Область техники, к которой относится изобретение
Изобретение относится к молекулярной биологии, генной инженерии, биотехнологиям и касается новых генетических конструкций, обеспечивающих экспрессию химерного белка, состоящего из N-концевого домена белка CTCF/N-концевого домена и двух ближайших к нему доменов типа «цинковый палец» белка CTCF/N-концевого домена и семи ближайших к нему доменов типа «цинковый палец» белка CTCF слитых с белками dCas9 и EGFP, пригодного к дальнейшим структурно-функциональным исследованиям, таким как создание линии трансгенных мутантных мышей с постоянной экспрессией данных химерных белков и gRNA к промотору белка, на экспрессию которого необходимо повлиять; электропорация данных плазмид и плазмид-носителей gRNA к онкосупрессору и близкому энхансеру в твердые опухоли с целью создания контакта между ними, начала экспрессии онкосупрессора и замедления развития рака; лентивирусная трансдукция предусмотренная строением векторной плазмиды в различные культуральные клеточные линии с плазмидой-носителем gRNA для различных фундаментальных эпигенетических исследований профилей контактов целевых локусов.
Уровень техники
Трехмерная организация хроматина является одним из ключевых факторов, влияющих на экспрессию генов. Различные формы его компактизации, такие как петли или топологически ассоциированные домены (ТАДы) позволяют генам взаимодействовать с регуляторными элементами. Нарушения нормального профиля таких контактов приводят к возникновению различных патологий, в частности, пороков развития и рака.
Известны множественные технологии по искусственному формированию хроматиновых контактов между двумя точками в геноме.
CLOuD9 (chromatin loop reorganization using CRISPR-dCas9). Используется плазмида, кодирующая два химерных белка - PYL1-dCas9 и ABI1-dCas9 (Morgan, S.L., Mariano, N.C., Bermudez, A., Arruda, N.L., Wu, F., Luo, Y., Shankar, G., Jia, L., Chen, H., Hu, J.-F., Hoffman, A.R., Huang, C.-C., Pitteri, S.J., Wang, K.C., 2017. Manipulation of nuclear architecture through CRISPR-mediated chromosomal looping. Nature Communications 8, 15993. https://doi.org/10.1038/ncomms15993). Две разные gRNA привлекают данные белки к двум точкам локуса. При добавлении 1мМ абсцизовой кислоты к культуре клеток, содержащих данные химерные белки и gRNAs, PYL1 и ABI1 димеризуются, замыкая хроматиновую петлю. При отмывке клеток от абсцизовой кислоты, PYL1 и ABI1 отсоединяются друг от друга и контакт исчезает. Однако, через данный метод образуются петли ограниченной длины, без участия нативных архитектурных белков, со специфическими условиями обработки, что делает систему непригодной для использования в моделях in vivo.
LADL (light-activated dynamic looping). Используются плазмида, кодирующая два химерных белка CIBN-dCas9, а также две гидовых РНК к ним и белок CRY2 (Kim, J.H., Rege, M., Valeri, J., Dunagin, M.C., Metzger, A., Titus, K.R., Gilgenast, T.G., Gong, W., Beagan, J.A., Raj, A., Phillips-Cremins, J.E., 2019. LADL: light-activated dynamic looping for endogenous gene expression control. Nature Methods 16, 633–639. https://doi.org/10.1038/s41592-019-0436-5). Под воздействием коротких световых импульсов с длиной волны 470 нм образуется тетрамер из двух белков CIBN и двух белков CRY2, замыкающий хроматиновую петлю. Аналогично варианту 1), через данный метод образуются петли ограниченной длины, без участия нативных архитектурных белков, со специфическими условиями обработки, что делает систему непригодной для использования в моделях in vivo.
SunTag-CTCF. Использование лентивирусов, несущих плазмиды, кодирующие химерные белки dCas9-10xGCN4-T2A-бластицидин и scFv-CTCF-Т2А-гидромицин (Oh, S., Shao, J., Mitra, J., Xiong, F., D’Antonio, M., Wang, R., Garcia-Bassets, I., Ma, Q., Zhu, X., Lee, J.-H., Nair, S.J., Yang, F., Ohgi, K., Frazer, K.A., Zhang, Z.D., Li, W., Rosenfeld, M.G., 2021. Enhancer release and retargeting activates disease-susceptibility genes. Nature 595, 735–740. https://doi.org/10.1038/s41586-021-03577-1). Привлечение CTCF через систему SunTag к промотору с разрушенным сайтом связывания CTCF способствует увеличению экспрессии генов данного локуса. Однако, в данной системе, используется полноразмерный вариант белка CTCF, содержащий 11 доменов вида «цинковый палец», а также С-концевой домен. Кроме того, используемая система SunTag привлекает к точке-мишени сразу 10 белков CTCF, что многократно приумножает негативные эффекты, описанные для системы 3).
