Изобретение относится к области экспериментальной медицины и фармакологии, в частности, химиотерапии рака и может быть использовано для усиления противоопухолевых эффектов известного фармпрепарата панобиностата (LBH589).
Панобиностат (он же фаридад или LBH589) – это одобренное в Великобритании и странах Евросоюза противоопухолевое средство, которое является производным гидроксамовой кислоты, а по механизму терапевтического действия – ингибитором ферментов гистон-деацетилаз (ГДАЦ). Панобиностат обладает высокой ингибирующей активностью против всех классов ГДАЦ, имеющих локализацию в ядре и цитоплазме (Mack, G.S. To selectivity and beyond. Nature Biotechnology 2010, 28, 1259–1266).
Ингибиторы ГДАЦ – это отдельный класс противоопухолевых фармпрепаратов, которые направленно воздействуют на эпигенетическую регуляцию в раковых клетках, подавляя деацетилирование гистоновых и негистоновых субстратов, таких как гистоны Н3 и Н4 хроматина, белок теплового шока 90 (HSP90), альфа-тубулин и др. (Revill, P., Mealy, N., Serradell, N., Bolos, J., Rosa, E. Panobinostat. Drugs Future. 2007, 32(4): 315. DOI: 10.1358/dof.2007.032.04.1094476).
В экспериментах in vitro (доклинические испытания) обнаружено, что панобиностат в наномолярных концентрациях подавляет пролиферацию и оказывает цитотоксическое действие в опухолевых клетках различных линий (Berdeja, J.G., Laubach, J.P., Richter, J., et al. Panobinostat From Bench to Bedside: Rethinking the Treatment Paradigm for Multiple Myeloma. Clin Lymphoma Myeloma Leuk. 2021, 21(11), 752-765. doi: 10.1016/j.clml.2021.06.020; Chen, E., Liu, NL., Zhao, Y., et al. Panobinostat reverses HepaCAM gene expression and suppresses proliferation by increasing histone acetylation in prostate cancer. Gene. 2022, 808, 145977. doi: 10.1016/j.gene.2021.145977; Wang, X., Xuzhi Yin, Y. Panobinostat inhibits breast cancer progression via Vps34-mediated exosomal pathway. Hum Cell. 2022, Nov 3. doi: 10.1007/s13577-022-00812-3. Online ahead of print).
На молекулярно-клеточном уровне противоопухолевые эффекты панобиностата обусловлены его мультитаргетным воздействием на эпигенетику и сигнальные пути раковой клетки. Ингибирование ГДАЦ панобиностатом усиливает ацетилирование гистонов Н3/Н4 в опухолевых клетках-мишенях. В результате такой модификации гистонов хроматина, панобиностат запускает экспрессию гена-супрессора опухоли p21 – ключевого медиатора остановки фазы G1 клеточного цикла и дифференцировки (Vallo, S., Mani, J., Stastny, M., et al. The prostate cancer blocking potential of the histone deacetylase inhibitor LBH589 is not enhanced by the multi receptor tyrosine kinase inhibitor TKI258. Invest. New Drugs 2013, 31(2), 265-272. doi: 10.1007/s10637-012-9851-5).
Важно отметить, что панобиностат ингибирует ГДАЦ6, тем самым вызывая ацетилирование и инактивацию белка теплового шока HSP90, что в свою очередь приводит к протеотоксическому стрессу и падению уровней или дисфункции онко-белков в раковой клетке-мишени, а также остановке HSP90-зависимых сигнальных путей, способствующих злокачественному росту, защите от апоптоза и устойчивости опухоли к терапевтическим воздействиям (Edwards, A., Li, J., Atadja, P., et al. Effect of the histone deacetylase inhibitor LBH589 against epidermal growth factor receptor-dependent human lung cancer cells. Mol. Cancer Ther. 2007, 6(9), 2515-2524. doi: 10.1158/1535-7163.MCT-06-0761; Kim, I.A., No, M., Lee, J.M., et al. Epigenetic modulation of radiation response in human cancer cells with activated EGFR or HER-2 signaling: potential role of histone deacetylase 6. Radiother. Oncol. 2009, 92(1), 125-132. doi: 10.1016/j.radonc.2009.03.008). Кроме того, ингибирование HDAC6 панобиностатом ведет к нарушению функции аггресом, формирующихся для регулируемой деградации внутриклеточных белков на протеасомах, что дополнительно понижает жизнеспособность и адаптивный потенциал раковой клетки (Richardson, P.G., Schlossman, R.L., Alsina, M., et al. PANORAMA 2: panobinostat in combination with bortezomib and dexamethasone in patients with relapsed and bortezomib-refractory myeloma. Blood. 2013, 122(14), 2331-2337. doi: 10.1182/blood-2013-01-481325).
Таким образом, результатами ингибирования ГДАЦ панобиностатом являются блокада клеточного цикла (т.е. подавление пролиферации), протеотоксический стресс и апоптоз в опухолевых клетках, а также падение их выживаемости после дополнительных терапевтических воздействий.
Однако в проведенных испытаниях панобиностат не показал высокой эффективности против многих других видов онкологических заболеваний, из-за чего этот фармпрепарат не стал широко применяемым противоопухолевым лекарством.
В онко-клинике панобиностат (он же фаридад или LBH589) применялся для моно- и комбинированной терапии множественной миеломы. «Фаридак» был разрешен для применения при рефрактерной или рецидивирующей множественной миеломе. Согласно инструкции, панобиностат применялся только после двух предшествующих курсах терапии. Препарат зарегистрирован в России в 2016 году, указано в государственном реестре. В 2015 получил ускоренное одобрение FDA для применения вместе с бортезомибом и дексаметазоном при множественной миеломе. В 2019 году права на препарат приобрела компания Secura Bio. Однако в ноябре 2021 года компания сообщила об отзыве панобиностата с рынка США из-за невозможности завершить необходимые клинические исследования, как этого требовали правила FDA после ускоренного одобрения. Панобиностат по-прежнему доступен в странах ЕС и Великобритании, указано на сайте препарата, принадлежащем компании Secura Bio, и официальном сайте Европейского агентства по лекарственным средствам EMA (https://pharmvestnik.ru/content/news/Minzdrav-otmenil-registraciu-originalnogo-protivoopuholevogo-preparata.html).
