ПЕРЕКРЕСТНЫЕ ССЫЛКИ
[0001] Настоящая заявка испрашивает приоритет на основании предварительной заявки на патент США №62/451819, поданной 30 января 2017 года; на патент США №62/504626, поданной 11 мая 2017 года; на патент США №62/512312, поданной 30 мая 2017 года, и на патент США №62/588985, поданной 21 ноября 2017 года, содержание которых полностью включено в настоящее описание посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и полностью включен в настоящее описание посредством ссылки. Копия ASCII, созданная 29 января 2018 года, называется INX00382_SL.txt и имеет размер 65784 байт.
УРОВЕНЬ ТЕХНИКИ
[0003] Соединение 2,3-бутандиол («2,3-BDO»), также известное как 2,3-бутиленгликоль, диметиленгликоль, диметилэтиленгликоль и бутан-2,3-диол (С4Н10О2; №CAS 513-85-9), представляет собой высокоценное химическое вещество, которое в настоящее время в основном получают из нефтяных источников. 2,3-BDO имеет широкий ряд промышленных применений. Например, 2,3-BDO можно применять в качестве предшественника для получения различных пластиков, пестицидов, синтетического каучука, печатных красок, ароматизирующих веществ, фумигантов, увлажняющих и смягчающих агентов, взрывчатых веществ, пластификаторов, пищевых продуктов и фармацевтических препаратов (Garg S.K. and Jain A. «Fermentative production of 2,3-butanediol», Bioresource Technology, p. 103-109 (1995)).
[0004] 2,3-BDO в настоящее время получают из сырой нефти. Тем не менее 2,3-BDO также продуцируется различными микроорганизмами, и его можно обнаружить в масле какао, в корнях руты душистой (Ruta graveolens), сладкой кукурузе и протухших мидиях. 2,3-BDO также является побочным продуктом спиртового брожения у дрожжей, и он обычно является одним из самых распространенных микрокомпонентов вина. Это происходит за счет уменьшения содержания ацетоина. (Romano, P. and Suzzi, G., «Origin and Production of Acetoin during Wine Yeast Fermentation)), Applied and Enviromental Microbiology, p. 309-315 (1996)).
[0005] В последние годы наблюдался некоторый интерес к получению 2,3-BDO путем ферментации. Ферментация обычно включает обеспечение источника углерода (обычно сахара) и его ферментацию с применением микроорганизма, который способен превращать указанный источник углерода в желаемый продукт.
[0006] Предпринимались многочисленные попытки сконструировать штаммы Saccharomyces cerevisiae с пониженным выходом ацетоина путем переориентации атомов углерода в направлении глицерина и 2,3-BDO с получением дрожжей, обеспечивающих низкое содержание спирта, с желаемыми органолептическими свойствами, позволяющими уменьшить содержание этанола в винах вплоть до 3°С. (Ehsani, М., et al., «Engineering of 2,3-butanediol dehydrogenase to reduce acetoin formation by glycerol-overproducing, low-alcohol Saccharomyces cerevisiae», Applied and Environmental Microrobiology, p. 3196-3205 (2009)).
[0007] Затраты на производство химических веществ, таких как 2,3-BDO, путем ферментации обычно зависят от применяемого источника углерода. В целом сахара являются дорогостоящими источниками углерода, и их применение также приводит к сокращению поставок продовольствия. В настоящее время одним из самых экономически эффективных и широко распространенных источников углерода является природный газ. Основным источником углерода в природном газе является метан (CH4), С1-углеродное соединение. Применяя такие дешевые источники углерода, как, например, метан, можно без экономического ущерба получать 2,3-BDO.
[0008] В настоящее время 2,3-BDO также получают с применением некоторых негенетически модифицированных микроорганизмов при очень малых титрах. При таких титрах стоимость ферментации была бы слишком велика, чтобы быть экономически целесообразной. Таким образом, для получения 2,3-BDO на экономически приемлемом уровне необходимо применение генной инженерии. Задача заключается в разработке способов ферментации и конструировании микроорганизмов для эффективного превращения дешевых источников углерода, таких как метан, в 2,3-BDO с применением способов ферментации.
[0009] Настоящее изобретение относится к микроорганизмам, таким как метанотрофы или дрожжи, генетически модифицированным для существенного улучшения биосинтеза 2,3-BDO.
ВКЛЮЧЕНИЕ ПОСРЕДСТВОМ ССЫЛКИ
[0010] Все публикации, патенты и патентные заявки, приведенные в настоящем документе, включены в него посредством ссылки в той же степени, как если бы каждая отдельная публикация, патент или патентная заявка были напрямую или косвенно указаны для включения посредством ссылки. В случае противоречия между термином в данном документе и термином во включенной ссылке, следует отдавать предпочтение терминам из настоящего документа.
[0011] Ссылка на любую публикацию предназначена для ее раскрытия до даты подачи, и ее не следует рассматривать как признание того, что настоящее изобретение не имеет права допустить такую публикацию в силу предшествующего изобретения. Кроме того, указанные даты публикации могут отличаться от фактических дат публикации, которые, возможно, должны подтверждаться отдельно.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0012] В настоящем описании предложены генетически модифицированные микроорганизмы, способные продуцировать желаемое органическое соединение, начиная с одноуглеродной молекулы углеводорода, такой как метан. Раскрыты различные способы получения указанного желаемого органического соединения, включая применение генетически модифицированных микроорганизмов.
[0013] Например, предложен генетически модифицированный микроорганизм, способный превращать С1-углеродное соединение в 2,3-бутандиол (2,3-BDO). Примерами С1-углеродных соединений, которые указанные микроорганизмы могут превращать, могут являться монооксид углерода (СО), диоксид углерода (СО2), метан (CH4) или любая их комбинация. Указанный генетически модифицированный микроорганизм может содержать один или более генов, кодирующих гетерологичные ферменты, такие как, например, ацетоинредуктаза, альфа-ацетолактатдекарбоксилаза (budA) и/или ацетолактатсинтаза (AlsS). Ген AlsS в этих микроорганизмах может экспрессироваться временно. Один или более указанных генов могут находиться под контролем переключателя, такого как, например, индуцируемый или репрессируемый промотор, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан.
[0014] Кодируемая AlsS может, например, содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична любой из последовательностей SEQ ID NO 1, 3 или 19. Кодируемая альфа-ацетолактатдекарбоксилаза (budA) может, например, содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична аминокислотной последовательности SEQ ID NO 7. Кодируемая ацетоинредуктаза может, например, содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична аминокислотной последовательности SEQ ID NO 9 Кодируемая ацетоинредуктаза может, например, представлять собой НАДФН-зависимую ацетоинредуктазу грамположительной бактерии, например, из рода Clostridium, например, Clostridium autoethanogenum. В некоторых случаях ацетоинредуктаза может быть НАДФН-зависимой. В некоторых случаях ацетоинредуктаза может быть НАДН-зависимой.
[0015] Генетически модифицированный организм может, например, быть метанотрофом, как, например, из рода Methylobacter, Methylomicrobium, Methylomonas, Methylocaldum, Methylococcus, Methylosoma, Methylosarcina, Methylothermus, Methylohalobius, Methylogaea, Methylovulum, Crenothrix, Clonothrix, Methylosphaera, Methylocapsa, Methylocella, Methylosinus, Methylocystis или Methyloacidophilum. Указанный метанотроф может быть из рода Methylococcus, например, из вида Methylococcus capsulatus.
[0016] Генетически модифицированный микроорганизм также может быть прокариотом. В некоторых случаях генетически модифицированный микроорганизм может представлять собой бактерию, дрожжи или водоросль.
[0017] В некоторых случаях генетически модифицированные микроорганизмы могут также продуцировать большее количество 2,3-BDO при 42°С по сравнению с теми же организмами при 37°С. В некоторых случаях генетически модифицированные микроорганизмы могут также продуцировать большее количество 2,3-BDO при 41°С по сравнению с теми же организмами при 37°С. В некоторых случаях генетически модифицированный микроорганизм может продуцировать большее количество 2,3-BDO при 42°С по сравнению с тем же организмом при 45°С. В некоторых случаях генетически модифицированный микроорганизм может продуцировать большее количество 2,3-BDO при 41°С по сравнению с тем же организмом при 45°С. В некоторых случаях генетически модифицированный микроорганизм может продуцировать большее количество 2,3-BDO при 37°С по сравнению с тем же организмом при 45°С.
[0018] В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, интегрируют при помощи вектора интеграции в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессируется в эписомальном векторе.
[0019] В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген.
[0020] Гетерологичные гены в векторе могут быть представлены в определенном порядке до, во время или после приведения в контакт с микроорганизмом. Например, ген может располагаться ближе к 5'-концу, чем любой другой гетерологичный ген в векторе до приведения в контакт с микроорганизмом. Однако после контакта с указанным микроорганизмом ген можно поместить в положение, или в вектор можно вставить другой ген, когда указанный ген уже будет располагаться ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген в данном векторе. Например, векторы можно модифицировать внутри микроорганизма таким образом, что будет изменен порядок генов. В некоторых случаях, определенного порядка генов можно достигнуть после того, как один или более гетерологичных генов будут вставлены в геном указанного микроорганизма. Например, для достижения определенного порядка генов в геноме микроорганизма можно применять различные векторы интеграции.
[0021] В настоящем описании также раскрыты векторы, которые содержат два или более из: гена ацетоинредуктазы (например, НАДФН-зависимой), гена альфа-ацетолактатдекарбоксилазы (budA) и гена AlsS. В некоторых случаях ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях различные гены могут находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Различные гены могут находиться под контролем разных промоторов, как, например, конститутивно экспрессируемого промотора или неконститутивно экспрессируемого промотора. Применяемые промоторы также могут быть активны внутри метанотрофа. Примеры таких векторов включают такие, которые содержат нуклеотидную последовательность, которая по меньшей мере на 90% идентична любой из последовательностей SEQ ID NO 15-18. В некоторых случаях указанный вектор представляет собой вектор интеграции, тогда как в других случаях указанный вектор представляет собой эписомально экспрессируемый вектор.
[0022] В настоящем описании также раскрыт способ получения генетически модифицированного микроорганизма, способного превращать С1-углеродное соединение в 2,3-BDO, включающий трансформацию микроорганизма при помощи нуклеиновой кислоты, которая экспрессирует по меньшей мере один гетерологичный ген, кодирующий i) ацетоинредуктазу (например, НАДФН-зависимую); ii) альфа-ацетолактатдекарбоксилазу (budA); iii) ацетолактатсинтазу (AlsS) или iv) любую их комбинацию. По меньшей мере один гетерологичный ген может, например, находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. В некоторых случаях при помощи указанного способа можно получать генетически модифицированные микроорганизмы, которые могут продуцировать большее количество 2,3-BDO при 42°С по сравнению с теми же организмами при 37°С. В некоторых случаях при помощи указанного способа можно получать генетически модифицированные микроорганизмы, которые могут продуцировать большее количество 2,3-BDO при 41°С по сравнению с теми же организмами при 37°С. В некоторых случаях при помощи указанного способа можно получать генетически модифицированные микроорганизмы, которые могут продуцировать большее количество 2,3-BDO при 42°С по сравнению с теми же организмами при 45°С. В некоторых случаях при помощи указанного способа можно получать генетически модифицированные микроорганизмы, которые могут продуцировать большее количество 2,3-BDO при 41°С по сравнению с теми же организмами при 45°С. В некоторых случаях при помощи указанного способа можно получать генетически модифицированные микроорганизмы, которые могут продуцировать большее количество 2,3-BDO при 37°С по сравнению с теми же организмами при 45°С.
[0023] В некоторых случаях указанные способы могут включать микроорганизмы, в которых ген(-ы), кодирующий гетерологичную ацетоинредуктазу, гетерологичную альфа ацетолактатдекарбоксилазу и/или гетерологичную ацетолактатсинтазу, интегрируют при помощи вектора интеграции в геном указанного микроорганизма. В некоторых случаях указанные способы могут включать микроорганизмы, в которых ген(-ы), кодирующий(-ие) гетерологичную ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессирует(-ют)ся в эписомальном векторе. В некоторых случаях указанный способ может включать микроорганизм, в котором ген(-ы), кодирующий(-ие) гетерологичную ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессирует(-ют)ся как в эписомальном векторе, так и интегрирован(-ы) в геном указанного микроорганизма (например, при помощи вектора интеграции).
[0024] В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген.
[0025] Также раскрыт способ получения 2,3-BDO, включающий (а) приведение генетически модифицированного микроорганизма в контакт с С1-углеродом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий (i) ацетоинредуктазу (например, НАДФН-зависимую); (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS или (iv) любую их комбинацию; и (b) выращивание указанного микроорганизма с получением 2,3-BDO. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, интегрируют при помощи вектора интеграции в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессируется в эписомальном векторе. В некоторых случаях указанный способ может включать микроорганизм, в котором ген(-ы), кодирующий(-ие) гетерологичную ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессирует(-ют)ся как в эписомальном векторе, так и интегрирован(-ы) в геном указанного микроорганизма (например, при помощи вектора интеграции). В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ может включать выращивание указанного микроорганизма при температуре от 32°С до 49°С. В некоторых случаях микроорганизм можно выращивать при температуре от 37°С до 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 41°С. В некоторых случаях по меньшей мере один гетерологичный ген может, например, находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемент, такого как лантан. Также указанный микроорганизм можно сначала вырастить в среде, которая содержит редкоземельный металл, такой как лантан (например, по меньшей мере 1 мкМ лантана), а затем впоследствии редкоземельный металл, такой как лантан, можно разбавить. Это может происходить до выращивания микроорганизма с получением 2,3-BDO. 2,3-BDO, полученный этим способом, можно выделить и, в некоторых случаях, он может быть по существу чистым.
[0026] Кроме того, раскрыт способ получения ацетоина, включающий (а) приведение генетически модифицированного микроорганизма в контакт с С1-углеродным соединением, при этом указанный микроорганизм содержит гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу (budA); и (b) выращивание указанного микроорганизма с получением ацетоина. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, интегрируют при помощи вектора интеграции в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, экспрессируется в эписомальном векторе. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, экспрессируется как в эписомальном векторе, так и интегрирован в геном указанного микроорганизма (например, при помощи вектора интеграции). Указанный способ может включать выращивание микроорганизма при температуре от 32°С до 49°С. В некоторых случаях микроорганизм можно выращивать при температуре от 37°С до 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 41°С. В некоторых случаях ген budA может, например, находиться под контролем переключателя, как, например, индуцируемого или репрессируемого переключателя, например, чувствительного к арабинозе или лантану переключателя. В некоторых случаях микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана). Впоследствии лантан можно использовать, удалить и/или разбавить. Это может происходить до выращивания микроорганизма с получением ацетоина. Ацетоин, полученный этим способом, можно выделить и, в некоторых случаях, он может быть по существу чистым. В случае, если ацетоин, полученный таким способом, не является по существу чистым, можно также выделить неацетоиновые побочные продукты, такие как 2,3-BDO.
[0027] После получения 2,3-BDO его можно превратить в другие желаемые продукты, например, такие как бутадиен или метилэтилкетон (MEK). Таким образом, в настоящем описании также раскрыт способ получения бутадиена, включающий (а) приведение генетически модифицированного микроорганизма в контакт с С1-углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий (i) НАДФН-зависимую ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS или (iv) любую их комбинацию; и (b) выращивание указанного микроорганизма с получением 2,3-BDO; и (с) приведение полученного на стадии (b) 2,3-BDO в контакт с катализатором с получением бутадиена. В некоторых случаях полученный на стадии (b) 2,3-BDO удаляют до стадии (с). В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, интегрируют при помощи вектора интеграции в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессируется в эписомальном векторе. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессируется как в эписомальном векторе, так и интегрирован в геном микроорганизма (например, при помощи вектора интеграции). В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ может включать выращивание микроорганизма при температуре от 32°С до 49°С. В некоторых случаях микроорганизм можно выращивать при температуре от 37°С до 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 41°С. Кроме того, катализатором может быть любой катализатор, который может дегидратировать 2,3-BDO, как, например, катализатор дигидрофосфат цезия (CsH2PO4), нанесенный на SiO2. Бутадиен, полученный этим способом, можно выделить и, в некоторых случаях, он может быть по существу чистым. Бутадиен в дальнейшем также можно переработать в синтетический каучук.
[0028] Также раскрыт способ получения MEK, включающий (а) приведение генетически модифицированного микроорганизма в контакт с С1-углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий (i) НАДФН-зависимую ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) ацетолактатсинтазу (AlsS) или (iv) любую их комбинацию; и (b) выращивание указанного микроорганизма с получением 2,3-BDO; и (с) приведение полученного на стадии (b) 2,3-BDO в контакт с катализатором с получением MEK. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, интегрируют при помощи вектора интеграции в геном указанного микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессируется в эписомальном векторе. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и/или ацетолактатсинтазу, экспрессируется как в эписомальном векторе, так и интегрирован в геном микроорганизма (например, при помощи вектора интеграции). В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ может включать выращивание микроорганизма при температуре от 32°С до 49°С. В некоторых случаях микроорганизм можно выращивать при температуре от 37°С до 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 42°С. В некоторых случаях микроорганизм можно выращивать при температуре примерно 41°С. В некоторых случаях катализатор представляет собой кислоту в виде твердого вещества. MEK, полученный этим способом, можно выделить, и в некоторых случаях он может быть по существу чистым. MEK в дальнейшем также можно переработать в пластики, ткани, парафиновый воск, лак, олифу, средство для удаления краски, клеи и/или чистящие средства. В некоторых случаях катализатор на стадии (с) может представлять собой диолдегидратазу (В12). В некоторых случаях ген диолдегидратазы может экспрессироваться одним и тем же или другим генетически модифицированным микроорганизмом. Таким образом, раскрытый в настоящем описании способ получения MEK включает (а) приведение генетически модифицированного микроорганизма в контакт с С1-углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий (i) НАДФН-зависимую ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS; (iv) диолдегидратазу или (v) любую их комбинацию; и (b) выращивание указанного микроорганизма с получением MEK.
[0029] Также раскрыты выделенные полинуклеиновые кислоты, содержащие нуклеотидные последовательности, которые по меньшей мере на 84% идентичны последовательности SEQ ID NO 2, по меньшей мере на 88% идентичны последовательности SEQ ID NO 4 или по меньшей мере на 60% идентичны последовательности SEQ ID NO 20. Эти нуклеотидные последовательности могут кодировать белок, обладающий ацетолактатсинтазной активностью. Также раскрыты выделенные полинуклеиновые кислоты, содержащие нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 6 или 8. Эти нуклеотидные последовательности могут кодировать белок, обладающий альфа-ацетолактатдекарбоксилазной активностью. Кроме того, раскрыты выделенные полинуклеиновые кислоты, которые по меньшей мере на 85% идентичны любой из последовательностей SEQ ID NO 10, 12 или 14. Эти нуклеотидные последовательности могут кодировать белок, обладающий бутандиолдегидрогеназной активностью.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0030] На ФИГ. 1 показан метаболический путь превращения метана (CH4) в 2,3-BDO.
[0031] На ФИГ. 2 показана эффективность сконструированных штаммов в эксперименте по ферментации клеток с высокой плотностью в течение 72 часов. Слева направо представлены следующие исследованные штаммы: XZ58 (MF1904), XZ06 (MF1905), XZ59 (MF1906) и XZ08 (MF1907). Образование 2,3-BDO и ацетоина исследовали в различные моменты времени.
[0032] На ФИГ. 3 показана нуклеотидная последовательность кассеты экспрессии гена пути 2,3-BDO в штамме XZ58 (SEQ ID NO 15). Следуя в направлении от 5' к 3', последовательности, состоящие из подчеркнутых заглавных букв, представляют собой промотор pBAD. Инициатор ATG и терминатор ТАА для гена g.Bsu AlsS (ацетолактатсинтаза) обозначены жирным шрифтом и заглавными буквами, а кодирующая область - строчными буквами. Сайт связывания с рибосомой rbsGTW001 обозначен заглавными буквами в обведенной области. Инициатор ATG и терминатор TGA для гена g.Kpn BudA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в курсиве. Терминатор rrnB обозначен заглавными буквами, за которым следует промотор pmxaF, обозначенный подчеркнутыми строчными буквами в курсиве. Инициатор ATG и терминатор TGA для гена g.Cau ButA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в жирном курсиве. Терминатор лямбда Т0 обозначен заглавными буквами в курсиве.
[0033] На ФИГ. 4 показана нуклеотидная последовательность кассеты экспрессии гена пути 2,3-BDO в штамме XZ59 (SEQ ID NO 16). Следуя в направлении от 5' к 3', последовательности, состоящие из подчеркнутых заглавных букв, представляют собой промотор pBAD. Инициатор ATG и терминатор ТАА для гена g.Bsu AlsS обозначены жирным шрифтом и заглавными буквами, а кодирующая область - строчными буквами. Сайт связывания с рибосомой rbsGTW001 обозначен заглавными буквами в обведенной области. Инициатор ATG и терминатор TGA для гена g.Kpn BudA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в курсиве. Терминатор rrnB обозначен заглавными буквами, за которым следует промотор pmxaF, обозначенный подчеркнутыми строчными буквами в курсиве. Инициатор ATG и терминатор TGA для гена g.Bsu ButA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в жирном курсиве. Терминатор лямбда Т0 обозначен заглавными буквами в курсиве.
[0034] На ФИГ. 5 показана нуклеотидная последовательность кассеты экспрессии гена пути 2,3-BDO в штамме XZ06 (SEQ ID NO 17). Следуя в направлении от 5' к 3', последовательности, состоящие из подчеркнутых заглавных букв, представляют собой промотор pBAD. Инициатор ATG и терминатор ТАА для гена g.Bsu AlsS обозначены жирным шрифтом и заглавными буквами, а кодирующая область - строчными буквами. Сайт связывания с рибосомой rbsGTW001 обозначен заглавными буквами в обведенной области. Инициатор ATG и терминатор TGA для гена g.Kpn BudA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в курсиве, за которой следует дополнительный сайт связывания с рибосомой rbsGTW001, обозначенный заглавными буквами в обведенной области. Инициатор ATG и терминатор TGA для гена g.Cau ButA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в жирном курсиве. Терминатор rrnB обозначен заглавными буквами.
[0035] На ФИГ. 6 показана нуклеотидная последовательность кассеты экспрессии гена пути 2,3-BDO в штамме XZ08 (SEQ ID NO 18). Следуя в направлении от 5' к 3', последовательности, состоящие из подчеркнутых заглавных букв, представляют собой промотор pBAD. Инициатор ATG и терминатор ТАА для гена g.Bsu AlsS обозначены жирным шрифтом и заглавными буквами, а кодирующая область - строчными буквами. Сайт связывания с рибосомой rbsGTW001 обозначен заглавными буквами в обведенной области. Инициатор ATG и терминатор TGA для гена g.Kpn BudA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в курсиве, за которой следует дополнительный сайт связывания с рибосомой rbsGTW001, обозначенный заглавными буквами в обведенной области. Инициатор ATG и терминатор TGA для гена g.Bsu ButA обозначены заглавными буквами в жирном курсиве, а кодирующая область - строчными буквами в жирном курсиве. Терминатор rrnB обозначен заглавными буквами.
[0036] ФИГ. 7А и 7В. На ФИГ. 7А показано образование ацетоина и 2,3-BDO 21 различным штаммом через 96 часов после разбавления лантансодержащей среды. Штаммы и генотипы этих штаммов перечислены в таблицах 3 и 4. Для штаммов с 1 по 21 полученные титры измеряли через 96 часов после разбавления культуры в соотношении 1:10 (10Х) свежей средой, тогда как для штаммов с 22 по 42 полученные титры измеряли после разбавления в соотношении 1:50 (50Х). Штаммы с 22 по 27 продуцировали высокие уровни 2,3-BDO по сравнению со штаммами, подвергавшимися меньшему разбавлению до фазы образования 2,3-BDO. На ФИГ. 7В показано образование ацетоина и 2,3-BDO 21 различным штаммом через 120 часов после разбавления лантансодержащей среды. Штаммы и генотипы этих штаммов перечислены в таблицах 3 и 4. Для штаммов с 1 по 21 полученные титры измеряли через 120 часов после разбавления культуры в соотношении 1:10 (10Х) свежей средой, тогда как для штаммов с 22 по 42 полученные титры измеряли после разбавления в соотношении 1:50 (50Х). Штаммы с 22 по 27 продуцировали высокие уровни 2,3-BDO по сравнению со штаммами, подвергавшимися меньшему разбавлению до фазы образования 2,3-BDO.
[0037] На ФИГ. 8 показано, что штаммы, экспрессирующие AlsS Bacillus licheniformis, демонстрируют значимо улучшенные титры 2,3-BDO. Для одного штамма, экспрессирующего AlsS Bacillus licheniformis (XZ562), титры 2,3-BDO увеличивались в среднем на 44,6% по сравнению со штаммом XZ58 (описаны на ФИГ. 3) в течение периода ферментации. Другой биологический репликат (XZ561) также продуцировал значимо более высокие средние титры 2,3-BDO по сравнению со штаммом XZ58.
[0038] На ФИГ. 9 показаны титры 2,3-BDO для 7 штаммов метанотрофов, генетически сконструированных для получения 2,3-BDO, при ферментации при температуре либо 42°С, либо 37°С. Видно, что все штаммы продуцировали более высокие титры 2,3-BDO при инкубации при 42°С. Штамм МВС2122 продуцировал примерно на 50% больше 2,3-BDO при 42°С по сравнению с 37°С.
[0039] На ФИГ. 10 показаны титры 2,3-BDO для 3 штаммов метанотрофов, генетически сконструированных для получения 2,3-BDO, при ферментации при температуре либо 45°С, либо 37°С. Видно, что все штаммы продуцировали более низкие титры 2,3-BDO при инкубации при 45°С. Большинство штаммов продуцировало примерно на 50% меньше 2,3-BDO при 45°С по сравнению с 37°С. Эписомальный штамм МВС1322 показал лучшие результаты при селективном давлении антибиотиков как при 37°С, так и при 45°С, что свидетельствует о проблеме со стабильностью. Интегрированные штаммы, напротив, были способны сохранять свою продуктивность без избирательного давления.
[0040] На ФИГ. 11 показана производительность (отношение 2,3-BDO/OD) штамма метанотрофа, генетически модифицированного для получения 2,3-BDO, при ферментации при 37°С, 41°С, 43°С, 45°С и 47°С. Наилучшую производительность наблюдали при температуре 41°С.
[0041] На ФИГ. 12 показано, что штаммы метанотрофов, которые содержали дополнительные копии ферментов пути 2,3-BDO (Штаммы В и С) в эписомальном векторе, продуцировали в 3 раза больше 2,3-BDO по сравнению с одиночно интегрированным штаммом (Штамм А) в экспериментах со встряхиваемыми флаконами. Штамм В и штамм С предварительно культивировали в среде с концентрацией лантана 10 мкМ в присутствии канамицина при 37°С в течение 48 часов. Штамм А предварительно культивировали в тех же условиях, за исключением того, что не добавляли канамицин. После 48 часов лантан разбавляли в 50 раз (50Х), а титры 2,3-BDO измеряли через 96 часов.
[0042] На ФИГ. 13 показано действие (в эксперименте со встряхиваемым флаконом) предварительного культивирования штамма метанотрофа, содержащего гены, кодирующие ферменты пути 2,3-BDO, в случае, когда эти гены экспрессируются эписомально (Штамм D). Этот штамм метанотрофа предварительно культивировали в среде с концентрацией лантана 10 мкМ, 3 мкМ и 1 мкМ. Среду для получения впоследствии разбавляли в 50 раз. Более высокие концентрации лантана приводили к более высоким титрам 2,3-BDO для эписомально экспрессируемых штаммов метанотрофов.
[0043] На ФИГ. 14 показано действие (в эксперименте со встряхиваемым флаконом) предварительного культивирования штамма метанотрофа, содержащего гены, кодирующие ферменты пути 2,3-BDO, в случае, когда эти гены интегрированы в геном указанного метанотрофа (Штамм А). Этот штамм предварительно культивировали в среде с концентрацией лантана 35 мкМ, 10 мкМ, 3 мкМ, 1 мкМ и 0 мкМ. Затем среду для получения разбавляли в 50 раз (50Х). Уровни образования 2,3-BDO увеличились в случае, когда указанный штамм предварительно культивировали в среде с более низкой концентрацией лантана.
[0044] На ФИГ. 15 показано действие предварительного культивирования при различных концентрациях лантана (10 мкМ, 3 мкМ и 1 мкМ) двух штаммов с ФИГ. 12 (Штаммы В и С) в эксперименте со встряхиваемым флаконом. Оба штамма содержали как интегрированную, так и эписомально экспрессируемую копию фермента пути 2,3-BDO. В целом штаммы, содержащие эписомально экспрессируемые ферменты пути 2,3-BDO, приводят к получению более высоких титров 2,3-BDO в случае, когда их предварительно культивируют в среде с концентрацией лантана 10 мкМ.
[0045] На ФИГ. 16 показаны краткие сведения о генотипах штаммов А, В, С и D.
ПОДРОБНОЕ ОПИАСНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
[0046] Как это было суммировано выше, аспекты настоящего изобретения включают генетически модифицированных микроорганизмов, которые могут превращать углеродные субстраты в химические продукты, такие как 2,3-BDO. Генетически модифицированные микроорганизмы включают метанотрофов, способные продуцировать 2,3-BDO с высоким титром из метанового источника. Также раскрыты способы получения и применения таких генетически модифицированных микроорганизмов.
[0047] Прежде чем настоящее изобретение будет описано более подробно, следует понимать, что указанное изобретение не ограничивается конкретными описанными случаями, поскольку такие, конечно же, могут варьироваться. Также следует понимать, что применяемая в настоящем описании терминология предназначена только для описания конкретных случаев и не предназначена для их ограничения, поскольку объем настоящего изобретения будет ограничен только прилагаемой формулой изобретения.
[0048] Если предоставлен диапазон значений, то следует понимать, что каждое промежуточное значение до десятой доли нижнего предела до тех пор, пока в контексте явно не указано иное, между верхним и нижним пределом этого диапазона и любым другим указанным или промежуточным значением в указанном диапазоне включено в настоящее изобретение. Верхние и нижние пределы этих меньших диапазонов могут независимым образом быть включены в меньшие диапазоны, и они также включены в настоящее изобретение с учетом любых, конкретных образом исключенных пределов в указанном диапазоне. В случае, если указанный диапазон включает одно или оба из этих пределов, диапазоны, исключающие один или оба из этих включенных пределов, также включены в настоящее изобретение.
I. ОПРЕДЕЛЕНИЯ
[0049] Термин «2,3-бутандиол» или «2,3-BDO» и его грамматические эквиваленты в контексте настоящего описания могут относиться ко всем энантиомерным и диастереомерным формам указанного соединения, включая (R,R), (S,S) и мезаформы в рацемической, частично стереоизомерно чистой и/или по существу стереоизомерно чистой формах.
[0050] Термин «бутен» или «бутилен» и его грамматические эквиваленты в контексте настоящего описания могут относиться ко всем структурным изомерам алкена, включая 2-бутен, бут-1-ен, 2-метилпропен и все стереоизомерные и геометрические изомерные формы указанного соединения, включая Z-бут-2-ен, Е-бут-2-ен, в смесях изомеров и чистых и/или по существу чистых формах.
[0051] Термин «бутадиен» и его грамматические эквиваленты в контексте настоящего описания могут относиться ко всем геометрическим изомерам диена, включая цис- и транс-1,3-бутадиен, в смесях изомеров и чистых и/или по существу чистых формах.
[0052] Термин «метилэтилкетон» или «MEK», или «бутанон» и его грамматические эквиваленты в контексте настоящего описания могут относиться ко всем изомерам кетона в частично чистых и/или по существу чистых формах.
[0053] Термин «примерно» по отношению к эталонному числовому значению и его грамматические эквиваленты в контексте настоящего описания могут включать само числовое значение и диапазон значений плюс или минус 10% от этого числового значения. Например, сумма «примерно 10» включает 10 и любые суммы от 9 до 11. Например, термин «примерно» по отношению к эталонному числовому значению может также включать диапазон значений плюс или минус 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% от этого значения. В некоторых случаях числовое значение, раскрытое в настоящем документе, может составлять «примерно» это числовое значение даже без конкретного упоминания термина «примерно».
[0054] Следует отметить, что в контексте настоящего описания и в прилагаемой формуле изобретения формы единственного числа включают множественные ссылки до тех пор, пока в контексте явно не указано иное. Также следует отметить, что формула изобретения может быть составлена таким образом, чтобы исключить любой необязательный элемент. По сути это утверждение предназначено для того, чтобы служить предшествующей основой для применения такой исключительной терминологии, как «исключительно», «только» и тому подобное в связи со ссылкой на элементы формулы изобретения, или применения «отрицательного» ограничения.
[0055] Термин «генетическая модификация» или «генетически модифицированный» и их грамматические эквиваленты в контексте настоящего описания могут относиться к одному или более изменениям нуклеиновой кислоты, например, нуклеиновой кислоты в геноме микроорганизма. Например, генетическая модификация может относиться к изменениям, дополнениям и/или делециям нуклеиновой кислоты (например, целых генов или фрагментов генов).
[0056] Термин «разрушение» и его грамматические эквиваленты в контексте настоящего описания могут относиться к процессу изменения гена, например, путем делеции, вставки, мутации, перегруппировки или любой их комбинации. Например, ген может быть разрушен в результате нокаута. Разрушение гена может представлять собой частичное уменьшение или полное подавление экспрессии (например, экспрессии мРНК и/или белка) гена. Разрушение может также включать ингибиторную технологию, как, например, с применением короткошпилечных РНК, миРНК, микроРНК, доминантно-отрицательного или любого другого способа ингибирования функциональности или экспрессии гена или белка.
[0057] Термин «редактирование гена» и его грамматические эквиваленты в контексте настоящего описания могут относиться к генетическому конструированию, при котором один или более нуклеотидов вставлен, заменен или удален из генома. Например, редактирование гена можно осуществить с применением нуклеазы (например, встречающейся в природе нуклеазы или искусственно сконструированной нуклеазы).
[0058] Термины «и/или» и «любая их комбинация» и их грамматические эквиваленты в контексте настоящего описания можно применять взаимозаменяемо. Эти термины могут указывать на то, что любая специальным образом предусмотрена любая комбинация. Исключительно в иллюстративных целях следующие фразы «А, В, и/или С» или «А, В, С или любая их комбинация» могут означать «А по отдельности; В по отдельности; С по отдельности; А и В; В и С; А и С; и А, В и С.
[0059] Как будет понятно специалистам в данной области техники после прочтения данного раскрытия, каждый из отдельных случаев, описанных и проиллюстрированных в настоящем документе, имеет отдельные компоненты и признаки, которые можно легко отделить или объединить с признаками любых других нескольких случаев без отступления от объема или сущности настоящего изобретения. Любой перечисленный способ осуществить в порядке перечисленных событий или в любом другом порядке, который возможен с логической точки зрения.
[0060] До тех пор, пока не указано иное, все технические и научные термины в контексте настоящего описания имеют то же значение, которое обычно понимается специалистом в данной области техники, к которой относится настоящее изобретение. Хотя любые способы и материалы, аналогичные или эквивалентные описанным в настоящем документе, также можно применять на практике или при исследовании в соответствии с настоящим описанием, в настоящее время описаны иллюстративные примеры способов и материалов.
[0061] Термин «полинуклеиновая кислота» и его грамматические эквиваленты в контексте настоящего описания могут относиться к органическому полимеру, состоящему из двух или более мономеров, включая нуклеотиды, нуклеозиды или их аналоги, включая, но не ограничиваясь ими, одноцепочечную или двухцепочечную, смысловую или антисмысловую дезоксирибонуклеиновую кислоту (ДНК) любой длины и, при необходимости, одноцепочечную или двухцепочечную, смысловую или антисмысловую рибонуклеиновую кислоту (РНК) любой длины, включая миРНК. Термин «нуклеотид» относится к любому из нескольких соединений, которые состоят из рибозного или дезоксирибозного сахара, присоединенного к пуриновому или пиримидиновому основанию и к фосфатной группе, и которые являются основными структурными единицами нуклеиновых кислот. Нуклеотиды могут встречаться в природе, быть искусственными и/или модифицированными нуклеотидами. Термин «нуклеозид» относится к соединению (как гуанозину или аденозину), которое состоит из пуриновой или пиримидиновой основы в сочетании с дезоксирибозой или рибозой, и которое часто обнаруживают в полинуклеиновых кислотах. Термин «аналог нуклеотида» или «аналог нуклеозида» относится соответственно к нуклеотиду или нуклеозиду, в котором один или более отдельных атомов были заменены другим атомом или другой функциональной группой. Термин «полинуклеиновая кислота» в контексте настоящего описания включает нуклеиновые кислоты любой длины, включая ДНК, РНК, открытые рамки считывания, аналоги и их фрагменты.
[0062] Примеры полинуклеиновых кислот включают олигонуклеотиды, длина которых обычно составляет от 2 нуклеотидов до примерно 100 нуклеотидов, и полинуклеотиды, длина которых обычно составляет примерно более 100 нуклеотидов. Будет понятно, что описанные в настоящем описании полинуклеиновые кислоты включают полинуклеотиды, такие как «гены», «промоторы», «опероны» и/или «векторы». В контексте настоящего описания термин «ген» и его грамматические эквиваленты относятся к полинуклеотиду, который кодирует определенную последовательность аминокислот, которая содержат все или часть одного или более белков или ферментов, и могут содержать регуляторные (нетранскрибируемые) последовательности ДНК, такие как промоторные последовательности, которые определяют, например, условия экспрессии гена. Транскрибируемая область гена может содержать нетранслируемые области, включая интроны, 5'-нетранслируемую область (UTR) и 3'-UTR, а также кодирующую последовательность.
[0063] Термин «промотор» и его грамматические эквиваленты в контексте настоящего описания могут относиться к последовательности нуклеиновой кислоты, способной контролировать экспрессию кодирующей последовательности или функциональной РНК. В целом кодирующая последовательность расположена на 3'-конце по отношению к промоторной последовательности. Промоторы можно полностью получить из нативного гена, или они могут состоять из различных элементов, полученных их различных промоторов, встречающихся в природе, или даже содержать сегменты синтетических нуклеиновых кислот. Специалистам в данной области техники будет понятно, что различные промоторы могут управлять экспрессией гена в разных тканях или типах клеток, или на разных стадиях развития, или в ответ на различные условия окружающей среды или физиологические условия. Промоторы, которые вызывают экспрессию гена в большинстве типов клеток в большинстве случаев, обычно называют «конститутивными промоторами». Кроме того, признано, что, поскольку в большинстве случаев точные границы регуляторных последовательностей определены неполностью, фрагменты ДНК различной длины могут иметь одинаковую активность промотора.
[0064] Термин «функционально связанный» и его грамматические эквиваленты в контексте настоящего описания могут относиться к совокупности последовательностей нуклеиновых кислот на одиночном фрагменте полинуклеиновой кислоты, так что функционирование одной из последовательностей зависит от функционирования другой. Например, промотор функционально связан с кодирующей последовательностью в случае, когда он способен воздействовать на экспрессию этой кодирующей последовательности (т.е. регуляция указанным промотором экспрессии указанной кодирующей последовательности происходит на уровне транскрипции). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в 5'-3' или 3'-5' направлении.
[0065] Термин «кодон-оптимизированный» и его грамматические эквиваленты в контексте настоящего описания в смысле того, что он относится к генам или кодирующим областям молекул нуклеиновых кислот (или к открытым рамкам считывания) для трансформации различных хозяев, могут относиться к изменению кодонов в указанном гене или кодирующих областях молекул нуклеиновой кислоты, указывая на обычное применение кодонов организмом хозяина без изменения полипептида, кодируемого ДНК.
[0066] Термин «открытая рамка считывания» («ОРС») и его грамматические эквиваленты в контексте настоящего описания могут относиться к последовательности полинуклеиновой кислоты или нуклеиновой кислоты (будь то встречающейся в природе, не встречающейся в природе или синтетической), содержащей непрерывную рамку считывания, состоящую из (i) инициирующего кодона, (ii) серии из двух (2) или более кодонов, представляющих аминокислоты, и (iii) терминирующего кодона, при этом чтение (или транслирование) ОРС происходит в направлении от 5'- к 3'-концу.
