Область техники, к которой относится изобретение
Настоящее изобретение относится к области антител. В частности, настоящее изобретение относится к биспецифическим антителам, к содержащим их фармацевтическим композициям и к их применениям.
Предпосылки создания изобретения
Биспецифические антитела (BsAbs) представляют собой антитела или антитело-подобные молекулы, имеющие две разные специфичности связывания. Биспецифические антитела широко используются в биомедицине, особенно в иммунотерапии опухолей.
Биспецифические антитела можно получить способами, такими как химическая инженерия, клеточная инженерия и генная инженерия. Преимуществом генной инженерии является то, что антитела легко можно модифицировать, таким образом, можно разработать и получить множество различных форм фрагментов биспецифических антител, включая биспецифические IgG (BsIgG), дополненные IgG, фрагменты биспецифических антител (фрагменты BsAb), биспецифические слитые белки и конъюгаты биспецифических антител (конъюгаты BsAb), (см. Christoph Spiess, Qianting Zhai, Paul J. Carter, Alternative molecular formats and therapeutic applications for bispecific antibodies. Molecular Immunology 67 (2015) 95-106).
Первый тип: биспецифические IgG, которые имеют такую же структуру и молекулярную массу, как у природных моноклональных антител, но имеют два разных Fab. (1) В ранее используемой технологии Triomab крысиные и мышиные гибридомы снова подвергают слиянию с образованием "гибрид-гибридомы" для продукции мышиных и крысиных химерных биспецифических антител. Очистку осуществляют в соответствии с селективностью крысиного и мышиного Fc к аффинному наполнителю, и затем получают представляющий интерес продукт. Эта технология имеет недостатки, такие как низкое экспрессируемое количество и низкий выход после очистки. (2) В дальнейшем для модификации Fc используется технология конструирования антител, например, “лейциновая молния”, “выступы-во-впадины” или электростатическое взаимодействие, таким образом, гетеродимер образуется из двух разных тяжелых цепей, а затем легкую цепь и тяжелую цепь спаривают должным образом с использованием общих легких цепей, технологии "выступы-во-впадины" или электростатического взаимодействия, или нужный представляющий интерес продукт получают путем конструирования различных подтипов κ и λ легкой цепи и очистки с использованием аффинного наполнителя, специфического к κ цепи и λ цепи. Биспецифические антитела такого типа имеют фармакокинетику, близкую к фармакокинетике природных антител, но их недостатком является то, что трудно идентифицировать и удалить примеси.
Второй тип: дополненные IgG, где природное моноклональное антитело IgG используется как каркас, а антигенсвязывающий домен (такой как фрагмент вариабельной области (Fv), одноцепочечный фрагмент вариабельной области (ScFv), антигенсвязывающий фрагмент (Fab) или полипептид и т.д.) слит на N-конце или C-конце тяжелой или легкой цепи антитела; биспецифические антитела такого типа имеют молекулярную массу больше чем 150кДа и преимущественно симметричную структуру. Фармакокинетика дополненных IgG подобна фармакокинетике природных антител, но дополненные IgG имеют больший период полужизни и удобны для очистки. Недостатками дополненных IgG являются низкое экспрессируемое количество и недостаточная стабильность и т.д.
Третий тип: фрагменты биспецифических антител, где фрагменты вариабельных областей антитела сохраняются, а часть или даже вся константная область антитела делетирована; фрагменты вариабельной области тяжелой цепи (VH) и фрагменты вариабельной области легкой цепи (VL), как правило, связаны с образованием ScFv посредством линкерного белка, или спонтанно спарены с образованием Fv. Молекулярная масса биспецифических антител такого типа не превышает 150кДа, их период полужизни короткий. Недостатки фрагментов биспецифических антител следующие: трудно достичь такого же уровня экспрессии фрагмента биспецифического антитела, как уровень экспрессии моноклонального антитела, молекула недостаточно стабильна, и низкий выход после очистки и т.д.
Четвертый тип: биспецифические слитые белки, где фрагменты вариабельных областей антитела сохраняются, а другие белки или фармацевтические молекулы (такие как константные области T-клеточного рецептора, человеческий сывороточный альбумин или белки токсинов и т.п.) слиты. Молекулярная масса биспецифических антител такого типа находится в пределах от 75кДа до 160кДа, процесс очистки затруднителен, а молекулярная стабильность также имеет определенные риски.
Пятый тип: конъюгаты биспецифических антител, где два разных моноклональных антитела или ScFv связаны вместе химическими методами. Способ получения антител такого типа является сложным и имеет низкий конечный выход, и поэтому антитела такого типа в настоящее время по существу нигде в мире не используются.
Все из вышеуказанных типов биспецифических антител имеют некоторые недостатки, что приводит к потенциальным рискам при клиническом применении таких продуктов. Для устранения этих рисков настоящее изобретение представляет новое биспецифическое антитело.
Сущность изобретения
Настоящее раскрытие представляет способ конструирования и получения нового биспецифического антитела. Конкретная структура биспецифического антитела показана на Фиг. 1. Все из антител в настоящем раскрытии имеют константную область IgG человека и относятся к дополненному IgG или фрагментам биспецифических антител.
В настоящем изобретении испытали аффинность, стабильность, биологическую активность и эффективность нового биспецифического антитела и сравнили с тремя существующими биспецифическими антителами. Результаты показывают, что новое биспецифическое антитело имеет лучшую стабильность, биологическую активность и эффективность, и способы его экспрессии и получения более удобны.
Технические решения
Биспецифическое антитело по настоящему изобретению имеет две идентичные гибридные тяжелые цепи и две идентичные гибридные легкие цепи, где две гибридные тяжелые цепи образуют пару, и гибридная легкая цепь и гибридная тяжелая цепь образуют пару.
В некоторых аспектах гибридные тяжелые цепи антитела связаны одной или несколькими дисульфидными связями.
В некоторых аспектах гибридная легкая цепь и гибридная тяжелая цепь антитела связаны одной или несколькими дисульфидными связями.
В некоторых аспектах гибридная тяжелая цепь антитела содержит вариабельную область тяжелой цепи антитела a (VHa), первую константную область (CH1), вариабельную область легкой цепи антитела b (VLb) и Fc фрагмент. Где VLb расположена между CH1 и Fc и связана при помощи линкера или пептидной связи. Гибридная легкая цепь антитела содержит вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи (CL) и вариабельную область тяжелой цепи антитела b (VHb). Где VHb расположена на C-конце CL и связана при помощи линкера или пептидной связи.
В некоторых аспектах гибридная тяжелая цепь антитела содержит вариабельную область тяжелой цепи антитела a (VHa), первую константную область (CH1), вариабельную область тяжелой цепи антитела b (VHb) и Fc фрагмент. Где VHb расположена между CH1 и Fc и связана при помощи линкера или пептидной связи. Гибридная легкая цепь антитела содержит вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи (CL) и вариабельную область легкой цепи антитела b (VLb). Где VLb расположена на C-конце CL и связана при помощи линкера или пептидной связи.
В некоторых аспектах VHa-VLa пара является специфической к антигену A, который включает, но не ограничивается этим, антигены поверхности опухолевых клеток, антигены поверхности иммунных клеток, вирусы, бактерии, эндотоксины, цитокины, такие как CD3, SLAMF7, CD38, BCMA, CD16a, CEA, PD-L1, PD-1, CTLA-4, TIGIT, LAG-3, VEGF, B7-H3, TGF-β, IL-10 и т.д.
В некоторых аспектах VHb-VLb пара является специфической к антигену B, который включает, но не ограничивается этим, антигены поверхности опухолевых клеток, антигены поверхности иммунных клеток, вирусы, бактерии, эндотоксины, цитокины, такие как CD3, SLAMF7, CD38, BCMA, CD16a, CEA, PD-L1, PD-1, CTLA-4, TIGIT, LAG-3, VEGF, B7-H3, TGF-β, IL-10 и т.д.
В частности, в одном аспекте настоящее изобретение обеспечивает биспецифическое антитело, включающее две идентичные гибридные тяжелые цепи и две идентичные гибридные легкие цепи, где две гибридные тяжелые цепи образуют пару, и гибридная легкая цепь и гибридная тяжелая цепь образуют пару, где гибридная тяжелая цепь включает вариабельную область тяжелой цепи антитела a (VHa), первую константную область CH1, вариабельную область 1 антитела b и Fc фрагмент, и вариабельная область 1 антитела b связана с CH1 через линкер 2 или пептидную связь, или связана с Fc через линкер 3 или пептидную связь. Гибридная легкая цепь биспецифического антитела включает вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи CL и вариабельную область 2 антитела b, где вариабельная область 2 антитела b связана с C-концом CL через линкер 1 или пептидную связь, где
(1) когда вариабельная область 1 антитела b представляет собой вариабельную область тяжелой цепи антитела b (VHb), вариабельная область 2 антитела b представляет собой вариабельную область легкой цепи антитела b (VLb);
или
(2) когда вариабельная область 1 антитела b представляет собой вариабельную область легкой цепи антитела b (VLb), вариабельная область 2 антитела b представляет собой вариабельную область тяжелой цепи антитела b (VHb),
где, VHa-VLa пара нацеливается на антиген A, а VHb-VLb пара нацеливается на антиген B.
В одном варианте осуществления структура биспецифического антитела по настоящему изобретению представляет собой F(ab)2-(Fv)2-Fc, где F(ab)2 включает две VHa, две CH1, две VLa и две CL; (Fv)2 включает две VHb и две VLb, где положения VHb и VLb являются взаимозаменяемыми; и Fc фрагмент включает шарнирную область и вторую константную область CH2 и третью константную область CH3.
В одном варианте осуществления биспецифическое антитело симметрично имеет структуру, показанную в формуле I от N-конца к C-концу:
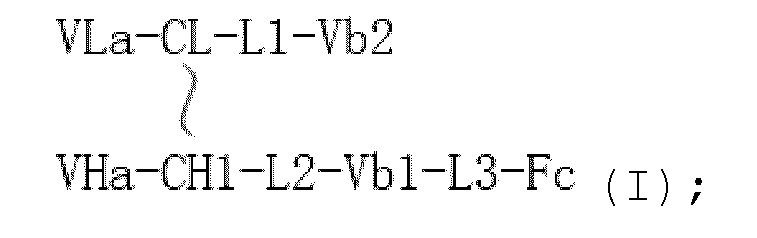
где
"  " представляет собой дисульфидную связь или ковалентную связь;
" представляет собой дисульфидную связь или ковалентную связь;
"-" представляет собой пептидную связь;
L1, L2 и L3 каждый независимо представляет собой пептидную связь или линкер или шарнир;
VHa представляет собой вариабельную область тяжелой цепи антитела a;
VLa представляет собой вариабельную область легкой цепи антитела a;
CH1 представляет собой первую константную область;
Fc представляет собой Fc фрагмент, содержащий шарнирную область;
CL представляет собой константную область легкой цепи;
Vb1 представляет собой вариабельную область 1 антитела b;
Vb2 представляет собой вариабельную область 2 антитела b.
В одном варианте осуществления биспецифическое антитело симметрично имеет структуру, показанную в формуле II или Формуле III от N-конца к C-концу:

где
" " представляет собой дисульфидную связь или ковалентную связь;
"-" представляет собой пептидную связь;
L1, L2 и L3 каждый независимо представляет собой пептидную связь или линкер;
VHa представляет собой вариабельную область тяжелой цепи антитела a;
VLa представляет собой вариабельную область легкой цепи антитела a;
CL представляет собой константную область легкой цепи;
H представляет собой шарнирную область;
CH1, CH2 и CH3 представляют собой первую константную область, вторую константную область и третью константную область, соответственно;
VHb представляет собой вариабельную область тяжелой цепи антитела b;
VLb представляет собой вариабельную область легкой цепи антитела b.
В одном варианте осуществления биспецифическое антитело включает:
(1) гибридную тяжелую цепь, где гибридная тяжелая цепь включает вариабельную область тяжелой цепи антитела a (VHa), первую константную область CH1, линкер 2 (или пептидную связь) и вариабельную область тяжелой цепи антитела b (VHb), линкер 3 (или пептидную связь) и Fc фрагмент в последовательности от N-конца к C-концу; и
(2) гибридную легкую цепь, где гибридная легкая цепь включает вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи CL, линкер 1 (или пептидную связь) и вариабельную область легкой цепи антитела b (VLb) в последовательности от N-конца к C-концу.
В одном варианте осуществления биспецифическое антитело включает:
(1) гибридную тяжелую цепь, где гибридная тяжелая цепь включает вариабельную область тяжелой цепи антитела a (VHa), первую константную область CH1, линкер 2 (или пептидную связь) и вариабельную область легкой цепи антитела b (VLb), линкер 3 (или пептидную связь) и Fc фрагмент в последовательности от N-конца к C-концу; и
(2) гибридную легкую цепь, где гибридная легкая цепь включает вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи CL, линкер 1 (или пептидную связь) и вариабельную область тяжелой цепи антитела b (VHb) в последовательности от N-конца к C-концу.
В одном варианте осуществления VHa-VLa пара образует одну или несколько межцепочечных дисульфидных связей, и VHb-VLb пара образует одну или несколько межцепочечных дисульфидных связей.
В одном варианте осуществления вариабельная область 1 антитела b представляет собой вариабельную область легкой цепи антитела b (VLb), а вариабельная область 2 антитела b представляет собой вариабельную область тяжелой цепи антитела b (VHb).
В одном варианте осуществления линкеры 1-3 могут быть одинаковыми или отличными друг от друга, и линкеры 1-3 каждый независимо выбран из группы, состоящей из SEQ ID NO: 69-90, предпочтительно выбран из SEQ ID NO: 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 82, 83, 85, 86, 87, 88, 89 или 90.
В одном варианте осуществления последовательность шарнирной области выбрана из группы, состоящей из SEQ ID NO: 91-103.
В одном варианте осуществления последовательность CL выбрана из группы, состоящей из SEQ ID NO: 104-110.
В одном варианте осуществления последовательность CH1 выбрана из группы, состоящей из SEQ ID NO: 111-114.
В одном варианте осуществления последовательность CH2 выбрана из группы, состоящей из SEQ ID NO: 115-119.
В одном варианте осуществления последовательность CH3 выбрана из группы, состоящей из SEQ ID NO: 120-128.
В одном варианте осуществления антиген A и антиген B могут быть одинаковыми или отличными друг от друга, предпочтительно антиген A или антиген B являются разными или представляют собой разные эпитопы на одном и том же антигене.
В одном варианте осуществления антиген A и антиген B каждый независимо выбран из группы, состоящей из: антигенов поверхности иммунных клеток, опухолевых антигенов, вирусов, бактерий, эндотоксинов, цитокинов или их комбинации.
В одном варианте осуществления антиген A и/или антиген B выбраны из PD-L1, PD-1, VEGFA, IL-10, IL-10R, BCMA, VEGF, TGF-β, CTLA-4, LAG-3, TIGIT, CEA, CD38, SLAMF7, B7-H3, Her2, EpCAM, CD19, CD20, CD30, CD33, CD47, CD52, CD133, EGFR, GD2, GD3, GM2, RANKL, CD3 и/или CD16a.
В одном варианте осуществления антиген A и/или антиген B выбраны из SEQ ID NO: 129-145.
В одном варианте осуществления антиген A и/или антиген B представляет собой PD-1; предпочтительно, один из антигена A и антигена B представляет собой PD-1, а другой выбран из группы, состоящей из PD-L1, PD-1, VEGFA, IL-10, IL-10R, BCMA, VEGF, TGF-β, CTLA-4, LAG-3, TIGIT, CEA, CD38, SLAMF7, B7-H3, HER2, CD3 или CD16a.
В одном варианте осуществления антиген A и антиген B выбраны из группы, состоящей из:
PD-L1 и VEGF,
PD-1 и VEGF,
PD-L1 и TGF-β,
PD-1 и TGF-β,
PD-1 и CTLA-4,
PD-1 и LAG-3,
PD-1 и TIGIT,
PD-1 и IL-10,
SLAMF7 и CD16a, и
Her2 и Her2.
В одном варианте осуществления VHa, VLa, VHb и/или VLb происходят из антитела, выбранного из группы, состоящей из антител животного происхождения (таких как мышиные антитела), химерных антител и гуманизированных антител; предпочтительно гуманизированные антитела включают полностью гуманизированные антитела и частично гуманизированные антитела.
В одном варианте осуществления VHa и/или VHb включают любую из следующих последовательностей:
a) аминокислотную последовательность, показанную в любой из SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 45, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 146, 148;
b) аминокислотную последовательность, имеющую идентичность последовательности 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более с аминокислотной последовательностью из a);
c) аминокислотную последовательность, которая имеет одну или несколько (предпочтительно одну или несколько, более предпочтительно 1, 2 или 3) аминокислотных модификаций по сравнению с аминокислотной последовательностью из a); и/или
VLa и/или VLb включают любую из следующих последовательностей:
d) аминокислотную последовательность, показанную в любой из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 147, 149;
e) аминокислотную последовательность, имеющую идентичность последовательности 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более с аминокислотной последовательностью из d);
f) аминокислотную последовательность, которая имеет одну или несколько (предпочтительно одну или несколько, более предпочтительно 1, 2, или 3) аминокислотных модификаций по сравнению с аминокислотной последовательностью из d).
В одном варианте осуществления биспецифическое антитело выбрано из группы, состоящей из Y100-A1, Y100-A2, Y100-A3, Y100-A4, Y100-A5, Y100-A6, Y100-A7, Y100-A8, Y100 -A9, Y100-A10, Y100-A11, Y100-AC1, Y100-AC2, Y100-AC3, Y101-A1, Y101-A2, Y101-A3, Y101-A4, Y101-A5, Y103-A1, Y103-A2 , Y103-A3, Y104-A1, Y104-A2, Y104-A3, Y105-A1, Y105-A2, Y105-A3, Y106-A1, Y106-A2, Y106-A3, Y106-A4, Y106-A5, Y110-A1, Y110-A2, Y110-A3, Y110-A4, Y116-A1, Y116-A2, Y116-A3, Y100-B1, Y100-B2, Y100-B3, Y100-B4, Y100-B5, Y100-B6, Y100-B7, Y100-B8, Y100-B9, Y100-B10, Y100-B11, Y100-B12, Y100-B13, Y100-BC1, Y100-BC2, Y100-BC3, Y101-B1, Y101-B2, Y101 -B3, Y101-B4, Y101-B5, Y101-B6, Y103-B1, Y103-B2, Y103-B3, Y104-B1, Y104-B2, Y104-B3, Y105-B1, Y105-B2, Y105-B3 , Y106-B1, Y106-B2, Y106-B3, Y106-B4, Y106-B5, Y110-B1, Y110-B2, Y110-B3, Y110-B4, Y110-B5, Y116-B1, Y116-B2, Y116 -B3, Y140-A1, Y140-A2, Y140-A3, Y140-A4, Y140-B1, Y140-B2, Y140-B3, Y140-B4, или их комбинации.
В одном варианте осуществления биспецифическое антитело выбрано из тех, которые показаны в любой из Таблицы 27 и Таблицы 34.
В одном варианте осуществления биспецифическое антитело выбрано из группы, состоящей из Y100-B6, Y100-B7, Y100-B8, Y100-B9, Y100-B10, Y100-B11, Y100-B12, Y100-BC1, Y100 -BC2, Y100-BC3, Y101-B1, Y101-B2, Y101-B3, Y101-B4, Y101-B5, Y101-B6, Y103-B1, Y103-B2, Y103-B3, Y104-B1, Y104-B2 , Y104-B3, Y105-B1, Y105-B2, Y105-B3, Y106-B1, Y106-B2, Y106-B3, Y106-B4, Y106-B5, Y110-B1, Y110-B2, Y110-B3, Y110 -B4, Y110-B5, Y116-B1, Y116-B2, Y116-B3, Y140-B1, Y140-B2, Y140-B3, Y140-B4, или их комбинации.
В одном варианте осуществления биспецифическое антитело выбрано из группы, состоящей из Y100-B7, Y100-B9, Y101-B2, Y101-B4, Y103-B2, Y104-B2, Y105-B2, Y106-B2, Y106 -B4, Y110-B2, Y110-B4, Y116-B2, или их комбинации.
В одном варианте осуществления биспецифическое антитело выбрано из группы, состоящей из следующих:
(1) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 82 и SEQ ID NO: 42; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 82, SEQ ID NO: 41, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(2) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 42; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 41, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(3) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 42; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 41, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(4) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 46; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 45, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(5) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 46; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 45, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(6) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(7) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(8) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(9) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(10) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 54; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 53, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(11) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(12) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 68; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 67, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(13) гибридная легкая цепь включает SEQ ID NO: 68, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 67, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(14) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(15) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(16) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(17) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(18) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(19) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 34; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 33, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(20) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 34; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 33, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(21) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 34; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 33, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(22) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 28; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 27, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(23) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 28; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 27, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(24) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 28; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 27, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(25) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 30; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 29, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(26) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 30; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 29, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(27) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 30; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 29, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(28) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(29) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(30) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(31) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(32) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(33) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 64; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 63, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(34) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 64; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 63, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(35) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 64; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 63, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(36) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 64; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 63, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(37) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 48; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 47, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(38) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 48; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 47, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(39) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 48; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 47, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(40) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 82 и SEQ ID NO: 41; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 82, SEQ ID NO: 42, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(41) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 41; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 42, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(42) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 41; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 42, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(43) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 45; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 46, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(44) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 45; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 46, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(45) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(46) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(47) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(48) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(49) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 53; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO:54, SEQ ID NO:74, SEQ ID NO:91, SEQ ID NO:117 и SEQ ID NO:120;
(50) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 84 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 84, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(51) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 86 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 86, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(52) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(53) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 67; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(54) гибридная легкая цепь включает SEQ ID NO: 68, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 67, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(55) гибридная легкая цепь включает SEQ ID NO: 68, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 67, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(56) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(57) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(58) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(59) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(60) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 84 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 84, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(61) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 86 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 86, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(62) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 33; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 34, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(63) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 86 и SEQ ID NO: 33; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 86, SEQ ID NO: 34, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(64) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 86 и SEQ ID NO: 33; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 86, SEQ ID NO: 34, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(65) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 27; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 28, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(66) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 27; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 28, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(67) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 27; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 28, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(68) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 29; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 30, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(69) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 29; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 30, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(70) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 29; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 30, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(71) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(72) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(73) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(74) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(75) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(76) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(77) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(78) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(79) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(80) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(80) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 89 и SEQ ID NO: 47; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 83, SEQ ID NO: 48, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(81) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 47; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 48, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(82) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 47; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 48, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(83) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 149; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 148, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(84) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 147; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 146, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(85) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 149; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 148, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(86) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 147; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 146, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(87) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 148; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 149, SEQ ID NO: 87, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(88) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 146; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 147, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(89) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 148; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 149, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120; или
(90) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 146; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 147, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120.
В одном варианте осуществления биспецифическое антитело обладает активностью одновременного связывания с антигеном A и антигеном B.
В одном варианте осуществления биспецифическое антитело представляет собой бипаратопное антитело.
В другом аспекте настоящее изобретение обеспечивает конъюгат или слитый белок, включающие биспецифические антитела по настоящему изобретению.
В конкретном варианте осуществления конъюгат или слитый белок включает вещество A, конъюгированное или слитое с антителом, и вещество A выбрано из группы, состоящей из терапевтических средств, пролекарств, белков (таких как ферменты), вирусов, липидов, модификаторов биологического ответа (таких как иммуномодуляторы), ПЭГ, гормонов, олигонуклеотидов, диагностических средств, цитотоксических средств, которые могут представлять собой лекарственные средства или токсины, усиливающих ультразвук агентов, нерадиоактивных маркеров, детектируемых маркеров, таких как хемилюминесцентные метящие соединения (такие как люминол, изолюминол, термолюминесцентный эфир акридиния, имидазол, соль акридиния и оксалат), или излучающие флуоресценцию металлы (такие как 152Eu, или лантанидные метки).
В одном варианте осуществления конъюгат или слитый белок является мономером, димером или мультимером.
В другом аспекте настоящее изобретение обеспечивает фармацевтическую композицию, включающую биспецифическое антитело по настоящему изобретению или конъюгат или слитый белок по настоящему изобретению.
В одном варианте осуществления фармацевтическая композиция также включает фармацевтически приемлемый носитель.
В одном варианте осуществления лекарственная форма фармацевтической композиции включает лекарственную форму для желудочно-кишечного введения или лекарственную форму для парентерального введения; предпочтительно лекарственная форма фармацевтической композиции представляет собой инъекцию, включая внутривенную инъекцию, внутривенное вливание, подкожную инъекцию, локальную инъекцию, внутримышечную инъекцию, интратуморальную инъекцию, интраперитонеальную инъекцию, интракраниальную инъекцию или внутриполостную инъекцию.
Еще в одном аспекте настоящее изобретение обеспечивает полинуклеотид, кодирующий биспецифическое антитело по настоящему изобретению.
В одном варианте осуществления полинуклеотид содержит первый полинуклеотид, кодирующий гибридную легкую цепь биспецифического антитела, и второй полинуклеотид, кодирующий гибридную тяжелую цепь. Предпочтительно соотношение первого полинуклеотида и второго полинуклеотида составляет 1:1.
В другом аспекте настоящее изобретение обеспечивает вектор, включающий полинуклеотид по настоящему изобретению; предпочтительно вектор включает: плазмиды, фаги, дрожжевые плазмиды, вирусы растений, вирусы млекопитающих, такие как аденовирусы, аденоассоциированные вирусы, ретровирусы или их комбинация.
Еще в одном аспекте настоящее изобретение обеспечивает клетку, включающую полинуклеотид по изобретению.
В одном варианте осуществления клетка включает вектор по настоящему изобретению, или геном клетки интегрирован с полинуклеотидом по настоящему изобретению.
В одном варианте осуществления клетка выбрана из группы, состоящей из Escherichia coli, Bacillus subtilis, дрожжевых клеток, клеток насекомых, клеток млекопитающих или их комбинации; предпочтительно представляет собой клетку млекопитающего, такую как CHO-S клетка или 293E клетка.
Еще в одном аспекте настоящее изобретение обеспечивает применение биспецифического антитела, или конъюгата, или слитого белка по настоящему изобретению для лечения опухолей (рака) или для получения лекарственных средств для лечения опухолей (рака).
В одном варианте осуществления заболевание представляет собой рак или опухоль. Предпочтительно опухоль выбрана из группы, состоящей из гематологических опухолей, солидных опухолей или их комбинации.
В другом аспекте настоящее изобретение обеспечивает применение биспецифического антитела, или конъюгата, или слитого белка по настоящему изобретению для получения реагентов или наборов для детекции опухолей (рака).
Еще в одном аспекте настоящее изобретение обеспечивает способ получения биспецифического антитела по настоящему изобретению, включающий следующие стадии:
(i) культивирование клеток по настоящему изобретению в подходящих условиях с получением смеси, содержащей биспецифическое антитело по настоящему изобретению; и
(ii) очистка и/или разделение смеси, полученной на стадии (i), с получением биспецифического антитела по настоящему изобретению.
В одном варианте осуществления очистка включает: аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, хроматографию с молекулярным ситом или их комбинацию.
В другом аспекте настоящее изобретение обеспечивает способ лечения заболеваний, включающий следующие стадии: введение терапевтически эффективного количества биспецифического антитела по настоящему изобретению, или конъюгата, или слитого белка по настоящему изобретению, или фармацевтической композиции по настоящему изобретению, или их комбинации субъекту, нуждающемуся в этом.
В одном варианте осуществления субъектом является человек или отличное от человека млекопитающее.
Краткое описание чертежей
Фиг. 1 схематически представляет структуру антитела, имеющего структуру 1 биспецифического антитела (A), и схематически представляет первичную структуру белка каждого компонента антитела (B), где вариабельная область 1 антитела b представлена как VL/Hb, а вариабельная область 2 антитела b представлена как VH/Lb.
Фиг. 2 схематически представляет структуру антитела, имеющего структуру 2 биспецифического антитела (A), и схематически представляет первичную структуру белка каждого компонента антитела (B).
Фиг. 3 схематически представляет структуру антитела, имеющего структуру 3 биспецифического антитела (A), и схематически представляет первичную структуру белка каждого компонента антитела (B).
Фиг. 4 схематически представляет структуру антитела, имеющего структуру 4 биспецифического антитела (A), и схематически представляет первичную структуру белка каждого компонента антитела (B).
Фиг. 5 показывает антитела со структурой 1 биспецифического антитела. (A) показывает уровень экспрессии антител серии Y100-A и серии Y100-B при транзиентной трансфекции в клетках CHO. (B) показывает уровни экспрессии антител серии Y101/Y103/Y104/Y105-A и серии Y101/Y103/Y104/Y105-B при транзиентной трансфекции в клетках CHO. (C) показывает уровень экспрессии антител серии Y106/Y110/Y116-A серии и серии Y106/Y110/Y116-B при транзиентной трансфекции в клетках CHO. Фиг. 5D показывает чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A, для антител серии Y100/Y101/Y103/Y104/Y105/Y106/Y110/Y116-B серии, которые экспрессируются на уровне больше чем 5 мг/л.
Фиг. 6 показывает уровень экспрессии антител серии Y200/Y201/Y203/Y204/Y205/Y206/Y210/Y216-A и B, имеющих структуру 2 биспецифического антитела, при транзиентной трансфекции в клетках CHO и чистоту, определенную методом эксклюзионной ВЭЖ, после аффинной хроматографии с белком A.
Фиг. 7 показывает уровни экспрессии антител Y300/Y304/Y316-A серии, B серии, C серии и D серии, имеющих структуру 3 биспецифического антитела, при транзиентной трансфекции в клетках CHO и чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A.
Фиг. 8 показывает уровни экспрессии антител Y400/Y404/Y416-A серии и B серии, имеющих структуру 4 биспецифического антитела, при транзиентной трансфекции в клетках CHO и чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A.
Фиг. 9 показывает сравнение уровня экспрессии и чистоты антител со структурой 1-4 биспецифического антитела, с одинаковой мишенью и последовательностью вариабельной области антитела. (A) Уровень экспрессии и чистота антител со структурой 1-4 биспецифического антитела с последовательностью вариабельной области антитела S70 (SEQ ID NO: 41 и 42) и с последовательностью вариабельной области антитела G631 (SEQ ID NO: 57 и 58). (B) Уровень экспрессии и чистота антител со структурой 1 и 2 биспецифического антитела с последовательностью вариабельной области антитела S70 (SEQ ID NO: 41 и 42) и с последовательностью вариабельной области антитела 3G12 (SEQ ID NO: 59 и 60). (C) Уровень экспрессии и чистота антител со структурой 1 и 2 биспецифического антитела с последовательностью вариабельной области антитела 12A4 (SEQ ID NO: 45 и 46) и с последовательностью вариабельной области антитела 3G12 (SEQ ID NO: 59 и 60). (D) Уровень экспрессии и чистота антител со структурой 1 и 2 биспецифического антитела с последовательностью вариабельной области антитела S70 (SEQ ID NO: 41 и 42) и с последовательностью вариабельной области антитела LAG35 (SEQ ID NO: 33 и 34). (E) Уровень экспрессии и чистота антител со структурой 1, 3 и 4 биспецифического антитела с последовательностью вариабельной области антитела 5C4 (SEQ ID NO: 37 и 38) и с последовательностью вариабельной области антитела Ервой (SEQ ID NO: 27 и 28). (F) Уровень экспрессии и чистота антител со структурой 1 и 2 биспецифического антитела с последовательностью вариабельной области антитела 5C4 (SEQ ID NO: 37 и 38) и с последовательностью вариабельной области антитела 10A7 (SEQ ID NO: 29 и 30). (G) Уровень экспрессии и чистота антител со структурой 1 и 2 биспецифического антитела с последовательностью вариабельной области антитела 5C4 (SEQ ID NO: 37 и 38) и с последовательностью вариабельной области антитела G631 (SEQ ID NO: 57 и 58). (H) Уровень экспрессии и чистота антител со структурой 1 и 2 биспецифического антитела с последовательностью вариабельной области антитела 5C4 (SEQ ID NO: 37 и 38) и с последовательностью вариабельной области антитела B-N10 (SEQ ID NO: 63 и 64). (I) Уровень экспрессии и чистота антител со структурой 1-4 биспецифического антитела с последовательностью вариабельной области антитела Элотузумаба (SEQ ID NO: 49 и 50) и с последовательностью вариабельной области антитела NM3E2 (SEQ ID NO: 47 и 48).
