Данная заявка представляет собой международную заявку согласно PCT, которая заявляет приоритет по предварительной заявке США с серийным №62/521287, поданной 16 июня 2017 г., которая включена в данный документ посредством ссылки в полном объеме.
Область техники
Данное раскрытие относится к пептидным иммуногенным конструкциям на основе C-терминального конца белка альфа-синуклеина (α-Syn) и составам на их основе для лечения синуклеинопатий.
Уровень техники
Синуклеиновые белки (рассматриваемые на веб-сайте: en.wikipedia.org/wiki/Synuclein) представляют собой семейство растворимых белков, характерных для позвоночных, которые экспрессируются главным образом в нервной ткани и в определенных опухолях. Семейство синуклеинов включает в себя три известных белка: альфа-синуклеин (рассматриваемый на веб-сайте: en.wikipedia.org/wiki/Alpha-synuclein), бета-синуклеин (веб-сайт: en.wikipedia.org/wiki/Beta-synuclein) и гамма-синуклеин. Для всех синуклеинов общим является наличие высококонсервативного альфа-спирального липид-связывающего мотива с аналогией с липид-связывающими доменами класса A2 взаимозаменяемых аполипопротеинов. Нормальные клеточные функции не были определены ни для какого из синуклеиновых белков, несмотря на то, что некоторые данные указывают на роль в регуляции стабильности и метаболизма мембран.
Полноразмерный белок альфа-синуклеин (α-Syn) представляет собой белок из 140 аминокислот (№доступа NP_000336) и кодируется геном SNCA. По меньшей мере три изоформы α-Syn образуются посредством альтернативного сплайсинга. Основной формой является полноразмерный белок. Другие изоформы представляют собой α-Syn-126, у которого отсутствуют остатки 41-54 в результате потери экзона 3; и α-Syn-112, у которого отсутствуют остатки 103-130 в результате потери экзона 5.
Первичная структура α-Syn обычно разделяется на три отдельных домена: (1) остатки 1-60: амфифильная N-терминальная область, в которой преобладают четыре повтора, состоящие из 11 остатков, содержащие консенсусную последовательность KTKEGV, которая имеет структурное свойство альфа-спирали, аналогичное связывающим доменам аполипопротеинов; (2) остатки 61-95: центральная гидрофобная область, которая содержит область неамилоидного β-компонента (NAC - non-amyloid-(component), которые участвуют в агрегации белка; и (3) остатки 96-140: сильнокислая и богатая пролином область, которая не имеет отдельного структурного свойства. Было обнаружено, что фрагмент α-Syn, состоящий из 35 аминокислот, области NAC, присутствует с A(в богатой амилоидом фракции. Позже было продемонстрировано, что NAC представляет собой фрагмент своего белка-предшественника, NACP (non-amyloid-(component protein), который, как было определено позже, представляет собой полноразмерный гомолог синуклеина от калифорнийского ската (Torpedo californica), обозначаемого в настоящее время как α-Syn человека.
Применение масс-спектрометрии ионной подвижности с высоким разрешением (IMS-MS - ion-mobility mass spectrometry) в отношении HPLC-очищенного α-Syn in vitro показало, что α-Syn является автопротеолитическим (самопротеолитическим), приводя к образованию ряда фрагментов с низкой молекулярной массой при инкубации. Было обнаружено, что полноразмерный белок с молекулярной массой 14,46 кДа приводит к образованию многочисленных меньших фрагментов, в том числе фрагмента с молекулярной массой 12,16 кДа (аминокислоты 14-133) и фрагмента с молекулярной массой 10,44 кДа (аминокислоты 40-140), образованных в результате C- и N-терминальных усечений, а также фрагмента с молекулярной массой 7,27 кДа (аминокислоты 72-140). Было продемонстрировано, что фрагмент с молекулярной массой 7,27 кДа, который содержит большую часть области NAC, агрегируется значительно быстрее, чем полноразмерный α-Syn. Возможно, что эти автопротеолитические продукты играют роль в качестве посредников или кофакторов в агрегации α-Syn.
α-Syn присутствует в большом количестве в головном мозге человека, составляя до 1% от всех белков в цитозоле клеток головного мозга и глиальных клеток. α-Syn в значительной степени экспрессируется в неокортексе, гиппокампе, зубчатой извилине, обонятельной луковице, полосатом теле, таламусе и мозжечке. Он также в значительной степени экспрессируется в гематопоэтических клетках, в том числе В-, T- и NK-клетках, а также моноцитах и тромбоцитах. Меньшие количества α-Syn встречаются в сердце, мышцах и других тканях. В головном мозге α-Syn встречается главным образом на кончиках нервных клеток (нейронов) в специализированных структурах, называемых пресинаптическими терминалями. В пределах этих структур α-Syn взаимодействует с фосфолипидами и белками. Пресинаптические терминали высвобождают химические посредники, называемые нейромедиаторами, такие как дофамин, из компартментов, известных как синаптические пузырьки. Высвобождение нейромедиаторов приводит к передаче сигналов между нейронами и является определяющим для нормальной функции головного мозга, в том числе когнитивной деятельности.
α-Syn в растворе считается внутренне неупорядоченным белком в том смысле, что у него отсутствует единственная стабильная 3D структура. Было продемонстрировано, что α-Syn в значительной степени взаимодействует с тубулином, а также что α-Syn может иметь активность в качестве потенциального белка, ассоциированного с микротрубочками, например, тау. α-Syn классически считался неструктурированным растворимым белком, немутированный α-Syn образует стабильно уложенный тетрамер, который является устойчивым к агрегации. Несмотря на это, α-Syn может агрегировать с образованием нерастворимых фибрилл в патологических условиях, характеризуемых тельцами Леви. Эти нарушения известны как синуклеинопатии (описаны на веб-сайте: en.wikipedia.org/wiki/Synucleinopathies).
Синуклеинопатии представляют собой разнообразную группу нейродегенеративных нарушений, которые имеют общую патологическую характеристику: при нейропатологических исследованиях характерные поражения, содержащие аномальные агрегаты нерастворимого α-Syn, присутствуют в выборочно восприимчивых популяциях нейронов и глиальных клеток. Наиболее распространенные синуклеинопатии включают в себя нарушения телец Леви (LBD - Lewy body disorders), например, болезнь Паркинсона (PD - Parkinson's disease), болезнь Паркинсона с деменцией (PDD - Parkinson's disease with dementia) и деменцию с тельцами Леви (DLB - dementia with Lewy bodies), а также множественную системную атрофию (MSA - Multiple System Atrophy) или нейродегенерацию с накоплением железа в головном мозге I типа (NBIA (Neurodegeneration with Brain Iron Accumulation) I типа). Существующие в настоящее время виды лечения этих заболеваний включают в себя симптоматические лекарственные препараты, такие как леводопа, антихолинергические лекарственные препараты, а также ингибиторы моноаминоксидазы. В то же время все существующие в настоящее время возможности лечения приводят только к симптоматической нормализации, но не индуцируют длительного модифицирующего эффекта на заболевание у пациентов.
LBD представляют собой прогрессирующие нейродегенеративные нарушения, характеризующиеся тремором, ригидностью, брадикинезией и потерей дофаминергических нейронов в головном мозге. В случае DLB и PDD симптомы также включают в себя когнитивное нарушение. У до 2% населения в возрасте старше 60 лет в западных странах развиваются типичные симптомы PD/LBD. По-видимому, генетическая предрасположенность и средовые факторы участвуют в развитии указанного заболевания. У пациентов, страдающих от указанного заболевания, развиваются характерные внутриклеточные включения, называемые тельцами Леви (LB - Lewy bodies), в корковых и подкорковых областях головного мозга, особенно в случае участков с высоким содержанием дофаминергических нейронов или отростков нейронов. При LBD α-Syn накапливается в LB по всем пораженным областям головного мозга. Кроме того, можно было продемонстрировать, что точечные мутации, а также дупликации или мультипликации в гене α-Syn ассоциированы с редкими семейными формами паркинсонизма.
Множественная системная атрофия (MSA - Multiple System Atrophy) представляет собой спорадическое нейродегенеративное нарушением, которое характеризуется симптомами устойчивого к леводопе паркинсонизма, мозжечковой атаксии и дисавтономии. Пациенты, страдающие от мультисистемной потери нейронов, будут поражены в различных областях головного мозга, в том числе полосатом теле, черной субстанции, мозжечке, варолиевом мосте, а также нижней оливе и спинном мозге. MSA характеризуется α-Syn-положительными глиальными цитоплазматическими (GCI - glial cytoplasmic) и редкими нейрональными включениями по всей центральной нервной системе.
Другие нервные нарушения, такие как различные нейроаксональные дистрофии, также имеют патологии α-Syn, в которых α-Syn представляет собой основной структурный компонент фибрилл телец Леви. Иногда тельца Леви содержат тау-белок; однако, α-Syn и тау представляют собой две различные подгруппы филаментов в одних и тех же тельцах включения. Патология α-Syn также встречается как при спорадических, так и при семейных случаях болезни Альцгеймера.
Механизм агрегации α-Syn является неясным. Мономерный α-Syn в природе является несвернутым в растворе, однако также может связываться с мембранами в α-спиральной форме. Несвернутый мономер может агрегировать сначала в небольшие олигомерные молекулы, которые можно стабилизировать с помощью взаимодействий, подобных β-складкам, и затем в нерастворимые молекулы с более высокой молекулярной массой. α-Syn существует в виде смеси неструктурированных конформеров с высоким содержанием альфа-спиралей и бета-складок в равновесии. Мутации или буферные условия, которые, как известно, нормализуют агрегацию, приводят к значительному повышению популяции бета-конформера, таким образом, указывая на то, что это может представлять собой конформацию, связанную с патогенной агрегацией. Имеются данные, указывающие на наличие структурированного посредника с высоким содержанием бета-структуры, который может представлять собой предшественника агрегации и в конечном итоге телец Леви.
Несколько физиологических факторов могут приводить к модификации α-Syn, приводя к образованию агрегатов, в том числе (1) фосфорилирование с помощью одной или нескольких киназ, (2) усечение с помощью протеазы, такой как кальпаины; и (3) нитрование с помощью оксида азота (NO) или других реактивных форм азота, которые присутствуют во время воспаления. Транспорт в ЭПС и аппарате Гольджи, синаптические пузырьки, митохондрии, лизосомы и другие протеолитический аппарат представляют собой некоторые из предложенных клеточных мишеней для токсичности, опосредованной α-Syn, в результате такой агрегации.
Среди стратегий лечения синуклеинопатий имеют место соединения, которые ингибируют агрегацию α-Syn. Было продемонстрировано, что малая молекула куминальдегид приводит к ингибированию фибриллирования α-Syn. Кроме терапии малыми молекулами, недавний отчет указывает на то, что на агрегаты α-Syn можно целенаправленно воздействовать иммунотерапией (описано Lee JS and Lee S-J, 2016). В то же время, указанный отчет указывает на некоторые потенциальные вопросы или проблемы, которые существуют при разработке иммунотерапии α-Syn, в том числе (1) потенциальное нежелательное воздействие на нормальную физиологическую функцию α-Syn; (2) сложности доставки лекарственного препарата, представляющего собой антитело, в паренхиму головного мозга; и (3) эффективность иммунотерапии.
На сегодняшний день еще имеет место неудовлетворенная потребность в разработке сайт-направленных пептидных иммуногенов и составов на их основе для малозатратного лечения пациентов, страдающих синуклеинопатиями.
Литературные источники:
1. "Alpha-synuclein," Wikipedia, The Free Encyclopedia, адрес веб-сайта: en.wikipedia.org/w/index.php?title=Alpha-synuclein&oldid=781366541 (доступно 30 мая 2017 г.).
2. "Synucleinopathies," Wikipedia, The Free Encyclopedia, адрес веб-сайта: en.wikipedia.org/w/index.php?title=Synucleinopathies&oldid=686287116 (доступно 30 мая 2017 г.).
3. "Beta-synuclein," Wikipedia, The Free Encyclopedia, адрес веб-сайта: en.wikipedia.org/w/index.php?title=Beta-synuclein&oldid=763171134 (доступно 30 мая 2017 г.).
4. "Synucleinopathies," Wikipedia, The Free Encyclopedia, адрес веб-сайта: en.wikipedia.org/w/index.php?title=Synucleinopathies&oldid=686287116 (доступно 30 мая 2017 г.).
5. LEE, J.S., et al., “Mechanism of Anti-α-Synuclein Immunotherapy”, J Mov Disord.; 9(1):14-19 (2016)
6. TRAGGIAI, E., et al. “An efficient method to make human monoclonal antibodies from memory B cells: potent neutralization of SARS coronavirus”, Nat Med.; 10(8):871-875 (2004)
7. WANG, C., et al. “Versatile Structures of α-Synuclein”, Front Mol Neurosci. 9:48 (2016)
Краткое описание СУЩНОСТИ изобретения
Данное раскрытие связано с пептидными иммуногенными конструкциями белка альфа-синуклеина (α-Syn). Данное раскрытие также связано с композициями, содержащими пептидные иммуногенные конструкции, способами получения и применения указанных пептидных иммуногенных конструкций и антителами, образуемыми указанными пептидными иммуногенными конструкциями.
Раскрытые пептидные иммуногенные конструкции содержат B-клеточный эпитоп из α-Syn, связанный с гетерологичным эпитопом T-хелперных клеток (Th) непосредственно или с помощью необязательного гетерологичного спейсера. Участок B-клеточного эпитопа пептидных иммуногенных конструкций содержит от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального участка α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn (SEQ ID NO: 1). Участок гетерологичного Th-эпитопа пептидных иммуногенных конструкций происходит из аминокислотных последовательностей, полученных из патогенных белков. Участки B-клеточного эпитопа и Th-эпитопа пептидных иммуногенных конструкций функционируют совместно при введении хозяину с целью стимуляции образования антител, которые специфически распознают и связываются с участком В-клеточного эпитопа α-Syn конструкций.
В некоторых вариантах осуществления пептидная иммуногенная конструкция α-Syn содержит: (a) B-клеточный эпитоп, содержащий от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального фрагмента α-Syn, соответствующего от приблизительно аминокислоты G111 до приблизительно аминокислоты D135 SEQ ID NO: 1; (b) T-хелперный эпитоп, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 70-98; и (c) необязательный гетерологичный спейсер, выбранный из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys и ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148), где B-клеточный эпитоп ковалентно связан с T-хелперным эпитопом непосредственно или с помощью необязательного гетерологичного спейсера. В конкретных вариантах осуществления пептидная иммуногенная конструкция α-Syn содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 107, 108, 111-113 и 115-147.
Данное раскрытие также связано с композициями, содержащими раскрытые пептидные иммуногенные конструкции, в том числе фармацевтические композиции. Раскрытые фармацевтические композиции способны вызывать иммунный ответ и продуцирование антител против раскрытых пептидных иммуногенных конструкций в организме хозяина. Раскрытые композиции могут содержать одну или смесь из более чем одной из раскрытых пептидных иммуногенных композиций. В некоторых вариантах осуществления композиции содержат раскрытые пептидные иммуногенные конструкции совместно с дополнительными компонентами, в том числе носителями, адъювантами, буферами и другими подходящими реагентами. В определенных вариантах осуществления композиции содержат раскрытые пептидные иммуногенные конструкции стабилизированного иммуностимулирующего комплекса с олигомером CpG, который необязательно дополняют адъювантом.
В некоторых вариантах осуществления композиции содержат пептидную иммуногенную конструкцию α-Syn, которая содержит аминокислотную последовательность, состоящую из SEQ ID NO: 107, 108, 111-113, 115-147. В определенных вариантах осуществления композиция представляет собой фармацевтическую композицию, содержащую пептидную иммуногенную конструкцию α-Syn, выбранную из группы, состоящей из SEQ ID NO: 107, 108, 111-113, 115-147, и фармацевтически приемлемого носителя или адъюванта.
Данное раскрытие также связано с антителами, которые продуцируются хозяином, которого иммунизируют раскрытыми пептидными иммуногенными конструкциями. Раскрытые антитела специфически распознают и связываются с участком В-клеточного эпитопа α-Syn пептидных иммуногенных конструкций. Раскрытые антитела к α-Syn имеют неожиданным образом высокую перекрестную реактивность по отношению к β-складке α-Syn в форме мономеров, олигомеров или фибрилл. Исходя из своих уникальных характеристик и свойств раскрытые антитела способны обеспечивать иммунотерапевтический подход к целенаправленному воздействию, идентификации и лечению синуклеинопатий.
В конкретных вариантах осуществления антитело или его эпитопсвязывающий фрагмент специфически связываются с B-клеточным эпитопом пептидной иммуногенной конструкции α-Syn, выбранной из группы, состоящей из SEQ ID NO: 107, 108, 111-113, 115-147.
Данное раскрытие также связано со способами получения и применения раскрытых пептидных иммуногенных конструкций, антител и композиций. Раскрытые способы предлагают малозатратное производство и контроль качества пептидных иммуногенных конструкций и композиций, содержащих указанные конструкции, которые могут быть использованы в способах предупреждения и лечения синопатий.
Данное раскрытие также включает в себя способы лечения и/или предупреждения синуклеинопатий с помощью пептидных иммуногенных конструкций и/или антител, направленных против пептидных иммуногенных конструкций. В некоторых вариантах осуществления способы лечения и/или предупреждения синуклеинопатий включают в себя введение хозяину раскрытой пептидной иммуногенной конструкции. В определенных вариантах осуществления композиции, используемые в способах, содержат раскрытую пептидную иммуногенную конструкцию в форме стабильного иммуностимулирующего комплекса отрицательно заряженными олигонуклеотидами, такими как олигомеры CpG, в результате электростатической ассоциации, указанные комплексы затем дополняются, необязательно, минеральными солями или маслом в качестве адъюванта, для введения пациентам с синуклеинопатиями. Раскрытые способы также включают в себя режимы дозирования, лекарственные формы и пути введения пептидных иммуногенных конструкций хозяину, имеющему риск развития или имеющему синуклеинопатии.
В различных вариантах осуществления описаны способы применения пептидной иммуногенной конструкции α-Syn и/или антител, образование которых вызывается пептидной иммуногенной конструкцией α-Syn. В конкретных вариантах осуществления описаны способы получения антител, ингибирования агрегации α-Syn, снижения количества агрегатов α-Syn и идентификации агрегатов α-Syn различных размеров. Различные способы включают в себя введение фармакологически эффективного количества пептидного иммуногена α-Syn хозяину, нуждающемуся в этом.
Краткое описание графических материалов
На Фиг. 1 продемонстрирован график, на котором представлен уровень агрегации α-Syn in vitro через 6 дней в присутствии антител, направленных против C-терминального конца α-Syn (образцы 1-4) или в присутствии контроля-носителя (образец 5). В частности, агрегацию α-Syn выполняли в присутствии анти-α-Syn антител, образование которых вызывается: α-Syn111-132 (образец 1); α-Syn121-135 (образец 2); α-Syn123-135 (образец 3); α-Syn126-135 (образец 4); или контролем-носителем (образец 5). Уровень агрегации α-Syn измеряли с помощью окрашивания агрегатов тиофлавином-T (ThT). Образцы 1-4 нормализовали по отношению к контролю-носителю образца 5. Планки погрешностей представляют SEM (стандартную ошибку среднего) каждого исследования в двух повторах.
На Фиг. 2 продемонстрирован график, на котором представлен уровень диссоциации предварительно образованных агрегатов α-Syn in vitro после инкубации агрегатов в течение 3 дней в присутствии антител, направленных против C-терминального конца α-Syn (образцы 1-3), или преимунного контроля-сыворотки (образец 4). В частности, предварительно образованные агрегаты α-Syn инкубировали с анти-α-Syn антителами, образование которых вызывается: α-Syn111-132 (образец 1); α-Syn126-135 (образец 2); комбинацией антител, образование которых вызывается α-Syn111-132 и α-Syn126-135 (образец 3); или преиммунным контролем-сывороткой (образец 4). Уровень агрегации α-Syn измеряли с помощью окрашивания агрегатов тиофлавином-T (ThT). Образцы 1-3 нормализовали по отношению к преимунному контролю-сыворотке образца 4. Планки погрешностей представляют SEM (стандартную ошибку среднего) каждого исследования в двух повторах.
На Фиг. 3 продемонстрирован график, на котором представлены уровни агрегации α-Syn и дезагреации α-Syn в клетках РС12, сверхэкспрессирующих α-Syn, инкубированных с фактором роста нервов (NGF) в присутствии антител, направленных против C-терминального конца α-Syn (образцы 1-4) или контроля-носителя (образец 5). В частности, клетки РС12 инкубировали с анти-α-Syn антителами, образование которых вызывается: α-Syn111-132 (образец 1); α-Syn121-135 (образец 2); α-Syn123-135 (образец 3); α-Syn126-135 (образец 4); или контролем-носителем (образец 5). Образцы 1-4 нормализовали по отношению к контролю-носителю образца 5. Планки погрешностей представляют SD (стандартное отклонение) каждого исследования в трех повторах.
На Фиг. 4 продемонстрирован график, на котором представлены уровни высвобождения TNF-(и IL-6, опосредованного агрегатами α-Syn из клеток, инкубированных в присутствии антител, направленных против C-терминального конца α-Syn (образцы 1-4) или контроля-носителя (образец 5). В частности, микроглиальные клетки инкубировали с анти-α-Syn антителами, образование которых вызывается: α-Syn111-132 (образец 1); α-Syn121-135 (образец 2); α-Syn123-135 (образец 3); α-Syn126-135 (образец 4); или контролем-носителем (образец 5). Образцы 1-4 нормализовали по отношению к контролю-носителю образца 5. Планки погрешностей представляют SD (стандартное отклонение) каждого исследования в трех повторах.
На Фиг. 5A-5C продемонстрированы графики, которые иллюстрируют влияние анти-α-Syn антител в модели нейродегенерации in vitro с использованием экзогенных предварительно образованных агрегатов α-Syn в индуцированных NGF клетках РС12, дифференцируемых в нейроны. На Фиг. 5A продемонстрирована оценка длины нейритов клеток PC12, обработанных только NGF (темная сплошная линия); NGF с экзогенными предварительно образованными агрегатами α-Syn (точечная линия); NGF с преиммунными сыворотками (светлая сплошная линия); и NGF, как с экзогенными предварительно образованными агрегатами α-Syn, так и преиммунными сыворотками (пунктирная линия). На Фиг. 5B продемонстрирована оценка длины нейритов клеток PC12, обработанных NGF наряду с носителем (темная сплошная линия); NGF с экзогенными предварительно образованными агрегатами α-Syn (точечная линия); NGF с анти-α-Syn антителами, образование которых вызывается α-Syn111-132 (SEQ ID NO:113) (светлая сплошная линия); и NGF, как с экзогенными предварительно образованными агрегатами α-Syn, так и анти-α-Syn антителами, образование которых вызывается α-Syn111-132 (SEQ ID NO:113) (пунктирная линия). На Фиг. 5С продемонстрирована оценка длины нейритов клеток PC12, обработанных только NGF с носителем (темная сплошная линия); NGF с экзогенными предварительно образованными агрегатами α-Syn (точечная линия); NGF с анти-α-Syn антителами, образование которых вызывается α-Syn126-135 (SEQ ID NO:112) (светлая сплошная линия); и NGF, как с экзогенными предварительно образованными агрегатами α-Syn, так и анти-α-Syn антителами, образование которых вызывается α-Syn126-135 (SEQ ID NO:112) (пунктирная линия).
На Фиг. 6A-6B продемонстрированы графики, которые иллюстрируют влияние анти-α-Syn антител на число клеток и длину нейритов в модели нейродегенерации in vitro с использованием индуцированных NGF клеток РС12 дикого типа, сверхэкспрессирующих α-Syn, дифференцируемых в нейроны. Клетки обрабатывали контролем-носителем (образец 1); анти-α-Syn антителами, образование которых вызывается α-Syn101-132 (образец 2), α-Syn111-132 (образец 3), α-Syn121-135 (образец 4), α-Syn123-135 (образец 5), α-Syn126-135 (образец 6), комбинацией анти-α-Syn антител, образование которых вызывается α-Syn111-132 и α-Syn126-135 (образец 7); или преимунным контролем-сывороткой (образец 8). На Фиг. 6A продемонстрирована оценка соответствующих защитных эффектов каждого образца в отношении восстановления количества клеток PC12. На Фиг. 6B продемонстрирована оценка длины нейритов клеток, обработанных каждым образцом. Образцы 1-8 нормализовали по отношению к индуцированным NGF клеткам дикого типа РС12, дифференцируемых в нейроны. t-Критерий использовали для исследования значимости (p-значение менее чем 0,05 определяли как статически значимое и обозначали звездочкой (*)).
На Фиг. 7A-7B продемонстрирована способность анти-α-Syn антител распознавать и связываться с агрегатами α-Syn различных размеров с помощью вестерн-блоттинга. На Фиг. 7A продемонстрировано изображение вестерн-блота, на котором сравнивают коммерчески доступное анти-α-Syn антитело, Syn211 (полоса 1); преиммунный контроль-сыворотка (полоса 2); анти-α-Syn антитело, вызванное Syn111-132 (полоса 3); анти-α-Syn антитело, вызванное Syn111-135 (полоса 4); анти-α-Syn антитело, вызванное Syn121-135 (полоса 5); анти-α-Syn антитело, образование которого вызывается Syn123-135 (полоса 6); и анти-α-Syn антитело, образование которого вызывается α-Syn126-135 (полоса 7). На Фиг. 7B продемонстрирована столбиковая диаграмма, на которой представлена относительная способность каждого антитела связываться с молекулярными комплексами α-Syn различных размеров (в том числе мономерами, димерами, тримерами, тетрамерами и олигомерами). Хемилюминесцентные сигналы полос вестерн-блоттинга, показанные на Фиг. 7A, оценивали количественно и обозначали на столбиковой диаграмме Фиг. 7B.
На Фиг. 8A-8C продемонстрированы дот-блоттинговые изображения, которые иллюстрируют, что антитела, направленные только против C-терминального конца α-Syn, распознают и связываются с различными молекулами α-Syn (т.е., α-спиральными мономерами, β-складчатыми мономерами, β-складчатыми олигомерами и β-складчатыми фибриллами), а не с такими же самыми молекулами других амилоидогенных белков (т.е., Aβ1-42 и Tau441). На Фиг. 8A продемонстрирован контрольный образец, демонстрирующий, что антитела, очищенные от преиммунной сыворотки от морских свинок, не проявляли детектируемого уровня по отношению к какому-либо из всех анализируемых белковых молекул. На Фиг. 8B продемонстрирована оценка способности анти-α-Syn антитела, вызванного α-Syn111-132 (SEQ ID NO:113), распознавать и связываться с различными молекулами α-Syn, белками Aβ1-42 и Tau441. На Фиг. 8С продемонстрирована оценка способности анти-α-Syn антитела, вызванного α-Syn126-135 (SEQ ID NO:112), распознавать и связываться с различными молекулами α-Syn, белками Aβ1-42 и Tau441.
На Фиг. 9 продемонстрирована таблица, в которой представлены сравнительные аффинности связывания антител, направленных против C-терминального конца α-Syn, по отношению к внутриклеточному α-Syn в различных линиях клеток PC12, измеряемые с помощью положительных сигналов в иммуногистохимическом (ИЦХ) исследовании. В частнсоти, сравнительные аффинности связывания анти-α-Syn антител, вызванных α-Syn111-132, α-Syn121-135, α-Syn126-135 или контрольным образцом преимунной сыворотки, оценивали в исходных клетках PC12, клетках PC12 пустого контроля, клетках РС12 дикого типа, сверхэкспрессирующих α-Syn, и клетках РС12, сверхэкспрессирующих α-Syn с мутацией A53T при обработке NGF.
На Фиг. 10A-10C продемонстрировано, что антитела, направленные только против C-терминального конца α-Syn, связываются с α-Syn в срезах головного мозга при PD, но не в срезах здорового головного мозга. На Фиг. 10A продемонстрировано, что антитела к α-Syn, вызванные пептидными иммуногенными конструкциями α-Syn, и преиммунные антитела не демонстрировали детектируемой иммунореактивности на панели нормальных тканей человека, в том числе срезах головного мозга. На Фиг. 10B продемонстрирована иммунореактивность антител, направленных против агрегатов α-Syn в срезах таламуса при PD, указанная с помощью стрелки. На Фиг. 10C продемонстрирована таблица, описывающая иммунореактивность антител, направленных против C-терминального конца α-Syn, и преиммунного контроля-сыворотки по отношению к агрегатам α-Syn в срезах головного мозга при PD, а также в срезах здорового головного, определенную с помощью подсчета положительных окрашивания при микроскопическом наблюдении.
На Фиг. 11A-11B продемонстрированы графики, представляющие уровень анти-α-Syn IgG в сыворотке крови в мышиных моделях PD после трех иммунизаций только адъювантом (незаштрихованный кружок) или пептидными иммуногенами, содержащими α-Syn111-132 (незаштрихованный прямоугольник); α-Syn126-135 (заштрихованный кружок); или комбинацией α-Syn111-132 и α-Syn126-135 (заштрихованный прямоугольник). На Фиг. 11A продемонстрированы уровни IgG в мышиной модели с индукцией MPP+. На Фиг. 11B продемонстрированы уровни IgG в мышиной модели с инокуляцией фибриллярного α-Syn.
На Фиг. 12A-12B продемонстрированы графики, представляющие уровень α-Syn в периферическом кровообращении в мышиных моделях PD после трех иммунизаций только адъювантом (незаштрихованный кружок) или пептидными иммуногенами, содержащими α-Syn111-132 (незаштрихованный прямоугольник); α-Syn126-135 (заштрихованный кружок); или комбинацией α-Syn111-132 и α-Syn126-135 (заштрихованный прямоугольник). На Фиг. 12A продемонстрированы уровни α-Syn в мышиной модели с индукцией MPP+. На Фиг. 12B продемонстрированы уровни α-Syn в мышиной модели с инокуляцией фибриллярного α-Syn.
На Фиг. 13A-13B продемонстрирован уровень олигомерного α-Syn в образцах головного мозга в модели необработанных здоровых мышей (полоса 1) или мышиных моделях при PD (полосы 2-3), получавших три иммунизации либо только с адъювантом (полоса 2), либо пептидными иммуногенами, содержащими α-Syn111-132 (полоса 3). Необработанные мыши Balb/c представляют собой модель здоровых мышей, в том время как мыши, индуцированные MPP+, представляют собой модели мышей с PD. На Фиг. 13A продемонстрирован вестерн-блот, демонстрирующий уровень олигомерного α-Syn, а также GAPDH в качестве контроля нагрузки белка в образцах. На Фиг. 13B продемонстрирован график, на котором сравнивают относительные уровни олигомерного α-Syn, показанные в вестерн-блоте Фиг. 13A, после того, как уровни белка нормализовали по отношению к уровню GAPDH, и отношение лизата в модели необработанных здоровых мышей дополнительно стандартизовали до уровня 1,00 для сравнения.
На Фиг. 14A-14G продемонстрирован уровень олигомерного α-Syn и тирозингидроксилазы в образцах головного мозга в модели необработанных здоровых мышей (полоса 1) или мышиных моделях при PD (полосы 2-4), получавших три иммунизации либо только с адъювантом (полоса 2), либо пептидными иммуногенами, содержащими α-Syn111-132 (полоса 3); или α-Syn126-135 (полоса 4). Необработанные мыши FVB представляют собой модель здоровых мышей, в том время как мыши, инокулированные фибриллярным α-Syn, представляют собой модели мышей с PD. На Фиг. 14A продемонстрирован вестерн-блот, демонстрирующий уровень олигомерного α-Syn и тирозингидроксилазы, а также GAPDH в качестве контроля нагрузки белка в лизатах черной субстанции ипсилатеральной стороны. На Фиг. 14B продемонстрирован график, на котором сравнивают относительные уровни олигомерного α-Syn, показанные в вестерн-блоте на Фиг. 14A, после того, как уровни белка нормализовали по отношению к уровню GAPDH. На Фиг. 14С продемонстрирован график, на котором сравнивают относительные уровни белка тирозингидроксилазы, показанные в вестерн-блоте на Фиг. 14A, после того, как уровни белка нормализовали по отношению к уровню GAPDH. На Фиг. 14D продемонстрирован вестерн-блот, демонстрирующий уровень олигомерного α-Syn, а также GAPDH в качестве контроля нагрузки белка в лизатах полосатого тела ипсилатеральной стороны. На Фиг. 14Е продемонстрирован график, на котором сравнивают относительные уровни олигомерного α-Syn, показанные в вестерн-блоте на Фиг. 14С, после того, как уровни белка нормализовали по отношению к уровню GAPDH. На Фиг. 14F продемонстрирован вестерн-блот, демонстрирующий уровень олигомерного α-Syn, а также GAPDH в качестве контроля нагрузки белка в лизатах полосатого тела контралатеральной стороны. На Фиг. 14G продемонстрирован график, на котором сравнивают относительные уровни олигомерного α-Syn, показанные в вестерн-блоте на Фиг. 14E, после того, как уровни белка нормализовали по отношению к уровню GAPDH.
На Фиг. 15A-15C продемонстрированы графики, на которых представлена оценка двигательной функции у мышей, измеряемая с помощью CatWalk™ XT в моделях здоровых мышей (полосы 1-2), обработанных солевым раствором (полоса 1) или только адъювантом (полоса 2); или моделях мышей при PD (полосы 3-5), иммунизированных либо только адъювантом (полоса 3), либо пептидными иммуногенами, содержащими α-Syn126-135 (полоса 4) или α-Syn111-132 (полоса 5). t-Критерий использовали для исследования значимости (p-значение менее чем 0,05 определяли как статически значимое и обозначали звездочкой «*»). На Фиг. 15A продемонстрирована оценка стойки(стоек) левой задней конечности у обработанных мышей, при этом необработанные мыши FVB представляют собой модель здоровых мышей, а мыши, инокулированные фибриллярным α-Syn, представляют собой модели мышей при PD. На Фиг. 15В продемонстрирована оценка продолжительности бега у обработанных мышей, при этом необработанные мыши FVB представляют собой модель здоровых мышей, а мыши, инокулированные фибриллярным α-Syn, представляют собой модели мышей при PD. На Фиг. 15С продемонстрирована оценка продолжительности бега у обработанных мышей, при этом необработанные мыши Balb/c представляют собой модель здоровых мышей, в то время как мыши, индуцированные МРР+, представляют собой модели мышей при PD.
Фиг. 16A-16H. На Фиг. 16A продемонстрировано, что PD-021514 (α-Syn85-140, нед. после иммуниз. 08) распознает фибриллы штамма α-Syn с максимальной аффинностью. Наблюдается высокое связывание с лентами и фибриллами-91 указанного штамма. Имеет место слабое связывание с олигомерами и фибриллами-65. Имеет место слабое связывание с мономером α-Syn и с фибриллами, не имеющими C-терминальные 30 аминокислотных остатков (Fib-110). На Фиг. 16B продемонстрировано, что PD-021522 (α-Syn85-140, нед. после иммуниз. 13) связывается со всеми штаммами/олигомерами, но не с мономерами, четко не наблюдается зависимое от концентрации повышение сигнала. Антитело связывается с фибриллами, не имеющими C-терминальные 30 аминокислотных остатков (Fib-110). Таким образом, эпитоп находится не в этом участке. На Фиг. 16C продемонстрировано, что PD-100806 (α-Syn126-135, нед. после иммуниз. 09) связывается со всеми штаммами, с максимальной аффинностью в отношении лент.Оно связывает нативный олигомерный α-Syn с более низкой эффективностью. Почти не наблюдалось связывания с глутаральдегидом, дофаминовыми перекрестно сшитыми олигомерами и мономерным a-syn. Антитело вероятно направлено против С-терминальных аминокислотных остатков a-syn 30, поскольку оно не связывает фибриллы, не имеющие C-терминальные 30 аминокислотных остатков (Fib-110). На Фиг. 16D продемонстрировано, что коммерческое антитело Syn1 (клон 42, BD bioscience) связывается со всеми штаммами α-Syn и с олигомерами, за исключением глутаральдегидовых перекрестных сшиваний. Оно также свуязывается с мономерным α-Syn. Описано, что его эпитоп находится на протяжении остатков 91-96/99. В соответствии с этим, оно связывает фибриллы, не имеющими C-терминальные 30 аминокислотных остатков (Fib-110). На Фиг. 16E продемонстрировано, что PRX002 распознает с немного более высокой аффинностью фибриллярный α-Syn по сравнению с мономерным α-Syn. На Фиг. 16F продемонстрирован контроль для фона антитела, образованных у морских свинок. На Фиг. 16G продемонстрирован контроль для фона антитела к Syn1. На Фиг. 16H продемонстрирован контроль для фона антитела к PRX002.
На Фиг. 17A-17D продемонстрирован ИГХ анализ специфичности антител UNS в отношении α-Syn в базальных ганглиях пациентов с деменцией с тельцами Леви (DLB). Средний процент площади агрегатов α-Syn, окрашенных каждым антителом (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в бледном шаре (Фиг. 17A), внутренней капсуле (Фиг. 17B) и островковой коре (Фиг. 17C). Иллюстративные микроскопические изображения, полученные в результате иммунологического окрашивания в бледном шаре с помощью каждого антитела, продемонстрированы на Фиг. 17D. Антитела UNS приводили к выявлению более высокого процента площади агрегатов α-Syn в бледном шаре (F(3,7)=1,550, p=0,284 с помощью ANOVA), внутренней капсуле (F(3,7)=1,356, p=0,332 с помощью ANOVA) и островковой коре (F(3,8)=2,050, p=0,195 с помощью ANOVA). P<0,05 (*); P<0,01 (**); P<0,001 (***). Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 18A-18D продемонстрирован ИГХ анализ специфичности антител UNS в отношении α-Syn в базальных ганглиях пациентов с деменцией с болезнью Паркинсона (PD). Средний процент площади агрегатов α-Syn, окрашенных каждым антителом (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в бледном шаре (Фиг. 18A), внутренней капсуле (Фиг. 18B) и островковой коре (Фиг. 18C) исходя из трех случаев PD. Иллюстративные микроскопические изображения, полученные в результате иммунологического окрашивания, продемонстрированы на Фиг. 18D для бледного шара. Антитела UNS приводили к выявлению более высокого процента площади агрегатов α-Syn в бледном шаре (F(3,18)=4,152, p=0,047 с помощью ANOVA), внутренней капсуле (F(3,8)=1,995, p=0,1934 с помощью ANOVA) и островковой коре (F(3,8)=0,4044, p=0,754 с помощью ANOVA). Значительно более высокий процент площади α-Syn выявляли в случае PD100806 по сравнению с NCL-L-ASYN (p=0,023 в случае PD100806 по сравнению с NCL-L-ASYN; n=3). P<0,05 (*); P<0,01 (**); P<0,001 (***). После однофакторного ANOVA использовали критерий Даннета. Данные продемонстрированы в виде среднего+SD (планки погрешностей).
Фиг. 19A-19C: ИГХ анализ специфичности антител UNS в отношении α-Syn в базальных ганглиях пациентов с множественной системной атрофией (MSA). Средний процент площади агрегатов α-Syn, окрашенных каждым антителом (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в бледном шаре (Фиг. 19A) и внутренней капсуле (Фиг. 19B) в трех случаях MSA. В островковой коре пациентов с MSA патологии не выявляли и, таким образом, ее количественно не оценивали. Антитела UNS приводили к выявлению более высокого процента площади агрегатов α-Syn в бледном шаре (F(3,8)=1,56, p=0,273 с помощью ANOVA) и внутренней капсуле (F(3,8)=1,126, p=0,395 с помощью ANOVA). Иллюстративные микроскопические изображения, полученные в результате иммунологического окрашивания каждым антителом, продемонстрированы на Фиг. 19C в случае бледного шара. P<0,05 (*); P<0,01 (**); P<0,001 (***). Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 20A-20E продемонстрирован ИГХ анализ специфичности антител UNS в отношении α-Syn в среднем мозге ганглиях пациентов с различными синуклеинопатиями. Средний процент площади агрегатов α-Syn, окрашенных каждым антителом (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в черной субстанции пациентов с PD (Фиг. 20A), DLB (Фиг. 20B) и MSA (Фиг. 20C). Процент площади, окрашенной каждым антителом, сравнивали с диагностическим антителом, NCL-L-ASYN. Антитела UNS приводили к выявлению более высокого процента площади агрегатов α-Syn в черной субстанции пациентов с MSA (F(3,8)=0,830, p=0,51 с помощью ANOVA); DLB (F(3,7)=2,493, p=0,144 с помощью ANOVA) и PD (F(3,7)=0,189, p=0,900 с помощью ANOVA). Иллюстративные микроскопические изображения, полученные в результате иммунологического окрашивания каждым антителом, продемонстрированы на Фиг. 20D (MSA) и Фиг. 20E (DLB). P<0,05 (*); P<0,01 (**); P<0,001 (***). Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 21A-21F продемонстрирован ИГХ анализ специфичности антител UNS в отношении α-Syn в белом и сером веществе височной коры пациентов с различными синуклеинопатиями. Средний процент площади агрегатов α-Syn, окрашенных каждым антителом (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в сером веществе коры головного мозга и белом веществе подкорки головного мозга пациентов с PD (Фиг. 21A и 21D), DLB (Фиг. 21В и 21Е) и MSA (Фиг. 21C и 21F). Процент площади, окрашенной каждым антителом, сравнивали с диагностическим антителом, NCL-L-ASYN. P<0,05 (*); P<0,01 (**); P<0,001 (***). После однофакторного ANOVA использовали критерий Даннета. Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 22A-22C продемонстрирован ИГХ анализ специфичности антител UNS в отношении α-Syn в мозжечке пациентов с различными синуклеинопатиями. Средний процент площади агрегатов α-Syn, окрашенных каждым антителом (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в белом веществе мозжечка пациентов с PD (Фиг. 22A), DLB (Фиг. 22B) и MSA (Фиг. 22C). Антитела UNS приводили к выявлению более высокого процента площади агрегатов α-Syn при MSA (F(3,8)=0,929, p=0,469 с помощью ANOVA); DLB (F(3,6)=1,426, p=0,325 с помощью ANOVA) и PD (F(3,6)=2,509, p=0,157 с помощью ANOVA). Процент площади, окрашенной каждым антителом, сравнивали с диагностическим антителом, NCL-L-ASYN. P<0,05 (*); P<0,01 (**); P<0,001 (***). Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 23A-23B продемонстрированы иллюстративные изображения иммунологического окрашивания каждым антителом черной субстанции (Фиг. 23A) и бледного шара (Фиг. 23B) из головного мозга контрольных пациентов без заболевания. Ни одно из антител UNS не приводило к выявлению какой-либо патологии α-Syn по сравнению с диагностическим антителом NCL-L-ASYN.
На Фиг. 24A-24D продемонстрирован ИГХ анализ специфичности антител UNS в отношении LB в островковой коре базальных ганглиев пациентов с DLB или PD. Средний процент площади иммуноположительных LB, выявленных с помощью каждого антитела (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в островковой коре пациентов с PD (Фиг. 24A) и DLB (Фиг. 24B). Процент площади LB представлен в виде доли всего α-Syn, выявленного с помощью каждого антитела. Антитела UNS приводили к выявлению более низкой доли LB (или более высокой доли LN (Lewy neurites)) в островковой коре пациентов с DLB (F(3,7)=0,836, p=0,516 с помощью ANOVA) и PD (F(3,4)=0,913, p=0,510 с помощью ANOVA). Процент площади, окрашенной каждым антителом, сравнивали с диагностическим антителом, NCL-L-ASYN. Иллюстративные микроскопические изображения, полученные в результате иммунологического окрашивания каждым антителом, продемонстрированы на Фиг. 24C (PD) и Фиг. 24D (DLB). P<0,05 (*); P<0,01 (**); P<0,001 (***). Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 25A-25D продемонстрирован ИГХ анализ специфичности антител UNS в отношении LB в сером веществе височной коры пациентов с DLB или PD. Средний процент площади иммуноположительных LB, выявленных с помощью каждого антитела (PD062220, PD062205, PD100806 и NCL-L-ASYN), определяли для общей площади 7,5 мм2 в сером веществе пациентов с PD (Фиг. 25A) и DLB (Фиг. 25B). Процент площади LB представлен в виде доли всего альфа-синуклеина, выявленного с помощью каждого антитела. Антитела UNS приводили к выявлению более низкой доли LB (или более высокой доли LN) в сером веществе пациентов с PD (F(2,3)=1,983, p=0,282 с помощью ANOVA) и DLB (F(3,7)=1,906, p=0,217 с помощью ANOVA). Процент площади, окрашенной каждым антителом, сравнивали с диагностическим антителом, NCL-L-ASYN. Иллюстративные микроскопические изображения, полученные в результате иммунологического окрашивания каждым антителом, продемонстрированы на Фиг. 25C (PD) и Фиг. 25D (DLB). P<0,05 (*); P<0,01 (**); P<0,001 (***). Данные продемонстрированы в виде среднего+SD (планки погрешностей).
