ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет по предварительной заявке США № 62/775194, поданной 4 декабря 2018 г., содержание которой полностью включено в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к векторам и их применению для разработки линий клеток-хозяев для продукции интересующих белков и, в частности, к векторам, в которых используется слабый промотор, для управления селектируемым маркером.
УРОВЕНЬ ТЕХНИКИ
Терапевтические белковые препараты являются важным классом лекарств, помогающих пациентам, которые больше всего нуждаются в новых видах терапии. Недавно были разработаны одобренные рекомбинантные белковые терапевтические средства для лечения широкого спектра клинических показаний, включая раковые заболевания, аутоиммунные/воспалительные заболевания, воздействие инфекционных агентов и генетические нарушения. Последние достижения в белково-инженерных технологиях позволяют разработчикам и производителям лекарственных средств точно регулировать и использовать желательные функциональные характеристики интересующих белков, сохраняя при этом (а в некоторых случаях и повышая) безопасность или эффективность продукта, либо и то, и другое.
Производство и продукция терапевтических белков являются очень сложными процессами. Например, типичный белковый препарат может включать в себя более 5000 критических стадий процесса, что во много раз превышает количество, необходимое для производства низкомолекулярного препарата.
Аналогичным образом, белковые терапевтические средства, которые содержат моноклональные антитела, а также крупные или слитые белки, могут быть на порядки больше по размеру, чем низкомолекулярные препараты, их молекулярные массы превышают 100 кДа. Кроме того, белковые терапевтические средства демонстрируют сложные вторичные и третичные структуры, которые необходимо поддерживать. Белковые терапевтические средства не могут быть полностью синтезированы химическими процессами и должны быть произведены в живых клетках или организмах; следовательно, выбор клеточной линии, видового происхождения и условий культивирования вместе влияют на характеристики конечного продукта. Более того, большинство биологически активных белков требуют посттрансляционных модификаций, которые могут быть нарушены при использовании гетерологичных систем экспрессии. Кроме того, поскольку продукты синтезируются клетками или организмами, вовлекаются сложные процессы очистки. Кроме того, для предотвращения серьезной проблемы для безопасности из-за вирусной контаминации белковых лекарственных субстанций применяются такие процессы очистки от вирусов, как удаление вирусных частиц с помощью фильтров или смол, а также стадии инактивации с использованием низких значений pH или детергентов. Учитывая сложность терапевтических белков с точки зрения их большого молекулярного размера, посттрансляционных модификаций и разнообразия биологических материалов, вовлеченных в процесс их производства, весьма желательна возможность повышения конкретных функциональных атрибутов при сохранении безопасности и эффективности продукта, достигаемых с помощью стратегий белковой инженерии.
Хотя интеграция новых стратегий и подходов к модификации белковых лекарственных средств не является тривиальной, потенциальные терапевтические выгоды способствовали более широкому использованию таких стратегий в процессе разработки лекарственных средств. В настоящее время используется ряд белково-инженерных платформенных технологий для увеличения периода полувыведения из кровотока, нацеливания и функциональности новых терапевтических белковых препаратов, а также для увеличения выхода продукции и повышения чистоты продукта. Например, в настоящее время для продления периода полувыведения препарата из кровотока используют способы конъюгации и дериватизации белков, включая Fc-слияние, альбумин-слияние и ПЭГилирование.
Продукция белкового фармацевтического препарата (биологического средства) является дорогостоящим и требует много времени. В данной области техники нужны более эффективные инструменты и процессы для производства этого важного класса лекарственных средств.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к векторам и их применению для разработки линий клеток-хозяев для продукции интересующих белков и, в частности, к векторам, в которых используется слабый промотор, для управления селектируемым маркером.
Соответственно, в некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается (предлагаются) вектор(-ы) для экспрессии интересующего белка, содержащий последовательность нуклеиновой кислоты, кодирующую селектируемый маркер в функциональной связи с первой промоторной последовательностью, которая была изменена для снижения активности промотора по сравнению с неизмененной версией или версией дикого типа первой промоторной последовательности, и последовательность нуклеиновой кислоты, кодирующая интересующий белок, функционально связана со второй промоторной последовательностью.
В некоторых предпочтительных вариантах осуществления первая промоторная последовательность, которая была изменена для снижения активности промотора по сравнению с неизмененной версией или версией дикого типа первой промоторной последовательности, представляет собой промоторную последовательность вирусного самоинактивирующегося (Self-Inactivating, SIN) длинного концевого повтора (Long Terminal Repeat, LTR). В некоторых предпочтительных вариантах осуществления промоторная последовательность SIN LTR является по меньшей мере на 95% идентичной SEQ ID NO:3. В некоторых предпочтительных вариантах осуществления промоторная последовательность SIN LTR представляет собой SEQ ID NO:3.
В некоторых предпочтительных вариантах осуществления селектируемый маркер представляет глутамин-синтетазу (Glutamine Synthetase, GS). В некоторых предпочтительных вариантах осуществления селектируемый маркер представляет дигидрофолат-редуктазу (Dihydrofolate Reductase, DHFR).
В некоторых вариантах осуществления вектор содержит одну сигнальную последовательность поли-A в функциональной связи с селектируемым маркером и нуклеиновой кислотой, кодирующей интересующий белок. В других вариантах осуществления вектор содержит первую сигнальную последовательность поли-A в функциональной связи с селектируемым маркером и вторую сигнальную последовательность поли-A в функциональной связи с нуклеиновой кислотой, кодирующей интересующий белок.
В некоторых предпочтительных вариантах осуществления интересующий белок выбран из группы, состоящей из Fc-слитого белка, фермента, слияния с альбумином, фактора роста, белкового рецептора, одноцепочечного антитела (scFv), одноцепочечного антитела-Fc (scFv-Fc), диатела и минитела (scFv-CH3), Fab, одноцепочечного Fab (scFab), тяжелой цепи иммуноглобулина и легкой цепи иммуноглобулина. В некоторых предпочтительных вариантах осуществления интересующий белок представляет собой Fc-слитый белок.
В некоторых вариантах осуществления вектор представляет собой плазмиду. В некоторых предпочтительных вариантах осуществления вектор представляет собой вирусный вектор. В некоторых предпочтительных вариантах осуществления вектор представляет собой ретровирусный вектор. В некоторых предпочтительных вариантах осуществления вектор представляет собой лентивирусный вектор.
В некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается (предлагаются) клетка-хозяин (клетки-хозяева), содержащая(-ие) описанный выше вектор. В некоторых предпочтительных вариантах осуществления линия клеток-хозяев представляет собой линию клеток с нокаутом GS. В некоторых предпочтительных вариантах осуществления линия клеток-хозяев представляет собой линию клеток с нокаутом DHFR. В некоторых предпочтительных вариантах осуществления линия клеток-хозяев представляет собой линию клеток яичника китайского хомячка (Chinese Hamster Ovary, CHO). В некоторых предпочтительных вариантах осуществления линия клеток-хозяев представляет собой HEK 293 или CAP линии клеток. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 1, 20, 50 до 1000 копий вектора. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 10 до 200 копий вектора. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 10 до 100 копий вектора. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 20 до 100 копий вектора.
В некоторых предпочтительных вариантах осуществления клетка-хозяин дополнительно содержит по меньшей мере второй вектор, который кодирует и обеспечивает экспрессию второго интересующего белка, и при этом указанный второй вектор не включает в себя селектируемый маркер. В некоторых предпочтительных вариантах осуществления клетка-хозяин дополнительно содержит по меньшей мере второй вектор, который кодирует и обеспечивает экспрессию второго интересующего белка, и при этом указанный второй вектор включает в себя селектируемый маркер, отличающийся от селектируемого маркера в первом векторе. В некоторых предпочтительных вариантах осуществления первый интересующий белок в первом векторе представляет собой одну из тяжелой или легкой цепи иммуноглобулина, и второй белок во втором векторе представляет собой другую из тяжелой или легкой цепи иммуноглобулина. В некоторых предпочтительных вариантах осуществления первый интересующий белок представляет собой тяжелую цепь иммуноглобулина, и второй интересующий белок представляет собой легкую цепь иммуноглобулина. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 1, 20, 50 или от 100 до 1000 копий второго вектора. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 10 до 200 копий второго вектора. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 10 до 100 копий второго вектора. В некоторых предпочтительных вариантах осуществления клетка-хозяин содержит от около 20 до 100 копий второго вектора.
В некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается культура клеток-хозяев, содержащая клетки-хозяева, описанные выше. В некоторых предпочтительных вариантах осуществления культура продуцирует от 1 до 50 грамм/литр/сутки интересующего белка. В некоторых предпочтительных вариантах осуществления культура продуцирует от 2 до 10 грамм/литр/сутки интересующего белка.
В некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается способ получения интересующего белка, включающий культивирование описанных выше клеток-хозяев и очистку интересующего белка от культуры клеток-хозяев.
В некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается инфекционная ретровирусная частица, содержащая следующие элементы в порядке от 5’ до 3’: 1) 5’ LTR; 2) ретровирусную упаковывающую область; 3) нуклеиновую кислоту, кодирующую селектируемый маркер; 4) внутренний промотор; 5) последовательность нуклеиновой кислоты, кодирующую интересующий белок, которая является функционально связанной с внутренним промотором; и 6) 3’ SIN LTR. В некоторых предпочтительных вариантах осуществления 5’LTR представляет собой LTR MoMuSV или SIN LTR. В некоторых вариантах осуществления 3’ LTR содержит сигнальную последовательность поли-A. В некоторых вариантах осуществления упаковывающая область содержит множество потенциальных сайтов инициации трансляции. В некоторых предпочтительных вариантах осуществления селектируемый маркер представляет собой GS. В некоторых предпочтительных вариантах осуществления внутренний промотор представляет собой промотор CMV. В некоторых вариантах осуществления частица содержит одну сигнальную последовательность поли-A после нуклеиновой кислоты, кодирующей интересующий белок. В некоторых вариантах осуществления частицы содержат первую сигнальную последовательность поли-A в функциональной связи с селектируемым маркером и вторую сигнальную последовательность поли-A в функциональной связи с нуклеиновой кислотой, кодирующей интересующий ген.
В дополнительных вариантах осуществления предлагается плазмида, содержащая следующие элементы в порядке от 5’ до 3’: 1) 5’ LTR (например, SIN LTR); 2) упаковывающую область; 3) селектируемый маркер (например, GS); 4) внутренний промотор (например, промотор CMV); 5) последовательность нуклеиновой кислоты, кодирующую интересующий белок, которая функционально связана с внутренним промотором; и 6) сигнальную последовательность поли-A. В некоторых вариантах осуществления плазмида содержит одну сигнальную последовательность поли-A после нуклеиновой кислоты, кодирующей интересующий белок.
В дополнительных вариантах осуществления предлагается система, содержащая a) первый вектор, содержащий последовательность нуклеиновой кислоты, кодирующую селектируемый маркер в функциональной связи с первой промоторной последовательностью, которая была изменена для снижения активности промотора по сравнению с неизмененной версией или версией дикого типа первой промоторной последовательности, и последовательность нуклеиновой кислоты, кодирующая первый интересующий белок, функционально связана со второй промоторной последовательностью; и b) второй вектор, содержащий последовательность нуклеиновой кислоты, кодирующую второй интересующий белок, функционально связанную с промоторной последовательностью, и при этом второй вектор не включает в себя селектируемый маркер.
В некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается способ получения интересующего белка, включающий: трансдукцию или трансфекцию клетки-хозяина или клетки с помощью инфекционной ретровирусной частицы, плазмиды или векторной системы, описанных выше, разработку линии клеток-хозяев, которая экспрессирует интересующий белок из этой клетки-хозяина или клеток; культивирование клеток-хозяев в таких условиях, чтобы интересующий белок продуцировался линией клеток-хозяев; и очистку интересующего белка от культуры клеток-хозяев.
В некоторых предпочтительных вариантах осуществления в настоящем изобретении предлагается инфекционная ретровирусная частица или плазмида, описанная выше, для применения при трансдукции клетки-хозяина или клеток для продукции интересующего белка.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГ. 1 - Карта полноразмерной конструкции MMLV.
ФИГ. 2 - Карта конструкции SIN LTR MMLV.
ФИГ. 3 - Последовательность полноразмерной конструкции MMLV (SEQ ID NO:1).
ФИГ. 4 - Последовательность конструкции SIN LTR MMLV (SEQ ID NO:2).
ФИГ. 5 - Сравнение объединенного титра линии клеток между полноразмерной конструкцией MMLV и конструкцией SIN LTR MMLV.
ФИГ. 6 - Сравнение объединенного титра линии клеток между процессами, с использованием полноразмерной конструкции MMLV и конструкции SIN LTR MMLV.
ФИГ. 7 - Последовательность SIN LTR (SEQ ID NO:3).
ФИГ. 8 - Карта провирусной плазмидной конструкции (SEQ ID NO: 4).
ФИГ. 9 - Последовательность провирусной плазмидной конструкции (SEQ ID NO: 4).
ОПРЕДЕЛЕНИЯ
Для облегчения понимания настоящего изобретения ниже приведены определения для ряда терминов.
Используемый в данном документе термин «клетки-хозяева» относится к любой эукариотической клетке (например, клеткам млекопитающих, клеткам птиц, клеткам амфибий, клеткам растений, клеткам рыб и клеткам насекомых), независимо от их локализации in vitro или in vivo.
Используемый в данном документе термин «клеточная культура» относится к любой культуре клеток in vitro. Этот термин включает непрерывные клеточные линии (например, с бессмертным фенотипом), первичные клеточные культуры, терминальные клеточные линии (например, нетрансформированные клетки) и любые другие популяции клеток, поддерживаемые in vitro, включая яйцеклетки и эмбрионы.
Используемый в данном документе термин «вектор» относится к любому генетическому элементу, такому как плазмида, фаг, транспозон, космида, хромосома, вирус, вирион и т.д., которые способны к репликации, когда они связаны с надлежащими регулирующими элементами и которые могут переносить генные последовательности между клетками. Таким образом, этот термин включает клонирующие и экспрессионные носители, а также вирусные векторы.
Используемый в данном документе термин «множественность заражения» или MOI относится к соотношению количеств интегрирующие векторы:хозяйские клетки, применяемому во время трансфекции или трансдукции клеток-хозяев. Например, если применяют 1000000 векторов для трансдукции 100000 клеток-хозяев, то множественность заражения составляет 10. Использование этого термина не ограничивается событиями, связанными с трансдукцией, а включает в себя введение вектора в хозяина с помощью таких методов, как липофекция, микроинъекция, осаждение фосфата кальция и электропорация.
Используемый в данном документе термин «геном» относится к генетическому материалу (например, хромосомам) организма.
Термин «интересующая нуклеотидная последовательность» относится к любой нуклеотидной последовательности (например, РНК или ДНК), манипуляция которой средним специалистом в данной области техники может считаться желаемой по какой-либо причине (например, лечение заболевания, приобретение улучшенных качеств, экспрессия интересующего белка в клетке-хозяине, экспрессия рибозима и т.д.). Такие нуклеотидные последовательностей включают в себя, но не ограничиваются ими, кодирующие последовательности структурных генов (например, репортерные гены, гены маркеров селекции, онкогены, гены резистентности к лекарственным веществам, факторы роста и т.д.), и некодирующие регуляторные последовательности, которые не кодируют мРНК или белковый продукт (например, промоторная последовательность, последовательность полиаденилирования, последовательность терминации, энхансерная последовательность и т.д.).
Используемый в данном документе термин «интересующий белок» относится к белку, кодируемому интересующей нуклеиновой кислотой.
Используемый в данном документе термин «кодирующая молекула нуклеиновой кислоты», «кодирующая последовательность ДНК», «кодирующая ДНК», «кодирующая последовательность РНК» и «кодирующая РНК» относится к порядку или последовательности дезоксирибонуклеотидов или рибонуклеотидов вдоль цепи дезоксирибонуклеиновой кислоты или рибонуклеиновой кислоты. Порядок этих дезоксирибонуклеотидов или рибонуклеотидов определяет порядок аминокислот вдоль полипептидной (белковой) цепи. Таким образом, последовательность ДНК или РНК кодирует аминокислотную последовательность.
Термин «промотор», «промоторный элемент» или «промоторная последовательность», используемый в данном документе, относится к последовательности ДНК, которую затем лигируют с интересующей нуклеотидной последовательностью, способна регулировать транскрипцию интересующей нуклеотидной последовательности в мРНК. Промотор, как правило, но не обязательно, расположен на 5' конце (т.е. перед) интересующей нуклеотидной последовательность, транскрипция которой в мРНК им регулируется, и предоставляет сайт для специфического связывания РНК-полимеразой и другими транскрипционными факторами для инициации транскрипции.
Используемый в данном документе термин «измененный», при применении касательно промотора, относится к промоторам, у которых имеются измененная последовательность нуклеиновой кислоты по сравнению с эталонной последовательностью дикого типа. Например, промотор последовательности может изменятся путем удаления определенных промоторных и/или энхансерных элементов.
Сигналы транскрипционной регуляции в эукариотах включает себя «промоторный» и «энхансерный» элементы. Промоторы и энхансеры состоят из коротких массивов последовательностей ДНК, которые специфически взаимодействуют с клеточными белками, участвующими в транскрипции (Maniatis et al., Science 236:1237 [1987]). Промоторные и энхансерные элементы были выделены из различных эукариотических источников, включая гены дрожжей, клеток насекомых и млекопитающих, и вирусов (аналогичные регулирующие элементы, т.е. промоторы, также обнаружены у прокариот). Выбор конкретного промотора и энхансера зависит от типа клетки, которую требуется применять для экспрессии интересующего белка. Некоторые эукариотические промоторы и энхансеры имеют широкий спектр хозяев, в то время как другие функционируют в ограниченном подмножестве типов клеток (обзор см., Voss et al., Trends Biochem. Sci., 11:287 [1986]; и Maniatis et al., выше). Например, ранний генный энхансер SV40 является очень активным в обширном наборе типов клеток во множестве видов млекопитающих и нашел широкое применение для экспрессии белков в клетках млекопитающих (Dijkema et al., EMBO J. 4:761 [1985]). Два других примера промоторных/энхансерных элементов, активных в широком спектре типов клеток млекопитающих, исходят от гена фактора 1α элонгации (Uetsuki et al., J. Biol. Chem., 264:5791 [1989]; Kim et al., Gene 91:217 [1990]; и Mizushima and Nagata, Nuc. Acids. Res., 18:5322 [1990]) и длинных концевых повторов вируса саркомы Рауса (Gorman et al., Proc. Natl. Acad. Sci. USA 79:6777 [1982]) и цитомегаловируса человека (Boshart et al., Cell 41:521 [1985]).