Исследуя влияние панобиностата на опухолевые клетки in vitro, нами обнаружился интересный и важный эффект, который сильно снижает противоопухолевую активность данного лекарственного препарата. Заявитель обращает внимание на один важный побочный эффект в молекулярно-клеточном механизме действия панобиностата, что этот побочный эффект существенно снижает цитотоксичность панобиностата для раковых клеток, но такого нежелательного снижения цитотоксичности можно избежать с помощью предлагаемого нами способа, который будет усиливать противоопухолевую активность панобиностата. Здесь имеется ввиду вызываемая панобиностатом инактивация белка теплового шока HSP90, что считается важным событием в механизме противоопухолевого действия этого фармпрепарата, т.к. благодаря этому происходит блокирование сразу нескольких HSP90-зависимых сигнальных путей, ответственных за пролиферацию злокачественных клеток и их устойчивость к цитотоксическим (терапевтическим) обработкам. Заявителю удалось выяснить, что дисфункция HSP90, вызванная панобиностатом, приводит к активации транскрипционного фактора HSF1 и быстрой (HSF1-опосредованной) индукции цитопротективных HSPs (HSP70, HSP27 и др.), накопление которых в раковых клетках делает их более устойчивой к панобиностату и, следовательно, помогает части раковых клеток пережить обработку этим лекарством. Чтобы справиться с этой проблемой, заявитель предлагает способ предотвращения активации HSF1 и индукции HSРs с помощью известных ингибиторов, что позволяет сенсибилизировать раковые клетки к действию панобиностата.
Для демонстрации действенности предлагаемого нами способа авторы использовали проникающие в клетку (т.е. низкомолекулярные) ингибиторы транскрипционной активности HSF1, такие как кверцетин (Hosokawa N, Hirayoshi K, Kudo H, Takeshi H, Aoike A, et al. Inhibition of the activation of heat shock factor in vivo and in vitro by flavonoids. Mol Cell Biol. 1992;12:3490–3498), триптолид (Yokota S, Kitahara M, Nagata K. Benzylidene lactam compound, KNK437, a novel inhibitor of acquisition of thermotolerance and heat shock protein induction in human colon carcinoma cells. Cancer Res. 2000;60:2942–2948), KNK437 (Westerheide SD, Kawahara TLA, Orton K, Morimoto RI. Triptolide, an inhibitor of the human heat shock response that enhances stress-induced cell death. J Biol Chem. 2006;281:9616–9622), NZ28 (Zaarur N, Gabai VL, Porco JA, Calderwood S, Sherman MY. Targeting heat shock response to sensitize cancer cells to proteasome and Hsp90 inhibitors. Cancer Res. 2006;66:1783–1791) и KRIBB11 (Yoon, Y.J., Kim, J.A., Shin, K.D, et al. KRIBB11 inhibits HSP70 synthesis through inhibition of heat shock factor 1 function by impairing the recruitment of positive transcription elongation factor b to the hsp70 promoter. J Biol Chem. 2011, 286(3), 1737-1747), или «нокдаун» технологию селективного подавления экспрессии HSF1 (или экспрессии основных HSPs) с помощью siRNA. Авторы показали, что комбинация с панобиностатом любого из этих ингибиторов или анти-HSF1/siRNA резко усиливает гибель обработанных раковых клеток на фоне подавления в них HSF1-зависимой экспрессии HSPs. Таким образом, суть способа заключается, во-первых, в выявлении феномена активации HSF1 и индукции HSPs в обработанных панобиностатом раковых клетках, что повышает их жизнеспособность и сопротивляемость данному фармпрепарату, и во-вторых, в доказательстве возможности устранять этот вредный побочный эффект с помощью специальных агентов, предотвращающих активацию HSF1 и накопление HSPs в раковых клетках-мишенях, что усиливает противоопухолевые эффекты панобиностата.
Самыми близкими (прототипами) являются следующие публикации:
- Известен способ усиления противоопухолевых эффектов ингибитора активности HSP90 – 17-allylamino-demethoxy geldanamycin (17AAG), который тоже активировал HSF1 (с последующей индукцией HSPs) в раковых клетках (Guo et al., Abrogation of heat shock protein 70 induction as a strategy to increase antileukemia activity of heat shock protein 90 inhibitor 17-allylamino-demethoxy geldanamycin. Cancer Res. 2005; 65(22):10536-44. doi: 10.1158/0008-5472.CAN-05-1799) (Guo, F., Rocha, K., Bali, P., Pranpat, M., et al. Abrogation of heat shock protein 70 induction as a strategy to increase antileukemia activity of heat shock protein 90 inhibitor 17-allylamino-demethoxy geldanamycin. Cancer Res. 2005; 65(22):10536-44. doi: 10.1158/0008-5472.CAN-05-1799). Это усиление достигалось с помощью предобработок клеток-мишеней либо известным ингибитором HSF1 – KNK437, либо генным вектором, экспрессирующим анти-HSP70 siRNA. Было показано, что в результате таких предобработок цитотоксичность 17AAG по отношению к раковым клеткам существенно возрастала.
- Известно исследование противоопухолевого (цитотоксическое) эффекта при обработках раковых клеток ингибиторами активности HSP90 (17AAG, радицикол) или ингибитором протеасом MG132, что в обоих случаях сопровождалось активацией HSF1 и индукцией HSPs в обработанных клетках (Zaarur et al., Targeting heat shock response to sensitize cancer cells to proteasome and Hsp90 inhibitors. Cancer Res. 2006; 66(3):1783-91. doi: 10.1158/0008-5472.CAN-05-3692). Исследователям удавалось значительно усилить цитотоксичность 17AAG, радицикола и MG132 путем подавления в раковых клетках-мишенях активации HSF1 (и, соответственно, накопления в них HSPs) с помощью низкомолекулярных ингибиторов HSF1 (кверцетин, NZ28) или генного вектора, экспрессирующего анти-HSF1 siRNA.
