Настоящее изобретение относится к получению олигосахаридов посредством микробной ферментации. Более конкретно, настоящее изобретение относится к применению гликозидаз для улучшения получения целевых олигосахаридов посредством микробной ферментации.
Предшествующий уровень техники
Человеческое грудное молоко содержит уникальную смесь разных олигосахаридов, называемых Олигосахаридами Грудного Молока (ОГМ). На сегодняшний день в человеческом грудном молоке идентифицировано более чем 150 структурно разных олигосахаридов. За очень редким исключением, ОГМ характеризуются лактозной группировкой на их восстанавливающем конце, и многие ОГМ содержат остаток фукозы и/или остаток N-ацетилнейраминовой кислоты на их невосстанавливающем конце. Обычно, остатки моносахаридов ОГМ происходят из D-глюкозы, D-галактозы, N-ацетилглюкозамина, L-фукозы и N-ацетилнейраминовой кислоты. Важность ОГМ для питания грудных детей прямо связано с их уникальными биологическими активностями, включая защиту новорожденного от патогенов, поддержку развития иммунной системы и познавательных способностей грудного ребенка. Таким образом, существует большой интерес к получению ОГМ в коммерческом масштабе.
Помимо химического синтеза отдельных ОГМ значительный прогресс был сделан в разработке получения ОГМ посредством микробной ферментации с использованием генетически модифицированных микроорганизмов, которые сверхэкспрессируют гетерологичную гликозилтрансферазу. При культивации таких микроорганизмов в среде и в условиях, являющихся пермиссивными в отношении экспрессии микроорганизмами указанной гетерологичной гликозилтрансферазы, ОГМ может быть продуцирован указанным микроорганизмом и выделен из культуральной среды или клеточного лизата.
Однако гликозилтрансферазы часто обладают ферментативными побочными активностями, таким образом, что их сверхэкспрессия для получения целевого олигосахарида обычно приводит к побочным продуктам, которые являются нежелательными. Обычно, данные побочные продукты также представляют собой олигосахариды, но которые должны быть удалены из препарата целевого олигосахарида для коммерческого применения продукта. Однако, удаление таких побочных продуктов из целевого олигосахарида является сложным и трудоемким. Один подход к удалению таких побочных продуктов включает применение гликозидаз, которых либо экзогенно добавляют к реакционной смеси/клеточной среде, содержащей желательные и нежелательные олигосахариды, либо продуцирует генетически сконструированный микроорганизм при индукции в конкретный момент времени в конце процесса ферментации для получения целевого олигосахарида.
Международная публикация WO 2015/032412 А1 относится к применению фукозы и в ней раскрыт способ, в котором генетически модифицированную клетку, экспрессирующую гетерологичную фукозилтрансферазу, культивируют в присутствии лактозы для продуцирования и секреции смеси 2'-фукозиллактозы (2'-FL- от англ. 2'-fucosyllactose) и дифукозиллактозы (DFL - от англ. difucosyllactose) во внеклеточное пространство культуральной среды с высоким выходом. Сахариды разделяют и подвергают гидролизу посредством кислоты или посредством фукозидазы с получением фукозы с высоким выходом.
В международной публикации №WO 2104/090261 А1 раскрыт способ образования смеси, содержащей по меньшей мере одну из 2'-FL и 3-фукозиллактозы β-FL), где DFL подвергается частичному гидролизу, например, ферментативному гидролизу или кислотному гидролизу. В ферментативном гидролизе DFL подвергают действию фукозидазы, которая может высвобождать один из остатков фукозы из DFL. DFL (10 мМ) инкубировали с 1,2-α-1_-фукозидазой из Xanthomonas manihotis при 37°С в инкубационном буфере, и гидролиз DFL отслеживали посредством ВЭЖХ (высокоэффективная жидкостная хроматография). Спустя 18 часов, DFL подвергали частичному гидролизу до 3-FL и фукозы. Лактозы не выявляли.
Европейская патентная заявка №ЕР 2845905 А1 относится к получению олигосахаридов и в ней раскрыто применение одного или более гликозидаз в способе получения и/или очистки олигосахарида. Способ включает а) культивирование микроорганизма-хозяина, подходящего для продукции целевого олигосахарида, в условиях и в среде, являющихся пермиссивными для продукции указанного целевого олигосахарида, в результате чего продуцируются данный олигосахарид и, где применимо, промежуточные соединения биосинтеза - сахариды и/или побочные продукты; b) применение гликозидазы в среде, в которой культивируется микроорганизм-хозяин, для деградации промежуточных соединений биосинтеза - сахаридов и/или побочных продуктов - сахаридов и/или неиспользованных субстратов сахаридов; и с) выделение целевого олигосахарида. В одном воплощении указанная гликозидаза эндогенно продуцируется в микроорганизме-хозяине, где гликозидаза представляет собой гликозидазу, которая не встречается в природе в микроорганизме-хозяине, и где экспрессия указанной гликозидазы в указанном микроорганизме-хозяине является индуцибельной, таким образом, что данная экспрессия может быть инициирована после того, как достаточное и/или по существу максимальное количество целевого олигосахарида было продуцировано во время культивации микроорганизмα-хозяина.
В итоге, в предшествующем уровне техники раскрыто применение гликозидаз для удаления нецелевых олигосахаридов из смеси целевых и нецелевых олигосахаридов посредством гидролиза нецелевых олигосахаридов в реакционной смеси/клеточной среде. Однако, данные подходы включают биосинтез нецелевых олигосахаридов микроорганизмом, включая применение субстратов и энергии, и данные подходы требуют удаления продуктов деградации нецелевых олигосахаридов из целевых олигосахаридов.
Таким образом, цель настоящего изобретения заключалась в предложении способа получения целевого олигосахарида посредством микробной ферментации без сопутствующей продукции/аккумуляции нецелевых побочных продуктов-сахаридов, а именно нецелевых олигосахаридов, в клеточной среде, содержащей микроорганизм, подлежащий ферментации.
Цель достигается посредством предложения генетически сконструированной микробной клетки-хозяина, способной продуцировать целевой олигосахарид, где указанная микробная клетка-хозяин экспрессирует гетерологичную гликозидазу, которая способна внутриклеточно деградировать побочные продукты метаболизма, которые образуются во время внутриклеточного биосинтеза целевого олигосахарида, таким образом, предотвращая образование смеси целевых и нецелевых сахаридов в культуральной среде. Указанные продукты деградации затем могут быть использованы в метаболизме микробной клетки-хозяина, например, для биосинтеза целевого олигосахарида.
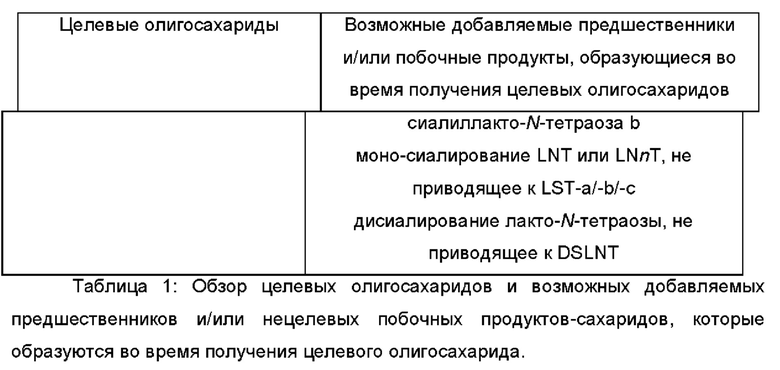
В Таблице 1 предоставлен подробный обзор целевых олигосахаридов и возможных предшественников, которые добавляются, и/или нецелевых побочных продуктов - сахаридов, которые образуются во время получения целевого олигосахарида.







Краткое изложение сущности изобретения
В первом аспекте раскрыт способ получения целевого олигосахарида с использованием генетически сконструированной микробной клетки-хозяина, которая способна продуцировать целевой олигосахарид, причем указанная микробная клетка-хозяин экспрессирует гетерологичную гликозидазу, способную осуществлять внутриклеточную деградацию побочных продуктов метаболизма - сахаридов, которые образуются в процессе внутриклеточного биосинтеза целевого олигосахарида.
Во втором аспекте раскрыта генетически сконструированная микробная клетка-хозяин для получения целевого олигосахарида, где указанная микробная клетка-хозяин способна продуцировать целевой олигосахарид, и где указанная микробная клетка-хозяин была генетически сконструирована с возможностью экспрессии гетерологичной гликозидазы, которая может осуществлять внутриклеточную деградацию побочных продуктов метаболизма - сахаридов, которые образуются в процессе внутриклеточного биосинтеза целевого олигосахарида.
В третьем аспекте раскрыто применение генетически сконструированной микробной клетки-хозяина согласно второму аспекту для получения целевого олигосахарида.
В четвертом аспекте раскрыты олигосахариды, а именно желательные олигосахариды, которые получают способом согласно первому аспекту и/или посредством использования генетически сконструированной микробной клетки-хозяина согласно второму аспекту.
В пятом аспекте раскрыто применение целевых олигосахаридов согласно четвертому аспекту для получения питательной композиции.
В шестом аспекте раскрыты питательные композиции, содержащие целевой олигосахарид согласно четвертому аспекту.
Краткое описание графических материалов
На Фиг. 1 показано схематичное представление воплощения микробной клетки-хозяина, экспрессирующей гетерологичную гликозидазу (например, альфа-1,3-фукозидазу), которая может осуществлять деградацию побочных продуктов метаболизма - сахаридов (например, 3-фукозиллактозы и 2'3-дифукозиллактозы), которые образуются во время внутриклеточного биосинтеза целевого олигосахарида (2'-фукозиллактоза), и где микробная клетка-хозяин может повторно использовать продукты деградации (например, фукозу и лактозу), являющиеся результатом ферментативной активности указанной гликозидазы, для продукции целевого олигосахарида.
Подробное описание
Согласно первому аспекту предложен способ получения целевого олигосахарида с использованием генетически сконструированной микробной клетки-хозяина, который включает следующие стадии:
(i) предоставление генетически сконструированной микробной клетки-хозяина, которая способна продуцировать целевой олигосахарид, где микробная клетка-хозяин генетически сконструирована с возможностью экспрессии гетерологичной гликозидазы, которая может осуществлять внутриклеточную деградацию побочных продуктов метаболизма - сахаридов, которые образуются в процессе внутриклеточного биосинтеза целевого олигосахарида, и где микробная клетка-хозяин способна повторно использовать продукты деградации, являющиеся результатом ферментативной активности указанной гликозидазы;
(ii) культивирование генетически сконструированной микробной клетки-хозяина в условиях и в среде, являющихся пермиссивными в отношении продукции целевого олигосахарида, с получением, таким образом, целевого олигосахарида; и
(iii) возможно выделение целевого олигосахарида.
Термин «целевой», в том виде, в котором он используется в данном документе в отношении олигосахаридов, относится к олигосахариду, который предназначен для того, чтобы быть продуцированным микробной клеткой-хозяином. Термин «целевой» используется для проведения различия между олигосахаридом, подлежащим целевой продукции, и другими олигосахаридами, которые может продуцировать микробная клетка-хозяин. Указанные другие олигосахариды считаются «нежелательными», независимо от того, обладают ли или нет данные другие олигосахариды биологической функцией, участвуют ли или нет в биосинтезе других соединений клетки, таких как гликолипиды, гликопротеины или полисахариды, или являются ли или нет продуктами метаболизма - сахаридами, которые образованы во время внутриклеточного биосинтеза целевого олигосахарида, или за счет запасных (нецелевых) ферментативных активностей одного или более ферментов, участвующих в биосинтезе целевого олигосахарида, или за счет ферментативной активности одного или более ферментов, которые не прямо участвуют в биосинтезе целевого олигосахарида, а используют олигосахарид в качестве субстрата, который образуется в виде промежуточного соединения в метаболическом пути, приводя к получению целевого олигосахарида.
Термин «олигосахарид», в том виде, в котором он используется в данном документе, относится к молекуле сахарида, состоящей из трех - двадцати остатков моносахаридов, где каждый из указанных остатков моносахаридов связан по меньшей мере с еще одним из указанных моносахаридных звеньев гликозидной связью. Олигосахарид может представлять собой линейную цепь моносахаридных остатков или разветвленную цепь моносахаридных остатков.
В дополнительном и/или альтернативном воплощении целевой олигосахарид представляет собой олигосахарид грудного молока (НМО).
В дополнительном и/или альтернативном воплощении целевой олигосахарид представляет собой ОГМ, выбранный из группы, состоящей из 2'-фукозиллактозы (2'-FL), 3-фукозиллактозы (3-FL), 2'3-дифукозиллактозы (DFL), лакто-N-триозы II, лакто-N-тетраозы (LNT), лакто-N-неотетраозы (LNnT), лакто-N-фукопентаозы I (LNFP-I), лакто-N-неофукопентаозы I (LNnFP-I), лакто-N-фукопентаозы II (LNFP-II), лакто-N-фукопентаозы III (LNFP-III), лакто-N-фукопентаозы V (LNFP-V), лакто-N-неофукопентаозы V (LNnFP-V), лакто-N-дифукогексаозы I, лакто-N-дифукозилгексаозы II, пара-Лакто-N-фукозилгексаозы, фукозил-лакто-N-сиалилпентаозы b, фукозил-лакто-N-сиалилпентаозы с, фукозил-лакто-N-сиалилпентаозы с, дисиалил-лакто-N-фукопентаозы, 3-фукозил-3'-сиалиллактозы, 3-фукозил-6'-сиалиллактозы, лакто-N-неодифукогексаозы I, 3'-сиалиллакозы β-SL), 6'-сиалиллактозы (6-SL), сиалиллакто-N-тетраозы a (LST-a), сиалиллакто-N-тетраозы b (LST-b), сиалиллакто-N-тетраозы с (LST-c) и дисиалиллакто-N-тетраозы.
Способ включает предоставление генетически сконструированной микробной клетки-хозяина, которая способна продуцировать целевой олигосахарид.
Термин «генетически сконструированный», в том виде, в котором он используется в данном документе, относится к модификации генетического состава клетки с использованием методов молекулярной биологии. Модификация генетического состава клетки может включать перенос генов в пределах и/или через видовые связи, вставку, удаление, замену и/или модификацию нуклеотидов, триплетов, генов, открытых рамок считывания, промоторов, энхансеров, терминаторов и других нуклеотидных последовательностей, опосредуя и/или контролируя экспрессию генов. Модификация генетического состава клетки нацелена на создание генетически модифицированного организма, обладающего конкретными, желательными свойствами. Генетически сконструированная микробная клетка-хозяин может содержать один или более генов, которые отсутствуют в нативной (не генетически сконструированной) форме клетки. Методики введения экзогенных молекул нуклеиновой кислоты и/или осуществления вставки экзогенных молекул нуклеиновой кислоты (рекомбинантных, гетерологичных) в наследуемую информацию клетки для вставки, удаления или изменения нуклеотидной последовательности генетической информации клетки известны квалифицированному специалисту. Генетически сконструированные клетки могут содержать один или более генов, которые находятся в нативной форме клетки, где указанные гены модифицированы и повторно вставлены в клетку искусственными средствами. Термин «генетически сконструированный» также охватывает клетки, которые содержат молекулу нуклеиновой кислоты, являющуюся эндогенной в отношении клетки, и которая модифицирована без удаления молекулы нуклеиновой кислоты из клетки. Такие модификации включают модификации, полученные в результате замены генов, сайт-специфичных мутаций, и родственные методики, включающие методики, обычно называемые «редактированием генома».
Генетически сконструированная микробная клетка-хозяин может представлять собой прокариотическую клетку или эукариотическую клетку. Подходящие микробные клетки-хозяева включают дрожжевые клетки, бактериальные клетки, клетки архебактерий и клетки грибов.
В дополнительном и/или альтернативном воплощении прокариотическая клетка представляет собой бактериальную клетку, предпочтительно бактериальную клетку, выбранную из бактерий рода, выбранного из группы, состоящей из Bacillus, Bifidobacterium, Clostridium, Corynebacterium, Enterococcus, Lactobacillus, Lactococcus, Micrococcus, Micromonospora, Pseudomonas, Rhodococcus и Sporolactobacillus. Подходящие виды бактерий представляют собой Bacillus subtilis, В, licheniformis, В. coagulans, В, thermophiius, В. laterosporus, В, megaterium, В. mycoides, В. pumilus, В. lentus, В. cereus, В. circulans, Bifidobacterium longum, B. infantis, B, bifidum, Citrobacter freundii, Clostridium cellulolyticum, C. Ijungdahiii, C, autoethanogenum, C. acetobutylicum, Corynebacterium glutamicum, Enterococcus faecium, E. thermophiles, Escherichia coli, Erwinia herbicoia (Pantoea agglomerans), Lactobacillus acidophilus, L. salivarius, L, plantarum, L, helveticus, L. deibrueckii, L, rhamnosus, L. buigaricus, L. crispatus, L. gasseri, L, casei, L. reuteri, L. jensenii, L. lactis, Pantoea citrea, Pectobacterium carotovorum, Proprionibacterium freudenreichii, Pseudomonas fluorescens, P. aeruginosa, Streptococcus thermophiles и Xanthomonas campestris.
В дополнительном и/или альтернативном воплощении эукариотическая клетка представляет собой дрожжевую клетку, предпочтительно, дрожжевую клетку, выбранную из группы, состоящей из Saccharomyces sp., в частности Saccharomyces cerevisiae, Saccharomycopsis sp., Pichia sp., в частности, Pichia pastoris, Hansenula sp., Kluyveromyces sp., Yarrowia sp., Rhodotorula sp, и Schizosaccharomyces sp.
Генетически сконструированная микробная клетка-хозяин может продуцировать целевой олигосахарид. Термин «может продуцировать» в том виде, в котором он используется в данном документе, относится к способности генетически сконструированной микробной клетки-хозяина продуцировать целевой олигосахарид, при условии, что микробную клетку-хозяина культивируют в условиях и в среде, которые являются пермиссивными в отношении продукции микробной клеткой-хозяином целевого олигосахарида. Таким образом, среда должна иметь значение рН в определенном интервале, состав ионов и питательных элементов, а также содержать соединения, требуемые для сохранения жизнеспособности и метаболической активности микробной клетки-хозяина. При необходимости для получения целевого олигосахарида, среда также должна содержать достаточные количества какого-либо предшественника, требуемого для биосинтеза целевого олигосахарида микробной клеткой-хозяином. Аналогично, условия (например, температура, рН, подача кислорода, перемешивание, снабжение питательными веществами и т.д.) для культивирования микробной клетки-хозяина для продукции целевого олигосахарида должны поддерживаться таким образом, чтобы микробная клетка-хозяин могла быть или оставаться метаболически активной для продуцирования целевого олигосахарида.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин, способная продуцировать целевой олигосахарид, представляет собой микробную клетку-хозяина, которая генетически сконструирована для того, чтобы иметь способность продуцировать целевой олигосахарид. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии гетерологичной гликозилтрансферазы. Гетерологичная гликозидаза экспрессируется в генетически сконструированной микробной клетке-хозяине во время ферментации, а именно во время продукции или биосинтеза целевого олигосахарида. В дополнительном и/или альтернативном воплощении экспрессия гетерологичной гликозидазы является конститутивной в генетически сконструированном хозяине - микробе.
Термин «гетерологичный», в том виде, в котором он используется в данном документе, относится к нуклеотидной последовательности, молекуле нуклеиновой кислоты или полипептиду, которые являются чужеродными в отношении клетки или организма, а именно, к нуклеотидной последовательности, молекуле нуклеиновой кислоты или полипептиду, которые в природе не встречаются в указанной клетке или организме. Термин «гетерологичная последовательность» или «гетерологичная нуклеиновая кислота» или «гетерологичный полипептид», в том виде, в котором он используется в данном документе, представляет собой последовательность или нуклеиновую кислоту или пептид, которые происходят из источника, являющегося чужеродным для конкретной клетки-хозяина (например, из другого вида), или, если из того же источника, модифицированы по сравнению со своей исходной формой. Таким образом, гетерологичная нуклеиновая кислота, функционально связанная с промотором, происходит из источника, отличного от источника, из которого происходит промотор, или, если из того же источника, является модифицированной в сравнении с ее исходной формой. Гетерологичная последовательность может быть стабильно введена, например, посредством трансфекции, трансформации, конъюгации или трансдукции, в геном микробной клетки-хозяина, таким образом, представляя генетически модифицированную клетку-хозяина. Можно применять методики, которые будут зависеть клетки-хозяина, последовательности, которая подлежит введению. Разные методики известны специалисту в данной области, и например, раскрыты в Sambrook ef al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989). Соответственно, «гетерологичный полипептид» представляет собой полипептид, который в природе не встречается в клетке дикого типа, от которой происходит генетически сконструированная клетка, и «гетерологичная гликозилтрансфераза» представляет собой гликозилтрансферазу, которая не встречается в природе в клетке дикого типа, от которой происходит генетически сконструированная клетка.
В дополнительном и/или альтернативном воплощении гетерологичная гликозилтрансфераза выбрана из группы, состоящей из фукозилтрансфераз, предпочтительно α-1,2-фукозилтрансфераз и α-1,3-фукозилтрансфераз, гликозилтрансфераз, галактозилтрансфераз, предпочтительно β-1,3-галактозилтрансфераз и β-1,4-галактозилтрансфераз, сиалилтрансфераз, предпочтительно α-2,3-сиалилтрансфераз и α-2,6-сиалилтрансфераз, и N-ацетил-глкжозаминилтрансфераз.
Фукозилтрансферазы катализируют перенос остатков фукозы от донора - гуанозиндифосфат-активированной L-фукозы (ГДФ-фукоза) к нескольким акцепторным молекулам. Фукозилтрансферазы экспрессируются в животных, растениях, грибах и бактериях, и они подразделяются на категории в соответствии со связью с фукозой на акцепторном субстрате. Таким образом, α-1,2-, α-1,3/4- и α-1,6-фукозилтрансферазы отличаются друг от друга. Подходящие фукозилтрансферазы для гетерологичной экспрессии в генетически сконструированной микробной клетке-хозяине раскрыты, например, в Европейской патентной заявке №17180176.
Сиалилтрансферазы катализируют перенос остатков N-ацетилнейраминовой кислоты (Neu5Ac) от донора CMP-Neu5Ac к акцепторным молекулам. Обнаружили, что сиалилтрансферазы экспрессируются в животных, растениях, грибах и бактериях. Сиалилтрансферазы подразделяются на категории в соответствии со связью, которая образуется между NeuSAc и акцепторной молекулой. Следовательно, α-2,3-, α-2,6- и α-2,8-сиалилтрансферазы отличаются друг от друга. Подходящие сиалилтрансферазы для гетерологичной экспрессии в генетически сконструированной микробной клетке-хозяине раскрыты, например, в Европейской патентной заявке №17183391.
Галактозилтрансферазы катализируют перенос остатка галактозы от донора, УДФ (уридиндифосфат)-галактозы, к акцепторным субстратам. Галактозилтрансферазы различают на основе связи между галактозой и акцепторной молекулой, которая образуется. Следовательно, β-1,3- и β-1,4-галактозилтрансферазы отличаются друг от друга. Подходящая β-1,3-галактозилтрансфераза для гетерологичной экспрессии в генетически сконструированной микробной клетке-хозяине кодируется геном wbdO Salmonella enterica. Подходящая β-1,4-галактозилтрансфераза для гетерологичной экспрессии в генетически сконструированной микробной клетке-хозяине кодируется геном lex1 Aggregatibacter aphrophilus.
Генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии гетерологичной гликозидазы, которая может осуществлять внутриклеточную деградацию побочных продуктов метаболизма - сахаридов, которые образуются во время внутриклеточного биосинтеза целевого олигосахарида. Подходящие гликозидазы представляют собой гликозидазы, которые являются специфичными в отношении гликозидной связи, которая гидролизуется под действием ферментативной активности, и/или в отношении субстрата, который гидролизуется гликозидазой. За счет указанной специфичности гликозидаза гидролизует нежелательные побочные продукты, а не целевой олигосахарид, подлежащий продуцированию. В дополнительном и/или альтернативном воплощении гликозидаза не гидролизует один или более предшественников, которые поглощаются или синтезируются микробной клеткой-хозяином для продуцирования целевого олигосахарида. Предпочтительно, гликозидаза представляет собой экзогликозидазу.
Экзогликозидазы представляют собой ферменты гликозид гидролазы, которые разрушают гликозидные связи на концевом остатке структуры олигосахарида.
В дополнительном и/или альтернативном воплощении гетерологичная гликозидаза выбрана из группы, состоящей из фукозидаз, включая α-1,2-фукозидазы и α-1,3-фукозидазы, сиалидаз, таких как α-2,3-сиалидазы, α-2,6-сиалидазы, α-2,8-сиалидазы, галактозидаз, таких как β-1,3-галактозидазы, β-1,4-галактозидазы и β-1,6-галактозидазы, β-N-ацетилгексозаминидаз и глюкозидаз, таких как β-1,3-глюкозидазы.
Подходящая фукозидаза представляет собой α-1,2-фукозидазу. α-1,2-фукозидаза представляет собой высокоспецифичную экзогликозидазу, которая катализирует гидролиз линейных альфа-1,2-связанных L-фукопиранозильных остатков от олигосахаридов. Предпочтительная α-1,2-фукозидаза представляет собой AfcA Bifidobacterium bifidum (SEQ ID NO: 2).
В дополнительном и/или альтернативном воплощении предложена генетически сконструированная микробная клетка-хозяин, которая может продуцировать 3-FL, где указанная генетически сконструированная микробная клетка-хозяин экспрессирует α-1,2-фукозидазу. Для того, чтобы мочь продуцировать 3-FL, генетически сконструированная микробная клетка-хозяин экспрессирует альфа-1,3-фукозилтрансферазу. Указанная альфа-1,3-фукозилтрансфераза может переносить остаток фукозы от ГДФ-фукозы на группировку глюкозы лактозы в качестве акцепторного субстрата, синтезируя, таким образом, 3-FL в качестве целевого олигосахарида. 2'-FL и 2'3-DFL представляют собой нежелательные побочные продукты - сахариды в получении 3-FL.
Посредством экспрессии гетерологичной α-1,2-фукозидазы в генетически сконструированной микробной клетке-хозяине, которая может продуцировать 3-FL, продукция побочных продуктов 2'-FL и 2'3-DFL может быть прекращена или по меньшей мере уменьшена, поскольку данные побочные продукты гидролизуются в пределах генетически-сконструированной микробной клетки-хозяина под действием гетерологичной α-1,2-фукозидазы. Полученные продукты деградации представляют собой фукозу и лактозу. Как фукоза, так и лактоза могут быть использованы генетически сконструированной микробной клеткой-хозяином для продукции целевой 3-FL.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии α-1,2-фукозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую α-1,2-фукозидазу для ее экспресии. Предпочтительно, нуклеотидная последовательность, кодирующая α-1,2-фукозидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, ка представлено SEQ ID NO: 1;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 1, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 1;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 2; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 2, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 2.
Термин «гибридизуется» или «гибридизация», в том виде, в котором он используется в данном документе, означает гибридизацию в общепринятых условиях, как описано в Sambrook et al. (1989) «Molecular Cloning, A Laboratory Manual» (Cold Spring Harbor Laboratory Press, New York), предпочтительно в жестких условиях. Жесткие условия гибридизации, например, выглядят следующим образом: гибридизация в 4 × SSC при 65°С и последующая множественная промывка в 0,1 × SSC при 65°С на протяжении в общей сложности примерно 1 часа. Менее жесткие условия гибридизации выглядят, например, следующим образом: гибридизация в 4 × SSC при 37°С и последующая множественная промывка в 1 × SSC при комнатной температуре (примерно 21°С). «Условия жесткой гибридизации» могут также означать: гибридизацию при 68°С в 0,25 М фосфате натрия, рН 7,2, 7% SDS (от англ. sodium dodecyl sulphate - додецилсульфат натрия), 1 мМ ЭДТА (Этилендиаминтетрауксусная кислота) и 1% БСА (Бычий сывороточный альбумин) в течение 16 часов и с последующими двумя промывками 2 × SSC и 0,1% SDS при 68°С.
Для экспрессии нуклеотидной последовательности, кодирующей α-1,2-фукозидазу или ее функциональный вариант, указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей α-1,2-фукозидазу или ее функциональный вариант в генетически сконструированной микробной клетке-хозяине.
«Последовательности контроля экспрессии» представляют собой регуляторные нуклеотидные последовательности, которые не являются частью нуклеотидной последовательности, кодирующей белок, а опосредуют экспрессию нуклеотидной последовательности, кодирующей белок. Нуклеотидные последовательности регуляторных элементов включают промоторы, цис-регуляторные элементы, энхансеры, интроны и терминаторы. В зависимости от типа регуляторного элемента, он находится на молекуле нуклеиновой кислоты перед нуклеотидной последовательностью, кодирующей белок (а именно 3'), или позади нуклеотидной последовательности, кодирующей белок (а именно 5'). Регуляторные элементы являются функциональными в микробной клетке-хозяине.
