Область техники, к которой относится изобретение
Описание настоящего изобретения относится к микроорганизму для продуцирования микоспорин-подобной аминокислоты и к способу получения микоспорин-подобной аминокислоты с использованием этого микроорганизма.
Предшествующий уровень техники
Излучаемый солнцем ультрафиолетовый свет состоит из УФ-А (ультрафиолета А, диапазон которого составляет от приблизительно 320 нм до 400 нм), УФ-В (ультрафиолета В, диапазон которого составляет от приблизительно 290 нм до 320 нм) и УФ-С (ультрафиолета С, диапазон которого составляет от приблизительно 100 нм до 280 нм). Среди солнечных лучей приблизительно 6% УФ-А и УФ-В достигает поверхности Земли, с другой стороны, УФ-С не достигает поверхности Земли, поскольку поглощается и рассеивается в озоновом слое и в атмосфере Земли.
Хотя эти УФ лучи могут обеспечивать полезные аспекты, такие как синтез витамина D в организме, лечение кожных заболеваний, эффекты стерилизации и т.д., они также несут в себе вредные аспекты, такие как солнечные ожоги, рак кожи, старение, заболевания, обусловленные светочувствительностью кожи, и мутагенез. Известно, что УФ-А проникает даже в слой дермы, что, в основном, вызывает пигментацию и старение кожи, и вовлечен в развитие заболеваний, обусловленных светочувствительностью кожи, а УФ-В связывают с солнечными ожогами, пигментацией и злокачественными новообразованиями кожи вследствие проникновения лучей высокой энергии в верхний слой эпидермиса и дерму.
Чтобы предотвратить эти побочные эффекты, вызываемые солнечными лучами, сделаны попытки блокирования солнечного света. Типы предназначенных для этой цели солнцезащитных агентов можно разделить на химические солнцезащитные агенты и физические солнцезащитные агенты. Химические солнцезащитные агенты прежде всего блокируют проникновение солнечного света посредством поглощения, при этом физические солнцезащитные агенты блокируют проникновение солнечного света посредством отражения и рассеивания солнечного света.
Химические солнцезащитные агенты содержат один или более компонентов, поглощающих УФ-свет, таких как пара-аминобензойная кислота (ПАБК), сложные эфиры ПАБК (амилдиметил-ПАБК, октилдиметил-ПАБК), циннаматы (циноксат), салицилаты (гомометилсалицилат), камфора и т.д., которые поглощают в основном УФ-В, и бензофенон (оксибензон, диоксибензон, сулизобензон), дибензоилметан, антранилат и т.д., поглощающие в основном УФ-А. Хотя эти химические солнцезащитные агенты могут обеспечить защитный эффект от УФ света за счет поглощения УФ лучей, некоторые из них могут вызывать раздражение кожи или глаз. В частности, известно, что ПАБК, сложные эфиры ПАБК, бензофеноны, циннаматы и т.д. вызывают контактный дерматит. Дополнительно сообщалось о проблемах, связанных с тем, что некоторые другие агенты обуславливают развитие реакций гиперчувствительности кожи, и поэтому применение химических солнцезащитных агентов и их используемое количество в некоторых странах ограничено.
Физические солнцезащитные агенты, которые содержат существующие в природе компоненты, защищают кожу за счет отражения и рассеивания проникающих в кожу УФ лучей. Например, такие физические солнцезащитные агенты, как диоксид титана, тальк (силикат магния), оксид магния, оксид цинка, каолин и т.д., могут обеспечить УФ-защитный эффект как от УФ-А, так и от УФ-В. Кроме того, их преимущество состоит в том, что они не обладают побочными эффектами, такими как контактный дерматит, и не смываются легко водой. Но они, наоборот, также имеют недостатки, связанные с тем, что при применении желаемой композиции физических солнцезащитных агентов поддержание их эффективного количества может быть затруднительным, и они оставляют белый оттенок при нанесении на кожу.
Микоспорин-подобные аминокислоты (МАК) представляют собой вещества, обнаруживаемые в природных организмах, и известные как эффективные поглотители УФ-А (320 нм - 400 нм) и УФ-В (290 нм - 320 нм). В природе существует 35 или более видов МАК в зависимости от типа аминокислот, циклогексенонового или циклогексениминового кольца, служащих в качестве предшественников. (Mar. Biol., 1991, 108: 157-166; Planta Med., 2015, 81: 813-820). Недавно в микроводорослях были обнаружены различные гликозилированные МАК, о которых сообщалось, что они играют роль превосходных антиоксидантов (Journal of Photochemistry and Photobiology, 2015, 142: 154-168). Дополнительно известно, что МАК не только обеспечивают защиту от УФ света, но также проявляют устойчивость к окислению, осмосу и тепловому стрессу (Comp. Biochem. Physiol. С Toxicol. Pharmacol., 2007, 146: 60-78; J. Photochem. Photobiol. В., 2007, 89: 29-35).
Тем не менее количество продуцируемой микроводорослями МАК слишком мало, на уровне нескольких мкг, а условия выделения, экстракции и очистки МАК путем культивирования микроводорослей сложны, и поэтому получить МАК на уровне крупномасштабного производства могло бы быть затруднительным.
Документация предшествующего уровня техники
Непатентная документация
Непатентный документ 1 Comp. Biochem. Physiol. В., 1995, 112: 105-114.
Непатентный документ 2 FEMS Microbiol Lett. 2007, 269: 1-10.
Непатентный документ 3 Ann. Rev. Physiol. 2002, 64: 223-262.
Непатентный документ 4 Mar. Biol. 1991, 108: 157 166.
Непатентный документ 5 Journal of Photochemistry and Photobiology B: Biology. 2015, 142: 154-168
Непатентный документ 6 Biol. Rev. 1999, 74: 311-345.
Непатентный документ 7 Mol. Biol. Evol. 2006, 23: 1437-1443.
Непатентный документ 8 Science, 2010, 329: 1653 1656.
Непатентный документ 9 Genomics 2010, 95: 120-128.
Непатентный документ 10 Geomicrobiol. J. 1997, 14: 231-241.
Непатентный документ 11 Comp. Biochem. Physiol. С Toxicol. Pharmacol. 2007. 146: 60-78.
Непатентный документ 12 Can. J. Bot. 2003. 81: 131-138.
Непатентный документ 13 J. Photochem. Photobiol. В., 2007, 89: 29-35.
Непатентный документ 14 J. Bacteriol. 2011. 193(21): 5923-5928.
Непатентный документ 15 PlantaMed. 2015. 81: 813-820
Непатентный документ 16 ACS Appl. Mater. Interfaces. 2015. 7: 16558-16564
Непатентный документ 17 Appl Environ Microbiol. 2016, 82(20): 6167-6173
Непатентный документ 18 ChemBioChem. 2015, 16: 320-327
Непатентный документ 19 Methods Mol Biol. 2013, 1073: 437
Непатентный документ 20 Enzyme Microb Technol., 2016, Jan, 82: 96-104
Непатентный документ 21 Nature Review, 2011, 9: 791-802
Описание
Техническая задача изобретения
Авторы настоящего изобретения приложили интенсивные усилия к повышению продукции МАК в микроорганизмах. В результате было подтверждено, что продукция МАК была повышена посредством различных исследований по инактивации активности 3-дегидрохиннатдегидратазы в микроорганизме, продуцирующем МАК, в результате чего было выполнено настоящее изобретение.
Техническое решение
Одним объектом настоящего изобретения является предоставление микроорганизма для продуцирования микроспорин-подобной аминокислоты, в котором активность 3-дегидрохиннатдегидратазы инактивирована по сравнению с немодифицированным микроорганизмом.
Другим объектом настоящего изобретения является предоставление способа продуцирования микроспорин-подобной аминокислоты, включающего:
культивирование микроорганизма; и
извлечение микроспорин-подобной аминокислоты из культивируемого микроорганизма или из среды.
Преимущественные эффекты изобретения
Микроорганизм по настоящему изобретению проявляет улучшенную способность к продуцированию микоспорин-подобной аминокислоты и, таким образом, может быть эффективно использован в получении микоспорин-подобной аминокислоты.
Подробное описание предпочтительных воплощений изобретения
Конкретные особенности настоящего изобретения могут быть пояснены ниже.
Тем не менее раскрытые в описании настоящего изобретения пояснения и воплощения могут быть применены к другим пояснениям и воплощениям соответственно. Таким образом, все комбинации различных элементов, раскрытых в настоящем документе, относятся к объему настоящего изобретения. Кроме того, объем настоящего изобретения не подлежит ограничению конкретными описаниями, приведенными в настоящем документе ниже. Кроме того, специалист среднего уровня в данной области техники с помощью лишь традиционных экспериментов сможет распознать или подтвердить множество эквивалентов конкретных аспектов изобретения, описанных в данной заявке. Более того, также подразумевают, что эти эквиваленты должны быть включены в настоящее изобретение.
Для достижения описанных выше целей в одном аспекте настоящего изобретения предложен микроорганизм для продуцирования микроспорин-подобной аминокислоты, в котором активность 3-дегидрохиннатдегидратазы инактивирована по сравнению с немодифицированным микроорганизмом.
При использовании в настоящем документе термин «3-дегидрохиннатдегидратаза» относится к ферменту, катализирующему обратимую реакцию, представленную на приведенной ниже схеме реакции, и может специфично преобразовывать 3-дегидрохиннат в 3-дегидрошикимат, но не ограничен этим.
Схема реакции
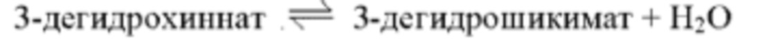
Используемый в настоящем документе термин «инактивация» относится к случаю, где активность белка-фермента, которой исходно обладает микроорганизм, ослаблена по сравнению с его эндогенной активностью или активностью до модификации; к случаю, где отсутствует какая-либо экспрессия белка; или к случаю, где белок экспрессируется, но не проявляет активности. Понятие инактивации включает в себя случай, где активность самого фермента ослаблена или отсутствует по сравнению с активностью фермента, которой исходно обладает микроорганизм, вследствие модификации в кодирующем этот фермент полинуклеотиде и т.д.; случай, где общий уровень активности фермента внутри клетки снижен или фермент удален по сравнению с микроорганизмом дикого типа вследствие ингибирования экспрессии кодирующего фермент гена или ингибирования его трансляции и т.д.; случай, где кодирующий фермент ген или его участок делетирован; и их комбинацию, но не ограничено этим. Термин «немодифицированный микроорганизм» относится к микроорганизму, обладающему активностью конкретного белка, которой исходно обладает родительский штамм до его модификации, при этом признаки микроорганизма модифицируют посредством генетической модификации в конкретном белке, которым обладает микроорганизм по сравнению с модификацией вследствие естественных или искусственных факторов. При использовании в настоящем документе термин «немодифицированный микроорганизм» можно использовать взаимозаменяемо с термином «микроорганизм, обладающий эндогенной активностью».
Инактивация ферментативной активности может быть достигнута различными способами, хорошо известными в данной области техники. Примеры этих способов включают 1) способ делетирования всего гена, кодирующего фермент на хромосоме, или его участка; 2) способ модифицирования регуляторной последовательности экспрессии таким образом, чтобы экспрессия гена, кодирующего фермент на хромосоме, была снижена; 3) способ модифицирования последовательности кодирующего белок гена на хромосоме таким образом, чтобы активность этого белка была устранена или ослаблена; 4) способ введения антисмыслового олигонуклеотида, который комплементарно связывается с транскриптом гена, кодирующего белок на хромосоме (например, антисмысловая РНК); 5) способ искусственного присоединения последовательности, комплементарной последовательности Шайна - Дальгарно гена, кодирующего белок на хромосоме, выше этой последовательности Шайна - Дальгарно с образованием вторичной структуры, в результате чего адгезия рибосомы становится невозможной; и 6) способ обратно-транскрипционного (ОТ) конструирования, при котором к 3'-концу открытой рамки считывания (ОРС) кодирующей белок полинуклеотидной последовательности присоединяют промотор, с которого должна идти обратная транскрипция, или комбинацию этих способов, но без конкретного ограничения этими ими.
Способ делетирования всего гена, кодирующего фермент на хромосоме, или его участка можно осуществлять путем замещения кодирующего эндогенный целевой белок полинуклеотида внутри хромосомы полинуклеотидом, имеющим частично делетированную нуклеиново-кислотную последовательность, или геном-маркером с помощью вектора для встраивания в хромосому микроорганизма. В качестве примера способа делетирования всего полинуклеотида или его участка можно, не ограничиваясь этим, использовать способ делетирования полинуклеотида посредством гомологичной рекомбинации.
Способ модифицирования регуляторной последовательности экспрессии можно осуществлять путем индукции модификации в регуляторной последовательности экспрессии посредством делеции, вставки, консервативной или неконсервативной замены, или их комбинации с целью дополнительного ослабления активности регуляторной последовательности экспрессии; или путем замены этой последовательности нуклеиново-кислотной последовательностью, обладающей ослабленной активностью. Регуляторная последовательность экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую домен связывания рибосомы, и последовательность регуляции терминации транскрипции и трансляции, но не ограничена ими.
Способ модифицирования последовательности гена на хромосоме можно осуществлять путем индукции модификации в последовательности гена посредством делеции, вставки, консервативной или неконсервативной замены, или их комбинации таким образом, чтобы дополнительно ослабить активность фермента; или путем замещения последовательности модифицированной последовательностью этого гена, обладающего более слабой активностью или модифицированной последовательностью этого гена, активность которого полностью отсутствует, но не ограничен этим.
Полинуклеотид может быть описан как ген в тех случаях, где он относится к комплексу полинуклеотидов, способному к выполнению функций. В описании настоящего изобретения термины «полинуклеотиды» и «гены» можно использовать взаимозаменяемо, и термины «полинуклеотидные последовательности» и «нуклеотидные последовательности» можно использовать взаимозаменяемо.
Используемый в настоящем документе термин «участок», хотя он может изменяться в зависимости от видов полинуклеотида, может конкретно относиться к участку длиной от 1 до 300 нуклеотидов, более конкретно от 1 до 100 нуклеотидов и еще более конкретно от 1 до 50 нуклеотидов, но в частности не ограничен ими.
В микроорганизме по настоящему изобретению активность по меньшей мере одного фермента, выбранного из группы, состоящей из 2-дегидро-3-дезоксифосфогептонатальдолазы, фосфоенолпируватсинтетазы, транскетолазы I/II и 3-дегидрохиннатсинтазы, в частности, активность одного или более, двух или более, трех или более, или всех этих ферментов, может быть усилена по сравнению с немодифицированным микроорганизмом.
2-Дегидро-3-дезоксифосфогептонатальдолаза относится к ферменту, катализирующему обратимую реакцию, представленную на приведенной ниже схеме реакции, и может специфично синтезировать 3-дезокси-арабино-гептулозонат-7-фосфат, но не ограниченному этим.
Схема реакции
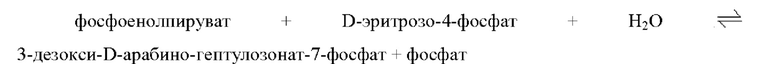
Фосфоенолпируватсинтетаза относится к ферменту, катализирующему обратимую реакцию, представленную на приведенной ниже схеме реакции, и может специфично синтезировать фосфоенолпируват, но не ограниченному этим.
Схема реакции
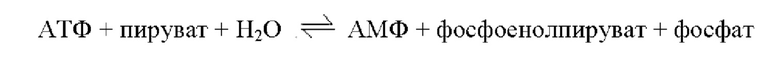
Транскетолаза I/II относится к ферменту, катализирующему обратимую реакцию, представленную на приведенной ниже схеме реакции.
Схема реакции
Седогептулозо-7-фосфат + D-глицеральдегид-3-фосфат = D-рибозо-5-фосфат + В-ксилулозо-5-фосфат
3-Дегидрохиннатсинтаза относится к ферменту, катализирующему обратимую реакцию, представленную на приведенной ниже схеме реакции, и может специфично синтезировать 3-дегидрохиннат (3-DHQ), но не ограничена этим.
Схема реакции
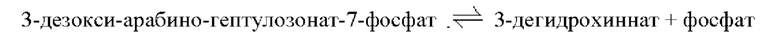
При использовании в настоящем документе термин «усиление активности» означает, что активность белка-фермента либо введена, либо усилена по сравнению с эндогенной активностью, которой обладает микроорганизм, или с активностью до модификации. «Введение» активности означает, что активность конкретного полипептида, которой микроорганизм исходно не обладал, экспрессируется естественным или искусственным путем. Например, усиление активности может включать в себя либо усиление за счет введения 2-дегидро-3-дезоксифосфогептонатальдолазы, фосфоенолпируватсинтетазы, транскетолазы VII и/или 3-дегидрохиннатсинтазы; либо усиление эндогенной активности 2-дегидро-3-дезоксифосфогептонатальдолазы, фосфоенолпируватсинтетазы, транскетолазы I/II и/или 3-дегидрохиннатсинтазы. В частности, усиление активности в настоящем изобретении можно осуществлять следующими способами:
1) способом увеличения числа копий полинуклеотида, кодирующего ферменты;
2) способом модифицирования регуляторной последовательности экспрессии таким образом, чтобы экспрессия полинуклеотида была повышена;
3) способом модифицирования полинуклеотидной последовательности на хромосоме таким образом, чтобы активность ферментов была усилена; и
4) способом модификации, усиливающей активность за счет комбинирования описанных выше способов, но не ограничиваясь ими.
Увеличение числа копий полинуклеотида в описанном выше способе 1) можно осуществить в такой форме, чтобы полинуклеотид был функционально связан с вектором, или путем встраивания полинуклеотида в хромосому клетки-хозяина, но конкретно не ограничиваясь этим. Дополнительно в аспекте увеличения числа копий его можно осуществлять путем введения в клетку-хозяина чужеродного полинуклеотида, проявляющего активность фермента, или модифицированного полинуклеотида, в котором кодоны чужеродного полинуклеотида оптимизированы. Чужеродный полинуклеотид можно использовать без ограничения его происхождением или последовательностью при условии, что он проявляет активность, идентичную или подобную активности фермента. Специалисты в данной области техники могут осуществить это введение, выбрав подходящий способ трансформации, известный в данной области техники, и в случае экспрессии введенных в клетку-хозяина полинуклеотидов может продуцироваться фермент, таким образом, повышая активность.
Кроме того, модификацию регуляторной последовательности экспрессии в способе 2) таким образом, чтобы экспрессия полинуклеотида была повышена, можно осуществить путем индукции модификации последовательности посредством делеции, вставки или неконсервативной или консервативной замены нуклеиново-кислотной последовательности, либо их комбинации таким образом, чтобы дополнительно усилить активность регуляторной последовательности экспрессии, либо путем замещения нуклеотидной последовательностью, обладающей более сильной активностью, но конкретно не ограничиваясь этим. Дополнительно регуляторная последовательность экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую домен связывания рибосомы, последовательность регуляции терминации транскрипции и трансляции и т.д., но без особых ограничений.
В частности, с областью, расположенной выше единицы экспрессии полинуклеотида, может быть связан сильный гетер о логичный промотор вместо исходного промотора. Примеры сильного промотора включают промотор CJ7, промотор lysCP1, промотор EF-Tu, промотор groEL, промотор асеА или асеВ и т.д. Более конкретно уровень экспрессии кодирующего фермент полинуклеотида можно повысить путем функционального связывания полинуклеотида с промотором lysCP1 (WO 2009/0966S9) или с промотором CJ7 (WO 2006/065095), но не ограничиваясь ими.
Кроме того, модификацию полинуклеотидной последовательности на хромосоме в способе 3) можно осуществить путем индукции модификации в регуляторной последовательности экспрессии посредством делеции, вставки или неконсервативной или консервативной замены нуклеиново-кислотной последовательности, либо их комбинации таким образом, чтобы дополнительно усилить активность полинуклеотидной последовательности, либо путем замещения этой полинуклеотидной последовательности модифицированной полинуклеотидной последовательностью, обладающей более сильной активностью, но конкретно не ограничиваясь этим.
Наконец, способ модификации с усилением активности посредством комбинирования способов 1) - 3) в способе 4) можно осуществить путем комбинированного применения по меньшей мере одного из следующих способов: увеличения числа копий полинуклеотида, кодирующего белок; модифицирования регуляторной последовательности экспрессии таким образом, чтобы экспрессия полинуклеотида была повышена; модифицирования полинуклеотидной последовательности на хромосоме и модифицирования чужеродного полинуклеотида, проявляющего активность фермента, или оптимизированного по кодонам полинуклеотида.
Используемый в настоящем документе термин «вектор» относится к конструкции ДНК, содержащей нуклеотидную последовательность кодирующего целевой белок полинуклеотида, которая функционально связана с подходящей регуляторной последовательностью так, чтобы целевой белок мог экспрессироваться в подходящей клетке-хозяине. Регуляторная последовательность включает в себя промотор, способный к инициации транскрипции, любую последовательность оператора для контроля транскрипции, последовательность, кодирующую домен связывания рибосомы соответствующей мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После того, как вектор трансформирован в подходящую клетку-хозяина, он может реплицироваться или функционировать независимо от генома клетки-хозяина, а может быть интегрирован в сам геном клетки-хозяина.
Используемый в настоящем изобретении вектор конкретно не ограничен при условии, что он может реплицироваться в клетке-хозяине, и можно использовать любой вектор, известный в данной области техники. Примеры традиционно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A, Charon21A и т.д., а в качестве плазмидного вектора можно использовать векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и т.д. В частности, можно использовать векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д., но вектор конкретно не ограничен ими.
Вектор, который можно использовать в настоящем изобретении, конкретно не ограничен, и можно использовать известный экспрессионный вектор. Дополнительно, кодирующий целевой белок полинуклеотид может быть вставлен в хромосому с помощью вектора для вставки в хромосому. Вставку полинуклеотида в хромосому можно осуществить любым способом, известным в данной области техники, например, посредством гомологической рекомбинации, но не ограничиваясь этим. Для подтверждения вставки в хромосому можно дополнительно включать селективный маркер. Селективный маркер используют для отбора трансформированной вектором клетки, т.е. для подтверждения вставки целевых молекул нуклеиновой кислоты, и можно использовать маркеры, способные обеспечивать селективные фенотипы, такие как устойчивость к лекарственным средствам, потребность в питательных веществах, устойчивость к цитотоксическим агентам или экспрессия поверхностных белков. В окружающей среде, обработанной этими селективными агентами, могут выживать или экспрессировать другие фенотипические признаки только клетки, экспрессирующие селективные маркеры, и таким образом можно легко отобрать трансформированные клетки.
Используемый в настоящем документе термин «трансформация» относится к процессу введения в клетку-хозяина вектора, включающего в себя полинуклеотид, кодирующий целевой полипептид, и, следовательно, обеспечивающего экспрессию кодируемого полинуклеотидом белка в клетке-хозяине. До тех пор, пока трансформированный полинуклеотид может экспрессироваться в клетке-хозяине, не имеет значения, встроен ли он в хромосому клетки-хозяина и расположен внутри нее, или расположен все хромосомы, и могут быть включены оба случая. Кроме того, полинуклеотид включает ДНК и РНК, которая кодирует целевой полипептид. Полинуклеотид может быть введен в любой форме при условии возможности его введения в клетку-хозяина и экспрессии в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой генную конструкцию, включающую в себя все необходимые элементы для самостоятельной экспрессии. Экспрессионная кассета традиционно может включать в себя промотор, функционально связанный с полинуклеотидом, терминатор, домен связывания рибосомы и стоп-кодон. Экспрессионная кассета может иметь форму экспрессионного вектора, способного к самостоятельной репликации. Кроме того, полинуклеотид можно вводить в клетку-хозяина как он есть и функционально связывать с последовательностью, необходимой для его экспрессии в клетке-хозяине, но без ограничений. Способ трансформации вектора включает любой способ введения нуклеиновой кислоты в клетку, и его можно осуществлять, выбирая подходящую стандартную методику, известную в данной области техники, в зависимости от клетки-хозяина. Например, трансформацию можно осуществлять с помощью методики электропорации, кальций-фосфатной (CaPO4) преципитации, кальций-хлоридной (CaCl2) преципитации, микроинъекции, методики, основанной на использовании полиэтиленгликоля (ПЭГ), методики, основанной на использовании ДЭАЭ-декстрана, катионно-липосомной методики, методики, основанной на использовании ацетата лития-ДМСО, и т.д., но способ не ограничен этими методиками.
Кроме того, используемый выше термин «функционально связанный» относится к функциональному соединению между описанной выше полинуклеотидной последовательностью и последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению. Функциональное связывание может быть получено с использованием известной в данной области методики рекомбинантных ДНК, а сайт-специфическое расщепление и лигирование ДНК можно осуществить с помощью известных в данной области техники ферментов, таких как лиазы, лигазы и т.д., но не ограничиваясь ими.
Генетическая информация о 3-дегидрохиннатдегидратазе, 2-дегидро-3-дезоксифосфогептонатальдолазе, фосфоенолпируватсинтетазе, транскетолазе и 3-дегидрохиннатсинтазе может быть получена из известной базы данных, и ее примеры включают GenBank Национального центра биотехнологической информации (National Center for Biotechnology Information; NCBI) и т.д., но известная база данных не ограничена ею.
Происхождение или последовательности 3-дегидрохиннатдегидратазы, 2-дегидро-3-дезоксифосфогептонатальдолазы, фосфоенолпируватсинтетазы, транскетолазы и 3-дегидрохиннатсинтазы могут быть не ограничены, поскольку в зависимости от видов бактерий или микроорганизмов могут существовать различия по аминокислотной последовательности белка, проявляющего эту активность.
В частности, 3-дегидрохиннатдегидратаза может представлять собой белок, включающий в себя аминокислотную последовательность SEQ ID NO: 72 или 80, а 2-дегидро-3-дезоксифосфогептонатальдолаза, фосфоенолпируватсинтетаза, транскетолаза и 3-дегидрохиннатсинтаза могут представлять собой белки, включающие в себя аминокислотные последовательности SEQ ID NO: 74, 76, 78 и 84 соответственно, но не ограниченные ими. В описании настоящего изобретения «белок, включающий в себя аминокислотную последовательность», можно использовать взаимозаменяемо с выражением «белок, имеющий аминокислотную последовательность» или «белок, состоящий из аминокислотной последовательности».
Дополнительно в описании настоящего изобретения ферменты могут включать не только белок, имеющий описанную выше аминокислотную последовательность SEQ ID, но также белок, имеющий гомологию 80% или более, в частности, 90% или более, более конкретно 95% или более, еще более конкретно 99% или более с описанными выше аминокислотными последовательностями при условии, что белок обладает биологической активностью, идентичной или соответствующей активности каждого из этих ферментов.
Кроме того, очевидно, что если аминокислотная последовательность имеет гомологию с описанными выше последовательностями, аминокислотная последовательность с делецией, модификацией, заменой или добавлением участка последовательности также включена в объем настоящего изобретения при условии, что эта аминокислотная последовательность обладает биологической активностью, по существу идентичной или соответствующей активности белка-фермента, имеющего описанную выше аминокислотную последовательность SEQ ID NO.
Полинуклеотид, кодирующий 3-дегидрохиннатдегидратазу, 2-дегидро-3-дезоксифосфогептонатальдолазу, фосфоенолпируватсинтетазу, транскетолазу и 3-дегидрохиннатсинтазу по настоящему изобретению, может включать полинуклеотид, кодирующий описанную выше аминокислотную последовательность SEQ ID NO, или полинуклеотид, кодирующий белок, имеющий гомологию 80% или более, в частности, 90% или более, более конкретно 95% или более, еще более конкретно 99% или более с описанными выше аминокислотными последовательностями при условии, что белок обладает биологической активностью, идентичной или соответствующей активности каждого из этих ферментов.
Дополнительно, в кодирующей области полинуклеотида, кодирующего 2-дегидро-3-дезоксифосфогептонатальдолазу, фосфоенолпируватсинтетазу, транскетолазу и 3-дегидрохиннатсинтазу, с учетом кодонов, предпочитаемых организмами для экспрессии белка вследствие вырожденности кодонов, могут быть выполнены различные модификации в пределах объема без изменения аминокислотной последовательности полипептида. Соответственно, полинуклеотид может включать любую полинуклеотидную последовательность, кодирующую каждый белок-фермент, без ограничений.
Кроме того, может быть без ограничений включен зонд, который может быть получен из последовательности известного гена, например, из любой последовательности, которая гибридизуется с последовательностью, комплементарной всей нуклеотидной последовательности или ее участку в жестких условиях гибридизации, кодирующей белок, обладающий активностью 3-дегидрохиннатдегидратазы, 2-дегидро-3-дезоксифосфогептонатальдолазы, фосфоенолпируватсинтетазы, транскетолазы и 3-дегидрохиннатсинтазы.
Используемый в настоящем документе термин «гомология» относится к степени соответствия данной аминокислотной последовательности или нуклеотидной последовательности и может быть выражен в процентах. В настоящем описании гомологичная последовательность, обладающая активностью, идентичной или подобной активности данной аминокислотной последовательности или нуклеотидной последовательности, может быть указана в виде «% гомологии». Например, гомология может быть подтверждена с помощью стандартного программного обеспечения для расчета таких показателей, как оценка в баллах, идентичность и подобие, в частности BLAST 2.0, или путем сравнения последовательностей с помощью экспериментов по гибридизации в определенных жестких условиях, и определенные подходящие жесткие условия гибридизации находятся в пределах компетенции специалистов в данной области техники и могут быть определены способом, известным специалистам в данной области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York). Термин «жесткие условия» относятся к условиям, дающим возможность для специфичной гибридизации между полинуклеотидами. Такие условия конкретно описаны в литературе (например, J. Sambrook et al.).
Используемый в настоящем документе термин «микоспорин-подобные аминокислоты (МАК)» относится к циклическим соединениям, поглощающим ультрафиолетовые лучи. В описание настоящего изобретения может быть включена любая микоспорин-подобная аминокислота без ограничений при условии, что она может поглощать УФ лучи, но более конкретно она может представлять собой соединение, содержащее центральное кольцо циклогексенона или циклогексенимина; или соединение, в котором различные вещества, такие как аминокислота и т.д., связаны с центральным кольцом. Более конкретно, она может представлять собой микоспорин-2-глицин, палитинол, палитеновую кислоту, дезоксигадузол, микоспорин-метиламин-треонин, микоспорин-глицин-валин, палитин, астерина-330 (asterina-330), шинорин (shinorine), порфиру-334, эугалотек-362 (euhalothece-362), микоспорин-глицин, микоспорин-орнитин, микоспорин-лизин, микоспорин-глутаминовая кислота-глицин, микоспорин-метиламин-серин, микоспорин-таурин, палитен, палитин-серин, палитин-серин-сульфат, палитинол, усуджирен (usujirene) или их комбинацию.
В описании настоящего изобретения микоспорин-подобные аминокислоты могут использоваться взаимозаменяемо с МАК и МАА (англ.).
Используемый в настоящем документе термин «микроорганизм для продуцирования микоспорин-подобной аминокислоты» относится к микроорганизму, включающему в себя ген фермента, вовлеченного в биосинтез микоспорин-подобных аминокислот, или кластер этих генов. Кроме того, используемый в настоящем документе термин «ген биосинтеза микоспорин-подобной аминокислоты» относится к гену, кодирующему фермент, вовлеченный в биосинтез микоспорин-подобных аминокислот, а также включает в себя кластер этих генов. Ген биосинтеза микоспорин-подобной аминокислоты включает чужеродные и/или эндогенные гены микроорганизма при условии, что микроорганизм, включающий в себя этот ген, может продуцировать микоспорин-подобные аминокислоты. Чужеродные гены могут быть гомологичными и/или гетерологичными.
Ген биосинтеза микоспорин-подобной аминокислоты может быть не ограничен видом микроорганизма, из которого получен этот ген, при условии, что включающий его микроорганизм продуцирует фермент, вовлеченный в биосинтез микоспорин-подобных аминокислот и, следовательно, продуцирует микоспорин-подобные аминокислоты. В частности, он может представлять собой Anabaena variabilis, Nostoc punctiforme, Nodularia spumigena, Cyanothece sp. PCC 7424, Lyngbya sp. PCC 8106, Microcystis aeruginosa, Microcoleus chthonoplastes, Cyanothece sp. АТСС 51142, Crocosphaera watsonii, Cyanothece sp. CCY 0110, Cylindrospermum stagnate sp. PCC 7417, Aphanothece halophytica или Trichodesmium erythraeum, которые являются видами цианобактерий, или Magnaporthe oryzae, Pyrenophora tritici-repentis, Aspergillus clavatus, Nectria haematococca, Aspergillus nidulans, Gibberella zeae, Verticillium albo-atrum, Botryotinia fuckeliana или Phaeosphaeria nodorum, являющиеся видами грибов, или Nematostella vectensis, Heterocapsa triquetra, Oxyrrhis marina, Karlodinium micrum, Actinosynnema mirum, etc., но не ограничен этими видами.
В соответствии с одним воплощением изобретения микроорганизм для продуцирования микоспорин-подобной аминокислоты по настоящему изобретению может включать в себя ген биосинтеза микоспорин-подобной аминокислоты.
В частности, ген биосинтеза микоспорин-подобной аминокислоты может быть не ограничен названием фермента или микроорганизмов, из которых они получены, при условии, что микроорганизм может продуцировать микоспорин-подобные аминокислоты, и может, в частности, включать по меньшей мере один, в частности, один или более, два или более, либо три или более, либо все белки-ферменты, выбранные из группы, состоящей из 2-деметил-4-дезоксигадузол-синтазы, О-метилтрансферазы и C-N лигазы; или ген, кодирующий белок-фермент, обладающий активностью, идентичной и/или подобной активности этих ферментов.
Например, 2-деметил-4-дезоксигадузолсинтаза представляет собой фермент, который преобразует седогептулозо-7-фосфат в 2-деметил-4-дезоксигадузол. О-метилтрансфераза представляет собой фермент, который преобразует 2-деметил-4-дезоксигадузол в 4-дезоксигадузол, а глицирование 4-дезоксигадузола катализирует C-N лигаза.