На Фиг. 26A-26B продемонстрированы иллюстративные изображения иммунологического окрашивания с помощью антител UNS и NCL-L-ASYN в черной субстанции среднего мозга пациентов с DLB (Фиг. 26A) и PD (Фиг. 26B). Имеет место более высокая детекция LN с помощью антител UNS по сравнению с NCL-L-ASYN.
На Фиг. 27A-27C продемонстрирована специфическая в отношении клеток агрегация α-Syn. Максимальная проекция приводила к наложению конфокальных изображений агрегатов α-Syn, полученных из базальных ганглиев и среднего мозга клинических случаев с PD (Фиг. 27A), DLB (Фиг. 27B) и MSA (Фиг. 27C). Показаны агрегаты α-Syn (PD062205, красный цвет) в нейронах (HuD, зеленый цвет) в случаях PD и DLB, но не MSA. α-Syn (PD062205) и HuD отмечены на фигурах в градациях серого цвета, которые подаются с данной заявкой; однако цветные копии доступны по требованию. Масштаб планок: 10 мкМ.
На Фиг. 28A-28C продемонстрирована специфическая в отношении клеток агрегация α-Syn. Максимальная проекция приводила к наложению конфокальных изображений агрегатов α-Syn, полученных на основании клинических случаев PD (Фиг. 28A), DLB (Фиг. 28B) и MSA (Фиг. 28C). Агрегаты α-Syn (PD062205, красный цвет) редко располагались в олигодендроцитах (Olig2, зеленый цвет) в случаях MSA, но не PD или DLB. α-Syn (PD062205) и Olig2 отмечены на фигурах в градациях серого цвета, которые подаются с данной заявкой; однако цветные копии доступны по требованию. Масштаб планок: 10 мкМ.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Данное раскрытие связано с пептидными иммуногенными конструкциями белка альфа-синуклеина (α-Syn). Данное раскрытие также связано с композициями, содержащими пептидные иммуногенные конструкции, способами получения и применения указанных пептидных иммуногенных конструкций и антителами, образуемыми пептидными иммуногенными конструкциями.
Раскрытые пептидные иммуногенные конструкции содержат B-клеточный эпитоп из α-Syn, связанный с гетерологичным эпитопом T-хелперных клеток (Th) непосредственно или с помощью необязательного гетерологичного спейсера. Участок B-клеточного эпитопа пептидных иммуногенных конструкций содержит от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn (SEQ ID NO: 1). Участок гетерологичного Th-эпитопа пептидных иммуногенных конструкций происходит из аминокислотных последовательностей, полученных из патогенных белков. Участки B-клеточного эпитопа и Th-эпитопа пептидных иммуногенных конструкций функционируют совместно при введении хозяину с целью стимуляции образования антител, которые специфически распознают и связываются с участком В-клеточного эпитопа α-Syn конструкций.
Данное раскрытие также связано с композициями, содержащими раскрытые пептидные иммуногенные конструкции, в том числе фармацевтические композиции. Раскрытые фармацевтические композиции способны вызывать иммунный ответ и продуцирование антител против раскрытых пептидных иммуногенных конструкций в организме хозяина. Раскрытые композиции могут содержать одну или смесь из более чем одной из раскрытых пептидных иммуногенных композиций. В некоторых вариантах осуществления композиции содержат раскрытые пептидные иммуногенные конструкции совместно с дополнительными компонентами, в том числе носителями, адъювантами, буферами и другими подходящими реагентами. В определенных вариантах осуществления композиции содержат раскрытые пептидные иммуногенные конструкции стабилизированного иммуностимулирующего комплекса с олигомером CpG, который необязательно дополняют адъювантом.
Данное раскрытие также связано с антителами, которые продуцируются хозяином, которого иммунизируют раскрытыми пептидными иммуногенными конструкциями. Раскрытые антитела специфически распознают и связываются с участком В-клеточного эпитопа α-Syn пептидных иммуногенных конструкций. Раскрытые антитела к α-Syn имеют неожиданным образом высокую перекрестную реактивность по отношению к β-складке α-Syn в форме мономеров, олигомеров или фибрилл. Исходя из своих уникальных характеристик и свойств раскрытые антитела способны обеспечивать иммунотерапевтический подход к целенаправленному воздействию, идентификации и лечению синуклеинопатий.
Данное раскрытие также связано со способами получения и применения раскрытых пептидных иммуногенных конструкций, антител и композиций. Раскрытые способы предлагают малозатратное производство и контроль качества пептидных иммуногенных конструкций и композиций, содержащих указанные конструкции, которые могут быть использованы в способах предупреждения и лечения синопатий.
Данное раскрытие также включает в себя способы лечения и/или предупреждения синуклеинопатий с помощью пептидных иммуногенных конструкций и/или антител, направленных против пептидных иммуногенных конструкций. В некоторых вариантах осуществления способы лечения и/или предупреждения синуклеинопатий включают в себя введение хозяину раскрытой пептидной иммуногенной конструкции. В определенных вариантах осуществления композиции, используемые в способах, содержат раскрытую пептидную иммуногенную конструкцию в форме стабильного иммуностимулирующего комплекса отрицательно заряженными олигонуклеотидами, такими как олигомеры CpG, в результате электростатической ассоциации, указанные комплексы затем дополняются, необязательно, минеральными солями или маслом в качестве адъюванта, для введения пациентам с синуклеинопатиями. Раскрытые способы также включают в себя режимы дозирования, лекарственные формы и пути введения пептидных иммуногенных конструкций хозяину, имеющему риск развития или имеющему синуклеинопатии.
Названия разделов, используемые в данном документе, представлены лишь для организационных целей, и не предполагают ограничивать описываемый предмет изобретения. Все ссылки или части ссылок, цитируемые в данной заявке, явно включены посредством ссылки в данный документ в полном объеме для любой цели.
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют такое же самое значение, которое обычно понятно специалисту в данной области техники, к которой относится данное изобретение. Формы существительного единственного числа включают в себя формы множественного числа, если контекст четко не указывает иное. Аналогично, выражение «или» предусматривает включение в себя «и», если контекст четко не указывает иное. Таким образом, «содержащий A или B» означает включение в себя А, или B, или A и B. Также необходимо понимать, что все размеры аминокислот и все значения молекулярного веса или молекулярной массы, приведенные для полипептидов, являются примерными, и предусмотрены для описания. Несмотря на то, что способы и материалы, аналогичные или эквивалентные способам или материалам, описанным в данном документе, могут быть применимы при практическом осуществлении или исследовании раскрытого способа, подходящие способы и материалы описаны ниже. Все публикации, заявки на патент, патенты и другие литературные источники, которые упоминаются в данном документе, включены посредством ссылки в полном объеме. В случае конфликта данное описание, в том числе объяснения терминов, будет иметь преимущественную силу. Кроме того, материалы, способы и примеры носят лишь иллюстративный характер и не предусматривают ограничения.
Пептидные иммуногенные конструкции α-Syn
В данном раскрытии предложены пептидные иммуногенные конструкции, содержащий B-клеточный эпитоп из α-Syn, ковалентно связанный с гетерологичным эпитопом T-хелперных клеток (Th) непосредственно или с помощью необязательного гетерологичного спейсера.
Фраза «пептидная иммуногенная конструкция α-Syn», используемая в данном документе, относится к пептиду, содержащему (a) B-клеточный эпитоп, имеющий от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn (SEQ ID NO: 1); (b) гетерологичный Th-эпитоп; и (c) необязательный гетерологичный спейсер.
В определенных вариантах осуществления пептидная иммуногенная конструкция может быть представлена формулами:
(Th)m-(A)n-(С-терминальный фрагмент α-Syn)-X
или
(С-терминальный фрагмент α-Syn)-(A)n-(Th)m-X,
где
Th представляет собой гетерологичный T-хелперный эпитоп;
A представляет собой гетерологичный спейсер;
(С-терминальный фрагмент α-Syn) представляет собой B-клеточный эпитоп, имеющий от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn;
X представляет собой α-COOH или α-CONH2 аминокислоты;
m составляет от 1 до приблизительно 4; и
n составляет от 0 до приблизительно 10.
Различные компоненты раскрытой пептидной иммуногенной конструкции α-Syn описаны ниже.
a. α-Syn и С-терминальный фрагменты α-Syn.
Термин «α-Syn», «альфа-синуклеин», «α-синуклеин» и т.п., используемые в данном документе, относится к (a) полноразмерному белку α-Syn и/или (b) его фрагментам из любого организма, который экспрессирует α-Syn. α-Syn характеризуется чрезвычайным конформационным разнообразием, которое адаптируется к различным условиям в состояниях связывания с мембраной, цитозоля и агрегации амилоида, и выполняет различные функции. В некоторых вариантах осуществления белок α-Syn происходит от человека. В определенных вариантах осуществления полноразмерный белок α-Syn человека имеет 140 аминокислот (№доступа NP_000336) (SEQ ID NO: 1).
Фраза «C-терминальный участок» или «C-терминальный конец» α-Syn, используемая в данном документе, относится к любой аминокислотной последовательности из карбокситерминальной части α-Syn. В определенных вариантах осуществления C-терминальный участок или C-терминальный конец α-Syn относится к аминокислотной последовательности между остатками 96-140, или их фрагментам, α-Syn. C-терминальный участок α-Syn имеет высокое содержание пролинов и отрицательно заряженных остатков, которые представляют собой общие характеристики, встречающиеся во внутренне неупорядоченных белках, с целью поддержания растворимости. C-терминальный участок α-Syn, как правило, присутствует в случайной спиральной структуре в связи со своей гидрофобностью и высоким суммарным отрицательным зарядом. Исследования in vitro показали, что агрегация α-Syn может быть индуцирована с помощью снижения pH, что приводит к нейтрализации указанных отрицательных зарядов.
Фраза «С-терминальный фрагмент α-Syn» или «B-клеточный эпитоп из C-терминального конца α-Syn», используемая в данном документе, относится к части полноразмерной последовательности α-Syn, которая содержит от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn. С-терминальный фрагмент α-Syn также обозначается в данном документе как пептид G111-D135 α-Syn и его фрагменты. Различные С-терминальные фрагменты α-Syn, описанные в данном документе, обозначаются с помощью положений своих аминокислот по отношению к полноразмерной последовательности α-Syn, представленной SEQ ID NO: 1.
Аминокислотные последовательности С-терминальных фрагментов α-Syn, используемые в пептидных иммуногенных конструкциях α-Syn, выбирали на основе количества принципов разработки. Некоторые из этих принципов включат в себя использование пептидной последовательности α-Syn, которая:
(i) не разделяет значительной гомологии последовательности с бета-синуклеином (β-Syn) с целью недопущения образования антител, которые перекрестно реагируют с β-Syn, поскольку β-Syn может связываться с α-Syn и предупреждать его агрегацию;
(ii) лишена аутологичного T-хелперного эпитопа в α-Syn с целью предупреждения активации аутологичных T-клеток, которые могут приводить к воспалению головного мозга, приводя к менингококковому энцефалиту, как ранее описывали в клинических исследования с помощью вакцины AN1792, целенаправленно воздействующей на Aβ1-42, с целью лечения болезни Альцгеймера;
(iii) содержится в участке α-Syn, который восприимчив к конформационным изменениями по отношению к своей нативной форме;
(iv) сама по себе является неиммуногенной, поскольку она представляет собой молекулу собственного организма;
(v) может быть выполнена иммуногенной с помощью белкового носителя или эффективного (эффективных) T-хелперного (Т-хелперных) эпитопа (эпитопов);
(vi) при выполнении иммуногенной и введении хозяину:
(a) вызывает образование высокого титра антител, направленных против пептидной последовательности α-Syn (B-клеточного эпитопа), а не против белкового носителя или эффективного (эффективных) Т-хелперного (Т-хелперных) эпитопа (эпитопов);
(b) вызывает образование высокого титра антител, которые реагируют с денатурированной β-складкой α-Syn, в форме мономеров, олигомеров или фибрилл, для того, чтобы позволить таким антителам предупреждать агрегацию α-Syn, вызывать дезагрегацию любых агрегатов α-Syn и приводить к удалению токсических олигомеров, агрегатов и/или фибрилл α-Syn, тем самым, снижая или предупреждая нагрузку агрегатами α-Syn в головном мозге;
(c) не вызывает образование антител, которые реагируют с нативным α-Syn, что будет представлять собой большую проблему для безопасности, поскольку нативный α-Syn представляет собой основной клеточной белок со значительным распределением в тканях.
С учетом указанных принципов разработки C-терминальный участок α-Syn выбирали в качестве мишени для разработки пептидных иммуногенов. Кроме того, C-терминальный участок α-Syn выбирали поскольку, исходя из его структурных характеристик, этот участок, по-видимому, является наиболее восприимчивым к модуляции антителом или другими физическими факторами по сравнению с другими участками α-Syn.
Оценка многочисленных пептидных последовательностей, происходящих из α-Syn, описанных далее в разделе Примеры, привела к идентификации и отбору многочисленных пептидов α-Syn, которые удовлетворяют принципам разработки, описанным выше. В частности, последовательности, которые удовлетворяют принципам разработки, включают в себя пептиды, имеющие от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального участка α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn.
В некоторых вариантах осуществления С-терминальный фрагмент α-Syn представляет собой пептид G111-D135 α-Syn из 25 аминокислот, представленный SEQ ID NO: 12. В других вариантах осуществления С-терминальный фрагмент α-Syn содержит приблизительно 10 смежных аминокислот пептида α-Syn G111-D135, представленного SEQ ID NO: 12. В определенных вариантах осуществления С-терминальный фрагмент α-Syn содержит 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 смежных аминокислот пептида G111-D135, представленного SEQ ID NO: 12. В конкретных вариантах осуществления С-терминальный фрагмент α-Syn имеет аминокислотную последовательность, представленную SEQ ID NO: 12-15, 17 или 49-64, как продемонстрировано в Табл. 1.
С-терминальный фрагмент α-Syn по данному раскрытию также включает в себя иммунологически функциональные аналоги или гомологи пептида G111-D135 α-Syn и их фрагменты. Функциональные иммунологические аналоги или гомологи пептида G111-D135 α-Syn и их фрагменты включают в себя варианты, которые сохраняют по сути такую же самую иммуногенность, что и исходный пептид. Иммунологически функциональные аналоги могут иметь консервативную замену в положении аминокислоты; изменение общего заряда; ковалентную связь с другим фрагментом; или добавления, вставки или делеции аминокислот; и/или любую их комбинацию.
Консервативные замены выполняют, когда один аминокислотный остаток замещают другим аминокислотным остатком с аналогичными химическими свойствами. Например, неполярные (гидрофобные) аминокислоты включают в себя аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагинин и глутамин; положительно заряженные (основные) аминокислоты включают в себя аргинин, лизин и гистидин; а отрицательно заряженные (кислотные) аминокислоты включают в себя аспарагиновую кислоту и глутаминовую кислоту.
Иммунологически функциональные аналоги включают в себя аминокислотные последовательности, которые содержат консервативные замены, добавления, делеции или вставки от одной до приблизительно четырех аминокислотных остатков, которые вызывают иммунные ответы, которые перекрестно реагируют с пептидом G111-D135 α-Syn. Консервативные замены, добавления и вставки можно выполнять с использованием природных или не встречающихся в природе аминокислот.Не встречающиеся в природе аминокислоты включают в себя, но не ограничиваясь ими, ε-N-лизин, β-аланин, орнитин, норлейцин, норвалин, гидроксипролин, тирозин, γ-аминомасляную кислоту, гомосерин, цитруллин, аминобензойную кислоту, 6-аминокапроновую кислоту (Aca; 6-аминогексановую кислоту), гидроксипролин, меркаптопропионовую кислоту (MPA), 3-нитротирозин, пироглутаминовую кислоту и т.п. Встречающиеся в природе аминокислоты включают в себя аланин, аргинин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, трипрофан, тирозин и валин.
В одном варианте осуществления функциональный иммунологический аналог определенного пептида содержит ту же самую аминокислотную последовательность, что и исходный пептид и дополнительно содержит три остатка лизина (Lys-Lys-Lys), добавленных к аминоконцу пептида G111-D135 α-Syn и его фрагментам В-клеточного эпитопа. В данном варианте осуществления включение трех остатков лизина в последовательность исходного пептида приводит к изменению общего заряда исходного пептида, но не приводит к изменению функции исходного пептида.
В определенных вариантах осуществления функциональный аналог С-терминального фрагмента α-Syn имеет по меньшей мере 50% идентичность по отношению к исходной аминокислотной последовательности. В других вариантах осуществления указанный функциональный аналог имеет по меньшей мере 80% идентичность по отношению к исходной аминокислотной последовательности. В еще одних вариантах осуществления указанный функциональный аналог имеет по меньшей мере 85% идентичность по отношению к исходной аминокислотной последовательности. В еще одних вариантах осуществления указанный функциональный аналог имеет по меньшей мере 90% или по меньшей мере 95% идентичность по отношению к исходной аминокислотной последовательности.
b. Гетерологичные T-хелперные эпитопы (Th-эпитопы)
В данном раскрытии предложены пептидные иммуногенные конструкции, содержащий B-клеточный эпитоп из α-Syn, ковалентно связанный с гетерологичным эпитопом T-хелперных клеток (Th) непосредственно или с помощью необязательного гетерологичного спейсера.
Гетерологичный Th-эпитоп в пептидной иммуногенной конструкции α-Syn приводит к усилению иммуногенности С-терминального фрагмента α-Syn, что облегчает получение высокого титра специфических антител, направленных против оптимизированного В-клеточного эпитопа (т.е., С-терминального фрагмента α-Syn) при рациональной разработке.
Термин «гетерологичный», используемый в данном документе, относится к аминокислотной последовательности, которая происходит из аминокислотной последовательности, которая не является частью или не является гомологичной последовательности дикого типа α-Syn. Таким образом, гетерологичный Th-эпитоп представляет собой Th-эпитоп, происходящий из аминокислотной последовательности, которая не встречается в природе в α-Syn (т.е., Th-эпитоп не является аналогичным α-Syn). Поскольку Th-эпитоп является гетерологичным по отношению к α-Syn, встречающаяся в природе аминокислотная последовательность α-Syn не продолжается ни в N-терминальных, ни в C-терминальных направлениях, если гетерологичный Th-эпитоп ковалентно связан с С-терминальным фрагментом α-Syn.
Гетерологичный Th-эпитоп по данному раскрытию может представлять собой любой Th-эпитоп, который не имеет аминокислотной последовательности, встречающей в природе в α-Syn. Th-эпитоп может иметь аминокислотную последовательность, происходящую от любого вида (например, человека, свиньи, крупного рогатого скота, собаки, крысы, мыши, морской свинки и т.д.). Th-эпитоп также может иметь неизбирательные связывающие мотив с молекулами МНС II класса многих видов. В определенных вариантах осуществления Th-эпитоп содержит многочисленные неизбирательные связывающие мотивы с MHC II класса с целью обеспечения максимальной активации T-хелперов, приводя к инициации и регуляции иммунных ответов. Th-эпитоп по своей природе предпочтительно является иммуномолчащим, т.е., небольшое количество, или вообще никакие антитела, полученные с помощью пептидных иммуногенных конструкций α-Syn, не будут направлены против Th-эпитопа, тем самым обеспечивая очень сфокусированный иммунный ответ, направленный на целевой B-клеточный эпитоп С-терминального фрагмента α-Syn.
Th-эпитопы по данному раскрытию содержат, но не ограничиваясь ими, аминокислотные последовательности, происходящие из чужеродных патогенов, как продемонстрировано в качестве примера в Табл. 2 (SEQ ID NO: 70-98). Кроме того, Th-эпитопы включают в себя идеализированные искусственные Th-эпитопы и комбинированные идеализированные Th-эпитопы (например, SEQ ID NO: 71 и 78-84). Пептиды гетерологичных Th-эпитопов, представленные в виде комбинаторной последовательности (например, SEQ ID NO: 79-82), содержат смесь аминокислотных остатков, представленных в определенных положениях в пептидном каркасе на основании вариабельных остатков гомологов для этого конкретного пептида. Сборка комбинаторных пептидов может происходить в одном процессе с помощью добавления смеси разработанных защитных аминокислот, вместо одной определенной аминокислоты, в определенном положении во время процесса синтеза. Сборка таких пептидов комбинаторных гетерологичных Th-эпитопов может обеспечивать широкий охват Th-эпитопов для животных, имеющих различный генетический фон. Иллюстративные комбинаторные последовательности пептидов гетерологичных Th-эпитопов включают в себя SEQ ID NO: 79-82, которые продемонстрированы в Табл. 2. Пептиды Th-эпитопов по данному изобретению обеспечивают значительную реактивность и иммуногенность в отношении животных и пациентов из генетически различных популяций.
Пептидные иммуногенные конструкции α-Syn, содержащие Th-эпитопы, образуются одновременно в одном твердофазном синтезе пептидов в тандеме с С-терминальным фрагментом α-Syn. Th-эпитопы также включают в себя иммунологические аналоги Th-эпитопов. Иммунологические Th-аналоги включают в себя иммуностимулирующие аналоги, перекрестно-реагирующие аналоги и сегменты любого из этих Th-эпитопов, которые являются достаточными для усиления или стимуляции иммунного ответа в отношении С-терминальных фрагментов α-Syn.
Функциональные иммунологические аналоги пептидов Th-эпитопов также являются эффективными и включены в виде части по данному изобретению. Функциональные иммунологические Th-аналоги могут включать в себя консервативные замены, добавления, делеции и вставки от одного до приблизительно пяти аминокислотных остатков в Th-эпитопе, которые необязательно приводят к модификации Th-стимулирующей функции Th-эпитопа. Консервативные замены, добавления и вставки можно выполнять с использованием природных или не встречающихся в природе аминокислот, как описано выше для С-терминальных фрагментов α-Syn. В Табл. 2 определена другая вариация функционального аналога пептида Th-эпитопа. В частности, SEQ ID NO: 71 и 78 MvF1 и Th MvF2 представляют собой функциональные аналоги SEQ ID NO: 81 и 83 MvF4 и MvF5 в том, что они отличаются в аминокислотном каркасе делецией (SEQ ID NO: 71 и 78) или включением (SEQ ID NO: 81 и 83) двух аминокислот, каждая из которых расположена на N- и C-концах. Различия между этими двумя сериями аналогичными последовательностей не будет влиять на функцию Th-эпитопов, содержащихся в указанных последовательностях. Таким образом, функциональные иммунологические Th-аналоги включают в себя несколько версий Th-эпитопа, происходящего из белка слияния вируса кори MvF1-4 Ths (SEQ ID NO: 71, 78, 79, 81 и 83) и из поверхностного белка вируса гепатита HBsAg 1-3 Ths (SEQ ID NO: 80, 82 и 84).
Th-эпитоп в пептидной иммуногенной конструкции α-Syn может быть ковалентно связан либо на N-, либо на C-терминальном конце С-терминального пептида α-Syn. В некоторых вариантах осуществления Th-эпитоп ковалентно связан с N-терминальным концом С-терминального пептида α-Syn. В других вариантах осуществления Th-эпитоп ковалентно связан с С-терминальным концом С-терминального пептида α-Syn. В определенных вариантах осуществления более чем один Th-эпитоп ковалентно связан с С-терминальным фрагментом α-Syn. Если более чем один Th-эпитоп связан с С-терминальным фрагментом α-Syn, каждый Th-эпитоп может иметь одинаковую аминокислотную последовательность или различные аминокислотные последовательности. Кроме того, если более чем один Th-эпитоп связан с С-терминальным фрагментом α-Syn, то Th-эпитопы могут быть упорядочены в любом порядке. Например, Th-эпитопы могут быть последовательно связаны с N-терминальным концом С-терминального фрагмента α-Syn, или последовательно связаны с C-терминальным концом С-терминального фрагмента α-Syn, или Th-эпитоп может быть ковалентно связан с N-терминальным концом С-терминального фрагмента α-Syn, в то время как отдельный Th-эпитоп ковалентно связан с C-терминальным концом С-терминального фрагмента α-Syn. Не существует ограничения в структуре Th-эпитопов в отношении С-терминального фрагмента α-Syn.
В некоторых вариантах осуществления Th-эпитоп ковалентно связан с С-терминальным фрагментом α-Syn непосредственно. В других вариантах осуществления Th-эпитоп ковалентно связан с С-терминальным фрагментом α-Syn с помощью гетерологичного спейсера, подробно описанного ниже.
c. Гетерологичный спейсер
Раскрытые пептидные иммуногенные конструкции α-Syn необязательно содержат гетерологичный спейсер, который ковалентно связывает B-клеточный эпитоп из α-Syn с гетерологичным Т-хелперным (Th) эпитопом.
Как обсуждается выше, термин «гетерологичный» относится к аминокислотной последовательности, которая происходит из аминокислотной последовательности, которая не является частью или не является гомологичной последовательности дикого типа α-Syn. Таким образом, встречающаяся в природе аминокислотная последовательность α-Syn не продолжается ни в N-терминальных, ни в C-терминальных направлениях, если гетерологичный спейсер ковалентно связан с B-клеточным эпитопом из α-Syn, поскольку спейсер является гетерологичным по отношению к последовательности α-Syn.
Спейсер представляет собой любую молекулу или химическую структуру, способные связывать две аминокислоты и/или пептиды вместе. Спейсер может варьировать по длине или полярности в зависимости от применения. Присоединение спейсера может осуществляться с помощью амидной или карбоксильной связи, однако другие функциональные группы также возможны. Спейсер может включать в себя химическое соединение, встречающуюся в природе аминокислоту или не встречающуюся в природе аминокислоту.
Спейсер может обеспечивать структурные свойства в отношении пептидной иммуногенной конструкции α-Syn. Структурно спейсер обеспечивает физическое разделение Th-эпитопа от B-клеточного эпитопа С-терминального фрагмента α-Syn. Физическое разделение с помощью спейсера может приводить к нарушению любых искусственных вторичных структур, созданных в результате связывания Th-эпитопа с B-клеточным эпитопом. Кроме того, физическое разделение эпитопов с помощью спейсера может приводить к устранению интерференции между Th-клеточными и/или B-клеточными ответами. Кроме того, спейсер может быть разработан для создания или модификации вторичной структуры пептидной иммуногенной конструкции. Например, спейсер может быть разработан для функционирования в качестве гибкого шарнира с целью усиления разделения Th-эпитопа и B-клеточного эпитопа. Гибкий шарнирный спейсер также может обеспечивать более эффективные взаимодействия между представленным пептидным иммуногеном и соответствующими Th-клетками и B-клетками с целью усиления иммунных ответов в отношении Th-эпитопа и B-клеточного эпитопа. Примеры последовательностей, кодирующих гибкие шарниры, встречаются в шарнирной области тяжелой цепи иммуноглобулинов, которые часто имеют высокое содержание пролина. Один особенно полезный гибкий шарнир, который может быть использован в качестве спейсера, представлен в виде последовательности Pro-Pro-Xaa-Pro-Xaa-Pro (SEQ ID NO: 148), где Xaa представляет собой любую аминокислоту, и предпочтительно аспарагиновую кислоту.
Спейсер также может обеспечивать функциональные свойства в отношении пептидной иммуногенной конструкции α-Syn. Например, спейсер может быть разработан с целью изменения общего заряда пептидной иммуногенной конструкции α-Syn, которая может влиять на растворимость пептидной иммуногенной конструкции. Кроме того, изменение суммарного заряда пептидной иммуногенной конструкции α-Syn может влиять на способность пептидной иммуногенной конструкции ассоциировать с другими соединениями и реагентами. Как обсуждается подробно ниже, пептидная иммуногенная конструкция α-Syn может быть образована в виде стабильного иммуностимулирующего комплекса с высокозаряженным олигонуклеотидом, таким как олигомеры CpG, с помощью электростатической ассоциации. Общий заряд пептидной иммуногенной конструкции α-Syn является важным для образования указанных стабильных иммуностимулирующих комплексов.
Химические соединения, которые могут быть использованы в качестве спейсера, включают в себя, но не ограничиваясь ими, (2-аминоэтокси)уксусную кислоту (AEA), 5-аминовалериановую кислоту (AVA), 6-аминокапроновую кислоту (Ahx), 8-амино-3,6-диоксаоктановую кислоту (AEEA, мини-PEG1), 12-амино-4,7,10-триоксадодекановую кислоту (мини-PEG2), 15-амино-4,7,10,13-тетраоксапентадекановую кислоту (мини-PEG3), триоксатридеканянтарную кислоту (Ttd), 12-аминододекановую кислоту, Fmoc-5-амино-3-оксапентановую кислоту (O1Pen) и т.п.
Встречающиеся в природе аминокислоты включают в себя аланин, аргинин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, трипрофан, тирозин и валин.
Не встречающиеся в природе аминокислоты включают в себя, но не ограничиваясь ими, ε-N-лизин, β-аланин, орнитин, норлейцин, норвалин, гидроксипролин, тирозин, γ-аминомасляную кислоту, гомосерин, цитруллин, аминобензойную кислоту, 6-аминокапроновую кислоту (Aca; 6-аминогексановую кислоту), гидроксипролин, меркаптопропионовую кислоту (MPA), 3-нитротирозин, пироглутаминовую кислоту и т.п.
Спейсер в пептидной иммуногенной конструкции α-Syn может быть ковалентно связан либо на N-, либо на C-терминальном конце Th-эпитопа и С-терминального пептида α-Syn. В некоторых вариантах осуществления спейсер ковалентно связан с С-терминальным концом Th-эпитопа и N-терминальным концом С-терминального пептида α-Syn. В других вариантах осуществления спейсер ковалентно связан с С-терминальным концом С-терминального пептида α-Syn и N-терминальным концом Th-эпитопа. В определенных вариантах осуществления может быть использован более чем один спейсер, например, если более чем один Th-эпитоп присутствует в пептидной иммуногенной конструкции. Если используется более чем один спейсер, то каждый спейсер может быть таким же самым, как и другие, или отличаться. Кроме того, если более чем один Th-эпитоп присутствует в пептидной иммуногенной конструкции, Th-эпитопы могут быть разделены спейсером, который может быть одинаковым или отличаться от спейсера, используемого для разделения Th-эпитопа от B-клеточного эпитопа. Не существует ограничения в структуре спейсера в отношении Th-эпитопа или С-терминального фрагмента α-Syn.
В определенных вариантах осуществления гетерологичный спейсер представляет собой встречающуюся в природе аминокислоту или не встречающуюся в природе аминокислоту. В других вариантах осуществления спейсер содержит более одной встречающейся в природе или не встречающейся в природе аминокислоты. В конкретных вариантах осуществления спейсер представляет собой Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys или ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148).
d. Конкретные варанты осуществления пептидной иммуногенной конструкции α-Syn
Пептидная иммуногенная конструкция α-Syn может быть представлена формулами:
(Th)m-(A)n-(С-терминальный фрагмент α-Syn)-X
или
(С-терминальный фрагмент α-Syn)-(A)n-(Th)m-X,
где
Th представляет собой гетерологичный T-хелперный эпитоп;
A представляет собой гетерологичный спейсер;
(С-терминальный фрагмент α-Syn) представляет собой B-клеточный эпитоп, имеющий от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn;
X представляет собой α-COOH или α-CONH2 аминокислоты;
m составляет от 1 до приблизительно 4; и
n составляет от 0 до приблизительно 10.
В определенных вариантах осуществления гетерологичный Th-эпитоп в пептидной иммуногенной конструкции α-Syn имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 70-98 или их комбинаций, показанных в Табл. 2. В конкретных вариантах осуществления Th-эпитоп имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 78-84. В определенных вариантах осуществления пептидная иммуногенная конструкция α-Syn содержит более чем один Th-эпитоп.
В определенных вариантах осуществления необязательный гетерологичный спейсер выбирают из любого из Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148) и их комбинаций. В конкретных вариантах осуществления гетерологичный спейсер представляет собой ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148).
В определенных вариантах осуществления С-терминальный фрагмент α-Syn имеет от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn. В конкретных вариантах осуществления С-терминальный фрагмент α-Syn имеет аминокислотную последовательность, представленную SEQ ID NO: 12-15, 17 или 49-64, как продемонстрировано в Табл. 1.
В определенных вариантах осуществления пептидная иммуногенная конструкция α-Syn имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 107-108, 111-113 и 115-147, как продемонстрировано в Табл. 3. В конкретных вариантах осуществления пептидная иммуногенная конструкция α-Syn имеет аминокислотную последовательность, выбранную из любой из SEQ ID NO: 107-108 и 111-113.
Композиции
В данном раскрытии также предложены композиции, содержащие раскрытую пептидную иммуногенную конструкцию α-Syn.
a. Пептидные композиции
Композиции, содержащие раскрытую пептидную иммуногенную конструкцию α-Syn, могут находится в жидкой или твердой форме. Жидкие композиции могут включать в себя воду, буферы, растворители, соли и/или любой другой приемлемый реагент, который не приводит к изменению структурных или функциональных свойств пептидной иммуногенной конструкции α-Syn. Пептидные композиции могут содержать одну или несколько из раскрытых пептидных иммуногенных конструкций α-Syn.
b. Фармацевтические композиции
Данное раскрытие также связано с фармацевтическими композициями, содержащими раскрытую пептидную иммуногенную конструкцию α-Syn.
Фармацевтические композиции могут содержать носители и/или другие вспомогательные вещества в фармацевтически приемлемой системе доставки. Соответственно, фармацевтические композиции могут содержать фармацевтически эффективное количество пептидной иммуногенной конструкции α-Syn совместно с фармацевтически приемлемым носителем, адъювантом и/или другими наполнителями, такими как разбавителя, вспомогательные вещества, стабилизирующие агенты, консерванты, солюбилизирующие агенты, буферы и т.п.
Фармацевтические композиции могут содержать один или несколько адъювантом, которые функционируют с целью ускорения, продления или усиления иммунного ответа в отношении пептидной иммуногенной конструкции α-Syn без оказания любого собственного специфического антигенного эффекта. Адъюванты, используемые в фармацевтической композиции, могут включать в себя масла, соли алюминия, виросомы, фосфат алюминия (например, ADJU-PHOS®), гидроксид алюминия (например, ALHYDROGEL®), липосин, сапонин, сквален, L121, Emulsigen®, монофосфорил-липид A (MPL), QS21, ISA 35, ISA 206, ISA50V, ISA51, ISA 720, а также другие адъюванты и эмульгаторы.
В некоторых вариантах осуществления фармацевтическая композиция содержит Montanide™ ISA 51 (композицию масляного адъюванта, состоящего из растительного масла и маннида олеата для получения эмульсий типа вода в масле), Tween(80 (также известный как полисорбат 80 или полиоксиэтилен (20) сорбитан моноолеат), олигонуклеотид CpG и/или любую их комбинацию. В других вариантах осуществления фармацевтическая композиция представляет собой эмульсию типа вода-в масле-в воде (т.е., w/o/w) с эмульсигеном или эмульсигеном D в качестве адъюванта.
Фармацевтические композиции могут быть составлены в виде составов с немедленным высвобождением или замедленным высвобождением. Кроме того, фармацевтические композиции могут быть составлены для индукции системного или локального слизистого иммунитета с помощью захвата иммуногена и совместного введения с микрочастицами. Такие системы доставки легко определяются специалистом в данной области техники.
Фармацевтические композиции могут быть получены в виде инъекционных препаратов, либо в виде жидких растворов, либо суспензий. Жидкие носители, содержащие пептидную иммуногенную конструкцию α-Syn, могут также быть получены перед инъекцией. Фармацевтическая композиция может быть введена с помощью любого подходящего способа применения, например, i.d., i.v., i.p., i.m., интраназально, перорально, подкожно и т.д., в любом подходящем устройстве доставки. В определенных вариантах осуществления фармацевтическую композицию составляют для внутривенного, подкожного, интрадермального или внутримышечного введения. Также могут быть получены фармацевтические композиции, пригодные для других способов введения, в том числе пероральных и интраназальных путей применения.
Фармацевтические композиции могут быть составлены в виде составов с немедленным высвобождением или замедленным высвобождением. Кроме того, фармацевтические композиции могут быть составлены для индукции системного или локального слизистого иммунитета с помощью захвата иммуногена и совместного введения с микрочастицами. Такие системы доставки легко определяются специалистом в данной области техники.
Фармацевтические композиции могут легко быть составлены в подходящей единичной дозированной форме. В некоторых вариантах осуществления фармацевтические композиции содержат от приблизительно 0,5 мкг до приблизительно 1 мг пептидной иммуногенной конструкции α-Syn на кг массы тела. Эффективные дозы фармацевтических композиций варьируют в зависимости от многих факторов, в том числе способов введения, целевого участка, физиологического состояния пациента, того, является ли пациент человеком или животным, других вводимых лекарственных препаратов, и того, является ли лечение профилактическим или терапевтическим. Обычно пациентом является человек, однако млекопитающие помимо человека, в том числе трансгенные млекопитающие, также могут получать лечение. При доставке в нескольких дозах фармацевтические композиции могут быть стандартным образом разделены на подходящее количество на единичную дозированную форму. Вводимая доза будет зависеть от возраста, веса и общего состояния здоровья субъекта, как хорошо известно в терапевтических областях.
В некоторых вариантах осуществления фармацевтическая композиция содержит более чем одну пептидную иммуногенную конструкцию α-Syn. Фармацевтическая композиция, содержащая смесь из более чем одной пептидной иммуногенной конструкции α-Syn может обеспечивать синергическое усиление иммунологической эффективности указанных конструкций. Фармацевтические композиции, содержащие более чем одну пептидную иммуногенную конструкцию α-Syn, могут быть более эффективными в более разнообразной генетической популяции в связи с широким охватом MHC класса II, тем самым, обеспечивая повышенный иммунный ответ в отношении пептидных иммуногенных конструкций α-Syn.
В некоторых вариантах осуществления фармацевтическая композиция содержит пептидную иммуногенную конструкцию α-Syn, выбранную из SEQ ID NO: 107-108, 111-113, 115-147, а также их гомологи, аналоги и/или комбинации. В конкретных вариантах осуществления фармацевтические композиции содержат пептидную иммуногенную конструкцию α-Syn, выбранную из SEQ ID NO: 107-108, 111-113, и любую их комбинацию.
Фармацевтические композиции, содержащие пептидную иммуногенную конструкцию α-Syn, могут быть использованы для того, чтобы вызывать иммунный ответ и приводить к продуцированию антител при введению хозяину.
c. Иммуностимулирующие комплексы
Данное раскрытие также связано с фармацевтическими композициями, содержащими пептидную иммуногенную конструкцию α-Syn в форме иммуностимулирующего комплекса с олигонуклеотидом CpG. Такие иммуностимулирующие комплексы специфически адаптированы с целью функционирования в качестве адъюванта и в качестве стабилизатора пептидного иммуногена. Иммуностимулирующие комплексы находятся в форме гранулы, которая может эффективно представлять пептидный иммуноген α-Syn клеткам иммунной системы с целью получения иммунного ответа. Иммуностимулирующие комплексы могут быть составлены в виде суспензии для парентерального введения. Иммуностимулирующие комплексы также могут быть составлены в форме эмульсий w/o, в виде суспензии в комбинации с минеральной солью или с гелеобразующим полимером in-situ для эффективной доставки пептидного иммуногена α-Syn в клетки иммунной системы хозяина после парентерального введения. Иммуностимулирующие комплексы способны приводить к иммунному ответу в отношении β-складки α-Syn (например, Фиг. 8A, 8B и 8C из Примера 13) с профилактическим/терапевтическим эффектом.
Стабилизированный иммуностимулирующий комплекс может быть образован в результате образования комплекса пептидной иммуногенной конструкции α-Syn с анионной молекулой, олигонуклеотидом, полинуклеотидом или их комбинациями с помощью электростатической ассоциации. Стабилизированный иммуностимулирующий комплекс может быть включен в фармацевтическую композицию в виде системы для доставки иммуногена.
В определенных вариантах осуществления пептидную иммуногенную конструкцию α-Syn разрабатывают с целью содержания катионной части, который является положительно заряженным при pH в диапазоне от 5,0 до 8,0. Суммарный заряд катионной части пептидной иммуногенной конструкции α-Syn или смеси конструкций рассчитывают с помощью присвоения заряда+1 в случае каждого лизина (K), аргинина (R) или гистидина (H), заряда -1 в случае каждой аспарагиновой кислоты (D) или глутаминовой кислоты (E) и заряда 0 в случае другой аминокислоты в последовательности. Заряды суммируются в катионной части пептидной иммуногенной конструкции α-Syn и выражаются в виде суммарного среднего заряда. Пригодный пептидный иммуноген имеет катионную часть с суммарным положительным зарядом+1. Предпочтительно пептидный иммуноген имеет суммарный положительный заряд в диапазоне, который составляет более чем +2. В некоторых вариантах осуществления катионная часть пептидной иммуногенной конструкции α-Syn представляет собой гетерологичный спейсер. В определенных вариантах осуществления катионная часть пептидной иммуногенной конструкции α-Syn имеет заряд +4, если последовательность спейсера представляет собой (α, ε-N)Lys, ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148).
Термин «анионная молекула», описанный в данном документе, относится к любой молекуле, которая является отрицательно заряженной при pH в диапазоне 5,0-8,0. В определенных вариантах осуществления анионная молекула представляет собой олигомер или полимер. Суммарный отрицательный заряд олигомера или полимера рассчитывают с помощью присвоения заряда -1 в случае каждой сложной фосфодиэферной или фосфоротиоатной группы в олигомере. Пригодный анионный олигонуклеотид представляет собой однонитевую молекулу ДНК с 8-64 нуклеотидными основаниями, при этом число повторов мотива CpG находится в диапазоне от 1 до 10. Предпочтительно иммуностимулирующие однонитевые молекулы ДНК в виде CpG содержат 18-48 нуклеотидных оснований, при этом число повторов мотива CpG находится в диапазоне от 3 до 8.
Более предпочтительно анионный олигонуклеотид представлен формулой: 5(X1CGX2 3', где C и G являются неметилированными; X1 выбирают из группы, состоящей из A (аденин), G (гуанин) и T (тимин); и X2 представляет собой C (цитозин) или T (тимин). Или же анионный олигонуклеотид представлен формулой: 5((X3)2CG(X4)2 3', где C и G являются неметилированными; X3 выбирают из группы, состоящей из A, T или G; и X4 представляет собой C или T.
Полученный в результате иммуностимулирующий комплекс находится в форме частиц с размером в типичном случае в диапазоне 1-50 микрон и представляет собой функцию от многих факторов, в том числе стехиометрии относительного заряда и молекулярной массы взаимодействующих молекул. Гранулированные иммуностимулирующий комплекс имеет преимущество обеспечения адъювантности и стимуляции иммунных ответов in vivo. Кроме того, стабилизированный иммуностимулирующий комплекс пригоден для получения фармацевтических композиций с помощью различных процессов, в том числе эмульсий типа вода в масле, суспензий минеральных солей и полимерных гелей.
Антитела
В данном раскрытии также предложены антитела, вызванные пептидной иммуногенной конструкцией α-Syn.
С-терминальные фрагменты α-Syn, имеющие от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального конца α-Syn, соответствующих последовательности от приблизительно глицина в положении аминокислоты 111 (G111) до приблизительно аспарагина в положении аминокислоты 135 (D135) полноразмерного α-Syn, по своей природе являются неиммуногенными или слабоиммуногенными. В то же время, раскрытые пептидные иммуногенные конструкции α-Syn, содержащие С-терминальный фрагмент α-Syn, гетерологичный Th-эпитоп и необязательный гетерологичный спейсер, способны вызывать иммунный ответ и приводить к образованию антител при введении хозяину. Разработка пептидных иммуногенных конструкций α-Syn может нарушать толерантность к собственному α-Syn и вызывать образование сайт-специфических антител, которые распознают конформационные, но не линейные, эпитопы.
Неожиданным образом, антитела, образованные с помощью пептидных иммуногенных конструкций α-Syn, не связываются с встречающейся в природе альфа-спиралью мономера α-Syn в своей нативной форме. В отличие от этого, антитела, образованные с помощью пептидных иммуногенных конструкций α-Syn, распознают и связываются с денатурированнй β-складкой α-Syn в формах мономеров, олигомеров и фибрилл. Кроме того, антитела, образованные с помощью пептидных иммуногенных конструкций α-Syn, не связываются с аналогичными структурами других амилоидогенных белков (т.е., Aβ1-42 и Tau441). Таким образом, определенная разработка пептидной иммуногенной конструкции α-Syn (содержащей С-терминальный фрагмент α-Syn, гетерологичный Th-эпитоп и необязательный гетерологичный спейсер), по-видимому, привела к изменению конформацию различных С-терминальных фрагментов α-Syn с тем, чтобы обеспечить конформацию, подобную β-складке.
Во многих функциональных анализах были выполнены разнообразные сравнения антител, происходящих из иммунных сывороток от животных, иммунизированных пептидными иммуногенными конструкциями α-Syn. Эти сравнения продемонстрировали, что антитела связываются с α-Syn в клетках РС12, обработанных фактором роста нервов (NGF), с высокой специфичностью только с β-складчатыми мономерами и олигомерами α-Syn, но не с другими молекулами амилоидогенных белков (см. Пример 9).
