ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет на основании предварительной заявки на патент США №63/033,514, поданной 2 июня 2020 года, содержание которой полностью включено в настоящую заявку посредством ссылки.
Настоящая заявка также испрашивает приоритет на основании предварительной заявки на патент США №63/033,516, поданной 2 июня 2020 года, содержание которой полностью включено в настоящую заявку посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к конструкциям нуклеиновых кислот и их применению для разработки линий клеток-хозяев для получения целевого белка и, в частности, к конструкциям нуклеиновых кислот, которые обеспечивают улучшенную селекцию для разработки высокопродуктивных линий клеток.
УРОВЕНЬ ТЕХНИКИ
Белковые терапевтические средства являются важным классом лекарственных средств, которые обеспечивают помощь пациентам, наиболее нуждающимся в новых видах терапии. Недавно одобренные рекомбинантные белковые терапевтические средства были разработаны для лечения широкого спектра клинических показаний, включая раковые заболевания, аутоиммунные нарушения/воспаление, воздействие инфекционных агентов и генетические нарушения. Последние достижения в области технологий конструирования белков позволили разработчикам и производителям лекарственных средств точно подбирать и использовать желаемые функциональные характеристики целевых белков при сохранении (и в некоторых случаях улучшении) безопасности или эффективности продукта или и того, и другого.
Производство и продуцирование терапевтических белков представляют собой очень сложные процессы. Например, процесс получения типичного белкового лекарственного средства может включать более 5000 критических стадий, что во много раз превышает число стадий, необходимых для получения низкомолекулярного лекарственного средства.
Аналогичным образом, белковые терапевтические средства, которые включают моноклональные антитела, а также большие или слитые белки, имеющие молекулярную массу свыше 100 кДа, могут быть на порядки больше по размеру, чем низкомолекулярные лекарственные средства. Кроме того, белковые терапевтические средства характеризуются сложными вторичными и третичными структурами, которые должны быть сохранены. Белковые терапевтические средства не могут быть синтезированы полностью с помощью химических способов, и требуется их получение в живых клетках или организмах; соответственно, выбор клеточной линии, природы вида и условий культивирования влияет на характеристики конечного продукта. Более того, большинство биологически активных белков нуждаются в посттрансляционных модификациях, которые могут нарушаться при использовании гетерологичных систем экспрессии. Кроме того, поскольку продукты синтезируются клетками или организмами, то задействованы сложные способы очистки. Кроме того, для предотвращения серьезной проблемы безопасности белковых лекарственных веществ, связанной с вирусной контаминацией, вводят процессы очистки от вирусов, такие как удаление вирусных частиц с помощью фильтров или смол, а также стадии инактивации путем использования низких значений рН или детергентов. С учетом сложности терапевтических белков, касающихся большого размера их молекул, посттрансляционных модификаций и разнообразия биологических материалов, включаемых в процесс их получения, весьма желательной является возможность улучшения конкретных функциональных характеристик продукта при сохранении его безопасности и эффективности, что обеспечивается с помощью стратегий конструирования белков.
В то время как внедрение новых стратегий и подходов к модификации белковых лекарственных продуктов не является рядовой задачей, потенциальные терапевтические преимущества обусловили более широкое применение таких стратегий в процессе разработки лекарственных средств. В настоящее время используют ряд платформенных технологий конструирования белков для увеличения периода полувыведения из циркуляции, усиления нацеливания и повышения функциональности новых белковых терапевтических средств, а также для повышения выхода и чистоты продукта. Например, для продления периода полувыведения лекарственного средства из циркуляции в настоящее время применяют подходы с конъюгированием и дериватизацией белков, включая слияние с Fc, слияние с альбумином и ПЭГилирование.
Получение белковых фармацевтических (биологических) препаратов является дорогостоящим и трудоемким. В данной области техники существует потребность в более эффективных инструментах и способах для получения этого важного класса лекарственных средств.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к конструкциям нуклеиновых кислот и их применению для разработки линий клеток-хозяев для получения целевого белка и, в частности, к конструкциям нуклеиновых кислот, которые обеспечивают улучшенную селекцию для разработки высокопродуктивных линий клеток.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложены конструкции нуклеиновых кислот для экспрессии целевого белка, содержащие следующие функционально связанные элементы по порядку от 5'- до 3'-конца: необязательно, первую промоторную последовательность; последовательность селектируемого маркера; вторую промоторную последовательность; последовательность нуклеиновой кислоты, кодирующую первый целевой белок, которая функционально связана со второй промоторной последовательностью; и сигнальную последовательность поли(А); при этом указанная конструкция нуклеиновой кислоты дополнительно содержит по меньшей мере один инсерционный элемент в положении или положениях, выбранных из группы, состоящей из 3'-конца относительно необязательной первой промоторной последовательности или последовательности селектируемого маркера, 3'-конца относительно сигнальной последовательности поли(А), положения между необязательной первой промоторной последовательностью и сигнальной последовательностью поли(А), положения между последовательностью селектируемого маркера и второй промоторной последовательностью, и одновременно 5'-конца относительно необязательной первой промоторной последовательности или последовательности селектируемого маркера и 3'-конца относительно сигнальной последовательности поли(А). В некоторых вариантах реализации конструкции нуклеиновых кислот содержат первую промоторную последовательность. В некоторых предпочтительных вариантах реализации конструкция не содержит сигнальной последовательности поли(А) между селектируемым маркером и вторым промотором. В некоторых предпочтительных вариантах реализации селектируемый маркер примыкает ко второму промотору. В некоторых предпочтительных вариантах реализации второй промотор примыкает к последовательности нуклеиновой кислоты, кодирующей первый целевой белок.
В некоторых предпочтительных вариантах реализации конструкция нуклеиновой кислоты содержит некодирующую область между первым промотором и селектируемым маркером. В некоторых предпочтительных вариантах реализации некодирующая область содержит несколько потенциальных последовательностей Козак и/или сайтов начала трансляции ATG. В некоторых предпочтительных вариантах реализации конструкция нуклеиновой кислоты содержит расширенную область упаковки (EPR) между первым промотором и селектируемым маркером. В некоторых предпочтительных вариантах реализации EPR содержит несколько потенциальных последовательностей Козак и/или сайтов начала трансляции ATG.
В некоторых предпочтительных вариантах реализации первая промоторная последовательность выбрана из группы, состоящей из промоторных последовательностей SIN-LTR, SV40, lac Е. coli, trp Е. coli, PL фага лямбда, PR фага лямбда, Т3, Т7, немедленно-раннего промотора цитомегаловируса (CMV), тимидинкиназы вируса простого герпеса (HSV), альфа-лактальбумина, фактора элонгации 1 альфа человека (hEF1 альфа) и металлотионеина-I мыши. В некоторых предпочтительных вариантах реализации первая промоторная последовательность не является ретровирусным промотором LTR.
В некоторых предпочтительных вариантах реализации последовательность селектируемого маркера представляет собой амплифицируемую последовательность селектируемого маркера, выбранную из группы, состоящей из последовательности глутаминсинтетазы (GS) и последовательности дигидрофолатредуктазы (DHFR). В некоторых предпочтительных вариантах реализации последовательность селектируемого маркера представляет собой последовательность маркера устойчивости к антибиотику, выбранную из группы, состоящей из последовательностей гена устойчивости к неомицину (neo), гена гигромицин В-фосфотрансферазы и гена пуромицин-М-ацетилтрансферазы.
В некоторых предпочтительных вариантах реализации вторая промоторная последовательность выбрана из группы, состоящей из промоторных последовательностей SV40, lac Е. coli, trp Е. coli, PL фага лямбда, PR фага лямбда, Т3, Т7, немедленно-раннего промотора цитомегаловируса (CMV), тимидинкиназы вируса простого герпеса (HSV), альфа-лактальбумина, фактора элонгации 1 альфа человека (hEF1 альфа) и металлотионеина-I мыши.
В некоторых предпочтительных вариантах реализации последовательность нуклеиновой кислоты, кодирующая целевой белок, кодирует белок, выбранный из группы, состоящей из последовательностей тяжелой и легкой цепей иммуноглобулина.
В некоторых предпочтительных вариантах реализации инсерционный элемент выбран из группы, состоящей из управляемого транспозоном инсерционного элемента, управляемого рекомбиназой инсерционного элемента и управляемого HDR инсерционного элемента. В некоторых предпочтительных вариантах реализации управляемый транспозоном инсерционный элемент представляет собой инвертированный концевой повтор. В некоторых предпочтительных вариантах реализации конструкция содержит два инвертированных концевых повтора, расположенных на 5'-конце относительно первого промотора и на 3'-конце относительно сигнальной последовательности поли(А). В некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент представляет собой сайт интеграции (att). В некоторых предпочтительных вариантах реализации сайт интеграции (att) представляет собой attB. В некоторых предпочтительных вариантах реализации управляемый HDR инсерционный элемент содержит последовательность генетически нейтрального ("safe harbor") локуса AAVS1. В некоторых предпочтительных вариантах реализации управляемый HDR инсерционный элемент представляет собой последовательность нуклеиновой кислоты, гомологичную сайту-мишени в хромосоме. В некоторых предпочтительных вариантах реализации последовательность нуклеиновой кислоты, гомологичная сайту-мишени в хромосоме, имеет длину от приблизительно 30 до 1000 оснований. В некоторых предпочтительных вариантах реализации конструкция содержит две последовательности нуклеиновой кислоты, гомологичные сайту-мишени в хромосоме, расположенные на 5'-конце относительно первого промотора и на 3'-конце относительно сигнальной последовательности поли(А). В некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент представляет собой сайт-мишень для распознавания Flp (FRT). В некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент представляет собой последовательность LoxP.
В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат элемент экспорта РНК. В некоторых предпочтительных вариантах реализации элемент экспорта РНК расположен на 3'- или 5'-конце относительно последовательности нуклеиновой кислоты, кодирующей целевой белок. В некоторых предпочтительных вариантах реализации элемент экспорта РНК представляет собой энхансер процессинга пре-мРНК (РРЕ). В некоторых предпочтительных вариантах реализации элемент экспорта РНК представляет собой посттранскрипционный регуляторный элемент (PRE). В некоторых предпочтительных вариантах реализации элемент экспорта РНК PRE представляет собой посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE).
В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат последовательность сигнального пептида, функционально связанную с первым целевым белком. В некоторых предпочтительных вариантах реализации последовательность сигнального пептида выбрана из группы, состоящей из последовательностей сигнальных пептидов тканевого активатора плазминогена, гормона роста человека, лактоферрина, альфа-казеина и альфа-лактальбумина.
В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат маркерную последовательность для очистки белка. В некоторых предпочтительных вариантах реализации маркерная последовательность для очистки белка представляет собой гексагистидиновую метку или гемагглютининовую (НА) метку.
В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат последовательность участка внутренней посадки рибосомы (IRES) и вторую последовательность нуклеиновой кислоты, кодирующую по меньшей мере второй целевой белок (например, в том числе третий, четвертый, пятый и т.д. целевой белок), расположенные на 3'-конце относительно последовательности нуклеиновой кислоты, кодирующей первый целевой белок. В некоторых предпочтительных вариантах реализации последовательность IRES выбрана из группы, состоящей из последовательностей IRES вируса ящура (FDV), вируса энцефаломиокардита и полиовируса.
В некоторых предпочтительных вариантах реализации конструкция нуклеиновой кислоты дополнительно содержит третий промотор, функционально связанный со второй последовательностью нуклеиновой кислоты, кодирующей второй целевой белок, расположенный на 3'-конце относительно последовательности нуклеиновой кислоты, кодирующей первый целевой белок. В некоторых предпочтительных вариантах реализации третья промоторная последовательность выбрана из группы, состоящей из промоторных последовательностей SV40, lac Е. coli, trp Е. coli, PL фага лямбда, PR фага лямбда, Т3, Т7, немедленно-раннего промотора цитомегаловируса (CMV), тимидинкиназы вируса простого герпеса (HSV), альфа-лактальбумина, фактора элонгации 1 альфа человека (hEF1 альфа) и металлотионеина-I мыши. В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат элемент экспорта РНК, функционально связанный со второй последовательностью нуклеиновой кислоты, кодирующей второй целевой белок. В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат сигнальную последовательность поли(А), функционально связанную со второй последовательностью нуклеиновой кислоты, кодирующей второй целевой белок.
В некоторых предпочтительных вариантах реализации первый целевой белок представляет собой одну из тяжелой и легкой цепей антитела, и второй целевой белок представляет собой другую из тяжелой и легкой цепей антитела.
В некоторых предпочтительных вариантах реализации конструкция нуклеиновой кислоты дополнительно содержит интрон, функционально связанный со второй последовательностью нуклеиновой кислоты, кодирующей второй целевой белок, расположенный на 3'-конце относительно последовательности нуклеиновой кислоты, кодирующей первый целевой белок. В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат элемент экспорта РНК, функционально связанный со второй последовательностью нуклеиновой кислоты, кодирующей второй целевой белок. В некоторых предпочтительных вариантах реализации конструкции дополнительно содержат сигнальную последовательность поли(А), функционально связанную со второй последовательностью нуклеиновой кислоты, кодирующей второй целевой белок. В некоторых предпочтительных вариантах реализации первый целевой белок представляет собой одну из тяжелой и легкой цепей антитела, и второй целевой белок представляет собой другую из тяжелой и легкой цепей антитела.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложен вектор, содержащий конструкцию нуклеиновой кислоты, описанную выше. В некоторых предпочтительных вариантах реализации вектор представляет собой плазмиду.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложена клетка-хозяин, содержащая конструкцию нуклеиновой кислоты, описанную выше, или вектор, описанный выше. В некоторых предпочтительных вариантах реализации клетка-хозяин выбрана из группы, состоящей из клеток яичника китайского хомячка (СНО), клеток HEK 293, клеток САР, эпителиальных клеток молочной железы крупного рогатого скота, линии клеток почки обезьяны CV1, трансформированной SV40, клеток почки детеныша хомяка, клеток Сертоли мыши, клеток почки обезьяны, клеток почки африканской зеленой мартышки, клеток карциномы шейки матки человека, клеток почки собаки, клеток печени крысы Buffalo, клеток легкого человека, клеток печени человека, клеток опухоли молочной железы мыши, клеток TRI, клеток MRC 5, клеток FS4, фибробластов крысы, клеток MDBK и линии клеток гепатомы человека. В некоторых предпочтительных вариантах реализации клетка-хозяин выбрана из группы, состоящей из клеток яичника китайского хомячка (СНО), клеток HEK 293 и клеток САР. В некоторых предпочтительных вариантах реализации линия клеток-хозяев представляет собой линию клеток с нокаутом GS. В некоторых предпочтительных вариантах реализации линия клеток-хозяев представляет собой линию клеток с нокаутом DHFR. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 1 до 1000 копий конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 10 до 200 копий конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 10 до 100 копий конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 20 до 100 копий конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от 50 до 500 копий конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от 50 до 250 копий конструкции нуклеиновой кислоты.
В некоторых предпочтительных вариантах реализации клетка-хозяин дополнительно содержит по меньшей мере вторую конструкцию нуклеиновой кислоты, которая кодирует второй целевой белок и обеспечивает его экспрессию. В некоторых предпочтительных вариантах реализации вторая конструкция нуклеиновой кислоты не содержит селектируемого маркера. В некоторых предпочтительных вариантах реализации вторая конструкция нуклеиновой кислоты содержит селектируемый маркер, отличный от селектируемого маркера в первой конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации первый целевой белок в первой конструкции нуклеиновой кислоты представляет собой одну из тяжелой или легкой цепей иммуноглобулина, и второй белок во второй конструкции нуклеиновой кислоты представляет собой другую из тяжелой или легкой цепей иммуноглобулина. В некоторых предпочтительных вариантах реализации первый целевой белок представляет собой тяжелую цепь иммуноглобулина, и второй целевой белок представляет собой легкую цепь иммуноглобулина. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 1 до 1000 копий второй конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 10 до 200 копий второй конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 10 до 100 копий второй конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от приблизительно 20 до 100 копий второй конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от 50 до 500 копий второй конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации клетка-хозяин содержит от 50 до 250 копий второй конструкции нуклеиновой кислоты.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложена культура клеток-хозяев, содержащая популяцию клеток-хозяев, описанную выше.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложены способы получения целевого белка, включающие культивирование клеток-хозяев, описанных выше, в условиях, при которых осуществляется экспрессия целевого(ых) белка(ов), и очистку целевого(ых) белка(ов) от культуры клеток-хозяев. В некоторых предпочтительных вариантах реализации клетки-хозяева выращивают в среде, содержащей ингибитор селектируемого маркера. В некоторых предпочтительных вариантах реализации селектируемый маркер представляет собой GS, и ингибитор представляет собой фосфинотрицин или метионинсульфоксимин (Msx). В некоторых предпочтительных вариантах реализации селектируемый маркер представляет собой DHFR, и ингибитор представляет собой метотрексат.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложен вектор, содержащий конструкцию нуклеиновой кислоты, описанную выше. В некоторых предпочтительных вариантах реализации вектор выбран из группы, состоящей из плазмидного вектора, ретровирусного вектора, лентивирусного вектора, вектора на основе AAV и транспозонного вектора.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложена система, содержащая: первую конструкцию нуклеиновой кислоты, описанную выше; и вторую конструкцию нуклеиновой кислоты, кодирующую фермент. В некоторых предпочтительных вариантах реализации конструкции обеспечены на различных векторах. В некоторых предпочтительных вариантах реализации конструкции обеспечены на одних и тех же векторах. В некоторых предпочтительных вариантах реализации фермент выбран из группы, состоящей из транспозазы, интегразы, рекомбиназы, нуклеазы и никазы. В некоторых предпочтительных вариантах реализации нуклеаза представляет собой нуклеазу Cas. В некоторых предпочтительных вариантах реализации никаза представляет собой никазу Cas. В некоторых предпочтительных вариантах реализации системы дополнительно содержат одну или более направляющих последовательностей РНК. В некоторых предпочтительных вариантах реализации фермент облегчает встраивание конструкции нуклеиновой кислоты или ее части в геном клетки-хозяина.
В некоторых предпочтительных вариантах реализации системы дополнительно содержат по меньшей мере третью конструкцию нуклеиновой кислоты, описанную выше, при этом указанная третья конструкция нуклеиновой кислоты кодирует целевой белок, который отличен от целевого белка в первой конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации третья конструкция нуклеиновой кислоты обеспечена в отдельном векторе. В некоторых предпочтительных вариантах реализации третья конструкция нуклеиновой кислоты обеспечена в том же векторе, что и первая и вторая конструкции нуклеиновых кислот.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложена система, содержащая по меньшей мере первую и вторую конструкции нуклеиновых кислот, описанные выше; при этом каждая из указанных первой и второй конструкций нуклеиновых кислот кодирует свой целевой белок. В некоторых предпочтительных вариантах реализации первая и вторая конструкции нуклеиновых кислот обеспечены в отдельных векторах. В некоторых предпочтительных вариантах реализации первая и вторая конструкции нуклеиновых кислот обеспечены в одном и том же векторе. В некоторых предпочтительных вариантах реализации системы дополнительно содержат третью конструкцию нуклеиновой кислоты, кодирующую фермент. В некоторых предпочтительных вариантах реализации фермент выбран из группы, состоящей из транспозазы, интегразы, рекомбиназы, нуклеазы и никазы. В некоторых предпочтительных вариантах реализации нуклеаза представляет собой нуклеазу Cas. В некоторых предпочтительных вариантах реализации никаза представляет собой никазу Cas. В некоторых предпочтительных вариантах реализации системы дополнительно содержат одну или более направляющих последовательностей РНК. В некоторых предпочтительных вариантах реализации фермент облегчает встраивание конструкции нуклеиновой кислоты или ее части в геном клетки-хозяина. В некоторых предпочтительных вариантах реализации третья конструкция нуклеиновой кислоты обеспечена в отдельном векторе. В некоторых предпочтительных вариантах реализации третья конструкция нуклеиновой кислоты обеспечена в том же векторе, что и первая и вторая конструкции нуклеиновых кислот.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложены способы получения целевого белка, включающие: введение конструкции нуклеиновой кислоты, вектора или системы, описанных выше, в клетку-хозяина в условиях, при которых конструкция нуклеиновой кислоты встраивается в геном клетки-хозяина; разработку линии клеток-хозяев, экспрессирующей целевой белок; культивирование клеток-хозяев из линии клеток-хозяев в условиях, при которых целевой белок продуцируется клетками-хозяевами; и очистку целевого белка от культуры клеток-хозяев.
В некоторых предпочтительных вариантах реализации клетка-хозяин выбрана из группы, состоящей из клеток яичника китайского хомячка (СНО), клеток HEK 293, клеток САР, эпителиальных клеток молочной железы крупного рогатого скота, линии клеток почки обезьяны CV1, трансформированной SV40, клеток почки детеныша хомяка, клеток Сертоли мыши, клеток почки обезьяны, клеток почки африканской зеленой мартышки, клеток карциномы шейки матки человека, клеток почки собаки, клеток печени крысы Buffalo, клеток легкого человека, клеток печени человека, клеток опухоли молочной железы мыши, клеток TRI, клеток MRC 5, клеток FS4, фибробластов крысы, клеток MDBK и линии клеток гепатомы человека. В некоторых предпочтительных вариантах реализации клетка-хозяин выбрана из группы, состоящей из клеток яичника китайского хомячка (СНО), клеток HEK 293 и клеток САР. В некоторых предпочтительных вариантах реализации линия клеток-хозяев представляет собой линию клеток с нокаутом GS. В некоторых предпочтительных вариантах реализации линия клеток-хозяев представляет собой линию клеток с нокаутом DHFR. В некоторых предпочтительных вариантах реализации клетки-хозяева выращивают в среде, содержащей ингибитор селектируемого маркера. В некоторых предпочтительных вариантах реализации селектируемый маркер представляет собой GS, и ингибитор представляет собой фосфинотрицин или метионинсульфоксимин (Msx). В некоторых предпочтительных вариантах реализации селектируемый маркер представляет собой DHFR, и ингибитор представляет собой метотрексат. В некоторых предпочтительных вариантах реализации культивирование клеток-хозяев из линии клеток-хозяев в условиях, при которых целевой белок продуцируется клетками-хозяевами, дополнительно включает культивирование в системе, выбранной из группы, состоящей из чашек Петри, луночных планшетов, роллерных флаконов, биореакторов, перфузионных систем и культур с периодическим добавлением питательной среды.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Сокращения, используемые на чертежах:
AmpR = Бактериальный ген устойчивости к ампициллину
attB = Бактериальный сайт интеграции
attP = Фаговый сайт интеграции
attR = Рекомбинированный вышележащий сайт интеграции
Остов = Остов плазмиды
CDS = Кодирующая последовательность
EPR = Расширенная область упаковки вируса лейкоза мышей Молони (MMLV)
GCI = Индекс копийности гена
GS = Глутаминсинтетаза
Н или НС = Тяжелая цепь
hCMV = Немедленно-ранний промотор цитомегаловируса человека
I = Интрон
L или LC = Легкая цепь
MoMuSV 5'LTR=5' - длинный концевой повтор вируса саркомы мышей Молони
Neo = Ген устойчивости к неомицину
РА или PolyA = Сигнал полиаденилирования
ProV SLNT-LTR = Провирусный самоинактивирующийся длинный концевой повтор
sCMV = Немедленно-ранний промотор цитомегаловируса обезьян
ДСН-ПААГ = Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия
SLN-3'LTR = Самоинактивирующийся 3'-длинный концевой повтор
SV40 = Вирус обезьян 40
TK = Тимидинкиназа
UTR = Нетранслируемая область
W или WPRE = Посттранскрипционный регуляторный элемент вируса гепатита сурков.
ФИГ. 1. Схема конструкции нуклеиновой кислоты для некоторых вариантов реализации настоящего изобретения.
ФИГ. 2. График кривых выживаемости клеток после трансфекции и селекции в отсутствие глутамина. Показаны средние значения для двух дублированных трансфекций.
ФИГ. 3. Таблица, представляющая анализ продуктивности и числа копий для объединенных клеточных линий, полученных с применением различных плазмид. Показаны средние значения для двух дублированных трансфекций.
ФИГ. 4. График кривых выживаемости клеток после трансфекции и селекции в отсутствие глутамина. Показаны средние значения для двух дублированных трансфекций.
ФИГ. 5. Карта плазмиды для экспрессии интегразы PhiC31.
ФИГ. 6. Последовательность плазмиды для экспрессии интегразы PhiC31.
ФИГ. 7. Карта плазмиды для экспрессии акцепторного сайта (Dock).
ФИГ. 8. Последовательность плазмиды для экспрессии акцепторного сайта (Dock).
ФИГ. 9. Карта плазмиды Dock-WPRE.
ФИГ. 10. Последовательность плазмиды Dock-WPRE.
ФИГ. 11. Карта плазмиды Transgene-Promoter-Any way. В этой плазмиде экспрессией GS управляет слабый провирусный самоинактивирующийся 5'-длинный концевой повтор вируса саркомы мышей Молони.
ФИГ. 12. Последовательность плазмиды Transgene-Promoter- Any way. В этой плазмиде и всех последующих плазмидах Transgene отсутствует промотор, который управлял бы экспрессией GS в плазмиде Transgene.
ФИГ. 13. Карта плазмиды Transgene-Any way.
ФИГ. 14. Последовательность плазмиды Transgene-Any way.
ФИГ. 15. Карта плазмиды Transgene-MCS.
ФИГ. 16. Последовательность плазмиды Transgene-MCS.
ФИГ. 17. Карта плазмиды Transgene-MCS-WPRE-hvtron-MCS.
ФИГ. 18. Последовательность плазмиды Transgene- MCS-WPRE-Intron-MCS.
ФИГ. 19. Карта плазмиды Transgene-MCS-WPRE-MCS-WPRE.
ФИГ. 20. Последовательность плазмиды Transgene-MCS-WPRE-MCS-WPRE.
ФИГ. 21. Карта плазмиды Transgene-Yourway-HWIL.
ФИГ. 22. Последовательность плазмиды Transgene-Yourway-HWIL.
ФИГ. 23. Карта плазмиды Transgene-Yourway-LWIH.
ФИГ. 24. Последовательность плазмиды Transgene-Yourway-LWIH.
ФИГ. 25. Карта плазмиды Transgene-Yourway-HWLW.
ФИГ. 26. Последовательность плазмиды Transgene-Yourway-HWLW.
ФИГ. 27. Карта плазмиды Transgene-Yourway-LWHW.
ФИГ. 28. Последовательность плазмиды Transgene-Yourway- LWHW.
ФИГ. 29. Диаграмма индекса копийности гена attR для не подвергнутых селекции пулов клеток с акцепторными сайтами, содержащих в среднем приблизительно 36 акцепторных сайтов на клетку, трансфицированных плазмидой Transgene-Promoter-Any way в указанных соотношениях.
ФИГ. 30. График процентного содержания жизнеспособных клеток с течением времени селекции в выбранных пулах согласно Фигуре 29.
ФИГ. 31. Таблица значений индекса копийности и числа копий гена attR для всех пулов согласно Фигуре 30.
ФИГ. 32. График процентного содержания жизнеспособных клеток с течением времени селекции в пулах клеток с акцепторными сайтами, содержащих в среднем приблизительно 135 акцепторных сайтов на клетку, трансфицированных не содержащей промотора плазмидой Transgene-Anyway и плазмидой Integrase в указанных соотношениях. Показано среднее значение для двух дублированных пулов.
ФИГ. 33. Таблица значений индекса копийности гена attR для пулов согласно Фигуре 32 после селекции. Показано среднее значение для двух дублированных пулов.
ФИГ. 34. График процентного содержания жизнеспособных клеток с течением времени селекции в клонах клеток с акцепторными сайтами, содержащих приблизительно 181 копию акцепторного сайта на клетку, трансфицированных плазмидой Transgene-Yourway-LWHW и плазмидой Integrase в указанных соотношениях. Показано среднее значение для двух дублированных пулов.
ФИГ. 35. Таблица значений индекса копийности гена attR для пулов согласно Фигуре 34 после селекции. Показано среднее значение для двух дублированных пулов.
ФИГ. 36. Таблица значений индексов копийности генов attR (заполненный акцепторный сайт) и attP (пустой акцепторный сайт), % заполненных акцепторных сайтов и конечного титра согласно анализу продуктивности при периодическом добавлении питательной среды для клонов, полученных из пулов с акцепторными сайтами, содержащих приблизительно 135 копий акцепторного сайта на клетку, трансфицированных плазмидой Transgene-Anyway и плазмидой Integrase.
ФИГ. 37. График зависимости титра согласно анализу продуктивности при периодическом добавлении среды Excell от индексов копийности гена attR для всех 25 клонов согласно Фигуре 36.
ФИГ. 38. График процентного содержания жизнеспособных клеток с течением времени селекции в клонах клеток с акцепторными сайтами, содержащих приблизительно 181 копию акцепторного сайта на клетку, трансфицированных плазмидами Transgene-Yourway-LWHW, Yourway-HWLW, Yourway-HWTL, Yourway-LWTH или Anyway (в отдельности) и плазмидой Integrase. Показано среднее значение для двух дублированных пулов.
ФИГ. 39. Таблица значений индекса копийности гена attR и конечного титра согласно анализу продуктивности при периодическом добавлении питательной среды для клонов, полученных из пулов с акцепторными сайтами, согласно Фигуре 38. Показано среднее значение для двух дублированных пулов.
ФИГ. 40. Анализ продуктов Transgene- Yourway и Transgene-Anyway посредством ДСН-ПААГ-электрофореза проводили как в невосстанавливающих (слева), так и в восстанавливающих (справа) условиях.
ФИГ. 41. График конечного титра через более чем 40 поколений согласно анализу продуктивности при периодическом добавлении питательной среды с применением двух различных сред/стратегий добавления питательной среды для 3 пулов, экспрессирующих Anyway.
ОПРЕДЕЛЕНИЯ
Для облегчения понимания настоящего изобретения ниже приведены определения ряда терминов.
В настоящей заявке термин «клетка-хозяин» относится к любой эукариотической клетке (например, клеткам млекопитающих, клеткам птиц, клеткам земноводных, клеткам растений, клеткам рыб и клеткам насекомых) независимо от того, находится ли клетка в условиях in vitro или in vivo.
В настоящей заявке термин «клеточная культура» относится к любой культуре клеток in vitro. Этот термин включает стабильные клеточные линии (например, имеющие иммортализованный фенотип), первичные клеточные культуры, перевиваемые клеточные линии (например, нетрансформированные клетки) и любую другую популяцию клеток, поддерживаемую in vitro, включая ооциты и эмбрионы.
В настоящей заявке термин «вектор» относится к любому генетическому элементу, такому как плазмида, фаг, транспозон, космида, хромосома, вирус, вирион и т.д., который способен к репликации, когда он связан с подходящими регуляторными элементами, и который может переносить последовательности генов между клетками. Таким образом, указанный термин включает переносчики для клонирования и экспрессии, а также вирусные векторы.
В настоящей заявке термин «геном» относится к генетическому материалу (например, хромосомам) организма.
Термин «целевая (представляющая интерес) нуклеотидная последовательность» относится к любой нуклеотидной последовательности (например, РНК или ДНК), манипуляции с которой могут считаться желательными по любой причине (например, для лечения заболевания, обеспечения улучшенных качеств, экспрессии целевого белка в клетке-хозяине, экспрессии рибозима и т.д.) специалистом в данной области техники. Такие нуклеотидные последовательности включают, не ограничиваясь указанными, кодирующие последовательности структурных генов (например, репортерных генов, генов селектируемых маркеров, онкогенов, генов устойчивости к лекарственным средствам, факторов роста и т.д.) и некодирующие регуляторные последовательности, которые не кодируют мРНК или белковый продукт (например, промоторную последовательность, последовательность полиаденилирования, последовательность терминации, энхансерную последовательность и т.д.).
В настоящей заявке термин «целевой белок» относится к белку, кодируемому целевой нуклеиновой кислотой.
В настоящей заявке термины «кодирующая молекула нуклеиновой кислоты», «кодирующая последовательность ДНК», «кодирующая ДНК», «кодирующая последовательность РНК» и «кодирующая РНК» относятся к расположению в определенном порядке или последовательности дезоксирибонуклеотидов или рибонуклеотидов вдоль цепи дезоксирибонуклеиновой кислоты или рибонуклеиновой кислоты. Порядок этих дезоксирибонуклеотидов или рибонуклеотидов определяет порядок аминокислот вдоль полипептидной (белковой) цепи. Таким образом, последовательность ДНК или РНК кодирует аминокислотную последовательность.
В настоящей заявке термин «промотор», «промоторный элемент» или «промоторная последовательность» относится к последовательности ДНК, которая, при лигировании с целевой нуклеотидной последовательностью, способна регулировать транскрипцию целевой нуклеотидной последовательности в мРНК. Промотор, как правило, хотя и не обязательно, расположен на 5'-конце относительно целевой нуклеотидной последовательности (то есть выше нее), транскрипцию которой в мРНК он регулирует, и обеспечивает сайт для специфичного связывания РНК-полимеразы и других факторов транскрипции для инициации транскрипции.
Сигналы контроля транскрипции у эукариот включают «промоторные» и «энхансерные» элементы. Промоторы и энхансеры состоят из коротких наборов последовательностей ДНК, которые специфично взаимодействуют с клеточными белками, участвующими в транскрипции (Maniatis el al, Science 236: 1237 [1987]). Промоторные и энхансерные элементы были выделены из различных эукариотических источников, включая гены в клетках дрожжей, насекомых и млекопитающих, а также вирусов (аналогичные регуляторные элементы, то есть промоторы, также обнаружены в прокариотах). Выбор конкретного промотора и энхансера зависит от типа клеток, который предполагается использовать для экспрессии целевого белка. Некоторые эукариотические промоторы и энхансеры имеют широкий спектр хозяев, тогда как другие являются функциональными в ограниченной субпопуляции типов клеток (см. обзор в источниках Voss et al., Trends Biochem. Sci., 11:287 [1986]; и Maniatis et al., см. выше). Например, энхансер раннего гена SV40 очень активен в широком спектре типов клеток из многих видов млекопитающих и широко используется для экспрессии белков в клетках млекопитающих (Dijkema et al., EMBO J. 4:761 [1985]). Два других примера промоторных/энхансерных элементов, которые активны в широком спектре типов клеток млекопитающих, представляют собой элементы из гена фактора элонгации человека la (Uetsuki et al., J. Biol. Chem., 264:5791 [1989]; Kim et al, Gene 91:217 [1990]; и Mizushima andNagata, Nuc. Acids. Res., 18:5322 [1990]) и длинные концевые повторы вируса саркомы Рауса (Gorman et al, Proc. Natl. Acad. Sci. USA 79:6777 [1982]) и цитомегаловируса человека (Boshart et al, Cell 41:521 [1985]).
В настоящей заявке термин «промотор/энхансер» обозначает сегмент ДНК, который содержит последовательности, способные обеспечивать как промоторные, так и энхансерные функции (то есть функции, обеспечиваемые промоторным элементом и энхансерным элементом; обсуждение этих функций см. выше). Например, длинные концевые повторы ретровирусов обладают как промоторными, так и энхансерными функциями. Энхансер/промотор может быть «эндогенным» или «экзогенным» или «гетерологичным». «Эндогенный» энхансер/промотор представляет собой энхансер/промотор, который природно связан с данным геном в геноме. «Экзогенный» или «гетерологичный» энхансер/промотор представляет собой энхансер/промотор, который помещают в смежное с геном положение посредством генетической манипуляции (то есть молекулярно-биологических методик, таких как клонирование и рекомбинация) таким образом, что транскрипцией этого гена управляет связанный с ним энхансер/промотор.
В настоящей заявке термин «длинный концевой повтор» или «LTR» относится к элементам контроля транскрипции, расположенным в 5'- и 3'-концевых участках области U3 ретро вирусного генома или выделенным из них.
Как известно в данной области техники, длинные концевые повторы могут применяться в качестве регуляторных элементов в ретровирусных векторах, либо они могут быть выделены из ретровирусного генома и применяться для регулирования экспрессии из векторов других типов.
В настоящей заявке термины «комплементарный» или «комплементарность» используются в отношении полинуклеотидов (то есть последовательности нуклеотидов), связанных в соответствии с правилами спаривания оснований. Например, последовательность «5'-A-G-T-3'» комплементарна последовательности «3'-T-C-A-5'». Комплементарность может быть «частичной», при которой только некоторые из оснований нуклеиновых кислот соответствуют правилам спаривания оснований. Либо, комплементарность между нуклеиновыми кислотами может быть «полной» или «абсолютной». Степень комплементарное™ между цепями нуклеиновых кислот оказывает значительное влияние на эффективность и силу гибридизации между цепями нуклеиновых кислот. Это особенно важно в реакциях амплификации, а также в способах детектирования, зависящих от связывания нуклеиновых кислот.
Термины «гомология» и «процент идентичности», используемые в отношении нуклеиновых кислот, относятся к степени комплементарности. Гомология может быть частичной (то есть частичная идентичность) или полной (то есть полная идентичность). Частично комплементарная последовательность представляет собой последовательность, которая по меньшей мере частично ингибирует гибридизацию полностью комплементарной последовательности с последовательностью нуклеиновой кислоты-мишени, и ее обозначают с помощью функционального термина «по существу гомологичная». Ингибирование гибридизации полностью комплементарной последовательности с последовательностью-мишенью может быть исследовано с применением анализа гибридизации (Саузерн-блоттинг или Нозерн-блоттинг, гибридизация в растворе и тому подобное) в условиях низкой жесткости. По существу гомологичные последовательность или зонд (то есть олигонуклеотид, который способен гибридизоваться с другим целевым олигонуклеотидом) будут конкурировать за последовательность-мишень и ингибировать ее связывание (то есть гибридизацию) с полностью гомологичной последовательностью в условиях низкой жесткости. Это не означает, что условия низкой жесткости допускают неспецифичное связывание; условия низкой жесткости требуют, чтобы связывание двух последовательностей друг с другом являлось специфичным (то есть селектируемым) взаимодействием. Отсутствие неспецифичного связывания может быть проверено путем использования второй мишени, у которой отсутствует даже частичная степень комплементарности (например, имеющей идентичность менее чем приблизительно 30%); в отсутствие неспецифичного связывания зонд не будет гибридизоваться со второй некомплементарной мишенью.
Термины «в функциональной комбинации», «в функциональном порядке» и «функционально связанный» в настоящей заявке относятся к такой связи между последовательностями нуклеиновых кислот, при которой молекула нуклеиновой кислоты способна управлять транскрипцией данного гена и/или обеспечивает синтез желаемой белковой молекулы. Указанный термин также относится к такой связи между аминокислотными последовательностями, при которой образуется функциональный белок.
В настоящей заявке термин «селектируемый маркер» относится к гену, который кодирует белок с ферментативной активностью или другой белок, обеспечивающий способность расти в среде, не содержащей элемента, который в противном случае являлся бы незаменимым питательным веществом; кроме того, селектируемый маркер может обеспечивать клетке, в которой он экспрессируется, устойчивость к антибиотику или лекарственному средству.
В настоящей заявке термин «ретровирус» относится к ретровирусной частице, способной проникать в клетку (то есть указанная частица содержит мембрано-ассоциированный белок, такой как белок оболочки или вирусный гликопротеин G, который может связываться с поверхностью клетки-хозяина и облегчать проникновение вирусной частицы в цитоплазму клетки-хозяина) и интегрировать ретровирусный геном (в виде двухцепочечного провируса) в геном клетки-хозяина. Термин "ретровирус" охватывает подсемейства Oncovirinae (например, вирус лейкоза мышей Молони (MoMLV), вирус саркомы мышей Молони (MoMSV) и вирус опухоли молочной железы мышей (MMTV)), Spumavirinae и Lentivirinae (например, вирус иммунодефицита человека, вирус иммунодефицита обезьян, вирус инфекционной анемии лошадей и вирус козьего артрита-энцефалита; см., например, патенты США №№5,994,136 и 6,013,516, оба из которых включены в настоящую заявку посредством ссылки).
