Приоритетная заявка
Данная заявка заявляет приоритет по предварительным заявкам США № 62/607850, поданной 19 декабря 2017 г., и 62/675070, поданной 22 мая 2018 г., которые явно включены в данный документ посредством ссылки в полном объеме, с конкретной ссылкой на фигуры, условные обозначения и формулу изобретений в них.
Перечень последовательностей
Данная заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и включен в данный документ посредством ссылки в полном объеме. Указанная копия ASCII, созданная 30 ноября 2018 г., имеет название 067461-5217-WO_ST25.txt и размер 605038 байт.
Уровень техники
Гомеостаз иммунной системы основан на тонком равновесии между различными популяциями иммунных клеток, в том числе CD8+ и CD4+ T-клеток (CD3+CD25-FOXP3-) и регуляторных T-клеток (Treg; CD3+CD4+CD25+FOXP3+). Нарушения этого равновесия могут привести к заболеваниям, таким как аутоиммунные заболевания, при которых Т-клетки остаются нерегулируемыми и поражают собственные ткани организма. В нормальном состоянии Treg модулируют дифференцировку Т-клеток и эффекторные и цитотоксические функции. Таким образом, основная предпосылка в этом отношении заключается в том, что нарушения числа и/или функции Treg-клеток являются сопутствующими факторами патологических состояний. В связи с этим, способность изменять равновесие между цитотоксичностью и регуляцией путем тонкой настройки Т-клеточного ответа имеет большой потенциал для лечения аутоиммунных и других заболеваний.
ИЛ-2 способствует пролиферации и дифференцировке В-клеток, Т-клеток и NK-клеток. ИЛ-2 также важен для функционирования и выживания Treg. ИЛ-2 осуществляет свою клеточную сигнальную функцию посредством связывания с высокоаффинным тримерным рецепторным комплексом, состоящим из трех отдельных белков: общей гамма-цепи (γc; CD132) и B-цепи рецептора ИЛ-2 (ИЛ-2Рß; CD122), которые являются общими с ИЛ-15, а также уникального рецептора альфа-цепи (ИЛ-2Рγ; CD25). ИЛ-2 также может проявлять свою клеточную сигнальную функцию посредством связывания с димерным рецепторным комплексом с промежуточной аффинностью, состоящим только из ИЛ-2Рß и γc (ИЛ-2Рßγ).
Вследствие низких концентраций ИЛ-2, которые в типичном случае существуют в тканях, ИЛ-2 преимущественно активирует клетки, которые экспрессируют высокоаффинный рецепторный комплекс (CD25:CD122:CD132; ИЛ-2Рγßγ), и поэтому оказывает предпочтительное действие в отношении FOXP3+ Treg, которые конститутивно экспрессируют CD25. В то же время ИЛ-2 может также активировать и индуцировать пролиферацию FOXP3- T-клеток, которые экспрессируют рецепторный комплекс с промежуточной аффинностью (CD122:CD132; ИЛ-2Рßγ). FOXP3- T-клетки, такие как CD4+ или CD8+ T-клетки, могут способствовать воспалению, аутоиммунной реакции, отторжению органного трансплантата или болезни «трансплантат против хозяина». Вследствие способности ИЛ-2 стимулировать или снижать активность как Т-клеток, так и Treg с ограниченной селективностью, существует значительная потребность в области создания более селективных модуляторов Treg. В качестве дополнения, в качестве потенциального лекарственного препарата, ИЛ-2 имеет недостаток, заключающийся в очень быстром клиренсе, при этом период полужизни измеряется в минутах, что затрудняет эффективное введение. Данное изобретение направлено на решение обеих указанных проблем в результате получения новых слитых белков ИЛ-2-Fc.
Соответственно, существует потребность в получении пригодных вариантов ИЛ-2 и слитых белков Fc.
Краткое описание сущности изобретения
Соответственно, в некоторых аспектах данное раскрытие относится к композициям, содержащим вариант белка ИЛ-2 человека (по сравнению с SEQ ID NO:2), в которых указанный вариант белка ИЛ-2 содержит замену (замены) аминокислоты, выбранную (выбранные) из группы T3A, R38A; R38D; R38E; R38F; R38G; R38H; R38I; R38K; R38L; R38M; R38N; R38P; R38Q; R38S; R38T; R38V; R38W; R38Y; T41A; T41D; T41E; T41F; T41G; T41H; T41I; T41K; T41L; T41M; T41N; T41P; T41Q; T41R; T41S; T41V; T41W; T41Y; F42A; F42D; F42E; F42G; F42H; F42I; F42K; F42L; F42M; F42N; F42P; F42Q; F42R; F42S; F42T; F42V; F42W; F42Y; R38Q/T41K; R38Q/T41Q; R38E/T41K; R38Q/T41R; R38N/T41Q; R38Q/T41V; R38N/T41V; R38Q/T41M; R38Q/T41S; R38Q/T41L; R38N/T41M; T41I/F42Y; T41E/F42Y’ T41D/F42Y; T41M/F42Y; 41Q/F42Y; T41E/F42H; T41E/F42L; T41E/F42P; R38Q/F42Y; R38N/T41R; R38N/T41K; R38V/T41R; R38P/T41R; T41E/F42K; T41D/F42K; T41M/F42K; T41Q/F42K; R38Q/F42K; T41I/F42K; R38N/F42K; T41H/F42K; R38Q/T41K/F42Y; R38Q/T41R/F42Y; R38Q/T41Q/F42Y; R38Q/T41V/F42Y; R38N/T41K/F42K; R38Q/T41H/F42K; R38Q/T41K/F42K; R38Q/T41Q/F42K; 38Q/T41V/F42K; R38Q/T41R/F42K; Q11E; L12D; Q13E; E15Q; H16Y; L19D; D20N; N29S/Y31H/K35R/T37A/R38L/K48E/V69A/N71R/Q74P/N88D/I89V/Q126T; Q22E; K35R; T37S; K43R; F44Y; Y45F; K48R; K49E; E61Q; E62Q; K64R; E68Q; V69L; L72I; R81D; D84N; S87T; N88D; V91L; I92L; E95Q; Y107F; E116R; N119D; R120D; T123S; C125S/Q126E; C125S/S127T; C125S/I129L; C125S/S130T; C125S/T133S; T3A; F42A/Y45A/L72G; N29S/Y31H/K35R/T37A/K48E/V69A/N71R/Q74P/N88D/I89V; V69A/Q74P/I128T; N29S/Y31H/K35R/T37A/K48E/V69A/N71R/Q74P/N88D/I89V/Q126T; C125S/Q126T; N88R; R38I; L80F/R81D/L85V/I92F; L18R/L80F/R81D/L85V/I92F/Q126T; L18R/L80F/R81D/L85V/I92F/Q126T/S130R; F42A/Y45A/L72G/N88R; F42A/Y45A/L72G/Q126T; F42A/Y45A/L72G/N88R/Q126T; L19D; D20N; N88D; N88K; N88R; N88R; N88R; F42A/Y45A/L72G; N29S/Y31H/K35R/T37A/K48E/N71R/N88D/I89V; L19D/N29S/Y31H/K35R/T37A/K48E/N71R; D20N/N29S/Y31H/K35R/T37A; K48E/N71R; L19D/N29S/Y31H/K35R/T37A/K48E; D20N/N29S/Y31H/K35R/T37A; K48E; L19D K35R; L19D/T37R; D20N/T37R; L19D/N71K; D20N/N71K; D20N/R38I; D20N/T37R; 38I; D20N/R38I/N71K; D20N/N71K; D20N; D20N/T37R; D20N/R38I; D20N/T37R R38I; D20N/R38I/N71K; D20N; D20N/T37R; D20N/N71K; D20N/R38I; D20N/T37R R38I; D20N/R38I/N71K; D20N; D20N/T37R; D20N/N71K; D20N/R38I; D20N/T37R/R38I; D20N/R38I/N71K; N29S/Y31H/K35R/T37A/K48E/V69A/N71R/Q74P/N88D/I89V/Q126T, R38A/C125S; R38D/C125S; R38E/C125S; R38F/C125S; R38G/C125S; R38H/C125S; R38I/C125S; R38K/C125S; R38L/C125S; R38M/C125S; R38N/C125S; R38P/C125S; R38Q/C125S; R38S/C125S; R38T/C125S; R38V/C125S; R38W/C125S; R38Y/C125S; T41A/C125S; T41D/C125S; T41E/C125S; T41F/C125S; T41G/C125S; T41H/C125S; T41I/C125S; T41K/C125S; T41L/C125S; T41M/C125S; T41N/C125S; T41P/C125S; T41Q/C125S; T41R/C125S; T41S/C125S; T41V/C125S; T41W/C125S; T41Y/C125S; F42A/C125S; F42D/C125S; F42E/C125S; F42G/C125S; F42H/C125S; F42I/C125S; F42K/C125S; F42L/C125S; F42M/C125S; F42N/C125S; F42P/C125S; F42Q/C125S; F42R/C125S; F42S/C125S; F42T/C125S; F42V/C125S; F42W/C125S; F42Y/C125S; R38Q/T41K/C125S; R38Q/T41Q/C125S; R38E/T41K/C125S; R38Q/T41R/C125S; R38N/T41Q/C125S; R38Q/T41V/C125S; R38N/T41V/C125S; R38Q/T41M/C125S; R38Q/T41S/C125S; R38Q/T41L/C125S; R38N/T41M/C125S; T41I/F42Y/C125S; T41E/F42Y/C125S’ T41D/F42Y/C125S; T41M/F42Y/C125S; 41Q/F42Y/C125S; T41E/F42H/C125S; T41E/F42L/C125S; T41E/F42P/C125S; R38Q/F42Y/C125S; R38N/T41R/C125S; R38N/T41K/C125S; R38V/T41R/C125S; R38P/T41R/C125S; T41E/F42K/C125S; T41D/F42K/C125S; T41M/F42K/C125S; T41Q/F42K/C125S; R38Q/F42K/C125S; T41I/F42K/C125S; R38N/F42K/C125S; T41H/F42K/C125S; R38Q/T41K/F42Y/C125S; R38Q/T41R/F42Y/C125S; R38Q/T41Q/F42Y/C125S; R38Q/T41V/F42Y/C125S; R38N/T41K/F42K/C125S; R38Q/T41H/F42K/C125S; R38Q/T41K/F42K/C125S; R38Q/T41Q/F42K/C125S; 38Q/T41V/F42K/C125S; R38Q/T41R/F42K/C125S; N29S/Y31H/K35R/T37A/R38L/K48E/V69A/N71R/Q74P/N88D/I89V/C125S/Q126T; Q11E/C125S; L12D/C125S; Q13E/C125S; E15Q/C125S; H16Y/C125S; L19D/C125S; D20N/C125S; Q22E/C125S; K35R/C125S; T37S/C125S; K43R/C125S; F44Y/C125S; Y45F/C125S; K48R/C125S; K49E/C125S; E61Q/C125S; E62Q/C125S; K64R/C125S; E68Q/C125S; V69L/C125S; L72I/C125S; R81D/C125S; D84N/C125S; S87T/C125S; N88D/C125S; V91L/C125S; I92L/C125S; E95Q/C125S; Y107F/C125S; E116R/C125S; N119D/C125S; R120D/C125S; T123S/C125S; C125S/Q126E; C125S/S127T; C125S/I129L; C125S/S130T; C125S/T133S; T3A/C125S; T3A/F42A/Y45A/L72G/C125A; N29S/Y31H/K35R/T37A/K48E/V69A/N71R/Q74P/N88D/I89V/C125S; V69A/Q74P/I128T/C125S; N29S/Y31H/K35R/T37A/K48E/V69A/N71R/Q74P/N88D/I89V/C125S/Q126T; C125S/Q126T; N88R/C125S; R38I/C125S; L80F/R81D/L85V/I92F/C125S; L18R/L80F/R81D/L85V/I92F/C125S/Q126T; L18R/L80F/R81D/L85V/I92F/C125S/Q126T/S130R; T3A/F42A/Y45A/L72G/N88R/C125A; T3A/F42A/Y45A/L72G/C125A/Q126T; T3A/F42A/Y45A/L72G/N88R/C125A/Q126T; T3A/L19D/C125S; T3A/D20N/C125S; T3A/N88D/C125S; T3A/N88K/C125S; N88R/C125S; N88R/C125S; N88R/C125S; T3A/F42A/Y45A/L72G/C125A; N29S/Y31H/K35R/T37A/K48E/N71R/N88D/I89V/C125S; L19D/N29S/Y31H/K35R/T37A/K48E/N71R/C125S; D20N/N29S/Y31H/K35R/T37A; K48E/N71R/C125S; L19D/N29S/Y31H/K35R/T37A/K48E/C125S; D20N/N29S/Y31H/K35R/T37A; K48E/C125S; T3A/L19D K35R/C125S; T3A/L19D/T37R/C125S; T3A/D20N/T37R/C125S; T3A/L19D/N71K/C125S; T3A/D20N/N71K/C125S; T3A/D20N/R38I/C125S; T3A/D20N/T37R; 38I/C125S; T3A/D20N/R38I/N71K/C125S; T3A/D20N/N71K/C125S; T3A/D20N/C125S; T3A/D20N/T37R/C125S; T3A/D20N/R38I/C125S; T3A/D20N/T37R R38I/C125S; T3A/D20N/R38I/N71K/C125S; T3A/D20N/C125S; T3A/D20N/T37R/C125S; T3A/D20N/N71K/C125S; T3A/D20N/R38I/C125S; T3A/D20N/T37R R38I/C125S; T3A/D20N/R38I/N71K/C125S; T3A/D20N/C125S; T3A/D20N/T37R/C125S; T3A/D20N/N71K/C125S; T3A/D20N/R38I/C125S; T3A/D20N/T37R/R38I/C125S; T3A/D20N/R38I/N71K/C125S; и N29S/Y31H/K35R/T37A/K48E/V69A/N71R/Q74P/N88D/I89V/C125S/Q126T.
В дополнительных аспектах в данном раскрытии представлены слитые димерные белки ИЛ-2-Fc, содержащие: а) первый мономер, содержащий от N- до С-конца: i) вариантный белок ИЛ-2 в соответствии с любым из пп. А1-А4; ii) линкер первого домена; и iii) первый вариантный Fс-домен; и b) второй мономер, содержащий второй вариантный Fc-домен.
В дополнительных аспектах в данном изобретении представлены слитые димерные белки ИЛ-2-Fc, в которых указанный второй мономер содержит от N- до С-конца: а) вариантный белок ИЛ-2 в соответствии с любым из пп. А1-А4; b) линкер второго домена; и с) указанный второй вариантный Fc-домен.
В дополнительных аспектах слитые димерные белки ИЛ-2-Fc имеют первый и второй вариантные Fc-домены, которые содержат гетеродимеризационные варианты, выбранные из группы, состоящей из L368D/K370S : S364K/E357Q; L368D/K370S : S364K; L368E/K370S : S364K; T411E/K360E/Q362E : D401K; и T366S/L368A/Y407V : T366W. В некоторых случаях слитые белки ИЛ-2-Fc дополнительно содержат абляционные варианты, в том числе P233P/L234V/L235A/G236_/S267K. В некоторых аспектах слитые белки также содержат Fc-домены с аминокислотными заменами M428L/N434S или M428L/N434A. В некоторых аспектах линкер доменов представляет собой шарнир IGG1, а в других аспектах он может включать в себя линкер, выбранный из (GS)n, (GSGGS)n, (GGGGS)n и (GGGS)n, где n представляет собой целое число, составляющее, по меньшей мере, один.
В дополнительных аспектах данное изобретение включает в себя полипептидную композицию, содержащую вариантный белок ИЛ-2 человека, в которой указанный вариантный белок ИЛ-2 содержит аминокислотную (аминокислотные) замену (замены) по сравнению с SEQ ID NO:2, выбранную (выбранные) из группы T3A/D20N/T37R и T3A/D20N/N71K. В некоторых случаях вариантный белок ИЛ-2 дополнительно содержит C125S вариант или C125A вариант.
В дополнительном аспекте полипептидная композиция представляет собой гомодимерный белковый комплекс, в котором каждый мономер белка содержит указанный вариантный белок ИЛ-2, ковалентно присоединенный к Fc-домену. В некоторых аспектах каждый из указанных Fc-доменов представляет собой вариантный Fc-домен.
В дополнительном аспекте полипептидная композиция представляет собой гетеродимерный белковый комплекс, содержащий первый мономер белка, содержащий указанный вариантный белок ИЛ-2, ковалентно присоединенный к первому вариантному Fc-домену, и второй мономер белка, содержащий второй вариантный Fc-домен.
В дополнительном аспекте полипептидная композиция имеет вариантные Fc-домены, которые представляют собой вариантные Fc-домены IgG1 человека, содержащие аминокислотные замены M428L/N434S.
В дополнительном аспекте полипептидная композиция имеет вариантные Fc-домены, которые представляют собой вариантные Fc-домены IgG1 человека, содержащие аминокислотные замены E233P/L234V/L235A/G236del/S267K.
В дополнительном аспекте полипептидная композиция имеет первый и второй вариантные Fc-домены, которые включают в себя совокупность гетеродимеризационных вариантов, выбранных из группы, состоящей из вариантов, изображенных на фиг. 2. В некоторых вариантах осуществления совокупность гетеродимеризационных вариантов выбирают из группы, состоящей из L368D/K370S : S364K/E357Q; L368D/K370S : S364K; L368E/K370S : S364K; T411E/K360E/Q362E : D401K; и T366S/L368A/Y407V : T366W.
В дополнительном аспекте полипептидную композицию выбирают из группы, состоящей из XENP27564 (SEQ ID NO:297 и 298), XENP27563 (SEQ ID NO:295 и 296), XENP26105 (SEQ ID NO:245 и 246) и XENP26109 (SEQ ID NO:249 и 250).
Также представлены композиции нуклеиновых кислот, содержащие: а) первую нуклеиновую кислоту, кодирующую указанный первый мономер белка по любому из пп. 6-13; и b) вторую нуклеиновую кислоту, кодирующую указанный второй мономер белка по любому из пп. 6-13 соответственно. В качестве дополнения представлены композиции экспрессионных векторов, содержащие а) первый экспрессионный вектор, содержащий указанную первую нуклеиновую кислоту; и b) второй экспрессионный вектор, содержащий указанную вторую нуклеиновую кислоту, и клетки-хозяева, содержащие композиции нуклеиновой кислоты и/или композиции экспрессионных векторов. Кроме того, представлены способы получения полипептидной композиции, включающие культивирование клетки-хозяина по данному изобретению в условиях, при которых указанная композиция образуется, и извлечение указанной композиции.
В дополнительном аспекте слитый димерный белок ИЛ-2-Fc выбирают из XENP24635; XENP24636; XENP24637; XENP24638; XENP24639; XENP24640; XENP24641; XENP24642; XENP24643; XENP24725; XENP24728; XENP24729; XENP24730; XENP24731; XENP24732; XENP25717; XENP25720; XENP25725; XENP25727; XENP25910; XENP25911; XENP25912; XENP26086; XENP26088; XENP26089; XENP26092; XENP26093; XENP26096; XENP26104; XENP26105; XENP26108; XENP26109; XENP26835; XENP26839; XENP26840; XENP26841; XENP26986; XENP26987; XENP26989; XENP26990, XENP26991, XENP25906, XENP25907; XENP25908; XENP25909; XENP26992; XENP26993; XENP26994; XENP26995; XENP26996; XENP27001; XENP27002; XENP27003; XENP27004; XENP27005; XENP27006 и XENP27007.
В дополнительных аспектах представлены способы активации CD25+ клеток, включающие приведение в контакт указанных клеток со слитым димерным белком ИЛ-2-Fc по данному изобретению, и способы лечения аутоиммунного заболевания, включающие введение пациенту, нуждающемуся в этом, белковой композиции, представленной в данном документе.
Краткое описание графических материалов
На фиг. 1 изображены аминокислотные последовательности (и номера доступа в GenBank) ИЛ-2 человека и его рецепторов: ИЛ-2Рα (также известного как CD25), ИЛ-2Рβ (также известного как CD122) и общей гамма-цепи (также известной как ИЛ-2Р или CD132).
На фиг. 2 изображены пригодные пары гетеродимеризационных вариантов Fc (в том числе искаженных и pI вариантов). На фиг. 2 изображены варианты, для которых отсутствуют соответствующие варианты «мономера 2»; они представляют собой pI варианты, которые могут быть использованы в отдельности в отношении любого мономера.
На фиг. 3 изображен перечень константных областей изостерических вариантных антител и их соответствующие замены. pI (-) обозначает варианты с более низким значением pI, в то время как pI_(+) обозначает варианты с более высоким значением pI. Они могут быть необязательно и независимо объединены с другими вариантами гетеродимеризации по данному изобретению (а также с другими типами вариантов, описанными в данном документе).
На фиг. 4 изображены пригодные абляционные варианты, которые приводят к устранению связывания FcγR (иногда обозначаемые как «нокауты» или «КО» варианты). Как правило, абляционные варианты встречаются на обоих мономерах, хотя в некоторых случаях они могут присутствовать только на одном мономере.
На фиг. 5 изображены особенно пригодные варианты осуществления «нецитокиновых» компонентов по данному изобретению.
На фиг. 6 изображены последовательности нескольких пригодных каркасов слитых форматов ИЛ-2-Fc на основе IgG человека, без цитокиновых последовательностей. Каркас 1 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих цепях, искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях. Каркас 2 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих цепях, искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях. Каркас 3 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих цепях, искаженные варианты S364K/E357Q: L368E/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368E/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях. Каркас 4 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих цепях, искаженные варианты D401K: K360E/Q362E/T411E, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами K360E/Q362E/T411E и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях. Каркас 5 основан на IgG1 человека (аллотип 356D/358L) и содержит C220S в обеих цепях, искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях. Каркас 6 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих цепях, искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях, а также вариант N297A в обеих цепях. Каркас 7 является идентичным 6, за исключением того, что мутация представляет собой N297S. Альтернативные форматы каркасов 6 и 7 могут исключать абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях. Каркас 8 основан на IgG4 человека и содержит искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S, а также вариант S228P (нумерация EU, т.е., S241P, как по Кабату) в обеих цепях, который приводит к устранению обмена Fab-фрагментами, как известно в данной области техники. Каркас 9 основан на IgG2 человека и содержит искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S. Каркас 10 основан на IgG2 человека и содержит искаженные варианты S364K/E357Q: L368D/K370S, pI варианты Q295E/N384D/Q418E/N421D в цепи с искаженными вариантами L368D/K370S, а также вариант S267K в обеих цепях. Каркас 11 идентичен каркасу 1, за исключением того, что содержит мутации Xtend M428L/N434S. Каркас 12 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих идентичных цепях и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих идентичных цепях. Каркас 13 основан на IgG1 человека (аллотип 356E/358M) и содержит C220S в обеих цепях, искаженные варианты S364K/E357Q: L368D/K370S, pI варианты P217R/P229R/N276K в цепи с искаженными вариантами S364K/E357Q и абляционные варианты E233P/L234V/L235A/G236del/S267K в обеих цепях.
Как будет понятно специалистам в данной области техники и изложено ниже, любые варианты ИЛ-2 могут быть включены в эти каркасы в любой комбинации. В каждый из этих каркасов включены последовательности, которые на 90, 95, 98 и 99% идентичны (как определено в данном документе) перечисленным последовательностям и/или содержат от 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 дополнительных аминокислотных замен (по сравнению с «родительской» из указанной фигуры, которая, как будет понятно специалистам в данной области техники, уже содержит ряд аминокислотных модификаций по сравнению с исходным IgG1 человека (или IgG2 или IgG4, в зависимости от каркаса). Таким образом, указанные каркасы могут содержать дополнительные аминокислотные модификации (как правило, аминокислотные замены) в дополнение к искаженным, pI и абляционным вариантам, содержащимся в каркасах указанной фигуры. В частности, также могут быть включены варианты FcRn, такие как M428L/N434S.
На фиг. 7 изображен ряд иллюстративных линкеров переменной длины. В некоторых вариантах осуществления указанные линкеры находят применение, связывая C-конец ИЛ-2 с N-концом Fc-области (которая в некоторых случаях включает шарнирный домен).
На фиг. 8А и 8В изображены А) структурная модель ИЛ-2, образующего комплекс с высокоаффинным рецептором ИЛ-2 (ИЛ-2Рαßγ), и В) положение трех остатков ИЛ-2, которые приводятся в контакт с ИЛ-2Рα, при этом замены предположительно ослабляют рН-зависимое связывание ИЛ-2 с ИЛ-2Рα.
На фиг. 9 изображены аминокислотные последовательности иллюстративных вариантов ИЛ-2, сконструированные для ослабления рН-зависимого связывания с ИЛ-2РRα. Важно отметить, что указанные варианты были созданы с использованием С-концевых меток полигистидина (Hisx8 или HHHHHHHH), которые были удалены из последовательностей, изображенных в данном документе.
На фиг. 10 изображены скорости ассоциации (ka), скорости диссоциации (Kd) и константы диссоциации (KD) вариантов ИЛ-2 по отношению к ИЛ-2Рα при рН 6,0, а также кратное повышение kd и KD по отношению к XENP14135 (ИЛ-2 дикого типа с мутацией C125S). СО указывает на отсутствие измеримого связывания.
На фиг. 11 изображены скорости ассоциации (ka), скорости диссоциации (Kd) и константы диссоциации (KD) вариантов ИЛ-2 по отношению к ИЛ-2Рα при рН 7,4, а также кратное повышение kd и KD по отношению к XENP14135 (ИЛ-2 дикого типа с мутацией C125S). СО указывает на отсутствие измеримого связывания.
На фиг. 12 изображена кратность повышения скорости диссоциации ИЛ-2 (kd) от ИЛ-2Рα при рН 6,0, опосредуемая различными точечными мутациями.
На фиг. 13 изображена кратность повышения скорости диссоциации ИЛ-2 (kd) из ИЛ-2Рα при рН 7,4, опосредуемая различными точечными мутациями.
На фиг. 14 изображены сенсограммы Biacore для А) XENP14135 (ИЛ-2 дикого типа с мутацией C125S) и В) XENP14142 (вариантного ИЛ-2 с R38I и C125S).
На фиг. 15 изображены аминокислотные последовательности варианта ИЛ-2 дополнительного предшествующего уровня техники (мутант 2-4 с Q126T, описанный в WO 2009/061853, опубликованном 14 мая 2009 г.) без R38L (XENP14277) и с R38L (XENP14381), включенные для ослабления рН-зависимого связывания с ИЛ-2Рα. Важно отметить, что указанные варианты были созданы с использованием С-концевых меток полигистидина (Hisx8 или HHHHHHHH), которые были удалены из последовательностей, изображенных в данном документе.
На фиг. 16 изображены скорости ассоциации (ka), скорости диссоциации (Kd) и константы диссоциации (KD) вариантов ИЛ-2, связывающихся с ИЛ-2Рα, с заменой или без замены R38L, при pH 7,4 и pH 6,0.
На фиг. 17 изображены аминокислотные последовательности иллюстративных вариантов ИЛ-2, сконструированные для изменения связывания с ИЛ-2Рα, ИЛ-2Рß, γc или ИЛ-2Рßγ. Важно отметить, что указанные варианты были созданы с использованием С-концевых меток полигистидина (Hisx8 или HHHHHHHH), которые были удалены из последовательностей, изображенных в данном документе.
На фиг. 18 изображен нормализованный ответ BLI (по отношению к XENP14135) иллюстративных вариантов ИЛ-2 для различных рецепторов ИЛ-2 рецепторов, определенный с помощью Octet. Цель заключается в том, чтобы либо увеличить связывание с ИЛ-2Рα, либо уменьшить связывание с ИЛ-2Рβ и ИЛ-2Рγ или с границей раздела ИЛ-2Рβγ, либо в том и другом.
На фиг. 19 изображено несколько форматов слияний ИЛ-2-Fc по данному изобретению. Моновалентный ИЛ-2-Fc или «моновИЛ-2-Fc» (фиг. 19A) содержит ИЛ-2, рекомбинантно слитый с N-концом гетеродимерной Fc-области, причем другая сторона молекулы представляет собой «только Fc» или «пустой Fc». Бивалентный ИЛ-2-Fc или «бивИЛ-2-Fc» (фиг. 19B) содержит ИЛ-2, рекомбинантно слитый с N-концом обеих сторон гомодимерной Fc-области. Моновалентный ИЛ-2-Fc с линкером или «моновИЛ-2-Fc (с линкером)» (фиг. 19C) содержит ИЛ-2, рекомбинантно слитый с N-концом гетеродимерной Fc-области с помощью линкера домена, при этом другая сторона молекулы представляет собой «только Fc» или «пустой Fc». Бивалентный ИЛ-2-Fc с линкером или «бивИЛ-2-Fc (с линкером)» (фиг. 19D) содержит ИЛ-2, рекомбинантно слитый с N-концом обеих сторон гомодимерной Fc-области с помощью линкера домена. Неограничивающие примеры линкеров доменов, подходящих для использования в форматах моновИЛ-2-Fc (с линкером) и бивИЛ-2-Fc (с линкером), изображены на фиг. 7.
На фиг. 20 изображены аминокислотные последовательности иллюстративных слияний моновИЛ-2-Fc по данному изобретению. Косые черты (/) указывают границу между ИЛ-2 и Fc-областями (в данном случае Fc-область содержит шарнир, а также вариант C220S).
На фиг. 21 изображены аффинность (KD), скорость ассоциации (ka) и скорость диссоциации (kd) иллюстративных слияний ИЛ-2-Fc для ИЛ-2Рα, ИЛ-2Рß и ИЛ-2Рßγ, определенные с помощью Octet.
На фиг. 22А-22F изображена индукция фосфорилирования STAT5 в CD4+CD45RA-Т-клетках, CD4+CD45RA+Т-клетках, CD8+CD45RA-Т-клетках, CD8+CD45RA+Т-клетках, а также Treg, с помощью А) XENP24636, B) XENP24638, C) XENP24641, D) XENP24642, E) XENP24643 и F) XENP24731.
На фиг. 23 изображены аминокислотные последовательности иллюстративных слияний бивИЛ-2-Fc по данному изобретению. Косые черты (/) указывают границу между ИЛ-2 и Fc-областями, в данном случае Fc-область также содержит домен шарнира IgG1 с вариантом C220S.
На фиг. 24А и 24В изображена индукция фосфорилирования STAT5 в CD4+CD45RA-Т-клетках, CD4+CD45RA+Т-клетках, CD8+CD45RA-Т-клетках, CD8+CD45RA+Т-клетках, а также Treg, с помощью А) XENP25906 и B) XENP25907.
На фиг. 25 изображены аминокислотные последовательности иллюстративных слияний ИЛ-2-Fc с линкерами доменов. Косые черты (/) указывают границу между ИЛ-2, линкерами и Fc-областями, в данном случае Fc-область также содержит домен шарнира IgG1 с вариантом C220S. Линкеры подчеркнуты двойной линией.
На фиг. 26А-26Е изображена индукция фосфорилирования STAT5 в CD4+CD45RA-Т-клетках, CD4+CD45RA+Т-клетках, CD8+CD45RA-Т-клетках, CD8+CD45RA+Т-клетках, а также Treg, с помощью А) XENP25908, B) XENP25909, C) XENP25910, D) XENP25911 и E) XENP25912.
На фиг. 27 изображены аминокислотные последовательности дополнительных слияний ИЛ-2-Fc с ИЛ-2, сконструированных для повышения аффинности к CD25 и снижения аффинности SEQ Figure \* ARABIC к CD122. Косые черты (/) указывают границу между ИЛ-2 и Fc-областями, в данном случае Fc-область также содержит домен шарнира IgG1 с вариантом C220S.
На фиг. 28А-28T изображено фосфорилирование STAT5 в CD8+ и CD4+ Т-клетках и Treg в качестве показателя активации с помощью A) XENP24635, B) XENP24636, С) XENP24637, D) XENP24638, Е) XENP24642, F) XENP25717, G) XENP25720, H) XENP25725, I) XENP25727, J) XENP26086, K) XENP26088, L) XENP26089, M) XENP26092, N) XENP26093, O) XENP2104, XENP26) Q26P26) Q2 R) XENP26108, S) XENP26109 и T) рекомбинантного ИЛ-2 человека.
На фиг. 29А и 29В изображено фосфорилирование STAT5 в А) Treg и В) CD4+ (CD45RA-) Т-клетках, в качестве показателя активации с помощью иллюстративных слияний ИЛ-2-Fc.
На фиг. 30 изображены аминокислотные последовательности дополнительных слияний ИЛ-2-Fc. Косые черты (/) указывают границу между ИЛ-2, линкерами доменов (с двойной подчеркнутой линией) и Fc-областями, в данном случае Fc-область также содержит домен шарнира IgG1 с вариантом C220S.
На фиг. 31 изображены некоторые предпочтительные варианты осуществления.
На фиг. 32 изображены некоторые предпочтительные варианты осуществления вариантов, сконструированных с A) заменами рН-переключателей, B) заменами селективности Treg и C) комбинацией замен с рН-переключателями и замен селективности Treg.
На фиг. 33 изображены аффинность (KD), скорость ассоциации (ka) и скорость диссоциации (kd) иллюстративных слияний ИЛ-2-Fc для ИЛ-2Рα, ИЛ-2Рß и ИЛ-2Рßγ, определенные с помощью Octet. СО указывает на отсутствие связывания.
На фиг. 34 изображена последовательность XENP27193, слияния Fc-ИЛ-2 (V91K/C125A), сконструированного для увеличения соотношения Treg к нерегуляторным Т-клеткам, описанного в WO 2014/153111. Косые черты (/) указывают границу между ИЛ-2, линкером доменов и Fc-областями (в данном случае Fc-область также содержит домен шарнира IgG1 с вариантом C220S); а линкер подчеркнут двойной линией.
На фиг. 35A-E изображена индукция фосфорилирования STAT5 в A) CD4+CD45RA- T-клетках, B) CD8+CD45RA- T-клетках, C) NK-клетках, D) γδ T-клетках и E) Treg с помощью вариантов слияний моновИЛ-2-Fc, XENP24638, XENP24642, XENP26105, XENP26109, XENP26835, XENP26839, XENP26991 и XENP25702. Данные демонстрируют, что вариантные слияния моновИЛ-2-Fc преимущественно индуцируют фосфорилирование STAT5 в Treg по отношению к CD4+ Т-клеткам памяти (CD45RA-), CD8+ Т-клеткам памяти (CD45RA-), NK-клеткам и γδ Т-клеткам по сравнению с рекомбинантным ИЛ-2 и моновалентным слиянием ИЛ-2-Fc ДТ (XENP24635), а также бивалентным слиянием ИЛ-2-Fc предшествующего уровня техники, описанным в WO 2014/153111 (XENP27193).
На фиг. 36 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами N88R/C125S, представленными в формате моновИЛ-2-Fc без линкера (XENP24642) и бивИЛ-2-Fc с линкером (XENP25908).
На фиг. 37 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами T3A/D20N/C125S, представленными в формате моновИЛ-2-Fc без линкера (XENP25720), бивИЛ-2-Fc без линкера (XENP26992) и бивИЛ-2-Fc с линкером (XENP27002).
На фиг. 38 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами T3A/D20N/T37R/C125S, представленными в формате моновИЛ-2-Fc без линкера (XENP26105), бивИЛ-2-Fc без линкера (XENP26993) и бивИЛ-2-Fc с линкером (XENP27003).
На фиг. 39 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами T3A/D20N/N71K/C125S, представленными в формате моновИЛ-2-Fc без линкера (XENP26109), бивИЛ-2-Fc без линкера (XENP26994) и бивИЛ-2-Fc с линкером (XENP27004).
На фиг. 40 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами T3A/D20N/R38I/C125S, представленными в формате моновИЛ-2-Fc без линкера (XENP26835), бивИЛ-2-Fc без линкера (XENP26995) и бивИЛ-2-Fc с линкером (XENP27005).
На фиг. 41 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами T3A/D20N/T37R/R38I/C125S, представленными в формате моновИЛ-2-Fc без линкера (XENP26839), бивИЛ-2-Fc без линкера (XENP26996) и бивИЛ-2-Fc с линкером (XENP27006).
На фиг. 42 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- T-клетках и Treg с помощью варианта ИЛ-2 с заменами T3A/D20N/R38I/N71K/C125S, представленными в формате моновИЛ-2-Fc с линкером (XENP26991), бивИЛ-2-Fc без линкера (XENP27001) и бивИЛ-2-Fc с линкером (XENP27007).
На фиг. 43 изображены аминокислотные последовательности иллюстративных бивалентных слияний ИЛ-2-Fc, содержащие Xtend (M428L/N434S) Fc по данному изобретению (в данном случае также содержащие шарнир и вариант C220S). Косые черты (/) указывают границу между ИЛ-2 и Fc-областями.
На фиг. 44 изображены аминокислотные последовательности иллюстративных моновалентных слияний ИЛ-2-Fc, содержащие Xtend (M428L/N434S) Fc по данному изобретению. Косые черты (/) указывают границу между ИЛ-2 и Fc-областями.
На фиг. 45A-E изображена индукция фосфорилирования STAT5 в различных популяциях лимфоцитов с помощью A) XENP26105, B) XENP26109, C) XENP24635, D) XENP25908 и E) XENP27193.
На фиг. 46 изображено иммунофенотипирование культур Treg с рапамицином, обработанных XENP27564 или рекомбинантным ИЛ-2. Данные демонстрируют более высокую экспрессию CD25 в результате лечения XENP27564.
На фиг. 47 Treg с рапамицином, обработанные XENP27564 или рекомбинантным ИЛ-2, изображены на гистограмме для оценки относительной экспрессии CD25. Данные демонстрируют, что культуры Treg с рапамицином, обработанные ИЛ-2-Fc XENP27564, демонстрируют более высокую экспрессию CD25.
На фиг. 48 изображены различные компартменты CD4+, в том числе Treg, после инкубации с рапамицином и XENP27564 или рекомбинантным ИЛ-2. Данные демонстрируют, что культуры, экспандированные с помощью XENP27564, демонстрируют более значительную популяцию эффекторных Treg (CD45RA-FoxP3mid-high) по сравнению с культурами, экспандированными с помощью рекомбинантного ИЛ-2.
На фиг. 49A и B изображена супрессия пролиферации А) CD8 респондерных Т-клеток и B) CD4 респондерных Т-клеток с помощью культур Treg с рапамицином, экспандированных либо с помощью XENP27564, либо с помощью рекомбинантного ИЛ-2. Данные предполагают, что Treg, экспандированные с помощью XENP27564, могут иметь повышенную супрессорную функцию.
На фиг. 50 изображена экспрессия CD25 на Treg в супрессорном анализе, изображенном на фиг. X, указанная с помощью А) СИФ CD25 на Treg и В) процента CD25+ Treg.
На фиг. 51А и В изображена экспрессия CD127 на Treg в супрессорном анализе, изображенном на фиг. X, указанная с помощью А) СИФ CD127 на Treg и В) процента CD127+ Treg.
На фиг. 52A-E изображена пролиферация (определенная с помощью разведения CFSE или Tag-it Violet) различных популяций лимфоцитов после инкубации или МКПК и Treg с помощью A) XENP27563, B) XENP27564, C) XENP24635, D) рекомбинантного ИЛ-2 и E) рекомбинантного ИЛ-15. Данные демонстрируют, что XENP27563 и XENP27564 демонстрируют селективность Treg.
На фиг. 53A-B изображена пролиферация CD8+ T-клеток, изображенная с помощью A) пролиферирующих клеток (определенных с помощью разведения CFSE или Tag-it Violet) и B) общего количества клеток после инкубации или МКПК и Treg с помощью XENP27563, XENP27564, XENP24635, рекомбинантного ИЛ-2, рекомбинантного ИЛ-15 и отрицательный контрольный анти-RSV мАТ XENP15074. Данные демонстрируют, что XENP27563 и XENP27564 гораздо менее эффективны в индукции пролиферации CD8+ T-клеток по сравнению с рекомбинантным ИЛ-2 и ИЛ-15, а также слиянием ИЛ-2-Fc, содержащим ИЛ-2 ДТ (с мутацией C125S).
На фиг. 54 изображена пролиферация CD4+ T-клеток, изображенная с помощью A) пролиферирующих клеток (определенных с помощью разведения CFSE или Tag-it Violet) и B) общего количества клеток после инкубации или МКПК и Treg с помощью XENP27563, XENP27564, XENP24635, рекомбинантного ИЛ-2, рекомбинантного ИЛ-15 и отрицательный контрольный анти-RSV мАТ XENP15074. Данные демонстрируют, что XENP27563 и XENP27564 гораздо менее эффективны в индукции пролиферации CD4+ T-клеток по сравнению с рекомбинантным ИЛ-2 и ИЛ-15, а также слиянием ИЛ-2-Fc, содержащим ИЛ-2 ДТ (с мутацией C125S).
На фиг. 55 изображены пролиферирующие CD8+ Т-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации CD8+ T-клеток.
На фиг. 56 изображены пролиферирующие CD4+ Т-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации CD4+ T-клеток.
На фиг. 57 изображены пролиферирующие CD45RA- CD8+ Т-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации CD8+CD45RA- T-клеток.
На фиг. 58 изображены пролиферирующие CD8+CD45RA+ Т-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации CD8+CD45RA+ T-клеток.
На фиг. 59 изображены пролиферирующие CD4+CD45RA- Т-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации CD4+CD45RA- T-клеток.
На фиг. 60 изображены пролиферирующие CD4+CD45RA+ Т-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации CD4+CD45RA+ T-клеток.
На фиг. 61 изображены пролиферирующие NK-клетки (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) имеют нарушение функции индукции пролиферации NK-клеток.
На фиг. 62 изображены пролиферирующие Treg (продемонстрированные с помощью процента клеток, экспрессирующих Ki67) после инкубации МКПК с указанными концентрациями указанных исследуемых препаратов и А) 5 нг/мл, B) 10 нг/мл или C) 20 нг/мл связанного с планшетом анти-CD3 (ОКТ3). Данные демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности CD25) индуцируют пролиферацию Treg.
На фиг. 63 изображены пролиферирующие CD8+ T-клетки, CD8+CD45RA- T-клетки, CD8+CD45RA+ T-клетки, CD4+ T-клетки, CD4+CD45RA- T-клетки, CD4+CD45RA+ T-клетки, NK-клетки и Treg (указанные в виде процента клеток, экспрессирующих Ki67) после обработки указанными концентрациями A) XENP27563, B) XENP27564, C) XENP24635, D) ИЛ-2 и E) ИЛ-15 и 5 нг/мл связанного с планшетом анти-CD3 (OKT3).
На фиг. 64 изображены пролиферирующие CD8+ T-клетки, CD8+CD45RA- T-клетки, CD8+CD45RA+ T-клетки, CD4+ T-клетки, CD4+CD45RA- T-клетки, CD4+CD45RA+ T-клетки, NK-клетки и Treg (указанные в виде процента клеток, экспрессирующих Ki67) после обработки указанными концентрациями A) XENP27563, B) XENP27564, C) XENP24635, D) ИЛ-2 и E) ИЛ-15 и 10 нг/мл связанного с планшетом анти-CD3 (OKT3).
На фиг. 65 изображены пролиферирующие CD8+ T-клетки, CD8+CD45RA- T-клетки, CD8+CD45RA+ T-клетки, CD4+ T-клетки, CD4+CD45RA- T-клетки, CD4+CD45RA+ T-клетки, NK-клетки и Treg (указанные в виде процента клеток, экспрессирующих Ki67) после обработки указанными концентрациями A) XENP27563, B) XENP27564, C) XENP24635, D) ИЛ-2 и E) ИЛ-15 и 20 нг/мл связанного с планшетом анти-CD3 (OKT3).
На фиг. 66 изображена экспансия А) CD4+CD45RA- Т-клеток, В) CD8+CD45RA- Т-клеток, C) CD8α-CD16+ NK-клеток и D) FoxP3+ Treg у яванских макак, которым вводили 3× дозу XENP27563 и 3× дозу XENP27564. Данные демонстрируют, что как XENP27563, так и XENP27564 приводят к селективной экспансии Treg, и что эти два исследуемых препарата вызывают аналогичную фармакологическую активность.
На фиг. 67 изображено изменение уровня концентрации в сыворотке крови с течением времени у яванских макак, которым вводили А) 3× дозу XENP27563 и В) 3× дозу XENP27564. Данные указывают на то, что эти два исследуемых препарата демонстрируют аналогичные фармакокинетические профили.
На фиг. 68 изображено изменение концентрации сывороточного альбумина у яванских макак, которым вводили 3× дозу XENP27563 и 3× дозу XENP27564. Данные демонстрируют, что у одного животного, которому вводили XENP27563, длительное снижение уровней альбумина было обнаружено после первого и второго введения; кроме того, у одного животного, которому вводили XENP27564, временное снижение уровня альбумина было обнаружено только после второго введения, при этом происходило быстрое восстановление до исходных уровней.
На фиг. 69 изображены телеметрические данные артериального давления для А) первого яванского макака, которому вводили XENP27563 в 0-й день и 15-й день, В) второго яванского макака, которому вводили XENP27563 в 0-й день и 15-й день, и С) третьего яванского макака, которому вводили XENP27564 в 0-й день и 15-й день. Данные демонстрируют резкое снижение кровяного давления у первой обезьяны через день после 2-й дозы и резкое снижение кровяного давления у второй обезьяны через день после 1-й дозы, в то время как у третьей обезьяны кровяное давление оставалось стабильным в течение всей продолжительности исследования.
На фиг. 70 изображена экспансия А) CD4+CD45RA- Т-клеток, В) CD8+CD45RA- Т-клеток, C) CD8α-CD16+ NK-клеток и D) Treg с течением времени у яванских макак в результате введения 1×, 2× и 3× дозы XENP27564.
На фиг. 71 изображена экспансия Treg в 7-й и 14-й дни у яванского макака, которому вводили 1× и 3× дозу XENP27564. Данные демонстрируют, что 1× и 3× дозы вызывают аналогичную фармакологическую активность у обезьян и предполагают, что максимальный эффект достигается при более низких дозах.
На фиг. 72 изображено изменение концентрации сывороточного альбумина (в качестве показателя пропотевания жидкости из сосудов) у яванских макак после введения 1×, 3× и 10× дозы XENP27564. Данные демонстрируют, что более высокие дозы XENP27564 повышали токсичность.
На фиг. 73 изображено изменение концентрации С-реактивного белка в сыворотке крови (в качестве показателя пропотевания жидкости из сосудов) у яванских макак после введения 1×, 3× и 10× дозы XENP27564. Данные демонстрируют, что более высокие дозы XENP27564 значительно повышали токсичность.
На фиг. 74 изображено изменение А) концентрации натрия, В) концентрация хлора, С) содержания эозинофилов и D) содержания базофилов у яванских макак, которым вводили 1×, 2× и 3× дозу XENP27564. Данные демонстрируют, что более высокие дозы XENP27564 повышали токсичность, в то время как меньшие дозы хорошо переносились яванскими макаками.
На фиг. 75 изображено изменение уровня концентрации в сыворотке крови с течением времени у яванских макак, которым вводили 1×, 2× и 3× дозу XENP27564. Данные демонстрируют устойчивую фармакокинетику в пределах нескольких дней у яванских макак.
На фиг. 76 изображено изменение А) содержания эозинофилов и В) содержания базофилов у яванских макак, которым вводили 3× дозу XENP27563 или 3× дозу XENP27563 в 0-й и 15-й дни. Данные демонстрируют, что повторное введение XENP27564 хорошо переносилось яванскими макаками.
На фиг. 77 изображена индукция фосфорилирования STAT5 в CD4+CD44hi клетках, CD8+CD44hi клетках и Treg (в спленоцитах от мышей В6) с помощью А) XENP26105, B) XENP26109 и С) рекомбинантного ИЛ-2 человека. Данные указывают на то, что сконструированные слияния ИЛ-2-Fc являются одинаково селективными и эффективными для Treg у мышей, что делает их пригодными для использования в доклинических моделях у мышей для исследования аутоиммунных заболеваний.
На фиг. 78 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- Т-клетках, CD4+CD45RA+ Т-клетках, CD8+CD45RA- Т-клетках, CD8+CD45RA+ Т-клетках, CD56+ NK-клетках, γδ T-клетках, а также Treg человека, с помощью А) XENP27563 и B) XENP27564.
На фиг. 79 изображена индукция фосфорилирования STAT5 в CD4+CD45RA- Т-клетках, CD4+CD45RA+ Т-клетках, CD8+CD45RA- Т-клетках, CD8+CD45RA+ Т-клетках, CD16+ NK-клетках, CD56+ NK-клетках, γδ T-клетках, а также Treg яванского макака, с помощью А) XENP27563 и B) XENP27564. Данные указывают на то, что сконструированные слияния ИЛ-2-Fc являются одинаково селективными и эффективными для Treg у яванских макак, что делает их пригодными для использования в доклинических моделях у яванских макак.
Подробное описание сущности изобретения
Введение
Данное изобретение направлено на композиции и способы, предназначенные для сконструированных слияний ИЛ-2-Fc для лечения аутоиммунных заболеваний. Аутоиммунные заболевания можно лечить с использованием механизмов, которые преимущественно активируют регуляторные Т-клетки, обычно называемые «Treg-клетками» или «Treg». Treg представляют собой субпопуляцию иммунных Т-клеток, которые являются иммуносупрессорными и модулируют иммунную систему в результате поддержания толерантности к аутоантигенам для предотвращения аутоиммунных заболеваний. Treg, как правило, подавляют пролиферацию эффекторных Т-клеток. Treg экспрессируют биомаркеры, в том числе CD4, FOXP3 и CD25 (CD25 также известен как белок ИЛ-2Рα).
Treg можно регулировать с помощью ИЛ-2, который необходим для функционирования и выживания Treg. Вследствие способности ИЛ-2 стимулировать или снижать активность как Т-клеток, так и Treg с ограниченной селективностью, существует значительная потребность в области создания более селективных модуляторов Treg. В качестве дополнения, в качестве потенциального лекарственного препарата, ИЛ-2 имеет недостаток, заключающийся в очень быстром клиренсе, при этом период полужизни измеряется в минутах, что затрудняет эффективное введение. Данное изобретение направлено на решение обеих указанных проблем в результате получения новых слитых белков ИЛ-2-Fc.
Таким образом, в данном изобретении представлены белки ИЛ-2, которые сконструированы двумя различными способами. Первый заключается в том, что варианты ИЛ-2 по данному изобретению предпочтительно активируют CD25+ клетки, такие как Treg, по сравнению с другими Т-клетками, которые являются CD25-, чтобы обеспечить повышенную селективность Treg по сравнению с другими Т-клетками и, таким образом, приводить к образованию композиций для подавления иммунной функции и, таким образом, способствовать лечению аутоиммунных заболеваний. Как правило, это осуществляют либо путем увеличения связывания с ИЛ-2Рα, либо путем уменьшения связывания с ИЛ-2Рβ (и/или ИЛ-2Рγ) или с границей разделе ИЛ-2Рβγ, либо и того, и другого.
В дополнение к разработке селективности, описанной выше, в данном изобретении также представлены белки ИЛ-2, которые имеют увеличенный период полужизни в сыворотке, что осуществляется с использованием слияний Fc. В этом случае добавление Fc-домена будет приводить к увеличению периода полужизни молекулы ИЛ-2, как общеизвестно в данной области техники. В то же время, в данном изобретении представлены два дополнительных способа увеличения периода полужизни в сыворотке крови.