Дополнительно микроорганизм для продуцирования микоспорин-подобной аминокислоты может включать в себя ген фермента, обладающего активностью присоединения к микоспорин-подобным аминокислотам дополнительных аминокислотных остатков, или кластер таких генов. Этот ген или кластер генов может быть не ограничен названием ферментов или микроорганизмов, из которых они получены, при условии, что микроорганизм для продуцирования микоспорин-подобной аминокислоты может продуцировать микоспорин-подобные аминокислоты, к которым присоединены два или более аминокислотных остатков, и может, в частности, включать ген по меньшей мере одного, в частности, одного или более, двух или более, трех или более, либо всех белков-ферментов, выбранных из группы, состоящей из синтетазы нерибосомных пептидов (NRPS), фермента, подобного синтетазе нерибосомных пептидов (NRPS-подобный фермент), и D-Ala-D-Ala лигазы (DDL); или ген, кодирующий белок-фермент, обладающий активностью, идентичной и/или подобной активности этих ферментов. Некоторые микоспорин-подобные аминокислоты могут включать второй аминокислотный остаток в микоспорин-глицине. По меньшей мере один фермент, выбранный из группы, состоящей из синтетазы нерибосомных пептидов, фермента, подобного синтетазе нерибосомных пептидов, и D-Ala-D-Ala лигазы, может присоединять второй аминокислотный остаток к микоспорин-глицину.
В соответствии с одним воплощением изобретения микроорганизм для продуцирования микоспорин-подобной аминокислоты может включать в себя любой фермент без ограничения названием ферментов или видов микроорганизмов, из которых они получены, при условии, что он обладает такой же активностью присоединения второй аминокислоты к микоспорин-глицину, как синтетаза нерибосомных пептидов, фермент, подобный синтетазе нерибосомных пептидов, и D-Ala-D-Ala лигаза.
В одном воплощении изобретения фермент, подобный синтетазе нерибосомных пептидов (Ava_3855), обнаруженный в Anabaena variabilis, или D-Ala-D-Ala лигаза (NpF5597), обнаруженная в Nostoc punctiforme, могут продуцировать шинорин (shinorine) путем присоединения остатка серина к микоспорин-глицину. В другом воплощении изобретения микоспорин-2-глицин может быть образован путем присоединения второго остатка глицина посредством гомолога D-Ala-D-Ala лигазы (Ар_3855), обнаруженного в Aphanothece halophytica. Аналогично, в Actinosynnema mirum серин или аланин может быть присоединен посредством D-Ala-D-Ala лигазы с образованием шинорина или микоспорин-глицин-аланина. Микроорганизм в соответствии с одним воплощением настоящего описания может включать в себя фермент, подходящий для продуцирования желаемых микоспорин-подобных аминокислот путем выбора из описанных выше ферментов или ферментов, обладающих активностью, идентичной и/иди подобной активности этих ферментов.
2-Деметил-4-дезоксигадузолсинтаза, О-метилтрансфераза, C-N лигаза, синтетаза нерибосомных пептидов, фермент, подобный синтетазе нерибосомных пептидов, и/или D-Ala-D-Ala лигаза, которые могут быть использованы в настоящем изобретении, могут быть не ограничены видами микроорганизмов, из которых они получены, и может быть включен любой фермент без ограничений, если известно, что он выполняет идентичные и/или подобные функции и обладает соответствующими активностями. Дополнительно диапазон числовых значений гомологии между этими ферментами может быть не ограничен. Например, My1A, My1B, My1D, My1E и My1C Cylindrospermum stagnate PCC 7417 гомологичны 2-деметил-4-дезоксигадузолсинтазе, О-метилтрансферазе, C-N лигазе и D-Ala-D-Ala лигазе, полученным из Anabaena variabilis и Nostoc punctiforme, и подобие между этими гомологами составляет от примерно 61 до 88% (Appl Environ Microbiol, 2016, 82(20), 6167-6173; J Bacteriol, 2011, 193(21), 5923-5928). Таким образом, фермент, который может быть использован в настоящем изобретении, может быть конкретно не ограничен видами микроорганизмов, из которых он получен, или гомологией последовательности, если известно, что он проявляет идентичные и/или подобные функции и эффекты. Далее, непатентная документация, раскрытая в предшествующем уровне техники, включена в настоящий документ посредством ссылки во всей своей полноте.
Дополнительно ген биосинтеза микоспорин-подобной аминокислоты может представлять собой полинуклеотид, кодирующий белок, включающий в себя аминокислотную последовательность SEQ ID NO: 2, 4, 86, 88, 90, 92, 94, 96, 98, 100, 102 или 104, но не ограниченный ими.
Дополнительно ген биосинтеза микоспорин-подобной аминокислоты может включать в себя нуклеотидную последовательность, кодирующую белок, включающий в себя аминокислотную последовательность, имеющую гомологию 50%, 60% или 70% или более, в частности 80% или более, более конкретно 90% или более, еще более конкретно 95% и еще более конкретно 99%, с аминокислотной последовательностью SEQ ID NO: 2, 4, 86, 88, 90, 92, 94, 96, 98, 100, 102 или 104, и может без ограничений включать в себя любую нуклеотидную последовательность, кодирующую белок, имеющий гомологию, отклоняющуюся от описанной выше, при условии, что микроорганизм продуцирует микоспорин-подобные аминокислоты. В частности, ген биосинтеза микоспорин-подобной аминокислоты может включать в себя без ограничений нуклеотидную последовательность SEQ ID NO: 1, 3, 85, 84, 89, 91, 93, 95, 97, 99, 101 или 103.
Кроме того, если аминокислотная последовательность имеет гомологию с описанной выше аминокислотной последовательностью, очевидно, что в объем настоящего изобретения также включена аминокислотная последовательность с делецией, модификацией, заменой или добавлением участка последовательности при условии, что эта аминокислотная последовательность обладает биологической активностью, идентичной или соответствующей активности белка, имеющего описанную выше аминокислотную последовательность SEQ ID NO.
Дополнительно, принимая во внимание кодоны, предпочитаемые организмами для экспрессии белка, вследствие вырожденности кодонов, могут быть выполнены различные модификации в кодирующей области в объеме, не измененяющем аминокислотную последовательность полипептида. Соответственно, ген биосинтеза микоспорин-подобной аминокислоты может включать в себя без ограничений любую нуклеотидную последовательность, кодирующую белок, вовлеченный в синтез микоспорин-подобных аминокислот.
Кроме того, может быть без ограничений включен зонд, который может быть получен из последовательности известного гена, например, из любой последовательности, которая гибридизуется в жестких условиях гибридизации с последовательностью, комплементарной всей нуклеотидной последовательности, кодирующей белок, вовлеченный в биосинтез микоспорин-подобных аминокислот.
В соответствии с одним воплощением изобретения микроорганизм для продуцирования микоспорин-подобной аминокислоты может включать в себя гены биосинтеза микоспорин-подобных аминокислот, полученных из источников различного происхождения.
В описании настоящего изобретения инактивацию белка, усиление активности белка и/или введение гена в настоящем документе можно осуществлять одновременно, последовательно или в обратном порядке.
Используемый в настоящем документе термин «микроорганизм для продуцирования микоспорин-подобной аминокислоты» обладает эндогенными и/или экзогенно введенными генами биосинтеза микоспорин-подобных аминокислот и, таким образом, может продуцировать микоспорин-подобные аминокислоты, и дополнительно он может представлять собой микроорганизм, у которого способность к продуцированию микоспорин-подобных аминокислот повышена за счет инактивации эндогенной активности 3-дегидрохиннатдегидратазы. Введение гена биосинтеза микоспорин-подобной аминокислоты и инактивацию 3-дегидрохиннатдегидратазы можно осуществлять одновременно, последовательно или в обратном порядке.
Дополнительно микроорганизм по настоящему изобретению может представлять собой природный микроорганизм, исходно имеющий ген биосинтеза микоспорин-подобной аминокислоты; и микроорганизм, в который введен гомологичный и/или гетерологичный ген биосинтеза микоспорин-подобной аминокислоты, но не ограничен ими.
Дополнительно микроорганизм по настоящему изобретению может представлять собой микроорганизм, обладающий усиленной активностью фермента, кодируемого эндогенным и/или введенным геном, связанным с биосинтезом микоспорин-подобной аминокислоты, но не ограничен ими.
Дополнительно микроорганизм по настоящему изобретению может не иметь ограничений при условии, что он обладает активностью 3-дегидрохиннатдегидратазы до модификации, и, в частности, он может представлять собой микроорганизм рода Corynebacterium, микроорганизм рода Escherichia или дрожжи.
Микроорганизм рода Corynebacterium может, в частности, представлять собой Corynebacterium glutamicum, Corynebacterium ammoniagenes, Brevibacterium lactofermentum, Brevibacterium flavum, Corynebacterium thermoaminogenes, Corynebacterium efficiens и т.д. и более конкретно он может представлять собой Corynebacterium glutamicum, но не ограничен ими.
Микроорганизм рода Escherichia может, в частности, представлять собой Escherichia albertii, Escherichia coli, Escherichia fergusonii, Escherichia hermannii, Escherichia vulneris, и т.д., и более конкретно он может представлять собой Escherichia coli, но не ограничен ими.
Дрожжи, в частности, могут представлять собой микроорганизм, принадлежащий к роду Saccharomycotina, Taphrinomycotina класса Ascomycota или Agaricomycotina, Pucciniomycotina класса Basidiomycota и т.д., и более конкретно он может представлять собой микроорганизм рода Saccharomyces, микроорганизм рода Schizosaccharomyces, микроорганизм рода Phaffia, микроорганизм рода Kluyveromyces, микроорганизм рода Pichia или микроорганизм рода Candida, и еще более конкретно он может представлять собой Saccharomyces cerevisiae, но не ограничен ими.
В настоящем изобретении в дрожжи для продуцирования микоспорин-подобной аминокислоты может быть введен ген, кодирующий 3-дегидрохиннатсинтазу, или может обладать усиленной активностью 3-дегидрохиннатсинтазы. Например, если в дрожжах весь ген ARO1 или его участок делетирован в целях инактивации активности 3-дегидрохиннатдегидратазы, функция 3-дегидрохиннатдегидратазы утрачивается, и, следовательно, синтез 3-DHQ может быть затруднительным. Таким образом, если в дрожжах делетирован весь ген ARO1 или его участок, в них может быть введен ген 3-дегидрохиннатсинтазы (например, ген aroB), но без ограничений.
В другом аспекте настоящего изобретения предложен способ продуцирования микроспорин-подобной аминокислоты, включающий:
культивирование микроорганизма по настоящему изобретению; и
извлечение микроспорин-подобной аминокислоты из культивируемого микроорганизма или из среды.
«Микроорганизм» и «микоспорин-подобные аминокислоты» являются такими, как определено выше.
Используемый в настоящем документе термин «культивирование» относится к культивированию микроорганизма в подобранной соответствующим образом окружающей среде. Процесс культивирования по настоящему изобретению может быть выполнен в соответствии с подходящей средой и условиями культивирования, известными в данной области техники. Процесс культивирования может быть легко скорректирован специалистами в данной области техники для применения в соответствии с выбранным микроорганизмом. Стадию культивирования микроорганизма можно осуществлять известным способом периодического культивирования, непрерывного культивирования, способа периодического культивирования с подпиткой и т.д., но в частности не ограничиваясь ими. Среда, используемая для культивирования микроорганизма по настоящему изобретению, и другие условия культивирования конкретно не ограничены, но можно использовать любую среду, применяемую для традиционного культивирования микроорганизма. В частности, микроорганизм по настоящему изобретению можно культивировать в традиционной среде, содержащей подходящие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины и т.д., в аэробных условиях при регулировании температуры, рН и т.д. В частности, рН культуры можно регулировать до подходящего рН (например, рН от 5 до 9, в частности, рН от 6 до 8 и наиболее конкретно рН 6,8), используя основное соединение (например, гидроксид натрия, гидроксид калия или аммиак) или кислое соединение (например, фосфорную кислоту или серную кислоту), но не ограничиваясь ими. Дополнительно для поддержания аэробного состояния культуры в культуру можно вводить кислород или кислородсодержащий газ; или для поддержания анаэробного или микроаэробного состояния можно вводить газообразный азот, газообразный водород или газообразный диоксид углерода или не вводить никакого газа, но без ограничений. Дополнительно температуру культуры можно поддерживать от 20 до 45°С, в частности, от 25 до 40°С, и культивирование можно осуществлять в течение от примерно 10 до 160 часов, но культивирование не ограничено этим. Дополнительно образование пены во время культивирования можно предотвращать, используя пеногаситель, такой как сложный полигликолевый эфир жирной кислоты и т.д., но не ограничиваясь им.
Дополнительно в качестве источника углерода для используемой культуральной среды сахара и углеводы (например, глюкозу, сахарозу, лактозу, фруктозу, мальтозу, мелассы, крахмал и целлюлозу), масла и жиры (например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло), жирные кислоты (например, пальмитиновую кислоту, стеариновую кислоту и линолевую кислоту), спирты (например, глицерин и этанол), органические кислоты (например, уксусную кислоту), и т.д., можно использовать по отдельности или в комбинации, но не ограничиваясь ими. В качестве источника азота можно использовать азотсодержащие органические соединения (например, пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину) или неорганические соединения (например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония) и т.д., по отдельности или в комбинации, но не ограничиваясь ими. В качестве источника фосфора можно использовать дигидрофосфат калия, вторичный кислый фосфат калия, соответствующие им натрийсодержащие соли и т.д., по отдельности или в комбинации, но не ограничиваясь ими. Кроме того, в среде могут содержаться незаменимые материалы, стимулирующие рост, такие как другие соли металлов (например, сульфат магния или сульфат железа), аминокислоты, витамины и т.д.
МАК, продуцируемые в культуре, могут секретироваться в среду или могут оставаться в клетках.
Используемый в настоящем документе термин «среда» относится к продукту, полученному в результате культивирования микроорганизма по настоящему изобретению. Понятие среды включает в себя как форму, содержащую микроорганизм, так и форму, где микроорганизм был удален из культурального раствора, содержащего микроорганизм, посредством центрифугирования, фильтрации и т.д.
На стадии извлечения МАК, продуцируемых на стадии культивирования по настоящему изобретению, желаемые МАК могут быть собраны из культурального раствора подходящим способом, известным в данной области техники. Например, можно использовать центрифугирование, фильтрацию, анионообменную хроматографию, кристаллизацию, высокоэффективную жидкостную хроматографию (ВЭЖХ) и т.д., и желаемые МАК могут быть выделены из культивируемого микроорганизма или из среды с использованием подходящего способа, известного в данной области техники. Кроме того, стадия выделения МАК может дополнительно включать процесс разделения и/или стадию очистки.
Способ осуществления изобретения
Настоящее изобретение будет описано более подробно с помощью примеров. Тем не менее эти примеры предназначены только для иллюстративных целей, и объем настоящего изобретения не предназначен для ограничения этими примерами.
Конструирование продуцирующих МАК рекомбинантных микроорганизмов, полученных из Е. coli., и получение МАК с их использованием
Пример 1 Конструирование векторов для гиперэкспрессии гена биосинтеза шинорина, полученного из микроводорослей
Генный кластер биосинтеза шинорина на основе A. variabilis состоит из четырех генов 2-деметил-4-дезоксигадузолсинтазы, О-метилтрансферазы, C-N лигазы и синтетазы нерибосомных пептидов, и один из видов цианобактерий Nostoc punctiforme также может продуцировать шинорин, используя эти гены. В геномной ДНК А. variabilis АТСС29413 и N. punctiforme АТСС29133 были идентифицированы генные кластеры биосинтеза шинорина. С использованием двух типов векторов pECCG 117_Ptrc_GFP_терминатор и pECCG 117_Pcj1_GFP_терминатор были сконструированы четыре вектора, каждый из которых содержал гены биосинтеза шинорина (Ava_ABCD и Npr_ABCD), полученные из A. variabilis АТСС29413 и N. punctiforme АТСС29133 соответственно. Названия четырех экспрессионных векторов гена биосинтеза шинорина и используемые для конструирования этих векторов матрицы и праймеры обобщены в таблице 1 ниже.


После получения фрагментов генов с использованием этих матриц и праймеров каждый фрагмент гена лигировали в векторы pECCG 117_Ptrc_GFP_терминатор и pECCG 117_Pcj1_GFP_терминатор, обработанные ферментами рестрикции EcoRV/XbaI, с помощью набора реактивов для клонирования In-Fusion® HD Cloning Kit (Clontech Laboratories, Inc.). Экспрессионные векторы были названы pECCG117_Ptrc_Ava_ABCD, pECCG117_Pcj1_Ava_ABCD, pECCG117_Ptrc_Npr_ABCD и pECCG117_Pcj1_Npr_ABCD соответственно, и каждый из этих экспрессионных векторов был подтвержден секвенированием. Нуклеотидные последовательности и аминокислотные последовательности Ava_ABCD и Npr_ABCD были описаны как SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 и SEQ ID NO: 4 соответственно.
Пример 2: Оценка способности к продуцированию шинорина штаммами, в которые введены экспрессионные векторы генов биосинтеза шинорина
Чтобы подтвердить способность к продуцированию МАК в Е. coli, четыре плазмиды, полученные в примере 1, были введены в штамм Е. coli W3110 дикого типа с получением штаммов с усиленным биосинтезом шинорина. Полученные таким образом штаммы высевали на плотную питательную среду Лурия-Бертани (LB), содержащую канамицин, а затем культивировали в течение ночи в термостате при 37°С. Одну платиновую петлю каждого штамма, культивированного в течение ночи на плотной питательной среде LB засевали в 25 мл среды для титрования (состав среды: 40 г/л глюкозы, 0,3 г/л KH2PO4, 0,6 г/л K2HPO4, 15 г/л (NH4)2SO4, 1 г/л MgSO4⋅7H2O, 2,5 г/л NaCl, 1,2 г/л цитрата натрия, 2,5 г/л дрожжевого экстракта, 40 г/л карбоната кальция: рН 7,0), а затем культивировали в термостате при 37°С со скоростью вращения 200 об/мин в течение 48 часов. Результаты представлены в таблице 2 ниже.

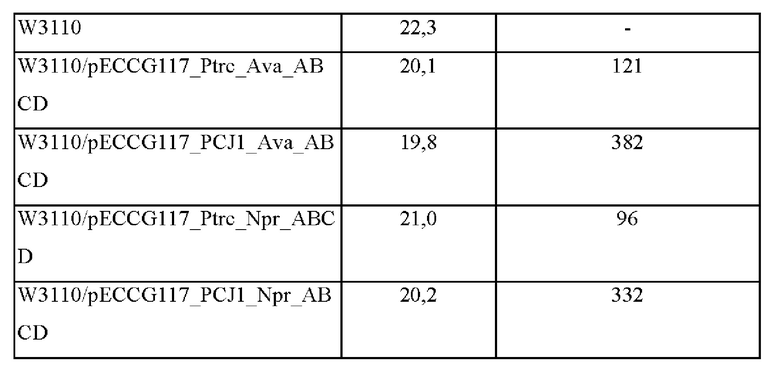
Как показано в таблице 2 выше, было подтверждено, что при введении в штамм W3110 генов биосинтеза шинорина продуцирование шинорина было возможно. Кроме того, было подтверждено, что продуцирование шинорина увеличивалось за счет повышения интенсивности промоторов (введен промотор PCJ1) посредством усиления пути биосинтеза.
Пример 3: Конструирование штаммов, в которых инактивирована 3 - дети дрохиннатдегидр атаза
Первый ген биосинтеза МАК Ava-A в микроводорослях имеет и использует общие субстраты DHQ (3-дегидрохиннат) в шикиматном пути и SH-7P (седогептулозо-7-фосфат) в пентозофосфатном пути. Чтобы получить штаммы, в которых 3-дегидрохиннатдегидратаза инактивирована в результате делеции гена aroD, применяли метод гомологической рекомбинации с использованием рекомбиназы Red фага лямбда. В качестве маркера вставки гена использовали ген резистентности к хлорамфениколу pKD3 и сконструировали кассету с делетированным геном aroD, включающую участок гена aroD и ген резистентности к хлорамфениколу плазмиды pKD3, с помощью полимеразной цепной реакции (ПЦР) с использованием праймеров SEQ ID NO: 9 (прямой) и 10 (обратный). После подготовки штаммов (Е. coli W3110 дикого типа), в которых должен быть делетирован ген aroD (SEQ ID NO: 71 и 72), эти штаммы трансформировали плазмидой pKD46, содержащей ген рекомбиназы Red фага лямбда, и впоследствии индуцировали экспрессию этого гена, используя арабинозу для подготовки компетентных клеток. Кассету с делетированным геном aroD вводили в компетентные клетки методом электропорации, а затем клетки высевали на плотную питательную среду LB, содержащую 30 мг/л хлорамфеникола. Полученные таким образом штаммы подвергали ПЦР, используя праймеры SEQ ID NO: 11 (прямой) и 12 (обратный), и делецию гена aroD подтверждали на основании выявления амплифицированных фрагментов 1300 п. о. (пар оснований).
Пример 4: Оценка способности к продуцированию шинорина штаммами, в которых инактивирована 3-дегидрохиннатдегидратаза
Две из четырех плазмид, полученных в примере 1, в которых экспрессия находится под контролем промотора PCJ1, вводили в штаммы, в которых был делетирован ген aroD, полученные в примере 3(W3110 ΔaraD/pECCG117_PCJ1_Ava_ABCD и W3110 ΔaroD/pECCG117 PCJ1_Npr_ABCD), а затем эти штаммы высевали на плотную питательную среду LB, содержащую канамицин. Впоследствии штаммы с делетированным aroD и штаммы без делеции aroD соответственно культивировали в течение ночи в термостате при 37°С, и одну платиновую петлю каждого штамма засевали в 25 мл среды для титрования (состав среды: 40 г/л глюкозы, 0,3 г/л KH2PO4, 0,6 г/л K2HPO4, 15 г/л (NH4)2SO4, 1 г/л MgSO4⋅7H2O, 2,5 г/л NaCl, 1,2 г/л цитрата натрия, 2,5 г/л дрожжевого экстракта, 40 г/л карбоната кальция: рН 7,0), а затем культивировали в термостате при 37°С со скоростью вращения 200 об/мин в течение 48 часов. Результаты представлены в таблице 3 ниже.
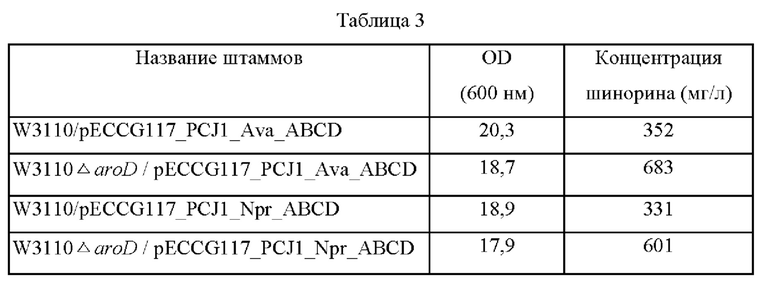
Как показано в таблице 3, концентрация шинорина, продуцируемого в штаммах с делетированным геном aroD, была повышена на 194% и 182% по сравнению с концентрацией шинорина, продуцируемого в штаммах без делеции aroD соответственно. Соответственно, штамм W3110 ΔaraD/pECCG117_PCJ1_Ava_ABCD и штамм W3110 ΔaraD/pECCG117_PCJ1_Npr_ABCD, представляющие собой штаммы с делетированным геном aroD, были названы СВ06-0017 и СВ06-0018 и депонированы в Корейском центре культур микроорганизмов (KССМ) в соответствии с Будапештским договором 26 июня 2017 г. под номерами доступа KCCM1204P и KCCM12045P соответственно.
Пример 5: Конструирование штаммов, обладающих повышенной активностью
2-дегидро-3-дезоксифосфогептонатальдолазы/фосфоенолпируватсинтетазы/транскетолазы I/II
Чтобы повысить способность продуцирующих МАК микроорганизмов к продуцированию МАК, была усилена активность 2-дегидро-3-дезоксифосфогептонатальдолазы/фосфоенолпируватсинтетазы/транскетолазы I/II. В частности, были дополнительно введены три гена, полученные из Е. coli W3110, а именно aroG (2-дегидро-3-дезоксифосфогептонатальдолаза; SEQ ID NO: 73 и 74), ppsA (фосфоенолпируватсинтетаза; SEQ ID NO: 75 и 76) и tktA (транскетолаза I/II; SEQ ID NO: 77 и 78). Для усиления генов aroG, ppsA и tktA сконструировали плазмиды pSKH130-ΔfluA-Pn-aroG-Pn-ppsA-Pn-tktA. Матрицы и праймеры, используемые при конструировании плазмид pSKH130-ΔfhuA-Pn-aroG-Pn-ppsA-Pn-tktA, представлены в таблице 4 ниже.
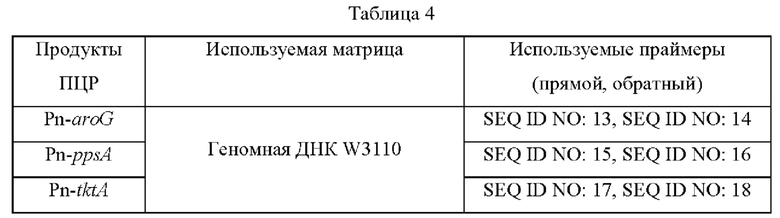
Фрагменты генов aroG, ppsA и tktA амплифицировали с помощью ПЦР с использованием описанных выше матрицы и праймеров, а затем соответственно вводили в вектор pSKH130- ΔfhuA, расщепленный ферментом рестрикции BamH1-Pst1. Для подтверждения клонирования и последовательностей генов векторов применяли секвенирование, а затем этими векторами трансформировали штамм Е. coli W3110 дикого типа и штамм Е. coli W3110 Δ aroD с делецией aroD методом электропорации. Трансформированные гены вводили в хромосому путем первичной рекомбинации (кроссинговера) с последующим исключением участка плазмиды из хромосомы путем вторичной рекомбинации (кроссинговера). Введение генов aroG, ppsA и tktA в трансформированные штаммы Е. coli, в которых была завершена вторая рекомбинация, подтверждали с использованием праймеров SEQ ID NO: 19 (прямой) и 20 (обратный).
Пример 6: Оценка способности к продуцированию шинорина штаммами, обладающими_повышенной_активностью 2-дегидро-3-дезоксифосфогептонатальдолазы/фосфоенолпируватсинтетазы/транскетолазы I/II
Две из четырех плазмид, полученных в примере 5, в которых экспрессия находится под контролем промотора PCJ1, соответственно вводили в штаммы, в которые были введены гены aroG, ppsA и tktA, полученные в примере 5, а затем эти штаммы высевали на плотную питательную среду LB. Впоследствии штаммы культивировали в течение ночи в термостате при 37°С, и одну платиновую петлю каждого штамма засевали в 25 мл среды для титрования, указанной в примере 4, а затем культивировали в термостате при 37°С со скоростью перемешивания 200 об/мин в течение 48 часов. Результаты представлены в таблице 5 ниже.
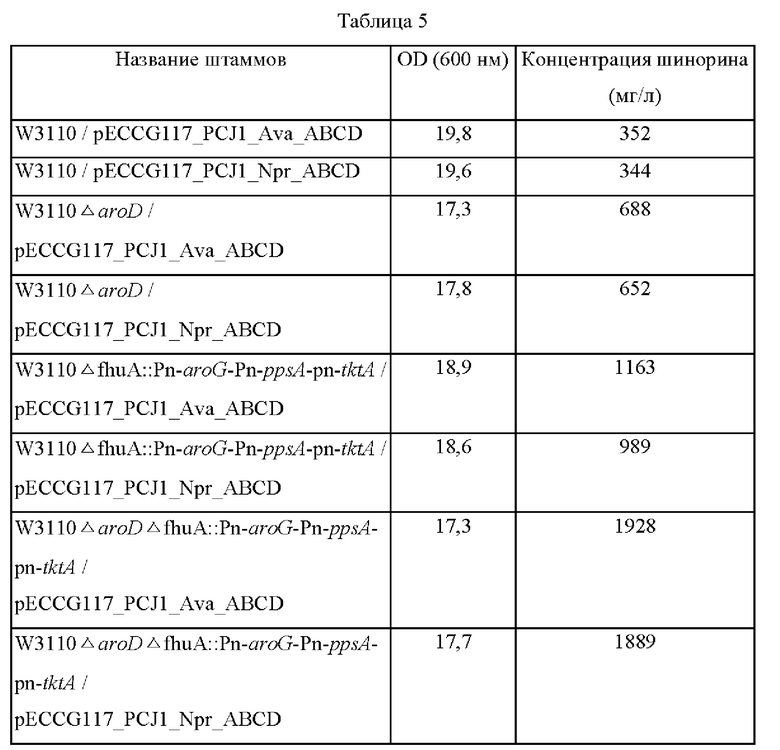
Как показано в таблице 5, концентрация шинорина, продуцируемого в штаммах, в которых была усилена экспрессия трех генов (aroG, ppsA и tktA), была повышена приблизительно на 300% по сравнению с контрольной группой.
Пример 7: Конструирование векторов и штаммов с хромосомной вставкой ava ABCD
Для введения генов биосинтеза шинорина в Е. coli были получены плазмиды pSKH130 ΔpinR::Ava-ABCD. Ava_ABCD подвергали ПЦР с использованием пары праймеров SEQ ID NO: 21 (прямой) и 22 (обратный) на основе pECCG117_Ptrc_Ava_ABCD в качестве матрицы. Чтобы получить pSKH130 ΔpirR::Ava_ABCD, фрагменты ПЦР около 7 кб (килобаз) лигировали в вектор pSKH130 ΔpinR, обработанный ферментами рестрикции BamHI и PstI, используя набор реактивов для клонирования In-Fusion® HD Cloning Kit (Clontech Laboratories, Inc.). Для контроля экспрессии Ava-ABCD фрагменты промотора Ptrc и PCJ1 впоследствии подвергали ПЦР, используя пары прямых и обратных праймеров SEQ ID NO: 23 24, SEQ ID NO: 25 и 26, и SEQ ID NO: 25 и 27, соответственно. Затем эти фрагменты лигировали в вектор pSKH130ΔpinR::Ava_ABCD, обработанный ферментом рестрикции ScaI, с получением pSKH130 ΔpinR::Ptrc-Ava-ABCD и pSKH130ΔpinR::PCJ1-Ava-ABCD, используя набор реактивов для клонирования In-Fusion® HD Cloning Kit (Clontech Laboratories, Inc.). Штаммы W3110 ΔaroΔfhuA::Pn-aroG-Pn-ppsA-Pn-tktA, полученные в примере 5, трансформировали рекомбинантными плазмидами методом электропорации и вводили их в хромосому штаммов посредством первичной рекомбинации (кроссинговера) с последующим исключением векторного участка из хромосомы за исключением целевого гена посредством вторичной рекомбинации (кроссинговера).
Введение гена Ava_ABCD в трансформированные штаммы Е. coli, в которых была завершена вторая рекомбинация, подтверждали с помощью ПЦР с использованием праймеров SEQ ID NO: 28 (прямой) и 29 (обратный).
Пример 8: Оценка способности к продуцированию шинорина штаммами с хромосомной вставкой ava_ABCD
Штаммы, полученные в примере 7, высевали на плотную среду LB, а затем культивировали в течение ночи в термостате при 37°С. Впоследствии одну платиновую петлю каждого штамма засевали в 25 мл среды для титрования, указанной в примере 4, а затем культивировали в термостате при 37°С со скоростью перемешивания 200 об/мин в течение 48 часов. Результаты представлены в таблице 6 ниже.
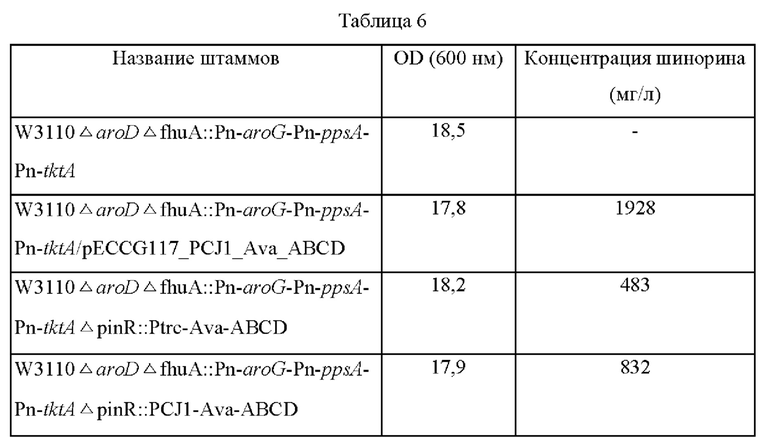
Как показано в таблице 6, было подтверждено, что шинорин продуцируется при введении в хромосому Ava-ABCD, и его концентрация была увеличена в соответствии с силой промоторов. Однако было также подтверждено, что продукция шинорина снижена по сравнению со штаммами, в которых биосинтез шинорина был усилен посредством плазмид. При дополнительном введении плазмиды pECCG117_PCJ1_Ava_ABCD в штаммы с введенной в хромосому вставкой Ava-ABCD продукция шинорина увеличивалась на 353% и 152% по сравнению со штаммом, в котором в хромосому была введена только вставка Ava-ABCD (на основе промотора СJ1), и со штаммом, в который была введена только плазмида, соответственно.
Пример 9: Конструирование векторов с гиперэкспрессией гена МАК и оценка их способности к продуцированию МАК
4-Дезоксигадузол и микоспорин-глицин являются промежуточными соединениями, образующимися в процессе биосинтеза шинорина, и представляют собой микоспорин-подобные аминокислоты, одновременно обладающие эффектом предотвращения воздействия УФ лучей. Чтобы подтвердить, могут ли эти вещества продуцироваться в штаммах Е. coli с делецией AroD, были сконструированы векторы. Результаты представлены в таблице 7 ниже.
Ptrc_Ava_АВ и Ptrc_Ava_ABC подвергали ПЦР, используя праймеры SEQ ID NO: 30 и 31 и SEQ ID NO: 30 и 32 на основе pECCG117_Ptrc_Ava_ABCD в качестве матрицы. pECCG117_Ptrc_Ava_AB и pECCG117_Ptrc_Ava_ABC были получены путем лигирования фрагментов ПЦР в вектор pECCG117 Pre GFP, обработанный ферментами рестрикции BamHI и SpeI. Таким же образом получили pECCG117_PCJ1_Ava_АВ и pECCG117_PCJ1_Ava_ABC путем лигирования фрагментов ПЦР, полученных с использованием пар праймеров SEQ ID NO: 30 и 31 и SEQ ID NO: 30 и 32 на основе pECCG117_PCJ1_Ava_ABCD в качестве матрицы, в вектор pECCG117 Pcj1 GFP, обработанный ферментами рестрикции BamHI и SpeI. Нуклеотидная последовательность и аминокислотная последовательность Ava_АВ и Ava_ABC были описаны как SEQ ID NO: 85-88.