В настоящей заявке термин «ретровирусный вектор» относится к ретровирусу, который был модифицирован для экспрессии целевого гена. Ретровирусные векторы могут применяться для эффективного переноса генов в клетки-хозяева путем использования вирусного инфекционного процесса. Чужеродные или гетерологичные гены, клонированные (то есть встроенные с использованием молекулярно-биологических методик) в ретровирусный геном, могут быть эффективно доставлены в клетки-хозяева, которые восприимчивы к инфицированию ретровирусом. С помощью хорошо известных генетических манипуляций может быть устранена способность ретровирусного генома к репликации. Полученные дефектные по репликации векторы могут применяться для введения нового генетического материала в клетку, но они не способны реплицироваться. Вирус-помощник или линия упаковывающих клеток могут применяться для обеспечения сборки векторных частиц и выхода их из клетки. Такие ретровирусные векторы содержат дефектный по репликации ретровирусный геном, содержащий последовательность нуклеиновой кислоты, кодирующую по меньшей мере один целевой ген (то есть полицистронная последовательность нуклеиновой кислоты может кодировать более одного целевого гена), 5'-ретровирусный длинный концевой повтор (5'-LTR) и 3'-ретровирусный длинный концевой повтор (3'-LTR).
В настоящей заявке термин «лентивирусный вектор» относится к ретровирусным векторам, полученным из семейства Lentiviridae (например, вируса иммунодефицита человека, вируса иммунодефицита обезьян, вируса инфекционной анемии лошадей и вируса козьего артрита-энцефалита), которые способны интегрироваться в неделящиеся клетки (см., например, патенты США №№5,994,136 и 6,013,516, оба из которых включены в настоящую заявку посредством ссылки).
В настоящей заявке термин «транспозон» относится к мобильным элементам (например, Tn5, Tn7 и TnlO), которые могут перемещаться или переставляться из одного положения в геноме в другое. Как правило, транспозиция находится под контролем транспозазы. В настоящей заявке термин «транспозонный вектор» относится к вектору, кодирующему целевую нуклеиновую кислоту, фланкированную концевыми участками транспозона. Примеры транспозонных векторов включают, не ограничиваясь указанными, векторы, описанные в патентах США №№6,027,722; 5,958,775; 5,968,785; 5,965,443 и 5,719,055, все из которых включены в настоящую заявку посредством ссылки.
В настоящей заявке термин «вектор на основе аденоассоциированного вируса (AAV)» относится к вектору, полученному из серотипа аденоассоциированного вируса, включая, но не ограничиваясь указанными, AAV-1, AAV-2, AAV-3, AAV-4, AAV-5, AAVX7 и т.д. Векторы на основе AAV могут иметь полную или частичную делецию одного или более генов AAV дикого типа, предпочтительно генов rep и/или cap, но сохраняют функциональные фланкирующие последовательности ITR.
Векторы на основе AAV могут быть сконструированы с использованием рекомбинантных методик, известных в данной области техники, таким образом, чтобы содержать одну или более гетерологичных нуклеотидных последовательностей, фланкированных на обоих концах (5' и 3') функциональными ITR AAV. При практическом применении настоящего изобретения вектор на основе AAV может содержать по меньшей мере одну ITR AAV и подходящую промоторную последовательность, расположенные выше гетерологичной нуклеотидной последовательности, и по меньшей мере одну ITR AAV, расположенную ниже гетерологичной последовательности. «Рекомбинантная векторная плазмида на основе AAV» относится к одному из типов рекомбинантных векторов на основе AAV, где указанный вектор содержит плазмиду. Как правило, как и в случае векторов на основе AAV, 5'- и 3'-ITR фланкируют выбранную гетерологичную нуклеотидную последовательность.
В настоящей заявке термин «аденовирусный вектор» относится к не имеющему оболочки двухцепочечному ДНК-вектору, содержащему аденовирусный остов.
В настоящей заявке термин «очищенный» относится к молекулам, последовательностям нуклеиновых кислот или аминокислот, которые извлечены из их природной среды, выделены или разделены. Таким образом, «последовательность выделенной нуклеиновой кислоты» представляет собой последовательность очищенной нуклеиновой кислоты. «По существу очищенные» молекулы по меньшей мере на 60%, предпочтительно по меньшей мере на 75% и более предпочтительно по меньшей мере на 90% не содержат других компонентов, с которыми они обычно связаны.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к конструкциям нуклеиновых кислот и их применению для разработки линий клеток-хозяев для получения целевого белка и, в частности, к конструкциям нуклеиновых кислот, которые обеспечивают улучшенную селекцию для разработки высокопродуктивных линий клеток.
В некоторых предпочтительных вариантах реализации настоящего изобретения предложены конструкции нуклеиновых кислот для применения при осуществлении экспрессии целевого белка или целевых белков в клетке-хозяине. В некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот содержат следующие функционально связанные элементы, наиболее предпочтительно по порядку от 5'- до 3'-конца:
первая промоторная последовательность - последовательность селектируемого маркера -вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - сигнальная последовательность поли(А).
В некоторых предпочтительных вариантах реализации конструкции согласно настоящему изобретению не содержат сигнальной последовательности поли(А) между последовательностью селектируемого маркера и второй промоторной последовательностью. Настоящее изобретение не ограничивается каким-либо конкретным механизмом действия.
Действительно, понимание механизма действия не является необходимым для практического применения настоящего изобретения. Тем не менее было обнаружено, что конструкции, не содержащие сигнальной последовательности поли(А) после селектируемого маркера, обеспечивали лучшие селекцию и продуцирование целевого белка в культурах клеток-хозяев. В других предпочтительных вариантах реализации селектируемый маркер примыкает ко второму промотору. В других предпочтительных вариантах реализации второй промотор примыкает к последовательности нуклеиновой кислоты, кодирующей первый целевой белок. В этом контексте термин «примыкает» означает, что между перечисленными компонентами нет промежуточного функционального элемента или интрона.
Конструкции нуклеиновых кислот могут применяться со многими различными векторами и векторными системами. Подходящие векторы и векторные системы включают, не ограничиваясь указанными, технологии встраивания генов с использованием вирусов, такие как ретровирусные, лентивирусные системы и системы на основе AAV, а также технологии встраивания генов без использования вирусов, такие как встраивание генов с помощью транспозазы, рекомбиназы, интегразы или CRISPR. Конкретные примеры технологий/ферментов, которые могут применяться с конструкциями нуклеиновых кислот согласно настоящему изобретению, включают системы транспозазы PiggyBac, системы транспозазы «Спящая красавица», системы транспозазы Mos1, системы транспозазы Tol2, системы транспозазы Leapin, системы рекомбиназы фага лямбда, системы FLP/FRT, системы Cre/Lox, системы интегразы MMLV, системы интегразы Rep 78 и системы CRISPR, которые могут содержать нуклеазы или никазы, а также направляющие последовательности. В некоторых предпочтительных вариантах реализации система представляет собой систему интеграции нуклеиновых кислот при условии, что эта система не является ретровирусной или лентивирусной системой, использующей ретровирусный или лентивирусный LTR.
В некоторых вариантах реализации конструкции являются подходящими для применения в клетках-хозяевах, содержащих интегрированные акцепторные сайты, описанных в предварительной заявке на патент США №63/033,516, содержание которой полностью включено в настоящую заявку посредством ссылки. Интегрированные акцепторные сайты предпочтительно содержат один или более инсерционных элементов (которые могут быть названы «инсерционный элемент акцепторного сайта»). Инсерционные элементы акцепторного сайта предпочтительно представляют собой последовательности нуклеиновых кислот, которые облегчают встраивание последовательности нуклеиновой кислоты, кодирующей целевой белок, в акцепторный сайт. Конструкции нуклеиновых кислот, которые могут быть встроены в акцепторные сайты в клетках-хозяевах согласно настоящему изобретению, подробно описаны ниже.
Например, в некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент акцепторного сайта содержит сайт интеграции (att). В некоторых особенно предпочтительных вариантах реализации сайт интеграции представляет собой attP. Эти сайты интеграции используются интегразой PhiC31, которая представляет собой фермент рекомбиназу и которая в предпочтительных вариантах реализации может быть обеспечена в клетке-хозяине с помощью вектора. Эти акцепторные сайты служат в качестве акцепторов для интеграции конструкций нуклеиновых кислот, содержащих сайт интеграции attB. В других предпочтительных вариантах реализации могут быть использованы сайты интеграции attR и attL.
В других предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент акцепторного сайта содержит сайт-мишень для распознавания Flp (FRT). Эти сайты используются ферментом флиппазой, которая представляет собой фермент рекомбиназу и которая в предпочтительных вариантах реализации может быть обеспечена в клетке-хозяине с помощью вектора. Эти акцепторные сайты служат в качестве акцепторов для интеграции конструкций нуклеиновых кислот, содержащих сайт FRT.
В других предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент акцепторного сайта содержит сайт LoxP. Эти сайты используются рекомбиназой Cre, которая в предпочтительных вариантах реализации может быть обеспечена в клетке-хозяине с помощью вектора. Эти акцепторные сайты служат в качестве акцепторов для интеграции конструкций нуклеиновых кислот, содержащих сайт LoxP.
В других предпочтительных вариантах реализации инсерционный элемент представляет собой управляемый HDR инсерционный элемент (направляемой гомологией репарации) акцепторного сайта. Управляемые HDR инсерционные элементы акцепторного сайта представляют собой последовательности нуклеиновых кислот, обеспечивающие область гомологии («плечо гомологии»), которая взаимодействует за счет спаривания оснований с соответствующими плечами гомологии на конструкции нуклеиновой кислоты, встраиваемой в этот сайт. Эти системы предпочтительно применяются с эндонуклеазами, которые вводят двухцепочечные разрывы в целевой сайт или целевые сайты, предпочтительно фланкированные плечами гомологии. В некоторых вариантах реализации управляемый HDR инсерционный элемент акцепторного сайта представляет собой генетически нейтральный локус AAVS1. В этих вариантах реализации применяют акцепторный сайт, используемый эндонуклеазой (никазой) Rep 78, которая может быть введена в клетку-хозяина с помощью вектора. Белок никаза Rep 78 способствует сайт-специфической интеграции последовательностей нуклеиновых кислот, несущих плечи гомологии, соответствующие генетически нейтральному локусу AAVS1.
В других предпочтительных вариантах реализации управляемый HDR инсерционный элемент акцепторного сайта содержит одно или более плеч гомологии, которые представляют собой экзогенные последовательности, имеющие длину от 30 до 1000 пар оснований. Эти акцепторные сайты предпочтительно применяются в сочетании с системами редактирования генов CRISPR. В некоторых вариантах реализации акцепторный сайт дополнительно содержит одну или более последовательностей, которые гомологичны последовательностям направляющей РНК. В этих вариантах реализации конструкция нуклеиновой кислоты, встраиваемая в акцепторный сайт, предпочтительно содержит плечи гомологии, которые гомологичны плечам гомологии в акцепторном сайте и взаимодействуют с ними за счет спаривания оснований. Для применения с системами редактирования генов CRISPR в клетку-хозяина вводят совместимую с системами редактирования генов CRISPR нуклеазу. Совместимая с системой редактирования генов CRISPR нуклеаза может представлять собой эндонуклеазу дикого типа, которая создает двухцепочечный разрыв в положении, определяемом направляющей РНК (и в пределах акцепторного сайта), или мутированную нуклеазу (то есть никазу), которая создает ступенчатые одноцепочечные разрывы в пределах акцепторного сайта в положениях, определяемых двумя направляющими РНК. Подходящие нуклеазы подробно описаны ниже при обсуждении конструкций для экспрессии нуклеиновых кислот.
В некоторых предпочтительных вариантах реализации акцепторный сайт предпочтительно может содержать подходящий промотор таким образом, что используется схема промоторной ловушки, когда в акцепторный сайт вводят подходящие конструкции нуклеиновых кислот. Подходящие промоторы включают, не ограничиваясь указанными, промоторные последовательности SIN-LTR, SV40, EF1a, lac Е. coli, trp Е. coli, PL фага лямбда, PR фага лямбда, Т3, Т7, немедленно-раннего промотора цитомегаловируса (CMV), тимидинкиназы вируса простого герпеса (HSV), альфа-лактальбумина и металлотионеина-I мыши. В некоторых предпочтительных вариантах реализации промоторная последовательность ориентирована в акцепторном сайте таким образом, что промотор будет обеспечивать экспрессию из встроенной конструкции нуклеиновой кислоты. В некоторых предпочтительных вариантах реализации промотор имеет 5'-ориентацию относительно акцепторного сайта. В некоторых особенно предпочтительных вариантах реализации промотор представляет собой SIN LTR. В этих вариантах реализации SIN-LTR и EPR расположены на 5'-конце относительно акцепторного сайта, и SIN LTR расположен на 3'-конце относительно акцепторного сайта.
Соответственно, в некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот содержат инсерционный элемент. Предпочтительно инсерционный элемент может быть расположен на 5'-конце относительно первого промотора, на 3'-конце относительно сигнальной последовательности поли(А), между первой промоторной последовательностью и сигнальной последовательностью поли(А), между последовательностью селектируемого маркера и второй промоторной последовательностью, и одновременно на 5'-конце относительно первого промотора и на 3'-конце относительно сигнальной последовательности поли(А). Подходящие конструкции представлены в следующих неограничивающих примерах:
инсерционный элемент экспрессионной конструкции - первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая (то есть внутренняя) промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - сигнальная последовательность поли(А).
первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - сигнальная последовательность поли(А) - инсерционный элемент экспрессионной конструкции.
инсерционный элемент экспрессионной конструкции - первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - сигнальная последовательность поли(А) - инсерционный элемент экспрессионной конструкции.
первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - инсерционный элемент экспрессионной конструкции - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - сигнальная последовательность поли(А).
В некоторых предпочтительных вариантах реализации конструкции могут содержать последовательности нуклеиновых кислот, кодирующие несколько целевых белков, например, 2, 3, 4 или 5 целевых белков. Подходящие конструкции для экспрессии двух целевых белков представлены в следующих неограничивающих примерах:
инсерционный элемент экспрессионной конструкции - первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая (то есть внутренняя) промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - третья промоторная последовательность или IRES - последовательность нуклеиновой кислоты, кодирующая второй целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А);
первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - третья промоторная последовательность - интрон (необязательно) - последовательность нуклеиновой кислоты, кодирующая второй целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - инсерционный элемент экспрессионной конструкции;
инсерционный элемент экспрессионной конструкции - первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - третья промоторная последовательность - интрон (необязательно) последовательность нуклеиновой кислоты, кодирующая второй целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - инсерционный элемент экспрессионной конструкции;
первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - инсерционный элемент экспрессионной конструкции - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - WPRE - сигнальная последовательность поли(А) - третья промоторная последовательность или IRES - последовательность нуклеиновой кислоты, кодирующая второй целевой белок - WPRE - сигнальная последовательность поли(А);
инсерционный элемент экспрессионной конструкции - первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - третья промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая второй целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - инсерционный элемент экспрессионной конструкции;
инсерционный элемент экспрессионной конструкции - первая промоторная последовательность (необязательно в зависимости от того, содержит ли уже акцепторный сайт экзогенную промоторную последовательность) - последовательность селектируемого маркера - вторая промоторная последовательность - последовательность нуклеиновой кислоты, кодирующая первый целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - третья промоторная последовательность - интрон - последовательность нуклеиновой кислоты, кодирующая второй целевой белок - WPRE (необязательно) - сигнальная последовательность поли(А) - инсерционный элемент экспрессионной конструкции.
В некоторых предпочтительных вариантах реализации первый целевой белок представляет собой одну из тяжелой и легкой цепей антитела, и второй целевой белок представляет собой другую из тяжелой и легкой цепей антитела.
Однако любые подходящие целевые белки могут быть экспрессированы с применением клеток-хозяев, конструкций и систем согласно настоящему изобретению. Примеры целевых белков включают иммуноглобулины, одноцепочечные антитела, антикоагулянтные белки, белки-факторы крови, костные морфогенетические белки, конструированные белковые каркасы, ферменты, содержащие Fc слитые белки, факторы роста, гормоны, интерфероны, интерлейкины, антигены и тромболитические белки. В других предпочтительных вариантах реализации конструкции согласно настоящему изобретению могут применяться для экспрессии вирусных векторов. В этих вариантах реализации последовательность целевого белка, описанная в приведенных выше типичных векторах, заменена на последовательность нуклеиновой кислоты, кодирующую остов вирусного вектора. Вирусные векторы, которые могут быть включены в конструкции согласно настоящему изобретению, включают, не ограничиваясь указанными, ретровирусные векторы, лентивирусные векторы, аденовирусные векторы и векторы на основе AAV. В некоторых предпочтительных вариантах реализации ретровирусные векторы сами по себе включают последовательность нуклеиновой кислоты, кодирующую целевой белок, описанный выше, которая экспрессируется вектором. В некоторых особенно предпочтительных вариантах реализации целевой белок, который экспрессируется вектором, представляет собой последовательность антигена для применения в вакцине.
В некоторых предпочтительных вариантах реализации инсерционные элементы представляют собой элементы, которые находят применение в сочетании с системами транспозонов, интеграз, рекомбиназ или CRISPR или распознаются указанными системами. Подходящие инсерционные элементы включают, не ограничиваясь указанными, инвертированные концевые повторы, сайты интеграции для интегразы (att) и плечи гомологичной рекомбинации, которые в контексте конструкций, описанных в настоящей заявке, могут быть описаны как инсерционные элементы гомологичной рекомбинации.
В некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот согласно настоящему изобретению содержат управляемые транспозоном инсерционные элементы, предпочтительно инвертированные концевые повторы, которые распознаются транспозонами. В некоторых предпочтительных вариантах реализации инвертированные концевые повторы расположены как на 5'-, так и на 3'-концах конструкции. Транспозоны представляют собой мобильные генетические элементы, которые могут перемещаться или переставляться из одного положения в геноме в другое. Транспозиция внутри генома находится под контролем фермента транспозазы, кодируемого транспозоном. В данной области техники известны многие примеры транспозонов, включая, но не ограничиваясь указанными, системы транспозаз Tn5 (см., например, de la Cruz et al., J. Bact. 175: 6932-38 [1993], Tn7 (см., например, Craig, Curr. Topics Microbiol. Immunol. 204: 27-48 [1996]) и TnlO (см., например, Morisato and Kleckner, Cell 51:101-111 [1987), а также системы транспозазы PiggyBac, системы транспозазы «Спящая красавица», системы транспозазы Mosl, системы транспозазы Tol2 и системы транспозазы Leapin. Способность транспозонов интегрироваться в геномы была использована для создания транспозонных векторов (описанных, например, в патентах США №№5,719,055; 5,968,785; 5,958,775 и 6,027,722; все из которых включены в настоящую заявку посредством ссылки, а также векторов, поставляемых компаниями System Biosciences (Пало-Альто, Калифорния; система PiggyBac), Creative Biolabs (Ширли, Нью-Йорк; система «Спящая красавица») и ATUM (Ньюарк, Калифорния; система Leapin)).
Транспозиция включает упорядоченную последовательность событий: (1) специфичное в отношении последовательности связывание транспозазы с концевыми инвертированными повторами (IR), присутствующими на концах транспозона, (2) расщепление обеих цепей ДНК на каждом конце транспозона, (3) синапсис концов за счет взаимодействий транспозаза-транспозаза, (4) захват ДНК-мишени и (5) перенос цепи со встраиванием элемента в мишень.
Транспозазы являются представителями суперсемейства белков интеграз ретровирусов. Несмотря на структурное сходство в своих каталитических доменах, эти белки осуществляют реакции переноса фосфорила с различной специфичностью. Некоторые из них расщепляют только одну цепь ДНК, в то время как РНКаза Н расщепляет одну цепь РНК в гибридном дуплексе РНК: ДНК. Другие создают двухцепочечные разрывы ДНК, задействуя различные механизмы. Транспозазы бактериальных транспозонов Tn5 и TnlO осуществляют расщепление первой цепи путем гидролиза с образованием 3'-гидроксила (3'-ОН) на каждом конце элемента, в то время как вторая цепь расщепляется путем трансэтерификации с использованием этого 3'-ОН в качестве атакующего нуклеофила. В результате этого на каждом конце элемента образуется шпилька ДНК, которая гидролизуется транспозазой с регенерацией 3'-ОН, необходимого для переноса цепи. V(В)J-рекомбинация и транспозиция эукариотического элемента Hermes, представителя семейства hAT, протекают по сходному механизму за исключением того, что порядок расщепления цепей является обратным, и шпилька образуется на фланкирующей, а не на вырезаемой ДНК. Другой бактериальный транспозон, Tn7, использует TnsB для осуществления расщепления первой цепи и рекрутирует второй белок, TnsA, для расщепления не перенесенной цепи.
Поскольку транспозоны не являются инфекционными, то транспозонные векторы вводят в клетки-хозяева с помощью способов, известных в данной области техники (например, электропорации, липофекции или микроинъекции). Таким образом, можно подбирать соотношение транспозонных векторов и клеток-хозяев для обеспечения желаемой множественности заражения для получения клеток-хозяев с большим числом копий. Транспозонные векторы, подходящие для применения в настоящем изобретении, обычно содержат нуклеиновую кислоту, кодирующую целевой белок, помещенную между двумя инсерционными последовательностями транспозонов. Некоторые векторы также содержат последовательность нуклеиновой кислоты, кодирующую фермент транспозазу. В этих векторах одна из инсерционных последовательностей расположена между ферментом транспозазой и нуклеиновой кислотой, кодирующей целевой белок, таким образом, что она не встраивается в геном клетки-хозяина в процессе рекомбинации. В качестве альтернативы, фермент транспозаза может быть обеспечен подходящим способом (например, липофекцией или микроинъекцией).
В некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот согласно настоящему изобретению содержат управляемый рекомбиназой инсерционный элемент, который распознается рекомбиназой. Подходящие управляемые рекомбиназой инсерционные элементы включают, не ограничиваясь указанными, сайты интеграции (att), сайты LoxP и последовательности LTR MMLV.
В некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент представляет собой attB и применяется в сочетании с интегразой phiC31 (BioCat GmbH, Гейдельберг, Германия или System Biosciences, Пало-Альто, Калифорния)). Интеграза phiC31 представляет собой специфичную в отношении последовательности рекомбиназу, кодируемую в геноме бактериофага phiC31. Интеграза phiC31 опосредует рекомбинацию между двумя последовательностями длиной 34 пары оснований, называемыми сайтами интеграции (att), одна из которых находится в фаге, а другая - в хозяине. Было показано, что эта сериновая интеграза эффективно функционирует во многих различных типах клеток, включая клетки млекопитающих. В присутствии интегразы phiC31 донорная плазмида, содержащая attB, может быть однонаправленно интегрирована в геном-мишень посредством рекомбинации в сайтах, имеющих сходство последовательностей с природным сайтом attP (называемых сайтами псевдо-attP). Интеграза phiC31 может интегрировать плазмиду любого размера в виде одной копии и не требует участия кофакторов. Интегрированные трансгены стабильно экспрессируются и наследуются.
В других предпочтительных вариантах реализации инсерционный элемент представляет собой последовательность нуклеиновой кислоты, гомологичную сайту-мишени в хромосоме, такой как хромосома в клетке-хозяине, и используемую в сочетании с рекомбиназой или такими системами как CRISPR. Подходящие системы на основе рекомбиназы включают системы CRE-Lox, FLP-FRT и рекомбиназы фага лямбда. Как правило, последовательность нуклеиновой кислоты, которая гомологична сайту-мишени в хромосоме, будет иметь длину от 30 до 1000 оснований.
В некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент представляет собой последовательность lox. Рекомбинация с участием Cre-Lox представляет собой сайт-специфическую технологию с применением рекомбиназы, используемую для введения делеций, инсерций, транслокаций и инверсий в специфические сайты в ДНК клеток. Это позволяет нацеливать модификацию ДНК на конкретный тип клеток или запускать ее с помощью специфического внешнего стимула. Указанную технологию реализуют как в эукариотической, так и в прокариотической системах. Система рекомбинации Cre-lox нашла применение, в частности, у нейробиологов при изучении головного мозга, в котором сложные типы клеток и нейронные сети вместе формируют когнитивные функции и поведение. Система состоит из одного фермента, Cre-рекомбиназы, который осуществляет рекомбинацию пары коротких последовательностей-мишеней, называемых последовательностями Lox. Эта система может быть применена на практике без встраивания каких-либо дополнительных вспомогательных белков или последовательностей. Фермент Cre и исходный сайт Lox, называемый последовательностью LoxP, получены из бактериофага PI. См., например, Targeted integration of DNA using mutant lox sites in embryonic stem cells. Araki, et al. Nucleic Acids Res, Feb 1997, vol. 25, Issue 4, pp. 868-872; High-Resolution Labeling and Functional Manipulation of Specific Neuron Types in Mouse Brain by Cre-Activated Viral Gene Expression. Kuhlman, et al. PLos One, Apr 2008, Vol.3, e2005; When reverse genetics meets physiology: the use of site-specific recombinases in mice. Tranche, et al. FEBS Letters, Aug 2002, Vol.529, Issue 1, pp. 116-121.
В некоторых предпочтительных вариантах реализации управляемый рекомбиназой инсерционный элемент представляет собой последовательность FRT. Система рекомбинации FLP-FRT представляет собой другую технологию сайт-направленной рекомбинации, принципиально очень похожую на Cre-lox, использующую флиппазу (Flp) и короткий сайт-мишень для распознавания флиппазой (FRT), которые являются аналогами Cre и loxP, соответственно. См., например, Candice et al., Cre/loxP, Flp/FRT Systems and Pluripotent Stem Cell Lines (2012) Topics in Current Genetics, vol 23. Технология FLP-FRT может являться эффективной альтернативой Cre-lox, и она также была использована в сочетании с Cre-lox, что обеспечило одновременный контроль двух отдельных событий рекомбинации.
В других предпочтительных вариантах реализации конструкции нуклеиновых кислот согласно настоящему изобретению могут применяться в сочетании с системами гомологичной рекомбинации (FFDR) CRISPR. В этих системах инсерционные элементы HDR содержат плечи гомологии, которые гомологичны последовательностям-мишеням в геноме или взаимодействуют с ними за счет спаривания оснований. FTDR инициируется присутствием двухцепочечных разрывов (DSB) в ДНК. Система CRISPR/Cas9 предпочтительно применяется для создания целевых двухцепочечных разрывов с помощью последовательности направляющей РНК таким образом, что может быть встроена конструкция нуклеиновой кислоты согласно настоящему изобретению. См., например, Zhang et al., Efficient precise knockin with a double cut HDR donor after CRISPR/Cas9-mediated double-stranded DNA cleavage (2017) Genome Biol. 18:35; Mali et al., Cas9 as a versatile tool for engineering biology. Nature MethodslO, 957-963 (2013); Mali et al., RNA-Guided Human Genome Engineering via Cas9. Science339(6121), 823-826 (2013); Ran et al., Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell, 155(2), 479-480(2013). Подходящие последовательности направляющих РНК (gPHK) могут быть сконструированы, как известно в данной области техники. В некоторых предпочтительных вариантах реализации в системах CRISPR для HDR применяются либо одна, либо две направляющие последовательности. В случае применения одной последовательности направляющей РНК предпочтительно использовать нуклеазу, такую как нуклеаза Cas9, которая создает один двухцепочечный разрыв под контролем последовательности направляющей РНК. В случае применения двух направляющих последовательностей предпочтительно использовать никазу, которая может представлять собой мутированную нуклеазу Cas9, которая создает только одноцепочечные разрывы в последовательности ДНК-мишени под контролем каждой из последовательностей направляющей РНК. Одноцепочечные разрывы предпочтительно расположены в ступенчатом порядке на разных цепях (то есть смысловой и антисмысловой цепях) последовательности ДНК-мишени. Такое расположение, как правило, улучшает эффективность HDR.
В целом, «система CRISPR» относится в совокупности к транскриптам и другим элементам, участвующим в экспрессии или управлении активностью CRISPR-ассоциированных («Cas») генов, включая последовательности, кодирующие ген Cas, последовательность tracr (трансактивирующую CRISPR) (например, tracrPHK или частично активную tracrPHK), последовательность tracr-mate (охватывающую «прямой повтор» и процессируемый tracrPHK частичный прямой повтор в контексте эндогенной системы CRISPR), направляющую последовательность (также называемую «спейсер» в контексте эндогенной системы CRISPR) или другие последовательности и транскрипты из локуса CRISPR. В некоторых вариантах реализации один или более элементов системы CRISPR получены из системы CRISPR I типа, II типа или III типа. В некоторых вариантах реализации один или более элементов системы CRISPR получены из конкретного организма, содержащего эндогенную систему CRISPR, такого как Streptococcus pyogenes. В целом, система CRISPR характеризуется элементами, которые способствуют образованию комплекса CRISPR в сайте последовательности-мишени (также называемой протоспейсером в контексте эндогенной системы CRISPR). В контексте образования комплекса CRISPR «последовательность-мишень» относится к последовательности, направляющую последовательность для которой создают таким образом, чтобы она была комплементарной данной последовательность-мишени, при этом гибридизация между последовательностью-мишенью и направляющей последовательностью способствует образованию комплекса CRISPR. Не обязательно требуется полная комплементарность, если имеющаяся комплементарность достаточна для того, чтобы вызывать гибридизацию и способствовать образованию комплекса CRISPR. Последовательность-мишень может содержать любой полинуклеотид, такой как полинуклеотиды ДНК или РНК. В некоторых вариантах реализации последовательность-мишень расположена в ядре или цитоплазме клетки. В некоторых вариантах реализации последовательность-мишень может находиться внутри органеллы эукариотической клетки, например, митохондрии или хлоропласта. Последовательность или матрицу, которая может применяться для рекомбинации в целевой локус, содержащий последовательности-мишени, называют «редактирующей матрицей» или «редактирующим полинуклеотидом» или «редактирующей последовательностью». В аспектах настоящего изобретения экзогенный матричный полинуклеотид может называться редактирующей матрицей. В одном из аспектов настоящего изобретения рекомбинация представляет собой гомологичную рекомбинацию.
Как правило, в контексте эндогенной системы CRISPR образование комплекса CRISPR (содержащего направляющую последовательность, гибридизованную с последовательностью-мишенью и находящуюся в комплексе с одним или более белками Cas) приводит к расщеплению одной или обеих цепей в последовательности-мишени или вблизи от нее (например, в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар оснований от последовательности-мишени). Без привязки к какой-либо теории, последовательность tracr, которая может содержать всю последовательность tracr дикого типа или ее часть или состоять из них (например, приблизительно или более чем приблизительно 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов последовательности tracr дикого типа), также может составлять часть комплекса CRISPR, например, путем гибридизации по меньшей мере части последовательности tracr со всей последовательностью tracr-mate или ее частью, которая функционально связана с направляющей последовательностью. В некоторых вариантах реализации последовательность tracr обладает достаточной комплементарностью в отношении последовательности tracr-mate для гибридизации и участия в образовании комплекса CRISPR. Как и в случае последовательности-мишени, полагают, что не требуется полная комплементарность, если имеющаяся комплементарность достаточна для функционирования. В некоторых вариантах реализации последовательность tracr имеет по меньшей мере 50%, 60%, 70%, 80%, 90%, 95% или 99% комплементарности последовательностей вдоль длины последовательности tracr-mate при оптимальном выравнивании. В некоторых вариантах реализации один или более векторов, обеспечивающих экспрессию одного или более элементов системы CRISPR, вводят в клетку-хозяина таким образом, что экспрессия элементов системы CRISPR определяет образование комплекса CRISPR в одном или более сайтах-мишенях. Например, фермент Cas, направляющая последовательность, связанная с последовательностью tracr-mate, и последовательность tracr могут быть функционально связаны с отдельными регуляторными элементами на отдельных векторах. В качестве альтернативы, два или более элементов, экспрессируемых с участием одного и того же или разных регуляторных элементов, могут быть объединены в один вектор с использованием при этом одного или более дополнительных векторов, обеспечивающих какие-либо компоненты системы CRISPR, не включенные в первый вектор. Элементы системы CRISPR, которые объединены в одном векторе, могут быть расположены в любой подходящей ориентации, например, один элемент может быть расположен на 5'-конце (то есть «выше») или на 3'-конце (то есть «ниже») относительно второго элемента. Кодирующая последовательность одного элемента может быть расположена на той же или противоположной цепи, что и кодирующая последовательность второго элемента, и ориентирована в том же или противоположном направлении. В некоторых вариантах реализации один промотор управляет экспрессией транскрипта, кодирующего фермент CRISPR, и одной или более из направляющей последовательности, последовательности tracr-mate (необязательно функционально связанной с направляющей последовательностью) и последовательности tracr, встроенных в одну или более интронных последовательностей (например, каждая в свой интрон, две или более в по меньшей мере один интрон или все в один интрон). В некоторых вариантах реализации фермент CRISPR, направляющая последовательность, последовательность tracr-mate и последовательность tracr функционально связаны и экспрессируются с одного и того же промотора.
Неограничивающие примеры белков Cas, подходящих для применения в настоящем изобретении, включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9 (также известный как Csn1 и Csxl2), CaslO, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csxl7, Csxl4, CsxlO, Csxl6, CsaX, Csx3, Csx1, Csxl5, Csf1, Csf2, Csf3, Csf4, их гомологи или их модифицированные варианты. Эти ферменты являются известными; например, аминокислотную последовательность белка Cas9 S. pyogenes можно найти в базе данных SwissProt под номером доступа Q99ZW2. В некоторых вариантах реализации немодифицированный фермент CRISPR обладает активностью расщепления ДНК, такой как активность Cas9. В некоторых вариантах реализации фермент CRISPR представляет собой Cas9 и может представлять собой Cas9 из S. pyogenes или S. pneumoniae. В некоторых вариантах реализации фермент CRISPR направляет расщепление одной или обеих цепей в положении последовательности-мишени, например, в пределах последовательности-мишени и/или в пределах комплемента последовательности-мишени. В некоторых вариантах реализации фермент CRISPR направляет расщепление одной или обеих цепей в пределах приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 50, 100, 200, 500 или более пар оснований от первого или последнего нуклеотида последовательности-мишени. В некоторых вариантах реализации вектор кодирует фермент CRISPR, который является мутированным по сравнению с соответствующим ферментом дикого типа таким образом, что мутированный фермент CRISPR не обладает способностью расщеплять одну или обе цепи целевого полинуклеотида, содержащего последовательность-мишень. Например, замена аспарагиновой кислоты на аланин (D10A) в каталитическом домене RuvC I Cas9 из S. pyogenes превращает Cas9 из нуклеазы, которая расщепляет обе цепи, до никазы (расщепляет одну цепь). Другие примеры мутаций, которые превращают Cas9 в никазу, включают, не ограничиваясь указанными, Н840А, N854A и N863A. В аспектах настоящего изобретения никазы могут применяться для редактирования генома посредством гомологичной рекомбинации.
В некоторых предпочтительных вариантах реализации управляемый HDR инсерционный элемент содержит генетически нейтральный локус AAVS1 и применяется в сочетании с интегразой Rep 78. В особенно предпочтительных вариантах реализации управляемый HDR инсерционный элемент содержит плечи гомологии, которые взаимодействуют за счет спаривания оснований с генетически нейтральным локусом AAVS1. Белок Rep 78 аденоассоциированного вируса серотипа 2 (AAV2) представляет собой специфичную в отношении цепи эндонуклеазу (никазу), которая способствует сайт-специфической интеграции трансгенных последовательностей, несущих плечи гомологии, соответствующие генетически нейтральному локусу AAVS1. См., например, Ramachandra et al., Efficient recombinase-mediated cassette exchange at the AAVS1 locus in human embryonic stem cells using baculoviral vectors (2011) Nucleic Acids Research, 39(16):e107; WO 1998027207).
Как указано выше, в некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот согласно настоящему изобретению содержат первую и вторую промоторные последовательности. Первая и вторая промоторные последовательности могут быть одинаковыми или различными. Подходящие первая и вторая промоторные последовательности включают, не ограничиваясь указанными, промотор LTR MMLV, промотор LTR MoMuSV, промотор LTR респираторно-синцитиального вируса (RSV), промотор SIN LTR, промотор SV40, немедленно-ранний промотор цитомегаловируса (CMV), промотор тимидинкиназы вируса простого герпеса (HSV), промотор альфа-лактальбумина, промотор металлотионеина-I мыши, промотор дигидрофолатредуктазы, промотор b-актина, промотор фосфоглицеринкиназы (PGK) и промоторные последовательности EF 1а и их комбинации. В некоторых предпочтительных вариантах реализации первая промоторная последовательность не является ретровирусным промотором LTR, то есть первый промотор представляет собой промоторную последовательность, отличную от ретровирусной промоторной последовательности LTR. Однако, когда промотор является ретровирусной промоторной последовательностью, он может представлять собой промоторную последовательность самоинактивирующегося (SIN) LTR. См., например, также находящуюся на рассмотрении заявку PCT/US2019/064423, которая полностью включена в настоящую заявку посредством ссылки. Подходящие промоторы Sin LTR известны в данной области техники и получены путем удаления всей области U3 LTR или ее части.
Как описано в PCT/US2019/064423, в некоторых предпочтительных вариантах реализации первый промотор, который управляет экспрессией селективного маркера, представляет собой слабый промотор. В некоторых предпочтительных вариантах реализации слабый промотор представляет собой промотор, предпочтительно конститутивный промотор, который, будучи функционально связанным с последовательностью селектируемого маркера, обладает активностью, равной активности или меньшей чем активность промотора SIN LTR, в целевом хозяине (например, клетке СНО). В других предпочтительных вариантах реализации слабый промотор представляет собой промотор, предпочтительно конститутивный промотор, который, будучи функционально связанным с последовательностью селектируемого маркера, обладает активностью, равной активности или меньшей чем активность промотора убиквитина С (UBC) человека, в целевом хозяине (например, клетке СНО). Подходящие способы оценки силы промоторов известны в данной области техники. См., например, Dandindorj et al. (2014) A Comparative Analysis of Constitutive Promoters Located in Adeno-Associated Viral Vectors, PLoS One 9(8): el06472; Zhang and Baum (2005) Evaluation of Viral and Mammalian Promoters for Use in Gene Delivery to Salivary Glands Mol. Ther. 12(3):528-536; Qin et al. (2010) Systematic Comparison of Constitutive Promoters and the Doxycycline-Inducible Promoter PLoS 5(5): el 0611; Jeyaseelan et al. (2001) Real-time detection of gene promoter activity: quantitation of toxin gene transcription, Nucleic Acids Research. 29 (12): 58e-58. В некоторых вариантах реализации слабые промоторы были изменены с целью снижения промоторной активности. Соответственно, в некоторых предпочтительных вариантах реализации настоящего изобретения предложен(ы) вектор(ы) для экспрессии целевого белка, содержащий(ие) последовательность нуклеиновой кислоты, кодирующую селектируемый маркер, функционально связанную с первой последовательностью слабого промотора или промоторной последовательностью, которая была изменена с целью снижения промоторной активности по сравнению с вариантом первой промоторной последовательности, не имеющим изменений, или дикого типа, и последовательность нуклеиновой кислоты, кодирующую целевой белок, функционально связанную со второй промоторной последовательностью. Одним из таких примеров является промоторная последовательность SIN LTR. Другие промоторные последовательности, описанные выше, также могут быть изменены с целью снижения активности и обеспечения слабого промотора, либо слабый промотор может представлять собой природный слабый промотор, такой как промотор UBC.
В некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот содержат селектируемый маркер. Подходящие селектируемые маркеры включают, не ограничиваясь указанными, глутаминсинтетазу (GS), дигидрофолатредуктазу (DHFR) и тому подобное. Эти гены описаны в патентах США №№5,770,359; 5,827,739; 4,399,216; 4,634,665; 5,149,636 и 6,455,275; все из которых включены в настоящую заявку посредством ссылки. В некоторых предпочтительных вариантах реализации используемый селектируемый маркер совместим с линией клеток-хозяев, которая является дефектной по продуцированию фермента, кодируемого последовательностью нуклеиновой кислоты селектируемого маркера. Подходящие линии клеток-хозяев более подробно описаны ниже. В других вариантах реализации селектируемый маркер представляет собой маркер устойчивости к антибиотику, то есть ген, продуцирующий белок, который обеспечивает экспрессирующие этот белок клетки устойчивостью к антибиотику. Подходящие маркеры устойчивости к антибиотикам включают гены, которые обеспечивают устойчивость кнеомицину (ген устойчивости к неомицину (neo)), гигромицину (ген фосфотрансферазы гигромицина В), пуромицину (N-ацетилтрансфераза пуромицина) и тому подобное.