Первый включает FcRn-рецептор. В IgG сайт в Fc между Cγ2- и Cγ3-доменами опосредует взаимодействие с неонатальным FcRn-рецептором. Связывание с FcRn приводит к рециркуляции эндоцитозированного антитела из эндосомы обратно в кровоток (Raghavan et al., 1996, Annu Rev Cell Dev Biol 12:181-220; Ghetie et al., 2000, Annu Rev Immunol 18:739-766, оба полностью включены посредством ссылки). Этот процесс, в сочетании с исключением фильтрации в почках вследствие большого размера полноразмерной молекулы приводит к предпочтительным периодам полужизни антител в сыворотке крови в пределах от одной до трех недель. Для того, чтобы увеличить удерживание Fc-белков in vivo, повышение аффинности связывания должно происходить на уровне около рН 6, в то время как поддержание более низкой аффинности должно происходить на уровне около рН 7,4. Несмотря на то, что вопрос все еще находится на изучении, считается, что Fc-области имеют более длительные периоды полужизни in vivo, поскольку связывание с FcRn при рН 6 в эндосоме приводит к секвестрированию Fc (Ghetie and Ward, 1997 Immunol Today. 18(12):592-598, включена в полном объеме посредством ссылки). Затем эндосомальный компартмент рециркулирует Fc на поверхность клетки. После того, как компартмент открывается во внеклеточное пространство, более высокое значение рН, ~7,4, способствует высвобождению Fc обратно в кровоток.
Соответственно, увеличенное время полужизни в сыворотке крови может способствовать использованию вариантов Fc, которые увеличивают связывание с FcRn и во многих случаях приводят к увеличению периода полужизни.
Дополнительный способ увеличения периода полужизни в сыворотке крови слитой молекулы ИЛ-2-Fc основан на рН-конструировании для рециркуляции из эндосомального пути сортировки. Как известно в данной области техники, эндоцитоз цитокинов, таких как ИЛ-2, в эндосому приводит к эндоцитотизной сортировке, при этом цитокин либо распадается, либо возвращается обратно в кровоток (см. Fallon et al., JBC 275(10):6790, 2000, тем самым включенная в данный документ посредством ссылки в полном объеме). После интернализации в эндосому ИЛ-2, ИЛ-2Рß и γc распадаются, в то время как ИЛ-2Р конститутивно рециркулируется к клеточной поверхности. Поскольку уровень pH крови составляет от приблизительно 7,2 до 7,4, а pH эндосомы составляет около 6, в результате конструирования ИЛ-2 для повышения связывания с лигандом ИЛ-2Рα при pH 6, ИЛ-2/ИЛ-2Рα рециркулируется, а не распадается, что приводит к увеличению периода полужизни в сыворотке крови.
В качестве дополнения, активность слитых молекул ИЛ-2-Fc по данному изобретению также может зависеть от других факторов. Например, в данном изобретении представлены бивалентные конструкции ИЛ-2, такие как изображенные на фиг. 19B, при этом образуются гомодимеры вариантов слияний ИЛ-2-Fc, приводя, тем самым, к образованию бивалентного связывания с рецепторами. В качестве альтернативы, в данном изобретении представлены моновалентные конструкции ИЛ-2, такие как изображенные на фиг. 19А, при этом образуются гетеродимеры, один из мономеров которых представляет собой вариант слияния ИЛ-2-Fc, а другой представляет собой Fc-мономер «с пустым фрагментом». В качестве дополнения, наличие дополнительных гибких линкеров в некоторых случаях может приводить к повышению активности, например, как показано на фиг. 19C для моновалентных конструкций и фиг. 19D для бивалентных конструкций.
Соответственно, в данном изобретении представлены сконструированные варианты ИЛ-2, а также сконструированные слитые белки ИЛ-2-Fc, которые демонстрируют предпочтительную активацию CD25+ клеток, таких как Treg, по сравнению с CD25- T-клетками, и которые демонстрируют увеличенный период полужизни в сыворотке крови.
Определения
С целью более полного понимания данной заявки, ниже приведены несколько определений. Такие определения предназначены для охвата грамматических эквивалентов.
В данном контексте под «абляцией» подразумевается снижение или устранение активности. Так, например, «устранение связывания FcγR» означает, что аминокислотный вариант Fc-области характеризуется менее 50% начального связывания по сравнению с Fc-областью, не содержащей специфического варианта, с потерей менее 70-80-90-95-98% активности, что является предпочтительным, и, как правило, активностью ниже уровня детектируемого связывания в анализе Biacore. Особенно применимыми при устранении связывания FcγR являются те, которые показаны на фиг. 4.
Под термином «ADCC» или «антителозависимая клеточно-опосредованная цитотоксичность», используемым в данном документе, подразумевается клеточно-опосредованная реакция, при которой неспецифические цитотоксические клетки, которые экспрессируют FcγR, распознают связанное антитело на клетке-мишени и впоследствии вызывают лизис клетки-мишени. ADCC коррелирует со связыванием Fc-области с FcγRIIIa; усиление связывания с FcγRIIIa приводит к увеличению активности ADCC. Как обсуждается в данном документе, многие варианты осуществления данного изобретения полностью приводят к устранению активности ADCC.
Под термином «модификация», используемым в данном документе, подразумевается аминокислотная замена, вставка и/или делеция в полипептидной последовательности или изменение фрагмента, химически связанного с белком. Например, модификация может представлять собой измененную углеводную или ПЭГ-структуру, присоединенную к белку. Под термином «аминокислотная модификация», используемым в данном документе, подразумевается аминокислотная замена, вставка и/или делеция в полипептидной последовательности. Для ясности, если не указано иное, аминокислотная модификация всегда относится к аминокислоте, кодируемой ДНК, например, к 20 аминокислотам, которые имеют кодоны в ДНК и РНК.
Под термином «аминокислотная замена» или «замена», используемым в данном документе, подразумевается замена аминокислоты в конкретном положении в родительской полипептидной последовательности другой аминокислотой. В частности, в некоторых вариантах осуществления данного изобретения замена представляет собой аминокислоту, которая не встречается в природе в конкретном положении, либо не встречается в природе в организме, либо в любом организме. Например, замена S364K относится к вариантному полипептиду, в данном случае к Fc варианту, в котором серин в положении 364 заменен лизином. Нумерация основана на нумерации родительского полипептида, например, R38W в контексте нумерации ИЛ-2. Для ясности, белок, который был сконструирован для изменения последовательности, кодирующей нуклеиновую кислоту, но не для замены исходной аминокислоты (например, обмен CGG (кодирующего аргинин) на CGA (все еще кодирующего аргинин) для повышения уровня экспрессии в организме хозяина), не является «аминокислотной заменой»; то есть, несмотря на создание нового гена, кодирующего тот же белок, если белок имеет ту же аминокислоту в конкретной позиции, с которой он начинался, это не аминокислотная замена.
Под термином «вставка аминокислоты» или «вставка», используемым в данном документ, подразумевается добавление аминокислотной последовательности в конкретном положении в последовательности родительского полипептида. Например, -233E или 233E обозначает вставку глутаминовой кислоты после положения 233 и перед положением 234. Кроме того, -233ADE или A233ADE обозначает вставку AlaAspGlu после положения 233 и перед положением 234.
Под термином «делеция аминокислоты» или «делеция», используемом в данном документе, подразумевается удаление аминокислотной последовательности в конкретном положении в последовательности родительского полипептида. Например, E233- или E233#, E233() или E233del обозначает делецию глутаминовой кислоты в положении 233. Кроме того, EDA233- или EDA233# обозначает делецию последовательности GluAspAla, которая начинается в положении 233.
Под термином «белок», используемым в данном документе, подразумевается по меньшей мере две ковалентно связанные аминокислоты, при этом указанный термин включает белки, полипептиды, олигопептиды и пептиды. Пептидильная группа может содержать встречающиеся в природе аминокислоты и пептидные связи или синтетические пептидомиметические структуры, то есть «аналоги», такие как пептоиды (см. Simon et al., PNAS USA 89(20):9367 (1992), которая включена в данный документ посредством ссылки в полном объеме). Аминокислоты могут быть либо встречающимися в природе, либо синтетическими (например, не быть аминокислотой, кодируемой ДНК); как будет понятно специалистам в данной области техники. Как правило, в белках по данному изобретению используются встречающиеся в природе аминокислоты. В качестве дополнения, вариантные полипептиды могут включать синтетическую дериватизацию одной или нескольких боковых цепей или концов, гликозилирование, пегилирование, циклическую перестановку, циклизацию, линкеры с другими молекулами, слияние с белками или доменами белков и добавление пептидных тегов или меток.
Под термином «остаток», используемым в данном документе, подразумевается положение в белке и ассоциированную с ним идентичность аминокислот. Например, аргинин 38 (также называемый Arg38 или R38) представляет собой остаток в положении 38 (нумерация от зрелой последовательности) в белке ИЛ-2 человека.
Под термином «вариантный белок» или «вариант белка» или «вариант», используемым в данном документе, подразумевается белок, который отличается от родительского белка, по меньшей мере, модификацией одной аминокислоты. Вариант белка может относиться к самому белку, композиции, содержащей белок, или к аминокислотной последовательности, которая его кодирует. Предпочтительно, вариант белка имеет, по меньшей мере, одну аминокислотную модификацию по сравнению с родительским белком, например, от около одной до около семидесяти аминокислотных модификаций, и предпочтительно от около одной до около пяти аминокислотных модификаций по сравнению с родительским белком. Как описано ниже, в некоторых вариантах осуществления данного изобретения родительский полипептид, например родительский полипептид Fc, представляет собой последовательность дикого типа человека, такую как Fc-область IgG1, IgG2, IgG3 или IgG4 человека. В контексте вариантов ИЛ-2, родительский полипептид представляет собой ИЛ-2 человека, зрелый последовательность которого показана на фиг. 1. Вариант последовательности белка в данном документе предпочтительно будет обладать, по меньшей мере, около 80% идентичностью с последовательностью исходного белка, и наиболее предпочтительно, по меньшей мере, около 90% идентичностью, более предпочтительно, по меньшей мере, около 95-98-99% идентичностью. Вариант белка может относиться к самому варианту белка, композициям, содержащим вариант белка, или последовательности ДНК, которая его кодирует.
Под термином «Fc» или «Fc-область» или «Fc-домен», используемым в данном документе, подразумевается полипептид, содержащий константную область антитела IgG, исключая первый константный домен домена иммуноглобулина и, в некоторых случаях, весь или часть шарнира. В IgG Fc-домен содержит иммуноглобулиновые домены Cγ2 и Cγ3 (CH2 и CH3) и шарнирную область между Cγ1 (CH1) и Cγ2 (CH2). В контексте антител IgG каждый изотип IgG имеет три СН-области. Соответственно, «CH»-домены в контексте IgG являются следующими: «CH1» относится к положениям 118-220 в соответствии с индексом EU, как по Кабату. «CH2» относится к положениям 237-340 в соответствии с индексом EU, как по Кабату, а «CH3» относится к положениям 341-447 в соответствии с индексом EU, как по Кабату. Если не указано иное, Fc-домены по данному изобретению содержат шарнир, начинающийся в положении 216 (нумерация EU) и заканчивающийся на С-конце СН3-домена в положении 447; это называется «шарнир-СН2-СН3» IgG. В некоторых случаях, таких как в случае Fc-слияний в данном документе, шарнир выступает в качестве линкера домена, описанного в данном документе. В некоторых вариантах осуществления, как более полно описано ниже, аминокислотные модификации осуществляются в Fc-области, например, для изменения связывания с одним или несколькими FcγR-рецепторами или с FcRn-рецептором или для облегчения гетеродимеризации Fc-доменов.
Соответственно, под термином «вариант Fc» или «вариантный Fc», используемым в данном документе, подразумевается белок, содержащий аминокислотную модификацию в Fc-домене. Варианты Fc по данному изобретению определены в соответствии с аминокислотными модификациями, которые их составляют. Таким образом, например, N434S или 434S представляет собой вариант Fc с замещенным серином в положении 434 относительно родительского Fc-полипептида, при этом нумерация соответствует индексу EU. Аналогично, M428L/N434S определяет вариант Fc с заменами M428L и N434S относительно исходного Fc-полипептида. Идентичность аминокислоты ДТ может быть неопределенной, и в этом случае вышеупомянутый вариант обозначается как 428L/434S. Следует отметить, что порядок, в котором представлены замены, является произвольным, т.е., например, 428L/434S представляет собой тот же самый вариант Fc, что и M428L/N434S, и т.д. Для всех положений, обсуждаемых в данном изобретении, которые связаны с антителами, если не указано иное, нумерация аминокислотных положений соответствует индексу EU. Индекс EU или EU-индекс, как по Кабату или схема нумерации EU, относится к нумерации антител EU (Edelman et al., 1969, Proc Natl Acad Sci USA 63:78-85, которая включена в данный документ посредством ссылки в полном объеме). Модификация может представлять собой добавление, делецию или замену. Замены могут включать встречающиеся в природе аминокислоты и, в некоторых случаях, синтетические аминокислоты. Примеры включают патенты США № 6586207; WO 98/48032; WO 03/073238; US2004-0214988A1; WO 05/35727A2; WO 05/74524A2; J. W. Chin et al., (2002), Journal of the American Chemical Society 124:9026-9027; J. W. Chin, & P. G. Schultz, (2002), ChemBioChem 11:1135-1137; J. W. Chin, et al., (2002), PICAS United States of America 99:11020-11024; и L. Wang & PG Schultz (2002), Chem. 1-10, все из которых включены в данный документ посредством ссылки в полном объеме.
Под термином «интерлейкин-2» или «ИЛ-2» в данном документе подразумевается ИЛ-2 человека, имеющий последовательность, показанную на фиг. 1.
Под термином «вариант ИЛ-2» или «вариантный ИЛ-2» в данном документе подразумевается белок, содержащий аминокислотную модификацию в последовательности зрелого ИЛ-2 человека, изображенную на фиг. 1. Варианты ИЛ-2 по данному изобретению определены в соответствии с аминокислотными модификациями, которые их составляют, как изложено выше, с помощью нумерации зрелой формы человека.
Под термином «слитый Fc-белок» или «иммуноадгезин» в данном документе подразумевается белок, содержащий Fc-область, как правило, связанную (необязательно с помощью линкерного фрагмента, описанного в данном документе, который может представлять собой шарнирную область IgG, такого как IgG1), с другим белком, таким как ИЛ-2. Таким образом, слитый белок ИЛ-2-Fc представляет собой белок, содержащий ИЛ-2 (в данном случае вариантный ИЛ-2) и Fc-домены, изложенные в данном документе (в данном случае также, как правило, варианты Fc). Они, как правило, имеют структуру ИЛ-2-шарнир-CH2-CH3. Как будет понятно из уровня техники, два Fc-домена будут подвергаться самосборке с образованием димерных слитых Fc-белков, изложенных в данном документе.
Под термином «положение», используемым в данном документе, подразумевается местоположение в последовательности белка. Положения могут быть пронумерованы последовательно или в соответствии с установленным форматом, например, индексом EU для нумерации антител.
Под термином «не встречающаяся в природе модификация», используемым в данном документе, подразумевается аминокислотная модификация, которая не является изотипической. Например, поскольку ни один из IgG не содержит серин в положении 434, замена 434S в IgG1, IgG2, IgG3 или IgG4 (или в их гибридах) считается не встречающейся в природе модификацией.
Под термином «аминокислота» и «идентичность аминокислоты», используемым в данном документе, подразумевается одна из 20 встречающихся в природе аминокислот, которые кодируются ДНК и РНК.
Под термином «эффекторная функция», используемым в данном документе, подразумевается биохимическое явление, которое возникает в результате взаимодействия Fc-области антитела с Fc-рецептором или лигандом. Эффекторные функции включают, но не ограничиваясь ими, ADCC, ADCP и CDC.
Под термином «Fc гамма-рецептор», «FcγR» или «Fc гамма Р», используемым в данном документе, подразумевается любой представитель семейства белков, который связывает Fc-область антитела IgG и кодируется геном FcγR. У людей это семейство включает, но не ограничиваясь ими, FcγRI (CD64), в том числе изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), в том числе изоформы FcγRIIa (в том числе аллотипы H131 и R131), FcγRIIb (в том числе FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16), в том числе изоформы FcγRIIIa (в том числе аллотипы V158 и F158) и FcγRIIIb (в том числе аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2) (Jefferis et al., 2002, Immunol Lett 82: 57-65, которая в полном объеме включена в данный документ посредством ссылки), а также любые неизвестные изоформы или аллотипы FcγRs или FcγR человека. FcγR может происходить из любого организма, в том числе, но не ограничиваясь ими, организма людей, мышей, крыс, кроликов и обезьян. FcγR мыши включают, но не ограничиваясь ими, FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (CD16-2), а также любые неизвестные изоформы или аллотипы FcγR или FcγR мыши.
Под термином «FcRn» или «неонатальный Fc-рецептор», используемым в данном документе, подразумевается белок, который связывает Fc-область антитела IgG и кодируется, по меньшей мере, отчасти, геном FcRn.
Под термином «родительский полипептид», используемым в данном документе, подразумевается исходный полипептид, который впоследствии модифицируют для получения варианта. Родительский полипептид может представлять собой встречающийся в природе полипептид или вариант, или сконструированный вариант встречающегося в природе полипептида. Родительский полипептид может относиться к самому полипептиду, композициям, которые содержат родительский полипептид, или к аминокислотной последовательности, которая его кодирует. Соответственно, под термином «родительский ИЛ-2», используемым в данном документе, подразумевается немодифицированный белок ИЛ-2 человека, который модифицирован для получения варианта, а под термином «родительский Fc» или «родительский Fc-домен», используемым в данном документе, подразумевается немодифицированный Fc-домен IgG человека, который модифицирован для получения вариантного Fc-домена.
Под «наличием цепей» в контексте мономеров гетеродимерных Fc-слияний по данному изобретению в данном документе подразумевается, что, подобно двум цепям ДНК, которые «совпадают», гетеродимеризационные варианты включены в каждый мономер с целью сохранения способности «совпадать» с образованием гетеродимеров. Например, если некоторые pI варианты сконструированы в мономер A (например, делая pI выше), тогда стерические варианты, которые представляют собой «пары зарядов», которые также могут использоваться, не влияют на pI варианты, например, варианты зарядов, которые делает pI выше, помещаются в одну и ту же «цепь» или «мономер» с тем, чтобы сохранить обе функциональные возможности. Аналогично, для «искаженных» вариантов, которые входят в пары совокупности, как более подробно изложено ниже, специалист в данной области техники будет учитывать pI при принятии решения, в какую цепь или мономер, которые включают одну совокупность пары, направляться с тем, чтобы сделать разделение pI максимальным также с использованием рI искаженных вариантов.
Под термином «дикий тип или ДТ», используемым в данном документе, подразумевается аминокислотная последовательность или нуклеотидная последовательность, которая встречается в природе, в том числе аллельные вариации. Белок ДТ имеет аминокислотную последовательность или нуклеотидную последовательность, которая не была намеренно модифицирована.
Белки по данному изобретению, как правило, являются выделенными или рекомбинантными. Термин «выделенный», при применении его для описания различных полипептидов, охарактеризованных в данном документе, означает полипептид, который был идентифицирован и отделен и/или извлечен из клетки или клеточной культуры, из которой он был экспрессирован. Обычно выделенный полипептид будет получен, по меньшей мере, в результате одного этапа очистки. Термин «рекомбинантный» означает, что антитела получают с помощью методик рекомбинантных нуклеиновых кислот в экзогенных клетках-хозяевах.
«Процент (%) идентичности аминокислотной последовательности» по отношению к последовательности белка определяется в виде процента аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в конкретной (родительской) последовательности после выравнивания последовательностей и введения гэпов, при необходимости, для достижения максимального процента идентичности последовательности, и без учета каких-либо консервативных замен в виде части идентичности последовательности. Выравнивание в целях определения процента идентичности аминокислотной последовательности может быть достигнуто различными способами, которые известны специалисту в данной области техники, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить соответствующие параметры для измерения выравнивания, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания по всей последовательностей, подлежащих сравнению. Одной из конкретных программ является программа ALIGN-2, описанная в параграфах [0279]-[0280] публикации США № 20160244525, включенная в данный документе посредством ссылки.
Степень идентичности между аминокислотной последовательностью по данному изобретению («последовательность по данному изобретению») и родительской аминокислотной последовательностью рассчитывают в виде количества точных совпадений в выравнивании двух последовательностей, деленное на длину «последовательности по данному изобретению» или длину родительской последовательности, в зависимости от того, какая из них является самой короткой. Результат выражается в проценте идентичности.
В некоторых вариантах осуществления две или более аминокислотных последовательности являются, по меньшей мере, на 50%, 60%, 70%, 80% или 90% идентичными. В некоторых вариантах осуществления две или более аминокислотных последовательности являются, по меньшей мере, на 95%, 97%, 98%, 99% или даже 100% идентичными.
Под термином «линкер» в данном документе подразумевается белковый линкер, который используется для соединения двух других белковых доменов (например, домена вариантного ИЛ-2 и вариантного Fc-домена). В некоторых случаях линкер представляет собой «линкер доменов», используемый для связывания любых двух доменов, как указано в данном документе, вместе. Несмотря на то, что может быть использован любой подходящий линкер, во многих вариантах осуществления используется глицин-сериновый полимер, в том числе, например, (GS)n, (GSGGS)n, (GGGGS)n и (GGGS)n, где n представляет собой целое число, из, по меньшей мере, одного (и, как правило, от 3 до 4 и до 5), а также любая пептидная последовательность, которая обеспечивает рекомбинантное присоединение двух доменов с достаточной длиной и гибкостью с тем, чтобы каждый домен сохранял свою биологическую функцию. В некоторых случаях и при уделении внимания «наличию цепей», как изложено ниже, могут использоваться заряженные линкеры доменов. Кроме того, шарнирный домен белка IgG1 человека также может представлять собой линкер доменов.
Под термином «регуляторные Т-клетки» или «Treg» в данном документе подразумеваются Т-клетки, которые представляют собой CD3+/CD4+/CD8-/CD25 +/FOXP3+.
Слитые белки ИЛ-2-Fc по данному изобретению
В данном изобретении представлены слитые белки (ИЛ-2)-(Fc-домен), продемонстрированные в данном документе и в целом на фиг. 19. Как будет понятно специалистам в данной области техники, слитые белки по данному изобретению фактически представляют собой два различных полипептида, которые подлежат самосборке в связи с наличием Fc-областей либо в гомодимерные белки фиг. 19В, либо в гетеродимерные белки (фиг. 19А). Белки по данному изобретению, как правило, имеют три различных домена: Fc-домен, один или несколько линкеров доменов и домен ИЛ-2.
Домены ИЛ-2 по данному изобретению
Слитые белки ИЛ-2-Fc по данному изобретению содержат домены ИЛ-2, которые представляют собой вариантные домены ИЛ-2 человека. Как обсуждается в данном документе, указанные домены сконструированы таким образом, чтобы включать варианты специфичности, которые приводят к повышенной активации Т-клеток, которые представляют собой CD25+, такие как Treg, по сравнению с популяциями CD25- Т-клеток, и необязательно также содержат аминокислотные замены, предназначенные для увеличения связывания ИЛ-2 с ИЛ-2Рα при рН 6, так что слитые Fc-белки варианта ИЛ-2 рециркулируют по эндоцитотическому пути, а не распадаются.
Экспрессионные варианты
Предварительно варианты ИЛ-2 по данному изобретению также содержат C125S вариант, который, как было показано ранее, увеличивает экспрессию ИЛ-2 человека. Таким образом, если не указано иное, все варианты, описанные в данном документе, содержат C125S вариант; в некоторых случаях также может использоваться C125A вариант.
В качестве дополнения, в некоторых случаях варианты ИЛ-2 по данному изобретению содержат вариант T3A, который приводит к удалению сайта O-гликозилирования с целью уменьшения сложности.
В качестве дополнения, варианты ИЛ-2 по данному изобретению содержат дополнительные мутации.
Варианты специфичности
Соответственно, в данном изобретении представлены вариантные белки ИЛ-2 с повышенной специфичностью к CD25+ Т-клеткам, в том числе Treg. Как правило, это осуществляют либо путем увеличения связывания с ИЛ-2Рα, либо путем уменьшения связывания с ИЛ-2Рβ и ИЛ-2Рγ или с границей раздела ИЛ-2Рβγ, либо и того, и другого.
В одном варианте осуществления вариантный ИЛ-2 содержит аминокислотную замену D20N, в качестве дополнения к варианту экспрессии C125S, и, таким образом, имеет набор аминокислот D20N/C125S. Следует отметить, что ранее сообщалось, что вариант D20N приводит к потере связывания как с высокоаффинным рецептором (ИЛ-2Рαβ), так и с рецептором с промежуточной аффинностью (ИЛ-2Рβ); см. Collins et al., PNAS USA 85:7709-7713 (1988), предполагая, что «дифференциальное связывание или активация между ИЛ-2Рβ или ИЛ-2Рαβγ не достигается с помощью замены Asp в положении 20», см. патент США № 6955807, Описание предшествующего уровня техники.
В одном варианте осуществления вариантный ИЛ-2 содержит аминокислотный вариант T3A, в качестве дополнения к аминокислотной замене D20N и варианту экспрессии C125S, и, таким образом, имеет набор аминокислот D20N/C125S.
В одном варианте осуществления вариантный ИЛ-2 содержит аминокислотный вариант T37R, в качестве дополнения к аминокислотному варианту T3A, аминокислотной замене D20N и варианту экспрессии C125S, и, таким образом, имеет набор аминокислот T3A/D20N/T37R/C125S.
В одном варианте осуществления вариантный ИЛ-2 содержит аминокислотный вариант N71K, в качестве дополнения к аминокислотному варианту T3A, аминокислотной замене D20N и варианту экспрессии C125S, и, таким образом, имеет набор аминокислот T3A/D20N/N71K/C125S.
В одном варианте осуществления вариантный ИЛ-2 содержит аминокислотные варианты N29S/Y31H/K35R/T37A/K48E/N71R/N88D/I89V, в качестве дополнения к варианту экспрессии C125S, и, таким образом, имеет набор аминокислот N29S/Y31H/K35R/T37A/K48E/N71R/N88D/I89V/C125S.
Анализы на специфичность Treg
Как известно в данной области техники, активация белков STAT5 (STAT5a и STAT5b) путем фосфорилирования является одним из самых ранних сигнальных событий, опосредованных ИЛ-2. Таким образом, рассматривая фосфорилирование STAT5 в различных популяциях Т-клеток с использованием конструкций по данному изобретению, можно оценить специфичность.
Как правило, как описано в Примерах, анализы фосфорилирования STAT5 проводят с использованием способов, изложенных в разделе Примеры. Как описано в разделе Примеры, как правило, исследуют 5 различных типов клеток, в том числе CD4+/CD45RA+, CD4+/CD45RA-, CD8+CD45RA+, CD8+CD45RA- и Treg (CD3+/CD4+/CD8-/CD25+/FOXP3+) для обеспечения отбора образцов других типов Т-клеток (например, CD45RA экспрессируется на Т-клетках памяти, а не на наивных Т-клетках).
Как правило, имеет место повышенная активность по сравнению с ИЛ-2 дикого типа.
pH варианты
Кроме того, в данном изобретении представлены вариантные белки ИЛ-2 с повышенной специфичностью к рН, в которых связывание при рН 6 (рН эндосомы) увеличивается.
В указанном варианте осуществления вариант ИЛ-2 может иметь одну или несколько аминокислотных замен, выбранных из R38A, R38D, R38E, R38F, R38G, R38H, R38I, R38K, R38L, R38M, R38N, R38P, R38Q, R38S, R38T, R38V, R38W, R38Y, T41A, T41D, T41E, T41F, T41G, T41H, T41I, T41K, T41L, T41M, T41N, T41P, T41Q, T41R, T41S, T41V, T41W, T41Y, F42A, F42D, F42E, F42G, F42G, F42I, F42K, F42L, F42M, F42N, F42P, F42Q, F42R, F42S, F42T, F42V, F42W и F42Y. В качестве дополнения, указанные мутации могут сочетаться с C125S.
В указанном варианте осуществления вариант ИЛ-2 может иметь аминокислотную (аминокислотные) замену (замены), выбранную (выбранные) из R38Q/T41K, R38Q/41Q, R38E/T41K, R38Q/T41R, R38N/T41Q, R38Q/T41V, R38N/T41V, R38Q/T41M, R38Q/T41S, R38Q/T41L, R38N/T41M, T41I/F42Y, T41E/F42Y, T41D/F42Y, T41M/F42Y, T41Q/F42Y, T41E/F42H, T41E/F42L, T41E/F42P, R38Q/F42Y, R38N/T41R, R38N/T41K, R38V/T41R, R38P/T41R, T41E/F42K, T41D/F42K, T41M/F42K, T41Q/F42K, R38Q/F42K, T41I/F42K, R38N/F42K, T41H/F42K, R38Q/T41K/F42Y, R38Q/T41R/F42Y, R38Q/T41Q/F42Y, R38Q/T41V/F42Y, R38N/T41K/F42K, R38Q/T41H/F42K, R38Q/T41K/F42K, R38Q/T41Q/F42K, R38Q/T41V/F42K и R38Q/T41R/F42K. В качестве дополнения, указанные мутации могут сочетаться с C125S.
Пригодные варианты ИЛ-2
В данном изобретении представлен ряд особенно пригодных вариантов ИЛ-2, которые имеют необходимые виды активности как отдельно, так и при слиянии с Fc-доменами, в том числе как Fc-доменами дикого типа, так и вариантными Fc-доменами, изложенными в данном документе. В качестве дополнения, указанные варианты ИЛ-2 могут быть использованы в моновалентных конструкциях (например, фиг. 19А) или в бивалентных конструкциях (например, фиг. 19В).
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены R38I/C125S и используется в бивалентной конструкции. В указанном варианте домен вариантного ИЛ-2 может быть слит с диким Fc-доменом, например, из IgG2 или IgG4. В качестве альтернативы, домен вариантного ИЛ-2 может быть слит с вариантным Fc-доменом, например, доменом, которые содержит абляционные варианты и варианты FcRn.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены R38I/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены R38L/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены R38L/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены D20N/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены D20N/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены N29S/Y31H/K35R/T37A/K48E/N71R/N88D/I89V/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены N29S/Y31H/K35R/T37A/K48E/N71R/N88D/I89V/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/T37R/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/T37R/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/N71K/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/N71K/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/T37R/R38I/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/T37R/R38I/C125S и используется в моновалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/R38I/N71K/C125S и используется в бивалентной конструкции.
В одном варианте осуществления домен вариантного ИЛ-2 имеет аминокислотные замены T3A/D20N/R38I/N71K/C125S и используется в моновалентной конструкции.
Особенно предпочтительные белки включают XENP14142, XENP14144, XENP23833, XENP25720, XENP26086, XENP26105, XENP26987, XENP27003, XENP26109, XENP26994, XENP26841, XENP27004, XENP26839, XENP26996, XENP26990, XENP27006, XENP26840, XENP27001, XENP26991, XENP27007, XENP27563, XENP26105, XENP27564 и XENP26109.
Fc-2 домены по данному изобретению
Как обсуждается в данном документе, в данном изобретении представлены слитые Fc-белки, содержащие два Fc-домена, при этом, по меньшей мере, один из Fc-доменов, содержащий слитый вариант ИЛ-2, необязательно содержит линкер доменов. Как показано на фиг. 19, димерные белки по данному изобретению могут быть сконфигурированы таким образом, чтобы иметь один вариант ИЛ-2, который иногда называется в данном документе как «моновалентный ИЛ-2», как показано на фиг. 19А, где один из Fc-доменов ковалентно связан с вариантом белка ИЛ-2, а другой является «пустым» или представляет собой «только Fc». Указанный вариант осуществления основан на гетеродимерных Fc-доменах, как обсуждается ниже. В качестве альтернативы, используют «бивалентные» конструкции ИЛ-2, такие, как показано на фиг. 19В, где каждый Fc-домен слит с вариантом ИЛ-2; в указанных вариантах осуществления используют гомодимерные Fc-домены, как обсуждается выше.
В обоих вариантах осуществления, независимо от того, используется ли гомо- или гетеродимерная слитая Fc-конструкция, Fc-домены, как правило, содержат некоторые конкретные аминокислотные варианты для нескольких функций.
Fc варианты для дополнительной функциональности
В качестве дополнения к pI аминокислотным вариантам существует ряд пригодных аминокислотных модификаций Fc, которые могут быть выполнены по ряду причин, в том числе, но не ограничиваясь ими, изменение связывания с одним или несколькими FcγR-рецепторами, изменение связывания с FcRn-рецепторами и т.д.
Соответственно, белки по данному изобретению могут содержать аминокислотные модификации, в том числе гетеродимеризационные варианты, изложенные в данном документе, которые включают pI варианты и стерические варианты. Каждая совокупность вариантов может быть независимо и необязательно включена или исключена из любого конкретного гетеродимерного белка.
FcγR варианты
Соответственно, существует ряд пригодных замен Fc, которые можно осуществить для изменения связывания с одним или несколькими FcγR-рецепторами. Могут быть пригодными замены, которые приводят к повышению связывания, а также к уменьшению связывания. Например, известно, что повышенное связывание с FcγRIIIa приводит к увеличению ADCC (антителозависимой клеточно-опосредованной цитотоксичности; клеточно-опосредованной реакции, при которой неспецифические цитотоксические клетки, которые экспрессируют FcγR, распознают связанное антитело на клетке-мишени и впоследствии вызывают лизис клетки-мишени). Аналогичным образом, в некоторых случаях также может быть пригодным уменьшение связывания с FcγRIIb (ингибирующий рецептор). Аминокислотные замены, которые находят применение в данном изобретении, включают замены, перечисленные в USSN 11/124620 (особенно на фиг. 41), 11/174287, 11/396495, 11/538406, все из которых явным образом включены в данный документ посредством ссылки в полном объеме и конкретно для описанных в них вариантов. Конкретные варианты, которые находят применение, включают, но не ограничиваясь ими, 236A, 239D, 239E, 332E, 332D, 239D/332E, 267D, 267E, 328F, 267E/328F, 236A/332E, 239D/332E/330Y, 239D, 332E/330L, 243A, 243L, 264A, 264V и 299T.
В качестве дополнения, существуют дополнительные замены Fc, которые находят применение в повышенном связывании с FcRn-рецептором и увеличенном периоде полужизни в сыворотке крови, как конкретно раскрыто в USSN 12/341769, полностью включенным в данный документ посредством ссылки, в том числе, но не ограничиваясь ими, 434S, 434A, 428L, 308F, 259I, 428L/434S, 428L/434A, 259I/308F, 436I/428L, 436I или V/434S, 436V/428L и 259I/308F/428L.
Абляционные варианты
Аналогичным образом, другая категория функциональных вариантов представляет собой «абляционные варианты FcγR» или «варианты Fc-нокаута (FcKO или KO)». В указанных вариантах осуществления для некоторых терапевтических применений желательно уменьшить или устранить нормальное связывание Fc-домена с одним или несколькими или всеми Fcγ-рецепторами (например, FcγR1, FcγRIIa, FcγRIIb, FcγRIIIa и т.д.), чтобы избежать дополнительных механизмов действия. Т.е., например, желательно устранять связывание с FcγRIIIa для избегания или значительного снижения активности ADCC, таким образом, что один из Fc-доменов содержит один или несколько абляционных вариантов Fcγ-рецепторов. Указанные абляционные варианты изображены на фиг. 4, и каждый из них может быть независимо и необязательно включен или исключен, причем в предпочтительных аспектах используются абляционные варианты, выбранные из группы, состоящей из G236R/L328R, E233P/L234V/L235A/G236del/S239K, E233P/L234V/L235A/G236del/S267K, E233P/L234V/L235A/G236del/S239K/A327G, E233P/L234V/L235A/G236del/S267K/A327G и E233P/L234V/L235A/G236del. Следует отметить, что абляционные варианты, упомянутые в данном документе, устраняют связывание с FcγR, но обычно не связывание с FcRn.
Гомодимерные Fc-домены
В некоторых вариантах осуществления в данном изобретении представлены бивалентные гомодимерные белки, содержащие гомодимерные Fc-домены, как в целом изображено на фиг. 19В. В указанном варианте осуществления каждый мономер является идентичным и, как правило, содержит вариантный ИЛ-2-линкер-Fc-домен, где линкер, как правило, представляет собой шарнир из IgG1.
В указанном варианте осуществления Fc-домены могут иметь абляционные варианты, как правило, изображенные на фиг. 4. Подходящие абляционные варианты изображены на фиг. 4, которые, как правило, приводят к устранению связывания с FcRI, FcγRIIb и FcγRIIIa. Особо применимым в указанном варианте осуществления IgG1 является набор абляционных аминокислот E233P/L234V/L235A/G236_/S267K («G236_» представляет собой делецию, описанную в данном документе).
В качестве дополнения, существуют дополнительные замены Fc, которые находят применение в повышенном связывании с FcRn-рецептором и увеличенном периоде полужизни в сыворотке крови, как конкретно раскрыто в USSN 12/341769, полностью включенным в данный документ посредством ссылки, в том числе, но не ограничиваясь ими, 434S, 434A, 428L, 308F, 259I, 428L/434S, 428L/434A, 259I/308F, 436I/428L, 436I или V/434S, 436V/428L и 259I/308F/428L.
Гетеродимерные Fc-домены
В качестве дополнения к гомодимерным бивалентным слитым белкам ИЛ-2 в альтернативных вариантах осуществления используются моновалентные слитые белки ИЛ-2, в которых один из Fc-доменов является «пустым», и данное изобретение основано на вариантах гетеродимеризации для объединения двух Fc-доменов, изображенных на фиг. 19В. Указанные варианты осуществления основаны на использовании двух различных вариантных последовательностей Fc, которые будут приводить к самосборке с образованием гетеродимерных Fc-доменов и гетеродимерных слитых Fc-белков.
Гетеродимерные белковые конструкции основаны на самособирающейся природе двух Fc-доменов тяжелых цепей антител, например, двух «мономеров», которые собираются в «димер». Гетеродимерные белки получают путем изменения аминокислотной последовательности каждого мономера, как более подробно обсуждается ниже. Таким образом, данное изобретение в целом направлено на создание гетеродимерных слитых Fc-белков, которые основаны на вариантах аминокислот в константных областях, которые различаются в каждой цепи, чтобы способствовать образованию гетеродимеров и/или обеспечить легкую очистку гетеродимеров по сравнению с гомодимерами.
Существует ряд механизмов, которые могут быть использованы для создания гетеродимеров по данному изобретению. В качестве дополнения, как будет понятно специалистам в данной области техники, указанные механизмы могут быть объединены для обеспечения высокой гетеродимеризации. Таким образом, аминокислотные варианты, которые приводят к получению гетеродимеров, называются «вариантами гетеродимеризации». Как обсуждается ниже, гетеродимеризационные варианты могут включать стерические варианты (например, варианты «выступы и впадины» или «искаженные» варианты, описанные ниже, и варианты «пары зарядов», описанные ниже), а также «pI варианты», что позволяет очищать гомодимеры от гетеродимеров. Как в целом описано в WO2014/145806, включенным в данный документ посредством ссылки в полном объеме и конкретно, как описано ниже для обсуждения «гетеродимеризационных вариантов», пригодные механизмы гетеродимеризации включают в себя «выступы и впадины» («KIH»; называемые иногда в данном документе, как «искаженные» варианты (см. обсуждение в WO2014/145806), «электростатическое взаимодействие» или «пары зарядов», описанные в WO2014/145806, pI варианты, описанные в WO2014/145806, и общие дополнительные варианты Fc, описанные в WO2014/145806 и ниже.
В данном изобретении существует несколько основных механизмов, которые могут приводить к облегчению очистки гетеродимерных белков; каждый из них основан на использовании pI вариантов таким образом, что каждый мономер имеет разное значение pI, что способствует изоэлектрической очистке димерных белков A-A, A-B и B-B. Кроме того, гетеродимерные белки по данному изобретению также способствуют разделению на основе размера. Как дополнительно изложено ниже, также возможно «искажать» образование гетеродимеров по сравнению с гомодимерами. Таким образом, комбинация стерических гетеродимеризационных вариантов и pI вариантов или вариантов пары зарядов находит конкретное применение в данном изобретении.
В целом, варианты осуществления, которые в частности используются в данном изобретении, основаны на наборах вариантов, которые включают искаженные варианты, которые способствуют образованию гетеродимеризации по сравнению с образованием гомодимеризации, в сочетании с pI вариантами, которые увеличивают разницу pI между двумя мономерами.
В качестве дополнения, как более подробно изложено в данном документе, pI варианты могут содержаться либо внутри константных доменов и/либо Fc-доменов мономера, или могут использоваться заряженные линкеры, такие как линкеры доменов.
В данном изобретении, в котором значение pI используется в качестве механизма разделения для очистки гетеродимерных белков, аминокислотные варианты могут быть введены в один или оба мономерных полипептида; т.е., значение pI одного из мономеров (для простоты называемого в данном документе «мономер A») может быть предусмотрено таким образом, что оно будет удалено от мономера B, или могут быть изменены как мономер A, так и мономер B, при этом увеличивается значение pI мономера A и уменьшается значение pI мономера B. Как обсуждается, изменения pI одного или обоих мономеров могут быть выполнены путем удаления или добавления заряженного остатка (например, нейтральная аминокислота заменяется положительно или отрицательно заряженным аминокислотным остатком, например, глицин заменяется на глутаминовую кислоту), приводя к изменению заряженного остатка с положительного или отрицательного на противоположный заряд (например, аспарагиновая кислота заменяется на лизин) или изменению заряженного остатка на нейтральный остаток (например, потеря заряда; лизин заменяется на серин). Несколько указанных вариантов изображены в разделе фигуры.
Соответственно, в указанном варианте осуществления данного изобретения представлено выполнение достаточного изменения pI, по меньшей мере, в одном из мономеров, таким образом, что гетеродимеры могут быть отделены от гомодимеров. Как будет понятно специалистам в данной области техники и как будет обсуждено ниже, это можно осуществить с помощью константной области тяжелой цепи «дикого типа» и вариантной области, которая была сконструирована для увеличения или уменьшения ее значения pI (A-+B дикого типа или A - -B дикого типа), или путем увеличения его в одной области и уменьшения в другой области (A+ -B- или A- B +).
Таким образом, в целом, компонент некоторых вариантов осуществления по данному изобретению представляет собой аминокислотные варианты в Fc-доменах, которые направлены на изменение изоэлектрической точки (pI), по меньшей мере, одного, если не обоих, мономеров димерного белка с помощью включения аминокислотных замен («pI вариантов» или «pI замен») в один или оба мономера. Как показано в данном документе, отделение гетеродимеров от двух гомодимеров может быть достигнуто, если значения pI указанных двух мономеров отличаются всего на 0,1 единицу рН, при этом все из значений 0,2, 0,3, 0,4 и 0,5 или больше находят применение в данном изобретении.
Гетеродимеризационные варианты
В данном изобретении представлены гетеродимерные белки, в том числе гетеродимерные антитела в различных форматах, которые используют гетеродимерные варианты для обеспечения образования гетеродимеров и/или очистки от гомодимеров. Несколько гетеродимеризационных вариантов изображены на фиг. 2.
Существует ряд подходящих пар наборов гетеродимеризационных искаженных гетеродимеризационных вариантов. Указанные варианты входят в «пары» «наборов». Т.е. один набор пары включен в первый мономер, а другой набор пары включен во второй мономер. Следует отметить, что указанные наборы не обязательно ведут себя как варианты «выступы во впадинах», с однозначным соответствием между остатком в одном мономере и остатком в другом; т.е. указанные пары наборов образуют границу раздела между двумя мономерами, которая стимулирует образование гетеродимеров и препятствует образованию гомодимеров, способствуя тому, что процент гетеродимеров, которые спонтанно образуются в биологических условиях, будет составлять более 90%, а не ожидаемые 50% (25% гомодимер A/A:50% гетеродимер A/B:25% гомодимер B/B).
Стерические варианты
В некоторых вариантах осуществления образование гетеродимеров может быть облегчено путем добавления стерических вариантов. Т.е. в результате изменения аминокислот в каждой тяжелой цепи, различные тяжелые цепи с большей вероятностью ассоциируются с образованием гетеродимерной структуры, чем с образованием гомодимеров с одинаковыми аминокислотными последовательностями Fc. Подходящие стерические варианты включены в раздел фигуры.
Один из механизмов обычно упоминается в данной области техники как «выступы во впадинах», обозначая конструирование аминокислот, которое приводит к созданию стерических влияний, способствующих образованию гетеродимеров, и препятствующих образованию гомодимеров; его иногда называют «выступы во впадинах», как описано в USSN 61/596846, Ridgway et al., Protein Engineering 9(7):617 (1996); Atwell et al., J. Mol. Biol. 1997 270:26; патент США № 8216805, все из которых включены в данный документ в полном объеме посредством ссылки. В разделе фигуры обозначено несколько пар «мономер A – мономер B», которые основаны на механизме «выступы во впадинах». В качестве дополнения, как описано в Merchant et al., Nature Biotech. 16:677 (1998), указанные мутации «выступы во впадинах» могут быть объединены с дисульфидными связями для образования искаженных гетеродимеризационных вариантов.
Дополнительный механизм, который находит применение в создании гетеродимеров, иногда называют «электростатическим влиянием», как описано в Gunasekaran et al., J. Biol. Chem. 285(25):19637 (2010), включенной в данный документ посредством ссылки в полном объеме. Его иногда обозначают в данном документе в качестве «пар зарядов». В указанном варианте осуществления электростатические силы используются для искажения образования вариантов в сторону гетеродимеризации. Специалистам в данной области техники будет понятно, что они также могут оказывать влияние на pI и, следовательно, на очистку, и, таким образом, в некоторых случаях также могут рассматриваться как pI варианты. В то же время, поскольку они были созданы для усиления гетеродимеризации и не были использованы в качестве инструментов очистки, они классифицируются как «стерические варианты». Они включают, но не ограничиваясь ими, D221E/P228E/L368E в паре с D221R/P228R/K409R (например, они представляют собой «наборы, соответствующие мономерам») и C220E/P228E/368E в паре с C220R/E224R/P228R/K409R.
Дополнительные варианты мономеров A и мономеров B, которые можно комбинировать с другими вариантами, необязательно и независимо в любом количестве, такими как pI варианты, описанные в данном документе, или другие стерические варианты, которые показаны на фиг. 37 US 2012/0149876, фигура и условные обозначения и SEQ ID NO которой явно включены в данный документ посредством ссылки.
В некоторых вариантах осуществления стерические варианты, изложенные в данном документе, могут быть необязательно и независимо включены с любым pI вариантом (или другими вариантами, такими как варианты Fc, варианты FcRn и т.д.) в один или оба мономера и могут быть независимо и необязательно включены или исключены из белков по данному изобретению.
Перечень подходящих искаженных вариантов находится на фиг. 2 на которой изображены некоторые пары, особенно пригодные в использовании во многих вариантах осуществления. Особенно пригодными в использовании во многих вариантах осуществления являются пары наборов, в том числе, но не ограничиваясь ими, S364K/E357Q : L368D/K370S; L368D/K370S : S364K; L368E/K370S : S364K; T411T/E360E/Q362E : D401K; L368D/K370S : S364K/E357L, K370S : S364K/E357Q и T366S/L368A/Y407V : T366W (необязательно содержащие связывающий дисульфид, T366S/L368A/Y407V/Y349C : T366W/S354C). С точки зрения номенклатуры, пара «S364K/E357Q: L368D/K370S» означает, что один из мономеров имеет набор с двойным вариантом S364K/E357Q, а другой имеет набор с двойным вариантом L368D/K370S; как указано выше, «наличие цепей» указанных пар зависит от исходного значения pI.
pI варианты (варианты с изоэлектрической точкой) для гетеродимеров
В целом, как будет понятно специалистам в данной области техники, существует две основные категории pI вариантов: варианты, которые приводят к повышению значения pI белка (основные изменения), и варианты, которые приводят к уменьшению значения pI белка (кислотные изменения). Как описано в данном документе, могут быть выполнены все комбинации указанных вариантов: один мономер может относиться к дикому типу или варианту, который не проявляет существенно отличающееся от дикого типа значение pI, а другой может быть либо более основным, либо более кислым. В качестве альтернативы, каждый мономер является измененным, один на более основной и один на более кислый.
Предпочтительные комбинации pI вариантов изображены на фиг. 5. Как изложено в данном документе и изображено в разделе фигуры, указанных изменения показаны относительно IgG1, но все изотипы могут быть изменены таким же образом, как и гибриды изотипов. В случае, когда константный домен тяжелой цепи происходит от IgG2-4, также могут быть использованы R133E и R133Q.
В одном варианте осуществления предпочтительная комбинация pI вариантов имеет один мономер (отрицательная сторона), содержащий варианты 295E/384D/418E/421D (Q295E/N384D/Q418E/N421D применительно к IgG1 человека), и второй мономер (положительная сторона), содержащий положительно заряженный линкер scFv, содержащий (GKPGS)4.
Изотипические варианты
В качестве дополнения, некоторые варианты осуществления по данному изобретению основаны на «импорте» pI аминокислот в определенных положениях из одного изотипа IgG в другой, тем самым уменьшая или исключая возможность внесения нежелательной иммуногенности в указанные варианты. Некоторые из них изображены на фиг. 21 публикации США 2014/0370013, включенной в данный документ посредством ссылки. Т.е. IgG1 представляет собой общий изотип для терапевтических антител по ряду причин, в том числе в связи с высокой эффекторной функцией. В то же время, константная область тяжелой цепи IgG1 имеет более высокое значение pI, чем у IgG2 (8,10 по сравнению с 7,31). В результате введения остатков IgG2 в определенных положениях в каркас IgG1 значение pI образованного в результате мономера снижается (или увеличивается) и дополнительно демонстрирует более длительный период полужизни в сыворотке крови. Например, IgG1 имеет глицин (pI 5,97) в положении 137, а IgG2 имеет глутаминовую кислоту (pI 3,22); импорт глутаминовой кислоты будет влиять на значение pI образованного в результате белка. Как описано ниже, как правило, требуется несколько аминокислотных замен для значительного влияния на pI вариантного антитела. В то же время, следует отметить, что, как обсуждается ниже, даже изменения в молекулах IgG2 способствуют увеличению периода полужизни в сыворотке крови.
В других вариантах осуществления выполняют замены неизотипных аминокислот либо для уменьшения общего заряженного состояния образованного в результате белка (например, путем изменения аминокислоты с более высоким значением pI на аминокислоту с более низким значением pI), либо для обеспечения приспособления в структуре с целью стабильности и т.д., как более подробно описано ниже.
Кроме того, с помощью pI-инженерии константных доменов как тяжелых, так и легких цепей можно наблюдать значительные изменения в каждом мономере гетеродимера. Как обсуждается в данном документе, наличие значений pI двух мономеров, отличающихся по меньшей мере на 0,5, может способствовать разделению с помощью ионообменной хроматографии или изоэлектрической фокусировки, или других способов, чувствительных к изоэлектрической точке.
Расчет значения pI
Значение pI каждого мономера может зависеть от pI домена вариантного ИЛ-2 и Fc-домена, изложенных в данном документе. Таким образом, в некоторых вариантах осуществления изменение pI рассчитывают на основе константного домена вариантной тяжелой цепи, используя диаграмму на фиг. 19 публикации США 2014/0370013. Как обсуждается в данном документе, мономер, подлежащий конструированию, как правило, определяется внутренним значением pI Fv и каркасных областей. В качестве альтернативы, значение pI каждого мономера можно сравнивать.
Комбинирование гетеродимерных вариантов и Fc вариантов
Как будет понятно специалистам в данной области техники, для использования в гетеродимерных Fc-доменах все из перечисленных гетеродимеризационных вариантов (в том числе искаженных и/или pI вариантов) могут быть необязательно и независимо объединены любым способом, при условии, что они сохраняют свое «наличие цепей» или «разделение мономеров». В качестве дополнения, все из указанных вариантов могут быть объединены в любой из форматов гетеродимеризации.
В случае pI вариантов, несмотря на то, что варианты осуществления, находящие особенное применение, изображены в разделе фигуры, могут быть созданы другие комбинации, на основе основного правила изменения разницы pI между двумя мономерами для облегчения очистки.
В качестве дополнения, любой из гетеродимеризационных вариантов, искаженных и pI вариантов также независимо и необязательно комбинируется с абляционными вариантами Fc, вариантами Fc, вариантами FcRn, как в целом изложено в данном документе.
