Изобретение относится к области биотехнологии и регенеративной медицины и может быть использовано при культивировании клеток костного мозга в условиях, требующих использования бессывороточной среды, а также для стимуляции пролиферативной активности клеток костного мозга при патологиях, связанных с нарушением кроветворения, например, при синдроме недостаточности костного мозга, при кровопотерях, после прохождения курсов химио- или лучевой терапии.
Кроветворение (гемопоэз) представляет собой сложный многостадийный процесс клеточных делений и дифференцировок, в результате которого образуются зрелые, функционально полноценные клетки крови. В постнатальный период основным кроветворным органом является костный мозг. В нем содержится основная масса стволовых кроветворных клеток и образуются все клетки крови. Высокая пролиферативная активность всех типов клеток является одной из важнейших характеристик ткани костного мозга. Регуляция кроветворения в костном мозге находится под контролем широкого спектра факторов, среди которых наиболее изученными являются гемопоэтические факторы роста. Главными источниками этих факторов являются макрофаги, эндотелиальные клетки, лимфоциты и клетки стромы.
В настоящее время одной из актуальных задач гематологии и регенеративной медицины является поиск способов и лекарственных средств, направленных на стимуляцию кроветворения в костном мозге с целью восстановления необходимого при заболеваниях системы крови и гематологических осложнениях, резистентных к современной медикаментозной терапии.
Известно использование средства Энтерол-250 для коррекции эритропоэза при инфекционной патологии. Указанное средство представляет собой лиофилизированный препарат культуры дрожжевых грибков Saccharomyces boulardii, содержащий в 1 г препарата следующие соединения: тиамин (60 мкг), рибофлавин (40 мкг), никотиновая кислота (480 мкг), пиридоксин (33 мкг), пантотеновая кислота (77 мкг), ферменты, аминокислоты, стимуляторы IgA и т.д. [патент на изобретение РФ №2189826 С1, Способ коррекции эритропоэза и лечения анемии при инфекционной патологии. 2001. МПК А61К 35/72 (2000.01), А61Р 7/06 (2000.01)]. Недостатком энтерола-250 является то, что его эффект ограничен увеличением количества эритроцитов на 17% по сравнению с исходным уровнем, при этом не установлено его влияние на другие клеточные линии костного мозга.
В качестве средства, стимулирующего гемопоэз при гипопластических состояниях костного мозга, вызванных цитостатиком, предложено использовать внутривенное введение 50 мг/кг N-ацетилнейраминовой (сиаловой) кислоты. Данное средство снижает токсическое действие цитостатиков на миелопоэз за счет стимуляции эритропоэза и грануломоноцитопоэза [патент на изобретение РФ №2058138 С1. Средство, стимулирующее гемопоэз. 1996. МПК А61К 31/7012 (2006.01), А61К 31/185 (2006.01), А61Р 7/00 (2006.01)]. Указанное средство позволяет увеличить количество эритроидных элементов, а также зрелых форм нейтрофильных лейкоцитов в костном мозге мышей, подвергнутых воздействию циклофосфана. Однако не установлено стимулирующее влияние указанного средства на количество лимфоцитов и лимфоцитарных клеток, а также ретикулярных клеток костного мозга.
Для восстановления гемопоэза после применения цитостатиков используют лекарственную композицию, в состав которой входит D-глюкуроновая кислота, милдронат и изотонический раствор в соотношении: Д-глюкуроновая кислота 0,133-0,267; 3(2,2,2-триметилгидрозиний)пропионат - 0,133-0,267; изотонический раствор - остальное [патент РФ №2033152 С1. Средство для стимуляции гемопоэза. 1995. МПК А61К 31/15, А61К 31/70]. Средство позволяет существенно повысить незрелых и зрелых форм нейтрофильных гранулоцитов и миелокариоцитов в костном мозге экспериментальных животных, получавших данный состав. В зависимости от соотношения ингредиентов содержание незрелых форм нейтрофильных гранулоцитов повышалось до 189-261%, зрелых нейтрофильных гранулоцитов - до 136-200%, общее количество миелокариоцитов у животных экспериментальной группы восстанавливалось до 106-118% от исходного уровня. Однако не установлено влияние указанного средства на количество лимфоцитов и лимфоцитарных клеток, а также ретикулярных клеток костного мозга.
Известно применение церулоплазмина для стимуляции гемопоэза при лучевой болезни у собак. Церуплазмин вводят капельно со скоростью 29-30 капель в минуту один раз в сутки в суточной дозе от 1,5 до 2,5 мг/кг массы тела животного. Курс лечения церуплазмином составляет 7 процедур с интервалом 47-48 часов. Установлено, что применение церулоплазмина на фоне лазерной остеоперфорации у собак происходит увеличение содержания в периферической крови эритроцитов в 1,29 раза от исходного уровня и снижение повышенного количества лейкоцитов в 1, 84 раза [патент РФ 2329791 С2. Способ стимуляции гемопоэза при лечении острой лучевой болезни у собак. 2006. МПК А61К 31/00 (2006.01)]. Недостатком известного средства является то, что его стимулирующее воздействие на гемопоэз ограничено увеличением эритроцитов и не распространяется на другие клеточные линии.
Для стимуляции миелопоэза вводят пегилированную гиалуронидазу в дозе 50 ЕД/кг 1 раз в сутки в течение 2 дней и дополнительно однократно внутривенно препарат, содержащий D-глюкуроновую кислоту, в дозе 2 мг/кг. Введение препарата на фоне моделирования цитостатической миелосупрессии сопровождалось стимуляцией процессов регенерации эритроидного и гранулоцитарного ростков кроветворения. Отмечалось увеличение содержания кроветворных прекурсоров (КОЕ-Э, КОЕ-ГМ) и морфологически распознаваемых клеточных элементов эритроидного и грануломоноцитарного ростков кроветворения в гемопоэтической ткани, а также количества зрелых клеток эритроидного и гранулоцитарного ряда в периферической крови. Способ позволяет ускорить регенеративные процессы гемопоэтической ткани, усиливая при этом миелопоэз [патент на изобретение РФ №2442589 С1. Способ стимуляции миелопоэза. 2012. МПК А61К 31/712 (2006.01), А61К 38/47 (2006.01), А61Р 43/00 (2006.01)]. Недостатком заявленного средства является его неэффективность для стимуляции пролиферации клеток лимфоцитарного звена.
Известно применение гиалуронидазы, иммобилизированной на низкомолекулярном полиэтиленгликоле, для стимуляции гемопоэза. Указанное средство вводят парентерально либо перорально 1 раз в сутки в течение 1-5 дней в дозе 10-1000 ЕД/кг [патент на изобретение РФ №2414926 С1. Гемостимулирующее средство и способ стимуляции гемопоэза. 2011. МПК А61К 38/43, А61К 41/00, А61К 47/48, А61Р 7/00]. Иммобилизированная гиалуронидаза оказывает выраженный стимулирующий эффект в отношении эритроидного, гранулоцитарного и мегакариоцитарного (тромбоцитарного) ростков кроветворения, однако не оказывает стимулирующего влияния на лимфоцитарное звено гемопоэза.
Недостатком перечисленных средств является также присутствие в препаратах компонентов ксеногенного происхождения.
Известно применение долихола и его эфиров в качестве активного компонента для стимулирования дифференцировки и пролиферации гемопоэтических стволовых клеток и улучшения гемопоэза при нарушении функций кроветворения [патент JP S6112622 (A). Agent for promoting differentiation and proliferation of hemopoietic stem cell. 1986. МПК A61K 31/045; A61P 7/00; C07C 27/00; C07C 33/02; C07C 67/00; C07C 69/145; (IPC1-7): A61K 31/045; С07С 33/02]. Эффективность средства ограничена его стимулирующим действием на эритроцитарное звено гемоэза, что обусловливает его использование для лечения старческой анемии.