В других вариантах реализации настоящего изобретения, где желательна секреция целевого белка, конструкции нуклеиновых кислот содержат последовательность сигнального пептида, функционально связанную с целевым белком. Специалистам в данной области техники известны последовательности некоторых подходящих сигнальных пептидов, включая, но не ограничиваясь указанными, последовательности, полученные из тканевого активатора плазминогена, гормона роста человека, лактоферрина, альфа-казеина и альфа-лактальбумина.
В других вариантах реализации настоящего изобретения конструкции нуклеиновых кислот содержат элемент экспорта РНК (см., например, патенты США №№5,914,267; 6,136,597 и 5,686,120; и WO 99/14310, все из которых включены в настоящую заявку посредством ссылки), расположенный либо на 3'-, либо на 5'-конце относительно последовательности нуклеиновой кислоты, кодирующей целевой белок. Предполагается, что применение элементов экспорта РНК обеспечивает высокие уровни экспрессии целевого белка без включения сигналов сплайсинга или интронов в последовательность нуклеиновой кислоты, кодирующую целевой белок.
В других вариантах реализации конструкции нуклеиновых кислот содержат по меньшей мере одну последовательность участка внутренней посадки рибосомы (IRES). Доступные последовательности некоторых подходящих IRES включают, не ограничиваясь указанными, последовательности, полученные из вируса ящура (FDV), вируса энцефаломиокардита и полиовируса. Последовательность IRES может быть помещена между двумя транскрипционными единицами (например, нуклеиновыми кислотами, кодирующими различные целевые белки или субъединицы мультисубъединичного белка, такого как антитело) с образованием полицистронной последовательности таким образом, что две транскрипционные единицы транскрибируются с одного и того же промотора.
Настоящее изобретение не ограничивается экспрессией какого-либо конкретного целевого белка. В некоторых предпочтительных вариантах реализации целевой белок выбран из группы, состоящей из содержащего Fc слитого белка, фермента, продукта слияния с альбумином, фактора роста, белкового рецептора, одноцепочечного антитела (scFv), одноцепочечного Fc (scFv-Fc), диатела и миниантитела (scFv-СН3), Fab, одноцепочечного Fab (scFab), тяжелой цепи иммуноглобулина и легкой цепи иммуноглобулина и других антигенсвязывающих белков.
В некоторых предпочтительных вариантах реализации конструкции нуклеиновых кислот встроены в вектор для экспрессии нуклеиновых кислот. Векторы включают, не ограничиваясь указанными, молекулы нуклеиновой кислоты, которые являются одноцепочечными, двухцепочечными или частично двухцепочечными; молекулы нуклеиновой кислоты, которые содержат один или более свободных концов, не содержат свободных концов (например, кольцевые); молекулы нуклеиновой кислоты, которые содержат ДНК, РНК или и то, и другое; и другие разновидности полинуклеотидов, известные в данной области техники. Один из типов векторов представляет собой «плазмиду», которая относится к кольцевой двухцепочечной ДНК в виде петли, в которую могут быть встроены дополнительные сегменты ДНК, например, с помощью стандартных методик молекулярного клонирования. Другой тип векторов представляет собой вирусный вектор, в котором присутствуют полученные из вирусов последовательности ДНК или РНК для упаковки в вирус (например, ретровирусы, дефектные по репликации ретровирусы, аденовирусы, дефектные по репликации аденовирусы и аденоассоциированные вирусы). Вирусные векторы также содержат полинуклеотиды, переносимые вирусом для трансфекции в клетку-хозяина. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, содержащие бактериальную точку начала репликации, и эписомальные векторы для экспрессии в клетках млекопитающих). Другие векторы (например, неэписомальные векторы для экспрессии в клетках млекопитающих) интегрируются в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. В настоящей заявке такие векторы обозначены термином «векторы экспрессии». Векторы экспрессии, широко используемые в методиках рекомбинантной ДНК, часто представлены в форме плазмид.
Соответственно, подходящие векторы для экспрессии нуклеиновых кислот включают, не ограничиваясь указанными, транспозонные векторы, описанные выше, а также плазмидные векторы, ретровирусные векторы, лентивирусные векторы, векторы на основе AAV, фаговые векторы и т.д.). Предполагается, что может быть использован любой вектор, если он способен к репликации и является жизнеспособным в организме-хозяине. В предпочтительных вариантах реализации векторы представляют собой векторы для экспрессии в клетках млекопитающих, которые содержат, наряду с другими элементами, описанными в настоящей заявке, точку начала репликации, подходящие промотор и энхансер, а также любые необходимые сайты связывания рибосом, сайты полиаденилирования, донорные и акцепторные сайты сплайсинга, последовательности терминации транскрипции и 5'-фланкирующие нетранскрибируемые последовательности.
Подходящие плазмидные векторы, которые могут быть адаптированы для встраивания конструкций нуклеиновых кислот согласно настоящему изобретению, включают специфические плазмидные системы для транспозонных векторов, системы FLP-FLT, системы Cre-lox, системы CRISPR-Cas9, системы рекомбиназ и системы интеграз, а также плазмидные векторы, полученные из pCIneo, pVAX1, рАСТ, плазмид Gateway, pAdvantage, pBIND, pG51uc, pTNT, pTarget, pCat3, pSI, pCMV, pSV и тому подобное.
В некоторых вариантах реализации векторы представляют собой ретровирусные векторы. Наиболее часто применяемые рекомбинантные ретровирусные векторы получают из амфотропного вируса лейкоза мышей Молони (MoMLV) (см., например, Miller and Baltimore Mol. Cell. Biol. 6:2895 [1986]). Система MoMLV имеет несколько преимуществ: 1) этот специфический ретровирус может инфицировать многие различные типы клеток, 2) для получения рекомбинантных вирусных частиц MoMLV доступны устойчивые линии упаковывающих клеток и 3) переносимые гены стабильно интегрированы в хромосому клетки-мишени. Устойчивые векторные системы на основе MoMLV содержат ДНК-вектор, содержащий небольшую часть ретровирусной последовательности (например, вирусный длинный концевой повтор, или «LTR», и сигнал упаковки «psi»), и линию упаковывающих клеток. Переносимый ген встраивают в ДНК-вектор. Вирусные последовательности, присутствующие на ДНК-векторе, обеспечивают сигналы, необходимые для встраивания или упаковки РНК вектора в вирусную частицу и для экспрессии встроенного гена. Линия упаковывающих клеток обеспечивает белки, необходимые для сборки частиц (Markowitz et ak, J. Virol. 62: 1120 [1988]).
В некоторых предпочтительных вариантах реализации ретровирусные векторы представляют собой псевдотипированные векторы, и в них используется, например, белок G вируса везикулярного стоматита (VSV) в качестве мембрано-ассоциированного белка. В отличие от ретровирусных белков оболочки, которые связываются со специфическим белковым рецептором на поверхности клетки для проникновения в клетку, белок G VSV взаимодействует с фосфолипидным компонентом плазматической мембраны (Mastromarino et al., J. Gen. Virol. 68:2359 [1977]). Поскольку проникновение VSV в клетку не зависит от наличия специфических белковых рецепторов, то VSV имеет чрезвычайно широкий спектр хозяев. Псевдотипированные ретровирусные векторы, несущие белок G VSV, имеют измененный спектр хозяев, характерный для VSV (то есть они могут инфицировать клетки почти всех видов позвоночных, беспозвоночных и насекомых). Важно отметить, что G-псевдотипированные ретровирусные векторы на основе VSV могут быть сконцентрированы в 2000 раз или больше посредством ультрацентрифугирования без значительной потери инфекционности (Bums et al. Proc. Natl. Acad. Sci. USA 90:8033 [1993]).
В некоторых предпочтительных вариантах реализации векторы представляют собой лентивирусные векторы. Лентивирусы (например, вирус инфекционной анемии лошадей, вирус козьего артрита-энцефалита, вирус иммунодефицита человека) представляют собой подсемейство ретровирусов, которые способны интегрироваться в неделящиеся клетки. Лентивирусный геном и провирусная ДНК содержат три гена, обнаруженные во всех ретровирусах: gag, pol и env, которые фланкированы двумя последовательностями LTR. Ген gag кодирует внутренние структурные белки (например, матричные, капсидные и нуклеокапсидные белки); ген pol кодирует белки обратную транскриптазу, протеазу и интегразу; и ген env кодирует гликопротеины вирусной оболочки. 5'- и 3'-LTR регулируют транскрипцию и полиаденилирование вирусных РНК. Дополнительные гены в лентивирусном геноме включают гены vif, vpr, tat, rev, vpu, nef и vpx.
Различные лентивирусные векторы и линии упаковывающих клеток известны в данной области техники и находят применение в настоящем изобретении (см., например, патенты США №№5,994,136 и 6,013,516, оба из которых включены в настоящую заявку посредством ссылки). Кроме того, белок G VSV также был использован для псевдотипирования ретровирусных векторов на основе вируса иммунодефицита человека (ВИЧ) (Naldini et al., Science 272:263 [1996]). Таким образом, белок G VSV может применяться для получения различных псевдотипированных ретровирусных векторов, и его использование не ограничивается векторами на основе MoMLV. Лентивирусные векторы также могут быть модифицированы, как описано выше, таким образом, что они будут содержать различные регуляторные последовательности (например, последовательности сигнальных пептидов, элементы экспорта РНК и TRES). После получения лентивирусных векторов они могут применяться для трансфекции клеток-хозяев, как описано выше для ретровирусных векторов.
В некоторых предпочтительных вариантах реализации векторы представляют собой векторы на основе аденоассоциированного вируса (AAV). Геном AAV состоит из линейной одноцепочечной молекулы ДНК, которая содержит приблизительно 4680 оснований. Геном содержит инвертированные концевые повторы (ITR) на каждом конце, которые функционируют в цис-положении как точки начала репликации ДНК и как сигналы упаковки для вируса. Внутренняя неповторяющаяся часть генома содержит две большие открытые рамки считывания, известные как области rep и cap AAV, соответственно. Эти области кодируют вирусные белки, участвующие в репликации и упаковке вириона. Из области rep AAV синтезируется семейство из по меньшей мере четырех вирусных белков, названных в соответствии с их кажущейся молекулярной массой, Rep 78, Rep 68, Rep 52 и Rep 40. Область cap AAV кодирует по меньшей мере три белка, VP1, VP2 и VP3 (подробное описание генома AAV см., например, в источниках Muzyczka, Current Topics Microbiol. Immunol. 158:97-129 [1992]; Kotin, Human Gene Therapy 5:793-801 [1994]).
AAV требует совместного инфицирования неродственным вирусом-помощником, таким как аденовирус, герпесвирус или вирус осповакцины, для того чтобы произошло продуктивное заражение. В отсутствие такой совместной инфекции AAV переходит в латентное состояние путем встраивания его генома в хромосому клетки-хозяина. Последующее инфицирование вирусом-помощником приводит к высвобождению интегрированной копии, которая затем может реплицироваться с образованием инфекционного вирусного потомства. В отличие от не псевдотипированных ретровирусов, AAV имеет широкий спектр хозяев и способен реплицироваться в клетках из любого вида при условии, что существует совместная инфекция с вирусом-помощником, который также будет размножаться в этом виде. Таким образом, например, AAV человека будет реплицироваться в клетках собаки, совместно инфицированных аденовирусом собак. Кроме того, в отличие от ретровирусов, AAV не ассоциирован с каким-либо заболеванием человека или животных; по-видимому, не изменяет биологические свойства клетки-хозяина при интеграции и способен интегрироваться в неделящиеся клетки. Недавно также было обнаружено, что AAV способен к сайт-специфической интеграции в геном клетки-хозяина.
В свете описанных выше свойств был разработан ряд рекомбинантных векторов на основе AAV для доставки генов (см., например, патенты США №№5,173,414; 5,139,941; источники WO 92/01070 и WO 93/03769, оба из которых включены в настоящую заявку посредством ссылки; Lebkowski et al., Molec. Cell. Biol. 8:3988-3996 [1988]; Carter, Current Opinion in Biotechnology 3:533-539 [1992]; Muzyczka, Current Topics in Microbiol and Immunol. 158:97-129 [1992]; Kotin, (1994) Human Gene Therapy 5:793-801; Shelling and Smith, Gene Therapy 1:165-169 [1994]; и Zhou et al., J. Exp. Med. 179:1867-1875 [1994]).
Рекомбинантные вирионы AAV могут быть получены в подходящей клетке-хозяине, трансфицированной как плазмидой-помощником на основе AAV, так и вектором на основе AAV. Плазмида-помощник на основе AAV обычно содержит области, кодирующие rep и cap AAV, но не содержит ITR AAV. Соответственно, плазмида-помощник самостоятельно не способна ни к репликации, ни к упаковке. Вектор на основе AAV, как правило, содержит выбранный целевой ген, ограниченный ITR AAV, которые обеспечивают функции репликации и упаковки вируса. И плазмиду-помощника, и вектор на основе AAV, несущий выбранный ген, вводят в подходящую клетку-хозяина посредством временной трансфекции. Затем трансфицированную клетку инфицируют вирусом-помощником, таким как аденовирус, который трансактивирует присутствующие на плазмиде-помощнике промоторы AAV, которые управляют транскрипцией и трансляцией областей rep и cap AAV. Образуются рекомбинантные вирионы AAV, несущие выбранный ген, которые могут быть очищены от препарата. После получения векторов на основе AAV они могут быть использованы для трансфекции (см., например, патент США №5,843,742, включенный в настоящую заявку посредством ссылки) клеток-хозяев при желаемой множественности заражения для получения клеток-хозяев с большим числом копий. Специалистам в данной области техники будет понятно, что векторы на основе AAV также могут быть модифицированы, как описано выше, таким образом, что они будут содержать различные регуляторные последовательности (например, последовательности сигнальных пептидов, элементы экспорта РНК и IRES).
В некоторых вариантах реализации настоящего изобретения предложены клетки-хозяева и культура клеток-хозяев, где клетки-хозяева экспрессируют целевой белок из конструкций нуклеиновых кислот, описанных выше. В предпочтительном варианте реализации клетки-хозяева представляют собой клетки-хозяева млекопитающих. Ряд линий клеток-хозяев млекопитающих известен в данной области техники. В целом, эти клетки-хозяева способны к росту и выживанию при помещении либо в монослойную культуру, либо в суспензионную культуру в среде, содержащей подходящие питательные вещества и факторы роста, как более подробно описано ниже. Как правило, указанные клетки способны экспрессировать и секретировать большие количества конкретного целевого белка в культуральную среду. Примеры подходящих клеток-хозяев млекопитающих включают, не ограничиваясь указанными, клетки яичника китайского хомячка (СНО-K1, АТСС СС1-61); эпителиальные клетки молочной железы крупного рогатого скота (АТСС CRL 10274; эпителиальные клетки молочной железы крупного рогатого скота); линию клеток почки обезьяны CV1, трансформированную SV40 (COS-7, АТСС CRL 1651); линию эмбриональных клеток почки человека (293 или клетки 293, субклонированные для роста в суспензионной культуре; см., например, Graham et al., J. Gen Virol., 36:59 [1977]); клетки почки детеныша хомяка (BHK, АТСС CCL 10); клетки Сертоли мыши (ТМ4, Mather, Biol. Reprod. 23:243-251 [1980]); клетки почки обезьяны (CV1 АТСС CCL 70); клетки почки африканской зеленой мартышки (VERO-76, АТСС CRL-1587); клетки карциномы шейки матки человека (HELA, АТСС CCL 2); клетки почки собаки (MDCK, АТСС CCL 34); клетки печени крысы Buffalo (BRL 3 А, АТСС CRL 1442); клетки легкого человека (W138, АТСС CCL 75); клетки печени человека (Hep G2, НВ 8065); клетки опухоли молочной железы мыши (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383:44-68 [1982]); клетки MRC 5; клетки FS4; фибробласты крысы (клетки 208F); клетки MDBK (клетки почки крупного рогатого скота); клетки CAP (CEVEC's Amniocyte Production); и линию клеток гепатомы человека (Hep G2).
В некоторых особенно предпочтительных вариантах реализации клетки-хозяева модифицированы таким образом, что они становятся дефектными, либо они по своей природе являются дефектными, по ферментативной активности, которая необходима для роста или выживания клеток в присутствии селектируемого агента и которая обеспечивается селектируемым маркером. Например, клетки яичника китайского хомячка (СНО) были модифицированы таким образом, что они стали дефектными по GS. В некоторых предпочтительных вариантах реализации, где вектор содержит селектируемый маркер GS, линия клеток-хозяев является дефектной по GS. В некоторых особенно предпочтительных вариантах реализации дефектная по GS линия клеток-хозяев представляет собой линию клеток CHOZN® GS, доступную от компании Merck KGaA. В других вариантах реализации, где селектируемый маркер представляет собой, например, DHFR, клеточная линия предпочтительно может быть дефектной по активности DHFR (то есть DHFR). Подходящие линии клеток DHFR- включают, не ограничиваясь указанными, CHO-DG44 и ее производные.
Конструкции нуклеиновых кислот и векторы согласно настоящему изобретению могут быть введены в клетки-хозяева любыми подходящими способами, такими как трансфекция, трансформация или трансдукция. В некоторых вариантах реализации после трансфекции или трансдукции клетки оставляют для размножения, а затем трипсинизируют и повторно высевают. Затем отбирают отдельные колонии для получения клонально отобранных клеточных линий. В других вариантах реализации клонально отобранные клеточные линии подвергают скринингу с помощью Саузерн-блоттинга или ПЦР-анализов для проверки того, что произошло желаемое количество событий интеграции. Также предполагается, что клональная селекция позволяет идентифицировать лучшие продуцирующие белок клеточные линии. В других вариантах реализации клетки не являются клонально отобранными после трансфекции.
В некоторых вариантах реализации клетки-хозяева трансфицируют векторами, кодирующими различные целевые белки. Векторы, кодирующие различные целевые белки, могут применяться для одновременной трансфекции клеток (например, клетки-хозяева подвергают воздействию раствора, содержащего векторы, кодирующие различные целевые белки), или трансфекция может быть последовательной (например, клетки-хозяева сначала трансфицируют вектором, кодирующим первый целевой белок, выжидают некоторый промежуток времени и затем клетки-хозяева трансфицируют вектором, кодирующим второй целевой белок). В некоторых предпочтительных вариантах реализации клетки-хозяева трансфицируют интегрирующим вектором, кодирующим первый целевой белок, при этом отбирают клеточные линии с высоким уровнем экспрессии, содержащие много интегрированных копий интегрирующего вектора (например, путем клонального отбора), и отобранную клеточную линию трансфицируют интегрирующим вектором, кодирующим второй целевой белок. Этот процесс можно повторять для введения нескольких целевых белков. В некоторых вариантах реализации можно регулировать множественность заражения (например, увеличивать или уменьшать) для повышения или снижения экспрессии целевого белка. Аналогичным образом, для изменения экспрессии целевых белков можно применять различные промоторы. Предполагается, что эти способы трансфекции могут применяться для конструирования линий клеток-хозяев, обеспечивающих полный экзогенный метаболический путь, или для получения клеток-хозяев с повышенной способностью к процессингу белков (например, клетки-хозяева могут быть обеспечены ферментами, необходимыми для посттрансляционной модификации).
В некоторых вариантах реализации настоящего изобретения, после трансформации подходящего штамма-хозяина и выращивания штамма-хозяина до требуемой плотности клеток в среде, целевой белок секретируется в процессе культивирования клеток-хозяев. В некоторых предпочтительных вариантах реализации, где применяются амплифицируемые маркеры, предусмотрена культура трансдуцированных клеток-хозяев в среде, содержащей ингибитор гена. Подходящие ингибиторы включают, не ограничиваясь указанными, метотрексат для ингибирования DHFR и метионинсульфоксимин (Msx) или фосфинотрицин для ингибирования GS. Предполагается, что по мере увеличения концентраций этих ингибиторов в системе с клеточной культурой отбираются клетки с большими числами копий амплифицируемого маркера (и, таким образом, генов или целевых генов) или клетки, содержащие инсерции, обеспечивающие более высокую продуктивность.
Соответственно, клетки-хозяева, содержащие векторы, описанные выше, предпочтительно культивируют в соответствии со способами, известными в данной области техники. Подходящие условия для культивирования клеток млекопитающих хорошо известны в данной области техники (см., например, J. Immunol. Methods (1983) 56:221-234 [1983], Animal Cell Culture: A Practical Approach 2nd Ed., Rickwood, D. and Hames, B.D., eds. Oxford University Press, New York [1992]).
Культуры клеток-хозяев согласно настоящему изобретению получают в среде, подходящей для конкретной культивируемой клетки. Коммерчески доступные среды, такие как среда ActiPro (HyClone), среда ExCell Advanced Fed Batch (SAFC), среда Хэма F10 (Sigma, Сент-Луис, Миссури), минимальная питательная среда (MEM, Sigma), RPMI-1640 (Sigma) и модифицированная по способу Дульбекко среда Игла (DMEM, Sigma), являются типичными питательными растворами. Подходящие среды также описаны в патентах США №№4,767,704; 4,657,866; 4,927,762; 5,122,469; 4,560,655; и источниках WO 90/03430 и WO 87/00195; раскрытия которых включены в настоящую заявку посредством ссылки. Любая из этих сред при необходимости может содержать в качестве добавок сыворотку, гормоны и/или другие факторы роста (такие как инсулин, трансферрин или эпидермальный фактор роста), соли (такие как хлорид натрия, кальций, магний и фосфат), буферы (такие как HEPES), нуклеозиды (такие как аденозин и тимидин), антибиотики (такие как гентамицин), микроэлементы (определяемые как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне), липиды (такие как линолевая кислота или другие жирные кислоты) и их подходящие носители и глюкозу или эквивалентный источник энергии. В некоторых предпочтительных вариантах реализации, где применяются селектируемые маркеры, такие как, например, GS, среда не будет содержать глутамина. Любые другие необходимые добавки также могут быть включены в подходящих концентрациях, которые известны специалистам в данной области техники.
Настоящее изобретение также предусматривает применение различных систем для культивирования трансфицированных клеток-хозяев (например, чашек Петри, 96-луночных планшетов, роллерных флаконов и биореакторов). Например, трансфицированные клетки-хозяева можно культивировать в перфузионной системе. Перфузионное культивирование относится к обеспечению непрерывного потока культуральной среды через культуру, в которой поддерживается высокая плотность клеток. Клетки являются суспендированными и не требуют твердой подложки для роста.
Как правило, должны непрерывно обеспечиваться свежие питательные вещества с одновременным удалением токсичных метаболитов и, в идеале, селектируемым удалением мертвых клеток. Способы фильтрования, улавливания и микрокапсулирования подходят для обновления культуральной среды с достаточной скоростью.
В качестве другого примера, в некоторых вариантах реализации может применяться процесс культивирования с периодическим добавлением питательной среды. В предпочтительном процессе культивирования клеток-хозяев млекопитающего с периодическим добавлением питательной среды клетки и культуральную среду сначала помещают в емкость для культивирования, а дополнительные питательные вещества для культивирования добавляют, непрерывно или с дискретными приращениями, в культуру в процессе культивирования с периодическим сбором клеток и/или продукта до окончания культивирования или без этого сбора. Культивирование с периодическим добавлением питательной среды может включать, например, полунепрерывное культивирование с периодическим добавлением питательной среды, при котором всю культуру (включая клетки и среду) периодически удаляют и заменяют свежей средой. Культивирование с периодическим добавлением питательной среды отличается от простого периодического культивирования, при котором все компоненты для культивирования клеток (включая клетки и все питательные вещества для культивирования) помещают в емкость для культивирования в начале процесса культивирования. Культивирование с периодическим добавлением питательной среды также можно отличить от перфузионного культивирования, поскольку супернатант не удаляют из емкости для культивирования в ходе процесса (при перфузионном культивировании клетки удерживают в культуре, например, посредством фильтрования, капсулирования, прикрепления к микроносителям и т.д., а культуральную среду непрерывно или периодически вводят в емкость для культивирования и удаляют из нее). В некоторых особенно предпочтительных вариантах реализации процессы культивирования с периодическим добавлением питательной среды проводят в роллерных флаконах.
Кроме того, клетки культуры могут быть размножены в соответствии с любой схемой или обычной процедурой, которые могут быть подходящими для конкретной клетки-хозяина и конкретного предполагаемого плана получения продукта. Таким образом, настоящее изобретение предусматривает одностадийный или многостадийный процесс культивирования. При одностадийном культивировании клетки-хозяева инокулируют в культуральную среду, и способы согласно настоящему изобретению применяют в течение единственной фазы получения клеточной культуры. В качестве альтернативы предусмотрено многостадийное культивирование. При многостадийном культивировании клетки можно культивировать в течение нескольких стадий или фаз. Например, клетки можно выращивать на первой стадии, или в фазе роста культуры, при этом клетки, предположительно извлеченные из хранилища, инокулируют в среду, подходящую для стимулирования роста и высокой жизнеспособности. Клетки можно поддерживать в фазе роста в течение подходящего периода времени путем добавления свежей среды к культуре клеток-хозяев.
Для усиления роста клеток млекопитающих в фазе роста клеточной культуры разрабатывают условия культивирования клеток с периодическим добавлением питательной среды или непрерывного культивирования клеток. В фазе роста клетки выращивают в условиях и в течение периода времени, которые обеспечивают максимальный рост. Условия культивирования, такие как температура, рН, растворенный кислород (dO2) и тому подобное, представляют собой условия, используемые с конкретным хозяином, и они будут понятны специалисту в данной области техники. Как правило, рН доводят до уровня между приблизительно 6,5 и 7,5 с использованием либо кислоты (например, СС), либо основания (например, Na2CO3 или NaOH). Подходящий температурный диапазон для культивирования клеток млекопитающих, таких как клетки СНО, составляет от приблизительно 30°С до 38°С, а подходящий dC составляет 5-90% насыщения воздуха.
После фазы получения полипептида целевой полипептид выделяют из культуральной среды с использованием методик, хорошо отработанных в данной области техники. Целевой белок предпочтительно выделяют из культуральной среды в виде секретируемого полипептида (например, секреция целевого белка направляется последовательностью сигнального пептида), хотя он также может быть выделен из лизатов клеток-хозяев. В качестве первой стадии культуральную среду или лизат центрифугируют для удаления частиц клеточного остатка. Затем полипептид очищают от загрязняющих растворимых белков и полипептидов, при этом следующие процедуры являются примерами подходящих процедур очистки: фракционирование на иммуноаффинных или ионообменных колонках; осаждение этанолом; обращенно-фазовая ВЭЖХ; хроматография на диоксиде кремния или на катионообменной смоле, такой как ДЭАЭ; хроматофокусирование; ДСН-ПААГ-электрофорез; осаждение сульфатом аммония; гель-фильтрации с использованием, например, сефадекса G-75; и колоночная хроматография на сефарозе с белком A (protein A Sepharose) для удаления загрязняющих веществ, таких как IgG. Ингибитор протеазы, такой как фенилметилсульфонилфторид (PMSF), также может быть подходящим для ингибирования протеолитической деградации в процессе очистки. Кроме того, целевой белок может присутствовать в слитом виде в рамке считывания с маркерной последовательностью, которая позволяет очищать целевой белок. Неограничивающие примеры маркерных последовательностей включают гексагистидиновую метку, которая может быть обеспечена с помощью вектора, предпочтительно вектора pQE-9, и гемагглютининовую (НА) метку. НА-метка соответствует эпитопу, полученному из белка гемагглютинина вируса гриппа (см., например, Wilson et al., Cell, 37:767 [1984]). Специалисту в данной области техники будет понятно, что способы очистки, подходящие для целевого полипептида, могут потребовать модификации для учета изменений в свойствах полипептида при экспрессии в культуре рекомбинантных клеток.
В некоторых предпочтительных вариантах реализации в системы включены конструкции нуклеиновых кислот. В некоторых вариантах реализации системы содержат многокомпонентные конструкции нуклеиновых кислот или векторы, описанные выше, которые предназначены для введения в клетку-хозяина. В других предпочтительных вариантах реализации системы содержат одну или более или один или более многокомпонентных конструкций нуклеиновых кислот или векторов, описанных выше, которые предназначены для введения в клетку-хозяина, в дополнение к нуклеиновой кислоте или вектору, кодирующим фермент, необходимый для встраивания конструкций нуклеиновых кислот в геном клетки-хозяина. Примеры ферментов включают, не ограничиваясь указанными, транспозазы для применения с транспозоиными векторными системами, интегразы для применения в системах, в которых используются интегрирующие последовательности, таких как система PhiC31, системы MMLV и тому подобное, рекомбиназы для применения в векторных системах, таких как Cre-loc, FLP-FRT и тому подобное, и нуклеазы Cas9 для применения в системах на основе CRISPR.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
В настоящем изобретении предложен уникальный способ комбинирования ретровирусной экспрессионной кассеты SIN-LTR с системой линии клеток СНО, имеющих нокаут гена глутаминсинтетазы (GS), для улучшения способов разработки клеточных линий с использованием случайной интеграции, который обеспечивает увеличение числа копий гена и повышение продуктивности на каждую копию. Указанный подход также обеспечивает улучшенный и неожиданный способ более строгой селекции пулов для дополнительного улучшения титра и обогащения пулов для клонов с более высокой продуктивностью. Он также обеспечивает быстрый и эффективный способ разработки высокопродуктивных линий клеток посредством нацеленной интеграции экспрессионных кассет (трансгенов) в заранее определенные сайты (акцепторные сайты) в пределах всего генома СНО.
Пример 1:
Три объединенные клеточные линии были получены в результате временной трансфекции пятью независимыми плазмидами (Фиг. 1), все из которых были сконструированы для экспрессии тест-белка «Anyway». Эти плазмиды названы в соответствии с промотором, используемым в них для управления экспрессией GS. Первая плазмида, SV40, представляет традиционный способ разработки клеточной линии плазмиду, содержащую ген селектируемого маркера (GS), управляемый сильным промотором SV40, а также содержащую интрон SV40 и сигнальную последовательность поли(А). Во второй плазмиде, WT-LTR, используется провирусный LTR дикого типа для обеспечения экспрессии GS, включенный в плазмиду аналогично тому, как выглядела бы его вставка в векторе GPEx. Несмотря на то, что этот промотор считается лишь относительно сильным, транскрипция с этого промотора не заканчивается после GS, а продолжается на протяжении последовательностей sCMV, Anyway и WPRE, используя последовательность поли(А) для ТК. Третья плазмида, SLN-LTR, идентична второй конструкции за исключением того, что она содержит усеченный вариант LTR, SIN-LTR (самоинактивирущийся LTR), который обладает более низкой промоторной активностью. Четвертая плазмида, pSLN, идентична первой плазмиде за исключением того, что вместо сильного промотора, управляющего экспрессией GS, в ней используется более слабый промоторный элемент из SIN-LTR. Пятая плазмида экспрессировала GFP, но не содержала гена GS и, таким образом, служила в качестве отрицательного контроля.
Пулы, полученные путем трансфекции этими плазмидами, отбирали по выживанию в отсутствие глутамина. Отобранные пулы были использованы для получения неспецифического продукта с периодическим добавлением питательной среды для определения их способности продуцировать белок Anyway.
Разработка клеточной линии CHOZN
Трансфекция клеток CHOZn: Объединенные клеточные линии, содержащие каждую из плазмид, интегрированную случайным образом, получали путем трансфекции клеток указанной плазмидой с использованием реагента для трансфекции Expifectamine СНО. К 1 мл среды OptiPro добавляли 20 мкг плазмиды. К 920 мкл OptiPro добавляли 80 мкл Expifectamine СНО. Эти два раствора перемешивали в течение 1 минуты, затем добавляли к 3 мл среды СНО-Gro, содержащей 30 миллионов клеток CHOZn. Клетки инкубировали в течение ночи при 37°С со встряхиванием при 250 об/мин. На следующее утро добавляли 15 мл среды Excell CD Fusion с добавлением 6 мМ глутамина. Клетки пересевали в этой среде до тех пор, пока они не восстанавливались после трансфекции.
Селекция клеток CHOZn: Как только жизнеспособность клеток достигала > 96%, их пересевали путем полной замены среды в среду Ex-Cell CD Fusion с добавлением 2% ClonaCell-СНО ACF, но без глутамина. Регулярно проводили мониторинг жизнеспособности клеток и плотности жизнеспособных клеток. Среду заменяли еженедельно до тех пор, пока концентрация клеток в культуре не достигала 1 миллиона клеток на мл, и клетки пересевали по стандартной методике.
Получение продукта с периодическим добавлением питательной среды: Перед получением продукта с периодическим добавлением питательной среды каждый пул адаптировали к среде ActiPro в течение по меньшей мере трех пересевов. Для получения продукта с периодическим добавлением питательной среды в центрифужные пробирки объемом 50 мл высевали клетки в концентрации 600000 клеток на мл в среде ActiPro (HyClone) и инкубировали в увлажненном (70-80%) встряхивателе-инкубаторе при 250 об/мин при 5% СО2 и температуре 37°С (34°С начиная с 5 дня). Культуры получали питание шесть раз в течение цикла получения с использованием двух различных питательных добавок. Ежедневно контролировали содержание глюкозы и добавляли ее, если уровень падал ниже 5 г/л. Культивирование прекращали, когда жизнеспособность составляла <70%.
Результаты
Как показано на Фиг. 2, пулы SV40, WT-LTR и SIN-LTR демонстрировали значительно различающиеся профили восстановления в процессе селекции. Пулы SV40 демонстрировали наиболее быстрое восстановление (>90% жизнеспособности), что указывало на то, что относительно большая доля клеток в не подвергнутом селекции пуле была устойчива к селекции. Пулы WT-LTR демонстрировали более медленное восстановление, что указывало на то, что устойчивой была меньшая доля в не подвергнутом селекции пуле. Пулы SIN-LTR демонстрировали заметно замедленное восстановление, что указывало на то, что устойчивой была очень небольшая доля в не подвергнутом селекции пуле.
Как показано на Фиг. 3, титр был значительно выше для пула STN-LTR по сравнению с пулами WT-LTR и SV40. Для сравнения, значения числа копий гена демонстрировали сходную тенденцию.
Эти данные указывают на то, что плазмида SIN-LTR обеспечивала селекцию большего числа копий и сайтов встраивания с более высокой активностью.
Неожиданно, в отдельном эксперименте, представленном на Фиг. 4, пулы pSTN имели время восстановления, сходное с таковым для пулов SV40 или WT-LTR. Следовательно, различия во времени восстановления не объясняются только промоторной активностью, поскольку pSTN содержит очень слабый промотор, но все еще демонстрировала быстрое восстановление. Другие элементы плазмиды SIN-LTR должны быть ответственными за более сильное селекционное давление. Без ограничения каким-либо конкретным механизмом действия, предполагается, что комбинация слабого промотора и длинного транскрипта, который также содержит вторую открытую рамку считывания, может влиять на эффективность транскрипции или трансляции GS. Аналогичным образом, без ограничения каким-либо конкретным механизмом, известное присутствие слабой последовательности Козак в EPR может приводить к аберрантной трансляции со снижением эффективности трансляции белка GS.
Пример 2:
Концепции GPEx Boost также могут применяться в комбинации с другими технологиями встраивания генов без использования вирусов, такими как встраивание генов с помощью транспозазы, рекомбиназы, интегразы или CRISPR. Технология GPEx может применяться для помещения множества копий последовательности распознавания для технологии встраивания без использования вирусов в высокоактивные сайты в пределах всего генома. Полученная линия клеток с акцепторными сайтами («Dock») затем может быть подвергнута временной совместной трансфекции с применением плазмиды, экспрессирующей транспозазу, рекомбиназу, интегразу или Cas9, в комбинации с плазмидой трансгена, которая содержит когнатную последовательность распознавания, селектируемый маркер GS и подлежащий экспрессии генный продукт. Транспозаза, рекомбиназа, интеграза или Cas9 будут опосредовать встраивание всей плазмиды трансгена или ее части в акцепторные сайты. Полученная клеточная линия будет содержать множество копий плазмиды трансгена, встроенных в высокоактивные акцепторные сайты в пределах всего генома. Некоторые примеры технологий/ферментов, которые можно применять, включают транспозазу PiggyBac, транспозазу «Спящая красавица», транспозазу Mos1, транспозазу Tol2, транспозазу Leapin, рекомбиназу фага лямбда, FLP/FRT, Cre/Lox, интегразу MMLV, интегразу Rep 78, интегразу Bxb1 и различные типы CRISPR. Авторы настоящего изобретения впервые проверили эту концепцию, применяя систему интегразы PhiC31 в комбинации с технологией GPEx.
Получение ретровектора и трансдукция для создания родительской линии клеток с акцепторными сайтами: Конструкцию для экспрессии акцепторного сайта, Фиг. 7 и 8, вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Также были проведены совместные трансфекции оболочкой вируса, содержащей экспрессионную плазмиду, и каждой из генетических конструкций. Совместная трансфекция приводила к получению дефектного по репликации ретровектора с высоким титром, который концентрировали посредством ультрацентрифугирования и использовали для трансдукции родительской линии клеток яичника китайского хомячка CHOZN (1,2). Проводили 5 последовательных циклов трансдукции, и клетки поддерживали в среде по стандартной методике с добавлением 6 мМ глутамина. Также успешно получали вторую объединенную линию клеток с акцепторными сайтами с применением тех же способов. Это было осуществлено с использованием немного другой генетической конструкции для экспрессии акцепторного сайта, представленной на Фиг. 9 и 10.
Трансфекция и селекция объединенной линии клеток с акцепторными сайтами: 1,5 миллиона клеток инкубировали с предварительно приготовленной смесью, содержащей 1 мкг (общее количество) ДНК плазмид Transgene и Integrase (Фиг. 11, 12, 5 и 6, соответственно) и 4 микролитра ExpiFectamine СНО™ (ThermoFisher Scientific) в конечном объеме 250 микролитров. Объединенные клеточные линии оставляли для восстановления в присутствии среды с добавлением 6 мМ глутамина до тех пор, пока жизнеспособность не возвращалась до уровня более 95%. Затем клетки переносили в среду, не содержащую глутамина. Проводили мониторинг жизнеспособности клеток, и среду заменяли еженедельно до тех пор, пока жизнеспособность полученных отобранных пулов клеток не возвращалась до уровня более 95%
Количественная оценка рекомбинации: Геномную ДНК выделяли из 3 миллионов клеток с использованием набора Qiagen DNEasy. attR является результатом рекомбинации между attP и attB. Количественную полимеразную цепную реакцию (кПЦР) с использованием красителя SYBR Green проводили для количественного определения attR в клетках с использованием прямого праймера в последовательности attP в акцепторном сайте и обратного праймера в последовательности attB в плазмиде трансгена. Амплификация с использованием этой пары праймеров будет детектировать плазмиду трансгена только в том случае, когда эта плазмида является рекомбинированной в акцепторный сайт и не является свободной, интегрированной случайным образом или интегрированной в сайт псевдо-attP плазмидой трансгена. Аналогичным образом, эта пара праймеров не будет детектировать последовательность нерекомбинированного (пустого) акцепторного сайта. Число циклов ПЦР, необходимых для пересечения порога интенсивности флуоресценции (значение Ct), определяли для этого набора праймеров, а также для набора праймеров для внутреннего референсного гена СНО. Индексы копийности гена (GCI) рассчитывали путем вычитания значения Ct для референсного гена из значения Ct для набора праймеров attR. Следует отметить, что значения GCI по своей природе являются логарифмическими, а не линейными, так что изменение на 1 единицу в нижней части шкалы, например, от GCI=1 до GCI=3, соответствует разнице, составляющей лишь несколько копий, тогда как изменение на одну единицу в верхней части шкалы, например, от GCI=6 до GCI=7, может соответствовать разнице, составляющей множество копий. В некоторых случаях плазмиду, содержащую желаемые ампликоны и имеющую известную концентрацию, также подвергали кПЦР, и выполняли линейный регрессионный анализ полученных данных для более точного определения числа присутствующих копий.
Результаты
Акцепторные сайты, содержащие последовательность распознавания attP для PhiC31, Фигуры 7+8, помещали в пределах всего генома клеток CHOZN с помощью 5 последовательных циклов трансдукции с использованием технологии GPEx, и полученный пул клеток содержал в среднем приблизительно 36 копий акцепторного сайта на клетку. Этот пул клеток с акцепторными сайтами был совместно трансфицирован плазмидами Transgene-Promoter- Any way и Integrase (Фиг. 11, 12, 5 и 6, соответственно) в соотношениях в диапазоне от 1:50 до 1:1, как было предложено в опубликованной литературе (Groth, 2000: Andreas, 2002: Farruggio, 2012). Эта плазмида Transgene-Prom oter-Any way со держит последовательность распознавания attB для PhiC31, ген глутаминсинтетазы (GS), управляемый слабым провирусным промотором STN-LTR (самоинактивирующегося длинного концевого повтора), и тест-продукт в виде слитого с Fc белка Anyway под контролем сильного промотора. Через 3 дня после трансфекции, но до селекции, проводили кПЦР для количественной оценки рекомбинации, но уровни attR (вышележащего продукта рекомбинации между attP и attB) были детектируемыми на уровне не выше фонового, при этом индексы копийности гена (GCI) составляли приблизительно -10. Когда трансфицированные клетки подвергали селекции путем удаления глутамина, они не восстанавливались за более чем 25 дней, что указывало на то, что интеграция и экспрессия GS в этих клетках не достигали достаточных уровней.