Используемый в данном документе термин «промотор/энхансер» обозначает сегмент ДНК, который содержит последовательности, способные обеспечивать как промоторные, так и энхансерные функции (т.е. функции, обеспечиваемые промоторным элементом и энхансерным элементом, см. выше обсуждение этих функций). Например, длинные концевые повторы ретровирусов содержат как промоторные, так и энхансерные функции. Энхансер/промотор может быть «эндогенным» или «экзогенным» или «гетерологичным». «Эндогенный» энхансер/промотор представляет собой такой энхансер/промотор, который естественным образом связан в геноме с данным геном. «Экзогенный» или «гетерологичный» энхансер/промотор представляет собой такой энхансер/промотор, который размещен рядом с геном посредством генетической манипуляции (т.е. молекулярно-биологических методов, таких как клонирование и рекомбинация), так что транскрипцией этого гена управляет связанный энхансер/промотор.
Используемый в данном документе термин «длинный концевой повтор» или LTR относится к регулирующим транскрипционным элементам, расположенным в области U3 направлений 5' и 3' ретровирусного генома или выделенным из нее. Как известно в данной области техники, длинные концевые повторы могут применять как регулирующие элементы в ретровирусных векторах, или выделять из ретровирусного генома и применять для регуляции экспрессии других типов векторов.
Используемые в данном документе термины «комплементарный» или «комплементарность» используют касательно полинуклеотидов (т.е. последовательности нуклеотидов), связанных по правилам спаривания оснований. Например, последовательность «5'-A-G-T-3'» является комплементарной с последовательностью «3'-T-C-A-5'». Комплементарность может быть «частичной», когда только некоторые основания нуклеиновых кислот соответствуют согласно правилам спаривания оснований. Или может быть «полная» или «тотальная» комплементарность между нуклеиновыми кислотами. Степень комплементарности между цепями нуклеиновых кислот оказывает значительное влияние на эффективность и силу гибридизации между цепями нуклеиновых кислот. Это особенно важно в реакциях амплификации, а также методах выявления, которые зависят от связывания между нуклеиновыми кислотами.
Термины «гомология» и «процент идентичности» при применении в отношении нуклеиновых кислот относится к степени комплементарности. Может существовать частичная гомология (т.е. частичная идентичность) или полная гомология (т.е. полная идентичность). Частично комплементарная последовательность представляет собой последовательность, которая по меньшей мере частично ингибирует полностью комплементарную последовательность при гибридизации с целевой последовательностью нуклеиновой кислоты, и относится к использованию функционального термина «по существу гомологичный». Ингибирование гибридизации полностью комплементарной последовательности с последовательностью-мишенью можно проверять с использованием гибридизационного анализа (Саузерн или нозерн блот, метод гибридизации в растворе и т.п.) в условия низкой жесткости. По существу гомологичная последовательность или зонд (т.е. олигонуклеотид, который способен гибридизироваться с другим интересующим олигонуклеотидом) будет конкурировать за связывание или ингибировать (т.е. гибридизироваться) полностью гомологичную последовательность к последовательности-мишени в условиях низкой жесткости. Это не означает, что условия низкой жесткости такие, что допускается неспецифическое связывание; условия низкой жесткости требуют, чтобы связывание двух последовательностей друг с другом было специфическим (т.е. селективным) взаимодействием. Отсутствие неспецифического связывания могут изучать применением второй мишени, у которой отсутствует даже частичная степень комплементарности (например, менее около 30% идентичности); в отсутствии неспецифического связывания зонд будет гибридизироваться со второй некомплементарной мишенью.
Термин «в функциональной комбинации», «в функциональном порядке» и «функционально связанный», используемый в данном документе, относится к связи последовательностей нуклеиновых кислот таким образом, что молекула нуклеиновой кислоты способна управлять транскрипцией данного гена и/или синтезом желаемой продуцируемой молекулы белка. Этот термин также относится к связи аминокислотных последовательностей таким образом, что продуцируется функциональный белок.
Используемый в данном документе термин «селектируемый маркер» относится к гену, кодирующему ферментативную активность или другой белок, который дает возможность расти в среде, не имея того, что в противном случае было бы необходимым питательным веществом; кроме того, селектируемый маркер может придавать устойчивость клетке к антибиотику или лекарственному веществу, в которой экспрессируется селектируемый маркер.
Используемый в данном документе термин «ретровирус» относится к ретровирусной частице, которая способна входить в клетку (т.е. частица содержит мембранно-ассоциированный белок, такой как белок оболочки или вирусный гликопротеин G, который может связываться с поверхностью клетки-хозяина и облегчать вход вирусных частиц в цитоплазму клетки-хозяина) и интегрировать ретровирусный геном (как двухцепочечный провирус) в геном клетки-хозяина. Термин «ретровирус» охватывает подсемейство Oncovirinae (например, вирус мышиного лейкоза Молони (MoMLV), вирус мышиной саркомы Молони (MoMSV) и вирус опухоли молочной железы мыши (MMTV), Spumavirinae и Lentivirinae (например, вирус иммунодефицита человека, вирус иммунодефицита обезьян, вирус инфекционной анемии у лошадей и вирус артрита-энцефалита коз и овец; см., например, патенты США №5994136 и 6013516, оба из которых включены в данный документ посредством ссылки).
Используемый в данном документе термин «ретровирусный вектор» относится к ретровирусу, который был модифицирован для экспрессии интересующего гена. Ретровирусные векторы могут использовать для эффективной передачи генов в клетки-хозяева, используя вирусный инфекционный процесс. Чужеродные или гетерологичные гены, клонированные (т.е. вставленные с использованием молекулярных биологических методов) в ретровирусный геном могут эффективно доставляться в клетки-хозяева, которые чувствительны к инфекции ретровируса. Благодаря хорошо известным генетическим манипуляциям может быть разрушена репликативная способность ретровирусного генома. Полученные в результате репликации дефектные векторы могут использовать для введения в клетку нового генетического материала, но они неспособны к репликации. Для обеспечения сборки и выхода из клетки векторных частиц могут использовать вирус-помощник или линия упаковывающих клеток. Такие ретровирусные векторы содержат дефектный по репликации ретровирусный геном содержащий последовательность нуклеиновой кислоты, кодирующую по меньшей мере один интересующий ген (т.е.полицистронная последовательность нуклеиновой кислоты может кодировать более одного интересующего гена), 5'-ретровирусный длинный концевой повтор (5' LTR); и 3'-ретровирусный длинный концевой повтор (3' LTR).
Термин «псевдотипированный ретровирусный вектор» относится к ретровирусному вектору, содержащему гетерологичный мембранный белок. Термин «мембранно-ассоциированный белок» относится к белку (например, гликопротеину вирусной оболочки или G-белкам вируса в семействе Rhabdoviridae, такому как ВВС, Пири, Чандипура и Мокола), который ассоциирован с мембраной, окружающей вирусную частицу; эти мембранно-ассоциированные белки опосредуют вход вирусных частиц в клетку-хозяина. Мембранно-ассоциированный белок может связываться со специфическими белковыми рецепторами клеточной поверхности, как в случае белков ретровирусной оболочки, или мембранно-ассоциированный белок может взаимодействовать с фосфолипидным компонентом плазматической мембраны клетки-хозяина, как в случае G-белков, происходящих из представителей семейства Rhabdoviridae.
Термин «гетерологичный мембранно-ассоциированный белок» относится к мембранно-ассоциированному белку, происходящему из вируса, который представляет не является представителем того же класса или семейства вирусов, от которого происходит нуклеокапсидный белок векторной частицы. «Класс или семейство вирусов» относится к таксономическому рангу класса или семейства, как определено Международным комитетом по таксономии вирусов.
Используемый в данном документе термин «лентивирусный вектор» относится к ретровирусному вектору, происходящему из семейства Lentiviridae (например, вируса иммунодефицита человека, вирусу иммунодефицита обезьян, вирус инфекционной анемии у лошадей и вирус артрита-энцефалита коз и овец), который способен интегрироваться в неделящиеся клетки (см., например, патенты США №5994136 и 6013516, оба из которых включены в данный документ посредством ссылки).
Термин «псевдотипированный лентивирусный вектор» относится к лентивирусному вектору, содержащему гетерологичный мембранный белок (например, гликопротеин вирусной оболочки или G-белки вирусов семейства Rhabdoviridae, таких как ВВС, Пири, Чандипура и Мокола).
Используемый в данном документе термин «транспозон» относится к мобильным элементам (например, Tn5, Tn7 и Tn10), которые могут перемещаться или переноситься в геноме из одного положения в другое. В целом, транспозиция контролируется транспозазой. Термин «транспозоновый вектор», используемый в данном документе, относится к вектору, кодирующему интересующую нуклеиновую кислоту, фланкированную терминальными концами транспозона. Примеры транспозоновых векторов включают в себя, но не ограничиваются ими, описанные в патентах США №6027722; 5958775; 5968785; 5965443 и 5719055, все из которых включены в данный документ посредством ссылки.
Используемый в данном документе термин «аденоассоциированный вирусный (AAV) вектор» относится к вектору, происходящему из серотипа аденоассоциированного вируса, включая без ограничений, AAV-1, AAV-2, AAV-3, AAV-4, AAV-5, AAVX7 и т.д., векторы AAV могут иметь один или более генов AAV дикого типа, удаленных полностью или частично, предпочтительно гены rep и/или cap, но сохранять функциональные фланкирующие последовательности ITR.
Векторы AAV вектор могут конструировать с использованием рекомбинантных методов, которые известны в данной области техники, для включения одного или более гетерологичных нуклеотидных последовательностей, фланкированных по обоим концам (5' и 3') функциональными ITR AAV. В практической реализации данного изобретения вектор AAV может включать в себя по меньшей мере один ITR AAV и подходящую промоторную последовательность, расположенную перед гетерологичной нуклеотидной последовательностью, и по меньшей мере один ITR AAV, расположенный после гетерологичной последовательности. «Плазмида рекомбинантого вектора AAV» относится к одному типу рекомбинантного вектора AAV, причем вектор содержит плазмиду. Как и с векторами AAV, в целом, 5' и 3' ITR фланкируют выбранную гетерологичную нуклеотидную последовательность.
Вектор AAV также может включать в себя транскрипционные последовательности, такие как сайт полиаденилирования, а также селектируемый маркер или репортерные гены, энхансерные последовательностей и другие регулирующие элементы, обеспечивающие индукцию транскрипции. Такие регулирующие элементы описаны выше.
Используемый в данном документе термин «вирион AAV» относится к полноценной вирусной частице. Вирион AAV может быть вирусной частицей AAV дикого типа (содержащей линейный одноцепочечный геном нуклеиновой кислоты AAV, ассоциированный с капсидом AAV, т.е. белковой оболочкой) или рекомбинантной вирусной частицей AAV (описанного ниже). В связи с этим, молекулы одноцепочечных нуклеиновых кислот AAV (либо смысловой/кодирующей цепи, либо антисмысловой/некодирующей цепи, как эти термины обычно определены) могут быть упакованы в вирион AAV; как смысловые, так и антисмысловые цепи в равной степени инфекционные.
Используемый в данном документе термин «рекомбинантный вирион AAV» или «rAAV» представляет собой инфекционный, дефектный по репликации вирус, состоящий из белкового окружения AAV, инкапсидирующего (т.е. окружающего белковой оболочкой) гетерологичную нуклеотидную последовательность, которая в свою очередь фланкирована по 5' и 3' с помощью ITR AAV. В данной области техники известен ряд методов конструирования рекомбинантных вирионов AAV (см., например, патент США №5173414; WO 92/01070; WO 93/03769; Lebkowski et al., Molec. Cell. Biol. 8:3988-3996 [1988]; Vincent et al., Vaccines 90 [1990] (Cold Spring Harbor Laboratory Press); Carter, Current Opinion in Biotechnology 3:533-539 [1992]; Muzyczka, Current Topics in Microbiol. and Immunol. 158:97-129 [1992]; Kotin, Human Gene Therapy 5:793-801 [1994]; Shelling and Smith, Gene Therapy 1:165-169 [1994]; и Zhou et al., J. Exp. Med. 179:1867-1875 [1994], все из которых включены в данный документ посредством ссылки.
Подходящие нуклеотидные последовательности для применения в векторах AAV (и, фактически, любые из описанных в данном документе векторов) включают в себя любую функциональную соответствующую нуклеотидную последовательность. Таким образом, векторы AAV по настоящему изобретению могут включать в себя любой желаемый ген, кодирующий белок, который является дефективным или отсутствующим по отношению к геному клетки-мишени или который кодирует ненативный белок, обладающий желаемым биологическим или терапевтическим действием (например, противовирусной функцией), или последовательность может соответствовать молекуле, обладающей антисмысловой или рибозимной функцией. Подходящие гены включают в себя гены, применяемые для лечения воспалительных заболеваний, аутоиммунных, хронических и инфекционных заболеваний, включая такие нарушения как СПИД, рак, неврологические заболевания, сердечно-сосудистое заболевание, гиперхолестеринемия; различные нарушения крови, включая различные анемии, талассемии и гемофилию; такие генетические дефекты, как муковисцидоз, болезнь Гоше, недостаточная активность аденозиндезаминазы (ADA), эмфизема и т.д. В данной области техники были описаны ряд антисмысловых олигонуклеотидов (например, короткие олигонуклеотиды, комплементарные последовательностям поблизости сайта инициации трансляции (кодон AUG) мРНК), которые используют в антисмысловой терапии раковых и вирусных заболеваний. (См., например, Han et al., Proc. Natl. Acad. Sci. USA 88:4313-4317 [1991]; Uhlmann et al., Chem. Rev. 90:543-584 [1990]; Helene et al., Biochim. Biophys. Acta. 1049:99-125 [1990]; Agarwal et al., Proc. Natl. Acad. Sci. USA 85:7079-7083 [1989]; и Heikkila et al., Nature 328:445-449 [1987]). Для обсуждения подходящих рибозимов см., например, Cech et al. (1992) J. Biol. Chem. 267:17479-17482 и патент США №5225347, которые включены в данный документ посредством ссылки.
Используемый в данном документе термин «очищенный» относится к молекулам как нуклеиновых, так и аминокислотных последовательностей, которые удалены их обычного окружения, выделены или отделены. Поэтому термин «выделенная последовательность нуклеиновой кислоты» представляет собой очищенную последовательность нуклеиновой кислоты. «По существу очищенные» молекулы являются по меньшей мере на 60% свободны, предпочтительно по меньшей мере на 75% свободны и предпочтительнее по меньшей мере на 90% свободны от других компонентов, с которыми они обычно ассоциированы.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к векторам и их применению для разработки линий клеток-хозяев для продукции интересующих белков и, в частности, к векторам, в которых используется слабый промотор, для управления селектируемым маркером.
В некоторых предпочтительных вариантах осуществления в экспрессионных системах по настоящему изобретению используется экспрессионный вектор, который включает в себя последовательность нуклеиновой кислоты, кодирующую интересующий белок (т.е. терапевтический белок или другой белок, которые требуется получать) в функциональной связи с дополнительными последовательностями нуклеиновых кислот, выполняющим различные функции. Таким образом, например, последовательность интересующей нуклеиновой кислоты может включаться в любой из множестве экспрессионных векторов для экспрессии полипептида. В некоторых вариантах осуществления по настоящему изобретению вектор включает в себя, но не ограничивается ими, ретровирусные векторы, хромосомные, нехромосомные последовательности и синтетические последовательности ДНК (например, производные SV40, бактериальные плазмиды, фаговую ДНК; бакуловирусные, дрожжевые плазмиды, векторы, происходящие из комбинаций плазмид и фаговых ДНК, и вирусные ДНК, такие как из вируса осповакцины, аденовируса, вируса оспы-дифтерита птиц и вирусов псевдобешенства). Подразумевается, что может использоваться любой вектор, при условии, что он способен реплицироваться и жизнеспособный в хозяине. В некоторых предпочтительных вариантах осуществления векторы являются ретровирусными векторами, как описано в патентах США №6852510 и 7332333, и публикациях патентов США. №200402335173 и 20030224415, содержание всех из которых в полном объеме включено в данный документ посредством ссылок. В некоторых особенно предпочтительных вариантах осуществления векторы представляют собой псевдотипированные ретровирусные векторы. В некоторых предпочтительных вариантах осуществления по настоящему изобретению экспрессионные векторы млекопитающих включают в себя точку начала репликации, подходящий промотор и энхансер, а также любые необходимые сайты связывания рибозимов, сайт полиаденилирования, донорные и акцепторные сайты сплайсинга, последовательности транскрипционной терминации и 5'-фланкирующие нетранскрибируемые последовательности. В других вариантах осуществления последовательности ДНК, происходящие из слайс-последовательности SV40, и сайт полиаденилирования могут применять для обеспечения требуемых нетранскрибируемых генетических элементов.
В некоторых предпочтительных вариантах осуществления векторы представляют собой ретровирусные векторы. В данной области техники хорошо известна архитектура геномов многочисленных ретровирусов, и это позволяет адаптировать ретровирусный геном для продукции ретровирусных векторов. Продукция рекомбинантного ретровирусного вектора, несущего интересующий ген, как правило достигается за две стадии.
