ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к биспецифическим антителам, которые связывают карциноэмбриональный антиген CEACAM5 (CEA) человека и CD3ε человека (анти-CEAxCD3 биспецифическое антитело). Кроме того, настоящее изобретение относится к полинуклеотидам, кодирующим такие биспецифические антитела, а также к векторам и клеткам-хозяевам, содержащим такие полинуклеотиды. Изобретение также относится к способам выбора и получения таких антител, и к способам применения таких антител в лечении заболеваний.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Успешное лечение прогрессирующего/метастатического солидного рака, например, рака поджелудочной железы, колоректального рака, рака желудка, рака легких, и так далее, все еще остается сложной задачей. В современной иммунотерапии рака разработаны способы/методики, помогающие иммунным клеткам организма более эффективно атаковать и уничтожать раковые клетки. Например, было разработано несколько методик/способов для усиления атаки опухолевых клеток Т-клетками. Примерами являются ингибиторы иммунных контрольных точек, такие как, например, моноклональные антитела, ингибирующие PD-1/PD-L1, Т-клеточные биспецифические антитела, связывающие опухоль-ассоциированный антиген (ОАА) и CD3 на Т-клетках или CAR-T-клетках. CAR-T-клетки и биспецифические антитела эффективны при гематологических злокачественных новообразованиях и одобрены, например, для лечения В-клеточных злокачественных новообразований или острого лимфоцитарного лейкоза ОЛЛ, но до настоящего времени применение этих методов в терапии прогрессирующего/метастатического солидного рака не привело к реальному успеху. Моноклональные антитела, а также биспецифические антитела, используемые в терапии, могут вызывать различные побочные эффекты. Важной разновидностью токсичности является синдром высвобождения цитокинов (СВЦ), который, например, был обнаружен при терапии алемтузумабом, муромонабом-CD3, ритуксимабом, тоситузумабом и биспецифическим анти-CD19xCD3 антителом блинатумомабом.
Tabernero с соавторами (J Clin Oncol 35, 2017 (suppl. abstr. 3002)) представили на ASCO 2017 данные клинического испытания фазы 1 по применению для пациентов с распространенным/метастатическим колоректальным раком анти-CEAxCD3 биспецифического антитела (RO 6958688, цибисатамаба, смотри ниже) в монотерапии и в сочетании с анти-PD-L1 антителом атезолизумабом. Цибисатамаб имеет так называемый формат 2+1: один Fab-фрагмент связывается с CD3 и два Fab-фрагмента связываются с CEA. Такие антитела описаны, например, в US20140242079 (WO2014131712) и US20140242080 (WO2014131711).
Используемый в настоящем документе термин «TCB2014» означает биспецифическое антитело, связывающее CEA и CD3 в формате 2+1, которое описано в US20140242080 (полное содержание которого включено в настоящий документ посредством ссылки), содержащее в качестве областей CDR области CDR с SEQ ID NO:270-276 и 290-296, приведенные в US20140242080 (смотри также области CDR с SEQ ID NO:4-10 и 24-30 в US20140242079, полное содержание которого включено в настоящий документ посредством ссылки). Используемый в настоящем документе термин «TCB2017» означает молекулу B в формате «2+1 IgG кросс-Fab, инвертированные» с модификациями заряда (обмен VH/VL в связывающем CD3 фрагменте, модификация заряда в CEA-связывающем фрагменте, гуманизированный CEA-связывающий фрагмент), описанную в WO2017055389 (содержание которого включено посредством ссылки в полном объеме), которая содержит в качестве областей CDR области CDR с SEQ ID NO:4-6, 8-10 и 14-19, описанные в WO2017055389.
Структура 2+1 довольно сильно отличается от нативных IgG антител. Структура также содержит искусственные аминокислотные (ак) мосты и две различные тяжелые цепи, объединенные ак последовательностями в Fc-части с помощью технологии «выступ во впадину» (смотри, например, US6737056, WO2013055958). Такие биспецифические антитела (например, RO6958688, цибисатамаб) являются иммуногенными и, вследствие этого, вызывают образование направленных против лекарственного средства антител (ADA) и потерю воздействия лекарственного средства вследствие нейтрализации лекарственного средства за счет ADA. Melero с соавторами сообщали о 50 или более % пациентов с ADA и о потере воздействия лекарственного средства у 45% пациентов при использовании доз 60-200 мг (Melero et al., ASCO 2017, Abstract 2549 и Poster No. 41, Abstract смотри Journal of Clinical Oncology 35, no. 15_suppl (May 20, 2017) 2549-2549). Потеря воздействия затрудняет контроль над практической терапией и значительно снижает вероятность успеха. Для минимизации образования ADA цибисатамаб, а именно, комбинация цибисатамаба и атезолизумаба, проходит клинические испытания в сочетании после предварительного лечения анти-CD20 антителом обинутузумабом (смотри ClinicalTrials.gov Trial NCT03866239). Предварительное лечение проводят для истощения В-клеток у пациентов с метастатическими колоректальными карциномами. Истощение В-клеток приводит к снижению уровня иммуноглобулинов у пациента и, следовательно, потенциальных ADA, но в то же время это приводит к ослаблению иммунной системы.
MEDI-565 (AMG211), следующее анти-CEAxCD3 биспецифическое антитело, одноцепочечное антитело, прошло стадию клинической разработки, результаты опубликованы (смотри, например, M. Pishvaian et al. Clin Colorectal Cancer. 2016 DEC; 15(4) 345-351). Сообщено о завершении исследования NCT01284231 (ClinicalTrials.gov), за последние годы не было начато ни одного нового исследования. Это одноцепочечное биспецифическое антитело (два scFv, соединенные ак линкером) имеет крайне низкий период полувыведения из организма от 2,2 до 6,5 часов (Pishvaian et al. Clin Colorectal Cancer. 2016 DEC; 15(4) 345-351) (включено в настоящий документ посредством ссылки).
Настоящее изобретение относится к менее иммуногенному анти-CEAxCD3 биспецифическому антителу с высокой эффективностью. Такое антитело имеет общую тяжелую цепь и в одном варианте осуществления легкую цепь каппа в CEA-связывающем фрагменте и легкую цепь лямбда в CD3-связывающем фрагменте.
О концепции использования общей тяжелой цепи для получения биспецифических антител упомянуто в Fischer et al., Nature Communications 6 (2015): 6113. https://doi.org/10.1038/ncomms7113 и Magistrelli G. et al., MABS 9 (2017) 231-239. Каппа-лямбда биспецифические антитела описаны, например, в WO2014087248 (включенном в настоящий документ посредством ссылки в полном объеме). Их структура практически неотличима от структуры естественного IgG, следствием чего является отсутствие, или в минимальной степени, образование ADA и, таким образом, низкая или минимальная потеря воздействия. Последовательность общей вариабельной области тяжелой цепи VH и последовательность VL анти-huCD3 1A4 по изобретению приведены в WO2019175658 (US2019/0284297) (включенном в настоящий документ посредством ссылки в полном объеме).
Как упомянуто выше, в WO2017055389 описаны анти-CEAxCD3 биспецифические антитела в формате 2+1, но связывающие иной домен, чем цибисатамаб. Одно из этих антител (RO7172508 или RG 6123) было протестировано в клиническом исследовании у пациентов с местнораспространенными и/или метастатическими CEA-положительными солидными опухолями (ClinicalTrials.gov; смотри RO7172508), а также с предварительным лечением обинутузумабом и в сочетании с атезолизумабом. Согласно описанию клинического исследования на сайте ClinicalTrials.gov, для некоторых когорт уровень сывороточного CEA (растворимого CEACAM5, sCEA) у пациентов, подлежащих лечению, должен был быть ниже определенного порога, чтобы они имели право на лечение, это позволяет предположить, что более высокий уровень растворимого CEACAM5 может снижать эффективность этого анти-CEAxCD3 биспецифического антитела. Для антител по настоящему изобретению характерно минимальное влияние растворимого CEA на эффективность уничтожения ими опухолевых клеток.
