Настоящее изобретение относится к сиалилтрансферазам, их применению в получении сиалированных олигосахаридов и к применению указанных сиалированных олигосахаридов в предоставлении питательных композиций.
Предшествующий уровень техники
На сегодняшний день идентифицировано более 150 структурно отличающихся олигосахаридов грудного молока (ОГМ). Несмотря на то, что ОГМ представляют только незначительное количество суммарных питательных веществ грудного молока, их высоко полезное действие на развитие детей, вскармливаемых грудью, стало очевидным за последние десятилетия.
Вплоть до 20% общего содержания ОГМ в грудном молоке является кислотным. Таким образом, данные молекулы ОГМ обладают по меньшей мере одной группировкой сиаловой кислоты. В то время как только 3% сиаловой кислоты, содержащейся в грудном молоке, доступно в свободной форме, 23% и 74% связаны с (глико-)протеинами и олигосахаридами, соответственно. Наиболее распространенным членом семейства сиаловых кислот является N-ацетилнейраминовая кислота (Neu5Ac). В качестве составной части олигомерного сахарида N-ацетилнейраминовая кислота часто обуславливает биологическую активность сахарида.
Наблюдалось, что сиалированные ОГМ (СОГМ) поддерживают устойчивость к патогенам, а также кишечной микрофлоре. Интересно, что недавние исследования дополнительно продемонстрировали защитное действие длинноцепочечных СОГМ от некротизирующего энтероколита, который является одним из наиболее распространенных и смертельных заболеваний у недоношенных младенцев. Кроме того, полагают, что СОГМ поддерживает развитие мозга младенца и его когнитивные способности.
Несмотря на то, что значительные варьирования в профиле ОГМ среди разных доноров препятствуют абсолютной количественной оценке кислотных олигосахаридов, особенно влияя на структурные изомеры сиалиллакто-N-тетраозы, наиболее распространенными кислотными ОГМ являются 3'-сиалиллактоза (3'-SL- от англ. sialyllactose), 6'-сиалиллактоза (6'-SL), сиалиллакто-N-тетраоза a (LST-a - от англ. sialyllacto-N-tetraose), сиалиллакто-N-тетраоза b (LST-b), сиалиллакто-N-тетраоза с (LST-c) и дисиалиллакто-N-тетраоза (DSLNT - от англ. disialyllacto-N-tetraose).
В отношении сложности структуры сиалированных ОГМ (Фиг. 1), их химические или химико-ферментативные синтезы являются проблематичными и ассоциированы с огромными сложностями, например, контролем стереохимии, образованием специфичных связей, доступностью сырья. Наконец, несмотря на то, что совокупность таких способов синтеза была успешной для некоторых СОГМ, их дороговизна и неудовлетворительные выходы ограничивают рентабельное получение сиалированных ОГМ в коммерческих целях.
В общем, метаболическое конструирование микроорганизмов с получением ОГМ представляет самый многообещающий подход к получению ОГМ в промышленном масштабе и оно уже было разработано для 2'-фукозиллактозы, 3-фукозиллактозы и 3'-сиалил-лактозы.
Тем не менее, конструирование путей биосинтеза для получения ОГМ часто ограничено специфичностью и активностью гликозилтрансфераз, которые вовлечены в биосинтез желательного ОГМ, например, фукозил-, галактозил-, N-ацетил-глюкозаминил- или сиалилтрансфераз, особенно в пределах гетерологичной экспрессионной системы, такой как рекомбинантная бактериальная клетка.
К сожалению, гены, кодирующие человеческие сиалилтрансферазы, едва экспрессируются в прокариотических микроорганизмах. Таким образом, данные гены и ферменты неприменимы в биотехнологических способах, использующих генетически модифицированные бактериальные штаммы, как например, Escherichia coli, для получения сиалированных ОГМ.
На настоящий момент идентифицировано и охарактеризовано несколько сиалилтрансфераз (SiaT) из видов бактерий, например, из Neisseria, Campylobacter, Pasteurella, Helicobacter и Photobacterium, а также из млекопитающих и вирусов. Сиалилтрансферазы обычно подразделяют на шесть семейств гликозилтрансфераз (GT - от англ. glycosyltransferase), в зависимости от сходства белковых последовательностей. При этом, все эукариотические и вирусные сиалилтрансферазы сгруппированы в семейство GT 29, тогда как бактериальные SiaT содержатся в группах GT4, GT38, GT42, GT52 или GT80. Кроме того, сиалилтрансферазы и полисиалилтрансферазы могут быть подразделены, благодаря связям, которые они образуют, например, на α-2,3-, α-2,6- и α-2,8-сиалилтрансферазы. Все данные сиалилтрансферазы переносят остаток сиаловой кислоты от цитидин 5'-монофосфат сиаловой кислоты (например, CMP-Neu5Ac) к множеству акцепторных молекул, обычно группировкам галактозы (Gal), N-ацетилгалактозамина (GalNAc) или N-ацетилглюкозамина (GlcNAc) или группировкам других сиаловых кислот (Sia).
Несколько бактериальных сиалилтрансфераз хорошо охарактеризованы ранее и, как уже доказано, подходят для получения 3'-SL или 6'-SL. Смогла быть получена ничтожно малая сумма знаний о сиалилтрансферазах, делающих возможным синтез сиалированных пента- или гексасахаридов, таких как LST-a, LST-b или DSLNT, ограничивая, таким образом, разработку способа получения для какого-либо из данных СОГМ. Вследствие этого, недоступность количеств данных желательных олигосахаридов высокой чистоты препятствует широкой научной оценке их лечебных свойств.
Таким образом, существует необходимость в экономически эффективных способах получения одного или боле СОГМ, особенно тетрасахаридов, пентасахаридов и гексасахаридов, обладающих одним или двумя остатками сиаловой кислоты, в больших количествах и высокой чистоты.
Краткое изложение сущности изобретения
Данная задача решается, помимо прочего, посредством идентификации и характеристики новых сиалилтрансфераз и их применения в получении сиалированных олигосахаридов грудного молока посредством цельноклеточной ферментации или биокатализа.
Согласно первому аспекту предложен способ получения сиалированных олигосахаридов, в котором для получения указанного сиалированного олигосахарида используется генетически модифицированная клетка. Указанная генетически модифицированная клетка содержит по меньшей мере одну гетерологичную сиалилтрансферазу, которая способна переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока.
Согласно второму аспекту предложена генетически модифицированная клетка для применения в способе получения сиалированных олигосахаридов, в котором указанная генетически модифицированная клетка была генетически модифицирована для экспрессии гетерологичной сиалилтрансферазы, которая способна переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока.
Согласно третьему аспекту предложена молекула рекомбинантной нуклеиновой кислоты для экспрессии гетерологичной сиалилтрансферазы при накоплении в клетке, где указанная сиалилтрансфераза способна переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока.
Согласно четвертому аспекту предложены сиалилтрансферазы, способные переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока.
Согласно пятому аспекту предложено применение сиалилтрансферазы, способной переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока, для получения сиалированных олигосахаридов.
Согласно шестому аспекту предложен способ получения сиалированных олигосахаридов посредством биокатализа in vitro, в котором используют сиалилтрансферазу, причем указанная сиалилтрансфераза способна переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока.
Согласно седьмому аспекту предложены сиалированные олигосахариды, получаемые способом согласно первому аспекту или способом согласно шестому аспекту.
Согласно восьмому аспекту предложено применение сиалированных олигосахаридов согласно седьмому аспекту для изготовления питательной композиции.
Согласно девятому аспекту предложена питательная композиция, содержащая по меньшей мере один сиалированный олигосахарид согласно седьмому аспекту.
Согласно десятому аспекту предложена детская смесь, содержащая по меньшей мере один сиалированный олигосахарид грудного молока.
Краткое описание графических материалов
На Фиг. 1 изображены химические структуры наиболее распространенных кислотных олигосахаридов в грудном молоке: 3'-SL (А), 6'-SL (В), LST-a (С), LST-b (D), LST-c (Е) и DSLNT (F).
На Фиг. 2 показаны карты плазмид pDEST14 и рЕТ11, сверхэкспрессирующих гены сиалилтрансфераз.
На Фиг. 3 показана продукция in vivo сиалиллакто-N-тетраозы-а и сиалиллакто-N-тетраозы-b (выделена стрелками) благодаря сверхэкспрессии подходящих сиалилтрансфераз, где изображено разделение внутриклеточной фракции siaT9- или siaT19-сверхэкспрессирующих клеток Е. coli BL21(DE3) #2130 посредством тонкослойной хроматографии.
Подробное описание
В попытке идентифицировать сиалилтрансферазы, которые подходят для применения в способе изготовления сиалированного ОГМ, исследовали базы данных нуклеиновых кислот и базы данных белков. Сто предположительных сиалилтрансфераз идентифицировали посредством сходства последовательностей с известными гликозилтрансферазами. Указанные предположительные сиалилтрансферазы оценивали в отношении активности сиалилтрансфераз.
Согласно первому аспекту предложен способ получения сиалированного олигосахарида, включающий следующие стадии:
a) предоставление по меньшей мере одной генетически модифицированной клетки, которая содержит гетерологичную сиалилтрансферазу, причем указанная гетерологичная сиалилтрансфераза способна обладать α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью для переноса остатка сиаловой кислоты от СМР-активированной формы в качестве донорного субстрата к акцепторной молекуле, выбранной из группы, состоящей из лактозы, LNT-II и олигосахаридов грудного молока;
b) культивирование по меньшей мере одной генетически модифицированной клетки в ферментационном бульоне и в условиях, пермиссивных для продукции указанного сиалированного олигосахарида; и
c) выделение указанного сиалированного олигосахарида.
Способ представляет собой способ получения сиалированного олигосахарида.
Термин «олигосахарид», в том виде, в котором он используется в данном документе, относится к полимерам из моносахаридных остатков, где указанные полимеры содержат по меньшей мере три моносахаридных остатка, но не больше чем 10 моносахаридных остатков, предпочтительно не больше чем 7 моносахаридных остатков. Олигосахариды представляют собой или линейную цепь моносахаридов или являются разветвленными. Кроме того, моносахаридные остатки олигосахаридов могут характеризоваться целым рядом химических модификаций. Соответственно, олигосахариды могу содержать одну или более несахаридных группировок.
Термин «сиалированный олигосахарид», в том виде, в котором он используется в данном документе, относится к олигосахаридам, содержащим один или более остатков сиаловой кислоты. В предпочтительном воплощении остаток сиаловой кислоты представляет собой остаток N-ацетилнейраминовой кислоты (Neu5Ac). Остаток N-ацетилнейраминовой кислоты обычно переносится с СМР-Neu5Ac в качестве донорного субстрата на акцепторную молекулу.
Способ получения сиалированного олигосахарида включает стадию предоставления генетически модифицированной клетки, содержащей гетерологичную сиалилтрансферазу, которая способна обладать α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью.
Генетически модифицированная клетка представляет собой прокариотическую клетку или эукариотическую клетку. Предпочтительно, генетически модифицированная клетка представляет собой микробную клетку. Соответствующие микробные клетки включают дрожжевые клетки, бактериальные клетки, клетки архебактерий, клетки водорослей и клетки грибов.
В дополнительном и/или альтернативном воплощении микробная клетка представляет собой прокариотическую клетку, предпочтительно бактериальную клетку, более предпочтительно бактериальную клетку, выбранную из группы, состоящей из Bacillus, Lactobacillus, Lactococcus, Enterococcus, Bifidobacterium, Sporolactobacillus spp., Micromomospora spp., Micrococcus spp., Rhodococcus spp., and Pseudomonas. Подходящие виды бактерий представляют собой Bacillus subtilis, Bacillus licheniformis, Bacillus coagulans, Bacillus thermophilus, Bacillus laterosporus, Bacillus megaterium, Bacillus mycoides, Bacillus pumilus, Bacillus lentus, Bacillus cereus, Bacillus circulans, Bifidobacterium longum, Bifidobacterium infantis, Bifidobacterium bifidum, Citrobacter freundii, Clostridium cellulolyticum, Clostridium ljungdahlii, Clostridium autoethanogenum, Clostridium acetobutylicum, Corynebacterium glutamicum, Enterococcus faecium, Enterococcus thermophiles, Escherichia coli, Erwinia herbicola (Pantoea agglomerans), Lactobacillus acidophilus, Lactobacillus salivarius, Lactobacillus plantarum, Lactobacillus helveticus, Lactobacillus delbrueckii, Lactobacillus rhamnosus, Lactobacillus bulgaricus, Lactobacillus crispatus, Lactobacillus gasseri, Lactobacillus casei, Lactobacillus reuteri, Lactobacillus jensenii, Lactococcus lactis, Pantoea citrea, Pectobacterium carotovorum, Proprionibacterium freudenreichii, Pseudomonas fluorescens, Pseudomonas aeruginosa, Streptococcus thermophiles и Xanthomonas campestris.
В альтернативном воплощении эукариотическая клетка представляет собой дрожжевую клетку, клетку насекомого, растительную клетку или клетку млекопитающего. Дрожжевая клетка предпочтительно выбрана из группы, состоящей из Saccharomyces sp., в частности, Saccharomyces cerevisiae, Saccharomycopsis sp., Pichia sp., в частности Pichia pastoris, Hansenula sp., Kluyveromyces sp., Yarrowia sp., Rhodotorula sp.и Schizosaccharomyces sp.
Генетически модифицированная клетка была генетически модифицирована с возможностью содержать гетерологичную сиалилтрансферазу.
Термин «генетически модифицированный», в том виде, в котором он используется в данном документе, относится к модификации организации генома клетки с использованием биологических способов. Модификация организации генома клетки может включать перенос генов в пределах и/или через видовые границы, осуществление вставки, осуществление делеции, осуществление замены и/или осуществление модификации нуклеотидов, триплетов, генов, открытых рамок считывания, промоторов, энхансеров, терминаторов и других нуклеотидных последовательностей, опосредующих и/или контролирующих экспрессию генов. Модификацая организации генома клетки нацелена на создание генетически модифицированного организма, обладающего конкретными, желательными свойствами. Генетически модифицированные клетки могут содержать один или более генов, которые отсутствуют в нативной (генетически немодифицированной) форме клетки. Методики введения экзогенных молекул нуклеиновых кислот и/или осуществления вставки экзогенных молекул нуклеиновых кислот (рекомбинантных. гетерологичных) в наследственную информацию клетки для осуществления вставки, осуществления делеции или изменения нуклеотидной последовательности генетической информации клетки хорошо известны специалисту в данной области. Генетически модифицированные клетки могут содержать один или более генов, которые находятся в нативной форме клетки, где указанные гены модифицируют и повторно вводят в клетку посредством искусственных средств. Термин «генетически модифицированный» также охватывает клетки, которые содержат молекулу нуклеиновой кислоты, являющуюся эндогенной в отношении данной клетки и которая была модифицирована без удаления молекулы нуклеиновой кислоты из данной клетки. Такие модификации включают модификации, полученные посредством замены гена, сайт-специфичных мутаций и родственных методик.
Генетически модифицированная клетка содержит гетерологичную сиалилтрансферазу.
Термин «сиалилтрансфераза», в том виде, в котором он используется в данном документе, относится к полипептидам, способным обладать сиалилтрансферазной активностью. «Сиалилтрансферазная активность» означает перенос остатка сиаловой кислоты, предпочтительно остатка N-ацетилнейраминовой кислоты (Neu5Ac), от донорного субстрата к акцепторной молекуле. Термин «сиалилтрансфераза» включает функциональные фрагменты сиалилтрансфераз, описанных в данном документе, функциональные варианты сиалилтрансфераз, описанных в данном документе, и функциональные фрагменты функциональных вариантов. «Функциональный» в данном отношении означает, что фрагменты и/или варианты обладают сиалилтрансферазной активностью. Функциональные фрагменты сиалилтрансферазы охватывают усеченные версии сиалилтрансферазы, как кодируется ее встречающимся в природе геном, усеченная версия которой способна обладать сиалилтрансферазной активностью. Примеры усеченных версий представляют собой сиалилтрансферазы, которые не содержат так называемой лидерной последовательности, которая обычно направляет полипептид в конкретную внутриклеточную локализацию. Обычно, такие лидерные последовательности удаляются из полипептида во время его внутриклеточной транспортировки и также отсутствуют в встречающейся в природе зрелой сиалилтрансферазе.
Гетерологичная сиалилтрансфераза способна переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле. Термин «способный к» в отношении гетерологичной сиалилтрансферазы относится к сиалилтрансферазной активности гетерологичной сиалилтрансферазы и условию, что подходящие условия реакции необходимы для того, чтобы гетерологичная сиалилтрансфераза обладала своей ферментативной активностью. При отсутствии подходящих условий реакции гетерологичная сиалилтрансфераза не обладает своей ферментативной активностью, а сохраняет свою ферментативную активность и обладает своей ферментативной активностью, когда подходящие условия реакции восстанавливаются. Подходящие условия реакции включают наличие подходящего донорного субстрата, наличие подходящей акцепторной молекулы, наличие важнейших кофакторов, таких как, например, одновалентные или двухвалентные ионы, значение рН в соответствующем диапазоне, подходящую температуру и тому подобное. Необязательно, чтобы были удовлетворены оптимальные значения для абсолютно всех факторов, воздействующих на ферментативную реакцию гетерологичной сиалилтрансферазы, но условия реакции должны быть такими, чтобы гетерологичная сиалилтрансфераза осуществляла свою ферментативную активность. Соответственно, термин «способный к» исключает какие-либо условия, при которых ферментативная активность данной гетерологичной сиалилтрансферазы была бы необратимо нарушена, и также исключал воздействие гетерологичной сиалилтрансферазы на любое такое условие. Напротив, термин «способный к» означает, что сиалилтрансфераза является ферментативно активной, то есть обладает своей сиалилтрансферазной активностью, если для сиалилтрансферазы обеспечиваются пермиссивные условия реакции (где все требования являются необходимыми для осуществления сиалилтрансферазой своей ферментативной активности).
Сиалилтрансферазы могут различаться по типу связи с сахаром, которую они образуют.В том виде, в котором они используются в данном документе, термины «α-2,3-сиалилтрансфераза» и «α-2,3-сиалилтрансферазная активность» относятся к полипептидам и их ферментативной активности, которые добавляют остаток сиаловой кислоты с альфα-2,3 связью к галактозе или остатку галактозы акцепторной молекулы. Аналогичным образом, термины «α-2,6-сиалилтрансфераза» и «α-2,6-сиалилтрансферазная активность» относятся к полипептидам и их ферментативной активности, которые добавляют остаток сиаловой кислоты с альфα-2,6 связью к галактозе, N-ацетилгалактозамину и/или N-ацетилглюкозамину, остатку галактозы или остатку N-ацетилгалактозамина и/или остатку N-ацетилгалактозамина и/или остатку N-ацетилглюкозамина акцепторной молекулы. Аналогичным образом, термины «α-2,8-сиалилтрансфераза» и «α-2,8-сиалилтрансферазная активность» относятся к полипептидам и их ферментативной активности, которые добавляют остаток сиаловой кислоты с альфа-2,8 связью к галактозе, N-ацетилгалактозамину и/или N-ацетилглюкозамину, остатку галактозы или остатку N-ацетилгалактозамина и/или остатку N-ацетилглюкозамина акцепторной молекулы.
Термин «гетерологичный», в том виде, в котором он используется в данном документе, относится к полипептиду, аминокислотной последовательности, молекуле нуклеиновой кислоты или нуклеотидной последовательности, которая является чужеродной для клетки или организма, то есть к полипептиду, аминокислотной последовательности, молекуле нуклеиновой кислоты или нуклеотидной последовательности, которая в природе не встречается в указанной клетке или организме. Термин «гетерологичная последовательность» или «гетерологичная нуклеиновая кислота» или «гетерологичный пептид», в том виде, в котором он используется в данном документе, представляет собой последовательность или нуклеиновую кислоту или пептид, который происходит из источника, чужеродного для конкретной клетки - хозяина (например, из другого вида), или, если из того же источника, является модифицированным, по сравнению с его исходной формой. Таким образом, гетерологичная нуклеиновая кислота, функционально связанная с промотором, происходит из источника, отличного от источника, из которого происходил промотор, или, если из того же источника, является модифицированной, по сравнению со своей исходной формой. Гетерологичная последовательность может стабильно вводиться, например, посредством трансфекции, трансформации, конъюгации или трансдукции, в геном микробной клетки-хозяина, таким образом, представляя генетически модифицированную клетку-хозяина. Можно применять методики, которые будут зависеть от клетки-хозяина, последовательности, которая подлежит вставке. Специалисту в данной области известны разные методики, и они раскрыты, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989). Соответственно, «гетерологичный пептид» представляет собой пептид, который в природе не встречается в клетке, и «гетерологичная сиалилтрансфераза» представляет собой сиалилтрансферазу, которая в природе не встречается в данной клетке.
Гетерологичная сиалилтрансфераза способна переносить остаток сиаловой кислоты, например, остаток N-ацетилнейраминовой кислоты (Neu5Ac), от донорного субстрата, например, CMP-Neu5Ac, к акцепторной молекуле. Акцепторная молекула представляет собой лактозу, лакто-N-триозу II (LNT-II) или олигосахарид, выбранный из группы, состоящей из олигосахаридов грудного молока.
В дополнительном и/или альтернативном воплощении акцепторная молекула представляет собой олигосахарид грудного молока, выбранный из группы, состоящей из трисахаридов, тетрасахаридов и пентасахаридов.
В дополнительном и/или альтернативном воплощении акцепторная молекула представляет собой олигосахарид грудного молока, выбранный из группы, состоящей из лакто-N-тетраозы, лакто-N-неотетраозы, LST-a и LST-b.
В одном воплощении гетерологичная сиалилтрансфераза выбрана из группы, состоящей из:
I. полипептидов, содержащих или состоящих из аминокислотной последовательности, как представлено любой из SEQ ID NO: 1-33;
II. полипептидов, содержащих или состоящих из аминокислотной последовательности, имеющей сходство последовательности, составляющее по меньшей мере 80%, с любой из аминокислотных последовательностей, как представлено любой из SEQ ID NO: 1-33; и
III. фрагментов любого из полипептидов I. и II.
В дополнительном и/или альтернативном воплощении генетически модифицированная клетка трансформирована с возможностью содержать молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую гетерологичную сиалилтрансферазу. Предпочтительно, нуклеотидная последовательность выбрана из группы, состоящей из:
i. нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 1-33;
ii. нуклеотидных последовательностей, как представлено любой из SEQ ID NO: 34-66;
iii. нуклеотидных последовательностей, имеющих по меньшей мере 80%-ое сходство последовательностей с одной из нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 1-33;
iv. нуклеотидных последовательностей, имеющих сходство последовательностей по меньшей мере 80% с любой из нуклеотидных последовательностей, представленных SEQ ID NO: 34-66;
v. нуклеотидных последовательностей, которые комплементарны любой из нуклеотидных последовательностей i., ii., iii. и iv; и
vi. фрагментов любой из нуклеотидных последовательностей i., ii., iii., iv. и v. Выражение «любая из SEQ ID NO: 1-33» относится к любой из группы,
состоящей из SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13. SEQ ID NO: 14. SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32 и SEQ ID NO: 33. Такой же принцип относится к выражению «любая из SEQ ID NO: 34-66». В общем, выражение «любая из SEQ ID NO: X - Z», где «X» и «Z» представляют натуральное число, относится ко всем последовательностям (нуклеотидным последовательностям или аминокислотным последовательностям), представленным любой из «SEQ ID NO», содержащим идентификационный номер от X до Z.
Кроме того, генетически модифицированную клетку генетически модифицировали для экспрессии нуклеотидной последовательности, кодирующей гетерологичную сиалилтрансферазу. В связи с этим, нуклеотидная последовательность, кодирующая гетерологичную сиалилтрансферазу, функционально связана с по меньшей мере одним контролем экспрессии, влияющим на транскрипцию и/или трансляцию указанной нуклеотидной последовательности, кодирующей гетерологичную сиалилтрансферазу, в генетически модифицированной клетке.
Термин «функционально связанный», в том виде, в котором он используется в данном документе, относится к функциональной связи между нуклеотидной последовательностью, кодирующей гетерологичную сиалилтрансферазу, и второй нуклеотидной последовательностью, нуклеотидной последовательностью контроля экспрессии (такой как промотор, оператор, энхансер, регулятор, целый ряд сайтов связывания факторов транскрипции, терминатор транскрипции, сайт связывания рибосомы), где последовательность контроля экспрессии влияет на транскрипцию и/или трансляцию нуклеиновой кислоты, соответствующей нуклеотидной последовательности, кодирующей гетерологичную сиалилтрансферазу. Соответственно, термин «промотор» обозначает последовательности ДНК, которые обычно «предшествуют» гену в полимере ДНК и предоставляют сайт для инициации транскрипции в мРНК. «Регуляторные» последовательности ДНК, также обычно «расположенные до» (то есть, предшествующие) гена в данном полимере ДНК, связывают белки, которые определяют частоту (или скорость) инициации транскрипции. Совместно называемые «промоторными/регуляторными» или «контрольными» последовательностями ДНК, данные последовательности, которые предшествуют выбранному гену (или серии генов) в функциональном полимере ДНК, содействуют определению того, будет ли происходить транскрипция (или возможная транскрипция) гена. Последовательности ДНК, которые «следуют за» геном в ДНК-полимере и обеспечивают сигнал для терминации транскрипции в мРНК, называются последовательностями, «терминирующими»транскрипцию.
В одном воплощении генетически модифицированная клетка содержит гетерологичную сиалилтрансферазу, способную обладать α-2,3-сиалилтрансферазной активностью, и олигосахарид грудного молока представляет собой LNT. Полученный таким образом сиалированный олигосахарид представляет собой LST-a.
В дополнительном и/или альтернативном воплощении гетерологичная сиалилтрансфераза, способная обладать α-2,3-сиалилтрансферазной активностью, выбрана из группы, состоящей из:
I. полипептидов, содержащих или состоящих из аминокислотной последовательности, как представлено любой из SEQ ID NO: 1-27;
II. полипептидов, содержащих или состоящих из аминокислотной последовательности, имеющей сходство по меньшей мере 80%, с любой из аминокислотных последовательностей, как представлено любой из SEQ ID NO: 1-27; и
III. фрагментов любого из полипептидов I. и II.
В дополнительном и/или альтернативном воплощении генетически модифицированная клетка содержит молекулу рекомбинантной или синтетической нуклеиновой кислоты, которая содержит по меньшей мере одну нуклеотидную последовательность, кодирующую указанную гетерологичную сиалилтрансферазу, способную обладать α-2,3-сиалилтрансферазной активностью, где указанная по меньшей мере одна нуклеотидная последовательность выбрана из группы, состоящей из:
i. нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 1-27;
ii. нуклеотидных последовательностей, как представлено любой из SEQ ID NO: 34-60;
iii. нуклеотидных последовательностей, имеющих по меньшей мере 80%-ое сходство последовательностей с одной из нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 1-27;
iv. нуклеотидных последовательностей, имеющих сходство последовательностей, составляющее по меньшей мере 80%, с любой из нуклеотидных последовательностей, представленных SEQ ID NO: 34-60;
v. нуклеотидных последовательностей, которые комплементарны любой из нуклеотидных последовательностей i., ii., iii. и iv; и
vi. фрагментов любой из нуклеотидных последовательностей i, ii, iii, iv и v.
В дополнительном и/или альтернативном воплощении гетерологичная сиалилтрансфераза, способная обладать α-2,3-сиалилтрансферазной активностью, обладает относительной эффективностью по меньшей мере 100-кратной, по меньшей мере 200-кратной, по меньшей мере 300-кратной, по меньшей мере 1000-кратной, по меньшей мере 10000-кратной, по сравнению с относительной эффективностью SiaT16, как представлено SEQ ID NO: 27, посредством количественного анализа сиалирования LNT с использованием ЖХ-МС/МС (жидкостная хроматография/масс-спектрометрия) в соответствии со способом, как описано в примере 5.
В другом воплощении гетерологичная сиалилтрансфераза может обладать α-2,6-сиалилтрансферазной активностью, и олигосахарид грудного молока представляет собой LNT. Полученный таким образом сиалированный олигосахарид представляет собой LST-b.
В дополнительном воплощении гетерологичная сиалилтрансфераза, способная обладать α-2,6-сиалилтрансферазной активностью, выбрана из группы, состоящей из:
I. полипептидов, содержащих или состоящих из аминокислотной последовательности, как представлено любой из SEQ ID NO: 28-33;
II. полипептидов, содержащих или состоящих из аминокислотной последовательности, имеющей сходство по меньшей 80%, с любой из аминокислотных последовательностей, как представлено любой из SEQ ID NO: 28-33; и
III. фрагментов любой из полипептидов I. и II.
В дополнительном и/или альтернативном воплощении генетически модифицированная клетка содержит молекулу рекомбинантной или синтетической нуклеиновой кислоты, которая содержит по меньшей мере одну нуклеотидную последовательность, кодирующую указанную гетерологичную сиалилтрансферазу, способную обладать α-2,6-сиалилтрансферазной активностью, когда указанная по меньшей мере одна нуклеотидная последовательность выбрана из группы, состоящей из:
i. нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 28-33;
ii. нуклеотидных последовательностей, как представлено любой из SEQ ID NOs: 61-66;
iii. нуклеотидных последовательностей, имеющих по меньшей мере 80%-ое сходство последовательностей с одной из нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 28-33;
iv. нуклеотидных последовательностей, имеющих сходство последовательностей, составляющее по меньшей мере 80%, с любой из нуклеотидных последовательностей, представленных SEQ ID NO: 61-66;
v. нуклеотидных последовательностей, которые комплементарны любой из нуклеотидных последовательностей i., ii., iii. и iv; и
vi. фрагментов любой из нуклеотидных последовательностей i., ii., iii., iv. и v.
В дополнительном и/или альтернативном воплощении гетерологичная сиалилтрансфераза, способная обладать α-2,3-сиалилтрансферазной активностью, обладает относительной эффективностью по меньшей мере 100-кратной, более предпочтительно по меньшей мере 200-кратной, наиболее предпочтительно по меньшей мере 300-кратной, по сравнению с относительной эффективностью SiaT5, как представлено SEQ ID NO: 33, посредством количественного анализа сиалирования LNT с использованием ЖХ-МС/МС в соответствии со способом, как описано в примере 5.
В дополнительном и/или альтернативном воплощении по меньшей мере одна генетически модифицированная клетка имеет повышенную продукцию одного или более нуклеотид-активированных сахаров, выбранных из группы, состоящей из CMP-N-ацетилнейраминовой кислоты (Neu5Ac), УДФ (уридиндифосфат)-N-ацетилглюкозамина, УДФ-галактозы и ГДФ (гуанозиндифосфат)-фукозы. Предпочтительно, по меньшей мере одна генетически модифицированная клетка дополнительно генетически модифицирована для возможности обладать увеличенной продукцией одного или более указанных нуклеотид-активированных сахаров. Продукция по меньшей мере одного из указанных нуклеотид-активированных сахаров увеличена в дополнительно генетически модифицированной клетке, по сравнению с продукцией того (тех) же нуклеотид-активированного(ых) сахара(ов) в клетке-предшественнике дополнительно генетически модифицированной клетки перед дополнительной генетической модификацией для того, чтобы обладать увеличенной продукцией по меньшей мере одного из указанных нуклеотид-активированных сахаров.
В дополнительном и/или альтернативном воплощении по меньшей мере одна клетка дополнительно генетически модифицирована для сверхэкспрессии одного или более генов, кодирующих полипептиды, способные обладать ферментативной активностью, выбранной из группы, состоящей из следующего: L-глутамин:D-фруктозо-6-фосфат аминотрансфераза, N-ацетилглюкозамин-1-фосфат уридилтрансфераза, глюкозамин-1-фосфат ацетилтрансфераза, фосфоглюкозаминмутаза, глюкозамин-6-фосфат-N-ацетилтрансфераза, N-ацетилглюкозамин-2-эпимераза, УДФ-N-ацетилглюкозамин-2-эпимераза, синтаза сиаловой кислоты, фосфоенолпируватсинтаза, синтетаза СМР-сиаловой кислоты, УДФ-галактозо-4-эпимераза, галактозо-1-фосфат уридилилтрансфераза, фосфоглюкомутаза, глюкозо-1-фосфат уридилилтрансфераза, фосфоманномутаза, маннозо-1-фосфат гуанозилтрансфераза, ГДФ-маннозо-4,6-дегидратаза, ГДФ-L-фукозосинтаза и фукозокиназа/L-фукозо-1-фосфат-гуанилтрансфераза. Указанная сверхэкспрессия одного или более генов представляет собой сверхэкспрессию, по сравнению с клеткой-предшественником дополнительно генетически модифицированной клетки перед дополнительной генетической модификацией для возможности обладания сверхэкспрессией указанного одного или более генов.
Сверхэкспрессия одного или более указанных генов увеличивает количество соответствующего(их) фермента(ов) в генетически модифицированной клетке и, следовательно, повышает соответствующую ферментативную активность в клетке с усилением внутриклеточной продукции по меньшей мере одного из указанных нуклеотид-активированных сахаров.
В дополнительном и/или альтернативном воплощении по меньшей мере одна генетически модифицированная клетка не обладает или обладает сниженной активностью одной или более ферментативных активностей, выбранных из группы, состоящей из β-галактозидазной активности, глюкозамин-6-фосфатдезаминазы, N-ацетилглюкозамин-6-фосфатдеацетилазы, N-ацетилманнозаминкиназы, N-ацетилманнозамин-6-фосфатэпимеразы и альдолазы N-ацетилнейраминовой кислоты, по сравнению с клеткой перед генетической модификацией.
В дополнительном и/или альтернативном воплощении один или более генов, кодирующих β-галактозидазу, глюкозамин-6-фосфатдезаминазу, N-ацетилглюкозамин-6-фосфатдезацетилазу, N-ацетилманнозаминкиназу, N-ацетилманнозамин-6-фосфатэпимеразу и альдолазу N-ацетилнейраминовой кислоты, был/были удален(удалены) из генома генетически модифицированной клетки или экспрессия одного или более генов, кодирующих β-галактозидазу, глюкозамин-6-фосфатдезаминазу, N-ацетилглюкозамин-6-фосфатдеацетилазу, N-ацетилманнозаминкиназу, N-ацетилманнозамин-6-фосфатэпимеразу и альдолазу N-ацетилнейраминовой кислоты, была инактивирована или по меньшей мере снижена в генетически модифицированной клетке посредством дополнительной генетической модификации клетки. Уровень экспрессии указанных генов снижается в дополнительно генетически модифицированной клетке, по сравнению с клеткой-предшественником дополнительно генетически модифицированной клетки перед дополнительной генетической модификацией для обладания сниженным уровнем экспрессии указанных генов.
В дополнительном и/или альтернативном воплощении по меньшей мере одна генетически модифицированная клетка содержит по меньшей мере одно, выбранное из группы, состоящей из функциональной лактозопермеазы, функциональной фукозопермеазы и функционального транспортера сиаловой кислоты (импортера), предпочтительно содержит и экспрессирует по меньшей мере одну нуклеотидную последовательность, кодирующую одно, выбранное из группы, состоящей из функциональной лактозопермеазы, функциональной фукозопермеазы и функционального транспортера сиаловой кислоты (импортера).
В дополнительном и/или альтернативном воплощении генетически модифицированная клетка обладает активностью по меньшей мере одной глюкозилтрансферазы, выбранной из группы, состоящей из β-1,3-N-ацетилглюкозаминилтрансферазы, β-1,3-галактозилтрансферазы, β-1,4-галактозилтрансферазы, α-2,3-сиалилтрансферазы и α-2,6-сиалилтрансферазы.
В дополнительном и/или альтернативном воплощении по меньшей мере одну генетически модифицированную клетку культивируют в ферментационном бульоне и в условиях, являющихся пермиссивными для продукции сиалированного олигосахарида.
Ферментационный бульон содержит по меньшей мере один источник углерода для генетически модифицированных клеток. По меньшей мере один источник углерода предпочтительно выбран из группы, состоящей из глюкозы, фруктозы, сахарозы, глицерина и их комбинаций.
В дополнительном и/или альтернативном воплощении ферментационный бульон содержит по меньшей мере одно, выбранное из группы, состоящей из N-ацетилглюкозамина, галактозы и сиаловой кислоты.
В дополнительном и/или альтернативном воплощении, где по меньшей мере одну генетически модифицированную клетку культивируют при отсутствии и/или без добавления одного или более, выбранных из группы, состоящей из N-ацетилглюкозамина, галактозы и сиаловой кислоты, по меньшей мере одну генетически модифицированную клетку культивируют в присутствии лактозы, лакто-N-триозы II (LNT-II) или по меньшей мере одного ОГМ, предпочтительно ОГМ, выбранного из группы, состоящей из трисахаридов, тетрасахаридов и пентасахаридов, более предпочтительно ОГМ, выбранного из группы, состоящей из LNT и LNnT.
Способ включает возможную стадию выделения сиалированного олигосахарида, который был продуцирован по меньшей мере одной генетически модифицированной клеткой во время ее культивирования в ферментационном бульоне. Сиалированный олигосахарид может быть выделен из ферментационного бульона после удаления генетически модифицированной клетки, например, посредством центрифугирования, и/или может быть выделен из клеток, например, в том отношении, что клетки собирают из ферментационного бульона посредством центрифугирования и подвергают стадии лизиса клеток. Затем, сиалированные олигосахариды могут быть дополнительно очищены из ферментационного бульона и/или клеточных лизатов подходящими методиками, известными специалисту в данной области. Подходящие методики включают микрофильтрацию, ультрафильтрацию, диафильтрацию, хроматографию с псевдодвижущимся слоем, электродиализ, обратный осмос, гель-фильтрацию, анионообменную хроматографию, катионообменную хроматографию и т.п.
Согласно второму аспекту предложена генетически модифицированная клетка для применения в способе получения сиалированных олигосахаридов. Указанная генетически модифицированная клетка и предпочтительные воплощения указанной генетически модифицированной клетки ранее описаны в данном документе в связи со способом. Следовательно, генетически модифицированная клетка содержит гетерологичную сиалилтрансферазу, причем указанная гетерологичная сиалилтрансфераза способна обладать α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью для переноса остатка сиаловой кислоты, например, остатка N-ацетилнейраминовой кислоты (Neu5Ac) от нуклеотид-активированной формы в качестве донорного субстрата, например, СМР-Neu5Ac, к акцепторной молекуле, где акцепторная молекула выбрана из группы, состоящей из лактозы, лакто-N-триозы II и олигосахаридов грудного молока.
Согласно третьему аспекту предложены молекулы рекомбинантной нуклеиновой кислоты для экспрессии сиалилтрансферазы при накоплении в клетке, причем указанная сиалилтрансфераза представляет собой гетерологичную сиалилтрансферазу при экспрессии в клетке. Молекула(ы) рекомбинантной нуклеиновой кислоты содержит(ат) нуклеотидную последовательность, кодирующую сиалилтрансферазу, которая способна переносить остаток сиаловой кислоты, например, остаток N-ацетилнейраминовой кислоты, от донорного субстрата к акцепторной молекуле, где указанная акцепторная молекула выбрана из группы, состоящей из лактозы, лакто-N-триозы II и олигосахаридов грудного молока.
Предпочтительные воплощения нуклеотидных последовательностей, кодирующих сиалилтрансферазу, которая способна переносить остаток сиаловой кислоты от донорного субстрата к акцепторной молекуле, где акцепторная молекула выбрана из группы, состоящей из лактозы, лакто-N-триозы II и олигосахаридов грудного молока, таких как предпочтительные нуклеотидные последовательности, раскрыты ранее в данном документе, в связи со способом получения сиалированных олигосахаридов. Например, сиалилтрансфераза способна переносить остаток N-ацетилнейраминовой кислоты от CMP-Neu5Ac к лактозе, лакто-N-триозе II или олигосахариду грудного молока.
Нуклеотидная последовательность, кодирующая сиалилтрансферазу, функционально связана с по меньшей мере одной последовательностью контроля экспрессии. Таким образом, в дополнительном и/или альтернативном воплощении молекула рекомбинантной нуклеиновой кислоты содержит по меньшей мере одну последовательность контроля экспрессии, опосредующую транскрипцию и/или трансляцию нуклеотидной последовательности, кодирующей сиалилтрансферазу, когда указанная молекула рекомбинантной нуклеиновой кислоты нарабатывается в клетке.
Согласно четвертому аспекту предложены сиалилтрансферазы, способные обладать α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью, переносящие остаток сиаловой кислоты, например, остаток N-ацетилнейраминовой кислоты, от донорного субстрата, например, CMP-Neu5Ac, к акцепторной молекуле, где указанная акцепторная молекула представляет собой лактозу, лакто-N-триозу II или олигосахарид грудного молока.
В одном воплощении акцепторная молекула выбрана из группы, состоящей из трисахаридов, тетрасахаридов и пентасахаридов. В дополнительном и/или альтернативном воплощении акцепторная молекула выбрана из группы, состоящей из LST-a и LST-b.
В дополнительном и/или альтернативном воплощении сиалилтрансфераза выбрана из группы, состоящей из:
I. полипептидов, содержащих или состоящих из аминокислотной последовательности, как представлено любой из SEQ ID NO: 1-33;
II. полипептидов, содержащих или состоящих из аминокислотной последовательности, обладающей сходством последовательности, составляющим по меньшей мере 80%, с любой из аминокислотных последовательностей, как представлено любой из SEQ ID NO: 1-33; и
III. фрагментов любого из полипептидов I. и II.
Согласно пятому аспекту предложено применение сиалилтрансфераз, ранее описанных в данном документе и способных переносить остаток сиаловой кислоты от донорного субстрата, например, остаток N-ацетилнейраминовой кислоты, от СМР Neu5AC к акцепторной молекуле, где указанная акцепторная молекула представляет собой лактозу, лакто-N-триозу II или олигосахарид грудного молока, для получения сиалированных олигосахаридов.
Указанные сиалилтрансферазы способны переносить остаток сиаловой кислоты к акцепторной молекуле, представляющей собой олигосахарид грудного молока, с получением, таким образом, сиалированного олигосахарида.
Олигосахарид грудного молока может представлять собой нейтральный олигосахарид или кислотный олигосахарид, то есть олигосахарид грудного молока, содержащий по меньшей мере один остаток сиаловой кислоты.
Сиалированный олигосахарид, полученный посредством применения сиалилтрансфераз, как описано ранее в данном документе, может представлять собой олигосахарид грудного молока или может представлять собой сиалированный олигосахарид, не обнаруженный во встречающемся в природе грудном молоке.
Согласно шестому аспекту предложен способ получения сиалированных олигосахаридов посредством биокатализа in vitro, где используется сиалилтрансфераза, причем указанная сиалилтрансфераза способна переносить остаток сиаловой кислоты от донорного субстрата, например, остаток N-ацетилнейраминовой кислоты от CMP-Neu5Ac к акцепторной молекуле, где указанная акцепторная молекула представляет собой олигосахарид грудного молока.
Способ включает следующие стадии:
- предоставление - в реакционной смеси - сиалилтрансферазы, способной переносить остаток сиаловой кислоты, предпочтительно N-ацетилнейраминовой кислоты, от донорного субстрата к акцепторной молекуле, донорного субстрата и акцепторной молекулы;
- обеспечение переноса сиалилтрансферазой остатка сиаловой кислоты от донорного субстрата к акцепторной молекуле с получением сиалированного олигосахарида; и
- выделение сиалированного олигосахарида из реакционной смеси. Согласно седьмому аспекту предложены сиалированные олигосахариды,
получаемые способом согласно первому аспекту или способом согласно шестому аспекту.
В одном воплощении сиалированный олигосахарид представляет собой олигосахарид грудного молока, предпочтительно, тетрасахарид, пентасахарид или гексасахарид, более предпочтительно, сиалированный олигосахарид, выбранный из группы, состоящей из LST-a, LST-b и DSLNT.
Согласно восьмому аспекту предложено применение сиалированного олигосахарида, получаемого посредством подхода на основе цельноклеточной ферментации или биокатализа in vitro, как описано ранее в данном документе, для изготовления питательной композиции. Указанная питательная композиция содержит по меньшей мере один сиалированный олигосахарид, который был получен способом, как ранее раскрыто в данном документе.
Таким образом, согласно девятому аспекту предложена питательная композиция, содержащая по меньшей мере один сиалированный олигосахарид, который был получен способом, как ранее раскрыто в данном документе. Предпочтительно, по меньшей мере один сиалированный олигосахарид представляет собой 3'-сиалиллактозу, 6'-сиалиллактозу, LST-a, LST-b, LST-c или DSLNT.
В дополнительном и/или альтернативном воплощении питательная композиция дополнительно содержит по меньшей мере один нейтральный ОГМ, предпочтительно 2'-FL.
В дополнительном и/или альтернативном воплощении питательная композиция содержит 3-SL, 6-SL и 2'-FL.
В дополнительном воплощении питательная композиция выбрана из группы, состоящей из лекарственных составов, детской смеси и биологически активных добавок.
Питательная композиция может быть представлена в жидкой форме или в твердой форме, включая порошки, гранулы, хлопья и пеллеты, но, не ограничиваясь ими.
Согласно десятому аспекту предложена детская смесь, содержащая по меньшей мере один сиалированный ОГМ. Указанный сиалированный ОГМ представляет собой ОГМ, выбранный из группы сиалированных олигосахаридов, которые были получены способом, как описано ранее в данном документе.
В одном воплощении по меньшей мере один сиалированный ОГМ, который содержится в детской питательной смеси, выбран из группы, состоящей из 3-SL, 6-SL, LST-a, LST-b, LST-c и DSLNT.
В дополнительном и/или альтернативном воплощении детская питательная смесь содержит по меньшей мере один сиалированный ОГМ и один или более нейтральных ОГМ.
В дополнительном и/или альтернативном воплощении детская питательная смесь содержит 3-SL, 6-SL и 2'-FL.