Антитела, вызванные пептидные иммуногенными конструкциями α-Syn, неожиданным образом могут предупреждать агрегацию α-Syn (антиагрегационная активность) и могут приводить к диссоциации предварительно образованных агрегатов α-Syn (дезагрегационная активность). Кроме того, неожиданным образом антитела могут приводить к снижению образованию TNF-альфа и IL6, индуцированных микроглиальными клетками, что указывает на то, что эти антитела могут приводить к эффективному снижению агрегатов α-Syn или микроглиальной активации, индуцированной фибриллами. Также было обнаружено, что эти антитела приводят к снижению нейродегенерации, вызванной как экзогенными агрегатами α-Syn, так и эндогенными агрегатами α-Syn в клетках, сверхэкспрессирующих α-Syn. Кроме того, такие антитела распознают и специфически связываются с патологическими олигомерными агрегатами или фибриллами α-Syn, но не реагируют с не являющимся патологическим α-Syn. В частности, антитела реагируют с тельцами Леви из срезов головного мозга, взятых от пациентов с болезнью Паркинсона из группы альфа-синуклеинопатий, но не с нормальными тканями человека.
Неожиданным образом было обнаружено, что две мышиные модели болезни Паркинсона (мышиная модель с индукцией MPP+ и мышиная модель с инокуляцией фибриллами α-Syn), которым вводили композиции, содержащие пептидные иммуногенные конструкции α-Syn, (a) приводили к образованию антител, которые были высоко перекрестно-реактивными в отношении β-складки α-Syn, (b) имели снижению уровней α-Syn в сыворотке крови, (c) имели снижение уровней олигомерного α-Syn в головном мозге, и (d) имели снижению нейропатологии, приводя к восстановлению двигательной функции.
Полученные в результате иммунные ответы от животных, иммунизированных пептидными иммуногенными конструкциями α-Syn по данному изобретению, продемонстрировали способность конструкций приводить к образованию эффективных сайт-направленных антител, которые реагируют с денатурированной β-складкой α-Syn в формах мономеров, олигомеров и фибрилл, но не случайной спиральной структуры C-терминального участка α-Syn в его нативной форме.
Функциональные анализы in vitro
Антитела, образуемые пептидными иммуногенными конструкциями α-Syn, могут быть использованы в функциональных анализах in vitro. Эти функциональные анализы включают в себе, но не ограничиваясь ими:
(a) ингибирование агрегации рекомбинантного α-Syn in vitro; и дезагрегацию предварительно образованных агрегатов рекомбинантного α-Syn (см. Пример 8);
(b) ингибирование агрегации клеточного α-Syn, и диссоциацию предварительно образованных агрегатов α-Syn в клетках (см. Пример 9);
(c) снижение секреции TNF-альфа и IL6 в микроглии (см. Пример 10);
(d) снижение нейродегенерации, вызываемой экзогенными агрегатами α-Syn (см. Пример 11);
(e) снижение нейродегенерации в клетках, сверхэкспрессирующих α-Syn (см. Пример 12);
(f) доказательство эффективности in vivo в модели болезни Паркинсона у мышей с инокуляцией фибриллярного α-Syn и индукцией MPP+, показывающее снижение уровня α-Syn в сыворотке крови, снижение уровня олигомерного α-Syn в головном мозге, снижение нейропатологии и восстановление двигательной активности (см. Пример 15).
Способы
Данное раскрытие также связано со способами получения и применения пептидных иммуногенных конструкций α-Syn, композиций и фармацевтических композиций.
a. Способы получения пептидной иммуногенной конструкции α-Syn
Пептидные иммуногенные конструкции α-Syn по данному раскрытию могут быть получены с помощью способов химического синтеза, хорошо известных специалисту в данной области техники (см., например, Fields et al., Chapter 3 in Synthetic Peptides: A User's Guide, ed. Grant, W. H. Freeman & Co., New York, NY, 1992, p.77). Пептидные иммуногенные конструкции α-Syn могут быть синтезированы с помощью автоматических методик по Меррифилду твердофазного синтеза с использованием α-NH2, защищенной либо химической структурой t-Boc, либо химической структурой F-moc, с помощью аминокислот с защищенной боковой цепью, например, в синтезаторе пептидном Applied Biosystems модели 430A или 431. Получение пептидных иммуногенных конструкций α-Syn, содержащих пептиды комбинаторных библиотек для Th-эпитопов, можно выполнять в результате получения смеси альтернативных аминокислот для связывания в определенном варьирующем положении.
После полной сборки необходимой пептидной иммуногенной конструкции α-Syn смолу можно обрабатывать в соответствии со стандартными процедурами с целью отделения пептида от смолы и функциональные группы на боковых цепях аминокислот могут быть разблокированы. Свободный пептид может быть очищен с помощью ВЭЖХ и охарактеризован биохимически, например, с помощью аминокислотного анализа или секвенирования. Способы очистки и характеристики пептидов хорошо известны специалисту в данной области техники.
Качество пептидов, образуемых в результате этого химического процесса, можно контролировать и определять, и в результате можно гарантировать воспроизводимость пептидных иммуногенных конструкций α-Syn, иммуногенность и выход. Подробное описание получения пептидной иммуногенной конструкции α-Syn с помощью твердофазного синтеза пептидов продемонстрировано в Примере 1.
Было обнаружено, что диапазон структурной изменчивости, который допускает сохранение предполагаемой иммунологической активности, является гораздо более подходящим, чем диапазон структурной изменчивости, допускаемый для сохранения специфической активности лекарственного средства с помощью низкомолекулярного лекарственного средства или необходимых видов активности и нежелательных видов токсичности, встречающихся в крупных молекулах, которые образуются совместно с лекарственными средствами, получаемыми биологически. Таким образом, аналоги пептидов, либо намеренно разработанные, либо неизбежно получаемые в результате ошибок процесса синтеза в виде смеси побочных продуктов удаленной последовательности, которые имеют хроматографические и иммунологические свойства, аналогичные предполагаемому пептиду, часто так же эффективны, как очищенный препарат необходимого пептида. Разработанные аналоги и непреднамеренные смеси аналогов эффективны до тех пор, пока процедура QC для распознания разрабатывают для мониторинга как производственного процесса, так и процесса оценки продукта, чтобы гарантировать воспроизводимость и эффективность конечного продукта, использующего эти пептиды.
Пептидные иммуногенные конструкции α-Syn также могут быть получены с использованием технологии рекомбинантной ДНК, в том числе молекул нуклеиновой кислоты, векторов и/или клеток-хозяев. Таким образом, молекулы нуклеиновой кислоты, кодирующие пептидную иммуногенную конструкцию α-Syn, и их иммунологически функциональные аналоги также охвачены данным раскрытием в виде части данного изобретения. Аналогичным образом, векторы, в том числе экспрессионные векторы, содержащие молекулы нуклеиновой кислоты, а также клетки-хозяева, содержащие указанные векторы, также охвачены данным раскрытием в виде части данного изобретения.
Различные иллюстративные варианты осуществления также охватывают способы получения пептидной иммуногенной конструкции α-Syn и иммунологически функциональных аналогов пептидных иммуногенных конструкций, происходящих из фрагмента G111-D135 α-Syn. Например, способы могут включать этап инкубации клетки-хозяина, содержащей экспрессионный вектор, содержащий молекулу нуклеиновой кислоты, кодирующую пептидную иммуногенную конструкцию α-Syn и/или ее иммунологически функциональный аналог, в таких условиях, когда экспрессируется пептид и/или аналог.Более длинные синтетические пептидные иммуногены могут быть синтезированы с помощью хорошо известных методик рекомбинантной ДНК. Такие методики представлены в известных стандартных руководствах с подробными протоколами. Для конструирования гена, кодирующего пептид по данному изобретению, аминокислотную последовательность подвергают обратной трансляции для получения последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность, предпочтительно с кодонами, которые являются оптимальными для организма, в которой ген подлежит экспрессии. Затем синтетический ген получают в типичном случае с помощью синтеза олигонуклеотидов, которые кодируют пептид и любые регуляторные элементы, при необходимости. Синтетический ген вставляется в подходящий клонирующий вектор и трансфицируют в клетку-хозяина. Затем пептид экспрессируется в соответствующих условиях, пригодных для выбранной системы экспрессии и хозяина. Пептид очищают и характеризуют с помощью стандартных способов.
b. Способы получения иммуностимулирующих комплексов
Различные иллюстративные варианты осуществления также охватывают способы получения иммуностимулирующих комплексов, содержащих пептидные иммуногенные конструкции α-Syn и молекулу олигодезоксинуклеотида (ODN) CpG. Стабилизированные иммуностимулирующие комплексы (ISC) происходят из катионной части пептидной иммуногенной конструкции α-Syn и полианионной молекулы CpG ODN. Самособирающаяся система активируется с помощью электростатической нейтрализации заряда. Стехиометрия молярного отношения заряда катионной части пептидной иммуногенной конструкции α-Syn по отношению к анионному олигомеру определяет степень ассоциации. Нековалентная электростатическая ассоциация пептидной иммуногенной конструкции α-Syn и CpG ODN представляет собой полностью воспроизводимый процесс.Агрегаты иммуностимулирующего комплекса пептид/CpG ODN, которые облегчают презентацию «профессиональным» антигенпрезентирующим клеткам (АРС) иммунной системы, таким образом, дополнительно усиливают иммуногенность комплексов. Эти комплексы легко характеризуются для контроля качества при получении. Пептид/CpG ISC хорошо переносится in vivo. Указанная новая гранулированная система, содержащая CpG ODN и пептидные иммуногенные конструкции, происходящие из фрагмента G111-D135 α-Syn, была разработана с тем, чтобы использовать преимущества обобщенной В-клеточной митогенности, ассоциированной с использованием CpG ODN, и в то же время способствовать сбалансированным ответам типа Th-1/Th-2.
CpG ODN в раскрытых фармацевтических композициях на 100% связан с иммуногеном в процессе, опосредованном электростатической нейтрализацией противоположного заряда, приводя к образованию частиц микронного размера. Гранулированная форма позволяет значительно снизить дозировку CpG по сравнению со стандартным применением адъювантов CpG, снизить вероятность неблагоприятных врожденных иммунных реакций и облегчить альтернативные пути процессинга иммуногена, в том числе антиген-презентирующие клетки (АПК). Следовательно, такие составы являются новыми концептуально и предлагают потенциальные преимущества, активируя стимуляцию иммунных реакций с помощью альтернативных механизмов.
с. Способы получения фармацевтических композиций
Различные иллюстративные варианты осуществления также охватывают фармацевтические композиции, содержащие пептидные иммуногенные конструкции α-Syn. В определенных вариантах осуществления в фармацевтических композициях используют эмульсии типа вода в масле и в суспензии с минеральными солями.
С целью применения фармацевтической композиции большим количеством населения и с целью профилактики агрегации α-Syn, также являющейся частью цели введения, безопасность становится другим важным фактором для рассмотрения. Несмотря на применение эмульсий типа масло в воде у людей для многих составов в клинических исследованиях, квасцы остаются основным адъювантом для применения в составах в связи с их безопасностью. Квасцы или их минеральные соли фосфат алюминия (ADJUPHOS), таким образом, часто используют в качестве адъювантов при получении для клинических областей применения.
d. Способы применения фармацевтических композиций
Данное раскрытие также включает в себя способы применения фармацевтических композиций, содержащих пептидные иммуногенные конструкции α-Syn.
В определенных вариантах осуществления фармацевтические композиции, содержащие пептидные иммуногенные конструкции α-Syn, могут быть использованы для:
(a) ингибирования агрегации α-Syn в организме хозяина;
(b) индукции дезагрегации предварительно образованных агрегатов α-Syn в организме хозяина;
(c) снижения секреции TNF-альфа и IL6 в микроглии в организме хозяина;
(d) снижения нейродегенерации, вызываемой экзогенными агрегатами α-Syn в организме хозяина;
(e) снижения нейродегенерации в клетках, сверхэкспрессирующих α-Syn;
(f) снижения уровней α-Syn в сыворотке крови в организме хозяина;
(g) снижения уровня олигомерного α-Syn в головном мозге хозяина;
(h) снижения нейропатологии и восстановления двигательной активности в организме хозяина и т.п.
Вышеописанные способы включают в себя введение фармацевтической композиции, содержащей фармакологически эффективное количество пептидной иммуногенной конструкции α-Syn хозяину, нуждающемуся в этом.
Конкретные варианты осуществления
Конкретные варианты осуществления данного изобретения включают в себя, но не ограничиваясь ими, следующие.
(1) Пептидная иммуногенная конструкция альфа-синуклеина (α-Syn), содержащая:
B-клеточный эпитоп, содержащий от приблизительно 10 до приблизительно 25 аминокислотных остатков из C-терминального фрагмента α-Syn, соответствующего от приблизительно аминокислоты G111 до приблизительно аминокислоты D135 SEQ ID NO: 1;
T-хелперный эпитоп, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 70-98; и
необязательный гетерологичный спейсер, выбранный из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys и ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148),
где B-клеточный эпитоп ковалентно связан с T-хелперным эпитопом непосредственно или с помощью необязательного гетерологичного спейсера.
(2) Пептидная иммуногенная конструкция α-Syn из (1), где B-клеточный эпитоп выбирают из группы, состоящей из SEQ ID NO: 12-15, 17 и 49-63.
(3) Пептидная иммуногенная конструкция α-Syn из (1), где Т-хелперный эпитоп выбирают из группы, состоящей из SEQ ID NO: 81, 83 и 84.
(4) Пептидная иммуногенная конструкция α-Syn из (1), где необязательный гетерологичный спейсер представляет собой (α, ε-N)Lys или ε-N-Lys-Lys-Lys-Lys (SEQ ID NO: 148).
(5) Пептидная иммуногенная конструкция α-Syn из (1), где T-хелперный эпитоп ковалентно связан с аминоконцом B-клеточного эпитопа.
(6) Пептидная иммуногенная конструкция α-Syn из (1), где T-хелперный эпитоп ковалентно связан с аминоконцом B-клеточного эпитопа с помощью необязательного гетерологичного спейсера.
(7) Пептидная иммуногенная конструкция α-Syn из (1), содержащая следующую формулу:
(Th)m-(A)n-(С-терминальный фрагмент α-Syn)-X
или
(С-терминальный фрагмент α-Syn)-(A)n-(Th)m-X,
где
Th представляет собой T-хелперный эпитоп;
A представляет собой гетерологичный спейсер;
(С-терминальный фрагмент α-Syn) представляет собой B-клеточный эпитоп;
X представляет собой α-COOH или α-CONH2 аминокислоты;
m составляет от 1 до приблизительно 4; и
n составляет от 1 до приблизительно 10.
(8) Пептидная иммуногенная конструкция α-Syn из (1), содержащая аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 107, 108, 111-113 и 115-147.
(9) Пептидная иммуногенная конструкция α-Syn из (1), содержащая аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 107, 108 и 111-113.
(10) Композиция, содержащая пептидную иммуногенную конструкцию α-Syn из (1).
(11) Композиция, содержащая более чем одну пептидную иммуногенную конструкцию α-Syn из (1).
(12) Композиция из (11), где пептидная иммуногенная конструкция α-Syn имеет аминокислотные последовательности SEQ ID NO: 112 и 113.
(13) Фармацевтическая композиция, содержащая пептидную иммуногенную конструкцию α-Syn из (1) и фармацевтически приемлемое средство доставки и/или адъювант.
(14) Фармацевтическая композиция из (13), где
a. пептидную иммуногенную конструкцию α-Syn выбирают из группы, состоящей из SEQ ID NO: 107, 108, 111-113 и 115-147; и
b. адъювант представляет собой минеральную соль алюминия, выбранную из группы, состоящей из Al(OH)3 или AlPO4.
(15) Фармацевтическая композиция из (13), где
a. пептидную иммуногенную конструкцию α-Syn выбирают из группы, состоящей из SEQ ID NO: 107, 108, 111-113 и 115-147; и
b. пептидную иммуногенную конструкцию α-Syn смешивают с олигодезоксинуклеотидом CpG (ODN) с образованием стабилизированного иммуностимулирующего комплекса.
(16) Выделенное антитело или его эпитопсвязывающий фрагмент, которые специфически связываются с B-клеточным эпитопом пептидной иммуногенной конструкции α-Syn из (1).
(17) Выделенное антитело или его эпитопсвязывающий фрагмент из (16), связанные с пептидной иммуногенной конструкцией α-Syn.
(18) Выделенное антитело или его эпитопсвязывающий фрагмент, которые специфически связываются с B-клеточным эпитопом пептидной иммуногенной конструкции α-Syn из (9).
(19) Композиция, содержащая выделенное антитело или его эпитопсвязывающий фрагмент из (16).
(20) Композиция, содержащая выделенное антитело или его эпитопсвязывающий фрагмент из (18).
(21) Композиция из (20), содержащая смесь
a. выделенного антитела или его эпитопсвязывающего фрагмента, которые специфически связываются с B-клеточным эпитопом SEQ ID NO: 112; и
b. выделенного антитела или его эпитопсвязывающего фрагмента, которые специфически связываются с B-клеточным эпитопом SEQ ID NO: 113.
(22) Способ получения антител, которые распознают α-Syn в организме хозяина, включающий в себя введение хозяину композиции, содержащей пептидный иммуноген α-Syn из (1) и средство доставки и/или адъювант.
(23) Способ ингибирования агрегации α-Syn у животного, включающий в себя введение фармакологически эффективного количества пептидного иммуногена α-Syn из (1) животному.
(24) Способ снижения количества агрегатов α-Syn у животного, включающий в себя введение фармакологически эффективного количества пептидного иммуногена α-Syn из (1) животному.
(25) Способ идентификации агрегатов α-Syn различных размеров в биологическом образце, включающий в себя:
a. воздействие на указанный биологический образец антитела или его эпитопсвязывающего фрагмента в соответствии с (16) в условиях, которые позволяют антителу или его эпитопсвязывающему фрагменту связываться с агрегатами α-Syn; и
b. выявление количества антитела или его эпитопсвязывающего фрагмента, связанных с агрегатами α-Syn, в биологическом образце.
Подробное описание используемых процедур предложено в следующем разделе Примеры.
Пример 1
Синтез пептидов, связанных с альфа-синуклеином, и получение составов на их основе
a. Синтез С-терминальных фрагментов α-Syn
Описаны способы синтеза разработанных С-терминальных фрагментов α-Syn, которые включали в программу разработки пептидных иммуногенных конструкций α-Syn. Пептиды синтезировали в небольших количествах, которые являются пригодными для серологических анализов, лабораторных пилотных и полевых исследований, а также в больших (килограммовых) количествах, которые являются пригодными для промышленного/коммерческого производства фармацевтических композиций. Большой репертуар антигенных пептидов, связанных с α-Syn, имеющих последовательности от примерно 10 до 40 аминокислот, разрабатывали для скрининга и отбора наиболее оптимальных пептидных конструкций для применения в эффективной пептидной иммуногенной конструкции α-Syn.
Иллюстративные полноразмерные α-Syn (SEQ ID NO:1) и β-Syn (SEQ ID No: 2), сегменты α-Syn, такие как α-Syn111-132, α-Syn126-135, 10-мерные пептиды и т.д., используемые для картирования эпитопов в различных серологических анализах, идентифицированы в Табл. 1 (SEQ ID NO: 1 и 3-69). Выбранные фрагменты α-Syn выполняли в пептидных иммуногенных конструкциях α-Syn с помощью синтетического связывания с тщательно разработанным T-хелперным (Th) эпитопом, происходящим из патогенных белков, в том числе белка слияния вируса кори (MVF), поверхностного антигенного белка вируса гепатита B (HBsAg), белка вируса гриппа, белка Clostridum tetani и белка вируса Эпштейн-Барра (EBV), идентифицированных в Табл. 2 (SEQ ID NO: 70-98). Th-эпитопы использовали либо в одиночных последовательностях (SEQ ID NO: 70-78 и 83-98), либо комбинаторной библиотеке (SEQ ID NO: 79-82) с целью усиления иммуногенности их соответствующих пептидных иммуногенных конструкций α-Syn.
Иллюстративные пептидные иммуногенные конструкции α-Syn, выбранные из свыше 100 пептидных конструкций, идентифицированы в Табл. 3 (SEQ ID NO: 99-147). Все пептиды, используемые для исследования иммуногенности или соответствующих серологических тестов для детекции и/или измерения анти-α-Syn антител, синтезировали в небольшом количестве с помощью химического синтеза F-moc с использованием пептидных синтезаторов Applied BioSystems Models 430A, 431 и/или 433. Каждый пептид получали с помощью независимого синтеза на твердофазной подложке, с защитой F-moc на N-конце и боковыми защитными группами трифункциональных аминокислот.Готовые пептиды отделяли от твердой подложки, а защитные группы боковых цепей удаляли с помощью 90% трифторуксусной кислоты (TFA). Синтетические пептидные препараты оценивали с помощью времяпролетной масс-спектрометрии с лазерной ионизацией и десорбцией из жидкой матрицы (MALDI-TOF) для обеспечения соответствующего содержания аминокислот.Каждый синтетический пептид также оценивали с помощью ВЭЖХ с обращенной фазой (ОФ-ВЭЖХ) с целью подтверждения синтетического профиля и концентрации препарата. Несмотря на жесткий контроль процесса синтеза (в том числе пошаговый мониторинг эффективности связывания), также получали пептидные аналоги в результате непредусмотренных событий во время циклов удлинения, в том числе вставки, делеции, замены аминокислот и преждевременной терминации. Таким образом, синтезированные препараты в типичном случае включали в себя многочисленные пептидные аналоги наряду с целевым пептидом. Несмотря на включение таких непредусмотренных пептидных аналогов, образующиеся в результате синтезированные пептидные препараты, тем не менее, были подходящими для применения в иммунологических областях применения, в том числе иммунодиагностике (в виде антител для для захвата антигенов) и фармацевтических композициях (в виде пептидных иммуногенов). В типичном случае такие пептидные аналоги, либо специально разработанные, либо образованные в процессе синтеза в виде смеси побочных продуктов, часто являются такими же эффективными, как и очищенный препарат необходимого пептида, в том случае, если процедуру QC для распознания разрабатывают для мониторинга как процесса производства, так и процесса оценки продукта с целью гарантирования воспроизводимости и эффективности конечного продукта, использующего указанные пептиды. Синтез пептидов в большом масштабе в количестве множества килограммов выполняли на изготовленном по индивидуальному заказу автоматическом синтезаторе пептидов UBI2003 или подобном на уровне от 15 ммоль до 50 ммоль. Для активных ингредиентов, используемых в конечной фармацевтической композиции для клинических исследований, пептидные конструкции α-Syn очищали с помощью препаративной ОФ-ВЭЖХ при поверхностном градиенте в элюенте и характеризовали с помощью масс-спектрометрии MALDI-TOF, аминокислотного анализа и ОФ-ВЭЖХ в отношении чистоты и идентичности.
b. Получение композиций, содержащих пептидные иммуногенные конструкции α-Syn
Готовили составы, использующие эмульсии и суспензии типа масло в воде с минеральными солями. С целью применения разработанной фармацевтической композиции большим количеством населения и с целью профилактики, также являющейся частью цели введения, безопасность становится другим важным фактором для рассмотрения. Несмотря на применение эмульсий типа масло в воде у людей для многих фармацевтических композиций в клинических исследованиях, квасцы остаются основным адъювантом для применения в фармацевтических композициях в связи с их безопасностью. Квасцы или их минеральные соли ADJUPHOS (фосфат алюминия), таким образом, часто используют в качестве адъювантов при получении для клинических областей применения.
Вкратце, составы, определенные в каждой из групп исследования, описанных ниже, как правило, содержали все типы разработанных пептидных иммуногенных конструкций α-Syn. Свыше 100 разработанных пептидных иммуногенных конструкций α-Syn изначально оценивали у морских свинок в отношении их относительной иммуногенности с соответствующим пептидным представителем α-Syn В-эпитопного пептида иммуногена, а также для оценки серологических видов перекрестной реактивности среди различных гомологичных пептидов с помощью ИФА с использованием планшетов, покрытых различными пептидами, выбранными из пептидов с SEQ ID NO: 1-153.
Пептидные иммуногенные конструкции α-Syn получали (i) в эмульсии типа масло в воде с Seppic Montanide™ ISA 51 в качестве одобренного масла для применения у человека, или (ii) смешанными с минеральными солями ADJUPHOS (фосфат алюминия) или ALHYDROGEL (квасцы), в различных количествах пептидных конструкций, как описано. Композиции в типичном случае получали в помощью растворения пептидных иммуногенных конструкций α-Syn в воде при от приблизительно 20 до 800 мкг/мл и составляли с Montanide™ ISA 51 в эмульсиях типа масло в воде (1:1 в объеме) или с минеральными солями или ALHYDROGEL (квасцы) (1:1 в объеме). Композиции хранили при комнатной температуре в течение приблизительно 30 минут и смешивали с помощью вортекса в течение от приблизительно от 10 до 15 секунд до иммунизации. Некоторых животных иммунизировали 2-3 дозами определенной композиции, которую вводили во время 0 (примирование) и через 3 недели после исходной иммунизации (нед. после иммуниз.) (бустинг), необязательно через 5 или 6 нед. после иммуниз. в случае второго бустинга, с помощью внутримышечного пути. Этих иммунизированных животных затем исследовали с выбранным (выбранными) В-эпитопным (В-эпитопными) пептидом (пептидами) для оценки иммуногенности различных пептидных иммуногенных конструкций α-Syn, присутствующих в составе, а также их перекрестной реактивности с соответствующими целевыми пептидами или белками. Пептидные иммуногенные конструкции α-Syn с высокой иммуногенностью при исходном скрининге у морских свинок затем дополнительно исследовали как в эмульсии типа вода в масле, минеральных солях, так и составах на основе квасцов у приматов в отношении режимов дозирования в течение определенного периода согласно протоколам иммунизации.
Только наиболее перспективные пептидные иммуногенные конструкции α-Syn дополнительно тщательно оценивали до включения в конечные составы для исследований иммуногенности, продолжительности применения, токсичности и эффективности в доклинических исследованиях под контролем GLP при подготовке заявки на регистрацию нового экспериментального лекарственного препарата и клинических исследований у пациентов с синуклеинопатиями.
Пример 2
Получение рекомбинантного белка альфа-синуклеина
Клонирование гена α-Syn в векторе pGEX-4T1 ранее описывали в Neurotoxicology and teratology 2004, 26 (3): 397-406. Целевую последовательность (SEQ ID NO: 1) включали в вектор pGEX-4T1 между сайтами рестрикции BamHI и XhoI. Указанный фрагмент получали с помощью полимеразной цепной реакции (ПЦР) с использованием ДНК-полимеразы KAPA HiFi (Kapa Biosystems, Inc., Вуберн, Массачусетс, США). Праймерные последовательности были следующими: прямой праймер, 5'-cgggatccgatgtgtttatgaaaggtctgag-3((SEQ ID NO: 149); обратный праймер, 5'-ggaattccgatgtgtttatgaaaggtctgag-3((SEQ ID NO: 150). Условие ПЦР было следующим: денатурация при 94°C в течение 1 минуты с последующими 30 циклами денатурации при 94°C в течение 15 секунд, отжиг при 60°C в течение 30 секунд и удлинение при 68°C в течение 2 минут, затем терминация еще через 5 минут при 68°C. Сайт-направленный мутагенез A53T α-Syn выполняли с помощью набора для сайт-направленного мутагенеза Q5 (New England BioLabs, Беверли, Массачусетс, США). Праймерные последовательности мутантного α-Syn были следующими: прямой праймер, 5'-tcatggtgtgaccaccgttgcag-3((SEQ ID NO: 151); обратный праймер, 5'-accacgccttctttggttttg-3((SEQ ID NO: 152).
α-Syn, клонированный в вектор pGEX-4T1 GST, трансформировали в E.coli BL21 (DE3) для экспрессии белка. E.coli культивировали в бульоне LB при 37°C и изопропил-β-D-1-тиогалактопиранозид (IPTG) добавляли до конечной концентрации 4 мМ, когда значение ОП600 достигало 0,8. Через 4 часа после инкубирования клетки собирали с помощью центрифугирования при 5000 x g в течение 20 минут при 4°C. Собранные клетки ресуспендировали в PBS, разрушали с помощью сонификации на льду и затем центрифугировали при 5000 x g в течение 20 минут.Фракцию супернатанта загружали в колонку с глутатионсефарозой-4B (GE Healthcare), уравновешенную с помощью PBS. После трех раз промывания с помощью PBS 1 мл тромбина (20 ЕД/мл в PBS) добавляли для переваривания в течение ночи при 4°C с высвобождением GST из слитого белка. Не содержащий тэга α-Syn затем элюировали, после чего удаляли тромбин с помощью колонки с бензамидином HiTrap FF (GE Healthcare). Диализированный α-Syn незамедлительно замораживали при -80°C. Очищенный α-Syn с ММ 14 кДа идентифицировали с помощью вестерн-блоттинга с использованием анти-α-Syn антитела (1:2000, Millipore, целенаправленно воздействующего на α-Syn111-131) после разделения с помощью 10% SDS-PAGE.
Пример 3
Серологические анализы и реагенты
Серологические анализы и реагенты для оценки функциональной иммуногенности синтетических пептидных конструкций и составов на их основе подробно описаны ниже.
a. ИФА пептидов для анализа специфичности антител
ИФА для оценки образцов иммунных сывороток, описанных в следующих Примерах, разрабатывали и описывали ниже. Лунки 96-луночных планшетов покрывали отдельно на 1 час при 37°C 100 мкл фрагментов целевого белка α-Syn пептидом A85-A140, A91-A140, A101-A140, A111-A140, D121-A140, E126-A140, K97-D135, G101-D135, G111-D135, D121-D135, E123-D135, E126-D135, G101-132 и G111-G132 (SEQ ID NO: 4-17) в концентрации 2 мкг/мл (если не указано иное), в 10 мМ буфере NaHCO3, pH 9,5 (если не указано иное).
b. Оценка реактивности антител в отношении пептида Th с помощью ИФА пептида Th
Лунки, покрытые пептидом (SEQ ID No: 70-98), инкубировали с 250 мкл 3% по массе желатина в PBS при 37°C в течение 1 часа с целью блокирования неспецифических сайтов связывания белка, после чего три раза промывали PBS, содержащим 0,05% по объему TWEEN(20, и высушивали. Сыворотки, подлежащие анализу, разбавляли 1:20 (если не указано иное) с помощью PBS, содержащего 20% по объему нормальной козьей сыворотки, 1% по массе желатина и 0,05% по объему TWEEN(20. Сто микролитров (100 мкл) разбавленных образцов (например, сыворотки крови, плазмы крови) добавляли в каждую из лунок и оставляли реагировать в течение 60 минут при 37°C. Затем лунки шесть раз промывали 0,05% по объему TWEEN(20 в PBS с целью удаления несвязанных антител. Конъюгированное с пероксидазой хрена (HRP) видоспецифическое (например, мышиное, морской свинки или человеческое) козье анти-IgG использовали в качестве меченого индикатора для связывания с антигенным комплексом антитело/пептид, образованным в положительных лунках. Сто микролитров меченого пероксидазой козьего анти-IgG в предварительно титрованном оптимальном разведении и в 1% по объему нормальной козьей сыворотке с 0,05% по объему TWEEN(20 в PBS добавляли в каждую лунку и инкубировали при 37°C еще в течение 30 минут.Лунки промывали шесть раз 0,05% по объему TWEEN(20 в PBS с целью удаления несвязанного антитела и проводили реакцию с 100 мкл смеси субстратов, содержащей 0,04% по массе 3’, 3’, 5’, 5-тетраметилбензидин (TMB) и 0,12% по объему перекись водорода в натриево-цитратном буфере еще в течение 15 минут.Эту субстратную смесь использовали для детекции пероксидазной метки путем образования окрашенного продукта. Реакции останавливали в результате добавления 100 мкл 1,0 M H2SO4 и определяли поглощение при 450 нм (A450). Для определения титров антител иммунизированных животных, которые получали различные пептидные иммуногены, происхоящие из α-Syn, исследовали 10-кратные серийные разведения сывороток от 1:100 до 1:10000 и титр исследуемой сыворотки, выраженный в виде Log10, рассчитывали с помощью линейного регрессионного анализа А450 с порогом А450, установленным на 0,5.
c. Детальный анализ специфичности и картирование эпитопов фрагментов α-Syn с помощью ИФА 10-мерных пептидов кластера В-клеточных эпитопов
Детальный анализ специфичности анти-α-Syn антител в организмах иммунизированных хозяев определяли с помощью картирования эпитопов. Вкратце, лунки 96-луночных планшетов покрывали отдельными 10-мерными пептидами α-Syn (SEQ ID NO: 18-69) в концентрации 0,5 мкг на 0,1 мл на лунку, а затем 100 мкл образцов сыворотки (разведение 1:100 в PBS) инкубировали в лунках 10-мерных планшетов в двойных повторах в соответствии с этапами способа ИФА, описанного выше. В-клеточный эпитоп пептидной иммуногенной конструкции α-Syn и анти-α-Syn антитела иммунных сывороток из соответствующих детальных анализов специфичности у иммунизированных хозяев также исследовали с соответствующими пептидами α-Syn (SEQ ID No: 99, 102, 108, 110, 112, 113) или его фрагментом без спейсера и последовательностей Th или с β-Syn (SEQ ID NO: 153) для подтверждения дополнительной реактивности и специфичности.
d. Оценка иммуногенности
Образцы преиммунной и иммунной сыворотки от животных собирали в соответствии с протоколами иммунизации и нагревали при 56°C в течение 30 минут с целью инактивации факторов комплемента сыворотки. После введения фармацевтической композиции образцы крови получали в соответствии с протоколами и оценивали их иммуногенность в отношении специфического (специфических) целевого (целевых) сайта (сайтов). Последовательно разведенные сыворотки исследовали и положительные титры выражали в виде Log10 реципрокного разведения. Иммуногенность определенной фармацевтической композиции оценивали по ее способности вызывать ответ В-клеточных антител с высоким титром, направленный против специфичности необходимого эпитопа в целевом антигене, при этом сохраняется низкая или незначительная реактивность антител по отношению к «Т-хелперным эпитопам», используемым для усиления необходимых В-клеточных ответов.
e. Иммунологический анализ уровня α-Syn в иммунных сыворотках мышей
Уровни сывороточного α-Syn у мышей, получавших пептидные иммуногены, происходящие из α-Syn, измеряли с помощью сэндвич-ИФА (Cloud-clon, SEB222Mu) с использованием анти-α-Syn антител в качестве антитела захвата и меченого биотином анти-α-Syn антитела в качестве детекторного антитела. Вкратце, антитело иммобилизировали на 96-луночных планшетах в концентрации 100 нг/лунка в покрывающем буфере (15 мМ Na2CO3, 35 мМ NaHCO3, pH 9,6) и инкубировали при 4°C в течение ночи. Покрытые лунки блокировали с помощью 200 мкг/лунка аналитических разбавителей (0,5% BSA, 0,05% TWEEN®-20, 0,02% ProClin 300 в PBS) при комнатной температуре в течение 1 часа. Планшеты промывали 3 раза с помощью 200 мкл/лунка промывочного буфера (PBS с 0,05% TWEEN®-20). Очищенный рекомбинантный α-Syn использовали для получения стандартной кривой (диапазон от 156 до 1250 нг/мл при 2-кратном серийном разведении) в аналитическом разбавителе с 5% сывороткой мыши. 50 мкл разведенных сывороток (1:20) и стандартов добавляли в покрытые лунки. Инкубацию проводили при комнатной температуре в течение 1 часа. Все лунки аспирировали и промывали 6 раз с помощью 200 мкл/лунка промывочного буфера. Захваченный α-Syn человека инкубировали с 100 мкл раствора детекторого антитела (50 нг/мл HP6029, меченого биотином, в аналитическом разбавителе) при комнатной температуре в течение 1 часа. Затем связанный биотин-HP6029 детектировали с использованием стрептавидина поли-HRP (разведение 1: 10000, Thermo Pierce) в течение 1 часа (100 мкл/лунка). Все лунки аспирировали и промывали 6 раз с помощью 200 мкл/лунка промывочного буфера, и реакцию останавливали добавлением 100 мкл/лунка 1 М H2SO4. Стандартную кривую получали с помощью программного обеспечения SoftMax Pro (Molecular Devices) с подбором четырехпараметрической логистической кривой и использовали для расчета концентраций α-Syn во всех исследуемых образцах. t-Тесты Стьюдента использовали для сравнения данных с использованием программного обеспечения Prism.
f. Получение агрегатов α-Syn с рекомбинантным α-Syn
Для получения агрегированного α-Syn очищенный α-Syn дикого типа или мутантный по A53T α-Syn [0,1 мкг/мкл в 100 мкл буфера для агрегации PBS/KCl (2,5 мМ MgCl2, 50 мМ HEPES и 150 мМ KCl в 1 х PBS, pH 7,4)] инкубировали при 37°С в 1,5 мл пробирках Eppendorf в течение 7 дней в термомиксере (Eppendorf) без встряхивания. Агрегированный α-Syn незамедлительно замораживали при -80°C для последующего применения.
g. Очистка анти-α-Syn антител
Ант-α-Syn антитела очищали от сывороток, собранных через 3-15 недель после инъекции (нед. после инъекц.) морских свинок, иммунизированных пептидными иммуногенными конструкциями α-Syn, содержащими пептиды различных последовательностей (SEQ ID NO: 99-121) с использованием аффинной колонки (Thermo Scientific, Рокфорд). Вкратце, после уравновешивания буфера (0,1 М фосфата и 0,15 М хлорида натрия, рН 7,2) 400 мкл сыворотки добавляли в спин-колонку с белком G Nab с последующим перемешиванием снизу вверх и сверху внизу в течение 10 минут и центрифугированием при 5800 x g в течение 1 минуты. Колонку трижды промывали буфером для связывания (400 мкл). Затем в спин-колонку добавляли элюирующий буфер (400 мкл, 0,1 М глицин, рН 2,0) с целью элюирования антител после центрифугирования при 5800 x g в течение 1 минуты. Элюированные антитела смешивали с нейтрализующим буфером (400 мкл, 0,1 М Tris, рН 8,0) и измеряли концентрации этих очищенных антител с помощью Nan-Drop при значении ОП280, при этом в качестве стандарта использовали BSA (бычий сывороточный альбумин).
h. Специфичность анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn различных размеров
Вестерн-блот использовали для скрининга анти-α-Syn антител из антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, в отношении специфичности связывания с молекулярным комплексом α-Syn различных размеров. 20 мкМ α-Syn разделяли на 12% Tris-глициновом SDS-PAGE и переносили на нитроцеллюлозную (NC) мембрану перед обработкой путем фотоиндуцированной перекрестной сшивки (PICUP). Мембрану инкубировали с анти-α-Syn антителами, очищенными от антисывороток морских свинок в концентрации 1 мкг/мл, и затем инкубировали с конъюгированным с HRP ослиным анти-антитело морской свинки (706-035-148, Jackson). Блот визуализировали с помощью хемилюминесцентного реагента ECL Pro Western Lightning (PerkinElmer). В результате мономерный α-Syn (ММ 14460 Да) подвергали блоттингу с размером около 14 кДа, в то время как димер, тример или олигомеры имели свои молекулярные массы, в несколько раз больше, чем мономерный α-Syn размером 14 кДа. В качестве положительного контроля использовали коммерческое антитело, которое способно приводить к детекции различных молекул олигомеров, таких как димеры, тримеры и более крупные олигомеры, Syn211 (Abcam).
i. Дот-блоттинг с использованием различных молекул амилоидогенных белков
Получение α-спиральных мономеров, β-складчатых мономеров, β-складчатых олигомеров и β-сладчатых фибрилл Aβ1-42, Tau и α-Syn описано далее.
1. α-Спиральные мономеры Aβ1-42: 20 мкг β-складчатых мономеров Aβ1-42 (50 мкл) добавляли в 1x PBS, содержащий 20% трифторуксусную кислоту и 20% гексафторизопропанол (10 мкл) и инкубировали при 4°C в течение 24 часов с образованием α-спиральных мономеров.
2. β-Складчатые мономеры Aβ1-42: 60 мкг Aβ1-42 в 120 мкл 1x PBS, содержащего 5% TFA, агрегированного при 37°C в течение 24 часов, переносили на 10 кДа отсекающий фильтр (Millipore) с извлечением β-складчатых мономеров.
3. β-Складчатые олигомеры Aβ1-42: 60 мкг Aβ1-42 в 120 мкл 1x PBS, агрегированного при 37°C в течение 3 дней, сонифицировали на льду и переносили на 10 и 30 кДа отсекающие фильтры (Millipore) с извлечением β-складчатых олигомерных фибрилл менее чем 35 кДа.
4. β-Складчатые фибриллы Aβ1-42: 60 мкг Aβ1-42 в 120 мкл 1x PBS, агрегированного при 37°C в течение 3 дней, сонифицировали на льду и переносили на 30 кДа отсекающие фильтры (Millipore) с выделением β-складчатых фибрилл.
5. α-Спиральные мономеры α-Syn: 40 мкг свежеприготовленного α-Syn растворяли в 100 мкл холодного 1x PBS при 4°C и незамедлительно переносили на 10 кДа отсекающий фильтр (Millipore) с извлечением α-спиральных мономеров.
6. β-Складчатые мономеры α-Syn: 40 мкг α-Syn, инкубированного в 100 мкл буфера PBS/KCl при 37°C в течение 24 часов, переносили на 10 кДа отсекающий фильтр (Millipore) с извлечением β-складчатых мономеров.
7. β-Складчатые олигомеры α-Syn: 40 мкг α-Syn, агрегированного в 100 мкл буфера PBS/KCl при 37°C в течение 8 дней, сонифицировали на льду и затем переносили на 30 и 100 кДа отсекающие фильтры (Millipore) с извлечением β-складчатых олигомеров.
8. β-Складчатые фибриллы α-Syn: 40 мкг α-Syn, агрегированного в 100 мкл буфера PBS/KCl при 37°C в течение 8 дней, сонифицировали на льду и затем переносили на 30 и 100 кДа отсекающие фильтры (Millipore) с выделением β-складчатых фибрилл.α-Спиральные мономеры Tau441: 60 мкг Tau в 100 мкл 1x PBS при 4°C и переносили на 100 кДа отсекающий фильтр (Millipore) с извлечением α-спиральных мономеров.
9. β-Складчатые мономеры Tau441: 60 мкг Tau, агрегированного в 100 мкл 1x PBS с 10 ЕД/мл гепарина при при 25°C в течение 48 часов, переносили на 100 кДа отсекающий фильтр (Millipore) при 4°C с извлечением β-складчатых мономеров.
10. β-Складчатые олигомеры Tau441: 60 мкг Tau, агрегированного в 100 мкл 1x PBS с 10 ЕД/мл гепарина при при 37°C в течение 48 часов, переносили на 100 и 300 кДа отсекающие фильтры (Pall) при 4°C с извлечением β-складчатых олигомеров.
11. β-Складчатые фибриллы Tau441: 60 мкг Tau, агрегированного в 100 мкл 1x PBS с 10 ЕД/мл гепарина при при 37°C в течение 6 дней, переносили на 300 кДа отсекающие фильтры (Pall) при 4°C с выделением β-складчатых фибрилл.
Указанные мономеры и олигомеры верифицировали с помощью флуоресценции тиофлавином-T (ThT, Sigma) или PAGE (электрофорез в полиакриламидном геле). Концентрации амилоидогенных белков измеряли с помощью Nano-Drop с использованием коммерческого амилоидогенного рабочего раствора Aβ1-42 в качестве стандарта. Указанные мономеры и олигомеры определяли индивидуально на мембранах PVDF с количеством 3 мкг в случае Aβ1-42, 4 мкг в случае α-Syn и 7 мкг в случае Tau. Мембраны инкубировали с анти-α-Syn антителами, очищенными от антисывороток морских свинок (разведение 1: 1000), в качестве первичного антитела, с последующей гибридизацией с конъюгированным с HRP вторичным анти-морской свинки антителом (1:5000; Vector Laboratories). Мембраны обрабатывали субстратами Luminata Western HRP (Bio-Rad, Геркуслес, Калифоврния, США) и сигналы регистрировали с помощью системы цифровых изображений ChemiDoc-It 810 (UVP Inc., Апленд, Калифорния, США).
j. Специфичность связывания с агрегированным α-Syn в клетках РС12, сверхэкспрессирующих α-Syn, при обработке фактором роста неровов (NGF)
Иммуноцитохимический анализ (ИЦХ) с помощью анти-α-Syn антител, очищенных от антисывороток морских свинок, собранных через 8 или 9 нед. после иммунзиц. в отношении исходных клеток PC12, клеток PC12 пустого контроля и клеток РС12, сверэкспрессирующих α-Syn после обработки NGF, проводили для оценки аффинности связывания антител, образование которых вызывается после иммунизации. Ядра клеток контрастно окрашивали DAPI (4',6-диамидино-2-фенилиндол). Фотографии выполняли с помощью флуоресцентного микроскопа, и отношение количества положительно окрашенных клеток к общему количеству клеток категорически оценивали с помощью -,+,++и+++, что составляет <1%, 1-15%, 16-50%, >50%.
