Изобретение относится к области препаративной биохимии, биофармакологии, биотехнологии и может быть использовано в медицине, иммунологии для получения вакцин и для исследовательских целей.
Белки теплового шока (heat shock proteins, HSPs) - обширное семейство высококонсервативных белков, представленных во всех организмах от бактерий до млекопитающих. HSPs играют важную роль в процессах сворачивания полипептидной цепи вновь синтезированных белков, репарации или элиминации неправильно свернутых или денатурированных белков, а также поддерживают стабильное состояние других внутриклеточных белков в условиях стресса. Согласно современной классификации, в основу которой положены различия в молекулярных массах, выделяют пять основных классов: HSP 100, HSP 90, HSP 70, HSP 60 и малые sHSP [J.G.Kiang and G.C.Tsokos. Heat Shock Protein 70 kDa: Molecular Biology, Biochemistry, and Physiology. Pharmacol. Then, 1998, 80(2): 183-201].
Наиболее изученным является семейство HSP70, белки которого локализуются в цитоплазме, ядре, эндоплазматическом ретикулуме и митохондриях. Идентифицировано 11 генов, кодирующих белки этого семейства [L.A.Sonna, J.Fujita, S.L.Gaffin, С.Lilly. Molecular Biology of Thermoregulation. Invited Review: Effects of heat and cold stress on mammalian gene expression. J Appl Physiol, 2002, 92: 1725-42].
В настоящее время большой интерес исследователей вызывают функции HSP70 как молекул межклеточной сигнализации, способных вызывать продукцию цитокинов клетками иммунной системы путем рецептор-опосредуемых взаимодействий [A.Asea, S.K.Kraeft, E.A.Kurt-Jones, M.A.Stevenson, L.B.Chen, R.W.Finberg, G.C.Koo, S.K.Calderwood. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med, 2000, 6: 435-42]. Другие исследования показали, что иммунизация препаратами HSP70, выделенными из опухолевых и инфицированных вирусами клеток, вызывает специфический антиопухолевый [Н.Udono, P.K.Srivastava. Heat shock protein 70-associated peptides elicit specific cancerimmunity. J Exp Med., 1993 Oct 1; 178(4): 1391-6] и антивирусный [А.Т.Ciupitu, M.Petersson, C.L.O′Donnell, K.Williams, S.Jindal, R.Kiessling, and R.M.Welsh. Immunization with a lymphocytic choriomeningitis virus peptide mixed with heat shock protein 70 results in protective antiviral immunity and specific cytotoxic lymphocytes. J. Exp. Med., 1998, 187: 685-91] клеточный иммунный ответ. Специфичность такого ответа обусловлена способностью HSP-молекул связывать различные белки и пептиды (в т.ч. онко- и вирусные пептиды) и доставлять их антиген-презентирующим клеткам [D.Arnold-Schild, D.Hanau, D.Spehner, С.Schmid, H.G.Rammensee, H. de la Salle, H.Schild. Receptor-mediated endocytosis of heat shock proteins by professional antigen-presenting cells. J Immunol., 1999, 162 (7): 3757-60]. Возможность образования комплексов HSP70 с белковыми антигенами in vitro позволила создавать на их основе противоопухолевые и противовирусные вакцины [N.E.Blachere, Z.Li, R.Y.Chandawarkar, R.Suto, N.S.Jaikaria, S.Basu, H.Udono, P.K.Srivastava. Heat-shock protein complexes, reconstituted in vitro, elicit peptide-specific cytotoxic T-lymphocyte response and tumor immunity. J. Exp.Med., 1997, 186(8): 1315-22].
Природные белковые препараты HSP70 млекопитающих были выделены из опухолевых тканей [Р.К.Srivastava. Purification of heat shock protein-peptide complexes for use in vaccination against cancers and intracellular pathogens. Methods, 1997 Jun, 12(2): 165-71]. Эти способы включали в себя накопление и гомогенизацию ткани, экстракцию целевого белка, а также фракционирование препарата с помощью нескольких хроматографических стадий. Основными недостатками получения HSP из природных источников являются необходимость накопления опухолевых тканей, трудоемкость процесса выделения и малый конечный выход.