Полученными таким образом векторами трансформировали штаммы W3110ΔaroDΔfhuA::Pn-aroG-Pn-ppsA-Pn-tktA, используемые в примере 8, с помощью общепринятого электроимпульсного метода, и каждый штамм высевали на плотную среду LB и культивировали в течение ночи в термостате при 37°С. Штаммы, культивированные в течение ночи на плотной среде LB, засевали в 25 мл среды для титрования, указанной в примере 4, а затем культивировали в термостате при 37°С со скоростью перемешивания 200 об/мин в течение 48 часов. После окончания культивирования культуры измеряли продукцию МАК методом жидкостной высокоскоростной хроматографии, и в таблице 8 ниже представлена концентрация МАК в культуре каждого штамма.

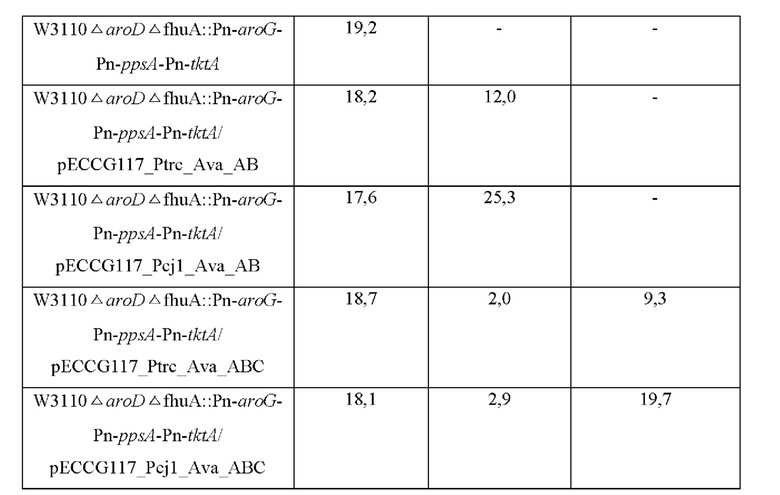
Как показано в таблице 8, было подтверждено, что при введении генов Ava_АВ и Ava_ABC продуцируются 4-дезоксигадузол и микоспорин-глицин, а их количество увеличивается по мере увеличения силы промотора.
Конструирование продуцирующих МАК рекомбинантных микроорганизмов, полученных из Corynebacterium glutamicum, и получение МАК с их использованием
Пример 10: Оценка способности к продуцированию шинорина штаммами, в которые введены векторы с гиперэкспрессией генов биосинтеза шинорина
Чтобы подтвердить способность Corynebacterium glutamicum к продуцированию МАК, четыре плазмиды, полученные в примере 1, вводили в штамм 13032 Corynebacterium glutamicum с получением штаммов, характеризующихся усиленным биосинтезом шинорина, и высевали эти штаммы на плотную питательную среду с добавлением сердечно-мозговой вытяжки (BHIS; Beef Heart Infusion Supplemented), содержащую канамицин, и культивировали в течение ночи в термостате при 30°С. Одну платиновую петлю каждого штамма, культивированного в течение ночи на плотной питательной среде BHIS, засевали в 25 мл среды для титрования (состав среды: 40 г/л глюкозы, 1 г/л KH2PO4, 10 г/л (NH4)2SO4, 5 г/л MgSO4⋅7H2O, 5 г/л NaCl, 5 г/л дрожжевого экстракта, 30 г/л карбоната кальция: рН 7,0), а затем культивировали в термостате при 37°С со скоростью вращения 200 об/мин в течение 48 часов. Результаты представлены в таблице 9 ниже.
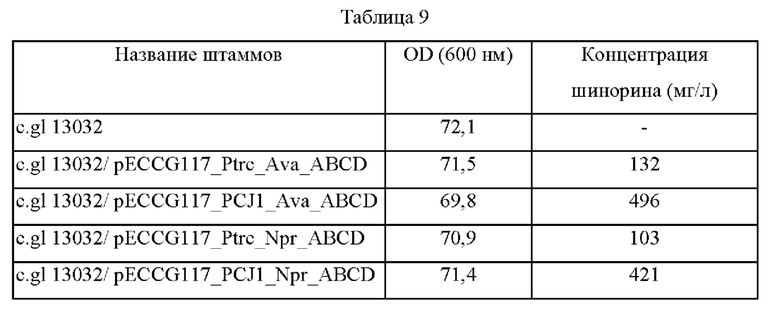
Как показано в таблице 9, было подтверждено, что продуцирование шинорина возможно при введении генов биосинтеза шинорина в Corynebacterium glutamicum 13032, и его продукция может быть повышена до 375% в зависимости от силы промоторов.
Пример 11: Конструирование векторов для введения гена биосинтеза шинорина в хромосому, и штаммов
Для введения генов биосинтеза шинорина в Corynebacterium glutamicum были получены плазмиды pDC AN1021_Ava_ABCD. Ava_ABCD подвергали ПЦР с использованием пары праймеров SEQ ID NO: 33 (прямой) и 34 (обратный) на основе pECCG117_Ptrc_Ava_ABCD в качестве матрицы. Чтобы получить pDC ΔN1021_Ava_ABCD, фрагменты ПЦР около 7 кб лигировали в вектор pDC ΔN1021, обработанный ферментом рестрикции NdeI, используя набор реактивов для клонирования In-Fusion® HD Cloning Kit (Clontech Laboratories, Inc.). Для контроля экспрессии Ava-ABCD фрагменты промотора CJ7, Lysc8 и O2 впоследствии подвергали ПЦР, используя пары прямых и обратных праймеров SEQ ID NO: 35 и 36, SEQ ID NO: 37 и 38 и SEQ ID NO: 39 и 40 соответственно. Затем эти фрагменты ПЦР лигировали в вектор pDC ΔN1021_Ava_ABCD, обработанный ферментом рестрикции NdeI, используя набор реактивов для клонирования In-Fusion® HD Cloning Kit (Clontech Laboratories, Inc.), для получения pDC ΔN1021_Pcj7_Ava_ABCD, pDC ΔN1021_Plysc8_Ava_ABCD и pDC ΔN1021_PO2_Ava_ABCD.
Этими рекомбинантными плазмидами трансформировали штамм 13032 Corynebacterium glutamicum методом электропорации (van der Rest et al. 1999) и вводили в хромосому путем первичной рекомбинации (кроссинговера) с последующим исключением плазмиды из хромосомы путем вторичной рекомбинации (кроссинговера).
Введение гена Ava_ABCD в трансформированные штаммы Corynebacterium glutamicum, в которых была завершена вторая рекомбинация, подтверждали с помощью ПЦР с использованием праймеров SEQ ID NO: 33 (прямой) и 34 (обратный).
Пример 12: Оценка способности к продуцированию шинорина штаммами, в хромосому которых введен ген биосинтеза шинорина
Все штаммы высевали на плотную среду BHIS, а затем культивировали в течение ночи в термостате при 30°С для подтверждения способности к продуцированию шинорина. Одну платиновую петлю каждого штамма, культивированного в течение ночи на плотной среде BHIS, засевали в 25 мл среды для титрования примера 11, а затем культивировали в термостате при 37°С со скоростью перемешивания 200 об/мин в течение 48 часов. Результаты представлены в таблице 10 ниже.
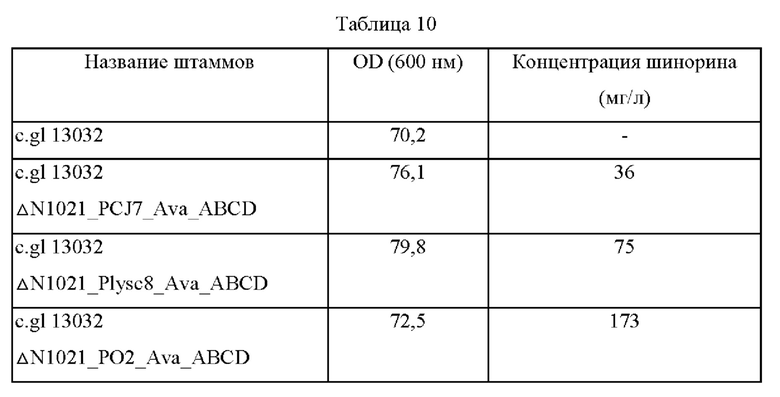
Как показано в таблице 10, было подтверждено, что при введении одной копии генов биосинтеза шинорина в штамм дикого типа Corynebacterium glutamicum шинорин может продуцироваться в количестве от 36 мг до 173 мг.
Пример 13: Конструирование векторов и штаммов Corynebacterium с делецией aroD (3-дегидрохиннатдегидратазы)
Как упоминалось в примере 3, были получены штаммы с делецией, чтобы подтвердить, что можно увеличить продукцию шинорина посредством делеции aroD (3-дегидрохиннатдегидратазы). Чтобы получить штаммы с сайт-специфической делецией гена aroD (SEQ ID NO: 79 и 80) Corynebacterium glutamicum, были сконструированы плазмиды pDC-ΔaroD, в которых была эндогенно делетирована открытая рамка считывания aroD. Эндогенная делеция гена pDC-ΔaroD была образована путем введения в вектор pDC фрагментов гена, образовавшихся в результате кроссоверной ПЦР с использованием прямых и обратных праймеров SEQ ID NO: 41 и 42 и SEQ ID ΔN1021_РО2_Ava_ABCD методом электропорации (van der Rest et al. 1999) и вводили плазмиды в хромосому путем первичной рекомбинации (кроссинговера) с последующим исключением плазмиды из хромосомы путем вторичной рекомбинации (кроссинговера).
Делецию гена aroD в трансформированных штаммах Corynebacterium glutamicum, в которых была завершена вторая рекомбинация, подтверждали с помощью ПЦР с использованием пары геноспецифичных праймеров SEQ ID NO: 41 и 44. Пример 14: Оценка делеции aroD (3-дегидрохиннатдегидратазы) Штамм Corynebacterium glutamicum 13032 ΔN1021_PO2_Ava_ABCD, в котором ожидалось накопление DHQ за счет делеции гена 3-дегидрохиннатдегидратазы, высевали на плотную среду BHIS и культивировали в течение ночи в термостате при 30°С. Одну платиновую петлю каждого штамма, культивированного в течение ночи на плотной среде BHIS, засевали в 25 мл среды для титрования (состав среды: 40 г/л глюкозы, 1 г/л KH2PO4, 10 г/л (NH4)2SO4, 5 г/л MgSO4-7H2O, 5 г/л NaCl, 5 г/л дрожжевого экстракта, 30 г/л карбоната кальция: рН 7,0), а затем кульивировали в термостате при 37°С со скоростью вращения 200 об/мин в течение 48 часов. Результаты представлены в таблице 11 ниже.
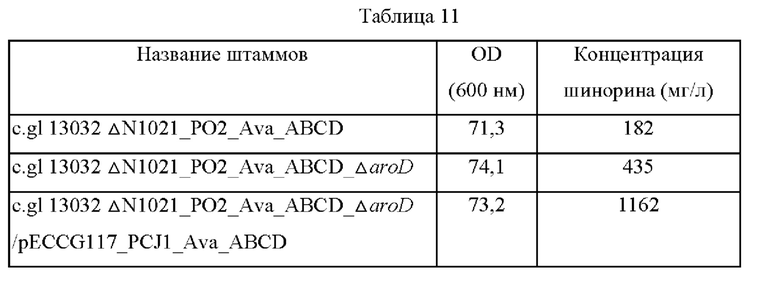
Как показано в таблице 11, было подтверждено, что при делетировании гена aroD концентрация шинорина повышалась на 239% по сравнению с контрольной группой, и за счет дополнительного усиления биосинтеза шинорина посредством pECCG117_PCJ1_Ava_ABCD концентрация шинорина повышалась. Соответственно, штамм с делецией aroD c.g1 13032 N1021_PO2_Ava_ABCD_ΔaroD был назван CB06-0019 и депонирован в Корейском центре культур микроорганизмов (KCCM) в соответствии с Будапештским договором от 26 июня 2017 г. с номером доступа KCCM12046P.
Конструирование продуцирующих МАК рекомбинантных микроорганизмов, полученных из дрожжей, и получение МАК с их использованием
Пример 15: Конструирование дрожжевых векторов с гиперэкспрессией гена биосинтеза шинорина, полученного из микроводорослей
Были сконструированы векторы S. cerevisiae, в которые были введены гены биосинтеза шинорина, основанные на геномной ДНК A. variabilis АТСС29413 и N. punctiforme АТСС29133. Для получения векторов использовали промоторы ADH, TEF и GPD S. cerevisiae, и в таблице 12 ниже представлены матрицы и праймеры, использованные для получения в общей сложности двадцати четырех экспрессионных векторов гена биосинтеза шинорина. Нуклеотидные последовательности и аминокислотные последовательности Ava_A, Ava_B, Ava_C, Ava_D, Npr_A, Npr_B, Npr_C и Npr_D обозначены номерами SEQ ID NO: 89-104 в порядке ферментов.
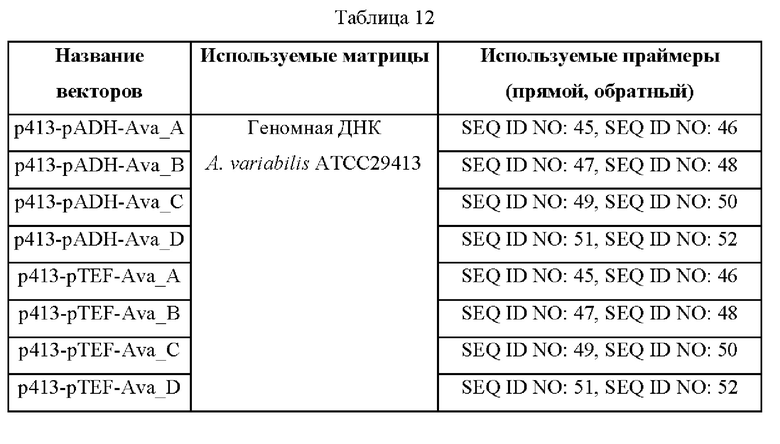
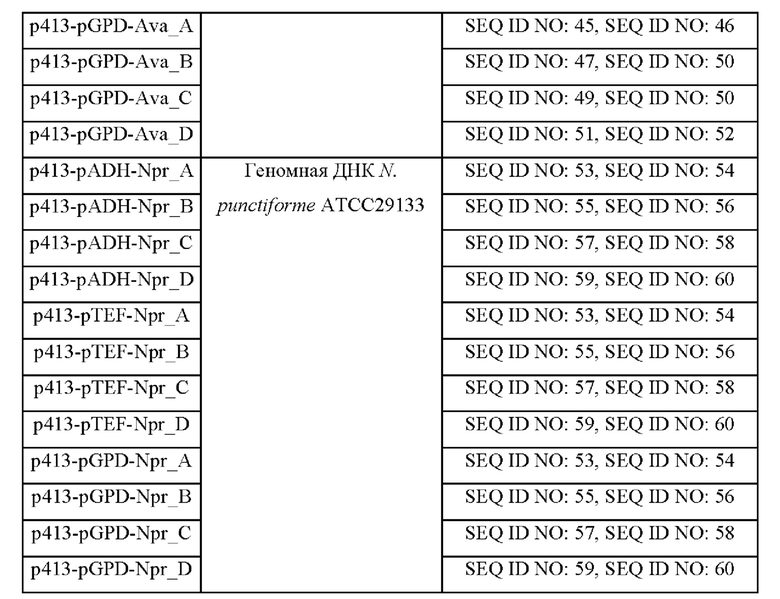
Фрагменты генов, полученные с помощью ПЦР с использованием описанных выше комбинаций матриц и праймеров, лигировали в вектор р413/414/415/416-pADH/pTEF/pGPD-CYC1_терминатор, обработанный ферментами рестрикции BamH1/XhoI, с использованием фермента лигазы Т4 (NEB) и, таким образом, получили двадцать четыре вектора p413/414/415/416-pADH/pTEF/pGPD-A, В, С, D. Конструирование каждого экспрессионного вектора и их последовательности генов подтверждали методом секвенирования. Полученные такие образом экспрессионные векторы вводили в штамм дикого типа S. cerevisiae CEN.PK-1D и, таким образом, получили штаммы, способные продуцировать шинорин.
Пример 16: Оценка способности к продуцированию шинорина штаммами, в которые введены векторы с гиперэкспрессией генов биосинтеза шинорина
Чтобы подтвердить способность дрожжей к продуцированию МАК, двадцать четыре плазмиды, полученные в примере 15, вводили в штамм Saccharomyces cerevisiae CEN.PK-1D (S. cerevisiae CEN.PK-1D) для получения штаммов, характеризующихся усиленным биосинтезом шинорина, и высевали эти штаммы на плотную питательную среду SC (полная синтетическая среда) с исключением Leu, Trp, Ura и His и культивировали в течение ночи в термостате при 30°С. Одну платиновую петлю каждого штамма, культивированного в течение ночи, засевали в 25 мл среды для титрования, показанной в таблице 13, а затем культивировали в термостате при 30°С со скоростью перемешивания 150 об/мин в течение 24 часов. Результаты представлены в таблице 14 ниже.
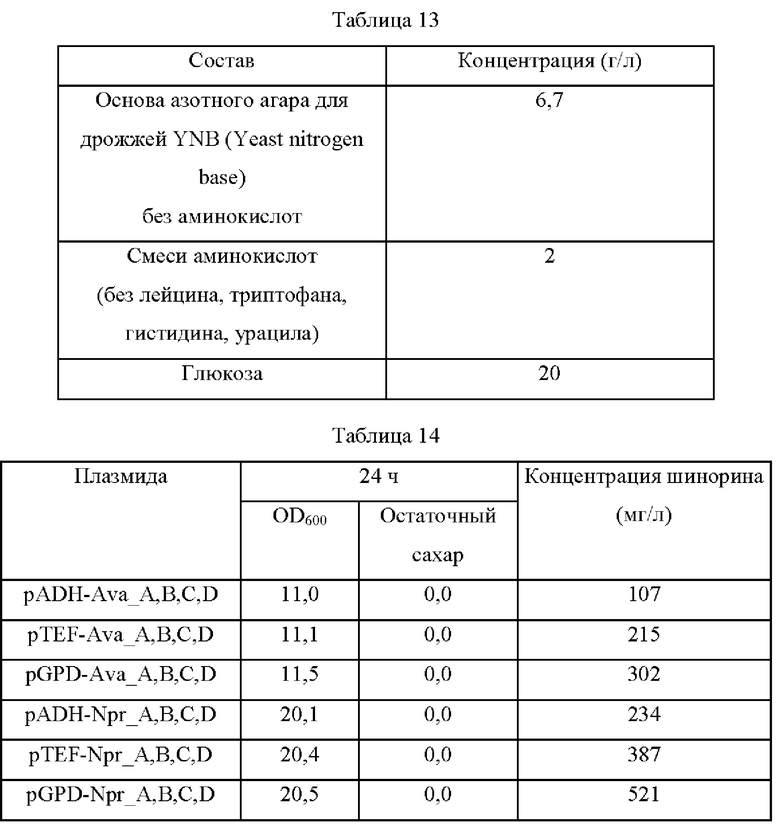
На основании приведенных выше результатов было подтверждено, что S. cerevisiae CEN.PK-1D (т.е. штамм дрожжей) показал высокую активность генов Npr А, В, С, D по сравнению с генами Ava А, В, С, D. Дополнительно было подтверждено, что уровень экспрессии генов регулируется в соответствии с силой промоторов, и в соответствии с этим продукция шинорина была изменена. В частности, было подтверждено, что самая высокая продукция шинорина (521 мг/л) была получена при введении вектора, несущего Npr А, В, С, D под контролем промотора GPD (глицеральдегид-3-фосфатдегидрогеназы).
Пример 17: Повышение продукции шинорина в результате делеции гена ARO1 в S. cerevisiae и введения гена aroB Е. coli
Чтобы определить, приводит ли инактивация дегидрохиннатдегидратазы к улучшению продукции шинорина в дрожжах, в S. cerevisiae CEN.PK-1D был делетирован tquARO1. TquARO1 S. cerevisiae представляет собой ген, выполняющий пять функций, и в результате делеции гена ARO1 функция 3-дегидрохиннатсинтазы, соответствующая гену aroB Е. coli, утрачивается, в результате чего синтез 3-DHQ становится невозможным. Поэтому после делеции гена ARO1 S. cerevisiae (SEQ ID NO: 81 и 82), который является гомологом гена aroB Е. coli на хромосоме, ген aroB Е. coli (SEQ ID NO: 83 и 84) был введен в том же положении под контролем промотора GPD. Использованные матрицы и праймеры показаны в таблице 15. Двадцать четыре плазмиды, полученные в примере 15, вводили в штамм S. cerevisiae CEN.PK-1D, в котором делетирован ген ARO1 и введен ген aroB Е. coli, и штаммы высевали на плотную среду SC (полную синтетическую) с исключением Leu, Trp, Ura и His и культивировали в течение ночи в термостате при 30°С. Одну платиновую петлю каждого штамма, культивированного в течение ночи, засевали в 25 мл среды для титрования, показанной в таблице 13, а затем культивировали в термостате при 30°С со скоростью перемешивания 150 об/мин в течение 24 часов. Результаты представлены в таблице 16 ниже.
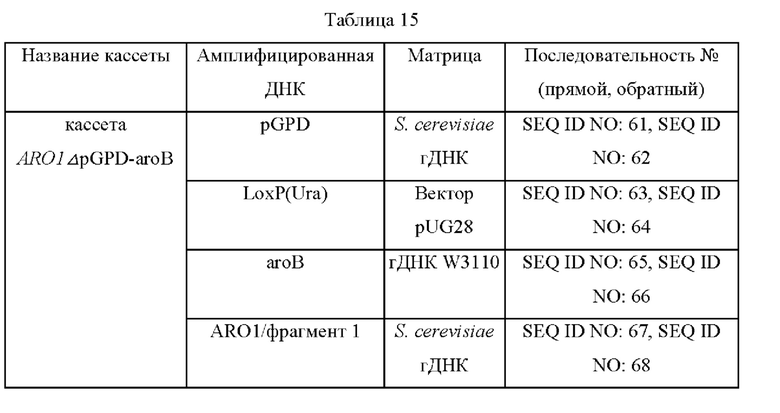
На основании результатов, представленных в таблице 16, было подтверждено что продукция шинорина повышалась в 3 раза в штаммах, в которых способность к продуцированию DHQ была повышена в результате делеции ARO1 и введения aroB Е. coli по сравнению со штаммами ДТ (дикого типа). Дополнительно было подтверждено, что эти штаммы проявляли более высокую активность генов Npr А, В, С, D по сравнению с генами Ava А, В, С, D, и продукция шинорина повышалась по мере увеличения силы промоторов. В частности, было подтверждено, что самая высокая продукция шинорина (1,6 г/л) была получена при введении вектора, несущего Npr А, В, С, D под контролем промотора GPD.
Специалистам в данной области техники будет понятно, что настоящее изобретение может быть воплощено в других конкретных формах без отклонения от его сущности или неотъемлемых характеристик. Описанные воплощения изобретения должны быть рассмотрены во всех аспектах только как иллюстративные, а не ограничивающие. Таким образом, объем настоящего изобретения в большей степени указан в прилагаемой формуле изобретения, чем в приведенном выше описании. Все изменения, которые следуют из значения и диапазона эквивалентности формулы изобретения, должны быть включены в объем настоящего изобретения.
--->
<110> CJ CHEILJEDANG CORPORATION
<120> МИКРООРГАНИЗМ ДЛЯ ПРОДУЦИРОВАНИЯ МИКРОСПОРИН-ПОДОБНОЙ АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ МИКРОСПОРИН-ПОДОБНОЙ АМИНОКИСЛОТЫ С ЕГО ИСПОЛЬЗОВАНИЕМ
<130> OPA18235-PCT
<150> KR10-2017-0103795
<151> 2017-08-16
<160> 104
<170> KopatentIn 2.0
<210> 1
<211> 6461
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 1
atgagtatcg tccaagcaaa gtttgaagct aaggaaacat cttttcatgt agaaggttac 60
gaaaagattg agtatgattt ggtgtatgta gatggtattt ttgaaatcca gaattctgca 120
ctagcagatg tatatcaagg ttttggacga tgcttggcga ttgtagatgc taacgtcagt 180
cggttgtatg gtaatcaaat tcaggcatat ttccagtatt atggtataga actgaggcta 240
tttcctatta ccattactga accagataag actattcaaa ctttcgagag agttatagat 300
gtctttgcag atttcaaatt agtccgcaaa gaaccagtat tagtcgtggg tggcggttta 360
attacagatg ttgtcggctt tgcttgttct acatatcgtc gcagcagcaa ttacatccgc 420
attcctacta cattgattgg attaattgat gccagtgtag caattaaggt agcagttaat 480
catcgcaaac tgaaaaaccg tttgggtgct tatcatgctt ctcgcaaagt atttttagat 540
ttctccttgt tgcgtactct ccctacagac caagtacgta acgggatggc ggaattggta 600
aaaatcgctg tagtagcgca tcaagaagtt tttgaattgt tggagaagta cggcgaagaa 660
ttactacgta ctcattttgg caatatagat gcaactccag agattaaaga aatagcccat 720
cgtttgactt acaaagctat ccataagatg ttggaattgg aagttcccaa cctgcatgag 780
ttagacctag atagggtgat tgcttacggt cacacttgga gtcccacctt ggaacttgcg 840
cctcgtctac ccatgttcca cggacacgcc gttaatgtag atatggcttt ctcggcaacg 900
atcgccgccc gtagaggata tattacaatt gcagaacgcg atcgtatttt aggattaatg 960
agtcgcgttg gtctatccct cgaccatccc atgttggata tagatatttt gtggcgtggt 1020
actgaatcta tcacattaac tcgtgatggt ttgttaagag ctgctatgcc aaaacccatt 1080
ggtgattgtg tcttcgtcaa tgacctgaca agagaagaat tagcagccgc attagctgac 1140
cacaaagaac tttgtaccag ttatccccgt ggtggtgaag gtgtggatgt gtatcccgtt 1200
tatcaaaaag aattaatcgg gagtgttaaa taatgacttt tttgaattca aaatgcaaaa 1260
tactccacgg atacactgcg cgagcgcggt agcatttctg ttcgcggagc gtcccgtagg 1320
gaaagagaag gctacgcaaa taatcggaca ctaattgtct ttaattttga attttgaatt 1380
ttgaattttg aattggagcg aagcgacttg acaaatgtga ttgtccaacc aacagctaga 1440
cctgttacac cattgggaat tttaaccaag cagttagaag ccatagtcca agaggttaag 1500
caacatccag atttacctgg ggaattgata gcaaacatcc atcaggcttg gcgtttagcc 1560
gcaggtatag acccttattt ggaagaatgc accactccag aatctcctga actcgctgca 1620
ttggcaaaaa ccacagccac cgaagcctgg ggagaacact tccacggagg tacaaccgtc 1680
cgtcctctag aacaagagat gctttctggt catatcgaag gacaaacctt aaagatgttt 1740
gttcacatga ccaaagctaa aaaagtctta gaaattggga tgtttaccgg ttattcggcg 1800
ctggcgatgg cggaagcatt accagaggat ggactgcttg tggcttgtga agttgaccct 1860
tacgcggcgg aaattggaca gaaagccttt caacaatctc cccacggtgg aaagattcgt 1920
gtggaattgg atgcagcctt agcaactctt gataagttag cagaagctgg ggagtctttt 1980
gacttggtat ttatcgacgc agataaaaaa gagtatgtag cctattttca caagttgcta 2040
ggtagcagtt tgttagcacc agatggcttt atttgtgtag ataacacctt attacaaggg 2100
gaagtttatc taccagcaga ggaacgtagc gtcaatggtg aagcgatcgc gcaatttaat 2160
catacagtag ctatagaccc ccgtgtagaa caggttttgt tgccgttgcg agatggttta 2220
acaattatcc gcagaataca accttaattg tccaatcgac tatggcacaa tcccttcccc 2280
tttcttccgc acctgctaca ccgtctcttc cttcccagac gaaaatagcc gcaattatcc 2340
aaaatatctg cactttggct ttgttattac tagcattgcc cattaatgcc accattgttt 2400
ttatatcctt gttagtcttc cgaccgcaaa aggtcaaagc agcaaacccc caaaccattc 2460
ttatcagtgg cggtaagatg accaaagctt tacaactagc aaggtcattc cacgcggctg 2520
gacatagagt tgtcttggtg gaaacccata aatactggtt gactggtcat cgtttttccc 2580
aagcagtgga taagttttac acagtccccg caccccagga caatccccaa gcttacattc 2640
aggctttggt agatatcgtc aaacaagaaa acatcgatgt ttatattccc gtcaccagtc 2700
cagtgggtag ctactacgac tcattagcca aaccagagtt atcccattat tgcgaagtgt 2760
ttcactttga cgcagatatt acccaaatgt tggatgataa atttgcgttg acacaaaaag 2820
cgcgatcgct tggtttatca gtacccaaat cctttaaaat tacctcacca gaacaagtca 2880
tcaacttcga tttttctgga gagacacgta aatacatcct caaaagcatt ccctacgact 2940
cagtgcggcg gttggactta accaaactcc cctgtgctac tccagaggaa acagcagcat 3000
tcgtcagaag tttgccaatt actcccgaaa aaccgtggat tatgcaggaa tttatccccg 3060
gtaaggaatt ctgcacccat agcaccgttc ggaatgggga actcagactg cattgctgtt 3120
gcgaatcttc agccttccaa gttaattatg agaatgtaaa taacccgcaa attaccgaat 3180
gggtacagca ttttgtcaag gaactgaaac tgacaggaca gatttccttt gactttatcc 3240
aagccgaaga cggaacagtt tacgccatcg agtgtaaccc ccgcacacat tcagcaatta 3300
ccacatttta cgaccacccc caggtagcag aagcgtactt gagtcaagca ccgacgactg 3360
aaaccataca accactaacg acaagcaagc ctacctattg gacttatcac gaagtttggc 3420
gtttaactgg tatccgttct ttcacccagt tgcaaagatg gctggggaat atttggcgcg 3480
ggactgatgc gatttatcag ccagatgacc ccttaccgtt tttgatggta catcattggc 3540
aaattcccct actgttattg aataatttgc gtcgtcttaa aggttggacg cggatagatt 3600
tcaatattgg gaagttggtg gaattggggg gagattagtt tttaaacgca gagggacgct 3660
gaggttagcg cagcgaaaag ttctggagga gggtttccct ccgtaggaaa cttttcaaga 3720
gagagggacg cggagtgtgt tttctctgcg tctctgcgtg agaaattttt tattattgag 3780
caaagttaga agatatgcag actatagatt ttaatattcg taagttactt gtagagtgga 3840
acgcgaccca cagagattat gatctttccc agagtttaca tgaactaatt gtagctcaag 3900
tagaacgaac acctgaggcg atcgctgtca cctttgacaa gcaacaacta acttatcaag 3960
aactaaatca taaagcaaac cagctaggac attatttaca aacattagga gtccagccag 4020
aaaccctggt aggcgtttgt ttagaacgtt ccttagaaat ggttatctgt cttttaggaa 4080
tcctcaaagc tgggggtgct tatgttccta ttgaccctga atatcctcaa gaacgcatag 4140
cttatatgct agaagattct caggtgaagg tactactaac tcaagaaaaa ttactcaatc 4200
aaattcccca ccatcaagca caaactatct gtgtagatag ggaatgggag aaaatttcca 4260
cacaagctaa taccaatccc aaaagtaata taaaaacgga taatcttgct tatgtaattt 4320
acacctctgg ttccactggt aaaccaaaag gtgcaatgaa cacccacaaa ggtatctgta 4380
atcgcttatt gtggatgcag gaagcttatc aaatcgattc cacagatagc attttacaaa 4440
aaaccccctt tagttttgat gtttccgttt gggagttctt ttggacttta ttaactggcg 4500
cacgtttggt aatagccaaa ccaggcggac ataaagatag tgcttacctc atcgatttaa 4560
ttactcaaga acaaatcact acgttgcatt ttgtcccctc aatgctgcaa gtgtttttac 4620
aaaatcgcca tgtaagcaaa tgcagctctc taaaaagagt tatttgtagc ggtgaagctt 4680
tatctataga tttacaaaat agatttttcc agcatttgca atgtgaatta cataacctct 4740
atggcccgac agaagcagca attgatgtca cattttggca atgtagaaaa gatagtaatt 4800
taaagagtgt acctattggt cgtcccattg ctaatactca aatttatatt cttgatgccg 4860
atttacaacc agtaaatatt ggtgtcactg gtgaaattta tattggtggt gtaggggttg 4920
ctcgtggtta tttgaataaa gaagaattga ccaaagaaaa atttattatt aatccctttc 4980
ccaattctga gtttaagcga ctttataaaa caggtgattt agctcgttat ttacccgatg 5040
gaaatattga atatcttggt agaacagatt atcaagtaaa aattcggggt tatagaattg 5100
aaattggcga gattgaaaat gttttatctt cacacccaca agtcagagaa gctgtagtca 5160
tagcgcggga tgataacgct caagaaaaac aaatcatcgc ttatattacc tataactcca 5220
tcaaacctca gcttgataat ctgcgtgatt tcctaaaagc aaggctacct gattttatga 5280
ttccagccgc ttttgtgatg ctggagcatc ttcctttaac tcccagtggt aaagtagacc 5340
gtaaggcatt acctaagcct gatttattta attatagtga acataattcc tatgtagcgc 5400
ctcggaatga agttgaagaa aaattagtac aaatctggtc gaatattctg catttaccta 5460
aagtaggtgt gacagaaaac tttttcgcta ttggtggtaa ttccctcaaa gctctacatt 5520
taatttctca aattgaagag ttatttgcta aagagatatc cttagcaaca cttttaacaa 5580
atccagtaat tgcagattta gccaaggtta ttcaagcaaa caaccaaatc cataattcac 5640
ccctagttcc aattcaacca caaggtaagc agcagccttt cttttgtata catcctgctg 5700
gtggtcatgt tttatgctat tttaaactcg cacaatatat aggaactgac caaccatttt 5760
atggcttaca agctcaagga ttttatggag atgaagcacc cttgacgcga gttgaagata 5820
tggctagtct ctacgtcaaa actattagag aatttcaacc ccaagggcct tatcgtgtcg 5880
gggggtggtc atttggtgga gtcgtagctt atgaagtagc acagcagtta catagacaag 5940
gacaagaagt atctttacta gcaatattag attcttacgt accgattctg ctggataaac 6000
aaaaacccat tgatgacgtt tatttagttg gtgttctctc cagagttttt ggcggtatgt 6060
ttggtcaaga taatctagtc acacctgaag aaatagaaaa tttaactgta gaagaaaaaa 6120
ttaattacat cattgataaa gcacggagcg ctagaatatt cccgcctggt gtagaacgtc 6180
aaaataatcg ccgtattctt gatgttttgg tgggaacttt aaaagcaact tattcctata 6240
taagacaacc atatccagga aaagtcactg tatttcgagc cagggaaaaa catattatgg 6300
ctcctgaccc gaccttagtt tgggtagaat tattttctgt aatggcggct caagaaatta 6360
agattattga tgtccctgga aaccattatt cgtttgttct agaaccccat gtacaggttt 6420
tagcacagcg tttacaagat tgtctggaaa ataattcata a 6461
<210> 2
<211> 2035
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 2
Met Ser Ile Val Gln Ala Lys Phe Glu Ala Lys Glu Thr Ser Phe His
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly
20 25 30
Ile Phe Glu Ile Gln Asn Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe
35 40 45
Gly Arg Cys Leu Ala Ile Val Asp Ala Asn Val Ser Arg Leu Tyr Gly
50 55 60
Asn Gln Ile Gln Ala Tyr Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu
65 70 75 80
Phe Pro Ile Thr Ile Thr Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu
85 90 95
Arg Val Ile Asp Val Phe Ala Asp Phe Lys Leu Val Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Ile Thr Asp Val Val Gly Phe Ala
115 120 125
Cys Ser Thr Tyr Arg Arg Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Ser Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Ser Arg Lys
165 170 175
Val Phe Leu Asp Phe Ser Leu Leu Arg Thr Leu Pro Thr Asp Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala His Gln
195 200 205
Glu Val Phe Glu Leu Leu Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr
210 215 220
His Phe Gly Asn Ile Asp Ala Thr Pro Glu Ile Lys Glu Ile Ala His
225 230 235 240
Arg Leu Thr Tyr Lys Ala Ile His Lys Met Leu Glu Leu Glu Val Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Arg Leu Pro Met Phe His Gly
275 280 285
His Ala Val Asn Val Asp Met Ala Phe Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ile Ala Glu Arg Asp Arg Ile Leu Gly Leu Met
305 310 315 320
Ser Arg Val Gly Leu Ser Leu Asp His Pro Met Leu Asp Ile Asp Ile
325 330 335
Leu Trp Arg Gly Thr Glu Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Asp Cys Val Phe Val Asn Asp
355 360 365
Leu Thr Arg Glu Glu Leu Ala Ala Ala Leu Ala Asp His Lys Glu Leu
