ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к областям молекулярной биологии и регенеративной медицины, конкретно, к способам модуляции пролиферации и/или дифференцировки клеток, в частности, в организме млекопитающего.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Клеточная трансплантационная терапия сопряжена с проблемами безопасности, связанными с риском нежелательной активации иммунной системы хозяина. Соответственно, в данной области остро ощущается потребность в биологической терапии, которая обеспечит функциональное восстановление системы организма реципиента и в то же время позволит избежать активации его иммунной системы.
КРАТКОЕ СОДЕРЖАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение связано с композициями и способами или применениями указанных композиций для профилактики, терапии или излечения заболеваний.
Композиции и способы согласно изобретению предлагаются в качестве замены традиционной клеточной терапии, которая сопряжена с риском для здоровья реципиента в результате нежелательной активации его иммунной системы и отторжения трансплантата. Композиции и способы согласно изобретению включают в себя препараты РНК или препараты тотальной РНК, выделенные из одного или более типов соматических клеток (включая регуляторные лимфоидные клетки и стволовые клетки). Описанные здесь композиции и способы, в силу своей неиммуногенности, дают возможность, в частности, отказаться от необходимости персонализированного консервирования клеток пуповинной крови.
Композиции и способы согласно изобретению предлагаются в качестве замены - в большом ряде случаев - терапевтической процедуры переливания крови, которая связана с определенным риском для здоровья реципиента.
Описанные здесь композиции, способы и применения способствуют структурной и функциональной репарации и восстановлению поврежденных тканей. Таким образом, композиции и способы согласно изобретению направлены не на симптоматическое лечение заболеваний, а дают возможность их излечения.
Композиции и способы согласно изобретению используются для модуляции нарушенной пролиферации и/или дифференцировки клеток, в частности, в организме млекопитающего. Более конкретно, композиции и способы согласно изобретению используются для модуляции численности и дифференцировки различных типов клеток (например, клеток млекопитающего) путем активации/нормализации регуляторной функции лимфоидных клеток. Композиции включают в себя один или более препаратов РНК, полученных из лимфоидных клеток селезенки, тимуса, лимфатических узлов, из лимфоцитов периферической крови, из костного мозга, из стволовых клеток (например, из пуповинной крови, из пупочного канатика и/или из плаценты) здоровых доноров, которые восстанавливают нормальную функцию ткани или клеточной популяции реципиента, для лечения, облегчения или профилактики заболевания, расстройства или состояния, ассоциированного с нарушением регуляции клеточной пролиферации и/или дифференцировки.
Композиции и способы согласно изобретению используются, в частности, для активации стволовых клеток и дальнейшей модуляции их стимулирующего действия. Композиции и способы согласно изобретению используются для лечения или профилактики нарушений гемопоэза, а также гематологических, дегенеративных, гиперпролиферативных или аутоиммунных заболеваний, расстройств или состояний. Композиции и способы согласно изобретению используются для коррекции ряда наследственных и врожденных дефектов.
Композиции и способы согласно изобретению используются для модуляции пролиферации и/или дифференцировки различных типов клеток млекопитающего in vivo или in vitro. Композиции включают в себя препараты РНК, полученные из одного или более органов, тканей или соматических клеток. Композиции согласно изобретению могут быть использованы в качестве дополнительного воздействия для лечения или профилактики нарушений гемопоэза, болезней крови, дегенеративных, опухолевых и аутоиммунных заболеваний, расстройств и состояний, а также для коррекции определенных наследственных и врожденных дефектов путем их компенсации.
В композициях и способах согласно изобретению используется морфогенетическая активность лимфоидных клеток для осуществления контроля над патологическими процессами в организме, в частности в организме млекопитающего, предпочтительно человека, при этом удается избежать и осложнений, связанных с нежелательной активацией иммунной системы реципиента, и осуществления трудоемких поисков наиболее подходящего донора. Композиции и способы согласно изобретению индуцируют клеточную пролиферацию и регенерацию в организме, в частности, в организме млекопитающего, предпочтительно человека. Композиции согласно изобретению включают в себя одну или более молекул РНК препарата тотальной РНК, полученного из одной или более клеток или из одной или более разновидностей клеток.
Препараты тотальной РНК лимфоидных клеток обладают морфогенетической активностью, сравнимой с таковой самих этих лимфоидных клеток. Под ʺморфогенетическойʺ активностью лимфоидных клеток здесь подразумевается их регуляторная активность, состоящая в осуществлении контроля пролиферации и/или дифференцировки различных типов клеток организма. Данные, приведенные в настоящем описании, свидетельствуют о морфогенетической активности препаратов тотальной РНК, полученных из лимфоидных клеток селезенки или тимуса, лимфоцитов периферической крови и из костного мозга. Кроме того, здесь показано, что препараты тотальной РНК, полученные из пуповинной крови, из самой пуповины или из плаценты, оказывают действие, подобное действию, оказываемому интактной пуповинной кровью, клетками пуповины или плаценты. Под ʺинтактнымиʺ лимфоидными клетками в настоящем документе подразумеваются ʺне подвергнутые активацииʺ лимфоидные клетки. Препараты тотальной РНК, полученные из любых других типов соматических клеток, оказывают такое же действие, какое обеспечивается самими клетками того же типа, из которых выделена указанная РНК. То есть РНК-содержащие препараты, полученные из любых других типов соматических клеток, эффективны в повышении функциональной активности и регенерационной способности гомологичных тканей, а кроме того, эффективны в связи с их трофической функцией, с пополнением недостаточности эндогенной РНК.
В композициях и способах согласно изобретению используются морфогенетические свойства препаратов тотальной РНК из лимфоидных клеток для модуляции пролиферативной активности клеток различных органов и тканей. Композиции, способы и применения согласно изобретению широко применимы в ветеринарии и в медицине, поскольку дают возможность заменить регуляторные клетки их функциональным аналогом в виде неиммуногенных препаратов тотальной РНК. Таким образом, в композициях и способах согласно изобретению используются неиммуногенные средства для переноса пролиферативных или антипролиферативных сигналов. Здесь термин «неиммуногенный» означает отсутствие у РНК индивидуальных и видоспецифических антигенных ограничений (см. также патент RU 2314814). Примером такого переноса могут служить результаты ксеногенного переноса тотальной РНК из лимфоидных клеток (см. пример 7). Поскольку препараты РНК согласно изобретению являются неиммуногенными, любые аллогенные и ксеногенные препараты тотальной РНК можно беспрепятственно вводить в организм млекопитающего, в частности человека. При этом регуляторные препараты тотальной РНК, выделенные из лимфоидных клеток и лимфоидных органов здоровых доноров, отличаются тем, что оказывают корригирующее действие не только в отношении пролиферативного статуса соматических клеток-мишеней, но и восстанавливают нарушенную регуляторную функцию самой системы лимфоидных клеток реципиента при самых разных патологических состояниях.
В одном из воплощений настоящего изобретения композиции и способы согласно изобретению включают в себя разновидности регуляторных препаратов тотальной РНК, выделенных из лимфоидных клеток селезенки, тимуса, лимфатических узлов, лимфоцитов периферической крови или из костного мозга здорового интактного донора, или же -альтернативно или дополнительно - из лимфоидных клеток селезенки, тимуса, лимфоцитов периферической крови или из костного мозга здорового донора, подвергнутого активации Т-клеточного звена иммунной системы, в фазе проявления ими стимулирующей (хелперной) или тормозящей (супрессорной) активности в отношении соматических клеток (например, соматических клеток определенного гистотипа).
Функциональные различия указанных регуляторных препаратов тотальной РНК определяются качественными различиями функционального состояния исходных донорских клеток, полученных в нормальных условиях, или донорских клеток, полученных на разных стадиях реализации их морфогенетической функции.
В другом воплощении настоящего изобретения композиции и способы согласно изобретению включают в себя разновидности препаратов тотальной РНК, полученные из клеток пуповинной крови или из цельной пуповинной крови, из клеток пупочного канатика или из цельной пуповины, или из плаценты здорового интактного донора.
Еще в одном из воплощений настоящего изобретения описанные здесь композиции и способы включают в себя препараты тотальной РНК, выделенные из любой разновидности клеток млекопитающего. Клетка млекопитающего может быть клеткой такого типа, которая необходима для восстановления структуры и функции ткани. Поскольку трансплантация клеток ассоциирована с возможными нежелательными эффектами и требует предварительной иммуносупрессии у реципиента, использование донорских клеток связано с дополнительным риском для здоровья и даже жизни пациента. Композиции и способы согласно изобретению позволяют избежать и реакцию трансплантат-против-хозяина (РТПХ), и необходимость подавления иммунной системы во избежание отторжения переносимых донорских клеток. Функциональное восстановление, наблюдаемое в организме реципиента после введения композиций согласно изобретению, содержащих препараты тотальной РНК, полученные из интактного и предварительно активированного костного мозга крыс-доноров, сравнимы с функциональным восстановлением, которое достигается после пересадки костного мозга (см. пример 6).
В одном из воплощений настоящего изобретения описанные здесь композиции и способы включают в себя препарат регуляторной тотальной РНК, выделенный из лимфоидных клеток или лимфоидных органов донора, который необязательно может содержать популяцию активированных (стимулирующих или супрессорных) Т-клеток, образующихся в ответ на активацию его иммунной системы. Активация Т-клеток здорового донора может быть предпринята in vivo, ex vivo или in vitro. Регуляторную тотальную РНК выделяют in vitro, предпочтительно из популяции донорских клеток (например, из лимфоидных клеток селезенки, тимуса, лимфатических узлов, лимфоцитов периферической крови или из костного мозга), включающей в себя по меньшей мере одну активированную Т-клетку, по стандартной методике с использованием тризольного реагента и фенол-хлороформной экстракции (Chomczynski P. BioTechniques, 1993, vol. 15, pp. 532-537). Предпочтительно, чтобы выделение РНК было предпринято на стадии, когда иммунные клетки проявляют свою стимулирующую или супрессорную активность в отношении клеток определенного гистотипа (гистотипов), с получением препарата регуляторной тотальной РНК, который обладает, соответственно, стимулирующей или супрессорной активностью в отношении таких же клеток реципиента.
В другом воплощении настоящего изобретения описанные здесь композиции и способы включают в себя регуляторные препараты тотальной РНК. Под «регуляторными» препаратами тотальной РНК подразумеваются препараты тотальной РНК, выделенные из интактных или активированных лимфоидных клеток селезенки, тимуса, лимфатических узлов, лимфоцитов периферической крови или из костного мозга здорового донора.
Препараты тотальной РНК могут быть выделены также из любой другой ткани или любых других соматических клеток здорового донора. В частности, препараты тотальной РНК могут быть получены из любой стволовой клетки здорового донора, включая клетки костного мозга, клетки пуповины (включая цельную пуповинную кровь и Вартонов студень (substantia gelatinea funiculi umbilicalis)) и плаценту. Здесь под ʺстволовыми клеткамиʺ подразумеваются клетки-предшественники соматических клеток, обладающие высоким пролиферативным потенциалом и тотипотентностью (то есть способностью дифференцироваться в любые соматические клетки организма). При этом под ʺсоматическими клеткамиʺ подразумеваются все клетки организма, кроме половых клеток.
В другом воплощении настоящего изобретения описанные здесь композиции и способы осуществляют модуляцию пролиферации и/или дифференцировки клеток, в частности, клеток млекопитающего. В одном из воплощений указанного способа композицию, содержащую препарат тотальной РНК или часть его, полученную из лимфоидных клеток и/или костного мозга здорового донора в нормальных условиях, вводят субъекту, в частности млекопитающему (предпочтительно человеку). Альтернативно или дополнительно, композицию, содержащую препарат тотальной РНК или часть его, полученную из лимфоидных клеток и/или костного мозга здорового донора в условиях активации Т-клеточного звена иммунной системы, во временных интервалах, когда исходные клетки in vivo или in vitro проявляют стимулирующую (приблизительно от 15 минут примерно до 48 часов после активации, в зависимости от ткани-мишени) или супрессорную (приблизительно от 48 часов примерно до 96 и более часов после активации, в зависимости от ткани-мишени) активности в отношении клеток того или иного гистотипа, вводят субъекту, в частности, млекопитающему (предпочтительно человеку).
В другом воплощении настоящего изобретения описанные здесь композиции и способы осуществляют модуляцию пролиферации и/или дифференцировки клеток для лечения или профилактики нарушений гемопоэза, болезней крови, дегенеративных, опухолевых и аутоиммунных заболеваний, расстройств и состояний, а также для коррекции определенных наследственных, врожденных или возрастных дефектов.
В другом воплощении настоящего изобретения описанные здесь композиции включают в себя комбинацию препаратов тотальной РНК, полученных из различных органов или соматических клеток, для лечения или профилактики нарушений гемопоэза, болезней крови, дегенеративных, опухолевых и аутоиммунных заболеваний, расстройств и состояний, а также для коррекции (устранения) ряда наследственных, врожденных или возрастных дефектов, включая, но не ограничиваясь остеопетрозом, ДЦП, нарушениями зрения, слуха (глухотой), и пр. Указанная комбинация может включать в себя препарат тотальной РНК, полученный из лимфоидных клеток и/или костного мозга, а также препарат тотальной РНК, полученный из различных органов, тканей, соматических клеток, в частности, из стволовых клеток.
В другом воплощении настоящего изобретения описанные здесь композиции и способы включают в себя стимулирующий препарат (препарат РНК или тотальной РНК, полученный из стимулированных, или активированных, лимфоидных клеток или клеток костного мозга) для модуляции морфогенетической функции лимфоцитов для воздействия на клетки различных тканей организма реципиента (необязательно млекопитающего).
В другом воплощении настоящего изобретения описанные здесь композиции и способы включают в себя тормозящий препарат (препарат РНК или тотальной РНК, полученный из стимулированных, или активированных, иммунных клеток (то есть лимфоидных клеток или клеток костного мозга) для модуляции морфогенетической функции лимфоцитов для воздействия на клетки различных тканей организма реципиента (необязательно млекопитающего).
В одном из воплощений композиции и способы согласно изобретению включают в себя применение препаратов тотальной РНК согласно изобретению в качестве замены переливания крови субъекту.
В другом воплощении композиции и способы согласно изобретению включают в себя применение препаратов тотальной РНК согласно изобретению в качестве замены клеточной терапии с использованием стволовых клеток у субъекта.
В другом воплощении композиции и способы согласно изобретению включают в себя применение препаратов тотальной РНК согласно изобретению в качестве замены одной или нескольких трансплантаций костного мозга у субъекта.
В одном из воплощений композиции и способы согласно изобретению включают в себя, но не ограничены препаратом тотальной РНК, полученным из любых интактных (то есть не подвергнутых активации Т-клеточной популяции) клеток здорового донора, и/или препаратом тотальной РНК, полученным из регуляторных препаратов тотальной РНК, полученных из лимфоидных клеток или лимфоидных органов здорового донора, подвергнутого активации Т-клеточного звена иммунной системы. Донорские клетки согласно изобретению могут быть получены у любого представителя позвоночных.
Донорские клетки согласно изобретению могут быть получены у любого здорового млекопитающего, предпочтительно у крупного рогатого скота. Альтернативно, донорские клетки могут быть получены у любого представителя позвоночных, не являющегося млекопитающим. Донорские клетки согласно изобретению могут быть получены из одной или нескольких тканей человека. При этом человек-донор может быть мужчиной или женщиной любого возраста. Предпочтительно, чтобы для получения препаратов тотальной РНК согласно изобретению были использованы ткани и/или клетки, полученные у молодых здоровых доноров.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы включают в себя препарат тотальной РНК, представляющий собой любое сочетание препаратов тотальной РНК, выбранных из группы, включающей в себя регуляторный препарат (препараты) тотальной РНК, выделенный из лимфоидных клеток селезенки, тимуса, лимфатических узлов, лимфоцитов периферической крови, из костного мозга интактного здорового донора и/или здорового донора, подвергнутого активации Т-клеточного звена иммунной системы, в фазе проявления стимулирующей (хелперной) или супрессорной активности в отношении клеток того же самого или другого гистотипа.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы осуществляют модуляцию пролиферации и/или дифференцировки клеток млекопитающего для лечения или профилактики иммунодефицита. Примеры иммунодефицитных состояний включают в себя, но не ограничены иммунодефицитами, при которых имеют место признаки аутоиммунных процессов, такими как атаксия-телеангиэктазия, тимома, сцепленная с полом гипогаммаглобулинемия, иммунодефицита с гиперпродукцией IgM, дефицит IgA, синдромы Nezelof и Вискотта-Олдрича; атрофический гастрит, Myasthenia gravis. Pemphigus vulgaris (пузырчатка обыкновенная); энцефаломиелит; коллагенозы, системная красная волчанка, ревматоидный артрит, синдром Шегрена; язвенный колит; синдром Эванса; иммунный тиреоидит; сахарный диабет 1 и 2 типа; иммунная тромбоцитопения, болезнь холодовых агглютининов, холодовая гемоглобинурия; гипертиреоз; бесплодие, обусловленное нарушением иммунных механизмов; симпатическая офтальмия; хронический активный гепатит; коагулопатия, обусловленная нарушением синтеза антител; первичный билиарный цирроз; факогенный увеит; идиопатическая форма болезни Аддисона; поствакцинальный энцефалит; идиопатический гипопаратиреоз; узелковый периартериит; дермато- или полимиозит; склеродермия; рассеянный склероз и пр.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы осуществляют модуляцию пролиферации и/или дифференцировки клеток млекопитающего для лечения или профилактики гематологического заболевания или расстройства у указанного млекопитающего. Гематологические заболевания или расстройства включают в себя, но не ограничены анемиями любой этиологии (включая наследственные формы анемии), такими, например, как постгеморрагическая анемия, гемолитическая анемия, средиземноморская анемия (талассемия), гипо- и апластическая анемия, железодефицитная анемия, В12-дефицитная анемия, фолиеводефицитная анемия, анемия смешанного генеза, гемофилия и пр. В определенных аспектах настоящего изобретения описанные здесь композиции и способы осуществляют модуляцию пролиферации и/или дифференцировки клеток млекопитающего для лечения или профилактики анемии, заменяя используемые в настоящее время методы лечения (например, переливание крови). Препараты РНК и препараты тотальной РНК согласно изобретению способствуют увеличению количества эритроцитов и повышению уровней гемоглобина как у здоровых, так и у анемичных индивидов.
Несмотря на огромные успехи в области трансфузиологии, становится все более очевидным, что эффективное лечение гематологических расстройств и заболеваний требует регенерации кроветворной ткани самого реципиента. Лечебный эффект от гемотрансфузии ограничен временем жизни перелитых элементов крови. В отличие от этого, композиции и способы согласно изобретению, свободные от ограничений, связанных с технологией переливания крови, способны стимулировать регенерацию кроветворной ткани пациента в течение значительного периода.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы, включающие в себя препараты РНК, выделенные из лимфоидных клеток, костного мозга, из пуповины, пуповинной крови и/или из плаценты, могут быть использованы для лечения гематологических заболеваний и расстройств, характеризующихся нарушением пролиферативных процессов, нарушением дифференцировки клеток в костном мозге, повреждением клеточных или тканевых мембран или функциональными нарушениями клеток. Примеры гематологических заболеваний и расстройств включают в себя, но не ограничены острыми или хроническими постгеморрагическими анемиями, наследственными или приобретенными дизэритропоэтическими анемиями, анемиями, связанными с нарушением выработки эритропоэтина или появлением ингибиторов эритропоэтина, аутоиммунными анемиями и панцитопениями; гемолитическими анемиями, возникающими при спленомегалии, при отравлении кислотами или тяжелыми металлами; врожденными анемиями, связанными с нарушением синтеза цепей гемоглобина (серповидноклеточная анемия, талассемии); наследственными гемолитическими анемиями, связанными с нарушением мембраны эритроцитов (наследственный микросфероцитоз, наследственный эллиптоцитоз, наследственный стоматоцитоз, наследственный акантоцитоз, анемия, связанная со снижением количества полиненасыщенных жирных кислот мембраны); наследственными гемолитическими анемиями, связанными с нарушением активности ферментов эритроцитов; врожденными мегалобластными анемиями, связанных с нарушением синтеза ДНК и РНК (в том числе при синдроме Роджерса, сопровождающемся глухотой, сахарным диабетом и мегалобластной анемией); симптоматическими анемиями у больных миелофиброзом, хроническим лимфолейкозом, инфекционным мононуклеозом, гематосаркомой, хроническим гепатитом, тимомой, хроническим миелолейкозом, лимфогранулематозом, системной красной волчанкой; симптоматическими анемиями, связанными с угнетением пролиферации клеток костного мозга после воздействия токсических или лекарственных веществ, цитостатиков или ионизирующей радиации; и врожденными или приобретенными тромбоцитопатиями и острым геморрагическим васкулитом.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для лечения заболеваний, связанных с нарушением кровотока в микроциркуляторном русле. Примеры заболеваний, ассоциированных с нарушением кровотока в микроциркуляторном русле, включают в себя, но не ограничены облитерирующим артериитом, ишемической болезнью сердца, атеросклерозом, сахарным диабетом 1 типа и 2 типа, а также синдромом длительного сдавления и восстановлением мышечной ткани после длительной иммобилизации конечностей. Введение препаратов РНК, выделенных из селезенки интактных или анемизированных животных, существенно увеличивает кровенаполнение многих органов, включая печень, селезенку, поджелудочную железу, почки (см. пример 9).
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для лечения или профилактики лучевого поражения или лучевой болезни у млекопитающего. Кроме того, композиции и способы согласно изобретению могут быть использованы для лечения или профилактики побочных эффектов лучевой терапии, например, у больных раком.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для лечения или профилактики побочных эффектов химиотерапии.
Еще в одном из предпочтительных воплощений способом модуляции пролиферации и/или дифференцировки клеток млекопитающего, согласно изобретению, является способ профилактики или лечения химического поражения костного мозга у указанного млекопитающего.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для лечения или профилактики химического поражения костного мозга.
Расстройства, заболевания и/или состояния, ассоциированные с нарушением регуляции клеточной пролиферации и/или дифференцировки, включают в себя, но не ограничены аутоиммунными расстройствами и заболеваниями (например, аутоиммунная гемолитическая анемия, аутоиммунная тромбоцитопеническая пурпура, болезнь Грейвса (диффузный токсический зоб), синдром Гудпасчера (геморрагический легочно-почечный синдром, системный капиллярит, наследственный), тиреоидит Хашимото, рассеянный склероз).
Расстройства, заболевания и/или состояния, ассоциированные с нарушением регуляции клеточной пролиферации и/или дифференцировки, включают в себя, но не ограничены дегенеративными заболеваниями (например, болезнь Альцгеймера, болезнь Паркинсона, амилоидоз).
Расстройства, заболевания и/или состояния, ассоциированные с нарушением регуляции клеточной пролиферации и/или дифференцировки, включают в себя, но не ограничены гиперпролиферативными или опухолевыми заболеваниями (например, аденома предстательной железы и рак предстательной железы, доброкачественная и злокачественная опухоль молочной железы).
Расстройства, заболевания и/или состояния, ассоциированные с нарушением регуляции клеточной пролиферации и/или дифференцировки, включают в себя, но не ограничены нейро-эндокринными расстройствами (например, поликистоз яичников; нарушения кроветворения и болезни крови).
Расстройства, заболевания и/или состояния, ассоциированные с нарушением регуляции клеточной пролиферации и/или дифференцировки, включают в себя, но не ограничены наследственными заболеваниями и дефектами, ассоциированными с нарушениями регуляции пролиферации или дифференцировки клеток (например, остеопетроз; ДЦП; нарушения слуха. Типовые нарушения слуха могут быть охарактеризованы ухудшением слуха, нейро-сенсорной тугоухостью, возрастной тугоухостью и глухотой (включая врожденную глухоту).
Расстройства, заболевания и/или состояния, ассоциированные с нарушением регуляции клеточной пролиферации и/или дифференцировки, включают в себя, но не ограничены ранозаживлением, псориазом, эрозией шейки матки, пародонтозом, альвеолитом, гингивитом, атеросклерозом, доброкачественной опухолью, злокачественной опухолью, опухолью, резистентной к химиотерапии; состояниями, требующими усиленной регенерации, такими как переломы костей, ожоги, язвы, гипертрофические рубцы, разрывы связок, травмы мягких тканей и внутренних органов и приживление кожных лоскутов.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для лечения или профилактики избыточной клеточной пролиферации. Избыточная клеточная пролиферация необязательно может быть вызвана модуляцией клеточной дифференцировки. Композиции и способы согласно изобретению могут предотвращать или ингибировать метастазирование злокачественных клеток. Типичные состояния, характеризующиеся избыточной клеточной пролиферацией, включают в себя, но не ограничены злокачественной опухолью, доброкачественной опухолью и гиперпролиферативными расстройствами (например, образование рубцов, псориаз и атеросклероз).
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для модуляции клеточной пролиферации и/или дифференцировки при всевозможных видах доброкачественных и злокачественных опухолей, включая опухоли, резистентные к химиотерапии или лучевой терапии.