[0067] Термин «оперон» и его грамматические эквиваленты в контексте настоящего описания могут относиться к двум или более генам, которые транскрибируются как единая транскрипционная единица с общего промотора. В некоторых случаях гены, полинуклеотиды или ОРС, содержащие оперон, являются смежными. Будет понятно, что транскрипцию всего оперона можно модифицировать (т.е. увеличить, уменьшить или исключить) путем модификации общего промотора. В качестве альтернативы любой ген, полинуклеотид или ОРС, или любую их комбинацию в опероне можно модифицировать для изменения функции или активности кодируемого полипептида. Модификация может привести к увеличению или снижению активности или функции кодируемого полипептида. Кроме того, модификация может придать кодируемому полипептиду новые функции.
[0068] Термин «вектор» и его грамматические эквиваленты в контексте настоящего описания могут относиться к любым средствам, с помощью которых нуклеиновая кислота может распространяться и/или передаваться между организмами, клетками или клеточными компонентами. Векторы включают вирусы, бактериофаги, провирусы, плазмиды, фагемиды, транспозоны и искусственные хромосомы, такие как YAC (искусственные дрожжевые хромосомы), ВАС (искусственные бактериальные хромосомы) и PLAC (искусственные хромосомы растений) и им подобные структуры, которые представляют собой «эписомы», которые реплицируются автономно или могут интегрироваться в хромосому микроорганизма-хозяина. Вектором также может быть полинуклеотид «оголенной» РНК, полинуклеотид «оголенной» ДНК, полинуклеотид, состоящий из ДНК и РНК в одной цепи, конъюгированная с полилизином ДНК или РНК, конъюгированная с пептидом ДНК или РНК, конъюгированная с липосомами ДНК или им подобные конструкции, которые не являются эписомами в природных условиях, или вектор может представлять собой организм, который содержит одну или более из указанных выше полинуклеотидных конструкций, таких как агробактерия или бактерия.
[0069] Термин «полипептид» и его грамматические эквиваленты в контексте настоящего описания могут относиться к любому органическому полимеру, содержащему две или более аминокислот, независимо от его размера. Хотя термин «белок» часто используют в отношении относительно больших полипептидов, а термин «пептид» часто используют в отношении небольших полипептидов, применение этих терминов в данной области техники пересекается и варьируется. Термин «полипептид» в контексте настоящего описания относится к пептидам, полипептидам и белкам до тех пора, пока не указано иное. В контексте настоящего описания термины «белок», «полипептид» и «пептид» применяются в настоящем документе взаимозаменяемо при ссылке на генный продукт. До тех пор, пока не указано иное, определенный полипептид также косвенно охватывает свои консервативно-замещенные варианты.
[0070] Термин «фермент» и его грамматические эквиваленты в контексте настоящего описания могут относиться к любому из многочисленных белков, которые действуют как биологический катализатор. Подобно традиционным химическим катализаторам, ферменты ускоряют скорость биологических реакций, создавая переходное состояние с меньшей энергией активации, чем в случае некатализированной реакции. Другими словами, ферменты являются белками, специализирующимися на реакциях, которые они катализируют. Примеры ферментов, описанных в настоящем документе, включают ацетолактатсинтазу (кодируемую геном AlsS), альфа-ацетолактатдекарбоксилазу (кодируемую геном BudA) и ацетоинредуктазу (кодируемую геном ButA).
[0071] Фразы «рекомбинантная клетка-хозяин», «генетически сконструированная клетка-хозяин», «сконструированная клетка-хозяин», «генетически модифицированная клетка-хозяин» и их грамматические эквиваленты в контексте настоящего описания могут применяться взаимозаменяемо и могут относиться к клеткам-хозяевам, которые были генетически модифицированы для: (а) экспрессии одной или более экзогенных полинуклеиновых кислот; (b) гиперэкспрессии одной или более эндогенных и/или одной или более экзогенных полинуклеиновых кислот, таких как те, которые включены в вектор, или у которых изменена экспрессия эндогенного гена; или (с) нокаута или подавляющей регуляции эндогенного гена. Кроме того, некоторые гены можно физически удалить из генома (например, нокаутом) или их можно сконструировать таким образом, что они будут обладать пониженной, измененной или повышенной активностью. Фразы «рекомбинантная клетка-хозяин», «генетически сконструированная клетка-хозяин», «сконструированная клетка-хозяин» и «генетически модифицированная клетка-хозяин» относятся не только к определенной исследуемой клетке-хозяину, но и к потомству или потенциальному потомству такой клетки. Поскольку определенные модификации могут происходить в последующих поколениях вследствие либо мутации, либо влияния окружающей среды, такое потомство, на самом деле, может не быть идентичным родительской клетке, однако оно все же находится в области действия термина(-ов) в разделе настоящего описания.
[0072] Термин «in vitro» и его грамматические эквиваленты в контексте настоящего описания могут относиться к внешнему окружению живой клетки, независимо от ее местоположения. Термин «in vivo» и его грамматические эквиваленты в контексте настоящего описания могут относиться к внутреннему окружению живой клетке независимо от ее местоположения.
[0073] Термины «конструировать», «генетически конструировать», «модифицировать», «генетически модифицировать» и их грамматические эквиваленты в контексте настоящего описания могут относиться к любым манипуляциям с микроорганизмом, которые приводят к обнаруживаемым изменениям в микроорганизме, при этом манипулирование включает, но не ограничивается этим, придание ненативной метаболической функциональности при помощи гетерологичных (экзогенных) полинуклеиновых кислот или исключение нативной функциональности через делеции, мутации или нокауты полинуклеиновых кислот. Термин «метаболически сконструированный» в целом включает рациональное проектирование путей и сборку биосинтетических генов (или открытых рамок считывания), связанных с оперонами генов и элементов контроля такими полинуклеиновыми кислотами для получения желаемого метаболита. «Метаболически сконструированный» может включать оптимизацию метаболического потока путем регуляции и оптимизации транскрипции, трансляции, стабильности белка и функциональности белка с применением методов генной инженерии и соответствующих условий культивирования, включая уменьшение, нарушение или нокаут конкурирующего метаболического пути, который конкурирует с промежуточным соединением, ведущим к желаемому пути.
[0074] Термин «переключатель» и его грамматические эквиваленты в контексте настоящего описания могут означать регуляторную единицу гена или генов, которая способна реагировать на конкретный стимул либо вызывая, либо подавляя экспрессию. Например, переключатели могут включать регуляторные единицы, которые реагируют на сахар (например, арабинозу) или редкоземельные металлы (например, лантан).
[0075] В контексте настоящего описания термины «генетическая модификация», «генетически модифицированный» и их грамматические эквиваленты могут относиться к любой модификации полинуклеиновой кислоты и/или полипептида, которая приводит к получению измененной нуклеиновой кислоты или полипептида (т.е. относительно последовательности нуклеиновой кислоты или полипептидной последовательности дикого типа). Генетическая модификация включает, например, точечные мутации, замены, делеции или вставки одного или нескольких остатков в полинуклеиновую кислоту (или кодируемый полипептид), которая включает изменения, возникающие в пределах кодирующей белок области гена, а также изменения в областях за пределами кодирующей белок последовательности, таких как, но не ограничиваясь ими, в регуляторных или промоторных последовательностях. Генетическая модификация может представлять собой изменение любого типа. Например, модификацией может быть удаление, вставка, мутация, перегруппировка или любая их комбинация. В некоторых случаях часть генетически модифицированного генома микроорганизма можно заменить одной или более гетерологичными (экзогенными) полинуклеиновыми кислотами. В некоторых случаях модификация происходит естественным путем. В других случаях модификация является результатом искусственного отбора. В других случаях модификация является результатом генной инженерии. Одной из форм генетической модификации является разрушение, как, например, путем нокаута. В контексте настоящего описания термин «введение» и его грамматические эквиваленты, применяемые в таких словосочетаниях, как «введение в клетку-хозяина» по меньшей мере одной полинуклеиновой кислоты включают методы, известные в данной области техники, позволяющих вводить в клетку полинуклеиновые кислоты, включая, но не ограничиваясь ими: трансформацию (например, хлоридом кальция, электропорацию), трансдукцию, трансфекцию, конъюгацию и тому подобное.
[0076] В контексте настоящего описания термины «экспрессия» или «экспрессируемый» и их грамматические эквиваленты в отношении последовательности гена, последовательности ОРС или последовательности полинуклеиновой кислоты могут относиться к транскрипции указанного гена, открытой рамки считывания или полинуклеиновой кислоты и, в случае, если это уместно, к трансляции полученного транскрипта мРНК в белок. Таким образом, как это будет ясно из контекста, экспрессия белка является результатом транскрипции и трансляции последовательности открытой рамки считывания. Уровень экспрессии желаемого конечного продукта в клетке-хозяине можно определить либо в зависимости от количества соответствующей мРНК, которая присутствует в указанной клетке-хозяине, либо от количества желаемого конечного продукта, кодируемого выбранной последовательностью. Например, содержание мРНК, транскрибируемой с выбранной последовательности, можно количественно определить при помощи ПНР или метода нозенр-гибридизации (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1989). Содержание белка, кодируемого выбранной последовательностью, можно количественно определить различными способами (например, при помощи ферментного иммуносорбентного анализа (ELISA), путем анализа биологической активности указанного белка или с использованием анализов, которые не зависят от такой активности, таких как вестерн-блоттинг или радиоиммуноанализ, с применением антител, которые распознают и связывают реагирующий белок).
[0077] В контексте настоящего описания термин «эндогенный» и его грамматические эквиваленты, применяемые в отношении полинуклеиновых кислот (и кодируемых ими полипептидов), относятся к полинуклеиновым кислотам и полипептидам, которые экспрессируются в организме, из которого они происходят (т.е. они присущи данному организму). Напротив, термины «гетерологичный» и «экзогенный», применяемые взаимозаменяемо и, как это определенно в настоящем описании, со ссылкой на полинуклеиновые кислоты (и кодируемые ими полипептиды), указывают на полинуклеиновые кислоты и полипептиды, которые экспрессируются в организме, отличном от того организма, из которого они (т.е. полинуклеотидные или полипептидные последовательности) происходят или были получены. В некоторых случаях термин «гетерологичный» и его грамматические эквиваленты могут означать «полученный из разных видов». Например, термин «гетерологичный ген» может означать ген, который получен из вида, отличного от эталонного вида. Например, метанотроф, содержащий «гетерологичный ген», содержит ген, полученный из другого метанотрофа. Указанный ген можно получить из другого микроорганизма, такого как дрожжи, или от другого вида, как, например, другого вида метанотрофов.
[0078] В контексте настоящего описания термин «субстрат» и его грамматические эквиваленты могут относиться к любому веществу или соединению, которое превращается или которое предназначено для превращения в другое соединение под действием фермента. Термин включает не только одиночное соединение, но и также комбинации соединений, как, например, растворы, смеси и другие материалы, которые содержат по меньшей мере один субстрат или его производные.
[0079] В контексте настоящего описания термины «С1-углеродное соединение», «С1-углеродные субстраты» и их грамматические эквиваленты могут относиться к любому органическому соединению, которое содержит один атом углерода. Примеры включают, но не ограничиваются ими, монооксид углерода (СО), метан (CH4) и диоксид углерода (СО2).
[0080] В контексте настоящего описания термин «ферментация» или «процесс ферментации» и его грамматические эквиваленты могут представлять собой процесс, в котором клетка-хозяин культивируется в среде для культивирования, содержащей сырье, такое как исходное сырье и питательные вещества, в которой клетка преобразует сырье, как, например, исходное сырье, в желаемые конечные продукты.
[0081] В контексте настоящего описания термин «гомолог» и его грамматические эквиваленты, применяемые в отношении исходного белка, полипептида, гена или полинуклеиновой кислоты (или их кодирующей ОРС) из семейства или вида с родством первой степени, могут относиться к различным белкам, генам или полинуклеиновым кислотам из семейства или вида с родством второй степени, которые (структурно, функционально и/или с точки зрения генома) соответствуют исходному белку, гену или полинуклеиновой кислоте из семейства или вида с родством первой степени. Чаще всего «гомологи» будут иметь функциональное, структурное сходство или сходство в геноме. Известны методы, с помощью которых гомологи белка, гена или полинуклеиновой кислоты можно легко клонировать с применением генетических зондов и метода ПЦР Идентичность клонированных последовательностей в качестве «гомологов» можно подтвердить с помощью функциональных анализов и/или путем геномного картирования генов.
[0082] Полипептид (или белок, или фермент) обладает «гомологией» или является «гомологичным» второму полипептиду в случае, если нуклеотидная последовательность, которая кодирует указанный полипептид, содержит последовательность, сходную с указанной нуклеотидной последовательностью, которая кодирует второй полипептид. В качестве альтернативы полипептид обладает гомологией со вторым полипептидом в случае, если два белка содержат «сходные» аминокислотные последовательности. Таким образом, термины «гомологичные белки» или «гомологичные полипептиды» и их грамматические эквиваленты могут относиться к двум полипептидам, содержащим сходные аминокислотные последовательности. В некоторых случаях согласно настоящему изобретению полинуклеотиды и полипептиды, гомологичные одному или более полинуклеотидам и/или полипептидам, указанным в таблице 1, можно легко идентифицировать с применением методов, известных в данной области техники для анализа последовательности и сравнения.
[0083] Гомологичную полинуклеотидную или полипептидную последовательность согласно настоящему изобретению также можно определить или идентифицировать с помощью BLAST-анализа (средство поиска основного локального выравнивания) или при помощи аналогичных средств биоинформатики, которые сравнивают запрашиваемую нуклеотидную или полипептидную последовательность с базой данных известных последовательностей. Например, поисковый анализ может быть выполнен с применением BLAST для определения идентичности или сходства последовательности с ранее опубликованными последовательностями, и в случае, если указанная последовательность еще не была опубликована, она может обеспечить соответствующее представление о функции последовательности ДНК или белка.
[0084] В контексте настоящего описания термин «по существу чистый» и его грамматические эквиваленты могут относиться к конкретному веществу, которое не содержит большую часть другого вещества. Например, «по существу чистый 2,3-BDO» может означать по меньшей мере 90% 2,3-BDO. В некоторых случаях «по существу чистый 2,3-BDO» может означать по меньшей мере 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9%, 99,99%, 99,999% или 99,9999% 2,3-BDO. Например, по существу чистый 2,3-BDO может означать по меньшей мере 70% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 75% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 80% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 85% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 90% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 91% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 92% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 93% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 94% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 95% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 96% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 97% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 98% 2,3-BDO. В некоторых случаях по существу чистый 2,3-BDO может означать по меньшей мере 99% 2,3-BDO.
[0085] В контексте настоящего описания термин «по существу схожий» и его грамматические эквиваленты при применении со ссылкой на сходство между последовательностью и эталонной последовательностью означают, что указанные последовательности являются по меньшей мере на 50% идентичными (но не на 100%). В некоторых случаях указанные последовательности на 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% являются идентичными. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная по меньшей мере на 50%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 55%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 60%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 65%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 70%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 75%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 80%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 81%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 82%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 83%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 84%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 85%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 86%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 87%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 88%. В других случаях термин «по существу схожий» относится к последовательности, которая идентичная на 89%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 90%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 91%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 92%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 93%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 94%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 95%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 96%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 97%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 98%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 99%. В некоторых случаях термин «по существу схожий» относится к последовательности, которая идентичная на 100%. Для определения процента идентичности между двумя последовательностями, указанные две последовательности выравнивают, применяя, например, способ выравнивания по Needleman and Wunsch (J. Mol Biol, 1970, 48: 443) в редакции Smith and Waterman (Adv. Appl Math., 1981, 2: 482), так что достигается совпадение наивысшего порядка между двумя указанными последовательностями, а количество идентичных аминокислот/нуклеотидов определяют между этими двумя последовательностями. Методы расчеты процента идентичности между двумя аминокислотными последовательностями в целом известны в данной области техники и включают, например, те методы, которые описаны в публикациях Carillo and Lipton (SIAM J. Applied Math., 1988, 48:1073) и Computational Molecular Biology, Lesk, e.d. Oxford University Press, New York, 1988, Biocomputing: Informatics и Genomics Projects. В целом такие вычисления проводятся при помощи компьютерных программ. Компьютерные программы, которые можно применять для этих целей, включают, но не ограничиваются ими, GCG (Devereux et al, Nucleic Acids Res., 1984, 12: 387) BLASTP, BLASTN и FASTA (Altschul et al., J. Molec. Biol, 1990:215:403). Особенно предпочтительный способ определения процента идентичности между двумя полипептидами включает алгоритм Clustal W (Thompson, J D, Higgines, D G and Gibson T J, 1994, Nucleic Acids Res., 22(22): 4673-4680 совместно с оценочной матрицей BLOSUM 62 (Hemkoff S & Hemkoff, J G, 1992, Proc. Natl. Acad. Sci. USA 89: 10915-10919), применяя штраф за открытие гэпа, составляющий 10, и штраф за продление гэпа, составляющий 0,1, достигая таким образом соответствия наивысшего порядка между двумя последовательностями, причем в выравнивании участвует по меньшей мере 50% от общей длины одной из двух последовательностей.
II. ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ МИКРООРГАНИЗМЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ
[0086] Настоящее изобретение частично относится к генетически модифицированным микроорганизмам, которые значительно улучшали скорости биосинтеза 2,3-BDO по сравнению с теми скоростями, которые наблюдаются при применении микроорганизмов дикого типа. В некоторых случаях скорости биосинтеза на порядки выше тех скоростей, которые обычно получают. В некоторых случаях микроорганизмы, которые естественным образом не продуцируют 2,3-BDO, были генетически модифицированы для синтеза 2,3-BDO, в том числе на достаточно высоких уровнях.
Микроорганизмы
[0087] В некоторых случаях микроорганизмы могут использовать С1-углеродные субстраты, такие как СО, СО2 и CH4, для синтеза желаемого конечного продукта. Тем не менее этот факт не означает, что указанные микроорганизмы используют исключительно С1-углеродные соединения. Некоторые из микроорганизмов можно получить для использования ими дополнительных углеродных субстратов, включая углеродные субстраты, которые этот микроорганизм естественным образом использует. Например, в случае, если микроорганизм естественным образом использует сахар в качестве углеродных субстратов, то такой микроорганизм можно получить для использования им другого источника углерода, такого как С1-углеродного соединения.
[0088] Микроорганизмы могут быть прокариотами или эукариотами. В некоторых случаях, например, указанные микроорганизмы могут быть бактериями, дрожжами или водорослями.
[0089] Микроорганизмы, которые могут превращать С1-углеродные субстраты в желаемые продукты, включают тех микроорганизмов, которые способны использовать природный газ в качестве углеродного субстрата. Например, микроорганизм может использовать метан, содержащийся в природном газе, в качестве источника углерода, для получения таких желаемых продуктов. К таким микроорганизмам могут относиться метанотрофы. Метанотрофы, которые могут быть особенно подходящими для применения, включают микроорганизмы из родов Methylobacter, Methylomicrobium, Methylomonas, Methylocaldum, Methylococcus, Methylosoma, Methylosarcina, Methylothermus, Methylohalobius, Methylogaea, Methylovulum, Crenothrix, Clonothrix, Methylosphaera, Methylocapsa, Methylocella, Methylosinus, Methylocystis, Methyloacidophilum или любой их комбинации. В некоторых случаях указанный метанотроф может быть представителем рода Methylococcus. В одном случае указанный метанотроф может быть из вида Methylococcus capsulatus.
[0090] Некоторые микроорганизмы способны использовать СО2 в качестве субстрата. К таким микроорганизмам относятся метаногены. Микроорганизмы, способные использовать СО2 в качестве субстрата, могут содержать хлорофилл. Примеры таких микроорганизмов включают водоросли и цианобактерии.
[0091] Некоторые микроорганизмы способны использовать СО в качестве субстрата. Примеры включают анаэробных микроорганизмов, таких как Clostridium. Эти микроорганизмы можно генетически модифицировать для получения с их помощью значительных количеств 2,3-BDO.
[0092] В некоторых случаях генетически модифицированные микроорганизмы, описанные в настоящем документе, могут продуцировать желаемый продукт при более высоких титрах при ферментации при более высокой температуре. Например, генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продуктов, таких как 2,3-BDO, бутадиен и/или MEK, при их инкубации при температуре выше 37°С (но не выше температуры 100°С). В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при их инкубации при 42°С по сравнению с 37°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при их инкубации при 41°С по сравнению с 37°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при его инкубации при 42°С по сравнению с 45°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при его инкубации при 41°С по сравнению с 45°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при их инкубации при 37°С по сравнению с 45°С. В некоторых случаях генетические модификации приводят к повышенной толерантности/ предпочтению к более высоким температурам.
Ферменты
[0093] Для того чтобы генетически сконструировать определенные микроорганизмы для получения определенных подходящих продуктов, таких как 2,3-BDO, бутадиен и/или MEK, микроорганизмы можно трансформировать при помощи одного или более генов, которые кодируют специфичные ферменты. Эти гены могут быть гетерологичными для данного микроорганизма.
[0094] Например, чтобы сконструировать микроорганизм, который может продуцировать 2,3-BDO, один или более генов (например, гетерологичные гены) можно трансформировать или трансфицировать в указанный микроорганизм либо на время, либо на постоянной основе. В некоторых случаях один или более из этих генов могут эписомально экспрессироваться. В некоторых случаях один или более из этих генов можно интегрировать в геном микроорганизма. В некоторых случаях один или более из этих генов могут экспрессироваться эписомально, тогда как один или более из этих генов также может быть интегрирован в геном микроорганизма. В некоторых случаях указанный сконструированный микроорганизм может использовать один или более из следующих ферментов: (i) ацетолактатсинтазу, (ii) альфа-ацетолактатдекарбоксилазу и/или (iii) ацетоинредуктазу. Ацетолактатсинтаза (кодируемая геном AlsS) превращает две молекулы пирувата в 2-ацетолактат.Альфа-ацетолактатдекарбоксилаза (кодируемая геном BudA) превращает 2-ацетолактат в ацетоин. Ацетоинредуктаза (кодируется геном ВшА) превращает ацетоин в 2,3-BDO, используя НАДФН или НАДН в качестве восстановленного кофактора. Ацетоинредуктазы, которые используют НАДФН в качестве кофактора, называются «НАДФН-зависимые ацетоинредуктазы». Ацетоинредуктазы, которые используют НАДН в качестве кофактора, называются «НАДН-зависимые ацетоинредуктазы». В некоторых случаях при применении вектора для экспрессии или интегрировании гена в микроорганизм, указанный ген, кодирующий ацетолактатсинтазу, может располагаться ближе к 5'-концу, чем любой другой ген в данном векторе. В некоторых случаях ген, кодирующий ацетоинредуктазу, может располагаться ближе к 3'-концу, чем любой другой ген в векторе. В некоторых случаях ген, кодирующий альфа-ацетолактатдекарбоксилазу, может быть расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой ген в векторе (например, существует по меньшей мере один ген, который расположен на 5'-конце, а также по меньшей мере один ген, расположенный на 3'-конце гена альфа-ацетолактатдекарбоксилазы). Гены в векторе могут быть представлены в определенном порядке до, во время или после контакта с микроорганизмом. Например, ген может располагаться ближе к 5'-концу, чем любой другой ген в векторе до контакта с микроорганизмом. После контакта с микроорганизмом ген так можно разместить в положении генома микроорганизма, что указанный ген будет располагаться ближе к 3'-концу, чем любой другой ген в векторе, или он уже не будет располагаться ни ближе к 3'-, ни ближе к 5'-концу, чем любой другой ген в векторе. В некоторых случаях, ген можно оставить в 5'-положении по отношению к любому другому гену. Например, векторы можно модифицировать внутри микроорганизма таким образом, что будет изменен порядок генов. В некоторых случаях после контакта с микроорганизмом ген можно вставить в вектор, при этом указанный ген будет располагаться ближе к 3'-концу, чем любой другой ген в векторе, или он уже не будет располагаться ни ближе к 3'-, ни ближе к 5'-концу, чем любой другой ген в векторе. В некоторых случаях определенного порядка генов можно достигнуть после того, как в геном микроорганизма будет вставлен один или более гетерологичных генов. Например, различные векторы интеграции можно применять для достижения определенного порядка генов в геноме микроорганизма. В некоторых случаях порядок генов определяют после того, как гетерологичный ген будет вставлен в геном микроорганизма.
[0095] В настоящем документе описаны микроорганизмы, применяемые для получения 2,3-BDO из С1-углеродного соединения (например, метана). В некоторых случаях указанный в настоящем документе микроорганизм можно трансформировать при помощи гена, кодирующего один или более следующих ферментов: (i) ацетоинредуктаза (НАДФН-зависимая и/или НАДН-зависимая); (ii) альфа-ацетолактатдекарбоксилаза и/или (iii) ацетолактатсинтаза (AlsS). Например, микроорганизм можно трансформировать при помощи гена, кодирующего НАДФН- или НАДН-зависимую ацетоинредуктазу. В качестве другого примера, микроорганизм можно трансформировать при помощи гена, кодирующего альфа-ацетолактатдекарбоксилазу. В еще одном примере, микроорганизм можно трансформировать при помощи гена, кодирующего ацетолактатсинтазу. Эти гены могут быть гетерологичными для указанного микроорганизма. В некоторых случаях эти гены могут эписомально экспрессироваться, быть интегрированы в геном микроорганизма (например, при помощи вектора интеграции) или представлять собой любую из таких комбинаций. В некоторых случаях ген, кодирующий гетерологичную ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, не располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген.
[0096] В некоторых случаях микроорганизм можно трансформировать при помощи двух или более генов, как, например, кодирующих НАДФН- и/или НАДН-зависимую ацетоинредуктазу и альфа-ацетолактатдекарбоксилазу. В качестве другого примера, микроорганизм также можно трансформировать при помощи генов, кодирующих НАДФН- или НАДН-зависимую ацетоинредуктазу и ацетолактатсинтазу. В еще одном примере, микроорганизм можно трансформировать при помощи генов, кодирующих альфа-ацетолактатдекарбоксилазу и ацетолактатсинтазу. Один или более генов могут быть гетерологичными для микроорганизма. В некоторых случаях эти гены могут эписомально экспрессироваться, быть интегрированы в геном микроорганизма или представлять собой любую из таких комбинаций.
[0097] В некоторых случаях микроорганизм можно трансформировать при помощи по меньшей мере трех или более генов, как, например, кодирующих НАДФН- и/или НАДН-зависимую ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и ацетолактатсинтазу. Один или более генов могут быть гетерологичными для микроорганизма. В некоторых случаях эти гены могут эписомально экспрессироваться, быть интегрированы в геном микроорганизма или представлять собой любую из таких комбинаций.
[0098] Ген, кодирующий НАДФН-зависимую ацетоинредуктазу, можно получить из бактерий (например, грамположительной или грамотрицательной бактерии). Бактерия, например, может быть из рода Clostridium, например, Clostridium autoethanogenum.
[0099] НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по существу схожа с SEQ ID NO 9. Например, НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичная последовательности SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая представляет собой последовательность SEQ ID NO 9. В некоторых случаях НАДФН-зависимая ацетоинредуктаза содержит аминокислотную последовательность SEQ ID NO 9.
[00100] Ген, кодирующий НАДН-зависимую ацетоинредуктазу, можно получить из бактерии (например, грамположительной или грамотрицательной бактерии). Примеры включают бактерии из рода Bacillus, например, Bacillus subtilis. Указанная бактерия может быть из рода Paenibacillus, например, Paenibacillus polymyxa.
[00101] В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по существу схожа с последовательностями SEQ ID NO 11 или 13. Например, НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза может содержать аминокислотную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 11 или 13. В некоторых случаях НАДН-зависимая ацетоинредуктаза содержит аминокислотную последовательность SEQ ID NO 11 или 13.
[00102] Ген, кодирующий альфа-ацетолактатдекарбоксилазу (budA), можно получить из бактерии (например, грамположительной бактерии или грамотрицательной бактерии). Примеры включают бактерии из рода Clostridium, например, Clostridium autoethanogenum. Другие примеры включают бактерии из рода Klebsiella, например, Klebsiella pneumoniae.
[00103] В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по существу схожа с SEQ ID NO 5 или 7. Например, альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 60% 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 5 или 7. Например, альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 5 или 7. Например, альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 5 или 7. Например, альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза может содержать аминокислотную последовательность, которая идентична SEQ ID NO 5 или 7. В некоторых случаях альфа-ацетолактатдекарбоксилаза содержит аминокислотную последовательность SEQ ID NO 5 или 7.
[00104] Ген, кодирующий ацетолактатсинтазу (AlsS), можно получить из бактерии (например, грамположительной бактерии). Примеры включают бактерии из рода Clostridium, например, Clostridium autoethanogenum. Другие примеры включают бактерии из рода Bacillus, например, Bacillus subtilis. Дополнительные примеры видов включают Bacillus licheniformis.
[00105] В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по существу схожа с одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 60% 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 60% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 65% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 70% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 75% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 80% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 85% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 90% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 91% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 92% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 93% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 94% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 95% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 96% идентична одной из последовательностей из SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 97% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 98% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая по меньшей мере на 99% идентична одной из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях ацетолактатсинтаза может содержать аминокислотную последовательность, которая идентична одной из последовательностей SEQ ID NO 1, 3 или 19.
[00106] В некоторых случаях микроорганизм можно обеспечить дополнительными ферментами, что приводит к получению других желаемых конечных продуктов путем ферментации.
[00107] В некоторых случаях аминокислотную последовательность можно оптимизировать в зависимости от микроорганизма, который будут обеспечивать генами или в котором будут экспрессироваться ферменты. В таких случаях консервативные замены аминокислотных кислот можно произвести в зависимости от того, использует ли указанный микроорганизм определенную аминокислоту или сколько этой определенной аминокислоты доступно для применения внутри микроорганизма.
Векторы
[00108] Полинуклеотидные конструкции, полученные для введения в прокариотического или эукариотического хозяина, могут как правило, но не всегда, содержать систему репликации (т.е. вектор), распознаваемую данным хозяином. Вектор содержит предполагаемый полинуклеотидный фрагмент, кодирующий желаемый полипептид и необязательно регуляторные последовательности инициации транскрипции и трансляции, функционально связанные с кодирующим полипептид сегментом. Системы экспрессии (как, например, векторы экспрессии) могут содержать, например, точку начала репликации или автономно реплицирующуюся последовательность (ARS), контрольные последовательности экспрессии, промотор, энхансер и необходимые сайты обработки информации, такие как сайты связывания с рибосомой, сайты сплайсинга РНК, сайты полиаденилирования, последовательности терминации транскрипции, мРНК-стабилизирующие последовательности, нуклеотидные последовательности, гомологичные хромосомной ДНК хозяина и/или сайт множественного клонирования. В случае необходимости также могут содержаться сигнальные пептиды, предпочтительно полученные из секретируемых полипептидов тех же или родственных видов, которые позволяют белку проходить и/или оседать в клеточных мембранах или секретироваться из клетки.
[00109] Векторы можно сконструировать с применением стандартных способов (см, например, Sambrook et al., Molecular Biology: A Laboratory Manual, Cold Spring Harbor, N.Y. 1989; и Ausubel, et al., Current Protocols in Molecular Biology, Greene Publishing, Co. N.Y, 1995).
[00110] Манипуляции с полинуклеотидами, которые кодируют ферменты, раскрытые в настоящем документе, обычно осуществляют в рекомбинантных векторах. Векторы, которые можно использовать, включают бактериальные плазмиды, бактериофаг, искусственные хромосомы, эписомальные векторы и векторы экспрессии генов. Векторы можно выбрать для размещения полинуклеотида, кодирующего белок желаемого размера. После получения выбранного вектора, подходящую клетку-хозяина (например, микроорганизмы, описанные в настоящем описании) трансфицируют или трансформируют при помощи указанного вектора. Каждый вектор содержит различные функциональные компоненты, которые в целом включают сайт клонирования, точку начала репликации и по меньшей мере один отбираемый маркерный ген. Вектор может дополнительно содержать один или более из следующих элементов: энхансер, промотор, последовательность терминации транскрипции и/или другие сигнальные последовательности. Такие элементы последовательности можно оптимизировать для выбранных видов хозяев. Такие элементы последовательности могут располагаться в непосредственной близости от сайта клонирования, так что они функционально связаны с геном, кодирующим предварительно отобранный фермент.
[00111] Векторы, включая векторы клонирования и экспрессии, могут содержать нуклеотидные последовательности, которые позволяют вектору реплицироваться в одном или более из выбранных микроорганизмов. Например, указанная последовательность может быть такой, которая позволяет вектору реплицироваться независимо от хромосомной ДНК хозяина, и которая может содержать точки начала репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для различных бактерий, дрожжей и вирусов. Например, точка начала репликации, полученная из плазмиды pBR322, является подходящей для большинства грамотрицательных бактерий, точка начала репликации, полученная из 2-микронной плазмиды, является подходящей для дрожжей, а полученные из различных вирусов точки начала репликации (например, из SV40, аденовируса) являются подходящими для векторов клонирования.
[00112] Вектор клонирования или экспрессии может содержать ген селекции, также называемый отбираемым маркером. Этот ген кодирует белок, необходимый для выживания или роста трансформированных микроорганизмов в селективной среде для культивирования. Микроорганизмы, не трансформированные при помощи вектора, содержащего ген селекции, таким образом, не выживут в указанной среде для культивирования. Типичные гены селекции кодируют белки, которые придают устойчивость к антибиотикам и другим токсинам, например, ампициллину, неомицину, метотрексату, гигромицину, тиострептону, апрамицину или тетрациклину, восполняют ауксотрофный дефицит или обеспечивают важными питательными веществами, которые отсутствуют в питательных средах.
[00113] Репликацию векторов можно осуществить в Е. coli. Примером отбираемого в E.coli маркера является ген [3-лактамазы, который придает устойчивость к антибиотику ампициллину. Эти отбираемые маркеры можно получить из плазмид Е. coli, таких как pBR322, или плазмиды pUC, как, например, pUC18 или pUC19, или pUC119.
[00114] Векторы согласно настоящему изобретению могут содержать один или более переключателей, как, например, индуцируемый или репрессируемый переключатель, например, переключатель, чувствительный к арабинозе или лантану. Векторы также могут содержать один или более разных/одинаковых промоторов.
Промоторы
[00115] Векторы могут содержать промотор, который распознается микроорганизмом-хозяином. Промотор может быть функционально связан с представляющей интерес кодирующей последовательностью. Такой промотор может быть индуцируемым или конститутивным. Полинуклеотиды считаются функционально связанными тогда, когда они находятся во взаимосвязи, позволяющей им функционировать по их назначению.
[00116] Для управления экспрессией генов можно применять различные промоторы. Например, в случае, если желательна временная экспрессия гена (т.е. неконститутивная экспрессия), экспрессия может быть опосредована индуцируемыми промоторами.
[00117] В некоторых случаях ген AlsS экспрессируется временно. Другими словами, ген AlsS экспрессируется неконститутивно. Экспрессия гена AlsS может быть опосредована индуцируемыми или репрессируемыми промоторами. Примеры индуцируемых или репрессируемых промоторов включают, но не ограничиваются ими, такие промоторы, которые индуцируются или репрессируются с помощью: (а) Сахаров, таких как арабиноза и лактоза (или неметаболизируемых аналогов, например, изопропил-β-D-1-тиогалактопиранозида (IPTG)); (b) металлов, таких как лантан, медь, кальций; (с) температуры; (d) азот-содержащего источника; (д) кислорода; (е) состояния клетки (состояния роста или стационарного состояния); (g) метаболитов, таких как фосфат; (h) CRISPRi; (i) jun; (j) fos, (k) металлотионеина и/или (l) теплового шока. Эти промоторы можно применять в системе метанотрофов. Примером индуцируемого промотора, которого можно применять в метанотрофах, является промотор pBAD или промотор pMxaF.
[00118] Индуцируемыми или репрессируемыми промоторами, которые могут быть особенно подходящими, являются переключатели в виде сахара и редкоземельных металлов. Например, промоторы, которые чувствительны к сахару арабинозе, можно применять в качестве индуцируемого переключателя. В некоторых случаях чувствительные к арабинозе переключатели можно применять для управления экспрессии одного или более генов. Например, в случае присутствия арабинозы, может быть «включен» механизм получения 2,3-BDO. Чувствительный к арабинозе переключатель может включать экспрессию ацетоинредуктазы. Чувствительный к арабинозе переключатель также может включать экспрессию альфа-ацетолактатдекарбоксилазы (budA). Чувствительный к арабинозе переключатель также может включать экспрессию ацетолактатсинтазы (AlsS).
[00119] Другими особенно подходящими переключателями могут быть переключатели, чувствительные к редкоземельным металлам, как, например, чувствительные к лантану переключатели (или переключатели, чувствительные к церию, празеодиму или неодиму). В некоторых случаях указанный переключатель, чувствительный к редкоземельному металлу (например, лантану, церию, празеодиму или неодиму) может представлять собой репрессируемый переключатель, который можно применять для подавления экспрессии одного или более генов до тех пор, пока указанный репрессор не будет удален, после чего происходит «включение» генов. Например, в случае присутствия металлического лантана, механизм получения 2,3-BDO может быть «выключен». Чувствительный к лантану переключатель можно выключить (и может быть индуцирована экспрессия генов) путем либо удаления лантана из среды, либо разбавлением лантана в среде до уровней, при которых его подавляющее воздействие уменьшено, минимизировано или устранено. В некоторых случаях переключатель, чувствительный к редкоземельному металлу (например, лантану, церию, празеодиму или неодиму), может контролировать экспрессию НАДФН-зависимой ацетоинредуктазы. Переключатель, чувствительный к редкоземельному металлу (например, лантану, церию, празеодиму или неодиму) можно применять для контроля экспрессии альфа-ацетолактатдекарбоксилазы (budA). Кроме того, переключатель, чувствительный к редкоземельному металлу (например, лантану, церию, празеодиму или неодиму), можно применять для контроля экспрессии ацетолактатсинтазы (AlsS).
[00120] Конститутивно экспрессируемые промоторы также можно применять в векторных системах согласно настоящему документу. Например, экспрессия НАДФН- или НАДН-зависимой ацетоинредуктазы и/или альфа-ацетолактатдекарбоксилазы может контролироваться конститутивно активными промоторами. Примеры таких промоторов включают, но не ограничиваются ими, pMxaF и p.Bba.J23111.
[00121] Промоторы, подходящие для применения в прокариотических хозяевах, могут включать, например, системы промоторов а-лактамазы и лактозы, щелочной фосфатазы, систему промотора триптофана (trp), промотор эритромицина, промотор апрамицина, промотор гигромицина, промотор метиленомицина и гибридные промоторы, такие как промотор tac. Промоторы для применения в бактериальных системах также в целом содержат последовательность Шайна-Дальгарно, функционально связанную с кодирующей последовательностью.
[00122] В целом для получения высокого уровня транскрипции и экспрессии желаемого продукта можно использовать сильный промотор.
[00123] Один или более промоторов единицы транскрипции могут являться индуцируемыми промоторами. Например, зеленый флуоресцентный белок (GFP) можно экспрессировать с помощью конститутивного промотора, в то время как индуцируемый промотор применяют для управления транскрипцией гена, кодирующего один или более ферментов, описанных в настоящем описании, и/или амплифицируемый отбираемый маркер.