- Похожие экспериментальные подходы применялись для демонстрации усиления радиосенсибилизирующих эффектов 17AAG и других ингибиторов активности HSP90 (гелданамицин, радицикол, NVP-AUY922) на культуры опухолевых клеток (Schilling et al., Sensitizing tumor cells to radiation by targeting the heat shock response. Cancer Lett. 2015, 360(2):294-301. doi: 10.1016/j.canlet.2015.02.033.; Kudryavtsev et al., Induction of Hsp70 in tumor cells treated with inhibitors of the Hsp90 activity: A predictive marker and promising target for radiosensitization. PLoS One, 2017, 12(3):e0173640. doi: 10.1371/journal.pone.0173640.) В тех работах тоже (как и в данной заявке) использовались такие ингибиторы активации HSF1 как кверцетин, NZ28, KNK43 и триптолид, которые блокировали HSF1-опосредованную индукцию цитопротективного HSP70 в обработанных клетках, тем самым усиливая искомый (радиосенсибилизирующий) эффект.
- Также известны способы усиления противоопухолевых и химио-сенсибилизирующих эффектов гипертермии с помощью предобработки раковых клеток-мишеней генным вектором, экспрессирующим анти-HSF1 si RNA (Rossi et al., Targeting the heat shock factor 1 by RNA interference: a potent tool to enhance hyperthermochemotherapy efficacy in cervical cancer. Cancer Res. 2006; 66(15):7678-85. doi: 10.1158/0008-5472.CAN-05-4282) или низкомолекулярными блокаторами HSF1 – кверцетином и NZ28 (Кудрявцев и др. Биомедицинская химия 2012) (Кудрявцев В.А., Макарова Ю.М., Кабаков А.Е. Термосенсибилизация опухолевых клеток ингибиторами активности и экспрессии шаперонов. Биомед. химия 2012, Т.58 (№6), с. 662-672).
Однако, следует отметить, что 17AAG не применяется в клинике, поэтому данные результаты не имеют большой практической ценности для медицины, при этом ни 17AAG, ни радицикол, ни MG132 не являются разрешенными к употреблению фармпрепаратами, что существенно снижает практическую значимость результатов.
Заявитель предлагает использовать аналогичный подход (т.е. подавление активации HSF1 и индукции HSPs в клетках-мишенях) для усиления противоопухолевых эффектов панобиностата, который подобно 17AAG или радициколу тоже ингибирует HSP90 и активирует HSF1, а именно, в виде вспомогательной обработки в комбинации с обработкой панобиностатом, но в отличие от 17AАG и радицикола, является известным фармпрепаратом, применявшимся для лечения множественной миеломы.
Техническим решением является предотвращение побочных эффектов панобиностата (связанных с индукцией HSРs и повышающих жизнеспособность и сопротивляемость опухолевых клеток) с помощью специальных агентов, ингибирующих активацию HSF1 и накопление HSPs в раковых клетках-мишенях, обработанных панобиностатом.
Технический результат достигается тем, что так же, как и в известных способах (прототипах) для усиления противоопухолевых эффектов используют ингибиторы HSF1 (кверетин, триптолид, KNK437, NZ28, KRIBB11) или siRNA-векторы для нокдауна HSF1.
Особенностью заявляемого способа является то, что опухолевые клетки перед обработкой панобиностатом в концентрации 2-400 нМ предварительно обрабатывают ингибитором HSF1, выбранным из кверцетина в концентрации 40 мкМ, триптолида в концентрации 20 нМ, KNK437 в концентрации 200 мкМ, NZ28 в концентрации 20 мкМ, KRIBB11 в концентрации 20 мкМ или миРНК-вектора для нокдауна HSF1.
Изобретение иллюстрируется подробным описанием, примерами и иллюстрациями, на которых изображено:
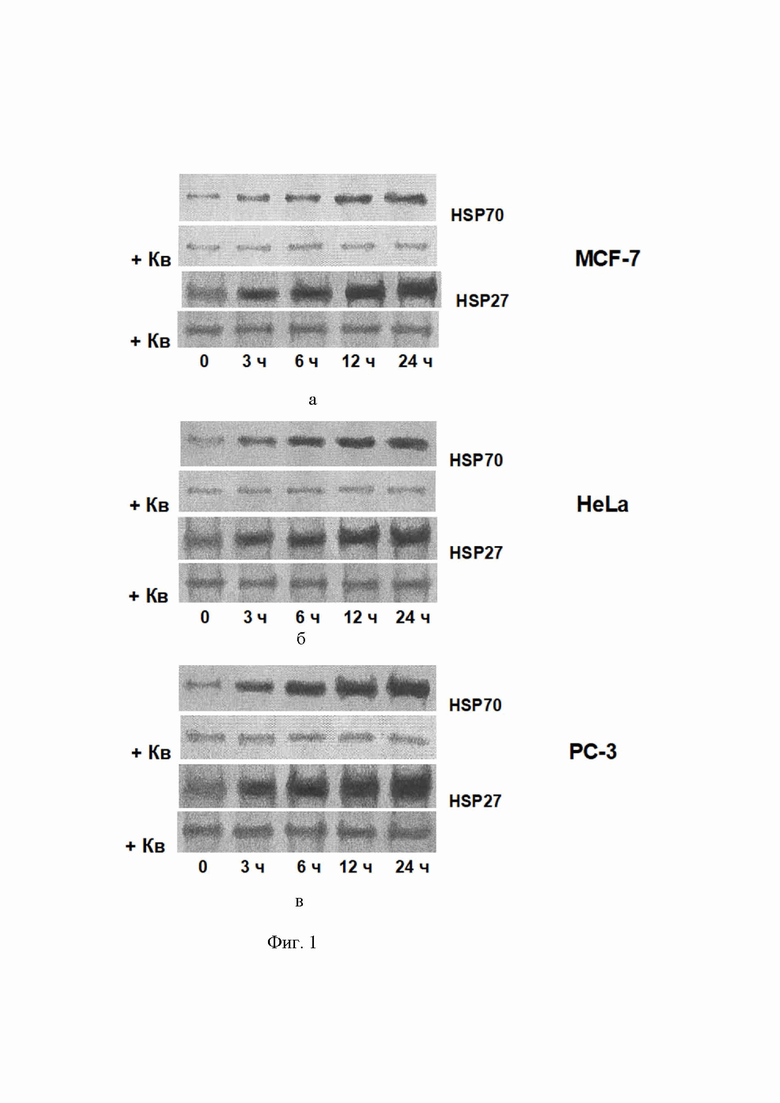
Фиг. 1 – фотоиллюстрация блотов, показывающая индукцию цитопротективных HSP70 и HSP27 в клетках опухолевых линий: а) опухолевые клетки линии MCF-7; б) опухолевые клетки линии HeLa; в) опухолевые клетки линии PC-3; обработанных 200 нМ панобиностатом. Время (в часах) инкубации клеток с реагентом отмечено цифрами по нижним краям блотов. Фотоиллюстрация также показывает ингибирование индукции 30 микроМ кверцетином (см. блоты, помеченные +Кв). Такой же результат получался при действии других ингибиторов (KNK437, NZ28, siRNA-векторы, триптолид и KRIBB11), см. в тексте.