370 375 380
Cys Thr Ser Tyr Pro Arg Gly Gly Glu Gly Val Asp Val Tyr Pro Val
385 390 395 400
Tyr Gln Lys Glu Leu Ile Gly Ser Val Lys Leu Thr Asn Val Ile Val
405 410 415
Gln Pro Thr Ala Arg Pro Val Thr Pro Leu Gly Ile Leu Thr Lys Gln
420 425 430
Leu Glu Ala Ile Val Gln Glu Val Lys Gln His Pro Asp Leu Pro Gly
435 440 445
Glu Leu Ile Ala Asn Ile His Gln Ala Trp Arg Leu Ala Ala Gly Ile
450 455 460
Asp Pro Tyr Leu Glu Glu Cys Thr Thr Pro Glu Ser Pro Glu Leu Ala
465 470 475 480
Ala Leu Ala Lys Thr Thr Ala Thr Glu Ala Trp Gly Glu His Phe His
485 490 495
Gly Gly Thr Thr Val Arg Pro Leu Glu Gln Glu Met Leu Ser Gly His
500 505 510
Ile Glu Gly Gln Thr Leu Lys Met Phe Val His Met Thr Lys Ala Lys
515 520 525
Lys Val Leu Glu Ile Gly Met Phe Thr Gly Tyr Ser Ala Leu Ala Met
530 535 540
Ala Glu Ala Leu Pro Glu Asp Gly Leu Leu Val Ala Cys Glu Val Asp
545 550 555 560
Pro Tyr Ala Ala Glu Ile Gly Gln Lys Ala Phe Gln Gln Ser Pro His
565 570 575
Gly Gly Lys Ile Arg Val Glu Leu Asp Ala Ala Leu Ala Thr Leu Asp
580 585 590
Lys Leu Ala Glu Ala Gly Glu Ser Phe Asp Leu Val Phe Ile Asp Ala
595 600 605
Asp Lys Lys Glu Tyr Val Ala Tyr Phe His Lys Leu Leu Gly Ser Ser
610 615 620
Leu Leu Ala Pro Asp Gly Phe Ile Cys Val Asp Asn Thr Leu Leu Gln
625 630 635 640
Gly Glu Val Tyr Leu Pro Ala Glu Glu Arg Ser Val Asn Gly Glu Ala
645 650 655
Ile Ala Gln Phe Asn His Thr Val Ala Ile Asp Pro Arg Val Glu Gln
660 665 670
Val Leu Leu Pro Leu Arg Asp Gly Leu Thr Ile Ile Arg Arg Ile Gln
675 680 685
Pro Met Ala Gln Ser Leu Pro Leu Ser Ser Ala Pro Ala Thr Pro Ser
690 695 700
Leu Pro Ser Gln Thr Lys Ile Ala Ala Ile Ile Gln Asn Ile Cys Thr
705 710 715 720
Leu Ala Leu Leu Leu Leu Ala Leu Pro Ile Asn Ala Thr Ile Val Phe
725 730 735
Ile Ser Leu Leu Val Phe Arg Pro Gln Lys Val Lys Ala Ala Asn Pro
740 745 750
Gln Thr Ile Leu Ile Ser Gly Gly Lys Met Thr Lys Ala Leu Gln Leu
755 760 765
Ala Arg Ser Phe His Ala Ala Gly His Arg Val Val Leu Val Glu Thr
770 775 780
His Lys Tyr Trp Leu Thr Gly His Arg Phe Ser Gln Ala Val Asp Lys
785 790 795 800
Phe Tyr Thr Val Pro Ala Pro Gln Asp Asn Pro Gln Ala Tyr Ile Gln
805 810 815
Ala Leu Val Asp Ile Val Lys Gln Glu Asn Ile Asp Val Tyr Ile Pro
820 825 830
Val Thr Ser Pro Val Gly Ser Tyr Tyr Asp Ser Leu Ala Lys Pro Glu
835 840 845
Leu Ser His Tyr Cys Glu Val Phe His Phe Asp Ala Asp Ile Thr Gln
850 855 860
Met Leu Asp Asp Lys Phe Ala Leu Thr Gln Lys Ala Arg Ser Leu Gly
865 870 875 880
Leu Ser Val Pro Lys Ser Phe Lys Ile Thr Ser Pro Glu Gln Val Ile
885 890 895
Asn Phe Asp Phe Ser Gly Glu Thr Arg Lys Tyr Ile Leu Lys Ser Ile
900 905 910
Pro Tyr Asp Ser Val Arg Arg Leu Asp Leu Thr Lys Leu Pro Cys Ala
915 920 925
Thr Pro Glu Glu Thr Ala Ala Phe Val Arg Ser Leu Pro Ile Thr Pro
930 935 940
Glu Lys Pro Trp Ile Met Gln Glu Phe Ile Pro Gly Lys Glu Phe Cys
945 950 955 960
Thr His Ser Thr Val Arg Asn Gly Glu Leu Arg Leu His Cys Cys Cys
965 970 975
Glu Ser Ser Ala Phe Gln Val Asn Tyr Glu Asn Val Asn Asn Pro Gln
980 985 990
Ile Thr Glu Trp Val Gln His Phe Val Lys Glu Leu Lys Leu Thr Gly
995 1000 1005
Gln Ile Ser Phe Asp Phe Ile Gln Ala Glu Asp Gly Thr Val Tyr Ala
1010 1015 1020
Ile Glu Cys Asn Pro Arg Thr His Ser Ala Ile Thr Thr Phe Tyr Asp
1025 1030 1035 1040
His Pro Gln Val Ala Glu Ala Tyr Leu Ser Gln Ala Pro Thr Thr Glu
1045 1050 1055
Thr Ile Gln Pro Leu Thr Thr Ser Lys Pro Thr Tyr Trp Thr Tyr His
1060 1065 1070
Glu Val Trp Arg Leu Thr Gly Ile Arg Ser Phe Thr Gln Leu Gln Arg
1075 1080 1085
Trp Leu Gly Asn Ile Trp Arg Gly Thr Asp Ala Ile Tyr Gln Pro Asp
1090 1095 1100
Asp Pro Leu Pro Phe Leu Met Val His His Trp Gln Ile Pro Leu Leu
1105 1110 1115 1120
Leu Leu Asn Asn Leu Arg Arg Leu Lys Gly Trp Thr Arg Ile Asp Phe
1125 1130 1135
Asn Ile Gly Lys Leu Val Glu Leu Gly Gly Asp Met Gln Thr Ile Asp
1140 1145 1150
Phe Asn Ile Arg Lys Leu Leu Val Glu Trp Asn Ala Thr His Arg Asp
1155 1160 1165
Tyr Asp Leu Ser Gln Ser Leu His Glu Leu Ile Val Ala Gln Val Glu
1170 1175 1180
Arg Thr Pro Glu Ala Ile Ala Val Thr Phe Asp Lys Gln Gln Leu Thr
1185 1190 1195 1200
Tyr Gln Glu Leu Asn His Lys Ala Asn Gln Leu Gly His Tyr Leu Gln
1205 1210 1215
Thr Leu Gly Val Gln Pro Glu Thr Leu Val Gly Val Cys Leu Glu Arg
1220 1225 1230
Ser Leu Glu Met Val Ile Cys Leu Leu Gly Ile Leu Lys Ala Gly Gly
1235 1240 1245
Ala Tyr Val Pro Ile Asp Pro Glu Tyr Pro Gln Glu Arg Ile Ala Tyr
1250 1255 1260
Met Leu Glu Asp Ser Gln Val Lys Val Leu Leu Thr Gln Glu Lys Leu
1265 1270 1275 1280
Leu Asn Gln Ile Pro His His Gln Ala Gln Thr Ile Cys Val Asp Arg
1285 1290 1295
Glu Trp Glu Lys Ile Ser Thr Gln Ala Asn Thr Asn Pro Lys Ser Asn
1300 1305 1310
Ile Lys Thr Asp Asn Leu Ala Tyr Val Ile Tyr Thr Ser Gly Ser Thr
1315 1320 1325
Gly Lys Pro Lys Gly Ala Met Asn Thr His Lys Gly Ile Cys Asn Arg
1330 1335 1340
Leu Leu Trp Met Gln Glu Ala Tyr Gln Ile Asp Ser Thr Asp Ser Ile
1345 1350 1355 1360
Leu Gln Lys Thr Pro Phe Ser Phe Asp Val Ser Val Trp Glu Phe Phe
1365 1370 1375
Trp Thr Leu Leu Thr Gly Ala Arg Leu Val Ile Ala Lys Pro Gly Gly
1380 1385 1390
His Lys Asp Ser Ala Tyr Leu Ile Asp Leu Ile Thr Gln Glu Gln Ile
1395 1400 1405
Thr Thr Leu His Phe Val Pro Ser Met Leu Gln Val Phe Leu Gln Asn
1410 1415 1420
Arg His Val Ser Lys Cys Ser Ser Leu Lys Arg Val Ile Cys Ser Gly
1425 1430 1435 1440
Glu Ala Leu Ser Ile Asp Leu Gln Asn Arg Phe Phe Gln His Leu Gln
1445 1450 1455
Cys Glu Leu His Asn Leu Tyr Gly Pro Thr Glu Ala Ala Ile Asp Val
1460 1465 1470
Thr Phe Trp Gln Cys Arg Lys Asp Ser Asn Leu Lys Ser Val Pro Ile
1475 1480 1485
Gly Arg Pro Ile Ala Asn Thr Gln Ile Tyr Ile Leu Asp Ala Asp Leu
1490 1495 1500
Gln Pro Val Asn Ile Gly Val Thr Gly Glu Ile Tyr Ile Gly Gly Val
1505 1510 1515 1520
Gly Val Ala Arg Gly Tyr Leu Asn Lys Glu Glu Leu Thr Lys Glu Lys
1525 1530 1535
Phe Ile Ile Asn Pro Phe Pro Asn Ser Glu Phe Lys Arg Leu Tyr Lys
1540 1545 1550
Thr Gly Asp Leu Ala Arg Tyr Leu Pro Asp Gly Asn Ile Glu Tyr Leu
1555 1560 1565
Gly Arg Thr Asp Tyr Gln Val Lys Ile Arg Gly Tyr Arg Ile Glu Ile
1570 1575 1580
Gly Glu Ile Glu Asn Val Leu Ser Ser His Pro Gln Val Arg Glu Ala
1585 1590 1595 1600
Val Val Ile Ala Arg Asp Asp Asn Ala Gln Glu Lys Gln Ile Ile Ala
1605 1610 1615
Tyr Ile Thr Tyr Asn Ser Ile Lys Pro Gln Leu Asp Asn Leu Arg Asp
1620 1625 1630
Phe Leu Lys Ala Arg Leu Pro Asp Phe Met Ile Pro Ala Ala Phe Val
1635 1640 1645
Met Leu Glu His Leu Pro Leu Thr Pro Ser Gly Lys Val Asp Arg Lys
1650 1655 1660
Ala Leu Pro Lys Pro Asp Leu Phe Asn Tyr Ser Glu His Asn Ser Tyr
1665 1670 1675 1680
Val Ala Pro Arg Asn Glu Val Glu Glu Lys Leu Val Gln Ile Trp Ser
1685 1690 1695
Asn Ile Leu His Leu Pro Lys Val Gly Val Thr Glu Asn Phe Phe Ala
1700 1705 1710
Ile Gly Gly Asn Ser Leu Lys Ala Leu His Leu Ile Ser Gln Ile Glu
1715 1720 1725
Glu Leu Phe Ala Lys Glu Ile Ser Leu Ala Thr Leu Leu Thr Asn Pro
1730 1735 1740
Val Ile Ala Asp Leu Ala Lys Val Ile Gln Ala Asn Asn Gln Ile His
1745 1750 1755 1760
Asn Ser Pro Leu Val Pro Ile Gln Pro Gln Gly Lys Gln Gln Pro Phe
1765 1770 1775
Phe Cys Ile His Pro Ala Gly Gly His Val Leu Cys Tyr Phe Lys Leu
1780 1785 1790
Ala Gln Tyr Ile Gly Thr Asp Gln Pro Phe Tyr Gly Leu Gln Ala Gln
1795 1800 1805
Gly Phe Tyr Gly Asp Glu Ala Pro Leu Thr Arg Val Glu Asp Met Ala
1810 1815 1820
Ser Leu Tyr Val Lys Thr Ile Arg Glu Phe Gln Pro Gln Gly Pro Tyr
1825 1830 1835 1840
Arg Val Gly Gly Trp Ser Phe Gly Gly Val Val Ala Tyr Glu Val Ala
1845 1850 1855
Gln Gln Leu His Arg Gln Gly Gln Glu Val Ser Leu Leu Ala Ile Leu
1860 1865 1870
Asp Ser Tyr Val Pro Ile Leu Leu Asp Lys Gln Lys Pro Ile Asp Asp
1875 1880 1885
Val Tyr Leu Val Gly Val Leu Ser Arg Val Phe Gly Gly Met Phe Gly
1890 1895 1900
Gln Asp Asn Leu Val Thr Pro Glu Glu Ile Glu Asn Leu Thr Val Glu
1905 1910 1915 1920
Glu Lys Ile Asn Tyr Ile Ile Asp Lys Ala Arg Ser Ala Arg Ile Phe
1925 1930 1935
Pro Pro Gly Val Glu Arg Gln Asn Asn Arg Arg Ile Leu Asp Val Leu
1940 1945 1950
Val Gly Thr Leu Lys Ala Thr Tyr Ser Tyr Ile Arg Gln Pro Tyr Pro
1955 1960 1965
Gly Lys Val Thr Val Phe Arg Ala Arg Glu Lys His Ile Met Ala Pro
1970 1975 1980
Asp Pro Thr Leu Val Trp Val Glu Leu Phe Ser Val Met Ala Ala Gln
1985 1990 1995 2000
Glu Ile Lys Ile Ile Asp Val Pro Gly Asn His Tyr Ser Phe Val Leu
2005 2010 2015
Glu Pro His Val Gln Val Leu Ala Gln Arg Leu Gln Asp Cys Leu Glu
2020 2025 2030
Asn Asn Ser
2035
<210> 3
<211> 4528
<212> ДНК
<213> Nostoc punctiforme ATCC29133
<400> 3
atgagtaatg ttcaagcatc gtttgaagca acggaagctg aattccgcgt ggaaggttac 60
gaaaaaattg agtttagtct tgtctatgta aatggtgcat ttgatatcag taacagagaa 120
attgcagaca gctatgagaa gtttggccgt tgtctgactg tgattgatgc taatgtcaac 180
agattatatg gcaagcaaat caagtcatat tttagacact atggtattga tctgaccgta 240
gttcccattg tgattactga gcctactaaa acccttgcaa cctttgagaa aattgttgat 300
gctttttctg actttggttt aatccgcaag gaaccagtat tagtagtggg tggtggttta 360
accactgatg tagctgggtt tgcgtgtgct gcttaccgtc gtaagagtaa ctatattcgg 420
gttccgacaa cgctgattgg tctgattgat gcaggtgtag cgattaaggt tgcagtcaac 480
catcgcaagt taaaaaatcg cttgggtgca tatcatgctc ctttgaaagt catcctcgat 540
ttctccttct tgcaaacatt accaacggct caagttcgta atgggatggc agagttggtc 600
aaaattgctg ttgtggcgaa ctctgaagtc tttgaattgt tgtatgaata tggcgaagag 660
ttgctttcca ctcactttgg atatgtgaat ggtacaaagg aactgaaagc gatcgcacat 720
aaactcaatt acgaggcaat aaaaactatg ctggagttgg aaactccaaa cttgcatgag 780
ttagacctcg atcgcgtcat tgcctacggt cacacttgga gtccgacctt agaattagct 840
ccgatgatac cgttgtttca cggtcatgcc gtcaatatag acatggcttt gtctgcaact 900
attgcggcaa gacgtggcta cattacatca ggagaacgcg atcgcatttt gagcttgatg 960
agtcgtatag gtttatcaat cgatcatcct ctactagatg gcgatttgct ctggtatgct 1020
acccaatcta ttagcttgac gcgagacggg aaacaacgcg cagctatgcc taaacccatt 1080
ggtgagtgct tctttgtcaa cgatttcacc cgtgaagaac tagatgcagc tttagctgaa 1140
cacaaacgtc tgtgtgctac ataccctcgt ggtggagatg gcattgacgc ttacatagaa 1200
actcaagaag aatccaaact attgggagtg tgaaaacatg accagtattt taggacgaga 1260
taccgcaaga ccaataacgc cacatagcat tctggtagca cagctacaaa aaaccctcag 1320
aatggcagag gaaagtaata ttccttcaga gatactgact tctctgcgcc aagggttgca 1380
attagcagca ggtttagatc cctatctgga tgattgcact acccctgaat cgaccgcatt 1440
gacagcacta gcgcagaaga caagcattga agactggagt aaacgcttca gtgatggtga 1500
aacagtgcgt caattagagc aagaaatgct ctcaggacat cttgaaggac aaacactaaa 1560
gatgtttgtg catatcacta aggctaaaag catcctagaa gtgggaatgt tcacaggata 1620
ttcagctttg gcaatggcag aggcgttacc agatgatggg cgactgattg cttgtgaagt 1680
agactcctat gtggccgagt ttgctcaaac ttgctttcaa gagtctcccc acggccgcaa 1740
gattgttgta gaagtggcac ctgcactaga gacgctgcac aagctggtgg ctaaaaaaga 1800
atcctttgat ctgatcttca ttgatgcgga taaaaaggag tatatagaat acttccaaat 1860
tatcttggat agccatttac tagctcccga cggattaatc tgtgtagata atactttgtt 1920
gcaaggacaa gtttacctgc catcagaaca gcgtacagcc aatggtgaag cgatcgctca 1980
attcaaccgc attgtcgccg cagatcctcg tgtagagcaa gttctgttac ccatacgaga 2040
tggtataacc ctgattagac gcttggtata agcggccgcg agctcctcga gatggcacaa 2100
tcaatctctt tatctcttcc tcaatccaca acgccatcaa agggtgtgag gctaaaaata 2160
gcagctctac tgaagactat cgggactcta atactactgc tgatagcctt gccgcttaat 2220
gctttgatag tattgatatc tctgatgtgt aggccgttta caaaaaaacc tgccgtagcc 2280
actcatcccc agaatatctt ggtcagtggc ggcaaaatga ccaaagcatt gcaacttgcc 2340
cgctccttcc atgcagccgg acacagagtt attctgattg aaggtcataa atactggtta 2400
tcagggcata gattctcaaa ttctgtgagt cgtttttata cagttcctgc accacaagac 2460
gacccagaag gctataccca agcgctattg gaaattgtca aacgagagaa gattgacgtt 2520
tatgtacccg tatgcagccc tgtagctagt tattacgact ctttggcaaa gtctgcacta 2580
tcagaatatt gtgaggtttt tcactttgat gctgatataa ccaagatgct ggatgataaa 2640
tttgccttta ccgatcgggc gcgatcgctt ggtttatcag ccccgaaatc ttttaaaatt 2700
accgatcctg aacaagttat caacttcgat tttagtaaag agacgcgcaa atatattctt 2760
aagagtattt cttacgactc agttcgccgc ttaaatttaa ccaaacttcc ttgtgatacc 2820
ccagaagaga cagcagcgtt tgtcaagagt ttacccatca gcccagaaaa accttggatt 2880
atgcaagaat ttattcctgg gaaagaatta tgcacccata gcacagtccg agacggcgaa 2940
ttaaggttgc attgctgttc aaattcttca gcgtttcaga ttaactatga aaatgtcgaa 3000
aatccccaaa ttcaagaatg ggtacaacat ttcgtcaaaa gtttacggct gactggacaa 3060
atatctcttg actttatcca agctgaagat ggtacagctt atgccattga atgtaatcct 3120
cgcacccatt cggcgatcac aatgttctac aatcacccag gtgttgcaga agcctatctt 3180
ggtaaaactc ctctagctgc acctttggaa cctcttgcag atagcaagcc cacttactgg 3240
atatatcacg aaatctggcg attgactggg attcgctctg gacaacaatt acaaacttgg 3300
tttgggagat tagtcagagg tacagatgcc atttatcgcc tggacgatcc gataccattt 3360
ttaactttgc accattggca gattacttta cttttgctac aaaatttgca acgactcaaa 3420
ggttgggtaa agatcgattt taatatcggt aaactcgtgg aattaggggg cgactaaaaa 3480
catgccagta cttaatatcc ttcatttagt tgggtctgca cacgataagt tttactgtga 3540
tttatcacgt ctttatgccc aagactgttt agctgcaaca gcagatccat cgctttataa 3600
ctttcaaatt gcatatatca cacccgatcg gcagtggcga tttcctgact ctctcagtcg 3660
agaagatatt gctcttacca aaccgattcc tgtgtttgat gccatacaat ttctaacagg 3720
ccaaaacatt gacatgatgt taccacaaat gttttgtatt cctggaatga ctcagtaccg 3780
tgccctattc gatctgctca agatccctta tataggaaat accccagata ttatggcgat 3840
cgcggcccac aaagccagag ccaaagcaat tgtcgaagca gcaggggtaa aagtgcctcg 3900
tggagaattg cttcgccaag gagatattcc aacaattaca cctccagcag tcgtcaaacc 3960
tgtaagttct gacaactctt taggagtagt cttagttaaa gatgtgactg aatatgatgc 4020
tgccttaaag aaagcatttg aatatgcttc ggaggtcatc gtagaagcat tcatcgaact 4080
tggtcgagaa gtcagatgcg gcatcattgt aaaagacggt gagctaatag gtttacccct 4140
tgaagagtat ctggtagacc cacacgataa acctatccgt aactatgctg ataaactcca 4200
acaaactgac gatggcgact tgcatttgac tgctaaagat aatatcaagg cttggatttt 4260
agaccctaac gacccaatca cccaaaaggt tcagcaagtg gctaaaaggt gtcatcaggc 4320
tttgggttgt cgccactaca gtttatttga cttccgaatc gatccaaagg gacaaccttg 4380
gttcttagaa gctggattat attgttcttt tgcccccaaa agtgtgattt cttctatggc 4440
gaaagcagcc ggaatccctc taaatgattt attaataacc gctattaatg aaacattggg 4500
tagtaataaa aaggtgttac aaaattga 4528
<210> 4
<211> 1496
<212> PRT
<213> Nostoc punctiforme ATCC29133
<400> 4
Met Ser Asn Val Gln Ala Ser Phe Glu Ala Thr Glu Ala Glu Phe Arg
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Phe Ser Leu Val Tyr Val Asn Gly
20 25 30
Ala Phe Asp Ile Ser Asn Arg Glu Ile Ala Asp Ser Tyr Glu Lys Phe
35 40 45
Gly Arg Cys Leu Thr Val Ile Asp Ala Asn Val Asn Arg Leu Tyr Gly
50 55 60
Lys Gln Ile Lys Ser Tyr Phe Arg His Tyr Gly Ile Asp Leu Thr Val
65 70 75 80
Val Pro Ile Val Ile Thr Glu Pro Thr Lys Thr Leu Ala Thr Phe Glu
85 90 95
Lys Ile Val Asp Ala Phe Ser Asp Phe Gly Leu Ile Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Thr Thr Asp Val Ala Gly Phe Ala
115 120 125
Cys Ala Ala Tyr Arg Arg Lys Ser Asn Tyr Ile Arg Val Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Gly Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Pro Leu Lys
165 170 175
Val Ile Leu Asp Phe Ser Phe Leu Gln Thr Leu Pro Thr Ala Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala Asn Ser
195 200 205
Glu Val Phe Glu Leu Leu Tyr Glu Tyr Gly Glu Glu Leu Leu Ser Thr
210 215 220
His Phe Gly Tyr Val Asn Gly Thr Lys Glu Leu Lys Ala Ile Ala His
225 230 235 240
Lys Leu Asn Tyr Glu Ala Ile Lys Thr Met Leu Glu Leu Glu Thr Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Met Ile Pro Leu Phe His Gly
275 280 285
His Ala Val Asn Ile Asp Met Ala Leu Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ser Gly Glu Arg Asp Arg Ile Leu Ser Leu Met
305 310 315 320
Ser Arg Ile Gly Leu Ser Ile Asp His Pro Leu Leu Asp Gly Asp Leu
325 330 335
Leu Trp Tyr Ala Thr Gln Ser Ile Ser Leu Thr Arg Asp Gly Lys Gln
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Glu Cys Phe Phe Val Asn Asp
355 360 365
Phe Thr Arg Glu Glu Leu Asp Ala Ala Leu Ala Glu His Lys Arg Leu
370 375 380
Cys Ala Thr Tyr Pro Arg Gly Gly Asp Gly Ile Asp Ala Tyr Ile Glu
385 390 395 400
Thr Gln Glu Glu Ser Lys Leu Leu Gly Val Met Thr Ser Ile Leu Gly
405 410 415
Arg Asp Thr Ala Arg Pro Ile Thr Pro His Ser Ile Leu Val Ala Gln
420 425 430
Leu Gln Lys Thr Leu Arg Met Ala Glu Glu Ser Asn Ile Pro Ser Glu
435 440 445
Ile Leu Thr Ser Leu Arg Gln Gly Leu Gln Leu Ala Ala Gly Leu Asp
450 455 460
Pro Tyr Leu Asp Asp Cys Thr Thr Pro Glu Ser Thr Ala Leu Thr Ala
465 470 475 480
Leu Ala Gln Lys Thr Ser Ile Glu Asp Trp Ser Lys Arg Phe Ser Asp
485 490 495
Gly Glu Thr Val Arg Gln Leu Glu Gln Glu Met Leu Ser Gly His Leu
500 505 510
Glu Gly Gln Thr Leu Lys Met Phe Val His Ile Thr Lys Ala Lys Ser
515 520 525
Ile Leu Glu Val Gly Met Phe Thr Gly Tyr Ser Ala Leu Ala Met Ala
530 535 540
Glu Ala Leu Pro Asp Asp Gly Arg Leu Ile Ala Cys Glu Val Asp Ser
545 550 555 560
Tyr Val Ala Glu Phe Ala Gln Thr Cys Phe Gln Glu Ser Pro His Gly
565 570 575
Arg Lys Ile Val Val Glu Val Ala Pro Ala Leu Glu Thr Leu His Lys
580 585 590
Leu Val Ala Lys Lys Glu Ser Phe Asp Leu Ile Phe Ile Asp Ala Asp
595 600 605
Lys Lys Glu Tyr Ile Glu Tyr Phe Gln Ile Ile Leu Asp Ser His Leu
610 615 620
Leu Ala Pro Asp Gly Leu Ile Cys Val Asp Asn Thr Leu Leu Gln Gly
625 630 635 640
Gln Val Tyr Leu Pro Ser Glu Gln Arg Thr Ala Asn Gly Glu Ala Ile
645 650 655
Ala Gln Phe Asn Arg Ile Val Ala Ala Asp Pro Arg Val Glu Gln Val
660 665 670
Leu Leu Pro Ile Arg Asp Gly Ile Thr Leu Ile Arg Arg Leu Val Met
675 680 685
Ala Gln Ser Ile Ser Leu Ser Leu Pro Gln Ser Thr Thr Pro Ser Lys
690 695 700
Gly Val Arg Leu Lys Ile Ala Ala Leu Leu Lys Thr Ile Gly Thr Leu
705 710 715 720
Ile Leu Leu Leu Ile Ala Leu Pro Leu Asn Ala Leu Ile Val Leu Ile
725 730 735
Ser Leu Met Cys Arg Pro Phe Thr Lys Lys Pro Ala Val Ala Thr His
740 745 750
Pro Gln Asn Ile Leu Val Ser Gly Gly Lys Met Thr Lys Ala Leu Gln
755 760 765
Leu Ala Arg Ser Phe His Ala Ala Gly His Arg Val Ile Leu Ile Glu
770 775 780
Gly His Lys Tyr Trp Leu Ser Gly His Arg Phe Ser Asn Ser Val Ser
785 790 795 800
Arg Phe Tyr Thr Val Pro Ala Pro Gln Asp Asp Pro Glu Gly Tyr Thr
805 810 815
Gln Ala Leu Leu Glu Ile Val Lys Arg Glu Lys Ile Asp Val Tyr Val
820 825 830
Pro Val Cys Ser Pro Val Ala Ser Tyr Tyr Asp Ser Leu Ala Lys Ser
835 840 845
Ala Leu Ser Glu Tyr Cys Glu Val Phe His Phe Asp Ala Asp Ile Thr
850 855 860
Lys Met Leu Asp Asp Lys Phe Ala Phe Thr Asp Arg Ala Arg Ser Leu
865 870 875 880
Gly Leu Ser Ala Pro Lys Ser Phe Lys Ile Thr Asp Pro Glu Gln Val
885 890 895
Ile Asn Phe Asp Phe Ser Lys Glu Thr Arg Lys Tyr Ile Leu Lys Ser
900 905 910
Ile Ser Tyr Asp Ser Val Arg Arg Leu Asn Leu Thr Lys Leu Pro Cys
915 920 925
Asp Thr Pro Glu Glu Thr Ala Ala Phe Val Lys Ser Leu Pro Ile Ser
930 935 940
Pro Glu Lys Pro Trp Ile Met Gln Glu Phe Ile Pro Gly Lys Glu Leu
945 950 955 960
Cys Thr His Ser Thr Val Arg Asp Gly Glu Leu Arg Leu His Cys Cys
965 970 975
Ser Asn Ser Ser Ala Phe Gln Ile Asn Tyr Glu Asn Val Glu Asn Pro
980 985 990
Gln Ile Gln Glu Trp Val Gln His Phe Val Lys Ser Leu Arg Leu Thr
995 1000 1005
Gly Gln Ile Ser Leu Asp Phe Ile Gln Ala Glu Asp Gly Thr Ala Tyr
1010 1015 1020
Ala Ile Glu Cys Asn Pro Arg Thr His Ser Ala Ile Thr Met Phe Tyr
1025 1030 1035 1040
Asn His Pro Gly Val Ala Glu Ala Tyr Leu Gly Lys Thr Pro Leu Ala
1045 1050 1055
Ala Pro Leu Glu Pro Leu Ala Asp Ser Lys Pro Thr Tyr Trp Ile Tyr
1060 1065 1070
His Glu Ile Trp Arg Leu Thr Gly Ile Arg Ser Gly Gln Gln Leu Gln
1075 1080 1085
Thr Trp Phe Gly Arg Leu Val Arg Gly Thr Asp Ala Ile Tyr Arg Leu
1090 1095 1100
Asp Asp Pro Ile Pro Phe Leu Thr Leu His His Trp Gln Ile Thr Leu
1105 1110 1115 1120
Leu Leu Leu Gln Asn Leu Gln Arg Leu Lys Gly Trp Val Lys Ile Asp
1125 1130 1135
Phe Asn Ile Gly Lys Leu Val Glu Leu Gly Gly Asp Met Pro Val Leu
1140 1145 1150
Asn Ile Leu His Leu Val Gly Ser Ala His Asp Lys Phe Tyr Cys Asp
1155 1160 1165
Leu Ser Arg Leu Tyr Ala Gln Asp Cys Leu Ala Ala Thr Ala Asp Pro
1170 1175 1180
Ser Leu Tyr Asn Phe Gln Ile Ala Tyr Ile Thr Pro Asp Arg Gln Trp
1185 1190 1195 1200
Arg Phe Pro Asp Ser Leu Ser Arg Glu Asp Ile Ala Leu Thr Lys Pro
1205 1210 1215
Ile Pro Val Phe Asp Ala Ile Gln Phe Leu Thr Gly Gln Asn Ile Asp
1220 1225 1230
Met Met Leu Pro Gln Met Phe Cys Ile Pro Gly Met Thr Gln Tyr Arg
1235 1240 1245
Ala Leu Phe Asp Leu Leu Lys Ile Pro Tyr Ile Gly Asn Thr Pro Asp
1250 1255 1260
Ile Met Ala Ile Ala Ala His Lys Ala Arg Ala Lys Ala Ile Val Glu
1265 1270 1275 1280
Ala Ala Gly Val Lys Val Pro Arg Gly Glu Leu Leu Arg Gln Gly Asp
1285 1290 1295
Ile Pro Thr Ile Thr Pro Pro Ala Val Val Lys Pro Val Ser Ser Asp
1300 1305 1310
Asn Ser Leu Gly Val Val Leu Val Lys Asp Val Thr Glu Tyr Asp Ala
1315 1320 1325
Ala Leu Lys Lys Ala Phe Glu Tyr Ala Ser Glu Val Ile Val Glu Ala
1330 1335 1340
Phe Ile Glu Leu Gly Arg Glu Val Arg Cys Gly Ile Ile Val Lys Asp
1345 1350 1355 1360
Gly Glu Leu Ile Gly Leu Pro Leu Glu Glu Tyr Leu Val Asp Pro His
1365 1370 1375
Asp Lys Pro Ile Arg Asn Tyr Ala Asp Lys Leu Gln Gln Thr Asp Asp
1380 1385 1390
Gly Asp Leu His Leu Thr Ala Lys Asp Asn Ile Lys Ala Trp Ile Leu
1395 1400 1405
Asp Pro Asn Asp Pro Ile Thr Gln Lys Val Gln Gln Val Ala Lys Arg
1410 1415 1420
Cys His Gln Ala Leu Gly Cys Arg His Tyr Ser Leu Phe Asp Phe Arg
1425 1430 1435 1440
Ile Asp Pro Lys Gly Gln Pro Trp Phe Leu Glu Ala Gly Leu Tyr Cys
1445 1450 1455
Ser Phe Ala Pro Lys Ser Val Ile Ser Ser Met Ala Lys Ala Ala Gly
1460 1465 1470
Ile Pro Leu Asn Asp Leu Leu Ile Thr Ala Ile Asn Glu Thr Leu Gly
1475 1480 1485
Ser Asn Lys Lys Val Leu Gln Asn
1490 1495
<210> 5
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_ABCD
<400> 5
acaatttcac acaggaaaga tatcatgagt atcgtccaag caaag 45
<210> 6
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_ABCD
<400> 6
ctcatccgcc aaaacagctc tagattatga attattttcc agaca 45
<210> 7
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Npr_ABCD
<400> 7
gatcgatatc atgagtaatg ttcaagcatc g 31
<210> 8
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Npr_ABCD
<400> 8
gatctctaga tcaattttgt aacacctttt tattac 36
<210> 9
<211> 71
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для кассеты с делецией aroD
<400> 9
tcatggggtt cggtgcctga caggctgacc gcgtgcagaa agggtaaaaa gctggagctg 60
cttcgaagtt c 71
<210> 10
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для кассеты с делецией aroD
<400> 10
atatattttt tagttcggcg gggagggtgt tcccgccgaa atattattgc gccatggtcc 60
atatgaatat cctccttag 79
<210> 11
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для делеции aroD
<400> 11
caaagatttc cctctggaat atg 23
<210> 12
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для делеции aroD
<400> 12
cagatgtgat tttccctacg c 21
<210> 13
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для aroG
<400> 13
aaggagctca ctagtggtac ggatgctcct gttatggtcg 40
<210> 14
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для aroG
<400> 14
ctctcttcga caatttccac ttacccgcga cgcgctttta c 41
<210> 15
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для ppsA
<400> 15
gtaaaagcgc gtcgcgggta agtggaaatt gtcgaagaga g 41
<210> 16
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для ppsA
<400> 16
gccctagtgg atctgatggg ttatttcttc agttcagcca gg 42
<210> 17
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для tktA
<400> 17
cctggctgaa ctgaagaaat aacccatcag atccactagg gc 42
<210> 18
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для tktA
<400> 18
cgacaccggt cagcagggtg ttacagcagt tcttttgctt tc 42
<210> 19