Генно-инженерные методы позволяют нарабатывать препаративные количества HSP, необходимые для исследования различных функций белка. Так, в клетках штамма E.coli BL21(DE3) был осуществлен биосинтез рекомбинантного HSP70 человека, соответствующий гену HSPA1L [S.Jindal, P.Murrey, S.Rosenberg, R.A.Young and K.P.Williams. Human stress protein hsp70: overexpression in E.coli, purification and characterization. Biotechnology, 1995, 13: 1105-9]. Для этого кДНК HSP70 была встроена по сайтам рестрикции NdeI-BamHI в экспрессионный вектор рЕТ-3а. После трансформации штамма E.coli BL21(DE3) полученной рекомбинантной плазмидой и индукции экспрессии гена изопропилтиогалактозидом целевой белок накапливался в клетках в виде телец включения. Выделение предварительно рефолдированного белка HSP70 было основано на аффинной хроматографии с использованием АТФ-агарозы. К недостаткам этого метода относится контаминация целевого белка DnaK - бактериальным аналогом эукариотического HSP70 и, как следствие, введение дополнительной стадии очистки посредством ионно-обменной хроматографии.
Задачей предлагаемого изобретения является создание эффективной системы экспрессии для получения по упрощенной технологии другого представителя семейства HSP70, кодируемого геном HSPA1B (Gene Bank NM005346).
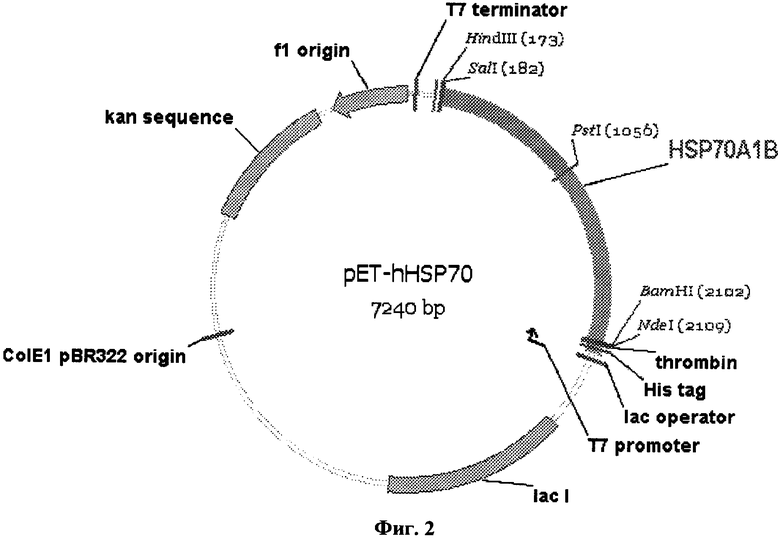
Техническим результатом заявленного изобретения является получение нового рекомбинантного белка теплового шока человека hHSP70A1B. Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pET-hHSP70, кодирующей рекомбинантный белок теплового шока человека hHSP70A1B с молекулярной массой 72,33 кДа и состоящей из NdeI-HindIII-фрагмента плазмиды рЕТ-28а(+) и HindIII-NdeI-фрагмента нуклеотидной последовательности, представленной на фиг.1, который кодирует hHSP70A1B и включает уникальные сайты узнавания рестрикционными эндонуклеазами со следующими координатами: SalI-182, PstI-1056. BamHI-2102 п.о., а также за счет получения штамма E.coli BL21(DE3)/pET-hHSP70-продуцента, обеспечивающего синтез рекомбинантного белка теплового шока человека hHSP70A1B размером 663 аминокислотных остатков (SEQ ID №1) с уровнем экспрессии 30-40% от суммарного содержания белка клеток.
