Перечень последовательностей
[0001] Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате с кодировкой ASCII и настоящим включен посредством ссылки во всей своей полноте. Копия указанного файла с кодировкой ASCII, созданная 30 апреля 2020 г., имеет название 022548_WO060_SL.txt, и ее размер составляет 7612 байт.
Предпосылки изобретения
[0002] Болезнь Альцгеймера (AD) характеризуется прогрессирующей нейродегенерацией, ведущей к потере памяти и снижению когнитивных функций. Ее патологические признаки включают накопление внеклеточных амилоидных бляшек и внутринейрональных фибрилл тау-белка. Терапевтически средства, нацеливающиеся на бета-амилоид (Aβ), активно изучались в течение многих лет вследствие его генетического и патологического влияния при AD (Tcw and Goate, Cold Spring Harb Perspect Med. (2017) 7(6): pii a024539). Хотя повышенные уровни белка-предшественника амилоида (АРР) и Aβ связаны с патогенезом AD, пептиды Aβ существуют в различных конформациях и фибриллярном статусе, и неясно, на какие разновидности необходимо нацеливаться для достижения терапевтической пользы (Benilova et al., Nat Neurosci. (2012) 15:349-57).
[0003] Несмотря на эту неопределенность, пассивная иммунотерапия против различных форм Aβ была тщательно протестирована в клинике; однако этим подходам препятствовали дополнительные проблемы. Во-первых, гематоэнцефалический барьер (BBB) ограничивает транспорт крупных биомолекул, что требует введения высоких доз на периферии для достижения терапевтически значимых уровней в головном мозге. В высоких дозах несколько антител к Aβ в клинических испытаниях вызывали нежелательные реакции, типичными для которых были аномалии визуализации, связанные с амилоидом (ARIA); считается, что данные нежелательные реакции вызваны накоплением антител в участках сосудистого амилоида, запускающих местное воспаление посредством Fc-зависимых эффекторных функций (Mo et al., Ann Clin Transl Neu. (2017) 4:931-42). Во-вторых, существует потребность в поддержании уровней выше минимальной терапевтической дозы, что требует длительной пассивной иммунотерапии, которая требует участия пациента и соблюдения режима лечения, а также характеризуется значительными затратами.
[0004] Перенос генов в центральную нервную систему (ЦНС) позволяет продуцировать терапевтический белок в нейрональных клетках и, следовательно, обходить BBB. Были произведены попытки AAV-опосредованной экспрессии либо цельных иммуноглобулинов (IgG), либо одноцепочечных вариабельных фрагментов (scFv) в ЦНС, но оба этих подхода имеют присущие им ограничения (Sudol et al., Mol Ther. (2009) 17:2031-40; Ryan et al., Mol Ther. (2010) 18:1471-81; Levites et al., J Neurosci. (2006) 26:11923-28; Levites et al., J Neurosci. (2015) 35:6265-76; Kou et al., JAD. (2011) 27:23-38; Fukuchi et al., Neurobio Dis. (2006) 23:502-11; Liu et al., J Neurosci. (2016) 36:12425-35). Экспрессия тяжелой и легкой цепей IgG в ЦНС была осуществлена только с применением саморасщепляющейся последовательности F2A для получения обеих цепей с кассеты с одним промотором. Пептид F2A остается прикрепленным либо к тяжелой, либо к легкой цепи и является потенциально иммуногенным (Saunders et al., J Vir. (2015) 89:8334-45). С другой стороны, доставка белков scFv с применением генов часто сопровождается существенной потерей аффинности вследствие потери валентности. Удаление Fc-области также приводит к потере связывания FcRn, что приводит к более короткому периоду полужизни на периферии и снижению оттока связанных с антигеном (Ag) scFv из головного мозга посредством обратного трансцитоза (Deane et al., J Neurosci. (2005) 25:11495-503; Boado, et al., Bioconjug Chem. (2007) 18:447-55; Zhang et al., J Neuroimm. (2001) 114:168-72; Schlachetzki et al., J Neurochem. (2002) 81:203-6). Следовательно, терапевтические средства на основе антител для лечения заболеваний ЦНС, таких как болезнь Альцгеймера, являются многообещающими, но были ограничены проблемами введения терапевтических белков в головной мозг, пораженный заболеванием. Соответственно, существует потребность в улучшении доступа терапевтических средств на основе антител к центральной нервной системе.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0005] Настоящее изобретение предусматривает способ экспрессии бивалентного связывающего элемента в клетке нервной системы, включающий введение в клетку кассеты экспрессии, кодирующей полипептид, содержащий вариабельный домен тяжелой цепи антитела (VH), вариабельный домен легкой цепи антитела (VL) и Fc-область IgG, где VH и VL образуют антигенсвязывающий сайт, который специфически связывается с целевым белком, и при этом в ходе экспрессии в клетке две молекулы полипептида образуют связанный дисульфидной связью гомодимерный бивалентный связывающий элемент, специфический в отношении целевого белка.
[0006] В некоторых вариантах осуществления клетка нервной системы представляет собой нейрон, глиальную клетку, эпендимальную клетку или эпителиальную клетку головного мозга. В дополнительных вариантах осуществления глиальная клетка выбрана из олигодендроцита, астроцита, перицита, шванновской клетки и клетки микроглии. В некоторых вариантах осуществления клетка представляет собой клетку человека, такую как клетка головного мозга пациента-человека.
[0007] В некоторых вариантах осуществления целевой белок представляет собой белок, экспрессируемый в головном мозге, и может представлять собой бета-амилоидный пептид (Aβ), тау-белок, SOD-1, TDP-43, ApoE или α-синуклеин.
[0008] В некоторых вариантах осуществления полипептид содержит от N-конца к C-концу (i) VH, пептидный линкер и VL, или VL, пептидный линкер и VH, а также (ii) Fc-область IgG. В дополнительных вариантах осуществления пептидный линкер содержит последовательность GGGGS (SEQ ID NO: 3); например, пептидный линкер имеет последовательность [G4S]3 (SEQ ID NO: 2).
[0009] В некоторых вариантах осуществления бивалентный связывающий элемент по настоящему изобретению связывается с неонатальным Fc-рецептором (FcRn), но не связывается с Fc-гамма-рецептором вследствие одной или более мутаций в Fc-области IgG.
[0010] В некоторых вариантах осуществления способ по настоящему изобретению включает введение вирусного вектора, содержащего кассету экспрессии. Вирусный вектор может представлять собой рекомбинантный вирус. В дополнительных вариантах осуществления рекомбинантный вирус вводится в головной мозг пациента посредством внутричерепной инъекции, интратекальной инъекции или инъекции в мостомозжечковую цистерну. Рекомбинантный вирус может представлять собой, например, рекомбинантный аденоассоциированный вирус (rAAV), например rAAV серотипа 1 или 2.
[0011] В некоторых вариантах осуществления экспрессия полипептида находится под транскрипционным контролем конститутивно активного промотора или индуцируемого промотора.
[0012] Способы по настоящему изобретению можно использовать для лечения пациента с нейродегенеративным заболеванием, например, болезнью Альцгеймера, церебральной амилоидной ангиопатией, синуклеопатией, таупатией или амиотрофическим латеральным склерозом (ALS).
[0013] В другом аспекте настоящее изобретение предусматривает способ лечения нейродегенеративного заболевания, включающий введение пациенту, нуждающемуся в этом, терапевтически эффективного количества композиции, содержащей вирусный вектор, раскрытый в данном документе, который экспрессирует бивалентный связывающий элемент по настоящему изобретению.
[0014] В другом аспекте настоящее изобретение предусматривает бивалентный связывающий элемент для применения в лечении пациента, нуждающегося в этом, и применение бивалентного связывающего элемента для изготовления лекарственного препарата для лечения пациента, нуждающегося в этом, при этом пациент страдает, например, нейродегенеративным заболеванием, таким как болезнь Альцгеймера, церебральная амилоидная ангиопатия, синуклеопатия, таупатия или ALS.
[0015] Другие признаки, цели и преимущества настоящего изобретения очевидны из приведенного далее подробного описания. Однако следует понимать, что подробное описание, несмотря на то, что в нем указаны варианты осуществления и аспекты настоящего изобретения, приведено лишь с целью иллюстрации, а не ограничения. Различные изменения и модификация в пределах объема настоящего изобретения станут очевидны специалисту в данной области техники из подробного описания.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0016] На фиг. 1A-C показаны конструкция и характеристика вектора AAV-IgG.
[0017] На фиг. 1А показана схема вектора для экспрессии полных тяжелой и легкой цепей. Указан размер генома.
[0018] На фиг. 1B, левая панель, показана устойчивая экспрессия и секреция AAV-αAβ IgG в головном мозге по сравнению с huIgG, измеренные у контрольных мышей, которым вводили PBS. Точки на графике представляют собой среднее значение +/- SEM, n=8 мышей на группу. На правой панели показана динамика AAV-опосредованной экспрессии AAV-αAβ IgG в головном мозге по сравнению с традиционным периферическим введением IgG к αAβ. На графиках показаны среднее значение +/- SEM. **p < 0,01, 1-факторный ANOVA через 7 недель после инъекции, n=5 мышей на момент времени.
[0019] На фиг. 1С показана цветная микрофотография нейронов, экспрессирующих трансген huIgG по всему гиппокампу (подробно показан CA2), с некоторыми GFAP+астроцитами поблизости, также экспрессирующими huIgG. Cc=мозолистое тело. Зеленый: человеческий IgG (huIgG). Красный: глиальный фибриллярный кислый белок (GFAP). Синий: DAPI.
[0020] На фиг. 2A и B показано связывание антигена посредством AAV-αAβ IgG на мышиной модели болезни Альцгеймера.
[0021] На фиг. 2А показана схема исследования для внутричерепного (AAV-αAβ IgG или контрольный AAV-IgG) и периферического введения доз (IgG к αAβ).
[0022] На фиг. 2В показана экспрессия AAV-αAβ IgG или контрольного AAV-IgG по всему гиппокампу и вышележащей коре головного мозга. Изображения на правой панели демонстрируют связывание IgG с бляшками во фронтальной коре. Масштабные отрезки=10 мкм. Синий: DAPI. Зеленый: huIgG. Красный: 4G8+GFAP.
[0023] На фиг. 3A-C показана оценка нейрональной экспрессии и нейротоксичности AAV-αAβ IgG.
[0024] На фиг. 3A, левая панель, показаны выявленные пептиды из тяжелой и легкой цепей huIgG из лизатов полушарий головного мозга мышей SCID, которым путем инъекции вводили AAV-αAβ IgG, по сравнению с животными, которым путем инъекции вводили PBS (симуляция), или из симулятивного гомогената головного мозга с добавлением эквивалентных уровней huIgG, как в группе AAV-αAβ IgG. Правая панель демонстрирует количественную оценку функционального huIgG по сравнению с общим huIgG, экспрессируемым у мышей SCID центрально или периферически. Данные представлены в виде среднего значения +/- SEM. **p < 0,01, непарный t-критерий Стьюдента.
[0025] На фиг. 3В, левая панель, показано окрашивание H&E гиппокампа головного мозга мышей C57BL/6 после экспрессии AAV-αAβ msIgG внутри гиппокампа в течение 16 недель по сравнению с контролем PBS. На вставке показаны детали, стрелки указывают на типичные гиалиновые включения. Масштабный отрезок=100 мкм. Краткое описание результатов показано в таблице на правой панели в виде количества животных с данной патологией или без нее.
[0026] На фиг. 3С показаны доказательства нейровоспаления по данным иммуногистохимического (IHC) анализа глиального фибриллярного кислого белка (GFAP) по сравнению с PBS. На левой панели показаны количественные данные (IHC) для GFAP+ области. На правой панели каждый круг представляет одну мышь. Столбцы указывают на групповое среднее значение +/- SEM GFAP+ площади, нормализованное к PBS. ***p < 0,001, непарный t-критерий Стьюдента, n=8 мышей на группу.
[0027] На фиг. 4A-C показаны конструкция и характеристика вектора AAV-scFv-IgG.
[0028] На фиг. 4A, левая панель, показана схема конструкции scFv-IgG. Средняя панель демонстрирует, что SDS-PAGE-анализ очищенного scFv-IgG в восстанавливающих или невосстанавливающих условиях демонстрировал чистоту и надлежащую дисульфид-зависимую димеризацию белка. В таблице на правой панели сравнивается аффинность связывания антигена (M) scFv-IgG с форматом IgG.
[0029] На фиг. 4B, левая панель, показана экспрессия AAV-scFv-IgG в сыворотке крови, измеренная посредством твердофазного иммуноферментного анализа антигена (ELISA) через один месяц после периферических внутривенных инъекций AAV мышам C57BL/6. На правой панели показана экспрессия AAV-scFv-IgG в головном мозге. ***p < 0,001, непарный t-критерий Стьюдента, n=5 мышей на группу для внутричерепной инъекции, 2 мыши на группу для внутривенной инъекции.
[0030] На фиг. 4С, левая панель, показаны нацеливание вектора на гиппокамп и трансдукция по всей гиппокампальной формации после IHC на сагиттальных срезах головного мозга мышей, взятых у тех же животных, что и на фиг. 4B, правая панель. На правой панели показано количественное определение scFv-IgG с применением ELISA в различных вырезанных областях головного мозга после двусторонней инъекции AAV-scFv-IgG в гиппокамп. Hipp=гиппокамп. Ctx=вышележащие области коры. Str=полосатое тело.
[0031] На фиг. 5A-C показаны экспрессия, диффузия и связывание с бляшками scFv-IgG к Aβ.
[0032] На фиг. 5A показано полное сканирование гиппокампа и вышележащей коры взрослых мышей через месяц после инъекции AAV-scFv-IgG к Aβ. Срезы подвергали иммуноокрашиванию в отношении бляшек Aβ (4G8, красный) и 6xHis (SEQ ID NO: 9) (зеленый). Масштабный отрезок=300 мкм. Cc=мозолистое тело. Изображения на правой панели демонстрируют ROI отдельных бляшек (пронумерованные в A) от проксимальной (1) до дистальной (6) от места инъекции. Обильное образование бляшек наблюдалось по всей коре (левая панель), и окрашивание с помощью антитела к His локализовано вместе с бляшками (правая панель). Представляющие интерес области (ROI) имеют диаметр 150 мкм. Красный: 4G8. Зеленый: антитело к HIS. Синий: DAPI.
[0033] На фиг. 5B, левая панель, показана схема исследования. Изображения на правой панели демонстрируют гиппокамп из корональных срезов мышей, которым путем инъекции вводили AAV. IHC-анализ позволил выявить присутствие метки по всему гиппокампу на стороне инъекции (красная стрелка) с дополнительной трансдукцией контралатерального гиппокампа. Головной мозг с введенным путем инъекции AAV без вставки не демонстрировал какого-либо присутствия метки к His. Масштабный отрезок=1 мм.