Наиболее близким к заявляемому методу, является использование плазмиды, кодирующей полноразмерный вариант белка CTCF слитый с белком dCas9 (Kubo, N., Ishii, H., Xiong, X., Bianco, S., Meitinger, F., Hu, R., Hocker, J.D., Conte, M., Gorkin, D., Yu, M., Li, B., Dixon, J.R., Hu, M., Nicodemi, M., Zhao, H., Ren, B., 2021. Promoter-proximal CTCF binding promotes distal enhancer-dependent gene activation. Nature Structural & Molecular Biology 28, 152–161. https://doi.org/10.1038/s41594-020-00539-5). Привлечение CTCF через dCas9 к промотору с разрушенным сайтом связывания CTCF способствует возрастанию частоты контактов с данной точкой, но не до исходного уровня. Однако, в данной системе используется полноразмерный вариант белка CTCF, содержащий 11 доменов вида «цинковый палец», а также С-концевой домен. Подобная структура негативно влияет на процесс формирования петлевых контактов, поскольку домены вида «цинковый палец» в CTCF – ДНК-связывающие и могут конкурировать с белком dCas9 за взаимодействие с ДНК в точке посадки. Кроме того, С-концевой домен CTCF имеет сторонние функции, не затрагивающие формирование и поддержание трехмерной организации генома. Взаимодействия С-концевого домена CTCF с другими белками, такими как CENP-E, могут оказывать нежелательное влияние на экспрессию генов в локусе привлечения.
Таким образом, техническая проблема, решаемая посредством заявляемого изобретения, заключается в необходимости преодоления недостатков, присущих аналогам и прототипу за счет создания химерного белка, обеспечивающего экспрессию данного белка вместе с соответствующими gRNAs в клетках млекопитающих способного устанавливать новые хроматиновые контакты в целевом локусе, либо обеспечивать восстановление тех контактов, что были разрушены мутацией и способствовали развитию патологий.
Раскрытие изобретения
Техническим результатом изобретения является получение химерных белков, состоящих из N-концевого домена белка CTCF/ N-концевого домена и двух ближайших к нему доменов типа «цинковый палец» белка CTCF/ N-концевого домена и семи ближайших к нему доменов типа «цинковый палец» белка CTCF слитого с белками dCas9 и EGFP, обеспечивающих повышение частоты контактов между точкой привлечения данных химерных белков и другими точками целевого локуса. Данный эффект был подтвержден на модельном локусе HoxD посредством трансфекции вышеописанных плазмид с таргетными gRNA с последующим детектированием контактов в локусе методом C-TALE (из семейства так называемых С-методов – методов захвата конформации хромосом); результаты C-TALE представлены в виде фрагментов тепловых карт с разрешением 5 т.п.н. (тысяч пар нуклеотидов) см. фиг. 8, повышение частоты контактов проявляется в виде пикселей в центре фрагментов по яркости превышающих окружение, в отличие от контроля.
Технический результат достигается генетической конструкцией, кодирующей N-концевой домен белка CTCF слитый с белками dCas9 и GFP/N-концевой домен и два ближайших к нему домена типа «цинковый палец» белка CTCF слитых с белками dCas9 и GFP/N-концевой домен и семь ближайших к нему доменов типа «цинковый палец» белка CTCF слитых с белками dCas9 и GFP, представляющей собой вектор, включающий нуклеотидные последовательности SEQ ID NO: 1 или SEQ ID NO: 2 или SEQ ID NO: 3.
При этом, вектор включает следующие функциональные фрагменты: ген dCas9 из Streptococcus pyogenes с инактивирующими рестриктазную активность Cas9 мутациями D10A и H840A; ген белка GFP, выделенного из Aequorea victoria; сигнал ядерной локализации большого Т-антигена вируса SV40; промотор гена фосфоглицерат киназы; ген устойчивости к пуромицину; компоненты ретровирусного генома, необходимые для трансдукции химерных белков в геном эукариотических клеток; участок ориджин репликации; бактериальный промотор гена устойчивости к ампицилину и ген устойчивости к ампицилину. В качестве компонентов ретровирусного генома, используют 3'-LTR, 5'-LTR, MESV Ψ, gag (trancated). В качестве вектора для сборки генетической конструкции используют коммерческий вектор, выбранный из pSLQ1658-dCas9-EGFP, pHAGE-TO-dCas9-3XmCherry, pHAGE-TO-dCas9-GFP, pdCas9:BFP-humanized, pHAGE-TO-dCas9-3XGFP, lentiCRISPR v2-dCas9, pLenti_dCas9-2xAM, pLenti_dCas9-2xAM_hIRF-1.
