Область техники
Группа изобретений относится к области биотехнологии, иммунологии и вирусологии. Предложенное иммунобиологическое средство может применяться для профилактики заболеваний, вызываемых вирусом клещевого энцефалита.
Уровень техники
Клещевой энцефалит (КЭ) является природно-очаговой трансмиссивной вирусной инфекцией с выраженной сезонностью. Заболеваемость зависит от периода активности иксодовых клещей - основных переносчиков инфекции. Течение заболевания характеризуется преимущественным поражением центральной нервной системы (ЦНС) и отличается полиморфизмом клинического течения, с высокой частотой остаточных явлений в виде параличей, а также летальностью.
С медицинской точки зрения вирус КЭ (ВКЭ) является наиболее важным представителем рода Flavivirus, семейства Flaviviridae. Вирусы семейства Flaviviridae - это РНК-содержащие сферические оболочечные вирусы. Вирион ВКЭ имеет сферическую форму, 45-50 нм в диаметре. Одноцепочечный несегментированный (+)РНК-геном вируса КЭ имеет длину около 11 000 нуклеотидов кодирует и 3 структурных (C, PrM, E) и 7 неструктурных генов (NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5). Белок Е является главным структурным белком вириона, который отвечает за сборку вириона, слияние с мембраной эукариотической клетки и связывание c рецептором. В организме иммунизированных животных непосредственно участвует в стимуляции синтеза вируснейтрализующих антител. На основании нуклеотидной последовательности гена, кодирующего белок Е, в настоящее время выделяют 4 генотипа ВКЭ: дальневосточный (генотип 1, прототипный штамм Софьин), европейский (центрально-европейский или западный, генотип 2, прототипный штамм Neudoerfl), сибирский (урало-сибирский, генотип 3, прототипные штаммы - Васильченко и Заусаев) и сравнительно недавно описанный байкальский (прототипный штамм 886-84). Для каждого из генотипов ВКЭ характерны ассоциация с определенным видом клеща-переносчика и кругом позвоночных хозяев, разный патогенный потенциал для человека и циркуляция в собственном ареале, в пределах которого отмечается его доминирование определённого генотипа.
ВКЭ является эндемичным для центральных регионов Европы и Азии, ареал циркуляции вируса достигает Франции, Японии и Албании. В настоящее время случаи КЭ официально зарегистрированы в 29 европейских странах и 6 стран Азии. В мире ежегодно регистрируется до 12000 случаев заболевания КЭ. На территории Евразии основным природным очагом ВКЭ является Россия. Факторы, оказывающие влияние на заболеваемость КЭ, включают продолжительность времени пребывания человека на территории природного очага (определяется профессиональной, хозяйственной деятельностью, проведением досуга), длительность присасывания клеща, а также экологические факторы (изменение климата, и как следствие увеличение численности клещей и расширение их ареалов), технологические (улучшение диагностики).
На сегодняшний день наиболее эффективным способом профилактики КЭ является вакцинация, тем не менее, несмотря на наличие вакцин, наблюдается рост заболеваемости КЭ, что связано с недостаточным охватом вакцинацией групп риска. На территории Российской Федерации для иммунизации населения против вируса КЭ разрешены к применению несколько инактивированных вакцин: «Вакцина клещевого энцефалита культуральная очищенная концентрированная инактивированная сухая» и «Клещ-Э-Вак» производства ФГАНУ «ФНЦИРИП им. М. П. Чумакова РАН» (Институт полиомиелита) (Москва, Россия), «ЭнцеВир®» и «ЭнцеВир® Нео детский» производства ФГУП «НПО «Микроген» Минздрава России (Россия), «ФСМЕ-Иммун®» и «ФСМЕ-Иммун® Джуниор» производства Пфайзер Инк (Австрия), «Энцепур®» и «Энцепур® детский» производства ГСК Вакцинс ГмбХ (Германия).
Данные о ежегодной заболеваемости КЭ среди вакцинированных лиц указывают на то, что современные вакцины не обеспечивают 100% защиты от ВКЭ. Неэффективность вакцинных препаратов может быть связана со свойствами самого препарата, а также особенностям этиологического агента или восприимчивого организма.
Среди направлений по разработке вакцин для профилактики инфицирования ВКЭ можно выделить производство аттенуированных или инактивированных вакцин (например, RU2070929C1; RU2288266C2; RU2541784C2), ДНК-вакцин (например RU2150294C1; RU2112038C1; RU2202612C2) или конструирование рекомбинантных вирусов, содержащих клонированные гены наиболее распространенных флавивирусов (например, RU2136312C1; RU2208635C2; RU2209082C2; RU2465326C2; EP0877086; US6497884; US8828687B2; RU2527891C2).
мРНК-вакцины представляют собой многообещающую альтернативу традиционным вакцинам, что обусловлено их высокой эффективностью, безопасностью применения, быстротой разработки и относительно недорогого производства. Последние технологические достижения дали возможность для развития многочисленных платформ мРНК-вакцин против инфекционных заболеваний и некоторых типов рака. Полученные препараты продемонстрировали обнадеживающие результаты как на животных моделях, так и на людях (например, RU2746406C2; RU2022106357; RU2768829C2; US10933127B2).