[00124] Некоторые векторы могут содержать прокариотические последовательности, которые способствуют размножению вектора в бактериях. Таким образом, векторы могут содержать другие компоненты, как, например, точку начала репликации (например, нуклеотидную последовательность, которая позволяет указанному вектору реплицироваться в одном или более из выбранных микроорганизмов), гены устойчивости к антибиотикам для отбора в бактериях и/или стоп-кодон amber, который может позволить трансляции проходить через кодон. Также могут быть включены дополнительные отбираемые гены. В целом в векторах клонирования точка начала репликации представляет собой точку, которая позволяет указанному вектору реплицироваться независимо от хромосомной ДНК хозяина, и включает точки начала репликации или автономно реплицирующиеся последовательности. Такие последовательности могут содержать точку начала репликации ColE1 в бактериальных или других известных последовательностях. Гены
[00125] Векторы, описанные в настоящем документе, могут содержать нуклеотидную последовательность одного или более генов, кодирующих: i) ацетоинредуктазу (НАДФН-зависимую и/или НАДН-зависимую); ii) альфа-ацетолактатдекарбоксилазу и/или iii) ацетолактатсинтазу (AlsS). Например, вектор может содержать ген НАДФН- или НАДН-зависимой ацетоинредуктазы. Вектор может содержать ген альфа-ацетолактатдекарбоксилазы. Вектор может содержать ген ацетолактатсинтазы. Эти векторы могут также содержать один или более регуляторных элементов (индуцируемых и/или репрессируемых промоторов), которые контролируют экспрессию генов в указанных векторах. В некоторых случаях переключатели, которые можно применять, включают, но не ограничиваются ими, индуцируемые или репрессируемые переключатели, например переключатели в виде арабинозы или лантана. Эти гены могут быть гетерологичными микроорганизму, с которым указанный вектор приводят в контакт (и в конечном итоге при помощи которого микроорганизм трансформируют). В некоторых случаях эти гены могут находиться в эписомальном векторе. В некоторых случаях вектор может представлять собой вектор, который можно применять для интегрирования одного или более таких векторов в геном микроорганизма. В некоторых случаях можно применять как эписомальный вектор, так и вектор интеграции. В некоторых случаях ген, кодирующий гетерологичную ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Гетерологичные гены в векторе могут быть представлены в определенном порядке до, во время или после приведения в контакт с микроорганизмом. Например, ген может быть расположен ближе к 5'-концу, чем любой другой ген в векторе до приведения в контакт с микроорганизмом. Однако после приведения в контакт с указанным микроорганизмом ген можно вставить в такое положение генома микроорганизма, когда указанный ген будет расположен ближе к 3'-концу, чем любой другой ген в векторе, или будет расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой ген в векторе. Например, векторы можно модифицировать внутри микроорганизма таким образом, что будет изменен порядок генов. В некоторых случаях, после приведения в контакт с микроорганизмом в вектор можно вставить ген, причем указанный вектор будет расположен ближе к 3'-концу, чем любой другой ген в векторе, или будет расположен ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой ген в векторе. В некоторых случаях определенного порядка генов можно достигнуть после того, как гетерологичные гены будут вставлены в геном микроорганизма. Например, можно применять различные векторы интеграции для достижения определенного порядка генов в геноме микроорганизма. В некоторых случаях порядок генов можно определить после того, как гетерологичный ген был вставлен в геном микроорганизма.
[00126] В некоторых случаях вектор может содержать два или более генов, кодирующих: i) ацетоинредуктазу (НАДФН-зависимую и/или НАДН-зависимую); ii) альфа-ацетолактатдекарбоксилазу и/или iii) ацетолактатсинтазу (AlsS). Например, вектор может содержать ген НАДФН- или НАДН-зависимой ацетоинредуктазы. В этом случае один или более генов могут быть гетерологичными микроорганизму, с которым указанный вектор приводят в контакт (и в конечном итоге при помощи которого микроорганизм трансформируют). В некоторых случаях вектор может представлять собой вектор, который можно применять для интегрирования одного или более таких векторов в геном микроорганизма. В некоторых случаях ген, кодирующий гетерологичную ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген.
[00127] В одном случае вектор может содержать по меньшей мере три или более генов, кодирующих НАДФН- и/или НАДН-зависимую ацетоинредуктазу, альфа-ацетолактатдекарбоксилазу и ацетолактатсинтазу. Один или более генов могут быть гетерологичными микроорганизму, с которым указанный вектор приводят в контакт (и в конечном итоге при помощи которого микроорганизм трансформируют). В некоторых случаях эти гены могут находиться в эписомальном векторе. В некоторых случаях вектор может представлять собой вектор, который можно применять для интегрирования одного или более таких векторов в геном микроорганизма. В некоторых случаях эти гены могут находиться как в эписомальном векторе, так и в векторе интеграции.
[00128] В одном случае ген ацетоинредуктазы получают из бактерии (например, грамположительной бактерии). Указанная бактерия может быть из рода Clostridium, например, вида Clostridium autoethanogenum.
[00129] Ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по существу схожа с SEQ ID NO 10. Например, ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 10. Например, ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере 80% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 10. В некоторых случаях ген НАДФН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая представляет собой последовательность SEQ ID NO 10.
[00130] В случае, когда желательна НАДН-зависимая ацетоинредуктаза, ген НАДН-зависимой ацетоинредуктазы можно получить из бактерии (например, грамположительной бактерии). Примеры включают бактерии из рода Bacillus, например, вида Bacillus subtilis, и рода Paenibacillus, например, вида Paenibacillus polymyxa.
[00131] Ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по существу схожа с SEQ ID NO 12 или 14. Например, ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 12 или 14. В некоторых случаях ген НАДН-зависимой ацетоинредуктазы может содержать нуклеотидную последовательность, которая идентична SEQ ID NO 12 или 14.
[00132] В случае, когда желателен ген альфа-ацетолактатдекарбоксилазы (budA), указанный ген альфа-ацетолактатдекарбоксилазы можно получить из бактерии. Примеры таких бактерий включают бактерии из рода Clostridium, например, вида Clostridium autoethanogenum, и бактерии из рода Klebsiella, например, вида Klebsiella pneumoniae. Ген альфа-ацетолактатдекарбоксилазы можно получить из грамположительной или грамотрицательной бактерии.
[00133] Ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по существу схожа с SEQ ID NO 6 или 8. Например, ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 6 или 8. В некоторых случаях ген альфа-ацетолактатдекарбоксилазы может содержать нуклеотидную последовательность, которая идентична SEQ ID NO 6 или 8.
[00134] В случае, когда желательна ацетолактатсинтаза (AlsS), указанную ацетолактатсинтазу можно получить из бактерии (например, грамположительной бактерии). Такие примеры включают бактерии из рода Clostridium, например, вида Clostridium autoethanogenum, и бактерии из рода Bacillus, например, вида Bacillus subtilis. Дополнительные примеры видов включают Bacillus licheniformis.
[00135] Ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по существу схожа с любой из последовательностей SEQ ID NO 2, 4 или 20. Например, ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. Ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по существу схожа с любой из последовательностей SEQ ID NO 2, 4 или 20. Например, ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях ген ацетолактатсинтазы может содержать нуклеотидную последовательность, которая идентична любой из последовательностей SEQ ID NO 2, 4 или 20.
[00136] Дополнительные гены можно разместить внутри микроорганизма для получения других желаемые конечных продуктов путем ферментации.
[00137] Нуклеотидную последовательность (или более конкретно кодоны, кодируемые нуклеотидной последовательностью) можно оптимизировать в зависимости от микроорганизма, в котором нуклеотидные последовательности будут экспрессироваться. Нуклеотидные последовательности могут быть кодон-оптимизированными в зависимости от количества тРНК, доступной в каждом отдельном микроорганизме. Другими словами, консервативные замены кодонов можно осуществить в зависимости от того, использует ли соответствующий микроорганизм специфичный кодон или сколько конкретной тРНК доступно в указанном микроорганизме.
Число копий гена
[00138] Любой из генов, описанных в настоящем документе, может иметь одну или более копий (интегрированных, эписомально экспрессируемых или их обоих). Например, каждая из ацетоинредуктазы (НАДФН-зависимой и/или НАДН-зависимой); ii) альфа-ацетолактатдекарбоксилазы и/или iii) ацетолактатсинтазы (AlsS) может иметь 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более копий гена. В некоторых случаях число копий генов равно 1. В некоторых случаях число копий гена равно 2. В некоторых случаях число копий гена равно 3. В некоторых случаях число копий гена равно 4. В некоторых случаях число копий гена равно 5. В некоторых случаях число копий гена равно 6. В некоторых случаях число копий гена равно 7. В некоторых случаях число копий гена равно 8. В некоторых случаях число копий гена равно 9. В некоторых случаях число копий гена равно 10. В некоторых случаях число копий гена равно 11. В некоторых случаях число копий гена равно 12. В некоторых случаях число копий гена равно 13. В некоторых случаях число копий гена равно 14. В некоторых случаях число копий гена равно 15. В некоторых случаях число копий гена равно 16. В некоторых случаях число копий гена равно 17. В некоторых случаях число копий гена равно 18. В некоторых случаях число копий гена равно 19. В некоторых случаях число копий гена равно 20. В некоторых случаях число копий гена равно 21. В некоторых случаях число копий гена равно 22. В некоторых случаях число копий гена равно 23. В некоторых случаях число копий гена равно 24. В некоторых случаях число копий гена равно 25. Обычно число копий гена меньше 25 копий, однако в некоторых случаях число может быть больше. Таким образом, в некоторых случаях число копий гена составляет более 25.
[00139] Кроме того, любой из описанных в настоящем документе генов (например, ген ацетоинредуктазы; альфа-ацетолактатдекарбоксилазы и/или ацетолактатсинтазы) может иметь одинаковое число копий генов. Например, ген, кодирующий ацетолактатсинтазу, может иметь от 1 до 25 копий, ген, кодирующий альфа-ацетолактатдекарбоксилазу, может иметь от 1 до 25 копий, а ген, кодирующий ацетоинредуктазу, может иметь от 1 до 25 копий, при этом все гены присутствуют в одном и том же количестве. В некоторых случаях любой из описанных в настоящем документе генов может иметь различное число копий генов. Например, ген, кодирующий ацетолактатсинтазу, может иметь от 1 до 25 копий, ген, кодирующий альфа ацетолактатдекарбоксилазу, может иметь от 1 до 25 копий, а ген, кодирующий ацетоинредуктазу, может иметь от 1 до 25 копий, при этом все гены присутствуют в разных количествах. В одном конкретном примере ген, кодирующий ацетолактатсинтазу, имеет от 5 до 10 копий, в то время как ген, кодирующий ацетоинредуктазу, может иметь от 1 до 6 копий, при этом ген, кодирующий альфа-ацетолактатдекарбоксилазу, может иметь от 3 до 8 копий. В этом же случае обычно число копий гена меньше 25 копий для любого рассматриваемого гена. Тем не менее, в некоторых случаях это число может быть больше.
[00140] Любое количество генов может экспрессироваться в векторе интеграции, эписомальном векторе или в них обоих. Что касается приведенного выше примера, ген, кодирующий ацетолактатсинтазу, может иметь от 5 до 10 копий, при этом от 2 до 5 копий могут быть интегрированы, а от 3 до 5 копий могут экспрессироваться в эписомальном векторе. Что касается приведенного выше примера, ген, кодирующий ацетоинредуктазу, может иметь от 1 до 6 копий, при этом от 1 до 4 копий могут быть интегрированы, а от 0 до 1 копий могут экспрессироваться в эписомальном векторе. Что касается приведенного выше примера, ген, кодирующий альфа-ацетолактатдекарбоксилазу, может иметь от 3 до 8 копий, при этом от 2 до 6 копий могут быть интегрированы, а от 1 до 2 копий могут экспрессироваться в эписомальном векторе.
[00141] В некоторых случаях копия гена может содержать частичную последовательность полного гена. В некоторых случаях указанная частичная последовательность полного гена может содержать активный сайт фермента, кодируемого геном. Подразумевается любая комбинация полного гена и частичной последовательности гена. Например, может быть 4 копии полного гена и 2 копии частичной последовательности гена. Считается, что это 6 копий гена.
Выделенные нуклеиновые кислоты
[00142] Гены, описанные в настоящем документе, могут быть представлены в форме выделенной полинуклеиновой кислоты. Другими словами, гены могут быть представлены в формах, которые не существуют в природе, в выделенной из хромосомы форме. Выделенные полинуклеиновые кислоты могут содержать последовательность одного или более генов, кодирующих: i) ацетоинредуктазу (НАДФН-зависимую и/или НАДН-зависимую); ii) альфа-ацетолактатдекарбоксилазу и/или iii) ацетолактатсинтазу (AlsS). Например, выделенная полинуклеиновая кислота может содержать ген НАДФН-зависимой и/или НАДН-зависимой ацетоинредуктазы. Выделенная полинуклеиновая кислота может содержать ген альфа ацетолактатдекарбоксилазы. Выделенная полинуклеиновая кислота может содержать ген ацетолактатсинтазы.
[00143] В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая кодирует ген НАДФН-зависимой ацетоинредуктазы. Например, выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по существу схожа с SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 81% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 82% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 83% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 84% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 86% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 87% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 88% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 89% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 10. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая представляет собой SEQ ID NO 10.
[00144] В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая кодирует НАДН-зависимую ацетоинредуктазу. Например, выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по существу схожа с SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может кодировать НАДН-зависимую ацетоинредуктазу. Например, выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по существу схожа с SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 81% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 82% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 83% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 84% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 86% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 87% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 88% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 89% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 12 или 14. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая идентична SEQ ID NO 12 или 14.
[00145] В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая кодирует альфа-ацетолактатдекарбоксилазу. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по существу схожа с последовательностями SEQ ГО NO 6 или 8. Например, выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична SEQ ID NO 6 или 8. Например, выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 81% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 82% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 83% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 84% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 86% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 87% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 88% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 89% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична SEQ ID NO 6 или 8. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая идентична SEQ ID NO 6 или 8.
[00146] В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая кодирует ацетолактатсинтазу. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по существу схожа с любой из последовательностей SEQ ID NO 2, 4 или 20. Например, выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. Выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 60% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 65% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 70% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 75% идентична любой из последовательностей SEQ ID NO 2, 4, или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 80% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 81% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 82% идентична любой из последовательностей SEQ ID NO 2, 4, или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 83% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 84% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 85% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 86% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 87% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 88% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 89% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 90% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 91% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 92% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 93% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 94% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 95% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 96% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 97% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 98% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая по меньшей мере на 99% идентична любой из последовательностей SEQ ID NO 2, 4 или 20. В некоторых случаях выделенная полинуклеиновая кислота может содержать нуклеотидную последовательность, которая идентична любой из последовательностей SEQ ID NO 2, 4 или 20.
Примеры векторных последовательностей
[00147] В настоящем документе также раскрыты векторы. Эти векторы можно интегрировать в различные микроорганизмы, как, например, метанотрофы, которые раскрыты в настоящем описании (например, в любую одну из последовательностей SEQ ID NO 15-18). В некоторых случаях эти векторы могут экспрессироваться эписомально. В некоторых случаях эти векторы могут быть как интегрированными, так и экспрессироваться эписомально. В некоторых случаях можно внести незначительные изменения в указанные векторы без существенных изменений в эффективности векторов или количества ферментов, которые эти векторы могут продуцировать. Таким образом, векторы могут быть по существу схожими с любой из последовательностей SEQ ID NO 15-18.
[00148] В некоторых случаях кассету экспрессии последовательности SEQ ID NO 15 или последовательности, которая по существу схожа с ней, можно привести в контакт с микроорганизмом (и вставить в него). Эта кассета экспрессии содержит промотор pBAD; ген g.Bsu AlsS (который содержит инициатор ATG и терминатор ТАА); сайт связывания с рибосомой rbsGTW001; ген g.Kpn BudA (который содержит инициатор ATG и терминатор TGA); терминатор rrnB; промотор pmxaF; ген g.Cau ButA (который содержит инициатор ATG и терминатор TGA) и терминатор лямбда Т0. Как только микроорганизм (такой как метанотроф) трансформируют при помощи этой кассеты экспрессии, то указанный микроорганизм называют штаммом XZ58. В некоторых случаях ген g.Bsu AlsS можно заменить на ген g.Blic_AlsS (например, SEQ ID NO 20).
[00149] В некоторых случаях кассету экспрессии последовательности SEQ ID NO 16 или последовательности, которая по существу схожа с ней, можно привести в контакт с микроорганизмом (и вставить в него). Эта кассета экспрессии содержит промотор pBAD; ген g.Bsu AlsS (который содержит инициатор ATG и терминатор ТАА); сайт связывания с рибосомой rbsGTW001; ген g.Kpn BudA (который содержит инициатор ATG и терминатор TGA); терминатор rrnB; промотор pmxaF; ген g.Bsu ButA (который содержит инициатор ATG и терминатор TGA) и терминатор лямбда Т0. Как только микроорганизм (такой как метанотроф) трансформируют при помощи этой кассеты экспрессии, то указанный микроорганизм называют штаммом XZ59. В некоторых случаях ген g.Bsu AlsS можно заменить на ген g.Blic_AlsS (например, SEQ ID NO 20).
[00150] В некоторых случаях кассету экспрессии последовательности SEQ ID NO 17 или последовательности, которая по существу схожа с ней, можно привести в контакт с микроорганизмом (и вставить в него). Эта кассета экспрессии содержит промотор pBAD; ген g.Bsu AlsS (который содержит инициатор ATG и терминатор ТАА); сайт связывания с рибосомой rbsGTW001; ген g.Kpn BudA (который содержит инициатор ATG и терминатор TGA); дополнительный сайт связывания с рибосомой rbsGTW001; терминатор rrnB; промотор pmxaF; ген g. Cau ButA (который содержит инициатор ATG и терминатор TGA) и терминатор rrnB. Как только микроорганизм (такой как метанотроф) трансформируют при помощи этой кассеты экспрессии, то указанный микроорганизм называют штаммом XZ06. В некоторых случаях ген g.Bsu AlsS можно заменить на ген g.Blic_AlsS (например, SEQ ID NO 20).
[00151] В некоторых случаях кассету экспрессии последовательности SEQ ID NO 18 или последовательности, которая по существу схожа с ней, можно привести в контакт с микроорганизмом (и вставить в него). Эта кассета экспрессии содержит промотор pBAD; ген g.Bsu AlsS (который содержит инициатор ATG и терминатор ТАА); сайт связывания с рибосомой rbsGTW001; ген g.Kpn BudA (который содержит инициатор ATG и терминатор TGA); дополнительный сайт связывания с рибосомой rbsGTWOOl; терминатор rrnB; промотор pmxaF; ген g.Bsu ButA (который содержит инициатор ATG и терминатор TGA) и терминатор rrnB. Как только микроорганизм (такой как метанотроф) трансформируют при помощи этой кассеты экспрессии, то указанный микроорганизм называют штаммом XZ08. В некоторых случаях ген g.Bsu AlsS можно заменить на ген g.Blic_AlsS (например, SEQ ID NO 20).
III. Способ получения генетически модифицированных микроорганизмов
[00152] Генетически модифицированные микроорганизмы, раскрытые в настоящем описании, можно получить различными способами. Микроорганизм можно модифицировать (например, сконструировать генетически) любым способом для того, чтобы указанный микроорганизм содержал и/или экспрессировал один или более полинуклеотидов, кодирующих ферменты в пути, которые катализируют превращение ферментируемого источника углерода (например, С1-углеродного соединения) в один или более промежуточных соединений в пути для получения 2,3-BDO, MEK и/или бутадиена. Такие ферменты могут включать ацетолактатсинтазу, альфа-ацетолактатдекарбоксилазу и ацетоинредуктазу. Например, один или более из любых указанных выше генов можно вставить в микроорганизм. Гены можно вставить при помощи вектора экспрессии. Гены также могут находиться под контролем одного или более разных/одних и тех же промоторов или один или более генов могут находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, например, чувствительного к арабинозе или лантану переключателя. Гены также можно стабильно интегрировать в геном микроорганизма. В некоторых случаях гены могут экспрессироваться в эписомальном векторе. В некоторых случаях гены можно интегрировать в геном микроорганизма. В некоторых случаях эти гены могут экспрессироваться в эписомальном векторе, а также быть интегрированы в геном микроорганизма. В некоторых случаях ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой ген в векторе. В некоторых случаях ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой ген в векторе. В некоторых случаях ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5', ни ближе к 3'-концу, чем любой другой ген в векторе. Гены в векторе могут быть представлены в определенном порядке до, во время или после приведения в контакт с микроорганизмом. Например, ген может располагаться ближе к 5'-концу, чем любой другой ген в векторе до приведения в контакт с микроорганизмом. Однако после приведения в контакт с указанным микроорганизмом ген можно вставить в положение, или в вектор может быть вставлен другой ген, когда указанный ген будет располагаться ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой ген в векторе. Например, векторы можно модифицировать внутри микроорганизма таким образом, что будет изменен порядок генов. В некоторых случаях определенного порядка генов можно достигнуть после того, как один или более гетерологичных генов будут вставлены в геном микроорганизма. Например, можно применять различные векторы интеграции для достижения определенного порядка генов в геноме микроорганизма. В некоторых случаях порядок генов можно определить после того, как гетерологичный ген был вставлен в геном микроорганизма.
[00153] Микроорганизм, применяемый в этом способе, может быть любым из описанных выше, включая, но не ограничиваясь ими, прокариот. Можно применять и другие микроорганизмы, такие как бактерии, дрожжи или водоросли. Микроорганизмы, представляющие особый интерес, включают метанотрофов, как, например, из родов Methylobacter, Methylomicrobium, Methylomonas, Methylocaldum, Methylococcus, Methylosoma, Methylosarcina, Methylothermus, Methylohalobius, Methylogaea, Methylovulum, Crenothrix, Clonothrix, Methylosphaera, Methylocapsa, Methylocella, Methylosinus, Methylocystis или Methyloacidophilum. Желаемый вид может включать Methylococcus capsulatus.
[00154] Примером способа получения генетически модифицированного микроорганизма, раскрытого в настоящем описании, является приведение в контакт (или трансформирование) микроорганизма с нуклеиновой кислотой, которая экспрессирует по меньшей мере один из гетерологичных генов, кодирующих i) ацетоинредуктазу (например, НАДФН- или НАДН-зависимую)); ii) альфа-ацетолактатдекарбоксилазу (budA); iii) ацетолактатсинтазу или iv) любую их комбинацию. Микроорганизм может представлять собой любой микроорганизм, способным превращать С1-углеродное соединение в продукт. В некоторых случаях указанный продукт представляет собой 2,3-BDO.
[00155] НАДФН- или НАДН-зависимая ацетоинредуктаза, альфа-ацетолактатдекарбоксилаза (budA) и/или ацетолактатсинтаза могут представлять собой любой из вариантов, описанных выше. Например, ацетоинредуктазу можно получить из грамположительной бактерии, такой как из рода Clostridium, как, например, вида Clostridium autoethanogenum.
[00156] Гены, которые вставляют в микроорганизм, могут быть гетерологичными для этого микроорганизма. Например, если указанный микроорганизм представляет собой метанотрофа, вставленные гены могут, например, быть получены из дрожжей, бактерии или различных видов метанотрофов. Кроме того, эти гены могут быть эндогенной частью генома микроорганизма.
[00157] Гены можно вставить в микроорганизм с применением векторов. В некоторых случаях гены можно вставить в геном микроорганизма. В некоторых случаях оба метода можно применять тогда, когда два или более генов вставляют в микроорганизм. Например, ген может вставить в геном микроорганизма, например, путем применения вектора интеграции. Впоследствии дополнительный ген можно трансформировать в микроорганизм через эписомальный вектор. В некоторых случаях вектор может представлять определенный порядок генов. Например, в некоторых случаях вектор может содержать ген, кодирующий ацетолактатсинтазу, при этом указанный ген расположен ближе к 5'-концу, чем любой другой ген в векторе. В некоторых случаях вектор может содержать ген, кодирующий ацетоинредуктазу, при этом указанный ген расположен ближе к 3'-концу, чем любой другой ген в векторе. В некоторых случаях вектор может содержать ген, кодирующий альфа-ацетолактатдекарбоксилазу, который располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой ген в векторе.
[00158] В некоторых случаях генетически модифицированные микроорганизмы, полученные способами, описанными в настоящем документе, могут продуцировать желаемый продукт при более высоких титрах при ферментации при более высокой температуре. Например, генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продуктов, таких как 2,3-BDO, бутадиена и/или МЕК, при их инкубации при температуре выше 37°С (но не выше температуры 100°С). В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при их инкубации при 42°С по сравнению с 37°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при их инкубации при 41°С по сравнению с 37°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при его инкубации при 42°С по сравнению с 45°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при его инкубации при 41°С по сравнению с 45°С. В некоторых случаях генетически модифицированные микроорганизмы можно получить для продуцирования ими более высоких титров продукта при их инкубации при 37°С по сравнению с 45°С. В некоторых случаях генетические модификации приводят к повышенной толерантности/предпочтению к более высоким температурам. Методы генной модификации
[00159] Микроорганизмы, раскрытые в настоящем описании, могут быть генетически сконструированы с применением классических микробиологических методов. Некоторые из таких методов в целом раскрыты, например, в публикации Sambrook et al., 1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Labs Press.
[00160] Генетически модифицированные микроорганизмы, раскрытые в настоящем описании, могут содержать полинуклеотид, который был вставлен, удален или модифицирован (т.е. мутирован; например, путем вставки, делеции, замены и/или инверсии нуклеотидов) таким образом, что такие модификации обеспечивают желаемый эффект экспрессии (например, гиперэкспрессии) одного или более предложенных в настоящем документе ферментов в микроорганизме. Генетические модификации, которые приводят к увеличению экспрессии или функции генов, могут упоминаться как амплификация, сверхсинтез, гиперэкспрессия, активация, усиление, добавление или повышенная регуляция гена. Добавление гена для увеличения экспрессии гена может включать поддержание гена(-ов) в реплицирующихся плазмидах или интеграцию указанного клонированного гена(-ов) в геном продуцирующего микроорганизма. Кроме того, увеличение экспрессии желаемого гена может включать оперативное связывание клонированного гена(-ов) с нативными или гетерологичными контрольными элементами транскрипции.
[00161] В случае, когда это желательно, экспрессия одного или более ферментов, предложенных в настоящем документе, находится под контролем регуляторной последовательности, которая напрямую или косвенно контролирует экспрессию фермента в зависимости от времени во время ферментации. В некоторых случаях для достижения этой цели можно применять индуцируемые промоторы.
[00162] В некоторых случаях микроорганизм трансформируют или трансфицируют при помощи генетического носителя, как, например, вектора экспрессии, содержащего гетерологичную полинуклеотидную последовательность, кодирующую ферменты, предложенные в настоящем документе В некоторых случаях вектор(-ы) может представлять собой эписомальный вектор, последовательность гена, которую можно интегрировать в геном микроорганизма, или любую их комбинацию. В некоторых случаях векторы, содержащие гетерологичную полинуклеотидную последовательность, кодирующую предложенные в настоящем документе ферменты, интегрируют в геном микроорганизма.
[00163] Для того, чтобы облегчить вставку и способствовать экспрессии различных генов, кодирующих ферменты, раскрытых в настоящем описании, из конструкций и векторов экспрессии, указанные конструкции можно спроектировать с применением по меньшей мере одного сайта клонирования для вставки любого гена, кодирующего любой фермент, раскрытый в настоящем описании. Сайт клонирования может представлять собой сайт множественного клонирования, например, содержащий множественные сайты рестрикции.
Трансфекция
[00164] Стандартные методы трансфекции можно применять для вставки генов в микроорганизм. В контексте настоящего описания термин «трансфекция» или «трансформация» и их грамматические эквиваленты могут относиться ко вставке экзогенной нуклеиновой кислоты или полинуклеотида в клетку-хозяина. Указанную экзогенную нуклеиновую кислоту или полинуклеотид можно сохранить в качестве неинтегрируемого вектора, например, плазмидного или эписомального вектора, или, в качестве альтернативы, их можно интегрировать в геном указанной клетки-хозяина. Термин «осуществление трансфекции» или «трансфекция» предназначен для охвата всех общепринятых методов введения нуклеиновой кислоты или полинуклеотида в микроорганизмы. Примеры методов трансфекции включают, но не ограничиваются ими, метод осаждения с фосфатом кальция, опосредованную DEAE-декстраном трансфекцию, липофекцию, электропорацию, микроинъекцию, трансфекцию, опосредованную хлоридом рубидия или поликатионами, слияние протопластов и обработку ультразвуком. Предпочтительным является способ трансфекции, который обеспечивает оптимальную частоту трансфекции и экспрессию конструкции в конкретной линии и типе клеток-хозяев. Для стабильных трансфектантов конструкции интегрируют таким образом, чтобы они стабильно сохранялись в хромосоме хозяина. В некоторых случаях предпочтительной трансфекцией является стабильная трансфекция. В некоторых случаях интеграция гена происходит в определенном локусе в геноме микроорганизма.
Трансформация
[00165] Векторы экспрессии или другие нуклеиновые кислоты можно вводить в выбранные микроорганизмы любым из множества подходящих способов. Например, векторные конструкции можно вводить в соответствующие клетки любым из ряда способов трансформации для плазмидных векторов. Для введения оголенной ДНК в бактерию до сих пор широко применяют стандартную, опосредованную кальций-хлоридом, бактериальную трансформацию (см., например, Sambrook et al., 1989, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.), однако также можно применять электропорацию и конъюгацию (см., например, Ausubel et al., 1988, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY, N.Y.).
[00166] Для введения векторных конструкций в клетки дрожжей или клетки других грибов можно применять химические способы трансформации (например, Rose et al., 1990, Methods in Yeast Genetics, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). Трансформированные клетки можно выделить на селективной среде, соответствующей применяемому отбираемому маркеру. В качестве альтернативы или помимо этого, планшеты или фильтры, снятые с планшетов, можно отсканировать на наличие флуоресценции GFP для идентификации трансформированных клонов.
[00167] Для введения векторов, содержащих дифференциально экспрессируемые последовательности, в определенные типы клеток, применяемый способ может зависеть от формы вектора. Плазмидные векторы можно ввести любым из множества способов трансфекции, включая, например, липид-опосредованную трансфекцию («липофекцию»), опосредованную DEAE-декстраном трансфекцию, электропорацию или осаждение с фосфатом кальция (см., например, Ausubel et al., 1988, Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY, NY.).
[00168] Агенты для липофекции и способы, подходящие для временной трансфекции широкого спектра трансформированных и нетрансформированных или первичных клеток, широко доступны, что делает липофекцию привлекательным способом введения конструкций в эукариотические клетки, в частности клетки млекопитающих, в культуре. Многие компании предлагают наборы и способы для этого вида трансфекции.
[00169] Клетка-хозяин может обладать способностью экспрессировать конструкцию, кодирующую желаемый белок, обрабатывать белок и транспортировать секретируемый белок к клеточной поверхности для секреции. Обработка включает совместную и посттрансляционную модификацию, такую как расщепление лидерного пептида, присоединение GPI, гликозилирование, убиквитинирование и образование дисульфидной связи.
[00170] Микроорганизмы можно трансформировать или трансфицировать при помощи описанных выше векторов экспрессии или полинуклеотидов, кодирующих один или более ферментов, описанных в настоящем документе, и культивировать в питательных средах, модифицированных для конкретного микроорганизма, индуцируя промоторы, выбирая трансформантов или амплифицируя гены, кодирующие желаемые последовательности. В некоторых случаях метод электропорация можно применять для доставки вектора экспрессии.
[00171] Экспрессию вектора (и гена, содержащегося в указанном векторе) можно подтвердить с помощью анализа экспрессии, например, кПЦР или путем измерения уровней РНК. Уровень экспрессии может также указывать на число копий. Например, в случае, если уровни экспрессии чрезвычайно высокие, это может свидетельствовать о том, что в геном интегрировали более одной копии гена. Альтернативным образом, высокая экспрессия может свидетельствовать о том, что ген был интегрирован в высоко транскрибируемую область, например, рядом с гиперкспрессируемым промотором. Экспрессию также можно подтвердить путем измерения уровня белка, например, при помощи вестерн-блоттинга. CRISPR/cas система
[00172] Способы, раскрытые в настоящем документе, могут включать точную вставку генов или делецию генов (или частей генов). В способах, описанных в настоящем документе, можно применять систему CRISPR/cas. Например, двухцепочечные разрывы (DSB) можно образовывать с применением системы CRISPR/cas, например, системы CRISPR/cas типа II. Фермент Cas, применяемый в раскрытом в настоящем документе способе, может представлять собой белок Cas9, который катализирует расщепление ДНК. Ферментативное действие Cas9, полученного из Streptococcus pyogenes или любого близкородственного Cas9 фермента, может образовывать двухцепочечные разрывы в последовательностях сайтов-мишеней, которые гибридизуются с 20 нуклеотидами гидовой последовательности и содержат примыкающий к протоспейсеру мотив (РАМ) после 20 нуклеотидов указанной последовательности-мишени.
[00173] Вектор может быть функционально связан с кодирующей фермент последовательностью, кодирующей фермент CRISPR, такой как белок Cas. Cas-белки, которые можно применять, включают класс 1 и класс 2. Неограничивающие примеры Cas-белков включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5d, Cas5t, Cas5h, Cas5a, Cas6, Cas7, Cas8, Cas9 (также известный как Csn1 или Csx12), Cas10, Csy1, Csy2, Csy3, Csy4, Cse1, Cse2, Cse3, Cse4, Cse5e, Csc1, Csc2, Csa5, Csn1, Csn2, Csm1, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx1S, Csf1, Csf2, CsO, Csf4, Csd1, Csd2, Cst1, Cst2, Csh1, Csh2, Csa1, Csa2, Csa3, Csa4, Csa5, C2c1, C2c2, C2c3, Cpf1, CARF, DinG, их гомологи или их модифицированные версии. Немодифицированный фермент CRISPR может обладать расщепляющей ДНК активностью, как, например, Cas9. Фермент CRISPR может управлять расщеплением одной или обеих цепей в последовательности-мишени, например, в последовательности-мишени и/или в комплименте последовательности-мишени. Например, фермент CRISPR может управлять расщеплением одной или обеих цепей в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 125, 150, 175, 200, 300, 400, 500 или более пар оснований в первом или последнем нуклеотиде последовательности-мишени. Можно применять вектор, который кодирует фермент CRISPR, который мутирован по сравнению с соответствующим ферментом дикого типа, так что указанный мутированный фермент CRISPR не способен расщеплять одну или обе цепи полинуклеотида-мишени, содержащего последовательность-мишень.
[00174] Можно применять вектор, который кодирует фермент CRISPR, содержащий одну или более последовательностей ядерной локализации (NLS). Например, можно применить 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 NLS. Фермент CRISPR может содержать NLS на или рядом с N-концом (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 NLS), или на или рядом с С-концом (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 NLS), или любую их комбинацию (например, одну или более NLS на N-конце и одну или более NLS на С-конце). В случае, когда имеется более одной NLS, каждую из них можно выбрать независимо от другой, так что одна NLS может присутствовать в более чем одной копии и/или в комбинации с одной или более другими NLS, представленными в одной или более копиях.
[00175] Применяемые в способах ферменты CRISPR могут содержать не более 6 NLS. Считается, что NLS расположена вблизи N- или С-конца в случае, когда ближайшая к NLS аминокислота находится в пределах 50 аминокислот вдоль полипептидной цепи от N- или С-конца, например, в пределах 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 40 или 50 аминокислот.
Гидовая РНК
[00176] В контексте настоящего описания термин «гидовая РНК» и его грамматические эквиваленты могут относиться к РНК, которая может быть специфичной для ДНК-мишени и может образовывать комплекс с белком Cas. Комплекс РНК/Cas может помочь «направить» белок Cas к ДНК-мишени.
[00177] Способ, раскрытый в настоящем описании, также может включать введение в клетку или эмбрион по меньшей мере одной гидовой РНК или нуклеиновой кислоты, например, ДНК, кодирующей по меньшей мере одну гидовую РНК. Гидовая РНК может взаимодействовать с направляемой РНК эндонуклеазой для направления указанной эндонуклеазы к определенному сайту-мишени, на котором 5'-конец нуклеотидов гидовой РНК образует пару со специфичной последовательностью протоспейсера в хромосомной последовательности.
[00178] Гидовая РНК может содержать две РНК, например, CRISPR РНК (crRNA) и трансактивирующую crRNA (tracrRNA). Гидовая РНК иногда может содержать одноцепочечную РНК или одиночную гидовую РНК (sgRNA), образованную слиянием части (например, функциональной части) crPHK и tracrPHK. Гидовая РНК также может представлять собой двойную РНК, содержащую crPHK и tracrHK. Кроме того, crPHK может гибридизоваться с ДНК-мишенью.
[00179] Как это обсуждалось выше, гидовая РНК может представлять собой продукт экспрессии. Например, ДНК, кодирующая гидовую РНК, может представлять собой вектор, содержащий последовательность, кодирующую указанную гидовую РНК. Гидовую РНК можно перенести в клетку или микроорганизм путем трансфекции указанной клетки или микроорганизма с помощью выделенной гидовой РНК или плазмидной ДНК, содержащей последовательность, кодирующую указанную гидовую РНК, и промотор. Гидовую РНК также можно перенести в клетку или микроорганизм другим способом, например, при помощи опосредованной вирусом доставки генов.
[00180] Гидовую РНК можно выделить. Например, гидовую РНК можно трансфицировать в форме выделенной РНК в клетку или микроорганизм. Гидовую РНК можно получить путем транскрипции in vitro с применением любой системы транскрипции in vitro. Гидовую РНК можно перенести в клетку в форме выделенной РНК, а не в форме плазмиды, содержащей кодирующую для гидовой РНК последовательность.
[00181] Гидовая РНК может содержать три области: первую область на 5'-конце, которая может быть комплементарна сайту-мишени в хромосомной последовательности, вторую внутреннюю область, которая может образовывать структуру типа «петля-на-стебле», и третью область на 3'-конце, которая может быть одноцепочечной. Первая область каждой гидовой РНК также может отличаться таким образом, что каждая гидовая РНК будет направлять белок слияния к специфичному сайту-мишени. Кроме того, вторая и третья области каждой гидовой РНК могут быть одинаковыми во всех гидовых РНК.
[00182] Первая область гидовой РНК может быть комплементарна последовательности в сайте-мишени в хромосомной последовательности, так что указанная первая область гидовой РНК может образовывать пары с нуклеотидами в сайте-мишени. В некоторых случаях первая область гидовой РНК может содержать от 10 нуклеотидов до 25 нуклеотидов (т.е. от 10 nt до 25 nt; или от 10 nt до 25 nt; или от 10 nt до 25 nt; или от 10 nt до 25 nt) или больше. Например, длина участка образования пар нуклеотидов между первой областью гидовой РНК и сайтом-мишенью в хромосомной последовательности может составлять 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 22, 23, 24, 25 или более нуклеотидов. Иногда длина первой области гидовой РНК может составлять 19, 20 или 21 нуклеотид.
[00183] Гидовая РНК также может содержать вторую область, которая образует вторичную структуру. Например, вторичная структура, образованная гидовой РНК, может содержать стебель (или шпильку) и петлю. Длина петли и стебля может варьироваться. Например, длина петли может составлять от 3 до 10 нуклеотидов, а длина ствола - от 6 до 20 пар оснований. Стебель может содержать одну или более выпуклостей от 1 до 10 нуклеотидов в длину. Общая длина второй области может составлять от 16 до 60 нуклеотидов в длину. Например, петля может состоять из 4 нуклеотидов в длину, а стебель может состоять из 12 пар оснований.
[00184] Гидовая РНК также может содержать третью область на 3'-конце, которая может быть по существу одноцепочечной. Например, третья область иногда не является комплементарной какой-либо хромосомной последовательности в представляющей интерес клетке, а иногда она не является комплементарной остальной части гидовой РНК. Кроме того, длина третьей области может варьироваться. Длина третьей области может составлять более 4 нуклеотидов. Например, длина третьей области может составлять от 5 до 60 нуклеотидов.
[00185] Гидовую РНК можно ввести в клетку или эмбрион в виде молекулы РНК. Например, молекула РНК может транскрибироваться in vitro и/или ее можно синтезировать химическим путем. РНК можно транскрибировать с синтетической молекулы ДНК, например, фрагмента гена gBlocks®. Гидовую РНК затем можно ввести в клетку или эмбрион в виде молекулы РНК. Гидовую РНК также можно ввести в клетку или эмбрион в форме молекулы нуклеиновой кислоты, не являющейся РНК, например, в форме молекулы ДНК. Например, ДНК, кодирующая гидовую РНК, может быть функционально связана с промоторной контрольной последовательностью для экспрессии указанной гидовой РНК в представляющей интерес клетке или эмбрионе. Кодирующая РНК последовательность может быть функционально связана с промоторной последовательностью, которая распознается РНК-полимеразой III (Pol III). Плазмидные векторы, которые можно применить для экспрессии гидовой РНК, включают, но не ограничиваются ими, векторы рх330 и векторы рх333. В некоторых случаях плазмидный вектор (например, вектор рх333) может содержать две последовательности ДНК, кодирующие гидовую РНК.
[00186] Последовательность ДНК, кодирующая гидовую РНК, также может быть частью вектора. Кроме того, вектор может содержать дополнительные контрольные последовательности экспрессии (например, энхансерные последовательности, последовательности Козака, последовательности полиаденилирования, последовательности терминации транскрипции и т.д.), отбираемые маркерные последовательности (например, гены устойчивости к антибиотикам), точки начала репликации и тому подобное. Молекула ДНК, кодирующая гидовую РНК, также может быть линейной. Молекула ДНК, кодирующая гидовую РНК, также может быть кольцевой.