В одном из воплощений настоящего изобретения описанные здесь композиции и способы могут быть использованы для модуляции пролиферации и/или дифференцировки клеток путем восстановления нормальных и/или характерных для здорового состояния уровней клеточной пролиферации и/или дифференцировки у субъекта с нарушенной пролиферацией и/или дифференцировкой.
Настоящее изобретение связано с фармацевтической композицией для лечения расстройства, заболевания или состояния, ассоциированного с нарушением клеточной пролиферации и/или дифференцировки, в организме субъекта, в частности млекопитающего, при этом указанная композиция включает в себя эффективное количество любого из описанных здесь препаратов РНК или их вариантов и фармацевтически приемлемый носитель, разбавитель или наполнитель.
В настоящем изобретении предусмотрена фармацевтическая композиция для восстановления нормальной функции, которая в условиях патологии нарушена, например, нарушенной клеточной пролиферации и/или дифференцировки, при этом указанная композиция включает в себя эффективное количество любого из описанных здесь препаратов РНК или их вариантов и фармацевтически приемлемый носитель, разбавитель или наполнитель. Типичные патологические состояния включают в себя, но не ограничены дегенеративными состояниями, гиперпролиферативными состояниями (опухоли) и аутоиммунными состояниями.
В настоящем изобретении предусмотрена фармацевтическая композиция для лечения расстройства, заболевания или состояния, ассоциированного с нарушенной клеточной пролиферацией и/или дифференцировкой в организме субъекта, в частности, млекопитающего, при этом указанная фармацевтическая композиция включает в себя эффективное количество любого из описанных здесь препаратов РНК или их вариантов, полученных из лимфоидных клеток и/или костного мозга, и, необязательно, препарат тотальной РНК, выделенный из одной или более клеток другого гистотипа, и фармацевтически приемлемый носитель, разбавитель или наполнитель.
Композиции и способы лечения, предлагаемые в настоящем изобретении, могут быть использованы как в практической медицине, так и в ветеринарии, например, для лечения сельскохозяйственных животных, домашнего и рабочего скота, а также домашних животных-любимцев, включая собак, кошек, грызунов и птиц.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Рис. 1 - процесс реконструкции эритропоэза в культуре ЭО (1 МЕ/мл эритропоэтина, 24 часа культивирования). На фото - инволюцирующие ЭО, в «короне» одного из которых находятся 2 проэритробласта).
Рис. 2 - типичная картина развития ЭО в культуре (0,5 МЕ/мл эритропоэтина, 24 часа культивирования). На фото - 2 ЭО 3 класса зрелости, содержащие в своей «короне» эритроидные клетки разной степени зрелости.
Рис. 3 - процесс созревания эритроидных клеток в культуре (1,5 МЕ/мл эритропоэтина, 96 часов культивирования). Слева на фото - инволюцирующий ЭО, «корона» которого состоит только из ретикулоцитов.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Являясь важнейшей и филогенетически более древней функциональной частью иммунной системы, чем та, которая обеспечивает развитие гуморального иммунитета и образование антител, морфогенетическая функция лимфоцитов осуществляет регуляцию пролиферативных процессов в организме. В норме регуляция означает своевременную стимуляцию и своевременное торможение пролиферации клеток любой ткани, обеспечивая таким образом постоянство клеточной численности и анатомической целостности всех органов и тканей в процессе роста и в процессе физиологической и репаративной регенерации. Морфогенетическая функция лимфоидных клеток обеспечивается путем реализации двухстадийной (двухфазной) программы регуляции пролиферации и дифференцировки клеток их тканей-мишеней, являясь при этом постоянной составляющей также и иммунных реакций и обеспечивая пролиферацию иммунокомпетентных клеток как при гуморальном, так и при клеточном иммунитете.
Настоящее изобретение позволяет получать регуляторные тканеспецифические средства с заданным направленным действием, корригирующие, стимулирующие и ингибирующие процессы клеточного деления и дифференцировки при различных патологических состояниях, - средства, которые могут быть использованы в медицинской практике в области гематологии, трансфузиологии, хирургии, онкологии, радиологии, гинекологии, в терапии дегенеративных, аутоиммунных, гипо- и гиперпролиферативных расстройств, заболеваний и состояний, в частности, опухолевых заболеваний, а также целого ряда наследственных болезней и дефектов.
Средства согласно изобретению получают путем выделения из лимфоидных клеток селезенки, тимуса, лимфатических узлов, из костного мозга, из лимфоцитов периферической крови здоровых доноров, в частности, из донорской крови человека, из пуповинной крови, из клеток пуповины, из цельной пуповины и из плаценты фракции тотальной РНК, различный характер действия которой определяется источником ее получения и/или вариацией функционального состояния исходных лимфоидных клеток в норме и на разных стадиях проявления их морфогенетической функции.
Изобретение связано со способом модуляции клеточной пролиферации и/или дифференцировки у пациента путем введения пациенту препаратов (средств) согласно настоящему изобретению. При этом указанный способ модуляции может быть использован для лечения заболеваний, расстройств или нарушений, выбранных из группы, не ограниченной перечисленным и включающей в себя ревматоидный артрит, артрит Лайма, системную красную волчанку, болезнь Крона, язвенный колит, воспалительное заболевание кишечника, инсулинзависимый сахарный диабет, инсулинонезависимый сахарный диабет, тиреоидит, астму, аллергические заболевания, псориаз, дерматит, склеродермию, болезнь трансплантат против хозяина, отторжение трансплантата любого органа или любой ткани, саркоидоз, атеросклероз, диссеминированное внутрисосудистое свертывание, болезнь Кавасаки, болезнь Грейвса, нефротический синдром, синдром хронической усталости, гранулематоз Вегенера, пурпуру Геноха-Шенлейна, микроскопический васкулит почек, хронический активный гепатит, кахексию, синдром приобретенного иммунодефицита, хорею Гентингтона, болезнь Паркинсона, болезнь Альцгеймера, инсульт, гемолитическую анемию, злокачественные заболевания, сердечную недостаточность, инфаркт миокарда, алопецию, артропатию любой этиологии, артериосклероз, атопическую аллергию, аутоиммунное буллезное заболевание, пернициозную анемию, гепатиты всех типов, синдром приобретенного иммунодефицита, общий транзиторный иммунодефицит, кардиомиопатию, женское бесплодие, преждевременное угасание функции яичников, фибротическое заболевание легких, интерстициальную болезнь легких, болезнь Шегрена, фиброз, гипопаратиреоз, острое и хроническое иммунное заболевание, ассоциированное с трансплантацией органа, остеоартроз, идиопатическую лейкопению, аутоиммунную нейтропению, гломерулонефрит, болезнь Лайма, идиопатическое мужское бесплодие, рассеянный склероз, симпатическую офтальмию, синдром Гудпасчера, спондилит, болезнь Стилла, системный склероз, синдром Шегрена, болезнь Такаясу, тромбоцитопению разного генеза, аутоиммунное заболевание щитовидной железы, гипертиреоз, гипотиреоз разного генеза, факогенный увеит, первичный васкулит, витилиго, хроническое заболевание печени, цирроз печени, психические нарушения (например, депрессию и шизофрению), опухолевые заболевания, такие как рак легкого, молочной железы, желудка, мочевого пузыря, ободочной кишки, поджелудочной железы, рак ободочной кишки, яичников, предстательной железы и прямой кишки; аллергический ринит, отторжение аллотрансплантата и/или ксенотрансплантата, амиотрофический боковой склероз, анемию, стенокардию, артериальную гипертонию, В-клеточную лимфому, отторжение трансплантата костного мозга (ВМТ), лимфому Беркитта, кардиомиопатию, ассоциированные с химиотерапией нарушения, хронический миелоцитарный лейкоз (CML), хронический алкоголизм, хронические воспалительные процессы, хронический лимфоцитарный лейкоз (CLL), хроническую обструктивную болезнь легких (COPD), застойную сердечную недостаточность, конъюнктивит, контактный дерматит, болезнь коронарных артерий, болезнь Крейтцфельдта-Якоба, муковисцидоз, демиелинизирующие заболевания, дерматологические состояния, несахарный диабет, диабетическое артериосклеротическое заболевание, синдром Дауна в младшем и среднем возрасте, экзему, энцефаломиелит, эндокардит, эндокринопатию, экстрапирамидные и церебеллярные нарушения, атаксию Фридрейха, функциональные периферические артериальные нарушения, язву желудка, гломерулонефрит, тромболитическую тромбоцитопеническую пурпуру, кровотечение, болезнь Ходжкина, астению, воздействие ионизирующей радиации, инсульт, спинально-мышечную атрофию, саркому Капоши, проказу, жировой отек, лимфатический отек, злокачественную лимфому, злокачественный гистиоцитоз, злокачественную меланому, множественную миелому, синдром миелодисплазии, нефроз, нейродегенеративные заболевания, неходжкинскую лимфому, органомегалию, остеопороз, периферическое атеросклеротическое заболевание, периферические васкулярные нарушения, перитонит, пернициозную анемию, лучевую терапию, болезнь Рейно, саркомы, сенильную деменцию, Т-клеточный или FAB ALL (острый лимфобластный лейкоз), телеангиэктазию, облитерирующий тромбоангиит, варикозные вены, васкулит, венозные заболевания, венозный тромбоз, болезнь Вильсона, отторжение ксенотрансплантата любого органа или любой ткани.
Настоящее изобретение связано также с фармацевтической композицией для лечения расстройства, заболевания или состояния, ассоциированного с нарушением пролиферации и/или дифференцировки клеток, включающей в себя эффективное количество любой из разновидностей регуляторных препаратов тотальной РНК согласно изобретению или любого их сочетания, полученных из лимфоидных клеток селезенки, тимуса, лимфатических узлов, из лимфоцитов периферической крови или из костного мозга здорового интактного донора или здорового донора, подвергнутого активации Т-клеточного звена иммунной системы, на стадии проявления ими стимулирующей (хелперной) или тормозящей (супрессорной) активности в отношении соматических клеток-мишеней того или иного гистотипа, и, необязательно, препарат(препараты) тотальной РНК, выделенной из пуповинной крови, из пупочного канатика и/или из указанных соматических клеток-мишеней, необязательно вместе с фармацевтически приемлемыми носителем, разбавителем или наполнителем.
Описанный здесь способ модуляции клеточной пролиферации и/или дифференцировки для лечения заболеваний, расстройств или нарушений может быть осуществлен путем введения субъекту композиции согласно изобретению по меньшей мере одним из способов, выбранных из парентерального, подкожного, внутримышечного, внутривенного, внутрисуставного, внутрибронхиального, внутрибрюшинного, интракапсулярного, внутрихрящевого, внутриполостного, интрацеребеллярного, интрацеребровентрикулярного введений, введения внутрь толстой/ободочной кишки, интрацервикального, внутрижелудочного, внутрипеченочного, интрамиокардиального, внутрикостного, внутритазового, интраперикардиального, внутрибрюшинного, внутриплеврального, внутрипростатического, внутрилегочного, интраректального, интраренального, интраретинального, интраспинального, интрасиновиального, внутригрудного, внутриматочного, вагинального, ректального, буккального, сублингвального, интраназального (например, ингаляционного) и трансдермального (например, местного), глазного, внутриглазного, ушного способов введения.
Следует отметить, что интраназальный путь введения в ряде случаев является особенно предпочтительным.
В одном из воплощений изобретения композицию составляют, в соответствии с общеизвестным процедурами, как фармацевтическую композицию, адаптированную для внутривенного, подкожного, внутримышечного, интраназального или местного введения человеку или животному. Обычно композиции для введения путем инъекции или инфузии представляют собой растворы в стерильном изотоническом водном буфере. Если необходимо (например, когда композиция представлена в лиофилизированной форме), к композиции может прилагаться также солюбилизирующий агент. В определенных случаях препараты РНК для парентерального введения требуют стерилизации. Стерильность легко достигается путем фильтрации через стерильные фильтрационные мембраны, например, до или после лиофилизации и восстановления. Парентеральный путь введения включает в себя известные способы, например, инъекцию или инфузию путем внутривенного, внутрибрюшинного, внутримышечного, внутриартериального, подкожного или внутриочагового введения. Способы получения фармацевтических композиций для парентерального, интраназального и внутриочагового введения, композиций в виде глазных и ушных капель хорошо известны в данной области и более подробно описаны в различных источниках, включая, например. Руководство Remington's Pharmaceutical Sciences and Introduction to Pharmaceutical Dosage Forms, 19 th ed., Mack Pub. Co., Easton, Pa. (1995).
В случае, если композиции согласно изобретению предназначены для местного введения, они могут быть составлены в виде мази, крема, трансдермального пластыря, лосьона, геля, шампуня, спрея, аэрозоля, раствора, эмульсии или других форм, известных специалистам в данной области (там же).
Приведенный в настоящей заявке разнообразный экспериментальный материал убедительно доказывает, что полученные препараты тотальной РНК воспроизводят определенные функциональные свойства исходных лимфоидных клеток и эффективно влияют на ход морфогенетических процессов, меняя их в нужном направлении; более того, введением препаратов согласно изобретению можно заменить перенос лимфоидных клеток, поскольку указанные препараты способны осуществлять коррекцию нарушенных функций соматических клеток любого гистотипа на основе естественных механизмов регуляции.
Изменчивость функциональных проявлений тотальной РНК под влиянием различных факторов, действующих на клетку, из которой она получена, дает основания предполагать, что ее использование в лечебной практике может найти очень широкое применение.
Как показали наши многолетние исследования, морфогенетическая функция лимфоидных клеток имеет ряд особенностей и закономерностей своего действия. В частности, морфогенетической функции лимфоидных клеток свойственна преимущественная органоспецифичность. Это означает, что при адоптивном переносе лимфоидных клеток они в организме реципиента обязательно влияют на пролиферацию клеток органа, гомологичного органу донора, подвергшемуся тому или иному повреждающему воздействию (например, оперативному вмешательству) или любому другому воздействию, активирующему Т-клеточное звено иммунной системы. При этом следует отметить, что реакция пролиферативной активности лимфоидных клеток реципиента всегда развивается в направлении, соответствующем перенесенному сигналу.
В действии активированных регенерационным процессом лимфоидных клеток, как и в самом регенерационном процессе, отмечается двухфазность. Сначала действуют лимфоциты, обладающие свойством стимулировать пролиферацию клеток органа-мишени, а затем, на высоте пролиферации, появляются лимфоциты, обладающие свойством тормозить клеточное деление в указанном органе. Эти лимфоциты не препятствуют завершению митотического цикла в клетках, вступивших в него, но препятствуют вступлению в цикл деления новых клеток. Тем самым они способствуют завершению пролиферативной волны и останавливают восстановительный процесс, предотвращая гиперрегенерацию. Таким образом, лимфоциты обеспечивают и начало, и завершение регенерационного процесса.
Функцию стимуляции и функцию ингибирования, как было нами показано, осуществляют разные популяции Т-лимфоцитов со свойствами Т-хелперов или Т-супрессоров. Однако было замечено, что в действии Т-супрессоров органоспецифичность выражена в меньшей степени, чем в действии Т-стимуляторов. Материалы по лимфоидной регуляции морфогенетических процессов обобщены в ряде монографий [Бабаева А.Г. Иммунологические механизмы регуляции восстановительных процессов. М., 1972, 150 с.; Бабаева А.Г. Регенерация и система иммуногенеза. М., 1985, 256 с.; Бабаева А.Г., Геворкян Н.М., Зотиков Е.А. Роль лимфоцитов в оперативном изменении программы развития тканей. М., Изд-во РАМН, 2009, 108 с.]. Здесь уместно подчеркнуть, что активированные лимфоциты со стимулирующими способностями индуцируют у реципиента модус ускоренной пролиферации, характерной для регенерационных процессов. Однако все описанные процессы реализуются лишь в сингенной системе. В аллогенной системе этот мощный собственный регулятор пролиферативных и восстановительных процессов до сих пор не мог быть использован из-за антигенного ограничения, связанного с иммунологической несовместимостью, и поэтому не получил применения в практической медицине.
В то же время успехи хирургии, развитие трансплантологии требовало решения проблемы несовместимости, что явилось мощным толчком к поиску средств для ее преодоления. Особенно остро эта проблема встала перед гематологами и трансфузиологами из-за возросшей потребности в трансплантации костного мозга и многократных переливаниях крови и все еще существующих серьезных проблем для беспрепятственного осуществления указанных процедур.
Вместе с тем до настоящего времени переливание крови, ее форменных элементов и других компонентов является одним из самых частых и востребованных инвазивных вмешательств в медицинской практике. На его счету миллионы спасенных жизней и столько же часов кропотливого упорного труда не одного поколения врачей-исследователей, чтобы сделать эту простую в техническом отношении процедуру безопасной для жизни пациента.
Но при колоссальных достижениях гемотрансфузии проблему ее безопасности нельзя считать окончательно решенной, что определяется как свойствами переливаемой ткани (кровь), так и особенностями реципиента - человека как представителя биологического вида. Причиной смертельной опасности является иммунологический конфликт при антигенной несовместимости донора и реципиента. Поэтому вся история развития трансфузиологии связана с необходимостью подбора совместимого донора, что и является краеугольной проблемой этой процедуры. Кровь человека является самой иммуногенной тканью для всех млекопитающих и особенно для человека. Форменные элементы крови являются носителями огромного числа антигенов: эритроциты - антигенов системы АВО, нескольких разновидностей резус-фактора и редких эритроцитарных антигенов М, N и некоторых других; на тромбоцитах обнаружено до 40 тромбоцитарных антигенов; и, наконец, лимфоциты являются носителями более сотни антигенов системы HLA (Human Leucocyte Antigens) - группы антигенов, определяемых главным комплексом гистосовместимости (МНС) у человека, число которых продолжает пополняться. В результате найти среди людей, не связанных родственными отношениями, совместимого донора (что определяется числом сочетаний разных антигенов) - задача весьма нелегкая (обычно это требует поиска среди тысяч вариантов донорской крови).
Для решения этой проблемы созданы банки типированных доноров. В настоящее время в целом можно считать решенной проблемой только подбор доноров по антигенам системы АВО и резус-фактору. Что же касается редких эритроцитарных антигенов, то сенсибилизация ими реципиентов не исключена. При первичном переливании крови такая несовместимость не проявляет себя, но повторное переливание, также, например, как последующая такая же по специфичности несовместимая беременность, приведут к иммунологическому конфликту, к развитию изоиммунной анемии и т.д. Такую же природу могут иметь и другие цитопении: тромбоцитопения, нейтропения, лимфоцитопения, иммунологическая тромбоцитарная рефрактерность.
В борьбе с этими явлениями, кроме тщательного подбора донора, пришли к практике перехода от переливания цельной крови к переливанию ее отдельных компонентов (эритроцитарной, тромбоцитарной и лейкоцитарной массы), что, безусловно, уменьшает возможность дополнительной сенсибилизации. Кроме того, при заведомо необходимых многократных гемотрансфузиях, так как перелитые форменные элементы имеют определенный срок жизни, перешли к практике пересадки костного мозга, более долговременного поставщика необходимой крови, с одной стороны, а с другой стороны, стали добиваться более быстрой регенерации кроветворной ткани самого реципиента, используя самые разнообразные стимуляторы регуляторных систем нормального и репаративного гемопоэза.
Среди систем регуляции восстановительных процессов лимфоидная регуляция занимает особое место.
Система иммуногенеза в целом контролирует пролиферацию большинства клеток организма, обеспечивая в норме, как уже было сказано, постоянство клеточной численности и анатомической целостности органов и тканей. Наличие такой морфогенетической функции было теоретически обосновано и экспериментально доказано в конце 60-х годов на модели адоптивного переноса лимфоцитами регенерационной информации от животных с частичной гепатэктомией сингенным неоперированным реципиентам [Бабаева А.Г. Иммунологические реакции в процессах нормального и восстановительного роста. В кн.: Регенерация и клеточное деление. М., 1968, с. 11-16]. Закономерный характер этого явления был подтвержден в опытах на ряде органов (почка, кишечник, кроветворная ткань, легкие, кожа) как самими авторами первой публикации, так и другими исследователями. Одной из самых важных особенностей этой системы регуляции является адресная направленность ее действия, ее преимущественная (хоть и не абсолютная) органо- и тканеспецифичность. Это означает, что лимфоидная регуляция реализует свои морфогенетические свойства главным образом в отношении тех органов и тканей, которые подвергаются воздействию патогенного фактора или оперативному вмешательству. Система иммуногенеза располагает возможностями идеальной естественной системы регуляции. Ее клетки и вырабатываемые ими биологически активные вещества (лимфокины) инициируют процессы деления клеток (Т-лимфоциты со свойствами Т-хелперов (Т-эффекторы)), останавливают их (Т-лимфоциты со свойствами Т-супрессоров (Т-регуляторы)) и уничтожают измененные клетки, например, изменившиеся под действием патогенных факторов (Т-киллеры). Вырабатываемые лимфоидными клетками факторы способствуют взаимодействию лимфоцитов друг с другом и с другими лимфоидными клетками и клетками органов-мишеней. При чрезвычайных обстоятельствах, но в естественных условиях (например, в процессе регенерации в ответ на повреждение), функция соответствующих популяций усиливается, что в результате приводит к успешному завершению процесса регенерации. В то же время в ситуациях, не имеющих место в природе (например, при трансплантации органа или адоптивном переносе чужеродных клеток), система иммуногенеза осуществляет защиту от вторжения в организм чужеродного начала.
Аллотрансплантация тканей, в том числе и костного мозга, а также гемотрансфузия относятся к числу воздействий, не встречающихся в природных условиях и отвергаемых иммунной системой, которая распознает чужеродность по многочисленным вариантам сочетаний антигенных различий (видовых и индивидуальных).
Чтобы обойти указанную проблему, в настоящее время, помимо подбора совместимых доноров и реципиентов, обычно у реципиента вызывают состояние иммуносупрессии, с тем, чтобы его иммунная система не в состоянии была отторгнуть чужеродную ткань. Однако иммуносупрессия сама по себе чревата целым рядом хорошо известных многочисленных сопутствующих ей нежелательных явлений.
В поисках решения этой проблемы авторы настоящего изобретения нашли более эффективный путь трансплантации или адоптивного переноса, используя вместо клеток такие их компоненты, которые сохраняли бы определенную функциональную активность клеток, но не имели бы присущих клеткам антигенных свойств. Таким компонентом оказался препарат тотальной РНК, полученный из клетки. По данным литературы, РНК практически не имеет не только индивидуальных, но и видовых антигенных детерминант. К примеру, ксеногенная дрожжевая РНК хорошо переносится млекопитающими, в том числе и человеком, и применяется в лечебной практике при ряде заболеваний - например, при глазных болезнях, при болезни Шегрена, дегенеративных заболеваниях нервно-мышечной системы, наследственных формах миопатий, последствиях нейроинфекций, спинальных амиотрофиях - в виде препарата для перорального введения и препарата для внутримышечных инъекций [Шабанова М.Е., Казаньев В.В., Баурина М.М., Красноштанова А.А., Крылов И.А. Способ усиления пролиферативной активности костного мозга. В сб.: «Нейроиммунопатология» (Тезисы докладов Четвертой российской конференции). Патогенез, 2006, №1, С. 71]. М.Е. Шабанова с соавт. (2006) в экспериментах на крысах установили, что костный мозг животных, которым вводили дрожжевую РНК, при его введении облученным крысам-реципиентам, в 2 раза увеличивает число селезеночных колоний (соответственно, эритроцитарного, гранулоцитарного и мегакариоцитарного рядов). Другие публикации, посвященные влиянию РНК на организм, в основном касаются усиления функциональных показателей и иммунологической функции: воздействия на реактивность и резистентность к инфекционным процессам, на иммунитет и повышение функциональной активности макрофагов и клеток лимфоцитарного ряда. И хотя в целом считается, что благотворное влияние РНК связано в основном с ее трофической функцией, на самом деле ее возможности значительно шире, так как ряд данных указывает на то, что в опытах с адоптивным переносом она способна передавать морфофункциональные особенности лимфоидных клеток, из которых она получена, вызванные изменениями внешней среды или повреждающими факторами.
Мы сосредоточили свое внимание на выяснении того, может ли препарат тотальной РНК лимфоидных клеток осуществлять присущую этим клеткам специфическую регуляторную функцию, то есть выступать в роли регулятора процессов пролиферации, заменяя функцию самих лимфоцитов, и сохраняется ли при этом свойственная лимфоцитам преимущественная органоспецифичность, например, у анемизированных животных. С этой целью мы решили проверить, влияет ли препарат тотальной РНК на эритропоэз и в целом на гемопоэз у реципиентов регуляторных РНК. Выяснить указанные вопросы нам удалось с использованием анемизированных животных и других in vitro и in vivo моделей изучения эритропоэза и гемопоэза в целом.
Для решения этого вопроса мы использовали несколько моделей, выявляющих регуляторные свойства лимфоидных клеток как в клеточной культуре, так и in vivo. Ранее в опытах на животных на модели с адоптивным переносом лимфоцитов нами было показано, что регенерационный процесс всегда сопровождается фазовыми изменениями морфогенетических свойств лимфоцитов, выражающимися в том, что способность этих клеток стимулировать клеточное деление сменяется способностью его угнетать и тем самым препятствовать возможности избыточного роста регенерирующего органа. В связи с этим в число первоочередных задач входило прежде всего выяснение вопроса о том, имеются ли сходные фазовые изменения функциональных свойств РНК, к примеру, при регенерации крови, то есть то, сохраняет ли препарат тотальной РНК способность лимфоидных клеток, из которых она получена, индуцировать модус ускоренной пролиферации и дифференцировки клеток.