Фиг. 10 показывает результаты ускоренного испытания на термостабильность различных структур биспецифических антител при 40°С. (A) Чистота Y100-B6, Y100-B7, Y200-A1, Y200-B1, Y200-B3, Y300-A1 и Y400-B1, определенная методом эксклюзионной ВЭЖХ после обработки при 40°C в течение 0, 7 и 14 дней; (B) чистота Y101-B1, Y101-B2, Y201-A1, Y201-B1, Y101-B3, Y101-B4, Y201-A2 и Y201-B2, определенная методом эксклюзионной ВЭЖХ после обработки при 40°C в течение 0, 7 и 14 дней; (C) чистота Y103-B1, Y103-B2, Y203-A1, Y203-B1, Y104-B1, Y104-B2, Y304-A1 и Y404-B1, определенная методом эксклюзионной ВЭЖХ после обработки при 40°C в течение 0, 7 и 14 дней; (D) чистота Y105-B1, Y105-B2, Y205-A2, Y205-B2, Y106-B1, Y106-B2, Y206-A2, Y206-B1 и Y206-B2, определенная методом эксклюзионной ВЭЖХ после обработки при 40°C в течение 0, 7 и 14 дней; (E) чистота Y110-B1, Y110-B2, Y210-A1, Y210-B1, Y116-B1, Y116-B2, Y216-A1, Y216-B1, Y316-A1, Y416-B1, определенная методом эксклюзионной ВЭЖХ после обработки при 40°C в течение 0, 7 и 14 дней.
Фиг. 11 показывает результаты испытания на кислотостойкость различных структур биспецифических антител. Значение pH используемого кислотного раствора составляет 3,5. Фиг. 11A показывает чистоту Y100-B6, Y100-B7, Y200-A1, Y200-B1, Y200-B3, Y300-A1 и Y400-B1, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0, 30 и 60 минут; Фиг. 11B показывает чистоту Y101-B1, Y101-B2, Y201-A1, Y201-B1, Y101-B3, Y101-B4, Y201-A2 и Y201-B2, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0, 30 и 60 минут; Фиг. 11C показывает чистоту Y103-B1, Y103-B2, Y203-A1, Y203-B1, Y104-B1, Y104-B2, Y304-A1 и Y404-B1, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0, 30 и 60 минут; Фиг. 11D показывает чистоту Y105-B1, Y105-B2, Y205-A2, Y205-B2, Y106-B1, Y106-B2, Y206-A2, Y206-B1 и Y206-B2, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0, 30 и 60 минут; Фиг. 11E показывает чистоту Y110-B1, Y110-B2, Y210-A1, Y210-B1, Y116-B1, Y116-B2, Y216-A1, Y216-B1, Y316-A1, Y416-B1, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0, 30 и 60 минут.
Фиг. 12 показывает результаты фармакодинамического эксперимента in vivo Y100-B7 со структурой 1 биспецифического антитела. Рак толстой кишки мыши MC38 используют в качестве опухолевой клеточной линии, и самок мышей C57BL/6 используют в качестве линии мышей.
Фиг. 13 показывает результаты фармакодинамического эксперимента in vivo Y101-B2 со структурой 1 биспецифического антитела. Рак толстой кишки мыши MC38 используют в качестве опухолевой клеточной линии, и самок мышей C57BL/6 используют в качестве линии мышей.
Определения
Следует отметить, что термин с артиклем "a" или "an" относится к одному или нескольким из таких объектов; например, "биспецифическое антитело" следует понимать как представляющее собой одно или несколько биспецифических антител. Таким образом, термины "a" (или "an"), "один или несколько" и "по меньшей мере один" могут использоваться взаимозаменяемо в настоящей заявке.
В контексте настоящей заявки термин "полипептид" предназначен для охвата одного "полипептида", а также нескольких "полипептидов", и относится к молекуле, состоящей из мономеров (аминокислот), линейно связанных амидными связями (также известными как пептидные связи). Термин "полипептид" относится к любой цепи или цепям из двух или более аминокислот, и не означает конкретную длину продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды, "белок", "аминокислотная цепь" или любой другой используемый термин, относящийся к цепи или цепям из двух или более аминокислот, все включены в определение "полипептид", и термин "полипептид" может использоваться вместо любого из этих терминов или взаимозаменяемо с ними. Термин "полипептид" также предназначен для обозначения продуктов постэкспрессионных модификаций полипептида, включая, без ограничения, гликозилирование, ацетилирование, фосфорилирование, амидирование, дериватизацию известными защитными/блокирующими группами, протеолитическое расщепление или модификацию не встречающимися в природе аминокислотами. Полипептид может быть получен из природного биологического источника или может быть получен с использованием рекомбинантной технологии, но не обязательно должен транслироваться из определенной нуклеиновокислотной последовательности. Его можно получить любым способом, в том числе путем химического синтеза.
В контексте настоящей заявки термин "рекомбинантный", когда он относится к полипептидам или полинуклеотидам, относится к форме полипептида или полинуклеотида, которая не существует в природе, неограничивающий пример которой может быть получен путем объединения полинуклеотидов или полипептидов, которые обычно не встречаются вместе.
"Гомология", или "идентичность", или "сходство" относится к сходству последовательностей между двумя пептидами или между двумя молекулами нуклеиновых кислот. Гомологию можно определить путем сравнения положений в каждой последовательности, которые могут быть выровнены для целей сравнения. Если положение в сравниваемой последовательности занято одним и тем же основанием или аминокислотой, то молекулы гомологичны в этом положении. Степень гомологии между последовательностями является функцией количества совпадающих или гомологичных положений, общих для последовательностей. "Неродственная" или "негомологическая" последовательность имеет менее 40% идентичности, хотя предпочтительно менее 25% идентичности, с одной из последовательностей по настоящему изобретению.
Полинуклеотид или полинуклеотидная область (или полипептид или полипептидная область), имеющие определенный процент (например, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99%) "идентичности последовательности" с другой последовательностью, означает, что при выравнивании такой процент оснований (или аминокислот) одинаков при сравнении двух последовательностей. Это выравнивание и процент гомологии или идентичности последовательностей могут быть определены с использованием компьютерных программ, известных в данной области техники, например программ, описанных в Ausubel et al. eds. (2007) Current Protocols in Molecular Biology. Предпочтительно для выравнивания используются параметры по умолчанию. Одной из программ выравнивания является BLAST, использующая параметры по умолчанию. В частности, программы BLASTN и BLASTP используют следующие параметры по умолчанию: Генетический код=стандартный; фильтр=нет; цепь=обе; отсечка=60; ожидание=10; Матрица=BLOSUM62; Описания=50 последовательностей; сортировка по=HIGH SCORE; Базы данных=неизбыточные, GenBank+EMBL+DDBJ+PDB+GenBank CDS translations+SwissProtein+SPupdate+PIR. Подробную информацию об этих программах можно найти по следующему адресу в Интернете: http://www.ncbi.nlm.nih.gov/blast/Blast.cgi, последний доступ 21 мая 2008 г. Биологически эквивалентными полинуклеотидами являются полинуклеотиды, имеющие вышеуказанные определенный процент гомологии и кодирующие полипептид, обладающий такой же или подобной биологической активностью.
Термин "кодировать" применительно к полинуклеотидам относится к полинуклеотиду, который "кодирует" полипептид и который в своем нативном состоянии или при манипулировании методами, хорошо известными специалистам в данной области, может транскрибироваться и/или транслироваться для продукции мРНК для полипептида и/или его фрагмента. Антисмысловая цепь является комплементом такой нуклеиновой кислоты, и из нее можно вывести кодирующую последовательность.
В контексте настоящей заявки термин "антитело" или "антигенсвязывающий полипептид" относится к полипептиду или полипептидному комплексу, который специфически распознает антиген и связывается с ним. Антитело может представлять собой интактное антитело и любой антигенсвязывающий фрагмент или одну его цепь. Таким образом, термин "антитело" включает любой белок или пептид, содержащий специфическую молекулу, где специфическая молекула включает по меньшей мере часть молекулы иммуноглобулина, обладающую биологической активностью связывания с антигеном. Примеры включают, но не ограничиваются этим, определяющую комплементарность область (CDR) тяжелой или легкой цепи или ее лигандсвязывающую часть, вариабельную область тяжелой или легкой цепи, константную область тяжелой или легкой цепи, каркасную (FR) область, или любую их часть, или по меньшей мере одну часть связывающего белка.
Термин "фрагмент антитела" или "антигенсвязывающий фрагмент" в контексте настоящей заявки означает часть антитела, такую как F(ab')2, F(ab)2, Fab', Fab, Fv, Fd, Fv, dAb, Fab/c, фрагменты определяющих комплементарность областей (CDR), связанные дисульфидной связью Fv (sdFv), одноцепочечные антитела (например, scFv), двухвалентные антитела или доменные антитела и т.п. Независимо от структуры фрагмент антитела связывается с тем же антигеном, который распознается интактным антителом. Термин "фрагмент антитела" включает аптамеры, шпигельмеры и диатела. Термин "фрагмент антитела" также включает любой синтетический или генетически сконструированный белок, который действует подобно антителу, связываясь со специфическим антигеном с образованием комплекса.
"Одноцепочечный вариабельный фрагмент", или "scFv", относится к слитому белку из вариабельных областей тяжелой (VH) и легкой (VL) цепей иммуноглобулинов. В некоторых аспектах области соединены коротким линкерным пептидом из от 10 до около 25 аминокислот. Линкер может быть обогащен глицином для гибкости, а также серином или треонином для растворимости, и может либо соединять N-конец VH с C-концом VL, либо наоборот. Этот белок сохраняет специфичность исходного иммуноглобулина, но константная область удалена и введен линкер. Молекулы ScFv известны в данной области и описаны, например, в патенте США 5892019.
Термин "антитело" охватывает широкий спектр полипептидов, которые могут биохимически распознаться. Специалистам в данной области техники должно быть понятно, что тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон (γ, μ, α, δ, ε) с некоторыми подклассами среди них (например, γ1-γ4). Именно природа этой цепи определяет "класс" антитела как IgG, IgM, IgA, IgG или IgE, соответственно. Подклассы (изотипы) иммуноглобулинов, например IgG1, IgG2, IgG3, IgG4, IgG5 и т.д., хорошо охарактеризованы и являются функционально специфическими. Модифицированные версии каждого из этих классов и изотипов легко распознаваемы для специалистов в данной области с учетом настоящего раскрытия и, соответственно, входят в объем настоящего раскрытия. Все классы иммуноглобулинов определенно входят в объем настоящего изобретения, последующее обсуждение в основном будет направлено на IgG класс молекул иммуноглобулина. Что касается IgG, то стандартная молекула иммуноглобулина включает два идентичных полипептида легкой цепи с молекулярной массой приблизительно 23000 дальтон и два идентичных полипептида тяжелой цепи с молекулярной массой 53000-70000. Эти четыре цепи обычно соединены дисульфидными связями в конфигурации "Y", в которой легкие цепи обрамляют тяжелые цепи, начинающиеся в устье "Y" и проходящие через вариабельную область.
Антитела, антигенсвязывающие полипептиды, их варианты или производные в настоящем изобретении включают, но не ограничиваются этим, поликлональные, моноклональные, мультиспецифические, человеческие, гуманизированные, приматизированные или химерные антитела, одноцепочечные антитела, антигенсвязывающие фрагменты, например Fab, Fab' и F(ab')2, Fd, Fv, одноцепочечные Fv (scFv), дисульфид-связанные одноцепочечные Fv (sdFv), фрагменты, включающие либо VL, либо VH домен, фрагменты, продуцируемые библиотекой экспрессии Fab, и антиидиотипические (анти-Id) антитела. Молекулы иммуноглобулинов или антител по настоящему изобретению могут относиться к любому типу (например, IgG, IgE, IgM, IgD, IgA и IgY), любому классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу молекул иммуноглобулина.
Легкие цепи классифицируются как каппа или лямбда (κ, λ). Каждый класс тяжелой цепи может быть связан с легкой либо каппа, либо лямбда цепью. Как правило, легкие и тяжелые цепи ковалентно связаны друг с другом, а "хвостовые" части двух тяжелых цепей связаны друг с другом ковалентными дисульфидными связями или нековалентными связями, когда иммуноглобулины генерируются гибридомами, B-клетками или генетически модифицированными клетками-хозяевами. В тяжелой цепи аминокислотные последовательности простираются от N-конца на разветвленных концах Y-конфигурации до С-конца в нижней части каждой цепи.
Как легкие, так и тяжелые цепи подразделяются на структурные области и области функциональной гомологии. Термины "константная" и "вариабельная" используются функционально. В этом отношении следует понимать, что вариабельные домены как легкой (VL), так и тяжелой (VH) цепи определяют распознавание антигена и специфичность в отношении антигена. Наоборот, константные домены легкой цепи (CL) и тяжелой цепи (СН1, СН2 или СН3) придают важные биологические свойства, такие как секреция, трансплацентарная подвижность, связывание с Fc рецептором, связывание комплемента и т.п. Как правило, количество доменов константной области увеличивается по мере того, как они становятся более удаленными от сайта связывания антигена или амино-конца антитела. N-концевая часть представляет собой вариабельную область, а С-концевая часть представляет собой константную область; CH3 и CL домены фактически включают карбокси-конец тяжелой и легкой цепи, соответственно.
Как указано выше, вариабельная область позволяет антителу селективно распознавать и специфически связываться с эпитопом на антигене. То есть VL домен и VH домен или подклассы определяющих комплементарность областей (CDR) антитела объединяются с образованием вариабельной области, которая определяет трехмерный антигенсвязывающий сайт. Эта структура четырехвалентного антитела образует сайт связывания антигена, который присутствует на конце каждого плеча в Y-конфигурации. Более конкретно, антигенсвязывающий сайт определяется тремя CDR на каждой из VH и VL цепей (т.е. CDR-H1, CDR-H2, CDR-H3, CDR-L1, CDR-L2 и CDR-L3). В некоторых примерах, например некоторых иммуноглобулинов, полученных из видов верблюдовых или сконструированных на основе иммуноглобулинов верблюдовых, их интактная молекула иммуноглобулина может состоять только из тяжелых цепей без легких цепей. См., например, Hamers-Casterman et al., Nature 363:446-448 (1993 г.).
В природных антителах шесть "определяющих комплементарность областей", или "CDR", присутствующих в каждом антигенсвязывающем домене, представляют собой короткие несмежные аминокислотные последовательности, которые специфически расположены с образованием антигенсвязывающего домена. Остальные аминокислоты в антигенсвязывающем домене, называемые "каркасной" областью, демонстрируют меньшую внутримолекулярную изменчивость. Каркасные области в основном принимают конфигурацию β-слоя, а CDR образуют петли, которые соединяют структуру β-слоя и иногда образуют часть структуры β-слоя. Поэтому каркасные области используются для формирования каркаса, который обеспечивает расположение CDR в правильном направлении за счет межцепочечных нековалентных взаимодействий. Антигенсвязывающий домен, образованный расположенной CDR, определяет поверхность, комплементарную эпитопу на иммунореактивном антигене. Эта комплементарная поверхность способствует нековалентному связыванию антитела с его гомологичным эпитопом. Специалисты в данной области могут легко идентифицировать аминокислоты CDR и каркасных областей для любой данной вариабельной области тяжелой или легкой цепи, поскольку они были четко определены (см. "Sequences of Proteins of Immunological Interest", Kabat, E., et al., US Department of Health and Human Services, (1983), Chothia and Lesk, J. MoI. Biol., 196:901-917 (1987), полный текст которого включен в настоящую заявку посредством ссылки).
В случае, когда имеется два или более определений термина, которые используются и/или приняты в данной области техники, предполагается, что определения термина, используемые в настоящей заявке, включают все значения, если прямо не указано иное. Конкретным примером является использование термина "определяющая комплементарность область" ("CDR") для описания несмежных антигенсвязывающих сайтов, присутствующих в вариабельных областях полипептидов как тяжелой, так и легкой цепи. Эта специфическая область была описана Kabat et al., U.S. Department of Health and Human Services, "Sequences of Proteins of Immunological Interest" (1983) и Chothia et al., J. MoI. Biol. 196:901-917 (1987), полный текст которого включен в настоящую заявку посредством ссылки. В соответствии с определением Kabat и Chothia, CDR включает перекрывающиеся аминокислотные остатки или аминокислотные субструктуры при сравнении друг с другом. Однако применение каждого определения для обозначения CDR антитела или его варианта будет находиться в пределах объема термина, как определено и используется в настоящей заявке. Соответствующие аминокислотные остатки, которые охватывают CDR, как определено в каждой из ссылок, приведенных выше, приведены в таблице ниже для сравнения. Точное количество остатков, которые охватывают конкретную CDR, будет варьироваться в зависимости от последовательности и размера CDR. Специалисты в данной области могут обычным способом определить, какие остатки составляют конкретную CDR, если предоставлена аминокислотная последовательность вариабельной области антитела.
Определение вариабельной области антитела
Kabat et al. также определили систему нумерации последовательностей вариабельных доменов, которая применима к любому типу антител. Специалисты в данной области техники могут однозначно использовать эту систему "нумерации по Kabat" для любой последовательности вариабельного домена, независимо от каких-либо экспериментальных данных, кроме самой последовательности. "Нумерации по Kabat", как используется в настоящей заявке, относится к системе нумерации, описанной Kabat et al., U.S. Department of Health and Human Services, "Sequences of Proteins of Immunological Interest" (1983).
В дополнение к приведенной выше таблице, CDR области, описанные по системе нумерации Kabat, являются следующими: CDR-H1 начинается с аминокислоты примерно в положении 31 (т.е. примерно через 9 остатков после первого цистеинового остатка) и включает примерно от 5 до 7 аминокислот и заканчивается на следующем триптофановом остатке. CDR-H2 начинается с пятнадцатого остатка после конца CDR-H1, включает приблизительно от 16 до 19 аминокислот и заканчивается на следующем аргининовом или лизиновом остатке. CDR-H3 начинается примерно с тридцать третьего аминокислотного остатка после конца CDR-H2; включает от 3 до 25 аминокислот; и заканчивается последовательностью W-G-X-G, где X представляет собой любую аминокислоту. CDR-L1 начинается с остатка примерно в положении 24 (т.е. после цистеинового остатка); включает приблизительно от 10 до 17 остатков; и заканчивается на следующем триптофановом остатке. CDR-L2 начинается примерно с шестнадцатого остатка после окончания CDR-L1 и включает примерно 7 остатков. CDR-L3 начинается примерно с тридцать третьего остатка после окончания CDR-L2 (т.е. после цистеинового остатка); включает примерно от 7 до 11 остатков и заканчивается последовательностью F или WGXG, где X представляет собой любую аминокислоту.
Описанные в настоящей заявке антитела могут быть любого животного происхождения, включая птиц и млекопитающих. Предпочтительно антитела представляют собой антитела человека, мыши, осла, кролика, козы, морской свинки, верблюда, ламы, лошади или цыпленка. В другом варианте осуществления вариабельная область может происходить от condricthoid (например, от акулы).
Термин "константная область тяжелой цепи", используемый в настоящей заявке, включает аминокислотные последовательности, полученные из тяжелых цепей иммуноглобулина. Полипептид, включающий константную область тяжелой цепи, включает по меньшей мере одно из следующего: СН1 домен, шарнирный домен (например, верхний шарнирный участок, средний шарнирный участок и/или нижний шарнирный участок), СН2 домен, СН3 домен, или вариант или фрагмент вышеуказанного. Например, антигенсвязывающий полипептид для применения в настоящем изобретении может включать полипептидную цепь, включающую CH1 домен; полипептид, включающий СН1 домен, по меньшей мере часть шарнирного домена и СН2 домен; полипептидную цепь, включающую СН1 домен и СН3 домен; полипептидную цепь, включающую СН1 домен, по меньшей мере часть шарнирного домена и СН3 домен, или полипептидную цепь, включающую СН1 домен, по меньшей мере часть шарнирного домена, СН2 домен и СН3 домен. В другом варианте осуществления полипептид по настоящему изобретению включает полипептидную цепь, включающую СН3 домен. Кроме того, в антителах, используемых в настоящем изобретении, может отсутствовать по меньшей мере часть CH2 домена (например, весь CH2 домен или его часть). Как указано выше, специалистам в данной области должно быть понятно, что константные области тяжелой цепи могут быть модифицированы таким образом, чтобы они отличались по аминокислотной последовательности от природных молекул иммуноглобулина.
Константные области тяжелой цепи раскрытого в настоящей заявке антитела могут быть получены из разных молекул иммуноглобулина. Например, константная область тяжелой цепи полипептида может включать домен CH1, полученный из молекулы IgG1, и шарнирную область, полученную из молекулы IgG3. В другом примере константная область тяжелой цепи может включать шарнирную область, которая частично происходит из молекулы IgG1 и частично из молекулы IgG3. В другом примере часть тяжелой цепи может включать химерный шарнир, который частично происходит из молекулы IgG1 и частично происходит из молекулы IgG4.
Термин "константная область легкой цепи", используемый в настоящей заявке, включает аминокислотную последовательность, полученную из легкой цепи антитела. Предпочтительно константная область легкой цепи включает по меньшей мере один из константного каппа-домена и константного лямбда-домена.
"Пара легкая цепь-тяжелая цепь" относится к объединению легкой цепи и тяжелой цепи, которые могут образовывать димер за счет дисульфидной связи между CL доменом легкой цепи и CH1 доменом тяжелой цепи.
Как указывалось ранее, известны субъединичные структуры и трехмерные структуры константных областей различных иммуноглобулинов. Используемый в настоящей заявке термин "VH домен" включает амино-концевой вариабельный домен тяжелой цепи иммуноглобулина, а термин "СН1 домен" включает первую (преимущественно амино-конец) константную область тяжелой цепи иммуноглобулина. CH1 домен является смежным с VH доменом и является амино-концом шарнирной области молекулы тяжелой цепи иммуноглобулина.
Используемый в настоящей заявке термин "CH2 домен" включает часть молекулы тяжелой цепи, например, от остатка примерно в положении 244 до остатка в положении 360 антитела в соответствии с общепринятой системой нумерации (остатки в положениях 244-360, в соответствии с системой нумерации Kabat; и остатки в положениях 231-340, в соответствии с системой нумерации ЕС; см. Kabat et al., U.S. Department of Health and Human Services, "Sequences of Proteins of Immunological Interest" (1983). CH2 домен уникален, поскольку он тесно не спаривается с другим доменом. Наоборот, между двумя CH2 доменами интактной природной молекулы IgG вставлены две N-связанные разветвленные углеводные цепи. Документально подтверждено, что CH3 домен простирается от CH2 домена к С-концу молекулы IgG и включает около 108 остатков.
Термин "шарнирная область", как он используется в настоящей заявке, включает часть молекулы тяжелой цепи, которая связывает СН1 домен с СН2 доменом. Шарнирная область включает около 25 остатков и является гибкой, что позволяет двум N-концевым антигенсвязывающим областям двигаться независимо. Шарнирные области можно разделить на три разных домена: верхний шарнирный домен, средний шарнирный домен и нижний шарнирный домен (Roux et al., J. Immunol 161:4083 (1998)).
Используемый в настоящей заявке термин "дисульфидная связь" включает ковалентную связь, образованную между двумя атомами серы. Аминокислота цистеин включает тиоловую группу, которая может образовывать дисульфидную связь или мостик со второй тиоловой группой. В большинстве природных IgG молекул CH1 и CL области связаны дисульфидной связью, а две тяжелые цепи связаны двумя дисульфидными связями в положениях, соответствующих положениям 239 и 242 по системе нумерации Kabat (положение 226 или 229 в соответствии с системой нумерации ЕС).
Термин "химерное антитело", как он используется в настоящей заявке, предназначен для обозначения любого антитела, в котором иммунореактивные области или сайты получены или происходят из первого вида, а константные области (которые могут быть интактными, частичными или модифицированными в соответствии с настоящим изобретением) получены из второго вида. В некоторых вариантах осуществления области или сайты связывания с мишенью будут происходить не от человека (например, от мыши или примата), а константная область будет происходить от человека.
В контексте настоящей заявки "процент гуманизации" рассчитывают путем определения количества различий аминокислот каркаса (т.е. различий, не относящихся к CDR) между гуманизированным доменом и доменом зародышевой линии и вычитания этого числа из общего количества аминокислот, что затем делят на общее количество аминокислот и умножают на 100.
Так называемое "специфическое связывание" или "специфическое к" обычно означает, что антитело связывается с эпитопом антигена через свой антигенсвязывающий домен, и что это связывание влечет за собой некоторую комплементарность между антигенсвязывающим доменом и эпитопом антигена. В соответствии с этим определением считается, что антитело "специфически связывается" с эпитопом антигена, когда оно связывается с эпитопом антигена, и это связывание через антигенсвязывающий домен легче, чем через связывание со случайным, неродственным эпитопом антигена. Термин "специфичность" используется в настоящей заявке для определения аффинности определенного антитела к связыванию с определенным эпитопом антигена. Например, можно считать, что антитело "А" обладает более высокой специфичностью в отношении данного антигенного эпитопа, чем антитело "В", или можно сказать, что антитело "А" обладает более высокой специфичностью при связывании с эпитопом "С", чем при связывании с родственным эпитопом "D".
Термин "осуществление лечения" ("лечить" или "лечение"), используемый в настоящей заявке, относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам, целью которых является предотвращение или замедление (уменьшение) нежелательного физиологического изменения или расстройства, такого как прогрессирование рака. Благоприятные или желаемые клинические результаты включают, но не ограничиваются этим, облегчение симптомов, уменьшение степени заболевания, стабилизацию (например, предотвращение его ухудшения) состояния заболевания, отсрочку или замедление прогрессирования заболевания, облегчение или паллиативную помощь при заболевании и облегчение (частичное или полное), независимо от того, поддается это обнаружению или нет. "Лечение" также может означать увеличение срока выживания по сравнению с ожидаемой выживаемостью без лечения. Нуждающиеся в лечении, включают тех, у кого уже есть заболевание или симптом, и тех, у кого есть предрасположенность к заболеванию или симптому, или тех, у кого заболевание или симптом необходимо предотвратить.
Варианты осуществления настоящего изобретения обеспечивают различные биспецифические антитела, которые включают две разные или идентичные антигенсвязывающие полипептидные единицы. Домен антитела, который связывается с антигеном, представляет собой Fab, или ScFv, или нековалентное спаривание (Fv) между вариабельной областью тяжелой цепи (VH) и вариабельной областью легкой цепи (VL). В частности, все эти биспецифические антитела имеют Fc-фрагмент константной области тяжелой цепи антитела, и Fc включает: (1) шарнир, (2) вторую константную область тяжелой цепи (CH2) и третью константную область тяжелой цепи (CH3).
Любые из вышеуказанных антител или полипептидов могут также включать дополнительные полипептиды, например, кодируемый полипептид, как описано в настоящей заявке, сигнальный пептид на N-конце антитела, используемый для направления секреции, или другие гетерологичные полипептиды, как описано в настоящей заявке.
Специалистам в данной области также будет понятно, что описанные в настоящей заявке антитела можно модифицировать таким образом, чтобы они отличались по аминокислотной последовательности от природных связывающих полипептидов, из которых они получены. Например, полипептидная или аминокислотная последовательность, полученная из указанного белка, может быть сходной с исходной последовательностью, например, может иметь определенный процент идентичности с исходной последовательностью, например, она может иметь идентичность 60%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% с исходной последовательностью.
Кроме того, можно осуществить нуклеотидные или аминокислотные замены, делеции или вставки, ведущие к консервативным заменам или изменениям в областях "заменимых" аминокислот. Например, полипептидная или аминокислотная последовательность, полученная из указанного белка, может быть идентична исходной последовательности, за исключением одной или нескольких независимых аминокислотных замен, вставок или делеций, например 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или более независимых аминокислотных замен, вставок или делеций. В некоторых вариантах осуществления полипептидная или аминокислотная последовательность, полученная из указанного белка, имеет от 1 до 5, от 1 до 10, от 1 до 15 или от 1 до 20 независимых аминокислотных замен, вставок или делеций по сравнению с исходной последовательностью.
В других вариантах осуществления антигенсвязывающие полипептиды по настоящему изобретению могут включать консервативные аминокислотные замены.
"Консервативная аминокислотная замена" представляет собой замену аминокислотного остатка аминокислотным остатком, имеющим аналогичную боковую цепь. В данной области определены семейства аминокислотных остатков, имеющих сходные боковые цепи, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), β-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Следовательно, заменимые аминокислотные остатки иммуноглобулиновых полипептидов предпочтительно заменяют другими аминокислотными остатками из того же семейства боковых цепей. В другом варианте осуществления цепочка аминокислот может быть заменена структурно аналогичной цепочкой аминокислот, которая отличается порядком и/или составом семейства боковых цепей.
Неограничивающие примеры консервативных аминокислотных замен представлены в таблице ниже, где оценка сходства 0 или выше указывает на консервативную замену между двумя аминокислотами.
Неограничивающий перечень консервативных аминокислотных замен
В некоторых вариантах осуществления антитело может быть конъюгировано с терапевтическими средствами, пролекарствами, пептидами, белками, ферментами, вирусами, липидами, модификаторами биологического ответа, фармацевтическими препаратами или ПЭГ.
Антитело может быть связано или объединено с терапевтическим средством, которое может включать детектируемые метки, такие как радиоактивные метки, иммуномодуляторы, гормоны, ферменты, олигонуклеотиды, фотоактивные терапевтические или диагностические средства, цитотоксические средства, которые могут представлять собой лекарственные средства или токсины, усиливающие ультразвук агенты, нерадиоактивные метки, их комбинацию и другие такие компоненты, известные в данной области техники.
Можно осуществить мечение антитела для возможности его детекции путем его связывания с хемилюминесцентными соединениями. Затем присутствие хемилюминесцентно-меченного антигенсвязывающего полипептида определяют путем детекции люминесценции, генерируемой в процессе химической реакции. Примерами особенно полезных хемилюминесцентных метящих соединений являются люминол, изолюминол, термолюминесцентный эфир акридиния, имидазол, соль акридиния и оксалатный эфир.
Также можно осуществить мечение антител для возможности их детекции с использованием излучающих флуоресценцию металлов, таких как 152Eu, или других лантанидных меток. Эти металлы можно присоединить к антителу с использованием следующих металл- хелатирующих групп, таких как диэтилентриаминпентауксусная кислота (DTPA) или этилендиаминтетрауксусная кислота (EDTA). Методы связывания различных фрагментов с антителами хорошо известны, см., например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", в Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (ред.), стр. 243-56 (Alan R. Liss, Inc. (1985); Hellstrom et al., "Antibodies For Drug Delivery", в Controlled Drug Delivery (второе издание), Robinson et al. (ред.), Marcel Dekker, Inc., стр.. 623-53 (1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", в Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (ред.), стр. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", в Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (ред.), Academic Press стр. 303-16 (1985), и Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev. (52:119-58 (1982))
Способ получения антител
Способы получения антител хорошо известны в данной области техники и описаны в настоящей заявке. В некоторых вариантах осуществления как вариабельные, так и константные области антигенсвязывающих полипептидов по настоящему изобретению являются полностью человеческими. Полностью человеческие антитела можно получить с использованием способов, описанных в предшествующем уровне техники, и как описано в настоящей заявке. Например, полностью человеческие антитела против конкретного антигена можно получить путем введения антигена трансгенному животному, которое было модифицировано для продукции такого антитела в ответ на стимуляцию антигеном, но чьи эндогенные локусы были отключены. Примеры способов, которые можно использовать для получения таких антител, описаны в патентах США №№: 6150584; 6458592; 6420140, полное содержание которых включено в настоящую заявку посредством ссылки.
Специфичность связывания антигенсвязывающего полипептида по настоящему изобретению можно измерить путем осуществления экспериментов in vitro, таких как иммунопреципитация, радиоиммуноанализ (RIA) или твердофазный иммуноферментный анализ (ELISA).
Альтернативно, способ, описанный для получения одноцепочечных структур (патент США № 4694778; Bird, Science 242:423-442 (1988); Huston et al. Proc. Natl. Acad. Sci. USA 55:5879-5883 (1988); и Ward et al., Nature 334:544-554 (1989)), можно использовать для получения одноцепочечных структур по настоящему изобретению. Одноцепочечные структуры получают путем связывания фрагментов тяжелой и легкой цепи Fv области через аминокислотный мостик, что приводит к одноцепочечному слитому пептиду. Также можно использовать способы синтеза функциональных Fv фрагментов в E. coli (Skerra et al., Science 242: 1038-1041 (1988)).
Примеры способов, которые можно использовать для получения одноцепочечных Fv (scFvs) и антител, включают способы, описанные в патентах США №№ 4946778 и 5258498; Huston et al., Methods in Enzymology 203:46-88 (1991); Shu et al., Proc. Natl. Sci. USA 90:1995-1999 (1993); и Skerra et al., Science 240:1038-1040 (1988). Для некоторых применений, включая применение антитела у людей in vivo и анализы детекци in vitro, предпочтительным может быть использование химерных, гуманизированных или человеческих антител. Химерные антитела представляют собой молекулы, в которых разные части антитела происходят от разных видов животных, такие как антитела, содержащие вариабельную область из мышиного моноклонального антитела и константную область человеческого иммуноглобулина. Способы получения химерных антител известны в данной области техники. См., например, Morrison, Science 229:1202 (1985); Oi et al., BioTechniques 4:214 (1986); Gillies et al., J. Immunol. Methods 125:191-202 (1989); патенты США №№ 5807715; 4816567 и 4816397, полное содержание которых включено в настоящую заявку посредством ссылки.