мРНК вакцины представляют собой препарат мРНК с модифицированными нуклеотидами, содержащей в своем составе последовательность антигена (АГ). Для синтеза мРНК обычно используется in vitro транскрипция. При этом используется РНК-полимераза бактериофага - T3, T7 или SP6, и молекула линеаризованной ДНК, содержащая последовательность целевого антигена. В ходе in vitro транскрипции не используются эукариотические клетки, поэтому получение мРНК таким способом проще и быстрее, чем крупномасштабное производство и очистка белка [Jain S, Venkataraman A, Wechsler ME, Peppas NA. Messenger RNA-based vaccines: Past, present, and future directions in the context of the COVID-19 pandemic. Adv Drug Deliv Rev. 2021 Dec;179:114000.].
Молекулы мРНК достигая клеток-мишеней, способны активировать гуморальные и клеточные иммунные реакции посредством экспрессии целевого гена в белковые продукты. Клеточные иммунные ответы, в которых основную защитную роль играют цитотоксические Т-клетки, обладают способностью воздействовать на инфицированные клетки, тогда как прямая нейтрализация или уничтожение патогена (вируса или бактерии) достигается за счет гуморального иммунитета. Для индукции клеточных иммунных ответов антигену необходимо быть доставленным в цитозоль, где происходит протеосомный процессинг. Вследствие этого, полученные пептиды доставляются в эндоплазматический ретикулум, где антигенные пептиды связываются с молекулами главного комплекса гистосовместимости класса I (MHC-I). Затем комплекс МНС-I / пептид транспортируются на поверхность клетки для распознавания CD8 + Т-лимфоцитами. Таким образом, доставка иммуногенов в пути процессинга антигена, является основной целью в современной разработке вакцин. Вакцины на основе нуклеиновых кислот особенно подходят для выработки мощных клеточных иммунных ответов, поскольку они обеспечивают экспрессию кодируемого белка в цитозоле антиген-презентирующей клетки-мишени [Pollard C, De Koker S, Saelens X, Vanham G, Grooten J. Challenges and advances towards the rational design of mRNA vaccines. Trends Mol Med. 2013 Dec;19(12):705-13. Jain S, Venkataraman A, Wechsler ME, Peppas NA. Messenger RNA-based vaccines: Past, present, and future directions in the context of the COVID-19 pandemic. Adv Drug Deliv Rev. 2021 Dec;179:114000].
За ближайший аналог может быть принят документ EP 3723796, из которого известна искусственная нуклеиновая кислота и полипептид, подходящие для применения при профилактике инфекции, вызванной флавивирусами, в частности инфекции, вызванной вирусом желтой лихорадки или вирусом денге, или расстройства, связанного с таким инфекционное заболевание. В качестве антигена для получения иммуногенной композиции используется нуклеотидная последовательность белка PrM-E вируса желтой лихорадки или вируса денге. Данная композиция может быть использована для профилактики инфицирования вирусом желтой лихорадкой или вируса денге, но не для профилактики клещевого энцефалита.
В качестве аналога также может быть принят документ US20200368343A1, из которого известна искусственная нуклеиновая кислота и полипептид, подходящие для применения при профилактике инфекции, вызванной флавивирусами, в частности инфекции, вызванной вирусом Зика, вирусом Чикунгунья или вирусом денге, или расстройства, связанного с таким инфекционное заболевание. В качестве антигена для получения иммуногенной композиции используется нуклеотидная последовательность белков PrM-E вирусов Зика, Чикунгунья или денге. Данная композиция может быть использована для профилактики инфицирования вирусом Зика, вирусом Чикунгунья или вируса денге, но не для профилактики клещевого энцефалита.
Таким образом, существует потребность в создании нового иммунно-биологического средства на основе мРНК ВКЭ, которое возможно использовать в качестве средства профилактики заболеваний, вызываемых ВКЭ.
Раскрытие сущности изобретения
Технической задачей заявленной группы изобретение является расширение арсенала иммунно-биологических средств на основе мРНК ВКЭ для профилактики заболеваний, вызываемых ВКЭ.
Технический результат заключается в создании иммуннобиологического средства на основе мРНК, кодирующей PrM-E белки ВКЭ, которое может быть использовано для индукции специфического иммунитета против заболеваний, вызываемых ВКЭ.
Указанный технический результат достигается тем, что создано иммунобиологическое средство для профилактики заболеваний, вызванных ВКЭ, в котором для in vitro транскрипции мРНК были использованы гены премембранного (PrM) и оболочечного (E) белков штамма Софьин ВКЭ. Синтезированная мРНК, кодирующая PrM-E белки ВКЭ, упакована в липидные наночастицы мРНК ВКЭ. В частном случае иммунобиологическое средство представляет собой кэпированную мРНК вируса клещевого энцефалита.
Также указанный технический результат достигается тем, что иммунобиологическое средство может применятся для получения вакцин для профилактики клещевого энцефалита.