[00187] В случае, когда последовательности ДНК, кодирующие направляемую РНК эндонуклеазу и гидовую РНК, вводят в клетку, каждая из последовательностей ДНК может представлять собой часть отдельной молекулы (например, один вектор, содержащий последовательность, кодирующую направляемую РНК эндонуклеазу, и второй вектор, содержащий последовательность, кодирующую гидовую РНК) или они обе могут быть частью одной и той же молекулы (например, один вектор, содержащий кодирующую (и регуляторную) последовательность как для направляемой РНК эндонуклеазы, так и для гидовой РНК).
Сайт-специфичные вставки
[00188] Вставка генов может быть сайт-специфичной. Например, один или более генов можно вставить рядом с промотором.
[00189] Модификацию целевого локуса микроорганизма можно произвести путем введения ДНК в микроорганизмы, при этом указанная ДНК обладает гомологией с локусом-мишенью. ДНК может содержать маркерный ген, позволяющий осуществлять отбор клеток, содержащих интегрированную конструкцию. Гомологичную ДНК в векторе-мишени можно рекомбинировать с ДНК в локусе-мишени. Маркерный ген может быть фланкирован с обеих сторон гомологичными последовательностями ДНК, 3'-плечом рекомбинации и 5'-плечом рекомбинации.
[00190] Разнообразные ферменты могут катализировать вставку чужеродной ДНК в геном микроорганизма. Например, сайт-специфические рекомбиназы можно сгруппировать в два семейства белков с различными биохимическими свойствами, а именно тирозин-рекомбиназы (в которых ДНК ковалентно связана с остатком тирозина) и серин-рекомбиназы (в которых ковалентное присоединение происходит по остатку серина). В некоторых случаях рекомбиназы могут содержать Cre, интегразу Ф31 (серин-рекомбиназу, полученную из фага Ф31 Streptomyces) или сайт-специфичные рекомбиназы, полученные из бактериофага (включая Flp, лямбда-интегразу, рекомбиназу бактериофага HK022, интегразу бактериофага R4 и интегразу фагаТР901-1).
[00191] Систему CRISPR/Cas можно применять для осуществления сайт-специфичной вставки. Например, при помощи CRISPR/cas можно получить одноцепочечный разрыв на сайте вставки в геноме для облегчения вставки трансгена в сайт вставки.
[00192] В способах, описанных в настоящем документе, можно использовать методы, которые можно применять для обеспечения возможности проникновения конструкции ДНК или РНК в клетку-хозяина, которые включают, но не ограничиваются ими, совместное осаждение ДНК с фосфатом кальция, микроинъекцию ДНК в ядро, электропорацию, слияние бактериальных протопластов с интактными клетками, трансфекцию, липофекцию, инфекцию, бомбардировку частицами, опосредованную спермой передачу генов или любой другой метод.
[00193] В определенных аспектах раскрытого в настоящем описании документа можно использовать векторы (в том числе описанные выше). Любые плазмиды и векторы можно применять до тех пор, пока они воспроизводятся и являются жизнеспособными в выбранном микроорганизме-хозяине. Векторы, известные в данной области техники и коммерчески доступные (и их варианты или производные), можно сконструировать так, что они будут содержать один или более сайтов рекомбинации для применения в указанных способах. Векторы, которые можно применять, включают, но не ограничиваются ими, эукариотические векторы экспрессии, такие как pFastBac, pFastBacHT, pFastBacDUAL, pSFV и pTet-Splice (Invitrogen), pEUK-C1, pPUR, pMAM, pMAMneo, pBI01, pBI121, pDR2, pCMVEBNA и pYACneo (Clontech), pSVK3, pSVL, pMSG, рСН110, и pKK232-8 (Pharmacia, Inc.), pXT1, pSG5, pPbac, pMbac, pMClneo и pOG44 (Stratagene, Inc.), и pYES2, pAC360, pBlueBa-cHis А, В, и С, pVL1392, pBlueBac111, pCDM8, pcDNA1, pZeoSV, pcDNA3, pREP4, pCEP4, и pEBVHis (Invitrogen, Corp.) и их варианты или производные.
[00194] Эти векторы можно применять для экспрессии гена или части гена, представляющего интерес. Часть гена или ген можно вставить с применением известных способов, таких как методов, основанных на ферменте рестриктазе.
IV. Другие способы
Способы получения подходящих химических соединений
[00195] Генетически модифицированные микроорганизмы, описанные в настоящем документе, можно применять для получения подходящих химических соединений, включая, но не ограничиваясь ими, ацетоин, 2,3-BDO, MEK, бутадиен и/или бутен.
[00196] Микроорганизм может представлять собой любой микроорганизм, описанный в настоящем документе, включая, но не ограничиваясь ими, прокариотов, таких как, например, метанотроф.
[00197] Углеродный субстрат может представлять собой любой углеродный субстрат, описанный в настоящем документе, включая, но не ограничиваясь ими, С1-углеродный субстрат, такой как метан.
[00198] Условия ферментации, применяемые во время получения подходящих химических соединений, могут быть любыми из описанных в настоящем документе, как, например, температура. Например, температуру ферментации можно поддерживать между 37°С и 45°С. В некоторых случаях температура ферментации может составлять 42°С. В некоторых случаях ферментацию можно проводить при температуре 41°С.
2-Ацетолактат
[00199] В настоящем документе раскрыт способ получения 2-ацетолактата, включающий приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, причем указанный микроорганизм содержит гетерологичный ген, кодирующий ацетолактатсинтазу (AlsS). В некоторых случаях гетерологичный ген интегрирован в геном микроорганизма. В некоторых случаях гетерологичный ген экспрессируется эписомально. В некоторых случаях гетерологичный ген является как эписомально эксрессируемым, так и интегрированным в геном микроорганизма. В некоторых случаях гетерологичный ген ацетолактатсинтазы расположен ближе к 5'-концу, чем любой другой гетерологичный ген (независимо от того, экспрессируется ли он в эписомальном векторе или интегрирован в геном микроорганизма). В некоторых случаях, когда применяемый микроорганизм содержит более одной хромосомы, термины «5'-конец» или «3'-конец» могут относиться к генам, содержащимся в одной хромосоме. Указанный способ может также включать выращивание микроорганизма с получением 2-ацетолактата. Последовательность AlsS может быть по существу схожа с любой из последовательностей SEQ ID NO 1, 3 или 19. В некоторых случаях AlsS может кодироваться нуклеиновой кислотой, которая по существу схожа с любой из последовательностей SEQ ID NO 2, 4 или 20. Ген AlsS может находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Также микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана), затем впоследствии лантан можно разбавить. Это может происходить до выращивания микроорганизма с получением 2-ацетолактата. Получаемый 2-ацетолактат может быть по существу чистым. Полученный 2-ацетолактат можно выделить. Кроме того, также можно выделить 2-ацетолактатные продукты (т.е. побочные продукты). Например, в качестве побочного продукта можно выделить диацетил.
[00200] 2-Ацетолактат также можно обработать при помощи того же микроорганизма, другого микроорганизма или вне микроорганизма (т.е. in vitro). Могут происходить различные ферментативные реакции или даже спонтанные реакции. Например, 2-ацетолактат может спонтанно превратиться в диацетил. 2-Ацетолактат также можно превратить в ацетоин путем применения альфа-ацетолактатдекарбоксилазы. Кроме того, 2-ацетолактат также можно превратить в 2,3-дигидрокси-2-метилбутановую кислоту посредством реакции восстановления редуктазой 3-кетокислоты.
[00201] Таким образом, один и тот же микроорганизм может содержать альфа-ацетолактатдекарбоксилазу и/или редуктазу 3-кетоки слоты. В других случаях, другой микроорганизм может содержать альфа-ацетолактатдекарбоксилазу и/или редуктазу 3-кетокислоты, или альфа-ацетолактатдекарбоксилазу и/или редуктазу 3-кетокислоты выделяют из клетки. В случае, если альфа-ацетолактатдекарбоксилаза и/или редуктаза 3-кетокислоты содержатся в другом микроорганизме или они выделены из клетки, указанный микроорганизм/выделенный фермент может превращать 2-ацетолактат, который находится в среде для культивирования (этого достигают либо путем дополнительного добавления, либо путем секреции микроорганизма, продуцирующего ацетоин). Превращение 2-ацетолактата при помощи альфа-ацетолактатдекарбоксилазы и/или редуктазы 3-кетокислоты может приводить к получению ацетоина и/или 2,3-дигидрокси-2-метилбутановой кислоты соответственно.
Ацетоин
[00202] В настоящем описании раскрыт способ получения ацетоина, включающий приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу (budA). В некоторых случаях гетерологичный ген интегрирован в геном микроорганизма. В некоторых случаях гетерологичный ген экспрессируется эписомально. В некоторых случаях гетерологичный ген является как эписомально эксрессируемым, так и интегрированным в геном микроорганизма. В некоторых случаях альфа-ацетолактатдекарбоксилаза располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген (независимо от того, экспрессируется ли он в эписомальном векторе, или интегрирован в геном микроорганизма). Например, что касается гена ацетолактатдекарбоксилазы, существует по меньшей мере один гетерологичный ген, который расположен в направлении 3'-5' (дополнительный 5'-конец) от гена ацетолактатдекарбоксилазы, а также по меньшей мере один дополнительный гетерологичный ген, который расположен в направлении 5'-3' (дополнительный 3'-конец) от гена ацетолактатдекарбоксилазы. В некоторых случаях, когда применяемый микроорганизм содержит более одной хромосомы, термины «5'-конец» или «3'-конец» могут относиться к генам, содержащимся в одной хромосоме. Указанный способ также может включать выращивание микроорганизма с получением ацетоина. Ген budA может находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который реагирует на присутствие или отсутствие компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Также микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана), а затем впоследствии лантан можно разбавить. Это может происходить до выращивания микроорганизма с получением ацетоина. Полученный ацетоин может быть по существу чистым. Полученный ацетоин можно выделить. Кроме того, неацетоиновые продукты (т.е. побочные продукты) также можно выделить.
[00203] Ацетоин также можно обработать при помощи одного и того же микроорганизма, другого микроорганизма или вне микроорганизма (т.е. in vitro) с применением ацетоинредуктазы (например, НАДН-зависимой или НАДФН-зависимой ацетоинредуктазы). Тот же микроорганизм может содержать ацетоинредуктазу (например, НАДН-зависимую или НАДФН-зависимую ацетоинредуктазу). В других случаях другой микроорганизм может содержать ацетоинредуктазу, или ацетоинредуктазу выделяют из клетки. В случае, если ацетоинредуктаза содержится в другом микроорганизме или она выделена из клетки, указанный микроорганизм/выделенный фермент может превращать ацетоинредуктазу, которая содержится в среде для культивирования (этого достигают либо путем дополнительного добавления, либо путем секреции микроорганизма, продуцирующего ацетоин). При выделении ацетоина с помощью ацетоинредуктазы может образовываться 2,3-BDO.
[00204] Кроме того, превращение 2,3-BDO в различные продукты может происходить посредством различных процессов ферментации или различных каталитических превращений.
2,3-BDO
[00205] В настоящем описании раскрыт способ получения 2,3-BDO, включающий приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий: (i) ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS или (iv) любую их комбинацию. В некоторых случаях по меньшей мере один гетерологичный ген интегрирован в геном микроорганизма. В некоторых случаях по меньшей мере один гетерологичный ген экспрессируется эписомально. В некоторых случаях по меньшей мере один гетерологичный ген является как эписомально эксрессируемым, так и интегрированным в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях, когда применяемый микроорганизм содержит более одной хромосомы, термины «5'-конец» или «3'-конец» могут относиться к генам, содержащимся в одной хромосоме. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, является гетерологичным геном, расположенным ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ также может включать выращивание микроорганизма с получением 2,3-BDO. По меньшей мере один гетерологичный ген может находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Также микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана), а затем впоследствии лантан можно разбавить. Это может происходить до выращивания микроорганизма с получением 2,3-BDO.
[00206] 2,3-BDO, который получают при помощи этих способов, может быть по существу чистым. Полученный 2,3-BDO можно выделить. Кроме того, также можно выделить продукты, не являющиеся 2,3-BDO (т.е. побочные продукты), такие как 2-ацетолактат и ацетоин.
MEK
[00207] В настоящем описании раскрыт способ получения MEK, включающий приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий: (i) ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS или (iv) любую их комбинацию. В некоторых случаях по меньшей мере один гетерологичный ген интегрирован в геном микроорганизма. В некоторых случаях по меньшей мере один гетерологичный ген экспрессируется эписомально. В некоторых случаях по меньшей мере один гетерологичный ген является как эписомально эксрессируемым, так и интегрированным в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях, когда применяемый микроорганизм содержит более одной хромосомы, термины «5'-конец» или «3'-конец» могут относиться к генам, содержащимся в одной хромосоме. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ также может включать выращивание микроорганизма с получением 2,3-BDO. Указанный способ также может включать приведение в контакт 2,3-BDO, полученного с применением катализатора с получением MEK. По меньшей мере один гетерологичный ген может находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Также микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана), а затем впоследствии лантан можно разбавить. Это может происходить до выращивания микроорганизма с получением 2,3-BDO. В некоторых случаях 2,3-BDO можно выделить или очистить перед приведением его в контакт с 2,3-BDO, полученным с применением катализатора для получения MEK.
[00208] Катализатор может представлять собой ферментативный катализатор или неферментативный катализатор. Катализатор может включать любой катализатор, при помощи которого можно получить MEK. Например, MEK можно получить методом прямой дегидратации 2,3-BDO на различных катализаторах, таких как оксид алюминия, путем осуществления прямой реакции с серной кислотой, Cu, AlO3 и/или цеолитом (или с другими твердыми кислотными катализаторами) (см., например, Emerson, R.R., et al., «Kinetics of dehydration of aqueous 2,3-butanediol to methyl ethyl ketone,» Ind. Eng. Chem. Prod. Res. Dev., p. 473-477 (1982)). Обычные кислотные катализаторы, такие как оксиды металлов и цеолиты, в основном приводят к получению MEK, IBA, бутенов и C1-С3-газообразных соединений. Например, применение 10% H3PO4/Силикагель 60 приводит к получению 43% 1,3-бутадиена, 41% MEK и 8% IBA. Применение цеолита ZSM-5 приводит к получению более 80% MEK. В некоторых случаях ZSM-5 производит к получению более 90% MEK. В некоторых случаях катализатор и/или применяемая смесь не содержит щелочных металлов, таких как калий (K), рубидий (Rb) и/или цезий (Cs).
[00209] Катализируемая кислотой реакция дегидратация одной молекулы воды от 2,3-BDO приводит к получению промежуточного соединения карбокатиона. Пинаколиновая перегруппировка производит к получению метилэтилкетона (MEK) и изобутилальдегида (IBA).
[00210] Кроме того, 2,3-BDO, получаемый при помощи микроорганизмов и способов, описанных в настоящем документе, можно также обработать диолдегидратазой (В12). Эта ферментативная реакция приводит к получению MEK (также известный как бутан-2-он). Таким образом, раскрыт способ получения MEK, включающий: (а) приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит гетерологичный ген, кодирующий диолдегидратазу; и (b) выращивание этого микроорганизма с получением МЕК. Микроорганизм может также содержать ацетолактатсинтазу (AlsS), альфа-ацетолактатдекарбоксилазу (budA) и/или ацетоинредуктазу (butA).
[00211] В некоторых случаях МЕК также можно обработать алкогольдегидрогеназой. Эта ферментативная реакция может приводить к получению бутан-2-ола (также известного как 2-бутанол). Таким образом, раскрыт способ получения бутан-2-ола, включающий: (а) приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит гетерологичный ген, кодирующий алкогольдегидрогиназу; и (b) выращивание микроорганизма с получением бутан-2-ола. Микроорганизм может также содержать ацетолактатсинтазу (AlsS), альфа-ацетолактатдекарбоксилазу (budA), ацетоинредуктазу (butA) и/или диолдегидратазу (В12).
1,3-бутадиен (бутадиен)
[00212] В настоящем описании раскрыт способ получения бутандиена, включающий приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий: (i) ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS или (iv) любую их комбинацию. В некоторых случаях по меньшей мере один гетерологичный ген интегрирован в геном микроорганизма. В некоторых случаях по меньшей мере один гетерологичный ген экспрессируется эписомально. В некоторых случаях по меньшей мере один гетерологичный ген является как эписомально эксрессируемым, так и интегрированным в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях, когда применяемый микроорганизм содержит более одной хромосомы, термины «5'-конец» или «3'-конец» могут относиться к генам, содержащимся в одной хромосоме. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ также может включать выращивание микроорганизма с получением 2,3-BDO. Указанный способ также может включать приведение в контакт 2,3-BDO, полученного с применением катализатора с получением MEK. По меньшей мере один гетерологичный ген может находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Также микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана), а затем впоследствии лантан можно разбавить. Это может происходить до выращивания микроорганизма с получением 2,3-BDO.
[00213] Катализатор может представлять собой ферментативный катализатор или неферментативный катализатор. Катализатор может включать любой катализатор, с помощью которого можно дегидратировать 2,3-BDO. Катализатор может включать любой катализатор, при помощи которого можно осуществить гидридный сдвиг, например, оксид алюминия, или с помощью осуществления непосредственной реакции с серной кислотой (см., например, Emerson, R.R., et al., «Kinetics of dehydration of aqueous 2,3-butanediol to methyl ethyl ketone,» Ind. Eng. Chem. Prod. Res. Dev., p. 473-477 (1982). Кроме того, более чем 80% превращение 2,3-BDO в бутадиен можно осуществить с применением катализатора дигидрофосфата цезия (CsH2PO4), нанесенного на SiO2. В целом можно применить 10% катализатора CsH2PO4. В некоторых случаях превращение может составлять более 90%. Примеры катализаторов, которые можно применять, включают, но не ограничиваются ими, 10% CsH2PO4/CARiACT Q6 и/или 10% CsH2PO4/CARiACT Q10. (Кроме того, применение катализатора, в котором содержится щелочной металл, такой как K, Rb или Cs, может привести к увеличению получения бутадиена и уменьшению получения MEK. (см., например, патент США №2016/0229765). Таким образом, в способе, раскрытом в настоящем описании, можно применять дополнительные щелочные металлы.
[00214] В качестве дополнительного примера, бутадиен также можно получить методом прямой дегидратации 2,3-BDO над ториевым катализатором, хотя большинство других катализаторов для дегидратации приводят к получению метилэтилкетон в качестве основного продукта. (Winfield, М.Е., «The Catalytic Dehydration of 2,3-Бутандиол to Butadiene. II. Adsorption Equilibri,» Australian Journal of Scientific Research, Series A: Physical Sciences, vol. 3, p. 290-305 (1945)). 2-Бутен (бутен)
[00215] В настоящем описании раскрыт способ получения бутена, включающий приведение генетически модифицированного микроорганизма в контакт с углеродным субстратом, при этом указанный микроорганизм содержит по меньшей мере один гетерологичный ген, кодирующий: (i) ацетоинредуктазу; (ii) альфа-ацетолактатдекарбоксилазу (budA); (iii) AlsS или (iv) любую их комбинацию. В некоторых случаях по меньшей мере один гетерологичный ген интегрирован в геном микроорганизма. В некоторых случаях по меньшей мере один гетерологичный ген экспрессируется эписомально. В некоторых случаях по меньшей мере один гетерологичный ген является как эписомально эксрессируемым, так и интегрированным в геном микроорганизма. В некоторых случаях гетерологичный ген, кодирующий ацетолактатсинтазу, расположен ближе к 5'-концу, чем любой другой гетерологичный ген. В некоторых случаях, когда применяемый микроорганизм содержит более одной хромосомы, термины «5'-конец» или «3'-конец» могут относиться к генам, содержащимся в одной хромосоме. В некоторых случаях гетерологичный ген, кодирующий ацетоинредуктазу, расположен ближе к 3'-концу, чем любой другой гетерологичный ген. В некоторых случаях гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, располагается ни ближе к 5'-, ни ближе к 3'-концу, чем любой другой гетерологичный ген. Указанный способ может также включать выращивание микроорганизма с получением 2,3-BDO. Указанный способ может также включать приведение в контакт 2,3-BDO, полученного с применением катализатора с получением бутена. По меньшей мере один гетерологичный ген может находиться под контролем переключателя, как, например, индуцируемого или репрессируемого промотора, который чувствителен к присутствию или отсутствию компонента в среде, например, сахара, такого как арабиноза, или редкоземельного элемента, такого как лантан. Также микроорганизм можно сначала вырастить в среде, содержащей лантан (например, по меньшей мере 1 мкМ лантана), а затем впоследствии лантан можно разбавить. Это может происходить до выращивания микроорганизма с получением 2,3-BDO.
[00216] Бутен можно получить из 2,3-BDO. Например, обработка dial HBr, а затем порошком Zn может привести в результате к получению бутена. Реакции отщепления брома происходят с высокой степенью антистереоспецифичности (House, Н.О, and Ro, R,S, "The Stereochemistry of Elimination Reactions Involving Halohydrin Derivatives and Metals," J. Am. Chem. Soc. 80(1), p. 182-187(1958); Gordon, M., and Hay, J.V., "Stereochemistry of vapor phase dehalogenation of meso-and DL-2,3-dibromobutane with zinc," J. Org. Chem. 33(1), p. 427-427 (1968)), мезо-изомер приводит к получению транс-бутена, а (+)-изомер - цис-бутена.
[00217] Бутен можно превратить в бутадиен. Например, можно осуществить каталитическую реакцию дегидратации бутена до 1,3-бутадиена в перегретом паре в качестве среды для разбавления и нагревания (Voloch, М., et al., «2,3-Butanediol,» Industrial Chemicals, Biochemicals and Fuels, Chapter 45, p. 933-947 (1985).
Способы получения продуктов, подходящих для коммерческого применения
[00218] Хотя 2,3-BDO сам по себе можно применять в некоторых подходящих с коммерческой точки зрения аспектах, многие из продуктов обработки 2,3-BDO можно применять для получения дополнительных продуктов, подходящих для коммерческого применения. Например, бутадиен, бутен и MEK можно впоследствии применять в различных способах для получения продуктов, подходящих для коммерческого применения.
[00219] Например, бутен можно применять для получения бензина и бутадиена. Бутен также можно применять в качестве предшественника или компонента для получения парафинов С12, как, например, изо-парафина, применяемого в качестве авиационного топлива (см., например, патент США №7338541).
[00220] МЕК можно применять для растворения многих веществ, и его можно применять, например, в качестве растворителя в способах с участием камеди, смол, целлюлозоацетата, нитроцеллюлозных покрытий и в виниловых пленках. Таким образом, МЕК может быть подходящим для получения пластмасс, тканей, парафинового воска, а также бытовых средств, таких как лак, средство для удаления краски и олифы, клеи, и в качестве чистящих средств. МЕК также можно применять в качестве денатурирующего агента для денатурированного спирта. МЕК также можно применять в маркерах на водной основе в качестве растворителя для стирающегося красителя. Помимо этого, MEK является предшественником пероксида метилэтилкетона, катализатора, применяемого в некоторых реакциях полимеризации. Кроме того, MEK можно превратить в 2-бутанол путем приведения в контакт MEK с катализатором, таким как рутений на углероде. МЕК также можно превратить в 2-бутнаол путем приведения в контакт с алкогольдегидрогеназой.
[00221] Одним из наиболее подходящих химических веществ, которые можно получить с помощью указанных в настоящем документе способов, является бутадиен. Бутадиен можно применять для получения различных, очень предпочтительных для применения продуктов, таких как синтетические каучуки и полимерные смолы. Хотя сам по себе полибутадиен является очень мягким, почти жидким материалом, полимеры, полученные из смесей бутадиена со стиролом или акрилонитрилом, как, например, ABS, являются жесткими и эластичными. Стирен-бутадиеновый каучук представляет собой материал, наиболее часто применяемый в производстве автомобильных шин. Бутадиен также можно применять для получения нейлона через промежуточное соединение адипонитрил, других синтетических каучуковых материалов, таких как хлорпрен, и растворителя сульфолана. Помимо этого, бутадиен можно применять в промышленном производстве 4-винилциклогексена через реакцию димеризации и циклододекатриена через реакцию тримеризации. Бутадиен также является подходящим для синтеза циклоалканов и циклоалкенов, поскольку он вступает в реакцию Дильса-Альдера через двойные и тройные углерод-углеродные связи. В качестве дополнительного примера, бутадиен можно применять для получения циклоалканов, циклоалкенов, додекандиоевой кислоты (DDDA), адипонитрила, капролактама, стирола, этилиденнорборнена, лауриллактама и 1,5-циклоооктадиена (COD).
[00222] Следует понимать, что способы согласно настоящему изобретению можно интегрировать или связать с одним или более способов получения продуктов обработки бутена, бутадиена и/или МЕК. Например, способы согласно настоящему изобретению могут напрямую или косвенно способствовать вступлению бутена, бутадиена и/или МЕК в химические процессы или реакции, достаточные для превращения или получения других подходящих химических продуктов. В некоторых случаях, как это отмечалось выше, 2,3-BDO можно превратить в один или более химических продуктов непосредственно через промежуточное соединения бутен, бутадиен и/или МЕК без необходимости выделения бутена, бутадиена и/или МЕК из способа перед последующим его применением для получения одного или более химических продуктов.
[00223] В конкретных случаях метан превращают в 2,3-BDO, который затем превращают в бутен, бутадиен и/или МЕК с помощью одного или более химических способов, который, в свою очередь, превращают в один или более химических продуктов при помощи одного или более химических способов. В конкретных случаях, один или более химических продуктов получают без выделения бутана, бутадиена и/или МЕК. В другом варианте реализации 2,3-BDO превращают в один или более химических продуктов в одном химическом способе с помощью одного или более промежуточных соединений бутана, бутадиена и/или MEK
V. Ферментация
[00224] В целом микроорганизмы, раскрытые в настоящем описании, следует применять в условиях ферментации, которые являются подходящими для превращения С1-углеродного соединения (такого как метан) в 2,3-BDO (или другой желаемый продукт). Условия реакции, которые следует учитывать, включают температуру, скорость потока среды, рН, окислительно-восстановительный потенциал среды, скорость перемешивания (при применении резервуарного реактора с непрерывным перемешиванием), уровень инокулюма, максимальные концентрации субстрата и скорости введения субстрата в биореактор во избежание того, чтобы уровень субстрата становился ограничивающим, и максимальные концентрации продукта во избежание ингибирования продукта.
[00225] Оптимальные условия реакции будут частично зависеть от конкретного применяемого микроорганизма. Однако в целом ферментацию можно проводить при давлении, превышающем давление окружающей среды. Работа при повышенных давлениях может позволить значительно увеличить скорость переноса С1-углеродного соединения из газовой фазы в жидкую фазу, где его может использовать указанный микроорганизмом в качестве источника углерода для получения 2,3-BDO. Это, в свою очередь, может означать, что время удерживания (определяемое как объем жидкости в биореакторе, деленный на входную скорость газового потока) можно уменьшить в случае, когда в биореакторах поддерживается повышенное давлении, а не атмосферное давление.
[00226] Применяемые системы под давлением могут значительным образом уменьшить требуемый объем биореактора и, следовательно, капитальные затраты на оборудование для ферментации. В некоторых случаях объем реактора можно уменьшить в линейной пропорции с увеличением рабочего давления реактора, т.е. объем биореакторов, работающих при давлении 10 атмосфер, должен составлять лишь одну десятую от объема биореакторов, работающих при давлении 1 атмосфера.
[00227] Также желательно, чтобы скорость введения газообразного субстрата, содержащего С1-углеродный субстрат (такой как метан), была такой, которая бы гарантировала, что концентрация С1-углеродного субстрата (такого как метан) в жидкой фазе не станет ограничивающей. Это связано с тем, что следствием ограниченных условий в отношении С1-углеродного субстрата (например, метана) может быть то, что продукт 2,3-BDO (или другой желаемый продукт) будет потребляться культурой.
Предварительное культивирование
[00228] Предварительное культивирование различных штаммов генетически модифицированных микроорганизмов, описанных в настоящем документе, может позволить увеличить титры многоуглеродистых продуктов, описанных в настоящем документе, таких как 2,3-BDO, бутадиен и/или MEK.
[00229] Предварительное культивирование может включать добавление в среду антибиотика, такого как канамицин, источника углерода, такого как метан или другое С1-углеродное соединение, и/или веществ для активации или подавления молекулярного переключателя.
[00230] Как правило, предварительное культивирование может быть проведено в течение менее 240 часов в зависимости от вида применяемого микроорганизма. Предварительное культивирование позволяет получить достаточное количество биомассы для эффективного увеличения титров продукта. В некоторых случаях время предварительного культивирования составляет менее 240 часов. В некоторых случаях время предварительного культивирования составляет менее 220 часов. В некоторых случаях время предварительного культивирования составляет менее 200 часов. В некоторых случаях время предварительного культивирования составляет менее 180 часов. В некоторых случаях время предварительного культивирования составляет менее 160 часов. В некоторых случаях время предварительного культивирования составляет менее 140 часов. В некоторых случаях время предварительного культивирования составляет менее 120 часов. В некоторых случаях время предварительного культивирования составляет менее 100 часов. В некоторых случаях время предварительного культивирования составляет менее 96 часов. В некоторых случаях время предварительного культивирования составляет менее 90 часов. В некоторых случаях время предварительного культивирования составляет менее 84 часов. В некоторых случаях время предварительного культивирования составляет менее 78 часов. В некоторых случаях время предварительного культивирования составляет менее 72 часов. В некоторых случаях время предварительного культивирования составляет менее 68 часов. В некоторых случаях время предварительного культивирования составляет менее 62 часов. В некоторых случаях время предварительного культивирования составляет менее 56 часов. В некоторых случаях время предварительного культивирования составляет менее 50 часов. В некоторых случаях время предварительного культивирования составляет менее 48 часов. В некоторых случаях время предварительного культивирования составляет менее 46 часов. В некоторых случаях время предварительного культивирования составляет менее 44 часов. В некоторых случаях время предварительного культивирования составляет менее 42 часов. В некоторых случаях время предварительного культивирования составляет менее 40 часов. В некоторых случаях время предварительного культивирования составляет менее 38 часов. В некоторых случаях время предварительного культивирования составляет менее 36 часов. В некоторых случаях время предварительного культивирования составляет менее 34 часов. В некоторых случаях время предварительного культивирования составляет менее 32 часов. В некоторых случаях время предварительного культивирования составляет менее 30 часов. В некоторых случаях время предварительного культивирования составляет менее 28 часов. В некоторых случаях время предварительного культивирования составляет менее 26 часов. В некоторых случаях время предварительного культивирования составляет менее 24 часов. В некоторых случаях время предварительного культивирования составляет менее 22 часов. В некоторых случаях время предварительного культивирования составляет менее 20 часов. В некоторых случаях время предварительного культивирования составляет менее 18 часов. В некоторых случаях время предварительного культивирования составляет менее 16 часов. В некоторых случаях время предварительного культивирования составляет менее 14 часов. В некоторых случаях время предварительного культивирования составляет менее 12 часов. В некоторых случаях время предварительного культивирования составляет менее 10 часов. В некоторых случаях время предварительного культивирования составляет менее 8 часов. В некоторых случаях время предварительного культивирования составляет менее 6 часов. В некоторых случаях время предварительного культивирования составляет менее 4 часов. В некоторых случаях время предварительного культивирования составляет менее 2 часов. В некоторых случаях время предварительного культивирования составляет менее 1 часа. В некоторых случаях предварительное культивирование не проводят.
[00231] В некоторых случаях время предварительного культивирования составляет от 240 часов до 1 часа. В некоторых случаях время предварительного культивирования составляет до 220 часов. В некоторых случаях время предварительного культивирования составляет от 200 часов до 2 часов. В некоторых случаях время предварительного культивирования составляет от 180 часов до 4 часов. В некоторых случаях время предварительного культивирования составляет от 160 часов до 6 часов. В некоторых случаях время предварительного культивирования составляет от 140 часов до 8 часов. В некоторых случаях время предварительного культивирования составляет от 120 часов до 10 часов. В некоторых случаях время предварительного культивирования составляет от 100 часов до 12 часов. В некоторых случаях время предварительного культивирования составляет от 96 часов до 18 часов. В некоторых случаях время предварительного культивирования составляет от 90 часов до 24 часов. В некоторых случаях время предварительного культивирования составляет от 84 часов до 36 часов. В некоторых случаях время предварительного культивирования составляет от 78 часов до 40 часов. В некоторых случаях время предварительного культивирования составляет от 72 часов до 42 часов. В некоторых случаях время предварительного культивирования составляет от 56 часов до 44 часов. В некоторых случаях время предварительного культивирования составляет от 50 часов до 46 часов.
[00232] В случае, когда вещество для активации или подавления молекулярного переключения добавляют в предварительную культуру, его количество может варьироваться. Например, в некоторых случаях, если вещество для активации или подавления молекулярного переключения представляет собой редкоземельный металл (такой как лантан), для генетически модифицированного микроорганизма, который содержит только интегрированные гены, не требуется какого-либо предварительного культивирования с редкоземельным металлом (таким как лантан). Однако в некоторых случаях для этих интегрированных штаммов требуется редкоземельный металл (такой как лантан), хотя и в небольших количествах. Например, для предварительного культивирования интегрированных штаммов количество требуемого редкоземельного металла (такого как лантан) может составлять менее 25 мкМ редкоземельного металла (такого как лантан). В некоторых случаях количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 20 мкМ. Количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 15 мкМ. Количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 10 мкМ. Количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 7,5 мкМ. Количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 5 мкМ. Количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 2,5 мкМ. Количество требуемого для предварительного культивирования интегрированных штаммов редкоземельного металла (такого как лантан) может составлять менее 1 мкМ.
[00233] Однако в случае, когда микроорганизм содержит эписомально экспрессируемые гены (независимо от того, присутствует ли уже в микроорганизме дополнительный интегрированный набор генов), во время предварительного культивирования требуется большее количество редкоземельного металла (такого как лантан) для оптимизации способа получения. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 1 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 2,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 5,0 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 7,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 10 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 12,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 15 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 17,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 20 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 25 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 30 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 35 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 40 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 45 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 50 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 75 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан) для предварительного культивирования этих эписомальных штаммов должно составлять более 100 мкМ.
[00234] В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, может составлять от 0,5 мкМ до 100 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 0,5 мкМ до 50 мкМ. В других случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 1 мкМ до 20 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 2 мкМ до 15 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 3 мкМ до 12,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 4 мкМ до 12 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 5 мкМ до 11,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 6 мкМ до 11 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 7 мкМ до 10,5 мкМ. В некоторых случаях количество редкоземельного металла (такого как лантан), требуемое для предварительного культивирования для оптимизации способа получения, составляет от 8 мкМ до 10 мкМ.
[00235] В некоторых других случаях можно применять другие редкоземельные металлы, такие как церий (Се), диспрозий (Dy), эрбий (Er), европий (Ей), гадолиний (Gd), гольмий (Но), лютеций (Lu), неодим (Nd), празеодим (Pr), прометий (Pm), самарий (Sm), скандий (Sc), тербий (Tb), тулий (Tm), иттербий (Yb), иттрий (Y) или любую их комбинацию. В других случаях можно применять сахара, такие как IPTG и арабиноза.
Условия ферментации
[00236] рН можно оптимизировать в зависимости от применяемого микроорганизма. Например, рН, применяемый во время ферментации метана метанотрофом в желаемый продукт, может составлять от 4 до 10. В других случаях рН может составлять от 5 до 9; от 6 до 8; от 6,1 до 7,9; от 6.2 до 7,8; от 6,3 до 7,7; от 6,4 до 7,6 или от 6,5 до 7,5. Например, рН может составлять от 6,6 до 7,4. В некоторых случаях рН может составлять от 5 до 9. В некоторых случаях рН может составлять от 6 до 8. В некоторых случаях рН может составлять от 6,1 до 7,9. В некоторых случаях рН может составлять от 6,2 до 7,8. В некоторых случаях рН может составлять от 6,3 до 7,7. В некоторых случаях рН может составлять от 6,4 до 7,6. В некоторых случаях рН может составлять от 6,5 до 7,5. В некоторых случаях рН, применяемый при ферментации метанотрофами, может быть больше 6.
[00237] Температуру также можно регулировать в зависимости от применяемого микроорганизма. Например, температура, применяемая во время ферментации метана метанотрофом в желаемый продукт, может составлять от 30 С° до 45 С°. В других случаях температура ферментации может составлять от 30 С° до 45 С°; от 31 С° до 44 С°; от 32 С° до 43 С°; от 33 С° до 42 С°; от 34 С° до 41 С°; от 35 С° до 40 С°. Например, температура может составлять от 36 С° до 39 С° (например, 36 С°, 37 С°, 38 С° или 39 С°). В некоторых случаях температура может составлять от 30 С° до 45 С° (например, 30 С°, 31 С°, 32 С°, 33 С°, 34 С°, 35 С°, 36 С°, 37 С°, 38 С°, 39 С°, 40 С°, 41 С°, 42 С°, 43 С°, 44 С° или 45 С°). В некоторых случаях температура может составлять от 31 С° до 44 С° (например, 31 С°, 32 С°, 33 С°, 34 С°, 35 С°, 36 С°, 37 С°, 38 С°, 39 С°, 40 С°, 41 С°, 42 С°, 43 С° или 44 С°). В некоторых случаях температура может составлять от 32 С° до 43 С°. В некоторых случаях температура может составлять от 33 С° до 42 С° (например, 33 С°, 34 С°, 35 С°, 36 С°, 37 С°, 38 С°, 39 С°, 40 С°, 41 С° или 42 С°). В некоторых случаях температура может составлять от 34 С° до 41 С° (например, 34 С°, 35 С°, 36 С°, 37 С°, 38 С°, 39 С°, 40 С° или 41 С°). В некоторых случаях температура может составлять от 35 С° до 40 С° (например, 35 С°, 36 С°, 37 С°, 38 С°, 39 С° или 40 С°).
[00238] В некоторых случаях температура может находиться в пределах одной десятой градуса. Например, в некоторых случаях температура ферментации может составлять 37,0 С°, 37,1 С°, 37,2 С°, 37,3 С°, 37,4 С°, 37,5 С°, 37,6 С°, 37,7 С°, 37,8 С°, 37,9 С°, 38,0 С°, 38,1 С°, 38,2 С°, 38,3 С°, 38,4 С°, 38,5 С°, 38,6 С°, 38,7 С°, 38,8 С°, 38,9 С°, 39,0 С°, 39,1 С°, 39,2 С°, 39,3 С°, 39,4 С°, 39,5 С°, 39,6 С°, 39,7 С°, 39,8 С°, 39,9 С°, 40,0 С°, 40,1 С°, 40,2 С°, 40,3 С°, 40,4 С°, 40,5 С°, 40,6 С°, 40,7 С°, 40,8 С°, 40,9 С°, 41,0 С°, 41,1 С°, 41,2 С°, 41,3 С°, 41,4 С°, 41,5 С°, 41,6 С°, 41,7 С°, 41,8 С°, 41,9 С°, 42,0 С°, 42,1 С°, 42,2 С°, 42,3 С°, 42,4 С°, 42,5 С°, 42,6 С°, 42,7 С°, 42,8 С°, 42,9 С°, 43,0 С°, 43,1 С°, 43,2 С°, 43,3 С°, 43,4 С°, 43,5 С°, 43,6 С°, 43,7 С°, 43,8 С°, 43,9 С°,44,0 С°, 44,1 С°, 44,2 С°, 44,3 С°, 44,4 С°, 44,5 С°, 44,6 С°, 44,7 С°, 44,8 С°, 44,9 С°,45,0 С°, 45,1 С°, 45,2 С°, 45,3 С°, 45,4 С°, 45,5 С°, 45,6 С°, 45,7 С°, 45,8 С°, 45,9 С°, 46,0 С°, 46,1 С°, 46,2 С°, 46,3 С°, 46,4 С°, 46,5 С°, 46,6 С°, 46,7 С°, 46,8 С°, 46,9 С°, 47,0 С°, 47,1 С°, 47,2 С°, 47,3 С°, 47,4 С°, 47,5 С°, 47,6 С°, 47,7 С°, 47,8 С° или 47,9 С°.