Гуманизированное антитело представляет собой молекулу антитела, которая получена от отличного от человека вида и связывается целевым антигеном, и такая молекула антитела содержит одну или несколько определяющих комплементарность областей (CDR) от отличного от человека вида и каркасную область из молекулы иммуноглобулина человека. Как правило, каркасные остатки в человеческой каркасной области будут изменяться путем замены соответствующими остатками из CDR донорного антитела, предпочтительно для повышения антигенсвязывающей способности. Эти замены в каркасной области идентифицируют способами, известными в данной области техники, например, путем моделирования взаимодействий между CDR и каркасными остатками для идентификации каркасных остатков, которые важны для связывания с антигеном и сравнения последовательностей, чтобы выявить аномальные каркасные остатки в определенных положениях. (См., например, патент США № 5585089 Queen et al.; Nature 332:323 (1988) Riechmann et al., полное содержание которых включено в настоящую заявку посредством ссылки). Антитела могут быть гуманизированы с использованием различных способов, известных в данной области техники, включая, например, CDR-прививку (EP 239400; публикация PCT № WO 91/09967; патенты США №№ 5225539; 5530101 и 5585089), маскировку поверхностных остатков или изменение поверхности вариабельных доменов (EP 592106; EP 519596; Padlan, Molecular Immunology 28(4/5):489-498 (1991); Studnicka et al., Protein Engineering 7(6):805-814 (1994); Roguska et al., Proc. Natl. Sci. USA 91:969-973 (1994)) и перестановку цепей (патент США № 5565332, полное содержание которых включено в настоящую заявку посредством ссылки).
Используя обычную технологию рекомбинантной ДНК, одна или несколько CDR антигенсвязывающего полипептида по настоящему изобретению могут быть встроены в каркасную область, например встроены в каркасную область человека для гуманизации нечеловеческих антител. Каркасная область может быть природной или гетерологичной каркасной областью и предпочтительно представляет собой каркасную область человека (см., например, Chothia et al., J. Mol. Biol. 278:457-479 (1998), перечень человеческих каркасных областей). Предпочтительно полинуклеотид, полученный комбинацией каркасной области и CDR, кодирует полипептид, который специфически связывается по меньшей мере с одним антигенным эпитопом желаемого полипептида, например, LIGHT. Предпочтительно в каркасной области можно осуществить одну или несколько аминокислотных замен, и предпочтительно аминокислотные замены улучшают связывание антитела с его антигеном. Кроме того, этот метод можно использовать для аминокислотных замен или делеций одного или нескольких цистеиновых остатков вариабельной области (цистеиновые остатки участвуют в образовании внутрицепочечных дисульфидных связей), тем самым создавая молекулы антител, лишенные одной или нескольких внутрицепочечных дисульфидных связей. Другие изменения, вносимые в полинуклеотиды, включены в объем настоящего изобретения и в объем предшествующего уровня техники.
Кроме того, можно использовать методы получения "химерных антител" путем сплайсинга генов из молекул мышиных антител (Morrison et al., Proc. Natl. Acad. Sci. USA: 851-855 (1984); Neuberger et al., Nature 37:604-608 (1984); Takeda et al. Nature 314:452-454 (1985)), и молекула с соответствующей антиген- специфичностью будет связана с геном молекулы человеческого антитела с соответствующей биологической активностью. Используемый в настоящей заявке термин "химерное антитело" означает молекулу, в которой разные части происходят от разных видов животных, например, антитела, содержащие вариабельную область мышиного моноклонального антитела и константную область человеческого иммуноглобулина.
Однако другой эффективный способ получения рекомбинантных антител раскрыт в Newman, Biotechnology 10: 1455-1460 (1992). В частности, эта методика привела к получению приматизированных антител, содержащих обезьяньи вариабельные домены и человеческие константные последовательности. Этот документ полностью включен в настоящую заявку посредством ссылки. Кроме того, этот метод также описан в принадлежащих одному и тому же правообладателю патентах США №№ 5658570, 5693780 и 5756096, каждый из которых включен в настоящую заявку посредством ссылки.
Альтернативно, клеточные линии, продуцирующие антитела, могут быть отобраны и культивированы с использованием способов, хорошо известных специалистам в данной области. Такие способы описаны в различных лабораторных руководствах и первичных публикациях. В этом отношении способы, подходящие для использования в настоящем изобретении, описаны, например, в Current Protocols in Immunology под редакцией Coligan et al., Green Publishing Associates and Wiley-Interscience, John Wiley and Sons, New York (1991), которые включены посредством ссылки во всей полноте, включая дополнительные ссылки.
Кроме того, для введения мутаций в нуклеотидные последовательности, кодирующие антитела по настоящему изобретению, можно использовать стандартные методы, известные специалистам в данной области, включая, но не ограничиваясь этим, сайт-направленный мутагенез и ПЦР-опосредованные мутации, которые приводят к аминокислотным заменам. Предпочтительно варианты (включая производные) по сравнению с эталонной вариабельной областью тяжелой цепи, CDR-H1, CDR-H2, CDR-H3, вариабельной областью легкой цепи, CDR-L1, CDR-L2 или CDR-L3, кодируют менее 50 аминокислотных замен, менее 40 аминокислотных замен, менее 30 аминокислотных замен, менее 25 аминокислотных замен, менее 20 аминокислотных замен, менее 15 аминокислотных замен и менее 10 аминокислотных замен, менее 5 аминокислотных замен, менее 4 аминокислотных замен, менее 3 аминокислотных замен или менее 2 аминокислотных замен. Альтернативно, мутации могут быть введены случайным образом вдоль всей или части кодирующей последовательности, например, путем насыщающего мутагенеза, и полученные мутанты можно скринировать на биологическую активность для выявления мутаций, сохраняющих активность.
Информация о структуре антител
Моноспецифические антитела представляют собой симметричные антитела, включающие две идентичные легкие цепи и две идентичные тяжелые цепи. Легкая цепь и тяжелая цепь связаны дисульфидной связью и нацелены на соответствующий антиген; тяжелая цепь и тяжелая цепь связаны дисульфидной связью; антитело в целом имеет "Y" конфигурацию. Легкая цепь включает вариабельную область легкой цепи (VL) и константную область легкой цепи (Lc), и тяжелая цепь включает вариабельную область тяжелой цепи (VH) и константную область тяжелой цепи, где константная область тяжелой цепи включает CH1 и Fc, и Fc включает шарнир, CH2 и CH3.
Структура 1 биспецифического антитела представляет собой F(ab)2-(Fv)2-Fc, которая представляет собой четырехвалентное симметричное биспецифическое антитело, включающее две идентичные гибридные тяжелые цепи и две идентичные гибридные легкие цепи, где две гибридные тяжелые цепи образуют пару, гибридная легкая цепь и гибридная тяжелая цепь образуют пару, и каждая пара образует одну или несколько межцепочечных дисульфидных связей. Гибридная тяжелая цепь антитела включает вариабельную область тяжелой цепи антитела a (VHa), первую константную область (CH1), вариабельную область 1 антитела b и Fc фрагмент. Вариабельная область 1 антитела b расположена между CH1 и Fc и связана линкером. Гибридная легкая цепь антитела включает вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи (CL) и вариабельную область 2 антитела b. Вариабельная область 2 антитела b расположена на C-конце CL и связана линкером; где (1) вариабельная область 1 антитела b представляет собой вариабельную область тяжелой цепи (VHb), вариабельная область 2 антитела b представляет собой вариабельную область легкой цепи (VLb), и антитело называется биспецифическим антителом серии A структуры 1, или (2) вариабельная область 1 антитела b представляет собой вариабельную область легкой цепи (VLb), вариабельная область 2 антитела b представляет собой вариабельную область тяжелой цепи (VHb), и антитело называется биспецифическим антителом серии B структуры 1. Где VHa-VLa пара нацелена на антиген A, а VHb-VLb пара нацелена на антиген B.
Фиг. 1A схематически представляет структуру биспецифического антитела структуры 1, и Фиг. 1B схематически представляет первичную структуру белка каждого компонента антитела.
Структура 2 биспецифического антитела представляет собой IgG(H)-ScFv, которая представляет собой четырехвалентное симметричное биспецифическое антитело, включающее две идентичные гибридные тяжелые цепи и две идентичные легкие цепи, где две гибридные тяжелые цепи образуют пару, легкая цепь и гибридная тяжелая цепь образуют пару, и каждая пара образует одну или несколько межцепочечных дисульфидных связей; гибридная тяжелая цепь включает вариабельную область тяжелой цепи (VHa), первую константную область тяжелой цепи (CH1), Fc и ScFv, где ScFv расположен на C-конце Fc и связан линкером. Пара легкая цепь-тяжелая цепь нацелена на антиген A, а ScFv нацелен на антиген B.
Фиг. 2A схематически представляет структуру биспецифического антитела структуры 2, и Фиг. 2B схематически представляет первичную структуру белка каждого компонента антитела.
Структура 3 биспецифического антитела представляет собой Тандем-ScFv-Fc, которая представляет собой четырехвалентное симметричное биспецифическое антитело, включающее два идентичных гибридных пептида. Гибридный пептид включает ScFv (ScFv-a), нацеливающийся на антиген A, (ScFv-b), нацеливающийся на антиген B, и Fc, где ScFv-b расположен между ScFv-a и Fc и связан линкером или шарниром.
Фиг. 3A схематически представляет структуру биспецифического антитела структуры 3, и Фиг. 3B схематически представляет первичную структуру белка каждого компонента антитела.
Структура 4 биспецифического антитела представляет собой DVD-IgG, которая представляет собой четырехвалентное симметричное биспецифическое антитело, включающее две идентичные гибридные легкие цепи и две идентичные гибридные тяжелые цепи, где две гибридные тяжелые цепи образуют пару, и две гибридные легкие цепи и гибридные тяжелые цепи образуют пару, и каждая пара образует одну или несколько межцепочечных дисульфидных связей; гибридная легкая цепь включает вариабельную область 1 антитела b и легкую цепь антитела a, которая связана линкером, где легкая цепь антитела a расположена на C-конце; гибридная тяжелая цепь включает вариабельную область 2 антитела b и тяжелую цепь антитела a, которая связана линкером, где тяжелая цепь антитела a расположена на C-конце. Где (1) вариабельная область 1 антитела b представляет собой вариабельную область легкой цепи (VLb), вариабельная область 2 антитела b представляет собой вариабельную область тяжелой цепи (VHb), или (2) вариабельная область 1 антитела b представляет собой вариабельную область тяжелой цепи (VHb), вариабельная область 2 антитела b представляет собой вариабельную область легкой цепи (VLb). VHa-VLa пара нацелена на антиген A, а VHb-VLb пара нацелена на антиген B.
Фиг. 4A схематически представляет структуру биспецифического антитела структуры 4, и Фиг. 4B схематически представляет первичную структуру белка каждого компонента антитела.
Все из вышеуказанных четырех структур биспецифических антител представляют собой симметричную структуру с Fc фрагментом, и все являются четырехвалентными, при этом два нацелены на антиген A, а другие два нацелены на антиген B.
Вариабельные области последовательностей антител
Последовательности вариабельных областей анти-CD3 антитела
(источник последовательности)
WVRQAPGKGLEWVARIRSKYNNYATYYADSVKD
RFTISRDDSKNTLYLQMNSLRAEDTAVYYCARHGN
FGNSYVSWFAYWGQGTLVTVSS
SNYANWVQQKPGQAPRGLIGGTNKRAPGV
PARFSGSLLGGKAALTLSGVQPEDEAEYYCA
LWYSNLWVFGGGTKVEIK
WVRQAPGKGLEWVARIRSKYNNYATYYADSVKD
RFTISRDDSKNTLYLQMNSLRAEDTAVYYCARHGN
FGNSYVSWAAYWGQGTLVTVSS
SNYANWFQQKPGQAPRGLIGGTNKRAPGVP
ARFSGSLLGGKAALTLSGVQPEDEAEYYCAL
WYSNLWVFGGGTKVEIK
(US8236308)
WVRQAPGKGLEWVARIRSKYNNYATYYADSVKDR
FTISRDDSKNTAYLQMNNLKTEDTAVYYCVRHGNF
GNSYISYWAYWGQGTLVTVSS
GNYPNWVQQKPGQAPRGLIGGTKFLAPGTP
ARFSGSLLGGKAALTLSGVQPEDEAEYYCVL
WYSNRWVFGGGTKLTVL
Последовательности вариабельных областей анти-B7-H3 антитела
(источник последовательности)
(Cancer Research 61, 4048-4054, 15 мая 2001 г.)
WVRQRPEQGLEWIGWIFPGDGSTQYNEKFKGKA
TLTTDTSSSTAYMQLSRLTSEDSAVYFCARQTTATW
FAYWGQGTLVTVSS
LHWYQQKSHESPRLLIKYASQSISGIPSRFSG
SGSGSDFTLSINSVEPEDVGVYYCQNGHSFP
LTFGAGTKLELK
(US20120294796A1)
QWVKQRPGQGLEWIGTIYPGDGDTRYTQKFKGK
ATLTADKSSSTAYMQLSSLASEDSAVYYCARRGIPR
LWYFDVWGAGTTVTVSS
LNWYQQKPDGTVKLLIYYTSRLHSGVPSRF
SGSGSGTDYSLTIDNLEQEDIATYFCQQGNTL
PPTFGGGTKLEIK
Последовательности вариабельных областей анти-CD38 антитела
(источник последовательности)
(US9040050)
WVRQAPGKGLEWVSAISGSGGGTYYADSVKGRF
TISRDNSKNTLYLQMNSLRAEDTAVYFCAKDKILW
FGEPVFDYWGQGTLVTVSS
LAWYQQKPGQAPRLLIYDASNRATGIPARFS
GSGSGTDFTLTISSLEPEDFAVYYCQQRSNW
PPTFGQGTKVEIK
(US8088896)
NWVRQAPGKGLEWVSGISGDPSNTYYADSVKGRF
TISRDNSKNTLYLQMNSLRAEDTAVYYCARDLPLV
YTGFAYWGQGTLVTVSS
VYWYQQKPGQAPVLVIYGDSKRPSGIPERFS
GSNSGNTATLTISGTQAEDEADYYCQTYTG
GASLVFGGGTKLTVLGQ
(US9040050)
WVRQAPGQGLEWMGRVIPFLGIANSAQKFQGRV
TITADKSTSTAYMDLSSLRSEDTAVYYCARDDIAAL
GPFDYWGQGTLVTVSS
LAWYQQKPEKAPKSLIYAASSLQSGVPSRFS
GSGSGTDFTLTISSLQPEDFATYYCQQYNSYP
RTFGQGTKVEIK
Последовательности вариабельных областей анти-EpCAM антитела
(источник последовательности)
WVRQAPGQGLEWMGGIIPIFGTANYAQKFQGRV
TITADESTSTAYMELSSLRSEDTAVYYCARGLLWN
YWGQGTLVTVSS
LAWYQQKPGQAPRLIIYGASTTASGIPARFS
ASGSGTDFTLTISSLQSEDFAVYYCQQYNNW
PPAYTFGQGTKLEIK
WVKQAPGKGLKWMGWINTETGEPTYADDFKGRF
AFSLETSASTAYLQINNLKNEDTATYFCARTAVYW
GQGTTVTVSS
LSWLQQEPDGTIKRLIYATSTLDSGVPKRFS
GSRSGSDYSLTISSLESEDFVDYYCLQYASYP
WTFGGGTKLEIK
Последовательности вариабельных областей анти-BCMA антитела
(источник последовательности)
(US9598500)
WVRQAPGQGLEWMGWIYFASGNSEYNQKFTGRV
TMTRDTSINTAYMELSSLTSEDTAVYFCASLYDYD
WYFDVWGQGTMVTVSS
NGNTYLHWYLQKPGQSPQLLIYKVSNRFSG
VPDRFSGSGSGTDFTLKISRVEAEDVGIYYCS
QSSIYPWTFGQGTKLEIK
(WO2016090320A1)
WVRQAPGQGLEWMGRIIPILGIANYAQKFQGRVTI
TADKSTSTAYMELSSLRSEDTAVYYCARGGYYSH
DMWSEDWGQGTLVTVSS
VNWYRQLPGAAPKLLIYSNNQRPPGVPVRFS
GSKSGTSASLAISGLQSEDEATYYCATWDDN
LNVHYVFGTGTKVTVLG
(US2017051068A1)
GWIRQPPGKGLEWIGSIYYSGITYYNPSLKSRVTIS
VDTSKNQFSLKLSSVTAADTAVYYCARHDGAVAG
LFDYWGQGTLVTVSS
VHWYQQPPGQAPVVVVYDDSDRPSGIPERF
SGNSNGNTATLTISRVEAGDEAVYYCQVWD
SSSDHVVFGGGTKLTVL
Последовательности вариабельных областей анти-CTLA-4 антитела
(источник последовательности)
WVRQAPGKGLEWVTFISYDGNNKYYADSVKGRFT
ISRDNSKNTLYLQMNSLRAEDTAIYYCARTGWLGP
FDYWGQGTLVTVSS
YLAWYQQKPGQAPRLLIYGAFSRATGIPDRF
SGSGSGTDFTLTISRLEPEDFAVYYCQQYGSS
PWTFGQGTKVEIK
Последовательности вариабельных областей анти-TIGIT антитела
(источник последовательности)
WVRQSPGKGLEWVAFIRSGSGIVFYADAVRGRFTI
SRDNAKNLLFLQMNDLKSEDTAMYYCARRPLGHN
TFDSWGQGTLVTVSS
SGVKENLLAWYQQKPGQSPKLLIYYASIRF
TGVPDRFTGSGSGTDYTLTITSVQAEDMGQY
FCQQGINNPLTFGDGTKLEIK
WGWIRQPPGKGLEWIGSIYYSGSTYYNPSLKSRATI
SVDTSKNQFSLKLSSVTAADTAVYYCARDGVLAL
NKRSFDIWGQGTMVTVSS
YLAWYQQKPGQAPRLLIYGASSRATGIPDRF
SGSGSGTDFTLTISRLEPEDFAVYYCQQHTV
RPPLTFGGGTKVEIK
Последовательности вариабельных областей анти-LAG-3 антитела
(источник последовательности)
NWIRQPPGKGLEWIGEINHRGSTNSNPSLKSRVTLS
LDTSKNQFSLKLRSVTAADTAVYYCAFGYSDYEY
NWFDPWGQGTLVTVSS
AWYQQKPGQAPRLLIYDASNRATGIPARFSG
SGSGTDFTLTISSLEPEDFAVYYCQQRSNWP
LTFGQGTNLEIK
HWVRQAPGQGLEWMGIINPSAGSTSYAQKFQGRV
TMTRDTSTSTVYMELSSLRSEDTAVYYCARELMAT
GGFDYWGQGTLVTVSS
NYVSWYQQHPGKAPKLMIYDVSNRPSGVS
NRFSGSKSGNTASLTISGLQAEDEANYYCSS
YTSSSTNVFGTGTKVTVL
Последовательности вариабельных областей анти-PD-1 антитела
(источник последовательности)
WVRQAPGKGLEWVAVIWYDGSKRYYADSVKGRF
TISRDNSKNTLFLQMNSLRAEDTAVYYCATNDDY
WGQGTLVTVSS
LAWYQQKPGQAPRLLIYDASNRATGIPARFS
GSGSGTDFTLTISSLEPEDFAVYYCQQSSNW
PRTFGQGTKVEIK
YWVRQAPGQGLEWMGGINPSNGGTNFNEKFKNR
VTLTTDSSTTTAYMELKSLQFDDTAVYYCARRDYR
FDMGFDYWGQGTTVTVSS
GYSYLHWYQQKPGQAPRLLIYLASYLESGV
PARFSGSGSGTDFTLTISSLEPEDFAVYYCQH
SRDLPLTFGGGTKVEIK
Последовательности вариабельных областей анти-PD-L1 антитела
(источник последовательности)
(WO2010077634A1)
WVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFT
ISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPG
GFDYWGQGTLVTVSS
AVAWYQQKPGKAPKLLIYSASFLYSGVPSRF
SGSGSGTDFTLTISSLQPEDFATYYCQQYLY
HPATFGQGTKVEIK
(WO2013079174A1)
WVRQAPGKGLEWVSSIYPSGGITFYADTVKGRFTI
SRDNSKNTLYLQMNSLRAEDTAVYYCARIKLGTVT
TVDYWGQGTLVTVSS
NYVSWYQQHPGKAPKLMIYDVSNRPSGVSN
RFSGSKSGNTASLTISGLQAEDEADYYCSSYT
SSSTRVFGTGTKVTVL
WVRQAPGQGLEWMGGIIPIFGKAHYAQKFQGRV
TITADESTSTAYMELSSLRSEDTAVYFCARKFHFVS
GSPFGMDVWGQGTTVTVSS
LAWYQQKPGQAPRLLIYDASNRATGIPARFS
GSGSGTDFTLTISSLEPEDFAVYYCQQRSNW
PTFGQGTKVEIK
Последовательности вариабельных областей анти-CD16 антитела
(источник последовательности)
WVRQAPGKGLEWVSGINWNGGSTGYADSVKGRF
TISRDNAKNSLYLQMNSLRAEDTAVYYCARGRSLL
FDYWGQGTLVTVSR
WYQQKPGQAPVLVIYGKNNRPSGIPDRFSGS
SSGNTASLTITGAQAEDEADYYCNSRDSSGN
HVVFGGGTKLTVL
Последовательность вариабельной области анти-SLAMF7 антитела
(источник последовательности)
(WO2004100898A2)
WVRQAPGKGLEWIGEINPDSSTINYAPSLKDKFIISR
DNAKNSLYLQMNSLRAEDTAVYYCARPDGNYWY
FDVWGQGTLVTVSS
AVAWYQQKPGKVPKLLIYWASTRHTGVPDR
FSGSGSGTDFTLTISSLQPEDVATYYCQQYSS
YPYTFGQGTKVEIK
Последовательности вариабельных областей анти-CEA антитела
(источник последовательности)
NWVRQAPGQGLEWMGWINTKTGEATYVEEFKG
RFVFSLDTSVSTAYLQISSLKADDTAVYYCARWDF
YDYVEAMDYWGQGTTVTVSS
NVAWYQQKPGKAPKLLIYSASYRYSGVPSR
FSGSGSGTDFTFTISSLQPEDIATYYCHQYYT
YPLFTFGQGTKVEIK
Последовательности вариабельных областей анти-VEGF антитела
(источник последовательности)
NWVRQAPGKGLEWVGWINTYTGEPTYAADFKRR
FTFSLDTSKSTAYLQMNSLRAEDTAVYYCAKYPHY
YGSSHWYFDVWGQGTLVTVSS
LNWYQQKPGKAPKVLIYFTSSLHSGVPSRFS
GSGSGTDFTLTISSLQPEDFATYYCQQYSTVP
WTFGQGTKVEIK
WVRQAPGKGLEWVGAIWPFGGYTHYADSVKGRF
TISADTSKNTAYLQMNSLRAEDTAVYYCARWGHS
TSPWAMDYWGQGTLVTVSS
LAWYQQKPGKAPKLLIYAASNLASGVPSRFS
GSGSGTDFTLTISSLQPEDFATYYCQQSNTSP
LTFGQGTKVEIK
WVRQAPGKGLEWVAGITPAGGYTYYADSVKGRF
TISADTSKNTAYLQMNSLRAEDTAVYYCARFVFFL
PYAMDYWGQGTLVTVSS
AVAWYQQKPGKAPKLLIYSASFLYSGVPSRF
SGSGSGTDFTLTISSLQPEDFATYYCQQGYG
NPFTFGQGTKVEIK
Последовательности вариабельных областей анти-TGF-β антитела
(источник последовательности)
WVRQAPGQGLEWMGGVIPIVDIANYAQRFKGRVT
ITADESTSTTYMELSSLRSEDTAVYYCASTLGLVLD
AMDYWGQGTLVTVSS
YLAWYQQKPGQAPRLLIYGASSRAPGIPDRF
SGSGSGTDFTLTISRLEPEDFAVYYCQQYAD
SPITFGQGTRLEIK
WVRQAPGQGLEWMGGVIPIVDIANYAQRFKGRVT
ITADESTSTTYMELSSLRSEDTAVYYCALPRAFVLD
AMDYWGQGTLVTVSS
YLAWYQQKPGQAPRLLIYGASSRAPGIPDRF
SGSGSGTDFTLTISRLEPEDFAVYYCQQYAD
SPITFGQGTRLEIK
Последовательности вариабельных областей анти-IL-10 антитела
(источник последовательности)
WVRQSPGKGLEWLGVIWRGGSTDYSAAFMSRLSI
TKDNSKSQVFFKMNSLQADDTAIYFCAKQAYGHY
MDYWGQGTSVTVSS
NGNTYLEWYLQKPGQSPKLLIYKVSNRFSG
VPDRFSGSGSGTDFTLKITRLEAEDLGVYYC
FQGSHVPWTFGGGTKLEIK
WVRQSPGKGLEWLGVIWRGGSTDYSAAFMSRLTI
SKDNSKNTVYLQMNSLRAEDTAVYFCAKQAYGH
YMDYWGQGTSVTVSS
NGNTYLEWYLQRPGQSPRLLIYKVSNRFSG
VPDRFSGSGSGTDFTLKISRVEAEDVGVYYC
FQGSHVPWTFGQGTKVEIK
Последовательности вариабельных областей антитела против люциферазы
(источник последовательности)
nwvrqspekglewvaqirnkpynyetyysdsvk
grftisrddskssvylqmnnlrvedmgiyyctgsy
ygmdywgqgtsvtvss
sngntylrwylqkpgqspkvliykvsnrfs
gvpdrfsgsgsgtdftlkisrveaedlgvyf
csqsthvpwtfgggtkleik
Последовательность других доменов
(1) Аминокислотные последовательности линкерного домена
Аминокислотные последовательности линкера
(2) Аминокислотные последовательности шарнирного домена
Аминокислотные последовательности шарнира
(3) Аминокислотная последовательность CL домена константной области легкой цепи
Аминокислотные последовательности CL
ID NO.
(4) Аминокислотные последовательности CH1 домена константной области тяжелой цепи
Аминокислотные последовательности CH1
ID NO.
Аминокислотные последовательности CH2 Fc
ID NO.
Аминокислотные последовательности CH3 Fc, образующие гетеродимер
ID NO.
Специфические последовательности антигенов
Аминокислотные последовательности опухолевого антигена
ID NO.
(источник: UniProtKB - P28907)
(источник: UniProtKB - Q02223)
(источник: UniProtKB - P16410)
(источник: UniProtKB - P18627)
(источник: UniProtKB - Q495A1)
(источник: UniProtKB - Q15116)
(источник: UniProtKB - Q9NZQ7)
(источник: UniProtKB - Q9NQ25)
(источник: UniProtKB - P06731)
(источник: UniProtKB - P07766)
(источник: UniProtKB - P08637)
(источник: UniProtKB - P01137)
(источник: UniProtKB - P61812)
(источник: UniProtKB - P10600)
(источник: UniProtKB - P15692)
(источник: UniProtKB - P22301)
(источник: UniProtKB - Q13651)
Получение антитела и детекция активности антитела
Пример 1. Получение биспецифического антитела
1. Способ конструирования плазмиды
pcDNA3.1 (приобретали у invitrogen) или pCHO1.0 (приобретали у Gibco) используют в качестве вектора.
1.1 Амплификация фрагмента ДНК, представляющего интерес, с использованием полимеразной цепной реакции (ПЦР)
Полученный продукт ПЦР представляет собой интересующий фрагмент ДНК.
1.2 Расщепление векторной плазмиды рестрикционным ферментом
Ферментативное расщепление осуществляют при оптимальной температуре в течение 4 часов.
Фермент-расщепленный продукт представляет собой фермент-расщепленную векторную ДНК.
*Векторы подразделяются на два типа: вектор экспрессии тяжелой цепи и вектор экспрессии легкой цепи. Вектор экспрессии тяжелой цепи содержит сигнальный пептид и последовательность ДНК константной области тяжелой цепи человеческого IgG1 (GenBank номер доступа MG920253.1), которая включает CH1, шарнир, CH2 и CH3; и сайт рестрикции расположен между 3' сигнального пептида и 5' концом CH1; при этом вектор экспрессии легкой цепи содержит сигнальный пептид и последовательность ДНК константной области каппа или лямбда легкой цепи человека, и сайт рестрикции расположен между 3' сигнального пептида и 5' концом константной области легкой цепи.
1.3 Очистка ПЦР продуктов или фермент-расщепленных продуктов
Использовали реагенты Tiangen для очистки ДНК. Что касается конкретных стадий, см. инструкции, прилагаемые к набору Tiangen. Полученный очищенный продукт представлял собой очищенный фрагмент ДНК, представляющий интерес, и очищенную фермент-расщепленную векторную ДНК.
(1) Рекомбинация фрагментов, представляющих интерес
Условия рекомбинации: 37°C в течение 30 минут. Полученный рекомбинантный продукт помещали на лед, и он был готов для трансформации.
Фрагмент тяжелой цепи был рекомбинирован в расщепленную ферментом векторную ДНК для экспрессии тяжелой цепи, а фрагмент легкой цепи был рекомбинирован в расщепленную ферментом векторную ДНК для экспрессии легкой цепи.
1.4 Трансформация методом теплового шока
10 мкл рекомбинантного продукта трансформировали в 100 мкл Транс10-компетентных клеток путем теплового шока в соответствии с общепринятым методом. Была выбрана единственная колония, которая была секвенирована Wuhan Genecreate. Плазмида, полученная путем рекомбинации фрагмента тяжелой цепи в фермент-расщепленную ДНК вектора экспрессии, называется плазмидой экспрессии тяжелой цепи, а плазмида, полученная путем рекомбинации фрагмента легкой цепи в фермент-расщепленную ДНК вектора экспрессии, называется плазмидой экспрессии легкой цепи, а плазмида, полученная путем рекомбинации гибридного пептидного фрагмента в фермент-расщепленную ДНК вектора экспрессии, называется плазмидой экспрессии гибридного пептида, плазмида, полученная путем рекомбинации фрагмента гибридной тяжелой цепи в фермент-расщепленную ДНК вектора экспрессии, называется плазмидой экспрессии гибридной тяжелой цепи, а плазмида, полученная путем рекомбинации фрагмента гибридной цепи в фермент-расщепленный вектор экспрессии, называется плазмидой экспрессии гибридной цепи.
Конструкция специфической плазмиды является следующей:
1) Структура 1 биспецифического антитела на Фиг. 1 включает конструкцию из двух плазмид. Эти две плазмиды представляют собой следующие: плазмида экспрессии гибридной легкой цепи (pFL) и плазмида экспрессии гибридной тяжелой цепи (pFH).
2) Структура 2 биспецифического антитела на Фиг. 2 включает конструкцию их двух плазмид. Эти две плазмиды представляют собой следующие: плазмида экспрессии легкой цепи (pL) и плазмида экспрессии гибридной тяжелой цепи 2 (pFH2).
3) Структура 3 биспецифического антитела на Фиг. 3 включает конструкцию из одной плазмиды. Эта плазмида представляет собой следующую: плазмида экспрессии гибридного пептида (pFP).
4) Структура 4 биспецифического антитела на Фиг. 4 включает конструкцию из двух плазмид. Эти две плазмиды представляют собой следующие: плазмида экспрессии гибридной легкой цепи 2 (pFL2) и плазмида экспрессии гибридной тяжелой цепи 3 (pFH3).
1.5 Способ экспрессии биспецифического антитела
Задействованы две системы экспрессии с транзиентной трансфекцией, CHAOS и 293E, и стадии подробно описаны ниже:
(1) Процедура транзиентной трансфекции CHO-S (100 мл общего объема трансфекции в качестве примера)
а) Пассаж клеток осуществляли за один день до трансфекции, например, для пассажа клеток можно использовать среду CD-CHO (приобретенную у Thermo Fisher); плотность клеток в суспензии доводили до 1×106 клеток/мл, а объем составлял 90 мл, чтобы гарантировать, что клетки находились в логарифмической фазе роста, и плотность клеток может достигать 2×106 клеток/мл на следующий день трансфекции;
b) клетки культивировали при 37°С, 125 об/мин, 5% CO2 в течение ночи на шейкере;
c) в день, определенный для трансфекции, трансфекционный реагент FectoPRO (приобретен у Polyplus transfection company) предварительно нагревали до комнатной температуры и осторожно смешивали; 50мкг плазмидной ДНК разбавляли 10мл бессывороточной среды, такой как opti PRO-SFM (приобретенной у Thermo Fisher), осторожно смешивали и добавляли в 100мкл трансфекционного реагента, тщательно смешивали и инкубировали при комнатной температуре в течение 10 минут с образованием трансфекционного комплекса; две или три плазмидные ДНК были котрансфицированы в процессе трансфекции.
d) трансфекционный комплекс равномерно добавляли в полученные 90 мл клеток в логарифмической фазе роста, встряхивали сразу после перемешивания и помещали на шейкер для культуры при 37°C, 125об/мин, 5% CO2;
e) через 2-4 часа после трансфекции добавляли 75мкл усилителя трансфекции Fecto PRO® Booster (приобретенного у Polyplus transfection company);
f) через 18-24 часа после трансфекции клетки охлаждали до 32°С и инкубировали;
g) подпитку осуществляли на 3, 5 и 7 день после трансфекции, и объем подпитки составлял 3,5% от общего объема клеток;
h) клетки собирали, когда жизнеспособность клеток была меньше чем 70%, и время экспрессии составляло 9-13 дней.
