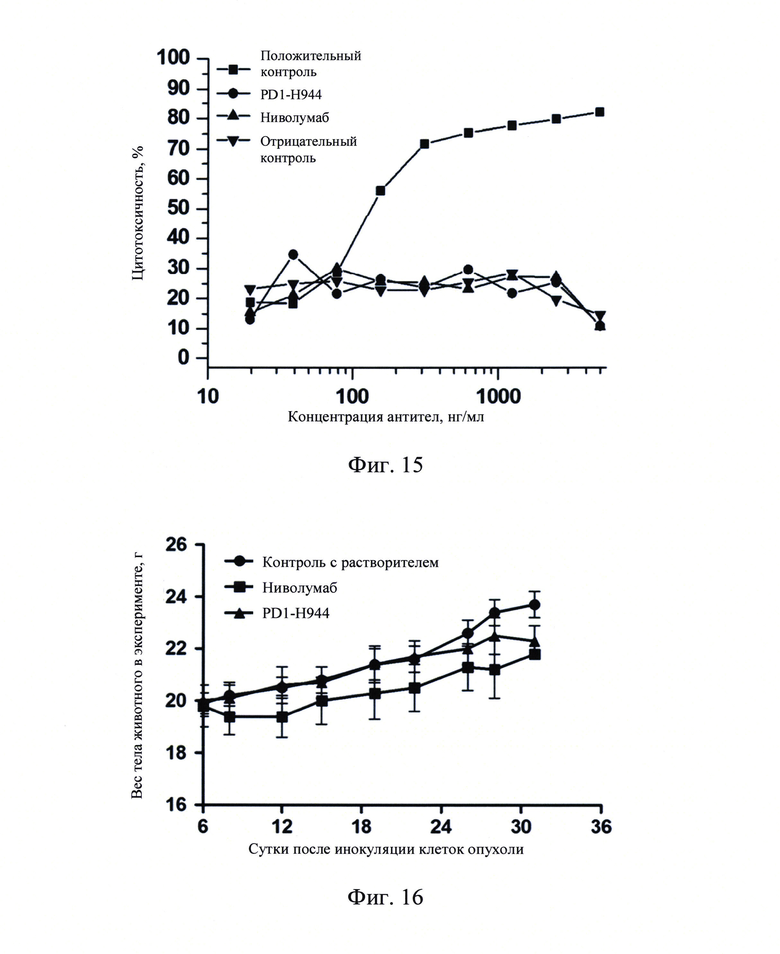
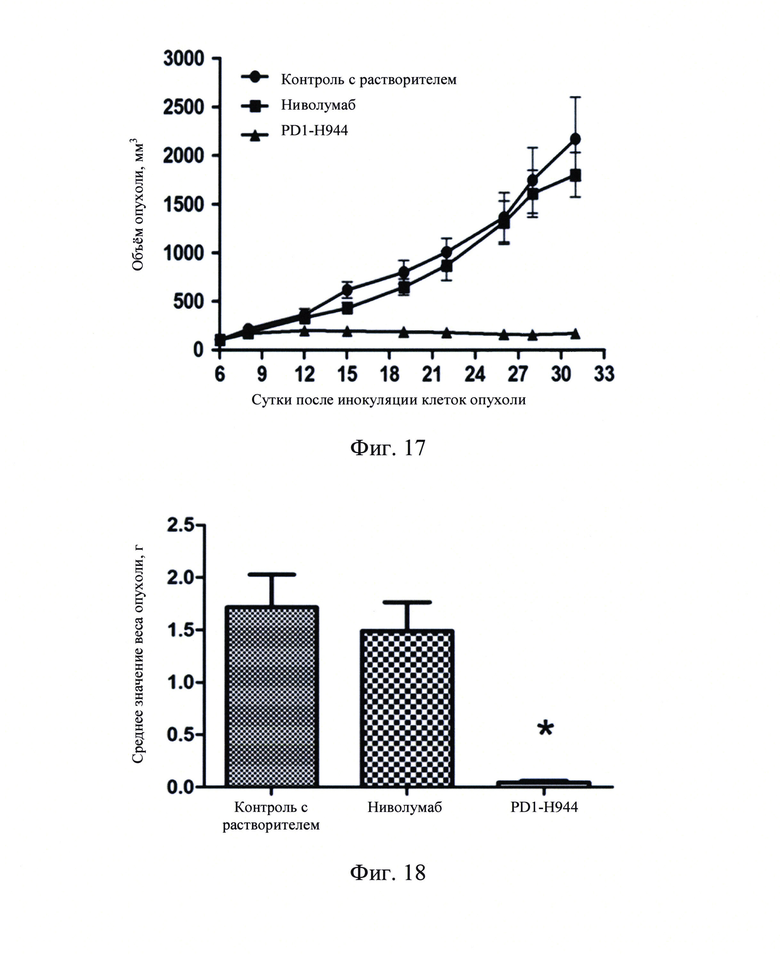
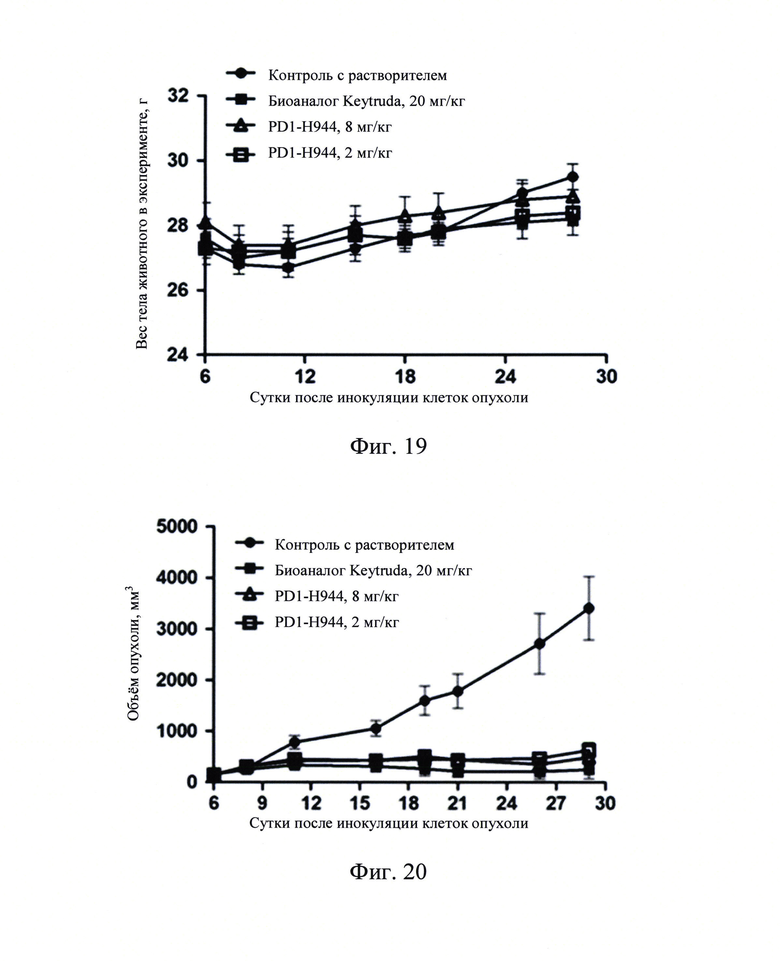
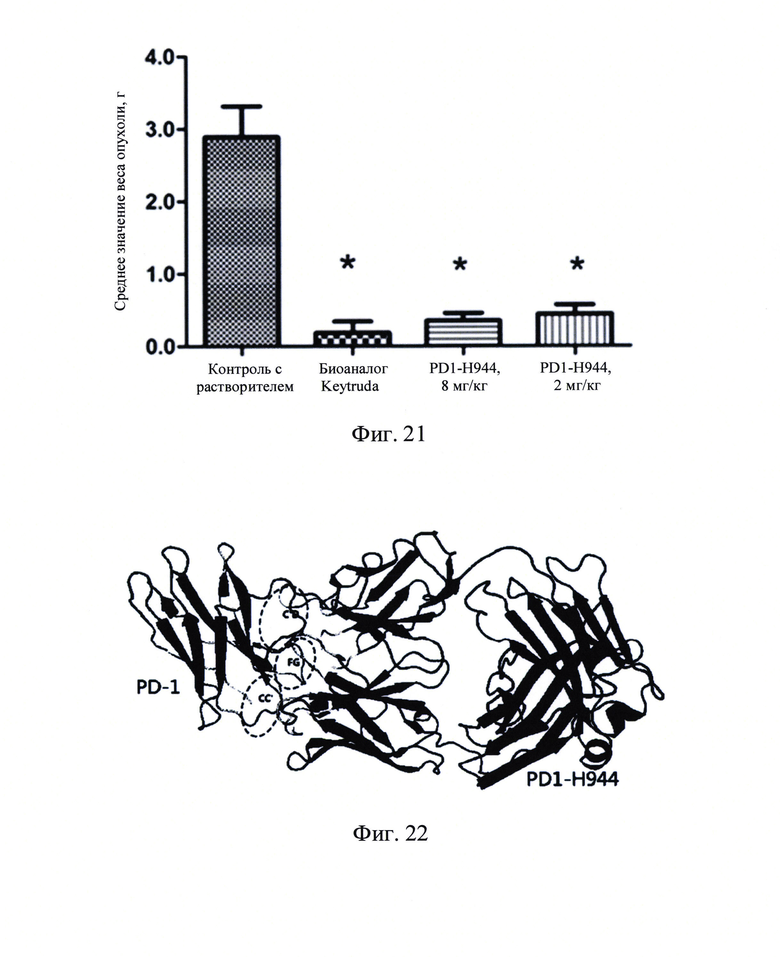
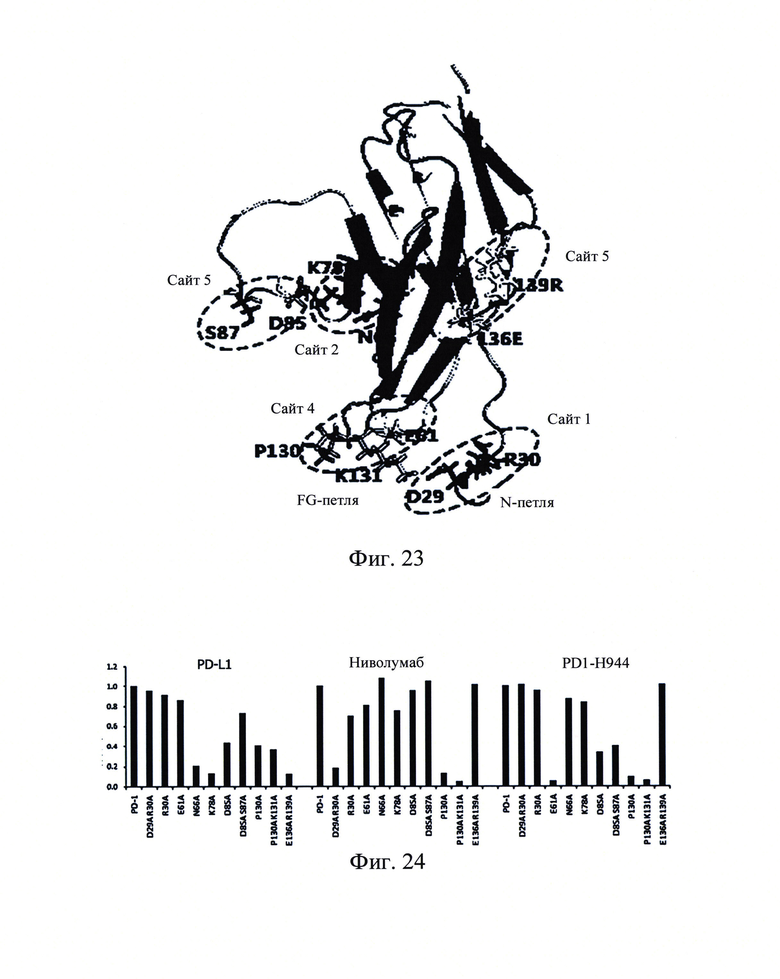
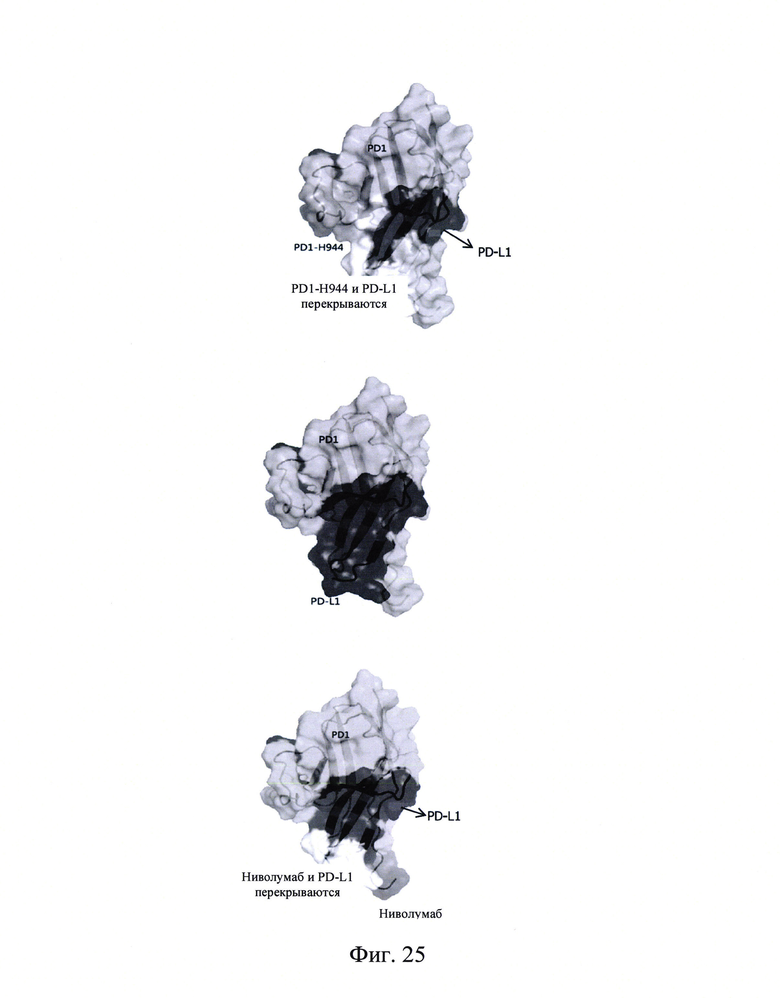
Изобретение относится к области биотехнологии, а именно к выделенному антителу против PD-1 или его антиген-связывающему фрагменту, а также к композиции, комбинации и набору, его содержащим. Также раскрыты выделенная нуклеиновая кислота, кодирующая вышеуказанное антитело или его фрагмент, а также вектор и клетка, ее содержащие. Изобретение также относится к способу получения выделенного антитела против PD-1 или его антигенсвязывающего фрагмента, предусматривающему выращивание вышеуказанной клетки. Изобретение эффективно для получения лекарственного препарата для лечения опухоли или рака, ассоциированного с PD-1, а также для лечения опухоли или рака, ассоциированного с PD-1. 11 н. и 11 з.п. ф-лы, 25 ил., 15 табл., 6 пр.