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для подтверждения плазмиды
pSKH130-deltafhuA-Pn-aroG-Pn-ppsA-Pn-tktA
<400> 19
gaaaatgccg atgggtaccg 20
<210> 20
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для подтверждения поазмиды
pSKH130-deltafhuA-Pn-aroG-Pn-ppsA-Pn-tktA
<400> 20
cgttagaacg cggctacaat taatac 26
<210> 21
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_ABCD
<400> 21
catctcacac cgagattatt ttagtactat gagtatcgtc caagcaaag 49
<210> 22
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_ABCD
<400> 22
caatttcgta tagagtctca ctagtaccgc aaaaaggcca tccgtc 46
<210> 23
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента промотора Ptrc
<400> 23
tctcacaccg agattatttt agtcgactgc acggtgcacc aat 43
<210> 24
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента промотора Ptrc
<400> 24
tttgcttgga cgatactcat agtctgtttc ctgtgtgaaa ttgttatccg 50
<210> 25
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента промотора PCJ1 или PCJ2.2
<400> 25
tctcacaccg agattatttt agtcaccgcg ggcttattcc att 43
<210> 26
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента промотора PCJ1 или PCJ2.2
<400> 26
ttgcttggac gatactcata gtatcttaat ctcctagatt gggtttcac 49
<210> 27
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента промотора PCJ2.2
<400> 27
tttgcttgga cgatactcat agtatcttaa tctcctagat ttggtttcac 50
<210> 28
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для подтверждения интеграции Ava-ABCD
<400> 28
cagaatgagt gaacaaccac gg 22
<210> 29
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для подтверждения интеграции Ava-ABCD
<400> 29
catatccaat aacccagaca aaacc 25
<210> 30
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ptrc/PCJ1-Ava_AB или Ptrc/PCJ1-Ava_ABC
<400> 30
gatcggatcc atgagtatcg tccaagcaaa g 31
<210> 31
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ptrc/PCJ1-Ava_AB
<400> 31
gatcactagt ttaaggttgt attctgcgg 29
<210> 32
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ptrc/PCJ1-Ava_ABC
<400> 32
gatcactagt ctaatctccc cccaattcc 29
<210> 33
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_ABCD
<400> 33
ccaaacacca acaaaaggct ctacccatat gatgagtatc gtccaagcaa agtttgaagc 60
60
<210> 34
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_ABCD
<400> 34
agagggtgac gctagtcaga actagtttat gaattatttt ccagacaatc ttgtaaacgc 60
60
<210> 35
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента промотора CJ7
<400> 35
ccaaacacca acaaaaggct ctacccatat gagaaacatc ccagcgctac taataggg 58
<210> 36
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента промотора CJ7
<400> 36
caaactttgc ttggacgata ctcatagtgt ttcctttcgt tgggtacg 48
<210> 37
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента промотора lysc8
<400> 37
caccaacaaa aggctctacc catatgaaca ctcctctggc taggtag 47
<210> 38
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента промотора lysc8
<400> 38
caaactttgc ttggacgata ctcatctttg tgcacctttc gatctacg 48
<210> 39
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента промотора O2
<400> 39
ccaaacacca acaaaaggct ctacccatat gcaataatcg tgaattttgg cagc 54
<210> 40
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента промотора O2
<400> 40
caaactttgc ttggacgata ctcattgttt tgatctcctc caataatcta tgc 53
<210> 41
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для генов aroD для конструирования pDC-дельта-aroD
<400> 41
gatcggatcc cactggacgt ttgggtgaga cc 32
<210> 42
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для генов aroD для конструирования pDC-дельта-aroD
<400> 42
gatcggatcc actagtttta ggttccattt ctaattg 37
<210> 43
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для генов aroD для конструирования pDC-дельта-aroD
<400> 43
gatcactagt atgaacgaca gtattctcc 29
<210> 44
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для генов aroD для конструирования pDC-дельта-aroD
<400> 44
gatcaagctt gttacatcct gacgttgtgg 30
<210> 45
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_A
<400> 45
cgcggatcca tgagtatcgt ccaagcaaag 30
<210> 46
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_A
<400> 46
cgcctcgagt tatttaacac tcccgattaa ttc 33
<210> 47
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_B
<400> 47
cgcggatcca tgacaaatgt gattgtccaa c 31
<210> 48
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_B
<400> 48
cgcctcgagt taaggttgta ttctgcggat aattg 35
<210> 49
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_C
<400> 49
cgcggatcca tggcacaatc ccttcccctt tc 32
<210> 50
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_C
<400> 50
cgcctcgagc taatctcccc ccaattccac c 31
<210> 51
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Ava_D
<400> 51
cgcggatcca tgcagactat agattttaat attc 34
<210> 52
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Ava_D
<400> 52
cgcctcgagt tatgaattat tttccagaca atc 33
<210> 53
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Npr_A
<400> 53
cgcggatcca tgagtaatgt tcaagcatcg 30
<210> 54
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Npr_A
<400> 54
cgcctcgagt cacactccca atagtttgg 29
<210> 55
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Npr_B
<400> 55
cgcggatcca tgaccagtat tttaggacga g 31
<210> 56
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Npr_B
<400> 56
cgcctcgagt tataccaagc gtctaatc 28
<210> 57
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Npr_C
<400> 57
cgcggatcca tggcacaatc aatctcttta tc 32
<210> 58
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Npr_C
<400> 58
cgcctcgagt tagtcgcccc ctaattccac g 31
<210> 59
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для Npr_D
<400> 59
cgcggatcca tgccagtact taatatcctt c 31
<210> 60
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для Npr_D
<400> 60
cgcctcgagt caattttgta acaccttttt attac 35
<210> 61
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для pGPD
<400> 61
acgtaagata attgtatatt acgcagttta tcattatcaa tactcgcc 48
<210> 62
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для pGPD
<400> 62
gtaacgacaa tcctctccat gtggtcttga atcaaagctg 40
<210> 63
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для LoxP(Ura)
<400> 63
ccgattgtca atcagcgtaa cagctgaagc ttcgtacgc 39
<210> 64
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для LoxP(Ura)
<400> 64
ttgttttggc cagtcggcaa gggcataggc cactagtgga tctg 44
<210> 65
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для aroB
<400> 65
cagctttgat tcaagaccac atggagagga ttgtcgttac 40
<210> 66
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для aroB
<400> 66
gcgtacgaag cttcagctgt tacgctgatt gacaatcgg 39
<210> 67
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента 1 ARO1
<400> 67
accattcaag agtatatgag agaaggaaaa ct 32
<210> 68
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента 1 ARO1
<400> 68
ggcgagtatt gataatgata aactgcgtaa tatacaatta tcttacg 47
<210> 69
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> прямой праймер для фрагмента 2 ARO1
<400> 69
cagatccact agtggcctat gcccttgccg actggccaaa acaa 44
<210> 70
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> обратный праймер для фрагмента 2 ARO1
<400> 70
caaatatcaa tggcacgttg 20
<210> 71
<211> 759
<212> ДНК
<213> Escherichia coli W3110
<400> 71
atgaaaaccg taactgtaaa agatctcgtc attggtacgg gcgcacctaa aatcatcgtc 60
tcgctgatgg cgaaagatat cgccagcgtg aaatccgaag ctctcgccta tcgtgaagcg 120
gactttgata ttctggaatg gcgtgtggac cactatgccg acctctccaa tgtggagtct 180
gtcatggcgg cagcaaaaat tctccgtgag accatgccag aaaaaccgct gctgtttacc 240
ttccgcagtg ccaaagaagg cggcgagcag gcgatttcca ccgaggctta tattgcactc 300
aatcgtgcag ccatcgacag cggcctggtt gatatgatcg atctggagtt atttaccggt 360
gatgatcagg ttaaagaaac cgtcgcctac gcccacgcgc atgatgtgaa agtagtcatg 420
tccaaccatg acttccataa aacgccggaa gccgaagaaa tcattgcccg tctgcgcaaa 480
atgcaatcct tcgacgccga tattcctaag attgcgctga tgccgcaaag taccagcgat 540
gtgctgacgt tgcttgccgc gaccctggag atgcaggagc agtatgccga tcgtccaatt 600
atcacgatgt cgatggcaaa aactggcgta atttctcgtc tggctggtga agtatttggc 660
tcggcggcaa cttttggtgc ggtaaaaaaa gcgtctgcgc cagggcaaat ctcggtaaat 720
gatttgcgca cggtattaac tattttacac caggcataa 759
<210> 72
<211> 252
<212> PRT
<213> Escherichia coli W3110
<400> 72
Met Lys Thr Val Thr Val Lys Asp Leu Val Ile Gly Thr Gly Ala Pro
1 5 10 15
Lys Ile Ile Val Ser Leu Met Ala Lys Asp Ile Ala Ser Val Lys Ser
20 25 30
Glu Ala Leu Ala Tyr Arg Glu Ala Asp Phe Asp Ile Leu Glu Trp Arg
35 40 45
Val Asp His Tyr Ala Asp Leu Ser Asn Val Glu Ser Val Met Ala Ala
50 55 60
Ala Lys Ile Leu Arg Glu Thr Met Pro Glu Lys Pro Leu Leu Phe Thr
65 70 75 80
Phe Arg Ser Ala Lys Glu Gly Gly Glu Gln Ala Ile Ser Thr Glu Ala
85 90 95
Tyr Ile Ala Leu Asn Arg Ala Ala Ile Asp Ser Gly Leu Val Asp Met
100 105 110
Ile Asp Leu Glu Leu Phe Thr Gly Asp Asp Gln Val Lys Glu Thr Val
115 120 125
Ala Tyr Ala His Ala His Asp Val Lys Val Val Met Ser Asn His Asp
130 135 140
Phe His Lys Thr Pro Glu Ala Glu Glu Ile Ile Ala Arg Leu Arg Lys
145 150 155 160
Met Gln Ser Phe Asp Ala Asp Ile Pro Lys Ile Ala Leu Met Pro Gln
165 170 175
Ser Thr Ser Asp Val Leu Thr Leu Leu Ala Ala Thr Leu Glu Met Gln
180 185 190
Glu Gln Tyr Ala Asp Arg Pro Ile Ile Thr Met Ser Met Ala Lys Thr
195 200 205
Gly Val Ile Ser Arg Leu Ala Gly Glu Val Phe Gly Ser Ala Ala Thr
210 215 220
Phe Gly Ala Val Lys Lys Ala Ser Ala Pro Gly Gln Ile Ser Val Asn
225 230 235 240
Asp Leu Arg Thr Val Leu Thr Ile Leu His Gln Ala
245 250
<210> 73
<211> 1053
<212> ДНК
<213> Escherichia coli W3110
<400> 73
atgaattatc agaacgacga tttacgcatc aaagaaatca aagagttact tcctcctgtc 60
gcattgctgg aaaaattccc cgctactgaa aatgccgcga atacggttgc ccatgcccga 120
aaagcgatcc ataagatcct gaaaggtaat gatgatcgcc tgttggttgt gattggccca 180
tgctcaattc atgatcctgt cgcggcaaaa gagtatgcca ctcgcttgct ggcgctgcgt 240
gaagagctga aagatgagct ggaaatcgta atgcgcgtct attttgaaaa gccgcgtacc 300
acggtgggct ggaaagggct gattaacgat ccgcatatgg ataatagctt ccagatcaac 360
gacggtctgc gtatagcccg taaattgctg cttgatatta acgacagcgg tctgccagcg 420
gcaggtgagt ttctcgatat gatcacccca caatatctcg ctgacctgat gagctggggc 480
gcaattggcg cacgtaccac cgaatcgcag gtgcaccgcg aactggcatc agggctttct 540
tgtccggtcg gcttcaaaaa tggcaccgac ggtacgatta aagtggctat cgatgccatt 600
aatgccgccg gtgcgccgca ctgcttcctg tccgtaacga aatgggggca ttcggcgatt 660
gtgaatacca gcggtaacgg cgattgccat atcattctgc gcggcggtaa agagcctaac 720
tacagcgcga agcacgttgc tgaagtgaaa gaagggctga acaaagcagg cctgccagca 780
caggtgatga tcgatttcag ccatgctaac tcgtccaaac aattcaaaaa gcagatggat 840
gtttgtgctg acgtttgcca gcagattgcc ggtggcgaaa aggccattat tggcgtgatg 900
gtggaaagcc atctggtgga aggcaatcag agcctcgaga gcggggagcc gctggcctac 960
ggtaagagca tcaccgatgc ctgcatcggc tgggaagata ccgatgctct gttacgtcaa 1020
ctggcgaatg cagtaaaagc gcgtcgcggg taa 1053
<210> 74
<211> 350
<212> PRT
<213> Escherichia coli W3110
<400> 74
Met Asn Tyr Gln Asn Asp Asp Leu Arg Ile Lys Glu Ile Lys Glu Leu
1 5 10 15
Leu Pro Pro Val Ala Leu Leu Glu Lys Phe Pro Ala Thr Glu Asn Ala
20 25 30
Ala Asn Thr Val Ala His Ala Arg Lys Ala Ile His Lys Ile Leu Lys
35 40 45
Gly Asn Asp Asp Arg Leu Leu Val Val Ile Gly Pro Cys Ser Ile His
50 55 60
Asp Pro Val Ala Ala Lys Glu Tyr Ala Thr Arg Leu Leu Ala Leu Arg
65 70 75 80
Glu Glu Leu Lys Asp Glu Leu Glu Ile Val Met Arg Val Tyr Phe Glu
85 90 95
Lys Pro Arg Thr Thr Val Gly Trp Lys Gly Leu Ile Asn Asp Pro His
100 105 110
Met Asp Asn Ser Phe Gln Ile Asn Asp Gly Leu Arg Ile Ala Arg Lys
115 120 125
Leu Leu Leu Asp Ile Asn Asp Ser Gly Leu Pro Ala Ala Gly Glu Phe
130 135 140
Leu Asp Met Ile Thr Pro Gln Tyr Leu Ala Asp Leu Met Ser Trp Gly
145 150 155 160
Ala Ile Gly Ala Arg Thr Thr Glu Ser Gln Val His Arg Glu Leu Ala
165 170 175
Ser Gly Leu Ser Cys Pro Val Gly Phe Lys Asn Gly Thr Asp Gly Thr
180 185 190
Ile Lys Val Ala Ile Asp Ala Ile Asn Ala Ala Gly Ala Pro His Cys
195 200 205
Phe Leu Ser Val Thr Lys Trp Gly His Ser Ala Ile Val Asn Thr Ser
210 215 220
Gly Asn Gly Asp Cys His Ile Ile Leu Arg Gly Gly Lys Glu Pro Asn
225 230 235 240
Tyr Ser Ala Lys His Val Ala Glu Val Lys Glu Gly Leu Asn Lys Ala
245 250 255
Gly Leu Pro Ala Gln Val Met Ile Asp Phe Ser His Ala Asn Ser Ser
260 265 270
Lys Gln Phe Lys Lys Gln Met Asp Val Cys Ala Asp Val Cys Gln Gln
275 280 285
Ile Ala Gly Gly Glu Lys Ala Ile Ile Gly Val Met Val Glu Ser His
290 295 300
Leu Val Glu Gly Asn Gln Ser Leu Glu Ser Gly Glu Pro Leu Ala Tyr
305 310 315 320
Gly Lys Ser Ile Thr Asp Ala Cys Ile Gly Trp Glu Asp Thr Asp Ala
325 330 335
Leu Leu Arg Gln Leu Ala Asn Ala Val Lys Ala Arg Arg Gly
340 345 350
<210> 75
<211> 2379
<212> ДНК
<213> Escherichia coli W3110
<400> 75
atgtccaaca atggctcgtc accgctggtg ctttggtata accaactcgg catgaatgat 60
gtagacaggg ttgggggcaa aaatgcctcc ctgggtgaaa tgattactaa tctttccgga 120
atgggtgttt ccgttccgaa tggtttcgcc acaaccgccg acgcgtttaa ccagtttctg 180
gaccaaagcg gcgtaaacca gcgcatttat gaactgctgg ataaaacgga tattgacgat 240
gttactcagc ttgcgaaagc gggcgcgcaa atccgccagt ggattatcga cactcccttc 300
cagcctgagc tggaaaacgc catccgcgaa gcctatgcac agctttccgc cgatgacgaa 360
aacgcctctt ttgcggtgcg ctcctccgcc accgcagaag atatgccgga cgcttctttt 420
gccggtcagc aggaaacctt cctcaacgtt cagggttttg acgccgttct cgtggcagtg 480
aaacatgtat ttgcttctct gtttaacgat cgcgccatct cttatcgtgt gcaccagggt 540
tacgatcacc gtggtgtggc gctctccgcc ggtgttcaac ggatggtgcg ctctgacctc 600
gcatcatctg gcgtgatgtt ctccattgat accgaatccg gctttgacca ggtggtgttt 660
atcacttccg catggggcct tggtgagatg gtcgtgcagg gtgcggttaa cccggatgag 720
ttttacgtgc ataaaccgac actggcggcg aatcgcccgg ctatcgtgcg ccgcaccatg 780
gggtcgaaaa aaatccgcat ggtttacgcg ccgacccagg agcacggcaa gcaggttaaa 840
atcgaagacg taccgcagga acagcgtgac atcttctcgc tgaccaacga agaagtgcag 900
gaactggcaa aacaggccgt acaaattgag aaacactacg gtcgcccgat ggatattgag 960
tgggcgaaag atggccacac cggtaaactg ttcattgtgc aggcgcgtcc ggaaaccgtg 1020
cgctcacgcg gtcaggtcat ggagcgttat acgctgcatt cacagggtaa gattatcgcc 1080
gaaggccgtg ctatcggtca tcgcatcggt gcgggtccgg tgaaagtcat ccatgacatc 1140
agcgaaatga accgcatcga acctggcgac gtgctggtta ctgacatgac cgacccggac 1200
tgggaaccga tcatgaagaa agcatctgcc atcgtcacca accgtggcgg tcgtacctgt 1260
cacgcggcga tcatcgctcg tgaactgggc attccggcgg tagtgggctg tggagatgca 1320
acagaacgga tgaaagacgg tgagaacgtc actgtttctt gtgccgaagg tgataccggt 1380
tacgtctatg cggagttgct ggaatttagc gtgaaaagct ccagcgtaga aacgatgccg 1440
gatctgccgt tgaaagtgat gatgaacgtc ggtaacccgg accgtgcttt cgacttcgcc 1500
tgcctaccga acgaaggcgt gggccttgcg cgtctggaat ttatcatcaa ccgtatgatt 1560
ggcgtccacc cacgcgcact gcttgagttt gacgatcagg aaccgcagtt gcaaaacgaa 1620
atccgcgaga tgatgaaagg ttttgattct ccgcgtgaat tttacgttgg tcgtctgact 1680
gaagggatcg cgacgctggg tgccgcgttt tatccgaagc gcgtcattgt ccgtctctct 1740
gattttaaat cgaacgaata tgccaacctg gtcggtggtg agcgttacga gccagatgaa 1800
gagaacccga tgctcggctt ccgtggcgcg ggccgctatg tttccgacag cttccgcgac 1860
tgtttcgcgc tggagtgtga agcagtgaaa cgtgtgcgca acgacatggg actgaccaac 1920
gttgagatca tgatcccgtt cgtgcgtacc gtagatcagg cgaaagcggt ggttgaagaa 1980
ctggcgcgtc aggggctgaa acgtggcgag aacgggctga aaatcatcat gatgtgtgaa 2040
atcccgtcca acgccttgct ggccgagcag ttcctcgaat atttcgacgg cttctcaatt 2100
ggctcaaacg atatgacgca gctggcgctc ggtctggacc gtgactccgg cgtggtgtct 2160
gaattgttcg atgagcgcaa cgatgcggtg aaagcactgc tgtcgatggc tatccgtgcc 2220
gcgaagaaac agggcaaata tgtcgggatt tgcggtcagg gtccgtccga ccacgaagac 2280
tttgccgcat ggttgatgga agaggggatc gatagcctgt ctctgaaccc ggacaccgtg 2340
gtgcaaacct ggttaagcct ggctgaactg aagaaataa 2379
<210> 76
<211> 792
<212> PRT
<213> Escherichia coli W3110
<400> 76
Met Ser Asn Asn Gly Ser Ser Pro Leu Val Leu Trp Tyr Asn Gln Leu
1 5 10 15
Gly Met Asn Asp Val Asp Arg Val Gly Gly Lys Asn Ala Ser Leu Gly
20 25 30
Glu Met Ile Thr Asn Leu Ser Gly Met Gly Val Ser Val Pro Asn Gly
35 40 45
Phe Ala Thr Thr Ala Asp Ala Phe Asn Gln Phe Leu Asp Gln Ser Gly
50 55 60
Val Asn Gln Arg Ile Tyr Glu Leu Leu Asp Lys Thr Asp Ile Asp Asp
65 70 75 80
Val Thr Gln Leu Ala Lys Ala Gly Ala Gln Ile Arg Gln Trp Ile Ile
85 90 95
Asp Thr Pro Phe Gln Pro Glu Leu Glu Asn Ala Ile Arg Glu Ala Tyr
100 105 110
Ala Gln Leu Ser Ala Asp Asp Glu Asn Ala Ser Phe Ala Val Arg Ser
115 120 125
Ser Ala Thr Ala Glu Asp Met Pro Asp Ala Ser Phe Ala Gly Gln Gln
130 135 140
Glu Thr Phe Leu Asn Val Gln Gly Phe Asp Ala Val Leu Val Ala Val
145 150 155 160
Lys His Val Phe Ala Ser Leu Phe Asn Asp Arg Ala Ile Ser Tyr Arg
165 170 175
Val His Gln Gly Tyr Asp His Arg Gly Val Ala Leu Ser Ala Gly Val
180 185 190
Gln Arg Met Val Arg Ser Asp Leu Ala Ser Ser Gly Val Met Phe Ser
195 200 205
Ile Asp Thr Glu Ser Gly Phe Asp Gln Val Val Phe Ile Thr Ser Ala
210 215 220
Trp Gly Leu Gly Glu Met Val Val Gln Gly Ala Val Asn Pro Asp Glu
225 230 235 240
Phe Tyr Val His Lys Pro Thr Leu Ala Ala Asn Arg Pro Ala Ile Val
245 250 255
Arg Arg Thr Met Gly Ser Lys Lys Ile Arg Met Val Tyr Ala Pro Thr
260 265 270
Gln Glu His Gly Lys Gln Val Lys Ile Glu Asp Val Pro Gln Glu Gln
275 280 285
Arg Asp Ile Phe Ser Leu Thr Asn Glu Glu Val Gln Glu Leu Ala Lys
290 295 300
Gln Ala Val Gln Ile Glu Lys His Tyr Gly Arg Pro Met Asp Ile Glu
305 310 315 320
Trp Ala Lys Asp Gly His Thr Gly Lys Leu Phe Ile Val Gln Ala Arg
325 330 335
Pro Glu Thr Val Arg Ser Arg Gly Gln Val Met Glu Arg Tyr Thr Leu
340 345 350
His Ser Gln Gly Lys Ile Ile Ala Glu Gly Arg Ala Ile Gly His Arg
355 360 365
Ile Gly Ala Gly Pro Val Lys Val Ile His Asp Ile Ser Glu Met Asn
370 375 380
Arg Ile Glu Pro Gly Asp Val Leu Val Thr Asp Met Thr Asp Pro Asp
385 390 395 400
Trp Glu Pro Ile Met Lys Lys Ala Ser Ala Ile Val Thr Asn Arg Gly
405 410 415
Gly Arg Thr Cys His Ala Ala Ile Ile Ala Arg Glu Leu Gly Ile Pro
420 425 430
Ala Val Val Gly Cys Gly Asp Ala Thr Glu Arg Met Lys Asp Gly Glu
435 440 445
Asn Val Thr Val Ser Cys Ala Glu Gly Asp Thr Gly Tyr Val Tyr Ala
450 455 460
Glu Leu Leu Glu Phe Ser Val Lys Ser Ser Ser Val Glu Thr Met Pro
465 470 475 480
Asp Leu Pro Leu Lys Val Met Met Asn Val Gly Asn Pro Asp Arg Ala
485 490 495
Phe Asp Phe Ala Cys Leu Pro Asn Glu Gly Val Gly Leu Ala Arg Leu
500 505 510
Glu Phe Ile Ile Asn Arg Met Ile Gly Val His Pro Arg Ala Leu Leu
515 520 525
Glu Phe Asp Asp Gln Glu Pro Gln Leu Gln Asn Glu Ile Arg Glu Met
530 535 540
Met Lys Gly Phe Asp Ser Pro Arg Glu Phe Tyr Val Gly Arg Leu Thr
545 550 555 560
Glu Gly Ile Ala Thr Leu Gly Ala Ala Phe Tyr Pro Lys Arg Val Ile
565 570 575
Val Arg Leu Ser Asp Phe Lys Ser Asn Glu Tyr Ala Asn Leu Val Gly
580 585 590
Gly Glu Arg Tyr Glu Pro Asp Glu Glu Asn Pro Met Leu Gly Phe Arg
595 600 605
Gly Ala Gly Arg Tyr Val Ser Asp Ser Phe Arg Asp Cys Phe Ala Leu
610 615 620
Glu Cys Glu Ala Val Lys Arg Val Arg Asn Asp Met Gly Leu Thr Asn
625 630 635 640
Val Glu Ile Met Ile Pro Phe Val Arg Thr Val Asp Gln Ala Lys Ala
645 650 655
Val Val Glu Glu Leu Ala Arg Gln Gly Leu Lys Arg Gly Glu Asn Gly
660 665 670
Leu Lys Ile Ile Met Met Cys Glu Ile Pro Ser Asn Ala Leu Leu Ala
675 680 685
Glu Gln Phe Leu Glu Tyr Phe Asp Gly Phe Ser Ile Gly Ser Asn Asp
690 695 700
Met Thr Gln Leu Ala Leu Gly Leu Asp Arg Asp Ser Gly Val Val Ser
705 710 715 720
Glu Leu Phe Asp Glu Arg Asn Asp Ala Val Lys Ala Leu Leu Ser Met
725 730 735
Ala Ile Arg Ala Ala Lys Lys Gln Gly Lys Tyr Val Gly Ile Cys Gly
740 745 750
Gln Gly Pro Ser Asp His Glu Asp Phe Ala Ala Trp Leu Met Glu Glu
755 760 765
Gly Ile Asp Ser Leu Ser Leu Asn Pro Asp Thr Val Val Gln Thr Trp
770 775 780
Leu Ser Leu Ala Glu Leu Lys Lys
785 790
<210> 77
<211> 1992
<212> ДНК
<213> Escherichia coli W3110
<400> 77
atgtcctcac gtaaagagct tgccaatgct attcgtgcgc tgagcatgga cgcagtacag 60
aaagccaaat ccggtcaccc gggtgcccct atgggtatgg ctgacattgc cgaagtcctg 120
tggcgtgatt tcctgaaaca caacccgcag aatccgtcct gggctgaccg tgaccgcttc 180
gtgctgtcca acggccacgg ctccatgctg atctacagcc tgctgcacct caccggttac 240
gatctgccga tggaagaact gaaaaacttc cgtcagctgc actctaaaac tccgggtcac 300
ccggaagtgg gttacaccgc tggtgtggaa accaccaccg gtccgctggg tcagggtatt 360
gccaacgcag tcggtatggc gattgcagaa aaaacgctgg cggcgcagtt taaccgtccg 420
ggccacgaca ttgtcgacca ctacacctac gccttcatgg gcgacggctg catgatggaa 480
ggcatctccc acgaagtttg ctctctggcg ggtacgctga agctgggtaa actgattgca 540
ttctacgatg acaacggtat ttctatcgat ggtcacgttg aaggctggtt caccgacgac 600
accgcaatgc gtttcgaagc ttacggctgg cacgttattc gcgacatcga cggtcatgac 660
gcggcatcta tcaaacgcgc agtagaagaa gcgcgcgcag tgactgacaa accttccctg 720
ctgatgtgca aaaccatcat cggtttcggt tccccgaaca aagccggtac ccacgactcc 780
cacggtgcgc cgctgggcga cgctgaaatt gccctgaccc gcgaacaact gggctggaaa 840
tatgcgccgt tcgaaatccc gtctgaaatc tatgctcagt gggatgcgaa agaagcaggc 900
caggcgaaag aatccgcatg gaacgagaaa ttcgctgctt acgcgaaagc ttatccgcag 960
gaagccgctg aatttacccg ccgtatgaaa ggcgaaatgc cgtctgactt cgacgctaaa 1020
gcgaaagagt tcatcgctaa actgcaggct aatccggcga aaatcgccag ccgtaaagcg 1080
tctcagaatg ctatcgaagc gttcggtccg ctgttgccgg aattcctcgg cggttctgct 1140
gacctggcgc cgtctaacct gaccctgtgg tctggttcta aagcaatcaa cgaagatgct 1200
gcgggtaact acatccacta cggtgttcgc gagttcggta tgaccgcgat tgctaacggt 1260
atctccctgc acggtggctt cctgccgtac acctccacct tcctgatgtt cgtggaatac 1320
gcacgtaacg ccgtacgtat ggctgcgctg atgaaacagc gtcaggtgat ggtttacacc 1380
cacgactcca tcggtctggg cgaagacggc ccgactcacc agccggttga gcaggtcgct 1440
tctctgcgcg taaccccgaa catgtctaca tggcgtccgt gtgaccaggt tgaatccgcg 1500
gtcgcgtgga aatacggtgt tgagcgtcag gacggcccga ccgcactgat cctctcccgt 1560
cagaacctgg cgcagcagga acgaactgaa gagcaactgg caaacatcgc gcgcggtggt 1620
tatgtgctga aagactgcgc cggtcagccg gaactgattt tcatcgctac cggttcagaa 1680
gttgaactgg ctgttgctgc ctacgaaaaa ctgactgccg aaggcgtgaa agcgcgcgtg 1740
gtgtccatgc cgtctaccga cgcatttgac aagcaggatg ctgcttaccg tgaatccgta 1800
ctgccgaaag cggttactgc acgcgttgct gtagaagcgg gtattgctga ctactggtac 1860
aagtatgttg gcctgaacgg tgctatcgtc ggtatgacca ccttcggtga atctgctccg 1920
gcagagctgc tgtttgaaga gttcggcttc actgttgata acgttgttgc gaaagcaaaa 1980
gaactgctgt aa 1992
<210> 78
<211> 663
<212> PRT
<213> Escherichia coli W3110
<400> 78
Met Ser Ser Arg Lys Glu Leu Ala Asn Ala Ile Arg Ala Leu Ser Met
1 5 10 15
Asp Ala Val Gln Lys Ala Lys Ser Gly His Pro Gly Ala Pro Met Gly
20 25 30
Met Ala Asp Ile Ala Glu Val Leu Trp Arg Asp Phe Leu Lys His Asn
35 40 45
Pro Gln Asn Pro Ser Trp Ala Asp Arg Asp Arg Phe Val Leu Ser Asn
50 55 60
Gly His Gly Ser Met Leu Ile Tyr Ser Leu Leu His Leu Thr Gly Tyr
65 70 75 80
Asp Leu Pro Met Glu Glu Leu Lys Asn Phe Arg Gln Leu His Ser Lys
85 90 95
Thr Pro Gly His Pro Glu Val Gly Tyr Thr Ala Gly Val Glu Thr Thr
100 105 110
Thr Gly Pro Leu Gly Gln Gly Ile Ala Asn Ala Val Gly Met Ala Ile
115 120 125
Ala Glu Lys Thr Leu Ala Ala Gln Phe Asn Arg Pro Gly His Asp Ile
130 135 140
Val Asp His Tyr Thr Tyr Ala Phe Met Gly Asp Gly Cys Met Met Glu
145 150 155 160
Gly Ile Ser His Glu Val Cys Ser Leu Ala Gly Thr Leu Lys Leu Gly
165 170 175
Lys Leu Ile Ala Phe Tyr Asp Asp Asn Gly Ile Ser Ile Asp Gly His
180 185 190
Val Glu Gly Trp Phe Thr Asp Asp Thr Ala Met Arg Phe Glu Ala Tyr
195 200 205
Gly Trp His Val Ile Arg Asp Ile Asp Gly His Asp Ala Ala Ser Ile
210 215 220
Lys Arg Ala Val Glu Glu Ala Arg Ala Val Thr Asp Lys Pro Ser Leu
225 230 235 240
Leu Met Cys Lys Thr Ile Ile Gly Phe Gly Ser Pro Asn Lys Ala Gly
245 250 255
Thr His Asp Ser His Gly Ala Pro Leu Gly Asp Ala Glu Ile Ala Leu
260 265 270
Thr Arg Glu Gln Leu Gly Trp Lys Tyr Ala Pro Phe Glu Ile Pro Ser
275 280 285
Glu Ile Tyr Ala Gln Trp Asp Ala Lys Glu Ala Gly Gln Ala Lys Glu
290 295 300
Ser Ala Trp Asn Glu Lys Phe Ala Ala Tyr Ala Lys Ala Tyr Pro Gln
305 310 315 320
Glu Ala Ala Glu Phe Thr Arg Arg Met Lys Gly Glu Met Pro Ser Asp
325 330 335
Phe Asp Ala Lys Ala Lys Glu Phe Ile Ala Lys Leu Gln Ala Asn Pro
340 345 350
Ala Lys Ile Ala Ser Arg Lys Ala Ser Gln Asn Ala Ile Glu Ala Phe
355 360 365
Gly Pro Leu Leu Pro Glu Phe Leu Gly Gly Ser Ala Asp Leu Ala Pro
370 375 380
Ser Asn Leu Thr Leu Trp Ser Gly Ser Lys Ala Ile Asn Glu Asp Ala
385 390 395 400
Ala Gly Asn Tyr Ile His Tyr Gly Val Arg Glu Phe Gly Met Thr Ala
405 410 415
Ile Ala Asn Gly Ile Ser Leu His Gly Gly Phe Leu Pro Tyr Thr Ser
420 425 430
Thr Phe Leu Met Phe Val Glu Tyr Ala Arg Asn Ala Val Arg Met Ala
435 440 445
Ala Leu Met Lys Gln Arg Gln Val Met Val Tyr Thr His Asp Ser Ile
450 455 460
Gly Leu Gly Glu Asp Gly Pro Thr His Gln Pro Val Glu Gln Val Ala
465 470 475 480
Ser Leu Arg Val Thr Pro Asn Met Ser Thr Trp Arg Pro Cys Asp Gln
485 490 495
Val Glu Ser Ala Val Ala Trp Lys Tyr Gly Val Glu Arg Gln Asp Gly
500 505 510
Pro Thr Ala Leu Ile Leu Ser Arg Gln Asn Leu Ala Gln Gln Glu Arg
515 520 525
Thr Glu Glu Gln Leu Ala Asn Ile Ala Arg Gly Gly Tyr Val Leu Lys
530 535 540
Asp Cys Ala Gly Gln Pro Glu Leu Ile Phe Ile Ala Thr Gly Ser Glu
545 550 555 560
Val Glu Leu Ala Val Ala Ala Tyr Glu Lys Leu Thr Ala Glu Gly Val
565 570 575
Lys Ala Arg Val Val Ser Met Pro Ser Thr Asp Ala Phe Asp Lys Gln
580 585 590
Asp Ala Ala Tyr Arg Glu Ser Val Leu Pro Lys Ala Val Thr Ala Arg
595 600 605
Val Ala Val Glu Ala Gly Ile Ala Asp Tyr Trp Tyr Lys Tyr Val Gly
610 615 620
Leu Asn Gly Ala Ile Val Gly Met Thr Thr Phe Gly Glu Ser Ala Pro
625 630 635 640
Ala Glu Leu Leu Phe Glu Glu Phe Gly Phe Thr Val Asp Asn Val Val
645 650 655
Ala Lys Ala Lys Glu Leu Leu
660
<210> 79
<211> 438
<212> ДНК
<213> Corynebacterium glutamicum ATCC 13032
<400> 79
atgcctggaa aaattctcct cctcaacggc ccaaacctga acatgctggg caaacgcgag 60
cctgacattt acggacacga caccttggaa gacgtcgtcg cgctggcaac cgctgaggct 120
gcgaagcacg gccttgaggt tgaggcgctg cagagcaatc acgaaggtga gctaatcgat 180
gcgctgcaca acgctcgcgg cacccacatc ggttgcgtga ttaaccccgg cggcctgact 240
cacacttcgg tggcgctttt ggatgctgtg aaggcgtctg agcttcctac cgttgaggtg 300
cacatttcca atccgcatgc ccgtgaagag ttccgccacc attcttacat ttccctcgcc 360
gcggtctccg ttatcgctgg cgctggcatc cagggttacc gtttcgcggt cgatatcctg 420
gcaaatctca aaaagtag 438
<210> 80
<211> 145
<212> PRT
<213> Corynebacterium glutamicum ATCC 13032
<400> 80
Met Pro Gly Lys Ile Leu Leu Leu Asn Gly Pro Asn Leu Asn Met Leu
1 5 10 15
Gly Lys Arg Glu Pro Asp Ile Tyr Gly His Asp Thr Leu Glu Asp Val
20 25 30
Val Ala Leu Ala Thr Ala Glu Ala Ala Lys His Gly Leu Glu Val Glu
35 40 45
Ala Leu Gln Ser Asn His Glu Gly Glu Leu Ile Asp Ala Leu His Asn
50 55 60
Ala Arg Gly Thr His Ile Gly Cys Val Ile Asn Pro Gly Gly Leu Thr
65 70 75 80
His Thr Ser Val Ala Leu Leu Asp Ala Val Lys Ala Ser Glu Leu Pro
85 90 95
Thr Val Glu Val His Ile Ser Asn Pro His Ala Arg Glu Glu Phe Arg
100 105 110
His His Ser Tyr Ile Ser Leu Ala Ala Val Ser Val Ile Ala Gly Ala
115 120 125
Gly Ile Gln Gly Tyr Arg Phe Ala Val Asp Ile Leu Ala Asn Leu Lys
130 135 140
Lys
145
<210> 81
<211> 4767
<212> ДНК
<213> Saccharomyces cerevisiae
<400> 81
atggtgcagt tagccaaagt cccaattcta ggaaatgata ttatccacgt tgggtataac 60
attcatgacc atttggttga aaccataatt aaacattgtc cttcttcgac atacgttatt 120
tgcaatgata cgaacttgag taaagttcca tactaccagc aattagtcct ggaattcaag 180
gcttctttgc cagaaggctc tcgtttactt acttatgttg ttaaaccagg tgagacaagt 240
aaaagtagag aaaccaaagc gcagctagaa gattatcttt tagtggaagg atgtactcgt 300
gatacggtta tggtagcgat cggtggtggt gttattggtg acatgattgg gttcgttgca 360
tctacattta tgagaggtgt tcgtgttgtc caagtaccaa catccttatt ggcaatggtc 420
gattcctcca ttggtggtaa aactgctatt gacactcctc taggtaaaaa ctttattggt 480
gcattttggc aaccaaaatt tgtccttgta gatattaaat ggctagaaac gttagccaag 540
agagagttta tcaatgggat ggcagaagtt atcaagactg cttgtatttg gaacgctgac 600
gaatttacta gattagaatc aaacgcttcg ttgttcttaa atgttgttaa tggggcaaaa 660
aatgtcaagg ttaccaatca attgacaaac gagattgacg agatatcgaa tacagatatt 720
gaagctatgt tggatcatac atataagtta gttcttgaga gtattaaggt caaagcggaa 780
gttgtctctt cggatgaacg tgaatccagt ctaagaaacc ttttgaactt cggacattct 840
attggtcatg cttatgaagc tatactaacc ccacaagcat tacatggtga atgtgtgtcc 900
attggtatgg ttaaagaggc ggaattatcc cgttatttcg gtattctctc ccctacccaa 960
gttgcacgtc tatccaagat tttggttgcc tacgggttgc ctgtttcgcc tgatgagaaa 1020
tggtttaaag agctaacctt acataagaaa acaccattgg atatcttatt gaagaaaatg 1080
agtattgaca agaaaaacga gggttccaaa aagaaggtgg tcattttaga aagtattggt 1140
aagtgctatg gtgactccgc tcaatttgtt agcgatgaag acctgagatt tattctaaca 1200
gatgaaaccc tcgtttaccc cttcaaggac atccctgctg atcaacagaa agttgttatc 1260
ccccctggtt ctaagtccat ctccaatcgt gctttaattc ttgctgccct cggtgaaggt 1320
caatgtaaaa tcaagaactt attacattct gatgatacta aacatatgtt aaccgctgtt 1380
catgaattga aaggtgctac gatatcatgg gaagataatg gtgagacggt agtggtggaa 1440
ggacatggtg gttccacatt gtcagcttgt gctgacccct tatatctagg taatgcaggt 1500
actgcatcta gatttttgac ttccttggct gccttggtca attctacttc aagccaaaag 1560
tatatcgttt taactggtaa cgcaagaatg caacaaagac caattgctcc tttggtcgat 1620
tctttgcgtg ctaatggtac taaaattgag tacttgaata atgaaggttc cctgccaatc 1680
aaagtttata ctgattcggt attcaaaggt ggtagaattg aattagctgc tacagtttct 1740
tctcagtacg tatcctctat cttgatgtgt gccccatacg ctgaagaacc tgtaactttg 1800
gctcttgttg gtggtaagcc aatctctaaa ttgtacgtcg atatgacaat aaaaatgatg 1860
gaaaaattcg gtatcaatgt tgaaacttct actacagaac cttacactta ttatattcca 1920
aagggacatt atattaaccc atcagaatac gtcattgaaa gtgatgcctc aagtgctaca 1980
tacccattgg ccttcgccgc aatgactggt actaccgtaa cggttccaaa cattggtttt 2040
gagtcgttac aaggtgatgc cagatttgca agagatgtct tgaaacctat gggttgtaaa 2100
ataactcaaa cggcaacttc aactactgtt tcgggtcctc ctgtaggtac tttaaagcca 2160
ttaaaacatg ttgatatgga gccaatgact gatgcgttct taactgcatg tgttgttgcc 2220
gctatttcgc acgacagtga tccaaattct gcaaatacaa ccaccattga aggtattgca 2280
aaccagcgtg tcaaagagtg taacagaatt ttggccatgg ctacagagct cgccaaattt 2340
ggcgtcaaaa ctacagaatt accagatggt attcaagtcc atggtttaaa ctcgataaaa 2400
gatttgaagg ttccttccga ctcttctgga cctgtcggtg tatgcacata tgatgatcat 2460
cgtgtggcca tgagtttctc gcttcttgca ggaatggtaa attctcaaaa tgaacgtgac 2520
gaagttgcta atcctgtaag aatacttgaa agacattgta ctggtaaaac ctggcctggc 2580
tggtgggatg tgttacattc cgaactaggt gccaaattag atggtgcaga acctttagag 2640
tgcacatcca aaaagaactc aaagaaaagc gttgtcatta ttggcatgag agcagctggc 2700
aaaactacta taagtaaatg gtgcgcatcc gctctgggtt acaaattagt tgacctagac 2760
gagctgtttg agcaacagca taacaatcaa agtgttaaac aatttgttgt ggagaacggt 2820
tgggagaagt tccgtgagga agaaacaaga attttcaagg aagttattca aaattacggc 2880
gatgatggat atgttttctc aacaggtggc ggtattgttg aaagcgctga gtctagaaaa 2940
gccttaaaag attttgcctc atcaggtgga tacgttttac acttacatag ggatattgag 3000
gagacaattg tctttttaca aagtgatcct tcaagacctg cctatgtgga agaaattcgt 3060
gaagtttgga acagaaggga ggggtggtat aaagaatgct caaatttctc tttctttgct 3120
cctcattgct ccgcagaagc tgagttccaa gctctaagaa gatcgtttag taagtacatt 3180
gcaaccatta caggtgtcag agaaatagaa attccaagcg gaagatctgc ctttgtgtgt 3240
ttaacctttg atgacttaac tgaacaaact gagaatttga ctccaatctg ttatggttgt 3300
gaggctgtag aggtcagagt agaccatttg gctaattact ctgctgattt cgtgagtaaa 3360
cagttatcta tattgcgtaa agccactgac agtattccta tcatttttac tgtgcgaacc 3420
atgaagcaag gtggcaactt tcctgatgaa gagttcaaaa ccttgagaga gctatacgat 3480
attgccttga agaatggtgt tgaattcctt gacttagaac taactttacc tactgatatc 3540
caatatgagg ttattaacaa aaggggcaac accaagatca ttggttccca tcatgacttc 3600
caaggattat actcctggga cgacgctgaa tgggaaaaca gattcaatca agcgttaact 3660
cttgatgtgg atgttgtaaa atttgtgggt acggctgtta atttcgaaga taatttgaga 3720
ctggaacact ttagggatac acacaagaat aagcctttaa ttgcagttaa tatgacttct 3780
aaaggtagca tttctcgtgt tttgaataat gttttaacac ctgtgacatc agatttattg 3840
cctaactccg ctgcccctgg ccaattgaca gtagcacaaa ttaacaagat gtatacatct 3900
atgggaggta tcgagcctaa ggaactgttt gttgttggaa agccaattgg ccactctaga 3960
tcgccaattt tacataacac tggctatgaa attttaggtt tacctcacaa gttcgataaa 4020
tttgaaactg aatccgcaca attggtgaaa gaaaaacttt tggacggaaa caagaacttt 4080
ggcggtgctg cagtcacaat tcctctgaaa ttagatataa tgcagtacat ggatgaattg 4140
actgatgctg ctaaagttat tggtgctgta aacacagtta taccattggg taacaagaag 4200
tttaagggtg ataataccga ctggttaggt atccgtaatg ccttaattaa caatggcgtt 4260
cccgaatatg ttggtcatac cgctggtttg gttatcggtg caggtggcac ttctagagcc 4320
gccctttacg ccttgcacag tttaggttgc aaaaagatct tcataatcaa caggacaact 4380
tcgaaattga agccattaat agagtcactt ccatctgaat tcaacattat tggaatagag 4440
tccactaaat ctatagaaga gattaaggaa cacgttggcg ttgctgtcag ctgtgtacca 4500
gccgacaaac cattagatga cgaactttta agtaagctgg agagattcct tgtgaaaggt 4560
gcccatgctg cttttgtacc aaccttattg gaagccgcat acaaaccaag cgttactccc 4620
gttatgacaa tttcacaaga caaatatcaa tggcacgttg tccctggatc acaaatgtta 4680
gtacaccaag gtgtagctca gtttgaaaag tggacaggat tcaagggccc tttcaaggcc 4740
atttttgatg ccgttacgaa agagtag 4767
<210> 82
<211> 1588
<212> PRT
<213> Saccharomyces cerevisiae
<400> 82
Met Val Gln Leu Ala Lys Val Pro Ile Leu Gly Asn Asp Ile Ile His
1 5 10 15
Val Gly Tyr Asn Ile His Asp His Leu Val Glu Thr Ile Ile Lys His
20 25 30
Cys Pro Ser Ser Thr Tyr Val Ile Cys Asn Asp Thr Asn Leu Ser Lys
35 40 45
Val Pro Tyr Tyr Gln Gln Leu Val Leu Glu Phe Lys Ala Ser Leu Pro
50 55 60
Glu Gly Ser Arg Leu Leu Thr Tyr Val Val Lys Pro Gly Glu Thr Ser
65 70 75 80
Lys Ser Arg Glu Thr Lys Ala Gln Leu Glu Asp Tyr Leu Leu Val Glu
85 90 95
Gly Cys Thr Arg Asp Thr Val Met Val Ala Ile Gly Gly Gly Val Ile
100 105 110
Gly Asp Met Ile Gly Phe Val Ala Ser Thr Phe Met Arg Gly Val Arg
115 120 125
Val Val Gln Val Pro Thr Ser Leu Leu Ala Met Val Asp Ser Ser Ile
130 135 140
Gly Gly Lys Thr Ala Ile Asp Thr Pro Leu Gly Lys Asn Phe Ile Gly
145 150 155 160
Ala Phe Trp Gln Pro Lys Phe Val Leu Val Asp Ile Lys Trp Leu Glu
165 170 175
Thr Leu Ala Lys Arg Glu Phe Ile Asn Gly Met Ala Glu Val Ile Lys
180 185 190
Thr Ala Cys Ile Trp Asn Ala Asp Glu Phe Thr Arg Leu Glu Ser Asn
195 200 205
Ala Ser Leu Phe Leu Asn Val Val Asn Gly Ala Lys Asn Val Lys Val
210 215 220
Thr Asn Gln Leu Thr Asn Glu Ile Asp Glu Ile Ser Asn Thr Asp Ile
225 230 235 240
Glu Ala Met Leu Asp His Thr Tyr Lys Leu Val Leu Glu Ser Ile Lys
245 250 255
Val Lys Ala Glu Val Val Ser Ser Asp Glu Arg Glu Ser Ser Leu Arg
260 265 270
Asn Leu Leu Asn Phe Gly His Ser Ile Gly His Ala Tyr Glu Ala Ile
275 280 285
Leu Thr Pro Gln Ala Leu His Gly Glu Cys Val Ser Ile Gly Met Val
290 295 300
Lys Glu Ala Glu Leu Ser Arg Tyr Phe Gly Ile Leu Ser Pro Thr Gln
305 310 315 320
Val Ala Arg Leu Ser Lys Ile Leu Val Ala Tyr Gly Leu Pro Val Ser
325 330 335
Pro Asp Glu Lys Trp Phe Lys Glu Leu Thr Leu His Lys Lys Thr Pro
340 345 350
Leu Asp Ile Leu Leu Lys Lys Met Ser Ile Asp Lys Lys Asn Glu Gly
355 360 365
Ser Lys Lys Lys Val Val Ile Leu Glu Ser Ile Gly Lys Cys Tyr Gly
370 375 380
Asp Ser Ala Gln Phe Val Ser Asp Glu Asp Leu Arg Phe Ile Leu Thr
385 390 395 400
Asp Glu Thr Leu Val Tyr Pro Phe Lys Asp Ile Pro Ala Asp Gln Gln
405 410 415
Lys Val Val Ile Pro Pro Gly Ser Lys Ser Ile Ser Asn Arg Ala Leu
420 425 430
Ile Leu Ala Ala Leu Gly Glu Gly Gln Cys Lys Ile Lys Asn Leu Leu
435 440 445
His Ser Asp Asp Thr Lys His Met Leu Thr Ala Val His Glu Leu Lys
450 455 460
Gly Ala Thr Ile Ser Trp Glu Asp Asn Gly Glu Thr Val Val Val Glu
465 470 475 480
Gly His Gly Gly Ser Thr Leu Ser Ala Cys Ala Asp Pro Leu Tyr Leu
485 490 495
Gly Asn Ala Gly Thr Ala Ser Arg Phe Leu Thr Ser Leu Ala Ala Leu
500 505 510
Val Asn Ser Thr Ser Ser Gln Lys Tyr Ile Val Leu Thr Gly Asn Ala
515 520 525
Arg Met Gln Gln Arg Pro Ile Ala Pro Leu Val Asp Ser Leu Arg Ala
530 535 540
Asn Gly Thr Lys Ile Glu Tyr Leu Asn Asn Glu Gly Ser Leu Pro Ile
545 550 555 560
Lys Val Tyr Thr Asp Ser Val Phe Lys Gly Gly Arg Ile Glu Leu Ala
565 570 575
Ala Thr Val Ser Ser Gln Tyr Val Ser Ser Ile Leu Met Cys Ala Pro
580 585 590
Tyr Ala Glu Glu Pro Val Thr Leu Ala Leu Val Gly Gly Lys Pro Ile
595 600 605
Ser Lys Leu Tyr Val Asp Met Thr Ile Lys Met Met Glu Lys Phe Gly
610 615 620
Ile Asn Val Glu Thr Ser Thr Thr Glu Pro Tyr Thr Tyr Tyr Ile Pro
625 630 635 640
Lys Gly His Tyr Ile Asn Pro Ser Glu Tyr Val Ile Glu Ser Asp Ala
645 650 655
Ser Ser Ala Thr Tyr Pro Leu Ala Phe Ala Ala Met Thr Gly Thr Thr
660 665 670
Val Thr Val Pro Asn Ile Gly Phe Glu Ser Leu Gln Gly Asp Ala Arg
675 680 685
Phe Ala Arg Asp Val Leu Lys Pro Met Gly Cys Lys Ile Thr Gln Thr
690 695 700
Ala Thr Ser Thr Thr Val Ser Gly Pro Pro Val Gly Thr Leu Lys Pro
705 710 715 720
Leu Lys His Val Asp Met Glu Pro Met Thr Asp Ala Phe Leu Thr Ala
725 730 735
Cys Val Val Ala Ala Ile Ser His Asp Ser Asp Pro Asn Ser Ala Asn
740 745 750
Thr Thr Thr Ile Glu Gly Ile Ala Asn Gln Arg Val Lys Glu Cys Asn
755 760 765
Arg Ile Leu Ala Met Ala Thr Glu Leu Ala Lys Phe Gly Val Lys Thr
770 775 780
Thr Glu Leu Pro Asp Gly Ile Gln Val His Gly Leu Asn Ser Ile Lys
785 790 795 800
Asp Leu Lys Val Pro Ser Asp Ser Ser Gly Pro Val Gly Val Cys Thr
805 810 815
Tyr Asp Asp His Arg Val Ala Met Ser Phe Ser Leu Leu Ala Gly Met
820 825 830
Val Asn Ser Gln Asn Glu Arg Asp Glu Val Ala Asn Pro Val Arg Ile
835 840 845
Leu Glu Arg His Cys Thr Gly Lys Thr Trp Pro Gly Trp Trp Asp Val
850 855 860
Leu His Ser Glu Leu Gly Ala Lys Leu Asp Gly Ala Glu Pro Leu Glu
865 870 875 880
Cys Thr Ser Lys Lys Asn Ser Lys Lys Ser Val Val Ile Ile Gly Met
885 890 895
Arg Ala Ala Gly Lys Thr Thr Ile Ser Lys Trp Cys Ala Ser Ala Leu
900 905 910
Gly Tyr Lys Leu Val Asp Leu Asp Glu Leu Phe Glu Gln Gln His Asn
915 920 925
Asn Gln Ser Val Lys Gln Phe Val Val Glu Asn Gly Trp Glu Lys Phe
930 935 940
Arg Glu Glu Glu Thr Arg Ile Phe Lys Glu Val Ile Gln Asn Tyr Gly
945 950 955 960
Asp Asp Gly Tyr Val Phe Ser Thr Gly Gly Gly Ile Val Glu Ser Ala
965 970 975
Glu Ser Arg Lys Ala Leu Lys Asp Phe Ala Ser Ser Gly Gly Tyr Val
980 985 990
Leu His Leu His Arg Asp Ile Glu Glu Thr Ile Val Phe Leu Gln Ser
995 1000 1005
Asp Pro Ser Arg Pro Ala Tyr Val Glu Glu Ile Arg Glu Val Trp Asn
1010 1015 1020
Arg Arg Glu Gly Trp Tyr Lys Glu Cys Ser Asn Phe Ser Phe Phe Ala
1025 1030 1035 1040
Pro His Cys Ser Ala Glu Ala Glu Phe Gln Ala Leu Arg Arg Ser Phe
1045 1050 1055
Ser Lys Tyr Ile Ala Thr Ile Thr Gly Val Arg Glu Ile Glu Ile Pro
1060 1065 1070
Ser Gly Arg Ser Ala Phe Val Cys Leu Thr Phe Asp Asp Leu Thr Glu
1075 1080 1085
Gln Thr Glu Asn Leu Thr Pro Ile Cys Tyr Gly Cys Glu Ala Val Glu
1090 1095 1100
Val Arg Val Asp His Leu Ala Asn Tyr Ser Ala Asp Phe Val Ser Lys
1105 1110 1115 1120
Gln Leu Ser Ile Leu Arg Lys Ala Thr Asp Ser Ile Pro Ile Ile Phe
1125 1130 1135
Thr Val Arg Thr Met Lys Gln Gly Gly Asn Phe Pro Asp Glu Glu Phe
1140 1145 1150
Lys Thr Leu Arg Glu Leu Tyr Asp Ile Ala Leu Lys Asn Gly Val Glu
1155 1160 1165
Phe Leu Asp Leu Glu Leu Thr Leu Pro Thr Asp Ile Gln Tyr Glu Val
1170 1175 1180
Ile Asn Lys Arg Gly Asn Thr Lys Ile Ile Gly Ser His His Asp Phe
1185 1190 1195 1200
Gln Gly Leu Tyr Ser Trp Asp Asp Ala Glu Trp Glu Asn Arg Phe Asn
1205 1210 1215
Gln Ala Leu Thr Leu Asp Val Asp Val Val Lys Phe Val Gly Thr Ala
1220 1225 1230
Val Asn Phe Glu Asp Asn Leu Arg Leu Glu His Phe Arg Asp Thr His
1235 1240 1245
Lys Asn Lys Pro Leu Ile Ala Val Asn Met Thr Ser Lys Gly Ser Ile
1250 1255 1260
Ser Arg Val Leu Asn Asn Val Leu Thr Pro Val Thr Ser Asp Leu Leu
1265 1270 1275 1280
Pro Asn Ser Ala Ala Pro Gly Gln Leu Thr Val Ala Gln Ile Asn Lys
1285 1290 1295
Met Tyr Thr Ser Met Gly Gly Ile Glu Pro Lys Glu Leu Phe Val Val
1300 1305 1310
Gly Lys Pro Ile Gly His Ser Arg Ser Pro Ile Leu His Asn Thr Gly
1315 1320 1325
Tyr Glu Ile Leu Gly Leu Pro His Lys Phe Asp Lys Phe Glu Thr Glu
1330 1335 1340
Ser Ala Gln Leu Val Lys Glu Lys Leu Leu Asp Gly Asn Lys Asn Phe
1345 1350 1355 1360
Gly Gly Ala Ala Val Thr Ile Pro Leu Lys Leu Asp Ile Met Gln Tyr
1365 1370 1375
Met Asp Glu Leu Thr Asp Ala Ala Lys Val Ile Gly Ala Val Asn Thr
1380 1385 1390
Val Ile Pro Leu Gly Asn Lys Lys Phe Lys Gly Asp Asn Thr Asp Trp
1395 1400 1405
Leu Gly Ile Arg Asn Ala Leu Ile Asn Asn Gly Val Pro Glu Tyr Val
1410 1415 1420
Gly His Thr Ala Gly Leu Val Ile Gly Ala Gly Gly Thr Ser Arg Ala
1425 1430 1435 1440
Ala Leu Tyr Ala Leu His Ser Leu Gly Cys Lys Lys Ile Phe Ile Ile
1445 1450 1455
Asn Arg Thr Thr Ser Lys Leu Lys Pro Leu Ile Glu Ser Leu Pro Ser
1460 1465 1470
Glu Phe Asn Ile Ile Gly Ile Glu Ser Thr Lys Ser Ile Glu Glu Ile
1475 1480 1485
Lys Glu His Val Gly Val Ala Val Ser Cys Val Pro Ala Asp Lys Pro
1490 1495 1500
Leu Asp Asp Glu Leu Leu Ser Lys Leu Glu Arg Phe Leu Val Lys Gly
1505 1510 1515 1520
Ala His Ala Ala Phe Val Pro Thr Leu Leu Glu Ala Ala Tyr Lys Pro
1525 1530 1535
Ser Val Thr Pro Val Met Thr Ile Ser Gln Asp Lys Tyr Gln Trp His
1540 1545 1550
Val Val Pro Gly Ser Gln Met Leu Val His Gln Gly Val Ala Gln Phe
1555 1560 1565
Glu Lys Trp Thr Gly Phe Lys Gly Pro Phe Lys Ala Ile Phe Asp Ala
1570 1575 1580
Val Thr Lys Glu
1585
<210> 83
<211> 1089
<212> ДНК
<213> Escherichia coli W3110
<400> 83
atggagagga ttgtcgttac tctcggggaa cgtagttacc caattaccat cgcatctggt 60
ttgtttaatg aaccagcttc attcttaccg ctgaaatcgg gcgagcaggt catgttggtc 120
accaacgaaa ccctggctcc tctgtatctc gataaggtcc gcggcgtact tgaacaggcg 180
ggtgttaacg tcgatagcgt tatcctccct gacggcgagc agtataaaag cctggctgta 240
ctcgataccg tctttacggc gttgttacaa aaaccgcatg gtcgcgatac tacgctggtg 300
gcgcttggcg gcggcgtagt gggcgatctg accggcttcg cggcggcgag ttatcagcgc 360
ggtgtccgtt tcattcaagt cccgacgacg ttactgtcgc aggtcgattc ctccgttggc 420
ggcaaaactg cggtcaacca tcccctcggt aaaaacatga ttggcgcgtt ctaccaacct 480
gcttcagtgg tggtggatct cgactgtctg aaaacgcttc ccccgcgtga gttagcgtcg 540
gggctggcag aagtcatcaa atacggcatt attcttgacg gtgcgttttt taactggctg 600
gaagagaatc tggatgcgtt gttgcgtctg gacggtccgg caatggcgta ctgtattcgc 660
cgttgttgtg aactgaaggc agaagttgtc gccgccgacg agcgcgaaac cgggttacgt 720
gctttactga atctgggaca cacctttggt catgccattg aagctgaaat ggggtatggc 780
aattggttac atggtgaagc ggtcgctgcg ggtatggtga tggcggcgcg gacgtcggaa 840
cgtctcgggc agtttagttc tgccgaaacg cagcgtatta taaccctgct caagcgggct 900
gggttaccgg tcaatgggcc gcgcgaaatg tccgcgcagg cgtatttacc gcatatgctg 960
cgtgacaaga aagtccttgc gggagagatg cgcttaattc ttccgttggc aattggtaag 1020
agtgaagttc gcagcggcgt ttcgcacgag cttgttctta acgccattgc cgattgtcaa 1080
tcagcgtaa 1089
<210> 84
<211> 362
<212> PRT
<213> Escherichia coli W3110
<400> 84
Met Glu Arg Ile Val Val Thr Leu Gly Glu Arg Ser Tyr Pro Ile Thr
1 5 10 15
Ile Ala Ser Gly Leu Phe Asn Glu Pro Ala Ser Phe Leu Pro Leu Lys
20 25 30
Ser Gly Glu Gln Val Met Leu Val Thr Asn Glu Thr Leu Ala Pro Leu
35 40 45
Tyr Leu Asp Lys Val Arg Gly Val Leu Glu Gln Ala Gly Val Asn Val
50 55 60
Asp Ser Val Ile Leu Pro Asp Gly Glu Gln Tyr Lys Ser Leu Ala Val
65 70 75 80
Leu Asp Thr Val Phe Thr Ala Leu Leu Gln Lys Pro His Gly Arg Asp
85 90 95
Thr Thr Leu Val Ala Leu Gly Gly Gly Val Val Gly Asp Leu Thr Gly
100 105 110
Phe Ala Ala Ala Ser Tyr Gln Arg Gly Val Arg Phe Ile Gln Val Pro
115 120 125
Thr Thr Leu Leu Ser Gln Val Asp Ser Ser Val Gly Gly Lys Thr Ala
130 135 140
Val Asn His Pro Leu Gly Lys Asn Met Ile Gly Ala Phe Tyr Gln Pro
145 150 155 160
Ala Ser Val Val Val Asp Leu Asp Cys Leu Lys Thr Leu Pro Pro Arg
165 170 175
Glu Leu Ala Ser Gly Leu Ala Glu Val Ile Lys Tyr Gly Ile Ile Leu
180 185 190
Asp Gly Ala Phe Phe Asn Trp Leu Glu Glu Asn Leu Asp Ala Leu Leu
195 200 205
Arg Leu Asp Gly Pro Ala Met Ala Tyr Cys Ile Arg Arg Cys Cys Glu
210 215 220
Leu Lys Ala Glu Val Val Ala Ala Asp Glu Arg Glu Thr Gly Leu Arg
225 230 235 240
Ala Leu Leu Asn Leu Gly His Thr Phe Gly His Ala Ile Glu Ala Glu
245 250 255
Met Gly Tyr Gly Asn Trp Leu His Gly Glu Ala Val Ala Ala Gly Met
260 265 270
Val Met Ala Ala Arg Thr Ser Glu Arg Leu Gly Gln Phe Ser Ser Ala
275 280 285
Glu Thr Gln Arg Ile Ile Thr Leu Leu Lys Arg Ala Gly Leu Pro Val
290 295 300
Asn Gly Pro Arg Glu Met Ser Ala Gln Ala Tyr Leu Pro His Met Leu
305 310 315 320
Arg Asp Lys Lys Val Leu Ala Gly Glu Met Arg Leu Ile Leu Pro Leu
325 330 335
Ala Ile Gly Lys Ser Glu Val Arg Ser Gly Val Ser His Glu Leu Val
340 345 350
Leu Asn Ala Ile Ala Asp Cys Gln Ser Ala
355 360
<210> 85
<211> 2247
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 85
atgagtatcg tccaagcaaa gtttgaagct aaggaaacat cttttcatgt agaaggttac 60
gaaaagattg agtatgattt ggtgtatgta gatggtattt ttgaaatcca gaattctgca 120
ctagcagatg tatatcaagg ttttggacga tgcttggcga ttgtagatgc taacgtcagt 180
cggttgtatg gtaatcaaat tcaggcatat ttccagtatt atggtataga actgaggcta 240
tttcctatta ccattactga accagataag actattcaaa ctttcgagag agttatagat 300
gtctttgcag atttcaaatt agtccgcaaa gaaccagtat tagtcgtggg tggcggttta 360
attacagatg ttgtcggctt tgcttgttct acatatcgtc gcagcagcaa ttacatccgc 420
attcctacta cattgattgg attaattgat gccagtgtag caattaaggt agcagttaat 480
catcgcaaac tgaaaaaccg tttgggtgct tatcatgctt ctcgcaaagt atttttagat 540
ttctccttgt tgcgtactct ccctacagac caagtacgta acgggatggc ggaattggta 600
aaaatcgctg tagtagcgca tcaagaagtt tttgaattgt tggagaagta cggcgaagaa 660
ttactacgta ctcattttgg caatatagat gcaactccag agattaaaga aatagcccat 720
cgtttgactt acaaagctat ccataagatg ttggaattgg aagttcccaa cctgcatgag 780
ttagacctag atagggtgat tgcttacggt cacacttgga gtcccacctt ggaacttgcg 840
cctcgtctac ccatgttcca cggacacgcc gttaatgtag atatggcttt ctcggcaacg 900
atcgccgccc gtagaggata tattacaatt gcagaacgcg atcgtatttt aggattaatg 960
agtcgcgttg gtctatccct cgaccatccc atgttggata tagatatttt gtggcgtggt 1020
actgaatcta tcacattaac tcgtgatggt ttgttaagag ctgctatgcc aaaacccatt 1080
ggtgattgtg tcttcgtcaa tgacctgaca agagaagaat tagcagccgc attagctgac 1140
cacaaagaac tttgtaccag ttatccccgt ggtggtgaag gtgtggatgt gtatcccgtt 1200
tatcaaaaag aattaatcgg gagtgttaaa taatgacttt tttgaattca aaatgcaaaa 1260
tactccacgg atacactgcg cgagcgcggt agcatttctg ttcgcggagc gtcccgtagg 1320
gaaagagaag gctacgcaaa taatcggaca ctaattgtct ttaattttga attttgaatt 1380
ttgaattttg aattggagcg aagcgacttg acaaatgtga ttgtccaacc aacagctaga 1440
cctgttacac cattgggaat tttaaccaag cagttagaag ccatagtcca agaggttaag 1500
caacatccag atttacctgg ggaattgata gcaaacatcc atcaggcttg gcgtttagcc 1560
gcaggtatag acccttattt ggaagaatgc accactccag aatctcctga actcgctgca 1620
ttggcaaaaa ccacagccac cgaagcctgg ggagaacact tccacggagg tacaaccgtc 1680
cgtcctctag aacaagagat gctttctggt catatcgaag gacaaacctt aaagatgttt 1740
gttcacatga ccaaagctaa aaaagtctta gaaattggga tgtttaccgg ttattcggcg 1800
ctggcgatgg cggaagcatt accagaggat ggactgcttg tggcttgtga agttgaccct 1860
tacgcggcgg aaattggaca gaaagccttt caacaatctc cccacggtgg aaagattcgt 1920
gtggaattgg atgcagcctt agcaactctt gataagttag cagaagctgg ggagtctttt 1980
gacttggtat ttatcgacgc agataaaaaa gagtatgtag cctattttca caagttgcta 2040
ggtagcagtt tgttagcacc agatggcttt atttgtgtag ataacacctt attacaaggg 2100
gaagtttatc taccagcaga ggaacgtagc gtcaatggtg aagcgatcgc gcaatttaat 2160
catacagtag ctatagaccc ccgtgtagaa caggttttgt tgccgttgcg agatggttta 2220
acaattatcc gcagaataca accttaa 2247
<210> 86
<211> 820
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 86
