Изобретение относится к фармакологии и биотехнологии и может быть использовано в медико-биологической промышленности и для терапии онкологических заболеваний.
Цель данного изобретения - получение химерного фермента, потенциально способного к таргетной терапии онкологических заболеваний.
Разработан химерный белок: L-метионин-гамма-лиаза (МГЛ) [1,2] сшитая с VHH антителом на ERBB3: MGL-BCD090-M456. Разработан способ получения препарата на его основе: биосинтеза, выделения, очистки. Основные физико-химические характеристики показывают наличие растворимой формы белка в препарате.
МГЛ (КФ 4.4.1.11) является пиридоксаль-5'-фосфат - зависимым ферментом, катализирующим реакцию γ-расщепления L-метионина с образованием метилмеркаптана, иона аммония и α-кетобутирата:
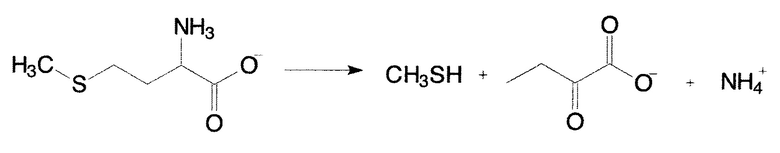
МГЛ также катализирует реакцию γ-расщепления L-цистеина и его S-замещенных производных, а также реакции α- и β-замещения в молекулах L-метионина, L-цистеина и их аналогов [3]. Фермент был обнаружен в бактериях Pseudomonas putida, Citrobacter freundii, Aeromonas sp., Clostridium sporogenes, Phophyromonas gingivalis, Brevivacterium linens BL2, а также у простейших эукариот Trichomonas vaginalis [4] и Entamoeba histolytica [5]. Однако стационарные кинетические параметры определены лишь для ферментов из P. putida [6, 7], Т. vaginalis [4] и С.freundii [8].
Известны штаммы, которые используют для получения рекомбинантной МГЛ из таких бактерий как P. putida и Т. vaginalis, имеется патент на получение рекомбинантной МГЛ из P. putida [9]. В патенте [10] заявлена возможность улучшения свойств МГЛ при помощи изучения структуры белка и основанной на этом белковой инженерии. В работе [11] обнаружен ген megL, кодирующий МГЛ в бактериях семейства Enterobacteriaceae. В патенте Soda, et al 1999 [9] заявлена возможность получения рекомбинантной МГЛ с использованием гибридной плазмиды pYH301, в которой ген megL из P. putida расположен под контролем Plac промотора. Способ выделения и очистки фермента не заявлен, стабилизирующие агенты не применяются. Согласно данного патента инкубация на 37°С в присутствии изопропил-β-D-1-тиогалактопиранозид (ИПТГ) 1 мМ в течение 17 часов штамма Escherichia coli JM109. содержащего плазмиду pYH301, приводит к получению биомассы, бесклеточный экстракт которой, обладает целевой активностью 0,81 ед/мг.
Противоопухолевая активность МГЛ показана в публикациях [2, 12-16].
В патенте Roger G Harrison 2004 г. [17] заявлена возможность получения химерно слитых белков МГЛ с коньюгатами, связывающимися с рецепторами ростовых факторов. В качестве лигандов для распознавания гиперэкспрессированых на опухолевых клетках рецепторов были предложены урокиназа, эпидермальный ростовой фактор, трансформирующий ростовой фактор альфа, инсулина-подобный фактор роста, интерлейкин-4, интерлейкин-6, фактор роста тромбоцитов, фактор роста фибробластов, ламинин, фактор роста эндотелий сосудов, аннексин V, различные антитела и фрагменты лигандов. Применение однодоменных антител на рецептор из ERBb семейства в данном документе не описано.
Гиперэкспрессия семейства рецепторов эпидермального фактора роста, в том числе EGFR, - очень распространенный маркер опухолевых клеток [18, 19].
В нашей работе был сделан химерный белок: МГЛ слитая с антителом BCD090-М456 [20] (далее MGL-456). Антитело BCD090-M456 специфично связывается ERBB3 [20, 21].