[00239] В некоторых случаях температура ферментации может составлять от 37,0 С° до 47,9 С°. В некоторых случаях температура ферментации может составлять от 37,1 С° до 47,8 С°. В некоторых случаях температура ферментации может составлять от 37,2 С° до 47,7 С°. В некоторых случаях температура ферментации может составлять от 37,3 С° до 47,6 С°. В некоторых случаях температура ферментации может составлять от 37,4 С° до 47,5 С°. В некоторых случаях температура ферментации может составлять от 37,5 С° до 47,4 С°. В некоторых случаях температура ферментации может составлять от 37,6 С° до 47,3 С°. В некоторых случаях температура ферментации может составлять от 37,7 С° до 47,2 С°. В некоторых случаях температура ферментации может составлять от 37,8 С° до 47,1 С°. В некоторых случаях температура ферментации может составлять от 37,9 С° до 47,0 С°. В некоторых случаях температура ферментации может составлять от 38,0 С° до 46,9 С°. В некоторых случаях температура ферментации может составлять от 38,1 С° до 46,8 С°. В некоторых случаях температура ферментации может составлять от 38,2 С° до 46,7 С°. В некоторых случаях температура ферментации может составлять от 38,3 С° до 46,6 С°. В некоторых случаях температура ферментации может составлять от 38,4 С° до 46,5 С°. В некоторых случаях температура ферментации может составлять от 38,5 С° до 46,4 С°. В некоторых случаях температура ферментации может составлять от 38,6 С° до 46,3 С°. В некоторых случаях температура ферментации может составлять от 38,7 С° до 46,2 С°. В некоторых случаях температура ферментации может составлять от 38,8 С° до 46,1 С°. В некоторых случаях температура ферментации может составлять от 38,9 С° до 46,0 С°. В некоторых случаях температура ферментации может составлять от 39,0 С° до 45,9 С°. В некоторых случаях температура ферментации может составлять от 39,1 С° до 45,8 С°. В некоторых случаях температура ферментации может составлять от 39,2 С° до 45,7 С°. В некоторых случаях температура ферментации может составлять от 39,3 С° до 45,6 С°. В некоторых случаях температура ферментации может составлять от 39,4 С° до 45,5 С°. В некоторых случаях температура ферментации может составлять от 39,5 С° до 45,4 С°. В некоторых случаях температура ферментации может составлять от 39,6 С° до 45,3 С°. В некоторых случаях температура ферментации может составлять от 39,7 С° до 45,2 С°. В некоторых случаях температура ферментации может составлять от 39,8 С° до 45,1 С°. В некоторых случаях температура ферментации может составлять от 39,9 С° до 45,0 С°. В некоторых случаях температура ферментации может составлять от 40,0 С° до 44,9 С°. В некоторых случаях температура ферментации может составлять от 40,1 С° до 44,8 С°. В некоторых случаях температура ферментации может составлять от 40,2 С° до 44,7 С°. В некоторых случаях температура ферментации может составлять от 40,3 С° до 44,6 С°. В некоторых случаях температура ферментации может составлять от 40,4 С° до 44,5 С°. В некоторых случаях температура ферментации может составлять от 40,5 С° до 44,4 С°. В некоторых случаях температура ферментации может составлять от 40,6 С° до 44,3 С°. В некоторых случаях температура ферментации может составлять от 40,7 С° до 44,2 С°. В некоторых случаях температура ферментации может составлять от 40,8 С° до 44,1 С°. В некоторых случаях температура ферментации может составлять от 40,9 С° до 44,0 С°. В некоторых случаях температура ферментации может составлять от 41,0 С° до 43,9 С°. В некоторых случаях температура ферментации может составлять от 41,1 С° до 43,8 С°. В некоторых случаях температура ферментации может составлять от 41,2 С° до 43,7 С°. В некоторых случаях температура ферментации может составлять от 41,3 С° до 43,6 С°. В некоторых случаях температура ферментации может составлять от 41,4 С° до 43,5 С°. В некоторых случаях температура ферментации может составлять от 41,5 С° до 43,4 С°. В некоторых случаях температура ферментации может составлять от 41,6 С° до 43,3 С°. В некоторых случаях температура ферментации может составлять от 41,7 С° до 43,2 С°. В некоторых случаях температура ферментации может составлять от 41,8 С° до 43,1 С°. В некоторых случаях температура ферментации может составлять от 41,9 С° до 43,0 С°. В некоторых случаях температура ферментации может составлять от 42,0 С° до 42,9 С°. В некоторых случаях температура ферментации может составлять от 42,1 С° до 42,8 С°. В некоторых случаях температура ферментации может составлять от 42,2 С° до 42,7 С°. В некоторых случаях температура ферментации может составлять от 42,3 С° до 42,6 С°. В некоторых случаях температура ферментации может составлять от 42,4 С° до 42,5 С°.
[00240] Доступность кислорода и других газов, таких как газообразные С1-углеродные субстраты (как, например, метан), может влиять на выход и скорость ферментации. Например, когда речь идет о доступности кислорода, процент растворенного кислорода (DO) в среде для ферментации может составлять от 1% до 40%. В некоторых случаях концентрация DO может составлять от 1,5% до 35%; от 2% до 30%; от 2,5% до 25%; от 3% до 20%; от 4% до 19%; от 5% до 18%; от 6% до 17%; от 7% до 16%; от 8% до 15%; от 9% до 14%; от 10% до 13% или от 11% до 12%. Например, в некоторых случаях концентрация DO может составлять от 2% до 30%. В других случаях DO может составлять от 3% до 20%. В некоторых случаях DO может составлять от 4% до 10%. В некоторых случаях DO может быть от 1,5% до 35%. В некоторых случаях DO может составлять от 2,5% до 25%. В некоторых случаях DO может составлять от 4% до 19%. В некоторых случаях DO может составлять от 5% до 18%. В некоторых случаях DO может составлять от 6% до 17%. В некоторых случаях DO может составлять от 7% до 16%. В некоторых случаях DO может составлять от 8% до 15%. В некоторых случаях DO может составлять от 9% до 14%. В некоторых случаях DO может быть от 10% до 13%. В некоторых случаях DO может составлять от 11% до 12%.
[00241] При применении метанотрофов тип метансодержащих соединений может влиять на выход и скорость ферментации. Например, можно применять природный газ, содержание метана в котором обычно составляет более 85% (например, более 90%). Другие компоненты природного газа могут включать, но не ограничиваться ими, этан, пропан, изо-бутан, нормальный бутан, изо-пентан, нормальный пентан, гексаны и другие углеводороды с числом атома больше 6, азот, диоксид углерода, кислород, водород и сероводород.
[00242] Также можно применять «чистый» метан. В этих случаях метан обычно поступает из резервуара. Содержание метана в этих резервуарах может варьироваться от 90% или более, а остальное количество газа составляют другие газы (такие как диоксид углерода). Например, в процессе ферментации можно применять газ с содержанием метана более 90%. В некоторых случаях концентрация метана может превышать 90%; 91%; 92%; 93%; 94%; 95%; 96%; 97%; 98%; 99% или 99,9%. В некоторых случаях концентрация метана может составлять 90%, а 10% составляют другие газы (такие как диоксид углерода). В других случаях концентрация метана может составлять 91%, а 9% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метана может составлять 92%, а 8% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метана может составлять 93%, а 7% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метана может составлять 94%, а 6% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метана может составлять 95%, а 5% составляют другие газы (такие как диоксид углерода). В других случаях концентрация метана может составлять 96%, а 4% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метана может составлять 97%, а 3% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метана может составлять 98%, а 2% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метанола может составлять 99%, а 1% составляют другие газы (такие как диоксид углерода). В некоторых случаях концентрация метанола может составлять 99,9%, а 0,1% составляют другие газы (такие как диоксид углерода).
[00243] В случаях, когда применяют переключатель, среда может содержать молекулу, которая индуцирует или подавляет указанный переключатель. Например, в случае, когда чувствительный к лантану переключатель применяют для подавления экспрессии одного или более генов, описанных в настоящем документе, среда может содержать лантан, который будет подавлять экспрессию одного или более генов под контролем указанного переключателя. В случае лантана любая из следующих концентраций может эффективно подавлять экспрессию одного или более генов: 0,1 мкМ; 0,5 мкМ; 1 мкМ; 2 мкМ; 3 мкМ; 4 мкМ; 5 мкМ; 6 мкМ; 7 мкМ; 8 мкМ; 9 мкМ; 10 мкМ; 12,5 мкМ; 15 мкМ; 17,5 мкМ; 20 мкМ; 25 мкМ; 50 мкМ; 100 мкМ или более. В одном случае можно применять 0,1 мкМ лантана для подавления экспрессии одного или более генов под контролем чувствительного к лантану переключателя. В других случаях можно применять по меньшей мере 0,5 мкМ лантана. В других случаях можно применять по меньшей мере 1 мкМ лантана. В других случаях можно применять по меньшей мере 2 мкМ лантана. В других случаях можно применять по меньшей мере 3 мкМ лантана. В других случаях можно применять по меньшей мере 4 мкМ лантана. В других случаях можно применять по меньшей мере 5 мкМ лантана. В других случаях можно применять по меньшей мере 6 мкМ лантана. В других случаях можно применять по меньшей мере 7 мкМ лантана. В других случаях можно применять по меньшей мере 8 мкМ лантана. В других случаях можно применять по меньшей мере 9 мкМ лантана. В других случаях можно применять по меньшей мере 10 мкМ лантана могут быть применены. В других случаях по меньшей мере 12,5 мкМ лантана. В других случаях можно применять по меньшей мере 15 мкМ лантана. В других случаях можно применять по меньшей мере 17,5 мкМ лантана. В других случаях можно применять по меньшей мере 20 мкМ лантана. В других случаях можно применять по меньшей мере 25 мкМ лантана. В других случаях можно применять по меньшей мере 50 мкМ лантана. В других случаях можно применять по меньшей мере 100 мкМ лантана. В некоторых случаях концентрация лантана от 0,5 мкМ до 100 мкМ будет эффективно подавлять экспрессию генов. В некоторых случаях концентрация лантана от 0,5 мкМ до 50 мкМ будет подавлять экспрессию генов. В других случаях концентрация лантана от 1 мкМ до 20 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 2 мкМ до 15 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 3 мкМ до 12,5 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 4 мкМ до 12 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 5 мкМ до 11,5 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 6 мкМ до 11 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 7 мкМ до 10,5 мкМ будет подавлять экспрессию генов. В некоторых случаях концентрация лантана от 8 мкМ до 10 мкМ будет подавлять экспрессию генов.
[00244] В некоторых случаях лантан в среде можно разбавить для «включения» экспрессии одного или более подавляемых лантаном генов. Например, в некоторых случаях разбавление среды, содержащей лантан, может составлять 1:1(1 часть среды, содержащей лантан, на 1 часть среды, не содержащей лантан). В некоторых случаях разбавление по меньшей мере может составлять 1:2; 1:3; 1:4; 1:5; 1:7,5; 1:10; 1:15; 1:20; 1:25; 1:30; 1:35; 1:40; 1:45; 1:50; 1:75; 1:100; 1: 200; 1:300; 1:400; 1:500; 1:1000 или 1:10000. Например, в некоторых случаях можно применять разбавление 1:2. В некоторых случаях можно применять разбавление по меньшей мере 1:3. В некоторых случаях можно применять разбавление по меньшей мере 1:4. В некоторых случаях можно применять разбавление по меньшей мере 1:5. В некоторых случаях можно применять разбавление по меньшей мере 1:7,5. В некоторых случаях можно применять разбавление по меньшей мере 1:10. В некоторых случаях можно применять разбавление по меньшей мере 1:15. В некоторых случаях можно применять разбавление по меньшей мере 1:20. В некоторых случаях можно применять разбавление по меньшей мере 1:25. В некоторых случаях можно применять разбавление по меньшей мере 1:30. В некоторых случаях можно применять разбавление по меньшей мере 1:35. В некоторых случаях можно применять разбавление по меньшей мере 1:40. В некоторых случаях можно применять разбавление по меньшей мере 1:45. В некоторых случаях можно применять разбавление по меньшей мере 1:50. В некоторых случаях можно применять разбавление по меньшей мере 1:75. В некоторых случаях можно применять разбавление по меньшей мере 1:100. В некоторых случаях можно применять разбавление по меньшей мере 1:200. В некоторых случаях можно применять разбавление по меньшей мере 1:300. В некоторых случаях можно применять разбавление по меньшей мере 1:400. В некоторых случаях можно применять разбавление по меньшей мере 1:500. В некоторых случаях можно применять разбавление по меньшей мере 1:1000. В некоторых случаях можно применять разбавление по меньшей мере 1:10000.
[00245] В некоторых случаях микроорганизм можно вырастить в среде, содержащей лантан. Указанную среду можно затем разбавить для эффективного «включения» экспрессии подавляемых лантаном генов. Микроорганизм затем можно вырастить в условиях, способствующих получению желаемых продуктов, таких как 2,3-BDO и ацетоин (или других продуктов, раскрытых в настоящем документе).
[00246] В некоторых случаях можно применять другие редкоземельные металлы. Например, для подавления или активации молекулярного переключателя можно применять другие редкоземельные металлы, такие как церий (Се), диспрозий (Dy), эрбий (Er), европий (Eu), гадолиний (Gd), гольмий (Но), лютеций (Lu), неодим (Nd), празеодим (Pr), прометий (Pm), самарий (Sm), скандий (Sc), тербий (Tb), тулий (Tm), иттербий (Yb), иттрий (Y) или любую их комбинацию.
Биореактор
[00247] Реакции ферментации можно осуществить в любом подходящем биореакторе. В некоторых случаях согласно настоящему изобретению биореактор может содержать первый реактор для роста, в котором культивируются микроорганизмы, и второй реактор для ферментации, в который подается бульон из указанного реактора для роста и в котором образуется большая часть продукта ферментации (например, 2,3-BDO).
Выделение продукта
[00248] При ферментации микроорганизмов, раскрытых в настоящем описании, может образовываться ферментативный бульон, содержащий желаемый продукт (например, 2,3-BDO, МЕК и/или бутадиен) и/или один или более побочных продуктов, а также микроорганизмы (например, генетически модифицированный метанотроф) в питательной среде.
[00249] При помощи описанных в настоящем документе микроорганизмов и способов можно получить 2,3-BDO с удивительно высокой эффективностью, которая выше, чем в других известных способах ферментации 2,3-BDO. Например, при помощи микроорганизмов и способов, раскрытых в настоящем документе, можно преобразовать С1-углеродный субстрат (такой как метан) со скоростью, превышающей 50%. Это означает, что по меньшей мере 50% С1-углеродных соединений в системах превращается в продукт, такой как 2,3-BDO. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 60%, 70%, 80%, 81%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 60%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 70%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 80%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 81%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 82%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 83%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 84%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 85%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 86%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 87%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 88%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 89%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 90%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 91%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 92%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 93%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 94%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 95%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 96%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 97%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 98%. В некоторых случаях превращение С1-углеродного субстрата в 2,3-BDO может происходить по меньшей мере на 99%.
[00250] В некоторых способах при получении 2,3-BDO концентрация 2,3-BDO в ферментативном бульоне может составлять по меньшей мере 1 г/л. Например, полученная концентрация 2,3-BDO в ферментативном бульоне, может составлять от 1 г/л до 5 г/л, от 2 г/л до 6 г/л, от 3 г/л до 7 г/л, от 4 г/л до 8 г/л, от 5 г/л до 9 г/л или от 6 г/л до 10 г/л. В некоторых случаях концентрация 2,3-BDO может составлять по меньшей мере 9 г/л. В некоторых случаях концентрация 2,3-BDO может составлять от 1 г/л до 5 г/л. В некоторых случаях концентрация 2,3-BDO может составлять от 2 г/л до 6 г/л. В некоторых случаях концентрация 2,3-BDO может составлять от 3 г/л до 7 г/л. В некоторых случаях концентрация 2,3-BDO может составлять от 4 г/л до 8 г/л. В некоторых случаях концентрация 2,3-BDO может составлять от 5 г/л до 9 г/л. В некоторых случаях концентрация 2,3-BDO может составлять от 6 г/л до 10 г/л.
[00251] В других случаях, когда применяют микроорганизмы, которые обычно продуцируют по меньшей мере 2,3-BDO, после генетической модификации и ферментации генетически модифицированный микроорганизм может продуцировать 2,3-BDO в концентрациях, которые по меньшей мере в 1,1 раза (1ДХ) превышают обычно получаемое количество. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 2Х, 3Х, 4Х, 5Х, 6Х, 7Х, 8Х, 9Х, 10Х, 15Х, 20Х, 25Х, 30Х, 35Х, 40Х, 45Х, 50Х, 60Х, 70Х, 80Х, 90Х или 100Х больше обычно получаемого количества (например, получаемого при помощи микроорганизма, который не является модифицированным, и представляет собой тот же вид, что и генетически модифицированный микроорганизм). В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 2Х, 3Х, 4Х, 5Х, 10Х, 25Х, 50Х и/или 100Х больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 2 раза больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 3 раза больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 4 раза больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 5 раз больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 10 раз больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 25 раз больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 50 раз больше обычно получаемого количества. В некоторых случаях генетически модифицированный микроорганизм может продуцировать по меньшей мере в 100 раз больше обычно получаемого количества.
[00252] Как это обсуждалось выше, в некоторых случаях 2,3-BDO, образующийся в реакции ферментации, превращается в МЕК, бутен и/или бутадиен (или другие продукты) непосредственно в ферментативном бульоне. В других случаях 2,3-BDO сначала выделяют из ферментативного бульона до превращения в МЕК, бутен и/или бутадиен.
[00253] В некоторых случаях 2,3-BDO можно постоянно удалять из части бульона и выделять его как очищенный 2,3-BDO. В определенных случаях, выделение 2,3-BDO включает пропускание удаленной части бульона, содержащего 2,3-BDO, через разделительную установку для отделения микроорганизмов (например, генетически модифицированных метанотрофов) от бульона с получением не содержащего клетки, но содержащего 2,3-BDO пермеата, и возврат микроорганизмов в биореактор. Не содержащий клетки, но содержащий 2,3-BDO можно затем хранить или применять для последующего превращения в бутен, МЕК и/или бутадиен (или другой желаемый продукт).
[00254] Выделение 2,3-BDO и/или одного или более других продуктов или побочных продуктов, образующихся в реакции ферментации, может включать постоянное удаление части бульона и отдельное выделение 2,3-BDO и одного или более других продуктов из удаленного часть бульона. В некоторых случаях выделение 2,3-BDO и/или одного или более других продуктов включает пропускание удаленной части бульона, содержащей 2,3-BDO и/или один или более других продуктов, через разделительную установку для отделения микроорганизмов от 2,3-BDO и/или один или более других продуктов с получением не содержащего клетки, но содержащего 2,3-BDO и один или более других продуктов пермеата, и возврат микроорганизмов в биореактор.
[00255] В указанных выше случаях выделение 2,3-BDO и одного или более других продуктов может включать сначала удаление 2,3-BDO из несодержащего клетки пермеата, а затем удаление одного или более других продуктов из несодержащего клетки пермеата. Несодержащий клетки пермеат также можно возвратить в биореактор.
[00256] 2,3-BDO или поток смешанного продукта, содержащий 2,3-BDO, можно выделить из ферментативного бульона. Например, способы могут включать, но не ограничиваться ими, фракционную дистилляцию или испарение, первапорацию и экстрактивную ферментацию. Дополнительные примеры включают: выделение с применением пара из цельного ферментативного бульона; обратный осмос в сочетании с дистилляцией; методы жидкостно-жидкостной экстракции, включающие экстракцию растворителем 2,3-BDO; водную двухфазную экстракцию 2,3-BDO в системе ПЭГ/декстран; экстракцию растворителем с применением спиртов или сложных эфиров, например, этилацетата, трибутилфосфата, диэтилового простого эфира, н-бутанола, додеканола, олеилового спирта и системы вида этанол/фосфат; двухфазных водных систем, состоящих из гидрофильных растворителей и неорганических солей. См. в целом Voloch, М., et al., (1985) и патент США №2012/0045807.
[00257] В некоторых случаях перед воздействием растворителя ферментативный бульон удаляют путем выпаривания или при помощи как метода микрофильтрации, так и обратного осмоса вследствие низкого коэффициента распределения и низкой селективности 2,3-BDO. Методы Repulsive экстракции или высаливания с применением хлорида калия (KCl) или дегидратированного K2CO3 также был исследованы на выделение 2,3-BDO (Syu 2001) по аналогии с высаливающим эффектом K2CO3 при выделении бутанола при ферментации ацетон-О-бутанол-этанола. Также исследовали удаление воды из ферментативного бульона перед высаливанием, поскольку концентрация 2,3-BDO в бульоне была слишком низкой для того, чтобы его высаливать даже из насыщенного раствора KCl или K2CO3. См. в целом патент США №2012/0045807.
[00258] Еще одним дополнительным примером способа выделения 2,3-BDO является вступление его в реакцию с формальдегидом с образованием формальдегида 2,3-BDO при катализе кислотой. Формальдегид 2,3-BDO собирается в верхней масляной фазе и позволяет метанолу вступить в реакцию с кислотой с образованием 2,3-BDO и метилаля. Метилаль можно гидролизовать до метанола и формальдегида. См. в целом патент США №2012/0045807.
[00259] Дополнительным примером может быть применение ионных жидкостей для экстракции этанола/2,3-BDO из осветленного бульона. Ионные жидкости можно подобрать различными способами для изменения физических свойств. Преимущество этого подхода заключается в том, что ионные жидкости не являются летучими. Некоторые реагируют на воду, а другие нет.
[00260] Методы первапорации или мембранной дистилляции под вакуумом, применяемые ранее при ферментации этанола и бутанола, можно применять для концентрации 2,3-BDO в воде в качестве экстракта, полученного из ферментативного бульона. В интегрированном способе применяют микропористую политетрафторэтиленовую (PTFE) мембрану, в то время как силиконовую мембрану обычно применяют для первапорации при ферментации этанола или бутанола. См. в целом патент США №2012/0045807.
[00261] Побочные продукты, такие как кислоты, включая ацетат и бутират, также можно выделить при ферментации, применяя способы, известные в данной области техники. Например, можно применить адсорбционную систему, включая фильтр с активированным углем, или электродиализ.
[00262] В некоторых случаях согласно настоящему изобретению 2,3-BDO и побочные продукты выделяют из ферментативного бульона путем постоянного удаления части бульона из биореактора, отделения микробных клеток от бульона (удобным методом, например, является фильтрация) и выделения 2,3-BDO и необязательно других спиртов и кислот из бульона. Спирт можно удобным образом выделить, например, методом дистилляции, а кислоты можно выделить, например, методом адсорбции на активированном угле. Отделенные микробные клетки можно возвратить в биореактор для ферментации. Несодержащий клетки пермеат, оставшийся после выделения спирта(-ов) и кислоты(-т), также можно возвратить в биореактор для ферментации. Дополнительные питательные вещества можно добавить к несодержащему клетки пермеату для пополнения питательной среды перед его возвратом в биореактор.
[00263] Кроме того, в случае, если рН бульона был отрегулирован во время выделения 2,3-BDO и/или побочных продуктов, то прежде, чем возвратить его в биореактор, рН следует повторно отрегулировать до значения, равного рН бульона в биореакторе для ферментации.
[00264] В некоторых случаях 2,3-BDO постоянно выделяют из ферментативного бульона или биореактора и подают непосредственно для его химического превращения в одно или более из бутена, бутадиена и метилэтилкетона. Например, 2,3-BDO можно подавать непосредственно через трубопровод в один или несколько сосудов, подходящих для химического синтеза одного или более из бутена, бутадиена и метилэтилкетона или других химических продуктов обработки.
[00265] Хотя в настоящем документе были показаны и описаны некоторые случаи, такие случаи представлены только в качестве примера. Специалист в данной области техники без отхода от настоящего изобретения может осуществить многочисленные вариации, изменения и замены. Следует понимать, что при практическом осуществлении настоящего изобретения будут использоваться различные альтернативы случаям, описанным в настоящем изобретении.
ПРИМЕРЫ
Пример 1: Генетическое конструирование метанотрофов
[00266] Для конструирования метанотрофа для получения 2,3-BDO авторы настоящего изобретения начали с М. capsulatus и исследовали несколько генов биосинтеза 2,3-BDO из Bacillus subtilis, Clostridium autoethanogenum, Klebsiella pneumoniae и Paenibacillus polymyxa. В приведенной ниже таблице 1 показаны исследованные гены и происхождение генов.
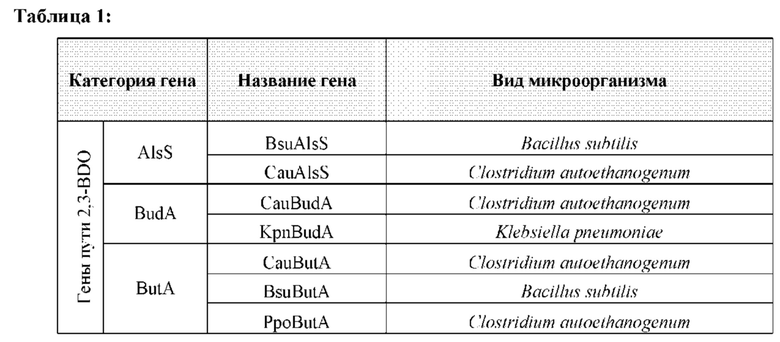
[00267] Для первого гена пути, AlsS, который кодирует фермент, превращающий две молекулы пирувата в 2-ацетолактат, авторы настоящего изобретения исследовали гомологи AlsS, полученные из Bacillus subtilis и Clostridium autoethanogenum. Для второго гена пути, BudA, который кодирует фермент, превращающий 2-ацетолактат в ацетоин, авторы настоящего изобретения исследовали гомологи BudA, полученные из Clostridium autoethanogenum и Klebsiella pneumonia. Для третьего гена пути, ButA, который кодирует фермент, превращающий ацетоин в 2,3-BDO с применением НАД(Ф)Н в качестве восстановленного кофактора, авторы настоящего изобретения исследовали гомологи ВшА, полученные из Clostridium autoethanogenum (НАДФН-зависимая), Bacillus subtilis (НАДН-зависимый) и Paenibacillus polymyxa (НАДН-зависимая).
[00268] Авторы настоящего изобретения создали 48 различных плазмид-хозяев, содержащих вариации в 3 генах пути 2,3-BDO, как это показано в таблице 2 (ниже в примере 2). В таблице 2 показаны генотипы полученных сконструированных штаммов.
Пример 2: производительность 2,3-BDO
[00269] Авторы настоящего изобретения трансформировали вышеупомянутые плазмиды (в примере 1) в отвечающий требованиям трансформации штамм метанотрофа, RL83A, и оценили 74 полученных штамма (включая биологически повторяющиеся штаммы) для получения 2,3-BDO в небольшом по размеру микротитровальном планшете для ферментации с применением метана в качестве источника углерода.
[00270] Как показано в таблице 2, конструкции, которые конститутивно экспрессируют гены AlsS, приводят либо к отсутствию трансформантов, либо к отсутствию образования 2,3-BDO, что свидетельствует о наличии негативного воздействия в случае, когда гены AlsS были сильно и конститутивно экспрессированы. Результаты показали, что штаммы, которые продуцировали наибольшее количество 2,3-BDO, содержали комбинации генов BsuAlsA-KpnBudA-CauButA или BsuAlsA-KpnBudA-BsuButA (таблица 2).
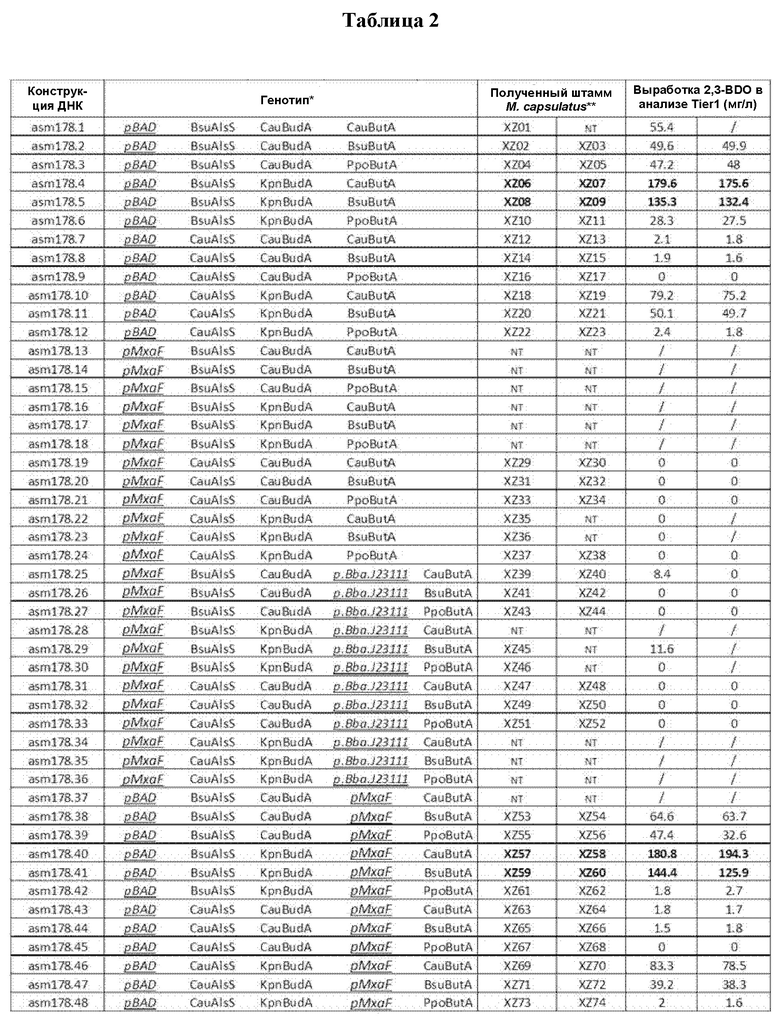
[00271] Для подтверждения результатов первичного микропланшетного анализа, авторы настоящего изобретения оценили четыре самых эффективных штамма, XZ58 и XZ06 (BsuAlsA-KpnBudA-CauButA), XZ59 и XZ08 (BsuAlsA-KpnBudA-BsuButA), в эксперименте по ферментации с высокой плотностью клеток в биореакторе на 1 л. Как это показано на ФИГ. 2, штаммы XZ58 и XZ06 продуцировали 9,3 г/л и 8,5 г/л 2,3-BDO соответственно без образования второстепенного продукта ацетоина (ФИГ. 2, левая панель). Данные показывают, что штаммы, экспрессирующие НАДФН-зависимую ацетоинредуктазу (CauButA), продуцируют только 2,3-BDO без сопутствующего продукта ацетоина. Напротив, штаммы, экспрессирующие НАДН-зависимую ацетоинредуктазу, продуцировали значительное количество ацетоина, предшественника 2,3-BDO (ФИГ. 2, правая панель). Неэффективное превращение ацетоина в 2,3-BDO в НАДН-зависимых путях может быть связано либо с недостаточной экспрессией фермента, либо с низкими пулами предшественников НАДН в М. capsulatus.
Пример 3: генетические переключатели, применяемые для получения 2,3-BDO
[00272] Для того, что эффективно контролировать получение 2,3-BDO в определенное время, была внедрена система генетического переключения. Метанотрофов трансформировали при помощи генов, находящихся под контролем чувствительного к лантану «переключателя». Чувствительные к лантану переключатели подавляют экспрессию генов в присутствии металла лантана. При удалении или разбавлении лантана в среде происходит «включение» подавленных генов.
[00273] Различные штаммы метанотрофов (как это показано в таблице 3 ниже, представлены штаммы и генотип штаммов) предварительно культивировали в присутствии 10 мкМ лантана.


[00274] После выращивания предварительных культур до значения плотности клеток ~ 3 при OD600, среду, содержащую лантан, разбавляли в соотношении 1:10 (среда, содержащая лантан: среда, не содержащая лантан) или 1:50. После 96 или 120 часов указанные культуры оценивали на образование 2,3-BDO и ацетоина (ФИГ. 7А, 96 часов) или (ФИГ. 7В, 120 часов). Ниже в таблице 4 показаны штаммы, уровни разбавления, полученные титры ацетоина после 96 часов, полученные титры 2,3-BDO через 96 часов, полученные титры ацетоина через 120 часов и полученные титры 2,3-BDO после 120 часов.
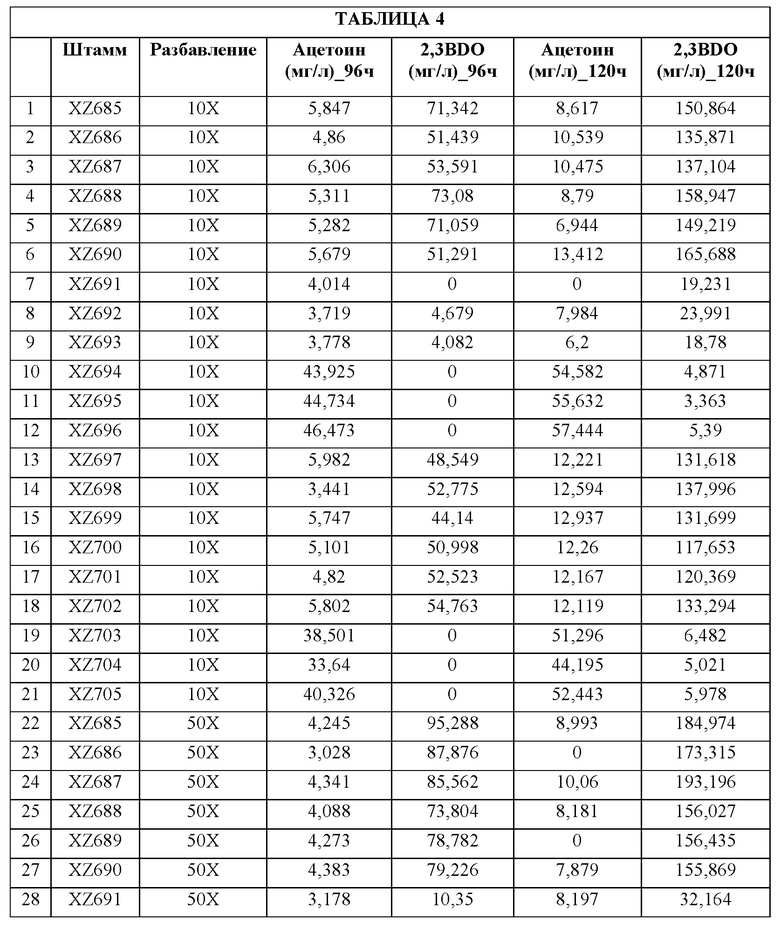
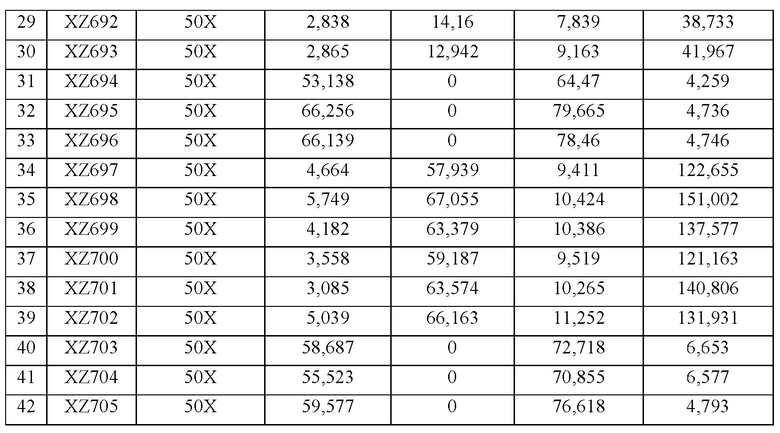
[00275] Как показано на ФИГ. 7А, 7В и в таблице 4, штаммы XZ685, XZ686, XZ687, XZ688, XZ689 и XZ690 (относящиеся к 22-27 соответственно на ФИГ. 7А и 7В) продуцировали самые высокие титры 2,3-BDO как при 96, так и при 120 часах при разбавлении содержащей лантан среды в соотношении 1:50 (50Х). Применение протокола разбавления 1:10 также приводило к значительному образованию 2,3-BDO, однако титры были ниже, чем титры с применением протокола разбавления 1:50 при 96 и 120 часах. Штаммы XZ697, XZ698, XZ699, XZ700, XZ701 и XZ702 (относящиеся к 34-39 соответственно на ФИГ. 7А и 7В) продуцировали более низкие титры при разбавлении 1:10 и 1:50 при 96 и 120 часах.
Пример 4: ацетолактатсинтаза
[00276] Плазмиды, описанные в таблице 5 (ниже), трансформировали в соответствующий требованиям трансформации штамм метанотрофов. Полученные штаммы исследовали на полученные титры 2,3-BDO в небольших по размеру микротитровальных планшетах с применением метана в качестве источника углерода.

[00277] Как показано в таблице 5 и на ФИГ. 8, штаммы, которые экспрессировали ген AlsS Bacillus licheniformis, показали более лучшие титры образования 2,3-BDO, чем штаммы, которые экспрессировали AlsS Bacillus subtilis (например, штаммы, описанные в примере 2). В одном примере штамм, который содержит замену только гена AlsS (например, штаммы XZ557), показал увеличение титра образования 2,3-BDO вплоть до 16,1% по сравнению со штаммом XZ58. В другом образце штамм XZ546, штамм, содержащий замену только сайта связывания с рибосомой на ген Kpn.BudA, практически не показал увеличения титров 2,3-BDO по сравнению со штаммом XZ58. Однако, что примечательно, штамм, который содержал rbs.Mca.MxaF для гена Kpn.BudA вместо rbs.GTW0001 и экспрессировал ген AlsS Bacillus licheniformis (например, штамм XZ562), показал значительное увеличение титров 2,3-BDO вплоть до 44,6% по сравнению со штаммом XZ58.
Пример 5: температура во время ферментации
[00278] Для предотвращения нестабильности, иногда присутствующей в экспрессирующих плазмиду штаммах, авторы настоящего изобретения создали штаммы с генами пути 2,3-BDO, интегрированными в хромосому в локусе Glgc. Всего был создан 21 штамм.
[00279] Семь штаммов были исследованы во встряхиваемых флаконах на их способность продуцировать 2,3-BDO при 37°С и при 42°С.Ферментацию проводили в течение 96 часов. Генотипы штаммов представлены ниже в ТАБЛИЦЕ 6.
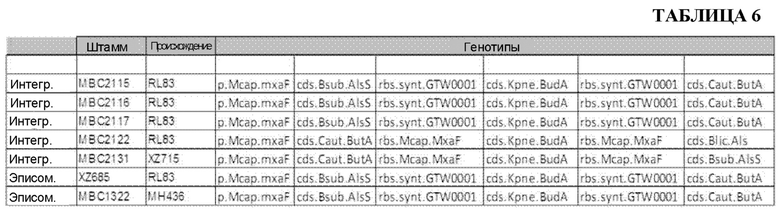
[00280] Как видно на ФИГ. 9, все исследованные штаммы продуцировали значимо большие количества 2,3-BDO в случае, когда ферментация происходила при 42°С. Некоторые штаммы, такие как МВС2122, продуцировали примерно в два раза большее количество 2,3-BDO при 42°С, а не при 37°С.
[00281] Три штамма дополнительно исследовали при их ферментации при 45°С и без селективного давления антибиотиков. По сравнению с тем же штаммом, инкубированным при 37°С, штаммы, которые ферментировали при 45°С, продуцировали значительно меньшее количество 2,3-BDO. См. ФИГ. 10. Эписомальный штамм МВС1322 показал лучшие результаты при селективном давлении антибиотиков как при 37°С, так и при 45°С, что свидетельствует о проблеме со стабильностью. Напротив, интегрированные штаммы обладали способностью сохранять свою продуктивность без селективного давления.
[00282] Дополнительные исследования проводили при 37°С, 41°С, 43°С, 45°С и 47°С.Были подсчитаны титры 2,3-BDO и соотношения OD. Как видно на ФИГ. 11 наилучшую производительность наблюдали при температуре 41°С.
Пример 6: дополнительные копии генов
[00283] Штамм метанотрофа, который содержал дополнительные копии ферментов пути 2,3-BDO (Штаммы В и С) в эписомальном векторе, продуцировал повышенное количество 2,3-BDO по сравнению с экспериментами с одиночно интегрированным штаммом (Штамм А) в эксперименте со встряхиваемым флаконом. Штамм В и штамм С предварительно культивировали в среде с 10 мкМ лантана в присутствии канамицина при 37°С в течение 48 часов. Штамм А предварительно культивировали в тех же условиях за исключением того, что не добавляли канамицин. Через 48 часов лантан разбавляли в 50 раз (50Х), и титр 2,3-BDO измеряли через 96 часов. Как видно на ФИГ. 12, штаммы, которые содержали дополнительные копии ферментов пути 2,3-BDO (Штаммы В и С) в эписомальном векторе, продуцировали в 3 раза большее количество 2,3-BDO по сравнению с одиночно интегрированным штаммом (Штамм А).