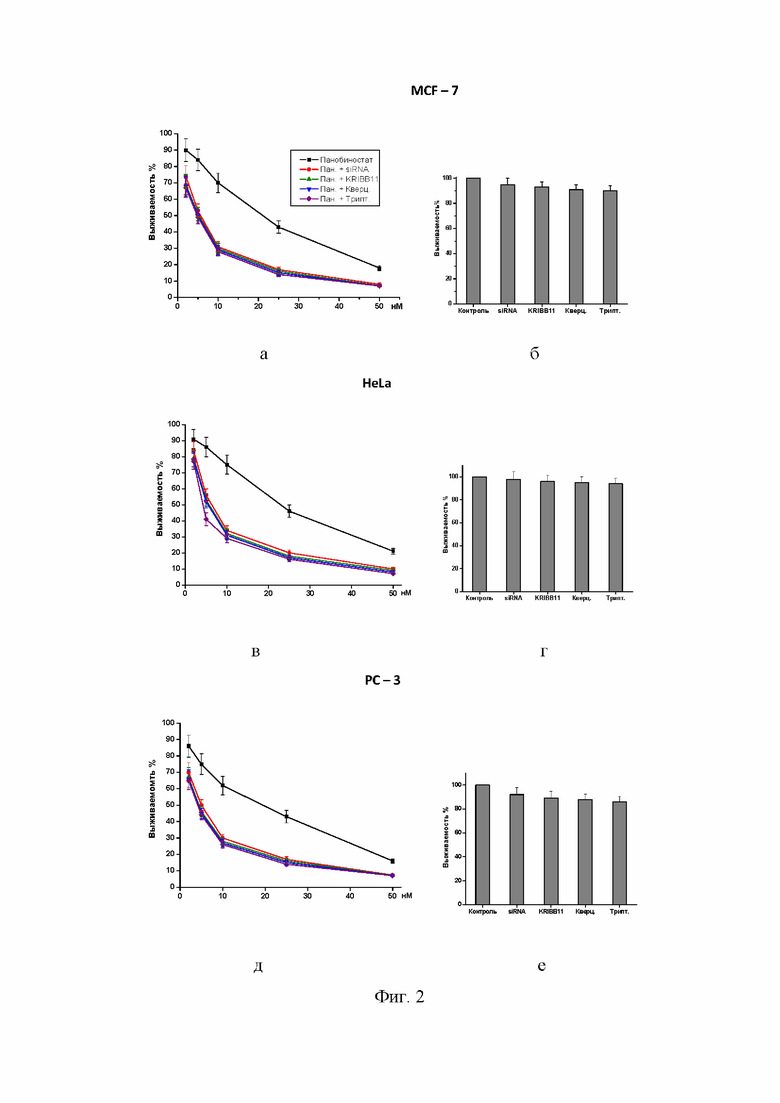
Фиг. 2 – диаграмма кривых выживания колоний, показывающая подавление клоногенности опухолевых клеток: а) опухолевые клетки линии MCF-7; в) опухолевые клетки линии HeLa; д) опухолевые клетки линии PC-3, инкубированных с разными концентрациями (10-50 нМ) панобиностата, а также резкое усиление цитотоксического эффекта в случае комбинирования панобиностата с siRNA-вектором, истощающим HSF1, KRIBB11 (15 микроМ), кверцетином (30 микроМ) или триптолидом (10 нМ). Все значения, полученные при комбинированной обработке, достоверно отличаются от значений при действии одного панобиностата (р < 0,05). Правая панель (столбчатая диаграмма) показывает нетоксичность примененных ингибиторов HSF1 для: б) клеток MCF-7; г) клеток HeLa; е) клеток PC-3.