Растворимый CEACAM5 является признанным опухолевым маркером. Уровни sCEA в плазме пациентов с раком могут превышать 1000 нг/мл, в то время как концентрации в плазме у здоровых людей составляют менее 10 нг/мл (например, Sandler B. et al. Anticancer Res 1999, 19(5B), 4229-33). Растворимый CEACAM5, таким образом, может конкурировать с мембраносвязанным CEA, находящимся на опухолевых клетках, за связывание с терапевтическими анти-CEA антителами и анти-CEA биспецифическими антителами, потенциально приводя к снижению уровней анти-CEA антитела или анти-CEAxCD3 антитела. TCB2017 и TCB2014 (смотри выше) были протестированы авторами изобретения in vitro в анализе опосредованного T-клетками лизиса CEA-положительных опухолевых клеток в присутствии растворимого CEA. Было обнаружено, что добавление sCEA в аналитическую смесь приводит к смещению кривой лизиса и, следовательно, значения EC50 для TCB2014 и TCB2017 к более высоким концентрациям, это указывает на то, что как TCB2014, так и TCB2017, в значительной степени связывают sCEA.
Семейство CEA человека включает 29 генов, из которых 18 экспрессируются: 7 принадлежат к подгруппе CEA, и 11 к подгруппе специфических для беременности гликопротеинов. Считается, что некоторые члены подгруппы CEA обладают свойствами клеточной адгезии. CEACAM5 экспрессируется не только клетками колоректального рака, но также клетками рака поджелудочной железы, рака желудка, рака легких и рака других видов. Считается, что CEACAM5 играет определенную роль во врожденном иммунитете (Hammarström S., Semin. Cancer Biol. 9(2):67-81 (1999)). Карциноэмбриональный антиген 5 (CEA, CEACAM5 или CD66e; UniProtKB - P06731) является членом семейства родственных карциноэмбриональным антигенам молекул клеточной адгезии (семейства CEACAM) и опухоль-ассоциированным антигеном (Gold and Freedman, J Exp. Med., 121:439-462, 1965; Berinstein N. L., J Clin Oncol., 20:2197-2207, 2002). Многие моноклональные антитела были получены против CEACAM5 в исследовательских целях, в качестве диагностических средств и в терапевтических целях (смотри, например, WO2012117002). Члены семейства карциноэмбриональных антигенов (CEACAM) широко экспрессируются и, в зависимости от ткани, способны регулировать различные функции, включая развитие опухоли, подавление опухоли, ангиогенез и активацию нейтрофилов. Четыре члена этого семейства, CEACAM1, CEACAM3, CEACAM6 и CEACAM8, экспрессируются и представлены в изобилии на человеческих нейтрофилах (http://www.proteinatlas.org). Учитывая механизм действия анти-CEAxCD3 биспецифических антител, перекрестная реактивность с другими CEACAM может приводить к истощению важных циркулирующих популяций здоровых клеток. Например, перекрестная реактивность с CEACAM8, который экспрессируется нейтрофилами или гемопоэтическими стволовыми клетками, может приводить к истощению таких клеточных популяций. Изобретение относится к анти-CEAxCD3 биспецифическому антителу с низкой перекрестной реактивностью с одним или более членами семейства CEACAM, CEACAM1, CEACAM3, CEACAM4, CEACAM6, CEACAM7, CEACAM8, CEACAM16, CEACAM18, CEACAM19, CEACAM20 и CEACAM21.
Мышиное моноклональное анти-CEACAM5 антитело PR1A3 было получено путем слияния клеток миеломы NS1 (P3/NS 1/I-Ag-4-1) с клетками селезенки мышей, иммунизированных нормальным эпителием толстой кишки. В публикации Richman P. I. and Bodmer W. F., Int. J. Cancer, 39:317-328, 1987 описано мышиное моноклональное антитело PR1A3. Картирование эпитопов PR1A3 показало, что антитело направлено на B3-домен и GPI-якорь молекулы CEA (Durbin H. et al., Proc. Natl. Acad. Sci. USA, 91:4313-4317, 1994). Эпитоп, который связывает PR1A3, представляет собой конформационный эпитоп, а не линейный эпитоп (Stewart et al., Cancer Immunol. Immunother., 47 (1999) 299-06). Гуманизированные антитела PR1A3 (hPR1A3) описаны, например, в публикации Conaghhan P. J., et al., Br. J. Cancer, 98 (2008)1217-1225 и WO2012117002. CEA-связывающий фрагмент, используемый в TCB2014 (называемый CH1A1A), является гуманизированным, с созревшей аффинностью, генетически модифицированным для стабильности вариантом, полученным из антитела PR1A3. M. Bacac et.al., Clin. Cancer Research 22(13);3286-97 (2016), Conaghan P, et al., Br J Cancer 2008;98:1217-25, и Durbin H, et al. Proc Natl Acad Sci USA 1994;91:4313-7).
Способ лечения рака сочетанием антагониста оси PD-1 человека и биспецифического анти-CEAxCD3 антитела описан в WO2017118657, и клинические результаты были опубликованы в материалах конференции ASCO 2017 года (Tabernero et al., J Clin Oncol 35, 2017 (suppl. abstr. 3002)). Способ лечения опухолей путем введения антагонистов иммунных контрольных точек, связывающих две или более разных мишеней пути иммунных контрольных точек, и перенаправляющего T-клетки средства, связывающего CEA и T-клеточный поверхностный антиген, описан в WO2015112534. Конъюгат, состоящий из однодоменного анти-CEACAM6 антитела и уриназы, в настоящее время проходит клинические испытания (NCT02309892; WO2016116907). Антитело класса I, связывающее CEACAM5, CEACAM6 и гранулоциты, описано в US20110064653. Биспецифические антитела, содержащие первую полипептидную цепь и вторую полипептидную цепь, ковалентно связанные друг с другом, описаны в WO2018053328.
Анти-CD3ε антителом, описанным в документах современного уровня техники, является SP34 (Yang SJ, The Journal of Immunology (1986) 137; 1097-1100). SP34 взаимодействует с CD3 как приматов, так и человека. SP34 доступно от компании BD Biosciences. Еще одним анти-CD3 антителом, описанным в документах современного уровня техники, является UCHT-1 (смотри WO2000041474). Еще одним анти-CD3 антителом, описанным в документах современного уровня техники, является BC-3 (Институт исследования рака имени Фреда Хатчинсона; используется в испытаниях I/II фазы у пациентов с РТПХ, Anasetti et al., Transplantation 54: 844 (1992)). SP34 отличается от UCHT-1 и BC-3 тем, что SP-34 узнает эпитоп, присутствующий исключительно на ε-цепи CD3 (смотри Salmeron et al., (1991) J. Immunol. 147: 3047), в то время как UCHT-1 и BC-3 узнают эпитоп, в формировании которого участвуют цепи ε и γ. Анти-CD3 антитела также описаны в WO2007042261, WO2008119565, WO2008119566, WO2008119567, WO2010037836, WO2010037837, WO2010037838 и US8236308. Биспецифические антитела, содержащие связывающий фрагмент, специфический для CEA, и связывающий фрагмент, специфический для CD3ε, например, описаны в US20140242079, WO2007071426, WO2013012414, WO2015112534, WO2017118675 и WO2017055389. Анти-CD3 антитело, содержащее последовательности второго связывающего фрагмента антитела по изобретению, описаны в US62/643095 и PCT/US2019/000232, полное содержание которых включено в настоящий документ посредством ссылки. В US2012321626 описан мультиспецифический слитый белок Fab, содержащий Fab-фрагмент, который связывает N-конец эпсилон-цепи CD3. В WO2018199593 описаны биспецифические антитела, которые связывают HER3 и CD3.