[0034] На фиг. 5C показана количественная оценка отложения бляшек в коре и гиппокампе животных из каждой соответствующей группы. N=10-13 животных на группу, по 3 среза на животное. *** p < 0,001, однофакторный ANOVA с множественными сравнениями. Планки погрешности представляют стандартную ошибку среднего значения (SEM).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0035] Настоящее изобретение предусматривает способ экспрессии бивалентного связывающего элемента в клетке нервной системы без побочных эффектов, наблюдаемых при использовании современных способов экспрессии. Клетки нервной системы в природе не экспрессируют антитела. Предыдущие исследования продемонстрировали, что экспрессия полных антител в головном мозге вызывает нейротоксичность. По сравнению с общепринятыми способами экспрессии IgG дикого типа в клетках головного мозга, способы экспрессии по настоящему изобретению обеспечивают более высокий выход (например, в два или более раза выше) и более низкую токсичность (например, на что указывает отсутствие выявляемого внутринейронального накопления белка гиалина в месте инъекции). Не ограничиваясь теорией, авторы настоящего изобретения предполагают, что клетки нервной системы не обеспечены эффективной системой экспрессии и сборки нативных антител, и что неспаренные цепи антител образуют тельца включения, токсичные для клеток; однако способы экспрессии по настоящему изобретению позволяют преодолеть эту проблему за счет снижения количества экспрессируемых полипептидных цепей с двух до одной. Способы экспрессии по настоящему изобретению также имеют преимущество перед предшествующими способами экспрессии scFv в головном мозге, поскольку способы по настоящему изобретению позволяют осуществлять экспрессию связывающей молекулы, которая характеризуется более высокой авидностью и лучшими фармакокинетическими профилями (например периодом полужизни).
Клетки нервной системы
[0036] Настоящее изобретение предусматривает способ экспрессии (например, включая секрецию) бивалентной молекулы клеткой нервной системы, которая является специфической в отношении целевого белка, экспрессируемого в нервной системе, такой как центральная нервная система, включая головной мозг и спинной мозг. Клетки нервной системы для экспрессии связывающего элемента по настоящему изобретению могут относиться к любому типу клеток в нервной системе, например к любому типу клеток в головном мозге. Например, посредством способа по настоящему изобретению можно экспрессировать связывающий элемент в нейрональной клетке (например, интернейроне, двигательном нейроне, чувствительном нейроне, нейроне головного мозга, дофаминергическом нейроне, холинергическом нейроне, глутаматергическом нейроне, ГАМКергическом нейроне или серотонинергическом нейроне); глиальной клетке (например, олигодендроците, астроците, периците, шванновской клетке или клетке микроглии); эпендимальной клетке или эпителиальной клетке головного мозга. В некоторых вариантах осуществления эти клетки представляют собой клетки человека. Клетки также могут представлять собой клетки, расположенные в любой целевой области головного мозга человека, такой как гиппокамп, кора, базальные ганглии, средний мозг или задний мозг.
Бивалентные связывающие элементы
[0037] Настоящее изобретение предусматривает бивалентный связывающий элемент, который экспрессируется в клетке нервной системы и связывает антиген-мишень, экспрессируемый в нервной системе, например в головном мозге. Антиген-мишень может представлять собой, например, белок, который опосредует неврологическое заболевание, такое как нейродегенеративное заболевание. Представляющие интерес антигены включают без ограничения бета-амилоидный пептид (Aβ), тау-белок, SOD-1, TDP-43, ApoE и α-синуклеин.
[0038] Бивалентный связывающий элемент представляет собой гомодимер полипептидной цепи, где полипептидная цепь содержит антигенсвязывающий домен и константную область антитела (например, шарнирную область, CH2-домен и CH3-домен IgG, такого как человеческий IgG). Таким образом, гомодимер содержит два антигенсвязывающих сайта и Fc-домен антитела.
[0039] В некоторых вариантах осуществления антигенсвязывающий домен полипептидной цепи представляет собой одноцепочечный Fv-домен (scFv). scFv-домены включают вариабельную область тяжелой цепи антитела (VH) и вариабельную область легкой цепи антитела (VL), где VH и VL необязательно разделены пептидным линкером и взаимодействуют с образованием антигенсвязывающего сайта. Способы получения полипептида scFv к интересующему антигену широко известны из уровня техники. Например, можно провести скрининг библиотеки фагового дисплея для получения комбинаций VH и VL, которые связываются с антигеном с высокой аффинностью, или можно получить последовательности VH и VL из уже существующего антитела, которое специфически связывается с антигеном.
[0040] Антигенсвязывающий домен, такой как scFv-домен, может быть слит, с пептидным линкером или без него (например таким, как приведенные в данном документе в качестве примеров, включая линкер с повторами 9-Gly (SEQ ID NO: 7)), с константной областью антитела, где константные области двух полипептидных цепей образуют Fc-домен антитела посредством одной или более дисульфидных связей. Используемый в данном документе термин "Fc-область" или "Fc-домен" относится к части нативного иммуноглобулина, образованной посредством димерной ассоциации одного или более константных доменов иммуноглобулина.
[0041] В некоторых вариантах осуществления каждая полипептидная последовательность Fc-домена может содержать часть тяжелой цепи одиночного иммуноглобулина (Ig), начинающуюся в шарнирной области непосредственно перед сайтом расщепления папаином и заканчивающуюся на С-конце тяжелой цепи Ig. Fc-домен может содержать шарнирную область, СН2 и СН3 иммуноглобулина. В зависимости от изотипа Ig, из которого получен Fc-домен, Fc-домен может содержать дополнительные константные домены (например CH4-домен IgE или IgM). Fc-домен может содержать мутации относительно последовательностей дикого типа, например, для повышения стабильности слитого димерного белка (например периода полужизни) и/или для модификации эффекторных функций слитых димерных белков. Мутации могут представлять собой добавления, делеции или замены одной или более аминокислот.
[0042] В некоторых вариантах осуществления Fc-домен получен из IgG, такого как человеческий IgG, и может относиться к любому подтипу IgG, например к подтипу человеческого IgG1, IgG2, IgG3 или IgG4. В таких случаях scFv-Fc по настоящему изобретению также называют scFv-IgG. Fc-домен может содержать всю шарнирную область или только ее часть из IgG, например, шарнирную область IgG1, IgG2, IgG3 или IgG4. В некоторых вариантах осуществления Fc-домен получен из человеческого IgG1 и содержит мутации L234A и L235A ("LALA") (нумерация согласно EU) таким образом, что Fc-домен не связывается с высокоаффинным Fc-гамма(γ)-рецептором(рецепторами) и характеризуется снижением эффекторных функций ADCC/CDC. Другие мутации Fc, которые могут быть введены в человеческий IgG1, включают без ограничения N297Q, N297A, N297G, C220S/C226S/C229S/P238S, C226S/C229S/E233P/L234V/L235A и L234F/L235E/P331S (нумерация EU). См., например, Wang et al., Protein Cell. (2018) 9(1):63-73; Strohl, Curr Opin Biotechnol. (2009) 20(6):685-91; Johnson et al., Nat Med. (2009) 15(8):901-6. В некоторых вариантах осуществления связывающий элемент содержит шарнирную область из человеческого IgG4, где шарнирная область содержит мутацию S228P (нумерация согласно EU) для снижения диссоциации двух полипептидных цепей связывающего элемента. В некоторых вариантах осуществления Fc-домен получен из человеческого IgG4 и содержит мутации S228P и L235E (нумерация согласно EU; соответствует S241P и L248E в нумерации согласно Kabat), которые снижают обмен полумолекул Fcγ и эффекторную функцию соответственно (Reddy et al., J Imm. (2000) 164:1925-33). Потеря или снижение эффекторных функций ADCC/CDC позволяет связывающему элементу связываться с антигеном-мишенью, не вызывая цитотоксичности или не индуцируя нежелательного воспаления в нервной системе. В дополнительных вариантах осуществления модифицированный Fc-домен сохраняет способность связываться с FcRn, неонатальным Fc-рецептором. Перенаправление способности связывания FcRn позволяет удалить связанный с антигеном связывающий элемент из нервной системы, например из головного мозга, посредством FcRn-опосредованного обратного трансцитоза.
[0043] В некоторых вариантах осуществления VH- и VL-домены связывающего элемента scFv-Fc и/или scFv- и Fc-домены связывающего элемента соединены посредством пептидного линкера. Подходящие пептидные линкеры широко известны из уровня техники. См., например, Bird et al., Science (1988) 242:423-26, и Huston et al., PNAS. (1988) 85:5879-83. Пептидный линкер может быть богат глицином и/или серином. Примерами пептидных линкеров являются G, GG, G3S (SEQ ID NO: 1), G4S (SEQ ID NO: 3) и [G4S]n (n=1, 2, 3 или 4; SEQ ID NO: 4). В некоторых вариантах осуществления линкер с повторами 9-Gly (SEQ ID NO: 7) используется для соединения scFv с частью IgG в формате scFv-IgG по настоящему изобретению.
[0044] В конкретных вариантах осуществления scFv-IgG по настоящему изобретению сконструирован таким образом, что он содержит вариабельные домены, соединенные посредством пептидного линкера с применением пептидного линкера типа [G4S]3 (SEQ ID NO: 2). Линкеры типа [G4S]3 (SEQ ID NO: 2) широко используются для соединения вариабельных доменов в структуре scFv (Huston, выше). Используемый в данном документе линкер типа [G4S]3 (SEQ ID NO: 2) относится к [G4S]3 (SEQ ID NO: 2) или его функциональному варианту (например пептидному линкеру, содержащему до четырех аминокислотных модификаций (например, вставок, делеций и/или замен) из [G4S]3 (SEQ ID NO: 2)). В качестве примеров функциональный вариант [G4S]3 (SEQ ID NO: 2) может представлять собой аминокислотную последовательность SGGGSGGGGSGGGGS (SEQ ID NO: 5) или аминокислотную последовательность GGGGSGGGGXGGGGYGGGGS (X=S, A или N, и Y=A или N; SEQ ID NO: 6).
[0045] В некоторых вариантах осуществления аминокислотная последовательность линкеров может быть модифицирована. Модификации могут включать делеции или вставки, которые изменяют длину линкера (например для корректировки гибкости), или аминокислотные замены, включая, например, с Gly на Ser или наоборот.
[0046] Полипептид scFv-Fc к Aβ показан ниже только для иллюстрации одного формата полипептида scFv-Fc. Следующая последовательность от N-конца к C-концу содержит сигнальный пептид (выделен курсивом), VL, линкер [G4S]3 (SEQ ID NO: 2) (подчеркнутый), VH, G9 (SEQ ID NO: 7) (в рамке), шарнир IgG1, Fc-домен и короткий линкер, прикрепленный к метке 6xHis (SEQ ID NO: 9) (выделен жирным шрифтом).
MDSKGSSQKG SRLLLLLVVS NLLLPQGVLA Seivmtqtpl slpvslgdra siscrsgqsl vhsngntylh wylqkpgqsp klliytvsnr fsgvpdrfsg sgsgsdftlt isrveaedlg vyfcsqntfv pwtfgggtkl eikrtssggg gsggggsggg gsevqlqqsg pevvkpgvsv kisckgsgyt ftdyamhwvk qspgkslewi gvistkygkt nynpsfqgqa tmtvdkssst aymelaslka sdsaiyycar gddgyswgqg tsvtvssast gggggggggs gvprdcgckp cictvpevss vfifppkpkd vltitltpkv tcvvvdiskd dpevqfswfv ddvevhtaqt qpreeqfast frsvselpim hqdwlngkef kcrvnsaafp apiektiskt kgrpkapqvy tipppkeqma kdkvsltcmi tdffpeditv ewqwngqpae nykntqpimd tdgsyfvysk lnvqksnwea gntftcsvlh eglhnhhtek slshspgsgs gsgshhhhhh (SEQ ID NO: 8).
Экспрессия связывающих элементов в нервной системе
[0047] Экспрессионная конструкция, содержащая кассету экспрессии для связывающего элемента, может быть введена в клетки нервной системы посредством широко известных способов. Например, для доставки in vivo или ex vivo можно использовать вирусный вектор. В некоторых вариантах осуществления вектор экспрессии остается в клетке в виде стабильной эписомы. В других вариантах осуществления вектор экспрессии интегрируется в геном клетки. Векторы экспрессии могут содержать последовательности для контроля экспрессии, такие как промоторы, энхансеры, последовательности сигналов транскрипции и последовательности терминации транскрипции, которые обеспечивают экспрессию кодирующей последовательности для связывающего элемента в клетках нервной системы. Подходящие промоторы включают без ограничения ретровирусный промотор RSV LTR (необязательно с энхансером RSV), промотор CMV (необязательно с энхансером CMV), немедленноранний промотор CMV, промотор SV40, промотор дигидрофолатредуктазы (DHFR), промотор β-актина, промотор фосфоглицераткиназы (PGK), промотор EF1α, LTR MoMLV, промотор CK6, промотор транстиретина (TTR), промотор TK, промотор, отвечающий на тетрациклин (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные промоторы, специфические в отношении печени (LSP), промотор E2F, промотор теломеразы (hTERT) и энхансер CMV/промотор β-актина курицы/β-глобина кролика (промотор CAG; Niwa et al., Gene (1991) 108(2):193-9). В некоторых вариантах осуществления промотор включает промотор β-глюкуронидазы человека или энхансер CMV, соединенный с промотором β-актина курицы (CBA). Промотор может представлять собой конститутивный, индуцируемый или репрессируемый промотор.
[0048] Может быть использован любой способ введения нуклеотидной последовательности в клетку, включая без ограничения электропорацию, осаждение фосфатом кальция, микроинъекцию, катионные или анионные липосомы, липосомы в сочетании с сигналом ядерной локализации, встречающиеся в природе липосомы (например экзосомы) или вирусная трансдукция.
[0049] Для in vivo доставки кассеты экспрессии для связывающего элемента можно использовать вирусную трансдукцию. Различные вирусные векторы, известные из уровня техники, могут быть адаптированы специалистом в данной области техники для применения в настоящем изобретении, например, рекомбинантные аденоассоциированные вирусы (rAAV), рекомбинантные аденовирусы, рекомбинантные ретровирусы, рекомбинантные поксвирусы, рекомбинантные лентивирусы и т. д. В некоторых вариантах осуществления используемый в данном документе вирусный вектор представляет собой вектор на основе rAAV. Векторы на основе AAV особенно подходят для доставки генов в ЦНС, поскольку они инфицируют как делящиеся, так и неделящиеся клетки, существуют в виде стабильных эписомальных структур для долговременной экспрессии и характеризуются очень низкой иммуногенностью (Hadaczek et al., Mol Ther. (2010) 18:1458-61; Zaiss, et al., Gene Ther. (2008) 15:808-16). Можно использовать любой подходящий серотип AAV. Например, можно использовать AAV серотипа 1, 2 или 9. AAV можно конструировать таким образом, чтобы его капсидные белки характеризовались сниженной иммуногенностью у людей. В некоторых вариантах осуществления AAV1 используется потому, что этот серотип демонстрирует превосходное паренхиматозное распространение, и хотя преобладает нейрональная трансдукция (как в случае большинства векторов на основе AAV), этот серотип также трансдуцирует астроциты, которые могут быть особенно подвержены высокому уровню экспрессии и секреции белка.