В качестве дополнения к абляционным вариантам, Fc-домены, как правило, также включают вариант C220S (например, поскольку Fc-домены по данному изобретению содержат шарнирную область, начинающуюся в положении 216, нумерация EU), который исключается, так как в него не включены легкие цепи, и указанный цистеин используется для образования дисульфида с легкой цепью.
В качестве дополнения, Fc-домены слитых белков по данному изобретению необязательно могут содержать аминокислотные замены с удлинением периода полужизни.
Недавно было высказано предположение, что антитела с вариабельными областями, которые имеют более низкие значения изоэлектрической точки, могут также иметь более длительный период полужизни в сыворотке крови (Igawa et al., 2010 PEDS. 23(5): 385-392, включенная в полном объеме посредством ссылки). В то же время, механизм этого до сих пор остается неясным. Кроме того, вариабельные области отличаются от антитела к антителу. Варианты константных областей с уменьшенным значением pI и увеличенным периодом полужизни будут обеспечивать более модульный подход к улучшению фармакокинетических свойств антител, описанных в данном документе.
Пригодные конструкции по данному изобретению
Как изложено в данном документе, в данном изобретении представлен ряд пригодных моновалентных и бивалентных конструкций.
Гетеродимерные моновалентные конструкции
В некоторых вариантах осуществления слитые белки ИЛ-2-Fc по данному изобретению представляют собой гетеродимерные моновалентные конструкции, такие как изображенные на фиг. 19A и 19C. В указанном варианте осуществления домен вариантного ИЛ-2 обычно сливают с Fc-доменом вариантного IgG1 человека, либо с помощью шарнира в качестве линкера домена (как правило, содержащего вариант C220S), либо с помощью дополнительного линкера, присоединенного к шарниру, при этом другой Fc-домен (содержащий шарнир) остается «пустым».
В некоторых вариантах осуществления домен вариантного ИЛ-2 присоединен к стороне «+» мономера (см. фиг. 5А), которая содержит Fc-домен вариантного IgG1 человека (в том числе шарнир с вариантом C220S), «искаженные варианты» S364K/E357Q» и абляционные варианты E233P/L234V/L235A/G236del/S267K, а «пустая Fc-сторона» представляет собой Fc-домен вариантного IgG1 человека (в том числе шарнир с вариантом C220S), «искаженные варианты» L368D/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K. В указанном варианте осуществления предпочтительные конструкции содержат домен вариантного ИЛ-2 с аминокислотными заменами, выбранными из группы, состоящей из T3A/D20N/T37R, T3A/D20N/T37R/C125S, T3A/D20N/T37R/C125A, T3A/D20N/N71K, T3A/D20N/N71K/C125S и T3A/D20N/N71K/C125A.
В некоторых вариантах осуществления домен вариантного ИЛ-2 присоединен к стороне «+» мономера (см. фиг. 5А), которая содержит Fc-домен вариантного IgG1 человека (в том числе шарнир с вариантом C220S), «искаженные варианты» S364K/E357Q» и абляционные варианты E233P/L234V/L235A/G236del/S267K, а также варианты FcRn M428L/N434S, а «пустая Fc-сторона» представляет собой Fc-домен вариантного IgG1 человека (в том числе шарнир с вариантом C220S), «искаженные варианты» L368D/K370S и абляционные варианты E233P/L234V/L235A/G236del/S267K, а также варианты FcRn M428L/N434S. В указанном варианте осуществления предпочтительные конструкции содержат домен вариантного ИЛ-2 с аминокислотными заменами, выбранными из группы, состоящей из T3A/D20N/T37R, T3A/D20N/T37R/C125S, T3A/D20N/T37R/C125A, T3A/D20N/N71K, T3A/D20N/N71K/C125S и T3A/D20N/N71K/C125A.
В одном варианте осуществления «мономер 1» содержит домен вариантного ИЛ-2, содержащий аминокислотные замены T3A/D20N/N71K/C125S (по сравнению с ИЛ-2 дикого типа, SEQ IN NO: 2), и «Fc-каркас мономера 1», выбранный из таковых на фиг. 6, в том числе SEQ ID NO: 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31 или 33. В указанном варианте осуществления «мономер 2» содержит «пустой Fc», выбранный из «Fc-каркасов мономера 2» из SEQ ID NO: 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32 и 34.
В одном варианте осуществления «мономер 1» содержит домен вариантного ИЛ-2, содержащий аминокислотные замены T3A/D20N/N71K/C125S (по сравнению с ИЛ-2 дикого типа, SEQ IN NO: 2), и «Fc-каркас мономера 1», выбранный из таковых на фиг. 6, в том числе SEQ ID NO: 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31 или 33, но с вариантами FcRn M428L/N434S. В указанном варианте осуществления «мономер 2» содержит «пустой Fc», выбранный из «Fc-каркасов мономера 2» из SEQ ID NO: 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32 и 34, но с вариантами FcRn M428L/N434S.
В одном варианте осуществления «мономер 1» содержит домен вариантного ИЛ-2, содержащий аминокислотные замены T3A/D20N/T37R/C125S (по сравнению с ИЛ-2 дикого типа, SEQ IN NO: 2), и «Fc-каркас мономера 1», выбранный из таковых на фиг. 6, в том числе SEQ ID NO: 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31 или 33. В указанном варианте осуществления «мономер 2» содержит «пустой Fc», выбранный из «Fc-каркасов мономера 2» из SEQ ID NO: 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32 и 34.
В одном варианте осуществления содержит домен вариантного ИЛ-2, содержащий аминокислотные замены T3A/D20N/T37R/C125S (по сравнению с ИЛ-2 дикого типа, SEQ IN NO: 2), и «Fc-каркас», выбранный из таковых на фиг. 6, в том числе SEQ ID NO: 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31 или 33, но с вариантами FcRn M428L/N434S. В указанном варианте осуществления «мономер 2» содержит «пустой Fc», выбранный из «Fc-каркасов мономера 2» из SEQ ID NO: 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32 и 34, но с вариантами FcRn M428L/N434S.
В указанных вариантах осуществления предпочтительные конструкции включают XENP26105, XENP27563, XENP26109 и XENP27564.
Гомодимерные бивалентные конструкции
В некоторых вариантах осуществления слитые белки ИЛ-2-Fc по данному изобретению представляют собой гомодимерные бивалентные конструкции, такие как изображенные на фиг. 19B и 19D. В указанном варианте осуществления каждый из доменов вариантного ИЛ-2 обычно сливают с Fc-доменом вариантного IgG1 человека, либо с помощью шарнира в качестве линкера домена (как правило, содержащего вариант C220S), либо с помощью дополнительного линкера, присоединенного к шарниру.
В некоторых вариантах осуществления домены вариантного ИЛ-2 присоединены к Fc-домену вариантного IgG1 человека (в том числе шарнир с вариантом C220S) и абляционным вариантам E233P/L234V/L235A/G236del/S267K. В указанном варианте осуществления предпочтительные конструкции содержат домен вариантного ИЛ-2 с аминокислотными заменами, выбранными из группы, состоящей из T3A/D20N/T37R, T3A/D20N/T37R/C125S, T3A/D20N/T37R/C125A, T3A/D20N/N71K, T3A/D20N/N71K/C125S и T3A/D20N/N71K/C125A.
В некоторых вариантах осуществления домены вариантного ИЛ-2 присоединены к Fc-домену вариантного IgG1 человека (в том числе шарнир с вариантом C220S), абляционным вариантам E233P/L234V/L235A/G236del/S267K и вариантам FcRn M428L/N434S. В указанном варианте осуществления предпочтительные конструкции содержат домен вариантного ИЛ-2 с аминокислотными заменами, выбранными из группы, состоящей из T3A/D20N/T37R, T3A/D20N/T37R/C125S, T3A/D20N/T37R/C125A, T3A/D20N/N71K, T3A/D20N/N71K/C125S и T3A/D20N/N71K/C125A.
Нуклеиновые кислоты по данному изобретению
В данном изобретении дополнительно представлены композиции нуклеиновых кислот, кодирующих гомодимерные бивалентные слитые белки ИЛ-2-Fc и гетеродимерные моновалентные слитые белки ИЛ-2.
Как будет понятно специалистам в данной области техники, композиции нуклеиновых кислот будут зависеть от формата гетеродимерного белка. Таким образом, например, когда для формата требуется две аминокислотные последовательности, например, в случае гетеродимерных моновалентных форматов из фиг. 19А, то две последовательности нуклеиновой кислоты могут быть включены в один или несколько экспрессионных векторов для экспрессии.
В качестве альтернативы, при использовании гомодимерных бивалентных форматов, изображенных на фиг. 19В, используются одна конструкции нуклеиновой кислоты и один экспрессионный вектор.
Как известно в данной области техники, нуклеиновые кислоты, кодирующие компоненты по данному изобретению, могут быть включены в экспрессионные векторы, известные в данной области техники, в зависимости от клеток-хозяев, используемых для получения гетеродимерных слитых Fc-белков по данному изобретению. Как правило, нуклеиновые кислоты функционально связаны с любым количеством регуляторных элементов (промоторов, точек репликации, селектируемых маркеров, сайтов связывания рибосом, индукторов и т.д.). Экспрессионные векторы могут представлять собой внехромосомные или интегрирующие векторы.
Затем нуклеиновые кислоты и/или экспрессионные векторы по данному изобретению трансформируют в любое количество различных типов клеток-хозяев, как хорошо известно в данной области техники, в том числе клетки млекопитающих, бактерий, дрожжей, насекомых и/или грибов, при этом клетки млекопитающих (например, клетки СНО) находят применение во многих вариантах осуществления.
В некоторых вариантах осуществления каждая из нуклеиновых кислот, кодирующая каждый мономер, в зависимости от формата, содержится в одном экспрессионном векторе, как правило, под контролем различных или одинаковых промоторов. В вариантах осуществления с особой применимостью в данном изобретении каждая из этих двух или трех нуклеиновых кислот содержится в разных экспрессионных векторах.
Гетеродимерный Fc-слитый белок по данному изобретению получают путем культивирования клеток-хозяев, содержащих экспрессионный (экспрессионные) вектор (векторы), как хорошо известно в данной области техники. После получения выполняются традиционные стадии очистки слитых белков или антител, в том числе стадия ионообменной хромотографии. Как обсуждается в данном документе, наличие значений pI двух мономеров, отличающихся по меньшей мере на 0,5, может способствовать разделению с помощью ионообменной хроматографии или изоэлектрической фокусировки, или других способов, чувствительных к изоэлектрической точке. Т.е. включение pI замен, которые изменяют изоэлектрическую точку (pI) каждого мономера, таким образом, что каждый мономер имеет другое значение pI, а гетеродимер также имеет другое значение pI, таким образом, облегчает изоэлектрическую очистку гетеродимера (например, анионообменные колонки, катионообменные колонки). Указанные замены также облегчают определение и мониторинг последующей очистки любых загрязняющих гомодимеров (например, гели IEF, cIEF и аналитические колонки IEX).
Составы
Составы композиций, используемые в соответствии с данным изобретением, готовят для хранения с помощью смешивания слитых белков, имеющих необходимую степень чистоты, с необязательными фармацевтически приемлемыми носителями, наполнителями или стабилизаторами (как в целом изложено в Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. [1980]), в форме лиофилизированных составов или водных растворов.
Лечение
Композиции слитых белков ИЛ-2-Fc по данному изобретению находят применение в лечении аутоиммунного заболевания, например, путем активации CD25+ клеток пациента с использованием димерных белков по данному изобретению.
Примеры
Пример 1. Конструирование ИЛ-2 для увеличения периода полужизни
Как обсуждалось выше, ИЛ-2 имеет недостаток, заключающийся в очень быстром клиренсе. Такой быстрый клиренс частично связан с интернализацией комплекса ИЛ-2:ИЛ-2Р. После интернализации в эндосому ИЛ-2, ИЛ-2Рß и γc распадаются, в то время как ИЛ-2Р конститутивно рециркулируется к клеточной поверхности. Вариант 2D1 ИЛ-2, который имеет мутации L18M/L19S, продемонстрировал более длительное время полужизни, чем ИЛ-2 дикого типа. Fallon et al. сообщили, что ИЛ-2 дикого типа имеет более низкую аффинность по отношению к ИЛ-2R при эндосомном значении pH, что указывает на рН-зависимое связывание между ИЛ-2 и ИЛ-2Рα. Кроме того, эта группа ученых обнаружила, что увеличенный период полужизни 2D1 является результатом рециркуляции с ИЛ-2Рα благодаря тому, что 2D1 имеет более высокую аффинность к ИЛ-2Рα при эндосомном значении pH, чем ИЛ-2 ДТ. Примечательно, было описано, что остатки L18 и L19 в ИЛ-2 связываются с ИЛ-2Рß и γc. В отличие от этого, было сделано предположение, что модифицирующие остатки на границе раздела ИЛ-2:ИЛ-2Рα будут более подходящими для ослабления рН-зависимого связывания, увеличения рециркуляцию ИЛ-2 с ИЛ-2Рα и, соответственно, увеличения периода полужизни.
Конструирование ИЛ-2 на границе раздела ИЛ-2/ИЛ-2Рα для ослабления рН-зависимого связывания
Изучив кристаллическую структуру границы раздела ИЛ-2 и ИЛ-2Рα (код 2ERJ в PDB), был обнаружен вероятный естественный «рН-переключатель» на границе раздела ИЛ-2/ИЛ-2Рα, состоящий из взаимодействия между Arg38, Thr41 и/или Phe42 ИЛ-2 и His120 ИЛ-2Рα (см. фиг. 8). Была выдвинута гипотеза о том, что при низком значении pH в эндосоме после интернализации комплекса ИЛ-2:ИЛ-2Рαßγ His20 станет протонированным, что приведет к высвобождению ИЛ-2 из ИЛ-2Рα и последующему лизосомальному разрушению оставшегося комплекса ИЛ-2:ИЛ-2Рßγ. Известно, что ИЛ-2Р рециркулирует к поверхности клетки, и повышение аффинности связывания ИЛ-2 с ИЛ-2Рα при рН 6,0 путем замены одного или нескольких из этих взаимодействующих остатков может повысить рециркуляцию ИЛ-2 и продлить период полужизни. С помощью вычислительные прогнозы с использованием технологии Protein Design Automation (см., например, WO 1998/047089, опубликованный 22 октября 1998 г.), создавали варианты для насыщения этих трех контактирующих остатков.
Плазмиду, кодирующую ИЛ-2, конструировали с помощью стандартного синтеза генов с последующим субклонированием в экспрессионный вектор pTT5. ИЛ-2 содержал С-концевую полигистидиновую метку (8xHis) для очистки и замену C125S для повышения экспрессии. Замены, прогнозируемые выше, вводили с помощью стандартных методик мутагенеза. Белки получали путем временной трансфекции в клетках HEK293E и очищали с помощью хроматографии Ni-NTA. Последовательности для иллюстративных вариантов изображены на фиг. 9, при этом полигистидиновую метку удаляли.
Аффинность ИЛ-2 в отношении ИЛ-2Рα при pH 7,4 и pH 6,0 определяли с использованием Biacore, технологии на основе поверхностного плазмонного резонанса (SPR). Экспериментальные этапы для Biacore, как правило, включали следующее: иммобилизацию (захват лиганда на сенсорном чипе); ассоциацию (пропускание различных концентраций аналита через сенсорный чип); и диссоциацию (пропускание буфера через сенсорные чипы) для определения аффинности исследуемых препаратов. Референтный поток только с буфером также включали в способ фоновой коррекции при обработке данных. В указанном конкретном скрининге CD25 человека (ИЛ-2Рα) сначала захватывали на сенсорных чипах, а затем различные концентрации вариантов ИЛ-2 пропускали через сенсорные чипы. Отдельные эксперименты проводили с использованием буфера при рН 7,4 для моделирования условий на поверхности клетки и с использованием буфера при рН 6,0 для моделирования эндосомальных условий. Полученные в результате константы диссоциации (KD), скорости ассоциации (ka) и скорости диссоциации (kd) изображены на фиг. 10 и 11. Аффинности связывания и кинетические константы скорости получали путем анализа обработанных данных с использованием модели связывания 1:1. На фиг. 12 изображена кратность повышения скорости диссоциации при рН 6,0, в то время как на фиг. 13 изображена кратность повышения скорости диссоциации при рН 7,4.
Большое количество вариантов с различными заменами при R38, T41 и F42 имели худшие или аналогичные скорости диссоциации при pH 6,0 по сравнению с ИЛ-2 ДТ; тем не менее, идентифицировали некоторые варианты со значительно повышенными скоростями диссоциации (т.е. варианты ИЛ-2, которые с большей вероятностью будут рециркулировать с ИЛ-2Рα). Предпочтительные варианты включают XENP14142 (R38I) и XENP14144 (R38L). На фиг. 14 изображены сенсограммы Biacore для XENP14135 (ИЛ-2 дикого типа с мутацией C125S) и XENP14142 (вариантного ИЛ-2 с R38I и C125S). Аналогичные кривые диссоциации для XENP14142 при рН 7,4 и рН 6,0 демонстрируют успешное ослабление рН-зависимого связывания.
Ослабление pH-зависимого связывания в контексте других вариантов ИЛ-2
Затем R38L объединяли с вариантом ИЛ-2 предшествующего уровня техники (мутант 2-4 с Q126T, описанный в WO 2009/061853, опубликованном 14 мая 2009 г.), чтобы исследовать, были ли снижены скорости диссоциации в контексте других вариантов ИЛ-2.
Как указано выше, замены вводили с помощью стандартных методик мутагенеза. Белки получали путем временной трансфекции в клетках HEK293E и очищали с помощью хроматографии Ni-NTA. Последовательности для варианта предшествующего уровня техники, описанного с R38L и без R38L изображены на фиг. 15, при этом полигистидиновую метку удаляли.
Аффинность ИЛ-2 в отношении ИЛ-2Рα при pH 7,4 и pH 6,0 определяли с использованием Biacore, как описано в Примере 1А. Полученные в результате константы диссоциации (KD), скорости ассоциации (ka) и скорости диссоциации (kd), а также отношение аффинности при рН 7,4 к аффинности при рН 6,0 изображены на фиг. 16. Данные демонстрируют, что отношение аффинностей увеличивалось с включением замены R38L, что указывало на успешное ослабление рН-зависимого связывания.
Пример 2. Конструирование ИЛ-2 для селективности Treg
Пролиферативная передача сигналов с помощью ИЛ-2 опосредуется ИЛ-2Рß и γc в виде части рецепторного комплекса ИЛ-2 с промежуточной аффинностью (ИЛ-2Рßγ) или в виде части высокоаффинного рецепторного комплекса ИЛ-2 (ИЛ-2Рαßγ). CD25 обеспечивает высокую аффинность связывания комплекса ИЛ-2Рαβγ с ИЛ-2, однако в остальном он сам по себе является недостаточным для передачи сигналов. В результате высокой аффинности связывания с комплексом ИЛ-2Рαβ, ИЛ-2 оказывает положительное влияние на Treg, которые конститутивно экспрессируют ИЛ-2Рα. Таким образом, было выдвинуто предположение, что увеличение аффинности ИЛ-2 в отношении ИЛ-2Рα могло привести к дополнительному искажению связывания в пользу комплекса ИЛ-2Рαßγ на Treg. В качестве альтернативы, снижение аффинности ИЛ-2 в отношении ИЛ-2Рß, γc или ИЛ-2Рßγ могло приводить к искажению связывания с CD25-отрицательными Т-клетками и NK-клетками.
Изучая кристаллическую структуру границы раздела между ИЛ-2 и его рецепторами, а также путем моделирования с использованием программного обеспечения MOE, прогнозировали остатки, которые могут быть замещены с целью повышения аффинности ИЛ-2 в отношении ИЛ-2Рα или уменьшения аффинности ИЛ-2 в отношении ИЛ-2Рß, γc и/или ИЛ-2Рßγ.
Плазмиды, кодирующие ИЛ-2, конструировали с помощью стандартного синтеза генов с последующим субклонированием в экспрессионный вектор pTT5. ИЛ-2 содержал С-концевую полигистидиновую метку (8xHis) для очистки и замену C125S для повышения экспрессии. Замены, прогнозируемые выше, вводили с помощью стандартных методик мутагенеза. Белки получали путем временной трансфекции в клетках HEK293E и очищали с помощью хроматографии Ni-NTA. Последовательности для иллюстративных вариантов изображены на фиг. 17, при этом полигистидиновую метку удаляли.
Связывание ИЛ-2 с его рецепторными компонентами определяли с помощью Octet, способа, основанного на биослойной интерферометрии (BLI). Экспериментальные этапы для Octet, как правило, включали следующее: иммобилизацию (захват лиганда в биосенсор); ассоциацию (погружение покрытых лигандом биосенсоров в лунки, содержащие серийные разведения аналита); и диссоциацию (возврат биосенсоров в лунку, содержащую буфер) для определения аффинности исследуемых препаратов. Референтную лунку, содержащую только буфер, также включали в способ фоновой коррекции при обработке данных. В частности, биосенсоры на основе анти-Fc человека (AHC) использовали для захвата бивалентного слияния CD25(ИЛ-2Рα)-Fc, бивалентного слияния CD122 (ИЛ-2Рß)-Fc или гетеродимерного слияния CD122:CD132 (ИЛ-2Рßγ)-Fc и погружали в несколько концентраций вариантов ИЛ-2. Образующийся в результате ответ BLI для вариантов ИЛ-2 нормализовали по отношению к ответу BLI для XENP14135 (ИЛ-2 дикого типа с C125S) и изображали на фиг. 18. Примечательно, что некоторые замены на границе раздела ИЛ-2:ИЛ-2Рß, такие как D20N и N88D, значительно снижали или устраняли связывание ИЛ-2 с ИЛ-2Рß.
Пример 3. Слитые белки ИЛ-2-Fc
Для дополнительного изучения проблемы короткого периода полужизни ИЛ-2 получали ИЛ-2 в виде Fc-слияния (далее называемого слияниями ИЛ-2-Fc) с целью облегчения производства и стимулирования FcRn-опосредованной рециркуляции комплекса и продления период полужизни.
Получения слияний ИЛ-2-Fc
Плазмиду, кодирующую ИЛ-2, конструировали с помощью стандартного синтеза генов с последующим субклонированием в экспрессионный вектор pTT5, содержащий партнеры Fc-слияний (например, константные области, изображенные на фиг. 6). ИЛ-2 может содержать замену C125S для повышения экспрессии и замену T3A для удаления сайта O-гликозилирования. Схемы иллюстративных форматов слияния ИЛ-2-Fc изображены на фиг. 19. Выбранные замены вводили с помощью стандартных методик мутагенеза.
Моновалентный формат ИЛ-2-Fc или «моновИЛ-2-Fc» (фиг. 19A) содержит ИЛ-2, слитый с N-концом первой гетеродимерной Fc-области (см., например, каркас 1 ИЛ-2-Fc - мономер 2, на фиг. 6), при этом другая сторона молекулы представляет собой гетеродимерный Fc типа «только Fc» или типа «пустой Fc» (см., например, каркас 1 ИЛ-2-Fc 1 - мономер 1, на фиг. 6). Последовательности для иллюстративных слияний моновИЛ-2-Fc изображены на фиг. 20. Бивалентный формат ИЛ-2-Fc или «бивИЛ-2-Fc» (фиг. 19B) содержит ИЛ-2, слитый с N-концом гомодимерной Fc-области (см., например, каркас 12 ИЛ-2-Fc, на фиг. 6). Последовательности для иллюстративных слияний бивИЛ-2-Fc изображены на фиг. 23. Слияния моновИЛ-2-Fc и слияния бивИЛ-2-Fc могут иметь линкер переменной длины между C-концом ИЛ-2 и N-концом Fc-области (см. неограничивающие примеры линкеров доменов на фиг. 7, которые могут найти применение в слияниях ИЛ-2-Fc, и изображений форматов на фиг. 19C-D). Последовательности для иллюстративных слияний ИЛ-2-Fc с линкерами переменной длины изображены на фиг. 25.
Белки получали путем временной трансфекции в клетках HEK293E и очищали с помощью двухстадийного способа очистки, включающего хроматографию с использованием белка А и анионообменную хроматографию.
Исследование вариантов ИЛ-2 предшествующего уровня техники, сконструированных в виде слияний моновИЛ-2-Fc
Для изучения надежности и эффективности формата слияния моновИЛ-2-Fc, получали ряд вариантов ИЛ-2 предшествующего уровня техники в указанном формате. Указанные слияния ИЛ-2-Fc включают XENP24637 (на основе варианта, описанного в WO 2012/107417, опубликованном 16 августа 2012 г.), XENP24638 (на основе мутанта 2-4, описанного в WO 2005/007121, опубликованном 27 января 2005 г.), XENP24639 (на основе мутанта M1, описанного в WO 2005/007121, опубликованном 27 января 2005 г.), XENP24640 (на основе мутанта 2-4 с Q126T, описанного в WO 2009/061853, опубликованном 14 мая 2009 г.), XENP24642 (на основе варианта, описанного в WO 1999/060128, опубликованного 25 ноября 1999 г.), XENP24728 (на основе H9-RET, описанного Mitra et al. 2015) и XENP24729 (на основе H9-RETR, описанного Mitra et al. 2015). Дополнительные варианты, такие как XENP24641, XENP24730, XENP24731 и XENP24732, были основаны на отдельных заменах или комбинациях замен, описанных в предшествующем уровне техники. Иллюстративные последовательности изображены на фиг. 20.
Скрининг аффинности слияний моновИЛ-2-Fc
Аффинность описанных выше слияний моновИЛ-2-Fc в отношении различных рецепторов ИЛ-2 определяли с помощью Octet, как в целом описано в Примере 2. В частности, для определения аффинности в отношении ИЛ-2Рα слияния CD25(ИЛ-2Рα)-Fc (R&D Systems, Миннеаполис, Миннесота) загружали на биосенсоры AR2G и погружали в несколько концентраций слияний ИЛ-2-Fc. Для определения аффинности в отношении ИЛ-2Рß и ИЛ-2Рßγ бивалентные слияния CD122(ИЛ-2Рß)-Fc-His или гетеродимерные слияния CD122:CD132(ИЛ-2Рßγ)-Fc-His загружали на биосенсоры HIS1K и погружали в несколько концентраций слияний ИЛ-2-Fc. Полученные в результате константы диссоциации (KD), скорости ассоциации (ka) и скорости диссоциации (kd) изображены на фиг. 21.
Индукция фосфорилирования STAT5 в результате слияния моновИЛ-2-Fc
После связывания цитокинов с их рецепторами киназы Janus (JAK) ассоциировали с рецепторами фосфорилирующих белков STAT, которые затем транслоцируются в ядро с целью дополнительной регуляции нисходящих процессов. Таким образом, фосфорилирование белков STAT (в частности, STAT5, который включает STAT5a и STAT5b) представляет собой одно из самых ранних сигнальных событий, запускаемых в результате связывания ИЛ-2 с высокоаффинными рецепторами ИЛ-2 или с рецепторами ИЛ-2 с промежуточной аффинностью (Lin and Leonard (2000); Wuest et al. (2008)). Соответственно, исследовали способность описанных выше слияний моновИЛ-2-Fc индуцировать фосфорилирование STAT5 в клетках различных типов, таких как CD8+ и CD4+ T-клетки, а также Treg.
Свежеотобранные МКПК инкубировали с указанными исследуемыми препаратами ИЛ-2-Fc в указанных концентрациях в течение 15 минут. После инкубации МКПК окрашивали анти-CD3-BV510 (UCHT1), анти-CD4-BV605 (RPA-T4) и анти-CD8-Alexa700 (SK1) в течение 30-45 минут при комнатной температуре. Клетки промывали и инкубировали с предварительно охлажденным (-20°C) 90% метанолом в течение 20-60 минут. После инкубации с метанолом клетки снова промывали и окрашивали анти-CD25-BV421 (M-A251), анти-CD45RA-PE (HI100), анти-FOXP3-Alexa488 (259D) и анти-pSTAT5-Alexa647 (pY687) с целью маркирования различных клеточных популяций и фосфорилирования STAT5. Данные, изображающие индукцию фосфорилирования STAT5 в CD8+ T-клетках (CD3+CD8+CD25-), CD4+ T-клетках (CD3+CD4+CD25-) и Treg (CD3+CD4+CD25+FOXP3+) показаны на фиг. 22. Примечательно, что слияния ИЛ-2-Fc XENP24638 и XENP24642 представляли собой сильные активаторы Treg (как указано с помощью фосфорилирования STAT5) с минимальной активацией CD8+ и CD4+ T-клеток, что согласуется с активностью, описанной для вариантов ИЛ-2, которые составляют слияния ИЛ-2-Fc.
Изучение вариантов ИЛ-2 предшествующего уровня техники, сконструированных в виде слияний бивИЛ-2-Fc
Для изучения надежности и эффективности формата слияния бивИЛ-2-Fc вариант ИЛ-2 предшествующего уровня техники (описанный в WO 1999/060128, опубликованном 25 ноября 1999 г.), а также контрольный ИЛ-2 (с заменами C125S и T3A) создавали в указанном формате, последовательности для которого изображены на фиг. 23.
Индукция фосфорилирования STAT5 с помощью слияний бивИЛ-2-Fc
Свежеотобранные МКПК инкубировали с указанными исследуемыми препаратами ИЛ-2-Fc в указанных концентрациях в течение 15 минут. После инкубации МКПК окрашивали анти-CD3-BUV395 (UCHT1), анти-CD4-BV605 (RPA-T4) и анти-CD8-Alexa700 (SK1) в течение 30-45 минут при комнатной температуре. Клетки промывали и инкубировали с предварительно охлажденным (-20°C) 90% метанолом в течение 20-60 минут. После инкубации с метанолом клетки снова промывали и окрашивали анти-CD25-BV510 (M-A251), анти-CD45RA-PE (HI100), анти-FOXP3-Alexa488 (259D) и анти-pSTAT5-Alexa647 (pY694) с целью маркирования различных клеточных популяций и фосфорилирования STAT5. Данные, изображающие индукцию фосфорилирования STAT5 в CD8+ T-клетках, CD4+ T-клетках и Treg показаны на фиг. 24.
Изучение вариантов ИЛ-2 предшествующего уровня техники, сконструированных в виде слияний ИЛ-2-Fc с линкерами доменов
Для изучения влияния включения линкеров между ИЛ-2 и Fc-областью варианты ИЛ-2 предшествующего уровня техники (как описано в WO 1999/060128, опубликованном 25 ноября 1999 г., и в WO 2012/107417, опубликованном 16 августа 2012 г.) получали в виде слияний моновИЛ-2-Fc или слияний бивИЛ-2-Fc с Gly-Ser линкерами. Последовательности для указанных слияний ИЛ-2-Fc изображены на фиг. 25.
Индукция фосфорилирования STAT5 с помощью слияний ИЛ-2-Fc с линкерами доменов
Свежеотобранные МКПК инкубировали с указанными исследуемыми препаратами ИЛ-2-Fc в указанных концентрациях в течение 15 минут. После инкубации МКПК окрашивали анти-CD3-BUV395 (UCHT1), анти-CD4-BV605 (RPA-T4) и анти-CD8-Alexa700 (SK1) в течение 30-45 минут при комнатной температуре. Клетки промывали и инкубировали с предварительно охлажденным (-20°C) 90% метанолом в течение 20-60 минут. После инкубации с метанолом клетки снова промывали и окрашивали анти-CD25-BV510 (M-A251), анти-CD45RA-PE (HI100), анти-FOXP3-Alexa488 (259D) и анти-pSTAT5-Alexa647 (pY694) с целью маркирования различных клеточных популяций и фосфорилирования STAT5. Данные, изображающие индукцию фосфорилирования STAT5 в CD8+ T-клетках, CD4+ T-клетках и Treg показаны на фиг. 26.
Пример 4. Конструирование вариантных слияний ИЛ-2-Fc с повышенной аффинностью в отношении CD25 и пониженной аффинностью в отношении CD122
Как обсуждалось в Примере 2, повышение аффинности ИЛ-2 в отношении ИЛ-2Рα могло приводить к дополнительному искажению связывания в пользу комплекса ИЛ-2Рαßγ на Treg, в то же время уменьшение аффинности ИЛ-2 в отношении ИЛ-2Рß, γc или ИЛ-2Рßγ могло приводить к искажению связывания с CD25-отрицательными Т-клетками и NK-клетками. В данном случае замены, которые повышали связывание с ИЛ-2Рα, комбинировали с заменами, которые уменьшали связывание с ИЛ-2Рβ в контексте слияний ИЛ-2-Fc с целью повышения селективности Treg.
Плазмиду, кодирующую ИЛ-2, конструировали с помощью стандартного синтеза генов с последующим субклонированием в экспрессионный вектор pTT5, содержащий партнеры Fc-слияний (например, константные области, изображенные на фиг. 6). ИЛ-2 содержал замену C125S для повышения экспрессии и замену T3A для удаления сайта O-гликозилирования. Выбранные замены, описанные в Примере 2, вводили с помощью стандартных методик мутагенеза. Белки получали путем временной трансфекции в клетках HEK293E и очищали с помощью двухстадийного способа очистки, включающего хроматографию с использованием белка А и анионообменную хроматографию. Последовательности для иллюстративных слияний ИЛ-2-Fc, сконструированных для повышения аффинности в отношении CD25 и/или снижения аффинности в отношении CD122, изображены на фиг. 27.
Скрининг аффинности вариантных слияний ИЛ-2-Fc, сконструированных для повышения аффинности в отношении CD25 и снижения аффинности в отношении CD122
Аффинность описанных выше вариантных слияний ИЛ-2-Fc в отношении различных рецепторов ИЛ-2 определяли с помощью Octet, как в целом описано в Примере 3В(а). В частности, для определения аффинности в отношении ИЛ-2Рα слияния CD25(ИЛ-2Рα)-Fc (R&D Systems, Миннеаполис, Миннесота) загружали на биосенсоры AR2G и погружали в несколько концентраций слияний ИЛ-2-Fc. Для определения аффинности в отношении ИЛ-2Рß и ИЛ-2Рßγ бивалентные слияния CD122(ИЛ-2Рß)-Fc-His или гетеродимерные слияния CD122:CD132(ИЛ-2Рßγ)-Fc-His загружали на биосенсоры HIS1K и погружали в несколько концентраций слияний ИЛ-2-Fc. Полученные в результате константы диссоциации (KD), скорости ассоциации (ka) и скорости диссоциации (kd) изображены на фиг. 33.
Фосфорилирование STAT5 с помощью вариантных слияний ИЛ-2-Fc, сконструированных для повышения аффинности в отношении CD25 и снижения аффинности в отношении CD122 в различных клеточных популяциях
Свежеотобранные МКПК инкубировали с указанными исследуемыми препаратами ИЛ-2-Fc в указанных концентрациях в течение 15 минут. После инкубации МКПК окрашивали анти-CD3-BV396 (UCHT1), анти-CD4-BV605 (RPA-T4) и анти-CD8-Alexa700 (SK1) в течение 30-45 минут при комнатной температуре. Клетки промывали и инкубировали с предварительно охлажденным (-20°C) 90% метанолом в течение 20-60 минут. После инкубации с метанолом клетки снова промывали и окрашивали анти-CD25-BV421 (M-A251), анти-CD45RA-PE (HI100), анти-FOXP3-Alexa488 (259D) и анти-pSTAT5-Alexa647 (pY687) с целью маркирования различных клеточных популяций и фосфорилирования STAT5. Данные, изображающие индукцию фосфорилирования STAT5 в различных клеточных популяциях, изображены на фиг. 28-29.
Данные демонстрируют, что по сравнению с контролями XENP24635 (ИЛ-2-Fc только с C125S) и рекомбинантным ИЛ-2 человека (рчИЛ-2), многие иллюстративные варианты представляли собой сильные активаторы Treg, при этом оказывали минимальную индукцию фосфорилирования STAT5 в CD25-отрицательных Т-клетках (CD8+ и CD4+).
Пример 5. Конструирование дополнительных вариантов слияния ИЛ-2-Fc
Дополнительные слияния ИЛ-2-Fc конструировали с комбинированием характеристик, описанных в предыдущих примерах, в том числе валентности, линкеров доменов, рН-переключателя и селективности Treg, и получали, как в целом описано в Примере 2. Иллюстративные последовательности изображены на фиг. 30. В качестве дополнения слияние Fc-ИЛ-2 (V91K/C125A), сконструированное для увеличения соотношения Treg к нерегуляторным Т-клеткам, описанного в WO 2014/153111, получали в качестве препарата сравнения (в данном документе называемого XENP27193; последовательность изображена на фиг. 34).
Индукция фосфорилирования STAT5 с помощью дополнительных моновалентных слияний ИЛ-2-Fc
Свежеотобранные МКПК инкубировали с указанными исследуемыми препаратами ИЛ-2-Fc в указанных концентрациях в течение 15 минут при 37°C. После инкубации МКПК сначала окрашивали анти-CD3-BUV395 (UCHT1), анти-CD4-BV605 (RPA-T4), анти-CD8-AF700 (SK1) и анти-CD56-PE антителами. После первого окрашивания клетки пермеабилизировали с использованием PerFix EXPOSE (Beckman Coulter, Индианаполис, Индиана). После пермеабилизации клетки окрашивали анти-CD25-BV421 (M-A251), анти-CD45RA-BV510 (HI100), анти-FoxP3-AF488 (259D) и анти-pSTAT5-AF647 (47/Stat5 (pY694)) антителами. После второго окрашивания клетки анализировали с помощью проточной цитометрии для изучения фосфорилирования STAT5 в различных популяциях лимфоцитов. Данные, изображающие СИФ pFIAT5 для различных популяций лимфоцитов, указывающие на передачу сигналов с помощью слияний ИЛ-2-Fc с участием рецепторов ИЛ-2, представлены на фиг. 35.
Данные демонстрируют, что каждый из вариантов индуцировал фосфорилирование STAT5 в Treg. Примечательно, что вариантные слияния ИЛ-2-Fc предпочтительно индуцировали Treg по отношению к CD4+ Т-клеткам памяти (CD45RA-), CD8+ Т-клеткам памяти (CD45RA-), NK-клеткам и γδ Т-клеткам по сравнению как с рекомбинантным ИЛ-2, так и моновалентным слиянием ИЛ-2-Fc ДТ (XENP24635). Для сравнения, вариантное слияние Fc-ИЛ-2 предшествующего уровня техники XENP27193 было сравнительно менее селективным в отношении Treg.
Бивалентные слияния ИЛ-2-Fc являются более эффективными, чем моновалентные аналоги.
Индукцию фосфорилирования STAT5 с помощью различных исследуемых препаратов ИЛ-2-Fc изучали, как описано в Примере 5А. Данные, изображающие СИФ pSTAT5 в Treg и CD4+ T-клетках памяти (CD45RA-), указывающие на передачу сигналов с помощью слияний ИЛ-2-Fc с участием рецепторов ИЛ-2, представлены на фиг. 36-42.
Данные демонстрируют, что для каждого варианта ИЛ-2 бивалентные версии ИЛ-2-Fc были более эффективными в индукции фосфорилирования STAT5 в Treg, чем соответствующие моновалентные версии ИЛ-2-Fc. В частности, конструирование линкеров доменов между компонентом ИЛ-2 и Fc-компонентом (например, в XENP27002, XENP27003, XENP27004, XENP27005, XENP27006 и XENP27007) приводит к дополнительному повышению активности бивалентных слияний ИЛ-2-Fc. Примечательно, что каждая из бивалентных слитых конструкций ИЛ-2-Fc (с линкерами и без них) сохраняла селективность в отношении Treg по сравнению с другими популяциями лимфоцитов, такими как CD4+CD45RA- T-клетки.
Пример 6. Сведение воздействия слияний ИЛ-2-Fc к максимуму
Включение Xtend Fc
Слияния ИЛ-2-Fc, описанные выше, конструировали с Xtend Fc (M428L/N434S) с повышенным связыванием с FcRn с целью дополнительной стимуляции FcRn-опосредованной рециркуляции слияний и последующего продления периода полужизни в кровотоке. Последовательности для иллюстративных бивалентных слияний ИЛ-2-Fc с Xtend Fc изображены на фиг. 43, а последовательности для иллюстративных моновалентных слияний ИЛ-2-Fc с Xtend Fc изображены на фиг. 44.
Отбор слияний ИЛ-2-Fc с балансом селективности и активности Treg
Также было доказано, что слияние ИЛ-2-Fc с более низкой активностью будет уменьшать антиген-зависимый клиренс и, следовательно, повышать период полужизни в кровотоке. С точки зрения данных в Примере 5B, демонстрирующих, что бивалентные слияния ИЛ-2-Fc и слияния ИЛ-2-Fc, имеющие линкеры доменов, обладают повышенной эффективностью, особый интерес вызывали моновалентные слияния ИЛ-2-Fc, в которых отсутствовали линкеры доменов. Соответственно, с целью идентификации слияний ИЛ-2-Fc с оптимальным балансом между селективностью и эффективностью, проводили сравнение эффективности in vitro (как показано с помощью индукции фосфорилирования STAT5 в различных популяциях лимфоцитов) моновалентных слияний ИЛ-2 (XENP26105 и XENP26109 соответственно, содержащих варианты D20N/T37R и D20N/N71K) с XENP24635 (моновалентное слияние ИЛ-2-Fc с мутацией C125S), а также с XENP25908 и XENP27193 (которые представляют собой препараты сравнения слияний ИЛ-2-Fc, продемонстрированные для лечения аутоиммунных заболеваний), в анализе фосфорилирования STAT5, данные для которого изображены на фиг. 45.
Примечательно, что как ХENP26105, так и XENP26109 были менее эффективными, чем XENP24635 и XENP27193 (соответственно EC50 1 нМ и 5 нМ по сравнению с 0,02 нМ и 0,01 нМ), однако были способны достигать аналогичных уровней активности в Treg при более высоких дозах, при этом сохранялась селективность в отношении Treg. Несмотря на то, что XENP26105 обладал эффективностью, сопоставимой с XENP25908 (соответственно EC50 1 нМ по сравнению с 0,7 нМ), данные демонстрируют, что XENP26105, а также XENP26109, были способны достигать гораздо более высоких уровней активности в Treg, чем XENP25908. Сниженная эффективность и селективность, наблюдаемые для XENP26105 и XENP26109, позволяют предположить, что они будут пригодны для селективной и устойчивой экспансии Treg в клинических условиях. Соответственно, дополнительно изучали потенциал XENP27563 и XENP27564, Xtend Fc-аналогов XENP26105 и XENP26109.
Пример 7. Характеристика in vitro XENP27563 и XENP27564
Культуры Treg, обработанные CD25-селективными слияниями ИЛ-2-Fc, демонстрируют более высокую экспрессию CD25
Ранее сообщалось, что рапамицин способствует пролиферации CD4+CD25+FOXP3+ Treg in vitro, и экспандированные в результате этого Treg подавляют пролиферацию CD4+ и CD8+ T-клеток (см., например, Battaglia et al. (2006). Rapamycin promotes expansion of functional CD4+CD25+FOXP3+ regulatory T cells of both healthy subjects and type 1 diabetic patients. J Immunol. 177(12) 8338-8347; и Strauss et al. (2007) Selective survival of naturally occurring human CD4+CD25+Foxp3+ regulatory T cells cultured with rapamycin. J Immunol. 178(1) 320-329).
CD4+ Т-клетки обогащали из МКПК человека путем отрицательной селекции с использованием набора для обогащения CD4+ Т-клеток человека EasySep™ (STEMCELL Technologies, Ванкувер, Канада). Treg экспандировали с помощью Human Treg Expander Dynabeads™ (Thermo Fisher Scientific, Уолтем, Массачусетс) в RPMI1640 + 10% фетальная бычья сыворотка + 0,1 мкг/мл рапамицина + 500 ЕД/мл ИЛ-2 в течение 1-4 дней. Treg переносили в колбы T75, покрытые 0,5 мкг/мл анти-CD3 (OKT3, Biolegend, Сан-Диего, Калифорния) и культивировали с RPMI1640 + 10% фетальная бычья сыворотка + 0,1 мкг/мл рапамицина + 100 ЕД/мл ИЛ-2 + 0,5 мкг/мл анти-CD28 мАт. Эксперименты проводили, по меньшей мере, через 8 дней после исходного обогащения CD4+ Т-клеток из МКПК. Treg, обогащенные и культивируемые таким образом, в дальнейшем называются Treg с рапамицином.
Treg с рапамицином дополнительно культивировали с 0,5 мкг/мл связанного с планшетом анти-CD3 (OKT3) в среде RPMI1640, содержащей 10% FBS, 0,5 мкг/мл анти-CD28 мАт, 100 нг/мл рапамицина и 10 нг/мл рекомбинантного ИЛ-2 или 10 мкг/мл XENP27564 (слияние ИЛ-2-Fc с вариантом ИЛ-2 (D20N/N71K/C125S)). Через 14 дней после культивирования Treg окрашивали анти-CD25-FITC (M-A251), анти-FoxP3-PE (PCH101), анти-CTLA-4-PE-Dazzle594 (L3D10), анти-PD-1-BB700 (EH12.1), анти-GITR-PE-Cy7 (108-17), анти-Ki67-Alexa647, анти-ICOS-Alexa700 (C398.4a), анти-TIGIT-BV421 (A15153G), анти-LAG-3 (11C3C65), анти-CCR4-BV605 (L291H4), анти-CD8-BV650 (SK1), анти-CD39-BV711 (A1), анти-TIM-3-BV785 (F38-2E2), анти-CD40BUV396 (SK3), анти-CD3-BUV496 (UCHT1), анти-CD45-BUV805 (HI30), анти-CD45RA-BUV737 (HI100) и Zombie NIR (APC-Cy7) и анализировали с помощью проточной цитометрии, данные которой изображены на фиг. 46-47. Данные демонстрируют, что Treg, обработанные CD25-селективным XENP27564, демонстрирует более высокую экспрессию CD25. Далее, как изображено на фиг. 48, XENP27564 продемонстрировал более высокую экспансию популяции эффекторных Treg (CD45RA-FoxP3mid-high).
7B: Культуры Treg, обработанные CD25-селективными слияниями ИЛ-2-Fc, демонстрируют более высокую супрессивную функцию
Treg с рапамицином, дополнительно культивируемые либо с ИЛ-2, либо с XENP27564, как описано в Примере 7A, оценивали в отношении их супрессорной функции на 15-й день. 1× 105 меченных CFSE МКПК инкубировали с указанным количеством меченых Tag-it Violet Treg в течение 4 дней и определяли экспансию CD8+ респондеров и CD4+ респондеров с помощью разведения CFSE. Популяции лимфоцитов окрашивали следующим образом: анти-CD8-PerCp-By5.5 (SK1), анти-CD3-PE-Cy7 (OKT3), анти-CD127-APC (A019D5), анти-CD25-APC-Fire750 (M-A251), анти-CD45RO-Alexa700 (UCHL1), анти-CD16-BV605 (3G6), анти-CD56-BV605 (HCD56), анти-CD45RA-BV785 (HI100), анти-CD4-BUV395 (SK3) и Zombie Aqua (BV510). Примечательно, что данные, изображенные на фиг. 49, указывают на то, что Treg, экспандированные с помощью Treg-селективных слияний ИЛ-2-Fc, могут иметь повышенную супрессорную функцию.
В качестве дополнения, изучали экспрессию CD25 и CD127 на Treg в супрессорном анализе, данные для которого изображены на фиг. 50-51. В соответствии с приведенными выше данными, экспандированные с помощью XENP27564 Treg демонстрировали более высокие уровни экспрессии CD25. Примечательно, что экспандированные с помощью XENP27564 Treg демонстрировали более низкую экспрессию CD127, маркера, который ранее, как было обнаружено, обратно коррелирует с супрессорной функцией Treg (Liu et al. (2006) CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4+ T reg cells. J Exp Med. 203(7): 1701-1711). Это может объяснить повышенную супрессорную функцию, наблюдаемую для экспандированных с помощью XENP27564 Treg.
CD25-селективные слияния ИЛ-2-Fc демонстрируют селективность Treg и супрессорный эффект в отношении других популяций лимфоцитов после активации анти-CD3
В супрессорном анализе меченные CFSE МКПК и меченные Tag-it Violet Treg с рапамицином инкубировали с указанными концентрациями указанных исследуемых препаратов в течение 4 дней с 100 нг/мл связанного с планшетом анти-CD3 (OKT3). Популяции лимфоцитов окрашивали следующим образом: анти-CD8-PerCp-Cy5.5 (SK1), анти-CD3-PE-Cy7 (OKT3), анти-CD25-APC-Cy7 (M-A251), анти-CD45RO-Alexa700 (UCHL1), анти-CD16-BV605 (3G6), анти-CD56-BV605 (HCD56), анти-CD45RA-BV785 (HI100), анти-CD4-BUV395 (SK3) и Zombie Aqua (BV510). Пролиферация различных популяций лимфоцитов (определяемая с помощью разведения CFSE или Tag-it Violet; краситель Zombie, используемый для исключения мертвых клеток) после обработки исследуемыми препаратами изображена на фиг. 52. Данные демонстрируют, что CD25-селективные слияния ИЛ-2-Fc XENP27563 и XENP27564 приводили к селективной экспансии Treg по сравнению с XENP24635 (ИЛ-2-Fc только с C125S), рекомбинантным ИЛ-2 и рекомбинантным ИЛ-15. Фактически, данные, изображенные на фиг. 53 и 54, демонстрируют, что CD25-селективные слияния ИЛ-2-Fc обладали значительно сниженной способностью индуцировать пролиферацию CD8+ и CD4+ T-клеток по сравнению с XENP24635, рекомбинантным ИЛ-2 и рекомбинантным ИЛ-15.
В анализе пролиферации МКПК инкубировали с указанными концентрациями указанных исследуемых препаратов с указанной концентрацией связанного с планшетом анти-CD3 мАт (ОКТ3). Популяции лимфоцитов окрашивали следующим образом: анти-FoxP3-PE (PCH101), анти-CD8-PerCP-Cy5.5 (SK1), анти-CD3-PE-Cy7 (OKT3), анти-Ki67-APC, анти-CD45RO-Alexa700 (UCHL1), анти-CD25-BV421 (M-A251), анти-CD16-BV605 (3G6), анти-CD56-BV605 (HCD56), анти-CD45RA-BV785 (HI100), анти-CD4-BUV396 (SK3) и Zombie NIR (APC-Cy7). Пролиферацию различных популяций лимфоцитов определяли на основе процента экспрессии Ki67, маркера пролиферации, данные для которого изображены на фиг. 55-65. В соответствии с данными, представленными выше в супрессорном анализе, данные анализа пролиферации, изображенные на фиг. 55-61, демонстрируют, что XENP27563 и XENP27564 (слияния ИЛ-2-Fc, сконструированные для селективности к CD25) имеют нарушенную функцию индукции пролиферации CD8+ Т-клеток, CD8+CD45RA- T-клеток, CD8+CD45RA+ T-клеток, CD4+ T-клеток, CD4+CD45RA- T-клеток, CD4+CD45RA+ T-клеток, NK-клеток; а данные, изображенные на фиг. 63-65, демонстрируют, что XENP27563 и XENP27564 приводят к селективной экспансии Treg по сравнению с другими популяциями лимфоцитов.
Пример 8. Слияния ИЛ-2-Fc активируют селективную и устойчивую экспансию Treg у яванских макак
С целью изучения клинического потенциала XENP27563 и XENP27564, изучали их активность у яванских макак. Перед введением препаратов животным подтверждали активность слияний ИЛ-2-Fc в отношении лимфоцитов яванских макак. Проводили два анализа, описанные ниже.
В первом анализе МКПК человека стимулировали различными концентрациями XENP27563 или XENP27564 в течение 15 минут при 37°C. Затем МКПК окрашивали анти-CD3-BUV395 (UCHT1), анти-CD4-BV605 (RPA-T4), анти-CD8-BV711 (RPA-T8), анти-CD25-BV421 (M-A251), анти-CD45RA-BV510 (HI100) и анти-CD56-PE. Затем клетки пермеабилизировали с использованием PerFix EXPOSE (Beckman Coulter, Индианаполис, Индиана). После пермеабилизации клетки окрашивали анти-CD16-AF700 (DJ130C), анти-FoxP3-AF488 (259D) и pSTAT5 (pY694) и анализировали с помощью проточной цитометрии в отношении фосфорилирования STAT5 в различных популяциях лимфоцитов, данные для которой изображены на фиг. 78.