Термин «функционально связанный» означает, что регуляторный элемент соединен таким путем с нуклеотидной последовательностью, кодирующей белок, а именно расположен таким образом относительно нуклеотидной последовательности, кодирующей белок, например, молекулы нуклеиновой кислоты, что экспрессия нуклеотидной последовательности, кодирующей белок, под контролем данного регуляторного элемента может происходить в живой клетке.
В целях настоящего изобретения «промотор» представляет собой нуклеотидную последовательность, регулирующую экспрессию гена, которая обычно находится на 5'-конце гена и посредством взаимодействия с конкретными ДНК-связывающими белками опосредует инициацию транскрипции посредством РНК-полимеразы.
Кроме того, подходящие промоторы включают синтетические промоторы. Существуют промоторы, которые были созданы посредством методик молекулярной биологии, которые не обнаруживаются в природе в данной конфигурации. Синтетический промотор представляет собой минималистичный промотор, содержащий только один или более выбранных определенных цис-элементов помимо минимального промотора. Данные цис-элементы представляют собой сайты связывания для ДНК-связывающих белков, таких как транскрипционные факторы, и выделены из природных промоторов, происходящих из ранее выделенных цис-элементов, или технически получены посредством методик случайной рекомбинации и выбраны соответствующими способами; по сравнению с природным промотором, за счет своей менее сложной конструкции синтетический промотор активируется только несколькими экзогенными и эндогенными факторами и, таким образом, более специфично регулируется.
«Минимальный промотор» или «коровый промотор» представляет собой нуклеотидную последовательность, которая содержит сайты связывания для комплекса базальных факторов транскрипции и делает возможной точную инициацию транскрипции посредством РНК-полимеразы II. Характеристические мотивы последовательности минимального промотора представляют собой ТАТА-бокс, инициаторный элемент (Inr), «элемент распознавания TFBII» (BRE) и «расположенный ниже элемент корового промотора» (ОРЕ). В минимальном промоторе данные элементы могут встречаться отдельно или в комбинации. Минимальный промотор или его мотивы последовательности доступны, например, из гена бактерии, гриба или вируса.
«Цис-элементы» представляют собой нуклеотидные последовательности, которые расположены на той же молекуле нуклеиновой кислоты, что и нуклеотидная последовательность, кодирующая белок, подлежащая экспрессии. Цис-элементы не должны кодировать РНК или белок и в направлении транскрипции могут быть расположены перед или после нуклеотидной последовательности, кодирующей белок, подлежащей экспрессии. Расположенные выше цис-элементы перед нуклеотидной последовательностью, кодирующей белок, подлежащей экспрессии, часто предоставляют необходимые мотивы связывания, в частности для транскрипционных факторов, которые участвуют как транс-действующие элементы (от лат.trans - за пределами), на молекулярном уровне, с другой стороны в регуляции транскрипции данного гена. Если, кроме того, цис-элементы приводят к ингибированию транскрипции, они называются сайленсерами. Цис-элементы, которые приводят к усилению транскрипции, называются энхансерами. Совокупность цис/транс-активностей в промоторе определяет интенсивность, с которой РНК-полимераза осуществляет транскрипцию.
Кроме того, промотор может представлять собой химерный промотор и/или промотор, который был модифицирован цис-элементами. Модификация промотора также может означать дополнительное включение цис-элемента в промотор, который, например, уже в природе имеет цис-элемент. Кроме того, модификация также включает мультимеризацию цис-элемента, в частности мультимеризацию существующего в природе цис-элемента. По сравнению с нативной версией, такой модифицированный промотор может, например, обладать измененными свойствами в отношении специфичности, уровня экспрессии или фоновой активности.
Терминаторы представляют собой нуклеотидные последовательности на ДНК, которые обычно отмечают конец гена и приводят к терминации транскрипции.
Другая подходящая фукозидаза представляет собой α-1,3-фукозидазу. α-1,3-фукозидаза представляет собой высоко специфичную гликозидазу, которая катализирует гидролиз α-1,3-связанных L-фукопиранозильных остатков от олигосахаридов. Предпочтительная α-1,3-фукозидаза представляет собой AfcB из Bifidobacterium bifidum (SEQ ID NO: 4).
В дополнительном и/или альтернативном воплощении предложена генетически сконструированная микробная клетка-хозяин, которая способна продуцировать 2'-FL, где указанный генетически сконструированный микроорганизм - хозяин экспрессирует α-1,3-фукозидазу. Для того, чтобы мочь продуцировать 2'-FL, генетически сконструированная микробная клетка-хозяин экспрессирует α-1,2-фукозилтрансферазу. Указанная альфа-1,2-фукозилтрансфераза может переносить остаток фукозы от ГДФ-фукозы к группировке галактозы лактозы в качестве акцепторного субстрата, синтезируя, таким образом, 2'-FL в качестве целевого олигосахарида. 3-FL и 2'3-DFL представляют собой нежелательные побочные продукты - сахариды в продукции 2-FL.
Посредством экспрессии гетерологичной α-1,3-фукозидазы в генетически сконструированной микробной клетке-хозяине, которая может продуцировать 2'-FL, продукция побочных продуктов 3-FL и 2'3-DFL может быть прекращена или по меньшей мере уменьшена, по той причине, что данные побочные продукты гидролизуются в генетически сконструированной микробной клетке-хозяине посредством гетерологичной α-1,3-фукозидазы. Полученные продукты деградации представляют собой фукозу и лактозу. Как фукоза, так и лактоза могут быть использованы генетически сконструированным микроорганизмом-хозяином для продукции целевой 2'-FL.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии α-1,3-фукозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую α-1,3-фукозидазу, для ее экспрессии. Предпочтительно, нуклеотидная последовательность, кодирующая α-1,3-фукозидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, как представлено SEQ ID NO: 3;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 3, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 3;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 4; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 4, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 4.
Для экспрессии нуклеотидной последовательности, кодирующей α-1,3-фукозидазу или ее функциональный вариант, указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей α-1,3-фукозидазу или ее функциональный вариант в генетически-сконструированной микробной клетке-хозяине.
В дополнительном и/или альтернативном воплощении предложена генетически сконструированная микробная клетка-хозяин, которая может продуцировать LNFP-I, где указанная генетически сконструированная микробная клетка-хозяин экспрессирует α-1,3-фукозидазу. Для того, чтобы мочь продуцировать LNFP-I, генетически сконструированная микробная клетка экспрессирует β-1,3-N-ацетилглюкозаминилтрансферазу, β-1,3-галактозилтрансферазу и α-1,2-фукозилтрансферазу. Указанная β-1,3-N-ацетилглюкозаминилтрансфераза может переносить остаток GlcNAc от УДФ-GlcNAc к галактозной группировке лактозы, синтезируя, таким образом, лакто-N-триозу-11 (LNT-II). Указанная β-1,3-галактозилтрансфераза может переносить остаток галактозы от УДФ-галактозы к группировке GlcNAc LNT-II, синтезируя, таким образом, лакто-N-тетраозу (LNT). Указанная α-1,2-фукозилтрансфераза может переносить остаток фукозы от ГДФ-фукозы к концевой группировке галактозы LNT, синтезируя, таким образом, LNFP-I. 3-FL и 2'3-DFL будут нежелательными побочными продуктами в получении LNFP-I. Посредством экспрессии α-1,3-фукозидазы в генетически сконструированной микробной клетке-хозяине, способной продуцировать LNFP-I, продукция побочных продуктов 3-FL и 2'3-DFL может быть прекращена или по меньшей мере уменьшена, по той причине, что данные побочные продукты гидролизуются под действием гетерологичной α-1,3-фукозидазы в пределах генетически-сконструированной микробной клетки-хозяина. Полученные продукты деградации представляют собой фукозу, лактозу и 2'-FL. Фукоза и лактоза могут использоваться генетически сконструированным микроорганизмом-хозяином для продукции целевой LNFP-I.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии α-1,3-фукозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую α-1,3-фукозидазу, для ее экспрессии. Предпочтительно, нуклеотидная последовательность, кодирующая α-1,3-фукозидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, как представлено SEQ ID NO: 3;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 3, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 3;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 4; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 4, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 4.
Для экспрессии нуклеотидной последовательности, кодирующей α-1,3-фукозидазу или ее функциональный вариант указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей α-1,3-фукозидазу или ее функциональный вариант в генетически сконструированной микробной клетке-хозяине.
Подходящая сиалидаза представляет собой α-2,3-сиалидазу. α-2,3-Сиалидаза представляет собой высоко специфичную экзогликозидазу, которая катализирует гидролиз линейных α-2,3-связанных L-сиалильных остатков от олигосахаридов. Предпочтительная α-2,3-сиалидаза представляет собой NanB Streptococcus pneumoniae (SEQ ID NO: 6).
В дополнительном и/или альтернативном воплощении предложена генетически сконструированная микробная клетка-хозяин, которая способна продуцировать 6'-SL, где указанная генетически сконструированная микробная клетка-хозяин экспрессирует α-2,3 сиалидазу. Для того чтобы мочь продуцировать 6'-SL, генетически сконструированная микробная клетка-хозяин экстрессирует α-2,6-сиалилтрансферазу. Указанная 2,6-сиалилтрансфераза может переносить остаток NeuSAc от CMP-Neu5Ac к галактозной группировке лактозы в качестве субстрата, синтезируя, таким образом, 6'-SL. 3'-SL представляет собой нецелевой побочный продукт в продукции 6'-SL.
Посредством экспрессии α-2,3-сиалидазы в генетически сконструированной микробной клетке-хозяине, которая может продуцировать 6'SL, продукция побочного продукта 3'-SL может быть прекращена или по меньшей мере уменьшена по той причине, что данный побочный продукт гидролизуется под действием α-2,3-сиалидазы в генетически модифицированной микробной клетке-хозяине. Полученные продукты деградации представляют собой N-ацетилнейарминовую кислоту и лактозу. Как N-ацетилнейраминовая кислота, так и лактоза могут быть использованы генетически сконструированным микроорганизмом-хозяином для продукции целевой 6'-SL.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии α-2,3-сиалидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую α-2,3-сиалидазу для ее экспрессии. Предпочтительно, нуклеотидная последовательность, кодирующая α-2,3-сиалидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, как представлено SEQ ID NO: 5;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 5, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 5;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 6; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 6, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 6.
Для экспрессии нуклеотидной последовательности, кодирующей α-2,3-сиалидазу или ее функциональный вариант, указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей α-2,3-сиалидазу или ее функциональный вариант, в генетически сконструированной микробной клетке-хозяине.
Подходящая галактозидаза представляет собой β-1,3-галактозидазу. β-1,3-Галактозидаза представляет собой фермент, который катализирует гидролиз остатка β-1,3-связанной галактозы от олигосахаридов. Предпочтительная β-1,3-галактозидаза представляет собой Bga42A Bifidobacterium longum (SEQ ID NO: 8).
В дополнительном и/или альтернативном воплощении предложена генетически сконструированная микробная клетка-хозяин, которая может продуцировать LNnT, где указанная генетически сконструированная микробная клетка-хозяин экспрессирует β-1,3-галактозидазу. Для того, чтобы иметь способность продуцировать LNnT, генетически сконструированная микробная клетка-хозяин экспрессирует β-1,3-N-ацетилглюкозаминилтрансферазу и β-1,4-галактозилтрансферазу. Указанная β-1,3-N-ацетилглюкозаминилтрансфераза может переносить остаток GlcNAc от УДФ-GlcNAc к группировке галактозы лактозы, синтезируя, таким образом, LNT-II. Указанная β-1,4-галактозилтрансфераза может переносить остаток галактозы от УДФ-галактозы на группировку GlcNAc LNT-II, синтезируя, таким образом, LNnT в качестве целевого олигосахарида.
LNT представляет собой нецелевой побочный продукт в продукции LNnT. Посредством экспрессии β-1,3-галактозидазы в генетически сконструированной микробной клетке-хозяине, способной продуцировать LNnT, продукция побочного продукта LNT может быть прекращена или по меньшей мере уменьшена по той причине, что данный побочный продукт гидролизуется в генетически сконструированной микробной клетке-хозяине под действием гетерологичной β-1,3-галактозидазы. Полученные продукты деградации представляют собой галактозу и LNT-II. Галактоза, а также LNT-II могут быть использованы генетически сконструированным микроорганизмом-хозяином для продукции целевой LNnT.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии β-1,3-галактозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую β-1,3-галактозидазу для ее экспрессии.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии β-1,3-галактозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструированная для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую β-1,3-галактозидазу для ее экспрессии. Предпочтительно, нуклеотидная последовательность, кодирующая β-1,3-галактозидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, как представлено SEQ ID NO: 7;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 7, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 7;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 8; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 8, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 8.
Для экспрессии нуклеотидной последовательности, кодирующей β-1,3-галактозидазу или ее функциональный вариант, указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей β-1,3-глюкозидазу или ее функциональный вариант в генетически сконструированной микробной клетке-хозяине.
Другая подходящая галактозидаза представляет собой галактан β-1,3-галактозидазу. Галактан β-1,3-галактозидаза представляет собой фермент, который катализирует гидролиз остатка β-1,3-связанной галактозы от олигосахаридных цепей, несущих галактозу. Предпочтительная галактан β-1,3-галактозидаза представляет собой Ct1,3Gal43A Clostridium thermocellum (SEQ ID NO: 10).
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии галактан β-1,3-галактозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую галактан β-1,3-галактозидазу для ее экспрессии. Предпочтительно, нуклеотидная последовательность, кодирующая галактан β-1,3-галактозидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, как представлено SEQ ID NO: 9;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 9, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 9;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 10; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 10, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 10.
Для экспрессии нуклеотидной последовательности, кодирующей галактан β-1,3-галактозидазу или ее функциональный вариант, указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей галактан β-1,3-глюкозидазу или ее функциональный вариант, в генетически сконструированной микробной клетке-хозяине.
Подходящая глюкозидаза представляет собой β-1,3-глюкозидазу. β-1,3-Глюкозидаза представляет собой высокоспецифичную экзогликозидазу, которая катализирует гидролиз остатка β-1,3-связанной глюкозы от олигосахаридов. Предпочтительная β-1,3-глюкозидаза представляет собой PgIA Paenibacillus sp.(SEQ ID NO: 12).
В дополнительном и/или альтернативном воплощении предложен генетически сконструированный микроорганизм-хозяин, который может продуцировать LNT или LNnT, где указанная генетически сконструированная микробная клетка-хозяин экспрессирует β1,3-глюкозидазу и/или β-1,3-галактозидазу. Для того, чтобы мочь продуцировать LNT, генетически сконструированная микробная клетка-хозяин экспрессирует β-1,3-N-ацетилглюкозаминилтрансферазу и β-1,3-галактозилтрансферазу. Указанная β-1,3-N-ацетилглюкозаминилтрансфераза может переносить остаток GlcNAc от УДФ-GlcNAc к группировке галактозы лактозы, синтезируя, таким образом, лакто-N-триозу-II (LNT-II). Указанная β-1,3-галактозилтрансфераза может переносить остаток галактозы от УДФ-галактозы к группировке GlcNAc LNT-II, синтезируя, таким образом, лакто-N-тетраозу (LNT). Для того, чтобы мочь продуцировать LNnT, генетически сконструированная микробная клетка-хозяин экспрессирует β-1,3-N-ацетилглюкозаминилтрансферазу и β-1,4-галактозилтрансферазу. Указанная β-1,3-N-ацетилглюкозаминилтрансфераза может синтезировать LNT-II. Указанная β-1,4-галактозилтрансфераза может переносить остаток галактозы от УДФ-галактозы к группировке GlcNAc LNT-II, синтезируя, таким образом, LNnT в качестве целевого олигосахарида.
Квалифицированному специалисту в данной области известно, что β-1,3-N-ацетилглюкозаминилтрансферазы, подобно LgtA Neisseria meningitides, акцептируют широкий спектр субстратов-доноров. Как правило, при переносе GlcNAc от УДФ-GlcNAc к соответствующему акцепторному сахариду, LgtA также может использовать УДФ-галактозу или УДФ-глюкозу в качестве субстратов-до но ров. Используя генетически сконструированный микроорганизм-хозяин, способный продуцировать LNT или LNnT, как описано, указанная β-1,3-N-ацетилглюкозаминилтрансфераза также может переносить остаток галактозы от УДФ-галактозы, а также остаток глюкозы от УДФ-глюкозы к группировке галактозы лактозы, синтезируя, таким образом, нежелательные побочные продукты Gal(β1,3)Gal(β1,4)Glc и Glc(β1,3)Gal(β1,4)Glc, соответственно.
Посредством экспрессии галактан β-1,3-галактозидазы и/или β-1,3-глюкозидазы в генетически сконструированной микробной клетке-хозяине, способной продуцировать LNT или LNnT, продукция побочных продуктов Gal(β1,3)Gal(β1,4)Glc и Glc(β1,3)Gal(β1,4)Glc может быть прекращена или по меньшей мере уменьшена по той причине, что данные побочный продукты гидролизуются в генетически сконструированной микробной клетке-хозяине под действием галактан β-1,3-галактозидазы и/или β1,3-глюкозидазы. Полученные продукты деградации представляют собой галактозу и/или глюкозу и лактозу. Оба моносахарида, а также лактоза могут быть использованы генетически сконструированной микробной клеткой-хозяином для продукции целевой LNT или LNnT.
В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована с возможностью экспрессии β-1,3-глюкозидазы. В дополнительном и/или альтернативном воплощении генетически сконструированная микробная клетка-хозяин генетически сконструирована для того, чтобы содержать молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую β-1,3-глюкозидазу для ее экспрессии. Предпочтительно, нуклеотидная последовательность, кодирующая β-1,3-глюкозидазу, представляет собой нуклеотидную последовательность, выбранную из группы, состоящей из:
- нуклеотидной последовательности, как представлено SEQ ID NO: 11;
- нуклеотидных последовательностей, которые комплементарны нуклеотидной последовательности, которая гибридизуется с нуклеотидной последовательностью, как представлено SEQ ID NO: 11, в жестких условиях;
- нуклеотидных последовательностей, обладающих идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с нуклеотидной последовательностью, как представлено SEQ ID NO: 11;
- нуклеотидных последовательностей, кодирующих полипептид, имеющий аминокислотную последовательность, как представлено SEQ ID NO: 12; и
- нуклеотидных последовательностей, кодирующих функциональный вариант полипептидных последовательностей, как представлено SEQ ID NO: 12, где аминокислотная последовательность функционального варианта обладает идентичностью последовательностей по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% с аминокислотной последовательностью, как представлено SEQ ID NO: 10.
Для экспрессии нуклеотидной последовательности, кодирующей β-1,3-глюкозидазу или ее функциональный вариант, указанная нуклеотидная последовательность функционально связана с последовательностями контроля экспрессии, которые опосредуют экспрессию нуклеотидной последовательности, кодирующей β-1,3-глюкозидазу или ее функциональный вариант, в генетически сконструированной микробной клетке-хозяине.
Генетически сконструированная микробная клетка-хозяин может повторно использовать по меньшей мере один из продуктов деградации, полученных в результате ферментативной активности гетерологичной гликозидазы в генетически сконструированной микробной клетке-хозяине. Таким образом, генетически сконструированная микробная клетка-хозяин может использовать по меньшей мере один из продуктов деградации, полученных за счет ферментативной активности гетерологичной гликозидазы для продукции целевого олигосахарида. Например, остаток моносахарида, высвобождающийся из нецелевого побочного продукта -сахарида под действием гетерологичной гликозидазы, может быть повторно активирован, а именно связан с нуклеотидом, перенесен от полученного нуклеотид-активированного моносахарида к субстрату-акцептору посредством соответствующей гликозилтрансферазы с получением целевого олигосахарида или предшественника целевого олигосахарида.
Способ включает стадию культивирования генетически сконструированной микробной клетки-хозяина в среде, которая является пермиссивной для продукции целевого олигосахарида указанным генетически сконструированным микроорганизмом-хозяином, и в условиях, которые являются пермиссивными для продукции целевого олигосахарида указанным генетически сконструированным микроорганизмом-хозяином.
Среда, которая является пермиссивной в отношении продукции целевого олигосахарида генетически сконструированной микробной клеткой-хозяином, содержит питательные вещества, по меньшей мере один источник энергии, незаменимые металлы и минеральные вещества и буферный агент. Среда возможно содержит предшественник целевого олигосахарида, причем указанный предшественник может поглощаться генетически сконструированной микробной клеткой-хозяином и использоваться для продукции целевого олигосахарида, при условии, что генетически сконструированная микробная клетка-хозяин не может синтезировать указанный предшественник самостоятельно. Затем, генетически сконструированная микробная клетка-хозяин поглощает предшественник и подвергает предшественник биосинтезу целевого олигосахарида. Например, лактозу можно считать предшественником 2'-фукозиллактозы.
Во время культивации генетически сконструированных микробных клеток-хозяев для продуцирования целевого олигосахарида поддерживаются пермиссивные условия. Условия являются «пермиссивными», если генетически сконструированные микробные клетки-хозяева, которые культивируют в данных условиях, остаются живыми и продуцируют целевой олигосахарид. Предпочтительно, пермиссивные условия культивирования делают возможным размножение генетически сконструированных микробных клеток-хозяев. Условия, которые нужно поддерживать при определенной величине или в пределах определенного диапазона, включают рН, температуру, кислород и концентрации питательных веществ, источников энергии и незаменимых металлов и минеральных веществ.
В дополнительном и/или альтернативном воплощении способ включает стадию выделения целевого олигосахарида. Целевой олигосахарид может быть выделен из ферментационного бульона и/или из генетически сконструированного микроорганизмα-хозяина.
Способ, как ранее описано в данном документе, является преимущественным в том отношении, что во время продукции целевого олигосахарида продуцируется меньше нецелевых побочных продуктов или они не продуцируются. Таким образом, менее трудоемко и затратно выделять и очищать целевой огилосахарид из ферментационного бульона или клеточного лизата.
Кроме того, гораздо больше субстрата специфично используется для продуцирования целевого олигосахарида вместо того, чтобы стать недоступным для продукции целевого олигосахарида, поскольку он включается в нежелательные побочные продукты, которые не могут быть метаболизированы микробной клеткой-хозяином.
Согласно второму аспекту предложены генетически сконструированные микробные клетки-хозяева для получения целевого олигосахарида, где микробная клетка-хозяин может продуцировать целевой олигосахарид, и где микробная клетка-хозяин генетически сконструирована с возможностью экспрессии гетерологичной гликозидазы, которая может осуществлять внутриклеточную деградацию побочных продуктов метаболизма, которые образуются во время внутриклеточного биосинтеза целевого олигосахарида.
Согласно третьему аспекту, генетически сконструированные микробные клетки-хозяева, как ранее описано в данном документе, используются для продуцирования целевого олигосахарида. Использование данных генетически сконструированных микробных клеток-хозяев для продуцирования целевого олигосахарида посредством ферментации является преимущественным, поскольку продуцирование нецелевых побочных продуктов-сахаридов предотвращается или даже прекращается. Таким образом, это сохраняет ресурсы и является менее трудоемким для выделения целевого олигосахарида из ферментационного бульона, поскольку можно избежать отделения целевого олигосахарида от нецелевых побочных продуктов - олигосахаридов. Кроме того, гораздо больше продукта извлечения и источников энергии, предоставленных генетически сконструированным микробным клеткам-хозяевам согласно настоящему изобретению, превращается в целевой продукт, по сравнению с нативной микробной клеткой-хозяином, которая не была генетически сконструирована с возможностью экспрессии гетерологичной гликозидазы.
Согласно четвертому аспекту, целевые олигосахариды, которые получены способом, и/или применением генетически сконструированных микробных клеток-хозяев, описанных в данном документе ранее, предпочтительно выбраны из группы ОГМ.
Целевые олигосахариды, которые получены способом, и/или применением генетически сконструированных микробных клеток-хозяев, описанных в данном документе, могут быть использованы для получения питательной композиции.
Питательная композиция представляет собой лечебную композицию, диетическую композицию, детскую питательную смесь или т.п.
Настоящее изобретение будет описано в отношении конкретных воплощений и со ссылкой на графические материалы, но данное изобретение не ограничивается ими, а только формулой изобретения. Кроме того, термины первый, второй и тому подобное в описании и в формуле изобретения используются для проведения различия между похожими элементами и не обязательно для описания последовательности, во времени, в пространстве, по рангу или любым другим образом. Следует понимать, что термины, используемые таким образом, являются взаимозаменяемыми в соответствующих обстоятельствах, и что воплощения изобретения, описанные в данном документе, способны работать в последовательностях, отличных от описанных или проиллюстрированных в данном документе.
Следует понимать, что термин «содержащий», используемый в формуле изобретения, не следует считать ограничивающимся средствами, перечисленными в дальнейшем; он не исключает других элементов или стадий. Таким образом, его следует считать определяющим наличие заявленных признаков, целых чисел, стадий или компонентов, на которые ссылаются, но он не исключает наличия или добавления одного или более других признаков, целых чисел, стадий или компонентов или их групп. Таким образом, объем выражения «устройство, содержащее средства А и В» не следует ограничивать устройствами, состоящими только из компонентов А и В. Оно означает, что в отношении настоящего изобретения, единственными релевантными компонентами устройства являются А и В.
Ссылка на всем протяжении данного описания изобретения на «одно воплощение» или «воплощение» означает, что конкретный признак, структура или характеристика, описанные в связи с данным воплощением, включены в по меньшей мере одно воплощение настоящего изобретения. Таким образом, появления фраз «в одном воплощении» или «в воплощении» в разных местах по всему объему данного описания изобретения не обязательно все относятся к одному и тому же воплощению, но могут. Кроме того, конкретные признаки, структуры или характеристики могут быть объединены любым подходящим образом, как будет очевидно обычному специалисту в данной области из данного раскрытия, в одном или более воплощениях.
Аналогично следует понимать, что в описании иллюстративных воплощений изобретения разные признаки изобретения иногда сгруппированы вместе в одном единственном воплощении, фигуре или его описании в целях упрощения раскрытия и помощи в понимании одного или более разных аспектов изобретения. Данный способ раскрытия, однако, не нужно считать отражающим мысль, что заявленное изобретение требует больше признаков, чем явным образом перечислены в каждом пункте. Скорее, как отражено в следующей формуле изобретения, аспекты изобретения заключаются меньше чем во всех признаках одного вышеизложенного раскрытого воплощения. Таким образом, формула изобретения после подробного описания явным образом включена тем самым в данное подробное описание, причем каждый пункт отдельно стоит в виде отдельного воплощения данного изобретения.
Кроме того, в то время как некоторые воплощения, описанные в данном документе, включают некоторые, но не все признаки, включенные в другие воплощения, подразумевается, что комбинации признаков разных воплощений находятся в объеме изобретения и образуют разные воплощения, как будет понятно специалистам в данной области. Например, в следующей формуле изобретения любое из заявленных воплощений можно использовать в любой комбинации.
Кроме того, некоторые из воплощений описаны в данном документе как способ или комбинация элементов способа, которые могут быть реализованы посредством процессора компьютерной системы или с помощью других средств выполнения функции. Таким образом, процессор с необходимыми инструкциями для осуществления такого способа или элемента способа образует средство осуществления способа или элемента способа. Кроме того, описанный в данном документе элемент воплощения аппарата представляет собой пример средства осуществления функции, выполняемой элементом, с целью осуществления изобретения.
В описании и графических материалах, предоставленных в данном документе, изложены многочисленные конкретные подробности. Однако, понятно, что воплощения изобретения можно осуществлять на практике без данных конкретных подробностей. В других примерах хорошо известные способы, структуры и методики не были показаны подробно для того, чтобы не затруднять понимание данного описания.
Теперь изобретение будет описано с помощью подробного описания нескольких воплощений изобретения. Ясно, что другие воплощения изобретения могут быть скомпонованы в соответствии со знаниями специалистов в данной области, не отклоняясь от истинной сущности или технической идеи изобретения, причем изобретение ограничено только условиями прилагаемой формулы изобретения.
Пример 1: Метаболическое конструирование штамма BL21(DE3) Е. coli для продукции 2'-фукозиллактозы
BL21(DE3) Е. coli (Novagen) использовали в качестве исходного штамма для конструирования штаммα-хозяина для продукции 2'-FL. Генетическое конструирование исходного штамма включало нарушение гена и события делеций и интеграцию гетерологичных генов.
Поскольку 2'-фукозиллактоза синтезируется из лактозы, ее применяют для бактериальной культуры, и из ГДФ-L-фукозы, которая продуцируется из живых клеток, сначала, копию дикого типа гена lacZ, кодирующего эндогенную β-галактозидазу, инактивировали посредством мутагенеза с использованием ошибки спаривания олигонуклеотидов (Ellis et al., «High efficiency mutagenesis, repair, and engineering of chromosomal DNA using single-stranded oligonucleotides”, Proc. Natl. Acad. Sci. USA 98: 6742-6746 (2001)). Используя тот же способ, нарушали ген арабинозоизомеразы araA.
Фрагмент гена lacZΩ вводили под контролем чувствительного к температуре транскрипционного репрессора cl857. Фрагмент гена lacZα экспрессируется под контролем промотора PgbA BL21 (DE3) Е, coli в данном штамме, обнаруживая LacZ+ штамм.