[0050] Описанные в данном документе вирусные векторы могут быть получены с применением способов, известных из уровня техники. Любые подходящие пермиссивные или упаковывающие клетки могут быть использованы для получения вирусных частиц. Например, клетки млекопитающих или насекомых можно использовать в качестве линии упаковывающих клеток.
[0051] Экспрессионные конструкции, такие как рекомбинантный вирус AAV, могут быть введены в головной мозг пациента посредством внутричерепной инъекции, интратекальной инъекции или инъекции в мостомозжечковую цистерну.
Пути применения
[0052] Способы экспрессии по настоящему изобретению можно использовать для доставки терапевтического связывающего элемента в нервную систему пациента. Затем связывающий элемент будет экспрессироваться и секретироваться трансфицированными/трансдуцированными клетками в нервной системе и проявлять свою терапевтическую активность локально в нервной системе, например в головном мозге. Эти способы можно использовать для нацеливания на патогенные антигены при нейродегенеративных заболеваниях, таких как болезнь Альцгеймера (например Aβ и ApoE), церебральная амилоидная ангиопатия, синуклеопатия (например α-синуклеин), таупатия (например тау-белок) или ALS (например SOD-1 и TDP-43 (Pozzi et al., JCI (2019) doi: 10.1172/JCI123931)), болезнь Паркинсона (например α-синуклеин), деменция (например тау-белок (Sigurdsson, J Alzheimers Dis. (2018) 66 (2): 855-6)), синдром, связанный с тельцами Леви (например α-синуклеин (Games et al., J Neurosci. (2014) 34 (28): 9441-54)), болезнь Хантингтона (например хантингтин (WO2016016278)) и множественная системная атрофия (например P25α и α-синуклеин (Games, выше)). В конкретном варианте осуществления нейродегенеративное заболевание представляет собой болезнь Альцгеймера. Связывающий элемент, экспрессируемый локально в нервной системе, будет нацеливаться на патогенный антиген и выводить его из нервной системы, например из головного мозга.
[0053] Соответственно, настоящее изобретение предусматривает способ лечения неврологического заболевания (например нейродегенеративного заболевания) у субъекта, такого как пациент-человек, нуждающегося в этом, включающий введение в нервную систему субъекта терапевтически эффективного количества (например количества, которое обеспечивает достаточную экспрессию связывающего элемента, чтобы вызвать требуемый терапевтический эффект) вирусного вектора (например rAAV), содержащего кодирующую последовательность связывающего элемента для антигена-мишени, функционально связанного с элементом(элементами), регулирующим транскрипцию, который активен в клетках нервной системы.
Фармацевтические композиции
[0054] В некоторых вариантах осуществления настоящее изобретение предусматривает фармацевтическую композицию, содержащую вирусный вектор, такой как рекомбинантный rAAV, рекомбинантный геном которого содержит кассету экспрессии для связывающего элемента scFv-Fc. Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый носитель, такой как вода, солевой раствор (например фосфатно-солевой буферный раствор), декстроза, глицерин, сахароза, лактоза, желатин, декстран, альбумин или пектин. Кроме того, композиция может содержать вспомогательные вещества, такие как смачивающие или эмульгирующие средства, средства для доведения pH, стабилизирующие средства или другие реагенты, которые повышают эффективность фармацевтической композиции. Фармацевтическая композиция может содержать среды-носители для доставки, такие как липосомы, нанокапсулы, микрочастицы, микросферы, липидные частицы и везикулы.
[0055] Доставка rAAV субъекту может осуществляться, например, посредством внутривенного введения. В определенных случаях может потребоваться доставка rAAV локально в ткань головного мозга, спинной мозг, спинномозговую жидкость (CSF), нейрональные клетки, глиальные клетки, мозговые оболочки, астроциты, олигодендроциты, интерстициальное пространство и т. п. В некоторых случаях рекомбинантные AAV могут быть доставлены непосредственно в ЦНС путем инъекции в желудочковую область, а также в полосатое тело и нервно-мышечное соединение или дольки мозжечка. AAV можно доставлять с помощью иглы, катетера или аналогичного устройства с применением нейрохирургических методик, известных из уровня техники, таких как стереотаксическая инъекция (См., например, Stein et al., J Vir. (1999) 73:3424-9; Davidson et al., PNAS. (2000) 97:3428-32; Davidson et al., Nat Genet. (1993) 3:219-23, и Alisky and Davidson, Hum. Gene Ther. (2000) 11:2315-29.)
[0056] Пути введения включают без ограничения интрацеребральное, интратекальное, внутричерепное, интрацеребральное, внутрижелудочковое, интратекальное, в мостомозжечковую цистерну, внутривенное, интраназальное или внутриглазное введение. В некоторых вариантах осуществления вирусный вектор распространяется по ткани ЦНС после прямого введения в спинномозговую жидкость (CSF), например посредством интратекальной и/или интрацеребральной инъекции или инъекции в мостомозжечковую цистерну. В других вариантах осуществления вирусные векторы преодолевают гематоэнцефалический барьер и достигают широкого распространения в ткани ЦНС субъекта после внутривенного введения. В некоторых аспектах вирусные векторы обладают различными способностями нацеливания на ткани ЦНС (например тропизмом к тканям ЦНС), что обеспечивает стабильный и нетоксичный перенос генов с высокой эффективностью.
[0057] В качестве примера фармацевтическая композиция может быть введена пациенту посредством внутрижелудочкового введения, например в желудочковую область переднего мозга пациента, такую как правый латеральный желудочек, левый латеральный желудочек, третий желудочек или четвертый желудочек. Фармацевтическая композиция может быть введена пациенту посредством интрацеребрального введения, например инъекции композиции в большой мозг, продолговатый мозг, варолиев мост, мозжечок, внутричерепную полость, мозговые оболочки, твердую оболочку, паутинную оболочку или мягкую оболочку головного мозга или рядом с ними. В некоторых случаях интрацеребральное введение может включать введение средства в спинномозговую жидкость (CSF) субарахноидального пространства, окружающего головной мозг.
[0058] В некоторых случаях интрацеребральное введение включает инъекцию с применением стереотаксических процедур. Стереотаксические процедуры широко известны из уровня техники и обычно включают применение компьютера и устройства для 3-мерного сканирования, которые используются совместно для направления инъекции в конкретную интрацеребральную область, например желудочковую область. Также можно использовать насосы для микроинъекций (например от World Precision Instruments). В некоторых случаях для доставки композиции, содержащей вирусный вектор, используется насос для микроинъекций. В некоторых случаях скорость инфузии композиции находится в диапазоне от 1 мкл/мин до 100 мкл/мин. Как будет понятно специалисту в данной области техники, скорость инфузии будет зависеть от множества факторов, включая, например, вид субъекта, возраст субъекта, вес/размер субъекта, серотип AAV, требуемую дозу и нацеливание на интрацеребральную область. Таким образом, другие значения скорости инфузии могут быть определены специалистом в данной области техники как подходящие в определенных обстоятельствах.
[0059] Если в данном документе не определено иное, научные и технические термины, используемые в контексте настоящего изобретения, должны иметь значения, которые обычно понятны специалистам в данной области техники. Иллюстративные способы и материалы описаны ниже, хотя при практическом осуществлении или при тестировании настоящего изобретения также можно использовать способы и материалы, аналогичные или эквивалентные описанным в данном документе. В случае противоречий настоящее описание, включая определения, будет иметь преимущественную силу. Как правило, используемая номенклатура, связанная с культивированием клеток и тканей, молекулярной биологией, иммунологией, микробиологией, генетикой, аналитической химией, синтетической органической химией, медицинской и фармацевтической химией и химией и гибридизацией белков и нуклеиновых кислот, а также соответствующие методики, описанные в данном документе, являются общеизвестными и широко используемыми в данной области техники. Ферментативные реакции и методики очистки выполняются в соответствии со спецификациями производителя, как обычно осуществляется в данной области техники или как описано в данном документе. Кроме того, если иное не требуется по контексту, термины в единственном числе включают множественное число, а термины во множественном числе включают единственное число. На протяжении всего настоящего описания и вариантов осуществления слова "иметь" и "содержать" или их варианты, такие как "имеет", "имеющий", "содержит" или "содержащий", следует понимать как подразумевающие включение указанного целого числа или группы целых чисел, но не исключение любого другого целого числа или группы целых чисел. "Приблизительно" следует понимать как в пределах 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,5%, 0,1%, 0,05% или 0,01% заявленного значения. Если иное не ясно из контекста, все числовые значения, представленные в данном документе, модифицированы термином "приблизительно". Понятно, что аспекты и варианты настоящего изобретения, описанные в данном документе, включают аспекты и вариации фраз "состоящий из" и/или "состоящий по сути из". Все публикации и другие литературные источники, упомянутые в данном документе, включены посредством ссылки во всей своей полноте. Хотя в данном документе цитируется ряд документов, данное цитирование не является признанием того, что любой из этих документов образует часть общеизвестных знаний в данной области техники. В случае противоречий настоящее описание, включая определения, будет иметь преимущественную силу. Кроме того, материалы, способы и примеры являются лишь иллюстративными и не предназначены для ограничения.
[0060] Для лучшего понимания настоящего изобретения приведены следующие примеры. Эти примеры предназначены только для целей иллюстрации и никоим образом не должны истолковываться как ограничивающие объем настоящего изобретения.
ПРИМЕРЫ
[0061] В приведенных ниже рабочих примерах авторы настоящего изобретения демонстрируют, что одноцепочечные антитела (Ab), слитые с Fc-доменом, сохраняющим связывание FcRn, но лишенным связывания Fc-гамма-рецептора (FcγR), называемые подвергнутым сайленсингу scFv-IgG, могут экспрессироваться и высвобождаться в ЦНС после переноса гена с помощью AAV. Посредством включения Fc в конструкцию scFv-IgG молекула восстанавливает бивалентность канонического IgG, что обеспечивает более высокую авидность в отношении мультимерных мишеней, таких как агрегированный амилоид, и обеспечивает возможность модулировать Fc-зависимую передачу сигнала, если это необходимо. Сохранение связывания Fc с FcRn на гематоэнцефалическом барьере может улучшить снижение проявления амилоидной патологии, наблюдаемое ранее при применении только scFv, за счет обеспечения клиренса комплекса антитело-антиген посредством опосредованного FcRn оттока из головного мозга. Хотя экспрессия канонического IgG в головном мозге приводила к признакам нейротоксичности, данное модифицированное антитело (Ab) эффективно секретировалось из нейрональных клеток и сохраняло специфичность в отношении мишени. Устойчивые уровни в головном мозге превышали пиковые уровни, полученные при внутривенной инъекции Ab. В модели прогрессирующего накопления амилоидных бляшек на трансгенных мышах ThyAPPmut экспрессия данного scFv-IgG в AAV снижает нагрузку бляшками в коре и гиппокампе по сравнению с контролем. Эти данные свидетельствуют о том, что доставка в ЦНС подвергнутого сайленсингу гена scFv-IgG к Aβ хорошо переносится, при это ген длительно экспрессируется и функционирует в соответствующей модели заболевания, демонстрируя потенциал данного способа для лечения болезни Альцгеймера и других неврологических заболеваний.
[0062] Материалы и способы, используемые в исследованиях, описанных в следующих примерах, описаны ниже.
Схема исследования
[0063] Данное исследование было инициировано для конструирования IgG к Aβ для AAV-опосредованной доставки в головной мозг для лечения болезни Альцгеймера. Данные конструкции IgG разрабатывали и первоначально тестировали in vitro 2-4 раза для подтверждения надлежащей экспрессии, сборки и активности связывания антигена перед экспериментами in vivo. Размеры выборки для исследований на животных C57BL/6 или SCID устанавливали на основании вариабельности, наблюдаемой в предшествующих экспериментах по экспрессии трансгенов in vivo с применением стереотаксической доставки AAV, и определяли для каждого эксперимента. Исследования по тестированию экспрессии in vivo проводили 2-3 раза. Размер выборки для мышей ThyAPPmut для количественной оценки нагрузки амилоидными бляшками устанавливали с учетом ожидаемой вариабельности образования бляшек у разных животных. Основываясь на предшествующих исследованиях с применением данной линии, исследование эффективности проводили один раз при n ≥ 10 на группу. Животных случайным образом распределяли в каждую группу для всех исследований. Идентификация ROI для автоматического анализа изображений была проведена исследователями, "ослепленными" в отношении экспериментальных условий. Все исследования на животных проводили согласно соответствующим руководствам.
Конструкции AAV-IgG
[0064] Вариабельные области получали из антитела к Aβ либо из исходного мышиного 13C3 (для AAV-αAβ msIgG), либо из гуманизированных последовательностей (для AAV-αAβ IgG) (Schupf et al., PNAS (2008) 105:14052-7), как описано в патентных заявках WO2009/065054 и WO2010/130946 соответственно. Вектор экспрессии huIgG получали посредством вставки кодирующих последовательностей тяжелой цепи человеческого IgG4, содержащих две аминокислотные замены, описанные для снижения полумолекул (S241P) и эффекторных функций (L248E) (Reddy et al., J Imm. (2000) 164:1925-33), и легкой каппа-цепи в кассету с двумя промоторами (без необходимости в расщеплении последовательности пептида 2A, показанной на фиг. 1A. Для экспериментов, требующих каркасной области мышиного IgG1, использовали исходное антитело 13C3 (Vandenberghe et al., Sci Rep (2016) 6:20958) с добавлением мутации N297A в тяжелой цепи для снижения эффекторной функции. Вектор AAV-контрольный IgG кодировал антитело для изотипического контроля huIgG4 PE, которое нацеливается на антиген, отличный от антигена млекопитающего.
Конструкция ScFv-IgG
[0065] Показана конструкция scFv-IgG (фиг. 4A; SEQ ID NO: 8). Вкратце, вариабельные области легкой цепи и вариабельные области тяжелой цепи исходного антитела 13C3 к бета-амилоиду соединяли 3 повторами гибкого линкера G4S (SEQ ID NO: 2) с образованием VL-VH scFv. За последовательностью scFv следовал дополнительный глициновый линкер из 9 повторов (SEQ ID NO: 7) (Balazs et al., Nature (2011) 481:81-4), при этом последовательность scFv содержала нативный шарнир мышиного IgG1 и CH2- и CH3-домены, составляющие Fc-область scFv-IgG. Как и в случае AAV-αAβ msIgG, аспарагин 297 в Fc подвергали мутации в аланин (N297A) с ослаблением эффекторной функции (Chao et al., Immunol Invest. (2009) 38:76-92); Jefferis et al., Immunol Rev. (1998) 163:59-76). C-концевую эпитопную метку 6xHis (SEQ ID NO: 9) включали, чтобы облегчить очистку in vitro и выявление in vivo у мышей. Экспрессией scFv-IgG управляли с помощью кассеты экспрессии с промотором hCMV/hEF1a с полиА Tbgh.