Как уже упоминалось выше, результаты первых клинических испытаний с T-клеточными биспецифическими антителами против ОААxCD3 (ОАА=опухоль-ассоциированный антиген) у пациентов с прогрессирующими солидными опухолями не оправдали ожиданий, но недавно были опубликованы предварительные результаты исследования фазы 1 для анти-CEAxCD3 биспецифического антитела цибисатамаба (RO6958688, смотри, например, Bacac et al. Clin. Cancer Res., 22(13), 3286-97 (2016); и US20140242079), которое при использовании у пациентов с прогрессирующим колоректальным раком в монотерапии и в сочетании с ингибированием PD-L1 привело к частичным ответам и стабилизации заболевания (J.Tabernero et.al., J. Clin. Oncol. 35, 2017 (suppl. Abstr. 3002)). Другой подход для получения лучших результатов может заключаться в добавлении к T-клеточным биспецифическим антителам не только ингибитора оси иммунной контрольной точки PD-1, но также в добавлении других ингибиторов иммунных контрольных точек или агонистов. Но пока считается, что нет перспективных клинических данных для такого комбинированного подхода.
Ограниченная доступность Т-клеток в прогрессирующих солидных опухолях, безусловно, является важным механизмом, ограничивающим эффективность, достижимую при использовании Т-клеточных биспецифических антител плюс ингибиторов оси PD-1.
Вместо добавления к сочетанию Т-клеточного биспецифического антитела и ингибитора оси PD-1 другого терапевтического средства с целью перенаправления Т-клеток на опухолевые клетки прогрессирующих солидных опухолей, более успешным может оказаться добавление терапевтического средства, перенаправляющего на опухолевые клетки другие иммунные клетки, в частности, макрофаги или макрофаги и клетки - естественные киллеры (NK).
Настоящее изобретение относится к новым анти-CEAxCD3 биспецифическим антителам, которые разработаны таким образом, что их можно применять параллельно с анти-CEAxCD47 биспецифическими антителами для перенаправления макрофагов, а также NK-клеток, на CEA-экспрессирующие солидные опухоли. Совместная атака Т-клеток, макрофагов и NK-клеток, направленная на СЕА-экспрессирующие опухоли, открывает широкие возможности для достижения превосходной эффективности/уничтожения и фагоцитоза СЕА-экспрессирующих опухолевых клеток.
Неутешительные результаты применения CAR-T-клеток при солидных опухолях могут иметь простое объяснение - количество CAR-T-клеток, проникающих в солидную опухоль и распределяющихся в ней, просто является недостаточным. Ситуация, безусловно, иная в случае большинства гематологических злокачественных новообразований; CAR-T-клетки имеют хороший доступ к опухолевым клеткам, что объясняет высокую эффективность при этих злокачественных новообразованиях в сравнении с неутешительной эффективностью при солидных опухолях. Кроме того, CAR-T-клетки могут быть сильно подавлены опухолевым микроокружением (TME) солидных опухолей, которое в большинстве случаев является в высокой степени иммуносупрессивным.
Настоящее изобретение относится к новым биспецифическим анти-CEAxCD3 антителам, отличающимся высокой эффективностью, слабым влиянием sCEA на эффективность, низкой или отсутствующей перекрестной реактивностью с другими CEACAM, кроме CEACAM5 (= CEA), и, следовательно, пониженной токсичностью, низкой иммуногенностью, возможностью применения в комбинированной терапии с анти-CEAxCD47 антителами, а также обладающим ценными фармакокинетическими свойствами.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном варианте осуществления изобретение относится к биспецифическому антителу (далее также называемому «анти-CEAxCD3 бсАт» или «анти-CEAxCD3 биспецифическое антитело»), содержащему первый связывающий фрагмент, специфически связывающий CEACAM5 человека (далее также называемый «анти-CEA»), и второй связывающий фрагмент, специфически связывающий CD3ε человека (далее также называемый «анти-CD3»).
В одном варианте осуществления биспецифическое антитело отличается тем, что указанное антитело является одновалентным для первого связывающего фрагмента и одновалентным для второго связывающего фрагмента.
В одном варианте осуществления биспецифическое антитело отличается тем, что каркасные последовательности константной и вариабельной областей являются человеческими.
В одном варианте осуществления биспецифическое антитело отличается тем, что каждый из первого и второго связывающих фрагментов содержит тяжелую цепь иммуноглобулина и легкую цепь иммуноглобулина.
В одном варианте осуществления биспецифическое антитело имеет первый связывающий фрагмент, содержащий тяжелую цепь, и второй связывающий фрагмент, содержащий тяжелую цепь, где тяжелая цепь в каждом связывающем фрагменте является одной и той же (то есть, общей тяжелой цепью). В одном варианте осуществления вариабельная область общей тяжелой цепи содержит в качестве областей CDR CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4. В одном варианте осуществления вариабельная область общей тяжелой цепи содержит SEQ ID NO:1. В одном варианте осуществления константная область общей тяжелой цепи содержит SEQ ID NO:30. В одном варианте осуществления общая тяжелая цепь содержит SEQ ID NO:43. В одном варианте осуществления общая тяжелая цепь содержит SEQ ID NO:44. В одном варианте осуществления общая тяжелая цепь содержит SEQ ID NO:45.
В одном варианте осуществления биспецифическое антитело отличается содержанием в первом связывающем фрагменте и втором связывающем фрагменте в качестве тяжелой цепи общей тяжелой цепи, содержанием в первом связывающем фрагменте в качестве легкой цепи легкой цепи каппа, и во втором связывающем фрагменте в качестве легкой цепи легкой цепи лямбда. В одном варианте осуществления легкая цепь второго связывающего фрагмента содержит SEQ ID NO:28, и тяжелая цепь второго связывающего фрагмента содержит SEQ ID NO:45 (например, полученные из AB1 и AB-1L3-1/N биспецифические антитела, такие как AB13L3-1/N, AB14L3-1/N, AB15L3-1/N, AB17L3-1/N, AB20L3-1/N, AB54L3-1/N, AB60L3-1/N, AB66L3-1N, AB71L3-1/N, AB72L3-1/N и AB73L3-1/N; для последовательностей CDR и VL смотри список последовательностей).
AB13,14,15, и так далее, обозначают первый связывающий фрагмент (плечи анти-CEACAM5 антитела) и L3-1 обозначает второй связывающий фрагмент (плечо анти-CD3 антитела, также называемое 1A4) биспецифических антител по настоящему изобретению. Любое ABXX анти-CEA плечо может быть объединено с L3-1 анти-CD3 плечом, с образованием биспецифического антитела: например, ABXXL3-1 обозначает анти-CEAxCD3 биспецифическое антитело по изобретению, содержащее Fc-фрагмент чIgG1 ДТ; ABXXL3-1/D обозначает анти-CEAxCD3 биспецифическое антитело по изобретению, содержащее Fc-фрагмент чIgG1, имеющий мутации L234A+L235A; ABXXL3-1/N обозначает анти-CEAxCD3 биспецифическое антитело по изобретению, содержащее Fc-фрагмент чIgG1, имеющий мутации L234A+L235A+P329A.