Геномные делеции осуществляли посредством Red-зависимой рекомбинации фага λ в соответствии с методом Datsenko и Warner («One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products», Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). Гены fuel и fucK, кодирующие L-фукозоизомеразу и L-фукулозокиназу, соответственно, удаляли для предотвращения деградации L-фукозы. Также удаляли гены wzxC-wcaJ. WcaJ возможно кодирует УДФ-глюкоза: ундекапренилфосфатглюкозо-1-фосфаттрансферазу, катализирующую первую стадию в синтезе колановой кислоты (Stevenson et al., «Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colonic acid», J. Bacterid. 178:4885-4893; (1996)); продуцирование колановой кислоты будет конкурировать за ГДФ-фукозу с реакцией фукозилтрансферазы.
Интеграцию в геном гетерологичных генов проводили посредством транспозиции. Большие кластеры генов интегрировали в геном при посредничестве гиперактивного С9-мутанта транспозазы mariner Himarl (Lampe et al., «Hyperactive transposase mutants of the Himarl mariner transposon», Proc. Natl. Acad. Sci. USA 96:11428-11433 (1999)), который вставляли в плазмиду pEcomar под транскрипционным контролем промотора Para. Для усиления de novo синтеза ГДФ-фукозы, гены, кодирующие фосфоманномутазу (manB), маннозо-1-фосфатгуанозилтрансферазу (manC), ГДФ-маннозо-4,6-дегидратазу (gmd) и ГДФ-L-фукозосинтазу (wcaG), из DH5α K12 Е, coli сверхэкспрессировали в штамме BL21(DE3) Е, coli; оперон manCB ставили под контролем конститутивного промотора Ptet, оперон gmd, wcaG транскрибируется с конститутивного промотора PT5-Транспозонную кассету <Ptet-manCB-PT5-gmd, wcaG-FRT-dhfr-FRT> (SEQ ID NO: 13), включающую ген дигидрофолатредуктазы для устойчивости к триметоприму, фланкированную инвертированными концевыми повторами, специфично узнаваемыми транспозазой mariner-подобных элементов Himar1, вставляли в геном Е, coli из pEcomar CQ-manCB-gmd, wcaG-dhfr.
Для хромосомной интеграции одиночных генов использовали транспозазу EZ-Tn5™ (Epicentre, США). Для получения транспосом EZ-Tn5 исследуемый ген вместе с FRT-сайт-фланкированной кассетой устойчивости к антибиотику амплифицировали с помощью праймеров, которые несли на обоих участках сайты распознавания 19 - п.н. Mosaic End (5'-CTGTCTCTTATACACATCT, SEQ ID NO: 21) для транспозазы EZ-Tn5. Используя транспозазу EZ-Tn5TM, ген импортера лактозы LacY от K12 TG1 Е, coli (учет. №ABN72583), ген 2-фукозилтрансферазы wbgL от Е. coli: O126 (учет. №ADN43847), и ген yberc0001_9420, кодирующий эффлюксный транспортер сахара суперсемейства мембранных транспортеров из Yersinia bercovieri АТСС 43970 (учет. №EEQ08298), интегрировали, используя соответствующие кассеты интеграции: <Ptet-lacY-FRT-aadA-FRT> (SEQ ID NO: 14), <Ptet-wbgL.co-FRT-neo-FRT> (SEQ ID NO: 15) и <Ptet-yberc0001_9420co-FRT-cat-FRT> (SEQ ID NO: 16). Гены wbgL и yberc0001_9420 синтетически синтезировали и осуществляли оптимизацию кодонов (со) GenScript Cooperation (США). После успешной интеграции гена lacY ген устойчивости устраняли из клонов, устойчивых к стрептомицину, посредством рекомбиназы FLP, кодируемой на плазмиде рСР20 (Datsenko and Warner, «One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products”, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)).
Поскольку BL21(DE3) E, coli не имеет функционального gal-оперона, нативно регулируемую копию оперона galETKM от Е, coli K интегрировали в штамм В посредством EZ-транспозиции с использованием интегрируемой кассеты <Pgal-galЕ-galT-galK-galM> (SEQ ID NO: 17). Интегранты выбирали из агара МакКонки, содержащего 1% галактозу, в виде красных колоний. Полученный штамм может метаболизировать моносахариды - глюкозу и галактозу, возникающие в результате гидролиза лактозы.
Дополнительного улучшения, касающегося синтеза 2'-фукозиллактозы штаммом Е, coli, достигали посредством делеции гена pfkA, кодирующего фосфофруктокиназу А. При культивировании Е, coli на глюконеогенном субстрате подобно глицерину, фосфорилирование фруктозо-6-фосфата под действием PfkA представляет собой тредмил-реакцию с высоким потреблением АТФ, и, кроме того, конкурирует с ManA за субстрат. Ген pfkA удаляли в результате гомологичной рекомбинации в соответствии с Datsenko and Wanner (2000) с использованием кассеты устойчивости к гентамицину (аасС1), которая фланкирована сайтами lox71/66 (Lambert, JM et al. (2007) Cre-lox-based system for multiple gene deletions and selectable - marker removal in Lactobacillus plantarum. Appl. Environ. Microbiol 73: 1126-1135). После удачной делеции гена pfkA ген устойчивости к антибиотику удаляли из генома Е, colic использованием рекомбиназы Cre (Abremski, K et al. (1983) Studies on the properties of P1 site-specific recombination: evidence for topologically unlinked products following recombination. Cell 32: 1301-1311), который клонировали под контролем промотора Para в каркас pKD46 (Datsenko and Wanner, 2000).
В случае разных фукозилтрансфераз помимо трансферазной активности была продемонстрирована ГДФ-L-фукозогидролазная активность. Кроме того, в случае wbgL, α-1,2-фукозилтрансферазы, используемой в данном документе для синтеза 2'-фукозиллактозы, была показана данная гидролитическая активность (см. ЕР 3 050 973 А1). Для сохранения свободной L-фукозы для продуцирования 2'-фукозиллактозы и для устранения загрязняющей L-фукозы из культурального бульона, ген fkp, кодирующий бифункциональную L-фукокиназо/L-фукозо-1-фосфатгуанилилтрансферазу Bacteroides fragilis, под транскрипционным контролем промотора Ptet, вместе с lox71/66-фланкированным геном аасС1 хромосомно интегрировали посредством транспозиции с использованием транспозазы EZ-Tn5™, <Ptet-fkp-lox-aacC1-lox> (SEQ ID NO: 18). После успешной интеграции ген устойчивости к гентамицину удаляли из генома, как описано выше.
Для усиления потока метаболизированного источника углерода-глицерина через глюконеогенный путь от триозофосфатов до фруктозо-6-фосфата для подпитки биосинтеза ГДФ-L-фукозы осуществляли сверхэкспрессию генов, кодирующих фруктозо-1,6-бисфосфатальдолазу (fbaB) и гетерологичную фруктозо-1,6-бисфосфатфосфотазу (fbpase), из Pisum sativum. Ген fbaB от BL21 (DE3) Е. coli сливали с промотором Ptet. Активность FBPase хлоропласта P. sativum аллостерически регулировали посредством замены дисульфид-дитиол за счет восстановления тиоредоксинами. Замена остатка цистеина 153 на серии приводит к конститутивно активному ферменту. Приобретали ген, кодирующий FBPase хлоропласта от P. sativum (учет. №AAD10213), осуществляли оптимизацию кодонов для экспрессии в Е. coli, метели N-концевой гексагистидиновой меткой и модифицировали для кодирования варианта C153S фермента из Genescript. Ген fbpase транскрибируется с промотора 17. Кассету <Ptet-fbaB-P17-His6-fbpase-lox-аасС1-lox> (SEQ ID NO: 19) использовали для транспозаза EZ-Tn5™ -опосредованной интеграции в штамме-хозяине. После удаления гена устойчивости к гентамицину из генома Е. coli штамм использовали для продукции 2'-фукозиллактозы. Впоследствии, данный штамм называется «штаммом А».
Пример 2: Конструирование штамма BL21(DE3) Е. coli для продукции 2'-фукозиллактозы с высокой чистотой
Культивирования с подпиткой с использованием штамма А для продукции 2'-фукозиллактозы обнаружили наличие побочных продуктов β-фукозиллактозы и 2'3-дифукозиллактозы) в культуральном бульоне. Для минимизации продукции данных побочных продуктов, а также для улучшения выхода углерода, α-1,3-фукозидазу субклонировали за конститутивным промотором и интегрировали в геном штамма А. Таким образом, ген afcB Bifidobacterium bifidum (учетн. № АВ474964) сливали с конститутивным промотором Pand и геном устойчивости к гентамицину. Полученную транспозонную кассету <Pand-afcS-lox-aacC7-lox> (SEQ ID NO: 20), фланкированную инвертированными концевыми повторами, специфично распознаваемыми транспозазой mariner-подобных элементов Himar1, вставляли в геном Е, coli из pEcomar afcB-aacC1, образуя «штамм В».
Пример 3: Анализ ВЭЖХ для выявления 2'-фукозиллактозы в супернатанте культуры
Анализы посредством ВЭЖХ проводили, используя рефрактометрический детектор (RID-10A) (Shimadzu, Германия) и колонку Waters XBridge Amide 3,5 мкм (250 × 4,6 мм) (Eschborn, Германия), соединенную с системой ВЭЖХ (Shimadzu, Германия). Элюирование проводили изократически с помощью 30% А: 50% (об./об.) ACN (от англ. acetonitrile - ацетонитрил) в ddH2O, 0,1% (об./об.) NH4OH и 70% В: 80% (об./об.) ACN в ddH2O, 0,1% (об./об.) NH4OH (об./об.) в качестве элюента при 35°С и при скорости потока 1,4 мл мин-1. Образцы для ВЭЖХ подвергали стерильной фильтрации (размер пор 0,22 мкм) и очищали посредством твердофазной экстракции на ионообменной матрице (Strata ABW, Phenomenex). 10 мкл образцов наносили на колонку, и концентрацию 2'-фукозиллактозы рассчитывали в соответствии со стандартной кривой. Другие сахара, такие как L-фукоза и/или другие моносахариды, лактоза и/или другие дисахариды, 3-фукозиллактоза и/или другие трисахариды, 2'3-дифукозиллактоза и/или другие тетрасахариды, а также глицерин, также могут быть выявлены с использованием данных условий анализа. Относительные количества выявленных Сахаров можно определять посредством сравнения AUC (от англ. area under the curve - площадь под кривой) всех пиков в хроматограмме. Пики, также присутствующие в водном контроле, исключают из данного расчета.
Пример 4: Получение 2'-фукозиллактозы в процессе ферментации
Ферментации проводили в 3л-ферментерах при 33°С (New Brunswick, Edison, США), начиная с 1000 мл среды с минеральными солями, содержащей 3 г/л KH2PO4, 12 г/л K2HPO4, 5 г/л (NH4)2SO4, 0,3 г/л лимонной кислоты, 2 г/л MgSO4 × 7⋅H2O, 0,1 г/л NaCl и 0,015 г/л CaCl2 × 6⋅H2O с добавлением 1 г/л раствора микроэлементов (54,4 г⋅л-1 цитрата железа (III)-аммония, 9,8 г/л MnCl2 × 4⋅H2O, 1,6 г/л CoCl2 × 6⋅Н2O, 1 г/л CuCl2 × 2⋅H2O, 1,9 г/л Н3ВО3, 9 г/л ZnSO4 × 7⋅H2O, 1,1 г/л Na2MoO4 × 2⋅H2O, 1,5 г/л Na2SeO3, 1.,5 г/л NiSO4 × 6⋅H2O) и содержащей 2% (об./об.) глицерина в качестве источника углерода, 60 мМ лактозу и антибиотик канамицин (25 мкг/мл). Аэрацию поддерживали на уровне 3 л/мин. Растворенный кислород поддерживали на уровне насыщения 20-30% посредством осуществления контроля скорости перемешивания. рН поддерживали на уровне 7,0 посредством добавления 25%-ного раствора аммония. Культивацию начинали с 2,5% (об./об.) инокулята из предварительной культуры, растущей в той же среде, содержащей глицерин, но не содержащей лактозы. После прохождения порционной фазы, на что указывало повышение уровня растворенного кислорода, осуществляли подачу глицерина (60% (об./об.) с добавлением 2 г/л MgSO4 × 7⋅H2O, 0,015 г/л CaCl2 × 6⋅H2O и 1 мл/л раствора микроэлементов) при скоростях потока 7,0-8,0 мл/ч, относительно исходного объема. Подачу лактозы (0,66 М) проводили на протяжении всей культивации и регулировали интуитивно для реализации постоянной подачи лактозы в культуральной бульоне. Подачу лактозы останавливали к концу ферментации, и культивацию продолжали до тех пор, пока лактоза полностью не превратится в 2'-фукозиллактозу. Через примерно 94 часа после засева ферментера титр 2'-фукозиллактозы в клеточной среде примерно 150 г/л достигался при использовании штамма, описанного в примере 1 (штамм А). Штамм, продуцирующий 2'-фукозиллактозу, генетически модифицированный, как описано в примере 2 (штамм В), культивировали аналогично, и он также приводил к получению титра 2'-фукозиллактозы примерно 150 г/л. Однако, количество побочных продуктов было значимо ниже, чем после культивирования штамма А (Таблица 2). Тогда как содержание сахарида в супернатанте культуры штамма А составляло только 94,22% 2'-фукозиллактозы, оно увеличивалось на 5,50% в супернатанте культуры штамма В, демонстрируя чистоту 99,72%.
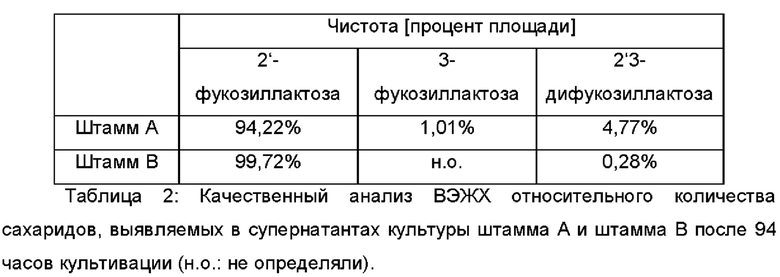
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Jennewein Biotechnologie GmbH
<120> ПРИМЕНЕНИЕ ГЛИКОЗИДАЗ В ПОЛУЧЕНИИ ОЛИГОСАХАРИДОВ
<130> P 1804 WO
<160> 20
<170> PatentIn version 3.5
<210> 1
<211> 5880
<212> ДНК
<213> Bifidobacterium bifidum
<400> 1
atgaaacata gagcgatgtc atcgcgtctg atgccactgg tggcgtcctg cgcgacggtc 60
ggcatgctgc tggccggact acctgtgtcg gccgtcgcgg tcggcacgac gagagcggca 120
gcgtccgacg cctcgtcctc caccacagca accatcaccc cctccgccga taccacgttg 180
cagacatgga cgagcgagaa gaattcctca atggcgtcca agccgtacat cggcacactg 240
caagggccct cgcaaggcgt gttcggcgag aagttcgagt ccacggatgc cgcggacacc 300
accgatctga agaccggcct gctgacgttc gacctgagcg cctacgacca tgcccccgat 360
tccgcaacgt tcgagatgac gtacctcggc taccgcggca acccgacggc caccgacacc 420
gacaccatca aggtgacccc cgtcgacacc accgtgtgca ccaataacgc cacagactgc 480
ggcgcgaatg tcgcgaccgg cgcgaccaag ccgaagttca gcatcaacga ctcctcattc 540
gtcgccgagt ccaagccgtt cgagtacggt acgacggttt acacgggcga cgccatcacc 600
gtggttcccg ccaataccaa gaaggtcacc gtagatgtga ccgaaatcgt gcgccagcag 660
ttcgccgaag gcaagaaggt catcaccctg gccgtgggcg agaccaagaa gaccgaggtt 720
cgtttcgcca gttccgaagg cacgacgtcc ctgaacggcg cgaccgcaga catggctccg 780
aagctgaccg tttccgtgtc caccaaggac gatctcaagc cctccgccga caccacgttg 840
caggcatggg ccagcgagaa gaacgagaag aagaacactg cggcctatgt cggcgcgctg 900
cagccggaag gcgattacgg cgacttcggt gagaagttca agtccaccga cgtccacgat 960
gtcacagacg ccaagatggg tctgatgacg ttcgacctgt ccgattacac cgcggcgccc 1020
gagcactcca tcctcacctt gacgtatctg ggctacgccg gtgcagacaa gaccgccacg 1080
gccaccgata aggtcaaggt ggtcgctgtt gacacgtcgc ggtgcaccgg caccgctccc 1140
tgcgacacca acaatgccac gtgggcgaac cgcccggact tcgaggtgac cgataccacg 1200
aagaccgcga cgtcccatgc gttcgcttat ggatctaaga agtattccga tggcatgacc 1260
gtcgaatcgg gcaacgccaa gaaggtcctg ctcgacgtgt ccgatgtcat caaggcagag 1320
ttcgccaagt tcagcgccgg cgccaccgag aagaagatca cgctggccct gggcgagctc 1380
aacaagtccg acatgcgttt cggcagcaag gaagtcacct cgctgaccgg cgccaccgaa 1440
gccatgcagc cgaccttgtc cgtcaccaag aagccgaagg catacacgct gagcatcgaa 1500
ggcccgacca aggtcaagta ccagaagggc gaggcgttcg acaaggccgg actcgtggtc 1560
aaggccacca gcacggctga cggcacggtc aagacgctga ccgaaggcaa cggtgaggat 1620
aactacacca tcgacaccag cgctttcgat agtgccagca tcggcgtata ccctgttacc 1680
gtgaagtaca acaaggaccc cgaaatcgcc gcttcgttca acgcctatgt catcgccagt 1740
gtcgaggacg gcggagacgg cgacaccagc aaagacgact ggctgtggta caagcagccc 1800
gcgtcgcaga ccgacgccac cgccaccgcc ggcggcaatt acggcaaccc cgacaacaac 1860
cgttggcagc agaccacctt gccgttcggc aacggcaaga tcggcggcac cgtctggggc 1920
gaggtcagcc gtgaacgcgt caccttcaac gaggagacgc tgtggaccgg cggccccgga 1980
tcctcgacca gctacaacgg cggcaacaac gagaccaagg gtcagaacgg cgccacgctg 2040
cgcgcgctca acaagcagct cgcgaacggc gccgagacgg tcaatcccgg caacctgacc 2100
ggcggcgaga acgcggccga gcagggcaac tacctgaact ggggcgacat ctacctcgac 2160
tacgggttca acgatacgac cgtcaccgaa taccgccgcg acctgaacct gagcaagggc 2220
aaggccgacg tcacgttcaa gcatgacggc gtcacctaca cgcgcgaata cttcgcgtcg 2280
aaccccgaca atgtcatggt cgcccgcctc acggccagca aagccggcaa gctgaacttc 2340
aacgtcagca tgccgaccaa cacgaactac tccaagaccg gcgaaaccac gacggtcaag 2400
ggtgacacgc tcaccgtcaa gggcgctctc ggcaacaacg gcctgctgta caactcgcag 2460
atcaaggtcg tcctcgacaa cggtgagggc acgctctccg aaggctccga cggcgcttcg 2520
ctgaaggtct ccgacgcgaa ggcggtcacg ctgtacatcg ccgccgcgac ggactacaag 2580
cagaagtatc cgtcctaccg caccggcgaa accgccgccg aggtgaacac ccgcgtcgcc 2640
aaggtcgtgc aggacgccgc caacaagggc tacaccgccg tcaagaaagc gcacatcgac 2700
gatcattccg ccatctacga ccgcgtgaag atcgatttgg gccagtccgg ccacagctcc 2760
gacggcgccg tcgccaccga cgcgctgctc aaggcgtacc agagaggctc cgcaaccacc 2820
gcgcagaagc gcgagctgga gacgctggtg tacaagtacg gccgctactt gaccatcggc 2880
tcctcccgtg agaacagcca gctgcccagc aacctgcagg gcatctggtc ggtcaccgcg 2940
ggcgacaacg cccacggcaa cacgccttgg ggctccgact tccacatgaa cgtgaacctc 3000
cagatgaact actggccgac ctattcggcc aacatgggag agctcgccga gccgctcatc 3060
gagtatgtgg agggtctggt caagcccggc cgtgtgaccg ccaaggtcta cgcgggcgcg 3120
gagacgacga accccgagac cacgccgatc ggcgagggcg agggctacat ggcccacacc 3180
gagaacaccg cctacggctg gaccgcaccc ggtcaatcgt tctcgtgggg ttggagcccg 3240
gccgccgtgc cgtggatcct gcagaacgtg tacgaggcgt acgagtactc cggcgaccct 3300
gccctgcttg atcgcgtgta cgcgctgctc aaggaggaat cgcacttcta cgtcaactac 3360
atgctgcaca aggccggctc cagctccggt gaccgcctga ctaccggcgt cgcgtactcg 3420
cccgaacagg gcccgctggg caccgacggc aacacgtacg agagctcgct cgtgtggcag 3480
atgctcaacg acgccatcga ggcggccaag gccaagggag atccggacgg tctggtcggc 3540
aataccaccg actgctcggc cgacaactgg gccaagaatg acagcggcaa cttcaccgat 3600
gcgaacgcca accgttcctg gagctgcgcc aagagcctgc tcaagccgat cgaggtcggc 3660
gactccggcc agatcaagga atggtacttc gaaggtgcgc tcggcaagaa gaaggatgga 3720
tccaccatca gcggctacca ggcggacaac cagcaccgtc acatgtccca cctgctcgga 3780
ctgttccccg gtgatttgat caccatcgac aactccgagt acatggatgc ggccaagacc 3840
tcgctgaggt accgctgctt caagggcaac gtgctgcagt ccaacaccgg ctgggccatt 3900
ggccagcgca tcaattcgtg ggctcgcacc ggcgacggca acaccacgta ccagctggtc 3960
gagctgcagc tcaagaacgc gatgtatgca aacctgttcg attaccatgc gccgttccag 4020
atcgacggca acttcggcaa cacctccggt gtcgacgaaa tgctgctgca gtccaactcc 4080
accttcaccg acaccgccgg caagaagtac gtgaactaca cgaacatcct gcccgccctg 4140
cccgatgcct gggcgggcgg ctcggtgagc ggcctcgtgg cccgcggcaa cttcaccgtc 4200
ggcacgacat ggaagaacgg caaggccacc gaagtcaggc tgacctccaa caagggcaag 4260
caggcggccg tcaagatcac cgccggcggc gcccagaact acgaggtcaa gaacggtgac 4320
accgccgtga acgccaaggt cgtgaccaac gcggacggcg cctcgctgct cgtgttcgat 4380
accaccgcag gcaccacgta cacgatcacg aagaaggcga gcgccaacgt gcccgtcacc 4440
ggcgtgaccg tgaccggcgc caacaccgcc accgcaggcg acaccgtcac tcttacggct 4500
accgtcgccc cggccaatgc gaccgacaag tccgtcacct ggtcgacctc cgacgccgcc 4560
gtagctacgg tcaacgccaa cggcgtggtg accacgaaga aggccggcaa ggtgaccatc 4620
accgccacgt cgaacggcga caagacgaag ttcggttcca tcgagatcac cgtctccgcc 4680
gcgaccgtgc ccgtcaccag cgtcaccgtt gccggcgacg ccgcgatgac cgtcgatgga 4740
gagcagaccc tgacggcgac cgtcgccccg gccactgcga ccgacaagac ggtcacgtgg 4800
aagtcctccg acgccactgt ggcgacggtt gacgccaacg gcaaggtcgt cgcgaagaag 4860
gccggcgaag tgacgatcac cgccacggcc ggtggcgtgt ccggcacgct gaagatcacg 4920
gtgagcgaca aggccccgac cgtcatcccg gtccagtccg tgaccgtgac aggcaagcag 4980
gagctcgtcg aaggcgcctc cacgaccctg acggcgaccg tcgccccggc tgacgcgacc 5040
gacaagacgg ttacgtggaa gtcgagcgac gagtccgtcg ccacggtcga caaggacggc 5100
gtcgtgaccg ccaagaaggc cggcacggtg accatcaccg ccacggccgg tggcgtgtcc 5160
ggcacgctcc acatcaccgt gacggccaag cccgtcgaga ccgtccccgt caccagcgtg 5220
gaggtcaccg tcgaggccgg caccaccgtc tccgtcggca agacactcca ggccaccgcg 5280
accgtcaagc ccggcaacgc caccaacaag aaggtgacgt ggaagtcgag cgacgaatcc 5340
atcgcgacgg tcgacgccaa cggcgtcatc accgcgaaga aggccggcaa ggtcgtcatc 5400
acggccacct cgaccgacgg cacggacaag tccggcagcg tcgagatcac cgtcgtggat 5460
gagaccaagc cgacgcccga ccacaagtcc gtcaaggccg ataccggcga cgtgaccgcc 5520
ggcaagaccg gtacggtcac cgagccgaag gacgtggcgg gctggaagag ccgctccatc 5580
atcaagcaag gcaagctcgg caaggccgaa atcgccgacg gcacgctcgt gtatgcggcc 5640
ggcgacaaga ccggtgacga cagcttcgtc gtgcagtaca cgatggccga cggcacggtc 5700
atcgacgtga cctacagcgt cacggtcaag gccgccgaaa ccggcaagaa cgacggcgac 5760
ggcaagggcg acggtgtcgc gaagaccggc gccgccgtcg gcgcgctcgc cggcctcggc 5820
ttgatgctgc tcgccgtcgg agtgagcgtg gtgatgattc gccgcaagca ctccgcctga 5880
<210> 2
<211> 1959
<212> ПРТ
<213> Bifidobacterium bifidum
<400> 2
Met Lys His Arg Ala Met Ser Ser Arg Leu Met Pro Leu Val Ala Ser
1 5 10 15
Cys Ala Thr Val Gly Met Leu Leu Ala Gly Leu Pro Val Ser Ala Val
20 25 30
Ala Val Gly Thr Thr Arg Ala Ala Ala Ser Asp Ala Ser Ser Ser Thr
35 40 45
Thr Ala Thr Ile Thr Pro Ser Ala Asp Thr Thr Leu Gln Thr Trp Thr