Иммунная толерантность
[0066] Для индукции иммунной толерантности мышам в дни 0, 2 и 10 путем инъекции вводили 7,5 мг/кг IP моноклонального антитела GK1.5 к CD4 (Bioxcell). Для подтверждения истощения CD4 Т-клеток на 12-й день отбирали кровь посредством ретроорбитального забора крови в пробирки, покрытые гепарином. Количество CD4+ Т-лимфоцитов определяли с применением FACS-анализа на устройстве BD Fortessa с применением стандартных протоколов с антителами CD45-FITC (клон 104 BD Pharmigen™), CD3e-AlexaFluor 647 (клон 17A2, eBioscience) и CD4-PE (клон RM4-4, BioLegend). У животных, обработанных с помощью GK1.5, наблюдалось снижение уровня CD4, о чем свидетельствует соотношение CD4+ лимфоциты/общее количество CD3+ лимфоцитов, составляющее 0,04 +/- 0,008 (среднее значение +/-SEM) у обработанных мышей по сравнению с 0,47 +/- 0,003 у необработанных мышей.
Культура клеток, экспрессия и очистка белков
[0067] Клетки Expi293™ (Life Tech) пассировали в среде Expi293T™ без сыворотки крови (Life Tech) и использовали для экспрессии белка. Плазмидами экспрессии трансфицировали клетки Expi293™ посредством липидной трансфекции (Fectopro, Polyplus) и среду для культивирования клеток, содержащую секретированный белок, собирали через 4 дня. После стерильной фильтрации меченные с помощью 6xHis (SEQ ID NO: 9) белки очищали посредством аффинной хроматографии с иммобилизованным металлом (IMAC). Вкратце, белки периодически адсорбировали на кобальтовой смоле (Thermo Scientific™) в течение ночи при 4°C, промывали 10 объемами колонки фосфатно-солевого буферного раствора, затем элюировали 500 мМ имидазолом. Белки подвергали диализу в забуференном с помощью HEPES солевом растворе в течение ночи, концентрировали (Centricon®) и замораживали при -80°C до применения.
ELISA
[0068] 96-луночные планшеты Immulon™ IIHB (Thermo) покрывали либо с помощью Aβ1-42 в концентрации 1 мкг/мл (Bachem H-1368) для ELISA антигена, либо мышиным поликлональным Ab к huIgG в концентрации 1 мкг/мл (Jackson 209-005-088) для захвата общего huIgG в карбонатном буфере в течение ночи при 25°C. Лунки промывали 5X с помощью TBS-0,5% tween (TBST) и блокировали в TBSTB (TBST+1,5% BSA) в течение 1 ч. Стандартные кривые с применением очищенного белка строили параллельно для образцов сыворотки крови или гомогенатов головного мозга для обеспечения количественного определения связанного scFv-IgG или huIgG. Образцы инкубировали в течение 2,5 ч, промывали 3X в TBST и затем инкубировали с вторичным антителом, конъюгированным с HRP, в течение 1 ч. После 5X промывки c помощью TBST лунки инкубировали с субстратом TMB в течение 5 мин перед гашением с помощью 0,5 М H2SO4. Сигнал связывания с планшетом количественно определяли по поглощению при 450 нм (Spectramax M5). Все образцы анализировали в трех повторностях.
LC-MS/MS
[0069] Эксперименты с применением LC/MS/MS проводили на масс-спектрометре Q Exactive™ (Thermo Scientific™) в сочетании с системой NanoAcQuity LC (Waters). IgG из гомогенатов тканей специфически обогащали и выделяли с применением аффинных смол CaptureSelect™ для HuIgG (Thermo Fisher). Обогащенные IgG расщепляли посредством инкубации с трипсином/Lys-C (1:100 вес/вес) в течение ночи при 37°C после восстановления с помощью DTT и алкилирования. Расщепление прекращали посредством добавления 1% муравьиной кислоты (FA). Полученные смеси триптических пептидов загружали в микрокапиллярную колонку (id 75 мкм, HSST3 15 см, 1,8 мкм, Waters) и разделяли. Данные получали в режиме PRM с разрешением 70000 (при соотношении масса/заряд 200), мишенью AGC 5 × 106 и максимальным временем ввода 500 мс. Запланированный перечень включения создавали на основании данных профилирования контрольных IgG. В способе PRM использовали выделение ионов-мишеней посредством окна выделения 2 Да, фрагментированных с нормализованной энергией соударения (NCE), составляющей 25. Сканы MS/MS получали с начальным диапазоном массы с соотношением масса/заряд, составляющим 100, и получали в виде типа данных профильного спектра. Определяли количество ионов-предшественников и фрагментов ионов с применением Skyline (MacCoss Lab Software).
Поверхностный плазмонный резонанс
[0070] Пептид Aβ1-42 (Bachem H-1368) инкубировали в 10 мМ HCl при 1 мг/мл в течение ночи при 37°C со встряхиванием при 600 об./мин. Полученный раствор фибрилл непосредственно подвергали иммобилизации на сенсорном чипе CM5 (GE Healthcare) с применением связывания с аминами. Растворы антител или scFv-IgG, полученные при 50, 30, 20, 10 и 5 нМ в буфере PBS-+P (GE Healthcare), вводили при относительно высокой скорости потока (50 мкл/мин) для ограничения эффектов авидности. Данные обрабатывали с применением программного обеспечения для оценки Biacore™ T200 и два раза сопоставляли посредством вычитания незанятой поверхности и введения только буфера перед общей подгонкой данных к модели связывания 1:1.
Плазмиды AAV ITR и получение аденоассоциированного вирусного вектора
[0071] Кассеты экспрессии для IgG или scFv-IgG субклонировали в плазмиду, содержащую AAV2-ITR, с ДНК-наполнителем A1AT, оставляли по мере необходимости для поддержания размера генома AAV для надлежащей упаковки. В случае плазмиды IgG ITR с двумя промоторами не включали ДНК-наполнитель, поскольку размер кассеты уже был максимально допустимым для эффективной упаковки. AAV-вектор без вставки состоял из промотора CBA, полиA Tbgh и ДНК-наполнителя A1AT. Вирус AAV2/1 получали посредством временной трансфекции. Вкратце, клетки HEK293 трансфицировали с применением PEI (полиэтиленимина) с соотношением трех плазмид (содержащих ITR, rep/cap AAV и хелперный элемент Ad), составляющим 1:1:1. Хелперную плазмиду Ad (pHelper) получали от Stratagene/Agilent Technologies (Санта-Клара, Калифорния). Очистку проводили с применением колоночной хроматографии, как описано ранее (Burnham et al., Hum Gene Ther Methods (2015) 26:228-42). Вирус титровали с применением qPCR против последовательности полиА и AAV хранили в 180 мМ хлориде натрия, 10 мМ фосфате натрия (5 мМ одноосновный+5 мМ двухосновный), 0,001% F68, pH 7,3, при -80°C до применения.
Животные
[0072] Применяемые животные представляли собой самцов C57BL/6, полученных из Jackson Labs (Бар-Харбор, США), в возрасте 2 месяцев, если не указано иное. Взрослых мышей SCID получали из Jackson Labs (B6.CB17-Prkdcscid/SzJ) в возрасте 2 месяцев. Трансгенные мыши ThyAPPmut, скрещенные с C57BL/6, описаны в Blanchard et al., Exp Neurol. (2003) 184:247-63. Хирургические группы размещали отдельно, чтобы обеспечить надлежащее восстановление после хирургических операций на головном мозге. Мышей содержали при 12-часовом цикле чередования света и темноты с пищей и водой, доступными в неограниченном количестве. Животных рандомизировали в разные группы и анализы проводили с операторами, ослепленными в отношении групп обработки.
Стереотаксические инъекции
[0073] Хирургическую операцию проводили в соответствии с процедурами, утвержденными комитетом по уходу за животными и их использованию. Мышей подвергали глубокой анестезии путем интраперитонеальной инъекции смеси (объем составляет 10 мл/кг) кетамина (100 мг/кг; Imalgene; Merial, Франция) и ксилазина (10 мг/кг; Rompun; Bayer, Франция). Перед помещением животного в стереотаксическую рамку (Kopf Instruments, США) кожу черепа мыши брили и дезинфицировали ветидином (Vetoquinol, Франция), местный анестетик бупивакаин (2 мг/кг в объеме, составляющем 5 мл/кг; Aguettant, Франция) вводили путем подкожной инъекции под кожу черепа и наносили Emla (Lidocaïne, Astrazeneca) на уши. Во время хирургической операции глаза защищали от света с помощью витамина A Dulcis и поддерживали постоянную температуру тела, составляющую 37°C, с помощью обогревающего одеяла.
[0074] Образцы вводили со скоростью 0,5 микролитров в минуту. Иглу оставляли на 2 мин, чтобы предотвратить обратный отток образца через тракт иглы, а затем медленно вынимали из головного мозга. Выполняли односторонние инъекции в гиппокамп мышей ThyAPPmut или двусторонние инъекции всем другим мышам. Координаты инъекций в гиппокамп представляли собой: AP -2,0, DV -2,0 и ML +/-1,5. Мышей держали в тепле, и они получали подкожную инъекцию карпрофена (5 мг/кг в объеме 5 мл/кг, Rimadyl®, Zoetis) после хирургической операции и их непрерывно наблюдали до восстановления. В конце исследования мышей подвергали эвтаназии посредством передозировки анестетика с Euthasol® (США) или кетамином/зилазином (Франция). После передозировки мышей держали в тепле до перфузии ледяным PBS.
Иммуногистохимический анализ
[0075] После перфузии холодным PBS ткань головного мозга фиксировали в 10% нейтральном забуференном формалине (NBF). Ткань, фиксированную в формалине, заключали в парафин, затем делали срезы толщиной 5 мкм в сагиттальной или коронарной плоскости. Все ткани окрашивали с применением красителя для автоматического окрашивания Leica BOND RX. Для иммунофлуоресцентного окрашивания выполняли опосредованное нагреванием демаскирование антигена с применением раствора 1 для демаскирования эпитопа (ER1; цитратный буфер, pH 6,0) в течение 10 мин. Затем ткань блокировали/повышали проницаемость ткани в козьей сыворотке крови+0,25% triton Х-100, затем инкубировали с первичными антителами в течение 1 ч при RT, промывали в TBST, затем инкубировали со вторичными антителами в течение 30 мин. Ядра выявляли с применением Spectral DAPI (Life). Для количественной оценки бляшек ткань подвергали иммуноокрашиванию с помощью конъюгированного с биотином антитела 4G8 (клон 4G8, BioLegend 800701) с применением набора Vectastain® ABC (PK-7100) согласно инструкциям производителя без демаскирования антигена или экстракции муравьиной кислотой.
Антитела
[0076] 6xHis (SEQ ID NO: 9) (Abcam Ab9108, 1:1000 IHC, Invitrogen™ R931-25, 1:1000 Western, ELISA) GFAP (Ebiosciences, 41-9892-82, 1:200, или Abcam Ab4674, 1:500 IHC) 4G8 (BioLegend 800701, 1:500 IHC). Вторичные антитела от Life Technologies: козье антитело Cy3 к мышиному антителу, козье антитело Alexa Fluor®647 к кроличьему антителу, козье антитело Alexa Fluor®488 к куриному антителу; все в соотношении 1:500. Для окрашивания амилоида с применением DAB: 4G8-биотин (BioLegend 800705, 1:250).
Анализ изображений
[0077] Предметные стекла для иммуногистохимического анализа сканировали при 20X увеличении с применением светлопольного сканера изображений Scanscope® XT (Aperio, Виста, Калифорния) или AxioScanZ1 (Carl Zeiss Microscopy GmBH, Германия). Полные изображения предметных стекол (WSI) после IHC GFAP просматривали и анализировали с применением программного обеспечения для анализа изображений HALO™ (Indica Labs, Корралес, Нью-Мексико, США). Для каждого WSI область гиппокампа вручную аннотировали и анализировали в отношении иммуноположительной области GFAP с применением алгоритма автоматического количественного анализа площади HALO. Для каждого образца GFAP-положительную площадь разделяли на общую площадь ткани для выбранной ROI, чтобы получить процент иммуноположительной площади. Для анализа бляшек у мышей ThyAPPmut в возрасте 6 месяцев собирали корональные срезы головного мозга толщиной 5 мкм на трех разных уровнях на расстоянии 50 мкм друг от друга. ROI коры и гиппокампа аннотировали вручную. Нагрузку амилоидными бляшками количественно определяли как % DAB+площадь ткани с применением специального алгоритма анализа изображений, разработанного с применением программного обеспечения ZEN 2 (Carl Zeiss Microscopy GmBH, Германия). Данные наносили на график с применением GraphPad Prism версии 6 (программное обеспечение GraphPad, Ла-Хойя, Калифорния, США).
Статистические анализы
[0078] Статистический анализ выполняли с помощью Graphpad Prism (v6 и v7) с применением 1-факторного ANOVA с множественными сравнениями (критерий Даннетта) для экспериментов с более чем двумя группами. Для сравнения двух групп использовали непарный t-критерий Стьюдента. *p < 0,05, **p < 0,01, ***p < 0,001. Размер выборки варьировался и был разным для каждого эксперимента.
Пример 1. Конструирование и определение характеристик вектора AAV-IgG, нацеливающегося на β-амилоид
[0079] Для осуществления экспрессии гена, кодирующего антитело, авторы настоящего изобретения использовали кассету экспрессии с двумя промоторами для экспрессии гуманизированного варианта антитела 13C3, которое связывает протофибриллярный и фибриллярный Aβ без аффинности в отношении мономерных форм, как описано у Schupf, выше. Тяжелая цепь IgG4 содержала мутации S228P и L248E, которые снижают эффекторную функцию Fcγ и обмен полумолекул (Yang et al., Curr Opin Biotechnol. (2014) 30:225-9; Reddy et al., J Imm. (2000) 164:1925-33).