Смысл применения антител в составе химерного белка с МГЛ состоит в том. что можно создать безметиониновую зону в организме вокруг раковых клеток в сóлидных (твердых) опухолях. В таких условиях отпадает необходимость транспорта МГЛ внутрь раковой клетки. Применение MGL-456 может синергично усилить эффект других химиотерапевтических препаратов. Например, известно, что химерная МГЛ с S3 лигандом на EGFR вызывает угнетение сигнального пути ERK и имеет синергию с гефитинибом (ингибитором EGFR) [22]. Кроме того, синергический эффект показан в патенте Манухова и др. 2020 г. [23], где изложен способ выделения и очистки МГЛ. Согласно этому документу для биосинтеза МГЛ (кодируется природным геном без модификаций в гетерологичной системе Е. coli) требуется добавление в среду сульфата аммония, очистка включает в себя стадию соосаждения спиртом, а применение в качестве антиракового препарата наиболее успешно в комбинации с доксорубицином. Модификация препарата с целью его таргетирования к раковым клеткам в данном патенте не рассматривается.
Ближайшим аналогом генетической конструкции является патент Soda et al 1996 г [22], где приведена последовательность рекомбинантной МГЛ из Pseudomonas putida. Ключевое отличие нашей генетической конструкции состоит в гомологичном гене МГЛ из С.s porogenus и следующим за ним в одной рамке считывания последовательности гена камелоидного антитела.
Ближайшим аналогом MGL-456 является патент [17], где было сделано коньюгирование L-метионин-гамма-лиазы из Pseudomonas putida с урокиназой. В отличии от [17] в нашей работе используется L-метионин-гамма-лиаза из С. sporogenus и однодоменное антитело с большей специфичностью по отношению к рецептору эпидермального фактора роста.
Задачей заявляемой группы изобретений является разработка нового химерного белка, перспективного для использования в качестве таргетного препарата в антираковой терапии,
Задача решена путем:
- разработки штаммов-продуцентов, несущих ген, кодирующий химерный белок-МГЛ слитый с VHH антителом М456 на С-конце и (вариант) с гистидиновой меткой (6 гистидинов), соединенной с геном МГЛ через последовательность TEV-протеазы на N-конце;
Проблема, решаемая данной группой изобретений, состоит в разработке таргетных препаратов, необходимых для снижения общей токсичности при химиотерапии. Техническим решением является создание плазмидных конструкций, содержащих заявляемую последовательность или ее гомолог, отличающийся вследствие вырожденности генетического кода, получение штамма продуцента на основе этих плазмид и демонстрация возможности выделения и очистки целевого белка.
Получение химерного белка МГЛ-456 в общем виде.
Получение генетической конструкции, кодирующей MGL-456 происходит путем амплификации гена megL, кодирующего L-метионин-гамма-лиазу из С. sporogenes [24], а также последовательности, кодирующей М456 [21] с последующим клонированием в плазмидном векторе. К 5'-концу megL гена с помощью ПЦР с последующей сборкой по Гибсону добавляется последовательность, кодирующая антитела.
Генетическая конструкции гибридной плазмиды, обеспечивающей биосинтез MGL-456, содержит ориджин репликации, ген устойчивости к канамицину и трансляционно слитую последовательность природной последовательности гена megL и VHH-антитела М456.
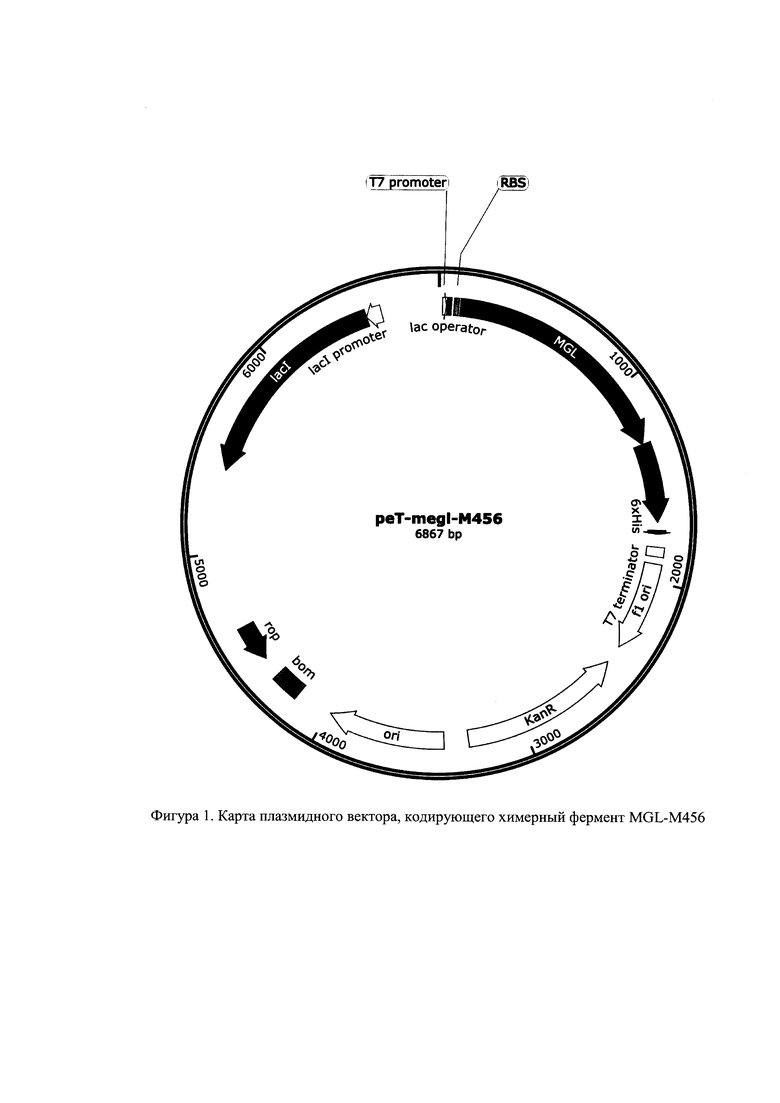
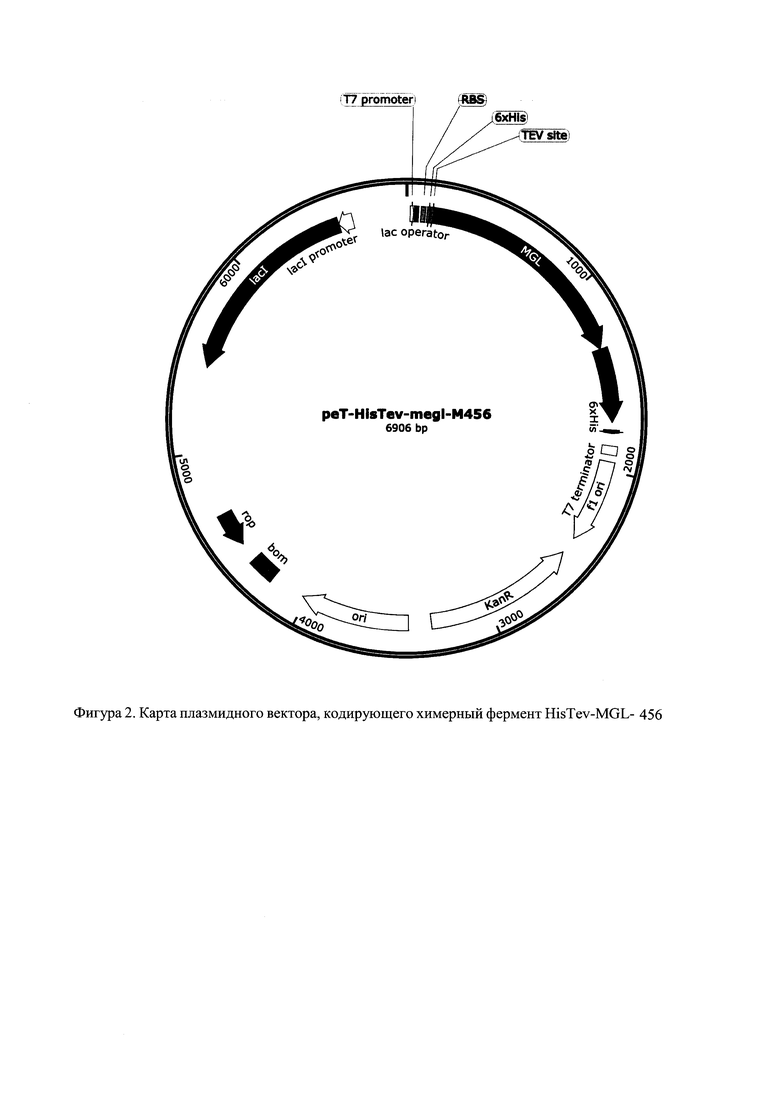
Изобретение проиллюстрировано фигурами 1 и 2 в виде карты гибридных плазмид, которые обеспечивают получение химерного белка для очистки с помощью ионно-обменной или никель-аффиной хроматографии соответственно. В этих плазмидах под контроль Т7 промотора клонированы гены megl-M456 и HisTev-megl-M456 соответственно.
Заключение.
В данной работе был разработаны химерный белок MGL-456, отработана технология его биосинтеза и хроматографической очистки. В результате достигнута цель - получена генетическая конструкция, кодирующая модифицированный фермент, обладающий потенциальным таргетным и цитостатическим свойством по отношению к опухолевым клеткам.
Пример 1.
Получение генетической конструкции, кодирующей MGL-456 на базе гибридных плазмид рЕТ 28a::megL_sporogenes, содержащий ген L-метионин-гамма-лиазы из С.sporogenes [24], а так же плазмиды pSol-SUMO-VHH456 [21].
Ген, кодирующий химерный белок - МГЛ слитый с VHH антителом получается путем клонирования. К 3'-концу megL гена с помощью кругового ПЦР с последующей сборкой по Гибсону добавляется последовательность антитела. Праймера пишутся так, чтобы получились перекрывающиеся концы. После этого две получившиеся цепочки ДНК лигируются с помощью сборки по Гибсону.
Генетическая конструкции гибридной плазмиды, обеспечивающей биосинтез MGL-456. содержит ориджин репликации, ген устойчивости к канамицину и трансляционно слитую последовательность природной последовательности гена megL и VHH-антитела М456. Карта гибридной плазмиды представлена на фигуре 1. Карта была построена с помощью программного обеспечения SnapGene Viewer (США).
Создание штамма-продуцента производилось путем химической трансформации клеток Е. coli BL21(DE 3) лигазной смесью, полученной на шаге, описанном выше. Получившиеся клоны были проверены с помощью определения последовательности по Сэнгеру.
Для биосинтеза используют среду, не содержащую NaCl (типа Rich medium) [25].
Очистка рекомбинантного белка MGL-456 из полученной биомассы проходит в соответствии с [24].
Полученный в результате раствор целевого белка, имеющий чистоту не менее 99% по МГЛ-456 может храниться как минимум 5 дней в темном месте при температуре 4°С.
Пример 2.
Получение генетической конструкции, кодирующей MGL-456 с HisTag-TEV меткой для никель-афинной очистки осуществляется с использованием гибридной плазмиды рЕТ 28a::megL_sporogenes, содержащий ген L-метионин-гамма-лиазы из С.sporogenes [24], а так же плазмиды pSol-SUMO-VHH456 [21].
Ген, кодирующий химерный белок - МГЛ слитый с VHH антителом и последовательнстью HisTag - TEV, получается путем клонирования. Сначала к гену megL с помощью лигирования по Гибсону и кругового ПЦР добавляется на N-конец последовательность HisTag-TEV, затем ставится два ПЦР - круговой на плазмиды с megL и для создания вставки в виде антитела на С-конец МГЛ. Праймера пишутся так, чтобы получились перекрывающиеся концы. После этого две получившиеся цепочки ДНК лигируется с помощью сборки по Гибсону. Карта гибридной плазмиды представлена на фигуре 2. Карта была построена с помощью программного обеспечения SnapGene Viewer (США).
Генетическая конструкции гибридной плазмиды, обеспечивающей биосинтез MGL-456, содержит ориджин репликации, ген устойчивости к канамицину и трансляционно слитую последовательность 6 His, сайта TEV протеазы, природной последовательности гена megL и VHH-антитела М456.
Создание штамма-продуцента производилось путем химической трансформации клеток Е. coli BL21(DE 3) лигазной смесью, полученной на шаге, описанном выше. Получившиеся клоны были проверены с помощью определения последовательности по Сэнгеру.
Для биосинтеза используют среду, не содержащую NaCl (типа Rich medium) [25].
Очистка рекомбинантного белка MGL-456 из полученной биомассы проходит в несколько стадий. Клетки биомассы лизируют в присутствии PMSF и пиродоксаль-фосфата на поточном дезинтеграторе. Полученный в результате лизиса клеточный дебрис, мембранную фракцию и нерастворимые белки штамма-продуцента отделяют осаждением с помощью ультрацентрифугирования (100000 g, 1 час). К осветленному лизату добавляют имидазола до 10 мМ и наносят batch-методом на NiNTA (Cytiva, США) смолу. Избавления от нецелевых белков проводят промывкой колонки буфером с 20 мМ имидазолом. Перед элюцией целевого белка проводят промывку сорбента с нанесенным белком с помощью Triton-X114 с целью очистки целевого белка от липополисахаридов штамма-продуцента. Элюцию целевого белка проводят в том же буфере с добавлением 400 мМ имидазола.
На следующем шаге концентрацию белка измеряют с помощью реактива Брэдфорда и добавляют TEV - протеазу из расчета 1 мг протеазы на 100 мг белка. Расщепление белка идет всю ночь на +4°С. На следующий день ставят диализ в мешках на 30 кДа против буфера с 10 мМ имидазола в течении 10 часов. После этого белок повторно наносится на NiNTA сорбент и элюируется. Полученная фракция белка без HisTag снова диализуется в течении ночи.
Метионазную активность MGL-456 определяют с помощью модифицированного метода Ристона [26] в последовательных ферментативной и колориметрической реакциях на реактив Несслера, дающего возможность определения концентрации аммиака, образующегося после распада метионина.
Полученный в результате раствор целевого белка, имеющий чистоту не менее 99% по MGL-456 и может храниться как минимум 5 дней в темном месте при температуре 4°С.




Использованные источники информации:
1. Anufrieva NV, Morozova ЕА, Kulikova VV, Bazhulina NP, Manukhov IV, Degtev DI, Gnuchikh EY, Rodionov AN, Zavilgelsky GB, Demidkina TV. Сульфоксиды - аналоги L-метионина и L-цистеина как пролекарства против грамположительных и грамотрицательных бактерий. Acta Naturae. 2015, 7(4): 128-35.
2. Hoffman R.M. Development of recombinant methioninase to target the general cancer-specific metabolic defect of methionine dependence: a 40-year odyssey. Expert opinion on biological therapy. - 2015. - T. 15. - №. 1. - C. 21-31.
3. Tanaka, H., Esaki, N. & Soda, K. A versatile bacterial enzyme: 1-methionine γ-lyase. Enzyme Microb. Technol. 1985, 7, 530-537.
4. McKie, A.E., Edlind, Т., Walker, J., Mottram, J.M. & Coombs, G.H. The Primitive protozoon trichomonas vaginalis contains two methionine g-lyase genes that encode members of the g-family of pyridoxal 5'-phosphate-dependent enzymes. J. Biol. Chem. 1998, 273, 5549-5556.
5. Tokoro, M., Asai, Т., Kobayashi, S., Takeuchi, Т., Nozaki, T. Identification and characterization of two isoenzymes of methionine gamma-lyase from Entamoeba histolytica: a key enzyme of sulfur-amino acid degradation in an anaerobic parasitic protist that lacks forward and reverse trans-sulfuration pathways. J. Biol. Chem. 2003, 278, 42717-42727.
6. Esaki, N., Nakayama, Т., Sawada, S., Tanaka, H., and Soda, K. Proton nmr studies of substrate hydrogen exchange reactions catalyzed by L-methionine.gamma.-lyase. Biochemistry. 1985, 24, 3857-3862.
7. Inoe, H., Inagaki, K., Adachi, N., Tamura, Т., Esaki, N., Soda, K. & Tanaka, H. Role of tyrosine 114 of L-methionine gamma-lyase from Pseudomonas putida. Biosci. Biotechnol. Biochem. 2000, 64, 2336-2343.
8. Манухов И.В., Мамаева Д.В., Морозова Е.А., Расторгуев С.М., Фалеев Н.Г., Демидкина Т.В., Завильгельский Г.Б. Биохимия. 2006 апрель; 71(4); 842-851. Манухов И.В., Мамаева Д.В., Морозова Е.А., Расторгуев С.М., Фалеев Н.Г., Демидкина Т.В., Завильгельский Г. Б. L-метионин-гамма-лиаза Citrobacter freundii: клонирование гена и кинетические параметры фермента. Биохимия. 2006, 71(4):842-851.
9. United States Patent 5,861,154. Inventors: Soda, et al. January 19, 1999. Recombinant L-methionine gamma.-lyase.
10. United States Patent 7,118,903. Inventors: Inagaki, et al. October 10, 2006. L-methionine gamma-lyase with modified function.
11. Manukhov IV, Mamaeva DV, Rastorguev SM, Faleev NG, Morozova EA, Demidkina TV, Zavilgelsky GB. A gene encoding L-methionine gamma-lyase is present in Enterobacteriaceae family genomes: identification and characterization of Citrobacter freundii L-methionine gamma-lyase. J Bacteriol. 2005, 187(11):3889-93.
12. Tan Y., Xu M., Tan X.-Z., Tan X., Wang X., Saikawa Y., Nagahama Т., Sun X., Lenz M., Hoffman R.M. Overexpression and large-scale production of recombinantl-methionine-a-deamino-y-mercaptomethane-lyase for novel anticancer therapy. 1997. Protein. Exp.Purif 9, 233-245.
13. Yoshioka Т., Wada Т., Uchida N., Maki H., Yoshida H., Ide N., Kasai H., Hojo K., Shono K., Maekawa R., Yagi S., Hoffman R.M. Anticancer efficacy in vivo and in vitro, synergy with 5-fluorouracil, and safety of recombinant methioninase. 1998. Cancer Res. 58, 2583-2587.