Краткое описание чертежей
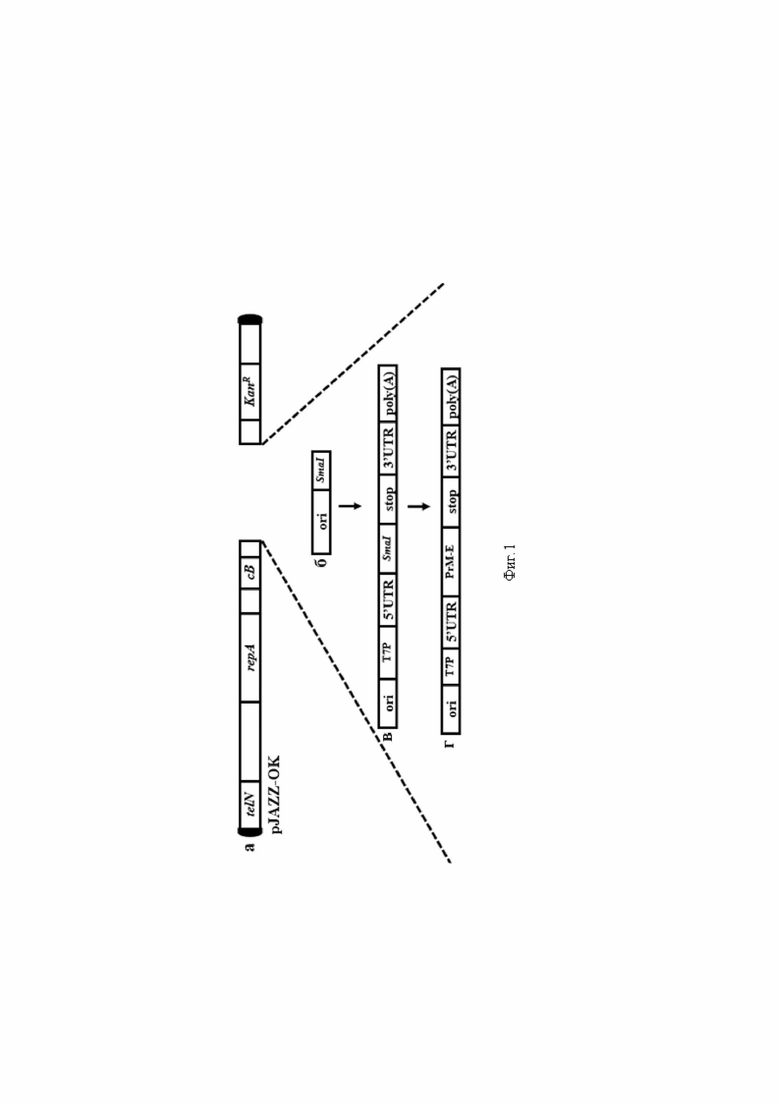
На фиг.1 представлена схема получения линейной плазмиды на основе линейного бактериального вектора pJAZZ-OK. а) Вектор pJAZZ-OK (telN - ген протеломеразы, repA - ориджин репликации, cB - репрессор репликации, KanR - ген устойчивости к канамицину, полусферы чёрного цвета - концевые протеломерные шпильки); b) ColE1 ориджин репликации с последующим сайтом рестрикции рестриктазы SmaI (ori - ориджин); c) Структурные компоненты для in vitro транскрипции (T7P - промотор для РНК-полимеразы бактериофага, 5′ и 3′ UTRs - 5’ и 3’некодирующие области мРНК, SmaI - сайт рестрикции, Stop - терминатор трансляции, poly(A) - сегментированный поли-A-хвост; d) Схематическое представление результирующей линейной плазмиды с открытой рамкой считывания целевого гена.
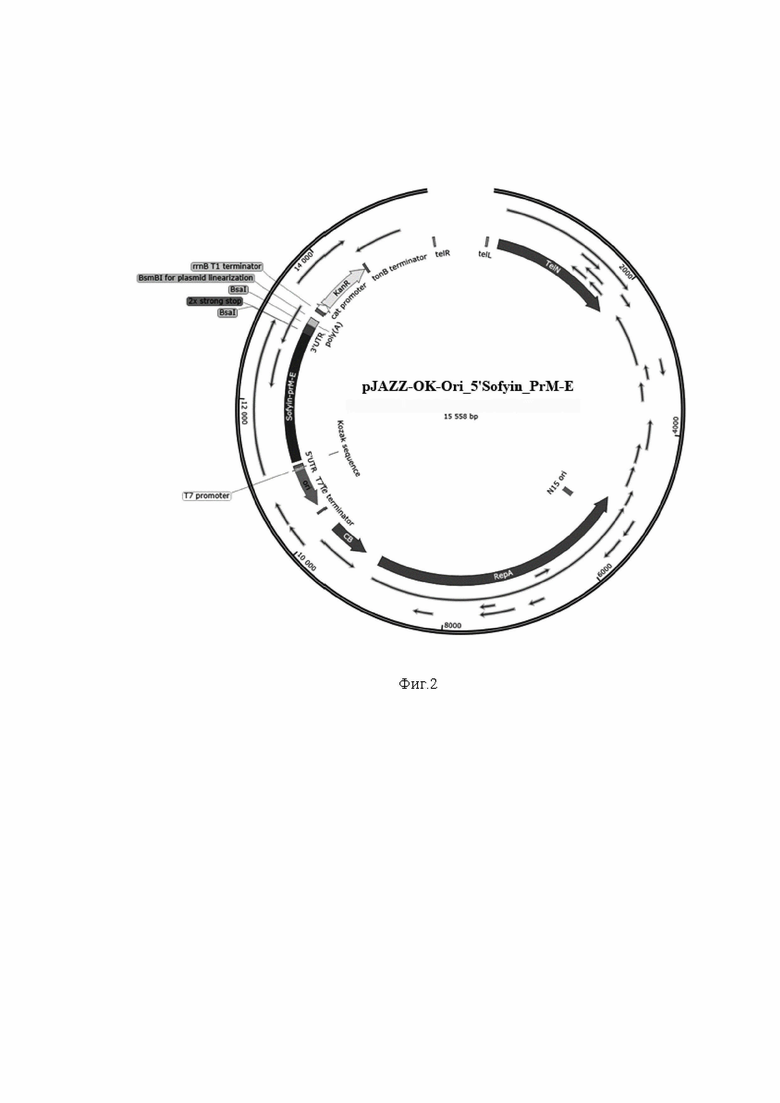
На фиг.2 представлена плазмида pJAZZ-OK, в состав которой включены ориджин репликации, структурные компоненты и гены, кодирующие PrM-E белки ВКЭ для in vitro транскрипции.