(2) Процедура транзиентной трансфекции 293E (20 мл общего объема трансфекции в качестве примера)
a) Пассаж клеток осуществляли за один день до трансфекции, например, для пассажа клеток можно использовать FreeStyle™ 293 (приобретенную у Thermo Fisher); плотность клеток в суспензии доводили до 0,6×106-0,8×106 клеток/мл, а объем составлял 20мл, чтобы гарантировать, что клетки находились в логарифмической фазе роста, и плотность клеток может достигать 1,2×106-1,6×106 клеток/мл на следующий день трансфекции;
b) клетки культивировали при 37°С, 125 об/мин, 5% CO2 в течение ночи на шейкере;
c) в день, определенный для трансфекции, LPEI предварительно нагревали до комнатной температуры и осторожно смешивали перед использованием;
d) 20мкг ДНК разбавляли 0,67 мл бессывороточной среды, такой как FreeStyle™ 293, и тщательно смешивали;
e) 40мкг LPEI разбавляли 0,67 мл бессывороточной среды, такой как FreeStyle™ 293, и тщательно смешивали;
f) разбавленную LPEI, полученную на стадии e), добавляли в разбавленную ДНК, полученную на стадии d), быстро тщательно смешивали и инкубировали при комнатной температуре в течение 15 минут с образованием комплекса;
g) трансфекционный комплекс, полученный на стадии f), равномерно добавляли в полученные 20 мл клеток в логарифмической фазе роста, встряхивали сразу после перемешивания и помещали на шейкер для культуры при 37°C, 125 об/мин, 5% CO2;
h) подпитку осуществляли в первый и третий день после трансфекции, и объем подпитки составлял 5% от общего объема клеток;
i) клетки экспрессировали вплоть до шестого дня и затем их собирали.
(3) Специфическая котрансфицированная плазмидная ДНК представляла собой следующую:
1) Для экспресии биспецифического антитела 1, показанного на Фиг. 1, требовались плазмиды pFL и pFH для котрансфекции в CHO-S или 293E клетках для экспрессии;
2) Для экспресии биспецифического антитела 2, показанного на Фиг. 2, требовались плазмиды pL и pFH2 для котрансфекции в CHO-S или 293E клетках для экспрессии;
3) Для экспресии биспецифического антитела 3, показанного на Фиг. 3, требовалась плазмида pFP для котрансфекции в CHO-S или 293E клетках для экспрессии;
4) Для экспресии биспецифического антитела 4, показанного на Фиг. 4, требовались плазмиды pFL2 и pFH3 для котрансфекции в CHO-S или 293E клетах для экспрессии.
Как правило, если две плазмиды котрансфицируют для экспрессии, молярное соотношение двух плазмид может быть 1:1 или любым другим соотношением.
2. Способ очистки биспецифического антитела:
Способ очистки антитела преимущественно включает аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию и молекулярные сита, и стадии являются следующими:
(1) культуральный раствор антитело-экспрессирующих клеток собирали и центрифугировали при 3000×g в течение 10 минут, затем супернатант собирали, фильтровали через 0,22мкм фильтр и хранили при 4°С для последующего использования;
(2) аффинная хроматография (MabSelect SuRe GE 17-5438-01, 18мл колоночного объема в качестве примера)
a) уравновешивание: колонку уравновешивали связывающим буфером (25 мМ Tris, pH 7,0-7,4) до тех пор, пока УФ детекция и значение проводимости не были стабильными или на исходном уровне, при этом использовали по меньшей мере пять колоночных объемов для уравновешивания;
b) загрузка образца: отфильтрованный супернатант, полученный на стадии (1), загружали при скорости потока 5мл/мин;
c) промывка и уравновешивание: пять колоночных объемов связывающего буфера использовали для промывки;
d) элюирование: образец элюировали элюирующим буфером (50мМ лимонной кислоты, pH 3,4±0,1) при скорости потока 5мл/мин, использовали 5 колоночных объемов для элюирования и элюируемый пик собирали;
e) нейтрализация: элюент нейтрализовали при помощи 1M Tris pH8,0 и pH образца доводили до 6,0±0,1.
(3) ионообменная хроматография (катионообменная хроматография в качестве примера, HiTrap SP-HP GE 17-1151-01 5мл колоночного объема)
a) подготовка образца: образец аффинной хроматографии подвергали микрофильтрации, а затем разбавляли сверхчистой водой, чтобы сделать проводимость меньше чем 5 мСм/см, и затем pH доводили до 6,0±0,1;
b) уравновешивание и загрузка образца: колонку сначала уравновешивали 5 колоночными объемами буфера B (25мМ лимонной кислоты+1M хлорида натрия, проводимость которого должна быть 80 ~ 90 мСм/см, pH 6,0±0,1), а затем уравновешивали по меньшей мере пятью колоночными объемами буфера A (25мМ лимонной кислоты, проводимость которого должна быть меньше чем 5 мСм/см, pH 6,0±0,1) до тех пор, пока проводимость, pH и исходный уровень УФ не стали стабильными, и затем загружали образец при скорости потока 3мл/мин;
c) промывка и уравновешивание: колонку промывали пятью колоночных объемами буфера A при скорости потока 5мл/мин;
d) элюирование: образец элюировали 20 колоночными объемами 0-30% буфера B, а затем 10 колоночными объемами 100% буфера B; скорость потока в течение всего процесса составляла 3мл/мин, и элюент собирали в отдельные пробирки и тестировали.
(4) гидрофобная хроматография (Capto phenyl ImpRes наполнитель GE XK16/20 11,5см/23мл)
a) обработка образца: 5M хлорида натрия добавляли в образец к 1M хлорида натрия и pH доводили до 6,0;
b) уравновешивание и загрузка образца: сначала колонку уравновешивали пятью колоночными объемами буфера A (25мМ цитрата+1M хлорида натрия, pH 6,0±0,1) при скорости потока 5мл/мин; и образец загружали при скорости потока 3,3мл/мин;
c) промывка и уравновешивание: для промывки использовали пять колоночных объемов буфера A при скорости потока 5мл/мин; для промывки использовали пять колоночных объемов 10% буфера B (25мМ цитрата, pH 6,0±0,1) при скорости потока 5мл/мин;
d) элюирование: для элюирования использовали 90% буфер B при скорости потока 5мл/мин, элюируемые пики собирали в отдельные пробирки и затем осуществляли детекцию;
(5) молекулярное сито (HiLoad Superdex 200pg GE 28989336 26/600)
a) уравновешивание и загрузка образца: колонку уравновешивали двумя колоночными объемами буфера (20мМ гистидина+0,15M хлорида натрия, pH6,0±0,1), а затем загружали образец при скорости потока 3мл/мин;
b) элюирование: для элюирования использовали два колоночных объема буфера, и элюируемые пики собирали в отдельные пробирки и затем осуществляли детекцию.
Коды антител некоторых специфически экспрессированных антител и соответствующие аминокислотные последовательности антител показаны в следующих таблицах:
Коды антител и аминокислотные последовательности некоторых антител со структурой 1 биспецифического антитела
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQR
SNWPTFGQGTKVEIK
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLRS
EDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQR
SNWPTFGQGTKVEIK
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLRS
EDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
KVLIYFTSSLHSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
YSTVPWTFGQGTKVEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
KGLEWVGWINTYTGEPTYAADFKRRFTFSLDTSKSTAYLQMNS
LRAEDTAVYYCAKYPHYYGSSHWYFDVWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
kpgqspkvliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedl
gvyfcsqsthvpwtfgggtkleik
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
kglewvaqirnkpynyetyysdsvkgrftisrddskssvylqm
nnlrvedmgiyyctgsyygmdywgqgtsvtvss
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
kpgqspkvliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedl
gvyfcsqsthvpwtfgggtkleik
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
kglewvaqirnkpynyetyysdsvkgrftisrddskssvylqm
nnlrvedmgiyyctgsyygmdywgqgtsvtvss
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
kpgqspkvliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedl
gvyfcsqsthvpwtfgggtkleik
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
kglewvaqirnkpynyetyysdsvkgrftisrddskssvylqm
nnlrvedmgiyyctgsyygmdywgqgtsvtvss
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPLTFGQGTNLEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTA
ADTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPLTFGQGTNLEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTA
ADTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPLTFGQGTNLEIK
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTA
ADTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDL
KSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDL
KSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDL
KSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKIRRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLRS
EDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQR
SNWPTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLRS
EDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQR
SNWPTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
KGLEWVGWINTYTGEPTYAADFKRRFTFSLDTSKSTAYLQMNS
LRAEDTAVYYCAKYPHYYGSSHWYFDVWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
KVLIYFTSSLHSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
YSTVPWTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
kglewvaqirnkpynyetyysdsvkgrftisrddskssvylqm
nnlrvedmgiyyctgsyygmdywgqgtsvtvss
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
kpgqspkvliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedl
gvyfcsqsthvpwtfgggtkleik
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
kpgqspkvliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedl
gvyfcsqsthvpwtfgggtkleik
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
kglewvaqirnkpynyetyysdsvkgrftisrddskssvylqm
nnlrvedmgiyyctgsyygmdywgqgtsvtvss
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
kpgqspkvliykvsnrfsgvpdrfsgsgsgtdftlkisrveaedl
gvyfcsqsthvpwtfgggtkleik
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
kglewvaqirnkpynyetyysdsvkgrftisrddskssvylqm
nnlrvedmgiyyctgsyygmdywgqgtsvtvss
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTA
ADTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPLTFGQGTNLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTA
ADTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPLTFGQGTNLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QYLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTA
ADTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
GLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPLTFGQGTNLEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDL
KSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDL
KSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQMNDL
KSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQ
QGYGNPFTFGQGTKVEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYADSPITFGQGTRLEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
SSNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
RLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQ
RSNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQA
DDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
GLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLR
SEDTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLG
VYYCFQGSHVPWTFGGGTKLEIK
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
VKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWL
NGKEYKCKVSNKALPAPIEKTISKAK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
Коды антител и аминокислотные последовательности некоторых антител со структурой 2 биспецифического антитела для сравнения
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQRSNWPTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGQGLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAY
MELSSLRSEDTAVYFCARKFHFVSGSPFGMDVWGQGTTV
TVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
APGKGLEWVGWINTYTGEPTYAADFKRRFTFSLDTSKSTA
YLQMNSLRAEDTAVYYCAKYPHYYGSSHWYFDVWGQGT
LVTVSS
KAPKVLIYFTSSLHSGVPSRFSGSGSGTDFTLTISSLQPEDFA
TYYCQQYSTVPWTFGQGTKVEIK
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
PGQGLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTY
MELSSLRSEDTAVYYCASTLGLVLDAMDYWGQGTLVTVS
S
GQAPRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDF
AVYYCQQYADSPITFGQGTRLEIK
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQRSNWPTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGQGLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAY
MELSSLRSEDTAVYFCARKFHFVSGSPFGMDVWGQGTTV
TVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
PGQGLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTY
MELSSLRSEDTAVYYCASTLGLVLDAMDYWGQGTLVTVS
S
GQAPRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDF
AVYYCQQYADSPITFGQGTRLEIK
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
PPGKGLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSL
KLRSVTAADTAVYYCAFGYSDYEYNWFDPWGQGTLVTV
SS
APRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAV
YYCQQRSNWPLTFGQGTNLEIK
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
APGKGLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTL
YLQMNSLRAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
GQAPRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDF
AVYYCQQYGSSPWTFGQGTKVEIK
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQ
MNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
WYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTIT
SVQAEDMGQYFCQQGINNPLTFGDGTKLEIK
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQ
MNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
WYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTIT
SVQAEDMGQYFCQQGINNPLTFGDGTKLEIK
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GKGLEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFK
MNSLQADDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
YLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITR
LEAEDLGVYYCFQGSHVPWTFGGGTKLEIK
KVPKLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDV
ATYYCQQYSSYPYTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQ
MNSLRAEDTAVYYCARPDGNYWYFDVWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
APGKGLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNS
LYLQMNSLRAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
APVLVIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEA
DYYCNSRDSSGNHVVFGGGTKLTVL
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQRSNWPTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGQGLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAY
MELSSLRSEDTAVYFCARKFHFVSGSPFGMDVWGQGTTV
TVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
KAPKVLIYFTSSLHSGVPSRFSGSGSGTDFTLTISSLQPEDFA
TYYCQQYSTVPWTFGQGTKVEIK
APGKGLEWVGWINTYTGEPTYAADFKRRFTFSLDTSKSTA
YLQMNSLRAEDTAVYYCAKYPHYYGSSHWYFDVWGQGT
LVTVSS
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQRSNWPTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGQGLEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAY
MELSSLRSEDTAVYFCARKFHFVSGSPFGMDVWGQGTTV
TVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GQAPRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDF
AVYYCQQYADSPITFGQGTRLEIK
PGQGLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTY
MELSSLRSEDTAVYYCASTLGLVLDAMDYWGQGTLVTVS
S
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GQAPRLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDF
AVYYCQQYADSPITFGQGTRLEIK
PGQGLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTY
MELSSLRSEDTAVYYCASTLGLVLDAMDYWGQGTLVTVS
S
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
APRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAV
YYCQQRSNWPLTFGQGTNLEIK
PPGKGLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSL
KLRSVTAADTAVYYCAFGYSDYEYNWFDPWGQGTLVTV
SS
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GQAPRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDF
AVYYCQQYGSSPWTFGQGTKVEIK
APGKGLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTL
YLQMNSLRAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQYLYHPATFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYL
QMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
WYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTIT
SVQAEDMGQYFCQQGINNPLTFGDGTKLEIK
GKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQ
MNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
WYQQKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTIT
SVQAEDMGQYFCQQGINNPLTFGDGTKLEIK
GKGLEWVAFIRSGSGIVFYADAVRGRFTISRDNAKNLLFLQ
MNDLKSEDTAMYYCARRPLGHNTFDSWGQGTLVTVSS
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
GKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDF
ATYYCQQGYGNPFTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
PGKGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAY
LQMNSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVS
S
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
QAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFA
VYYCQQSSNWPRTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNT
LFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
YLQKPGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITR
LEAEDLGVYYCFQGSHVPWTFGGGTKLEIK
GKGLEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFK
MNSLQADDTAIYFCAKQAYGHYMDYWGQGTSVTVSS
KVPKLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDV
ATYYCQQYSSYPYTFGQGTKVEIK
KVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKH
KVYACEVTHQGLSSPVTKSFNRGEC
APGKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQ
MNSLRAEDTAVYYCARPDGNYWYFDVWGQGTLVTVSS
NSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYIC
NVNHKPSNTKVDKKVEPKSC
PEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVH
QDWLNGKEYKCKVSNKGLPAPIEKTISKTK
ESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGN
VFSCSVMHEALHNHYTQKSLSLSPG
APVLVIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEA
DYYCNSRDSSGNHVVFGGGTKLTVL
APGKGLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNS
LYLQMNSLRAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
Коды антител и аминокислотные последовательности некоторых антител со структурой 3 биспецифического антитела для сравнения
KGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMN
SLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQYLYHPATFGQGTKVEIK
KGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQM
NSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQGYGNPFTFGQGTKVEIK
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
GKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQ
MNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
PRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYY
CQQSSNWPRTFGQGTKVEIK
GKGLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQ
MNSLRAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
APRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVY
YCQQYGSSPWTFGQGTKVEIK
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
GKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNS
LRAEDTAVYYCARPDGNYWYFDVWGQGTLVTVSS
VPKLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATY
YCQQYSSYPYTFGQGTKVEIK
GKGLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQ
MNSLRAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
VLVIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYY
CNSRDSSGNHVVFGGGTKLTVL
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
KGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMN
SLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQYLYHPATFGQGTKVEIK
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQGYGNPFTFGQGTKVEIK
KGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQM
NSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
GKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQ
MNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
PRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYY
CQQSSNWPRTFGQGTKVEIK
APRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVY
YCQQYGSSPWTFGQGTKVEIK
GKGLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQ
MNSLRAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
GKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNS
LRAEDTAVYYCARPDGNYWYFDVWGQGTLVTVSS
VPKLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATY
YCQQYSSYPYTFGQGTKVEIK
VLVIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYY
CNSRDSSGNHVVFGGGTKLTVL
GKGLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQ
MNSLRAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQYLYHPATFGQGTKVEIK
KGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMN
SLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
KGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQM
NSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQGYGNPFTFGQGTKVEIK
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
PRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYY
CQQSSNWPRTFGQGTKVEIK
GKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQ
MNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
GKGLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQ
MNSLRAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
APRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVY
YCQQYGSSPWTFGQGTKVEIK
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
GKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNS
LRAEDTAVYYCARPDGNYWYFDVWGQGTLVTVSS
VPKLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATY
YCQQYSSYPYTFGQGTKVEIK
GKGLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQ
MNSLRAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
VLVIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYY
CNSRDSSGNHVVFGGGTKLTVL
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQYLYHPATFGQGTKVEIK
KGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMN
SLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSS
APKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYY
CQQGYGNPFTFGQGTKVEIK
KGLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQM
NSLRAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
PRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYY
CQQSSNWPRTFGQGTKVEIK
GKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQ
MNSLRAEDTAVYYCATNDDYWGQGTLVTVSS
APRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVY
YCQQYGSSPWTFGQGTKVEIK
GKGLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQ
MNSLRAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
VPKLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATY
YCQQYSSYPYTFGQGTKVEIK
GKGLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNS
LRAEDTAVYYCARPDGNYWYFDVWGQGTLVTVSS
VLVIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYY
CNSRDSSGNHVVFGGGTKLTVL
GKGLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQ
MNSLRAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
VQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQD
WLNGKEYKCKVSNKGLPAPIEKTISKTK
NGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSC
SVMHEALHNHYTQKSLSLSPGK
Коды антител и аминокислотные последовательности некоторых антитела со структурой 4 биспецифического антитела для сравнения
KLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
YLYHPATFGQGTKVEIK
KLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
GYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRA
EDTAVYYCARRHWPGGFDYWGQGTLVTVSS
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQS
SNWPRTFGQGTKVEIK
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
LEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRA
EDTAVYYCARRHWPGGFDYWGQGTLVTVSS
KLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
GYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
KLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
YLYHPATFGQGTKVEIK
GLEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSL
RAEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
PRLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQ
QYGSSPWTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQS
SNWPRTFGQGTKVEIK
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSL
RAEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQ
QYSSYPYTFGQGTKVEIK
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
ALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKP
SNTKVDKKVEPKSC
QFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLN
GKEYKCKVSNKGLPAPIEKTISKTK
GQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSV
MHEALHNHYTQKSLSLSPGK
Коды антител и аминокислотные последовательности моноклональных антител
KLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
YLYHPATFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRA
EDTAVYYCARRHWPGGFDYWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQR
SNWPTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWMGGIIPIFGKAHYAQKFQGRVTITADESTSTAYMELSSLRSE
DTAVYFCARKFHFVSGSPFGMDVWGQGTTVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
RLLIYGASSRAPGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQ
YADSPITFGQGTRLEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWMGGVIPIVDIANYAQRFKGRVTITADESTSTTYMELSSLRS
EDTAVYYCASTLGLVLDAMDYWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
LIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRS
NWPLTFGQGTNLEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWIGEINHRGSTNSNPSLKSRVTLSLDTSKNQFSLKLRSVTAA
DTAVYYCAFGYSDYEYNWFDPWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
LLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQS
SNWPRTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSL
RAEDTAVYYCATNDDYWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
QKPGQSPKLLIYYASIRFTGVPDRFTGSGSGTDYTLTITSVQAED
MGQYFCQQGINNPLTFGDGTKLEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
PGQSPKLLIYKVSNRFSGVPDRFSGSGSGTDFTLKITRLEAEDLGV
YYCFQGSHVPWTFGGGTKLEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWLGVIWRGGSTDYSAAFMSRLSITKDNSKSQVFFKMNSLQAD
DTAIYFCAKQAYGHYMDYWGQGTSVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
KLLIYWASTRHTGVPDRFSGSGSGTDFTLTISSLQPEDVATYYCQQ
YSSYPYTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWIGEINPDSSTINYAPSLKDKFIISRDNAKNSLYLQMNSLRAE
DTAVYYCARPDGNYWYFDVWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
KLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQ
GYGNPFTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
LEWVAGITPAGGYTYYADSVKGRFTISADTSKNTAYLQMNSLR
AEDTAVYYCARFVFFLPYAMDYWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
RLLIYGAFSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQ
YGSSPWTFGQGTKVEIK
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVTFISYDGNNKYYADSVKGRFTISRDNSKNTLYLQMNSLR
AEDTAIYYCARTGWLGPFDYWGQGTLVTVSS
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
VIYGKNNRPSGIPDRFSGSSSGNTASLTITGAQAEDEADYYCNSR
DSSGNHVVFGGGTKLTVL
NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACE
VTHQGLSSPVTKSFNRGEC
GLEWVSGINWNGGSTGYADSVKGRFTISRDNAKNSLYLQMNSL
RAEDTAVYYCARGRSLLFDYWGQGTLVTVSR
LTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSN
TKVDKKVEPKSC
KFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLN
GKEYKCKVSNKALPAPIEKTISKAK
QPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMH
EALHNHYTQKSLSLSPGK
Уровни экспрессии некоторых антител со структурой 1 биспецифического антитела по настоящему изобретению показаны на Фиг. 5A-D. Фиг. 5A показывает уровни экспрессии при транзиентной трансфекции антител серии Y100-A серии и серии Y100-B в клетках CHO. Фиг. 5B показывает уровни экспрессии при транзиентной трансфекции антител серии Y101/Y103/Y104/Y105-A и серии Y101/Y103/Y104/Y105-B в клетках CHO. Фиг. 5C показывает уровни экспрессии при транзиентной трансфекции антител серии Y106/Y110/Y116-A и серии Y106/Y110/Y116-B в клетках CHO. Фиг. 5D показывает чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A для антител серии Y100/Y101/Y103/Y104/Y105/Y106/Y110/Y116-A и B, которые экспрессируются на уровне больше чем 5 мг/л.
На основании Фиг. 5A-C, можно видеть, что уровень экспрессии при транзиентной трансфекции двух серий биспецифических антител структуры 1 (т.е. серии A и серии B) существенно различается, при этом уровень экспрессии серии A редко превышает 10 мг/л, а уровень экспрессии серии B выше. Кроме того, как можно видеть из Фиг. 5D, чистота биспецифического антитела структуры 1 серии B составляет более 70%, и чистота биспецифического антитела структуры 1 серии A является по существу такой же, как у серии B. Чистоту антител, уровни экспрессии которых меньше чем 5 мг/л, не определяют.
Фиг. 6 показывает уровень экспрессии некоторых антител со структурой 2 биспецифического антитела для сравнения. В частности, Фиг. 6 показывает уровни экспрессии при транзиентной трансфекции антител Y200/Y201/Y203/Y204/Y205/Y206/Y210/Y216-A серии и B серии в клетках CHO, и чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A.
Как можно видеть из Фиг. 6, две серии антител со структурой 2 биспецифического антитела (т.е. серии A и серии B) не имеют существенной разницы в уровне транзиентной экспрессии, и уровень экспрессии не превышает 20 мг/л. Кроме того, что касается чистоты биспецифического антитела структуры 2, определенной методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A, результат показывает, что чистота разных молекул сильно варьируется от 30% до 90%. Уровень экспрессии некоторых антител слишком низкий для определения чистоты.
Фиг. 7 показывает уровень экспрессии некоторых антител со структурой 3 биспецифического антитела для сравнения. В частности, Фиг. 7 показывает уровни экспрессии при транзиентной трансфекции антител Y300/Y304/Y316 серии A, серии B, серии C и серии D в клетках CHO и чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A.
Как можно видеть из Фиг. 7, уровни экспрессии при транзиентной трансфекции четырех серий биспецифических антител структуры 3 (т.е. серии A, серии B, серии C серии и серии D) не являются высокими, и уровни экспрессии не превышают 12 мг/л. Кроме того, что касается чистоты антител со структурой 3 биспецифического антитела, определенной методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A, результат показывает, что чистота является низкой и не более чем 50%. Уровень экспрессии некоторых антител слишком низкий для определения чистоты.
Фиг. 8 показывает уровень экспрессии некоторых антител со структурой 4 биспецифического антитела для сравнения. В частности, Фиг. 8 показывает уровень экспрессии при транзиентной трансфекции антител Y400/Y404/Y416 серии A и серии B в клетках CHO и чистоту, определенную методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A.
Как можно видеть из Фиг. 8, уровень экспрессии двух серий биспецифических антител структуры 4 (т.е. серии A и серии B) не является высоким, и уровень экспрессии не превышает 6мг/л. Кроме того, что касается чистоты биспецифического антитела структуры 4, определенной методом эксклюзионной ВЭЖХ после аффинной хроматографии с белком A, результат показывает, что чистота сильно варьируется от 30% до 90%. Уровень экспрессии некоторых антител слишком низкий для определения чистоты.
Сравнивали уровень экспрессии и чистоту между антителами структур 1-4 биспецифического антитела с одинаковой мишенью и последовательностью вариабельной области антитела, и результаты показаны на Фиг. 9A-I. Фиг. 9A показывает уровень экспрессии и чистоту антител со структурой 1-4 биспецифического антитела с последовательностями вариабельных областей антитела S70 (SEQ ID NO: 41 и 42) и последовательностями вариабельных областей антитела G631 (SEQ ID NO: 57 и 58). Фиг. 9B показывает уровень экспрессии и чистоту антител со структурой 1 и 2 биспецифического антитела с последовательностями вариабельных областей антитела S70 (SEQ ID NO: 41 и 42) и последовательностями вариабельных областей антитела 3G12 (SEQ ID NO: 59 и 60). Фиг. 9C показывает уровень экспрессии и чистоту антител со структурой 1 и 2 биспецифического антитела с последовательностями вариабельных областей антитела 12A4 (SEQ ID NO: 45 и 46) и последовательностями вариабельных областей антитела 3G12 (SEQ ID NO: 59 и 60). Фиг. 9D показывает уровень экспрессии и чистоту антител со структурой 1 и 2 биспецифического антитела с последовательностями вариабельных областей антитела S70 (SEQ ID NO: 41 и 42) и последовательностями вариабельных областей антитела LAG35 (SEQ ID NO: 33 и 34). Фиг. 9E показывает уровень экспрессии и чистоту антител со структурой 1, 3 и 4 биспецифического антитела с последовательностями вариабельных областей антитела 5C4 (SEQ ID NO: 37 и 38) и последовательностями вариабельных областей антитела Ервой (SEQ ID NO: 27 и 28). Фиг. 9F показывает уровень экспрессии и чистоту антител со структурой 1 и 2 биспецифического антитела с последовательностями вариабельных областей антитела 5C4 (SEQ ID NO: 37 и 38) и последовательностями вариабельных областей антитела 10A7 (SEQ ID NO: 29 и 30). Фиг. 9G показывает уровень экспрессии и чистоту антител со структурой 1 и 2 биспецифического антитела с последовательностями вариабельных областей антитела 5C4 (SEQ ID NO: 37 и 38) и последовательностями вариабельных областей антитела G631 (SEQ ID NO: 57 и 58). Фиг. 9H показывает уровень экспрессии и чистоту антител со структурой 1 и 2 биспецифического антитела с последовательностями вариабельных областей антитела 5C4 (SEQ ID NO: 37 и 38) и последовательностями вариабельных областей антитела B-N10 (SEQ ID NO: 63 и 64). Фиг. 9I показывает уровень экспрессии и чистоту антител со структурой 1-4 биспецифического антитела с последовательностями вариабельных областей антитела Элотузумаба (SEQ ID NO: 49 и 50) и последовательностями вариабельных областей антитела NM3E2 (SEQ ID NO: 47 и 48).
Как можно видеть из Фиг. 9A, антитело со структурой 1 биспецифического антитела имеет более высокий уровень экспрессии и чистоту по сравнению с другими структурами, при этом Y100-B6 и Y100-B7 имеют лучший уровень экспрессии и/или чистоту по сравнению с антителами других структур, а Y100-B7 имеет самый лучший уровень экспрессии и чистоту; Фиг. 9B показывает, что антитела со структурой 1 биспецифического антитела имеют более высокий уровень экспрессии и чистоту по сравнению с другими структурами, при этом Y101-B1 и Y101-B2 имеют лучший уровень экспрессии и чистоту по сравнению с антителами других структур, а Y101-B2 имеет самый лучший уровень экспрессии и чистоту; Фиг. 9C показывает, что антитела со структурой 1 биспецифического антитела имеют более высокий уровень экспрессии и/или чистоту антител по сравнению с другими структурами, при этом Y101-B3 и Y101-B4 имеют лучший уровень экспрессии и чистоту, чем антитела со структурой 2 биспецифического антитела, а Y101-B4 имеет самый лучший уровень экспрессии и чистоту; Фиг. 9D показывает, что антитела со структурой 1 биспецифического антитела имеют лучший уровень экспрессии и чистоту по сравнению с другими структурами, при этом Y103-B1 и Y103-B2 имеют лучший уровень экспрессии и чистоту, чем антитела со структурой 2 биспецифического антитела, а Y103-B2 имеет самый лучший уровень экспрессии и чистоту; Фиг. 9E показывает, что антитела со структурой 1 биспецифического антитела имеют лучший уровень экспрессии и/или чистоту антител по сравнению с другими структурами, при этом Y104-B1 и Y104-B2 имеют лучший уровень экспрессии и чистоту, чем антитела со структурой 3 и структурой 4 биспецифического антитела, а Y104-B2 имеет самый лучший уровень экспрессии и чистоту; Фиг. 9F показывает, что антитела со структурой 1 биспецифического антитела имеют лучший уровень экспрессии и/или чистоту антител по сравнению с другими структурами, при этом Y105-B1 и Y105-B2 имеют лучший уровень экспрессии и чистоту, чем антитела со структурой 2 биспецифического антитела, а Y105-B2 имеет самый лучший уровень экспрессии и чистоту; как можно видеть из Фиг. 9G, антитела со структурой 1 биспецифического антитела имеют лучший уровень экспрессии и чистоту антител по сравнению с другими структурами, при этом Y106-B1 и Y106-B2 имеют лучший уровень экспрессии и/или чистоту, чем антитела со структурой 2 биспецифического антитела, а Y106-B2 имеет самый лучший уровень экспрессии и чистоту; как можно видеть из Фиг. 9H, антитела со структурой 1 биспецифического антитела имеют более высокий уровень экспрессии и чистоту по сравнению с другими структурами, при этом Y110-B1 и Y110-B2 имеют лучший уровень экспрессии и чистоту, чем антитела со структурой 2 биспецифического антитела, а Y110-B2 имеет самый лучший уровень экспрессии и чистоту; Фиг. 9I показывает, что антитела со структурой 1 биспецифического антитела имеют более высокий уровень экспрессии и/или чистоту антител по сравнению с другими структурами, при этом Y116-B1 и Y116-B2 имеют лучший уровень экспрессии и чистоту по сравнению с антителами других структур биспецифических антител, а Y116-B2 имеет самый лучший уровень экспрессии и чистоту.
Если шарнирная область, CH2 домен и CH3 домен каждого из вышеуказанных антител были заменены соответствующими последовательностями из IgG2 (GenBank номер доступа MH025834.2), IgG3 (GenBank номер доступа AJ390278) или IgG4 (GenBank номер доступа KJ901516), а последовательность вариабельной области антитела и линкер остались такими же, с получением замещенного антитела, не было никакой существенной разницы в уровне экспрессии и чистоте между полученным замещенным антителом и исходным антителом. На основании представленных выше данных можно видеть, что структура 1 биспецифического антитела имеет существенные преимущества по сравнению с другими структурами биспецифических антител, что касается уровня экспрессии и чистоты.
Кроме того, антитела, такие как Y100-B7, Y101-B2 и Y101-B4, экспрессируются в пуле клеток CHO со стабильной экспрессией с уровнем экспрессии более чем 700 мг/л и с чистотой более чем 85%. Антитело подвергали аффинной хроматографии с белком A, а затем способам очистки моноклональных антител, таким как традиционная анионо/катионообменная хроматография, с получением конечного продукта, представляющего интерес, с чистотой более 95% и общим выходом 40%, что согласуется с данными для моноклональных антител при их экспрессии и очистке.