Пример 4
Клетки и животные, используемые в исследованиях иммуногенности и эффективности
a. Клетки РС12, сверхэкпрессирующие α-Syn
Плазмиду pZD/XOL-L-α-Syn конструировали с помощью вставки последовательности кДНК, кодирующей полноразмерный α-Syn человека дикого типа или α-Syn с мутацией по A53T, в вектор pZD/XOL-L с промотором CMV. Конструкции трансфицировали в клетки РС12 с использованием реагента для трансфекции липофектамина LTX (Invitrogen, Карлсбад, Калифорния, США) в соответствии с процедурой производителя. 2,5 мкл смеси для трансфекции, 500 мкл среды Opti-MEM, 2,5 мкл реагента PLUS и 8,75 мкл липофектамина LTX смешивали и затем инкубировали в течение 25 минут при комнатной температуре. После замены среды для культивирования 1,5 мл среды для выращивания клеток RMPI 1640 500 мкл смеси для трансфекции добавляли непосредственно в каждую лунку с последующей инкубацией при 37°С в течение одного дня. Эффективность трансфекции подтверждали с помощью ПЦР и вестерн-блоттинга.
b. Морские свинки
Исследования иммуногенности проводили на зрелых, наивных, взрослых самцах и самках морских свинок Дункана-Хартли (300-350 г/массу тела). В экспериментах использовали не менее 3 морских свинок на группу. Протоколы с использованием морских свинок Дункана-Хартли (в возрасте 8-12 недель; Covance Research Laboratories, Денвер, Пенсильвания, США) выполняли в соответствии с утвержденными заявками IACUC в договорном виварии, а также в UBI в качестве спонсора.
c. Модель мышей с болезнью Паркинсона, инокулированных фибриллярным α-Syn
Самок мышей FVB (масса, варьирующая в диапазоне 25-30 г) поддерживали с циклом 12 часов света: 12 часов темноты, а уход за животными осуществляли в соответствии с одобренными руководствами AAALAC. Фибриллярный α-Syn получали с помощью инкубации пептидов α-Syn (5 мг/мл) при 37°С в 0,1% PBS, содержащим NaN3/высококонцентрированном KCl буфере без встряхивания в течение 7 дней. Фибриллизацию контролировали с помощью измерения флуоресценции ThT и подтверждение выполняли, когда сигнал увеличился более чем в 3 раза по сравнению с исходным мономером α-Syn. Вестерн-блоттинг также использовали для подтверждения агрегации α-Syn до инокуляции в черную субстанцию с одной стороны (передне-задний; -3,0 мм; медиально-латеральный: -1,3 мм; дорсально-вентральный: -4,7 мм от брегмы и твердой мозговой оболочки) и дорсального неостриатума (передне-задний;+0,2 мм; медиально-латеральный: -2 мм; дорсально-вентральный: -3,2 мм от брегмы и твердой мозговой оболочки) животных, анестезированных изофлураном.
d. Мышиная модель болезни Паркинсона с индукцией MPP+
Самок мышей Balb/c (масса, варьирующая в диапазоне 18-20 г) поддерживали с циклом 12 часов света: 12 часов темноты, а уход за животными осуществляли в соответствии с одобренными руководствами AAALAC. MPP+йоидид (Sigma, Сент-Луис, Миссури) растворяли в солевом растворе и инъецировали с 10 мкл раствора, содержащего 18 мкг MPP+йодида (0,8 мг/кг) в желудочек с одной стороны анестезированного животного. Стереотаксические координаты участка инъекции составляли: брегма -1,0 мм, латеральный 1,0 мм, глубина 2,0 мм.
Пример 5
Принцип разработки, скрининг, идентификация и оптимизация многокомпонентных фармацевтических композиций, включающих пептидные иммуногенные конструкции альфа-синуклеина
a. История разработки
Каждая пептидная иммуногенная конструкция или иммунотерапевтический продукт α-Syn требует своей индивидуальной направленности и подхода на основании конкретного механизма заболевания и целевого (целевых) белка (белков), необходимых для вмешательства. Мишени, разработки которых моделируются впоследствии, могут включать в себя клеточные белки, участвующие в пути заболевания, или инфекционный агент, в который могут быть вовлечены несколько белков из патогена. Процесс от исследования до коммерциализации является очень длительным, в типичном случае для завершения занимает одно или несколько десятилетий.
После выбора целевой молекулы требуется значительный процесс серологической проверки. Идентификация и распределение В-клеточных и Т-клеточных эпитопов в целевой молекуле важны для разработки пептидной иммуногенной конструкции α-Syn. После распознавания целевого В-клеточного эпитопа проводят последовательные пилотные исследования иммуногенности на мелких животных для оценки функциональных свойств антител, образование которых вызывается фармацевтическими композициями разработанных пептидов. Такое серологическое применение затем проводят на животных целевых видов для дальнейшей проверки иммуногенности пептидной иммуногенной конструкции α-Syn и функциональных свойств антител, образование которых вызывается. Все исследования проводятся в нескольких параллельных группах с сыворотками, взятыми у иммунизированных хозяев для оценки. Ранние исследования иммуногенности на целевых видах или приматах, не относящихся к человеку, в случае фармацевтических композиций для применения у человека, также проводят для дальнейшей проверки иммуногенности и направления разработки. Затем целевые пептиды получают в различных смесях для оценки незначительного различия функционального свойства, связанного с соответствующими взаимодействиями между пептидными конструкциями, при использовании в комбинациях для получения соответствующих разработок составов. После дополнительных оценок устанавливают конечные пептидные конструкции, пептидные композиции и составы на их основе, наряду с соответствующими физическими параметрами составов, приводящие к процессу разработки конечного продукта.
b. Разработка и валидация пептидных иммуногенных конструкций, происходящих из α-Syn, для фармацевтических композиций с возможностью лечения пациентов с синуклеинопатиями
С целью получения наиболее эффективных пептидных конструкций для включения в фармацевтические композиции, значительный репертуар неизбирательных Т-хелперных эпитопов, происходящих из различных патогенов или искусственных Т-хелперных эпитопов, дополнительно разработанных из последовательности белка слияния вируса кори (MVF - Measles Virus Fusion protein) или белка поверхностного антигена вируса гепатита В (HBsAg - Hepatitis B Surface Antigen protein), включали в исследования иммуногенности на морских свинках. Иллюстративное исследование пептидных конструкций, происходящих из α-Syn126-140, α-Syn121-140, α-Syn111-140, α-Syn101-140, α-Syn91-140, α-Syn85-140, α-Syn121-135, α-Syn111-135, α-Syn101-135, α-Syn97-135, α-Syn123-135, α-Syn126-135, α-Syn111-132 и α-Syn101-132, продемонстрировано в Табл. 3 (SEQ ID NO: 99-121), где пептид α-Syn связывали с помощью εK и/или KKK в качестве спейсера (спейсеров) с отдельными неизбирательными T-хелперными эпитопами.
i) Выбор C-терминальной части α-Syn в качестве мишени для разработки пептидных иммуногенов.
α-Syn представляет собой внутренне неупорядоченный белок. Он состоит из 140 аминокислот и разделен на три участка. N-терминальный участок (остатки 1-60) способен образовывать амфипатическую спираль, которая представляет собой типичную конформацию для распознавания и ассоциации мембран. Центральный участок, содержащий остатки 61-95, хорошо известен как неамилоидный β-компонент (NAC), впервые идентифицированный в сенильных бляшках при AD. Этот участок обладает высокой склонностью к образованию конформации с высоким содержанием β-складок и крайне подвержен агрегации. Различные типы посттрансляционных модификаций в этом участке демонстрируют различные эффекты на модулирование агрегации α-Syn. С-терминальный участок с остатками 96-140 имеет высокое содержание пролина и отрицательно заряженных остатков, которые представляют собой общую характеристику, встречающуюся во внутренне неупорядоченных белках, с целью поддержания растворимости. Указанный C-терминальный домен присутствует в случайной спиральной структуре в связи со своей гидрофобностью и высоким суммарным отрицательным зарядом. Исследования in vitro показали, что агрегацию α-Syn можно индуцировать с помощью снижения pH, который нейтрализует эти отрицательные заряды. α-Syn отличается чрезвычайным конформационным разнообразием, которое адаптируется к различным условиям в состояниях связывания с мембраной, цитозоля и амилоидной агрегации и выполняет разнообразные функции. При значительном рассмотрении С-терминальную случайную петлю и внутренне неупорядоченный участок, важные для поддержания растворимости белка, выбирали в качестве мишени для разработки пептидного иммуногена, поскольку этот участок был наиболее восприимчивым к модуляции антителом или другими физическими факторами, по сравнению с N-терминальной амфипатической спиралью и центральными областями с высоким содержанием β-складок.
ii) Идентификация аутологичных Th-эпитопов для исключения в разработке В-эпитопа α-Syn.
Предварительный анализ иммуногенности подтвердил наличие особенности структуры Т-хелперного (Т-хелперных) эпитопа (эпитопов) на С-конце α-Syn, где делеция пептидной последовательности с N-конца последовательности α-Syn делала пептиды α-Syn126-140 (SEQ ID NO: 9), α-Syn121-140 (SEQ ID NO: 8), α-Syn111-140 (SEQ ID NO: 7) полностью неиммуногенными, в то время как некоторую наиболее умеренную иммуногенность наблюдали в случае пептидов α-Syn101-140 (SEQ ID NO: 6), α-Syn91-140 (SEQ ID NO: 5) и α-Syn85-140 (SEQ ID NO: 4) (Табл. 4), указывая на присутствие потенциальной аутологичной Th-подобной структуры в C-терминальной последовательности. Включение такой последовательности при разработке B-эпитопа (В-эпитопов) может потенциально вызывать воспаление головного мозга при бустерной иммунизации в результате активации аутологичных Т-клеток, как в предыдущем патенте AN1792 для вакцины против болезни Альцгеймера. Таким образом, этот результат требует разработки пептидных иммуногенных конструкций α-Syn с В-клеточным (В-клеточными) эпитопом (эпитопами), начинающимся с аминокислотного остатка G111, с тем чтобы исключить любую возможность включения аутологичного Т-клеточного (Т-клеточных) эпитопа (эпитопов) при разработке В-эпитопа.
iii) Ранжирование гетерологичных T-хелперных эпитопов и их включение в пептидные иммуногенные конструкции α-Syn для восстановления и усиления иммуногенности выбранного пептида В-эпитопа α-Syn.
В Табл. 2 представлено в общей сложности 29 гетерологичных Th-эпитопов (SEQ ID NO: 70-98), которые исследовали в нашей группе в отношении их относительной эффективности у многих видах, таких как мыши, крысы, морские свинки, бабуины, макаки и т.д., для усиления иммуногенности B-клеточных эпитопов. Как продемонстрировано в Табл. 5, оба Т-клеточные эпитопы UBITh1 (SEQ ID NO: 83) и UBITh2 (SEQ ID NO: 84), происходящие из белка MvF, могут потенциировать неиммуногенный пептид α-Syn101-140 (SEQ ID NO: 6) до значительной и умеренной иммуногенности соответственно. Значительное исследование многочисленных пептидных иммуногенных конструкций, происходящих из α-Syn, выполняли с целью обеспечения ранжирования относительной иммуногенности среди указанных иммуногенных конструкций. Аналогичная иммунопотенцирующая активность встречается у UBITh3 (SEQ ID NO: 81), если он ковалентно связывается с помощью спейсера с различными C-терминальными пептидами α-Syn (SEQ ID NO: 4-9), как продемонстрировано в Табл. 6, при исследовании в ИФА с использованием планшета, покрытого длинным пептидом α-Syn A91-A140 (SEQ ID NO: 5).
iv) Оценка иммуногенности C-терминальных пептидных иммуногенных конструкций α-Syn в отношении реактивности их антител с соответствующими α-Syn и β-Syn.
Семейство синуклеинов включает в себя три известных белка: α-Syn, β-Syn и гамма-синуклеин. Все синуклеины имеют в качестве общего свойства высококонсервативный альфа-спиральный липидсвязывающий мотив со сходством с липидсвязывающими доменами класса А2 взаимозаменяемых аполипопротеинов. β-Syn является высокогомологичным по отношению к α-Syn. Предполагается, что β-Syn представляет собой ингибитор агрегации α-Syn, которая встречается при нейродегенеративных заболеваниях, таких как болезнь Паркинсона. Таким образом, β-Syn может защищать центральную нервную систему от нейротоксических эффектов α-Syn. Таким образом, предпочтительно, чтобы пептидные иммуногенные конструкции α-Syn вызывали образование антител, которые предпочтительно реагируют с α-синуклеином, а не с соответствующим защищающим от агрегации β-Syn. При исследовании шести пептидных иммуногенных конструкций, все их которых имели С-терминальное окончание с А140, все из антител, происходящих из иммунных сывороток указанных конструкций, показали значительную перекрестную реактивность в отношении β-Syn соответствующего размера, как продемонстрировано в Табл. 6. После тщательного изучения гомологии последовательностей между α-Syn и β-Syn (SEQ ID NO: 1 и 2) было продемонстрировано, что последовательность, соответствующая С-терминальным пяти аминокислотам YEPEA, была идентичной для двух белков. Таким образом, желательно разработать В-эпитоп (В-эпитопы), исключая последовательность, содержащую указанные пять аминокислот YEPEA. Таким образом, результат на основании исследований иммуногенности, показанных в Табл. 6, привел к делеции YEPEA (от Y136 до A140) при разработке B-эпитопа (В-эпитопов). При включении спейсерной последовательности и, например, искусственного T-хелперного пептида UBITh1 (SEQ ID NO: 83) в пептидную иммуногенную конструкцию α-Syn с использованием последовательностей B-клеточных эпитопов, за исключением хвоста YEPEA, как продемонстрировано с помощью пептидных иммуногенов α-Syn (SEQ ID NO: 107-114) в Табл. 7, все они стали высокоиммуногенными при оценке длинного пептида α-Syn K97-A140 (SEQ ID NO: 110). Ни одна из иммунных сывороток не реагировала с β-Syn. Исходя из данных, полученных из Табл. 6 и 7, разработка B-эпитопа для пептидных иммуногенных конструкций, таким образом, будет ограничена G111-D135 α-Syn и его фрагментами.
v) Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-Syn, реагировали исключительно с мономером бета-складчатым мономером, олигомером или фибриллой, но не α-спиральным мономером.
Несмотря на то, что мы использовали обоснованные принципы при нашей разработке пептидных иммуногенов α-Syn, неожиданным было обнаружение того, что антитела, полученные из разработанных пептидных иммуногенных конструкций α-Syn с В-эпитопами, имеющие свои последовательности, начинающиеся в G111 и заканчивающиеся в D135, или их фрагменты, антитела, образование которых вызывали, специфически реагируют с β-складкой мономера, олигомера и фибриллы α-Syn; и не реагирует с β-складкой Aβ1-42 или Tau1-441, таким образом, предлагая идеальные кандидаты в пептидную иммуногенную конструкцию α-Syn, как продемонстрировано иллюстративно с помощью пептидных иммуногенных конструкций α-Syn (SEQ ID NO: 112 и 113) на Фиг. 8.
vi) Расширение охвата MHC с помощью пептидных иммуногенных конструкций α-Syn с различными неизбирательными T-хелперными эпитопами.
При разработке фармацевтической композиции для лечения пациентов с различным генетическим фоном важно, чтобы эта разработка охватывала максимальную популяцию с различным генетическим фоном. В связи с этим исследовали синергетическую иммуногенность пептидных иммуногенных конструкций, происходящих из α-Syn, для такой комбинации. Поскольку неизбирательные T-хелперные эпитопы, происходящие из MVF или HBsAg, представляют собой одни из наиболее эффективных для обеспечения такого усиления иммуногенности, для такого исследования разрабатывали комбинацию пептидных конструкций, содержащих T-хелперный эпитоп.Было обнаружено, что смесь двух пептидных иммуногенных конструкций с одинаковыми B-эпитопами вызывает значительный иммунный ответ по сравнению с тем, который вызывается соответствующей отдельной пептидной конструкцией.
Пример 6
Направленный ответ антител, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, только в отношении целевого B-клеточного эпитопа
Хорошо известно, что все белки-носители (например, гемоцианин лимфы улитки (KLH) или другие белки-носители, такие как белки дифтерийного анатоксина (DT) и столбнячного анатоксина (TT)), используемые для усиления иммунного ответа, направленного против целевого пептида эпитопа В-клеток, с помощью химического конъюгирования такого пептида В-клеточного эпитопа с соответствующим белком-носителем будет вызывать образование более 90% антител, направленных против потенцирующего белка-носителя, и менее чем 10% антител, вновь направленных против целевого В-клеточного эпитопа у иммунизированных хозяев. В связи с этим представляет интерес оценка специфичности пептидных иммуногенных конструкций α-Syn по данному изобретению. Для оценки иммуногенности получали серию из восьми пептидных иммуногенных конструкций α-Syn (SEQ ID NO: 107-114) с B-клеточными эпитопами различной длины, которые связаны с помощью спейсерной последовательности с гетерологичным T-клеточным эпитопом UBITh1 (SEQ ID NO: 83). UBITh1 (T-хелперный пептид, используемый для иммунопотенцирования B-эпитопа) наносили на планшеты, а иммунные сыворотки морских свинок использовали для исследования перекрестных реактивностей с пептидом UBITh1, используемым для иммунопотенцирования. В отличие от высокой иммуногенности этих конструкций по отношению к соответствующим целевым B-эпитопам, о чем свидетельствовали высокие титры антител, образуемых по отношению к B-эпитопу (В-эпитопам), как продемонстрировано в Табл. 6 и 7, было обнаружено, что большинство, если не все, из иммунных сывороток были не реагируют с пептидом UBITh1, как продемонстрировано в Табл. 8.
Таким образом, простая разработка иммуногена, включающая целевой В-клеточный эпитоп, связанный с тщательно отобранным Т-хелперным эпитопом, позволяет получать направленный и чистый иммунный ответ, целенаправленной воздействующий только на В-клеточный эпитоп α-Syn. В случае разработки фармацевтической композиции, чем более специфичным является иммунный ответ, к которому она приводит, тем более высокий профиль безопасности требуется в отношении композиции. Таким образом, пептидные иммуногенные конструкции α-Syn по данному изобретению являются высокоспецифичными, но в то же время высокоэффективными в отношении своей мишени.
Пример 7
Картирование эпитопов для детального анализа специфичности с помощью иммунных сывороток (через 9 нед. после иммуниз.) против различных пептидных иммуногенных конструкций альфа-синуклеина
В исследовании детального картирования эпитопов (Табл. 9) для определения связывающего (связывающих) сайта (сайтов) антитела со специфическими остатками в С-терминальном участке α-Syn, синтезировали 52 перекрывающихся 10-мера (SEQ ID NO: 18-69), которые охватывают аминокислотную последовательность α-Syn (K80-A140). Два более длинных пептида (97-135, SEQ ID No: 10) и (111-132, SEQ ID No: 17) использовали в качестве положительного контроля. Указанные 10-мерные пептиды и два более длинных пептида отдельно наносили на лунки 96-луночных микротитрационных планшетов в виде твердофазных иммуноабсорбентов. Объединенные антисыворотки морских свинок добавляли с разведением 1:100 в буфере для разведения образцов в лунки планшета, покрытые 10-мерным пептидом в концентрации 2,0 мкг/мл, и затем инкубировали в течение одного часа при 37°С. После промывки лунок планшета промывочным буфером добавляли белок A/G, конъюгированный с пероксидазой хрена, и инкубировали в течение 30 минут. После повторного промывания PBS субстрат добавляли в лунки для измерения оптической плотности при 450 нм с помощью планшет-ридера для ИФА, образцы которого анализируют в двух повторах. Связывание антисывороток с соответствующим длинным пептидом α-Syn иммуногенной конструкции B-эпитопа представляет собой максимальное связывание.
Как продемонстрировано в Табл. 9, объединенные сыворотки морских свинок через 9 нед. после иммуниз., полученные соответственно из шести пептидных иммуногенных конструкций α-Syn [(K97-D135, SEQ ID No: 110), (G111-D135, SEQ ID No: 108), (G111-G132, SEQ ID No: 113), (E126-D135, SEQ ID No: 112), (G101-A140, SEQ ID No: 104) и (E126-A140, SEQ ID No: 99)] выбирали для детального картирования эпитопов. Эти шесть фрагментов B-эпитопов различной длины полностью охватывают последовательность 97-140 С-терминального участка α-синуклеина. Результаты ИФА показали, что все шесть иммунных сывороток в значительной степени реагировали с иллюстративным длинным пептидом α-Syn (97-135, SEQ ID No: 10). В случае исследования по детальному картированию эпитопов 10-меров результаты показали охват иммуногенного эпитопа возле участка от AA114 до 125 (пептиды 114-123, 115-124, 116-125 SEQ ID No: 52, 53 и 54) и высокоиммуногенного участка на C-терминальном конце, представленном пептидом 131-140 (SEQ ID NO: 69). Интересно, что большинство иммунных сывороток, происходящих из C-терминальных пептидных иммуногенных конструкций α-Syn, вызывали образвоание антител, которые распознают не линейные, а конформационные эпитопы, за исключением одного, который расположен на C-конце α-Syn с последовательностью EGYQDYEPEA (SEQ ID NO: 69) и отвечает за перекрестную реактивность с белком β-Syn.
Обнаружение указанного эпитопа было менее ожидаемым, но высоко коррелировало с обнаружением того, что указанные антитела, происходящие из пептидных иммуногенных конструкций α-Syn, представленных 111-132 α-Syn (SEQ ID NO: 113) и 126-135 α-Syn (SEQ ID NO: 112) из C-терминального случайного петлевого участка α-Syn, которые связаны со структурой гетерологичных Th-эпитопов, приводили к образованию конформационной структуры, напоминающей денатурированную β-складку α-Syn, и не реагировали перекрестно с нативной α-спиралью α-Syn.
Пример 8
Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, и составами на их основе: антиагрегационные и дезагрегационные эффекты в отношении рекомбинантного белка альфа-синуклеина
Оценили влияние пептидных иммуногенных конструкций α-Syn в антиагрегационных и дезагрегационных анализах in vitro с использованием анти-α-Syn антител, очищенных от антисывороток морских свинок, в отношении рекомбинантного α-Syn.
a. Ингибирование агрегации α-Syn
Исходный скрининговый анализ различных анти-α-Syn антител, очищенных от морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn в отношении потенциальной антиагрегационной способности, проводили с помощью количественной оценки уровня изменений агрегации α-Syn с помощью измерения тиофлавина Т, как описано в Примере 3. Рекомбинантный α-Syn, полученный в PBS в концентрации 100 мкМ, затем инкубировали в 40 мкл буфера PBS/KCl (2,5 мМ MgCl2, 50 мМ HEPES и 150 мМ KCl в 1х PBS, pH 7,4) в концентрации 5 мкМ в 384-луночном планшете в течение 6 дней с тем, чтобы вызвать агрегацию. Различные концентрации (0,05, 0,5 или 5 мкг/мл) анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, собранных в различные временные точки, добавляли в инкубационную смесь для оценки соответственно эффектов на ингибирование агрегации α-Syn. К концу инкубации уровень агрегации определяли с использованием анализов ThT. Показания, полученные при каждом отрезке исследования, нормализовали, принимая уровень агрегации в контроле-носителе за 100% и принимая показания, полученные в отсутствие α-Syn, за 0%.
Как продемонстрировано в Табл. 10, три анти-α-Syn антитела, образование которых вызывали α-Syn111-132, α-Syn121-135 или α-Syn126-135, собранные через 9 нед. после иммуниз. и более, показали более эффективное и зависимое от концентрации ингибирование агрегации α-Syn. Из всех анализируемых анти-α-Syn антител четыре выбранных антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO:113), α-Syn121-135 (SEQ ID NO:107), α-Syn123-135 (SEQ ID NO:111) или α-Syn126-135 (SEQ ID NO:112) (собранные через 9 нед. после иммуниз.), демонстрировали ингибирующий эффект в отношении агрегации α-Syn почти на 40% (Фиг. 1) по сравнению с агрегацией контролем-носителем в виде 100%.
b. Диссоциация предварительно образованных агрегатов α-Syn
На основании вышеупомянутых исследований было отмечено, что анти-α-Syn антитела, очищенные от антисывороток морских свинок, иммунизированных определенными пептидными иммуногенными конструкциями α-Syn, оказывали эффекты на ингибирование агрегации α-Syn. Для дополнительной оценки того, остаются ли антитела, образование которых вызывали пептидными иммуногенными конструкциями α-Syn, эффективными при диссоциации предварительно образованных агрегатов α-Syn, проводили анализы дезагрегации in vitro с использованием анти-α-Syn антител, очищенных от антисывороток морских свинок.
α-Syn агрегировали в 200 мкл буфера PBS/KCl в концентрации 5 мкМ в течение 3 дней. После центрифугирования (13000 x g, 4°С, 30 минут) агрегаты α-Syn собирали и подтверждали с помощью анализов ThT. Предварительно образованные агрегаты α-Syn затем инкубировали в 100 мкл буфера PBS/KCl с или без анти-α-Syn антител, очищенных от антисывороток морских свинок (5 мкг/мл), в течение 3 дней. После инкубации агрегаты собирали после центрифугирования 13000 x g при 4°С в течение 30 минут и затем количественно оценивали с помощью анализа ThT, как описано в Примере 3. Остаточные агрегаты α-Syn после спонтанных диссоциаций в контроле-носителе нормализовали по отношению к 100%.
Два выбранных анти-α-Syn антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO:113) или α-Syn126-135 (SEQ ID NO:112), и комбинацией α-Syn111-132 (SEQ ID NO:113), образование которых вызывали, α-Syn126-135 (SEQ ID NO:112), образование которых вызывали анти-α-Syn антителами, исследовали указанном дезагрегационном анализе in vitro. В результате анти-α-Syn антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) и α-Syn111-132 (SEQ ID NO: 113), продемонстрировали диссоциирующий эффект на предварительно образованные агрегаты α-Syn почти на 50% по сравнению с контролем-носителем в виде 100%, в то время как другие анти-α-Syn антитела и антитела, очищенные от предварительно иммунизированных животных, не продемонстрировали сопоставимых эффектов (Фиг. 2).
Пример 9
Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, и составами на их основе: антиагрегационные и дезагрегационные эффекты в отношении кинетики агрегации α-SYN в клетках, сверхэкспрессирующих α-SYN
Известно, что агрегация α-Syn ускоряется во время дифференцировки нейронов. С целью оценки влияния пептидных иммуногенных конструкций α-Syn либо на ингибирование агрегации α-Syn, либо на диссоциацию предварительно образованных агрегатов α-Syn в условиях на клетках, анти-α-Syn антитела, очищенные от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, оценивали с помощью антиагегационных анализов и дезаграгационных анализов в обработанных NGF клетках РС12, сверхэкспрессирующих α-Syn, дифференцируемых в нейроны.
a. Ингибирование агрегации α-Syn
Клетки РС12, сверхэкспрессирующие α-Syn, высевали на предварительно покрытые поли-D-лизином 96-луночные планшеты и затем обрабатывали фактором роста нервов (NGF) (100 нг/мл) наряду с анти-α-Syn антителами, очищенными от морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn (0 или 0,5 мкг/мл), в течение 4 дней для подтверждения активности против агрегации.
Обработанные клетки лизировали и 20 мкг клеточных лизатов отделяли с помощью SDS-PAGE и затем детектировали с помощью антитела к α-Syn (BD). Количество детектируемых сигналов α-Syn в области с более высокой молекулярной массой оценивали количественно и затем нормировали по отношению к контролю-носителю в виде 100%. Как продемонстрировано на Фиг. 3, ингибирующие эффекты на количество агрегированного α-Syn до 80-90% наблюдали среди всех четырех выбранных анти-α-Syn антител, образование которых вызывали пептидными иммуногенными конструкциями α-Syn α-Syn111-132 (SEQ ID NO:113), α-Syn121-135 (SEQ ID NO:107), α-Syn123-135 (SEQ ID NO:111) или α-Syn126-135 (SEQ ID NO:112), по сравнению с количеством агрегированного α-Syn в контроле-носителе.
b. Диссоциация предварительно образованных агрегатов α-Syn
С целью подтверждения дезагрегацинной активности в отношении предварительно образованых агрегатов α-Syn клетки PC12, сверхэкспрессирующие α-Syn, обрабатывали NGF (100 нг/мл) в течение 3 дней для нейрональной дифференцировки, чтобы инициировать агрегацию α-Syn, перед тем, как дополнительно обрабатывать анти-α-Syn антителами, очищенными от морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn (0 или 0,5 мкг/мл), в течение еще 4 дней.
Обработанные клетки лизировали и 20 мкг клеточных лизатов отделяли с помощью SDS-PAGE и затем детектировали с помощью антитела к α-Syn (BD). Количество детектируемых сигналов α-Syn в области с более высокой молекулярной массой оценивали количественно и затем нормировали по отношению к контролю-носителю в виде 100%. Как также продемонстрировано на Фиг. 3, снижение количества агрегированного α-Syn на 50-60% наблюдали при использовании анти-α-Syn антител, образование которых вызывали пептидными иммуногенными конструкциями α-Syn121-135 (SEQ ID NO:107), α-Syn123-135 (SEQ ID O:111) или α-Syn126-135 (SEQ ID NO:112), в то время как анти-α-Syn антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113), продемонстрировали более 90% снижение количества агрегированного α-Syn.
Пример 10
Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, и составами на их основе: влияние снижения секреции TNF-(и IL-6 в микроглии
Считается, что повреждение нигральных нейронов приводит к высвобождению агрегированного α-Syn в черную субстанцию, что активирует микроглию, при этом продуцируются провоспалительные медиаторы, тем самым приводя к стойкой и прогрессирующей нигральной нейродегенерации при PD. Для оценки эффектов снижения активации микроглии анти-α-Syn антителами, очищенными от морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, измеряли количество провоспалительных медиаторов, TNF-((фактор некроза опухоли альфа) и IL-6 (интерлейкин -6), высвобождаемых микроглией при обработке агрегатами α-Syn в присутствии или отсутствии различных анти-α-Syn антител.
Мышиные клетки BV2 или человеческие клетки SVG p12 высевали по 5000 клеток/лунка в среде RPMI 1640 с добавлением 1% FBS. Клетки обрабатывали 1 мкМ α-Syn и инкубировали при 37°C, 5% CO2 в увлажненной атмосфере в течение 24 часов. После этого ксреду для культивирования клеток собирали, центрифугировали и супернатанты выделяли. Концентрации IL-6, секретируемого клетками BV2, и TNF-α, секретируемого клетками SVG p12 в супернатантах, анализировали в трех повторах, используя наборы ИФА для IL-6 мыши или TNF-(человека (Thermofisher) соответственно. Сигнал нормализовали до контроля-носителя в виде 100%.
Данные показали, что анти-α-Syn антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113) и α-Syn123-135 (SEQ ID NO: 111), снижали опосредованное агрегатами α-Syn высвобождение TNF-(клетками SVG p12 на 30-50%, тогда как анти-α-Syn-антитела, образование которых вызывали α-Syn123-135 (SEQ ID NO: 111), снижали высвобождение IL-6 клетками SVGP12 примерно на 30% (Фиг. 4). Результаты указывали на то, что анти-α-Syn-антитела, образование которых вызывали α-Syn123-135 (SEQ ID NO: 111), были более эффективными, чем другие анти-α-Syn-антитела, исследуемые при снижении опосредованной агрегатами α-Syn активации микроглии.
Пример 11
Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, и составами на их основе: влияние на снижение нейродегенерации, вызванной экзогенным альфа-синуклеином
С целью оценки нейропротекторных эффектов анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, принимали модель нейродегенерации in vitro с использованием экзогенных, предварительно образованных агрегатов α-Syn в отношении обработанных NGF клеток РС12, дифференцируемых в нейроны.
Клетки РС12 обрабатывали NGF (100 нг/мл) в течение 6 дней с целью индукции дифференцировки нейронов. Морфологию дифференцируемых в нейроны клеток подтверждали и анализировали с помощью системы анализа изображений InCell с высоким содержанием (GE Healthcare). Нейротрофические эффекты NGF отражались на разрастании нейритов, и количество дифференцируемых в нейроны клеток оценивали количественно. Уровни разрастания нейритов и количество дифференцируемых клеток показывали в процентах (среднее (SEM) после нормализации. Длину нейритов клеток РС12 с обработкой и без обработки NGF принимали за 100% и 0% соответственно. Количество дифференцируемых в нейроны клеток РС12 через 6 дней после обработки NGF нормализовали до 100%.
Нейродегенерацию наблюдали с помощью добавления экзогенных, предварительно образованных агрегатов α-Syn к дифференцируемым в нейроны клеткам PC12. В присутствии предварительно образованных агрегатов α-Syn длина нейрита укорачивалась, а количество клеток в дифференцируемых в нейроны клетках РС12 уменьшалось. Указанная нейродегенерация, вызванная агрегатами α-Syn, была пропорциональна количеству добавленных экзогенных агрегатов α-Syn и могла блокироваться куркумином, широко известным своими нейропротективными эффектами по отношению к нейротоксичности агрегатов α-Syn, зависимым от концентрации образом. Коммерчески доступные анти-α-Syn антитела (BD bioscience), но не антитела, очищенные от наивных морских свинок, ослабляли нейродегенерацию, вызванную агрегатами α-Syn. Указанную модель принимали в качестве платформы для скрининга, чтобы определить, какие анти-α-Syn-антитела, очищенные от антисывороток морских свинкок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, обладали нейропротекторными эффектами в восстановлении роста нейритов, а также выживания нейронов в зависимости от концентрации (Табл. 11 и 12).
В результате анти-α-Syn-антитела, очищенные от антисывороток морских свинок, иммунизированных более чем половиной различных пептидных иммуногенных конструкций α-Syn, восстанавливали концентрацию нейритов в зависимости от концентрации (Табл. 11), а анти-α-Syn-антитела, очищенные от антисывороток морских свинок, иммунизированных почти всеми различными пептидными иммуногенными конструкциями α-Syn, защищали клетки РС12, дифференцируемые в нейроны, от гибели нейронов, вызванной агрегатами α-Syn (Табл. 12). Учитывая указанные два различные параметра вместе, обнаружили, что почти треть проанализированных анти-α-Syn-антител обладает эффектами как в отношении длины нейритов, так и выживаемости клеток против нейротоксичности агрегатов α-Syn. Антинейродегенеративные эффекты анти-α-Syn антител, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113), α-Syn126-135 (SEQ ID NO: 112), и преиммунными антителами от наивных морских свинок, наблюдали и количественно оценивали длину нейритов и количество клеток с кальцеином AM (Life Technologies), флуоресцентным красителем для маркировки живых клеток. Было продемонстрировано, что в клетках РС12, дифференцируемых в нейроны, с высоким содержанием нейритов, анти-α-Syn-антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113) (Фиг. 5B) и α-Syn126-135 (SEQ ID NO: 112) (Фиг. 5С), но не преиммунные антитела, очищенные от наивных морских свинок (Фиг. 5А), продемонстрировали защитные эффекты в отношении опосредованного агрегатами α-Syn укорочения длины нейритов.
Пример 12
Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, и составами на их основе: влияние на снижение нейродегенерации в клетках, сверхэкспрессирующих α-SYN
С целью оценки нейропротекторных эффектов анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, принимали модели нейродегенерации in vitro с использованием клеток РС12 дикого типа, сверхэспрессирующих α-Syn, и клеток РС12, мутантных по A53T, сверхэспрессирующих α-Syn.
После инкубации с NGF в клетках пустого контроля (трансфицированные плазмидным вектором) происходило удлинение длинных нейритов и увеличивалось количество клеток, аналогичных исходным клеткам РС12 дикого типа, тогда как в клетках РС12 дикого типа, сверхэкспрессирующих α-Syn, и клетках РС12, сверхэкспрессирующих α-Syn, с мутацией по A53T, не смогло происходить сопоставимое удлинение нейритов или увеличение количества клеток, подтверждая нейродегенеративные эффекты, сопровождаемые агрегированным α-Syn при обработке NGF. При характеристике сверхэкспрессированного α-Syn в клетках РС12 дикого типа, сверхэкспрессирующих α-Syn при обработке NGF, проводили вестерн-блоттинг и анализы ThT с использованием клеточных лизатов клеток РС12 дикого типа, сверхэкспрессирующих α-Syn после обработки NGF. Результаты вестерн-блоттинга продемонстрировали, что сверхэкспрессированный α-Syn в клеточных лизатах клеток РС12 дикого типа, сверхэкспрессирующих α-Syn при обработке NGF,был олигомерным, а результаты анализа ThT показали, что α-Syn в клеточном лизате клеток РС12 дикого типа, сверхэкспрессирующих α-Syn при обработке NGF, имел β-складчатую структуру (т.е., повышенные сигналы флуоресценции ThT). По сравнению с результатами вестерн-блоттинга и анализа ThT клеток РС12 дикого типа, сверхэкспрессирующими α-Syn без обработки NGF, предполагали, что структурный переход α-спирали в β-складку сверхэкспрессированного α-Syn происходит индуцированной NGF нейрональной дифференцировке, которая могла впоследствии вызывать нейродегенеративные эффекты β-складки олигомерного α-Syn. Кроме того, по сравнению с клетками РС12 дикого типа, сверхэкспрессирующими α-Syn, сверхэкспрессируемый мутантный по A53T α-Syn приводил к более сильным нейродегенеративным эффектам, отраженным как укороченной длиной нейритов, так и уменьшением количества клеток при обработке NGF, что указывало на то, что мутантный по A53T α-Syn вызывал более сильные нейродегенеративные эффекты, чем α-Syn дикого типа в клетках РС12, сверхэкспрессирующих α-Syn.
Анти-α-Syn антитела, образование которых вызывали α-Syn101-132 (SEQ ID NO:114), α-Syn111-132 (SEQ ID NO:113), α-Syn121-135 (SEQ ID NO:107), α-Syn123-135 (SEQ ID NO:111) или α-Syn126-135 (SEQ ID NO:112), и комбинацию анти-α-Syn антител, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113) и (-Syn126-135 (SEQ ID NO: 112), анализировали с помощью модели нейродегенерации in vitro с использованием клеток РС12 дикого типа, сверхэкспрессирующих α-Syn, для оценки их индивидуальных защитных эффектов против нейродегенерации. Клетки PC12 дикого типа, сверхэкспрессирующие α-Syn, обрабатывали NGF в течение 3 дней для инициации нейрональной дифференцировки, перед инкубацией как с анти-α-Syn антителами (конечной концентрации 5 мкг/мл), так и с NGF еще в течение 3 дней. Микроскопическое исследование клеток к концу инкубационного периода показало восстановленную длину нейритов и увеличенное количество клеток при совместной инкубации с выбранными анти-α-Syn антителами по сравнению с контролем-носителем. Количественную оценку длины нейритов и количества клеток выполняли по показаниям исходных клеток РС12, обработанных NGF в течение 6 дней, нормализованных до 100%. В результате анти-α-Syn, образование которых вызывали α-Syn101-132 (SEQ ID NO:114), α-Syn111-132 (SEQ ID NO:113) или α-Syn123-135 (SEQ ID NO:111), и комбинация анти-α-Syn, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113) и α-Syn126-135 (SEQ ID NO: 112), показали значительно большее количество клеток, в то время как анти-(-Syn антитела, образование которых вызывали α-Syn101-132 (SEQ ID NO:114), α-Syn111-132 (SEQ ID NO:113), α-Syn123-135 (SEQ ID NO:111) или α-Syn126-135 (SEQ ID NO:112), и комбинация анти-α-Syn антител, образование которых вызывали α-Syn111-132 (SEQ ID NO: 115) и α-Syn126-135 (SEQ ID NO: 114), показали значительную более длинную длину нейритов по сравнению с контролем-носителем (Фиг.6A и 6B).
Пример 13
Антитела, образование которых вызывали пептидными иммуногенными конструкциями α-SYN, и составами на их основе: специфичность по отношению к бета-складчатому олигомерному и фибриллярному белку альфа-синуклеину
С целью более эффективной характеристики специфичности анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, проводили серию анализов in vitro молекулярного комплекса α-Syn различных размеров, различных амилоидогенных белков, в том числе α-Syn, A(и тау-белка, и агрегированного α-Syn в клетках РС12, сверхэкспрессирующих α-Syn при обработке NGF.
a. Специфичность в отношении более крупных молекулярных комплексов α-Syn
Вестерн-блоттинг молекулярных комплексов α-Syn разных размеров проводили с использованием анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn в качестве первичных антител. Результаты показали, что все анти-α-Syn в значительной степени реагировали с молекулярными комплексами α-Syn более крупных размеров, в том числе димерами, тримерами, тетрамерами и олигомерами, в дополнение к мономерному α-Syn меньшего размера. По сравнению с коммерчески доступным анти-α-Syn-антителом, Syn211 (Abcam), анти-α-Syn-антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO:113), α-Syn121-135 (SEQ ID NO:107), α-Syn123-135 (SEQ ID NO:111) и α-Syn126-135 (SEQ ID NO:112), продемонстрировали более высокое отношение сигнала молекулярных комплексов α-Syn более крупных размеров (в том числе димеров, тримеров, тетрамеров и олигомеров) к сигналу мономерного α-Syn меньшего размера (Фиг. 7A и 7B), указывая на то, что анти-α-Syn антитела обладали специфичностью по отношению к более крупным молекулярным комплексам α-Syn.
b. Специфичность по отношению к α-Syn среди различных амилоидогенных белков
Дот-блоттинг с использованием различных молекул (т.е., α-спиральных мономеров, β-складчатых мономеров, β-складчатых олигомеров и β-складчатых фибрилл) различных амилоидогенных белков (т.е., α-Syn, Aβ1-42 и Tau441), полученных, как описано в Примере 3, проводили с использованием анти-α-Syn, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn в качестве первичных антител. Полученные результаты показали, что анти-α-Syn антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) и α-Syn111-132 (SEQ ID NO: 113), специфически реагировали на все β-складчатые формы α-Syn (мономерные, олигомерные и фибриллярные молекулы), но не на α-спиральные мономеры (Фиг.8A, 8B и 8C). Кроме того, анти-α-Syn-антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) и α-Syn111-132 (SEQ ID NO: 113), сильнее реагировали на β-складчатые фибриллы и β-складчатые олигомеры α-Syn, чем на β-складчатые мономеры α-Syn. В отличие от этого, анти-α-Syn-антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) и α-Syn111-132 (SEQ ID NO: 113), не демонстрировали детектируемую реактивность по отношению к β-Syn или различным молекулам (т.е., α-спиральным мономерам, β-складчатым мономерам, β-складчатым олигомерам и β-складчатым фибриллам) амилоидогенных белков Aβ1-42 и Tau441 (Фиг. 8A, 8B и 8C). Полученные результаты указывали на то, что анти-α-Syn-антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) и α-Syn111-132 (SEQ ID NO: 113), обладали специфичностью по отношению к α-Syn β-складчатых мономерных, β-складчатых олигомерных и β-складчатых фибриллярных форм.
c. Специфичность связывания с агрегированным α-Syn в клетках РС12, сверхэкспрессирующих α-Syn, при обработке NGF
Иммуноцитохимический анализ (ИЦХ) с использованием анти-α-Syn антител, очищенных от антисывороток морских свинок, иммунизированных различными пептидными иммуногенными конструкциями α-Syn, проводили на исходных клетках PC12, клетках PC12 пустого контроля, клетках PC12, сверхэкспрессирующих α-Syn дикого типа, и клетках РС12, мутантных по A53T, сверхэкспрессирующих α-Syn, для оценки аффинности связывания антител с агрегированным α-Syn при обработке NGF, как описано в Примере 3. Как показал результат количественного определения на Фиг. 9, анти-α-Syn антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113), α-Syn121-135 (SEQ ID NO: 107) или α-Syn126-135 (SEQ ID NO: 112), продемонстрировали более сильную реактивность в клетках РС12 дикого типа, сверхэкспрессирующих α-Syn, и в клетках РС12, мутантных по A53T, сверхэкспрессирующих α-Syn, чем в исходных клетках PC12 или клетках PC12 пустого контроля, при обработке NGF. Поскольку агрегацию сверхэкспрессированного α-Syn индуцировали при обработке NGF, полученные результаты указывали на то, что анти-α-Syn антитела, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113), α-Syn121-135 (SEQ ID NO: 107) или α-Syn126-135 (SEQ ID NO: 112), обладали специфичностью к агрегированному α-Syn в клетках PC12 дикого типа, сверхэкспрессирующих α-Syn, и клетках РС12, мутантных по A53T, сверхэкспрессирующих α-Syn, при обработке NGF.