50 55 60
Ser Glu Lys Asn Ser Ser Met Ala Ser Lys Pro Tyr Ile Gly Thr Leu
65 70 75 80
Gln Gly Pro Ser Gln Gly Val Phe Gly Glu Lys Phe Glu Ser Thr Asp
85 90 95
Ala Ala Asp Thr Thr Asp Leu Lys Thr Gly Leu Leu Thr Phe Asp Leu
100 105 110
Ser Ala Tyr Asp His Ala Pro Asp Ser Ala Thr Phe Glu Met Thr Tyr
115 120 125
Leu Gly Tyr Arg Gly Asn Pro Thr Ala Thr Asp Thr Asp Thr Ile Lys
130 135 140
Val Thr Pro Val Asp Thr Thr Val Cys Thr Asn Asn Ala Thr Asp Cys
145 150 155 160
Gly Ala Asn Val Ala Thr Gly Ala Thr Lys Pro Lys Phe Ser Ile Asn
165 170 175
Asp Ser Ser Phe Val Ala Glu Ser Lys Pro Phe Glu Tyr Gly Thr Thr
180 185 190
Val Tyr Thr Gly Asp Ala Ile Thr Val Val Pro Ala Asn Thr Lys Lys
195 200 205
Val Thr Val Asp Val Thr Glu Ile Val Arg Gln Gln Phe Ala Glu Gly
210 215 220
Lys Lys Val Ile Thr Leu Ala Val Gly Glu Thr Lys Lys Thr Glu Val
225 230 235 240
Arg Phe Ala Ser Ser Glu Gly Thr Thr Ser Leu Asn Gly Ala Thr Ala
245 250 255
Asp Met Ala Pro Lys Leu Thr Val Ser Val Ser Thr Lys Asp Asp Leu
260 265 270
Lys Pro Ser Ala Asp Thr Thr Leu Gln Ala Trp Ala Ser Glu Lys Asn
275 280 285
Glu Lys Lys Asn Thr Ala Ala Tyr Val Gly Ala Leu Gln Pro Glu Gly
290 295 300
Asp Tyr Gly Asp Phe Gly Glu Lys Phe Lys Ser Thr Asp Val His Asp
305 310 315 320
Val Thr Asp Ala Lys Met Gly Leu Met Thr Phe Asp Leu Ser Asp Tyr
325 330 335
Thr Ala Ala Pro Glu His Ser Ile Leu Thr Leu Thr Tyr Leu Gly Tyr
340 345 350
Ala Gly Ala Asp Lys Thr Ala Thr Ala Thr Asp Lys Val Lys Val Val
355 360 365
Ala Val Asp Thr Ser Arg Cys Thr Gly Thr Ala Pro Cys Asp Thr Asn
370 375 380
Asn Ala Thr Trp Ala Asn Arg Pro Asp Phe Glu Val Thr Asp Thr Thr
385 390 395 400
Lys Thr Ala Thr Ser His Ala Phe Ala Tyr Gly Ser Lys Lys Tyr Ser
405 410 415
Asp Gly Met Thr Val Glu Ser Gly Asn Ala Lys Lys Val Leu Leu Asp
420 425 430
Val Ser Asp Val Ile Lys Ala Glu Phe Ala Lys Phe Ser Ala Gly Ala
435 440 445
Thr Glu Lys Lys Ile Thr Leu Ala Leu Gly Glu Leu Asn Lys Ser Asp
450 455 460
Met Arg Phe Gly Ser Lys Glu Val Thr Ser Leu Thr Gly Ala Thr Glu
465 470 475 480
Ala Met Gln Pro Thr Leu Ser Val Thr Lys Lys Pro Lys Ala Tyr Thr
485 490 495
Leu Ser Ile Glu Gly Pro Thr Lys Val Lys Tyr Gln Lys Gly Glu Ala
500 505 510
Phe Asp Lys Ala Gly Leu Val Val Lys Ala Thr Ser Thr Ala Asp Gly
515 520 525
Thr Val Lys Thr Leu Thr Glu Gly Asn Gly Glu Asp Asn Tyr Thr Ile
530 535 540
Asp Thr Ser Ala Phe Asp Ser Ala Ser Ile Gly Val Tyr Pro Val Thr
545 550 555 560
Val Lys Tyr Asn Lys Asp Pro Glu Ile Ala Ala Ser Phe Asn Ala Tyr
565 570 575
Val Ile Ala Ser Val Glu Asp Gly Gly Asp Gly Asp Thr Ser Lys Asp
580 585 590
Asp Trp Leu Trp Tyr Lys Gln Pro Ala Ser Gln Thr Asp Ala Thr Ala
595 600 605
Thr Ala Gly Gly Asn Tyr Gly Asn Pro Asp Asn Asn Arg Trp Gln Gln
610 615 620
Thr Thr Leu Pro Phe Gly Asn Gly Lys Ile Gly Gly Thr Val Trp Gly
625 630 635 640
Glu Val Ser Arg Glu Arg Val Thr Phe Asn Glu Glu Thr Leu Trp Thr
645 650 655
Gly Gly Pro Gly Ser Ser Thr Ser Tyr Asn Gly Gly Asn Asn Glu Thr
660 665 670
Lys Gly Gln Asn Gly Ala Thr Leu Arg Ala Leu Asn Lys Gln Leu Ala
675 680 685
Asn Gly Ala Glu Thr Val Asn Pro Gly Asn Leu Thr Gly Gly Glu Asn
690 695 700
Ala Ala Glu Gln Gly Asn Tyr Leu Asn Trp Gly Asp Ile Tyr Leu Asp
705 710 715 720
Tyr Gly Phe Asn Asp Thr Thr Val Thr Glu Tyr Arg Arg Asp Leu Asn
725 730 735
Leu Ser Lys Gly Lys Ala Asp Val Thr Phe Lys His Asp Gly Val Thr
740 745 750
Tyr Thr Arg Glu Tyr Phe Ala Ser Asn Pro Asp Asn Val Met Val Ala
755 760 765
Arg Leu Thr Ala Ser Lys Ala Gly Lys Leu Asn Phe Asn Val Ser Met
770 775 780
Pro Thr Asn Thr Asn Tyr Ser Lys Thr Gly Glu Thr Thr Thr Val Lys
785 790 795 800
Gly Asp Thr Leu Thr Val Lys Gly Ala Leu Gly Asn Asn Gly Leu Leu
805 810 815
Tyr Asn Ser Gln Ile Lys Val Val Leu Asp Asn Gly Glu Gly Thr Leu
820 825 830
Ser Glu Gly Ser Asp Gly Ala Ser Leu Lys Val Ser Asp Ala Lys Ala
835 840 845
Val Thr Leu Tyr Ile Ala Ala Ala Thr Asp Tyr Lys Gln Lys Tyr Pro
850 855 860
Ser Tyr Arg Thr Gly Glu Thr Ala Ala Glu Val Asn Thr Arg Val Ala
865 870 875 880
Lys Val Val Gln Asp Ala Ala Asn Lys Gly Tyr Thr Ala Val Lys Lys
885 890 895
Ala His Ile Asp Asp His Ser Ala Ile Tyr Asp Arg Val Lys Ile Asp
900 905 910
Leu Gly Gln Ser Gly His Ser Ser Asp Gly Ala Val Ala Thr Asp Ala
915 920 925
Leu Leu Lys Ala Tyr Gln Arg Gly Ser Ala Thr Thr Ala Gln Lys Arg
930 935 940
Glu Leu Glu Thr Leu Val Tyr Lys Tyr Gly Arg Tyr Leu Thr Ile Gly
945 950 955 960
Ser Ser Arg Glu Asn Ser Gln Leu Pro Ser Asn Leu Gln Gly Ile Trp
965 970 975
Ser Val Thr Ala Gly Asp Asn Ala His Gly Asn Thr Pro Trp Gly Ser
980 985 990
Asp Phe His Met Asn Val Asn Leu Gln Met Asn Tyr Trp Pro Thr Tyr
995 1000 1005
Ser Ala Asn Met Gly Glu Leu Ala Glu Pro Leu Ile Glu Tyr Val
1010 1015 1020
Glu Gly Leu Val Lys Pro Gly Arg Val Thr Ala Lys Val Tyr Ala
1025 1030 1035
Gly Ala Glu Thr Thr Asn Pro Glu Thr Thr Pro Ile Gly Glu Gly
1040 1045 1050
Glu Gly Tyr Met Ala His Thr Glu Asn Thr Ala Tyr Gly Trp Thr
1055 1060 1065
Ala Pro Gly Gln Ser Phe Ser Trp Gly Trp Ser Pro Ala Ala Val
1070 1075 1080
Pro Trp Ile Leu Gln Asn Val Tyr Glu Ala Tyr Glu Tyr Ser Gly
1085 1090 1095
Asp Pro Ala Leu Leu Asp Arg Val Tyr Ala Leu Leu Lys Glu Glu
1100 1105 1110
Ser His Phe Tyr Val Asn Tyr Met Leu His Lys Ala Gly Ser Ser
1115 1120 1125
Ser Gly Asp Arg Leu Thr Thr Gly Val Ala Tyr Ser Pro Glu Gln
1130 1135 1140
Gly Pro Leu Gly Thr Asp Gly Asn Thr Tyr Glu Ser Ser Leu Val
1145 1150 1155
Trp Gln Met Leu Asn Asp Ala Ile Glu Ala Ala Lys Ala Lys Gly
1160 1165 1170
Asp Pro Asp Gly Leu Val Gly Asn Thr Thr Asp Cys Ser Ala Asp
1175 1180 1185
Asn Trp Ala Lys Asn Asp Ser Gly Asn Phe Thr Asp Ala Asn Ala
1190 1195 1200
Asn Arg Ser Trp Ser Cys Ala Lys Ser Leu Leu Lys Pro Ile Glu
1205 1210 1215
Val Gly Asp Ser Gly Gln Ile Lys Glu Trp Tyr Phe Glu Gly Ala
1220 1225 1230
Leu Gly Lys Lys Lys Asp Gly Ser Thr Ile Ser Gly Tyr Gln Ala
1235 1240 1245
Asp Asn Gln His Arg His Met Ser His Leu Leu Gly Leu Phe Pro
1250 1255 1260
Gly Asp Leu Ile Thr Ile Asp Asn Ser Glu Tyr Met Asp Ala Ala
1265 1270 1275
Lys Thr Ser Leu Arg Tyr Arg Cys Phe Lys Gly Asn Val Leu Gln
1280 1285 1290
Ser Asn Thr Gly Trp Ala Ile Gly Gln Arg Ile Asn Ser Trp Ala
1295 1300 1305
Arg Thr Gly Asp Gly Asn Thr Thr Tyr Gln Leu Val Glu Leu Gln
1310 1315 1320
Leu Lys Asn Ala Met Tyr Ala Asn Leu Phe Asp Tyr His Ala Pro
1325 1330 1335
Phe Gln Ile Asp Gly Asn Phe Gly Asn Thr Ser Gly Val Asp Glu
1340 1345 1350
Met Leu Leu Gln Ser Asn Ser Thr Phe Thr Asp Thr Ala Gly Lys
1355 1360 1365
Lys Tyr Val Asn Tyr Thr Asn Ile Leu Pro Ala Leu Pro Asp Ala
1370 1375 1380
Trp Ala Gly Gly Ser Val Ser Gly Leu Val Ala Arg Gly Asn Phe
1385 1390 1395
Thr Val Gly Thr Thr Trp Lys Asn Gly Lys Ala Thr Glu Val Arg
1400 1405 1410
Leu Thr Ser Asn Lys Gly Lys Gln Ala Ala Val Lys Ile Thr Ala
1415 1420 1425
Gly Gly Ala Gln Asn Tyr Glu Val Lys Asn Gly Asp Thr Ala Val
1430 1435 1440
Asn Ala Lys Val Val Thr Asn Ala Asp Gly Ala Ser Leu Leu Val
1445 1450 1455
Phe Asp Thr Thr Ala Gly Thr Thr Tyr Thr Ile Thr Lys Lys Ala
1460 1465 1470
Ser Ala Asn Val Pro Val Thr Gly Val Thr Val Thr Gly Ala Asn
1475 1480 1485
Thr Ala Thr Ala Gly Asp Thr Val Thr Leu Thr Ala Thr Val Ala
1490 1495 1500
Pro Ala Asn Ala Thr Asp Lys Ser Val Thr Trp Ser Thr Ser Asp
1505 1510 1515
Ala Ala Val Ala Thr Val Asn Ala Asn Gly Val Val Thr Thr Lys
1520 1525 1530
Lys Ala Gly Lys Val Thr Ile Thr Ala Thr Ser Asn Gly Asp Lys
1535 1540 1545
Thr Lys Phe Gly Ser Ile Glu Ile Thr Val Ser Ala Ala Thr Val
1550 1555 1560
Pro Val Thr Ser Val Thr Val Ala Gly Asp Ala Ala Met Thr Val
1565 1570 1575
Asp Gly Glu Gln Thr Leu Thr Ala Thr Val Ala Pro Ala Thr Ala
1580 1585 1590
Thr Asp Lys Thr Val Thr Trp Lys Ser Ser Asp Ala Thr Val Ala
1595 1600 1605
Thr Val Asp Ala Asn Gly Lys Val Val Ala Lys Lys Ala Gly Glu
1610 1615 1620
Val Thr Ile Thr Ala Thr Ala Gly Gly Val Ser Gly Thr Leu Lys
1625 1630 1635
Ile Thr Val Ser Asp Lys Ala Pro Thr Val Ile Pro Val Gln Ser
1640 1645 1650
Val Thr Val Thr Gly Lys Gln Glu Leu Val Glu Gly Ala Ser Thr
1655 1660 1665
Thr Leu Thr Ala Thr Val Ala Pro Ala Asp Ala Thr Asp Lys Thr
1670 1675 1680
Val Thr Trp Lys Ser Ser Asp Glu Ser Val Ala Thr Val Asp Lys
1685 1690 1695
Asp Gly Val Val Thr Ala Lys Lys Ala Gly Thr Val Thr Ile Thr
1700 1705 1710
Ala Thr Ala Gly Gly Val Ser Gly Thr Leu His Ile Thr Val Thr
1715 1720 1725
Ala Lys Pro Val Glu Thr Val Pro Val Thr Ser Val Glu Val Thr
1730 1735 1740
Val Glu Ala Gly Thr Thr Val Ser Val Gly Lys Thr Leu Gln Ala
1745 1750 1755
Thr Ala Thr Val Lys Pro Gly Asn Ala Thr Asn Lys Lys Val Thr
1760 1765 1770
Trp Lys Ser Ser Asp Glu Ser Ile Ala Thr Val Asp Ala Asn Gly
1775 1780 1785
Val Ile Thr Ala Lys Lys Ala Gly Lys Val Val Ile Thr Ala Thr
1790 1795 1800
Ser Thr Asp Gly Thr Asp Lys Ser Gly Ser Val Glu Ile Thr Val
1805 1810 1815
Val Asp Glu Thr Lys Pro Thr Pro Asp His Lys Ser Val Lys Ala
1820 1825 1830
Asp Thr Gly Asp Val Thr Ala Gly Lys Thr Gly Thr Val Thr Glu
1835 1840 1845
Pro Lys Asp Val Ala Gly Trp Lys Ser Arg Ser Ile Ile Lys Gln
1850 1855 1860
Gly Lys Leu Gly Lys Ala Glu Ile Ala Asp Gly Thr Leu Val Tyr
1865 1870 1875
Ala Ala Gly Asp Lys Thr Gly Asp Asp Ser Phe Val Val Gln Tyr
1880 1885 1890
Thr Met Ala Asp Gly Thr Val Ile Asp Val Thr Tyr Ser Val Thr
1895 1900 1905
Val Lys Ala Ala Glu Thr Gly Lys Asn Asp Gly Asp Gly Lys Gly
1910 1915 1920
Asp Gly Val Ala Lys Thr Gly Ala Ala Val Gly Ala Leu Ala Gly
1925 1930 1935
Leu Gly Leu Met Leu Leu Ala Val Gly Val Ser Val Val Met Ile
1940 1945 1950
Arg Arg Lys His Ser Ala
1955
<210> 3
<211> 4482
<212> ДНК
<213> Bifidobacterium bifidum
<400> 3
atgctacaca cagcatcaag aggatgctcg cgttcgtggc tgcgcagact caccgcattg 60
atagcggtct cggcgctcgc gttcgtggca ttgccgaacg tcgcggtggc ggcggatccg 120
atggaatacc tcgatgtgtc gttcggcggc acgttcgctg cagacaccta caccacaggt 180
ggcgacgagg tggcgaaggg ccccgtgacc aagcacggca gcataccgac caagcttgac 240
ggcggcggca tcaccctcgc tggcggcacc aacggcgtga cattcacctc gaccgcgagc 300
ttcagcgaga gtgggaaggt gaacaaggga ttccgcgccg aaatggagta ccgtacgacg 360
cagacgccca gcaacctcgc cacattgttc tccgccatgg gcaacatctt cgtgcgggcg 420
aacggcagca acctcgaata cggcttctcc acgaaccctt ccggcagtac atggaacgac 480
tacacaaagt ccgtgacgct gccttccaac aatgtgaagc acatcatcca gctgacatat 540
ctgccgggag ccgacggcgc tgcctcgacg ttgcagttgt cggtggatgg cgtggccggc 600
gagaccgcca cctccgcggc cggcgagctc gcggccgtca gcgattccgt cgggaacaag 660
ttcgggatcg gctacgaggt gaaccccgct tccggcgcgg cgagccgcgg tcttgccggt 720
gacgtgttcc gcgcgcgtgt cgccgattcg gacgccccgt gggagattct tgacgcatcc 780
cagctgctgc atgtcaattt caacggcacg ttcagcggca cctcatatac cgcggcgagc 840
ggcgagcaga tgctgggctc gctggtgtcg cgctcggcca atccgtccat ctcgaactcc 900
gccgtcacgc tgggcggcgg cacggccgga ttcgatttca cgcccacgga cttcaccctc 960
ggtgacaacg aggccatcac ccgcccgctg gtcgcggagc tgcgcttcac cccgacgcag 1020
accggcgaca accagaccct gttcggcgcg ggcggcaacc tgttcctgcg ctacgagtcg 1080
aacaagctcg tgttcggcgc ctccaccaag tccggcgata attggaccga ccacaagatc 1140
gagtccgcgg ccgccacggg tgcggagcac gtcgtgtcgg tggcgtacgt gcccaataag 1200
gccggcaccg gcgcgaagct tgtcatgcgc gtggatggcg gcgacgccca gaccaaggac 1260
atcactggtc tggcttacct gaattcgagc atcaagggca aggtcggctt cggcaacgac 1320
gtgcataccg acgcgctcag ccgcggcttc gtcggctcgc tgagcgagat ccgcctggcc 1380
gaaacctccg cgaacttcac caccaacgaa ttcaagctgg tctactctca ggtcagctgc 1440
gacacgtcgg gcatcaagga ggcgaatacc ttcgacgtgg agcccgccga gtgcgaggcc 1500
gcgcttaaga ccaagctgtc caagctgcgt ccgaccgaag ggcaggccga ctacatcgac 1560
tggggtcaga tcggattcct ccattacggc atcaacacgt actacaacca ggagtggggt 1620
cacggtaacg aggatccctc ccgcatcaac ccgaccggcc tcgacaccga ccagtgggcg 1680
aagtccttcg ccgacggtgg cttcaagatg atcatggtga cggtcaagca ccatgacggt 1740
ttcgagctgt acgactcgcg gtacaacacc gagcacgact gggcaaacac cgccgtcgcc 1800
aagcgcacgg gggagaagga cctgttccgc aagattgtcg cctcggcgaa gaaatacggc 1860
ctgaaggtcg gcatctacta ttcgccggcc gattcctaca tggagaggaa gggcgtctgg 1920
ggcaacaact ccgcacgcgt cgagcgcacg atccccacgc tggtggagaa cgacgaccgc 1980
gccggcaagg tggcttccgg caaactgccc acgttcaagt acaaggccac ggattacggc 2040
gcctacatgc tcaaccagct ctatgagctg ctgactgagt acggcgacat ctccgaggtc 2100
tggttcgacg gtgcccaagg caacaccgca ggcactgagc attacgacta tggcgtgttc 2160
tacgagatga tccgccggct tcagccccag gcaattcagg ccaacgccgc atacgatgcc 2220
cgatgggtgg gcaacgagga cggctgggcc cgtcagaccg agtggagccc gcaggcggca 2280
tacaacgacg gcgtggacaa ggtgtcgctc aagcctggcc agatggcccc cgacggtaag 2340
cttggcagca tgtcgagcgt gctgtccgag atccgcagcg gcgccgccaa ccagctgcac 2400
tggtatccgg ccgaagtcga cgccaagaac cggcccggat ggttctaccg tgccagccaa 2460
tcgccggcgt ccgtagccga agtcgtgaag tactacgagc agtccacggg acgcaactcg 2520
cagtatctgc tgaacgtccc accgtccgat accggcaagc tcgccgatgc ggatgccgcg 2580
ggacttaagg ggctgggcga ggagctcgcc cgacgctacg gcaccgatct tgccctgggc 2640
aagagcgcga ccgtcgccgc gtccgcgaac gacactgcgg tagcggcccc gaagctgacc 2700
gacggttcga agctctcctc cgacaaggcc gtgggcaata cgccgacgta caccatcgat 2760
ctgggcagca ctgtcgccgt ggatgcagtg aagatctccg aggacgtgcg caatgccggc 2820
cagcagatcg aaagcgccac tctgcaggga cgagtcaatg gaacatggac gaatctggcg 2880
actatgacga cggtcgggca gcagcgcgac cttcgcttca cgtcccagaa catcgatgcc 2940
atccgtctgg tggtcaactc ctcccgcggt ccggtgcgtc tgagccgtct tgaggtgttc 3000
cacaccgaat ccgagattca gaccggcgcc cgcgcctact acatcgatcc gacggcgcag 3060
accgcgggag atggattcac gaaggacaag cccatgacgt cgatcgagca gctgcacgat 3120
gtgaccgtcg cgccaggctc cgtgatcttc gtcaaggcgg gcaccgagct gaccggggac 3180
ttcgccgtct tcggctacgg caccaaggac gagcccatca ccgtgacgac atacggcgaa 3240
agcgacaaag ccaccaccgc gagcttcgac ggcatgaccg ccgggctgac gctgaagcag 3300
gcgctgaagg cgctcggcaa ggacgacgcc ggctgggtcg tggccgattc cgccactgca 3360
ccggcctccc gcgtgtatgt cccgcaggat gagatcagcg tgcacgccca gtcgtcgcag 3420
aactccggcg cagaggcggc gagggcgctc gacggcgact cgtcgacgag ctggcactcc 3480
cagtacagcc cgaccaccgc gtctgctccg cattgggtga ctctcgatct cggcaaatcg 3540
cgtgagaacg tcgcctactt cgactacctc gcccgtatcg acggcaacaa taacggtgcc 3600
gccaaggatt acgaggtgta tgtctccgac gatcccaacg attttggagc ccctgtggcc 3660
tcgggcacgt tgaagaacgt cgcctacacg cagcgcatca agctgacccc caagaacgga 3720
cggtacgtca agttcgtcat caagaccgat tattccggat cgaacttcgg ctccgcggcg 3780
gaaatgaatg tcgagttgct gcccacggcc gtagaggagg acaaggtcgc caccccgcag 3840
aagccgacag tggacgatga tgccgataca tacaccatcc ccgacatcga gggagtcgtg 3900
tacaaggtcg acggcaaggt gttggccgct ggttccgtag tgaacgtggg cgatgaggac 3960
gtgaccgtca cggtcaccgc cgagcccgcc gacggatacc gcttcccgga tggtgtgacg 4020
tccccagtca cgtatgagct gacgttcacc aagaagggtg gcgagaagcc tccgaccgaa 4080
gtcaacaagg acaagctgca cgccacgatc accaaggctc aggcgatcga ccgttccgcc 4140
tatacggacg agtcgctcaa ggtgcttgat gacaagctcg ccgcagcgct caaggtctat 4200
gacgatgaca aggtgagcca ggatgatgtc gatgccgccg aggcggctct gtctgcggcg 4260
atcgacgcgc tgaagaccaa gccgacgacc cccggcggtg aaggtgagaa gcctggtgaa 4320
ggtgaaaagc ccggtgacgg caacaagccc ggtgacggca agaagcccgg cgacgtgatc 4380
gcaaagaccg gcgcctccac aatgggcgtt gtcttcgctg cactcgcgat ggtagcgggt 4440
gcggtcgtga cgcttgaagc caagcgtaag tccaaccggt aa 4482
<210> 4
<211> 1493
<212> PRT
<213> Bifidobacterium bifidum
<400> 4
Met Leu His Thr Ala Ser Arg Gly Cys Ser Arg Ser Trp Leu Arg Arg
1 5 10 15
Leu Thr Ala Leu Ile Ala Val Ser Ala Leu Ala Phe Val Ala Leu Pro
20 25 30
Asn Val Ala Val Ala Ala Asp Pro Met Glu Tyr Leu Asp Val Ser Phe
35 40 45
Gly Gly Thr Phe Ala Ala Asp Thr Tyr Thr Thr Gly Gly Asp Glu Val
50 55 60
Ala Lys Gly Pro Val Thr Lys His Gly Ser Ile Pro Thr Lys Leu Asp
65 70 75 80
Gly Gly Gly Ile Thr Leu Ala Gly Gly Thr Asn Gly Val Thr Phe Thr
85 90 95
Ser Thr Ala Ser Phe Ser Glu Ser Gly Lys Val Asn Lys Gly Phe Arg
100 105 110
Ala Glu Met Glu Tyr Arg Thr Thr Gln Thr Pro Ser Asn Leu Ala Thr
115 120 125
Leu Phe Ser Ala Met Gly Asn Ile Phe Val Arg Ala Asn Gly Ser Asn
130 135 140
Leu Glu Tyr Gly Phe Ser Thr Asn Pro Ser Gly Ser Thr Trp Asn Asp
145 150 155 160
Tyr Thr Lys Ser Val Thr Leu Pro Ser Asn Asn Val Lys His Ile Ile
165 170 175
Gln Leu Thr Tyr Leu Pro Gly Ala Asp Gly Ala Ala Ser Thr Leu Gln
180 185 190
Leu Ser Val Asp Gly Val Ala Gly Glu Thr Ala Thr Ser Ala Ala Gly
195 200 205
Glu Leu Ala Ala Val Ser Asp Ser Val Gly Asn Lys Phe Gly Ile Gly
210 215 220
Tyr Glu Val Asn Pro Ala Ser Gly Ala Ala Ser Arg Gly Leu Ala Gly
225 230 235 240
Asp Val Phe Arg Ala Arg Val Ala Asp Ser Asp Ala Pro Trp Glu Ile
245 250 255
Leu Asp Ala Ser Gln Leu Leu His Val Asn Phe Asn Gly Thr Phe Ser
260 265 270
Gly Thr Ser Tyr Thr Ala Ala Ser Gly Glu Gln Met Leu Gly Ser Leu
275 280 285
Val Ser Arg Ser Ala Asn Pro Ser Ile Ser Asn Ser Ala Val Thr Leu
290 295 300
Gly Gly Gly Thr Ala Gly Phe Asp Phe Thr Pro Thr Asp Phe Thr Leu
305 310 315 320
Gly Asp Asn Glu Ala Ile Thr Arg Pro Leu Val Ala Glu Leu Arg Phe
325 330 335
Thr Pro Thr Gln Thr Gly Asp Asn Gln Thr Leu Phe Gly Ala Gly Gly
340 345 350
Asn Leu Phe Leu Arg Tyr Glu Ser Asn Lys Leu Val Phe Gly Ala Ser
355 360 365
Thr Lys Ser Gly Asp Asn Trp Thr Asp His Lys Ile Glu Ser Ala Ala
370 375 380
Ala Thr Gly Ala Glu His Val Val Ser Val Ala Tyr Val Pro Asn Lys
385 390 395 400
Ala Gly Thr Gly Ala Lys Leu Val Met Arg Val Asp Gly Gly Asp Ala
405 410 415
Gln Thr Lys Asp Ile Thr Gly Leu Ala Tyr Leu Asn Ser Ser Ile Lys
420 425 430
Gly Lys Val Gly Phe Gly Asn Asp Val His Thr Asp Ala Leu Ser Arg
435 440 445
Gly Phe Val Gly Ser Leu Ser Glu Ile Arg Leu Ala Glu Thr Ser Ala
450 455 460
Asn Phe Thr Thr Asn Glu Phe Lys Leu Val Tyr Ser Gln Val Ser Cys
465 470 475 480
Asp Thr Ser Gly Ile Lys Glu Ala Asn Thr Phe Asp Val Glu Pro Ala
485 490 495
Glu Cys Glu Ala Ala Leu Lys Thr Lys Leu Ser Lys Leu Arg Pro Thr
500 505 510
Glu Gly Gln Ala Asp Tyr Ile Asp Trp Gly Gln Ile Gly Phe Leu His
515 520 525
Tyr Gly Ile Asn Thr Tyr Tyr Asn Gln Glu Trp Gly His Gly Asn Glu
530 535 540
Asp Pro Ser Arg Ile Asn Pro Thr Gly Leu Asp Thr Asp Gln Trp Ala
545 550 555 560
Lys Ser Phe Ala Asp Gly Gly Phe Lys Met Ile Met Val Thr Val Lys
565 570 575
His His Asp Gly Phe Glu Leu Tyr Asp Ser Arg Tyr Asn Thr Glu His
580 585 590
Asp Trp Ala Asn Thr Ala Val Ala Lys Arg Thr Gly Glu Lys Asp Leu
595 600 605
Phe Arg Lys Ile Val Ala Ser Ala Lys Lys Tyr Gly Leu Lys Val Gly
610 615 620
Ile Tyr Tyr Ser Pro Ala Asp Ser Tyr Met Glu Arg Lys Gly Val Trp
625 630 635 640
Gly Asn Asn Ser Ala Arg Val Glu Arg Thr Ile Pro Thr Leu Val Glu
645 650 655
Asn Asp Asp Arg Ala Gly Lys Val Ala Ser Gly Lys Leu Pro Thr Phe
660 665 670
Lys Tyr Lys Ala Thr Asp Tyr Gly Ala Tyr Met Leu Asn Gln Leu Tyr
675 680 685
Glu Leu Leu Thr Glu Tyr Gly Asp Ile Ser Glu Val Trp Phe Asp Gly
690 695 700
Ala Gln Gly Asn Thr Ala Gly Thr Glu His Tyr Asp Tyr Gly Val Phe
705 710 715 720
Tyr Glu Met Ile Arg Arg Leu Gln Pro Gln Ala Ile Gln Ala Asn Ala
725 730 735
Ala Tyr Asp Ala Arg Trp Val Gly Asn Glu Asp Gly Trp Ala Arg Gln
740 745 750
Thr Glu Trp Ser Pro Gln Ala Ala Tyr Asn Asp Gly Val Asp Lys Val
755 760 765
Ser Leu Lys Pro Gly Gln Met Ala Pro Asp Gly Lys Leu Gly Ser Met
770 775 780
Ser Ser Val Leu Ser Glu Ile Arg Ser Gly Ala Ala Asn Gln Leu His
785 790 795 800
Trp Tyr Pro Ala Glu Val Asp Ala Lys Asn Arg Pro Gly Trp Phe Tyr
805 810 815
Arg Ala Ser Gln Ser Pro Ala Ser Val Ala Glu Val Val Lys Tyr Tyr
820 825 830
Glu Gln Ser Thr Gly Arg Asn Ser Gln Tyr Leu Leu Asn Val Pro Pro
835 840 845
Ser Asp Thr Gly Lys Leu Ala Asp Ala Asp Ala Ala Gly Leu Lys Gly
850 855 860
Leu Gly Glu Glu Leu Ala Arg Arg Tyr Gly Thr Asp Leu Ala Leu Gly
865 870 875 880
Lys Ser Ala Thr Val Ala Ala Ser Ala Asn Asp Thr Ala Val Ala Ala
885 890 895
Pro Lys Leu Thr Asp Gly Ser Lys Leu Ser Ser Asp Lys Ala Val Gly
900 905 910
Asn Thr Pro Thr Tyr Thr Ile Asp Leu Gly Ser Thr Val Ala Val Asp
915 920 925
Ala Val Lys Ile Ser Glu Asp Val Arg Asn Ala Gly Gln Gln Ile Glu