14. Tan Y., Xu M., Hoffman R.M. Broad selective efficacy of rMETase and PEG-rMETase on cancer cells in vitro. Anticancer Res. 2010, 30, 793-798.
15. Pokrovsky VS, Yu Anisimova N, Zh Davydov D, Bazhenov SV, Bulushova NV, Zavilgelsky GB, Kotova VY, Manukhov IV. Methionine gamma lyase from Clostridium sporogenes increases the anticancer effect of doxorubicin in A549 cells and human cancer xenografts. Invest New Drugs. 2019 Jun 15. doi: 10.1007/s10637-018-0619-4.
16. Pokrovsky V.S., Anisimova N.Yu., Davydov D.Zh., Bazhenov S.V., Bulushova N.V., Zavilgelsky G.B., Kotova V.Y., I.V. Manukhov. Methionine dependence of cancer and aging: methods and protocols. R.M. Hoffman, Editor. Chapter 18. Methionine gamma lyase from Clostridium sporogenes increases the anticancer effect of doxorubicin in A549 cancer cells in vitro and human cancer xenografts. Springer Nature. Springer Science+Business Media, LLC, Humana Press. ISBN 978-1-4939-8795-5. 2019.
17. WO 2004112717A3 2004. Conjugate for the specific targeting of anticancer agents to cancer cells and production thereof. Inventors: Roger G Harrison, Thomas J Pento.
18. Yarden, Y., Pines, G. The ERBB network: at last, cancer therapy meets systems biology. Nat Rev Cancer 2012, 12, 553-563. https://doi.org/10.1038/nrc3309.
19. Ciardiello F., Tortora G. EGFR antagonists in cancer treatment. New England Journal of Medicine. 2008. 358(11) 1160-1174.
20. Eliseev, I.E.; Ukrainskaya, V.M.; Yudenko, A.N.; Mikushina, A.D.; Shmakov, S.V.; Afremova, A.I.; Ekimova, V.M.; Vronskaia, A.A.; Knyazev, N.A.; Shamova, O.V. Targeting ErbB3 Receptor in Cancer with Inhibitory Antibodies from Llama. Biomedicines 2021, 9, 1106. https://doi.org/10.3390/biomedicines9091106.
21. 'Однодоменное антитело, специфически связывающее рецептор ErbB3 человека, без канонической дисульфидной связи' Патент РФ №272185 от 25.05.2020 Юденко А.Н., Елисеев И.Е., Украинская В.М.
22. Japan patent JPH09182592A Inventors: Kenji Soda, Hidehiko Tanaka, Hiroyuki Inoue, Manabu Sugimoto, Kenji Inagaki, Nobuyoshi Ezaki. 1996. Recombinant 1-methionine-gammay-lyase'
23. Bondarev N. et al. MGL S3 Chimeric Enzyme Drives Apoptotic Death of EGFR-Dependent Cancer Cells through ERK Downregulation. International Journal of Molecular Sciences. 2022. - T. 23. - №. 21. - C. 12807.
24. Патент РФ №2733440 от 04.06.2020 «Способ получения фермента метионин-гамма-лиазы, противоопухолевое лекарственное средство ГЛФ МГЛ на основе этого фермента и применение этого средства для торможения роста опухоли (варианты)».
Авторы: Манухов И.В., Баженов С.В., Гнучих Е.Ю., Покровский B.C., Давыдов Д.Ж., Котова В.Ю., Булушова Н.В., Завильгельский Г.Б.
25. Studier F.W. Protein production by auto-induction in high-density shaking cultures. Protein Expr. Purif. 2005, 41, 207-234.