Краткое описание чертежей
Сущность изобретения поясняется следующими иллюстративными материалами.
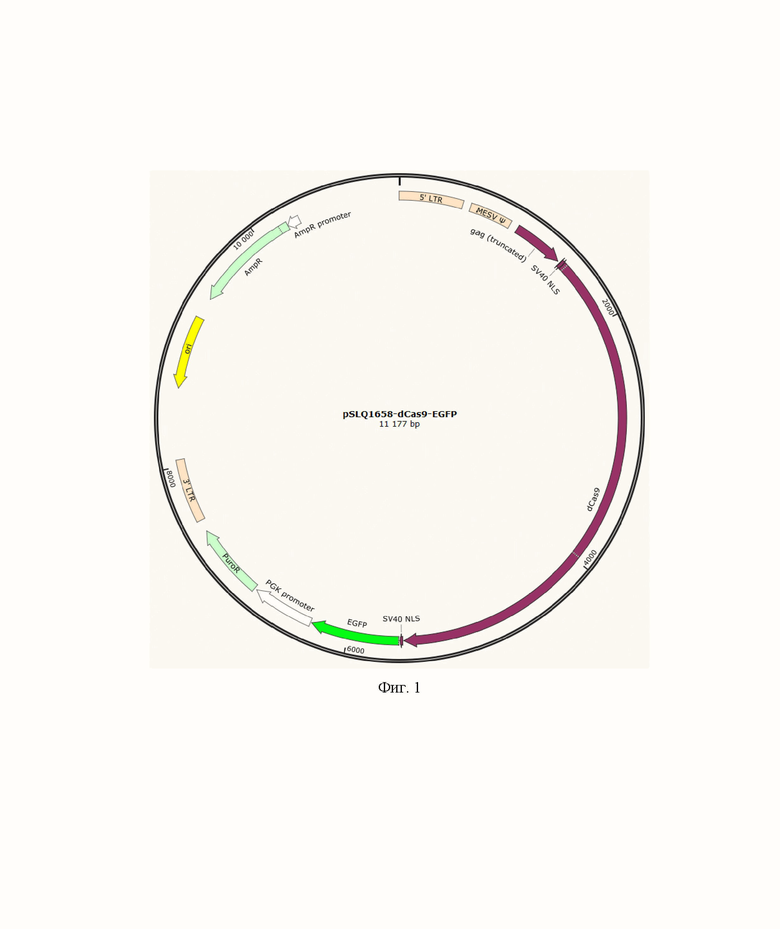
На фиг. 1 представлено схематичное изображение коммерческой плазмиды pSLQ1658-dCas9-EGFP, в которую осуществлялось клонирование фрагментов гена CTCF.
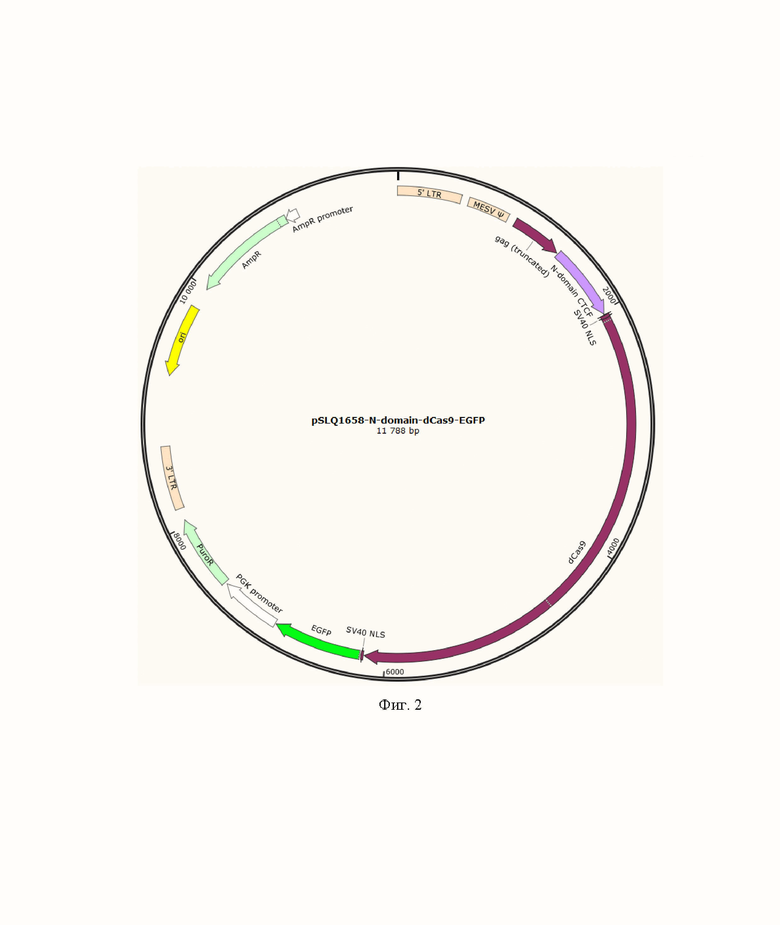
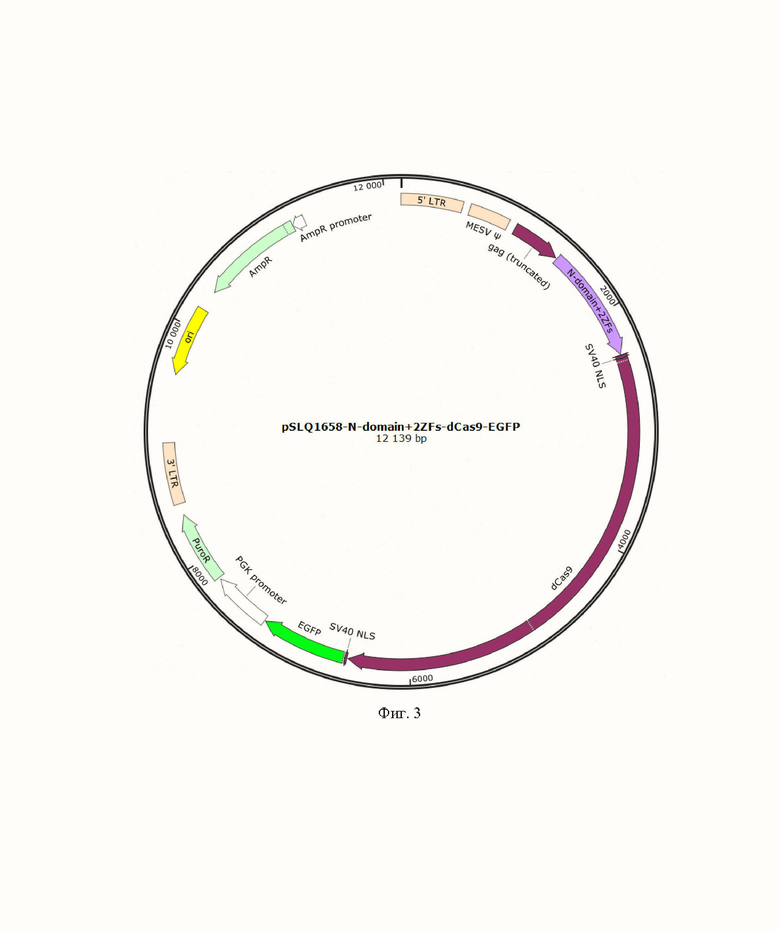
На фиг. 2, 3 и 4 представлены схематичные изображения плазмид, кодирующих химерные белки состоящих из N-концевого домена белка CTCF / N-концевого домена и двух ближайших к нему доменов типа «цинковый палец» белка CTCF/ N-концевого домена и семи ближайших к нему доменов типа «цинковый палец» белка CTCF слитых с белками dCas9 и EGFP.