Во втором анализе МКПК яванского макака стимулировали различными концентрациями XENP27563 или XENP27564 в течение 15 минут при 37°C. МКПК затем окрашивали анти-CD3-BV421 (SP34), анти-CD4-BV785 (OKT4), анти-CD8-BUV395 (RPA-T8), анти-CD25-BV510 (M-A251), анти-CD45RA-APC/H7 (HI100) и анти-CD56-PE. Затем клетки пермеабилизировали с использованием PerFix EXPOSE (Beckman Coulter, Индианаполис, Индиана). После пермеабилизации клетки окрашивали анти-CD16-AF700 (DJ130C), анти-FoxP3-AF488 (259D) и pSTAT5 (pY694) и анализировали с помощью проточной цитометрии в отношении фосфорилирования STAT5 в различных популяциях лимфоцитов, данные для которой изображены на фиг. 79.
Данные демонстрируют, что XENP27563 и XENP27564 являются в равной степени селективными и эффективными в отношении как Treg человека, так и Treg яванского макака.
8А: Сравнение ФД и ФК для XENP27563 и XENP27564
В первом исследовании на яванских макаках животным (n = 2) внутривенно вводили либо 3× дозу XENP27563, либо 3× дозу XENP27564 в 0-й и 15-й дни. С течением времени собирали кровь с целью изучения экспансии различных популяций лимфоцитов и изучения концентрации исследуемых препаратов в сыворотке крови. Концентрации сывороточного альбумина и кровяное давление животных также учитывали для изучения переносимости исследуемых препаратов.
На фиг. 66 изображена экспансия различных популяций лимфоцитов с течением времени. Данные демонстрируют, что оба исследуемых препарата были способны к экспансии Treg, при этом сохранялись близкие к исходным уровни CD8+CD45RA- T-клеток, CD4+CD45RA- T-клеток и CD16+ NK-клеток. Кроме того, данные демонстрируют, что два исследуемых препарата вызывают аналогичную фармакологическую активность у яванских макак. На фиг. 67 изображена концентрация в сыворотке крови исследуемых препаратов. Данные демонстрируют, что эти два исследуемых препарата демонстрировали аналогичные фармакокинетические профили, при этом в случае XENP27564 период полужизни составлял 1,5 дня. В совокупности это подтверждает, что слияния ИЛ-2-Fc, сконструированные для селективности к CD25 и уменьшения эффективности, обеспечивают селективную и устойчивую экспансию Treg.
Синдром капиллярной утечки представляет собой характерный токсический побочный эффект, ассоциированный с лечением цитокинами, такими как ИЛ-2. Одним из признаков утечки жидкости из сосудов является гипоальбуминемия, снижение концентрации сывороточного альбумина. Соответственно, изучали изменение концентрации сывороточного альбумина у животных, данные для которого изображены на фиг. 68. Примечательно, что у одного животного, которому вводили XENP27563, длительное снижение уровня альбумина обнаруживали как после первого, так и после второго введения. У одного животного, которому вводили XENP27564, длительное снижение уровня альбумина обнаруживали после второго введения, однако концентрация быстро возвращалась к исходным уровням. Эти данные свидетельствуют о том, что XENP27564, обладающий меньшей эффективностью, чем XENP27563, может способствовать более высокой переносимости и терапевтическому индексу.
Еще одним показателем утечки жидкости из сосудов является значительное снижение кровяного давления. Соответственно, фиксировали кровяное давление для животных в 0-й, 1-й, 3-й, 5-й, 9-й, 16-й, 18-й, 20-й и 24-й дни, данные для которого изображены на фиг. 69. Примечательно, что первая обезьяна, которой вводили XENP27563, испытывала снижение кровяного давления в 1-й день (1-й день после 1-й дозы), а вторая обезьяна, которой вводили XENP27563, испытывала снижение артериального давления в 16-й день (1-й день после 2-й дозы), в то время как стабильное артериальное давление наблюдали во все дни для обезьяны, которой вводили XENP27564 (телеметрические данные были повреждены для 2-й обезьяны, которой вводили XENP27564). Это также свидетельствует о том, что слияния ИЛ-2-Fc с более низкой эффективностью могут способствовать более высокой переносимости и терапевтическому индексу.
В конечном итоге, также изучали содержание эозинофилов и базофилов в качестве дополнительных показателей переносимости, данные для которых изображены на фиг. 76. В совокупности данные демонстрируют повторное введение XENP27564.
8B: Исследование с повышением дозы XENP27564
В первом исследовании на яванских макаках животным (n = 3) внутривенно вводили либо 1×, 3×, либо 10× дозу XENP27564. С течением времени собирали кровь с целью изучения экспансии различных популяций лимфоцитов, а также концентрации сывороточного альбумина и С-реактивного белка (CRP).
Экспансия различных популяций лимфоцитов изображена на фиг. 70-71. В соответствии с данными первого исследования на яванских макаках, XENP27564 обеспечивает селективную и устойчивую экспансию Treg. Кроме того, данные демонстрируют, что 1× и 3× дозы вызывают аналогичную фармакологическую активность (как показано с помощью экспансии Treg) у обезьян. Примечательно, что более высокая доза (10× доза) XENP27564 не приводила к усилению фармакодинамики. В соответствии с данными, представленными выше, на фиг. 75 изображена устойчивая фармакокинетика в пределах нескольких дней у яванских макак при всех исследуемых дозах XENP27564.
Как и в случае первого исследования, изучали снижение уровня альбумина в качестве показателя пропотевания жидкости из сосудов и переносимости, данные для которых изображены на фиг. 72. Кроме того, изучали концентрацию CRP в сыворотке крови, белка острой фазы, ассоциированного с воспалением, в качестве еще одного показателя переносимости, данные для которой изображены на фиг. 73. Также изучали концентрация натрия, концентрация хлора, содержание эозинофилов и содержание базофилов в качестве дополнительных показателей переносимости (данные для которых изображены на фиг. 74). Примечательно, что данные демонстрируют, что более высокие дозы XENP27564 приводили к повышению токсичности, о чем свидетельствовало как снижение уровня альбумина, так и повышение концентрации CRP в сыворотке крови (а также концентрация натрия, концентрация хлоридов, содержание эозинофилов и содержание базофилов), в то же время более низкие дозы, которые все еще приводили к значительному повышению Treg, были более переносимыми.
Пример 9. Слияния ИЛ-2-Fc в равной степени селективны в отношении Treg у мышей
Спленоциты от мышей B6 инкубировали со слияниями ИЛ-2-Fc и рекомбинантным ИЛ-2 в течение 15 минут. После инкубации клетки окрашивали анти-CD4-PE (GK1.5), анти-CD25-BV605 (PC61) и анти-CD44-BV510 (IM7). Затем клетки пермеабилизировали с использованием PerFix EXPOSE (Beckman Coulter, Индианаполис, Индиана). После пермеабилизации клетки окрашивали анти-CD3-AF700 (2C11), анти-CD8-AF488 (53-6.7), анти-FoxP3-eF450 (FJK-16S) и анти-pSTAT5 (pY694) и анализировали с помощью проточной цитометрии в отношении фосфорилирования STAT5 в различных популяциях лимфоцитов, данные для которой изображены на фиг. 77. Данные указывают на то, что сконструированные слияния ИЛ-2-Fc являются одинаково селективными и эффективными для Treg у мышей, что делает их пригодными для использования в доклинических моделях у мышей для исследования аутоиммунных заболеваний.
| название | год | авторы | номер документа |
|---|---|---|---|
| Гетеродимерные слитые белки ИЛ-15/ИЛ-15Рα - Fc и их применение | 2019 |
|
RU2827796C2 |
| МУТЕИНЫ ИЛ-2 ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ И ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ | 2021 |
|
RU2833295C1 |
| СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ | 2010 |
|
RU2598719C2 |
| ОХ40-СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2802070C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К CD25 И Fc ГАММА-РЕЦЕПТОРУ ДЛЯ ЭЛИМИНАЦИИ ОПУХОЛЕСПЕЦИФИЧЕСКИХ КЛЕТОК | 2017 |
|
RU2759970C2 |
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
| БИОЛОГИЧЕСКИ АКТИВИРУЕМЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА НА ОСНОВЕ ЦИТОКИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2819307C2 |
| ВАРИАНТЫ РЕЦЕПТОРА ИНТЕРЛЕЙКИНА-2 (IL2R) И ИНТЕРЛЕЙКИНА-2 (IL2) ДЛЯ СПЕЦИФИЧЕСКОЙ АКТИВАЦИИ ИММУННЫХ ЭФФЕКТОРНЫХ КЛЕТОК | 2020 |
|
RU2832154C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА-ИНГИБИТОРЫ КОНТРОЛЬНЫХ ТОЧЕК | 2017 |
|
RU2767357C2 |
| ПОЛИВАЛЕНТНЫЕ МОДУЛЯТОРЫ РЕГУЛЯТОРНЫХ T-КЛЕТОК | 2017 |
|
RU2769871C2 |










































































































































































































































































































































































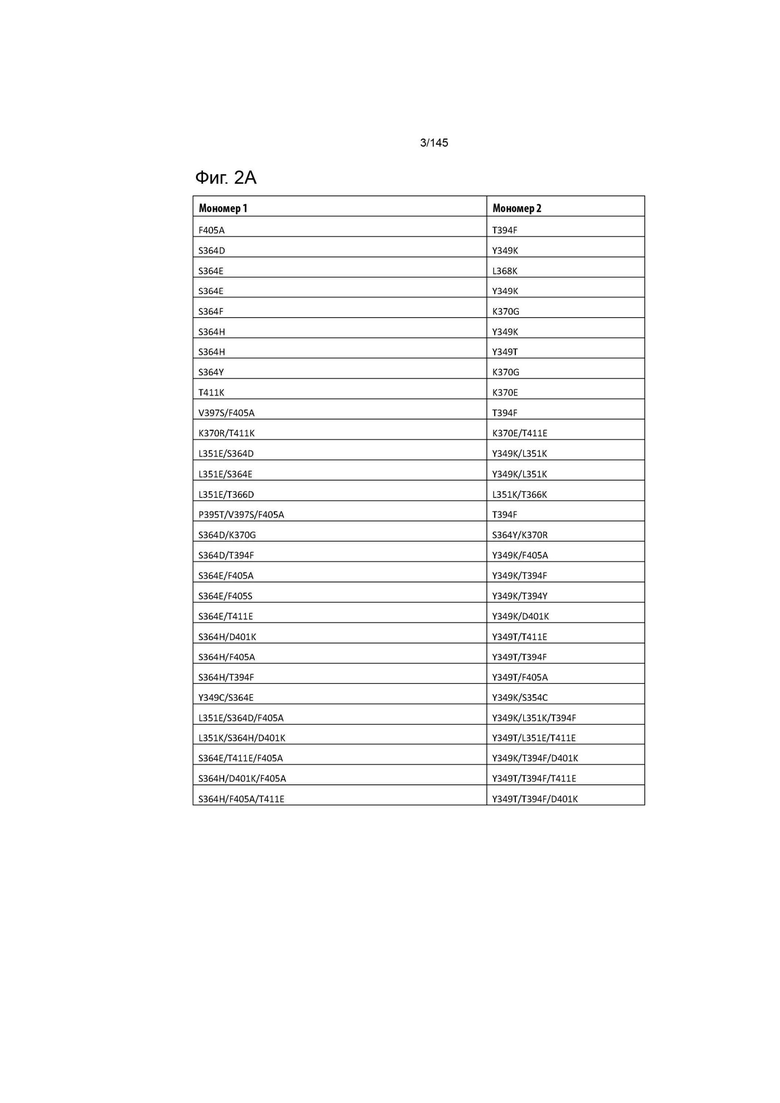












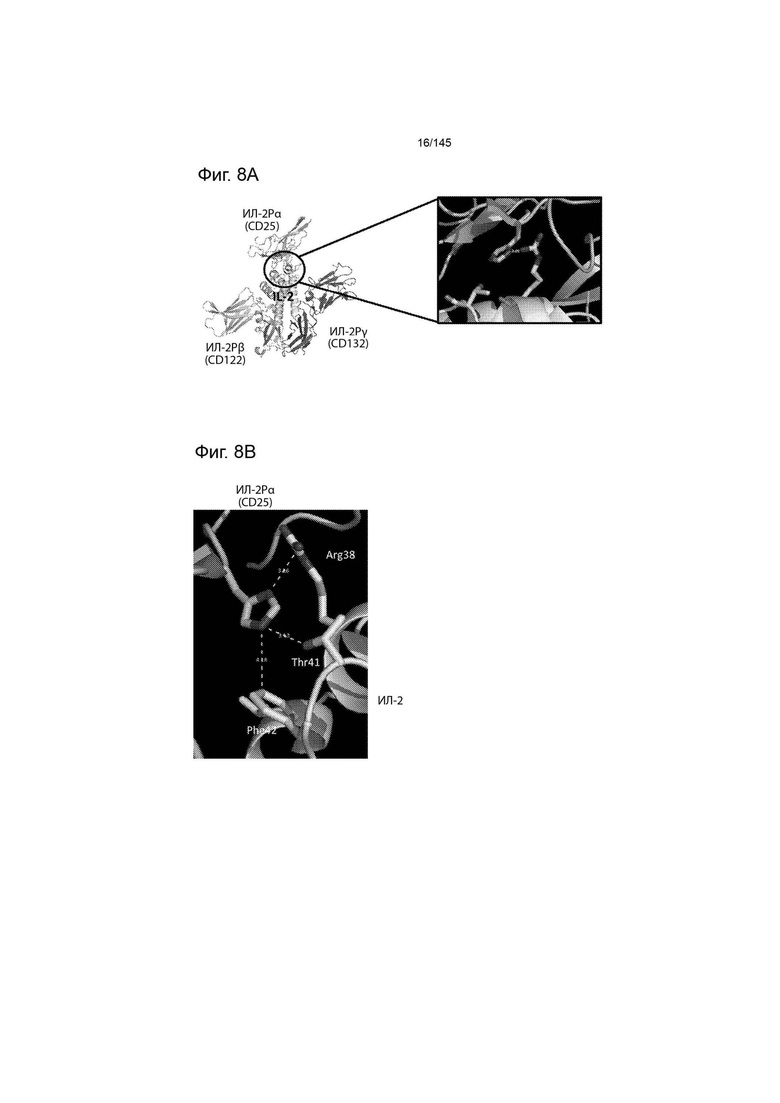








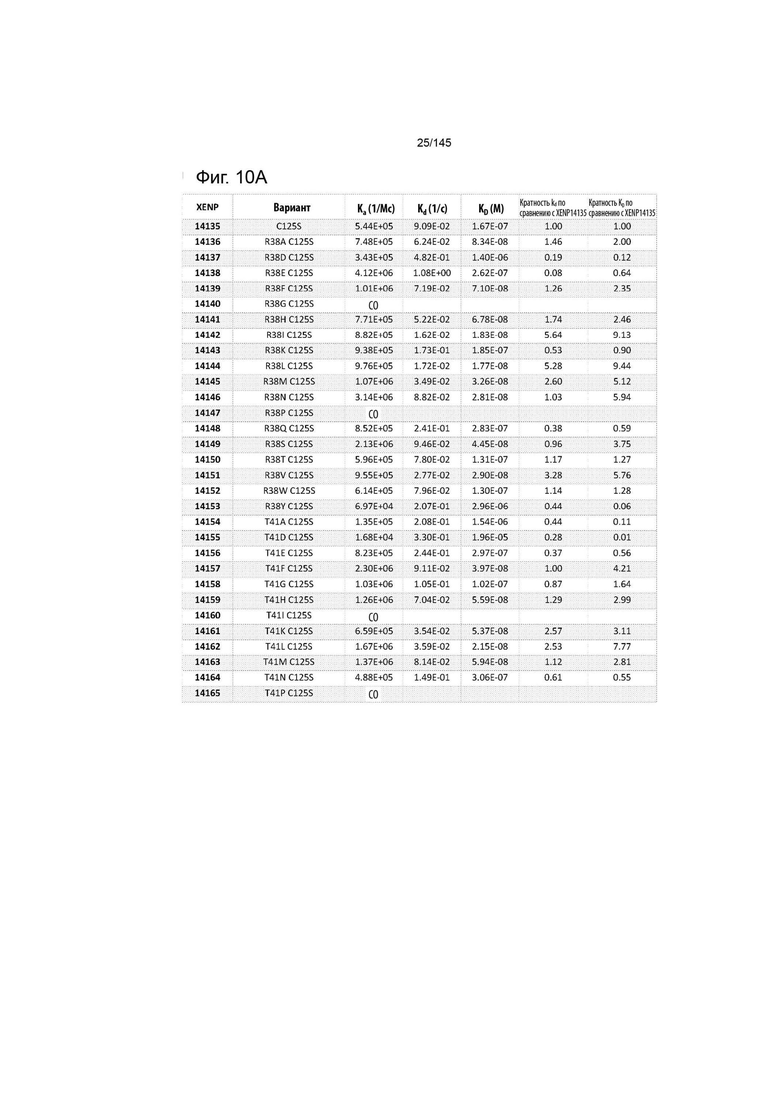
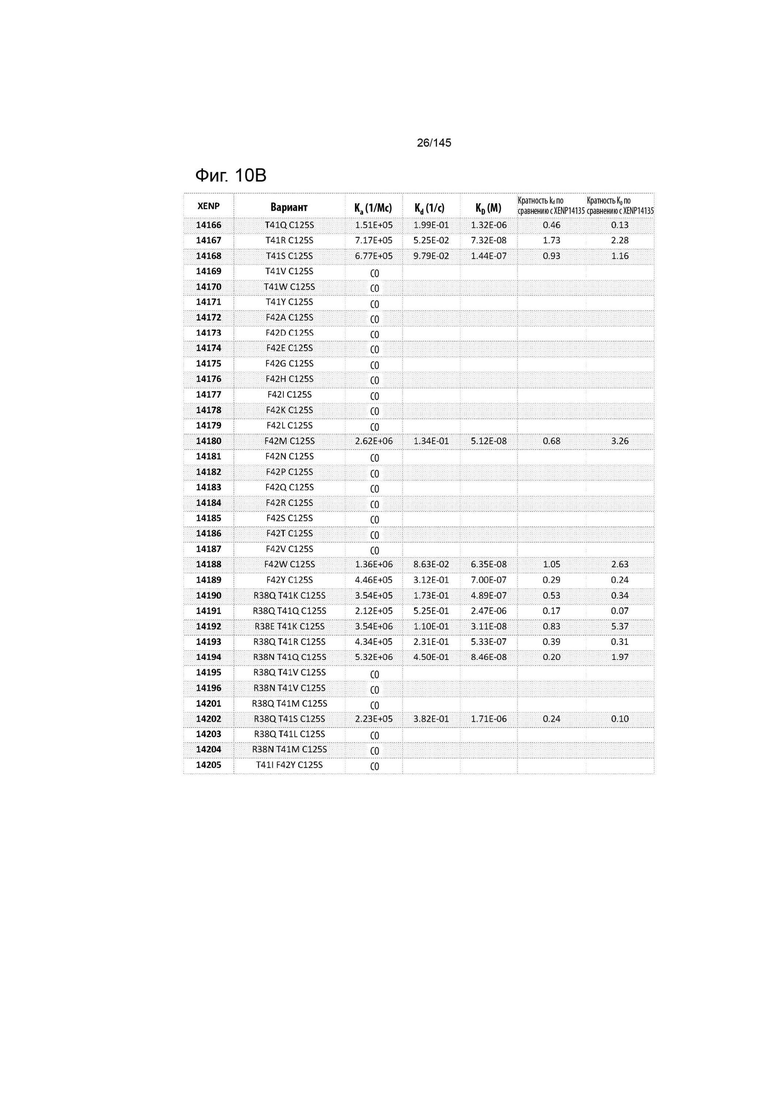
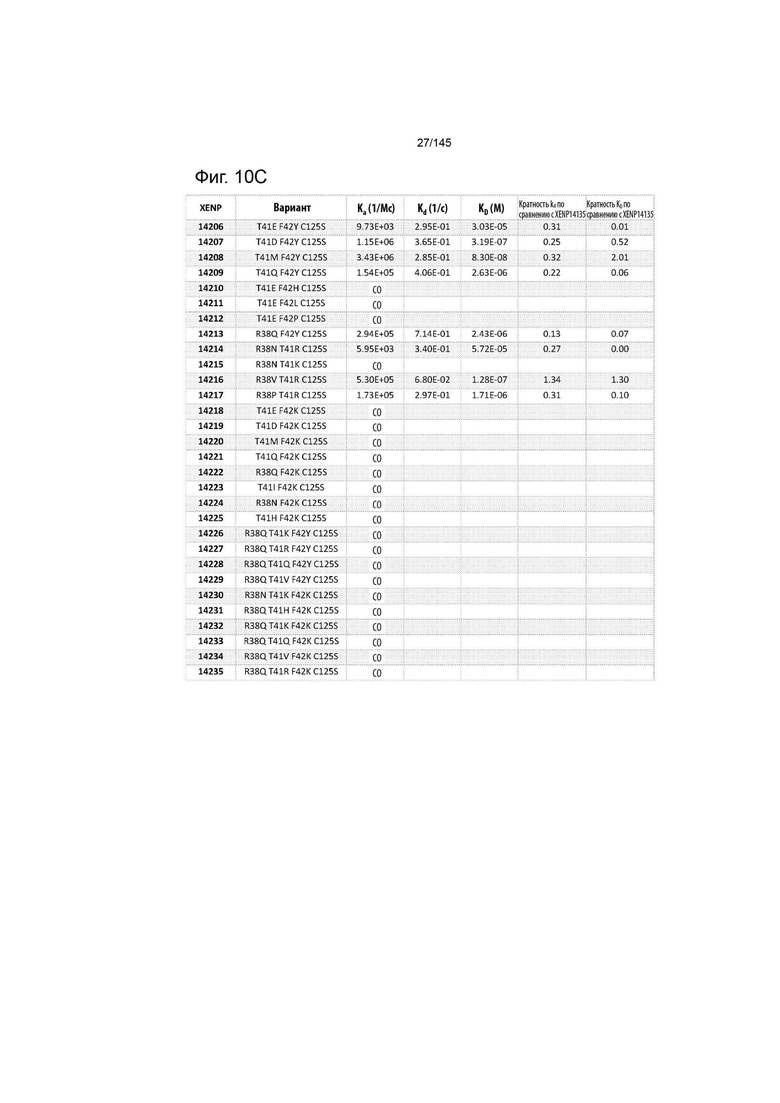
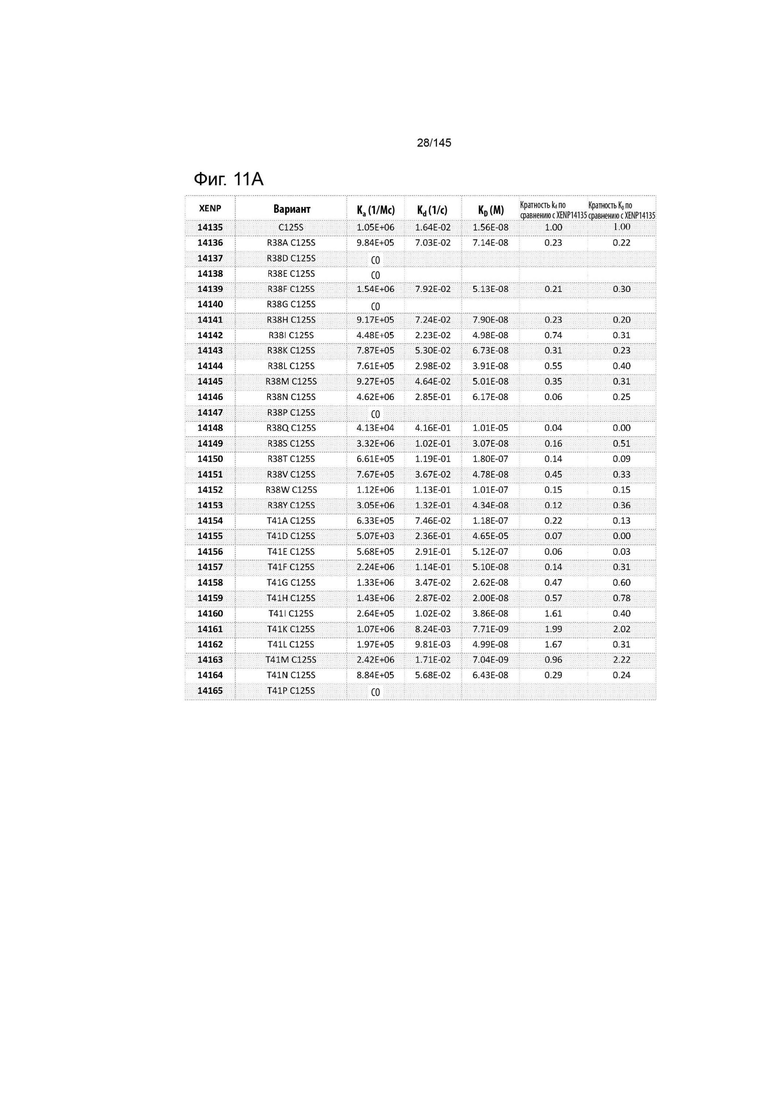
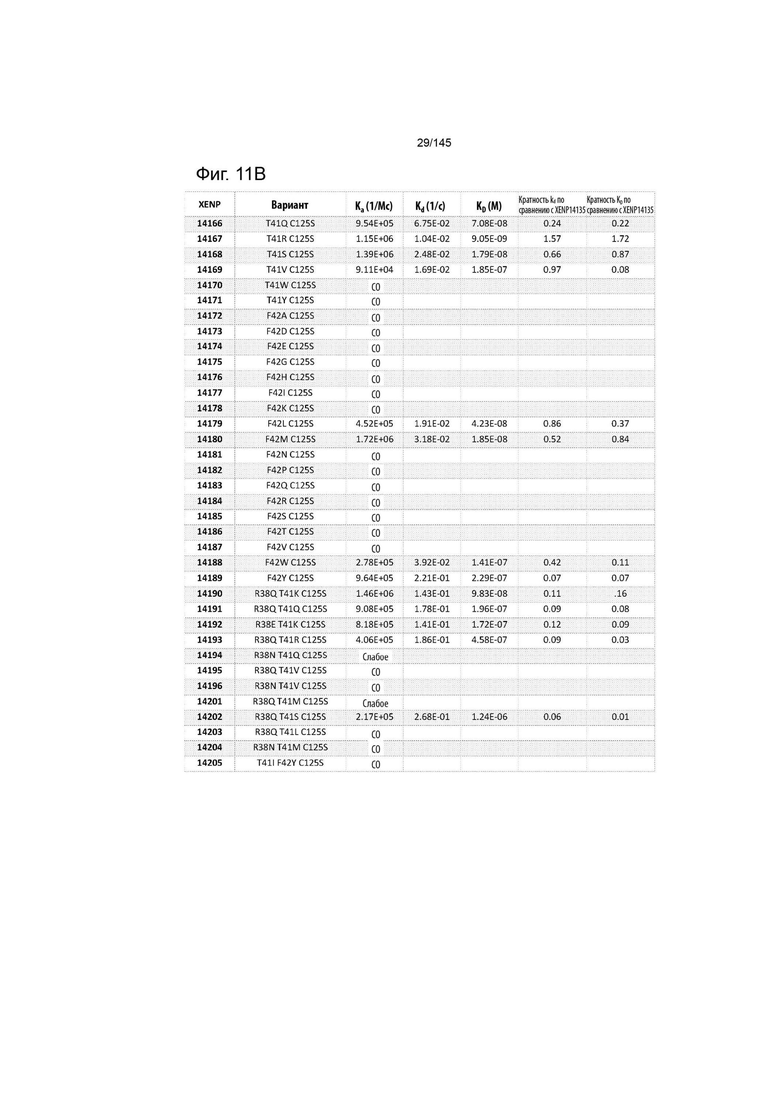
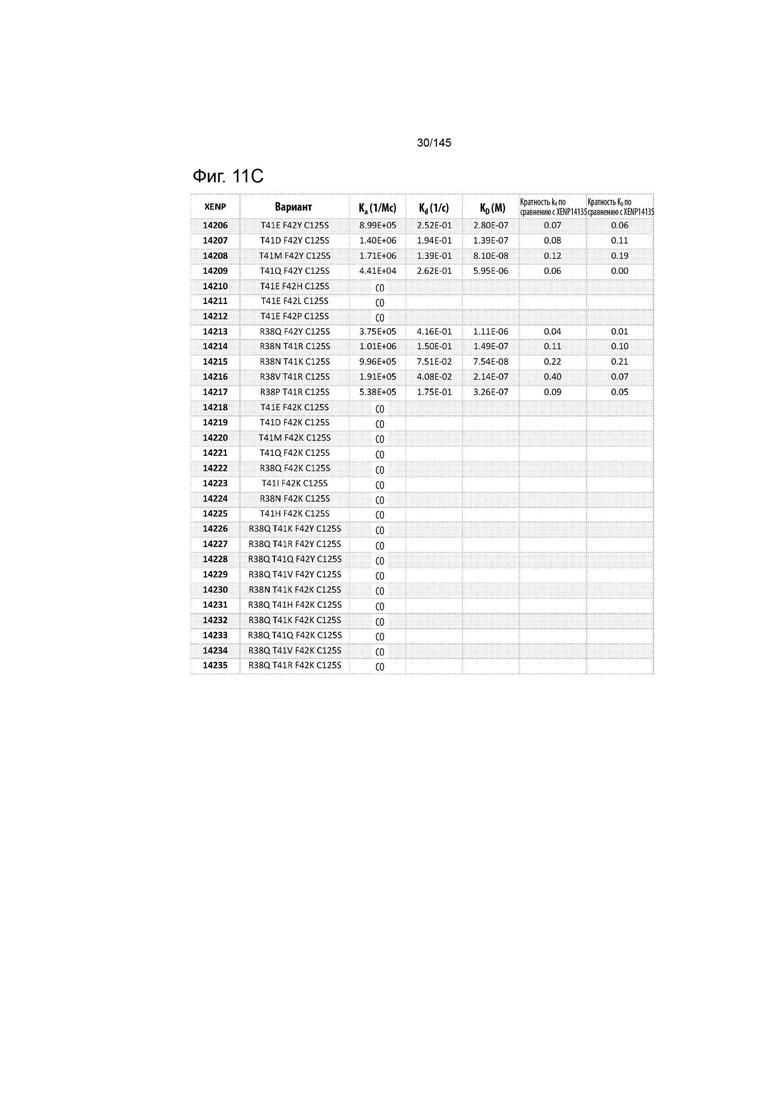
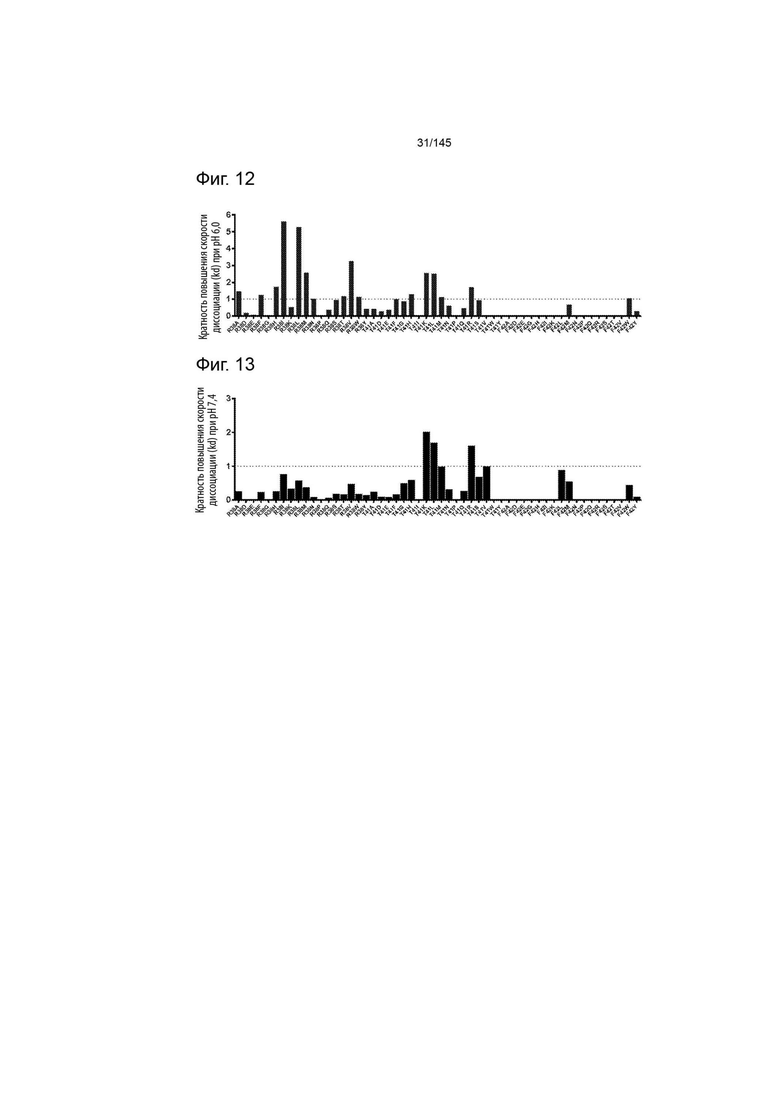
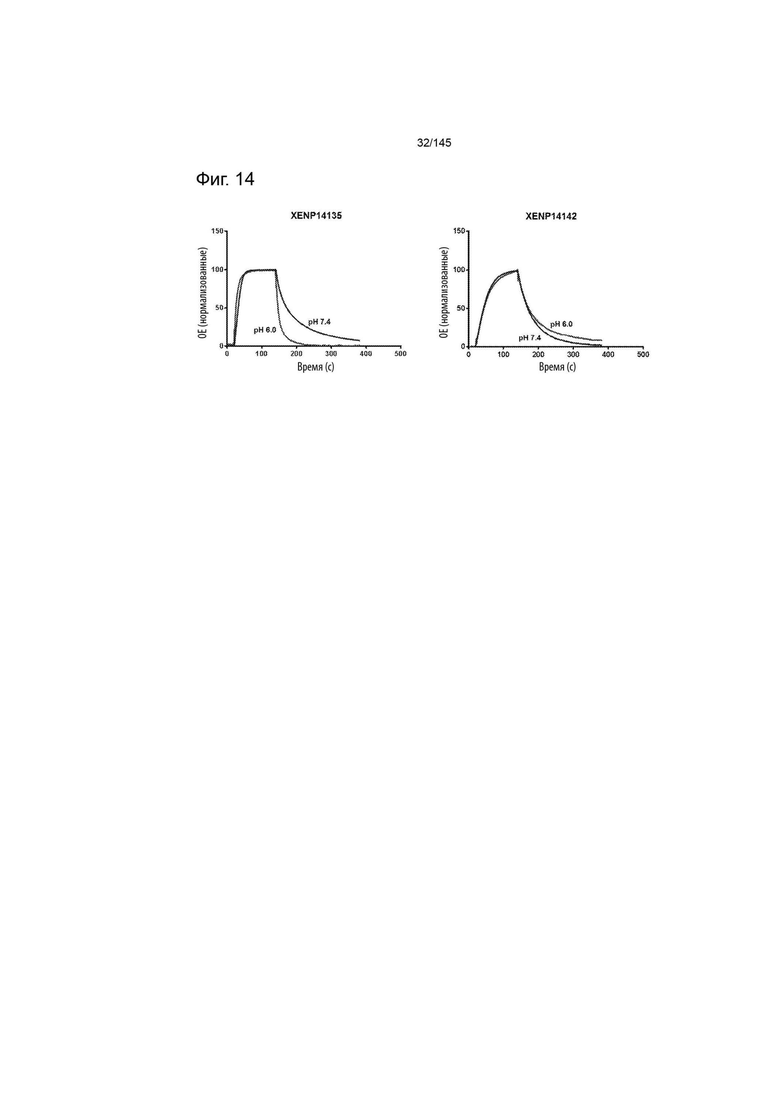

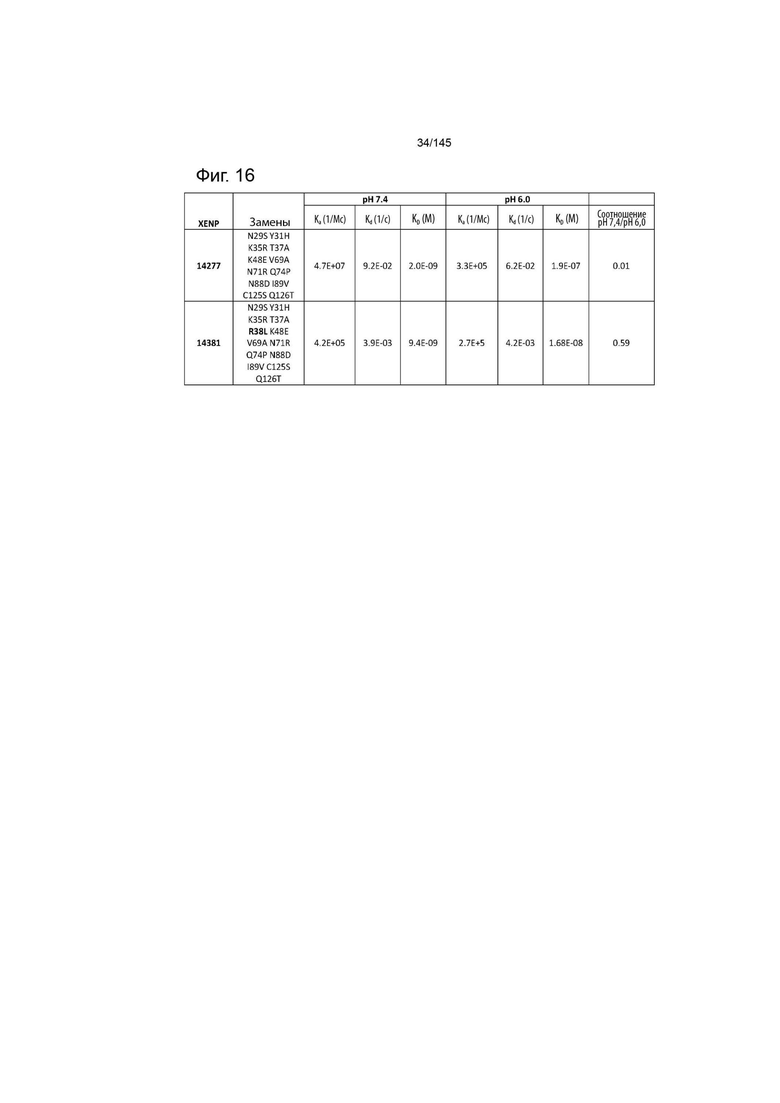




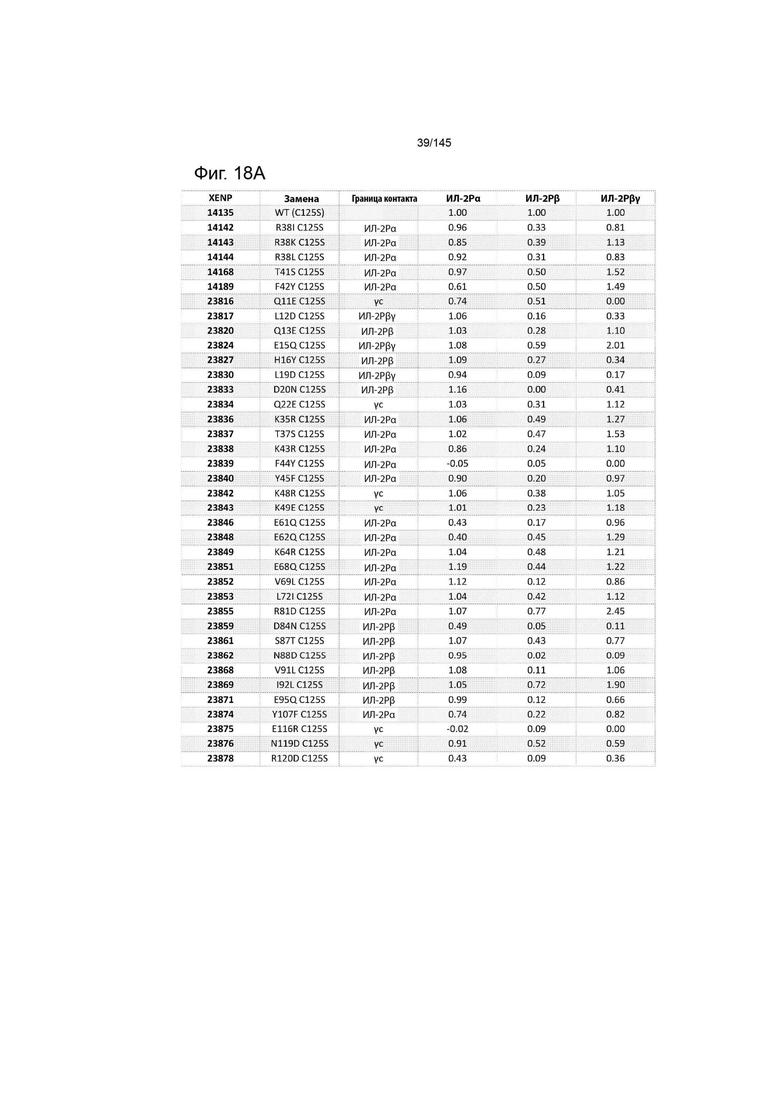
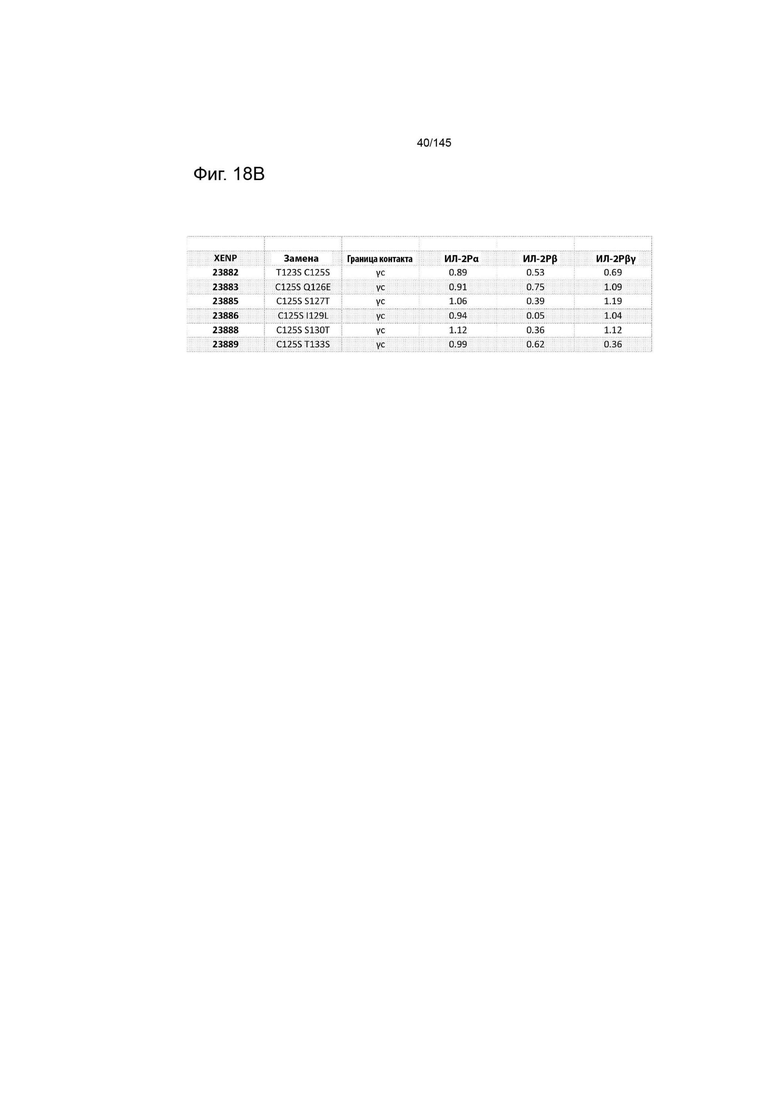
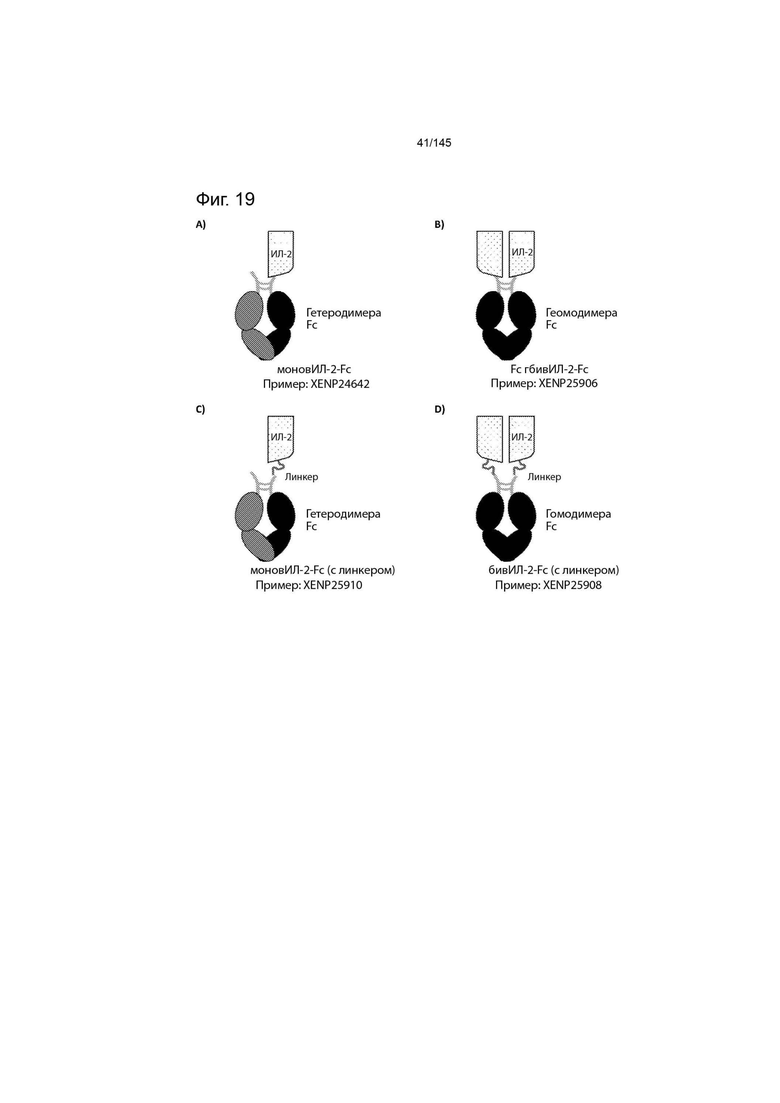





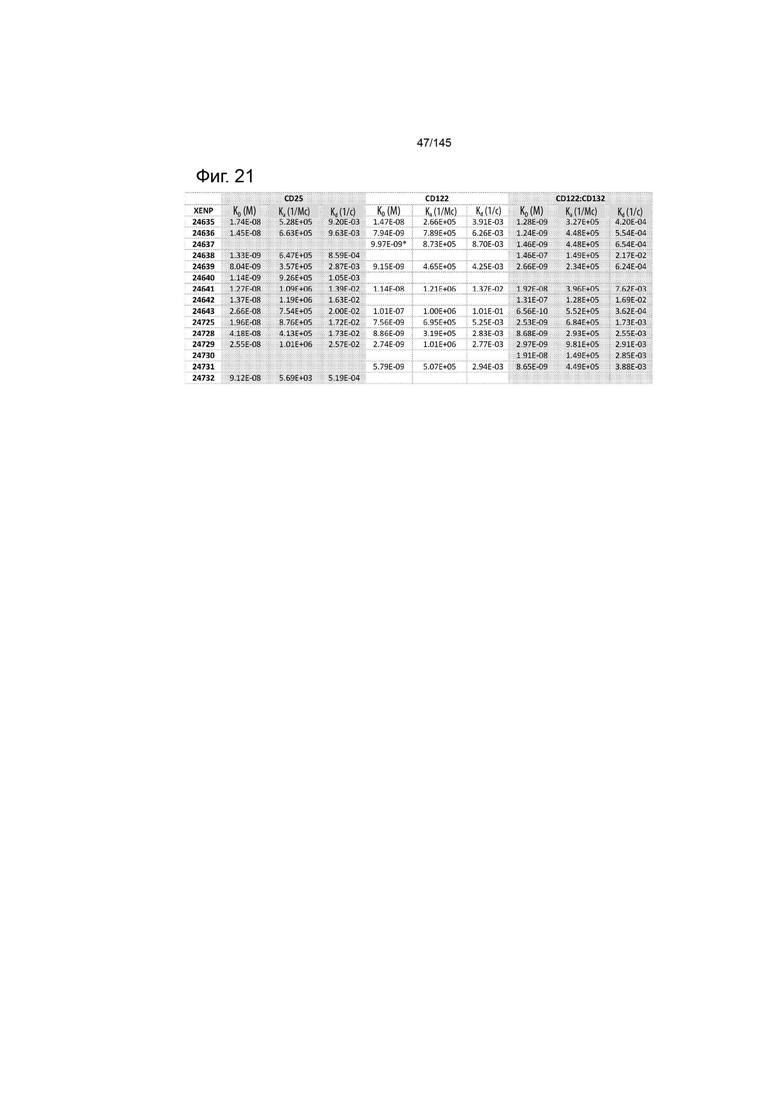
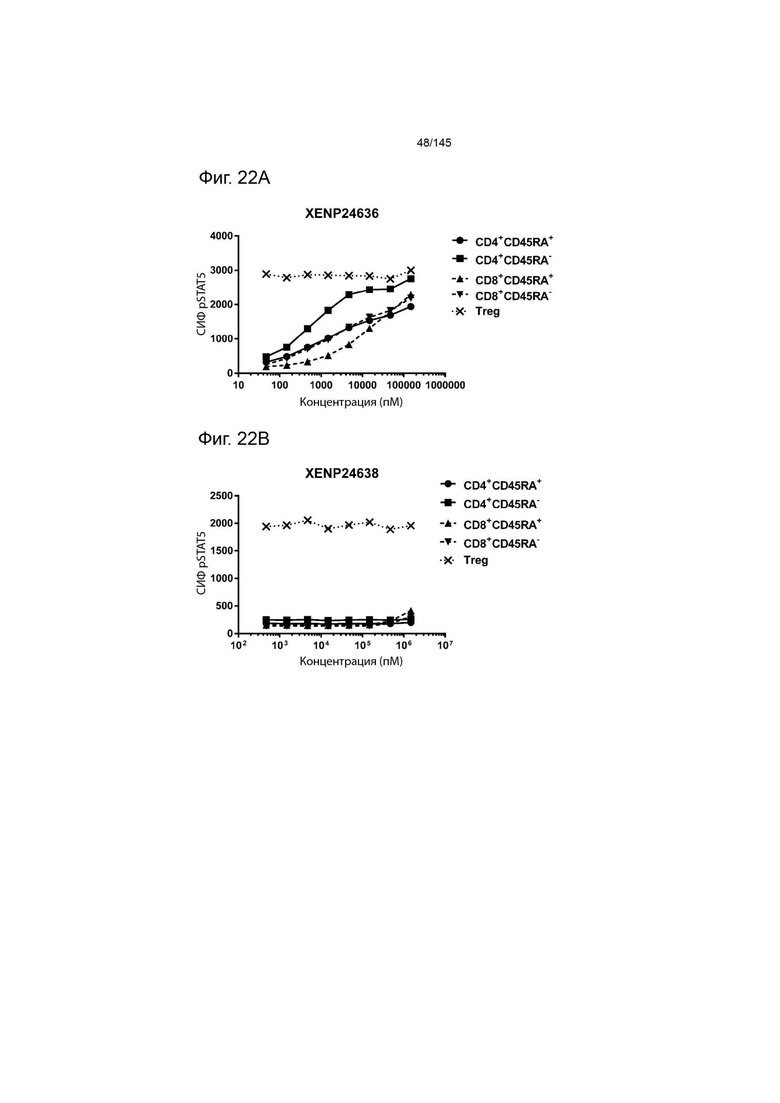
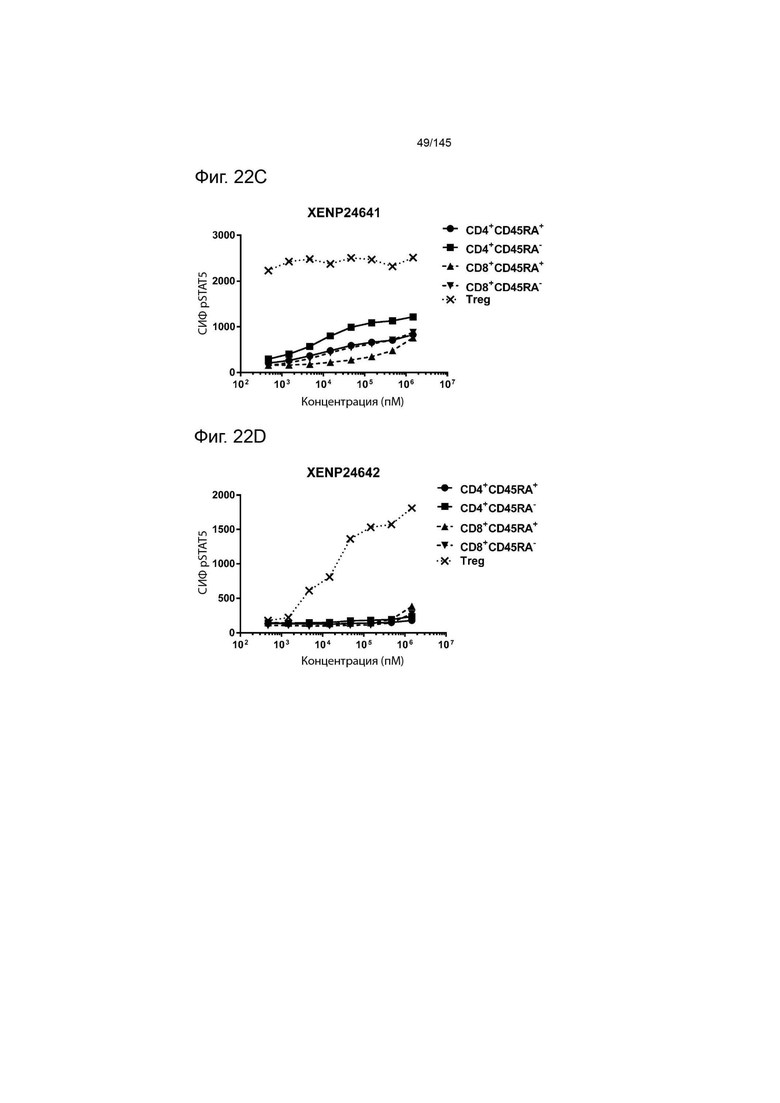
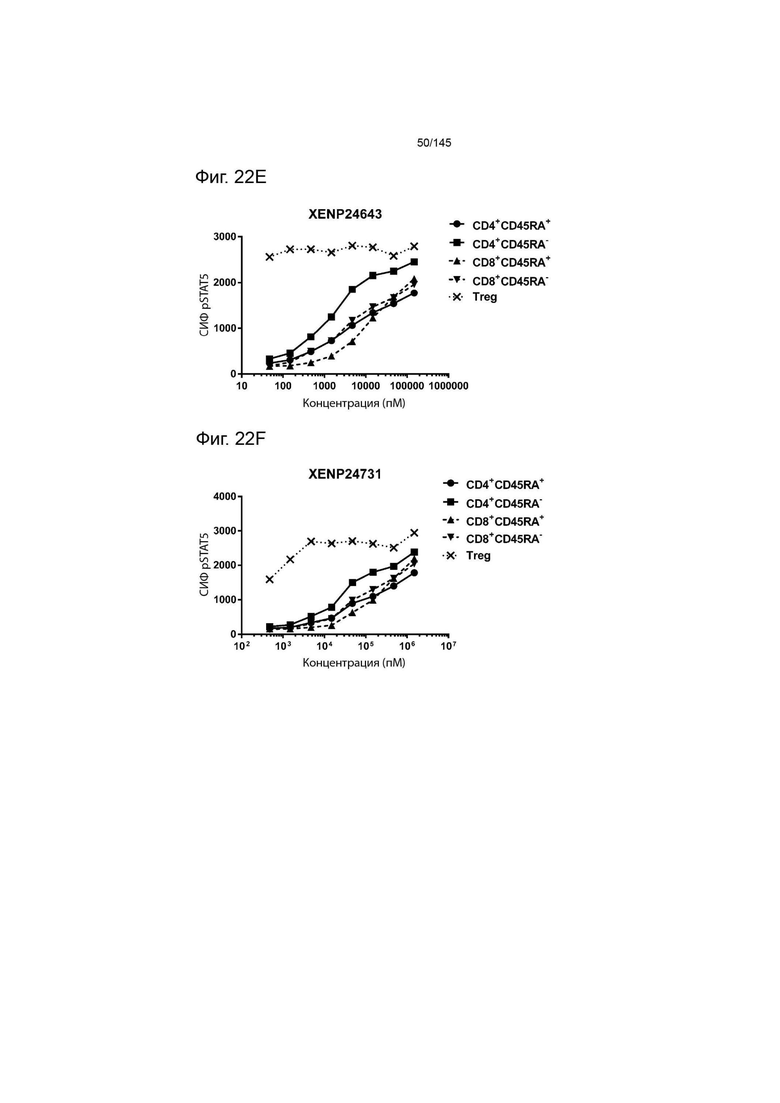

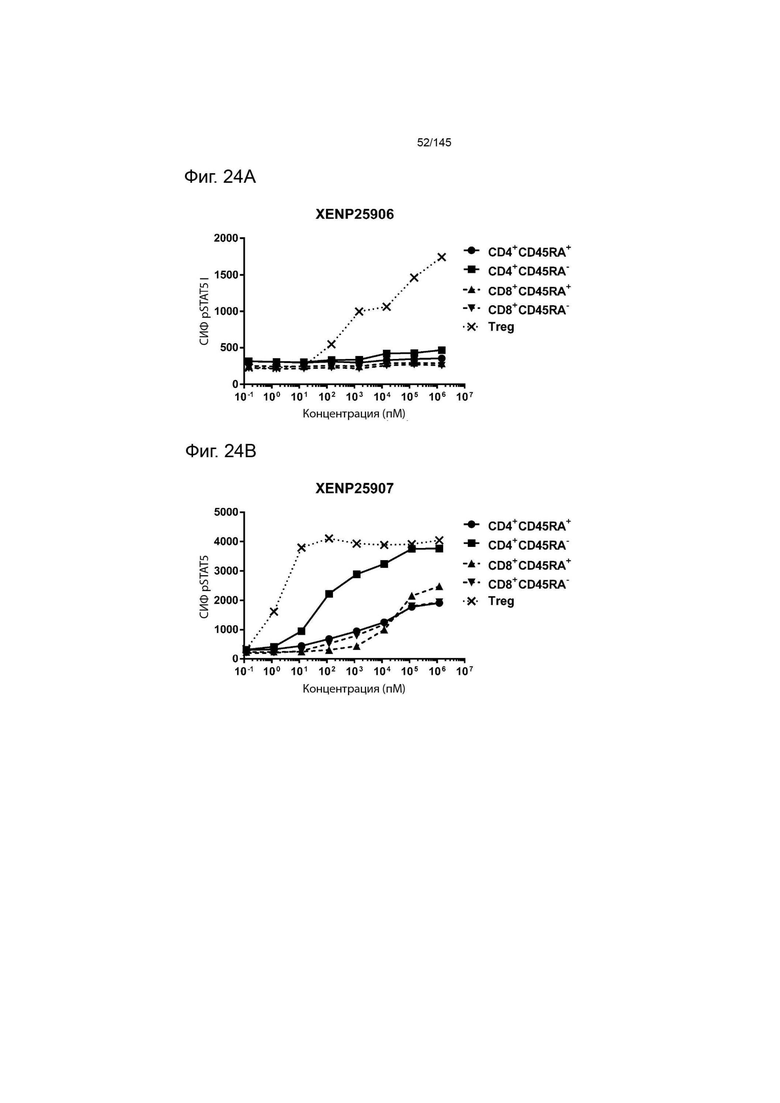


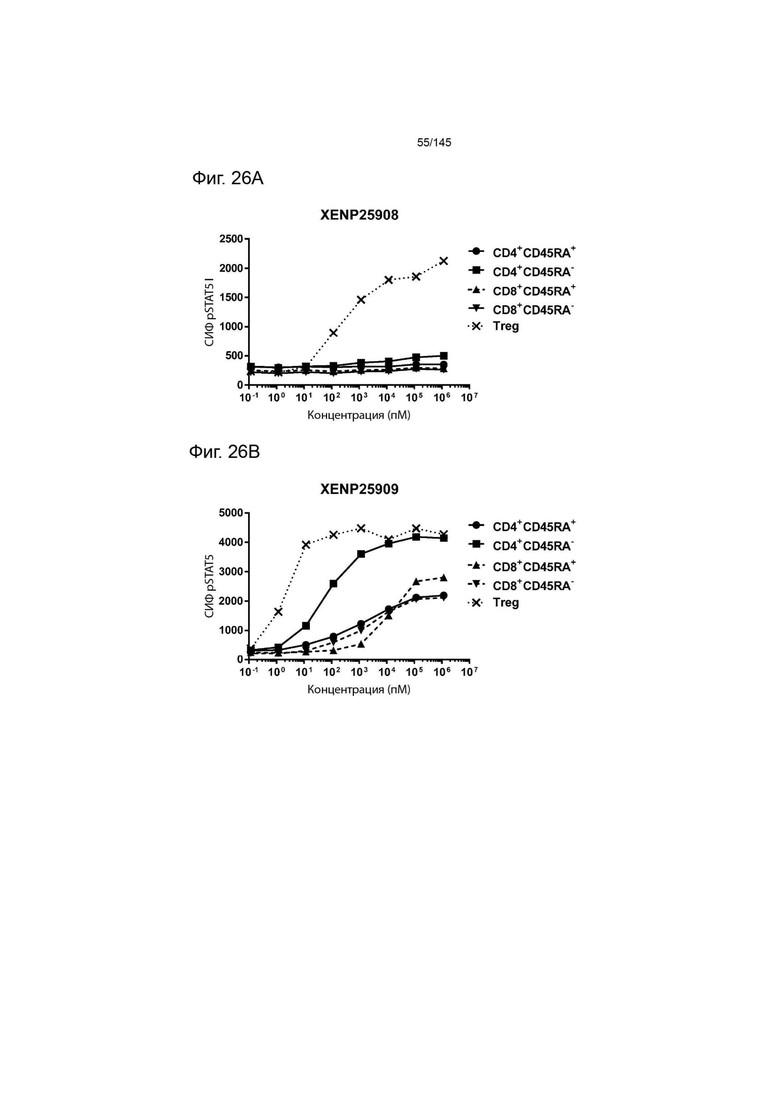
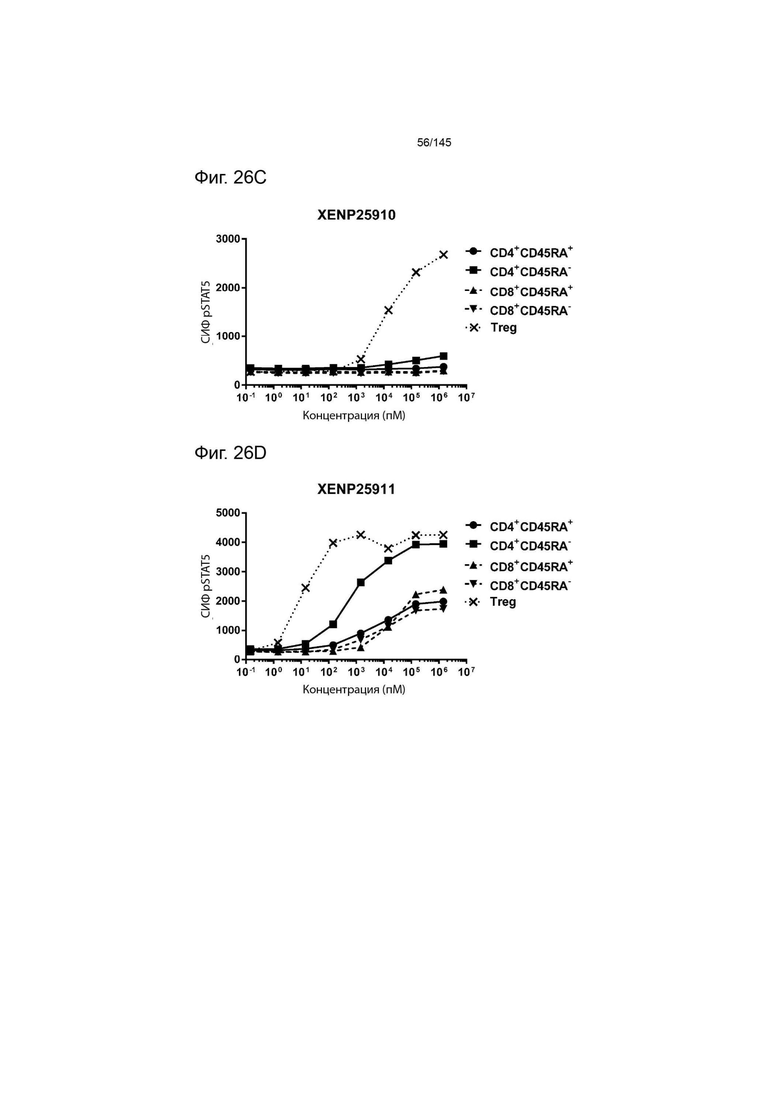
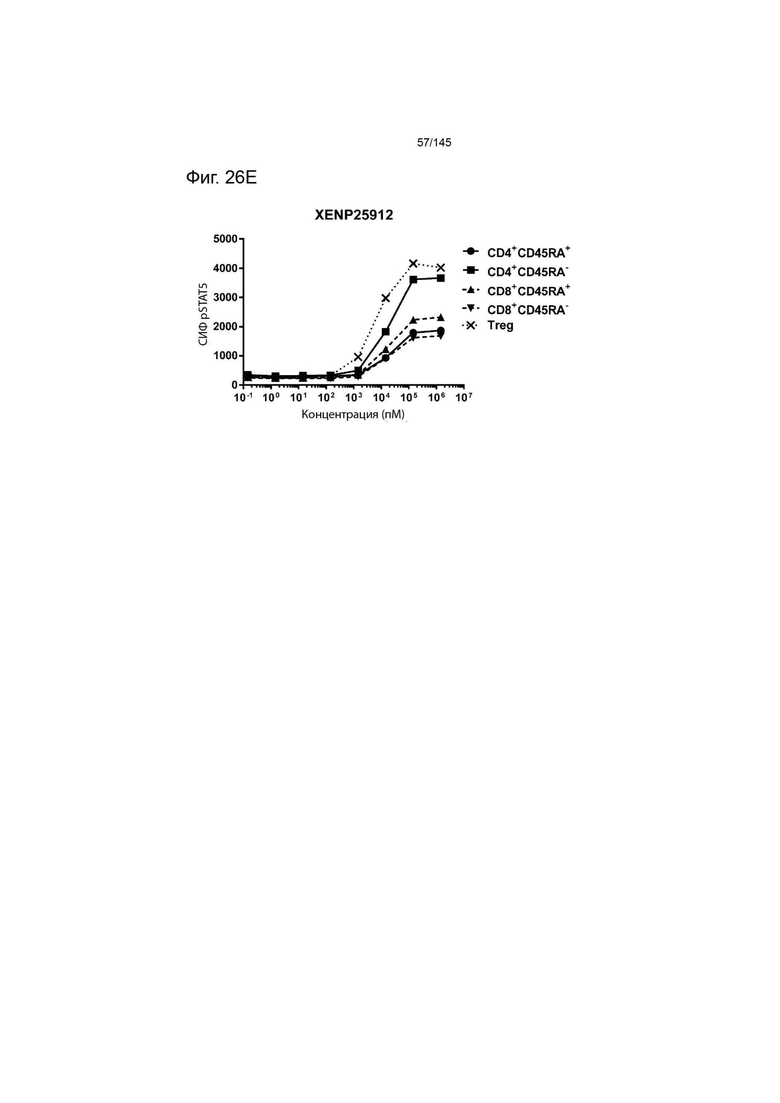





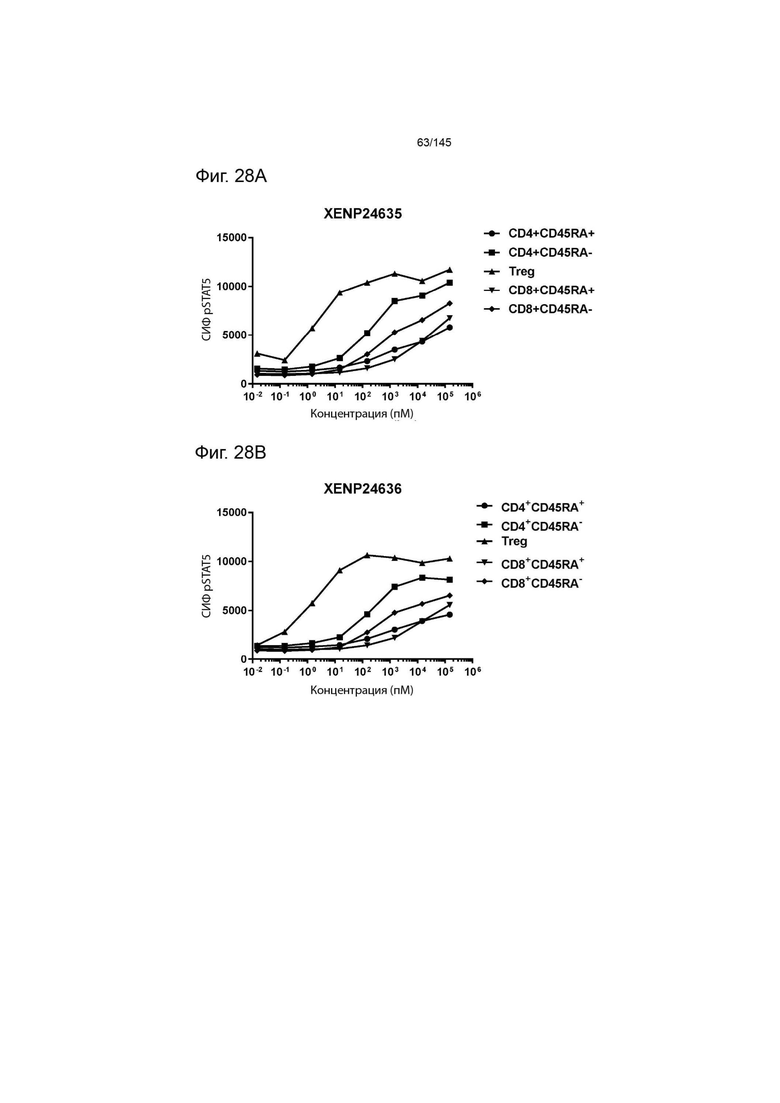
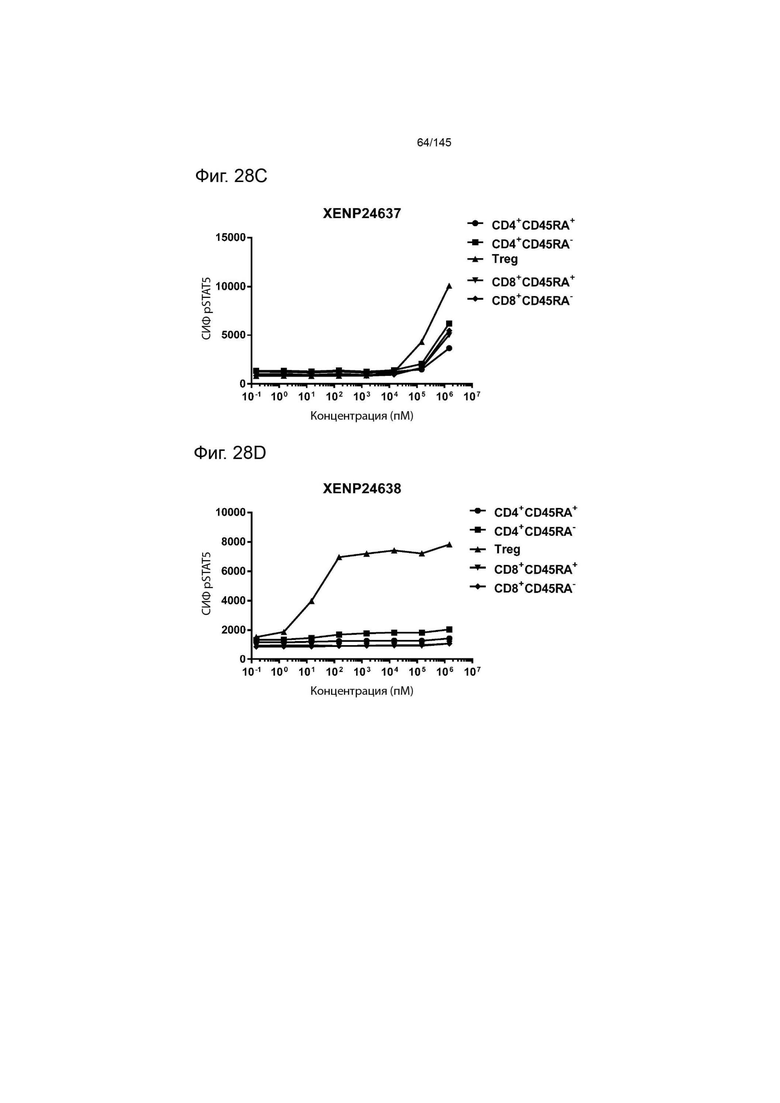
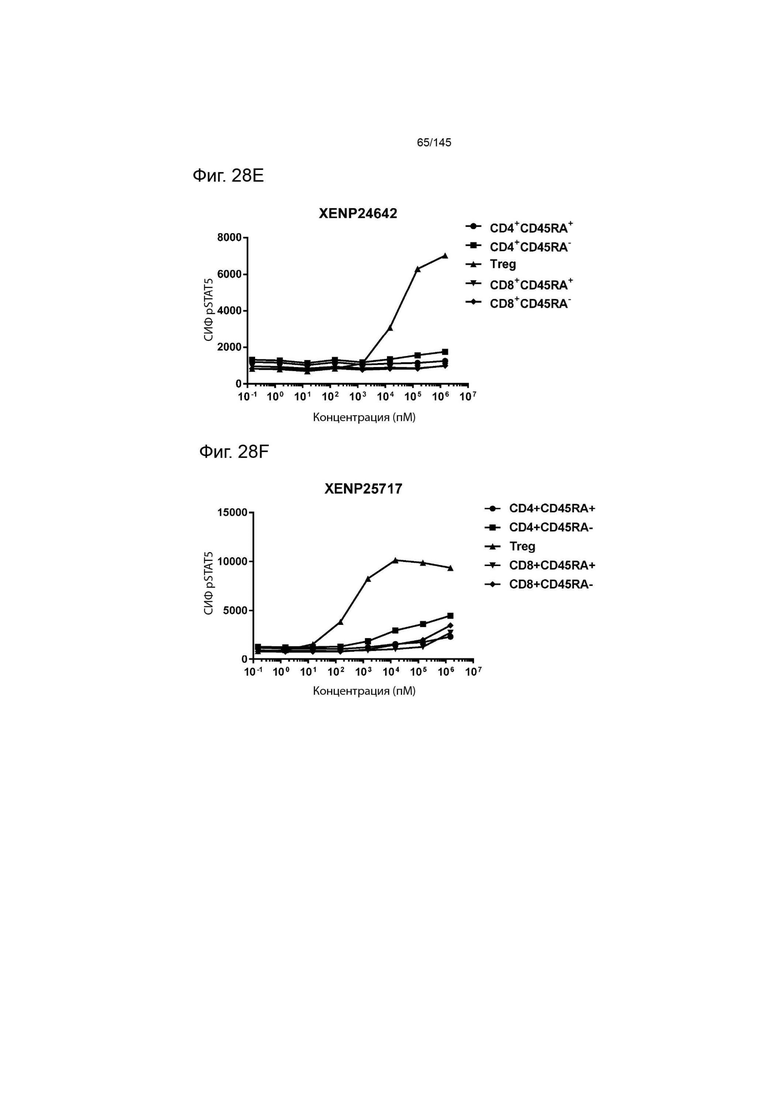
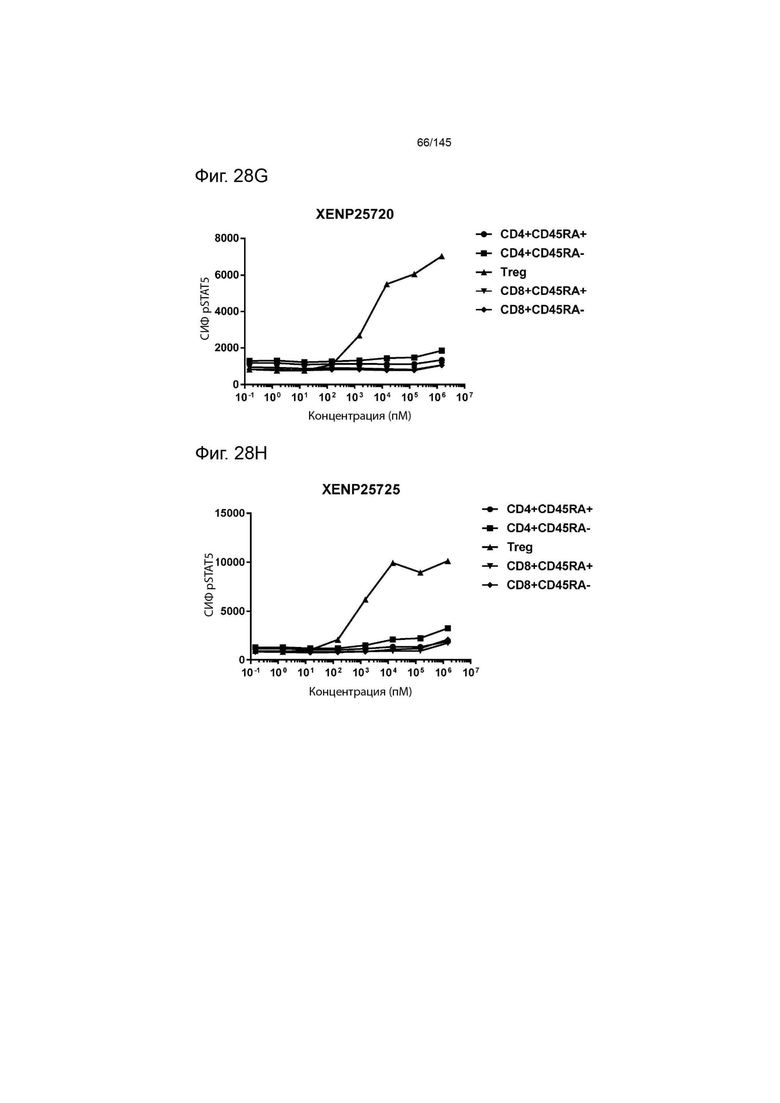
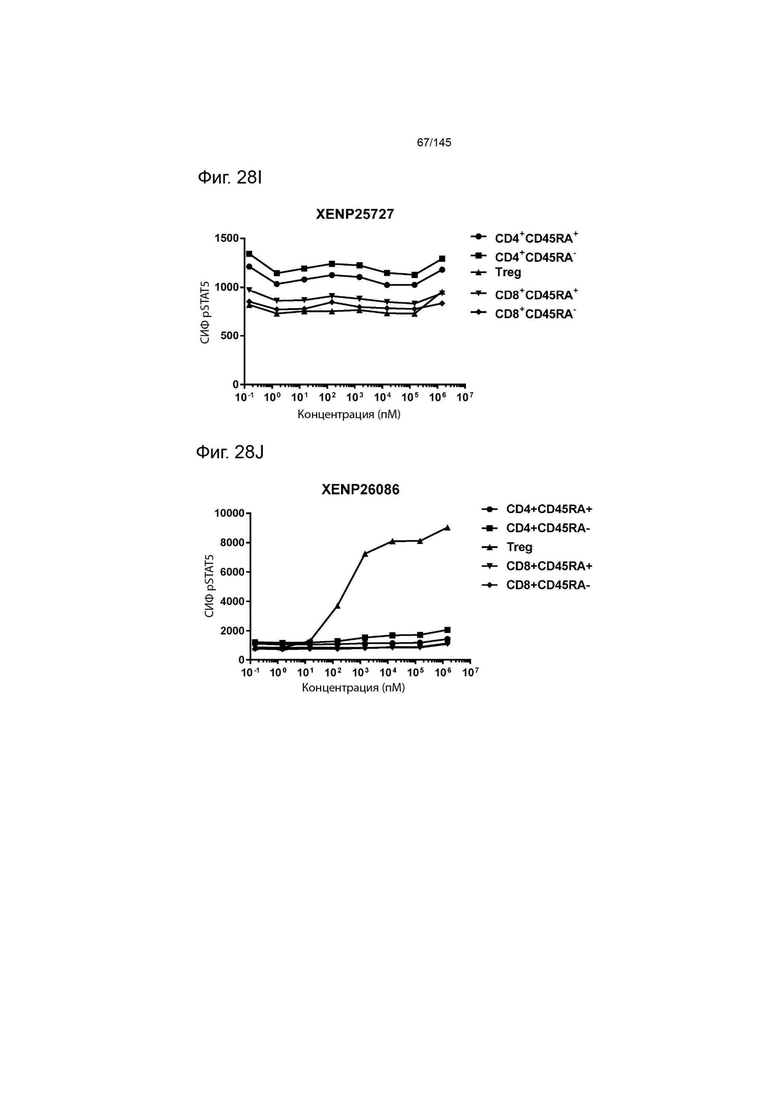
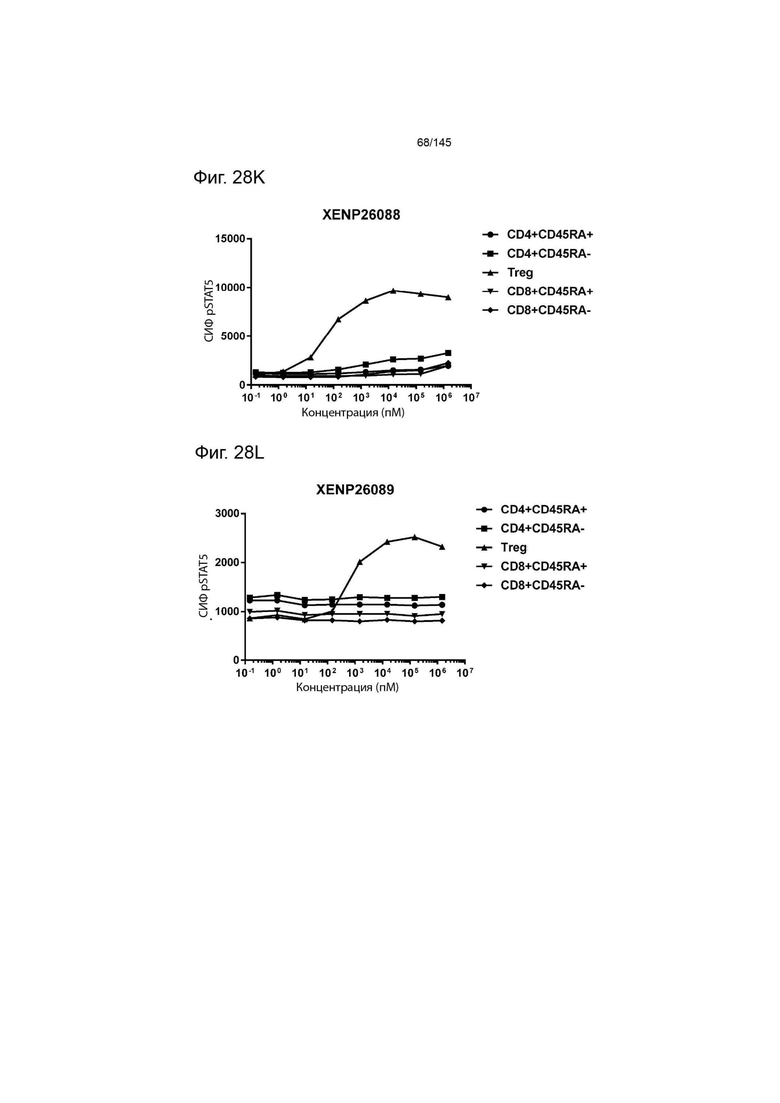
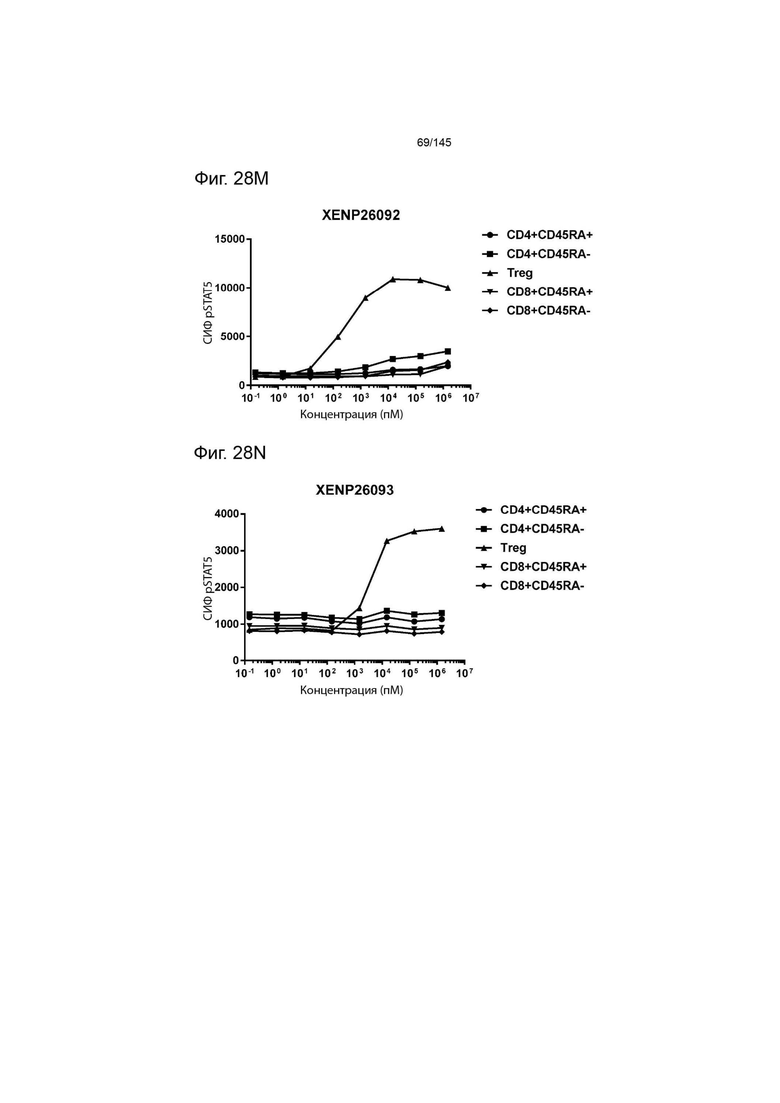
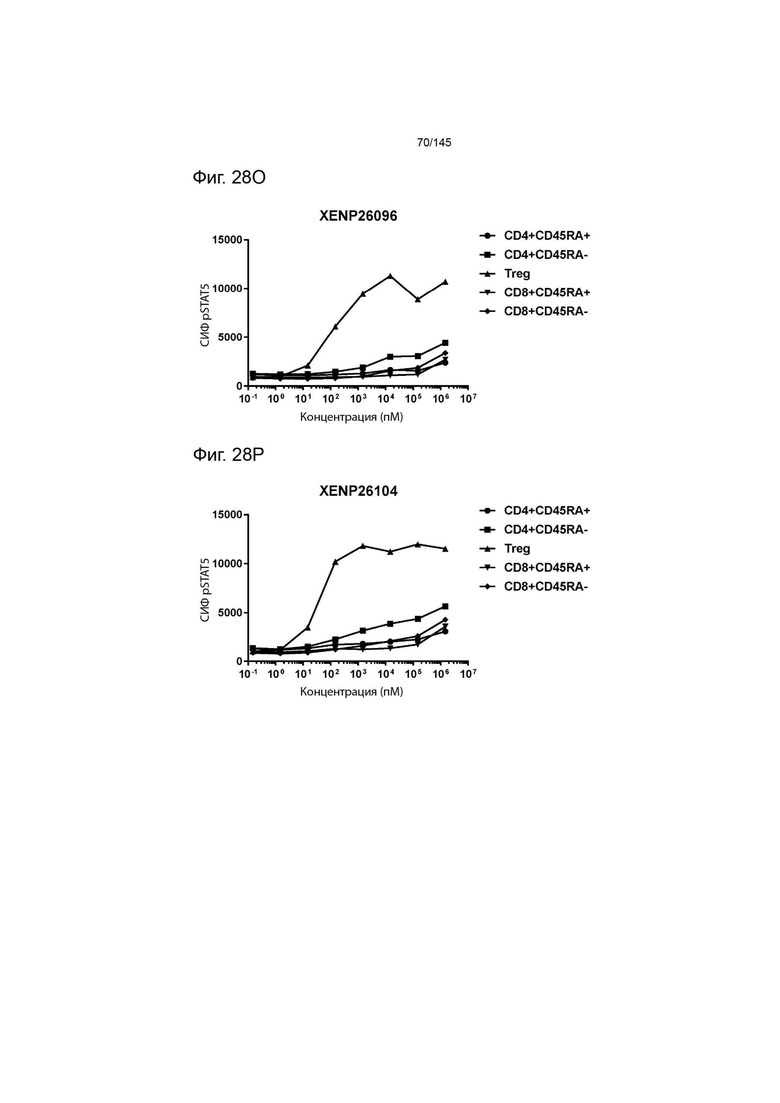
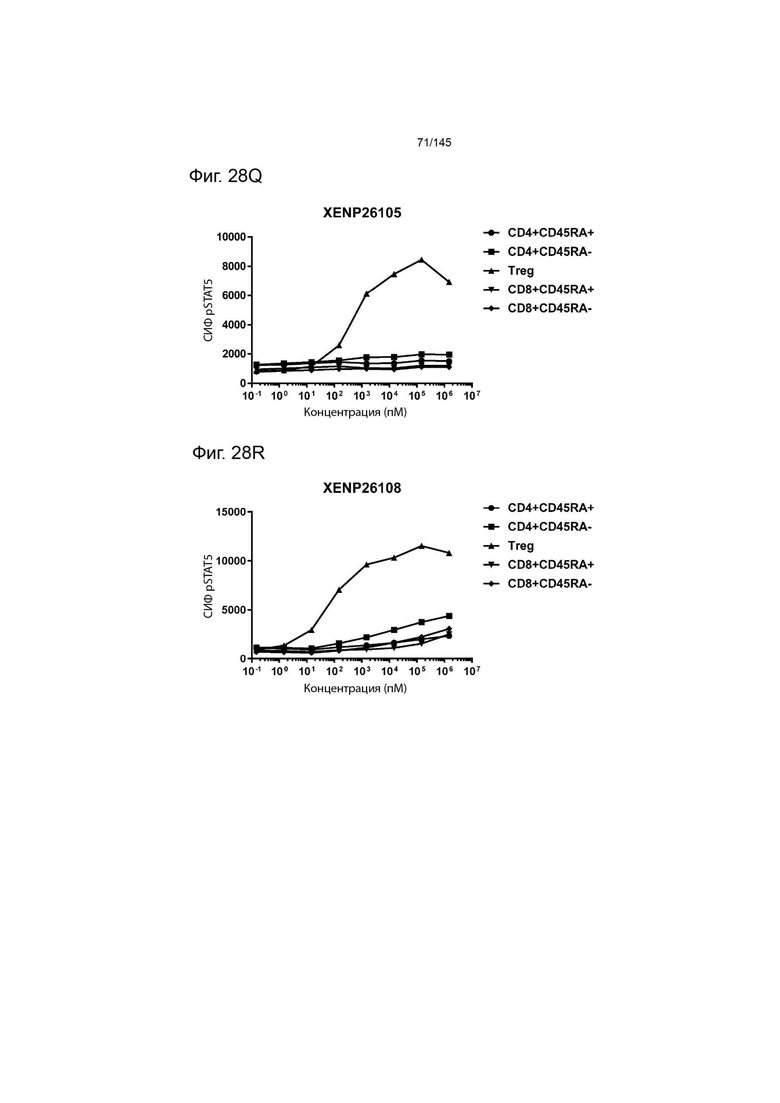
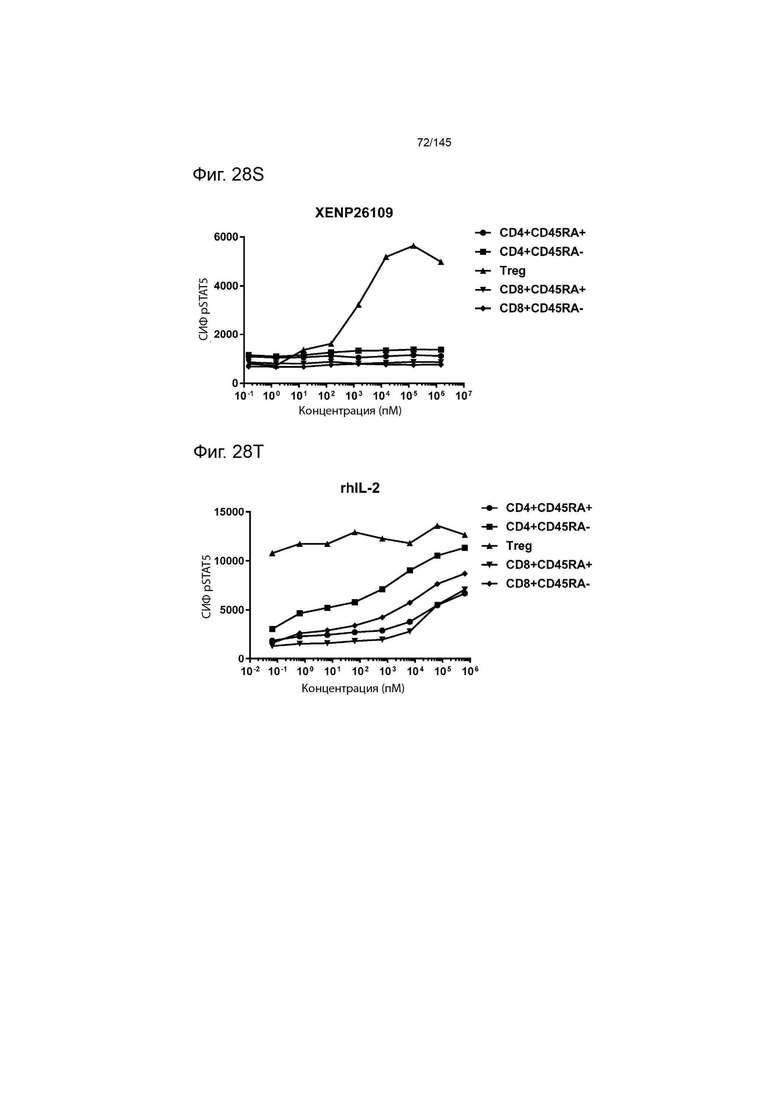
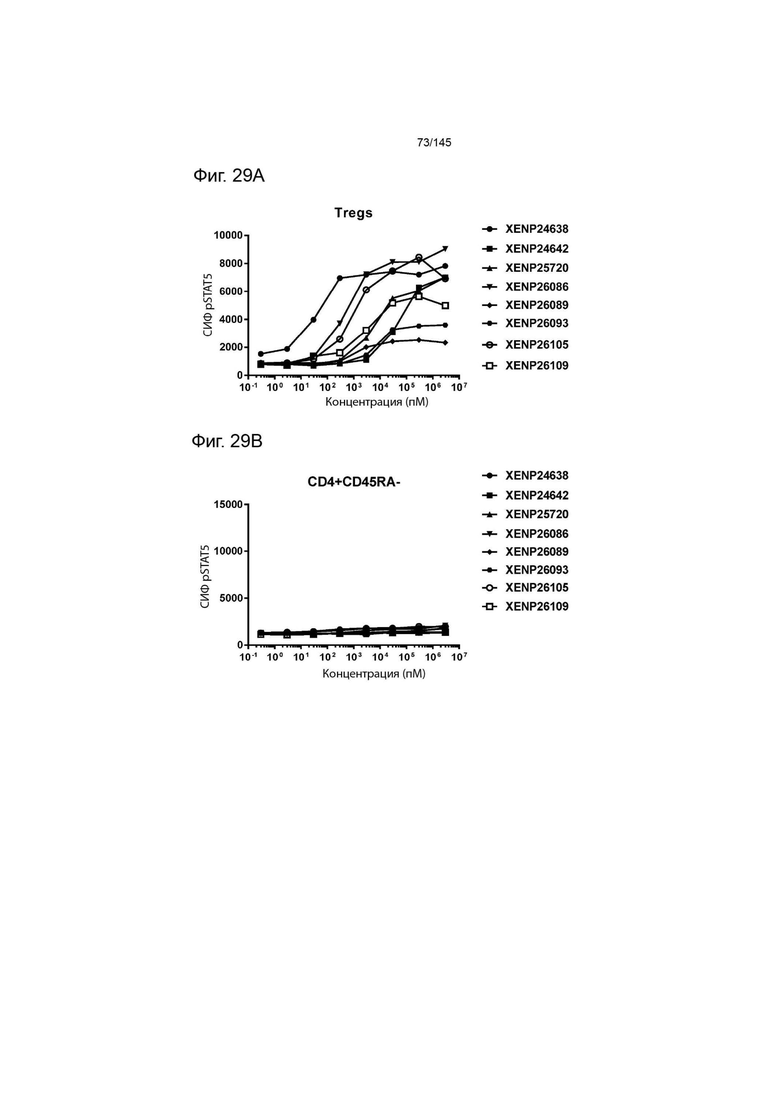





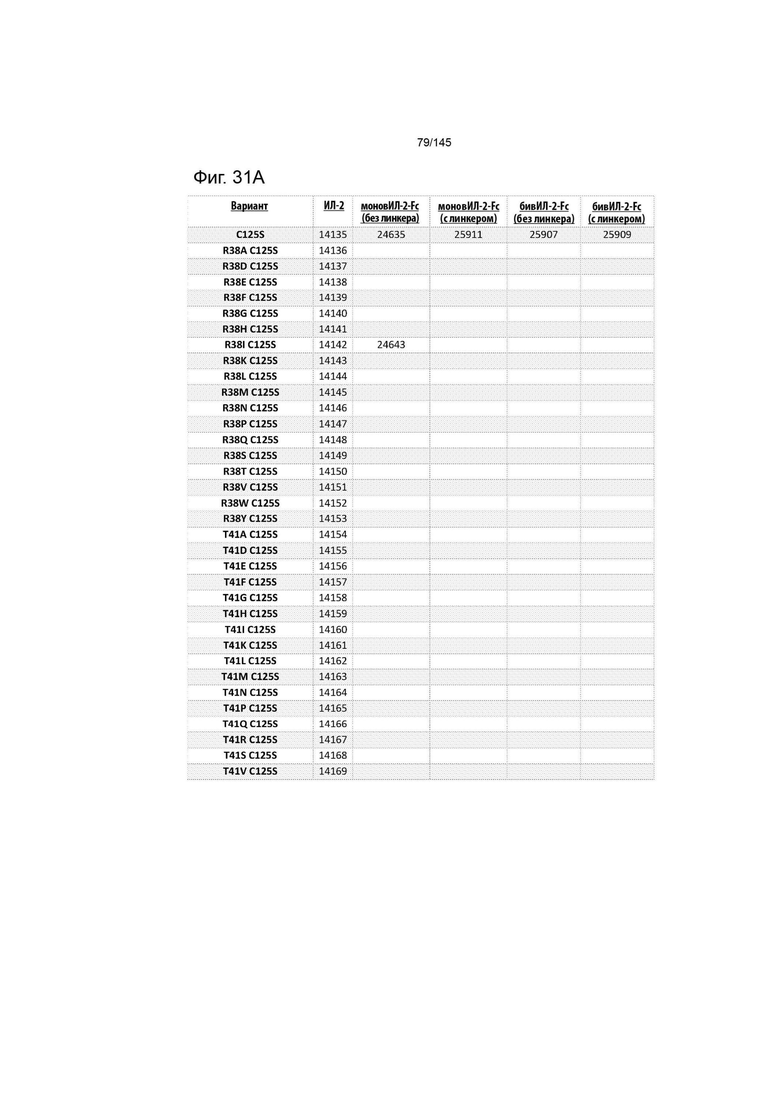
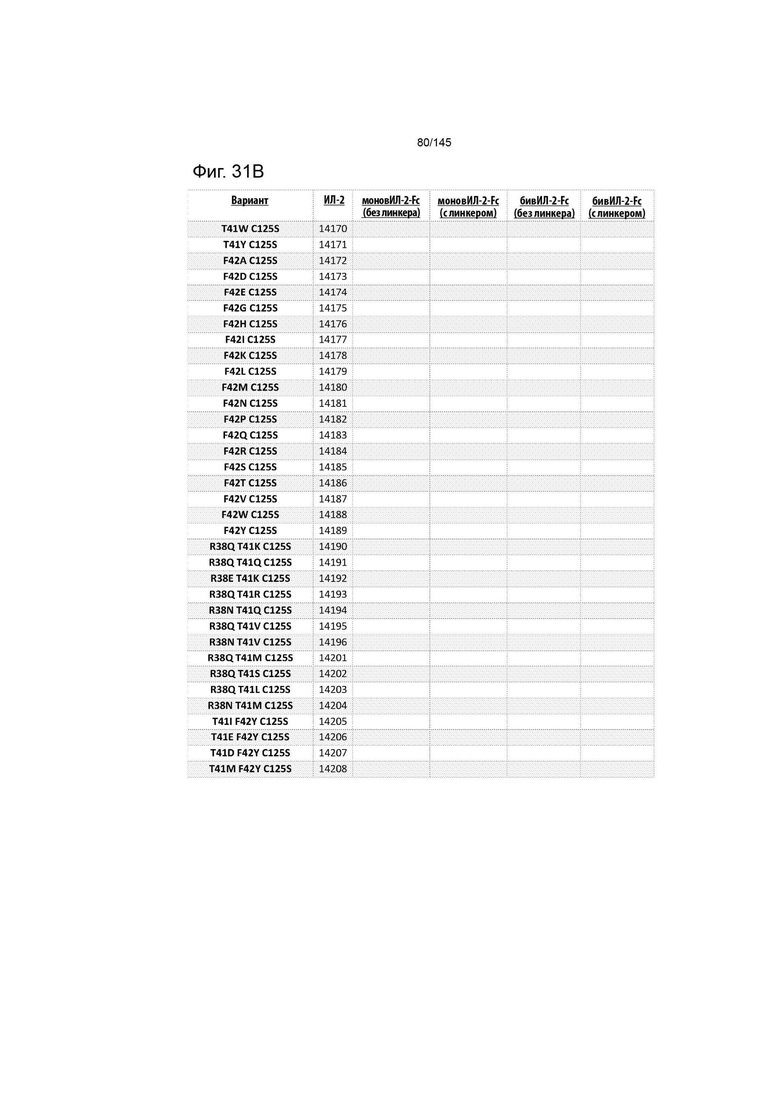
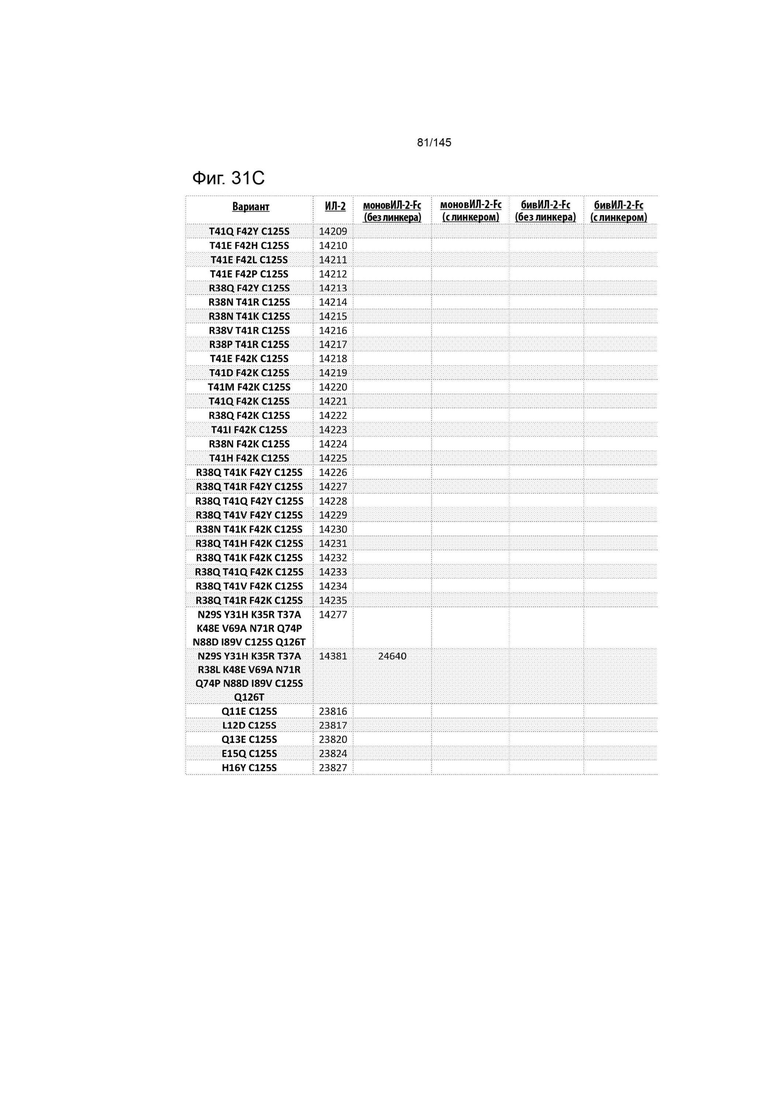
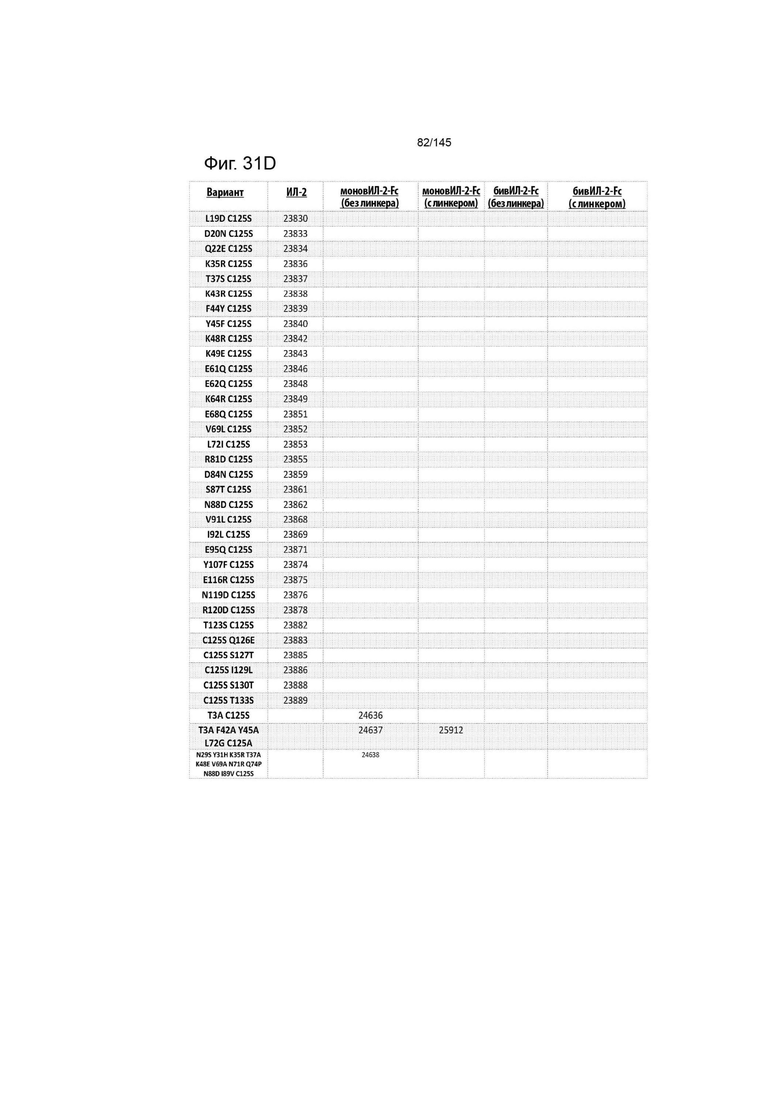
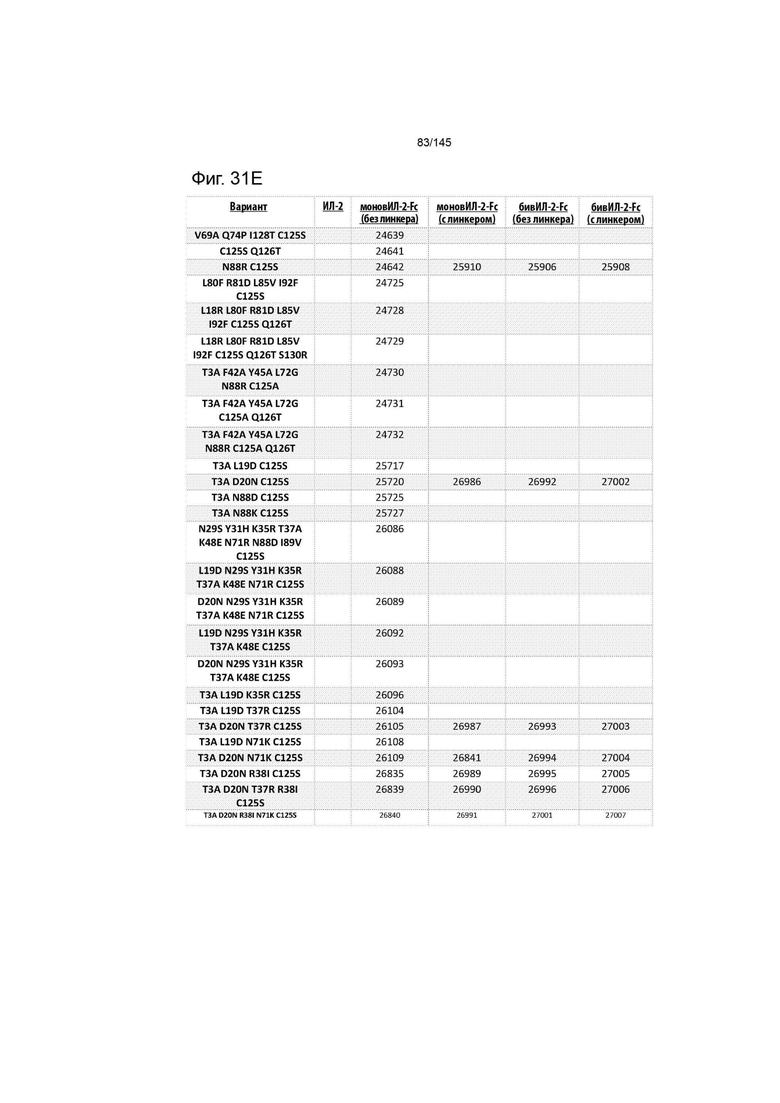

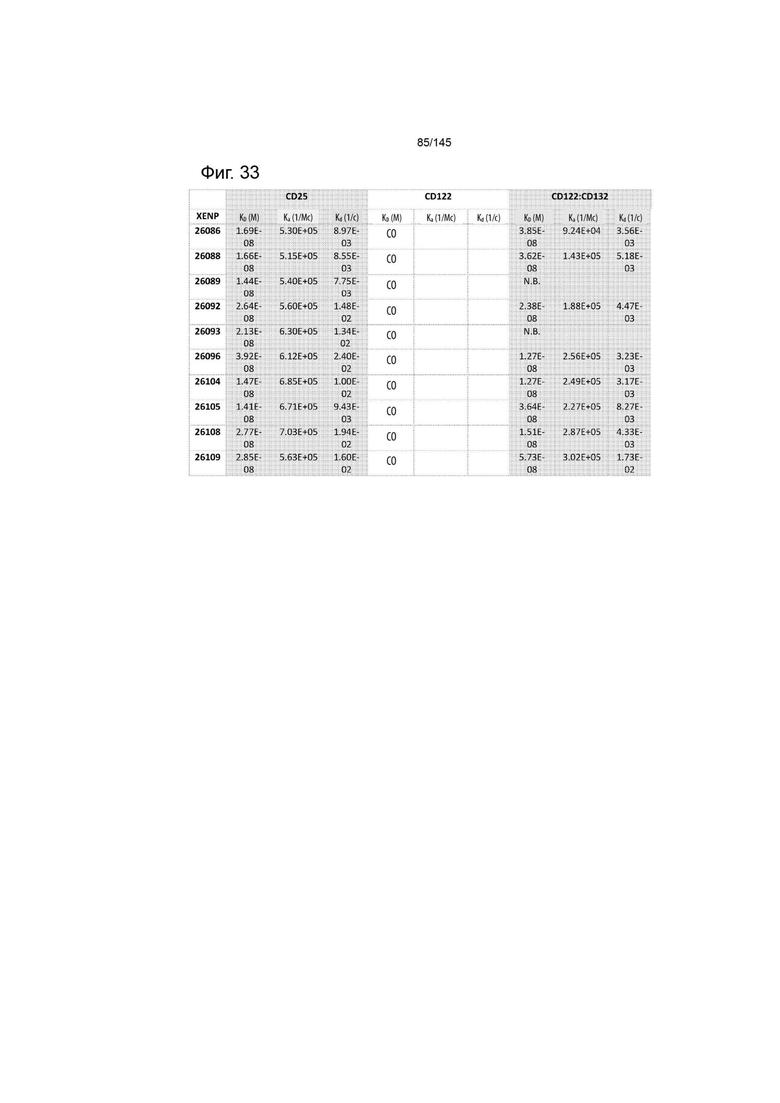

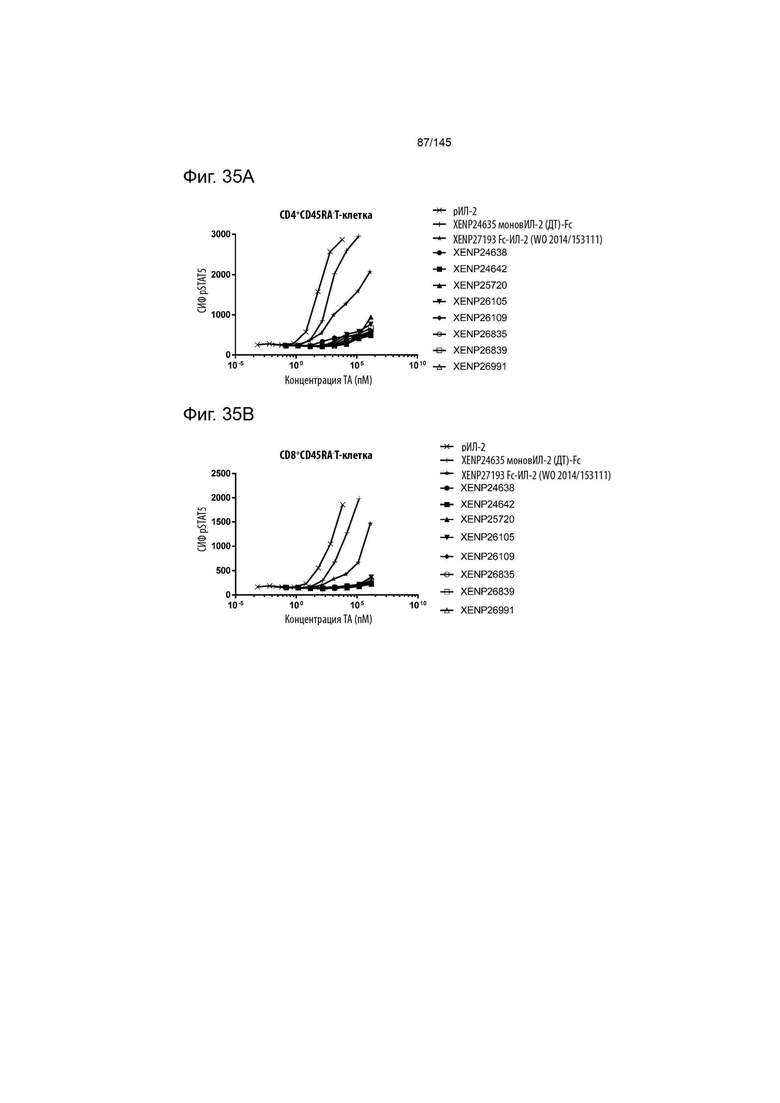
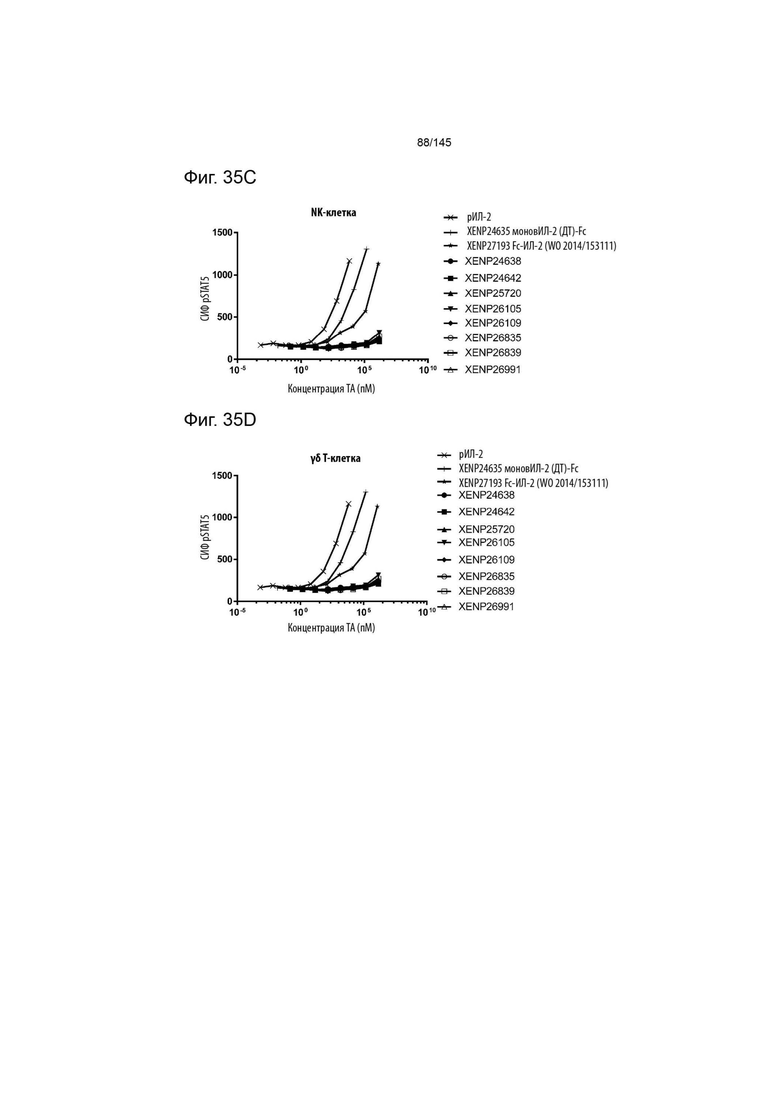
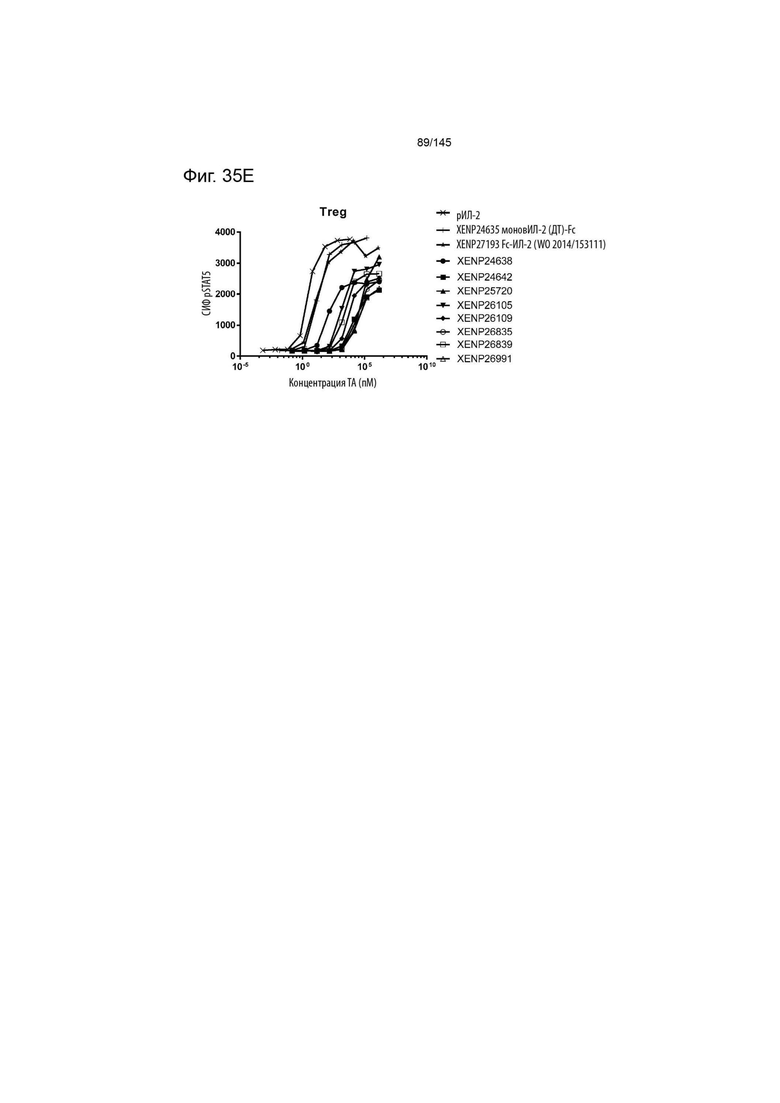