[0080] Тяжелые и легкие цепи экспрессировали с разных промоторов и всю кассету конструировали таким образом, чтобы не превысить предел упаковки генома AAV (фиг. 1A). Используемая в данном документе конструкция с двойным промотором позволяет избежать потенциальных недостатков, связанных с иммуногенностью и экспрессией, индуцируемых другими конструкциями, в которых используется один промотор, но требует применения расщепляющейся последовательности F2A или внутреннего сайта посадки рибосомы для бицистронной экспрессии (Saunders, выше; Mizuguchi et al., Mol Ther. (2000) 1:376-82). Эту кассету упаковывали в капсид AAV1 (AAV-αAβ IgG) для прямой инъекции в головной мозг, так как этот серотип демонстрирует превосходное паренхиматозное распространение, и хотя преобладает нейрональная трансдукция (как в случае большинства векторов на основе AAV), этот серотип также трансдуцирует астроциты, которые могут быть более подвержены высокому уровню экспрессии и секреции белка. Для тестирования экспрессии AAV-αAβ IgG использовали мышей C57BL/6-SCID (SCID) для предотвращения иммунных ответов против huIgG, которые могут препятствовать экспрессии трансгена. Антитело активно транспортируется из головного мозга посредством обратного трансцитоза. Поэтому авторы настоящего изобретения контролировали экспрессию AAV-αAβ IgG в головном мозге с применением сбора сыворотки крови один раз в две недели. Образцы сыворотки крови отбирали с 2-недельными интервалами в течение 16 недель после двусторонней инъекции AAV-αAβ IgG в гиппокамп (2E10 на сторону, с применением GC) мышей SCID. Иммуноанализ связывания с фибриллами Aβ1-42 использовали для измерения уровней экспрессируемого функционального антитела после двусторонней инъекции 2E10 AAV-αAβ IgG в гиппокамп с применением GC.
[0081] Вектор демонстрировал стабильную экспрессию до 16 недель (фиг. 1B, слева). Чтобы получить представление о том, как экспрессия AAV-опосредованного антитела в головном мозге сравнивается с уровнями, наблюдаемыми при стандартном подходе пассивной иммунотерапии, уровни huIgG в гиппокампе мышей SCID измеряли в разные моменты времени параллельно с отдельной группой, которая получала одну внутривенную (IV) болюсную инъекцию 20 мг/кг αAβ IgG. Мышам SCID один раз путем инъекции вводили 2E10 AAV-αAβ IgG с применением GC двусторонне в гиппокамп или один раз внутривенно вводили 20 мг/кг очищенного IgG перед сбором ткани в указанные моменты времени с получением динамики воздействия IgG на головной мозг. Образцы ипсилатерального гиппокампа гомогенизировали и анализировали в отношении huIgG посредством ELISA антигена. Вектор AAV-αAβ IgG поддерживал экспрессию в гиппокампе на уровне почти 300 нг/г в течение периода времени, как измерено с помощью ELISA антигена (фиг. 1B, справа). Уровни IgG в гиппокампе через 24 ч после внутривенной инъекции приближались к 200 нг/г, но эти уровни снижались по мере того, как IgG выводился из головного мозга (в соответствии с известным периодом полужизни в сыворотке крови), что приводило к 11-кратному снижению по сравнению с AAV-αAβ IgG через 7 недель.
[0082] На фиг. 1C показано, что внутринейрональная и глиальная экспрессия AAV-IgG выявлялась в гиппокампе. В частности, экспрессию как в нейронах, так и в астроцитах подтверждали посредством IHC в отношении продукта экспрессии huIgG, при этом нейроны легко идентифицировали по морфологии в CA2 гиппокампа и совместной локализации с GFAP, указывающей на экспрессию в астроцитах (фиг. 1C).
[0083] Эти данные демонстрируют, что вектор AAV-αAβ IgG способен поддерживать стабильные уровни антитела в головном мозге, значительно превышающие уровни, которые могут быть достигнуты с применением традиционных протоколов пассивной иммунотерапии.
Пример 2. Связывание антигена посредством AAV-αAβ IgG на мышиной модели болезни Альцгеймера
[0084] Затем авторы настоящего изобретения экспрессировали AAV-αAβ IgG на мышиной модели амилоидных бляшек с экспрессией мутантного белка-предшественника амилоида (ThyAPPmut), чтобы оценить степень трансдукции головного мозга и определить, секретируется ли антитело во внеклеточное пространство для связывания бляшек in vivo. Данная модель демонстрирует прогрессирующее накопление амилоидных бляшек в коре, начиная с возраста примерно 2-3 месяца (Blanchard et al., Exp Neurol. (2003) 184:247-63). Чтобы предотвратить ответы против антитела huIgG животным повышали иммунотолерантность с применением CD4-истощающего антитела до и после введения вектора (фиг. 2A). Вкратце, чтобы легко выявить IgG у мышей, авторы настоящего изобретения самцам мышей ThyAPPmut в возрасте 2 месяца внутрь гиппокампа вводили AAV-αAβ IgG или AAV, экспрессирующий изотипический контрольный IgG (контрольный AAV-IgG). Мышам ThyAPPmut повышали иммунотолерантность за счет истощения CD4 Т-клеток между днями 2-10. AAV-αAβ IgG или вектор AAV-IgG для изотипического контроля путем инъекции двусторонне вводили в гиппокамп (2E10 на инъекцию, с применением GC) в дни 4-5. Отдельной группе один раз в неделю интраперитонеально путем инъекции вводили очищенный αAβ huIgG при 10 мг/кг в течение всего исследования в качестве положительного контроля активности связывания бляшек. Через 8 недель собирали и подвергали иммуноокрашиванию сагиттальные срезы головного мозга толщиной 5 мкм. Ранее было показано, что эта доза αAβ IgG и парадигма интраперитонеальной доставки приводят к связыванию бляшек in vivo у животных ThyAPPmut (Pradier et al., Alzheimer's & Dementia (2013) 9(4):P808-P809).
[0085] Через два месяца после инъекции, в возрасте, когда у этих животных наблюдается отложение бляшек в лобной коре, сагиттальные срезы головного мозга обрабатывали для IHC. В частности, IHC-окрашивание huIgG позволило выявить экспрессию по всему гиппокампу и вышележащей коре, окружающей след иглы. Увеличенные представляющие интерес области (ROI) (ширина 500 мкм) демонстрируют детальную экспрессию huIgG в нейронах и в нейропиле гиппокампа. Напротив, группа, которой интраперитонеально вводили αAβ IgG с окрашиванием, ограниченным амилоидными бляшками, не проявляла никакой экспрессии в телах клеток (фиг. 2B, слева). Флуоресцентный IHC-анализ в отношении huIgG, бляшек Aβ и GFAP демонстрировал совместную локализацию huIgG с кортикальными бляшками как в группах AAV-αAβ IgG, так и в группах с внутривенным введением αAβ IgG, но не в контрольной группе AAV-IgG. В частности, AAV-αAβ IgG и периферически доставленный IgG к αAβ демонстрировали точно подтвержденное связывание с отложениями 4G8+ амилоида, в то время как контрольный AAV-IgG не демонстрировал выявляемого связывания (фиг. 2B, справа).
[0086] Эти данные демонстрируют, что AAV-αAβ IgG секретировался во внеклеточное пространство и был способен связываться с бляшками Aβ в областях головного мозга, отдаленных от места инъекции.
Пример 3. Оценка нейрональной экспрессии и нейротоксичности AAV-αAβ IgG
[0087] Нейрональные клетки являются высокоспециализированными в отношении секреции факторов, связанных с нейротрансмиссией, а не крупных макромолекул, таких как IgG. Неизвестно, могут ли в этих клетках происходить эффективный процессинг и секреция IgG. Чтобы определить имеет ли место ненадлежащий процессинг нейронально экспрессируемого IgG, авторы настоящего изобретения выполняли масс-спектрометрический анализ с измерением общих уровней тяжелых и легких цепей в образцах головного мозга через 1 месяц экспрессии AAV-αAβ IgG у мышей SCID. Экспрессия AAV-αAβ IgG в гиппокампе была связана с ожидаемыми уровнями тяжелой цепи - аналогично лизатам головного мозга, введенным в головной мозг с физиологическим раствором с добавлением очищенного αAβ IgG, но с низким количеством когнатной легкой цепи по сравнению с добавленным контролем (фиг. 3A). Это открытие свидетельствует о том, что экспрессия AAV-αAβ IgG в клетках головного мозга приводит к недостаточному продуцированию легких цепей, что приводит к дисбалансу в соотношении тяжелых и легких цепей.
[0088] Авторы настоящего изобретения также использовали ELISA для количественного определения общего IgG (H+L-цепи) по сравнению с процентом в данной группе, которая способна связывать антиген (Ag). В частности, уровни функционального антитела к Aβ в экстрактах головного мозга мышей SCID, экспрессирующих AAV-αAβ IgG, количественно оценивали посредством ELISA антигена и сравнивали параллельно с ELISA пан-huIgG. Авторы настоящего изобретения обнаружили, что ~20% общего IgG, экспрессируемого в головном мозге (всего 2E10 при инъекции в гиппокамп с применением GC), были функциональными, в то время как AAV-αAβ IgG, экспрессируемый в периферических тканях после внутривенной инъекции вектора (всего 1E12 при внутривенной инъекции с применением GC), не характеризовался дисбалансом соотношения общий IgG/функциональный IgG. В частности, уровни huIgG, связанного с антигеном, составляли только 21% от общего huIgG при экспрессии в головном мозге, тогда как это несоответствие не было выявлено в образцах сыворотки крови через один месяц после периферической экспрессии вектора (фиг. 3A, справа).
[0089] Затем авторы настоящего изобретения исследовали, есть ли доказательства нейротоксичности в результате экспрессии IgG. Для начальных процедур определения характеристик вектора AAV-αAβ IgG авторы настоящего изобретения использовали вариант huIgG данного антитела, который характеризуется более прямым потенциалом трансляции в случае людей и позволяет осуществлять точное выявление у мышей. Однако для тестирования в отношении любой токсичности или нейровоспаления, которые могут быть связаны с экспрессией IgG в головном мозге без вклада переменной воздействия ксеногенного huIgG, авторы настоящего изобретения использовали вектор на основе AAV, названный AAV-αAβ msIgG, который экспрессирует исходный мышиный вариант αAβ IgG (Schupf, выше; Pradier, выше; Vandenberghe et al., Sci Rep. (2016) 6:20958). Данный вектор путем инъекции вводили в гиппокамп мышей C57BL/6 и через один месяц ткань головного мозга обрабатывали для гистологического анализа. Гистопатологический анализ позволил выявить высокую частоту образования гиалиновых/эозинофильных цитоплазматических отложений в нейрональных клетках в гиппокампе, что напоминает сверхэкспрессию гликопротеина (фиг. 3B). Нейрональные, эозинофильные или гиалин-подобные включения, напоминающие накопление гликопротеина, наблюдались только в образцах головного мозга после инъекции вектора для экспрессии антитела. Эти структуры также наблюдали в гиппокампе мышей, которым путем инъекции вводили контрольный AAV-IgG (6/12 мышей), что указывает на то, что данная токсичность не была специфической в отношении экспрессии IgG к αAβ. Эти гиалиновые отложения никогда не наблюдались в гиппокампе мышей, которым путем инъекции вводили вектор на основе AAV1 без вставки или только PBS (фиг. 3B).
[0090] Авторы настоящего изобретения также наблюдали доказательства нейровоспаления посредством иммуногистохимического анализа GFAP по сравнению с PBS. В данном эксперименте мышам C57BL/6 путем инъекции вводили либо PBS, либо AAV-αAβ msIgG (2E10, с применением GC, в гиппокамп), и через 16 недель собирали сагиттальные срезы головного мозга толщиной 5 мкм (фиг. 3C). Вектор на основе AAV1 без вставки также не вызывал значительного глиоза по сравнению с PBS (1,11 +/- 0,12, 5 мышей, среднее значение +/- SEM, нормализованное к площади GFAP+ PBS), что позволяет предположить, что нейровоспаление было вызвано экспрессией IgG.
[0091] Эти данные демонстрируют, что хотя клетки головного мозга способны экспрессировать и секретировать IgG, только подгруппа из приблизительно 20% данного IgG является функциональной и способна связывать антиген, и данная экспрессия индуцирует выявляемое нейровоспаление по всей трансдуцированной области.
Пример 4. Конструирование и определение характеристик вектора AAV-scFv-IgG
[0092] В то время как IgG, доставленный с помощью вектора, разработанного авторами настоящего изобретения, секретировался и связывал амилоидные бляшки in vivo, авторы настоящего изобретения выдвинули гипотезу, что альтернативный формат Ig способен минимизировать неправильное спаривание и нейротоксичность, индуцированные AAV-IgG. На основе того же мышиного антитела к αAβ (Schupf, выше) авторы настоящего изобретения синтезировали модифицированный одноцепочечный Fv-фрагмент с вариабельной областью легкой цепи IgG, слитой с вариабельной областью тяжелой цепи, COOH-конец (C-конец) которой был соединен с шарниром мышиного IgG1, CH2- и CH3-доменами (фиг. 4A; scFv-IgG). Чтобы минимизировать провоспалительные эффекты Fc-области Fc-домен мышиного IgG1 подвергали мутации для устранения гликозилирования по аспарагину 297 (N297A), что предотвращает связывание со всеми FcγR (Johnson, выше; Chao, выше). В частности, scFv-IgG конструировали таким образом, чтобы он содержал вариабельные области мышиного IgG к Aβ, связанные 3 повторами гибкой линкерной последовательности GGGGS (SEQ ID NO: 3). ScFv соединяли с Fc мышиного IgG1 N297A посредством линкера с 9 повторами Gly (SEQ ID NO: 7). Метку 6xHis (SEQ ID NO: 9) добавляли к C-концу. ScFv-IgG экспрессировали в клетках Expi293 и очищали посредством аффинной хроматографии с иммобилизованным металлом (IMAC) с применением последовательности С-концевой гистидиновой (His) метки.
[0093] Анализ посредством SDS-PAGE позволил подтвердить, что данный белок эффективно собирается в димер, связанный дисульфидной связью (фиг. 4A). Данный scFv-IgG проявлял связывание с фибриллярным Aβ1-42 по результатам поверхностного плазмонного резонанса (SPR), сравнимое с исходным антителом. Аффинность (M) определяли посредством SPR, пропуская scFv-IgG или IgG через иммобилизованные фибриллы Aβ1-42 при различных молярных концентрациях для анализа кинетики связывания. Исходный IgG проявлял кажущуюся константу диссоциации (KD), составляющую 1,3×10-10 М, по сравнению с несколько более низкой аффинностью связывания с scFv-IgG, составляющей 5,2×10-10 (фиг. 4A, таблица).
[0094] Данную кассету экспрессии вставляли в вектор на основе AAV1, чтобы определить, можно ли синтезировать модифицированный IgG in vivo. Внутривенную инъекцию AAV использовали в качестве положительного контроля активности вируса, разработанного авторами настоящего изобретения, поскольку периферические ткани тщательно проверены в отношении экспрессии и секреции молекул IgG (Saunders, выше; Shimada et al., PloS ONE (2013) 8:e57606; Hicks et al., Sci Transl Med. (2012) 4:140ra187; Chen et al., Sci Rep. (2017) 7:46301; Balazs et al., Nature (2011) 481:81-4; Balazs et al., Nat Biotech. (2013) 31:647-52; Balazs et al., Nat Med. (2014) 20:296-300). Через месяц после внутривенной инъекции AAV-scFv-IgG (всего 1E12, с применением GC) уровни в сыворотке крови достигли 63 мкг/мл, демонстрируя устойчивую активность вектора AAV в периферических тканях (фиг. 4B, слева).