Во-первых, интересующий ген вставляют в ретровирусный вектор, который содержит последовательности, необходимые для эффективной экспрессии интересующего гена (включая промоторный и/или энхансерный элементы, которые могут быть обеспечены вирусными длинными концевыми повторами (LTR) или внутренним промотором/энхансером и соответствующими сигналами сплайсинга), последовательности, необходимые для эффективной упаковки вирусной РНК в инфекционные вирионы (например, упаковывающий сигнал (Psi), сайт связывания праймера тРНК (-PBS), 3'-регуляторные последовательности, требуемые для обратной транскрипции (+PBS)) и вирусные LTR. LTR содержат последовательности, требуемые для связи вирусной геномной РНК, функций обратной транскриптазы и интегразы, и последовательности, вовлеченные в управлении экспрессии геномной РНК, требующей упаковки в вирусные частицы. По соображениям безопасности, у многих рекомбинантных ретровирусных векторов отсутствуют функциональные копии генов, которые необходимы для вирусной репликации (эти необходимые гены либо удалены, либо неактивны); поэтому говорят, что получаемый вирус является репликационно дефективным.
Во-вторых, после конструирования рекомбинантного вектора в упаковывающую линию клеток вводят векторную ДНК. Упаковывающие линии клеток предоставляют белки, требуемые в положении транс для упаковки вирусной геномной РНК в вирусные частицы, имеющие желаемый диапазон хозяев (т.е. кодируемые вирусом белки gag, pol и env). Диапазоном хозяев управляют, в частности, типом генного продукта оболочки, экспрессируемого на поверхности вирусной частицы. Упаковывающие линии клеток могут экспрессировать экотропные, амфотропные или ксенотропные генные продукты оболочки. Альтернативно в упаковывающей линии клеток могут отсутствовать последовательности, кодирующие белок вирусной оболочки (env). В этом случае упаковывающая линия клеток будет паковать вирусный геном в частицы, у которых отсутствует мембранно-ассоциированный белок (например, белок env). Чтобы продуцировать вирусные частицы, содержащие мембранно-ассоциированный белок, который позволит проникать вирусу в клетку, упаковывающую линию клеток, содержащую ретровирусные последовательности, трансфицируют последовательностями, кодирующими мембранно-ассоциированный белок (например, G-белок вируса везикулярного стоматита (ВВС)). Трансфицированная упаковывающая клетка затем будет продуцировать вирусные частицы, которые содержат мембранно-ассоциированный белок, экспрессируемый трансфицированной упаковывающей линией клеток; эти вирусные частицы, которые содержат вирусную геномную РНК, происходящую от одного вируса, инкапсидированные белками оболочки другого вируса, называют псевдотипированными вирусными частицами.
Ретровирусный вектор по настоящему изобретению может дополнительно модифицироваться для включения в него дополнительных регуляторных последовательностей. Как описано выше, ретровирусные векторы по настоящему изобретению включает в себя следующие элементы в функциональной связи: a) 5' LTR; b) упаковывающий сигнал; c) 3' LTR и d) нуклеиновую кислоту, кодирующую интересующий белок, расположенный между 5' и 3' LTR. В некоторых вариантах осуществления по настоящему изобретению интересующую нуклеиновую кислоту могут размещать в противоположной ориентации к 5' LTR, когда желательна транскрипция от внутреннего промотора. Подходящий внутренний промотор включает в себя, но не ограничивается ими, промотор альфа-лактальбумина, промотор CMV (человека или обезьян) и промотор тимидинкиназы.
В других вариантах осуществления по настоящему изобретению, в которых желательна секреция интересующего белка, векторы модифицированы путем включения сигнальной пептидной последовательности в функциональной связи с интересующим белком. Последовательности некоторых подходящих сигнальных пептидов известны специалистам в данной области техники, включая, но ограничиваясь ими, происходящие из тканевого активатора плазминогена, гормона роста человека, лактоферрина, альфа-казеина и альфа-лактальбумина.
В других вариантах осуществления по настоящему изобретению векторы модифицируют введением элемента экспорта РНК (см., например, патенты США №5914267; 6136597 и 5686120; и WO99/14310, все из которых включены в данный документ посредством ссылки) либо в направлении 3', либо 5' по отношению к последовательности нуклеиновой кислоты, кодирующей интересующий белок. Подразумевается, что применение элемента экспорта РНК обеспечивает высокие уровни экспрессии интересующего белка без введения сплайс-сигналов или интронов в последовательность нуклеиновой кислоты, кодирующую интересующий белок.
В еще других вариантах осуществления вектор дополнительно содержит по меньшей мере одну последовательность сайт внутренней посадки рибосомы (IRES). Доступны последовательности нескольких подходящих IRES, включающие, но ограничивающиеся ими, происходящие от вируса ящура (FDV), вируса энцефаломиокардита и полиовируса. Последовательность IRES могут вставлять между двух транскрипционных единиц (например, нуклеиновых кислот, кодирующих различные интересующие белки или субъединицы мультисубъединичного белка, такого как антитело) с целью образования полицистронной последовательности так, что две транскрипционные единицы транскрибируются из того же промотора.
Наиболее широко применяемые рекомбинантные ретровирусные векторы походят от амфотропного вируса мышиного лейкоза Молони (MoMLV) (см. например, Miller and Baltimore Mol. Cell. Biol. 6:2895 [1986]). У системы MoMLV имеется несколько преимуществ: 1) этот специфический ретровирус может инфицировать множество различных типов клеток, 2) доступны устойчивые упаковывающие линии клеток для продукции рекомбинантных вирусных частиц MoMLV и 3) перенесенные гены постоянно интегрированы в хромосому клетки-мишени. Устойчивые векторные системы MoMLV включают в себя вектор ДНК, содержащий небольшую часть ретровирусной последовательности (например, вирусный длинный концевой повтор или LTR, и упаковывающий или psi сигнал) и упаковывающую линию клеток. В ДНК-вектор вставляют ген, требующий переноса. Вирусные последовательности, находящиеся на ДНК-векторе, предоставляют сигнал, необходимый для вставки или упаковки векторной РНК в вирусную частицу и для экспрессии вставленного гена. Упаковывающая линия клеток предоставляет белки, требуемые для сборки частиц (Markowitz et al., J. Virol. 62:1120 [1988]).
В некоторых предпочтительных вариантах осуществления ретровирусные векторы являются псевдотипированными и, например, используют G-белок ВВС в качестве мембранно-ассоциированного белка. В отличии от ретровирусных белков оболочки, которые связываются со специфическим белковым рецептором поверхности клеток для обеспечения проникновения в клетку, G-белок ВВС взаимодействует с фосфолипидным компонентом плазматической мембраны (Mastromarino et al., J. Gen. Virol. 68:2359 [1977]). Поскольку проникновение ВВС в клетку не зависит от наличия специфических белковых рецепторов, ВВС имеет чрезвычайно широкий диапазон хозяев. Псевдотипированные ретровирусные векторы, несущие G-белок ВВС, имеют измененные в отношении диапазона хозяев свойства ВВС (т.е. они могут инфицировать большинство из всех видов клеток позвоночных, беспозвоночных и насекомых). Важно отметить, что G-псевдотипированные по ВВС ретровирусные векторы могут концентрировать в 2000 или более раз путем ультрацентрифугирования без значительной потери инфекционности (Burns et al. Proc. Natl. Acad. Sci. USA 90:8033 [1993]).
Настоящее изобретение не ограничивается применением G-белка ВВС, при котором вирусный G-белок применяют как гетерологичный мембранно-ассоциированный белок у вирусной частицы (см., например, патент США №5512421, который включен в данный документ посредством ссылки). G-белки вирусов рода Vesiculovirus, отличающиеся от ВВС, такие как вирусы Пири и Чандипура, которые высокогомологичны G-белку ВВС и, подобно G-белку ВВС, содержат ковалентно связанную пальмитиновую кислоту (Brun et al. Intervirol. 38:274 [1995] и Masters et al., Virol. 171:285 (1990]). Таким образом, G-белок вирусов Пири и Чандипура могут использовать вместо G-белка ВВС для псевдотипирования вирусных частиц. Кроме того, G-белок ВВС вирусов в пределах рода Lyssa вирусов, таких как вирусы бешенства и Мокола, демонстрируют высокую степень консервативности (консервативность аминокислотной последовательности, а также функциональную консервативность) с G-белками ВВС. Например, показано, что G-белок вируса Мокола функционирует подобным G-белку ВВС образом (т.е. опосредует слияние с мембраной) и, поэтому, его могут использовать вместо G-белка ВВС для псевдотипирования вирусных частиц (Mebatsion et al., J. Virol. 69:1444 [1995]). Вирусные частицы могут псевдотипировать с использованием любого из G-белков Пири, Чандипура или Мокола, как описано в примере 2, з исключением того, что применяют содержащие плазмиду последовательности, кодирующие любой из G-белков Пири, Чандипура или Мокола, находятся под транскрипционным контролем подходящего промоторного элемента (например, немедленно-ранний промотор CMV; доступны многочисленные экспрессионные векторы, содержащие промотор IE CMV, такие как вектор pcDNA3.1 (Invitrogen)) вместо pHCMV-G. Могут применять последовательности, кодирующие другие G-белки, производные от других представителей семейства Rhabdoviridae; последовательности, кодирующие многочисленные рабдовирусные G-белки, доступные в базе данных Genbank.
В некоторых предпочтительных вариантах осуществления векторы представляют собой лентивирусные векторы. Лентивирусы (например, вирус инфекционной анемии у лошадей, вирус артрита-энцефалита коз и овец, вирус иммунодефицита человека) представляют собой подсемейство ретровирусов, которые способны к интеграции в неделящиеся клетки. Лентивирусный геном и провирусная ДНК обычно содержат три гена, обнаруженные в ретровирусах: gag, pol и env, которые фланкированы двумя последовательностями LTR. Ген gag кодирует внутренние структурные (матриксный, капсидный и нуклеокапсидный) белки; ген pol кодирует обратную транскриптазу, протеазу и интегразу; и ген env кодирует гликопротеины вирусной оболочки. 5' и 3' LTR контролируют транскрипцию и полиаденилирование вирусной РНК. Дополнительные гены в лентивирусном геноме включают гены vif, vpr, tat, rev, vpu, nef и vpx.
Разнообразные лентивирусные векторы и упаковывающие линии клеток известны в данной области техники и находят применение в настоящем документе (см., например, патенты США №5994136 и 6013516, оба из которых включены в данный документ посредством ссылки). Кроме того, G-белок ВВС также применяется для псевдотипирования ретровирусных векторов, на основе вируса иммунодефицита человека (ВИЧ) (Naldini et al., Science 272:263 [1996]). Таким образом, G-белок ВВС могут применять для создания разнообразных псевдотипированных ретровирусных векторов, и они не ограничиваются векторами, основанными на MoMLV. Лентивирусные векторы также могут также модифицировать, как описано выше, с целью заключения в них различных регуляторных последовательностей (например, сигнальных пептидных последовательностей, элементов экспорта РНК и IRES). После продуцирования лентивирусных векторов, их могут применять для трансфекции клеток-хозяева, как описано выше для ретровирусных векторов.
В некоторых предпочтительных вариантах осуществления векторы представляют собой аденоассоциированные векторы (AAV). AAV представляет собой ДНК-парвовирус человека, который принадлежит роду Dependovirus. Геном AAV состоит из линейной одноцепочечной молекулы ДНК, которая содержит приблизительно 4680 оснований. Геном включает в себя инвертированные концевые повторы (ITR) на каждом конце, которые функционируют в положении цис, как точки начала репликации ДНК и как упаковывающие сигналы для вируса. Внутреняя часть генома без повторов включает в себя две большие открытые рамки считывания, известные соответственно как области rep и cap AAV. Эти области кодируют вирусные белки, участвующие в репликации и упаковке вириона. Семейство из по меньшей мере четырех вирусных белков синтезируется в области rep AAV, Rep 78, Rep 68, Rep 52 и Rep 40, называемые в соответствии с их кажущейся молекулярной массой. Область cap AAV, кодирует по меньшей мере три белка, VP1, VP2 и VP3 (для получения подробного описания генома AAV, см., например, Muzyczka, Current Topics Microbiol. Иммун. 158:97-129 [1992]; Kotin, Человека Ген Therapy 5:793-801 [1994]).
AAV требует коинфекции с неродственным вирусом-помощником, таким как аденовирус, герпесвирус или вирус осповакцины, с целью прохождения продуктивной инфекции. В отсутствии такой коинфекции AAV переходит в латентное состояние путем вставки своего генома в хромосому клетки-хозяина. Последующая инфекция вирусом-помощником разблокирует интегрированную копию, которая потом может реплицироваться для продуцирования инфекционного вирусного потомства. В отличие от непсевдотипированных ретровирусов AAV имеет широкий спектр хозяев и способен реплицироваться в клетках от любых видов, при условии, что существует коинфекция с вирусом-помощником, который будет также мультиплицироваться в таких биологических видах. Таким образом, например, AAV человека будет реплицироваться в клетках псовых, коинфицированных аденовирусом псовых. Кроме того, в отличие от ретровирусов, AAV не ассоциирован с любым заболеванием человека или животного, по-видимому не изменяет биологические свойства клетки-хозяина после интеграции и способен интегрироваться в неделящиеся клетки. Недавно также обнаружено, что AAV способен к сайт-специфической интеграции в геном клетки-хозяина.
В свете описанных выше свойств разработали ряд рекомбинантных векторов AAV для доставки генов (см., например, патенты США №5173414; 5139941; WO 92/01070 и WO93/03769, оба вида включены в данный документ посредством ссылки; Lebkowski et al., Molec. Cell. Biol. 8:3988-3996 [1988]; Carter, Current Opinion in Biotechnology 3:533-539 [1992]; Muzyczka, Current Topics in Microbiol. and Immunol. 158:97-129 [1992]; Kotin, (1994) Human Gene Therapy 5:793-801; Shelling and Smith, Gene Therapy 1:165-169 [1994]; и Zhou et al., J. Exp. Med. 179:1867-1875 [1994]).
Рекомбинантные вирионы AAV могут продуцировать в подходящей клетке-хозяине, которая была трансфицирована как плазмидой-помощником AAV, так и вектором AAV. Плазмида-помощник AAV обычно включает в себя кодирующие rep и cap AAV области, но у нее отсутствуют ITR AAV. Соответственно, плазмида-помощник не может самостоятельно ни реплицироваться, ни упаковываться. Вектор AAV обычно включает в себя выбранный интересующий ген, связанный ITR AAV, которые обеспечивают функции вирусной репликации и упаковки. И плазмиду-помощник, и вектор AAV, несущий выбранный ген, вводят в подходящую клетку-хозяина, путем транзиентной трансфекции. Затем трансфицированную клетку инфицируют вирусом-помощником, таким как аденовирус, который трансактивирует промотор AAV, находящийся на плазмиде-помощнике, контролирующей транскрипцию и трансляцию областей rep и cap AAV. Формируются рекомбинантные вирионы AAV, содержащие выбранный ген, и их могут очищать от среды получения. Сразу после продуцирования векторов AAV, их могут применять для трансфекции (см., например, патент США 5843742, в данном документе включены в данный документ посредством ссылки) клеток-хозяев с требуемой множественностью инфекции для получения клеток-хозяев с высоким числом копий. Специалисту в данной области техники будет понятно, что векторы AAV могут также модифицировать, как описано выше, с целью заключения в них различных регуляторных последовательностей (например, сигнальных пептидных последовательностей, элементов экспорта РНК и IRES).
В некоторых предпочтительных вариантах осуществления векторы представляют собой векторы на основе транспозонов. Транспозоны представляют собой мобильные генетические элементы, которые могут перемещаться или переноситься в геноме из одного положения в другое. Транспозиция в пределах генома управляется ферментом транспозазой, который кодируется этим транспозоном. Множество примеров транспозонов известны в данной области техники, включая, но ограничиваясь ими, Tn5 (см. например, de la Cruz et al., J. Bact. 175: 6932-38 [1993], Tn7 (см. например, Craig, Curr. Topics Microbiol. Immunol. 204: 27-48 [1996]), и Tn10 (см., например, Morisato and Kleckner, Cell 51:101-111 [1987]). Способность транспозонов интегрироваться в геном использована для создания транспозонных векторов (см., например, патенты США №5719055; 5968785; 5958775; и 6027722; все из которых включены в данный документ посредством ссылки). Поскольку транспозоны не являются инфекционными, транспозонные векторы вводят в клетки-хозяева посредством известной в данной области техники методов (например, электропорацией, липофекцией или микроинъекцией). Поэтому соотношение транспозонных векторов в клетки-хозяева могут корректировать для обеспечения желаемой множественности инфекции с целью получения клеток-хозяев по настоящему изобретению с высоким числом копий.
Транспозонные векторы, подходящие для применения в настоящем изобретении, обычно включают в себя нуклеиновую кислоту, кодирующую интересующий белок, вставленный между двух последовательностей вставки транспозона. Некоторые векторы также включают в себя последовательность нуклеиновой кислоты, кодирующую фермент транспозазу. В этих векторах, одна из последовательностей вставки расположена между ферментом транспозазой и нуклеиновой кислотой, кодирующей интересующий белок таким образом, что он не включается в геном клетки-хозяина во время рекомбинации. Альтернативно, фермент транспозазу могут предоставлять подходящим методом (например, липофекцией или микроинъекцией). Специалисту в данной области техники будет понятно, что транспозонные векторы могут также модифицировать, как описано выше, с целью заключения в них различных регуляторных последовательностей (например, сигнальных пептидных последовательностей, элементов экспорта РНК и IRES).