Предложен препарат (3-интерферона для внутримышечного введения с целью стимуляции эритропоэза при нарушениях, связанных с недостаточным созреванием клеток предшественников крови в эритроциты [патент США 5104653. Use of human interferon-beta for stimulation of erythropoiesis. 1992]. Недостатком указанного средства является то, что оно стимулирует эритроцитарное звено гемопоэза и не оказывает более широкого стимулирующего действия на другие клеточные линии гемопоэза.
Известно использование ингибитора сфингозин киназы 2 (SphK2) для восстановления системы костно-мозгового гемопоза после лучевой терапии и химиотерапии [патент AU 2019101780 (А4). Use of SPHK2 inhibitor as drug for repairing bone marrow hematopoietic system injury or treating bone marrow hematopoietic dysfunction. 2021. МПК A01K 67/027; A61K 31/4409; A61K 31/7088; G01N 33/50; A61P 37/00; A61P 43/00; А61Р 7/00]. На экспериментальных животных (мыши) показано, что инъекция ингибитора SPHK2 АВС294640 позволяет значительно ускорить восстановление количества лейкоцитов и тромбоцитов после радиационной стимуляции, тромбоцитов после воздействия химиотерапевтических препаратов, восстановление количества лейкоцитов и тромбоцитов после трансплантации мышам гемопоэтических стволовых клеток костного мозга. Однако указанное средство не обеспечивает увеличение количества моноцитарных и лимфоцитарных клеток, лимфоцитов.
Предложено средство, стимулирующее гемопоэз у млекопитающих in vivo или в культуре стволовых клеток млекопитающих in vitro, включающее мутантный рекомбинантный гемоглобин либо композицию, включающую указанный гемоглобин и один или более гемопоэтических ростовых факторов (ГМКСФ, ГКСФ, интерлейкины 1-13 и др.) [патент США №5631219. Method of stimulating hematopoiesis with hemoglobin. 1997. МПК A61K 38/42 (20060101); A61K 38/41 (20060101); C12N 5/06 (20060101); A61K 038/16; A01N 061/00; A01N 001/02; G01N 033/20]. Недостатком известного средства является то, что его эффект ограничен действием гемоглобина или его комбинации с эритропоэтином на эритроидные клетки.
Известно средство, стимулирующее гемопоэз, включающее гибридный рекомбинантный белок, содержащий гранулоцитарный макрофагальный колониестимулирующий фактор (GM-CSF) и эритропоэтин (ЕРО), слитые с помощью линкера. Установлено, что гибридный белок проявляет более выраженный эффект на дифференцировку эритроидного ростка в культуре гематопоэтических стволовых клеток костного мозга по сравнению с эквимолярной смесью несвязанных молекул GM-CSF и ЕРО [патент США 5916773. Recombinant production of fusion proteins comprising erythropoietin and GM-CSF components. 1999. МПК C07K 14/535; C07K 14/435; C07K 14/505; A61K 38/00; C07K 019/00; C12N 015/62; A61K 038/19]. Недостатком известного средство является то, что его эффект ограничен стимуляцией эритропоэза в культуре стволовых клеток.
Известен препарат, содержащий пептидные фрагменты а 1-антитрипсина, стимулирующие образование ретикулоцитов. Этот фактор можно выделить и очистить из человеческой плазмы. Он обладает высокой способностью стимулировать генерацию ретикулоцитов у мышей. Фактор роста ретикулоцитов можно применять для изготовления лекарственного средства, предназначенного для лечения тяжелой анемии [патент РФ 2703176 С2. Фактор роста ретикулоцитов, способ его получения и применения. 2014. МПК С07К 14/475 (2006.01), С07К 14/505 (2006.01), А61Р 7/06 (2006.01)]. Недостатком данного средства является то, что оно обладает избирательным действием на генерацию ретикулоцитов и не оказывает комплексного воздействия на другие гемопоэтические ростки.
Известно, что сывороточные липопротеины высокой плотности (ЛПВП), помимо участия в обратном транспорте холестерина из периферических тканей в печень, оказывают важное регуляторное влияние на функции клеток разных органов и тканей организма [White R., Giordano S. and Datta G. Role of HDL-Associated Proteins and Lipids in the Regulation of Inflammation. In.: Advances in Lipoprotein Research., 2017, http://dx.doi.org/10.5772/67141]. Механизм регуляторного действия ЛПВП связывают с наличием на поверхности липопротеиновой частицы белкового компонента - аполипопротеина A-I (АпоА-I) [Nofer J.R. Signal transduction by HDL: agonists, receptors, and signaling cascades. Handb Exp Pharmacol. 2015, 224, pp. 229-256. doi: 10.1007/978-3-319-09665-0 6; Mangara M et al., Apolipoprotein A-I: A Molecule of Diverse Function. Ind J Clin Biochem, 2016. 31(3), pp. 253-259. DOI 10.1007/s12291-015-0513-l]. Основная часть данного белка находится в связанном с липидами состоянии в составе ЛПВП.
Известно, что как ЛПВП, так и изолированный АпоА-I стимулируют пролиферацию клеток костного мозга in vitro, но по сравнению с исходными частицами ЛПВП изолированный апоА-I оказывает более выраженный стимулирующий эффект in vitro [Usynin I.F., Dudarev A.N., Gorodetskaya A. Yu., Miroshnichenko S. M., Tkachenko T. A., Tkachenko V. I. Apolipoprotein A-I Stimulates Cell Proliferation in Bone Marrow Cell Culture // Bulletin of Experimental Biology and Medicine. 2018. Vol. 164, P. 308-311. https://doi.org/10.1007/s10517-018-3978-0.]. В культуре клеток костного мозга апоА-1 стимулирует пролиферацию моноцитарных (монобласты, промоноциты) и гранулоцитарных (миелобласты, промиелоциты) прогениторных клеток, а также стромальных клеток костного мозга. Однако пролиферативный эффект апоА-1 недостаточно выражен, в том числе в отношении полихроматофильных и оксифильных нормоцитов, клеток моноцитарного ростка (монобласты, промоноциты и Ring-клетки) и клеток лимфоцитарного ростка (лимфобласт, пролимфоцит, лимфоцит) и не показан его эффект на пролиферацию клеток костного мозга in vivo. Данное средство выбрано в качестве прототипа.
Задачей, на решение которой направлено изобретение, является повышение эффективности средства, стимулирующего пролиферативную активность клеток костного мозга in vitro и in vivo.
Решение изобретательской задачи достигается тем, что предложено средство, включающее комплекс АпоА-I и тетрагидрокортизола (ТГК), полученный путем совместной инкубации белка АпоА-I в изотоническом фосфатно-солевом буфере (рН 7.4) в присутствии гормона ТГК в течение 15 мин при температуре 22±2°С в соотношении белка и гормона, выраженное в мкг как АпоА-I :ТГК=20:1.
Техническим результатом является повышение пролиферативной активности клеток костного мозга in vivo и повышение пролиферативной активности клеток костного мозга в отношении полихроматофильных и оксифильных нормоцитов клеток моноцитарного ростка (монобласты, промоноциты и Ring-клетки) и клеток лимфоцитарного ростка (лимфобласт, пролимфоцит, лимфоцит) in vitro.