Пример 2. Детекция биологической активности биспецифических антител
1. Аффинность к антигену
1) Получение антигена: для антигенных белков VEGFA, TGF-β1, IL-10, PD-1, CTLA-4, TIGIT, SLAMF7, LAG-3 и CD16a (последовательности, соответственно, см. в Таблице 26) и т.п. была сконструирована плазмида экспрессии pcDNA3.1 (приобретена у invitrogen) с His-меткой, где для конструкции были выбраны внеклеточные домены пяти белков PD-1, CTLA-4, TIGIT, SLAMF7 и CD16a, затем осуществляли транзиентную трансфекцию в 293E клетки для экспрессии и очистки; способ очистки разделяли на две стадии, т.е. очистка колоночной никель-хелатной хроматографией и очистка с использованием молекулярного сита, и в результате чистота антигенного белка, определенная методом SDS-PAGE, была не менее 95%; концентрацию каждого антигенного белка доводили до 2 мкг/мл, и микротитровальный планшет покрывали антигенным белком при 100 мкл/лунка, 4°С в течение ночи; супернатант сливали, в каждую лунку добавляли 250 мкл блокирующего раствора (3% BSA в PBS);
2) добавление антитела: в соответствии с планом экспериментa, процедуру осуществляли при комнатной температуре и антитело разбавляли в градиенте с 1% BSA в PBS. Например, исходная концентрация антитела для разведения составляла 3000 нМ, и антитело разбавляли 3-кратно с 11 градиентами. Разбавленное антитело добавляли в лунки микротитровального планшета при 200 мкл/лунка, инкубировали при комнатной температуре в течение 2 ч и супернатант затем сливали;
3) промывка: планшет промывали PBST (PBS, содержащий 0,1% Tween20) при 200 мкл/лунка 3 раза;
4) инкубация вторичного антитела: разбавленное вторичное антитело HRP-меченное античеловеческое IgG (Sigma, A8792) добавляли в объеме 100 мкл/лунка и инкубировали при комнатной температуре в течение 1ч, где вторичное антитело было разбавлено при 1:40000 и разбавитель представлял собой 1% BSA в PBS, и лунки покрывали только антигеном, а вторичное антитело использовали в качестве контроля;
5) промывка: планшет промывали PBST (PBS, содержащий 0,1% Tween20) при 200мкл/лунка 5 раз;
6) проявление цвета: цветопроявляющий раствор TMB (полученный из цветопроявляющих растворов A и B, приобретенных у Wuhan Boster Company, и смешанный в соответствии с A:B=1:1, готовый к использованию) добавляли при 100мкл/лунка и проявление цвета осуществляли при 37°С в течение 5 мин;
7) 2M HCl стоп-раствор добавляли при 100мкл/лунка, а затем обязательно нужно было использовать считывающее устройство для микропланшетов для считывания при 450нм в течение 30 минут, и данные представляли графически, используя программу Graphpad Prism 5, с концентрацией антитела (нМ) по оси абсцисс и средней интенсивностью флуоресценции по оси ординат; EC50 значение, которое показывает аффинность антитела к соответствующему антигену-мишени, рассчитывали методом односайтового специфического связывания. Результаты показаны в Таблице 32. Результаты показывают, что в случае антител с одинаковыми последовательностями вариабельных областей, аффинность биспецифического антитела по настоящему изобретению (т.е. биспецифическое антитело структуры 1) к соответствующему антигену-мишени значительно выше, чем у биспецифических антител структур 2-4. Кроме того, биспецифические антитела по настоящему изобретению (биспецифические антитела структуры 1) имеют самую сильную антигенсвязывающую активность в отношении мишени.
2. Клеточная аффинность
1) Подготовка клеток: PD-L1-положительные клетки H358 (приобретенные у Cell Resource Center of Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences) использовали для определения связывающей активности молекул биспецифических антител против PD-L1 клеток на терминальной стадии дифференцировки; достаточное количество клеток центрифугировали при 300×g в течение 5 минут, супернатант сливали; клетки ресуспендировали в 1% FBS-PBS, доводили до плотности 4×106/мл; 50 мкл клеток отбирали для каждой лунки и высевали при 2×105 на лунку; при этом центрифугирование осуществляли при 300×g при 4°С в течение 5 минут, супернатант сливали и посев клеток осуществляли на льду;
2) Добавление антитела: в соответствии с планом экспериментa, антитела разбавляли в градиенте и разбавление антител осуществляли на льду; например, исходная концентрация антитела для разведения составляла 3000 нМ, и антитело разбавляли 3-кратно с 11 градиентами концентрации; разбавленное антитело добавляли в лунку при 50мкл на лунку, тщательно смешивали при осторожной продувке и инкубировали при 4°С с встряхиванием при 1100 об/мин в течение 2ч;
3) промывка: клетки ресуспендировали с 150мкл 1% FBS-PBS и центрифугировали при 300×g при 4°C в течение 5 минут; супернатант сливали; и клетки промывали еще один раз;
4) инкубация вторичного антитела: разбавленное вторичное антитело PE античеловеческое IgG FC (Biolegend, 409304) добавляли в объеме 50мкл/лунка, при этом конечная концентрация вторичного антитела составляла 8 мкг/мл, тщательно смешивали при осторожной продувке и инкубировали при 4°С в темноте с встряхиванием при 1100об/мин в течение 1ч; при этом лунку, в которую добавляли только клетки и вторичное антитело, использовали в качестве контроля;
5) промывка: клетки ресуспендировали с 150мкл 1% FBS-PBS и центрифугировали при 300g при 4°C в течение 5 минут, супернатант сливали; клетки промывали еще один раз;
6) фиксация: 2% раствор параформальдегида добавляли при 200мкл/лунка для ресуспендирования и фиксации клеток при комнатной температуре в течение 20 минут и затем клетки центрифугировали при 300×g в течение 5 минут; супернатант сливали;
7) ресуспендирование клеток: клетки ресуспендировали с 200мкл 1% FBS-PBS, центрифугировали при 300×g в течение 5 минут, и супернатант сливали;
8) загрузка для проточной цитометрии: клетки ресуспендировали с 150мкл 1% FBS-PBS и осуществляли детекцию на проточном цитрометре;
9) анализ данных: данные анализировали с использованием программы для проточного анализа FlowJo 7.6 с получением средней интенсивности флуоресценции при конкретной концентрации антител, и представляли графически, используя программу Graphpad Prism 5, с концентрацией антитела (нМ) по оси абсцисс и средней интенсивностью флуоресценции по оси ординат; метод односайтового специфического связывания использовали для расчета EC50 значения, которое показывает клеточную аффинность антитела к соответствующим антигенам-мишеням; результаты показаны в Таблице 32.
Активность связывания некоторых антител, имеющих структуру 1-4 биспецифического антитела
(пМ)
Y100-B7
Y200-A1
Y200-B1
Y200-B3
Y300-A1
Y400-B1
169,60
156,24
173,00
207,15
180,95
254,02
35,19
237,93
188,41
47,33
307,96
268,35
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y101-B2
Y201-A1
Y201-B1
191,23
173,80
192,18
NA
NA
NA
3,09
25,05
28,46
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y101-B4
Y201-A2
Y201-B2
119,07
143,63
124,05
NA
NA
NA
1,86
31,18
28,01
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y103-B2
Y203-A1
Y203-B1
169,09
143,09
187,96
NA
NA
NA
NA
NA
NA
12,68
65,70
89,44
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y104-B2
Y304-A1
Y404-B1
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
93,78
464,65
388,51
69,68
468,46
330,93
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y105-B2
Y205-A2
Y205-B2
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
114,70
306,23
155,11
NA
NA
NA
86,97
484,29
409,16
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y106-B2
Y206-A2
Y206-B1
Y206-B2
NA
NA
NA
NA
53,91
63,03
212,78
59,64
NA
NA
NA
NA
NA
NA
NA
NA
112,44
449,79
164,52
311,52
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
Y110-B2
Y210-A1
Y210-B1
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
113,27
148,67
198,43
NA
NA
NA
NA
NA
NA
66,62
409,05
234,03
NA
NA
NA
NA
NA
NA
Y116-B2
Y216-A1
Y216-B1
Y316-A1
Y416-B1
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
NA
648,23
714,64
499,21
9176,07
5590,07
436,73
3257,47
3220,97
3842,87
3557,75
Из данных Таблицы 32 можно видеть, что когда последовательности вариабельных областей разных антител идентичны, связывающая активность таких антител, имеющих разные структуры биспецифического антитела, с антигеном-мишенью существенно различается, при этом для любых мишеней антитела со структурой 1 биспецифического антитела имеют самую сильную антигенсвязывающую активность. Если шарнирная область, CH2 домен и CH3 домен каждого из вышеуказанных антител заменяются соответствующими последовательностями из IgG2, IgG3 или IgG4 (supra), а последовательности вариабельной области антитела и линкера остаются такими же, с получением замещенного антитела, нет никакой существенной разницы в антигенсвязывающей активности между полученным замещенным антителом и исходным антителом.
Пример 3. Испытание антител на стабильность
Экспериментальная процедура:
1. Конкретные стадии ускоренного испытания на термостабильность при 40°С являются следующими:
1) образец переносили в специфический буфер, компонент буфера представляет собой 20мМ лимонной кислоты, pH5,5, и концентрацию образца доводили до 1 мг/мл;
2) каждый образец разделяли по 500мкл на пробирку (всего 6 пробирок) и пробирки герметично закрывали и помещали в 40°С водяную баню. В день 0, день 3, день 5, день 7, день 10 и день 14 брали образцы для эксклюзионной ВЭЖХ. Водяную баню поддерживали в общей сложности 14 дней.
2. Испытание на кислотостойкость, также известное как испытание на стабильность в условиях низкого pH, представляет собой испытание для определения, может ли молекула антитела сохранять свое исходное состояние после обработки в кислотных условиях в течение определенного периода времени с последующей нейтрализацией до физиологических условий. Конкретные стадии являются следующими:
когда молекулы антител подвергали аффинной хроматографии с белком A, раствор элюируемого антитела не нейтрализовали на стадии кислотного элюирования (использовали цитратный буфер при pH 3,5); после хранения в указанном буфере в течение определенного периода времени образцы отбирали через 30 минут и 60 минут, добавляли 1/10 объема 1M Tris-HCl (pH 8,0) для нейтрализации и образцы подвергали детекции методом эксклюзионной ВЭЖХ.
3.1 Испытание на стабильность биспецифического антитела со структурой 1
(1) Ускоренное испытание на термостабильность при 40°С биспецифических антител с различными структурами
Результаты испытаний показаны на Фиг. 10A-E. Фиг. 10A показывает чистоту Y100-B6, Y100-B7, Y200-A1, Y200-B1, Y200-B3, Y300-A1 и Y400-B1, определенную методом эксклюзионной ВЭЖХ в день 0, день 7 и день 14, после того как их обрабатывали при 40°C; Фиг. 10B показывает чистоту Y101-B1, Y101-B2, Y201-A1, Y201-B1, Y101-B3, Y101-B4, Y201-A2 и Y201-B2, определенную методом эксклюзионной ВЭЖХ в день 0, день 7 и день 14, после того как их обрабатывали при 40°C; Фиг. 10C показывает чистоту Y103-B1, Y103-B2, Y203-A1, Y203-B1, Y104-B1, Y104-B2, Y304-A1 и Y404-B1, определенную методом эксклюзионной ВЭЖХ в день 0, день 7 и день 14, после того как их обрабатывали при 40°C; Фиг. 10D показывает чистоту Y105-B1, Y105-B2, Y205-A2, Y205-B2, Y106-B1, Y106-B2, Y206-A2, Y206-B1 и Y206-B2, определенную методом эксклюзионной ВЭЖХ в день 0, день 7 и день 14, после того как их обрабатывали при 40°C; Фиг. 10E показывает чистоту Y110-B1, Y110-B2, Y210-A1, Y210-B1, Y116-B1, Y116-B2, Y216-A1, Y216-B1, Y316-A1, Y416-B1, определенную методом эксклюзионной ВЭЖХ в день 0, день 7 и день 14, после того как их обрабатывали при 40°C.
Как можно видеть из Фиг. 10, чистота разных антител со структурой 1 биспецифического антитела сохранялась на уровне больше чем 90% после обработки при 40°C в течение 14 дней, без большого количества агрегации или разложения, и это указывает на то, что биспецифическое антитело структуры 1 имеет хорошую термостабильность. Чистота биспецифических антител с другими структурами существенно снижается после обработки при 40°C в течение 14 дней.
(2) Испытание на кислотостойкость различных структур биспецифических антител
Результаты испытаний показаны на Фиг. 11A-E. Фиг. 11A показывает чистоту Y100-B6, Y100-B7, Y200-A1, Y200-B1, Y200-B3, Y300-A1 и Y400-B1, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0 минут, 30 минут и 60 минут; Фиг. 11B показывает чистоту Y101-B1, Y101-B2, Y201-A1, Y201-B1, Y101-B3, Y101-B4, Y201-A2 и Y201-B2, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0 минут, 30 минут и 60 минут; Фиг. 11C показывает чистоту Y103-B1, Y103-B2, Y203-A1, Y203-B1, Y104-B1, Y104-B2, Y304-A1 и Y404-B1, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0 минут, 30 минут и 60 минут; Фиг. 11D показывает чистоту Y105-B1, Y105-B2, Y205-A2, Y205-B2, Y106-B1, Y106-B2, Y206-A2, Y206-B1 и Y206-B2, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0 минут, 30 минут и 60 минут; Фиг. 11E показывает чистоту Y110-B1, Y110-B2, Y210-A1, Y210-B1, Y116-B1, Y116-B2, Y216-A1, Y216-B1, Y316-A1 и Y416-B1, определенную методом эксклюзионной ВЭЖХ после обработки в условиях низкого pH в течение 0 минут, 30 минут и 60 минут.
Как можно видеть из Фиг. 11A-E, когда антитела со структурой 1 биспецифического антитела обрабатывали в течение 60 минут в условиях низкого pH, чистота антител существенно не изменялась, указывая на то, что биспецифические антитела, имеющие структуру 1, обладают хорошей кислотостойкостью. Что касается других структур биспецифических антител, чистота некоторых антител значительно снижалась после 60 минут обработки в условиях низкого pH.
На основании Фиг. 10 и 11 можно видеть, что антитело со структурой 1 биспецифического антитела имеет лучшую термостабильность и кислотостойкость по сравнению с другими структурами биспецифических антител.
Когда шарнирная область, CH2 домен и CH3 домен каждого из вышеуказанных антител заменены соответствующей последовательностью из IgG2, IgG3 или IgG4 (supra), а последовательности вариабельной области антитела и линкера остаются такими же, с получением замещенного антитела, нет никакой существенной разницы в стабильности между полученным замещенным антителом и исходным антителом.
Пример 4. Эксперимент 1 по определению фармакодинамики биспецифических антител in vivo
1). Используемые для экспериментa материалы:
Клетки: MC38 (мышиная клеточная линия рака толстой кишки, приобретенная у ATCC);
Мыши: C57BL/6 мыши, самки, приобретенные у Beijing Vital River;
Способ инокуляции: MC38 клетки культивировали и собирали, регулировали концентрацию клеток, затем клетки подкожно инокулировали в область спины при 1*106 клеток/мышь (0,1мл/мышь). Когда опухоль вырастала до 100-200мм3, мышей разделяли на группы введения, по 8 мышей в каждой группе:
Испытываемое лекарственное средство: Y100-B7;
Отрицательный контроль: физиологический раствор;
Положительный контроль: Tecentriq (моноклональное антитело к PD-L1, Roche);
Способ введения: были установлены разные дозы (13,3мг/кг и 4мг/кг) для Y100-B7, а вводимая доза Tecentriq составляла 10мг/кг; лекарственное средство вводили интраперитонеально, начиная в день 0, 3 раза в неделю в течение одной недели.
Объем опухоли: длину и ширину опухоли измеряли через каждые 2-3 дня. Когда объем опухоли группы приближался к 2000мм3 или объем опухоли отдельной мыши достигал 3000мм3, группа прекращала испытание.
2). Результаты экспериментa
Результаты испытания фармакодинамики in vivo для Y100-B7 показаны на Фиг. 12. Фиг. 12 показывает фармакодинамику биспецифического антитела Y100-B7 in vivo в мышиной модели опухоли и объем опухоли. Результаты показывают, что группа, получавшая высокую дозу Y100-B7 (13,3 мг/кг), показала существенный противоопухолевый эффект, и этот эффект лучше, чем у Tecentriq.
Пример 5. Эксперимент 2 по определению фармакодинамики биспецифических антител in vivo
1). Используемые для экспериментa материалы:
Клетки: MC38 (мышиная клеточная линия рака толстой кишки, приобретенная у ATCC);
Мыши: C57BL/6 мыши, самки, приобретенные у Beijing Vital River;
Способ инокуляции: MC38 клетки культивировали, собирали, регулировали концентрацию клеток, затем клетки подкожно инокулировали в область спины при 1*106 клеток/мышь (0,1мл/мышь). Когда опухоль вырастала до 100-200мм3, мышей разделяли на группы введения, по 8 мышей в каждой группе:
Испытываемое лекарственное средство: Y101-B2;
Отрицательный контроль: физиологический раствор;
Положительный контроль: Tecentriq (моноклональное антитело к PD-L1, Roche);
Способ введения: были установлены разные дозы (13,3мг/кг и 4мг/кг) для Y101-B2, а вводимая доза Tecentriq составляла 10мг/кг; лекарственное средство вводили интраперитонеально, начиная в день 0, 3 раза в неделю в течение 3 недель.
Объем опухоли: длину и ширину опухоли измеряли 3 раза в неделю. Когда объем опухоли группы приближался к 2000мм3 или объем опухоли отдельной мыши достигал 3000мм3, группа прекращала испытание.
2). Результаты экспериментa
Результаты испытания фармакодинамики in vivo для Y101-B2 показаны на Фиг. 13. Фиг. 13 показывает фармакодинамику биспецифического антитела Y101-B2 in vivo в мышиной модели опухоли и объем опухоли. Результаты показывают, что группа, получавшая высокую дозу Y101-B2 (13,3 мг/кг), показала существенный противоопухолевый эффект, и этот эффект лучше, чем у Tecentriq.
Пример 6: Получение и определение биологической активности бипаратопного антитела
Бипаратопное антитело является разновидностью биспецифического антитела и может связываться с двумя разными эпитопами одного и того же антигена.
Конструкция плазмиды экспрессии, транзиентная трансфекция 293E клеток и способ получения антитела были такими же, как описанные выше способы.
Рассматриваемый антиген представлял собой внеклеточный домен Her2 человека (источник последовательности: UniProtKB-P04626).
Соответствующие последовательности вариабельных областей антитела показаны в таблице ниже:
Последовательности вариабельных областей анти-Her2 антитела
(источник последовательности)
WVRQAPGKGLEWVARIYPTNGYTRYADSVKGRFT
ISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDG
FYAMDYWGQGTLVTVSS
AVAWYQQKPGKAPKLLIYSASFLYSGVPSRF
SGSRSGTDFTLTISSLQPEDFATYYCQQHYTT
PPTFGQGTKVEIK
WVRQAPGKGLEWVADVNPNSGGSIYNQRFKGRFT
LSVDRSKNTLYLQMNSLRAEDTAVYYCARNLGPSF
YFDYWGQGTLVTVSS
VAWYQQKPGKAPKLLIYSASYRYTGVPSRFS
GSGSGTDFTLTISSLQPEDFATYYCQQYYIYP
YTFGQGTKVEIK
Последовательности сконструированных антител показаны в следующих таблицах:
Коды антител и аминокислотные последовательности некоторых антител со структурой 1 биспецифического антитела
VARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVY
YCSRWGGDGFYAMDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
WYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC
KVSNKALPAPIEKTISKAK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
WYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC
KVSNKALPAPIEKTISKAK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
WYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKC
KVSNKGLPAPIEKTISKTK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
WYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKC
KVSNKGLPAPIEKTISKTK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
WYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC
KVSNKALPAPIEKTISKAK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPPTFGQGTKVEIK
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
WYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC
KVSNKALPAPIEKTISKAK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
WYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKC
KVSNKGLPAPIEKTISKTK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
WYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKC
KVSNKGLPAPIEKTISKTK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
WYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC
KVSNKALPAPIEKTISKAK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
YSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYYIYPYT
FGQGTKVEIK
QSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLS
SPVTKSFNRGEC
WVARIYPTNGYTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTA
VYYCSRWGGDGFYAMDYWGQGTLVTVSS
WVADVNPNSGGSIYNQRFKGRFTLSVDRSKNTLYLQMNSLRAEDTA
VYYCARNLGPSFYFDYWGQGTLVTVSS
GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDK
KVEPKSC
IYSASFLYSGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCQQHYTTPP
TFGQGTKVEIK
WYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKC
KVSNKALPAPIEKTISKAK
NNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNH
YTQKSLSLSPGK
Уровень экспрессии при транзиентной трансфекции обоих Герцептина mAb и Перьета mAb составляет 30 мг/л; уровень экспрессии при транзиентной трансфекции биспецифического антитела Y140-B1/B2/B3/B4 не меньше чем 25мг/л; уровень экспрессии биспецифического антитела Y140-A1/A2/A3/A4 составляет 7 мг/л - 20мг/л, и чистота всех антител по данным эксклюзионной хроматографии не меньше чем 80%.
2. Детекция биологической активности
Клеточная аффинность
1) Подготовка клеток: Her2-положительные BT-474 клетки (приобретенные у the Cell Bank of the Type Culture Collection Committee of the Chinese Academy of Sciences) использовали для определения аффинности молекулы биспецифического антитела к Her2-концу; достаточное количество клеток центрифугировали при 300×g в течение 5 минут, супернатант сливали; клетки ресуспендировали в 1% FBS-PBS, доводили до плотности 2×106/мл, и отбирали 50 мкл клеток на лунку и затем высевали при 1×105 на лунку; при этом клетки центрифугировали при 300×g при 4°С в течение 5 минут, супернатант сливали, а посев клеток осуществляли на льду;
2) Добавление антитела: в соответствии с планом экспериментa, антитело, представленное в Таблице 34, разбавляли в градиенте и разбавление антитела осуществляли на льду; например исходная концентрация антитела для разведения составляла 3000 нМ, и антитело разбавляли 3-кратно с 11 градиентами концентрации; разбавленное антитело добавляли в лунку при 50мкл на лунку, тщательно смешивали при осторожной продувке и инкубировали при 4°С с встряхиванием при 1100 об/мин в течение 2ч;
3) промывка: клетки ресуспендировали с 150мкл 1% FBS-PBS и центрифугировали при 300×g при 4°C в течение 5 минут; супернатант сливали; и клетки промывали еще один раз;
4) инкубация вторичного антитела: разбавленное вторичное антитело PE античеловеческое IgG FC (Biolegend, 409304) добавляли при объеме 50мкл/лунка, при этом конечная концентрация вторичного антитела составляла 8 мкг/мл, тщательно смешивали при осторожной продувке и инкубировали при 4°С в темноте с встряхиванием при 1100об/мин в течение 1ч; при этом лунку, в которую добавляли только клетки и вторичное антитело, использовали в качестве контроля;
5) промывка: клетки ресуспендировали с 150мкл 1% FBS-PBS и центрифугировали при 300g при 4°C в течение 5 минут, супернатант сливали; клетки промывали еще один раз;
6) фиксация: 2% раствор параформальдегида добавляли при 200мкл/лунка для ресуспендирования и фиксации клеток при комнатной температуре в течение 20 минут и затем клетки центрифугировали при 300×g в течение 5 минут; супернатант сливали;
7) ресуспендирование клеток: клетки ресуспендировали с 200мкл 1% FBS-PBS, центрифугировали при 300×g в течение 5 минут, и супернатант сливали;
8) загрузка для проточной цитометрии: клетки ресуспендировали с 150мкл 1% FBS-PBS и осуществляли детекцию на проточном цитрометре;
9) анализ данных: данные анализировали с использованием программы для проточного анализа FlowJo 7.6 с получением средней интенсивности флуоресценции при конкретной концентрации антител, и представляли графически, используя программу Graphpad Prism 5, с концентрацией антитела (нМ) по оси абсцисс и средней интенсивностью флуоресценции по оси ординат; метод односайтового специфического связывания использовали для расчета EC50 значения, которое показывает клеточную аффинность антитела к соответствующим антигенам-мишеням.
Связывающая активность некоторых биспецифических антител
Из приведенной выше таблицы видно, что аффинность биспецифических антител, сформированных при помощи двух антител, которые нацелены на один и тот же антиген, но разные эпитопы, значительно улучшена по сравнению с аффинностью двух моноклональных антител, а аффинность биспецифических антител серии В значительно сильнее, чем у антител серии А.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> УХАНЬ АйЗиАй БАЙОФАРМА КО., ЛТД.