Пример 14
Иммуногистохимическое окрашивание головного мозга человека при болезни Паркинсона для оценки тканевой специфичности пептидных иммуногенных конструкций α-SYN и составов на их основе
Иммуногистопатологическое исследование с использованием преиммунных анти-α-Syn антител, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113), и комбинацией 1:1 обоих анти-α-Syn антител выполняли на нормальных тканях человека, чтобы контролировать специфичность и нежелательную аутореактивность антител. Панель тканей человека (Pantomics) депарафинизировали ксилолом, регидратировали в этаноле и затем обрабатывали 0,25% раствором трипсина с 0,5% CaCl2 в PBS в течение 30 минут и инкубировали в 1% перекиси водорода в метаноле для блокирования активности эндогенной пероксидазы, а затем инкубировали с 10% Block Ace (Sigma) в PBS перед тем, как использовали анти-α-Syn антитела от морских свинок, иммунизированных α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113), и комбинацией 1:1 обоих антител (разведение 1:300). Срезы визуализировали с использованием 3-3'-диаминобензидина (DAB) и были контрастно окрашивали гематоксилином перед микроскопическим исследованием. В отличие от положительной реактивности коммерческого анти-α-Syn (BD, 610708), анти-α-Syn антитела, очищенные от морских свинок, иммунизированных α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113) и комбинацией 1:1 обоих антител, показали отрицательную реактивность к нормальным тканям человека, что было сопоставимо с паттерном преиммунных антител от наивных морских свинок (Фиг. 10А).
Другое иммуногистопатологическое исследование с использованием преиммунных анти-α-Syn антител, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113), и комбинации 1:1 обоих анти-α-Syn антител, проводили для проверки их реактивности в отношении головного мозга человека при болезни Паркинсона. Анализировали срезы тканей трех областей (т.е., мозжечка, мозолистого тела и таламуса) (BioChain). В результате анти-α-Syn антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113), и комбинация 1:1 обоих анти-α-Syn антител, продемонстрировали положительную реактивность (с указанием стрелок) по отношению к срезам головного мозга при PD во всех трех областях по сравнению с отрицательной реактивностью в нормальных срезах головного мозга (Фиг. 10B и 10C). Количественное определение реактивности к агрегатам α-Syn в срезах головного мозга при PD проводили путем подсчета положительных пятен при микроскопическом наблюдении. Результаты показали, что анти-α-Syn антитела, образование которых вызывали α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113), и комбинация 1: 1 обоих анти-α-Syn антител обладали сильной положительностью в срезах головного мозга при PD по сравнению с срезами головного мозга здорового человека. Все их трех различных проанализированных анти-α-Syn антител, образование которых вызывали α-Syn111-132 (SEQ ID NO: 113), обладали наиболее сильной иммунореактивностью по отношению к агрегатам α-Syn в срезах головного мозга при PD.
Пример 15
Доказательство эффективности пептидных иммуногенных конструкций α-SYN и составов на их основе в животных моделях
a. Иммунизация и забор крови/ткани головного мозга
Мышиные модели болезни Паркинсона (PD) получали, как описано в Примере 4. Через две недели после инъекции MPP+ или через 7 недель после инокуляции фибриллярного α-Syn мышей случайным образом разделяли на три группы, включая пептид, в том числе связанный с UBITh1 пептид α-Syn111-132 (SEQ ID NO: 113), связанный с UBITh1 пептид α-Syn126-135 (SEQ ID NO: 112) и комбинация обоих пептидов, помимо группы адъюванта (иммунизированной адъювантами и растворителем, используемыми при получении композиций (ISA 51 VG, CpG3, 0,2% TWEEN®-80)). Внутримышечную (IM) иммунизацию вводили три раза с интервалом в 3 недели в дозе 40 мкг.Схемы введения и забора крови выполняли в соответствии с Табл. 13.
В каждой временной точке 200 мкл крови извлекали с помощью забора образца крови из лицевой вены. Кровь, капающую из проколотой поднижнечелюстной вены, собирали в микропробирку, а сыворотку крови получали в результате центрифугирования при 300 об./мин. в течение 10 минут.После умерщвления животных образцы ткани головного мозга собирали для вестерн-блоттинга.
b. Иммунные ответы в мышиных моделях PD, получавших композиции, содержащие пептидные иммуногенные конструкции α-Syn111-132 (SEQ ID NO: 113) или/и α-Syn126-135 (SEQ ID NO: 112)
Объединенные образцы сыворотки каждой группы обработки разводили в 1% BSA (в PBST) и затем наносили на планшет для ИФА, покрытый 200 мкл полноразмерного пептида α-Syn (Cloud-clone) в 0,1 М бикарбонате натрия (концентрация α-Syn 4,4 мкг/мкл, рН 9,6). Через 2 часа после инкубации при комнатной температуре и после трех промывок PBST добавляли 100 мкл конъюгированного с HRP анти-мышиного IgG антитела, разведенного в 1:3000 с 1% BSA, для реагирования в течение 2 часов при комнатной температуре. После этого планшеты трижды промывали PBST и инкубировали с 100 мкл 3,3,5,5-тетраметилбензидина (TMB) в течение 10 минут в темноте. Затем наносили 100 мкл 2M H2SO4 и инкубировали в течение 15-30 минут, после чего измеряли значение оптической плотности (ОП) при 450 нм с помощью многорежимной детекции с использованием SpectraMax i3x (Molecular Devices).
Две мышиные модели PD, иммунизированные составленным α-Syn111-132 (SEQ ID NO: 113), составленным α-Syn126-135 (SEQ ID NO: 112) или комбинацией обеих пептидных иммуногенных конструкций, имели значение оптической плотности (ОП) анти-α-Syn антител более 3,0 после второй иммунизации, которое оставалось повышенным к моменту окончания исследования через 15 или 19 недель после исходной иммунизации, в модели с индукцией MPP+(Фиг. 11A) или в модели с инокуляцией фибриллярного α-Syn (Фиг. 11B) соответственно, в то время как животные, которым вводили адъювант, не вызывали измеримого иммунного ответа в отношении анти-α-Syn антител.
Отмечено, что в модели с инокуляцией фибриллярного α-Syn конструкция α-Syn111-132 вызвала более сильный иммунный ответ, чем конструкция α-Syn126-135 (Фиг. 11B), в то время как различие в иммуногенности не наблюдали в модели с индукцией MPP+(Фиг. 11А).
c. Снижение уровня α-Syn в сыворотке крови
Уровни α-Syn объединенной сыворотки крови от животных из каждой группы оценивали с помощью набора для ИФА (SEB222Mu, USCN), который мог выявлять как альфа-спиральный, так и β-складчатый α-Syn, описанные в Примере 3.
Количественный ИФА α-Syn предназначался для исследования того, был ли ответ анти-α-Syn антител иммунизированных групп ассоциирован со сниженным количеством периферического α-Syn по сравнению с необработанными животными. Было продемонстрировано, что иммунизация α-Syn126-135 (SEQ ID NO: 112), α-Syn111-132 (SEQ ID NO: 113) или комбинацией этих конструкций приводила к снижению значений оптической плотности (ОП) уровней α-Syn по сравнению с животными, которым вводили адъювант, как в модели с индукцией MPP+(Фиг. 12A), так и в модели с инокуляцией фибриллярного α-Syn (Фиг. 12B). Результаты указывали на то, что в случае получения ответа анти-α-Syn антител при иммунизации пептидными иммуногенными конструкциями α-Syn количество α-Syn в периферической крови соответственно уменьшалось.
d. Снижение уровня олигомерного α-Syn в головном мозге
После умерщвления животных образцы ткани головного мозга собирали для вестерн-блоттинга. У мышей, индуцированных MPP+, головной мозг удаляли и гомогенизировали, в то время как у мышей, инокулированных фибриллярным α-Syn, сначала выделяли области полосатого тела и черной субстанции, а затем гомогенизировали. Лизат ткани головного мозга получали в результате добавления в гомогенат буфера для лизиса (Amresco) и 1х ингибитора протеиназы (Roche). Затем лизат разделяли с помощью 10% SDS-PAGE (электрофорез в додецилсульфат-полиакриламидном геле натрия), переносили на мембрану из поливинилидендифторида (PVDF) и инкубировали в течение ночи с 5% молоком в PBS. Для детекции высокого содержания дофаминергических нейронов, мембраны инкубировали с анти-тирозингидроксилаза антителом (разведение 1:1000, Abcam), а затем гибридизировали с вторичным антителом, конъюгированным с HRP козьим анти-кроличьим IgG (H+L) антителом (разведение 1:5000, Jackson Immunoresearch). Для визуализации использовали субстраты Luminata Western HRP и полученный сигнал захватывали цифровой системой изображений ChemiDoc-It 810. Количественное определение уровня олигомерного α-Syn осуществляли путем нормализации к уровню GAPDH, и соотношение ненарушенного лизата дополнительно стандартизировали до 100% для сравнения.
В модели с индукцией MPP+снижение доли олигомерного α-Syn показывали у животных, иммунизированных пептидной иммуногенной конструкцией α-Syn111-132 (Фиг. 13A). Аналогичным образом у мышей, инокулированных фибриллярным α-Syn, вестерн-блоттинг с использованием лизатов черной субстанции, а также полосатого тела ипсилатеральной стороны при инокуляции фибриллярным α-Syn (Фиг. 14A и 14D) и с использованием лизатов полосатого тела контралатеральной стороны при инокуляции фибриллярным α-Syn (Фиг. 14F) показал, что повышение уровня олигомерного α-Syn в 2-3 раза, наблюдаемое у контрольных мышей с использование адъюванта, снижалось после обработки составленным α-Syn111-132 (SEQ ID NO:113) и конструкции α-Syn126-135 (SEQ ID NO:112). Количественную оценку результатов вестерн-блоттинга представляли на Фиг. 13B, 14B, 14C, 14D и 14G.
e. Снижение нейропатологии
У мфшей, инокулированных фибриллярным α-Syn, сначала выделяли участки черной субстанции, а затем их гомогенизировали. Лизат ткани получали в результате добавления в гомогенат буфера для лизиса (Amresco) и 1х ингибитора протеиназы (Roche). Затем лизат разделяли с помощью 10% SDS-PAGE (электрофорез в додецилсульфат-полиакриламидном геле натрия), переносили на мембрану из поливинилидендифторида (PVDF) и инкубировали в течение ночи с 5% молоком в PBS. Для детекции высокого содержания дофаминергических нейронов, мембраны инкубировали с анти-тирозингидроксилаза антителом (разведение 1:1000, Abcam), а затем гибридизировали с вторичным антителом, конъюгированным с HRP козьим анти-кроличий IgG (H+L) антителом (разведение 1:5000, Jackson Immunoresearch). Для визуализации использовали субстраты Luminata Western HRP и полученный сигнал захватывали цифровой системой изображений ChemiDoc-It 810. Уровень экспрессии α-Syn стандартизировали по отношению к GAPDH (глицеральдегид-3-фосфатдегидрогеназа), используемому в качестве контроля нагрузки белка.
Результаты продемонстрировали, что иммунизация конструкцией α-Syn111-132 восстановливала количество тирозин-гидроксилазы до уровня, эквивалентного уровню у не пораженных нормальных животных (Фиг. 14C-14D), что свидетельствовало о нейропротекторном эффекте пептидных иммуногенных конструкций α-Syn против нейротоксичности, ассоциированной с агрегированным α-Syn, инокулированным мышам.
f. Восстановление двигательной активности
CatWalk™ XT (Noldus information Technology, Вагенинген, Нидерланды) представляет собой систему анализа на основе видео, используемую для объективного измерения различных аспектов походки в динамическом режиме на основании положения, давления и площади поверхности каждого шага. Всех мышей учили последовательно пересекать дорожку, по меньшей мере, три раза в день перед экспериментом. Успешную пробежку определяли, когда животное пробегало по дорожке без перерыва или колебаний, а мышей, которые не прошли обучение, исключали из исследования.
В среднем анализировали 5 пересечений каждой мыши. Поскольку инокуляцию фибриллярным α-Syn выполняли на правом стороне головного мозга, время стояния на левой задней лапе считалось эталонным параметром, наряду с продолжительностью пробега.
В модели с инокуляцией фибриллярного α-Syn, существенное различие в измерении времени стояния на левой задней конечности наблюдали после обработки композициями, содержащими α-Syn126-135 (SEQ ID NO: 112) или α-Syn111-132 (SEQ ID NO: 113) (Фиг. 15А). Между тем, как в модели с инокуляцией фибриллярного α-Syn, так и в модели с индукцией MPP+, значительную разницу в измерении продолжительности пробега наблюдали после обработки композициями, содержащими α-Syn111-132 (SEQ ID NO:113) (Фиг. 15B и 15C). Результаты указывали на ассоциацию обработки пептидными иммуногенными конструкциями α-Syn составленного α-Syn126-135 (SEQ ID NO:112) или составленного α-Syn111-132 (SEQ ID NO:113) и улучшением двигательных функций в двух мышиных моделях PD.
Пример 16
Реактивности антител, образованных с помощью пептидных иммуногенных конструкций α-SYN с различными видами α-SYN, встречающимися в нейродегенеративных заболеваниях
α-Syn вызывает болезнь Паркинсона и другие синуклеинопатии. Белок α-Syn способен образовывать различные типы агрегатов, которые имеют различные размеры и структуры, и разные эффекты на клетки, таким образом, что каждое из этих заболеваний вызывается одним или несколькими различными типами агрегатов. Агрегаты α-Syn различной формы могут вызывать различные паттерны повреждения в головном мозге и даже могут вызывать различные заболевания головного мозга. Данное исследование разрабатывали для оценки того, как антитела, образуемые пептидными иммуногенными конструкциями α-Syn, взаимодействуют с различными видами α-Syn, встречающимися при нейродегенеративных заболеваниях.
Доктор Рональд Мелки (Dr. Ronald Melki) участвовал в данном исследовании. В лаборатории получали различные типы агрегатов α-Syn, которые включают в себя (а) фибриллу, длинную, скрученную, соединенную вместе нить из белков α-Syn; (b) ленту, более широкую, более плоскую структуру, и (c) олигомеры α-Syn (O550), стабилизированные дофамином (ODA) и глутаральдегидом (OGA).
Антитела, образующиеся у морских свинок с помощью различных пептидных иммуногенных конструкций α-Syn, раскрытых в данном документе, исследовали в отношении их относительных аффиностей. Иллюстративные образцы из PD-021514 (α-Syn85-140, нед. после иммуниз. 08), PD-021522 (α-Syn85-140, нед. после иммуниз. 13), PD-100806 (α-Syn126-135, нед. после иммуниз. 09), PRX002 и коммерческого моноклонального антитела Syn1 (клон 42) исследовали в отношении отдельных структур α-Syn, в том числе: фибрилл, лент, фибрилл 65, фибрилл 91, фибрилл 110, в отношении олигомеров α-Syn фибриллярного пути сборки (O550), стабилизированных дофамином (ODA) и стабилизированных глутаральдегидом (OGA) олигомеров, наряду с контрольным мономером с использованием анализа ловушки на фильтре.
Способы и материалы
a. Сборка α-Syn в фибриллы и ленты
Для образования фибрилл растворимый α-Syn ДТ инкубировали в буфере A (50 мМ Tris-HCl, pH 7,5, 150 мМ KCl) при 37°C при непрерывном встряхивании в термомиксере Eppendorf, установленном при 600 об./мин. Сборку непрерывно контролировали в спектрофлуориметре Cary Eclipse (Varian Inc., Пало-Альто, Калифорния, США) в присутствии тиофлавина Т (15 мкМ) в кюветах размером 1 (1 см при перемешивании (100 об./мин.) с использованием магнитной мешалки (6 (3 мм) с длиной волны возбуждения, установленной на 440 нм, и длиной волны излучения, установленной на 440 нм и 480 нм, и временем усреднения, равным 1 с.Для образования ленты α-Syn Дт диализовали в течение 16 часов против 1000 объема буфера B (5 мМ Tris-HCl, pH 7,5) при 4°C, затем инкубировали при 37°C при непрерывном встряхивании в термомиксере Eppendorf, установленном при 600 об./мин. Сборку контролировали с помощью измерения рассеянного света при 440 нм. В качестве альтернативы количество белка, оставшегося в супернатанте после осаждения при 35000 x g, определяли путем измерения оптической плотности при 280 нм на спектрофотометре с диодной матрицей Hewlett Packard 8453. Природу олигомерных молекул оценивали с использованием TEM Jeol 1400 (Jeol Ltd.) после адсорбции образцов на покрытых углеродом сетках 200 меш и отрицательного окрашивания 1% уранилацетатом. Изображения записывали с помощью CCD-камеры Gatan Orius (Gatan). Способность сструктур α-Syn связывать конго красный оценивали следующим образом: фибриллы и ленты α-Syn инкубировали в течение 1 часа с 100 мкМ конго красного (Sigma-Aldrich, Сент-Луис, Миссури, США) в 20 мМ буфере Tris (рН 7,5). Затем полимеры осаждали при 20°С в ультрацентрифуге TL100 Tabletop Beckman (Beckman Instruments, Inc., Фуллертон, Калифорния, США) при 25000 g в течение 30 минут.Гранулы промывали четыре раза, используя равный объем воды. После ресуспендирования гранул аликвоту помещали на стеклянное покровное стекло и немедленно визуализировали или давали высохнуть. Образцы просматривали в светлом поле и кроссполяризационном свете с помощью поляризационной микроскопии с использованием микроскопа Leica (MZ12.5), оснащенного кроссполяризаторами (Leica Microsystems, Ltd., Хербруг, Швейцария).
b. Определение концентраций фибрилл и лент α-Syn
Гетерогенность длины фибрилл и лент α-Syn снижали с помощью соникации в течение 20 минут на льду в 2-мл пробирках Eppendorf в VialTweeter, работающем на ультразвуковом процессоре UIS250v (250 Вт, 2,4 кГц; Hielscher Ultrasonic, Тетлоу, Германия), установленном на 75% амплитуде, с импульсами 0,5 с.Измеряли скорости седиментации фибрилл и лент α-Syn. Границы седиментации анализировали с помощью программного обеспечения Sedfit, используя моделирование методом наименьших квадратов ls - g* (s), которое лучше всего подходит для гетерогенных смесей крупных частиц. Это приводило к распределению частиц с коэффициентами седиментации от 50 до 150 S для лент α-Syn, от 100 до 1000 S для фибрилл α-Syn, с сосредоточением на молекулах, которые имеют коэффициент седиментации ~ 90 S и 375 S для лент и фибрилл α-Syn соответственно, соответствующие частицам, которые имеют молекулярную массу ~ 11500 кДа, например, составленные из ~ 800 молекул α-Syn (12000 кДа/14,5 кДа) для лент α-Syn, ~ 102000 кДа, например, составленные из ~ 7000 молекул α-Syn (102000 кДа/14,5 кДа) для фибрилл α-Syn. Таким образом, при рабочей концентрации 20 мкМ общие концентрации частиц лент и фибрилл α-Syn составляют 20 мкМ/~800=~0,02 мкМ, 20 мкМ/~7000=~ 0,003 мкМ для лент и фибрилл α-Syn соответственно, учитывая, что 100% α-Syn собирается в лентах или фибриллах в устойчивом состоянии, так как 100% белка находится во фракции гранул при центрифугировании образцов.
c. Оценка аффинности эндотел в отношени различных фибрилл и лент α-Syn
Аффинность антител, образуемых с помощью пептидных иммуногенных конструкций α-Syn, раскрытых в данном документе, оценивали для различных структур α-Syn с использованием анализа ловушки на фильтре с антителом в качестве эталона. Структуры α-Syn (фибриллы, ленты, фибриллы 65, фибриллы 91, фибриллы 110, олигомеры фибриллярного пути сборки α-Syn (O550), стабилизированные дофамином (ODA) и стабилизированные глутаральдегидом (OGA) олигомеры) описаны в Bousset L. et al., 2013 Nat Commun 4:2575; Makky A. et al., 2016 Sci Rep 6:37970; and Pieri L. et al, 2016 Sci. Rep 6:24526. Также использовали контрольный мономерный α-Syn.
Возрастающие количества фибриллярного, олигомерного или мономерного α-Syn в диапазоне от 20 пг до 200 нг обнаруживали на нитроцеллюлозных фильтрах с использованием устройства для щелевой фильтрации. Затем фильтры блокировали с обезжиренным молоком, инкубировали с антителом PRX002 или Syn1 или исследуемыми антителами GP по данному изобретению в указанном разведении. После тщательной промывки для детекции профилей связывания первичных антител использовали второе анти-человека или анти-морской свинки IgG-HRP. Также исследовали контроль с использованием только вторичного антитела. На блотах использовали Super Signal ECL (Pierce №34096), а затем блоты визуализировали на сканере BioRad (система визуализации Chemidoc MP/программное обеспечение BioRad imagelab). Время экспозиции и динамический диапазон указаны на Фиг. 16A-16H. В этом наборе измерений на мембране обнаруживали гомогенат головного мозга человека со случаем DLB.
d. Результаты
Аффинность антител морских свинок (GP) PD-021514 (α-Syn85-140, нед. после иммуниз. 08), PD-021522 (α-Syn85-140, нед. после иммуниз. 13), PD-100806 (α-Syn126-135, нед. после иммуниз. 09) из иммунизированных GP, PRX002 и коммерческих антител Syn1 (клон 42) сравнивали для различных структур α-Syn с использованием анализа ловушки на фильтре. Используемые структуры α-Syn включали в себя фибриллы, ленты, фибриллы 65, фибриллы 91, фибриллы 110, олигомеры фибриллярного пути сборки α-Syn (O550), стабилизированные дофамином (ODA) и стабилизированные глутаральдегидом (OGA) олигомеры, а также контрольный мономерный α-Syn.
На Фиг. 16А-16Н продемонстрировано, что эталонное антитело, PRX002, распознает с немного более высокой аффинностью фибриллярный α-Syn по сравнению с мономерным α-Syn; тогда как PD-100806 и PD-021514, оба направленные против пептидной конструкции α-Syn126-135 по данному раскрытию, имеют гораздо более высокую аффиность по отношению к фибриллярному α-Syn по сравнению с мономерным α-Syn, что указывает на то, что оба имеют преимущественное связывание с фибриллярным (-Syn. Было обнаружено, что аффиности PRX002 по отношению к олигомерному и фибриллярному α-Syn являются аналогичными. Моноклональное антитело Syn1 связывалось с фибриллярным α-Syn, а также с олигомерным и мономерным α-Syn без особого дифференциального предпочтения.
Пример 17
Иммуногистохимическое исследование антител, происходящих из пептидных иммуногенных конструкций α-SYN, с использованием срезов головного мозга пациентов с болезнью Паркинсона (PD), множественной системной атромфией (MSA) и деменцией с тельцами Леви (DLB)
Антитела, полученные при иммунизации морских свинок иллюстративной пептидной иммуногенной конструкцией α-Syn126-135 по данному изобретению, использовали в иммунохимическом исследовании для характеристики их способности связываться с α-Syn, присутствующим в срезах головного мозга, полученных от пациентов с альфа-синуклеинопатиями. Исследование проводили в сотрудничестве с профессором Роксаной Караре (Prof. Roxana Carare). Оценивали способность антител связывать α-Syn, присутствующий в срезах головного мозга, полученных от пациентов с PD, LBD и MSA. Здоровые ткани включали в исследование в качестве отрицательного контроля. NCL-L-ASYN, коммерчески доступное моноклональное антитело, используемое для посмертной диагностики альфа-синуклеинопатий, включали в качестве положительного контроля. В данном исследовании предложено доказательство положительной иммунореактивности антител, направленных против пептидной иммуногенной конструкции α-Syn126-135 на срезах тканей головного мозга, полученных от пациентов с PD, LBD и MSA. Связывание было особенно заметно в головном мозге пациентов с синуклеопатиями, но не в головном мозге лиц, не являющихся пациентами, при этом связывание было более выраженным при использовании исследуемых антител, чем в случае коммерческого диагностического антитела.
Способы и материалы
a. Описание используемых реагентов и их поставщиков
Антитела, полученные в результате иммунизации морских свинок иллюстративной пептидной иммуногенной конструкцией α-Syn126-135, использовали в разведении 1:100. PD062220-09-1-2-Syn; PD062205-09-1-2-Syn; PD100806-09-1-2-Syn предоставляли United NeuroScience (UNS), NCL-L-ASYN (мышиное моноклональное антитело, используемое в разведении 1:100) предоставляли Leica Biosystems, HuD (EI) (мышиное моноклональное антитело в разведении 1:100) предоставляли Santa Cruz Biotechnology, Olig2 (кроличьи антитела в разведении 1:100) предоставляли Millipore, Alexa Flour 594 (козье-анти-морской свинки в разведении 1:200), Alexa Flour 488 (козье-анти-мышиное в 1:200) и Alexa Flour 488 (козье-кроличье в разведении 1: 200) предоставляли Molecular Probes life technologies.
b. Ткань головного мозга человека
Срезы мкм толщины получали из банка головного мозга UCL, используемого в данном исследовании. Все образцы собирали и получали в соответствии с утвержденными протоколами Национальной службы этики исследований.
Ткань получали от субъектов (Табл. 15) с первичной патологией α-Syn, включающей множественную системную атрофию (MSA; n=3), деменцию с тельцами Леви (DLB; n=3) и болезнь Паркинсона (PD; n=3). Субъектов диагностировали посмертно в соответствии с опубликованными критериями**.
c. Иммуногистохимический анализ субъектов-людей с синуклеинопатиями
Иммуногистохимический анализ (ИГХ) субъектов-людей с тремя различными синуклеинопатиями (MSA, DLB и PD) проводили с целью количественного сравнения специфичности в отношении агрегатов α-Syn трех антител, полученных United Neuroscience (UNS). Специфичность антител UNS (PD062220, PD062205 и PD100806) для агрегатов α-Syn сравнивали со специфичностью коммерчески доступного диагностического антитела (NCL-L-ASYN). Специфичность антител анализировали в следующих четырех областях головного мозга у каждого пациента и в каждом типе заболевания (1) бледный шар, внутренняя капсула и островковая кора; (2) средний мозг: черная субстанция; (3) височная кора: серое вещество коры головного мозга; и (4) мозжечок: белое вещество подкорки; белое вещество мозжечка.
Известно, что эти области головного мозга подвержены агрегации α-Syn в различной степени и на разных стадиях прогрессирования заболевания при каждом типе заболевания. Как правило, базальные ганглии и средний мозг поражаются на ранних стадиях DLB, PD и MSA, а также имеют наибольшую нагрузку агрегатами. Височная кора и мозжечок поражаются на более поздних стадиях заболевания с очень небольшим количеством агрегатов в мозжечке, присутствующих при PD и DLB. Отрицательный контроль (без использования первичного антитела) использовали наряду с каждым протоколом ИГХ, чтобы подтвердить отсутствие неспецифического связывания вторичного антитела. Заключенные в парафин микропрепараты депарафинировали в печи при 60°С в течение 15-20 минут и затем погружали в ксилол I и II, на 5 минут в каждый. Ткань регидратировали в 4 разведениях IMS от 100% до 50%, в течение 5 минут в каждом. Ткань промывали 3 раза в течение 5 минут в 1x PBS и затем инкубировали в течение 3 минут в 100% муравьиной кислоте для извлечения антигена. Ткань тщательно промывали 1x PBS перед гашением активностью эндогенной пероксидазы с 3% H2O2 в течение 10 минут.Ткани давали остыть и еще 3 раза промывали в 1x PBS (по 5 минут каждый) перед микроволновой обработкой в цитратном буфере (15 мМ Tris-цитрат натрия, TWEEN, pH 6) при средней температуре в течение 25 минут, чтобы обеспечить равномерную обработки в микроволновке на цикл, 3 стойки микропрепаратов в 3 контейнерах включали каждый раз. Микропрепараты оставляли для охлаждения и трижды промывали в 1x PBS (5 минут) перед блокированием неспецифических сайтов связывания с 15% нормальной козьей сывороткой. Ткань инкубировали в первичном антителе (1:100 в 0,1% TBS/t) в течение ночи при 4°С.Ткань промывали 3 (5 минут в 1x PBS и инкубировали в течение 1 часа (КТ) в биотинилированном вторичном антителе. Раствор ABC получали за 30 минут до его применения. После промывки ткани 3×5 мин 1x PBS ее инкубировали в ABC в течение 1 часа при КТ. VIP субстрат пероксидазы готовили с использованием набора пероксидазы ImmPACTVIP, как подробно описано в инструкциях производителя. VIP субстрат пероксидазы добавляли в течение 7 минут при КТ и промывали в dH2O. Перед установкой в DPX ткань обезвоживали в течение 2 минут каждый раз в IMS 50%, 70%, 95%, 100%, 100% и ксилолах I и II. Для двойного иммунофлуоресцентного окрашивания ткань не гасили 3% H2O2 до нанесения первичного антитела. После нанесения первого первичного и эквивалентного вторичного антител ткань блокировали 15% нормальной козьей сывороткой в течение 30 минут и инкубировали со вторым первичным и вторичным антителом, как описано ранее. После окончательного нанесения вторичных антител, меченых флуоресцентно, ткань инкубировали в 1% судановом черном в течение 5 минут, чтобы погасить аутофлуоресценцию, промывали 0,1% TBS/T и незамедлительно помещали в Mowiol Cituflour. Флуоресцентно окрашенную ткань хранили при 4°С до получения изображения.
d. Анализ изображений и статистический анализ
Микропрепараты сканировали для анализа с объективом x20 с использованием высокопроизводительной системы виртуальной микроскопии Olympus VS110 или системы виртуальной микроскопии слайдов Olympus dot. Тридцать изображений (каждое 500 мкм2) получали с отсканированного изображения с использованием программного обеспечения Olympus VS из эквивалентных областей каждого участка из каждого объекта (см. Фиг. 17A-17D, 18A-18D, 19A-19C, 20A-20E, 21A-21F, 22A- 22C, 24A-24D и 25A-25D). Это позволило проанализировать общую площадь 7,5 мм2 в каждой области головного мозга. Программное обеспечение версии ImageJ для Fiji windows-64 использовали для количественного анализа иммунореактивности α-Syn каждого изображения.
Для анализа общего количества α-Syn, выявленного с помощью каждого антитела, иммунореактивность описывали в виде процента от общей площади изображения. Пороговое значение, применяемое для выбора положительной в отношении α-Syn иммунореактивности, корректировали для каждой анализируемой области головного мозга, чтобы учесть различия в фоновом окрашивании фона, которые могли повлиять на результаты. Средний процент площади, покрытой положительными в отношении α-Syn агрегатами, рассчитывали для каждого анализируемого антитела и области головного мозга.
Для анализа относительной специфичности каждого антитела в отношении LB или LN использовали программное обеспечение Fiji для количественной оценки иммунореактивности LB на основе параметров размера и округлости, чтобы отличать их от LN (см. Фиг. 24A-24D, 25A-25D и 26A-26В). Области головного мозга с четкой морфологией LB и LN отбирали для этого анализа, чтобы избежать ложноположительных результатов, и включали островковую кору базальных ганглиев и корковое серое вещество височной коры. Иммунореактивность LB выражали в процентах от общей иммунореактивности α-Syn.
Статистический анализ проводили с использованием программного обеспечения GraphPad Prism v7.01 и представляли в виде среднего значения+SD (если не указано иное). Результаты анализировали с помощью однофакторного дисперсионного анализа (ANOVA) с последующим апостериорным анализом с поправками Даннетта, при необходимости. Различия считали значимыми при р<0,05 (*). Количества (n) относятся к количеству субъектов, используемых для каждого эксперимента.
Качественный анализ местоположения α-Syn в нейронах или глие выполняли с помощью двойного иммунофлуоресцентного окрашивания, как описано ранее. Микропрепараты просматривали с помощью лазерного сканирующего конфокального микроскопа Leica SP8. Изображения с наложением максимальных проекций получали при объективе х40 последовательно. Эти изображения содержали серию изображений z-слайдов, сложенных вместе с наложенными обоими цветовыми каналами с тем, чтобы показать их относительные положения.
e. Антитела к α-Syn126-135 приводили к детекции отличного паттерна агрегатов α-Syn по сравнению с NCL-L-ASYN
Тип клеток и субклеточная локализация агрегатов α-Syn варьируют при различных синуклеинопатиях. MSA характеризуется глиальными цитоплазматическими включениями (GCI), в то время как при DLB и PD агрегация α-Syn происходит в телах нейронов (LB) и аксональных отростках (LN). Анализ процента окрашенной площади позволил количественно оценить суммарные агрегаты α-Syn, выявленные с помощью каждого антитела. В то же время при этом не учитывали различия в типе или субклеточном расположении выявленных агрегатов. Отчетливый паттерн агрегатов α-Syn в клеточных телах и нейритах в случаях PD и DLB позволил определить относительную чувствительность антител UNS по данному раскрытию в отношении этих различных типов агрегатов α-Syn.
С целью исследования этого, долю агрегатов, выявляемых в клеточных телах, оценивали для каждого антитела в случаях DLB и PD. С помощью программного обеспечения FIJI агрегаты в клеточных телах выбирали на основе их размера и округлости. Средний процент площади агрегатов клеточных тел затем рассчитывали в виде доли от общего выявленного α-Syn, а результаты продемонстрированы на Фиг. 24A-24D и 25A-25D. Разницу в проценте площади общего α-Syn и α-Syn клеточных тел связывали с аксональными агрегатами α-Syn (LN) на основании качественного анализа ткани. Снижение доли детекции α-Syn клеточных тел отвечает повышению детекции LN. Указанный анализ проводили в сером веществе височной коры и островковой коры, поскольку этих области демонстрировали как LB, так и LN-подобную патологию. LN были очень редкими и неравномерно распределялись по бледному шару и капсуле, и, следовательно, эти области базальных ганглиев не выбирали для данного анализа. Аналогичную корреляцию наблюдали в черной субстанции среднего мозга (Фиг. 26A и 26B) в случае антител UNS по данному раскрытию, которые приводили к детекции более высоких уровней LN по сравнению с NCL-L-ASYN при DLB и PD. В то же время из-за сложной морфологии LN и LB было невозможным надежно различить и количественно оценить их с помощью одного и того же способа.
Результаты на Фиг. 24A-24D показывают, что из общего α-Syn, выявленного с помощью каждого антитела, доля агрегатов, выявленных в клеточных телах, была снижена с помощью антител UNS по сравнению с NCL-L-ASYN. Это означает, что отношение включений в клеточных телах по отношению к LN было снижено, и более высокую долю LN выявляли в случае антителам UNS. Из антител UNS PD062205 было устойчивым при DLB и PD при выявлении высоких долей LN в островковой коре (Фиг. 17A-17D и 18A-18D). В отличие от этого, все антитела к α-Syn126-135 приводили к детекции более высоких долей агрегатов клеточных тел по сравнению с NCL-L-ASYN в сером веществе височной коры случаев DLB и PD (Фиг. 25A-25B).
f. Агрегация α-Syn является специфической в отношении типа клеток
Агрегаты, содержащие α-Syn, представляют собой характерный патогенный признак синуклеинопатий, в том числе MSA, DLB и PD. Несмотря на то, что агрегация α-Syn связана с основным белков, являющимся причиной синуклеинопатий, паттерн агрегации и типы клеток, которые являются восприимчивыми а образования агрегатов, отличаются между конкретными подтипами заболеваний. Клиническая характеристика MSA, DLB и PD описана в виде накопления α-Syn в клеточных телах и отростках нейритов нейронов как в DLB, так и в PD, но при MSA оно встречается в основном в глиальных клетках и олигодендроцитах.
С целью определения селективности антител к α-Syn126-135 в отношении специфических в отношении клеток агрегатов α-Syn, проводили двойной иммунофлуоресцентный анализ с использованием PD062205 и маркеров либо для нейронов (HuD), либо для олигодендроцитов (Olig2).
Результаты на Фиг. 27A-27C показывают, что α-Syn, выявленный с помощью PD062205, локализуется совместно с клеточными телами нейронов в базальных ганглиях и среднем мозге (областях высокой патологии) при PD и DLB, но не при MSA. Использование маркеров олигодендроцитов (Olig2) на Фиг. 28A-28C показывает, что при MSA, но не при PD или DL, α-Syn агрегирует в клетках глии. Эти результаты демонстрируют, что антитела к α-Syn126-135 согласуются с клинической характеристикой этих синуклеинопатий и подтверждают специфичность этих антител в случае патологических агрегатов α-Syn.
Результаты
a. Количественный анализ антител, образующихся в результате иммунизации у морских свинок иллюстративной пептидной иммуногенной конструкцией α-Syn126-135 с целью иммунотерапии
С целью исследования применения новых анти-α-Syn антител с целью иммунотерапии проводили количественный анализ относительной специфичности каждого антитела к α-Syn с помощью иммуногистохимического анализа (ИГХ) в случаях трех синуклеинопатий (MSA, DLB и PD) у человека.
b. Антитела, образующиеся в результате иммунизации у морских свинок иллюстративной пептидной иммуногенной конструкцией α-Syn126-135, являются более чувствительными, чем коммерчески используемые диагностические антитела при связывании с агрегатами α-Syn
С целью исследования относительной антигенности раскрытых антител к α-Syn126-135, нагрузку α-Syn, выявляемую с помощью каждого антитела, сравнивали с коммерчески доступным диагностическим антителом для синуклеинопатий (NCL-L-ASYN). Изначально изучив общую картину результатов, показанных на Фиг. 17A-D-22A-22C, можно увидеть, что наблюдается заметное увеличение среднего процента площади α-Syn, детектируемого с помощью антител к α-Syn126-135, по сравнению с NCL- L-ASYN. Эта тенденция является последовательной для каждой области головного мозга и типа заболевания и предполагает, что раскрытые антитела к α-Syn126-135 являются более чувствительными или селективными при связывании с агрегированным α-Syn, чем NCL-L-ASYN. Хотя размер выборки в данном исследовании был относительно небольшим (n=3), все еще можно заметить отчетливую тенденцию в данных. Специфичность раскрытых антител к α-Syn126-135 в отношении α-Syn подтверждали в тех же областях головного мозга, что и в головном мозге контрольных пациентов без заболевания. Эти результаты, которые продемонстрированы на Фиг. 23A-23B, показывают отсутствие какого-либо иммуноположительного окрашивания с помощью каждого антитела, в том числе NCL-L-ASYN. Эти данные указывают на то, что раскрытые антитела к α-Syn126-135 являются специфическими в отношении патологических форм α-Syn.
c. Более высокий уровень α-Syn, выявленный с помощью антител к α-Syn126-135, указывает на их повышенную чувствительность и специфичность по сравнению с коммерческими антителами
Антитела к α-Syn126-135 по данному раскрытию приводят к детекции большего количества α-Syn по сравнению с NCL-L-ASYN, что указывает на то, что раскрытые антитела являются более предпочтительными для применения в иммунотерапии для облегчения выведения указанных агрегатов α-Syn.
Первым этап выбора подходящего антитела для применения в качестве реагента для иммунотерапии заключается в определении селективности антител в отношении целевого антигена (α-Syn) в ткани головного мозга человека с первичной патологией α-Syn. Различные синуклеинопатии различаются по механизмам и нейроанатомическому типу агрегации α-Syn, а также по восприимчивости определенных типов клеток к агрегации.
Важно оценить селективность антител к α-Syn126-135 по отношению к α-Syn при различных синуклеинопатиях с различной невропатологией, чтобы исследовать использование реагента в качестве иммунотерапии для синуклеинопатий в целом. Для этой цели выбирали клинически подтвержденные случаи PD, DLB и MSA. PD и DLB представляю собой вторые вторыми наиболее распространенные формы деменции и главным образом вызваны накоплением α-Syn в нейронах (LB и LN). В отличие от PD, патологии амилоида бета и тау, как известно, способствуют нейродегенерации при DLB2. Другой паттерн агрегации α-Syn наблюдается при MSA, где агрегаты главным образом образуются в глиальных клетках, а не в нейронах (Фиг. 27A-27C и 28A-28B). Кроме того, прогрессирование патологии α-Syn варьируется между типами заболеваний, при этом средний мозг и базальные ганглии являются распространенными областями ранней патологии. Исследование антигенности каждого антитела в областях головного мозга, пораженных на разных стадиях заболевания, позволит понять, какое антитело может быть более эффективным для лечения ранних стадий заболевания.
d. Антитела к α-Syn126-135 (PD062220, PD062205 и PD100806) споосбны специфически связыаться с патологическими агрегатами α-Syn в ткани головного мозга человека, полученных при PD, DLB и MSA (Фиг. 17A-D-22A-22C) без детекции какой-либо патологии синуклеинов в здоровом контроле (Фиг. 23A-23B).
Детекцию α-Syn с помощью раскрытых антител к α-Syn126-135 выполняли с той же специфичностью в отношении типа клеток, которая была описана при клинической нейропатологии (Фиг. 27A-27B и 28A-28B). Важно, что раскрытые антитела к α-Syn126-135 не продемонстрировали одинаковую антигенность в отношении всех форм α-Syn человека.
Специфичность PD062205 и PD100806 дополнительно подтверждали способностью каждого антитела приводить к детекции более высокой долю LN, чем NCL-L-ASYN в базальных ганглиях (Фиг. 24A-24D). Это также наблюдали визуально в среднем мозге (Фиг. 26А-26 В). В совокупности при более высоком проценте площади α-Syn, выявленного с помощью PD062205 и PD100806, эти результаты указывают на то, что дополнительный α-Syn, выявленный с помощью раскрытых антител к α-Syn126-135, может быть частично связан с повышенной специфичностью этих антител в отношении LN. Эти результаты пригодны для иммунотерапии, поскольку на ранних стадиях заболевания LN представляют собой преобладающую форму агрегации α-Syn в базальных ганглиях. Другие реагенты для лечения синуклеинопатий, которые находятся в доклинической разработке, не обеспечивают ИГХ детекцию LN. Таким образом, раскрытые пептидные иммуногенные конструкции и антитела к α-Syn126-135, образованные из пептидных иммуногенных конструкций, имеют уникальные свойства и характеристики по сравнению с другими коммерчески доступными продуктами.
В данном исследовании использовали ИГХ для анализа чувствительности антител к α-Syn126-135, образование которых вызывали раскрытыми пептидными иммуногенными конструкциями, с помощью измерения среднего количества агрегатов α-Syn в пораженных областях головного мозга. В данном исследовании, в котором количественно определяют средний процент площади α-Syn в образцах головного мозга, продемонстрировано, что раскрытые антитела к α-Syn126-135 были очень чувствительны к выявлению α-Syn ранее при прогрессировании заболевания по сравнению с MSA, DLB и PD по сравнению с коммерчески доступным антителом.
Более высокая чувствительность, обнаруженная в этом исследовании, может быть связана с более высокой специфичностью раскрытых антител по отношению к LN по сравнению с диагностическим антителом NCL-L-ASYN. Эти результаты позволяют предположить, что раскрытые антитела к α-Syn126-135, вероятно, являются наиболее эффективными кандидатами для исследования опосредованного антителами выведения агрегатов α-Syn при синуклеинопатиях.