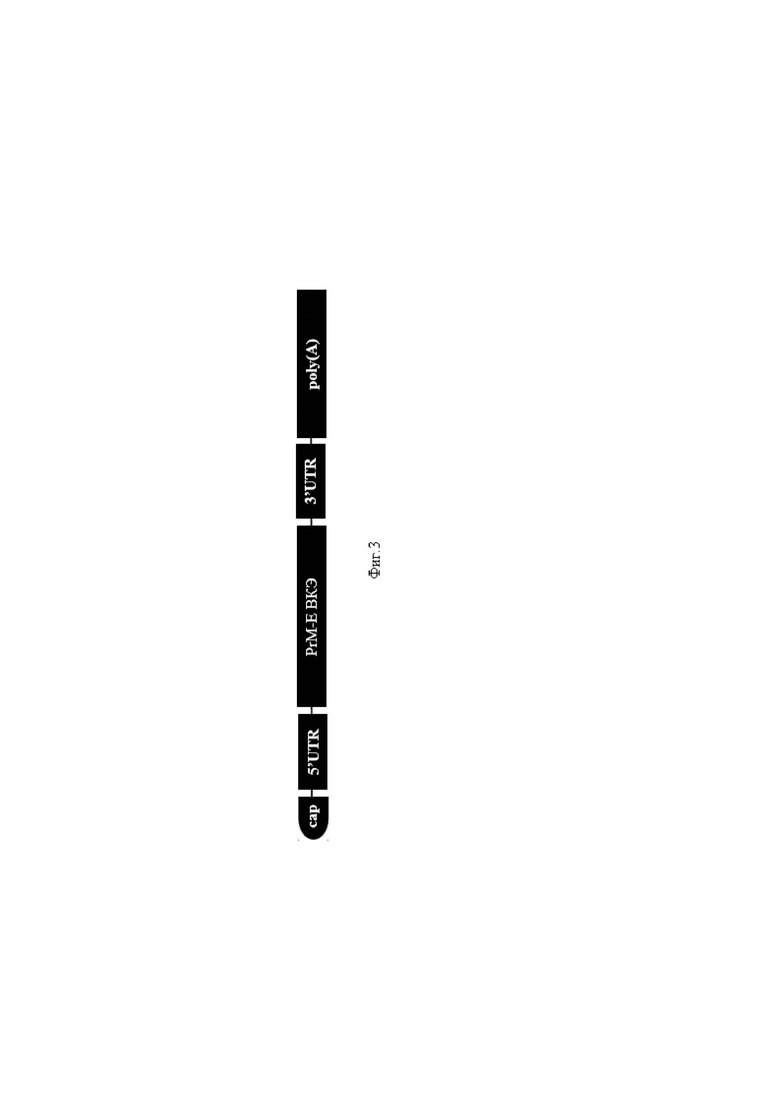
На фиг. 3 представлено строение молекулы мРНК-компонента иммунобиологического средства: cap - модификация 5'-конца мРНК, осуществляемая ферментом гуанилилтрансферазой, 5′ и 3′ UTRs - 5’- и 3’-некодирующие области мРНК соответственно; PrM-E ВКЭ - последовательность, кодирующая PrM-E белки ВКЭ, poly(A) - сегментированный поли-A-хвост.
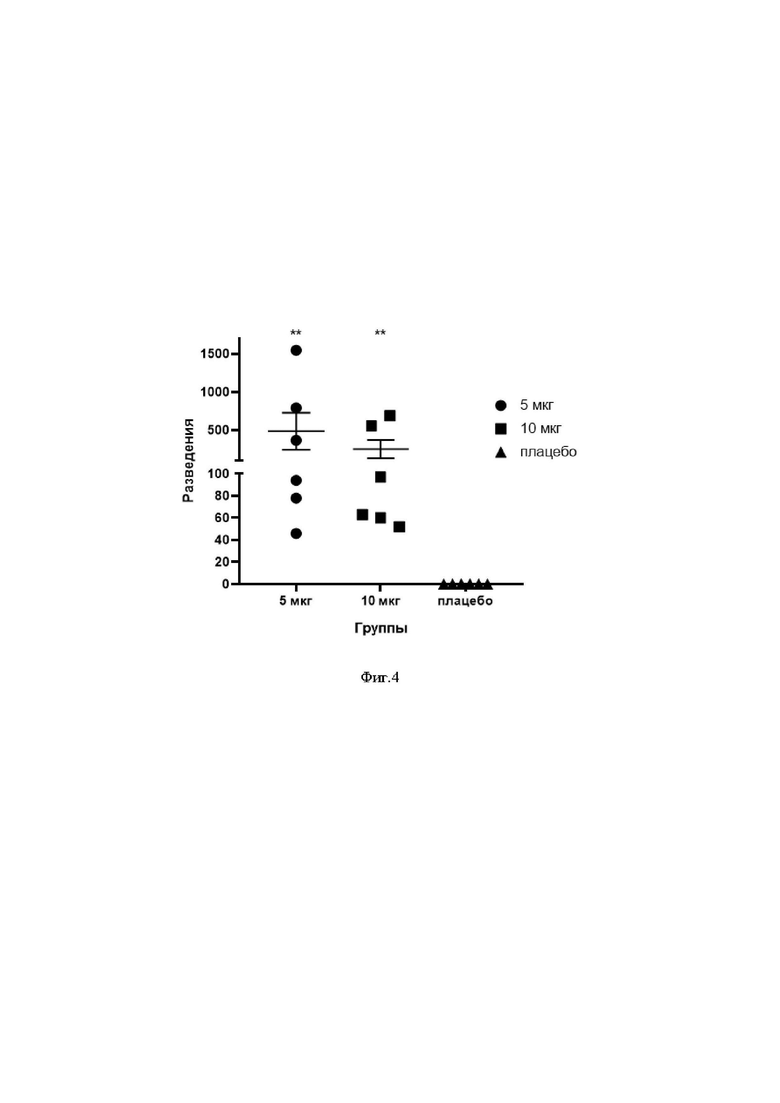
На фиг.4 представлены результаты оценки уровня вируснейтрализующих антител у модельных животных, иммунизированных разными дозами препарата мРНК, кодирующего PrM-E белки ВКЭ. ** - уровень вируснейтрализующих антител статистически достоверно отличается от контрольной группы.