<120> ЧЕТЫРЕХВАЛЕНТНЫЕ СИММЕТРИЧНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА
<130> FP200576MX
<160> 149
<170> PatentIn version 3.3
<210> 1
<211> 125
<212> Белок
<213> Искусственная последовательность
<400> 1
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 2
<211> 109
<212> Белок
<213> Искусственная последовательность
<400> 2
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 3
<211> 125
<212> Белок
<213> Искусственная последовательность
<400> 3
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Ala
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 4
<211> 109
<212> Белок
<213> Искусственная последовательность
<400> 4
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Val Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 5
<211> 125
<212> Белок
<213> Искусственная последовательность
<400> 5
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 6
<211> 109
<212> Белок
<213> Искусственная последовательность
<400> 6
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 7
<211> 118
<212> Белок
<213> Искусственная последовательность
<400> 7
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Phe Pro Gly Asp Gly Ser Thr Gln Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Thr Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Arg Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gln Thr Thr Ala Thr Trp Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 8
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 8
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1 5 10 15
Asp Arg Val Ser Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Asp Tyr
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Ser Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Pro
65 70 75 80
Glu Asp Val Gly Val Tyr Tyr Cys Gln Asn Gly His Ser Phe Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 9
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 9
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Met Gln Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Tyr Pro Gly Asp Gly Asp Thr Arg Tyr Thr Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Ile Pro Arg Leu Trp Tyr Phe Asp Val Trp Gly Ala
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 10
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 10
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Asp Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 11
<211> 122
<212> Белок
<213> Искусственная последовательность
<400> 11
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Phe Asn Ser Phe
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Lys Asp Lys Ile Leu Trp Phe Gly Glu Pro Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 12
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 12
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 13
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 13
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Asp Pro Ser Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Pro Leu Val Tyr Thr Gly Phe Ala Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 14
<211> 109
<212> Белок
<213> Искусственная последовательность
<400> 14
Asp Ile Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Ser Cys Ser Gly Asp Asn Leu Arg His Tyr Tyr Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Thr Gly Gly Ala Ser Leu
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105
<210> 15
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Phe Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Phe Leu Gly Ile Ala Asn Ser Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Asp Ile Ala Ala Leu Gly Pro Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 16
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 16
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Glu Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 115
<212> Белок
<213> Искусственная последовательность
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Leu Trp Asn Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 18
<211> 109
<212> Белок
<213> Искусственная последовательность
<400> 18
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Ile Ile
35 40 45
Tyr Gly Ala Ser Thr Thr Ala Ser Gly Ile Pro Ala Arg Phe Ser Ala
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asn Asn Trp Pro Pro
85 90 95
Ala Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 19
<211> 113
<212> Белок
<213> Искусственная последовательность
<400> 19
Glu Val Gln Leu Val Glu Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Met His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Thr Ala Val Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
100 105 110
Ser
<210> 20
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 20
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Glu Ile Ser Val Ser
20 25 30
Leu Ser Trp Leu Gln Gln Glu Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Thr Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Val Asp Tyr Tyr Cys Leu Gln Tyr Ala Ser Tyr Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 21
<211> 119
<212> Белок
<213> Искусственная последовательность
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Pro Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Phe Ala Ser Gly Asn Ser Glu Tyr Asn Gln Lys Phe
50 55 60
Thr Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Asn Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Asp Tyr Asp Trp Tyr Phe Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 22
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 22
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Ser Gln Ser
85 90 95
Ser Ile Tyr Pro Trp Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 23
<211> 121
<212> Белок
<213> Искусственная последовательность
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Tyr Tyr Ser His Asp Met Trp Ser Glu Asp Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 24
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 24
Leu Pro Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Arg Ser Ser Asn Ile Gly Ser Asn
20 25 30
Ser Val Asn Trp Tyr Arg Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Pro Gly Val Pro Val Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Thr Tyr Tyr Cys Ala Thr Trp Asp Asp Asn Leu
85 90 95
Asn Val His Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 25
<211> 121
<212> Белок
<213> Искусственная последовательность
<400> 25
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Ser Tyr Phe Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ile Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg His Asp Gly Ala Val Ala Gly Leu Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 26
<211> 108
<212> Белок
<213> Искусственная последовательность
<400> 26
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Pro Pro Gly Gln Ala Pro Val Val Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Asn
50 55 60
Ser Asn Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Val Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 27
<211> 118
<212> Белок
<213> Искусственная последовательность
<400> 27
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Thr Phe Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Trp Leu Gly Pro Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 28
<211> 108
<212> Белок
<213> Искусственная последовательность
<400> 28
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Phe Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 29
<211> 119
<212> Белок
<213> Искусственная последовательность
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Thr Gln Pro Gly Lys
1 5 10 15
Ser Leu Lys Leu Ser Cys Glu Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Thr Met His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Ser Gly Ser Gly Ile Val Phe Tyr Ala Asp Ala Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Leu Leu Phe
65 70 75 80
Leu Gln Met Asn Asp Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Pro Leu Gly His Asn Thr Phe Asp Ser Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 30
<211> 113
<212> Белок
<213> Искусственная последовательность
<400> 30
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ala Val Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Lys Ser Ser Gln Ser Leu Tyr Tyr Ser
20 25 30
Gly Val Lys Glu Asn Leu Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Tyr Ala Ser Ile Arg Phe Thr Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr
65 70 75 80
Ile Thr Ser Val Gln Ala Glu Asp Met Gly Gln Tyr Phe Cys Gln Gln
85 90 95
Gly Ile Asn Asn Pro Leu Thr Phe Gly Asp Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 31
<211> 123
<212> Белок
<213> Искусственная последовательность
<400> 31
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Glu Ser Gly
20 25 30
Leu Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Ala Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Asp Gly Val Leu Ala Leu Asn Lys Arg Ser Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 32
<211> 109
<212> Белок
<213> Искусственная последовательность
<400> 32
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Thr Val Arg Pro
85 90 95
Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 33
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 33
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Asp Tyr
20 25 30
Tyr Trp Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Arg Gly Ser Thr Asn Ser Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Leu Ser Leu Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Phe Gly Tyr Ser Asp Tyr Glu Tyr Asn Trp Phe Asp Pro Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 34
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 34
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Asn Leu Glu Ile Lys
100 105
<210> 35
<211> 119
<212> Белок
<213> Искусственная последовательность
<400> 35
Glu Val Gln Leu Leu Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Ala Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Met Ala Thr Gly Gly Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 36
<211> 110
<212> Белок
<213> Искусственная последовательность
<400> 36
Gln Ser Val Leu Thr Gln Pro Ala Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asn Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Asn Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 37
<211> 113
<212> Белок
<213> Искусственная последовательность
<400> 37
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser
<210> 38
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 38
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 39
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 39
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Phe Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 40
<211> 111
<212> Белок
<213> Искусственная последовательность
<400> 40
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Gly Val Ser Thr Ser
20 25 30
Gly Tyr Ser Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Leu Ala Ser Tyr Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Ser Arg
85 90 95
Asp Leu Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 41
<211> 118
<212> Белок
<213> Искусственная последовательность
<400> 41
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 42
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 42
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 43
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 43
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ile Met Met Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Pro Ser Gly Gly Ile Thr Phe Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Lys Leu Gly Thr Val Thr Thr Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 44
<211> 110
<212> Белок
<213> Искусственная последовательность
<400> 44
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Arg Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 45
<211> 123
<212> Белок
<213> Искусственная последовательность
<400> 45
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Thr Ser Gly Asp Thr Phe Ser Thr Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Lys Ala His Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Lys Phe His Phe Val Ser Gly Ser Pro Phe Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 46
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 46
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 47
<211> 117
<212> Белок
<213> Искусственная последовательность
<400> 47
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Asn Trp Asn Gly Gly Ser Thr Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Arg Ser Leu Leu Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Arg
115
<210> 48
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 48
Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln Thr
1 5 10 15
Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala Ser
20 25 30
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Gly
35 40 45
Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser Ser
50 55 60
Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser Ser Gly Asn His Val
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 49
<211> 119
<212> Белок
<213> Искусственная последовательность
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asp Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Lys Phe Ile Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Asp Gly Asn Tyr Trp Tyr Phe Asp Val Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 50
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 50
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Gly Ile Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg His Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 51
<211> 121
<212> Белок
<213> Искусственная последовательность
<400> 51
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Val Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Tyr Asp Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 52
<211> 108
<212> Белок
<213> Искусственная последовательность
<400> 52
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 53
<211> 123
<212> Белок
<213> Искусственная последовательность
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Ala Asp Phe
50 55 60
Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Pro His Tyr Tyr Gly Ser Ser His Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 54
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Phe Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Val Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 55
<211> 121
<212> Белок
<213> Искусственная последовательность
<400> 55
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Ile Asn Gly Ser
20 25 30
Trp Ile Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Ala Ile Trp Pro Phe Gly Gly Tyr Thr His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly His Ser Thr Ser Pro Trp Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 56
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 56
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Val Ile Arg Arg Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Thr Ser Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 57
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Ile Ser Asp Tyr
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gly Ile Thr Pro Ala Gly Gly Tyr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Val Phe Phe Leu Pro Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 58
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 58
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Tyr Gly Asn Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 59
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 59
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn
20 25 30
Val Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Ile Pro Ile Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Thr Leu Gly Leu Val Leu Asp Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 60
<211> 108
<212> Белок
<213> Искусственная последовательность
<400> 60
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 61
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Ser Asn
20 25 30
Val Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Ile Pro Ile Val Asp Ile Ala Asn Tyr Ala Gln Arg Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Thr Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Pro Arg Ala Phe Val Leu Asp Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 62
<211> 108
<212> Белок
<213> Искусственная последовательность
<400> 62
Glu Thr Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Leu Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Pro Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ala Asp Ser Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 63
<211> 117
<212> Белок
<213> Искусственная последовательность
<400> 63
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Leu Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Ser Cys Thr Val Ser Gly Phe Ser Leu Ala Thr Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Ser Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Phe Cys Ala
85 90 95
Lys Gln Ala Tyr Gly His Tyr Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 64
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 64
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Thr Arg Leu Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 65
<211> 117
<212> Белок
<213> Искусственная последовательность
<400> 65
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ala Thr Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Tyr Ser Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Phe Cys Ala
85 90 95
Lys Gln Ala Tyr Gly His Tyr Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 66
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 66
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 67
<211> 118
<212> Белок
<213> Искусственная последовательность
<400> 67
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 68
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 68
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Arg Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Val Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 69
<211> 2
<212> Белок
<213> Искусственная последовательность
<400> 69
Ser Ser
1
<210> 70
<211> 2
<212> Белок
<213> Искусственная последовательность
<400> 70
Ala Ser
1
<210> 71
<211> 5
<212> Белок
<213> Искусственная последовательность
<400> 71
Gly Gly Gly Gly Ser
1 5
<210> 72
<211> 7
<212> Белок
<213> Искусственная последовательность
<400> 72
Gly Gly Gly Ser Ala Ala Ala
1 5
<210> 73
<211> 7
<212> Белок
<213> Искусственная последовательность
<400> 73
Gly Gly Gly Gly Ser Ala Ser
1 5
<210> 74
<211> 8
<212> Белок
<213> Искусственная последовательность
<400> 74
Gly Tyr Pro Gly Gly Gly Gly Ser
1 5
<210> 75
<211> 10
<212> Белок
<213> Искусственная последовательность
<400> 75
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 76
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 76
Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro
1 5 10
<210> 77
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 77
Ala Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Ala
1 5 10
<210> 78
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 78
Gly Lys Ser Ser Gly Ser Gly Ser Glu Ser Lys Ser
1 5 10
<210> 79
<211> 14
<212> Белок
<213> Искусственная последовательность
<400> 79
Gly Ser Thr Ser Gly Ser Gly Lys Ser Ser Glu Gly Lys Gly
1 5 10
<210> 80
<211> 14
<212> Белок
<213> Искусственная последовательность
<400> 80
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Ser Pro Pro Ser
1 5 10
<210> 81
<211> 14
<212> Белок
<213> Искусственная последовательность
<400> 81
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Ser Pro Pro Ser
1 5 10
<210> 82
<211> 15
<212> Белок
<213> Искусственная последовательность
<400> 82
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 83
<211> 14
<212> Белок
<213> Искусственная последовательность
<400> 83
Gly Ala Ala Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 84
<211> 15
<212> Белок
<213> Искусственная последовательность
<400> 84
Glu Ala Ala Ala Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 85
<211> 15
<212> Белок
<213> Искусственная последовательность
<400> 85
Gly Gly Gly Gly Ser Glu Ala Ala Ala Lys Gly Gly Gly Gly Ser
1 5 10 15
<210> 86
<211> 15
<212> Белок
<213> Искусственная последовательность
<400> 86
Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys Gly Gly Gly Gly Ser
1 5 10 15
<210> 87
<211> 17
<212> Белок
<213> Искусственная последовательность
<400> 87
Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
Gly
<210> 88
<211> 18
<212> Белок
<213> Искусственная последовательность
<400> 88
Gly Ser Thr Ser Gly Ser Gly Lys Ser Ser Glu Gly Ser Gly Ser Thr
1 5 10 15
Lys Gly
<210> 89
<211> 19
<212> Белок
<213> Искусственная последовательность
<400> 89
Gly Ala Ala Ala Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Gly Ser
<210> 90
<211> 20
<212> Белок
<213> Искусственная последовательность
<400> 90
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 91
<211> 7
<212> Белок
<213> Искусственная последовательность
<400> 91
Asp Lys Thr His Thr Cys Pro
1 5
<210> 92
<211> 9
<212> Белок
<213> Искусственная последовательность
<400> 92
Glu Arg Lys Cys Cys Val Glu Cys Pro
1 5
<210> 93
<211> 60
<212> Белок
<213> Искусственная последовательность
<400> 93