Преимуществом заявленного технического решения является, во-первых, тот факт, что рекомбинантный белок накапливается в клетках как в виде телец включения, так и в растворимой форме (примерно в равных количествах), что обусловлено сочетанием свойств аминокислотной последовательности белка hHSP70A1B и свойств применяемой экспрессионной системы, в частности наличие оптимальных регуляторных элементов: Т7-lac-промотора для предотвращения базальной экспрессии гена до момента начала индукции и высокого уровня транскрипции соответствующей мРНК при индукции, высокоэффективного терминатора транскрипции Т7. Во-вторых, введение в состав полипептидной цепи 6 остатков гистидина, значительно упрощающее выделение рекомбинантного белка, и наличие сайта узнавания тромбином для удаления (при необходимости) гистидиновых остатков из конечного белкового продукта. Также преимуществом предлагаемой генно-инженерной конструкции является наличие С- и N-концевых сайтов рестрикции SalI и BamHI для возможности создания гибридных (fusion) белков, имитирующих комплексы hНSР70А1В-антиген. Преимущество предлагаемого штамма E.coli заключается в использовании бактерий с пониженным уровнем протеолитического расщепления рекомбинантного белка и загрязнения выделяемого белка наиболее активными протеазами E.coli.
Конструирование новой последовательности ДНК, кодирующей белок hHSP70A1B, осуществляют на основе векторной плазмиды рЕТ28-а(+). кДНК белка теплового шока hHSP70A1B человека, соответствующую гену HSPA1B (GenBank NM_005346) синтезируют на мРНК с помощью обратной транскриптазы мышиного вируса лейкемии Молони с использованием праймера на 3′-нетранслируемую область мРНК. Транслируемую область гена HSPA1B человека, фланкированную сайтами рестрикции NdeI и HindIII, получали промежуточным клонированием в плазмиде pUC19.
Рекомбинантная плазмидная ДНК pET-hHSP70 характеризуется следующими признаками:
- имеет размер 7240 п.о.;
- кодирует белок теплового шока человека hHSP70A1B;
- состоит из NdeI-HindIII-фрагмента плазмиды рЕТ-28а(+) и HindIII-NdeI-фрагмента приведенной на фиг.1 нуклеотидной последовательности длиной 1941 п.о., кодирующего hHSP70A1B;
- содержит генетический маркер Kan, детерминирующий устойчивость трансформированных плазмидой pET-hHSP70 клеток E.coli к канамицину, а также уникальные сайты узнавания рестрикционными эндонуклеазами со следующими координатами: SalI-182, PstI-1056, BamHI-2102 п.о.
Для получения штамма-продуцента белка теплового шока hHSP70 компетентные клетки E.coli BL21(DE3) трансформировали рекомбинантной плазмидной ДНК рЕТ-hHSP70 с последующей селекцией на агаризованной среде LB с канамицином. Отобранные клоны наращивали для оценки продуктивности, устойчивости к антибиотику и стабильности трансформации.
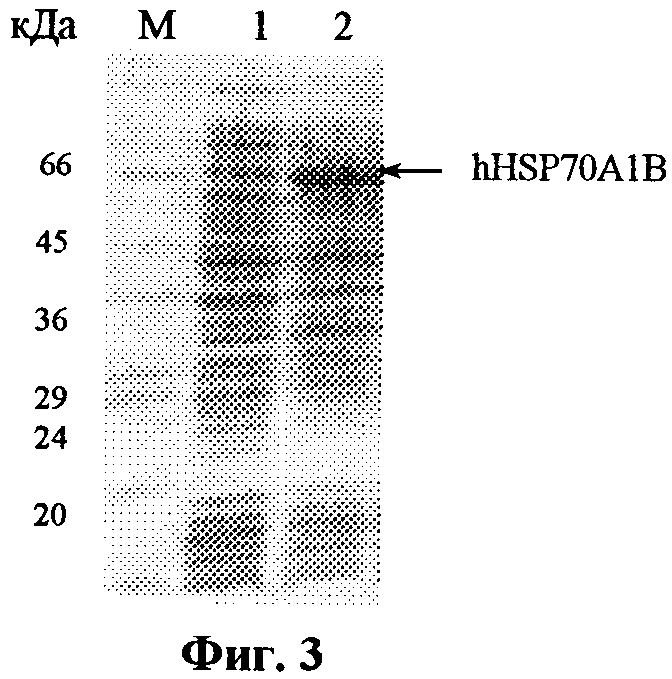