В попытке улучшить частоту рекомбинации авторы настоящего изобретения пришли к выводу, что отношение плазмиды, экспрессирующей интегразу, к плазмиде трансгена может являться критическим параметром для эффективной рекомбинации. Для исследования этой возможности авторы настоящего изобретения подвергли совместной трансфекции пул клеток с акцепторными сайтами с использованием диапазона соотношений плазмид Transgene-Promoter-Anyway и Integrase. Через 3 дня после трансфекции, но до селекции, проводили кПЦР для количественной оценки рекомбинации, Фиг. 29. Диапазон соотношений, содержащий низкие соотношения Transgene:Integrase (1:20-100), часто используемые в литературе, соответствовал GCI для attR около фонового уровня, составляющего -10. Авторы настоящего изобретения неожиданно обнаружили, что высокие соотношения Transgene:Integrase (5-100:1) соответствовали GCI для attR, составляющим -3, что означало число копий, которое приблизительно в 200 раз превышало число копий для самых низких соотношений.
Затем проводили селекцию путем удаления глутамина на образцах с наибольшими GCI для attR согласно предварительной селекции, Фиг. 30. Эти пулы начинали восстанавливаться с 9 дня селекции. После полного восстановления авторы настоящего изобретения провели кПЦР, Фиг. 31, и обнаружили, что исследуемые пулы содержали вплоть до приблизительно 28 копий трансгена на клетку. Эти данные указывают на то, что путем использования более высоких соотношений Transgene:Integrase авторы настоящего изобретения смогли достичь эффективной интеграции в среднем вплоть до 28 трансгенов на клетку в пуле, который содержал в среднем приблизительно 36 акцепторных сайтов на клетку. Кроме того, уровень этой рекомбинации был приблизительно на 2 порядка выше, чем уровень рекомбинации, наблюдаемый с использованием более низких соотношений, описанных в литературе.
Пример 3:
После обнаружения заполнения приблизительно на 80% пула с акцепторными сайтами, содержащего приблизительно 36 копий на клетку, авторы настоящего изобретения далее попытались определить, могли ли они дополнительно увеличить число интегрированных плазмид Transgene путем использования пула с акцепторными сайтами, который содержал более 36 акцепторных сайтов на клетку. Авторы настоящего изобретения также попытались определить, можно ли было также применять в этой системе плазмиду Transgene, не содержащую промотора GS. Такая плазмида способна экспрессировать GS и таким образом обеспечивать устойчивость только в том случае, если она является рекомбинированной в акцепторный сайт, а не интегрированной случайным образом или интегрированной в сайты псевдо-attP.
Получение ретровектора и трансдукция для создания родительской линии клеток с акцепторными сайтами:
Конструкцию для экспрессии акцепторного сайта (Фиг. 7 и 8) вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Также были проведены совместные трансфекции оболочкой вируса, содержащей экспрессионную плазмиду, и каждой из генетических конструкций. Совместная трансфекция приводила к получению дефектного по репликации ретровектора с высоким титром, который концентрировали посредством ультрацентрифугирования и использовали для трансдукции родительской линии клеток яичника китайского хомячка CHOZN (1,2). Проводили 9 последовательных циклов трансдукции, и клетки поддерживали в среде по стандартной методике с добавлением 6 мМ глутамина.
Трансфекция и селекция объединенной линии клеток с акцепторными сайтами: 3 миллиона клеток инкубировали с предварительно приготовленной смесью, содержащей 2 мкг (общее количество) плазмидной ДНК плазмид Transgene и Integrase и 8 микролитров ExpiFectamine СНО™ (ThermoFisher Scientific) в конечном объеме 500 микролитров. Объединенные клеточные линии оставляли для восстановления в присутствии среды с добавлением 6 мМ глутамина до тех пор, пока жизнеспособность не возвращалась до уровня более 95%. Затем клетки переносили в среду, не содержащую глутамина. Проводили мониторинг жизнеспособности клеток, среду заменяли еженедельно, и клетки пересевали до тех пор, пока жизнеспособность полученных отобранных пулов клеток не возвращалась до уровня более 95%
Количественная оценка рекомбинации: Геномную ДНК выделяли из 3 миллионов клеток с использованием набора Qiagen DNEasy. attR является результатом рекомбинации между attP и attB. Количественную полимеразную цепную реакцию (кПЦР) с использованием красителя SYBR Green проводили для количественного определения attR в клетках с использованием прямого праймера в последовательности attP в акцепторном сайте и обратного праймера в последовательности attB в трансгене. Амплификация с использованием этой пары праймеров будет детектировать плазмиду трансгена только в том случае, когда эта плазмида является рекомбинированной в акцепторный сайт и не является свободной, интегрированной случайным образом или интегрированной в сайт псевдо-attP плазмидой трансгена. Аналогичным образом, эта пара праймеров не будет детектировать последовательности нерекомбинированных (пустых) акцепторных сайтов. Число циклов ПНР, необходимых для пересечения порога интенсивности флуоресценции (значение Ct), определяли для этого набора праймеров, а также для набора праймеров для внутреннего референсного гена СНО. Индексы копийности гена (GCI) рассчитывали путем вычитания значения Ct для референсного гена из значения Ct для набора праймеров attR. Следует отметить, что значения GCI по своей природе являются логарифмическими, а не линейными, так что изменение на 1 единицу в нижней части шкалы, например, от GCI=1 до GCI=3, соответствует разнице, составляющей лишь несколько копий, тогда как изменение на одну единицу в верхней части шкалы, например, от GCI=6 до GCI=7, может соответствовать разнице, составляющей множество копий. В некоторых случаях плазмиду, содержащую желаемые ампликоны и имеющую известную концентрацию, также подвергали кПЦР, и выполняли линейный регрессионный анализ полученных данных для более точного определения числа присутствующих копий.
Результаты
Акцепторные сайты, содержащие последовательность распознавания attP для PhiC31, помещали в пределах всего генома клеток CHOZN с помощью 9 последовательных циклов трансдукции с использованием технологии GPEx, и полученный пул клеток имел GCI для EPR, составляющий 6,7, и содержал в среднем приблизительно 135 копий акцепторного сайта на клетку. Этот пул клеток с акцепторными сайтами был совместно трансфицирован плазмидами Transgene-Anyway и Integrase (Фиг. 13, 14, 5 и 6, соответственно) в соотношениях в диапазоне от 50:1 до 400:1 с последующей селекцией путем удаления глутамина. Наименьшая (минимальная) жизнеспособность, согласно Фиг. 32, была выше, чем в линии с акцепторными сайтами, содержащей приблизительно 36 копий на клетку. GCI для attR, Фиг. 33, также были выше, чем для линии клеток, содержащей приблизительно 36 копий акцепторного сайта, и увеличивались при более высоких соотношениях Transgene:Integrase, что позволяет предположить, что может быть возможным дальнейшее улучшение числа интегрированных трансгенов при более высоких соотношениях Transgene: Integrase и большем числе копий акцепторного сайта. Кроме того, авторы настоящего изобретения также продемонстрировали устойчивое восстановление после селекции при использовании плазмиды Transgene-Any way, Фиг. 13 и 14, которая не содержит промотора для GS и, таким образом, должна зависеть от слабого промотора SIN-LTR в конструкции для экспрессии акцепторного сайта.
Пример 4:
После улучшения числа интегрированных плазмид Transgene с использованием большего количества акцепторных сайтов и более высоких соотношений Transgene:Integrase, авторы настоящего изобретения далее попытались определить, могли ли они еще больше увеличить число интегрированных плазмид Transgene путем выделения клона с более высоким числом копий акцепторного сайта из пула, содержащего 135 копий акцепторного сайта, и путем проверки еще более высоких соотношений плазмид Transgene:Integrase. Авторы настоящего изобретения также попытались определить, можно ли с помощью этой технологии встраивать плазмиды с большими размерами.
Клонирование родительской линии клеток с акцепторными сайтами: Пул клеток с акцепторными сайтами, полученный с помощью 9 последовательных циклов трансдукции, клонировали с использованием прибора Berkeley Lights, Beacon. Клоны размножали, подвергали скринингу посредством кПЦР, и отбирали клон с наибольшим числом инсерций акцепторного сайта.
Трансфекция и селекция объединенной линии клеток с акцепторными сайтами: 3 миллиона клеток инкубировали с предварительно приготовленной смесью, содержащей 2 мкг (общее количество) ДНК плазмид Transgene и Integrase и 8 микролитров ExpiFectamine СНО™ (ThermoFisher Scientific) в конечном объеме 500 микролитров. Объединенные клеточные линии оставляли для восстановления в присутствии среды с добавлением 6 мМ глутамина до тех пор, пока жизнеспособность не возвращалась до уровня более 95%. Затем клетки переносили в среду, не содержащую глутамина. Проводили мониторинг жизнеспособности клеток, и среду заменяли еженедельно до тех пор, пока жизнеспособность полученных отобранных пулов клеток не возвращалась до уровня более 95%.
Количественная оценка рекомбинации: Геномную ДНК выделяли из 3 миллионов клеток с использованием набора Qiagen DNEasy. attR является результатом рекомбинации между attP и attB. Количественную полимеразную цепную реакцию (кПЦР) с использованием красителя SYBR Green проводили для количественного определения attR в клетках с использованием прямого праймера в последовательности attP в акцепторном сайте и обратного праймера в последовательности attB в трансгене. Амплификация с использованием этой пары праймеров будет детектировать плазмиду трансгена только в том случае, когда эта плазмида является рекомбинированной в акцепторный сайт и не является свободной, интегрированной случайным образом или интегрированной в сайт псевдо-attP плазмидой трансгена. Аналогичным образом, эта пара праймеров не будет детектировать последовательность нерекомбинированного (пустого) акцепторного сайта. Число циклов ПЦР, необходимых для пересечения порога интенсивности флуоресценции (значение Ct), определяли для этого набора праймеров, а также для набора праймеров для внутреннего референсного гена СНО. Праймеры, специфичные к части EPR акцепторного сайта (Фиг. 5 и 6), были использованы для классификации клонов на основе GCI для EPR. Значения индекса копийности гена рассчитывали путем вычитания значения Ct для референсного гена из значения Ct для набора праймеров attR. Следует отметить, что значения GCI по своей природе являются логарифмическими, а не линейными, так что изменение на 1 единицу в нижней части шкалы, например, от GCI=1 до GCI=3, соответствует разнице, составляющей лишь несколько копий, тогда как изменение на одну единицу в верхней части шкалы, например, от GCI=6 до GCI=7, может соответствовать разнице, составляющей множество копий. В некоторых случаях плазмиду, содержащую желаемые ампликоны и имеющую известную концентрацию, также подвергали кПЦР, и выполняли линейный регрессионный анализ полученных данных для более точного определения числа присутствующих копий.
Получение продукта с периодическим добавлением питательной среды: Для получения продукта с периодическим добавлением питательной среды в центрифужные пробирки объемом 50 мл высевали клетки в концентрации 600000 клеток на мл в 20 мл среды Ex-Cell Advanced СНО Fed-Batch™ (MilliporeSigma) и инкубировали в увлажненном (70-80%) встряхивателе-инкубаторе при 250 об/мин при 5% СО2 и температуре 37°С (34°С начиная с 4 дня). Культуры получали дополнительное питание через день, начиная со 2 дня с использованием 6,25% (об./об.) питательной смеси, содержащей 66% среды Ex-cell Advanced СНО Feed 1™ и 33% среды Cellvento 4Feed (MilliporeSigma). Ежедневно контролировали содержание глюкозы и добавляли ее, если уровень падал ниже 5 г/л. Культивирование прекращали, когда жизнеспособность составляла <70% или в конце 20 дня.
Результаты
Для выделения клона с высоким числом копий акцепторного сайта пул клеток с акцепторными сайтами, полученный с помощью 9 циклов трансдукции, подвергали процедуре клонирования одиночных клеток с использованием прибора Berkely Lights, Beacon®. Клоны выделяли, размножали и подвергали кПЦР с использованием праймеров, специфичных для области EPR акцепторного сайта. Клон 1F7 содержал приблизительно 181 копию плазмиды для экспрессии акцепторного сайта на клетку и был выбран для дальнейших экспериментов. Клон 1F7 с акцепторными сайтами совместно трансфицировали плазмидой Transgene-Yourway-LWHW, экспрессирующей обе легкую и тяжелую цепи антитела, и плазмидой Integrase, Фигуры 27+28 и 5+6, в соотношениях в диапазоне от 50:1 до 8000:1. Полученные пулы подвергали селекции путем удаления глутамина, Фигура 34. Пулы с соотношениями 4000:1 и 8000:1 не выжили в процессе селекции. кПЦР-анализ содержания attR в выживших пулах, Фигура 35, показал, что более крупные плазмиды с размером вплоть до по меньшей мере 9,8 т.п.н. могли быть эффективно интегрированы с помощью применяемой технологии, и оптимальное соотношение плазмид Transgen:Inetegrase этого размера составляло 500:1.
Пример 5:
После обнаружения относительно высокой эффективности интеграции в клоне 1F7 с акцепторными сайтами, авторы настоящего изобретения далее попытались определить, могли ли клоны, полученные из пулов с высокими уровнями интегрированных трансгенов, обеспечивать еще более высокие уровни интеграции трансгенов, и определить эффективность этих клонов в качестве продуцентов.
Трансфекция и селекция объединенной линии клеток с акцепторными сайтами: 3 миллиона клеток инкубировали с предварительно приготовленной смесью, содержащей 2 мкг (общее количество) ДНК плазмид Transgene и Integrase (Фиг. 13, 14, 5 и 6, соответственно) и 8 микролитров ExpiFectamine СНО™ (ThermoFisher Scientific) в конечном объеме 500 микролитров. Объединенные клеточные линии оставляли для восстановления в присутствии среды с добавлением 6 мМ глутамина до тех пор, пока жизнеспособность не возвращалась до уровня более 95%. Затем клетки переносили в среду, не содержащую глутамина. Проводили мониторинг жизнеспособности клеток, и среду заменяли еженедельно до тех пор, пока жизнеспособность полученных отобранных пулов клеток не возвращалась до уровня более 95%
Клонирование пулов с интегрированными плазмидами Transgene: Пулы с интегрированными плазмидами Transgene клонировали с использованием прибора Berkeley Lights, Beacon. Для определения относительной продуктивности этих клонов использовали анализ Spotlight®. Клоны с наибольшей продуктивностью извлекали из устройства и размножали.
Количественная оценка рекомбинации: Геномную ДНК выделяли из 3 миллионов клеток с использованием набора Qiagen DNEasy. attR является результатом рекомбинации между attP и attB. Количественную полимеразную цепную реакцию (кПЦР) с использованием красителя SYBR Green проводили для количественного определения attR в клетках с использованием прямого праймера в последовательности attP в акцепторном сайте и обратного праймера в последовательности attB в трансгене. Амплификация с использованием этой пары праймеров будет детектировать плазмиду трансгена только в том случае, когда эта плазмида является рекомбинированной в акцепторный сайт и не является свободной, интегрированной случайным образом или интегрированной в сайт псевдо-attP плазмидой трансгена. Аналогичным образом, эта пара праймеров не будет детектировать последовательность нерекомбинированного (пустого) акцепторного сайта. Число циклов ПЦР, необходимых для пересечения порога интенсивности флуоресценции (значение Ct), определяли для этого набора праймеров, а также для набора праймеров для внутреннего референсного гена СНО. Для оценки доли заполненных акцепторных сайтов были использованы праймеры, специфичные в отношении attP, который присутствовал только в не интегрировавших акцепторных сайтах. Значения индекса копийности гена рассчитывали путем вычитания значения Ct для референсного гена из значения Ct для набора праймеров attR. Следует отметить, что значения GCI по своей природе являются логарифмическими, а не линейными, так что изменение на 1 единицу в нижней части шкалы, например, от GCI=1 до GCI=3, соответствует разнице, составляющей лишь несколько копий, тогда как изменение на одну единицу в верхней части шкалы, например, от GCI=6 до GCI=7, может соответствовать разнице, составляющей множество копий. В некоторых случаях плазмиду, содержащую желаемые ампликоны и имеющую известную концентрацию, также подвергали кПЦР, и выполняли линейный регрессионный анализ полученных данных для более точного определения числа присутствующих копий.
Результаты
Клон 1F7 с акцепторными сайтами совместно трансфицировали плазмидами Transgene-Anyway и Integrase (Фиг. 13, 14, 5 и 6, соответственно), и полученные пулы подвергали селекции путем удаления глутамина. GCI для attR для выбранного пула составлял 6,9. Этот пул подвергали процедуре клонирования одиночных клеток с использованием прибора Berkely Lights, Beacon. Клоны классифицировали и извлекали на основании относительной экспрессии Anyway, определенной с использованием анализа Spotlight®. Были размножены 27 клонов, и GCI для attR в этих клонах, Фиг. 36, находились в диапазоне от 5,2 до 7,5. Для этих клонов также определяли GCI для attP, который являлся мерой числа пустых акцепторных сайтов, что позволило авторам настоящего изобретения оценить долю заполненных акцепторных сайтов в каждом клоне, Фиг. 36. В этих клонах средний процент заполнения составлял 65%. Это соответствовало приблизительно 118 копиям интегрированной плазмиды Transgene. Клон 1В7 имел GCI для attR, составляющий 7,5, что было эквивалентно GCI для attP (пустой акцепторный сайт) для родительского клона 1F7 с акцепторными сайтами. Авторы настоящего изобретения неожиданно не смогли детектировать attP в этом клоне при использовании двух разных пар праймеров. Эти данные указывают на то, что, неожиданно, после только одной трансфекции авторы настоящего изобретения смогли получить клон со всеми приблизительно 181 акцепторными сайтами, заполненными трансгеном.
Для определения эффективности всех этих клонов в качестве продуцентов белка проводили анализ их продуктивности при получении неспецифического продукта с периодическим добавлением питательной среды, конечные титры также показаны на Фиг. 36. Высокие уровни GCI для attR были связаны с высоким конечным титром, Фиг. 37, что указывало на то, что, как и ожидалось, увеличение количества событий нацеленной интеграции трансгенов в высокоактивные акцепторные сайты приводило к повышению эффективности клеточной линии в качестве продуцента белка. Эти данные также позволяют предположить, что авторами настоящего изобретения еще не было достигнуто насыщение эффективности этих клеток в качестве продуцентов даже при 181 интегрировавшей копии.
Пример 6:
После обнаружения относительно высокой эффективности интеграции и экспрессии слитого белка в клоне 1F7 с акцепторными сайтами, авторы настоящего изобретения далее попытались определить, могли ли они также применять эту систему для интеграции и экспрессии моноклональных антител с обеими тяжелой и легкой цепями на одной и той же плазмиде Transgene.
Трансфекция и селекция объединенной линии клеток с акцепторными сайтами: 3 миллиона клеток инкубировали с предварительно приготовленной смесью, содержащей 2 мкг (общее количество) ДНК плазмид Transgene и Integrase и 8 микролитров ExpiFectamine СНО™ (ThermoFisher Scientific) в конечном объеме 500 микролитров. Объединенные клеточные линии оставляли для восстановления в присутствии среды с добавлением 6 мМ глутамина до тех пор, пока жизнеспособность не возвращалась до уровня более 95%. Затем клетки переносили в среду, не содержащую глутамина. Проводили мониторинг жизнеспособности клеток, и среду заменяли еженедельно до тех пор, пока жизнеспособность полученных отобранных пулов клеток не возвращалась до уровня более 95%.
Количественная оценка рекомбинации: Геномную ДНК выделяли из 3 миллионов клеток с использованием набора Qiagen DNEasy. attR и attL представляют собой результат рекомбинации между attP и attB. Количественную полимеразную цепную реакцию (кПЦР) с использованием красителя SYBR Green проводили для количественного определения attR в клетках с использованием прямого праймера в последовательности attP в акцепторном сайте и обратного праймера в последовательности attB в трансгене. Амплификация с использованием этой пары праймеров будет детектировать плазмиду трансгена только в том случае, когда эта плазмида является рекомбинированной в акцепторный сайт и не является свободной, интегрированной случайным образом или интегрированной в сайт псевдо-attP плазмидой трансгена. Аналогичным образом, эта пара праймеров не будет детектировать последовательность нерекомбинированного (пустого) акцепторного сайта. Число циклов ПЦР, необходимых для пересечения порога интенсивности флуоресценции (значение Ct), определяли для этого набора праймеров, а также для набора праймеров для внутреннего референсного гена СНО. Для оценки доли заполненных акцепторных сайтов были использованы праймеры, специфичные в отношении attP, который присутствовал только в не интегрировавших акцепторных сайтах. Значения индекса копийности гена рассчитывали путем вычитания значения Ct для референсного гена из значения Ct для набора праймеров attR. Следует отметить, что значения GCI по своей природе являются логарифмическими, а не линейными, так что изменение на 1 единицу в нижней части шкалы, например, от GCI=1 до GCI=3, соответствует разнице, составляющей лишь несколько копий, тогда как изменение на одну единицу в верхней части шкалы, например, от GCI=6 до GCI=7, может соответствовать разнице, составляющей множество копий. В некоторых случаях плазмиду, содержащую желаемые ампликоны и имеющую известную концентрацию, также подвергали кПЦР, и выполняли линейный регрессионный анализ полученных данных для более точного определения числа присутствующих копий.
Получение продукта с периодическим добавлением питательной среды: В центрифужные пробирки объемом 50 мл высевали клетки в концентрации 600000 клеток на мл в 20 мл среды Ex-Cell Advanced СНО FED-BATCH™ (MilliporeSigma) и инкубировали в увлажненном (70-80%) встряхивателе-инкубаторе при 250 об/мин при 5% СО2 и температуре 37°С (34°С начиная с 4 дня). Культуры получали дополнительное питание через день начиная со 2 дня с использованием 6,25% (об./об.) питательной смеси, содержащей 66% среды Ex-Cell Advanced СНО Feed 1™ и 33% среды Cellvento 4Feed (MilliporeSigma). Ежедневно контролировали содержание глюкозы и добавляли ее, если уровень падал ниже 5 г/л. Культивирование прекращали, когда жизнеспособность составляла <70% или в конце 20 дня.
Электрофорез белков в геле. Супернатанты от получения продукта с периодическим добавлением питательной среды (см. выше) собирали и делали прозрачными. 3 мкг каждого антитела или содержащего Fc слитого белка смешивали с буфером для нанесения образцов LDS с добавлением или без добавления денатурирующего агента. Перед электрофорезом денатурированные образцы также нагревали до 70°С в течение 10 минут. Все образцы наносили на гель NuPAGE Novex 4-12% Bis-Tris (Invitrogen) и подвергали электрофорезу в буфере IX MES в течение 15 минут при 60 В и затем 105 минут при 100 В. Затем гель промывали деионизированной водой и окрашивали с помощью красителя SYPRO-Ruby. Окрашенные гели были визуализированы, и на Фиг. 40 представлено «негативное» изображение (с обращением цветов) окрашенного геля.
Результаты
Хорошо известно, что и экспрессия, и очистка моноклональных антител чувствительны к относительным количествам экспрессируемых легкой цепи и тяжелой цепи. Система, предложенная авторами настоящего изобретения, предназначена для интеграции легкой цепи и тяжелой цепи при соотношении генов 1:1. С целью оптимизации относительной экспрессии каждой цепи авторы настоящего изобретения разработали и проверили четыре различные экспрессионные конструкции, которые содержат гены, расположенные в различном порядке, и энхансерные элементы (см. Фиг. 21, 22, 23, 24, 25, 26, 27 и 28). Все исследованные конструкции не содержат промотора GS и содержат сильные промоторы и последовательности поли(А) для генов и тяжелой цепи, и легкой цепи. В первой конструкции, называемой HWIL (для подчеркивания различий между конструкциями), кодирующая последовательность тяжелой цепи (Н) экспрессируется с вышележащего промотора, и за ней следует посттранскрипционный регуляторный элемент вируса гепатита сурков (W или WPRE). Кодирующая последовательность легкой цепи (L) экспрессируется с нижележащего промотора, и ей предшествует интронная последовательность (I). Остальные три экспрессионные конструкции следуют той же номенклатуре. Клон 1F7 с акцепторными сайтами, содержащий приблизительно 181 копию акцепторного сайта, был совместно трансфицирован всеми четырьмя плазмидами Transgene-Yourway или плазмидой Transgene-Any way (в отдельности) и плазмидой Integrase (Фиг. 5+6, 21+22, 23+24, 25+26, 27+28, 13+14), и полученные пулы подвергали селекции путем удаления глутамина, Фиг. 38. Интересно, что пулы, трансфицированные плазмидой LWIH, после селекции восстанавливались медленнее, чем при трансфекции другими плазмидами. кПЦР-анализ полученных пулов, Фиг. 38, показал, что был достигнут высокий уровень интеграции трансгена, аналогичный таковому для предыдущих примеров, несмотря на больший размер этих плазмид. Также проводили анализ продуктивности при периодическом добавлении питательной среды для определения эффективности этих пулов в качестве продуцентов белка, Фиг. 39.
Три из четырех экспрессионных плазмид демонстрировали стабильную экспрессию, при этом HWTL и LWHW обеспечивали самые высокие титры. Полученные белки подвергали анализу посредством ДСН-ПААГ-электрофореза как в невосстанавливающих, так и в восстанавливающих условиях, Фиг. 40, для оценки относительной экспрессии тяжелой и легкой цепей и сборки зрелого антитела. Все четыре экспрессионные плазмиды демонстрировали высокую долю образовавшихся зрелых антител с молекулярной массой 150 кДа относительно свободных легких и тяжелых цепей. Все четыре плазмиды для экспрессии антител также демонстрировали незначительный избыток экспрессии легкой цепи, что является желательным для очистки с помощью белка А, чтобы свести к минимуму очистку свободной тяжелой цепи. Аналогичным образом, экспрессия одноцепочечного слитого белка, Anyway, демонстрировала как высокий титр, Фигура 39, так и высокую долю зрелого, димеризованного белка прогнозируемого размера.
Пример 7:
Далее авторы настоящего изобретения хотели определить стабильность продуцирования в пулах, полученных с применением этой технологии, поскольку это является необходимой характеристикой для производства.
Получение ретровектора и трансдукция для создания родительской линии клеток с акцепторными сайтами: Конструкцию для экспрессии акцепторного сайта, Фиг. 7 и 8, вводили в линию клеток HEK 293, которая конститутивно продуцирует белки gag, pro и pol MLV. Также были проведены совместные трансфекции оболочкой вируса, содержащей экспрессионную плазмиду, и каждой из генетических конструкций. Совместная трансфекция приводила к получению дефектного по репликации ретровектора с высоким титром, который концентрировали посредством ультрацентрифугирования и использовали для трансдукции родительской линии клеток яичника китайского хомячка CHOZN (1,2). Проводили 5 последовательных циклов трансдукции, и клетки поддерживали в среде по стандартной методике с добавлением 6 мМ глутамина.
Трансфекция и селекция объединенной линии клеток с акцепторными сайтами: 3 миллиона клеток инкубировали с предварительно приготовленной смесью, содержащей 2 мкг (общее количество) ДНК плазмид Transgene и Integrase и 8 микролитров ExpiFectamine СНО™ (ThermoFisher Scientific) в конечном объеме 500 микролитров. Объединенные клеточные линии оставляли для восстановления в присутствии среды с добавлением 6 мМ глутамина до тех пор, пока жизнеспособность не возвращалась до уровня более 95%. Затем клетки переносили в среду, не содержащую глутамина. Проводили мониторинг жизнеспособности клеток, и среду заменяли еженедельно до тех пор, пока жизнеспособность полученных отобранных пулов клеток не возвращалась до уровня более 95%.
Получение продукта с периодическим добавлением питательной среды - среда Ex-cell: В центрифужные пробирки объемом 50 мл высевали клетки в концентрации 600000 клеток на мл в 20 мл среды Ex-Cell Advanced СНО FED-BATCH™ (MilliporeSigma) и инкубировали в увлажненном (70-80%) встряхивателе-инкубаторе при 250 об/мин при 5% СО2 и температуре 37°С (34°С начиная с 4 дня). Культуры получали дополнительное питание через день начиная со 2 дня с использованием 6,25% (об./об.) питательной смеси, содержащей 66% среды Ex-Cell Advanced СНО Feed 1™ и 33% среды Cellvento 4Feed (MilliporeSigma). Ежедневно контролировали содержание глюкозы и добавляли ее, если уровень падал ниже 5 г/л. Культивирование прекращали, когда жизнеспособность составляла <70% или в конце 20 дня.
Получение продукта с периодическим добавлением питательной среды - среда ActiPro: В центрифужные пробирки объемом 50 мл высевали клетки в концентрации 600000 клеток на мл в 20 мл среды Hyclone ActiPro™ (Activa Life Sciences) и инкубировали в увлажненном (70-80%) встряхивателе-инкубаторе при 250 об/мин при 5% СО2 и температуре 37°С (34°С начиная с 4 дня). Культуры получали дополнительное питание через день начиная со 2 дня с использованием 3% (об./об.) среды Hyclone Cell Boost 7А и 0,3% среды Hyclone Cell Boost 7b (Activa Life Sciences). Ежедневно контролировали содержание глюкозы и добавляли ее, если уровень падал ниже 5 г/л. Культивирование прекращали, когда жизнеспособность составляла<70% или в конце 20 дня.
Результаты
Для определения стабильности продуцирования в пулах, экспрессирующих гибридный белок Anyway, пул клеток с акцепторными сайтами, полученный с помощью 9 циклов трансдукции, был совместно трансфицирован плазмидами Transgene-Anyway и Integrase, Фигуры 13+14 и 5+6. Полученные пулы подвергали селекции путем удаления глутамина. Три пула непрерывно пересевали, и аликвоты замораживали еженедельно для более чем 40 поколений. После получения 40 поколений для всех пулов размораживали флаконы с ранее замороженными поколениями и проводили анализ продуктивности при периодическом добавлении питательной среды с применением двух различных сред/стратегий добавления питательной среды. Конечные титры согласно анализам продуктивности при периодическом добавлении питательной среды, Фиг. 41, показывают, что даже после непрерывного культивирования в течение более 40 поколений титры белка оставались стабильными во всех трех пулах, что указывало как на устойчивую генетическую стабильность интегрированных плазмид трансгенов, так и на стабильную экспрессию из интегрированных плазмид трансгенов, и оба эти параметра являются критическими характеристиками для применения этой технологии в производстве лекарственных веществ.