Раскрытие сущности изобретения
Средство для стимуляции пролиферативной активности костного мозга представляет собой комплекс АпоА-I и ТГК, полученный путем совместной инкубации в изотоническом фосфатно-солевом буфере (рН 7.4) белка АпоА-I и гормона ТГК в течение 15 мин при температуре 22±2°С в соотношении АпоА-I :ТГК=20:1, выраженном в мкг.
Для получения АпоА-I используют фракцию липопротеинов высокой плотности (ЛПВП), выделенную из плазмы крови здоровых доноров. Способ получения АпоА-I описан в примере 1.
В качестве второго компонента комплекса может быть использован коммерчески доступный препарат тетрагидрокортизол (ТГК), полученный путем синтеза с высокой степенью очистки от примесей, например, ТГК производства компании Research Plus, Inc., (США). ТГК является метаболитом кортизола и выявляется у здоровых людей в плазме крови и моче. Он имеет молекулярную массу 366,5 и эмпирическую формулу С21Н34О5.
Образование комплекса ТГК с АпоА-I путем совместной инкубации в изотоническом фосфатно-солевом буфере подтверждают методом тушения флуоресценции триптофана, как описано в примере 2.
Для стимуляции пролиферативной активности клеток костного мозга in vivo данный комплекс вводят в терапевтически эффективных дозах внутривенно для получения быстрых терапевтических эффектов, как показано в примере 3, однако возможно и подкожное введение для получения долгосрочных терапевтических эффектов.
Для стимуляции пролиферативной активности клеток костного мозга in vitro заявленный комплекс вводят в первичную культуру клеток костного мозга в бессыворотной питательной среде, как описано в примерах 4-6, что позволяет также определить пролиферацию отдельных клеточных линий костного мозга.
Перечень фигур
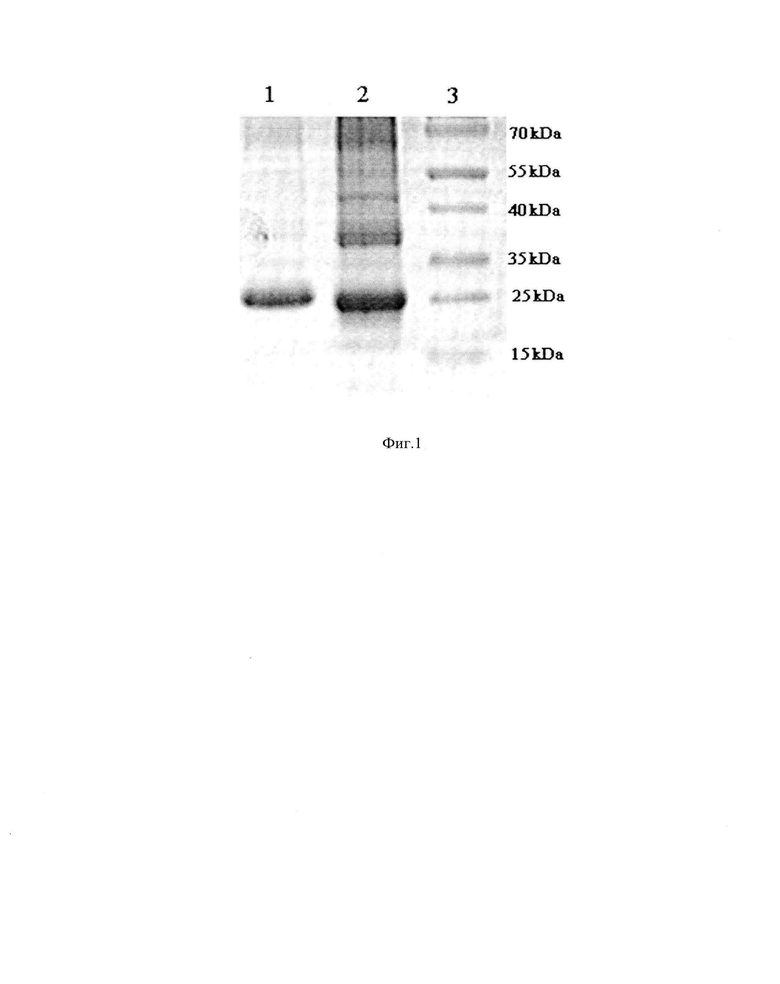
Фиг. 1. Электрофореграмма суммарной фракции ЛПВП и АпоА-I, выделенных из сыворотки крови человека с помощью процедуры, описанной в примере 1. Обозначения: 1 - АпоА-I ; 2 - ЛПВП; 3 - белки-стандарты.
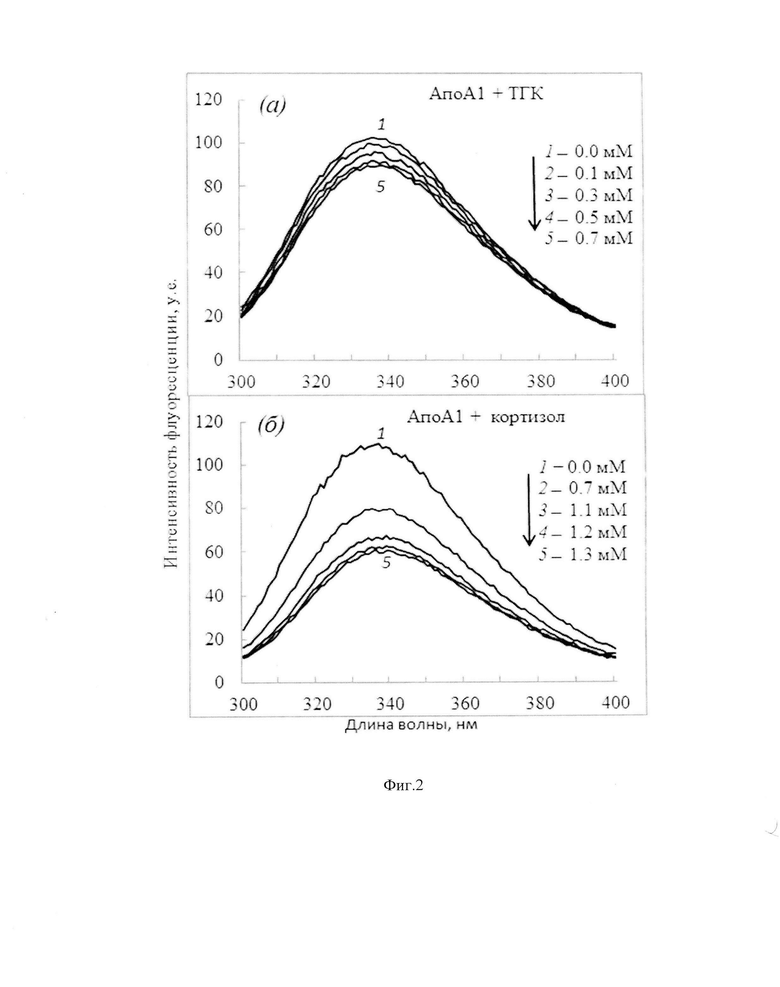
Фиг. 2. Тушение собственной триптофановой флуоресценцию АпоА-I (0,1 мг/мл) при образовании комплексов с ТГК (а) и кортизолом (б).