В некоторых предпочтительных вариантах осуществления векторы включают в себя селектируемый маркер. Подходящие селектируемые маркеры, включают в себя, но не ограничиваются ими, глутамин-синтазу (GS), дигидрофолат-редуктазу (DHFR) и т.п. Эти гены описаны в патентах США №5770359; 5827739; 4399216; 4634665; 5149636 и 6455275; все из которых включены в данный документ посредством ссылки. В некоторых особенно предпочтительных вариантах осуществления селектируемый маркер представляет собой GS и имеет последовательность, которая разделяет по меньшей мере 80%, 90%, 95%, 99% или 100% идентичности с последовательностью по положениям от 1789 до 2910 из SEQ ID NO:2. В некоторых предпочтительных вариантах осуществления используемый селектируемый маркер является совместимым с линией клеток-хозяев, которая дефектна по продукции фермента, кодируемого последовательностью нуклеиновой кислоты селектируемого маркера. Подходящие линии клеток-хозяев более подробно описаны ниже. В других вариантах осуществления селектируемый маркер представляет собой маркер резистентности к антибиотику, т.е. ген, продуцирующий белок, который позволяет клеткам экспрессировать этот белок с резистентностью к антибиотику. Подходящие маркеры резистентности к антибиотикам включают в себя гены, которые обеспечивают резистентность к неомицину (ген резистентности к неомицину (neo)), гигромицину (ген фосфатрансферазы гигромицина B), пуромицину (пуромицин N-ацетил-трансфераза) и подобные.
В некоторых вариантах осуществления последовательность нуклеиновой кислоты, кодирующая селектируемый маркер, функционально связана с промоторной последовательностью. В некоторых особенно предпочтительных вариантах осуществления промоторная последовательность была изменена, например путем мутации, чтобы уменьшить промоторную активность по сравнению с неизмененной версией промотора. Эти промоторы могут описывать как слабые промоторы, в том отношении, что экспрессия гена, связанного с промотором, происходит на низком уровне, в противоположность высокому уровню. Гены, регулируемые сильными промоторами, дают больше мРНК и, поэтому, больше продуцируют белка, чем гены, регулируемые слабыми промоторами. Таким образом, в настоящем изобретении предпочтительно используется промотор для селектируемого маркера, который был изменен так, чтобы давать меньше мРНК, чем сравнимый неизмененный промотор.
В некоторых предпочтительных вариантах осуществления промотор, функционально связанный с селектируемым маркером, представляет собой длинный концевой повтор (LTR) из ретровируса или лентивируса. В некоторых особенно предпочтительных вариантах осуществления промотор представляет собой LTR, который был изменен путем удаления либо всей, либо части области U3 LTR. В некоторых вариантах осуществления промотор, функционально связанный с селектируемым маркером, является самоинактивирующимся (SIN) LTR. Подходящие SIN LTR известны в данной области техники. В некоторых предпочтительных вариантах осуществления SIN LTR по настоящему изобретению предпочтительно имеют по меньшей мере 80%, 90%, 95%, 99% или 100% идентичности с SEQ ID NO:3, и SIN LTR наиболее предпочтительно содержит в себе делецию в области U3, которая снижает промоторную активность по сравнению с промоторной активностью, если в этой области U3 нет делеции.
Легко понять, что когда вектор представляет собой ретровирусный вектор, то данный вектор сконструирован таким образом, что SIN LTR представляет собой 3’ LTR последовательности ретровирусного вектора. Благодаря тому факту, что делеция U3 копируется на 5′ и 3′ LTR во время обратной транскрипции, интегрированные векторы SIN содержат только LTR с удаленными U3. Карта типового ретровирусного вектора по настоящему изобретению представлена как ФИГ. 2. Как видно, вектор содержит 3’ SIN LTR. После обратной траскрипции LTR hCMV-MOMuSV, изображенный на карте вектора, заменяется 3’ SIN LTR, изображенным на карте вектора таким образом, что когда вектор интегрируется в хромосому клетки-хозяина, SIN LTR с ней функционально связывается и направляет экспрессию описанной кДНК GS.
В некоторых предпочтительных вариантах осуществления по настоящему изобретению последовательность нуклеиновой кислоты, кодирующая интересующий белок в векторе экспрессии, функционально связана с соответствующей(-ими) последовательностью(-ями) регуляции экспрессии (промотор) для управления синтезом мРНК. Промотор, используемый в настоящем изобретении, включает в себя, но не ограничивается ими, промотор LTR или SV40, lac или trp E. coli, PL и PR фага лямбда, промотор T3 и T7 и немедленно-ранний промотор цитомегаловируса (CMV), промоторы тимидин-киназы вируса простого герпеса (HSV) и металлотионеина-I мыши и другие промоторы, которые, как известно, регулируют экспрессию гена в прокариотических или эукариотических клетках или их вирусах.
Как указано выше, типовая карта вектора по настоящему изобретению представлена на ФИГ. 3 и соответствует SEQ ID NO:2. В некоторых предпочтительных вариантах осуществления векторы включают в себя следующий элемент в направлении от 5’ до 3’:
- 5’ LTR (на примере LTR hCMV-MoMuSV);
- ретровирусную упаковывающую область;
- нуклеиновую кислоту, кодирующую селектируемый маркер (например, кДНК GS);
- внутренний промотор (на примере промотора CMV обезьян (sCMV));
- последовательность нуклеиновой кислоты, кодирующую интересующий белок (на примере последовательности «anyway»), которая функционально связана с внутренним промотором;
- последовательность WPRE;
- 3’ SIN LTR.
В некоторых вариантах осуществления вектор представляет собой плазмиду, содержащую следующие элементы в порядке от 5’ до 3’: 1) 5’ LTR (например, SIN LTR); 2) упаковывающую область; 3) селектируемый маркер (например, GS); 4) внутренний промотор (например, CMV); 5) последовательность нуклеиновой кислоты, кодирующую интересующий белок, которая функционально связана с внутренним промотором.
В некоторых вариантах осуществления векторы содержат одну сигнальную последовательность поли-A после нуклеиновой кислоты, кодирующей интересующий белок. Например, в некоторых вариантах осуществления вектор содержит следующие компоненты в порядке от 5’ до 3’: Промотор (например, SIN) селектируемый маркер - стоп-кодон - промотор (например, CMV) - интересующий белок - стоп-кодон - поли-A.
В некоторых вариантах осуществления в настоящем изобретении предлагаются клетки-хозяева и культура клеток-хозяев, причем клетки-хозяева экспрессируют интересующий белок из описанных выше векторов. В предпочтительном варианте осуществления клетки-хозяева являются клетками-хозяевами млекопитающих. Ряд линий клеток-хозяев млекопитающих известен в данной области техники. В целом, эти клетки-хозяева способны расти и выживать при выращивании либо в монослойной культуре, либо в суспензионной культуре в среде, содержащей соответствующие питательные вещества и факторы роста, как более подробно описано ниже. Как правило, клетки способны экспрессировать и секретировать большие количества конкретного интересующего белка в культуральную среду. Примеры подходящих клеток-хозяев млекопитающих включают в себя, но не ограничиваются ими, клетки яичника китайского хомячка (CHO-K1, ATCC CCl-61); эпителиальные клетки молочной железы крупного рогатого скота (ATCC CRL 10274; эпителиальные клетки молочной железы крупного рогатого скота); линия CV1 почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); эмбриональная линия почки человека (293 или клетки 293, субклонированные для выращивания в суспензионной культуре; см., например, Graham et al., J. Gen Virol., 36:59 [1977]); клетки почек детенышей хомяка (BHK, ATCC CCL 10); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23:243-251 [1980]); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы линии Buffalo (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383:44-68 [1982]); клетки MRC 5; клетки FS4; фибробласты крысы (клетки 208F); клетки MDBK (клетки почки крупного рогатого скота); клетки CAP (CEVEC от Amniocyte Production) и линия гепатомы человека (Hep G2).
В некоторых особенно предпочтительных вариантах осуществления клетки-хозяева модифицированы таким образом, что они являются дефектными, или они дефектны естественным путем, по ферментативной активности, которая требуется для роста или выживаемости клеток в присутствии агента селекции, предоставляемого селектируемым маркером. Например, клетки яичника китайского хомячка (CHO) были модифицированы для обеспечения дефектности по GS. В некоторых предпочтительных вариантах осуществления, в которых вектор включает в себя селектируемый маркер GS, линия клеток-хозяев дефектна по GS. В некоторых особенно предпочтительных вариантах осуществления дефектная по GS линия клеток-хозяев представляет собой CHOZN® GS-/- линию клеток, предлагаемую Merck KGaA. В других вариантах осуществления, в которых селектируемый маркер представляет собой, например, DHFR, линия клеток предпочтительно может быть дефектна по активности DHFR (т.е. является DHFR-). Подходящие DHFR- линии клеток включают в себя, но не ограничиваются ими, CHO-DG44 и их производные.
Векторы могут вводить в клетки-хозяева путем любых подходящих средств. В некоторых предпочтительных вариантах осуществления получали векторы (например, ретровирусные векторы), кодирующие интересующий белок, их можно применять для трансфекции или трансдукции клеток-хозяев. Предпочтительно, клетки-хозяева трансфицируют или трансдуцируют интегрирующимися векторами со множественностью инфекции, достаточной для получения интеграции по меньшей мере 1 и предпочтительно по меньшей мере 2 или более ретровирусных векторов. В некоторых вариантах осуществления могут использовать значения множественности инфекции от 10 до 1000000, так что геномы инфицированных клеток-хозяев содержат от 2 до 1000 копий интегрированных векторов, предпочтительно от 5 до 1000 копий интегрированных векторов и наиболее предпочтительно от 20 до 500 копий интегрированных векторов. В других вариантах осуществления применяют множественность инфекции от 10 до 10000. Когда для инфекции используют непсевдотипированные ретровирусные векторы, клетки-хозяева инкубируют с культуральной средой от продуцирующих ретровирусы клеток, содержащей желаемый титр (т.е. количество колониеобразующих единиц, КОЕ) инфекционных векторов. Когда применяют псевдотипированные ретровирусные векторы, эти векторы концентрируют для получения соответствующего титра путем ультрацентрифугирования, а затем добавляют к культуре клеток-хозяев. Альтернативно, концентрированные векторы могут разбавлять в культуральной среде, подходящей для данного типа клеток. Кроме того, когда желательна экспрессия более одного интересующего белка клетки-хозяева, клетки-хозяева могут трансфицировать множеством векторов, каждый из которых содержит нуклеиновую кислоту, кодирующую отличающийся интересующий белок.
В каждом случае клетки-хозяева помещают в среду, содержащую инфекционные ретровирусные векторы, в течение достаточного периода времени для прохождения инфекции и последующей интеграции векторов. В целом, количество среды, используемой для покрытия клеток, необходимо поддерживать на уровне минимально возможного объема с целью способствования максимального количества интеграционных событий на клетку. Как общее руководство, количество колониеобразующих единиц (КОЕ) на миллилитр должно составлять приблизительно от 105 до 107 КОЕ/мл, в зависимости от количества желаемых интеграционных событий.
В некоторых вариантах осуществления после трансфекции или трансдукции, клеткам позволяют размножиться, и затем их трипсинизируют и пересевают. Затем отбирают отдельные колонии для получения клонально отобранных линий клеток. В еще дополнительных вариантах осуществления клонально отобранные линии клеток подвергают скринингу Саузерн блоттингом или анализами ПЦР для проверки прохождения желаемого количества интеграционных событий. Подразумевается также, что клональный отбор позволяет идентифицировать линии клеток, продуцирующие наибольшие количества белка. В других вариантах осуществления клетки клонально не отбирают после трансфекции.
В некоторых вариантах осуществления клетки-хозяева трансфицируют с помощью векторов, кодирующих различные интересующие белки. Векторы, кодирующие различные интересующие белки, могут применять для трансфекции клеток одновременно (например, клетки-хозяева подвергают действию раствора, содержащего векторы, кодирующие различные интересующие белки), или трансфекция может быть последовательной (например, клетки-хозяева сначала трансфицируют вектором, кодирующим первый интересующий белок, выдерживают определенный период времени и затем клетки-хозяева трансфицируют вектором, кодирующим второй интересующий белок). В некоторых предпочтительных вариантах осуществления клетки-хозяева трансфицируют интегрирующим вектором, кодирующим первый интересующий белок, отбирают линии клеток с высокой экспрессией (например, клонально отбирают), содержащие множество интегрированных копий интегрирующегося вектора, и отобранные линии клеток трансфицируют интегрирующимся вектором, кодирующим второй интересующий белок. Это процесс может повторяться для введения множества интересующих белков. В некоторых вариантах осуществления могут манипулировать множественностью инфекции (например, увеличивать или уменьшать ее) для увеличения или уменьшения экспрессии интересующего белка. Аналогичным образом, могут применять различные промоторы для изменения экспрессии интересующих белков. Подразумевается, что эти способы могут применять для конструирования линий клеток-хозяева, содержащих целые экзогенные метаболические пути, или для предоставления клеткам-хозяевам повышенной способности процессировать белки (например, клетки-хозяева могут обеспечивать ферментами, необходимыми для посттрансляционной модификации).
В еще дополнительных вариантах осуществления линии клеток последовательно трансфицируют векторами, кодирующими одинаковый ген. В некоторых предпочтительных вариантах осуществления клетки-хозяева трансфицируют (например, при MOI приблизительно от 10 до 1000000, предпочтительно от 100 до 10000) интегрирующим вектором, кодирующим интересующий белок, отбирают линии клеток (например, клонально отбирают), содержащие одну или множество интегрированных копии интегрирующегося вектора или экспрессирующие высокие уровни желаемого белка, и отобранные линии клеток ретрансфицируют тем же вектором (например, при MOI приблизительно от 10 до 1000000, предпочтительно от 100 до 10000). В некоторых вариантах осуществления идентифицируют и отбирают линии клеток содержат по меньшей мере две интегрированные копии вектора. Это процесс может повторяться множество раз до получения желаемого уровня экспрессии белка и может также повторяться для введения векторов, кодирующих множество интересующих белков. Неожиданно обнаружено, что последовательная трансфекция одинаковым геном приводит к увеличениям продукции белков из получаемых клеток, что не является просто аддитивным эффектом.
В настоящем изобретении подразумевается разнообразные последовательные процедуры трансфекции. В некоторых вариантах осуществления, в которых применяют ретровирусные векторы, предлагаются последовательные процедуры трансдукции. В предпочтительных вариантах осуществления последовательная трансдукция выполняется на пуле клеток. В этих вариантах осуществления исходный пул клеток-хозяев приводят в контакт с ретровирусными векторами, предпочтительно при множественности инфекции в диапазоне от около 0,5 до около 1000 векторов/клетку-хозяина. Затем клетки культивируют в течение нескольких дней в подходящей среде (например, с агентом селекции). Потом отбирают аликвоту клеток для определения количества интегрированных векторов и замораживают для будущего возможного использования. Затем оставшиеся клетки повторно приводят в контакт с ретровирусными векторами, снова предпочтительно при множественности инфекции в диапазоне от около 0,5 до около 1000 векторов/клетку-хозяина. Этот процесс повторяют пока не получат желаемое количество интегрированных векторов. Например, процесс могут повторять от 10 до 20 или более раз. В некоторых вариантах осуществления клетки могут клонально отбирать после любого конкретного этапа трансдукция, если так требуется, однако применение пула клеток в отсутствии результатов трансдукции приводит к увеличению времени для получения желаемого количества интегрированных копий вектора.
В некоторых вариантах осуществления по настоящему изобретению после трансформации подходящего хозяйского штамма и выращивания хозяйского штамма в среде до соответствующей плотности клеток, интересующий белок секретируется во время культивирования клеток-хозяев. В предпочтительных вариантах осуществления, в которых применяют амплифицируемые маркеры, подразумевается, что культура трансдуцированных клеток-хозяев находится в среде, содержащей ингибитор данного гена. Подходящие ингибиторы включают в себя, но не ограничиваются ими, метотрексат для ингибирования DHFR и метионинсульфоксимин (Msx) или фосфинотрицин для ингибирования GS. Подразумевается, что по мере увеличения концентраций этих ингибиторов в системе клеточной культуры, отбираются клетки с большим количеством копий амплифицируемого маркера (и, таким образом, генов или интересующих генов) или клетки, которые содержат более высоко-продуцирующие инсерции.
Соответственно, клетки-хозяева, содержащие векторы, как описано выше, культивируют в соответствии со способами, известными в данной области техники. Подходящие условия культивирования для клеток млекопитающих хорошо известны в данной области техники (см. например, J. Immunol. Methods (1983) 56:221-234 [1983], Animal Cell Culture: A Practical Approach 2nd Ed., Rickwood, D. and Hames, B. D., eds. Oxford University Press, New York [1992]).
Культуры клеток-хозяев по настоящему изобретению готовят в среде, подходящей для конкретной культивируемой клетки. Примерами питательных растворов являются коммерчески доступные среды, такие как среда ActiPro (HyClone), усовершенствованная среда для периодической подпитки ExCell (SAFC), среда Хэма F10 (Sigma, г. Сент-Луис, штат Миссури), минимальная питательная среда (MEM, Sigma), RPMI-1640 (Sigma) и модифицированная Дульбекко среда Игла (DMEM, Sigma). Подходящие среды также описаны в патентах США №4767704; 4657866; 4927762; 5122469; 4560655; и WO 90/03430 и WO 87/00195; описания которых включены в данный документ посредством ссылки. Любые из этих сред могут дополнять, при необходимости, сывороткой, гормонами и/или другими факторами роста (такие как инсулин, трансферрин или эпидермальным фактором роста), солями (такими как хлорид натрия, соли кальция, магния и фосфата), буферами (такими как HEPES), нуклеозидами (такими как аденозин и тимин), антибиотиками (такими как гентамицин (гентамицин), микроэлементами (известными как неорганические соединения, обычно присутствующими в конечных концентрациях в микромолярном диапазоне), липидами (такими как линолевая кислота или другие жирные кислоты) и их подходящими носителями, и глюкозу или эквивалентный источник энергии. В некоторых предпочтительных вариантах осуществления, в которых применяют селектируемый маркер, такой как GS, например в среде будет отсутствовать глутамин. В соответствующих концентрациях могут включать любые другие необходимые добавки, которые могут быть известны специалистам в данной области техники.