Met Ser Ile Val Gln Ala Lys Phe Glu Ala Lys Glu Thr Ser Phe His
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly
20 25 30
Ile Phe Glu Ile Gln Asn Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe
35 40 45
Gly Arg Cys Leu Ala Ile Val Asp Ala Asn Val Ser Arg Leu Tyr Gly
50 55 60
Asn Gln Ile Gln Ala Tyr Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu
65 70 75 80
Phe Pro Ile Thr Ile Thr Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu
85 90 95
Arg Val Ile Asp Val Phe Ala Asp Phe Lys Leu Val Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Ile Thr Asp Val Val Gly Phe Ala
115 120 125
Cys Ser Thr Tyr Arg Arg Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Ser Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Ser Arg Lys
165 170 175
Val Phe Leu Asp Phe Ser Leu Leu Arg Thr Leu Pro Thr Asp Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala His Gln
195 200 205
Glu Val Phe Glu Leu Leu Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr
210 215 220
His Phe Gly Asn Ile Asp Ala Thr Pro Glu Ile Lys Glu Ile Ala His
225 230 235 240
Arg Leu Thr Tyr Lys Ala Ile His Lys Met Leu Glu Leu Glu Val Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Arg Leu Pro Met Phe His Gly
275 280 285
His Ala Val Asn Val Asp Met Ala Phe Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ile Ala Glu Arg Asp Arg Ile Leu Gly Leu Met
305 310 315 320
Ser Arg Val Gly Leu Ser Leu Asp His Pro Met Leu Asp Ile Asp Ile
325 330 335
Leu Trp Arg Gly Thr Glu Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Asp Cys Val Phe Val Asn Asp
355 360 365
Leu Thr Arg Glu Glu Leu Ala Ala Ala Leu Ala Asp His Lys Glu Leu
370 375 380
Cys Thr Ser Tyr Pro Arg Gly Gly Glu Gly Val Asp Val Tyr Pro Val
385 390 395 400
Tyr Gln Lys Glu Leu Ile Gly Ser Val Lys Met Ser Ile Val Gln Ala
405 410 415
Lys Phe Glu Ala Lys Glu Thr Ser Phe His Val Glu Gly Tyr Glu Lys
420 425 430
Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly Ile Phe Glu Ile Gln Asn
435 440 445
Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe Gly Arg Cys Leu Ala Ile
450 455 460
Val Asp Ala Asn Val Ser Arg Leu Tyr Gly Asn Gln Ile Gln Ala Tyr
465 470 475 480
Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu Phe Pro Ile Thr Ile Thr
485 490 495
Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu Arg Val Ile Asp Val Phe
500 505 510
Ala Asp Phe Lys Leu Val Arg Lys Glu Pro Val Leu Val Val Gly Gly
515 520 525
Gly Leu Ile Thr Asp Val Val Gly Phe Ala Cys Ser Thr Tyr Arg Arg
530 535 540
Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr Leu Ile Gly Leu Ile Asp
545 550 555 560
Ala Ser Val Ala Ile Lys Val Ala Val Asn His Arg Lys Leu Lys Asn
565 570 575
Arg Leu Gly Ala Tyr His Ala Ser Arg Lys Val Phe Leu Asp Phe Ser
580 585 590
Leu Leu Arg Thr Leu Pro Thr Asp Gln Val Arg Asn Gly Met Ala Glu
595 600 605
Leu Val Lys Ile Ala Val Val Ala His Gln Glu Val Phe Glu Leu Leu
610 615 620
Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr His Phe Gly Asn Ile Asp
625 630 635 640
Ala Thr Pro Glu Ile Lys Glu Ile Ala His Arg Leu Thr Tyr Lys Ala
645 650 655
Ile His Lys Met Leu Glu Leu Glu Val Pro Asn Leu His Glu Leu Asp
660 665 670
Leu Asp Arg Val Ile Ala Tyr Gly His Thr Trp Ser Pro Thr Leu Glu
675 680 685
Leu Ala Pro Arg Leu Pro Met Phe His Gly His Ala Val Asn Val Asp
690 695 700
Met Ala Phe Ser Ala Thr Ile Ala Ala Arg Arg Gly Tyr Ile Thr Ile
705 710 715 720
Ala Glu Arg Asp Arg Ile Leu Gly Leu Met Ser Arg Val Gly Leu Ser
725 730 735
Leu Asp His Pro Met Leu Asp Ile Asp Ile Leu Trp Arg Gly Thr Glu
740 745 750
Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu Arg Ala Ala Met Pro Lys
755 760 765
Pro Ile Gly Asp Cys Val Phe Val Asn Asp Leu Thr Arg Glu Glu Leu
770 775 780
Ala Ala Ala Leu Ala Asp His Lys Glu Leu Cys Thr Ser Tyr Pro Arg
785 790 795 800
Gly Gly Glu Gly Val Asp Val Tyr Pro Val Tyr Gln Lys Glu Leu Ile
805 810 815
Gly Ser Val Lys
820
<210> 87
<211> 3638
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 87
atgagtatcg tccaagcaaa gtttgaagct aaggaaacat cttttcatgt agaaggttac 60
gaaaagattg agtatgattt ggtgtatgta gatggtattt ttgaaatcca gaattctgca 120
ctagcagatg tatatcaagg ttttggacga tgcttggcga ttgtagatgc taacgtcagt 180
cggttgtatg gtaatcaaat tcaggcatat ttccagtatt atggtataga actgaggcta 240
tttcctatta ccattactga accagataag actattcaaa ctttcgagag agttatagat 300
gtctttgcag atttcaaatt agtccgcaaa gaaccagtat tagtcgtggg tggcggttta 360
attacagatg ttgtcggctt tgcttgttct acatatcgtc gcagcagcaa ttacatccgc 420
attcctacta cattgattgg attaattgat gccagtgtag caattaaggt agcagttaat 480
catcgcaaac tgaaaaaccg tttgggtgct tatcatgctt ctcgcaaagt atttttagat 540
ttctccttgt tgcgtactct ccctacagac caagtacgta acgggatggc ggaattggta 600
aaaatcgctg tagtagcgca tcaagaagtt tttgaattgt tggagaagta cggcgaagaa 660
ttactacgta ctcattttgg caatatagat gcaactccag agattaaaga aatagcccat 720
cgtttgactt acaaagctat ccataagatg ttggaattgg aagttcccaa cctgcatgag 780
ttagacctag atagggtgat tgcttacggt cacacttgga gtcccacctt ggaacttgcg 840
cctcgtctac ccatgttcca cggacacgcc gttaatgtag atatggcttt ctcggcaacg 900
atcgccgccc gtagaggata tattacaatt gcagaacgcg atcgtatttt aggattaatg 960
agtcgcgttg gtctatccct cgaccatccc atgttggata tagatatttt gtggcgtggt 1020
actgaatcta tcacattaac tcgtgatggt ttgttaagag ctgctatgcc aaaacccatt 1080
ggtgattgtg tcttcgtcaa tgacctgaca agagaagaat tagcagccgc attagctgac 1140
cacaaagaac tttgtaccag ttatccccgt ggtggtgaag gtgtggatgt gtatcccgtt 1200
tatcaaaaag aattaatcgg gagtgttaaa taatgacttt tttgaattca aaatgcaaaa 1260
tactccacgg atacactgcg cgagcgcggt agcatttctg ttcgcggagc gtcccgtagg 1320
gaaagagaag gctacgcaaa taatcggaca ctaattgtct ttaattttga attttgaatt 1380
ttgaattttg aattggagcg aagcgacttg acaaatgtga ttgtccaacc aacagctaga 1440
cctgttacac cattgggaat tttaaccaag cagttagaag ccatagtcca agaggttaag 1500
caacatccag atttacctgg ggaattgata gcaaacatcc atcaggcttg gcgtttagcc 1560
gcaggtatag acccttattt ggaagaatgc accactccag aatctcctga actcgctgca 1620
ttggcaaaaa ccacagccac cgaagcctgg ggagaacact tccacggagg tacaaccgtc 1680
cgtcctctag aacaagagat gctttctggt catatcgaag gacaaacctt aaagatgttt 1740
gttcacatga ccaaagctaa aaaagtctta gaaattggga tgtttaccgg ttattcggcg 1800
ctggcgatgg cggaagcatt accagaggat ggactgcttg tggcttgtga agttgaccct 1860
tacgcggcgg aaattggaca gaaagccttt caacaatctc cccacggtgg aaagattcgt 1920
gtggaattgg atgcagcctt agcaactctt gataagttag cagaagctgg ggagtctttt 1980
gacttggtat ttatcgacgc agataaaaaa gagtatgtag cctattttca caagttgcta 2040
ggtagcagtt tgttagcacc agatggcttt atttgtgtag ataacacctt attacaaggg 2100
gaagtttatc taccagcaga ggaacgtagc gtcaatggtg aagcgatcgc gcaatttaat 2160
catacagtag ctatagaccc ccgtgtagaa caggttttgt tgccgttgcg agatggttta 2220
acaattatcc gcagaataca accttaattg tccaatcgac tatggcacaa tcccttcccc 2280
tttcttccgc acctgctaca ccgtctcttc cttcccagac gaaaatagcc gcaattatcc 2340
aaaatatctg cactttggct ttgttattac tagcattgcc cattaatgcc accattgttt 2400
ttatatcctt gttagtcttc cgaccgcaaa aggtcaaagc agcaaacccc caaaccattc 2460
ttatcagtgg cggtaagatg accaaagctt tacaactagc aaggtcattc cacgcggctg 2520
gacatagagt tgtcttggtg gaaacccata aatactggtt gactggtcat cgtttttccc 2580
aagcagtgga taagttttac acagtccccg caccccagga caatccccaa gcttacattc 2640
aggctttggt agatatcgtc aaacaagaaa acatcgatgt ttatattccc gtcaccagtc 2700
cagtgggtag ctactacgac tcattagcca aaccagagtt atcccattat tgcgaagtgt 2760
ttcactttga cgcagatatt acccaaatgt tggatgataa atttgcgttg acacaaaaag 2820
cgcgatcgct tggtttatca gtacccaaat cctttaaaat tacctcacca gaacaagtca 2880
tcaacttcga tttttctgga gagacacgta aatacatcct caaaagcatt ccctacgact 2940
cagtgcggcg gttggactta accaaactcc cctgtgctac tccagaggaa acagcagcat 3000
tcgtcagaag tttgccaatt actcccgaaa aaccgtggat tatgcaggaa tttatccccg 3060
gtaaggaatt ctgcacccat agcaccgttc ggaatgggga actcagactg cattgctgtt 3120
gcgaatcttc agccttccaa gttaattatg agaatgtaaa taacccgcaa attaccgaat 3180
gggtacagca ttttgtcaag gaactgaaac tgacaggaca gatttccttt gactttatcc 3240
aagccgaaga cggaacagtt tacgccatcg agtgtaaccc ccgcacacat tcagcaatta 3300
ccacatttta cgaccacccc caggtagcag aagcgtactt gagtcaagca ccgacgactg 3360
aaaccataca accactaacg acaagcaagc ctacctattg gacttatcac gaagtttggc 3420
gtttaactgg tatccgttct ttcacccagt tgcaaagatg gctggggaat atttggcgcg 3480
ggactgatgc gatttatcag ccagatgacc ccttaccgtt tttgatggta catcattggc 3540
aaattcccct actgttattg aataatttgc gtcgtcttaa aggttggacg cggatagatt 3600
tcaatattgg gaagttggtg gaattggggg gagattag 3638
<210> 88
<211> 1099
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 88
Met Ser Ile Val Gln Ala Lys Phe Glu Ala Lys Glu Thr Ser Phe His
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly
20 25 30
Ile Phe Glu Ile Gln Asn Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe
35 40 45
Gly Arg Cys Leu Ala Ile Val Asp Ala Asn Val Ser Arg Leu Tyr Gly
50 55 60
Asn Gln Ile Gln Ala Tyr Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu
65 70 75 80
Phe Pro Ile Thr Ile Thr Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu
85 90 95
Arg Val Ile Asp Val Phe Ala Asp Phe Lys Leu Val Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Ile Thr Asp Val Val Gly Phe Ala
115 120 125
Cys Ser Thr Tyr Arg Arg Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Ser Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Ser Arg Lys
165 170 175
Val Phe Leu Asp Phe Ser Leu Leu Arg Thr Leu Pro Thr Asp Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala His Gln
195 200 205
Glu Val Phe Glu Leu Leu Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr
210 215 220
His Phe Gly Asn Ile Asp Ala Thr Pro Glu Ile Lys Glu Ile Ala His
225 230 235 240
Arg Leu Thr Tyr Lys Ala Ile His Lys Met Leu Glu Leu Glu Val Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Arg Leu Pro Met Phe His Gly
275 280 285
His Ala Val Asn Val Asp Met Ala Phe Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ile Ala Glu Arg Asp Arg Ile Leu Gly Leu Met
305 310 315 320
Ser Arg Val Gly Leu Ser Leu Asp His Pro Met Leu Asp Ile Asp Ile
325 330 335
Leu Trp Arg Gly Thr Glu Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Asp Cys Val Phe Val Asn Asp
355 360 365
Leu Thr Arg Glu Glu Leu Ala Ala Ala Leu Ala Asp His Lys Glu Leu
370 375 380
Cys Thr Ser Tyr Pro Arg Gly Gly Glu Gly Val Asp Val Tyr Pro Val
385 390 395 400
Tyr Gln Lys Glu Leu Ile Gly Ser Val Lys Met Ser Ile Val Gln Ala
405 410 415
Lys Phe Glu Ala Lys Glu Thr Ser Phe His Val Glu Gly Tyr Glu Lys
420 425 430
Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly Ile Phe Glu Ile Gln Asn
435 440 445
Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe Gly Arg Cys Leu Ala Ile
450 455 460
Val Asp Ala Asn Val Ser Arg Leu Tyr Gly Asn Gln Ile Gln Ala Tyr
465 470 475 480
Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu Phe Pro Ile Thr Ile Thr
485 490 495
Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu Arg Val Ile Asp Val Phe
500 505 510
Ala Asp Phe Lys Leu Val Arg Lys Glu Pro Val Leu Val Val Gly Gly
515 520 525
Gly Leu Ile Thr Asp Val Val Gly Phe Ala Cys Ser Thr Tyr Arg Arg
530 535 540
Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr Leu Ile Gly Leu Ile Asp
545 550 555 560
Ala Ser Val Ala Ile Lys Val Ala Val Asn His Arg Lys Leu Lys Asn
565 570 575
Arg Leu Gly Ala Tyr His Ala Ser Arg Lys Val Phe Leu Asp Phe Ser
580 585 590
Leu Leu Arg Thr Leu Pro Thr Asp Gln Val Arg Asn Gly Met Ala Glu
595 600 605
Leu Val Lys Ile Ala Val Val Ala His Gln Glu Val Phe Glu Leu Leu
610 615 620
Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr His Phe Gly Asn Ile Asp
625 630 635 640
Ala Thr Pro Glu Ile Lys Glu Ile Ala His Arg Leu Thr Tyr Lys Ala
645 650 655
Ile His Lys Met Leu Glu Leu Glu Val Pro Asn Leu His Glu Leu Asp
660 665 670
Leu Asp Arg Val Ile Ala Tyr Gly His Thr Trp Ser Pro Thr Leu Glu
675 680 685
Leu Ala Pro Arg Leu Pro Met Phe His Gly His Ala Val Asn Val Asp
690 695 700
Met Ala Phe Ser Ala Thr Ile Ala Ala Arg Arg Gly Tyr Ile Thr Ile
705 710 715 720
Ala Glu Arg Asp Arg Ile Leu Gly Leu Met Ser Arg Val Gly Leu Ser
725 730 735
Leu Asp His Pro Met Leu Asp Ile Asp Ile Leu Trp Arg Gly Thr Glu
740 745 750
Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu Arg Ala Ala Met Pro Lys
755 760 765
Pro Ile Gly Asp Cys Val Phe Val Asn Asp Leu Thr Arg Glu Glu Leu
770 775 780
Ala Ala Ala Leu Ala Asp His Lys Glu Leu Cys Thr Ser Tyr Pro Arg
785 790 795 800
Gly Gly Glu Gly Val Asp Val Tyr Pro Val Tyr Gln Lys Glu Leu Ile
805 810 815
Gly Ser Val Lys Met Thr Asn Val Ile Val Gln Pro Thr Ala Arg Pro
820 825 830
Val Thr Pro Leu Gly Ile Leu Thr Lys Gln Leu Glu Ala Ile Val Gln
835 840 845
Glu Val Lys Gln His Pro Asp Leu Pro Gly Glu Leu Ile Ala Asn Ile
850 855 860
His Gln Ala Trp Arg Leu Ala Ala Gly Ile Asp Pro Tyr Leu Glu Glu
865 870 875 880
Cys Thr Thr Pro Glu Ser Pro Glu Leu Ala Ala Leu Ala Lys Thr Thr
885 890 895
Ala Thr Glu Ala Trp Gly Glu His Phe His Gly Gly Thr Thr Val Arg
900 905 910
Pro Leu Glu Gln Glu Met Leu Ser Gly His Ile Glu Gly Gln Thr Leu
915 920 925
Lys Met Phe Val His Met Thr Lys Ala Lys Lys Val Leu Glu Ile Gly
930 935 940
Met Phe Thr Gly Tyr Ser Ala Leu Ala Met Ala Glu Ala Leu Pro Glu
945 950 955 960
Asp Gly Leu Leu Val Ala Cys Glu Val Asp Pro Tyr Ala Ala Glu Ile
965 970 975
Gly Gln Lys Ala Phe Gln Gln Ser Pro His Gly Gly Lys Ile Arg Val
980 985 990
Glu Leu Asp Ala Ala Leu Ala Thr Leu Asp Lys Leu Ala Glu Ala Gly
995 1000 1005
Glu Ser Phe Asp Leu Val Phe Ile Asp Ala Asp Lys Lys Glu Tyr Val
1010 1015 1020
Ala Tyr Phe His Lys Leu Leu Gly Ser Ser Leu Leu Ala Pro Asp Gly
1025 1030 1035 1040
Phe Ile Cys Val Asp Asn Thr Leu Leu Gln Gly Glu Val Tyr Leu Pro
1045 1050 1055
Ala Glu Glu Arg Ser Val Asn Gly Glu Ala Ile Ala Gln Phe Asn His
1060 1065 1070
Thr Val Ala Ile Asp Pro Arg Val Glu Gln Val Leu Leu Pro Leu Arg
1075 1080 1085
Asp Gly Leu Thr Ile Ile Arg Arg Ile Gln Pro
1090 1095
<210> 89
<211> 1233
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 89
atgagtatcg tccaagcaaa gtttgaagct aaggaaacat cttttcatgt agaaggttac 60
gaaaagattg agtatgattt ggtgtatgta gatggtattt ttgaaatcca gaattctgca 120
ctagcagatg tatatcaagg ttttggacga tgcttggcga ttgtagatgc taacgtcagt 180
cggttgtatg gtaatcaaat tcaggcatat ttccagtatt atggtataga actgaggcta 240
tttcctatta ccattactga accagataag actattcaaa ctttcgagag agttatagat 300
gtctttgcag atttcaaatt agtccgcaaa gaaccagtat tagtcgtggg tggcggttta 360
attacagatg ttgtcggctt tgcttgttct acatatcgtc gcagcagcaa ttacatccgc 420
attcctacta cattgattgg attaattgat gccagtgtag caattaaggt agcagttaat 480
catcgcaaac tgaaaaaccg tttgggtgct tatcatgctt ctcgcaaagt atttttagat 540
ttctccttgt tgcgtactct ccctacagac caagtacgta acgggatggc ggaattggta 600
aaaatcgctg tagtagcgca tcaagaagtt tttgaattgt tggagaagta cggcgaagaa 660
ttactacgta ctcattttgg caatatagat gcaactccag agattaaaga aatagcccat 720
cgtttgactt acaaagctat ccataagatg ttggaattgg aagttcccaa cctgcatgag 780
ttagacctag atagggtgat tgcttacggt cacacttgga gtcccacctt ggaacttgcg 840
cctcgtctac ccatgttcca cggacacgcc gttaatgtag atatggcttt ctcggcaacg 900
atcgccgccc gtagaggata tattacaatt gcagaacgcg atcgtatttt aggattaatg 960
agtcgcgttg gtctatccct cgaccatccc atgttggata tagatatttt gtggcgtggt 1020
actgaatcta tcacattaac tcgtgatggt ttgttaagag ctgctatgcc aaaacccatt 1080
ggtgattgtg tcttcgtcaa tgacctgaca agagaagaat tagcagccgc attagctgac 1140
cacaaagaac tttgtaccag ttatccccgt ggtggtgaag gtgtggatgt gtatcccgtt 1200
tatcaaaaag aattaatcgg gagtgttaaa taa 1233
<210> 90
<211> 410
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 90
Met Ser Ile Val Gln Ala Lys Phe Glu Ala Lys Glu Thr Ser Phe His
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly
20 25 30
Ile Phe Glu Ile Gln Asn Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe
35 40 45
Gly Arg Cys Leu Ala Ile Val Asp Ala Asn Val Ser Arg Leu Tyr Gly
50 55 60
Asn Gln Ile Gln Ala Tyr Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu
65 70 75 80
Phe Pro Ile Thr Ile Thr Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu
85 90 95
Arg Val Ile Asp Val Phe Ala Asp Phe Lys Leu Val Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Ile Thr Asp Val Val Gly Phe Ala
115 120 125
Cys Ser Thr Tyr Arg Arg Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Ser Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Ser Arg Lys
165 170 175
Val Phe Leu Asp Phe Ser Leu Leu Arg Thr Leu Pro Thr Asp Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala His Gln
195 200 205
Glu Val Phe Glu Leu Leu Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr
210 215 220
His Phe Gly Asn Ile Asp Ala Thr Pro Glu Ile Lys Glu Ile Ala His
225 230 235 240
Arg Leu Thr Tyr Lys Ala Ile His Lys Met Leu Glu Leu Glu Val Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Arg Leu Pro Met Phe His Gly
275 280 285
His Ala Val Asn Val Asp Met Ala Phe Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ile Ala Glu Arg Asp Arg Ile Leu Gly Leu Met
305 310 315 320
Ser Arg Val Gly Leu Ser Leu Asp His Pro Met Leu Asp Ile Asp Ile
325 330 335
Leu Trp Arg Gly Thr Glu Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Asp Cys Val Phe Val Asn Asp
355 360 365
Leu Thr Arg Glu Glu Leu Ala Ala Ala Leu Ala Asp His Lys Glu Leu
370 375 380
Cys Thr Ser Tyr Pro Arg Gly Gly Glu Gly Val Asp Val Tyr Pro Val
385 390 395 400
Tyr Gln Lys Glu Leu Ile Gly Ser Val Lys
405 410
<210> 91
<211> 840
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 91
ttgacaaatg tgattgtcca accaacagct agacctgtta caccattggg aattttaacc 60
aagcagttag aagccatagt ccaagaggtt aagcaacatc cagatttacc tggggaattg 120
atagcaaaca tccatcaggc ttggcgttta gccgcaggta tagaccctta tttggaagaa 180
tgcaccactc cagaatctcc tgaactcgct gcattggcaa aaaccacagc caccgaagcc 240
tggggagaac acttccacgg aggtacaacc gtccgtcctc tagaacaaga gatgctttct 300
ggtcatatcg aaggacaaac cttaaagatg tttgttcaca tgaccaaagc taaaaaagtc 360
ttagaaattg ggatgtttac cggttattcg gcgctggcga tggcggaagc attaccagag 420
gatggactgc ttgtggcttg tgaagttgac ccttacgcgg cggaaattgg acagaaagcc 480
tttcaacaat ctccccacgg tggaaagatt cgtgtggaat tggatgcagc cttagcaact 540
cttgataagt tagcagaagc tggggagtct tttgacttgg tatttatcga cgcagataaa 600
aaagagtatg tagcctattt tcacaagttg ctaggtagca gtttgttagc accagatggc 660
tttatttgtg tagataacac cttattacaa ggggaagttt atctaccagc agaggaacgt 720
agcgtcaatg gtgaagcgat cgcgcaattt aatcatacag tagctataga cccccgtgta 780
gaacaggttt tgttgccgtt gcgagatggt ttaacaatta tccgcagaat acaaccttaa 840
840
<210> 92
<211> 410
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 92
Met Ser Ile Val Gln Ala Lys Phe Glu Ala Lys Glu Thr Ser Phe His
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Tyr Asp Leu Val Tyr Val Asp Gly
20 25 30
Ile Phe Glu Ile Gln Asn Ser Ala Leu Ala Asp Val Tyr Gln Gly Phe
35 40 45
Gly Arg Cys Leu Ala Ile Val Asp Ala Asn Val Ser Arg Leu Tyr Gly
50 55 60
Asn Gln Ile Gln Ala Tyr Phe Gln Tyr Tyr Gly Ile Glu Leu Arg Leu
65 70 75 80
Phe Pro Ile Thr Ile Thr Glu Pro Asp Lys Thr Ile Gln Thr Phe Glu
85 90 95
Arg Val Ile Asp Val Phe Ala Asp Phe Lys Leu Val Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Ile Thr Asp Val Val Gly Phe Ala
115 120 125
Cys Ser Thr Tyr Arg Arg Ser Ser Asn Tyr Ile Arg Ile Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Ser Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Ser Arg Lys
165 170 175
Val Phe Leu Asp Phe Ser Leu Leu Arg Thr Leu Pro Thr Asp Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala His Gln
195 200 205
Glu Val Phe Glu Leu Leu Glu Lys Tyr Gly Glu Glu Leu Leu Arg Thr
210 215 220
His Phe Gly Asn Ile Asp Ala Thr Pro Glu Ile Lys Glu Ile Ala His
225 230 235 240
Arg Leu Thr Tyr Lys Ala Ile His Lys Met Leu Glu Leu Glu Val Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Arg Leu Pro Met Phe His Gly
275 280 285
His Ala Val Asn Val Asp Met Ala Phe Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ile Ala Glu Arg Asp Arg Ile Leu Gly Leu Met
305 310 315 320
Ser Arg Val Gly Leu Ser Leu Asp His Pro Met Leu Asp Ile Asp Ile
325 330 335
Leu Trp Arg Gly Thr Glu Ser Ile Thr Leu Thr Arg Asp Gly Leu Leu
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Asp Cys Val Phe Val Asn Asp
355 360 365
Leu Thr Arg Glu Glu Leu Ala Ala Ala Leu Ala Asp His Lys Glu Leu
370 375 380
Cys Thr Ser Tyr Pro Arg Gly Gly Glu Gly Val Asp Val Tyr Pro Val
385 390 395 400
Tyr Gln Lys Glu Leu Ile Gly Ser Val Lys
405 410
<210> 93
<211> 840
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 93
ttgacaaatg tgattgtcca accaacagct agacctgtta caccattggg aattttaacc 60
aagcagttag aagccatagt ccaagaggtt aagcaacatc cagatttacc tggggaattg 120
atagcaaaca tccatcaggc ttggcgttta gccgcaggta tagaccctta tttggaagaa 180
tgcaccactc cagaatctcc tgaactcgct gcattggcaa aaaccacagc caccgaagcc 240
tggggagaac acttccacgg aggtacaacc gtccgtcctc tagaacaaga gatgctttct 300
ggtcatatcg aaggacaaac cttaaagatg tttgttcaca tgaccaaagc taaaaaagtc 360
ttagaaattg ggatgtttac cggttattcg gcgctggcga tggcggaagc attaccagag 420
gatggactgc ttgtggcttg tgaagttgac ccttacgcgg cggaaattgg acagaaagcc 480
tttcaacaat ctccccacgg tggaaagatt cgtgtggaat tggatgcagc cttagcaact 540
cttgataagt tagcagaagc tggggagtct tttgacttgg tatttatcga cgcagataaa 600
aaagagtatg tagcctattt tcacaagttg ctaggtagca gtttgttagc accagatggc 660
tttatttgtg tagataacac cttattacaa ggggaagttt atctaccagc agaggaacgt 720
agcgtcaatg gtgaagcgat cgcgcaattt aatcatacag tagctataga cccccgtgta 780
gaacaggttt tgttgccgtt gcgagatggt ttaacaatta tccgcagaat acaaccttaa 840
840
<210> 94
<211> 279
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 94
Met Thr Asn Val Ile Val Gln Pro Thr Ala Arg Pro Val Thr Pro Leu
1 5 10 15
Gly Ile Leu Thr Lys Gln Leu Glu Ala Ile Val Gln Glu Val Lys Gln
20 25 30
His Pro Asp Leu Pro Gly Glu Leu Ile Ala Asn Ile His Gln Ala Trp
35 40 45
Arg Leu Ala Ala Gly Ile Asp Pro Tyr Leu Glu Glu Cys Thr Thr Pro
50 55 60
Glu Ser Pro Glu Leu Ala Ala Leu Ala Lys Thr Thr Ala Thr Glu Ala
65 70 75 80
Trp Gly Glu His Phe His Gly Gly Thr Thr Val Arg Pro Leu Glu Gln
85 90 95
Glu Met Leu Ser Gly His Ile Glu Gly Gln Thr Leu Lys Met Phe Val
100 105 110
His Met Thr Lys Ala Lys Lys Val Leu Glu Ile Gly Met Phe Thr Gly
115 120 125
Tyr Ser Ala Leu Ala Met Ala Glu Ala Leu Pro Glu Asp Gly Leu Leu
130 135 140
Val Ala Cys Glu Val Asp Pro Tyr Ala Ala Glu Ile Gly Gln Lys Ala
145 150 155 160
Phe Gln Gln Ser Pro His Gly Gly Lys Ile Arg Val Glu Leu Asp Ala
165 170 175
Ala Leu Ala Thr Leu Asp Lys Leu Ala Glu Ala Gly Glu Ser Phe Asp
180 185 190
Leu Val Phe Ile Asp Ala Asp Lys Lys Glu Tyr Val Ala Tyr Phe His
195 200 205
Lys Leu Leu Gly Ser Ser Leu Leu Ala Pro Asp Gly Phe Ile Cys Val
210 215 220
Asp Asn Thr Leu Leu Gln Gly Glu Val Tyr Leu Pro Ala Glu Glu Arg
225 230 235 240
Ser Val Asn Gly Glu Ala Ile Ala Gln Phe Asn His Thr Val Ala Ile
245 250 255
Asp Pro Arg Val Glu Gln Val Leu Leu Pro Leu Arg Asp Gly Leu Thr
260 265 270
Ile Ile Arg Arg Ile Gln Pro
275
<210> 95
<211> 2667
<212> ДНК
<213> Anabaena variabilis ATCC29413
<400> 95
atgcagacta tagattttaa tattcgtaag ttacttgtag agtggaacgc gacccacaga 60
gattatgatc tttcccagag tttacatgaa ctaattgtag ctcaagtaga acgaacacct 120
gaggcgatcg ctgtcacctt tgacaagcaa caactaactt atcaagaact aaatcataaa 180
gcaaaccagc taggacatta tttacaaaca ttaggagtcc agccagaaac cctggtaggc 240
gtttgtttag aacgttcctt agaaatggtt atctgtcttt taggaatcct caaagctggg 300