Таблица 1
Аминокислотные последовательности α-Syn и их фрагменты, используемые в серологических анализах
Таблица 1 (продолжение)
NO:
Таблица 2
Аминокислотные последовательности Th-эпитопов, происходящих из патогенных белков, в том числе идеализированные искусственные Th-эпитопы для использования в разработке пептидных иммуногенных конструкций α-Syn
NO:
T RT TR T
RRRIKII RII I L IR
VRVV VV V I V
F FF FF F V F
F
T RT TR
FFLL L ITTI
Таблица 3
Аминокислотные последовательности пептидных иммуногенных конструкций α-Syn
NO:
Таблица 3 (продолжение)
NO:
T RT TR T
RRRIKII RII I L IR
VRVV VV V I V
F FF FF F V F
F
FFLL L ITTI
Таблица 4
Оценка иммуногенности C-терминальных пептидных фрагментов α-Syn для идентификации аутологичных Th-эпитопов у морских свинок
Таблица 5
Ранжирование иммуногенности пептидных иммуногенных конструкций α-Syn у морских свинок
Таблица 6
Оценка иммуногенности пептидных иммуногенных конструкций α-Syn у морских свинок
Таблица 7
Оценка иммуногенности пептидных иммуногенных конструкций α-Syn у морских свинок
Таблица 8
Оценка иммуногенности против участка Th-эпитопа пептидных иммуногенных конструкций α-Syn у морских свинок
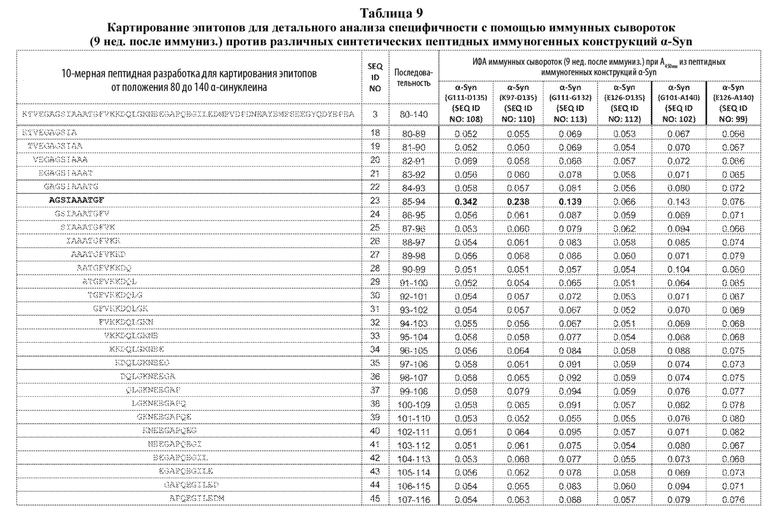
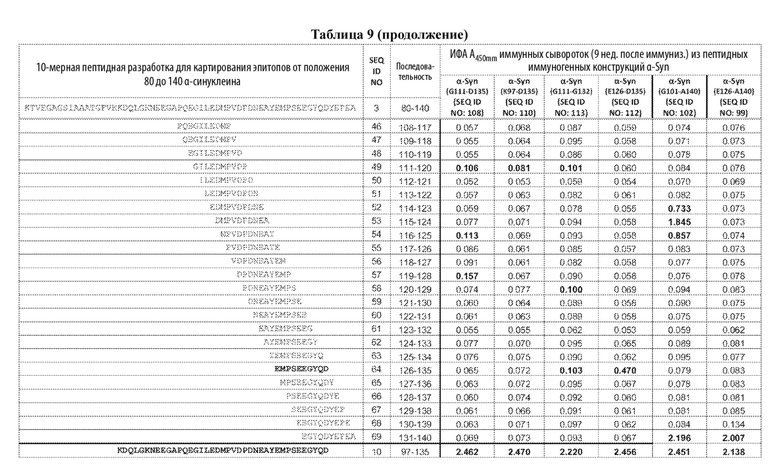
Таблица 10
Ингибирование агрегации α-Syn антителами от животных, получающих пептидных иммуногенные конструкции α-Syn
Таблица 11
Оценка нейропротекторной способности в отношении нейродегенерации, вызываемой агрегатами α-Syn, с помощью количественной оценки длины нейритов посредством многопараметрического скрининга с использованием антител от животных, получающих пептидные иммуногенные конструкции α-Syn
Таблица 12
Нейропротекторная оценка в нейродегенеративных нейронах, образование которых вызывается агрегатами α-Syn, с помощью количественной количества нейронов посредством многопараметрического скрининга с использованием антител от животных, получающих пептидные иммуногенные конструкции α-Syn
Таблица 15
Перечень случаев, полученных из UCL, и их диагноз после вскрытия
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>United Neuroscience
Wang, Chang Yi
<120>ПЕПТИДНЫЕ ИММУНОГЕНЫ ИЗ C-ТЕРМИНАЛЬНОГО КОНЦА БЕЛКА АЛЬФА-СИНУКЛЕИНА И СОСТАВЫ НА ИХ ОСНОВЕ ДЛЯ ЛЕЧЕНИЯ СИНУКЛЕИНОПАТИЙ
<130>UNS1001-US
<140>TBD
<141>2018-06-15
<150>US 62/521,287
<151>2017-06-16
<160>153
<170>PatentIn версия 3.5
<210>1
<211>140
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(140)
<223>альфа-Синуклеин 1-140
<400>1
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210>2
<211>134
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(134)
<223>бета-Синуклеин 1-134
<400>2
Met Asp Val Phe Met Lys Gly Leu Ser Met Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Thr Glu Ala Ala Glu Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Arg Glu Gly Val
35 40 45
Val Gln Gly Val Ala Ser Val Ala Glu Lys Thr Lys Glu Gln Ala Ser
50 55 60
His Leu Gly Gly Ala Val Phe Ser Gly Ala Gly Asn Ile Ala Ala Ala
65 70 75 80
Thr Gly Leu Val Lys Arg Glu Glu Phe Pro Thr Asp Leu Lys Pro Glu
85 90 95
Glu Val Ala Gln Glu Ala Ala Glu Glu Pro Leu Ile Glu Pro Leu Met
100 105 110
Glu Pro Glu Gly Glu Ser Tyr Glu Asp Pro Pro Gln Glu Glu Tyr Gln
115 120 125
Glu Tyr Glu Pro Glu Ala
130
<210>3
<211>61
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(61)
<223>альфа-Синуклеин 80-140
<400>3
Lys Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val
1 5 10 15
Lys Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly
20 25 30
Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met
35 40 45
Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
50 55 60
<210>4
<211>56
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(56)
<223>альфа-Синуклеин 85-140
<400>4
Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys Lys Asp Gln Leu
1 5 10 15
Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp Met
20 25 30
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
Tyr Gln Asp Tyr Glu Pro Glu Ala
50 55
<210>5
<211>50
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(50)
<223>альфа-Синуклеин 91-140
<400>5
Ala Thr Gly Phe Val Lys Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly
1 5 10 15
Ala Pro Gln Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn
20 25 30
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro
35 40 45
Glu Ala
50
<210>6
<211>40
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(40)
<223>альфа-Синуклеин 101-140
<400>6
Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp Met
1 5 10 15
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
20 25 30
Tyr Gln Asp Tyr Glu Pro Glu Ala
35 40
<210>7
<211>30
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(30)
<223>альфа-Синуклеин 111-140
<400>7
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
1 5 10 15
Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
20 25 30
<210>8
<211>20
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(20)
<223>альфа-Синуклеин 121-140
<400>8
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr
1 5 10 15
Glu Pro Glu Ala
20
<210>9
<211>15
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<400>9
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
1 5 10 15
<210>10
<211>39
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(39)
<223>альфа-Синуклеин 97-135
<400>10
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
1 5 10 15
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
20 25 30
Ser Glu Glu Gly Tyr Gln Asp
35
<210>11
<211>35
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(35)
<223>альфа-Синуклеин 101-135
<400>11
Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp Met
1 5 10 15
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
20 25 30
Tyr Gln Asp
35
<210>12
<211>25
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(25)
<223>альфа-Синуклеин 111-135
<400>12
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
1 5 10 15
Met Pro Ser Glu Glu Gly Tyr Gln Asp
20 25
<210>13
<211>15
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>альфа-Синуклеин 121-135
<400>13
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
1 5 10 15
<210>14
<211>13
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(13)
<223>альфа-Синуклеин 123-135
<400>14
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
1 5 10
<210>15
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 126-135
<400>15
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
1 5 10
<210>16
<211>32
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(32)
<223>альфа-Синуклеин 101-132
<400>16
Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp Met
1 5 10 15
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
20 25 30
<210>17
<211>22
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(22)
<223>альфа-Синуклеин 111-132
<400>17
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
1 5 10 15
Met Pro Ser Glu Glu Gly
20
<210>18
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 80-89
<400>18
Lys Thr Val Glu Gly Ala Gly Ser Ile Ala
1 5 10
<210>19
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 81-90
<400>19
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala
1 5 10
<210>20
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 82-91
<400>20
Val Glu Gly Ala Gly Ser Ile Ala Ala Ala
1 5 10
<210>21
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 83-92
<400>21
Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr
1 5 10
<210>22
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 84-93
<400>22
Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly
1 5 10
<210>23
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 85-94
<400>23
Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe
1 5 10
<210>24
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 86-95
<400>24
Gly Ser Ile Ala Ala Ala Thr Gly Phe Val
1 5 10
<210>25
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 87-96
<400>25
Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
1 5 10
<210>26
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 88-97
<400>26
Ile Ala Ala Ala Thr Gly Phe Val Lys Lys
1 5 10
<210>27
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 89-98
<400>27
Ala Ala Ala Thr Gly Phe Val Lys Lys Asp
1 5 10
<210>28
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 90-99
<400>28
Ala Ala Thr Gly Phe Val Lys Lys Asp Gln
1 5 10
<210>29
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 91-100
<400>29
Ala Thr Gly Phe Val Lys Lys Asp Gln Leu
1 5 10
<210>30
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 92-101
<400>30
Thr Gly Phe Val Lys Lys Asp Gln Leu Gly
1 5 10
<210>31
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 93-102
<400>31
Gly Phe Val Lys Lys Asp Gln Leu Gly Lys
1 5 10
<210>32
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 94-103
<400>32
Phe Val Lys Lys Asp Gln Leu Gly Lys Asn
1 5 10
<210>33
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 95-104
<400>33
Val Lys Lys Asp Gln Leu Gly Lys Asn Glu
1 5 10
<210>34
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 96-105
<400>34
Lys Lys Asp Gln Leu Gly Lys Asn Glu Glu
1 5 10
<210>35
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 97-106
<400>35
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly
1 5 10
<210>36
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 98-107
<400>36
Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala
1 5 10
<210>37
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 99-108
<400>37
Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro
1 5 10
<210>38
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 100-109
<400>38
Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln
1 5 10
<210>39
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 101-110
<400>39
Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu
1 5 10
<210>40
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 102-111
<400>40
Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly
1 5 10
<210>41
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 103-112
<400>41
Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
1 5 10
<210>42
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 104-113
<400>42
Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu
1 5 10
<210>43
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 105-114
<400>43
Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu
1 5 10
<210>44
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 106-115
<400>44
Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp
1 5 10
<210>45
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 107-116
<400>45
Ala Pro Gln Glu Gly Ile Leu Glu Asp Met
1 5 10
<210>46
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 108-117
<400>46
Pro Gln Glu Gly Ile Leu Glu Asp Met Pro
1 5 10
<210>47
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 109-118
<400>47
Gln Glu Gly Ile Leu Glu Asp Met Pro Val
1 5 10
<210>48
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 110-119
<400>48
Glu Gly Ile Leu Glu Asp Met Pro Val Asp
1 5 10
<210>49
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 111-120
<400>49
Gly Ile Leu Glu Asp Met Pro Val Asp Pro
1 5 10
<210>50
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 112-121
<400>50
Ile Leu Glu Asp Met Pro Val Asp Pro Asp
1 5 10
<210>51
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа1-Синуклеин 113-122
<400>51
Leu Glu Asp Met Pro Val Asp Pro Asp Asn
1 5 10
<210>52
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 114-123
<400>52
Glu Asp Met Pro Val Asp Pro Asp Asn Glu
1 5 10
<210>53
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 115-124
<400>53
Asp Met Pro Val Asp Pro Asp Asn Glu Ala
1 5 10
<210>54
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 116-125
<400>54
Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
1 5 10
<210>55
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 117-126
<400>55
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
1 5 10
<210>56
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 118-127
<400>56
Val Asp Pro Asp Asn Glu Ala Tyr Glu Met
1 5 10
<210>57
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 119-128
<400>57
Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
1 5 10
<210>58
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 120-129
<400>58
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser
1 5 10
<210>59
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 121-130
<400>59
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu
1 5 10
<210>60
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 122-131
<400>60
Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu
1 5 10
<210>61
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 123-132
<400>61
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
1 5 10
<210>62
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 124-133
<400>62
Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr
1 5 10
<210>63
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 125-134
<400>63
Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln
1 5 10
<210>64
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 126-135
<400>64
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
1 5 10
<210>65
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 127-136
<400>65
Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr
1 5 10
<210>66
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 128-137
<400>66
Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu
1 5 10
<210>67
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 129-138
<400>67
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro
1 5 10
<210>68
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 130-139
<400>68
Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu
1 5 10
<210>69
<211>10
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(10)
<223>альфа-Синуклеин 131-140
<400>69
Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
1 5 10
<210>70
<211>17
<212>БЕЛОК
<213>Clostridium tetani
<220>
<221>ПЕПТИД
<222>(1)..(17)
<223>Clostridium tetani 1 Th
<400>70
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu
<210>71
<211>15
<212>БЕЛОК
<213>Вирус кори
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>MvF1 Th
<400>71
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val
1 5 10 15
<210>72
<211>24
<212>БЕЛОК
<213>Bordetella pertussis
<220>
<221>ПЕПТИД
<222>(1)..(24)
<223>Bordetella pertussis Th
<400>72
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu
20
<210>73
<211>17
<212>БЕЛОК
<213>Clostridium tetani
<220>
<221>ПЕПТИД
<222>(1)..(17)
<223>Clostridium tetani 2 Th
<400>73
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr
<210>74
<211>23
<212>БЕЛОК
<213>дифтерийная палочка
<220>
<221>ПЕПТИД
<222>(1)..(23)
<223>Diphtheria Th
<400>74
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys
20
<210>75
<211>21
<212>БЕЛОК
<213>Plasmodium falciparum
<220>
<221>ПЕПТИД
<222>(1)..(21)
<223>Plasmodium falciparum Th
<400>75
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser
20
<210>76
<211>17
<212>БЕЛОК
<213>Schistosoma mansoni
<220>
<221>ПЕПТИД
<222>(1)..(17)
<223>Schistosoma mansoni Th
<400>76
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His
<210>77
<211>25
<212>БЕЛОК
<213>Холерный токсин
<220>
<221>ПЕПТИД
<222>(1)..(25)
<223>Холерный токсин Th
<400>77
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser
20 25
<210>78
<211>15
<212>БЕЛОК
<213>Вирус кори
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>MvF 2 Th
<400>78
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile
1 5 10 15
<210>79
<211>22
<212>БЕЛОК
<213>Вирус кори
<220>
<221>ПЕПТИД
<222>(1)..(22)
<223>KKKMvF 3 Th
<220>
<221>САЙТ
<222>(7)..(7)
<223>S или T
<220>
<221>САЙТ
<222>(10)..(10)
<223>K или R
<220>
<221>САЙТ
<222>(11)..(11)
<223>G или T
<220>
<221>САЙТ
<222>(15)..(15)
<223>H или T
<220>
<221>САЙТ
<222>(16)..(16)
<223>K или R
<220>