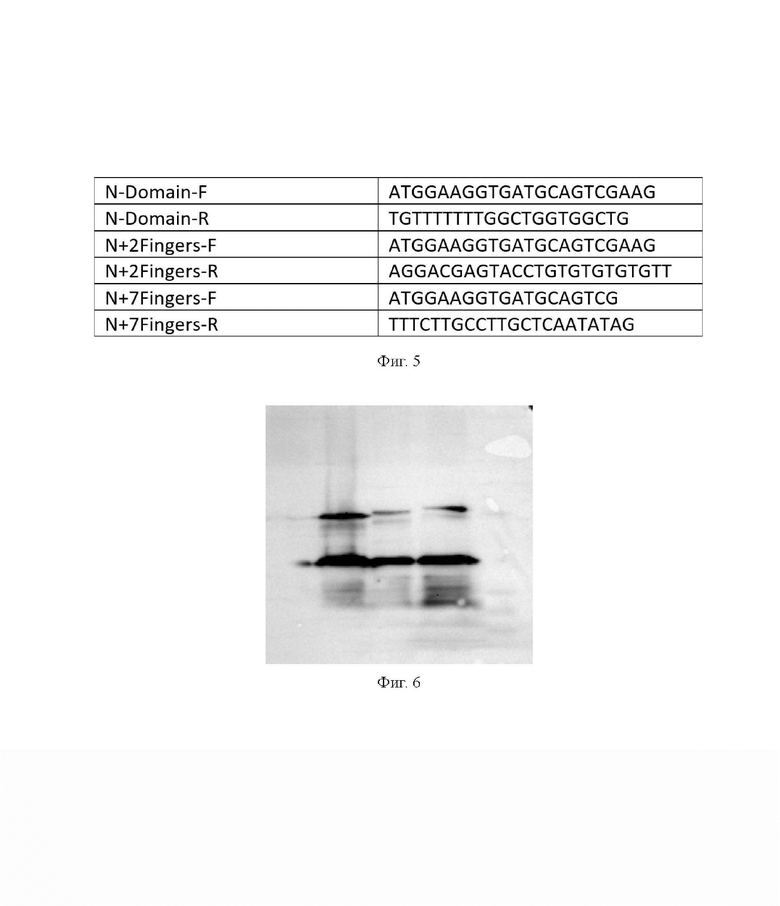
На фиг. 5 представлена таблица специализированных праймеров для синтеза фрагментов гена CTCF для вставки в коммерческую плазмиду pSLQ1658-dCas9-EGFP. Праймеры для синтеза ДНК N-концевого домена белка CTCF обозначены как N-Domain-F и N-Domain-R. Праймеры для синтеза ДНК N-концевого домена и двух ближайших к нему доменов типа «цинковый палец» белка CTCF обозначены как N+2Fingers-F и N+2Fingers-R. Праймеры для синтеза ДНК N-концевого домена и семи ближайших к нему доменов типа «цинковый палец» белка CTCF обозначены как N+7Fingers-F и N+7Fingers-R.