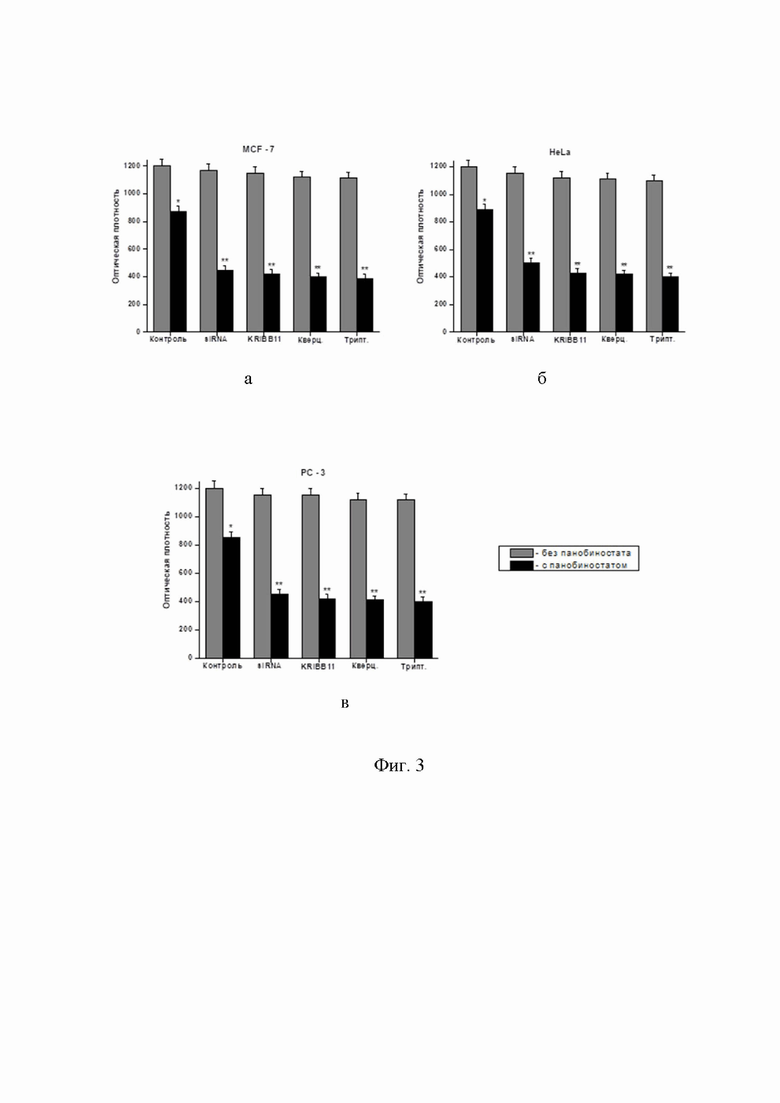
Фиг. 3 – диаграмма результатов МТТ-теста, демонстрирующая резкое снижение жизнеспособности обработанных опухолевых клеток в случае комбинирования панобиностата с любым из примененных ингибиторов: а) HSF1MCF-7; б) HeLa; в) PC-3. Клетки в течение 24 часов инкубировались с панобиностатом (100 нМ) и/или одним из ингибиторов HSF1: siRNA-вектором, KRIBB11 (15 микроМ), кверцетином (30 микроМ) или триптолидом (10 нМ), затем жизнеспособность клеток оценивалась в МТТ-тесте по стандартной методике (Kudryavtsev, V.A., Khokhlova, A.V., Mosina, V.A., et al. Induction of Hsp70 in tumor cells treated with inhibitors of the Hsp90 activity: A predictive marker and promising target for radiosensitization. PLoS One. 2017, 12(3):e0173640. doi: 10.1371/journal.pone.0173640; Kabakov, A.E., Kudryavtsev, V.A., Gabai, V.L. Determination of cell survival or death. Methods Mol Biol. 2011, 787, 231-244. doi: 10.1007/978-1-61779-295-3_17). Аналогичный результат, а именно усиление снижения жизнеспособности клеток после их комбинированной обработки, получались и с другими примененными авторами ингибиторами HSF1: KNK43 (100 микроМ) и NZ28 (10 микроМ) (данные не приводятся).
* - достоверное отличие (ANOVA) от контроля, р < 0,05;
** - достоверное отличие (ANOVA) от контроля и от значений, помеченных *, p < 0,01.
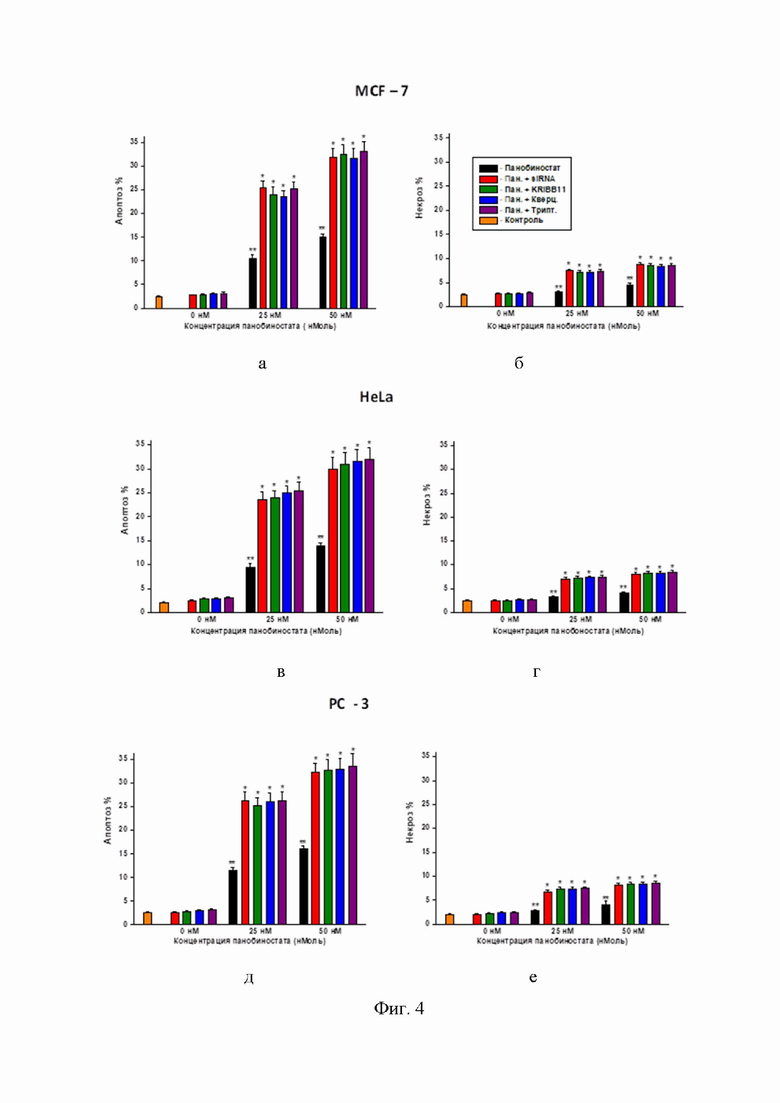
Фиг. 4 – диаграмма, показывающая усиление клеточной гибели - апоптоза (а, в, д) и некроза (б, г, е) в опухолевых клетках MCF-7 (а, б), HeLa (в, г) и PC-3 (д, е), обработанных только панобиностатом или панобиностатом в комбинации с ингибиторами HSF1;
* - достоверное отличие (ANOVA) от контроля и от значений, помеченных *, р < 0,01;
** - достоверное отличие (ANOVA) от контроля, p < 0,05.
Способ осуществляют следующим образом.