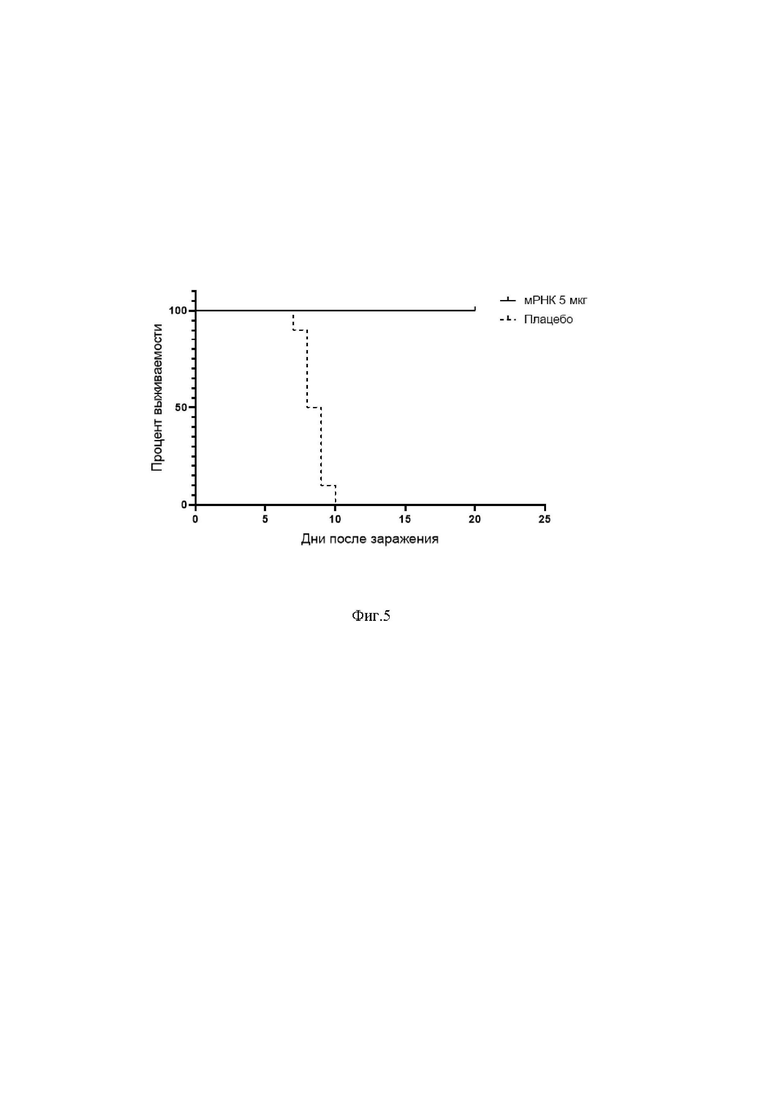
На фиг.5 представлена динамика выживаемости иммунизированных препаратом мРНК животных после экспериментального инфицирования ВКЭ.
Осуществление изобретения
Первым этапом в разработке иммунобиологического средства против коронавируса ВКЭ являлся выбор вакцинного антигена. Проведённый анализ данных литературы показал, что наиболее перспективным антигеном для создания кандидатной мРНК вакцины являются белки PrM и Е оболочки вируса. Это трансмембранный белки гликопротеин I типа, который отвечает за связывание, слияние и проникновение вирусных частиц в клетку. Было показано, что он является индуктором нейтрализующих антител (Zimna М et al, Functional characterization and immunogenicity of a novel vaccine candidate against tick-borne encephalitis virus based on Leishmania-derived virus-like particles. Antiviral Res. 2023 Jan:209:105511. doi: 10.1016/j.antiviral.2022.105511. Epub 2022 Dec 27.).