На фиг. 6 представлена электронная фотография результатов анализа химерных белков, получаемых с вышеописанных плазмид методом Western Blot с антителами к N-концевому домену белка CTCF. Съемка велась на приборе iBright FL1500 Imaging System (Invitrogen, США). Анализ проводился на клетках HeLa трансфецированных описываемыми плазмидами. На представленной фотографии наверху слева направо химерные белки: N-концевой домен белка CTCF слитый с dCas9 и EGFP; N-концевой домен и два ближайших к нему домена типа «цинковый палец» белка CTCF, слитых с dCas9 и EGFP; N-концевой домен и семь ближайших к нему доменов типа «цинковый палец» белка CTCF, слитых с dCas9 и EGFP. Вес химерных белков: 209, 223 и 240 кДа соответственно. Искомые полосы химерных белков видны рядом с полосой маркера PageRuler (ThermoFisher, США) в 250 кДа, а сильный сигнал ниже – нативный белок CTCF клеток HeLa весом в 120 кДа, расположенный около полосы в 130 кДа.
На фиг. 7 представлена электронная фотография результатов анализа химерных белков, получаемых с вышеописанных плазмид методом Western Blot с антителами антителами к белку EGFP. Съемка велась на приборе iBright FL1500 Imaging System (Invitrogen, США). Анализ проводился на клетках HeLa трансфецированных описываемыми плазмидами. На представленной фотографии наверху слева направо: N-концевой домен белка CTCF слитый с dCas9 и EGFP; контрольный продукт коммерческой плазмиды pSLQ1658-dCas9-EGFP без вставок; N-концевой домен и два ближайших к нему домена типа «цинковый палец» белка CTCF, слитых с dCas9 и EGFP; N-концевой домен и семь ближайших к нему доменов типа «цинковый палец» белка CTCF, слитых с dCas9 и EGFP. Контрольный химерный белок без вставки (dCas9-EGFP) весит 187 кДа, а нижние полосы – деградировавшие фрагменты.
На фиг. 8 представлены фрагменты тепловых C-TALE карт локуса chr2: 176,204kb X 176,344kb с привлечением химерных белков, состоящих из N-концевого домена белка CTCF/ N-концевого домена и двух ближайших к нему доменов типа «цинковый палец» белка CTCF/ N-концевого домена и семи ближайших к нему доменов типа «цинковый палец» белка CTCF слитых с белками dCas9 и EGFP в сравнении с контрольной картой. Из результатов C-TALE следует, что наиболее сильные хроматиновые контакты формируются при использовании плазмиды, кодирующей N-концевой домена и два ближайших к нему домена типа «цинковый палец» белка CTCF слитых с белками dCas9 и GFP.
Осуществление изобретения
Ниже представлено более подробное описание заявляемого изобретения. Настоящее изобретение может подвергаться различным изменениям и модификациям, понятным специалисту на основе прочтения данного описания. Такие изменения не ограничивают объем притязаний.