В настоящем изобретении также подразумевается использование различных систем культивирования (например, чашки Петри, 96-луночные планшеты, роллерные бутыли и биореакторы) для трансфицированных клеток-хозяев. Например, трансфицированные клетки-хозяева могут культивировать в перфузионной системе. Перфузионная культура относится к обеспечению непрерывного потока культуральной среды через культуру, поддерживаемую при высокой плотности клеток. Клетки суспендированы и не требуют твердой подложки для роста на ней. В целом, свежие питательные вещества могут поставляться непрерывно с сопутствующим удалением токсических метаболитов и, в идеале, избирательным удалением мертвых клеток. Для обновления окружения культуры в достаточных соотношениях пригодны все способы фильтрования, захвата и микрокапсулирования.
В качестве другого примера, в некоторых вариантах осуществления может применяться процедура периодического культивирования с подпиткой. При предпочтительном периодическом культивировании с подпиткой клеток хозяина млекопитающего первоначально в культуральный сосуд добавляют клетки и среду для культивирования, и непрерывно или дискретными добавлениями подпитывают дополнительными питательными веществами для культуры в процессе культивирования с периодическим сбором клеток и/или продуктов или без сбора до прекращения культивирования. Периодическая культура с подпиткой может включать, например, полунепрерывную периодическую культуру с подпиткой, в которой периодически цельную культуру (включающую в себя клетки и среду) удаляют и заменяют свежей средой. Периодическая культура с подпиткой отличается от простой периодической культуры, в которой все компоненты для культивирования клеток (включая клетки и все питательные вещества для культуры) добавляют в культуральный сосуд в начале процесса культивирования. Периодическая культура с подпиткой может отличаться от перфузионного культивирования в той мере, что супернатант не удаляют из культурального сосуда во время данного процесса (при перфузионном культивировании клетки удерживаются в культуре, например путем фильтрации, инкапсулирования, прикрепления к микроносителям и т.д., и культуральную среду непрерывно или периодически вводят и удаляют из культурального сосуда). В некоторых особенно предпочтительных вариантах осуществления периодические культуры поддерживают в роллерных бутылях.
Дополнительно клетки культуры могут размножать в соответствии с любой схемой или практикой, которые могут быть подходящими для конкретной клетки-хозяина и конкретного предполагаемого плана получения. Поэтому настоящее изобретение предусматривает одностадийную или многостадийную процедуру культивирования. При одностадийной культуре клетки-хозяева инокулируют в культуральное окружение, и применяют процессы по настоящему изобретению во время единственной фазы получения культуры клеток. Альтернативно, предусмотрена многостадийная культура. При многостадийной культуре клетки могут культивировать в ряде стадий или фаз. К примеру, клетки могут выращивать в первую стадию или фазу роста культуры, причем клетки зачастую извлекают из места хранения, их инокулируют в среду, пригодную для стимулирования роста и высокой жизнеспособности. Клетки могут поддерживать в фазе роста в течение подходящего периода времени путем добавления свежей среды к культуре клеток-хозяев.
Разрабатывают условия периодической культуры с подпиткой или непрерывной культуры клеток для усиления роста клеток млекопитающих в фазе роста культуры клеток. В фазе роста клетки выращивают в условиях и в течение периода времени, которые максимально подходят для роста. Условия культивирования, такие как температура, pH, растворенный кислород (dO2) и т.п., соответствуют применяемым с конкретным хозяином и будут очевидны среднему специалист в данной области техники. В целом, корректируют pH на уровне приблизительно между 6,5 и 7,5 с использованием либо кислоты (например, CO2), либо основания (например, Na2CO3 или NaOH). Подходящий диапазон температур для культивирования млекопитающих клеток, таких как клетки CHO составляет между приблизительно между 30o и 38o C и подходящее значение dO2 составляет между 5-90% насыщения воздухом.
После фазы продуцирования полипептида интересующий полипептид выделяют из культуральной среды с использованием методик, которые хорошо установлены в данной области техники. Интересующий белок предпочтительно выделяют из культуральной среды в виде секретируемого полипептида (например, секреция интересующего белка управляется сигнальной пептидной последовательностью), хотя его также могут выделять из лизатов клеток-хозяев. Не первой стадии культуральную среду или лизат центрифугируют для удаления дебриса фрагментов клеток. После чего полипептид очищают от загрязняющих растворимых белков и полипептидов, с последующими процедурами, являющимися типовыми для подходящих процедур очистки: фракционирования на иммунноаффинных или ионообменных колонках; осаждения этанолом; обращенно-фазной ВЭЖХ; хроматографии на силикагеле или на катионообменной смоле, такой как ДЭАЭ; хроматофокусирования; ДСН-ПААГ-электрофореза; осаждения сульфатом аммония; гель-фильтрации с использованием, например, Сефадекс G-75; и колонок с сефарозой и белком A для удаления таких контаминантов, как IgG. Для ингибирования протеолитического разрушения во время очистки также может быть полезен ингибитор протеаз, такой как фенилметилсульфофторид (PMSF). Кроме того, интересующий белок могут сливать в рамке считывания с маркерной последовательностью, которая обеспечивает очистку данного интересующего белка. Неограничивающий пример маркерных последовательностей включает в себя гексагистидиновым маркером, который может поставляться вектором, предпочтительно вектором pQE-9, и гемагглютининовым (HA) маркером. Маркер HA соответствует эпитопу, происходящему от белка-гемагглютинина вируса гриппа (см., например, Wilson et al., Cell, 37:767 [1984]). Специалист в данной области поймет, что методы очистки, подходящие для интересующего полипептида, могут потребовать модификации с учетом изменений в природе полипептида при экспрессии в культуре рекомбинантных клеток.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
В настоящем изобретении предлагается уникальный путь комбинирования ретровирусной трансдукции (называемой в данном документе технология GPEx®) в комбинации с системой линии клеток CHO с нокаутом глутаминсинтазы (GS), которая дала неожиданное улучшение способности клеток в отношении продуцирования высоких титров и удельных продуктивностей интересующего белка. Линия клеток с нокаутом GS, использованная в этих исследованиях, была линией клеток CHOZN, предлагаемой MilliporeSigma. Применена на практике технология GPEx® аналогичным образом, как она была применена раннее для нормальных клеток CHO. Однако в вирусном векторе экспрессии ген GS, используемый для селекции линии клеток с нокаутом GS, ослаблялся очень слабым промотором из SIN (самоинактивирующегося) LTR, присутствующем в одной из версий вектора экспрессии GPEx, который авторы изобретения использовали в компании Catalent. Комбинация такого вектора, процесс получения из нормальной линии клеток GPEx и линия клеток CHO с нокаутом GS дали уровни продуцирования до 7 раз больше, чем традиционный процесс с GPEx в немодифицированной линии клеток CHO.
Эксперимент 1
С использованием различных вирусных векторов получали две объединенные линии клеток. Обе линии получали с использованием нормальных процессов трансдукции GPEx и линии клеток CHOZN с нокаутом GS. Оба экспрессионые векторы сконструированы для экспрессии тестового белка «Anyway». Anyway представляет собой Fc-слитый белок. Единственное отличие между этими двумя экспрессионными векторами заключалось в том, что один вектор после вставки в линию клеток мог получить управление экспрессией гена GS полноразмерным LTR вирусом мышиного лейкоза Молони (MMLV), а другой вектор могу получить управление экспрессией GS посредством SIN LTR MMLV. Генные конструкции, используемые для продуцирования ретровекторных частиц, представлены на Фиг. 1 и 2. Последовательности для генных конструкций предложены на Фиг. 3 и 4.
Разработка линии клеток CHOZN
Продукция ретровекторов: отмеченные выше экспрессионные конструкции вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Содержащую ген оболочки экспрессионную плазмиду также ко-трансфекцировали с каждой из генных конструкций. Ко-трансфекция привела к продукции репликационно некомпетентного ретровектора с высокими титрами, которого концентрировали путем ультрацентрифугирования и использовали для трансдукций клеток (1, 2). Использование трансдукции ретровекторами, продуцированными с применением вышеуказанных конструкций, на клетках CHO приводит к дуплицированию последовательности 3’ LTR к 5’-концу последовательности, заменяя 5’LTR hCMV-MoMuSV, и впоследствии управляя экспрессией гена GS в объединенной линии клеток.
Трансдукция клеток CHOZN с помощью ретровектора: создавали объединенные линии клеток, содержащие ретровекторные вставки от каждой из вышеуказанных конструкций, путем выполнения трансдукции исходной линии клеток яичника китайского хомячка CHOZN с помощью ретровектора, полученного из генных конструкций, разработанных для экспрессии белка Anyway. Выполняли один цикл трансдукции с целью создания объединенной линии клеток для каждой из двух конструкций. После завершения трансдукций клеток линии клеток помещали в бесглутаминовую среду для осуществления селекции по GS.
Периодическая продукция Anyway с подпиткой из двух объединенных популяций клеток: после трансдукции объединенные линии клеток масштабировали для продуцирования в исследуемой периодической культуре с подпиткой в двух повторах в колбах для встряхивания на 250 мл. Каждую колбу для встряхивания засевали 300 000 жизнеспособных клеток на мл в рабочем объеме 50 мл усовершенствованной среды для периодической подпитки ExCell (SAFC) и инкубировали в увлажняемом инкубаторе (70-80%) со встряхиванием при 110 об/мин с 5% CO2 и температуре 37 °C. Во время продукции культуры подпитывали через сутки, начиная с дня 3, используя одно добавление подпитки. Ежесуточно отслеживали концентрацию глюкозы и добавляли ее, если уровень падал ниже 4 г/л. Культивирования прекращали, когда уровни жизнеспособности становились ≤ 50%.
Полученные результаты
Результаты представлены в таблице 1 и на ФИГ. 5. Эти результаты оказываются неожиданными. При аналогичном среднем числе копий гена для двух пулов, отмечено значительную разницу титров между этими двумя объединенным линиями клеток. Так даже при немного большем среднем числе копий генов генная конструкция полноразмерного LTR или дикого типа давали значительно меньшие титры, и это превращалось в значительно меньшую удельную продуктивность клетки на генную вставку. Происходит отбор или конкурентное преимущество для высоко экспрессирующих клеток, содержащих версию SIN генной конструкции.
Таблица 1. Сравнение объединенных линий клеток для двух генных конструкций
Эксперимент 2
На основе предыдущих результатов авторы изобретения спланировали эксперимент для сравнения пути, которым авторы традиционно осуществляли на практике технологию GPEx для практического осуществления технологии точно таким же путем, но с использованием вектора SIN-GS, описанного выше, в комбинации с линией клеток CHOZN с нокаутом GS. Процессы сохраняли максимально подобными только с двумя основными отличиями. Первое заключалось в том, что в традиционной GPEx используется исходная линия клеток китайского хомячка (GCHO) GPEx®, а в новой версии используется линия клеток CHOZN. Второе отличие заключалось в том, что были отличающиеся генные конструкции, использованные для создания ретровектора. В традиционной GPEx конструкция не содержит ген GS и в новой версии она не содержит ген GS. Все другие компоненты генной конструкции были аналогичными. Каждая из конструкций снова экспрессирует белок Anyway. После завершения получения объединенных линий клеток для каждого из методов, их сравнивали по продукции в анализе периодического культивирования с подпиткой.
Разработка линии клеток GCHO и CHOZN
Продукция ретровекторов: экспрессионные конструкции вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Содержащую ген оболочки экспрессионную плазмиду также ко-трансфекцировали с каждой из генных конструкций. Ко-трансфекция привела к продукции репликационно некомпетентного ретровектора с высокими титрами, которого концентрировали путем ультрацентрифугирования и использовали для трансдукций клеток (1, 2).
Трансдукция клеток CHOZN с помощью ретровектора: создавали объединенные линии клеток, содержащие ретровектроные вставки от каждой из конструкций, путем выполнения трансдукции либо исходной линии клеток яичника китайского хомячка CHOZN, либо исходной линии клеток яичника китайского хомячка GCHO с помощью ретровектора, полученного из либо генной конструкции, содержащей GS, либо генной конструкции, не содержащей GS, обе разработаны для экспрессии белка Anyway. Выполняли три цикла трансдукции с целью создания объединенной линии клеток для каждой из двух конструкций в соответствующей линии клеток. Дополнительные циклы, как правило, увеличивали количество вставок в линию клеток и, впоследствии, число копий генов. После завершения каждого цикла трансдукции пулы на основе клеток CHOZN помещали в бесглутаминовую среду для селекции по GS. Клетки традиционной GPEx не подвергали селекции как в нормальной процедуре.
Полученные результаты
Так как ожидаемое число копий возросло для обоих способов с повторными циклами трансдукции. Однако в каждом из новых пулов клеток GPEx наблюдали большие числа копий генов по сравнению с традиционной GPEx, как показано в таблице 2.
Таблица 2. Сравнение чисел копий генов объединенных линий клеток для двух различных процессов
Периодическая продукция Anyway с подпиткой из двух объединенных популяций клеток (3 цикла трансдукции): после трансдукции объединенные линии клеток масштабировали для продуцирования в исследуемой периодической культуре с подпиткой в двух повторах в колбах для встряхивания на 250 мл. Каждую колбу для встряхивания засевали 300 000 жизнеспособных клеток на мл в рабочем объеме 50 мл среды ActiPro (HyClone) и инкубировали в увлажняемом инкубаторе (70-80%) со встряхиванием при 120 об/мин с 5% CO2 и температуре 37 °C (34 °C, начиная с дня 6). Культуры подпитывали шесть раз во время анализа продукции с использованием двух различных подпитывающих добавок. Ежесуточно отслеживали концентрацию глюкозы и добавляли ее, если уровень падал ниже 4 г/л. Культивирования прекращали, когда уровни жизнеспособности становились ≤ 50%.
Результаты представлены в таблице 3 и на ФИГ. 6. И снова наблюдали очень неожиданные результаты. Клональные линии клеток по оптимизированной традиционной GPEx, продуцирующие продукт Anyway, экспрессировали максимум 1,8 г/л, а новый пул GPEx превзошел эту экспрессию более чем двухкратно до какого-либо клонального отбора. Новый пул GPEx содержит и большее число копий, а также обеспечивает большую продукцию на генную вставку, как показано в эксперименте №1. Вместе эти факторы привели к значительной разнице в клеточной удельной продуктивности с почти 3-кратно большим значением пг/клетку/сутки для нового пула GPEx. Общая плотность жизнеспособных клеток была также выше для нового пула GPEx по сравнению с традиционным пулом, также способствуя значительной наблюдавшейся разнице в титрах.
Таблица 3. Сравнение объединенных линий клеток для двух различных процессов для получения Anyway
Эксперименты 3 и 4
Приведенные выше данные касаются Fc-слияния, продуцированного из одного гена. В экспериментах 3 и 4 применяется трансдукция с отдельными векторами для тяжелой и легкой цепей (2 трансдукции для легкой цепи и 3 трансдукции для тяжелой цепи в обоих экспериментах) с двумя различными антителами «Peelaway» и «Yourway» с как технологии GPEx®, так и нового процесса с использованием SIN LTR для контроля экспрессии GS.
Продукция ретровекторов: экспрессионные конструкции вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Содержащую ген оболочки экспрессионную плазмиду также ко-трансфекцировали с каждой из генных конструкций. Ко-трансфекция привела к продукции репликационно некомпетентного ретровектора с высокими титрами, которого концентрировали путем ультрацентрифугирования и использовали для трансдукций клеток (1, 2).
Трансдукция клеток CHOZN с помощью ретровектора: создавали объединенные линии клеток, содержащие ретровектроные вставки от каждой из конструкций, путем выполнения трансдукции либо исходной линии клеток яичника китайского хомячка CHOZN, либо исходной линии клеток яичника китайского хомячка GCHO с помощью ретровектора, полученного из генных конструкций тяжелой и легкой цепей, содержащих GS, либо генных конструкций, не содержащих GS, разработанных для экспрессии двух различных антител. Выполняли два цикла трансдукции для легкой цепи и три цикла трансдукции для тяжелой цепи с целью создания объединенной линии клеток для каждой из четырех конструкций в каждой из линии клеток соответствующего антитела. Дополнительные циклы, как правило, увеличивали количество вставок в линию клеток и, впоследствии, число копий генов. После завершения каждого цикла трансдукции пулы на основе клеток CHOZN помещали в бесглутаминовую среду для селекции по GS. Клетки традиционной GPEx не подвергали селекции как в нормальной процедуре.
Периодическая продукция Peelaway и Yourway с подпиткой из объединенных популяций линий клеток (2 цикла трансдукции для легкой цепи и 3 цикла трансдукции для тяжелой цепи): после трансдукции объединенные линии клеток масштабировали для продуцирования в исследуемой периодической культуре с подпиткой в двух повторах в колбах для встряхивания на 250 мл. Каждую колбу для встряхивания засевали 300 000 жизнеспособных клеток на мл в рабочем объеме 50 мл среды ActiPro (HyClone) и инкубировали в увлажняемом инкубаторе (70-80%) со встряхиванием при 120 об/мин с 5% CO2 и температуре 37 °C (34 °C, начиная с дня 6). Культуры подпитывали шесть раз во время анализа продукции с использованием двух различных подпитывающих добавок. Ежесуточно отслеживали концентрацию глюкозы и добавляли ее, если уровень падал ниже 4 г/л. Культивирования прекращали, когда уровни жизнеспособности становились ≤ 50%.
Полученные результаты
Результаты представлены в таблицах 4 и 5. Наблюдали значительные преимущества для значений титра с новым вектором и процессом.
Таблица 4. Сравнение объединенных линий клеток для двух различных процессов для получения антитела Peelaway
Таблица 5. Сравнение объединенных линий клеток для двух различных процессов для получения антитела Yourway
На 4 пулах клеток Yourway выполняли дополнительный анализ числа копий. Пулы клеток, созданных после 1 цикла трансдукции легкой цепи и 2 циклов трансдукции тяжелой цепи, сравнивали с пулами клеток, созданными после 2 циклов трансдукции легкой цепи и 3 циклов трансдукции тяжелой цепи. Каждая из этих объединенных линий клеток либо находилась в условиях непрерывного роста в содержащей глутамин среде, либо помещалась в среду без глутамина. Для каждой из четырех объединенных линий клеток рассчитывали количество копий гена к общему числу генов, число гена тяжелой цепи и число гена легкой цепи.