РНК ВКЭ содержит одну открытую рамку считывания (open reading frame (ORF)), которая транслируется в большой полипротеин, затем полипротеин ко- и посттрансляционно расщепляется клеточными и вирусными протеазами с образованием трех структурных (E, C и M) и семи неструктурных белков, участвующих в цикл репликации вируса внутри клетки (Barrows NJ et al. Biochemistry and molecular Biology of flaviviruses Chem. Rev., 118 (8) (2018), pp. 4448-4482). Два вирусных белка (гликопротеин Е и малый мембранный белок М) играют важную роль в проникновении вируса в клетки-мишени. Гликопротеин оболочки E, как наиболее экспонируемый структурный элемент вирионов, участвует в сборке инфекционных частиц и играет роль в проникновении вируса, поскольку он обеспечивает взаимодействие со специфическими рецепторами клеточной поверхности и индуцирует слияние вирусной оболочки и мембраны клетки-хозяина. Он состоит из трех структурных доменов и трансмембранного домена, который необходим для закрепления белка в липидной мембране. Домен I содержит сайт N-гликозилирования, а пептид слитой петли расположен в домене II (Lattová E et al. Comprehensive N-glycosylation mapping of envelope glycoprotein from tick-borne encephalitis virus grown in human and tick cells. Sci. Rep. [Internet], 10 (1) (2020), pp. 1-10, 10.1038/s41598-020-70082-2). Домен I и II вместе ответственны за димеризацию белка E. Иммуноглобулинподобный домен III является наиболее вероятным кандидатом на взаимодействие с клеточными рецепторами. Также было показано, что при инфекции большинство нейтрализующих антител направлено против домена III гликопротеина Е (Zhang X et al. Structures and functions of the envelope glycoprotein in flavivirus infections. Viruses, 9 (11) (2017), pp. 1-14). Гликопротеин PrM/M представляет собой небольшой мембранный белок, который расщепляется до пептида Pr и белка M, присутствующих в зрелых вирионах во время созревания вирусных частиц. В Pr-фрагменте присутствует один N-гликосайт. Точная роль белка PrM во флавивирусах полностью не определена, но считается, что он является шапероноподобным белком, способствующим правильному сворачиванию гликопротеина E. Этот белок также необходим для pH-зависимых перестроек во время созревания вирионов и защиты от преждевременного слияния с клеточными мембранами (Roby J.A. et al. Post-translational regulation and modifications of flavivirus structural proteins. J. Gen. Virol., 96 (7) (2015), pp. 1551-1569, 2015).
С целью получения мРНК, содержащую гены белков PrM-E ВКЭ, в качестве матрицы для последующей in vitro транскрипции использовали линейную плазмиду, созданную на основе вектора pJAZZ-OK, который представляет собой линейную плазмидную систему на основе генома бактериофага N15. С целью получения плазмиды, пригодной для последующей in vitro транскрипции, провели последовательно несколько клонирований с сборок промежуточных плазмид. Схема последовательного клонинга и сборок представлена на фиг. 1а-г. Схема конечной линейной плазмиды pJAZZ-OK-Ori_5’Sofyin_PrM-E, в состав которой включены ориджин репликации, структурные компоненты и гены, кодирующие PrM-E белки ВКЭ для in vitro транскрипции представлена на фиг 2.
Для последующей in vitro транскрипции и котранскрипционного кэпирования использовали полученную плазмидную ДНК. Полученная мРНК затем была инкапсулирована в липидные наночастицы с использованием процесса самосборки, при котором водный раствор мРНК при pH 4,0 быстро смешивается с раствором липидов, растворенных в этаноле.
Оценку способности упакованной в липидные наночастицы мРНК ВКЭ стимулировать образование вируснейтрализующих антител против клещевого энцефалита проводили с использованием модельных животных. Было установлено, что сыворотка от иммунизированных животных специфически нейтрализует вирус КЭ штамм Софьин в разведении от 1/20 до 1/1500, что позволяет сделать вывод о появлении у животных вируснейтрализующих антител.
мРНК ВКЭ может быть использована для профилактики заболевания вызванного ВКЭ. Осуществление изобретения подтверждается следующими примерами.
Пример 1. С целью получения плазмиды, пригодной для последующей in vitro транскрипции, сначала к источнику репликации ColE1 добавляли сайт рестрикции SmaI на одном конце и вставляли в pJAZZ-OK для увеличения количества плазмиды. Полученную промежуточную плазмиду затем расщепляли SmaI, и собирали с помощью метода Гибсона (Gibson Assembly) продукты полимеразной цепной реакции, содержащие элементы мРНК (промотор Т7, 5'-UTR, 3'-UTR и поли(A) хвост. Затем перед следующей сборкой сайт SmaI был вставлен между 5'- и 3'-UTR, после обработки полученной плазмиды SmaI PrM-E клонировали непосредственно между 5'- и 3'-UTR, также используя сборку Гибсона, с одновременным удалением сайта SmaI. Схема последовательного клонинга и сборок представлена на фиг. 1а-г.
Пример 2. Схема конечной линейной плазмиды pJAZZ-OK-Ori_5’Sofyin_PrM-E, в состав которой включены ориджин репликации, структурные компоненты и гены, кодирующие PrM-E белки ВКЭ для in vitro транскрипции представлена на фиг 2.
Пример 3. ДНК плазмиды pJAZZ-OK-Ori_5’Sofyin_PrM-E для in vitro транскрипции выделяли и очищали из культуры E. coli с использованием набора Plasmid Maxi Kit (QIAGEN). Для in vitro готовили реакционную смесь, содержащую 3 мкг ДНК-матрицы (pJAZZ-OK-Ori_5’Sofyin_PrM-E), 3 мкл РНК-полимеразы Т7 (Biolabmix) и 10×буфера (TriLink), 4 мМ аналога тринуклеотидного кэпа 1 ((3'-OMe-m7G)-5'-ppp-5). ′-(2′-OMeA)pG)) (Biolabmix), 5 мМ m1ΨTP (Biolabmix) и 5 мМ GTP, АТФ и CTP. После 2 ч инкубации при 37°С добавляли 6 мкл ДНКазы I (Thermo Fisher Scientific) еще на 15 мин с последующим осаждением мРНК 2М LiCl (инкубация 1 ч во льду и центрифугирование 10 мин при 14 000 g, 4°). в) и тщательно промывали 80% этанолом. Целостность РНК оценивали электрофорезом в 8% денатурирующем ПААГ. В результате in vitro транскрипции получали мРНК, схематичное изображение которой представлено на фиг.3.