Все публикации и патенты, упомянутые в приведенном выше описании, включены в настоящую заявку посредством ссылки. Специалистам в данной области техники будут понятны различные модификации и варианты описанных способа и системы согласно настоящему изобретению без отступления от объема и сущности настоящего изобретения. Несмотря на то, что настоящее изобретение было описано в связи с конкретными предпочтительными вариантами реализации, следует понимать, что заявленное изобретение не должно быть неоправданно ограничено такими конкретными вариантами реализации. Действительно, предполагается, что различные модификации описанных способов практического осуществления настоящего изобретения, понятные для специалистов в области техники, к которой относится настоящее изобретение, находятся в пределах объема приведенной ниже формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Catalent Pharma Solutions, LLC
<120> КОНСТРУКЦИИ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ ПОЛУЧЕНИЯ БЕЛКОВ
<130> CATA-38549.601
<150> US 63/033,514
<151> 2020-06-02
<150> US 63/033,516
<151> 2020-06-02
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 7516
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 1
tcaatattgg ccattagcca tattattcat tggttatata gcataaatca atattggcta 60
ttggccattg catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc 120
aatatgaccg ccatgttggc attgattatt gactagttat taatagtaat caattacggg 180
gtcattagtt catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc 240
gcctggctga ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat 300
agtaacgcca atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc 360
ccacttggca gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga 420
cggtaaatgg cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg 480
gcagtacatc tacgtattag tcatcgctat taccatggtg atgcggtttt ggcagtacac 540
caatgggcgt ggatagcggt ttgactcacg gggatttcca agtctccacc ccattgacgt 600
caatgggagt ttgttttggc accaaaatca acgggacttt ccaaaatgtc gtaacaactg 660
cgatcgcccg ccccgttgac gcaaatgggc ggtaggcgtg tacggtggga ggtctatata 720
agcagagctc gtttagtgaa ccgtcagatc actagaagct ttattgcggt agtttatcac 780
agttaaattg ctaacgcagt cagtgcttct gacacaacag tctcgaactt aagctgcagt 840
gactctctta aggtagcctt gcagaagttg gtcgtgaggc actgggcagg taagtatcaa 900
ggttacaaga caggtttaag gagaccaata gaaactgggc ttgtcgagac agagaagact 960
cttgcgtttc tgataggcac ctattggtct tactgacatc cactttgcct ttctctccac 1020
aggtgtccac tcccagttca attacagctc ttaaaaattg gatctccatt cgccattcag 1080
gctgcgcaac tgctgggaag gacgatcaga gcgggcctct tcgctattac gccagctggc 1140
gaaagggacg tggcaagcaa ggcgattaag ttgagttacg ccaggatttt cccagtcacg 1200
acgttgtaaa acgacggcca gagaattata atacgactca ctatagggcg aattcggatc 1260
cgccgccacc atggtgacct acgccggagc ctacgacaga cagagccggg agagagagaa 1320
cagcagcgcc gccagccccg ccacccagag aagcgccaac gaggccaagg ccgccgccct 1380
gcagagagag atcgagaggg ccggcggcag attcagattt gtgggccact tcagcgaggc 1440
ccctggcacc agcgccttcg gcaccgccga gagacccgag ttcgagagaa tcctgaacga 1500
gtgtagggcc ggcaggctga acatgatcat cgtgtacgac gtgtcccggt tcagcaggct 1560
gaaggtgatg gacgccatcc ctatcgtgtc cgagctgctg gccctgggcg tgaccatcgt 1620
gtccacccag gaaggcgtct ttagacaggg caacgtgatg gacctgatcc acctgatcat 1680
gaggctggac gccagccaca aggagagcag cctgaagagc gccaagatcc tggacaccaa 1740
gaacctgcag agggagctgg gcggctatgt gggcggcaag gccccctacg gcttcgagct 1800
ggtgtccgag accaaggaga tcacccggaa cggcaggatg gtgaacgtgg tgatcaacaa 1860
gctggcccac agcaccaccc ccctgaccgg ccccttcgag tttgagcccg acgtgatcag 1920
gtggtggtgg cgggagatca agacccacaa gcacctgcct ttcaagcccg gcagccaggc 1980
cgccatccac cccggcagca tcaccggcct gtgtaagaga atggacgccg acgccgtgcc 2040
caccagaggc gagaccatcg gcaagaaaac cgccagcagc gcctgggacc ccgccaccgt 2100
gatgagaatc ctgagggacc ctaggatcgc cggcttcgcc gccgaggtga tctacaagaa 2160
gaagcccgac ggcaccccca ccaccaagat cgagggctac agaatccaga gagaccccat 2220
caccctgaga cctgtggagc tggactgtgg ccctatcatc gagcctgccg agtggtacga 2280
gctgcaggcc tggctggacg gcagaggcag aggcaagggc ctgagcagag gccaggccat 2340
cctgagcgcc atggacaagc tgtactgtga gtgtggcgcc gtgatgacca gcaagagagg 2400
cgaggagagc atcaaggaca gctaccggtg ccggagaaga aaggtggtgg accccagcgc 2460
ccctggccag cacgagggca cctgtaatgt gagcatggcc gccctggaca agttcgtggc 2520
cgagcggatc ttcaacaaga tccggcacgc cgagggcgac gaggagaccc tggccctgct 2580
gtgggaggcc gccagaagat tcggcaagct gaccgaggcc cccgagaaga gcggcgagag 2640
ggccaacctg gtggccgaga gagccgacgc cctgaacgcc ctggaggagc tgtacgagga 2700
cagagccgcc ggagcctatg acggccctgt gggcaggaag cacttcagaa agcagcaggc 2760
cgccctgacc ctgagacagc agggcgccga ggaaagactg gccgagctgg aggccgccga 2820
ggcccctaag ctgcccctgg atcagtggtt ccccgaggat gccgacgccg accccaccgg 2880
ccccaagtcc tggtggggca gagccagcgt ggacgacaag agggtgttcg tgggcctgtt 2940
cgtggataag atcgtggtga ccaagagcac caccggcagg ggccagggca cccccatcga 3000
gaagagagcc agcatcacct gggccaagcc tcccaccgac gacgacgagg atgacgccca 3060
ggacggcacc gaggacgtgg ccgcccctaa gaaaaagcgg aaagtgtgac tcgagacgcg 3120
tgatatcttt cccgggggta ccgtcgactg cggccgcgaa ttccaagctt gagtattcta 3180
tcgtgtcacc taaataactt ggcgtaatca tggtcatatc tgtttcctgt gtgaaattgt 3240
tatccgctca caattccaca caacatacga gccggaagca taaagtgtaa agcctggggt 3300
gcctaatgag tgagctaact cacattaatt gcgttgcgcg atgcttccat tttgtgaggg 3360
ttaatgcttc gagaagacat gataagatac attgatgagt ttggacaaac cacaacaaga 3420
atgcagtgaa aaaaatgctt tatttgtgaa atttgtgatg ctattgcttt atttgtaacc 3480
attataagct gcaataaaca agttaacaac aacaattgca ttcattttat gtttcaggtt 3540
cagggggaga tgtgggaggt tttttaaagc aagtaaaacc tctacaaatg tggtaaaatc 3600
cgataaggat cgattccgga gcctgaatgg cgaatggacg cgccctgtag cggcgcatta 3660
agcgcggcgg gtgtggtggt tacgcgcacg tgaccgctac acttgccagc gccctagcgc 3720
ccgctccttt cgctttcttc ccttcctttc tcgccacgtt cgccggcttt ccccgtcaag 3780
ctctaaatcg ggggctccct ttagggttcc gatttagtgc tttacggcac ctcgacccca 3840
aaaaacttga ttagggtgat ggttcacgta gtgggccatc gccctgatag acggtttttc 3900
gccctttgac gttggagtcc acgttcttta atagtggact cttgttccaa actggaacaa 3960
cactcaaccc tatctcggtc tattcttttg atttataagg gattttgccg atttcggcct 4020
attggttaaa aaatgagctg atttaacaaa aatttaacgc gaattttaac aaaatattaa 4080
cgcttacaat ttcgcctgtg taccttctga ggcggaaaga accagctgtg gaatgtgtgt 4140
cagttagggt gtggaaagtc cccaggctcc ccagcaggca gaagtatgca aagcatgcat 4200
ctcaattagt cagcaaccag gtgtggaaag tccccaggct ccccagcagg cagaagtatg 4260
caaagcatgc atctcaatta gtcagcaacc atagtcccgc ccctaactcc gcccatcccg 4320
cccctaactc cgcccagttc cgcccattct ccgccccatg gctgactaat tttttttatt 4380
tatgcagagg ccgaggccgc ctcggcctct gagctattcc agaagtagtg aggaggcttt 4440
tttggaggcc taggcttttg caaaaagctt gattcttctg acacaacagt ctcgaactta 4500
aggctagagc caccatgatt gaacaagatg gattgcacgc aggttctccg gccgcttggg 4560
tggagaggct attcggctat gactgggcac aacagacaat cggctgctct gatgccgccg 4620
tgttccggct gtcagcgcag gggcgcccgg ttctttttgt caagaccgac ctgtccggtg 4680
ccctgaatga actgcaggac gaggcagcgc ggctatcgtg gctggccacg acgggcgttc 4740
cttgcgcagc tgtgctcgac gttgtcactg aagcgggaag ggactggctg ctattgggcg 4800
aagtgccggg gcaggatctc ctgtcatctc accttgctcc tgccgagaaa gtatccatca 4860
tggctgatgc aatgcggcgg ctgcatacgc ttgatccggc tacctgccca ttcgaccacc 4920
aagcgaaaca tcgcatcgag cgagcacgta ctcggatgga agccggtctt gtcgatcagg 4980
atgatctgga cgaagagcat caggggctcg cgccagccga actgttcgcc aggctcaagg 5040
cgcgcatgcc cgacggcgag gatctcgtcg tgacccatgg cgatgcctgc ttgccgaata 5100
tcatggtgga aaatggccgc ttttctggat tcatcgactg tggccggctg ggtgtggcgg 5160
accgctatca ggacatagcg ttggctaccc gtgatattgc tgaagagctt ggcggcgaat 5220
gggctgaccg cttcctcgtg ctttacggta tcgccgctcc cgattcgcag cgcatcgcct 5280
tctatcgcct tcttgacgag ttcttctgag cgggactctg gggttcgaaa tgaccgacca 5340
agcgacgccc aacctgccat cacgatggcc gcaataaaat atctttattt tcattacatc 5400
tgtgtgttgg ttttttgtgt gcgtacgaag atccgcgtat ggtgcactct cagtacaatc 5460
tgctctgatg ccgcatagtt aagccagccc cgacacccgc caacacccgc tgacgcgccc 5520
tgacgggctt gtctgctccc ggcatccgct tacagacaag ctgtgaccgt ctccgggagc 5580
tgcatgtgtc agaggttttc accgtcatca ccgaaacgcg cgagacgaaa gggcctcgtg 5640
atacgcctat ttttataggt taatgtcatg ataataatgg tttcttagac gtcaggtggc 5700
acttttcggg gaaatgtgcg cggaacccct atttgtttat ttttctaaat acattcaaat 5760
atgtatccgc tcatgagaca ataaccctga taaatgcttc aataatattg aaaaaggaag 5820
agtatgagta ttcaacattt ccgtgtcgcc cttattccct tttttgcggc attttgcctt 5880
cctgtttttg ctcacccaga aacgctggtg aaagtaaaag atgctgaaga tcagttgggt 5940
gcacgagtgg gttacatcga actggatctc aacagcggta agatccttga gagttttcgc 6000
cccgaagaac gttttccaat gatgagcact tttaaagttc tgctatgtgg cgcggtatta 6060
tcccgtattg acgccgggca agagcaactc ggtcgccgca tacactattc tcagaatgac 6120
ttggttgagt actcaccagt cacagaaaag catcttacgg atggcatgac agtaagagaa 6180
ttatgcagtg ctgccataac catgagtgat aacactgcgg ccaacttact tctgacaacg 6240
atcggaggac cgaaggagct aaccgctttt ttgcacaaca tgggggatca tgtaactcgc 6300
cttgatcgtt gggaaccgga gctgaatgaa gccataccaa acgacgagcg tgacaccacg 6360
atgcctgtag caatggcaac aacgttgcgc aaactattaa ctggcgaact acttactcta 6420
gcttcccggc aacaattaat agactggatg gaggcggata aagttgcagg accacttctg 6480
cgctcggccc ttccggctgg ctggtttatt gctgataaat ctggagccgg tgagcgtggg 6540
tctcgcggta tcattgcagc actggggcca gatggtaagc cctcccgtat cgtagttatc 6600
tacacgacgg ggagtcaggc aactatggat gaacgaaata gacagatcgc tgagataggt 6660
gcctcactga ttaagcattg gtaactgtca gaccaagttt actcatatat actttagatt 6720
gatttaaaac ttcattttta atttaaaagg atctaggtga agatcctttt tgataatctc 6780
atgaccaaaa tcccttaacg tgagttttcg ttccactgag cgtcagaccc cgtagaaaag 6840
atcaaaggat cttcttgaga tccttttttt ctgcgcgtaa tctgctgctt gcaaacaaaa 6900
aaaccaccgc taccagcggt ggtttgtttg ccggatcaag agctaccaac tctttttccg 6960
aaggtaactg gcttcagcag agcgcagata ccaaatactg tccttctagt gtagccgtag 7020
ttaggccacc acttcaagaa ctctgtagca ccgcctacat acctcgctct gctaatcctg 7080
ttaccagtgg ctgctgccag tggcgataag tcgtgtctta ccgggttgga ctcaagacga 7140
tagttaccgg ataaggcgca gcggtcgggc tgaacggggg gttcgtgcac acagcccagc 7200
ttggagcgaa cgacctacac cgaactgaga tacctacagc gtgagctatg agaaagcgcc 7260
acgcttcccg aagggagaaa ggcggacagg tatccggtaa gcggcagggt cggaacagga 7320
gagcgcacga gggagcttcc agggggaaac gcctggtatc tttatagtcc tgtcgggttt 7380
cgccacctct gacttgagcg tcgatttttg tgatgctcgt caggggggcg gagcctatgg 7440
aaaaacgcca gcaacgcggc ctttttacgg ttcctggcct tttgctggcc ttttgctcac 7500
atggctcgac agatct 7516
<210> 2
<211> 5043
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 2
gaattaattc ataccagatc accgaaaact gtcctccaaa tgtgtccccc tcacactccc 60
aaattcgcgg gcttctgcct cttagaccac tctaccctat tccccacact caccggagcc 120
aaagccgcgg cccttccgtt tctttgctgt ccggccatta gccatattat tcattggtta 180
tatagcataa atcaatattg gctattggcc attgcatacg ttgtatccat atcataatat 240
gtacatttat attggctcat gtccaacatt accgccatgt tgacattgat tattgactag 300
ttattaatag taatcaatta cggggtcatt agttcatagc ccatatatgg agttccgcgt 360
tacataactt acggtaaatg gcccgcctgg ctgaccgccc aacgaccccc gcccattgac 420
gtcaataatg acgtatgttc ccatagtaac gccaataggg actttccatt gacgtcaatg 480
ggtggagtat ttacggtaaa ctgcccactt ggcagtacat caagtgtatc atatgccaag 540
tacgccccct attgacgtca atgacggtaa atggcccgcc tggcattatg cccagtacat 600
gaccttatgg gactttccta cttggcagta catctacgta ttagtcatcg ctattaccat 660
ggtgatgcgg ttttggcagt acatcaatgg gcgtggatag cggtttgact cacggggatt 720
tccaagtctc caccccattg acgtcaatgg gagtttgttt tggcaccaaa atcaacggga 780
ctttccaaaa tgtcgtaaca actccgcccc attgacgcaa atgggcggta ggcgtgtacg 840
gtgggaggtc tatataagca gagctcaata aaagagccca caacccctca ctcggcgcgc 900
cagtcttccg atagactgcg tcgcccgggt acccgtattc ccaataaagc ctcttgctgt 960
ttgcatccga atcgtggtct cgctgttcct tgggagggtc tcctctgagt gattgactac 1020
ccacgacggg ggtctttcat ttgggggctc gtccgggatt tggagacccc tgcccaggga 1080
ccaccgaccc accaccggga ggtaagctgg ccagcaactt atctgtgtct gtccgattgt 1140
ctagtgtcta tgtttgatgt tatgcgcctg cgtctgtact agttagctaa ctagctctgt 1200
atctggcgga cccgtggtgg aactgacgag ttctgaacac ccggccgcaa ccctgggaga 1260
cgtcccaggg actttggggg ccgtttttgt ggcccgacct gaggaaggga gtcgatgtgg 1320
aatccgaccc cgtcaggata tgtggttctg gtaggagacg agaacctaaa acagttcccg 1380
cctccgtctg aatttttgct ttcggtttgg aaccgaagcc gcgcgtcttg tctgctgcag 1440
cgctgcagca tcgttctgtg ttgtctctgt ctgactgtgt ttctgtattt gtctgaaaat 1500
tagggccaga ctgttaccac tcccttaagt ttgaccttag gtcactggaa agatgtcgag 1560
cggatcgctc acaaccagtc ggtagatgtc aagaagagac gttgggttac cttctgctct 1620
gcagaatggc caacctttaa cgtcggatgg ccgcgagacg gcacctttaa ccgagacctc 1680
atcacccagg ttaagatcaa ggtcttttca cctggcccgc atggacaccc agaccaggtc 1740
ccctacatcg tgacctggga agccttggct tttgaccccc ctccctgggt caagcccttt 1800
gtacacccta agcctccgcc tcctcttcct ccatccgccc cgtctctccc ccttgaacct 1860
cctcgttcga ccccgcctcg atcctccctt tatccagccc tcactccttc tctaggcgcc 1920
ggaattgacg cgcgcgtagg cctgcggccg cagtactgac ggacacaccg aagccccggc 1980
ggcaaccctc agcggatgcc ccggggcttc acgttttccc aggtcagaag cggttttcgg 2040
gagtagtgcc ccaactgggg taacctttga gttctctcag ttgggggcgt agggtcgccg 2100
acatgacaca aggggttgtg accggggtgg acacgtacgc gggtgcttac gaccgtcagt 2160
cgcgcgagcg cgatagtctc gagttcgaca tcgataaaat aaaagatttt atttagtctc 2220
cagaaaaagg ggggaatgaa agaccccacc tgtaggtttg gcaagctagc ttaagtaacg 2280
ccattttgca aggcatggaa aaatacataa ctgagaatag aaaagttcag atcaaggtca 2340
ggaacagatg gaacagggtc gaccggtcga ccggtcgacc ctagagaacc atcagatgtt 2400
tccagggtgc cccaaggacc tgaaatgacc ctgtgcctta tttgaactaa ccaatcagtt 2460
cgcttctcgc ttctgttcgc gcgcttctgc tccccgagct caataaaaga gcccacaacc 2520
cctcactcgg ggcgccagtc ctccgattga ctgagtcgcc cgggtacccg tgtatccaat 2580
aaaccctctt gcagttgcat ccgacttgtg gtctcgctgt tccttgggag ggtctcctct 2640
gagtgattga ctacccgtca gcgggggtct ttcatttggg ggctcgtccg ggatcgggag 2700
acccctgccc agggaccacc gacccaccac cgggaggtaa gctggctgcc tcgcgcgttt 2760
cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca cagcttgtct 2820
gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg ttggcgggtg 2880
tcggggcgca gccatgaccc agtcacgtag cgatagcgga gtgtatactg gcttaactat 2940
gcggcatcag agcagattgt actgagagtg caccatatgc ggtgtgaaat accgcacaga 3000
tgcgtaagga gaaaataccg catcaggcgc tcttccgctt cctcgctcac tgactcgctg 3060
cgctcggtcg ttcggctgcg gcgagcggta tcagctcact caaaggcggt aatacggtta 3120
tccacagaat caggggataa cgcaggaaag aacatgtatg catgagcaaa aggccagcaa 3180
aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct ccgcccccct 3240
gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac aggactataa 3300
agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc gaccctgccg 3360
cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc tcatagctca 3420
cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg tgtgcacgaa 3480
ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga gtccaacccg 3540
gtaagacacg acttatcgcc actggcagca gccactggta acaggattag cagagcgagg 3600
tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta cactagaaga 3660
acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag agttggtagc 3720
tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg caagcagcag 3780
attacgcgca gaaaaaaagg atctcaagaa gatcctttga tcttttctac ggggtctgac 3840
gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc aaaaaggatc 3900
ttcacctaga tccttttaaa ttaaaaatga agttttaaat caatctaaag tatatatgag 3960
taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc agcgatctgt 4020
ctatttcgtt catccatagt tgcctgactc cccgtcgtgt agataactac gatacgggag 4080
ggcttaccat ctggccccag tgctgcaatg ataccgcgag acccacgctc accggctcca 4140
gatttatcag caataaacca gccagccgga agggccgagc gcagaagtgg tcctgcaact 4200
ttatccgcct ccatccagtc tattaattgt tgccgggaag ctagagtaag tagttcgcca 4260
gttaatagtt tgcgcaacgt tgttgccatt gctgcaggca tcgtggtgtc acgctcgtcg 4320
tttggtatgg cttcattcag ctccggttcc caacgatcaa ggcgagttac atgatccccc 4380
atgttgtgca aaaaagcggt tagctccttc ggtcctccga tcgttgtcag aagtaagttg 4440
gccgcagtgt tatcactcat ggttatggca gcactgcata attctcttac tgtcatgcca 4500
tccgtaagat gcttttctgt gactggtgag tactcaacca agtcattctg agaatagtgt 4560
atgcggcgac cgagttgctc ttgcccggcg tcaacacggg ataataccgc gccacatagc 4620
agaactttaa aagtgctcat cattggaaaa cgttcttcgg ggcgaaaact ctcaaggatc 4680
ttaccgctgt tgagatccag ttcgatgtaa cccactcgtg cacccaactg atcttcagca 4740
tcttttactt tcaccagcgt ttctgggtga gcaaaaacag gaaggcaaaa tgccgcaaaa 4800
aagggaataa gggcgacacg gaaatgttga atactcatac tcttcctttt tcaatattat 4860
tgaagcattt atcagggtta ttgtctcatg agcggataca tatttgaatg tatttagaaa 4920
aataaacaaa taggggttcc gcgcacattt ccccgaaaag tgccacctga cgtctaagaa 4980
accattatta tcatgacatt aacctataaa aataggcgta tcacgaggcc ctttcgtctt 5040
caa 5043
<210> 3
<211> 5638
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 3
gaattaattc ataccagatc accgaaaact gtcctccaaa tgtgtccccc tcacactccc 60
aaattcgcgg gcttctgcct cttagaccac tctaccctat tccccacact caccggagcc 120
aaagccgcgg cccttccgtt tctttgctgt ccggccatta gccatattat tcattggtta 180
tatagcataa atcaatattg gctattggcc attgcatacg ttgtatccat atcataatat 240
gtacatttat attggctcat gtccaacatt accgccatgt tgacattgat tattgactag 300
ttattaatag taatcaatta cggggtcatt agttcatagc ccatatatgg agttccgcgt 360
tacataactt acggtaaatg gcccgcctgg ctgaccgccc aacgaccccc gcccattgac 420
gtcaataatg acgtatgttc ccatagtaac gccaataggg actttccatt gacgtcaatg 480
ggtggagtat ttacggtaaa ctgcccactt ggcagtacat caagtgtatc atatgccaag 540
tacgccccct attgacgtca atgacggtaa atggcccgcc tggcattatg cccagtacat 600
gaccttatgg gactttccta cttggcagta catctacgta ttagtcatcg ctattaccat 660
ggtgatgcgg ttttggcagt acatcaatgg gcgtggatag cggtttgact cacggggatt 720
tccaagtctc caccccattg acgtcaatgg gagtttgttt tggcaccaaa atcaacggga 780
ctttccaaaa tgtcgtaaca actccgcccc attgacgcaa atgggcggta ggcgtgtacg 840
gtgggaggtc tatataagca gagctcaata aaagagccca caacccctca ctcggcgcgc 900
cagtcttccg atagactgcg tcgcccgggt acccgtattc ccaataaagc ctcttgctgt 960
ttgcatccga atcgtggtct cgctgttcct tgggagggtc tcctctgagt gattgactac 1020
ccacgacggg ggtctttcat ttgggggctc gtccgggatt tggagacccc tgcccaggga 1080
ccaccgaccc accaccggga ggtaagctgg ccagcaactt atctgtgtct gtccgattgt 1140
ctagtgtcta tgtttgatgt tatgcgcctg cgtctgtact agttagctaa ctagctctgt 1200
atctggcgga cccgtggtgg aactgacgag ttctgaacac ccggccgcaa ccctgggaga 1260
cgtcccaggg actttggggg ccgtttttgt ggcccgacct gaggaaggga gtcgatgtgg 1320
aatccgaccc cgtcaggata tgtggttctg gtaggagacg agaacctaaa acagttcccg 1380
cctccgtctg aatttttgct ttcggtttgg aaccgaagcc gcgcgtcttg tctgctgcag 1440
cgctgcagca tcgttctgtg ttgtctctgt ctgactgtgt ttctgtattt gtctgaaaat 1500
tagggccaga ctgttaccac tcccttaagt ttgaccttag gtcactggaa agatgtcgag 1560
cggatcgctc acaaccagtc ggtagatgtc aagaagagac gttgggttac cttctgctct 1620
gcagaatggc caacctttaa cgtcggatgg ccgcgagacg gcacctttaa ccgagacctc 1680
atcacccagg ttaagatcaa ggtcttttca cctggcccgc atggacaccc agaccaggtc 1740
ccctacatcg tgacctggga agccttggct tttgaccccc ctccctgggt caagcccttt 1800
gtacacccta agcctccgcc tcctcttcct ccatccgccc cgtctctccc ccttgaacct 1860
cctcgttcga ccccgcctcg atcctccctt tatccagccc tcactccttc tctaggcgcc 1920
ggaattgacg cgcgcgtagg cctgcggccg cagtactgac ggacacaccg aagccccggc 1980
ggcaaccctc agcggatgcc ccggggcttc acgttttccc aggtcagaag cggttttcgg 2040
gagtagtgcc ccaactgggg taacctttga gttctctcag ttgggggcgt agggtcgccg 2100
acatgacaca aggggttgtg accggggtgg acacgtacgc gggtgcttac gaccgtcagt 2160
cgcgcgagcg cgatagtctc gagttcgaca tcgataatca acctctggat tacaaaattt 2220
gtgaaagatt gactggtatt cttaactatg ttgctccttt tacgctatgt ggatacgctg 2280
ctttaatgcc tttgtatcat gctattgctt cccgtatggc tttcattttc tcctccttgt 2340
ataaatcctg gttgctgtct ctttatgagg agttgtggcc cgttgtcagg caacgtggcg 2400
tggtgtgcac tgtgtttgct gacgcaaccc ccactggttg gggcattgcc accacctgtc 2460
agctcctttc cgggactttc gctttccccc tccctattgc cacggcggaa ctcatcgccg 2520
cctgccttgc ccgctgctgg acaggggctc ggctgttggg cactgacaat tccgtggtgt 2580
tgtcggggaa atcatcgtcc tttccttggc tgctcgcctg tgttgccacc tggattctgc 2640
gcgggacgtc cttctgctac gtcccttcgg ccctcaatcc agcggacctt ccttcccgcg 2700
gcctgctgcc ggctctgcgg cctcttccgc gtcttcgcct tcgccctcag acgagtcgga 2760
tctccctttg ggccgcctcc ccgcatcgat aaaataaaag attttattta gtctccagaa 2820
aaagggggga atgaaagacc ccacctgtag gtttggcaag ctagcttaag taacgccatt 2880
ttgcaaggca tggaaaaata cataactgag aatagaaaag ttcagatcaa ggtcaggaac 2940
agatggaaca gggtcgaccg gtcgaccggt cgaccctaga gaaccatcag atgtttccag 3000
ggtgccccaa ggacctgaaa tgaccctgtg ccttatttga actaaccaat cagttcgctt 3060
ctcgcttctg ttcgcgcgct tctgctcccc gagctcaata aaagagccca caacccctca 3120
ctcggggcgc cagtcctccg attgactgag tcgcccgggt acccgtgtat ccaataaacc 3180
ctcttgcagt tgcatccgac ttgtggtctc gctgttcctt gggagggtct cctctgagtg 3240
attgactacc cgtcagcggg ggtctttcat ttgggggctc gtccgggatc gggagacccc 3300
tgcccaggga ccaccgaccc accaccggga ggtaagctgg ctgcctcgcg cgtttcggtg 3360
atgacggtga aaacctctga cacatgcagc tcccggagac ggtcacagct tgtctgtaag 3420
cggatgccgg gagcagacaa gcccgtcagg gcgcgtcagc gggtgttggc gggtgtcggg 3480
gcgcagccat gacccagtca cgtagcgata gcggagtgta tactggctta actatgcggc 3540
atcagagcag attgtactga gagtgcacca tatgcggtgt gaaataccgc acagatgcgt 3600
aaggagaaaa taccgcatca ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc 3660
ggtcgttcgg ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac 3720
agaatcaggg gataacgcag gaaagaacat gtatgcatga gcaaaaggcc agcaaaaggc 3780
caggaaccgt aaaaaggccg cgttgctggc gtttttccat aggctccgcc cccctgacga 3840
gcatcacaaa aatcgacgct caagtcagag gtggcgaaac ccgacaggac tataaagata 3900
ccaggcgttt ccccctggaa gctccctcgt gcgctctcct gttccgaccc tgccgcttac 3960
cggatacctg tccgcctttc tcccttcggg aagcgtggcg ctttctcata gctcacgctg 4020
taggtatctc agttcggtgt aggtcgttcg ctccaagctg ggctgtgtgc acgaaccccc 4080
cgttcagccc gaccgctgcg ccttatccgg taactatcgt cttgagtcca acccggtaag 4140
acacgactta tcgccactgg cagcagccac tggtaacagg attagcagag cgaggtatgt 4200
aggcggtgct acagagttct tgaagtggtg gcctaactac ggctacacta gaagaacagt 4260
atttggtatc tgcgctctgc tgaagccagt taccttcgga aaaagagttg gtagctcttg 4320
atccggcaaa caaaccaccg ctggtagcgg tggttttttt gtttgcaagc agcagattac 4380
gcgcagaaaa aaaggatctc aagaagatcc tttgatcttt tctacggggt ctgacgctca 4440
gtggaacgaa aactcacgtt aagggatttt ggtcatgaga ttatcaaaaa ggatcttcac 4500
ctagatcctt ttaaattaaa aatgaagttt taaatcaatc taaagtatat atgagtaaac 4560
ttggtctgac agttaccaat gcttaatcag tgaggcacct atctcagcga tctgtctatt 4620
tcgttcatcc atagttgcct gactccccgt cgtgtagata actacgatac gggagggctt 4680
accatctggc cccagtgctg caatgatacc gcgagaccca cgctcaccgg ctccagattt 4740
atcagcaata aaccagccag ccggaagggc cgagcgcaga agtggtcctg caactttatc 4800
cgcctccatc cagtctatta attgttgccg ggaagctaga gtaagtagtt cgccagttaa 4860
tagtttgcgc aacgttgttg ccattgctgc aggcatcgtg gtgtcacgct cgtcgtttgg 4920
tatggcttca ttcagctccg gttcccaacg atcaaggcga gttacatgat cccccatgtt 4980
gtgcaaaaaa gcggttagct ccttcggtcc tccgatcgtt gtcagaagta agttggccgc 5040
agtgttatca ctcatggtta tggcagcact gcataattct cttactgtca tgccatccgt 5100
aagatgcttt tctgtgactg gtgagtactc aaccaagtca ttctgagaat agtgtatgcg 5160
gcgaccgagt tgctcttgcc cggcgtcaac acgggataat accgcgccac atagcagaac 5220
tttaaaagtg ctcatcattg gaaaacgttc ttcggggcga aaactctcaa ggatcttacc 5280
gctgttgaga tccagttcga tgtaacccac tcgtgcaccc aactgatctt cagcatcttt 5340
tactttcacc agcgtttctg ggtgagcaaa aacaggaagg caaaatgccg caaaaaaggg 5400
aataagggcg acacggaaat gttgaatact catactcttc ctttttcaat attattgaag 5460
catttatcag ggttattgtc tcatgagcgg atacatattt gaatgtattt agaaaaataa 5520
acaaataggg gttccgcgca catttccccg aaaagtgcca cctgacgtct aagaaaccat 5580
tattatcatg acattaacct ataaaaatag gcgtatcacg aggccctttc gtcttcaa 5638
<210> 4
<211> 8348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 4
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgaatt catttaaatg aaagacccca 420
cctgtaggtt tggcaagcta gcttaagtaa cgccattttg caaggcatgg aaaaatacat 480
aactgagaat agaaaagttc agatcaaggt caggaacaga tggaacaggg tcgaccggtc 540
gaccggtcga ccctagagaa ccatcagatg tttccagggt gccccaagga cctgaaatga 600
ccctgtgcct tatttgaact aaccaatcag ttcgcttctc gcttctgttc gcgcgcttct 660
gctccccgag ctcaataaaa gagcccacaa cccctcactc ggggcgccag tcttccgata 720
gactgcgtcg cccgggtacc cgtattccca ataaagcctc ttgctgtttg catccgaatc 780
gtggtctcgc tgttccttgg gagggtctcc tctgagtgat tgactaccca cgacgggggt 840
ctttcatttg ggggctcgtc cgggatttgg agacccctgc ccagggacca ccgacccacc 900
accgggaggt aagctggcca gcaacttatc tgtgtctgtc cgattgtcta gtgtctatgt 960
ttgatgttat gcgcctgcgt ctgtactagt tagctaacta gctctgtatc tggcggaccc 1020
gtggtggaac tgacgagttc tgaacacccg gccgcaaccc tgggagacgt cccagggact 1080
ttgggggccg tttttgtggc ccgacctgag gaagggagtc gatgtggaat ccgaccccgt 1140
caggatatgt ggttctggta ggagacgaga acctaaaaca gttcccgcct ccgtctgaat 1200
ttttgctttc ggtttggaac cgaagccgcg cgtcttgtct gctgcagcgc tgcagcatcg 1260
ttctgtgttg tctctgtctg actgtgtttc tgtatttgtc tgaaaattag ggccagactg 1320
ttaccactcc cttaagtttg accttaggtc actggaaaga tgtcgagcgg atcgctcaca 1380
accagtcggt agatgtcaag aagagacgtt gggttacctt ctgctctgca gaatggccaa 1440
cctttaacgt cggatggccg cgagacggca cctttaaccg agacctcatc acccaggtta 1500
agatcaaggt cttttcacct ggcccgcatg gacacccaga ccaggtcccc tacatcgtga 1560
cctgggaagc cttggctttt gacccccctc cctgggtcaa gccctttgta caccctaagc 1620
ctccgcctcc tcttcctcca tccgccccgt ctctccccct tgaacctcct cgttcgaccc 1680
cgcctcgatc ctccctttat ccagccctca ctccttctct aggcgccgga attgccttcc 1740
accatggcca cctcagcaag ttcccacttg aacaaaaaca tcaagcaaat gtacttgtgc 1800
ctgccccagg gtgagaaagt ccaagccatg tatatctggg ttgatggtac tggagaagga 1860
ctgcgctgca aaacccgcac cctggactgt gagcccaagt gtgtagaaga gttacctgag 1920
tggaattttg atggctctag tacctttcag tctgagggct ccaacagtga catgtatctc 1980
agccctgttg ccatgtttcg ggaccccttc cgcagagatc ccaacaagct ggtgttctgt 2040
gaagttttca agtacaaccg gaagcctgca gagaccaatt taaggcactc gtgtaaacgg 2100
ataatggaca tggtgagcaa ccagcacccc tggtttggaa tggaacagga gtatactctg 2160
atgggaacag atgggcaccc ttttggttgg ccttccaatg gctttcctgg gccccaaggt 2220
ccgtattact gtggtgtggg cgcagacaaa gcctatggca gggatatcgt ggaggctcac 2280
taccgcgcct gcttgtatgc tggggtcaag attacaggaa caaatgctga ggtcatgcct 2340
gcccagtggg agttccaaat aggaccctgt gaaggaatcc gcatgggaga tcatctctgg 2400
gtggcccgtt tcatcttgca tcgagtatgt gaagactttg gggtaatagc aacctttgac 2460
cccaagccca ttcctgggaa ctggaatggt gcaggctgcc ataccaactt tagcaccaag 2520