В одном варианте осуществления биспецифическое антитело отличается содержанием в первом связывающем фрагменте и втором связывающем фрагменте в качестве тяжелой цепи общей тяжелой цепи, содержанием в первом связывающем фрагменте в качестве вариабельной области легкой цепи область лямбда-типа и в качестве константной области легкой цепи область каппа-типа («легкая цепь гибридного формата»), и во втором связывающем фрагменте в качестве легкой цепи легкую цепь лямбда (например, L3-1AB8 H-CK5/D, смотри Фигуру 2 и описание Фигуры 2).
В одном варианте осуществления биспецифическое антитело отличается содержанием в первом связывающем фрагменте и втором связывающем фрагменте в качестве тяжелой цепи общей тяжелой цепи, содержанием в первом связывающем фрагменте в качестве вариабельной области легкой цепи области лямбда-типа и в качестве константной области легкой цепи области лямбда-типа, и во втором связывающем фрагменте в качестве вариабельной области легкой цепи области лямбда-типа и в качестве константной области легкой цепи области каппа-типа («легкая цепь гибридного формата»); например, AB8L3-1 H-CK5/D.
Биспецифические антитела по изобретению проявляют низкую связывающую/перекрестную реактивность с членами семейства CEACAM, отличными от CEACAM5. В одном варианте осуществления биспецифическое антитело отличается тем, что значение СИФ для связывания клеток PEAKrapid (ATCC® CRL-2828™), экспрессирующих CEACAM, выбранный из группы, состоящей из CEACAM1, CEACAM3, CEACAM4, CEACAM6, CEACAM7, CEACAM8, CEACAM16, CEACAM18, CEACAM19, CEACAM20 и CEACAM21, не более чем в 2 раза выше в сравнении со значением СИФ для связывания клеток PEAK ДТ (то есть, не трансфицированных клеток PEAK) в одних и тех же экспериментальных условиях. В одном варианте осуществления биспецифическое антитело отличается тем, что значение СИФ для связывания клеток PEAKrapid, экспрессирующих CEACAM, выбранный из группы, состоящей из CEACAM1, CEACAM3, CEACAM4, CEACAM6 и CEACAM8, не более чем в 2 раза выше в сравнении со значением СИФ для связывания клеток PEAK ДТ в одних и тех же экспериментальных условиях. В одном варианте осуществления биспецифическое антитело отличается тем, что значение СИФ для связывания клеток PEAKrapid, экспрессирующих CEACAM8, не более чем в 2 раза выше в сравнении со значением СИФ для связывания клеток PEAK ДТ в одних и тех же экспериментальных условиях. Экспериментальный метод трансфекции клеток PEAK и количественного определения связывания антител с этими клетками PEAK описан в примерах 1 и 5.
В одном варианте осуществления биспецифическое антитело связывает клетки MKN-45 (DSMZ No: ACC 409) со значением EC50, составляющим от 0,5 нМ до 50 нМ. В одном варианте осуществления биспецифическое антитело связывает клетки MKN-45 со значением EC50, составляющим от 0,5 нМ до 30 нМ. В одном варианте осуществления биспецифическое антитело по изобретению отличается тем, что значение СИФ для связывания клеток MKN-45 при концентрации 200 нМ, 1000 нМ и 5000 нМ по меньшей мере в два раза больше значения СИФ, полученного для TCB2014. Анализ связывания описан в примере 7a. В одном варианте осуществления значение EC50 для уничтожения опухолевой линии клеток MKN-45, измеренное в анализе с использованием человеческих МКПК, для биспецифических антител по настоящему изобретению на 40% или более ниже, чем значение EC50, измеренное для TCB2014. В одном варианте осуществления значение EC50 для уничтожения опухолевой линии клеток LS-174T, измеренное в анализе с использованием человеческих МКПК, для биспецифических антител по настоящему изобретению на 40% или более ниже, чем значение EC50, измеренное для TCB2014.
В одном варианте осуществления биспецифическое антитело уничтожает клетки LS-174T в анализе с использованием человеческих МКПК зависимым от концентрации образом со значением EC50, составляющим 0,01-10 нМ. В одном варианте осуществления биспецифическое антитело уничтожает клетки LS-174T в анализе с использованием человеческих МКПК зависимым от концентрации образом со значением EC50, составляющим 0,01-1 нМ.
Анализ для количественного определения T-клеточного перенаправленного лизиса/уничтожения CEA-положительных клеток описан в примере 8.
В одном варианте осуществления биспецифическое антитело отличается тем, что значение EC50 в том же анализе (уничтожение CEA-положительных опухолевых клеток LS-174T) не увеличивается более чем в 20 раз, в одном варианте осуществления не более чем в 15 раз, и в одном варианте осуществления не более чем в 10 раз в присутствии 5 мкг/мл растворимого CEACAM5, в сравнении со значением EC50 для лизиса без растворимого CEACAM5 в одних и тех же экспериментальных условиях.
В другом варианте осуществления биспецифическое антитело отличается тем, что значение EC50 в том же анализе не увеличивается более чем в 10, в одном варианте осуществления не более чем в 5 раз в присутствии 1 мкг/мл растворимого CEACAM5, в сравнении со значением EC50 для лизиса без растворимого CEACAM5 в одних и тех же экспериментальных условиях.
В одном варианте осуществления биспецифическое антитело отличается тем, что указанное биспецифическое антитело ингибирует рост объема опухоли в модели HPAF-II до дня 18 на 25% или более, в сравнении с ростом объема опухоли в группе введения растворителя в одних и тех же экспериментальных условиях. В одном варианте осуществления биспецифическое антитело отличается тем, что указанное биспецифическое антитело ингибирует рост объема опухоли в модели HPAF-II до дня 18 аналогичным, и статистически не отличающимся образом, в сравнении с TCB2014 в одних и тех же экспериментальных условиях. Мышиная модель опухоли описана в примере 9a.
В одном варианте осуществления биспецифическое антитело имеет в каждой субъединице Fc-домена аминокислотные замены, уменьшающие связывание с активирующим Fc-рецептором и/или снижающие эффекторную функцию, где указанные аминокислотные замены представляют собой L234A и L235A и/или замену P329, выбранную из группы, состоящей из P329A, P329G и P329R (нумерация Kabat EU-индекс). В одном варианте осуществления биспецифическое антитело имеет в каждой субъединице Fc-домена аминокислотные замены L234A и L235A, и P329A (нумерация Kabat EU-индекс). L234A и L235A (LALA) означает, что аминокислота лейцин в положении 234/235 заменена на аланин. P329A (PA) означает, что аминокислота пролин в положении 329 заменена на аланин.
В одном варианте осуществления биспецифическое антитело содержит общую тяжелую цепь. В одном варианте осуществления биспецифическое антитело содержит общую тяжелую цепь, содержащую в качестве областей CDR CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4. В одном варианте осуществления биспецифическое антитело содержит во втором связывающем фрагменте область легкой цепи, содержащую в качестве областей CDR CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
В одном варианте осуществления биспецифическое антитело содержит в первом связывающем фрагменте в качестве константной области легкой цепи область с SEQ ID NO:39. В одном варианте осуществления биспецифическое антитело содержит в первом связывающем фрагменте в качестве константной области легкой цепи область с SEQ ID NO:41. В одном варианте осуществления биспецифическое антитело содержит в первом связывающем фрагменте в качестве константной области легкой цепи область с SEQ ID NO:58. В одном варианте осуществления биспецифическое антитело содержит общую тяжелую цепь с SEQ ID NO:43 или общую тяжелую цепь с SEQ ID NO:44, или общую тяжелую цепь с SEQ ID NO:45. В одном варианте осуществления биспецифическое антитело содержит в первом связывающем фрагменте в качестве константной области легкой цепи область с SEQ ID NO:39 и общую тяжелую цепь с SEQ ID NO:45.