Заявляемые плазмиды были получены путем модификации коммерческой плазмиды pSLQ1658-dCas9-EGFP (#51023 в каталоге Addgene, США). Данная плазмида имеет функциональные элементы, важные для получения плазмид, представленными на фиг. 1:
1) dCas9 – ген dCas9 из Streptococcus pyogenes с инактивирующими рестриктазную активность Cas9 мутациями D10A и H840A;
2) SV40 NLS - сигнал ядерной локализации большого Т-антигена вируса SV40, важен для доставки химерных белков в ядро клетки для связывания с ДНК;
3) EGFP (enhanced GFP) – ген белка GFP, выделенного из Aequorea victoria со следующими модификациями M1_S2insV/F64L/S65T/H231L; позволяет детектировать химерные белки в клетке под лазером с λ=488 нм;
4) PGK promoter – промотор гена фосфоглицерат киназы из Saccharomyces cerevisiae, необходим для обеспечения высококопийной экспрессии гена устойчивости к пуромицину;
5) PuroR – ген устойчивости к пуромицину, может быть использован как селективный маркер для эукатиотических клеток.
6) 3'-LTR, 5'-LTR, MESV Ψ, gag (trancated) – компоненты ретровирусного генома, необходимые для трансдукции химерных белков в геном эукариотических клеток.
7) ori - ориджин репликации, важен для наработки плазмид в клетках Escherichia coli
8) AmpR promoter – бактериальный промотор гена устойчивости к ампицилину, важен для наработки плазмид в клетках Escherichia coli
9) AmpR – ген устойчивости к ампицилину, важен для наработки плазмид в клетках Escherichia coli
Эти фрагменты можно функционально разделить на три блока. Фрагменты (участки) 7, 8 и 9 должны быть расположены вместе и в той же очередности, что и в плазмиде pSLQ1658-dCas9-EGFP. Это необходимо для возможности наращивания больших объемов заявляемых плазмид (генетических конструкций) в клетках Escherichia coli. Фрагменты (участки) 6, 1, 2, 3 также должны быть расположены вместе и в той же очередности, что и в плазмиде pSLQ1658-dCas9-EGFP. Это необходимо для возможности вставки в начало рамки считывания, а также для возможности вирусной трансдукции фрагментов плазмид, кодирущих химерные белки. Фрагменты (участки) 4 и 5 также должны расположены вместе и в той же очередности, что и в плазмиде pSLQ1658-dCas9-EGFP. Это необходимо для возможности пуромициновой селекции клеток, трансфецированных заявляемыми плазмидами.
Равноценной заменой вектору pSLQ1658-dCas9-EGFP могут служить следующие плазмиды из каталога Addgene: 1) pHAGE-TO-dCas9-3XmCherry (#64108), 2) pHAGE-TO-dCas9-GFP (#64106), 3) pdCas9::BFP-humanized (#44247), 4) pHAGE-TO-dCas9-3XGFP (#64107), 5) lentiCRISPR v2-dCas9 (#112233) 6) pLenti_dCas9-2xAM (#92220) 7) pLenti_dCas9-2xAM_hIRF-1 (#92221). Данные плазмиды имеют схожий с pSLQ1658-dCas9-EGFP функционал, оставляющий при этом доступным для вставок начало химерного продукта: dCas9 из Streptococcus pyogenes с мутациями D10A и H840A, возможность детекции продукта (mcherry для 1, GFP для 2 и 4, BFP для 3, FLAG для 5, 2xAM tag для 6 и 7), а также компоненты для лентивирусной трансдукции. Плазмиды 5, 6 и 7 также несут остов gRNA, что может быть актуально при нежелательности использования котрансфекции с отдельной плазмидой-носителем gRNA.
Особенно важным свойством является свободное для клонирования начало рамки считывания dCas9 (и N-конец получаемого химерного белка), а также расположение сигнала для детектирования в конце рамки считывания (С-конец получаемого химерного белка), поскольку для правильного функционирования получаемых химерных белков необходимо, чтобы N-концевой домен белка CTCF был открыт и функционально доступен.