Штамм E.coli BL21(DE3), трансформированный плазмидой pET-hHSP70, выращивают в среде LB в аэробных условиях с добавлением канамицина до 50 мкг/мл в течение 12-14 часов (посевной материал), инокулируют новую порцию питательной среды в соотношении 1:50, растят культуру до достижения оптической плотности A600 0,6-0,8, индуцируют изопропилтио-β-D-галактозидом (IPTG) и растят еще 3-4 часа. Клетки E.coli BL21(DE3)/pET-hHSP70 являются суперпродуцентом. При индукции IPTG происходит эффективный биосинтез рекомбинантного hHSP70A1B, который накапливается в клетках, как в растворимой форме, так и в виде телец включения, в количестве более 30% суммарного белка бактерий (фиг.3). Получение из клеток продуцента рекомбинантного hHSP70A1B включает несколько стадий. Бактерии отделяют от культуральной среды и разрушают одним из обычно применяемых способов. Центрифугированием отделяют растворимую клеточную фракцию от телец включения. Последние отмывают буферным раствором от водорастворимых компонентов клетки, проводят солюбилизацию, восстановление и рефолдинг рекомбинантного белка. Окончательную очистку целевого белка проводят методом металло-хелатной хроматографии. Выход полученного таким способом рекомбинантного hHSP70A1B составляет примерно 10 мг с 1 л бактериальной культуры. Выделение рекомбинантного hHSP70A1B из растворимой клеточной фракции проводят в одну стадию - методом металло-хелатной хроматографии. При этом выход целевого белка составляет примерно 30 мг с 1 л бактериальной культуры.
Полученный штамм-продуцент E.coli BL21(DE3)/pET-hHSP70 характеризуется следующими признаками.
Морфологические признаки: клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки: при росте на агаризованной среде LB колонии круглые, гладкие, блестящие, с ровными краями. Диаметр колоний 1-3 мм. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением, осадок легко седиментирует.
Физиолого-биохимические признаки: клетки растут при температуре 4-42°С, температурный оптимум для роста клеток 37°С, оптимум рН 6,8-7,5. В качестве источника азота используют как минеральные соли азота, так и органические соединения в виде аминокислот, пептона, триптона, дрожжевого экстракта. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам: клетки проявляют устойчивость к канамицину, обусловленную наличием в плазмиде гена kan. Устойчивость к канамицину у штамма не ниже 120 мкг/мл на твердых агаризованных средах, при наращивании в жидких полноценных средах - не менее 60 мкг/мл.
Генотипические признаки: основной генотипический признак штамма E.coli BL21(DE3)/pET-hHSP70 (F-, ompT, hsdS β(r- βm- β), gal dcm, λ(DE3)) состоит в наличии под регуляцией Т7 промотора нуклеотидной последовательности кодирующей области гена белка теплового шока человека HSPA1B, интегрированной в состав экспрессионного вектора, содержащего в качестве селективного маркера ген kan.
Стабильность плазмиды pET-hHSP70 в штамме E.coli BL21(DE3)/pET-hHSP70: при хранении клеток на агаризованной среде (сроком до 1 месяца) и в процессе культивирования в жидкой среде с антибиотиком не происходит потери и перестройки плазмиды pET-hHSP70. Штамм сохраняет плазмиду в процессе не менее 5 пересевов на бесселективной среде.
Изобретение иллюстрируется следующими фигурами графических изображений.