В качестве объекта для выяснения функциональных свойств полученных нами препаратов тотальной РНК лимфоидных клеток мы выбрали прежде всего кровь, с тем, чтобы на примере гемопоэза изучить влияние указанных препаратов РНК, полученных в нормальных условиях, то есть из интактных лимфоидных клеток (препарат РНК-1), и в условиях регенерации кроветворной ткани - как в фазе проявления стимулирующих кроветворение свойств лимфоидных клеток (препарат РНК-2), так и в фазе проявления их ингибирующей активности в отношении кроветворения (препарат РНК-3). Степень выраженности активности препаратов РНК определяли также при разных функциональных состояниях кроветворной ткани реципиента: a) в условиях физиологического гемопоэза; b) в условиях усиленного кроветворения; и c) в условиях острого и хронического угнетения кроветворения.
Весь комплекс проведенных нами экспериментов без исключения однозначно показал, что препарат тотальной РНК, выделенной из лимфоидных клеток, обладает теми из указанных выше морфогенетических свойств, которые присущи и самим исходным лимфоидным клеткам.
Для того чтобы настоящее изобретение легче было понять и внедрить в практику, далее с помощью примеров будут описаны некоторые из предпочтительных воплощений, со ссылкой на сопроводительные материалы.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проводили на молодых половозрелых белых беспородных крысах обоего пола и на культурах эритробластических островков (ЭО) костного мозга (КМ). В отдельных опытах была использована линия мышей C57BL/RsJYLeprdb/+ с выраженной формой сахарного диабета 2 типа (питомник Светлые Горы). Организация работы соответствует международным этическим нормам, согласно Конвенции Европейского Союза и Европейской Конвенции о защите позвоночных животных, используемых для экспериментальных или иных научных целей (Страсбург, 1986), в международных правилах Good Laboratory Practice for Nonclinical Laboratory Studies от 04.03.2002 г.
Использованных в работе крыс массой 180-220 г размещали в стандартных пластиковых клетках и содержали на стандартном рационе вивария при свободном доступе к воде в помещении с температурой воздуха 18-25°C.
Активация лимфоидных клеток для получения из них препаратов тотальной РНК осуществлялась путем кровопускания в объеме 2% от массы тела животных. Для получения лимфоидных клеток селезенки, обладающих стимулирующим свойством, крыс умерщвляли (эфирным наркозом) через 17 час. после кровопускания. А для получения лимфоидных клеток селезенки с ингибирующим свойством - через 96 часов после потери крови. Такие конкретно сроки (то есть именно 17 часов и 96 часов после кровопускания), иначе называемые «донорским интервалом», в довольно широких пределах стимулирующего и тормозящего интервалов, в рамках которых была доказана ранее высокая экспрессия указанных активностей, были выбраны исключительно из соображений удобства планирования экспериментов. На самом деле эффективные периоды стимулирующей и ингибирующей активностей достаточно продолжительные (приблизительно от 15 минут до 48 часов составляет длительность периода стимуляции и приблизительно от 48 часов до 96 и более часов - периода торможения). Их продолжительность зависит от вида и возраста животного, масштаба оперативного вмешательства или повреждающего воздействия, а также от конкретного органа- или ткани-мишени, подвергшегося указанному воздействию [Бабаева А.Г. Регенерация и система иммуногенеза. М., Медицина, 1985, 256 с.]. У крыс и мышей эти периоды практически одинаковы.
Источником для получения регуляторной тотальной РНК служили лимфоидные клетки селезенки, тимуса, нефракционированный костный мозг молодых крыс обоего пола весом 80-130 г, лимфоциты периферической крови человека (донорская кровь); кроме того, препараты тотальной РНК были получены из пуповинной крови, пупочного канатика и плаценты человека и крысы.
С использованием методики определения количества РНК и ДНК в полученных нами препаратах [Трудолюбова М.Г. Количественное определение РНК и ДНК в субклеточных фракциях клеток животных (модификация метода Шмидта-Тангаузера). В сб.: Современные методы в биохимии. Под ред. академика АМН СССР В.Н. Ореховича. М., Медицина, 1977, с. 313-316] было показано, что содержание ДНК в них равно нулю. В них также не содержится белок, что было показано микробиуретовым методом.
Тотальную РНК выделяли по стандартной методике с использованием тризольного реагента и фенол-хлороформной экстракции [Chomczynski, P.A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. BioTechniques, 1993, vol. 15. pp. 532-537].
Клетки, выделенные из разных органов, не смешивали, и определяли их функциональные свойства по отдельности.
Препараты тотальной РНК получены из разных лимфоидных органов интактных животных, а также из лимфоидных органов анемизированных животных на разных сроках после кровопотери. При этом сохранение функциональной активности полученных препаратов при введении их в организм реципиентов дает основания считать, что получено три принципиальных разновидности препарата: РНК-1 (от интактных животных, т.е. из неактивированных лимфоидных клеток), РНК-2 со стимулирующими свойствами и РНК-3 с ингибирующими свойствами. Активность указанных препаратов изучали в культуре in vitro и в опытах на животных in vivo. Следует также отметить, что препарат тотальной РНК, полученный из лимфоидных клеток на стадии проявления ими хелперной или супрессорной активности, является препаратом, обладающим, соответственно, хелперной или супрессорной активностью в отношении клеток того же самого и/или другого гистотипа как в организме реципиента, так и в культуре соответствующих клеток-мишеней in vitro.
С использованием электрофореза в 1,5% агарозе было обнаружено, в частности, что регуляторные препараты тотальной РНК, выделенные из лимфоидных клеток того или иного лимфоидного органа и в той или иной фазе процесса морфогенеза (регенерации), могут содержать молекулы РНК размером приблизительно от 50 примерно до 50000 и более нуклеотидов, например, приблизительно от 50 примерно до 400 и более нуклеотидов, приблизительно от 50 примерно до 3000 и более нуклеотидов, приблизительно от 50 примерно до 10000 и более нуклеотидов. Таким образом, заявленные регуляторные свойства могут быть связаны с молекулами РНК, имеющими молекулярную массу в интервале приблизительно от 15 кДа примерно до 18000 кДа и выше, или насчитывающими приблизительно от 50 нуклеотидов примерно до 50000 нуклеотидов и более, например, приблизительно 136000 нуклеотидов (в частности, например, 135639 нуклеотидов), и даже более высокомолекулярные образцы РНК.
Препараты согласно изобретению для культуральных исследований добавляли из расчета 2 мкг/мл культуральной среды или 4 мкг/мл культуральной среды. В опытах in vivo препараты вводили животным парентерально в дозе от 5 до 45 мкг на 100 г веса животного. Следует отметить, что такая доза в десятки и сотни раз меньше, чем доза, используемая другими авторами [Шабанова М.Е., Казаньев В.В., Баурина М.М., Красноштанова А.А., Крылов И.А. Способ усиления пролиферативной активности костного мозга. В сб.: «Нейроиммунопатология» (Тезисы докладов Четвертой российской конференции). Патогенез, 2006, №1, С. 71; Патент РФ 2238756 (2003)].
Эритропоэтическую активность препаратов определяли на культуре эритробластических островков (ЭО) костного мозга (КМ) крыс с использованием модели in vitro физиологического эритропоэза [Тишевская Н.В., Захаров Ю.М., Тишевской И.А. Влияние эритропоэтина в различных концентрациях на культуру эритробластических островков. Российский физиологический журнал им. И.М. Сеченова, 1998, Т. 84, №12, с. 1412-1419]. Авторы указанной модели, отрабатывая метод поддержания эритропоэза в культуре ЭО КМ на уровне физиологического, показали, что физиологический уровень гемопоэза достигается в культуре при добавлении 0,5 МЕ/мл эритропоэтина. Это означает, что если сравнивать качественный состав эритробластических островков, то у интактных животных in vivo и в культуре ЭО КМ с 0,5 МЕ/мл эритропоэтина он будет одинаковым.
В полученных нами культурах ЭО определяли общее количество островков на 1 см2 поверхности культурального сосуда и их распределение по классам зрелости, пользуясь классификацией, предложенной Ю.М. Захаровым и соавт. (1990), учитывая число и стадию дифференцировки эритроидных клеток, входящих в «корону», окружающую центральный макрофаг. В соответствии с классификацией Ю.М. Захарова с соавт. [Захаров Ю.М., Мельников И.Ю., Рассохин А.Г. Классификация эритробластических островков костного мозга с учетом изменения их клеточного состава. Арх. анат., гистол. и эмбриол., 1990, №5, с. 38-42], различают следующие классы эритробластических островков: 1) к ЭО 1 класса зрелости относят островки, «корона» которых представлена проэритробластами, эритробластами и базофильными нормобластами в количестве от 2 до 8 клеток; 2) ЭО 2 класса содержат в «короне» базофильные и ранние полихроматофильные нормобласты в количестве от 9 до 16 клеток; 3) ЭО 3 класса содержат в «короне» средние и поздние полихроматофильные нормобласты, оксифильные нормобласты и ретикулоциты в количестве 17-32 клеток; 4) ЭО инволюцирующие содержат в «короне» поздние полихроматофильные, оксифильные нормобласты и ретикулоциты с числом ядросодержащих клеток в количестве менее 16; 5) ЭО реконструирующиеся представляют собой инволюцирующие ЭО с присоединившими к их ʺкоронеʺ молодыми способными к делению эритроидными клетками - проэритробластами, эритробластами и/или базофильными нормобластами (как в ЭО 1-го и 2-го классов) (рис. 1-3).
Для оценки интенсивности формирования ЭО и развития их эритроидной «короны» пользовались расчетными показателями [Воргова Л.В., Захаров Ю.М. Об изменении эритробластических островков костного мозга у животных при сочетании тепловых и мышечных нагрузок. Физиол. журнал СССР им. И.М. Сеченова, 1990, Т. 76, №2, с. 200-206].
1) «показатель интенсивности вовлечения КОЕ(э) в дифференцировку» (где КОЕ(э) - колониеобразующая единица эритроидного ряда):
ЭО 1-го класса + ЭО рек.
2) «показатель созревания эритроидных клеток в ЭО» - отношение суммы ЭО со зрелыми клетками в «короне» к сумме ЭО пролиферирующих классов:
ЭО 3-го класса + ЭО инв./ЭО 1-го класса + ЭО 2-го класса + ЭО рек.
3) «показатель повторного вовлечения макрофагов в эритропоэз»:
ЭО рек./ЭО инв.
Показателем стимуляции эритропоэза служило достоверное увеличение числа ЭО 1, 2 классов и ЭО реконструирующихся - и уменьшение числа инволюцирующих ЭО.
Показателем угнетения эритропоэза служило, наоборот, уменьшение числа ЭО 1 и 2 классов, реконструирующихся ЭО - и увеличение числа инволюцирующих ЭО.
Испытываемые препараты добавляли в среду культивирования в чашках Петри с одинаковым количеством ЭО (1500 ЭО на 3 мл культуральной среды на чашку).
Для изучения активности препаратов РНК-1, РНК-2 и РНК-3 в отношении эритропоэза было поставлено несколько серий опытов.
Для получения препаратов РНК прежде всего были использованы лимфоидные клетки селезенки, поскольку первоначально морфогенетическая функция лимфоцитов была обнаружена в опытах с адоптивным переносом именно лимфоцитов селезенки [Бабаева А.Г. Иммунологические механизмы регуляции восстановительных процессов. М., 1972, 150 с.; Бабаева А.Г. Регенерация и система иммуногенеза. М., 1985, 256 с.]. В соответствии с поставленной задачей, сначала было получено три препарата РНК: РНКс-1 - из лимфоидных клеток селезенки интактных крыс; РНКс-2 - из лимфоидных клеток селезенки крыс через 17 часов после кровопотери (кровопускание в объеме 2% от массы тела животного), обладающих стимулирующим эритропоэз действием (этот срок наиболее полно охарактеризован нами в предыдущих исследованиях); РНКс-3 - из лимфоидных клеток селезенки крыс через 96 часов после кровопотери (2% от массы тела), обладающих ингибирующим действием в отношении эритропоэза. Позже тем же способом и на тех же стадиях регенерационного процесса нами были получены аналогичные варианты препаратов тотальной РНК из костного мозга (КМ) и тимуса анемизированных крыс, (РНКкм-2), (РНКкм-3) и (РНКт-2). (РНКт-3), соответственно, а также препараты тотальной РНК из КМ и тимуса интактных животных (РНКкм-1) и (РНКт-1). Таким же образом могут быть получены и использованы варианты препаратов тотальной РНК из лимфатических узлов интактных животных и животных, претерпевающих процесс регенерации. Наконец, точно так же можно получать препараты тотальной РНК из любых клеток организма, включая стволовые клетки. Согласно нашим данным, препараты тотальной РНК воспроизводят (способствуют восполнению) в организме реципиента функциональные свойства тех клеток, из которых они получены.
В первой серии экспериментов изучали влияние препаратов согласно изобретению на физиологический эритропоэз in vitro, то есть на изменение уровня эритропоэза под влиянием указанных препаратов в культурах ЭО КМ, полученных от интактных крыс.
Культивирование эритробластических островков костного мозга крысы производили в газопроточном мультигазовом инкубаторе (SANYO, Япония) с системой самодеконтаминации и автоматической регулировкой скорости подачи CO2. Относительная влажность атмосферы в инкубаторе поддерживалась на уровне 95%. Погрешность поддержания заданной температуры при 37°C составляла ±0,15°C, градиент температуры в рабочем объеме камеры колебался на уровне ±0,3°C.
Все манипуляции по приготовлению компонентов культуральной среды, выделению и суспендированию ЭО, заполнению чашек Петри с адгезировавшимися островками средой, подготовленной для культивирования, производили в стерильном ламинарном шкафу СШЛ-0.5/130 (ЗАО «Асептические медицинские системы», г. Миасс) с вертикальным нисходящим малотурбулентным потоком воздуха. Степень очистки подаваемого внутрь воздуха от взвешенных частиц размером более 0,5 мкм составляла 99,95%.
Для текущего контроля жизнеспособности культуры с помощью фазово-контрастной микроскопии использовали инвертированный микроскоп Биолам П-1 с объективом 10×0,22 и бинокулярной насадкой АУ-12 с увеличением 1,5. Исследование клеточного состава культур после их фиксации и окраски осуществляли с помощью бинокулярного лабораторного микроскопа TS-136 («Tenso», Германия).
Эритробластические островки культивировали в отдельных стерильных пластиковых чашках Петри диаметром 35 мм (производитель «Coming-Costar»). В качестве основной составляющей среды для культивирования ЭО использовали среду RPMI-1640. Отдельно в среду добавляли L-глутамин в количестве 146 мг/л и 7,5% раствор бикарбоната натрия из расчета 26,7 мл/л [Гольдберг Е.Д., Дыгай A.M., Шахов В.П. Методы культуры тканей в гематологии. Томск, 1992, 272 с.]. В качестве антиоксиданта и восстановителя сульфгидрильных групп использовали β-меркаптоэтанол. Чтобы обеспечить развивающиеся эритроидные клетки белками, липидами, ферментами и ростовыми факторами, культуральную среду готовили с эмбриональной телячьей сывороткой, тестированной на цитотоксичность и отсутствие микоплазмы (отборная сыворотка совместного производства Германии и Франции, показавшая наилучший рост культур при тестировании в компании «ПанЭко»). Непосредственно перед началом эксперимента в культуральную среду добавляли разведенную средой RPMI-1640 смесь антибиотиков, проявляющих токсичность в отношении грамположительных и грамотрицательных бактерий, - калиевую соль бензилпенициллина (C16H17N2O4SK., М.м. 372,5 Да) в количестве 50 ЕД/мл (токсическая доза для кроветворных клеток составляет 10000 ЕД/мл) и стрептомицин C21H39N7O12×1.5H2SO4, М.м. 728,7 Да) в количестве 0,05 мг/мл (угнетение синтеза нуклеиновых кислот в клетках наблюдается при превышении дозы 1 мг/мл). Для стимуляции функциональной активности центральных макрофагов ЭО и создания своеобразного «депо» цитокинов вокруг развивающихся клеток в культуральную среду добавляли гепарин, повышающий адгезивную способность клеток в культуре и активирующий процессы пролиферации эритроидных, миелоидных и моноцитарных клеток [Luikart S.D., Sackrison J.L., Manglia C.A. Bone marrow matrix modulation of HL-60 phenotype. Blood, 1987, vol. 70, pp. 1119-1126; Luikart S.D., Manglia L.T., Furch J.B. A heparan sulfate fraction of bone marrow induces maturation of HL60 cells in vitro. Cancer Res., 1990, vol. 50, pp. 3781-3791; Юшков Б.Г., Попов Г.К., Северин М.В. и др. Гликозаминокгликаны и эритропоэз. Екатеринбург, Уральское отд. РАН, 1994, 127 с.].
Полный состав готовой культуральной среды (100 мл):
- Среда RPMI-1640 - 62 мл
- Эмбриональная телячья сыворотка - 30 мл
- Гепарин - 1,3 мл (6500 ЕД)
- Бензилпенициллина калиевая соль - 1 мл (5000 ЕД)
- Стрептомицин - 1 мл (5 мг)
- 2-меркаптоэтанол - 1 мл маточного раствора
- L-глутамин - 1 мл (14,6 мг)
- Бикарбонат натрия - 2,7 мл 7,5% раствора
Компоненты культуральной среды смешивали в стерильных условиях непосредственно перед началом эксперимента, и подготовленную среду фильтровали через ацетатный фильтр с диаметром пор 0,22 мкм.
Для успешного роста культур эритробластических островков в культуральную среду добавляли рекомбинантный эритропоэтин человека «Рекормон» («Boehringer Mannheim GmbH», Германия) в дозе 0,5 ME на 1 мл готовой культуральной среды.
Выделение ЭО из костного мозга бедренных костей производили по методике, предложенной Ю.М.Захаровым с соавт. [Захаров Ю.М., Рассохин А.Г., Мельников И.Ю. Исследование эритропоэза модифицированным методом выделения эритробластических островков костного мозга. Гематология и трансфузиология, 1984, Т. 29, №4, с. 52-54] и являющейся модификацией методических приемов, впервые разработанных в 1975 году Y.Le Charpentier и M. Prenant [Charpentier Y., Prenant M. Isolement de l'ilots erythroblastique. Etude en microscopie optique et electronique a balayage. Nouv. Rev. Franc. Hemat., 1975, vol. 15, pp. 119-140]. Костный мозг получали в результате промывания канала бедренной кости 1,5 мл препаративной среды, аналогичной по составу среде для культивирования, но без 2-меркаптоэтанола и эритропоэтина. Полученную суспензию ЭО и отдельных клеток костного мозга размещали с помощью дозатора на поверхностях чашек Петри.
Для разделения ЭО и взвеси костномозговых клеток чашки Петри на 30 мин помещали в газопроточный инкубатор при температуре 37°C, относительной влажности 95% и содержании CO2 - 4,5%. По окончании инкубации с помощью шприца отмывали монослой островков от неадгезировавшихся элементов костномозговой ткани, используя для этого среду RPMI-1640. Далее чашки Петри заполняли культуральной средой, с помощью микродозатора в них добавляли исследуемые вещества и помещали в газопроточный термостат, работающий в указанном выше режиме. Культивирование производили в течение 24-х часов.
Каждый из трех полученных препаратов, РНКс-1, РНКс-2 и РНКс-3, добавляли в чашки Петри, полностью подготовленные к культивированию, непосредственно перед постановкой их в газовопроточный инкубатор, в количестве 2 мкг или 4 мкг препарата на 1 мл культуральной среды. Каждый препарат был протестирован на 30 культурах ЭО. В качестве контроля, одновременно с опытными образцами культивировали также ЭО КМ интактных крыс без добавления препаратов (по 10 культур). Такое же количество культур было использовано для определения «фона», то есть состояния культур непосредственно перед началом культивирования. Всего было использовано 40 беспородных белых крыс-самцов в возрасте 4-5 месяцев (с массой тела 140-160 г).
Во второй и третьей сериях экспериментов влияние препаратов РНКс-1 и РНКс-2 тестировали на культурах ЭО КМ, полученного от крыс с эритропоэзом, подавленным экспериментально вызванной полицитемией (модель (I) посттрансфузионной полицитемии).
Модель (I) посттрансфузионной полицитемии. Для создания модели угнетенного эритропоэза у крыс-доноров (массой 250-300 г) из нижней полой вены забирали кровь, и путем трехкратного центрифугирования с 0,9% раствором NaCl получали 80%-ную эритроцитарную взвесь, затем эту взвесь однократно внутрибрюшинно вводили крысам-реципиентам (массой 90-100 г) из расчета 7 мл на 100 г массы животного. ЭО КМ из бедренной кости полицитемичных крыс выделяли на 5 сутки после трансфузии эритроцитарной взвеси, когда количество ретикулоцитов в крови доноров КМ уменьшалось вдвое.
Крысы с экспериментальной полицитемией были использованы и как доноры ЭО для изучения их реакции на препараты РНК в культуре in vitro, и как реципиенты этих препаратов в опытах in vivo. Оба варианта этой модели служили для изучения степени выраженности стимулирующего влияния РНК лимфоидных клеток селезенки на развитие эритроидных клеток в костном мозге у крыс в условиях исходно угнетенного эритропоэза (то есть в условиях, когда для проявления стимулирующей эритропоэз активности указанным препаратам РНК нужно было сначала преодолеть эффект торможения эритропоэза, вызванный полицитемией).
На 5-е сутки после трансфузии эритромассы у крыс определяли гематокрит, количество эритроцитов, гемоглобин и число ретикулоцитов в периферической крови. В эксперимент включали животных, у которых гематокрит превышал 60%, количество эритроцитов было не менее 9×1012/л, количество ретикулоцитов снижалось не менее чем в 2 раза по сравнению с исходным значением. В этот же день (на пятые сутки после трансфузии) в опытах in vivo 5 крысам внутривенно однократно вводили РНКс-1 и 5 крысам - РНКс-2 (соответственно, от интактных доноров и доноров, подвергнутых 2% кровопусканию (то есть анемизированных животных), из расчета 15 мкг/100 г массы тела (группы «РНКс-1» и «РНКс-2»). Группу контроля составили 5 крыс с посттрансфузионной полицитемией, которых забивали на 5-е сутки после трансфузии эритромассы для определения «фона» полицитемии. Ранее нами и другими исследователями было показано, что полномасштабное угнетение эритроидного ростка кроветворения у лабораторных животных (в частности, у крыс), начинающееся сразу после однократного введения 80% взвеси гомологичных эритроцитов, достигает максимума к 5-м суткам после трансфузии и сохраняется вплоть до 10 дня эксперимента [Мельников И.Ю. Исследование эритробластических островков костного мозга при различных функциональных состояниях эритропоэза: дис.… канд. мед. наук. Челябинск, 1987, 200 с.; Рассохин А.Г. Состояние эритропоэза и функциональные характеристики эритробластических островков костного мозга при стимуляции и ингибиции эритропоэза. Вестник РАМН, 2000, №2, с. 9-14; Тишевская Н.В. Динамика состава гликозаминогликанов при различных состояних эритропоэза в эритробластических островках: дис.… канд. мед. наук. Челябинск, 1995, 112 с.; Моисеева О.И. Физиологические механизмы регуляции эритропоэза. Л.: Наука, 1985, 183 с.; Филимонов В.И. Механизмы торможения эритропоэза. Патологическая физиология и экспериментальная терапия, 1972, Т. 16, №5, с. 33-37; Гительзон И.И., Сидько С.Ф. Популяционно-клеточные механизмы торможения эритропоэза при полицитемии. Цитология, 1977, Т. 19, №6, с. 632-638; Рассохин А.Г. О влиянии эритропоэтина на состояние эритропоэза в эритробластических островках костного мозга при полицитемии. Материалы II съезда физиологов Урала, 1990, с. 29-30]. В эти дни в костном мозге отмечается резкое уменьшение общего количества ЭО, снижение числа островков пролиферирующих классов, образование суперостровков, содержащих в своей короне более 64 эритроидных клеток, уменьшение показателей интенсивности вовлечения КОЕэ (колониеобразующая единица эритроцитарная) в дифференцировку и повторного вовлечения макрофагов в эритропоэз, увеличение количества резидентных макрофагов; в периферической крови в этот момент наблюдается двух-трехкратное снижение числа ретикулоцитов. Далее ежедневно, вплоть до 10-х суток, у опытных и контрольных животных общепринятыми методами исследовали указанные выше показатели периферической крови. На 10-е сутки животных выводили из эксперимента, и получали костный мозг с целью изучения эритропоэза в эритробластических островках (ЭО) костного мозга. Для качественной оценки эритропоэза, протекающего в ЭО КМ, использовали уже упомянутую классификацию Ю.М. Захарова с соавт. [Захаров Ю.М., Мельников И.Ю., Рассохин А.Г. Классификация эритробластических островков костного мозга с учетом изменения их клеточного состава. Арх. анат., гистол. и эмбриол., 1990, №5, с. 38-42].