26. Wriston, J.C. Asparaginase. Methods in Enzymology. 1970, 17A, 732-742.
--->
Перечень последовательностей
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="МГЛ-VHH456.xml"
softwareName="WIPO Sequence" softwareVersion="2.2.0"
productionDate="2022-12-09">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate></FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>-</ApplicantFileReference>
<ApplicantName languageCode="ru">Российский биотехнологический
университет (РОСБИОТЕХ)</ApplicantName>
<ApplicantNameLatin>BIOTECH University</ApplicantNameLatin>
<InventorName languageCode="ru">Бондарев Николай
Антонович</InventorName>
<InventorNameLatin>Bondarev Nikolai Antonovich</InventorNameLatin>
<InventionTitle languageCode="ru">Химерный фермент на основе L-
метионин-гамма-лиазы слитой с VHH антителом 456, фрагмент ДНК,
кодирующий указанный фермент</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>532</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..532</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Escherichia coli</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MENIEKMGFATKAIHGGHIGDKQFGSLATPIYQTSTFIFDSAEQGGRRF
AGEESGYIYSRLGNPTSTEVENKLALLEGGEAAVVAASGMGAIAASLWSALKSGDHVVASDTLYGCTFAL
LNHGLTRYGVEVTFVDVSNLDEVKNALKPNTKVVYLETPANPTLKVTDIRKISNMVHESNKECFVFVDNT
FCTPYIQRPLELGADVVVHSATKYLNGHGDVIAGFAVGKEEFINQVKLFGIKDMTGSVTGPFESFLIIRG
MKTLQLRMEKHCSNAMEVAKFLESHPAVEKVYYPGLESFEYYQLAREQMKLPGAMISFELKGGVEEGKIV
MNNVKLATLAVSLGDSETLIQHPASMTHSPYTAEERKAAGISDGLVRLSVGLEDAEDIIDDLKQALDLIV
KQVQLVQSGGGLVQSGGSLKLSCVASGGPFSTYLMGWFRQAPGKEREFVTAISRSGLNTYYADSVKGRFT
ISRDNAKNTVYLQMHSLKPEDTAVYHCAARRGGTNSGSYYFDRPAVSDEYDLWGQGTLVTVSS</INSDS
eq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>1599</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1599</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Escherichia coli</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggagaatattgaaaaaatggggtttgccacaaaggctatacatggag
gacatataggagataaacagtttggttcattagcaacaccaatatatcaaacatctacatttatatttga
ttctgcagagcaggggggaagaagatttgcaggagaagaaagtggatatatatattcaaggttaggtaat
cctacatctacagaagtagagaataaattggctttgttagaaggtggagaagcagctgttgtagcagcat
caggtatgggtgctatagcagcatctttatggtcagctttaaagtcaggagatcatgtagttgcatcaga
tactttatatgggtgcacatttgcactattaaatcatggattaacaagatatggagtagaggttacattt
gtagatgtatctaatttagatgaggtaaagaatgcattaaaaccaaatactaaagtagtttatttggaaa
ctccagctaatccaacactaaaggttactgatattagaaagatatcaaatatggttcatgagagcaataa
agagtgttttgtttttgttgataatactttttgtacgccatacatacaaagaccattagaattaggtgct
gatgtggttgtacattctgctactaaatatttaaatggtcatggtgatgttatagctggatttgcagtag
gaaaagaagaatttataaatcaagttaaattatttggcataaaagatatgacaggatcagttacaggacc
tttcgagtcattcttaataataagaggaatgaaaacattacaattaagaatggaaaaacattgcagtaat
gctatggaagtagcaaagtttttagaatcacatccagcagttgaaaaagtttattatccaggattggaaa
gctttgaatactatcagcttgctagagagcagatgaaattacctggtgctatgatttcctttgagttaaa
gggtggagtagaagaaggaaaaatagttatgaataatgttaaattagcaactcttgcagtaagtcttgga
gattcagaaacactaattcagcatccagcatcaatgacacattctccttatacagcagaagaaagaaaag
cagctggtataagtgatggattagtaagattatctgtaggtcttgaagatgcagaagatataatagatga
tttaaaacaagctttagatttaatagttaagcaggtgcagctggtgcagtctgggggaggattggtgcag
tctgggggctctctgaaactctcctgtgtagcctctggaggccccttcagtacgtatttgatgggctggt
tccgccaggctccagggaaggagcgtgagtttgtcacagctattagcaggagtggccttaacacatacta
tgcagactccgtgaagggccgattcaccatctccagagacaacgccaagaacacggtgtatctgcaaatg
cacagcctgaaacctgaggacacggccgtttatcactgtgcagcccgtcgaggtggcactaatagtggta
gttactacttcgaccgccccgcggtctcagatgagtatgacttgtggggccaggggaccctggtcaccgt
ctcctcataa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Химерный фермент МГЛ-S3 - метионин-гамма-лиаза, слитая с S3 доменом белка VGF из Vaccinia virus, способ получения МГЛ-S3 и противоопухолевый препарат на основе этого фермента | 2022 |
|
RU2816486C2 |