930 935 940
Ser Ala Thr Leu Gln Gly Arg Val Asn Gly Thr Trp Thr Asn Leu Ala
945 950 955 960
Thr Met Thr Thr Val Gly Gln Gln Arg Asp Leu Arg Phe Thr Ser Gln
965 970 975
Asn Ile Asp Ala Ile Arg Leu Val Val Asn Ser Ser Arg Gly Pro Val
980 985 990
Arg Leu Ser Arg Leu Glu Val Phe His Thr Glu Ser Glu Ile Gln Thr
995 1000 1005
Gly Ala Arg Ala Tyr Tyr Ile Asp Pro Thr Ala Gln Thr Ala Gly
1010 1015 1020
Asp Gly Phe Thr Lys Asp Lys Pro Met Thr Ser Ile Glu Gln Leu
1025 1030 1035
His Asp Val Thr Val Ala Pro Gly Ser Val Ile Phe Val Lys Ala
1040 1045 1050
Gly Thr Glu Leu Thr Gly Asp Phe Ala Val Phe Gly Tyr Gly Thr
1055 1060 1065
Lys Asp Glu Pro Ile Thr Val Thr Thr Tyr Gly Glu Ser Asp Lys
1070 1075 1080
Ala Thr Thr Ala Ser Phe Asp Gly Met Thr Ala Gly Leu Thr Leu
1085 1090 1095
Lys Gln Ala Leu Lys Ala Leu Gly Lys Asp Asp Ala Gly Trp Val
1100 1105 1110
Val Ala Asp Ser Ala Thr Ala Pro Ala Ser Arg Val Tyr Val Pro
1115 1120 1125
Gln Asp Glu Ile Ser Val His Ala Gln Ser Ser Gln Asn Ser Gly
1130 1135 1140
Ala Glu Ala Ala Arg Ala Leu Asp Gly Asp Ser Ser Thr Ser Trp
1145 1150 1155
His Ser Gln Tyr Ser Pro Thr Thr Ala Ser Ala Pro His Trp Val
1160 1165 1170
Thr Leu Asp Leu Gly Lys Ser Arg Glu Asn Val Ala Tyr Phe Asp
1175 1180 1185
Tyr Leu Ala Arg Ile Asp Gly Asn Asn Asn Gly Ala Ala Lys Asp
1190 1195 1200
Tyr Glu Val Tyr Val Ser Asp Asp Pro Asn Asp Phe Gly Ala Pro
1205 1210 1215
Val Ala Ser Gly Thr Leu Lys Asn Val Ala Tyr Thr Gln Arg Ile
1220 1225 1230
Lys Leu Thr Pro Lys Asn Gly Arg Tyr Val Lys Phe Val Ile Lys
1235 1240 1245
Thr Asp Tyr Ser Gly Ser Asn Phe Gly Ser Ala Ala Glu Met Asn
1250 1255 1260
Val Glu Leu Leu Pro Thr Ala Val Glu Glu Asp Lys Val Ala Thr
1265 1270 1275
Pro Gln Lys Pro Thr Val Asp Asp Asp Ala Asp Thr Tyr Thr Ile
1280 1285 1290
Pro Asp Ile Glu Gly Val Val Tyr Lys Val Asp Gly Lys Val Leu
1295 1300 1305
Ala Ala Gly Ser Val Val Asn Val Gly Asp Glu Asp Val Thr Val
1310 1315 1320
Thr Val Thr Ala Glu Pro Ala Asp Gly Tyr Arg Phe Pro Asp Gly
1325 1330 1335
Val Thr Ser Pro Val Thr Tyr Glu Leu Thr Phe Thr Lys Lys Gly
1340 1345 1350
Gly Glu Lys Pro Pro Thr Glu Val Asn Lys Asp Lys Leu His Ala
1355 1360 1365
Thr Ile Thr Lys Ala Gln Ala Ile Asp Arg Ser Ala Tyr Thr Asp
1370 1375 1380
Glu Ser Leu Lys Val Leu Asp Asp Lys Leu Ala Ala Ala Leu Lys
1385 1390 1395
Val Tyr Asp Asp Asp Lys Val Ser Gln Asp Asp Val Asp Ala Ala
1400 1405 1410
Glu Ala Ala Leu Ser Ala Ala Ile Asp Ala Leu Lys Thr Lys Pro
1415 1420 1425
Thr Thr Pro Gly Gly Glu Gly Glu Lys Pro Gly Glu Gly Glu Lys
1430 1435 1440
Pro Gly Asp Gly Asn Lys Pro Gly Asp Gly Lys Lys Pro Gly Asp
1445 1450 1455
Val Ile Ala Lys Thr Gly Ala Ser Thr Met Gly Val Val Phe Ala
1460 1465 1470
Ala Leu Ala Met Val Ala Gly Ala Val Val Thr Leu Glu Ala Lys
1475 1480 1485
Arg Lys Ser Asn Arg
1490
<210> 5
<211> 2094
<212> ДНК
<213> Streptococcus pneumoniae
<400> 5
atgaataaaa gaggtcttta ttcaaaacta ggaatttctg ttgtaggcat tagtctttta 60
atgggagtcc ccactttgat tcatgcgaat gaattaaact atggtcaact gtccatatct 120
cctatttttc aaggaggttc atatcaactg aacaataaga gtatagatat cagctctttg 180
ttattagata aattgtctgg agagagtcag acagtagtaa tgaaatttaa agcagataaa 240
ccaaactctc ttcaagcttt gtttggccta tctaatagta aagcaggctt taaaaataat 300
tacttttcaa ttttcatgag agattctggt gagataggtg tagaaataag agacgcccaa 360
gagggaataa attatttatt ttctagacca gcttcattat ggggaaagca taaaggacag 420
gcagttgaaa atacactagt atttgtatct gattctaaag ataaaacata cacaatgtat 480
gttaatggaa tagaagtgtt ctctgaaaca gttgatacat ttttgccaat ttcaaatata 540
aatggtatag ataaggcaac actaggagct gttaatcgtg aaggtaagga acattacctc 600
gcaaaaggaa gtattggtga aatcagtcta tttaacaaag caattagtga tcaggaagtt 660
tcaaatattc ccttgtcaaa tccatttcag ttaattttcc aatcaggaga ttctactcaa 720
gctaactatt ttagaatacc gacactatat acattaagta gtggaagagt tctatcaagt 780
attgatgcac gttatggtgg gactcatgat tctaaaagta agattaatat tgccacttct 840
tatagtgatg ataatgggaa aacgtggagt gagccaattt ttgctatgaa gtttaatgac 900
tatgaggagc agttagttta ctggccacga gataataaat taaagaatag tcaaattagt 960
ggaagtgctt cattcataga ttcatccatt gttgaagata aaaaatctgg gaaaacgata 1020
ttactagctg atgttatgcc tgcgggtatt ggaaataata atgcaaataa agccgactca 1080
ggttttaaag aaataaatgg tcattattat ttaaaactaa agaagaatgg agataacgat 1140
ttccgttata cagttagaga aaatggtgtc gtttatgatg aaacaactaa taaacctaca 1200
aattatacta taaatgataa gtatgaagtt ttggagggag gaaagtcttt aacagtcgaa 1260
caatattcgg ttgattttga tagtggctct ttaagagaaa ggcataatgg aaaacaggtt 1320
cctatgaatg ttttctacaa agattcgtta tttaaagtga ctcctactaa ttatatagca 1380
atgacaacta gtcagaatag aggagagagt tgggaacaat ttaagttgtt gcctccgttc 1440
ttaggagaaa aacataatgg aacttacttg tgtcctggac aaggtttagc attaaaatca 1500
agtaacagat tgatttttgc aacatatact agtggagaac taacctatct catttcggat 1560
gatagtggtc aaacatggaa gaaatcctca gcttcaattc cgtttaaaaa tgcaacagca 1620
gaagcacaaa tggttgaact gagagatggt gtgattagaa cattctttag aaccactaca 1680
ggtaagatag cttatatgac tagtagagat tctggagaaa catggtcgaa agtttcgtat 1740
attgatggaa ttcaacaaac ttcatatggc acacaagtat ctgcaattaa atactctcaa 1800
ttaattgatg gaaaagaagc agtcattttg agtacaccaa attctagaag tggccgtaag 1860
ggaggccaat tagttgtcgg tttggtcaat aaagaagatg atagtattga ttggagatac 1920
cactatgata ttgatttgcc ttcgtatggt tatgcctatt ctgcgattac agaattgcca 1980
aatcatcaca taggtgtact gtttgaaaaa tatgattcgt ggtcgagaaa tgaattgcat 2040
ttaagcaatg tagttcagta tatagatttg gaaattaatg atttaacaaa ataa 2094
<210> 6
<211> 697
<212> ПРТ
<213> Streptococcus pneumoniae
<400> 6
Met Asn Lys Arg Gly Leu Tyr Ser Lys Leu Gly Ile Ser Val Val Gly
1 5 10 15
Ile Ser Leu Leu Met Gly Val Pro Thr Leu Ile His Ala Asn Glu Leu
20 25 30
Asn Tyr Gly Gln Leu Ser Ile Ser Pro Ile Phe Gln Gly Gly Ser Tyr
35 40 45
Gln Leu Asn Asn Lys Ser Ile Asp Ile Ser Ser Leu Leu Leu Asp Lys
50 55 60
Leu Ser Gly Glu Ser Gln Thr Val Val Met Lys Phe Lys Ala Asp Lys
65 70 75 80
Pro Asn Ser Leu Gln Ala Leu Phe Gly Leu Ser Asn Ser Lys Ala Gly
85 90 95
Phe Lys Asn Asn Tyr Phe Ser Ile Phe Met Arg Asp Ser Gly Glu Ile
100 105 110
Gly Val Glu Ile Arg Asp Ala Gln Lys Gly Ile Asn Tyr Leu Phe Ser
115 120 125
Arg Pro Ala Ser Leu Trp Gly Lys His Lys Gly Gln Ala Val Glu Asn
130 135 140
Thr Leu Val Phe Val Ser Asp Ser Lys Asp Lys Thr Tyr Thr Met Tyr
145 150 155 160
Val Asn Gly Ile Glu Val Phe Ser Glu Thr Val Asp Thr Phe Leu Pro
165 170 175
Ile Ser Asn Ile Asn Gly Ile Asp Lys Ala Thr Leu Gly Ala Val Asn
180 185 190
Arg Glu Gly Lys Glu His Tyr Leu Ala Lys Gly Ser Ile Asp Glu Ile
195 200 205
Ser Leu Phe Asn Lys Ala Ile Ser Asp Gln Glu Val Ser Thr Ile Pro
210 215 220
Leu Ser Asn Pro Phe Gln Leu Ile Phe Gln Ser Gly Asp Ser Thr Gln
225 230 235 240
Ala Asn Tyr Phe Arg Ile Pro Thr Leu Tyr Thr Leu Ser Ser Gly Arg
245 250 255
Val Leu Ser Ser Ile Asp Ala Arg Tyr Gly Gly Thr His Asp Ser Lys
260 265 270
Ser Lys Ile Asn Ile Ala Thr Ser Tyr Ser Asp Asp Asn Gly Lys Thr
275 280 285
Trp Ser Glu Pro Ile Phe Ala Met Lys Phe Asn Asp Tyr Glu Glu Gln
290 295 300
Leu Val Tyr Trp Pro Arg Asp Asn Lys Leu Lys Asn Ser Gln Ile Ser
305 310 315 320
Gly Ser Ala Ser Phe Ile Asp Ser Ser Ile Val Glu Asp Lys Lys Ser
325 330 335
Gly Lys Thr Ile Leu Leu Ala Asp Val Met Pro Ala Gly Ile Gly Asn
340 345 350
Asn Asn Ala Asn Lys Ala Asp Ser Gly Phe Lys Glu Ile Asn Gly His
355 360 365
Tyr Tyr Leu Lys Leu Lys Lys Asn Gly Asp Asn Asp Phe Arg Tyr Thr
370 375 380
Val Arg Glu Asn Gly Val Val Tyr Asn Glu Thr Thr Asn Lys Pro Thr
385 390 395 400
Asn Tyr Thr Ile Asn Asp Lys Tyr Glu Val Leu Glu Gly Gly Lys Ser
405 410 415
Leu Thr Val Glu Gln Tyr Ser Val Asp Phe Asp Ser Gly Ser Leu Arg
420 425 430
Glu Arg His Asn Gly Lys Gln Val Pro Met Asn Val Phe Tyr Lys Asp
435 440 445
Ser Leu Phe Lys Val Thr Pro Thr Asn Tyr Ile Ala Met Thr Thr Ser
450 455 460
Gln Asn Arg Gly Glu Ser Trp Glu Gln Phe Lys Leu Leu Pro Pro Phe
465 470 475 480
Leu Gly Glu Lys His Asn Gly Thr Tyr Leu Cys Pro Gly Gln Gly Leu
485 490 495
Ala Leu Lys Ser Ser Asn Arg Leu Ile Phe Ala Thr Tyr Thr Ser Gly
500 505 510
Glu Leu Thr Tyr Leu Ile Ser Asp Asp Ser Gly Gln Thr Trp Lys Lys
515 520 525
Ser Ser Ala Ser Ile Pro Phe Lys Asn Ala Thr Ala Glu Ala Gln Met
530 535 540
Val Glu Leu Arg Asp Gly Val Ile Arg Thr Phe Phe Arg Thr Thr Thr
545 550 555 560
Gly Lys Ile Ala Tyr Met Thr Ser Arg Asp Ser Gly Glu Thr Trp Ser
565 570 575
Lys Val Ser Tyr Ile Asp Gly Ile Gln Gln Thr Ser Tyr Gly Thr Gln
580 585 590
Val Ser Ala Ile Lys Tyr Ser Gln Leu Ile Asp Gly Lys Glu Ala Val
595 600 605
Ile Leu Ser Thr Pro Asn Ser Arg Ser Gly Arg Lys Gly Gly Gln Leu
610 615 620
Val Val Gly Leu Val Asn Lys Glu Asp Asp Ser Ile Asp Trp Lys Tyr
625 630 635 640
His Tyr Asp Ile Asp Leu Pro Ser Tyr Gly Tyr Ala Tyr Ser Ala Ile
645 650 655
Thr Glu Leu Pro Asn His His Ile Gly Val Leu Phe Glu Lys Tyr Asp
660 665 670
Ser Trp Ser Arg Asn Glu Leu His Leu Ser Asn Val Val Gln Tyr Ile
675 680 685
Asp Leu Glu Ile Asn Asp Leu Thr Lys
690 695
<210> 7
<211> 2076
<212> ДНК
<213> Bifidobacterium longum
<400> 7
atggaacata gagcgttcaa gtggccgcag ccacttgcgg gcaacaagcc ccgcatctgg 60
tacggcggcg attacaaccc cgaccaatgg cctgaggaag tgtgggacga agatgtagcc 120
ctcatgcagc aggccggcgt caacctcgtc tccgtagcca tcttctcctg ggccaagctt 180
gagcccgaag aaggcgtgta cgacttcgat tggctcgacc gcgtcatcga caagctcggc 240
aaggccggca tcgccgtcga tctcgcctcc ggcaccgcat ccccgccgat gtggatgacc 300
caggcccacc cggagatcct ctgggtcgac taccgcggcg acgtctgcca gcccggtgcc 360
cgccagcact ggcgcgccac cagcccggtc ttccttgact acgcgctcaa cctgtgccgc 420
aagatggccg agcactacaa ggacaacccc tatgtggtct cttggcatgt gagcaacgag 480
tacggctgcc acaaccgctt cgactattcc gaagacgccg agcgcgcctt ccagaagtgg 540
tgcgagaaga agtacggcac catcgacgct gtcaacgacg cctggggcac cgccttctgg 600
gcgcagcgca tgaacaattt ctccgagatc atcccgccgc gattcatcgg cgacggcaac 660
ttcatgaacc cgggcaagct gcttgattgg aagcgtttca gctccgacgc gctgctggac 720
ttctacaagg ccgagcgcga cgccctgctc gagatcgccc ccaagccgca gaccaccaac 780
ttcatggtct ccgcgggctg caccgtcctc gactacgaca agtggggtca tgacgtggac 840
ttcgtgtcca acgaccatta cttctcgccc ggcgaggccc acttcgacga gatggcctac 900
gcggcctgcc tcaccgacgg catcgcccgc aagaacccgt ggttcctcat ggaacattcc 960
acgtccgccg tcaactggcg cccgaccaac taccggctcg agcccggcga gctggtgcgc 1020
gactccctgg cccatctggc catgggcgcc gacgccatct gctacttcca gtggcgtcag 1080
tccaaggccg gcgccgagaa gtggcattcc gccatggtgc cccacgcagg ccccgactcc 1140
cagatcttcc gcgatgtgtg cgagctgggt gccgacctca acaagcttgc tgacgagggc 1200
ctgctgagca ccaagctggt caagtccaag gtcgccatcg tcttcgacta cgagtcccag 1260
tgggccaccg agcacaccgc cacccccacg caggaggtgc gccactggac cgagccgctg 1320
gactggttcc gcgcgctggc ggacaatggc ctgaccgccg acgtggtgcc ggtccgcggt 1380
ccttgggatg agtacgaggc cgtcgtgttg ccgagcctgg ccatcctgtc cgagcagacc 1440
acgcgccgcg tgcgcgagta tgtggcgaac ggcggcaagc tgttcgtgac ctactacacc 1500
ggtctggtgg acgacaggga tcacgtctgg ctgggcggct accccggctc cattcgcgac 1560
gtggtgggcg tgcgcgtcga ggaattcgcc ccgatgggca ccgacgcccc cggcaccatg 1620
gaccaccttg acttggacaa cggaaccgtg gcgcacgatt tcgccgacgt gatcacctcc 1680
gtggccgata ccgctcacgt ggtcgcctcc ttcaaggcag ataagtggac cggtttcgac 1740
ggcgctcccg ccatcaccgt caacgacttc ggcgacggca aggccgcata cgtcggtgcc 1800
cgtctcgggc gtgagggctt ggccaagagc ctgcccgcgc tgctggagga actcggcatc 1860
gagacttcgg ctgaggacga tcgtggtgaa gtgctgcgcg tcgagcgtgc ggacgaaact 1920
ggcgagaacc acttcgtgtt cctgttcaac cgcacccacg atgttgcggt cgtggacgtg 1980
gaaggcgaac cgctggtcgc ctcgctggcc caggtcaacg agtccgagca cacggccgcc 2040
atccagccca acggcgtact cgtcgtcaag ctgtaa 2076
<210> 8
<211> 691
<212> ПРТ
<213> Bifidobacterium longum
<400> 8
Met Glu His Arg Ala Phe Lys Trp Pro Gln Pro Leu Ala Gly Asn Lys
1 5 10 15
Pro Arg Ile Trp Tyr Gly Gly Asp Tyr Asn Pro Asp Gln Trp Pro Glu
20 25 30
Glu Val Trp Asp Glu Asp Val Ala Leu Met Gln Gln Ala Gly Val Asn
35 40 45
Leu Val Ser Val Ala Ile Phe Ser Trp Ala Lys Leu Glu Pro Glu Glu
50 55 60
Gly Val Tyr Asp Phe Asp Trp Leu Asp Arg Val Ile Asp Lys Leu Gly
65 70 75 80
Lys Ala Gly Ile Ala Val Asp Leu Ala Ser Gly Thr Ala Ser Pro Pro
85 90 95
Met Trp Met Thr Gln Ala His Pro Glu Ile Leu Trp Val Asp Tyr Arg
100 105 110
Gly Asp Val Cys Gln Pro Gly Ala Arg Gln His Trp Arg Ala Thr Ser
115 120 125
Pro Val Phe Leu Asp Tyr Ala Leu Asn Leu Cys Arg Lys Met Ala Glu
130 135 140
His Tyr Lys Asp Asn Pro Tyr Val Val Ser Trp His Val Ser Asn Glu
145 150 155 160
Tyr Gly Cys His Asn Arg Phe Asp Tyr Ser Glu Asp Ala Glu Arg Ala
165 170 175
Phe Gln Lys Trp Cys Glu Lys Lys Tyr Gly Thr Ile Asp Ala Val Asn
180 185 190
Asp Ala Trp Gly Thr Ala Phe Trp Ala Gln Arg Met Asn Asn Phe Ser
195 200 205
Glu Ile Ile Pro Pro Arg Phe Ile Gly Asp Gly Asn Phe Met Asn Pro
210 215 220
Gly Lys Leu Leu Asp Trp Lys Arg Phe Ser Ser Asp Ala Leu Leu Asp
225 230 235 240
Phe Tyr Lys Ala Glu Arg Asp Ala Leu Leu Glu Ile Ala Pro Lys Pro
245 250 255
Gln Thr Thr Asn Phe Met Val Ser Ala Gly Cys Thr Val Leu Asp Tyr
260 265 270
Asp Lys Trp Gly His Asp Val Asp Phe Val Ser Asn Asp His Tyr Phe
275 280 285
Ser Pro Gly Glu Ala His Phe Asp Glu Met Ala Tyr Ala Ala Cys Leu
290 295 300
Thr Asp Gly Ile Ala Arg Lys Asn Pro Trp Phe Leu Met Glu His Ser
305 310 315 320
Thr Ser Ala Val Asn Trp Arg Pro Thr Asn Tyr Arg Leu Glu Pro Gly
325 330 335
Glu Leu Val Arg Asp Ser Leu Ala His Leu Ala Met Gly Ala Asp Ala
340 345 350
Ile Cys Tyr Phe Gln Trp Arg Gln Ser Lys Ala Gly Ala Glu Lys Trp
355 360 365
His Ser Ala Met Val Pro His Ala Gly Pro Asp Ser Gln Ile Phe Arg
370 375 380
Asp Val Cys Glu Leu Gly Ala Asp Leu Asn Lys Leu Ala Asp Glu Gly
385 390 395 400
Leu Leu Ser Thr Lys Leu Val Lys Ser Lys Val Ala Ile Val Phe Asp
405 410 415
Tyr Glu Ser Gln Trp Ala Thr Glu His Thr Ala Thr Pro Thr Gln Glu
420 425 430
Val Arg His Trp Thr Glu Pro Leu Asp Trp Phe Arg Ala Leu Ala Asp
435 440 445
Asn Gly Leu Thr Ala Asp Val Val Pro Val Arg Gly Pro Trp Asp Glu
450 455 460
Tyr Glu Ala Val Val Leu Pro Ser Leu Ala Ile Leu Ser Glu Gln Thr
465 470 475 480
Thr Arg Arg Val Arg Glu Tyr Val Ala Asn Gly Gly Lys Leu Phe Val
485 490 495
Thr Tyr Tyr Thr Gly Leu Val Asp Asp Arg Asp His Val Trp Leu Gly
500 505 510
Gly Tyr Pro Gly Ser Ile Arg Asp Val Val Gly Val Arg Val Glu Glu
515 520 525
Phe Ala Pro Met Gly Thr Asp Ala Pro Gly Thr Met Asp His Leu Asp
530 535 540
Leu Asp Asn Gly Thr Val Ala His Asp Phe Ala Asp Val Ile Thr Ser
545 550 555 560
Val Ala Asp Thr Ala His Val Val Ala Ser Phe Lys Ala Asp Lys Trp
565 570 575
Thr Gly Phe Asp Gly Ala Pro Ala Ile Thr Val Asn Asp Phe Gly Asp
580 585 590
Gly Lys Ala Ala Tyr Val Gly Ala Arg Leu Gly Arg Glu Gly Leu Ala
595 600 605
Lys Ser Leu Pro Ala Leu Leu Glu Glu Leu Gly Ile Glu Thr Ser Ala
610 615 620
Glu Asp Asp Arg Gly Glu Val Leu Arg Val Glu Arg Ala Asp Glu Thr
625 630 635 640
Gly Glu Asn His Phe Val Phe Leu Phe Asn Arg Thr His Asp Val Ala
645 650 655
Val Val Asp Val Glu Gly Glu Pro Leu Val Ala Ser Leu Ala Gln Val
660 665 670
Asn Glu Ser Glu His Thr Ala Ala Ile Gln Pro Asn Gly Val Leu Val
675 680 685
Val Lys Leu
690
<210> 9
<211> 1626
<212> ДНК
<213> Clostridium thermocellum
<400> 9
atggcagaag gggttatagt caacggaact cagtttaaag acacatcggg aaatgtgata 60
catgcccatg ggggaggcat gttaaagcat ggtgactatt attactggta cggtgaatac 120
cgggacgact ccaacttgtt tttgggtgta agttgctaca ggtcaaaaga tcttgtaaac 180
tgggaataca gaggagaagt gctgagccga aattccgctc ctgaactgaa tcactgcaat 240
attgaaagac cgaaagtcat gtacaacgca tcaaccggtg aatttgtcat gtggatgcac 300
tgggagaacg gcataaacta cggtcaggca agagcagctg ttgcgtattc caaaacgccc 360
gacggcaaat tcacatacat tcgaagcttt cgtcccatgc aggataccgg cgttatggat 420
catggccttc cgggatatat gtcaagggac tgcaatgtat ttgtggacac tgacggcaag 480
ggatatttta tatccgcagc caatgagaac atggacctgc acctttatga gctgacacct 540
gactataaaa atattgcatc ccttaaggca aagctgtttg tcggacagca gagggaagca 600
ccatgcctta taaagagaaa cggctactat taccttatta cttccggttg tacaggttgg 660
aacccgaatc aggctaaata cgcatattcc aaagatttgg ccagtggctg gtcccagctt 720
tacaatcttg gtaattcaac cacctacagg tcacagccga cttttatcat tcccgttcag 780
ggaagctcgg gaaccagtta tctttatatg ggtgaccgtt gggccggtgc ctggggagga 840
aaggttaatg actcccaata tgtatggctt cccttaaact tcatatccga tacaacactt 900
gaactgccct attatgactc tgtaaagatt gatgcttctt caggaataat ttccgagtac 960
ataccggaca ctacacgcta caagctggta aacaaaaaca gcggaaaagt cctggatgtt 1020
cttgacggtt ctgtcgataa tgcagcccag atagtccaat ggaccgataa cgggtctttg 1080
agtcaacagt ggtaccttgt ggacgtgggc ggtggttata aaaagattgt aaatgtaaag 1140
agcggaagag ccttggatgt aaaagacgaa tccaaggaag acggtggagt attaatacaa 1200
tataccagca acggcggata taatcagcac tggaaattca cagacatagg tgacgggtat 1260
tacaagattt ccagccgcca ctgcggaaaa cttatagatg tgcgaaaatg gtcaacggaa 1320
gacggcggaa taattcagca gtggtccgat gccggaggaa caaatcagca ttggaagctg 1380
gtgcttgtat caagtcccga gccttcacca tcaccttctc cccaagtggt taaaggagat 1440
gtaaacggcg acttgaaagt aaattcaacg gatttttcca tgttaagaag atatttactt 1500
aaaaccattg acaattttcc gacagaaaac ggaaaacagg ctgccgattt gaacggagac 1560
ggcagaataa actcttcgga tcttacaatg ctgaaaagat acttgcttat ggaagtggat 1620
ttgtaa 1626
<210> 10
<211> 541
<212> ПРТ
<213> Clostridium thermocellum
<400> 10
Met Ala Glu Gly Val Ile Val Asn Gly Thr Gln Phe Lys Asp Thr Ser
1 5 10 15
Gly Asn Val Ile His Ala His Gly Gly Gly Met Leu Lys His Gly Asp
20 25 30
Tyr Tyr Tyr Trp Tyr Gly Glu Tyr Arg Asp Asp Ser Asn Leu Phe Leu
35 40 45
Gly Val Ser Cys Tyr Arg Ser Lys Asp Leu Val Asn Trp Glu Tyr Arg
50 55 60
Gly Glu Val Leu Ser Arg Asn Ser Ala Pro Glu Leu Asn His Cys Asn
65 70 75 80
Ile Glu Arg Pro Lys Val Met Tyr Asn Ala Ser Thr Gly Glu Phe Val
85 90 95
Met Trp Met His Trp Glu Asn Gly Ile Asn Tyr Gly Gln Ala Arg Ala
100 105 110
Ala Val Ala Tyr Ser Lys Thr Pro Asp Gly Lys Phe Thr Tyr Ile Arg
115 120 125
Ser Phe Arg Pro Met Gln Asp Thr Gly Val Met Asp His Gly Leu Pro
130 135 140
Gly Tyr Met Ser Arg Asp Cys Asn Val Phe Val Asp Thr Asp Gly Lys
145 150 155 160
Gly Tyr Phe Ile Ser Ala Ala Asn Glu Asn Met Asp Leu His Leu Tyr
165 170 175
Glu Leu Thr Pro Asp Tyr Lys Asn Ile Ala Ser Leu Lys Ala Lys Leu
180 185 190
Phe Val Gly Gln Gln Arg Glu Ala Pro Cys Leu Ile Lys Arg Asn Gly
195 200 205
Tyr Tyr Tyr Leu Ile Thr Ser Gly Cys Thr Gly Trp Asn Pro Asn Gln
210 215 220
Ala Lys Tyr Ala Tyr Ser Lys Asp Leu Ala Ser Gly Trp Ser Gln Leu
225 230 235 240
Tyr Asn Leu Gly Asn Ser Thr Thr Tyr Arg Ser Gln Pro Thr Phe Ile
245 250 255
Ile Pro Val Gln Gly Ser Ser Gly Thr Ser Tyr Leu Tyr Met Gly Asp
260 265 270
Arg Trp Ala Gly Ala Trp Gly Gly Lys Val Asn Asp Ser Gln Tyr Val
275 280 285
Trp Leu Pro Leu Asn Phe Ile Ser Asp Thr Thr Leu Glu Leu Pro Tyr
290 295 300
Tyr Asp Ser Val Lys Ile Asp Ala Ser Ser Gly Ile Ile Ser Glu Tyr
305 310 315 320
Ile Pro Asp Thr Thr Arg Tyr Lys Leu Val Asn Lys Asn Ser Gly Lys
325 330 335
Val Leu Asp Val Leu Asp Gly Ser Val Asp Asn Ala Ala Gln Ile Val
340 345 350
Gln Trp Thr Asp Asn Gly Ser Leu Ser Gln Gln Trp Tyr Leu Val Asp
355 360 365
Val Gly Gly Gly Tyr Lys Lys Ile Val Asn Val Lys Ser Gly Arg Ala
370 375 380
Leu Asp Val Lys Asp Glu Ser Lys Glu Asp Gly Gly Val Leu Ile Gln
385 390 395 400
Tyr Thr Ser Asn Gly Gly Tyr Asn Gln His Trp Lys Phe Thr Asp Ile
405 410 415
Gly Asp Gly Tyr Tyr Lys Ile Ser Ser Arg His Cys Gly Lys Leu Ile
420 425 430
Asp Val Arg Lys Trp Ser Thr Glu Asp Gly Gly Ile Ile Gln Gln Trp
435 440 445
Ser Asp Ala Gly Gly Thr Asn Gln His Trp Lys Leu Val Leu Val Ser
450 455 460
Ser Pro Glu Pro Ser Pro Ser Pro Ser Pro Gln Val Val Lys Gly Asp
465 470 475 480
Val Asn Gly Asp Leu Lys Val Asn Ser Thr Asp Phe Ser Met Leu Arg
485 490 495
Arg Tyr Leu Leu Lys Thr Ile Asp Asn Phe Pro Thr Glu Asn Gly Lys
500 505 510
Gln Ala Ala Asp Leu Asn Gly Asp Gly Arg Ile Asn Ser Ser Asp Leu
515 520 525
Thr Met Leu Lys Arg Tyr Leu Leu Met Glu Val Asp Leu
530 535 540
<210> 11
<211> 2631
<212> ДНК
<213> Paenibacillus sp.