На фиг.1 представлена нуклеотидная последовательность фрагмента плазмиды pET-hHSP70 и кодируемая им аминокислотная последовательность рекомбинантного белка hHSP70A1B; на фиг.2 - физическая карта плазмиды pET-hHSP70. Указаны промотор РНК-полимеразы фага Т7, участок инициации репликации (ori) и терминатор транскрипции Т7, генетический маркер Kan устойчивости к канамицину, уникальные сайты рестрикции; на фиг.3 - электрофореграмма лизатов клеток штамма-продуцента E.coli BL21(DE3)/pET-hHSP70 до индукции (дорожка 1), лизатов клеток штамма-продуцента E.coli BL21(DE3)/pET-hHSP70 после индукции изопропилтио-β-D-галактозидом (дорожка 2) в 12%-ном полиакриламидном геле в присутствии SDS (М-белковые маркеры молекулярной массы; стрелкой указан белок hHSP70A1B).
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование промежуточной рекомбинантной плазмидной ДНК pUC-hHSP70A1B.
кДНК белка теплового шока hHSP70A1B человека синтезируют на мРНК с помощью обратной транскриптазы мышиного вируса лейкемии Молони с использованием праймера на 3'-нетранслируемую область мРНК 5'-GGC CCC TAA TCT ACC-3'.
Амплификацию ДНК проводят с использованием 2 пар праймеров: прямой праймер 5'-CAT ATG GGA TCC GCC AAA GCC GCG G-3', обратный праймер 5'-G AAG TCC TGC AGC AGC TTC TGC-3' (фрагмент I, 1068 п.о.); прямой праймер 5'-GCA GAA GCT GCT GCA GGA CTT C-3', обратный праймер 5'-AAG CTT TTA GTC GAC CTC CTC AAT GC-3'(фрагмент II, 895 п.о.). Для синтеза фрагментов I и II проводят последовательно 5 (94°С, 30 с; 44°С, 30 с; 72°С, 30 с) и 30 (94°С, 30 с; 67°С, 30 с; 72°С, 30 с) циклов амплификации. Реакционная смесь в объеме 50 мкл содержит 1 мкл кДНК, по 20 пмоль/мкл каждого праймера для амплификации соответствующих фрагментов, 5 мкл буфера для ПЦР, 200 мкМ dNTPs, 5 ед Taq-полимеразы. Продукты амплификации очищают электрофорезом в 1,2%-ном агарозном геле, полосы величиной 1068 п.о. (фрагмент I) и 895 п.о (фрагмент II) выделяют из 1%-ной легкоплавкой агарозы.
2 мкг плазмидной ДНК pUC19 (Fermentas) обрабатывают рестриктазой SmaI (Fermentas) (10 ед) в 50 мкл буферного раствора Y+/tango (33 мМ Tris-ацетат (рН 7,9 при 37°С); 10 мМ Mg-ацетат, 66 мМ К-ацетат, 0,1 мг/мл БСА) в течение 1,5 ч при 30°С, переосаждают ДНК из раствора этанолом и дефосфорилируют щелочной фосфотазой CIAP (0,07 ед) в течение 30 мин при 37°С в 50 мкл буфера для CIAP (0,1 М Tris-HCl (рН 7,5 при 37°C); 0,1 M MgCl2).
180 нг (0,26 пмоль) выделенного из 1% легкоплавкой агарозы фрагмента I длиной 1068 п.о. (предварительно кинированного полинуклеотидкиназой фага Т4 (10 ед) в 20 мкл буфера, содержащего 500 мМ Tris-HCl (рН 7,6 при 25°С); 100 мМ MgCl2; 50 мМ ДТТ; 1 мМ спермидин; 1 мМ ЭДТА, в течение 30 мин при 37°С) и 150 нг (0,08 пмоль) линеаризованной дефосфорилированной плазмиды pUC19 лигируют по сайту рестрикции SmaI при помощи 2 ед. Т4 ДНК-лигазы в 10 мкл буфера для лигирования (400 мМ Tris-HCl (рН 7,8 при 25°С); 100 мМ MgCl2; 100 мМ ДТТ; 5 мМ АТФ) в течение 12 ч при 8°С. 10 мкл реакционной смеси используют для трансформации 100 мкл компетентных клеток штамма E.coli JM109. Трансформированные клетки высевают на агаризованную среду LB, содержащую 50 мкг/мл ампициллина. Отбор положительных клонов проводят при помощи ПЦР с использованием универсальных праймеров для pUC-векторов. Из отобранных клонов выделяют плазмидную ДНК pUC19-фрагмент I. Нуклеотидную последовательность встроенного фрагмента I определяют методом секвенирования по Сенгеру.