gccatgcggg aggagaatgg tctgaagcac atcgaggagg ccatcgagaa actaagcaag 2580
cggcaccggt accacattcg agcctacgat cccaaggggg gcctggacaa tgcccgtcgt 2640
ctgactgggt tccacgaaac gtccaacatc aacgactttt ctgctggtgt cgccaatcgc 2700
agtgccagca tccgcattcc ccggactgtc ggccaggaga agaaaggtta ctttgaagac 2760
cgccgcccct ctgccaactg tgaccccttt gcagtgacag aagccatcgt ccgcacatgc 2820
cttctcaatg agactggcga cgagcccttc caatacaaaa actaaagatc cctatggcta 2880
ttggccaggt tcaatactat gtattggccc tatgccatat agtattccat atatgggttt 2940
tcctattgac gtagatagcc cctcccaatg ggcggtccca tataccatat atggggcttc 3000
ctaataccgc ccatagccac tcccccattg acgtcaatgg tctctatata tggtctttcc 3060
tattgacgtc atatgggcgg tcctattgac gtatatggcg cctcccccat tgacgtcaat 3120
tacggtaaat ggcccgcctg gctcaatgcc cattgacgtc aataggacca cccaccattg 3180
acgtcaatgg gatggctcat tgcccattca tatccgttct cacgccccct attgacgtca 3240
atgacggtaa atggcccact tggcagtaca tcaatatcta ttaatagtaa cttggcaagt 3300
acattactat tggaagtacg ccagggtaca ttggcagtac tcccattgac gtcaatggcg 3360
gtaaatggcc cgcgatggct gccaagtaca tccccattga cgtcaatggg gaggggcaat 3420
gacgcaaatg ggcgttccat tgacgtaaat gggcggtagg cgtgcctaat gggaggtcta 3480
tataagcaat gctcgtttag ggaaccgcca ttctgcctgg ggacgtcgga ggagctcgaa 3540
agcttctaga caattgccgc caccatgatg tcctttgtct ctctgctcct ggttggcatc 3600
ctattccatg ccacccaggc cagtgataca ggtagacctt tcgtagagat gtacagtgaa 3660
atccccgaaa ttatacacat gactgaagga agggagctcg tcattccctg ccgggttacg 3720
tcacctaaca tcactgttac tttaaaaaag tttccacttg acactttgat ccctgatgga 3780
aaacgcataa tctgggacag tagaaagggc ttcatcatat caaatgcaac gtacaaagaa 3840
atagggcttc tgacctgtga agcaacagtc aatgggcatt tgtataagac aaactatctc 3900
acacatcgac aaaccaatac aatcatagat gtcgttctga gtccgtctca tggaattgaa 3960
ctatctgttg gagaaaagct tgtcttaaat tgtacagcaa gaactgaact aaatgtgggg 4020
attgacttca actgggaata cccttcttcg aagcatcagc ataagaaact tgtaaaccga 4080
gacctaaaaa cccagtctgg gagtgagatg aagaagtttt tgagcacctt aactatagat 4140
ggtgtaaccc ggagtgacca aggattgtac acctgtgcag catccagtgg gctgatgacc 4200
aagaaaaaca gcacatttgt cagggtccat gaaaaagaca aaactcacac atgcccaccg 4260
tgcccagcac ctgaactcct ggggggaccc tcagtcttcc tcttcccccc aaaacccaag 4320
gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac 4380
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 4440
acaaagccac gggaggagca gtacaacagc acatatcgtg tggtcagcgt cctcaccgtc 4500
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 4560
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 4620
tacaccctgc ccccatcccg ggatgagctg accaagaacc aggtcagcct gacctgcctg 4680
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 4740
aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctctacagc 4800
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 4860
catgaggctc tgcacaacca ctacacgcag aagagcctct ccctgtctcc cgggaaatga 4920
tgagatctcg agttcgacat cgataatcaa cctctggatt acaaaatttg tgaaagattg 4980
actggtattc ttaactatgt tgctcctttt acgctatgtg gatacgctgc tttaatgcct 5040
ttgtatcatg ctattgcttc ccgtatggct ttcattttct cctccttgta taaatcctgg 5100
ttgctgtctc tttatgagga gttgtggccc gttgtcaggc aacgtggcgt ggtgtgcact 5160
gtgtttgctg acgcaacccc cactggttgg ggcattgcca ccacctgtca gctcctttcc 5220
gggactttcg ctttccccct ccctattgcc acggcggaac tcatcgccgc ctgccttgcc 5280
cgctgctgga caggggctcg gctgttgggc actgacaatt ccgtggtgtt gtcggggaaa 5340
tcatcgtcct ttccttggct gctcgcctgt gttgccacct ggattctgcg cgggacgtcc 5400
ttctgctacg tcccttcggc cctcaatcca gcggaccttc cttcccgcgg cctgctgccg 5460
gctctgcggc ctcttccgcg tcttcgcctt cgccctcaga cgagtcggat ctccctttgg 5520
gccgcctccc cgcatcgatg ggggaggcta actgaaacac ggaaggagac aataccggaa 5580
ggaacccgcg ctatgacggc aataaaaaga cagaataaaa cgcacgggtg ttgggtcgtt 5640
tgttcataaa cgcggggttc ggtcccaggg ctggcactct gtcgataccc caccgagacc 5700
ccattggggc caatacgccc gcgtttcttc cttttcccca ccccaccccc caagttcggg 5760
tgaaggccca gggctcgcag ccaacgtcgg ggcggcaggc cctgccatag cggtcgacga 5820
tgtaggtcac ggtctcgaag ccgcggtgcg ggtgccaggg cgtgcccttg ggctccccgg 5880
gcgcgtactc cacctcaccc atctggtcca tcatgatgaa cgggtcgagg tggcggtagt 5940
tgatcccggc gaacgcgcgg cgcaccggga agccctcgcc ctcgaaaccg ctgggcgcgg 6000
tggtcacggt gagcacggga cgtgcgacgg cgtcggcggg tgcggatacg cggggcagcg 6060
tcagcgggtt ctcgacggtc acggcgggca tgatttccac tgtacgcgta gcttggcgta 6120
atcatggtca tagctgtttc ctgtgtgaaa ttgttatccg ctcacaattc cacacaacat 6180
acgagccgga agcataaagt gtaaagcctg gggtgcctaa tgagtgagct aactcacatt 6240
aattgcgttg cgctcactgc ccgctttcca gtcgggaaac ctgtcgtgcc agctgcatta 6300
atgaatcggc caacgcgcgg ggagaggcgg tttgcgtatt gggcgctctt ccgcttcctc 6360
gctcactgac tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag ctcactcaaa 6420
ggcggtaata cggttatcca cagaatcagg ggataacgca ggaaagaaca tgtgagcaaa 6480
aggccagcaa aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct 6540
ccgcccccct gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac 6600
aggactataa agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc 6660
gaccctgccg cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc 6720
tcatagctca cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg 6780
tgtgcacgaa ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga 6840
gtccaacccg gtaagacacg acttatcgcc actggcagca gccactggta acaggattag 6900
cagagcgagg tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta 6960
cactagaaga acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag 7020
agttggtagc tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg 7080
caagcagcag attacgcgca gaaaaaaagg atctcaagaa gatcctttga tcttttctac 7140
ggggtctgac gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc 7200
aaaaaggatc ttcacctaga tccttttaaa ttaaaaatga agttttaaat caatctaaag 7260
tatatatgag taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc 7320
agcgatctgt ctatttcgtt catccatagt tgcctgactc cccgtcgtgt agataactac 7380
gatacgggag ggcttaccat ctggccccag tgctgcaatg ataccgcgag acccacgctc 7440
accggctcca gatttatcag caataaacca gccagccgga agggccgagc gcagaagtgg 7500
tcctgcaact ttatccgcct ccatccagtc tattaattgt tgccgggaag ctagagtaag 7560
tagttcgcca gttaatagtt tgcgcaacgt tgttgccatt gctacaggca tcgtggtgtc 7620
acgctcgtcg tttggtatgg cttcattcag ctccggttcc caacgatcaa ggcgagttac 7680
atgatccccc atgttgtgca aaaaagcggt tagctccttc ggtcctccga tcgttgtcag 7740
aagtaagttg gccgcagtgt tatcactcat ggttatggca gcactgcata attctcttac 7800
tgtcatgcca tccgtaagat gcttttctgt gactggtgag tactcaacca agtcattctg 7860
agaatagtgt atgcggcgac cgagttgctc ttgcccggcg tcaatacggg ataataccgc 7920
gccacatagc agaactttaa aagtgctcat cattggaaaa cgttcttcgg ggcgaaaact 7980
ctcaaggatc ttaccgctgt tgagatccag ttcgatgtaa cccactcgtg cacccaactg 8040
atcttcagca tcttttactt tcaccagcgt ttctgggtga gcaaaaacag gaaggcaaaa 8100
tgccgcaaaa aagggaataa gggcgacacg gaaatgttga atactcatac tcttcctttt 8160
tcaatattat tgaagcattt atcagggtta ttgtctcatg agcggataca tatttgaatg 8220
tatttagaaa aataaacaaa taggggttcc gcgcacattt ccccgaaaag tgccacctga 8280
cgtctaagaa accattatta tcatgacatt aacctataaa aataggcgta tcacgaggcc 8340
ctttcgtc 8348
<210> 5
<211> 7011
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 5
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaaagctt ctagacaatt gccgccacca 2520
tgatgtcctt tgtctctctg ctcctggttg gcatcctatt ccatgccacc caggccagtg 2580
atacaggtag acctttcgta gagatgtaca gtgaaatccc cgaaattata cacatgactg 2640
aaggaaggga gctcgtcatt ccctgccggg ttacgtcacc taacatcact gttactttaa 2700
aaaagtttcc acttgacact ttgatccctg atggaaaacg cataatctgg gacagtagaa 2760
agggcttcat catatcaaat gcaacgtaca aagaaatagg gcttctgacc tgtgaagcaa 2820
cagtcaatgg gcatttgtat aagacaaact atctcacaca tcgacaaacc aatacaatca 2880
tagatgtcgt tctgagtccg tctcatggaa ttgaactatc tgttggagaa aagcttgtct 2940
taaattgtac agcaagaact gaactaaatg tggggattga cttcaactgg gaataccctt 3000
cttcgaagca tcagcataag aaacttgtaa accgagacct aaaaacccag tctgggagtg 3060
agatgaagaa gtttttgagc accttaacta tagatggtgt aacccggagt gaccaaggat 3120
tgtacacctg tgcagcatcc agtgggctga tgaccaagaa aaacagcaca tttgtcaggg 3180
tccatgaaaa agacaaaact cacacatgcc caccgtgccc agcacctgaa ctcctggggg 3240
gaccctcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc tcccggaccc 3300
ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc aagttcaact 3360
ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccacgggag gagcagtaca 3420
acagcacata tcgtgtggtc agcgtcctca ccgtcctgca ccaggactgg ctgaatggca 3480
aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag aaaaccatct 3540
ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca tcccgggatg 3600
agctgaccaa gaaccaggtc agcctgacct gcctggtcaa aggcttctat cccagcgaca 3660
tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc acgcctcccg 3720
tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac aagagcaggt 3780
ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac aaccactaca 3840
cgcagaagag cctctccctg tctcccggga aatgatgaga tctcgagttc gacatcgata 3900
atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac tatgttgctc 3960
cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt gcttcccgta 4020
tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat gaggagttgt 4080
ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca acccccactg 4140
gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc cccctcccta 4200
ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg gctcggctgt 4260
tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct tggctgctcg 4320
cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct tcggccctca 4380
atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt ccgcgtcttc 4440
gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgcat cgatggggga 4500
ggctaactga aacacggaag gagacaatac cggaaggaac ccgcgctatg acggcaataa 4560
aaagacagaa taaaacgcac gggtgttggg tcgtttgttc ataaacgcgg ggttcggtcc 4620
cagggctggc actctgtcga taccccaccg agaccccatt ggggccaata cgcccgcgtt 4680
tcttcctttt ccccacccca ccccccaagt tcgggtgaag gcccagggct cgcagccaac 4740
gtcggggcgg caggccctgc catagcccta gcagcttggc gtaatcatgg tcatagctgt 4800
ttcctgtgtg aaattgttat ccgctcacaa ttccacacaa catacgagcc ggaagcataa 4860
agtgtaaagc ctggggtgcc taatgagtga gctaactcac attaattgcg ttgcgctcac 4920
tgcccgcttt ccagtcggga aacctgtcgt gccagctgca ttaatgaatc ggccaacgcg 4980
cggggagagg cggtttgcgt attgggcgct cttccgcttc ctcgctcact gactcgctgc 5040
gctcggtcgt tcggctgcgg cgagcggtat cagctcactc aaaggcggta atacggttat 5100
ccacagaatc aggggataac gcaggaaaga acatgtgagc aaaaggccag caaaaggcca 5160
ggaaccgtaa aaaggccgcg ttgctggcgt ttttccatag gctccgcccc cctgacgagc 5220
atcacaaaaa tcgacgctca agtcagaggt ggcgaaaccc gacaggacta taaagatacc 5280
aggcgtttcc ccctggaagc tccctcgtgc gctctcctgt tccgaccctg ccgcttaccg 5340
gatacctgtc cgcctttctc ccttcgggaa gcgtggcgct ttctcatagc tcacgctgta 5400
ggtatctcag ttcggtgtag gtcgttcgct ccaagctggg ctgtgtgcac gaaccccccg 5460
ttcagcccga ccgctgcgcc ttatccggta actatcgtct tgagtccaac ccggtaagac 5520
acgacttatc gccactggca gcagccactg gtaacaggat tagcagagcg aggtatgtag 5580
gcggtgctac agagttcttg aagtggtggc ctaactacgg ctacactaga agaacagtat 5640
ttggtatctg cgctctgctg aagccagtta ccttcggaaa aagagttggt agctcttgat 5700
ccggcaaaca aaccaccgct ggtagcggtg gtttttttgt ttgcaagcag cagattacgc 5760
gcagaaaaaa aggatctcaa gaagatcctt tgatcttttc tacggggtct gacgctcagt 5820
ggaacgaaaa ctcacgttaa gggattttgg tcatgagatt atcaaaaagg atcttcacct 5880
agatcctttt aaattaaaaa tgaagtttta aatcaatcta aagtatatat gagtaaactt 5940
ggtctgacag ttaccaatgc ttaatcagtg aggcacctat ctcagcgatc tgtctatttc 6000
gttcatccat agttgcctga ctccccgtcg tgtagataac tacgatacgg gagggcttac 6060
catctggccc cagtgctgca atgataccgc gagacccacg ctcaccggct ccagatttat 6120
cagcaataaa ccagccagcc ggaagggccg agcgcagaag tggtcctgca actttatccg 6180
cctccatcca gtctattaat tgttgccggg aagctagagt aagtagttcg ccagttaata 6240
gtttgcgcaa cgttgttgcc attgctacag gcatcgtggt gtcacgctcg tcgtttggta 6300
tggcttcatt cagctccggt tcccaacgat caaggcgagt tacatgatcc cccatgttgt 6360
gcaaaaaagc ggttagctcc ttcggtcctc cgatcgttgt cagaagtaag ttggccgcag 6420
tgttatcact catggttatg gcagcactgc ataattctct tactgtcatg ccatccgtaa 6480
gatgcttttc tgtgactggt gagtactcaa ccaagtcatt ctgagaatag tgtatgcggc 6540
gaccgagttg ctcttgcccg gcgtcaatac gggataatac cgcgccacat agcagaactt 6600
taaaagtgct catcattgga aaacgttctt cggggcgaaa actctcaagg atcttaccgc 6660
tgttgagatc cagttcgatg taacccactc gtgcacccaa ctgatcttca gcatctttta 6720
ctttcaccag cgtttctggg tgagcaaaaa caggaaggca aaatgccgca aaaaagggaa 6780
taagggcgac acggaaatgt tgaatactca tactcttcct ttttcaatat tattgaagca 6840
tttatcaggg ttattgtctc atgagcggat acatatttga atgtatttag aaaaataaac 6900
aaataggggt tccgcgcaca tttccccgaa aagtgccacc tgacgtctaa gaaaccatta 6960
ttatcatgac attaacctat aaaaataggc gtatcacgag gccctttcgt c 7011
<210> 6
<211> 5680
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 6
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgcaagcttc aattgaggcc 2520
tcctaggtta attaagttta aacagatctc tcgagttcga catcgataat caacctctgg 2580
attacaaaat ttgtgaaaga ttgactggta ttcttaacta tgttgctcct tttacgctat 2640
gtggatacgc tgctttaatg cctttgtatc atgctattgc ttcccgtatg gctttcattt 2700
tctcctcctt gtataaatcc tggttgctgt ctctttatga ggagttgtgg cccgttgtca 2760
ggcaacgtgg cgtggtgtgc actgtgtttg ctgacgcaac ccccactggt tggggcattg 2820
ccaccacctg tcagctcctt tccgggactt tcgctttccc cctccctatt gccacggcgg 2880
aactcatcgc cgcctgcctt gcccgctgct ggacaggggc tcggctgttg ggcactgaca 2940
attccgtggt gttgtcgggg aaatcatcgt cctttccttg gctgctcgcc tgtgttgcca 3000
cctggattct gcgcgggacg tccttctgct acgtcccttc ggccctcaat ccagcggacc 3060
ttccttcccg cggcctgctg ccggctctgc ggcctcttcc gcgtcttcgc cttcgccctc 3120
agacgagtcg gatctccctt tgggccgcct ccccgcatcg atgggggagg ctaactgaaa 3180
cacggaagga gacaataccg gaaggaaccc gcgctatgac ggcaataaaa agacagaata 3240
aaacgcacgg gtgttgggtc gtttgttcat aaacgcgggg ttcggtccca gggctggcac 3300
tctgtcgata ccccaccgag accccattgg ggccaatacg cccgcgtttc ttccttttcc 3360
ccaccccacc ccccaagttc gggtgaaggc ccagggctcg cagccaacgt cggggcggca 3420
ggccctgcca tagccctagc agcttggccg taatcatggt catagctgtt tcctgtgtga 3480
aattgttatc cgctcacaat tccacacaac atacgagccg gaagcataaa gtgtaaagcc 3540
tggggtgcct aatgagtgag ctaactcaca ttaattgcgt tgcgctcact gcccgctttc 3600
cagtcgggaa acctgtcgtg ccagctgcat taatgaatcg gccaacgcgc ggggagaggc 3660
ggtttgcgta ttgggcgctc ttccgcttcc tcgctcactg actcgctgcg ctcggtcgtt 3720
cggctgcggc gagcggtatc agctcactca aaggcggtaa tacggttatc cacagaatca 3780
ggggataacg caggaaagaa catgtgagca aaaggccagc aaaaggccag gaaccgtaaa 3840
aaggccgcgt tgctggcgtt tttccatagg ctccgccccc ctgacgagca tcacaaaaat 3900
cgacgctcaa gtcagaggtg gcgaaacccg acaggactat aaagatacca ggcgtttccc 3960
cctggaagct ccctcgtgcg ctctcctgtt ccgaccctgc cgcttaccgg atacctgtcc 4020
gcctttctcc cttcgggaag cgtggcgctt tctcatagct cacgctgtag gtatctcagt 4080
tcggtgtagg tcgttcgctc caagctgggc tgtgtgcacg aaccccccgt tcagcccgac 4140
cgctgcgcct tatccggtaa ctatcgtctt gagtccaacc cggtaagaca cgacttatcg 4200
ccactggcag cagccactgg taacaggatt agcagagcga ggtatgtagg cggtgctaca 4260
gagttcttga agtggtggcc taactacggc tacactagaa gaacagtatt tggtatctgc 4320
gctctgctga agccagttac cttcggaaaa agagttggta gctcttgatc cggcaaacaa 4380
accaccgctg gtagcggtgg tttttttgtt tgcaagcagc agattacgcg cagaaaaaaa 4440
ggatctcaag aagatccttt gatcttttct acggggtctg acgctcagtg gaacgaaaac 4500
tcacgttaag ggattttggt catgagatta tcaaaaagga tcttcaccta gatcctttta 4560
aattaaaaat gaagttttaa atcaatctaa agtatatatg agtaaacttg gtctgacagt 4620
taccaatgct taatcagtga ggcacctatc tcagcgatct gtctatttcg ttcatccata 4680
gttgcctgac tccccgtcgt gtagataact acgatacggg agggcttacc atctggcccc 4740
agtgctgcaa tgataccgcg agacccacgc tcaccggctc cagatttatc agcaataaac 4800
cagccagccg gaagggccga gcgcagaagt ggtcctgcaa ctttatccgc ctccatccag 4860
tctattaatt gttgccggga agctagagta agtagttcgc cagttaatag tttgcgcaac 4920
gttgttgcca ttgctacagg catcgtggtg tcacgctcgt cgtttggtat ggcttcattc 4980
agctccggtt cccaacgatc aaggcgagtt acatgatccc ccatgttgtg caaaaaagcg 5040
gttagctcct tcggtcctcc gatcgttgtc agaagtaagt tggccgcagt gttatcactc 5100
atggttatgg cagcactgca taattctctt actgtcatgc catccgtaag atgcttttct 5160
gtgactggtg agtactcaac caagtcattc tgagaatagt gtatgcggcg accgagttgc 5220
tcttgcccgg cgtcaatacg ggataatacc gcgccacata gcagaacttt aaaagtgctc 5280
atcattggaa aacgttcttc ggggcgaaaa ctctcaagga tcttaccgct gttgagatcc 5340
agttcgatgt aacccactcg tgcacccaac tgatcttcag catcttttac tttcaccagc 5400
gtttctgggt gagcaaaaac aggaaggcaa aatgccgcaa aaaagggaat aagggcgaca 5460
cggaaatgtt gaatactcat actcttcctt tttcaatatt attgaagcat ttatcagggt 5520
tattgtctca tgagcggata catatttgaa tgtatttaga aaaataaaca aataggggtt 5580
ccgcgcacat ttccccgaaa agtgccacct gacgtctaag aaaccattat tatcatgaca 5640
ttaacctata aaaataggcg tatcacgagg ccctttcgtc 5680
<210> 7
<211> 8085
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 7
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgccctaggg tttaaacaga 2520
tctatcgata atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac 2580
tatgttgctc cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt 2640
gcttcccgta tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat 2700
gaggagttgt ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca 2760
acccccactg gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc 2820
cccctcccta ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg 2880
gctcggctgt tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct 2940
tggctgctcg cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct 3000
tcggccctca atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt 3060
ccgcgtcttc gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgcat 3120
cgattactaa tcagccatac cacatttgta gaggttttac ttgctttaaa aaacctccca 3180
cacctccccc tgaacctgaa acataaaatg aatgcaattg ttgttgttaa cttgtttatt 3240
gcagcttata atggttacaa ataaagcaat agcatcacaa atttcacaaa taaagcattt 3300
ttttcactgc attctagttg tggtttgtcc aaactcatca atgtatctta tcatgtctgg 3360
aattcggacg gtgactgcag tgaataataa aatgtgtgtt tgtccgaaat acgcgttttg 3420
agatttctgt cgccgactaa attcatgtcg cgcgatagtg gtgtttatcg ccgatagaga 3480
tggcgatatt ggaaaaatcg atatttgaaa atatggcata ttgaaaatgt cgccgatgtg 3540
agtttctgtg taactgatat cgccattttt ccaaaagtga tttttgggca tacgcgatat 3600
ctggcgatag cgcttatatc gtttacgggg gatggcgata gacgactttg gtgacttggg 3660
cgattctgtg tgtcgcaaat atcgcagttt cgatataggt gacagacgat atgaggctat 3720
atcgccgata gaggcgacat caagctggca catggccaat gcatatcgat ctatacattg 3780
aatcaatatt ggccattagc catattattc attggttata tagcataaat caatattggc 3840
tattggccat tgcatacgtt gtatccatat cataatatgt acatttatat tggctcatgt 3900
ccaacattac cgccatgttg acattgatta ttgactagtt attaatagta atcaattacg 3960
gggtcattag ttcatagccc atatatggag ttccgcgtta cataacttac ggtaaatggc 4020
ccgcctggct gaccgcccaa cgacccccgc ccattgacgt caataatgac gtatgttccc 4080
atagtaacgc caatagggac tttccattga cgtcaatggg tggagtattt acggtaaact 4140
gcccacttgg cagtacatca agtgtatcat atgccaagta cgccccctat tgacgtcaat 4200
gacggtaaat ggcccgcctg gcattatgcc cagtacatga ccttatggga ctttcctact 4260
tggcagtaca tctacgtatt agtcatcgct attaccatgg tgatgcggtt ttggcagtac 4320
atcaatgggc gtggatagcg gtttgactca cggggatttc caagtctcca ccccattgac 4380
gtcaatggga gtttgttttg gcaccaaaat caacgggact ttccaaaatg tcgtaacaac 4440
tccgccccat tgacgcaaat gggcggtagg cgtgtacggt gggaggtcta tataagcaga 4500
gctcgtttag tgaaccgtca gatcgcctgg agacgccatc cacgctgttt tgacctccat 4560
agaagacacc gggaccgatc cagcctccgc ggccgggaac ggtgcattgg aacgcggatt 4620
ccccgtgcca agagtgacgt aagtaccgcc tatagagtct ataggcccac ccccttggct 4680
tcttatgcat gctatactgt ttttggcttg gggtctatac acccccgctt cctcatgtta 4740
taggtgatgg tatagcttag cctataggtg tgggttattg accattattg accactcccc 4800
tattggtgac gatactttcc attactaatc cataacatgg ctctttgcca caactctctt 4860
tattggctat atgccaatac actgtccttc agagactgac acggactctg tatttttaca 4920
ggatggggtc tcatttatta tttacaaatt cacatataca acaccaccgt ccccagtgcc 4980
cgcagttttt attaaacata acgtgggatc tccacgcgaa tctcgggtac gtgttccgga 5040
catgggctct tctccggtag cggcggagct tctacatccg agccctgctc ccatgcctcc 5100
agcgactcat ggtcgctcgg cagctccttg ctcctaacag tggaggccag acttaggcac 5160
agcacgatgc ccaccaccac cagtgtgccg cacaaggccg tggcggtagg gtatgtgtct 5220
gaaaatgagc tcggggagcg ggcttgcacc gctgacgcat ttggaagact taaggcagcg 5280
gcagaagaag atgcaggcag ctgagttgtt gtgttctgat aagagtcaga ggtaactccc 5340
gttgcggtgc tgttaacggt ggagggcagt gtagtctgag cagtactcgt tgctgccgcg 5400
cgcgccacca gacataatag ctgacagact aacagactgt tcctttccat gggtcttttc 5460
tgcagtcacc gtccttgaca cgaagttcga atcaggataa gggcgaattc cgacgtaggc 5520
ctattcgaag tctacttaat taaaagcttt ctagagcctc gagcgatggg ggaggctaac 5580
tgaaacacgg aaggagacaa taccggaagg aacccgcgct atgacggcaa taaaaagaca 5640
gaataaaacg cacgggtgtt gggtcgtttg ttcataaacg cggggttcgg tcccagggct 5700
ggcactctgt cgatacccca ccgagacccc attggggcca atacgcccgc gtttcttcct 5760
tttccccacc ccacccccca agttcgggtg aaggcccagg gctcgcagcc aacgtcgggg 5820
cggcaggccc tgccatagcc ctagcagctt ggccgtaatc atggtcatag ctgtttcctg 5880
tgtgaaattg ttatccgctc acaattccac acaacatacg agccggaagc ataaagtgta 5940
aagcctgggg tgcctaatga gtgagctaac tcacattaat tgcgttgcgc tcactgcccg 6000
ctttccagtc gggaaacctg tcgtgccagc tgcattaatg aatcggccaa cgcgcgggga 6060
gaggcggttt gcgtattggg cgctcttccg cttcctcgct cactgactcg ctgcgctcgg 6120
tcgttcggct gcggcgagcg gtatcagctc actcaaaggc ggtaatacgg ttatccacag 6180
aatcagggga taacgcagga aagaacatgt gagcaaaagg ccagcaaaag gccaggaacc 6240
gtaaaaaggc cgcgttgctg gcgtttttcc ataggctccg cccccctgac gagcatcaca 6300
aaaatcgacg ctcaagtcag aggtggcgaa acccgacagg actataaaga taccaggcgt 6360
ttccccctgg aagctccctc gtgcgctctc ctgttccgac cctgccgctt accggatacc 6420
tgtccgcctt tctcccttcg ggaagcgtgg cgctttctca tagctcacgc tgtaggtatc 6480
tcagttcggt gtaggtcgtt cgctccaagc tgggctgtgt gcacgaaccc cccgttcagc 6540
ccgaccgctg cgccttatcc ggtaactatc gtcttgagtc caacccggta agacacgact 6600
tatcgccact ggcagcagcc actggtaaca ggattagcag agcgaggtat gtaggcggtg 6660
ctacagagtt cttgaagtgg tggcctaact acggctacac tagaagaaca gtatttggta 6720
tctgcgctct gctgaagcca gttaccttcg gaaaaagagt tggtagctct tgatccggca 6780
aacaaaccac cgctggtagc ggtggttttt ttgtttgcaa gcagcagatt acgcgcagaa 6840
aaaaaggatc tcaagaagat cctttgatct tttctacggg gtctgacgct cagtggaacg 6900
aaaactcacg ttaagggatt ttggtcatga gattatcaaa aaggatcttc acctagatcc 6960
ttttaaatta aaaatgaagt tttaaatcaa tctaaagtat atatgagtaa acttggtctg 7020
acagttacca atgcttaatc agtgaggcac ctatctcagc gatctgtcta tttcgttcat 7080
ccatagttgc ctgactcccc gtcgtgtaga taactacgat acgggagggc ttaccatctg 7140
gccccagtgc tgcaatgata ccgcgagacc cacgctcacc ggctccagat ttatcagcaa 7200
taaaccagcc agccggaagg gccgagcgca gaagtggtcc tgcaacttta tccgcctcca 7260
tccagtctat taattgttgc cgggaagcta gagtaagtag ttcgccagtt aatagtttgc 7320
gcaacgttgt tgccattgct acaggcatcg tggtgtcacg ctcgtcgttt ggtatggctt 7380
cattcagctc cggttcccaa cgatcaaggc gagttacatg atcccccatg ttgtgcaaaa 7440
aagcggttag ctccttcggt cctccgatcg ttgtcagaag taagttggcc gcagtgttat 7500
cactcatggt tatggcagca ctgcataatt ctcttactgt catgccatcc gtaagatgct 7560
tttctgtgac tggtgagtac tcaaccaagt cattctgaga atagtgtatg cggcgaccga 7620
gttgctcttg cccggcgtca atacgggata ataccgcgcc acatagcaga actttaaaag 7680
tgctcatcat tggaaaacgt tcttcggggc gaaaactctc aaggatctta ccgctgttga 7740
gatccagttc gatgtaaccc actcgtgcac ccaactgatc ttcagcatct tttactttca 7800
ccagcgtttc tgggtgagca aaaacaggaa ggcaaaatgc cgcaaaaaag ggaataaggg 7860
cgacacggaa atgttgaata ctcatactct tcctttttca atattattga agcatttatc 7920
agggttattg tctcatgagc ggatacatat ttgaatgtat ttagaaaaat aaacaaatag 7980
gggttccgcg cacatttccc cgaaaagtgc cacctgacgt ctaagaaacc attattatca 8040
tgacattaac ctataaaaat aggcgtatca cgaggccctt tcgtc 8085
<210> 8
<211> 7683
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 8
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgccctaggg tttaaacaga 2520
tctatcgata atcaacctct ggattacaaa atttgtgaaa gattgactgg tattcttaac 2580
tatgttgctc cttttacgct atgtggatac gctgctttaa tgcctttgta tcatgctatt 2640
gcttcccgta tggctttcat tttctcctcc ttgtataaat cctggttgct gtctctttat 2700
gaggagttgt ggcccgttgt caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca 2760
acccccactg gttggggcat tgccaccacc tgtcagctcc tttccgggac tttcgctttc 2820
cccctcccta ttgccacggc ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg 2880
gctcggctgt tgggcactga caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct 2940
tggctgctcg cctgtgttgc cacctggatt ctgcgcggga cgtccttctg ctacgtccct 3000
tcggccctca atccagcgga ccttccttcc cgcggcctgc tgccggctct gcggcctctt 3060
ccgcgtcttc gccttcgccc tcagacgagt cggatctccc tttgggccgc ctccccgcat 3120
cgattactaa tcagccatac cacatttgta gaggttttac ttgctttaaa aaacctccca 3180
cacctccccc tgaacctgaa acataaaatg aatgcaattg ttgttgttaa cttgtttatt 3240
gcagcttata atggttacaa ataaagcaat agcatcacaa atttcacaaa taaagcattt 3300
ttttcactgc attctagttg tggtttgtcc aaactcatca atgtatctta tcatgtctgg 3360
aattcggacg gtgactgcag tgaataataa aatgtgtgtt tgtccgaaat acgcgttttg 3420
agatttctgt cgccgactaa attcatgtcg cgcgatagtg gtgtttatcg ccgatagaga 3480
tggcgatatt ggaaaaatcg atatttgaaa atatggcata ttgaaaatgt cgccgatgtg 3540
agtttctgtg taactgatat cgccattttt ccaaaagtga tttttgggca tacgcgatat 3600
ctggcgatag cgcttatatc gtttacgggg gatggcgata gacgactttg gtgacttggg 3660
cgattctgtg tgtcgcaaat atcgcagttt cgatataggt gacagacgat atgaggctat 3720
atcgccgata gaggcgacat caagctggca catggccaat gcatatcgat ctatacattg 3780
aatcaatatt ggccattagc catattattc attggttata tagcataaat caatattggc 3840
tattggccat tgcatacgtt gtatccatat cataatatgt acatttatat tggctcatgt 3900
ccaacattac cgccatgttg acattgatta ttgactagtt attaatagta atcaattacg 3960
gggtcattag ttcatagccc atatatggag ttccgcgtta cataacttac ggtaaatggc 4020
ccgcctggct gaccgcccaa cgacccccgc ccattgacgt caataatgac gtatgttccc 4080
atagtaacgc caatagggac tttccattga cgtcaatggg tggagtattt acggtaaact 4140
gcccacttgg cagtacatca agtgtatcat atgccaagta cgccccctat tgacgtcaat 4200
gacggtaaat ggcccgcctg gcattatgcc cagtacatga ccttatggga ctttcctact 4260
tggcagtaca tctacgtatt agtcatcgct attaccatgg tgatgcggtt ttggcagtac 4320
atcaatgggc gtggatagcg gtttgactca cggggatttc caagtctcca ccccattgac 4380
gtcaatggga gtttgttttg gcaccaaaat caacgggact ttccaaaatg tcgtaacaac 4440