Модификацией, проведенной для получения плазмид, служило клонирование в плазмиду pSLQ1658-dCas9-EGFP фрагментов кДНК транскрипта 1 гена CTCF клеточной линии К562. Из общей массы всех РНК культуры клеток К562 методов обратной транскрипции была получена совокупность кДНК всех транскриптов клеток. Из него методом ПЦР с специфическими праймерами (фиг. 5) получены последовательности, кодирующие N-концевой домен белка CTCF/ N-концевой домен и два ближайших к нему домена типа «цинковый палец» белка CTCF/ N-концевой домен и семь ближайших к нему доменов типа «цинковый палец» белка CTCF. Вышеупомянутые праймеры были подобраны к началу гена CTCF и к фрагменту его ДНК кодирующему либо конец N- концевой домен, либо конец второго домена типа «цинковый палец», либо к линкерной ДНК после седьмого домена типа «цинковый палец». Последовательность ДНК гена CTCF была взята из общедоступной базы данных NCBI, а карта расположения доменов типа «цинковый палец» из статьи Pugacheva, E.M., Kubo, N., Loukinov, D., Tajmul, M., Kang, S., Kovalchuk, A.L., Strunnikov, A.V., Zentner, G.E., Ren, B., Lobanenkov, V.V., 2020. CTCF mediates chromatin looping via N-terminal domain-dependent cohesin retention. Proc Natl Acad Sci USA 117, 2020–2031. https://doi.org/10.1073/pnas.1911708117 . Праймеры были синтезированы компанией «ДНК-Синтез». Для обеспечения цельной рамки считывания между вставленными фрагментами и последовательностью dCas9-EGFP, плазмиды были разрезаны рестриктазой NcoΙ, концы ДНК в месте разреза заполнены нуклеотидами с помощью фрагмента Кленова (NEB, Великобритания), после чего вновь соединены воедино Т4 ДНК лигазой (NEB, Великобритания). При трансфекции полученных плазмид в эукариотические клетки, вес производимых химерных белков составляет 209, 223 и 240 кДа соответственно. Следует особо подчеркнуть, что в получаемых химерных белках именно N-концевой домен белка CTCF является свободным для взаимодействия, в отличие от аналогов 3) и 4), где С-концевой домен является свободным и открытым для взаимодействия, а N-концевой домен слит с другими белками и не является свободным. Данное различие очень важно, поскольку, известно, что именно через свой N-концевой домен белок CTCF связывается с белком когезином и участвует в процессах образовании и регуляции хроматиновых контактов (Pugacheva, E.M., Kubo, N., Loukinov, D., Tajmul, M., Kang, S., Kovalchuk, A.L., Strunnikov, A.V., Zentner, G.E., Ren, B., Lobanenkov, V.V., 2020. CTCF mediates chromatin looping via N-terminal domain-dependent cohesin retention. Proc Natl Acad Sci U S A 117, 2020–2031. https://doi.org/10.1073/pnas.1911708117).
Для проверки правильности вставки, полученные плазмиды с фрагментами гена CTCF были просеквенированы по Сэнгеру в компании Евроген (Россия). Для проверки цельности получаемых химерных белков использовался метод Western Blot с антителами к N-концевому домену CTCF (Active Motif, США), а также к белку GFP (любезно предоставленными лабораторией биоорганической химии ферментов ИХБФМ СО РАН) (фиг. 6 и фиг. 7).
Получение вышеописанных химерных белков дает возможность редактирования пространственной организации генома без изменений в первичной последовательности ДНК. Трехмерная организация хроматина является одним из ключевых факторов, влияющих на экспрессию генов. Различные формы его компактизации, такие как петли или топологически ассоциированные домены (ТАДы) позволяют генам взаимодействовать с регуляторными элементами. Нарушения нормального профиля таких контактов приводят к возникновению различных патологий, в частности, пороков развития и рака. Белок CTCF – один из наиболее важных участников процесса образования хроматиновых петель. Связываясь с когезином, этот белок образует петлевые контакты и тем самым определяет границы ТАДов и зоны действия регуляторных элементов. Недавние работы (1. Pugacheva, E.M., Kubo, N., Loukinov, D., Tajmul, M., Kang, S., Kovalchuk, A.L., Strunnikov, A.V., Zentner, G.E., Ren, B., Lobanenkov, V.V., 2020. CTCF mediates chromatin looping via N-terminal domain-dependent cohesin retention. Proc Natl Acad Sci U S A 117, 2020–2031. https://doi.org/10.1073/pnas.1911708117; 2. Li, Y., Haarhuis, J.H.I., Cacciatore, Á.S., Oldenkamp, R., van Ruiten, M.S., Willems, L., Teunissen, H., Muir, K.W., de Wit, E., Rowland, B.D., Panne, D., 2020. The structural basis for cohesin-CTCF anchored loops. Nature 578, 472–476. https://doi.org/10.1038/s41586-019-1910-z) показывают особенную важность N-концевого домена CTCF в установлении подобных контактов. Использование различных форм химерных белков, описанных выше позволяет создавать хроматиновые контакты разной частоты и силы.