Влияние препарата РНКс-3 на эритропоэз в культуре ЭО КМ изучали на двух моделях: в культуре ЭО КМ при физиологическом эритропоэзе (модель (II) физиологического эритропоэза) и в культуре ЭО КМ, стимулированного 2% кровопотерей (модель (III) компенсационного эритропоэза). Всего было исследовано 30 культур (по 5 в каждой группе).
Для выяснения вопроса о том, эффективен ли препарат, стимулирующий эритропоэз, также и в системе организма в условиях патологии, и какова его эффективность, были использованы две экспериментальные модели: модель индуцированной бензолом гипопластической анемии и сублетальное облучение крыс.
Модель (IV) хронической гипопластической бензольной анемии. Для создания бензольной анемии интактным крысам весом 130-250 г с нормальными показателями периферической крови трехкратно подкожно вводили равную по объему смесь бензола и стерильного растительного масла, при этом доза бензола составляла 0,05 мл/100 г массы тела животного. Инъекции бензола вводили с интервалом в 7 суток.
Для контроля состояния периферического отдела кроветворной системы у крыс с бензольной анемией еженедельно определяли количество эритроцитов, ретикулоцитов, лейкоцитов, тромбоцитов. Через 4 недели после последней инъекции бензола у животных отмечалось достоверное снижение в 4,5 раза количества лейкоцитов и тромбоцитов и в 7 раз - число ретикулоцитов.
Таковым был исходный фон кроветворения, когда через 4 недели после последней инъекции бензола начинали вводить испытываемые препараты РНК-1 и РНК-2, трижды, с интервалом в 10 дней. Животным контрольной группы в эти же сроки вводили 0,9% раствор NaCl.
В дополнительном опыте, в тех же условиях развившейся бензольной анемии, через 4 недели после последней инъекции бензола животным экспериментальной группы начинали вводить препарат РНК из костного мозга анемизированных крыс, полученный на стадии стимуляции гемопоэза (в нашем случае через 17 часов после 2% кровопускания), из расчета 15 мкг/100 г массы тела, трижды, с интервалом в 10 дней. Животным контрольной группы в эти же сроки вводили 0,9% раствор NaCl.
Каждое животное получило суммарную дозу 45 мкг/100 г веса тела. Каждые 10 дней осуществляли контроль за состоянием указанных выше показателей периферической крови.
Модель (V). Сублетальное облучение. 15 беспородных крыс-самок массой от 140 до 170 граммов подвергли однократному γ-облучению в сублетальной дозе 6 Гр. Далее животные были разделены на 3 равные группы, одна из которых стала контрольной, а 2 - опытными. Через 2 часа после облучения животным одной из опытных групп внутривенно вводили по 30 мкг препарата РНК костного мозга интактных крыс (РНКкм-1), другой - ту же дозу препарата РНК костного мозга, стимулированного кровопотерей в количестве 2% от массы тела (РНКкм-2) (забор костного мозга для дальнейшего выделения РНК осуществляли, как и в других экспериментах со стимуляцией, через 17 часов после указанной манипуляции). Через 7 суток после облучения опытным животным повторно внутривенно вводили соответствующий препарат РНКкм в дозе 20 мкг на 1 крысу. На 14 сутки после начала эксперимента осуществляли третье введение препарата РНК в дозе 30 мкг на 1 крысу, но на этот раз вводили препарат, полученный не из костного мозга, а из лимфоидных клеток тимуса интактных (РНКт-1) и анемизированных (РНКт-2) крыс по описанной ранее схеме. Контрольным животным вместо препаратов РНК в те же сроки и в том же объеме внутривенно вводили 0,9% раствор NaCl.
На 3, 5, 7, 10, 15, 20, 25, 31 и 40 сутки после начала эксперимента у опытных и контрольных животных общепринятыми методами подсчитывали количество форменных элементов крови (число ретикулоцитов, эритроцитов, лейкоцитов и тромбоцитов). На 45-е сутки животные были выведены из эксперимента с целью оценки состояния костномозгового кроветворения. Клеточный состав костного мозга в миелограммах анализировали по известной методике [Филимонов В.И. Механизмы торможения эритропоэза / Патологическая физиология и экспериментальная терапия, 1972, Т. 16, №5, с. 33-37]. Для изучения особенностей восстановления эритропоэза в эритробластических островках (ЭО) в суспензии костного мозга определяли абсолютное количество ЭО (103/бедр. кость), а после адгезии островков к поверхности чашки Петри - распределение их по классам зрелости, пользуясь классификацией Захарова Ю.М. с соавт. [Захаров Ю.М., Мельников И.Ю., Рассохин А.Г. Классификация эритробластических островков костного мозга с учетом изменения их клеточного состава. Арх. анат., гистол. и эмбриол., 1990, №5, с. 38-42].
Полученные результаты обрабатывали стандартными методами описательной статистики с расчетом средних значений, ошибки среднего, доверительных интервалов, стандартного отклонения. Сравнения групп проводили методами непараметрической статистики с использованием критериев Колмогорова-Смирнова, Манна-Уитни и Краскела-Уоллеса. Различия считались достоверными при вероятности ошибки 1 рода <0,05.
Выделение лимфоцитов проводили из гепаринизированной крови (10-15 единиц гепарина на 1 мл крови), разведенной 1:1 физиологическим раствором и наслоенной на 2 мл градиента плотности (смесь фикола и верографина) с удельной плотностью 1,077 г/мл. Центрифугирование проводили в течение 25 минут при 1500 об/мин на центрифуге ОПН-3 (400 g). Образующееся в градиенте плотности «кольцо» лимфоцитов собирали в отдельные пробирки. Двойное отмывание клеток проводили физиологическим раствором в объеме 10 мл при следующем режиме центрифугирования: один раз - при 400 g в течение 10 минут, второй раз (для освобождения от тромбоцитов) при 350 g в течение 6-7 минут.
Модель (VI) аллоксанового сахарного диабета (СД) 1 типа
В работе была использована модель аллоксанового диабета. Аллоксан характеризуется трехкетонной структурой, сходной по своему строению с глюкозой. Он избирательно захватывается переносчиком глюкозы GLUT-2 и транспортируется в панкреатические β-эндокриноциты. В процессе окисления в цитоплазме β-эндокриноцитов аллоксан продуцирует цитотоксичные свободнорадикальные метаболиты, которые вызывают массивный некроз β-клеток островков Лангерганса с последующим развитием абсолютной недостаточности инсулина.
Воспроизведение модели СД 1 типа:
1. Интактным крысам массой 180-220 г однократно п/к вводили полный адъювант Фрейнда в дозе 0,5 мл/крысу (см. патент РФ №2400822).
2. Через 24 часа (на фоне 24-часового голодания, но свободного доступа к воде) животным однократно п/к вводили аллоксан (аллоксана тригидрат «La Chema», Чехия) в дозе 200 мг/кг (4% раствор в 0,9% NaCl) [Волчегорский И.А., Долгушин О.Л., Колесников В.Е. и др. Экспериментальное моделирование и лабораторная оценка адаптивных реакций организма. Челябинск/ Челябинский гос. педагогический университет, 2000, с. 167; Волчегорский И.А., Тишевская Н.В., Дементьева Е.В. Антианемическое действие реамберина при остром аллоксановом диабете у крыс. Экспериментальная и клиническая фармакология, 2008, Т. 71, No. 6, с. 23-27].
Для предотвращения фатального кетоацидоза, с 3-х суток после введения аллоксана все крысы получали базисную инсулинотерапию [Federiuk, I.F., Casey H.M., Quinn M.J., Wood M.D., Ward W.K. Induction of type-1 diabetes mellitus in laboratory rats by use of alloxan: Route of administration, pitfalls, and insulin treatment. Сотр. Med.,2004, vol. 54, no. 3, pp. 252-257]. Для этого животным 1 раз в сутки подкожно вводили раствор инсулина аспарата двухфазного (НовоМикс 30 Пенфилл, «Novo nordisk», Дания) в дозе 3 МЕд/кг. При снижении уровня глюкозы в крови ниже 10 ммоль/л введение инсулина прекращали.
Через 14 дней после введения аллоксана, перед введением препаратов РНК, уровень глюкозы в крови крыс составлял от 19 до 24 ммоль/л.
Пример 1
Стимуляция костномозгового эритропоэза в культуре ЭО под влиянием препарата тотальной РНК (препарат РНКс-2), полученного из лимфоидных клеток селезенки.
Работа выполнена на 36 белых беспородных крысах (24 донора эритроцитарной массы обоего пола массой 280-315 граммов, 12 самок-реципиентов массой 100-120 граммов). Создание модели угнетенного эритропоэза (см. модель (I) посттрансфузионной полицитемии) подробно изложена в разделе «Материалы и методы исследования».
Как показал анализ культур ЭО КМ интактных крыс (модель (II) физиологического эритропоэза), добавление в среду культивирования препаратов РНКс-1 и РНКс-2 в количестве 2 мкг/мл культуральной среды не так существенно отразилось на количестве ЭО, находящихся на чашках Петри. Их число через 24 часа культивирования достоверно не отличалось от фона и от контроля. Однако качественный состав культур ЭО при внесении в культуральную среду препарата РНКс-2, полученного из лимфоидных клеток селезенки анемизированных крыс, обладающих свойством стимулировать эритропоэз (в данном случае через 17 часов после кровопускания (донорский интервал)), изменился. Препарат РНКс-2, в отличие от препарата РНКс-1 (из лимфоидных клеток селезенки интактных крыс), достоверно стимулировал в культуре процесс формирования реконструирующихся островков, что свидетельствует об активации эритропоэза de repeto в условиях in vitro. По данным авторов этого метода, при стимуляции красного ростка кроветворения как в условиях in vitro, так и в условиях in vivo, именно процесс реконструкции эритропоэза реагирует на стимулирующее воздействие в первую очередь (Таблица 1).
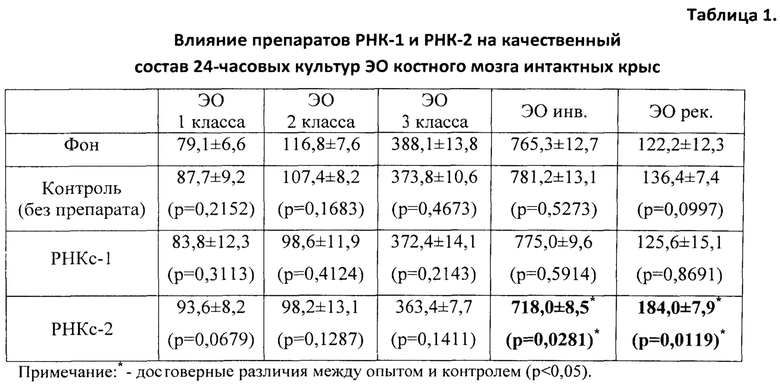
Одновременно с увеличением числа реконструирующихся ЭО в культурах, содержащих РНКс-2 лимфоидных клеток крыс с активированным эритропоэзом, достоверно уменьшалось число инволюцирующих ЭО за счет вовлечения их центральных макрофагов в процесс повторного эритропоэза и превращения ЭО инволюцирующих в ЭО реконструирующиеся. Процесс созревания существенно ускоряется, о чем свидетельствуют расчетные данные по интенсивности эритропоэза (Таблица 2). При этом увеличение абсолютного числа реконструирующихся островков составляет 33%, а уменьшение числа инволюцирующих островков составляет немногим менее 10%, так что полученные данные с несомненностью указывают на стимулирующее действие препарата РНКс-2, полученного из лимфоидных клеток селезенки крыс, подвергшихся кровопусканию, на стадии проявления ими стимулирующих свойств в отношении гемопоэза (то есть в первой фазе ответа на кровопотерю). Достоверным усилением эритропоэза отвечали только культуры с препаратом РНКс-2, в то время как препарат РНКс-1, полученный из лимфоидных клеток селезенок интактных крыс, никаких изменений не вызывал.
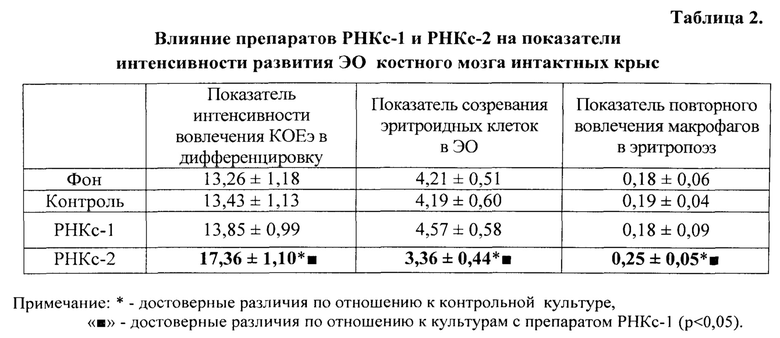
Для проверки этого результата дополнительно были поставлены опыты с увеличенной дозой препаратов до 4 мкг на 1 мл культуральной среды. Кроме того, была предпринята попытка испытать возможности стимулирующего препарата на культурах ЭО КМ крыс с угнетенной пролиферацией кроветворных клеток в результате экспериментально вызванной полицитемии (см. раздел «Модель (I) посттрансфузионной полицитемии»). В контрольные культуры добавляли только эритропоэтин.
Результаты опытов представлены в таблице 3, из которой видно, что полицитемичные культуры ЭО оказались довольно чувствительными к внесению 4 мкг препаратов РНКс-1 или РНКс-2 на 1 мл культуральной среды, поскольку уже через 24 часа культивирования качественный состав ЭО практически не отличался от состава культур интактных животных, то есть в данном случае в опытных культурах наблюдалось нормальное физиологическое течение эритропоэза. Более того, число реконструирующихся ЭО в культурах с препаратом РНКс-2 оказалось достоверно выше их количества в культурах интактных животных, что свидетельствует об истинной стимуляции эритропоэза, поскольку сопровождается увеличением контактов центральных макрофагов с КОЕэ.

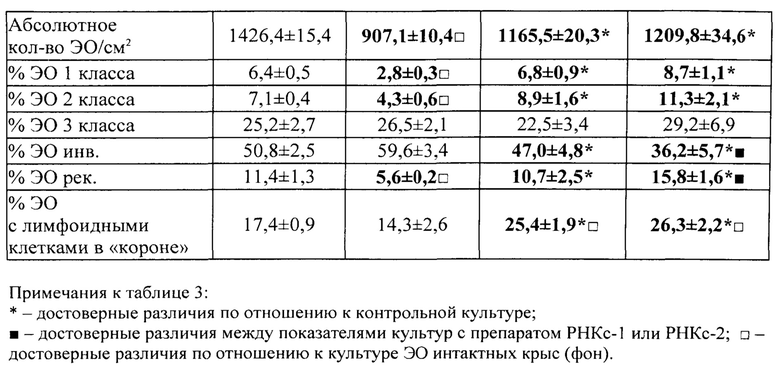
Таким образом, показано, что препараты РНКс-1 и РНКс-2 обладают стимулирующими эритропоэз свойствами, и степень выраженности этих свойств особенно заметна на модели (I) посттрансфузионной полицитемии, когда для стимуляции процесса кроветворения препаратам согласно изобретению удается преодолеть даже вызванное в этой модели интенсивное торможение эритропоэза.
Результаты, полученные в опытах с культурой ЭО, получили подтверждение и дальнейшее развитие и в опытах in vivo.
Пример 2
Влияние препаратов РНКс селезенки in vivo на эритропоэз у полицитемичных крыс.
После воспроизведения полицитемии животные имели достоверно более высокие (по сравнению с исходными) количества эритроцитов, уровни гемоглобина и гематокрита, одновременно с трехкратным снижением в данном опыте количества ретикулоцитов в периферической крови, что свидетельствует об эффективности созданной модели угнетения эритропоэза (Таблица 4) и полностью совпадает с данными, полученными ранее нами и другими исследователями [Мельников И.Ю. Исследование эритробластических островков костного мозга при различных функциональных состояниях эритропоэза: дис.… канд. мед. наук. Челябинск, 1987, 200 с.; Рассохин А.Г., Круглов Д.Г., Захаров Ю.М. Состояние эритропоэза и функциональные характеристики эритробластических островков костного мозга при стимуляции и ингибиции эритропоэза. Вестник РАМП, 2000, №2, с. 9-14; Тишевская Н.В. Динамика состава гликозаминогликанов при различных состояних эритропоэза в эритробластических островках: дис.… канд. мед. наук. Челябинск, 1995, 112 с.; Моисеева О.И. Физиологические механизмы регуляции эритропоэза. Л.: Наука, 1985, 183 с.; Филимонов В.И. Механизмы торможения эритропоэза. Патологическая физиология и экспериментальная терапия, 1972, Т. 16, №5, с. 33-37; Гительзон И.И., Сидько С.Ф. Популяционно-клеточные механизмы торможения эритропоэза при полицитемии. Цитология, 1977, Т. 19, №6, с. 632-638; Рассохин А.Г. О влиянии эритропоэтина на состояние эритропоэза в эритробластических островках костного мозга при полицитемии. В сб.: Материалы II съезда физиологов Урала. 1990, с. 29-30].
Внутривенное (в/в) ведение крысам с посттрансфузионной полицитемией препарата РНКс-2 способствовало достоверному снижению указанных показателей периферической крови по сравнению с контрольной группой: на 3 сутки после инъекции РНКс-2 снизились количество гемоглобина и гематокрит, а на 4 и 5 сутки, помимо указанных параметров, уменьшилось и число эритроцитов.
Отдельного внимания заслуживают данные о влиянии препаратов РНК согласно изобретению на количество ретикулоцитов в периферической крови. Так, очень быстро, начиная со 2 суток после введения препарата РНКс-2, наблюдался достоверный прирост числа ретикулоцитов в кровяном русле, а в последующие дни наблюдения уже оба препарата, РНКс-1 и РНКс-2, вызывали достоверное повышение данного показателя.
Следует отметить и то, что параллельно увеличению числа ретикулоцитов в периферической крови в последующие дни происходит неуклонное уменьшение гемоглобина и гематокрита, в конечном итоге с достоверным снижением этих показателей, и сопровождается это достоверным уменьшением численности эритроцитов в группе крыс, получавших препарат РНКс-2. Этот факт указывает на то, что стимуляция эритропоэза, обусловленная активацией макрофагов эритробластических островков, отражает общую активацию всей лимфоидно-макрофагальной системы организма, что приводит к усиленному разрушению и фагоцитозу перелитых аллогенных эритроцитов. И, по-видимому, показатель количества эритроцитов здесь отражает главным образом численность клеток собственного производства.

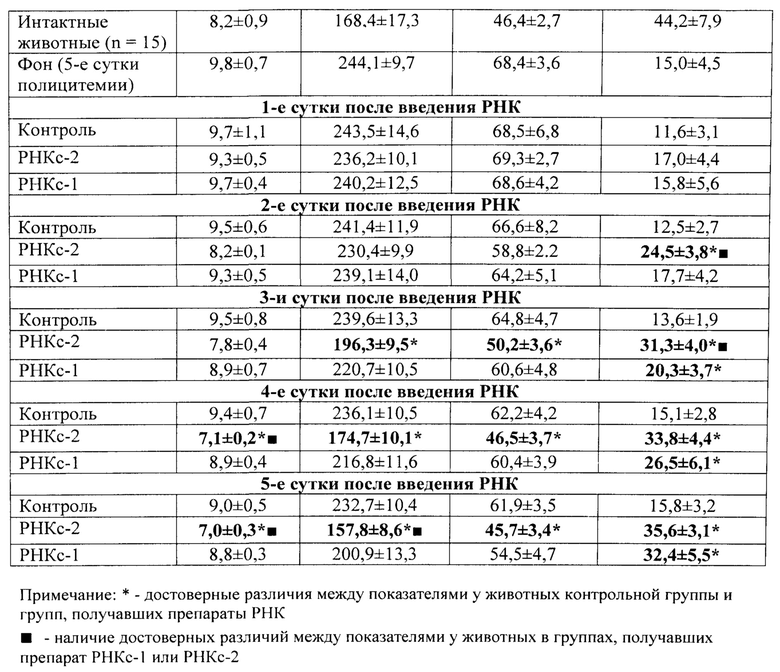
Явная активация эритроидного ростка на периферии сопровождалась изменениями процессов развития эритроидной ткани в костном мозге (таблица 5 (10-е сутки после трансфузии эритромассы, 5-е сутки после трансфузии препаратов). Оба исследуемых препарата РНК оказывали стимулирующее действие в отношении эритропоэза в ЭО: и в группе «РНКс-1», и в группе «РНКс-2» через 5 суток после инъекции препаратов достоверно увеличивалось абсолютное количество островков в костном мозге, и появлялись островки 1 класса зрелости, формирующиеся на основе контакта свободных костномозговых макрофагов с КОЕэ. Кроме того, под влиянием препаратов РНК ускорялся процесс реконструкции эритропоэза в ЭО: достоверно увеличивалось количество островков с повторной «волной» эритропоэза (реконструирующиеся ЭО). Помимо стимуляции процессов пролиферации, препараты РНК согласно изобретению способствовали увеличению в костном мозге количества зрелых островков 3 класса (что также свидетельствует об интенсификации развития красного ростка костномозгового кроветворения) и достоверному уменьшению количества инволюцирующих ЭО, что также свидетельствует об интенсификации эритропоэза.
Несмотря на то, что оба препарата РНК демонстрировали однонаправленное действие в отношении формирования и развития ЭО в костном мозге, они все же отличались по интенсивности влияния на ЭО. Препарат РНКс-2, полученный из селезенки анемизированных животных, оказывал более выраженный эффект стимуляции развития эритроидной ткани в костном мозге, и в целом под его влиянием эритропоэз в костном мозге достигает практически исходного уровня.
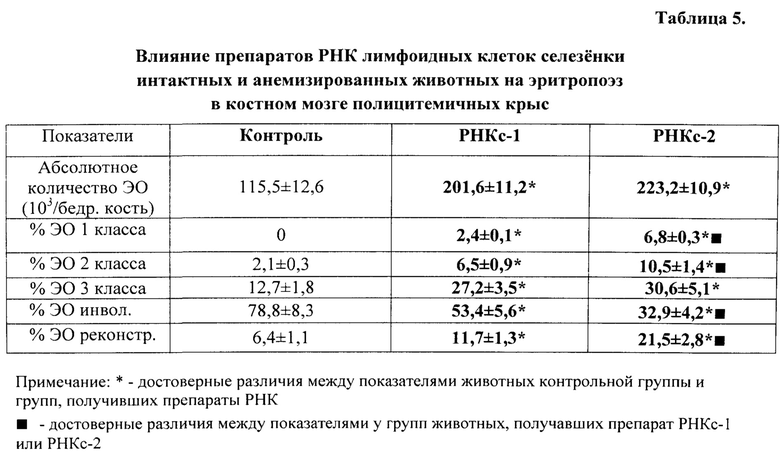
Таким образом, на основании полученных данных можно сделать вывод, что исследуемые препараты РНК лимфоидных клеток селезенки оказывают стимулирующее влияние не только на физиологический, но и на угнетенный эритропоэз.
Пример 3
Свойство РНК лимфоидных клеток селезенки, выделенных на определенной стадии после острой кровопотери, подавлять эритропоэз.
Доказательство наличия супрессорного свойства у препарата тотальной РНК лимфоидных клеток на определенной стадии восстановительных процессов, а именно во второй их фазе, очень важно не только для раскрытия механизма регуляции морфогенетической функции лимфоцитов и их способности передавать регенерационную информацию, но и для отработки принципа РНК-терапии тяжелых аутоиммунных заболеваний и так называемых болезней пролиферации, или гиперпролиферативных состояний.
Эффективность супрессорного действия препарата РНК согласно изобретению (препарат РНКс-3), полученного через 4 суток после массивной кровопотери из лимфоцитов селезенки, которые, как известно, на этой стадии обладают способностью подавлять кроветворение, мы проверяли в опытах на культуре ЭО КМ как интактных крыс (модель (II) физиологического эритропоэза), так и крыс со стимулированным кроветворением, вызванным острой кровопотерей (модель (III) компенсационного эритропоэза); кроме того, для полноты представлений о масштабах возможностей такого препарата был исследован уровень супрессорной активности препарата РНКс-3 в опытах на культуре ЭО КМ крыс не только со стимулированным кроветворением после острой кровопотери, но еще и в условиях повышенного содержания эритропоэтина в культуре в качестве дополнительного стимулятора эритропоэза (оба этих воздействия являются разновидностями модели (III) компенсационного эритропоэза). Иными словами, в этом случае для проявления супрессорной активности препарату РНКс-3 необходимо было преодолеть стимуляцию кроветворения, вызванную, во-первых, острой кровопотерей in vivo, а во-вторых, дополнительным воздействием in vitro, стимулирующим созревание эритробластических островков пролиферирующих классов (ЭО 1-го, 2-го классов и ЭО реконструирующихся).
Помимо этого, ингибирующие свойства препарата РНКс-3 проверяли и в опытах на животных в условиях усиленного кроветворения, вызванного острой кровопотерей. Препарат вводили крысам внутривенно (в/в), однократно, через 1 час после кровопотери, в дозе 30 мкг/100 г массы тела.