tccgccccat tgacgcaaat gggcggtagg cgtgtacggt gggaggtcta tataagcaga 4500
gctcgtttag tgaaccggat ccttaaccgt gaaagcttag gccttctaga gcctcgagtt 4560
cgacatcgat aatcaacctc tggattacaa aatttgtgaa agattgactg gtattcttaa 4620
ctatgttgct ccttttacgc tatgtggata cgctgcttta atgcctttgt atcatgctat 4680
tgcttcccgt atggctttca ttttctcctc cttgtataaa tcctggttgc tgtctcttta 4740
tgaggagttg tggcccgttg tcaggcaacg tggcgtggtg tgcactgtgt ttgctgacgc 4800
aacccccact ggttggggca ttgccaccac ctgtcagctc ctttccggga ctttcgcttt 4860
ccccctccct attgccacgg cggaactcat cgccgcctgc cttgcccgct gctggacagg 4920
ggctcggctg ttgggcactg acaattccgt ggtgttgtcg gggaaatcat cgtcctttcc 4980
ttggctgctc gcctgtgttg ccacctggat tctgcgcggg acgtccttct gctacgtccc 5040
ttcggccctc aatccagcgg accttccttc ccgcggcctg ctgccggctc tgcggcctct 5100
tccgcgtctt cgccttcgcc ctcagacgag tcggatctcc ctttgggccg cctccccgca 5160
tcgatggggg aggctaactg aaacacggaa ggagacaata ccggaaggaa cccgcgctat 5220
gacggcaata aaaagacaga ataaaacgca cgggtgttgg gtcgtttgtt cataaacgcg 5280
gggttcggtc ccagggctgg cactctgtcg ataccccacc gagaccccat tggggccaat 5340
acgcccgcgt ttcttccttt tccccacccc accccccaag ttcgggtgaa ggcccagggc 5400
tcgcagccaa cgtcggggcg gcaggccctg ccatagccct agcagcttgg ccgtaatcat 5460
ggtcatagct gtttcctgtg tgaaattgtt atccgctcac aattccacac aacatacgag 5520
ccggaagcat aaagtgtaaa gcctggggtg cctaatgagt gagctaactc acattaattg 5580
cgttgcgctc actgcccgct ttccagtcgg gaaacctgtc gtgccagctg cattaatgaa 5640
tcggccaacg cgcggggaga ggcggtttgc gtattgggcg ctcttccgct tcctcgctca 5700
ctgactcgct gcgctcggtc gttcggctgc ggcgagcggt atcagctcac tcaaaggcgg 5760
taatacggtt atccacagaa tcaggggata acgcaggaaa gaacatgtga gcaaaaggcc 5820
agcaaaaggc caggaaccgt aaaaaggccg cgttgctggc gtttttccat aggctccgcc 5880
cccctgacga gcatcacaaa aatcgacgct caagtcagag gtggcgaaac ccgacaggac 5940
tataaagata ccaggcgttt ccccctggaa gctccctcgt gcgctctcct gttccgaccc 6000
tgccgcttac cggatacctg tccgcctttc tcccttcggg aagcgtggcg ctttctcata 6060
gctcacgctg taggtatctc agttcggtgt aggtcgttcg ctccaagctg ggctgtgtgc 6120
acgaaccccc cgttcagccc gaccgctgcg ccttatccgg taactatcgt cttgagtcca 6180
acccggtaag acacgactta tcgccactgg cagcagccac tggtaacagg attagcagag 6240
cgaggtatgt aggcggtgct acagagttct tgaagtggtg gcctaactac ggctacacta 6300
gaagaacagt atttggtatc tgcgctctgc tgaagccagt taccttcgga aaaagagttg 6360
gtagctcttg atccggcaaa caaaccaccg ctggtagcgg tggttttttt gtttgcaagc 6420
agcagattac gcgcagaaaa aaaggatctc aagaagatcc tttgatcttt tctacggggt 6480
ctgacgctca gtggaacgaa aactcacgtt aagggatttt ggtcatgaga ttatcaaaaa 6540
ggatcttcac ctagatcctt ttaaattaaa aatgaagttt taaatcaatc taaagtatat 6600
atgagtaaac ttggtctgac agttaccaat gcttaatcag tgaggcacct atctcagcga 6660
tctgtctatt tcgttcatcc atagttgcct gactccccgt cgtgtagata actacgatac 6720
gggagggctt accatctggc cccagtgctg caatgatacc gcgagaccca cgctcaccgg 6780
ctccagattt atcagcaata aaccagccag ccggaagggc cgagcgcaga agtggtcctg 6840
caactttatc cgcctccatc cagtctatta attgttgccg ggaagctaga gtaagtagtt 6900
cgccagttaa tagtttgcgc aacgttgttg ccattgctac aggcatcgtg gtgtcacgct 6960
cgtcgtttgg tatggcttca ttcagctccg gttcccaacg atcaaggcga gttacatgat 7020
cccccatgtt gtgcaaaaaa gcggttagct ccttcggtcc tccgatcgtt gtcagaagta 7080
agttggccgc agtgttatca ctcatggtta tggcagcact gcataattct cttactgtca 7140
tgccatccgt aagatgcttt tctgtgactg gtgagtactc aaccaagtca ttctgagaat 7200
agtgtatgcg gcgaccgagt tgctcttgcc cggcgtcaat acgggataat accgcgccac 7260
atagcagaac tttaaaagtg ctcatcattg gaaaacgttc ttcggggcga aaactctcaa 7320
ggatcttacc gctgttgaga tccagttcga tgtaacccac tcgtgcaccc aactgatctt 7380
cagcatcttt tactttcacc agcgtttctg ggtgagcaaa aacaggaagg caaaatgccg 7440
caaaaaaggg aataagggcg acacggaaat gttgaatact catactcttc ctttttcaat 7500
attattgaag catttatcag ggttattgtc tcatgagcgg atacatattt gaatgtattt 7560
agaaaaataa acaaataggg gttccgcgca catttccccg aaaagtgcca cctgacgtct 7620
aagaaaccat tattatcatg acattaacct ataaaaatag gcgtatcacg aggccctttc 7680
gtc 7683
<210> 9
<211> 10196
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 9
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgccgccacc atgatgtcct 2520
ttgtctctct gctcctggtt ggcatcctat tccatgccac ccaggcccag gtgcagctgg 2580
tggagtccgg cggcggcgtc gtgcagcccg gccggtccct gcggctgtcc tgcgccgcct 2640
ccggcttcac cttctcctcc tacaccatgc actgggtgcg gcaggccccc ggcaagggcc 2700
tggagtgggt gactttcatc tcctacgacg gcaacaacaa gtactacgcc gactccgtga 2760
agggccggtt caccatctcc cgcgacaact ccaagaacac cctgtacctg cagatgaact 2820
ccctgcgggc cgaggacacc gccatctact actgcgcccg gaccggctgg ctgggcccct 2880
tcgactactg gggccagggc accctggtga ccgtgtcctc cgcctccacc aagggcccat 2940
cggtcttccc cctggcaccc tctagcaaga gcacctctgg gggcacagcg gccctgggct 3000
gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca ggcgccctga 3060
ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac tccctcagca 3120
gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc aacgtgaatc 3180
acaagcccag caacaccaag gtggacaagc gggttgagcc caaatcttgt gacaaaactc 3240
acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc ttcctcttcc 3300
ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca tgcgtggtgg 3360
tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac ggcgtggagg 3420
tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac cgtgtggtca 3480
gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag tgcaaggtct 3540
ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa gggcagcccc 3600
gagaaccaca ggtgtacacc ctgcctccat cccgcgatga gctgaccaag aaccaggtca 3660
gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag tgggagagca 3720
atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc gacggctcct 3780
tcttcctcta tagcaagctc accgtggaca agagcaggtg gcagcagggg aacgtcttct 3840
catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc ctctccctgt 3900
ctcctgggaa atgatgagat ctatcgataa tcaacctctg gattacaaaa tttgtgaaag 3960
attgactggt attcttaact atgttgctcc ttttacgcta tgtggatacg ctgctttaat 4020
gcctttgtat catgctattg cttcccgtat ggctttcatt ttctcctcct tgtataaatc 4080
ctggttgctg tctctttatg aggagttgtg gcccgttgtc aggcaacgtg gcgtggtgtg 4140
cactgtgttt gctgacgcaa cccccactgg ttggggcatt gccaccacct gtcagctcct 4200
ttccgggact ttcgctttcc ccctccctat tgccacggcg gaactcatcg ccgcctgcct 4260
tgcccgctgc tggacagggg ctcggctgtt gggcactgac aattccgtgg tgttgtcggg 4320
gaaatcatcg tcctttcctt ggctgctcgc ctgtgttgcc acctggattc tgcgcgggac 4380
gtccttctgc tacgtccctt cggccctcaa tccagcggac cttccttccc gcggcctgct 4440
gccggctctg cggcctcttc cgcgtcttcg ccttcgccct cagacgagtc ggatctccct 4500
ttgggccgcc tccccgcatc gattactaat cagccatacc acatttgtag aggttttact 4560
tgctttaaaa aacctcccac acctccccct gaacctgaaa cataaaatga atgcaattgt 4620
tgttgttaac ttgtttattg cagcttataa tggttacaaa taaagcaata gcatcacaaa 4680
tttcacaaat aaagcatttt tttcactgca ttctagttgt ggtttgtcca aactcatcaa 4740
tgtatcttat catgtctgga attcggacgg tgactgcagt gaataataaa atgtgtgttt 4800
gtccgaaata cgcgttttga gatttctgtc gccgactaaa ttcatgtcgc gcgatagtgg 4860
tgtttatcgc cgatagagat ggcgatattg gaaaaatcga tatttgaaaa tatggcatat 4920
tgaaaatgtc gccgatgtga gtttctgtgt aactgatatc gccatttttc caaaagtgat 4980
ttttgggcat acgcgatatc tggcgatagc gcttatatcg tttacggggg atggcgatag 5040
acgactttgg tgacttgggc gattctgtgt gtcgcaaata tcgcagtttc gatataggtg 5100
acagacgata tgaggctata tcgccgatag aggcgacatc aagctggcac atggccaatg 5160
catatcgatc tatacattga atcaatattg gccattagcc atattattca ttggttatat 5220
agcataaatc aatattggct attggccatt gcatacgttg tatccatatc ataatatgta 5280
catttatatt ggctcatgtc caacattacc gccatgttga cattgattat tgactagtta 5340
ttaatagtaa tcaattacgg ggtcattagt tcatagccca tatatggagt tccgcgttac 5400
ataacttacg gtaaatggcc cgcctggctg accgcccaac gacccccgcc cattgacgtc 5460
aataatgacg tatgttccca tagtaacgcc aatagggact ttccattgac gtcaatgggt 5520
ggagtattta cggtaaactg cccacttggc agtacatcaa gtgtatcata tgccaagtac 5580
gccccctatt gacgtcaatg acggtaaatg gcccgcctgg cattatgccc agtacatgac 5640
cttatgggac tttcctactt ggcagtacat ctacgtatta gtcatcgcta ttaccatggt 5700
gatgcggttt tggcagtaca tcaatgggcg tggatagcgg tttgactcac ggggatttcc 5760
aagtctccac cccattgacg tcaatgggag tttgttttgg caccaaaatc aacgggactt 5820
tccaaaatgt cgtaacaact ccgccccatt gacgcaaatg ggcggtaggc gtgtacggtg 5880
ggaggtctat ataagcagag ctcgtttagt gaaccgtcag atcgcctgga gacgccatcc 5940
acgctgtttt gacctccata gaagacaccg ggaccgatcc agcctccgcg gccgggaacg 6000
gtgcattgga acgcggattc cccgtgccaa gagtgacgta agtaccgcct atagagtcta 6060
taggcccacc cccttggctt cttatgcatg ctatactgtt tttggcttgg ggtctataca 6120
cccccgcttc ctcatgttat aggtgatggt atagcttagc ctataggtgt gggttattga 6180
ccattattga ccactcccct attggtgacg atactttcca ttactaatcc ataacatggc 6240
tctttgccac aactctcttt attggctata tgccaataca ctgtccttca gagactgaca 6300
cggactctgt atttttacag gatggggtct catttattat ttacaaattc acatatacaa 6360
caccaccgtc cccagtgccc gcagttttta ttaaacataa cgtgggatct ccacgcgaat 6420
ctcgggtacg tgttccggac atgggctctt ctccggtagc ggcggagctt ctacatccga 6480
gccctgctcc catgcctcca gcgactcatg gtcgctcggc agctccttgc tcctaacagt 6540
ggaggccaga cttaggcaca gcacgatgcc caccaccacc agtgtgccgc acaaggccgt 6600
ggcggtaggg tatgtgtctg aaaatgagct cggggagcgg gcttgcaccg ctgacgcatt 6660
tggaagactt aaggcagcgg cagaagaaga tgcaggcagc tgagttgttg tgttctgata 6720
agagtcagag gtaactcccg ttgcggtgct gttaacggtg gagggcagtg tagtctgagc 6780
agtactcgtt gctgccgcgc gcgccaccag acataatagc tgacagacta acagactgtt 6840
cctttccatg ggtcttttct gcagtcaccg tccttgacac gaagttcgaa tcaggataag 6900
ggcgaattcc gacgtaggcc tattcgaagt ctacttaatt aaaagcttgc cgccaccatg 6960
atgtcctttg tctctctgct cctggttggc atcctattcc atgccaccca ggccgagatc 7020
gtgctgaccc agtcccccgg caccctgtcc ctgtcccccg gcgagcgggc caccctgtcc 7080
tgccgggcct cccagtccgt gggctcctcc tacctggcct ggtaccagca gaagcccggc 7140
caggcccccc ggctgctgat ctacggcgcc ttctcccgcg ccaccggcat ccccgaccgg 7200
ttctccggct ccggctccgg caccgacttc accctgacca tctcccggct ggagcccgag 7260
gacttcgccg tgtactactg ccagcagtac ggctcctccc cctggacctt cggccagggc 7320
accaaggtgg agatcaagcg aactgtggct gcaccatctg tcttcatctt cccgccatct 7380
gatgagcagc ttaagtccgg aactgctagc gttgtgtgcc tgctgaataa cttctatccc 7440
agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag 7500
agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg 7560
agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg 7620
agctcgcccg tcacaaagag cttcaacagg ggagagtgtt agtgagatct cgagcgatgg 7680
gggaggctaa ctgaaacacg gaaggagaca ataccggaag gaacccgcgc tatgacggca 7740
ataaaaagac agaataaaac gcacgggtgt tgggtcgttt gttcataaac gcggggttcg 7800
gtcccagggc tggcactctg tcgatacccc accgagaccc cattggggcc aatacgcccg 7860
cgtttcttcc ttttccccac cccacccccc aagttcgggt gaaggcccag ggctcgcagc 7920
caacgtcggg gcggcaggcc ctgccatagc cctagcagct tggccgtaat catggtcata 7980
gctgtttcct gtgtgaaatt gttatccgct cacaattcca cacaacatac gagccggaag 8040
cataaagtgt aaagcctggg gtgcctaatg agtgagctaa ctcacattaa ttgcgttgcg 8100
ctcactgccc gctttccagt cgggaaacct gtcgtgccag ctgcattaat gaatcggcca 8160
acgcgcgggg agaggcggtt tgcgtattgg gcgctcttcc gcttcctcgc tcactgactc 8220
gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct cactcaaagg cggtaatacg 8280
gttatccaca gaatcagggg ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa 8340
ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc cataggctcc gcccccctga 8400
cgagcatcac aaaaatcgac gctcaagtca gaggtggcga aacccgacag gactataaag 8460
ataccaggcg tttccccctg gaagctccct cgtgcgctct cctgttccga ccctgccgct 8520
taccggatac ctgtccgcct ttctcccttc gggaagcgtg gcgctttctc atagctcacg 8580
ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc 8640
ccccgttcag cccgaccgct gcgccttatc cggtaactat cgtcttgagt ccaacccggt 8700
aagacacgac ttatcgccac tggcagcagc cactggtaac aggattagca gagcgaggta 8760
tgtaggcggt gctacagagt tcttgaagtg gtggcctaac tacggctaca ctagaagaac 8820
agtatttggt atctgcgctc tgctgaagcc agttaccttc ggaaaaagag ttggtagctc 8880
ttgatccggc aaacaaacca ccgctggtag cggtggtttt tttgtttgca agcagcagat 8940
tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc ttttctacgg ggtctgacgc 9000
tcagtggaac gaaaactcac gttaagggat tttggtcatg agattatcaa aaaggatctt 9060
cacctagatc cttttaaatt aaaaatgaag ttttaaatca atctaaagta tatatgagta 9120
aacttggtct gacagttacc aatgcttaat cagtgaggca cctatctcag cgatctgtct 9180
atttcgttca tccatagttg cctgactccc cgtcgtgtag ataactacga tacgggaggg 9240
cttaccatct ggccccagtg ctgcaatgat accgcgagac ccacgctcac cggctccaga 9300
tttatcagca ataaaccagc cagccggaag ggccgagcgc agaagtggtc ctgcaacttt 9360
atccgcctcc atccagtcta ttaattgttg ccgggaagct agagtaagta gttcgccagt 9420
taatagtttg cgcaacgttg ttgccattgc tacaggcatc gtggtgtcac gctcgtcgtt 9480
tggtatggct tcattcagct ccggttccca acgatcaagg cgagttacat gatcccccat 9540
gttgtgcaaa aaagcggtta gctccttcgg tcctccgatc gttgtcagaa gtaagttggc 9600
cgcagtgtta tcactcatgg ttatggcagc actgcataat tctcttactg tcatgccatc 9660
cgtaagatgc ttttctgtga ctggtgagta ctcaaccaag tcattctgag aatagtgtat 9720
gcggcgaccg agttgctctt gcccggcgtc aatacgggat aataccgcgc cacatagcag 9780
aactttaaaa gtgctcatca ttggaaaacg ttcttcgggg cgaaaactct caaggatctt 9840
accgctgttg agatccagtt cgatgtaacc cactcgtgca cccaactgat cttcagcatc 9900
ttttactttc accagcgttt ctgggtgagc aaaaacagga aggcaaaatg ccgcaaaaaa 9960
gggaataagg gcgacacgga aatgttgaat actcatactc ttcctttttc aatattattg 10020
aagcatttat cagggttatt gtctcatgag cggatacata tttgaatgta tttagaaaaa 10080
taaacaaata ggggttccgc gcacatttcc ccgaaaagtg ccacctgacg tctaagaaac 10140
cattattatc atgacattaa cctataaaaa taggcgtatc acgaggccct ttcgtc 10196
<210> 10
<211> 10196
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 10
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgccgccacc atgatgtcct 2520
ttgtctctct gctcctggtt ggcatcctat tccatgccac ccaggccgag atcgtgctga 2580
cccagtcccc cggcaccctg tccctgtccc ccggcgagcg ggccaccctg tcctgccggg 2640
cctcccagtc cgtgggctcc tcctacctgg cctggtacca gcagaagccc ggccaggccc 2700
cccggctgct gatctacggc gccttctccc gcgccaccgg catccccgac cggttctccg 2760
gctccggctc cggcaccgac ttcaccctga ccatctcccg gctggagccc gaggacttcg 2820
ccgtgtacta ctgccagcag tacggctcct ccccctggac cttcggccag ggcaccaagg 2880
tggagatcaa gcgaactgtg gctgcaccat ctgtcttcat cttcccgcca tctgatgagc 2940
agcttaagtc cggaactgct agcgttgtgt gcctgctgaa taacttctat cccagagagg 3000
ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag gagagtgtca 3060
cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg ctgagcaaag 3120
cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc 3180
ccgtcacaaa gagcttcaac aggggagagt gttagtgaga tctatcgata atcaacctct 3240
ggattacaaa atttgtgaaa gattgactgg tattcttaac tatgttgctc cttttacgct 3300
atgtggatac gctgctttaa tgcctttgta tcatgctatt gcttcccgta tggctttcat 3360
tttctcctcc ttgtataaat cctggttgct gtctctttat gaggagttgt ggcccgttgt 3420
caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca acccccactg gttggggcat 3480
tgccaccacc tgtcagctcc tttccgggac tttcgctttc cccctcccta ttgccacggc 3540
ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg gctcggctgt tgggcactga 3600
caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct tggctgctcg cctgtgttgc 3660
cacctggatt ctgcgcggga cgtccttctg ctacgtccct tcggccctca atccagcgga 3720
ccttccttcc cgcggcctgc tgccggctct gcggcctctt ccgcgtcttc gccttcgccc 3780
tcagacgagt cggatctccc tttgggccgc ctccccgcat cgattactaa tcagccatac 3840
cacatttgta gaggttttac ttgctttaaa aaacctccca cacctccccc tgaacctgaa 3900
acataaaatg aatgcaattg ttgttgttaa cttgtttatt gcagcttata atggttacaa 3960
ataaagcaat agcatcacaa atttcacaaa taaagcattt ttttcactgc attctagttg 4020
tggtttgtcc aaactcatca atgtatctta tcatgtctgg aattcggacg gtgactgcag 4080
tgaataataa aatgtgtgtt tgtccgaaat acgcgttttg agatttctgt cgccgactaa 4140
attcatgtcg cgcgatagtg gtgtttatcg ccgatagaga tggcgatatt ggaaaaatcg 4200
atatttgaaa atatggcata ttgaaaatgt cgccgatgtg agtttctgtg taactgatat 4260
cgccattttt ccaaaagtga tttttgggca tacgcgatat ctggcgatag cgcttatatc 4320
gtttacgggg gatggcgata gacgactttg gtgacttggg cgattctgtg tgtcgcaaat 4380
atcgcagttt cgatataggt gacagacgat atgaggctat atcgccgata gaggcgacat 4440
caagctggca catggccaat gcatatcgat ctatacattg aatcaatatt ggccattagc 4500
catattattc attggttata tagcataaat caatattggc tattggccat tgcatacgtt 4560
gtatccatat cataatatgt acatttatat tggctcatgt ccaacattac cgccatgttg 4620
acattgatta ttgactagtt attaatagta atcaattacg gggtcattag ttcatagccc 4680
atatatggag ttccgcgtta cataacttac ggtaaatggc ccgcctggct gaccgcccaa 4740
cgacccccgc ccattgacgt caataatgac gtatgttccc atagtaacgc caatagggac 4800
tttccattga cgtcaatggg tggagtattt acggtaaact gcccacttgg cagtacatca 4860
agtgtatcat atgccaagta cgccccctat tgacgtcaat gacggtaaat ggcccgcctg 4920
gcattatgcc cagtacatga ccttatggga ctttcctact tggcagtaca tctacgtatt 4980
agtcatcgct attaccatgg tgatgcggtt ttggcagtac atcaatgggc gtggatagcg 5040
gtttgactca cggggatttc caagtctcca ccccattgac gtcaatggga gtttgttttg 5100
gcaccaaaat caacgggact ttccaaaatg tcgtaacaac tccgccccat tgacgcaaat 5160
gggcggtagg cgtgtacggt gggaggtcta tataagcaga gctcgtttag tgaaccgtca 5220
gatcgcctgg agacgccatc cacgctgttt tgacctccat agaagacacc gggaccgatc 5280
cagcctccgc ggccgggaac ggtgcattgg aacgcggatt ccccgtgcca agagtgacgt 5340
aagtaccgcc tatagagtct ataggcccac ccccttggct tcttatgcat gctatactgt 5400
ttttggcttg gggtctatac acccccgctt cctcatgtta taggtgatgg tatagcttag 5460
cctataggtg tgggttattg accattattg accactcccc tattggtgac gatactttcc 5520
attactaatc cataacatgg ctctttgcca caactctctt tattggctat atgccaatac 5580
actgtccttc agagactgac acggactctg tatttttaca ggatggggtc tcatttatta 5640
tttacaaatt cacatataca acaccaccgt ccccagtgcc cgcagttttt attaaacata 5700
acgtgggatc tccacgcgaa tctcgggtac gtgttccgga catgggctct tctccggtag 5760
cggcggagct tctacatccg agccctgctc ccatgcctcc agcgactcat ggtcgctcgg 5820
cagctccttg ctcctaacag tggaggccag acttaggcac agcacgatgc ccaccaccac 5880
cagtgtgccg cacaaggccg tggcggtagg gtatgtgtct gaaaatgagc tcggggagcg 5940
ggcttgcacc gctgacgcat ttggaagact taaggcagcg gcagaagaag atgcaggcag 6000
ctgagttgtt gtgttctgat aagagtcaga ggtaactccc gttgcggtgc tgttaacggt 6060
ggagggcagt gtagtctgag cagtactcgt tgctgccgcg cgcgccacca gacataatag 6120
ctgacagact aacagactgt tcctttccat gggtcttttc tgcagtcacc gtccttgaca 6180
cgaagttcga atcaggataa gggcgaattc cgacgtaggc ctattcgaag tctacttaat 6240
taaaagcttg ccgccaccat gatgtccttt gtctctctgc tcctggttgg catcctattc 6300
catgccaccc aggcccaggt gcagctggtg gagtccggcg gcggcgtcgt gcagcccggc 6360
cggtccctgc ggctgtcctg cgccgcctcc ggcttcacct tctcctccta caccatgcac 6420
tgggtgcggc aggcccccgg caagggcctg gagtgggtga ctttcatctc ctacgacggc 6480
aacaacaagt actacgccga ctccgtgaag ggccggttca ccatctcccg cgacaactcc 6540
aagaacaccc tgtacctgca gatgaactcc ctgcgggccg aggacaccgc catctactac 6600
tgcgcccgga ccggctggct gggccccttc gactactggg gccagggcac cctggtgacc 6660
gtgtcctccg cctccaccaa gggcccatcg gtcttccccc tggcaccctc tagcaagagc 6720
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 6780
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 6840
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 6900
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagcgg 6960
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 7020
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 7080
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 7140
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag 7200
cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg 7260
aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa 7320
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcctccatcc 7380
cgcgatgagc tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 7440
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 7500
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 7560
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 7620
cactacacgc agaagagcct ctccctgtct cctgggaaat gatgagatct cgagcgatgg 7680
gggaggctaa ctgaaacacg gaaggagaca ataccggaag gaacccgcgc tatgacggca 7740
ataaaaagac agaataaaac gcacgggtgt tgggtcgttt gttcataaac gcggggttcg 7800
gtcccagggc tggcactctg tcgatacccc accgagaccc cattggggcc aatacgcccg 7860
cgtttcttcc ttttccccac cccacccccc aagttcgggt gaaggcccag ggctcgcagc 7920
caacgtcggg gcggcaggcc ctgccatagc cctagcagct tggccgtaat catggtcata 7980
gctgtttcct gtgtgaaatt gttatccgct cacaattcca cacaacatac gagccggaag 8040
cataaagtgt aaagcctggg gtgcctaatg agtgagctaa ctcacattaa ttgcgttgcg 8100
ctcactgccc gctttccagt cgggaaacct gtcgtgccag ctgcattaat gaatcggcca 8160
acgcgcgggg agaggcggtt tgcgtattgg gcgctcttcc gcttcctcgc tcactgactc 8220
gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct cactcaaagg cggtaatacg 8280
gttatccaca gaatcagggg ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa 8340
ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc cataggctcc gcccccctga 8400
cgagcatcac aaaaatcgac gctcaagtca gaggtggcga aacccgacag gactataaag 8460
ataccaggcg tttccccctg gaagctccct cgtgcgctct cctgttccga ccctgccgct 8520
taccggatac ctgtccgcct ttctcccttc gggaagcgtg gcgctttctc atagctcacg 8580
ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc 8640
ccccgttcag cccgaccgct gcgccttatc cggtaactat cgtcttgagt ccaacccggt 8700
aagacacgac ttatcgccac tggcagcagc cactggtaac aggattagca gagcgaggta 8760
tgtaggcggt gctacagagt tcttgaagtg gtggcctaac tacggctaca ctagaagaac 8820
agtatttggt atctgcgctc tgctgaagcc agttaccttc ggaaaaagag ttggtagctc 8880
ttgatccggc aaacaaacca ccgctggtag cggtggtttt tttgtttgca agcagcagat 8940
tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc ttttctacgg ggtctgacgc 9000
tcagtggaac gaaaactcac gttaagggat tttggtcatg agattatcaa aaaggatctt 9060
cacctagatc cttttaaatt aaaaatgaag ttttaaatca atctaaagta tatatgagta 9120
aacttggtct gacagttacc aatgcttaat cagtgaggca cctatctcag cgatctgtct 9180
atttcgttca tccatagttg cctgactccc cgtcgtgtag ataactacga tacgggaggg 9240
cttaccatct ggccccagtg ctgcaatgat accgcgagac ccacgctcac cggctccaga 9300
tttatcagca ataaaccagc cagccggaag ggccgagcgc agaagtggtc ctgcaacttt 9360
atccgcctcc atccagtcta ttaattgttg ccgggaagct agagtaagta gttcgccagt 9420
taatagtttg cgcaacgttg ttgccattgc tacaggcatc gtggtgtcac gctcgtcgtt 9480
tggtatggct tcattcagct ccggttccca acgatcaagg cgagttacat gatcccccat 9540
gttgtgcaaa aaagcggtta gctccttcgg tcctccgatc gttgtcagaa gtaagttggc 9600
cgcagtgtta tcactcatgg ttatggcagc actgcataat tctcttactg tcatgccatc 9660
cgtaagatgc ttttctgtga ctggtgagta ctcaaccaag tcattctgag aatagtgtat 9720
gcggcgaccg agttgctctt gcccggcgtc aatacgggat aataccgcgc cacatagcag 9780
aactttaaaa gtgctcatca ttggaaaacg ttcttcgggg cgaaaactct caaggatctt 9840
accgctgttg agatccagtt cgatgtaacc cactcgtgca cccaactgat cttcagcatc 9900
ttttactttc accagcgttt ctgggtgagc aaaaacagga aggcaaaatg ccgcaaaaaa 9960
gggaataagg gcgacacgga aatgttgaat actcatactc ttcctttttc aatattattg 10020
aagcatttat cagggttatt gtctcatgag cggatacata tttgaatgta tttagaaaaa 10080
taaacaaata ggggttccgc gcacatttcc ccgaaaagtg ccacctgacg tctaagaaac 10140
cattattatc atgacattaa cctataaaaa taggcgtatc acgaggccct ttcgtc 10196
<210> 11
<211> 9778
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 11
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgccgccacc atgatgtcct 2520
ttgtctctct gctcctggtt ggcatcctat tccatgccac ccaggcccag gtgcagctgg 2580
tggagtccgg cggcggcgtc gtgcagcccg gccggtccct gcggctgtcc tgcgccgcct 2640
ccggcttcac cttctcctcc tacaccatgc actgggtgcg gcaggccccc ggcaagggcc 2700
tggagtgggt gactttcatc tcctacgacg gcaacaacaa gtactacgcc gactccgtga 2760
agggccggtt caccatctcc cgcgacaact ccaagaacac cctgtacctg cagatgaact 2820
ccctgcgggc cgaggacacc gccatctact actgcgcccg gaccggctgg ctgggcccct 2880
tcgactactg gggccagggc accctggtga ccgtgtcctc cgcctccacc aagggcccat 2940
cggtcttccc cctggcaccc tctagcaaga gcacctctgg gggcacagcg gccctgggct 3000
gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca ggcgccctga 3060
ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac tccctcagca 3120
gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc aacgtgaatc 3180
acaagcccag caacaccaag gtggacaagc gggttgagcc caaatcttgt gacaaaactc 3240
acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc ttcctcttcc 3300
ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca tgcgtggtgg 3360
tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac ggcgtggagg 3420
tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac cgtgtggtca 3480
gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag tgcaaggtct 3540
ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa gggcagcccc 3600
gagaaccaca ggtgtacacc ctgcctccat cccgcgatga gctgaccaag aaccaggtca 3660
gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag tgggagagca 3720
atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc gacggctcct 3780
tcttcctcta tagcaagctc accgtggaca agagcaggtg gcagcagggg aacgtcttct 3840
catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc ctctccctgt 3900
ctcctgggaa atgatgagat ctatcgataa tcaacctctg gattacaaaa tttgtgaaag 3960
attgactggt attcttaact atgttgctcc ttttacgcta tgtggatacg ctgctttaat 4020
gcctttgtat catgctattg cttcccgtat ggctttcatt ttctcctcct tgtataaatc 4080
ctggttgctg tctctttatg aggagttgtg gcccgttgtc aggcaacgtg gcgtggtgtg 4140
cactgtgttt gctgacgcaa cccccactgg ttggggcatt gccaccacct gtcagctcct 4200
ttccgggact ttcgctttcc ccctccctat tgccacggcg gaactcatcg ccgcctgcct 4260
tgcccgctgc tggacagggg ctcggctgtt gggcactgac aattccgtgg tgttgtcggg 4320
gaaatcatcg tcctttcctt ggctgctcgc ctgtgttgcc acctggattc tgcgcgggac 4380
gtccttctgc tacgtccctt cggccctcaa tccagcggac cttccttccc gcggcctgct 4440
gccggctctg cggcctcttc cgcgtcttcg ccttcgccct cagacgagtc ggatctccct 4500
ttgggccgcc tccccgcatc gattactaat cagccatacc acatttgtag aggttttact 4560
tgctttaaaa aacctcccac acctccccct gaacctgaaa cataaaatga atgcaattgt 4620
tgttgttaac ttgtttattg cagcttataa tggttacaaa taaagcaata gcatcacaaa 4680
tttcacaaat aaagcatttt tttcactgca ttctagttgt ggtttgtcca aactcatcaa 4740
tgtatcttat catgtctgga attcggacgg tgactgcagt gaataataaa atgtgtgttt 4800
gtccgaaata cgcgttttga gatttctgtc gccgactaaa ttcatgtcgc gcgatagtgg 4860
tgtttatcgc cgatagagat ggcgatattg gaaaaatcga tatttgaaaa tatggcatat 4920
tgaaaatgtc gccgatgtga gtttctgtgt aactgatatc gccatttttc caaaagtgat 4980
ttttgggcat acgcgatatc tggcgatagc gcttatatcg tttacggggg atggcgatag 5040
acgactttgg tgacttgggc gattctgtgt gtcgcaaata tcgcagtttc gatataggtg 5100