1. Выделенное антитело против PD-1 или его антиген-связывающий фрагмент, содержащий вариабельный участок легкой цепи или его часть и вариабельный участок тяжелой цепи или его часть;
при этом указанный вариабельный участок легкой цепи или его часть содержит CDR1 легкой цепи, имеющий последовательность аминокислот SEQ ID NO: 10, CDR2 легкой цепи, имеющий последовательность аминокислот SEQ ID NO: 11, и CDR3 легкой цепи, имеющий последовательность аминокислот SEQ ID NO: 12; и указанный вариабельный участок тяжелой цепи или его часть содержит CDR1 тяжелой цепи, имеющий последовательность аминокислот SEQ ID NO: 13, CDR2 тяжелой цепи, имеющий последовательность аминокислот SEQ ID NO: 14, и CDR3 тяжелой цепи, имеющий последовательность аминокислот SEQ ID NO: 15.
2. Выделенное антитело против PD-1 или его антиген-связывающий фрагмент по п. 1, отличающееся тем, что указанное антитело или его антиген-связывающий фрагмент содержит, состоит из или по существу состоит из: последовательности аминокислот, имеющей по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью SEQ ID NO: 23 вариабельного участка легкой цепи антитела против PD-1, и/или последовательности аминокислот, имеющей по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью SEQ ID NO: 22 вариабельного участка тяжелой цепи антитело против PD-1.
3. Выделенное антитело против PD-1 по п. 1 или 2, отличающееся тем, что указанное антитело содержит константный участок легкой цепи и константный участок тяжелой цепи, при этом предпочтительно указанный константный участок легкой цепи содержит последовательность аминокислот, имеющую по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью SEQ ID NO: 25 константного участка легкой цепи каппа, и/или указанный константный участок тяжелой цепи содержит последовательность аминокислот, имеющую по меньшей мере 90%, 92%, 95%, 98% или 100% идентичность с последовательностью SEQ ID NO: 24 константного участка тяжелой цепи антитела IgG4.
4. Выделенное антитело против PD-1 по любому из пп. 1-3, которое является антителом IgG, предпочтительно антителом IgG4.
5. Выделенное антитело против PD-1 по любому из пп. 1-4, являющееся моноклональным антителом.
6. Выделенное антитело против PD-1 или его антиген-связывающий фрагмент по любому из пп. 1-5, которое связывается с рекомбинантным белком PD-1 человека с аффинностью со средним KD в диапазоне 20-200 пМ, предпочтительно 60-70 пМ, более предпочтительно 64,8 пМ.
7. Выделенное антитело против PD-1 или его антиген-связывающий фрагмент по любому из пп. 1-6, которое специфически связывается с молекулой белка PD-1, содержащего последовательность аминокислот SEQ ID NO: 1 или с молекулой белка, имеющей последовательности аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 100% идентичной последовательности SEQ ID NO: 1.