Результаты этих опытов представлены в таблицах 6, 7, 8.
В опытах на культуре ЭО были использованы разные дозы препарата РНКс-3 и эритропоэтина, а именно было исследовано
5 культур с 0,5 МЕ/мл эритропоэтина и 4 мкг/мл препарата
5 культур с 0,5 МЕ/мл эритропоэтина и 2 мкг/мл препарата
5 культур с 1,5 МЕ/мл эритропоэтина и 4 мкг/мл препарата
5 культур с 1,5 МЕ/мл эритропоэтина и 2 мкг/мл препарата
контрольные культуры с 0,5 МЕ/мл эритропоэтина
контрольные культуры с 1,5 МЕ/мл эритропоэтина.
Из таблицы 6 следует, что присутствие препарата РНКс-3 в культуре эритробластических островков (ЭО) приводило к подавлению развития эритроидных клеток. Тормозящий эффект исследуемого препарата в модели (II) физиологического эритропоэза был наиболее выражен, когда в культуральную среду, наряду с 0,5 МЕ/мл эритропоэтина, было добавлено не 2, а 4 мкг/мл РНКс-3 (Таблица 6). При анализе количества ЭО различных классов зрелости было обнаружено, что при развитии торможения эритропоэза происходит подавление процесса присоединения КОЕэ и проэритробластов к мембране центральных макрофагов, о чем свидетельствует снижение количества ЭО пролиферирующих классов в культуре (ЭО 1-го, 2-го классов и ЭО рек.). Кроме того, о торможении процесса дифференцировки и созревания эритроидных клеток говорит также и увеличение количества ЭО 3 класса зрелости, рост числа которых, вероятно, обусловлен замедлением созревания оксифильных нормобластов, торможением их денуклеации и задержкой выхода ретикулоцитов из «короны» ЭО.
Увеличение дозы препарата РНКс-3 вызвало эффект более отчетливый, но существенно не отличающийся от эффекта, вызванного меньшей дозой.
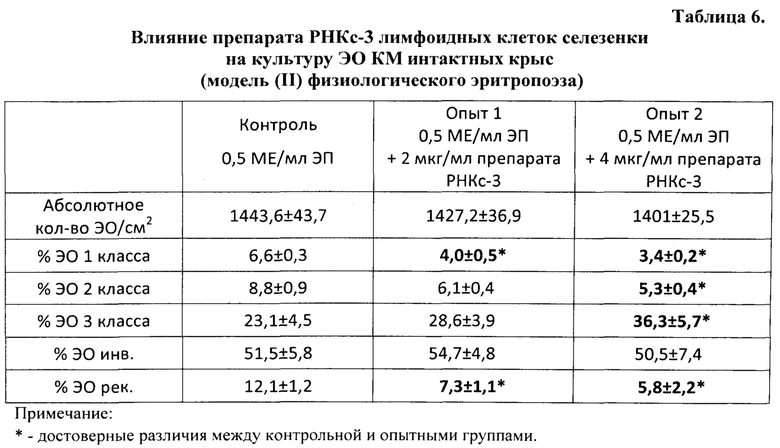
В модели (III) компенсационного эритропоэза также наблюдалось угнетение развития эритроидных клеток, но только при добавлении большего количества РНКс (Таблица 7). Присутствие 4 мкг/мл препарата РНКс-3 в культуральной среде уменьшало способность центральных макрофагов формировать новые ЭО как de novo, так и de repeto, о чем свидетельствует уменьшение количества островков 1 класса зрелости и реконструирующихся ЭО в культуре. Меньшая выраженность тормозящего действия РНКс-3 на компенсационный эритропоэз по сравнению с физиологическим, вероятно, связана с возникающим дисбалансом стимулирующих и угнетающих эритроидный росток факторов, поскольку при добавлении в культуральную среду большого количества эритропоэтина равновесие смещается в сторону стимуляции эритропоэза.
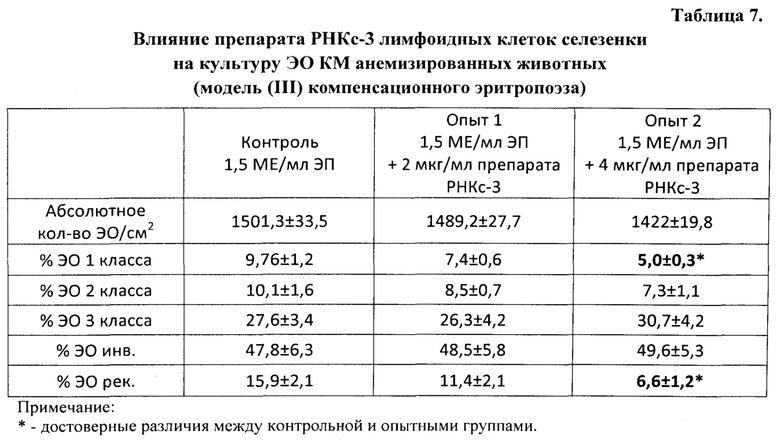
При микроскопировании культур ЭО с 0,5 МЕ/мл эритропоэтина (модель (II) физиологического эритропоэза) и 4 мкг/мл препарата РНКс-3 в культуральной среде мы обратили внимание на необычайно частую встречаемость контактов ЭО 3 класса с лимфоидными клетками, в связи с чем нами был проведен анализ числа контактов лимфоидных клеток с «короной» островков различных классов зрелости. Ранее подобные подсчеты проводились нами при изучении влияния разных доз эритропоэтина на интенсивность эритропоэза in vitro, и было установлено, что наибольшее число контактов с лимфоидными клетками имеют островки с активно пролиферирующими клетками в «короне» - ЭО 1 класса и ЭО 2 класса. Однако в данном случае при добавлении в культуру препарата РНКс-3 наибольшее число контактов стало наблюдаться у ЭО 3 класса, что, безусловно, заслуживает особого внимания.

Результаты опытов in vivo полностью согласуются с результатами, полученными на культурах ЭО костного мозга. Внутривенное введение крысам препарата РНКс-3, в отличие от введения контрольным крысам такого же количества физиологического раствора (0,1 мл) через 1 час после кровопотери (модель (III) компенсационного эритропоэза), приводило к достоверному уменьшению количества ретикулоцитов в периферической крови (Таблица 9) и к достоверному уменьшению в костном мозге количества ЭО 1 класса, ЭО реконструирующихся, а также к увеличению количества инволюцирующих эритробластических островков (Таблица 10).



Таким образом, все без исключения опыты, приведенные в 1, 2 и 3 примерах, несомненно, указывают на то, что препараты тотальной РНК из лимфоидных клеток полностью сохраняют основные свойства самих лимфоидных клеток и, в полном соответствии с ними, выполняют присущую им регуляторную функцию в отношении процессов пролиферации и дифференцировки кроветворных клеток.
В задачу последующих исследований входило выяснение следующих вопросов:
1) способен ли стимулирующий препарат РНКс-2 вылечить тяжелую хроническую анемию и тем самым заменить используемые для этой цели систематические гемотрансфузии?
2) затрагивают ли восстановительные свойства препарата РНКс-2 другие ростки гемопоэза, или же его действие касается только эритропоэза?
3) обладают ли такими же свойствами препараты тотальной РНК, полученные из других лимфоидных органов (тимуса, костного мозга) и периферической крови?
4) обладает ли лечебными свойствами препарат РНК, полученный из костного мозга интактных крыс?
Для решения вопроса о силе воздействия препаратов тотальных РНК-1 и РНК-2 были использованы две экспериментальные модели: модель (IV) хронической гипопластической бензольной анемии и модель (V) сублетального облучения.
Пример 4
Влияние препаратов тотальной РНКс-1 и РНКс-2 на эритропоэз у крыс с хронической гипопластической бензольной анемией (модель IV)
Бензольную хроническую гипопластическую анемию вызывали путем трехкратного подкожного введения смеси бензола со стерильным растительным маслом. Контрольным крысам вводили в таком же количестве только растительное масло (см. подробности в разделе «Материалы и методы исследования»).
В опыте было использовано 18 крыс. Препараты РНКс-1 и РНКс-2 начинали вводить через 28 суток после последней инъекции бензола и вводили трижды внутривенно, с интервалом в 10 суток, в дозе 15 мкг/100 г веса тела (общая доза составила 45 мкг/100 г веса тела).
Динамика изменений показателей численности форменных элементов периферической крови представлена в таблице 11. Из этой таблицы следует, что под влиянием бензола достоверно снизились показатели всех ростков кроветворения: ретикулоциты - в 7 раз, лейкоциты и тромбоциты - в 4,5 раза.
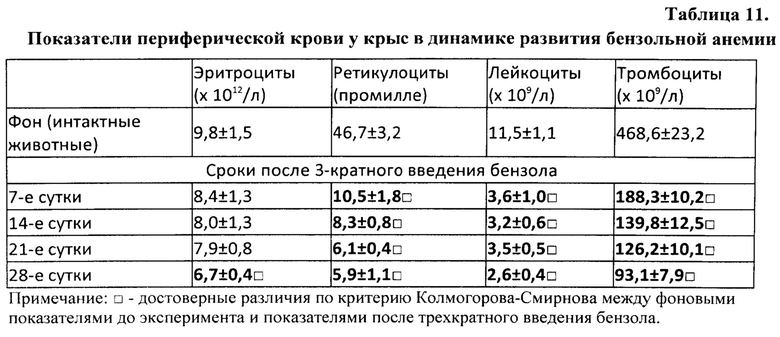
Животные были разбиты на 3 равные группы:
контрольная группа (бензольная анемия без РНК);
группа «РНКс-1» - животные, которым вводили препарат тотальной РНК лимфоидных клеток селезенки интактных крыс;
группа «РНК-2» - животные, которым вводили препарат тотальной РНК, полученный из лимфоидных клеток селезенки через 17 часов после кровопотери.
Препараты РНК вводили внутривенно трижды с интервалом в 10 дней. Животным контрольной группы в эти же сроки вводили равное по объему количество 0,9% раствора NaCl. Исходно концентрация препарата тотальной РНК лимфоидных клеток интактных и анемизированных животных составляла 3 мкг/мкл.
Каждые 9-10 дней определяли характер влияния РНК лимфоидных клеток селезенки на состояние периферической крови животных с бензольной анемией (Таблица 12). Первая же инъекция РНКс-2 на 10-й день привела к достоверному 2-кратному увеличению числа ретикулоцитов в крови. После второй инъекции этого препарата количество ретикулоцитов возросло в 3 раза, а также в 1,4 раза увеличилось число тромбоцитов. После 3-й инъекции РНКс-2 продолжало увеличиваться число ретикулоцитов - этот показатель в опытной группе в 5 раз превышал контрольное значение. К 18-м суткам после последнего введения препарата РНКс-2 стимуляция выхода ретикулоцитов из костного мозга отразилась и на количестве эритроцитов - их число в опытной группе достоверно превысило контрольный показатель.
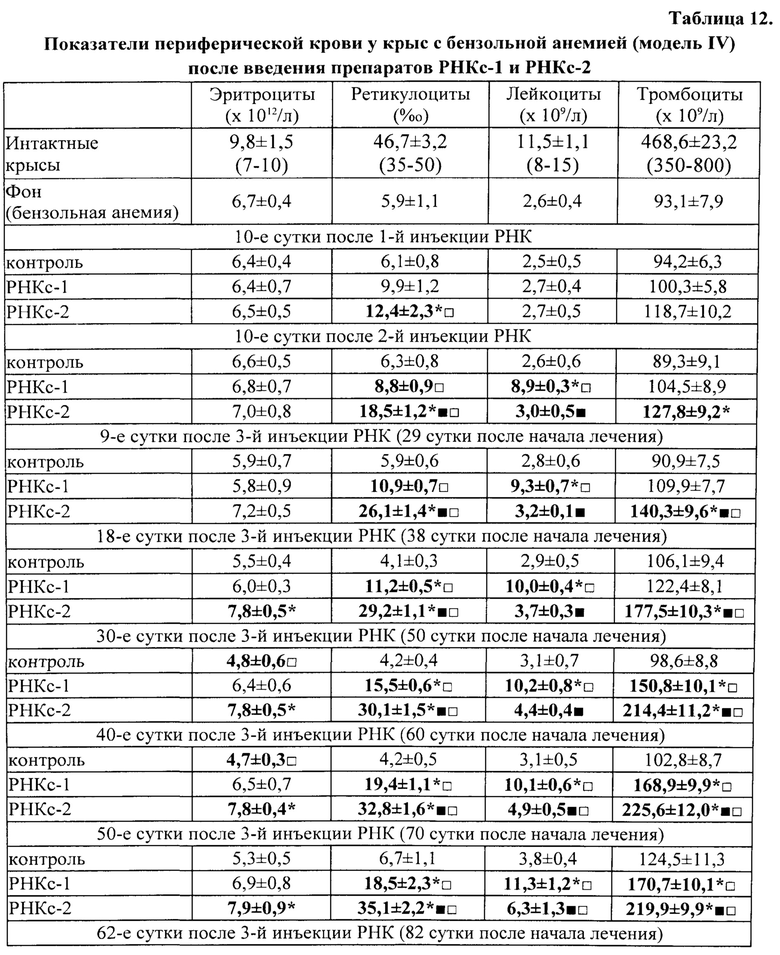
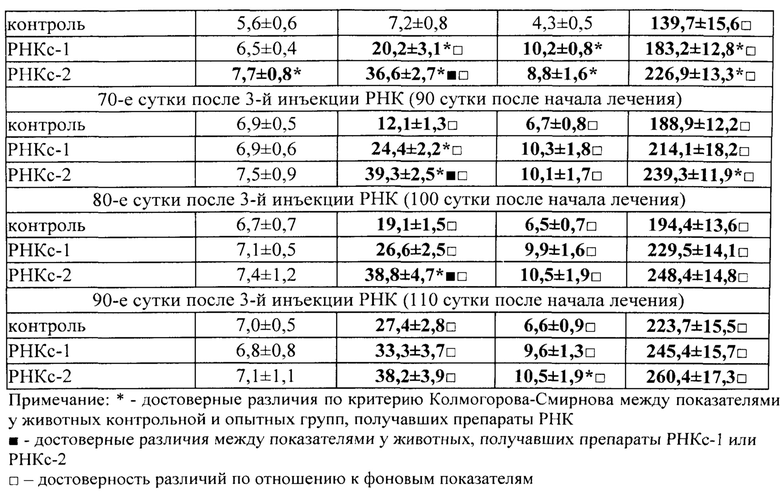
Через 40 суток после третьей инъекции препарата РНКс-2 животным с бензольной анемией количество ретикулоцитов у них увеличилось в 8 раз. число тромбоцитов - более чем в 2 раза, количество эритроцитов - в 1,7 раза. При этом в контроле к этому сроку количество ретикулоцитов и эритроцитов продолжает падать, а количество тромбоцитов едва достигает значений, которые получены в эксперименте уже на 10-й день после первого введения препаратов РНК согласно изобретению.
Введение животным с бензольной анемией препарата РНКс-1 в первую очередь отразилось на количестве лейкоцитов в периферической крови - этот показатель достоверно вырос уже к 10 дню после 2-ой инъекции препарата, и к 40-м суткам воздействия превысил контрольное значение в 3 раза. Кроме того, уже к 30-м суткам после трехкратной инъекции РНКс-1 наблюдалось отчетливое улучшение картины периферической крови по всем трем росткам кроветворения: почти в 4 раза возросло число ретикулоцитов, в 3 раза увеличилось количество лейкоцитов ив 1,5 раза повысилось количество тромбоцитов, что дает полное основание говорить о препарате тотальной РНК лимфоидных клеток селезенки интактных животных как о средстве, обладающем стимулирующей активностью в отношении гемопоэза в целом.
Получив данные о частичном восстановлении количества лейкоцитов в крови животных с бензольной анемией после введения им РНКс-1, мы выяснили, за счет каких форм лейкоцитов происходят изменения в периферической части «белого» ростка кроветворения (Таблица 13). Полученные результаты свидетельствуют о том, что РНК интактных лимфоидных клеток селезенки активирует прежде всего лимфоидную ткань организма, подвергшегося бензольной интоксикации, вследствие чего уже собственные лимфоциты, «усиленные» экзогенной РНКс-1, ускоряют (интенсифицируют) процессы восстановления кроветворения.
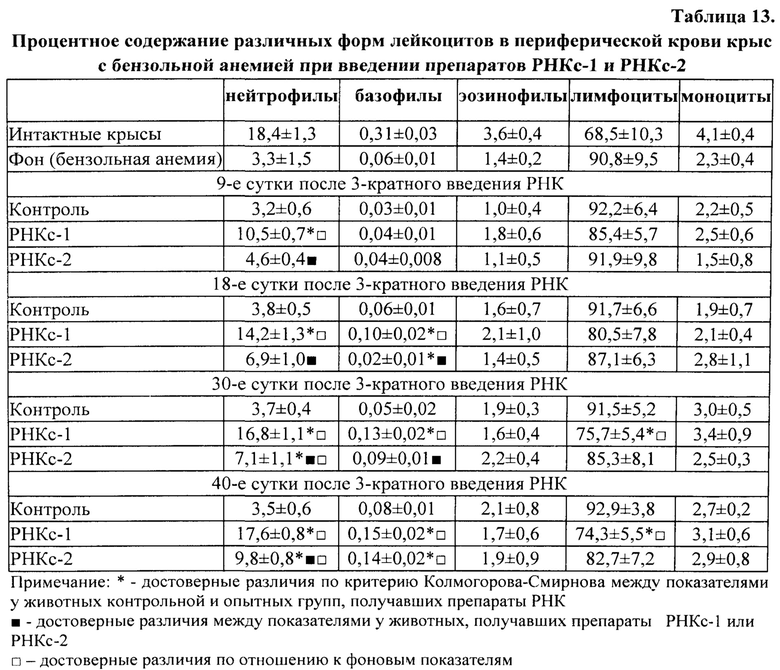
Доказано (Захаров В.Н., Караулов А.В. Соколов В.В. и др. Изменения системы крови при воздействии радиации и бензола (под ред. И.И. Гительзона). Новосибирск: Наука. Сибирское отделение, 1990, 241 с.), что именно лимфоидная ткань обеспечивает нормальное течение процессов клеточной дифференцировки в гемопоэзе, поэтому любые нарушения межклеточных взаимодействий между элементами лимфоидного ряда и другими видами кроветворных клеток могут служить пусковым механизмом гематотропного действия бензола. Не исключено также, что регулирующее влияние препаратов РНК лимфоидных клеток на кроветворение животных с бензольной анемией опосредовано макрофагами костного мозга.
В целом опыты с моделью (IV) хронической гипопластической бензольной анемии дают основание утверждать следующее:
Препараты РНК (как РНКс-1, так и РНКс-2) обладают выраженной не только эритропоэтической, но и в целом гемопоэтической активностью. Препарат РНКс-2 имеет более высокую активность, что выражается в его более быстром и более интенсивном воздействии на уровни ретикулоцитов и тромбоцитов во все сроки наблюдения, включая и конечный срок. Оба препарата обладают пролонгированным действием: через 40 суток после последней инъекции препаратов показатели периферической крови продолжают улучшаться.
По своему действию препараты РНКс-1 и РНКс-2 отличаются не только уровнем активности, но и преимущественной избирательностью своего действия. Это особенно заметно на начальных сроках стимуляции гемопоэза. Стимулированный кровопотерей, препарат РНКс-2 способствует прежде всего восстановлению эритропоэза, во вторую очередь - восстановлению уровня тромбоцитов, и только в последнюю очередь - уровня лейкоцитов. Препарат РНКс-1, в отличие от препарата РНКс-2, в данном случае в первую очередь действует на гомологичную ткань, проявляя значительно большую корригирующую способность в отношении белого ростка кроветворения. В связи с эти мы полагаем, что в случаях, когда необходимо восстанавливать все ростки кроветворения, для полноты регенерации следует вводить оба препарата, РНКс-1 и РНКс-2.
Через 2 месяца после начала инъекций препаратов РНК получено весьма значительное восстановление гемопоэза притом, что были использованы минимальные дозы препарата. Следовательно, совершенно очевидно, что для повышения эффективности используемых в данном опыте препаратов можно увеличить их дозу. Это было учтено нами в последующих экспериментах, в частности, в опытах с облучением.
Пример 5
Влияние препаратов РНК из костного мозга анемизированных животных на эритропоэз у крыс с хронической гипопластической бензольной анемией (модель IV)
В продолжение предыдущего опыта, в попытке добиться полного восстановления показателей периферической крови и костномозгового кроветворения, мы решили попробовать повысить эффективность регенерации крови не дозой вводимого препарата, а источником его получения.
С этой целью препарат тотальной РНК тем же самым способом выделили из костного мозга анемизированных крыс на стадии стимуляции гемопоэза (через 17 часов после 2% кровопускания) (РНКкм-2) и изучали его влияние на гемопоэз у крыс с хронической гипопластической бензольной анемией (модель IV). Для этого через 4 недели после последней инъекции бензола трехкратно, с интервалом в 10 дней, 6 животным экспериментальной группы вводили указанный препарат РНК из расчета 15 мкг/100 г массы тела. Животным контрольной группы в эти же сроки вводили 0,9% раствор NaCl.
В опыте было использовано 11 крыс. Препарат РНКкм-2 начинали вводить через 28 суток после последней инъекции бензола и вводили трижды внутривенно, с интервалом в 10 суток, в дозе 15 мкг/100 г веса тела животного (общая доза составила 45 мкг/100 г веса тела). За время наблюдения каждые 7 дней производили подсчет количества форменных элементов крови. Через 10 суток после последнего введения препарата РНКкм-2 опытные и контрольные животные были выведены из эксперимента для изучения состояния эритропоэза в костном мозге.
В этом случае уже после первой инъекции РНКкм-2 в периферической крови в 3 раза возросло количество ретикулоцитов (Таблица 14), а измерения, проведенные через 7 дней после второй инъекции, обнаружили достоверное увеличение содержания ретикулоцитов, лейкоцитов и тромбоцитов.
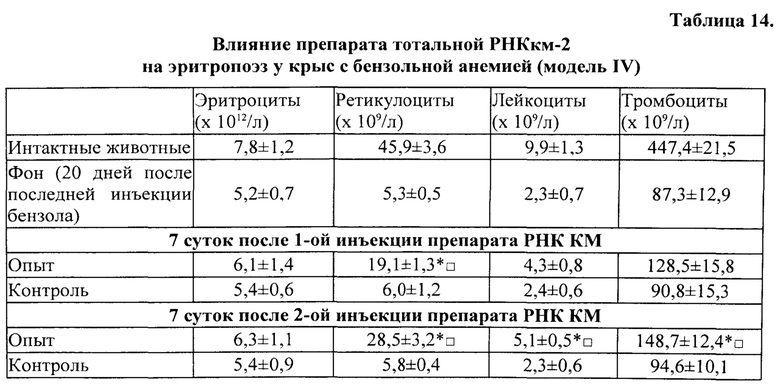

При исследовании количественного и качественного состава ЭО костного мозга были получены данные, свидетельствующие о том, что введение животным с бензольной анемией препарата РНКкм-2, полученного из костного мозга анемизированных крыс, полностью восстанавливает физиологический темп и характер развития эритроидных клеток в ЭО костного мозга (Таблица 15).
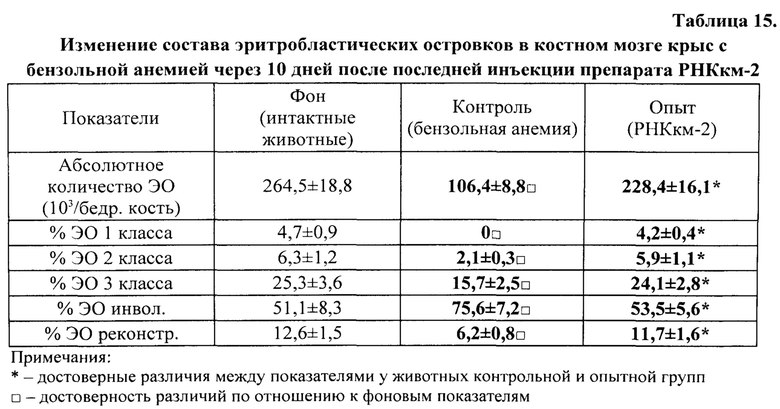
Из таблицы 20 видно, что через 10 дней после последней инъекции препарата тотальной РНКкм-2 согласно изобретению эритробластические островки ни по количеству, ни по качественным характеристикам не отличались от показателей у интактных животных.
Пример 6
Влияние препаратов тотальной РНК костного мозга и тимуса на процессы восстановления показателей периферической крови и костномозгового кроветворения после острого лучевого воздействия
В опытах с облучением (см. описание модели V в разделе «Материалы и методы исследования») для нормализации кроветворной функции были использованы для первого и второго введений более высокие дозы препаратов тотальной РНК (составляющие, соответственно, 30 мкг и 20 мкг/100 г массы животного, выделенных из костного мозга), и 30 мкг/100 г массы животного препарата тотальной РНК, выделенного из тимуса (вместо дозы в 15 мкг/100 г массы тела, используемой в Примерах 1-5). Препараты тотальной РНК тем же самым способом выделяли из костного мозга и лимфоидных клеток тимуса, с целью сравнения характера и эффективности действия препаратов тотальной РНК согласно изобретению, полученных из регуляторных клеток разных лимфоидных органов.