Пример 7: предварительное культивирование
[00284] Действие предварительного культивирования исследовали как в эписомальных, так и на интегрированных штаммах, а также на штаммах, содержащих как эписомально экспрессируемые гены пути 2,3-BDO, так и интегрированные копии генов.
[00285] На ФИГ. 13 показано действие предварительного культивирования штамма метанотрофа (во встряхиваемом флаконе), в котором гены, кодирующие ферменты пути 2,3-BDO, экспрессируются эписомально (Штамм D). Указанный штамм метанотрофа предварительно культивировали в среде с концентрацией лантана 10 мкМ, 3 мкМ и 1 мкМ во время предварительного культивирования. Среду для получения впоследствии разбавляли в 50 раз. Более высокие концентрации лантана приводили к более высоким титрам 2,3-BDO для эписомально экспрессируемых штаммов.
[00286] На ФИГ. 14 показано действие предварительного культивирования штамма метанотрофа (во встряхиваемом флаконе), в котором гены, кодирующие ферменты пути 2,3-BDO, были интегрированы в геном микроорганизма (Штамм А). Указанный штамм метанотрофа (Штамм А) предварительно культивировали в среде с концентрацией лантана 35 мкМ, 10 мкМ, 3 мкМ, 1 мкМ и 0 мкМ. Среду для получения впоследствии разбавляли в 50 раз (50Х). Уровни образования увеличивались в случае, когда указанный штамм предварительно культивировали в среде с более низкими концентрациями лантана.
[00287] На ФИГ. 15 показан эффект предварительного культивирования при различных концентрациях лантана (10 мкМ, 3 мкМ и 1 мкМ) двух штаммов с ФИГ. 12 (Штаммы В и С) в эксперименте со встряхиваемым флаконом. Оба штамма содержат одну интегрированную копию и эписомально экспрессированные копии ферментов пути 2,3-BDO. В целом штаммы, содержащие эписомально экспрессируемые ферменты пути 2,3-BDO, позволяют получить более высокие титры 2,3-BDO при предварительном культивировании в среде с концентрацией лантана 10 мкМ.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Intrexon Corporation
<120> СПОСОБЫ И МИКРООРГАНИЗМЫ ДЛЯ ПОЛУЧЕНИЯ 2,3-БУТАНДИОЛА И
ЕГО ПРОИЗВОДНЫХ ИЗ C1-УГЛЕРОДНЫХ СОЕДИНЕНИЙ
<130> INX00382US
<160> 20
<170> PatentIn, версия 3.5
<210> 1
<211> 571
<212> Белок
<213> Bacillus subtilis
<400> 1
Met Leu Thr Lys Ala Thr Lys Glu Gln Lys Ser Leu Val Lys Asn Arg
1 5 10 15
Gly Ala Glu Leu Val Val Asp Cys Leu Val Glu Gln Gly Val Thr His
20 25 30
Val Phe Gly Ile Pro Gly Ala Lys Ile Asp Ala Val Phe Asp Ala Leu
35 40 45
Gln Asp Lys Gly Pro Glu Ile Ile Val Ala Arg His Glu Gln Asn Ala
50 55 60
Ala Phe Met Ala Gln Ala Val Gly Arg Leu Thr Gly Lys Pro Gly Val
65 70 75 80
Val Leu Val Thr Ser Gly Pro Gly Ala Ser Asn Leu Ala Thr Gly Leu
85 90 95
Leu Thr Ala Asn Thr Glu Gly Asp Pro Val Val Ala Leu Ala Gly Asn
100 105 110
Val Ile Arg Ala Asp Arg Leu Lys Arg Thr His Gln Ser Leu Asp Asn
115 120 125
Ala Ala Leu Phe Gln Pro Ile Thr Lys Tyr Ser Val Glu Val Gln Asp
130 135 140
Val Lys Asn Ile Pro Glu Ala Val Thr Asn Ala Phe Arg Ile Ala Ser
145 150 155 160
Ala Gly Gln Ala Gly Ala Ala Phe Val Ser Phe Pro Gln Asp Val Val
165 170 175
Asn Glu Val Thr Asn Thr Lys Asn Val Arg Ala Val Ala Ala Pro Lys
180 185 190
Leu Gly Pro Ala Ala Asp Asp Ala Ile Ser Ala Ala Ile Ala Lys Ile
195 200 205
Gln Thr Ala Lys Leu Pro Val Val Leu Val Gly Met Lys Gly Gly Arg
210 215 220
Pro Glu Ala Ile Lys Ala Val Arg Lys Leu Leu Lys Lys Val Gln Leu
225 230 235 240
Pro Phe Val Glu Thr Tyr Gln Ala Ala Gly Thr Leu Ser Arg Asp Leu
245 250 255
Glu Asp Gln Tyr Phe Gly Arg Ile Gly Leu Phe Arg Asn Gln Pro Gly
260 265 270
Asp Leu Leu Leu Glu Gln Ala Asp Val Val Leu Thr Ile Gly Tyr Asp
275 280 285
Pro Ile Glu Tyr Asp Pro Lys Phe Trp Asn Ile Asn Gly Asp Arg Thr
290 295 300
Ile Ile His Leu Asp Glu Ile Ile Ala Asp Ile Asp His Ala Tyr Gln
305 310 315 320
Pro Asp Leu Glu Leu Ile Gly Asp Ile Pro Ser Thr Ile Asn His Ile
325 330 335
Glu His Asp Ala Val Lys Val Glu Phe Ala Glu Arg Glu Gln Lys Ile
340 345 350
Leu Ser Asp Leu Lys Gln Tyr Met His Glu Gly Glu Gln Val Pro Ala
355 360 365
Asp Trp Lys Ser Asp Arg Ala His Pro Leu Glu Ile Val Lys Glu Leu
370 375 380
Arg Asn Ala Val Asp Asp His Val Thr Val Thr Cys Asp Ile Gly Ser
385 390 395 400
His Ala Ile Trp Met Ser Arg Tyr Phe Arg Ser Tyr Glu Pro Leu Thr
405 410 415
Leu Met Ile Ser Asn Gly Met Gln Thr Leu Gly Val Ala Leu Pro Trp
420 425 430
Ala Ile Gly Ala Ser Leu Val Lys Pro Gly Glu Lys Val Val Ser Val
435 440 445
Ser Gly Asp Gly Gly Phe Leu Phe Ser Ala Met Glu Leu Glu Thr Ala
450 455 460
Val Arg Leu Lys Ala Pro Ile Val His Ile Val Trp Asn Asp Ser Thr
465 470 475 480
Tyr Asp Met Val Ala Phe Gln Gln Leu Lys Lys Tyr Asn Arg Thr Ser
485 490 495
Ala Val Asp Phe Gly Asn Ile Asp Ile Val Lys Tyr Ala Glu Ser Phe
500 505 510
Gly Ala Thr Gly Leu Arg Val Glu Ser Pro Asp Gln Leu Ala Asp Val
515 520 525
Leu Arg Gln Gly Met Asn Ala Glu Gly Pro Val Ile Ile Asp Val Pro
530 535 540
Val Asp Tyr Ser Asp Asn Ile Asn Leu Ala Ser Asp Lys Leu Pro Lys
545 550 555 560
Glu Phe Gly Glu Leu Met Lys Thr Lys Ala Leu
565 570
<210> 2
<211> 1713
<212> ДНК
<213> Bacillus subtilis
<400> 2
atgaccaagg ccaccaagga acagaaaagc ctggtcaaga accgcggtgc tgaactggtt 60
gtggactgcc tcgtggaaca gggcgtgacc catgtcttcg gcatcccggg cgccaagatc 120
gacgccgtct tcgacgccct gcaggataaa ggtccggaaa tcatcgtggc acgccatgag 180
cagaacgcag ccttcatggc ccaggccgtc ggtcggctga cgggtaagcc cggcgtggtg 240
ctggtcacct ccggtccggg agcctcgaac ctggccacgg gactgctcac cgccaacacc 300
gaaggcgacc cggtggtcgc cctggccggt aatgtcatcc gggcggatcg cctgaagcgc 360
acccatcagt ccctggataa cgcggccctg ttccagccaa tcaccaaata tagtgtcgaa 420
gtgcaggatg tgaagaacat cccggaagcc gtcaccaatg cgttccgaat cgcgtccgcc 480
ggccaagcag gggcagcatt cgtgagcttc ccccaggacg tggtcaatga agtgaccaac 540
accaaaaacg tcagagccgt agccgccccg aagctgggcc ctgcagcaga tgacgccatc 600
tccgctgcca tcgcgaagat ccagaccgca aagctgccgg tcgtgctggt cggaatgaag 660
ggcggacgcc cggaggccat caaggccgtg cgtaaactgc tgaagaaggt gcagctaccg 720
ttcgtggaaa cctaccaggc cgccggcacc ctgagtcggg acttggaaga ccagtatttc 780
ggccgtatcg gcctgttccg caaccagccg ggcgacctgc tcctggaaca agccgatgtg 840
gtgctgacca tcggctacga cccgatcgaa tatgacccga agttctggaa catcaatggc 900
gaccgcacga tcatccatct ggacgaaatc atcgccgaca tcgaccatgc ctatcagccg 960
gacctggaac tgatcggcga catcccgagc accatcaacc acatcgaaca cgatgccgtg 1020
aaggtggaat ttgccgaacg cgaacagaag atcctgtcgg acctgaagca gtatatgcat 1080
gagggcgaac aggtgcctgc cgactggaag tcggacagag cccatccgct ggaaatcgtg 1140
aaggaactgc gtaacgccgt cgacgaccat gtcaccgtca cctgcgatat cggcagccat 1200
gccatttgga tgagccgcta cttccggagc tatgaaccgc tgaccctgat gatctccaac 1260
ggtatgcaga ccctcggcgt cgccctcccg tgggccatcg gcgcaagtct ggtgaagccg 1320
ggcgaaaaag tggtcagcgt gtccggcgac ggcggcttcc tgttctccgc tatggaactg 1380
gaaaccgcgg tccgcctgaa ggccccgatc gtgcatatcg tgtggaacga cagcacctac 1440
gacatggtcg ccttccagca gctgaaaaag tacaaccgca ccagcgccgt ggacttcggc 1500
aatatcgaca tcgtgaagta tgccgaatcc ttcggagcca ccggactgcg cgtggaatcc 1560
ccggaccagc tggcggacgt tctgcgtcag ggcatgaatg ccgaaggtcc cgtgattatc 1620
gatgtgcccg tcgactacag cgacaacatc aacctggcct cggacaaatt gccgaaggag 1680
ttcggcgaac tgatgaaaac aaaagcacta taa 1713
<210> 3
<211> 556
<212> Белок
<213> Clostridium autoethanogenum
<400> 3
Met Asn Arg Asp Ile Lys Lys Glu Val Gln Leu Asn Thr Ala Gln Met
1 5 10 15
Leu Val Lys Cys Leu Glu Ala Glu Gly Val Lys Tyr Ile Phe Gly Ile
20 25 30
Pro Gly Glu Glu Asn Leu Glu Ile Met Asn Ala Ile Ser Asp Ser Thr
35 40 45
Ile Glu Phe Ile Thr Thr Arg His Glu Gln Gly Ala Ala Phe Met Ala
50 55 60
Asp Val Tyr Gly Arg Leu Thr Gly Lys Ala Gly Val Cys Leu Ser Thr
65 70 75 80
Leu Gly Pro Gly Ala Thr Asn Leu Val Thr Gly Val Ala Asp Ala Asp
85 90 95
Ser Asp Gly Ala Pro Val Val Ala Ile Thr Gly Gln Val Gly Thr Glu
100 105 110
Arg Met His Ile Thr Ser His Gln Phe Leu Asp Leu Cys Lys Met Phe
115 120 125
Glu Pro Ile Thr Lys Arg Ser Lys Gln Ile Val Arg Pro Asp Thr Val
130 135 140
Ser Glu Ile Ile Arg Leu Val Phe Lys Tyr Ala Glu Ser Glu Lys Pro
145 150 155 160
Gly Ala Cys His Ile Asp Leu Pro Val Asn Ile Ala Lys Met Pro Val
165 170 175
Gly Ala Leu Glu Lys Pro Leu Glu Lys Lys Ile Pro Pro Lys Glu His
180 185 190
Ala Asp Leu Ser Thr Ile Glu Glu Ala Ala Ser Glu Ile Phe Lys Ala
195 200 205
Lys Asn Pro Ile Ile Leu Ala Gly Ser Gly Ala Ile Arg Gly Asn Ser
210 215 220
Ser Lys Ala Val Thr Glu Phe Ala Thr Lys Leu Lys Ile Pro Val Ile
225 230 235 240
Asn Thr Met Met Ala Lys Gly Ile Ile Pro Met Asp Asn Lys Tyr Ser
245 250 255
Met Trp Thr Ile Gly Ile Pro Gln Lys Asp Tyr Val Asn Lys Ile Ile
260 265 270
Glu Glu Ala Asp Leu Val Ile Thr Ile Gly Tyr Asp Ile Val Glu Tyr
275 280 285
Ala Pro Ser Lys Trp Asn Ile Asn Gly Asp Ile Lys Ile Val His Ile
290 295 300
Asp Ala Arg Pro Ser His Ile Asn Lys Leu Tyr Gln Pro Ile Val Glu
305 310 315 320
Val Val Gly Asp Ile Ser Asp Ala Leu Tyr Asn Ile Leu Arg Arg Thr
325 330 335
Ser Ser Lys Asp Glu Pro Val Lys Ala Leu Glu Ile Lys Ser Glu Met
340 345 350
Leu Ala Glu His Glu Ser Tyr Ala Asn Asp Asn Ala Phe Pro Met Lys
355 360 365
Pro Gln Arg Ile Leu Asn Asp Val Arg Lys Val Met Gly Pro His Asp
370 375 380
Ile Val Ile Ser Asp Val Gly Ala His Lys Met Trp Ile Ala Arg His
385 390 395 400
Tyr Asn Cys Tyr Glu Pro Asn Thr Cys Ile Ile Ser Asn Gly Phe Ala
405 410 415
Thr Met Gly Ile Gly Val Pro Gly Ala Ile Ala Ala Lys Leu Ile Asn
420 425 430
Pro Asp Lys Lys Val Leu Ala Ile Val Gly Asp Gly Gly Phe Met Met
435 440 445
Asn Asn Gln Glu Leu Glu Thr Ala Leu Arg Ile Lys Thr Pro Ile Val
450 455 460
Val Leu Ile Phe Asn Asp Ser Asn Tyr Gly Leu Ile Lys Trp Lys Gln
465 470 475 480
Glu Glu His Tyr Gly Lys Ser Cys Tyr Val Asp Phe Thr Asn Pro Asp
485 490 495
Phe Val Lys Leu Ala Glu Ser Met Tyr Ala Lys Gly Tyr Arg Val Glu
500 505 510
Lys Ala Glu Asp Leu Ile Pro Thr Leu Glu Glu Ala Phe Lys Gln Asn
515 520 525
Val Pro Ala Val Ile Asp Cys Gln Val Asp Tyr Gly Glu Asn Ile Lys
530 535 540
Leu Thr Lys His Leu Lys Glu Val Tyr Glu Asn Met
545 550 555
<210> 4
<211> 1671
<212> ДНК
<213> Clostridium autoethanogenum
<400> 4
atgaatcggg atatcaagaa agaggtgcag ctcaacacgg cccagatgct ggtcaagtgt 60
ctggaagccg agggcgtcaa gtatatcttc ggcatcccgg gcgaggagaa tctcgaaatc 120
atgaacgcca tctcggattc cacgatcgag ttcatcacca cccgccatga acagggcgcg 180
gccttcatgg ccgacgtgta cggccggctg accggcaagg cgggcgtgtg tctgagcacc 240
ctcggccccg gcgcgaccaa cctggtcacc ggcgtggccg acgccgactc cgacggcgcc 300
cccgtggtcg cgatcaccgg ccaggtgggc acggagcgga tgcacatcac ctcccatcag 360
ttcctcgacc tctgcaagat gttcgagccg atcaccaagc ggagcaagca gatcgtccgc 420
ccggacacgg tgtcggagat catccgcctg gtgttcaagt acgccgaaag cgaaaagccc 480
ggcgcctgtc atatcgacct gccggtcaac atcgccaaga tgcccgtcgg cgccctggag 540
aagccgctgg agaaaaaaat cccgccgaag gaacacgcgg acctgtccac catcgaggaa 600
gcggcgtccg agatcttcaa ggccaaaaac cccatcatcc tggccggcag cggcgccatc 660
cgcggcaaca gcagcaaggc ggtcaccgag ttcgccacca agctgaagat ccccgtcatc 720
aacacgatga tggccaaggg catcatcccg atggacaaca agtatagcat gtggaccatc 780
ggcatccccc agaaggacta tgtgaacaag atcatcgaag aggccgacct ggtcatcacc 840
atcggctacg acatcgtgga atatgccccg tcgaaatgga acatcaacgg cgacatcaag 900
atcgtccata tcgacgcccg cccctcgcac atcaacaaac tctaccagcc catcgtggag 960
gtggtcggcg acatcagcga cgcgctgtat aacatcctgc gccgcaccag ctcgaaagac 1020
gagccggtca aggcgctgga gatcaagtcg gaaatgctgg cggagcacga gtcctacgcg 1080
aacgacaatg cgttcccgat gaagccgcag cgcatcctca acgatgtgcg caaagtcatg 1140
ggcccgcacg acatcgtgat ctccgatgtg ggcgcccata aaatgtggat cgcccgccac 1200
tataactgct acgagccgaa tacctgcatc atctcgaacg gcttcgccac gatgggcatc 1260
ggcgtcccgg gcgcgatcgc cgccaaactc atcaacccgg ataagaaggt cctggccatc 1320
gtcggcgacg gcggcttcat gatgaataac caggaactgg agacggcgct gcgcatcaaa 1380
acgcccatcg tggtcctcat cttcaacgac tccaattacg gcctcatcaa gtggaagcag 1440
gaggagcatt atggcaaatc gtgctatgtg gacttcacca acccggactt cgtgaagctg 1500
gccgagagca tgtacgccaa aggctatcgc gtggagaaag ccgaggatct gatcccgacc 1560
ctcgaagagg ccttcaagca gaatgtcccg gcggtcatcg actgccaggt ggactatggc 1620
gagaatatca agctcaccaa gcacctcaag gaggtctatg aaaacatgtg a 1671
<210> 5
<211> 239
<212> Белок
<213> Clostridium autoethanogenum
<400> 5
Met Asp Asp Glu Val Lys Val Pro Asn His Ile Tyr Gln Met Ser Thr
1 5 10 15
Ile Asn Ala Leu Val Ser Gly Leu Tyr Asp Gly Cys Val Ser Leu Ser
20 25 30
Lys Leu Leu Lys Lys Gly Asn Phe Gly Ile Gly Thr Phe Lys Gly Leu
35 40 45
Asp Gly Glu Leu Thr Leu Leu Asn Gly Thr Phe Tyr Arg Thr Lys Pro
50 55 60
Asp Gly Ser Val Tyr Val Cys Ser Lys Asn Val Ser Val Pro Phe Ala
65 70 75 80
Val Val Thr Glu Leu Glu Asn Tyr Asn Thr Tyr Asn Ile Gln Asn Arg
85 90 95
Thr Ser Tyr Glu Asp Ile Arg Lys Glu Leu Asp Ser Phe Ile Glu Ser
100 105 110
Lys Asn Ile Phe Tyr Ala Phe Tyr Met Glu Gly Lys Phe Asn Tyr Val
115 120 125
Lys Thr Arg Thr Val Val Lys Gln Asn Met Pro Tyr Lys Pro Met Ala
130 135 140
Glu Val Val Lys Asp Gln Pro Met Phe Glu Tyr Asn Gly Val Asp Gly
145 150 155 160
Tyr Val Val Gly Phe Arg Cys Pro Asp Tyr Val Glu Gly Leu Asn Val
165 170 175
Pro Gly Tyr His Phe His Phe Ile Asn Lys Asp Lys Lys Phe Gly Gly
180 185 190
His Ile Ser Glu Phe Ser Ile Glu Asn Ala Lys Val Tyr Val Gln Asn
195 200 205
Cys Ser Cys Phe Arg Met Glu Leu Pro Lys Asn Glu Ser Phe Tyr Asn
210 215 220
Met Glu Val Gln Asp Arg Asn Asp Glu Ile Thr Ser Val Glu Lys
225 230 235
<210> 6
<211> 720
<212> ДНК
<213> Clostridium autoethanogenum
<400> 6
atggatgatg aggtgaaagt cccgaaccac atctaccaga tgtcgaccat caatgccctg 60
gtcagcggcc tctacgacgg ctgtgtgtcg ctctcgaagc tcctgaaaaa gggcaatttc 120
ggcatcggca cgttcaaggg cctggatggc gagctgaccc tcctgaacgg cacgttctat 180
cgcaccaaac cggatggctc cgtgtacgtg tgcagcaaga acgtgagcgt ccccttcgcg 240
gtcgtcaccg agctggagaa ctacaatacc tataacatcc agaatcgcac ctcctatgag 300
gacatccgca aggagctgga ctcgttcatc gagtcgaaga acatcttcta tgccttctat 360
atggaaggca aattcaacta cgtcaaaacc cgcaccgtcg tgaagcagaa catgccgtac 420
aagccgatgg ccgaggtggt caaagaccag ccgatgttcg aatacaacgg cgtcgatggc 480
tacgtcgtcg gcttccggtg cccggattat gtggaaggcc tcaatgtgcc cggctaccat 540
ttccacttca tcaacaagga caaaaagttc ggcggccaca tctccgagtt ctcgatcgag 600
aacgccaaag tctacgtcca gaactgctcc tgtttccgca tggagctccc gaagaatgag 660
agcttctaca acatggaggt ccaggaccgc aacgacgaaa tcacgtccgt ggagaaatga 720
<210> 7
<211> 259
<212> Белок
<213> Klebsiella pneumoniae
<400> 7
Met Asn His Ser Ala Glu Cys Thr Cys Glu Glu Ser Leu Cys Glu Thr
1 5 10 15
Leu Arg Ala Phe Ser Ala Gln His Pro Glu Ser Val Leu Tyr Gln Thr
20 25 30
Ser Leu Met Ser Ala Leu Leu Ser Gly Val Tyr Glu Gly Ser Thr Thr
35 40 45
Ile Ala Asp Leu Leu Lys His Gly Asp Phe Gly Leu Gly Thr Phe Asn
50 55 60
Glu Leu Asp Gly Glu Leu Ile Ala Phe Ser Ser Gln Val Tyr Gln Leu
65 70 75 80
Arg Ala Asp Gly Ser Ala Arg Lys Ala Gln Pro Glu Gln Lys Thr Pro
85 90 95
Phe Ala Val Met Thr Trp Phe Gln Pro Gln Tyr Arg Lys Thr Phe Asp
100 105 110
His Pro Val Ser Arg Gln Gln Leu His Glu Val Ile Asp Gln Gln Ile
115 120 125
Pro Ser Asp Asn Leu Phe Cys Ala Leu Arg Ile Asp Gly His Phe Arg
130 135 140
His Ala His Thr Arg Thr Val Pro Arg Gln Thr Pro Pro Tyr Arg Ala
145 150 155 160
Met Thr Asp Val Leu Asp Asp Gln Pro Val Phe Arg Phe Asn Gln Arg
165 170 175
Glu Gly Val Leu Val Gly Phe Arg Thr Pro Gln His Met Gln Gly Ile
180 185 190
Asn Val Ala Gly Tyr His Glu His Phe Ile Thr Asp Asp Arg Lys Gly
195 200 205
Gly Gly His Leu Leu Asp Tyr Gln Leu Asp His Gly Val Leu Thr Phe
210 215 220
Gly Glu Ile His Lys Leu Met Ile Asp Leu Pro Ala Asp Ser Ala Phe
225 230 235 240
Leu Gln Ala Asn Leu His Pro Asp Asn Leu Asp Ala Ala Ile Arg Ser
245 250 255
Val Glu Ser
<210> 8
<211> 780
<212> ДНК
<213> Klebsiella pneumoniae
<400> 8
atgaaccact cggccgaatg cacctgcgaa gagagcctct gcgaaaccct ccgggccttc 60
tcggcccagc acccggagag cgtcctgtac cagacgagcc tgatgtcggc gctgctgtcg 120
ggcgtgtatg aaggctcgac gaccatcgcc gacctgctga agcatggcga cttcggcctg 180
ggcaccttca atgaactgga cggcgagctc atcgccttca gctcgcaggt gtatcagctc 240
cgggccgatg gctccgcccg gaaggcccag cccgaacaga agaccccgtt cgccgtgatg 300
acctggttcc agccgcagta tcggaagacc ttcgaccacc ccgtgagccg ccagcagctc 360
cacgaggtga tcgaccagca gatcccgagc gacaacctct tctgcgccct gcgcatcgac 420
ggccatttcc gccacgcgca tacccgcacc gtcccgcggc agaccccgcc ctaccgcgcc 480
atgaccgatg tcctggatga ccagccggtc ttccggttca accagcgcga gggcgtcctg 540
gtcggcttcc gcaccccgca gcacatgcag ggcatcaacg tcgcgggcta tcatgaacac 600
ttcatcaccg atgatcgcaa gggcggcggc cacctcctcg actaccagct ggaccacggc 660
gtcctgacct tcggcgaaat ccataagctg atgatcgacc tccccgccga cagcgccttc 720
ctgcaggcga atctgcatcc ggacaacctc gatgccgcca tccgctccgt cgagtcgtga 780
<210> 9
<211> 357
<212> Белок
<213> Clostridium autoethanogenum
<400> 9
Met Lys Ala Val Leu Trp Tyr Asp Lys Lys Asp Val Arg Val Glu Glu
1 5 10 15
Ile Glu Glu Pro Lys Val Lys Glu Asn Ala Val Lys Ile Lys Val Lys
20 25 30
Trp Cys Gly Ile Cys Gly Ser Asp Leu His Glu Tyr Leu Gly Gly Pro
35 40 45
Ile Phe Ile Pro Val Gly Thr Pro His Pro Leu Ser Lys Ser Thr Ala
50 55 60
Pro Val Val Leu Gly His Glu Phe Ser Gly Glu Val Val Glu Ile Gly
65 70 75 80
Ser Lys Val Thr Lys Phe Lys Ala Gly Asp Arg Val Ile Val Glu Pro
85 90 95
Ile Val Ala Cys Gly Lys Cys Pro Ala Cys Leu Glu Gly Lys Tyr Asn
100 105 110
Leu Cys Glu Ala Leu Gly Phe His Gly Leu Cys Gly Ser Gly Gly Gly
115 120 125
Phe Ala Glu Tyr Thr Val Phe Pro Glu Asp Phe Val His Lys Ile Pro
130 135 140
Asp Thr Met Asp Tyr Glu Gln Ala Ala Leu Val Glu Pro Met Ala Val
145 150 155 160
Ala Leu His Ser Leu Arg Val Gly Asn Phe Thr Thr Gly Asn Thr Ala
165 170 175
Leu Val Leu Gly Ala Gly Pro Ile Gly Leu Ala Thr Ile Gln Cys Leu
180 185 190
Lys Ala Ser Gly Ala Arg Ile Val Ile Val Phe Gln Arg Lys Ser Val
195 200 205
Arg Gln Glu Tyr Ala Lys Lys Phe Gly Ala Asp Val Val Leu Asp Pro
210 215 220
Asn Glu Val Asp Val Ile Glu Glu Ile Lys Lys Leu Thr Gly Gly Val
225 230 235 240
Gly Val Asp Thr Ser Phe Glu Thr Thr Gly Ala Asn Val Gly Ile Asn
245 250 255
Thr Ala Ile Gln Ala Leu Lys Tyr Glu Gly Thr Ala Val Ile Thr Ser
260 265 270
Val Trp Glu Lys Asn Ala Glu Ile Asn Pro Asn Asp Leu Val Phe Thr
275 280 285
Glu Lys Lys Val Val Gly Thr Leu Ala Tyr Arg His Glu Phe Pro Ser
290 295 300
Thr Ile Ala Leu Met Asn Asp Gly Arg Ile Lys Thr Asp Gly Tyr Ile
305 310 315 320
Thr Lys Arg Ile Ala Leu Glu Asp Ile Val Lys Glu Gly Phe Glu Thr
325 330 335
Leu Thr Gly Pro Glu Lys Lys Lys His Val Lys Ile Ile Val Thr Pro
340 345 350
Asp Lys Ser Leu Leu
355
<210> 10
<211> 1074
<212> ДНК
<213> Clostridium autoethanogenum
<400> 10
atgaaggccg tcctgtggta cgacaaaaag gatgtccgcg tggaagaaat cgaggaaccg 60
aaggtgaaag aaaacgccgt gaagatcaaa gtcaagtggt gcggcatctg cggctcggac 120
ctgcatgagt atctcggcgg cccgatcttc atcccggtcg gcacccccca cccgctgtcg 180
aagagcaccg cgcccgtcgt gctgggccac gagttctcgg gcgaagtggt ggagatcggc 240
agcaaagtga ccaagttcaa ggcgggcgac cgcgtcatcg tggaaccgat cgtcgcctgc 300
ggcaaatgcc cggcctgcct ggaaggcaag tacaatctgt gcgaggcgct gggcttccac 360
ggcctgtgcg gcagcggcgg cggcttcgcc gagtacacgg tgttcccgga agatttcgtg 420
cacaagatcc ccgacacgat ggattatgaa caggccgcgc tggtggagcc gatggcggtc 480
gcgctgcact ccctgcgggt gggcaacttc accacgggca acaccgccct ggtcctgggc 540
gcgggcccga tcggcctggc caccatccag tgcctcaaag cgtcgggcgc ccggatcgtc 600
atcgtcttcc agcgcaaatc ggtgcggcag gaatacgcca agaagttcgg cgcggacgtg 660
gtcctcgacc cgaatgaggt ggacgtgatc gaggaaatca aaaagctgac cggcggcgtg 720
ggcgtggaca cgagcttcga aaccaccggc gccaacgtcg gcatcaacac cgcgatccag 780
gcgctgaaat atgagggcac cgccgtcatc acctccgtct gggagaagaa cgccgagatc 840
aatccgaacg acctggtctt caccgaaaag aaggtcgtcg gcaccctcgc gtaccggcac 900
gagttcccgt cgaccatcgc cctgatgaac gacggccgca tcaagaccga tggctatatc 960
accaagcgga tcgccctgga agacatcgtc aaggagggct tcgaaaccct gaccggcccg 1020
gagaagaaaa agcacgtcaa aatcatcgtc acgcccgata aaagcctcct gtga 1074
<210> 11
<211> 350
<212> Белок
<213> Bacillus subtilis
<400> 11
Met Lys Ala Leu Leu Trp His Asn Gln Arg Asp Val Arg Val Glu Glu
1 5 10 15
Val Pro Glu Pro Ala Val Arg Ser Gly Ala Val Lys Ile Lys Val Lys
20 25 30
Trp Cys Gly Ile Cys Gly Thr Asp Leu His Glu Tyr Leu Ala Gly Pro
35 40 45
Ile Phe Ile Pro Thr Glu Glu His Pro Leu Thr His Val Lys Ala Pro
50 55 60
Val Ile Leu Gly His Glu Phe Ser Gly Glu Val Val Glu Ile Gly Glu
65 70 75 80
Gly Val Thr Asn His Lys Val Gly Asp Arg Val Val Val Glu Pro Ile
85 90 95
Tyr Ser Cys Gly Lys Cys Glu Ala Cys Lys His Gly His Tyr Asn Val
100 105 110
Cys Glu Gln Leu Val Phe His Gly Leu Gly Gly Asp Gly Gly Gly Phe
115 120 125
Ser Glu Tyr Thr Val Val Pro Ala Asp Met Val His His Ile Pro Asp
130 135 140
Glu Met Thr Tyr Glu Gln Gly Ala Leu Val Glu Pro Ala Ala Val Ala
145 150 155 160
Val His Ala Val Arg Gln Ser Lys Leu Lys Glu Gly Glu Ala Val Ala
165 170 175
Val Phe Gly Cys Gly Pro Ile Gly Leu Leu Val Ile Gln Ala Ala Lys
180 185 190
Ala Ala Gly Ala Thr Pro Val Ile Ala Val Glu Leu Ser Lys Glu Arg
195 200 205
Gln Glu Leu Ala Lys Leu Ala Gly Ala Asp Tyr Val Leu Asn Pro Ala
210 215 220
Glu Gln Asp Val Val Ala Glu Ile Arg Asn Leu Thr Asn Gly Leu Gly
225 230 235 240
Val Asn Val Ser Phe Glu Val Thr Gly Val Glu Val Val Leu Arg Gln
245 250 255
Ala Ile Glu Ser Thr Ser Phe Glu Gly Gln Thr Val Ile Val Ser Val
260 265 270
Trp Glu Lys Asp Ala Thr Ile Thr Pro Asn Asn Leu Val Leu Lys Glu
275 280 285
Lys Glu Val Val Gly Ile Leu Gly Tyr Arg His Ile Phe Pro Ser Val
290 295 300
Ile Lys Leu Ile Ser Ser Gly Gln Ile Gln Ala Glu Lys Leu Ile Thr
305 310 315 320
Lys Lys Ile Thr Val Asp Gln Val Val Glu Glu Gly Phe Glu Ala Leu
325 330 335
Val Lys Asp Lys Lys Gln Val Lys Ile Leu Val Ser Pro Lys
340 345 350
<210> 12
<211> 1053
<212> ДНК
<213> Bacillus subtilis
<400> 12
atgaaagccc tgctgtggca taaccagcgc gacgtgcggg tggaagaggt cccggagccc 60
gccgtccgca gcggcgcggt gaaaatcaaa gtgaaatggt gcggcatctg tggcaccgac 120
ctgcatgaat atctggccgg ccccatcttc atcccgacgg aggaacatcc gctgacgcac 180
gtcaaggccc cggtcatcct cggccatgag ttcagcggcg aggtggtgga gatcggcgaa 240
ggcgtcacca atcacaaagt cggcgatcgc gtggtcgtcg aaccgatcta ctcgtgcggc 300
aagtgtgagg cgtgcaagca cggccactat aatgtctgcg agcagctggt gttccacggc 360
ctgggcggcg acggcggcgg cttctcggag tacaccgtgg tgccggcgga tatggtccac 420
cacatcccgg atgaaatgac ctacgagcag ggcgccctgg tcgagccggc cgccgtggcg 480
gtgcacgcgg tgcgccagag caaactcaag gagggcgaag ccgtggccgt cttcggctgc 540
ggcccgatcg gcctgctggt catccaggcg gccaaagcgg cgggcgcgac ccccgtcatc 600
gcggtcgagc tgtcgaagga acgccaggag ctcgccaagc tggcgggcgc ggattatgtc 660
ctgaaccccg ccgaacagga cgtggtggcg gaaatccgga acctgaccaa cggcctgggc 720
gtcaacgtct ccttcgaggt caccggcgtg gaagtcgtcc tgcggcaggc gatcgaatcg 780
acctcgttcg agggccagac ggtcatcgtg tcggtctggg agaaggacgc caccatcacg 840
cccaataatc tggtcctgaa agagaaggaa gtggtcggca tcctcggcta ccggcatatc 900
ttcccgtccg tcatcaagct gatctcgtcg ggccagatcc aggccgagaa actcatcacc 960
aagaagatca cggtggacca ggtggtcgaa gaaggcttcg aagcgctggt caaggataag 1020
aagcaggtga agatcctcgt gtcgccgaag tga 1053
<210> 13
<211> 350
<212> Белок
<213> Paenibacillus polymyxa
<400> 13
Met Gln Ala Leu Arg Trp His Gly Ile Lys Asp Leu Arg Leu Glu Asn
1 5 10 15
Ile Glu Gln Pro Ala Ala Leu Pro Gly Lys Val Lys Ile Lys Val Glu
20 25 30
Trp Cys Gly Ile Cys Gly Ser Asp Leu His Glu Tyr Val Ala Gly Pro
35 40 45
Ile Phe Ile Pro Glu Asn Ala Gln His Pro Leu Thr Gly Glu Lys Ser
50 55 60
Pro Ile Val Met Gly His Glu Phe Ser Gly Gln Phe Phe Asp Phe Gly
65 70 75 80
Glu Gly Val Thr Lys Ile Gln Val Gly Asp Arg Glu Val Val Glu Pro
85 90 95
Val Phe Ala Cys Gly Glu Cys Asp Ala Cys Arg Gln Gly Lys Tyr Asn
100 105 110
Leu Cys Asp Lys Met Gly Phe Leu Gly Leu Ala Gly Gly Gly Gly Gly
115 120 125
Phe Ser Glu Tyr Val Ala Ala Asp Glu His Met Val His Lys Ile Pro
130 135 140
Glu Ser Val Ser Phe Glu Gln Gly Ala Leu Val Glu Pro Ser Ala Val
145 150 155 160
Ala Leu Tyr Ala Val Arg Gln Ile Gln Leu Lys Val Asp Asp Lys Ala
165 170 175
Val Val Phe Gly Ala Gly Pro Ile Gly Leu Leu Val Ile Glu Ala Leu
180 185 190
Asn Ala Ser Gly Ala Ser Glu Ile Tyr Ala Glu Glu Leu Ser Glu Glu
195 200 205
Arg Thr Ala Lys Ala Glu Asp Leu Gly Ala Ile Val Leu Asp Pro Asn
210 215 220
Thr Tyr Asp Val Val Glu Glu Leu His Lys Arg Thr Asn Gly Gly Val
225 230 235 240
Tyr Val Pro Tyr Glu Val Thr Glu Val Pro Pro Val Leu Thr Gln Ala
245 250 255
Ile Glu Ser Ala Lys Ile Ser Gly Glu Ile Met Ile Val Ile Ile Phe
260 265 270
Glu Lys Glu Ala Leu Ile Lys Pro Asn Asn Ile Val Met Asn Glu Arg
275 280 285
Asn Leu Thr Gly Leu Ile Cys Tyr Asp Asp Val Phe Pro Ala Leu Ile
290 295 300
Ser Leu Met Glu Asn Gly Tyr Phe Pro Ala Asp Lys Leu Val Ile Lys
305 310 315 320
Arg Ile Lys Leu Val Asp Val Ile Glu Ala Ala Phe Glu Ser Leu Leu
325 330 335
Ile Glu Glu Tyr Gln Val Thr Ile Leu Val Ser Pro His Ala
340 345 350
<210> 14
<211> 1053
<212> ДНК
<213> Paenibacillus polymyxa
<400> 14
atgcaggcgc tgcgctggca cggcatcaag gacctgcggc tggagaacat cgagcagccc 60
gccgccctcc cgggcaaggt gaagatcaag gtggaatggt gcggcatctg cggcagcgac 120
ctgcatgaat atgtcgccgg cccgatcttc atccccgaaa acgcgcagca tccgctcacg 180
ggcgagaagt cgcccatcgt gatgggccat gagttctccg gccagttctt cgacttcggc 240
gaaggcgtga cgaaaatcca ggtgggcgac cgcgaagtgg tggagccggt cttcgcgtgt 300
ggcgaatgcg atgcgtgccg gcagggcaaa tataacctgt gcgataagat gggcttcctg 360
ggcctggccg gcggcggcgg cggcttctcg gaatatgtcg ccgcggatga gcatatggtg 420
cacaaaatcc ccgagtccgt gtccttcgaa cagggcgccc tggtcgagcc gtccgccgtc 480
gccctctacg cggtccgcca gatccagctg aaggtcgatg acaaggcggt ggtcttcggc 540
gccggcccca tcggcctgct cgtcatcgaa gcgctgaacg ccagcggcgc gagcgaaatc 600
tatgcggaag agctcagcga agagcgcacc gccaaagccg aagacctggg cgccatcgtg 660
ctcgacccca acacgtacga tgtcgtcgag gaactccata agcgcacgaa tggcggcgtc 720
tacgtcccct atgaggtcac ggaagtcccg cccgtgctga cccaggccat cgagtccgcc 780
aagatctccg gcgaaatcat gatcgtcatc atcttcgaaa aggaggccct catcaagccg 840
aacaacatcg tcatgaatga acggaacctg acgggcctga tctgctacga cgatgtgttc 900
ccggccctga tctccctcat ggagaatggc tacttccccg ccgacaagct ggtcatcaaa 960
cggatcaagc tggtggatgt catcgaagcg gccttcgagt cgctcctgat cgaggagtac 1020
caggtgacca tcctcgtgtc gccgcacgcc tga 1053
<210> 15
<211> 4672
<212> ДНК
<213> Штамм XZ58
<400> 15
aagaaaccaa ttgtccatat tgcatcagac attgccgtca ctgcgtcttt tactggctct 60
tctcgctaac caaaccggta accccgctta ttaaaagcat tctgtaacaa agcgggacca 120
aagccatgac aaaaacgcgt aacaaaagtg tctataatca cggcagaaaa gtccacattg 180
attatttgca cggcgtcaca ctttgctatg ccatagcatt tttatccata agattagcgg 240
atcctacctg acgcttttta tcgcaactct ctactgtttc tccatacccg tttttttggg 300
ctaacaggag gaattaacca tgaccaaggc caccaaggaa cagaaaagcc tggtcaagaa 360
ccgcggtgct gaactggttg tggactgcct cgtggaacag ggcgtgaccc atgtcttcgg 420
catcccgggc gccaagatcg acgccgtctt cgacgccctg caggataaag gtccggaaat 480
catcgtggca cgccatgagc agaacgcagc cttcatggcc caggccgtcg gtcggctgac 540
gggtaagccc ggcgtggtgc tggtcacctc cggtccggga gcctcgaacc tggccacggg 600
actgctcacc gccaacaccg aaggcgaccc ggtggtcgcc ctggccggta atgtcatccg 660
ggcggatcgc ctgaagcgca cccatcagtc cctggataac gcggccctgt tccagccaat 720
caccaaatat agtgtcgaag tgcaggatgt gaagaacatc ccggaagccg tcaccaatgc 780
gttccgaatc gcgtccgccg gccaagcagg ggcagcattc gtgagcttcc cccaggacgt 840
ggtcaatgaa gtgaccaaca ccaaaaacgt cagagccgta gccgccccga agctgggccc 900
tgcagcagat gacgccatct ccgctgccat cgcgaagatc cagaccgcaa agctgccggt 960
cgtgctggtc ggaatgaagg gcggacgccc ggaggccatc aaggccgtgc gtaaactgct 1020
gaagaaggtg cagctaccgt tcgtggaaac ctaccaggcc gccggcaccc tgagtcggga 1080
cttggaagac cagtatttcg gccgtatcgg cctgttccgc aaccagccgg gcgacctgct 1140
cctggaacaa gccgatgtgg tgctgaccat cggctacgac ccgatcgaat atgacccgaa 1200
gttctggaac atcaatggcg accgcacgat catccatctg gacgaaatca tcgccgacat 1260
cgaccatgcc tatcagccgg acctggaact gatcggcgac atcccgagca ccatcaacca 1320
catcgaacac gatgccgtga aggtggaatt tgccgaacgc gaacagaaga tcctgtcgga 1380
cctgaagcag tatatgcatg agggcgaaca ggtgcctgcc gactggaagt cggacagagc 1440
ccatccgctg gaaatcgtga aggaactgcg taacgccgtc gacgaccatg tcaccgtcac 1500
ctgcgatatc ggcagccatg ccatttggat gagccgctac ttccggagct atgaaccgct 1560
gaccctgatg atctccaacg gtatgcagac cctcggcgtc gccctcccgt gggccatcgg 1620
cgcaagtctg gtgaagccgg gcgaaaaagt ggtcagcgtg tccggcgacg gcggcttcct 1680
gttctccgct atggaactgg aaaccgcggt ccgcctgaag gccccgatcg tgcatatcgt 1740
gtggaacgac agcacctacg acatggtcgc cttccagcag ctgaaaaagt acaaccgcac 1800
cagcgccgtg gacttcggca atatcgacat cgtgaagtat gccgaatcct tcggagccac 1860
cggactgcgc gtggaatccc cggaccagct ggcggacgtt ctgcgtcagg gcatgaatgc 1920
cgaaggtccc gtgattatcg atgtgcccgt cgactacagc gacaacatca acctggcctc 1980
ggacaaattg ccgaaggagt tcggcgaact gatgaaaaca aaagcactat aaaaaggagg 2040
tacgtatgaa ccactcggcc gaatgcacct gcgaagagag cctctgcgaa accctccggg 2100
ccttctcggc ccagcacccg gagagcgtcc tgtaccagac gagcctgatg tcggcgctgc 2160
tgtcgggcgt gtatgaaggc tcgacgacca tcgccgacct gctgaagcat ggcgacttcg 2220
gcctgggcac cttcaatgaa ctggacggcg agctcatcgc cttcagctcg caggtgtatc 2280
agctccgggc cgatggctcc gcccggaagg cccagcccga acagaagacc ccgttcgccg 2340
tgatgacctg gttccagccg cagtatcgga agaccttcga ccaccccgtg agccgccagc 2400
agctccacga ggtgatcgac cagcagatcc cgagcgacaa cctcttctgc gccctgcgca 2460
tcgacggcca tttccgccac gcgcataccc gcaccgtccc gcggcagacc ccgccctacc 2520
gcgccatgac cgatgtcctg gatgaccagc cggtcttccg gttcaaccag cgcgagggcg 2580
tcctggtcgg cttccgcacc ccgcagcaca tgcagggcat caacgtcgcg ggctatcatg 2640
aacacttcat caccgatgat cgcaagggcg gcggccacct cctcgactac cagctggacc 2700
acggcgtcct gaccttcggc gaaatccata agctgatgat cgacctcccc gccgacagcg 2760
ccttcctgca ggcgaatctg catccggaca acctcgatgc cgccatccgc tccgtcgagt 2820
cgtgatttca gcctgataca gattaaatca gaacgcagaa gcggtctgat aaaacagaat 2880
ttgcctggcg gcagtagcgc ggtggtccca cctgacccca tgccgaactc agaagtgaaa 2940
cgccgtagcg ccgatggtag tgtggggtct ccccatgcga gagtagggaa ctgccaggca 3000
tcaaataaaa cgaaaggctc agtcgaaaga ctgggccttt cgttttatct gttgtttgtc 3060
ggtgaacgct ctcctgagta ggacaaatcc gccgggagcg gatttgaacg ttgcgaagca 3120
acggcccgga gggtggcggg caggacgccc gccataaact gccaggcatc aaattaagca 3180
gaaggccatc ctgacggatg gcctttttgc gtttcgaggt tcaggcgaaa ccgcagactc 3240
aagggcgctt gctcccggga aagatcgtat tagtttgcct cgatcggcgg tccttgtgac 3300
agggagatat tcccgacgga tccggggcat tcgagcggaa ccgcccgccg tgggagtttt 3360
tccagcgagc attcgagagt ttttcaaggc ggcttcgagg ggttattccg taacgccgcc 3420
gacatgatct gtcccagaat ctccgccgct gttcgtagag cgccgatgca gggtcggcat 3480
caatcattct tggaggagac acatgaaggc cgtcctgtgg tacgacaaaa aggatgtccg 3540
cgtggaagaa atcgaggaac cgaaggtgaa agaaaacgcc gtgaagatca aagtcaagtg 3600
gtgcggcatc tgcggctcgg acctgcatga gtatctcggc ggcccgatct tcatcccggt 3660
cggcaccccc cacccgctgt cgaagagcac cgcgcccgtc gtgctgggcc acgagttctc 3720
gggcgaagtg gtggagatcg gcagcaaagt gaccaagttc aaggcgggcg accgcgtcat 3780
cgtggaaccg atcgtcgcct gcggcaaatg cccggcctgc ctggaaggca agtacaatct 3840
gtgcgaggcg ctgggcttcc acggcctgtg cggcagcggc ggcggcttcg ccgagtacac 3900
ggtgttcccg gaagatttcg tgcacaagat ccccgacacg atggattatg aacaggccgc 3960
gctggtggag ccgatggcgg tcgcgctgca ctccctgcgg gtgggcaact tcaccacggg 4020
caacaccgcc ctggtcctgg gcgcgggccc gatcggcctg gccaccatcc agtgcctcaa 4080
agcgtcgggc gcccggatcg tcatcgtctt ccagcgcaaa tcggtgcggc aggaatacgc 4140
caagaagttc ggcgcggacg tggtcctcga cccgaatgag gtggacgtga tcgaggaaat 4200
caaaaagctg accggcggcg tgggcgtgga cacgagcttc gaaaccaccg gcgccaacgt 4260
cggcatcaac accgcgatcc aggcgctgaa atatgagggc accgccgtca tcacctccgt 4320
ctgggagaag aacgccgaga tcaatccgaa cgacctggtc ttcaccgaaa agaaggtcgt 4380
cggcaccctc gcgtaccggc acgagttccc gtcgaccatc gccctgatga acgacggccg 4440
catcaagacc gatggctata tcaccaagcg gatcgccctg gaagacatcg tcaaggaggg 4500
cttcgaaacc ctgaccggcc cggagaagaa aaagcacgtc aaaatcatcg tcacgcccga 4560
taaaagcctc ctgtgagact cctgttgata gatccagtaa tgacctcaga actccatctg 4620
gatttgttca gaacgctcgg ttgccgccgg gcgtttttta ttggtgagaa tc 4672
<210> 16
<211> 4651
<212> ДНК
<213> Штамм XZ59
<400> 16
aagaaaccaa ttgtccatat tgcatcagac attgccgtca ctgcgtcttt tactggctct 60
tctcgctaac caaaccggta accccgctta ttaaaagcat tctgtaacaa agcgggacca 120
aagccatgac aaaaacgcgt aacaaaagtg tctataatca cggcagaaaa gtccacattg 180
attatttgca cggcgtcaca ctttgctatg ccatagcatt tttatccata agattagcgg 240
atcctacctg acgcttttta tcgcaactct ctactgtttc tccatacccg tttttttggg 300
ctaacaggag gaattaacca tgaccaaggc caccaaggaa cagaaaagcc tggtcaagaa 360
ccgcggtgct gaactggttg tggactgcct cgtggaacag ggcgtgaccc atgtcttcgg 420
catcccgggc gccaagatcg acgccgtctt cgacgccctg caggataaag gtccggaaat 480
catcgtggca cgccatgagc agaacgcagc cttcatggcc caggccgtcg gtcggctgac 540
gggtaagccc ggcgtggtgc tggtcacctc cggtccggga gcctcgaacc tggccacggg 600
actgctcacc gccaacaccg aaggcgaccc ggtggtcgcc ctggccggta atgtcatccg 660
ggcggatcgc ctgaagcgca cccatcagtc cctggataac gcggccctgt tccagccaat 720
caccaaatat agtgtcgaag tgcaggatgt gaagaacatc ccggaagccg tcaccaatgc 780
gttccgaatc gcgtccgccg gccaagcagg ggcagcattc gtgagcttcc cccaggacgt 840
ggtcaatgaa gtgaccaaca ccaaaaacgt cagagccgta gccgccccga agctgggccc 900
tgcagcagat gacgccatct ccgctgccat cgcgaagatc cagaccgcaa agctgccggt 960
cgtgctggtc ggaatgaagg gcggacgccc ggaggccatc aaggccgtgc gtaaactgct 1020
gaagaaggtg cagctaccgt tcgtggaaac ctaccaggcc gccggcaccc tgagtcggga 1080
cttggaagac cagtatttcg gccgtatcgg cctgttccgc aaccagccgg gcgacctgct 1140
cctggaacaa gccgatgtgg tgctgaccat cggctacgac ccgatcgaat atgacccgaa 1200
gttctggaac atcaatggcg accgcacgat catccatctg gacgaaatca tcgccgacat 1260
cgaccatgcc tatcagccgg acctggaact gatcggcgac atcccgagca ccatcaacca 1320
catcgaacac gatgccgtga aggtggaatt tgccgaacgc gaacagaaga tcctgtcgga 1380
cctgaagcag tatatgcatg agggcgaaca ggtgcctgcc gactggaagt cggacagagc 1440
ccatccgctg gaaatcgtga aggaactgcg taacgccgtc gacgaccatg tcaccgtcac 1500
ctgcgatatc ggcagccatg ccatttggat gagccgctac ttccggagct atgaaccgct 1560
gaccctgatg atctccaacg gtatgcagac cctcggcgtc gccctcccgt gggccatcgg 1620
cgcaagtctg gtgaagccgg gcgaaaaagt ggtcagcgtg tccggcgacg gcggcttcct 1680
gttctccgct atggaactgg aaaccgcggt ccgcctgaag gccccgatcg tgcatatcgt 1740
gtggaacgac agcacctacg acatggtcgc cttccagcag ctgaaaaagt acaaccgcac 1800
cagcgccgtg gacttcggca atatcgacat cgtgaagtat gccgaatcct tcggagccac 1860
cggactgcgc gtggaatccc cggaccagct ggcggacgtt ctgcgtcagg gcatgaatgc 1920
cgaaggtccc gtgattatcg atgtgcccgt cgactacagc gacaacatca acctggcctc 1980
ggacaaattg ccgaaggagt tcggcgaact gatgaaaaca aaagcactat aaaaaggagg 2040
tacgtatgaa ccactcggcc gaatgcacct gcgaagagag cctctgcgaa accctccggg 2100
ccttctcggc ccagcacccg gagagcgtcc tgtaccagac gagcctgatg tcggcgctgc 2160
tgtcgggcgt gtatgaaggc tcgacgacca tcgccgacct gctgaagcat ggcgacttcg 2220
gcctgggcac cttcaatgaa ctggacggcg agctcatcgc cttcagctcg caggtgtatc 2280
agctccgggc cgatggctcc gcccggaagg cccagcccga acagaagacc ccgttcgccg 2340
tgatgacctg gttccagccg cagtatcgga agaccttcga ccaccccgtg agccgccagc 2400
agctccacga ggtgatcgac cagcagatcc cgagcgacaa cctcttctgc gccctgcgca 2460
tcgacggcca tttccgccac gcgcataccc gcaccgtccc gcggcagacc ccgccctacc 2520
gcgccatgac cgatgtcctg gatgaccagc cggtcttccg gttcaaccag cgcgagggcg 2580
tcctggtcgg cttccgcacc ccgcagcaca tgcagggcat caacgtcgcg ggctatcatg 2640
aacacttcat caccgatgat cgcaagggcg gcggccacct cctcgactac cagctggacc 2700
acggcgtcct gaccttcggc gaaatccata agctgatgat cgacctcccc gccgacagcg 2760
ccttcctgca ggcgaatctg catccggaca acctcgatgc cgccatccgc tccgtcgagt 2820
cgtgatttca gcctgataca gattaaatca gaacgcagaa gcggtctgat aaaacagaat 2880
ttgcctggcg gcagtagcgc ggtggtccca cctgacccca tgccgaactc agaagtgaaa 2940
cgccgtagcg ccgatggtag tgtggggtct ccccatgcga gagtagggaa ctgccaggca 3000
tcaaataaaa cgaaaggctc agtcgaaaga ctgggccttt cgttttatct gttgtttgtc 3060
ggtgaacgct ctcctgagta ggacaaatcc gccgggagcg gatttgaacg ttgcgaagca 3120
acggcccgga gggtggcggg caggacgccc gccataaact gccaggcatc aaattaagca 3180
gaaggccatc ctgacggatg gcctttttgc gtttcgaggt tcaggcgaaa ccgcagactc 3240
aagggcgctt gctcccggga aagatcgtat tagtttgcct cgatcggcgg tccttgtgac 3300
agggagatat tcccgacgga tccggggcat tcgagcggaa ccgcccgccg tgggagtttt 3360
tccagcgagc attcgagagt ttttcaaggc ggcttcgagg ggttattccg taacgccgcc 3420
gacatgatct gtcccagaat ctccgccgct gttcgtagag cgccgatgca gggtcggcat 3480
caatcattct tggaggagac acatgaaagc cctgctgtgg cataaccagc gcgacgtgcg 3540
ggtggaagag gtcccggagc ccgccgtccg cagcggcgcg gtgaaaatca aagtgaaatg 3600
gtgcggcatc tgtggcaccg acctgcatga atatctggcc ggccccatct tcatcccgac 3660
ggaggaacat ccgctgacgc acgtcaaggc cccggtcatc ctcggccatg agttcagcgg 3720
cgaggtggtg gagatcggcg aaggcgtcac caatcacaaa gtcggcgatc gcgtggtcgt 3780
cgaaccgatc tactcgtgcg gcaagtgtga ggcgtgcaag cacggccact ataatgtctg 3840
cgagcagctg gtgttccacg gcctgggcgg cgacggcggc ggcttctcgg agtacaccgt 3900
ggtgccggcg gatatggtcc accacatccc ggatgaaatg acctacgagc agggcgccct 3960
ggtcgagccg gccgccgtgg cggtgcacgc ggtgcgccag agcaaactca aggagggcga 4020
agccgtggcc gtcttcggct gcggcccgat cggcctgctg gtcatccagg cggccaaagc 4080
ggcgggcgcg acccccgtca tcgcggtcga gctgtcgaag gaacgccagg agctcgccaa 4140
gctggcgggc gcggattatg tcctgaaccc cgccgaacag gacgtggtgg cggaaatccg 4200
gaacctgacc aacggcctgg gcgtcaacgt ctccttcgag gtcaccggcg tggaagtcgt 4260
cctgcggcag gcgatcgaat cgacctcgtt cgagggccag acggtcatcg tgtcggtctg 4320
ggagaaggac gccaccatca cgcccaataa tctggtcctg aaagagaagg aagtggtcgg 4380
catcctcggc taccggcata tcttcccgtc cgtcatcaag ctgatctcgt cgggccagat 4440
ccaggccgag aaactcatca ccaagaagat cacggtggac caggtggtcg aagaaggctt 4500
cgaagcgctg gtcaaggata agaagcaggt gaagatcctc gtgtcgccga agtgagactc 4560
ctgttgatag atccagtaat gacctcagaa ctccatctgg atttgttcag aacgctcggt 4620
tgccgccggg cgttttttat tggtgagaat c 4651
<210> 17
<211> 4302
<212> ДНК
<213> Штамм XZ06
<400> 17
aagaaaccaa ttgtccatat tgcatcagac attgccgtca ctgcgtcttt tactggctct 60
tctcgctaac caaaccggta accccgctta ttaaaagcat tctgtaacaa agcgggacca 120
aagccatgac aaaaacgcgt aacaaaagtg tctataatca cggcagaaaa gtccacattg 180
attatttgca cggcgtcaca ctttgctatg ccatagcatt tttatccata agattagcgg 240
atcctacctg acgcttttta tcgcaactct ctactgtttc tccatacccg tttttttggg 300
ctaacaggag gaattaacca tgaccaaggc caccaaggaa cagaaaagcc tggtcaagaa 360
ccgcggtgct gaactggttg tggactgcct cgtggaacag ggcgtgaccc atgtcttcgg 420
catcccgggc gccaagatcg acgccgtctt cgacgccctg caggataaag gtccggaaat 480
catcgtggca cgccatgagc agaacgcagc cttcatggcc caggccgtcg gtcggctgac 540
gggtaagccc ggcgtggtgc tggtcacctc cggtccggga gcctcgaacc tggccacggg 600
actgctcacc gccaacaccg aaggcgaccc ggtggtcgcc ctggccggta atgtcatccg 660
ggcggatcgc ctgaagcgca cccatcagtc cctggataac gcggccctgt tccagccaat 720
caccaaatat agtgtcgaag tgcaggatgt gaagaacatc ccggaagccg tcaccaatgc 780
gttccgaatc gcgtccgccg gccaagcagg ggcagcattc gtgagcttcc cccaggacgt 840
ggtcaatgaa gtgaccaaca ccaaaaacgt cagagccgta gccgccccga agctgggccc 900
tgcagcagat gacgccatct ccgctgccat cgcgaagatc cagaccgcaa agctgccggt 960
cgtgctggtc ggaatgaagg gcggacgccc ggaggccatc aaggccgtgc gtaaactgct 1020
gaagaaggtg cagctaccgt tcgtggaaac ctaccaggcc gccggcaccc tgagtcggga 1080
cttggaagac cagtatttcg gccgtatcgg cctgttccgc aaccagccgg gcgacctgct 1140
cctggaacaa gccgatgtgg tgctgaccat cggctacgac ccgatcgaat atgacccgaa 1200
gttctggaac atcaatggcg accgcacgat catccatctg gacgaaatca tcgccgacat 1260
cgaccatgcc tatcagccgg acctggaact gatcggcgac atcccgagca ccatcaacca 1320
catcgaacac gatgccgtga aggtggaatt tgccgaacgc gaacagaaga tcctgtcgga 1380
cctgaagcag tatatgcatg agggcgaaca ggtgcctgcc gactggaagt cggacagagc 1440
ccatccgctg gaaatcgtga aggaactgcg taacgccgtc gacgaccatg tcaccgtcac 1500
ctgcgatatc ggcagccatg ccatttggat gagccgctac ttccggagct atgaaccgct 1560
gaccctgatg atctccaacg gtatgcagac cctcggcgtc gccctcccgt gggccatcgg 1620
cgcaagtctg gtgaagccgg gcgaaaaagt ggtcagcgtg tccggcgacg gcggcttcct 1680
gttctccgct atggaactgg aaaccgcggt ccgcctgaag gccccgatcg tgcatatcgt 1740
gtggaacgac agcacctacg acatggtcgc cttccagcag ctgaaaaagt acaaccgcac 1800
cagcgccgtg gacttcggca atatcgacat cgtgaagtat gccgaatcct tcggagccac 1860
cggactgcgc gtggaatccc cggaccagct ggcggacgtt ctgcgtcagg gcatgaatgc 1920
cgaaggtccc gtgattatcg atgtgcccgt cgactacagc gacaacatca acctggcctc 1980
ggacaaattg ccgaaggagt tcggcgaact gatgaaaaca aaagcactat aaaaaggagg 2040
tacgtatgaa ccactcggcc gaatgcacct gcgaagagag cctctgcgaa accctccggg 2100