8. Выделенное антитело или его антиген-связывающий фрагмент по любому из пп. 1-7, которое специфически связывается с последовательностью пептида, выбранного из по меньшей мере одного из внеклеточных участков 60SESFV64, 78KLAAFPEDRSQP89, 128LAPKAQI134 белка PD-1; предпочтительно специфически связывается с остатками аминокислот, выбранных из по меньшей мере одного из Е61, K78, D85 и Р130 внеклеточных участков белка PD-1.
9. Выделенное антитело против PD-1 или его антиген-связывающий фрагмент по любому из пп. 1-8, отличающееся тем, что указанный антиген-связывающий фрагмент представлен в форме Fv, Fab, Fab', Fab'-SH, F(ab')2, фрагмента Fd, фрагмента Fd', молекулы одноцепочечного антитела или единичного домена антитела; отличающееся тем, что указанной молекулой одноцепочечного антитела предпочтительно является scFv, di-scFv, tri-scFv, диатело или scFab.
10. Выделенная нуклеиновая кислота, содержащая, состоящая из, или по существу состоящая из последовательности нуклеотидов, кодирующая выделенное антитело против PD-1 или его антиген-связывающий фрагмент по любому из пп. 1-9, предпочтительно выделенная нуклеиновая кислота, содержащая, состоящая из, или по существу состоящая из последовательности нуклеотидов SEQ ID NO: 26 и 27.
11. Вектор экспрессии, содержащий выделенную нуклеиновую кислоту по п. 10.
12. Выделенная клетка-хозяин для получения антитела против PD-1 или его антиген-связывающего фрагмента по любому из пп.1-9, содержащая выделенную нуклеиновую кислоту по п. 10, и/или содержащая вектор по п. 11.
13. Способ получения выделенного антитело против PD-1 или его антигенсвязывающего фрагмента по любому из пп. 1-9, включающий выращивание выделенных клеток-хозяев по п. 12 и очистки антитела против PD-1 или его антигенсвязывающий фрагмента.
14. Применение выделенного антитела против PD-1 или его антиген-связывающего фрагмента по любому из пп. 1-9 для получения лекарственного препарата для лечения опухоли или рака, ассоциированного с PD-1.
15. Применение по п. 14, отличающееся тем, что указанная опухоль или рак является раком толстой кишки.
16. Применение выделенного антитела против PD-1 или его антиген-связывающего фрагмента по любому из пп. 1-9 для лечения опухоли или рака, ассоциированного с PD-1.
17. Применение по п. 16, отличающееся тем, что указанная опухоль или рак является раком толстой кишки.
18. Фармацевтическая композиция для лечения опухоли или рака, ассоциированного с PD-1, содержащая эффективное количество выделенного антитела против PD-1 или его антиген-связывающий фрагмент по любому из пп. 1-9.
19. Фармацевтическая комбинация для лечения опухоли или рака, ассоциированного с PD-1, содержащая эффективное количество фармацевтической композиции по п. 18 с одним или более другим терапевтическими агентами.
20. Набор для лечения опухоли или рака, ассоциированного с PD-1, содержащий выделенное антитело против PD-1 или его антигенсвязывающий фрагмент по любому из пп. 1-9, фармацевтическую композицию по п. 18 или фармацевтическую комбинацию по п. 19.
21. Набор по п. 20, дополнительно содержащий устройство для введения.
22. Способ лечения опухоли или рака, ассоциированного с PD-1, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества выделенного антитела против PD-1 или его антиген-связывающего фрагмента по любому из пп. 1-9, или фармацевтической композиции по п. 18, или фармацевтической комбинации по п. 19, или набора по п. 20 для лечения указанной опухоли или рака, предпочтительно отличающееся тем, что указанной опухолью или раком является рак толстой кишки.
| WO2018113258 A1, 28.06.2018 | |||
| WO2017011580 A2, 19.01.2017 | |||
| ANNA MARIA VALENTINI et al., PD-L1 expression in colorectal cancer defines three subsets of tumor immune microenvironments, Oncotarget, 2018, Vol.9, No.9, pp | |||
| Приспособление для зажима и подачи к круглой пиле кокосовых орехов | 1927 |
|
SU8584A1 |
| US8735553 B1, 27.05.2014 | |||
| АНТИТЕЛА К PD-L1 И ИХ ПРИМЕНЕНИЕ ДЛЯ УСИЛЕНИЯ ФУНКЦИИ Т-КЛЕТОК | 2009 |
|
RU2636023C2 |