180 нг (0,31 пмоль) выделенного из 1% легкоплавкой агарозы и кинированного фрагмента II длиной 895 п.о. встраивают по сайту рестрикции SmaI в линеаризованную дефосфорилированную плазмиду pUC19 (0,1 мкг, 0,08 пмоль) в 10 мкл буфера для лигирования с 2 ед. Т4 ДНК-лигазы. 10 мкл реакционной смеси трансформируют 100 мкл компетентных клеток штамма E.coli JM109. 200 мкл трансформированных клеток высевают на агаризованную среду LB, содержащую 50 мкг/мл ампициллина. Отбор положительных клонов проводят при помощи ПЦР с использованием специфических праймеров для pUC-векторов. Из отобранных клонов выделяют плазмидную ДНК pUC19-фрагмент II. Последовательность ДНК, кодирующую фрагмент II, определяют секвенированием по методу Сенгера.
2 мкг плазмидных ДНК pUC19-фрагмент I и рUС19-фрагмент II обрабатывают рестриктазами PstI (5 ед) и HindIII (10 ед) (Fermentas) в 50 мкл буферного раствора Y+/tango (33 мМ Tris-ацетат (рН 7,9 при 37°С); 10 мМ Mg-ацетат, 66 мМ К-ацетат, 0,1 мг/мл БСА) в течение 1,5 ч при 37°С и выделяют в 1%-ном геле легкоплавкой агарозы фрагмент II (895 п.о.).
0,1 мкг (0,04 пмоль) векторной части плазмиды рUС19-фрагмент I и 0,09 мкг (0,15 пмоль) рестриктированного по PstI и HindIII фрагмента II длиной 895 п.о. лигируют при помощи 1,5 ед. Т4 ДНК-лигазы в 10 мкл буфера для лигирования. 5-10 мкл реакционной смеси используют для трансформации 100 мкл компетентных клеток штамма E.coli JM109. Трансформированные клетки высевают на агаризованную среду LB, содержащую 50 мкг/мл ампициллина. Из положительных клонов выделяют промежуточную плазмидную ДНК pUC19-hHSP70, содержащую полноразмерную транслируемую область гена HSPA1B длиной 1941 п.о.
Пример 2. Конструирование рекомбинантной плазмидной ДНК pET-hHSP70.
1 мкг плазмидной ДНК рЕТ-28а(+) (Novagen) обрабатывают рестриктазами NdeI и HindIII (Fermentas) (по 5 ед) в 50 мкл буферного раствора R+ (10 мМ Tris-HCl (рН 8,5 при 37°С); 10 мМ MgCl2, 100 мМ KCl, 0,1 мг/мл БСА) в течение 30 мин при 37°С и после инактивации реакции 65°С (20 мин) переосаждают ДНК из раствора этанолом.
2 мкг плазмидной ДНК pUC-hHSP70 обрабатывают рестриктазами NdeI и HindIII (по 10 ед) в 50 мкл буферного раствора R+ (10 мМ Tris-HCl (pH 8,5 при 37°С); 10 мМ MgCl2, 100 мМ KCl, 0,1 мг/мл БСА) в течение 30 мин при 37°С и из полученного гидролизата выделяют в 1%-ном геле легкоплавкой агарозы фрагмент длиной 1941 п.о. Полученный фрагмент гена HSPA1B длиной 1941 п.о. (72 нг, 0,058 пмоль) и линеаризованную плазмидную ДНК рЕТ-28а(+) (90 нг, 0,025 пмоль) соединяют при помощи лигазной реакции в 10 мкл буфера для лигирования, содержащего 2 ед. Т4 ДНК-лигазы. 10 мкл реакционной смеси используют для трансформации 100 мкл компетентных клеток BL21(DE3). Трансформированные клетки высевают на агаризованную среду LB, содержащую 50 мкг/мл канамицина. Из отобранных методом ПЦР (с помощью универсальных праймеров для рЕТ-векторов) клонов выделяют целевую плазмидную ДНК pET-hHSP70 (фиг.2). Нуклеотидную последовательность встроенного фрагмента подтверждают секвенированием по Сенгеру.