В одном варианте осуществления биспецифическое антитело содержит во втором связывающем фрагменте в качестве легкой цепи легкую цепь, выбранную из группы, состоящей из SEQ ID NO:25, 26, 27, 28 и 29, или из группы легких цепей гибридного формата (LC) с SEQ ID NO:67, 68, 69, 70 и 71.
В одном варианте осуществления биспецифическое антитело содержит в первом связывающем фрагменте в качестве константной области легкой цепи область с SEQ ID NO:39, общую тяжелую цепь с SEQ ID NO:45, и во втором связывающем фрагменте легкую цепь с SEQ ID NO:28.
В одном варианте осуществления биспецифическое антитело конкурирует с анти-CEA антителом, выбранным из группы, состоящей из анти-CEA антитела, содержащего в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:48 и 49 (анти-CEA антитело MEDI), содержащего в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:46 и 47 (антитело SM3E), содержащего в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:56 и 57 (лабетузумаб (Lab)), содержащего в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:50 и 51 (SAR), содержащего в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:54 и 55 (T86.66) и содержащего в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:52 и 53 (CH1A1A). Смотри также Фигуру 1 и пример 5c).
Примерами антител, которые можно использовать в качестве VL или CL-областей для CEA в биспецифических антителах по изобретению, и конкурирующих с MEDI за связывание рекомбинантного CEA, являются анти-CEA антитело AB1 и антитела, полученные путем оптимизации направленности из AB1 (смотри пример 11 для экспериментального метода). Анти-CEA антитела AB13, 14, 15, 17, 20, 54, 60, 66, 71, 72, 73 и соответствующие биспецифические анти-CEAxCD3 антитела AB13L3-1, AB14L3-1, AB15L3-1, AB17L3-1, AB20L3-1, AB54L3-1, AB60L3-1, AB66L3-1, AB71L3-1, AB72L3-1 и AB73L3-1 конкурируют так же, как антитело AB1, соответствующее AB1L3-1. Примерами антител, которые можно использовать в качестве VL или CL-областей для CEA в биспецифических антителах по изобретению, и конкурирующих с SM3E за связывание рекомбинантного CEA, являются анти-CEA антитело AB8 и антитела, полученные путем олигонуклеотид-направленного мутагенеза из AB8 с использованием вырожденных олигонуклеотидов. Примерами антител, которые можно использовать в качестве VL или CL-областей для CEA в биспецифических антителах по изобретению, и конкурирующих с T84.66 за связывание рекомбинантного CEA, являются анти-CEA антитело 1B4 и антитела, полученные путем олигонуклеотид-направленного мутагенеза из 1B4 с использованием вырожденных олигонуклеотидов.
Примерами антител, которые можно использовать в качестве VL или CL-областей для CEA в биспецифических антителах по изобретению, но не конкурирующих ни с одним из эталонных антител за связывание рекомбинантного CEA, являются анти-CEA антитело C11 и антитела, полученные путем олигонуклеотид-направленного мутагенеза из C11 с использованием вырожденных олигонуклеотидов.
AB1 представляет собой анти-CEA антитело с HC SEQ ID NO:43 и каппа LC SEQ ID NO:40, закодированное нуклеотидными последовательностями, приведенными в SEQ ID NO:80 и 78, соответственно.
AB8 представляет собой анти-CEA антитело с HC SEQ ID NO:43 и лямбда LC SEQ ID NO:42, закодированное нуклеотидными последовательностями, приведенными в SEQ ID NO:80 и 79, соответственно.
1B4 представляет собой анти-CEA антитело с HC SEQ ID NO:43 и лямбда LC SEQ ID NO:74, закодированное нуклеотидными последовательностями, приведенными в SEQ ID NO:80 и 77, соответственно.
C11 представляет собой анти-CEA антитело с HC SEQ ID NO:43 и каппа LC SEQ ID NO:73, закодированное нуклеотидными последовательностями, приведенными в SEQ ID NO:80 и 76, соответственно.
Легкая цепь для CEA, которую можно использовать в качестве легкой цепи каппа, содержит SEQ ID NO:40. Легкая цепь для CEA, которую можно использовать в качестве легкой цепи каппа, содержит SEQ ID NO:73. Легкая цепь для CEA, которую можно использовать в качестве легкой цепи лямбда, содержит SEQ ID NO:74. Легкая цепь для CEA, которую можно использовать в качестве гибридной легкой цепи каппа, содержит SEQ ID NO:75.
В одном варианте осуществления биспецифическое антитело имеет в каждой субъединице Fc-домена до трех аминокислотных замен, которые уменьшают связывание с активирующим Fc-рецептором и/или эффекторную функцию, где указанные аминокислотные замены представляют собой L234A, L235A и замену P329, выбранную из группы, состоящей из P329A, P329G и P329R (нумерация Kabat EU-индекс). В одном варианте осуществления общая тяжелая цепь антитела по изобретению содержит SEQ ID NO:43, 44 или 45. В одном варианте осуществления общая тяжелая цепь антитела по изобретению содержит SEQ ID NO:45 (L234A, L235A и P329A).
В одном варианте осуществления биспецифическое антитело проявляет одно или более свойств, выбранных из группы, включающей: a) связывание клеток MKN-45 со значением EC50, составляющим от 0,5 нМ до 50 нМ; b1) конкуренцию с анти-CEA антителом, содержащим в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:48 и 49 (MEDI), или b2) конкуренцию с анти-CEA антителом, содержащим в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:46 и 47 (SM3E), или b3) конкуренцию с анти-CEA антителом, содержащим в качестве VL и VH-доменов VL и CH с последовательностями SEQ ID NO:54 и 55 (T84.66), или b4) отсутствие конкуренции с любым из инструментальных антител (смотри пример 5c для инструментальных антител); c) наличие в каждой субъединице Fc-домена аминокислотных замен, которые уменьшают связывание с активирующим Fc-рецептором и/или эффекторную функцию, где указанные аминокислотные замены представляют собой L234A и L235A и замену P329, выбранную из группы, состоящей из P329A, P329G и P329R (нумерация Kabat EU-индекс); d) уничтожение клеток MKN-45, HPAF-II и/или LS-174T в анализе с использованием человеческих МКПК зависимым от концентрации образом со значением EC50, составляющим 0,01-10 нМ.
В одном варианте осуществления биспецифическое антитело проявляет одно или более свойств, выбранных из группы, включающей:
a) связывание клеток MKN-45 со значением EC50, составляющим от 0,5 нМ до 50 нМ,
b) уничтожение клеток MKN-45, HPAF-II или LS-174T в анализе с использованием человеческих МКПК зависимым от концентрации образом со значением EC50, составляющим 0,01-10 нМ,
c) связывание CEACAM5-экспрессирующих клеток PEAK, но без перекрестного реагирования с CEACAM8-экспрессирующими клетками PEAK,
d) EC50 уничтожения в анализе TDCC (пример 8) при использовании опухолевых клеток LS-174T в качестве клеток-мишеней не увеличивается более чем в 5 раз в присутствии 1 мкг/мл sCEA,
e) ингибирование роста опухоли в модели HPAF-II на 25% или более в сравнении с контрольной группой (только растворитель),
f) конкуренцию с анти-CEA антителом, содержащим в качестве VL и VH-доменов VL и VH с последовательностями SEQ ID NO:48 и 49 (MEDI), и
g) наличие в каждой субъединице Fc-домена аминокислотных замен L234A, L235A и P329A (нумерация Kabat EU-индекс).
В одном варианте осуществления биспецифическое антитело проявляет свойства a) - d). В одном варианте осуществления биспецифическое антитело проявляет все свойства из a) - f). В одном варианте осуществления биспецифическое антитело проявляет свойства a) - d) и g). В одном варианте осуществления биспецифическое антитело проявляет свойства a) - d) и f), и g). В одном варианте осуществления биспецифическое антитело проявляет все свойства из a) - g).