Пример 4. Транскрибированную in vitro мРНК, кодирующую PrM-E белки ВКЭ, инкапсулировали в липидные наночастицы с использованием процесса самосборки, при котором водный раствор мРНК при pH 4,0 быстро смешивается с раствором липидов, растворенных в этаноле. С этой целью липиды растворяли в этаноле при молярном соотношении 46,3:9:42,7:1,6 (ионизируемый липид:дистеароил-ФХ:холестерин:ПЭГ-липид). Липидную смесь объединяли с подкисляющим буфером из 10 мМ цитрата натрия (рН 4,0), содержащим мРНК (0,2 мг/мл) в объемном соотношении 3:1 (водный раствор: этанол), используя устройство Nanoassemblr Spark (Precision NanoSystems). Отношение ионизируемых атомов азота в ионизируемом липиде к числу фосфатных групп в мРНК (соотношение N:P) было установлено равным 6 для каждого состава. Составы диализовали против PBS (рН 7,2) в диализных кассетах Slide-A-Lyzer (Thermo Fisher Scientific) в течение по меньшей мере 24 часов. Составы концентрировали с использованием ультрацентробежных фильтров Amicon (EMD Millipore), затем пропускали через фильтр с размером пор 0,22 мкм и хранили при 4°C (PBS) до использования. Состав тестировали на размер частиц, зета-потенциал, инкапсуляцию РНК. Основные физико-химические характеристики липидных наночастиц представлены в таблице 1.
Таблица 1. Основные физико-химические характеристики частиц
± SD (стандартное отклонение)
Пример 4. Оценку способности полученного препарат мРНК стимулировать образование вируснейтрализующих антител против вируса клещевого энцефалита проводили с использованием инбредных мышей линии BALB/c и двукратной схемы иммунизации.
Предварительно, самок BALB/c внутримышечно иммунизировали препаратом мРНК в дозе 5 мкг или 10 мкг. Вторую (бустирующей) эквивалентную дозу препарата вводили животных спустя две недели после первой иммунизации. Сыворотку крови животных собирали через 2 недели после второй иммунизации, для последующего определения вируснейтрализующей активности. Контрольной группе животных в качестве плацебо вводили физраствор. В таблице 2 представлены группы животных и схема иммунизации.
Таблица 2. Группы животных и схема иммунизации
Реакцию вируснейтрализации проводили в 96-луночных планшетах (SPL, США). Для этого готовили серийные двукратные разведения сывороток животных и смешивали со 100 TCID50 вируса клещевого энцефалита (штамм Софьин). Планшеты инкубировали 1 ч при 37°С. Затем, смесь разведений сывороток с вирусом переносили на монослой клеток Spev. Ингибирование вирус-индуцированного цитопатического эффекта под действием разведений сывороток оценивали спустя 4 дня с помощью МТТ теста. 50%-й нейтрализующий титр сыворотки (НТ50) рассчитывали с помощью регрессионного анализа с использованием GraphPad Prism 9 (GraphPad Software Inc., США).
В реакции нейтрализации установлено, что сыворотка от иммунизированных животных специфически нейтрализует вирус КЭ штамм Софьин со значениями НТ50 от 1/40 до 1/1600, что позволяет сделать вывод о появлении у животных вируснейтрализующих антител. Результаты сравнения уровня вируснейтрализующих антител в сыворотке иммунизированных животных представлены на фиг 4.
Пример 5. Оценку способности полученного препарат мРНК защищать животных от инфицирования ВКЭ проводили с использованием инбредных мышей линии BALB/c и двукратной схемы иммунизации.
Предварительно самок BALB/c внутримышечно иммунизировали 5 мкг препарата мРНК на животное. Второй (бустирующей) дозой препарата животных иммунизировали через 2 недели, в той же дозе. В контрольной группе в качестве плацебо для иммунизации использовали физраствор. В таблице 3 представлены группы животных и схема иммунизации.
Таблица 3. Группы животных и схема иммунизации
Через 4 недели после первой иммунизации животных внутрибрюшинно инфицировали штаммом Софьин ВКЭ в дозе 100 LD50 и наблюдали за состоянием животных в течении 4 недель после инфицирования. За время наблюдения отмечали гибель животных только в группе плацебо. Выживаемость животных в группе мРНК составила 100% (фиг. 5).