Таблица 6
Эти данные показывают, что даже несмотря на то, что каждая из клеток в этих пулах содержит многочисленные копии гена GS, ослабляемого SIN LTR, существует значительная селекция, осуществляемая для клеток с большим количеством копий и, согласно вышеприведенных данных, таких копий, которые обеспечивают высокие уровни экспрессии.
Эксперимент 5
Для этого эксперимента применяли продукт-антитело Yourway. Генная конструкция тяжелой цепи, использованная для создания линии клеток, была такой же как в эксперименте 4. Генная конструкция легкой цепи была идентичной генной конструкции тяжелой цепи во всех аспектах, за исключением того, что она содержит кодирующую легкую цепь последовательностью и у нее отсутствует ген GS. Выполняли единственную трансдукцию для каждого содержащего легкую цепь (-GS) и тяжелую цепь (+GS) с целью создания объединенной линии клеток.
Продукция ретровекторов: экспрессионные конструкции вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Содержащую ген оболочки экспрессионную плазмиду также ко-трансфекцировали с каждой из генных конструкций. Ко-трансфекция привела к продукции репликационно некомпетентного ретровектора с высокими титрами, которого концентрировали путем ультрацентрифугирования и использовали для трансдукций клеток (1, 2).
Трансдукция клеток CHOZN с помощью ретровектора: создавали объединенные линии клеток, содержащие ретровектроные вставки от каждой из вышеуказанных конструкций, путем выполнения трансдукции исходной линии клеток яичника китайского хомячка CHOZN с помощью ретровектора, полученного из генных конструкций легкой цепи без GS и конструкций тяжелой цепи, содержащих GS. Выполняли один цикл трансдукции для легкой цепи и один цикл трансдукции для тяжелой цепи с целью создания объединенной линии клеток. После завершения обоих циклов трансдукции разделяли пул на основе клеток CHOZN, половину его помещали в бесглутаминовую среды, а другую половину постоянно подпитывали глутамином во время наращивание клеток и последующей оценки продуктивности.
Периодическая продукция Yourway с подпиткой из объединенных популяций линий клеток (1 трансдукция для легкой цепи и 1 трансдукция для тяжелой цепи): объединенные линии клеток масштабировали для продуцирования в исследуемой периодической культуре с подпиткой в колбах для встряхивания на 250 мл. Колбы для встряхивания засевали 300 000 жизнеспособных клеток на мл в рабочем объеме 50 мл среды ActiPro (HyClone) и инкубировали в увлажняемом инкубаторе (70-80%) со встряхиванием при 120 об/мин с 5% CO2 и температуре 37 °C (34 °C, начиная с дня 6). Культуры подпитывали шесть раз во время анализа продукции с использованием двух различных подпитывающих добавок. Ежесуточно отслеживали концентрацию глюкозы и добавляли ее, если уровень падал ниже 4 г/л. Культуры с добавлениями глутамина ежедневно отслеживали на содержание глутамина и подпитывали, когда его уровень падал ниже 2 мМ. Культивирования прекращали, когда уровни жизнеспособности становились ≤ 50%.
Полученные результаты
Результаты представлены в таблице 7. Селекция путем удаления глутамина значительно увеличила количество копий гена тяжелой цепи в пуле клеток. Несколько удивительно оказалось то, что также увеличилось число копий легкой цепи, но не в той же мере, что и тяжелой цепи. Общая продукция антитела также была значительно выше, чем у культуры, которая не подвергалась отбору, достигая уровней, показанных в таблице 5, при выполнении только двух трансдукций вместо пяти, осуществленных в этом эксперименте.
Таблица 7. Сравнение объединенных линий клеток с глутаминовой селекцией и без нее для получения антитела Yourway. Объединенную линию клеток продуцировали в результате одной трансдукции легкой цепи (-GS) и одной трансдукции тяжелой цепи трансдукция (+GS)
Эксперимент 6
Генную конструкцию разрабатывали и получали для исследования того, будет ли технология работать аналогичным образом с использованием традиционных методологий трансфекции клеток, по сравнению с трансдукцией ретровекторами. Один пример созданной традиционной плазмидной конструкции для трансфекции показан на ФИГ. 8 (карта плазмиды) и ФИГ. 9 (действительная плазмидная последовательность; SEQ ID NO:4).
1. Bleck, G.T. 2005 An alternative method for the rapid generation of stable, high-expressing mammalian cell lines (A Technical Review). Bioprocessing J. Sep/Oct. pp 1-7.
2. Bleck, G.T., 2010. GPEx® A Flexible Method for the Rapid Generation of Stable, High Expressing, Antibody Producing Mammalian Cell Lines Chapter 4 In: Current Trends in Monoclonal Antibody Development and Manufacturing, Biotechnology: Pharmaceutical Aspects, Edited by: S.J. Shire et al. © 2010 American Association of Pharmaceutical Scientists, DOI 10.1007/978-0-387-76643-0_4.
Все публикации и патенты, упомянутые в приведенном выше описании, включены в данный документ посредством ссылки. Различные модификации и вариации описанных способа и системы по данному изобретению будут очевидны для специалистов в данной области техники без отступления от объема и сущности изобретения. Хотя изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявляемое изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. Действительно, подразумевается, что различные модификации описанных способов осуществления данного изобретения, которые очевидны специалистам в данной области техники, входят в объем следующей формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CATALENT PHARMA SOLUTIONS, LLC
<120> ВЕКТОРЫ ДЛЯ ПРОИЗВОДСТВА БЕЛКОВ
<130> GALA-37375.601
<150> US 62/775 194
<151> 2018-12-04
<160> 4
<170> PatentIn версия 3.5
<210> 1
<211> 6217
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 1
gtccggccat tagccatatt attcattggt tatatagcat aaatcaatat tggctattgg
60
ccattgcata cgttgtatcc atatcataat atgtacattt atattggctc atgtccaaca
120
ttaccgccat gttgacattg attattgact agttattaat agtaatcaat tacggggtca
180
ttagttcata gcccatatat ggagttccgc gttacataac ttacggtaaa tggcccgcct
240
ggctgaccgc ccaacgaccc ccgcccattg acgtcaataa tgacgtatgt tcccatagta
300
acgccaatag ggactttcca ttgacgtcaa tgggtggagt atttacggta aactgcccac
360
ttggcagtac atcaagtgta tcatatgcca agtacgcccc ctattgacgt caatgacggt
420
aaatggcccg cctggcatta tgcccagtac atgaccttat gggactttcc tacttggcag
480
tacatctacg tattagtcat cgctattacc atggtgatgc ggttttggca gtacatcaat
540
gggcgtggat agcggtttga ctcacgggga tttccaagtc tccaccccat tgacgtcaat
600
gggagtttgt tttggcacca aaatcaacgg gactttccaa aatgtcgtaa caactccgcc
660
ccattgacgc aaatgggcgg taggcgtgta cggtgggagg tctatataag cagagctcaa
720
taaaagagcc cacaacccct cactcggcgc gccagtcttc cgatagactg cgtcgcccgg
780
gtacccgtat tcccaataaa gcctcttgct gtttgcatcc gaatcgtggt ctcgctgttc
840
cttgggaggg tctcctctga gtgattgact acccacgacg ggggtctttc atttgggggc
900
tcgtccggga tttggagacc cctgcccagg gaccaccgac ccaccaccgg gaggtaagct
960
ggccagcaac ttatctgtgt ctgtccgatt gtctagtgtc tatgtttgat gttatgcgcc
1020
tgcgtctgta ctagttagct aactagctct gtatctggcg gacccgtggt ggaactgacg
1080
agttctgaac acccggccgc aaccctggga gacgtcccag ggactttggg ggccgttttt
1140
gtggcccgac ctgaggaagg gagtcgatgt ggaatccgac cccgtcagga tatgtggttc
1200
tggtaggaga cgagaaccta aaacagttcc cgcctccgtc tgaatttttg ctttcggttt
1260
ggaaccgaag ccgcgcgtct tgtctgctgc agcgctgcag catcgttctg tgttgtctct
1320
gtctgactgt gtttctgtat ttgtctgaaa attagggcca gactgttacc actcccttaa
1380
gtttgacctt aggtcactgg aaagatgtcg agcggatcgc tcacaaccag tcggtagatg
1440
tcaagaagag acgttgggtt accttctgct ctgcagaatg gccaaccttt aacgtcggat
1500
ggccgcgaga cggcaccttt aaccgagacc tcatcaccca ggttaagatc aaggtctttt
1560
cacctggccc gcatggacac ccagaccagg tcccctacat cgtgacctgg gaagccttgg
1620
cttttgaccc ccctccctgg gtcaagccct ttgtacaccc taagcctccg cctcctcttc
1680
ctccatccgc cccgtctctc ccccttgaac ctcctcgttc gaccccgcct cgatcctccc
1740
tttatccagc cctcactcct tctctaggcg ccggaattgc cttccaccat ggccacctca
1800
gcaagttccc acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag
1860
aaagtccaag ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc
1920
cgcaccctgg actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc
1980
tctagtacct ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg
2040
tttcgggacc ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac
2100
aaccggaagc ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg
2160
agcaaccagc acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg
2220
cacccttttg gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt
2280
gtgggcgcag acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg
2340
tatgctgggg tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc
2400
caaataggac cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc
2460
ttgcatcgag tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct
2520
gggaactgga atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag
2580
aatggtctga agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac
2640
attcgagcct acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac
2700
gaaacgtcca acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc
2760
attccccgga ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc
2820
aactgtgacc cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact
2880
ggcgacgagc ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat
2940
actatgtatt ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga
3000
tagcccctcc caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata
3060
gccactcccc cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg
3120
ggcggtccta ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc
3180
gcctggctca atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg
3240
ctcattgccc attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc
3300
ccacttggca gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa
3360
gtacgccagg gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga
3420
tggctgccaa gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt
3480
tccattgacg taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg
3540
tttagggaac cgccattctg cctggggacg tcggaggagc tcgaaagctt ctagacaatt
3600
gccgccacca tgatgtcctt tgtctctctg ctcctggttg gcatcctatt ccatgccacc
3660
caggccagtg atacaggtag acctttcgta gagatgtaca gtgaaatccc cgaaattata
3720
cacatgactg aaggaaggga gctcgtcatt ccctgccggg ttacgtcacc taacatcact
3780
gttactttaa aaaagtttcc acttgacact ttgatccctg atggaaaacg cataatctgg
3840
gacagtagaa agggcttcat catatcaaat gcaacgtaca aagaaatagg gcttctgacc
3900
tgtgaagcaa cagtcaatgg gcatttgtat aagacaaact atctcacaca tcgacaaacc
3960
aatacaatca tagatgtcgt tctgagtccg tctcatggaa ttgaactatc tgttggagaa
4020
aagcttgtct taaattgtac agcaagaact gaactaaatg tggggattga cttcaactgg
4080
gaataccctt cttcgaagca tcagcataag aaacttgtaa accgagacct aaaaacccag
4140
tctgggagtg agatgaagaa gtttttgagc accttaacta tagatggtgt aacccggagt
4200
gaccaaggat tgtacacctg tgcagcatcc agtgggctga tgaccaagaa aaacagcaca
4260
tttgtcaggg tccatgaaaa agacaaaact cacacatgcc caccgtgccc agcacctgaa
4320
ctcctggggg gaccctcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc
4380
tcccggaccc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc
4440
aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccacgggag
4500
gagcagtaca acagcacata tcgtgtggtc agcgtcctca ccgtcctgca ccaggactgg
4560
ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag
4620
aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca
4680
tcccgggatg agctgaccaa gaaccaggtc agcctgacct gcctggtcaa aggcttctat
4740
cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc
4800
acgcctcccg tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac
4860
aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac
4920
aaccactaca cgcagaagag cctctccctg tctcccggga aatgatgaga tctcgagttc
4980
gacatcgata atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac
5040
tatgttgctc cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt
5100
gcttcccgta tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat
5160
gaggagttgt ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca
5220
acccccactg gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc
5280
cccctcccta ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg
5340
gctcggctgt tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct
5400
tggctgctcg cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct
5460
tcggccctca atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt
5520
ccgcgtcttc gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgcat
5580
cgataaaata aaagatttta tttagtctcc agaaaaaggg gggaatgaaa gaccccacct
5640
gtaggtttgg caagctagct taagtaacgc cattttgcaa ggcatggaaa aatacataac
5700
tgagaataga gaagttcaga tcaaggtcag gaacagatgg aacagctgaa tatgggccaa
5760
acaggatatc tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gatggaacag
5820
ctgaatatgg gccaaacagg atatctgtgg taagcagttc ctgccccggc tcagggccaa
5880
gaacagatgg tccccagatg cggtccagcc ctcagcagtt tctagagaac catcagatgt
5940
ttccagggtg ccccaaggac ctgaaatgac cctgtgcctt atttgaacta accaatcagt