Настоящее изобретение будет описано в отношении конкретных воплощений и со ссылкой на графические материалы, но изобретение не ограничивается ими, а только формулой изобретения. Кроме того, термины первый, второй и т.д. в описании и в формуле изобретения используются для проведения различия между похожими элементами и не обязательно для описания последовательности, или во времени, или в пространстве, или в расположении или любым другим образом. Следует понимать, что термины, используемые таким образом, являются взаимозаменяемыми в соответствующих обстоятельствах, и что воплощения изобретения, описанные в данном документе, способны действовать в последовательностях, отличных от последовательностей, описанных или проиллюстрированных в данном документе.
Следует отметить, что термин «содержащий», используемый в формуле изобретения, не следует считать ограниченным средствами, соответственно перечисленными; он не исключает других элементов или стадий. Таким образом, его следует понимать как точно определяющий наличие установленных признаков, целых чисел, стадий или компонентов, на которые ссылаются, но он не исключает наличия или добавления одного или более признаков, целых чисел, стадий или компонентов или их групп. Таким образом, объем выражения «устройство, содержащее средства А и В», не должен ограничиваться устройствами, состоящими только из компонентов А и В. Это означает, что в отношении настоящего изобретения А и В являются лишь релевантными компонентами устройства.
Ссылка на всем протяжении данного описания изобретения на «одно воплощение» или «воплощение» означает, что конкретный признак, структура или характеристика, описанная в связи с воплощением, включена в по меньшей мере одно воплощение настоящего изобретения. Таким образом, появления фраз «в одном воплощении» или «в воплощении» в разных местах на всем протяжении данного описания изобретения не обязательно все относятся к одному и тому же воплощению, но могут относиться к одному и тому же воплощению. Кроме того, конкретные признаки, структуры или характеристики могут объединяться любым подходящим образом, как будет очевидно обычному специалисту в данной области на основе данного раскрытия, в одном или более воплощениях.
Аналогично, следует понимать, что в описании иллюстративных воплощений изобретения разные признаки изобретения иногда группируют вместе в одном единственном воплощении, фигуре или их его описании в целях упрощения раскрытия и оказания помощи в понимании одного или более разных аспектов изобретения. Данный способ раскрытия, однако, не должен восприниматься как свидетельствование о намерении, что заявленное изобретение требует больше признаков, чем явным образом перечислены в каждом пункте. Напротив, о чем свидетельствует нижеприведенная формула изобретения, аспекты изобретения заключаются меньше чем во всех признаках одного единственного вышеприведенного раскрытого воплощения. Таким образом, формула изобретения, следующая после подробного описания, тем самым явным образом включена в данное подробное описание, причем каждый пункт формулы изобретения выступает сам по себе в качестве отдельного воплощения данного изобретения.
Кроме того, в то время как некоторые воплощения, описанные в данном документе, включают некоторые, но не все признаки, включенные в другие воплощения, подразумевается, что комбинации признаков разных воплощений находятся в пределах объема изобретения и образуют разные воплощения, как будет понятно специалистам в данной области. Например, в нижеследующей формуле изобретения любые из заявленных воплощений могут быть использованы в любой комбинации.
Кроме того, некоторые воплощения описаны в данном документе как способ или комбинация элементов способа, который может осуществляться посредством процессора компьютерной системы или посредством других средств осуществления функции. Таким образом, процессор с необходимыми инструкциями для осуществления такого способа или элемента способа образует средство осуществления способа или элемента способа. Кроме того, описанный в данном документе элемент воплощения устройства представляет собой пример средства осуществления функции, выполняемой элементом, с целью осуществления изобретения.
В описании и графических материалах, предложенных в данном документе, изложено множество конкретных подробностей. Однако, понятно, что воплощения изобретения могут быть осуществлены без данных конкретных подробностей. В других примерах хорошо известные способы, структуры и методики не показаны подробно для того, чтобы не мешать пониманию данного описания.
Теперь изобретение будет описано посредством подробного описания нескольких воплощений изобретения. Ясно, что другие воплощения изобретения могут быть сконфигурированы в соответствии со знанием специалистов в данной области без отступления от истинной сущности или технической идеи изобретения, причем изобретение ограничивается только терминами прилагаемой формулы изобретения.
Примеры
Пример 1: Разработка Neu5Ac-продуцирующего штамма Е. coli, делающего возможным скрининг in vivo сиалилтрансфераз, использующих лактозу в качестве акцептора
Метаболическое конструирование включало мутагенез и осуществление делеций конкретных генов, соответственно, и геномные интеграции гетерологичных генов. Гены lacZ и araA инактивировали посредством мутагенеза, используя ошибочно спаренные олигонуклеотиды, как описано Ellis et al., (Proc. Natl. Acad. Sci. USA 98: 6742-6746 (2001)).
Геномные делеции осуществляли в соответствии со способом Datsenko и Wanner (Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). Для предотвращения внутриклеточной деградации N-ацетилнейраминовой кислоты осуществляли делецию генов, кодирующих N-ацетилглюкозамин-6-фосфатдеацетилазу (nagA) и глюкозамин-6-фосфатдезаминазу (nagB), а также целый кластер генов катаболизма N-ацетилнейраминовой кислоты, кодирующих N-ацетилманнозаминкиназу (nanK), N-ацетилманнозамин-6-фосфатэпимеразу (nanE), альдолазу N-ацетилнейраминовой кислоты (nanA) и пермеазу сиаловой кислоты (nanT), из генома штамма Е. coli BL21 (DE3). Также осуществляли делецию генов wzxC-wcaJ. WcaJ кодирует УДФ-глюкозо:ундекапренил-фосфат-глюкозо-1-фосфат трансферазу, катализирующую первую стадию в синтезе колановой кислоты (Stevenson et al., J. Bacteriol. 1996, 178:4885-4893). Кроме того, удаляли гены fuel и fucK, кодирующие L-фукозоизомеразу и L-фукулозокиназу, соответственно.
Геномную интеграцию гетерологичных генов проводили посредством транспозиции. Одну из транспозаз EZ-Tn5™ (Epicentre, США) использовали для интеграции линейных ДНК-фрагментов, или для транспозиции использовали геперактивный С9-мутант mariner транспозазы Himar1 (Lampe et al., Proc. Natl. Acad. Sci. 1999, USA 96:11428-11433). Для получения транспосом EZ-Tn5 исследуемый ген вместе с маркерным геном устойчивости к антибиотику, фланкированным FRT-сайтами, амплифицировали с помощью праймера 1119 и 1120 (все используемые праймеры перечислены ниже в таблице 3); полученный ПЦР-продукт нес на обоих сайтах 19-п.н. сайты распознавания Mosaic End для транспозазы EZ-Tn5. Для интеграции с использованием тарспозазы Himar1 исследуемые экспрессионные конструкции (опероны) аналогично клонировали вместе с маркерным геном устойчивости к антибиотику, фланкированным FRT-сайтами, в вектор pEcomar. Вектор pEcomar кодирует гиперактивный С9-мутант mariner транспозазы Himar1 под контролем промотора, индуцируемого арабинозой ParaB. Экспрессионный фрагмент <Ptet-lacY-FRT-aadA-FRT> (SEQ ID NO: 67) интегрировали посредством использования транспозазы EZ-Tn5. После удачной интеграции гена для лактозного импортера LacY из Е. coli К12 TG1 (номер доступа ABN72583) ген устойчивости исключали из клонов, устойчивых к стрептомицину, посредством FLP рекомбиназы, кодируемой на плазмиде рСР20 (Datsenko and Wanner, Proc. Natl. Acad. Sci. 2000, USA 97:6640-6645), с образованием штамма #534. Кроме того, кластер csc-генов Е. coli W (номер доступа СР002185.1), содержащий гены сахарозопермеазы, фруктокиназы, сахарозогидролазы и репрессора транскрипции (гены cscB, cscK, cscA и cscR, соответственно), которые позволяют штамму расти на сахарозе в качестве единственного источника углерода, вставляли в геном. Данный csc-кластер интегрировали в геном штамма Е. coli BL21(DE3) посредством транспозиции с использованием плазмиды pEcomar-cscABKR Для усиления синтеза de novo УДФ-N-ацетилглюкозамина осуществляли оптимизацию кодонов генов, кодирующих L-глутамин:D-фруктозо-6-фосфат-аминотрансферазу (glmS), фосфоглюкозаминмутазу из подштамма Е. coli К-12 MG1655 (glmM) и N-ацетилглюкозамин-1-фосфат уридилтрансферазу/глюкозамин-1-фосфат ацетилтрансферазу (glmU) из подштамма Е. coli К-12 MG1655 (номер доступа NP_418185, NP_417643, NP_418186, соответственно), и их получали посредством синтеза генов. Оперон glmUM клонировали под контролем конститутивного тетрациклинового промотора Ptet, в то время как glmS клонировали под контролем конститутивного промотора РТ5. Кассету транспозонов <Ptet-glmUM-PT5-glmS-FRT-dhfr-FRT> (SEQ ID NO: 68), фланкированную инвертированными концевыми повторами, специфично распознаваемыми транспозазой mariner-подобного элемента Himar1, вставляли из pEcomar-glmUM-glmS. В итоге, описанные модификации генома приводили к получению штамма Е. coli BL21(DE3) #942, который представляет каркас для разработки штамма. В Таблицах 1, 2 и 3 содержатся все штаммы, олигонуклеотиды, используемые для клонирования, а также общие плазмиды, используемые в данном исследовании, соответственно.
Штамм #942 модифицировали для продукции сиаловой кислоты посредством геномной интеграции экспрессионных кассет <Ptet-glmSm-gna1-FRT-aacC1-FRT> (SEQ ID NO: 69), <Ptet-slr1975-FRT-caf-FRT> (SEQ ID NO: 70), <Ptet-neuBC-FRT-kan-FRT> (SEQ ID NO: 71) и <Ptet-ppsA-FRT-aad1-FRT> (SEQ ID NO: 72). Для всех генов осуществляли оптимизацию кодонов для экспрессии в Е. coli и их получали синтетическим способом посредством кооперации GenScript. GlmSm представляет подвергнутую мутагенезу версию GlmS, что, таким образом, исключает ингибирование глюкозамин-6-фосфатом по типу обратной связи. Ген gna1 кодирует глюкозамин-6-фосфат-ацетилтрансферазу, происходящую из Saccharomyces cerevisiae. Гены субклонировали в виде оперона позади конститутивного промотора Ptet и сливали с геном устойчивости к гентамицину, фланкированным FRT-сайтами, используя праймеры glmSm/gna1_1-8. Аналогично, гены neuB (номер доступа AF305571), кодирующий синтазу сиаловой кислоты, и neuC (номер доступа AF305571), кодирующий УДФ-N-ацетилглюкозамин-2-эпимеразу, оба из которых происходят из Campylobacter jejuni, субклонировали в виде оперона позади конститутивного промотора Ptet и сливали с геном устойчивости к канамицину, фланкированным FRT-сайтами, с использованием праймеров neuBC_1-6. Ген slr1975 (номер доступа BAL35720), также клонируемый позади конститутивного промотора Ptet и слитый с геном устойчивости к хлорамфениколу, фланкированным FRT-сайтами, используя праймеры slr_1-4, кодирует N-ацетилглюкозамин 2-эпимеразу из Synechocystis sp. РСС6803. Ген ppsA (номер доступа АСТ43527), кодирующий фосфоенолпируватсинтазу Е. coli BL21(DE3), аналогично клонировали для конститутивной экспрессии и сливали с геном устойчивости к стрептомицину, фланкированным FRT-сайтами, используя праймеры ppsA_1-4. Геномные интеграции, в конечном итоге, приводили к Neu5Ac-продуцирующему штамму #1363, который использовали для скрининга сиалилтрансфераз 1-26.






Пример 2: Разработка штамма Е. coli, делающего возможным скрининг in vivo сиалилтрансфераз, использующих лактозу в качестве акцептора, но требующих внешнего добавления сиаловой кислоты
Штамм Escherichia coli BL21(DE3) #942 использовали для создания штамма для скрининга кодируемых плазмидой сиалилтрансфераз 27-100. Таким образом, для обеспечения поглощения и активации сиаловой кислоты под действием нуклеотида интегрировали гены nanT и neuA, соответственно. Ген nanT (номер доступа В21_03035), кодирующий главный транспортер суперсемейства мембранных транспортеров Neu5Ac Е. coli, амплифицировали из геномной ДНК Е. coli BL21(DE3) и осуществляли оптимизацию кодонов гена neuA, происходящего из Campylobacter jejuni (номер доступа AF305571), и его получали посредством синтеза. Гены клонировали в виде оперона под контролем конститутивного тетрациклинового промотора Ptet, и полученный экспрессионный фрагмент <Pter-neuA-nanT-lox66-kan-lox72> (SEQ ID NO: 73) интегрировали посредством использования транспозазы EZ-Tn5 с получением штамма скрининга #1730.


Пример 3: Получение коллекции плазмид, кодирующих сиалилтрансферазы
Последовательности генов охарактеризованных или предполагаемых сиалилтрансфераз получали из литературы и общедоступных баз данных. Поскольку часто описывают, что сиалилтрансферазы демонстрируют высокую активность, когда удален их сигнальный пептид, авторы изобретения проанализировали соответствующие белковые последовательности посредством инструмента прогнозирования on-line SignalP (Petersen et al., Nature Methods, 2011 Sep 29; 8(10):785-6). Гены синтетическим способом синтезировали посредством кооперации GenScript, или, как аннотировано, в полноразмерной форме или когда предсказан сигнальный пептид, в виде усеченного варианта, не обладающего N-концевым сигнальным пептидом (Таблица 4).
Каждую из сиалилтрансфераз 1-26 субклонировали в виде оперона с neuA в pDEST14 посредством SLIC (от англ. Sequence- and ligation-independent cloning - сиквенс-независимое безлигазное клонирование) с использованием ген-специфичных праймеров (Таблица 2) с получением плазмид общего вида: pDEST14-siaT-neuA. Оставшиеся сиалилтрансферазы 27-100 прямо субклонировали посредством кооперации GenScript в плазмиду рЕТ11а с использованием сайтов рестрикции Ndel и SamHI. Обе экспрессионные системы обеспечивают IPTG (IsoPropyl-β-D-ThioGalactoside - изопропил-β-D-тиогалактозид)-индуцибельную экспрессию генов (Фиг. 2). Для осуществления скрининга активности in vivo плазмидами трансформировали или штамм #1363 или #1730, тогда как Е. coli BL21(DE3) дикого типа или его вариант с отсутствием lacZ (штамм #287) использовали для анализов in vitro.


Сиалилтрансферазы 1-11, 13-21 и 24 клонировали в виде оперона с neuA (номер доступа AY102622) Campylobacter jejuni. Клонирование сиалилтрансфераз 30-32, 34, 37, 39, 41, 42, 51 и 73 происходило через сайты Ndel и BamHI. Гены сиалилтрансфераз либо клонировали в виде полноразмерных конструкций (FL), либо без предсказанного сигнального пептида (Δ). Число, стоящее после Δ, показывает N-концевые аминокислоты, удаленные из соответствующей последовательности.
Пример 4: Идентификация и характеристика α-2,3- и α-2,6-сиалилтрансфераз, использующих лактозу в качестве акцепторного субстрата
Штаммы Escherichia coli BL21(DE3) #1363 и #1730, несущие плазмиды, кодирующие 100 сиалилтрансфераз, выращивали при 30°С в 100 мл встряхиваемых колбах, наполненных 20 мл среды на основе минеральных солей (Samain et al., J. Biotech. 1999, 72:33-47) с добавлением 2% (масс./об.) глюкозы, 100 мкг/мл ампициллина, 15 мкг/мл канамицина и 40 мкг/мл зеоцина. Когда культуры достигали OD600 (от англ. optical density - оптическая плотность) 0,1-0,3, экспрессию генов индуцировали добавлением 0,3 мМ IPTG. После инкубации в течение 1 часа 1,5 мМ лактозу добавляли к культурам #1363, тогда как к культурам #1730 добавляли 1,5 мМ лактозу, а также 1,5 мМ сиаловую кислоту. Инкубация продолжалась на протяжении 72-96 часов. Затем, клетки собирали посредством центрифугирования и механически разрушали в определенном объеме с использованием стеклянных шариков. Далее, к образцам применяли тонкослойную хроматографию (ТСХ) на силикагеле 60 F254 (Merck KGaA, Дармштадт, Германия). Смесь бутанол:ацетон:уксусная кислота:H2O (35/35/7/23 (об./об./об./об.)) использовали в качестве подвижной фазы. Для выявления разделенных веществ ТСХ-пластину вымачивали в тимоловом реагенте (0,5 г тимол, растворенный в 95 мл этаноле, с добавлением 5 мл серной кислоты) и нагревали.
Результаты обобщены в Таблице 5. При полном скрининге тридцать два гена, как было идентифицировано, кодируют α-2,3-сиалилтрансферазы, таким образом, продуцируя 3'-SL. 19 Ферментов синтезировали 6'-SL и были изображены как α-2,6-сиалилтрансферазы. α-2,3-, а также α-2,6-сиалилтрансферазную активность можно было наблюдать только в случае 3 ферментов. Соответственно, экспрессия 46 ферментов не приводила к образованию сиалиллактозы. Скрининг оказался высокоточным, поскольку смогли быть подтверждены описанные активности широко охарактеризованных сиалилтрансфераз, например, SiaT1 (Gilbert et al., J Biol Chem. 1996 Nov 8; 271(45):28271-6; Gilbert et al., Eur J Biochem. 1997 Oct 1;249(1): 187-94), SiaT6 (Tsukamoto et al., J Biochem. 2008 Feb; 143(2):187-97) и SiaT11 (Yamamoto et al., J Biochem. 1996 Jul; 120(1):104-10). Относительно образования продукта, условия проведения эксперимента сделали возможным полуколичественное сравнение ферментов, подвергающихся скринингу. Но для достижения более глубокого понимания их кинетических свойств, 3 α-2,3- и α-2,6-сиалилтранферазы с возможно наилучшей эффективностью применяли для анализов in vitro.
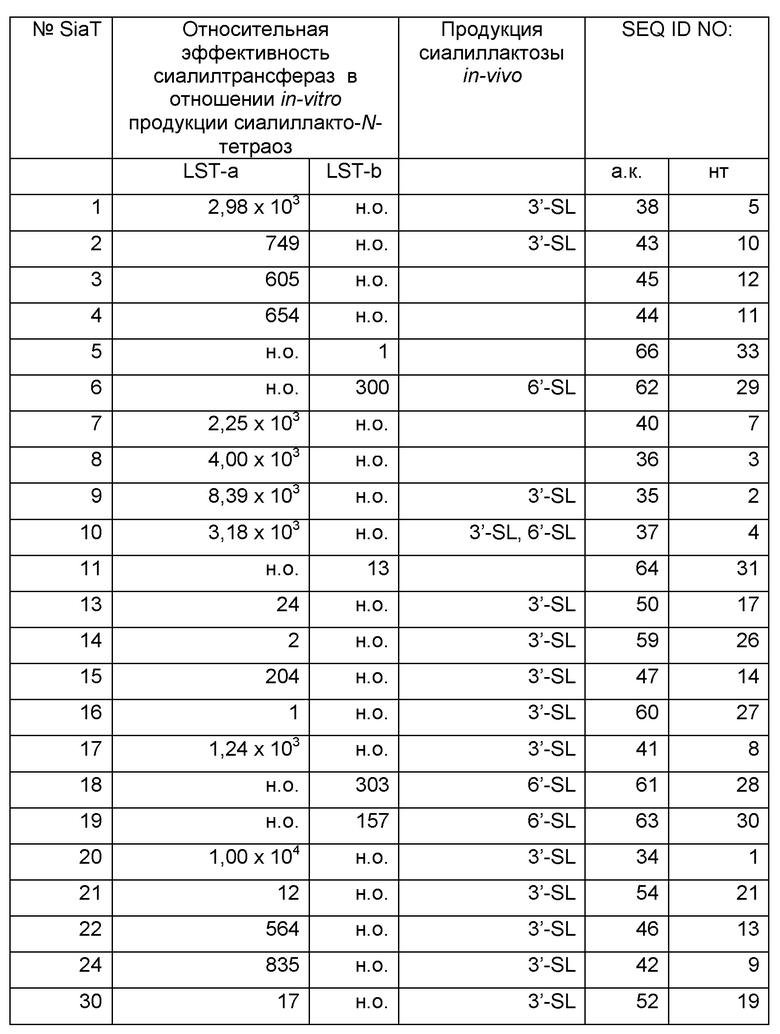
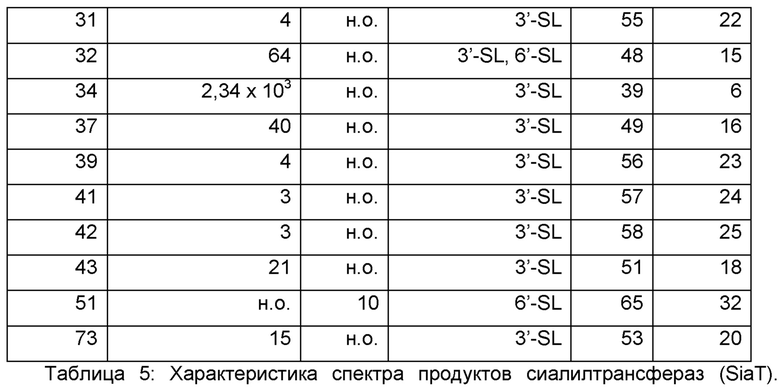
Активности определяли посредством in vivo или in vitro экспериментов с использованием лактозы или LNT в качестве субстратов на основе гликанов, соответственно. Идентификация сиалированной лактозы осуществлялась количественно посредством тонкослойной хроматографии. Продукты сиалирования LNT количественно оценивали посредством ЖХ-МС/МС. Изображено относительное количество LST-a или -b относительно фермента с наилучшей эффективностью, (н.о: неопределяемый).
Пример 5: Идентификация и характеристика α-2,3- и α-2,6-сиалилтрансфераз с использованием лакто-N-тетраозы в качестве акцепторного субстрата
Escherichia coli BL21(DE3), несущие плазмиды, кодирующие 100 сиалилтрансфераз, выращивали при 30°С в 100 мл встряхиваемых колбах, наполненных 20 мл среды 2YT с добавлением 100 мкг/мл ампициллина. Когда OD600 культур достигала 0,1-0,3, экспрессию генов индуцировали добавлением 0,3 мМ IPTG, и инкубацию продолжали на протяжении 12-16 часов. Клетки собирали посредством центрифугирования и механически разрушали в определенном объеме 50 мМ Tris-HCl, рН 7,5, используя стеклянные шарики. Белковый экстракт выдерживали на льду до начала анализа. Анализ in vitro проводили в общем объеме 25 мкл, включающем 50 мМ Tris-HCl рН 7,5, 5 мМ MgCl2, 10 мМ CMP-Neu5Ac и 5 мМ LNT. Анализ начинался с добавления 3 мкл белкового экстракта и продолжался в течение 16 часов. Образование продукта определяли посредством масс-спектрометрии.
Масс-спектрометрический анализ проводили посредством MRM (от англ. multiple reaction monitoring - мониторинг множественных реакций) с использованием трехквадрупольной системы обнаружения ЖХ-МС. Ионы-предшественники отбирали и анализировали в квадруполе 1, фрагментация происходит в столкновительной ячейке с использованием аргона в качестве газа CID (от англ. collision-induced dissociation - индуцированная столкновениями диссоциация), отбор фрагментарных ионов проводят в квадруполе 3. Хроматографическое разделение лактозы, 3'-сиалиллактозы и 6'-сиалиллактозы после разведения супернатанта культуры 1:100 H2O (степень чистоты для ЖХ/МС), проводили на колонке ВЭЖХ (высоко-эффективная жидкостная хроматография) XBridge Amide (3,5 мкм, 2,1 × 50 мм (Waters, США) с защитным картриджем XBridge Amide (3,5 мкм, 2,1 × 10 мм) (Waters, США). Температура термостата колонки системы ВЭЖХ составляла 50°С. Подвижная фаза состояла из ацетонитрила:H2O с 10 мМ ацетатом аммония. 1 мкл образец инъецировали в прибор, прогон проводили в течение 3,60 мин со скоростью потока 400 мкл/мин. 3'-Сиалиллактозу и 6'-Сиалиллактозу анализировали посредством MRM в режиме положительной ионизации ИЭР (ионизация электрораспылением). Масс-спектрометр работал при единичном разрешении. Сиалиллактоза образует ион m/z 656,2 [М+Na]. Ион-предшественник сиалиллактозы дополнительно фрагментировали в столкновительной ячейке до фрагментарных ионов m/z 612,15, m/z 365,15 и m/z 314,15. Энергию столкновения, предварительную систематическую погрешность измерения Q1 и Q3 оптимизировали для каждого аналита отдельно. Хроматографическое разделение лактозы, LNT-II, LNT и LST-a или -b после разведения не содержащей частиц реакционной смеси биокатализа или неочищенного экстракта, соответственно, 1:50 H2O (степень чистоты для ЖХ/МС) проводили на колонке ВЭЖХ XBridge Amide (3,5 мкм, 2,1 × 50 мм (Waters, США) с защитным картриджем XBridge Amide (3,5 мкм, 2,1 × 10 мм) (Waters, США). Термостат колонки запускали при 35°С. Подвижная фаза состояла из ацетонитрила:H2O с 0,1% гидроксидом аммония. 1 Мкл образец инъецировали в прибор, прогон проводили в течение 3,50 мин со скоростью потока 300 мкл/мин. Лактозу, LNT-II, LNT, а также LST-a и -b анализировали посредством MRM в режиме отрицательной ионизации ИЭР. Масс-спектрометр функционировал при единичном разрешении. Лактоза образует ион m/z 341,00 [М-Н]. Ион-предшественник лактозы дополнительно фрагментировали в столкновительной ячейке до фрагментарных ионов m/z 179,15, m/z 161,15 и m/z 101,05. LNT-II образует ион m/z 544,20 [М-Н]. Ион-предшественник LNT-II дополнительно фрагментировали до фрагментарных ионов m/z 382,10, m/z 161,00 и m/z 112,90. LNT образует ион m/z 706,20 [М-Н]. Ион-предшественник LNT дополнительно фрагментировали до фрагментарных ионов m/z 382,10, m/z 202,10 и m/z 142,00. LST-a и -b образует ион m/z 997,20 [М-Н]. Ион-предшественник LST-a и -b дополнительно фрагментировали до фрагментарных ионов m/z 290,15, m/z 202,15 и m/z 142,15. Энергию столкновения, предварительную систематическую погрешность измерения Q1 и Q3 оптимизировали отдельно для каждого аналита. Способы количественной оценки устанавливали, используя имеющиеся в продаже стандарты (Carbosynth, Compton, UK).
Результаты скрининга in vitro обобщенно приведены в Таблице 5. Идентифицировали, что двадцать восемь генов продуцируют LST-a, тогда как только 6 ферментов синтезировали LST-b. Соответственно, экспрессия 66 ферментов не приводила к образованию как LST-a, так и LST-b. Анализ считается точным, поскольку активность SiaT1, которая, как уже было описано, сиалирует LNT (Gilbert et al., J Biol Chem. 1996 Nov 8; 271(45):28271-6; Gilbert et al., Eur J Biochem. 1997 Oct 1; 249(1): 187-94), могла быть подтверждена. Безотносительно уровня сверхэкспрессии белка, сиалилтрансферазы, которые наилучшим образом продуцировались, выбирали для определения Km и Vmax.
Пример 6: Характеристика кинетических свойств выбранных сиалилтрансфераз
Для ранжирования сиалилтрансфераз с наилучшей эффективностью их значения Km для донорных и акцепторных субстратов определяли in vitro. Штамм Escherichia coli BL21(DE3) #287 использовали для чрезмерной продукции ферментов. Клетки инкубировали в 100 мл среды 2YT во встряхиваемых колбах с добавлением 100 мкг/мл ампициллина при 30°С до тех пор, пока OD600 не достигнет 0,3. Затем добавляли 0,3 мМ IPTG, и инкубацию продолжали в течение 12-16 часов. Клетки собирали посредством центрифугирования и механически разрушали в определенном объеме 50 мМ Tris-HCl рН 7,5, используя стеклянные шарики. Белковый экстракт держали на льду до начала анализа. Анализ in vitro проводили в общем объеме 50 мл, включающем 50 мМ Tris-HCl, рН7,5, 5 мМ MgCl2 и варьирующие концентрации CMP-Neu5Ac (0,05-30 мМ), а также лактозы или LNT (0,1-50 мМ). Анализ начинался с добавления 35-750 мкг белкового экстракта. После 1-10 минут инкубации при 30°С анализ инактивировали при 95°С в течение 5 минут. Образование продукта определяли посредством масс-спектрометрии. Данные оценивали, используя модуль ферментативной кинетики SigmaPlot v12.5 для расчета Km и Vmax.
Во время скрининга наиболее эффективные α-2,3-сиалилтрансферазы для получения LST-a, по-видимому, представляют собой SiaT8, SiaT9 и SiaT20. Для сравнения, SiaT6, SiaT18 и SiaT19, как наблюдали, сиалируют LNT наиболее эффективно среди анализируемых α-2,6-сиалилтрансфераз. Их кинетические параметры для CMP-Neu5Ac и LNT, а также лактозы, изображены в Таблице 6. Только SiaT20 не соответствует кинетике Михаэлиса-Ментен.
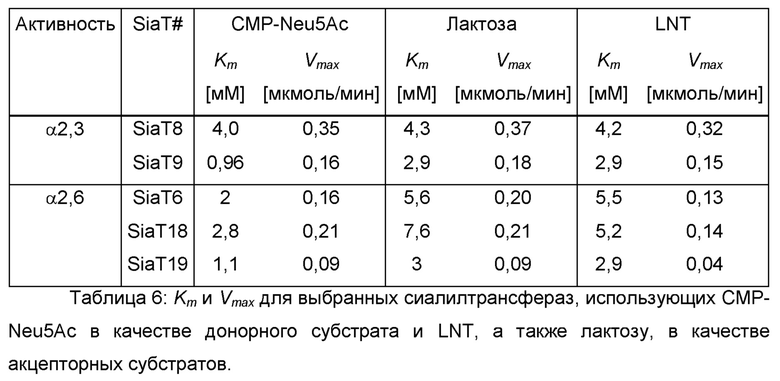
Пример 7: Получение штамма, продуцирующего лакто-N-тетраозу, для скрининга активности in vivo сиалилтрансфераз, использующих LNT в качестве акцепторного субстрата
Штамм Escherichia coli BL21(DE3) #534 использовали для конструирования штамма, продуцирующего лакто-N-тетраозу (LNT). Осуществляли оптимизацию кодонов гена β-1,3-N-ацетилглюкозаминилтрансферазы IgtA из Neisseria meningitidis МС58 (номер доступа NP_274923) для экспрессии в Е. coli и его получали синтетическим способом посредством синтеза генов. Вместе с геном galT, кодирующим галактозо-1-фосфат уридинилтрансферазу, из подштамма Е. coli К-12 MG1655 (номер доступа NP_415279), который аналогично получали посредством синтеза генов, посредством транспозиции вставляли IgtA (SEQ ID NO: 188) с использованием плазмиды pEcomar-IgtA-gaI7. Для усиления синтеза de novo УДФ-N-ацетилглюкозамина осуществляли оптимизацию кодонов генов, кодирующих L-глутамин:D-фруктозо-6-фосфат аминотрансферазу (glmS), фосфоглюкозаминмутазу из подштамма Е. coli К-12 MG1655 (glmM) и N-ацетилглюкозамин-1-фосфат уридилтрансферазу/глюкозамин-1-фосфат-ацетилтрансферазу (glmU), из подштамма Е. coli К-12 MG1655 (номер доступа NP_418185, NP_417643, NP_418186, соответственно), и их получали посредством синтеза генов. Оперон glmUM клонировали под контролем конститутивного тетрациклинового промотора Ptet, в то время как glmS клонировали под контролем конститутивного промотора РТ5- Кассету транспозонов <Ptet-glmUM-PT5-glmS-FRT-dhfr-FRT>, фланкированную инвертированными концевыми повторами, специфично распознаваемыми транспозазой mariner-подобного элемента Himar1, вставляли из pEcomar-glmUM-glmS, обнаруживая штамм, продуцирующий лакто-N-триозу II. Метаболическое конструирование дополнительно включало геномную интеграцию кассет транспозонов <Ptet-wbdO-PT5-galE-FRT-caf-FRT> (SEQ ID NO: 187), фланкированных инвертированными концевыми повторами, специфично распознаваемыми транспозазой mariner-подобного элемента Himar1, которую вставляли из pEcomar-wbdO-galE. Для предупреждения внутриклеточной деградации N-ацетилнейраминовой кислоты кластер генов nanKETA удаляли из генома штамма Е. coli BL21(DE3) в соответствии со способом Datsenko и Wanner (Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). Для предоставления достаточного донорного субстрата (CMP-Neu5Ac) для сиалирования LNT механизм поглощения сиаловой кислоты, а так же способность ее активации под действием нуклеотида внедряли в штамм Е. coli. Как описано выше, гены nanT и neuA клонировали в виде оперона (с использованием праймеров neuA/nanT_1-6) под контролем конститутивного тетрациклинового промотора Ptet, и полученный экспрессионный фрагмент <Pter-neuA-nanT-lox66-kan-lox72> интегрировали посредством применения транспозазы EZ-Tn5, получая, в конечном итоге, штамм #2130.
Пример 8: Периодическая ферментация штамма Е. coli BL21(DE3) #2130, экспрессирующего разные сиалилтрансферазы
Клетки Escherichia coli BL21(DE3) #2130, несущие экспрессионные плазмиды, кодирующие сиалилтрансферазы SiaT9 или SiaT19, выращивали при 30°С в 100 мл встряхиваемых колбах, наполненных 25 мл среды на основе минеральных солей (Samain et al., J. Biotech. 1999, 72:33-47) с добавлением 2% (масс./об.) глюкозы, 5 г/л NH4Cl, 100 мкг/мл ампициллина, 15 мкн/мл канамицина и 5 мкг/мл гентамицина. Когда культуры достигали OD600 0,5-1, добавляли 3 мМ лактозу. Спустя 24 часа инкубации, экспрессию гена сиалилтрансферазы индуцировали добавлением 0,3 мМ IPTG. Одновременно, к культурам добавляли 3 мМ сиаловую кислоту. Инкубация продолжалась в течение 48 часов. Затем клетки собирали посредством центрифугирования и механически разрушали в определенном объеме, используя стеклянные шарики. Далее, тонкослойную хроматографию (ТСХ) осуществляли для подтверждения образования внутри клетки сиалиллакто-N-тетраоз-а и -b. Как показано на Фиг. 3, экспрессия siaT9 или siaT19 в штамме #2130 приводила к образованию LST-a и LST-b, соответственно. Результаты подтверждали посредством масс-спектрометрии.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Jennewein Biotechnologie GmbH
<120> СИАЛИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ В ПОЛУЧЕНИИ СИАЛИРОВАННЫХ ОЛИГОСАХАРИДОВ
<130> P 1703 WO
<160> 188
<170> PatentIn version 3.5
<210> 1
<211> 1410
<212> ДНК
<213> Campylobacter coli
<400> 1
atgcaaaacg tcattatcgc tggtaacggt ccgagcctgc aatcaatcaa ctatcaacgc 60
ctgccgaaag aatacgacat cttccgctgc aaccagttct acttcgaaga taaatactac 120
ctgggcaaaa acatcaaagc ggcctttttc aatccgtatc cgttcctgca gcaataccat 180
accgcgaaac agctggtgtt caacaacgaa tacaaaatcg aaaacatctt ttgtagcacg 240
ttcaatctgc cgttcatcga aaaagataac ttcatcaaca aattttacga tttctttccg 300
gacgctaaac tgggtcacaa aatcatcgaa aacctgaaag aattttacgc gtacatcaaa 360
tacaacgaaa tctacctgaa caaacgtatt accagcggca tctatatgtg cgcaattgct 420
atcgcgctgg gttataaaaa catttacctg tgtggcatcg atttctatga aggtgaaacg 480
atctacccgt tcaaagccat gtctaaaaac attaagaaaa tttttccgtg gatcaaagat 540
ttcaacccga gtaacttcca ttccaaagaa tacgacatcg aaatcctgaa actgctggaa 600
tcaatctaca aagttaacat ctacgcactg tgcgataact cggccctggc aaattacttc 660
ccgctgctgg tgaacaccga caattcattt gttctggaaa acaaatcgga tgactgtatc 720
aacgatatcc tgctgaccaa caatacgccg ggcattaact tctataaaag ccagatccaa 780
gtcaacaata ccgaaattct gctgctgaac tttcagaata tgatcagcgc caaagaaaac 840
gaaatttcta acctgaacaa aatcctgcaa gactcataca aaaccatcaa cacgaaagaa 900
aacgaaatta gtaatctgaa taaaatcctg caggattcct ataaaacgat taataccaaa 960
gaaaatgaaa tttcgaatct gaacaaaatc ctgcaggata aagacaaact gctgatcgtt 1020
aaagaaaacc tgctgaattt caaaagccgt catggtaaag ccaaatttcg cattcagaac 1080
caactgtctt ataaactggg ccaggcaatg atggtcaata gcaaatctct gctgggttat 1140
atccgtatgc cgtttgtgct gagttacatc aaagacaaac acaaacagga acaaaaaatc 1200
tatcaggaaa aaattaagaa agatccgagc ctgaccctgc cgccgctgga agattatccg 1260
gactacaaag aagctctgaa agaaaaagaa tgcctgacct atcgcctggg ccagacgctg 1320
attaaagcgg atcaagaatg gtacaaaggt ggctatgtga aaatgtggtt cgaaatcaaa 1380
aaactgaaga aagaatacaa aaagaaataa 1410
<210> 2
<211> 1146
<212> ДНК
<213> Vibrio sp.
<400> 2
atgaacaacg acaactccac gaccaccaac aataacgcta ttgaaatcta tgtggatcgt 60
gcgaccctgc cgacgatcca gcaaatgacc aaaattgtta gccagaaaac gtctaacaaa 120
aaactgatct catggtcgcg ctacccgatt accgataaaa gcctgctgaa gaaaattaac 180
gcggaatttt tcaaagaaca atttgaactg acggaaagcc tgaaaaacat catcctgtct 240
gaaaacatcg ataacctgat cattcatggc aataccctgt ggagtattga tgtggttgac 300
attatcaaag aagtcaacct gctgggcaaa aatattccga tcgaactgca cttttatgat 360
gacggttccg ccgaatacgt tcgtatctac gaatttagta aactgccgga atccgaacag 420
aaatacaaaa ccagcctgtc taaaaacaac atcaaattct caatcgatgg caccgactcg 480
ttcaaaaaca cgatcgaaaa catctacggt ttcagccaac tgtatccgac cacgtaccac 540
atgctgcgtg cagatatctt cgacaccacg ctgaaaatta acccgctgcg cgaactgctg 600
tcaaacaaca tcaaacagat gaaatgggat tacttcaaag acttcaacta caaacaaaaa 660
gatatctttt actcactgac caacttcaac ccgaaagaaa tccaggaaga cttcaacaaa 720
aactcgaaca aaaacttcat cttcatcggc agtaactccg cgaccgccac ggcagaagaa 780
caaatcaata ttatcagcga agcgaagaaa gaaaacagca gcattatcac caattcaatt 840
tcggattatg acctgttttt caaaggtcat ccgtctgcca cgtttaacga acagattatc 900
aatgcacacg atatgatcga aatcaacaac aaaatcccgt tcgaagctct gatcatgacc 960
ggcattctgc cggatgccgt tggcggtatg ggtagttccg tctttttcag tatcccgaaa 1020
gaagtcaaaa acaaattcgt gttctataaa agtggtacgg atatcgaaaa taactccctg 1080
attcaggtga tgctgaaact gaatctgatt aaccgcgata atattaaact gatctctgac 1140
atttaa 1146
<210> 3
<211> 1173
<212> ДНК
<213> Photobacterium sp.
<400> 3
atgggctgta atagcgactc caaccacaac aactccgacg gcaacatcac caaaaacaaa 60
acgatcgaag tttatgtcga tcgtgcaacc ctgccgacga ttcagcaaat gacccagatc 120
atcaacgaaa atagcaacaa caaaaaactg atttcatggt cgcgctaccc gatcaatgat 180
gaagaactgc tggaatcaat taacggctcg tttttcaaaa acaactctga actgatcaaa 240
agtctggatt ccatgattct gaccaatgac attaagaaag tgatcatcaa cggtaacacg 300
ctgtgggcgg ccgatgtggt taacatcatc aaatcaatcg aagcgttcgg caagaaaacc 360
gaaatcgaac tgaactttta tgatgacggt tcggccgaat atgtgcgtct gtacgacttt 420
agcaaactgc cggaatctga acaggaatac aaaattagcc tgtctaaaga taacattctg 480
agcagcatca acggcaccca gccgttcgaa aacgtcgtgg aaaacatcta cggtttcagt 540
caactgtacc cgaccacgta ccacatgctg cgtgccgata tctttgaaac caatctgccg 600
ctgcgcagtc tgaaaggcgt tctgtccaac aacatcaaac agatgaaatg ggattacttc 660
aaaaccttca acagccagca aaaagacaaa ttctacaact tcacgggttt taacccggat 720
gaaattatgg aacaatacaa agcaagcccg aacaaaaatt ttatcttcgt cggcaccaat 780
tctggcaccg caacggctga acagcaaatt gatatcctga ccgaagctaa aaacccgaac 840
agcccgatta tcacgaaatc gatccagggc ttcgacctgt ttttcaaagg tcatccgtct 900
gcaacctaca acaaacaaat catcgatgct cacaacatga tcgaaatcta caacaaaatc 960
ccgttcgaag cgctgatcat gaccgatgcc ctgccggatg cggtgggcgg tatgggcagc 1020
agcgtgtttt tcagcctgcc gaataccgtg gaaaacaaat tcattttcta taaatccgat 1080
acggacattg aaaacaatgc cctgatccag gttatgattg aactgaatat cgtgaaccgt 1140
aatgatgtga aactgatctc ggacctgcaa taa 1173
<210> 4
<211> 1167
<212> ДНК
<213> Pasteurella multocida
<400> 4
atgaaaacga ttaccctgta tctggacccg gcgtccctgc cggcactgaa ccaactgatg 60
gattttacgc agaacaatga agacaaaacc catccgcgta tctttggcct gtctcgcttc 120
aaaattccgg ataacattat cacccaatat cagaatatcc actttgttga actgaaagac 180
aatcgtccga cggaagccct gttcaccatt ctggatcagt acccgggtaa cattgaactg 240
gacatccatc tgaatattgc tcacagcgtc cagctgattc gtccgatcct ggcgtatcgc 300
tttaaacatc tggatcgtgt gtccatccag cgcctgaacc tgtatgatga cggctcaatg 360
gaatacgttg atctggaaaa agaagaaaac aaagacatct cggcagaaat taaacaagct 420
gaaaaacagc tgagccatta tctgctgacg ggtaaaatca aattcgataa cccgaccatt 480
gcgcgctacg tttggcagtc tgcctttccg gtcaaatatc acttcctgag tacggactac 540
tttgaaaaag cagaatttct gcaaccgctg aaagaatatc tggcggaaaa ttaccagaaa 600
atggattgga cggcctatca gcaactgacc ccggaacagc aagcatttta cctgaccctg 660
gttggcttca acgacgaagt caaacagagt ctggaagtgc agcaagcgaa atttattttc 720
acgggcacca cgacctggga aggtaatacc gatgttcgtg aatattacgc ccagcaacag 780
ctgaacctgc tgaatcattt tacccaggcg ggcggcgacc tgtttattgg tgaccattac 840
aaaatttact tcaaaggtca cccgcgcggc ggtgaaatca acgattacat cctgaacaac 900
gcaaaaaaca tcacgaatat cccggctaat atctctttcg aagtgctgat gatgaccggc 960
ctgctgccgg ataaagtcgg cggtgtggct agctctctgt acttcagtct gccgaaagaa 1020
aaaattagtc acatcatctt caccagcaac aaacaggtca aatcaaaaga agatgccctg 1080
aacaatccgt acgtgaaagt tatgcgtcgc ctgggtatta tcgatgaatc gcaagtgatc 1140
ttttgggaca gcctgaaaca gctgtaa 1167
<210> 5
<211> 1116
<212> ДНК
<213> Neisseria meningitidis
<400> 5
atgggcctga aaaaagcctg cctgaccgtg ctgtgtctga tcgtgttttg cttcggcatc 60
ttttatacgt tcgatcgtgt gaaccagggt gaacgcaatg cagttagtct gctgaaagaa 120
aaactgttta acgaagaagg cgaaccggtg aatctgatct tctgttacac cattctgcaa 180
atgaaagttg ccgaacgtat tatggcacag catccgggtg aacgctttta tgtggttctg 240
atgagcgaaa accgtaacga aaaatacgat tactacttca accagatcaa agataaagcg 300
gaacgcgcct atttctttca cctgccgtac ggcctgaaca aaagttttaa tttcattccg 360
acgatggcgg aactgaaagt gaaaagcatg ctgctgccga aagttaaacg tatctatctg 420
gcaagcctgg aaaaagtgtc tattgcggcc tttctgagca cctacccgga tgcggaaatc 480
aaaaccttcg atgatggcac gggtaatctg attcagagct ctagttatct gggcgatgaa 540
ttttctgtta acggtacgat caaacgtaat ttcgcccgca tgatgatcgg tgattggtct 600
attgcgaaaa cccgcaacgc cagtgatgaa cattacacga tcttcaaagg cctgaaaaac 660
atcatggatg atggtcgtcg caaaatgacc tacctgccgc tgttcgatgc gtctgaactg 720
aaaacgggcg atgaaaccgg cggtacggtg cgtattctgc tgggtagccc ggataaagaa 780
atgaaagaaa tctctgaaaa agcagcgaaa aacttcaaaa tccagtatgt tgccccgcac 840
ccgcgtcaga cctacggcct gagtggtgtg accacgctga acagcccgta tgttattgaa 900
gattacatcc tgcgtgaaat taagaaaaac ccgcataccc gctatgaaat ctacacgttt 960
ttcagcggcg ccgcactgac catgaaagat tttccgaacg tgcacgttta tgcactgaaa 1020
ccggcgtctc tgccggaaga ttattggctg aaaccggtgt acgcgctgtt tacccagagt 1080
ggtattccga tcctgacgtt cgatgataaa aattaa 1116
<210> 6
<211> 852
<212> ДНК
<213> Pasteurella multocida
<400> 6
atggataaat ttgcagaaca tgaaattccg aaagcagtga tcgttgctgg caacggtgaa 60
agtctgtccc agattgatta tcgtctgctg ccgaaaaact acgacgtctt ccgttgcaac 120
caattctact tcgaagaacg ctacttcctg ggcaataaaa tcaaagccgt gtttttcacc 180
ccgggtgttt ttctggaaca gtattacacg ctgtatcatc tgaaacgcaa caatgaatac 240
tttgtcgata acgtgattct gagctctttc aatcacccga ccgtggacct ggaaaaatca 300
cagaaaatcc aagcactgtt catcgatgtt atcaacggct acgaaaaata cctgtcgaaa 360
ctgaccgctt tcgatgttta tctgcgttac aaagaactgt atgaaaatca gcgcattacg 420
agcggtgttt acatgtgcgc tgtcgcgatc gccatgggct ataccgatat ttacctgacg 480
ggtatcgact tttatcaagc gtctgaagaa aactacgcct tcgataacaa aaaaccgaat 540
attatccgtc tgctgccgga ctttcgcaaa gaaaaaaccc tgttcagcta tcattctaaa 600
gatattgacc tggaagcgct gtcatttctg cagcaacatt accacgtgaa cttctactca 660
atctcgccga tgagtccgct gtccaaacat tttccgatcc cgacggttga agatgactgt 720
gaaaccacgt tcgtcgcccc gctgaaagaa aactatatta atgacatcct gctgccgccg 780
cactttgtct atgaaaaact gggcgtggat aaactggcgg ccgcactgga acatcaccat 840
caccatcact aa 852
<210> 7
<211> 1158
<212> ДНК
<213> Pasteurella dagmatis
<400> 7
atgaccattt acctggaccc ggcgtctctg ccgaccctga accaactgat gcattttacg 60
aaagaaagcg aagacaaaga aaccgcacgt atttttggct tctctcgctt taaactgccg 120
gaaaaaatca cggaacagta caacaacatc catttcgtgg aaatcaaaaa caatcgtccg 180
acggaagata ttttcaccat cctggaccag tacccggaaa aactggaact ggatctgcat 240
ctgaacattg cacacagcat ccagctgttt catccgattc tgcaatatcg tttcaaacac 300
ccggatcgca ttagtatcaa atccctgaac ctgtatgatg acggcaccat ggaatacgtt 360
gatctggaaa aagaagaaaa caaagacatc aaaagtgcga tcaaaaaagc cgaaaaacag 420
ctgtccgatt atctgctgac gggtaaaatt aactttgaca atccgaccct ggcacgctac 480
gtttggcagt cacaatatcc ggtcaaatac catttcctgt cgacggaata ttttgaaaaa 540
gctgaattcc tgcagccgct gaaaacctat ctggcgggca aataccaaaa aatggattgg 600
tcagcctatg aaaaactgtc gccggaacag caaacgtttt acctgaaact ggtcggtttc 660
agtgatgaaa ccaaacagct gtttcacacg gaacaaacca aatttatttt cacgggcacc 720
acgacctggg agggtaacac cgatatccgt gaatattacg cgaaacagca actgaatctg 780
ctgaaacatt ttacccacag cgaaggcgac ctgtttatcg gtgaccagta caaaatctac 840
ttcaaaggcc atccgcgcgg cggtgatatt aacgactata tcctgaaaca cgcaaaagat 900
attacgaaca tcccggctaa tattagcttc gaaatcctga tgatgaccgg tctgctgccg 960
gacaaagtcg gcggtgtggc gagctctctg tacttctctc tgccgaaaga aaaaatcagc 1020
cacattatct tcacctctaa caagaaaatt aaaaacaaag aagatgccct gaatgacccg 1080
tacgtgcgtg ttatgctgcg tctgggtatg attgacaaaa gccaaattat cttctgggat 1140
tctctgaaac aactgtaa 1158
<210> 8
<211> 1173
<212> ДНК
<213> Photobacterium phosphoreum
<400> 8
atgggctgta actccgatag caaacacaat aacagtgatg gcaatattac caaaaacaaa 60
acgatcgaag tctatgtgga ccgtgcgacc ctgccgacga ttcagcaaat gacccagatc 120
atcaacgaaa atagcaacaa caaaaaactg atttcatggt cgcgttaccc gatcaatgat 180
gaaacgctgc tggaatcaat taatggctcg tttttcaaaa accgcccgga actgatcaaa 240
agtctggatt ccatgattct gaccaacgaa attaagaaag tgatcatcaa cggtaacacg 300
ctgtgggcag ttgacgtggt taatattatc aaaagcattg aagctctggg caagaaaacc 360
gaaatcgaac tgaacttcta tgatgacggt tctgcggaat atgtgcgtct gtacgatttt 420
agccgcctgc cggaatctga acaggaatac aaaattagcc tgtctaaaga taacattcag 480
agcagcatca acggcaccca accgttcgac aacagcatcg aaaacatcta cggtttctct 540
cagctgtatc cgaccacgta ccacatgctg cgtgccgata tctttgaaac caatctgccg 600
ctgacgagtc tgaaacgcgt tatctccaac aacatcaaac agatgaaatg ggattacttc 660
accacgttca attcccagca gaaaaacaaa ttttacaact tcaccggctt caacccggaa 720
aaaatcaaag aacaatacaa agcgagtccg cacgaaaatt ttattttcat tggcaccaac 780
tccggcaccg ccaccgcaga acagcaaatt gatatcctga ccgaagccaa aaaaccggac 840
tcaccgatta tcaccaacag cattcagggc ctggacctgt ttttcaaagg tcatccgtct 900
gcgacctata accagcaaat tatcgacgcc cacaacatga tcgaaatcta caacaaaatc 960
ccgttcgaag cactgatcat gaccgatgca ctgccggacg ctgttggcgg tatgggtagt 1020
tccgtctttt tctcactgcc gaataccgtc gaaaacaaat tcattttcta taaatcggat 1080
acggacattg aaaacaatgc tctgatccag gttatgatcg aactgaatat cgtgaaccgc 1140
aatgatgtga aactgattag tgacctgcaa taa 1173
<210> 9
<211> 1254
<212> ДНК
<213> Avibacterium paragallinarum
<400> 9
atgcgtaaaa tcatcacctt cttcagcctg ttcttctcga tctcagcgtg gtgtcaaaaa 60
atggaaatct acctggacta tgcgtcgctg ccgagcctga acatgatcct gaacctggtt 120
gaaaacaaaa acaacgaaaa agtcgaacgt attatcggct tcgaacgctt tgatttcaac 180
aaagaaattc tgaatagctt ctctaaagaa cgtatcgaat ttagtaaagt ctccattctg 240
gatatcaaag aattttcaga caaactgtac ctgaacattg aaaaatcgga tacgccggtg 300
gacctgatta tccataccaa tctggatcac tcagttcgtt cgctgctgag catctttaaa 360
accctgagtc cgctgttcca taaaatcaac atcgaaaaac tgtacctgta cgatgacggc 420
agcggtaact atgttgatct gtaccagcac cgccaagaaa atatttctgc gattctgatc 480
gaagcccaga aaaaactgaa agacgcgctg gaaaatcgtg aaacggatac cgacaaactg 540
catagcctga cgcgctatac ctggcacaaa atctttccga cggaatatat cctgctgcgt 600
ccggattacc tggatattga cgaaaaaatg caaccgctga aacatttcct gagcgatacc 660
atcgtgtcta tggacctgtc tcgctttagt catttctcca aaaaccagaa agaactgttt 720
ctgaaaatca cgcacttcga tcaaaacatc ttcaacgaac tgaacatcgg caccaaaaac 780
aaagaataca aaacgttcat cttcaccggc accacgacct gggaaaaaga taagaaaaaa 840
cgtctgaaca acgcgaaact gcagacggaa attctggaat cttttatcaa accgaacggc 900
aaattctacc tgggtaacga tatcaaaatc tttttcaaag gccacccgaa aggtgatgac 960
attaacgact acattatccg caaaaccggc gcagaaaaaa ttccggctaa catcccgttt 1020
gaagttctga tgatgacgaa tagtctgccg gattatgtcg gcggtattat gagtaccgtg 1080
tacttttccc tgccgccgaa aaatattgat aaagtggttt tcctgggttc cgaaaaaatc 1140
aaaaacgaaa acgacgccaa atcacagacc ctgtcgaaac tgatgctgat gctgaacgtc 1200
atcacgccgg aacagatttt ctttgaagaa atgccgaacc cgattaactt ttaa 1254
<210> 10
<211> 1293
<212> ДНК
<213> Campylobacter jejuni
<400> 10
atgacccgca cccgtatgga aaacgaactg attgtgagca aaaacatgca gaacattatt 60
atcgccggta acggtccgag cctgaaaaat attaactata aacgtctgcc gcgcgaatac 120
gatgtgttcc gttgcaacca gttctacttc gaagacaaat actacctggg caagaaaatt 180
aaagccgtgt ttttcaatcc gggcgtgttt ctgcaacaat atcataccgc aaaacagctg 240
attctgaaaa acgaatacga aatcaaaaac atcttttgta gcaccttcaa tctgccgttt 300
atcgaatcta acgatttcct gcaccaattt tataactttt tcccggacgc taaactgggc 360
tacgaagtca tcgaaaacct gaaagaattt tacgcgtaca tcaaatacaa cgaaatctac 420
ttcaacaaac gcatcacctc tggcgtgtat atgtgcgcga ttgccatcgc actgggttat 480
aaaacgattt acctgtgtgg catcgatttc tatgaaggtg acgttattta cccgtttgaa 540
gcaatgagta ccaacattaa aacgatcttc ccgggtatca aagatttcaa accgagtaac 600
tgccattcca aagaatatga catcgaagcg ctgaaactgc tgaaaagcat ctacaaagtt 660
aacatctacg ccctgtgtga tgacagtatt ctggcaaatc atttcccgct gtccattaac 720
atcaacaaca acttcaccct ggaaaacaaa cacaacaact caatcaacga tattctgctg 780
accgacaata cgccgggcgt ctcgttttat aaaaatcagc tgaaagccga taacaaaatc 840
atgctgaact tctacaacat cctgcatagc aaagataacc tgatcaaatt cctgaacaaa 900
gaaatcgctg ttctgaaaaa acagaccacg caacgtgcta aagcgcgcat tcagaaccac 960
ctgagctata aactgggcca agccctgatt atcaatagca aatctgtcct gggtttcctg 1020
tctctgccgt ttattatcct gtcaattgtg atctcgcaca aacaggaaca aaaagcgtat 1080
aaattcaaag tgaagaaaaa cccgaacctg gcactgccgc cgctggaaac ctatccggat 1140
tacaacgaag ccctgaaaga aaaagaatgc ttcacgtaca aactgggcga agaatttatc 1200
aaagcaggta aaaactggta tggcgaaggt tacatcaaat ttatcttcaa agatgttccg 1260
cgtctgaaac gtgaatttga aaaaggcgaa taa 1293
<210> 11
<211> 1188
<212> ДНК
<213> Heliobacter acinonychis
<400> 11
atgaataaga aaccgctgat tattgctggc aacgggccaa gcatcaaaga cttagattat 60
gcgttgttcc cgaaagactt tgatgtattc cgatgtaatc aattctactt cgaggacaaa 120
tactatttag ggcgggaaat aaaaggggtg ttctttaacg cgcacgtctt cgatctccaa 180
atgaagatca ctaaagccat agtcaaaaac ggggaatatc acccggacca catatattgc 240
acacatgtcg aaccgtacgg ttacgttaac ggaaaccagc aactcatgca agagtacctg 300
gaaaaacatt ttgtgggagt ccgaagcacg tacgcatacc tgaaagatct agagccattc 360
tttattctgc acagtaagta tcgcaacttc tacgaccagc acttcacaac gggcatcatg 420
atgctactgg tggccatcca attgggatac aaagaaatat acctgtgcgg aatagacttc 480
tacgaaaacg gattcggaca tttctacgag aaccaagggg gattctttga agaggatagc 540
gatccgatgc acgataagaa catagacatc caagcactgg aactggcaaa gaaatacgcg 600
aaaatctacg cactggtacc gaacagcgcc ctagtgaaaa tgattccgtt gagcagccaa 660
aaaggagttc tggaaaaggt gaaggaccgg atcgggttgg gcgagtttaa gagagagaaa 720
ttcgggcaaa aagaattgga aagacagaag gaattagaac gacaaaaaga gctcgaacgc 780
caaaaggagc ttgaacgtca aaaggaactt gaacgacaaa aagagttgga gaggcagaaa 840
gaactcgaac gccaaaaaga attagagaga cagaaggaat tagagcgcca aaaggagctt 900
gagcgtcaaa aagaattaga gaggcagaag gagttagaaa ggcagaaaga actggagaga 960
cagaaagaac tcgaaaggca gaaggagttg gaacgccaaa aagaactaga attagaacga 1020
tccttaaaag cacgattgaa agcggtactc gcgagcaaag gcatccgcgg cgacaacctg 1080
ataatcgtaa gtttaaaaga cacctaccga ctgtttaaag ggggatttgc gttactcttg 1140
gacctgaagg cgctaaagtc aatcattaaa gcattcctga agagataa 1188
<210> 12
<211> 783
<212> ДНК
<213> Campylobacter jejuni
<400> 12
atgggcaaaa aagtgattat tgcgggcaac ggcccgagcc tgaaagaaat tgattatagc 60
cgtctgccga acgattttga tgtgtttcgc tgcaaccagt tttatttcga agataaatat 120
tacctgggca aaaaatgcaa agcggtgttc tataatccga tcctgttctt cgaacagtat 180
tacaccctga aacatctgat tcagaaccag gaatatgaaa ccgaactgat catgtgcagc 240
aactataacc aggcgcatct ggaaaacgaa aactttgtga aaaccttcta cgattatttt 300
ccggatgcgc atctgggcta tgattttttc aaacagctga aagatttcaa cgcgtacttc 360
aaattccacg aaatctattt caaccagcgt attaccagcg gcgtgtatat gtgcgcggtg 420
gcgattgcgc tgggctataa agaaatttat ctgagcggca tcgattttta tcagaacggc 480
agcagctatg cgtttgatac caaacagaaa aacctgctga aactggcccc gaactttaaa 540
aacgataaca gccactatat tggccatagc aaaaacaccg atatcaaagc gctggaattt 600
ctggaaaaaa cctataaaat caaactgtat tgcctgtgcc cgaacagcct gctggccaac 660
tttattgaac tggcaccgaa tctgaacagc aacttcatca tccaggaaaa aaacaactat 720
accaaagata ttctgattcc gagcagcgaa gcgtatggca aattcagcaa aaacatcaac 780
taa 783
<210> 13
<211> 897
<212> ДНК
<213> Streptococcus entericus
<400> 13
atgaagaaag tctacttctg ccatacggtc taccatctgc tgattaccct gtgcaaaatt 60
agcgttgaag aacaagttga aattattgtg ttcgataccg ttagtaatca tgaactgatt 120
gtccagaaaa tccgcgacgt gtttgttaac accacggtgc tgttcgcaga acaaaatacc 180
gatttttcca ttctggaaat cgatcgcgct acggacattt atgtgttcaa cgactggacc 240
ccgatcggcg cgtatctgcg taaaaacaaa ctgttttacc atctgatcga agatggttat 300
aactaccacg aatataacgt ttacgcgaat gccctgacca tgaaacgtcg cctgctgaac 360
ttcgtgctgc gtcgcgaaga accgtcaggc ttttcgcgtt atgttcgcag cattgaagtt 420
aaccgtgtca aatacctgcc gaatgattgc cgcaaaagca aatgggttga aaaaccgcgt 480
tctgccctgt tcgaaaatct ggtcccggaa cataaacaga aaatcatcac gatcttcggc 540
ctggaaaact atcaagatag cctgcgcggt gtcctggtgc tgacccagcc gctggtgcaa 600
gactactggg atcgcgacat taccacggaa gaagaacagc tggaatttta tcgtcaaatc 660
gtggaatctt acggcgaagg tgaacaggtg tttttcaaaa ttcacccgcg tgataaagtt 720
gactatagct ctctgaccaa cgtcattttt ctgaagaaaa acgtcccgat ggaagtgtac 780
gaactgattg ccgattgtca ttttaccaaa ggtatcacgc acagttccac cgcactggac 840
ttcctgtcct gtgtggataa gaaaatcacc ctgaaacaaa tgaaagcaaa tagttaa 897
<210> 14
<211> 888
<212> ДНК
<213> Haemophilus ducreyi
<400> 14
atgaaagaaa tcgccatcat ctccaaccaa cgcatgttct tcctgtactg tctgctgacc 60
aataaaaatg tcgaagacgt gttcttcatt tttgaaaaag gcgcgatgcc gaacaatctg 120
accagcattt ctcatttcat cgtgctggat cacagtaaat ccgaatgcta tgactttttc 180
tacttcaact tcatcagttg taaatatcgt ctgcgcggcc tggatgttta cggtgcagac 240
catatcaaag gcgctaaatt tttcctggaa cgtcaccgct ttttcgtggt tgaagatggt 300
atgatgaact acagcaaaaa catgtacgca ttctctctgt tccgtacccg caatccggtg 360
attctgccgg gcggttttca tccgaacgtt aaaaccatct tcctgacgaa agataatccg 420
attccggacc agatcgctca caaacgtgaa atcatcaaca tcaaaaccct gtggcaagcg 480
aaaaccgcca cggaaaaaac gaaaattctg agctttttcg aaatcgatat gcaggaaatt 540
tcagttatca aaaaccgctc gtttgtcctg tatacccaac cgctgtcaga agataaactg 600
ctgacggaag cggaaaaaat tgacatctat cgtaccattc tgacgaaata caaccattcg 660
cagaccgtta tcaaaccgca cccgcgcgat aaaacggact ataaacaact gtttccggat 720
gcctatgtca tgaaaggcac ctacccgagt gaactgctga cgctgctggg tgtcaacttc 780
aacaaagtga tcaccctgtt ttccacggcg gtcttcgatt atccgaaaga aaaaatcgac 840
ttctacggca ccgcggtgca tccgaaactg ctggatttct ttgactaa 888
<210> 15
<211> 1467
<212> ДНК
<213> Alistipes sp.
<400> 15
atggccctgc tgagcggtac cgccgcatgc tcagatgacg aagtctcgca gaacctgatc 60
gtgattaatg gcggtgaaca ttttctgagc ctggatggtc tggcccgtgc aggtaaaatt 120
agcgtgctgg caccggctcc gtggcgtgtt acgaaagcag ctggtgatac ctggtttcgc 180
ctgagcgcaa ccgaaggtcc ggctggttac agcgaagtgg aactgtctct ggatgaaaat 240
ccgggtgccg cacgtagcgc acagctggcg tttgcctgtg gtgatgcgat tgtgccgttc 300
cgcctgagtc aaggcgcact gtccgctggt tatgattcac cggactatta cttttacgtt 360
accttcggca cgatgccgac cctgtatgcc ggtatccatc tgctgagcca cgataaaccg 420
ggctatgtct tttactcacg ttcgaaaacg tttgacccgg ccgaattccc ggcacgtgct 480
gaagttacca ccgcagctga tcgtaccgcc gatgcaaccc aggccgaaat ggaagcaatg 540
gctcgcgaaa tgaaacgtcg catcctggaa attaactctg cggatccgac cgccgtgttt 600
ggcctgtatg ttgatgacct gcgttgccgc attggctacg attggttcgt ggcgcagggt 660
atcgacagtg cccgtgtcaa agtgagcatg ctgtctgatg gcaccggcac gtacaacaat 720
ttttataact acttcggtga cgcggccacg gcggaacaaa attgggaaag ttatgcgtcc 780
gaagttgaag ccctggattg gaatcacggc ggtcgttatc cggaaacccg ctcgctgccg 840
gaatttgaaa gctacacgtg gccgtattac ctgtctaccc gtccggatta tcgcctggtg 900
gttcaggacg gcagtctgct ggaaagctct tgtccgttta ttaccgaaaa actgggtgaa 960
atggaaatcg aatccattca accgtatgaa atgctgtcag ccctgccgga aagttcccgt 1020
aaacgctttt atgatatggc aggcttcgat tacgacaaat ttgcagctct gttcgatgcg 1080
tccccgaaga aaaacctgat tatcattggt acctctcatg cggatgatgc cagtgcacgt 1140
ctgcagcgtg attacgttgc acgcatcatg gaacagtatg gcgctcaata cgatgtcttt 1200
ttcaaaccgc acccggcaga caccacgtca gctggttatg aaacggaatt tccgggcctg 1260
accctgctgc cgggtcaaat gccgtttgaa atcttcgttt ggtccctgat tgatcgtgtc 1320
gacatgatcg gcggttatcc gtcaacggtc tttctgaccg ttccggtcga taaagtgcgc 1380
tttatttttg ccgcggatgc agcttctctg gtgcgtccgc tgaatatcct gttccgcgat 1440
gcgaccgacg ttgaatggat gcagtaa 1467
<210> 16
<211> 876
<212> ДНК
<213> Campylobacter jejuni
<400> 16
atgaagaaag tgattatcgc cggcaatggt ccgagcctga aagaaattga ttattctcgt 60
ctgccgaatg atttcgacgt ctttcgctgc aaccagttct actttgaaga caaatattac 120
ctgggcaaaa aatgtaaagc cgtgttttat accccgaact ttttctttga acagtattac 180
acgctgaaac atctgattca gaaccaagaa tatgaaaccg aactgatcat gtgctcaaac 240
tacaatcaag cacatctgga aaacgaaaac ttcgtcaaaa cgttctacga ttacttcccg 300
gacgctcacc tgggttacga tttctttaaa cagctgaaag aattcaacgc gtacttcaaa 360
ttccacgaaa tctacttcaa ccaacgtatc acctcaggcg tgtatatgtg tgcggttgcc 420
attgcactgg gttataaaga aatttacctg tcgggcatcg atttttatca gaatggtagc 480
tcttacgcct tcgacacgaa acaagaaaat ctgctgaaac tggcaccgga ttttaaaaac 540
gaccgctcac attatattgg ccactcgaaa aacaccgata tcaaagctct ggaattcctg 600
gaaaaaacgt acaaaatcaa actgtactgc ctgtgtccga atagtctgct ggctaacttt 660
atcgaactgg cgccgaacct gaattccaac ttcatcatcc aggagaaaaa caactacacc 720
aaagatatcc tgatcccgag ttccgaagcg tacggcaaat ttagcaaaaa catcaacttc 780
aagaaaatta aaatcaaaga aaacgtgtat tacaaactga ttaaagatct gctgcgtctg 840
ccgtctgaca tcaaacatta ttttaaaggt aaataa 876
<210> 17
<211> 939
<212> ДНК
<213> Streptococcus agalactiae
<400> 17
atgacgaatc gcaaaatcta tgtctgccac accctgtacc atctgctgat ctgcctgtat 60
aaagaagaaa tctactcaaa tctggaaatt atcctgagca gcagcattcc ggatgtggac 120
aacctggaga aaaaactgaa aagcaaaacc atcaacatcc atattctgga agaatcctca 180
ggcgaatctg aagaactgct gagtgttctg aaagatgcag gtctgtctta cagtaaattc 240
gatagcaact gcttcatctt caacgacgct accccgattg gccgtacgct gatcaaacac 300
ggtatttatt acaatctgat cgaagatggc ctgaactgtt ttacctactc gattttcagc 360
cagaaactgt ggaaatacta cgtgaaaaaa tacatcctgc ataaaattca accgcacggc 420
ttttcccgct actgcctggg tatcgaagtg aacagtctgg ttaatctgcc gaaagatccg 480
cgttacaaaa aattcatcga agtcccgcgc aaagaactgt tcgacaatgt tacggaatac 540
cagaaagaaa tggcgatcaa cctgtttggc gccgtccgtg tgtctattaa atccccgtca 600
gttctggtcc tgacccagcc gctgtccatc gataaagaat ttatgtcata caacaacaaa 660
atcgaaacgt cggaagaaca attcaacttc tacaaaagca tcgtgaacga atacatcaac 720
aaaggttaca acgtctacct gaaagtgcat ccgcgtgatg tggttgacta ttctaaactg 780
ccggttgaac tgctgccgag taacgtcccg atggaaatta tcgaactgat gctgaccggc 840
cgctttgaat gcggtattac ccatagcagc accgccctgg atttcctgac ctgtgtggac 900
aagaaaatta cgctggttga tctgaaagac attaaataa 939
<210> 18
<211> 1233
<212> ДНК
<213> Bibersteinia trehalosi
<400> 18
atggaattct gcaaaatggc aacgacgcaa aaaatctgtg tctacctgga ctatgctacg 60
atcccgagcc tgaactacat cctgcacttt gcgcaacatt tcgaagatca ggaaaccatt 120
cgtctgtttg gcctgtcccg cttccacatt ccggaatcag tcatccagcg ctatccgaaa 180
ggtgtggttc aattttaccc gaaccaggaa aaagacttca gcgcgctgct gctggccctg 240
aaaaacatcc tgatcgaagt taaacagcaa cagcgtaaat gcgaaatcga actgcatctg 300
aacctgtttc actatcagct gctgctgctg ccgttcctga gtctgtatct ggatacccag 360
gactactgtc atctgacgct gaaattttac gatgacggct ctgaagcgat tagtgccctg 420
caggaactgg cactggctcc ggatctggcg gcccaaatcc agtttgaaaa acaacagttc 480
gacgaactgg tcgtgaaaaa atcgtttaaa ctgtcgctgc tgagccgcta tttttggggt 540
aaactgttcg aaagcgaata catttggttc aatcaagcaa tcctgcagaa agctgaactg 600
caaattctga aacaggaaat cagctctagt cgtcagatgg attttgcaat ttatcaacag 660
atgtccgacg aacaaaaaca gctggtgctg gaaattctga acatcgatct gaataaagtt 720
gcttacctga aacaactgat ggaaaaccag ccgtcttttc tgttcctggg caccacgctg 780
tttaatatta cccaggaaac caaaacgtgg ctgatgcaga tgcatgtgga tctgatccaa 840
cagtattgcc tgccgagcgg ccagtttttc aacaataaag ccggctatct gtgtttttac 900
aaaggtcacc cgaacgaaaa agaaatgaac caaatgatcc tgtctcagtt caaaaacctg 960
atcgcgctgc cggatgacat tccgctggaa atcctgctgc tgctgggcgt tattccgagt 1020
aaagtcggcg gttttgcatc ctcagctctg tttaacttca ccccggcgca gatcgaaaat 1080
attatctttt tcacgccgcg ttatttcgaa aaagataatc gcctgcacgc cacgcaatac 1140
cgtctgatgc agggcctgat tgaactgggt tatctggacg ctgaaaaatc tgtgacccac 1200
tttgaaatca tgcaactgct gacgaaagaa taa 1233
<210> 19
<211> 1221
<212> ДНК
<213> Haemophilus parahaemolyticus
<400> 19
atgaccgaac agtacatcaa aaacgtggaa gtttacctgg attacgcgac catcccgacg 60
ctgaactact tctaccattt caccgaaaac aaagatgaca tcgccacgat tcgtctgttt 120
ggcctgggtc gcttcaacat cagtaaatcc atcatcgaaa gctacccgga aggcattatc 180
cgttactgcc cgattatctt tgaagatcaa accgcatttc agcaactgtt cattaccctg 240
ctgacggaag acagtttttg tcagtatcgc tttaacttcc atattaacct gtttcactcc 300
tggaaaatgc tgatcccgct gctgcatatt atctggcagt ttaaacacaa agtcctggat 360
attaaactga acttctatga tgacggcagt gaaggtctgg tgacgctgtc caaaatcgaa 420
cagaactaca gctctgaaat cctgcaaaaa atcatcgata tcgactcaca gtcgttttat 480
gcagataaac tgtctttcct ggatgaagac attgctcgtt acctgtggaa cagtctgttt 540
gaatcccatt attacctgct gaacgacttc ctgctgaaaa acgaaaaact gtcactgctg 600
aaaaactcga tcaaatactg ccacatcatg gatctggaac gctacctgca gtttacccaa 660
gaagaaaaag actttttcaa cgaactgctg ggcatcaaca tccagagtct ggaagataaa 720
atcaaaatct tccagcagaa gaaaaccttt attttcacgg gtaccacgat cttcagcctg 780
ccgaaagaag aagaagaaac cctgtatcgt ctgcatctga acgcaatcct gaattatatt 840
cacccgaacg gcaaatactt tattggcgat ggtttcacgc tggttatcaa aggtcatccg 900
caccagaaag aaatgaacag ccgcctggaa aaatcttttg aaaaagctgt catgctgccg 960
gataatatcc cgttcgaaat tctgtatctg atcggctgca aaccggacaa aattggcggt 1020
tttgtgagca cctcttactt cagctgtgat aagaaaaaca ttgcggacct gctgtttatc 1080
tctgcccgtc aagaagaagt tcgcaaaaac gattacctgt ttaacatcca gtaccaactg 1140
cgtgacatga tgattaaaac cggttttatc caggaagaaa aaacgcactt ctactcagat 1200
atcccgatct tcatctcgta a 1221
<210> 20
<211> 903
<212> ДНК
<213> Haemophilus somnus
<400> 20
atgaaatata acatcaaaat taaagctatc gtcatcgtgt cgagcctgcg tatgctgctg 60
atcttcctga tgctgaataa ataccacctg gatgaagttc tgtttgtctt caacgaaggc 120
ttcgaactgc ataaaaaata caaaatcaaa cactatgtgg cgattaaaaa gaaaattacc 180
aaattctggc gtctgtacta caaactgtac ttctaccgtt tcaaaattga ccgcatcccg 240
gtttatggcg cagatcatct gggttggacc gactattttc tgaaatactt cgatttctac 300
ctgattgaag acggcatcgc taacttctcc ccgaaacgtt acgaaattaa cctgacgcgc 360
aatatcccgg tctttggttt ccataaaacc gtgaagaaaa tttacctgac gagtctggaa 420
aatgttccgt ccgatattcg tcataaagtc gaactgatca gcctggaaca cctgtggaaa 480
acccgcacgg cgcaggaaca acacaacatc ctggatttct ttgcctttaa tctggacagc 540
ctgatctctc tgaaaatgaa aaaatacatc ctgttcaccc agtgcctgtc agaagatcgc 600
gtcatttcgg aacaggaaaa aatcgcgatc taccaacata tcatcaaaaa ctacgatgaa 660
cgtctgctgg ttatcaaacc gcacccgcgc gaaaccacgg actatcagaa atactttgaa 720
aatgtcttcg tgtaccaaga tgtggttccg agcgaactgt ttgaactgct ggacgtgaac 780
ttcgaacgtg ttattaccct gttttctacg gccgtgttca aatatgatcg caatatcgtt 840
gacttctacg gtacgcgcat ccacgacaaa atctatcaat ggttcggcga catcaaattc 900
taa 903
<210> 21
<211> 1146
<212> ДНК
<213> Vibrio harveyi
<400> 21
atggattctt cgccggaaaa caccagctct acgctggaaa tttacatcga ttcagcaacc 60
ctgccgtcgc tgcagcacat ggtgaaaatt atcgacgaac aaagtggcaa caaaaaactg 120
atcaactgga aacgttatcc gatcgatgac gaactgctgc tggataaaat caacgctctg 180
agcttttctg ataccacgga cctgacccgt tatatggaaa gtattctgct gatcggcgat 240
attaaacgcg tggttattaa cggtaatagt ctgtccaact acaatattgt cggcgtgatg 300
cgctccatca acgccctggg tctggatctg gacgttgaaa tcaattttta tgatgacggt 360
tcagcagaat atgtccgtct gtacaacttc tcgcagctgc cggaagctga acgcgaactg 420
ctggtgtcaa tgtcgaaaaa caatattctg gcggccgtta acggcatcgg ttcttatgat 480
agcggctctc cggaaaatat ttacggtttt gcgcagattt atccggccac ctaccacatg 540
ctgcgtgcgg acattttcga tacggacctg gaaatcggcc tgattcgcga tatcctgggt 600
gacaacgtca aacagatgaa atggggccaa tttctgggtt tcaacgaaga acagaaagaa 660
ctgttttatc aactgaccag cttcaacccg gataaaatcc aggcgcaata caaagaatct 720
ccgaacaaaa acttcgtttt cgtcggcacc aacagtcgtt ccgcaacggc tgaacagcaa 780
atcaacatca tcaaagaagc caaaaaactg gatagcgaaa ttatcccgaa cagcatcgat 840
ggctatgacc tgtttttcaa aggtcatccg agcgcgacct acaaccagca aattgttgat 900
gcccacgaca tgaccgaaat ctataatcgc acgccgtttg aagtcctggc aatgacgagt 960
tccctgccgg atgctgtggg cggtatgggc tcatcgctgt ttttctcact gccgaaaacc 1020
gtggaaacga aattcatttt ctataaaagt ggcaccgata ttgaatccaa tgcgctgatc 1080
caggttatgc tgaaactggg tatcattacg gacgaaaaag tgcgctttac gacggacatc 1140
aaataa 1146
<210> 22
<211> 1452
<212> ДНК
<213> Alistipes sp.
<400> 22
atggccagct gttctgatga cgataaagaa cagacgggtt ttcaaatcga cgatggctct 60
ggtttcctga gtctggatgc agctgcgcgt agtggctcca ttgccatcac cgcaaacaat 120
tcatggtcgg tgacgcagga taaagacagc gaatggctga ccctgagcac cacgtctggt 180
gcagcaggtc gtaccgaaat tggtatcatg ctggaagcga acccgggcga agctcgtaat 240
gcgggtctga cctttaactc tggcggtcgc acgtatccgt tcgtgattac ccagagtgcc 300
catgttacgg cagattttga cgatgctgac cactgctttt atatcacctt tggtaccctg 360
ccgaccctgt atgcaggtct gcatgtgctg tcccacgata aaccgtcata tgtgtttttc 420
cagcgttccc aaacctttcg cccggaagaa ttcccggccc atgcagaagt tacgattgct 480
gcggatccgt cagctaatgc gaccgatgaa gacatggaac gtatgcgcac ggccatgaaa 540
cagcaaattc tgaaaatcaa cgttgaagat ccgaccgcag tttttggcct gtatgtcgac 600
gatctgcgtt gtggcattgg ttacgattgg ttcgtcgccc agggtatcga cagtacccgc 660
gtgaaagtta gtatgctgtc cgatggcacc ggcacgtaca acaacttcta caactacttc 720
ggcgatccgg ccaccgcaga acaaaactgg gaaaattacg ccgcacaggt ggaagcgctg 780
gattggcaac acggcggtcg ttttccggaa acccgcatgc cggatggttt tgacttctat 840
gaatggccgt attacctggc aacgcgtccg aactaccgcc tggttctgca ggacgatgac 900
ctgctggaag cgacgtctcc gtttatgacc gaacgtctgc agcaaatgcg caccgaatcg 960
aaacagccgt atgaactgct ggccagcctg ccggctgaag cccgtcaacg ctttttccgt 1020
atggctggct ttgattacga cgcgtttgct gcgctgttcg atgccagccc gaagaaaaac 1080
ctggtcatta tcggcacgtc acatacctcg gaagaaagcg aagcacagca agccgcatat 1140
gtggaacgta ttatcggcga ttatggtacc gcctacgaca ttttctttaa accgcacccg 1200
gcagatagct ctagttccaa ctacgaagaa cgctttgaag gtctgaccct gctgccgggt 1260
cagatgccgt ttgaaatttt cgtctggtcg ctgctggata aagtggacct gatcggcggt 1320
tattcatcga cggtgtttct gaccgtcccg gtggaaaaaa ccggctttat tttcgctgcg 1380
aatgctgaaa gcctgccgcg cccgctgaac gttctgttcc gtaatgcgga acatgtccgc 1440
tggatccagt aa 1452
<210> 23
<211> 1452
<212> ДНК
<213> Alistipes shahii
<400> 23
atggacgatg gcaccccgag tgtcagcatc aacggcggca ccgacttcct gagcctggac 60
cacctggcac gcagcggcaa aatcacggtc aacgcaccgg ctccgtggtc tgtgaccctg 120
gccccggaaa attacggcca ggatgaaaaa ccggactggc tgaccctgag cgccgaagaa 180
ggcccggcag gttatagcga aatcgatgtt acctttgcgg aaaacccggg tccggcccgt 240
tccgcatcac tgctgttcag ctgcgatggt aaaaccctgg cctttacggt ttcgcagagc 300
gcaggcggta cgggtttcga tgctccggac tattactttt atatttcggt cggcaccatg 360
ccgacgctgt actcgggtct gcatctgctg agccacgata aaccgtctta tgttagttac 420
gaacgtgcga gcacctttga tgcggccgaa ttcccggacc gcgcgtttgt ctatccggtg 480
gccgatccga ccggtcatgc aaccaacgaa gaactgcgtg cgatgagcga agccatgaaa 540
cgtcgcatcc tggaaattaa tgcagaagat ccgaccgctg ttttcggtct gtgggtcgat 600
gacctgcgtt gccgcctggg ctacgattgg tttgtggctc aaggtatcga ctctgcgcgc 660
gtgaaagtta cgatgctgag tgatggcacc gcgacgtata acaattttca taactacttc 720
ggtgacgcag ctaccgccga acagaactgg aatgattatg cggccgaagt tgaagcactg 780
gactggaatc atggcggtcg ttatccggaa acccgtgccc cggaagaatt cgcctcctac 840
acctggccgt attacctgtc aacgcgtccg gattatcgcc tgatgctgca aaacagctct 900
ctgatggaaa gttcctgtcc gtttatcgca gatcgcctgg cagctatgaa aatggaatcc 960
gtgcagccgt atgaactgct gacggcactg ccggaagctt caaaacagca attctatcgt 1020
atggccaaat ttgattacgc acgctttgct ggcctgttcg acctgtctcc gaagaaaaac 1080
ctgattatca ttggtacctc tcattcatcg gcggccagtg aacagcaaca ggcagcttac 1140
gtcgaacgta tcattcaaca gtatggcagt gattacgaca ttttctttaa accgcacccg 1200
gcagatagct ctagtgctgg ttatccggac cgctttgaag gtctgaccct gctgccgggt 1260
cagatgccgt ttgaaatctt cgtttgggcg ctgctggata aaatcgacat gattggcggt 1320
tatccgtcca ccacgtttat ttcagtgccg ctggataaag ttggctttct gttcgcggcc 1380
gatgccgacg gtctggtccg cccgctgaat atcctgttcc gtgacgctgc aaatgtcgaa 1440
tggattcaat aa 1452
<210> 24
<211> 1206
<212> ДНК
<213> Actinobacillus suis
<400> 24
atggaacgca cgccgcaact gcaagcggtg gacatttaca ttgacttcgc aacgatcccg 60
agcctgagct actttctgca ctttctgaaa cataaacacg atgatcagcg tctgcgtctg 120
ttcagcctgg cccgttttga aatgccgcaa accctgattg aacagtatga aggcattatc 180
cagttctcgc gcaacgtgga acataatgtt gaaccgctgc tggaacagct gcaaacgatc 240
ctgtcacaag aaggtaaaca gtttgaactg catctgcacc tgaacctgtt tcattcgttc 300
gaaatgtttc tgaatctgag cccgacctac acgcagtaca aagaaaaaat ctctaaaatc 360
gttctgcacc tgtatgatga cggcagtgaa ggtgtcatga aacagtacca actgcagaaa 420
agctctagtc tggtgcagga tctggcggcc accaaagcat ctctggttag cctgttcgaa 480
aacggcgaag gttcgtttag ccagattgat ctgatccgtt atgtctggaa tgctgtgctg 540
gaaacccatt attacctgct gtctgatcac tttctgctgg acgaaaaact gcagccgctg 600
aaagcagaac tgggccatta ccaactgctg aacctgagtg cttatcagta cctgtcctca 660
gaagatctgc tgtggctgaa acagattctg aaaatcgaca ccgaactgga aagcctgatg 720
caaaaactga cggcgcagcc ggtgtatttc tttagcggta ccacgttttt caacatcagt 780
ttcgaagata aacaacgtct ggcgaatatc catgccattc tgatccgcga acacctggac 840
ccgaactccc agctgtttat tggcgaaccg tacctgtttg tcttcaaagg tcatccgaac 900
tcaccggaaa ttaatcaggc cctgcgtgaa tattacccga acgttatctt cctgccggaa 960
aatattccgt ttgaaatcct gaccctgctg ggcttctccc cgcaaaaaat tggcggtttt 1020
gcgtcaacga tccacgttaa ttccgaacag tcaaaactgg ccaaactgtt tttcctgacc 1080
tcgacggatg aacaagaacg ccagctgagc gacggttata ttaaacaata cgcactggct 1140
caggctatgc tggaaatgca actggtctcg caagaacaag tctattactg ctcgctgtcg 1200
tcgtaa 1206
<210> 25
<211> 1206
<212> ДНК
<213> Actinobacillus capsulatus
<400> 25
atggaacgca tcccgcaact gcaagctgtc gatatttaca ttgacttcgc cacgatcccg 60
agcctgtcct actttctgca ctttctgaaa cataaacacg atcatcagcg tctgcgcctg 120
ttcagcctgg cgcgttttga aatgccgcag accgtcattg aacaatatga aggcattatc 180
cagttctcac gcaacgtgga acacaatgtt gaacaactgc tggaacagct gcaaacgatc 240
ctgtcgcagg aaggtaaaca atttgaactg cacctgcatc tgaacctgtt tcacagtttc 300
gaaatgtttc tgaatctgtc cccgacctac acgaaataca aagaaaaaat ctcaaaaatc 360
gttctgcatc tgtatgatga cggctcggaa ggtgtcatga aacagtacca actgcagcaa 420
agtaactccc tggcacagga tctggctagc accaaagcgt cactggtttc gctgttcaaa 480
aacggcgaag gtgccttttc tcagattgat ctgatccgtt atgtctggaa tgcagtgctg 540
gaaacccact attacctgct gtcagaccac tttctggccc atgaaaaact gcagccgctg 600
aaaattgaac tgggccatta ccagctgctg aatctgtctg cctatcaata cctgagctct 660
gaagatctgc tgtggctgaa acaaattctg aaaatcgacg cagaactgga aagtctgatg 720
cataaactga ccacgcagcc ggtgtatttc tttagcggta ccacgttttt caacatttcg 780
ttcgaagata aacagcgtct ggccaatatc cacgcaattc tgatccgcga acatctggac 840
ccgaacagtc agctgtttat cggcgaaccg tacctgtttg ttttcaaagg tcacccgaac 900
tccccggaaa ttaatcaggc tctgcgcgaa tattacccga acgcgatctt cctgccggaa 960
aatattccgt ttgaaatcct gaccctgctg ggcttcagcc cgcagaaaat tggcggtttt 1020
gcttctacga tccatgtgaa cagcgaacaa tctaaactgg cgaaactgtt tttcctgacc 1080
agtacggatg aacaggaacg taatcgctcc gacggttata ttaaacagta cgcgctggcc 1140
caagcaatgc tggaaatgca actggtctcg caagaacaag tctactactg ctcgctgtcg 1200
tcgtaa 1206
<210> 26
<211> 936
<212> ДНК
<213> Haemophilus somnus
<400> 26
atgttccgtg aagacaatat gaacctgatt atctgctgta cgccgctgca agtgattatc 60
gccgaaaaaa ttatcgaacg ctatccggaa cagaaatttt atggcgttat gctggaatca 120
ttctacaacg ataaattcga cttctacgaa aacaaactga aacatctgtg ccacgaattt 180
ttctgtatca aaatcgcacg tttcaaactg gaacgctata aaaacctgct gtcactgctg 240
aaaatcaaaa acaaaacctt cgatcgtgtc ttcctggcta acatcgaaaa acgctacatc 300
catatcatcc tgtcgaacat tttctttaaa gaactgtaca ccttcgatga cggcacggcg 360
aacatcgccc cgaatagtca tctgtatcaa gaatacgatc actccctgaa aaaacgtatt 420
accgacatcc tgctgccgaa ccattacaac agcaacaaag tgaaaaacat cagcaaactg 480
cactactcta tctaccgctg caaaaacaac atcatcgata acatcgaata catgccgctg 540
tttaacctgg agaaaaaata cacggcacag gataaaagta tttccatcct gctgggtcaa 600
ccgattttct atgacgaaga gaaaaacatt cgtctgatca aagaagtcat cgccaaattc 660
aaaatcgatt actacttccc gcacccgcgc gaagattact acatcgacaa cgtgtcttac 720
atcaaaaccc cgctgatctt tgaagaattt tacgcggaac gttcaatcga aaattcgatc 780
aaaatctata cctttttcag ctctgccgtg ctgaacatcg ttacgaaaga aaatattgat 840
cgcatctacg cactgaaacc gaaactgacg gaaaaagcgt atctggattg ttacgacatc 900
ctgaaagatt tcggtatcaa agttatcgac atctaa 936
<210> 27
<211> 1200
<212> ДНК
<213> Haemophilus ducreyi
<400> 27
atgctgattc aacagaacct ggaaatctac ctggactacg caaccatccc gagcctggcc 60
tgctttatgc acttcattca acacaaagat gacgtcgata gtattcgtct gtttggcctg 120
gcacgcttcg atatcccgca gtccattatc gaccgttacc cggctaacca cctgttttat 180
cacaacatcg ataatcgcga cctgaccgca gtgctgaacc agctggcgga tattctggcc 240
caggaaaata aacgttttca aatcaacctg catctgaacc tgtttcacag cattgacctg 300
tttttcgcta tttatccgat ctaccagcaa tatcagcata aaatttctac catccagctg 360
caactgtacg atgacggcag cgaaggtatt gttacgcagc attctctgtg caaaattgcg 420
gatctggaac agctgatcct gcaacacaaa aacgtgctgc tggaactgct gaccaaaggc 480
acggccaacg ttccgaatcc gaccctgctg cgttatctgt ggaacaatat tatcgattca 540
cagtttcatc tgatctcgga ccattttctg caacacccga aactgcaacc gctgaaacgt 600
ctgctgaaac gctacaccat tctggatttt acgtgttatc cgcgcttcaa tgccgaacag 660
aaacaactgc tgaaagaaat tctgcatatc tcaaacgaac tggaaaatct gctgaaactg 720
ctgaaacagc acaacacctt tctgttcacg ggcaccacgg cgtttaatct ggatcaggaa 780
aaactggacc tgctgaccca actgcatatc ctgctgctga acgaacacca gaatccgcat 840
tcaacgcact acattggcaa caattatctg ctgctgatca aaggtcatgc aaactcgccg 900
gctctgaatc ataccctggc gctgcacttt ccggatgcga ttttcctgcc ggccaatatt 960
ccgtttgaaa tcttcgcgat gctgggcttt acgccgaaca aaatgggcgg tttcgccagc 1020
acctcttaca ttaattatcc gacggaaaac atcaatcacc tgtttttcct gaccagtgat 1080
cagccgtcca ttcgcacgaa atggctggac tacgaaaaac aatttggtct gatgtattcc 1140
ctgctggcaa tgcagaaaat caacgaagat caggcgttta tgtgcaccat tcacaattaa 1200
<210> 28
<211> 1494
<212> ДНК
<213> Photobacterium leiognathi
<400> 28
atgtgtaacg ataatcaaaa tacggtcgat gttgttgtga gcaccgttaa cgataacgtc 60
atcgaaaaca acacgtacca agttaaaccg atcgataccc cgaccacgtt tgacagttac 120
tcctggattc agacgtgcgg caccccgatc ctgaaagatg acgaaaaata ttcactgtcg 180
tttgatttcg tcgccccgga actggatcag gacgaaaaat tctgtttcga atttaccggc 240
gatgttgacg gtaaacgtta tgtcacgcag accaacctga cggtggttgc accgaccctg 300
gaagtttacg tcgatcatgc tagtctgccg tccctgcagc aactgatgaa aatcatccag 360
cagaaaaacg aatactcaca gaatgaacgt ttcatttcgt ggggccgcat cggtctgacg 420
gaagataacg cggaaaaact gaatgcccat atttatccgc tggcaggcaa caatacctca 480
caggaactgg tggatgcagt gatcgattac gctgactcga aaaaccgtct gaatctggaa 540
ctgaacacga ataccgcgca cagctttccg aacctggccc cgattctgcg cattatcagc 600
tctaaaagca acatcctgat ctctaacatc aacctgtacg atgacggcag tgctgaatat 660
gtgaacctgt acaattggaa agataccgaa gacaaatccg tgaaactgag cgattctttc 720
ctggttctga aagactactt taacggtatt agttccgaaa aaccgagcgg catctatggt 780
cgctacaact ggcatcaact gtataatacg tcttattact tcctgcgtaa agattacctg 840
accgttgaac cgcagctgca cgacctgcgc gaatatctgg gcggtagtct gaaacaaatg 900
tcctgggatg gcttttcaca gctgtcgaaa ggtgacaaag aactgttcct gaacattgtc 960
ggctttgatc aggaaaaact gcagcaagaa taccagcaat cagaactgcc gaatttcgtg 1020
tttacgggca ccacgacctg ggcaggcggt gaaaccaaag aatattacgc tcagcaacag 1080
gtgaacgtcg tgaacaatgc gattaatgaa accagcccgt attacctggg ccgtgaacat 1140
gacctgtttt tcaaaggtca cccgcgcggc ggtattatca atgatattat cctgggcagt 1200
ttcaacaata tgattgacat cccggccaaa gtgtcctttg aagttctgat gatgacgggt 1260
atgctgccgg ataccgtggg cggtattgcg tcatcgctgt attttagcat cccggccgaa 1320
aaagtctctt tcattgtgtt taccagctct gatacgatca ccgatcgtga agacgcgctg 1380
aaatctccgc tggtgcaggt tatgatgacc ctgggcattg ttaaagaaaa agatgtgctg 1440
ttctggtcgg atctgccgga ttgttcctcg ggtgtttgta ttgctcagta ttaa 1494
<210> 29
<211> 1497
<212> ДНК
<213> Photobacterium sp.
<400> 29
atgagtgaag aaaacaccca gtccattatt aaaaacgaca tcaacaaaac catcatcgat 60
gaagaatacg ttaacctgga accgatcaac cagtctaaca tcagttttac caaacatagc 120
tgggtccaga cctgcggtac gcagcaactg ctgacggaac aaaacaaaga atcaatttcg 180
ctgagcgtgg ttgcgccgcg tctggatgac gatgaaaaat actgtttcga tttcaacggt 240
gttagtaata aaggcgaaaa atacatcacc aaagtcacgc tgaatgtcgt ggcaccgtct 300
ctggaagttt atgtggatca tgctagtctg ccgaccctgc aacaactgat ggatattatc 360
aaatcggaag aagaaaaccc gaccgcacag cgttacattg cttggggccg catcgtgccg 420
acggacgaac agatgaaaga actgaatatt accagctttg cgctgatcaa caatcacacg 480
ccggccgatc tggttcagga aattgtcaaa caggcgcaaa ccaaacatcg tctgaacgtg 540
aaactgagca gcaatacggc ccactcgttt gacaatctgg ttccgattct gaaagaactg 600
aacagcttca acaatgtgac cgttacgaat atcgatctgt atgacgatgg cagcgcggaa 660
tatgttaacc tgtacaattg gcgcgacacc ctgaacaaaa cggataatct gaaaattggc 720
aaagactatc tggaagatgt cattaacggt atcaatgaag ataccagcaa caccggcacg 780
agttccgtgt acaattggca gaaactgtat ccggctaact accattttct gcgtaaagat 840
tatctgaccc tggaaccgtc cctgcacgaa ctgcgcgact acattggtga ttcactgaaa 900
cagatgcaat gggacggctt caaaaaattc aactcgaaac agcaagaact gtttctgagc 960
atcgtgaatt tcgataaaca gaaactgcaa aacgaataca attcatcgaa cctgccgaat 1020
tttgtgttca ccggtaccac ggtttgggca ggcaaccacg aacgcgaata ctacgctaaa 1080
cagcaaatca acgttatcaa caacgccatc aacgaaagct ctccgcatta tctgggtaat 1140
tcctacgacc tgtttttcaa aggccacccg ggcggtggca ttatcaacac cctgatcatg 1200
cagaattatc cgtcaatggt cgatattccg tccaaaatct catttgaagt gctgatgatg 1260
accgacatgc tgccggatgc cgtggcaggt attgcgagtt ccctgtactt cacgatcccg 1320
gccgaaaaaa tcaaattcat cgttttcacc tctacggaaa ccattacgga tcgtgaaacc 1380
gccctgcgta gtccgctggt ccaggtgatg attaaactgg gcatcgtgaa agaagaaaat 1440
gtgctgttct gggcggacct gccgaattgc gaaacgggtg tctgtattgc tgtctga 1497
<210> 30
<211> 1449
<212> ДНК
<213> Photobacterium leiognathi
<400> 30
atgaacgata atcaaaatac ggtggacgtg gtggtctcaa ccgtcaacga taacgtgatc 60
gaaaacaaca cgtaccaagt caaaccgatc gataccccga ccacgttcga ctcatactcg 120
tggattcaga cgtgcggcac cccgatcctg aaagatgacg aaaaatatag cctgtctttt 180
gatttcgttg ccccggaact ggatcaagac gaaaaattct gtttcgaatt taccggcgat 240
gtggatggta aacgttatgt gacgcagacc aacctgacgg tggttgcacc gaccctggaa 300
gtttacgtcg atcatgcttc actgccgtcg ctgcagcaac tgatgaaaat catccagcag 360
aaaaacgaat acagccagaa tgaacgcttt atttcttggg gccgtatccg cctgacggaa 420
gataacgcgg aaaaactgaa tgcccatatt tatccgctgg caggcaacaa taccagccag 480
gaactggtgg acgcagttat cgattacgct gactctaaaa accgtctgaa tctggaactg 540
aacacgaata ccggccacag tttccgtaac attgcgccga tcctgcgcgc caccagctct 600
aaaaacaaca tcctgatctc caacatcaac ctgtacgatg acggtagtgc tgaatatgtg 660
tccctgtaca actggaaaga taccgacaat aaatcacaga aactgagtga ttcctttctg 720
gttctgaaag actacctgaa tggcatcagt tccgaaaaac cgaacggtat ttatagcatc 780
tacaattggc atcagctgta tcactcatcg tattacttcc tgcgtaaaga ttacctgacg 840
gtggaaacca aactgcacga cctgcgcgaa tatctgggcg gttcactgaa acaaatgtcg 900
tgggatacct ttagccagct gtctaaaggc gacaaagaac tgttcctgaa cattgttggt 960
tttgatcagg aaaaactgca gcaagaatac cagcaaagcg aactgccgaa tttcgtcttt 1020
acgggcacca cgacctgggc aggcggtgaa accaaagaat attacgctca gcaacaggtg 1080
aacgtcgtga acaatgcgat taatgaaacc tctccgtatt acctgggccg tgaacatgac 1140
ctgtttttca aaggtcaccc gcgcggcggt attatcaatg atattatcct gggctcattc 1200
aacaatatga ttgacatccc ggccaaagtt tcgtttgaag tcctgatgat gacgggtatg 1260
ctgccggata ccgttggcgg tattgcgagc agcctgtatt ttagtatccc ggccgaaaaa 1320
gtgtccttca ttgtttttac cagttccgat acgatcaccg atcgcgaaga cgcgctgaaa 1380
agtccgctgg tccaagtgat gatgaccctg ggcattgtga aagaaaaaga tgtgctgttc 1440
tggtgctaa 1449
<210> 31
<211> 2028
<212> ДНК
<213> Photobacterium damsela
<400> 31
atgaaaaaga tcctgaccgt cctgagcatc tttatcctga gcgcctgtaa tagcgacaac 60
acctctctga aagaaaccgt ctccagcaac agcgcggatg tggttgaaac ggaaacctat 120
cagctgaccc cgattgacgc cccgagcagc tttctgagcc attcttggga acagacgtgc 180
ggcaccccga tcctgaatga aagtgataaa caagcgattt cctttgactt cgtggccccg 240
gaactgaaac aggatgaaaa atactgtttc acgttcaaag gcatcaccgg tgaccaccgc 300
tacattacga acaccaccct gaccgttgtg gcaccgacgc tggaagtgta tatcgatcat 360
gctagtctgc cgagcctgca acaactgatt cacattatcc aggcgaaaga tgaatacccg 420
tcaaaccaac gctttgtttc gtggaaacgt gttaccgtcg atgcggacaa cgccaataaa 480
ctgaatattc atacctatcc gctgaaaggc aacaatacgt caccggaaat ggttgcggcc 540
atcgatgaat atgcacaatc gaaaaaccgc ctgaatattg aattttacac gaataccgct 600
catgtcttca acaatctgcc gccgattatc cagccgctgt acaacaacga aaaagtcaaa 660
atttcacaca tctcgctgta cgatgacggt agttccgaat atgtgagtct gtaccagtgg 720
aaagataccc cgaacaaaat tgaaacgctg gaaggcgaag tgagcctgct ggcaaattat 780
ctggctggca ccagcccgga tgcaccgaaa ggcatgggta accgttataa ttggcataaa 840
ctgtacgata ccgactatta ctttctgcgc gaagattatc tggacgtgga agcgaacctg 900
cacgatctgc gtgactacct gggttcatcg gcaaaacaga tgccgtggga tgaatttgct 960
aaactgagtg actcccagca aaccctgttt ctggatatcg ttggcttcga caaagaacag 1020
ctgcaacaac agtattcaca atcgccgctg ccgaatttta tttttaccgg caccaccacc 1080
tgggcgggcg gtgaaacgaa agaatattac gcccaacagc aagtgaacgt tattaacaat 1140
gccatcaatg aaaccagccc gtattacctg ggcaaagatt acgacctgtt tttcaaaggt 1200
catccggcag gcggtgtgat caacgatatt atcctgggca gttttccgga catgattaat 1260
atcccggcta aaatttcctt cgaagtgctg atgatgaccg atatgctgcc ggacacggtt 1320
gcaggtatcg ctagctctct gtattttacc attccggcgg ataaagtgaa ctttatcgtt 1380
ttcacgagtt ccgatacgat taccgaccgt gaagaagccc tgaaaagccc gctggtccag 1440
gtgatgctga ccctgggcat cgtcaaagaa aaagatgtgc tgttctgggc agaccacaaa 1500
gttaatagca tggaagtcgc gattgatgaa gcctgcaccc gcattatcgc aaaacgtcag 1560
ccgacggctt ctgatctgcg cctggtgatt gcgattatca aaacgatcac cgatctggaa 1620
cgtattggcg acgttgccga atctattgcg aaagtcgcgc tggaatcttt ttctaacaaa 1680
cagtacaatc tgctggttag cctggaatct ctgggtcaac ataccgtgcg catgctgcac 1740
gaagttctgg atgcattcgc tcgtatggac gtcaaagcag ctatcgaagt gtatcaggaa 1800
gatgaccgca tcgatcaaga atacgaaagt attgtccgtc agctgatggc ccacatgatg 1860
gaagatccgt catcgattcc gaacgttatg aaagtcatgt gggcggcccg ttccatcgaa 1920
cgcgttggtg atcgttgcca gaatatttgt gaatacatca tctacttcgt gaaaggcaaa 1980
gatgttcgcc acaccaaacc ggatgacttc ggtacgatgc tggactaa 2028
<210> 32
<211> 1533
<212> ДНК
<213> Photobacterium damsela
<400> 32
atgaaaaaga tcctgaccgt cctgagcatc tttatcctga gcgcctgtaa tagcgacaac 60
acctctctga aagaaaccgt ctccagcaac agcgcggatg tggttgaaac ggaaacctat 120
cagctgaccc cgattgacgc cccgagcagc tttctgagcc attcttggga acagacgtgc 180
ggcaccccga tcctgaatga aagtgataaa caagcgattt cctttgactt cgtggccccg 240
gaactgaaac aggatgaaaa atactgtttc acgttcaaag gcatcaccgg tgaccaccgc 300
tacattacga acaccaccct gaccgttgtg gcaccgacgc tggaagtgta tatcgatcat 360
gctagtctgc cgagcctgca acaactgatt cacattatcc aggcgaaaga tgaatacccg 420
tcaaaccaac gctttgtttc gtggaaacgt gttaccgtcg atgcggacaa cgccaataaa 480
ctgaatattc atacctatcc gctgaaaggc aacaatacgt caccggaaat ggttgcggcc 540
atcgatgaat atgcacaatc gaaaaaccgc ctgaatattg aattttacac gaataccgct 600
catgtcttca acaatctgcc gccgattatc cagccgctgt acaacaacga aaaagtcaaa 660
atttcacaca tctcgctgta cgatgacggt agttccgaat atgtgagtct gtaccagtgg 720
aaagataccc cgaacaaaat tgaaacgctg gaaggcgaag tgagcctgct ggcaaattat 780
ctggctggca ccagcccgga tgcaccgaaa ggcatgggta accgttataa ttggcataaa 840
ctgtacgata ccgactatta ctttctgcgc gaagattatc tggacgtgga agcgaacctg 900
cacgatctgc gtgactacct gggttcatcg gcaaaacaga tgccgtggga tgaatttgct 960
aaactgagtg actcccagca aaccctgttt ctggatatcg ttggcttcga caaagaacag 1020
ctgcaacaac agtattcaca atcgccgctg ccgaatttta tttttaccgg caccaccacc 1080
tgggcgggcg gtgaaacgaa agaatattac gcccaacagc aagtgaacgt tattaacaat 1140
gccatcaatg aaaccagccc gtattacctg ggcaaagatt acgacctgtt tttcaaaggt 1200
catccggcag gcggtgtgat caacgatatt atcctgggca gttttccgga catgattaat 1260
atcccggcta aaatttcctt cgaagtgctg atgatgaccg atatgctgcc ggacacggtt 1320
gcaggtatcg ctagctctct gtattttacc attccggcgg ataaagtgaa ctttatcgtt 1380
ttcacgagtt ccgatacgat taccgaccgt gaagaagccc tgaaaagccc gctggtccag 1440
gtgatgctga ccctgggcat cgtcaaagaa aaagatgtgc tgttctgggc agacctgccg 1500
gactgctcgt ctggtgtgtg tatcgacaaa taa 1533
<210> 33
<211> 1269
<212> ДНК
<213> Heliobacter acinonychis
<400> 33
atggggacca ttaaaaagcc cttaatcata gcaggaaatg gtccatcaat taaggaccta 60
gactatgctt tatttccaaa agacttcgat gtctttcgct gcaaccagtt ttacttcgag 120
gataaatatt acctaggacg cgaaataaaa ggagtgttct ttaacccttg tgtattaagc 180
agtcaaatgc aaacagtgca ataccttatg gacaatggcg aatatagcat agaacgcttc 240
ttttgcagtg tttcaacaga tcgccacgat tttgatgggg attaccaaac gattttaccg 300
gtagacggtt atttaaaagc acactatccg ttcgtctgcg atacattcag cttattcaaa 360
ggtcacgaag aaatcttaaa acacgtgaaa taccacctga aaacgtacag caaagaactt 420
agtgcgggtg tcttaatgtt attgagtgca gtggtattag gatacaaaga aatataccta 480
gtaggaatcg acttcggcgc ctcatcttgg gggcacttct atgacgaaag ccaatcccaa 540
cactttagca atcacatggc agattgtcac aatatctatt acgacatgct gactatttgt 600
ctctgtcaaa agtatgcaaa attgtacgca ttagcaccca attcaccatt atcacatttg 660
cttacactaa atccacaggc caaataccca tttgaactat tagataaacc tatcgggtat 720
actagcgacc taattattag tagcccgttg gaagagaagt tgctcgaatt taagaatatc 780
gaagagaagt tgcttgagtt caaaaacata gaagagaaac tcttagagtt caagaatatt 840
gaagagaaac tattagaatt taaaaacatc gaggaaaaac ttttggagtt caaaaatata 900
gaagagaaac tcctagagtt caagaacatt gaggaaaagt tgcttgagtt caaaaatatt 960
gaggaaaagt tgctcgaatt taagaatatc gaggaaaaac ttttggaatt taagaacata 1020
gaagaaaagt tactcgaatt taaaaacatt gaagagaaac tattggaatt taaaaatata 1080
gaggaaaagt tacttgagtt caaaaacata gaggaaaagt tacttgaatt taagaacata 1140
gaagagaaac ttctcgcaag ccgactgaac aacattctac gtaaaatcaa gcggaaaata 1200
cttccattct tttggggcgg aggtgtaacc ccaacattaa aagttagttt ccgttgggga 1260
gctgcataa 1269
<210> 34
<211> 469
<212> ПРТ
<213> Campylobacter coli
<400> 34
Met Gln Asn Val Ile Ile Ala Gly Asn Gly Pro Ser Leu Gln Ser Ile
1 5 10 15
Asn Tyr Gln Arg Leu Pro Lys Glu Tyr Asp Ile Phe Arg Cys Asn Gln
20 25 30
Phe Tyr Phe Glu Asp Lys Tyr Tyr Leu Gly Lys Asn Ile Lys Ala Ala
35 40 45
Phe Phe Asn Pro Tyr Pro Phe Leu Gln Gln Tyr His Thr Ala Lys Gln
50 55 60
Leu Val Phe Asn Asn Glu Tyr Lys Ile Glu Asn Ile Phe Cys Ser Thr
65 70 75 80
Phe Asn Leu Pro Phe Ile Glu Lys Asp Asn Phe Ile Asn Lys Phe Tyr
85 90 95
Asp Phe Phe Pro Asp Ala Lys Leu Gly His Lys Ile Ile Glu Asn Leu
100 105 110
Lys Glu Phe Tyr Ala Tyr Ile Lys Tyr Asn Glu Ile Tyr Leu Asn Lys
115 120 125
Arg Ile Thr Ser Gly Ile Tyr Met Cys Ala Ile Ala Ile Ala Leu Gly
130 135 140
Tyr Lys Asn Ile Tyr Leu Cys Gly Ile Asp Phe Tyr Glu Gly Glu Thr
145 150 155 160
Ile Tyr Pro Phe Lys Ala Met Ser Lys Asn Ile Lys Lys Ile Phe Pro
165 170 175
Trp Ile Lys Asp Phe Asn Pro Ser Asn Phe His Ser Lys Glu Tyr Asp
180 185 190
Ile Glu Ile Leu Lys Leu Leu Glu Ser Ile Tyr Lys Val Asn Ile Tyr
195 200 205
Ala Leu Cys Asp Asn Ser Ala Leu Ala Asn Tyr Phe Pro Leu Leu Val
210 215 220
Asn Thr Asp Asn Ser Phe Val Leu Glu Asn Lys Ser Asp Asp Cys Ile
225 230 235 240
Asn Asp Ile Leu Leu Thr Asn Asn Thr Pro Gly Ile Asn Phe Tyr Lys
245 250 255
Ser Gln Ile Gln Val Asn Asn Thr Glu Ile Leu Leu Leu Asn Phe Gln
260 265 270
Asn Met Ile Ser Ala Lys Glu Asn Glu Ile Ser Asn Leu Asn Lys Ile
275 280 285
Leu Gln Asp Ser Tyr Lys Thr Ile Asn Thr Lys Glu Asn Glu Ile Ser
290 295 300
Asn Leu Asn Lys Ile Leu Gln Asp Ser Tyr Lys Thr Ile Asn Thr Lys
305 310 315 320
Glu Asn Glu Ile Ser Asn Leu Asn Lys Ile Leu Gln Asp Lys Asp Lys
325 330 335
Leu Leu Ile Val Lys Glu Asn Leu Leu Asn Phe Lys Ser Arg His Gly
340 345 350
Lys Ala Lys Phe Arg Ile Gln Asn Gln Leu Ser Tyr Lys Leu Gly Gln
355 360 365
Ala Met Met Val Asn Ser Lys Ser Leu Leu Gly Tyr Ile Arg Met Pro
370 375 380
Phe Val Leu Ser Tyr Ile Lys Asp Lys His Lys Gln Glu Gln Lys Ile
385 390 395 400
Tyr Gln Glu Lys Ile Lys Lys Asp Pro Ser Leu Thr Leu Pro Pro Leu
405 410 415
Glu Asp Tyr Pro Asp Tyr Lys Glu Ala Leu Lys Glu Lys Glu Cys Leu
420 425 430
Thr Tyr Arg Leu Gly Gln Thr Leu Ile Lys Ala Asp Gln Glu Trp Tyr
435 440 445
Lys Gly Gly Tyr Val Lys Met Trp Phe Glu Ile Lys Lys Leu Lys Lys
450 455 460
Glu Tyr Lys Lys Lys
465
<210> 35
<211> 381
<212> ПРТ
<213> Vibrio sp.
<400> 35
Met Asn Asn Asp Asn Ser Thr Thr Thr Asn Asn Asn Ala Ile Glu Ile
1 5 10 15
Tyr Val Asp Arg Ala Thr Leu Pro Thr Ile Gln Gln Met Thr Lys Ile
20 25 30
Val Ser Gln Lys Thr Ser Asn Lys Lys Leu Ile Ser Trp Ser Arg Tyr
35 40 45
Pro Ile Thr Asp Lys Ser Leu Leu Lys Lys Ile Asn Ala Glu Phe Phe
50 55 60
Lys Glu Gln Phe Glu Leu Thr Glu Ser Leu Lys Asn Ile Ile Leu Ser
65 70 75 80
Glu Asn Ile Asp Asn Leu Ile Ile His Gly Asn Thr Leu Trp Ser Ile
85 90 95
Asp Val Val Asp Ile Ile Lys Glu Val Asn Leu Leu Gly Lys Asn Ile
100 105 110
Pro Ile Glu Leu His Phe Tyr Asp Asp Gly Ser Ala Glu Tyr Val Arg
115 120 125
Ile Tyr Glu Phe Ser Lys Leu Pro Glu Ser Glu Gln Lys Tyr Lys Thr
130 135 140
Ser Leu Ser Lys Asn Asn Ile Lys Phe Ser Ile Asp Gly Thr Asp Ser
145 150 155 160
Phe Lys Asn Thr Ile Glu Asn Ile Tyr Gly Phe Ser Gln Leu Tyr Pro
165 170 175
Thr Thr Tyr His Met Leu Arg Ala Asp Ile Phe Asp Thr Thr Leu Lys
180 185 190
Ile Asn Pro Leu Arg Glu Leu Leu Ser Asn Asn Ile Lys Gln Met Lys
195 200 205
Trp Asp Tyr Phe Lys Asp Phe Asn Tyr Lys Gln Lys Asp Ile Phe Tyr
210 215 220
Ser Leu Thr Asn Phe Asn Pro Lys Glu Ile Gln Glu Asp Phe Asn Lys
225 230 235 240
Asn Ser Asn Lys Asn Phe Ile Phe Ile Gly Ser Asn Ser Ala Thr Ala
245 250 255
Thr Ala Glu Glu Gln Ile Asn Ile Ile Ser Glu Ala Lys Lys Glu Asn
260 265 270
Ser Ser Ile Ile Thr Asn Ser Ile Ser Asp Tyr Asp Leu Phe Phe Lys
275 280 285
Gly His Pro Ser Ala Thr Phe Asn Glu Gln Ile Ile Asn Ala His Asp
290 295 300
Met Ile Glu Ile Asn Asn Lys Ile Pro Phe Glu Ala Leu Ile Met Thr
305 310 315 320
Gly Ile Leu Pro Asp Ala Val Gly Gly Met Gly Ser Ser Val Phe Phe
325 330 335
Ser Ile Pro Lys Glu Val Lys Asn Lys Phe Val Phe Tyr Lys Ser Gly
340 345 350
Thr Asp Ile Glu Asn Asn Ser Leu Ile Gln Val Met Leu Lys Leu Asn
355 360 365
Leu Ile Asn Arg Asp Asn Ile Lys Leu Ile Ser Asp Ile
370 375 380
<210> 36
<211> 390
<212> ПРТ
<213> Photobacterium sp.
<400> 36
Met Gly Cys Asn Ser Asp Ser Asn His Asn Asn Ser Asp Gly Asn Ile
1 5 10 15
Thr Lys Asn Lys Thr Ile Glu Val Tyr Val Asp Arg Ala Thr Leu Pro
20 25 30
Thr Ile Gln Gln Met Thr Gln Ile Ile Asn Glu Asn Ser Asn Asn Lys
35 40 45
Lys Leu Ile Ser Trp Ser Arg Tyr Pro Ile Asn Asp Glu Glu Leu Leu
50 55 60
Glu Ser Ile Asn Gly Ser Phe Phe Lys Asn Asn Ser Glu Leu Ile Lys
65 70 75 80
Ser Leu Asp Ser Met Ile Leu Thr Asn Asp Ile Lys Lys Val Ile Ile
85 90 95
Asn Gly Asn Thr Leu Trp Ala Ala Asp Val Val Asn Ile Ile Lys Ser
100 105 110
Ile Glu Ala Phe Gly Lys Lys Thr Glu Ile Glu Leu Asn Phe Tyr Asp
115 120 125
Asp Gly Ser Ala Glu Tyr Val Arg Leu Tyr Asp Phe Ser Lys Leu Pro
130 135 140
Glu Ser Glu Gln Glu Tyr Lys Ile Ser Leu Ser Lys Asp Asn Ile Leu
145 150 155 160
Ser Ser Ile Asn Gly Thr Gln Pro Phe Glu Asn Val Val Glu Asn Ile
165 170 175
Tyr Gly Phe Ser Gln Leu Tyr Pro Thr Thr Tyr His Met Leu Arg Ala
180 185 190
Asp Ile Phe Glu Thr Asn Leu Pro Leu Arg Ser Leu Lys Gly Val Leu
195 200 205
Ser Asn Asn Ile Lys Gln Met Lys Trp Asp Tyr Phe Lys Thr Phe Asn
210 215 220
Ser Gln Gln Lys Asp Lys Phe Tyr Asn Phe Thr Gly Phe Asn Pro Asp
225 230 235 240
Glu Ile Met Glu Gln Tyr Lys Ala Ser Pro Asn Lys Asn Phe Ile Phe
245 250 255
Val Gly Thr Asn Ser Gly Thr Ala Thr Ala Glu Gln Gln Ile Asp Ile
260 265 270
Leu Thr Glu Ala Lys Asn Pro Asn Ser Pro Ile Ile Thr Lys Ser Ile
275 280 285
Gln Gly Phe Asp Leu Phe Phe Lys Gly His Pro Ser Ala Thr Tyr Asn
290 295 300
Lys Gln Ile Ile Asp Ala His Asn Met Ile Glu Ile Tyr Asn Lys Ile
305 310 315 320
Pro Phe Glu Ala Leu Ile Met Thr Asp Ala Leu Pro Asp Ala Val Gly
325 330 335
Gly Met Gly Ser Ser Val Phe Phe Ser Leu Pro Asn Thr Val Glu Asn
340 345 350
Lys Phe Ile Phe Tyr Lys Ser Asp Thr Asp Ile Glu Asn Asn Ala Leu
355 360 365
Ile Gln Val Met Ile Glu Leu Asn Ile Val Asn Arg Asn Asp Val Lys
370 375 380
Leu Ile Ser Asp Leu Gln
385 390
<210> 37
<211> 388
<212> ПРТ
<213> Pasteurella multocida
<400> 37
Met Lys Thr Ile Thr Leu Tyr Leu Asp Pro Ala Ser Leu Pro Ala Leu
1 5 10 15
Asn Gln Leu Met Asp Phe Thr Gln Asn Asn Glu Asp Lys Thr His Pro
20 25 30
Arg Ile Phe Gly Leu Ser Arg Phe Lys Ile Pro Asp Asn Ile Ile Thr
35 40 45
Gln Tyr Gln Asn Ile His Phe Val Glu Leu Lys Asp Asn Arg Pro Thr
50 55 60
Glu Ala Leu Phe Thr Ile Leu Asp Gln Tyr Pro Gly Asn Ile Glu Leu
65 70 75 80
Asp Ile His Leu Asn Ile Ala His Ser Val Gln Leu Ile Arg Pro Ile
85 90 95
Leu Ala Tyr Arg Phe Lys His Leu Asp Arg Val Ser Ile Gln Arg Leu
100 105 110
Asn Leu Tyr Asp Asp Gly Ser Met Glu Tyr Val Asp Leu Glu Lys Glu
115 120 125
Glu Asn Lys Asp Ile Ser Ala Glu Ile Lys Gln Ala Glu Lys Gln Leu
130 135 140
Ser His Tyr Leu Leu Thr Gly Lys Ile Lys Phe Asp Asn Pro Thr Ile
145 150 155 160
Ala Arg Tyr Val Trp Gln Ser Ala Phe Pro Val Lys Tyr His Phe Leu
165 170 175
Ser Thr Asp Tyr Phe Glu Lys Ala Glu Phe Leu Gln Pro Leu Lys Glu
180 185 190
Tyr Leu Ala Glu Asn Tyr Gln Lys Met Asp Trp Thr Ala Tyr Gln Gln
195 200 205
Leu Thr Pro Glu Gln Gln Ala Phe Tyr Leu Thr Leu Val Gly Phe Asn
210 215 220
Asp Glu Val Lys Gln Ser Leu Glu Val Gln Gln Ala Lys Phe Ile Phe
225 230 235 240
Thr Gly Thr Thr Thr Trp Glu Gly Asn Thr Asp Val Arg Glu Tyr Tyr
245 250 255
Ala Gln Gln Gln Leu Asn Leu Leu Asn His Phe Thr Gln Ala Gly Gly
260 265 270
Asp Leu Phe Ile Gly Asp His Tyr Lys Ile Tyr Phe Lys Gly His Pro
275 280 285
Arg Gly Gly Glu Ile Asn Asp Tyr Ile Leu Asn Asn Ala Lys Asn Ile
290 295 300
Thr Asn Ile Pro Ala Asn Ile Ser Phe Glu Val Leu Met Met Thr Gly
305 310 315 320
Leu Leu Pro Asp Lys Val Gly Gly Val Ala Ser Ser Leu Tyr Phe Ser
325 330 335
Leu Pro Lys Glu Lys Ile Ser His Ile Ile Phe Thr Ser Asn Lys Gln
340 345 350
Val Lys Ser Lys Glu Asp Ala Leu Asn Asn Pro Tyr Val Lys Val Met
355 360 365
Arg Arg Leu Gly Ile Ile Asp Glu Ser Gln Val Ile Phe Trp Asp Ser
370 375 380
Leu Lys Gln Leu
385
<210> 38
<211> 371
<212> ПРТ
<213> Neisseria meningitidis
<400> 38
Met Gly Leu Lys Lys Ala Cys Leu Thr Val Leu Cys Leu Ile Val Phe
1 5 10 15
Cys Phe Gly Ile Phe Tyr Thr Phe Asp Arg Val Asn Gln Gly Glu Arg
20 25 30
Asn Ala Val Ser Leu Leu Lys Glu Lys Leu Phe Asn Glu Glu Gly Glu
35 40 45
Pro Val Asn Leu Ile Phe Cys Tyr Thr Ile Leu Gln Met Lys Val Ala
50 55 60
Glu Arg Ile Met Ala Gln His Pro Gly Glu Arg Phe Tyr Val Val Leu
65 70 75 80
Met Ser Glu Asn Arg Asn Glu Lys Tyr Asp Tyr Tyr Phe Asn Gln Ile
85 90 95
Lys Asp Lys Ala Glu Arg Ala Tyr Phe Phe His Leu Pro Tyr Gly Leu
100 105 110
Asn Lys Ser Phe Asn Phe Ile Pro Thr Met Ala Glu Leu Lys Val Lys
115 120 125
Ser Met Leu Leu Pro Lys Val Lys Arg Ile Tyr Leu Ala Ser Leu Glu
130 135 140
Lys Val Ser Ile Ala Ala Phe Leu Ser Thr Tyr Pro Asp Ala Glu Ile
145 150 155 160
Lys Thr Phe Asp Asp Gly Thr Gly Asn Leu Ile Gln Ser Ser Ser Tyr
165 170 175
Leu Gly Asp Glu Phe Ser Val Asn Gly Thr Ile Lys Arg Asn Phe Ala
180 185 190
Arg Met Met Ile Gly Asp Trp Ser Ile Ala Lys Thr Arg Asn Ala Ser
195 200 205
Asp Glu His Tyr Thr Ile Phe Lys Gly Leu Lys Asn Ile Met Asp Asp
210 215 220
Gly Arg Arg Lys Met Thr Tyr Leu Pro Leu Phe Asp Ala Ser Glu Leu
225 230 235 240
Lys Thr Gly Asp Glu Thr Gly Gly Thr Val Arg Ile Leu Leu Gly Ser
245 250 255
Pro Asp Lys Glu Met Lys Glu Ile Ser Glu Lys Ala Ala Lys Asn Phe
260 265 270
Lys Ile Gln Tyr Val Ala Pro His Pro Arg Gln Thr Tyr Gly Leu Ser
275 280 285
Gly Val Thr Thr Leu Asn Ser Pro Tyr Val Ile Glu Asp Tyr Ile Leu
290 295 300
Arg Glu Ile Lys Lys Asn Pro His Thr Arg Tyr Glu Ile Tyr Thr Phe
305 310 315 320
Phe Ser Gly Ala Ala Leu Thr Met Lys Asp Phe Pro Asn Val His Val
325 330 335
Tyr Ala Leu Lys Pro Ala Ser Leu Pro Glu Asp Tyr Trp Leu Lys Pro
340 345 350
Val Tyr Ala Leu Phe Thr Gln Ser Gly Ile Pro Ile Leu Thr Phe Asp
355 360 365
Asp Lys Asn
370
<210> 39
<211> 283
<212> ПРТ
<213> Pasteurella multocida
<400> 39
Met Asp Lys Phe Ala Glu His Glu Ile Pro Lys Ala Val Ile Val Ala
1 5 10 15
Gly Asn Gly Glu Ser Leu Ser Gln Ile Asp Tyr Arg Leu Leu Pro Lys
20 25 30
Asn Tyr Asp Val Phe Arg Cys Asn Gln Phe Tyr Phe Glu Glu Arg Tyr
35 40 45
Phe Leu Gly Asn Lys Ile Lys Ala Val Phe Phe Thr Pro Gly Val Phe
50 55 60
Leu Glu Gln Tyr Tyr Thr Leu Tyr His Leu Lys Arg Asn Asn Glu Tyr
65 70 75 80
Phe Val Asp Asn Val Ile Leu Ser Ser Phe Asn His Pro Thr Val Asp
85 90 95
Leu Glu Lys Ser Gln Lys Ile Gln Ala Leu Phe Ile Asp Val Ile Asn
100 105 110
Gly Tyr Glu Lys Tyr Leu Ser Lys Leu Thr Ala Phe Asp Val Tyr Leu
115 120 125
Arg Tyr Lys Glu Leu Tyr Glu Asn Gln Arg Ile Thr Ser Gly Val Tyr
130 135 140
Met Cys Ala Val Ala Ile Ala Met Gly Tyr Thr Asp Ile Tyr Leu Thr
145 150 155 160
Gly Ile Asp Phe Tyr Gln Ala Ser Glu Glu Asn Tyr Ala Phe Asp Asn
165 170 175
Lys Lys Pro Asn Ile Ile Arg Leu Leu Pro Asp Phe Arg Lys Glu Lys
180 185 190
Thr Leu Phe Ser Tyr His Ser Lys Asp Ile Asp Leu Glu Ala Leu Ser
195 200 205
Phe Leu Gln Gln His Tyr His Val Asn Phe Tyr Ser Ile Ser Pro Met
210 215 220
Ser Pro Leu Ser Lys His Phe Pro Ile Pro Thr Val Glu Asp Asp Cys
225 230 235 240
Glu Thr Thr Phe Val Ala Pro Leu Lys Glu Asn Tyr Ile Asn Asp Ile
245 250 255
Leu Leu Pro Pro His Phe Val Tyr Glu Lys Leu Gly Val Asp Lys Leu
260 265 270
Ala Ala Ala Leu Glu His His His His His His
275 280
<210> 40
<211> 385
<212> ПРТ
<213> Pasteurella dagmatis
<400> 40
Met Thr Ile Tyr Leu Asp Pro Ala Ser Leu Pro Thr Leu Asn Gln Leu
1 5 10 15
Met His Phe Thr Lys Glu Ser Glu Asp Lys Glu Thr Ala Arg Ile Phe
20 25 30
Gly Phe Ser Arg Phe Lys Leu Pro Glu Lys Ile Thr Glu Gln Tyr Asn
35 40 45
Asn Ile His Phe Val Glu Ile Lys Asn Asn Arg Pro Thr Glu Asp Ile
50 55 60
Phe Thr Ile Leu Asp Gln Tyr Pro Glu Lys Leu Glu Leu Asp Leu His
65 70 75 80
Leu Asn Ile Ala His Ser Ile Gln Leu Phe His Pro Ile Leu Gln Tyr
85 90 95
Arg Phe Lys His Pro Asp Arg Ile Ser Ile Lys Ser Leu Asn Leu Tyr
100 105 110
Asp Asp Gly Thr Met Glu Tyr Val Asp Leu Glu Lys Glu Glu Asn Lys
115 120 125
Asp Ile Lys Ser Ala Ile Lys Lys Ala Glu Lys Gln Leu Ser Asp Tyr
130 135 140
Leu Leu Thr Gly Lys Ile Asn Phe Asp Asn Pro Thr Leu Ala Arg Tyr
145 150 155 160
Val Trp Gln Ser Gln Tyr Pro Val Lys Tyr His Phe Leu Ser Thr Glu
165 170 175
Tyr Phe Glu Lys Ala Glu Phe Leu Gln Pro Leu Lys Thr Tyr Leu Ala
180 185 190
Gly Lys Tyr Gln Lys Met Asp Trp Ser Ala Tyr Glu Lys Leu Ser Pro
195 200 205
Glu Gln Gln Thr Phe Tyr Leu Lys Leu Val Gly Phe Ser Asp Glu Thr
210 215 220
Lys Gln Leu Phe His Thr Glu Gln Thr Lys Phe Ile Phe Thr Gly Thr
225 230 235 240
Thr Thr Trp Glu Gly Asn Thr Asp Ile Arg Glu Tyr Tyr Ala Lys Gln
245 250 255
Gln Leu Asn Leu Leu Lys His Phe Thr His Ser Glu Gly Asp Leu Phe
260 265 270
Ile Gly Asp Gln Tyr Lys Ile Tyr Phe Lys Gly His Pro Arg Gly Gly
275 280 285
Asp Ile Asn Asp Tyr Ile Leu Lys His Ala Lys Asp Ile Thr Asn Ile
290 295 300
Pro Ala Asn Ile Ser Phe Glu Ile Leu Met Met Thr Gly Leu Leu Pro
305 310 315 320
Asp Lys Val Gly Gly Val Ala Ser Ser Leu Tyr Phe Ser Leu Pro Lys
325 330 335
Glu Lys Ile Ser His Ile Ile Phe Thr Ser Asn Lys Lys Ile Lys Asn
340 345 350
Lys Glu Asp Ala Leu Asn Asp Pro Tyr Val Arg Val Met Leu Arg Leu
355 360 365
Gly Met Ile Asp Lys Ser Gln Ile Ile Phe Trp Asp Ser Leu Lys Gln
370 375 380
Leu
385
<210> 41
<211> 390
<212> ПРТ
<213> Photobacterium phosphoreum
<400> 41
Met Gly Cys Asn Ser Asp Ser Lys His Asn Asn Ser Asp Gly Asn Ile
1 5 10 15
Thr Lys Asn Lys Thr Ile Glu Val Tyr Val Asp Arg Ala Thr Leu Pro
20 25 30
Thr Ile Gln Gln Met Thr Gln Ile Ile Asn Glu Asn Ser Asn Asn Lys
35 40 45
Lys Leu Ile Ser Trp Ser Arg Tyr Pro Ile Asn Asp Glu Thr Leu Leu
50 55 60
Glu Ser Ile Asn Gly Ser Phe Phe Lys Asn Arg Pro Glu Leu Ile Lys
65 70 75 80
Ser Leu Asp Ser Met Ile Leu Thr Asn Glu Ile Lys Lys Val Ile Ile
85 90 95
Asn Gly Asn Thr Leu Trp Ala Val Asp Val Val Asn Ile Ile Lys Ser
100 105 110
Ile Glu Ala Leu Gly Lys Lys Thr Glu Ile Glu Leu Asn Phe Tyr Asp
115 120 125
Asp Gly Ser Ala Glu Tyr Val Arg Leu Tyr Asp Phe Ser Arg Leu Pro
130 135 140
Glu Ser Glu Gln Glu Tyr Lys Ile Ser Leu Ser Lys Asp Asn Ile Gln
145 150 155 160
Ser Ser Ile Asn Gly Thr Gln Pro Phe Asp Asn Ser Ile Glu Asn Ile
165 170 175
Tyr Gly Phe Ser Gln Leu Tyr Pro Thr Thr Tyr His Met Leu Arg Ala
180 185 190
Asp Ile Phe Glu Thr Asn Leu Pro Leu Thr Ser Leu Lys Arg Val Ile
195 200 205
Ser Asn Asn Ile Lys Gln Met Lys Trp Asp Tyr Phe Thr Thr Phe Asn
210 215 220
Ser Gln Gln Lys Asn Lys Phe Tyr Asn Phe Thr Gly Phe Asn Pro Glu
225 230 235 240
Lys Ile Lys Glu Gln Tyr Lys Ala Ser Pro His Glu Asn Phe Ile Phe
245 250 255
Ile Gly Thr Asn Ser Gly Thr Ala Thr Ala Glu Gln Gln Ile Asp Ile
260 265 270
Leu Thr Glu Ala Lys Lys Pro Asp Ser Pro Ile Ile Thr Asn Ser Ile
275 280 285
Gln Gly Leu Asp Leu Phe Phe Lys Gly His Pro Ser Ala Thr Tyr Asn
290 295 300
Gln Gln Ile Ile Asp Ala His Asn Met Ile Glu Ile Tyr Asn Lys Ile
305 310 315 320
Pro Phe Glu Ala Leu Ile Met Thr Asp Ala Leu Pro Asp Ala Val Gly
325 330 335
Gly Met Gly Ser Ser Val Phe Phe Ser Leu Pro Asn Thr Val Glu Asn
340 345 350
Lys Phe Ile Phe Tyr Lys Ser Asp Thr Asp Ile Glu Asn Asn Ala Leu
355 360 365
Ile Gln Val Met Ile Glu Leu Asn Ile Val Asn Arg Asn Asp Val Lys
370 375 380
Leu Ile Ser Asp Leu Gln
385 390
<210> 42
<211> 417
<212> ПРТ
<213> Avibacterium paragallinarum
<400> 42
Met Arg Lys Ile Ile Thr Phe Phe Ser Leu Phe Phe Ser Ile Ser Ala
1 5 10 15
Trp Cys Gln Lys Met Glu Ile Tyr Leu Asp Tyr Ala Ser Leu Pro Ser
20 25 30
Leu Asn Met Ile Leu Asn Leu Val Glu Asn Lys Asn Asn Glu Lys Val
35 40 45
Glu Arg Ile Ile Gly Phe Glu Arg Phe Asp Phe Asn Lys Glu Ile Leu
50 55 60
Asn Ser Phe Ser Lys Glu Arg Ile Glu Phe Ser Lys Val Ser Ile Leu
65 70 75 80
Asp Ile Lys Glu Phe Ser Asp Lys Leu Tyr Leu Asn Ile Glu Lys Ser
85 90 95
Asp Thr Pro Val Asp Leu Ile Ile His Thr Asn Leu Asp His Ser Val
100 105 110
Arg Ser Leu Leu Ser Ile Phe Lys Thr Leu Ser Pro Leu Phe His Lys
115 120 125
Ile Asn Ile Glu Lys Leu Tyr Leu Tyr Asp Asp Gly Ser Gly Asn Tyr
130 135 140
Val Asp Leu Tyr Gln His Arg Gln Glu Asn Ile Ser Ala Ile Leu Ile
145 150 155 160
Glu Ala Gln Lys Lys Leu Lys Asp Ala Leu Glu Asn Arg Glu Thr Asp
165 170 175
Thr Asp Lys Leu His Ser Leu Thr Arg Tyr Thr Trp His Lys Ile Phe
180 185 190
Pro Thr Glu Tyr Ile Leu Leu Arg Pro Asp Tyr Leu Asp Ile Asp Glu
195 200 205
Lys Met Gln Pro Leu Lys His Phe Leu Ser Asp Thr Ile Val Ser Met
210 215 220
Asp Leu Ser Arg Phe Ser His Phe Ser Lys Asn Gln Lys Glu Leu Phe
225 230 235 240
Leu Lys Ile Thr His Phe Asp Gln Asn Ile Phe Asn Glu Leu Asn Ile
245 250 255
Gly Thr Lys Asn Lys Glu Tyr Lys Thr Phe Ile Phe Thr Gly Thr Thr
260 265 270
Thr Trp Glu Lys Asp Lys Lys Lys Arg Leu Asn Asn Ala Lys Leu Gln
275 280 285
Thr Glu Ile Leu Glu Ser Phe Ile Lys Pro Asn Gly Lys Phe Tyr Leu
290 295 300
Gly Asn Asp Ile Lys Ile Phe Phe Lys Gly His Pro Lys Gly Asp Asp
305 310 315 320
Ile Asn Asp Tyr Ile Ile Arg Lys Thr Gly Ala Glu Lys Ile Pro Ala
325 330 335
Asn Ile Pro Phe Glu Val Leu Met Met Thr Asn Ser Leu Pro Asp Tyr
340 345 350
Val Gly Gly Ile Met Ser Thr Val Tyr Phe Ser Leu Pro Pro Lys Asn
355 360 365
Ile Asp Lys Val Val Phe Leu Gly Ser Glu Lys Ile Lys Asn Glu Asn
370 375 380
Asp Ala Lys Ser Gln Thr Leu Ser Lys Leu Met Leu Met Leu Asn Val
385 390 395 400
Ile Thr Pro Glu Gln Ile Phe Phe Glu Glu Met Pro Asn Pro Ile Asn
405 410 415
Phe
<210> 43
<211> 430
<212> ПРТ
<213> Campylobacter jejuni
<400> 43
Met Thr Arg Thr Arg Met Glu Asn Glu Leu Ile Val Ser Lys Asn Met
1 5 10 15
Gln Asn Ile Ile Ile Ala Gly Asn Gly Pro Ser Leu Lys Asn Ile Asn
20 25 30
Tyr Lys Arg Leu Pro Arg Glu Tyr Asp Val Phe Arg Cys Asn Gln Phe
35 40 45
Tyr Phe Glu Asp Lys Tyr Tyr Leu Gly Lys Lys Ile Lys Ala Val Phe
50 55 60
Phe Asn Pro Gly Val Phe Leu Gln Gln Tyr His Thr Ala Lys Gln Leu
65 70 75 80
Ile Leu Lys Asn Glu Tyr Glu Ile Lys Asn Ile Phe Cys Ser Thr Phe
85 90 95
Asn Leu Pro Phe Ile Glu Ser Asn Asp Phe Leu His Gln Phe Tyr Asn
100 105 110
Phe Phe Pro Asp Ala Lys Leu Gly Tyr Glu Val Ile Glu Asn Leu Lys
115 120 125
Glu Phe Tyr Ala Tyr Ile Lys Tyr Asn Glu Ile Tyr Phe Asn Lys Arg
130 135 140
Ile Thr Ser Gly Val Tyr Met Cys Ala Ile Ala Ile Ala Leu Gly Tyr
145 150 155 160
Lys Thr Ile Tyr Leu Cys Gly Ile Asp Phe Tyr Glu Gly Asp Val Ile
165 170 175
Tyr Pro Phe Glu Ala Met Ser Thr Asn Ile Lys Thr Ile Phe Pro Gly
180 185 190
Ile Lys Asp Phe Lys Pro Ser Asn Cys His Ser Lys Glu Tyr Asp Ile
195 200 205
Glu Ala Leu Lys Leu Leu Lys Ser Ile Tyr Lys Val Asn Ile Tyr Ala
210 215 220
Leu Cys Asp Asp Ser Ile Leu Ala Asn His Phe Pro Leu Ser Ile Asn
225 230 235 240
Ile Asn Asn Asn Phe Thr Leu Glu Asn Lys His Asn Asn Ser Ile Asn
245 250 255
Asp Ile Leu Leu Thr Asp Asn Thr Pro Gly Val Ser Phe Tyr Lys Asn
260 265 270
Gln Leu Lys Ala Asp Asn Lys Ile Met Leu Asn Phe Tyr Asn Ile Leu
275 280 285
His Ser Lys Asp Asn Leu Ile Lys Phe Leu Asn Lys Glu Ile Ala Val
290 295 300
Leu Lys Lys Gln Thr Thr Gln Arg Ala Lys Ala Arg Ile Gln Asn His
305 310 315 320
Leu Ser Tyr Lys Leu Gly Gln Ala Leu Ile Ile Asn Ser Lys Ser Val
325 330 335
Leu Gly Phe Leu Ser Leu Pro Phe Ile Ile Leu Ser Ile Val Ile Ser
340 345 350
His Lys Gln Glu Gln Lys Ala Tyr Lys Phe Lys Val Lys Lys Asn Pro
355 360 365
Asn Leu Ala Leu Pro Pro Leu Glu Thr Tyr Pro Asp Tyr Asn Glu Ala
370 375 380
Leu Lys Glu Lys Glu Cys Phe Thr Tyr Lys Leu Gly Glu Glu Phe Ile
385 390 395 400
Lys Ala Gly Lys Asn Trp Tyr Gly Glu Gly Tyr Ile Lys Phe Ile Phe
405 410 415
Lys Asp Val Pro Arg Leu Lys Arg Glu Phe Glu Lys Gly Glu
420 425 430
<210> 44
<211> 395
<212> ПРТ
<213> Heliobacter acinonychis
<400> 44
Met Asn Lys Lys Pro Leu Ile Ile Ala Gly Asn Gly Pro Ser Ile Lys
1 5 10 15
Asp Leu Asp Tyr Ala Leu Phe Pro Lys Asp Phe Asp Val Phe Arg Cys
20 25 30
Asn Gln Phe Tyr Phe Glu Asp Lys Tyr Tyr Leu Gly Arg Glu Ile Lys
35 40 45
Gly Val Phe Phe Asn Ala His Val Phe Asp Leu Gln Met Lys Ile Thr
50 55 60
Lys Ala Ile Val Lys Asn Gly Glu Tyr His Pro Asp His Ile Tyr Cys
65 70 75 80
Thr His Val Glu Pro Tyr Gly Tyr Val Asn Gly Asn Gln Gln Leu Met
85 90 95
Gln Glu Tyr Leu Glu Lys His Phe Val Gly Val Arg Ser Thr Tyr Ala
100 105 110
Tyr Leu Lys Asp Leu Glu Pro Phe Phe Ile Leu His Ser Lys Tyr Arg
115 120 125
Asn Phe Tyr Asp Gln His Phe Thr Thr Gly Ile Met Met Leu Leu Val
130 135 140
Ala Ile Gln Leu Gly Tyr Lys Glu Ile Tyr Leu Cys Gly Ile Asp Phe
145 150 155 160
Tyr Glu Asn Gly Phe Gly His Phe Tyr Glu Asn Gln Gly Gly Phe Phe
165 170 175
Glu Glu Asp Ser Asp Pro Met His Asp Lys Asn Ile Asp Ile Gln Ala
180 185 190
Leu Glu Leu Ala Lys Lys Tyr Ala Lys Ile Tyr Ala Leu Val Pro Asn
195 200 205
Ser Ala Leu Val Lys Met Ile Pro Leu Ser Ser Gln Lys Gly Val Leu
210 215 220
Glu Lys Val Lys Asp Arg Ile Gly Leu Gly Glu Phe Lys Arg Glu Lys
225 230 235 240
Phe Gly Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys
245 250 255
Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg
260 265 270
Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu
275 280 285
Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys
290 295 300
Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg
305 310 315 320
Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu Glu Arg Gln Lys Glu Leu
325 330 335
Glu Leu Glu Arg Ser Leu Lys Ala Arg Leu Lys Ala Val Leu Ala Ser
340 345 350
Lys Gly Ile Arg Gly Asp Asn Leu Ile Ile Val Ser Leu Lys Asp Thr
355 360 365
Tyr Arg Leu Phe Lys Gly Gly Phe Ala Leu Leu Leu Asp Leu Lys Ala
370 375 380
Leu Lys Ser Ile Ile Lys Ala Phe Leu Lys Arg
385 390 395
<210> 45
<211> 260
<212> ПРТ
<213> Campylobacter jejuni
<400> 45
Met Gly Lys Lys Val Ile Ile Ala Gly Asn Gly Pro Ser Leu Lys Glu
1 5 10 15
Ile Asp Tyr Ser Arg Leu Pro Asn Asp Phe Asp Val Phe Arg Cys Asn
20 25 30
Gln Phe Tyr Phe Glu Asp Lys Tyr Tyr Leu Gly Lys Lys Cys Lys Ala
35 40 45
Val Phe Tyr Asn Pro Ile Leu Phe Phe Glu Gln Tyr Tyr Thr Leu Lys
50 55 60
His Leu Ile Gln Asn Gln Glu Tyr Glu Thr Glu Leu Ile Met Cys Ser
65 70 75 80
Asn Tyr Asn Gln Ala His Leu Glu Asn Glu Asn Phe Val Lys Thr Phe
85 90 95
Tyr Asp Tyr Phe Pro Asp Ala His Leu Gly Tyr Asp Phe Phe Lys Gln
100 105 110
Leu Lys Asp Phe Asn Ala Tyr Phe Lys Phe His Glu Ile Tyr Phe Asn
115 120 125
Gln Arg Ile Thr Ser Gly Val Tyr Met Cys Ala Val Ala Ile Ala Leu
130 135 140
Gly Tyr Lys Glu Ile Tyr Leu Ser Gly Ile Asp Phe Tyr Gln Asn Gly
145 150 155 160
Ser Ser Tyr Ala Phe Asp Thr Lys Gln Lys Asn Leu Leu Lys Leu Ala
165 170 175
Pro Asn Phe Lys Asn Asp Asn Ser His Tyr Ile Gly His Ser Lys Asn
180 185 190
Thr Asp Ile Lys Ala Leu Glu Phe Leu Glu Lys Thr Tyr Lys Ile Lys
195 200 205
Leu Tyr Cys Leu Cys Pro Asn Ser Leu Leu Ala Asn Phe Ile Glu Leu
210 215 220
Ala Pro Asn Leu Asn Ser Asn Phe Ile Ile Gln Glu Lys Asn Asn Tyr
225 230 235 240
Thr Lys Asp Ile Leu Ile Pro Ser Ser Glu Ala Tyr Gly Lys Phe Ser
245 250 255
Lys Asn Ile Asn
260
<210> 46
<211> 298
<212> ПРТ
<213> Streptococcus entericus
<400> 46
Met Lys Lys Val Tyr Phe Cys His Thr Val Tyr His Leu Leu Ile Thr
1 5 10 15
Leu Cys Lys Ile Ser Val Glu Glu Gln Val Glu Ile Ile Val Phe Asp
20 25 30
Thr Val Ser Asn His Glu Leu Ile Val Gln Lys Ile Arg Asp Val Phe
35 40 45
Val Asn Thr Thr Val Leu Phe Ala Glu Gln Asn Thr Asp Phe Ser Ile
50 55 60
Leu Glu Ile Asp Arg Ala Thr Asp Ile Tyr Val Phe Asn Asp Trp Thr
65 70 75 80
Pro Ile Gly Ala Tyr Leu Arg Lys Asn Lys Leu Phe Tyr His Leu Ile
85 90 95
Glu Asp Gly Tyr Asn Tyr His Glu Tyr Asn Val Tyr Ala Asn Ala Leu
100 105 110
Thr Met Lys Arg Arg Leu Leu Asn Phe Val Leu Arg Arg Glu Glu Pro
115 120 125
Ser Gly Phe Ser Arg Tyr Val Arg Ser Ile Glu Val Asn Arg Val Lys
130 135 140
Tyr Leu Pro Asn Asp Cys Arg Lys Ser Lys Trp Val Glu Lys Pro Arg
145 150 155 160
Ser Ala Leu Phe Glu Asn Leu Val Pro Glu His Lys Gln Lys Ile Ile
165 170 175
Thr Ile Phe Gly Leu Glu Asn Tyr Gln Asp Ser Leu Arg Gly Val Leu
180 185 190
Val Leu Thr Gln Pro Leu Val Gln Asp Tyr Trp Asp Arg Asp Ile Thr
195 200 205
Thr Glu Glu Glu Gln Leu Glu Phe Tyr Arg Gln Ile Val Glu Ser Tyr
210 215 220
Gly Glu Gly Glu Gln Val Phe Phe Lys Ile His Pro Arg Asp Lys Val
225 230 235 240
Asp Tyr Ser Ser Leu Thr Asn Val Ile Phe Leu Lys Lys Asn Val Pro
245 250 255
Met Glu Val Tyr Glu Leu Ile Ala Asp Cys His Phe Thr Lys Gly Ile
260 265 270
Thr His Ser Ser Thr Ala Leu Asp Phe Leu Ser Cys Val Asp Lys Lys
275 280 285
Ile Thr Leu Lys Gln Met Lys Ala Asn Ser
290 295
<210> 47
<211> 295
<212> ПРТ
<213> Haemophilus ducreyi
<400> 47
Met Lys Glu Ile Ala Ile Ile Ser Asn Gln Arg Met Phe Phe Leu Tyr
1 5 10 15
Cys Leu Leu Thr Asn Lys Asn Val Glu Asp Val Phe Phe Ile Phe Glu
20 25 30
Lys Gly Ala Met Pro Asn Asn Leu Thr Ser Ile Ser His Phe Ile Val
35 40 45
Leu Asp His Ser Lys Ser Glu Cys Tyr Asp Phe Phe Tyr Phe Asn Phe
50 55 60
Ile Ser Cys Lys Tyr Arg Leu Arg Gly Leu Asp Val Tyr Gly Ala Asp
65 70 75 80
His Ile Lys Gly Ala Lys Phe Phe Leu Glu Arg His Arg Phe Phe Val
85 90 95
Val Glu Asp Gly Met Met Asn Tyr Ser Lys Asn Met Tyr Ala Phe Ser
100 105 110
Leu Phe Arg Thr Arg Asn Pro Val Ile Leu Pro Gly Gly Phe His Pro
115 120 125
Asn Val Lys Thr Ile Phe Leu Thr Lys Asp Asn Pro Ile Pro Asp Gln
130 135 140
Ile Ala His Lys Arg Glu Ile Ile Asn Ile Lys Thr Leu Trp Gln Ala
145 150 155 160
Lys Thr Ala Thr Glu Lys Thr Lys Ile Leu Ser Phe Phe Glu Ile Asp
165 170 175
Met Gln Glu Ile Ser Val Ile Lys Asn Arg Ser Phe Val Leu Tyr Thr
180 185 190
Gln Pro Leu Ser Glu Asp Lys Leu Leu Thr Glu Ala Glu Lys Ile Asp
195 200 205
Ile Tyr Arg Thr Ile Leu Thr Lys Tyr Asn His Ser Gln Thr Val Ile
210 215 220
Lys Pro His Pro Arg Asp Lys Thr Asp Tyr Lys Gln Leu Phe Pro Asp
225 230 235 240
Ala Tyr Val Met Lys Gly Thr Tyr Pro Ser Glu Leu Leu Thr Leu Leu
245 250 255
Gly Val Asn Phe Asn Lys Val Ile Thr Leu Phe Ser Thr Ala Val Phe
260 265 270
Asp Tyr Pro Lys Glu Lys Ile Asp Phe Tyr Gly Thr Ala Val His Pro
275 280 285
Lys Leu Leu Asp Phe Phe Asp
290 295
<210> 48
<211> 488
<212> ПРТ
<213> Alistipes sp.
<400> 48
Met Ala Leu Leu Ser Gly Thr Ala Ala Cys Ser Asp Asp Glu Val Ser
1 5 10 15
Gln Asn Leu Ile Val Ile Asn Gly Gly Glu His Phe Leu Ser Leu Asp
20 25 30
Gly Leu Ala Arg Ala Gly Lys Ile Ser Val Leu Ala Pro Ala Pro Trp
35 40 45
Arg Val Thr Lys Ala Ala Gly Asp Thr Trp Phe Arg Leu Ser Ala Thr
50 55 60
Glu Gly Pro Ala Gly Tyr Ser Glu Val Glu Leu Ser Leu Asp Glu Asn
65 70 75 80
Pro Gly Ala Ala Arg Ser Ala Gln Leu Ala Phe Ala Cys Gly Asp Ala
85 90 95
Ile Val Pro Phe Arg Leu Ser Gln Gly Ala Leu Ser Ala Gly Tyr Asp
100 105 110
Ser Pro Asp Tyr Tyr Phe Tyr Val Thr Phe Gly Thr Met Pro Thr Leu
115 120 125
Tyr Ala Gly Ile His Leu Leu Ser His Asp Lys Pro Gly Tyr Val Phe
130 135 140
Tyr Ser Arg Ser Lys Thr Phe Asp Pro Ala Glu Phe Pro Ala Arg Ala
145 150 155 160
Glu Val Thr Thr Ala Ala Asp Arg Thr Ala Asp Ala Thr Gln Ala Glu
165 170 175
Met Glu Ala Met Ala Arg Glu Met Lys Arg Arg Ile Leu Glu Ile Asn
180 185 190
Ser Ala Asp Pro Thr Ala Val Phe Gly Leu Tyr Val Asp Asp Leu Arg
195 200 205
Cys Arg Ile Gly Tyr Asp Trp Phe Val Ala Gln Gly Ile Asp Ser Ala
210 215 220
Arg Val Lys Val Ser Met Leu Ser Asp Gly Thr Gly Thr Tyr Asn Asn
225 230 235 240
Phe Tyr Asn Tyr Phe Gly Asp Ala Ala Thr Ala Glu Gln Asn Trp Glu
245 250 255
Ser Tyr Ala Ser Glu Val Glu Ala Leu Asp Trp Asn His Gly Gly Arg
260 265 270
Tyr Pro Glu Thr Arg Ser Leu Pro Glu Phe Glu Ser Tyr Thr Trp Pro
275 280 285
Tyr Tyr Leu Ser Thr Arg Pro Asp Tyr Arg Leu Val Val Gln Asp Gly
290 295 300
Ser Leu Leu Glu Ser Ser Cys Pro Phe Ile Thr Glu Lys Leu Gly Glu
305 310 315 320
Met Glu Ile Glu Ser Ile Gln Pro Tyr Glu Met Leu Ser Ala Leu Pro
325 330 335
Glu Ser Ser Arg Lys Arg Phe Tyr Asp Met Ala Gly Phe Asp Tyr Asp
340 345 350
Lys Phe Ala Ala Leu Phe Asp Ala Ser Pro Lys Lys Asn Leu Ile Ile
355 360 365
Ile Gly Thr Ser His Ala Asp Asp Ala Ser Ala Arg Leu Gln Arg Asp
370 375 380
Tyr Val Ala Arg Ile Met Glu Gln Tyr Gly Ala Gln Tyr Asp Val Phe
385 390 395 400
Phe Lys Pro His Pro Ala Asp Thr Thr Ser Ala Gly Tyr Glu Thr Glu
405 410 415
Phe Pro Gly Leu Thr Leu Leu Pro Gly Gln Met Pro Phe Glu Ile Phe
420 425 430
Val Trp Ser Leu Ile Asp Arg Val Asp Met Ile Gly Gly Tyr Pro Ser
435 440 445
Thr Val Phe Leu Thr Val Pro Val Asp Lys Val Arg Phe Ile Phe Ala
450 455 460
Ala Asp Ala Ala Ser Leu Val Arg Pro Leu Asn Ile Leu Phe Arg Asp
465 470 475 480
Ala Thr Asp Val Glu Trp Met Gln
485
<210> 49
<211> 291
<212> ПРТ
<213> Campylobacter jejuni
<400> 49
Met Lys Lys Val Ile Ile Ala Gly Asn Gly Pro Ser Leu Lys Glu Ile
1 5 10 15
Asp Tyr Ser Arg Leu Pro Asn Asp Phe Asp Val Phe Arg Cys Asn Gln
20
Phe Tyr Phe Glu Asp Lys Tyr Tyr Leu Gly Lys Lys Cys Lys Ala Val
35 40 45
Phe Tyr Thr Pro Asn Phe Phe Phe Glu Gln Tyr Tyr Thr Leu Lys His
50 55 60
Leu Ile Gln Asn Gln Glu Tyr Glu Thr Glu Leu Ile Met Cys Ser Asn
65 70 75 80
Tyr Asn Gln Ala His Leu Glu Asn Glu Asn Phe Val Lys Thr Phe Tyr
85 90 95
Asp Tyr Phe Pro Asp Ala His Leu Gly Tyr Asp Phe Phe Lys Gln Leu
100 105 110
Lys Glu Phe Asn Ala Tyr Phe Lys Phe His Glu Ile Tyr Phe Asn Gln
115 120 125
Arg Ile Thr Ser Gly Val Tyr Met Cys Ala Val Ala Ile Ala Leu Gly
130 135 140
Tyr Lys Glu Ile Tyr Leu Ser Gly Ile Asp Phe Tyr Gln Asn Gly Ser
145 150 155 160
Ser Tyr Ala Phe Asp Thr Lys Gln Glu Asn Leu Leu Lys Leu Ala Pro
165 170 175
Asp Phe Lys Asn Asp Arg Ser His Tyr Ile Gly His Ser Lys Asn Thr
180 185 190
Asp Ile Lys Ala Leu Glu Phe Leu Glu Lys Thr Tyr Lys Ile Lys Leu
195 200 205
Tyr Cys Leu Cys Pro Asn Ser Leu Leu Ala Asn Phe Ile Glu Leu Ala
210 215 220
Pro Asn Leu Asn Ser Asn Phe Ile Ile Gln Glu Lys Asn Asn Tyr Thr
225 230 235 240
Lys Asp Ile Leu Ile Pro Ser Ser Glu Ala Tyr Gly Lys Phe Ser Lys
245 250 255
Asn Ile Asn Phe Lys Lys Ile Lys Ile Lys Glu Asn Val Tyr Tyr Lys
260 265 270
Leu Ile Lys Asp Leu Leu Arg Leu Pro Ser Asp Ile Lys His Tyr Phe
275 280 285
Lys Gly Lys
290
<210> 50
<211> 312
<212> ПРТ
<213> Streptococcus agalactiae
<400> 50
Met Thr Asn Arg Lys Ile Tyr Val Cys His Thr Leu Tyr His Leu Leu
1 5 10 15
Ile Cys Leu Tyr Lys Glu Glu Ile Tyr Ser Asn Leu Glu Ile Ile Leu
20 25 30
Ser Ser Ser Ile Pro Asp Val Asp Asn Leu Glu Lys Lys Leu Lys Ser
35 40 45
Lys Thr Ile Asn Ile His Ile Leu Glu Glu Ser Ser Gly Glu Ser Glu
50 55 60
Glu Leu Leu Ser Val Leu Lys Asp Ala Gly Leu Ser Tyr Ser Lys Phe
65 70 75 80
Asp Ser Asn Cys Phe Ile Phe Asn Asp Ala Thr Pro Ile Gly Arg Thr
85 90 95
Leu Ile Lys His Gly Ile Tyr Tyr Asn Leu Ile Glu Asp Gly Leu Asn
100 105 110
Cys Phe Thr Tyr Ser Ile Phe Ser Gln Lys Leu Trp Lys Tyr Tyr Val
115 120 125
Lys Lys Tyr Ile Leu His Lys Ile Gln Pro His Gly Phe Ser Arg Tyr
130 135 140
Cys Leu Gly Ile Glu Val Asn Ser Leu Val Asn Leu Pro Lys Asp Pro
145 150 155 160
Arg Tyr Lys Lys Phe Ile Glu Val Pro Arg Lys Glu Leu Phe Asp Asn
165 170 175
Val Thr Glu Tyr Gln Lys Glu Met Ala Ile Asn Leu Phe Gly Ala Val
180 185 190
Arg Val Ser Ile Lys Ser Pro Ser Val Leu Val Leu Thr Gln Pro Leu
195 200 205
Ser Ile Asp Lys Glu Phe Met Ser Tyr Asn Asn Lys Ile Glu Thr Ser
210 215 220
Glu Glu Gln Phe Asn Phe Tyr Lys Ser Ile Val Asn Glu Tyr Ile Asn
225 230 235 240
Lys Gly Tyr Asn Val Tyr Leu Lys Val His Pro Arg Asp Val Val Asp
245 250 255
Tyr Ser Lys Leu Pro Val Glu Leu Leu Pro Ser Asn Val Pro Met Glu
260 265 270
Ile Ile Glu Leu Met Leu Thr Gly Arg Phe Glu Cys Gly Ile Thr His
275 280 285
Ser Ser Thr Ala Leu Asp Phe Leu Thr Cys Val Asp Lys Lys Ile Thr
290 295 300
Leu Val Asp Leu Lys Asp Ile Lys
305 310
<210> 51
<211> 410
<212> ПРТ
<213> Bibersteinia trehalosi
<400> 51
Met Glu Phe Cys Lys Met Ala Thr Thr Gln Lys Ile Cys Val Tyr Leu
1 5 10 15
Asp Tyr Ala Thr Ile Pro Ser Leu Asn Tyr Ile Leu His Phe Ala Gln
20 25 30
His Phe Glu Asp Gln Glu Thr Ile Arg Leu Phe Gly Leu Ser Arg Phe
35 40 45
His Ile Pro Glu Ser Val Ile Gln Arg Tyr Pro Lys Gly Val Val Gln
50 55 60
Phe Tyr Pro Asn Gln Glu Lys Asp Phe Ser Ala Leu Leu Leu Ala Leu
65 70 75 80
Lys Asn Ile Leu Ile Glu Val Lys Gln Gln Gln Arg Lys Cys Glu Ile
85 90 95
Glu Leu His Leu Asn Leu Phe His Tyr Gln Leu Leu Leu Leu Pro Phe
100 105 110
Leu Ser Leu Tyr Leu Asp Thr Gln Asp Tyr Cys His Leu Thr Leu Lys
115 120 125
Phe Tyr Asp Asp Gly Ser Glu Ala Ile Ser Ala Leu Gln Glu Leu Ala
130 135 140
Leu Ala Pro Asp Leu Ala Ala Gln Ile Gln Phe Glu Lys Gln Gln Phe
145 150 155 160
Asp Glu Leu Val Val Lys Lys Ser Phe Lys Leu Ser Leu Leu Ser Arg
165 170 175
Tyr Phe Trp Gly Lys Leu Phe Glu Ser Glu Tyr Ile Trp Phe Asn Gln
180 185 190
Ala Ile Leu Gln Lys Ala Glu Leu Gln Ile Leu Lys Gln Glu Ile Ser
195 200 205
Ser Ser Arg Gln Met Asp Phe Ala Ile Tyr Gln Gln Met Ser Asp Glu
210 215 220
Gln Lys Gln Leu Val Leu Glu Ile Leu Asn Ile Asp Leu Asn Lys Val
225 230 235 240
Ala Tyr Leu Lys Gln Leu Met Glu Asn Gln Pro Ser Phe Leu Phe Leu
245 250 255
Gly Thr Thr Leu Phe Asn Ile Thr Gln Glu Thr Lys Thr Trp Leu Met
260 265 270
Gln Met His Val Asp Leu Ile Gln Gln Tyr Cys Leu Pro Ser Gly Gln
275 280 285
Phe Phe Asn Asn Lys Ala Gly Tyr Leu Cys Phe Tyr Lys Gly His Pro
290 295 300
Asn Glu Lys Glu Met Asn Gln Met Ile Leu Ser Gln Phe Lys Asn Leu
305 310 315 320
Ile Ala Leu Pro Asp Asp Ile Pro Leu Glu Ile Leu Leu Leu Leu Gly
325 330 335
Val Ile Pro Ser Lys Val Gly Gly Phe Ala Ser Ser Ala Leu Phe Asn
340 345 350
Phe Thr Pro Ala Gln Ile Glu Asn Ile Ile Phe Phe Thr Pro Arg Tyr
355 360 365
Phe Glu Lys Asp Asn Arg Leu His Ala Thr Gln Tyr Arg Leu Met Gln
370 375 380
Gly Leu Ile Glu Leu Gly Tyr Leu Asp Ala Glu Lys Ser Val Thr His
385 390 395 400
Phe Glu Ile Met Gln Leu Leu Thr Lys Glu
405 410
<210> 52
<211> 406
<212> ПРТ
<213> Haemophilus parahaemolyticus
<400> 52
Met Thr Glu Gln Tyr Ile Lys Asn Val Glu Val Tyr Leu Asp Tyr Ala
1 5 10 15
Thr Ile Pro Thr Leu Asn Tyr Phe Tyr His Phe Thr Glu Asn Lys Asp
20 25 30
Asp Ile Ala Thr Ile Arg Leu Phe Gly Leu Gly Arg Phe Asn Ile Ser
35 40 45
Lys Ser Ile Ile Glu Ser Tyr Pro Glu Gly Ile Ile Arg Tyr Cys Pro
50 55 60
Ile Ile Phe Glu Asp Gln Thr Ala Phe Gln Gln Leu Phe Ile Thr Leu
65 70 75 80
Leu Thr Glu Asp Ser Phe Cys Gln Tyr Arg Phe Asn Phe His Ile Asn
85 90 95
Leu Phe His Ser Trp Lys Met Leu Ile Pro Leu Leu His Ile Ile Trp
100 105 110
Gln Phe Lys His Lys Val Leu Asp Ile Lys Leu Asn Phe Tyr Asp Asp
115 120 125
Gly Ser Glu Gly Leu Val Thr Leu Ser Lys Ile Glu Gln Asn Tyr Ser
130 135 140
Ser Glu Ile Leu Gln Lys Ile Ile Asp Ile Asp Ser Gln Ser Phe Tyr
145 150 155 160
Ala Asp Lys Leu Ser Phe Leu Asp Glu Asp Ile Ala Arg Tyr Leu Trp
165 170 175
Asn Ser Leu Phe Glu Ser His Tyr Tyr Leu Leu Asn Asp Phe Leu Leu
180 185 190
Lys Asn Glu Lys Leu Ser Leu Leu Lys Asn Ser Ile Lys Tyr Cys His
195 200 205
Ile Met Asp Leu Glu Arg Tyr Leu Gln Phe Thr Gln Glu Glu Lys Asp
210 215 220
Phe Phe Asn Glu Leu Leu Gly Ile Asn Ile Gln Ser Leu Glu Asp Lys
225 230 235 240
Ile Lys Ile Phe Gln Gln Lys Lys Thr Phe Ile Phe Thr Gly Thr Thr
245 250 255
Ile Phe Ser Leu Pro Lys Glu Glu Glu Glu Thr Leu Tyr Arg Leu His
260 265 270
Leu Asn Ala Ile Leu Asn Tyr Ile His Pro Asn Gly Lys Tyr Phe Ile
275 280 285
Gly Asp Gly Phe Thr Leu Val Ile Lys Gly His Pro His Gln Lys Glu
290 295 300
Met Asn Ser Arg Leu Glu Lys Ser Phe Glu Lys Ala Val Met Leu Pro
305 310 315 320
Asp Asn Ile Pro Phe Glu Ile Leu Tyr Leu Ile Gly Cys Lys Pro Asp
325 330 335
Lys Ile Gly Gly Phe Val Ser Thr Ser Tyr Phe Ser Cys Asp Lys Lys
340 345 350
Asn Ile Ala Asp Leu Leu Phe Ile Ser Ala Arg Gln Glu Glu Val Arg
355 360 365
Lys Asn Asp Tyr Leu Phe Asn Ile Gln Tyr Gln Leu Arg Asp Met Met
370 375 380
Ile Lys Thr Gly Phe Ile Gln Glu Glu Lys Thr His Phe Tyr Ser Asp
385 390 395 400
Ile Pro Ile Phe Ile Ser
405
<210> 53
<211> 300
<212> ПРТ
<213> Haemophilus somnus
<400> 53
Met Lys Tyr Asn Ile Lys Ile Lys Ala Ile Val Ile Val Ser Ser Leu
1 5 10 15
Arg Met Leu Leu Ile Phe Leu Met Leu Asn Lys Tyr His Leu Asp Glu
20 25 30
Val Leu Phe Val Phe Asn Glu Gly Phe Glu Leu His Lys Lys Tyr Lys
35 40 45
Ile Lys His Tyr Val Ala Ile Lys Lys Lys Ile Thr Lys Phe Trp Arg
50 55 60
Leu Tyr Tyr Lys Leu Tyr Phe Tyr Arg Phe Lys Ile Asp Arg Ile Pro
65 70 75 80
Val Tyr Gly Ala Asp His Leu Gly Trp Thr Asp Tyr Phe Leu Lys Tyr
85 90 95
Phe Asp Phe Tyr Leu Ile Glu Asp Gly Ile Ala Asn Phe Ser Pro Lys
100 105 110
Arg Tyr Glu Ile Asn Leu Thr Arg Asn Ile Pro Val Phe Gly Phe His
115 120 125
Lys Thr Val Lys Lys Ile Tyr Leu Thr Ser Leu Glu Asn Val Pro Ser
130 135 140
Asp Ile Arg His Lys Val Glu Leu Ile Ser Leu Glu His Leu Trp Lys
145 150 155 160
Thr Arg Thr Ala Gln Glu Gln His Asn Ile Leu Asp Phe Phe Ala Phe
165 170 175
Asn Leu Asp Ser Leu Ile Ser Leu Lys Met Lys Lys Tyr Ile Leu Phe
180 185 190
Thr Gln Cys Leu Ser Glu Asp Arg Val Ile Ser Glu Gln Glu Lys Ile
195 200 205
Ala Ile Tyr Gln His Ile Ile Lys Asn Tyr Asp Glu Arg Leu Leu Val
210 215 220
Ile Lys Pro His Pro Arg Glu Thr Thr Asp Tyr Gln Lys Tyr Phe Glu
225 230 235 240
Asn Val Phe Val Tyr Gln Asp Val Val Pro Ser Glu Leu Phe Glu Leu
245 250 255
Leu Asp Val Asn Phe Glu Arg Val Ile Thr Leu Phe Ser Thr Ala Val
260 265 270
Phe Lys Tyr Asp Arg Asn Ile Val Asp Phe Tyr Gly Thr Arg Ile His
275 280 285
Asp Lys Ile Tyr Gln Trp Phe Gly Asp Ile Lys Phe
290 295 300
<210> 54
<211> 381
<212> ПРТ
<213> Vibrio harveyi
<400> 54
Met Asp Ser Ser Pro Glu Asn Thr Ser Ser Thr Leu Glu Ile Tyr Ile
1 5 10 15
Asp Ser Ala Thr Leu Pro Ser Leu Gln His Met Val Lys Ile Ile Asp
20 25 30
Glu Gln Ser Gly Asn Lys Lys Leu Ile Asn Trp Lys Arg Tyr Pro Ile
35 40 45
Asp Asp Glu Leu Leu Leu Asp Lys Ile Asn Ala Leu Ser Phe Ser Asp
50 55 60
Thr Thr Asp Leu Thr Arg Tyr Met Glu Ser Ile Leu Leu Ile Gly Asp
65 70 75 80
Ile Lys Arg Val Val Ile Asn Gly Asn Ser Leu Ser Asn Tyr Asn Ile
85 90 95
Val Gly Val Met Arg Ser Ile Asn Ala Leu Gly Leu Asp Leu Asp Val
100 105 110
Glu Ile Asn Phe Tyr Asp Asp Gly Ser Ala Glu Tyr Val Arg Leu Tyr
115 120 125
Asn Phe Ser Gln Leu Pro Glu Ala Glu Arg Glu Leu Leu Val Ser Met
130 135 140
Ser Lys Asn Asn Ile Leu Ala Ala Val Asn Gly Ile Gly Ser Tyr Asp
145 150 155 160
Ser Gly Ser Pro Glu Asn Ile Tyr Gly Phe Ala Gln Ile Tyr Pro Ala
165 170 175
Thr Tyr His Met Leu Arg Ala Asp Ile Phe Asp Thr Asp Leu Glu Ile
180 185 190
Gly Leu Ile Arg Asp Ile Leu Gly Asp Asn Val Lys Gln Met Lys Trp
195 200 205
Gly Gln Phe Leu Gly Phe Asn Glu Glu Gln Lys Glu Leu Phe Tyr Gln
210 215 220
Leu Thr Ser Phe Asn Pro Asp Lys Ile Gln Ala Gln Tyr Lys Glu Ser
225 230 235 240
Pro Asn Lys Asn Phe Val Phe Val Gly Thr Asn Ser Arg Ser Ala Thr
245 250 255
Ala Glu Gln Gln Ile Asn Ile Ile Lys Glu Ala Lys Lys Leu Asp Ser
260 265 270
Glu Ile Ile Pro Asn Ser Ile Asp Gly Tyr Asp Leu Phe Phe Lys Gly
275 280 285
His Pro Ser Ala Thr Tyr Asn Gln Gln Ile Val Asp Ala His Asp Met
290 295 300
Thr Glu Ile Tyr Asn Arg Thr Pro Phe Glu Val Leu Ala Met Thr Ser
305 310 315 320
Ser Leu Pro Asp Ala Val Gly Gly Met Gly Ser Ser Leu Phe Phe Ser
325 330 335
Leu Pro Lys Thr Val Glu Thr Lys Phe Ile Phe Tyr Lys Ser Gly Thr
340 345 350
Asp Ile Glu Ser Asn Ala Leu Ile Gln Val Met Leu Lys Leu Gly Ile
355 360 365
Ile Thr Asp Glu Lys Val Arg Phe Thr Thr Asp Ile Lys
370 375 380
<210> 55
<211> 483
<212> ПРТ
<213> Alistipes sp.
<400> 55
Met Ala Ser Cys Ser Asp Asp Asp Lys Glu Gln Thr Gly Phe Gln Ile
1 5 10 15
Asp Asp Gly Ser Gly Phe Leu Ser Leu Asp Ala Ala Ala Arg Ser Gly
20 25 30
Ser Ile Ala Ile Thr Ala Asn Asn Ser Trp Ser Val Thr Gln Asp Lys
35 40 45
Asp Ser Glu Trp Leu Thr Leu Ser Thr Thr Ser Gly Ala Ala Gly Arg
50 55 60
Thr Glu Ile Gly Ile Met Leu Glu Ala Asn Pro Gly Glu Ala Arg Asn
65 70 75 80
Ala Gly Leu Thr Phe Asn Ser Gly Gly Arg Thr Tyr Pro Phe Val Ile
85 90 95
Thr Gln Ser Ala His Val Thr Ala Asp Phe Asp Asp Ala Asp His Cys
100 105 110
Phe Tyr Ile Thr Phe Gly Thr Leu Pro Thr Leu Tyr Ala Gly Leu His
115 120 125
Val Leu Ser His Asp Lys Pro Ser Tyr Val Phe Phe Gln Arg Ser Gln
130 135 140
Thr Phe Arg Pro Glu Glu Phe Pro Ala His Ala Glu Val Thr Ile Ala
145 150 155 160
Ala Asp Pro Ser Ala Asn Ala Thr Asp Glu Asp Met Glu Arg Met Arg
165 170 175
Thr Ala Met Lys Gln Gln Ile Leu Lys Ile Asn Val Glu Asp Pro Thr
180 185 190
Ala Val Phe Gly Leu Tyr Val Asp Asp Leu Arg Cys Gly Ile Gly Tyr
195 200 205
Asp Trp Phe Val Ala Gln Gly Ile Asp Ser Thr Arg Val Lys Val Ser
210 215 220
Met Leu Ser Asp Gly Thr Gly Thr Tyr Asn Asn Phe Tyr Asn Tyr Phe
225 230 235 240
Gly Asp Pro Ala Thr Ala Glu Gln Asn Trp Glu Asn Tyr Ala Ala Gln
245 250 255
Val Glu Ala Leu Asp Trp Gln His Gly Gly Arg Phe Pro Glu Thr Arg
260 265 270
Met Pro Asp Gly Phe Asp Phe Tyr Glu Trp Pro Tyr Tyr Leu Ala Thr
275 280 285
Arg Pro Asn Tyr Arg Leu Val Leu Gln Asp Asp Asp Leu Leu Glu Ala
290 295 300
Thr Ser Pro Phe Met Thr Glu Arg Leu Gln Gln Met Arg Thr Glu Ser
305 310 315 320
Lys Gln Pro Tyr Glu Leu Leu Ala Ser Leu Pro Ala Glu Ala Arg Gln
325 330 335
Arg Phe Phe Arg Met Ala Gly Phe Asp Tyr Asp Ala Phe Ala Ala Leu
340 345 350
Phe Asp Ala Ser Pro Lys Lys Asn Leu Val Ile Ile Gly Thr Ser His
355 360 365
Thr Ser Glu Glu Ser Glu Ala Gln Gln Ala Ala Tyr Val Glu Arg Ile
370 375 380
Ile Gly Asp Tyr Gly Thr Ala Tyr Asp Ile Phe Phe Lys Pro His Pro
385 390 395 400
Ala Asp Ser Ser Ser Ser Asn Tyr Glu Glu Arg Phe Glu Gly Leu Thr
405 410 415
Leu Leu Pro Gly Gln Met Pro Phe Glu Ile Phe Val Trp Ser Leu Leu
420 425 430
Asp Lys Val Asp Leu Ile Gly Gly Tyr Ser Ser Thr Val Phe Leu Thr
435 440 445
Val Pro Val Glu Lys Thr Gly Phe Ile Phe Ala Ala Asn Ala Glu Ser
450 455 460
Leu Pro Arg Pro Leu Asn Val Leu Phe Arg Asn Ala Glu His Val Arg
465 470 475 480
Trp Ile Gln
<210> 56
<211> 483
<212> ПРТ
<213> Alistipes shahii
<400> 56
Met Asp Asp Gly Thr Pro Ser Val Ser Ile Asn Gly Gly Thr Asp Phe
1 5 10 15
Leu Ser Leu Asp His Leu Ala Arg Ser Gly Lys Ile Thr Val Asn Ala
20 25 30
Pro Ala Pro Trp Ser Val Thr Leu Ala Pro Glu Asn Tyr Gly Gln Asp
35 40 45
Glu Lys Pro Asp Trp Leu Thr Leu Ser Ala Glu Glu Gly Pro Ala Gly
50 55 60
Tyr Ser Glu Ile Asp Val Thr Phe Ala Glu Asn Pro Gly Pro Ala Arg
65 70 75 80
Ser Ala Ser Leu Leu Phe Ser Cys Asp Gly Lys Thr Leu Ala Phe Thr
85 90 95
Val Ser Gln Ser Ala Gly Gly Thr Gly Phe Asp Ala Pro Asp Tyr Tyr
100 105 110
Phe Tyr Ile Ser Val Gly Thr Met Pro Thr Leu Tyr Ser Gly Leu His
115 120 125
Leu Leu Ser His Asp Lys Pro Ser Tyr Val Ser Tyr Glu Arg Ala Ser
130 135 140
Thr Phe Asp Ala Ala Glu Phe Pro Asp Arg Ala Phe Val Tyr Pro Val
145 150 155 160
Ala Asp Pro Thr Gly His Ala Thr Asn Glu Glu Leu Arg Ala Met Ser
165 170 175
Glu Ala Met Lys Arg Arg Ile Leu Glu Ile Asn Ala Glu Asp Pro Thr
180 185 190
Ala Val Phe Gly Leu Trp Val Asp Asp Leu Arg Cys Arg Leu Gly Tyr
195 200 205
Asp Trp Phe Val Ala Gln Gly Ile Asp Ser Ala Arg Val Lys Val Thr
210 215 220
Met Leu Ser Asp Gly Thr Ala Thr Tyr Asn Asn Phe His Asn Tyr Phe
225 230 235 240
Gly Asp Ala Ala Thr Ala Glu Gln Asn Trp Asn Asp Tyr Ala Ala Glu
245 250 255
Val Glu Ala Leu Asp Trp Asn His Gly Gly Arg Tyr Pro Glu Thr Arg
260 265 270
Ala Pro Glu Glu Phe Ala Ser Tyr Thr Trp Pro Tyr Tyr Leu Ser Thr
275 280 285
Arg Pro Asp Tyr Arg Leu Met Leu Gln Asn Ser Ser Leu Met Glu Ser
290 295 300
Ser Cys Pro Phe Ile Ala Asp Arg Leu Ala Ala Met Lys Met Glu Ser
305 310 315 320
Val Gln Pro Tyr Glu Leu Leu Thr Ala Leu Pro Glu Ala Ser Lys Gln
325 330 335
Gln Phe Tyr Arg Met Ala Lys Phe Asp Tyr Ala Arg Phe Ala Gly Leu
340 345 350
Phe Asp Leu Ser Pro Lys Lys Asn Leu Ile Ile Ile Gly Thr Ser His
355 360 365
Ser Ser Ala Ala Ser Glu Gln Gln Gln Ala Ala Tyr Val Glu Arg Ile
370 375 380
Ile Gln Gln Tyr Gly Ser Asp Tyr Asp Ile Phe Phe Lys Pro His Pro
385 390 395 400
Ala Asp Ser Ser Ser Ala Gly Tyr Pro Asp Arg Phe Glu Gly Leu Thr
405 410 415
Leu Leu Pro Gly Gln Met Pro Phe Glu Ile Phe Val Trp Ala Leu Leu
420 425 430
Asp Lys Ile Asp Met Ile Gly Gly Tyr Pro Ser Thr Thr Phe Ile Ser
435 440 445
Val Pro Leu Asp Lys Val Gly Phe Leu Phe Ala Ala Asp Ala Asp Gly
450 455 460
Leu Val Arg Pro Leu Asn Ile Leu Phe Arg Asp Ala Ala Asn Val Glu
465 470 475 480
Trp Ile Gln
<210> 57
<211> 401
<212> ПРТ
<213> Actinobacillus suis
<400> 57
Met Glu Arg Thr Pro Gln Leu Gln Ala Val Asp Ile Tyr Ile Asp Phe
1 5 10 15
Ala Thr Ile Pro Ser Leu Ser Tyr Phe Leu His Phe Leu Lys His Lys
20 25 30
His Asp Asp Gln Arg Leu Arg Leu Phe Ser Leu Ala Arg Phe Glu Met
35 40 45
Pro Gln Thr Leu Ile Glu Gln Tyr Glu Gly Ile Ile Gln Phe Ser Arg
50 55 60
Asn Val Glu His Asn Val Glu Pro Leu Leu Glu Gln Leu Gln Thr Ile
65 70 75 80
Leu Ser Gln Glu Gly Lys Gln Phe Glu Leu His Leu His Leu Asn Leu
85 90 95
Phe His Ser Phe Glu Met Phe Leu Asn Leu Ser Pro Thr Tyr Thr Gln
100 105 110
Tyr Lys Glu Lys Ile Ser Lys Ile Val Leu His Leu Tyr Asp Asp Gly
115 120 125
Ser Glu Gly Val Met Lys Gln Tyr Gln Leu Gln Lys Ser Ser Ser Leu
130 135 140
Val Gln Asp Leu Ala Ala Thr Lys Ala Ser Leu Val Ser Leu Phe Glu
145 150 155 160
Asn Gly Glu Gly Ser Phe Ser Gln Ile Asp Leu Ile Arg Tyr Val Trp
165 170 175
Asn Ala Val Leu Glu Thr His Tyr Tyr Leu Leu Ser Asp His Phe Leu
180 185 190
Leu Asp Glu Lys Leu Gln Pro Leu Lys Ala Glu Leu Gly His Tyr Gln
195 200 205
Leu Leu Asn Leu Ser Ala Tyr Gln Tyr Leu Ser Ser Glu Asp Leu Leu
210 215 220
Trp Leu Lys Gln Ile Leu Lys Ile Asp Thr Glu Leu Glu Ser Leu Met
225 230 235 240
Gln Lys Leu Thr Ala Gln Pro Val Tyr Phe Phe Ser Gly Thr Thr Phe
245 250 255
Phe Asn Ile Ser Phe Glu Asp Lys Gln Arg Leu Ala Asn Ile His Ala
260 265 270
Ile Leu Ile Arg Glu His Leu Asp Pro Asn Ser Gln Leu Phe Ile Gly
275 280 285
Glu Pro Tyr Leu Phe Val Phe Lys Gly His Pro Asn Ser Pro Glu Ile
290 295 300
Asn Gln Ala Leu Arg Glu Tyr Tyr Pro Asn Val Ile Phe Leu Pro Glu
305 310 315 320
Asn Ile Pro Phe Glu Ile Leu Thr Leu Leu Gly Phe Ser Pro Gln Lys
325 330 335
Ile Gly Gly Phe Ala Ser Thr Ile His Val Asn Ser Glu Gln Ser Lys
340 345 350
Leu Ala Lys Leu Phe Phe Leu Thr Ser Thr Asp Glu Gln Glu Arg Gln
355 360 365
Leu Ser Asp Gly Tyr Ile Lys Gln Tyr Ala Leu Ala Gln Ala Met Leu
370 375 380
Glu Met Gln Leu Val Ser Gln Glu Gln Val Tyr Tyr Cys Ser Leu Ser
385 390 395 400
Ser
<210> 58
<211> 401
<212> ПРТ
<213> Actinobacillus capsulatus
<400> 58
Met Glu Arg Ile Pro Gln Leu Gln Ala Val Asp Ile Tyr Ile Asp Phe
1 5 10 15
Ala Thr Ile Pro Ser Leu Ser Tyr Phe Leu His Phe Leu Lys His Lys
20 25 30
His Asp His Gln Arg Leu Arg Leu Phe Ser Leu Ala Arg Phe Glu Met
35 40 45
Pro Gln Thr Val Ile Glu Gln Tyr Glu Gly Ile Ile Gln Phe Ser Arg
50 55 60
Asn Val Glu His Asn Val Glu Gln Leu Leu Glu Gln Leu Gln Thr Ile
65 70 75 80
Leu Ser Gln Glu Gly Lys Gln Phe Glu Leu His Leu His Leu Asn Leu
85 90 95
Phe His Ser Phe Glu Met Phe Leu Asn Leu Ser Pro Thr Tyr Thr Lys
100 105 110
Tyr Lys Glu Lys Ile Ser Lys Ile Val Leu His Leu Tyr Asp Asp Gly
115 120 125
Ser Glu Gly Val Met Lys Gln Tyr Gln Leu Gln Gln Ser Asn Ser Leu
130 135 140
Ala Gln Asp Leu Ala Ser Thr Lys Ala Ser Leu Val Ser Leu Phe Lys
145 150 155 160
Asn Gly Glu Gly Ala Phe Ser Gln Ile Asp Leu Ile Arg Tyr Val Trp
165 170 175
Asn Ala Val Leu Glu Thr His Tyr Tyr Leu Leu Ser Asp His Phe Leu
180 185 190
Ala His Glu Lys Leu Gln Pro Leu Lys Ile Glu Leu Gly His Tyr Gln
195 200 205
Leu Leu Asn Leu Ser Ala Tyr Gln Tyr Leu Ser Ser Glu Asp Leu Leu
210 215 220
Trp Leu Lys Gln Ile Leu Lys Ile Asp Ala Glu Leu Glu Ser Leu Met
225 230 235 240
His Lys Leu Thr Thr Gln Pro Val Tyr Phe Phe Ser Gly Thr Thr Phe
245 250 255
Phe Asn Ile Ser Phe Glu Asp Lys Gln Arg Leu Ala Asn Ile His Ala
260 265 270
Ile Leu Ile Arg Glu His Leu Asp Pro Asn Ser Gln Leu Phe Ile Gly
275 280 285
Glu Pro Tyr Leu Phe Val Phe Lys Gly His Pro Asn Ser Pro Glu Ile
290 295 300
Asn Gln Ala Leu Arg Glu Tyr Tyr Pro Asn Ala Ile Phe Leu Pro Glu
305 310 315 320
Asn Ile Pro Phe Glu Ile Leu Thr Leu Leu Gly Phe Ser Pro Gln Lys
325 330 335
Ile Gly Gly Phe Ala Ser Thr Ile His Val Asn Ser Glu Gln Ser Lys
340 345 350
Leu Ala Lys Leu Phe Phe Leu Thr Ser Thr Asp Glu Gln Glu Arg Asn
355 360 365
Arg Ser Asp Gly Tyr Ile Lys Gln Tyr Ala Leu Ala Gln Ala Met Leu
370 375 380
Glu Met Gln Leu Val Ser Gln Glu Gln Val Tyr Tyr Cys Ser Leu Ser
385 390 395 400
Ser
<210> 59
<211> 311
<212> ПРТ
<213> Haemophilus somnus
<400> 59
Met Phe Arg Glu Asp Asn Met Asn Leu Ile Ile Cys Cys Thr Pro Leu
1 5 10 15
Gln Val Ile Ile Ala Glu Lys Ile Ile Glu Arg Tyr Pro Glu Gln Lys
20 25 30
Phe Tyr Gly Val Met Leu Glu Ser Phe Tyr Asn Asp Lys Phe Asp Phe
35 40 45
Tyr Glu Asn Lys Leu Lys His Leu Cys His Glu Phe Phe Cys Ile Lys
50 55 60
Ile Ala Arg Phe Lys Leu Glu Arg Tyr Lys Asn Leu Leu Ser Leu Leu
65 70 75 80
Lys Ile Lys Asn Lys Thr Phe Asp Arg Val Phe Leu Ala Asn Ile Glu
85 90 95
Lys Arg Tyr Ile His Ile Ile Leu Ser Asn Ile Phe Phe Lys Glu Leu
100 105 110
Tyr Thr Phe Asp Asp Gly Thr Ala Asn Ile Ala Pro Asn Ser His Leu
115 120 125
Tyr Gln Glu Tyr Asp His Ser Leu Lys Lys Arg Ile Thr Asp Ile Leu
130 135 140
Leu Pro Asn His Tyr Asn Ser Asn Lys Val Lys Asn Ile Ser Lys Leu
145 150 155 160
His Tyr Ser Ile Tyr Arg Cys Lys Asn Asn Ile Ile Asp Asn Ile Glu
165 170 175
Tyr Met Pro Leu Phe Asn Leu Glu Lys Lys Tyr Thr Ala Gln Asp Lys
180 185 190
Ser Ile Ser Ile Leu Leu Gly Gln Pro Ile Phe Tyr Asp Glu Glu Lys
195 200 205
Asn Ile Arg Leu Ile Lys Glu Val Ile Ala Lys Phe Lys Ile Asp Tyr
210 215 220
Tyr Phe Pro His Pro Arg Glu Asp Tyr Tyr Ile Asp Asn Val Ser Tyr
225 230 235 240
Ile Lys Thr Pro Leu Ile Phe Glu Glu Phe Tyr Ala Glu Arg Ser Ile
245 250 255
Glu Asn Ser Ile Lys Ile Tyr Thr Phe Phe Ser Ser Ala Val Leu Asn
260 265 270
Ile Val Thr Lys Glu Asn Ile Asp Arg Ile Tyr Ala Leu Lys Pro Lys
275 280 285
Leu Thr Glu Lys Ala Tyr Leu Asp Cys Tyr Asp Ile Leu Lys Asp Phe
290 295 300
Gly Ile Lys Val Ile Asp Ile
305 310
<210> 60
<211> 399
<212> ПРТ
<213> Haemophilus ducreyi
<400> 60
Met Leu Ile Gln Gln Asn Leu Glu Ile Tyr Leu Asp Tyr Ala Thr Ile
1 5 10 15
Pro Ser Leu Ala Cys Phe Met His Phe Ile Gln His Lys Asp Asp Val
20 25 30
Asp Ser Ile Arg Leu Phe Gly Leu Ala Arg Phe Asp Ile Pro Gln Ser
35 40 45
Ile Ile Asp Arg Tyr Pro Ala Asn His Leu Phe Tyr His Asn Ile Asp
50 55 60
Asn Arg Asp Leu Thr Ala Val Leu Asn Gln Leu Ala Asp Ile Leu Ala
65 70 75 80
Gln Glu Asn Lys Arg Phe Gln Ile Asn Leu His Leu Asn Leu Phe His
85 90 95
Ser Ile Asp Leu Phe Phe Ala Ile Tyr Pro Ile Tyr Gln Gln Tyr Gln
100 105 110
His Lys Ile Ser Thr Ile Gln Leu Gln Leu Tyr Asp Asp Gly Ser Glu
115 120 125
Gly Ile Val Thr Gln His Ser Leu Cys Lys Ile Ala Asp Leu Glu Gln
130 135 140
Leu Ile Leu Gln His Lys Asn Val Leu Leu Glu Leu Leu Thr Lys Gly
145 150 155 160
Thr Ala Asn Val Pro Asn Pro Thr Leu Leu Arg Tyr Leu Trp Asn Asn
165 170 175
Ile Ile Asp Ser Gln Phe His Leu Ile Ser Asp His Phe Leu Gln His
180 185 190
Pro Lys Leu Gln Pro Leu Lys Arg Leu Leu Lys Arg Tyr Thr Ile Leu
195 200 205
Asp Phe Thr Cys Tyr Pro Arg Phe Asn Ala Glu Gln Lys Gln Leu Leu
210 215 220
Lys Glu Ile Leu His Ile Ser Asn Glu Leu Glu Asn Leu Leu Lys Leu
225 230 235 240
Leu Lys Gln His Asn Thr Phe Leu Phe Thr Gly Thr Thr Ala Phe Asn
245 250 255
Leu Asp Gln Glu Lys Leu Asp Leu Leu Thr Gln Leu His Ile Leu Leu
260 265 270
Leu Asn Glu His Gln Asn Pro His Ser Thr His Tyr Ile Gly Asn Asn
275 280 285
Tyr Leu Leu Leu Ile Lys Gly His Ala Asn Ser Pro Ala Leu Asn His
290 295 300
Thr Leu Ala Leu His Phe Pro Asp Ala Ile Phe Leu Pro Ala Asn Ile
305 310 315 320
Pro Phe Glu Ile Phe Ala Met Leu Gly Phe Thr Pro Asn Lys Met Gly
325 330 335
Gly Phe Ala Ser Thr Ser Tyr Ile Asn Tyr Pro Thr Glu Asn Ile Asn
340 345 350
His Leu Phe Phe Leu Thr Ser Asp Gln Pro Ser Ile Arg Thr Lys Trp
355 360 365
Leu Asp Tyr Glu Lys Gln Phe Gly Leu Met Tyr Ser Leu Leu Ala Met
370 375 380
Gln Lys Ile Asn Glu Asp Gln Ala Phe Met Cys Thr Ile His Asn
385 390 395
<210> 61
<211> 497
<212> ПРТ
<213> Photobacterium leiognathi
<400> 61
Met Cys Asn Asp Asn Gln Asn Thr Val Asp Val Val Val Ser Thr Val
1 5 10 15
Asn Asp Asn Val Ile Glu Asn Asn Thr Tyr Gln Val Lys Pro Ile Asp
20 25 30
Thr Pro Thr Thr Phe Asp Ser Tyr Ser Trp Ile Gln Thr Cys Gly Thr
35 40 45
Pro Ile Leu Lys Asp Asp Glu Lys Tyr Ser Leu Ser Phe Asp Phe Val
50 55 60
Ala Pro Glu Leu Asp Gln Asp Glu Lys Phe Cys Phe Glu Phe Thr Gly
65 70 75 80
Asp Val Asp Gly Lys Arg Tyr Val Thr Gln Thr Asn Leu Thr Val Val
85 90 95
Ala Pro Thr Leu Glu Val Tyr Val Asp His Ala Ser Leu Pro Ser Leu
100 105 110
Gln Gln Leu Met Lys Ile Ile Gln Gln Lys Asn Glu Tyr Ser Gln Asn
115 120 125
Glu Arg Phe Ile Ser Trp Gly Arg Ile Gly Leu Thr Glu Asp Asn Ala
130 135 140
Glu Lys Leu Asn Ala His Ile Tyr Pro Leu Ala Gly Asn Asn Thr Ser
145 150 155 160
Gln Glu Leu Val Asp Ala Val Ile Asp Tyr Ala Asp Ser Lys Asn Arg
165 170 175
Leu Asn Leu Glu Leu Asn Thr Asn Thr Ala His Ser Phe Pro Asn Leu
180 185 190
Ala Pro Ile Leu Arg Ile Ile Ser Ser Lys Ser Asn Ile Leu Ile Ser
195 200 205
Asn Ile Asn Leu Tyr Asp Asp Gly Ser Ala Glu Tyr Val Asn Leu Tyr
210 215 220
Asn Trp Lys Asp Thr Glu Asp Lys Ser Val Lys Leu Ser Asp Ser Phe
225 230 235 240
Leu Val Leu Lys Asp Tyr Phe Asn Gly Ile Ser Ser Glu Lys Pro Ser
245 250 255
Gly Ile Tyr Gly Arg Tyr Asn Trp His Gln Leu Tyr Asn Thr Ser Tyr
260 265 270
Tyr Phe Leu Arg Lys Asp Tyr Leu Thr Val Glu Pro Gln Leu His Asp
275 280 285
Leu Arg Glu Tyr Leu Gly Gly Ser Leu Lys Gln Met Ser Trp Asp Gly
290 295 300
Phe Ser Gln Leu Ser Lys Gly Asp Lys Glu Leu Phe Leu Asn Ile Val
305 310 315 320
Gly Phe Asp Gln Glu Lys Leu Gln Gln Glu Tyr Gln Gln Ser Glu Leu
325 330 335
Pro Asn Phe Val Phe Thr Gly Thr Thr Thr Trp Ala Gly Gly Glu Thr
340 345 350
Lys Glu Tyr Tyr Ala Gln Gln Gln Val Asn Val Val Asn Asn Ala Ile
355 360 365
Asn Glu Thr Ser Pro Tyr Tyr Leu Gly Arg Glu His Asp Leu Phe Phe
370 375 380
Lys Gly His Pro Arg Gly Gly Ile Ile Asn Asp Ile Ile Leu Gly Ser
385 390 395 400
Phe Asn Asn Met Ile Asp Ile Pro Ala Lys Val Ser Phe Glu Val Leu
405 410 415
Met Met Thr Gly Met Leu Pro Asp Thr Val Gly Gly Ile Ala Ser Ser
420 425 430
Leu Tyr Phe Ser Ile Pro Ala Glu Lys Val Ser Phe Ile Val Phe Thr
435 440 445
Ser Ser Asp Thr Ile Thr Asp Arg Glu Asp Ala Leu Lys Ser Pro Leu
450 455 460
Val Gln Val Met Met Thr Leu Gly Ile Val Lys Glu Lys Asp Val Leu
465 470 475 480
Phe Trp Ser Asp Leu Pro Asp Cys Ser Ser Gly Val Cys Ile Ala Gln
485 490 495
Tyr
<210> 62
<211> 498
<212> ПРТ
<213> Photobacterium sp.
<400> 62
Met Ser Glu Glu Asn Thr Gln Ser Ile Ile Lys Asn Asp Ile Asn Lys
1 5 10 15
Thr Ile Ile Asp Glu Glu Tyr Val Asn Leu Glu Pro Ile Asn Gln Ser
20 25 30
Asn Ile Ser Phe Thr Lys His Ser Trp Val Gln Thr Cys Gly Thr Gln
35 40 45
Gln Leu Leu Thr Glu Gln Asn Lys Glu Ser Ile Ser Leu Ser Val Val
50 55 60
Ala Pro Arg Leu Asp Asp Asp Glu Lys Tyr Cys Phe Asp Phe Asn Gly
65 70 75 80
Val Ser Asn Lys Gly Glu Lys Tyr Ile Thr Lys Val Thr Leu Asn Val
85 90 95
Val Ala Pro Ser Leu Glu Val Tyr Val Asp His Ala Ser Leu Pro Thr
100 105 110
Leu Gln Gln Leu Met Asp Ile Ile Lys Ser Glu Glu Glu Asn Pro Thr
115 120 125
Ala Gln Arg Tyr Ile Ala Trp Gly Arg Ile Val Pro Thr Asp Glu Gln
130 135 140
Met Lys Glu Leu Asn Ile Thr Ser Phe Ala Leu Ile Asn Asn His Thr
145 150 155 160
Pro Ala Asp Leu Val Gln Glu Ile Val Lys Gln Ala Gln Thr Lys His
165 170 175
Arg Leu Asn Val Lys Leu Ser Ser Asn Thr Ala His Ser Phe Asp Asn
180 185 190
Leu Val Pro Ile Leu Lys Glu Leu Asn Ser Phe Asn Asn Val Thr Val
195 200 205
Thr Asn Ile Asp Leu Tyr Asp Asp Gly Ser Ala Glu Tyr Val Asn Leu
210 215 220
Tyr Asn Trp Arg Asp Thr Leu Asn Lys Thr Asp Asn Leu Lys Ile Gly
225 230 235 240
Lys Asp Tyr Leu Glu Asp Val Ile Asn Gly Ile Asn Glu Asp Thr Ser
245 250 255
Asn Thr Gly Thr Ser Ser Val Tyr Asn Trp Gln Lys Leu Tyr Pro Ala
260 265 270
Asn Tyr His Phe Leu Arg Lys Asp Tyr Leu Thr Leu Glu Pro Ser Leu
275 280 285
His Glu Leu Arg Asp Tyr Ile Gly Asp Ser Leu Lys Gln Met Gln Trp
290 295 300
Asp Gly Phe Lys Lys Phe Asn Ser Lys Gln Gln Glu Leu Phe Leu Ser
305 310 315 320
Ile Val Asn Phe Asp Lys Gln Lys Leu Gln Asn Glu Tyr Asn Ser Ser
325 330 335
Asn Leu Pro Asn Phe Val Phe Thr Gly Thr Thr Val Trp Ala Gly Asn
340 345 350
His Glu Arg Glu Tyr Tyr Ala Lys Gln Gln Ile Asn Val Ile Asn Asn
355 360 365
Ala Ile Asn Glu Ser Ser Pro His Tyr Leu Gly Asn Ser Tyr Asp Leu
370 375 380
Phe Phe Lys Gly His Pro Gly Gly Gly Ile Ile Asn Thr Leu Ile Met
385 390 395 400
Gln Asn Tyr Pro Ser Met Val Asp Ile Pro Ser Lys Ile Ser Phe Glu
405 410 415
Val Leu Met Met Thr Asp Met Leu Pro Asp Ala Val Ala Gly Ile Ala
420 425 430
Ser Ser Leu Tyr Phe Thr Ile Pro Ala Glu Lys Ile Lys Phe Ile Val
435 440 445
Phe Thr Ser Thr Glu Thr Ile Thr Asp Arg Glu Thr Ala Leu Arg Ser
450 455 460
Pro Leu Val Gln Val Met Ile Lys Leu Gly Ile Val Lys Glu Glu Asn
465 470 475 480
Val Leu Phe Trp Ala Asp Leu Pro Asn Cys Glu Thr Gly Val Cys Ile
485 490 495
Ala Val
<210> 63
<211> 482
<212> ПРТ
<213> Photobacterium leiognathi
<400> 63
Met Asn Asp Asn Gln Asn Thr Val Asp Val Val Val Ser Thr Val Asn
1 5 10 15
Asp Asn Val Ile Glu Asn Asn Thr Tyr Gln Val Lys Pro Ile Asp Thr
20 25 30
Pro Thr Thr Phe Asp Ser Tyr Ser Trp Ile Gln Thr Cys Gly Thr Pro
35 40 45
Ile Leu Lys Asp Asp Glu Lys Tyr Ser Leu Ser Phe Asp Phe Val Ala
50 55 60
Pro Glu Leu Asp Gln Asp Glu Lys Phe Cys Phe Glu Phe Thr Gly Asp
65 70 75 80
Val Asp Gly Lys Arg Tyr Val Thr Gln Thr Asn Leu Thr Val Val Ala
85 90 95
Pro Thr Leu Glu Val Tyr Val Asp His Ala Ser Leu Pro Ser Leu Gln
100 105 110
Gln Leu Met Lys Ile Ile Gln Gln Lys Asn Glu Tyr Ser Gln Asn Glu
115 120 125
Arg Phe Ile Ser Trp Gly Arg Ile Arg Leu Thr Glu Asp Asn Ala Glu
130 135 140
Lys Leu Asn Ala His Ile Tyr Pro Leu Ala Gly Asn Asn Thr Ser Gln
145 150 155 160
Glu Leu Val Asp Ala Val Ile Asp Tyr Ala Asp Ser Lys Asn Arg Leu
165 170 175
Asn Leu Glu Leu Asn Thr Asn Thr Gly His Ser Phe Arg Asn Ile Ala
180 185 190
Pro Ile Leu Arg Ala Thr Ser Ser Lys Asn Asn Ile Leu Ile Ser Asn
195 200 205
Ile Asn Leu Tyr Asp Asp Gly Ser Ala Glu Tyr Val Ser Leu Tyr Asn
210 215 220
Trp Lys Asp Thr Asp Asn Lys Ser Gln Lys Leu Ser Asp Ser Phe Leu
225 230 235 240
Val Leu Lys Asp Tyr Leu Asn Gly Ile Ser Ser Glu Lys Pro Asn Gly
245 250 255
Ile Tyr Ser Ile Tyr Asn Trp His Gln Leu Tyr His Ser Ser Tyr Tyr
260 265 270
Phe Leu Arg Lys Asp Tyr Leu Thr Val Glu Thr Lys Leu His Asp Leu
275 280 285
Arg Glu Tyr Leu Gly Gly Ser Leu Lys Gln Met Ser Trp Asp Thr Phe
290 295 300
Ser Gln Leu Ser Lys Gly Asp Lys Glu Leu Phe Leu Asn Ile Val Gly
305 310 315 320
Phe Asp Gln Glu Lys Leu Gln Gln Glu Tyr Gln Gln Ser Glu Leu Pro
325 330 335
Asn Phe Val Phe Thr Gly Thr Thr Thr Trp Ala Gly Gly Glu Thr Lys
340 345 350
Glu Tyr Tyr Ala Gln Gln Gln Val Asn Val Val Asn Asn Ala Ile Asn
355 360 365
Glu Thr Ser Pro Tyr Tyr Leu Gly Arg Glu His Asp Leu Phe Phe Lys
370 375 380
Gly His Pro Arg Gly Gly Ile Ile Asn Asp Ile Ile Leu Gly Ser Phe
385 390 395 400
Asn Asn Met Ile Asp Ile Pro Ala Lys Val Ser Phe Glu Val Leu Met
405 410 415
Met Thr Gly Met Leu Pro Asp Thr Val Gly Gly Ile Ala Ser Ser Leu
420 425 430
Tyr Phe Ser Ile Pro Ala Glu Lys Val Ser Phe Ile Val Phe Thr Ser
435 440 445
Ser Asp Thr Ile Thr Asp Arg Glu Asp Ala Leu Lys Ser Pro Leu Val
450 455 460
Gln Val Met Met Thr Leu Gly Ile Val Lys Glu Lys Asp Val Leu Phe
465 470 475 480
Trp Cys
<210> 64
<211> 675
<212> ПРТ
<213> Photobacterium damsela
<400> 64
Met Lys Lys Ile Leu Thr Val Leu Ser Ile Phe Ile Leu Ser Ala Cys
1 5 10 15
Asn Ser Asp Asn Thr Ser Leu Lys Glu Thr Val Ser Ser Asn Ser Ala
20 25 30
Asp Val Val Glu Thr Glu Thr Tyr Gln Leu Thr Pro Ile Asp Ala Pro
35 40 45
Ser Ser Phe Leu Ser His Ser Trp Glu Gln Thr Cys Gly Thr Pro Ile
50 55 60
Leu Asn Glu Ser Asp Lys Gln Ala Ile Ser Phe Asp Phe Val Ala Pro
65 70 75 80
Glu Leu Lys Gln Asp Glu Lys Tyr Cys Phe Thr Phe Lys Gly Ile Thr
85 90 95
Gly Asp His Arg Tyr Ile Thr Asn Thr Thr Leu Thr Val Val Ala Pro
100 105 110
Thr Leu Glu Val Tyr Ile Asp His Ala Ser Leu Pro Ser Leu Gln Gln
115 120 125
Leu Ile His Ile Ile Gln Ala Lys Asp Glu Tyr Pro Ser Asn Gln Arg
130 135 140
Phe Val Ser Trp Lys Arg Val Thr Val Asp Ala Asp Asn Ala Asn Lys
145 150 155 160
Leu Asn Ile His Thr Tyr Pro Leu Lys Gly Asn Asn Thr Ser Pro Glu
165 170 175
Met Val Ala Ala Ile Asp Glu Tyr Ala Gln Ser Lys Asn Arg Leu Asn
180 185 190
Ile Glu Phe Tyr Thr Asn Thr Ala His Val Phe Asn Asn Leu Pro Pro
195 200 205
Ile Ile Gln Pro Leu Tyr Asn Asn Glu Lys Val Lys Ile Ser His Ile
210 215 220
Ser Leu Tyr Asp Asp Gly Ser Ser Glu Tyr Val Ser Leu Tyr Gln Trp
225 230 235 240
Lys Asp Thr Pro Asn Lys Ile Glu Thr Leu Glu Gly Glu Val Ser Leu
245 250 255
Leu Ala Asn Tyr Leu Ala Gly Thr Ser Pro Asp Ala Pro Lys Gly Met
260 265 270
Gly Asn Arg Tyr Asn Trp His Lys Leu Tyr Asp Thr Asp Tyr Tyr Phe
275 280 285
Leu Arg Glu Asp Tyr Leu Asp Val Glu Ala Asn Leu His Asp Leu Arg
290 295 300
Asp Tyr Leu Gly Ser Ser Ala Lys Gln Met Pro Trp Asp Glu Phe Ala
305 310 315 320
Lys Leu Ser Asp Ser Gln Gln Thr Leu Phe Leu Asp Ile Val Gly Phe
325 330 335
Asp Lys Glu Gln Leu Gln Gln Gln Tyr Ser Gln Ser Pro Leu Pro Asn
340 345 350
Phe Ile Phe Thr Gly Thr Thr Thr Trp Ala Gly Gly Glu Thr Lys Glu
355 360 365
Tyr Tyr Ala Gln Gln Gln Val Asn Val Ile Asn Asn Ala Ile Asn Glu
370 375 380
Thr Ser Pro Tyr Tyr Leu Gly Lys Asp Tyr Asp Leu Phe Phe Lys Gly
385 390 395 400
His Pro Ala Gly Gly Val Ile Asn Asp Ile Ile Leu Gly Ser Phe Pro
405 410 415
Asp Met Ile Asn Ile Pro Ala Lys Ile Ser Phe Glu Val Leu Met Met
420 425 430
Thr Asp Met Leu Pro Asp Thr Val Ala Gly Ile Ala Ser Ser Leu Tyr
435 440 445
Phe Thr Ile Pro Ala Asp Lys Val Asn Phe Ile Val Phe Thr Ser Ser
450 455 460
Asp Thr Ile Thr Asp Arg Glu Glu Ala Leu Lys Ser Pro Leu Val Gln
465 470 475 480
Val Met Leu Thr Leu Gly Ile Val Lys Glu Lys Asp Val Leu Phe Trp
485 490 495
Ala Asp His Lys Val Asn Ser Met Glu Val Ala Ile Asp Glu Ala Cys
500 505 510
Thr Arg Ile Ile Ala Lys Arg Gln Pro Thr Ala Ser Asp Leu Arg Leu
515 520 525
Val Ile Ala Ile Ile Lys Thr Ile Thr Asp Leu Glu Arg Ile Gly Asp
530 535 540
Val Ala Glu Ser Ile Ala Lys Val Ala Leu Glu Ser Phe Ser Asn Lys
545 550 555 560
Gln Tyr Asn Leu Leu Val Ser Leu Glu Ser Leu Gly Gln His Thr Val
565 570 575
Arg Met Leu His Glu Val Leu Asp Ala Phe Ala Arg Met Asp Val Lys
580 585 590
Ala Ala Ile Glu Val Tyr Gln Glu Asp Asp Arg Ile Asp Gln Glu Tyr
595 600 605
Glu Ser Ile Val Arg Gln Leu Met Ala His Met Met Glu Asp Pro Ser
610 615 620
Ser Ile Pro Asn Val Met Lys Val Met Trp Ala Ala Arg Ser Ile Glu
625 630 635 640
Arg Val Gly Asp Arg Cys Gln Asn Ile Cys Glu Tyr Ile Ile Tyr Phe
645 650 655
Val Lys Gly Lys Asp Val Arg His Thr Lys Pro Asp Asp Phe Gly Thr
660 665 670
Met Leu Asp
675
<210> 65
<211> 510
<212> ПРТ
<213> Photobacterium damsela
<400> 65
Met Lys Lys Ile Leu Thr Val Leu Ser Ile Phe Ile Leu Ser Ala Cys
1 5 10 15
Asn Ser Asp Asn Thr Ser Leu Lys Glu Thr Val Ser Ser Asn Ser Ala
20 25 30
Asp Val Val Glu Thr Glu Thr Tyr Gln Leu Thr Pro Ile Asp Ala Pro
35 40 45
Ser Ser Phe Leu Ser His Ser Trp Glu Gln Thr Cys Gly Thr Pro Ile
50 55 60
Leu Asn Glu Ser Asp Lys Gln Ala Ile Ser Phe Asp Phe Val Ala Pro
65 70 75 80
Glu Leu Lys Gln Asp Glu Lys Tyr Cys Phe Thr Phe Lys Gly Ile Thr
85 90 95
Gly Asp His Arg Tyr Ile Thr Asn Thr Thr Leu Thr Val Val Ala Pro
100 105 110
Thr Leu Glu Val Tyr Ile Asp His Ala Ser Leu Pro Ser Leu Gln Gln
115 120 125
Leu Ile His Ile Ile Gln Ala Lys Asp Glu Tyr Pro Ser Asn Gln Arg
130 135 140
Phe Val Ser Trp Lys Arg Val Thr Val Asp Ala Asp Asn Ala Asn Lys
145 150 155 160
Leu Asn Ile His Thr Tyr Pro Leu Lys Gly Asn Asn Thr Ser Pro Glu
165 170 175
Met Val Ala Ala Ile Asp Glu Tyr Ala Gln Ser Lys Asn Arg Leu Asn
180 185 190
Ile Glu Phe Tyr Thr Asn Thr Ala His Val Phe Asn Asn Leu Pro Pro
195 200 205
Ile Ile Gln Pro Leu Tyr Asn Asn Glu Lys Val Lys Ile Ser His Ile
210 215 220
Ser Leu Tyr Asp Asp Gly Ser Ser Glu Tyr Val Ser Leu Tyr Gln Trp
225 230 235 240
Lys Asp Thr Pro Asn Lys Ile Glu Thr Leu Glu Gly Glu Val Ser Leu
245 250 255
Leu Ala Asn Tyr Leu Ala Gly Thr Ser Pro Asp Ala Pro Lys Gly Met
260 265 270
Gly Asn Arg Tyr Asn Trp His Lys Leu Tyr Asp Thr Asp Tyr Tyr Phe
275 280 285
Leu Arg Glu Asp Tyr Leu Asp Val Glu Ala Asn Leu His Asp Leu Arg
290 295 300
Asp Tyr Leu Gly Ser Ser Ala Lys Gln Met Pro Trp Asp Glu Phe Ala
305 310 315 320
Lys Leu Ser Asp Ser Gln Gln Thr Leu Phe Leu Asp Ile Val Gly Phe
325 330 335
Asp Lys Glu Gln Leu Gln Gln Gln Tyr Ser Gln Ser Pro Leu Pro Asn
340 345 350
Phe Ile Phe Thr Gly Thr Thr Thr Trp Ala Gly Gly Glu Thr Lys Glu
355 360 365
Tyr Tyr Ala Gln Gln Gln Val Asn Val Ile Asn Asn Ala Ile Asn Glu
370 375 380
Thr Ser Pro Tyr Tyr Leu Gly Lys Asp Tyr Asp Leu Phe Phe Lys Gly
385 390 395 400
His Pro Ala Gly Gly Val Ile Asn Asp Ile Ile Leu Gly Ser Phe Pro
405 410 415
Asp Met Ile Asn Ile Pro Ala Lys Ile Ser Phe Glu Val Leu Met Met
420 425 430
Thr Asp Met Leu Pro Asp Thr Val Ala Gly Ile Ala Ser Ser Leu Tyr
435 440 445
Phe Thr Ile Pro Ala Asp Lys Val Asn Phe Ile Val Phe Thr Ser Ser
450 455 460
Asp Thr Ile Thr Asp Arg Glu Glu Ala Leu Lys Ser Pro Leu Val Gln
465 470 475 480
Val Met Leu Thr Leu Gly Ile Val Lys Glu Lys Asp Val Leu Phe Trp
485 490 495
Ala Asp Leu Pro Asp Cys Ser Ser Gly Val Cys Ile Asp Lys
500 505 510
<210> 66
<211> 422
<212> ПРТ
<213> Heliobacter acinonychis
<400> 66
Met Gly Thr Ile Lys Lys Pro Leu Ile Ile Ala Gly Asn Gly Pro Ser
1 5 10 15
Ile Lys Asp Leu Asp Tyr Ala Leu Phe Pro Lys Asp Phe Asp Val Phe
20 25 30
Arg Cys Asn Gln Phe Tyr Phe Glu Asp Lys Tyr Tyr Leu Gly Arg Glu
35 40 45
Ile Lys Gly Val Phe Phe Asn Pro Cys Val Leu Ser Ser Gln Met Gln
50 55 60
Thr Val Gln Tyr Leu Met Asp Asn Gly Glu Tyr Ser Ile Glu Arg Phe
65 70 75 80
Phe Cys Ser Val Ser Thr Asp Arg His Asp Phe Asp Gly Asp Tyr Gln
85 90 95
Thr Ile Leu Pro Val Asp Gly Tyr Leu Lys Ala His Tyr Pro Phe Val
100 105 110
Cys Asp Thr Phe Ser Leu Phe Lys Gly His Glu Glu Ile Leu Lys His
115 120 125
Val Lys Tyr His Leu Lys Thr Tyr Ser Lys Glu Leu Ser Ala Gly Val
130 135 140
Leu Met Leu Leu Ser Ala Val Val Leu Gly Tyr Lys Glu Ile Tyr Leu
145 150 155 160
Val Gly Ile Asp Phe Gly Ala Ser Ser Trp Gly His Phe Tyr Asp Glu
165 170 175
Ser Gln Ser Gln His Phe Ser Asn His Met Ala Asp Cys His Asn Ile
180 185 190
Tyr Tyr Asp Met Leu Thr Ile Cys Leu Cys Gln Lys Tyr Ala Lys Leu
195 200 205
Tyr Ala Leu Ala Pro Asn Ser Pro Leu Ser His Leu Leu Thr Leu Asn
210 215 220
Pro Gln Ala Lys Tyr Pro Phe Glu Leu Leu Asp Lys Pro Ile Gly Tyr
225 230 235 240
Thr Ser Asp Leu Ile Ile Ser Ser Pro Leu Glu Glu Lys Leu Leu Glu
245 250 255
Phe Lys Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu
260 265 270
Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys
275 280 285
Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu Lys Leu
290 295 300
Leu Glu Phe Lys Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys Asn Ile
305 310 315 320
Glu Glu Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu Lys Leu Leu Glu
325 330 335
Phe Lys Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu
340 345 350
Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys
355 360 365
Asn Ile Glu Glu Lys Leu Leu Glu Phe Lys Asn Ile Glu Glu Lys Leu
370 375 380
Leu Ala Ser Arg Leu Asn Asn Ile Leu Arg Lys Ile Lys Arg Lys Ile
385 390 395 400
Leu Pro Phe Phe Trp Gly Gly Gly Val Thr Pro Thr Leu Lys Val Ser
405 410 415
Phe Arg Trp Gly Ala Ala
420
<210> 67
<211> 2851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Фрагмент экспрессии < Ptet-lacY-FRT-add1-FRT>
<400> 67
tggccagatg attaattcct aatttttgtt gacactctat cattgataga gttattttac 60
cactccctat cagtgataga gaaaagtgaa atgaatagtt cgacaaaaat ctagaaataa 120
ttttgtttaa ctttaagaag gagatataca aatgtactat ttaaaaaaca caaacttttg 180
gatgttcggt ttattctttt tcttttactt ttttatcatg ggagcctact tcccgttttt 240
cccgatttgg ctacatgaca tcaaccatat cagcaaaagt gatacgggta ttatttttgc 300
cgctatttct ctgttctcgc tattattcca accgctgttt ggtctgcttt ctgacaaact 360
cgggctgcgc aaatacctgc tgtggattat taccggcatg ttagtgatgt ttgcgccgtt 420
ctttattttt atcttcgggc cactgttaca atacaacatt ttagtaggat cgattgttgg 480
tggtatttat ctaggctttt gttttaacgc cggtgcgcca gcagtagagg catttattga 540
gaaagtcagc cgtcgcagta atttcgaatt tggtcgcgcg cggatgtttg gctgtgttgg 600
ctgggcgctg tgtgcctcga ttgtcggcat catgttcacc atcaataatc agtttgtttt 660
ctggctgggc tctggctgtg cactcatcct cgccgtttta ctctttttcg ccaaaacgga 720
tgcgccctct tctgccacgg ttgccaatgc ggtaggtgcc aaccattcgg catttagcct 780
taagctggca ctggaactgt tcagacagcc aaaactgtgg tttttgtcac tgtatgttat 840
tggcgtttcc tgcacctacg atgtttttga ccaacagttt gctaatttct ttacttcgtt 900
ctttgctacc ggtgaacagg gtacgcgggt atttggctac gtaacgacaa tgggcgaatt 960
acttaacgcc tcgattatgt tctttgcgcc actgatcatt aatcgcatcg gtgggaaaaa 1020
cgccctgctg ctggctggca ctattatgtc tgtacgtatt attggctcat cgttcgccac 1080
ctcagcgctg gaagtggtta ttctgaaaac gctgcatatg tttgaagtac cgttcctgct 1140
ggtgggctgc tttaaatata ttaccagcca gtttgaagtg cgtttttcag cgacgattta 1200
tctggtctgt ttctgcttct ttaagcaact ggcgatgatt tttatgtctg tactggcggg 1260
caatatgtat gaaagcatcg gtttccaggg cgcttatctg gtgctgggtc tggtggcgct 1320
gggcttcacc ttaatttccg tgttcacgct tagcggcccc ggcccgcttt ccctgctgcg 1380
tcgtcaggtg aatgaagtcg ctgggagcta agcggccgcg tcgacacgca aaaaggccat 1440
ccgtcaggat ggccttctgc ttaatttgat gcctggcagt ttatggcggg cgtcctgccc 1500
gccaccctcc gggccgttgc ttcgcaacgt tcaaatccgc tcccggcgga tttgtcctac 1560
tcaggagagc gttcaccgac aaacaacaga taaaacgaaa ggcccagtct ttcgactgag 1620
cctttcgttt tatttgatgc ctggcagttc cctactctcg catggggaga ccccacacta 1680
ccatcatgta tgaatatcct ccttagttcc tattccgaag ttcctattct ctagaaagta 1740
taggaacttc ggcgcgtcct acctgtgaca cgcgtgccgc agtctcacgc ccggagcgta 1800
gcgaccgagt gagctagcta tttgtttatt tttctaaata cattcaaata tgtatccgct 1860
catgagacaa taaccctgat aaatgcttca ataatattga aaaaggaaga gtatgaggga 1920
agcggtgatc gccgaagtat cgactcaact atcagaggta gttggcgtca tcgagcgcca 1980
tctcgaaccg acgttgctgg ccgtacattt gtacggctcc gcagtggatg gcggcctgaa 2040
gccacacagt gatattgatt tgctggttac ggtgaccgta aggcttgatg aaacaacgcg 2100
gcgagctttg atcaacgacc ttttggaaac ttcggcttcc cctggagaga gcgagattct 2160
ccgcgctgta gaagtcacca ttgttgtgca cgacgacatc attccgtggc gttatccagc 2220
taagcgcgaa ctgcaatttg gagaatggca gcgcaatgac attcttgcag gtatcttcga 2280
gccagccacg atcgacattg atctggctat cttgctgaca aaagcaagag aacatagcgt 2340
tgccttggta ggtccagcgg cggaggaact ctttgatccg gttcctgaac aggatctatt 2400
tgaggcgcta aatgaaacct taacgctatg gaactcgccg cccgactggg ctggcgatga 2460
gcgaaatgta gtgcttacgt tgtcccgcat ttggtacagc gcagtaaccg gcaaaatcgc 2520
gccgaaggat gtcgctgccg actgggcaat ggagcgcctg ccggcccagt atcagcccgt 2580
catacttgaa gctagacagg cttatcttgg acaagaagaa gatcgcttgg cctcgcgcgc 2640
agatcagttg gaagaatttg tccactacgt gaaaggcgag atcaccaagg tagtcggcaa 2700
ataatgtcta acaattcgtt caagccgagg ggccgcaaga tccggccacg atgacccggt 2760
cgtcgggtac cggcagggcg gggcgtaagg cgcgccattt aaatgaagtt cctattccga 2820
agttcctatt ctctagaaag tataggaact t 2851
<210> 68
<211> 6521
<212> ДНК
<213> Искусственная ПОследовательность
<220>
<223> Кассета транспонзонов <Ptet-glmUM-PT5-glmS-FRT-dhfr-FRT>
<400> 68
acaggttggc tgataagtcc ccggtctagc ttgcatgcag attgcagcat tacacgtctt 60
gagcgattgt gtaggctgga gctgcttcga aattaatacg actcactata ggggaattga 120
ttctggtacc aaatgagtcg accggccaga tgattaattc ctaatttttg ttgacactct 180
atcattgata gagttatttt accactccct atcagtgata gagaaaagtg aaatgaatag 240
ttcgacaaaa atctagaaat aattttgttt aactttaaga aggagatata caaatgctga 300
acaacgcgat gtctgttgtt atcctggcgg cgggtaaagg tacccgtatg tactctgacc 360
tgccgaaagt tctgcacacc ctggcgggta aagcgatggt tcagcacgtt atcgacgcgg 420
cgaacgaact gggtgcggcg cacgttcacc tggtttacgg tcacggtggt gacctgctga 480
aacaggcgct gaaagacgac aacctgaact gggttctgca ggcggaacag ctgggtaccg 540
gtcacgcgat gcagcaggcg gcgccgttct tcgcggacga cgaagacatc ctgatgctgt 600
acggtgacgt tccgctgatc tctgttgaaa ccctgcagcg tctgcgtgac gcgaaaccgc 660
agggtggtat cggtctgctg accgttaaac tggacgaccc gaccggttac ggtcgtatca 720
cccgtgaaaa cggtaaagta accggtatcg ttgaacacaa agacgcgacc gacgaacagc 780
gtcagatcca ggagatcaac accggtatcc tgatcgcgaa cggtgcagac atgaaacgtt 840
ggctggcgaa actgaccaac aacaacgcgc agggtgaata ctacatcacc gacatcatcg 900
cgctggcgta ccaggaaggt cgtgaaatcg ttgcggttca cccgcagcgt ctgtctgaag 960
ttgaaggtgt taacaaccgt ctgcagctgt ctcgtctgga acgtgtttac cagtctgaac 1020
aggcggaaaa actgctgctg gcgggtgtta tgctgcgtga cccggcgcgt ttcgacctgc 1080
gtggtaccct gacccacggt cgtgacgttg aaatcgacac caacgttatc atcgaaggta 1140
acgttaccct gggtcaccgt gtaaaaatcg gcaccggttg cgttatcaaa aactctgtta 1200
tcggtgacga ctgcgaaatc tctccgtaca ccgttgttga agacgcgaac ctggcggcgg 1260
cgtgcaccat cggtccgttc gcgcgtctgc gtccgggtgc ggaactgctg gaaggtgcgc 1320
acgttggtaa cttcgttgaa atgaaaaaag cgcgtctggg taaaggttct aaagcgggtc 1380
acctgaccta cctgggtgac gcggaaatcg gtgacaacgt taacatcggt gcgggtacca 1440
tcacctgcaa ctacgacggt gcgaacaaat tcaaaaccat catcggtgac gacgttttcg 1500
ttggttctga cacccagctg gttgcgccgg ttaccgttgg taaaggtgcg accatcgcgg 1560
cgggtaccac cgttacccgt aacgttggtg aaaacgcgct ggcgatctct cgtgttccgc 1620
agacccagaa agaaggttgg cgtcgtccgg ttaaaaaaaa ataacgaagg agatagaacc 1680
atgtccaacc gtaaatactt cggtacggac ggtatccgtg gtcgtgtagg tgatgctccg 1740
attacgccgg atttcgtcct gaaactcggt tgggcagcgg gtaaagttct cgcacgtcac 1800
ggctctcgta aaatcatcat cggtaaagac acccgtatct ctggttacat gctcgaatct 1860
gcactggaag cgggtctggc tgcagctggt ctgtctgcac tgttcacggg tccgatgcca 1920
accccagctg tagcgtacct gactcgcact ttccgtgcag aagcaggtat cgtgatctct 1980
gcctctcaca acccgttcta cgacaacggt atcaaattct tcagcatcga tggtaccaaa 2040
ctcccagacg cggttgaaga ggctatcgaa gcggaaatgg agaaagaaat ctcttgtgta 2100
gactctgccg aactcggtaa agcgtctcgt atcgttgatg cagcgggtcg ttacatcgag 2160
ttctgcaaag ccacctttcc gaacgaactg agcctgtctg agctgaaaat cgtcgtagac 2220
tgtgccaacg gtgcgactta ccacattgcc ccaaacgtac tgcgtgagct gggtgctaac 2280
gtcatcgcga tcggttgtga accgaacggt gtcaacatca acgcggaagt aggtgcgacc 2340
gatgttcgtg cactgcaggc tcgtgtactc gcggagaaag cggatctcgg tatcgccttt 2400
gacggtgatg gtgaccgtgt tatcatggtt gaccacgaag gtaacaaagt ggatggtgac 2460
cagatcatgt acatcattgc ccgtgaaggt ctgcgtcagg gtcagctgcg tggtggtgca 2520
gtaggtaccc tcatgagcaa catgggtctg gaactggccc tgaaacagct gggtatccca 2580
ttcgctcgtg ctaaagtagg cgaccgttac gttctggaga aaatgcagga gaaaggttgg 2640
cgtatcggtg ccgaaaactc tggtcacgtc atcctgctgg acaaaaccac taccggtgac 2700
ggtatcgtag caggtctgca ggtactcgcc gctatggccc gtaaccacat gtccctccat 2760
gacctctgct ctggtatgaa aatgttcccg cagatcctgg ttaacgttcg ttacaccgca 2820
ggttctggtg atccgctgga acacgagtct gtgaaagccg ttaccgcaga agtggaagcg 2880
gccctgggta accgtggtcg tgtactgctg cgtaaatccg gtactgagcc actgatccgt 2940
gttatggttg agggcgaaga tgaagcccag gtcaccgaat ttgcgcaccg tattgccgac 3000
gcagtcaaag cggtttaatt tcgtcgacac acaggaaaca tattaaaaat taaaacctgc 3060
aggagtttaa acgcggccgc gatatcgttg taaaacgacg gccagtgcaa gaatcataaa 3120
aaatttattt gctttcagga aaatttttct gtataataga ttcataaatt tgagagagga 3180
gtttttgtga gcggataaca attccccatc ttagtatatt agttaagtat aaatacacaa 3240
ggagatatac atatgtgcgg tatcgttggt gctatcgcac agcgtgatgt agcggagatc 3300
ctcctggaag gtctgcgtcg tctcgaatac cgtggttacg actctgccgg tctggcagta 3360
gtggatgcag aaggtcacat gactcgtctg cgtcgtctgg gtaaagtgca gatgctcgcg 3420
caggcggcgg aagaacaccc actccacggt ggtacgggta tcgcacacac tcgttgggca 3480
acccacggtg aaccgtctga ggtcaacgca cacccgcatg ttagcgagca catcgtagtc 3540
gttcacaacg gtatcatcga gaaccacgaa ccactccgtg aggaactcaa agcccgtggt 3600
tacaccttcg taagcgaaac cgacacggaa gttatcgccc acctcgttaa ctgggaactc 3660
aaacagggtg gtactctgcg tgaagcagtt ctgcgtgcca ttccacagct gcgtggtgca 3720
tacggtaccg tgatcatgga ctctcgtcat ccggataccc tgctcgccgc acgttctggt 3780
tctccactcg ttatcggtct gggtatgggt gagaacttca tcgcctctga tcagctggcc 3840
ctgctcccag ttacccgtcg cttcatcttc ctggaagagg gtgacatcgc cgaaatcacc 3900
cgtcgttccg ttaacatctt cgacaaaacg ggtgcggaag ttaaacgtca ggacatcgag 3960
tctaacctgc agtatgacgc tggtgacaaa ggcatctacc gtcactacat gcagaaagag 4020
atctacgaac agccgaacgc gatcaaaaac accctgaccg gtcgtatctc tcacggtcag 4080
gttgacctgt ctgagctggg tccaaacgcg gacgaactcc tgtccaaagt cgagcacatc 4140
cagatcctgg cttgtggtac ctcttacaac tccggtatgg tttctcgtta ctggttcgaa 4200
tctctggcag gtatcccatg cgacgttgaa atcgcctccg aattccgtta tcgtaaatct 4260
gcggtacgtc gtaactccct catgatcacc ctgtctcagt ctggtgaaac cgctgatact 4320
ctggcaggtc tgcgtctcag caaagaactg ggttacctgg gttctctggc catctgcaac 4380
gttccgggtt ctagcctggt tcgtgagtct gacctggctc tgatgaccaa cgcgggtacg 4440
gagatcggtg ttgcctctac caaagcgttc actacccagc tcactgtcct gctgatgctg 4500
gttgccaaac tgtctcgtct caaaggcctc gacgctagca tcgaacacga catcgtacac 4560
ggtctgcagg ccctcccatc tcgtatcgag cagatgctgt ctcaggacaa acgtatcgaa 4620
gcactggcag aagacttcag cgacaaacac cacgcgctgt ttctgggtcg tggtgaccag 4680
tacccaattg cgctggaagg tgccctgaaa ctgaaagaga tcagctacat ccatgcagag 4740
gcatacgcag cgggtgagct gaaacatggt ccactggccc tgatcgacgc agatatgccg 4800
gttattgtgg ttgctccgaa caacgaactg ctggagaaac tgaaatccaa catcgaggaa 4860
gtacgtgcgc gtggtggtca gctgtacgtg tttgctgacc aggacgcggg tttcgtttcc 4920
agcgacaaca tgcacatcat cgaaatgccg catgttgaag aggtaatcgc gccaatcttc 4980
tacaccgtac cgctgcagct gctggcgtac catgtagccc tgatcaaagg tacggacgtt 5040
gaccagccgc gtaacctggc gaaatccgtg accgtggaat aacgcggagg cgcgccattt 5100
aaatcaacct cagcggtcat agctgtttcc tgtgactgag caataactag cataacccct 5160
tggggcctct aaacgggtct tgaggggttt tttgctgaaa ccaatttgcc tggcggcagt 5220
agcgcggtgg tcccacctga ccccatgccg aactcagaag tgaaacgccg tagcgccgat 5280
ggtagtgtgg ggtctcccca tgcgagagta gggaactgcc aggcatcaaa taaaacgaaa 5340
ggctcagtcg aaagactggg cctttcggga tccaggccgg cctgttaacg aattaatctt 5400
ccgcggcggt atcgataagc ttgatatcga attccgaagt tcctattctc tagaaagtat 5460
aggaacttca ggtctgaaga ggagtttacg tccagccaag ctagcttggc tgcaggtcgt 5520
cgaaattcta ccgggtaggg gaggcgcttt tcccaaggca gtctggagca tgcgctttag 5580
cagccccgct gggcacttgg cgctacacaa gtggcctctg gcctcgcaca cattccacat 5640
ccaccggtag gcgccaaccg gctccgttct ttggtggccc cttcgcgcca ccttctactc 5700
ctcccctagt caggaagttc ccccccgccc cgcagctcgc gtcgtgcagg acgtgacaaa 5760
tggaagtagc acgtctcact agtctcgtgc agatggacag caccgctgag caatggaagc 5820
gggtaggcct ttggggcagc ggccaatagc agctttgctc cttcgctttc tgggctcaga 5880
ggctgggaag gggtgggtcc gggggcgggc tcaggggcgg gctcaggggc ggggcgggcg 5940
cccgaaggtc ctccggaggc ccggcattct gcacgcttca aaagcgcacg tctgccgcgc 6000
tgttctcctc ttcctcatct ccgggccttt cgacctgcag cctgttgaca attaatcatc 6060
ggcatagtat atcggcatag tataatacga caaggtgagg aactaaacca tgggtcaaag 6120
tagcgatgaa gccaacgctc ccgttgcagg gcagtttgcg cttcccctga gtgccacctt 6180
tggcttaggg gatcgcgtac gcaagaaatc tggtgccgct tggcagggtc aagtcgtcgg 6240
ttggtattgc acaaaactca ctcctgaagg ctatgcggtc gagtccgaat cccacccagg 6300
ctcagtgcaa atttatcctg tggctgcact tgaacgtgtg gcctaatgag gggatcaatt 6360
ctctagagct cgctgatcag aagttcctat tctctagaaa gtataggaac ttcgatggcg 6420
cctcatccct gaagccaata caacaaaaat taggaattaa tcatctggcc aatttcaggt 6480
ggcacttttc gggcagaccg gggacttatc agccaacctg t 6521
<210> 69
<211> 3919
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Экспрессионная кассета <Ptet-glmSm-gna1-FRT-aacC1-FRT>
<400> 69
ggtacccaaa tatgcataat cgaaattaat acgactcact ataggggaat tgattctggt 60
accaaatgag tcgaccggcc agatgattaa ttcctaattt ttgttgacac tctatcattg 120
atagagttat tttaccactc cctatcagtg atagagaaaa gtgaaatgaa tagttcgaca 180
aaaatctaga aataattttg tttaacttta agaaggagat atacaaatgt gcggtatcgt 240
tggtgctatc gcacagcgtg atgtagcgaa aatcctcctg gaaggtctgc gtcgtctcga 300
ataccgtggt tacgactctg ccggtctggc agtagtggat gcagaaggtc acatgactcg 360
tctgcgtcgt ctgggtaaag tgcagatgct cgcgcaggcg gcggaagaac acccactcca 420
cggtggtacg ggtatcgcac acactcgttg ggcaacccac ggtgaaccgt ctgaggtcaa 480
cgcacacccg catgttagcg agcacatcgt agtcgttcac aacggtatca tcgagaacca 540
cgaaccactc cgtgaggaac tcaaagcccg tggttacacc ttcgtaagcg aaaccgacac 600
ggaagttatc gcccacctcg ttaactggga actcaaacag ggtggtactc tgcgtgaagc 660
agttctgcgt gccattccac agctgcgtgg tgcatacggt accgtgatca tggactctcg 720
tcatccggat accctgctcg ccgcacgttc tggttctcca ctcgttatcg gtctgggtat 780
gggtgagaac ttcatcgcct ctgatcagct ggccctgctc ccagttaccc gtcgcttcat 840
cttcctggaa gagggtgaca tcgccgaaat cacccgtcgt tccgttaaca tcttcgacaa 900
aacgggtgcg gaagttaaac gtcaggacat cgagtctaac ctgcagtatg acgctggtga 960
caaaggcatc taccgtcact acatgcagaa agagatctac gaacagccga acgcgatcaa 1020
aaacaccctg accggtcgta tctctcacgg tcaggttgac ctgtctgagc tgggtccaaa 1080
cgcggacgaa ctcctgtcca aagtcgagca catccagatc ctggcttgtg gtacctctta 1140
caactccggt atggtttctc gttactggtt cgaatctctg gcaggtatcc catgcgacgt 1200
tgaaatcgcc tccgaattcc gttatcgtaa atctgcggta cgtcgtaact ccctcatgat 1260
caccctgtct cagtctggtg aaaccgctga tactctggca ggtctgcgtc tcagcaaaga 1320
actgggttac ctgggttctc tggccatctg caacgttccg ggttctagcc tggttcgtga 1380
gtctgtgctg gctctgatga ccaacgcggg tacggagatc ggtgttgcct ctaccaaagc 1440
gttcactacc cagctcactg tcctgctgat gctggttgcc aaactgtctc gtctcaaagg 1500
cctcgacgct agcatcgaac acgacatcgt acacggtctg caggccctcc catctcgtat 1560
cgagcagatg ctgccgcagg acaaacgtat cgaagcactg gcagaagact tcagcgacaa 1620
acaccacgcg ctgtttctgg gtcgtggtga ccagtaccca attgcgctgg aaggtgccct 1680
gaaactgaaa gagatcagct acatccatgc agaggcatac gcagcgggtg agctgaaaca 1740
tggtccactg gccctgatcg acgcagatat gccggttatt gtggttgctc cgaacaacgg 1800
cctgctggag aaactgaaat ccaacatcga ggaagtacgt gcgcgtggtg gtcagctgta 1860
cgtgtttgct gaccaggacg cgggtttcgt ttccagcgac aacatgcaca tcatcgaaat 1920
gccgcatgtt gaagaggtaa tcgcgccaat cttctacacc gtaccgctgc agctgctggc 1980
gtaccatgta gccctgatca aaggtacgga cgttgaccag ccgcgtaacc tggcgaaatc 2040
cgtgaccgtg gaataacgaa ggagatagaa ccatgagctt acccgatgga ttttatataa 2100
ggcgaatgga agagggggat ttggaacagg tcactgagac gctaaaggtt ttgaccaccg 2160
tgggcactat tacccccgaa tccttcagca aactcataaa atactggaat gaagccacag 2220
tatggaatga taacgaagat aaaaaaataa tgcaatataa ccccatggtg attgtggaca 2280
agcgcaccga gacggttgcc gctacgggga atatcatcat cgaaagaaag atcattcatg 2340
aactggggct atgtggccac atcgaggaca ttgcagtaaa ctccaagtat cagggccaag 2400
gtttgggcaa gctcttgatt gatcaattgg taactatcgg ctttgactac ggttgttata 2460
agattatttt agattgcgat gagaaaaatg tcaaattcta tgaaaaatgt gggtttagca 2520
acgcaggcgt ggaaatgcaa attagaaaat agaataacta gcataacccc ttggggcctc 2580
taaacgggtc ttgaggggtt ttttgctgaa accaatttgc ctggcggcag tagcgcggtg 2640
gtcccacctg accccatgcc gaactcagaa gtgaaacgcc gtagcgccga tggtagtgtg 2700
gggtctcccc atgcgagagt agggaactgc caggcatcaa ataaaacgaa aggctcagtc 2760
gaaagactgg gcctttcggg atccaggccg gcctgttaac gaattaatct tccgcggcgg 2820
tatcgataag cttgatatcg aattccgaag ttcctattct ctagaaagta taggaacttc 2880
aggtctgaag aggagtttac gtccagccaa gctagcttgg ctgcaggtcg tcgaaattct 2940
acgatctcgg cttgaacgaa ttgttaggtg gcggtacttg ggtcgatatc aaagtgcatc 3000
acttcttccc gtatgcccaa ctttgtatag agagccactg cgggatcgtc accgtaatct 3060
gcttgcacgt agatcacata agcaccaagc gcgttggcct catgcttgag gagattgatg 3120
agcgcggtgg caatgccctg cctccggtgc tcgccggaga ctgcgagatc atagatatag 3180
atctcactac gcggctgctc aaacctgggc agaacgtaag ccgcgagagc gccaacaacc 3240
gcttcttggt cgaaggcagc aagcgcgatg aatgtcttac tacggagcaa gttcccgagg 3300
taatcggagt ccggctgatg ttgggagtag gtggctacgt ctccgaactc acgaccgaaa 3360
agatcaagag cagcccgcat ggatttgact tggtcagggc cgagcctaca tgtgcgaatg 3420
atgcccatac ttgagccacc taactttgtt ttagggcgac tgccctgctg cgtaacatcg 3480
ttgctgctgc gtaacatcgt tgctgctcca taacatcaaa catcgaccca cggcgtaacg 3540
cgcttgctgc ttggatgccc gaggcataga ctgtacaaaa aaacagtcat aacaagccat 3600
gaaaaccgcc actgcgccgt taccaccgct gcgttcggtc aaggttctgg accagttgcg 3660
tgagcgcata cgctacttgc attacagttt acgaaccgaa caggcttatg tcaactgggt 3720
tcgtgccttc atccgtttcc acggtgtgcg ctgcacttga acgtgtggcc taatgagggg 3780
atcaattctc tagagctcgc tgatcagaag ttcctattct ctagaaagta taggaacttc 3840
gatggcgcct catccctgaa gccaataggg ataacagggt aatgatcgga tcccgggccc 3900
gtcgactgca gaggcctgc 3919
<210> 70
<211> 2850
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Экспрессионная кассета <Ptet-slr1975-FRT-cat-FRT>
<400> 70
atgcataatc gaaattaata cgactcacta taggggaatt gattctggta ccaaatgagt 60
cgaccggcca gatgattaat tcctaatttt tgttgacact ctatcattga tagagttatt 120
ttaccactcc ctatcagtga tagagaaaag tgaaatgaat agttcgacaa aaatctagaa 180
ataattttgt ttaactttaa gaaggagata tacaaatgat cgctcaccgt cgtcaggaac 240
tggctcaaca gtattatcag gctctgcacc aagatgtgct gccgttctgg gaaaagtatt 300
cgctggatcg tcaaggcggt ggctatttta cctgcctgga ccgcaagggt caggtttttg 360
atacggacaa gttcatttgg ctgcaaaacc gtcaagtgtg gcaatttgcg gttttctaca 420
atcgcctgga accgaaaccg cagtggctgg aaatcgctcg tcatggtgcg gattttctgg 480
cacgtcacgg tcgtgatcag gacggtaact ggtatttcgc cctggatcag gaaggcaaac 540
cgctgcgcca accgtacaat gtgttttccg actgtttcgc ggcgatggcg tttagccagt 600
atgcactggc ttctggtgct caagaagcga aggccattgc actgcaagcg tataacaatg 660
ttctgcgtcg ccagcataac ccgaaaggtc aatatgaaaa gagttacccg ggtacccgtc 720
cgctgaaatc cctggcagtg ccgatgatcc tggctaatct gacgctggaa atggaatggc 780
tgctgccgcc gaccacggtc gaagaagtgc tggcccagac cgttcgtgaa gtcatgacgg 840
attttctgga cccggaaatt ggcctgatgc gcgaagcagt taccccgacg ggtgaatttg 900
tcgattcatt cgaaggccgc ctgctgaacc cgggtcatgg cattgaagcg atgtggttta 960
tgatggatat tgcccagcgt tcgggtgacc gccagctgca agaacaggct attgcggtgg 1020
ttctgaatac cctggaatat gcatgggatg aagaatttgg tggcatcttt tacttcctgg 1080
accgtcaagg tcacccgccg cagcaactgg aatgggatca gaaactgtgg tgggtccatc 1140
tggaaaccct ggtggccctg gcaaaaggtc accaggcgac gggccaagaa aagtgctggc 1200
agtggtttga acgcgtgcat gattatgcat ggagccactt tgctgacccg gaatatggtg 1260
aatggttcgg ctacctgaac cgtcgcggtg aagtgctgct gaatctgaaa ggtggcaaat 1320
ggaagggctg cttccacgtt ccgcgtgcgc tgtggctgtg tgccgaaacc ctgcaactgc 1380
cggtctctta aaataactag cataacccct tggggcctct aaacgggtct tgaggggttt 1440
tttgctgaaa ccaatttgcc tggcggcagt agcgcggtgg tcccacctga ccccatgccg 1500
aactcagaag tgaaacgccg tagcgccgat ggtagtgtgg ggtctcccca tgcgagagta 1560
gggaactgcc aggcatcaaa taaaacgaaa ggctcagtcg aaagactggg cctttcggga 1620
tccaggccgg cctgttaacg aattaatctt ccgcggcggt atcgataagc ttgatatcga 1680
ggctgacatg ggaattagcc atggtccata tgaatatcct ccttagttcc tattccgaag 1740
ttcctattct ctagaaagta taggaacttc ggcgcgccta cctgtgacgg aagatcactt 1800
cgcagaataa ataaatcctg gtgtccctgt tgataccggg aagccctggg ccaacttttg 1860
gcgaaaatga gacgttgatc ggcacgtaag aggttccaac tttcaccata atgaaataag 1920
atcactaccg ggcgtatttt ttgagttgtc gagattttca ggagctaagg aagctaaaat 1980
ggagaaaaaa atcactggat ataccaccgt tgatatatcc caatggcatc gtaaagaaca 2040
ttttgaggca tttcagtcag ttgctcaatg tacctataac cagaccgttc agctggatat 2100
tacggccttt ttaaagaccg taaagaaaaa taagcacaag ttttatccgg cctttattca 2160
cattcttgcc cgcctgatga atgctcatcc ggaattacgt atggcaatga aagacggtga 2220
gctggtgata tgggatagtg ttcacccttg ttacaccgtt ttccatgagc aaactgaaac 2280
gttttcatcg ctctggagtg aataccacga cgatttccgg cagtttctac acatatattc 2340
gcaagatgtg gcgtgttacg gtgaaaacct ggcctatttc cctaaagggt ttattgagaa 2400
tatgtttttc gtctcagcca atccctgggt gagtttcacc agttttgatt taaacgtggc 2460
caatatggac aacttcttcg cccccgtttt caccatgggc aaatattata cgcaaggcga 2520
caaggtgctg atgccgctgg cgattcaggt tcatcatgcc gtttgtgatg gcttccatgt 2580
cggcagatgc ttaatgaata caacagtact gcgatgagtg gcagggcggg gcgtaaggcg 2640
cgccatttaa atgaagttcc tattccgaag ttcctattct ctagaaagta taggaacttc 2700
gaagcagctc cagcctacac aatcgctcaa gacgtgtaat gctgcaatct gcatgcaagc 2760
ttggcactgg cgatggcgcc tcatccctga agccaatagg gataacaggg taatgatcgg 2820
atcccgggcc cgtcgactgc agaggcctgc 2850
<210> 71
<211> 4360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Экспрессионная кассета <Ptet-neuBC-FRT-kan-FRT>
<400> 71
ggtacccaaa tatgcataat cgaaattaat acgactcact ataggggaat tgattctggt 60
accaaatgag tcgaccggcc agatgattaa ttcctaattt ttgttgacac tctatcattg 120
atagagttat tttaccactc cctatcagtg atagagaaaa gtgaaatgaa tagttcgaca 180
aaaatctaga aataattttg tttaacttta agaaggagat atacaaatga aagaaatcaa 240
aatccagaac atcatcatca gcgaagaaaa agcgccgctg gttgtgccgg aaatcggcat 300
taaccataat ggtagtctgg aactggcaaa aatcatggtg gatgcggcct ttagcgccgg 360
tgcaaaaatc attaaacatc agacccacat tgtggaagat gaaatgtcta aagcagcgaa 420
aaaagttatc ccgggcaacg cgaaaatcag tatctacgaa atcatgcaga aatgcgcgct 480
ggattacaaa gatgaactgg ccctgaaaga atataccgaa aaactgggtc tggtgtacct 540
gtctaccccg tttagtcgtg cgggtgcaaa ccgtctggaa gatatgggtg ttagtgcgtt 600
caaaatcggc agcggtgaat gtaacaatta tccgctgatc aaacatattg ccgcatttaa 660
aaaaccgatg attgttagca ccggcatgaa tagcatcgaa tctattaaac cgacggtgaa 720
aatcctgctg gataacgaaa ttccgtttgt tctgatgcat accacgaatc tgtacccgac 780
cccgcacaac ctggtgcgtc tgaatgccat gctggaactg aaaaaagaat tctcttgcat 840
ggttggtctg agtgatcaca ccacggataa tctggcatgc ctgggtgcag tggttctggg 900
tgcgtgtgtg ctggaacgtc atttcaccga tagcatgcac cgctctggtc cggatattgt 960
ttgtagtatg gatacgaaag cactgaaaga actgatcatt cagagcgaac agatggcgat 1020
cattcgcggc aacaatgaat ctaaaaaagc ggccaaacag gaacaggtga ccatcgattt 1080
tgcattcgcg agtgtggtta gcatcaaaga tatcaaaaaa ggcgaagtgc tgagcatgga 1140
taatatttgg gttaaacgtc cgggtctggg cggtatctct gcagcggaat ttgaaaacat 1200
tctgggcaaa aaagcactgc gcgatattga aaatgatgcg cagctgtctt atgaagattt 1260
cgcctaatcg aaggagatac aaccatgaag aaaattctgt ttatcaccgg cagccgtgca 1320
gattacagta aaatcaaaag cctgatgtac cgcgtgcaga acagctctga atttgaactg 1380
tatattttcg cgaccggcat gcatctgagc aaaaacttcg gttacacggt taaagaactg 1440
tacaaaaacg gtttcaaaaa catctacgaa ttcatcaact acgataaata ctaccagacc 1500
gataaagcgc tggccaccac gatcgatggt ttcagccgtt acgccaatga actgaaaccg 1560
gatctgattg tggttcacgg cgatcgtatt gaaccgctgg cggcggcaat tgtgggtgca 1620
ctgaacaata ttctggttgc gcatatcgaa ggcggtgaaa tttctggtac gatcgatgat 1680
agtctgcgtc acgcaattag caaactggcg catatccacc tggtgaacga tgaatttgcg 1740
aaacgtcgcc tgatgcagct gggcgaagat gaaaaatcta tcttcatcat cggtagtccg 1800
gatctggaac tgctgaacga taataaaatc agcctgtctg aagcgaaaaa atactacgat 1860
atcaactacg aaaactacgc cctgctgatg tttcatccgg ttaccacgga aattaccagt 1920
atcaaaaacc aggccgataa tctggtgaaa gcactgatcc agagcaacaa aaactacatc 1980
gttatctacc cgaacaacga tctgggcttt gaactgattc tgcagtctta tgaagaattc 2040
aaaaacaatc cgcgttttaa actgttcccg agtctgcgct ttgaatactt cattaccctg 2100
ctgaaaaacg ccgattttat tatcggtaat agtagctgca tcctgaaaga agcgctgtat 2160
ctgaaaacgg ccggcattct ggtgggtagc cgtcagaatg gtcgtctggg taacgaaaat 2220
accctgaaag ttaacgccaa ctctgatgaa atcctgaaag caatcaacac gatccacaaa 2280
aaacaggatc tgtttagcgc aaaactggaa attctggatt ctagtaaact gtttttcgaa 2340
tatctgcaga gcggcgattt ctttaaactg tctacccaga aagtgttcaa agatattaaa 2400
taaaataact agcataaccc cttggggcct ctaaacgggt cttgaggggt tttttgctga 2460
aaccaatttg cctggcggca gtagcgcggt ggtcccacct gaccccatgc cgaactcaga 2520
agtgaaacgc cgtagcgccg atggtagtgt ggggtctccc catgcgagag tagggaactg 2580
ccaggcatca aataaaacga aaggctcagt cgaaagactg ggcctttcgg gatccaggcc 2640
ggcctgttaa cgaattaatc ttccgcggcg gtatcgataa gcttgatatc gaggctgaca 2700
tgggaattag ccatggtcca tatgaatatc ctccttagtt cctattccga agttcctatt 2760
ctctagaaag tataggaact tcggcgcgtc ctacctgtga cacgcgtcaa gatcccctca 2820
cgctgccgca agcactcagg gcgcaagggc tgctaaagga agcggaacac gtagaaagcc 2880
agtccgcaga aacggtgctg accccggatg aatgtcagct actgggctat ctggacaagg 2940
gaaaacgcaa gcgcaaagag aaagcaggta gcttgcagtg ggcttacatg gcgatagcta 3000
gactgggcgg ttttatggac agcaagcgaa ccggaattgc cagctggggc gccctctggt 3060
aaggttggga agccctgcaa agtaaactgg atggctttct tgccgccaag gatctgatgg 3120
cgcaggggat caagatctga tcaagagaca ggatgaggat cgtttcgcat gattgaacaa 3180
gatggattgc acgcaggttc tccggccgct tgggtggaga ggctattcgg ctatgactgg 3240
gcacaacaga caatcggctg ctctgatgcc gccgtgttcc ggctgtcagc gcaggggcgc 3300
ccggttcttt ttgtcaagac cgacctgtcc ggtgccctga atgaactgca ggacgaggca 3360
gcgcggctat cgtggctggc cacgacgggc gttccttgcg cagctgtgct cgacgttgtc 3420
actgaagcgg gaagggactg gctgctattg ggcgaagtgc cggggcagga tctcctgtca 3480
tctcaccttg ctcctgccga gaaagtatcc atcatggctg atgcaatgcg gcggctgcat 3540
acgcttgatc cggctacctg cccattcgac caccaagcga aacatcgcat cgagcgagca 3600
cgtactcgga tggaagccgg tcttgtcgat caggatgatc tggacgaaga gcatcagggg 3660
ctcgcgccag ccgaactgtt cgccaggctc aaggcgcgca tgcccgacgg cgaggatctc 3720
gtcgtgaccc atggcgatgc ctgcttgccg aatatcatgg tggaaaatgg ccgcttttct 3780
ggattcatcg actgtggccg gctgggtgtg gcggaccgct atcaggacat agcgttggct 3840
acccgtgata ttgctgaaga gcttggcggc gaatgggctg accgcttcct cgtgctttac 3900
ggtatcgccg ctcccgattc gcagcgcatc gccttctatc gccttcttga cgagttcttc 3960
tgagcgggac tctggggttc gaaatgaccg accaagcgac gcccaacctg ccatcacgag 4020
atttcgattc caccgccgcc ttctatgaaa ggttgggctt cggaatcgtt ttccgggacg 4080
ccggctggat gatcctccag cgcggggatc tcatgctgga gttcttcgcc caccccagct 4140
tcaaaagcgc tctcggtacc ggcagggcgg ggcgtaaggc gcgccattta aatgaagaag 4200
ttcctattcc gaagttccta ttctctagaa agtataggaa cttcgaagca gctccagcct 4260
acacaatcgc tcaagacgtg taatgctgca atctgcatgc aagcttggca ctggcgatgg 4320
cgcctcatcc ctgaagccaa tagggataac agggtaatga 4360
<210> 72
<211> 3981
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Экспрессионная кассета <Ptet-ppsA-FRT-aad1-FRT>
<400> 72
attctggtac caaatgagtc gaccggccag atgattaatt cctaattttt gttgacactc 60
tatcattgat agagttattt taccactccc tatcagtgat agagaaaagt gaaatgaata 120
gttcgacaaa aatctagaaa taattttgtt tggcgtcgag aaggagatag aaccatgtcc 180
aacaatggct cgtcaccgct ggtgctttgg tataaccaac tcggcatgaa tgatgtagac 240
agggttgggg gcaaaaatgc ctccctgggt gaaatgatta ctaacctttc cggaatgggt 300
gtttccgttc cgaatggttt cgccacaacc gccgacgcgt ttaaccagtt tctggaccaa 360
agcggcgtaa accagcgcat ttatgaactg ctggataaaa cggatattga cgatgttact 420
cagcttgcga aagcgggcgc gcaaatccgc cagtggatta tcgacactcc cttccagcct 480
gagctggaaa acgccatcag cgaagcctat gcacagcttt ctgccgatga cgaaaacgcc 540
tcttttgcgg tgcgctcctc cgccaccgca gaagatatgc cggacgcttc ttttgccggt 600
cagcaggaaa ccttcctcaa cgttcagggt tttgacgccg ttctcgtggc agtgaaacat 660
gtatttgctt ctctgtttaa cgatcgcgcc atctcttatc gtgtgcacca gggttacgat 720
caccgtggtg tggcgctctc cgccggtgtt caacggatgg tgcgctctga cctcgcatca 780
tctggcgtga tgttctccat tgataccgaa tccggctttg accaggtggt gtttatcact 840
tccgcatggg gccttggtga gatggtcgtg cagggtgcgg ttaacccgga tgagttttac 900
gtgcataaac cgacactggc ggcgaatcgc ccggctatcg tgcgccgcac catggggtcg 960
aaaaaaatcc gcatggttta cgcgccgacc caggagcacg gcaagcaggt taaaatcgaa 1020
gacgtaccgc aggaacagcg tgacatcttc tcgctgacca acgaagaagt gcaggaactg 1080
gcaaaacagg ccgtacaaat tgagaaacac tacggtcgcc cgatggatat tgagtgggcg 1140
aaagatggcc acaccggtaa actgttcatt gtgcaggcgc gtccggaaac cgtgcgctca 1200
cgcggtcagg tcatggagcg ttatacgctg cattcacagg gtaagattat cgccgaaggc 1260
cgtgctatcg gtcatcgcat cggtgcgggt ccggtgaaag tcatccatga tatcagcgaa 1320
atgaaccgca tcgaacctgg tgacgtgctg gtcactgaca tgaccgaccc ggactgggaa 1380
ccgatcatga agaaagcatc tgccatcgtc accaaccgtg gcggtcgtac ctgtcacgcg 1440
gcgatcatcg ctcgtgaact gggcattccg gcggtagtgg gctgtggtga tgcaacagaa 1500
cggatgaaag acggtgagaa cgtcactgtt tcttgtgccg aaggtgatac cggttacgtc 1560
tatgcggagt tgctggaatt tagcgtgaaa agctccagcg tagaaacgat gccggatctg 1620
ccgttgaaag tgatgatgaa cgtcggtaac ccggaccgag ctttcgactt cgcctgtctg 1680
ccgaacgaag gcgtgggact tgcgcgtctg gaatttatca tcaaccgtat gattggcgtc 1740
cacccacgcg cactgcttga gtttgacgat caggaaccgc agttgcaaaa cgaaatccgc 1800
gagatgatga aaggttttga ttctccgcgt gaattttacg ttggtcgtct gactgaaggg 1860
atcgcgacgc tgggtgccgc gttttatccg aagcgcgtca ttgtccgtct ctctgatttt 1920
aaatcgaacg aatatgccaa cctggtcggt ggtgagcgtt acgagccaga tgaagagaac 1980
ccgatgctcg gcttccgtgg cgcgggacgc tatatttccg acagcttccg cgactgtttc 2040
gcgctggagt gcgaagcagt gaaacgtgtg cgcaacgaca tggggctgac caacgttgag 2100
atcatgatcc cgttcgtgcg aaccgtagat caggcgaaag cggtggttga ggaactggcg 2160
cgtcaggggc tgaaacgtgg tgagaacggg ctgaaaatca tcatgatgtg tgaaatcccg 2220
tccaacgcct tgctggccga gcagttcctc gaatatttcg acggcttctc aattggctca 2280
aacgacatga cgcagctggc gctcggtctg gatcgtgact ccggcgtggt gtctgaactg 2340
ttcgatgagc gcaacgatgc ggtgaaagca ctgctgtcga tggcgattcg tgccgcgaag 2400
aaacagggca aatatgtcgg gatttgcggt cagggtccgt ccgaccacga agactttgcc 2460
gcatggttga tggaagaggg gatcgatagc ctgtctctga acccggacac cgtggtgcaa 2520
acctggttaa gcctggctga actgaagaaa taaaataact agcataaccc cttggggcct 2580
ctaaacgggt cttgaggggt tttttgctga aaccaatttg cctggcggca gtagcgcggt 2640
ggtcccacct gaccccatgc cgaactcaga agtgaaacgc cgtagcgccg atggtagtgt 2700
ggggtctccc catgcgagag tagggaactg ccaggcatca aataaaacga aaggctcagt 2760
cgaaagactg ggcctttcgg gatccaggcc ggcctgttaa cgaattaatc ttccgcggcg 2820
gtatcgataa gcttgatatc gaggctgaca tgggaattag ccatggtcca tatgaatatc 2880
ctccttagtt cctattccga agttcctatt ctctagaaag tataggaact tccgagctct 2940
agagaatgat ccctaaatgc ttcaataata ttgaaaaagg aagagtatga gggaagcggt 3000
gatcgccgaa gtatcgactc aactatcaga ggtagttggc gtcatcgagc gccatctcga 3060
accgacgttg ctggccgtac atttgtacgg ctccgcagtg gatggcggcc tgaagccaca 3120
cagtgatatt gatttgctgg ttacggtgac cgtaaggctt gatgaaacaa cgcggcgagc 3180
tttgatcaac gaccttttgg aaacttcggc ttcccctgga gagagcgaga ttctccgcgc 3240
tgtagaagtc accattgttg tgcacgacga catcattccg tggcgttatc cagctaagcg 3300
cgaactgcaa tttggagaat ggcagcgcaa tgacattctt gcaggtatct tcgagccagc 3360
cacgatcgac attgatctgg ctatcttgct gacaaaagca agagaacata gcgttgcctt 3420
ggtaggtcca gcggcggagg aactctttga tccggttcct gaacaggatc tatttgaggc 3480
gctaaatgaa accttaacgc tatggaactc gccgcccgac tgggctggcg atgagcgaaa 3540
tgtagtgctt acgttgtccc gcatttggta cagcgcagta accggcaaaa tcgcgccgaa 3600
ggatgtcgct gccgactggg caatggagcg cctgccggcc cagtatcagc ccgtcatact 3660
tgaagctaga caggcttatc ttggacaaga agaagatcgc ttggcctcgc gcgcagatca 3720
gttggaagaa tttgtccact acgtgaaagg cgagatcacc aaggtagtcg gcaaataata 3780
gcgggactct gggaatttcg acgacctgca gccaagcgaa gttcctattc cgaagttcct 3840
attctctaga aagtatagga acttcgaagc agctccagcc tacacaatcg ctcaagacgt 3900
gtaatgctgc aatctgcatg caagcttggc actggcgatg gcgcctcatc cctgaagcca 3960
atagggataa cagggtaatg a 3981
<210> 73
<211> 4097
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Экспрессионный фрагмент <Ptet-neuA-nanT-lox66-kan-lox72>
<400> 73
tgagcgattg tgtaggctgg agctgcttgg ccagatgatt aattcctaat ttttgttgac 60
actctatcat tgatagagtt attttaccac tccctatcag tgatagagaa aagtgaaatg 120
aatagttcga caaaaatcta gaaataattt tgtttaactt taagaaggag atatacatga 180
gcctggccat tatcccggca cgtggcggtt ctaaaggcat caaaaacaaa aacctggttc 240
tgctgaacaa taaaccgctg atttattaca ccatcaaagc ggccctgaac gccaaaagta 300
ttagcaaagt ggttgtgagc tctgattctg atgaaatcct gaactacgca aaaagtcaga 360
acgttgatat cctgaaacgt ccgatcagtc tggcacagga tgataccacg agcgataaag 420
tgctgctgca tgcgctgaaa ttctacaaag attacgaaga tgttgtgttc ctgcagccga 480
ccagcccgct gcgtacgaat attcacatca acgaagcgtt caacctgtac aaaaacagca 540
acgcaaacgc gctgatttct gttagtgaat gcgataacaa aatcctgaaa gcgtttgtgt 600
gcaatgattg tggcgatctg gccggtattt gtaacgatga atacccgttc atgccgcgcc 660
agaaactgcc gaaaacctat atgagcaatg gtgccatcta catcctgaaa atcaaagaat 720
tcctgaacaa cccgagcttc ctgcagtcta aaacgaaaca tttcctgatg gatgaaagta 780
gctctctgga tattgattgc ctggaagatc tgaaaaaagt ggaacagatc tggaaaaaat 840
aagagctcga gtcgaaggag atagaaccat gagtactaca acccagaata tcccgtggta 900
tcgccatctc aaccgtgcac aatggcgcgc attttccgct gcctggttgg gatatctgct 960
tgacggtttt gatttcgttt taatcgccct ggtactcacc gaagtacaag gtgaattcgg 1020
gctgacgacg gtgcaggcgg caagtctgat ctctgcagcc tttatctctc gctggttcgg 1080
cggcctgatg ctcggcgcta tgggtgaccg ctacgggcgt cgtctggcaa tggtcaccag 1140
catcgttctc ttctcggccg ggacgctggc ctgcggcttt gcgccaggct acatcaccat 1200
gtttatcgct cgtctggtca tcggcatggg gatggcgggt gaatacggtt ccagcgccac 1260
ctatgtcatt gaaagctggc caaaacatct gcgtaacaaa gccagtggtt ttttgatttc 1320
aggcttctct gtgggggccg tcgttgccgc tcaggtctat agcctggtgg ttccggtctg 1380
gggctggcgt gcgctgttct ttatcggcat tttgccaatc atctttgctc tctggctgcg 1440
taaaaacatc ccggaagcgg aagactggaa agagaaacac gcaggtaaag caccagtacg 1500
cacaatggtg gatattctct accgtggtga acatcgcatt gccaatatcg taatgacact 1560
ggcggcggct actgcgctgt ggttctgctt cgccggtaac ctgcaaaatg ccgcgatcgt 1620
cgctgttctt gggctgttat gcgccgcaat ctttatcagc tttatggtgc agagtgcagg 1680
caaacgctgg ccaacgggcg taatgctgat ggtggtcgtg ttgtttgctt tcctctactc 1740
atggccgatt caggcgctgc tgccaacgta tctgaaaacc gatctggctt ataacccgca 1800
tactgtagcc aatgtgctgt tctttagtgg ctttggcgcg gcggtgggat gctgcgtagg 1860
tggcttcctc ggtgactggc tgggaacccg caaagcgtac gtttgtagcc tgctggcctc 1920
gcagctgctg attattccgg tatttgcgat tggcggcgca aacgtctggg tgctcggtct 1980
gttactgttc ttccagcaaa tgcttggaca agggatcgcc gggatcttac caaaactgat 2040
tggcggttat ttcgataccg accagcgtgc agcgggcctg ggctttacct acaacgttgg 2100
cgcattgggc ggtgcactgg ccccaatcat cggcgcgttg atcgctcaac gtctggatct 2160
gggtactgcg ctggcatcgc tctcgttcag tctgacgttc gtggtgatcc tgctgattgg 2220
gctggatatg ccttctcgcg ttcagcgttg gttgcgcccg gaagcgttgc gtactcatga 2280
cgctatcgac ggtaaaccat tcagcggtgc cgtgccgttt ggcagcgcca aaaacgattt 2340
agtcaaaacc aaaagttaat aaatcgatac tagcataacc ccttggggcc tctaaacgcg 2400
tcgacacgca aaaaggccat ccgtcaggat ggccttctgc ttaatttgat gcctggcagt 2460
ttatggcggg cgtcctgccc gccaccctcc gggccgttgc ttcgcaacgt tcaaatccgc 2520
tcccggcgga tttgtcctac tcaggagagc gttcaccgac aaacaacaga taaaacgaaa 2580
ggcccagtct ttcgactgag cctttcgttt tatttgatgc ctggcagttc cctactctcg 2640
catggggaga ccccacacta ccatccggta tcgataagct tgatggcgaa agggggatgt 2700
gctgcaaggc gattaagttg ggtaacgcca gggttttccc agtcacgacg ttgtaaaacg 2760
acggccagtg aattcgagct cggtacctac cgttcgtata atgtatgcta tacgaagtta 2820
tcgagctcta gagaatgatc ccctccctca cgctgccgca agcactcagg gcgcaagggc 2880
tgctaaagga agcggaacac gtagaaagcc agtccgcaga aacggtgctg accccggatg 2940
aatgtcagct actgggctat ctggacaagg gaaaacgcaa gcgcaaagag aaagcaggta 3000
gcttgcagtg ggcttacatg gcgatagcta gactgggcgg ttttatggac agcaagcgaa 3060
ccggaattgc cagctggggc gccctctggt aaggttggga agccctgcaa agtaaactgg 3120
atggctttct tgccgccaag gatctgatgg cgcaggggat caagatctga tcaagagaca 3180
ggatgaggat cgtttcgcat gattgaacaa gatggattgc acgcaggttc tccggccgct 3240
tgggtggaga ggctattcgg ctatgactgg gcacaacaga caatcggctg ctctgatgcc 3300
gccgtgttcc ggctgtcagc gcaggggcgc ccggttcttt ttgtcaagac cgacctgtcc 3360
ggtgccctga atgaactgca ggacgaggca gcgcggctat cgtggctggc cacgacgggc 3420
gttccttgcg cagctgtgct cgacgttgtc actgaagcgg gaagggactg gctgctattg 3480
ggcgaagtgc cggggcagga tctcctgtca tctcaccttg ctcctgccga gaaagtatcc 3540
atcatggctg atgcaatgcg gcggctgcat acgcttgatc cggctacctg cccattcgac 3600
caccaagcga aacatcgcat cgagcgagca cgtactcgga tggaagccgg tcttgtcgat 3660
caggatgatc tggacgaaga gcatcagggg ctcgcgccag ccgaactgtt cgccaggctc 3720
aaggcgcgca tgcccgacgg cgaggatctc gtcgtgaccc atggcgatgc ctgcttgccg 3780
aatatcatgg tggaaaatgg ccgcttttct ggattcatcg actgtggccg gctgggtgtg 3840
gcggaccgct atcaggacat agcgttggct acccgtgata ttgctgaaga gcttggcggc 3900
gaatgggctg accgcttcct cgtgctttac ggtatcgccg ctcccgattc gcagcgcatc 3960
gccttctatc gccttcttga cgagttcttc tgagcgggac tctgggaatt tcgacgacct 4020
gcagccaagc ataacttcgt ataatgtatg ctatacgaac ggtaggatcc tctagagtcg 4080
acctgcaggc atgcaag 4097
<210> 74
<211> 58
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 74
ctgtctctta tacacatctc ctgaaattgg ccagatgatt aattcctaat ttttgttg 58
<210> 75
<211> 50
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 75
ctgtctctta tacacatctc agcattacac gtcttgagcg attgtgtagg 50
<210> 76
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 76
cctgacgacg gtgagcgatc atttgtatat ctccttctta aagttaaaca aaattatttc 60
<210> 77
<211> 59
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 77
aaccctgcaa ctgccggtct cttaaaataa ctagcataac cccttggggc ctctaaacg 59
<210> 78
<211> 59
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 78
aactttaaga aggagatata caaatgatcg ctcaccgtcg tcaggaactg gctcaacag 59
<210> 79
<211> 57
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 79
aggccccaag gggttatgct agttatttta agagaccggc agttgcaggg tttcggc 57
<210> 80
<211> 62
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 80
gccccaaggg gttatgctag ttattttatt ccacggtcac ggatttcgcc aggttacgcg 60
gc 62
<210> 81
<211> 62
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 81
agcaccaacg ataccgcaca tttgtatatc tccttcttaa agttaaacaa aattatttct 60
ag 62
<210> 82
<211> 62
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 82
gcgaaatccg tgaccgtgga ataaaataac tagcataacc ccttggggcc tctaaacggg 60
tc 62
<210> 83
<211> 58
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 83
ctttaagaag gagatataca aatgtgcggt atcgttggtg ctatcgcaca gcgtgatg 58
<210> 84
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 84
cgtggaaatg caaattagaa aatagaataa ctagcataac cccttggggc ctctaaacgg 60
<210> 85
<211> 64
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 85
tataaaatcc atcgggtaag ctcatggttc tatctccttc gttattccac ggtcacggat 60
ttcg 64
<210> 86
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 86
atccgtgacc gtggaataac gaaggagata gaaccatgag cttacccgat ggattttata 60
taagg 65
<210> 87
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 87
ccgtttagag gccccaaggg gttatgctag ttattctatt ttctaatttg catttccacg 60
<210> 88
<211> 59
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 88
ctggttaagc ctggctgaac tgaagaaata aaataactag cataacccct tggggcctc 59
<210> 89
<211> 59
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 89
tagaggcccc aaggggttat gctagttatt ttatttcttc agttcagcca ggcttaacc 59
<210> 90
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 90
tttgtttggc gtcgagaagg agatagaacc atgtccaaca atggctcgtc accgctggtg 60
<210> 91
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 91
aagcaccagc ggtgacgagc cattgttgga catggttcta tctccttctc gacgccaaac 60
<210> 92
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 92
ctacccagaa agtgttcaaa gatattaaat aaaataacta gcataacccc ttggggcctc 60
<210> 93
<211> 61
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 93
tttagaggcc ccaaggggtt atgctagtta ttttatttaa tatctttgaa cactttctgg 60
g 61
<210> 94
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 94
gatgatgatg ttctggattt tgatttcttt catttgtata tctccttctt aaagttaaac 60
<210> 95
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 95
tttgtttaac tttaagaagg agatatacaa atgaaagaaa tcaaaatcca gaacatcatc 60
<210> 96
<211> 70
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 96
agctgtctta tgaagatttc gcctaatcga aggagataca accatgaaga aaattctgtt 60
tatcaccggc 70
<210> 97
<211> 70
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 97
tgccggtgat aaacagaatt ttcttcatgg ttgtatctcc ttcgattagg cgaaatcttc 60
ataagacagc 70
<210> 98
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 98
taattttgtt taactttaag aaggagatat acatgagcct ggccattatc ccggcacgtg 60
<210> 99
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 99
tgccgggata atggccaggc tcatgtatat ctccttctta aagttaaaca aaattatttc 60
<210> 100
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 100
aaaataagag ctcgagtcga aggagataga accatgagta ctacaaccca gaatatcccg 60
<210> 101
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 101
tagtactcat ggttctatct ccttcgactc gagctcttat tttttccaga tctgttccac 60
<210> 102
<211> 60
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 102
aaaacgattt agtcaaaacc aaaagttaat aaatcgatac tagcataacc ccttggggcc 60
<210> 103
<211> 56
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 103
aaggggttat gctagtatcg atttattaac ttttggtttt gactaaatcg tttttg 56
<210> 104
<211> 73
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 104
ggggacaagt ttgtacaaaa aagcaggcta gaaggaggta tacaaatggg cctgaaaaaa 60
gcctgcctga ccg 73
<210> 105
<211> 75
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 105
ggggacaagt ttgtacaaaa aagcaggcta gaaggagata tacaaatgac ccgcacccgt 60
atggaaaacg aactg 75
<210> 106
<211> 62
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 106
ggggaccact ttgtacaaga aagctgggtt tattttttcc agatctgttc cacttttttc 60
ag 62
<210> 107
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 107
aaaaagcagg ctagaaggag gtatacaaat gggcaaaaaa gtgattattg cgggcaacgg 60
cccgagcc 68
<210> 108
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 108
aggctcataa ttgtacctcc ttcgaggttt agttgatgtt tttgctgaat ttgccatacg 60
cttcgc 66
<210> 109
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 109
tggcaaattc agcaaaaaca tcaactaaac ctcgaaggag gtacaattat gagcctggcc 60
attatccc 68
<210> 110
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 110
tgcccgcaat aatcactttt ttgcccattt gtatacctcc ttctagcctg cttttttgta 60
caaacttg 68
<210> 111
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 111
aaaagcaggc tagaaggagg tatacaaatg aataagaaac cgctgattat tgctggcaac 60
gggcc 65
<210> 112
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 112
aggctcataa ttgtacctcc ttcgaggttt atctcttcag gaatgcttta atgattgact 60
ttagcgcc 68
<210> 113
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 113
atcattaaag cattcctgaa gagataaacc tcgaaggagg tacaattatg agcctggcca 60
ttatc 65
<210> 114
<211> 69
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 114
agcaataatc agcggtttct tattcatttg tatacctcct tctagcctgc ttttttgtac 60
aaacttgtg 69
<210> 115
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 115
aaaagcaggc tagaaggagg tatacaaatg gggaccatta aaaagccctt aatcatagca 60
ggaaatgg 68
<210> 116
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 116
aggctcataa ttgtacctcc ttcgaggttt atgcagctcc ccaacggaaa ctaactttta 60
atgttggg 68
<210> 117
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 117
tagtttccgt tggggagctg cataaacctc gaaggaggta caattatgag cctggccatt 60
atcccggc 68
<210> 118
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 118
attaagggct ttttaatggt ccccatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 119
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 119
aaaagcaggc tagaaggagg tatacaaatg agtgaagaaa acacccagtc cattattaaa 60
aacgac 66
<210> 120
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 120
aggctcataa ttgtacctcc ttcgaggttc agacagcaat acagacaccc gtttcgcaat 60
tcggcagg 68
<210> 121
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 121
aaacgggtgt ctgtattgct gtctgaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 122
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 122
atggactggg tgttttcttc actcatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 123
<211> 64
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 123
aaaagcaggc tagaaggagg tatacaaatg accatttacc tggacccggc gtctctgccg 60
accc 64
<210> 124
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 124
aggctcataa ttgtacctcc ttcgaggttt acagttgttt cagagaatcc cagaagataa 60
tttggc 66
<210> 125
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 125
ttctgggatt ctctgaaaca actgtaaacc tcgaaggagg tacaattatg agcctggcca 60
ttatccc 67
<210> 126
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 126
acgccgggtc caggtaaatg gtcatttgta tacctccttc tagcctgctt ttttgtacaa 60
acttgtg 67
<210> 127
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 127
aaagcaggct agaaggaggt atacaaatgg gctgtaatag cgactccaac cacaacaact 60
ccgacgg 67
<210> 128
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 128
aggctcataa ttgtacctcc ttcgaggttt attgcaggtc cgagatcagt ttcacatcat 60
tacgg 65
<210> 129
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 129
tgaaactgat ctcggacctg caataaacct cgaaggaggt acaattatga gcctggccat 60
tatccc 66
<210> 130
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 130
tggttggagt cgctattaca gcccatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 131
<211> 64
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 131
aaagcaggct agaaggaggt atacaaatga acaacgacaa ctccacgacc accaacaata 60
acgc 64
<210> 132
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 132
ctcataattg tacctccttc gaggtttaaa tgtcagagat cagtttaata ttatcgcggt 60
taatcag 67
<210> 133
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 133
atattaaact gatctctgac atttaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 134
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 134
tggtcgtgga gttgtcgttg ttcatttgta tacctccttc tagcctgctt ttttgtacaa 60
acttgtg 67
<210> 135
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 135
aaagcaggct agaaggaggt atacaaatga aaacgattac cctgtatctg gacccggcgt 60
ccctgcc 67
<210> 136
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 136
aggctcataa ttgtacctcc ttcgaggttt acagctgttt caggctgtcc caaaagatca 60
cttgcg 66
<210> 137
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 137
tttgggacag cctgaaacag ctgtaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 138
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 138
tccagataca gggtaatcgt tttcatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 139
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 139
aaagcaggct agaaggaggt atacaaatga aaaagatcct gaccgtcctg agcatcttta 60
tcctgagc 68
<210> 140
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 140
aggctcataa ttgtacctcc ttcgaggttt agtccagcat cgtaccgaag tcatccggtt 60
tggtgtgg 68
<210> 141
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 141
atgacttcgg tacgatgctg gactaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 142
<211> 70
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 142
tgctcaggac ggtcaggatc tttttcattt gtatacctcc ttctagcctg cttttttgta 60
caaacttgtg 70
<210> 143
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 143
aaagcaggct agaaggaggt atacaaatga cgaatcgcaa aatctatgtc tgccacaccc 60
tgtacc 66
<210> 144
<211> 70
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 144
aggctcataa ttgtacctcc ttcgaggttt atttaatgtc tttcagatca accagcgtaa 60
ttttcttgtc 70
<210> 145
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 145
tggttgatct gaaagacatt aaataaacct cgaaggaggt acaattatga gcctggccat 60
tatccc 66
<210> 146
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 146
agacatagat tttgcgattc gtcatttgta tacctccttc tagcctgctt ttttgtacaa 60
acttgtg 67
<210> 147
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 147
aaagcaggct agaaggaggt atacaaatgt tccgtgaaga caatatgaac ctgattatct 60
gctgtacg 68
<210> 148
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 148
aggctcataa ttgtacctcc ttcgaggttt agatgtcgat aactttgata ccgaaatctt 60
tcagg 65
<210> 149
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 149
tcggtatcaa agttatcgac atctaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 150
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 150
aggttcatat tgtcttcacg gaacatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 151
<211> 69
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 151
aaagcaggct agaaggaggt atacaaatga aagaaatcgc catcatctcc aaccaacgca 60
tgttcttcc 69
<210> 152
<211> 69
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 152
aggctcataa ttgtacctcc ttcgaggttt agtcaaagaa atccagcagt ttcggatgca 60
ccgcggtgc 69
<210> 153
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 153
tccgaaactg ctggatttct ttgactaaac ctcgaaggag gtacaattat gagcctggcc 60
attatccc 68
<210> 154
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 154
ttggagatga tggcgatttc tttcatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 155
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 155
aaagcaggct agaaggaggt atacaaatgc tgattcaaca gaacctggaa atctacctgg 60
actacgc 67
<210> 156
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 156
aggctcataa ttgtacctcc ttcgaggttt aattgtgaat ggtgcacata aacgcctgat 60
cttcgttg 68
<210> 157
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 157
aggcgtttat gtgcaccatt cacaattaaa cctcgaagga ggtacaatta tgagcctggc 60
cattatcc 68
<210> 158
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 158
atttccaggt tctgttgaat cagcatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 159
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 159
aaagcaggct agaaggaggt atacaaatgg gctgtaactc cgatagcaaa cacaataaca 60
gtgatggc 68
<210> 160
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 160
aggctcataa ttgtacctcc ttcgaggttt attgcaggtc actaatcagt ttcacatcat 60
tgcgg 65
<210> 161
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 161
tgaaactgat tagtgacctg caataaacct cgaaggaggt acaattatga gcctggccat 60
tatcccgg 68
<210> 162
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 162
tgtttgctat cggagttaca gcccatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 163
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 163
aaagcaggct agaaggaggt atacaaatgt gtaacgataa tcaaaatacg gtcgatgttg 60
ttgtgagc 68
<210> 164
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 164
aggctcataa ttgtacctcc ttcgaggttt aatactgagc aatacaaaca cccgaggaac 60
aatccggc 68
<210> 165
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 165
tcgggtgttt gtattgctca gtattaaacc tcgaaggagg tacaattatg agcctggcca 60
ttatccc 67
<210> 166
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 166
accgtatttt gattatcgtt acacatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 167
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 167
aaaagcaggc tagaaggagg tatacaaatg aacgataatc aaaatacggt ggacgtggtg 60
gtctc 65
<210> 168
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 168
aggctcataa ttgtacctcc ttcgaggttt agcaccagaa cagcacatct ttttctttca 60
caatgcc 67
<210> 169
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 169
aaaaagatgt gctgttctgg tgctaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 170
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 170
tccaccgtat tttgattatc gttcatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 171
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 171
aaaagcaggc tagaaggagg tatacaaatg caaaacgtca ttatcgctgg taacggtccg 60
agcctgc 67
<210> 172
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 172
aggctcataa ttgtacctcc ttcgaggttt atttcttttt gtattctttc ttcagttttt 60
tgatttcg 68
<210> 173
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 173
tgaagaaaga atacaaaaag aaataaacct cgaaggaggt acaattatga gcctggccat 60
tatcccgg 68
<210> 174
<211> 68
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 174
ttaccagcga taatgacgtt ttgcatttgt atacctcctt ctagcctgct tttttgtaca 60
aacttgtg 68
<210> 175
<211> 64
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 175
aaaagcaggc tagaaggagg tatacaaatg gattcttcgc cggaaaacac cagctctacg 60
ctgg 64
<210> 176
<211> 66
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 176
aggctcataa ttgtacctcc ttcgaggttt atttgatgtc cgtcgtaaag cgcacttttt 60
cgtccg 66
<210> 177
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 177
tgcgctttac gacggacatc aaataaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 178
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 178
tggtgttttc cggcgaagaa tccatttgta tacctccttc tagcctgctt ttttgtacaa 60
acttgtg 67
<210> 179
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 179
aaaagcaggc tagaaggagg tatacaaatg aagaaagtct acttctgcca tacggtctac 60
catctgc 67
<210> 180
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 180
aggctcataa ttgtacctcc ttcgaggttt aactatttgc tttcatttgt ttcagggtga 60
ttttc 65
<210> 181
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 181
tgaaacaaat gaaagcaaat agttaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 182
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 182
tatggcagaa gtagactttc ttcatttgta tacctccttc tagcctgctt ttttgtacaa 60
acttgtg 67
<210> 183
<211> 65
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 183
aaagcaggct agaaggaggt atacaaatgc gtaaaatcat caccttcttc agcctgttct 60
tctcg 65
<210> 184
<211> 70
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 184
aggctcataa ttgtacctcc ttcgaggttt aaaagttaat cgggttcggc atttcttcaa 60
agaaaatctg 70
<210> 185
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 185
aaatgccgaa cccgattaac ttttaaacct cgaaggaggt acaattatga gcctggccat 60
tatcccg 67
<210> 186
<211> 67
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Праймер
<400> 186
tgaagaaggt gatgatttta cgcatttgta tacctccttc tagcctgctt ttttgtacaa 60
acttgtg 67
<210> 187
<211> 3856
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Экспрессионная кассета <Ptet-wbdO-PT5-galE-FRT-cat-FRT>
<400> 187
acaggttggc tgataagtcc ccggtctgcc cgaaaagtgc cacctgaaat tggccagatg 60
attaattcct aatttttgtt gattctggta ccaaatgagt cgaccggcca gatgattaat 120
tcctaatttt tgttgacact ctatcattga tagagttatt ttaccactcc ctatcagtga 180
tagagaaaag tgaaatgaat agttcgacaa aaatctagaa ataattttgt ttaactttaa 240
gaaggagata tacaaatgct gacggaagtg cgcccggtct ctacgacgaa accgctggtg 300
tctgtgattc tgccggtgaa caaattcaac ccgtatctgg atcgtgcaat tcattcaatc 360
ctgagtcagt cctatccgtc gattgaactg attatcattg caaacaattg caccaatgac 420
tttttcgatg ctctgaaaaa acgtgaatgt gaaaccatta aagtgctgcg cacgaacatc 480
gcgtatctgc cgtactgcct gaataaaggc ctggatctgt gtaacggtga ctttgttgcc 540
cgcatggatt cagatgacat ttcgcacccg gaacgtatcg atcgccaggt cgacttcctg 600
attaacaatc cggacatcga tgtggttggc accaatgcag tctatattga tgaagatgac 660
atcgaactgg aaaaaagcaa cctgccggtg aacaataacg ctattcgtaa aatgctgccg 720
tataaatgct gtctggtgca tccgtctgtt atgtttcgca aaaatgtcgt gatcaccagc 780
ggcggttaca tgttcgcgaa ttattctgaa gattacgaac tgtggaaccg tctggccgtt 840
gaaggccgca atttttataa cctgagcgaa tacctgctgt attaccgtct gcacaataac 900
caatcaacgt cgaaaaataa cctgtttatg gtgatggcga acgatgtcgc cattaaagtg 960
aaatatttcc tgctgaccaa gaaaattagc tacctgctgg gtatcattcg cacggtcttt 1020
tctgtgttct attgcaaata catcaaatga tttcgtcgac acacaggaaa catattaaaa 1080
attaaaacct gcaggagttt aaacgcggcc gcgatatcgt tgtaaaacga cggccagtgc 1140
aagaatcata aaaaatttat ttgctttcag gaaaattttt ctgtataata gattcataaa 1200
tttgagagag gagtttttgt gagcggataa caattcccca tcttagtata ttagttaagt 1260
ataaatacac cgcggaggcg tcgaaggaga tacaaccatg agagttctgg ttaccggtgg 1320
tagcggttac attggaagtc atacctgtgt gcaattactg caaaacggtc atgatgtcat 1380
cattcttgat aacctctgta acagtaagcg cagcgtactg cctgttatcg agcgtttagg 1440
cggcaaacat ccaacgtttg ttgaaggcga tattcgtaac gaagcgttga tgaccgagat 1500
cctgcacgat cacgctatcg acaccgtgat ccacttcgcc gggctgaaag ccgtgggcga 1560
atcggtacaa aaaccgctgg aatattacga caacaatgtc aacggcactc tgcgcctgat 1620
tagcgccatg cgcgccgcta acgtcaaaaa ctttattttt agctcctccg ccaccgttta 1680
tggcgatcag cccaaaattc catacgttga aagcttcccg accggcacac cgcaaagccc 1740
ttacggcaaa agcaagctga tggtggaaca gatcctcacc gatctgcaaa aagcccagcc 1800
ggactggagc attgccctgc tgcgctactt caacccggtt ggcgcgcatc cgtcgggcga 1860
tatgggcgaa gatccgcaag gcattccgaa taacctgatg ccatacatcg cccaggttgc 1920
tgtaggccgt cgcgactcgc tggcgatttt tggtaacgat tatccgaccg aagatggtac 1980
tggcgtacgc gattacatcc acgtaatgga tctggcggac ggtcacgtcg tggcgatgga 2040
aaaactggcg aacaagccag gcgtacacat ctacaacctc ggcgctggcg taggcaacag 2100
cgtgctggac gtggttaatg ccttcagcaa agcctgcggc aaaccggtta attatcattt 2160
tgcaccgcgt cgcgagggcg accttccggc ctactgggcg gacgccagca aagccgaccg 2220
tgaactgaac tggcgcgtaa cgcgcacact cgatgaaatg gcgcaggaca cctggcactg 2280
gcagtcacgc catccacagg gatatcccga ttaacgccat ttaaatcaac ctcagcggtc 2340
atagctgttt cctgtgactg agcaataact agcataaccc cttggggcct ctaaacgggt 2400
cttgaggggt tttttgctga aaccaatttg cctggcggca gtagcgcggt ggtcccacct 2460
gaccccatgc cgaactcaga agtgaaacgc cgtagcgccg atggtagtgt ggggtctccc 2520
catgcgagag tagggaactg ccaggcatca aataaaacga aaggctcagt cgaaagactg 2580
ggcctttcgg gatccaggcc ggcctgttaa cgaattaatc ttccgcggcg gtatcgataa 2640
gcttgatatc gaggctgaca tgggaattag ccatggtcca tatgaatatc ctccttagtt 2700
cctattccga agttcctatt ctctagaaag tataggaact tcggcgcgcc tacctgtgac 2760
ggaagatcac ttcgcagaat aaataaatcc tggtgtccct gttgataccg ggaagccctg 2820
ggccaacttt tggcgaaaat gagacgttga tcggcacgta agaggttcca actttcacca 2880
taatgaaata agatcactac cgggcgtatt ttttgagttg tcgagatttt caggagctaa 2940
ggaagctaaa atggagaaaa aaatcactgg atataccacc gttgatatat cccaatggca 3000
tcgtaaagaa cattttgagg catttcagtc agttgctcaa tgtacctata accagaccgt 3060
tcagctggat attacggcct ttttaaagac cgtaaagaaa aataagcaca agttttatcc 3120
ggcctttatt cacattcttg cccgcctgat gaatgctcat ccggaattac gtatggcaat 3180
gaaagacggt gagctggtga tatgggatag tgttcaccct tgttacaccg ttttccatga 3240
gcaaactgaa acgttttcat cgctctggag tgaataccac gacgatttcc ggcagtttct 3300
acacatatat tcgcaagatg tggcgtgtta cggtgaaaac ctggcctatt tccctaaagg 3360
gtttattgag aatatgtttt tcgtctcagc caatccctgg gtgagtttca ccagttttga 3420
tttaaacgtg gccaatatgg acaacttctt cgcccccgtt ttcaccatgg gcaaatatta 3480
tacgcaaggc gacaaggtgc tgatgccgct ggcgattcag gttcatcatg ccgtttgtga 3540
tggcttccat gtcggcagat gcttaatgaa tacaacagta ctgcgatgag tggcagggcg 3600
gggcgtaagg cgcgccattt aaatgaagtt cctattccga agttcctatt ctctagaaag 3660
tataggaact tcgaagcagc tccagcctac acaatcgctc aagacgtgta atgctgcaat 3720
ctgcatgcaa gcttggcact ggcgatggcg cctcatccct gaagccaata agcagctcca 3780
gcctacacaa tcgctcaaga cgtgtaatgc tgcaatctgc atgcaagcta gaccggggac 3840
ttatcagcca acctgt 3856
<210> 188
<211> 4568
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Экспрессионная кассета <Ptet-lgtA-PT5-galT-FRT-kan-FRT>.
<400> 188
acaggttggc tgataagtcc ccggtctagc ttgcatgcag attgcagcat tacacgtctt 60
gagcgattgt gtaggctgga gctgcttcga agttcctata ctttctagag aataggaact 120
tcggaatagg aacttcattt aaatggcgcg ccttacgccc cgccctgccg gtaccgagag 180
cgcttttgaa gctggggtgg gcgaagaact ccagcatgag atccccgcgc tggaggatca 240
tccagccggc gtcccggaaa acgattccga agcccaacct ttcatagaag gcggcggtgg 300
aatcgaaatc tcgtgatggc aggttgggcg tcgcttggtc ggtcatttcg aaccccagag 360
tcccgctcag aagaactcgt caagaaggcg atagaaggcg atgcgctgcg aatcgggagc 420
ggcgataccg taaagcacga ggaagcggtc agcccattcg ccgccaagct cttcagcaat 480
atcacgggta gccaacgcta tgtcctgata gcggtccgcc acacccagcc ggccacagtc 540
gatgaatcca gaaaagcggc cattttccac catgatattc ggcaagcagg catcgccatg 600
ggtcacgacg agatcctcgc cgtcgggcat gcgcgccttg agcctggcga acagttcggc 660
tggcgcgagc ccctgatgct cttcgtccag atcatcctga tcgacaagac cggcttccat 720
ccgagtacgt gctcgctcga tgcgatgttt cgcttggtgg tcgaatgggc aggtagccgg 780
atcaagcgta tgcagccgcc gcattgcatc agccatgatg gatactttct cggcaggagc 840
aaggtgagat gacaggagat cctgccccgg cacttcgccc aatagcagcc agtcccttcc 900
cgcttcagtg acaacgtcga gcacagctgc gcaaggaacg cccgtcgtgg ccagccacga 960
tagccgcgct gcctcgtcct gcagttcatt cagggcaccg gacaggtcgg tcttgacaaa 1020
aagaaccggg cgcccctgcg ctgacagccg gaacacggcg gcatcagagc agccgattgt 1080
ctgttgtgcc cagtcatagc cgaatagcct ctccacccaa gcggccggag aacctgcgtg 1140
caatccatct tgttcaatca tgcgaaacga tcctcatcct gtctcttgat cagatcttga 1200
tcccctgcgc catcagatcc ttggcggcaa gaaagccatc cagtttactt tgcagggctt 1260
cccaacctta ccagagggcg ccccagctgg caattccggt tcgcttgctg tccataaaac 1320
cgcccagtct agctatcgcc atgtaagccc actgcaagct acctgctttc tctttgcgct 1380
tgcgttttcc cttgtccaga tagcccagta gctgacattc atccggggtc agcaccgttt 1440
ctgcggactg gctttctacg tgttccgctt cctttagcag cccttgcgcc ctgagtgctt 1500
gcggcagcgt gaggggatct tgacgcgtgt cacaggtagg acgcgccgaa gttcctatac 1560
tttctagaga ataggaactt cggaatagga actaaggagg atattcatac atgatggtag 1620
tgttcgaaat taatacgact cactataggg gaattgattc tggtaccaaa tgagtcgacc 1680
ggccagatga ttaattccta atttttgttg acactctatc attgatagag ttattttacc 1740
actccctatc agtgatagag aaaagtgaaa tgaatagttc gacaaaaatc tagaaataat 1800
tttgtttaac tttaagaagg agatatacaa atgccgtccg aagcattccg tcgtcaccgt 1860
gcttatcgcg aaaacaaact gcagccactg gtctctgtcc tgatctgcgc atacaacgtt 1920
gagaaatact tcgcacagtc tctggcagct gtagttaacc agacctggcg taacctggat 1980
atcctgatcg tagatgacgg ctctacggat ggtacgctgg cgatcgcaca gcgtttccag 2040
gaacaggacg gtcgtatccg cattctcgct cagccgcgta actctggtct gatcccgtct 2100
ctgaacatcg gtctggacga actggccaaa tctggtggtg gtggcgaata catcgcccgt 2160
actgacgccg acgacattgc ggccccggat tggatcgaaa aaatcgtagg tgaaatggag 2220
aaagaccgct ctatcatcgc gatgggtgct tggctggaag ttctgtccga agagaaagac 2280
ggtaaccgtc tggcccgtca ccatgaacac ggcaaaatct ggaaaaaacc gacccgtcac 2340
gaagatatcg cggacttctt cccgttcggt aacccgatcc ataacaacac catgatcatg 2400
cgtcgtagcg taatcgacgg tggtctgcgt tacaacaccg aacgtgattg ggcagaagac 2460
taccagtttt ggtatgacgt gtctaaactg ggtcgtctgg cttactaccc agaagcgctg 2520
gttaaatacc gtctgcacgc caaccaggtt agctccaaat actccatccg tcagcacgaa 2580
atcgcacagg gtatccagaa aacggctcgt aacgacttcc tgcagtccat gggtttcaaa 2640
acccgtttcg actctctgga gtaccgtcag atcaaagcgg ttgcgtatga gctgctggag 2700
aaacacctgc cggaagagga ctttgaacgt gcgcgtcgtt tcctgtacca gtgcttcaaa 2760
cgtaccgaca ctctgccggc gggtgcatgg ctcgactttg cagcggatgg tcgtatgcgt 2820
cgtctgttta ccctgcgtca gtacttcggt atcctgcatc gtctcctgaa aaaccgctaa 2880
tgatttcgtc gacacacagg aaacatatta aaaattaaaa cctgcaggag tttaaacgcg 2940
gccgcgatat cgttgtaaaa cgacggccag tgcaagaatc ataaaaaatt tatttgcttt 3000
caggaaaatt tttctgtata atagattcat aaatttgaga gaggagtttt tgtgagcgga 3060
taacaattcc ccatcttagt atattagtta agtataaata cacaaggaga tataccatga 3120
cgcaatttaa tcccgttgat catccacatc gccgctacaa cccgctcacc gggcaatgga 3180
ttctggtttc accgcaccgc gctaagcgcc cctggcaggg ggcgcaggaa acgccagcca 3240
aacaggtgtt acctgcgcac gatccagatt gcttcctctg cgcaggtaat gtgcgggtga 3300
caggcgataa aaaccccgat tacaccggga cttacgtttt cactaatgac tttgcggctt 3360
tgatgtctga cacgccagat gcgccagaaa gtcacgatcc gctgatgcgt tgccagagcg 3420
cgcgcggcac cagccgggtg atctgctttt caccggatca cagtaaaacg ctgccagagc 3480
tcagcgttgc agcattgacg gaaatcgtca aaacctggca ggagcaaacc gcagaactgg 3540
ggaaaacgta cccatgggtg caggtttttg aaaacaaagg cgcggcgatg ggctgctcta 3600
acccgcatcc gcacggtcag atttgggcaa atagcttcct gcctaacgaa gctgagcgcg 3660
aagaccgcct gcaaaaagaa tattttgccg aacagaaatc accaatgctg gtggattatg 3720
ttcagcgcga gctggcagac ggtagccgta ccgttgtcga aaccgaacac tggttagccg 3780
tcgtgcctta ctgggctgcc tggccgttcg aaacgctact gctgcccaaa gcccacgttt 3840
tacggatcac cgatttgacc gacgcccagc gcagcgatct ggcgctggcg ttgaaaaagc 3900
tgaccagtcg ttatgacaac ctcttccagt gctccttccc ctactctatg ggctggcacg 3960
gcgcgccatt taatggcgaa gagaatcaac actggcagct gcacgcgcac ttttatccgc 4020
ctctgctgcg ctccgccacc gtacgtaaat ttatggttgg ttatgaaatg ctggcagaga 4080
cccagcgaga cctgaccgca gaacaggcag cagagcgttt gcgcgcagtc agcgatatcc 4140
attttcgcga atccggagtg taacgcggag gcgcgccatt taaatcaacc tcagcggtca 4200
tagctgtttc ctgtgactga gcaataacta gcataacccc ttggggcctc taaacgggtc 4260
ttgaggggtt ttttgctgaa accaatttgc ctggcggcag tagcgcggtg gtcccacctg 4320
accccatgcc gaactcagaa gtgaaacgcc gtagcgccga tggtagtgtg gggtctcccc 4380
atgcgagagt agggaactgc caggcatcaa ataaaacgaa aggctcagtc gaaagactgg 4440
gcctttcggg atccaggccg gcctgttaac gaattaatct tccgcggcaa caaaaattag 4500
gaattaatca tctggccaat ttcaggtggc acttttcggg cagaccgggg acttatcagc 4560
caacctgt 4568
<---
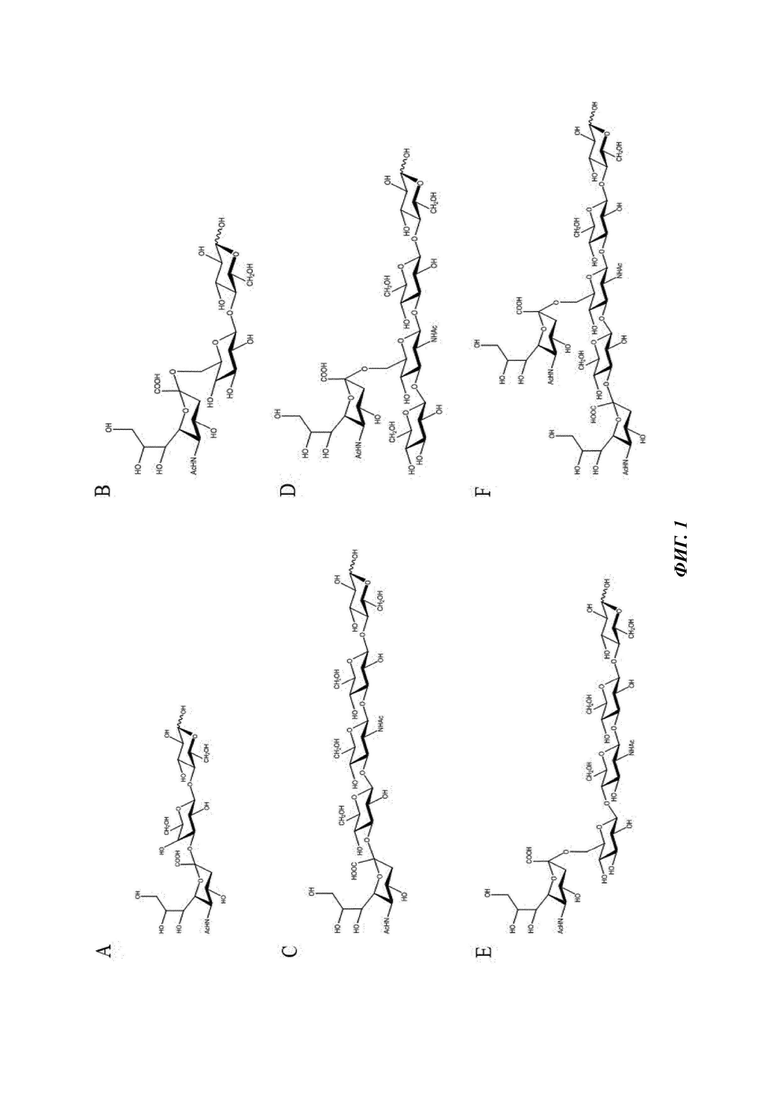
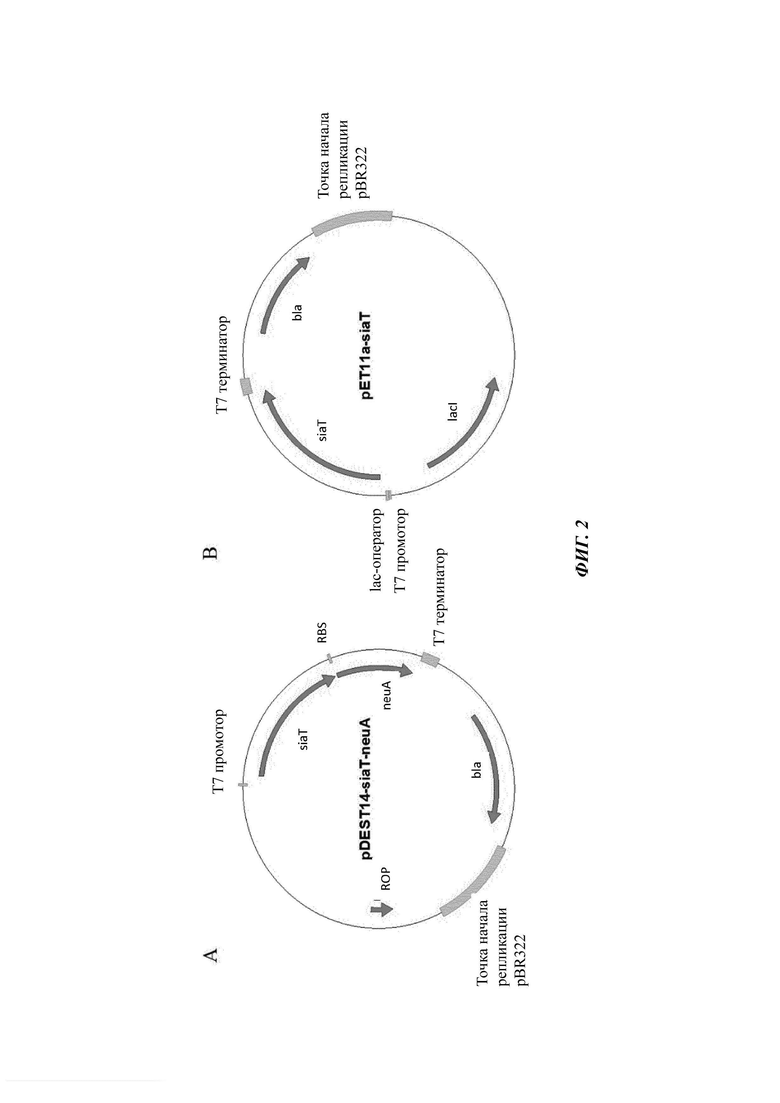

Изобретение относится к биотехнологии. Предложены варианты способа получения сиалированного олигосахарида с использованием генетически модифицированной клетки, которая содержит гетерологичную сиалилтрансферазу, причем указанная гетерологичная сиалилтрансфераза обладает α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью. При этом сиалированный олигосахарид выбран из группы, состоящей из 3’-сиалиллактозы, 6’-сиалиллактозы, сиалиллакто-N-тетраозы a (LST-a), сиалиллакто-N-тетраозы b (LST-b), сиалиллакто-N-тетраозы с (LST-c) и дисиалиллакто-N-тетраозы (DSLNT). Изобретение обеспечивает эффективное получение сиалированного олигосахарида. 2 н. и 11 з.п. ф-лы, 3 ил., 6 табл., 8 пр.
1. Способ получения сиалированного олигосахарида, где сиалированный олигосахарид выбран из группы, состоящей из 3’-сиалиллактозы, 6’-сиалиллактозы, сиалиллакто-N-тетраозы a (LST-a), сиалиллакто-N-тетраозы b (LST-b), сиалиллакто-N-тетраозы с (LST-c) и дисиалиллакто-N-тетраозы (DSLNT), где способ включает следующие стадии:
а) предоставление по меньшей мере одной генетически модифицированной клетки, которая содержит гетерологичную сиалилтрансферазу, причем указанная гетерологичная сиалилтрансфераза обладает α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью для переноса остатка сиаловой кислоты от ее нуклеотид-активированной формы в качестве донорного субстрата к акцепторной молекуле, при этом акцепторная молекула выбрана из группы, состоящей из лактозы, лакто-N-триозы II, лакто-N-тетраозы и лакто-N-неотетраозы, причем гетерологичная сиалилтрансфераза выбрана из группы, состоящей из полипептидов, содержащих или состоящих из аминокислотной последовательности, как представлено любой из SEQ ID NO: 34-37, 39-48 и 50-66;
b) культивирование по меньшей мере одной клетки в ферментационном бульоне и в условиях, являющихся пермиссивными для продукции указанного сиалированного олигосахарида; и, возможно,
в) выделение указанного сиалированного олигосахарида.
2. Способ по п. 1, в котором ферментационный бульон содержит по меньшей мере один источник углерода, при этом указанный по меньшей мере один источник углерода предпочтительно выбран из группы, состоящей из глюкозы, фруктозы, сахарозы, глицерина и их комбинаций.
3. Способ по п. 1 или 2, в котором ферментационный бульон содержит по меньшей мере одно, выбранное из группы, состоящей из N-ацетилглюкозамина, галактозы и сиаловой кислоты.
4. Способ по любому из пп. 1-3, в котором указанную по меньшей мере одну клетку культивируют при отсутствии и/или без добавления одного или более, выбранных из группы, состоящей из N-ацетилглюкозамина, галактозы и сиаловой кислоты.
5. Способ по любому из пп. 1-4, в котором указанную по меньшей мере одну клетку культивируют в присутствии по меньшей мере одного, выбранного из группы, состоящей из лактозы, лакто-N-триозы II, LNT и LNnT.
6. Способ по любому из пп. 1-5, где генетически модифицированная клетка содержит молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, которая кодирует гетерологичную сиалилтрансферазу, причем указанная нуклеотидная последовательность выбрана из группы, состоящей из:
i. нуклеотидных последовательностей, кодирующих полипептид, как представлено любой из SEQ ID NO: 34-37, 39-48 и 50-66;
ii. нуклеотидных последовательностей, как представлено любой из SEQ ID NO: 1-4, 6-11, 13-15 и 17-33; и
iii. нуклеотидных последовательностей, которые комплементарны любой из нуклеотидных последовательностей i. и ii.;
где указанная нуклеотидная последовательность функционально связана с по меньшей мере одной последовательностью контроля экспрессии нуклеиновой кислоты, влияющей на транскрипцию и/или трансляцию указанной нуклеотидной последовательности, кодирующей гетерологичную сиалилтрансферазу в генетически модифицированной клетке.
7. Способ по любому из пп. 1-6, где генетически модифицированная клетка обладает повышенной продукцией одного или более нуклеотид-активированных сахаров, выбранных из группы, состоящей из CMP-N-ацетилнейраминовой кислоты, УДФ-N- ацетилглюкозамина, УДФ-галактозы и ГДФ-фукозы, по сравнению с клеткой перед генетической модификацией.
8. Способ по любому из пп. 1-7, где генетически модифицированная клетка обладает сверхэкспрессией, по сравнению с клеткой перед генетической модификацией, одного или более генов, кодирующих полипептид, способный обладать ферментативной активностью, выбранный из группы, состоящей из следующего: L-глутамин:D-фруктозо-6-фосфат аминотрансфераза, N-ацетилглюкозамин-1-фосфат уридилтрансфераза, глюкозамин-1-фосфат ацетилтрансфераза, фосфоглюкозаминмутаза, глюкозамин-6-фосфат-N-ацетилтрансфераза, N-ацетилглюкозамин-2-эпимераза, УДФ-N-ацетилглюкозамин-2-эпимераза, синтаза сиаловой кислоты, фосфоенолпируватсинтаза, синтаза CMP-сиаловой кислоты, УДФ-галактозо-4-эпимераза, галактозо-1-фосфат уридилилтрансфераза, фосфоглюкомутаза, глюкозо-1-фосфат уридилилтрансфераза, фосфоманномутаза, маннозо-1-фосфат гуанозилтрансфераза, ГДФ-маннозо-4,6-дегидратаза, ГДФ-L-фукозосинтаза и фукозокиназа/L-фукозо-1-фосфат-гуанилтрансфераза.
9. Способ по любому из пп. 1-8, где клетка не обладает или обладает сниженной активностью, по сравнению с клеткой перед генетической модификацией, одной или более ферментативных активностей, выбранных из группы, состоящей из β-галактозидазы, глюкозамин-6-фосфатдезаминазы, N-ацетилглюкозамин-6-фосфатдеацетилазы, N-ацетилманнозаминкиназы, N-ацетилманнозамин-6-фосфатэпимеразы и альдолазы N-ацетилнейраминовой кислоты.
10. Способ по любому из пп. 1-9, где один или более генов, кодирующих β-галактозидазу, глюкозамин-6-фосфатдезаминазу, N-ацетилглюкозамин-6-фосфатдеацетилазу, N-ацетилманнозаминкиназу, N-ацетилманнозамин-6- фосфатэпимеразу и альдолазу N-ацетилнейраминовой кислоты, были удалены или их экспрессия была инактивирована в генетически модифицированной клетке.
11. Способ по любому из пп. 1-10, где указанная по меньшей мере одна клетка содержит по меньшей мере одно, выбранное из группы, состоящей из функциональной лактозопермеазы, функциональной фукозопермеазы и функционального транспортера сиаловой кислоты (импортера), предпочтительно содержит и экспрессирует по меньшей мере одну нуклеотидную последовательность, кодирующую одно, выбранное из группы, состоящей из функциональной лактозопермеазы, функциональной фукозопермеазы и функционального транспортера сиаловой кислоты (импортера).
12. Способ по любому из пп. 1-11, где клетка обладает активностью по меньшей мере одной гликозилтрансферазы, выбранной из группы, состоящей из β-1,3-N-ацетилглюкозаминилтрансферазы, β-1,3-галактозилтрансферазы, β-1,4-галактозилтрансферазы, α-2,3-сиалилтрансферазы и α-2,6-сиалилтрансферазы.
13. Способ получения сиалированного олигосахарида, где сиалированный олигосахарид выбран из группы, состоящей из 3’-сиалиллактозы, 6’-сиалиллактозы, LST-a, LST-b, LST-c и DSLNT, где способ включает:
а) предоставление сиалилтрансферазы в реакционной смеси, причем указанная сиалилтрансфераза обладает α-2,3-сиалилтрансферазной активностью и/или α-2,6-сиалилтрансферазной активностью для переноса остатка сиаловой кислоты от ее нуклеотид-активированной формы в качестве донорного субстрата к акцепторной молекуле, при этом акцепторная молекула выбрана из группы, состоящей из лактозы, лакто-N-триозы II, лакто-N-тетраозы и лакто-N-неотетраозы, и причем сиалилтрансфераза выбрана из группы, состоящей из полипептидов, содержащих или
состоящих из аминокислотной последовательности, как представлено любой из SEQ ID NO: 34-37, 39-48 и 50-66; и
b) приведение указанной сиалилтрансферазы в контакт с донорным субстратом и акцепторной молекулой, где акцепторная молекула выбрана из группы, состоящей из лактозы, лакто-N-триозы II, лакто-N-тетраозы и лакто-N-неотетраозы.
| WO2014153253A1, 25.09.2014 | |||
| US 20050089956 А1, 28.04.2005 | |||
| US 2012070863 A, 22.03.2012 | |||
| US 7968310 B2, 28.06.2011 | |||
| УСТАНОВКА ДЛЯ ОБРАБОТКИ ТАБАЧНЫХ ЛИСТБЕВ ПЕРЕД РЕЗАНИЕМ | 0 |
|
SU177314A1 |
| ЛИФТОВАЯ УСТАНОВКА | 2007 |
|
RU2441832C2 |
| US 8187838 B2, 29.05.2012 | |||
| US 8187853 B2, 29.05.2012 | |||
| WO 2017101958 A1, 22.06.2017 | |||
| CHEN X | |||
| Human Milk Oligosaccharides (HMOS): Structure, Function, and Enzyme-Catalyzed | |||