Как показали проведенные опыты, острое лучевое воздействие в сублетальной дозе привело к значительным изменениям количественного состава форменных элементов в периферической крови. На 3-й сутки после облучения в крови животных контрольной группы полностью отсутствовали ретикулоциты, встречались лишь единичные лейкоциты, и отмечалось восьмикратное уменьшение числа тромбоцитов по сравнению с указанными показателями у интактных крыс (Таблица 16). Одна крыса из контрольной группы погибла уже к началу 3-х суток эксперимента.
В опытных же группах животных картина периферической крови выглядела иначе: в группе, получившей препарат костного мозга интактных крыс (РНКкм-1), уже на 3-й сутки обнаруживались ретикулоциты, число лейкоцитов в 3 раза превышало этот показатель в контрольной группе, а количество тромбоцитов было на 18,5% выше, чем в контроле. В группе животных, получивших препарат РНК анемизированных крыс (РНКкм-2), отмечалась еще более выраженная положительная динамика: число ретикулоцитов и лейкоцитов было в 2 раза больше, а количество тромбоцитов на 12,5% выше, чем в группе, получившей препарат РНКкм-1. Таким образом, можно утверждать, что уже на 3-й сутки после облучения препараты РНКкм вызывают оживление эритропоэза, поскольку появление ретикулоцитов в периферической крови при повреждении костного мозга γ-лучами является самым ранним и достоверным признаком начала нормализации кроветворения [Внутренние болезни. Военно-полевая терапия (под ред. Ракова А.Л., Сосюкина А.Е.) С.-Петербург: Фолиант, 2003, 253 с.].
Подобный темп восстановления периферического звена эритроидного, миелоидного и мегакариоцитарного ростков кроветворения сохранялся и в последующие сроки наблюдения (на 5-е и 7-е сутки эксперимента). Так, ретикулоцитарный ответ у животных, получивших препараты РНК, существенно превышал таковой в контрольной группе, причем введение препарата РНКкм-2 по сравнению с препаратом РНКкм-1 приводило к большему увеличению количества ретикулоцитов. В контрольной группе животных первые ретикулоциты появлялись в периферической крови лишь к 7-м суткам. Количество лейкоцитов у животных, получивших препарат РНКкм-2, на 5-е сутки превышало этот показатель в группе, получившей препарат РНКкм-1, в 3 раза, а на 7-е сутки - в 2,4 раза. Число тромбоцитов в опытных группах на эти сроки наблюдения было выше контрольных значений почти в 2 раза. Важно отметить, что, помимо улучшения показателей периферической крови, у крыс опытных групп наблюдалось значительное улучшение общего состояния. Если контрольные крысы вплоть до 7-х суток эксперимента не проявляли практически никакой двигательной и исследовательской активности, а также не занимались грумингом, то поведение крыс опытных групп уже на 3-и сутки после облучения ничем не отличалось от поведения интактных животных.
На 7 день после облучения опытным животным была сделана повторная инъекция препаратов РНК (РНКкм-1 или РНКкм-2 соответственно) в дозе 20 мкг/100 г массы тела.
К 10-м суткам в контрольной группе погибла еще одна крыса, и, таким образом, летальность в контроле составила 40%. В опытных группах на фоне превышающих контрольное значение уровней ретикулоцитов, лейкоцитов и тромбоцитов к этому сроку наблюдения достоверно возросло и количество эритроцитов по сравнению с показателем у контрольной группы животных.
Убедившись в положительном влиянии препаратов РНК костного мозга на темп восстановления числа форменных элементов периферической крови сублетально облученных животных, мы предположили, что позитивную ответную реакцию со стороны системы кроветворения может поддержать и усилить препарат РНК лимфоидных клеток тимуса. Ранее другими исследователями было показано, что экранирование тимуса при облучении или введение тимозина в первые сутки после острого лучевого воздействия вызывает стимуляцию регенерации лимфоидных тканей и кроветворных органов [Москалев Ю.И. Отдаленные последствия воздействия ионизирующих излучений. М.: Медицина, 1991, 464 с.]. Для выявления возможного стимулирующего действия РНК лимфоидных клеток тимуса на кроветворение в нашем эксперименте крысам опытных групп, помимо введения препаратов РНК костного мозга, на 14 сутки после сублетального облучения было введено по 30 мкг препарата РНК тимуса интактных (РНКт-1) или анемизированных (РНКт-2) животных. Такое последовательное введение препаратов будет обозначаться далее через запятую, то есть (РНКкм-1), (РНКт-1) или (РНКкм-2), (РНКт-2).
На 15-е сутки после начала эксперимента (через 24 часа после дополнительного введения препарата РНК вилочковой железы) количество ретикулоцитов в группе животных, получавших препараты РНК из лимфоидных органов интактных крыс, в 3,1 раза превысило контрольный уровень, а в группе, получавшей препараты РНК из лимфоидных органов анемизированных крыс - в 4,7 раза. Таким образом, к 15-м суткам после γ-облучения в сублетальной дозе количество ретикулоцитов в периферической крови под влиянием препаратов РНК анемизированных животных достигло нормального уровня, поскольку этот показатель к этому моменту уже достоверно не отличался от фонового значения. В этой же группе животных заметно увеличилось число лейкоцитов и тромбоцитов: количество лейкоцитов по сравнению с контрольным значением возросло в 3,7 раза, количество тромбоцитов - в 2,1 раза.
Далее вплоть до 31-х суток наблюдения скорость восстановления показателей периферической крови в группе животных, получивших препараты РНК костного мозга и лимфоидных клеток тимуса крыс, подвергшихся предварительному кровопусканию, превышала темп нарастания числа форменных элементов в крови животных, получивших препараты РНК, выделенные из лимфоидных органов интактных крыс.
К 40-м суткам эксперимента ретикулоцитарный ответ системы кроветворения у всех облученных животных (и контрольной, и опытной групп) достиг нормальных значений, количество эритроцитов также не отличалось от фонового уровня. Количество лейкоцитов, хотя и не достигло исходного, но находилось в границах нормальных значений этого показателя для данного вида животных. Число тромбоцитов в контрольной группе и у животных, получавших препараты РНК из лимфоидных органов интактных крыс, было несколько меньше исходного значения, а в группе, получавшей препараты РНК из лимфоидных органов анемизированных животных, достоверно его превысило.
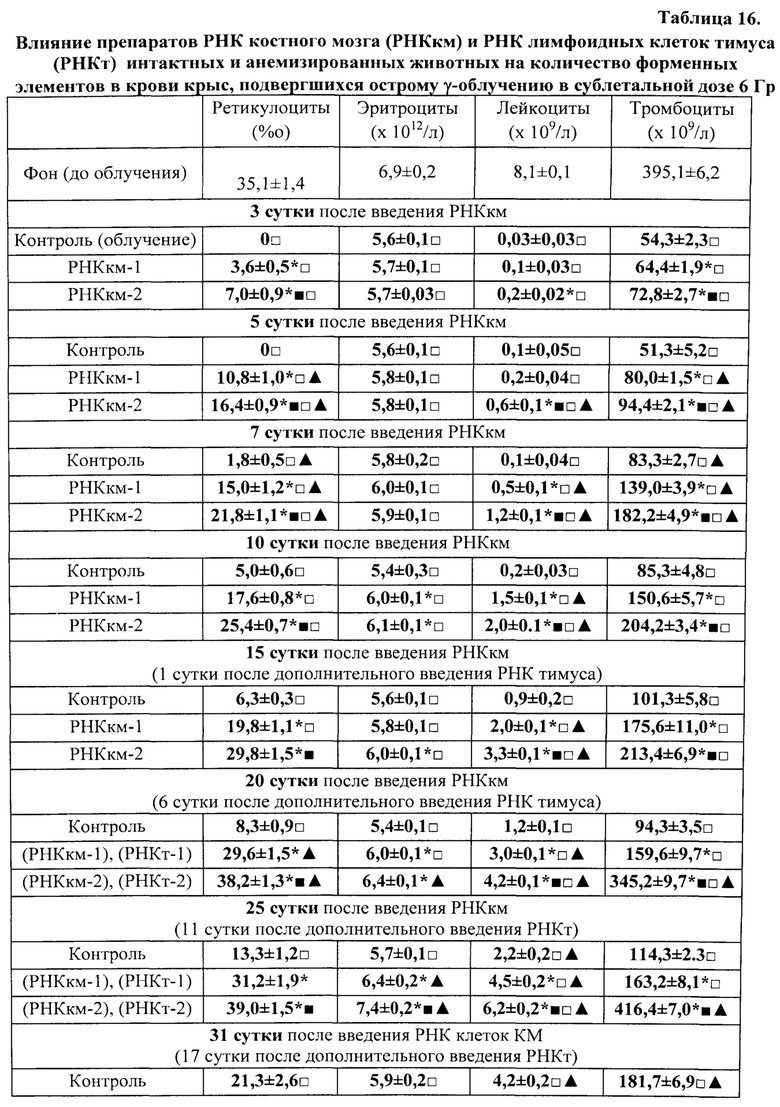
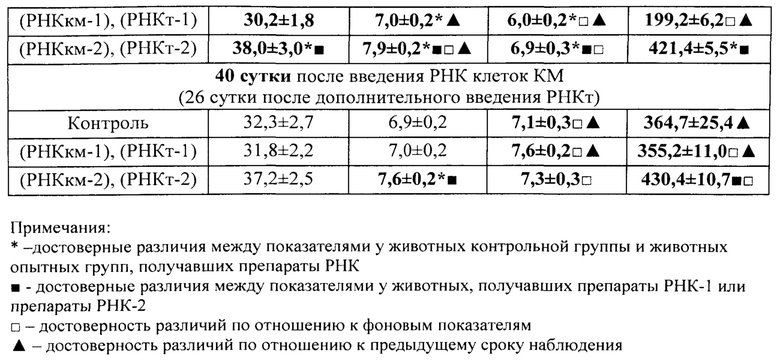
При анализе клеточного состава костного мозга животных на 45 сутки после облучения было установлено, что, несмотря на почти полную нормализацию состава периферической крови, гемопоэз контрольных крыс отличался от интактного кроветворения полным отсутствием молодых бластных элементов белого и красного ростков и значительным уменьшением количества промежуточных форм (Таблица 17). Большая часть клеток костного мозга контрольных животных была представлена дифференцированными элементами - палочко- и сегментоядерными нейтрофилами, эозинофилами и оксифильными эритробластами. В костном мозге животных, получавших препараты РНК из лимфоидных органов интактных крыс, процентное соотношение клеток отличалось от нормы только по числу промиелоцитов, количество которых было в 1,4 раза меньше. Введение же препаратов РНК согласно изобретению из лимфоидных органов анемизированных крыс в организм сублетально облученных животных привело к стимуляции кроветворения, особенно выраженной в направлении развития эритроидного ростка.

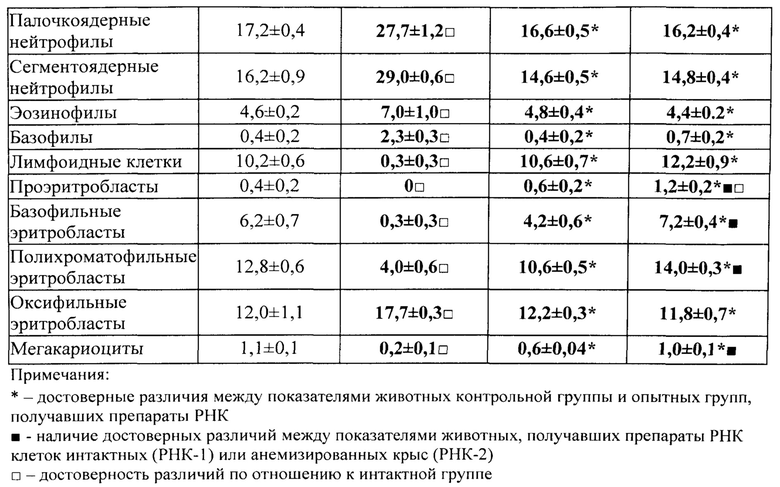
С целью более детального изучения процессов пролиферации, дифференцировки и созревания эритроидных клеток в условиях нашего эксперимента мы проанализировали количественный и качественный состав ЭО костного мозга животных контрольной и опытной групп (Таблица 18). В результате было выявлено, что у облученных крыс через 45 дней после воздействия радиации в костном мозге достоверно увеличивается доля пролиферирующих островков и количество зрелых ЭО 3 класса. Важно отметить, что влияние препарата РНК из лимфоидных органов анемизированных крыс по характеру воздействия на эритропоэз в ЭО отличалось от эффекта препарата РНК из лимфоидных органов интактных животных. Если в группе, получавшей препарат РНК из лимфоидных органов интактных крыс, стимуляция эритропоэза осуществлялась только за счет формирования новых островков, то у животных, получавших препарат РНК из лимфоидных органов анемизированных крыс, ЭО активно образовывались и путем реконструкции эритропоэза. Эта стимуляция процесса комплексирования КОЕэ как со свободными костномозговыми макрофагами, так и с макрофагами, ранее участвовавшими в эритропоэзе, в данной группе животных и привела к достоверному увеличению абсолютного количества ЭО в костном мозге.

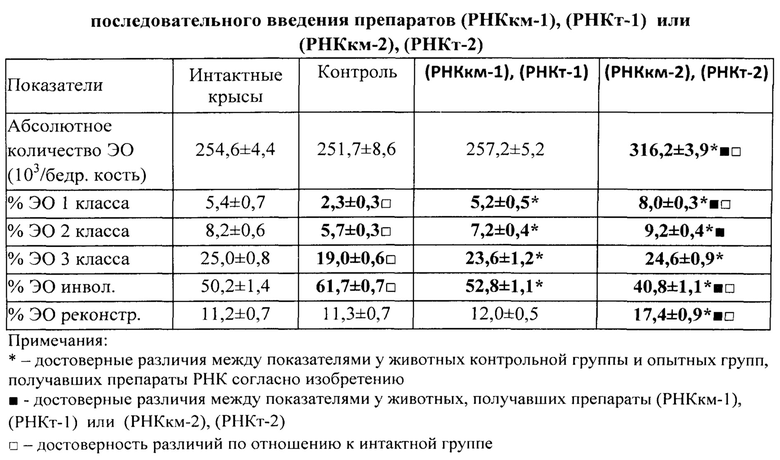
О выраженном эритропоэз-активирующем влиянии препаратов РНК из лимфоидных органов анемизированных животных свидетельствует и изменение показателей интенсивности развития ЭО (Таблица 19). На основании выполненных расчетов можно утверждать, что указанные препараты РНК достоверно повышают интенсивность процесса вовлечения КОЕэ в эритропоэз ЭО, скорость созревания эритроидных клеток в островках и стимулируют повторное вовлечение макрофагов в процесс формирования ЭО, то есть активируют реконструкцию эритропоэза в костном мозге.


Более быстрое и более полное восстановление численности форменных элементов в периферической крови облученных животных под влиянием препарата РНК стимулирующих клеток обусловлено, как оказалось, не только большей дозой вводимого препарата (по сравнению с дозой в 15 мкг/100 г веса тела животного), но и особенностями исходного клеточного материала, из которого выделен препарат тотальной РНК согласно изобретению. Обращает на себя внимание особенно высокая активность препарата РНК из костного мозга анемизированных животных. Вероятно, это обусловлено наличием в костном мозге стволовых клеток, что требует проверки и будет проанализировано далее.
Данные, представленные в настоящем разделе, с несомненностью указывают на то, что под влиянием одного и того же стимулирующего воздействия активация лимфоидных клеток происходит одновременно и однонаправленно. Показано, что действие полученных из них препаратов тотальной РНК согласно изобретению взаимозаменяемо и вполне совместимо.
Важно, что препараты тотальной РНК из лимфоидных органов интактных животных также обладают стимулирующими свойствами, хотя и менее выраженными. Следует также подчеркнуть, что в наших опытах было достигнуто полное восстановление гемопоэза, то есть всех ростков кроветворения, в течение периода немногим более одного месяца.
Результаты данного раздела работы ставят вопрос о возможной целесообразности сочетанного действия препаратов тотальной РНК, полученной из разных источников. Одним из путей проверки такой возможности могут явиться опыты по изучению процесса восстановления кроветворной ткани после хронического бензольного отравления (модель IV) под влиянием препарата тотальной РНК согласно изобретению, полученного из лимфоцитов периферической крови (РНКлпк-1) здорового донора, на том основании, что в периферической крови смешение лимфоцитов из разных лимфоидных органов, по законам гомеостаза, должно происходить самым оптимальным образом.
Результаты такого опыта представляют особый интерес.
Пример 7
Влияние препаратов тотальной РНК из лимфоцитов периферической крови здоровых доноров на процессы восстановления показателей периферической крови и костномозгового кроветворения у крыс с хронической гипопластической бензольной анемией
Особенность данного эксперимента заключается в том, что на этот раз препарат тотальной РНК был получен из чистой фракции нестимулированных лимфоцитов периферической крови здоровых людей (РНКлпкч-1). В результате удалось получить весьма интересные факты.
Во-первых, как оказалось, активность препарата тотальной РНК лимфоцитов периферической крови здорового человека нисколько не ниже, чем активность препарата тотальной РНК суммарных (то есть неразделенных) лимфоидных клеток.
Во-вторых, представленные здесь результаты опыта еще раз убедительно подтверждают отсутствие видового ограничения в отношении действенности и возможности применения препаратов ксеногенной тотальной РНК.
Эксперимент выполнен на 10 белых беспородных крысах-самках массой 120-140 граммов. Гипопластическая анемия у животных была вызвана четырехкратным введением бензола, как описано выше (модель IV). Через 35 дней после последней инъекции бензола в периферической крови крыс отмечалось значительное снижение количества форменных элементов, что свидетельствовало о развитии гипоплазии всех ростков кроветворения в костном мозге. После анализа показателей периферической крови животные были разделены на две группы. Опытной группе однократно внутривенно ввели препарат РНК лимфоцитов здорового донора (группа РНКлпкч-1) из расчета 30 мкг/100 г веса тела животного. Животным контрольной группы также внутривенно было введено по 0,1 мл 0,9% раствора NaCl.
В данном опыте всего одна инъекция препарата тотальной РНК, полученной из лимфоцитов периферической крови человека, приводит к полной нормализации кроветворения у крыс с бензольной анемией уже на 16-21 сутки после инъекции (Таблица 20).
Через 5 суток после введения препарата РНК в периферической крови крыс опытной группы отмечалось достоверное по сравнению с контролем увеличение количества ретикулоцитов (таблица 20), к 10-м суткам, помимо возросшего числа ретикулоцитов, в крови опытных животных начался рост количества лейкоцитов. Через 21 день после введения препарата РНК у животных опытной группы начали регистрироваться даже изменения числа эритроцитов. Важно отметить, что в промежутке между 16 и 30 сутками в периферической крови опытных животных отмечались наиболее выраженные скачки числа ретикулоцитов, лейкоцитов и тромбоцитов: в этот период через каждые 5 дней регистрируемые показатели достоверно превышали значения, полученные в предыдущий срок наблюдения.


Достаточно высокая активность препарата тотальной РНК, полученного из лимфоцитов периферической крови здоровых доноров, освобождает от многих затруднений в плане получения эффективного в конечном итоге препарата для стимуляции гемопоэза.
Тем не менее, как следует из предыдущих опытов, по степени активности препараты, полученные из стимулированных лимфоцитов, безусловно, превосходят препараты, полученные из нестимулированных лимфоидных клеток интактных животных. В связи с этим в определенных случаях, когда необходима срочная стимуляция кроветворения, мы полагаем, что в качестве доноров активированного кроветворения могут выступать, например, жители высокогорных районов. Кроме того, аллогенные варианты препаратов тотальной РНК с еще большей стимулирующей или ингибирующей активностью можно получить также и из разделенных на клеточном сортере фракций Т-хелперов и Т-супрессоров.
Поскольку известно, что активность Т-супрессоров, в отличие от Т-хелперов, отличается значительно менее выраженной тканеспецифичностью, подавляя пролиферацию далеко не только той ткани, которая индуцировала их появление, это еще более расширяет возможности использования регуляторных лимфоидных клеток и полученных из них препаратов тотальной РНК согласно изобретению.
Описанные выше результаты, свидетельствующие о реальных возможностях лечения анемии, заставили нас задуматься о том, эффективны ли препараты согласно изобретению - и насколько они эффективны - при других путях их введения в организм. Этот вопрос встал, в частности, в связи с поиском менее инвазивных и неинвазивных путей введения, и в этом отношении особый интерес представляет, в частности, интраназальное (и/н) введение. Результаты указанного поиска описаны в следующем примере 8.
Пример 8
Эффективность препаратов тотальной РНК в зависимости от пути их введения
С целью выяснения сравнительной эффективности препаратов тотальной РНК согласно изобретению при разных путях введения, была предпринята отдельная серия экспериментов, в которой, используя крыс с хронической гипопластической бензольной анемией (модель (IV)), лечение разных групп животных осуществляли, используя разные пути введения стимулирующего препарата тотальной РНКс-2.
В эксперименте использовали 30 крыс-самцов массой 190-210 г.
Для подкожного (п/к), внутримышечного (в/м), внутрибрюшинного (в/б) и внутривенного (в/в) введений была выбрана следующая схема ведений:
Первую инъекцию препарата РНКс-2 осуществляли в дозе 30 мкг/100 г массы, вторую (через 7 дней) - в дозе 10 мкг/100 г массы животного.
Интраназальное (и/н) введение: закапывание в обе ноздри 3 дня подряд в дозе 10 мкг/100 г массы, 4-й раз - через 7 дней после последнего введения - в дозе 10 мкг/100 г массы животного.
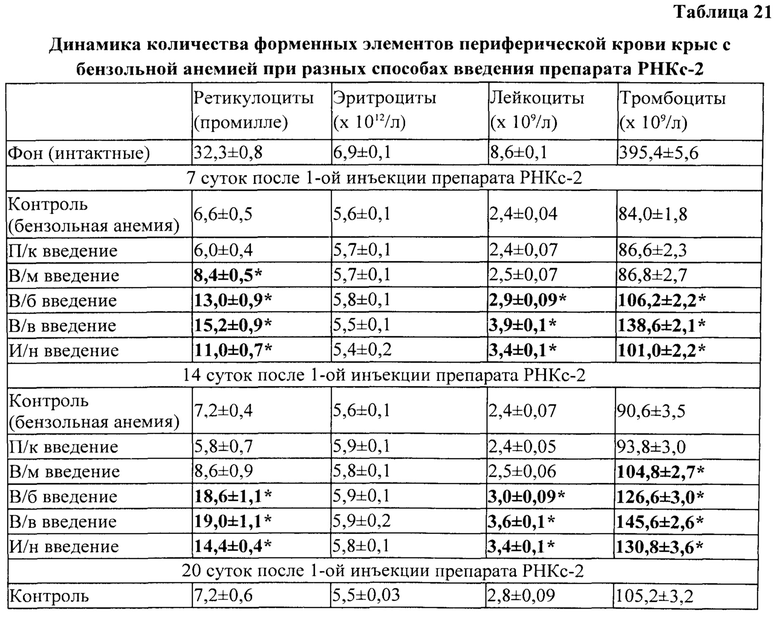

Помимо указанной выше статистической обработки полученных данных, для определения наиболее удачных способов введения препарата РНКс-2 был предпринят также кластерный анализ, который позволил выявить, что полученные результаты образуют 2 кластера. В первый кластер вошли показатели контрольной группы и групп животных, получивших препарат подкожно или внутримышечно. Второй кластер образован данными, полученными в группах животных, которым исследуемое вещество вводили внутривенно, внутрибрюшинно или интраназально. Эти данные свидетельствуют о том, что, несмотря на полученные в данном опыте отличия, внутрибрюшинный и интраназальный пути введения являются не менее эффективными и перспективными, чем внутривенное введение препарата РНКс-2. На основании таких оценок интраназальный путь был использован нами для введения и других препаратов согласно изобретению. В частности, в опытах по лечению сахарного диабета интраназальный путь введения, наряду с внутривенным и внутрибрюшинным, оказался не только приемлемым, но и достаточно эффективным.
В этой связи, есть все основания полагать, что не менее эффективным окажется и ректальный путь введения препаратов согласно изобретению.
Следует, однако, отметить, что результаты подкожного и внутримышечного введения, полученные в данном примере, означают только то, что такие пути введения не являются эффективными для реализации эффекта препаратов на уровне целого организма, или то, что для реализации эффекта в этих случаях требуются более высокие дозы препаратов. Тем не менее, как станет очевидно из примера 13. даже наружное (местное) применение препаратов согласно изобретению оказывается достаточно эффективным.
Из основной используемой здесь концепции авторов настоящего изобретения следует, что система иммуногенеза как система общей регуляции на уровне целого организма должна оказывать модулирующее действие не только на лимфоидные ткани и не только на гемопоэтические клетки, но и на клетки других гистотипов. Кроме того, на основании данных других авторов, препараты РНК, полученные из клеток того или иного органа, оказывают положительное влияние на клетки гомологичных органов и тканей других организмов.