<400> 11
atgaatcgac acgtcctgct tcatccgtat ctccaccgga aggcgttgcc tctgctcctg 60
gccttgacgc tgctgacggg catcgccctg ttcccggcct ccaccgcgca ggcggcgacg 120
accgtgacgt cgatgacgta cttctctgcc aatgacggtc ccgtcatctc caaatccggc 180
gtcgggcaag ccagctacgg tttcgtcatg ccgatcttca acggaggcgc tgcgacctgg 240
aacgatgtcg ccgatgacgt cggcgttcgc gtcaaggtcg gcggcagctg ggtcgacatt 300
gacagcgttg gcggctatgt gtacaaccag aactggggcc attggaacga cagcggcacc 360
tatggctact ggttcaccct ctccgccacg accgagctgc agctctactc caaggcgaac 420
agcagcgtca cactcaacta cacgctcgtc ttccagaatg tcaatgaaac gaccattacc 480
tcgatgacac cgacccaggg cccgcaattg accgcagggt ataccggcgg cgcaggcttc 540
acctatccgg tcttcaacaa cgatccctcc atcccgtatg cagccgtagc cggcgatctg 600
aaggtgtacg tcaagccagt cgccagcagt acctggatcg atatcgacaa caacgcggcg 660
agcggctgga tctacgacag caacttcggc cagttcaccg aaggcggcgg cggctactgg 720
ttcaccgtca ccgagtcgat caacgtcaag ctcgagtcca ggacgtcctc ggccaacgtc 780
gtctatacga tcaacttccc gcagccgacg cgcagcagct acacactctc cgcctatgac 840
ggcacgacct acagcgccga tgcgagcggc gcgatcggta tcccgctgcc gcggatcgac 900
ggcaccccgg cgatcggcag cgagctcggc aacttcgtct accagatcta ccggaacggc 960
cagtgggtcg agatgagcaa ctcggcgcag agcagcttcg tctactcggc caatggctac 1020
aacaacatgt ccgacgccaa tcaatggggc tactgggccg actacatcta cggcctctgg 1080
ttccggccga tccaggagga tatgcagatc cgcatcggct atccgctgaa tggccagtcc 1140
ggcggcagcg tcggcagcaa cttcgtcacc tatacgctga tcggcaaccc gaacgcgccg 1200
cgacccgatg tgagcgacca gggcgacgtc gagatcggca cgcccaccga tccggccatc 1260
gcaggatgga atctgtattg gcaggatgaa ttcgccggca gcgcgctcga tctgaacaag 1320
tggaactacg agaccggcta ctacatcggc aacgacccca atctgtgggg ctggggcaac 1380
gccgagatgc agcactatac gacgagcacg caaaatgtct tcgtcgctga cggcaaactc 1440
aacatccgag cgctccacga ttaccaatcg ttcccgcagg acccgaaccg ctacgcgacc 1500
tactcctccg gcaagatcaa caccaaggac aacatgtcgc tgcagtacgg ccgcgtcgat 1560
atccgcgcca agctgccgac tggcgatggc gtctggccgg cactgtggat gctgccggag 1620
gactccgtct acggcgcatg ggcggcatca ggagagatcg acatcatgga ggcgaagggc 1680
cgtctgcccg gcacgacgag cggcgcgatc cactacggcg gccaatggcc ggtcaaccgc 1740
tacctcgccg gagaatgcta cctcccgcaa ggtacgacat tcgccgacga ctttaatgtg 1800
tacacgatga tctgggaaga ggacaacatg aagtggtacg ttaacggtga gtttttcttc 1860
aaggtgacgc gcgagcagtg gtactccgtc gccgccccca acaatccgga cgcgccgttc 1920
gaccagccgt tctatctgat catgaacctg gcggtcggcg gccacttcga cggcgggcgt 1980
acgcccgacc cgtccgacat cccggcgacg atgcagatcg actacgtgcg ggtgtacaaa 2040
gagggcgcgg gcggcggtcc gggcaacccg ggcggcaacg tcgcggtgac cggcgttagc 2100
gtgaccccgg caacggcgca ggtgcaggtc ggtcagaccg tctcgctgag cgccaacgtc 2160
gcgccagcca atgcaacgaa caagcaagtg acctggtcag tcgccaatgg cagcatcgcc 2220
tcggtgagcg ccagcggcgt cgtcagtgga ctcgctgctg gcacgacgac cgtaaccgcc 2280
acgaccgcag acggcaaccg caccgcctcg gcgacgatca ccgtcgtgcc gccaccgacg 2340
acgaccgtca tcatcggcga tagcgtgcgc ggcatccgaa agaccggcga caacctgctc 2400
ttctacgtca acggcgcaac ctacgccgac ctgcactaca aggtgaacgg cggcggtcag 2460
cctaatgtcg cgatgacgca cacaggaggc ggcaactaca cctacccggt gcatggcctc 2520
caacaaggcg ataccgtcga atacttcttc acctacaacc ccggcaacgg cgcgctagac 2580
acgccttggc agacttatgt gcatggggta acacaaggtg ttgttgagta a 2631
<210> 12
<211> 876
<212> ПРТ
<213> Paenibacillus sp.
<400> 12
Met Asn Arg His Val Leu Leu His Pro Tyr Leu His Arg Lys Ala Leu
1 5 10 15
Pro Leu Leu Leu Ala Leu Thr Leu Leu Thr Gly Ile Ala Leu Phe Pro
20 25 30
Ala Ser Thr Ala Gln Ala Ala Thr Thr Val Thr Ser Met Thr Tyr Phe
35 40 45
Ser Ala Asn Asp Gly Pro Val Ile Ser Lys Ser Gly Val Gly Gln Ala
50 55 60
Ser Tyr Gly Phe Val Met Pro Ile Phe Asn Gly Gly Ala Ala Thr Trp
65 70 75 80
Asn Asp Val Ala Asp Asp Val Gly Val Arg Val Lys Val Gly Gly Ser
85 90 95
Trp Val Asp Ile Asp Ser Val Gly Gly Tyr Val Tyr Asn Gln Asn Trp
100 105 110
Gly His Trp Asn Asp Ser Gly Thr Tyr Gly Tyr Trp Phe Thr Leu Ser
115 120 125
Ala Thr Thr Glu Leu Gln Leu Tyr Ser Lys Ala Asn Ser Ser Val Thr
130 135 140
Leu Asn Tyr Thr Leu Val Phe Gln Asn Val Asn Glu Thr Thr Ile Thr
145 150 155 160
Ser Met Thr Pro Thr Gln Gly Pro Gln Leu Thr Ala Gly Tyr Thr Gly
165 170 175
Gly Ala Gly Phe Thr Tyr Pro Val Phe Asn Asn Asp Pro Ser Ile Pro
180 185 190
Tyr Ala Ala Val Ala Gly Asp Leu Lys Val Tyr Val Lys Pro Val Ala
195 200 205
Ser Ser Thr Trp Ile Asp Ile Asp Asn Asn Ala Ala Ser Gly Trp Ile
210 215 220
Tyr Asp Ser Asn Phe Gly Gln Phe Thr Glu Gly Gly Gly Gly Tyr Trp
225 230 235 240
Phe Thr Val Thr Glu Ser Ile Asn Val Lys Leu Glu Ser Arg Thr Ser
245 250 255
Ser Ala Asn Val Val Tyr Thr Ile Asn Phe Pro Gln Pro Thr Arg Ser
260 265 270
Ser Tyr Thr Leu Ser Ala Tyr Asp Gly Thr Thr Tyr Ser Ala Asp Ala
275 280 285
Ser Gly Ala Ile Gly Ile Pro Leu Pro Arg Ile Asp Gly Thr Pro Ala
290 295 300
Ile Gly Ser Glu Leu Gly Asn Phe Val Tyr Gln Ile Tyr Arg Asn Gly
305 310 315 320
Gln Trp Val Glu Met Ser Asn Ser Ala Gln Ser Ser Phe Val Tyr Ser
325 330 335
Ala Asn Gly Tyr Asn Asn Met Ser Asp Ala Asn Gln Trp Gly Tyr Trp
340 345 350
Ala Asp Tyr Ile Tyr Gly Leu Trp Phe Arg Pro Ile Gln Glu Asp Met
355 360 365
Gln Ile Arg Ile Gly Tyr Pro Leu Asn Gly Gln Ser Gly Gly Ser Val
370 375 380
Gly Ser Asn Phe Val Thr Tyr Thr Leu Ile Gly Asn Pro Asn Ala Pro
385 390 395 400
Arg Pro Asp Val Ser Asp Gln Gly Asp Val Glu Ile Gly Thr Pro Thr
405 410 415
Asp Pro Ala Ile Ala Gly Trp Asn Leu Tyr Trp Gln Asp Glu Phe Ala
420 425 430
Gly Ser Ala Leu Asp Leu Asn Lys Trp Asn Tyr Glu Thr Gly Tyr Tyr
435 440 445
Ile Gly Asn Asp Pro Asn Leu Trp Gly Trp Gly Asn Ala Glu Met Gln
450 455 460
His Tyr Thr Thr Ser Thr Gln Asn Val Phe Val Ala Asp Gly Lys Leu
465 470 475 480
Asn Ile Arg Ala Leu His Asp Tyr Gln Ser Phe Pro Gln Asp Pro Asn
485 490 495
Arg Tyr Ala Thr Tyr Ser Ser Gly Lys Ile Asn Thr Lys Asp Asn Met
500 505 510
Ser Leu Gln Tyr Gly Arg Val Asp Ile Arg Ala Lys Leu Pro Thr Gly
515 520 525
Asp Gly Val Trp Pro Ala Leu Trp Met Leu Pro Glu Asp Ser Val Tyr
530 535 540
Gly Ala Trp Ala Ala Ser Gly Glu Ile Asp Ile Met Glu Ala Lys Gly
545 550 555 560
Arg Leu Pro Gly Thr Thr Ser Gly Ala Ile His Tyr Gly Gly Gln Trp
565 570 575
Pro Val Asn Arg Tyr Leu Ala Gly Glu Cys Tyr Leu Pro Gln Gly Thr
580 585 590
Thr Phe Ala Asp Asp Phe Asn Val Tyr Thr Met Ile Trp Glu Glu Asp
595 600 605
Asn Met Lys Trp Tyr Val Asn Gly Glu Phe Phe Phe Lys Val Thr Arg
610 615 620
Glu Gln Trp Tyr Ser Val Ala Ala Pro Asn Asn Pro Asp Ala Pro Phe
625 630 635 640
Asp Gln Pro Phe Tyr Leu Ile Met Asn Leu Ala Val Gly Gly His Phe
645 650 655
Asp Gly Gly Arg Thr Pro Asp Pro Ser Asp Ile Pro Ala Thr Met Gln
660 665 670
Ile Asp Tyr Val Arg Val Tyr Lys Glu Gly Ala Gly Gly Gly Pro Gly
675 680 685
Asn Pro Gly Gly Asn Val Ala Val Thr Gly Val Ser Val Thr Pro Ala
690 695 700
Thr Ala Gln Val Gln Val Gly Gln Thr Val Ser Leu Ser Ala Asn Val
705 710 715 720
Ala Pro Ala Asn Ala Thr Asn Lys Gln Val Thr Trp Ser Val Ala Asn
725 730 735
Gly Ser Ile Ala Ser Val Ser Ala Ser Gly Val Val Ser Gly Leu Ala
740 745 750
Ala Gly Thr Thr Thr Val Thr Ala Thr Thr Ala Asp Gly Asn Arg Thr
755 760 765
Ala Ser Ala Thr Ile Thr Val Val Pro Pro Pro Thr Thr Thr Val Ile
770 775 780
Ile Gly Asp Ser Val Arg Gly Ile Arg Lys Thr Gly Asp Asn Leu Leu
785 790 795 800
Phe Tyr Val Asn Gly Ala Thr Tyr Ala Asp Leu His Tyr Lys Val Asn
805 810 815
Gly Gly Gly Gln Pro Asn Val Ala Met Thr His Thr Gly Gly Gly Asn
820 825 830
Tyr Thr Tyr Pro Val His Gly Leu Gln Gln Gly Asp Thr Val Glu Tyr
835 840 845
Phe Phe Thr Tyr Asn Pro Gly Asn Gly Ala Leu Asp Thr Pro Trp Gln
850 855 860
Thr Tyr Val His Gly Val Thr Gln Gly Val Val Glu
865 870 875
<210> 13
<211> 6783
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Транспозонная кассета
<400> 13
gccagatgat taattcctaa tttttgttga cactctatca ttgatagagt tattttacca 60
ctccctatca gtgatagaga aaagtgaaat gaatagttcg acaaaaatct agaaataatt 120
ttgtttaact ttaagaagga gatatacaat ttcgtcgaca cacaggaaac atattaaaaa 180
ttaaaacctg caggagtttg aaggagatag aaccatggcg cagtcgaaac tctatccagt 240
tgtgatggca ggtggctccg gtagccgctt atggccgctt tcccgcgtac tttatcccaa 300
gcagttttta tgcctgaaag gcgatctcac catgctgcaa accaccatct gccgcctgaa 360
cggcgtggag tgcgaaagcc cggtggtgat ttgcaatgag cagcaccgct ttattgtcgc 420
ggaacagctg cgtcaactga acaaacttac cgagaacatt attctcgaac cggcagggcg 480
aaacacggca cctgccattg cgctggcggc gctggcggca aaacgtcata gcccggagag 540
cgacccgtta atgctggtat tggcggcgga tcatgtgatt gccgatgaag acgcgttccg 600
tgccgccgtg cgtaatgcca tgccatatgc cgaagcgggc aagctggtga ccttcggcat 660
tgtgccggat ctaccagaaa ccggttatgg ctatattcgt cgcggtgaag tgtctgcggg 720
tgagcaggat atggtggcct ttgaagtggc gcagtttgtc gaaaaaccga atctggaaac 780
cgctcaggcc tatgtggcaa gcggcgaata ttactggaac agcggtatgt tcctgttccg 840
cgccggacgc tatctcgaag aactgaaaaa atatcgcccg gatatcctcg atgcctgtga 900
aaaagcgatg agcgccgtcg atccggatct caattttatt cgcgtggatg aagaagcgtt 960
tctcgcctgc ccggaagagt cggtggatta cgcggtcatg gaacgtacgg cagatgctgt 1020
tgtggtgccg atggatgcgg gctggagcga tgttggctcc tggtcttcat tatgggagat 1080
cagcgcccac accgccgagg gcaacgtttg ccacggcgat gtgattaatc acaaaactga 1140
aaacagctat gtgtatgctg aatctggcct ggtcaccacc gtcggggtga aagatctggt 1200
agtggtgcag accaaagatg cggtgctgat tgccgaccgt aacgcggtac aggatgtgaa 1260
aaaagtggtc gagcagatca aagccgatgg tcgccatgag catcgggtgc atcgcgaagt 1320
gtatcgtccg tggggcaaat atgactctat cgacgcgggc gaccgctacc aggtgaaacg 1380
catcaccgtg aaaccgggcg agggcttgtc ggtacagatg caccatcacc gcgcggaaca 1440
ctgggtggtt gtcgcgggaa cggcaaaagt caccattgat ggtgatatca aactgcttgg 1500
tgaaaacgag tccatttata ttccgctggg ggcgacgcat tgcctggaaa acccggggaa 1560
aattccgctc gatttaattg aagtgcgctc cggctcttat ctcgaagagg atgatgtggt 1620
gcgtttcgcg gatcgctacg gacgggtgta aacgtcgcat caggcaatga atgcgaaacc 1680
gcggtgtaaa taacgacaaa aataaaattg gccgcttcgg tcagggccaa ctattgcctg 1740
aaaaagggta acgatatgaa aaaattaacc tgctttaaag cctatgatat tcgcgggaaa 1800
ttaggcgaag aactgaatga agatatcgcc tggcgcattg gtcgcgccta tggcgaattt 1860
ctcaaaccga aaaccattgt gttaggcggt gatgtccgcc tcaccagcga aaccttaaaa 1920
ctggcgctgg cgaaaggttt acaggatgcg ggcgttgacg tgctggatat tggtatgtcc 1980
ggcaccgaag agatctattt cgccacgttc catctcggcg tggatggcgg cattgaagtt 2040
accgccagcc ataatccgat ggattataac ggcatgaagc tggttcgcga gggggctcgc 2100
ccgatcagcg gagataccgg actgcgcgac gtccagcgtc tggctgaagc caacgacttt 2160
cctcccgtcg atgaaaccaa acgcggtcgc tatcagcaaa tcaacctgcg tgacgcttac 2220
gttgatcacc tgttcggtta tatcaatgtc aaaaacctca cgccgctcaa gctggtgatc 2280
aactccggga acggcgcagc gggtccggtg gtggacgcca ttgaagcccg ctttaaagcc 2340
ctcggcgcgc ccgtggaatt aatcaaagtg cacaacacgc cggacggcaa tttccccaac 2400
ggtattccta acccactact gccggaatgc cgcgacgaca cccgcaatgc ggtcatcaaa 2460
cacggcgcgg atatgggcat tgcttttgat ggcgattttg accgctgttt cctgtttgac 2520
gaaaaagggc agtttattga gggctactac attgtcggcc tgttggcaga agcattcctc 2580
gaaaaaaatc ccggcgcgaa gatcatccac gatccacgtc tctcctggaa caccgttgat 2640
gtggtgactg ccgcaggtgg cacgccggta atgtcgaaaa ccggacacgc ctttattaaa 2700
gaacgtatgc gcaaggaaga cgccatctat ggtggcgaaa tgagcgccca ccattacttc 2760
cgtgatttcg cttactgcga cagcggcatg atcccgtggc tgctggtcgc cgaactggtg 2820
tgcctgaaag ataaaacgct gggcgaactg gtacgcgacc ggatggcggc gtttccggca 2880
agcggtgaga tcaacagcaa actggcgcaa cccgttgagg cgattaaccg cgtggaacag 2940
cattttagcc gtgaggcgct ggcggtggat cgcaccgatg gcatcagcat gacctttgcc 3000
gactggcgct ttaacctgcg cacctccaat accgaaccgg tggtgcgcct gaatgtggaa 3060
tcgcgcggtg atgtgccgct gatggaagcg cgaacgcgaa ctctgctgac gttgctgaac 3120
gagtaaaaac gcggccgcga tatcgttgta aaacgacggc cagtgcaaga atcataaaaa 3180
atttatttgc tttcaggaaa atttttctgt ataatagatt cataaatttg agagaggagt 3240
ttttgtgagc ggataacaat tccccatctt agtatattag ttaagtataa atacaccgcg 3300
gaggacgaag gagatagaac catgtcaaaa gtcgctctca tcaccggtgt aaccggacaa 3360
gacggttctt acctggcaga gtttctgctg gaaaaaggtt acgaggtgca tggtattaag 3420
cgtcgcgcat cgtcattcaa caccgagcgc gtggatcaca tttatcagga tccgcacacc 3480
tgcaacccga aattccatct gcattatggc gacctgagtg atacctctaa cctgacgcgc 3540
attttgcgtg aagtacagcc ggatgaagtg tacaacctgg gcgcaatgag ccacgttgcg 3600
gtctcttttg agtcaccaga atataccgct gacgtcgacg cgatgggtac gctgcgcctg 3660
ctggaggcga tccgcttcct cggtctggaa aagaaaactc gtttctatca ggcttccacc 3720
tctgaactgt atggtctggt gcaggaaatt ccgcagaaag agaccacgcc gttctacccg 3780
cgatctccgt atgcggtcgc caaactgtac gcctactgga tcaccgttaa ctaccgtgaa 3840
tcctacggca tgtacgcctg taacggaatt ctcttcaacc atgaatcccc gcgccgcggc 3900
gaaaccttcg ttacccgcaa aatcacccgc gcaatcgcca acatcgccca ggggctggag 3960
tcgtgcctgt acctcggcaa tatggattcc ctgcgtgact ggggccacgc caaagactac 4020
gtaaaaatgc agtggatgat gctgcagcag gaacagccgg aagatttcgt tatcgcgacc 4080
ggcgttcagt actccgtgcg tcagttcgtg gaaatggcgg cagcacagct gggcatcaaa 4140
ctgcgctttg aaggcacggg cgttgaagag aagggcattg tggtttccgt caccgggcat 4200
gacgcgccgg gcgttaaacc gggtgatgtg attatcgctg ttgacccgcg ttacttccgt 4260
ccggctgaag ttgaaacgct gctcggcgac ccgaccaaag cgcacgaaaa actgggctgg 4320
aaaccggaaa tcaccctcag agagatggtg tctgaaatgg tggctaatga cctcgaagcg 4380
gcgaaaaaac actctctgct gaaatctcac ggctacgacg tggcgatcgc gctggagtca 4440
taagcatgag taaacaacga gtttttattg ctggtcatcg cgggatggtc ggttccgcca 4500
tcaggcggca gctcgaacag cgcggtgatg tggaactggt attacgcacc cgcgacgagc 4560
tgaacctgct ggacagccgc gccgtgcatg atttctttgc cagcgaacgt attgaccagg 4620
tctatctggc ggcggcgaaa gtgggcggca ttgttgccaa caacacctat ccggcggatt 4680
tcatctacca gaacatgatg attgagagca acatcattca cgccgcgcat cagaacgacg 4740
tgaacaaact gctgtttctc ggatcgtcct gcatctaccc gaaactggca aaacagccga 4800
tggcagaaag cgagttgttg cagggcacgc tggagccgac taacgagcct tatgctattg 4860
ccaaaatcgc cgggatcaaa ctgtgcgaat catacaaccg ccagtacgga cgcgattacc 4920
gctcagtcat gccgaccaac ctgtacgggc cacacgacaa cttccacccg agtaattcgc 4980
atgtgatccc agcattgctg cgtcgcttcc acgaggcgac ggcacagaat gcgccggacg 5040
tggtggtatg gggcagcggt acaccgatgc gcgaatttct gcacgtcgat gatatggcgg 5100
cggcgagcat tcatgtcatg gagctggcgc atgaagtctg gctggagaac acccagccga 5160
tgttgtcgca cattaacgtc ggcacgggcg ttgactgcac tatccgcgag ctggcgcaaa 5220
ccatcgccaa agtggtgggt tacaaaggcc gggtggtttt tgatgccagc aaaccggatg 5280
gcacgccgcg caaactgctg gatgtgacgc gcctgcatca gcttggctgg tatcacgaaa 5340
tctcactgga agcggggctt gccagcactt accagtggtt ccttgagaat caagaccgct 5400
ttcggggggg gagctaacgc gccatttaaa tcaacctcag cggtcatagc tgtttcctgt 5460
gactgagcaa taactagcat aaccccttgg ggcctctaaa cgggtcttga ggggtttttt 5520
gctgaaacca atttgcctgg cggcagtagc gcggtggtcc cacctgaccc catgccgaac 5580
tcagaagtga aacgccgtag cgccgatggt agtgtggggt ctccccatgc gagagtaggg 5640
aactgccagg catcaaataa aacgaaaggc tcagtcgaaa gactgggcct ttcgggatcc 5700
aggccggcct gttaacgaat taatcttccg cggcggtatc gataagcttg atatcgaatt 5760
ccgaagttcc tattctctag aaagtatagg aacttcaggt ctgaagagga gtttacgtcc 5820
agccaagcta gcttggctgc aggtcgtcga aattctaccg ggtaggggag gcgcttttcc 5880
caaggcagtc tggagcatgc gctttagcag ccccgctggg cacttggcgc tacacaagtg 5940
gcctctggcc tcgcacacat tccacatcca ccggtaggcg ccaaccggct ccgttctttg 6000
gtggcccctt cgcgccacct tctactcctc ccctagtcag gaagttcccc cccgccccgc 6060
agctcgcgtc gtgcaggacg tgacaaatgg aagtagcacg tctcactagt ctcgtgcaga 6120
tggacagcac cgctgagcaa tggaagcggg taggcctttg gggcagcggc caatagcagc 6180
tttgctcctt cgctttctgg gctcagaggc tgggaagggg tgggtccggg ggcgggctca 6240
ggggcgggct caggggcggg gcgggcgccc gaaggtcctc cggaggcccg gcattctgca 6300
cgcttcaaaa gcgcacgtct gccgcgctgt tctcctcttc ctcatctccg ggcctttcga 6360
cctgcagcct gttgacaatt aatcatcggc atagtatatc ggcatagtat aatacgacaa 6420
ggtgaggaac taaaccatgg gtcaaagtag cgatgaagcc aacgctcccg ttgcagggca 6480
gtttgcgctt cccctgagtg ccacctttgg cttaggggat cgcgtacgca agaaatctgg 6540
tgccgcttgg cagggtcaag tcgtcggttg gtattgcaca aaactcactc ctgaaggcta 6600
tgcggtcgag tccgaatccc acccaggctc agtgcaaatt tatcctgtgg ctgcacttga 6660
acgtgtggcc taatgagggg atcaattctc tagagctcgc tgatcagaag ttcctattct 6720
ctagaaagta taggaacttc gatggcgcct catccctgaa gccaataggg ataacagggt 6780
aat 6783
<210> 14
<211> 2851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета интеграции
<400> 14
tggccagatg attaattcct aatttttgtt gacactctat cattgataga gttattttac 60
cactccctat cagtgataga gaaaagtgaa atgaatagtt cgacaaaaat ctagaaataa 120
ttttgtttaa ctttaagaag gagatataca aatgtactat ttaaaaaaca caaacttttg 180
gatgttcggt ttattctttt tcttttactt ttttatcatg ggagcctact tcccgttttt 240
cccgatttgg ctacatgaca tcaaccatat cagcaaaagt gatacgggta ttatttttgc 300
cgctatttct ctgttctcgc tattattcca accgctgttt ggtctgcttt ctgacaaact 360
cgggctgcgc aaatacctgc tgtggattat taccggcatg ttagtgatgt ttgcgccgtt 420
ctttattttt atcttcgggc cactgttaca atacaacatt ttagtaggat cgattgttgg 480
tggtatttat ctaggctttt gttttaacgc cggtgcgcca gcagtagagg catttattga 540
gaaagtcagc cgtcgcagta atttcgaatt tggtcgcgcg cggatgtttg gctgtgttgg 600
ctgggcgctg tgtgcctcga ttgtcggcat catgttcacc atcaataatc agtttgtttt 660
ctggctgggc tctggctgtg cactcatcct cgccgtttta ctctttttcg ccaaaacgga 720
tgcgccctct tctgccacgg ttgccaatgc ggtaggtgcc aaccattcgg catttagcct 780
taagctggca ctggaactgt tcagacagcc aaaactgtgg tttttgtcac tgtatgttat 840
tggcgtttcc tgcacctacg atgtttttga ccaacagttt gctaatttct ttacttcgtt 900
ctttgctacc ggtgaacagg gtacgcgggt atttggctac gtaacgacaa tgggcgaatt 960
acttaacgcc tcgattatgt tctttgcgcc actgatcatt aatcgcatcg gtgggaaaaa 1020
cgccctgctg ctggctggca ctattatgtc tgtacgtatt attggctcat cgttcgccac 1080
ctcagcgctg gaagtggtta ttctgaaaac gctgcatatg tttgaagtac cgttcctgct 1140
ggtgggctgc tttaaatata ttaccagcca gtttgaagtg cgtttttcag cgacgattta 1200
tctggtctgt ttctgcttct ttaagcaact ggcgatgatt tttatgtctg tactggcggg 1260
caatatgtat gaaagcatcg gtttccaggg cgcttatctg gtgctgggtc tggtggcgct 1320
gggcttcacc ttaatttccg tgttcacgct tagcggcccc ggcccgcttt ccctgctgcg 1380
tcgtcaggtg aatgaagtcg ctgggagcta agcggccgcg tcgacacgca aaaaggccat 1440
ccgtcaggat ggccttctgc ttaatttgat gcctggcagt ttatggcggg cgtcctgccc 1500
gccaccctcc gggccgttgc ttcgcaacgt tcaaatccgc tcccggcgga tttgtcctac 1560
tcaggagagc gttcaccgac aaacaacaga taaaacgaaa ggcccagtct ttcgactgag 1620
cctttcgttt tatttgatgc ctggcagttc cctactctcg catggggaga ccccacacta 1680
ccatcatgta tgaatatcct ccttagttcc tattccgaag ttcctattct ctagaaagta 1740
taggaacttc ggcgcgtcct acctgtgaca cgcgtgccgc agtctcacgc ccggagcgta 1800
gcgaccgagt gagctagcta tttgtttatt tttctaaata cattcaaata tgtatccgct 1860
catgagacaa taaccctgat aaatgcttca ataatattga aaaaggaaga gtatgaggga 1920
agcggtgatc gccgaagtat cgactcaact atcagaggta gttggcgtca tcgagcgcca 1980
tctcgaaccg acgttgctgg ccgtacattt gtacggctcc gcagtggatg gcggcctgaa 2040
gccacacagt gatattgatt tgctggttac ggtgaccgta aggcttgatg aaacaacgcg 2100
gcgagctttg atcaacgacc ttttggaaac ttcggcttcc cctggagaga gcgagattct 2160