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro Arg Cys
1 5 10 15
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
20 25 30
Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro Glu
35 40 45
Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
50 55 60
<210> 94
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 94
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro
1 5 10
<210> 95
<211> 5
<212> Белок
<213> Искусственная последовательность
<400> 95
Asp Lys Thr His Thr
1 5
<210> 96
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 96
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 97
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 97
Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro
1 5 10
<210> 98
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 98
Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro
1 5 10
<210> 99
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 99
Arg Gly Arg Gly Ser Asp Lys Thr His Thr Cys Pro
1 5 10
<210> 100
<211> 12
<212> Белок
<213> Искусственная последовательность
<400> 100
Asp Gly Asp Gly Ser Asp Lys Thr His Thr Cys Pro
1 5 10
<210> 101
<211> 13
<212> Белок
<213> Искусственная последовательность
<400> 101
Gly Arg Gly Arg Gly Ser Asp Lys Thr His Thr Cys Pro
1 5 10
<210> 102
<211> 15
<212> Белок
<213> Искусственная последовательность
<400> 102
Ala Ser Thr Arg Gly Arg Gly Ser Asp Lys Thr His Thr Cys Pro
1 5 10 15
<210> 103
<211> 15
<212> Белок
<213> Искусственная последовательность
<400> 103
Gly Gln Pro Asp Gly Asp Ala Ser Asp Lys Thr His Thr Cys Pro
1 5 10 15
<210> 104
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 104
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 105
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 105
Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 106
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 106
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 107
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 107
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Ala Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 108
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 108
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Lys Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Asn Thr Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Ala Glu Cys Ser
100 105
<210> 109
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 109
Gly Gln Pro Lys Ala Ala Pro Thr Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Lys Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Ala Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 110
<211> 105
<212> Белок
<213> Искусственная последовательность
<400> 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
1 5 10 15
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
20 25 30
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
35 40 45
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
50 55 60
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
65 70 75 80
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
85 90 95
Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 111
<211> 103
<212> Белок
<213> Искусственная последовательность
<400> 111
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys
100
<210> 112
<211> 98
<212> Белок
<213> Искусственная последовательность
<400> 112
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val
<210> 113
<211> 98
<212> Белок
<213> Искусственная последовательность
<400> 113
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 114
<211> 98
<212> Белок
<213> Искусственная последовательность
<400> 114
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 115
<211> 113
<212> Белок
<213> Искусственная последовательность
<400> 115
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
1 5 10 15
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
20 25 30
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
35 40 45
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
50 55 60
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
65 70 75 80
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
85 90 95
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
100 105 110
Lys
<210> 116
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 116
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
1 5 10 15
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
20 25 30
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn
35 40 45
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
50 55 60
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
65 70 75 80
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
85 90 95
Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
100 105 110
<210> 117
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 117
Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro
1 5 10 15
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
20 25 30
Cys Val Val Val Asp Val Ser His Glu Ala Pro Glu Val Gln Phe Asn
35 40 45
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
50 55 60
Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
65 70 75 80
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
85 90 95
Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
100 105 110
<210> 118
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 118
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
1 5 10 15
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
20 25 30
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys
35 40 45
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
50 55 60
Glu Glu Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val
65 70 75 80
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
85 90 95
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
100 105 110
<210> 119
<211> 110
<212> Белок
<213> Искусственная последовательность
<400> 119
Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 120
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 120
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
100 105
<210> 121
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 121
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
100 105
<210> 122
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 122
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Ile Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
100 105
<210> 123
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 123
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
100 105
<210> 124
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 124
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 125
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 125
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 126
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 126
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Ile Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 127
<211> 106
<212> Белок
<213> Искусственная последовательность
<400> 127
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu
1 5 10 15
Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
100 105
<210> 128
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 128
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Ala
100 105
<210> 129
<211> 258
<212> Белок
<213> Искусственная последовательность
<400> 129
Val Pro Arg Trp Arg Gln Gln Trp Ser Gly Pro Gly Thr Thr Lys Arg
1 5 10 15
Phe Pro Glu Thr Val Leu Ala Arg Cys Val Lys Tyr Thr Glu Ile His
20 25 30
Pro Glu Met Arg His Val Asp Cys Gln Ser Val Trp Asp Ala Phe Lys
35 40 45
Gly Ala Phe Ile Ser Lys His Pro Cys Asn Ile Thr Glu Glu Asp Tyr
50 55 60
Gln Pro Leu Met Lys Leu Gly Thr Gln Thr Val Pro Cys Asn Lys Ile
65 70 75 80
Leu Leu Trp Ser Arg Ile Lys Asp Leu Ala His Gln Phe Thr Gln Val
85 90 95
Gln Arg Asp Met Phe Thr Leu Glu Asp Thr Leu Leu Gly Tyr Leu Ala
100 105 110
Asp Asp Leu Thr Trp Cys Gly Glu Phe Asn Thr Ser Lys Ile Asn Tyr
115 120 125
Gln Ser Cys Pro Asp Trp Arg Lys Asp Cys Ser Asn Asn Pro Val Ser
130 135 140
Val Phe Trp Lys Thr Val Ser Arg Arg Phe Ala Glu Ala Ala Cys Asp
145 150 155 160
Val Val His Val Met Leu Asn Gly Ser Arg Ser Lys Ile Phe Asp Lys
165 170 175
Asn Ser Thr Phe Gly Ser Val Glu Val His Asn Leu Gln Pro Glu Lys
180 185 190
Val Gln Thr Leu Glu Ala Trp Val Ile His Gly Gly Arg Glu Asp Ser
195 200 205
Arg Asp Leu Cys Gln Asp Pro Thr Ile Lys Glu Leu Glu Ser Ile Ile
210 215 220
Ser Lys Arg Asn Ile Gln Phe Ser Cys Lys Asn Ile Tyr Arg Pro Asp
225 230 235 240
Lys Phe Leu Gln Cys Val Lys Asn Pro Glu Asp Ser Ser Cys Thr Ser
245 250 255
Glu Ile
<210> 130
<211> 54
<212> Белок
<213> Искусственная последовательность
<400> 130
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala
50
<210> 131
<211> 124
<212> Белок
<213> Искусственная последовательность
<400> 131
Met His Val Ala Gln Pro Ala Val Val Leu Ala Ser Ser Arg Gly Ile
1 5 10 15
Ala Ser Phe Val Cys Glu Tyr Ala Ser Pro Gly Lys Ala Thr Glu Val
20 25 30
Arg Val Thr Val Leu Arg Gln Ala Asp Ser Gln Val Thr Glu Val Cys
35 40 45
Ala Ala Thr Tyr Met Met Gly Asn Glu Leu Thr Phe Leu Asp Asp Ser
50 55 60
Ile Cys Thr Gly Thr Ser Ser Gly Asn Gln Val Asn Leu Thr Ile Gln
65 70 75 80
Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile Cys Lys Val Glu Leu
85 90 95
Met Tyr Pro Pro Pro Tyr Tyr Leu Gly Ile Gly Asn Gly Thr Gln Ile
100 105 110
Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser Asp
115 120
<210> 132
<211> 422
<212> Белок
<213> Искусственная последовательность
<400> 132
Val Pro Val Val Trp Ala Gln Glu Gly Ala Pro Ala Gln Leu Pro Cys
1 5 10 15
Ser Pro Thr Ile Pro Leu Gln Asp Leu Ser Leu Leu Arg Arg Ala Gly
20 25 30
Val Thr Trp Gln His Gln Pro Asp Ser Gly Pro Pro Ala Ala Ala Pro
35 40 45
Gly His Pro Leu Ala Pro Gly Pro His Pro Ala Ala Pro Ser Ser Trp
50 55 60
Gly Pro Arg Pro Arg Arg Tyr Thr Val Leu Ser Val Gly Pro Gly Gly
65 70 75 80
Leu Arg Ser Gly Arg Leu Pro Leu Gln Pro Arg Val Gln Leu Asp Glu
85 90 95
Arg Gly Arg Gln Arg Gly Asp Phe Ser Leu Trp Leu Arg Pro Ala Arg
100 105 110
Arg Ala Asp Ala Gly Glu Tyr Arg Ala Ala Val His Leu Arg Asp Arg
115 120 125
Ala Leu Ser Cys Arg Leu Arg Leu Arg Leu Gly Gln Ala Ser Met Thr
130 135 140
Ala Ser Pro Pro Gly Ser Leu Arg Ala Ser Asp Trp Val Ile Leu Asn
145 150 155 160
Cys Ser Phe Ser Arg Pro Asp Arg Pro Ala Ser Val His Trp Phe Arg
165 170 175
Asn Arg Gly Gln Gly Arg Val Pro Val Arg Glu Ser Pro His His His
180 185 190
Leu Ala Glu Ser Phe Leu Phe Leu Pro Gln Val Ser Pro Met Asp Ser
195 200 205
Gly Pro Trp Gly Cys Ile Leu Thr Tyr Arg Asp Gly Phe Asn Val Ser
210 215 220
Ile Met Tyr Asn Leu Thr Val Leu Gly Leu Glu Pro Pro Thr Pro Leu
225 230 235 240
Thr Val Tyr Ala Gly Ala Gly Ser Arg Val Gly Leu Pro Cys Arg Leu
245 250 255
Pro Ala Gly Val Gly Thr Arg Ser Phe Leu Thr Ala Lys Trp Thr Pro
260 265 270
Pro Gly Gly Gly Pro Asp Leu Leu Val Thr Gly Asp Asn Gly Asp Phe
275 280 285
Thr Leu Arg Leu Glu Asp Val Ser Gln Ala Gln Ala Gly Thr Tyr Thr
290 295 300
Cys His Ile His Leu Gln Glu Gln Gln Leu Asn Ala Thr Val Thr Leu
305 310 315 320
Ala Ile Ile Thr Val Thr Pro Lys Ser Phe Gly Ser Pro Gly Ser Leu
325 330 335
Gly Lys Leu Leu Cys Glu Val Thr Pro Val Ser Gly Gln Glu Arg Phe
340 345 350
Val Trp Ser Ser Leu Asp Thr Pro Ser Gln Arg Ser Phe Ser Gly Pro
355 360 365
Trp Leu Glu Ala Gln Glu Ala Gln Leu Leu Ser Gln Pro Trp Gln Cys
370 375 380
Gln Leu Tyr Gln Gly Glu Arg Leu Leu Gly Ala Ala Val Tyr Phe Thr
385 390 395 400
Glu Leu Ser Ser Pro Gly Ala Gln Arg Ser Gly Arg Ala Pro Gly Ala
405 410 415
Leu Pro Ala Gly His Leu
420
<210> 133
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 133
Met Met Thr Gly Thr Ile Glu Thr Thr Gly Asn Ile Ser Ala Glu Lys
1 5 10 15
Gly Gly Ser Ile Ile Leu Gln Cys His Leu Ser Ser Thr Thr Ala Gln
20 25 30
Val Thr Gln Val Asn Trp Glu Gln Gln Asp Gln Leu Leu Ala Ile Cys
35 40 45
Asn Ala Asp Leu Gly Trp His Ile Ser Pro Ser Phe Lys Asp Arg Val
50 55 60
Ala Pro Gly Pro Gly Leu Gly Leu Thr Leu Gln Ser Leu Thr Val Asn
65 70 75 80
Asp Thr Gly Glu Tyr Phe Cys Ile Tyr His Thr Tyr Pro Asp Gly Thr
85 90 95
Tyr Thr Gly Arg Ile Phe Leu Glu Val Leu Glu Ser Ser Val Ala Glu
100 105 110
His Gly Ala Arg Phe Gln Ile Pro
115 120
<210> 134
<211> 150
<212> Белок
<213> Искусственная последовательность
<400> 134
Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp Asn Pro Pro Thr
1 5 10 15
Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp Asn Ala Thr Phe
20 25 30
Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val Leu Asn Trp Tyr
35 40 45
Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala Ala Phe Pro Glu
50 55 60
Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg Val Thr Gln Leu
65 70 75 80
Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg Ala Arg Arg Asn
85 90 95
Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu Ala Pro Lys Ala
100 105 110
Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val Thr Glu Arg Arg
115 120 125
Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro Arg Pro Ala Gly
130 135 140
Gln Phe Gln Thr Leu Val
145 150
<210> 135
<211> 220
<212> Белок
<213> Искусственная последовательность
<400> 135
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser Tyr Arg
50 55 60
Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg
210 215 220
<210> 136
<211> 204
<212> Белок
<213> Искусственная последовательность
<400> 136
Ser Gly Pro Val Lys Glu Leu Val Gly Ser Val Gly Gly Ala Val Thr
1 5 10 15
Phe Pro Leu Lys Ser Lys Val Lys Gln Val Asp Ser Ile Val Trp Thr
20 25 30
Phe Asn Thr Thr Pro Leu Val Thr Ile Gln Pro Glu Gly Gly Thr Ile
35 40 45
Ile Val Thr Gln Asn Arg Asn Arg Glu Arg Val Asp Phe Pro Asp Gly
50 55 60
Gly Tyr Ser Leu Lys Leu Ser Lys Leu Lys Lys Asn Asp Ser Gly Ile
65 70 75 80
Tyr Tyr Val Gly Ile Tyr Ser Ser Ser Leu Gln Gln Pro Ser Thr Gln
85 90 95
Glu Tyr Val Leu His Val Tyr Glu His Leu Ser Lys Pro Lys Val Thr
100 105 110
Met Gly Leu Gln Ser Asn Lys Asn Gly Thr Cys Val Thr Asn Leu Thr
115 120 125
Cys Cys Met Glu His Gly Glu Glu Asp Val Ile Tyr Thr Trp Lys Ala
130 135 140
Leu Gly Gln Ala Ala Asn Glu Ser His Asn Gly Ser Ile Leu Pro Ile
145 150 155 160
Ser Trp Arg Trp Gly Glu Ser Asp Met Thr Phe Ile Cys Val Ala Arg
165 170 175
Asn Pro Val Ser Arg Asn Phe Ser Ser Pro Ile Leu Ala Arg Lys Leu
180 185 190
Cys Glu Gly Ala Ala Asp Asp Pro Asp Ser Ser Met
195 200
<210> 137
<211> 651
<212> Белок
<213> Искусственная последовательность
<400> 137
Lys Leu Thr Ile Glu Ser Thr Pro Phe Asn Val Ala Glu Gly Lys Glu
1 5 10 15
Val Leu Leu Leu Val His Asn Leu Pro Gln His Leu Phe Gly Tyr Ser
20 25 30
Trp Tyr Lys Gly Glu Arg Val Asp Gly Asn Arg Gln Ile Ile Gly Tyr
35 40 45
Val Ile Gly Thr Gln Gln Ala Thr Pro Gly Pro Ala Tyr Ser Gly Arg
50 55 60
Glu Ile Ile Tyr Pro Asn Ala Ser Leu Leu Ile Gln Asn Ile Ile Gln
65 70 75 80
Asn Asp Thr Gly Phe Tyr Thr Leu His Val Ile Lys Ser Asp Leu Val
85 90 95
Asn Glu Glu Ala Thr Gly Gln Phe Arg Val Tyr Pro Glu Leu Pro Lys
100 105 110
Pro Ser Ile Ser Ser Asn Asn Ser Lys Pro Val Glu Asp Lys Asp Ala
115 120 125
Val Ala Phe Thr Cys Glu Pro Glu Thr Gln Asp Ala Thr Tyr Leu Trp
130 135 140
Trp Val Asn Asn Gln Ser Leu Pro Val Ser Pro Arg Leu Gln Leu Ser
145 150 155 160
Asn Gly Asn Arg Thr Leu Thr Leu Phe Asn Val Thr Arg Asn Asp Thr
165 170 175
Ala Ser Tyr Lys Cys Glu Thr Gln Asn Pro Val Ser Ala Arg Arg Ser
180 185 190
Asp Ser Val Ile Leu Asn Val Leu Tyr Gly Pro Asp Ala Pro Thr Ile
195 200 205
Ser Pro Leu Asn Thr Ser Tyr Arg Ser Gly Glu Asn Leu Asn Leu Ser
210 215 220
Cys His Ala Ala Ser Asn Pro Pro Ala Gln Tyr Ser Trp Phe Val Asn
225 230 235 240
Gly Thr Phe Gln Gln Ser Thr Gln Glu Leu Phe Ile Pro Asn Ile Thr
245 250 255
Val Asn Asn Ser Gly Ser Tyr Thr Cys Gln Ala His Asn Ser Asp Thr
260 265 270
Gly Leu Asn Arg Thr Thr Val Thr Thr Ile Thr Val Tyr Ala Glu Pro
275 280 285
Pro Lys Pro Phe Ile Thr Ser Asn Asn Ser Asn Pro Val Glu Asp Glu
290 295 300
Asp Ala Val Ala Leu Thr Cys Glu Pro Glu Ile Gln Asn Thr Thr Tyr
305 310 315 320
Leu Trp Trp Val Asn Asn Gln Ser Leu Pro Val Ser Pro Arg Leu Gln
325 330 335
Leu Ser Asn Asp Asn Arg Thr Leu Thr Leu Leu Ser Val Thr Arg Asn
340 345 350
Asp Val Gly Pro Tyr Glu Cys Gly Ile Gln Asn Lys Leu Ser Val Asp
355 360 365
His Ser Asp Pro Val Ile Leu Asn Val Leu Tyr Gly Pro Asp Asp Pro
370 375 380
Thr Ile Ser Pro Ser Tyr Thr Tyr Tyr Arg Pro Gly Val Asn Leu Ser
385 390 395 400
Leu Ser Cys His Ala Ala Ser Asn Pro Pro Ala Gln Tyr Ser Trp Leu
405 410 415
Ile Asp Gly Asn Ile Gln Gln His Thr Gln Glu Leu Phe Ile Ser Asn
420 425 430
Ile Thr Glu Lys Asn Ser Gly Leu Tyr Thr Cys Gln Ala Asn Asn Ser
435 440 445
Ala Ser Gly His Ser Arg Thr Thr Val Lys Thr Ile Thr Val Ser Ala
450 455 460
Glu Leu Pro Lys Pro Ser Ile Ser Ser Asn Asn Ser Lys Pro Val Glu
465 470 475 480
Asp Lys Asp Ala Val Ala Phe Thr Cys Glu Pro Glu Ala Gln Asn Thr
485 490 495
Thr Tyr Leu Trp Trp Val Asn Gly Gln Ser Leu Pro Val Ser Pro Arg
500 505 510
Leu Gln Leu Ser Asn Gly Asn Arg Thr Leu Thr Leu Phe Asn Val Thr
515 520 525
Arg Asn Asp Ala Arg Ala Tyr Val Cys Gly Ile Gln Asn Ser Val Ser
530 535 540
Ala Asn Arg Ser Asp Pro Val Thr Leu Asp Val Leu Tyr Gly Pro Asp
545 550 555 560
Thr Pro Ile Ile Ser Pro Pro Asp Ser Ser Tyr Leu Ser Gly Ala Asn
565 570 575
Leu Asn Leu Ser Cys His Ser Ala Ser Asn Pro Ser Pro Gln Tyr Ser
580 585 590
Trp Arg Ile Asn Gly Ile Pro Gln Gln His Thr Gln Val Leu Phe Ile
595 600 605
Ala Lys Ile Thr Pro Asn Asn Asn Gly Thr Tyr Ala Cys Phe Val Ser
610 615 620
Asn Leu Ala Thr Gly Arg Asn Asn Ser Ile Val Lys Ser Ile Thr Val
625 630 635 640
Ser Ala Ser Gly Thr Ser Pro Gly Leu Ser Ala
645 650
<210> 138
<211> 104
<212> Белок
<213> Искусственная последовательность
<400> 138
Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys Val
1 5 10 15
Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Pro Gln Tyr Pro Gly
20 25 30
Ser Glu Ile Leu Trp Gln His Asn Asp Lys Asn Ile Gly Gly Asp Glu
35 40 45
Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp His Leu Ser Leu Lys Glu
50 55 60
Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg Gly
65 70 75 80
Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu Tyr Leu Arg Ala Arg Val
85 90 95
Cys Glu Asn Cys Met Glu Met Asp
100
<210> 139
<211> 192
<212> Белок
<213> Искусственная последовательность
<400> 139
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
1 5 10 15
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
20 25 30
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
35 40 45
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
50 55 60
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
65 70 75 80
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
85 90 95
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
100 105 110
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
115 120 125
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
130 135 140
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Phe
145 150 155 160
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
165 170 175
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
180 185 190
<210> 140
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 140
Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser Thr Glu Lys Asn Cys Cys
1 5 10 15
Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys Asp Leu Gly Trp Lys Trp
20 25 30
Ile His Glu Pro Lys Gly Tyr His Ala Asn Phe Cys Leu Gly Pro Cys
35 40 45
Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr Ser Lys Val Leu Ala Leu
50 55 60
Tyr Asn Gln His Asn Pro Gly Ala Ser Ala Ala Pro Cys Cys Val Pro
65 70 75 80
Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr Tyr Val Gly Arg Lys Pro
85 90 95
Lys Val Glu Gln Leu Ser Asn Met Ile Val Arg Ser Cys Lys Cys Ser
100 105 110
<210> 141
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 141
Ala Leu Asp Ala Ala Tyr Cys Phe Arg Asn Val Gln Asp Asn Cys Cys
1 5 10 15
Leu Arg Pro Leu Tyr Ile Asp Phe Lys Arg Asp Leu Gly Trp Lys Trp
20 25 30
Ile His Glu Pro Lys Gly Tyr Asn Ala Asn Phe Cys Ala Gly Ala Cys
35 40 45
Pro Tyr Leu Trp Ser Ser Asp Thr Gln His Ser Arg Val Leu Ser Leu
50 55 60
Tyr Asn Thr Ile Asn Pro Glu Ala Ser Ala Ser Pro Cys Cys Val Ser
65 70 75 80
Gln Asp Leu Glu Pro Leu Thr Ile Leu Tyr Tyr Ile Gly Lys Thr Pro
85 90 95
Lys Ile Glu Gln Leu Ser Asn Met Ile Val Lys Ser Cys Lys Cys Ser
100 105 110
<210> 142
<211> 112
<212> Белок
<213> Искусственная последовательность
<400> 142
Ala Leu Asp Thr Asn Tyr Cys Phe Arg Asn Leu Glu Glu Asn Cys Cys
1 5 10 15
Val Arg Pro Leu Tyr Ile Asp Phe Arg Gln Asp Leu Gly Trp Lys Trp
20 25 30
Val His Glu Pro Lys Gly Tyr Tyr Ala Asn Phe Cys Ser Gly Pro Cys
35 40 45
Pro Tyr Leu Arg Ser Ala Asp Thr Thr His Ser Thr Val Leu Gly Leu
50 55 60
Tyr Asn Thr Leu Asn Pro Glu Ala Ser Ala Ser Pro Cys Cys Val Pro
65 70 75 80
Gln Asp Leu Glu Pro Leu Thr Ile Leu Tyr Tyr Val Gly Arg Thr Pro
85 90 95
Lys Val Glu Gln Leu Ser Asn Met Val Val Lys Ser Cys Lys Cys Ser
100 105 110
<210> 143
<211> 206
<212> Белок
<213> Искусственная последовательность
<400> 143
Ala Pro Met Ala Glu Gly Gly Gly Gln Asn His His Glu Val Val Lys
1 5 10 15
Phe Met Asp Val Tyr Gln Arg Ser Tyr Cys His Pro Ile Glu Thr Leu
20 25 30
Val Asp Ile Phe Gln Glu Tyr Pro Asp Glu Ile Glu Tyr Ile Phe Lys
35 40 45
Pro Ser Cys Val Pro Leu Met Arg Cys Gly Gly Cys Cys Asn Asp Glu
50 55 60
Gly Leu Glu Cys Val Pro Thr Glu Glu Ser Asn Ile Thr Met Gln Ile
65 70 75 80
Met Arg Ile Lys Pro His Gln Gly Gln His Ile Gly Glu Met Ser Phe
85 90 95