ccttctcggc ccagcacccg gagagcgtcc tgtaccagac gagcctgatg tcggcgctgc 2160
tgtcgggcgt gtatgaaggc tcgacgacca tcgccgacct gctgaagcat ggcgacttcg 2220
gcctgggcac cttcaatgaa ctggacggcg agctcatcgc cttcagctcg caggtgtatc 2280
agctccgggc cgatggctcc gcccggaagg cccagcccga acagaagacc ccgttcgccg 2340
tgatgacctg gttccagccg cagtatcgga agaccttcga ccaccccgtg agccgccagc 2400
agctccacga ggtgatcgac cagcagatcc cgagcgacaa cctcttctgc gccctgcgca 2460
tcgacggcca tttccgccac gcgcataccc gcaccgtccc gcggcagacc ccgccctacc 2520
gcgccatgac cgatgtcctg gatgaccagc cggtcttccg gttcaaccag cgcgagggcg 2580
tcctggtcgg cttccgcacc ccgcagcaca tgcagggcat caacgtcgcg ggctatcatg 2640
aacacttcat caccgatgat cgcaagggcg gcggccacct cctcgactac cagctggacc 2700
acggcgtcct gaccttcggc gaaatccata agctgatgat cgacctcccc gccgacagcg 2760
ccttcctgca ggcgaatctg catccggaca acctcgatgc cgccatccgc tccgtcgagt 2820
cgtgaaaagg aggtacgtat gaaggccgtc ctgtggtacg acaaaaagga tgtccgcgtg 2880
gaagaaatcg aggaaccgaa ggtgaaagaa aacgccgtga agatcaaagt caagtggtgc 2940
ggcatctgcg gctcggacct gcatgagtat ctcggcggcc cgatcttcat cccggtcggc 3000
accccccacc cgctgtcgaa gagcaccgcg cccgtcgtgc tgggccacga gttctcgggc 3060
gaagtggtgg agatcggcag caaagtgacc aagttcaagg cgggcgaccg cgtcatcgtg 3120
gaaccgatcg tcgcctgcgg caaatgcccg gcctgcctgg aaggcaagta caatctgtgc 3180
gaggcgctgg gcttccacgg cctgtgcggc agcggcggcg gcttcgccga gtacacggtg 3240
ttcccggaag atttcgtgca caagatcccc gacacgatgg attatgaaca ggccgcgctg 3300
gtggagccga tggcggtcgc gctgcactcc ctgcgggtgg gcaacttcac cacgggcaac 3360
accgccctgg tcctgggcgc gggcccgatc ggcctggcca ccatccagtg cctcaaagcg 3420
tcgggcgccc ggatcgtcat cgtcttccag cgcaaatcgg tgcggcagga atacgccaag 3480
aagttcggcg cggacgtggt cctcgacccg aatgaggtgg acgtgatcga ggaaatcaaa 3540
aagctgaccg gcggcgtggg cgtggacacg agcttcgaaa ccaccggcgc caacgtcggc 3600
atcaacaccg cgatccaggc gctgaaatat gagggcaccg ccgtcatcac ctccgtctgg 3660
gagaagaacg ccgagatcaa tccgaacgac ctggtcttca ccgaaaagaa ggtcgtcggc 3720
accctcgcgt accggcacga gttcccgtcg accatcgccc tgatgaacga cggccgcatc 3780
aagaccgatg gctatatcac caagcggatc gccctggaag acatcgtcaa ggagggcttc 3840
gaaaccctga ccggcccgga gaagaaaaag cacgtcaaaa tcatcgtcac gcccgataaa 3900
agcctcctgt gatttcagcc tgatacagat taaatcagaa cgcagaagcg gtctgataaa 3960
acagaatttg cctggcggca gtagcgcggt ggtcccacct gaccccatgc cgaactcaga 4020
agtgaaacgc cgtagcgccg atggtagtgt ggggtctccc catgcgagag tagggaactg 4080
ccaggcatca aataaaacga aaggctcagt cgaaagactg ggcctttcgt tttatctgtt 4140
gtttgtcggt gaacgctctc ctgagtagga caaatccgcc gggagcggat ttgaacgttg 4200
cgaagcaacg gcccggaggg tggcgggcag gacgcccgcc ataaactgcc aggcatcaaa 4260
ttaagcagaa ggccatcctg acggatggcc tttttgcgtt tc 4302
<210> 18
<211> 4281
<212> ДНК
<213> Штамм XZ08
<400> 18
aagaaaccaa ttgtccatat tgcatcagac attgccgtca ctgcgtcttt tactggctct 60
tctcgctaac caaaccggta accccgctta ttaaaagcat tctgtaacaa agcgggacca 120
aagccatgac aaaaacgcgt aacaaaagtg tctataatca cggcagaaaa gtccacattg 180
attatttgca cggcgtcaca ctttgctatg ccatagcatt tttatccata agattagcgg 240
atcctacctg acgcttttta tcgcaactct ctactgtttc tccatacccg tttttttggg 300
ctaacaggag gaattaacca tgaccaaggc caccaaggaa cagaaaagcc tggtcaagaa 360
ccgcggtgct gaactggttg tggactgcct cgtggaacag ggcgtgaccc atgtcttcgg 420
catcccgggc gccaagatcg acgccgtctt cgacgccctg caggataaag gtccggaaat 480
catcgtggca cgccatgagc agaacgcagc cttcatggcc caggccgtcg gtcggctgac 540
gggtaagccc ggcgtggtgc tggtcacctc cggtccggga gcctcgaacc tggccacggg 600
actgctcacc gccaacaccg aaggcgaccc ggtggtcgcc ctggccggta atgtcatccg 660
ggcggatcgc ctgaagcgca cccatcagtc cctggataac gcggccctgt tccagccaat 720
caccaaatat agtgtcgaag tgcaggatgt gaagaacatc ccggaagccg tcaccaatgc 780
gttccgaatc gcgtccgccg gccaagcagg ggcagcattc gtgagcttcc cccaggacgt 840
ggtcaatgaa gtgaccaaca ccaaaaacgt cagagccgta gccgccccga agctgggccc 900
tgcagcagat gacgccatct ccgctgccat cgcgaagatc cagaccgcaa agctgccggt 960
cgtgctggtc ggaatgaagg gcggacgccc ggaggccatc aaggccgtgc gtaaactgct 1020
gaagaaggtg cagctaccgt tcgtggaaac ctaccaggcc gccggcaccc tgagtcggga 1080
cttggaagac cagtatttcg gccgtatcgg cctgttccgc aaccagccgg gcgacctgct 1140
cctggaacaa gccgatgtgg tgctgaccat cggctacgac ccgatcgaat atgacccgaa 1200
gttctggaac atcaatggcg accgcacgat catccatctg gacgaaatca tcgccgacat 1260
cgaccatgcc tatcagccgg acctggaact gatcggcgac atcccgagca ccatcaacca 1320
catcgaacac gatgccgtga aggtggaatt tgccgaacgc gaacagaaga tcctgtcgga 1380
cctgaagcag tatatgcatg agggcgaaca ggtgcctgcc gactggaagt cggacagagc 1440
ccatccgctg gaaatcgtga aggaactgcg taacgccgtc gacgaccatg tcaccgtcac 1500
ctgcgatatc ggcagccatg ccatttggat gagccgctac ttccggagct atgaaccgct 1560
gaccctgatg atctccaacg gtatgcagac cctcggcgtc gccctcccgt gggccatcgg 1620
cgcaagtctg gtgaagccgg gcgaaaaagt ggtcagcgtg tccggcgacg gcggcttcct 1680
gttctccgct atggaactgg aaaccgcggt ccgcctgaag gccccgatcg tgcatatcgt 1740
gtggaacgac agcacctacg acatggtcgc cttccagcag ctgaaaaagt acaaccgcac 1800
cagcgccgtg gacttcggca atatcgacat cgtgaagtat gccgaatcct tcggagccac 1860
cggactgcgc gtggaatccc cggaccagct ggcggacgtt ctgcgtcagg gcatgaatgc 1920
cgaaggtccc gtgattatcg atgtgcccgt cgactacagc gacaacatca acctggcctc 1980
ggacaaattg ccgaaggagt tcggcgaact gatgaaaaca aaagcactat aaaaaggagg 2040
tacgtatgaa ccactcggcc gaatgcacct gcgaagagag cctctgcgaa accctccggg 2100
ccttctcggc ccagcacccg gagagcgtcc tgtaccagac gagcctgatg tcggcgctgc 2160
tgtcgggcgt gtatgaaggc tcgacgacca tcgccgacct gctgaagcat ggcgacttcg 2220
gcctgggcac cttcaatgaa ctggacggcg agctcatcgc cttcagctcg caggtgtatc 2280
agctccgggc cgatggctcc gcccggaagg cccagcccga acagaagacc ccgttcgccg 2340
tgatgacctg gttccagccg cagtatcgga agaccttcga ccaccccgtg agccgccagc 2400
agctccacga ggtgatcgac cagcagatcc cgagcgacaa cctcttctgc gccctgcgca 2460
tcgacggcca tttccgccac gcgcataccc gcaccgtccc gcggcagacc ccgccctacc 2520
gcgccatgac cgatgtcctg gatgaccagc cggtcttccg gttcaaccag cgcgagggcg 2580
tcctggtcgg cttccgcacc ccgcagcaca tgcagggcat caacgtcgcg ggctatcatg 2640
aacacttcat caccgatgat cgcaagggcg gcggccacct cctcgactac cagctggacc 2700
acggcgtcct gaccttcggc gaaatccata agctgatgat cgacctcccc gccgacagcg 2760
ccttcctgca ggcgaatctg catccggaca acctcgatgc cgccatccgc tccgtcgagt 2820
cgtgaaaagg aggtacgtat gaaagccctg ctgtggcata accagcgcga cgtgcgggtg 2880
gaagaggtcc cggagcccgc cgtccgcagc ggcgcggtga aaatcaaagt gaaatggtgc 2940
ggcatctgtg gcaccgacct gcatgaatat ctggccggcc ccatcttcat cccgacggag 3000
gaacatccgc tgacgcacgt caaggccccg gtcatcctcg gccatgagtt cagcggcgag 3060
gtggtggaga tcggcgaagg cgtcaccaat cacaaagtcg gcgatcgcgt ggtcgtcgaa 3120
ccgatctact cgtgcggcaa gtgtgaggcg tgcaagcacg gccactataa tgtctgcgag 3180
cagctggtgt tccacggcct gggcggcgac ggcggcggct tctcggagta caccgtggtg 3240
ccggcggata tggtccacca catcccggat gaaatgacct acgagcaggg cgccctggtc 3300
gagccggccg ccgtggcggt gcacgcggtg cgccagagca aactcaagga gggcgaagcc 3360
gtggccgtct tcggctgcgg cccgatcggc ctgctggtca tccaggcggc caaagcggcg 3420
ggcgcgaccc ccgtcatcgc ggtcgagctg tcgaaggaac gccaggagct cgccaagctg 3480
gcgggcgcgg attatgtcct gaaccccgcc gaacaggacg tggtggcgga aatccggaac 3540
ctgaccaacg gcctgggcgt caacgtctcc ttcgaggtca ccggcgtgga agtcgtcctg 3600
cggcaggcga tcgaatcgac ctcgttcgag ggccagacgg tcatcgtgtc ggtctgggag 3660
aaggacgcca ccatcacgcc caataatctg gtcctgaaag agaaggaagt ggtcggcatc 3720
ctcggctacc ggcatatctt cccgtccgtc atcaagctga tctcgtcggg ccagatccag 3780
gccgagaaac tcatcaccaa gaagatcacg gtggaccagg tggtcgaaga aggcttcgaa 3840
gcgctggtca aggataagaa gcaggtgaag atcctcgtgt cgccgaagtg atttcagcct 3900
gatacagatt aaatcagaac gcagaagcgg tctgataaaa cagaatttgc ctggcggcag 3960
tagcgcggtg gtcccacctg accccatgcc gaactcagaa gtgaaacgcc gtagcgccga 4020
tggtagtgtg gggtctcccc atgcgagagt agggaactgc caggcatcaa ataaaacgaa 4080
aggctcagtc gaaagactgg gcctttcgtt ttatctgttg tttgtcggtg aacgctctcc 4140
tgagtaggac aaatccgccg ggagcggatt tgaacgttgc gaagcaacgg cccggagggt 4200
ggcgggcagg acgcccgcca taaactgcca ggcatcaaat taagcagaag gccatcctga 4260
cggatggcct ttttgcgttt c 4281
<210> 19
<211> 572
<212> Белок
<213> Bacillus licheniformis
<400> 19
Met Asn Asn Val Ala Ala Lys Asn Glu Thr Leu Thr Val Arg Gly Ala
1 5 10 15
Glu Leu Val Val Asp Ser Leu Ile Gln Gln Gly Val Thr His Val Phe
20 25 30
Gly Ile Pro Gly Ala Lys Ile Asp Ala Val Phe Asp Val Leu Lys Asp
35 40 45
Lys Gly Pro Glu Leu Ile Val Cys Arg His Glu Gln Asn Ala Ala Phe
50 55 60
Met Ala Ala Ala Val Gly Arg Leu Thr Gly Lys Pro Gly Val Cys Leu
65 70 75 80
Val Thr Ser Gly Pro Gly Ala Ser Asn Leu Ala Thr Gly Leu Val Thr
85 90 95
Ala Asn Thr Glu Gly Asp Pro Val Val Ala Leu Ala Gly Ala Val Lys
100 105 110
Arg Ala Asp Arg Leu Lys Lys Thr His Gln Ser Met Asp Asn Ala Ala
115 120 125
Leu Phe Gln Pro Ile Thr Lys Tyr Ser Ala Glu Val Glu Asp Ala Asn
130 135 140
Asn Ile Pro Glu Ala Val Thr Asn Ala Phe Arg Ala Ala Ala Ser Gly
145 150 155 160
Gln Ala Gly Ala Ala Phe Leu Ser Phe Pro Gln Asp Val Thr Ala Gly
165 170 175
Pro Ala Thr Ala Lys Pro Val Lys Thr Met Pro Ala Pro Lys Leu Gly
180 185 190
Ala Ala Ser Asp Glu Gln Ile Ser Ala Ala Ile Ala Lys Ile His Asn
195 200 205
Ala Asn Leu Pro Val Val Leu Val Gly Met Lys Gly Gly Arg Pro Glu
210 215 220
Ala Ile Glu Ala Val Arg Arg Leu Leu Arg Lys Val Lys Leu Pro Phe
225 230 235 240
Val Glu Thr Tyr Gln Ala Ala Gly Thr Leu Ser His Asp Leu Glu Asp
245 250 255
Gln Tyr Phe Gly Arg Ile Gly Leu Phe Arg Asn Gln Pro Gly Asp Met
260 265 270
Leu Leu Glu Lys Ala Asp Val Val Leu Thr Val Gly Tyr Asp Pro Ile
275 280 285
Glu Tyr Asp Pro Val Phe Trp Asn Gly Lys Gly Glu Arg Ser Val Ile
290 295 300
His Leu Asp Glu Ile Gln Ala Asp Ile Asp His Asp Tyr Gln Pro Glu
305 310 315 320
Ile Glu Leu Ile Gly Asp Ile Ala Glu Thr Leu Asn His Ile Glu His
325 330 335
Asp Ser Leu Pro Val Ser Ile Asp Glu Ser Phe Ala Pro Val Leu Asp
340 345 350
Tyr Leu Lys Lys Ala Leu Glu Glu Gln Ser Glu Pro Pro Lys Glu Thr
355 360 365
Lys Thr Asp Leu Val His Pro Leu Gln Ile Val Arg Asp Leu Arg Glu
370 375 380
Leu Leu Ser Asp Asp Ile Thr Val Thr Cys Asp Ile Gly Ser His Ala
385 390 395 400
Ile Trp Met Ser Arg Tyr Phe Arg Thr Tyr Arg Pro His Gly Leu Leu
405 410 415
Ile Ser Asn Gly Met Gln Thr Leu Gly Val Ala Leu Pro Trp Ala Ile
420 425 430
Ala Ala Thr Leu Val Asn Pro Gly Gln Lys Val Val Ser Val Ser Gly
435 440 445
Asp Gly Gly Phe Leu Phe Ser Ala Met Glu Leu Glu Thr Ala Val Arg
450 455 460
Leu Lys Ala Pro Ile Val His Ile Val Trp Asn Asp Ser Thr Tyr Asp
465 470 475 480
Met Val Ala Phe Gln Gln Glu Met Lys Tyr Lys Arg Thr Ser Gly Val
485 490 495
Asp Phe Gly Gly Ile Asp Ile Val Lys Tyr Ala Glu Ser Phe Gly Ala
500 505 510
Lys Gly Leu Arg Val Asn Ser Pro Asp Glu Leu Ala Glu Val Leu Lys
515 520 525
Ala Gly Leu Asp Ala Glu Gly Pro Val Val Ile Asp Ile Pro Val Asp
530 535 540
Tyr Ser Asp Asn Ile His Leu Ala Asp Gln Arg Phe Pro Lys Lys Phe
545 550 555 560
Glu Glu His Phe Asn Lys Glu Ala Ser Lys Gln Ser
565 570
<210> 20
<211> 1719
<212> ДНК
<213> Bacillus licheniformis
<400> 20
atgaataacg tcgcggccaa gaacgaaacc ctgaccgtcc ggggcgccga actcgtggtg 60
gatagcctga tccagcaggg cgtgacccat gtcttcggca tcccgggcgc caaaatcgac 120
gcggtcttcg acgtgctgaa ggataagggc cccgaactga tcgtctgccg ccatgagcag 180
aacgcggcct tcatggccgc cgccgtcggc cgcctgacgg gcaagccggg cgtctgcctg 240
gtcacctccg gcccgggcgc ctcgaatctc gcgaccggcc tggtcaccgc gaacacggaa 300
ggcgacccgg tggtcgccct ggcgggcgcc gtgaagcggg cggatcggct gaagaagacg 360
caccagtcga tggataacgc cgccctgttc cagcccatca cgaagtacag cgcggaggtg 420
gaagacgcga acaacatccc ggaggccgtg acgaacgcct tccgcgccgc ggcgtccggc 480
caggccggcg cggccttcct cagcttcccc caggatgtca ccgccggccc ggccaccgcc 540
aagccggtca aaaccatgcc cgccccgaag ctgggcgccg cgagcgatga acagatctcc 600
gccgcgatcg cgaagatcca caacgcgaat ctgccggtgg tcctcgtggg catgaagggc 660
ggccggccgg aagccatcga agccgtgcgc cgcctgctcc gcaaggtcaa gctcccgttc 720
gtggaaacct accaggcggc cggcacgctg tcgcacgatc tggaggatca gtacttcggc 780
cggatcggcc tgttccggaa ccagccgggc gacatgctcc tggaaaaggc cgacgtggtc 840
ctgaccgtgg gctacgaccc gatcgagtac gatccggtgt tctggaatgg caaaggcgaa 900
cgctcggtca tccacctcga cgaaatccag gccgatatcg atcacgacta ccagcccgag 960
atcgaactca tcggcgacat cgcggaaacc ctcaatcaca tcgagcatga ctcgctgccg 1020
gtgtccatcg acgaatcctt cgcgcccgtg ctcgactatc tcaagaaggc gctcgaagaa 1080
cagtcggagc ccccgaagga aacgaagacc gatctggtcc acccgctcca gatcgtgcgc 1140
gacctgcgcg agctgctctc cgatgacatc accgtcacct gcgacatcgg cagccacgcc 1200
atctggatgt cccgctattt ccgcacctat cgcccgcatg gcctcctgat ctccaacggc 1260
atgcagacgc tgggcgtcgc cctgccgtgg gcgatcgccg cgaccctggt gaacccgggc 1320
cagaaggtgg tgtcggtcag cggcgatggc ggcttcctct tctccgcgat ggaactcgaa 1380
accgccgtcc gcctcaaggc gccgatcgtg cacatcgtgt ggaacgactc cacgtacgac 1440
atggtcgcgt tccagcagga aatgaagtac aagcgcacct ccggcgtcga tttcggcggc 1500
atcgacatcg tcaagtatgc ggaatccttc ggcgccaaag gcctccgcgt gaatagcccc 1560
gatgaactgg ccgaggtcct gaaggccggc ctcgacgcgg agggcccggt ggtcatcgac 1620
atccccgtcg actactcgga taacatccac ctggccgacc agcgcttccc gaagaagttc 1680
gaggagcact tcaacaagga agcgtcgaag cagtcctga 1719
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРИМЕНЕНИЕ ГЛИКОЗИДАЗ В ПОЛУЧЕНИИ ОЛИГОСАХАРИДОВ | 2019 |
|
RU2810730C2 |
| ИНСЕКТИЦИДНЫЕ ПОЛИПЕПТИДЫ С ШИРОКИМ СПЕКТРОМ АКТИВНОСТИ И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2750459C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НЕВИРУСНОЙ ГЕННОЙ ТЕРАПИИ | 2019 |
|
RU2834625C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2019 |
|
RU2810220C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2819144C2 |
| АBC-ТРАНСПОРТЕРЫ ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2020 |
|
RU2795855C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| ПРИМЕНЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ, ЭКСПРЕССИРУЮЩИХ ФАКТОР VIII | 2019 |
|
RU2803163C2 |
| ФЕРМЕНТАТИВНЫЙ СИНТЕЗ N-АЦЕТИЛНЕЙРАМИНОВОЙ КИСЛОТЫ | 2018 |
|
RU2809787C2 |
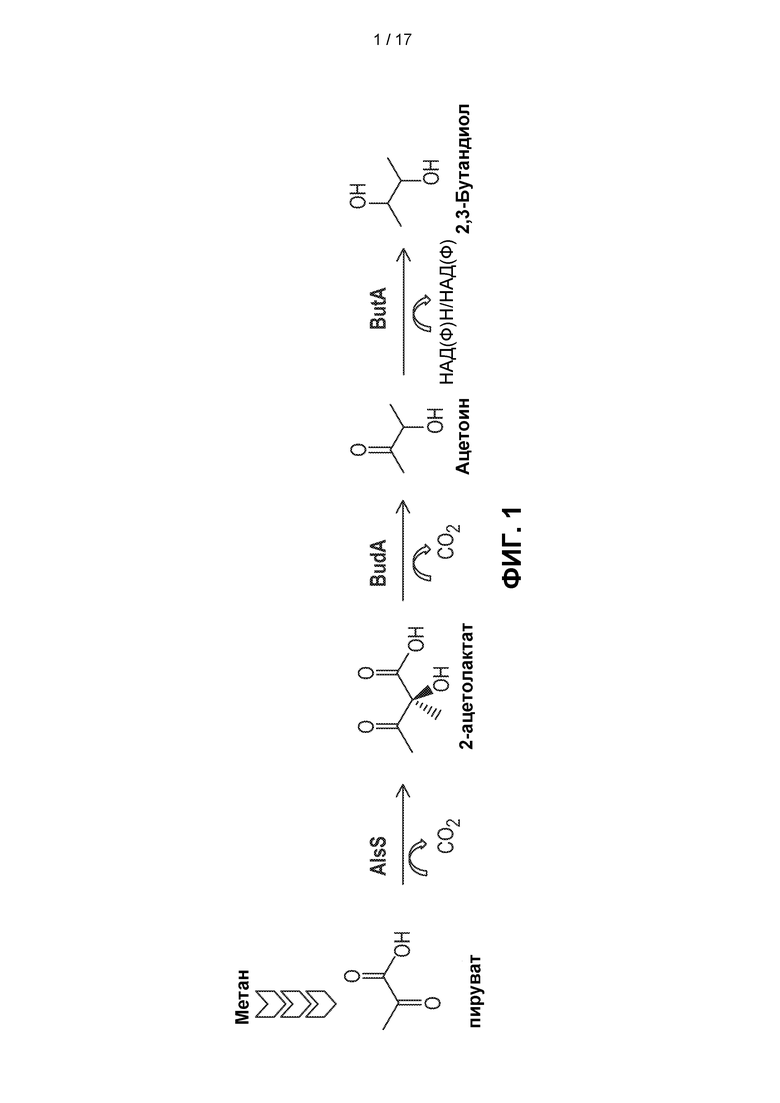





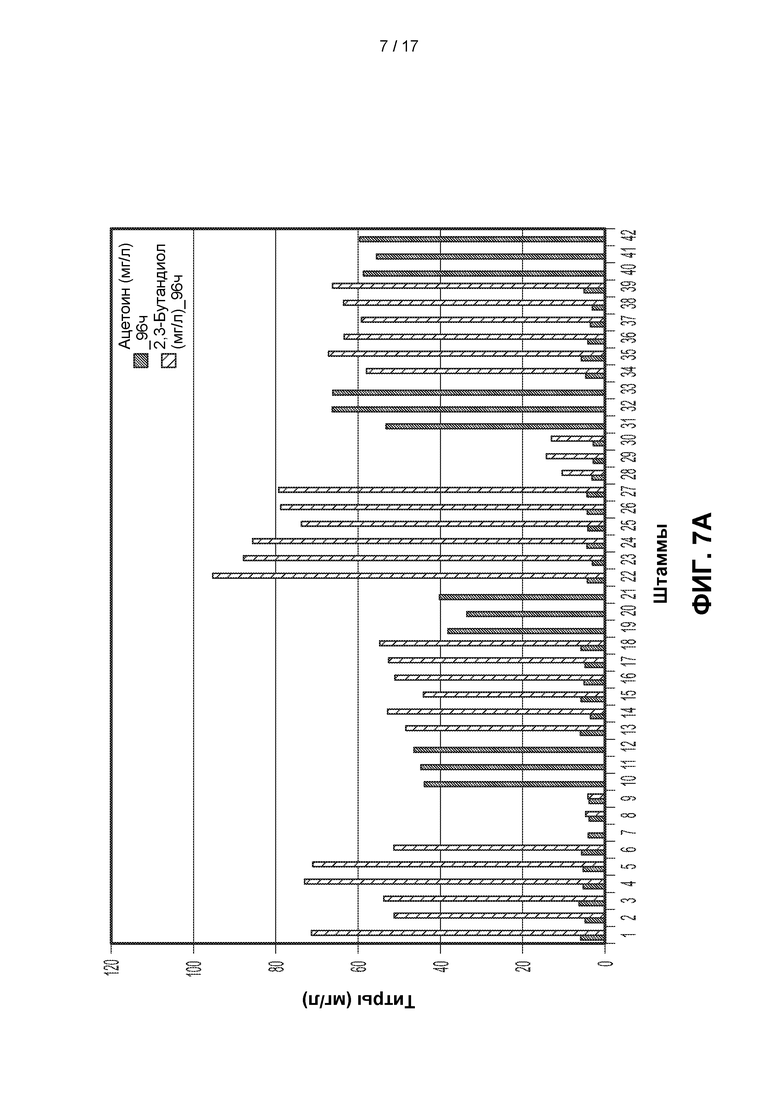
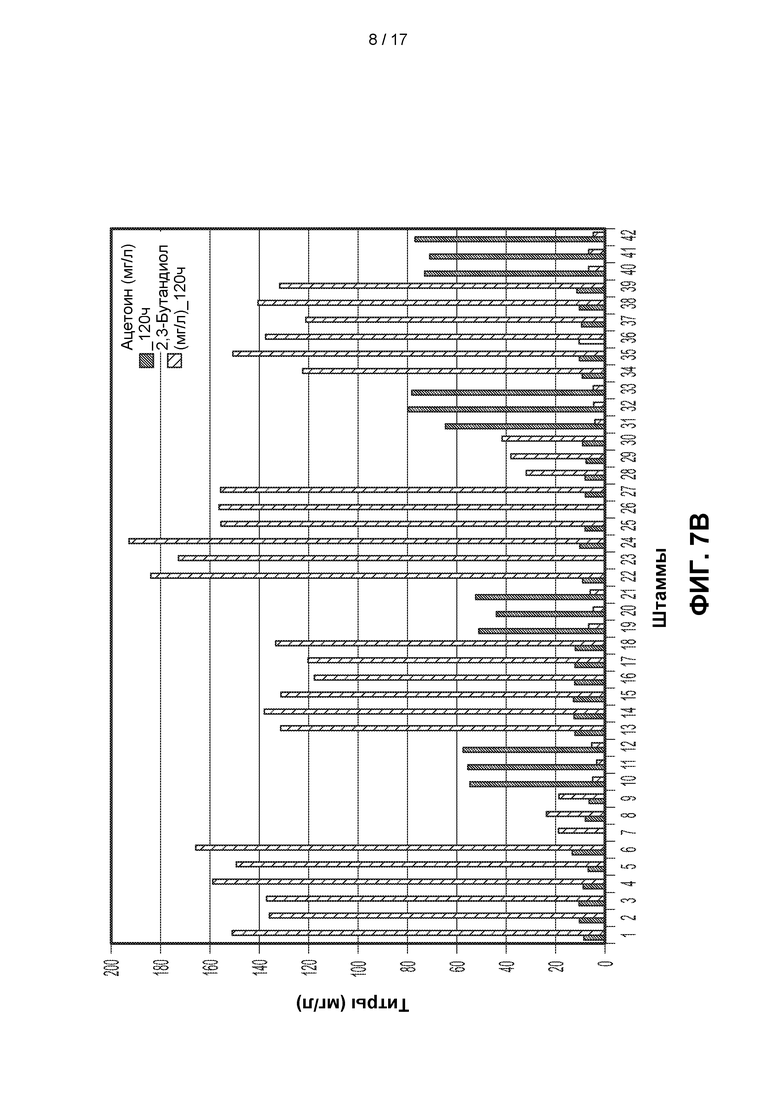
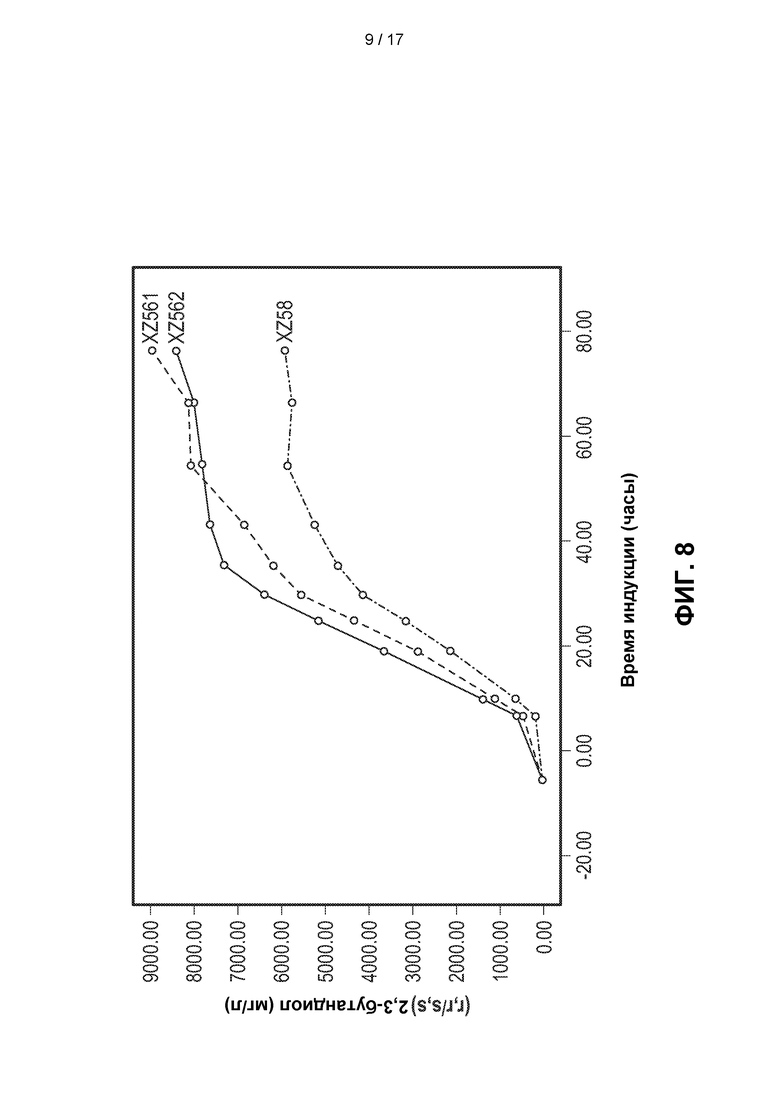
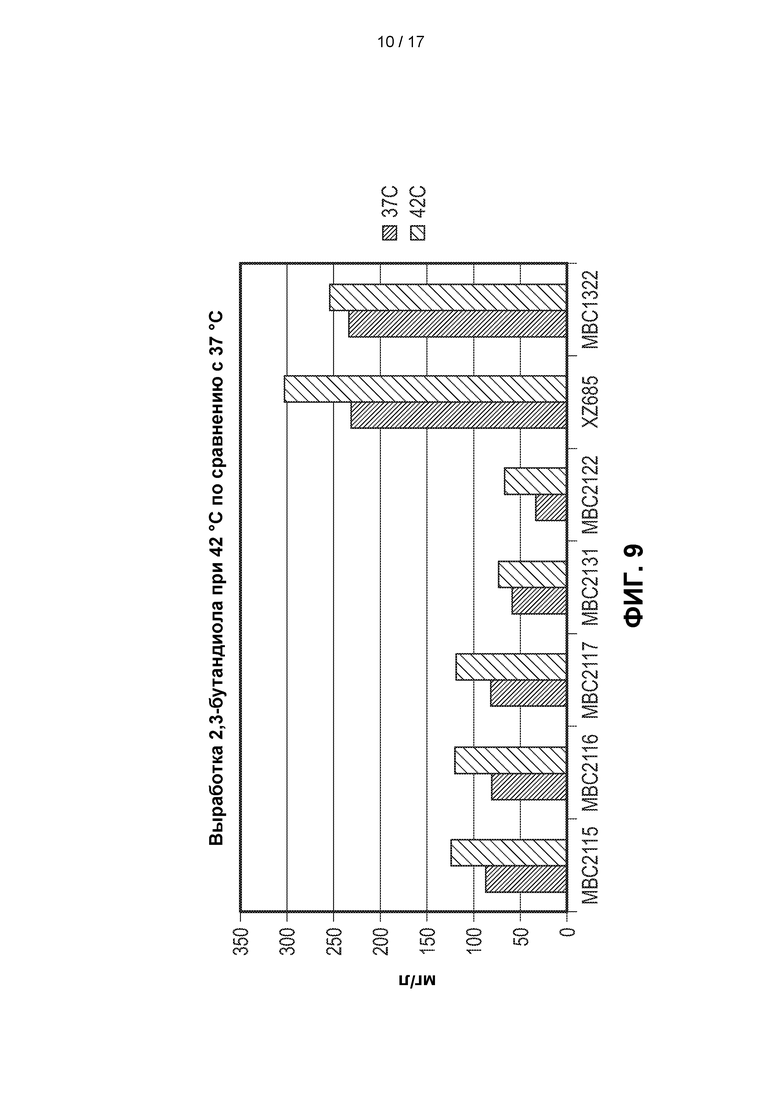
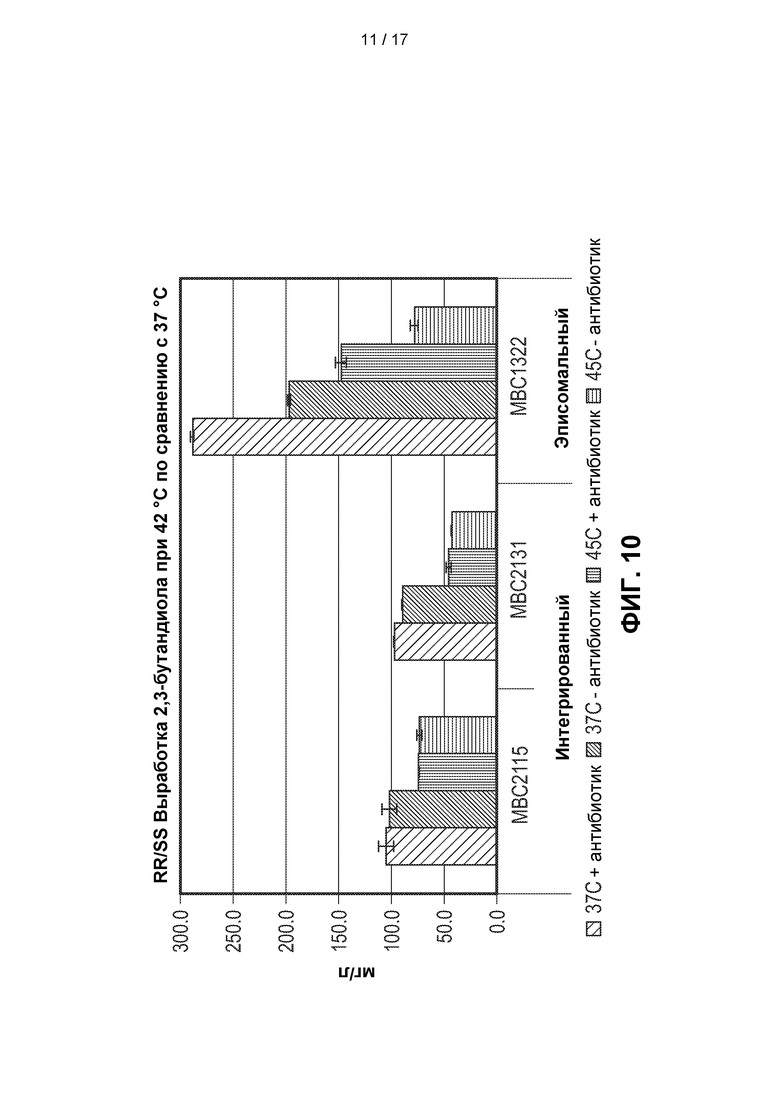
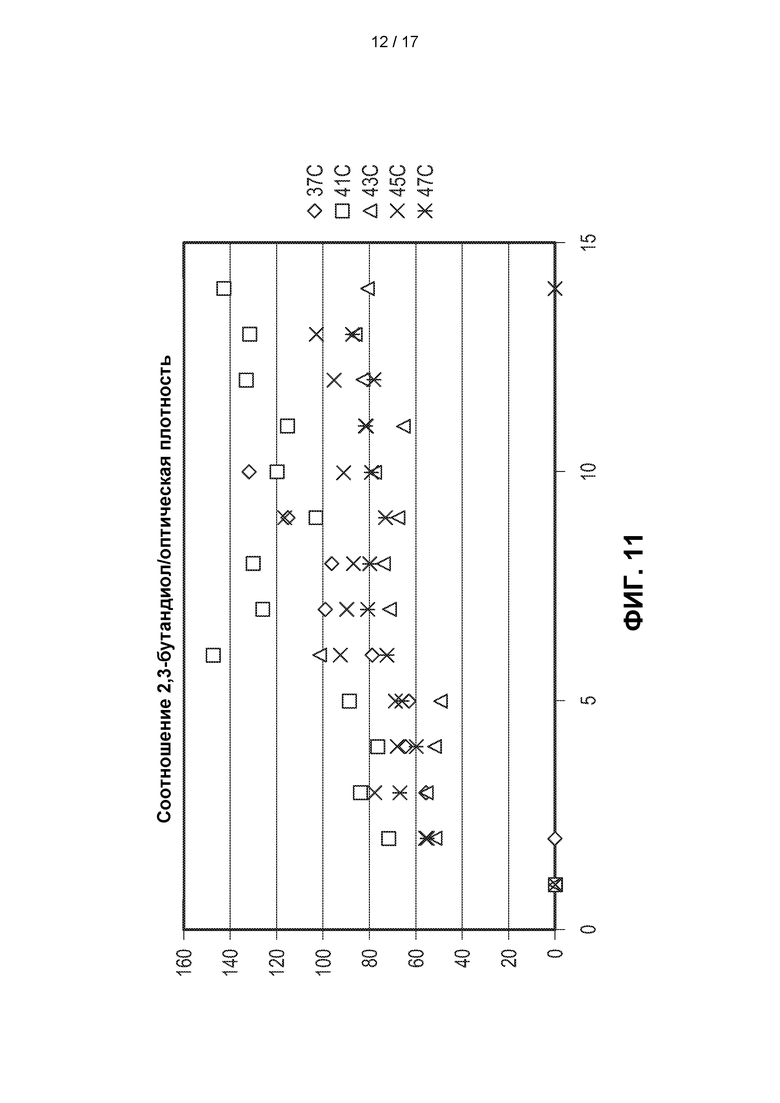
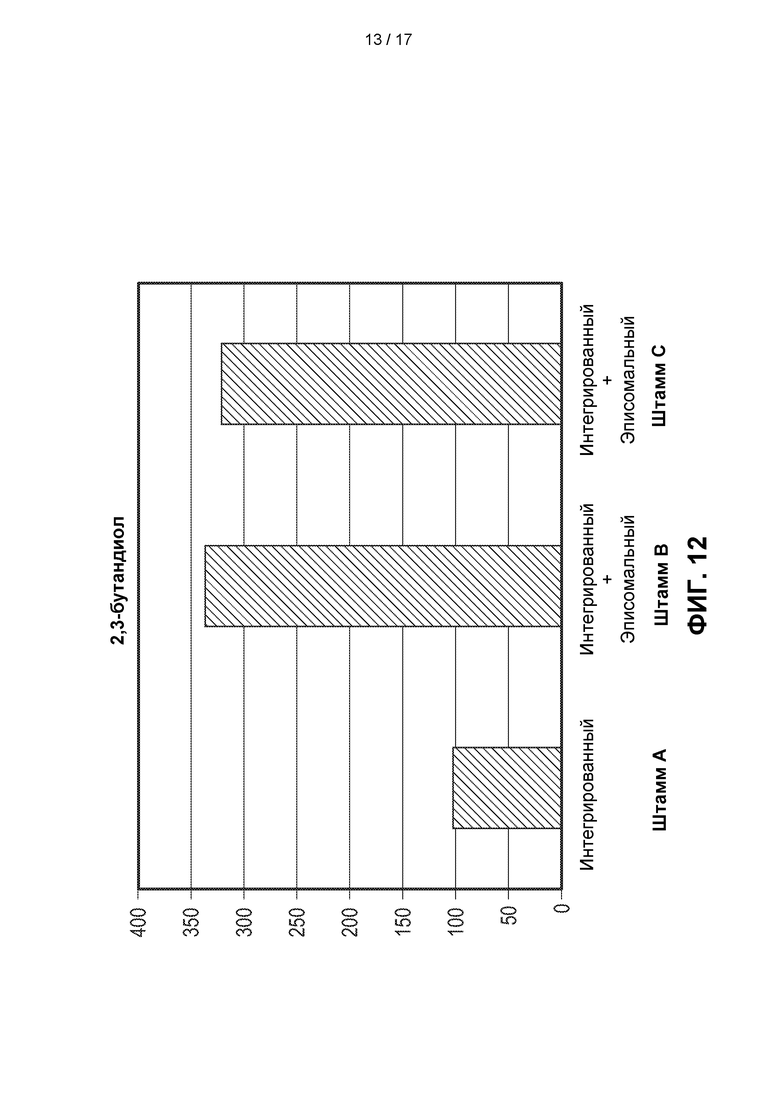
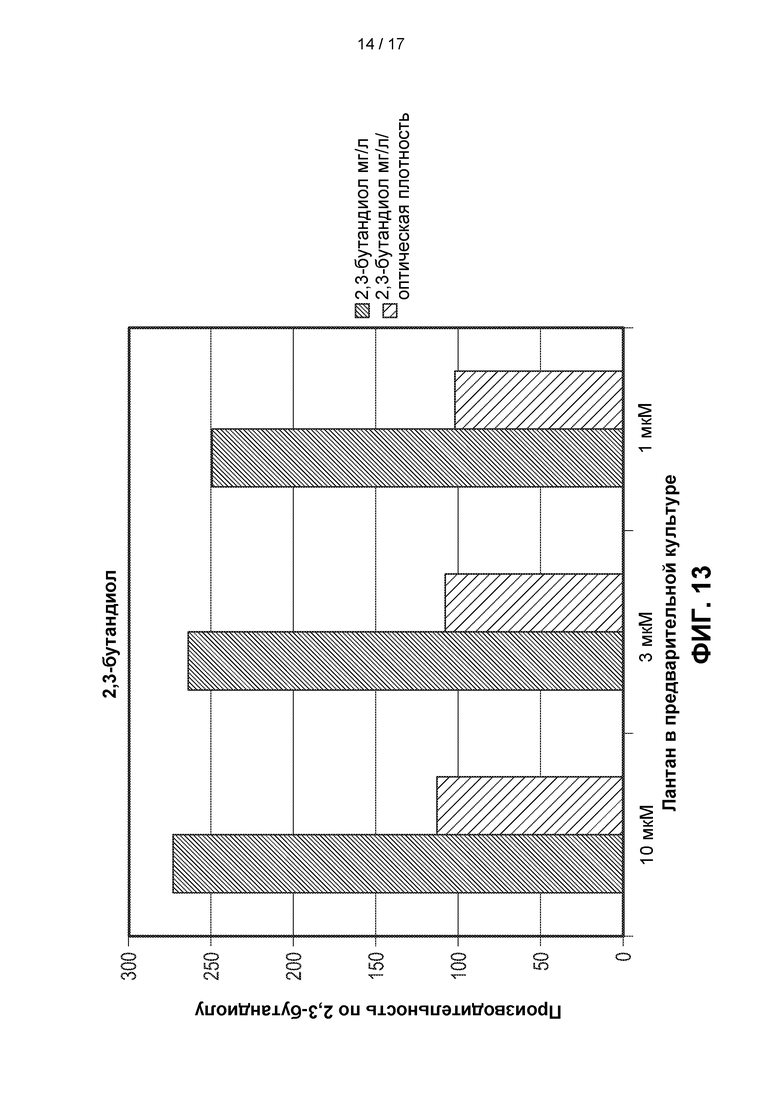
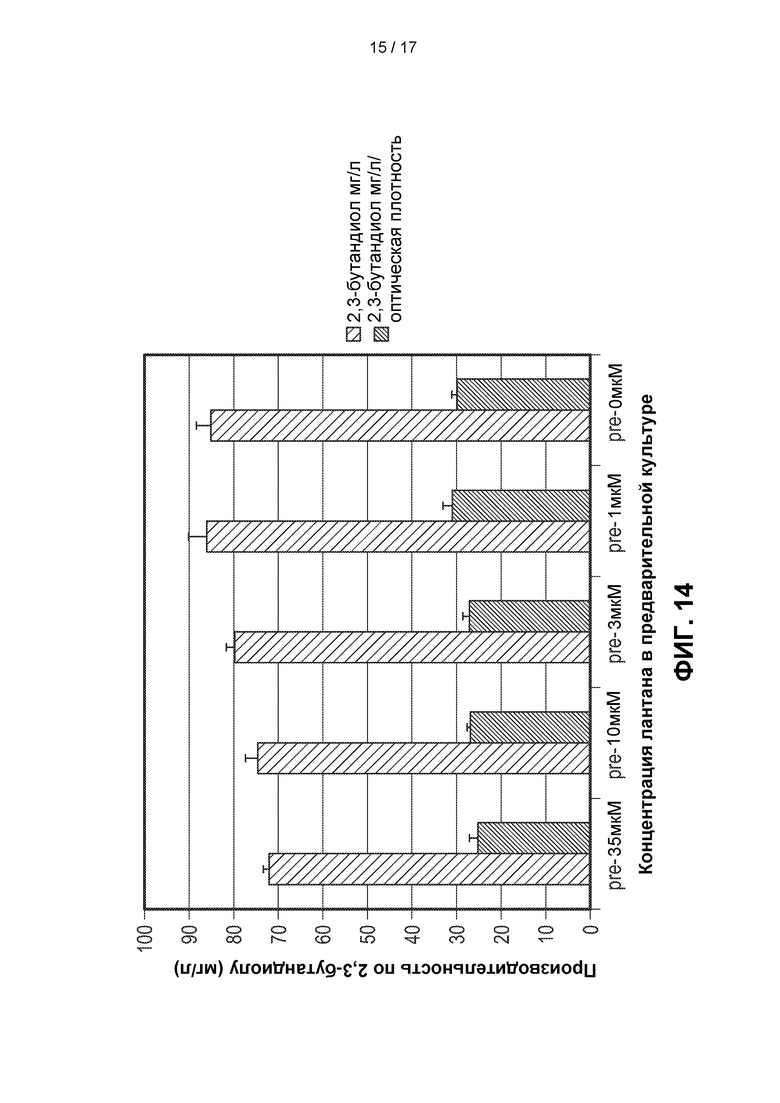
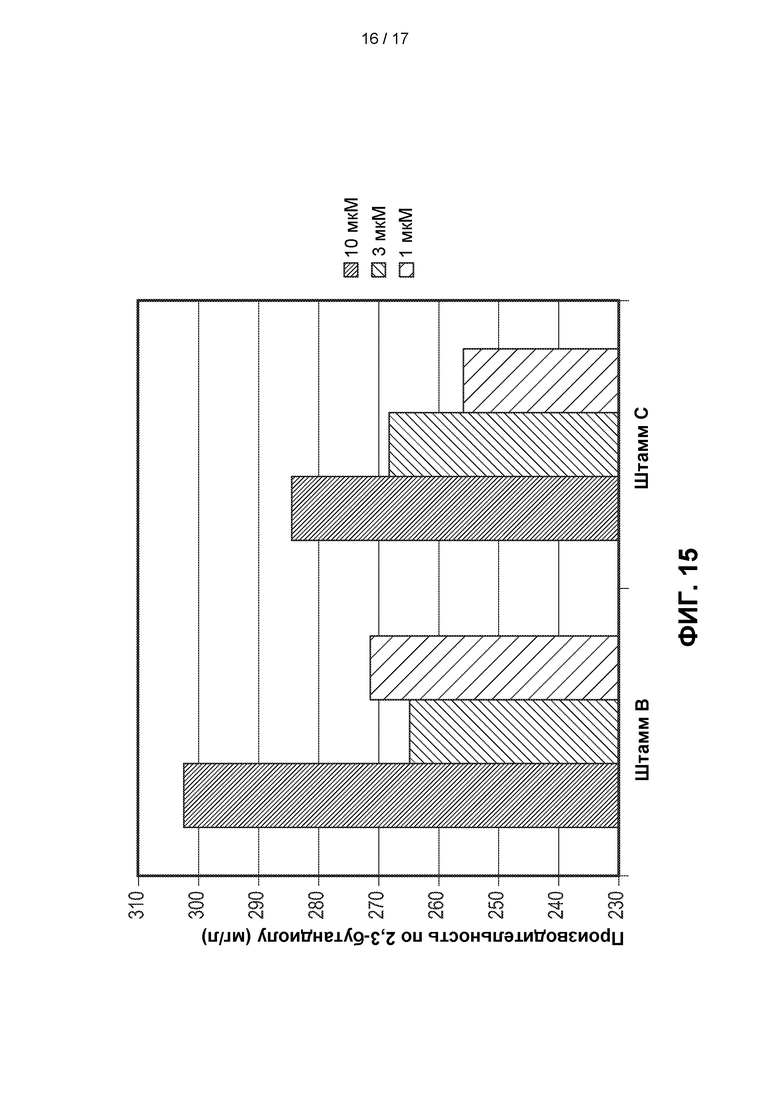
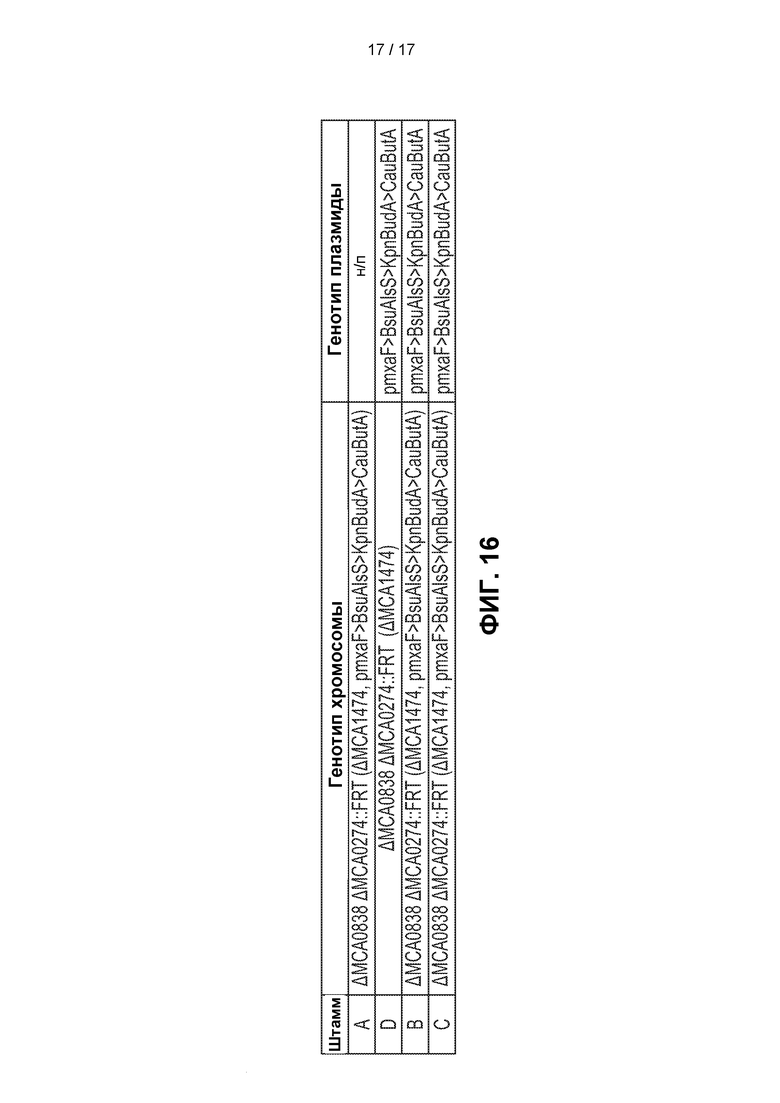
Изобретение относится к биотехнологии и представляет собой генетически модифицированный микроорганизм, способный превращать C1-углеродное соединение в 2,3-бутандиол, причем указанный микроорганизм содержит гетерологичный ген, кодирующий ацетоин редуктазу, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO:9. Изобретение позволяет эффективно получать 2,3-бутандиол и в дальнейшем бутадиен или метилэтилкетон с использованием указанного микроорганизма. 4 н. и 13 з.п. ф-лы, 16 ил., 6 табл., 7 пр.
1. Генетически модифицированный микроорганизм, способный превращать C1-углеродное соединение в 2,3-бутандиол, где указанный микроорганизм содержит гетерологичный ген, кодирующий ацетоин редуктазу, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 9.
2. Микроорганизм по п. 1, отличающийся тем, что указанный микроорганизм является метанотрофом, например, метанотрофом из рода Methylobacter, Methylomicrobium, Methylomonas, Methylocaldum, Methylococcus, Methylosoma, Methylosarcina, Methylothermus, Methylohalobius, Methylogaea, Methylovulum, Crenothrix, Clonothrix, Methylosphaera, Methylocapsa, Methylocella, Methylosinus, Methylocystis или Methyloacidophilum.
3. Микроорганизм по п. 1 или 2, отличающийся тем, что указанный микроорганизм принадлежит к виду Methylococcus capsulatus.
4. Микроорганизм по любому из пп. 1-3, где указанный микроорганизм продуцирует большее количество 2,3-бутандиола при 42°C по сравнению с тем же микроорганизмом при 37°C,
большее количество 2,3-бутандиола при 42°C по сравнению с тем же микроорганизмом при 45°C, или большее количество 2,3-бутандиола при 37°C по сравнению с тем же микроорганизмом при 45°C.
5. Микроорганизм по любому из пп. 1-4, в котором гетерологичный ген, кодирующий ацетоин редуктазу, интегрирован в геном указанного микроорганизма или экспрессируется в эписомальном векторе.
6. Микроорганизм по любому из пп. 1-5, отличающийся тем, что гетерологичный ген, кодирующий ацетоин редуктазу, расположен ближе к 3'-концу по отношению к любому другому гетерологичному гену, и при этом указанный микроорганизм дополнительно содержит гетерологичный ген, кодирующий ацетолактатсинтазу, который расположен ближе к 5'-концу по отношению к любому другому гетерологичному гену,
и/или
гетерологичный ген, кодирующий альфа-ацетолактатдекарбоксилазу, который расположен ни ближе к 5'-, ни ближе к 3'-концу по отношению к любому другому гетерологичному гену.
7. Микроорганизм по любому из пп. 1-6, отличающийся тем, что указанный гетерологичный ген, кодирующий ацетоин редуктазу, находится под контролем переключателя.
8. Микроорганизм по любому из пп. 1-7, отличающийся тем, что указанный гетерологичный ген, кодирующий ацетоин редуктазу, находится под контролем неконститутивного промотора, например:
(a) индуцируемого переключателя, такого как переключатель, чувствительный к арабинозе; или
(b) репрессируемого переключателя, такого как переключатель, чувствительный к редкоземельным металлам.
9. Микроорганизм по любому из пп. 1-8, отличающийся тем, что указанный гетерологичный ген, кодирующий ацетоин редуктазу, находится под контролем переключателя, чувствительного к лантану.
10. Микроорганизм по любому из пп. 1-9, отличающийся тем, что указанный микроорганизм экспрессирует:
(a) ацетоинредуктазу, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 9; и
(b) альфа-ацетолактатдекарбоксилазу, содержащую аминокислотную последовательность, которая по меньшей мере на 90% идентична SEQ ID NO: 7 и/или ацетолактатсинтазу, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 1 или 19.
11. Способ получения 2,3-бутандиола, включающий:
a) приведение микроорганизма по любому из пп. 1-10 в контакт с С1-углеродным соединением; и
b) выращивание указанного микроорганизма с получением 2,3-бутандиола.
12. Способ получения бутадиена или метилэтилкетона (MEK), включающий:
a) приведение микроорганизма по любому из пп. 1-10 в контакт с C1-углеродным соединением;
b) выращивание указанного микроорганизма с получением 2,3-бутандиола;
c) необязательно выделение 2,3-бутандиола; и
d) приведение указанного 2,3-бутандиола в контакт с катализатором с получением бутадиена или MEK.
13. Способ по п. 12, в котором указанный катализатор:
(a) способен дегидратировать 2,3-бутандиол, например, дигидрофосфат цезия (CsH2PO4), нанесенный на SiO2; или
(b) представляет собой катализатор на основе твердой кислоты.
14. Способ по любому из пп. 11-13, отличающийся тем, что экспрессия гетерологичного гена происходит под контролем переключателя, чувствительного к лантану, причем указанный способ дополнительно включает:
(a) приведение микроорганизма в контакт со средой, содержащей лантан, например, по меньшей мере 1 мкМ лантана; и
(b) необязательно, разбавление указанного лантана до выращивания микроорганизма с получением 2,3-бутандиола.
15. Способ по любому из пп. 11-14, отличающийся тем, что указанное C1-углеродное соединение представляет собой монооксид углерода, диоксид углерода и/или метан.
16. Способ по любому из пп. 11-15, в котором указанный микроорганизм выращивают при температуре от 32°C до 49°C.
17. Способ получения генетически модифицированного микроорганизма по любому из пп. 1-10, включающий трансформацию микроорганизма при помощи гетерологичной нуклеиновой кислоты, содержащей ген, кодирующий ацетоин редуктазу, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO: 9.
| WO 2014092562 A1, 19.06.2014 | |||
| US 20070292927 A1, 20.12.2007 | |||
| Вертикальная печь для сушки и сухой перегонки угля и др. сыпучих материалов | 1926 |
|
SU18720A1 |