ccgcgctgta gaagtcacca ttgttgtgca cgacgacatc attccgtggc gttatccagc 2220
taagcgcgaa ctgcaatttg gagaatggca gcgcaatgac attcttgcag gtatcttcga 2280
gccagccacg atcgacattg atctggctat cttgctgaca aaagcaagag aacatagcgt 2340
tgccttggta ggtccagcgg cggaggaact ctttgatccg gttcctgaac aggatctatt 2400
tgaggcgcta aatgaaacct taacgctatg gaactcgccg cccgactggg ctggcgatga 2460
gcgaaatgta gtgcttacgt tgtcccgcat ttggtacagc gcagtaaccg gcaaaatcgc 2520
gccgaaggat gtcgctgccg actgggcaat ggagcgcctg ccggcccagt atcagcccgt 2580
catacttgaa gctagacagg cttatcttgg acaagaagaa gatcgcttgg cctcgcgcgc 2640
agatcagttg gaagaatttg tccactacgt gaaaggcgag atcaccaagg tagtcggcaa 2700
ataatgtcta acaattcgtt caagccgagg ggccgcaaga tccggccacg atgacccggt 2760
cgtcgggtac cggcagggcg gggcgtaagg cgcgccattt aaatgaagtt cctattccga 2820
agttcctatt ctctagaaag tataggaact t 2851
<210> 15
<211> 2858
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета интеграции
<400> 15
ggccagatga ttaattccta atttttgttg acactctatc attgatagag ttattttacc 60
actccctatc agtgatagag aaaagtgaaa tgaatagttc gacaaaaatc tagaaataat 120
tttgtttaac tttaagaagg agatatacaa atgggcagca ttattcgtct gcagggtggt 180
ctgggtaatc agctgtttca gtttagcttt ggttatgccc tgagcaaaat taatggtaca 240
ccgctgtatt tcgacattag ccattatgcc gaaaacgatg atcatggtgg ttatcgtctg 300
aataatctgc agattccgga agaatatctg cagtattata ccccgaaaat taataatatt 360
tataaactgc tggtgcgtgg cagccgtctg tatccggata tttttctgtt tctgggcttt 420
tgcaacgaat ttcatgccta tggctacgat tttgaatata ttgcccagaa atggaaaagc 480
aaaaaataca ttggctactg gcagagcgaa cacttttttc ataaacatat tctggacctg 540
aaagaatttt ttattccgaa aaatgtgagc gaacaggcaa atctgctggc agcaaaaatt 600
ctggaaagcc agagcagcct gagcattcat attcgtcgtg gcgattatat taaaaacaaa 660
accgcaaccc tgacacatgg tgtttgtagc ctggaatatt ataaaaaagc cctgaacaaa 720
atccgcgatc tggcaatgat tcgtgatgtg tttatcttta gcgacgatat cttctggtgc 780
aaagaaaata ttgaaaccct gctgagcaaa aaatataata tttattatag cgaagatctg 840
agccaagaag aggatctgtg gctgatgagc ctggcaaatc atcatattat tgccaatagc 900
agctttagtt ggtggggtgc atatctgggt agcagcgcaa gccagattgt tatttatccg 960
accccgtggt atgatattac cccgaaaaac acctatatcc cgattgtgaa ccattggatc 1020
aacgttgata aacatagcag ctgctaagcg gccgcgtcga cacgcaaaaa ggccatccgt 1080
caggatggcc ttctgcttaa tttgatgcct ggcagtttat ggcgggcgtc ctgcccgcca 1140
ccctccgggc cgttgcttcg caacgttcaa atccgctccc ggcggatttg tcctactcag 1200
gagagcgttc accgacaaac aacagataaa acgaaaggcc cagtctttcg actgagcctt 1260
tcgttttatt tgatgcctgg cagttcccta ctctcgcatg gggagacccc acactaccat 1320
catgtatgaa tatcctcctt agttcctatt ccgaagttcc tattctctag aaagtatagg 1380
aacttcggcg cgtcctacct gtgacacgcg tcaagatccc ctcacgctgc cgcaagcact 1440
cagggcgcaa gggctgctaa aggaagcgga acacgtagaa agccagtccg cagaaacggt 1500
gctgaccccg gatgaatgtc agctactggg ctatctggac aagggaaaac gcaagcgcaa 1560
agagaaagca ggtagcttgc agtgggctta catggcgata gctagactgg gcggttttat 1620
ggacagcaag cgaaccggaa ttgccagctg gggcgccctc tggtaaggtt gggaagccct 1680
gcaaagtaaa ctggatggct ttcttgccgc caaggatctg atggcgcagg ggatcaagat 1740
ctgatcaaga gacaggatga ggatcgtttc gcatgattga acaagatgga ttgcacgcag 1800
gttctccggc cgcttgggtg gagaggctat tcggctatga ctgggcacaa cagacaatcg 1860
gctgctctga tgccgccgtg ttccggctgt cagcgcaggg gcgcccggtt ctttttgtca 1920
agaccgacct gtccggtgcc ctgaatgaac tgcaggacga ggcagcgcgg ctatcgtggc 1980
tggccacgac gggcgttcct tgcgcagctg tgctcgacgt tgtcactgaa gcgggaaggg 2040
actggctgct attgggcgaa gtgccggggc aggatctcct gtcatctcac cttgctcctg 2100
ccgagaaagt atccatcatg gctgatgcaa tgcggcggct gcatacgctt gatccggcta 2160
cctgcccatt cgaccaccaa gcgaaacatc gcatcgagcg agcacgtact cggatggaag 2220
ccggtcttgt cgatcaggat gatctggacg aagagcatca ggggctcgcg ccagccgaac 2280
tgttcgccag gctcaaggcg cgcatgcccg acggcgagga tctcgtcgtg acccatggcg 2340
atgcctgctt gccgaatatc atggtggaaa atggccgctt ttctggattc atcgactgtg 2400
gccggctggg tgtggcggac cgctatcagg acatagcgtt ggctacccgt gatattgctg 2460
aagagcttgg cggcgaatgg gctgaccgct tcctcgtgct ttacggtatc gccgctcccg 2520
attcgcagcg catcgccttc tatcgccttc ttgacgagtt cttctgagcg ggactctggg 2580
gttcgaaatg accgaccaag cgacgcccaa cctgccatca cgagatttcg attccaccgc 2640
cgccttctat gaaaggttgg gcttcggaat cgttttccgg gacgccggct ggatgatcct 2700
ccagcgcggg gatctcatgc tggagttctt cgcccacccc agcttcaaaa gcgctctcgg 2760
taccggcagg gcggggcgta aggcgcgcca tttaaatgaa gttcctattc cgaagttcct 2820
attctctaga aagtatagga acttcgaagc agctccag 2858
<210> 16
<211> 2631
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета интеграции
<400> 16
ggccagatga ttaattccta atttttgttg acactctatc attgatagag ttattttacc 60
actccctatc agtgatagag aaaagtgaaa tgaatagttc gacaaaaatc tagaaataat 120
tttgtttaac tttaagaagg agatatacaa atgaagtcgg cactgacctt ttcccgtcgc 180
atcaatccgg tgtttctggc gttctttgtc gttgcttttc tgagcggtat cgcaggcgca 240
ctgcaggctc cgaccctgag tctgtttctg tccacggaag tgaaagttcg tccgctgtgg 300
gttggtctgt tctataccgt caacgcaatc gctggcatta cggttagctt tatcctggcg 360
aaacgttcag attcgcgcgg tgaccgtcgc aagctgatta tggtgtgcta tctgatggcg 420
gttggcaact gtctgctgtt tgccttcaat cgtgattacc tgaccctgat cacggcaggt 480
gtgctgctgg cgagcgttgc caacaccgca atgccgcaga ttttcgcgct ggcccgtgaa 540
tatgccgaca gctctgcacg cgaagtggtt atgtttagtt ccatcatgcg cgctcaactg 600
agtctggcat gggtgattgg tccgccgctg tcctttatgc tggcgctgaa ttacggtttt 660
accctgatgt tctcaatcgc ggccggcatt ttcgttctgt cggccctggt cgtgtggttt 720
atcctgccga gtgtcccgcg tgcagaaccg gttgtcgatg caccggtggt tgtccagggt 780
tcactgttcg cagacaaaaa cgttctgctg ctgtttatcg cgtcgatgct gatgtggacc 840
tgcaatacga tgtatattat cgatatgccg ctgtacatta ccgcaagcct gggtctgccg 900
gaacgtctgg ctggtctgct gatgggtacc gcagctggcc tggaaattcc gatcatgctg 960
ctggcgggtt attctgtgcg ttactttggc aaacgcaaga ttatgctgtt cgctgttctg 1020
gcgggtgtcc tgttttatac cggcctggtt ctgtttaaat tcaagacggc cctgatgctg 1080
ctgcagatct ttaacgcaat tttcatcggt attgtggctg gcattggtat gctgtacttc 1140
caagatctga tgccgggtcg tgcaggtgca gcaaccacgc tgtttaccaa tagcatctct 1200
acgggtgtca ttctggcagg cgtgctgcaa ggcggtctga ccgaaacgtg gggccatgac 1260
agcgtctatg tgatggcgat ggtcctgtct attctggccc tgattatctg tgcacgtgtg 1320
cgcgaagctt aaatcgatac tagcataacc ccttggggcc tctaaacgcg tcgacacgca 1380
aaaaggccat ccgtcaggat ggccttctgc ttaatttgat gcctggcagt ttatggcggg 1440
cgtcctgccc gccaccctcc gggccgttgc ttcgcaacgt tcaaatccgc tcccggcgga 1500
tttgtcctac tcaggagagc gttcaccgac aaacaacaga taaaacgaaa ggcccagtct 1560
ttcgactgag cctttcgttt tatttgatgc ctggcagttc cctactctcg catggggaga 1620
ccccacacta ccatcatgta tgaatatcct ccttagttcc tattccgaag ttcctattct 1680
ctagaaagta taggaacttc ggcgcgtcct acctgtgacg gaagatcact tcgcagaata 1740
aataaatcct ggtgtccctg ttgataccgg gaagccctgg gccaactttt ggcgaaaatg 1800
agacgttgat cggcacgtaa gaggttccaa ctttcaccat aatgaaataa gatcactacc 1860
gggcgtattt tttgagttgt cgagattttc aggagctaag gaagctaaaa tggagaaaaa 1920
aatcactgga tataccaccg ttgatatatc ccaatggcat cgtaaagaac attttgaggc 1980
atttcagtca gttgctcaat gtacctataa ccagaccgtt cagctggata ttacggcctt 2040
tttaaagacc gtaaagaaaa ataagcacaa gttttatccg gcctttattc acattcttgc 2100
ccgcctgatg aatgctcatc cggaattacg tatggcaatg aaagacggtg agctggtgat 2160
atgggatagt gttcaccctt gttacaccgt tttccatgag caaactgaaa cgttttcatc 2220
gctctggagt gaataccacg acgatttccg gcagtttcta cacatatatt cgcaagatgt 2280
ggcgtgttac ggtgaaaacc tggcctattt ccctaaaggg tttattgaga atatgttttt 2340
cgtctcagcc aatccctggg tgagtttcac cagttttgat ttaaacgtgg ccaatatgga 2400
caacttcttc gcccccgttt tcaccatggg caaatattat acgcaaggcg acaaggtgct 2460
gatgccgctg gcgattcagg ttcatcatgc cgtttgtgat ggcttccatg tcggcagatg 2520
cttaatgaat acaacagtac tgcgatgagt ggcagggcgg ggcgtaaggc gcgccattta 2580
aatgaagttc ctattccgaa gttcctattc tctagaaagt ataggaactt c 2631
<210> 17
<211> 4259
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета интеграции
<400> 17
ttactcagca ataaactgat attccgtcag gctggaatac tcttcgccag gacgcaggaa 60
gcagtccggt tgcggccatt cagggtggtt cgggctgtcc ggtagaaact cgctttccag 120
agccagccct tgccagtcgg cgtaaggttc ggttccccgc gacggtgtgc cgccgaggaa 180
gttgccggag tagaattgca gagccggagc ggtggtgtag accttcagct gcaatttttc 240
atctgctgac cagacatgcg ccgccacttt cttgccatcg cctttggcct gtaacaagaa 300
tgcgtgatcg taacctttca ctttgcgctg atcgtcgtcg gcaagaaact cactggcgat 360
gattttggcg ctgcggaaat caaaagacgt tccggcgaca gatttcaggc cgtcgtgcgg 420
aatgccgcct tcatcaaccg gcagatattc gtccgccaga atctgcaact tgtgattgcg 480
cacgtcagac tgctcgccgt caagattgaa atagacgtga ttagtcatat tcaccgggca 540
aggtttatca actgtggcgc gataagtaat ggagatacgg ttatcgtcgg tcagacgata 600
ttgcaccgtc gcgccgagat tacccgggaa gccctgatca ccatcatctg aactcagggc 660
aaacagcacc tgacgatcgt tctggttcac aatctgccag cgacgtttgt cgaacccttc 720
cggcccgccg tgcagctggt taacgccctg acttggcgaa agcgtcacgg tttcaccgtc 780
aaaggtataa cggctattgg cgatacggtt ggcataacga ccaatagagg cccccagaaa 840
cgcggcctga tcctgatagc attccgggct ggcacagccg agcagcgcct cgcggacgct 900
gccatcggaa agcggaatac gggcggaaag taaagtcgca ccccagtcca tcagcgtgac 960
taccatccct gcgttgttac gcaaagttaa cagtcggtac ggctgaccat cgggtgccag 1020
tgcgggagtt tcgttcagca ctgtcctgct ccttgtgatg gtttacaaac gtaaaaagtc 1080
tctttaatac ctgtttttgc ttcatattgt tcagcgacag cttgctgtac ggcaggcacc 1140
agctcttccg ggatcagcgc gacgatacag ccgccaaatc cgccgccggt catgcgtacg 1200
ccacctttgt cgccaatcac agctttgacg atttctacca gagtgtcaat ttgcggcacg 1260
gtgatttcga aatcatcgcg catagaggca tgagactccg ccatcaactc gcccatacgt 1320
ttcaggtcgc cttgctccag cgcgctggca gcttcaacgg tgcgggcgtt ttcagtcagt 1380
atatgacgca cgcgttttgc cacgatcggg tccagttcat gcgcaacagc gttgaactct 1440
tcaatggtga catcacgcag ggctggctgc tggaagaaac gcgcaccggt ttcgcactgt 1500
tcacgacggg tgttgtattc gctgccaacc agggtacgtt tgaagttact gttgatgatg 1560
acgacagcca cacctttggg catggaaact gctttggtcc ccagtgagcg gcaatcgatc 1620
agcaaggcat gatctttctt gccgagcgcg gaaattagct gatccatgat cccgcagtta 1680
cagcctacaa actggttttc tgcttcctga ccgttaagcg cgatttgtgc gccgtccagc 1740
ggcagatgat aaagctgctg caatacggtt ccgaccgcga cttccagtga agcggaagaa 1800
cttaacccgg caccctgcgg cacattgccg ctgatcacca tgtccacgcc gccgaagctg 1860
ttgttacgca gttgcagatg tttcaccacg ccacgaacgt agttagccca ttgatagttt 1920
tcatgtgcga caatgggcgc atcgagggaa aactcgtcga gctgattttc ataatcggct 1980
gccatcacgc gaactttacg gtcatcgcgt ggtgcacaac tgatcacggt ttgataatca 2040
atcgcgcagg gcagaacgaa accgtcgttg tagtcggtgt gttcaccaat caaattcacg 2100
cggccaggcg cctgaatggt gtgagtggca gggtagccaa atgcgttggc aaacagagat 2160
tgtgtttttt ctttcagact catttcttac actccggatt cgcgaaaatg gatatcgctg 2220
actgcgcgca aacgctctgc tgcctgttct gcggtcaggt ctcgctgggt ctctgccagc 2280
atttcataac caaccataaa tttacgtacg gtggcggagc gcagcagagg cggataaaag 2340
tgcgcgtgca gctgccagtg ttgattctct tcgccattaa atggcgcgcc gtgccagccc 2400
atagagtagg ggaaggagca ctggaagagg ttgtcataac gactggtcag ctttttcaac 2460
gccagcgcca gatcgctgcg ctgggcgtcg gtcaaatcgg tgatccgtaa aacgtgggct 2520
ttgggcagca gtagcgtttc gaacggccag gcagcccagt aaggcacgac ggctaaccag 2580
tgttcggttt cgacaacggt acggctaccg tctgccagct cgcgctgaac ataatccacc 2640
agcattggtg atttctgttc ggcaaaatat tctttttgca ggcggtcttc gcgctcagct 2700
tcgttaggca ggaagctatt tgcccaaatc tgaccgtgcg gatgcgggtt agagcagccc 2760
atcgccgcgc ctttgttttc aaaaacctgc acccatgggt acgttttccc cagttctgcg 2820
gtttgctcct gccaggtttt gacgatttcc gtcaatgctg caacgctgag ctctggcagc 2880
gttttactgt gatccggtga aaagcagatc acccggctgg tgccgcgcgc gctctggcaa 2940
cgcatcagcg gatcgtgact ttctggcgca tctggcgtgt cagacatcaa agccgcaaag 3000
tcattagtga aaacgtaagt cccggtgtaa tcggggtttt tatcgcctgt cacccgcaca 3060
ttacctgcgc agaggaagca atctggatcg tgcgcaggta acacctgttt ggctggcgtt 3120
tcctgcgccc cctgccaggg gcgcttagcg cggtgcggtg aaaccagaat ccattgcccg 3180
gtgagcgggt tgtagcggcg atgtggatga tcaacgggat taaattgcgt catggtcgtt 3240
ccttaatcgg gatatccctg tggatggcgt gactgccagt gccaggtgtc ctgcgccatt 3300
tcatcgagtg tgcgcgttac gcgccagttc agttcacggt cggctttgct ggcgtccgcc 3360
cagtaggccg gaaggtcgcc ctcgcgacgc ggtgcaaaat gataattaac cggtttgccg 3420
caggctttgc tgaaggcatt aaccacgtcc agcacgctgt tgcctacgcc agcgccgagg 3480
ttgtagatgt gtacgcctgg cttgttcgcc agtttttcca tcgccacgac gtgaccgtcc 3540
gccagatcca ttacgtggat gtaatcgcgt acgccagtac catcttcggt cggataatcg 3600
ttaccaaaaa tcgccagcga gtcgcgacgg cctacagcaa cctgggcgat gtatggcatc 3660
aggttattcg gaatgccttg cggatcttcg cccatatcgc ccgacggatg cgcgccaacc 3720
gggttgaagt agcgcagcag ggcaatgctc cagtccggct gggctttttg cagatcggtg 3780
aggatctgtt ccaccatcag cttgcttttg ccgtaagggc tttgcggtgt gccggtcggg 3840
aagctttcaa cgtatggaat tttgggctga tcgccataaa cggtggcgga ggagctaaaa 3900
ataaagtttt tgacgttagc ggcgcgcatg gcgctaatca ggcgcagagt gccgttgaca 3960
ttgttgtcgt aatattccag cggtttttgt accgattcgc ccacggcttt cagcccggcg 4020
aagtggatca cggtgtcgat agcgtgatcg tgcaggatct cggtcatcaa cgcttcgtta 4080
cgaatatcgc cttcaacaaa cgttggatgt ttgccgccta aacgctcgat aacaggcagt 4140
acgctgcgct tactgttaca gaggttatca agaatgatga catcatgacc gttttgcagt 4200
aattgcacac aggtatgact tccaatgtaa ccgctaccac cggtaaccag aactctcat 4259
<210> 18
<211> 4223
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета интеграции
<400> 18
tggccagatg attaattcct aatttttgtt gacactctat cattgataga gttattttac 60
cactccctat cagtgataga gaaaagtgaa atgaatagtt cgacaaaaat ctagaaataa 120
ttttgtttaa ctttaagaag gagatataca aatgcaaaaa ctactatctt taccgtccaa 180
tctggttcag tcttttcatg aactggagag ggtgaatcgt accgattggt tttgtacttc 240
cgacccggta ggtaagaaac ttggttccgg tggtggaaca tcctggctgc ttgaagaatg 300
ttataatgaa tattcagatg gtgctacttt tggagagtgg cttgaaaaag aaaaaagaat 360
tcttcttcat gcgggtgggc aaagccgtcg tttacccggc tatgcacctt ctggaaagat 420
tctcactccg gttcctgtgt tccggtggga gagagggcaa catctgggac aaaatctgct 480
ttctctgcaa cttcccctat atgaaaaaat catgtctttg gctccggata aactccatac 540
actgattgcg agtggtgatg tctatattcg ttcggagaaa cctttgcaga gtattcccga 600
agcggatgtg gtttgttatg gactgtgggt agatccgtct ctggctaccc atcatggcgt 660
gtttgcttcc gatcgcaaac atcccgaaca actcgacttt atgcttcaga agccttcgtt 720
ggcagaattg gaatctttat cgaagaccca tttgttcctg atggacatcg gtatatggct 780
tttgagtgac cgtgccgtag aaatcttgat gaaacgttct cataaagaaa gctctgaaga 840
actaaagtat tatgatcttt attccgattt tggattagct ttgggaactc atccccgtat 900
tgaagacgaa gaggtcaata cgctatccgt tgctattctg cctttgccgg gaggagagtt 960
ctatcattac gggaccagta aagaactgat ttcttcaact ctttccgtac agaataaggt 1020
ttacgatcag cgtcgtatca tgcaccgtaa agtaaagccc aatccggcta tgtttgtcca 1080
aaatgctgtc gtgcggatac ctctttgtgc cgagaatgct gatttatgga tcgagaacag 1140
tcatatcgga ccaaagtgga agattgcttc acgacatatt attaccgggg ttccggaaaa 1200
tgactggtca ttggctgtgc ctgccggagt gtgtgtagat gtggttccga tgggtgataa 1260
gggctttgtt gcccgtccat acggtctgga cgatgttttc aaaggagatt tgagagattc 1320
caaaacaacc ctgacgggta ttccttttgg tgaatggatg tccaaacgcg gtttgtcata 1380
tacagatttg aaaggacgta cggacgattt acaggcagtt tccgtattcc ctatggttaa 1440
ttctgtagaa gagttgggat tggtgttgag gtggatgttg tccgaacccg aactggagga 1500
aggaaagaat atctggttac gttccgaaca tttttctgcg gacgaaattt cggcaggtgc 1560
caatctgaag cgtttgtatg cacaacgtga agagttcaga aaaggaaact ggaaagcatt 1620
ggccgttaat catgaaaaaa gtgtttttta tcaacttgat ttggccgatg cagctgaaga 1680
ttttgtacgt cttggtttgg atatgcctga attattgcct gaggatgctc tgcagatgtc 1740
acgcatccat aaccggatgt tgcgtgcgcg tattttgaaa ttagacggga aagattatcg 1800
tccggaagaa caggctgctt ttgatttgct tcgtgacggc ttgctggacg ggatcagtaa 1860
tcgtaagagt accccaaaat tggatgtata ttccgatcag attgtttggg gacgtagccc 1920
cgtgcgcatc gatatggcag gtggatggac cgatactcct ccttattcac tttattcggg 1980
aggaaatgtg gtgaatctag ccattgagtt gaacggacaa cctcccttac aggtctatgt 2040
gaagccgtgt aaagacttcc atatcgtcct gcgttctatc gatatgggtg ctatggaaat 2100
agtatctacg tttgatgaat tgcaagatta taagaagatc ggttcacctt tctctattcc 2160
gaaagccgct ctgtcattgg caggctttgc acctgcgttt tctgctgtat cttatgcttc 2220
attagaggaa cagcttaaag atttcggtgc aggtattgaa gtgactttat tggctgctat 2280
tcctgccggt tccggtttgg gcaccagttc cattctggct tctaccgtac ttggtgccat 2340
taacgatttc tgtggtttag cctgggataa aaatgagatt tgtcaacgta ctcttgttct 2400
tgaacaattg ctgactaccg gaggtggatg gcaggatcag tatggaggtg tgttgcaggg 2460
tgtgaagctt cttcagaccg aggccggctt tgctcaaagt ccattggtgc gttggctacc 2520
cgatcattta tttacgcatc ctgaatacaa agactgtcac ttgctttatt ataccggtat 2580
aactcgtacg gcaaaaggga tcttggcaga aatagtcagt tccatgttcc tcaattcatc 2640
gttgcatctc aatttacttt cggaaatgaa ggcgcatgca ttggatatga atgaagctat 2700
acagcgtgga agttttgttg agtttggccg tttggtagga aaaacctggg aacaaaacaa 2760
agcattggat agcggaacaa atcctccggc tgtggaggca attatcgatc tgataaaaga 2820
ttataccttg ggatataaat tgccgggagc cggtggtggc gggtacttat atatggtagc 2880
gaaagatccg caagctgctg ttcgtattcg taagatactg acagaaaacg ctccgaatcc 2940
gcgggcacgt tttgtcgaaa tgacgttatc tgataaggga ttccaagtat cacgatcata 3000
actgaaacca atttgcctgg cggcagtagc gcggtggtcc cacctgaccc catgccgaac 3060
tcagaagtga aacgccgtag cgccgatggt agtgtggggt ctccccatgc gagagtaggg 3120
aactgccagg catcaaataa aacgaaaggc tcagtcgaaa gactgggcct ttcgggatcc 3180
aggccggcct gttaagacgg ccagtgaatt cgagctcggt acctaccgtt cgtataatgt 3240
atgctatacg aagttatcga gctctagaga atgatcccct cattaggcca cacgttcaag 3300
tgcagcgcac accgtggaaa cggatgaagg cacgaaccca gttgacataa gcctgttcgg 3360
ttcgtaaact gtaatgcaag tagcgtatgc gctcacgcaa ctggtccaga accttgaccg 3420
aacgcagcgg tggtaacggc gcagtggcgg ttttcatggc ttgttatgac tgtttttttg 3480
tacagtctat gcctcgggca tccaagcagc aagcgcgtta cgccgtgggt cgatgtttga 3540
tgttatggag cagcaacgat gttacgcagc agcaacgatg ttacgcagca gggcagtcgc 3600
cctaaaacaa agttaggtgg ctcaagtatg ggcatcattc gcacatgtag gctcggccct 3660
gaccaagtca aatccatgcg ggctgctctt gatcttttcg gtcgtgagtt cggagacgta 3720
gccacctact cccaacatca gccggactcc gattacctcg ggaacttgct ccgtagtaag 3780
acattcatcg cgcttgctgc cttcgaccaa gaagcggttg ttggcgctct cgcggcttac 3840
gttctgccca ggtttgagca gccgcgtagt gagatctata tctatgatct cgcagtctcc 3900
ggcgagcacc ggaggcaggg cattgccacc gcgctcatca atctcctcaa gcatgaggcc 3960
aacgcgcttg gtgcttatgt gatctacgtg caagcagatt acggtgacga tcccgcagtg 4020
gctctctata caaagttggg catacgggaa gaagtgatgc actttgatat cgacccaagt 4080
accgccacct aacaattcgt tcaagccgag atcgtagaat ttcgacgacc tgcagccaag 4140
cataacttcg tataatgtat gctatacgaa cggtaggatc ctctagagtc gacctgcagg 4200
catgagatgt gtataagaga cag 4223
<210> 19
<211> 3792
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета интеграции
<400> 19
gggaattgat tctggtacca aatgagtcga ccggccagat gattaattcc taatttttgt 60
tgacactcta tcattgatag agttatttta ccactcccta tcagtgatag agaaaagtga 120
aatgaatagt tcgacaaaaa tctagaaata attttgttta actttaagaa ggagatatac 180
aaatgattac ccgcaaaagg cgggccagga caatccatag ccgatatcca atcggaattt 240
acgggagcat agtaatgaca gatattgcac agttgcttgg caaagacgcc gacaaccttt 300
tacagcaccg ttgtatgact attccttctg accagcttta tctccccgga catgactacg 360
tagaccgcgt gatgattgac aataatcgcc cgccagcggt gttacgtaat atgcagacgt 420
tgtacaacac tgggcgtctg gctggcacag gatatctttc tattctgccg gttgaccagg 480
gcgttgagca ctctgccgga gcttcatttg ctgctaaccc gctctacttt gacccgaaaa 540
acattgttga actggcgatc gaagcgggct gtaactgtgt ggcatcaact tacggcgtgt 600
tggcgtcggt atcgcggcgc tatgcgcatc gcattccatt cctcgtcaaa cttaatcaca 660
acgagacgct aagttacccg aacacctacg atcaaacgct gtatgccagc gtggagcagg 720
ccttcaacat gggcgcggtg gcggttggtg cgactatcta ttttggttcg gaagagtcac 780
gtcgccagat tgaagaaatt tctgcggctt ttgaacgtgc gcacgagctg ggcatggtga 840
cagtgctgtg ggcctatttg cgtaactccg cctttaagaa agatggcgtt gattaccatg 900
tttccgccga cctgaccggt caggcaaacc atctggcggc gaccataggt gcagatatcg 960
tcaaacaaaa aatggcggaa aataacggcg gctataaagc aattaattac ggttataccg 1020
acgatcgcgt gtacagcaag ttaaccagcg aaaacccgat tgatctggtg cgttatcagt 1080
tagctaactg ctatatgggc cgggccgggt tgataaactc cggcggtgct gcaggcggtg 1140
aaactgacct cagcgatgca gtgcgtactg cggttatcaa caaacgcgct ggcggaatgg 1200
ggctgattct tggacgtaag gcgttcaaga aatcgatggc tgacggcgtg aaactgatta 1260
acgccgtgca ggatgtttat ctcgatagca aaattactat cgcctaagag gatcgagatc 1320
tcgatcccgc gaaattaata cgactcacta taggggaatt gtgagcggat aacaattccc 1380
ctctagaaat aattttgttt aactttaaga aggagatata ccatgggcca tcatcatcat 1440
catcatcatc atcatcacag cagcggccat atcgaaggtc gtcatatggc ggtgaaagaa 1500
gcgaccagcg agaccaagaa gcgtagcggt tacgagatca ttaccctgac cagctggctg 1560
ctgcaacaag aacagaaggg tatcattgac gcggaactga ccatcgttct gagcagcatt 1620
agcatggcgt gcaaacagat cgcgagcctg gtgcaacgtg cgaacattag caacctgacc 1680
ggtacccaag gcgcggttaa catccagggt gaagaccaaa agaaactgga tgttattagc 1740
aacgaggtgt tcagcaactg cctgcgtagc agcggtcgta ccggcatcat tgcgagcgag 1800
gaagaggacg tggcggttgc ggtggaagag agctacagcg gtaactatat cgtggttttt 1860
gacccgctgg atggcagcag caacctggat gcggctgtga gcaccggtag catcttcggc 1920
atttacagcc cgaacgacga gagcctgccg gattttggtg acgatagcga cgataacacc 1980
ctgggcaccg aagagcaacg ttgcatcgtt aacgtgtgcc aaccgggtag caacctgctg 2040
gcggcgggct actgcatgta tagcagcagc gttgcgttcg tgctgaccat tggcaagggc 2100
gttttcgtgt ttaccctgga cccgctgtac ggtgaattcg tgctgaccca ggagaacctg 2160
caaatcccga agagcggtga aatttacagc tttaacgagg gcaactataa actgtgggat 2220
gaaaacctga agaaatatat cgacgatctg aaggaaccgg gtccgagcgg taaaccgtac 2280
agcgcgcgtt atatcggtag cctggttggc gacttccacc gtaccctgct gtacggtggc 2340
atttacggtt atccgcgtga taagaaaagc aagaacggca aactgcgtct gctgtatgaa 2400
tgcgcgccga tgagctttat tgttgagcag gcgggtggca aaggtagcga cggccaccag 2460
cgtgtgctgg atatccaacc gaccgaaatt caccagcgtg ttccgctgta cattggtagc 2520
accgaagagg ttgaaaaagt tgaaaagtat ctggcgtaat cgagtctggt aaagaaaccg 2580
ctgctgcgaa atttgaacgc cagcacatgg actcgtctac tagcgcagct taattaacct 2640
aggctgctgc caccgctgag caataactag cataacccct tggggcctct aaacgggtct 2700
tgaggggttt tttgctgaaa ggaggaacta tatccggatt ggcgaatggg acgcgccctg 2760
tagcggcgca ttaagcgcgg cgggtggacg gccagtgaat tcgagctcgg tacctaccgt 2820
tcgtataatg tatgctatac gaagttatcg agctctagag aatgatcccc tcattaggcc 2880
acacgttcaa gtgcagcgca caccgtggaa acggatgaag gcacgaaccc agttgacata 2940
agcctgttcg gttcgtaaac tgtaatgcaa gtagcgtatg cgctcacgca actggtccag 3000
aaccttgacc gaacgcagcg gtggtaacgg cgcagtggcg gttttcatgg cttgttatga 3060
ctgttttttt gtacagtcta tgcctcgggc atccaagcag caagcgcgtt acgccgtggg 3120
tcgatgtttg atgttatgga gcagcaacga tgttacgcag cagcaacgat gttacgcagc 3180
agggcagtcg ccctaaaaca aagttaggtg gctcaagtat gggcatcatt cgcacatgta 3240
ggctcggccc tgaccaagtc aaatccatgc gggctgctct tgatcttttc ggtcgtgagt 3300
tcggagacgt agccacctac tcccaacatc agccggactc cgattacctc gggaacttgc 3360
tccgtagtaa gacattcatc gcgcttgctg ccttcgacca agaagcggtt gttggcgctc 3420
tcgcggctta cgttctgccc aggtttgagc agccgcgtag tgagatctat atctatgatc 3480
tcgcagtctc cggcgagcac cggaggcagg gcattgccac cgcgctcatc aatctcctca 3540
agcatgaggc caacgcgctt ggtgcttatg tgatctacgt gcaagcagat tacggtgacg 3600
atcccgcagt ggctctctat acaaagttgg gcatacggga agaagtgatg cactttgata 3660
tcgacccaag taccgccacc taacaattcg ttcaagccga gatcgtagaa tttcgacgac 3720
ctgcagccaa gcataacttc gtataatgta tgctatacga acggtaggat cctctagagt 3780
cgacctgcag gc 3792
<210> 20
<211> 5917
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Транспозонная кассета
<400> 20
acaggttggc tgataagtcc ccggtctagc ttgcatgcag attgcagcat tacacgtctt 60
gatttgacgg ctagctcagt cctaggtaca gtgctagcac tgctttgtgg aaggagatag 120
acttatggcg gatccgatgg aatacctcga tgtgtcgttc ggcggcacgt tcgctgcaga 180
cacctacacc acaggtggcg acgaggtggc gaagggcccc gtgaccaagc acggcagcat 240
accgaccaag cttgacggcg gcggcatcac cctcgctggc ggcaccaacg gcgtgacatt 300
cacctcgacc gcgagcttca gcgagagtgg gaaggtgaac aagggattcc gcgccgaaat 360
ggagtaccgt acgacgcaga cgcccagcaa cctcgccaca ttgttctccg ccatgggcaa 420
catcttcgtg cgggcgaacg gcagcaacct cgaatacggc ttctccacga acccttccgg 480
cagtacatgg aacgactaca caaagtccgt gacgctgcct tccaacaatg tgaagcacat 540
catccagctg acatatctgc cgggagccga cggcgctgcc tcgacgttgc agttgtcggt 600
ggatggcgtg gccggcgaga ccgccacctc cgcggccggc gagctcgcgg ccgtcagcga 660
ttccgtcggg aacaagttcg ggatcggcta cgaggtgaac cccgcttccg gcgcggcgag 720
ccgcggtctt gccggtgacg tgttccgcgc gcgtgtcgcc gattcggacg ccccgtggga 780
gattcttgac gcatcccagc tgctgcatgt caatttcaac ggcacgttca gcggcacctc 840
atataccgcg gcgagcggcg agcagatgct gggctcgctg gtgtcgcgct cggccaatcc 900
gtccatctcg aactccgccg tcacgctggg cggcggcacg gccggattcg atttcacgcc 960
cacggacttc accctcggtg acaacgaggc catcacccgc ccgctggtcg cggagctgcg 1020
cttcaccccg acgcagaccg gcgacaacca gaccctgttc ggcgcgggcg gcaacctgtt 1080
cctgcgctac gagtcgaaca agctcgtgtt cggcgcctcc accaagtccg gcgataattg 1140
gaccgaccac aagatcgagt ccgcggccgc cacgggtgcg gagcacgtcg tgtcggtggc 1200
gtacgtgccc aataaggccg gcaccggcgc gaagcttgtc atgcgcgtgg atggcggcga 1260
cgcccagacc aaggacatca ctggtctggc ttacctgaat tcgagcatca agggcaaggt 1320
cggcttcggc aacgacgtgc ataccgacgc gctcagccgc ggcttcgtcg gctcgctgag 1380
cgagatccgc ctggccgaaa cctccgcgaa cttcaccacc aacgaattca agctggtcta 1440
ctctcaggtc agctgcgaca cgtcgggcat caaggaggcg aataccttcg acgtggagcc 1500
cgccgagtgc gaggccgcgc ttaagaccaa gctgtccaag ctgcgtccga ccgaagggca 1560
ggccgactac atcgactggg gtcagatcgg attcctccat tacggcatca acacgtacta 1620
caaccaggag tggggtcacg gtaacgagga tccctcccgc atcaacccga ccggcctcga 1680
caccgaccag tgggcgaagt ccttcgccga cggtggcttc aagatgatca tggtgacggt 1740
caagcaccat gacggtttcg agctgtacga ctcgcggtac aacaccgagc acgactgggc 1800
aaacaccgcc gtcgccaagc gcacggggga gaaggacctg ttccgcaaga ttgtcgcctc 1860
ggcgaagaaa tacggcctga aggtcggcat ctactattcg ccggccgatt cctacatgga 1920
gaggaagggc gtctggggca acaactccgc acgcgtcgag cgcacgatcc ccacgctggt 1980
ggagaacgac gaccgcgccg gcaaggtggc ttccggcaaa ctgcccacgt tcaagtacaa 2040
ggccacggat tacggcgcct acatgctcaa ccagctctat gagctgctga ctgagtacgg 2100
cgacatctcc gaggtctggt tcgacggtgc ccaaggcaac accgcaggca ctgagcatta 2160
cgactatggc gtgttctacg agatgatccg ccggcttcag ccccaggcaa ttcaggccaa 2220
cgccgcatac gatgcccgat gggtgggcaa cgaggacggc tgggcccgtc agaccgagtg 2280
gagcccgcag gcggcataca acgacggcgt ggacaaggtg tcgctcaagc ctggccagat 2340
ggcccccgac ggtaagcttg gcagcatgtc gagcgtgctg tccgagatcc gcagcggcgc 2400
cgccaaccag ctgcactggt atccggccga agtcgacgcc aagaaccggc ccggatggtt 2460
ctaccgtgcc agccaatcgc cggcgtccgt agccgaagtc gtgaagtact acgagcagtc 2520
cacgggacgc aactcgcagt atctgctgaa cgtcccaccg tccgataccg gcaagctcgc 2580
cgatgcggat gccgcgggac ttaaggggct gggcgaggag ctcgcccgac gctacggcac 2640
cgatcttgcc ctgggcaaga gcgcgaccgt cgccgcgtcc gcgaacgaca ctgcggtagc 2700
ggccccgaag ctgaccgacg gttcgaagct ctcctccgac aaggccgtgg gcaatacgcc 2760
gacgtacacc atcgatctgg gcagcactgt cgccgtggat gcagtgaaga tctccgagga 2820
cgtgcgcaat gccggccagc agatcgaaag cgccactctg cagggacgag tcaatggaac 2880
atggacgaat ctggcgacta tgacgacggt cgggcagcag cgcgaccttc gcttcacgtc 2940
ccagaacatc gatgccatcc gtctggtggt caactcctcc cgcggtccgg tgcgtctgag 3000
ccgtcttgag gtgttccaca ccgaatccga gattcagacc ggcgcccgcg cctactacat 3060
cgatccgacg gcgcagaccg cgggagatgg attcacgaag gacaagccca tgacgtcgat 3120
cgagcagctg cacgatgtga ccgtcgcgcc aggctccgtg atcttcgtca aggcgggcac 3180
cgagctgacc ggggacttcg ccgtcttcgg ctacggcacc aaggacgagc ccatcaccgt 3240
gacgacatac ggcgaaagcg acaaagccac caccgcgagc ttcgacggca tgaccgccgg 3300
gctgacgctg aagcaggcgc tgaaggcgct cggcaaggac gacgccggct gggtcgtggc 3360
cgattccgcc actgcaccgg cctcccgcgt gtatgtcccg caggatgaga tcagcgtgca 3420
cgcccagtcg tcgcagaact ccggcgcaga ggcggcgagg gcgctcgacg gcgactcgtc 3480
gacgagctgg cactcccagt acagcccgac caccgcgtct gctccgcatt gggtgactct 3540
cgatctcggc aaatcgcgtg agaacgtcgc ctacttcgac tacctcgccc gtatcgacgg 3600
caacaataac ggtgccgcca aggattacga ggtgtatgtc tccgacgatc ccaacgattt 3660
tggagcccct gtggcctcgg gcacgttgaa gaacgtcgcc tacacgcagc gcatcaagct 3720
gacccccaag aacggacggt acgtcaagtt cgtcatcaag accgattatt ccggatcgaa 3780
cttcggctcc gcggcggaaa tgaatgtcga gttgctgccc acggccgtag aggaggacaa 3840
ggtcgccacc ccgcagaagc cgacagtgga cgatgatgcc gatacataca ccatccccga 3900
catcgaggga gtcgtgtaca aggtcgacgg caaggtgttg gccgctggtt ccgtagtgaa 3960
cgtgggcgat gaggacgtga ccgtcacggt caccgccgag cccgccgacg gataccgctt 4020
cccggatggt gtgacgtccc cagtcacgta tgagctgacg ttcaccaaga agggtggcga 4080
gaagcctccg accgaagtca acaaggacaa gctgcacgcc acgatcacca aggctcaggc 4140
gatcgaccgt tccgcctata cggacgagtc gctcaaggtg cttgatgaca agctcgccgc 4200
agcgctcaag gtctatgacg atgacaaggt gagccaggat gatgtcgatg ccgccgaggc 4260
ggctctgtct gcggcgatcg acgcgctgaa gaccaagccg acgacccccg gcggtgaagg 4320
tgagaagcct ggtgaaggtg aaaagcccgg tgacggcaac aagcccggtg acggcaagaa 4380
gcccggcgac gtgatcgcaa agaccggcgc ctccacaatg taactagcat aaccccttgg 4440
ggcctctaaa cgggtcttga ggggtttttt gctgaaacca atttgcctgg cggcagtagc 4500
gcggtggtcc cacctgaccc catgccgaac tcagaagtga aacgccgtag cgccgatggt 4560
agtgtggggt ctccccatgc gagagtaggg aactgccagg catcaaataa aacgaaaggc 4620
tcagtcgaaa gactgggcct ttcgggatcc aggccggcct gttaacgaat taatcttccg 4680
cggcggtatc gataagcttg atatcgaatt ccgaagttcc tattctctag acgccattca 4740
ggctgcgcaa ctgttgggaa gggcgatcgg tgcgggcctc ttcgctatta cgccagctgg 4800
cgaaaggggg atgtgctgca aggcgattaa gttgggtaac gccagggttt tcccagtcac 4860
gacgttgtaa aacgacggcc agtgaattcg agctcggtac ctaccgttcg tataatgtat 4920
gctatacgaa gttatcgagc tctagagaat gatcccctca ttaggccaca cgttcaagtg 4980
cagcgcacac cgtggaaacg gatgaaggca cgaacccagt tgacataagc ctgttcggtt 5040
cgtaaactgt aatgcaagta gcgtatgcgc tcacgcaact ggtccagaac cttgaccgaa 5100
cgcagcggtg gtaacggcgc agtggcggtt ttcatggctt gttatgactg tttttttgta 5160
cagtctatgc ctcgggcatc caagcagcaa gcgcgttacg ccgtgggtcg atgtttgatg 5220
ttatggagca gcaacgatgt tacgcagcag caacgatgtt acgcagcagg gcagtcgccc 5280
taaaacaaag ttaggtggct caagtatggg catcattcgc acatgtaggc tcggccctga 5340
ccaagtcaaa tccatgcggg ctgctcttga tcttttcggt cgtgagttcg gagacgtagc 5400
cacctactcc caacatcagc cggactccga ttacctcggg aacttgctcc gtagtaagac 5460
attcatcgcg cttgctgcct tcgaccaaga agcggttgtt ggcgctctcg cggcttacgt 5520
tctgcccaga tttgagcagc cgcgtagtga gatctatatc tatgatctcg cagtctccgg 5580
cgagcaccgg aggcagggca ttgccaccgc gctcatcaat ctcctcaagc atgaggccaa 5640
cgcgcttggt gcttatgtga tctacgtgca agcagattac ggtgacgatc ccgcagtggc 5700
tctctataca aagttgggca tacgggaaga agtgatgcac tttgatatcg acccaagtac 5760
cgccacctaa caattcgttc aagccgagat cgtagaattt cgacgacctg cagccaagca 5820
taacttcgta taatgtatgc tatacgaacg gtaggatcct ctagagtcga ccaggtggca 5880
cttttcgggc agaccgggga cttatcagcc aacctgt 5917
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ СИАЛИЛИРОВАННЫХ САХАРИДОВ | 2019 |
|
RU2819876C2 |
| СИАЛИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ В ПОЛУЧЕНИИ СИАЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2018 |
|
RU2822039C2 |
| АBC-ТРАНСПОРТЕРЫ ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2020 |
|
RU2795855C2 |
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ КУРОНЫ, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2822800C2 |
| СПОСОБ СНИЖЕНИЯ ОШИБОЧНОГО ВКЛЮЧЕНИЯ НЕКАНОНИЧЕСКИХ АМИНОКИСЛОТ С РАЗВЕТВЛЕННОЙ ЦЕПЬЮ | 2019 |
|
RU2816654C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2741103C2 |
| Рекомбинантный микроорганизм для получения L-глутаминовой кислоты и способ получения L-глутаминовой кислоты с его использованием | 2021 |
|
RU2817194C1 |
| ГЕНЕТИЧЕСКИ СКОНСТРУИРОВАННЫЕ КЛЕТКИ, ЧУВСТВИТЕЛЬНЫЕ К КЛОСТРИДИАЛЬНЫМ НЕЙРОТОКСИНАМ | 2020 |
|
RU2824389C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |
| Новый вариант белка и способ получения L-лизина с его применением | 2021 |
|
RU2793405C1 |
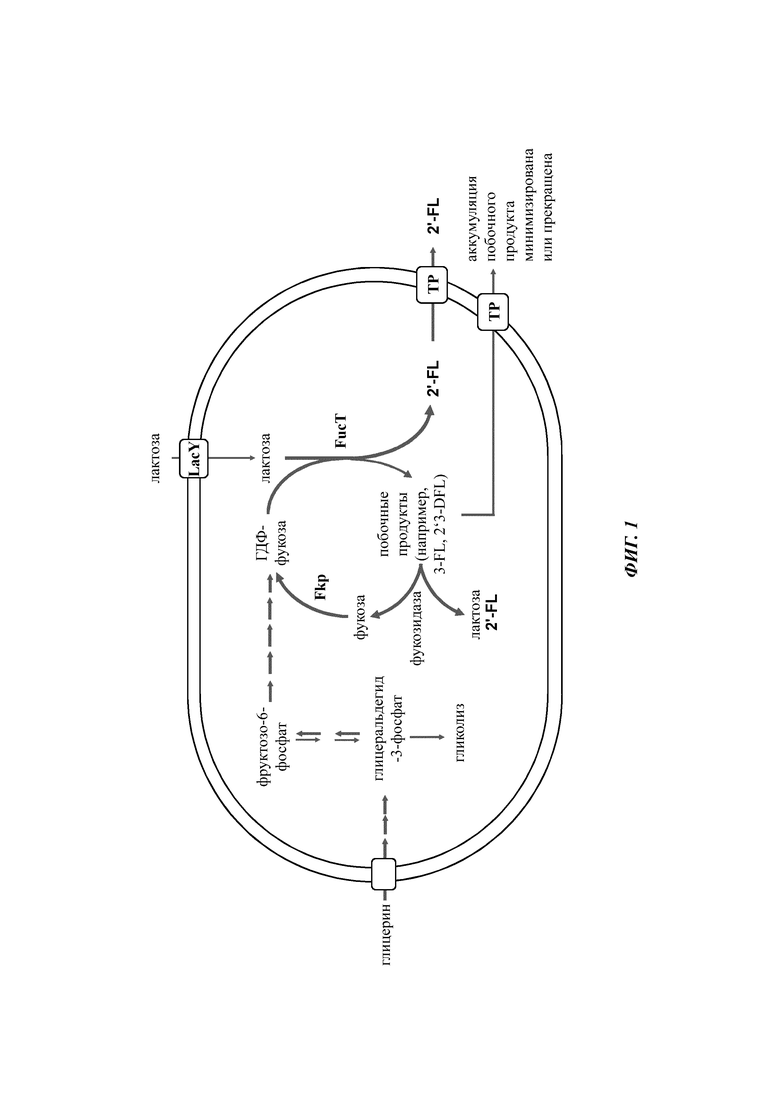
Изобретение относится к получению олигосахаридов. Предложенный способ получения 2'-фукозиллактозы с использованием генетически сконструированной клетки Escherichia coli включает предоставление генетически сконструированной клетки Escherichia coli, которая продуцирует 2'-фукозиллактозу. Причем клетка Escherichia coli генетически сконструирована для экспрессии по меньшей мере одной гетерологичной α-1,3-фукозидазы, осуществляющей внутриклеточную деградацию 3-фукозиллактозы и 2'3-дифукозиллактозы, которые образуются в процессе внутриклеточного биосинтеза 2'-фукозиллактозы. При этом клетка Escherichia coli повторно использует продукты деградации – лактозу и фукозу, полученные в результате ферментативной активности указанной α-1,3-фукозидазы, для продуцирования 2'-фукозиллактозы, причем генетически сконструированная клетка Escherichia coli содержит бифункциональную L-фукокиназо/L-фукозо-1-фосфатгуанилил-трансферазу для сохранения фукозы для продуцирования 2'-фукозиллактозы. Осуществляют культивирование генетически сконструированной клетки Escherichia coli в условиях и в среде, являющихся пермиссивными в отношении продуцирования 2'-фукозиллактозы, с получением, таким образом, 2'-фукозиллактозы и возможно выделение 2'-фукозиллактозы. Также предложена генетически сконструированная клетка Escherichia coli для получения 2'-фукозиллактозы и ее применение для получения 2'-фукозиллактозы. Изобретение направлено на получение 2'-фукозиллактозы с меньшим количеством нецелевых побочных продуктов. 3 н. и 5 з.п. ф-лы, 1 ил., 2 табл., 4 пр.
1. Способ получения 2'-фукозиллактозы с использованием генетически сконструированной клетки Escherichia coli, который включает следующие стадии:
(i) предоставление генетически сконструированной клетки Escherichia coli, которая продуцирует 2'-фукозиллактозу, причем клетка Escherichia coli генетически сконструирована для экспрессии по меньшей мере одной гетерологичной α-1,3-фукозидазы, осуществляющей внутриклеточную деградацию 3-фукозиллактозы и 2'3-дифукозиллактозы, которые образуются в процессе внутриклеточного биосинтеза 2'-фукозиллактозы, и при этом клетка Escherichia coli повторно использует продукты деградации – лактозу и фукозу, полученные в результате ферментативной активности указанной α-1,3-фукозидазы, для продуцирования 2'-фукозиллактозы, причем генетически сконструированная клетка Escherichia coli содержит бифункциональную L-фукокиназо/L-фукозо-1-фосфатгуанилил-трансферазу для сохранения фукозы для продуцирования 2'-фукозиллактозы;
(ii) культивирование генетически сконструированной клетки Escherichia coli в условиях и в среде, являющихся пермиссивными в отношении продуцирования 2'-фукозиллактозы, с получением, таким образом, 2'-фукозиллактозы; и
(iii) возможно выделение 2'-фукозиллактозы.
2. Генетически сконструированная клетка Escherichia coli для получения 2'-фукозиллактозы, где клетка Escherichia coli:
a) внутриклеточно продуцирует 2'-фукозиллактозу;
b) генетически сконструирована для экспрессии по меньшей мере одной гетерологичной α-1,3-фукозидазы, которая осуществляет внутриклеточную деградацию 3-фукозиллактозы и 2'3-дифукозиллактозы, которые образуются в процессе внутриклеточного биосинтеза 2'-фукозиллактозы;
c) генетически сконструирована с содержанием бифункциональной L-фукокиназо/L-фукозо-1-фосфатгуанилил-трансферазы для сохранения свободной фукозы для продуцирования 2'-фукозиллактозы; и
d) повторно использует продукты деградации – лактозу и фукозу, полученные в результате ферментативной активности указанной α-1,3-фукозидазы, для продуцирования 2'-фукозиллактозы.
3. Способ по п. 1 или генетически сконструированная клетка Escherichia coli по п. 2, где генетически сконструированная клетка Escherichia coli генетически сконструирована для экспрессии гетерологичной гликозилтрансферазы, предпочтительно гликозилтрансферазы, выбранной из группы, состоящей из фукозилтрансфераз, сиалилтрансфераз, галактозилтрансфераз, N-ацетил-глюкозаминилтрансфераз и глюкозилтрансфераз.
4. Способ по любому из пп. 1 и 3 или генетически сконструированная клетка Escherichia coli по любому из пп. 2, 3, где генетически сконструированная клетка Escherichia coli генетически сконструирована для экспрессии гетерологичной α-1,2-фукозилтрансферазы и гетерологичной α-1,3-фукозидазы.
5. Способ по любому из пп. 1 и 3 или генетически сконструированная клетка Escherichia coli по любому из пп. 2, 3, где генетически сконструированная клетка Escherichia coli генетически сконструирована для экспрессии α-1,2-фукозилтрансферазы гена 2-фукозилтрансферазы wbgL от E. coli:O126.
6. Способ или генетически сконструированная клетка Escherichia coli по любому из пп. 4 или 5, где генетически сконструированная клетка Escherichia coli генетически сконструирована для экспрессии α-1,3-фукозидазы AfcB из Bifidobacterium bifidum.
7. Способ по любому из пп. 1, 3-6 или генетически сконструированная клетка Escherichia coli по любому из пп. 2-6, где генетически сконструированная клетка Escherichia coli генетически сконструирована с содержанием гена fkp, кодирующего бифункциональную L-фукокиназо/L-фукозо-1-фосфатгуанилилтрансферазу, предпочтительно кодирующего бифункциональную L-фукокиназо/L-фукозо-1-фосфатгуанилилтрансферазу Bacteroides fragilis.
8. Применение генетически сконструированной клетки Escherichia coli по любому из пп. 2-7 для получения 2'-фукозиллактозы.
| EP 2845905 A1, 11.03.2015 | |||
| EP 3131912 A1, 22.02.2017 | |||
| JP 8140691 A, 04.06.1996 | |||
| ПРОДУКТЫ ПИТАНИЯ, СОДЕРЖАЩИЕ ОЛИГОСАХАРИДЫ ЧЕЛОВЕЧЕСКОГО МОЛОКА, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2012 |
|
RU2591123C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЛИКОЗИДОВ АКРИЛАТНЫХ ПРОИЗВОДНЫХ С ИСПОЛЬЗОВАНИЕМ ПОЛИСАХАРИДОВ И ГЛИКОЗИДАЗ ИЛИ ГЛИКОЗИЛТРАНСФЕРАЗ | 2011 |
|
RU2570556C2 |