[0095] Для оценки экспрессии вектора в головном мозге уровни scFv-IgG количественно определяли в экстрактах, полученных из одной сагиттальной половины головного мозга, называемой полушарием, через один месяц после инъекции в гиппокамп всего 2E10 AAV мышам C57BL6 с применением GC. Уровни экспрессии достигали среднего значения ~ 600 нг/г (фиг. 4B, справа). Примечательно, что данная концентрация была более чем в 3 раза выше, чем наблюдаемая через 24 ч после внутривенной инъекции 20 мг/кг IgG, и в 2,5 раза выше, чем наблюдаемая для AAV-αAβ IgG (фиг. 1B). Гистологический анализ продемонстрировал, что несмотря на более высокие уровни экспрессии в головном мозге, чем у вектора AAV-αAβ IgG, трансдукция с помощью AAV-scFv-IgG не вызывала какого-либо выявляемого внутринейронального накопления белка гиалина в гиппокампе при введении путем инъекции (0/5 мышей), что позволяет предположить, что scFv-IgG более эффективно процессировался нейрональными клетками, чем IgG.
[0096] Для определения распределения в головном мозге клеток, трансдуцированных с помощью scFv-IgG, выполняли IHC с применением DAB-6xHis ("6xHis" раскрыта под SEQ ID NO: 9) на сагиттальных срезах через один месяц после инъекции в гиппокамп с применением антитела к His-метке. Вектор AAV-scFv-IgG трансдуцировал весь гиппокамп с редкой трансдукцией в области коры, лежащей над гиппокампом, вокруг следа иглы и субикулума (фиг. 4C). Образцы головного мозга, трансдуцированного векторами на основе AAV без вставки (контрольный AAV) в качестве отрицательного контроля, не демонстрировали выявляемого иммуноокрашивания в отношении His (фиг. 4C). Следует отметить, что IHC в отношении His у мышей C57BL6 позволяет выявить только внутриклеточную экспрессию, поскольку любой секретируемый внеклеточный scFv-IgG, вероятно, вымывается вследствие отсутствия доступного антигена.
[0097] Экспрессию как внутриклеточного, так и внеклеточного scFv-IgG биохимически оценивали в ипсилатеральных областях головного мозга как проксимальнее, так и дистальнее места инъекции. Через месяц после инъекции AAV области головного мозга 3 мышей вырезали и определяли количество экспрессированного белка посредством ELISA антигена для каждой области головного мозга с применением гомогената головного мозга после инъекции PBS для вычитания фонового сигнала. В частности, гиппокамп, вышележащую кору и полосатое тело вырезали и гомогенизировали для количественного определения scFv-IgG посредством ELISA антигена (фиг. 4C, справа). Наблюдали градиент концентрации, при этом самые высокие уровни выявлялись в месте инъекции (гиппокамп), и постепенно более низкие уровни наблюдались в более дистальных областях головного мозга (фиг. 4C, справа). Несмотря на более низкие уровни по сравнению с уровнями в месте инъекции, концентрация scFv-IgG в ткани полосатого тела оставалась на уровне около 200 нг/г - стабильные уровни в головном мозге, которые обычно не достигаются при пассивной инфузии IgG.
Пример 5. Связывание антигена с помощью scFv-IgG на мышиной модели β-амилоидоза
[0098] Затем авторы настоящего изобретения определили, секретируется ли доставленный с помощью AAV scFv-IgG во внеклеточное пространство и способен ли он связываться с антигеном in vivo. Вектор AAV-scFv-IgG путем инъекции вводили в гиппокамп самок мышей ThyAPPmut в возрасте 5 месяцев (Blanchard, выше), в возрасте, когда у них уже образовались бляшки по всей новой коре. Сагиттальные срезы образцов головного мозга толщиной 5 мкм обрабатывали для IHC через один месяц после односторонней инъекции 1 мкл (всего 1E10, с применением GC) вектора AAV-scFv-IgG и окрашивали в отношении реактивности по отношению к His-метке и бляшкам Aβ. Изображения справа демонстрируют ROI отдельных бляшек (пронумерованные в A) от проксимальной (1) до дистальной (6) от места инъекции. Изображения накладывали на изображения иммуноокрашивания с применением 6xHis (SEQ ID NO: 9) (зеленый) и DAPI (синий) (фиг. 5A). Как и ожидалось, обильное образование бляшек наблюдалось по всей коре (фиг. 5A, слева), и окрашивание с помощью антитела к His локализовано вместе с бляшками (фиг. 5A, справа). Обратите внимание на прогрессирующее, но достигающее дальних областей снижение интенсивности мечения с применением 6xHis (SEQ ID NO: 9) на бляшках, которые расположены более дистально по отношению к гиппокампу и затылочным зонам коры с экспрессией AAV-scFv-IgG, что указывает на наличие четкого градиента концентрации связанного с бляшками scFv-IgG, при этом бляшки дистальнее гиппокампа прогрессивно демонстрируют более низкие уровни связанного scFv-IgG, чем бляшки, находящиеся ближе к месту инъекции. Эти данные указывают на то, что scFv-IgG к Aβ экспрессировался и секретировался из клеток в гиппокампе, что позволяло ему связываться с бляшками, находящимися дистальнее от места инъекции.
[0099] Эти данные свидетельствуют о том, что scFv-IgG, доставленный с помощью вирусного вектора, взаимодействует со своей физиологически релевантной мишенью in vivo. Затем авторы настоящего изобретения определили, может ли длительная экспрессия на данной мышиной модели амилоидоза снижать частоту образования бляшек. План схемы исследования. Четырем группам самцов мышей ThyAPPmut в возрасте 2 месяцев (приблизительный возраст, когда начинается образование бляшек) путем инъекции вводили в гиппокамп односторонне либо AAV-scFv-IgG, контрольный вектор на основе AAV, IgG к Aβ, либо контрольный изотип IgG. Данные группы сравнивали с животными, обработанными посредством пассивной иммунотерапии с еженедельными интраперитонеальными инъекциями мышиного антитела к Aβ (IgG к Aβ) или антитела для изотипического контроля при 10 мг/кг (фиг. 5B, слева). Образцы головного мозга собирали через 16 недель (4 месяца) лечения и корональные срезы подвергали иммуноокрашиванию в отношении амилоидных бляшек или 6xHis (SEQ ID NO: 9) и анализировали в отношении экспрессии трансгена. AAV-scFv-IgG экспрессировался по всему гиппокампу после инъекции, и также наблюдался четкий транспорт вектора в контралатеральный субикулум, о чем свидетельствует окрашивание тел клеток с применением αHis (фиг. 5B, справа). Нагрузку бляшками Aβ в коре и гиппокампе количественно определяли посредством IHC в отношении His в корональных срезах головного мозга. ROI из обоих полушарий объединяли для количественной оценки и нагрузку бляшками выражали в виде DAB-положительного окрашивания в процентах от площади ROI в ткани. По сравнению с их соответствующими контролями, однократная инъекция AAV-scFv-IgG вызывала снижение частоты образования бляшек в гиппокампе на такую же величину, что и IgG к αAβ в качестве стандарта, несмотря на различия в нагрузке бляшками между контрольными группами (фиг. 5C). Снижение частоты образования бляшек также было значительно снижено в коре (фиг. 5C), что согласуется с доказательствами того, что scFv-IgG диффундирует из места экспрессии, чтобы связываться с дистальными бляшками.
[00100] Эти результаты демонстрировали, что однократная инъекция AAV-scFv-IgG в амилоидной мышиной модели приводила к длительной экспрессии и секреции из места инъекции для связывания с бляшками по всему головному мозгу. Типичный режим пассивной иммунотерапии при 10 мг/кг IgG к Aβ в течение 16 недель вызывал значительное снижение частоты образования амилоидных бляшек у животных ThyAPPmut. Напротив, однократная внутричерепная инъекция AAV-scFv-IgG приводила к сопоставимой эффективности через 4 месяца экспрессии.
[00101] Подводя итог, в вышеупомянутом исследовании scFv-IgG получали из антитела, специфического в отношении протофибриллярных и фибриллярных разновидностей Aβ, которое снижало нагрузку амилоидными бляшками in vivo. scFv-IgG, разработанный авторами настоящего изобретения, хорошо экспрессируется in vitro, что позволяет проводить очистку и последующий анализ аффинности связывания антигена посредством SPR. По сравнению с формой IgG, scFv-IgG связывался с антигеном в сходной степени. AAV1 был выбран в качестве серотипа для данного показания, поскольку его капсид способствует распространению вектора в ЦНС после паренхиматозной инъекции. Данный серотип инфицирует преимущественно нейрональные клетки, но действительно трансдуцирует некоторые типы клеток, отличных от нейрональных, расширяя потенциальный репертуар клеток, доступных для экспрессии трансгена. При относительно высокой дозе (1E10 в гиппокамп, с применением GC) стабильные уровни в месте инъекции были в 3-4 раза выше, чем уровни, которые могли быть максимально достигнуты с применением пассивного IgG в качестве стандарта, который авторы настоящего изобретения выбрали для сравнения (уровни антител в головном мозге через 24 ч после внутривенной инъекции 20 мг/кг очищенного IgG). Для достижения уровней, достигаемых при применении вектора AAV-scFv-IgG, на периферии потребуется доза, составляющая ~ 60 мг/кг внутривенно.
[00102] После однократной инъекции экспрессия в гиппокампе сохранялась в течение по меньшей мере 4 месяцев, при этом концентрации белка превышали концентрации при пассивной иммунотерапии в качестве стандарта даже в областях головного мозга на несколько миллиметров дистальнее места инъекции. Маловероятно, что трансдуцированные клетки мигрируют из места инъекции, чтобы секретировать белок в областях, дистальных по отношению к гиппокампу, так как клетки, положительные в отношении 6xHis (SEQ ID NO: 9), не были обнаружены далеко за пределами места инъекции или тракта иглы (данные не показаны). Длительная экспрессия данного вектора у мышей ThyAPPmut вызывала снижение частоты образования бляшек как в коре (снижение на 52%), так и в гиппокампе (снижение на 87%). Это было более эффективное снижение, чем наблюдаемое в других исследованиях с применением scFv, где снижение частоты образования бляшек составляло от 0 до 60% (Levites, 2006, выше; Levites, 2015, выше; Kou, выше; Fukuchi, выше, и Wang et al., Brain, Behavior, and Immunity (2010) 24:1281-93). Наблюдаемая величина снижения частоты образования бляшек у животных, обработанных посредством однократной внутричерепной инъекции AAV-scFv-IgG, была сходной с величиной у животных, обработанных посредством внутривенных инъекций 10 мг/кг антитела к Aβ один раз в неделю в течение 4 месяцев, что подчеркивает ценность доставки генов в случае парадигм долговременного лечения.