Все три полученные плазмиды со вставой фрагментов гена CTCF, дают химерные белки способные влиять на топологию целевых локусов. Данная функция была проверена через трансфекцию клеток К562 каждой из трех плазмид и gRNA в точку chr2: 176,204 kb локуса HoxD. Образование петлевого контакта между точкой-мишенью и другим местом в локусе (приблизительно 176,344 kb) наблюдалось посредством метода C-TALE (см. фиг. 8). В результате C-TALE были получены тепловые карты контактов ДНК внутри локуса HoxD с разрешением 5 т.п.н. (тысяч пар нуклеотидов). Увеличение яркости сигнала в центре приведенных фрагментов карт свидетельствует о повышении частоты контактов между таргетной точкой chr2: 176,204 kb и точкой 176,344 kb локуса HoxD, в отличие от контроля.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННАЯ ДРОЖЖЕВАЯ ДВУГИБРИДНАЯ СИСТЕМА ДЛЯ ЭФФЕКТИВНОГО ИССЛЕДОВАНИЯ ВЗАИМОДЕЙСТВИЯ МЕЖДУ БЕЛКАМИ И ИХ ДОМЕНАМИ. | 2012 |
|
RU2529356C2 |
| Применение домена белка S-слоя из Lactobacillus brevis в качестве компонента системы для экспонирования слитых белков на поверхности клеток молочнокислых бактерий | 2016 |
|
RU2639246C1 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2712833C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2568056C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К КАТАЛИТИЧЕСКОМУ ДОМЕНУ FOKI НУКЛЕАЗЫ, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2830090C2 |
| БИОМОЛЕКУЛЯРНЫЕ КОМПЛЕКСЫ ДЛЯ СВЯЗЫВАНИЯ С МОЛЕКУЛАМИ ДНК НА ОСНОВЕ CAS-БЕЛКОВ И ХИМЕРНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ РНК, СОДЕРЖАЩИХ ДИМЕРИЗУЮЩИЕСЯ РНК-ПОСЛЕДОВАТЕЛЬНОСТИ | 2023 |
|
RU2835928C1 |
| ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ОПЕРАТИВНО СЛИТЫЙ ИНТРАМОЛЕКУЛЯРНЫЙ ДИМЕР БЕЛКА KillerRed, КАССЕТА ЭКСПРЕССИИ, КЛЕТКА, ПРОДУЦИРУЮЩАЯ ХИМЕРНЫЙ БЕЛОК И СОДЕРЖАЩАЯ КАССЕТУ ЭКСПРЕССИИ, ВЫДЕЛЕННЫЙ ХИМЕРНЫЙ БЕЛОК | 2011 |
|
RU2491342C1 |
| Универсальный эпигеномный репрессор транскрипции целевых генов в клетках человека | 2020 |
|
RU2748997C1 |
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |

Изобретение относится к области биотехнологии, конкретно к генетической конструкции для экспрессии N-концевого домена белка CTCF, слитого с белками dCas9 и GFP. Данные химерные белки могут быть использованы для модификации топологии геномных локусов и образования хроматиновых петель. Предложенные генетические конструкции имеют оптимизированные нуклеотидные последовательности, кодирующие части нативного человеческого архитектурного белка CTCF, но при этом минимизирующие сторонние эффекты С-концевого домена данного белка. Котрансфекция данных генетических конструкций вместе с gRNA в эукариотическую систему позволяет влиять на регуляторные контакты любого локуса генома. 3 з.п. ф-лы, 8 ил.
1. Генетическая конструкция для экспрессии N-концевого домена белка CTCF, слитого с белками dCas9 и GFP, характеризующаяся тем, что представляет собой вектор, включающий нуклеотидную последовательность SEQ ID NO: 1.
2. Генетическая конструкция по п. 1, характеризующаяся тем, что вектор включает следующие функциональные фрагменты: ген dCas9 из Streptococcus pyogenes с инактивирующими рестриктазную активность Cas9 мутациями D10A и H840A; ген белка GFP, выделенного из Aequorea victoria; сигнал ядерной локализации большого Т-антигена вируса SV40; промотор гена фосфоглицерат киназы; ген устойчивости к пуромицину; компоненты ретровирусного генома, необходимые для трансдукции химерных белков в геном эукариотических клеток; участок ориджин репликации; бактериальный промотор гена устойчивости к ампицилину и ген устойчивости к ампицилину.
3. Генетическая конструкция по п. 2, характеризующаяся тем, что в качестве компонентов ретровирусного генома используют 3'-LTR, 5'-LTR, MESV Ψ, gag truncated.
4. Генетическая конструкция по п. 2, характеризующаяся тем, что в качестве вектора для сборки генетической конструкции используют вектор, выбранный из pSLQ1658-dCas9-EGFP, pHAGE-TO-dCas9-3XmCherry, pHAGE-TO-dCas9-GFP, pdCas9:BFP-humanized, pHAGE-TO-dCas9-3XGFP, lentiCRISPR v2-dCas9, pLenti_dCas9-2xAM, pLenti_dCas9-2xAM_hIRF-1.
| US 11293018 B2, 05.04.2022 | |||
| NAOKI KUBO et al., Promoter-proximal CTCF binding promotes distal enhancer-dependent gene activation, Nat | |||
| Struct | |||
| & Mol | |||
| Biol., 2021, vol | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Способ образования азокрасителей на волокнах | 1918 |
|
SU152A1 |
| STEFANIE L MORGAN et al., CRISPR-Mediated Reorganization of Chromatin Loop Structure, J Vis Exp., 2018, vol | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| САЛЬНИКОВ П.А | |||
| Увеличение | |||