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь (cHC) и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит
i) константную область легкой цепи каппа (CL) и вариабельную область легкой цепи (VL), содержащую CDRL1, CDRL2 и CDRL3; CDRL1 с SEQ ID NO:32, CDRL2 с SEQ ID NO:33 и CDRL3 с SEQ ID NO:34, или вариабельную область легкой цепи, полученную из SEQ ID NO:31 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов,
c) второй связывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
d) второй связывающий фрагмент содержит константную область легкой цепи лямбда.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:25, 26, 27, 28 и 29.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи тяжелую цепь, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит вариабельную область легкой цепи (VL), содержащую в качестве областей CDR CDRL1, полученную из SEQ ID NO:32 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов и имеющую до четырех аминокислотных замен, CDRL2, полученную из SEQ ID NO:33 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов и имеющую до четырех аминокислотных замен, CDRL3, полученную из SEQ ID NO:34 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов и имеющую до четырех аминокислотных замен,
c) второй связывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3; CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
Такие биспецифические антитела включают, но не ограничиваются ими, биспецифические анти-CEAxCD3 антитела AB13L3-1, AB14L3-1, AB15L3-1, AB17L3-1, AB20L3-1, AB54L3-1, AB60L3-1, AB66L3-1, AB71L3-1, AB72L3-1 и AB73L3-1.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что:
a) первый связывающий фрагмент содержит вариабельную область тяжелой цепи VH, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит в качестве вариабельной области легкой цепи вариабельную область легкой цепи, содержащую набор CDRL, выбранный из группы, состоящей из
b1) CDRL1 с SEQ ID NO:32, CDRL2 с SEQ ID NO:33 и CDRL3 с SEQ ID NO:34,
b2) CDRL1 с SEQ ID NO:81, CDRL2 с SEQ ID NO:82 и CDRL3 с SEQ ID NO:83,
b3) CDRL1 с SEQ ID NO:84, CDRL2 с SEQ ID NO:85 и CDRL3 с SEQ ID NO:86,
b4) CDRL1 с SEQ ID NO:87, CDRL2 с SEQ ID NO:88 и CDRL3 с SEQ ID NO:89,
b5) CDRL1 с SEQ ID NO:90, CDRL2 с SEQ ID NO:91 и CDRL3 с SEQ ID NO:92,
b6) CDRL1 с SEQ ID NO:93, CDRL2 с SEQ ID NO:94 и CDRL3 с SEQ ID NO:95,
b7) CDRL1 с SEQ ID NO:96, CDRL2 с SEQ ID NO:97 и CDRL3 с SEQ ID NO:98,
b8) CDRL1 с SEQ ID NO:99, CDRL2 с SEQ ID NO:100 и CDRL3 с SEQ ID NO:101,
b9) CDRL1 с SEQ ID NO:102, CDRL2 с SEQ ID NO:103 и CDRL3 с SEQ ID NO:104,
b10) CDRL1 с SEQ ID NO:105, CDRL2 с SEQ ID NO:106 и CDRL3 с SEQ ID NO:107,
b11) CDRL1 с SEQ ID NO:108, CDRL2 с SEQ ID NO:109 и CDRL3 с SEQ ID NO:110, и
b12) CDRL1 с SEQ ID NO:111, CDRL2 с SEQ ID NO:112 и CDRL3 с SEQ ID NO:113, и
c) второй связывающий фрагмент содержит вариабельную область тяжелой цепи VH, содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4, и вариабельную область легкой цепи VL, содержащую CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что:
a) первый связывающий фрагмент содержит вариабельную область тяжелой цепи VH, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит в качестве вариабельной области легкой цепи вариабельную область легкой цепи, содержащую набор CDRL, выбранный из группы, состоящей из
b1) CDRL1 с SEQ ID NO:90, CDRL2 с SEQ ID NO:91 и CDRL3 с SEQ ID NO:92,
b2) CDRL1 с SEQ ID NO:96, CDRL2 с SEQ ID NO:97 и CDRL3 с SEQ ID NO:98,
b3) CDRL1 с SEQ ID NO:99, CDRL2 с SEQ ID NO:100 и CDRL3 с SEQ ID NO:101,
b4) CDRL1 с SEQ ID NO:102, CDRL2 с SEQ ID NO:103 и CDRL3 с SEQ ID NO:104,
b5) CDRL1 с SEQ ID NO:105, CDRL2 с SEQ ID NO:106 и CDRL3 с SEQ ID NO:107, и
b6) CDRL1 с SEQ ID NO:111, CDRL2 с SEQ ID NO:112 и CDRL3 с SEQ ID NO:113, и
c) второй связывающий фрагмент содержит вариабельную область тяжелой цепи VH, содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4, и вариабельную область легкой цепи VL, содержащую CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что:
a) первый связывающий фрагмент содержит вариабельную область тяжелой цепи VH, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит в качестве вариабельной области легкой цепи вариабельную область легкой цепи, содержащую набор CDRL, выбранный из группы, состоящей из
b1) CDRL1 с SEQ ID NO:90, CDRL2 с SEQ ID NO:91 и CDRL3 с SEQ ID NO:92,
b2) CDRL1 с SEQ ID NO:105, CDRL2 с SEQ ID NO:106 и CDRL3 с SEQ ID NO:107, и
b3) CDRL1 с SEQ ID NO:111, CDRL2 с SEQ ID NO:112 и CDRL3 с SEQ ID NO:113, и
c) второй связывающий фрагмент содержит вариабельную область тяжелой цепи VH, содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4, и вариабельную область легкой цепи VL, содержащую CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
В одном варианте осуществления изобретение относится к биспецифическому антителу по изобретению, отличающемуся содержанием
a) в первом связывающем фрагменте вариабельной области тяжелой цепи VH, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:1, и содержащей CDR1 с SEQ ID NO:2, CDR2 с SEQ ID NO:3 и CDR3 с SEQ ID NO:4, и
b) вариабельной области легкой цепи VL, выбранной из группы, состоящей из
b1) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:31, и содержащей CDRL1 с SEQ ID NO:32, CDRL2 с SEQ ID NO:33 и CDRL3 с SEQ ID NO:34,
b2) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:114, и содержащей CDRL1 с SEQ ID NO:81, CDRL2 с SEQ ID NO:82 и CDRL3 с SEQ ID NO:83,
b3) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:115, и содержащей CDRL1 с SEQ ID NO:84, CDRL2 с SEQ ID NO:85 и CDRL3 с SEQ ID NO:86,
b4) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:116, и содержащей CDRL1 с SEQ ID NO:87, CDRL2 с SEQ ID NO:88 и CDRL3 с SEQ ID NO:89,
b5) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:117, и содержащей CDRL1 с SEQ ID NO:90, CDRL2 с SEQ ID NO:91 и CDRL3 с SEQ ID NO:92,
b6) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:118, и содержащей CDRL1 с SEQ ID NO:93, CDRL2 с SEQ ID NO:94 и CDRL3 с SEQ ID NO:95,
b7) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:119, и содержащей CDRL1 с SEQ ID NO:96, CDRL2 с SEQ ID NO:97 и CDRL3 с SEQ ID NO:98,
b8) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:120, и содержащей CDRL1 с SEQ ID NO:99, CDRL2 с SEQ ID NO:100 и CDRL3 с SEQ ID NO:101,
b9) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:121, и содержащей CDRL1 с SEQ ID NO:102, CDRL2 с SEQ ID NO:103 и CDRL3 с SEQ ID NO:104,
b10) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:122, и содержащей CDRL1 с SEQ ID NO:105, CDRL2 с SEQ ID NO:106 и CDRL3 с SEQ ID NO:107, и
b11) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:123, и содержащей CDRL1 с SEQ ID NO:108, CDRL2 с SEQ ID NO:109 и CDRL3 с SEQ ID NO:110, и
b12) вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:124, и содержащей CDRL1 с SEQ ID NO:111, CDRL2 с SEQ ID NO:112 и CDRL3 с SEQ ID NO:113,
c) во втором связывающем фрагменте вариабельной области тяжелой цепи VH, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:1, и содержащей CDR1 с SEQ ID NO:2, CDR2 с SEQ ID NO:3 и CDR3 с SEQ ID NO:4, и вариабельной области легкой цепи VL, имеющей 97%, 98%, 99% или 100% идентичности аминокислотной последовательности с SEQ ID NO:17, и содержащей CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
В одном варианте осуществления изобретение относится к биспецифическому антителу по изобретению, отличающемуся содержанием
a) в первом связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и
b) вариабельной области легкой цепи VL, выбранной из группы, состоящей из
b1) вариабельной области легкой цепи VL с SEQ ID NO:31,
b2) вариабельной области легкой цепи VL с SEQ ID NO:114,
b3) вариабельной области легкой цепи VL с SEQ ID NO:115,
b4) вариабельной области легкой цепи VL с SEQ ID NO:116,
b5) вариабельной области легкой цепи VL с SEQ ID NO:117,
b6) вариабельной области легкой цепи VL с SEQ ID NO:118,
b7) вариабельной области легкой цепи VL с SEQ ID NO:119,
b8) вариабельной области легкой цепи VL с SEQ ID NO:120,
b9) вариабельной области легкой цепи VL с SEQ ID NO:121,
b10) вариабельной области легкой цепи VL с SEQ ID NO:122,
b11) вариабельной области легкой цепи VL с SEQ ID NO:123 и
b12) вариабельной области легкой цепи VL с SEQ ID NO:124, и
c) во втором связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и вариабельной области легкой цепи VL с SEQ ID NO:17.