ПОСЛЕДОВАТЕЛЬНОСТИ
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> САНОФИ
<120> ЭКСПРЕССИЯ АНТИГЕНСВЯЗЫВАЮЩИХ БЕЛКОВ В НЕРВНОЙ СИСТЕМЕ
<130> 022548.WO060
<140>
<141>
<150> EP 19306310.4
<151> 2019-10-08
<150> 62/848,659
<151> 2019-05-16
<160> 9
<170> PatentIn version 3.5
<210> 1
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 1
Gly Gly Gly Ser
1
<210> 2
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 2
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10 15
<210> 3
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 3
Gly Gly Gly Gly Ser
1. 5
<210> 4
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> САЙТ
<222> (1)..(20)
<223> /примечание="Эта последовательность может охватывать 1-4 повторяющихся звена
'Gly Gly Gly Gly Ser'"
<400> 4
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1. 5 10 15
Gly Gly Gly Ser
20
<210> 5
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 5
Ser Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10 15
<210> 6
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<220>
<221> ВАРИАНТ
<222> (10)..(10)
<223> /замена="Ala" или "Asn"
<220>
<221> ВАРИАНТ
<222> (15)..(15)
<223> /замена="Asn"
<220>
<221> SITE
<222> (1)..(20)
<223> /примечание="Вариантные остатки, приведенные в последовательности,
не являются предпочтительными по отношению остаткам, к указанным в
аннотациях для вариантных положений"
<400> 6
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ala Gly
1. 5 10 15
Gly Gly Gly Ser
20
<210> 7
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический пептид"
<400> 7
Gly Gly Gly Gly Gly Gly Gly Gly Gly
1. 5
<210> 8
<211> 530
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический полипептид"
<400> 8
Met Asp Ser Lys Gly Ser Ser Gln Lys Gly Ser Arg Leu Leu Leu Leu
1. 5 10 15
Leu Val Val Ser Asn Leu Leu Leu Pro Gln Gly Val Leu Ala Ser Glu
20 25 30
Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly Asp
35 40 45
Arg Ala Ser Ile Ser Cys Arg Ser Gly Gln Ser Leu Val His Ser Asn
50 55 60
Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser Pro
65 70 75 80
Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro Asp
85 90 95
Arg Phe Ser Gly Ser Gly Ser Gly Ser Asp Phe Thr Leu Thr Ile Ser
100 105 110
Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Asn Thr
115 120 125
Phe Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
130 135 140
Thr Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
145 150 155 160
Gly Ser Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Val Val Lys Pro
165 170 175
Gly Val Ser Val Lys Ile Ser Cys Lys Gly Ser Gly Tyr Thr Phe Thr
180 185 190
Asp Tyr Ala Met His Trp Val Lys Gln Ser Pro Gly Lys Ser Leu Glu
195 200 205
Trp Ile Gly Val Ile Ser Thr Lys Tyr Gly Lys Thr Asn Tyr Asn Pro
210 215 220
Ser Phe Gln Gly Gln Ala Thr Met Thr Val Asp Lys Ser Ser Ser Thr
225 230 235 240
Ala Tyr Met Glu Leu Ala Ser Leu Lys Ala Ser Asp Ser Ala Ile Tyr
245 250 255
Tyr Cys Ala Arg Gly Asp Asp Gly Tyr Ser Trp Gly Gln Gly Thr Ser
260 265 270
Val Thr Val Ser Ser Ala Ser Thr Gly Gly Gly Gly Gly Gly Gly Gly
275 280 285
Gly Ser Gly Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr
290 295 300
Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp
305 310 315 320
Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp
325 330 335
Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp
340 345 350
Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Ala
355 360 365
Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp
370 375 380
Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro
385 390 395 400
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala
405 410 415
Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp
420 425 430
Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile
435 440 445
Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn
450 455 460
Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys
465 470 475 480
Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys
485 490 495
Ser Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu
500 505 510
Ser His Ser Pro Gly Ser Gly Ser Gly Ser Gly Ser His His His His
515 520 525
His His
530
<210> 9
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности: синтетический
6xHis tag"
<400> 9
His His His His His His
1. 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ДОСТАВКИ ТЕРАПЕВТИЧЕСКОГО БЕЛКА | 2019 |
|
RU2818779C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИНТЕРНАЛИЗАЦИИ ФЕРМЕНТОВ | 2018 |
|
RU2806021C2 |
| АНТИТЕЛА, СТАБИЛИЗИРУЮЩИЕ TREM2 | 2019 |
|
RU2807996C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2809246C2 |
| СРЕДСТВА, ПУТИ ПРИМЕНЕНИЯ И СПОСОБЫ ЛЕЧЕНИЯ СИНУКЛЕОПАТИИ | 2016 |
|
RU2765303C2 |
| СКОНСТРУИРОВАННЫЕ АДЕНОАССОЦИИРОВАННЫЕ (AAV) ВЕКТОРЫ ДЛЯ ЭКСПРЕССИИ ТРАНСГЕНА | 2020 |
|
RU2839774C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ ПОСРЕДСТВОМ ЭКСПРЕССИИ ФЕРМЕНТА, ОБЛАДАЮЩЕГО ДЕЗОКСИРИБОНУКЛЕАЗНОЙ (ДНКазной) АКТИВНОСТЬЮ, В ПЕЧЕНИ | 2019 |
|
RU2773691C2 |
| ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ TLR2-ОПОСРЕДОВАННЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2019 |
|
RU2791022C2 |
| СЛИТАЯ МОЛЕКУЛА, СПОСОБНАЯ ИНДУЦИРОВАТЬ НЕВОСПАЛИТЕЛЬНЫЙ ФАГОЦИТОЗ | 2022 |
|
RU2823919C1 |
| ИНТРАТЕКАЛЬНОЕ ВВЕДЕНИЕ ВЕКТОРОВ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ГЕННОЙ ТЕРАПИИ | 2016 |
|
RU2775138C2 |
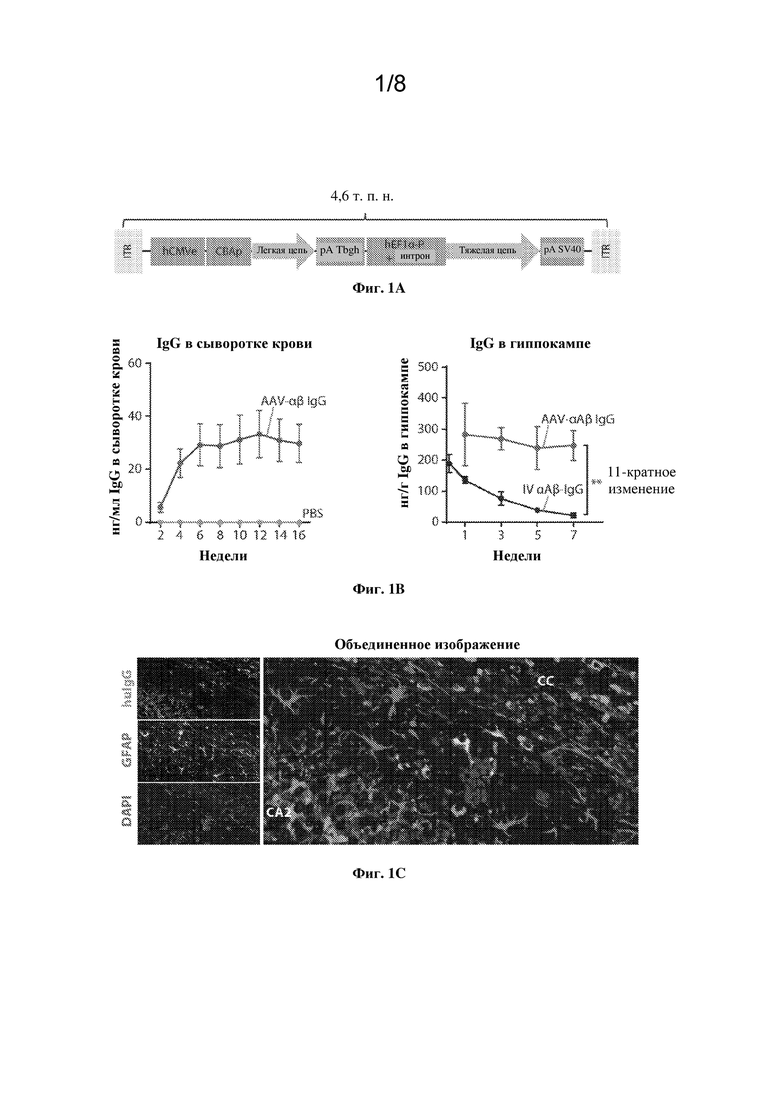
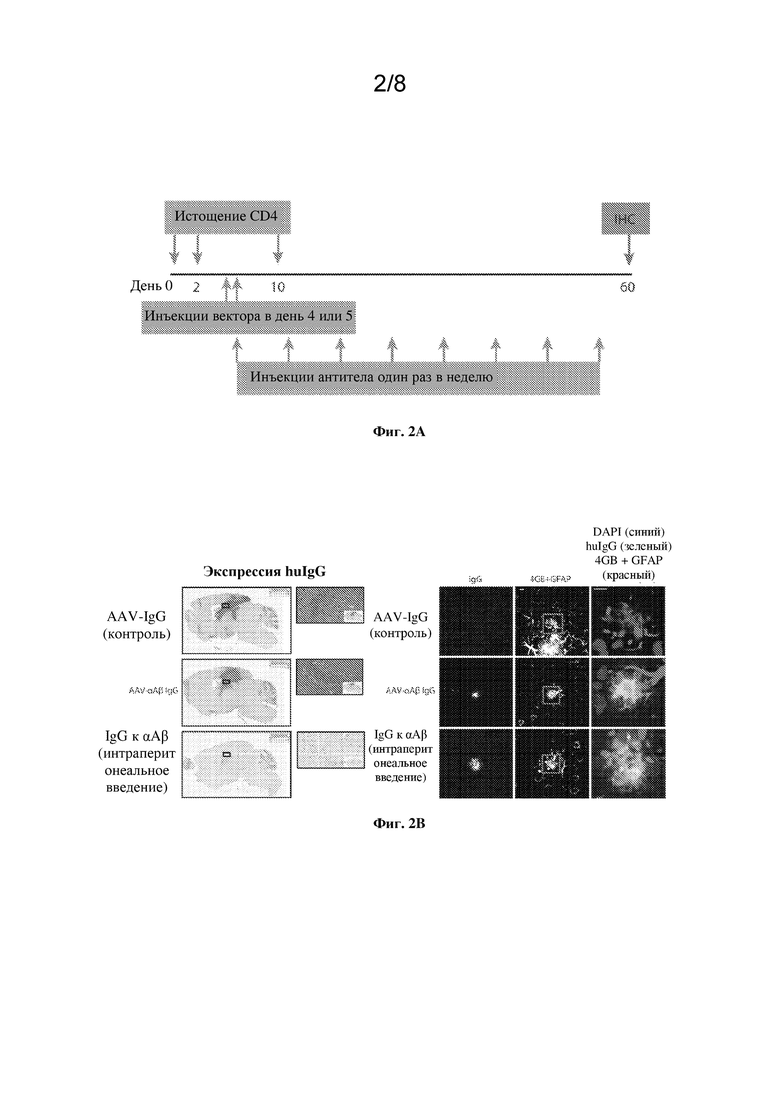
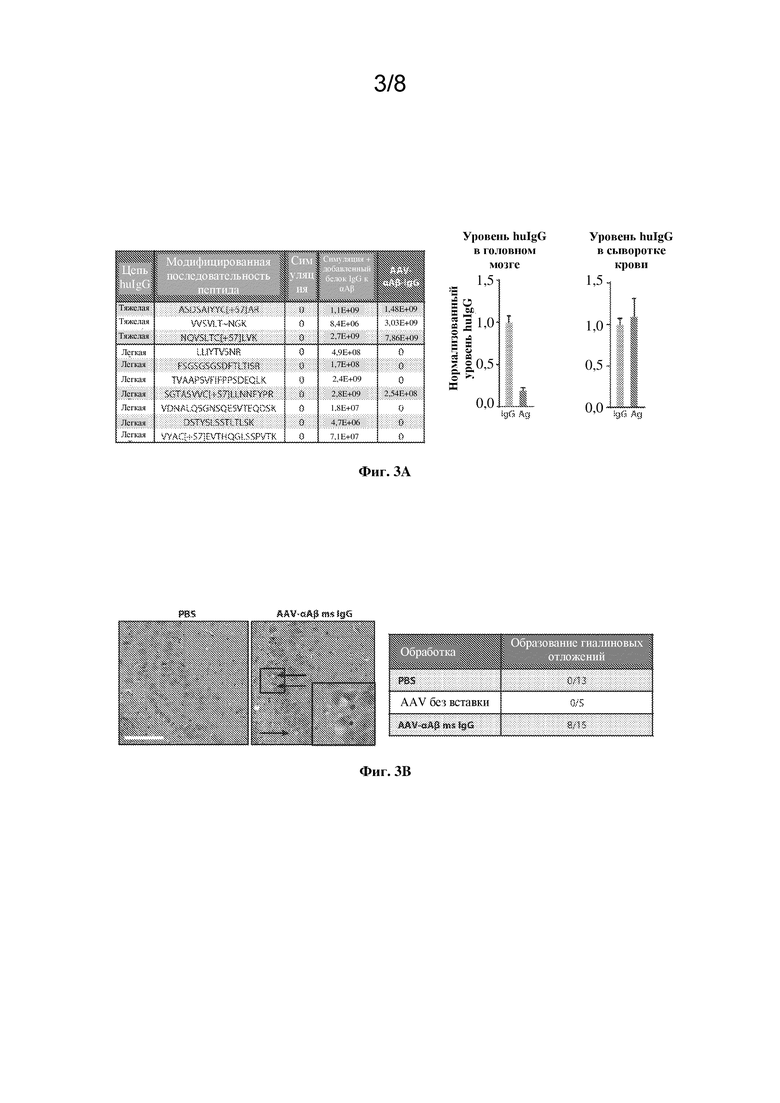
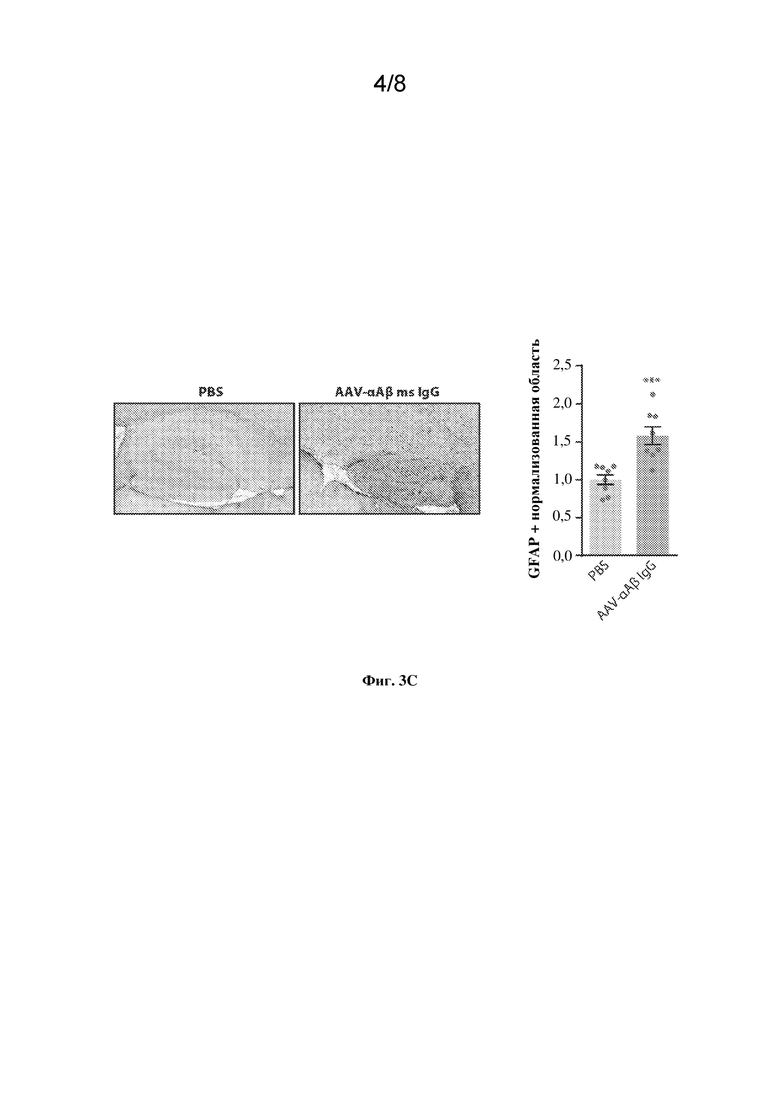
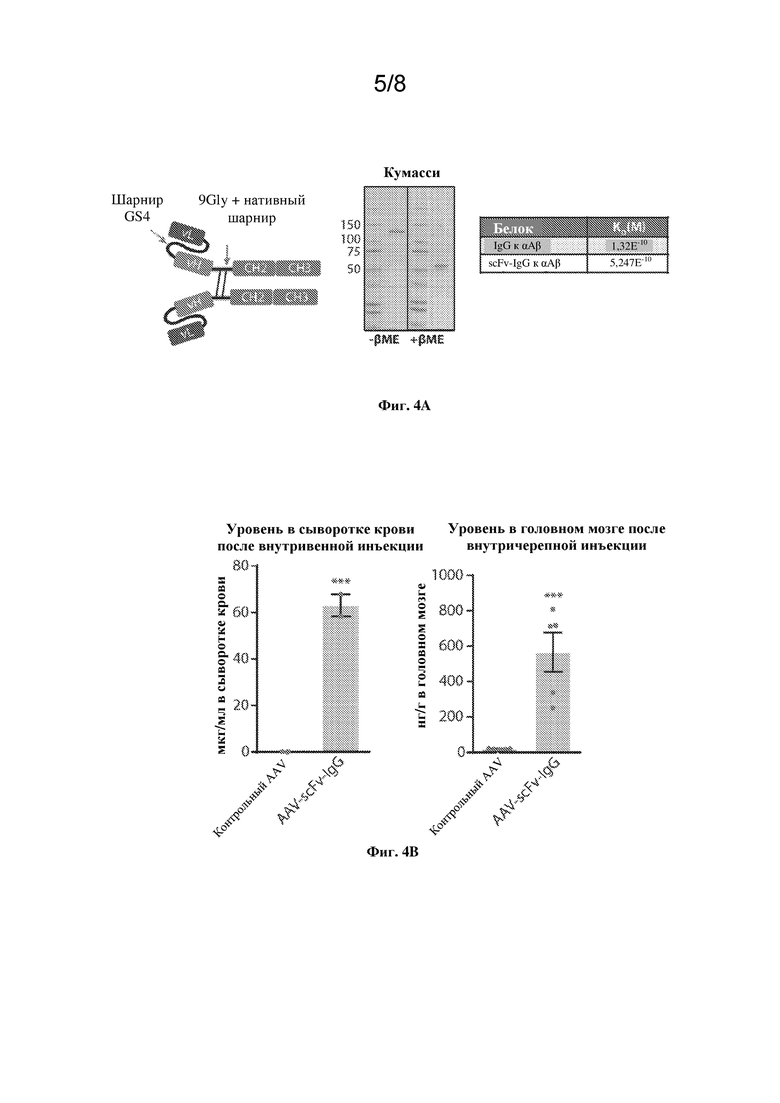
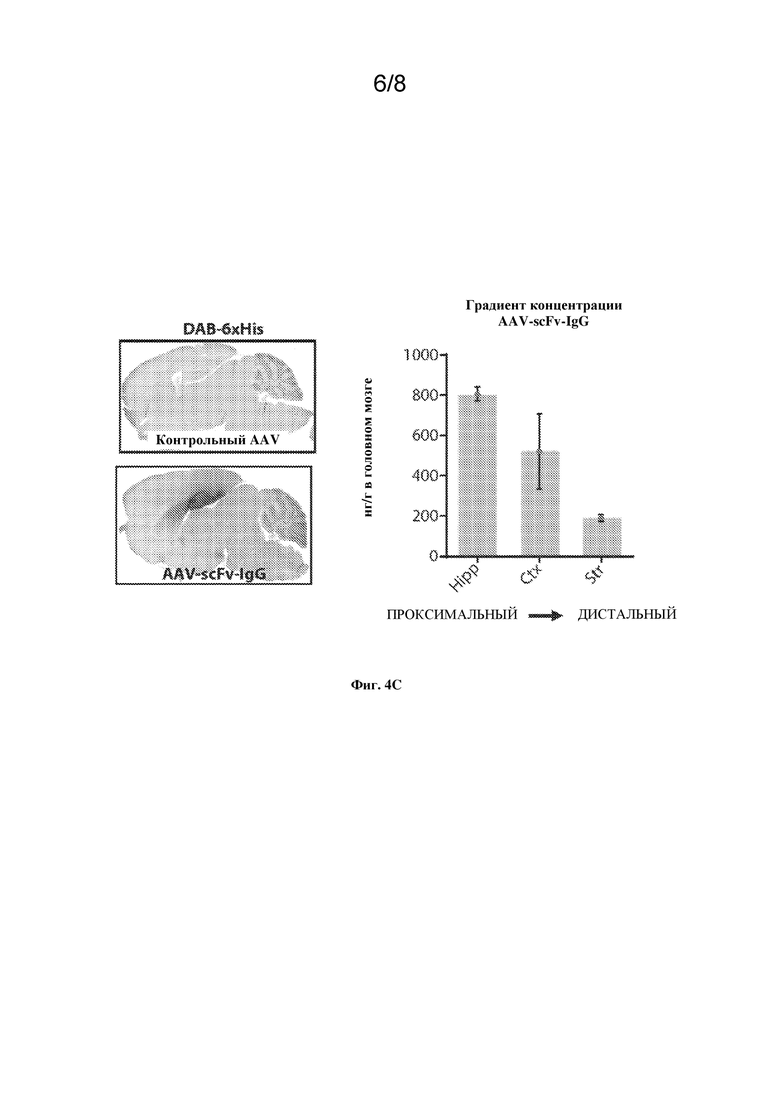
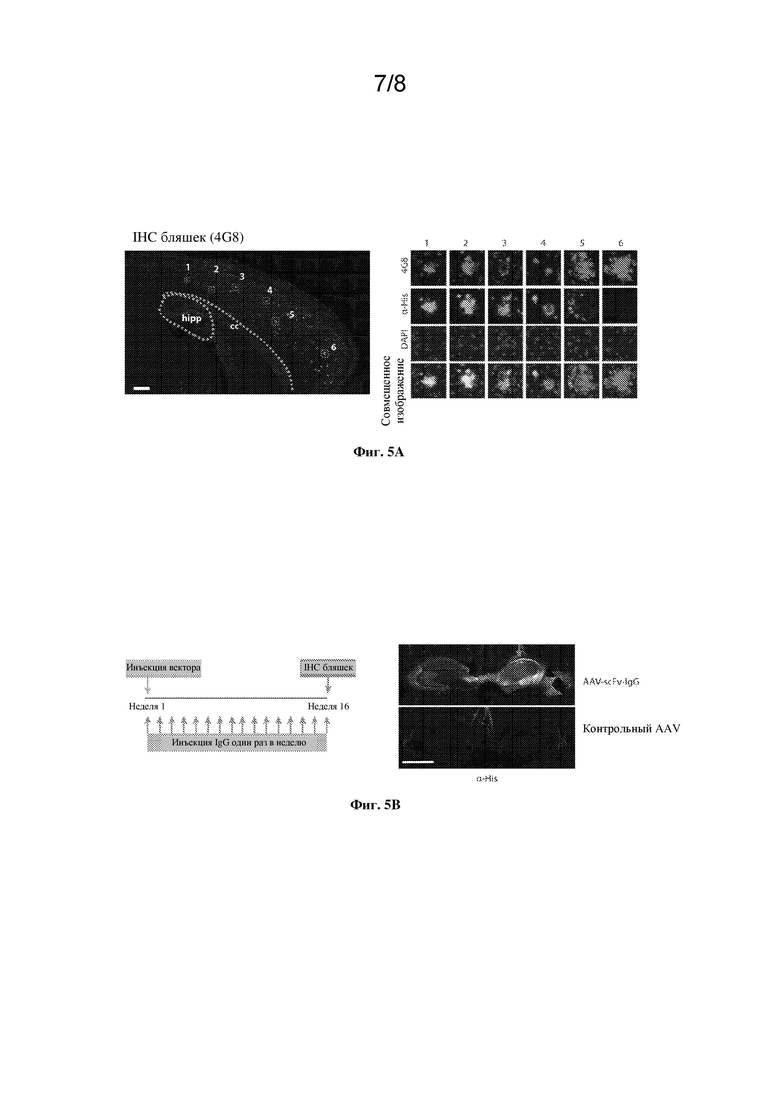
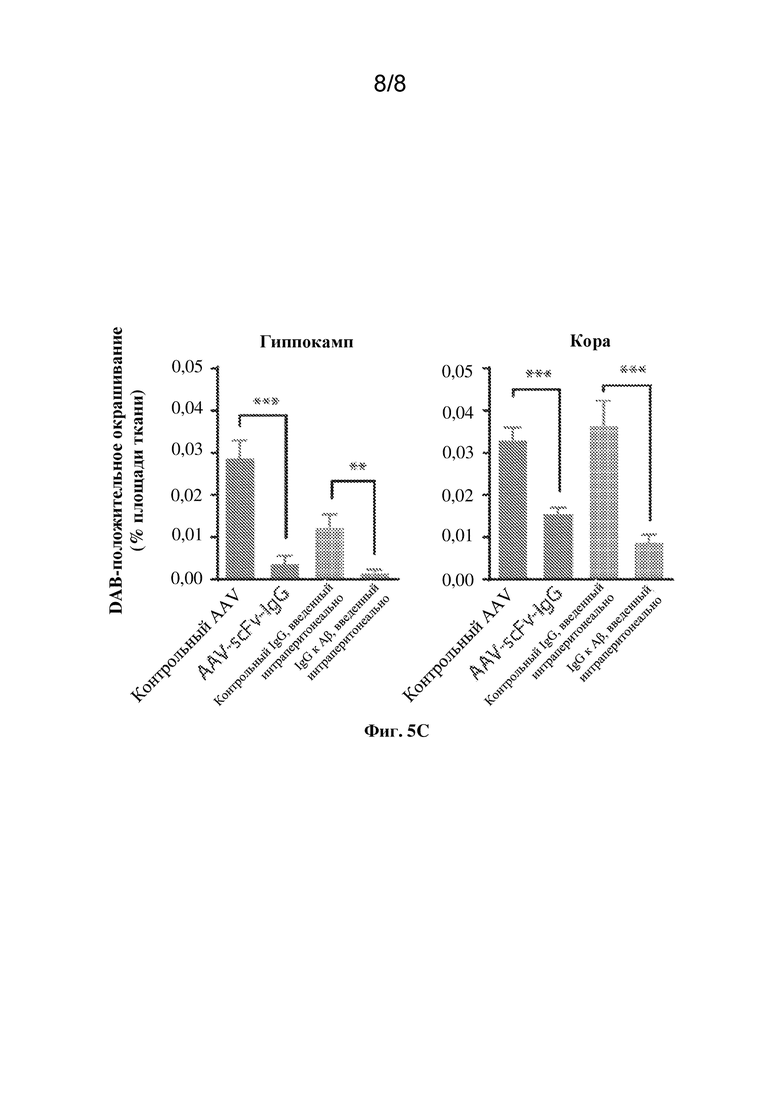