| Способ получения фермента метионин-гамма-лиазы, противоопухолевое лекарственное средство ГЛФ МГЛ на основе этого фермента и применение этого средства для торможения роста опухоли (варианты) | 2018 |
|
RU2733440C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИАЛКИЛ ТИОСУЛЬФИНАТОВ С ПОМОЩЬЮ ДВУХКОМПОНЕНТНОЙ СИСТЕМЫ МЕТИОНИН-γ-ЛИАЗА + СУЛЬФОКСИДЫ S-АЛКИЛ-L-ЦИСТЕИНА | 2021 |
|
RU2814985C2 |
| ГЕН ПЕПТИДООБРАЗУЮЩЕГО ФЕРМЕНТА, ПЕПТИДООБРАЗУЮЩИЙ ФЕРМЕНТ И СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА | 2002 |
|
RU2280077C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЭРИТРОЦИТЫ, В КОТОРЫХ ЗАКЛЮЧЕН ФЕРМЕНТ, ЗАВИСИМЫЙ ОТ ПИРИДОКСАЛЬФОСФАТА, И ЕГО КОФАКТОР | 2015 |
|
RU2697086C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЭРИТРОЦИТЫ, В КОТОРЫХ ЗАКЛЮЧЕН ФЕРМЕНТ, ЗАВИСИМЫЙ ОТ ПИРИДОКСАЛЬФОСФАТА, И ЕГО КОФАКТОР | 2015 |
|
RU2744659C2 |
| МОДИФИЦИРОВАННЫЕ ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ ЭКСПРЕССИИ N-КОНЦЕВОГО ДОМЕНА БЕЛКА CTCF/N-КОНЦЕВОГО ДОМЕНА И ДВУХ БЛИЖАЙШИХ К НЕМУ ДОМЕНОВ ТИПА "ЦИНКОВЫЙ ПАЛЕЦ" БЕЛКА CTCF/N-КОНЦЕВОГО ДОМЕНА И СЕМИ БЛИЖАЙШИХ К НЕМУ ДОМЕНОВ ТИПА "ЦИНКОВЫЙ ПАЛЕЦ" БЕЛКА CTCF, СЛИТЫХ С БЕЛКАМИ DCAS9 И EGFP | 2023 |
|
RU2836135C1 |
| Рекомбинантный слитый белок, специфически связывающий и нейтрализующий вирус бешенства, способ его получения и применение | 2023 |
|
RU2829507C1 |
| Рекомбинантная плазмидная ДНК pET-15b_T1_RL, обеспечивающая синтез рекомбинантного слитого белка, состоящего из опухоль-специфического пептида и противоопухолевого пептида RL2, и рекомбинантный слитый белок, обладающий цитотоксической активностью по отношению к раковым клеткам и таргетными свойствами к опухолевой ткани | 2016 |
|
RU2619050C1 |
| Клеточная линия PSCA-CAR-YT, обладающая поверхностной экспрессией химерных антигенных рецепторов и проявляющая цитотоксическую активность по отношению к PSCA-позитивным раковым клеткам человека | 2018 |
|
RU2712901C1 |
Группа изобретений относится к биотехнологии. Предложен химерный рекомбинантный белок MGL-M456 для терапии онкологических заболеваний, состоящий из L-метионин-гамма-лиазы, слитой с VHH антителом BCD090-М456. Также предложена генетическая конструкция, кодирующая указанный химерный белок. Группа изобретений может быть эффективно использована для терапии онкологических заболеваний. 2 н.п. ф-лы, 2 ил., 2 пр.
1. Химерный рекомбинантный белок MGL-M456 для терапии онкологических заболеваний, состоящий из L-метионин-гамма-лиазы, слитой с VHH антителом BCD090-М456, характеризующийся аминокислотной последовательностью SEQ ID №1, соответствующей аминокислотной последовательности МГЛ из Clostridium sporogenes, к С-концу которой добавлена последовательность VHH антитела BCD090-M456.
2. Генетическая конструкция, кодирующая химерный белок по п. 1, характеризующаяся тем, что имеет нуклеотидную последовательность, указанную в списке последовательностей под номером SEQ ID NO: 2, в которой последовательность гена megL С. sporogenes трансляционным слиянием соединена с последовательностью, кодирующей VHH антитело BCD090-M456, или последовательность, отличающуюся от последовательности SEQ ID NO: 2, вследствие вырожденности генетического кода.
| WO 2004112717 A3, 29.12.2004 | |||
| Способ получения фермента метионин-гамма-лиазы, противоопухолевое лекарственное средство ГЛФ МГЛ на основе этого фермента и применение этого средства для торможения роста опухоли (варианты) | 2018 |
|
RU2733440C2 |
| Однодоменное антитело, специфически связывающее рецептор ErbB3 человека, без канонической дисульфидной связи | 2018 |
|
RU2721854C1 |
| BONDAREV N | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Int J Mol Sci | |||
| Способ получения продуктов конденсации фенолов с формальдегидом | 1924 |
|
SU2022A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| ZANG X.P | |||
| et al | |||
| Targeting a methioninase-containing fusion protein to | |||