В одном варианте осуществления изобретение относится к биспецифическому антителу по изобретению, отличающемуся содержанием
a) в первом связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и
b) вариабельной области легкой цепи VL, выбранной из группы, состоящей из
b1) вариабельной области легкой цепи VL с SEQ ID NO:117,
b2) вариабельной области легкой цепи VL с SEQ ID NO:119,
b3) вариабельной области легкой цепи VL с SEQ ID NO:120,
b4) вариабельной области легкой цепи VL с SEQ ID NO:121,
b5) вариабельной области легкой цепи VL с SEQ ID NO:122 и
b6) вариабельной области легкой цепи VL с SEQ ID NO:124, и
c) во втором связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и вариабельной области легкой цепи VL с SEQ ID NO:17.
В одном варианте осуществления изобретение относится к биспецифическому антителу по изобретению, отличающемуся содержанием
a) в первом связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и
b) вариабельной области легкой цепи VL, выбранной из группы, состоящей из
b1) вариабельной области легкой цепи VL с SEQ ID NO:117,
b2) вариабельной области легкой цепи VL с SEQ ID NO:122 и
b3) вариабельной области легкой цепи VL с SEQ ID NO:124, и
c) во втором связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и вариабельной области легкой цепи VL с SEQ ID NO:17.
В одном варианте осуществления изобретение относится к биспецифическому антителу по изобретению, отличающемуся содержанием в первом связывающем фрагменте
a) вариабельной области тяжелой цепи VH с SEQ ID NO:1 и
b) легкой цепи, выбранной из группы, состоящей из
b1) легкой цепи с SEQ ID NO:40
b2) легкой цепи с SEQ ID NO:125,
b3) легкой цепи с SEQ ID NO:126,
b4) легкой цепи с SEQ ID NO:127,
b5) легкой цепи с SEQ ID NO:128,
b6) легкой цепи с SEQ ID NO:129,
b7) легкой цепи с SEQ ID NO:130,
b8) легкой цепи с SEQ ID NO:131
b9) легкой цепи с SEQ ID NO:132,
b10) легкой цепи с SEQ ID NO:133 и
b11) легкой цепи с SEQ ID NO:134 и
b12) легкой цепи с SEQ ID NO:135, и
c) во втором связывающем фрагменте вариабельной области тяжелой цепи VH с SEQ ID NO:1 и легкой цепи с SEQ ID NO:28.
В одном варианте осуществления изобретение относится к биспецифическому антителу по изобретению, отличающемуся содержанием в первом связывающем фрагменте
a) общей тяжелой цепи, выбранной из группы, состоящей из
a1) тяжелой цепи с SEQ ID NO:43,
a2) тяжелой цепи с SEQ ID NO:44 или
a3) тяжелой цепи с SEQ ID NO:45 и
b) легкой цепи, выбранной из группы, состоящей из
b1) легкой цепи с SEQ ID NO:40
b2) легкой цепи с SEQ ID NO:125,
b3) легкой цепи с SEQ ID NO:126,
b4) легкой цепи с SEQ ID NO:127,
b5) легкой цепи с SEQ ID NO:128,
b6) легкой цепи с SEQ ID NO:129,
b7) легкой цепи с SEQ ID NO:130,
b8) легкой цепи с SEQ ID NO:131
b9) легкой цепи с SEQ ID NO:132,
b10) легкой цепи с SEQ ID NO:133
b11) легкой цепи с SEQ ID NO:134 или
b12) легкой цепи с SEQ ID NO:135, и
c) во втором связывающем фрагменте легкой цепи с SEQ ID NO:28.
В одном варианте осуществления биспецифическое антитело по изобретению (AB17L3-1/N) содержит общую тяжелую цепь с SEQ ID NO:45 (/N) и во втором связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:28 (1A4 LC, соответственно, L3-1), и в первом связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:128 (AB17). В одном варианте осуществления биспецифическое антитело по изобретению (AB71L3-1/N) содержит общую тяжелую цепь с SEQ ID NO:45 (/N) и во втором связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:28 (L3-1), и в первом связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:133 (AB71). В одном варианте осуществления биспецифическое антитело по изобретению (AB73L3-1/N) содержит общую тяжелую цепь с SEQ ID NO:45 (/N) и во втором связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:28 (L3-1), и в первом связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:135 (AB73).
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит
i) константную область легкой цепи лямбда (CL) и
ii) вариабельную область легкой цепи (VL), содержащую CDRL1, CDRL2 и CDRL3; CDRL1 с SEQ ID NO:36, CDRL2 с SEQ ID NO:37 и CDRL3 с SEQ ID NO:38, или вариабельную область легкой цепи, полученную из SEQ ID NO:35 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов,
c) второй связывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
d) второй связывающий фрагмент содержит константную область гибридной цепи каппа.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:67, 68, 69, 70 и 71.
В одном варианте осуществления второй связывающий фрагмент может содержать в качестве константной области легкой цепи константную область легкой цепи лямбда с SEQ ID NO:41; в этом случае первый связывающий фрагмент содержит, в одном варианте осуществления, в качестве константной области легкой цепи область гибридной легкой цепи каппа с SEQ ID NO:58. В одном варианте осуществления выбор плеча, содержащего константную область гибридной легкой цепи, основан на общих свойствах бсАт, включая, но без ограничения, стабильность и продуктивность.