6000
tcgcttctcg cttctgttcg cgcgcttctg ctccccgagc tcaataaaag agcccacaac
6060
ccctcactcg gggcgccagt cctccgattg actgagtcgc ccgggtaccc gtgtatccaa
6120
taaaccctct tgcagttgca tccgacttgt ggtctcgctg ttccttggga gggtctcctc
6180
tgagtgattg actacccgtc agcgggggtc tttcatt
6217
<210> 2
<211> 6065
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 2
gtccggccat tagccatatt attcattggt tatatagcat aaatcaatat tggctattgg
60
ccattgcata cgttgtatcc atatcataat atgtacattt atattggctc atgtccaaca
120
ttaccgccat gttgacattg attattgact agttattaat agtaatcaat tacggggtca
180
ttagttcata gcccatatat ggagttccgc gttacataac ttacggtaaa tggcccgcct
240
ggctgaccgc ccaacgaccc ccgcccattg acgtcaataa tgacgtatgt tcccatagta
300
acgccaatag ggactttcca ttgacgtcaa tgggtggagt atttacggta aactgcccac
360
ttggcagtac atcaagtgta tcatatgcca agtacgcccc ctattgacgt caatgacggt
420
aaatggcccg cctggcatta tgcccagtac atgaccttat gggactttcc tacttggcag
480
tacatctacg tattagtcat cgctattacc atggtgatgc ggttttggca gtacatcaat
540
gggcgtggat agcggtttga ctcacgggga tttccaagtc tccaccccat tgacgtcaat
600
gggagtttgt tttggcacca aaatcaacgg gactttccaa aatgtcgtaa caactccgcc
660
ccattgacgc aaatgggcgg taggcgtgta cggtgggagg tctatataag cagagctcaa
720
taaaagagcc cacaacccct cactcggcgc gccagtcttc cgatagactg cgtcgcccgg
780
gtacccgtat tcccaataaa gcctcttgct gtttgcatcc gaatcgtggt ctcgctgttc
840
cttgggaggg tctcctctga gtgattgact acccacgacg ggggtctttc atttgggggc
900
tcgtccggga tttggagacc cctgcccagg gaccaccgac ccaccaccgg gaggtaagct
960
ggccagcaac ttatctgtgt ctgtccgatt gtctagtgtc tatgtttgat gttatgcgcc
1020
tgcgtctgta ctagttagct aactagctct gtatctggcg gacccgtggt ggaactgacg
1080
agttctgaac acccggccgc aaccctggga gacgtcccag ggactttggg ggccgttttt
1140
gtggcccgac ctgaggaagg gagtcgatgt ggaatccgac cccgtcagga tatgtggttc
1200
tggtaggaga cgagaaccta aaacagttcc cgcctccgtc tgaatttttg ctttcggttt
1260
ggaaccgaag ccgcgcgtct tgtctgctgc agcgctgcag catcgttctg tgttgtctct
1320
gtctgactgt gtttctgtat ttgtctgaaa attagggcca gactgttacc actcccttaa
1380
gtttgacctt aggtcactgg aaagatgtcg agcggatcgc tcacaaccag tcggtagatg
1440
tcaagaagag acgttgggtt accttctgct ctgcagaatg gccaaccttt aacgtcggat
1500
ggccgcgaga cggcaccttt aaccgagacc tcatcaccca ggttaagatc aaggtctttt
1560
cacctggccc gcatggacac ccagaccagg tcccctacat cgtgacctgg gaagccttgg
1620
cttttgaccc ccctccctgg gtcaagccct ttgtacaccc taagcctccg cctcctcttc
1680
ctccatccgc cccgtctctc ccccttgaac ctcctcgttc gaccccgcct cgatcctccc
1740
tttatccagc cctcactcct tctctaggcg ccggaattgc cttccaccat ggccacctca
1800
gcaagttccc acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag
1860
aaagtccaag ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc
1920
cgcaccctgg actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc
1980
tctagtacct ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg
2040
tttcgggacc ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac
2100
aaccggaagc ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg
2160
agcaaccagc acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg
2220
cacccttttg gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt
2280
gtgggcgcag acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg
2340
tatgctgggg tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc
2400
caaataggac cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc
2460
ttgcatcgag tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct
2520
gggaactgga atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag
2580
aatggtctga agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac
2640
attcgagcct acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac
2700
gaaacgtcca acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc
2760
attccccgga ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc
2820
aactgtgacc cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact
2880
ggcgacgagc ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat
2940
actatgtatt ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga
3000
tagcccctcc caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata
3060
gccactcccc cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg
3120
ggcggtccta ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc
3180
gcctggctca atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg
3240
ctcattgccc attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc
3300
ccacttggca gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa
3360
gtacgccagg gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga
3420
tggctgccaa gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt
3480
tccattgacg taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg
3540
tttagggaac cgccattctg cctggggacg tcggaggagc tcgaaagctt ctagacaatt
3600
gccgccacca tgatgtcctt tgtctctctg ctcctggttg gcatcctatt ccatgccacc
3660
caggccagtg atacaggtag acctttcgta gagatgtaca gtgaaatccc cgaaattata
3720
cacatgactg aaggaaggga gctcgtcatt ccctgccggg ttacgtcacc taacatcact
3780
gttactttaa aaaagtttcc acttgacact ttgatccctg atggaaaacg cataatctgg
3840
gacagtagaa agggcttcat catatcaaat gcaacgtaca aagaaatagg gcttctgacc
3900
tgtgaagcaa cagtcaatgg gcatttgtat aagacaaact atctcacaca tcgacaaacc
3960
aatacaatca tagatgtcgt tctgagtccg tctcatggaa ttgaactatc tgttggagaa
4020
aagcttgtct taaattgtac agcaagaact gaactaaatg tggggattga cttcaactgg
4080
gaataccctt cttcgaagca tcagcataag aaacttgtaa accgagacct aaaaacccag
4140
tctgggagtg agatgaagaa gtttttgagc accttaacta tagatggtgt aacccggagt
4200
gaccaaggat tgtacacctg tgcagcatcc agtgggctga tgaccaagaa aaacagcaca
4260
tttgtcaggg tccatgaaaa agacaaaact cacacatgcc caccgtgccc agcacctgaa
4320
ctcctggggg gaccctcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc
4380
tcccggaccc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc
4440
aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccacgggag
4500
gagcagtaca acagcacata tcgtgtggtc agcgtcctca ccgtcctgca ccaggactgg
4560
ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag
4620
aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca
4680
tcccgggatg agctgaccaa gaaccaggtc agcctgacct gcctggtcaa aggcttctat
4740
cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc
4800
acgcctcccg tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac
4860
aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac
4920
aaccactaca cgcagaagag cctctccctg tctcccggga aatgatgaga tctcgagttc
4980
gacatcgata atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac
5040
tatgttgctc cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt
5100
gcttcccgta tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat
5160
gaggagttgt ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca
5220
acccccactg gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc
5280
cccctcccta ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg
5340
gctcggctgt tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct
5400
tggctgctcg cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct
5460
tcggccctca atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt
5520
ccgcgtcttc gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgcat
5580
cgataaaata aaagatttta tttagtctcc agaaaaaggg gggaatgaaa gaccccacct
5640
gtaggtttgg caagctagct taagtaacgc cattttgcaa ggcatggaaa aatacataac
5700
tgagaataga gaagttcaga tcaaggtcag gaacagatgg aacagggtcg accggtcgac
5760
cggtcgaccc tagagaacca tcagatgttt ccagggtgcc ccaaggacct gaaatgaccc
5820
tgtgccttat ttgaactaac caatcagttc gcttctcgct tctgttcgcg cgcttctgct
5880
ccccgagctc aataaaagag cccacaaccc ctcactcggg gcgccagtcc tccgattgac
5940
tgagtcgccc gggtacccgt gtatccaata aaccctcttg cagttgcatc cgacttgtgg
6000
tctcgctgtt ccttgggagg gtctcctctg agtgattgac tacccgtcag cgggggtctt
6060
tcatt
6065
<210> 3
<211> 442
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 3
aatgaaagac cccacctgta ggtttggcaa gctagcttaa gtaacgccat tttgcaaggc
60
atggaaaaat acataactga gaatagagaa gttcagatca aggtcaggaa cagatggaac
120
agggtcgacc ggtcgaccgg tcgaccctag agaaccatca gatgtttcca gggtgcccca
180
aggacctgaa atgaccctgt gccttatttg aactaaccaa tcagttcgct tctcgcttct
240
gttcgcgcgc ttctgctccc cgagctcaat aaaagagccc acaacccctc actcggggcg
300
ccagtcctcc gattgactga gtcgcccggg tacccgtgta tccaataaac cctcttgcag
360
ttgcatccga cttgtggtct cgctgttcct tgggagggtc tcctctgagt gattgactac
420
ccgtcagcgg gggtctttca tt
442
<210> 4
<211> 8072
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 4
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca
60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg
120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc
180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc
240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat
300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt
360
tttcccagtc acgacgttgt aaaacgacgg ccagtgaatt catttaaatg aaagacccca
420
cctgtaggtt tggcaagcta gcttaagtaa cgccattttg caaggcatgg aaaaatacat
480
aactgagaat agaaaagttc agatcaaggt caggaacaga tggaacaggg tcgaccggtc
540
gaccggtcga ccctagagaa ccatcagatg tttccagggt gccccaagga cctgaaatga
600
ccctgtgcct tatttgaact aaccaatcag ttcgcttctc gcttctgttc gcgcgcttct
660
gctccccgag ctcaataaaa gagcccacaa cccctcactc ggggcgccag tcttccgata
720
gactgcgtcg cccgggtacc cgtattccca ataaagcctc ttgctgtttg catccgaatc
780
gtggtctcgc tgttccttgg gagggtctcc tctgagtgat tgactaccca cgacgggggt
840
ctttcatttg ggggctcgtc cgggatttgg agacccctgc ccagggacca ccgacccacc
900
accgggaggt aagctggcca gcaacttatc tgtgtctgtc cgattgtcta gtgtctatgt
960
ttgatgttat gcgcctgcgt ctgtactagt tagctaacta gctctgtatc tggcggaccc
1020
gtggtggaac tgacgagttc tgaacacccg gccgcaaccc tgggagacgt cccagggact
1080
ttgggggccg tttttgtggc ccgacctgag gaagggagtc gatgtggaat ccgaccccgt
1140
caggatatgt ggttctggta ggagacgaga acctaaaaca gttcccgcct ccgtctgaat
1200
ttttgctttc ggtttggaac cgaagccgcg cgtcttgtct gctgcagcgc tgcagcatcg
1260
ttctgtgttg tctctgtctg actgtgtttc tgtatttgtc tgaaaattag ggccagactg
1320
ttaccactcc cttaagtttg accttaggtc actggaaaga tgtcgagcgg atcgctcaca
1380
accagtcggt agatgtcaag aagagacgtt gggttacctt ctgctctgca gaatggccaa
1440
cctttaacgt cggatggccg cgagacggca cctttaaccg agacctcatc acccaggtta
1500
agatcaaggt cttttcacct ggcccgcatg gacacccaga ccaggtcccc tacatcgtga
1560
cctgggaagc cttggctttt gacccccctc cctgggtcaa gccctttgta caccctaagc
1620
ctccgcctcc tcttcctcca tccgccccgt ctctccccct tgaacctcct cgttcgaccc
1680
cgcctcgatc ctccctttat ccagccctca ctccttctct aggcgccgga attgccttcc
1740
accatggcca cctcagcaag ttcccacttg aacaaaaaca tcaagcaaat gtacttgtgc
1800
ctgccccagg gtgagaaagt ccaagccatg tatatctggg ttgatggtac tggagaagga
1860
ctgcgctgca aaacccgcac cctggactgt gagcccaagt gtgtagaaga gttacctgag
1920
tggaattttg atggctctag tacctttcag tctgagggct ccaacagtga catgtatctc
1980
agccctgttg ccatgtttcg ggaccccttc cgcagagatc ccaacaagct ggtgttctgt
2040
gaagttttca agtacaaccg gaagcctgca gagaccaatt taaggcactc gtgtaaacgg
2100
ataatggaca tggtgagcaa ccagcacccc tggtttggaa tggaacagga gtatactctg
2160
atgggaacag atgggcaccc ttttggttgg ccttccaatg gctttcctgg gccccaaggt
2220
ccgtattact gtggtgtggg cgcagacaaa gcctatggca gggatatcgt ggaggctcac
2280
taccgcgcct gcttgtatgc tggggtcaag attacaggaa caaatgctga ggtcatgcct
2340
gcccagtggg agttccaaat aggaccctgt gaaggaatcc gcatgggaga tcatctctgg
2400
gtggcccgtt tcatcttgca tcgagtatgt gaagactttg gggtaatagc aacctttgac
2460
cccaagccca ttcctgggaa ctggaatggt gcaggctgcc ataccaactt tagcaccaag
2520
gccatgcggg aggagaatgg tctgaagcac atcgaggagg ccatcgagaa actaagcaag
2580
cggcaccggt accacattcg agcctacgat cccaaggggg gcctggacaa tgcccgtcgt
2640
ctgactgggt tccacgaaac gtccaacatc aacgactttt ctgctggtgt cgccaatcgc
2700
agtgccagca tccgcattcc ccggactgtc ggccaggaga agaaaggtta ctttgaagac
2760
cgccgcccct ctgccaactg tgaccccttt gcagtgacag aagccatcgt ccgcacatgc
2820
cttctcaatg agactggcga cgagcccttc caatacaaaa actaaagatc cctatggcta
2880
ttggccaggt tcaatactat gtattggccc tatgccatat agtattccat atatgggttt
2940
tcctattgac gtagatagcc cctcccaatg ggcggtccca tataccatat atggggcttc
3000
ctaataccgc ccatagccac tcccccattg acgtcaatgg tctctatata tggtctttcc
3060
tattgacgtc atatgggcgg tcctattgac gtatatggcg cctcccccat tgacgtcaat
3120
tacggtaaat ggcccgcctg gctcaatgcc cattgacgtc aataggacca cccaccattg
3180
acgtcaatgg gatggctcat tgcccattca tatccgttct cacgccccct attgacgtca
3240
atgacggtaa atggcccact tggcagtaca tcaatatcta ttaatagtaa cttggcaagt
3300
acattactat tggaagtacg ccagggtaca ttggcagtac tcccattgac gtcaatggcg
3360
gtaaatggcc cgcgatggct gccaagtaca tccccattga cgtcaatggg gaggggcaat
3420
gacgcaaatg ggcgttccat tgacgtaaat gggcggtagg cgtgcctaat gggaggtcta
3480
tataagcaat gctcgtttag ggaaccgcca ttctgcctgg ggacgtcgga ggagctcgaa
3540
agcttctaga caattgccgc caccatgatg tcctttgtct ctctgctcct ggttggcatc
3600
ctattccatg ccacccaggc cagtgataca ggtagacctt tcgtagagat gtacagtgaa
3660
atccccgaaa ttatacacat gactgaagga agggagctcg tcattccctg ccgggttacg
3720
tcacctaaca tcactgttac tttaaaaaag tttccacttg acactttgat ccctgatgga
3780
aaacgcataa tctgggacag tagaaagggc ttcatcatat caaatgcaac gtacaaagaa
3840
atagggcttc tgacctgtga agcaacagtc aatgggcatt tgtataagac aaactatctc