Опухолевые клетки 3-х линий из карцином молочной железы (линия MCF-7), шейки матки (линия HeLa) и предстательной железы (линия PC-3) человека обрабатывают панобиностатом в наномолярной концентрации 2-400 наноМ после предварительной обработки ингибиторами HSF1 в следующих концентрациях: 40 микроМ кверцетин, 20 наноМ триптолид, 100-200 микроМ KNK437, 10-20 микроМ NZ28 и 15-20 микроМ KRIBB11, которые блокировали HSF1-опосредованную индукцию HSPs, или с генными векторами, экспрессирующими siRNA для истощения (нокдауна) HSF1, что тоже блокировало HSF1-опосредованную индукцию HSPs.
Проверялось влияние именно наномолярных концентраций панобиностата, т.е. концентраций, которые возможно создать в тканях пациента с помощью инъекций.
Исследовались следующие комбинации.
Панобиностат (2 - 400 наноМ) в комбинации с ингибитором активации HSF1 кверцетином (30-40 микроМ). Полезный (сенсибилизирующий) эффект наблюдался: в тесте на клоногенность – для диапазона концентраций панобиностата 2–50 нМ в комбинации с 30-40 микроМ кверцетином (Фиг. 2 а, в, д); в МТТ-тесте – в сочетании: панобиностат (50-400 нМ) + 30-40 микроМ кверцетин (Фиг. 3); при определении клеточной гибели (апоптоза и некроза) – в сочетании панобиностат (20-100 наноМ) + 30-40 микроМ кверцетин (Фиг. 4). Как видно на Фиг. 1, именно такая концентрация (30 микоМ) блокировала индукцию цитопротективную индукцию HSP70 и HSP27 (см. в Фиг.1 блоты, помеченные +Кв). При концентрациях панобиностата > 100 наноМ (200, 300 и 400 наноМ) во всех исследованных комбинациях с кверцетином наблюдалась 100% цитотоксичность (колонии не вырастали, отсутствие жизнеспособных клеток в МТТ-тесте и 100% апоптоз + некроз), поэтому эти эффекты не показаны на графиках.
Панобиностат (2–400 наноМ) в комбинации с ингибитором активации HSF1 триптолидом (5–20 наноМ). Полезный (сенсибилизирующий) эффект наблюдался: в тесте на клоногенность – для диапазона концентраций панобиностата 2–50 наноМ в комбинации с 10-20 наноМ триптолидом (Фиг. 2 а, в, д); в МТТ-тесте – в сочетании: панобиностат (50-400 наноМ) + 10-20 наноМ триптолид (Фиг. 3); при определении клеточной гибели (апоптоза и некроза) – в сочетании панобиностат (20-100 наноМ) + 10-20 наноМ триптолид (Фиг. 4). При концентрациях панобиностата > 100 наноМ (200, 300 и 400 наноМ) во всех исследованных комбинациях с триптолидом наблюдалась 100% цитотоксичность (колонии не вырастали, отсутствие жизнеспособных клеток в МТТ-тесте и 100% апоптоз + некроз), поэтому эти эффекты не показаны на графиках.
Панобиностат (2–400 наноМ) в комбинации с ингибитором активации HSF1 KNK437 (100-200 микроМ). Полезный (сенсибилизирующий) эффект наблюдался: в тесте на клоногенность – для диапазона концентраций панобиностата 2–50 наноМ в комбинации с 100 микроМ KNK437 (данные не приводятся); в МТТ-тесте – в сочетании: панобиностат (50-400 наноМ) + 100 микроМ KNK437 (данные не приводятся); при определении клеточной гибели (апоптоза и некроза) – в сочетании панобиностат (20-100 наноМ) + 100 микроМ KNK437 (данные не приводятся). При концентрациях панобиностата > 100 наноМ (200, 300 и 400 наноМ) во всех исследованных комбинациях с KNK437 наблюдалась 100% цитотоксичность (колонии не вырастали, отсутствие жизнеспособных клеток в МТТ-тесте и 100% апоптоз + некроз), поэтому эти эффекты не показаны на графиках.
Панобиностат (2–400 наноМ) в комбинации с ингибитором активации HSF1 NZ28 (10-20 микроМ). Полезный (сенсибилизирующий) эффект наблюдался: в тесте на клоногенность – для диапазона концентраций панобиностата 2–50 наноМ в комбинации с 10 микроМ NZ28 (данные не приводятся); в МТТ-тесте – в сочетании: панобиностат (50-400 наноМ) + 10 микроМ NZ28 (данные не приводятся); при определении клеточной гибели (апоптоза и некроза) – в сочетании панобиностат (20-100 наноМ) + 10 микроМ NZ28 (данные не приводятся). При концентрациях панобиностата > 100 наноМ (200, 300 и 400 наноМ) во всех исследованных комбинациях с NZ28 наблюдалась 100% цитотоксичность (колонии не вырастали, отсутствие жизнеспособных клеток в МТТ-тесте и 100% апоптоз + некроз), поэтому эти эффекты не показаны на графиках.