Пример 3. Получение штамма-продуцента E.coli BL21(DE3)/pET-hHSP70 и определение его продуктивности.
Штамм-продуцент E.coli BL21(DE3)/pET-hHSP70 получают методом кальциевой трансформации компетентных клеток E.coli BL21(DE3) плазмидой pET-hHSP70. Эффективность трансформации составляет около 105 клонов на 1 мкг плазмидной ДНК. Трансформанты отбирают на среде с канамицином, изменение генотипа определяют методом ПЦР с помощью праймеров для рЕТ-векторов. Для определения продуктивности отобранные клоны выращивают при 37°С в 3,5 мл LB с 50 мкг/мл канамицина в течение 2,5-3 ч на качалке со скоростью вращения 250 об/мин до мутности А600˜0,6, добавляют изопропилтио-β-D-галактозид до концентрации 0,5 мМ и продолжают процесс еще 4 ч. Отбирают пробы клеточных суспензий до и после индукции в количестве, соответствующем 0,25 о.е., центрифугируют 5 мин при 6000 об/мин, после чего суспендируют в 50 мкл лизирующего буфера (30 мМ Трис-HCl, рН 6,8, 10% глицерин, 1% додецилсульфат натрия, 3% меркаптоэтанол, 0,005% бромфеноловый синий), нагревают 5 мин при 95°С и образцы объемом 20 мкл анализируют электрофорезом в 12% полиакриламидном геле с додецилсульфатом натрия. Гель прокрашивают кумасси R-250 (фиг.3) по стандартной методике и сканируют для определения относительного количества белка в полосе целевого белка. По данным сканирования содержание рекомбинантного hHSP70A1B составляет 30-40% от всех клеточных белков.

Изобретение относится к области биотехнологии и может быть использовано в медико-биологической промышленности. На основе вектора рЕТ28а(+) сконструирована плазмидная ДНК pET-hHSP70, которая содержит нуклеотидную последовательность, кодирующую белок теплового шока человека hHSP70A1B. Путем трансформации клеток Е.coli плазмидной ДНК pET-hHSP70 получен штамм Е.coli BL21(DE3)/pET-hHSP70 - продуцент рекомбинантной формы hHSP70. Использование предложенного изобретения позволяет получить рекомбинантный белок hHSP70 с высоким выходом (в количестве 30-40% от суммарного содержания белка клеток) и упростить его выделение и очистку. 3 ил.
Штамм E.coli BL21(DE3)/pET-hHSP70 - продуцент рекомбинантного белка теплового шока человека hHSP70 с аминокислотной последовательностью SEQ ID № 1.
| JINDAL et al | |||
| Biotechnology, 1995, 13, 1105-1109 http://www.merckbiosciences.co.uk/NVG/pET Table | |||
| КОМПОЗИЦИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ, СПОСОБ ПОЛУЧЕНИЯ ТАКОЙ КОМПОЗИЦИИ, ФАРМАЦЕВТИЧЕСКИЙ НАБОР РЕАГЕНТОВ ДЛЯ ИММУНОТЕРАПИИ И ПРОФИЛАКТИЧЕСКОЙ ВАКЦИНАЦИИ ОПУХОЛЕВЫХ ЗАБОЛЕВАНИЙ АНОГЕНИТАЛЬНОЙ СФЕРЫ, СПОСОБ ИММУНОТЕРАПИИ И ПРОФИЛАКТИЧЕСКОЙ ВАКЦИНАЦИИ НА ЕГО ОСНОВЕ | 2002 |
|
RU2229307C1 |