Таким образом, можно сделать вывод, что разработанное иммунобиологическое средство может быть использовано для профилактики инфицирования ВКЭ.
| название | год | авторы | номер документа |
|---|---|---|---|
| Иммунобиологическое средство для профилактики заболеваний, вызванных вирусом клещевого энцефалита на основе рекомбинантного вируса рода Flavivirus | 2022 |
|
RU2795800C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PC-NS3, ОБЕСПЕЧИВАЮЩАЯ ИНТЕГРАЦИЮ КОМПЛЕКСА ГЕНОВ C, PRM, E, NS1, NS2A, NS2B, NS3 ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА (ВКЭ) В ГЕНОМ ВИРУСА ОСПОВАКЦИНЫ (ВОВ) И РЕКОМБИНАНТНЫЙ ШТАММ ВИРУСА ОСПОВАКЦИНЫ, ЭКСПРЕССИРУЮЩИЙ В КЛЕТКАХ ИММУНИЗИРОВАННОГО ОРГАНИЗМА КОМПЛЕКС ГЕНОВ C, PRM, E, NS1, NS2A, NS2B, NS3 ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА | 1995 |
|
RU2112038C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ТРЕХ ГИБРИДНЫХ БЕЛКОВ ОБОЛОЧКИ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ОПРЕДЕЛЯЮЩИХ ПРИНАДЛЕЖНОСТЬ К СИБИРСКОМУ (DBD2-D3S), ЕВРОПЕЙСКОМУ (DBD2-D3E) И ДАЛЬНЕВОСТОЧНОМУ (DBD2-D3D) ПОДТИПАМ ВИРУСА; РЕКОМБИНАНТНЫЕ ПЛАЗМИДЫ pDBD2-D3S, pDBD2-D3E И pDBD2-D3D; ШТАММЫ-ПРОДУЦЕНТЫ Escherichia coli M15 [pREP4]; ХИМЕРНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2560588C1 |
| Иммунобиологическое средство для индукции комплексного иммунного ответа против вируса SARS-CoV-2 и вируса гриппа и его применение | 2024 |
|
RU2836687C1 |
| Иммунобиологическое средство для индукции иммунного ответа против вируса гриппа и его применение | 2024 |
|
RU2838904C1 |
| Рекомбинантная плазмида pHis6-flagG-protE, обеспечивающая синтез рекомбинантного химерного белка, включающего эпитопы гликопротеина Е вируса клещевого энцефалита и флагеллин G S.typhii и используемого в качестве основы для вакцины против вируса клещевого энцефалита | 2018 |
|
RU2702716C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pSC13D6, СОДЕРЖАЩАЯ ГЕН ОДНОЦЕПОЧЕЧНОГО АНТИТЕЛА ПРОТИВ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3)/pSC13D6 - ПРОДУЦЕНТ ОДНОЦЕПОЧЕЧНЫХ АНТИТЕЛ ПРОТИВ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ОБЛАДАЮЩИХ ВИРУСНЕЙТРАЛИЗУЮЩИМИ СВОЙСТВАМИ | 2008 |
|
RU2378378C1 |
| Иммунобиологическое средство для терапии онкологических заболеваний на основе мРНК вектора. | 2024 |
|
RU2828631C1 |
| ЧЕТЫРЕХВАЛЕНТНАЯ ВЕКТОРНАЯ ВАКЦИНА И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ ДЛЯ ПРОФИЛАКТИКИ ЛИХОРАДКИ ДЕНГЕ | 2023 |
|
RU2812251C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PTLS, СОДЕРЖАЩАЯ ГЕН ОДНОЦЕПОЧЕЧНОГО АНТИТЕЛА ПРОТИВ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI-ПРОДУЦЕНТ ОДНОЦЕПОЧЕЧНЫХ АНТИТЕЛ ПРОТИВ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА | 1998 |
|
RU2158309C2 |
Группа изобретений относится к области биотехнологии, иммунологии и вирусологии. Создано иммунобиологическое средство для профилактики заболеваний, вызванных вирусом клещевого энцефалита, на основе мРНК вируса клещевого энцефалита, содержащей область, кодирующую антигенные премембранный (PrM) и оболочечный (E) белки оболочки вируса клещевого энцефалита. Изобретение расширяет арсенал средств для борьбы с энцефалитом. 2 н. и 3 з.п. ф-лы, 5 ил., 3 табл., 5 пр.
1. Иммунобиологическое средство для профилактики заболеваний, вызванных вирусом клещевого энцефалита, на основе мРНК вируса клещевого энцефалита, содержащей область, кодирующую антигенные премембранный (PrM) и оболочечный (E) белки оболочки вируса клещевого энцефалита.
2. Иммунобиологическое средство по п.1, в котором для синтеза мРНК были использованы гены премембранного (PrM) и оболочечного (E) белков штамма Софьин вируса клещевого энцефалита.
3. Иммунобиологическое средство по пп.1, 2, которое представляет собой кэпированную мРНК вируса клещевого энцефалита.
4. Иммунобиологическое средство по пп.1-3, которое представляет собой упакованную в липидные наночастицы мРНК вируса клещевого энцефалита.
5. Применение иммунобиологического средства по пп.1-4 для получения вакцин для профилактики клещевого энцефалита.
| НУКЛЕИНОВАЯ КИСЛОТА, СОДЕРЖАЩАЯ ИЛИ КОДИРУЮЩАЯ СТРУКТУРУ "СТЕБЕЛЬ-ПЕТЛЯ" ГИСТОНОВ И ПОСЛЕДОВАТЕЛЬНОСТЬ ПОЛИ(А) ИЛИ СИГНАЛ ПОЛИАДЕНИЛИРОВАНИЯ, ДЛЯ ПОВЫШЕНИЯ ЭКСПРЕССИИ КОДИРУЕМОГО ПАТОГЕННОГО АНТИГЕНА | 2013 |
|
RU2639528C2 |
| Реактивный струйный двигатель | 1928 |
|
SU16490A1 |
| WO 2008137163 A1, 13.11.2008. | |||