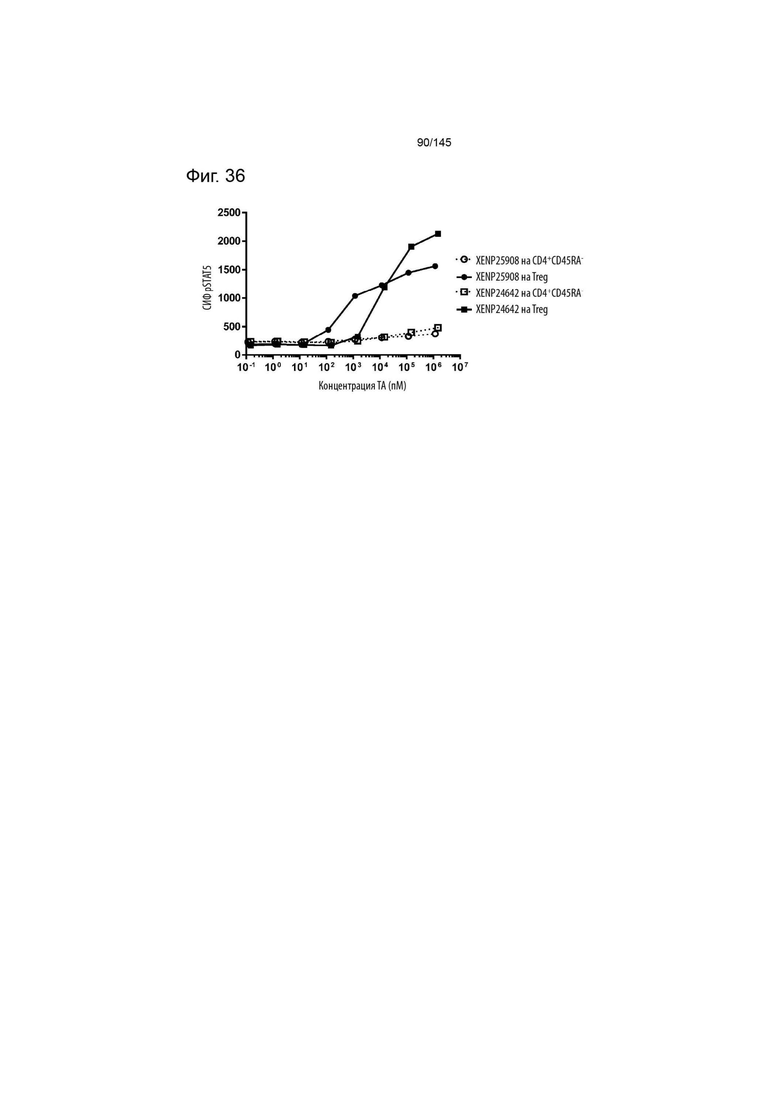
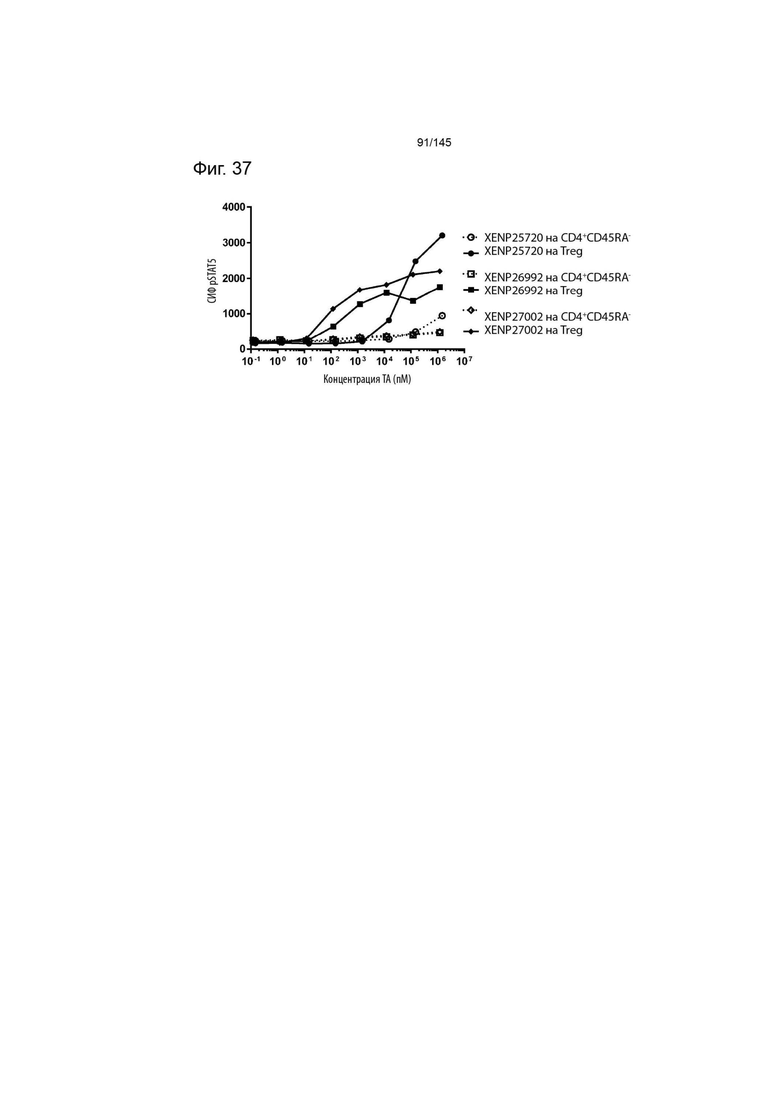
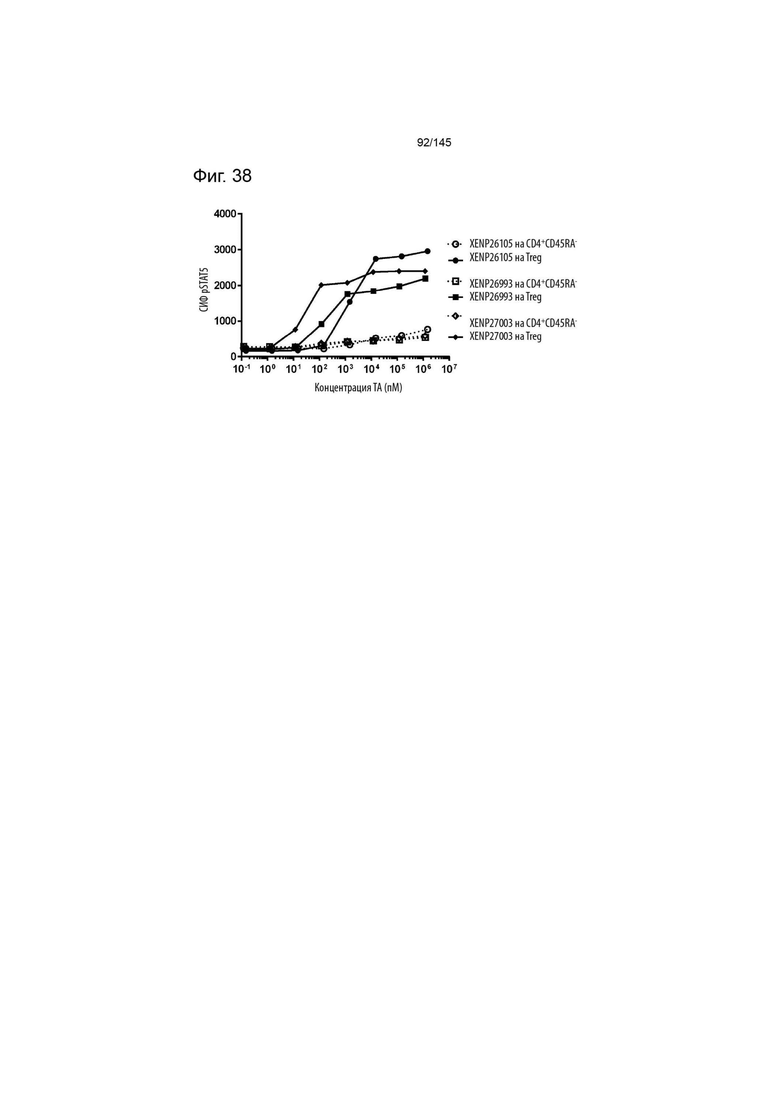
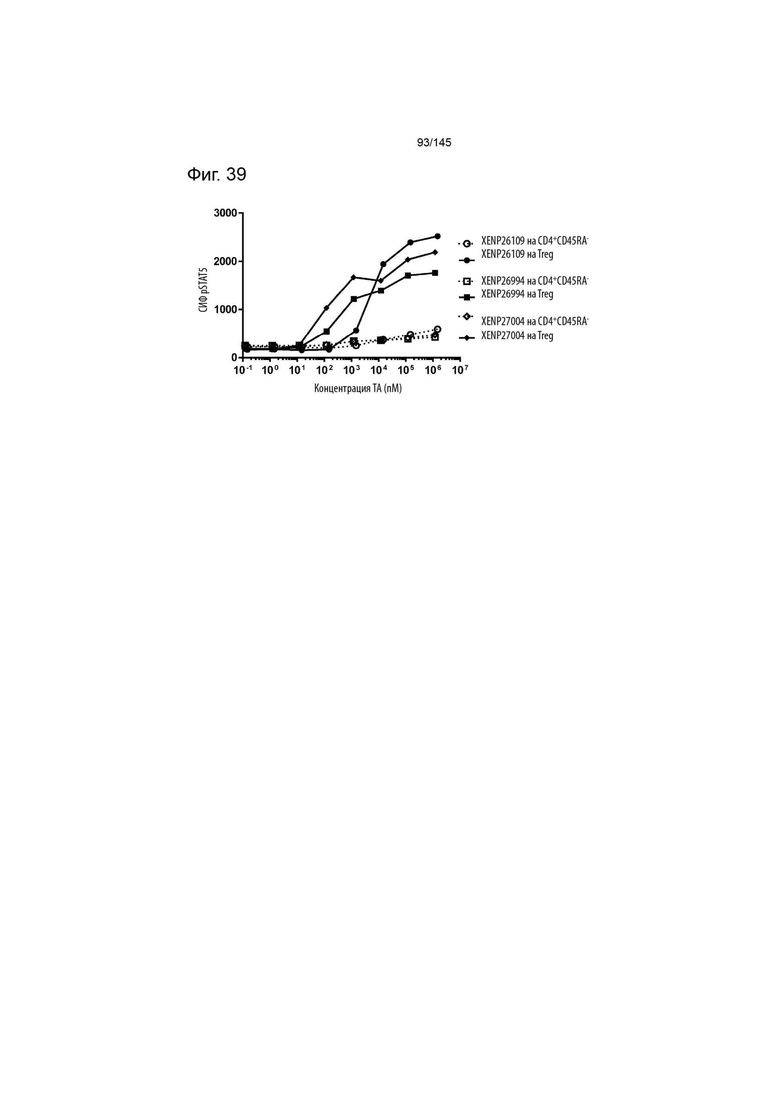
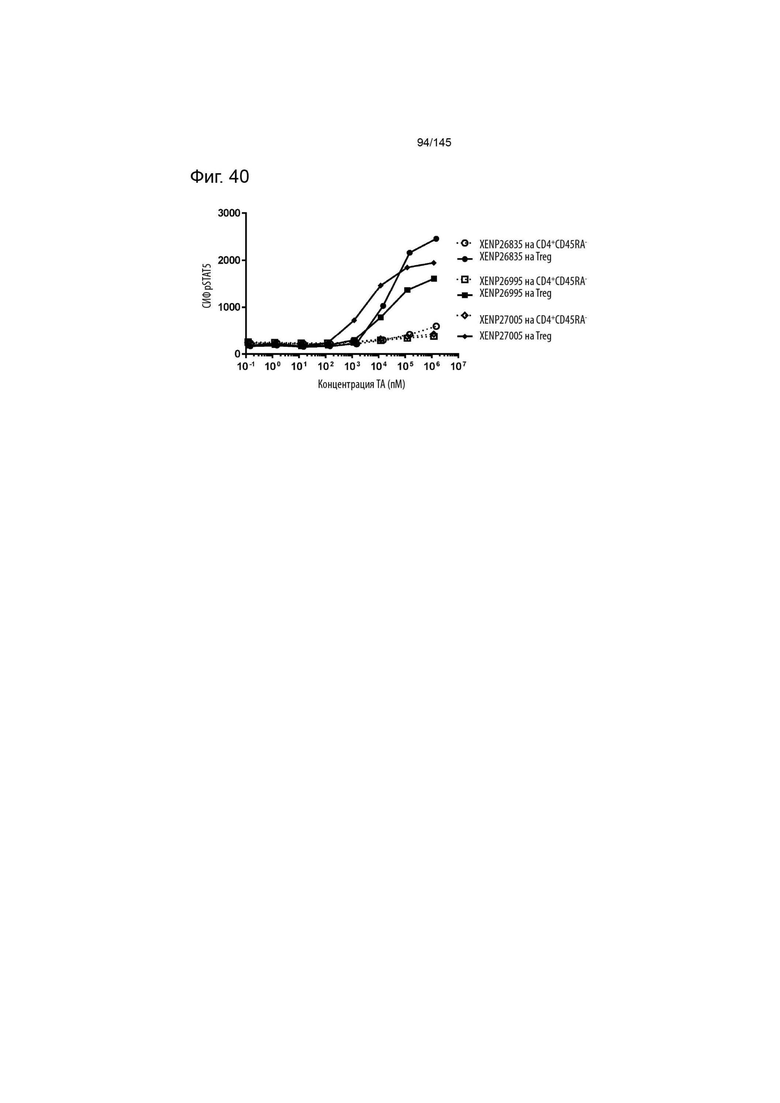
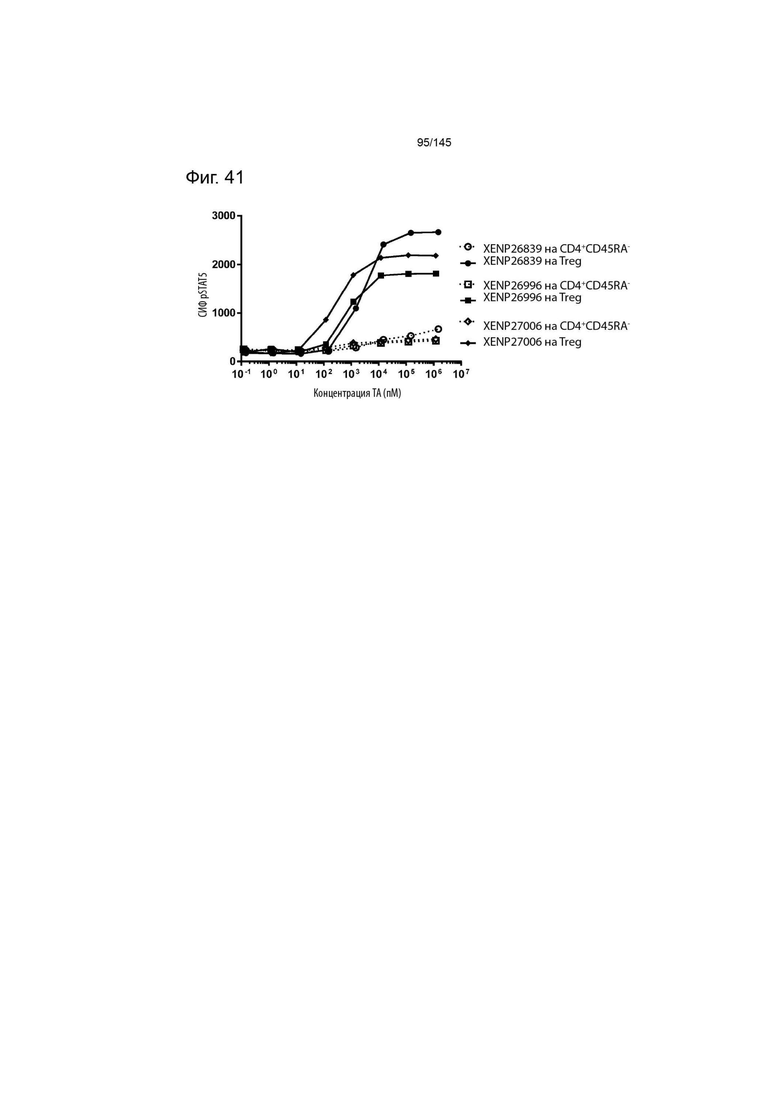
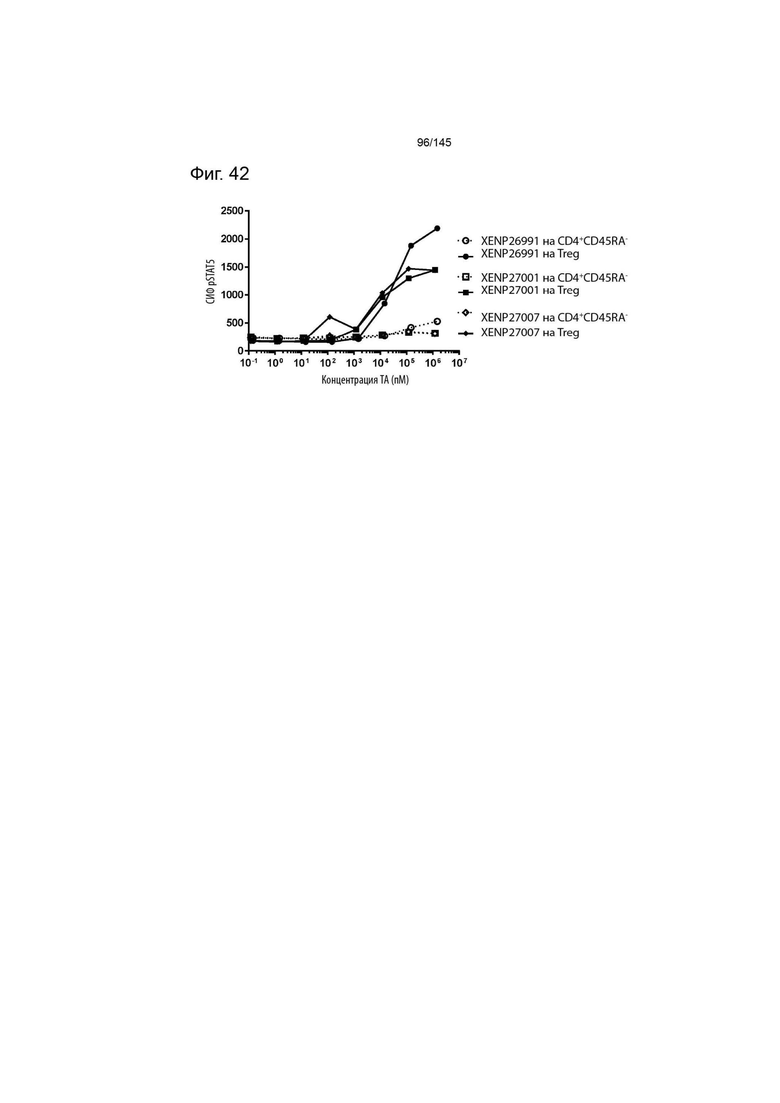






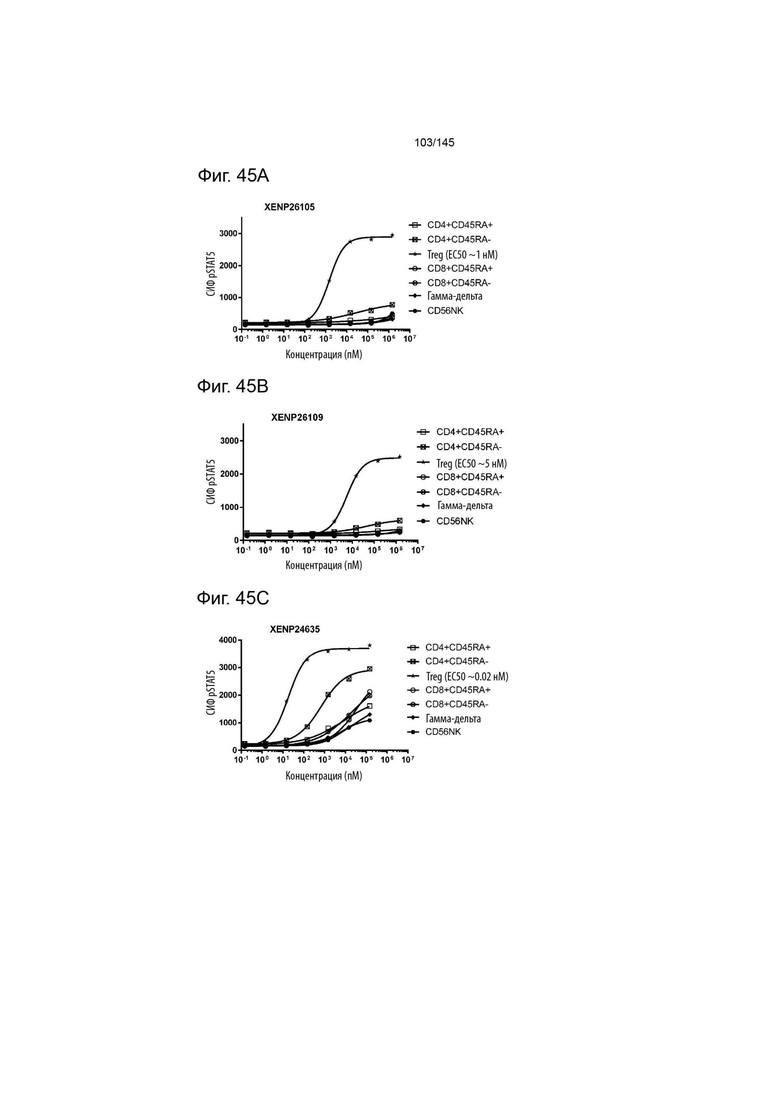
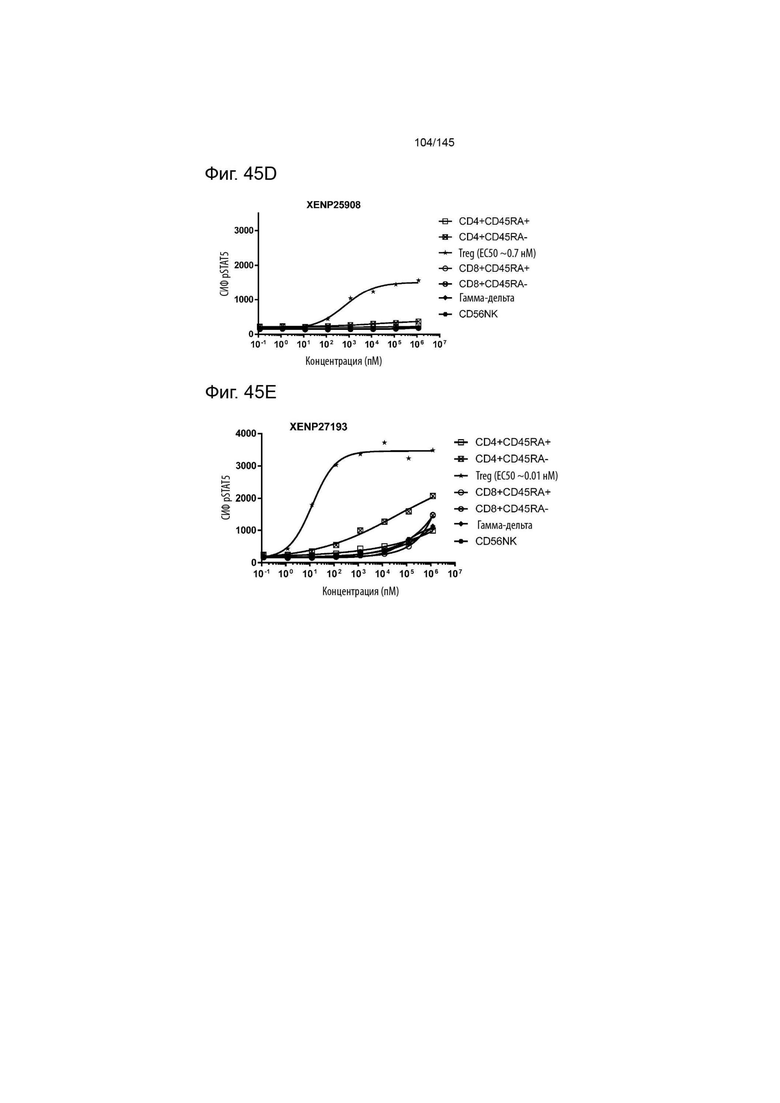
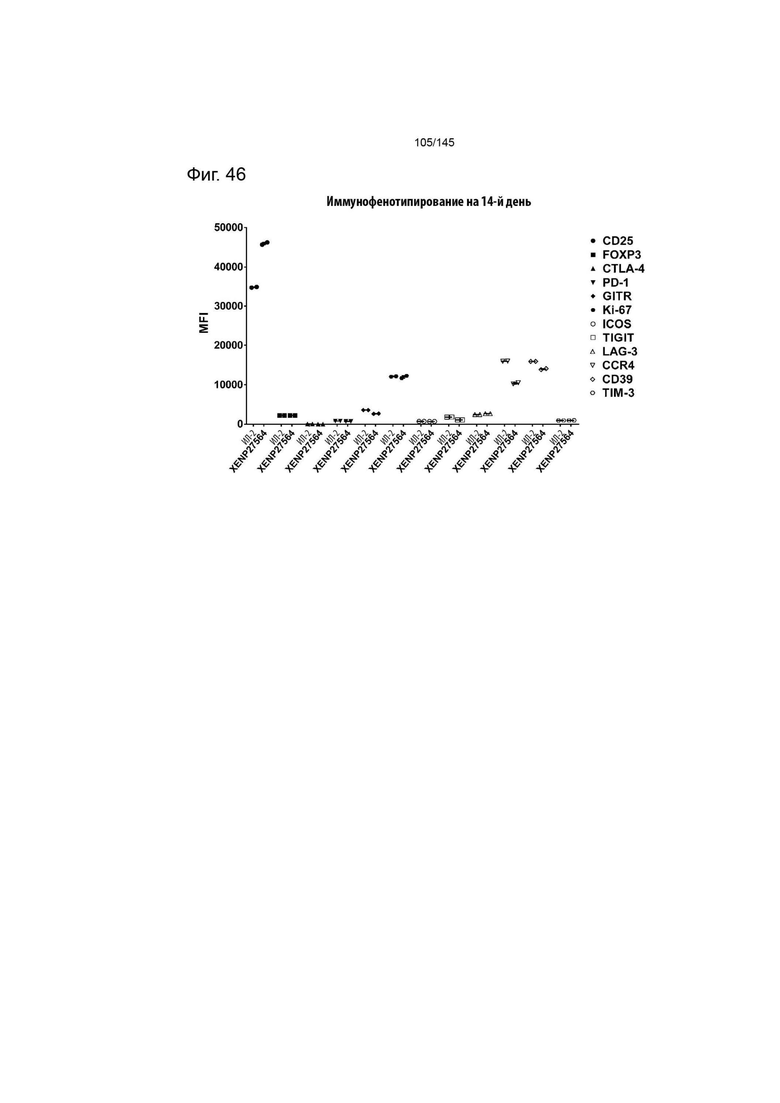
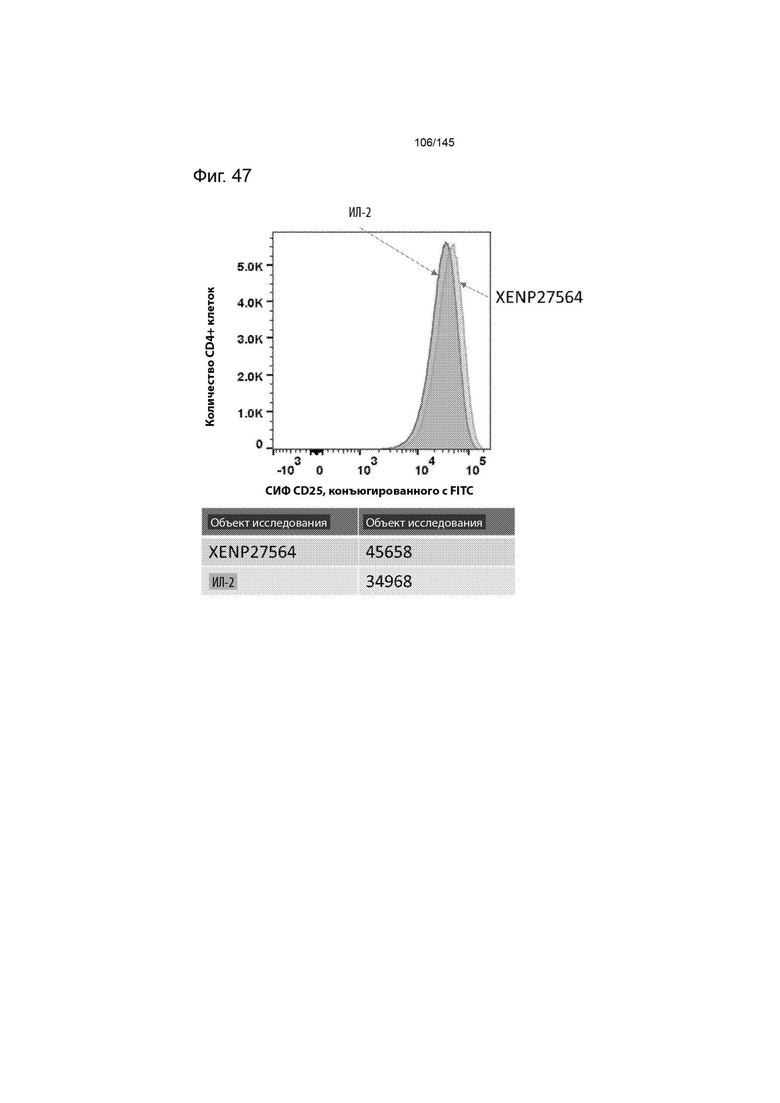
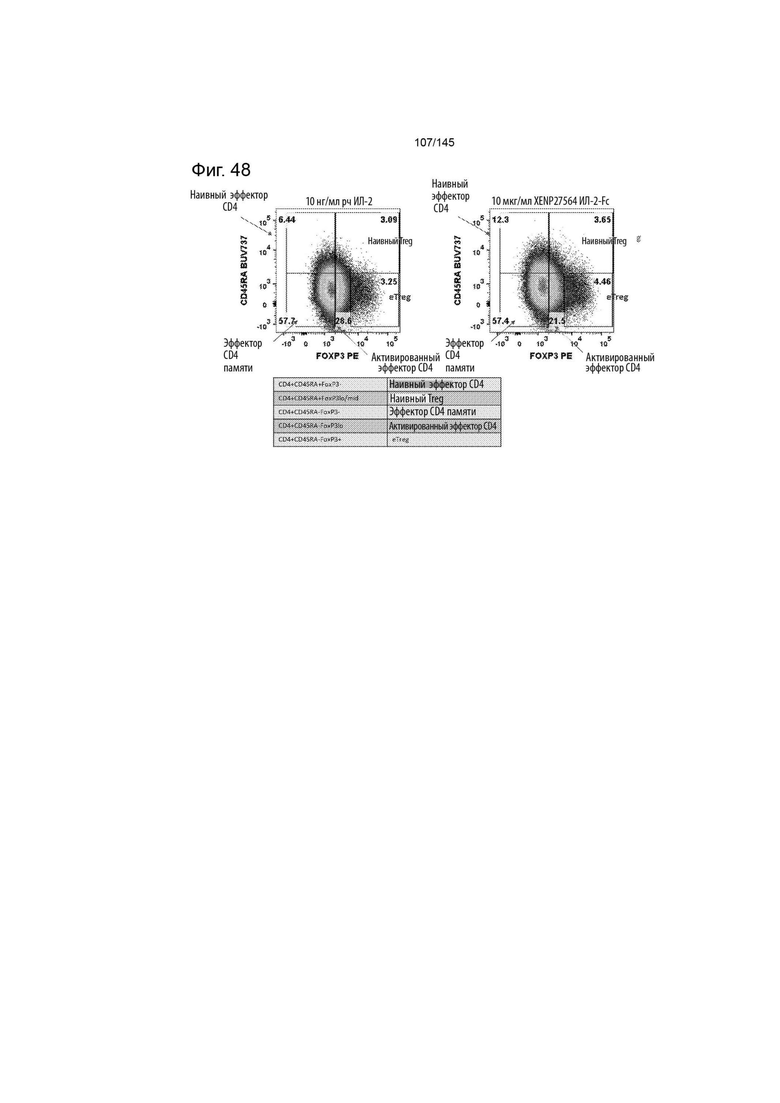
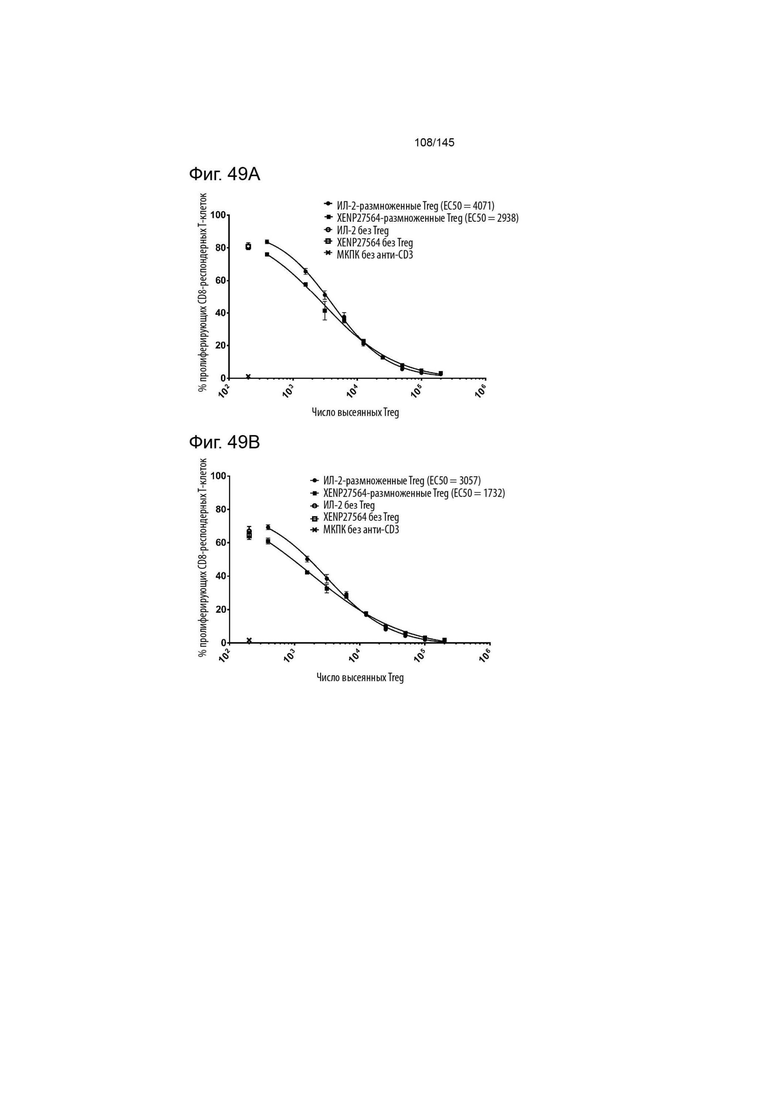
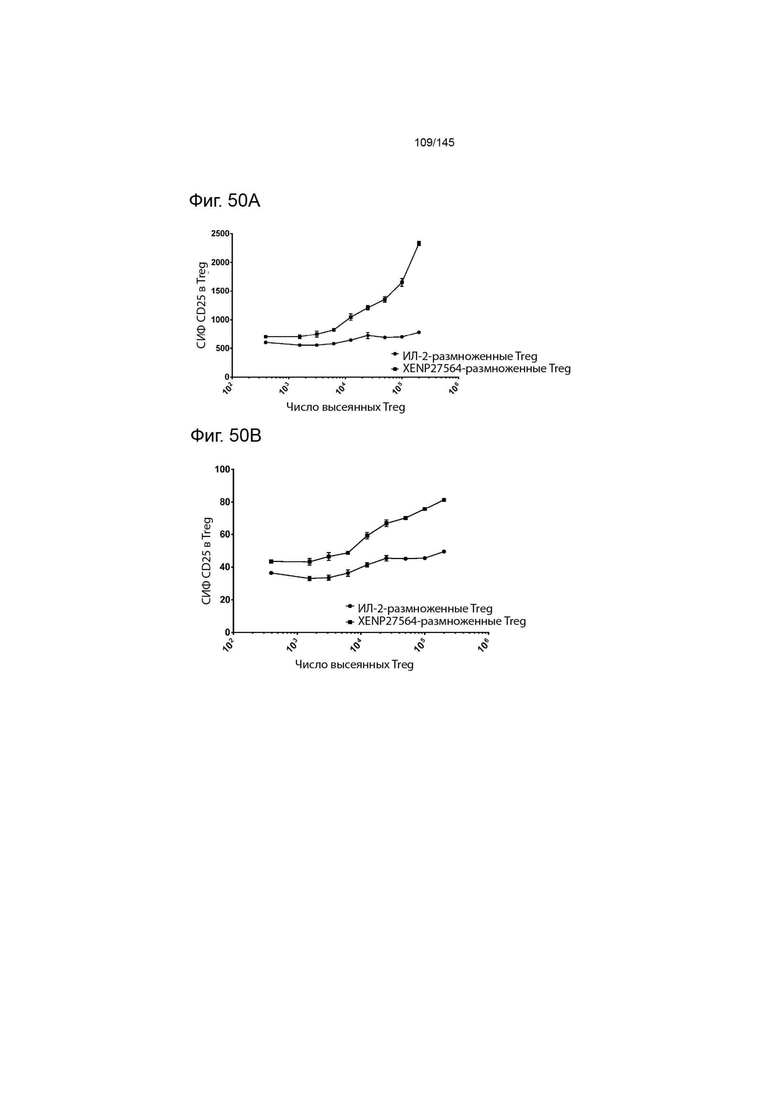
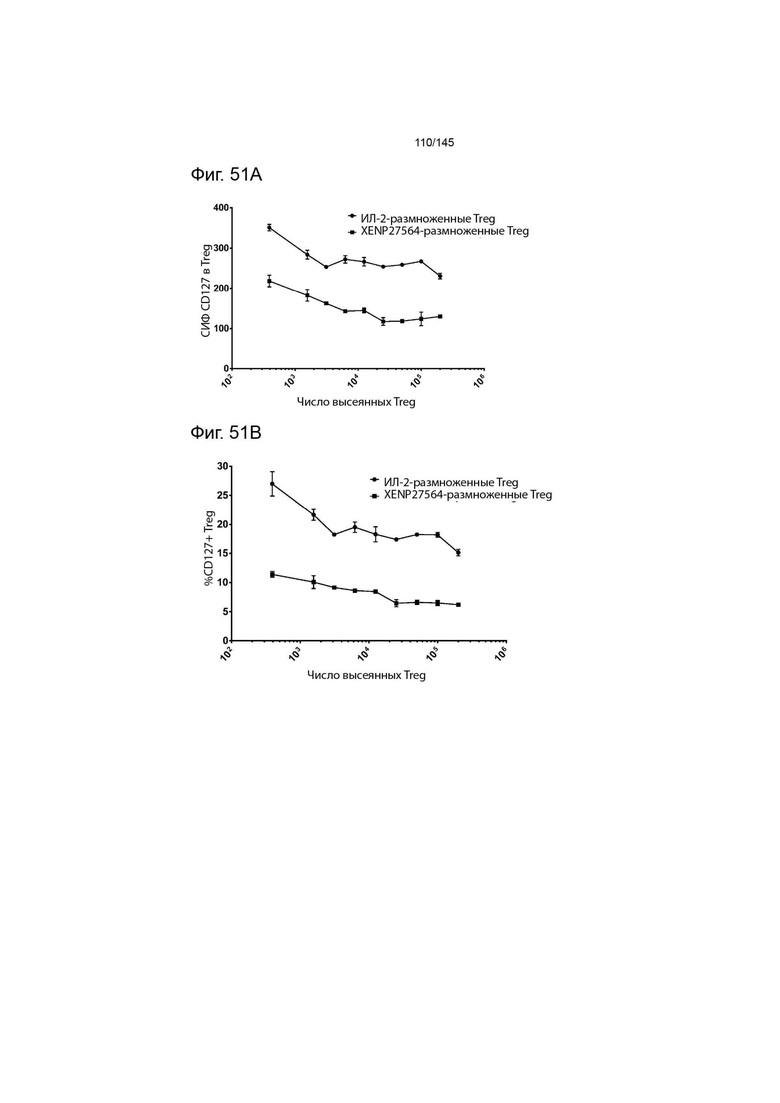
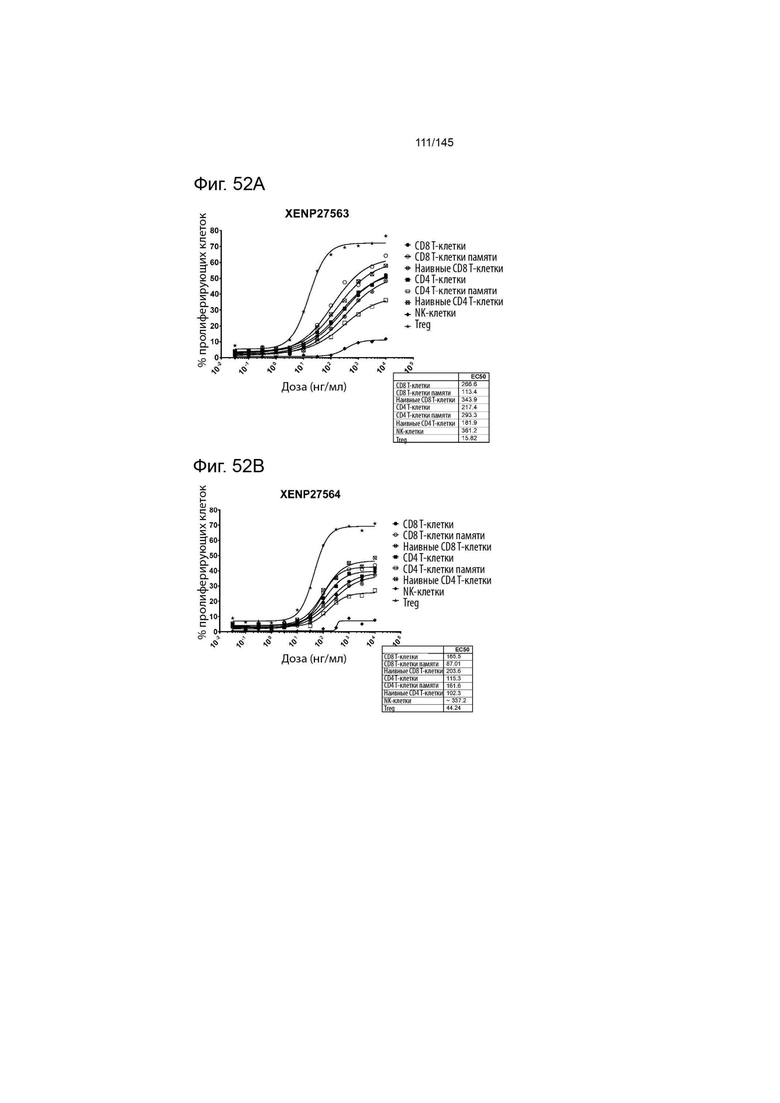
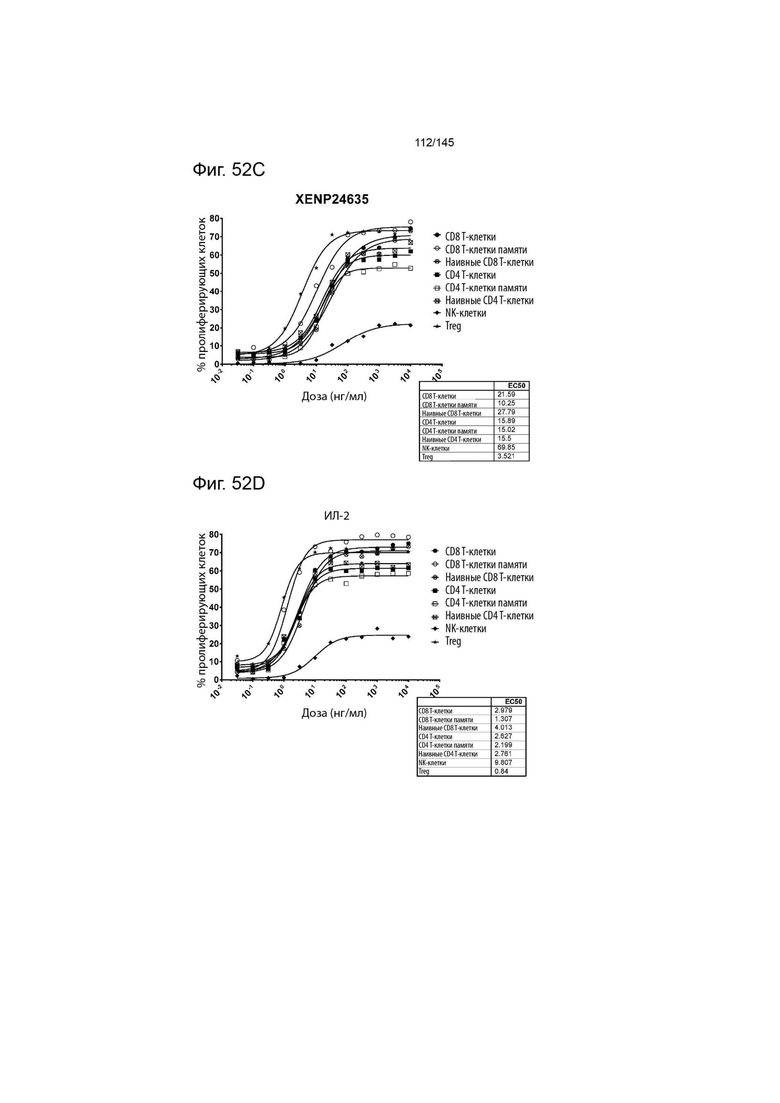
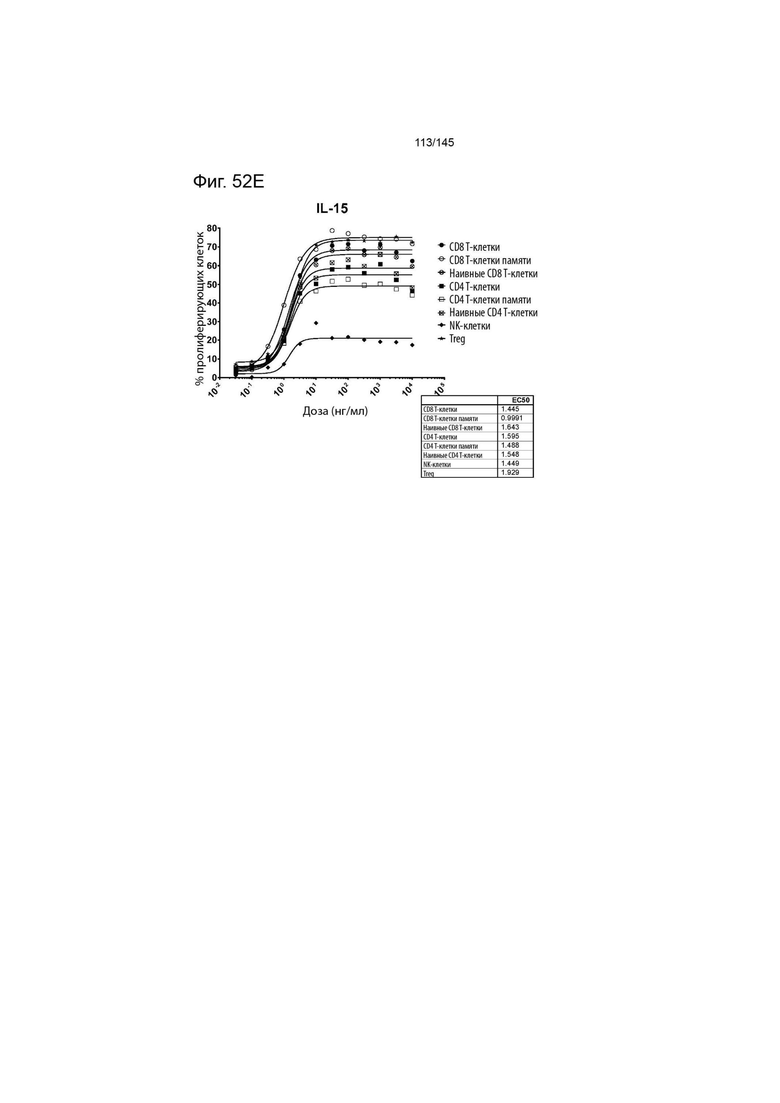
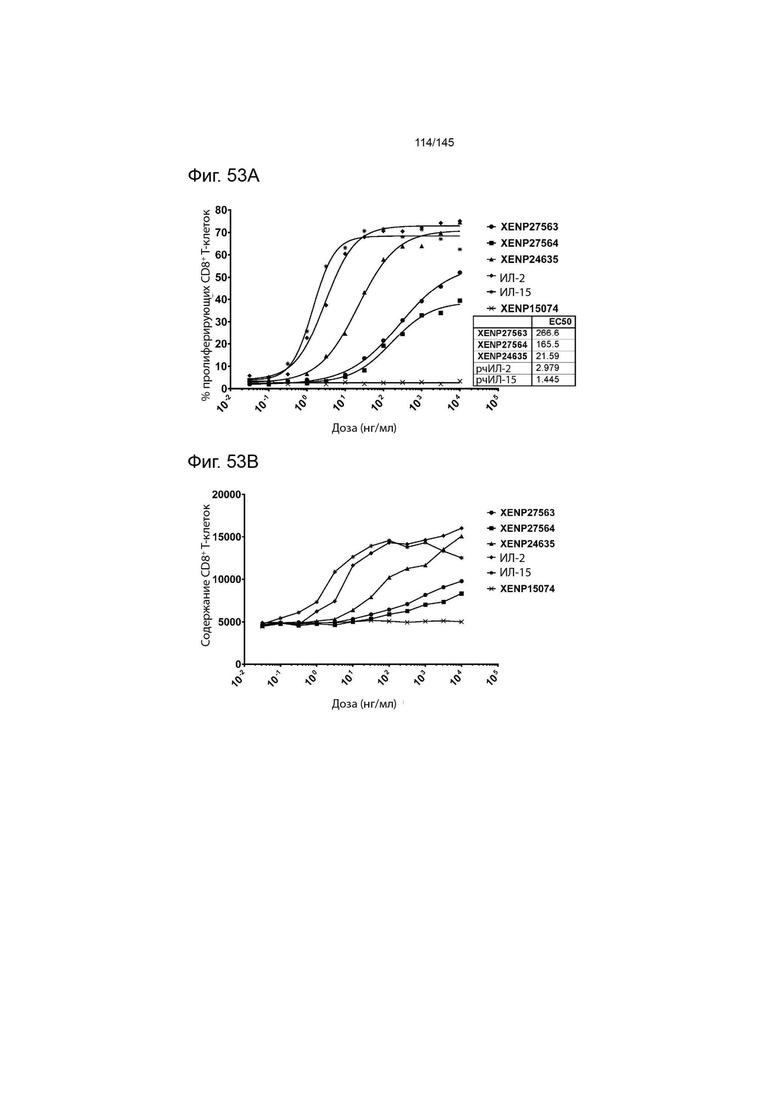
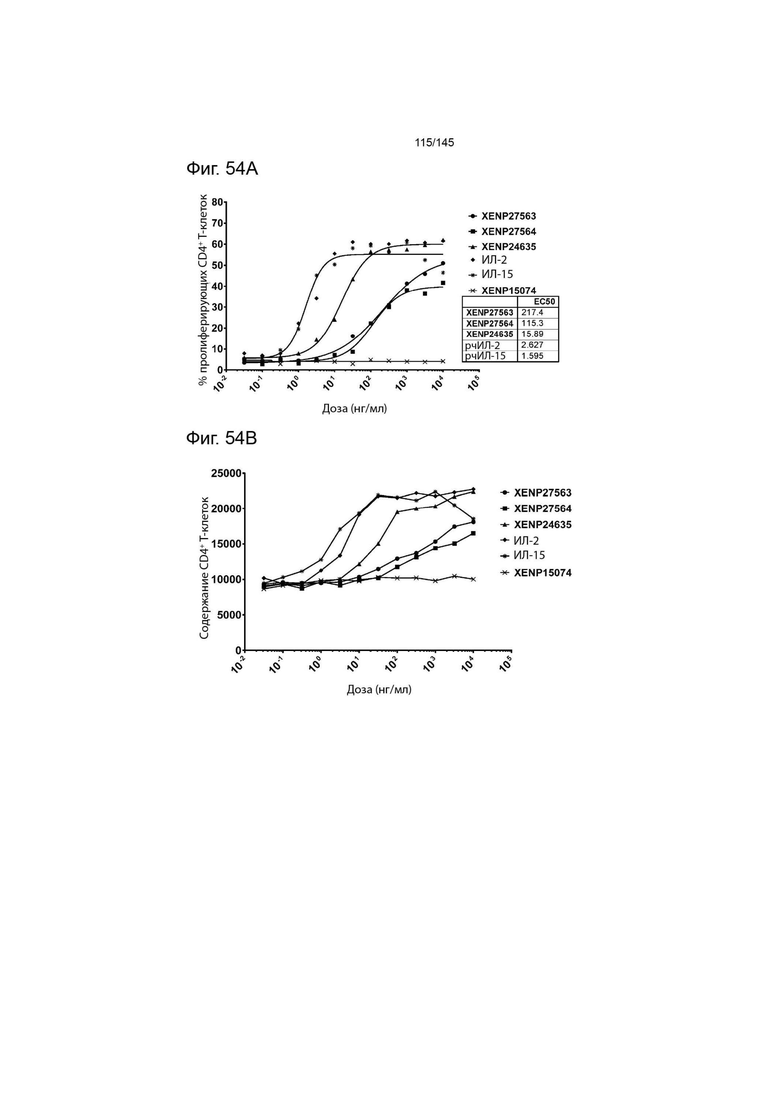
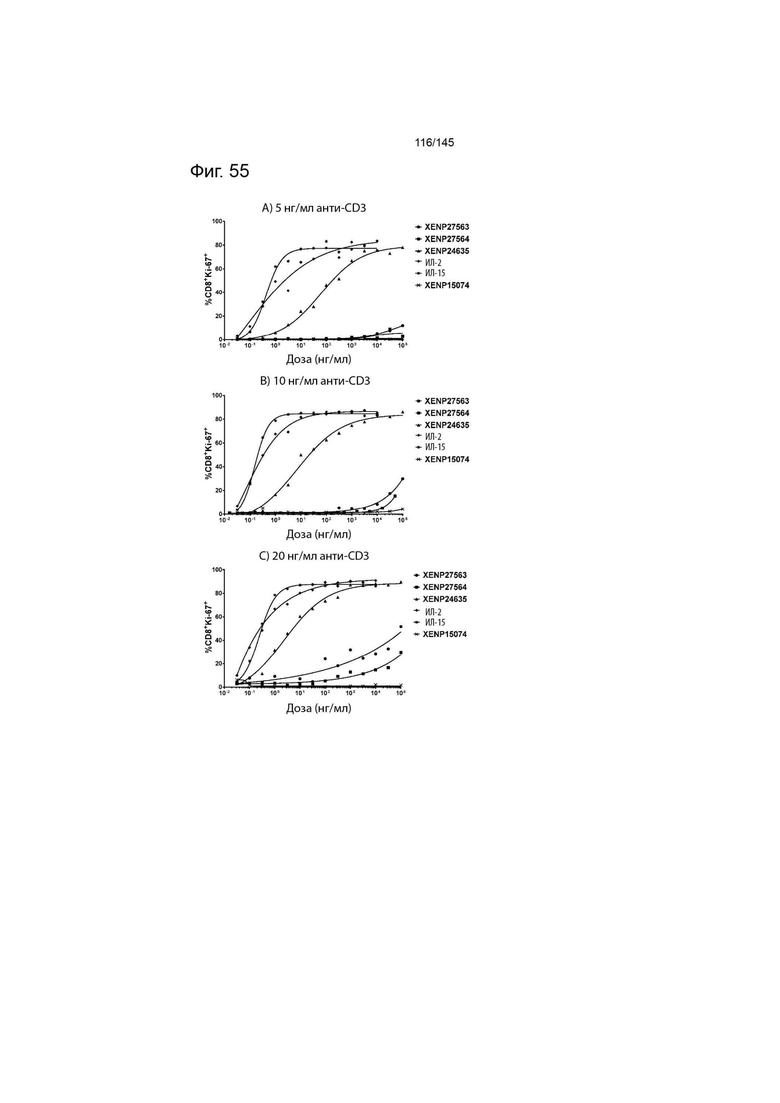
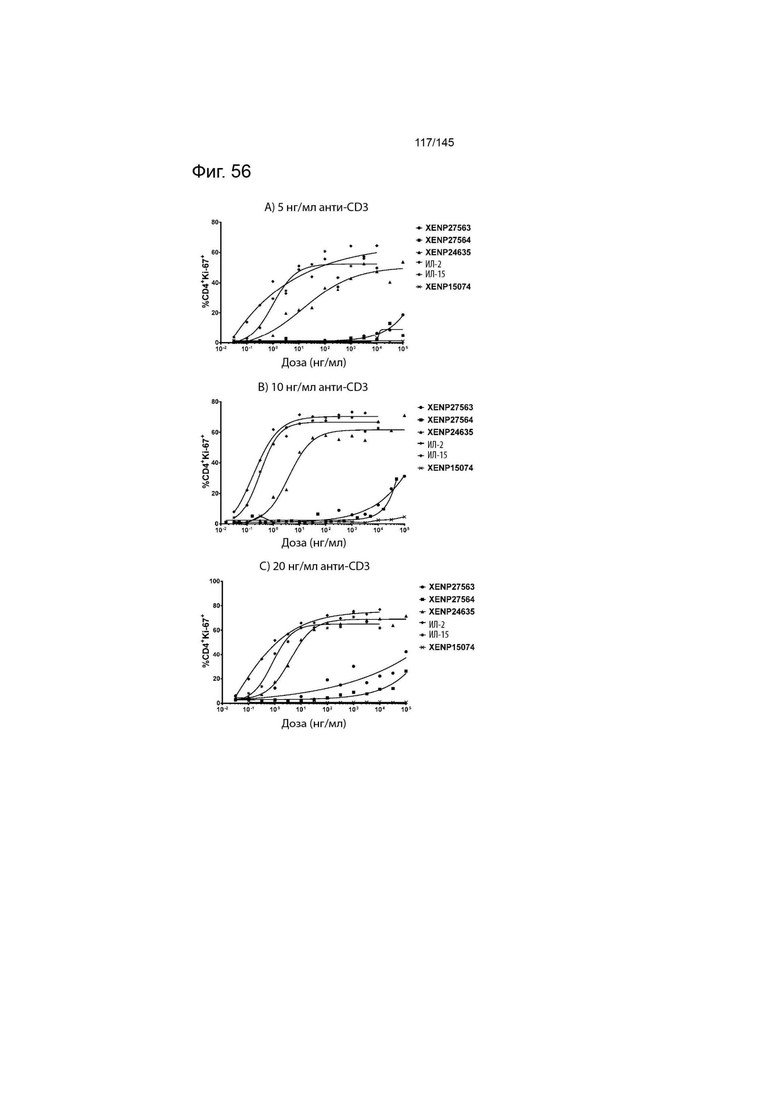
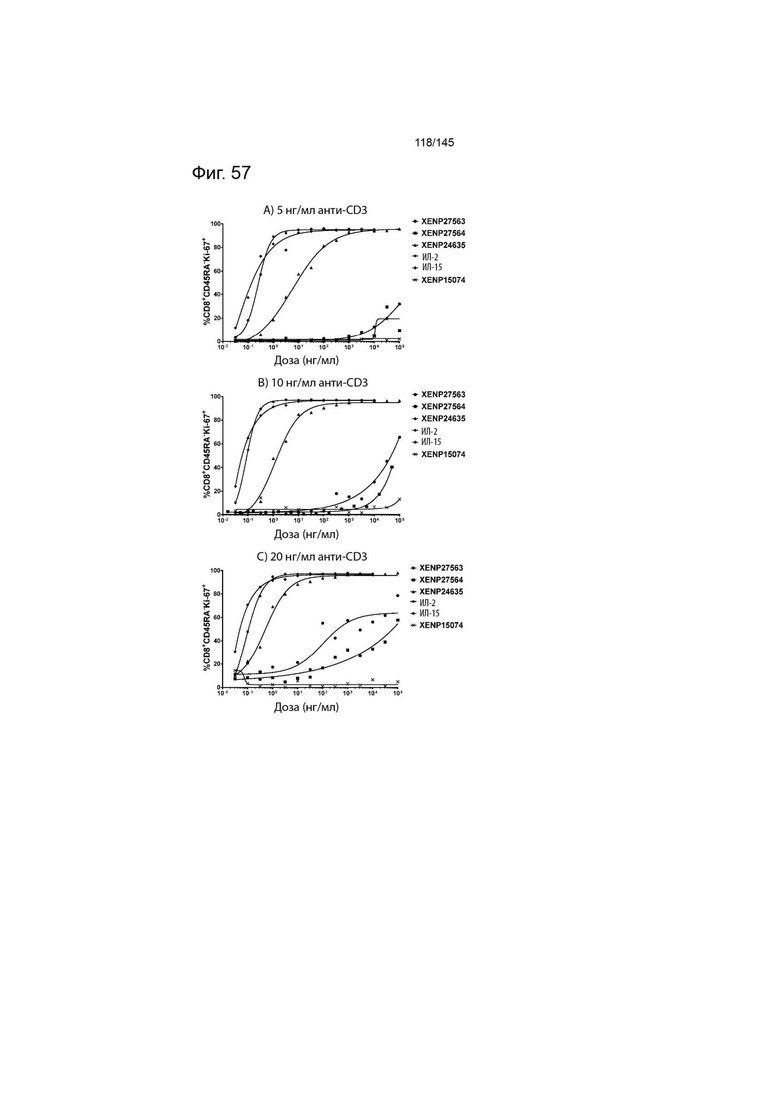
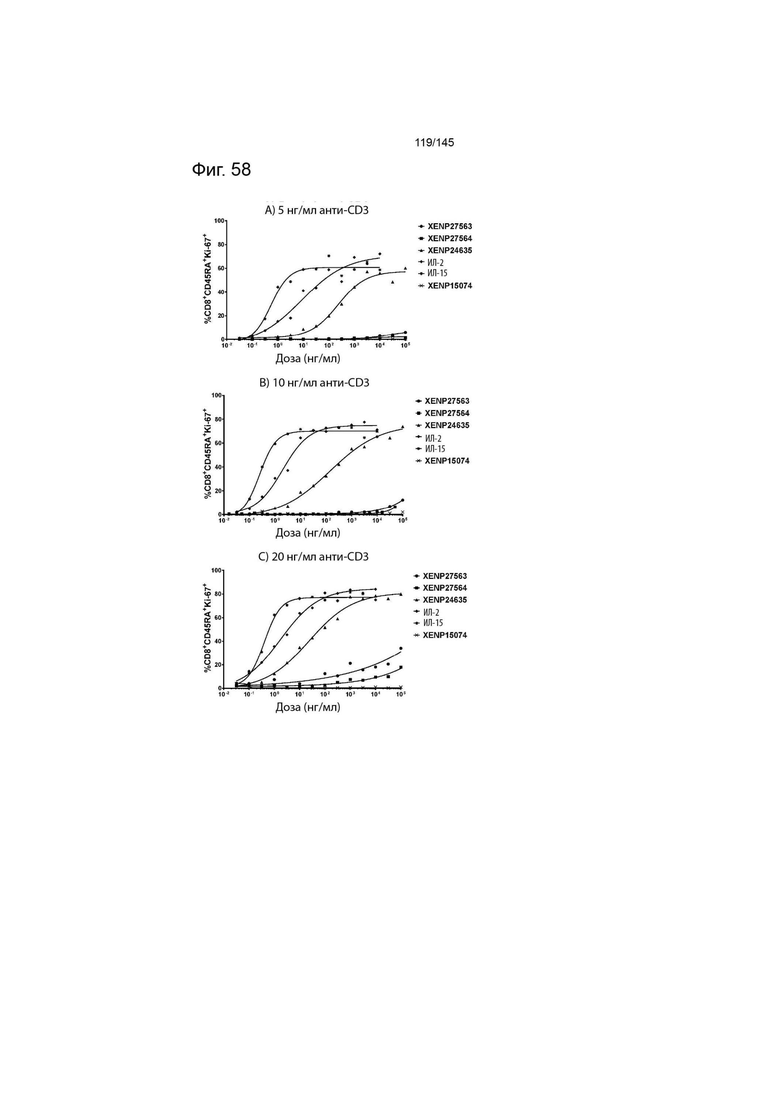
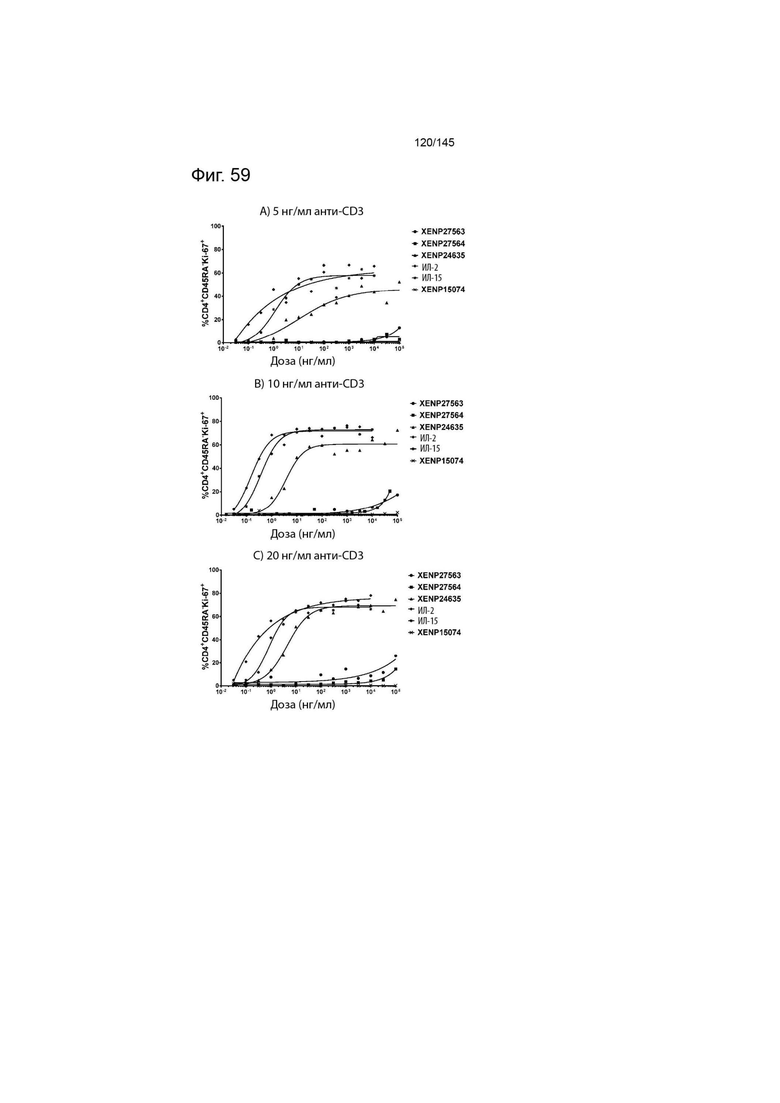
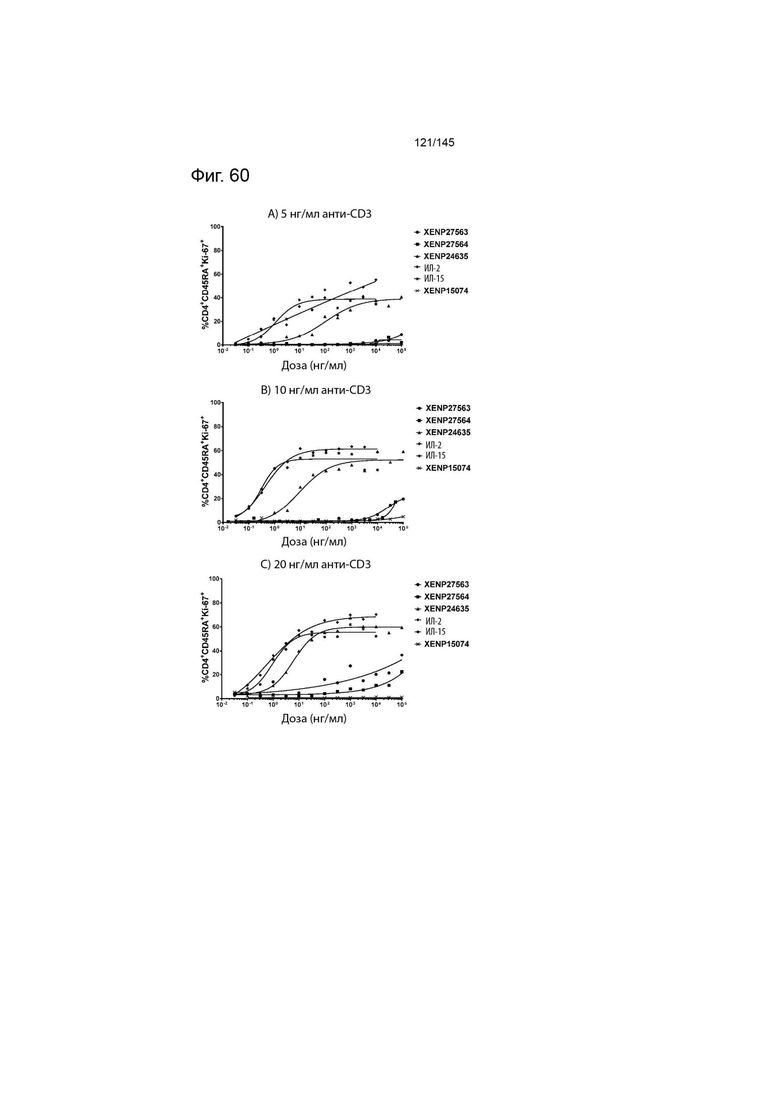
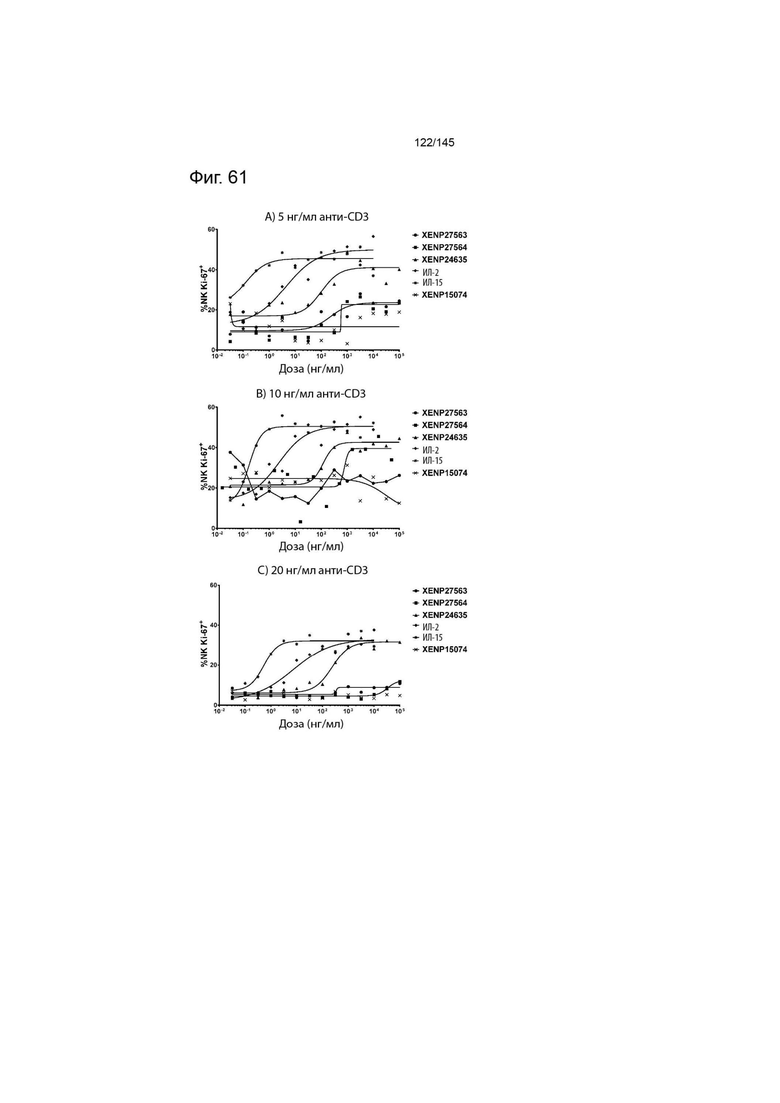
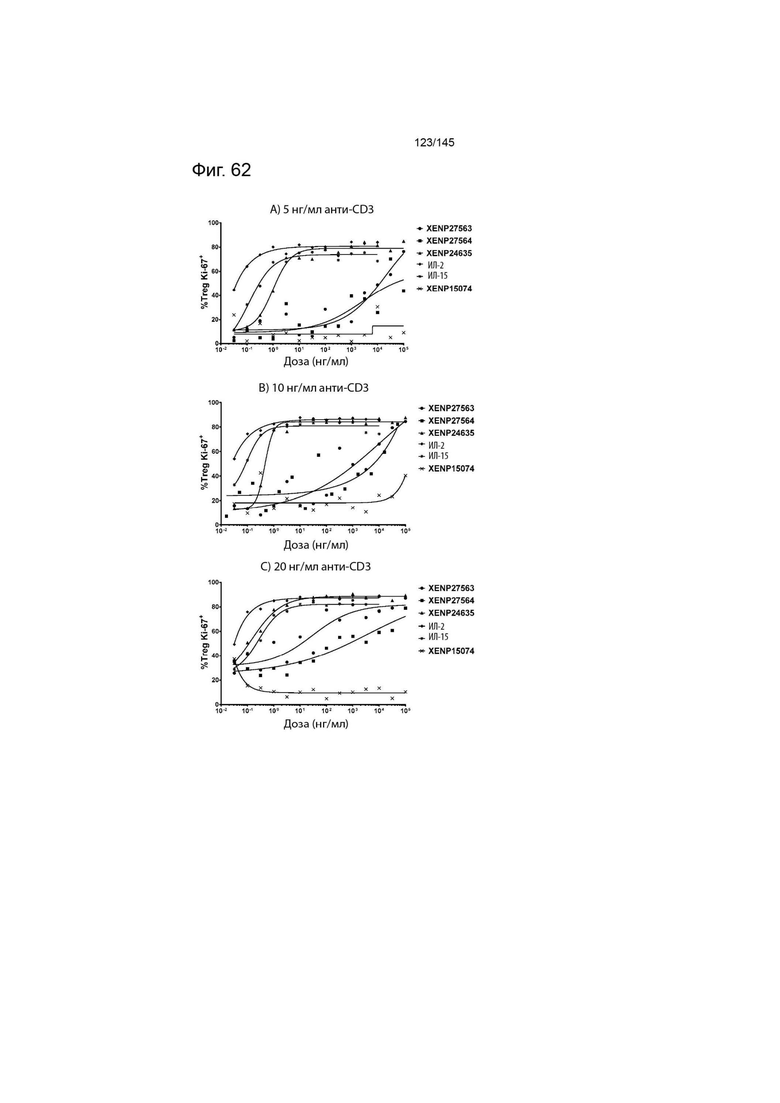
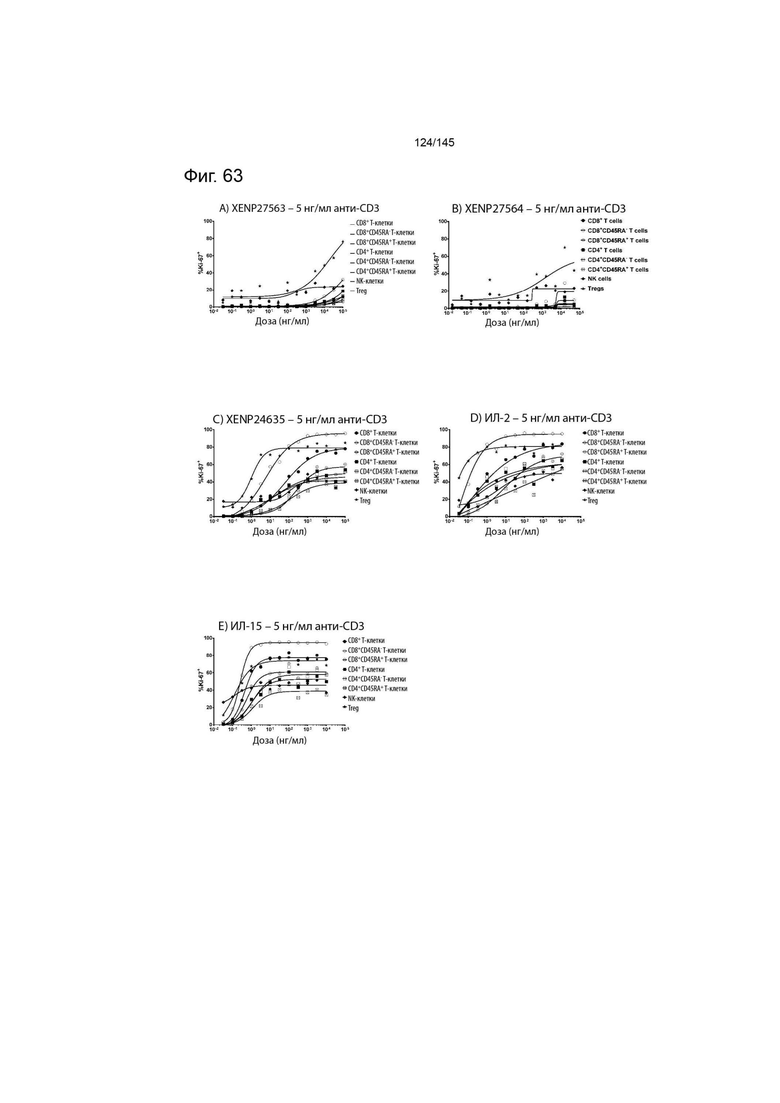
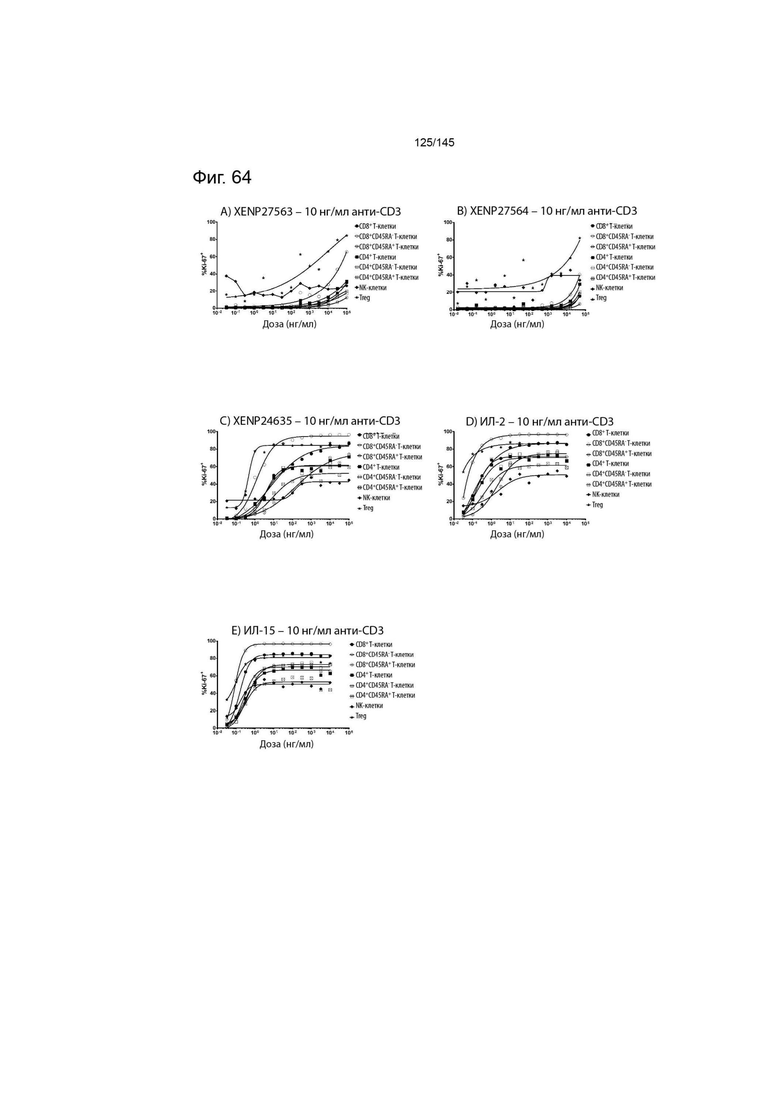
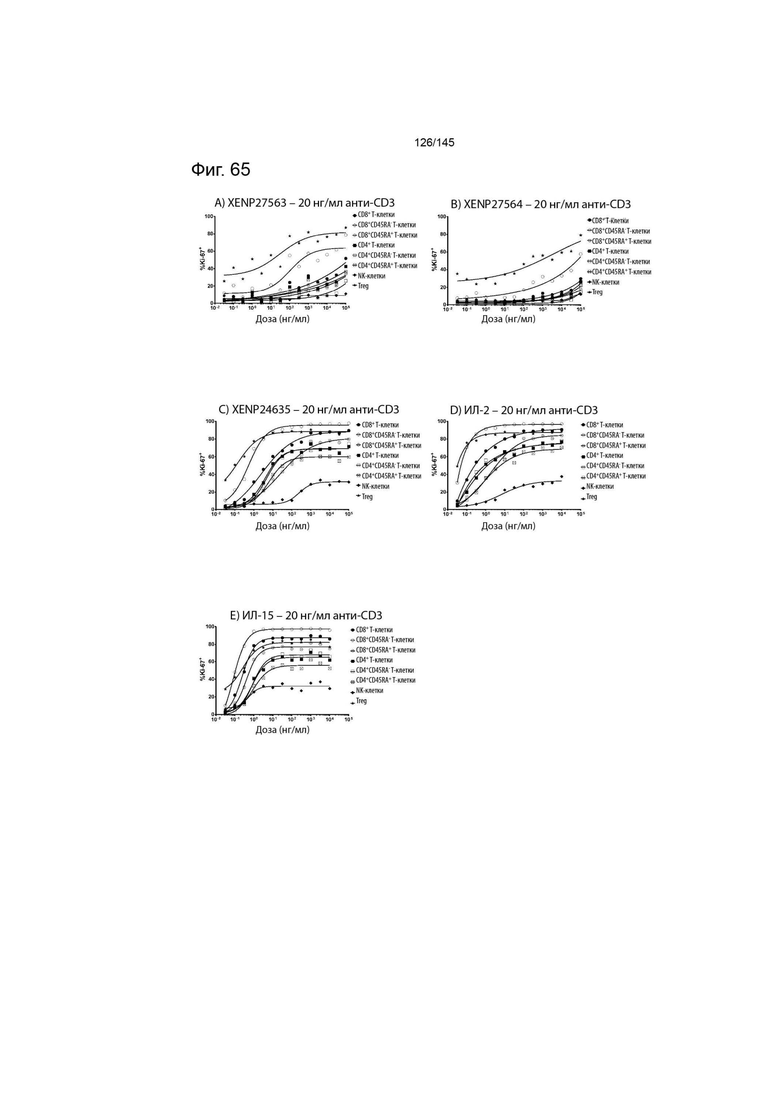
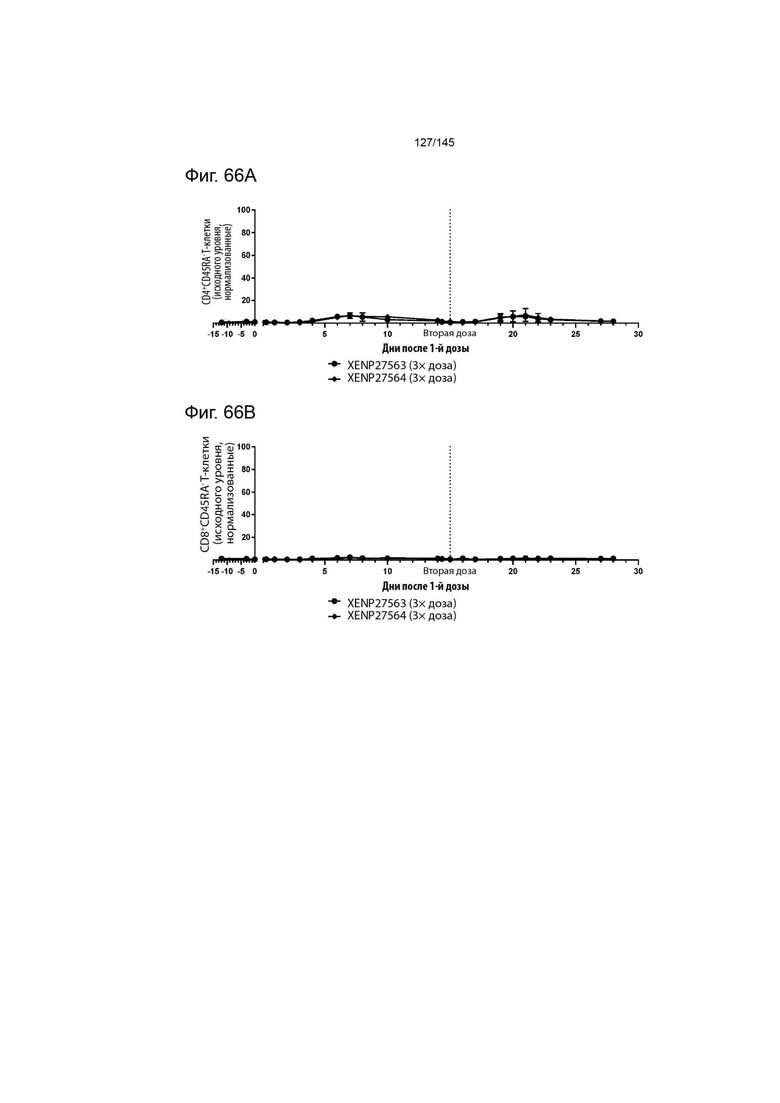
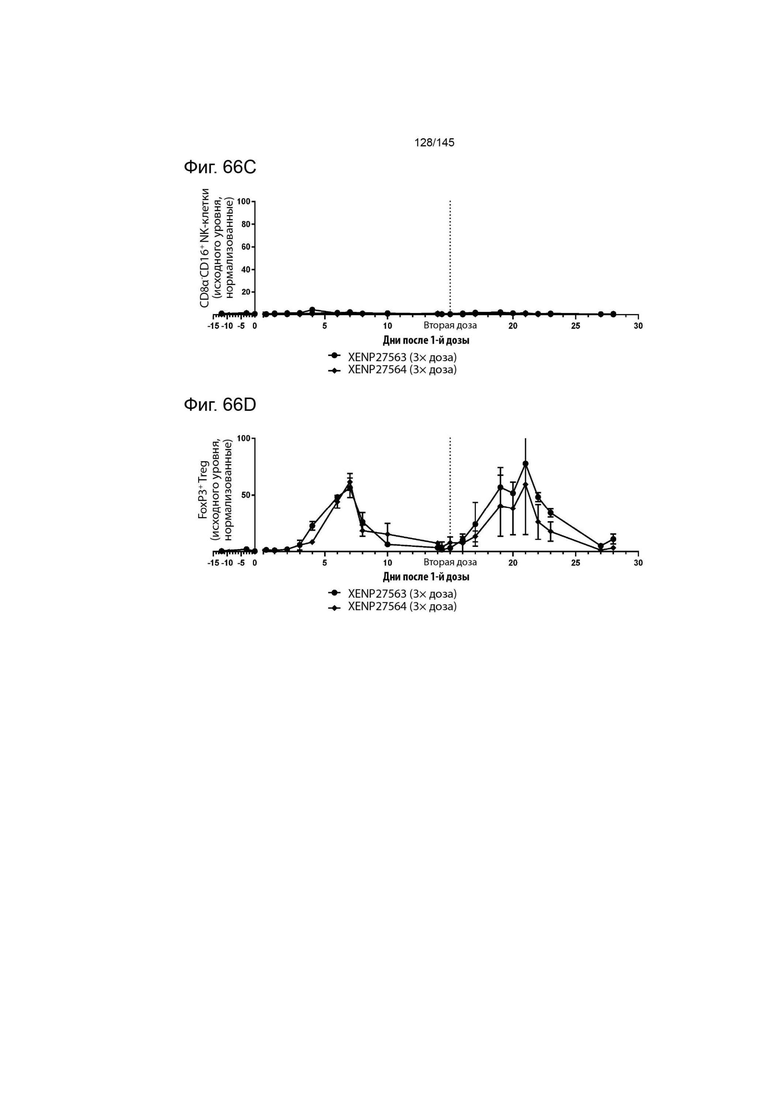
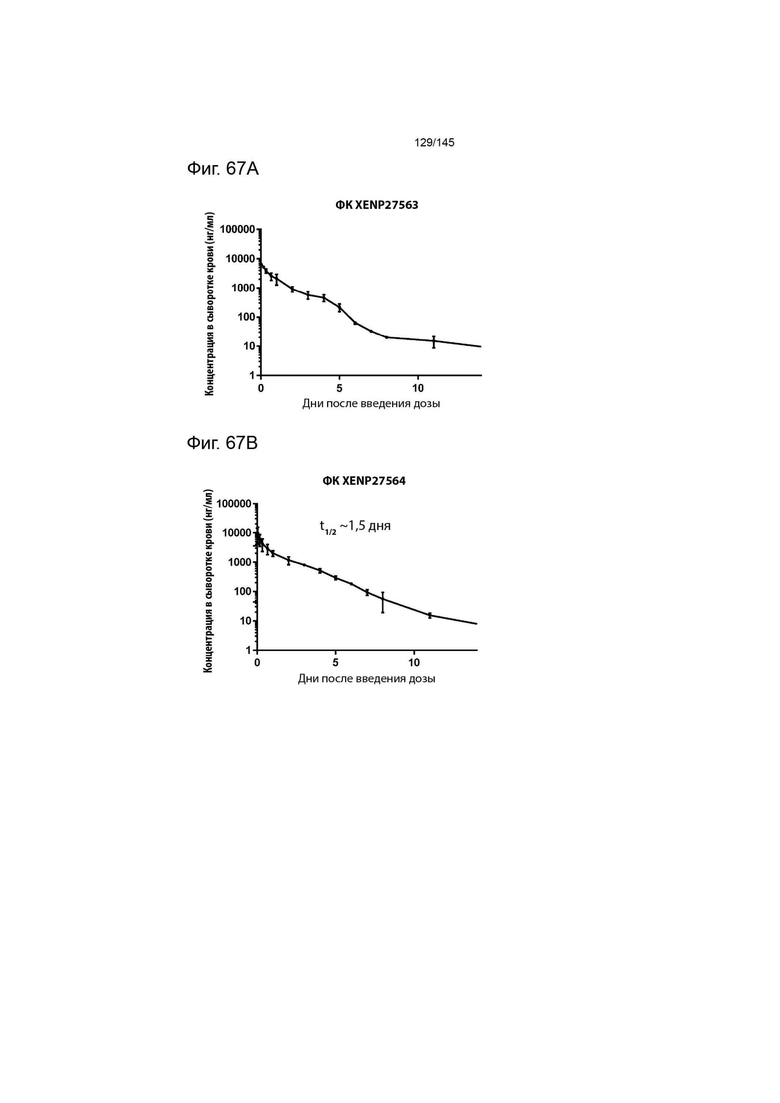
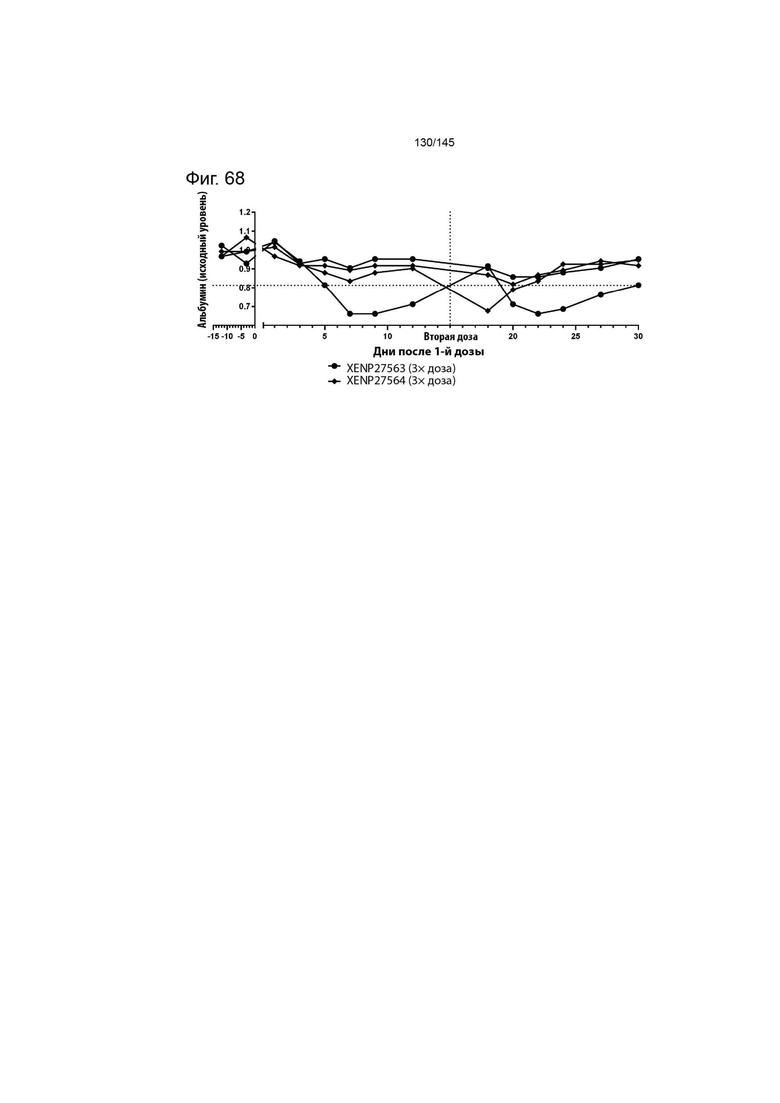

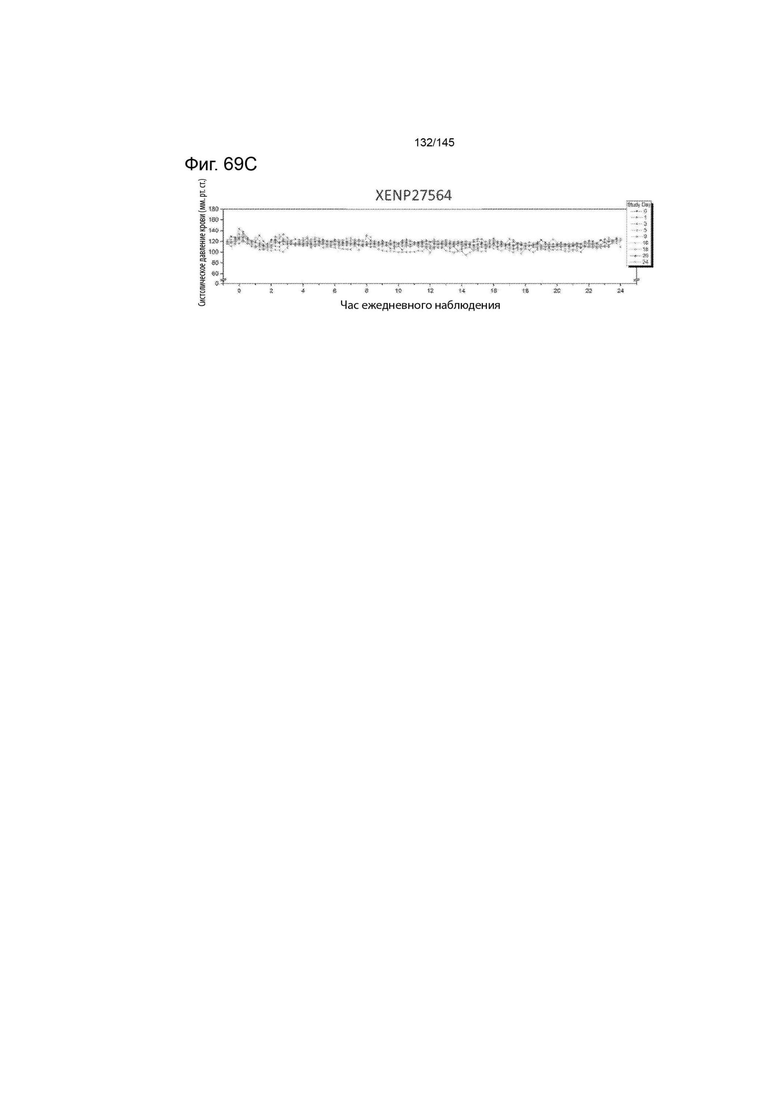
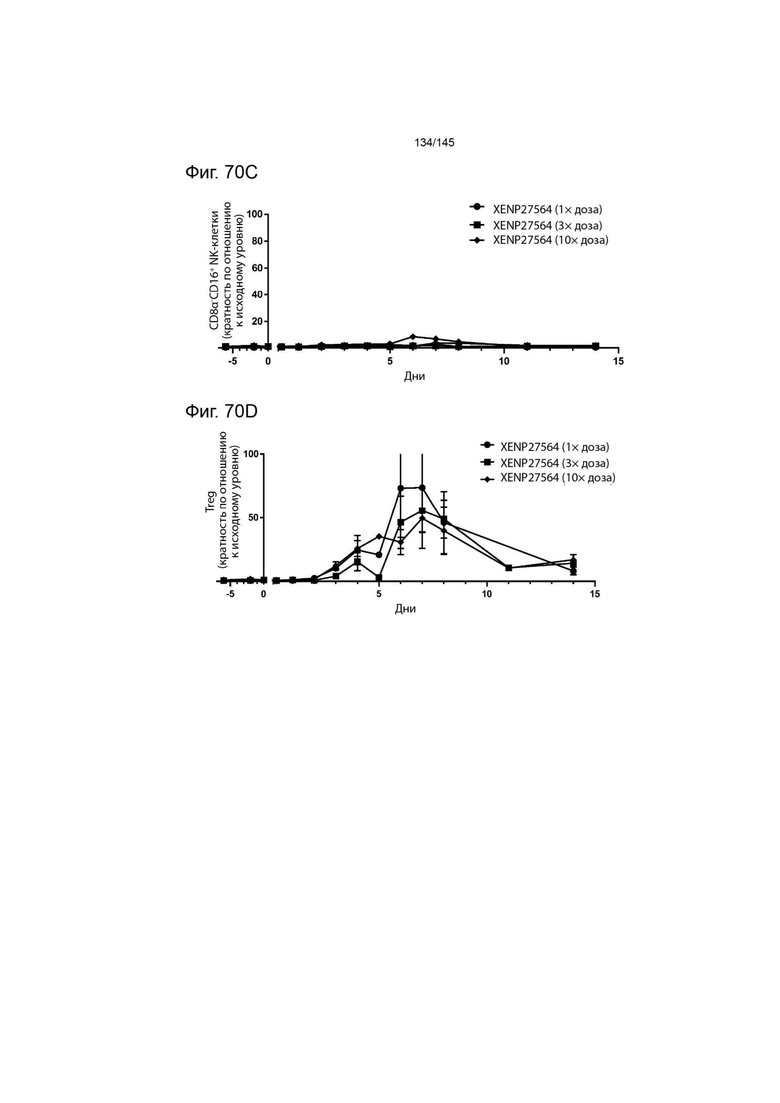
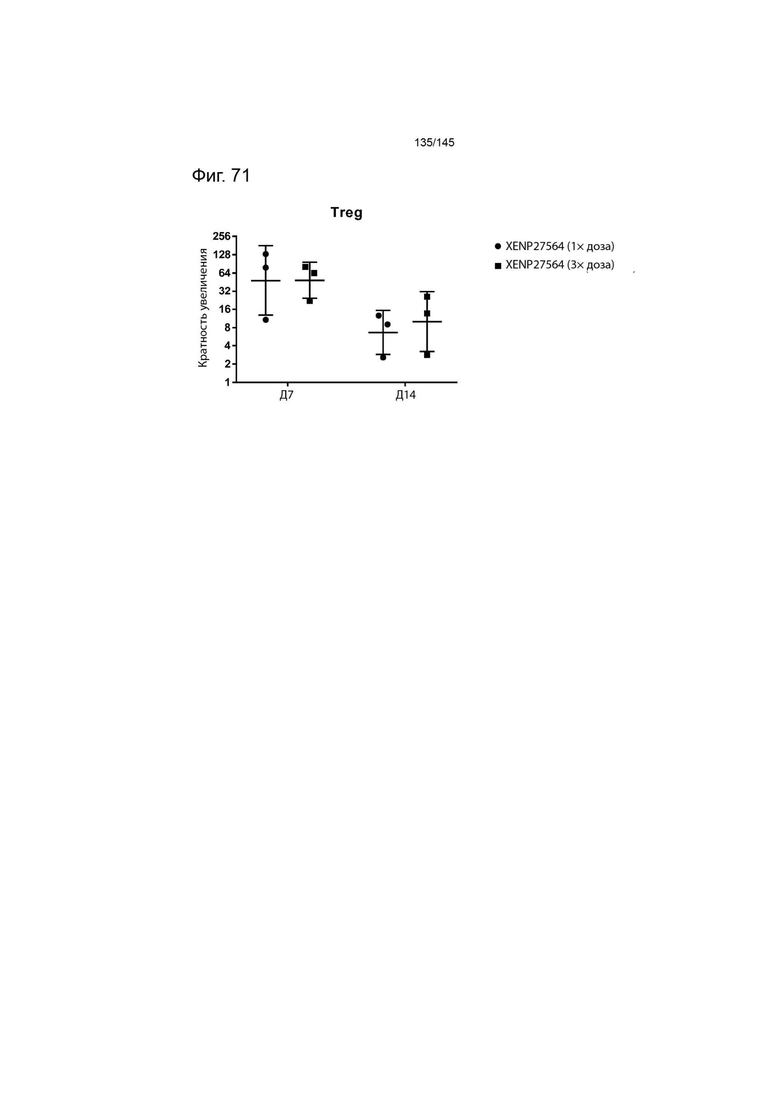
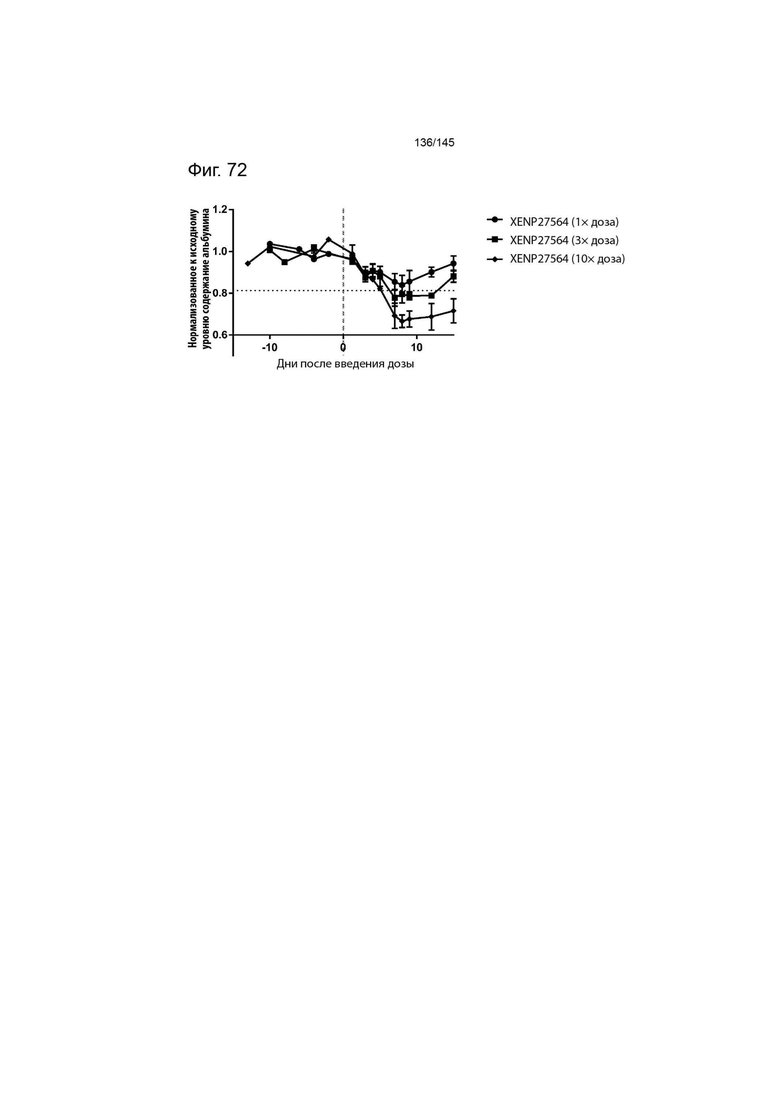
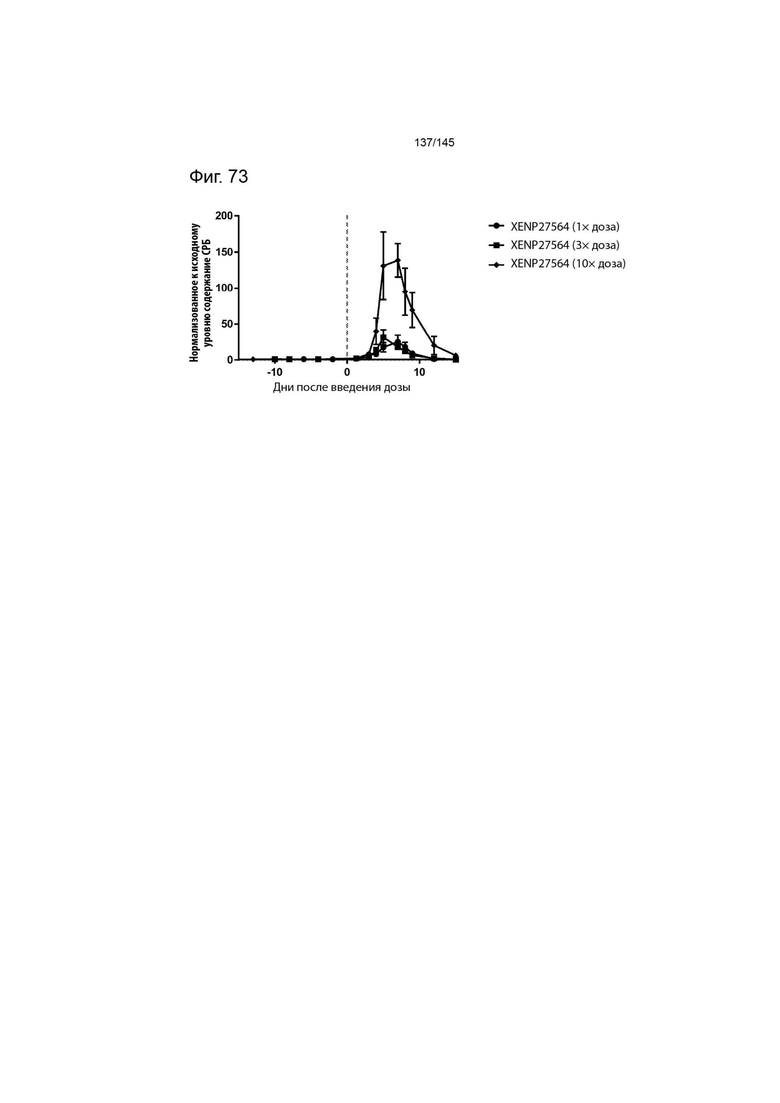
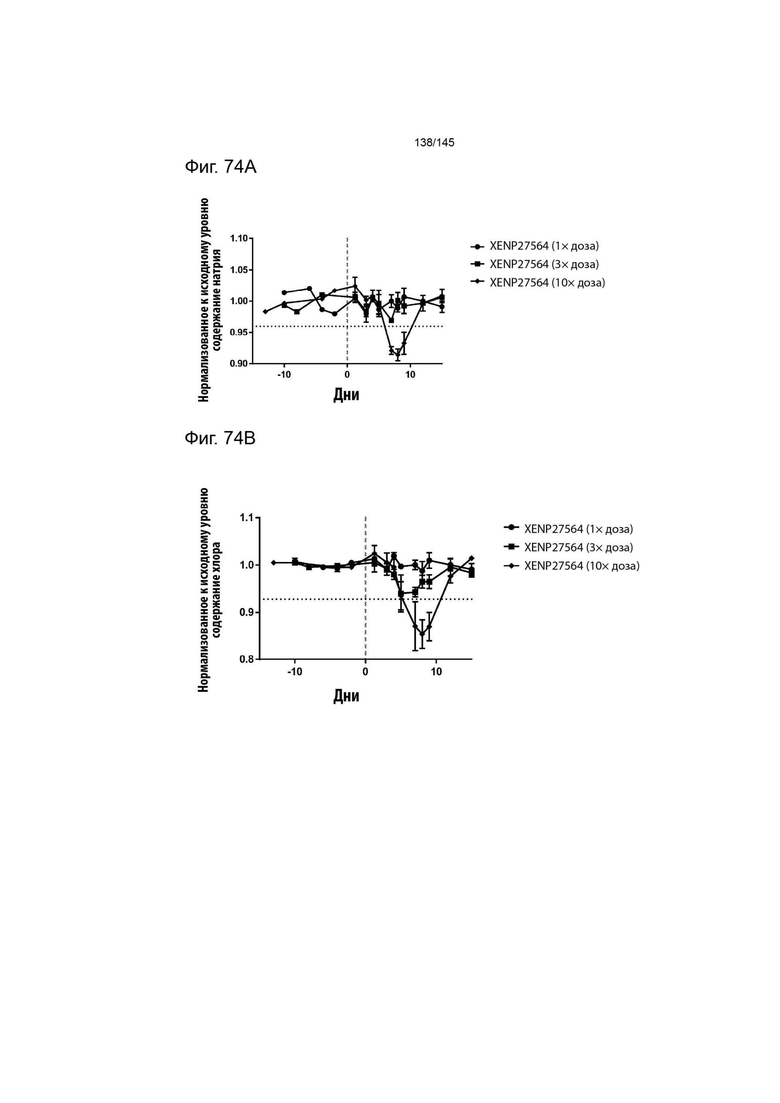
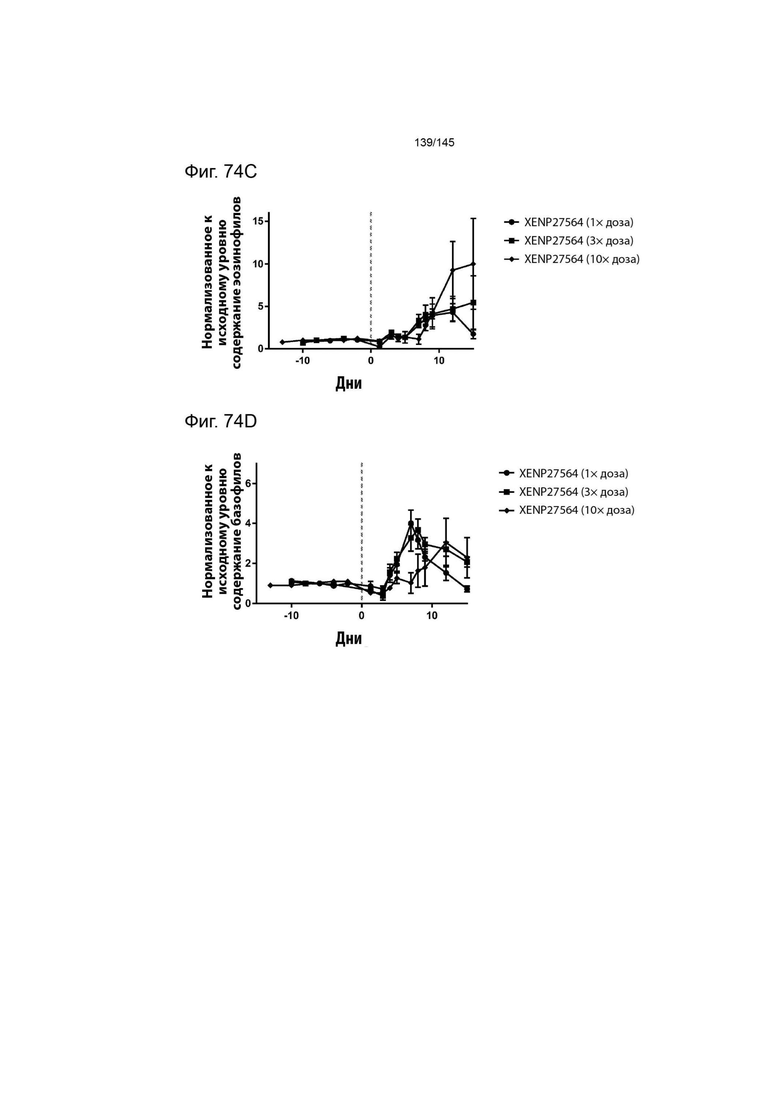
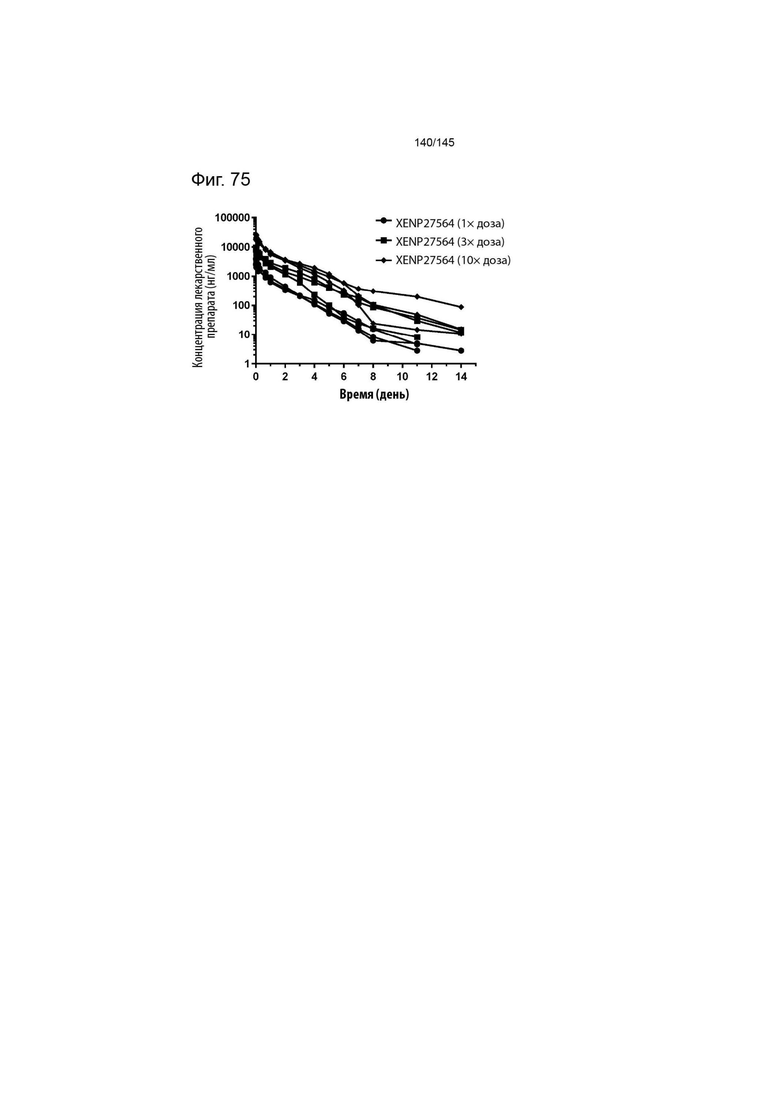
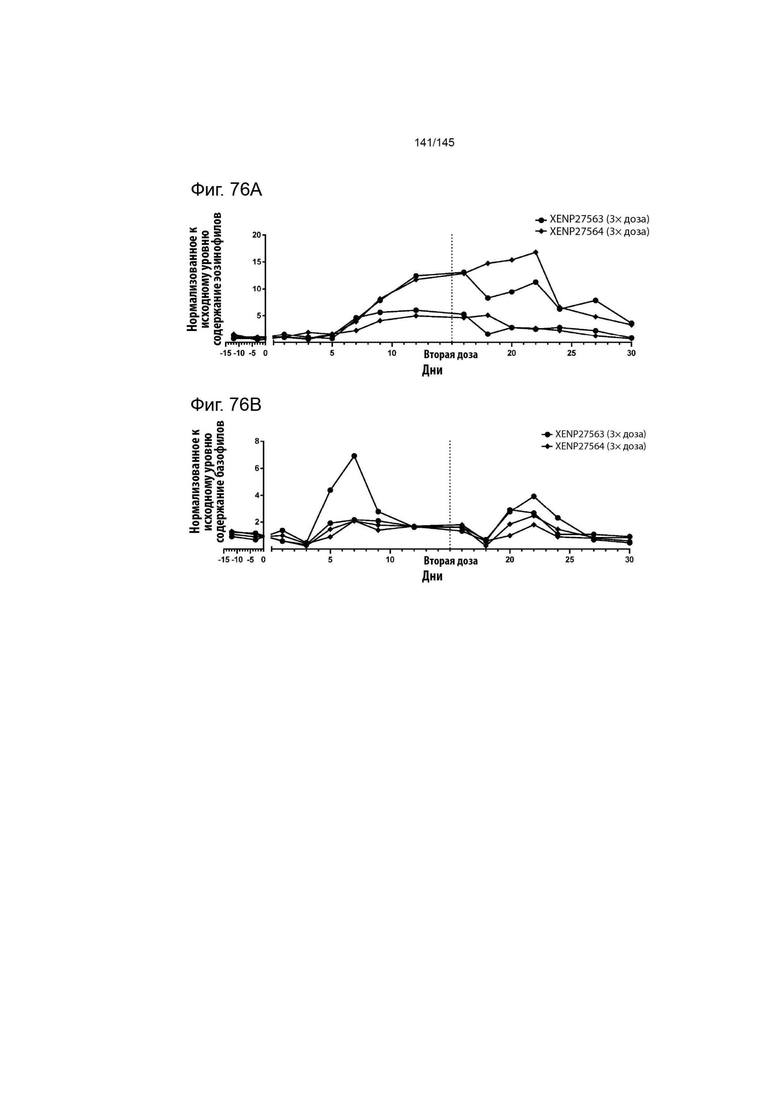
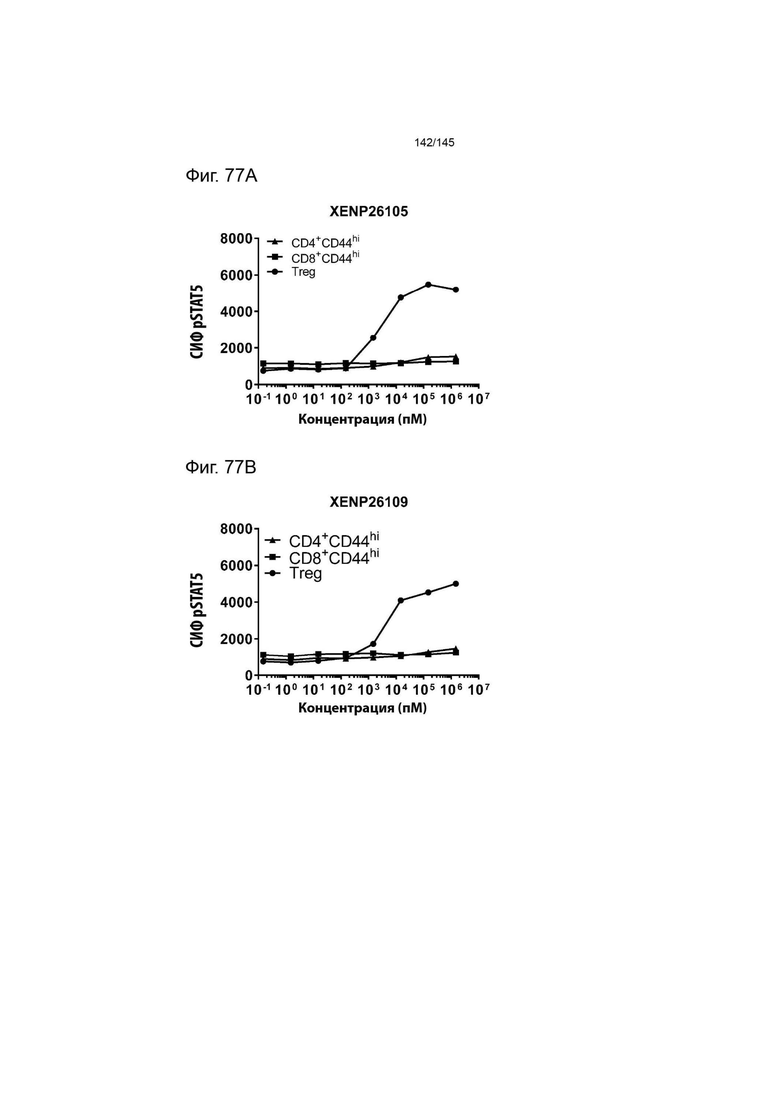
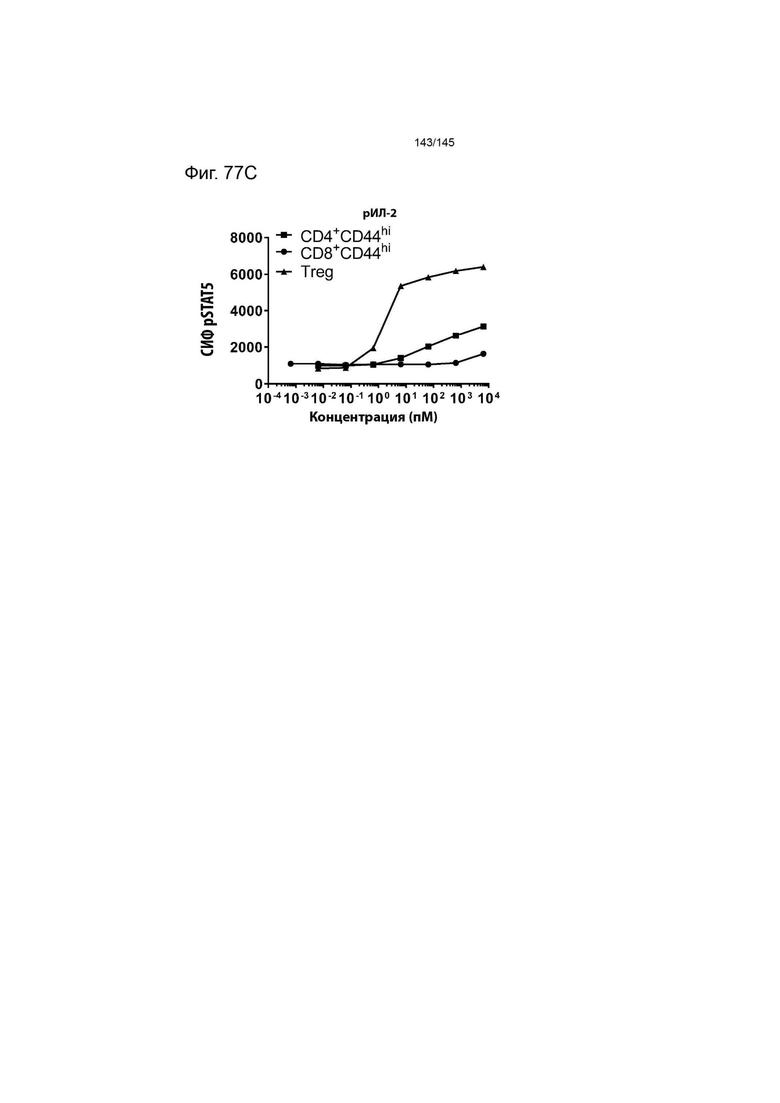
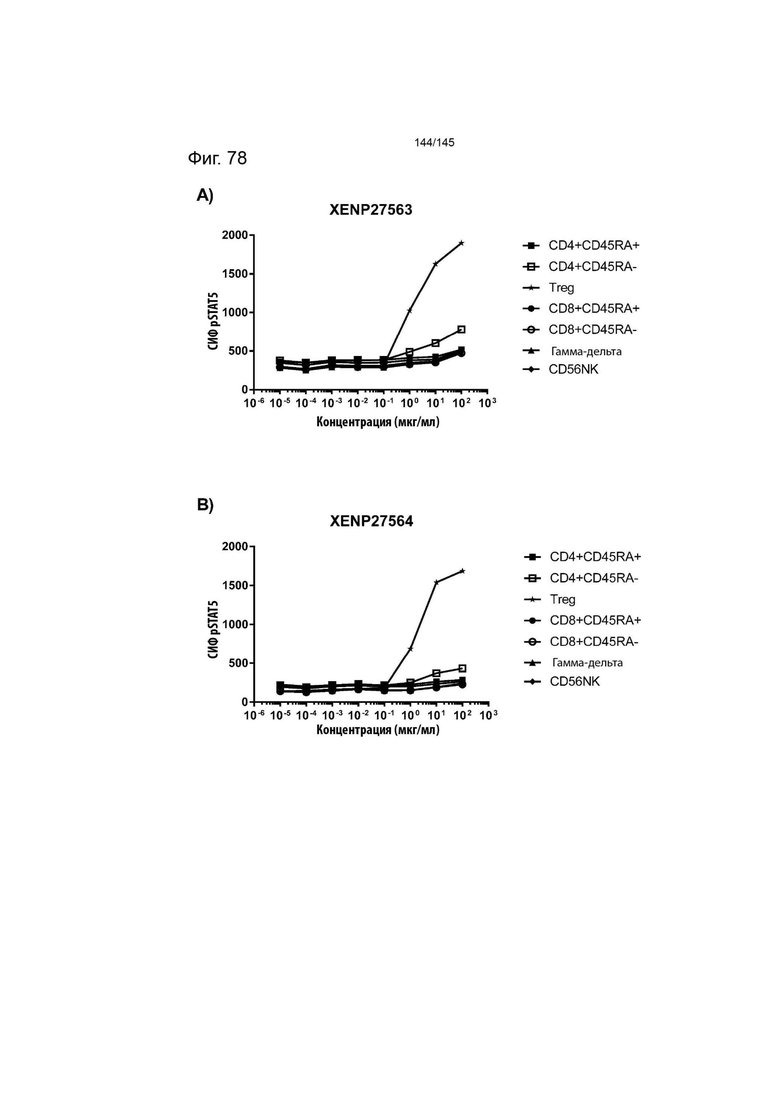
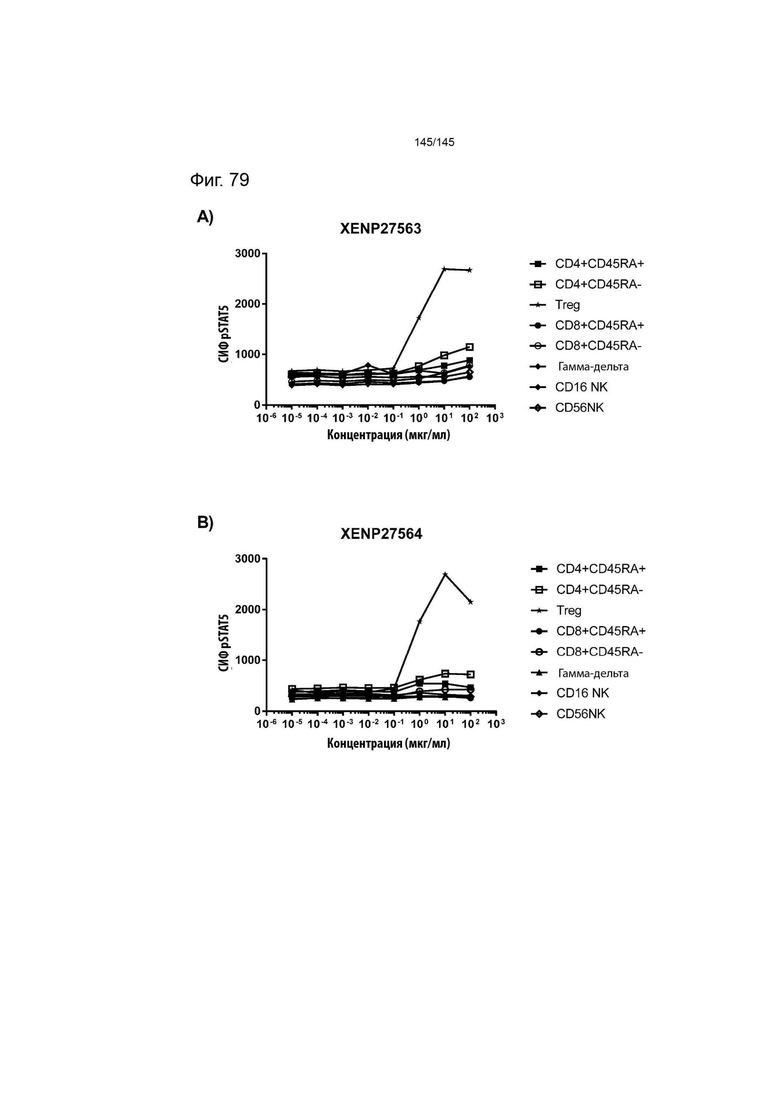
Группа изобретений относится к биотехнологии. Представлены полипептидные композиции для экспансии Tregs, одна из которых содержит вариантный белок ИЛ-2 человека, в которой указанный вариантный белок ИЛ-2 содержит одну или несколько аминокислотных замен по сравнению с SEQ ID NO:2, выбранных из группы T3A/D20N/T37R, T3A/D20N/N71K, T3A/D20N/T37R/C125S, T3A/D20N/N71K/C125S, T3A/D20N/T37R/C125A и T3A/D20N/N71K/C125A. Также раскрыты полипептидные композиции для экспансии Tregs, содержащие гетеродимерный белковый комплекс для экспансии Tregs. Изобретение обеспечивает селективную и устойчивую экспансию Treg. 5 н. и 8 з. п. ф-лы, 145 ил., 9 пр.
1. Полипептидная композиция для экспансии Tregs, отличающаяся тем, что содержит вариантный белок ИЛ-2 человека, причем указанный вариантный белок ИЛ-2 содержит одну или несколько аминокислотных замен по сравнению с SEQ ID NO:2, выбранных из группы T3A/D20N/T37R, T3A/D20N/N71K, T3A/D20N/T37R/C125S, T3A/D20N/N71K/C125S, T3A/D20N/T37R/C125A и T3A/D20N/N71K/C125A.
2. Полипептидная композиция, содержащая гомодимерный белковый комплекс для экспансии Tregs, в котором гомодимерный белковый комплекс содержит первый и второй белковые мономеры, где каждый белковый мономер содержит вариантный белок ИЛ-2, ковалентно присоединенный к Fc-домену, и где вариантный белок ИЛ-2 содержит одну или несколько аминокислотных замен по сравнению с SEQ ID NO:2, выбранных из группы T3A/D20N/T37R, T3A/D20N/N71K, T3A/D20N/T37R/C125S, T3A/D20N/N71K/C125S, T3A/D20N/T37R/C125A и T3A/D20N/N71K/C125A.