3900
acacatcgac aaaccaatac aatcatagat gtcgttctga gtccgtctca tggaattgaa
3960
ctatctgttg gagaaaagct tgtcttaaat tgtacagcaa gaactgaact aaatgtgggg
4020
attgacttca actgggaata cccttcttcg aagcatcagc ataagaaact tgtaaaccga
4080
gacctaaaaa cccagtctgg gagtgagatg aagaagtttt tgagcacctt aactatagat
4140
ggtgtaaccc ggagtgacca aggattgtac acctgtgcag catccagtgg gctgatgacc
4200
aagaaaaaca gcacatttgt cagggtccat gaaaaagaca aaactcacac atgcccaccg
4260
tgcccagcac ctgaactcct ggggggaccc tcagtcttcc tcttcccccc aaaacccaag
4320
gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac
4380
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag
4440
acaaagccac gggaggagca gtacaacagc acatatcgtg tggtcagcgt cctcaccgtc
4500
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc
4560
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg
4620
tacaccctgc ccccatcccg ggatgagctg accaagaacc aggtcagcct gacctgcctg
4680
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag
4740
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctctacagc
4800
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg
4860
catgaggctc tgcacaacca ctacacgcag aagagcctct ccctgtctcc cgggaaatga
4920
tgagatctcg agttcgacat cgataatcaa cctctggatt acaaaatttg tgaaagattg
4980
actggtattc ttaactatgt tgctcctttt acgctatgtg gatacgctgc tttaatgcct
5040
ttgtatcatg ctattgcttc ccgtatggct ttcattttct cctccttgta taaatcctgg
5100
ttgctgtctc tttatgagga gttgtggccc gttgtcaggc aacgtggcgt ggtgtgcact
5160
gtgtttgctg acgcaacccc cactggttgg ggcattgcca ccacctgtca gctcctttcc
5220
gggactttcg ctttccccct ccctattgcc acggcggaac tcatcgccgc ctgccttgcc
5280
cgctgctgga caggggctcg gctgttgggc actgacaatt ccgtggtgtt gtcggggaaa
5340
tcatcgtcct ttccttggct gctcgcctgt gttgccacct ggattctgcg cgggacgtcc
5400
ttctgctacg tcccttcggc cctcaatcca gcggaccttc cttcccgcgg cctgctgccg
5460
gctctgcggc ctcttccgcg tcttcgcctt cgccctcaga cgagtcggat ctccctttgg
5520
gccgcctccc cgcatcgatg ggggaggcta actgaaacac ggaaggagac aataccggaa
5580
ggaacccgcg ctatgacggc aataaaaaga cagaataaaa cgcacgggtg ttgggtcgtt
5640
tgttcataaa cgcggggttc ggtcccaggg ctggcactct gtcgataccc caccgagacc
5700
ccattggggc caatacgccc gcgtttcttc cttttcccca ccccaccccc caagttcggg
5760
tgaaggccca gggctcgcag ccaacgtcgg ggcggcaggc cctgccatag cggatccttt
5820
ccactgtacg cgtagcttgg cgtaatcatg gtcatagctg tttcctgtgt gaaattgtta
5880
tccgctcaca attccacaca acatacgagc cggaagcata aagtgtaaag cctggggtgc
5940
ctaatgagtg agctaactca cattaattgc gttgcgctca ctgcccgctt tccagtcggg
6000
aaacctgtcg tgccagctgc attaatgaat cggccaacgc gcggggagag gcggtttgcg
6060
tattgggcgc tcttccgctt cctcgctcac tgactcgctg cgctcggtcg ttcggctgcg
6120
gcgagcggta tcagctcact caaaggcggt aatacggtta tccacagaat caggggataa
6180
cgcaggaaag aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta aaaaggccgc
6240
gttgctggcg tttttccata ggctccgccc ccctgacgag catcacaaaa atcgacgctc
6300
aagtcagagg tggcgaaacc cgacaggact ataaagatac caggcgtttc cccctggaag
6360
ctccctcgtg cgctctcctg ttccgaccct gccgcttacc ggatacctgt ccgcctttct
6420
cccttcggga agcgtggcgc tttctcatag ctcacgctgt aggtatctca gttcggtgta
6480
ggtcgttcgc tccaagctgg gctgtgtgca cgaacccccc gttcagcccg accgctgcgc
6540
cttatccggt aactatcgtc ttgagtccaa cccggtaaga cacgacttat cgccactggc
6600
agcagccact ggtaacagga ttagcagagc gaggtatgta ggcggtgcta cagagttctt
6660
gaagtggtgg cctaactacg gctacactag aagaacagta tttggtatct gcgctctgct
6720
gaagccagtt accttcggaa aaagagttgg tagctcttga tccggcaaac aaaccaccgc
6780
tggtagcggt ggtttttttg tttgcaagca gcagattacg cgcagaaaaa aaggatctca
6840
agaagatcct ttgatctttt ctacggggtc tgacgctcag tggaacgaaa actcacgtta
6900
agggattttg gtcatgagat tatcaaaaag gatcttcacc tagatccttt taaattaaaa
6960
atgaagtttt aaatcaatct aaagtatata tgagtaaact tggtctgaca gttaccaatg
7020
cttaatcagt gaggcaccta tctcagcgat ctgtctattt cgttcatcca tagttgcctg
7080
actccccgtc gtgtagataa ctacgatacg ggagggctta ccatctggcc ccagtgctgc
7140
aatgataccg cgagacccac gctcaccggc tccagattta tcagcaataa accagccagc
7200
cggaagggcc gagcgcagaa gtggtcctgc aactttatcc gcctccatcc agtctattaa
7260
ttgttgccgg gaagctagag taagtagttc gccagttaat agtttgcgca acgttgttgc
7320
cattgctaca ggcatcgtgg tgtcacgctc gtcgtttggt atggcttcat tcagctccgg
7380
ttcccaacga tcaaggcgag ttacatgatc ccccatgttg tgcaaaaaag cggttagctc
7440
cttcggtcct ccgatcgttg tcagaagtaa gttggccgca gtgttatcac tcatggttat
7500
ggcagcactg cataattctc ttactgtcat gccatccgta agatgctttt ctgtgactgg
7560
tgagtactca accaagtcat tctgagaata gtgtatgcgg cgaccgagtt gctcttgccc
7620
ggcgtcaata cgggataata ccgcgccaca tagcagaact ttaaaagtgc tcatcattgg
7680
aaaacgttct tcggggcgaa aactctcaag gatcttaccg ctgttgagat ccagttcgat
7740
gtaacccact cgtgcaccca actgatcttc agcatctttt actttcacca gcgtttctgg
7800
gtgagcaaaa acaggaaggc aaaatgccgc aaaaaaggga ataagggcga cacggaaatg
7860
ttgaatactc atactcttcc tttttcaata ttattgaagc atttatcagg gttattgtct
7920
catgagcgga tacatatttg aatgtattta gaaaaataaa caaatagggg ttccgcgcac
7980
atttccccga aaagtgccac ctgacgtcta agaaaccatt attatcatga cattaaccta
8040
taaaaatagg cgtatcacga ggccctttcg tc
8072
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНСТРУКЦИИ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ ПОЛУЧЕНИЯ БЕЛКОВ | 2021 |
|
RU2840923C1 |
| Нуклеиновая кислота, имеющая промоторную активность, и ее применение | 2022 |
|
RU2818112C2 |
| СПОСОБ ВРЕМЕННОЙ ТРАНСФЕКЦИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСА | 2016 |
|
RU2749717C2 |
| КОМПОЗИЦИИ ПРОМОТОРОВ | 2014 |
|
RU2742435C2 |
| СТАБИЛЬНЫЕ КЛЕТОЧНЫЕ ЛИНИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСОВ | 2016 |
|
RU2752498C2 |
| ВЕКТОРНАЯ СИСТЕМА НА ОСНОВЕ ASLV | 2010 |
|
RU2566563C2 |
| КОМПОЗИЦИЯ И СПОСОБЫ ВЫСОКОЭФФЕКТИВНОГО ПЕРЕНОСА ГЕНОВ С ПОМОЩЬЮ ВАРИАНТОВ КАПСИДА AAV | 2013 |
|
RU2683497C2 |
| АНТИГЕННЫЕ ПЕПТИДЫ NEISSERIA | 2000 |
|
RU2281956C2 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ ГЛИКОПРОТЕИНОВ С МОДИФИЦИРОВАННЫМ ГЛИКОЗИЛИРОВАНИЕМ | 2015 |
|
RU2704830C2 |

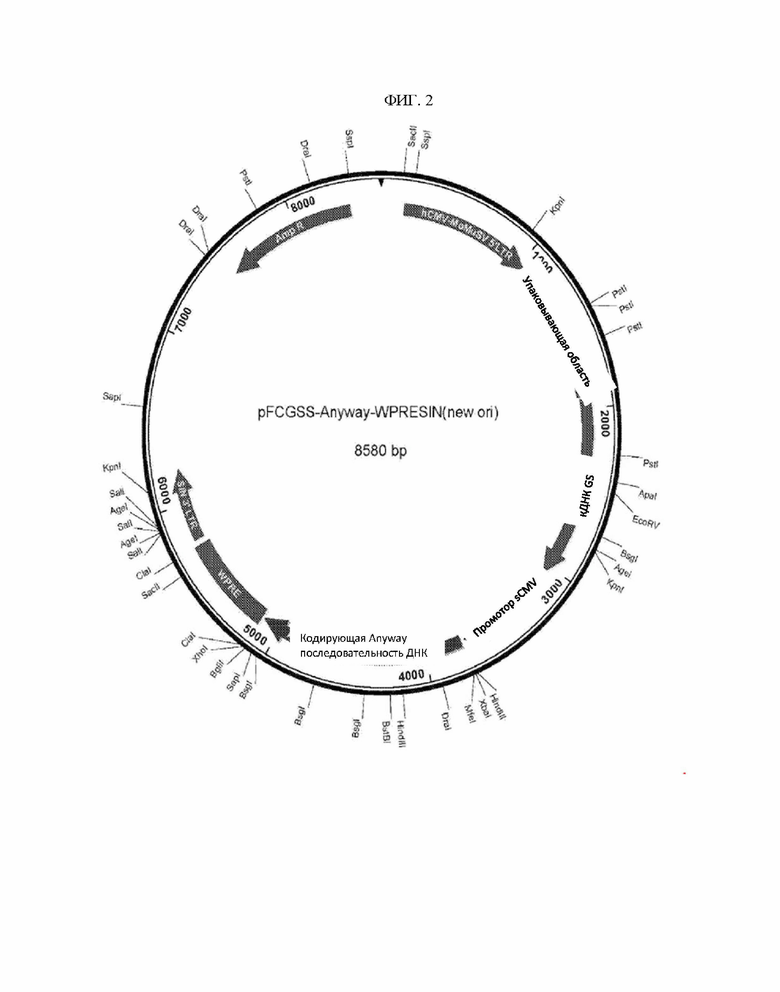




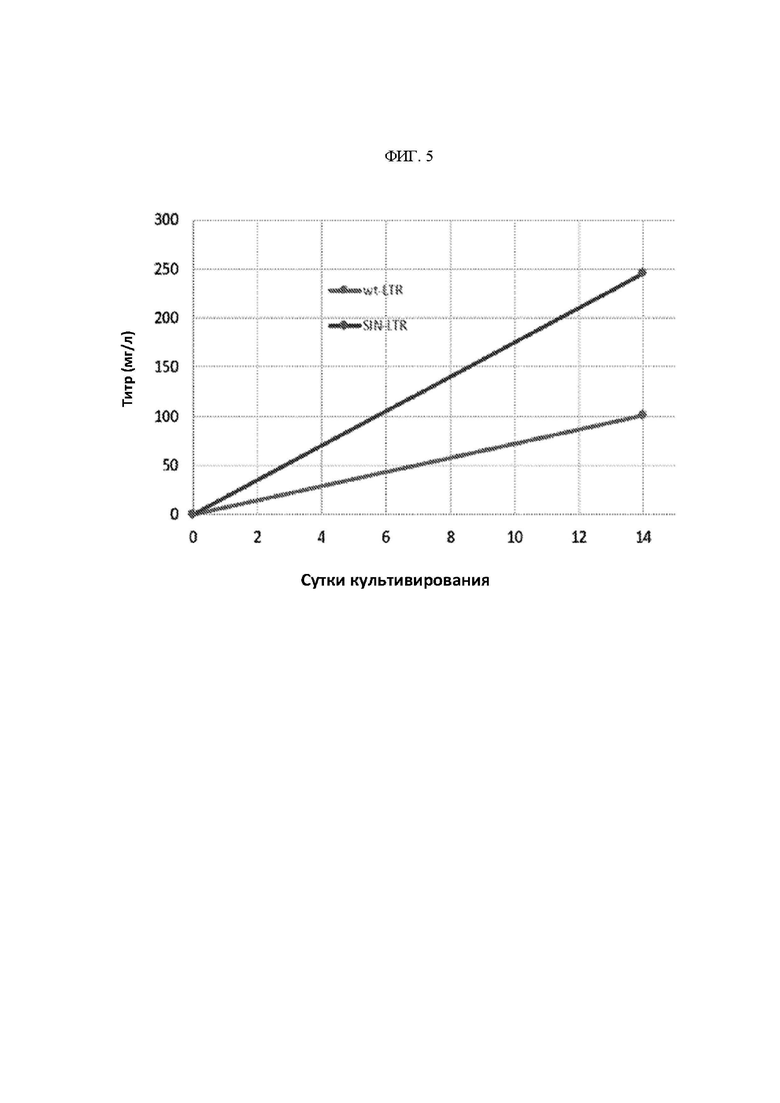
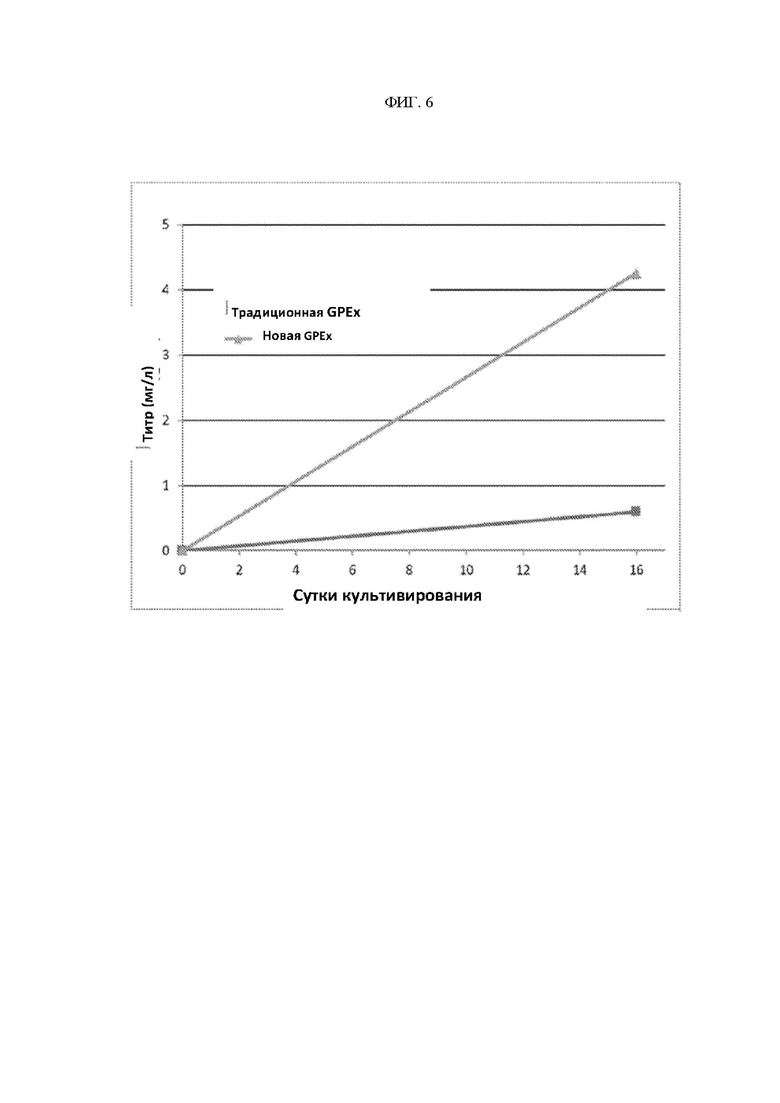

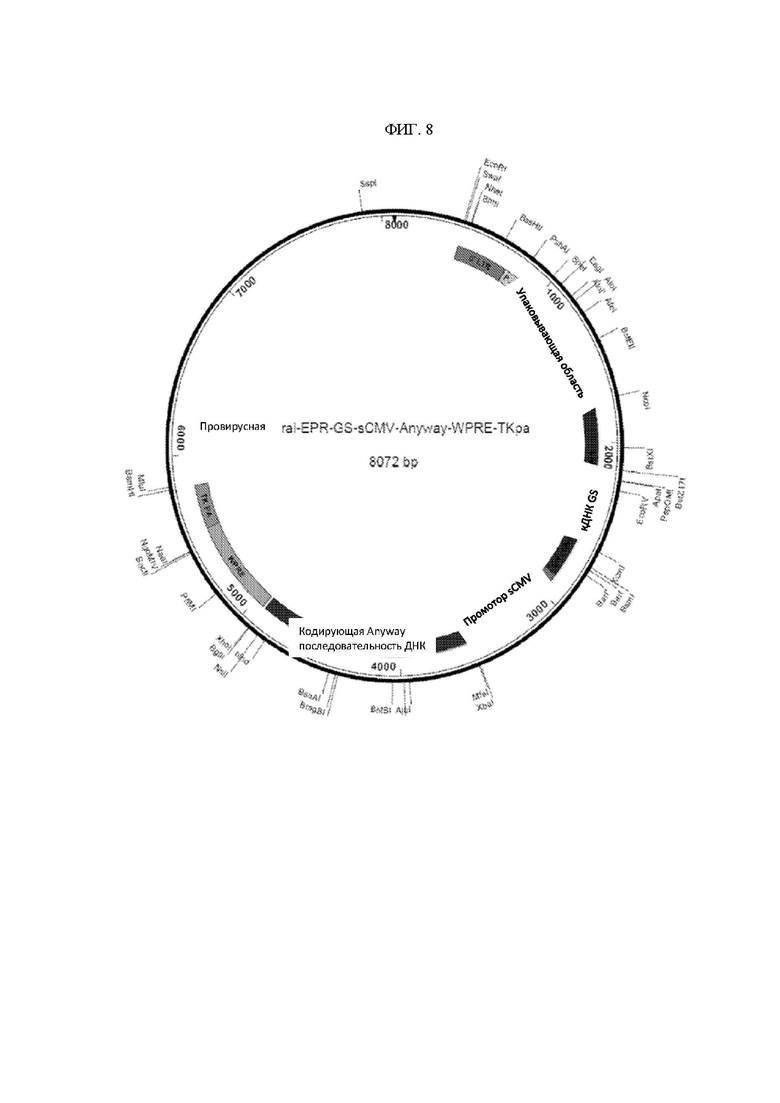

Настоящее изобретение относится к области биотехнологии, в частности к векторам экспрессии для продукции представляющих интерес белков. Изобретение раскрывает экспрессионный вектор со слабым промотором SIN LTR, управляющим экспрессией селектируемого маркера. Наличие в структуре вектора экспрессии такого слабого промотора, находящегося в функциональной связи с селектируемым маркером, улучшает способность клеточной культуры продуцировать более высокие титры и обеспечивает высокую удельную продуктивность представляющего интерес белка. 5 н. и 9 з.п. ф-лы, 9 ил., 7 табл., 6 пр.
1. Вектор для экспрессии представляющего интерес белка, содержащий последовательность нуклеиновой кислоты, кодирующую селектируемый маркер в функциональной связи с первой промоторной последовательностью, которая представляет собой промоторную последовательность вирусного самоинактивирующегося (Self-Inactivating, SIN) длинного концевого повтора (Long Terminal Repeat, LTR), и последовательность нуклеиновой кислоты, кодирующую представляющий интерес белок, функционально связанную со второй промоторной последовательностью.
2. Вектор по п. 1, причем указанная промоторная последовательность SIN LTR является по меньшей мере на 95% идентичной SEQ ID NO:3, при этом предпочтительно указанная промоторная последовательность SIN LTR представляет собой SEQ ID NO:3.
3. Вектор по любому из пп. 1, 2, в котором селектируемый маркер представляет собой глутамин-синтетазу (Glutamine Synthetase, GS) или дигидрофолат-редуктазу (Dihydrofolate Reductase, DHFR).
4. Вектор по любому из пп. 1-3, причем указанный вектор содержит одну сигнальную последовательность поли-A в функциональной связи с указанным селектируемым маркером и указанной нуклеиновой кислотой, кодирующей представляющий интерес белок, или причем указанный вектор содержит первую сигнальную последовательность поли-A в функциональной связи с указанным селектируемым маркером и вторую сигнальную последовательность поли-A в функциональной связи с указанной нуклеиновой кислотой, кодирующей представляющий интерес белок.
5. Вектор по любому из пп. 1-4, в котором представляющий интерес белок выбран из группы, состоящей из Fc-слитого белка, фермента, слияния с альбумином, фактора роста, белкового рецептора, одноцепочечного антитела (scFv), одноцепочечного антитела-Fc (scFv-Fc), диатела и минитела (scFv-CH3), Fab, одноцепочечного Fab (scFab), тяжелой цепи иммуноглобулина и легкой цепи иммуноглобулина, при этом предпочтительно указанный представляющий интерес белок представляет собой Fc-слитый белок.
6. Вектор по любому из пп. 1-5, причем указанный вектор представляет собой вирусный вектор, предпочтительно ретровирусный вектор и/или плазмиду.
7. Клетка-хозяин для экспрессии представляющего интерес белка, содержащая вектор по любому из пп. 1-6, при этом предпочтительно указанная клетка-хозяин дополнительно содержит по меньшей мере второй вектор, который кодирует и обеспечивает экспрессию второго представляющего интерес белка, и при этом i) указанный второй вектор не включает в себя селектируемый маркер, или ii) указанный второй вектор включает в себя селектируемый маркер, отличающийся от селектируемого маркера в первом векторе.
8. Клетка-хозяин по п. 7, причем линия клеток-хозяев выбрана из группы, состоящей из линии клеток с нокаутом GS, линии клеток яичника китайского хомячка (CHO), линии клеток HEK 293 и линии клеток CAP, предпочтительно линии клеток с нокаутом DHFR.
9. Клетка-хозяин по любому из пп. 7, 8, причем указанная клетка-хозяин содержит приблизительно от 1 дo 1000 копий вектора, предпочтительно, приблизительно от 10 дo 200 копий вектора, более предпочтительно приблизительно от 10 дo 100 копий вектора, наиболее предпочтительно приблизительно от 20 дo 100 копий вектора.
10. Клетка-хозяин по любому из пп. 7-9, в которой первый представляющий интерес белок в первом векторе представляет собой одну из тяжелой или легкой цепи иммуноглобулина, и второй белок во втором векторе представляет собой другую из тяжелой или легкой цепи иммуноглобулина, при этом предпочтительно первый представляющий интерес белок представляет собой тяжелую цепь иммуноглобулина, и второй представляющий интерес белок представляет собой легкую цепь иммуноглобулина.
11. Клетка-хозяин по любому из пп. 7-10, причем указанная клетка-хозяин содержит приблизительно от 1 дo 1000 копий второго вектора, предпочтительно приблизительно от 10 дo 200 копий второго вектора, более предпочтительно приблизительно от 10 дo 100 копий второго вектора, наиболее предпочтительно приблизительно от 20 дo 100 копий второго вектора.
12. Культура клеток-хозяев для продукции представляющего интерес белка, содержащая клетки-хозяева по любому из пп. 7-11, причем предпочтительно указанная культура продуцирует от 1 до 150 пикограмм/клетку/сутки представляющего интерес белка, более предпочтительно указанная культура продуцирует от 2 до 50 пикограмм/клетку/сутки представляющего интерес белка.
13. Система экспрессии, содержащая:
первый вектор, содержащий последовательность нуклеиновой кислоты, кодирующую селектируемый маркер в функциональной связи с первой промоторной последовательностью, которая представляет собой промоторную последовательность вирусного самоинактивирующегося (Self-Inactivating, SIN) длинного концевого повтора (Long Terminal Repeat, LTR), и последовательность нуклеиновой кислоты, кодирующую первый представляющий интерес белок, функционально связанную со второй промоторной последовательностью, при этом предпочтительно указанный первый представляющий интерес белок представляет собой одну из тяжелой или легкой цепи иммуноглобулина; и
второй вектор, содержащий последовательность нуклеиновой кислоты, кодирующую второй представляющий интерес белок, функционально связанную с промоторной последовательностью, при этом предпочтительно указанный второй представляющий интерес белок представляет собой другую из тяжелой или легкой цепи иммуноглобулина и при этом указанный второй вектор не включает в себя селектируемый маркер.
14. Способ получения представляющего интерес белка, включающий:
трансдукцию или трансфекцию клетки-хозяина или клеток-хозяев системой по п. 13;
разработку линии клеток-хозяев из указанных трансдуцированных или трансфецированных клетки-хозяина или клеток-хозяев, которые экспрессируют представляющий интерес белок;
культивирование клеток-хозяев в таких условиях, чтобы представляющий интерес белок продуцировался указанной линией клеток-хозяев; и
очистку представляющего интерес белка от культуры клеток-хозяев.
| POLLOK K.E | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Gene Ther., 1999, v | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| LOGAN A.C | |||
| et al | |||
| "Integrated self-inactivating lentiviral vectors produce full-length genomic transcripts competent for encapsidation and | |||