Таким образом, исходя из указанной совокупности знаний, мы вплотную подошли к исследованию возможностей лечения заболеваний, патогенез которых связан с нарушениями в системе лимфоидных клеток. В этой связи необходимо отметить, что особого внимания заслуживают полученные нами данные по использованию препарата тотальной РНК, полученного из лимфоцитов периферической крови человека (см. пример 7). Такой препарат, не требующий подбора по группе крови и антигенам гистосовместимости, к тому же при условии интраназального введения в организм, может быть использован в медицинской практике уже в ближайшее время.
Пример 9
Влияние препаратов РНК, полученных из лимфоидных клеток селезенки, на состояние мутантных мышей линии CSPBL/RsJYLeprdb/+ с сахарным диабетом 2 типа.
В самых предварительных опытах мы изучали влияние однократного введения препаратов РНКс-1, РНКс-2 и РНКс-3 из лимфоидных клеток селезенки крыс на состояние мутантных мышей линии C57BL/KsJYLeprdb/+, которая, как было показано, может быть использована в качестве адекватной модели сахарного диабета 2 типа в эксперименте (Степанова О.И., Каркищенко, В.Н., Баранова, О.В. и др. Мутантные мыши линии C57BL/KsJYLeprdb/+ как генетическая модель сахарного диабета 2-го типа Бюл. экспер. биол. и мед., 2007, №12, с. 664-667). Хотя численные экспериментальные данные здесь не приведены, однако в этих опытах нами были получены результаты, свидетельствующие о благотворном, но качественно различном влиянии указанных препаратов РНК. согласно изобретению на общее состояние животных и на уровни глюкозы в их крови.
Так, уже к шестому дню после однократного внутрибрюшинного введения препаратов РНКс-1 или РНКс-2 приблизительно у 40% животных наблюдался положительный эффект в отношении уровня глюкозы крови. У двух животных из 28 с первоначальным уровнем сахара в крови >33, в ответ на однократное введение препарата РНКс-2, уровень глюкозы в крови снизился соответственно на 35,2% и 25,1%, после чего в течение 49 дней (вплоть до забоя) у первого из них и в течение 35 дней у второго уровень сахара в крови неизменно снижался.
Весьма интересным результатом является и то, что после однократного внутрибрюшинного введения препаратов РНКс-1 наблюдалось постепенное и полное заживление мацераций кожных покровов у всех животных (у 8 из 28), у которых такие явления имели место. Здесь уместно отметить, что, согласно литературным данным [Степанова О.И. Дис.… канд. биол. наук «Трансплантация клеток костного мозга для коррекции патогенетических нарушений при сахарном диабете 2 типа», Москва, 2009], у 20-25% мышей-диабетиков db/db в возрасте 120-158 дней в области холки развивается мацерация кожных покровов, которая в течение последующих 5-14 дней становится обширной незаживающей раной и остается у животного вплоть до его гибели. В наших опытах обращает на себя внимание также и тот факт, что эффект ранозаживления, достигнутый в результате однократного введения препарата РНКс-1, был достаточно длительным (более одного месяца). По прошествии указанного срока у некоторых животных стали появляться участки с мацерацией кожи, что четко коррелировало с ослаблением эффекта однократного введения препаратов РНК, поскольку совпадало и с некоторым повышением уровня глюкозы в крови. Под действием препаратов РНКс-1 и РНКс-2 у большинства животных наблюдалась нормализация массы тела и снижение диуреза, и в целом животные стали потреблять меньше воды и пищи.
При сравнении результатов гистологического исследования, проведенного через 2 месяца после начала лечения препаратами РНК согласно изобретению, с результатами гистологического анализа контрольных мышей-диабетиков db/db указанной линии того же 4-6-месячного возраста [Степанова О.И. Дис.… канд. биол. наук «Трансплантация клеток костного мозга для коррекции патогенетических нарушений при сахарном диабете 2 типа», Москва, 2009], было обнаружено следующее. У 4-6-месячных мышей линии C57BL/KsJYLeprdb/+, служащих экспериментальной моделью сахарного диабета 2 типа, в поджелудочной железе отмечались явления выраженного перидуктального и интралобулярного склероза с атрофией паренхимы железы, а также интра- и перилобулярного липоматоза. Между прослойками соединительной и жировой ткани определялись очень мелкие атрофичные панкреатические островки, представленные скоплением небольшого количества базофильных клеток. Селезенка у этих мышей подвергалась прогрессирующей гипоплазии. В лимфоидных фолликулах селезенки были отмечены явления гипоплазии и атрофии. Площадь лимфоидных фолликулов в селезенке и в регионарном лимфатическом узле была уменьшена более чем в 2 раза по сравнению с нормальным контролем.
По прошествии 2-месячного срока после начала лечения препаратами РНК согласно изобретению в поджелудочной железе леченых животных количество панкреатических островков в пределах нормы, панкреатические островки среднего размера, правильной овальной или округлой формы, с четкими контурами, все островки клеточного строения. Отмечалось выраженное полнокровие стромальных сосудов. В селезенке определялись признаки умеренной гиперплазии лимфоидной ткани, с образованием единичных лимфоидных фолликулов, признаки выраженного полнокровия красной пульпы. Вообще особого внимания заслуживает тот факт, что на фоне лечения препаратами согласно изобретению кровенаполнение отмечалось также и в печени, и в почечной ткани.
Итак, результаты проведенных нами к настоящему времени предварительных исследований на экспериментальной модели сахарного диабета 2 типа определенно свидетельствуют о повышении регенерационной способности не только железистого эпителия поджелудочной железы, но и кожного эпителия под действием препарата РНКс-1.
Недостатком генетических моделей СД, как считают авторы патента RU 2400822, является то, что они воспроизводятся у животных, наследственно предрасположенных к СД, что искажает характер компенсаторных механизмов и регенерацию островковой ткани поджелудочной железы вследствие исходно измененного реагирования адаптивных систем организма. В то же время аутоиммунная модель СД, получаемая при сочетанием введении адъюванта Фрейнда и аллоксана (см. патент RU 2400822) здоровым крысам, позволила разработать схемы адекватной терапии данного заболевания у животных без генетических дефектов развития иммунной системы.
К этому можно добавить, что генетическая модель, хоть и является наиболее приближенной популяционной моделью, однако, в силу ее нестандартизованности и больших индивидуальных различий даже в группах животных одинакового возраста, достаточно проблематична для экспериментальных исследований и статистической обработки результатов.
Пример 10
Лечение аллоксановой модели сахарного диабета (СД) 1 типа (модель (VI))
Способ 1
Приступая к лечению аллоксановой модели сахарного диабета 1 типа, мы учитывали, в частности, ставшую известной из medicalnewstoday.com информацию о том, что у больных сахарным диабетом обнаружено наличие микроангиопатии, поражения мелких кровеносных сосудов, костного мозга (http:/www.medkurs.ru/news/39325.html). Это обстоятельство, наряду с описанными выше результатами наших опытов, свидетельствующими о высокой активности регуляторных препаратов тотальной РНК из костного мозга, послужило дополнительным аргументом в пользу применения препаратов РНКкм в комплексном лечении аллоксанового диабета. Не менее важным компонентом в лечении данной патологии мы считали разновидности препаратов РНК лимфоидных клеток селезенки. Наконец, таким же точно способом, что и остальные препараты согласно изобретению, мы получили препарат тотальной РНК из гомогената поджелудочной железы (п/ж) крыс и использовали его во всех схемах лечения аллоксанового диабета. При этом мы исходили из того, что для нормализации процессов, протекающих в организме при данной патологии, важно нормализовать не только регуляторные процессы с помощью препаратов РНК из лимфоидных клеток здоровых животных, но и поддержать функциональный резерв поджелудочной железы реципиента. Кроме того, в ряде публикаций предшествующего уровня техники сообщалось о благотворном влиянии экзогенных РНК, полученных из разных органов донора, на функцию гомологичных органов в организме реципиента.
Животные были разделены на группы (по 5 крыс в каждой группе). Группам контрольных животных с экспериментальным СД 1 типа внутрибрюшинно вводили 0,9% раствор NaCl в те же сроки и в том же объеме, что и исследуемые препараты животным опытных групп.
В общей сложности в двух сериях опытов было использовано 60 животных. При этом в первой серии на 35 крысах, по 5 крыс в разных экспериментальных группах и в группе контрольных животных, изучали поэтапное действие трех разновидностей препаратов РНК согласно изобретению в разных сочетаниях и в разной последовательности: РНКкм-1; РНКкм-2; РНКс-1; РНКс-2; РНКт-3; РНКп/ж. На 30 крысах опытных групп были испробованы разные режимы и схемы последовательного введения препаратов РНК, и в каждом случае препараты вводили внутрибрюшинно один раз в 7 дней в дозе 15 мкг/100 г веса животного.
Во второй серии опытов на 25 крысах (по 5 в разных экспериментальных группах и в группе контрольных животных), изучали некоторые нюансы отработанных в первой серии схем введения путем варьирования интервалов между введениями тех или иных препаратов. Помимо этого, во второй серии экспериментов мы изучали возможность суммарного введения препаратов РНК, оказавшихся, по результатам первой серии опытов, оптимальными для достижения полного восстановления уровня глюкозы в крови экспериментальных животных. В этом же опыте варьировали количества и пути введения оптимального набора средств согласно изобретению из тех, что указаны выше.
Постановка опыта в трех экспериментальных группах, в которых выбранные схемы лечения оказались самыми оптимальными, отражена в таблице 22.
В указанных трех экспериментальных группах схемы введения препаратов РНК были следующими:
1 - контроль (аллоксановый диабет)
2 - (РНКкм-1, РНКс-1, РНКп/ж) по отдельности, внутрибрюшинно
3 - (РНКкм-1+РНКс-1+РНКп/ж) в смеси, внутрибрюшинно
4 - (РНКкм-1+РНКс-1+РНКп/ж) в смеси, интраназально
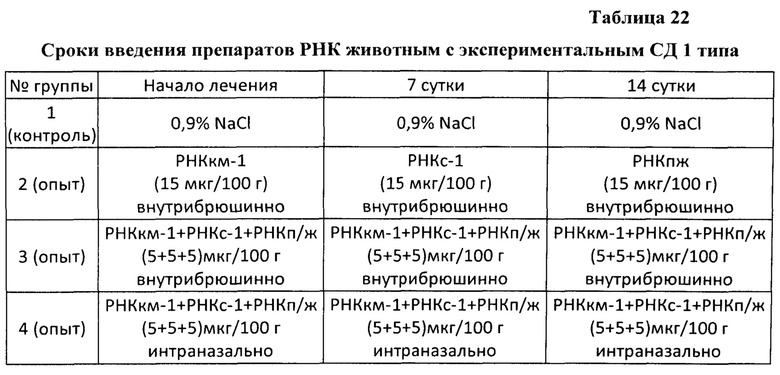
В таблице 23 представлены результаты, полученные в ходе лечения в указанных трех экспериментальных группах с самыми оптимальными из изученных нами схемами лечения.
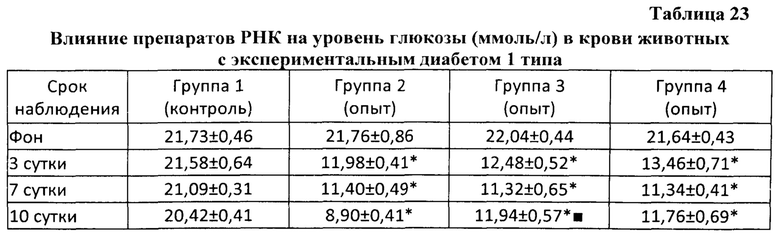
Еще одной группе крыс с аллоксановым диабетом однократно в/б вводили одновременно все три указанных препарата, РНКкм-1+РНКс-1+РНКп/ж (15+15+15) мкг/100 г массы тела животного, чтобы проверить, достаточно ли эффективны указанные три препарата согласно изобретению при их совместном однократном введении, чтобы нормализовать уровень глюкозы в крови больных животных. Результаты данного опыта, представленные в таблице 24, свидетельствуют о том, что совместное введение всех трех компонентов в надлежащей дозе (15 мкг/100 г массы тела) приводит даже к еще более быстрому темпу восстановления функции поджелудочной железы, чем в наиболее эффективной опытной группе 2 (см. таблицу 23).
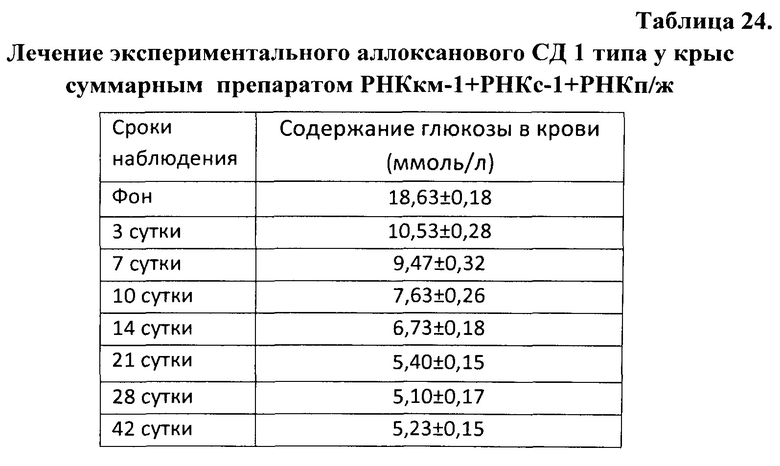
Итак, на основании результатов, полученных в предпринятых нами сериях исследований, можно считать установленным, что полного восстановления функции островкового аппарата поджелудочной железы у крыс с аллоксановым диабетом 1 типа можно добиться уже за три недели (21 день) в результате еженедельного последовательного в/б введения трех препаратов тотальной РНК согласно изобретению, РНКкм-1, РНКс-1 и РНКп/ж, в количестве 15 мкг/100 г веса тела (см. опыты 3 и 4 в таблицах 22 и 23). При сочетанном введении равных количеств всех трех указанных препаратов РНК, суммарная доза которых на одно введение составляла 15 мкг/100 г веса тела животного, полное восстановление функции островкового аппарата поджелудочной железы наступало на 42-е сутки лечения. Важно подчеркнуть, что при такой схеме лечения препаратами согласно изобретению внутрибрюшинный и интраназальный пути введения оказались практически одинаковыми по эффективности. Это открывает широкие возможности для неинвазивного лечения диабета 1 типа с помощью предлагаемых в настоящей заявке препаратов на основе естественных и в то же время неиммуногенных компонентов клеток здорового организма.
Весьма существенным результатом наших исследований данной модели явилось и то, что абсолютно все предпринятые нами схемы лечения оказались всего лишь по срокам излечения менее оптимальными, чем три приведенные выше схемы. При этом любое промежуточное воздействие любым из шести указанных выше препаратов согласно настоящему изобретению, которое в процессе проводимого нами лечения оказывалось менее эффективным по сравнению с самыми оптимальными, без каких-либо осложнений корректировалось последующим введением оптимального варианта РНК согласно изобретению. Это, собственно, и являлось в каждом случае единственной причиной удлинения срока полного восстановления уровня глюкозы в крови.
Важно отметить еще некоторые особенности процессов восстановления функции островкового аппарата поджелудочной железы, которые мы наблюдали при изучении данной модели сахарного диабета 1 типа. Каждый из использованных нами препаратов (РНКкм-1, РНКс-1 и РНКп/ж) требовал вполне определенного времени для реализации восстановительных процессов, происходящих с его участием. По прошествии необходимого периода падение уровня глюкозы в крови прекращалось, после чего дальнейшего падения не происходило, но и достигнутый уровень удерживался системой организма достаточно стабильно. Дальнейшее восстановление функции островкового аппарата поджелудочной железы под воздействием следующего препарата из указанной триады тоже требовало вполне определенного времени для реализации своей регенераторной функции и приводило к дальнейшему снижению уровня глюкозы в крови, но тоже до определенных значений, ниже которых этот показатель не опускался даже при длительном интервале между введениями (14 и более суток). И только под действием всех трех указанных препаратов, вводимых в любой последовательности, обеспечивалось полное восстановление функции инсулинпродуцирующего аппарата поджелудочной железы.
Из этого следует, что использованные три препарата РНК качественно различны и, что важно, не являются взаимозаменяемыми. По-видимому, каждый из них имеет свою мишень.
Наконец, здесь уместно добавить информацию, свидетельствующую об отсутствии токсичности описанных в настоящей заявке регуляторных препаратов. Так, когда была сделана попытка лечения мышей-генетических моделей СД 2 типа (пример 9), эффективные количества препаратов еще не были отработаны, поэтому животным вводили заведомо большие дозы препаратов (однократно в/б приблизительно от 45 до 100 мкг/100 г веса тела). Поскольку в дальнейшем опыты на крысах, поставленные на разных описанных здесь моделях, показали, что эффективными являются уже количества от 5 до 15 мкг/ 100 г массы животного и поскольку у мышей, получивших 6-7-кратные дозы препаратов, никаких необычных признаков поведения отмечено не было, можно смело утверждать, что предлагаемые препараты в пределах указанных доз никакой токсичностью не обладают.
Примеры 11 и 12
Лечение аллоксановой модели сахарного диабета (СД) 1 типа (модель (VI))
Способ 2 и Способ 3
В поисках дальнейших путей лечения, не связанных с введением ксеногенных для человека препаратов, которые позволили бы в максимально сжатые сроки реализовать способы согласно настоящему изобретению в медицинской практике, были предприняты еще две попытки лечения аллоксанового диабета на избыточных контрольных животных, у которых за 70 дней до этого срока был вызван аллоксановый диабет. С этой целью указанным животным однократно в/б вводили либо препарат тотальной РНК из клеток стромы пупочного канатика человека (РНКмск) (в количестве 10,0 мкг/100 г массы крысы) совместно с регуляторным препаратом тотальной РНК из лимфоцитов периферической крови человека (РНКлпкч-1) (в количестве 23,4 мкг/100 г массы), либо только РНКмск из стромы пупочного канатика человека (в количестве 10,7 мкг/100 г массы крысы).
Полученные результаты представлены в таблице 25.

Таким образом, как видно из таблицы, эффективны оба варианта лечения, в одном из которых используется препарат тотальной РНК из стволовых клеток стромы пуповины человека, а в другом - суммарный препарат тотальной РНК из стволовых клеток стромы пуповины человека и тотальной РНК из лимфоцитов периферической крови человека.
Данный пример является, помимо прочего, еще и свидетельством того, что препарат тотальной РНК, полученный из стволовых клеток, может быть эффективной заменой самих стволовых клеток при лечении заболеваний, поддающихся лечению стволовыми клетками. К таковым в настоящее время относят заболевания, выбранные из группы, включающей в себя, но не ограниченной боковым амиотрифическим склерозом (БАС), детским церебральным параличом (ДЦП), эпилепсией, травмой спинного мозга, травмой головного мозга и травматической инфекцией мозга, инсультом, болезнью Паркинсона, множественной системной атрофией, рассеянным склерозом, системной красной волчанкой, болезнью Девика, аутоиммунными заболеваниями, дистрофией сетчатки, пигментным ретинитом, глаукомой и другими глазными болезнями и нарушениями зрения, сахарным диабетом, диабетической стопой, мышечной дистрофией, аутизмом и глубокой задержкой развития, прогрессивным надъядерным параличом, кортикобазальной дегенерацией, болезнью Альцгеймера, болезнью Хантингтона, болезнью Баттена, наследственной атаксией, спиноцеребеллярной атаксией, атаксией Фридрейха, кардиомиопатией, хронической сердечной недостаточностью, инфарктом миокарда (осложнения), алопецией, артритом, хронической почечной недостаточностью, циррозом печени, ишемией нижних конечностей, остеопорозом и некрозом головки бедренной кости, ретинопатией недоношенных, нейро-сенсорной тугоухостью, врожденным амаврозом Лебера.
Несмотря на высокую эффективность воздействия препарата тотальной РНК, полученного из стволовых клеток здоровых доноров, мы полагаем, что сочетанное введение препаратов, полученных из стволовых клеток, с препаратами регуляторной РНК, полученными, в частности, из лимфоцитов периферической крови здорового человека, повышают не только эффективность (см. таблицу 25), но также надежность, и безопасность лечения по сравнению с лечением стволовыми клетками как таковыми.
Пример 13
Влияние препаратов РНКт-1 и РНКт-3, полученных из лимфоидных клеток тимуса, на рост волос.
В связи с предполагаемыми возможностями применения препаратов согласно изобретению также и в косметических целях - в данном случае для проверки возможной стимуляции и торможения роста волос - была исследована регуляторная активность препаратов РНКт-1 и РНКт-3, полученных, соответственно, из лимфоидных клеток тимуса интактных крыс и из лимфоидных клеток тимуса крыс через четверо суток после острой кровопотери, то есть в фазе торможения.
В соответствии с поставленной задачей, у беспородных крыс черного окраса осуществляли тримминг на симметричных участках кожи на спине по обе стороны позвоночника. Площадь участка, лишенного волосяного покрова, составляла в среднем 4 см2. На левый участок каждой из крыс ежедневно наружно наносили 50 мкл раствора, содержащего либо 10 мкг РНКт-1, либо 10 мкг РНКт-3, а на правый - такое же количество дистиллированной воды.
На 17-е сутки такой обработки длина волоса у крысы с удаленным волосяным покровом на участке, который обрабатывали препаратом РНКт-1, составила 6 мм, тогда как на контрольном участке кожи у той же крысы она составляла 3-4 мм.
В этот же срок, на 17-е сутки обработки, длина волоса у другой крысы с удаленным волосяным покровом на участке, который обрабатывали препаратом РНКт-3, составила 3 мм, тогда как на контрольном участке кожи у этой же самой крысы она составляла 6-8 мм.
Следует отметить, что крысы, которые были взяты в опыт, заметно отличались друг от друга густотой и длиной волосяного покрова. В связи с этим для опыта по торможению роста волос было специально выбрано животное с очень густо растущей и длинной шерстью, у которого, по нашим представлениям, труднее было затормозить рост волос, а для опыта по стимуляции роста волос - наоборот, крыса со значительно менее пышным волосяным покровом.
Таким образом, мы показали, в подтверждение изложенной здесь концепции, что препараты согласно настоящему изобретению способны даже при местном введении осуществлять регуляцию роста - в данном случае регуляцию роста волос. Так проявляется общерегуляторное действие РНК, полученной из клеток системы общей регуляции, к которой, как мы полагаем, в каждом случае для усиления эффекта в отношении конкретной ткани-мишени следует добавлять органоспецифический компонент, предпочтительно полученный у молодого здорового донора.
Теперь, возвращаясь к вопросу о путях введения препаратов согласно изобретению (см. пример 8), можно смело утверждать, что, говоря о том, что подкожное и внутримышечное введения, в отличие от внутривенного, внутрибрюшинного и интраназального, малоэффективны, надо помнить, что речь идет о слабой эффективности этих путей введения только на уровне целого организма, но что при локальном введении они могут быть исключительно результативны.
Далее приведены некоторые иллюстративные воплощения настоящего изобретения:
1. Композиция, содержащая препарат тотальной РНК, выделенный из интактной (не подвергнутой активации) лимфоидной клетки или ткани костного мозга здорового донора и/или из донорских лимфоидных клеток или ткани костного мозга здорового донора, подвергнутых активации Т-клеточной популяции.
2. Композиция согласно воплощению 1, где указанная композиция или препарат тотальной РНК модулируют пролиферацию и/или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа.
3. Композиция согласно воплощениям 1 или 2, где указанная композиция или препарат тотальной РНК, выделенный из интактной лимфоидной клетки или ткани костного мозга здорового донора и/или из донорских лимфоидных клеток или ткани костного мозга здорового донора, подвергнутых активации Т-клеточной популяции, являются регуляторными.
4. Композиция согласно воплощению 1, где указанная активация Т-клеточной популяции осуществляется in vivo, ex vivo или in vitro.
5. Композиция согласно воплощению 2, где указанная модуляция гомологичной ткани или клетки и/или соматической клетки другого гистотипа осуществляется in vivo, ex vivo или in vitro.
6. Композиция согласно воплощениям 2 или 5, где модуляция пролиферации и/или дифференцировки осуществляется в организме реципиента при введении ему композиции препарата тотальной РНК.
7. Композиция согласно воплощениям 1, 2 или 6, где модуляция пролиферации и/или дифференцировки представляет собой стимуляцию пролиферации или дифференцировки гомологичной ткани или клетки и/или соматической клетки другого гистотипа.
8. Композиция согласно воплощению 7, где препарат тотальной РНК выделяют в фазе проявления у клеток донора способности стимулировать пролиферацию или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа.
9. Композиция согласно воплощению 7, где указанная способность стимулировать пролиферацию или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа осуществляется в интервале приблизительно от 15 минут примерно до 48 часов после активации Т-клеточной популяции.
10. Композиция согласно воплощениям 1, 3 или 6, где модуляция пролиферации и/или дифференцировки представляет собой ингибирование пролиферации и/или дифференцировки гомологичной ткани или клетки и/или соматической клетки другого гистотипа.