acagacgata tgaggctata tcgccgatag aggcgacatc aagctggcac atggccaatg 5160
catatcgatc tatacattga atcaatattg gccattagcc atattattca ttggttatat 5220
agcataaatc aatattggct attggccatt gcatacgttg tatccatatc ataatatgta 5280
catttatatt ggctcatgtc caacattacc gccatgttga cattgattat tgactagtta 5340
ttaatagtaa tcaattacgg ggtcattagt tcatagccca tatatggagt tccgcgttac 5400
ataacttacg gtaaatggcc cgcctggctg accgcccaac gacccccgcc cattgacgtc 5460
aataatgacg tatgttccca tagtaacgcc aatagggact ttccattgac gtcaatgggt 5520
ggagtattta cggtaaactg cccacttggc agtacatcaa gtgtatcata tgccaagtac 5580
gccccctatt gacgtcaatg acggtaaatg gcccgcctgg cattatgccc agtacatgac 5640
cttatgggac tttcctactt ggcagtacat ctacgtatta gtcatcgcta ttaccatggt 5700
gatgcggttt tggcagtaca tcaatgggcg tggatagcgg tttgactcac ggggatttcc 5760
aagtctccac cccattgacg tcaatgggag tttgttttgg caccaaaatc aacgggactt 5820
tccaaaatgt cgtaacaact ccgccccatt gacgcaaatg ggcggtaggc gtgtacggtg 5880
ggaggtctat ataagcagag ctcgtttagt gaaccggatc cttaaccgtg aaagcttgcc 5940
gccaccatga tgtcctttgt ctctctgctc ctggttggca tcctattcca tgccacccag 6000
gccgagatcg tgctgaccca gtcccccggc accctgtccc tgtcccccgg cgagcgggcc 6060
accctgtcct gccgggcctc ccagtccgtg ggctcctcct acctggcctg gtaccagcag 6120
aagcccggcc aggccccccg gctgctgatc tacggcgcct tctcccgcgc caccggcatc 6180
cccgaccggt tctccggctc cggctccggc accgacttca ccctgaccat ctcccggctg 6240
gagcccgagg acttcgccgt gtactactgc cagcagtacg gctcctcccc ctggaccttc 6300
ggccagggca ccaaggtgga gatcaagcga actgtggctg caccatctgt cttcatcttc 6360
ccgccatctg atgagcagct taagtccgga actgctagcg ttgtgtgcct gctgaataac 6420
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccctcca atcgggtaac 6480
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 6540
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 6600
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgtta gtgagatctc 6660
gagttcgaca tcgataatca acctctggat tacaaaattt gtgaaagatt gactggtatt 6720
cttaactatg ttgctccttt tacgctatgt ggatacgctg ctttaatgcc tttgtatcat 6780
gctattgctt cccgtatggc tttcattttc tcctccttgt ataaatcctg gttgctgtct 6840
ctttatgagg agttgtggcc cgttgtcagg caacgtggcg tggtgtgcac tgtgtttgct 6900
gacgcaaccc ccactggttg gggcattgcc accacctgtc agctcctttc cgggactttc 6960
gctttccccc tccctattgc cacggcggaa ctcatcgccg cctgccttgc ccgctgctgg 7020
acaggggctc ggctgttggg cactgacaat tccgtggtgt tgtcggggaa atcatcgtcc 7080
tttccttggc tgctcgcctg tgttgccacc tggattctgc gcgggacgtc cttctgctac 7140
gtcccttcgg ccctcaatcc agcggacctt ccttcccgcg gcctgctgcc ggctctgcgg 7200
cctcttccgc gtcttcgcct tcgccctcag acgagtcgga tctccctttg ggccgcctcc 7260
ccgcatcgat gggggaggct aactgaaaca cggaaggaga caataccgga aggaacccgc 7320
gctatgacgg caataaaaag acagaataaa acgcacgggt gttgggtcgt ttgttcataa 7380
acgcggggtt cggtcccagg gctggcactc tgtcgatacc ccaccgagac cccattgggg 7440
ccaatacgcc cgcgtttctt ccttttcccc accccacccc ccaagttcgg gtgaaggccc 7500
agggctcgca gccaacgtcg gggcggcagg ccctgccata gccctagcag cttggccgta 7560
atcatggtca tagctgtttc ctgtgtgaaa ttgttatccg ctcacaattc cacacaacat 7620
acgagccgga agcataaagt gtaaagcctg gggtgcctaa tgagtgagct aactcacatt 7680
aattgcgttg cgctcactgc ccgctttcca gtcgggaaac ctgtcgtgcc agctgcatta 7740
atgaatcggc caacgcgcgg ggagaggcgg tttgcgtatt gggcgctctt ccgcttcctc 7800
gctcactgac tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag ctcactcaaa 7860
ggcggtaata cggttatcca cagaatcagg ggataacgca ggaaagaaca tgtgagcaaa 7920
aggccagcaa aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct 7980
ccgcccccct gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac 8040
aggactataa agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc 8100
gaccctgccg cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc 8160
tcatagctca cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg 8220
tgtgcacgaa ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga 8280
gtccaacccg gtaagacacg acttatcgcc actggcagca gccactggta acaggattag 8340
cagagcgagg tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta 8400
cactagaaga acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag 8460
agttggtagc tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg 8520
caagcagcag attacgcgca gaaaaaaagg atctcaagaa gatcctttga tcttttctac 8580
ggggtctgac gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc 8640
aaaaaggatc ttcacctaga tccttttaaa ttaaaaatga caatctaaag tatatatgag 8700
taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc agcgatctgt 8760
ctatttcgtt catccatagt tgcctgactc cccgtcgtgt agataactac gatacgggag 8820
ggcttaccat ctggccccag tgctgcaatg ataccgcgag acccacgctc accggctcca 8880
gatttatcag caataaacca gccagccgga agggccgagc gcagaagtgg tcctgcaact 8940
ttatccgcct ccatccagtc tattaattgt tgccgggaag ctagagtaag tagttcgcca 9000
gttaatagtt tgcgcaacgt tgttgccatt gctacaggca tcgtggtgtc acgctcgtcg 9060
tttggtatgg cttcattcag ctccggttcc caacgatcaa ggcgagttac atgatccccc 9120
atgttgtgca aaaaagcggt tagctccttc ggtcctccga tcgttgtcag aagtaagttg 9180
gccgcagtgt tatcactcat ggttatggca gcactgcata attctcttac tgtcatgcca 9240
tccgtaagat gcttttctgt gactggtgag tactcaacca agtcattctg agaatagtgt 9300
atgcggcgac cgagttgctc ttgcccggcg tcaatacggg ataataccgc gccacatagc 9360
agaactttaa aagtgctcat cattggaaaa cgttcttcgg ggcgaaaact ctcaaggatc 9420
ttaccgctgt tgagatccag ttcgatgtaa cccactcgtg cacccaactg atcttcagca 9480
tcttttactt tcaccagcgt ttctgggtga gcaaaaacag gaaggcaaaa tgccgcaaaa 9540
aagggaataa gggcgacacg gaaatgttga atactcatac tcttcctttt tcaatattat 9600
tgaagcattt atcagggtta ttgtctcatg agcggataca tatttgaatg tatttagaaa 9660
aataaacaaa taggggttcc gcgcacattt ccccgaaaag tgccacctga cgtctaagaa 9720
accattatta tcatgacatt aacctataaa aataggcgta tcacgaggcc ctttcgtc 9778
<210> 12
<211> 9788
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтетическая
<400> 12
tcgcgcgttt cggtgatgac ggtgaaaacc tctgacacat gcagctcccg gagacggtca 60
cagcttgtct gtaagcggat gccgggagca gacaagcccg tcagggcgcg tcagcgggtg 120
ttggcgggtg tcggggctgg cttaactatg cggcatcaga gcagattgta ctgagagtgc 180
accatatgcg gtgtgaaata ccgcacagat gcgtaaggag aaaataccgc atcaggcgcc 240
attcgccatt caggctgcgc aactgttggg aagggcgatc ggtgcgggcc tcttcgctat 300
tacgccagct ggcgaaaggg ggatgtgctg caaggcgatt aagttgggta acgccagggt 360
tttcccagtc acgacgttgt aaaacgacgg ccagtgggat ccccgtcgac gatgtaggtc 420
acggtctcga agccgcggtg cgggtgccag ggcgtgccct tgggctcccc gggcgcgtac 480
tccacctcac ccatctggtc catcatgatg aacgggtcga ggtggcggta gttgatcccg 540
gcgaacgcgc ggcgcaccgg gaagccctcg ccctcgaaac cgctgggcgc ggtggtcacg 600
gtgagcacgg gacgtgcgac ggcgtcggcg ggtgcggata cgcggggcag cgtcagcggg 660
ttctcgacgg tcacggcggg catgtcgacc cttccaccat ggccacctca gcaagttccc 720
acttgaacaa aaacatcaag caaatgtact tgtgcctgcc ccagggtgag aaagtccaag 780
ccatgtatat ctgggttgat ggtactggag aaggactgcg ctgcaaaacc cgcaccctgg 840
actgtgagcc caagtgtgta gaagagttac ctgagtggaa ttttgatggc tctagtacct 900
ttcagtctga gggctccaac agtgacatgt atctcagccc tgttgccatg tttcgggacc 960
ccttccgcag agatcccaac aagctggtgt tctgtgaagt tttcaagtac aaccggaagc 1020
ctgcagagac caatttaagg cactcgtgta aacggataat ggacatggtg agcaaccagc 1080
acccctggtt tggaatggaa caggagtata ctctgatggg aacagatggg cacccttttg 1140
gttggccttc caatggcttt cctgggcccc aaggtccgta ttactgtggt gtgggcgcag 1200
acaaagccta tggcagggat atcgtggagg ctcactaccg cgcctgcttg tatgctgggg 1260
tcaagattac aggaacaaat gctgaggtca tgcctgccca gtgggagttc caaataggac 1320
cctgtgaagg aatccgcatg ggagatcatc tctgggtggc ccgtttcatc ttgcatcgag 1380
tatgtgaaga ctttggggta atagcaacct ttgaccccaa gcccattcct gggaactgga 1440
atggtgcagg ctgccatacc aactttagca ccaaggccat gcgggaggag aatggtctga 1500
agcacatcga ggaggccatc gagaaactaa gcaagcggca ccggtaccac attcgagcct 1560
acgatcccaa ggggggcctg gacaatgccc gtcgtctgac tgggttccac gaaacgtcca 1620
acatcaacga cttttctgct ggtgtcgcca atcgcagtgc cagcatccgc attccccgga 1680
ctgtcggcca ggagaagaaa ggttactttg aagaccgccg cccctctgcc aactgtgacc 1740
cctttgcagt gacagaagcc atcgtccgca catgccttct caatgagact ggcgacgagc 1800
ccttccaata caaaaactaa agatccctat ggctattggc caggttcaat actatgtatt 1860
ggccctatgc catatagtat tccatatatg ggttttccta ttgacgtaga tagcccctcc 1920
caatgggcgg tcccatatac catatatggg gcttcctaat accgcccata gccactcccc 1980
cattgacgtc aatggtctct atatatggtc tttcctattg acgtcatatg ggcggtccta 2040
ttgacgtata tggcgcctcc cccattgacg tcaattacgg taaatggccc gcctggctca 2100
atgcccattg acgtcaatag gaccacccac cattgacgtc aatgggatgg ctcattgccc 2160
attcatatcc gttctcacgc cccctattga cgtcaatgac ggtaaatggc ccacttggca 2220
gtacatcaat atctattaat agtaacttgg caagtacatt actattggaa gtacgccagg 2280
gtacattggc agtactccca ttgacgtcaa tggcggtaaa tggcccgcga tggctgccaa 2340
gtacatcccc attgacgtca atggggaggg gcaatgacgc aaatgggcgt tccattgacg 2400
taaatgggcg gtaggcgtgc ctaatgggag gtctatataa gcaatgctcg tttagggaac 2460
cgccattctg cctggggacg tcggaggagc tcgaagcggc cgccgccacc atgatgtcct 2520
ttgtctctct gctcctggtt ggcatcctat tccatgccac ccaggccgag atcgtgctga 2580
cccagtcccc cggcaccctg tccctgtccc ccggcgagcg ggccaccctg tcctgccggg 2640
cctcccagtc cgtgggctcc tcctacctgg cctggtacca gcagaagccc ggccaggccc 2700
cccggctgct gatctacggc gccttctccc gcgccaccgg catccccgac cggttctccg 2760
gctccggctc cggcaccgac ttcaccctga ccatctcccg gctggagccc gaggacttcg 2820
ccgtgtacta ctgccagcag tacggctcct ccccctggac cttcggccag ggcaccaagg 2880
tggagatcaa gcgaactgtg gctgcaccat ctgtcttcat cttcccgcca tctgatgagc 2940
agcttaagtc cggaactgct agcgttgtgt gcctgctgaa taacttctat cccagagagg 3000
ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag gagagtgtca 3060
cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg ctgagcaaag 3120
cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc 3180
ccgtcacaaa gagcttcaac aggggagagt gttagtgaga tctatcgata atcaacctct 3240
ggattacaaa atttgtgaaa gattgactgg tattcttaac tatgttgctc cttttacgct 3300
atgtggatac gctgctttaa tgcctttgta tcatgctatt gcttcccgta tggctttcat 3360
tttctcctcc ttgtataaat cctggttgct gtctctttat gaggagttgt ggcccgttgt 3420
caggcaacgt ggcgtggtgt gcactgtgtt tgctgacgca acccccactg gttggggcat 3480
tgccaccacc tgtcagctcc tttccgggac tttcgctttc cccctcccta ttgccacggc 3540
ggaactcatc gccgcctgcc ttgcccgctg ctggacaggg gctcggctgt tgggcactga 3600
caattccgtg gtgttgtcgg ggaaatcatc gtcctttcct tggctgctcg cctgtgttgc 3660
cacctggatt ctgcgcggga cgtccttctg ctacgtccct tcggccctca atccagcgga 3720
ccttccttcc cgcggcctgc tgccggctct gcggcctctt ccgcgtcttc gccttcgccc 3780
tcagacgagt cggatctccc tttgggccgc ctccccgcat cgattactaa tcagccatac 3840
cacatttgta gaggttttac ttgctttaaa aaacctccca cacctccccc tgaacctgaa 3900
acataaaatg aatgcaattg ttgttgttaa cttgtttatt gcagcttata atggttacaa 3960
ataaagcaat agcatcacaa atttcacaaa taaagcattt ttttcactgc attctagttg 4020
tggtttgtcc aaactcatca atgtatctta tcatgtctgg aattcggacg gtgactgcag 4080
tgaataataa aatgtgtgtt tgtccgaaat acgcgttttg agatttctgt cgccgactaa 4140
attcatgtcg cgcgatagtg gtgtttatcg ccgatagaga tggcgatatt ggaaaaatcg 4200
atatttgaaa atatggcata ttgaaaatgt cgccgatgtg agtttctgtg taactgatat 4260
cgccattttt ccaaaagtga tttttgggca tacgcgatat ctggcgatag cgcttatatc 4320
gtttacgggg gatggcgata gacgactttg gtgacttggg cgattctgtg tgtcgcaaat 4380
atcgcagttt cgatataggt gacagacgat atgaggctat atcgccgata gaggcgacat 4440
caagctggca catggccaat gcatatcgat ctatacattg aatcaatatt ggccattagc 4500
catattattc attggttata tagcataaat caatattggc tattggccat tgcatacgtt 4560
gtatccatat cataatatgt acatttatat tggctcatgt ccaacattac cgccatgttg 4620
acattgatta ttgactagtt attaatagta atcaattacg gggtcattag ttcatagccc 4680
atatatggag ttccgcgtta cataacttac ggtaaatggc ccgcctggct gaccgcccaa 4740
cgacccccgc ccattgacgt caataatgac gtatgttccc atagtaacgc caatagggac 4800
tttccattga cgtcaatggg tggagtattt acggtaaact gcccacttgg cagtacatca 4860
agtgtatcat atgccaagta cgccccctat tgacgtcaat gacggtaaat ggcccgcctg 4920
gcattatgcc cagtacatga ccttatggga ctttcctact tggcagtaca tctacgtatt 4980
agtcatcgct attaccatgg tgatgcggtt ttggcagtac atcaatgggc gtggatagcg 5040
gtttgactca cggggatttc caagtctcca ccccattgac gtcaatggga gtttgttttg 5100
gcaccaaaat caacgggact ttccaaaatg tcgtaacaac tccgccccat tgacgcaaat 5160
gggcggtagg cgtgtacggt gggaggtcta tataagcaga gctcgtttag tgaaccggat 5220
ccttaaccgt gaaagcttgc cgccaccatg atgtcctttg tctctctgct cctggttggc 5280
atcctattcc atgccaccca ggcccaggtg cagctggtgg agtccggcgg cggcgtcgtg 5340
cagcccggcc ggtccctgcg gctgtcctgc gccgcctccg gcttcacctt ctcctcctac 5400
accatgcact gggtgcggca ggcccccggc aagggcctgg agtgggtgac tttcatctcc 5460
tacgacggca acaacaagta ctacgccgac tccgtgaagg gccggttcac catctcccgc 5520
gacaactcca agaacaccct gtacctgcag atgaactccc tgcgggccga ggacaccgcc 5580
atctactact gcgcccggac cggctggctg ggccccttcg actactgggg ccagggcacc 5640
ctggtgaccg tgtcctccgc ctccaccaag ggcccatcgg tcttccccct ggcaccctct 5700
agcaagagca cctctggggg cacagcggcc ctgggctgcc tggtcaagga ctacttcccc 5760
gaaccggtga cggtgtcgtg gaactcaggc gccctgacca gcggcgtgca caccttcccg 5820
gctgtcctac agtcctcagg actctactcc ctcagcagcg tggtgaccgt gccctccagc 5880
agcttgggca cccagaccta catctgcaac gtgaatcaca agcccagcaa caccaaggtg 5940
gacaagcggg ttgagcccaa atcttgtgac aaaactcaca catgcccacc gtgcccagca 6000
cctgaactcc tggggggacc gtcagtcttc ctcttccccc caaaacccaa ggacaccctc 6060
atgatctccc ggacccctga ggtcacatgc gtggtggtgg acgtgagcca cgaagaccct 6120
gaggtcaagt tcaactggta cgtggacggc gtggaggtgc ataatgccaa gacaaagccg 6180
cgggaggagc agtacaacag cacgtaccgt gtggtcagcg tcctcaccgt cctgcaccag 6240
gactggctga atggcaagga gtacaagtgc aaggtctcca acaaagccct cccagccccc 6300
atcgagaaaa ccatctccaa agccaaaggg cagccccgag aaccacaggt gtacaccctg 6360
cctccatccc gcgatgagct gaccaagaac caggtcagcc tgacctgcct ggtcaaaggc 6420
ttctatccca gcgacatcgc cgtggagtgg gagagcaatg ggcagccgga gaacaactac 6480
aagaccacgc ctcccgtgct ggactccgac ggctccttct tcctctatag caagctcacc 6540
gtggacaaga gcaggtggca gcaggggaac gtcttctcat gctccgtgat gcatgaggct 6600
ctgcacaacc actacacgca gaagagcctc tccctgtctc ctgggaaatg atgagatctc 6660
gagttcgaca tcgataatca acctctggat tacaaaattt gtgaaagatt gactggtatt 6720
cttaactatg ttgctccttt tacgctatgt ggatacgctg ctttaatgcc tttgtatcat 6780
gctattgctt cccgtatggc tttcattttc tcctccttgt ataaatcctg gttgctgtct 6840
ctttatgagg agttgtggcc cgttgtcagg caacgtggcg tggtgtgcac tgtgtttgct 6900
gacgcaaccc ccactggttg gggcattgcc accacctgtc agctcctttc cgggactttc 6960
gctttccccc tccctattgc cacggcggaa ctcatcgccg cctgccttgc ccgctgctgg 7020
acaggggctc ggctgttggg cactgacaat tccgtggtgt tgtcggggaa atcatcgtcc 7080
tttccttggc tgctcgcctg tgttgccacc tggattctgc gcgggacgtc cttctgctac 7140
gtcccttcgg ccctcaatcc agcggacctt ccttcccgcg gcctgctgcc ggctctgcgg 7200
cctcttccgc gtcttcgcct tcgccctcag acgagtcgga tctccctttg ggccgcctcc 7260
ccgcatcgat gggggaggct aactgaaaca cggaaggaga caataccgga aggaacccgc 7320
gctatgacgg caataaaaag acagaataaa acgcacgggt gttgggtcgt ttgttcataa 7380
acgcggggtt cggtcccagg gctggcactc tgtcgatacc ccaccgagac cccattgggg 7440
ccaatacgcc cgcgtttctt ccttttcccc accccacccc ccaagttcgg gtgaaggccc 7500
agggctcgca gccaacgtcg gggcggcagg ccctgccata gccctagcag cttggccgta 7560
atcatggtca tagctgtttc ctgtgtgaaa ttgttatccg ctcacaattc cacacaacat 7620
acgagccgga agcataaagt gtaaagcctg gggtgcctaa tgagtgagct aactcacatt 7680
aattgcgttg cgctcactgc ccgctttcca gtcgggaaac ctgtcgtgcc agctgcatta 7740
atgaatcggc caacgcgcgg ggagaggcgg tttgcgtatt gggcgctctt ccgcttcctc 7800
gctcactgac tcgctgcgct cggtcgttcg gctgcggcga gcggtatcag ctcactcaaa 7860
ggcggtaata cggttatcca cagaatcagg ggataacgca ggaaagaaca tgtgagcaaa 7920
aggccagcaa aaggccagga accgtaaaaa ggccgcgttg ctggcgtttt tccataggct 7980
ccgcccccct gacgagcatc acaaaaatcg acgctcaagt cagaggtggc gaaacccgac 8040
aggactataa agataccagg cgtttccccc tggaagctcc ctcgtgcgct ctcctgttcc 8100
gaccctgccg cttaccggat acctgtccgc ctttctccct tcgggaagcg tggcgctttc 8160
tcatagctca cgctgtaggt atctcagttc ggtgtaggtc gttcgctcca agctgggctg 8220
tgtgcacgaa ccccccgttc agcccgaccg ctgcgcctta tccggtaact atcgtcttga 8280
gtccaacccg gtaagacacg acttatcgcc actggcagca gccactggta acaggattag 8340
cagagcgagg tatgtaggcg gtgctacaga gttcttgaag tggtggccta actacggcta 8400
cactagaaga acagtatttg gtatctgcgc tctgctgaag ccagttacct tcggaaaaag 8460
agttggtagc tcttgatccg gcaaacaaac caccgctggt agcggtggtt tttttgtttg 8520
caagcagcag attacgcgca gaaaaaaagg atctcaagaa gatcctttga tcttttctac 8580
ggggtctgac gctcagtgga acgaaaactc acgttaaggg attttggtca tgagattatc 8640
aaaaaggatc ttcacctaga tccttttaaa ttaaaaatga agttttaaat caatctaaag 8700
tatatatgag taaacttggt ctgacagtta ccaatgctta atcagtgagg cacctatctc 8760
agcgatctgt ctatttcgtt catccatagt tgcctgactc cccgtcgtgt agataactac 8820
gatacgggag ggcttaccat ctggccccag tgctgcaatg ataccgcgag acccacgctc 8880
accggctcca gatttatcag caataaacca gccagccgga agggccgagc gcagaagtgg 8940
tcctgcaact ttatccgcct ccatccagtc tattaattgt tgccgggaag ctagagtaag 9000
tagttcgcca gttaatagtt tgcgcaacgt tgttgccatt gctacaggca tcgtggtgtc 9060
acgctcgtcg tttggtatgg cttcattcag ctccggttcc caacgatcaa ggcgagttac 9120
atgatccccc atgttgtgca aaaaagcggt tagctccttc ggtcctccga tcgttgtcag 9180
aagtaagttg gccgcagtgt tatcactcat ggttatggca gcactgcata attctcttac 9240
tgtcatgcca tccgtaagat gcttttctgt gactggtgag tactcaacca agtcattctg 9300
agaatagtgt atgcggcgac cgagttgctc ttgcccggcg tcaatacggg ataataccgc 9360
gccacatagc agaactttaa aagtgctcat cattggaaaa cgttcttcgg ggcgaaaact 9420
ctcaaggatc ttaccgctgt tgagatccag ttcgatgtaa cccactcgtg cacccaactg 9480
atcttcagca tcttttactt tcaccagcgt ttctgggtga gcaaaaacag gaaggcaaaa 9540
tgccgcaaaa aagggaataa gggcgacacg gaaatgttga atactcatac tcttcctttt 9600
tcaatattat tgaagcattt atcagggtta ttgtctcatg agcggataca tatttgaatg 9660
tatttagaaa aataaacaaa taggggttcc gcgcacattt ccccgaaaag tgccacctga 9720
cgtctaagaa accattatta tcatgacatt aacctataaa aataggcgta tcacgaggcc 9780
ctttcgtc 9788
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЕКТОРЫ ДЛЯ ПРОИЗВОДСТВА БЕЛКОВ | 2019 |
|
RU2820218C2 |
| ВЕКТОРНАЯ СИСТЕМА НА ОСНОВЕ ASLV | 2010 |
|
RU2566563C2 |
| НУКЛЕАЗА PaCas9 | 2018 |
|
RU2706298C1 |
| ГЕННОЕ РЕДАКТИРОВАНИЕ ГЛУБОКИХ ИНТРОННЫХ МУТАЦИЙ | 2016 |
|
RU2759335C2 |
| РЕДАКТИРОВАНИЕ ГЕНОВ С ИСПОЛЬЗОВАНИЕМ МОДИФИЦИРОВАННОЙ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2018 |
|
RU2811724C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| Нуклеиновая кислота, имеющая промоторную активность, и ее применение | 2022 |
|
RU2818112C2 |
| КОМПОНЕНТЫ СИСТЕМЫ CRISPR-CAS, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2701662C2 |
| МУТАЦИИ ФЕРМЕНТА CRISPR, УМЕНЬШАЮЩИЕ НЕЦЕЛЕВЫЕ ЭФФЕКТЫ | 2016 |
|
RU2752834C2 |
| ДОСТАВКА И ПРИМЕНЕНИЕ СИСТЕМ CRISPR-CAS, ВЕКТОРОВ И КОМПОЗИЦИЙ ДЛЯ ЦЕЛЕНАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ И ТЕРАПИИ В ПЕЧЕНИ | 2014 |
|
RU2716420C2 |

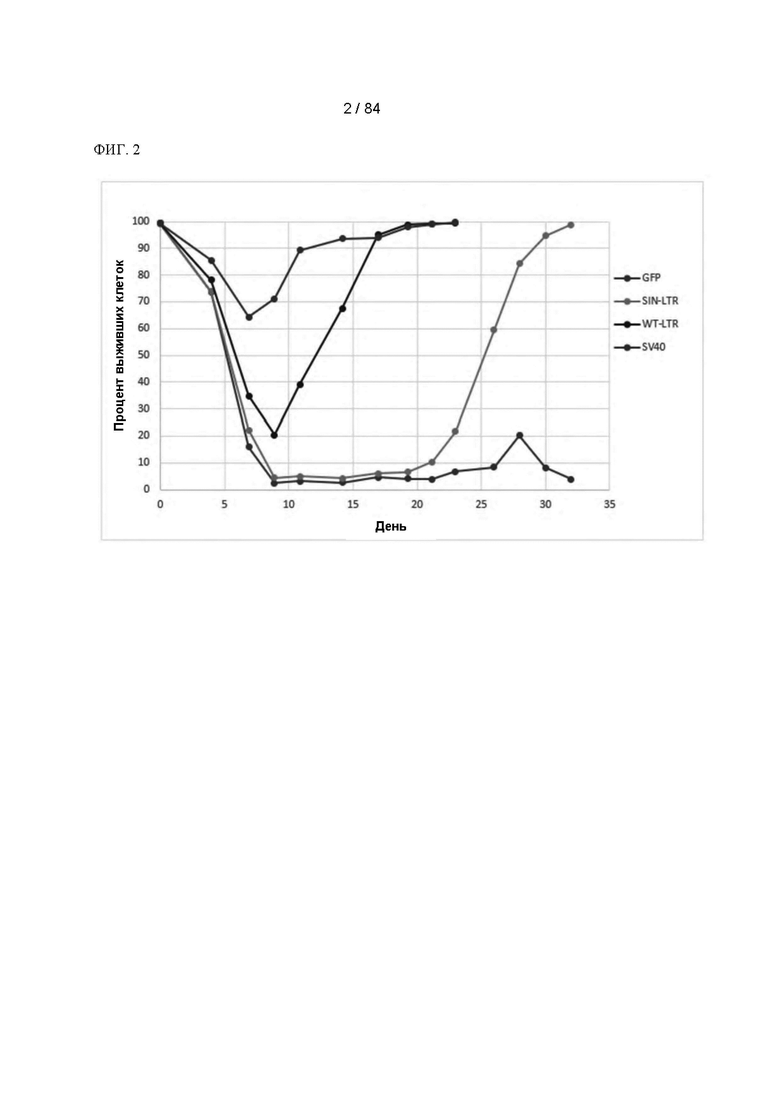
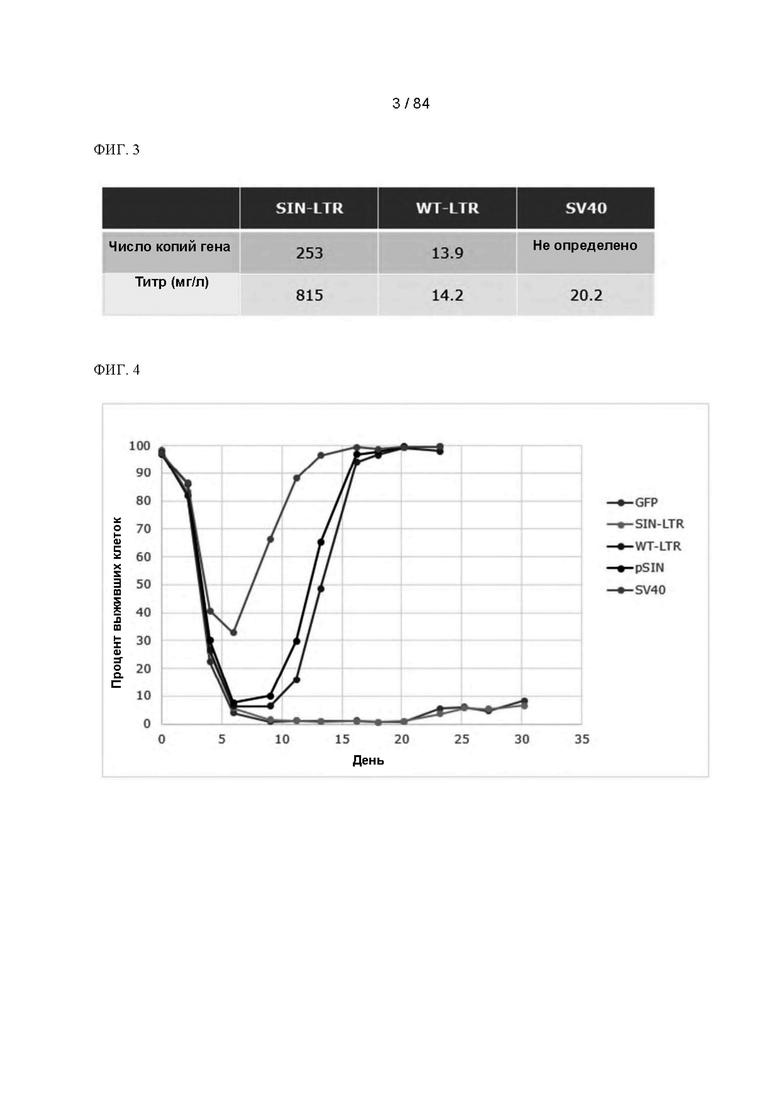
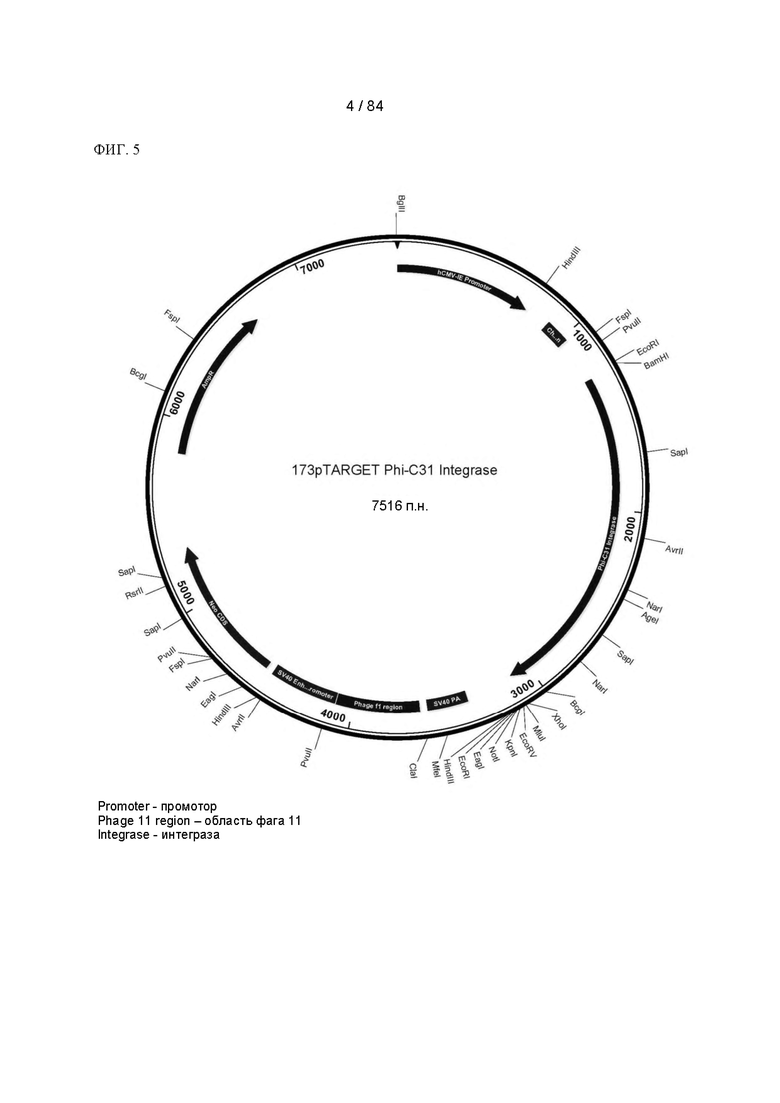





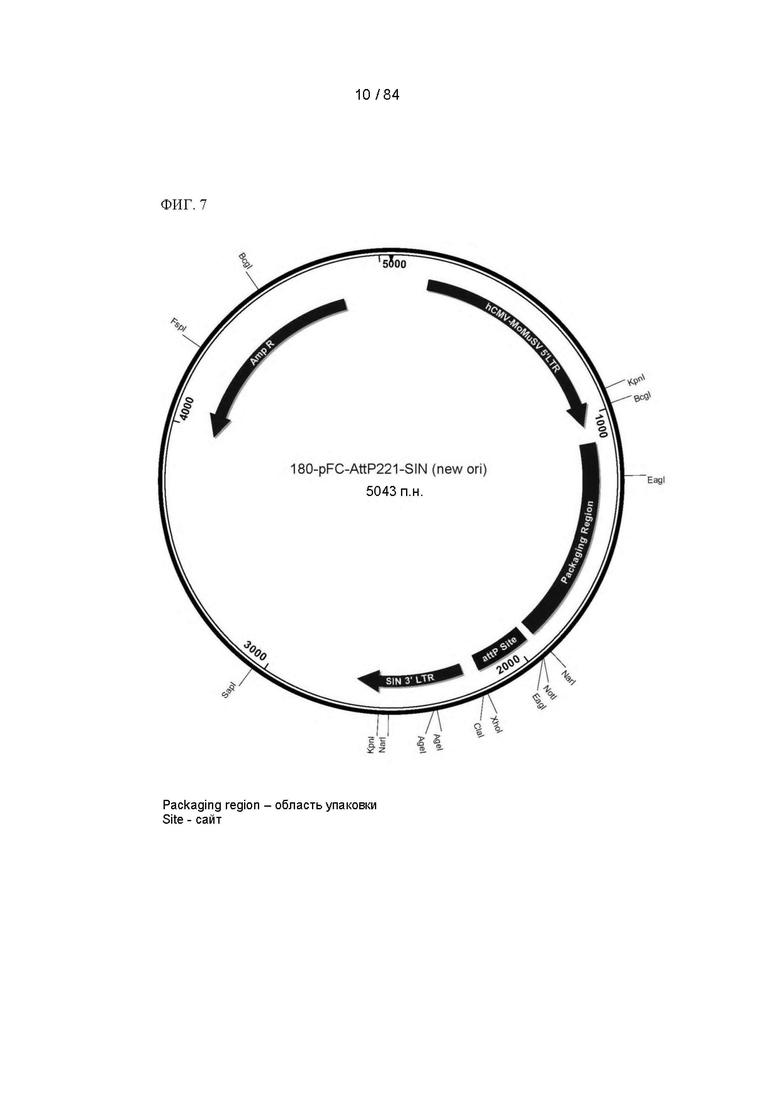

























































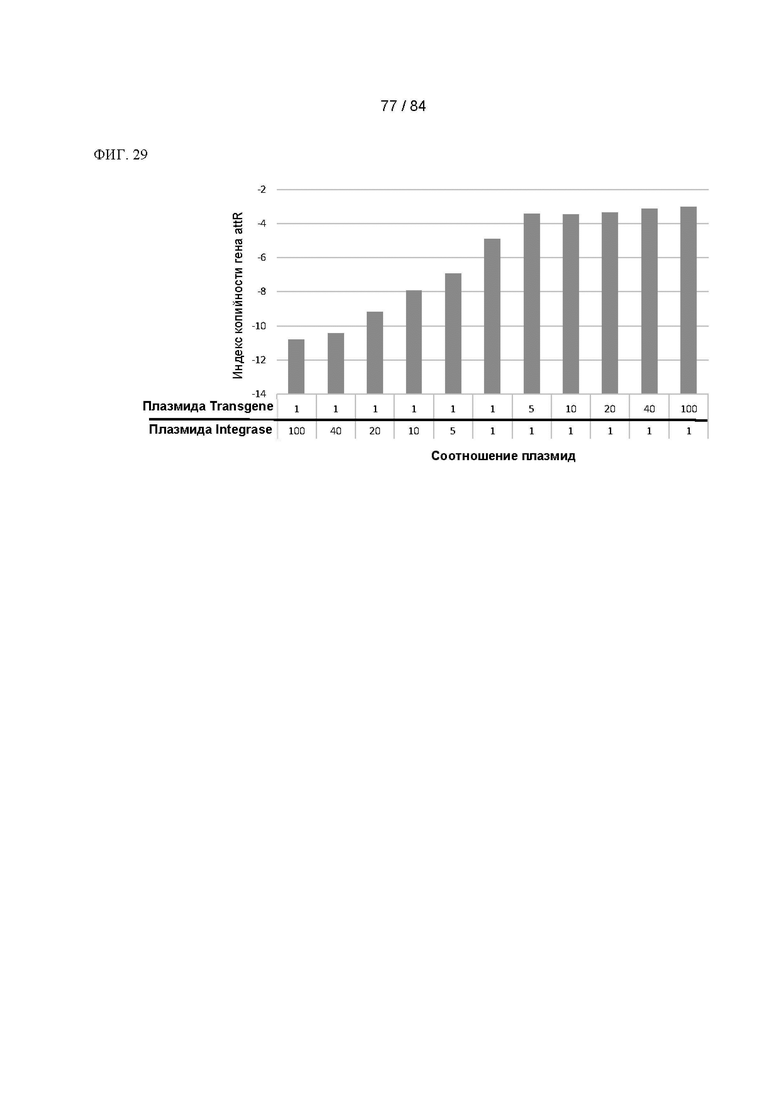
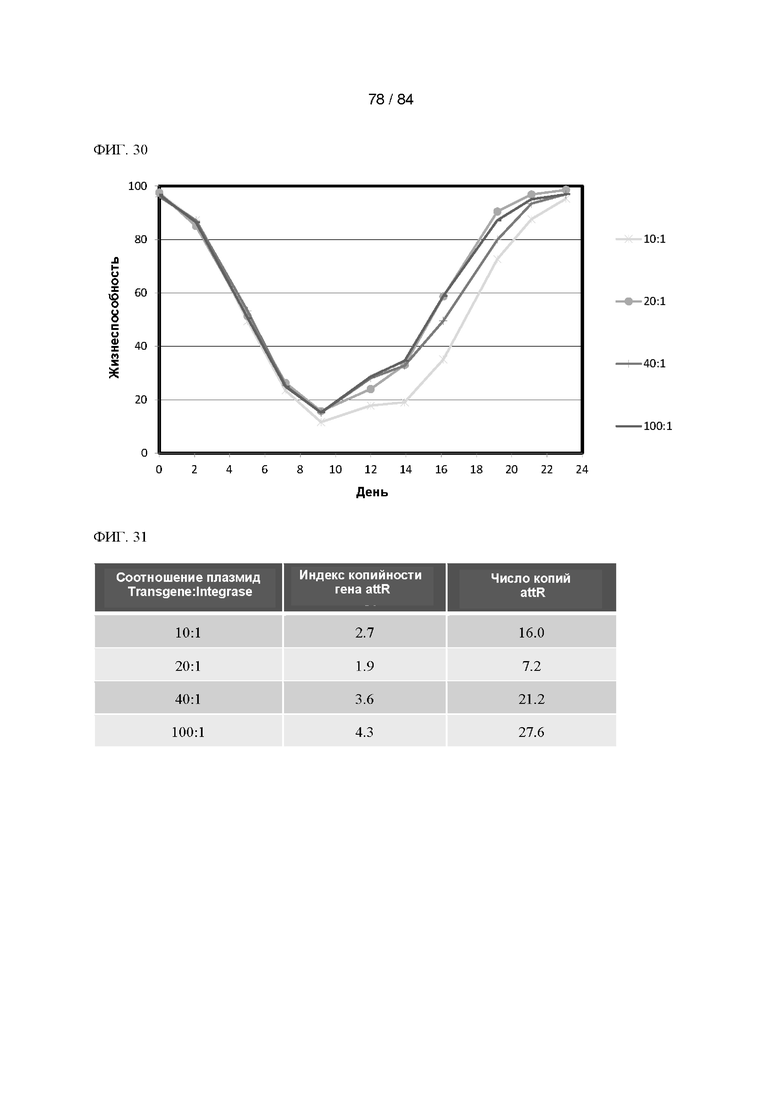
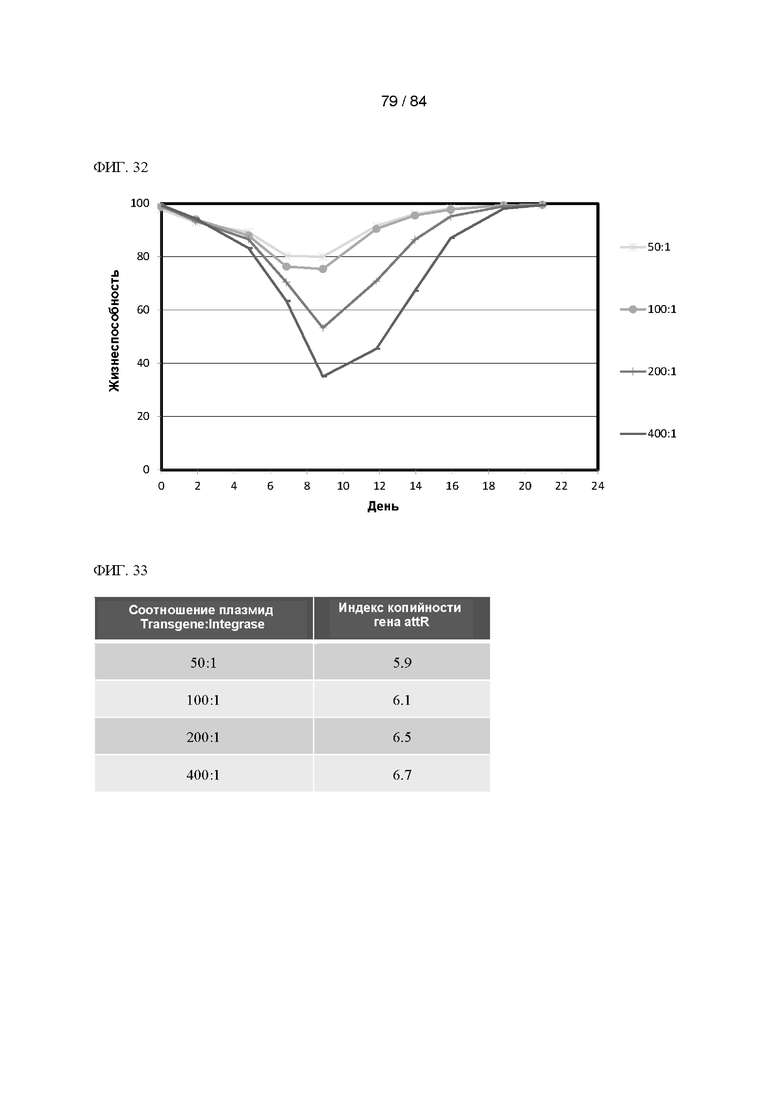
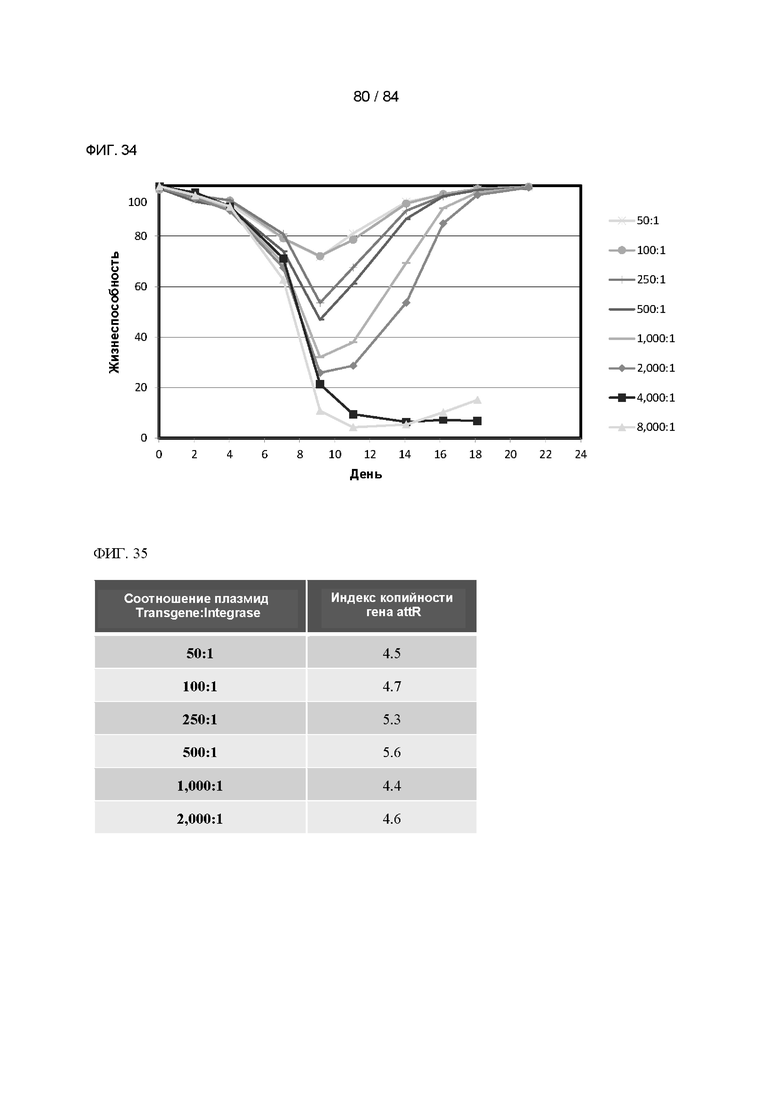
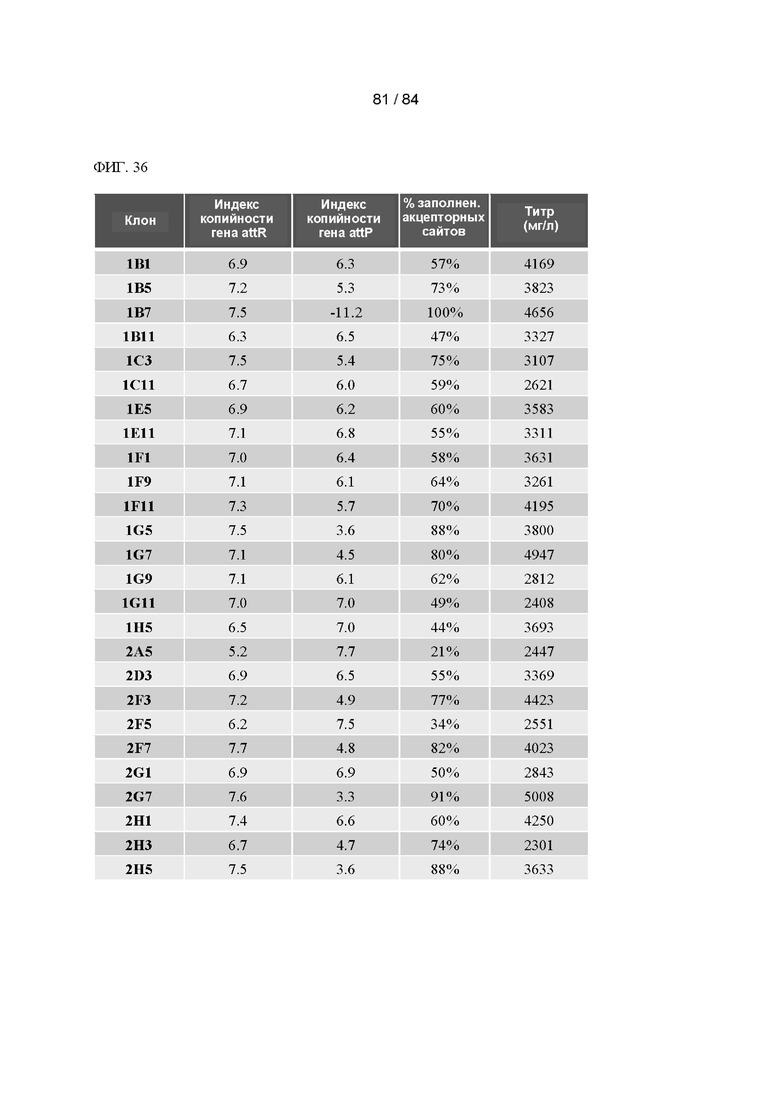
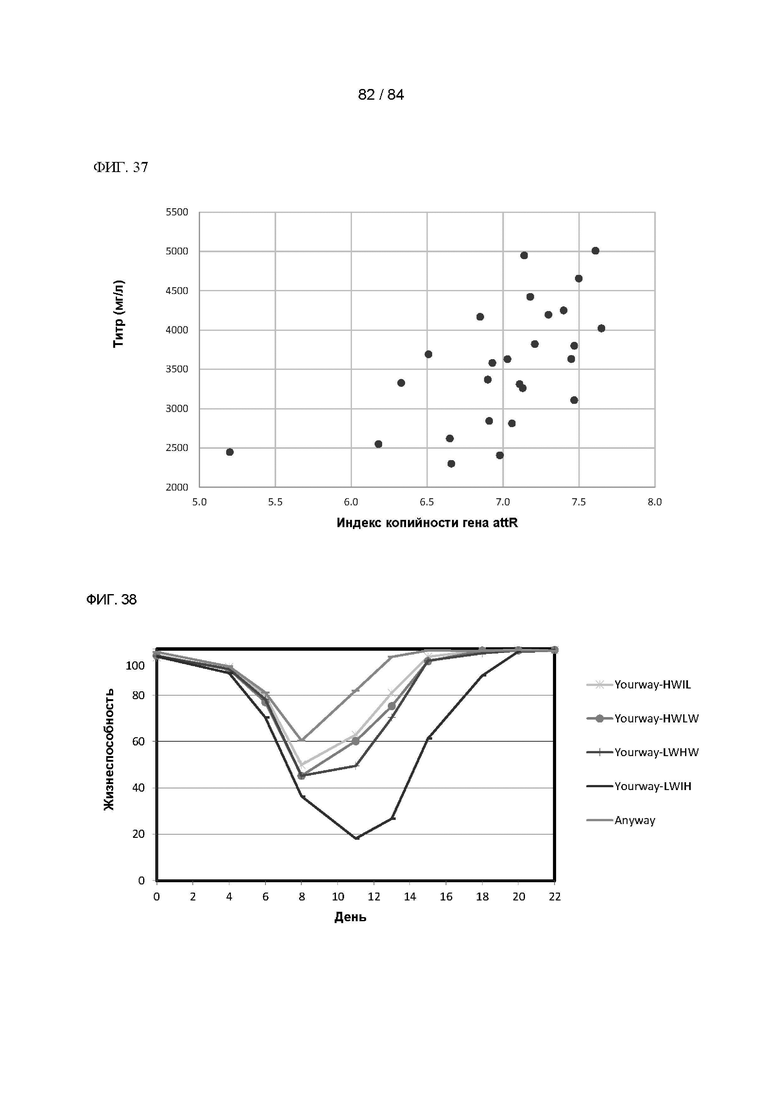
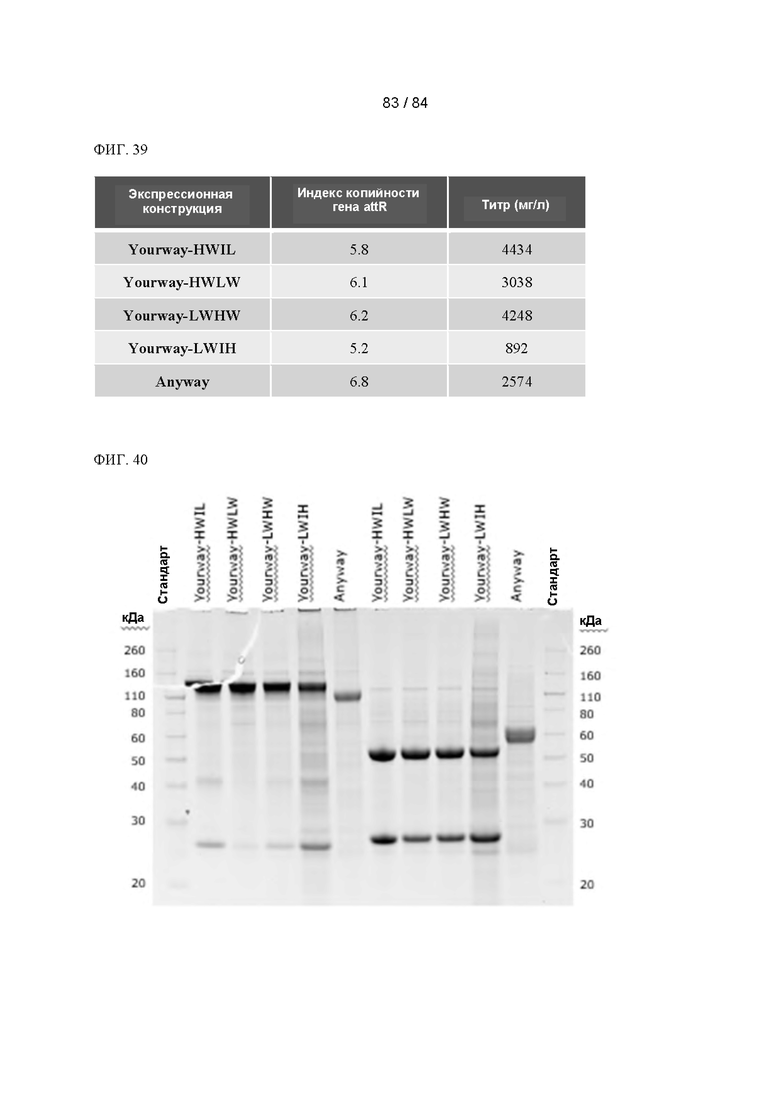
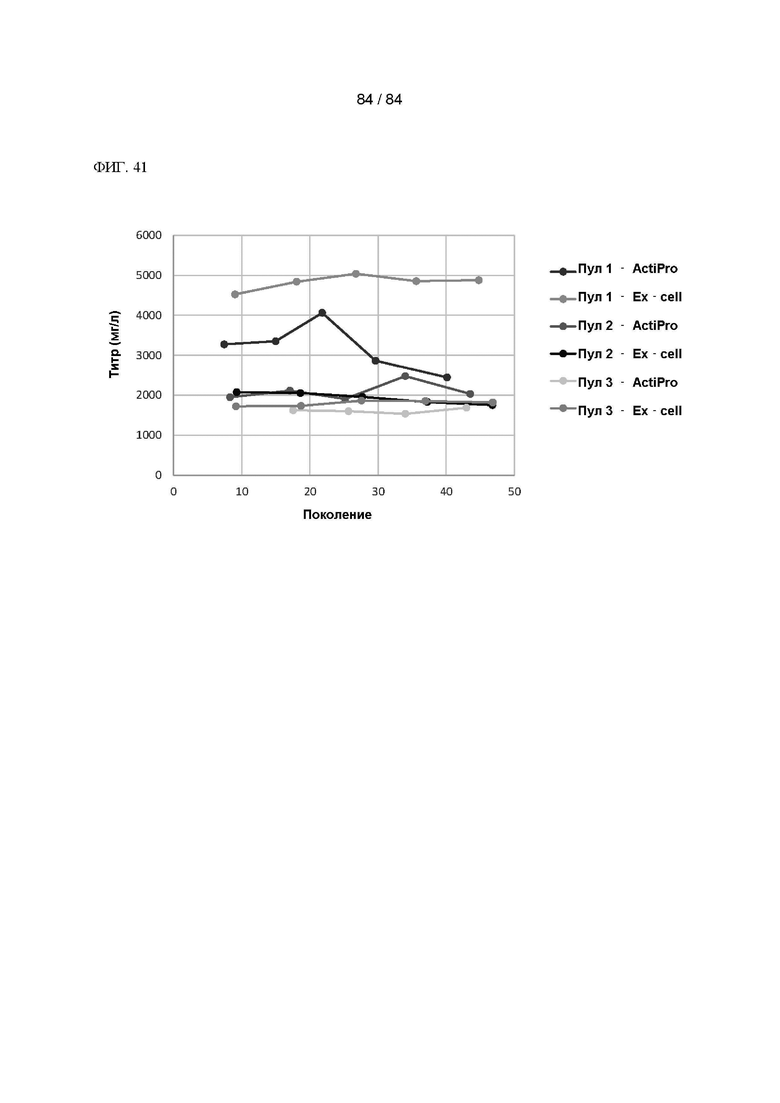
Изобретение относится к биотехнологии и молекулярной биологии, в частности к конструкциям нуклеиновых кислот, и их применению для разработки линий клеток-хозяев для получения целевого белка. Конструкции по настоящему изобретению обеспечивают улучшенную селекцию для разработки высокопродуктивных линий клеток. 4 н. и 10 з.п. ф-лы, 41 ил., 7 пр.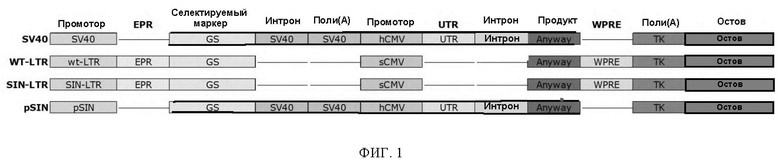
1. Конструкция нуклеиновой кислоты для экспрессии целевого белка, содержащая следующие функционально связанные элементы по порядку от 5'- до 3'-конца:
необязательно, первую промоторную последовательность;
последовательность селектируемого маркера, которая функционально связана с указанной первой промоторной последовательностью в случае её присутствия;
вторую промоторную последовательность;
последовательность нуклеиновой кислоты, кодирующую целевой белок, которая функционально связана с указанной второй промоторной последовательностью; и
сигнальную последовательность поли(А);
при этом указанная конструкция нуклеиновой кислоты дополнительно содержит по меньшей мере один инсерционный элемент, выбранный из группы, состоящей из управляемого транспозоном инсерционного элемента и управляемого рекомбиназой инсерционного элемента, в положении или положениях, выбранных из группы, состоящей из 5'-конца относительно указанной необязательной первой промоторной последовательности или указанной последовательности селектируемого маркера, 3'-конца относительно указанной сигнальной последовательности поли(А), положения между указанной необязательной первой промоторной последовательностью и указанной сигнальной последовательностью поли(А), положения между указанной последовательностью селектируемого маркера и указанной второй промоторной последовательностью, и одновременно 5'-конца относительно указанной необязательной первой промоторной последовательности или указанной последовательности селектируемого маркера и 3'-конца относительно указанной сигнальной последовательности поли(А).
2. Конструкция нуклеиновой кислоты по п. 1, отличающаяся тем, что указанная конструкция нуклеиновой кислоты не содержит сигнальной последовательности поли(А) между указанным селектируемым маркером и указанным вторым промотором.
3. Конструкция нуклеиновой кислоты по любому из пп. 1-2, отличающаяся тем, что указанный селектируемый маркер примыкает к указанному второму промотору.
4. Конструкция нуклеиновой кислоты по любому из пп. 1-3, отличающаяся тем, что указанный второй промотор примыкает к последовательности нуклеиновой кислоты, кодирующей указанный первый целевой белок.
5. Конструкция нуклеиновой кислоты по любому из пп. 1-4, отличающаяся тем, что указанная конструкция нуклеиновой кислоты содержит расширенную область упаковки (EPR) между указанным первым промотором и указанным селектируемым маркером, при этом предпочтительно указанная EPR содержит несколько потенциальных последовательностей Козак и/или сайтов начала трансляции ATG.
6. Конструкция нуклеиновой кислоты по любому из пп. 1-5, отличающаяся тем, что указанная первая промоторная последовательность представляет собой последовательность слабого промотора, при этом предпочтительно указанная первая промоторная последовательность выбрана из группы, состоящей из промоторных последовательностей SIN-LTR, SV40, lac E. coli, trp E. coli, PL фага лямбда, PR фага лямбда, T3, T7, немедленно-раннего промотора цитомегаловируса (CMV), тимидинкиназы вируса простого герпеса (HSV), альфа-лактальбумина, фактора элонгации 1 альфа человека (hEF1 альфа) и металлотионеина-I мыши.
7. Конструкция нуклеиновой кислоты по любому из пп. 1-6, отличающаяся тем, что указанная последовательность селектируемого маркера представляет собой амплифицируемую последовательность селектируемого маркера, выбранную из группы, состоящей из последовательности глутаминсинтетазы (GS) и последовательности дигидрофолатредуктазы (DHFR), при этом предпочтительно указанная последовательность селектируемого маркера представляет собой последовательность маркера устойчивости к антибиотику, выбранную из группы, состоящей из последовательностей гена устойчивости к неомицину (neo), гена гигромицин B-фосфотрансферазы и гена пуромицин-N-ацетилтрансферазы.
8. Конструкция нуклеиновой кислоты по любому из пп. 1-7, отличающаяся тем, что указанная вторая промоторная последовательность выбрана из группы, состоящей из промоторных последовательностей SV40, lac E. coli, trp E. coli, PL фага лямбда, PR фага лямбда, T3, T7, немедленно-раннего промотора цитомегаловируса (CMV), тимидинкиназы вируса простого герпеса (HSV), альфа-лактальбумина, фактора элонгации 1 альфа человека (hEF1 альфа) и металлотионеина-I мыши.
9. Конструкция нуклеиновой кислоты по любому из пп. 1-8, отличающаяся тем, что указанная последовательность нуклеиновой кислоты, кодирующая целевой белок, кодирует белок, выбранный из группы, состоящей из последовательностей тяжелой и легкой цепей иммуноглобулина.
10. Конструкция нуклеиновой кислоты по п. 1, отличающаяся тем, что указанный управляемый транспозоном инсерционный элемент представляет собой инвертированный концевой повтор, предпочтительно при этом указанный конструкт содержит два инвертированных концевых повтора, расположенных на 5'-конце относительно указанного первого промотора и на 3'-конце относительно указанной сигнальной последовательности поли(А).
11. Конструкция нуклеиновой кислоты по п. 1, отличающаяся тем, что указанный управляемый рекомбиназой инсерционный элемент представляет собой сайт интеграции (att), при этом предпочтительно указанный сайт интеграции (att) представляет собой attB.
12. Клетка-хозяин для экспрессии целевого белка, содержащая конструкцию нуклеиновой кислоты по любому из пп. 1-11, при этом предпочтительно указанная клетка-хозяин содержит от приблизительно 1 до 1000 копий указанной конструкции нуклеиновой кислоты.
13. Способ получения целевого белка, включающий культивирование клеток-хозяев по п. 12 и очистку указанного целевого белка от указанной культуры клеток-хозяев.
14. Комбинация конструкций нуклеиновых кислот для обеспечения экспрессии целевого белка в клетке, содержащая:
первую конструкцию нуклеиновой кислоты по любому из пп. 1-11; и
вторую конструкцию нуклеиновой кислоты, кодирующую фермент, при этом предпочтительно указанный фермент выбран из группы, состоящей из транспозазы, интегразы, рекомбиназы, нуклеазы и никазы.
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| SEKTAS M | |||
| et al., Expression plasmid with a very tight two-step control: Int/att-mediated gene inversion with respect to the stationary promoter, Gene, 2001, v | |||
| Тепловой измеритель силы тока | 1921 |
|
SU267A1 |
| Кулиса для фотографических трансформаторов и увеличительных аппаратов | 1921 |
|
SU213A1 |
| HICKMAN A.B., DYDA F., Mechanisms of DNA Transposition, Microbiol Spectr, 2015, v | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