<221>САЙТ
<222>(19)..(19)
<223>G или T
<400>79
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe
20
<210>80
<211>18
<212>БЕЛОК
<213>Вирус гепатита В
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg 1 Th
<220>
<221>САЙТ
<222>(1)..(1)
<223>K или R
<220>
<221>САЙТ
<222>(2)..(2)
<223>K или R
<220>
<221>САЙТ
<222>(3)..(3)
<223>K или R
<220>
<221>САЙТ
<222>(4)..(4)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(5)..(5)
<223>F или K или R
<220>
<221>САЙТ
<222>(6)..(6)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(7)..(7)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(9)..(9)
<223>K или R
<220>
<221>САЙТ
<222>(10)..(10)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(11)..(11)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(13)..(13)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(15)..(15)
<223>Q или L или I или V или F
<220>
<221>САЙТ
<222>(17)..(17)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(18)..(18)
<223>D или R
<400>80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa
<210>81
<211>19
<212>БЕЛОК
<213>Вирус кори
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<400>81
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210>82
<211>18
<212>БЕЛОК
<213>Вирус гепатита В
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg 2 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>I или F
<220>
<221>САЙТ
<222>(5)..(5)
<223>I или F
<220>
<221>САЙТ
<222>(6)..(6)
<223>T или L
<220>
<221>САЙТ
<222>(7)..(7)
<223>I или L
<220>
<221>САЙТ
<222>(11)..(11)
<223>I или L
<220>
<221>САЙТ
<222>(14)..(14)
<223>P или I
<220>
<221>САЙТ
<222>(15)..(15)
<223>Q или T
<220>
<221>САЙТ
<222>(16)..(16)
<223>S или T
<220>
<221>САЙТ
<222>(17)..(17)
<223>L или I
<400>82
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp
<210>83
<211>19
<212>БЕЛОК
<213>Вирус кори
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 5 Th
<400>83
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe
<210>84
<211>18
<212>БЕЛОК
<213>Вирус гепатита В
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg 3 Th
<400>84
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp
<210>85
<211>11
<212>БЕЛОК
<213>Вирус гриппа
<220>
<221>ПЕПТИД
<222>(1)..(11)
<223>Белок матрикса 1 вируса гриппа _1 Th
<220>
<221>ПЕПТИД
<222>(1)..(11)
<223>Белок матрикса 1 вируса гриппа_1 Th
<400>85
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg
1 5 10
<210>86
<211>15
<212>БЕЛОК
<213>Вирус гриппа
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>Белок матрикса 1 вируса гриппа_2 Th
<400>86
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val
1 5 10 15
<210>87
<211>9
<212>БЕЛОК
<213>Вирус гриппа
<220>
<221>ПЕПТИД
<222>(1)..(9)
<223>Неструктурный белок 1 вируса гриппа Th
<400>87
Asp Arg Leu Arg Arg Asp Gln Lys Ser
1 5
<210>88
<211>19
<212>БЕЛОК
<213>Вирус Эпштейна-Бар (EBV)
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>EBV BHRF1 Th
<400>88
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly
<210>89
<211>15
<212>БЕЛОК
<213>Clostridium tetani
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>Clostridium tetani TT1 Th
<400>89
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu
1 5 10 15
<210>90
<211>20
<212>БЕЛОК
<213>Вирус Эпштейна-Бар
<220>
<221>ПЕПТИД
<222>(1)..(20)
<223>EBV EBNA-1 Th
<400>90
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile
20
<210>91
<211>21
<212>БЕЛОК
<213>Clostridium tetani
<220>
<221>ПЕПТИД
<222>(1)..(21)
<223>Clostridium tetani TT2 Th
<400>91
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu
20
<210>92
<211>16
<212>БЕЛОК
<213>Clostridium tetani
<220>
<221>ПЕПТИД
<222>(1)..(16)
<223>Clostridium tetani TT3 Th
<400>92
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
<210>93
<211>16
<212>БЕЛОК
<213>Clostridium tetani
<220>
<221>ПЕПТИД
<222>(1)..(16)
<223>Clostridium tetani TT4 Th
<400>93
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
<210>94
<211>18
<212>БЕЛОК
<213>Вирус Эпштейна-Бар
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>EBV CP Th
<400>94
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu
<210>95
<211>14
<212>БЕЛОК
<213>Цитомегаловирус человека (HCMV)
<220>
<221>ПЕПТИД
<222>(1)..(14)
<223>HCMV IE1 Th
<400>95
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His
1 5 10
<210>96
<211>15
<212>БЕЛОК
<213>Вирус Эпштейна-Бар
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>EBV GP340 Th
<400>96
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr
1 5 10 15
<210>97
<211>13
<212>БЕЛОК
<213>Вирус Эпштейна-Бар
<220>
<221>ПЕПТИД
<222>(1)..(13)
<223>EBV BPLF1 Th
<400>97
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln
1 5 10
<210>98
<211>11
<212>БЕЛОК
<213>Вирус Эпштейна-Бар
<220>
<221>ПЕПТИД
<222>(1)..(11)
<223>EBV EBNA-2 Th
<400>98
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu
1 5 10
<210>99
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(38)
<223>альфа-Синуклеин 126-140
<400>99
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Glu Met Pro Ser Glu Glu Gly Tyr Gln
20 25 30
Asp Tyr Glu Pro Glu Ala
35
<210>100
<211>43
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(43)
<223>альфа-Синуклеин 121-140
<400>100
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Asp Asn Glu Ala Tyr Glu Met Pro Ser
20 25 30
Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
35 40
<210>101
<211>53
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(53)
<223>альфа-Синуклеин 111-140
<400>101
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
35 40 45
Tyr Glu Pro Glu Ala
50
<210>102
<211>63
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(63)
<223>альфа-Синуклеин 126-140
<400>102
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Lys Asn Glu Glu Gly Ala Pro Gln
20 25 30
Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
35 40 45
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
50 55 60
<210>103
<211>63
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(63)
<223>альфа-Синуклеин 101-140
<400>103
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Lys Asn Glu Glu Gly Ala Pro Gln
20 25 30
Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
35 40 45
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
50 55 60
<210>104
<211>62
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg3 Th
<220>
<221>САЙТ
<222>(19)..(19)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(19)..(22)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(23)..(62)
<223>альфа-Синуклеин 101-140
<400>104
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Lys Lys Lys Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu
20 25 30
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
35 40 45
Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
50 55 60
<210>105
<211>73
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(73)
<223>альфа-Синуклеин 91-140
<400>105
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Ala Thr Gly Phe Val Lys Lys Asp Gln
20 25 30
Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp
35 40 45
Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu
50 55 60
Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
65 70
<210>106
<211>79
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(79)
<223>альфа-Синуклеин 85-140
<400>106
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Ala Gly Ser Ile Ala Ala Ala Thr Gly
20 25 30
Phe Val Lys Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln
35 40 45
Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
50 55 60
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
65 70 75
<210>107
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(38)
<223>альфа-Синуклеин 121-135
<400>107
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Asp Asn Glu Ala Tyr Glu Met Pro Ser
20 25 30
Glu Glu Gly Tyr Gln Asp
35
<210>108
<211>48
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(48)
<223>альфа-Синуклеин 111-135
<400>108
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
35 40 45
<210>109
<211>58
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(58)
<223>альфа-Синуклеин 101-135
<400>109
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Lys Asn Glu Glu Gly Ala Pro Gln
20 25 30
Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
35 40 45
Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
50 55
<210>110
<211>62
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(62)
<223>альфа-Синуклеин 97-135
<400>110
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Lys Asp Gln Leu Gly Lys Asn Glu Glu
20 25 30
Gly Ala Pro Gln Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp
35 40 45
Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
50 55 60
<210>111
<211>36
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(36)
<223>альфа-Синуклеин 123-135
<400>111
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Glu Ala Tyr Glu Met Pro Ser Glu Glu
20 25 30
Gly Tyr Gln Asp
35
<210>112
<211>33
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(33)
<223>альфа-Синуклеин 126-135
<400>112
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Glu Met Pro Ser Glu Glu Gly Tyr Gln
20 25 30
Asp
<210>113
<211>45
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(45)
<223>альфа-Синуклеин 111-132
<400>113
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>114
<211>55
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(55)
<223>альфа-Синуклеин 101-132
<400>114
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Lys Asn Glu Glu Gly Ala Pro Gln
20 25 30
Glu Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
35 40 45
Glu Met Pro Ser Glu Glu Gly
50 55
<210>115
<211>45
<212>БЕЛОК
<213>Mus musculus
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(45)
<223>Mouse альфа-Синуклеин 111-132
<400>115
Ile Ser Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Gly Ser Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>116
<211>33
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(33)
<223>альфа-Синуклеин 126-135
<400>116
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Glu Met Pro Ser Glu Glu Gly Tyr Gln
20 25 30
Asp
<210>117
<211>45
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF 4 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>S или T
<220>
<221>САЙТ
<222>(7)..(7)
<223>K или R
<220>
<221>САЙТ
<222>(8)..(8)
<223>G или T
<220>
<221>САЙТ
<222>(12)..(12)
<223>H или T
<220>
<221>САЙТ
<222>(13)..(13)
<223>K или R
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон-К
<220>
<221>ПЕПТИД
<222>(20)..(23)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(45)
<223>альфа-Синуклеин 111-132
<400>117
Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Lys Lys Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>118
<211>30
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(21)..(30)
<223>альфа-Синуклеин 126-135
<400>118
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
20 25 30
<210>119
<211>42
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>MvF5 Th
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(21)..(42)
<223>альфа-Синуклеин 111-132
<400>119
Ile Ser Ile Thr Glu Ile Lys Gly Val Ile Val His Arg Ile Glu Thr
1 5 10 15
Ile Leu Phe Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn
20 25 30
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>120
<211>29
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg3 Th
<220>
<221>САЙТ
<222>(19)..(19)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(20)..(29)
<223>альфа-Синуклеин 126-135
<400>120
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Glu Met Pro Ser Glu Glu Gly Tyr Gln Asp
20 25
<210>121
<211>41
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg3 Th
<220>
<221>САЙТ
<222>(19)..(19)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(20)..(41)
<223>альфа-Синуклеин 111-132
<400>121
Lys Lys Lys Ile Ile Thr Ile Thr Arg Ile Ile Thr Ile Ile Thr Thr
1 5 10 15
Ile Asp Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>122
<211>40
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(17)
<223>Clostridium tetani1 Th
<220>
<221>САЙТ
<222>(18)..(18)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(19)..(40)
<223>альфа-Синуклеин 111-132
<400>122
Lys Lys Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu
1 5 10 15
Leu Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala
20 25 30
Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>123
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>MvF1 Th
<220>
<221>САЙТ
<222>(16)..(16)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(17)..(38)
<223>альфа-Синуклеин 111-132
<400>123
Leu Ser Glu Ile Lys Gly Val Ile Val His Arg Leu Glu Gly Val Lys
1 5 10 15
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
20 25 30
Met Pro Ser Glu Glu Gly
35
<210>124
<211>47
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(24)
<223>Bordetella pertussis Th
<220>
<221>САЙТ
<222>(25)..(25)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(26)..(47)
<223>альфа-Синуклеин 111-132
<400>124
Gly Ala Tyr Ala Arg Cys Pro Asn Gly Thr Arg Ala Leu Thr Val Ala
1 5 10 15
Glu Leu Arg Gly Asn Ala Glu Leu Lys Gly Ile Leu Glu Asp Met Pro
20 25 30
Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>125
<211>40
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(17)
<223>Clostridium tetani2 Th
<220>
<221>САЙТ
<222>(18)..(18)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(19)..(40)
<223>альфа-Синуклеин 111-132
<400>125
Trp Val Arg Asp Ile Ile Asp Asp Phe Thr Asn Glu Ser Ser Gln Lys
1 5 10 15
Thr Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala
20 25 30
Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>126
<211>46
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(23)
<223>Diphtheria Th
<220>
<221>САЙТ
<222>(24)..(24)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(25)..(46)
<223>альфа-Синуклеин 111-132
<400>126
Asp Ser Glu Thr Ala Asp Asn Leu Glu Lys Thr Val Ala Ala Leu Ser
1 5 10 15
Ile Leu Pro Gly His Gly Cys Lys Gly Ile Leu Glu Asp Met Pro Val
20 25 30
Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>127
<211>44
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(21)
<223>Plasmodium falciparum Th
<220>
<221>САЙТ
<222>(22)..(22)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(23)..(44)
<223>альфа-Синуклеин 111-132
<400>127
Asp His Glu Lys Lys His Ala Lys Met Glu Lys Ala Ser Ser Val Phe
1 5 10 15
Asn Val Val Asn Ser Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro
20 25 30
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>128
<211>40
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(17)
<223>Schistosoma mansoni Th
<220>
<221>САЙТ
<222>(18)..(18)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(19)..(40)
<223>альфа-Синуклеин 111-132
<400>128
Lys Trp Phe Lys Thr Asn Ala Pro Asn Gly Val Asp Glu Lys His Arg
1 5 10 15
His Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala
20 25 30
Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>129
<211>48
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(25)
<223>Холерный токсин Th
<220>
<221>САЙТ
<222>(26)..(26)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(27)..(48)
<223>альфа-Синуклеин 111-132
<400>129
Ala Leu Asn Ile Trp Asp Arg Phe Asp Val Phe Cys Thr Leu Gly Ala
1 5 10 15
Thr Thr Gly Tyr Leu Lys Gly Asn Ser Lys Gly Ile Leu Glu Asp Met
20 25 30
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>130
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>MvF2 Th
<220>
<221>САЙТ
<222>(16)..(16)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(17)..(38)
<223>альфа-Синуклеин 111-132
<400>130
Ile Ser Glu Ile Lys Gly Val Ile Val His Lys Ile Glu Gly Ile Lys
1 5 10 15
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
20 25 30
Met Pro Ser Glu Glu Gly
35
<210>131
<211>45
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(22)
<223>KKKMvF3 Th
<220>
<221>САЙТ
<222>(7)..(7)
<223>S или T
<220>
<221>САЙТ
<222>(10)..(10)