11. Композиция согласно воплощению 10, где препарат тотальной РНК выделяют в фазе проявления у клеток донора способности ингибировать пролиферацию и/или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа.
12. Композиция согласно воплощениям 10 или 11, где указанная способность ингибировать пролиферацию и/или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа осуществляется в интервале приблизительно от 48 часов примерно до 96 часов после активации Т-клеточной популяции.
13. Композиция согласно любому из указанных выше воплощений, дополнительно содержащая препарат тотальной РНК, выделенный из соматической клетки здорового донора.
14. Композиция согласно воплощению 13, где указанная соматическая клетка является стволовой клеткой.
15. Композиция согласно любому из указанных выше воплощений, где лимфоидная клетка является лимфоидной клеткой, выделенной из селезенки, тимуса, лимфатического узла или из популяции лимфоцитов периферической крови.
16. Композиция согласно любому из указанных выше воплощений, где донором является млекопитающее.
17. Композиция согласно воплощению 16, где донором-млекопитающим является аллогенный донор.
18. Композиция согласно воплощению 16, где донором-млекопитающим является человек.
19. Композиция согласно воплощению 15, где донором-млекопитающим является ксеногенный донор.
20. Композиция согласно воплощению 1, где донор не является млекопитающим.
21. Композиция согласно любому из указанных выше воплощений, где указанную композицию вводят реципиенту-млекопитающему.
22. Композиция согласно воплощению 21, где реципиентом-млекопитающим является человек.
23. Композиция согласно воплощениям 1-16, 18, 20, где реципиент не является млекопитающим.
24. Композиция согласно воплощению 3, где регуляторный препарат РНК выделен из интактной лимфоидной клетки или интактной ткани костного мозга.
25. Композиция согласно любому из воплощений 1-24, где указанный здоровый донор является молодым донором.
26. Композиция согласно любому из воплощений 1-12, 15-25, где у донора получена фракция регуляторной РНК.
27. Композиция согласно воплощению 26, где фракция регуляторной РНК имеет среднюю длину в интервале приблизительно от 50 примерно до 50000 и более нуклеотидов.
28. Композиция согласно воплощению 1, содержащая препарат тотальной РНК и фармацевтически приемлемый носитель, разбавитель и/или наполнитель.
29. Композиция согласно воплощению 28, представленная в жидкой, лиофилизированной или твердой форме.
30. Композиция согласно воплощениям 1 или 28, составленная для системного или локального введения.
31. Композиция согласно воплощениям 1, 28 или 30, где под введением подразумевается интраназальное, парентеральное, внутриочаговое или местное введение.
32. Способ получения препарата тотальной РНК согласно воплощениям 1-27, где из лимфоидных клеток или из ткани костного мозга донора, подвергнутых активации Т-клеточной популяции,
а) в фазе проявления у клеток донора способности стимулировать пролиферацию или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа или
б) в фазе проявления у клеток донора способности ингибировать пролиферацию и/или дифференцировку гомологичной ткани или клетки и/или соматической клетки другого гистотипа,
или
из интактных (не подвергнутых активации) лимфоидных клеток или ткани костного мозга донора выделяют препарат тотальной РНК.
33. Способ согласно воплощению 32, где указанная активация осуществляется in vivo, ex vivo или in vitro.
34. Способ согласно воплощению 32, где стадию активации осуществляют in vivo.
35. Способ согласно воплощению 32, где стадию активации осуществляют in vitro.
36. Способ согласно воплощению 32, где стадию активации осуществляют ех vivo.
37. Способ согласно воплощению 32, где способность стимулировать осуществляется в интервале приблизительно от 15 минут примерно до 48 часов после активации Т-клеточной популяции.
38. Способ согласно воплощению 32, где способность ингибировать осуществляется в интервале приблизительно от 48 часов примерно до 96 часов после активации Т-клеточной популяции.
39. Способ согласно любому из воплощений 32-38, где лимфоидная клетка является лимфоидной клеткой селезенки, тимуса, лимфатического узла или клеткой из популяции лимфоцитов периферической крови.
40. Способ согласно любому из воплощений 32-38, где донором является млекопитающее.
41. Способ согласно воплощению 40, где донором-млекопитающим является молодое здоровое млекопитающее.
42. Способ модуляции пролиферации и/или дифференцировки соматических клеток-мишеней в организме реципиента, предусматривающий введение указанному реципиенту терапевтически эффективного количества композиции согласно воплощениям 1 или 28 или препарата тотальной РНК согласно воплощению 1.
43. Способ согласно воплощению 42, согласно которому клетка-мишень отличается нарушенной пролиферативной активностью и/или дифференцировкой.
44. Способ согласно воплощениям 42 или 43, согласно которому клетка-мишень является соматической клеткой любого гистотипа.
45. Способ согласно любому из воплощений 42-44, где реципиентом является млекопитающее.
46. Способ согласно воплощению 45, где указанным млекопитающим является человек.
47. Способ согласно любому из воплощений 43-44, где реципиент не является млекопитающим.
48. Способ лечения заболевания или расстройства, ассоциированного с нарушенной пролиферацией и/или дифференцировкой соматической клетки-мишени (клеток-мишеней) того или иного гистотипа (гистотипов), предусматривающий введение пациенту терапевтически эффективного количества композиции согласно воплощениям 1 или 28 или препарата тотальной РНК согласно воплощению 1.
49. Способ согласно воплощению 48, где заболевание или расстройство, ассоциированное с нарушенной пролиферацией и/или дифференцировкой соматической клетки-мишени, является дегенеративным заболеванием или расстройством, нейродегенеративным заболеванием или расстройством; аутоиммунным заболеванием или расстройством, гипопролиферативным заболеванием или расстройством, гиперпролиферативным заболеванием или расстройством, доброкачественным неопластическим расстройством, злокачественным неопластическим расстройством; наследственным или врожденным дефектом, формой сахарного диабета или расстройством, поддающимся лечению на основе стволовых клеток.
50. Способ согласно воплощению 49, где неопластическим заболеванием или расстройством является аденома предстательной железы.
51. Способ лечения и профилактики гематологического заболевания или расстройства, требующего переливания крови или ее форменных элементов, предусматривающий введение пациенту терапевтически эффективного количества композиции согласно воплощениям 1 или 28 или препарата тотальной РНК согласно воплощению 1 в качестве полной или частичной замены гемотрансфузии.
52. Способ согласно воплощению 51, где гематологическим заболеванием или расстройством является анемия.
53. Способ согласно воплощению 51, где указанный пациент был подвергнут облучению.
54. Способ согласно воплощению 53, где облучение обусловлено лечением опухолевого заболевания.
55. Способ согласно воплощениям 53 или 54, где указанное гематологическое заболевание или расстройство обусловлено облучением.
56. Способ согласно воплощению 51, где указанный пациент был подвергнут химиотерапии.
57. Способ согласно воплощению 56, где указанное гематологическое заболевание или расстройство обусловлено химиотерапией.
58. Способ согласно воплощениям 53-57, где указанное введение пациенту терапевтически эффективного количества композиции согласно воплощениям 1 или 28 или препарата тотальной РНК согласно воплощению 1 предпринимают в интервале от 15 минут до 3 часов перед облучением или химиотерапией.
59. Способ согласно воплощению 49, где расстройством, поддающимся лечению на основе стволовых клеток, является боковой амиотрифический склероз (БАС), детский церебральный паралич (ДЦП), эпилепсия, травма спинного мозга, травма головного мозга, травматическая инфекция мозга, инсульт, болезнь Паркинсона, множественная системная атрофия, рассеянный склероз, системная красная волчанка, болезнь Девика, аутоиммунное заболевание, дистрофия сетчатки, пигментный ретинит, глаукома, глазная болезнь, нарушение зрения, сахарный диабет, мышечная дистрофия, аутизм, задержка развития, прогрессивный надъядерный паралич, кортикобазальная дегенерация, болезнь Альцгеймера, болезнь Хантингтона, болезнь Баттена, наследственная атаксия, спиноцеребеллярная атаксия, атаксия Фридрейха, кардиомиопатия, хроническая сердечная недостаточность, инфаркт миокарда, алопеция, артрит, хроническая почечная недостаточность, цирроз печени, ишемия нижних конечностей, остеопороз, некроз головки бедренной кости, ретинопатия недоношенных, нейро-сенсорная тугоухость, врожденный амавроз Лебера.
60. Способ согласно воплощению 59, где указанным расстройством, поддающимся лечению на основе стволовых клеток, является сахарный диабет.
61. Способ согласно любому из воплощений 42, 48 или 51, предусматривающий совместное или последовательное введение пациенту терапевтически эффективного количества препарата тотальной РНКлпкч-1 и препарата тотальной РНК из клеток или ткани пуповины и/или из пуповинной крови.
62. Способ согласно воплощению 61, где пуповиной является пуповина человека.
63. Способ лечения и профилактики заболевания или расстройства, требующего пересадки костного мозга, предусматривающий введение пациенту терапевтически эффективного количества композиции согласно воплощениям 1 или 28 или препарата тотальной РНК согласно воплощению 1 в качестве полной или частичной замены пересадки костного мозга.
64. Способ согласно воплощению 63, где препарат тотальной РНК выделен из ткани костного мозга здорового донора.
65. Способ коррекции или реверсии признака (признаков) или симптома (симптомов) возрастных изменений, предусматривающий введение субъекту эффективного количества композиции согласно воплощениям 1 или 28 или препарата тотальной РНК согласно воплощению 1.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИС-(2-ЭТИЛГЕКСАНОАТО)БИС(ТРИЭТИЛЕНТЕТРАМИН) КОБАЛЬТ (II), ПРОЯВЛЯЮЩИЙ СВОЙСТВА СТИМУЛЯТОРА ГЕМОПОЭЗА | 1994 |
|
RU2078767C1 |
| Средство для стимуляции пролиферативной активности клеток костного мозга | 2023 |
|
RU2811901C1 |
| ГЕМОСТИМУЛЯТОР | 1994 |
|
RU2094045C1 |
| СПОСОБ ЛЕЧЕНИЯ ДЕПРЕССИИ ЭРИТРОИДНОГО РОСТКА КРОВЕТВОРЕНИЯ ПРИ ЦИТОСТАТИЧЕСКОЙ МИЕЛОСУПРЕССИИ | 2008 |
|
RU2364398C1 |
| СПОСОБ ЛЕЧЕНИЯ ИММУННОЙ ДИСФУНКЦИИ, ТАКОЙ КАК РЕАКЦИЯ "ТРАНСПЛАНТАТ ПРОТИВ ХОЗЯИНА" ИЛИ "ХОЗЯИН ПРОТИВ ТРАНСПЛАНТАТА" | 2006 |
|
RU2497530C2 |
| СРЕДСТВО, ПРЕПЯТСТВУЮЩЕЕ РАЗВИТИЮ ДЕПРЕССИИ ЭРИТРОИДНОГО РОСТКА КРОВЕТВОРЕНИЯ ПРИ ЦИТОСТАТИЧЕСКИХ МИЕЛОДЕПРЕССИЯХ | 2007 |
|
RU2351333C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЭФФЕКТИВНОСТИ ГЕМОСТИМУЛЯТОРОВ ПРИ ЦИТОСТАТИЧЕСКОЙ МИЕЛОСУПРЕССИИ | 2009 |
|
RU2421720C2 |
| СРЕДСТВО, СТИМУЛИРУЮЩЕЕ ЭРИТРОПОЭЗ | 1985 |
|
RU2061496C1 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ РОСТА И РЕГЕНЕРАЦИИ КЛЕТОК, А ТАКЖЕ СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ | 2006 |
|
RU2341270C2 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ СТИМУЛЯЦИИ ПРОДУКЦИИ СТВОЛОВЫХ КЛЕТОК В КОСТНОМ МОЗГЕ | 2007 |
|
RU2410116C2 |
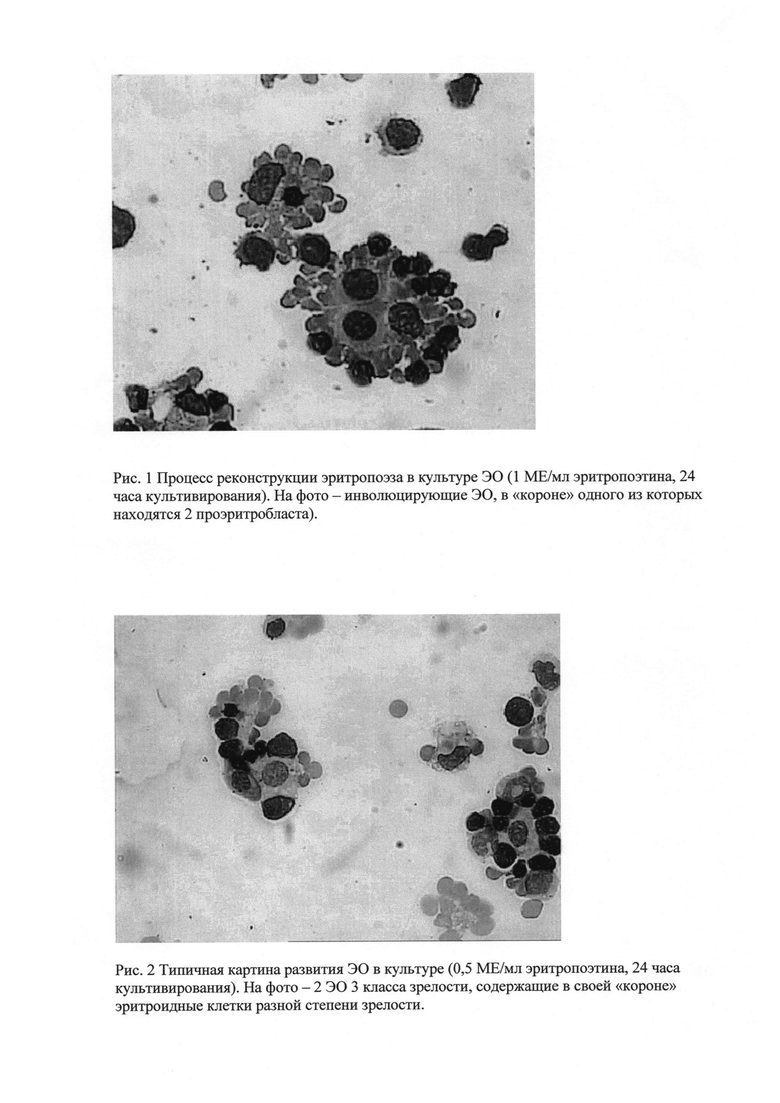
Изобретение относится к области фундаментальной биологии, практической регенеративной медицины, ветеринарии, клеточной биологии и может быть использовано для лечения и профилактики заболеваний, расстройств или состояний, связанных с нарушением процессов пролиферации и дифференцировки клеток разных органов и тканей, для активации регенерационного потенциала органов и тканей человека и животных при возрастных изменениях и после экстремальных воздействий, а также для медико-биологических исследований. Настоящее изобретение может найти широкое применение в области гемотрансфузии, пересадки органов, а также послужить общим подходом к разработкам надежных способов коррекции возрастных изменений у пожилых людей. Изобретение может быть использовано также в косметической промышленности для изготовления активных добавок для усиления регенерации и улучшения состояния кожи головы, лица и тела, в частности для изготовления активных добавок для борьбы с морщинами, для устранения кожных дефектов, для стимуляции и ускорения роста волос, для борьбы с избыточным оволосением. 9 н. и 41 з.п. ф-лы, 3 ил., 25 табл., 13 пр.
1. Регуляторный препарат тотальной РНК, полученный из интактных лимфоидных клеток или костного мозга здорового донора и/или из донорских лимфоидных клеток или костного мозга, подвергнутых активации Т-клеточного звена иммунной системы, обладающий модулирующей активностью в отношении пролиферации и/или дифференцировки клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа.
2. Регуляторный препарат тотальной РНК по п. 1, где под активацией лимфоидных клеток или костного мозга донора подразумевается in vivo активация Т-клеточного звена иммунной системы.
3. Препарат тотальной РНК по п. 1, где указанная модулирующая активность в отношении пролиферации и/или дифференцировки клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа осуществляется в организме реципиента при введении ему указанного препарата.
4. Препарат тотальной РНК по п. 1, где указанная активация лимфоидных Т-клеток или костного мозга осуществляется in vitro.
5. Препарат тотальной РНК по п. 1, где под лимфоидными клетками подразумеваются лимфоидные клетки селезенки, тимуса, лимфатических узлов, периферической крови донора.
6. Препарат тотальной РНК по п. 5, где указанный донор является млекопитающим.
7. Препарат тотальной РНК по п. 6, где указанный донор-млекопитающее является аллогенным донором.
8. Препарат тотальной РНК по п. 7, где указанным донором-млекопитающим является человек.
9. Препарат тотальной РНК по п. 6, где указанный донор-млекопитающее является ксеногенным донором.
10. Препарат тотальной РНК по п. 5, где указанный донор не является млекопитающим.
11. Препарат тотальной РНК по п. 3, где указанный реципиент является млекопитающим.
12. Препарат тотальной РНК по п. 11, где указанным реципиентом-млекопитающим является человек.
13. Препарат тотальной РНК по п. 3, где указанный реципиент не является млекопитающим.
14. Регуляторный препарат тотальной РНК по любому из пп. 1, 3, 5-13, полученный из интактных лимфоидных клеток или костного мозга здорового донора.
15. Регуляторный препарат тотальной РНК по п. 1, полученный в фазе проявления стимулирующей активности в отношении пролиферации и/или дифференцировки клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа.
16. Регуляторный препарат тотальной РНК по п. 15, полученный в фазе проявления стимулирующей активности в отношении пролиферации и/или дифференцировки клеток одноименной ткани и/или ткани какого-либо иного гистотипа в организме реципиента при введении ему указанного препарата.
17. Регуляторный препарат тотальной РНК по п. 1, полученный в фазе проявления тормозящей активности в отношении пролиферации и/или дифференцировки клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа.
18. Регуляторный препарат тотальной РНК по п. 17, полученный в фазе проявления тормозящей активности в отношении пролиферации и/или дифференцировки клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа в организме реципиента при введении ему указанного препарата.
19. Регуляторный препарат тотальной РНК по любому из пп. 1-13, 15-18, дополнительно содержащий препарат тотальной РНК, полученный из любого типа соматических клеток здорового донора.
20. Регуляторный препарат тотальной РНК по п. 14, дополнительно содержащий препарат тотальной РНК, полученный из любого типа соматических клеток здорового донора.
21. Препарат тотальной РНК по п. 19, где указанными соматическими клетками являются стволовые клетки.
22. Препарат тотальной РНК по п. 19, где указанным донором является молодой здоровый донор.
23. Препарат тотальной РНК по п. 20, где указанным донором является молодой здоровый донор.
24. Способ получения препарата тотальной РНК по пп. 1-23, согласно которому из лимфоидных клеток или из костного мозга интактного донора, или же из донорских лимфоидных клеток или костного мозга, которые сначала подвергают активации Т-клеточного звена иммунной системы, по стандартной методике с использованием тризольного реагента и фенол-хлороформной экстракции, в фазе проявления стимулирующей или супрессорной активности в отношении клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа, выделяют препарат регуляторной тотальной РНК, обладающий, соответственно, стимулирующей или супрессорной активностью в отношении клеток одноименной ткани и/или соматических клеток какого-либо иного гистотипа.
25. Способ по п. 24, где под активацией Т-клеточного звена иммунной системы подразумевается активация Т-клеточного звена in vivo.
26. Способ по п. 24, где указанная активация лимфоидных Т-клеток или костного мозга осуществляется in vitro.
27. Способ по п. 24, согласно которому указанная фаза проявления стимулирующей активности реализуется, в зависимости от ткани-мишени, в интервале приблизительно от 30 и менее минут примерно до 48 часов после активации Т-клеточного звена иммунной системы, а фаза проявления супрессорной активности реализуется, в зависимости от ткани-мишени, в интервале приблизительно от 48 часов примерно до 96 и более часов после активации Т-клеточного звена иммунной системы.
28. Способ по любому из пп. 24-27, где указанные лимфоидные клетки получены из селезенки, тимуса, лимфатических узлов или периферической крови донора.
29. Способ по п. 28, где указанным донором является млекопитающее.
30. Способ по п. 29, где указанным млекопитающим является молодое и здоровое млекопитающее.
31. Способ по п. 28, где указанный донор не является млекопитающим.
32. Композиция, содержащая регуляторный препарат тотальной РНК по любому из пп. 1-23 и фармацевтически приемлемый носитель, разбавитель и/или наполнитель.
33. Композиция по п. 32, представленная в жидкой, лиофилизированной или твердой форме.
34. Композиция по пп. 32, 33, предназначенная для интраназального, парентерального, внутриочагового или местного введения.
35. Способ модуляции пролиферации и/или дифференцировки соматических клеток с нарушенной пролиферацией и/или дифференцировкой в организме реципиента, предусматривающий введение указанному реципиенту регуляторного препарата тотальной РНК по любому из пп. 1-23 в эффективном количестве и сочетании.
36. Способ модуляции по п. 35, где указанным реципиентом является млекопитающее.
37. Способ модуляции по п. 36, где указанным млекопитающим является человек.
38. Способ модуляции по п. 35, где указанный реципиент не является млекопитающим.
39. Способ лечения заболеваний или расстройств, ассоциированных с нарушением пролиферации и/или дифференцировки соматических клеток (клетки-мишени) того или иного гистотипа, предусматривающий введение пациенту регуляторных препаратов тотальной РНК по любому из пп. 1-23 в эффективном количестве и сочетании.
40. Способ по п. 39, где заболевание или расстройство, ассоциированное с нарушением пролиферации и/или дифференцировки соматических клеток, выбрано из дегенеративных заболеваний или состояний, включая нейродегенеративные заболевания или состояния; аутоиммунных, гипо- и гиперпролиферативных расстройств, включая доброкачественные и злокачественные опухолевые заболевания; а также наследственных или врожденных дефектов.
41. Способ по п. 40, где опухолевым заболеванием является аденома предстательной железы.
42. Способ лечения или профилактики гематологических заболеваний или расстройств гемопоэза при заболеваниях другого генеза, требующих переливания крови или ее форменных элементов, заключающийся в замене (полной или частичной) гемотрансфузии введением препаратов тотальной РНК по любому из пп. 1-23 в эффективном количестве и сочетании.
43. Способ по п. 42, где указанным расстройством гемопоэза является анемия любого генеза.
44. Способ по п. 42, где указанное расстройство гемопоэза вызвано облучением.
45. Способ по п. 44, где указанное облучение обусловлено рентгенотерапией.
46. Способ по п. 42, где указанное расстройство гемопоэза вызвано химиотерапией.
47. Способ лечения заболеваний, поддающихся лечению стволовыми клетками и выбранных из группы, включающей в себя боковой амиотрофический склероз (БАС), детский церебральный паралич (ДЦП), эпилепсию, травму спинного мозга, травму головного мозга и травматическую инфекцию мозга, инсульт, болезнь Паркинсона, множественную системную атрофию, рассеянный склероз, системную красную волчанку, болезнь Девика, аутоиммунное заболевание, дистрофию сетчатки, пигментный ретинит, глаукому, глазную болезнь, нарушение зрения, сахарный диабет, мышечную дистрофию, аутизм, задержку развития, прогрессивный надъядерный паралич, кортикобазальную дегенерацию, болезнь Альцгеймера, болезнь Хантингтона, болезнь Баттена, наследственную атаксию, спиноцеребеллярную атаксию, атаксию Фридрейха, кардиомиопатию, хроническую сердечную недостаточность, инфаркт миокарда, алопецию, артрит, хроническую почечную недостаточность, цирроз печени, ишемию нижних конечностей, остеопороз и некроз головки бедренной кости, ретинопатию недоношенных, нейросенсорную тугоухость, врожденный амавроз Лебера, заключающийся в замене введения стволовых клеток здоровых доноров введением препаратов регуляторной РНК по пп. 1-23 в эффективном количестве и сочетании.
48. Способ лечения заболеваний, состояний или расстройств, требующих пересадки костного мозга, заключающийся в замене (полной или частичной) пересадки костного мозга введением препаратов регуляторной РНК по любому из пп. 1-16, 19-23 в эффективном количестве и сочетании.
49. Способ по п. 48, где указанное введение осуществляют внутривенным, внутрибрюшинным или интраназальным путем.
50. Способ коррекции у субъекта признака (признаков) или симптома (симптомов) возрастных изменений, предусматривающий введение указанному субъекту регуляторного препарата РНК по любому из пп. 1-16, 19-23 в эффективном количестве и сочетании.
| БАБАЕВА А.Г | |||
| Иммунологические механизмы регуляции восстановительных процессов, М., 1972 г., 150 с. | |||
| БАБАЕВА А.Г | |||
| и др | |||
| Роль лимфоцитов в оперативном изменении программы развития тканей, М., изд | |||
| РАМН, 2009 г., 108 с. | |||
| КСЕНОГЕННЫЕ ОЛИГО- ИЛИ/И ПОЛИРИБОНУКЛЕОТИДЫ В КАЧЕСТВЕ СРЕДСТВ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2002 |
|
RU2314814C2 |