ggtgcttatg ttcctattga ccctgaatat cctcaagaac gcatagctta tatgctagaa 360
gattctcagg tgaaggtact actaactcaa gaaaaattac tcaatcaaat tccccaccat 420
caagcacaaa ctatctgtgt agatagggaa tgggagaaaa tttccacaca agctaatacc 480
aatcccaaaa gtaatataaa aacggataat cttgcttatg taatttacac ctctggttcc 540
actggtaaac caaaaggtgc aatgaacacc cacaaaggta tctgtaatcg cttattgtgg 600
atgcaggaag cttatcaaat cgattccaca gatagcattt tacaaaaaac cccctttagt 660
tttgatgttt ccgtttggga gttcttttgg actttattaa ctggcgcacg tttggtaata 720
gccaaaccag gcggacataa agatagtgct tacctcatcg atttaattac tcaagaacaa 780
atcactacgt tgcattttgt cccctcaatg ctgcaagtgt ttttacaaaa tcgccatgta 840
agcaaatgca gctctctaaa aagagttatt tgtagcggtg aagctttatc tatagattta 900
caaaatagat ttttccagca tttgcaatgt gaattacata acctctatgg cccgacagaa 960
gcagcaattg atgtcacatt ttggcaatgt agaaaagata gtaatttaaa gagtgtacct 1020
attggtcgtc ccattgctaa tactcaaatt tatattcttg atgccgattt acaaccagta 1080
aatattggtg tcactggtga aatttatatt ggtggtgtag gggttgctcg tggttatttg 1140
aataaagaag aattgaccaa agaaaaattt attattaatc cctttcccaa ttctgagttt 1200
aagcgacttt ataaaacagg tgatttagct cgttatttac ccgatggaaa tattgaatat 1260
cttggtagaa cagattatca agtaaaaatt cggggttata gaattgaaat tggcgagatt 1320
gaaaatgttt tatcttcaca cccacaagtc agagaagctg tagtcatagc gcgggatgat 1380
aacgctcaag aaaaacaaat catcgcttat attacctata actccatcaa acctcagctt 1440
gataatctgc gtgatttcct aaaagcaagg ctacctgatt ttatgattcc agccgctttt 1500
gtgatgctgg agcatcttcc tttaactccc agtggtaaag tagaccgtaa ggcattacct 1560
aagcctgatt tatttaatta tagtgaacat aattcctatg tagcgcctcg gaatgaagtt 1620
gaagaaaaat tagtacaaat ctggtcgaat attctgcatt tacctaaagt aggtgtgaca 1680
gaaaactttt tcgctattgg tggtaattcc ctcaaagctc tacatttaat ttctcaaatt 1740
gaagagttat ttgctaaaga gatatcctta gcaacacttt taacaaatcc agtaattgca 1800
gatttagcca aggttattca agcaaacaac caaatccata attcacccct agttccaatt 1860
caaccacaag gtaagcagca gcctttcttt tgtatacatc ctgctggtgg tcatgtttta 1920
tgctatttta aactcgcaca atatatagga actgaccaac cattttatgg cttacaagct 1980
caaggatttt atggagatga agcacccttg acgcgagttg aagatatggc tagtctctac 2040
gtcaaaacta ttagagaatt tcaaccccaa gggccttatc gtgtcggggg gtggtcattt 2100
ggtggagtcg tagcttatga agtagcacag cagttacata gacaaggaca agaagtatct 2160
ttactagcaa tattagattc ttacgtaccg attctgctgg ataaacaaaa acccattgat 2220
gacgtttatt tagttggtgt tctctccaga gtttttggcg gtatgtttgg tcaagataat 2280
ctagtcacac ctgaagaaat agaaaattta actgtagaag aaaaaattaa ttacatcatt 2340
gataaagcac ggagcgctag aatattcccg cctggtgtag aacgtcaaaa taatcgccgt 2400
attcttgatg ttttggtggg aactttaaaa gcaacttatt cctatataag acaaccatat 2460
ccaggaaaag tcactgtatt tcgagccagg gaaaaacata ttatggctcc tgacccgacc 2520
ttagtttggg tagaattatt ttctgtaatg gcggctcaag aaattaagat tattgatgtc 2580
cctggaaacc attattcgtt tgttctagaa ccccatgtac aggttttagc acagcgttta 2640
caagattgtc tggaaaataa ttcataa 2667
<210> 96
<211> 888
<212> PRT
<213> Anabaena variabilis ATCC29413
<400> 96
Met Gln Thr Ile Asp Phe Asn Ile Arg Lys Leu Leu Val Glu Trp Asn
1 5 10 15
Ala Thr His Arg Asp Tyr Asp Leu Ser Gln Ser Leu His Glu Leu Ile
20 25 30
Val Ala Gln Val Glu Arg Thr Pro Glu Ala Ile Ala Val Thr Phe Asp
35 40 45
Lys Gln Gln Leu Thr Tyr Gln Glu Leu Asn His Lys Ala Asn Gln Leu
50 55 60
Gly His Tyr Leu Gln Thr Leu Gly Val Gln Pro Glu Thr Leu Val Gly
65 70 75 80
Val Cys Leu Glu Arg Ser Leu Glu Met Val Ile Cys Leu Leu Gly Ile
85 90 95
Leu Lys Ala Gly Gly Ala Tyr Val Pro Ile Asp Pro Glu Tyr Pro Gln
100 105 110
Glu Arg Ile Ala Tyr Met Leu Glu Asp Ser Gln Val Lys Val Leu Leu
115 120 125
Thr Gln Glu Lys Leu Leu Asn Gln Ile Pro His His Gln Ala Gln Thr
130 135 140
Ile Cys Val Asp Arg Glu Trp Glu Lys Ile Ser Thr Gln Ala Asn Thr
145 150 155 160
Asn Pro Lys Ser Asn Ile Lys Thr Asp Asn Leu Ala Tyr Val Ile Tyr
165 170 175
Thr Ser Gly Ser Thr Gly Lys Pro Lys Gly Ala Met Asn Thr His Lys
180 185 190
Gly Ile Cys Asn Arg Leu Leu Trp Met Gln Glu Ala Tyr Gln Ile Asp
195 200 205
Ser Thr Asp Ser Ile Leu Gln Lys Thr Pro Phe Ser Phe Asp Val Ser
210 215 220
Val Trp Glu Phe Phe Trp Thr Leu Leu Thr Gly Ala Arg Leu Val Ile
225 230 235 240
Ala Lys Pro Gly Gly His Lys Asp Ser Ala Tyr Leu Ile Asp Leu Ile
245 250 255
Thr Gln Glu Gln Ile Thr Thr Leu His Phe Val Pro Ser Met Leu Gln
260 265 270
Val Phe Leu Gln Asn Arg His Val Ser Lys Cys Ser Ser Leu Lys Arg
275 280 285
Val Ile Cys Ser Gly Glu Ala Leu Ser Ile Asp Leu Gln Asn Arg Phe
290 295 300
Phe Gln His Leu Gln Cys Glu Leu His Asn Leu Tyr Gly Pro Thr Glu
305 310 315 320
Ala Ala Ile Asp Val Thr Phe Trp Gln Cys Arg Lys Asp Ser Asn Leu
325 330 335
Lys Ser Val Pro Ile Gly Arg Pro Ile Ala Asn Thr Gln Ile Tyr Ile
340 345 350
Leu Asp Ala Asp Leu Gln Pro Val Asn Ile Gly Val Thr Gly Glu Ile
355 360 365
Tyr Ile Gly Gly Val Gly Val Ala Arg Gly Tyr Leu Asn Lys Glu Glu
370 375 380
Leu Thr Lys Glu Lys Phe Ile Ile Asn Pro Phe Pro Asn Ser Glu Phe
385 390 395 400
Lys Arg Leu Tyr Lys Thr Gly Asp Leu Ala Arg Tyr Leu Pro Asp Gly
405 410 415
Asn Ile Glu Tyr Leu Gly Arg Thr Asp Tyr Gln Val Lys Ile Arg Gly
420 425 430
Tyr Arg Ile Glu Ile Gly Glu Ile Glu Asn Val Leu Ser Ser His Pro
435 440 445
Gln Val Arg Glu Ala Val Val Ile Ala Arg Asp Asp Asn Ala Gln Glu
450 455 460
Lys Gln Ile Ile Ala Tyr Ile Thr Tyr Asn Ser Ile Lys Pro Gln Leu
465 470 475 480
Asp Asn Leu Arg Asp Phe Leu Lys Ala Arg Leu Pro Asp Phe Met Ile
485 490 495
Pro Ala Ala Phe Val Met Leu Glu His Leu Pro Leu Thr Pro Ser Gly
500 505 510
Lys Val Asp Arg Lys Ala Leu Pro Lys Pro Asp Leu Phe Asn Tyr Ser
515 520 525
Glu His Asn Ser Tyr Val Ala Pro Arg Asn Glu Val Glu Glu Lys Leu
530 535 540
Val Gln Ile Trp Ser Asn Ile Leu His Leu Pro Lys Val Gly Val Thr
545 550 555 560
Glu Asn Phe Phe Ala Ile Gly Gly Asn Ser Leu Lys Ala Leu His Leu
565 570 575
Ile Ser Gln Ile Glu Glu Leu Phe Ala Lys Glu Ile Ser Leu Ala Thr
580 585 590
Leu Leu Thr Asn Pro Val Ile Ala Asp Leu Ala Lys Val Ile Gln Ala
595 600 605
Asn Asn Gln Ile His Asn Ser Pro Leu Val Pro Ile Gln Pro Gln Gly
610 615 620
Lys Gln Gln Pro Phe Phe Cys Ile His Pro Ala Gly Gly His Val Leu
625 630 635 640
Cys Tyr Phe Lys Leu Ala Gln Tyr Ile Gly Thr Asp Gln Pro Phe Tyr
645 650 655
Gly Leu Gln Ala Gln Gly Phe Tyr Gly Asp Glu Ala Pro Leu Thr Arg
660 665 670
Val Glu Asp Met Ala Ser Leu Tyr Val Lys Thr Ile Arg Glu Phe Gln
675 680 685
Pro Gln Gly Pro Tyr Arg Val Gly Gly Trp Ser Phe Gly Gly Val Val
690 695 700
Ala Tyr Glu Val Ala Gln Gln Leu His Arg Gln Gly Gln Glu Val Ser
705 710 715 720
Leu Leu Ala Ile Leu Asp Ser Tyr Val Pro Ile Leu Leu Asp Lys Gln
725 730 735
Lys Pro Ile Asp Asp Val Tyr Leu Val Gly Val Leu Ser Arg Val Phe
740 745 750
Gly Gly Met Phe Gly Gln Asp Asn Leu Val Thr Pro Glu Glu Ile Glu
755 760 765
Asn Leu Thr Val Glu Glu Lys Ile Asn Tyr Ile Ile Asp Lys Ala Arg
770 775 780
Ser Ala Arg Ile Phe Pro Pro Gly Val Glu Arg Gln Asn Asn Arg Arg
785 790 795 800
Ile Leu Asp Val Leu Val Gly Thr Leu Lys Ala Thr Tyr Ser Tyr Ile
805 810 815
Arg Gln Pro Tyr Pro Gly Lys Val Thr Val Phe Arg Ala Arg Glu Lys
820 825 830
His Ile Met Ala Pro Asp Pro Thr Leu Val Trp Val Glu Leu Phe Ser
835 840 845
Val Met Ala Ala Gln Glu Ile Lys Ile Ile Asp Val Pro Gly Asn His
850 855 860
Tyr Ser Phe Val Leu Glu Pro His Val Gln Val Leu Ala Gln Arg Leu
865 870 875 880
Gln Asp Cys Leu Glu Asn Asn Ser
885
<210> 97
<211> 1233
<212> ДНК
<213> Nostoc punctiforme ATCC29133
<400> 97
atgagtaatg ttcaagcatc gtttgaagca acggaagctg aattccgcgt ggaaggttac 60
gaaaaaattg agtttagtct tgtctatgta aatggtgcat ttgatatcag taacagagaa 120
attgcagaca gctatgagaa gtttggccgt tgtctgactg tgattgatgc taatgtcaac 180
agattatatg gcaagcaaat caagtcatat tttagacact atggtattga tctgaccgta 240
gttcccattg tgattactga gcctactaaa acccttgcaa cctttgagaa aattgttgat 300
gctttttctg actttggttt aatccgcaag gaaccagtat tagtagtggg tggtggttta 360
accactgatg tagctgggtt tgcgtgtgct gcttaccgtc gtaagagtaa ctatattcgg 420
gttccgacaa cgctgattgg tctgattgat gcaggtgtag cgattaaggt tgcagtcaac 480
catcgcaagt taaaaaatcg cttgggtgca tatcatgctc ctttgaaagt catcctcgat 540
ttctccttct tgcaaacatt accaacggct caagttcgta atgggatggc agagttggtc 600
aaaattgctg ttgtggcgaa ctctgaagtc tttgaattgt tgtatgaata tggcgaagag 660
ttgctttcca ctcactttgg atatgtgaat ggtacaaagg aactgaaagc gatcgcacat 720
aaactcaatt acgaggcaat aaaaactatg ctggagttgg aaactccaaa cttgcatgag 780
ttagacctcg atcgcgtcat tgcctacggt cacacttgga gtccgacctt agaattagct 840
ccgatgatac cgttgtttca cggtcatgcc gtcaatatag acatggcttt gtctgcaact 900
attgcggcaa gacgtggcta cattacatca ggagaacgcg atcgcatttt gagcttgatg 960
agtcgtatag gtttatcaat cgatcatcct ctactagatg gcgatttgct ctggtatgct 1020
acccaatcta ttagcttgac gcgagacggg aaacaacgcg cagctatgcc taaacccatt 1080
ggtgagtgct tctttgtcaa cgatttcacc cgtgaagaac tagatgcagc tttagctgaa 1140
cacaaacgtc tgtgtgctac ataccctcgt ggtggagatg gcattgacgc ttacatagaa 1200
actcaagaag aatccaaact attgggagtg tga 1233
<210> 98
<211> 410
<212> PRT
<213> Nostoc punctiforme ATCC29133
<400> 98
Met Ser Asn Val Gln Ala Ser Phe Glu Ala Thr Glu Ala Glu Phe Arg
1 5 10 15
Val Glu Gly Tyr Glu Lys Ile Glu Phe Ser Leu Val Tyr Val Asn Gly
20 25 30
Ala Phe Asp Ile Ser Asn Arg Glu Ile Ala Asp Ser Tyr Glu Lys Phe
35 40 45
Gly Arg Cys Leu Thr Val Ile Asp Ala Asn Val Asn Arg Leu Tyr Gly
50 55 60
Lys Gln Ile Lys Ser Tyr Phe Arg His Tyr Gly Ile Asp Leu Thr Val
65 70 75 80
Val Pro Ile Val Ile Thr Glu Pro Thr Lys Thr Leu Ala Thr Phe Glu
85 90 95
Lys Ile Val Asp Ala Phe Ser Asp Phe Gly Leu Ile Arg Lys Glu Pro
100 105 110
Val Leu Val Val Gly Gly Gly Leu Thr Thr Asp Val Ala Gly Phe Ala
115 120 125
Cys Ala Ala Tyr Arg Arg Lys Ser Asn Tyr Ile Arg Val Pro Thr Thr
130 135 140
Leu Ile Gly Leu Ile Asp Ala Gly Val Ala Ile Lys Val Ala Val Asn
145 150 155 160
His Arg Lys Leu Lys Asn Arg Leu Gly Ala Tyr His Ala Pro Leu Lys
165 170 175
Val Ile Leu Asp Phe Ser Phe Leu Gln Thr Leu Pro Thr Ala Gln Val
180 185 190
Arg Asn Gly Met Ala Glu Leu Val Lys Ile Ala Val Val Ala Asn Ser
195 200 205
Glu Val Phe Glu Leu Leu Tyr Glu Tyr Gly Glu Glu Leu Leu Ser Thr
210 215 220
His Phe Gly Tyr Val Asn Gly Thr Lys Glu Leu Lys Ala Ile Ala His
225 230 235 240
Lys Leu Asn Tyr Glu Ala Ile Lys Thr Met Leu Glu Leu Glu Thr Pro
245 250 255
Asn Leu His Glu Leu Asp Leu Asp Arg Val Ile Ala Tyr Gly His Thr
260 265 270
Trp Ser Pro Thr Leu Glu Leu Ala Pro Met Ile Pro Leu Phe His Gly
275 280 285
His Ala Val Asn Ile Asp Met Ala Leu Ser Ala Thr Ile Ala Ala Arg
290 295 300
Arg Gly Tyr Ile Thr Ser Gly Glu Arg Asp Arg Ile Leu Ser Leu Met
305 310 315 320
Ser Arg Ile Gly Leu Ser Ile Asp His Pro Leu Leu Asp Gly Asp Leu
325 330 335
Leu Trp Tyr Ala Thr Gln Ser Ile Ser Leu Thr Arg Asp Gly Lys Gln
340 345 350
Arg Ala Ala Met Pro Lys Pro Ile Gly Glu Cys Phe Phe Val Asn Asp
355 360 365
Phe Thr Arg Glu Glu Leu Asp Ala Ala Leu Ala Glu His Lys Arg Leu
370 375 380
Cys Ala Thr Tyr Pro Arg Gly Gly Asp Gly Ile Asp Ala Tyr Ile Glu
385 390 395 400
Thr Gln Glu Glu Ser Lys Leu Leu Gly Val
405 410
<210> 99
<211> 834
<212> ДНК
<213> Nostoc punctiforme ATCC29133
<400> 99
atgaccagta ttttaggacg agataccgca agaccaataa cgccacatag cattctggta 60
gcacagctac aaaaaaccct cagaatggca gaggaaagta atattccttc agagatactg 120
acttctctgc gccaagggtt gcaattagca gcaggtttag atccctatct ggatgattgc 180
actacccctg aatcgaccgc attgacagca ctagcgcaga agacaagcat tgaagactgg 240
agtaaacgct tcagtgatgg tgaaacagtg cgtcaattag agcaagaaat gctctcagga 300
catcttgaag gacaaacact aaagatgttt gtgcatatca ctaaggctaa aagcatccta 360
gaagtgggaa tgttcacagg atattcagct ttggcaatgg cagaggcgtt accagatgat 420
gggcgactga ttgcttgtga agtagactcc tatgtggccg agtttgctca aacttgcttt 480
caagagtctc cccacggccg caagattgtt gtagaagtgg cacctgcact agagacgctg 540
cacaagctgg tggctaaaaa agaatccttt gatctgatct tcattgatgc ggataaaaag 600
gagtatatag aatacttcca aattatcttg gatagccatt tactagctcc cgacggatta 660
atctgtgtag ataatacttt gttgcaagga caagtttacc tgccatcaga acagcgtaca 720
gccaatggtg aagcgatcgc tcaattcaac cgcattgtcg ccgcagatcc tcgtgtagag 780
caagttctgt tacccatacg agatggtata accctgatta gacgcttggt ataa 834
<210> 100
<211> 277
<212> PRT
<213> Nostoc punctiforme ATCC29133
<400> 100
Met Thr Ser Ile Leu Gly Arg Asp Thr Ala Arg Pro Ile Thr Pro His
1 5 10 15
Ser Ile Leu Val Ala Gln Leu Gln Lys Thr Leu Arg Met Ala Glu Glu
20 25 30
Ser Asn Ile Pro Ser Glu Ile Leu Thr Ser Leu Arg Gln Gly Leu Gln
35 40 45
Leu Ala Ala Gly Leu Asp Pro Tyr Leu Asp Asp Cys Thr Thr Pro Glu
50 55 60
Ser Thr Ala Leu Thr Ala Leu Ala Gln Lys Thr Ser Ile Glu Asp Trp
65 70 75 80
Ser Lys Arg Phe Ser Asp Gly Glu Thr Val Arg Gln Leu Glu Gln Glu
85 90 95
Met Leu Ser Gly His Leu Glu Gly Gln Thr Leu Lys Met Phe Val His
100 105 110
Ile Thr Lys Ala Lys Ser Ile Leu Glu Val Gly Met Phe Thr Gly Tyr
115 120 125
Ser Ala Leu Ala Met Ala Glu Ala Leu Pro Asp Asp Gly Arg Leu Ile
130 135 140
Ala Cys Glu Val Asp Ser Tyr Val Ala Glu Phe Ala Gln Thr Cys Phe
145 150 155 160
Gln Glu Ser Pro His Gly Arg Lys Ile Val Val Glu Val Ala Pro Ala
165 170 175
Leu Glu Thr Leu His Lys Leu Val Ala Lys Lys Glu Ser Phe Asp Leu
180 185 190
Ile Phe Ile Asp Ala Asp Lys Lys Glu Tyr Ile Glu Tyr Phe Gln Ile
195 200 205
Ile Leu Asp Ser His Leu Leu Ala Pro Asp Gly Leu Ile Cys Val Asp
210 215 220
Asn Thr Leu Leu Gln Gly Gln Val Tyr Leu Pro Ser Glu Gln Arg Thr
225 230 235 240
Ala Asn Gly Glu Ala Ile Ala Gln Phe Asn Arg Ile Val Ala Ala Asp
245 250 255
Pro Arg Val Glu Gln Val Leu Leu Pro Ile Arg Asp Gly Ile Thr Leu
260 265 270
Ile Arg Arg Leu Val
275
<210> 101
<211> 1386
<212> ДНК
<213> Nostoc punctiforme ATCC29133
<400> 101
atggcacaat caatctcttt atctcttcct caatccacaa cgccatcaaa gggtgtgagg 60
ctaaaaatag cagctctact gaagactatc gggactctaa tactactgct gatagccttg 120
ccgcttaatg ctttgatagt attgatatct ctgatgtgta ggccgtttac aaaaaaacct 180
gccgtagcca ctcatcccca gaatatcttg gtcagtggcg gcaaaatgac caaagcattg 240
caacttgccc gctccttcca tgcagccgga cacagagtta ttctgattga aggtcataaa 300
tactggttat cagggcatag attctcaaat tctgtgagtc gtttttatac agttcctgca 360
ccacaagacg acccagaagg ctatacccaa gcgctattgg aaattgtcaa acgagagaag 420
attgacgttt atgtacccgt atgcagccct gtagctagtt attacgactc tttggcaaag 480
tctgcactat cagaatattg tgaggttttt cactttgatg ctgatataac caagatgctg 540
gatgataaat ttgcctttac cgatcgggcg cgatcgcttg gtttatcagc cccgaaatct 600
tttaaaatta ccgatcctga acaagttatc aacttcgatt ttagtaaaga gacgcgcaaa 660
tatattctta agagtatttc ttacgactca gttcgccgct taaatttaac caaacttcct 720
tgtgataccc cagaagagac agcagcgttt gtcaagagtt tacccatcag cccagaaaaa 780
ccttggatta tgcaagaatt tattcctggg aaagaattat gcacccatag cacagtccga 840
gacggcgaat taaggttgca ttgctgttca aattcttcag cgtttcagat taactatgaa 900
aatgtcgaaa atccccaaat tcaagaatgg gtacaacatt tcgtcaaaag tttacggctg 960
actggacaaa tatctcttga ctttatccaa gctgaagatg gtacagctta tgccattgaa 1020
tgtaatcctc gcacccattc ggcgatcaca atgttctaca atcacccagg tgttgcagaa 1080
gcctatcttg gtaaaactcc tctagctgca cctttggaac ctcttgcaga tagcaagccc 1140
acttactgga tatatcacga aatctggcga ttgactggga ttcgctctgg acaacaatta 1200
caaacttggt ttgggagatt agtcagaggt acagatgcca tttatcgcct ggacgatccg 1260
ataccatttt taactttgca ccattggcag attactttac ttttgctaca aaatttgcaa 1320
cgactcaaag gttgggtaaa gatcgatttt aatatcggta aactcgtgga attagggggc 1380
gactaa 1386
<210> 102
<211> 461
<212> PRT
<213> Nostoc punctiforme ATCC29133
<400> 102
Met Ala Gln Ser Ile Ser Leu Ser Leu Pro Gln Ser Thr Thr Pro Ser
1 5 10 15
Lys Gly Val Arg Leu Lys Ile Ala Ala Leu Leu Lys Thr Ile Gly Thr
20 25 30
Leu Ile Leu Leu Leu Ile Ala Leu Pro Leu Asn Ala Leu Ile Val Leu
35 40 45
Ile Ser Leu Met Cys Arg Pro Phe Thr Lys Lys Pro Ala Val Ala Thr
50 55 60
His Pro Gln Asn Ile Leu Val Ser Gly Gly Lys Met Thr Lys Ala Leu
65 70 75 80
Gln Leu Ala Arg Ser Phe His Ala Ala Gly His Arg Val Ile Leu Ile
85 90 95
Glu Gly His Lys Tyr Trp Leu Ser Gly His Arg Phe Ser Asn Ser Val
100 105 110
Ser Arg Phe Tyr Thr Val Pro Ala Pro Gln Asp Asp Pro Glu Gly Tyr
115 120 125
Thr Gln Ala Leu Leu Glu Ile Val Lys Arg Glu Lys Ile Asp Val Tyr
130 135 140
Val Pro Val Cys Ser Pro Val Ala Ser Tyr Tyr Asp Ser Leu Ala Lys
145 150 155 160
Ser Ala Leu Ser Glu Tyr Cys Glu Val Phe His Phe Asp Ala Asp Ile
165 170 175
Thr Lys Met Leu Asp Asp Lys Phe Ala Phe Thr Asp Arg Ala Arg Ser
180 185 190
Leu Gly Leu Ser Ala Pro Lys Ser Phe Lys Ile Thr Asp Pro Glu Gln
195 200 205
Val Ile Asn Phe Asp Phe Ser Lys Glu Thr Arg Lys Tyr Ile Leu Lys
210 215 220
Ser Ile Ser Tyr Asp Ser Val Arg Arg Leu Asn Leu Thr Lys Leu Pro
225 230 235 240
Cys Asp Thr Pro Glu Glu Thr Ala Ala Phe Val Lys Ser Leu Pro Ile
245 250 255
Ser Pro Glu Lys Pro Trp Ile Met Gln Glu Phe Ile Pro Gly Lys Glu
260 265 270
Leu Cys Thr His Ser Thr Val Arg Asp Gly Glu Leu Arg Leu His Cys
275 280 285
Cys Ser Asn Ser Ser Ala Phe Gln Ile Asn Tyr Glu Asn Val Glu Asn
290 295 300
Pro Gln Ile Gln Glu Trp Val Gln His Phe Val Lys Ser Leu Arg Leu
305 310 315 320
Thr Gly Gln Ile Ser Leu Asp Phe Ile Gln Ala Glu Asp Gly Thr Ala
325 330 335
Tyr Ala Ile Glu Cys Asn Pro Arg Thr His Ser Ala Ile Thr Met Phe
340 345 350
Tyr Asn His Pro Gly Val Ala Glu Ala Tyr Leu Gly Lys Thr Pro Leu
355 360 365
Ala Ala Pro Leu Glu Pro Leu Ala Asp Ser Lys Pro Thr Tyr Trp Ile
370 375 380
Tyr His Glu Ile Trp Arg Leu Thr Gly Ile Arg Ser Gly Gln Gln Leu
385 390 395 400
Gln Thr Trp Phe Gly Arg Leu Val Arg Gly Thr Asp Ala Ile Tyr Arg
405 410 415
Leu Asp Asp Pro Ile Pro Phe Leu Thr Leu His His Trp Gln Ile Thr
420 425 430
Leu Leu Leu Leu Gln Asn Leu Gln Arg Leu Lys Gly Trp Val Lys Ile
435 440 445
Asp Phe Asn Ile Gly Lys Leu Val Glu Leu Gly Gly Asp
450 455 460
<210> 103
<211> 1047
<212> ДНК
<213> Nostoc punctiforme ATCC29133
<400> 103
atgccagtac ttaatatcct tcatttagtt gggtctgcac acgataagtt ttactgtgat 60
ttatcacgtc tttatgccca agactgttta gctgcaacag cagatccatc gctttataac 120
tttcaaattg catatatcac acccgatcgg cagtggcgat ttcctgactc tctcagtcga 180
gaagatattg ctcttaccaa accgattcct gtgtttgatg ccatacaatt tctaacaggc 240
caaaacattg acatgatgtt accacaaatg ttttgtattc ctggaatgac tcagtaccgt 300
gccctattcg atctgctcaa gatcccttat ataggaaata ccccagatat tatggcgatc 360
gcggcccaca aagccagagc caaagcaatt gtcgaagcag caggggtaaa agtgcctcgt 420
ggagaattgc ttcgccaagg agatattcca acaattacac ctccagcagt cgtcaaacct 480
gtaagttctg acaactcttt aggagtagtc ttagttaaag atgtgactga atatgatgct 540
gccttaaaga aagcatttga atatgcttcg gaggtcatcg tagaagcatt catcgaactt 600
ggtcgagaag tcagatgcgg catcattgta aaagacggtg agctaatagg tttacccctt 660
gaagagtatc tggtagaccc acacgataaa cctatccgta actatgctga taaactccaa 720
caaactgacg atggcgactt gcatttgact gctaaagata atatcaaggc ttggatttta 780
gaccctaacg acccaatcac ccaaaaggtt cagcaagtgg ctaaaaggtg tcatcaggct 840
ttgggttgtc gccactacag tttatttgac ttccgaatcg atccaaaggg acaaccttgg 900
ttcttagaag ctggattata ttgttctttt gcccccaaaa gtgtgatttc ttctatggcg 960
aaagcagccg gaatccctct aaatgattta ttaataaccg ctattaatga aacattgggt 1020
agtaataaaa aggtgttaca aaattga 1047
<210> 104
<211> 348
<212> PRT
<213> Nostoc punctiforme ATCC29133
<400> 104
Met Pro Val Leu Asn Ile Leu His Leu Val Gly Ser Ala His Asp Lys
1 5 10 15
Phe Tyr Cys Asp Leu Ser Arg Leu Tyr Ala Gln Asp Cys Leu Ala Ala
20 25 30
Thr Ala Asp Pro Ser Leu Tyr Asn Phe Gln Ile Ala Tyr Ile Thr Pro
35 40 45
Asp Arg Gln Trp Arg Phe Pro Asp Ser Leu Ser Arg Glu Asp Ile Ala
50 55 60
Leu Thr Lys Pro Ile Pro Val Phe Asp Ala Ile Gln Phe Leu Thr Gly
65 70 75 80
Gln Asn Ile Asp Met Met Leu Pro Gln Met Phe Cys Ile Pro Gly Met
85 90 95
Thr Gln Tyr Arg Ala Leu Phe Asp Leu Leu Lys Ile Pro Tyr Ile Gly
100 105 110
Asn Thr Pro Asp Ile Met Ala Ile Ala Ala His Lys Ala Arg Ala Lys
115 120 125
Ala Ile Val Glu Ala Ala Gly Val Lys Val Pro Arg Gly Glu Leu Leu
130 135 140
Arg Gln Gly Asp Ile Pro Thr Ile Thr Pro Pro Ala Val Val Lys Pro
145 150 155 160
Val Ser Ser Asp Asn Ser Leu Gly Val Val Leu Val Lys Asp Val Thr
165 170 175
Glu Tyr Asp Ala Ala Leu Lys Lys Ala Phe Glu Tyr Ala Ser Glu Val
180 185 190
Ile Val Glu Ala Phe Ile Glu Leu Gly Arg Glu Val Arg Cys Gly Ile
195 200 205
Ile Val Lys Asp Gly Glu Leu Ile Gly Leu Pro Leu Glu Glu Tyr Leu
210 215 220
Val Asp Pro His Asp Lys Pro Ile Arg Asn Tyr Ala Asp Lys Leu Gln
225 230 235 240
Gln Thr Asp Asp Gly Asp Leu His Leu Thr Ala Lys Asp Asn Ile Lys
245 250 255
Ala Trp Ile Leu Asp Pro Asn Asp Pro Ile Thr Gln Lys Val Gln Gln
260 265 270
Val Ala Lys Arg Cys His Gln Ala Leu Gly Cys Arg His Tyr Ser Leu
275 280 285
Phe Asp Phe Arg Ile Asp Pro Lys Gly Gln Pro Trp Phe Leu Glu Ala
290 295 300
Gly Leu Tyr Cys Ser Phe Ala Pro Lys Ser Val Ile Ser Ser Met Ala
305 310 315 320
Lys Ala Ala Gly Ile Pro Leu Asn Asp Leu Leu Ile Thr Ala Ile Asn
325 330 335
Glu Thr Leu Gly Ser Asn Lys Lys Val Leu Gln Asn
340 345
1
<---
Изобретение относится к области биотехнологии. Описан микроорганизм для продуцирования микоспорин-подобной аминокислоты, в котором активность 3-дегидрохиннатдегидратазы инактивирована, а также способ получения микоспорин-подобной аминокислоты с использованием этого микроорганизма. Изобретение позволяет увеличить продуцирование микоспорин-подобных аминокислот. 2 н. и 6 з.п. ф-лы, 16 табл., 17 пр.
1. Микроорганизм для продуцирования микроспорин-подобной аминокислоты, в котором активность 3-дегидрохиннатдегидратазы инактивирована по сравнению с немодифицированным микроорганизмом.
2. Микроорганизм по п. 1, содержащий ген, кодирующий по меньшей мере один белок, выбранный из группы, состоящей из 2-деметил-4-дезоксигадузолсинтазы, О-метилтрансферазы и C-N лигазы.
3. Микроорганизм по п. 1, содержащий ген, кодирующий по меньшей мере один белок, выбранный из группы, состоящей из синтетазы нерибосомных пептидов, фермента, подобного синтетазе нерибосомных пептидов (NRPS-подобного фермента), и D-Ala-D-Ala лигазы.
4. Микроорганизм по п. 1, в котором активность по меньшей мере одного белка, выбранного из группы, состоящей из 2-дегидро-3-дезоксифосфогептонатальдолазы, фосфоенолпируватсинтетазы, транскетолазы I/II и 3-дегидрохиннатсинтазы, дополнительно усилена по сравнению с немодифицированным микроорганизмом.
5. Микроорганизм по п. 1, представляющий собой микроорганизм рода Corynebacterium, микроорганизм рода Escherichia или дрожжи.
6. Микроорганизм по п. 5, где в дрожжи введен ген, кодирующий 3-дегидрохиннатсинтазу.
7. Микроорганизм по п. 1, где микоспорин-подобная аминокислота представляет собой по меньшей мере одну аминокислоту, выбранную из группы, состоящей из микоспорин-2-глицина, палитинола, палитеновой кислоты, дезоксигадузола, микоспорин-метиламин-треонина, микоспорин-глицин-валина, палитина, астерина-330 (asterina-330), шинорина (shinorine), порфира-334, эугалотек-362 (euhalothece-362), микоспорин-глицина, микоспорин-орнитина, микоспорин-лизина, микоспорин-глутаминовой кислоты, микоспорин-метиламин-серина, микоспорин-таурина, палитена, палитин-серина, палитин-серин-сульфата, палитинола и усуджирена (usujirene).
8. Способ получения микоспорин-подобной аминокислоты, включающий:
культивирование микроорганизма по любому одному из пп. 1-7 и
извлечение микроспорин-подобной аминокислоты из культивируемого микроорганизма или из среды.
| US 20170202762 А1, 20.07.2017 | |||
| Герметичный кабельный ввод | 1975 |
|
SU663011A1 |
| WO 2014025938 A1, 13.02.2014 | |||
| ЗЛОБИН А.А | |||
| Основы биотехнологии | |||
| Методические указания к лекционному курсу, Киров, 2014, С.8-10. | |||