<223>K или R
<220>
<221>САЙТ
<222>(11)..(11)
<223>G или T
<220>
<221>САЙТ
<222>(15)..(15)
<223>H или T
<220>
<221>САЙТ
<222>(16)..(16)
<223>K или R
<220>
<221>САЙТ
<222>(19)..(19)
<223>G или T
<220>
<221>САЙТ
<222>(23)..(23)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(24)..(45)
<223>альфа-Синуклеин 111-132
<400>131
Lys Lys Lys Ile Ser Ile Xaa Glu Ile Xaa Xaa Val Ile Val Xaa Xaa
1 5 10 15
Ile Glu Xaa Ile Leu Phe Lys Gly Ile Leu Glu Asp Met Pro Val Asp
20 25 30
Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40 45
<210>132
<211>41
<212>БЕЛОК
<213>Вирус гепатита В
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg 1 Th
<220>
<221>САЙТ
<222>(1)..(1)
<223>K или R
<220>
<221>САЙТ
<222>(2)..(2)
<223>K или R
<220>
<221>САЙТ
<222>(3)..(3)
<223>K или R
<220>
<221>САЙТ
<222>(4)..(4)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(5)..(5)
<223>F или K или R
<220>
<221>САЙТ
<222>(6)..(6)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(7)..(7)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(9)..(9)
<223>K или R
<220>
<221>САЙТ
<222>(10)..(10)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(11)..(11)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(13)..(13)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(15)..(15)
<223>Q или L или I или V или F
<220>
<221>САЙТ
<222>(17)..(17)
<223>L или I или V или F
<220>
<221>САЙТ
<222>(18)..(18)
<223>D или R
<220>
<221>САЙТ
<222>(19)..(19)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(20)..(41)
<223>альфа-Синуклеин 111-132
<400>132
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Xaa Xaa Thr Xaa Pro Xaa Ser
1 5 10 15
Xaa Xaa Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>133
<211>41
<212>БЕЛОК
<213>Вирус гепатита В
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>HBsAg 2 Th
<220>
<221>САЙТ
<222>(4)..(4)
<223>I или F
<220>
<221>САЙТ
<222>(5)..(5)
<223>I или F
<220>
<221>САЙТ
<222>(6)..(6)
<223>T или L
<220>
<221>САЙТ
<222>(7)..(7)
<223>I или L
<220>
<221>САЙТ
<222>(11)..(11)
<223>I или L
<220>
<221>САЙТ
<222>(14)..(14)
<223>P или I
<220>
<221>САЙТ
<222>(15)..(15)
<223>Q или T
<220>
<221>САЙТ
<222>(16)..(16)
<223>S или T
<220>
<221>САЙТ
<222>(17)..(17)
<223>L или I
<220>
<221>САЙТ
<222>(19)..(19)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(20)..(41)
<223>альфа-Синуклеин 111-132
<400>133
Lys Lys Lys Xaa Xaa Xaa Xaa Thr Arg Ile Xaa Thr Ile Xaa Xaa Xaa
1 5 10 15
Xaa Asp Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>134
<211>34
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(11)
<223>Вирус гриппа MP1_1 Th
<220>
<221>САЙТ
<222>(12)..(12)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(13)..(34)
<223>альфа-Синуклеин 111-132
<400>134
Phe Val Phe Thr Leu Thr Val Pro Ser Glu Arg Lys Gly Ile Leu Glu
1 5 10 15
Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu
20 25 30
Glu Gly
<210>135
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>Вирус гриппа MP1_2 Th
<220>
<221>САЙТ
<222>(16)..(16)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(17)..(38)
<223>альфа-Синуклеин 111-132
<400>135
Ser Gly Pro Leu Lys Ala Glu Ile Ala Gln Arg Leu Glu Asp Val Lys
1 5 10 15
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
20 25 30
Met Pro Ser Glu Glu Gly
35
<210>136
<211>32
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(9)
<223>Вирус гриппа NSP1 Th
<220>
<221>САЙТ
<222>(10)..(10)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(11)..(32)
<223>альфа-Синуклеин 111-132
<400>136
Asp Arg Leu Arg Arg Asp Gln Lys Ser Lys Gly Ile Leu Glu Asp Met
1 5 10 15
Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
20 25 30
<210>137
<211>42
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(19)
<223>EBV BHRF1 Th
<220>
<221>ПЕПТИД
<222>(20)..(20)
<223>эпсилон К в качестве спейсера
<220>
<221>САЙТ
<222>(20)..(20)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(21)..(42)
<223>альфа-Синуклеин 111-132
<400>137
Ala Gly Leu Thr Leu Ser Leu Leu Val Ile Cys Ser Tyr Leu Phe Ile
1 5 10 15
Ser Arg Gly Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn
20 25 30
Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>138
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>Clostridium tetani TT1 Th
<220>
<221>САЙТ
<222>(16)..(16)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(17)..(38)
<223>альфа-Синуклеин 111-132
<400>138
Gln Tyr Ile Lys Ala Asn Ser Lys Phe Ile Gly Ile Thr Glu Leu Lys
1 5 10 15
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
20 25 30
Met Pro Ser Glu Glu Gly
35
<210>139
<211>43
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(20)
<223>EBV EBNA-1 Th
<220>
<221>САЙТ
<222>(21)..(21)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(22)..(43)
<223>альфа-Синуклеин 111-132
<400>139
Pro Gly Pro Leu Arg Glu Ser Ile Val Cys Tyr Phe Met Val Phe Leu
1 5 10 15
Gln Thr His Ile Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp
20 25 30
Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>140
<211>44
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(21)
<223>Clostridium tetani TT2 Th
<220>
<221>САЙТ
<222>(22)..(22)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(23)..(44)
<223>альфа-Синуклеин 111-132
<400>140
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro
20 25 30
Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>141
<211>39
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(16)
<223>Clostridium tetani TT3 Th
<220>
<221>САЙТ
<222>(17)..(17)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(18)..(39)
<223>альфа-Синуклеин 111-132
<400>141
Lys Phe Ile Ile Lys Arg Tyr Thr Pro Asn Asn Glu Ile Asp Ser Phe
1 5 10 15
Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
20 25 30
Glu Met Pro Ser Glu Glu Gly
35
<210>142
<211>39
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(16)
<223>Clostridium tetani TT4 Th
<220>
<221>САЙТ
<222>(17)..(17)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(18)..(39)
<223>альфа-Синуклеин 111-132
<400>142
Val Ser Ile Asp Lys Phe Arg Ile Phe Cys Lys Ala Leu Asn Pro Lys
1 5 10 15
Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr
20 25 30
Glu Met Pro Ser Glu Glu Gly
35
<210>143
<211>41
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(18)
<223>EBV CP Th
<220>
<221>САЙТ
<222>(19)..(19)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(20)..(41)
<223>альфа-Синуклеин 111-132
<400>143
Val Pro Gly Leu Tyr Ser Pro Cys Arg Ala Phe Phe Asn Lys Glu Glu
1 5 10 15
Leu Leu Lys Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu
20 25 30
Ala Tyr Glu Met Pro Ser Glu Glu Gly
35 40
<210>144
<211>37
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(14)
<223>HCMV IE1 Th
<220>
<221>САЙТ
<222>(15)..(15)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(16)..(37)
<223>альфа-Синуклеин 111-132
<400>144
Asp Lys Arg Glu Met Trp Met Ala Cys Ile Lys Glu Leu His Lys Gly
1 5 10 15
Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met
20 25 30
Pro Ser Glu Glu Gly
35
<210>145
<211>38
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(15)
<223>EBV GP340 Th
<220>
<221>САЙТ
<222>(16)..(16)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(17)..(38)
<223>альфа-Синуклеин 111-132
<400>145
Thr Gly His Gly Ala Arg Thr Ser Thr Glu Pro Thr Thr Asp Tyr Lys
1 5 10 15
Gly Ile Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu
20 25 30
Met Pro Ser Glu Glu Gly
35
<210>146
<211>36
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(13)
<223>EBV BPLF1 Th
<220>
<221>САЙТ
<222>(14)..(14)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(15)..(36)
<223>альфа-Синуклеин 111-132
<400>146
Lys Glu Leu Lys Arg Gln Tyr Glu Lys Lys Leu Arg Gln Lys Gly Ile
1 5 10 15
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
20 25 30
Ser Glu Glu Gly
35
<210>147
<211>34
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(11)
<223>EBV EBNA-2 Th
<220>
<221>САЙТ
<222>(12)..(12)
<223>эпсилон К в качестве спейсера
<220>
<221>ПЕПТИД
<222>(13)..(34)
<223>альфа-Синуклеин 111-132
<400>147
Thr Val Phe Tyr Asn Ile Pro Pro Met Pro Leu Lys Gly Ile Leu Glu
1 5 10 15
Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro Ser Glu
20 25 30
Glu Gly
<210>148
<211>4
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(4)
<223>эпсилон K-KKK в качестве спейсера
<220>
<221>САЙТ
<222>(1)..(1)
<223>эпсилон-К
<400>148
Lys Lys Lys Lys
1
<210>149
<211>31
<212>ДНК
<213>Homo sapiens
<220>
<221>primer_bind
<222>(1)..(31)
<223>последовательность прямого праймера для альфа-синуклеина
<400>149
cgggatccga tgtgtttatg aaaggtctga g 31
<210>150
<211>31
<212>ДНК
<213>Homo sapiens
<220>
<221>primer_bind
<222>(1)..(31)
<223>обратный праймер для альфа-синуклеина
<400>150
ggaattccga tgtgtttatg aaaggtctga g 31
<210>151
<211>23
<212>ДНК
<213>Homo sapiens
<220>
<221>primer_bind
<222>(1)..(23)
<223>Последовательности праймеров для мутантного альфа-синуклеина
<400>151
tcatggtgtg accaccgttg cag 23
<210>152
<211>21
<212>ДНК
<213>Homo sapiens
<220>
<221>primer_bind
<222>(1)..(21)
<223>обратный праймер для мутантного альфа-синуклеина
<400>152
accacgcctt ctttggtttt g 21
<210>153
<211>32
<212>БЕЛОК
<213>Homo sapiens
<220>
<221>ПЕПТИД
<222>(1)..(32)
<223>бета-синуклеин 103-134
<400>153
Ala Glu Glu Pro Leu Ile Glu Pro Leu Met Glu Pro Glu Gly Glu Ser
1 5 10 15
Tyr Glu Asp Pro Pro Gln Glu Glu Tyr Gln Glu Tyr Glu Pro Glu Ala
20 25 30
<---
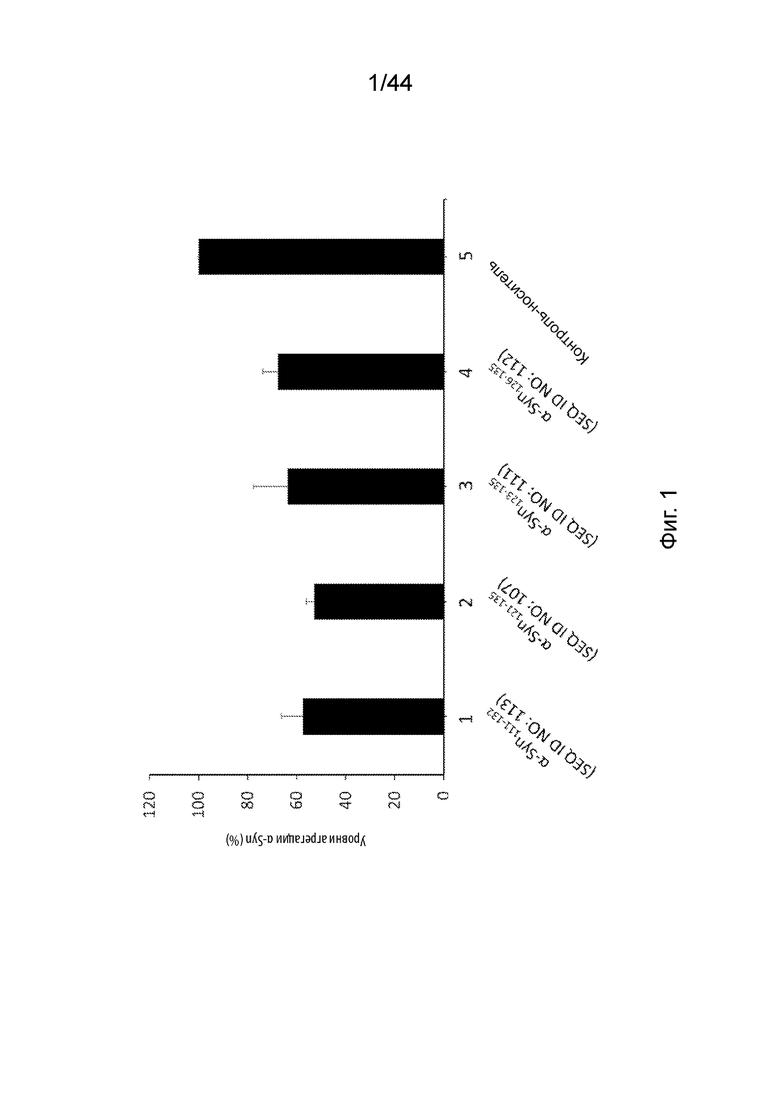
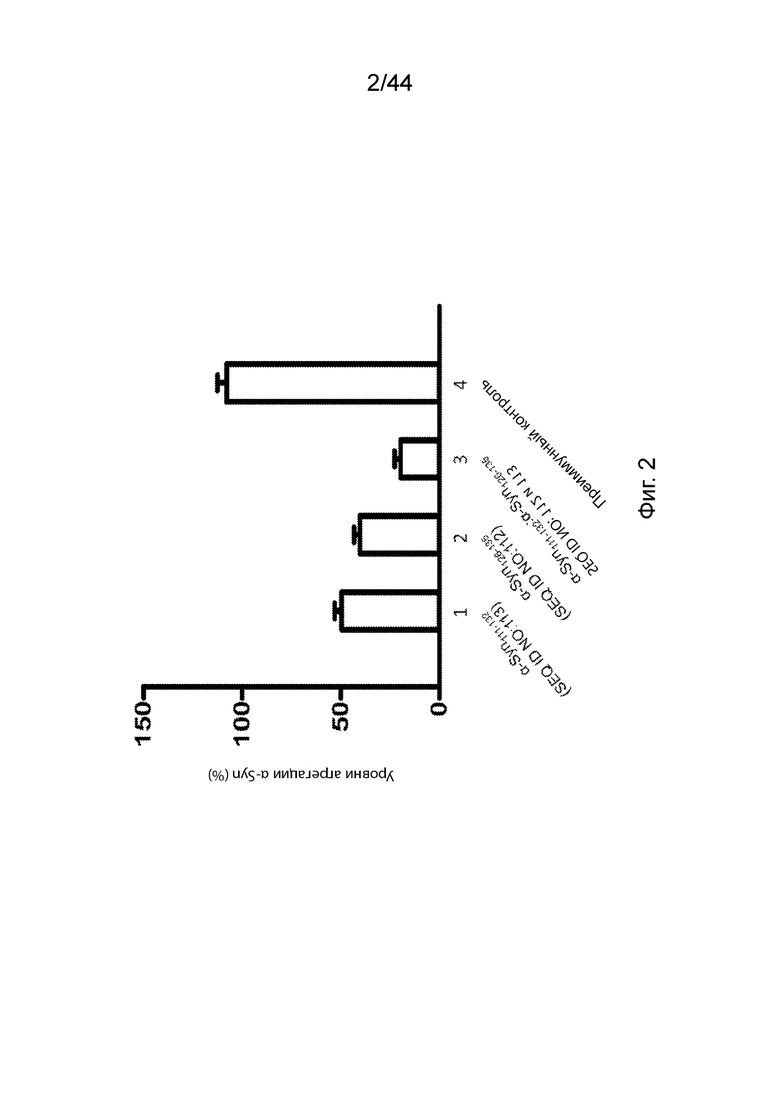
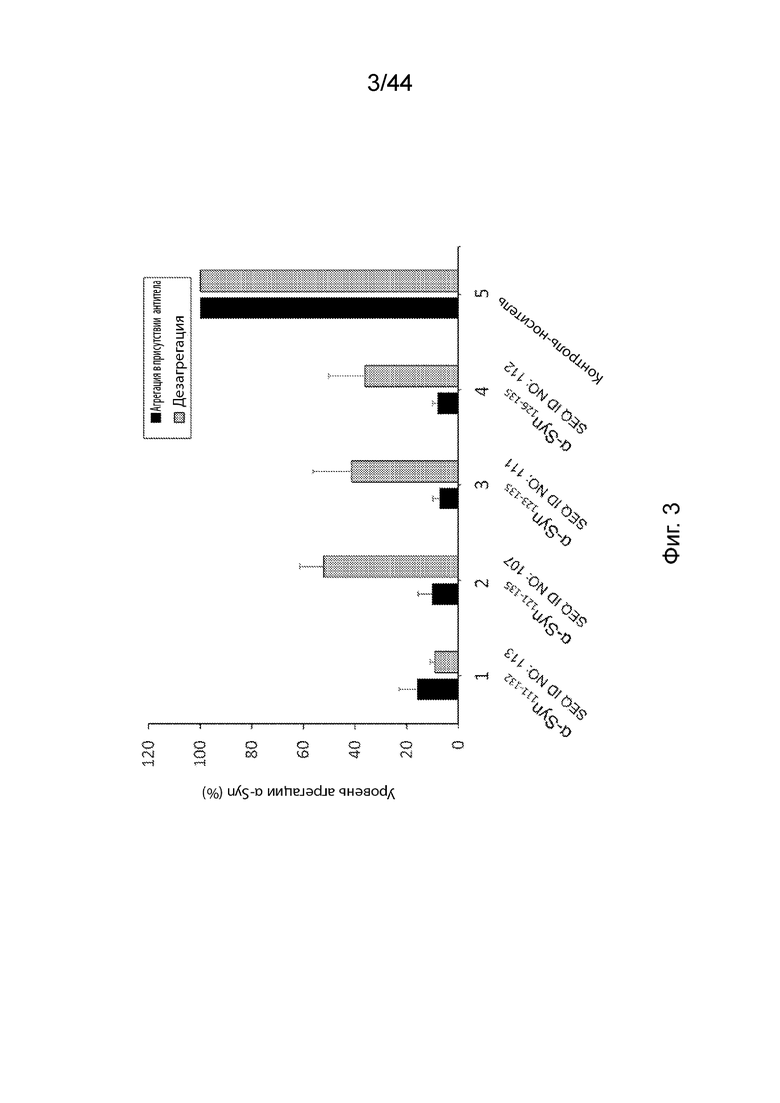
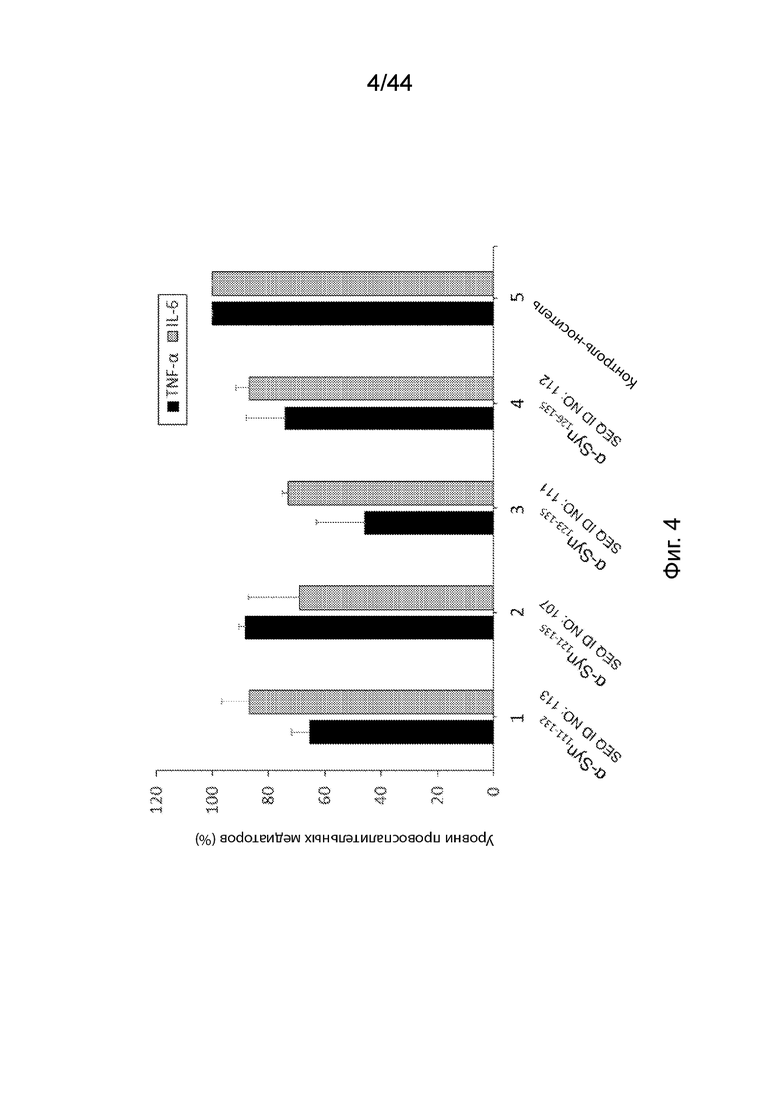
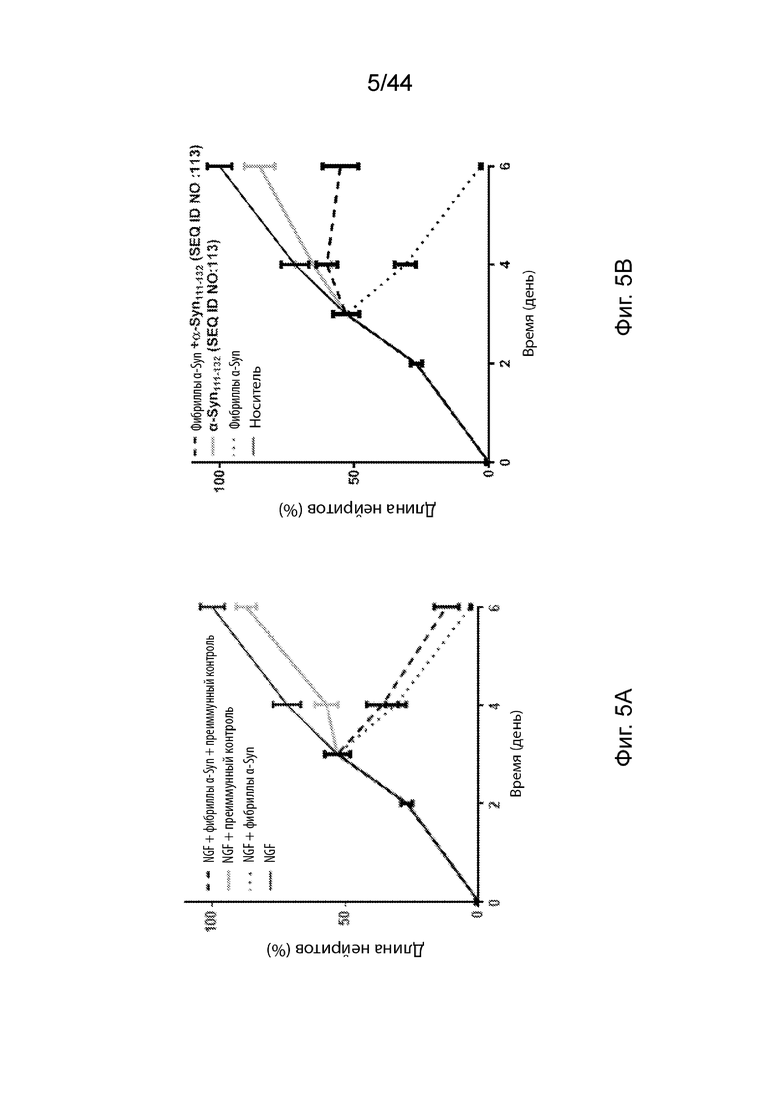
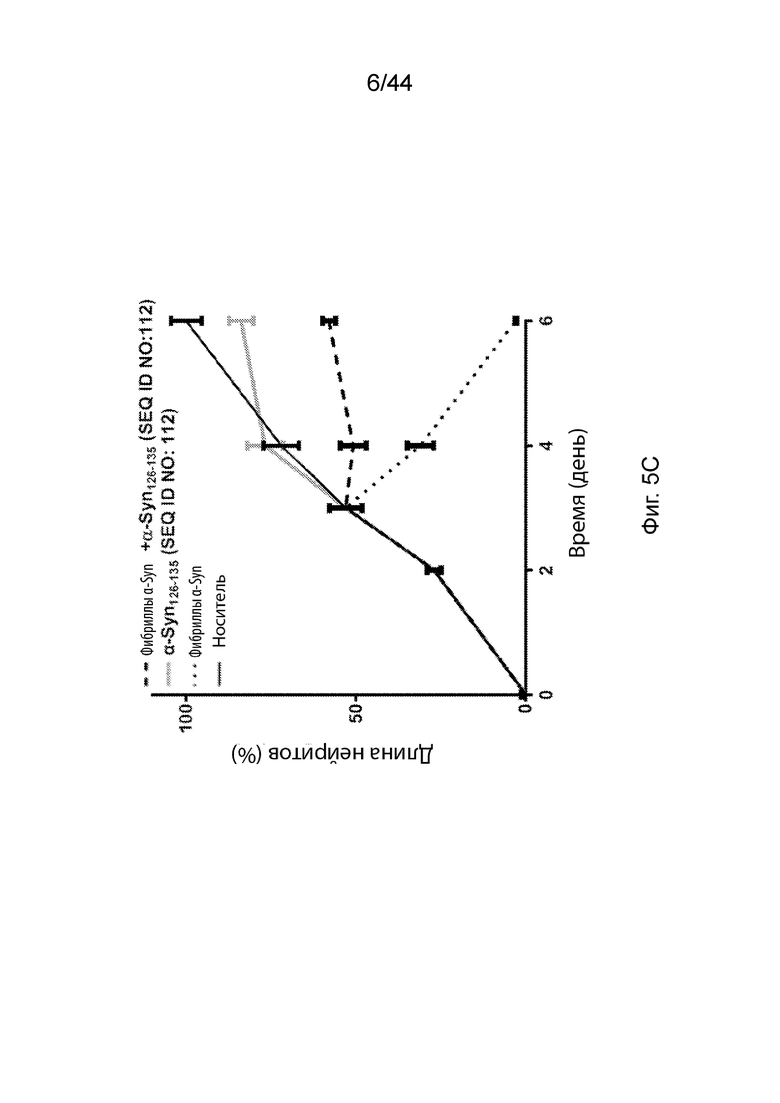
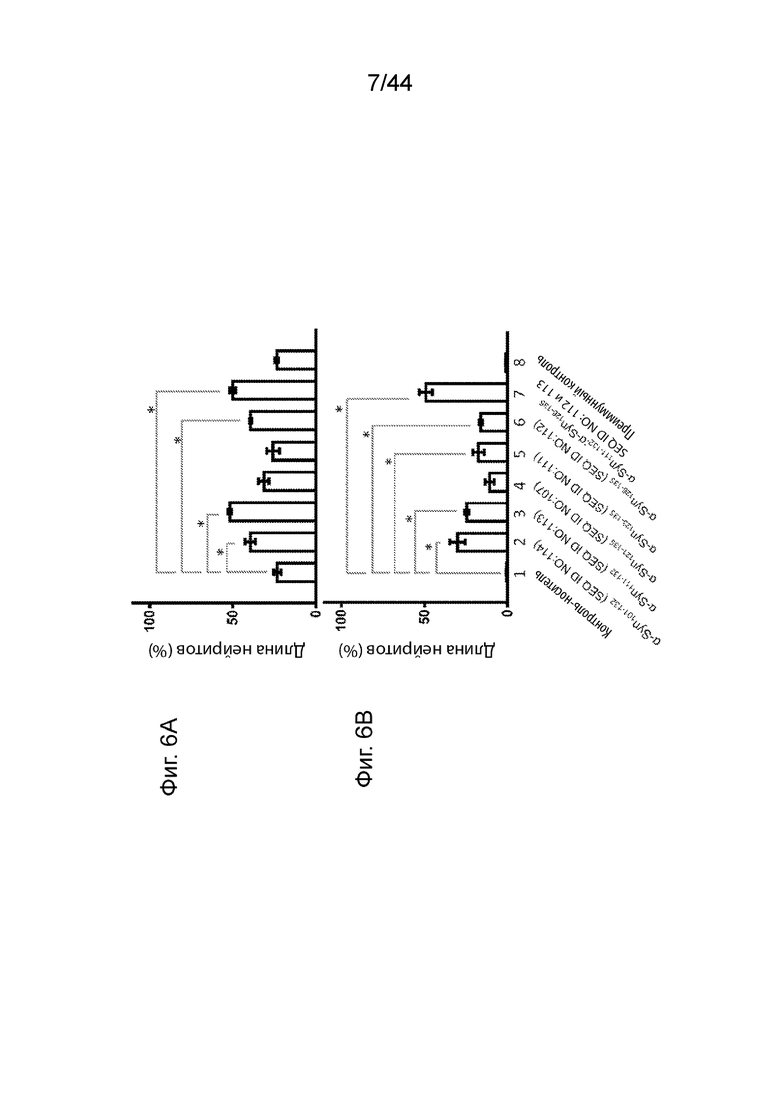
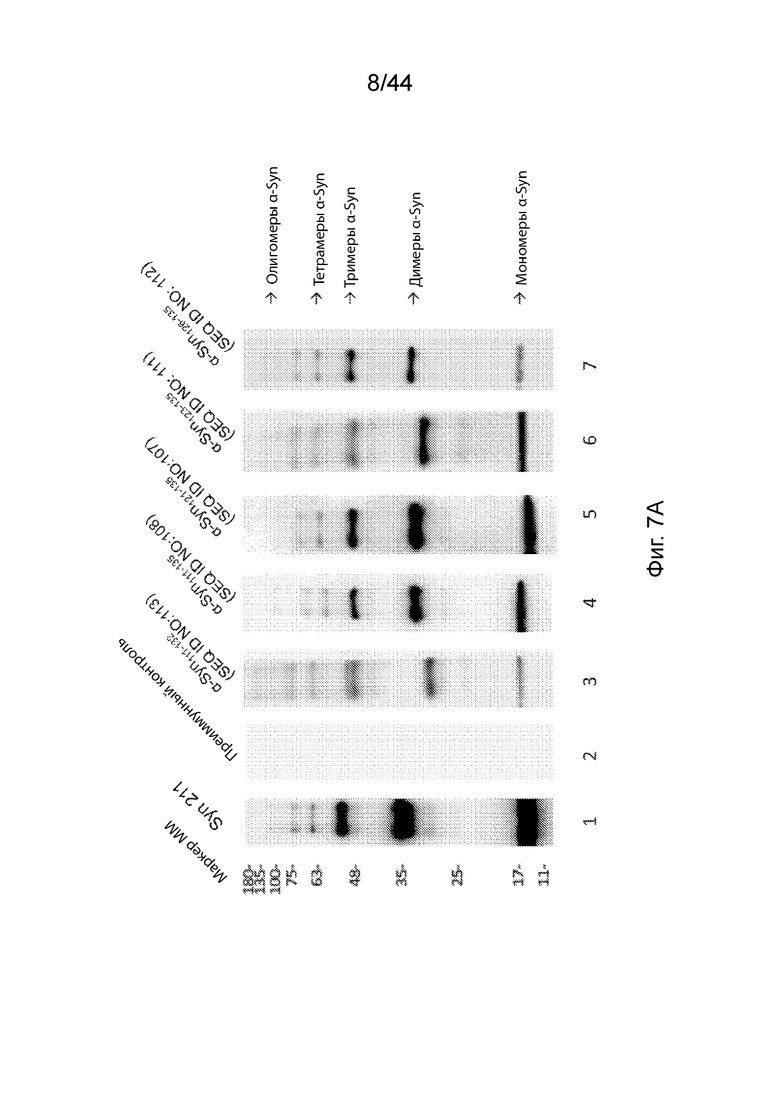
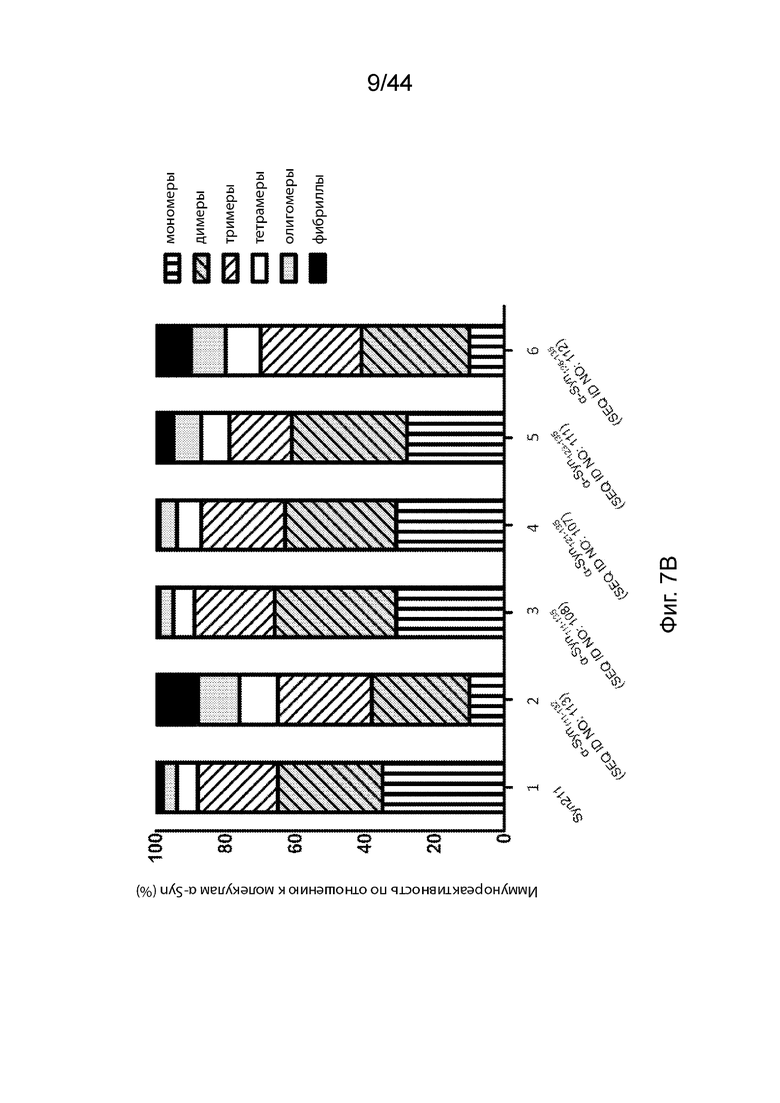
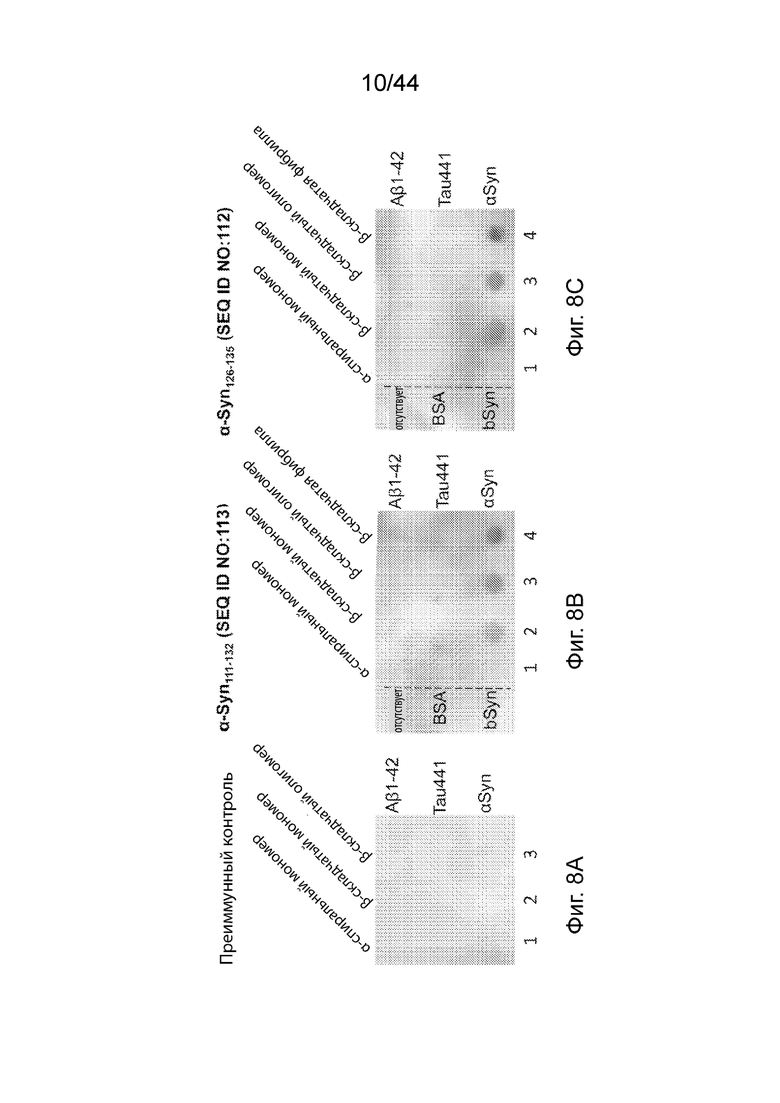
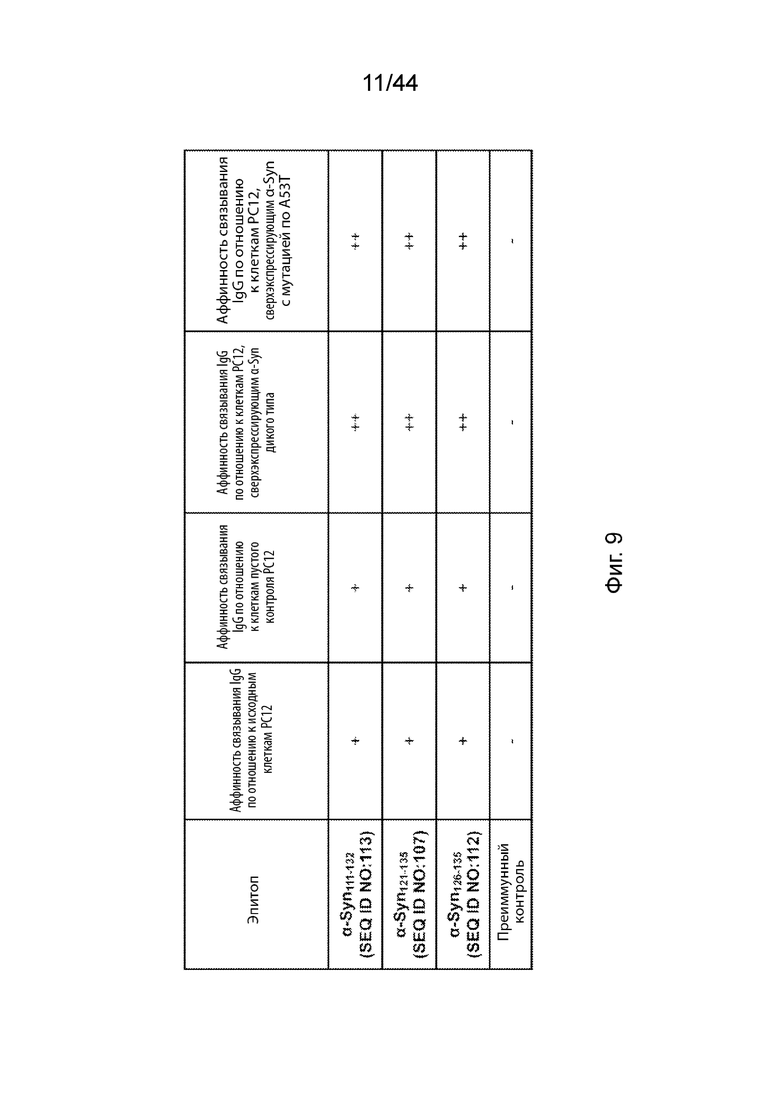

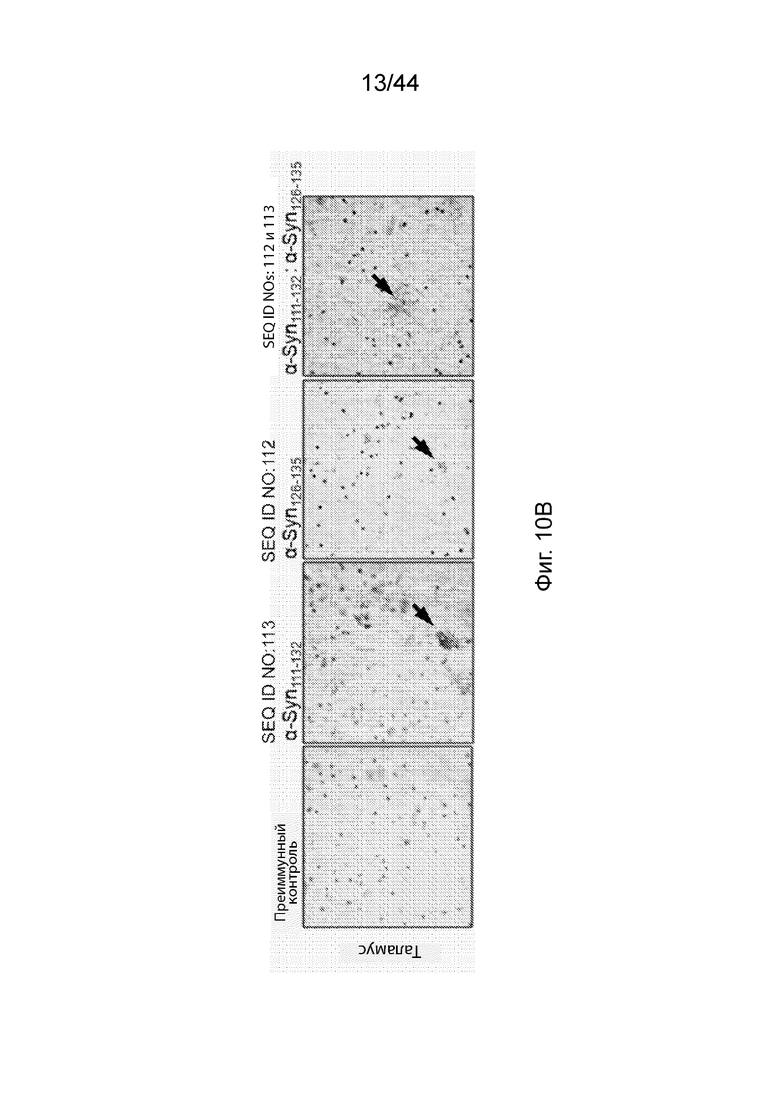
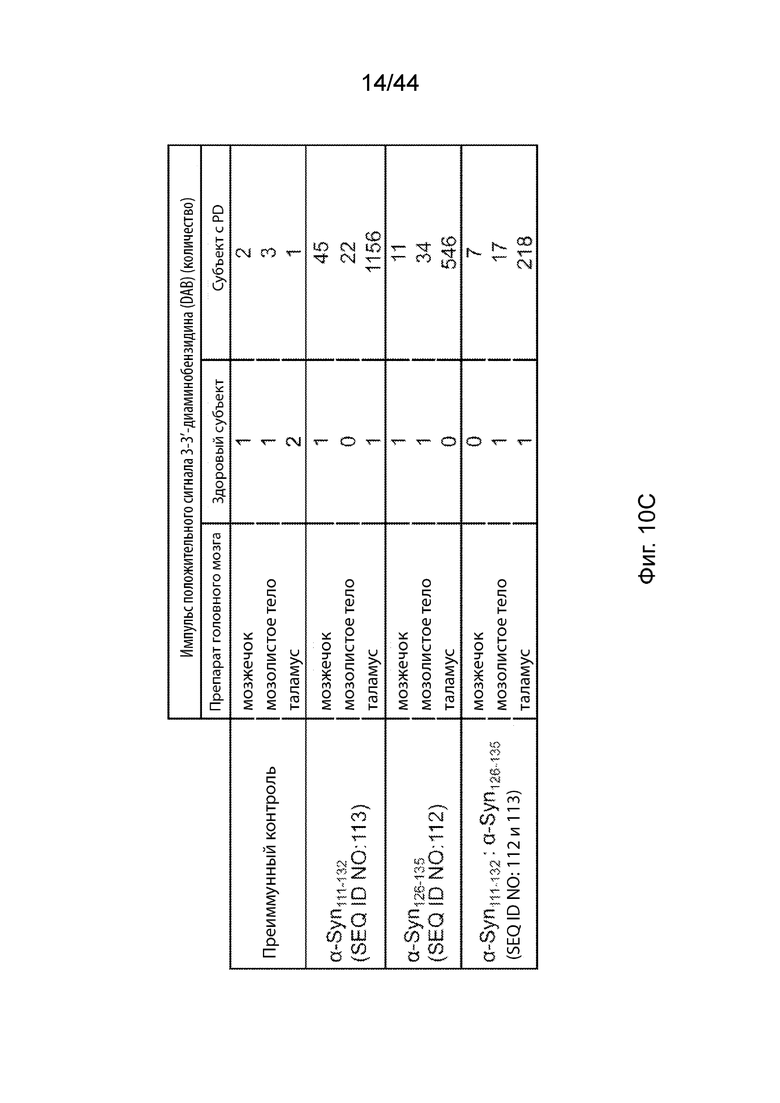
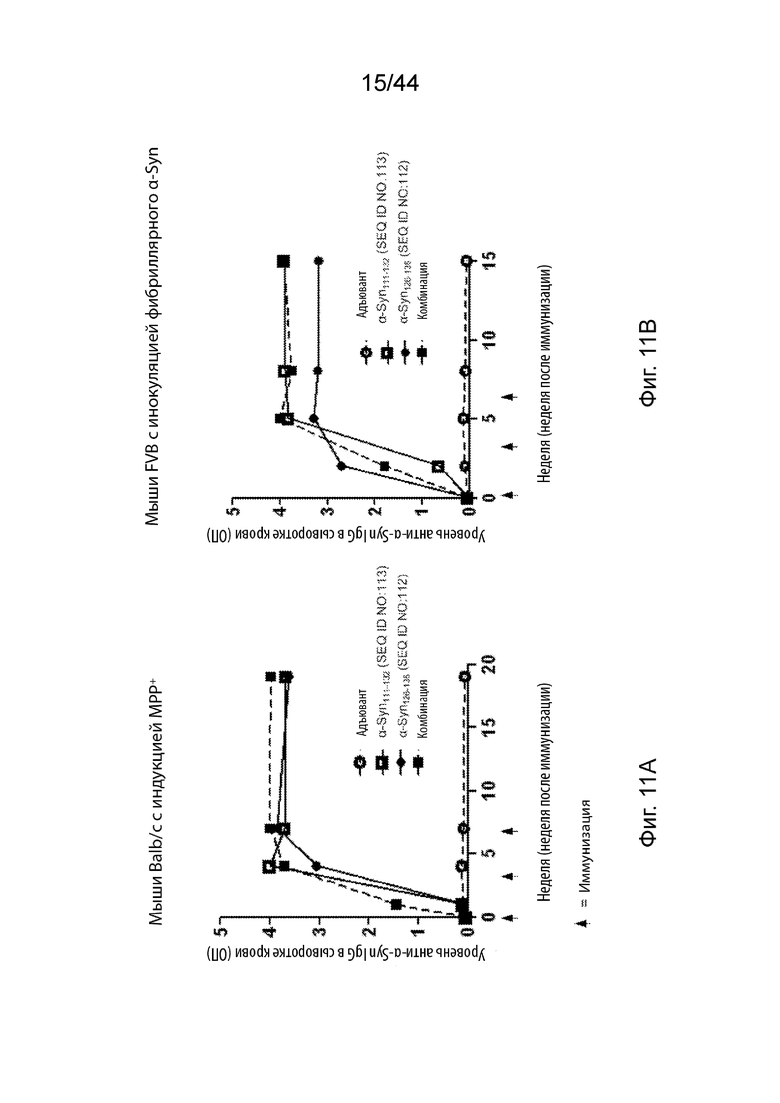
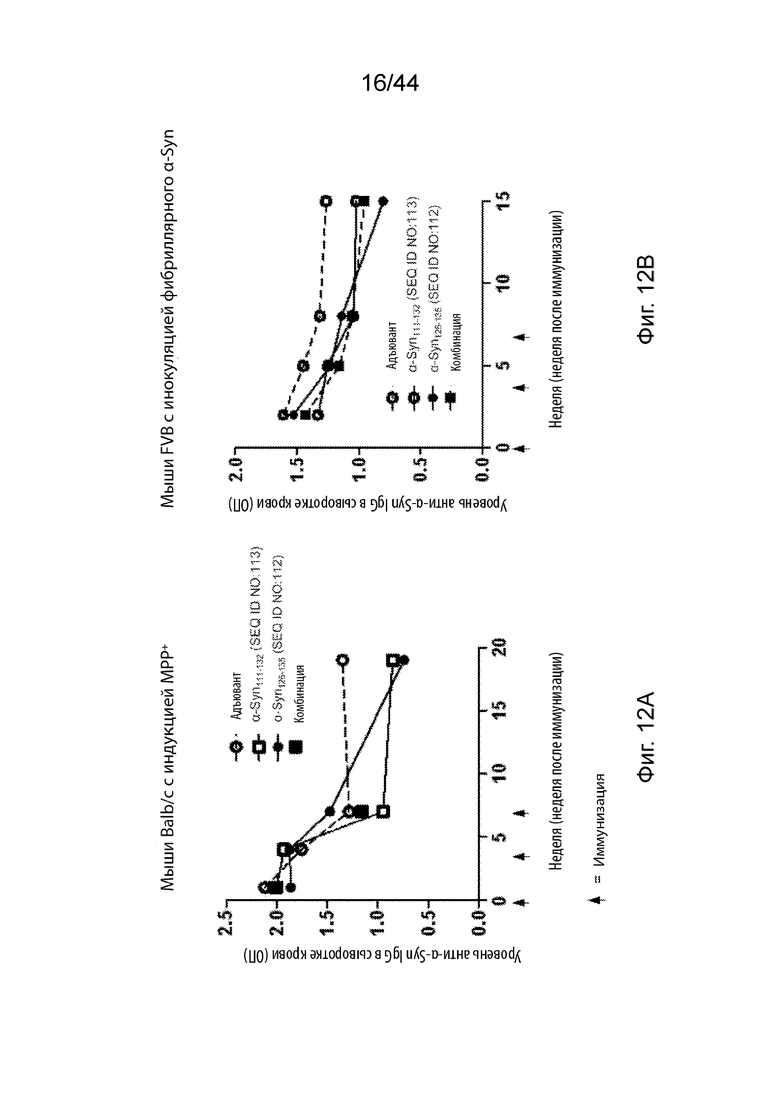
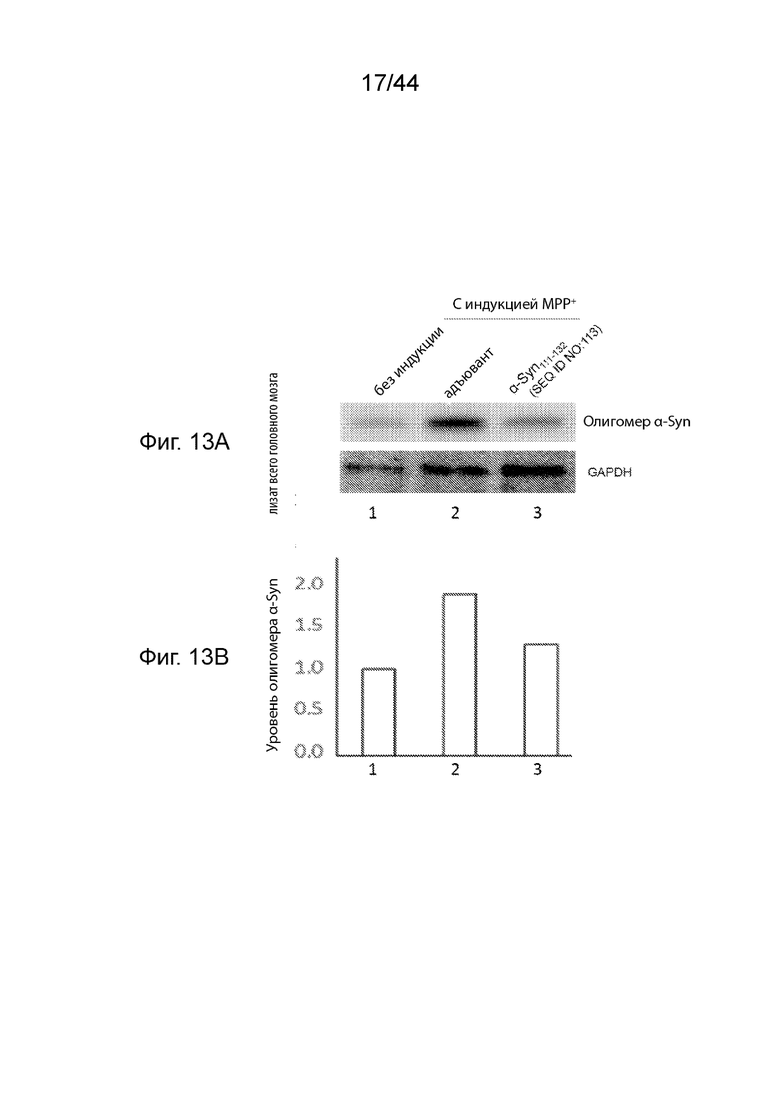
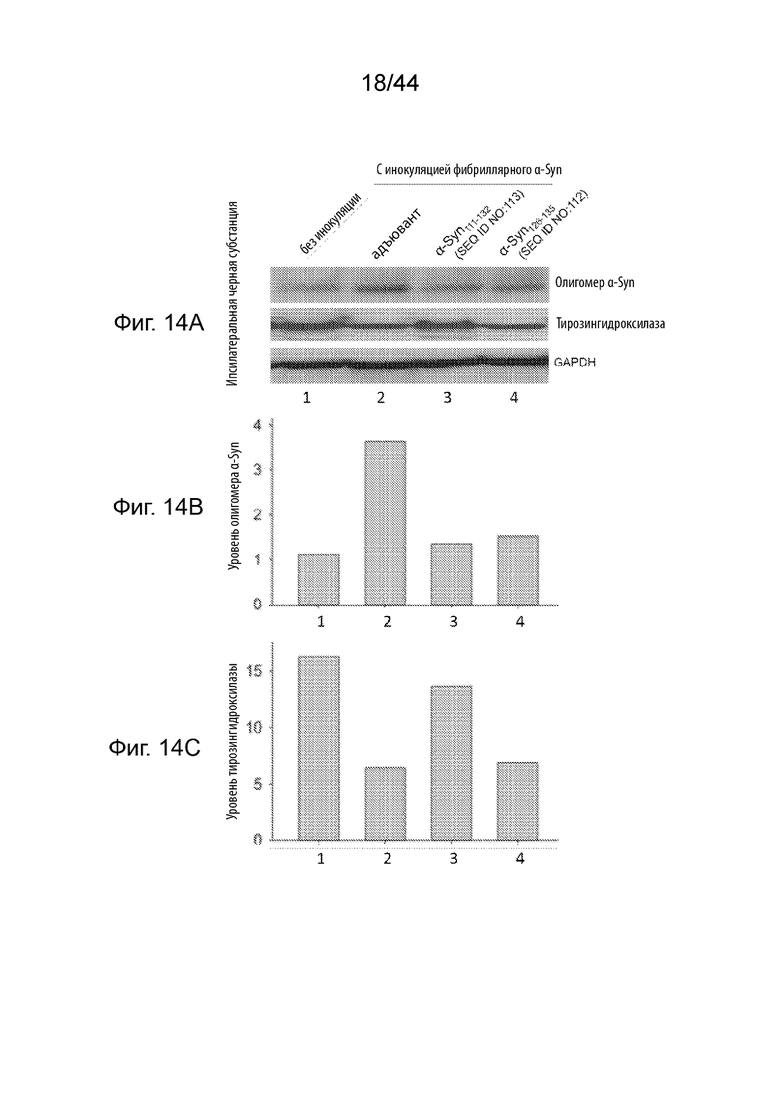
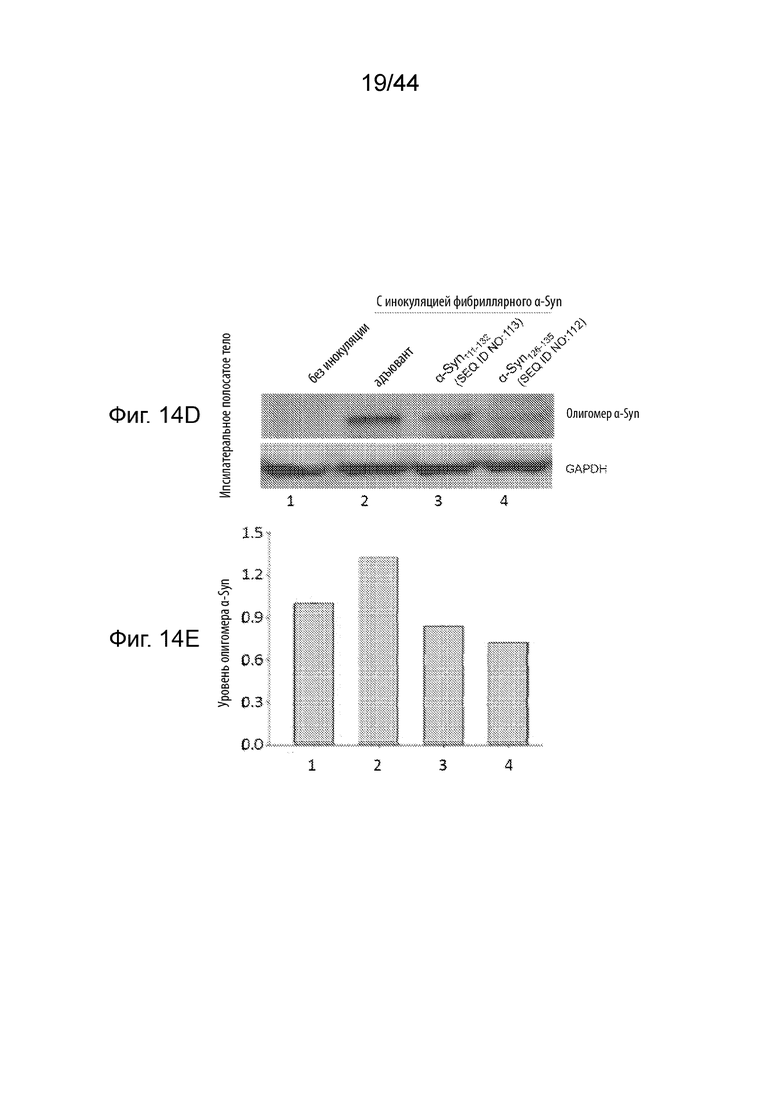
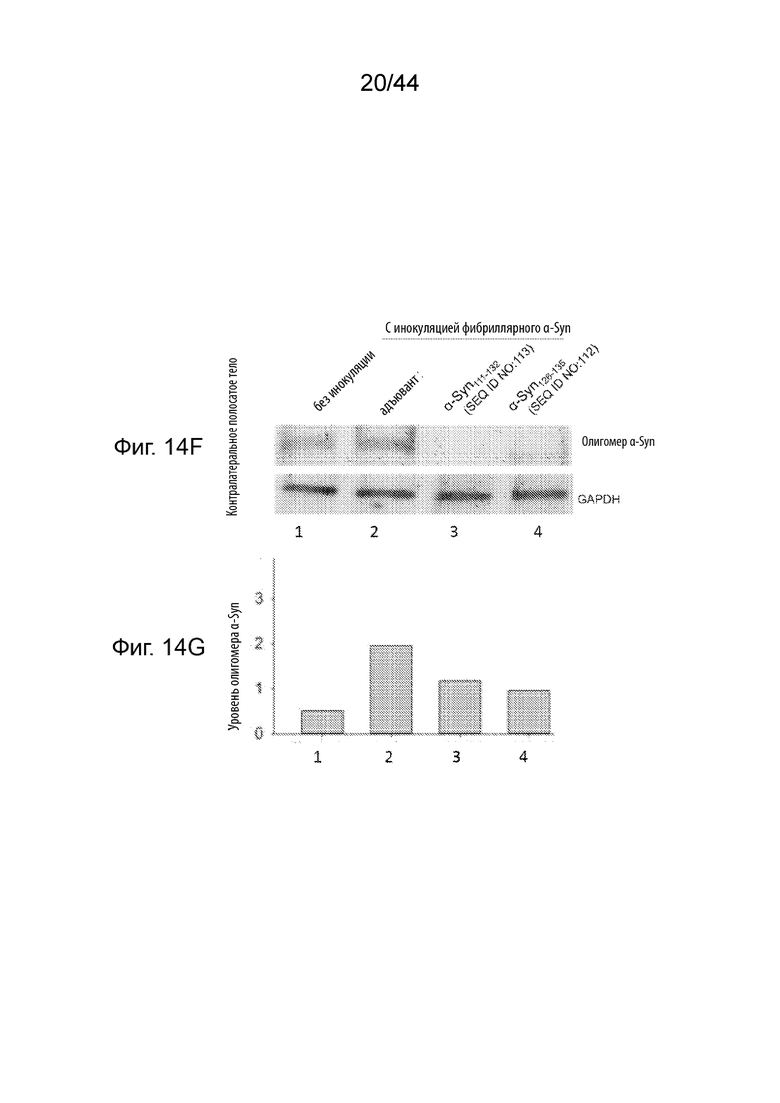
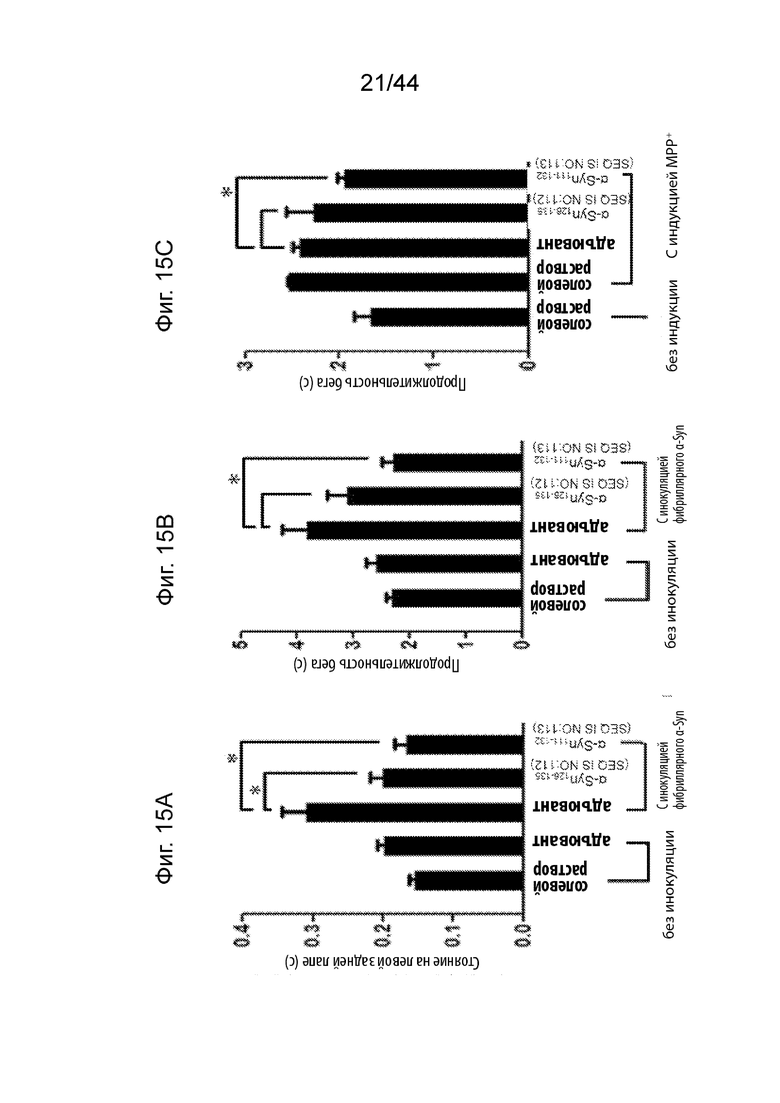
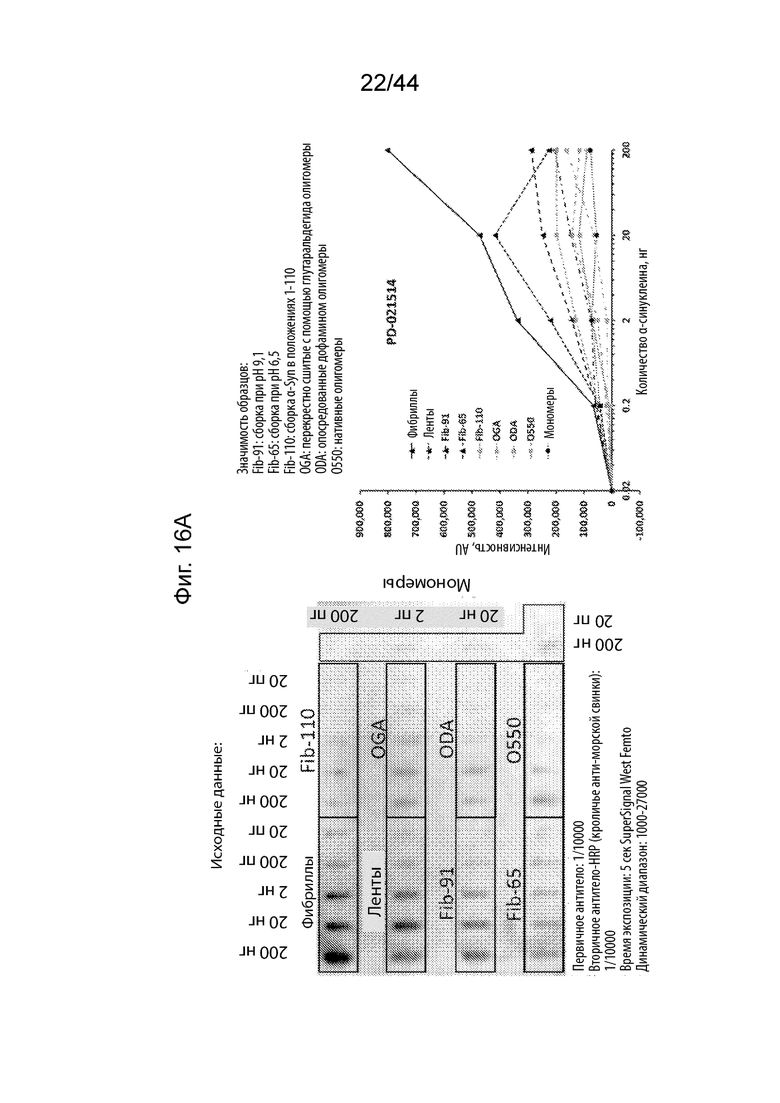
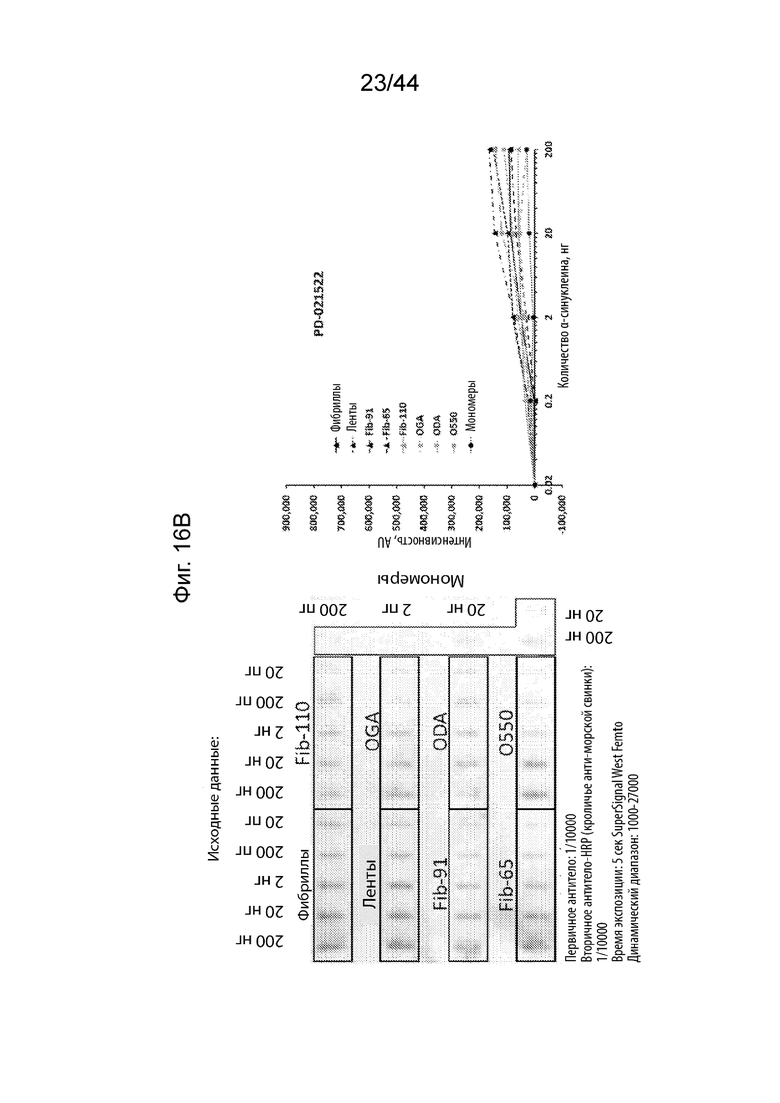
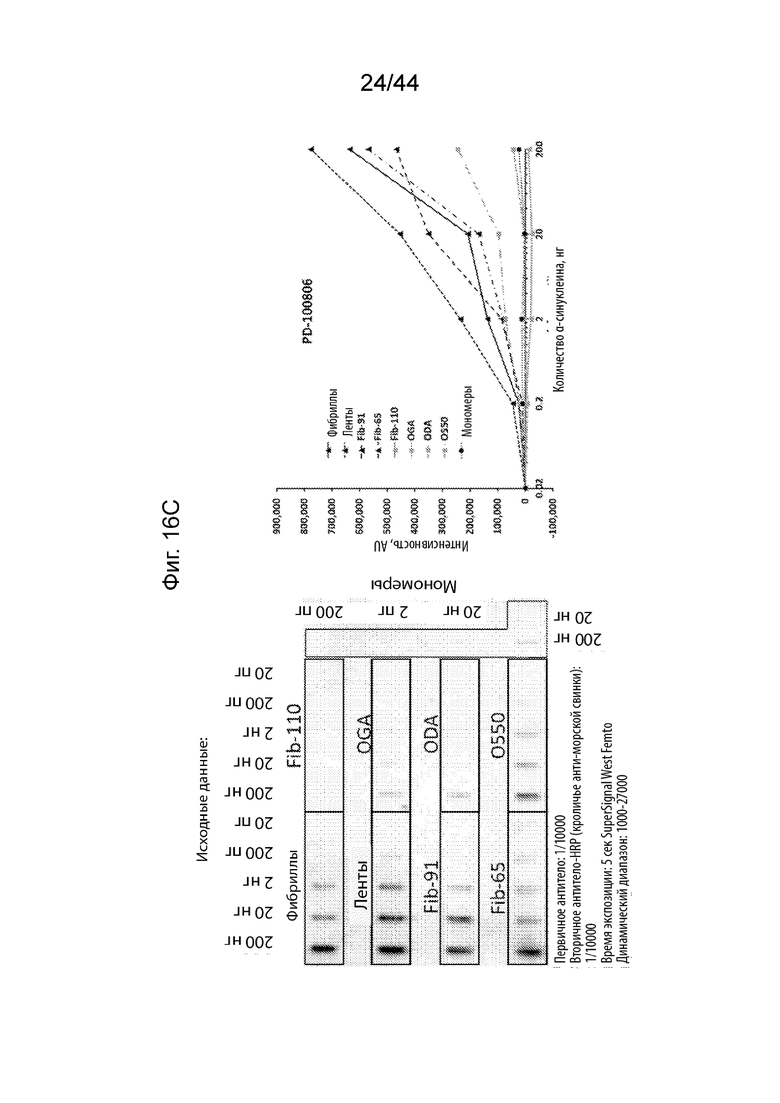
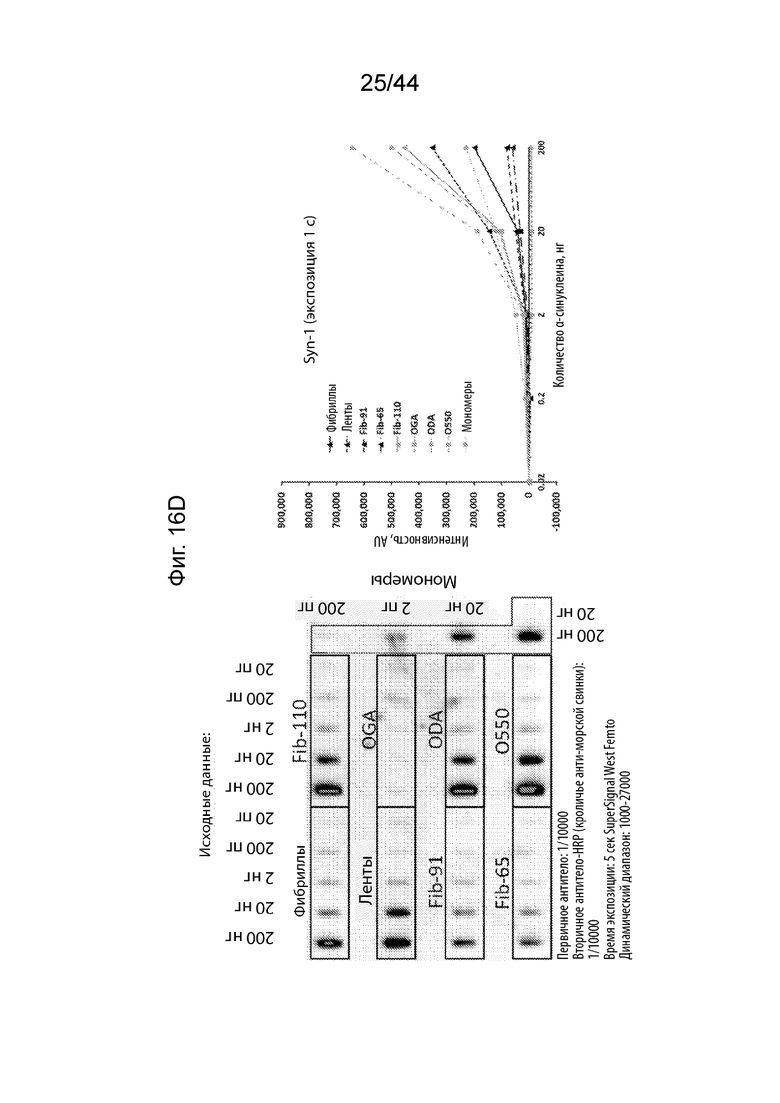



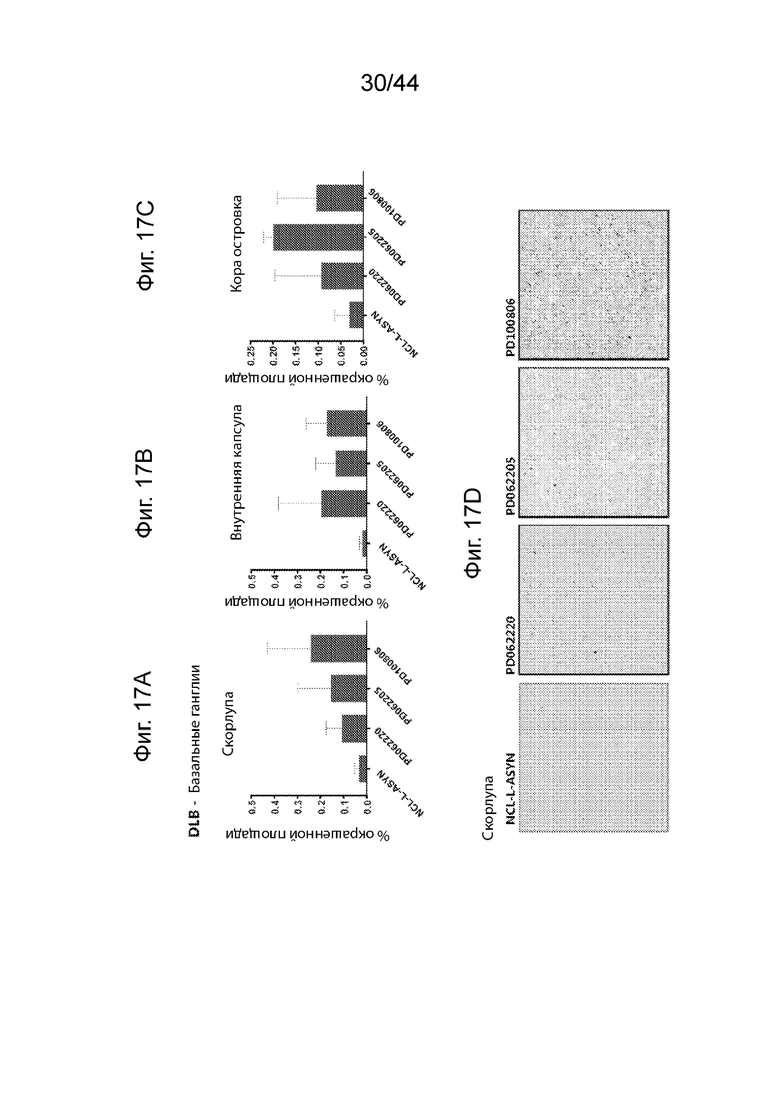
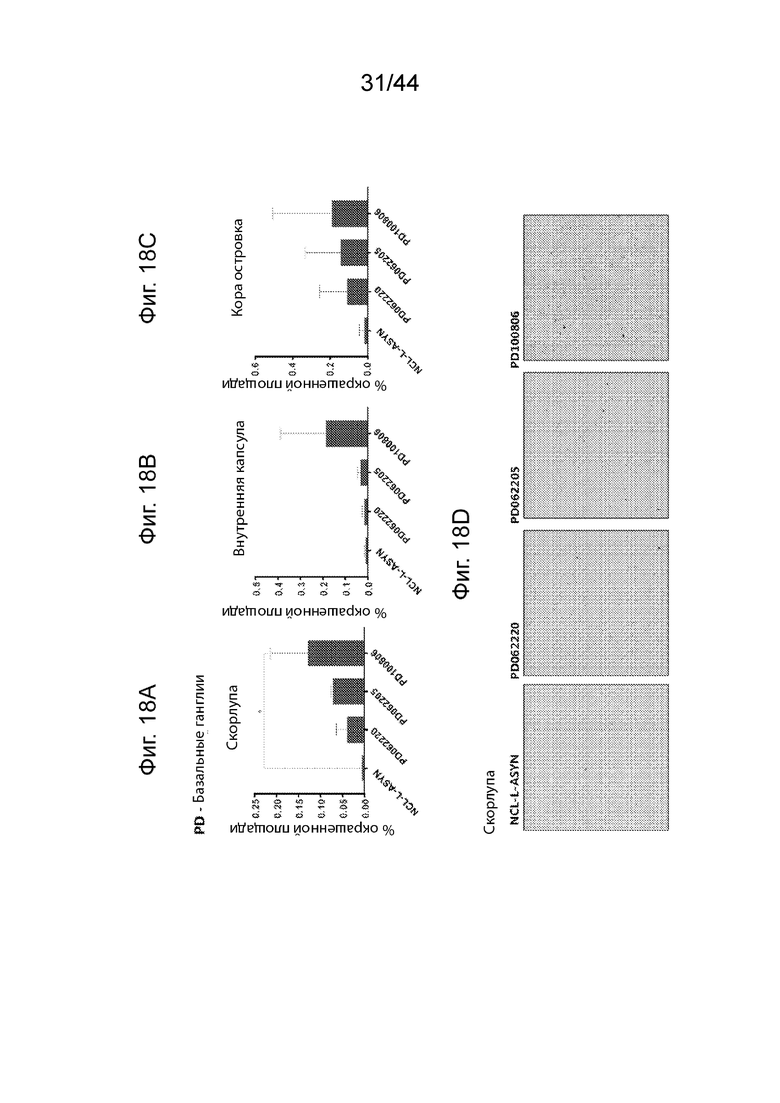
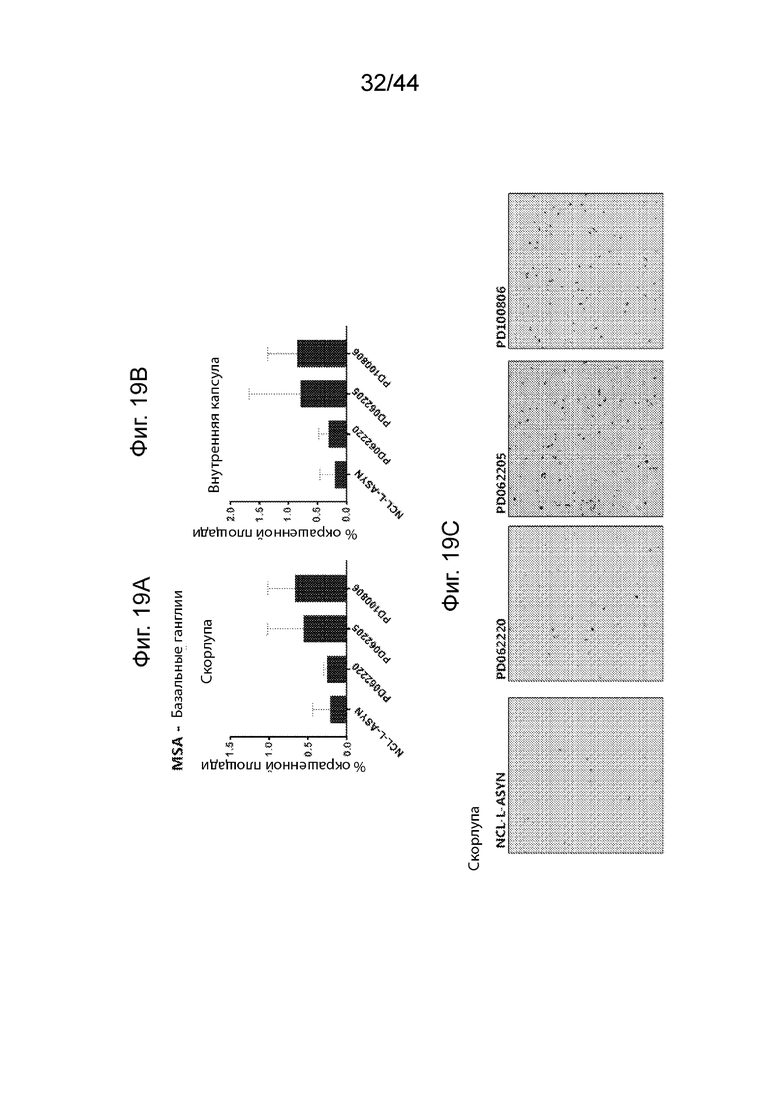
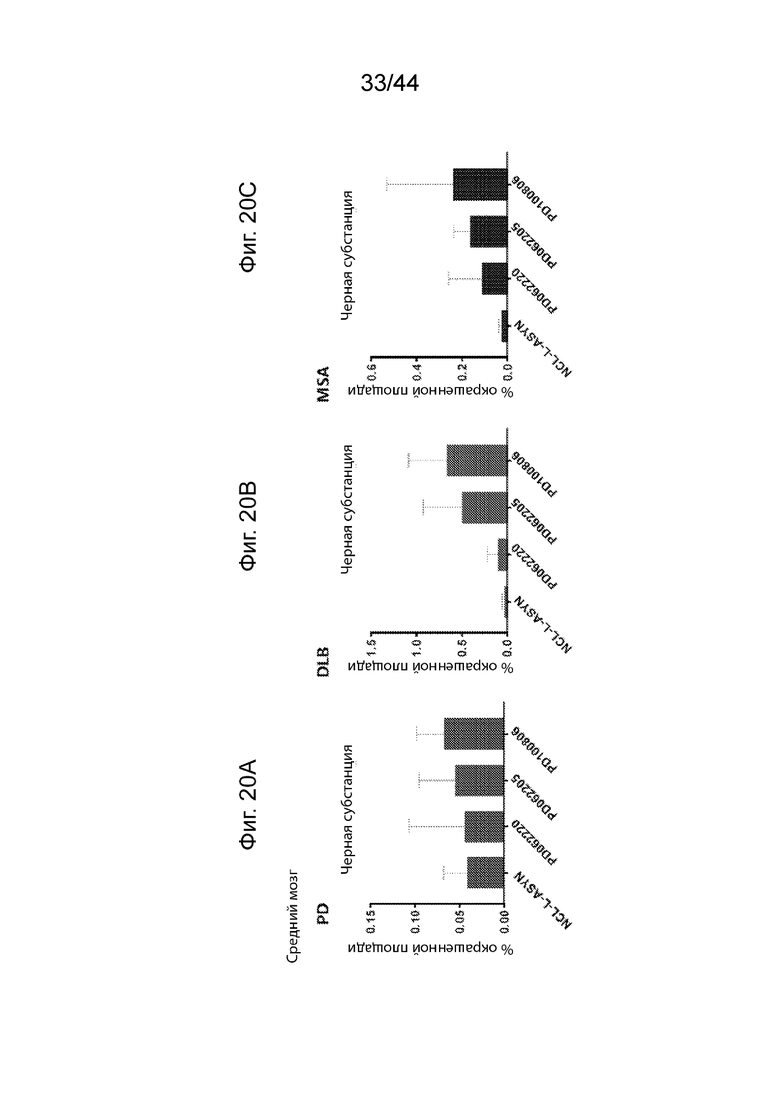

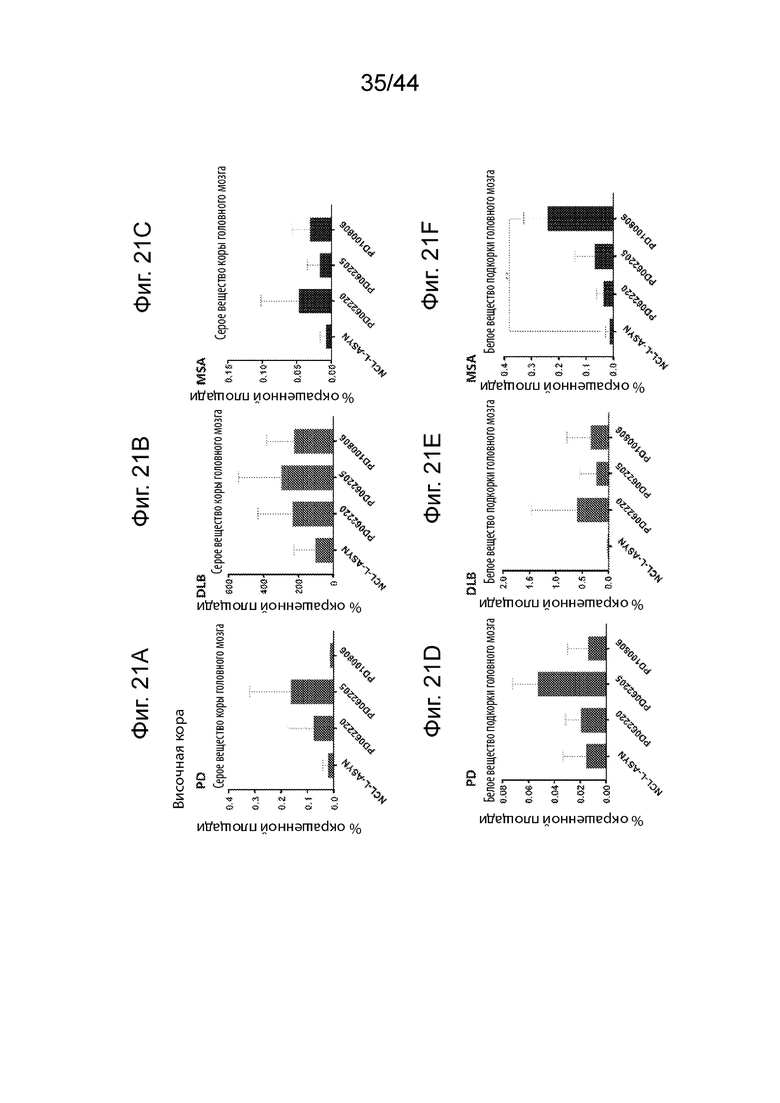
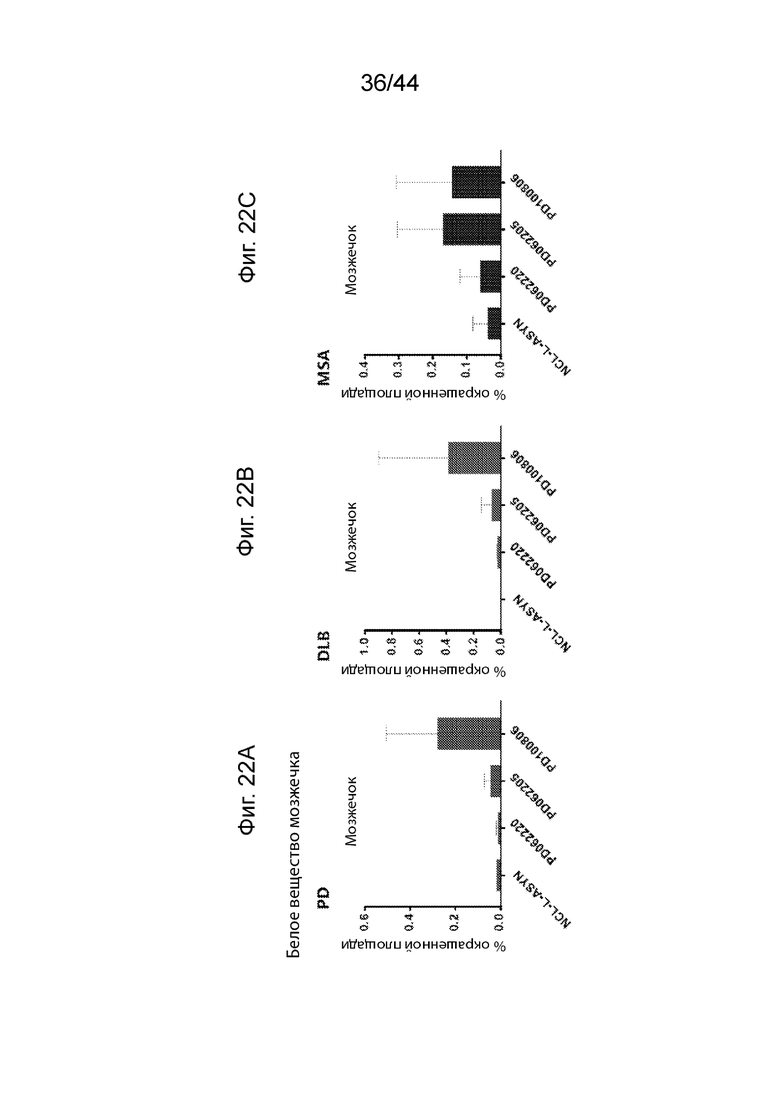

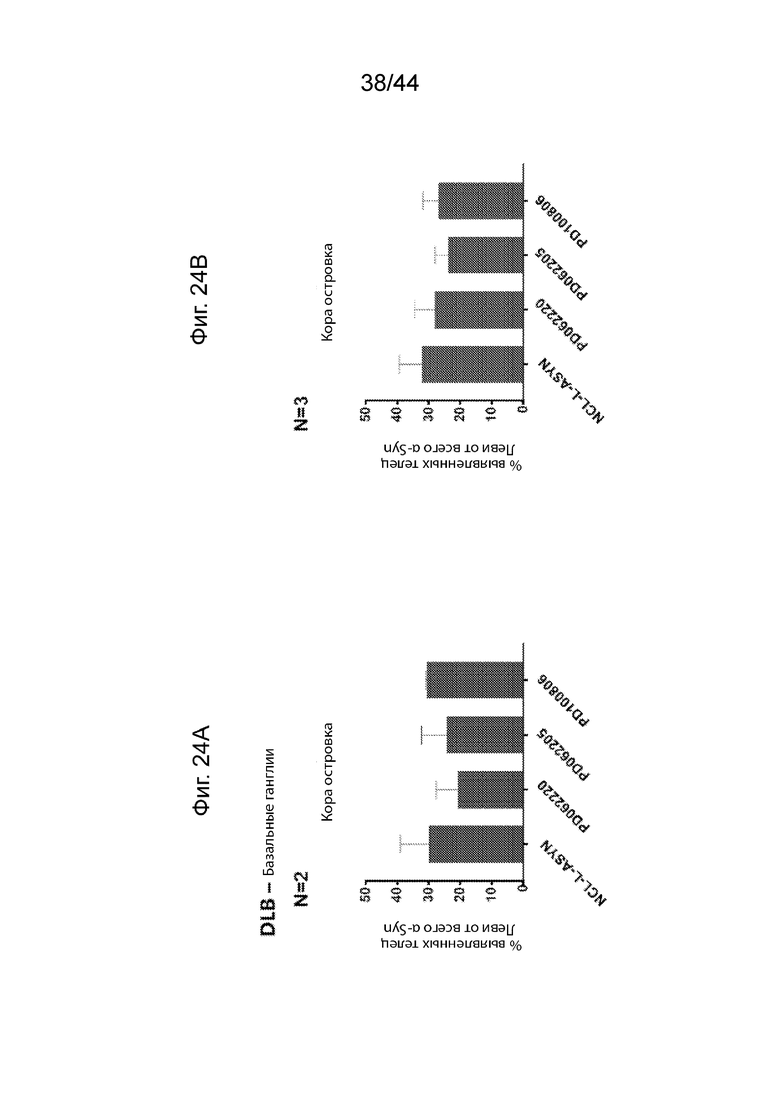

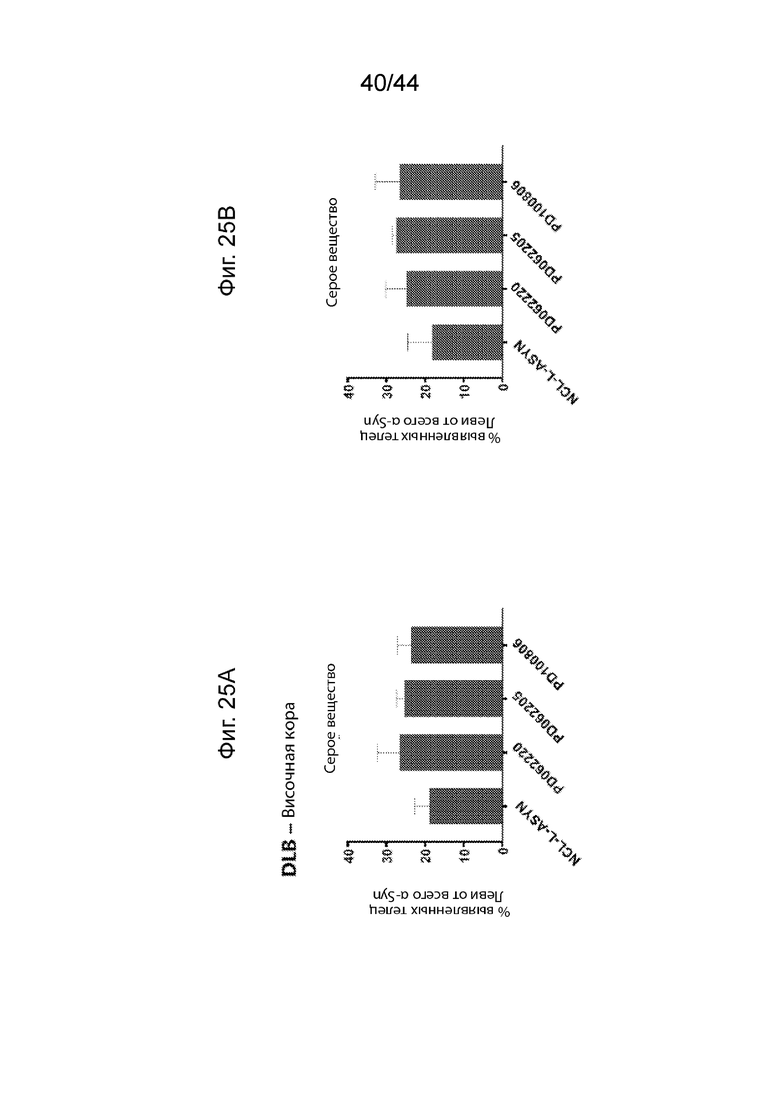


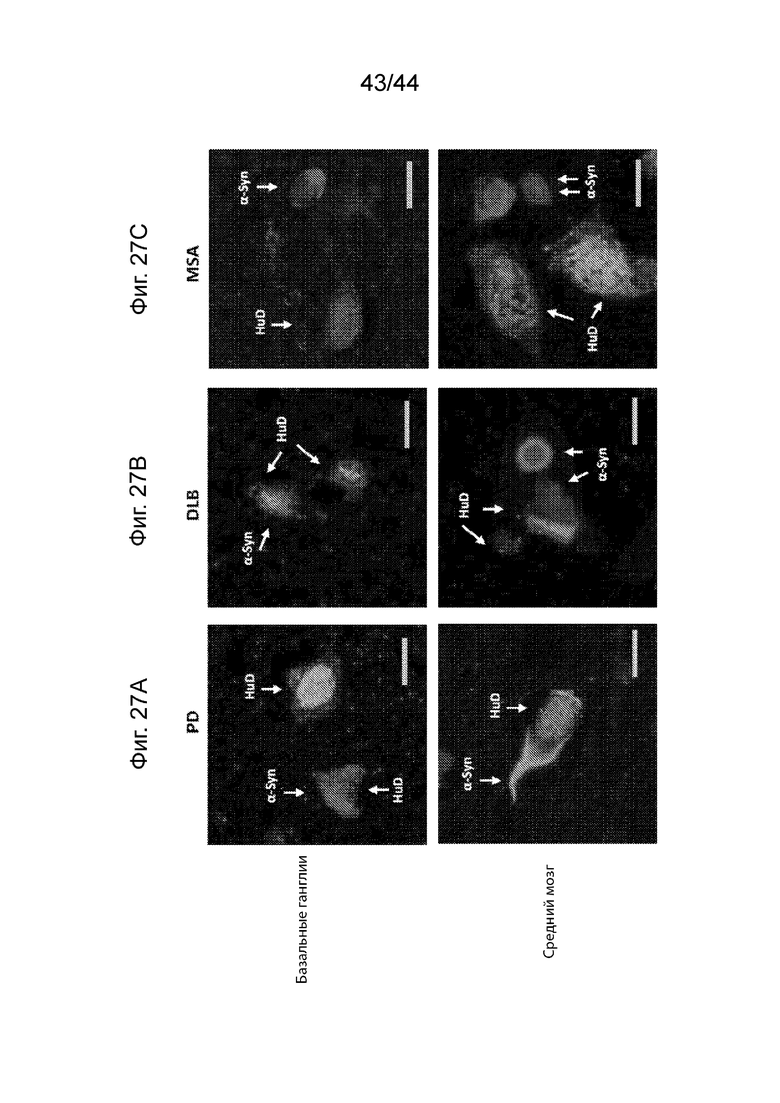
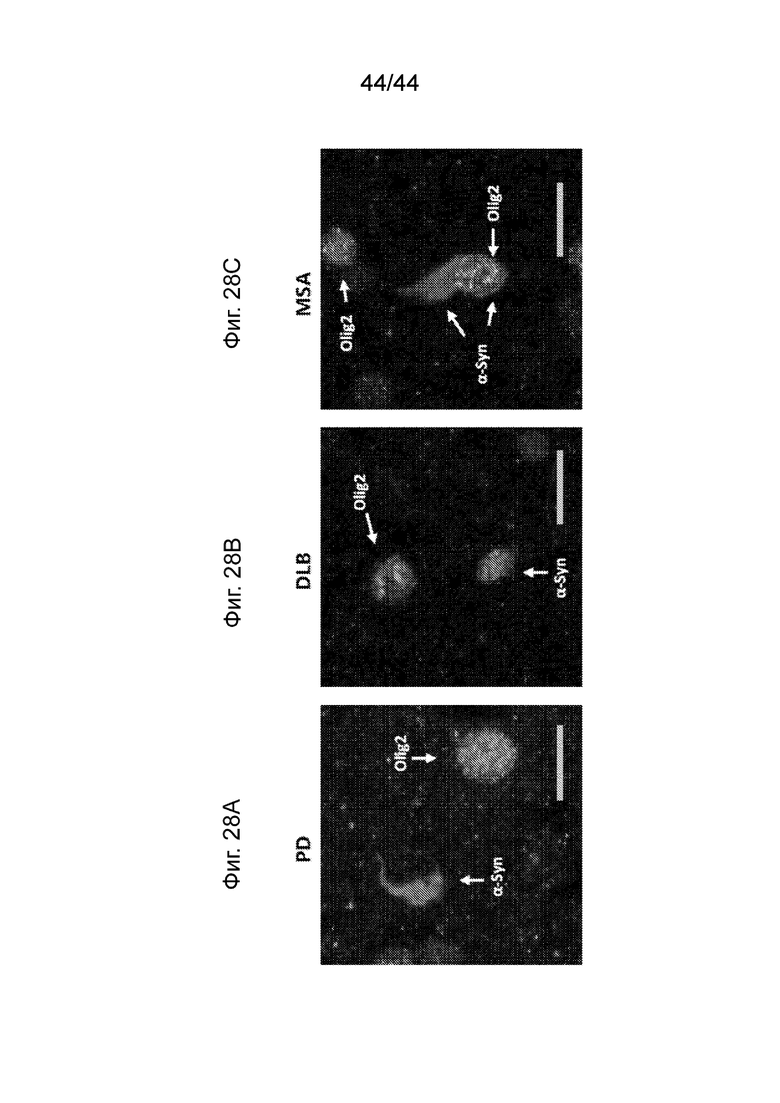
Изобретение относится к области биотехнологии, конкретно к пептидным иммуногенным конструкциям альфа-синуклеина (α-Syn), и может быть использовано в медицине для лечения заболевания головного мозга у индивида с синуклеинопатией. Предложенные пептидные иммуногенные конструкции α-Syn содержат B-клеточный эпитоп из α-Syn, связанный с гетерологичным эпитопом T-хелперных клеток (Th) непосредственно или с помощью гетерологичного спейсера. Изобретение обеспечивает получение пептидных иммуногенных конструкций α-Syn, которые стимулируют образование высокоспецифических антител, перекрестно реагирующих с β-листом α-Syn в виде мономеров, олигомеров и фибрилл, но не с встречающейся в природе α-спиралью α-Syn, обеспечивая терапевтические иммунные ответы при риске развития синуклеинопатий. 7 н. и 10 з.п. ф-лы, 28 ил., 15 табл., 17 пр.
1. Пептидная иммуногенная конструкция альфа-синуклеина (α-Syn) для лечения заболевания головного мозга у индивида c синуклеинопатией, содержащая следующую формулу:
(Th)m-(A)n-(С-терминальный фрагмент α-Syn)-X
или
(С-терминальный фрагмент α-Syn)-(A)n-(Th)m-X,
где (С-терминальный фрагмент α-Syn) представляет собой B-клеточный эпитоп, содержащий фрагмент из аминокислот G111-D135 последовательности SEQ ID NO:1;
Th представляет собой T-хелперный эпитоп, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:70-98; и
А представляет собой необязательный гетерологичный спейсер, выбранный из группы, состоящей из аминокислоты, Lys-, Gly-, Lys-Lys-Lys-, (α, ε-N)Lys и ε-N-Lys-Lys-Lys-Lys (SEQ ID NO:148),
X представляет собой α-COOH или α-CONH2 аминокислоты;
m равно от 1 до около 4; и
n равно от 1 до около 10;
где B-клеточный эпитоп ковалентно связан с T-хелперным эпитопом непосредственно или с помощью необязательного гетерологичного спейсера.
2. Пептидная иммуногенная конструкция α-Syn по п. 1, где B-клеточный эпитоп выбран из группы, состоящей из SEQ ID NO:12-15, 17 и 49-63.
3. Пептидная иммуногенная конструкция α-Syn по п. 1, где Т-хелперный эпитоп выбран из группы, состоящей из SEQ ID NO:81, 83 и 84.
4. Пептидная иммуногенная конструкция α-Syn по п. 1, где необязательный гетерологичный спейсер представляет собой (α, ε-N)Lys или ε-N-Lys-Lys-Lys-Lys (SEQ ID NO:148).
5. Пептидная иммуногенная конструкция α-Syn по п. 1, где T-хелперный эпитоп ковалентно связан с аминоконцом B-клеточного эпитопа.
6. Пептидная иммуногенная конструкция α-Syn по п. 1, где T-хелперный эпитоп ковалентно связан с аминоконцом B-клеточного эпитопа с помощью необязательного гетерологичного спейсера.
7. Пептидная иммуногенная конструкция α-Syn по п. 1, содержащая аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:107, 108, 111-113 и 115-147.
8. Пептидная иммуногенная конструкция α-Syn по п. 1, содержащая аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:107, 108 и 111-113.
9. Композиция для лечения заболевания головного мозга у индивида с синуклеинопатией, содержащая эффективное количество пептидной иммуногенной конструкции α-Syn по п. 1 в качестве активного средства композиции.
10. Композиция для лечения заболевания головного мозга у индивида с синуклеинопатией, содержащая эффективное количество более чем одной пептидной иммуногенной конструкции α-Syn по п. 1 в качестве активного средства композиции.
11. Композиция по п. 10, где пептидная иммуногенная конструкция α-Syn имеет аминокислотные последовательности SEQ ID NO:112 и 113.
12. Фармацевтическая композиция для лечения заболевания головного мозга у индивида с синуклеинопатией, содержащая пептидную иммуногенную конструкцию α-Syn по п. 1 и фармацевтически приемлемое средство доставки и/или адъювант.
13. Фармацевтическая композиция по п. 13, где
a. пептидная иммуногенная конструкция α-Syn выбрана из группы, состоящей из SEQ ID NO:107, 108, 111-113 и 115-147; и
b. адъювант представляет собой минеральную соль алюминия, выбранную из группы, состоящей из Al(OH)3 или AlPO4.
14. Фармацевтическая композиция по п. 12, где
a. пептидная иммуногенная конструкция α-Syn выбрана из группы, состоящей из SEQ ID NO:107, 108, 111-113 и 115-147; и
b. пептидная иммуногенная конструкция α-Syn смешана с олигодезоксинуклеотидом CpG (ODN) с образованием стабилизированного иммуностимулирующего комплекса.
15. Способ получения антител, которые распознают α-Syn в организме животного, включающий введение животному композиции, содержащей пептидный иммуноген α-Syn по п. 1 и средство доставки и/или адъювант.
16. Способ ингибирования агрегации α-Syn у животного, включающий введение фармакологически эффективного количества пептидного иммуногена α-Syn по п. 1 животному.
17. Способ снижения количества агрегатов α-Syn у животного, включающий введение фармакологически эффективного количества пептидного иммуногена α-Syn по п. 1 животному.
| СПОСОБ ПОЛУЧЕНИЯ СУХОЙ СМЕСИ ДЛЯ ДЕТСКОГО ПИТАНИЯ | 2003 |
|
RU2272539C2 |
| ET AL., Effects of alpha-synuclein immunization in a mouse model of Parkinsons disease, NEURON, CELL PRESS, 2005, v | |||
| Способ изготовления звездочек для французской бороны-катка | 1922 |
|
SU46A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |