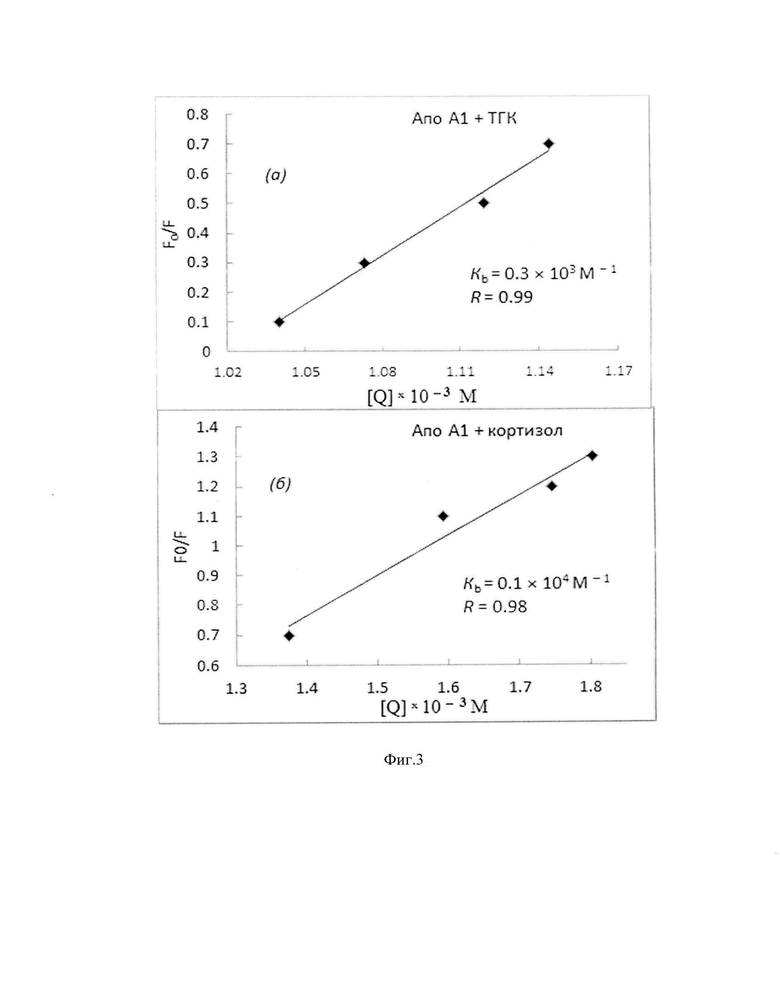
Фиг. 3. Графики Штерна-Фольмера для вычисления констант связывания (Kb) на основании концентрационного тушения триптофановой флуоресценции АпоА-I при добавлении возрастающих концентраций ТГК (а) и кортизола (б). Примечание: По оси ординат - отношение максимальной флуоресценции белка в отсутствие лиганда [F0] к интенсивности флуоресценции белка после добавления лиганда [F]. По оси абсцисс -концентрация лигандов [Q]. R - коэффициент корреляции Пирсона.
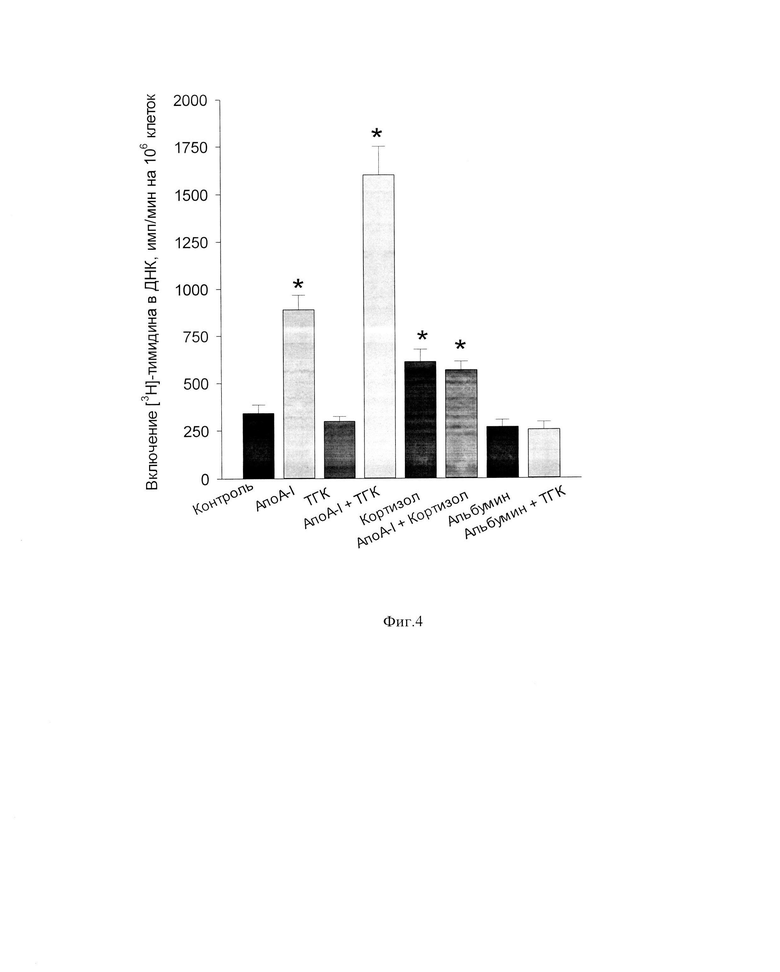
Фиг. 4. Влияние заявленного средства (комплекс АпоА-I -ТГК) на пролиферативную активность клеток костного мозга, культивируемых в бессывороточной среде в течение 24 ч. Примечание: Для сравнения представлены результаты влияния на данный показатель исходных компонентов комплекса (АпоА-I и ТГК), а также кортизола и комплекса АпоА-I-кортизол, альбумина и комплекса альбумина с ТГК. * - Статистически значимые различия по сравнению с контролем (р<0,05).
Осуществление изобретения
Пример 1. Получение белка АпоА-I из сыворотки крови человека и его биохимическая характеристика.
Из 50-100 мл сыворотки крови человека выделяют фракцию липопротеинов высокой плотности (ЛПВП) методом изоплотностного ультрацентрифугирования в растворе KBr при 105000 g [Mills G.L., Lane P.A., Weech P.K. In: Laboratory techniques in biochemistry and molecular biology: a guidebook to lipoprotein technique / Eds. R.H. Burdon, R.H. Knippenberg P.H. Elsevier, Amsterdam, 1984. - P. 18-116]. Из полученной фракции ЛПВП удаляют липиды смесью бутанола с диизопропиловым эфиром [Cham В.Е., Knowlee B.R. A solvent system for delipidation of plasma or serum without protein precipitation / J. Lipid Res. 17. - 1976. - P. 176-181]. Из полученной белковой фракции выделяют основной белок ЛПВП, используя метод высаливания сульфатом аммония с последующим растворением белка в 6 М мочевине и его ренатурацией с помощью диализа против фосфатно-солевого буфера (рН 7.4) в течение 24 ч [Jiang L., Неа L., Fountoulakis М. Comparison of protein precipitation methods for sample preparation prior to proteomic analysis / Journal of Chromatography A, 1023. - 2004. - P. 317-320; Пыхтина М.Б., Иванов И.Д., Беклемишев А.Б. Разработка эффективных способов выделения аполипопротеина A-I из плазмы крови человека. Биофарм. журн. - 2012. - Т. 4. - №6. - С. 37-45.]. Полученный концентрированный белок фильтруют через одноразовые стерильные фильтры с диаметром пор 0,22 мкм, что позволяет отделить не только потенциально содержащиеся микроорганизмы, но и агрегаты белка. Чистоту выделенного белка и его молекулярную массу оценивают с помощью электрофореза в ПААГ по методу Лэмли, используя набор белковых маркеров. На основании литературных данных о молекулярной массе белков, входящих в состав ЛПВП [Sakata N., Hoshii Y., Nakamura Т., Kiyama M., Arai H., Omoto M., Morimatsu M., Ishihara Т. Colocalization of apolipoprotein AI in various kinds of systemic amyloidosis / J. Histochem Cytochem. 2005. - Feb; 53. - P. 237-42.], делают вывод о соответствии выделенного белка аполипопротеину A-I (АпоА-I). Как видно из фиг. 1, используемый метод позволяет очистить АпоА-I от альбумина и минорных белков ЛПВП. С помощью данного способа из 100 мл сыворотки крови человека можно получить не менее 40 мг очищенного АпоА-I .
Пример 2. Получение комплекса АпоА-I с ТГК и определение константы связывания ТГК с АпоА-I .
Для получения заявленного средства очищенный АпоА-I, полученный как указано в примере 1, разводят в изотоническом фосфатно-солевом буфере (ФСБ) (рН 7.4) до концентрации 2-5 мг/мл.
На основе коммерческого препарата ТГК готовят концентрированный раствор ТГК в смеси органических растворителей - спирта и диметилсульфоксида в соотношении 1:1 и хранят при -20°С. Перед приготовлением комплекса АпоА-I с ТГК концентрированный раствор ТГК разводят до рабочей концентрации 100 мкг/мл в изотоническом фосфатно-солевом буфере (рН 7.4).
Указанные растворы АпоА-I и ТГК в фосфатно-солевом буфере смешивают в пропорциях, обеспечивающих соотношение АпоА-I :ТГК, равное 20:1, выраженное в мкг, и инкубируют в течение 15 мин при температуре 22±2°С.
Окончательное содержание органических растворителей при инкубации смеси АпоА-I и ТГК не превышает 0,5%.
Для подтверждения образования комплекса АпоА-I с ТГК оценивают взаимодействие ТГК с АпоА-I методом тушения (снижения) флуоресценции триптофана [Lakowicz J.R. // Principles of Fluorescence Spectroscopy, 3rd ed. Springer, Boston, MA, 2006]. Для сравнения в аналогичных условиях анализируют взаимодействие кортизола (активная форма стероидного гормона) с АпоА-I. Для этой цели измеряют интенсивность флуоресценции раствора АпоА-I в отсутствие и в присутствии возрастающих концентраций гормонов (кортизола или ТГК), выступающих в качестве тушителей флуоресценции. Титрование проводят в термостатируемой (37°С) кварцевой кювете, содержащей 100 мкг АпоА-I в 1 мл фосфатно-солевого буфера (рН 7.4). После добавления каждой аликвоты гормона к раствору АпоА-I содержимое кюветы интенсивно перемешивают и проводят измерение несколько раз до достижения максимального снижения флуоресценции. Флуоресценцию триптофановых остатков АпоА-I регистрируют на спектрофлуориметре RF-5301 (Shimadzu, Япония) при длине волны возбуждения 280 нм и испускания (эмиссии) в диапазоне 300-500 нм. Вклад растворителя без гормона в интенсивность флуоресценции составляет не более 1.5% и не изменяется в исследуемом диапазоне волн 300-500 нм.
Как известно, аминокислотная последовательность АпоА-I (28 кДа) состоит из 243 аминокислот, включая триптофан [Gursky О., Atkinson D. Thermal unfolding of human high-density apolipoprotein A-l: implications for a lipid-free molten globular state // Proc. Natl. Acad. Sci. USA. 1996. V. 93. P. 2991-2995]. Анализ спектральных свойств показал, что исходный спектр флуоресценции АпоА-I имеет максимум при 336 нм (фиг. 2). При регистрации спектров в присутствии возрастающих концентраций гормонов максимум спектра флуоресценции АпоА-I не изменялся (фиг. 2). Наблюдаемое снижение интенсивности флуоресценции АпоА-I в ответ на увеличение концентрации гормонов свидетельствует о формировании комплексов АпоА-I -ТГК или АпоА-I - кортизол.
В данном примере наибольшее снижение флуоресценции триптофана обнаружено у АпоА-I под влиянием кортизола. Различия, обнаруженные между влиянием гормонов на спектральные характеристики белков, можно объяснить разной способностью гормонов проникать во внутренние области белка. Менее выраженное снижение флуоресценции АпоА-I под влиянием ТГК, вероятно, связано с тушением остатков триптофана преимущественно на поверхности данного белка и отражает неспособность ТГК проникать во внутренние области белка.
Расчет констант связывания (Kb) гормонов проводят с помощью уравнения Штерна-Фольмера: F0 / F=1 - Kb × [Q]. Для определения Kb измеряют интенсивность флуоресценции АпоА-I в отсутствие (F0) и в присутствии (F) различных концентраций [Q] кортизола или ТГК, но при постоянной концентрации АпоА-I [Lakowicz J.R. Principles of Fluorescence Spectroscopy, 3rd ed. Springer, Boston, MA, 2006.]. На основании кривых тушения флуоресценции триптофана рассчитывают константы связывания, которые составили для комплекса АпоА-I - кортизол - (1.0±0.1) × 103 м-1 и для комплекса АпоА-I -ТГК-(3.1±0.4) × 102 М-1 (фиг. 3).
Как известно, взаимодействие гормонов с белком может сопровождаться конформационными изменениями последнего. Например, изменение вторичной структуры человеческого альбумина в сторону снижения доли α-спиралей зарегистрировано при его взаимодействии с тестостероном [Chanphai P., Vesper A.R., Bariyanga J.,  Tajmir-Riahi H.A. Review on the delivery of steroids by carrier proteins // J. Photochem. Photobiol. B. 2016. V. 161. P. 184-191], дексаметазоном [Naik P.N., Chimatadar S.A., Nandibewoor S.T. Interaction between a potent corticosteroid drug -Dexamethasone with bovine serum albumin and human serum albumin: A fluorescence quenching and fourier transformation infrared spectroscopy study // J. Photochem. Photobiol. B. 2010. V. 100. P. 147-159.] и прогестероном [Abu Teir M.M., Ghithan J.H., Darwish S.M., Abu-Hadidal M.M. Study of Progesterone interaction with Human Serum Albumin: Spectroscopic approach // J. Appl. Biol. Sci. 2011. V. 5. P. 35-47].
Tajmir-Riahi H.A. Review on the delivery of steroids by carrier proteins // J. Photochem. Photobiol. B. 2016. V. 161. P. 184-191], дексаметазоном [Naik P.N., Chimatadar S.A., Nandibewoor S.T. Interaction between a potent corticosteroid drug -Dexamethasone with bovine serum albumin and human serum albumin: A fluorescence quenching and fourier transformation infrared spectroscopy study // J. Photochem. Photobiol. B. 2010. V. 100. P. 147-159.] и прогестероном [Abu Teir M.M., Ghithan J.H., Darwish S.M., Abu-Hadidal M.M. Study of Progesterone interaction with Human Serum Albumin: Spectroscopic approach // J. Appl. Biol. Sci. 2011. V. 5. P. 35-47].
Используя инфракрасную Фурье-спектроскопию, ранее нами было показано, что при инкубации АпоА-I в присутствии ТГК происходит дозозависимое снижение содержания α-спиралей и увеличение содержания β-поворотов в АпоА-I. Под влиянием кортизола, напротив, количество а-спиралей в АпоА-I возрастало, а β-поворотов - снижалось [Дударев А. Н., Городецкая А.Ю., Ткаченко Т.А., Усынин И.Ф. Влияние кортизола и тетрагидрокортизола на вторичную структуру АпоА-I по данным инфракрасной Фурье-спектроскопии // Биоорганическая химия, 2021, том 47, №6, с. 785-794]. Эти результаты позволяют предполагать, что конформационные изменения АпоА-I, происходящие под влиянием стероидных гормонов в процессе комплексообразования, могут приводить к изменению акцепторных и регуляторных свойств данного белка.
Пример 3. Оценка влияние заявленного комплекса АпоА-I -ТГК на пролиферативную активность клеток костного мозга у интактных животных и в разные сроки после острой кровопотери.
Острая кровопотеря является одной из причин смертности при техногенных и природных катастрофах, во время военных конфликтов, а также при проведении оперативных вмешательств. Эффективность лечения острой кровопотери зависит как от средств ее возмещения, так и от объема трансфузионной терапии. Возмещение утраченной крови адекватным количеством донорской крови в настоящее время считается патогенетически и патофизиологически необоснованным. Показаниями к проведению гемотрансфузии при острой кровопотере являются лишь угрожающие жизни анемия и гипопротеанемия. Во всех остальных случаях предпочтение отдается кровезаменителям, компонентам и препаратам крови [Кузнецов Н.Л. Современные технологии лечения острой кровопотери. 2003, Consilium Medicum. Т. 5., №6. С. 347-357]. К числу последних могут быть отнесены стимуляторы пролиферации клеток костного мозга. В связи с этим, демонстрация эффективности влияния заявленного комплекса АпоА-I - ТГК на пролиферативную активность клеток костного мозга in vivo осуществлена на модели острой кровопотери.
Описание модели острой кровопотери
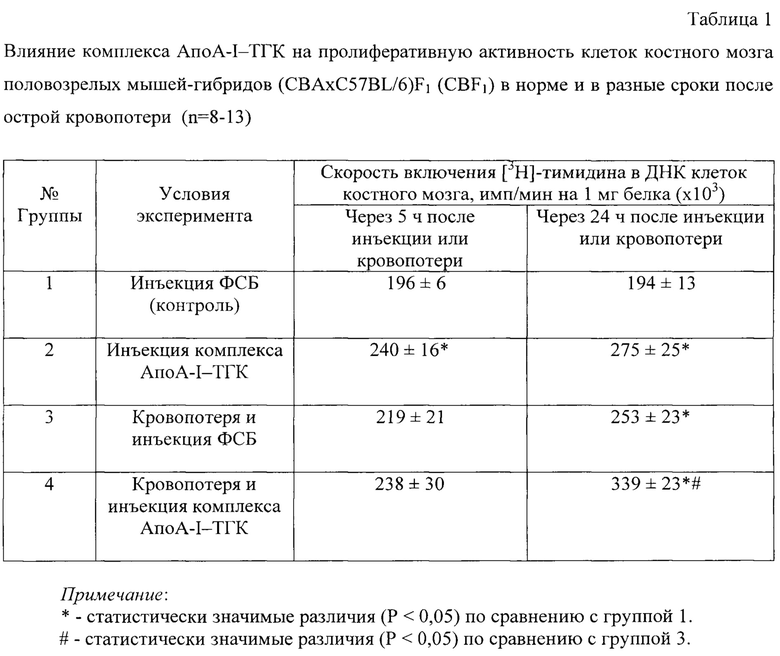
Для доказательства влияния заявленного комплекса на пролиферативную активность костного мозга in vivo используют модель острой кровопотери. Для этого под общим эфирным наркозом из ретроорбитального синуса животного с помощью гематокритного капилляра или пастеровской пипетки однократно отбирают 0,5 мл крови [Sadahira Y. et al. Impaired splenic erythropoiesis in phlebotomized mice injected with CL2MDP-liposome: an experimental model for studying the role of stromal macrophages in erythropoiesis. 2000. J. Leukoc. Biol. 68: 464÷470].
Испытания проводят в несколько серий опытов, каждая из которых имеет свой контроль (таблица 1). В работе используют здоровых половозрелых мышей-гибридов (CBA×C57BL/6) F1 (CBF1), 10-12- недельного возраста, массой 25±2 г. Контрольные и опытные животные содержатся в виварии в стандартных пластиковых клетках на стандартном рационе. Все исследования проводятся в одно и то же время суток. Хирургические манипуляции с экспериментальными животными проводят под ингаляционным эфирным наркозом в соответствии с правилами, принятыми Европейской конвенцией по защите животных (Страсбург, 1986) и одобренные комитетом по биомедицинской этике ФИЦ ФТМ. Оценку различий между выборками проводят с использованием непараметрического U-критерия Манна-Уитни [Гланц С. Медико-биологическая статистика. М., Практика, 1998. - 459 с.].
Заявленный комплекс готовят согласно примеру 2 и вводят в хвостовую вену мышей из расчета 15 мг АпоА-I в составе указанного средства на 1 кг массы тела животного сразу после проведения кровопотери (группа 2 и 4) (таблица 1). Одновременно контрольным животным (группа 1 и 3) вводят внутривенно эквивалентный объем фосфатно-солевого буфера (ФСБ).
Метод оценки влияния заявленного средства на пролиферативную активность клеток костного мозга животных при кровопотере
Пролиферативную активность костного мозга оценивают через 5 и 24 ч после проведения кровопотери. Для этого за 60 мин до забоя животным внутрибрюшинно вводят меченый предшественник [3Н]-тимидин («Изотоп», Россия) из расчета 1,5 мкКю на 1 г массы тела. Клетки костного мозга получают из бедренной кости стандартным методом [Гольдберг Е.Д., Дыгай A.M., Шахов В.П. Методы культуры ткани в гематологии. Томск. - 1992.]. После лизиса клеток полученный гомогенат наносят на фильтры, которые проходят многоэтапную отмывку от не связавшейся метки. Радиоактивность проб измеряют с помощью жидкостного сцинтилляционного счетчика [Остерман Л. А. Методы исследования белков и нуклеиновых кислот: Электрофорез и ультрацентрифугирование. М.: Наука, 1981. 288 с.]. Скорость включения [3Н]-тимидина в ДНК клеток костного мозга оценивают в имп/мин на 1 мг белка гомогената.
Результаты оценки влияния заявленного средства на пролиферативную активность клеток костного мозга in vivo при кровопотере
Как видно из таблицы 1, внутривенное введение мышам заявленного комплекса ТГК-АпоА-I сопровождается возрастанием скорости включения [3Н]-тимидина в ДНК клеток костного мозга через 5 и 24 ч на 22% и 42%, соответственно. Скорость биосинтеза ДНК в течение первых 5 ч после кровопотери достоверно не изменялась, а через 24 ч возрастала на 30%. Инъекция комплекса на фоне кровопотери через 24 ч приводила к увеличению пролиферативной активности клеток костного мозга на 74% по сравнению с интактными животными и на 34% по сравнению с животными, которым не вводили комплекс (группа 3).
Таким образом, использование комплекса АпоА-I -ТГК позволяет повысить пролиферативную активность клеток костного мозга in vivo.
Пример 4. Получение культуры клеток костного мозга для оценки влияния заявленного средства на пролиферативную активность клеток костного мозга in vitro.
Для демонстрации эффективности заявленного средства на пролиферативную активность клеток костного мозга используют первичную культуру клеток костного мозга экспериментальных животных. Клетки получают из бедренной кости крыс линии Wistar стандартным методом [Гольдберг Е.Д., Дыгай A.M., Шахов В.П. Методы культуры ткани в гематологии. Томск. - 1992.]. Очистку костного мозга от эритроцитов и лейкоцитов проводят с помощью противоточного центрифугирования в элютриаторном роторе JE-5.0 центрифуги Avanti J-26XP («Beckman Coulter», США) при 2500 об/мин и скорости тока жидкости 14 мл/мин.
Полученную суспензию клеток, содержащую не более 10% эритроцитов и лимфоцитов, культивируют в 24-луночных планшетах (2-5×106 клеток на 1 лунку) в бессывороточной среде RPMI-1640 в присутствии заявленного средства. Культивирование клеток проводят в СО2 - инкубаторе («Cole Panner», США) в атмосфере, содержащей 5% СО2 и 95% воздуха, при температуре 37°С.
Пример 5. Метод оценки влияния заявленного средства на пролиферативную активность клеток костного мозга в культуральной среде
Пролиферативную активность клеток костного мозга определяют по включению радиоактивно меченого предшественника [Н]-тимидина («Изотоп», Россия) в ДНК [Dawson C.W., Young L.S. In vitro assays to study epithelial cell growth / Methods Mol Biol. -2001. - Vol. 174. - P. 165-172.]. Данный показатель отражает скорость биосинтеза ДНК, что лежит в основе пролиферативной активности клеток, т.к. [3Н]-тимидин включается в ДНК в синтетической фазе (S-фазе) клеточного цикла. Для оценки данного показателя за 2 ч до окончания инкубации клеток костного мозга в культуральную среду вносят по 2,0 мкКю/мл меченого предшественника [3Н]-тимидина. После лизиса клеток костного мозга гомогенат клеток наносят на фильтры, которые проходят многоэтапную отмывку от не связавшейся метки. Радиоактивность проб измеряют с помощью жидкостного сцинтилляционного счетчика [Остерман Л.А. Методы исследования белков и нуклеиновых кислот: Электрофорез и ультрацентрифугирование. М.: Наука, 1981. 288 с.]. Скорость включения меченого предшественника оценивают в импульсах (имп) в 1 мин на 106 клеток или в % по отношению к контролю. Результаты представляют как среднее значение ± стандартное отклонение от трех независимых экспериментов, выполненных в трех параллелях. Оценку различий между выборками проводят с использованием непараметрического U-критерия Манна-Уитни [Гланц С. Медико-биологическая статистика. М., Практика, 1998. - 459 с.].
Пример 6. Влияние заявленного средства на пролиферативную активность клеток костного мозга при их культивировании в бессывороточной среде
На фиг. 4 дано сравнение пролиферативной активности клеток костного мозга при культивировании в присутствии заявленного средства (комплекс АпоА-I -ТГК) и компонентов заявленного средства раздельно (АпоА-I и ТГК). Для сравнения представлены данные о влиянии в этих условиях сывороточного человеческого альбумина и его комплекса с ТГК, а также кортизола и его комплекса с АпоА-I. Комплексы приготовлены согласно примеру 2. Культура клеток костного мозга получена согласно примеру 4. Оценка пролиферативной активности клеток костного мозга выполнена согласно примеру 5. Для доказательства наиболее выраженного эффекта заявленного средства по сравнению с эффектами его компонентов использовали эквивалентное содержание всех компонентов в культуральной среде.
Как видно из фиг. 4, культивирование клеток костного мозга в бессывороточной питательной среде RPMI-1640 в присутствии альбумина (10 мкг/мл) или ТГК (0,5 мкг/мл) или их комплекса не оказывало статистически значимого влияния на пролиферативную активность клеток костного мозга. Добавление в культуральную среду кортизола (0,5 мкг/мл) или АпоА-I (10 мкг/мл) приводило к повышению скорости синтеза ДНК в 1,8 и 2,6 раза, соответственно. Еще более выраженный эффект на данный показатель обнаружен при культивировании клеток костного мозга в присутствии комплекса АпоА-I -ТГК (10 мкг/мл АпоА-I и 0,5 мкг/мл ТГК): скорость включения [3Н]-тимидина в ДНК в присутствии данного комплекса была в 1,8 раз выше, чем в присутствии одного АпоА-I и в 4,7 раз выше, чем в контроле (без добавок). Следует отметить, что в отличие от комплекса АпоА-I -ТГК, инкубация клеток костного мозга в присутствии комплекса АпоА-I -кортизол (10 мкг/мл АпоА-I и 0,5 мкг/мл кортизола) приводила к снижению скорости биосинтеза ДНК в 1,6 раза по сравнению с эффектом АпоА-I (фиг. 4).
Механизм влияния стероидных гормонов на конформацию АпоА-I не изучен, и поэтому полученный максимальный эффект комплекса сывороточного АпоА-I -ТГК на функциональную активность клеток костного мозга с очевидностью не вытекает из уровня техники.
Пример 7. Влияние заявленного средства на изменение клеточного состава клеток костного мозга при их культивировании в бессывороточной среде.
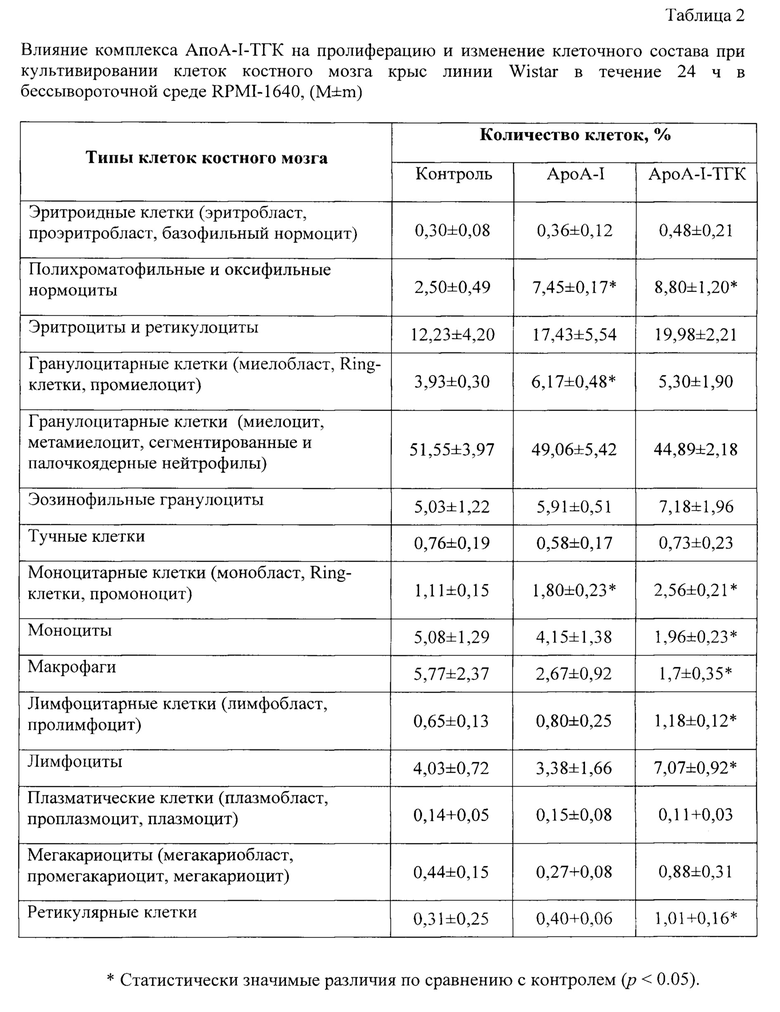
Костный мозг состоит из чрезвычайно гетерогенной популяции клеток, включающей клетки ретикулярной стромы, кроветворные клетки костного мозга и клетки крови на различных этапах дифференцировки [Гаврилов О.К., Козинец Г.И., Черняк Н.Б. Клетки костного мозга и периферической крови. М.: Медицина, 1985. 288 с.]. Для идентификации разных типов клеток костного мозга готовят мазки клеток и окрашивают по методу Романовского-Гимза [Гольдберг Е.Д., Дыгай A.M., Шахов В.П. Методы культуры ткани в гематологии. Томск. - 1992]. Учитывая морфологические особенности некоторых типов клеток костного мозга крысы, проводят дифференциальный анализ клеточного состава культивируемых клеток [Bolliger А.P. Cytologic evaluation of bone marrow in rats: indications, methods, and normal morphology. // Vet. Clin Pathol. 2004. Vol. 33, N 2. P. 58-67]. Подсчитывают не менее 1000 клеток и вычисляют процент каждого вида клеток костного мозга для составления миелограммы.
Культуру клеток костного мозга готовят, как описано в примере 4. Комплекс АпоА-I -ТГК готовят согласно примеру 2. После культивирования клеток в течение 24 ч в присутствии заявленного средства готовят мазки клеток костного мозга и анализируют миелограмму.
Как видно из таблицы 2, культивирование клеток костного мозга в присутствии заявленного средства сопровождается достоверно более значительным в сравнении с прототипом пролиферативным эффектом, заключающимся в достоверном увеличении количества полихроматофильных и оксифильных нормоцитов, клеток моноцитарного ростка (монобласты, промоноциты и Ring-клетки), клеток лимфоцитарного ростка (лимфобласт, пролимфоцит, лимфоцит), ретикулярных клеток, а также в некотором увеличении количества эритроцитов и ретикулоцитов, эозинофильных гранулоцитов, мегакариоцитов. При этом возрастает в 3,5 раза процентное содержание полихроматофильных и оксифильных нормоцитов, в 2,3 раза - клеток моноцитарного ростка (монобласты, промоноциты и Ring-клетки) и в 1,8 раза - клеток лимфоцитарного ростка (лимфобласт, пролимфоцит, лимфоцит).
Добавление в культуру клеток костного мозга только АпоА-I вызывало подобные, но менее выраженные изменения (табл. 2). Напротив, в этих условиях относительное содержание моноцитов и макрофагов снижалось, а содержание эритроцитов, клеток гранулоцитарного ростка, тучных клеток и мегакариоцитов не изменялось. Вероятно, для созревания клеток этих ростков необходимо присутствие в культуральной среде специфических ростовых факторов. Например, известно, что для формирования эритроидных островков и предотвращения апоптоза эритроидных предшественников необходимо постоянное присутствие в окружающей микросреде эритропоэтина [Sadahira Y., Mori М. Role of the macrophage in erythropoiesis. Pathol. Inter., 1999, 49, 841-848].
Таким образом, использование комплекса АпоА-I - ТГК позволяет повысить пролиферативную активность клеток костного мозга in vitro при культивировании клеток в бессывороточной среде, содержащей данный комплекс, а также in vivo с помощью внутривенного введения комплекса в организм на фоне нестимулированного или стимулированного кроветворения в костном мозге. Учитывая, что стимулирующий эффект комплекса АпоА-I - ТГК достигается при небольшой концентрации сывороточного АпоА-I, это позволяет использовать в составе комплекса АпоА-I, выделенный из сыворотки крови донора, одновременно являющегося реципиентом, что обеспечит практически безопасную персонифицированную терапию у больных при кровопотерях, после проведения химиотерапии и в других случаях, когда возникает необходимость стимуляции пролиферации клеток костного мозга различных линий.
Установленный эффект заявленного средства открывает также перспективы для применения культуры клеток костного мозга в бессывороточной питательной среде для клеточной терапии, при этом предпочтительным является включение в состав питательной среды комплекса АпоА-I -ТГК, полученного с использованием АпоА-I донора, одновременно являющегося реципиентом клеток костного мозга.
Работа выполнена в рамках выполнения государственного задания Министерства науки и высшего образования Российской Федерации (регистрационный номер 1021050400914-1-1.6.4) с использованием оборудования ЦКП «Спектрометрические измерения» и ЦКП «Протеомный анализ», поддержанного финансированием Минобрнауки России (соглашение №075-15-2021-691).
| название | год | авторы | номер документа |
|---|---|---|---|
| Среда для культивирования клеток костного мозга, предназначенных для клеточной терапии | 2020 |
|
RU2756926C1 |
| ВЕЩЕСТВО И СПОСОБ МОДУЛЯЦИИ ПРОЛИФЕРАЦИИ И ДИФФЕРЕНЦИРОВКИ РЕГУЛЯТОРНЫХ, СТВОЛОВЫХ И ДРУГИХ СОМАТИЧЕСКИХ КЛЕТОК | 2014 |
|
RU2620069C2 |
| Способ лечения и профилактики гипергликемии и инсулинорезистентности и связанных с ними заболеваний и функциональных нарушений органов и систем и средство для его осуществления | 2023 |
|
RU2823334C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЭФФЕКТИВНОСТИ ГЕМОСТИМУЛЯТОРОВ ПРИ ЦИТОСТАТИЧЕСКОЙ МИЕЛОСУПРЕССИИ | 2009 |
|
RU2421720C2 |
| СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2006 |
|
RU2322264C1 |
| ГЕМОСТИМУЛЯТОР | 1994 |
|
RU2094045C1 |
| МЕТАДИХОЛ® - ЖИДКИЕ И ГЕЛЕВЫЕ НАНОЛЕКАРСТВЕННЫЕ ПРЕПАРАТЫ | 2014 |
|
RU2631600C2 |
| СПОСОБ СТИМУЛЯЦИИ ЭНДОГЕННОЙ ПРОДУКЦИИ ЦИТОКИНОВ И ГЕМОПОЭТИЧЕСКИХ ФАКТОРОВ | 2012 |
|
RU2498821C1 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2018 |
|
RU2675233C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ИММУНОСТИМУЛИРУЮЩИМ И ГЕМОСТИМУЛИРУЮЩИМ ДЕЙСТВИЕМ | 2009 |
|
RU2414223C1 |
Изобретение относится к области биотехнологии и может быть использовано в медицине. Предложено применение комплекса сывороточного белка аполипопротеин A-I с тетрагидрокортизолом для стимуляции пролиферативной активности клеток костного мозга. Изобретение может применяться при культивировании клеток костного мозга в условиях, требующих использования бессывороточной среды, а также в регенеративной медицине для стимуляции пролиферативной активности клеток костного мозга in vivo при патологиях, связанных с нарушением кроветворения, например, при синдроме недостаточности костного мозга или при кровопотерях, наступивших после прохождения курсов химио- или лучевой терапии. 2 н.п. ф-лы, 4 ил., 2 табл., 7 пр.
1. Применение комплекса аполипопротеина A-I и тетрагидрокортизола в качестве средства для стимуляции пролиферативной активности клеток костного мозга.
2. Способ получения комплекса аполипопротеина A-I и тетрагидрокортизола по п.1, включающий совместную инкубацию в изотоническом фосфатно-солевом буфере с рН 7.4 аполипопротеина A-I и тетрагидрокортизола в соотношении 20:1, выраженном в мкг, при температуре 22±2°С в течение 15 мин.
| ПАНИН Л.Е | |||
| и др.: "Роль аполипопротеина A-I в реализации анаболического действия стероидных гормонов", Проблемы Эндокринологии, 2002, т | |||
| Приспособление для автоматической односторонней разгрузки железнодорожных платформ | 1921 |
|
SU48A1 |
| PANIN L.E | |||
| et al.: "Effect of Complexes of Apolipoprotein A-I with Tetrahydrocortisol and Pregnenolone on Protein Biosynthesis in Rat Hepatocytes Culture", Bulletin of Experimental Biology and | |||