Изобретение относится к медицинской биотехнологии. Предложен способ экспрессии гомодимерного бивалентного связывающего элемента в клетке нервной системы. Способ включает введение в клетку кассеты экспрессии для экспрессии полипептида, содержащего вариабельный домен тяжелой цепи антитела (VH), вариабельный домен легкой цепи антитела (VL) и Fc-область IgG, где VH и VL образуют антигенсвязывающий сайт, который специфически связывается с целевым белком, и где указанная Fc-область IgG содержит часть тяжелой цепи одиночного иммуноглобулина (Ig), начинающуюся в шарнирной области непосредственно перед сайтом расщепления папаином и заканчивающуюся на С-конце тяжелой цепи Ig, и при экспрессии в клетке две молекулы полипептида образуют связанный дисульфидной связью гомодимерный бивалентный связывающий элемент, специфический в отношении целевого белка. Предложено применение экспрессионной конструкции для производства лекарственного средства для лечения нуждающегося в этом пациента-человека. Способ позволяет модифицировать клетки нервной системы для экспрессии связывающих элементов в головном мозге пациентов, страдающих неврологическим заболеванием, таким как нейродегенеративное заболевание. 2 н. и 14 з.п. ф-лы, 14 ил., 5 пр.
1. Способ экспрессии гомодимерного бивалентного связывающего элемента в клетке нервной системы, включающий введение в клетку кассеты экспрессии для экспрессии полипептида, содержащего вариабельный домен тяжелой цепи антитела (VH), вариабельный домен легкой цепи антитела (VL) и Fc-область IgG, где VH и VL образуют антигенсвязывающий сайт, который специфически связывается с целевым белком, и где указанная Fc-область IgG содержит часть тяжелой цепи одиночного иммуноглобулина (Ig), начинающуюся в шарнирной области непосредственно перед сайтом расщепления папаином и заканчивающуюся на С-конце тяжелой цепи Ig, и при экспрессии в клетке две молекулы полипептида образуют связанный дисульфидной связью гомодимерный бивалентный связывающий элемент, специфический в отношении целевого белка.
2. Способ по п. 1, где клетка представляет собой клетку человека.
3. Применение экспрессионной конструкции для производства лекарственного средства для лечения пациента-человека, нуждающегося в этом, где указанная экспрессионная конструкция содержит кассету экспрессии для экспрессии полипептида, содержащего вариабельный домен тяжелой цепи антитела (VH), вариабельный домен легкой цепи антитела (VL) и Fc-область IgG, где VH и VL образуют антигенсвязывающий сайт, который специфически связывается с целевым белком, и где указанная Fc-область IgG содержит часть тяжелой цепи одиночного иммуноглобулина (Ig), начинающуюся в шарнирной области непосредственно перед сайтом расщепления папаином и заканчивающуюся на С-конце тяжелой цепи Ig, и, при экспрессии в клетке нервной системы указанного пациента-человека, две молекулы полипептида образуют связанный дисульфидной связью гомодимерный бивалентный связывающий элемент, специфический в отношении целевого белка, где указанная клетка находится в головном мозге указанного пациента.
4. Способ по п. 1 или 2 или применение по п. 3, где клетка нервной системы представляет собой нейрон, глиальную клетку, олигодендроцит, астроцит, перицит, шванновскую клетку, клетку микроглии, эпендимальную клетку или эпителиальную клетку головного мозга.
5. Способ по любому из пп. 1, 2 или 4 или применение по п. 3 или 4, где целевой белок представляет собой белок, экспрессируемый в головном мозге.
6. Способ по любому из пп. 1, 2, 4 или 5 или применение по любому из пп. 3-5, где белок представляет собой бета-амилоидный пептид (Aβ), тау-белок, SOD-1, TDP-43, ApoE или α-синуклеин.
7. Способ по любому из пп. 1, 2 или 4-6 или применение по любому из пп. 3-6, где полипептид содержит от N-конца к C-концу
(i) VH, пептидный линкер и VL, или VL, пептидный линкер и VH, и
(ii) Fc-область IgG.
8. Способ или применение по п. 7, где пептидный линкер содержит последовательность GGGGS (SEQ ID NO: 3).
9. Способ по любому из пп. 1, 2 или 4-8 или применение по любому из пп. 3-8, где бивалентный связывающий элемент связывается с неонатальным Fc-рецептором (FcRn), но не связывается с Fc-гамма-рецептором вследствие одной или более мутаций в Fc-области IgG.
10. Способ по любому из пп. 1, 2 или 4-9 или применение по любому из пп. 3-9, где стадия введения включает введение рекомбинантного вируса, геном которого содержит кассету экспрессии.
11. Способ или применение по п. 10, где рекомбинантный вирус вводят в головной мозг пациента посредством внутричерепной инъекции, интратекальной инъекции или инъекции в мостомозжечковую цистерну.
12. Способ или применение по п. 10 или 11, где рекомбинантный вирус представляет собой рекомбинантный аденоассоциированный вирус (AAV).
13. Способ или применение по п. 12, где рекомбинантный AAV характеризуется серотипом 1 или 2.
14. Способ по любому из пп. 1, 2 или 4-13 или применение по любому из пп. 3-13, где экспрессия полипептида находится под транскрипционным контролем конститутивно активного промотора или индуцируемого промотора.
15. Применение по любому из пп. 3-14, где пациент страдает нейродегенеративным заболеванием.
16. Применение по п. 15, где пациент страдает болезнью Альцгеймера, церебральной амилоидной ангиопатией, синуклеопатией, таупатией или амиотрофическим латеральным склерозом.
| WO 2016086320 A1, 09.06.2016 | |||
| WO 2009026274 A1, 26.02.2009 | |||
| БЕЛКОВЫЙ КОМПЛЕКС, ПОЛУЧЕННЫЙ С ИСПОЛЬЗОВАНИЕМ ФРАГМЕНТА ИММУНОГЛОБУЛИНА, И СПОСОБ ПОЛУЧЕНИЯ ТАКОГО КОМПЛЕКСА | 2004 |
|
RU2356909C2 |
| Elmer B.M | |||
| et al | |||
| Gene delivery of a modified antibody to Aβ reduces progression of murine Alzheimer’s disease // PLoS One, 2019, 14(12), p | |||
| e0226245, формула, описание | |||
| АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2607038C2 |
| US 20120231006 A, 13.09.2012. | |||