3. Полипептидная композиция по п. 2, отличающаяся тем, что каждый из указанных Fc-доменов представляет собой вариантный Fc-домен.
4. Полипептидная композиция, содержащая гетеродимерный белковый комплекс для экспансии Tregs, где гетеродимерный белковый комплекс содержит первый белковый мономер и второй белковый мономер, где первый белковый мономер содержит вариантный белок ИЛ-2, ковалентно присоединенный к первому вариантному Fc-домену, и где вариантный белок ИЛ-2 содержит одну или несколько аминокислотных замен по сравнению с SEQ ID NO:2, выбранных из группы T3A/D20N/T37R, T3A/D20N/N71K, T3A/D20N/T37R/C125S, T3A/D20N/N71K/C125S, T3A/D20N/T37R/C125A и T3A/D20N/N71K/C125A, и где второй белковый мономер содержит второй вариантный Fc-домен.
5. Полипептидная композиция по п. 3 или 4, отличающаяся тем, что указанные вариантные Fc-домены, первый вариантный Fc-домен и/или второй вариантный Fc-домен, представляют собой вариантные Fc-домены IgG1 человека, содержащие аминокислотные замены M428L/N434S.
6. Полипептидная композиция по п. 3 или 4, отличающаяся тем, что указанные вариантные Fc-домены, первый вариантный Fc-домен и/или второй вариантный Fc-домен, представляют собой вариантные Fc-домены IgG1 человека, содержащие аминокислотные замены E233P/L234V/L235A/G236del/S267K.
7. Полипептидная композиция по пп. 4, 5 или 6, отличающаяся тем, что указанные первый и второй вариантные Fc-домены, которые включают в себя совокупность гетеродимеризационных вариантов, выбранных из группы, состоящей из вариантов, изображенных на фиг. 2.
8. Полипептидная композиция по п. 7, отличающаяся тем, что указанную совокупность гетеродимеризационных вариантов выбирают из группы, состоящей из L368D/K370S : S364K/E357Q; L368D/K370S : S364K; L368E/K370S : S364K; T411E/K360E/Q362E : D401K; и T366S/L368A/Y407V : T366W.
9. Полипептидная композиция по пп. 4, 5, 6, 7, или 8, отличающаяся тем, что один из указанных вариантных Fc-доменов включает в себя совокупность pI вариантов, выбранных из группы, состоящей из вариантов, изображенных на фиг. 3.
10. Полипептидная композиция по п. 9, отличающаяся тем, что указанная совокупность pI вариантов содержит Q295D/N384D/Q418E/N421D.
11. Полипептидная композиция по п. 4, отличающаяся тем, что указанную полипептидную композицию выбирают из группы, состоящей из XENP27564 (SEQ ID NO: 297 и 298), XENP27563 (SEQ ID NO: 295 и 296), XENP26105 (SEQ ID NO: 245 и 246) и XENP26109 (SEQ ID NO: 249 и 250).
12. Полипептидная композиция для экспансии Tregs, содержащая гетеродимерный белковый комплекс для экспансии Tregs, где гетеродимерный белковый комплекс содержит:
первый мономер, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 297; и
второй мономер, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 298.
13. Полипептидная композиция для экспансии Tregs, содержащая гетеродимерный белковый комплекс для экспансии Tregs, где гетеродимерный белковый комплекс содержит:
первый мономер, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 295; и
второй мономер, содержащий аминокислотную последовательность, приведенную в SEQ ID NO: 296.
| WO 2016025385 A1, 18.02.2016 | |||
| КАНАЛЬНЫЙ ВЕНТИЛЯТОР | 2016 |
|
RU2639241C1 |
| WO 2005007121 A2, 27.01.2005; | |||
| WO 201207417 Al, 16.08.2012 | |||
| WO 9960128 Al, 25.11.1999 | |||
| WO 2009061853 A2, 14.05.2009 | |||
| COLLINS L | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