Панобиностат (2–400 наноМ) в комбинации с ингибитором активации HSF1 KRIBB11 (15-20 микроМ). Полезный (сенсибилизирующий) эффект наблюдался: в тесте на клоногенность – для диапазона концентраций панобиностата 2–50 наноМ в комбинации с 15 микроМ KRIBB11 (Фиг. 2 а, в, д); в МТТ-тесте – в сочетании: панобиностат (50-400 наноМ) + 15 микроМ KRIBB11 (Фиг. 3); при определении клеточной гибели (апоптоза и некроза) – в сочетании панобиностат (20-100 наноМ) + 15 микроМ KRIBB11 (Фиг. 4). При концентрациях панобиностата > 100 наноМ (200, 300 и 400 наноМ) во всех исследованных комбинациях с кверцетином наблюдалась 100% цитотоксичность (колонии не вырастали, отсутствие жизнеспособных клеток в МТТ-тесте и 100% апоптоз + некроз), поэтому эти эффекты не показаны на графиках.
Панобиностат (2–400 наноМ) в комбинации с генными векторами siRNA для истощения (нокдауна) HSF1 в клетках-мишенях. Культуры опухолевых клеток MCF-7, HeLa и PC-3 трансфицировали RNAi-Ready pSIREN-RetroQ вектором (BD Biosciences, San Jose, CA, USA) с устойчивостью к пуромицину (вектор был предоставлен Dr. V.L. Gabai, University of Boston, Boston, MA, USA) Процедура трансфекции опухолевых клеток проводилась согласно инструкции производителя (BD Biosciences) и как описано в ссылке (Zaarur et al., Cancer Res., 2006, Targeting heat shock response to sensitize cancer cells to proteasome and Hsp90 inhibitors. 66(3):1783-91. doi: 10.1158/0008-5472.CAN-05-3692.). После селекции в среде с пуромицином выжившие клетки не реагировали на тепловой шок экспрессией индуцибельного HSP70 (данные не приводятся), что доказывало произведенное siRNA истощение HSF1 в этих клетках. Полезный (сенсибилизирующий) эффект наблюдался в трансфектантах при тесте на клоногенность – для диапазона концентраций панобиностата 2 – 50 наноМ (Фиг. 2 а, в, д); в МТТ-тесте – для 50-400 наноМ панобиностата (Фиг. 3); при определении клеточной гибели (апоптоза и некроза) – для 20-100 наноМ панобиностата (Фиг. 4). При концентрациях панобиностата > 100 наноМ (200, 300 и 400 наноМ) во всех культурах клеток с истощением (нокдауном) HSF1 наблюдалась 100% цитотоксичность (колонии не вырастали, отсутствие жизнеспособных клеток в МТТ-тесте и 100% апоптоз + некроз), поэтому эти эффекты не показаны на графиках.
Экспериментальная часть, подтверждающая эффективность усиления цитотоксических эффектов панобиностата на опухолевые клетки.
Синтетический ингибитор NZ28, а также siRNA-векторы для подавления экспрессии HSF1 и контрольные векторы были получены из Университета Бостона (Бостон, США). Коммерчески доступные ингибиторы активации HSF1: кверцетин, KNK437 и KRIBB11 были приобретены у фирмы Calbiochem.
Для демонстрации полезных эффектов в экспериментах использовались предварительно подобранные (минимальные и нетоксичные) концентрации и дозы ингибиторов (включая siRNA-векторы), которые полностью блокировали HSF1-зависимую индукцию HSPs в исследуемых опухолевых клетках 3-х линий (Фиг. 1-4).
Факт индукции HSPs (HSP70 и HSP27) в обработанных панобиностатом опухолевых клетках, а также предотвращение этой индукции кверцетином (Фиг. 1 а, б, в) демонстрировались с помощью иммуноблоттинга.
Цитотоксические эффекты (и их усиление) выявлялись посредством определения клоногенности (Фиг. 2 а, б, в), клеточной жизнеспособности в МТТ-тесте (Фиг. 3 а, б, в) и уровней апоптоза/некроза (Фиг. 4 а, б, в, г, д, е). Статистический анализ различий между результатами, полученными для разных групп образцов проводился по описанным ранее схемам (Кудрявцев В.А., Макарова Ю.М., Кабаков А.Е. Термосенсибилизация опухолевых клеток ингибиторами активности и экспрессии шаперонов. Биомед. химия 2012, Т.58 (6), с. 662-672).
Результаты
Добавленный в наномолярной концентрации 2-400 наноМ панобиностат действительно индуцирует цитопротективные HSPs (HSP70 и HSP27) в опухолевых клетках 3-х линий: MCF-7, HeLa, PC-3 (Фиг. 1 а, б, в). Здесь важно отметить, что 40 микроМ кверцетин (см. блоты, помеченные +Кв) полностью блокировал эту нежелательную индукцию НSPs (см. нижний сегмент в каждой паре блотов на Фигуре 1). Точно такие же (блокирующие индукцию HSPs) эффекты получались и с другими использованными авторами ингибиторами HSF1: KNK437 - 200 микроМ, NZ28 - 20 микроМ, триптолид - 20 наноМ и KRIBB11 - 20 наноМ, а также после трансфекции siRNA-векторами (данные здесь не приводятся).
Демонстрируется резкое усиление цитотоксичности, если опухолевые клетки (MCF-7, HeLa, PC-3) обрабатывались панобиностатом в комбинации с ингибиторами HSF1 (siRNA-векторы, KRIBB11, кверцетин или триптолид) (Фиг. 2 а, в, д). Аналогичные эффекты (усиленное подавление клоногенности) также наблюдались при комбинировании панобиностата с 100 микроМ KNK437 или 10 микроМ NZ28 (данные не приводятся). Из кривых клоногенности (относительное выживание колоний) видно, что комбинирование панобиностата с ингибиторами HSF1 уменьшает LD50 в 4-5 раз. При этом сами по себе (без панобиностата) примененные в указанных концентрациях ингибиторы HSF1 не оказывали цитотоксического эффекта (Фиг. 2 б, г, е).
Данные МТТ-теста (Фиг. 3 а, б, в), тоже доказывают, что комбинирование панобиностата с ингибиторами HSF1 усиливает цитотоксичность обработки опухолевых клеток. По сравнению с эффектами моно-обработки панобиностатом, жизнеспособность опухолевых клеток 3-х линий (MCF-7, HeLa и PC-3) достоверно снижалась еще примерно в 2 раза после комбинирования панобиностата с ингибиторами HSF1, что блокировало индукцию HSPs в обработанных клетках (для примера см. эффект кверцетина в Фиг. 1 - блоты, помеченные +Кв). Как и в случае оценки клоногенности (Фиг. 2 а, в, д), МТТ-тест здесь демонстрирует нетоксичность использованных концентраций ингибиторов HSF1 в отсутствие панобиностата.
Резкое усиление цитотоксичности, наблюдается при комбинировании панобиностата с ингибиторами HSF1 (Фиг. 4 а, в, г, д, е), вероятно, что это обусловлено существенным (приблизительно двукратным) повышением уровней апоптоза и некроза в опухолевых клетках, подвергнутых комбинированной обработке.
Важно добавить, что описываемое усиление цитотоксичеких эффектов при комбинировании панобиностата с каким-либо из описанных ингибиторов HSF1 наблюдалось только при концентрациях (дозах) ингибитора, которые предотвращали индукцию HSP70 и HSP27 в опухолевых клетках, обработанных панобиностатом (Фиг. 1, эффект кверцетина (+Кв)).
Предложенный способ показал, что комбинация с панобиностатом любого из перечисленных ингибиторов активации HSF1 или анти-HSF1/siRNA резко усиливает гибель обработанных раковых клеток на фоне подавления в них HSF1-зависимой экспрессии HSPs. Таким образом выявлен феномен активации HSF1 и индукции HSPs в обработанных панобиностатом раковых клетках, что повышает их жизнеспособность и сопротивляемость данному фармпрепарату, и во-вторых, в доказательстве возможности устранять этот вредный побочный эффект с помощью специальных агентов, предотвращающих активацию HSF1 и накопление HSPs в раковых клетках-мишенях, что усиливает противоопухолевые эффекты панобиностата.
| название | год | авторы | номер документа |
|---|---|---|---|
| Терапевтическая вакцина против рака на основе стрессовых белков в качестве иммуногенов | 2014 |
|
RU2694761C2 |
| ИНДУЦИРОВАНИЕ ГИБЕЛИ КЛЕТОК ПУТЕМ ИНГИБИРОВАНИЯ АДАПТИВНОГО ТЕПЛОВОГО ШОКОВОГО ОТВЕТА | 2009 |
|
RU2474612C2 |
| СПОСОБ СНИЖЕНИЯ КОЛИЧЕСТВА СТВОЛОВЫХ КЛЕТОК АДЕНОКАРЦИНОМЫ МОЛОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА | 2022 |
|
RU2798550C2 |
| Использование производного соединения тиохромено[2,3-с]хинолин-12-она для лечения немелкоклеточного рака легких | 2016 |
|
RU2663929C2 |
| ПЕПТИД, ПОЛУЧЕННЫЙ ИЗ HSP70, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ РАКА, СОДЕРЖАЩАЯ ТАКОЙ ПЕПТИД, ИНДУКТОР ИММУННОГО ОТВЕТА И СПОСОБ ПОЛУЧЕНИЯ АНТИГЕН-ПРЕЗЕНТИРУЮЩЕЙ КЛЕТКИ | 2015 |
|
RU2684911C2 |
| Композиция, ингибирующая рост и выживаемость опухолевых клеток | 2019 |
|
RU2707554C1 |
| Средство, обладающее противоклоногенной активностью в отношении опухолевых клеток человека | 2016 |
|
RU2648820C2 |
| СПОСОБ ИНГИБИРОВАНИЯ РАДИАЦИОННО-ИНДУЦИРОВАННОГО УВЕЛИЧЕНИЯ КОЛИЧЕСТВА СТВОЛОВЫХ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА | 2022 |
|
RU2800366C2 |
| СПОСОБ СНИЖЕНИЯ КЛОНОГЕННОЙ АКТИВНОСТИ СТВОЛОВЫХ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2019 |
|
RU2700695C2 |
| ШТАММ E.coli BL21(DE3)/pET-hHSP70 - ПРОДУЦЕНТ БЕЛКА ТЕПЛОВОГО ШОКА ЧЕЛОВЕКА HSP70 | 2006 |
|
RU2333956C1 |
Изобретение относится к области фармацевтики и медицины, а именно, к способу усиления цитотоксического эффекта панобиностата на опухолевые клетки линий MCF-7, HeLa, PC-3, характеризующийся тем, что опухолевые клетки перед обработкой панобиностатом в концентрации 2-400 нМ предварительно обрабатывают ингибитором HSF1, выбранным из кверцетина в концентрации 40 мкМ, триптолида в концентрации 20 нМ, KNK437 в концентрации 200 мкМ, NZ28 в концентрации 20 мкМ, KRIBB11 в концентрации 20 мкМ или миРНК-вектора для нокдауна HSF1. Технический результат заключается в предотвращении побочных эффектов панобиностата с помощью агентов, ингибирующих активацию HSF1 и накопление HSPs в раковых клетках-мишенях, обработанных панобиностатом. 4 ил., 1 пр.
Способ усиления цитотоксического эффекта панобиностата на опухолевые клетки линий MCF-7, HeLa, PC-3, характеризующийся тем, что опухолевые клетки перед обработкой панобиностатом в концентрации 2-400 нМ предварительно обрабатывают ингибитором HSF1, выбранным из кверцетина в концентрации 40 мкМ, триптолида в концентрации 20 нМ, KNK437 в концентрации 200 мкМ, NZ28 в концентрации 20 мкМ, KRIBB11 в концентрации 20 мкМ или миРНК-вектора для нокдауна HSF1.
| WASIM L., CHOPRA M | |||
| Synergistic anticancer effect of panobinostat and topoisomerase inhibitors through ROS generation and intrinsic apoptotic pathway induction in cervical cancer cells // Cellular Oncology | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| - Vol | |||
| Механический грохот | 1922 |
|
SU41A1 |
| - P | |||
| Питательное приспособление к трепальным машинам для лубовых растений | 1922 |
|
SU201A1 |
| PACHECO M | |||
| B | |||
| et al | |||
| Hydralazine and panobinostat attenuate malignant properties of prostate cancer cell | |||