Leu Gln His Asn Lys Cys Glu Cys Arg Pro Lys Lys Asp Arg Ala Arg
100 105 110
Gln Glu Lys Lys Ser Val Arg Gly Lys Gly Lys Gly Gln Lys Arg Lys
115 120 125
Arg Lys Lys Ser Arg Tyr Lys Ser Trp Ser Val Tyr Val Gly Ala Arg
130 135 140
Cys Cys Leu Met Pro Trp Ser Leu Pro Gly Pro His Pro Cys Gly Pro
145 150 155 160
Cys Ser Glu Arg Arg Lys His Leu Phe Val Gln Asp Pro Gln Thr Cys
165 170 175
Lys Cys Ser Cys Lys Asn Thr Asp Ser Arg Cys Lys Ala Arg Gln Leu
180 185 190
Glu Leu Asn Glu Arg Thr Cys Arg Cys Asp Lys Pro Arg Arg
195 200 205
<210> 144
<211> 159
<212> Белок
<213> Искусственная последовательность
<400> 144
Pro Gly Gln Gly Thr Gln Ser Glu Asn Ser Cys Thr His Phe Pro Gly
1 5 10 15
Asn Leu Pro Asn Met Leu Arg Asp Leu Arg Asp Ala Phe Ser Arg Val
20 25 30
Lys Thr Phe Phe Gln Met Lys Asp Gln Leu Asp Asn Leu Leu Leu Lys
35 40 45
Glu Ser Leu Leu Glu Asp Phe Lys Gly Tyr Leu Gly Cys Gln Ala Leu
50 55 60
Ser Glu Met Ile Gln Phe Tyr Leu Glu Glu Val Met Pro Gln Ala Glu
65 70 75 80
Asn Gln Asp Pro Asp Ile Lys Ala His Val Asn Ser Leu Gly Glu Asn
85 90 95
Leu Lys Thr Leu Arg Leu Arg Leu Arg Arg Cys His Arg Phe Leu Pro
100 105 110
Cys Glu Asn Lys Ser Lys Ala Val Glu Gln Val Lys Asn Ala Phe Asn
115 120 125
Lys Leu Gln Glu Lys Gly Ile Tyr Lys Ala Met Ser Glu Phe Asp Ile
130 135 140
Phe Ile Asn Tyr Ile Glu Ala Tyr Met Thr Met Lys Ile Arg Asn
145 150 155
<210> 145
<211> 214
<212> Белок
<213> Искусственная последовательность
<400> 145
His Gly Thr Glu Leu Pro Ser Pro Pro Ser Val Trp Phe Glu Ala Glu
1 5 10 15
Phe Phe His His Ile Leu His Trp Thr Pro Ile Pro Asn Gln Ser Glu
20 25 30
Ser Thr Cys Tyr Glu Val Ala Leu Leu Arg Tyr Gly Ile Glu Ser Trp
35 40 45
Asn Ser Ile Ser Asn Cys Ser Gln Thr Leu Ser Tyr Asp Leu Thr Ala
50 55 60
Val Thr Leu Asp Leu Tyr His Ser Asn Gly Tyr Arg Ala Arg Val Arg
65 70 75 80
Ala Val Asp Gly Ser Arg His Ser Asn Trp Thr Val Thr Asn Thr Arg
85 90 95
Phe Ser Val Asp Glu Val Thr Leu Thr Val Gly Ser Val Asn Leu Glu
100 105 110
Ile His Asn Gly Phe Ile Leu Gly Lys Ile Gln Leu Pro Arg Pro Lys
115 120 125
Met Ala Pro Ala Asn Asp Thr Tyr Glu Ser Ile Phe Ser His Phe Arg
130 135 140
Glu Tyr Glu Ile Ala Ile Arg Lys Val Pro Gly Asn Phe Thr Phe Thr
145 150 155 160
His Lys Lys Val Lys His Glu Asn Phe Ser Leu Leu Thr Ser Gly Glu
165 170 175
Val Gly Glu Phe Cys Val Gln Val Lys Pro Ser Val Ala Ser Arg Ser
180 185 190
Asn Lys Gly Met Trp Ser Lys Glu Glu Cys Ile Ser Leu Thr Arg Gln
195 200 205
Tyr Phe Thr Val Thr Asn
210
<210> 146
<211> 120
<212> Белок
<213> Искусственная последовательность
<400> 146
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 147
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 147
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 148
<211> 119
<212> Белок
<213> Искусственная последовательность
<400> 148
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 149
<211> 107
<212> Белок
<213> Искусственная последовательность
<400> 149
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ, НАЦЕЛЕННЫЕ НА СИСТЕМУ СЕКРЕЦИИ ТИПА III | 2016 |
|
RU2759949C2 |
| МУЛЬТИВАЛЕНТНЫЕ И МУЛЬТИСПЕЦИФИЧЕСКИЕ ГИБРИДНЫЕ БЕЛКИ, СВЯЗЫВАЮЩИЕСЯ С DR5 | 2016 |
|
RU2748620C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| ГИБРИДНЫЕ БЕЛКИ ВАРИАНТА sPD-1—FC | 2019 |
|
RU2785993C2 |
| ПОЛИФУНКЦИОНАЛЬНЫЕ БЕЛКОВЫЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ ДЕКОРИН, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2823357C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| МОНОВАЛЕНТНЫЕ АНТИТЕЛА К ПРОПЕРДИНУ И ФРАГМЕНТЫ АНТИТЕЛ | 2018 |
|
RU2790103C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| НОВЫЙ МОДИФИЦИРОВАННЫЙ СЛИТЫЙ БЕЛОК FC-ФРАГМЕНТА ИММУНОГЛОБУЛИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800919C2 |
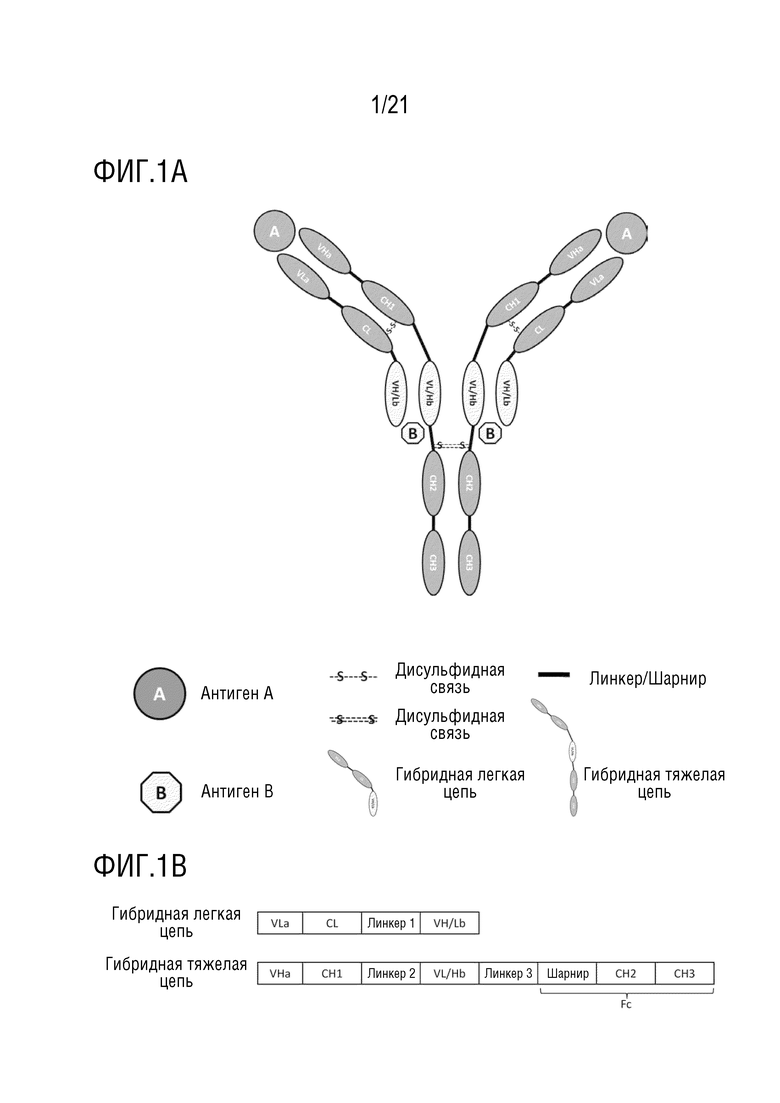
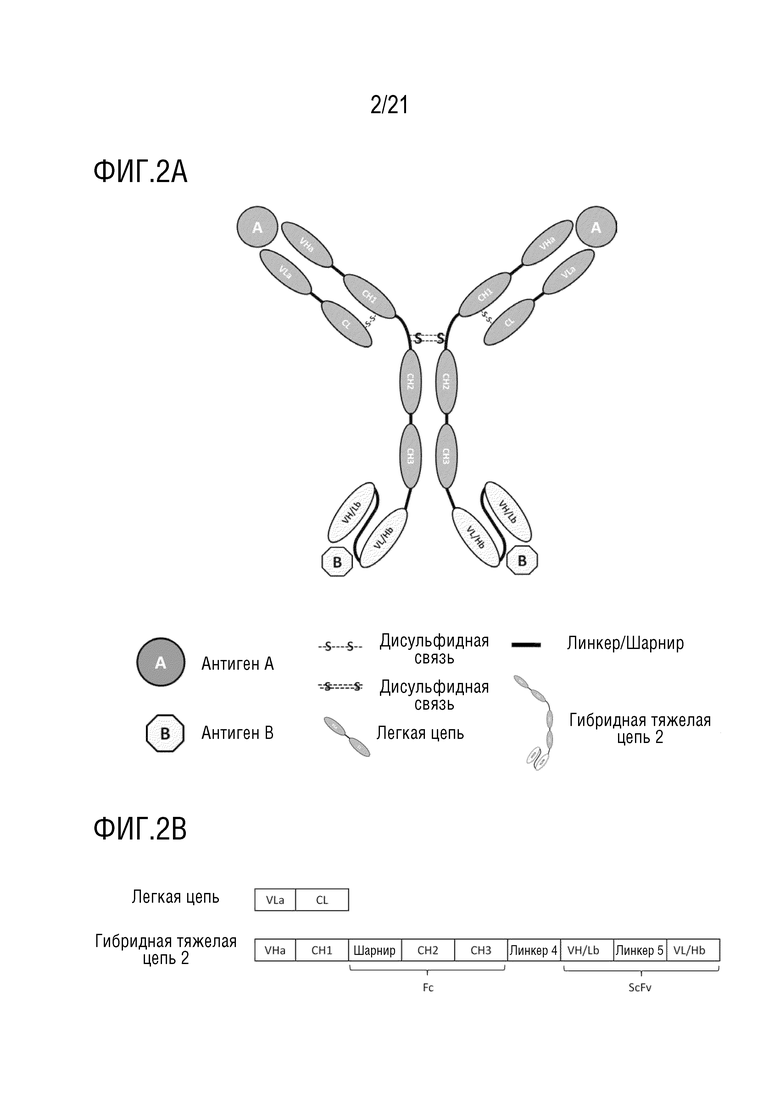
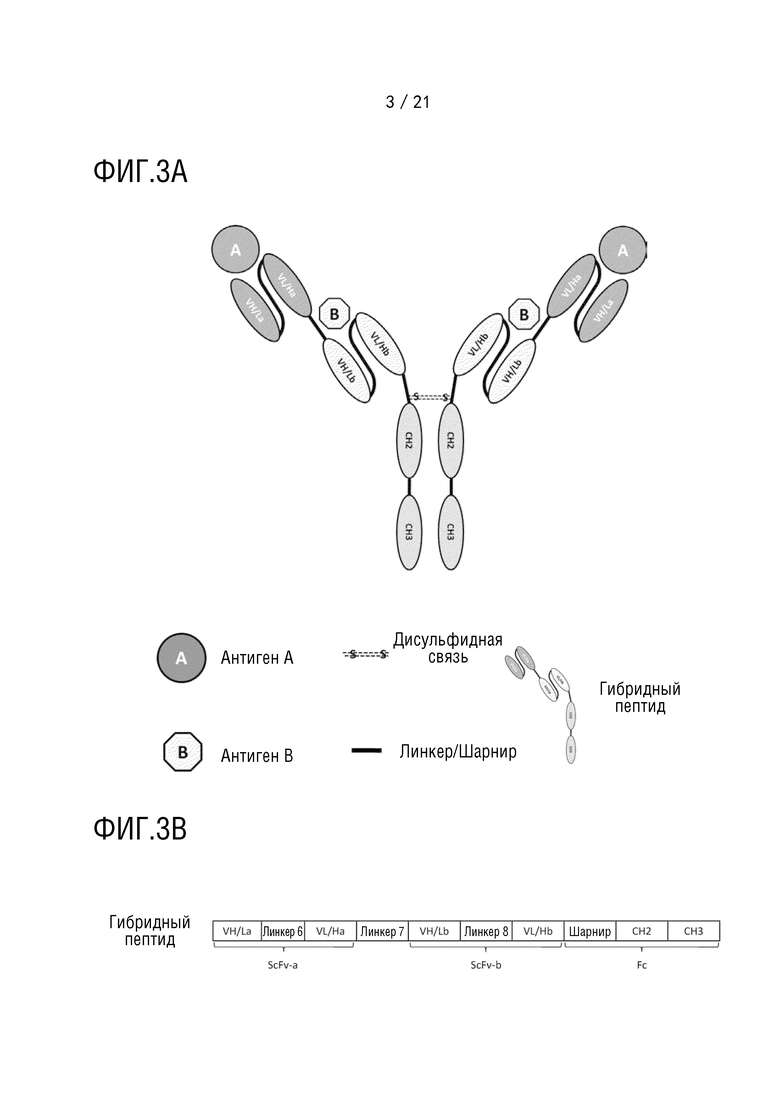
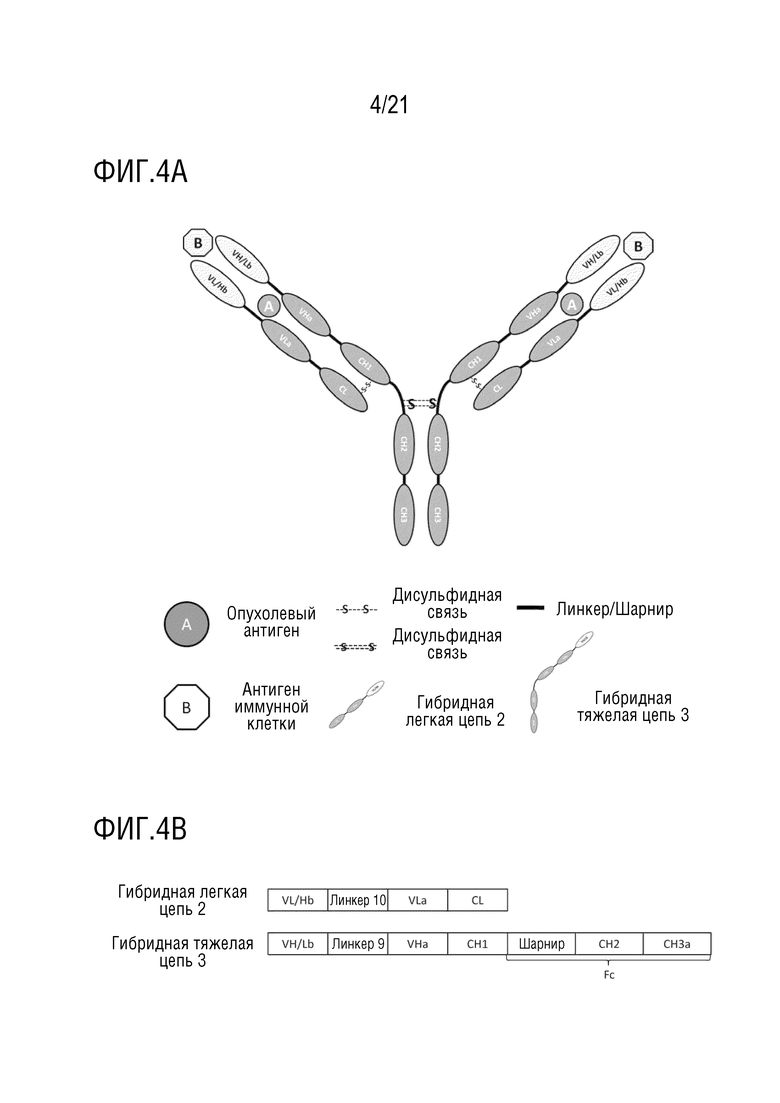
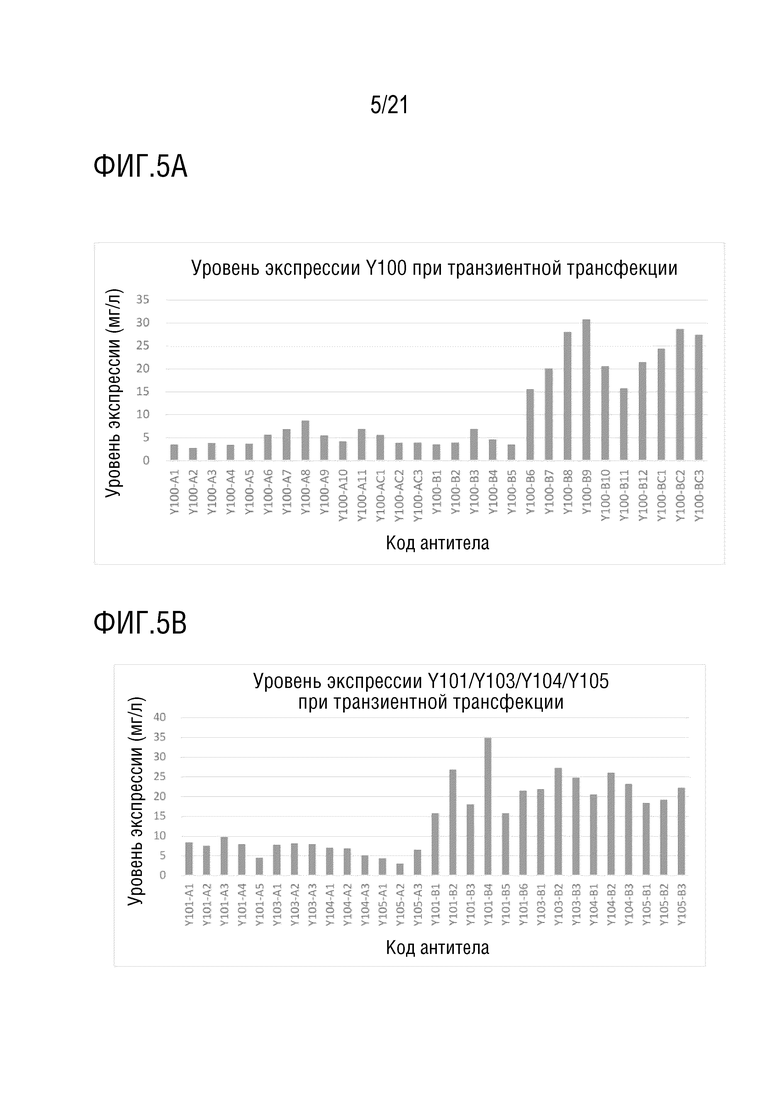
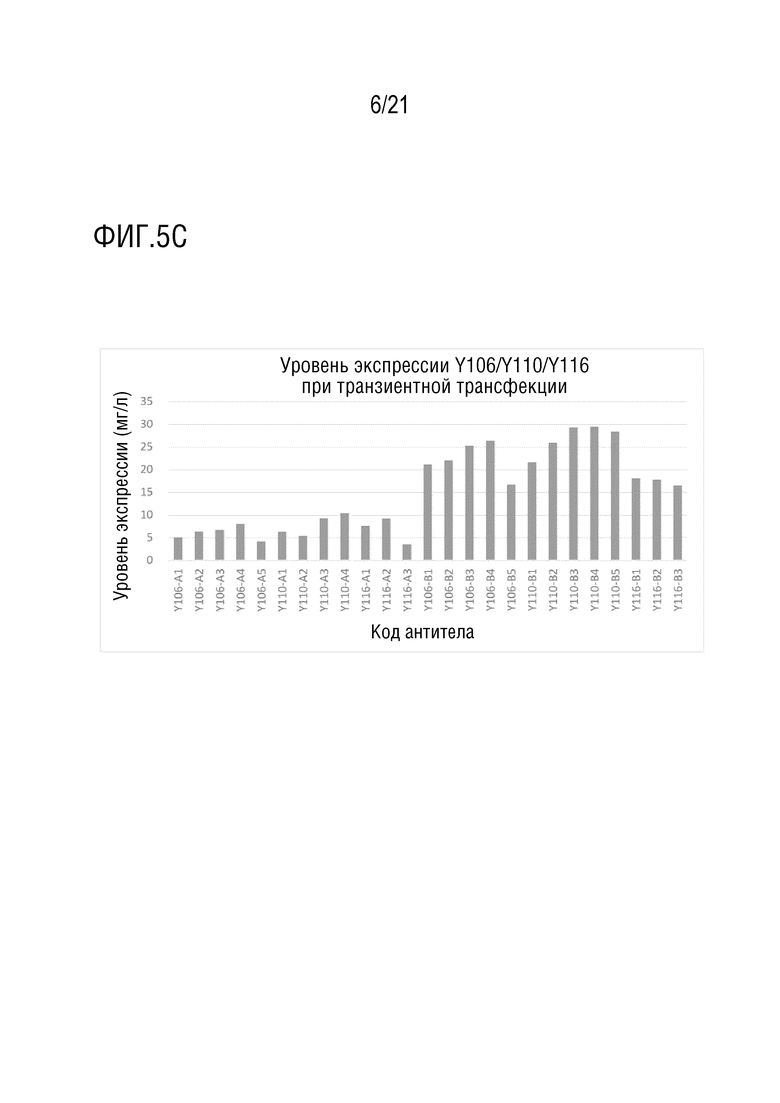

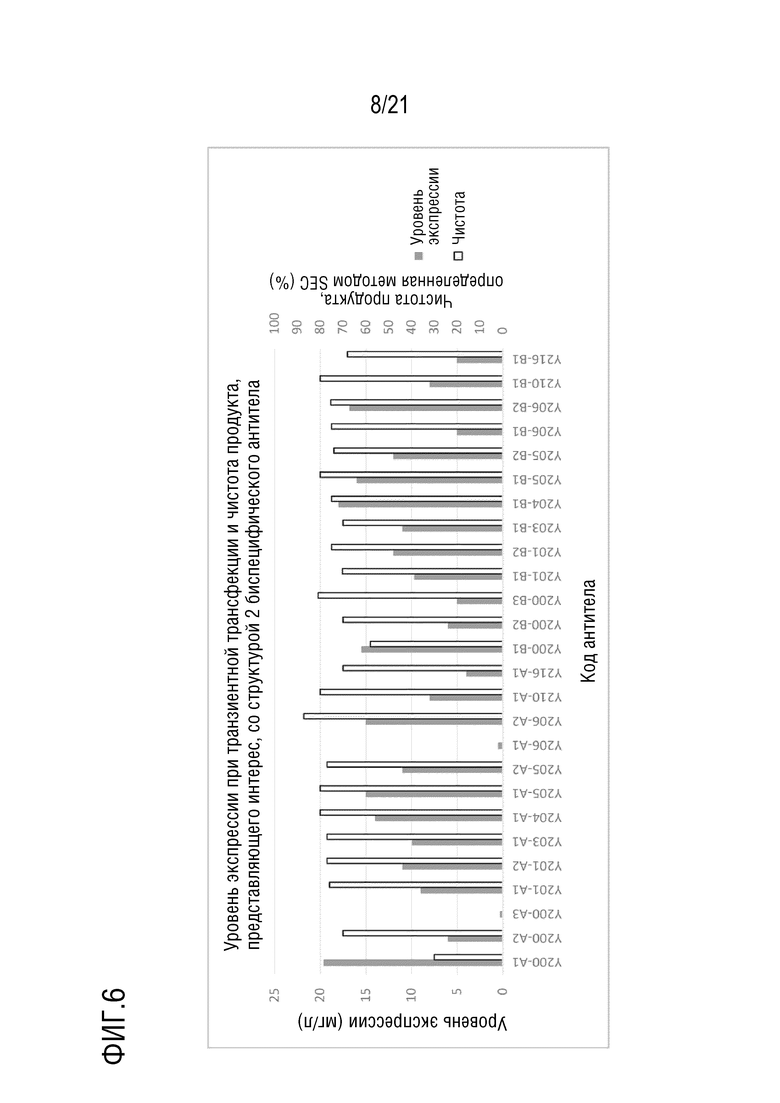
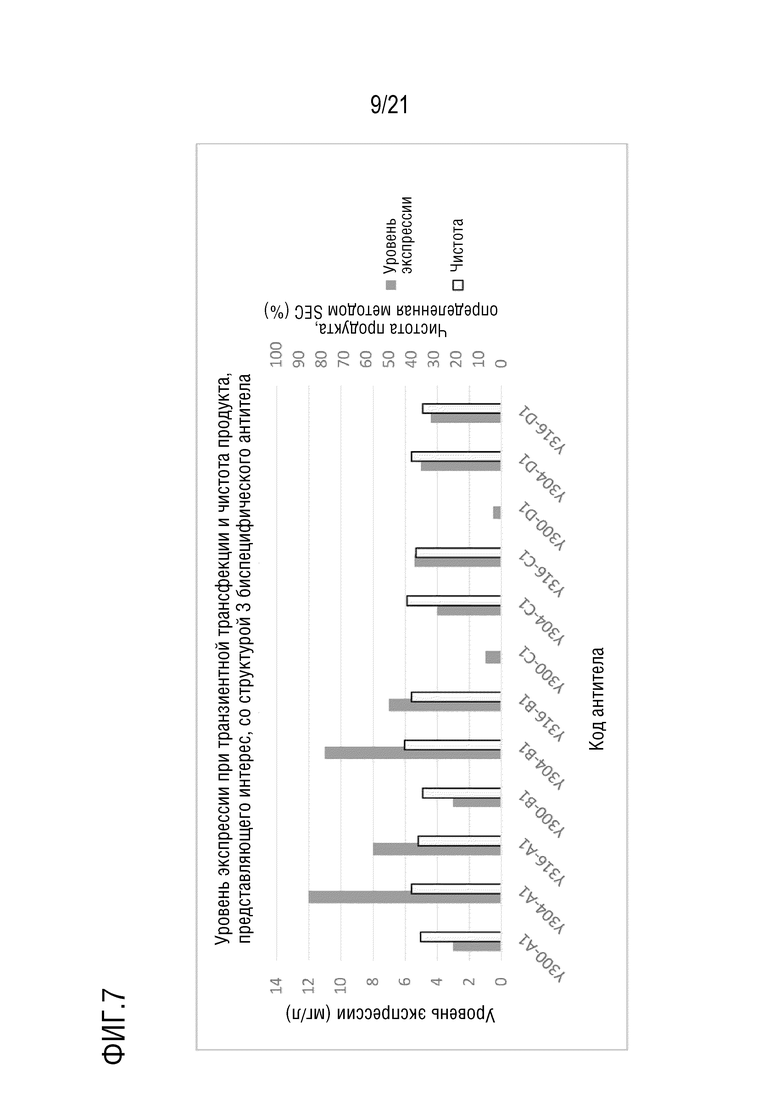
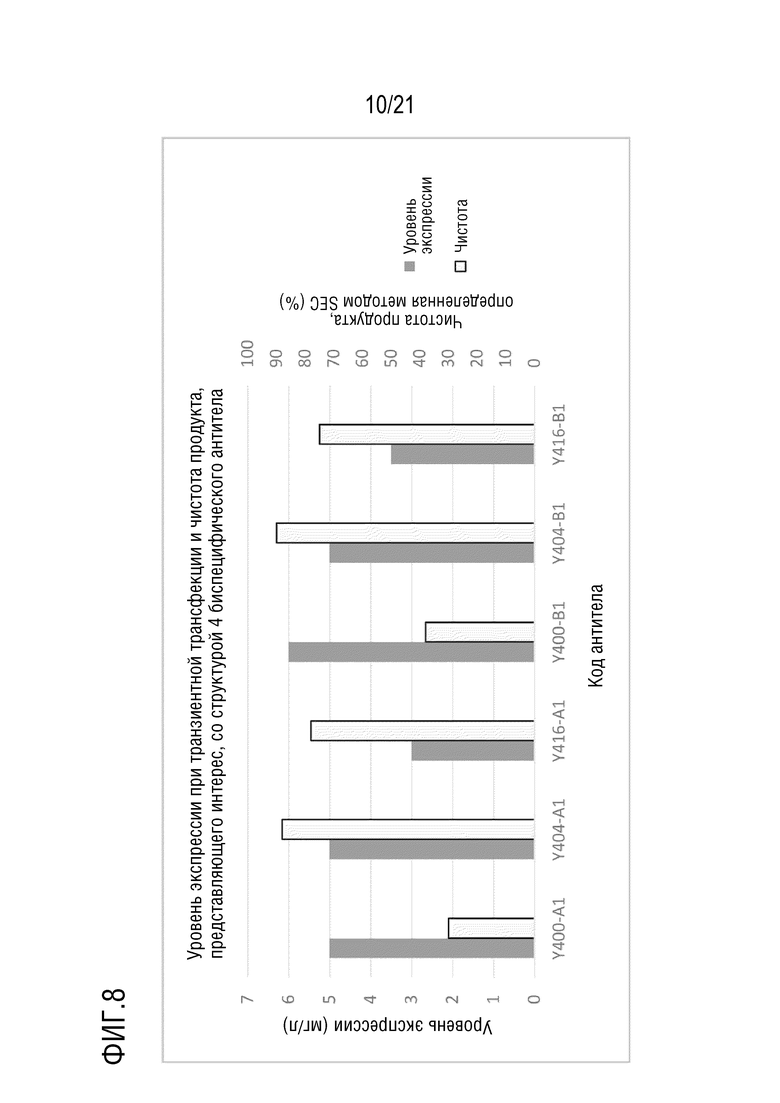
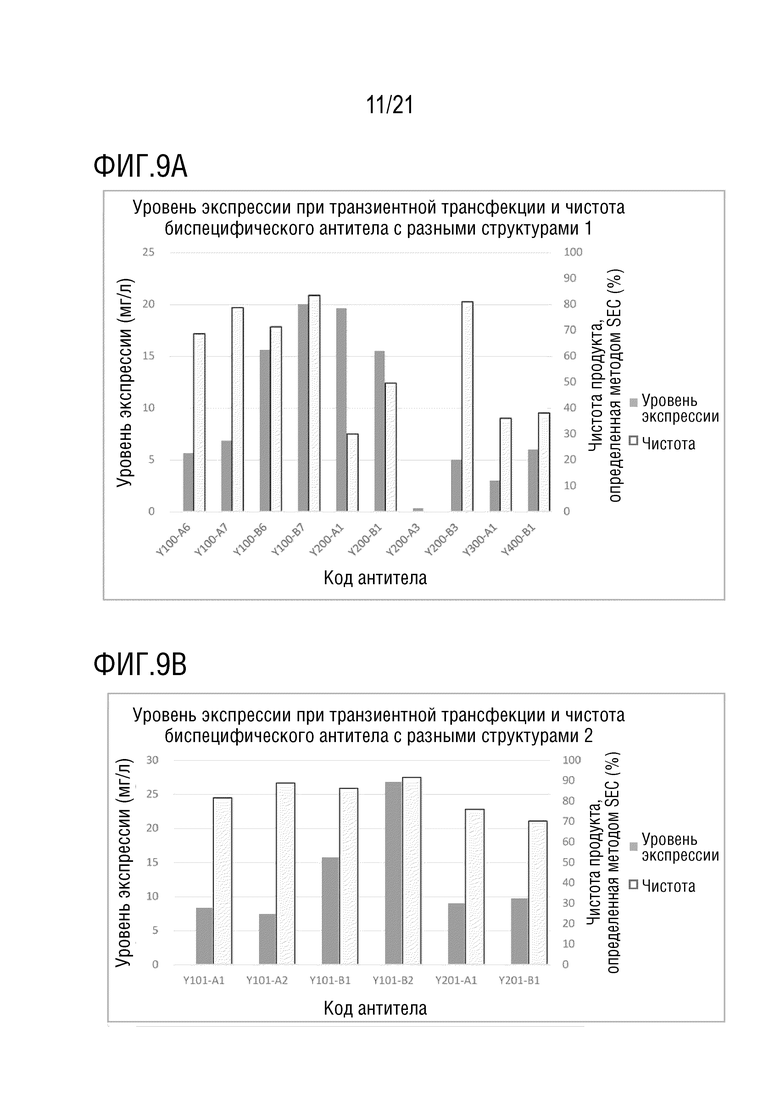
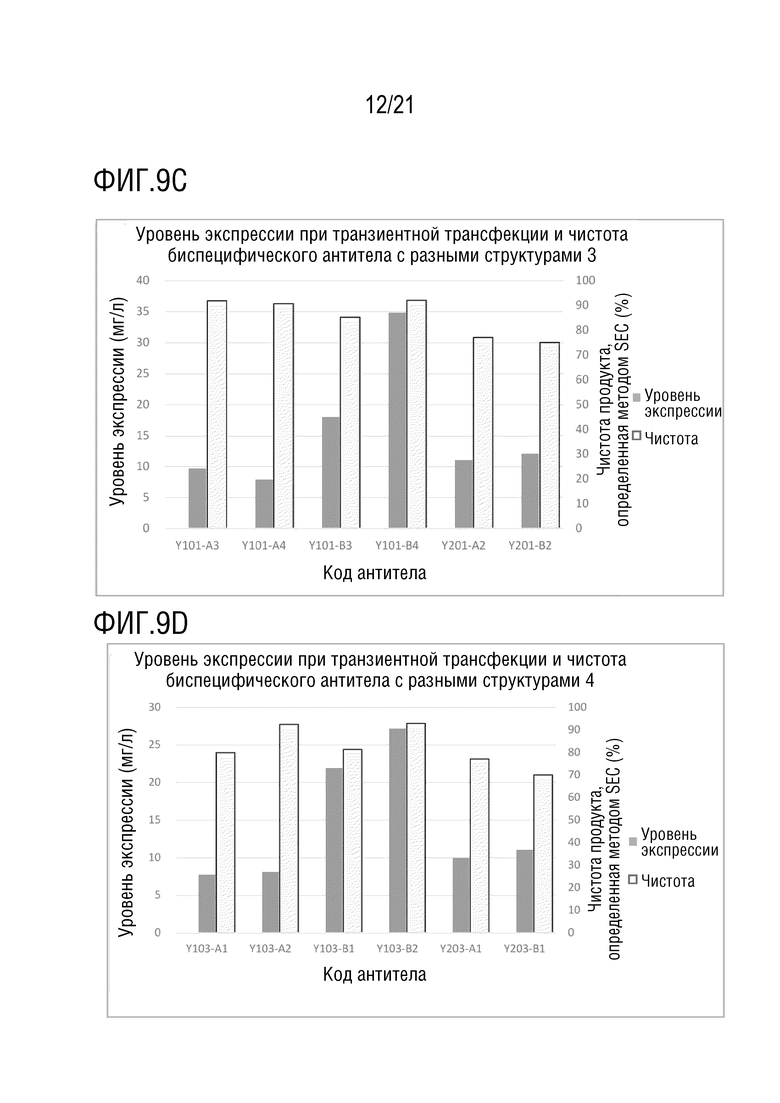
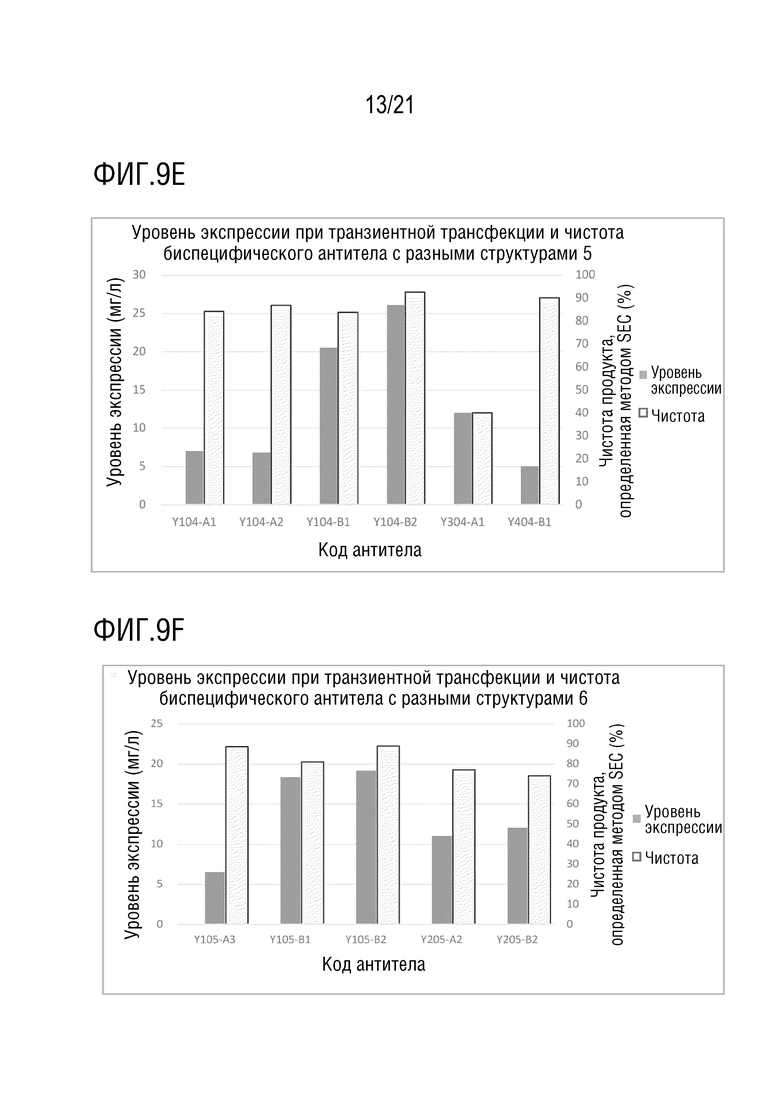
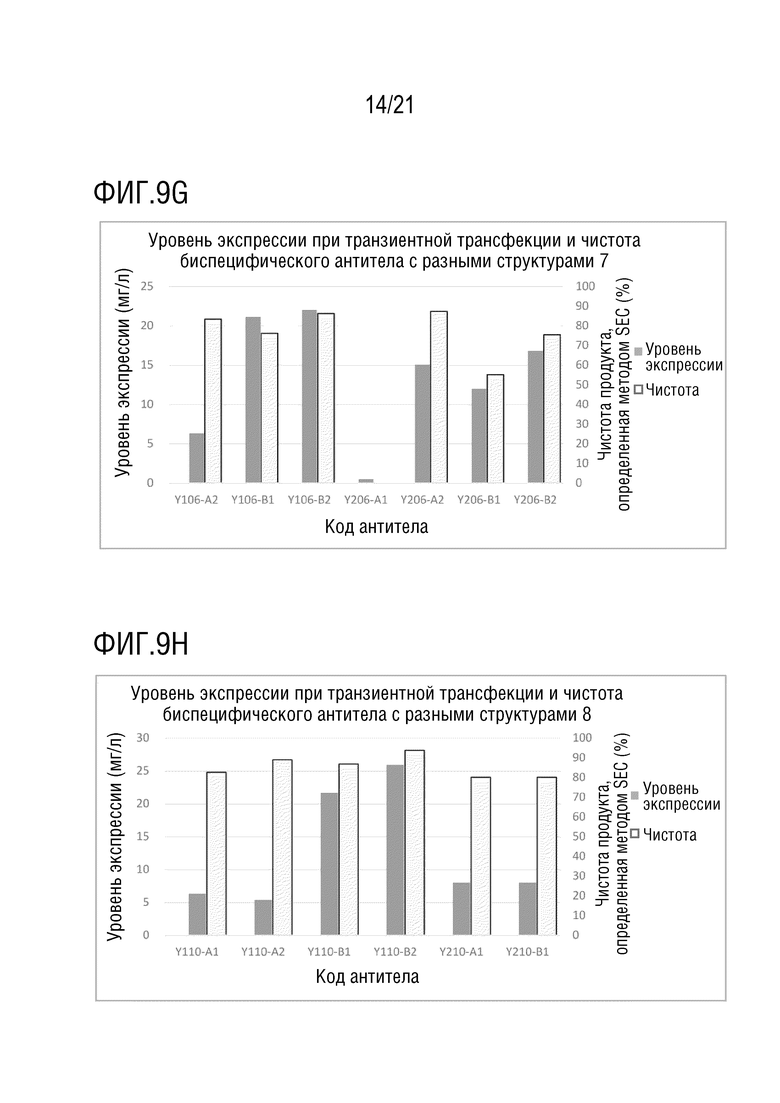
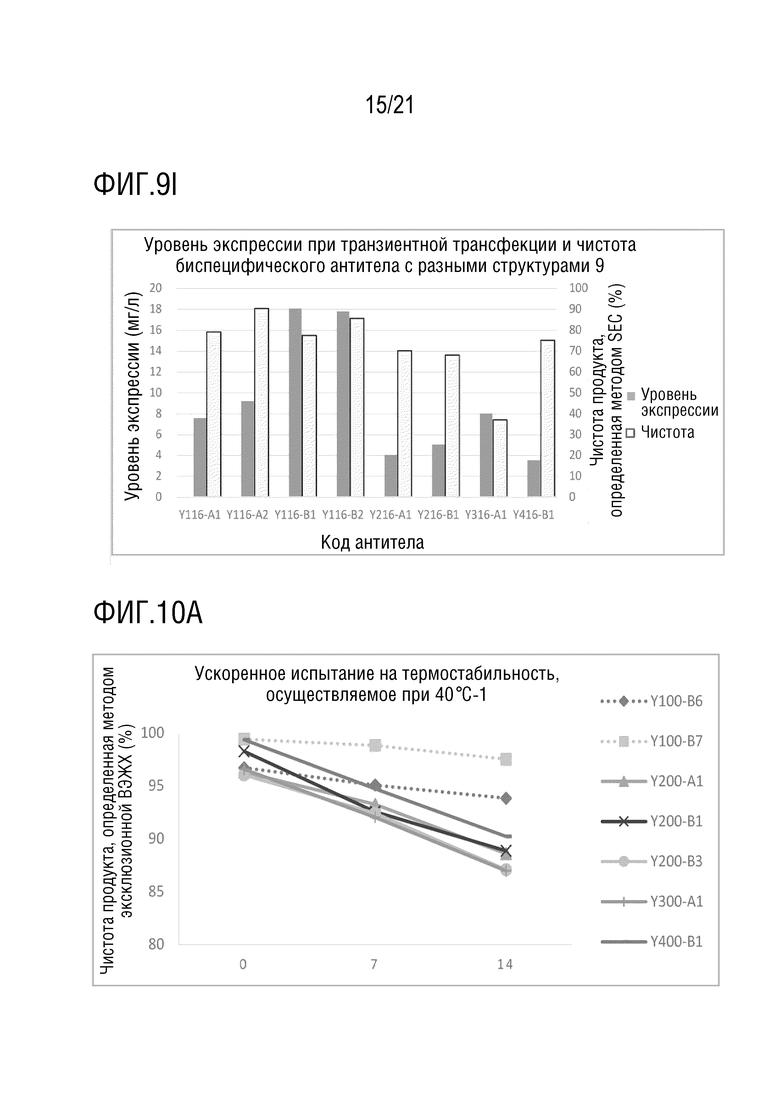
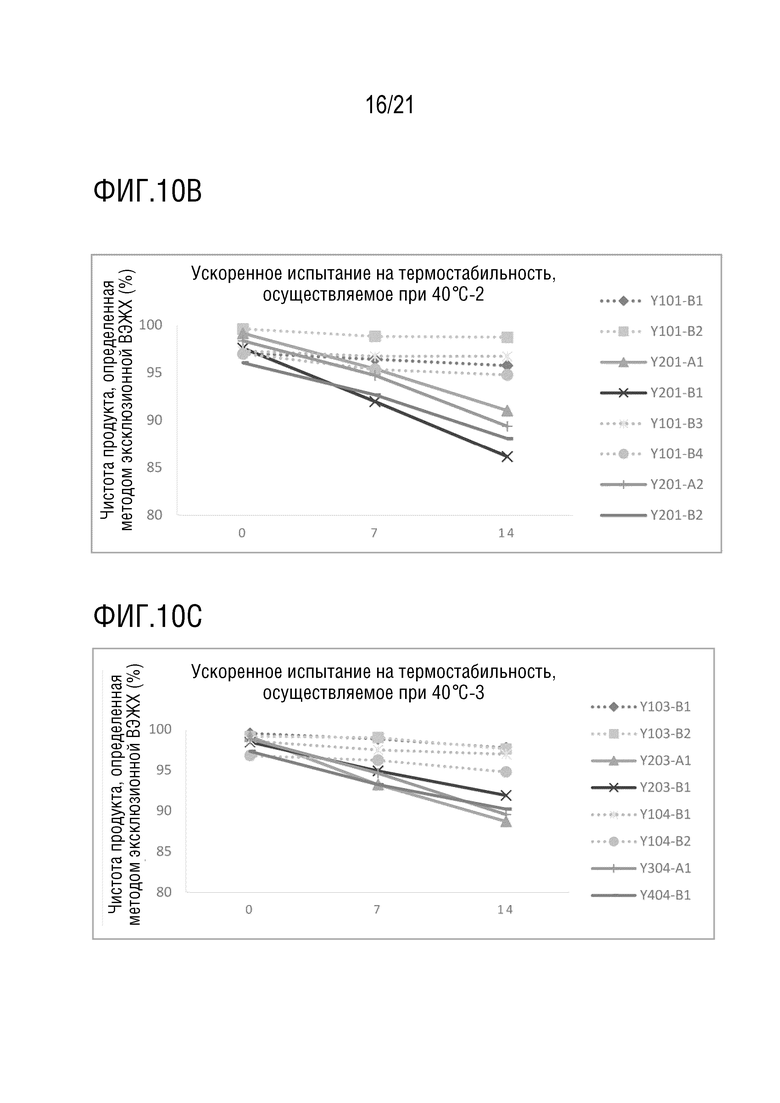
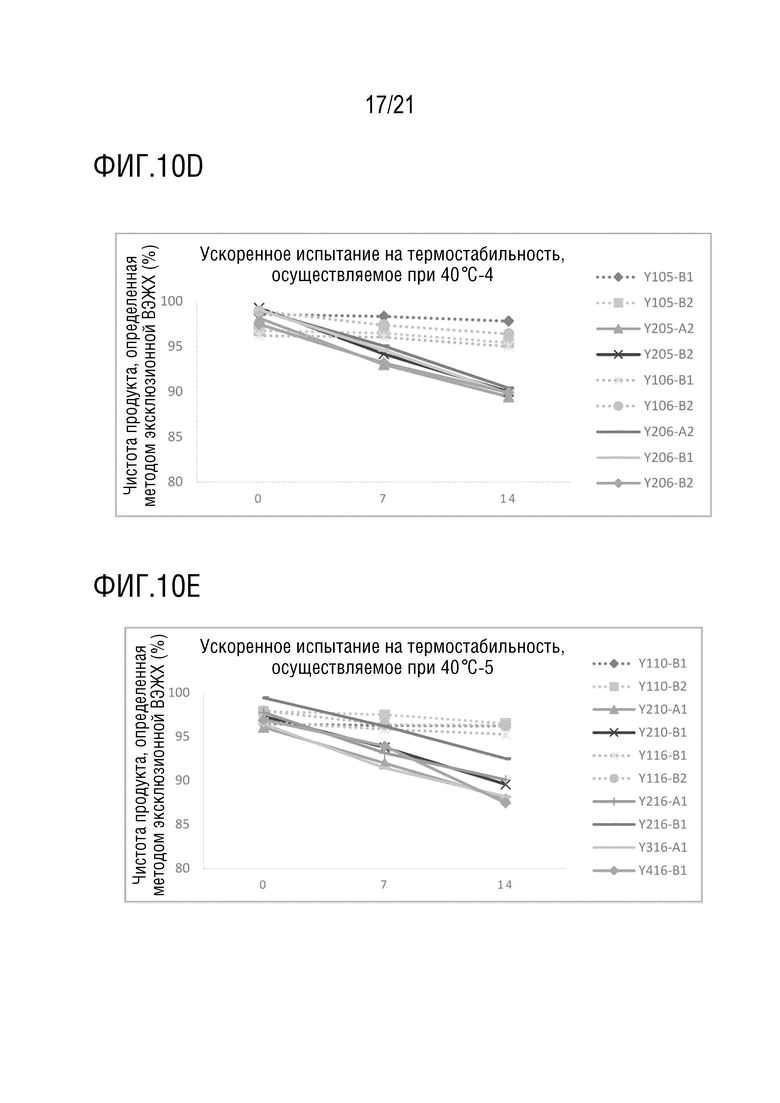
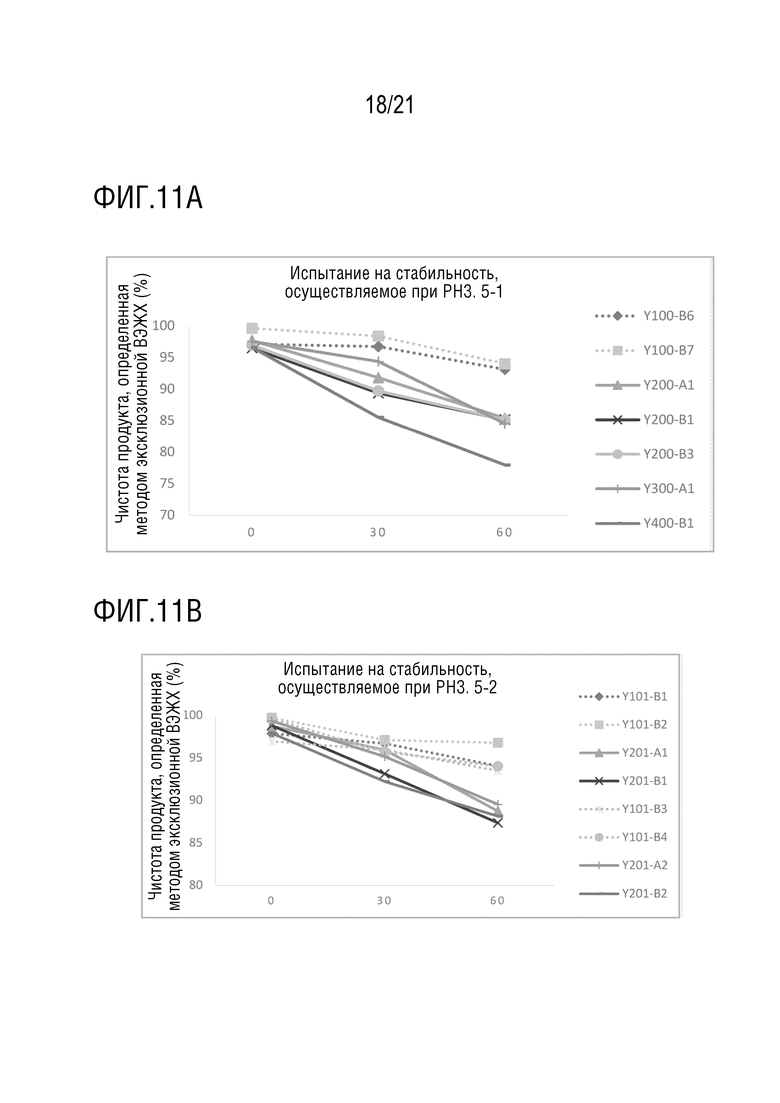
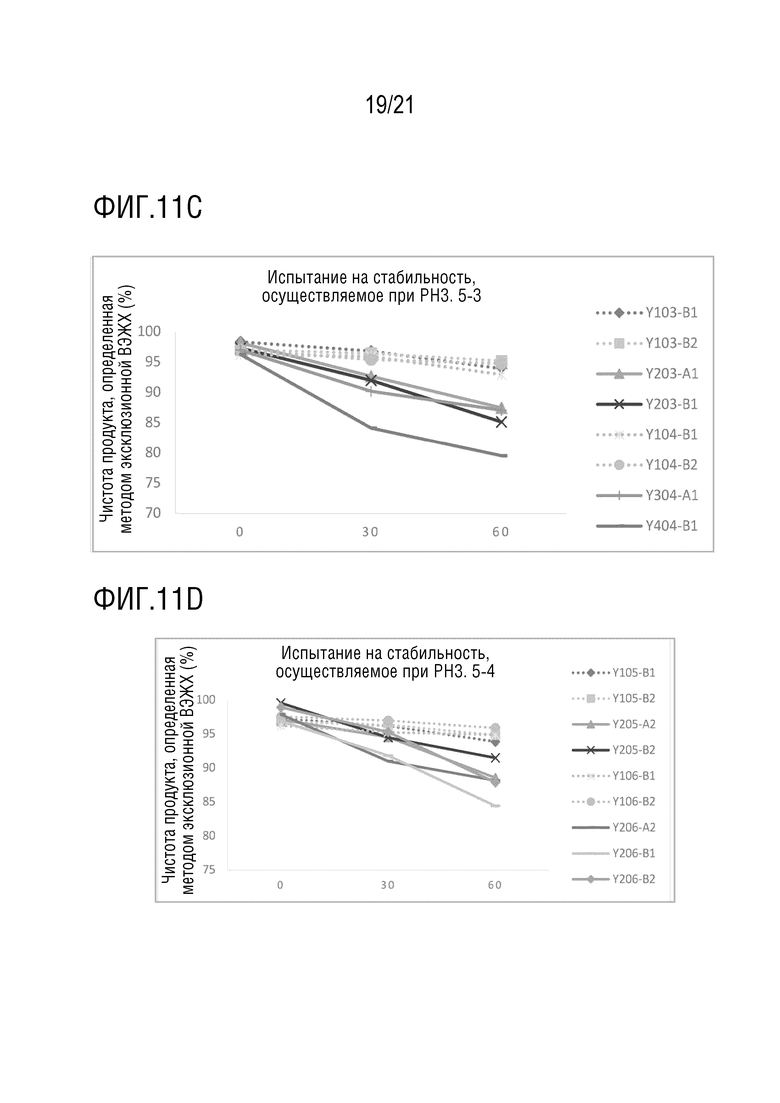
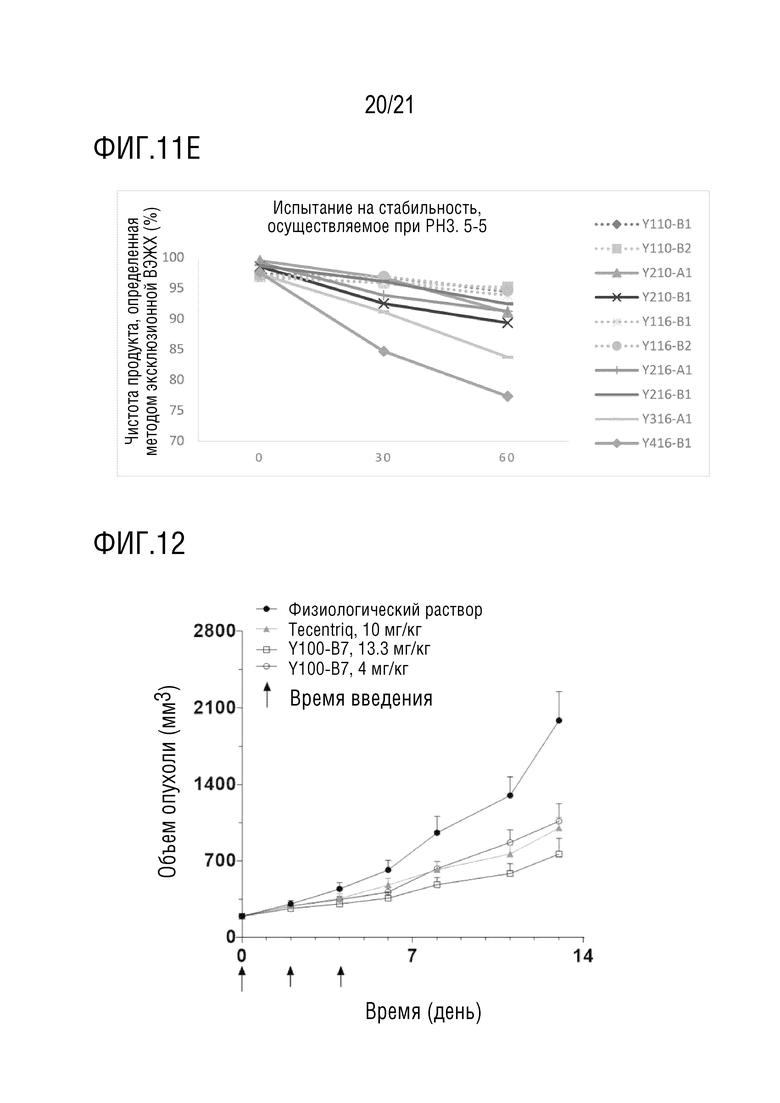
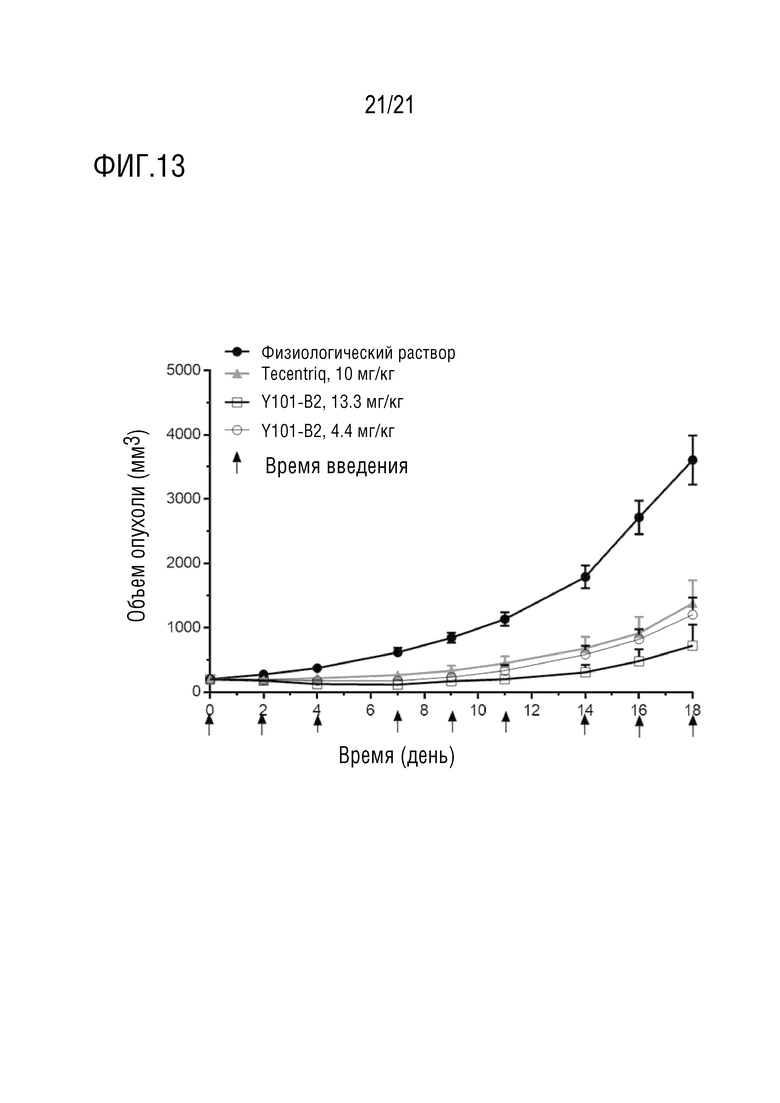
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая биспецифическое антитело, связывающее два разных эпитопа, фармацевтическую композицию для лечения опухоли, связанной с первым эпитопом и вторым эпитопом, включающую вышеуказанное биспецифическое антитело, конъюгат, связывающий первый эпитоп и второй эпитоп, слитый белок, связывающий первый эпитоп и второй эпитоп, полинуклеотид, кодирующий биспецифическое антитело, клетку, экспрессирующую биспецифическое антитело, применение биспецифического антитела или конъюгата или слитого белка для лечения опухоли, связанной с первым и вторым эпитопом и применение биспецифического антитела или конъюгата или слитого белка для получения реагента или набора для детекции опухоли, связанной с первым эпитопом и вторым эпитопом. Изобретение расширяет арсенал средств, связывающих два разных эпитопа. 8 н. и 22 з.п. ф-лы, 36 ил., 35 табл., 6 пр.
1. Биспецифическое антитело, связывающее два разных эпитопа, содержащее две тяжелые цепи и две легкие цепи, отличающееся тем, что две тяжелые цепи и две легкие цепи представляют собой две идентичные гибридные тяжелые цепи и две идентичные гибридные легкие цепи, где две гибридные тяжелые цепи образуют пару и гибридная легкая цепь и гибридная тяжелая цепь образуют пару; при этом гибридная тяжелая цепь включает вариабельную область тяжелой цепи антитела a (VHa), первую константную область CH1, вариабельную область 1 антитела b и Fc-фрагмент, вариабельная область 1 антитела b связана с CH1 линкером 2 или пептидной связью, а с Fc линкером 3 или пептидной связью; гибридная легкая цепь биспецифического антитела включает вариабельную область легкой цепи антитела a (VLa), константную область легкой цепи CL и вариабельную область 2 антитела b, где вариабельная область 2 антитела b связана с C-концом CL через линкер 1 или пептидную связь, где
(1) вариабельная область 1 антитела b представляет собой вариабельную область тяжелой цепи антитела b (VHb), а вариабельная область 2 антитела b представляет собой вариабельную область легкой цепи антитела b (VLb);
или
(2) вариабельная область 1 антитела b представляет собой вариабельную область легкой цепи антитела b (VLb), а вариабельная область 2 антитела b представляет собой вариабельную область тяжелой цепи антитела b (VHb),
при этом VHa-VLa пара нацелена на первый эпитоп, а VHb-VLb пара нацелена на второй эпитоп, и
где линкер 1 и линкер 2 представляют собой SEQ ID NO: 85.
2. Биспецифическое антитело по п. 1, где биспецифическое антитело представляет собой четырехвалентное симметричное биспецифическое антитело со структурой F(ab)2-(Fv)2-Fc, где F(ab)2 включает две VHa, две CH1, две VLa и две CL; (Fv)2 включает две VHb и две VLb, где положения VHb и VLb являются взаимозаменяемыми; и Fc-фрагмент включает шарнирную область, вторую константную область CH2 и третью константную область CH3.
3. Биспецифическое антитело по п. 1, где VHa-VLa пара образует одну или несколько межцепочечных дисульфидных связей и VHb-VLb пара образует одну или несколько межцепочечных дисульфидных связей.
4. Биспецифическое антитело по любому из пп. 1-3, где вариабельная область 1 антитела b представляет собой вариабельную область легкой цепи антитела b (VLb), а вариабельная область 2 антитела b представляет собой вариабельную область тяжелой цепи антитела b (VHb).
5. Биспецифическое антитело по любому из пп. 1-4, где линкеры 1-3 могут быть одинаковыми или отличными друг от друга.
6. Биспецифическое антитело по любому из пп. 1-5, где последовательность CL выбрана из группы, состоящей из SEQ ID NO: 104-110.
7. Биспецифическое антитело по любому из пп. 1-6, где последовательность CH1 выбрана из группы, состоящей из SEQ ID NO: 111-114.
8. Биспецифическое антитело по любому из пп. 1-7, где последовательность CH2 выбрана из группы, состоящей из SEQ ID NO: 115-119.
9. Биспецифическое антитело по любому из пп. 1-8, где последовательность CH3 выбрана из группы, состоящей из SEQ ID NO: 120-128.
10. Биспецифическое антитело по любому из пп. 1-9, где первый эпитоп и второй эпитоп выбраны, каждый независимо, из группы, состоящей из: антигенов поверхности иммунных клеток, опухолевых антигенов, вирусов, бактерий, эндотоксинов, цитокинов или их комбинации.
11. Биспецифическое антитело по любому из пп. 1-10, где первый эпитоп и/или второй эпитоп выбран из PD-L1, PD-1, VEGFA, IL-10, IL-10R, BCMA, VEGF, TGF-β, CTLA-4, LAG-3, TIGIT, CEA, CD38, SLAMF7, B7-H3, Her2, EpCAM, CD19, CD20, CD30, CD33, CD47, CD52, CD133, EGFR, GD2, GD3, GM2, RANKL, CD3 и/или CD16a.
12. Биспецифическое антитело по любому из пп. 1-11, где первый эпитоп и/или второй эпитоп выбран из SEQ ID NO: 129-145.
13. Биспецифическое антитело по любому из пп. 1-12, где первый эпитоп и второй эпитоп выбраны из группы, состоящей из:
PD-L1 и VEGF,
PD-1 и VEGF,
PD-L1 и TGF-β,
PD-1 и TGF-β,
PD-1 и CTLA-4,
PD-1 и LAG-3,
PD-1 и TIGIT,
PD-1 и IL-10,
SLAMF7 и CD16a, и
Her2 и Her2.
14. Биспецифическое антитело по любому из пп. 1-11, которое выбрано из группы, состоящей из:
(1) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 42; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 41, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(2) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 46; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 45, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(3) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(4) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(5) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 54; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 53, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(6) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(7) гибридная легкая цепь включает SEQ ID NO: 68, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 67, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(8) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(9) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(10) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(11) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 34; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 33, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(12) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 34; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 33, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(13) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 28; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 27, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(14) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 28; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 27, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(15) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 30; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 29, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(16) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 30; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 29, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(17) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 58; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 57, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(18) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(19) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 60; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 59, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(20) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 64; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 63, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(21) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 64; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 63, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(22) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 48; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 47, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(23) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 48; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 47, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(24) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 41; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 42, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(25) гибридная легкая цепь включает SEQ ID NO: 58, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 45; гибридная тяжелая цепь включает SEQ ID NO: 57, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 46, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(26) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(27) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(28) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 53; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 54, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(29) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(30) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 67; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 68, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(31) гибридная легкая цепь включает SEQ ID NO: 68, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 67, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(32) гибридная легкая цепь включает SEQ ID NO: 68, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 67, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(33) гибридная легкая цепь включает SEQ ID NO: 42, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 41, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(34) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(35) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 27; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 28, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(36) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 27; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 28, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(37) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 29; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 30, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(38) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 29; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 30, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(39) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 57; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 58, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(40) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(41) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 59; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 60, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(42) гибридная легкая цепь включает SEQ ID NO: 38, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 37, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(43) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(44) гибридная легкая цепь включает SEQ ID NO: 46, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 63; гибридная тяжелая цепь включает SEQ ID NO: 45, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 64, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(45) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 47; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 48, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(46) гибридная легкая цепь включает SEQ ID NO: 50, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 47; гибридная тяжелая цепь включает SEQ ID NO: 49, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 48, SEQ ID NO: 74, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(47) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 149; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 148, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(48) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 147; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 146, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(49) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 149; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 148, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(50) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 147; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 146, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120;
(51) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 148; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 149, SEQ ID NO: 87, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(52) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 146; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 147, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 117 и SEQ ID NO: 120;
(53) гибридная легкая цепь включает SEQ ID NO: 147, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 148; гибридная тяжелая цепь включает SEQ ID NO: 146, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 149, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120; или
(54) гибридная легкая цепь включает SEQ ID NO: 149, SEQ ID NO: 104, SEQ ID NO: 85 и SEQ ID NO: 146; гибридная тяжелая цепь включает SEQ ID NO: 148, SEQ ID NO: 111, SEQ ID NO: 85, SEQ ID NO: 147, SEQ ID NO: 82, SEQ ID NO: 91, SEQ ID NO: 115 и SEQ ID NO: 120.
15. Фармацевтическая композиция для лечения опухоли, связанной с первым эпитопом и вторым эпитопом, включающая биспецифическое антитело по любому из пп. 1-14.
16. Фармацевтическая композиция по п. 15, которая представляет собой лекарственную форму для желудочно-кишечного введения или лекарственную форму для парентерального введения.
17. Фармацевтическая композиция по п. 16, где лекарственная форма фармацевтической композиции представляет собой инъекцию, включая внутривенную инъекцию, внутривенное вливание, подкожную инъекцию, локальную инъекцию, внутримышечную инъекцию, интратуморальную инъекцию, интраперитонеальную инъекцию, интракраниальную инъекцию или внутриполостную инъекцию.
18. Конъюгат, связывающий первый эпитоп и второй эпитоп, включающий биспецифическое антитело по любому из пп. 1-14 и терапевтическое средство, пролекарство, белок, вирус, липид, модификатор биологического ответа, ПЭГ, гормон, олигонуклеотид, диагностическое средство или цитотоксическое средство.
19. Слитый белок, связывающий первый эпитоп и второй эпитоп, включающий биспецифическое антитело по любому из пп. 1-14 и терапевтическое средство, пролекарство, белок, модификатор биологического ответа, гормон, диагностическое средство или цитотоксическое средство.
20. Конъюгат или слитый белок по п.18 или 19, где белок представляет собой фермент и модификатор биологического ответа представляет собой иммуномодулятор.
21. Конъюгат или слитый белок по п. 18 или 19, где диагностическое средство выбрано из усиливающего ультразвук агента, нерадиоактивного маркера, детектируемого маркера.
22. Конъюгат или слитый белок по п. 21, где детектируемый маркер представляет собой хемилюминесцентное метящее соединение или излучающий флуоресценцию металл.
23. Конъюгат или слитый белок по п. 22, где хемилюминесцентное метящее соединение выбрано из люминола, изолюминола, термолюминесцентного эфира акридиния, имидазола, соли акридиния и оксалата, а излучающий флуоресценцию металл представляет собой 152Eu или лантанидную метку.
24. Конъюгат или слитый белок по п. 18 или 19, где цитотоксическое средство выбрано из лекарственного средства и токсина.
25. Конъюгат или слитый белок по п. 18 или 19, где белок представляет собой фермент и модификатор биологического ответа является иммуномодулятором.
26. Полинуклеотид, кодирующий биспецифическое антитело, связывающее два разных эпитопа, по любому из пп. 1-14.
27. Клетка, экспрессирующая биспецифическое антитело по любому из пп. 1-14, включающая полинуклеотид по п. 26.
28. Применение биспецифического антитела по любому из пп. 1-14 или конъюгата или слитого белка по любому из пп. 18-25 для лечения опухоли, связанной с первым эпитопом и вторым эпитопом.
29. Применение биспецифического антитела по любому из пп. 1-14 или конъюгата или слитого белка по любому из пп. 18-25 для получения реагента или набора для детекции опухоли, связанной с первым эпитопом и вторым эпитопом.
30. Применение по п. 28 или 29, где опухоль является раком.
| RU 2015144098 A, 21.04.2017 | |||
| Устройство для размывания льда | 1933 |
|
SU33766A1 |
| WO 1995009917 A1, 13.04.1995 | |||
| WO 2003025018 A2, 27.03.2003 | |||
| WO 2010028795 A1, 18.03.2010 | |||
| О.Н | |||
| Солопова, В.А | |||
| Мисюрин, Биспецифические антитела в клинике и клинических исследованиях (обзор литературы), Клиническая онкогематология | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Плуг с фрезерным барабаном для рыхления пласта | 1922 |
|
SU125A1 |