В одном варианте осуществления вариабельная область легкой цепи первого связывающего фрагмента получена из SEQ ID NO:35 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов, общая тяжелая цепь содержит SEQ ID NO:45, и вариабельная область общей тяжелой цепи содержит SEQ ID NO:1.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит
i) константную область легкой цепи каппа (CL) и
ii) вариабельную область легкой цепи (VL), содержащую CDRL1, CDRL2 и CDRL3; CDRL1 с SEQ ID NO:64, CDRL2 с SEQ ID NO:65 и CDRL3 с SEQ ID NO:66, или вариабельную область легкой цепи, полученную из SEQ ID NO:63 путем олигонуклеотид-направленного мутагенеза,
c) второй связывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
d) второй связывающий фрагмент содержит константную область легкой цепи лямбда.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:25, 26, 27, 28 и 29.
В одном варианте осуществления изобретения вариабельная область легкой цепи первого связывающего фрагмента получена из SEQ ID NO:63 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов, общая тяжелая цепь содержит SEQ ID NO:45.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит константную область легкой цепи лямбда (CL) и вариабельную область легкой цепи (VL), содержащую CDRL1, CDRL2 и CDRL3; CDRL1 с SEQ ID NO:60, CDRL2 с SEQ ID NO:61 и CDRL3 с SEQ ID NO:62, или вариабельную область легкой цепи, полученную из SEQ ID NO:59 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов,
c) второй связывающий фрагмент содержит вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24,
d) второй связывающий фрагмент содержит константную область гибридной цепи каппа.
В следующем варианте осуществления изобретения второй связывающий фрагмент содержит в качестве константной области легкой цепи константную область легкой цепи лямбда с SEQ ID NO:41; в этом случае первый связывающий фрагмент содержит, в одном варианте осуществления, в качестве константной области легкой цепи константную область гибридной легкой цепи каппа с SEQ ID NO:58. Выбор плеча, содержащего константную область гибридной легкой цепи, основан на общих свойствах конечного бсАт, включая, но без ограничения, стабильность и продуктивность.
В одном варианте осуществления вариабельная область легкой цепи первого связывающего фрагмента получена из SEQ ID NO:59 путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит константную область легкой цепи человеческого типа каппа и вариабельную область легкой цепи человеческого типа каппа, содержащую CDRL1, CDRL2 и CDRL3; CDRL1 с SEQ ID NO:32 с заменой 0, 1, 2, 3 или 4 аминокислот, CDRL2 с SEQ ID NO:33 с заменой 0, 1, 2, 3 или 4 аминокислот и CDRL3 с SEQ ID NO:34 с заменой 0, 1, 2, 3, 4 или 5 аминокислот,
c) второй связывающий фрагмент содержит константную область легкой цепи человеческого типа лямбда и вариабельную область легкой цепи человеческого типа лямбда, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:25, 26, 27, 28 и 29.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит константную область легкой цепи человеческого типа лямбда и вариабельную область легкой цепи человеческого типа лямбда, содержащую CDRL1, CDRL2 и CDRL3, набор CDRL, выбранный из группы, состоящей из CDRL1 с SEQ ID NO:36 с заменой 0, 1, 2, 3, 4 или 5 аминокислот, CDRL2 с SEQ ID NO:37 с заменой 0, 1, 2, 3, 4 или 5 аминокислот и CDRL3 с SEQ ID NO:38, с заменой 0, 1, 2, 3, 4 или 5 аминокислот,
c) второй связывающий фрагмент содержит константную область гибридной легкой цепи каппа и вариабельную область легкой цепи человеческого типа лямбда, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:67, 68, 69, 70 и 71.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит константную область легкой цепи человеческого типа лямбда и вариабельную область легкой цепи человеческого типа лямбда, содержащую CDRL1, CDRL2 и CDRL3, набор CDRL, выбранный из группы, состоящей из CDRL1 с SEQ ID NO:60 с заменой 0, 1, 2, 3 или 4 аминокислот, CDRL2 с SEQ ID NO:61 с заменой 0, 1, 2, 3 или 4 аминокислот и CDRL3 с SEQ ID NO:62 с заменой 0, 1, 2, 3, 4 или 5 аминокислот, и
c) второй связывающий фрагмент содержит константную область легкой цепи человеческого типа каппа и вариабельную область легкой цепи человеческого типа лямбда, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:67, 68, 69, 70 и 71.
В одном варианте осуществления биспецифическое антитело отличается содержанием первого связывающего фрагмента, специфически связывающего CEACAM5 человека, и второго связывающего фрагмента, специфически связывающего CD3ε человека, и отличается тем, что
a) каждый из первого связывающего фрагмента и второго связывающего фрагмента содержит в качестве тяжелой цепи общую тяжелую цепь, и содержит в качестве вариабельной области вариабельную область, содержащую CDRH1, CDRH2 и CDRH3; CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит константную область легкой цепи человеческого типа каппа и вариабельную область легкой цепи человеческого типа каппа, содержащую CDRL1, CDRL2 и CDRL3, набор CDRL, выбранный из группы, состоящей из CDRL1 с SEQ ID NO:64 с заменой 0, 1, 2, 3, 4 или 5 аминокислот, CDRL2 с SEQ ID NO:65 с заменой 0, 1, 2, 3, 4 или 5 аминокислот и CDRL3 с SEQ ID NO:66 и с заменой 0, 1, 2, 3, 4 или 5 аминокислот
c) второй связывающий фрагмент содержит константную область легкой цепи человеческого типа лямбда и вариабельную область легкой цепи человеческого типа лямбда, содержащую CDRL1, CDRL2 и CDRL3, группу областей CDR, выбранных из группы, состоящей из
I) CDRL1 с SEQ ID NO:6, CDRL2 с SEQ ID NO:7 и CDRL3 с SEQ ID NO:8,
II) CDRL1 с SEQ ID NO:10, CDRL2 с SEQ ID NO:11 и CDRL3 с SEQ ID NO:12,
III) CDRL1 с SEQ ID NO:14, CDRL2 с SEQ ID NO:15 и CDRL3 с SEQ ID NO:16,
IV) CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20, и
V) CDRL1 с SEQ ID NO:22, CDRL2 с SEQ ID NO:23 и CDRL3 с SEQ ID NO:24.
В одном варианте осуществления второй связывающий фрагмент в c) содержит легкую цепь, выбранную из группы, состоящей из SEQ ID NO:25, 26, 27, 28 и 29.
В одном варианте осуществления биспецифическое антитело содержит первый связывающий фрагмент, специфически связывающий CEACAM5 человека, и второй связывающий фрагмент, специфически связывающий CD3ε человека, где:
a) первый связывающий фрагмент содержит вариабельную область тяжелой цепи (VH), содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит вариабельную область легкой цепи (VL), содержащую набор CDRL, в котором CDRL1 имеет консенсусную последовательность SEQ ID NO:136, CDRL2 имеет консенсусную последовательность SEQ ID NO:137 и CDRL3 имеет консенсусную последовательность SEQ ID NO:138, и
c) второй связывающий фрагмент содержит VH, содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4.
В одном варианте осуществления первый связывающий фрагмент содержит в качестве каркасной последовательности вариабельной области легкой цепи каркасную последовательность SEQ ID NO:31. В одном варианте осуществления первый связывающий фрагмент содержит в качестве каркасной последовательности вариабельной области легкой цепи каркасную последовательность SEQ ID NO:35. В одном варианте осуществления первый связывающий фрагмент содержит в качестве каркасной последовательности вариабельной области легкой цепи каркасную последовательность SEQ ID NO:59. В одном варианте осуществления первый связывающий фрагмент содержит в качестве каркасной последовательности вариабельной области легкой цепи каркасную последовательность SEQ ID NO:63.
В одном варианте осуществления первый связывающий фрагмент специфически связывает CEA, содержит в качестве вариабельной области тяжелой цепи вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:1, и в качестве вариабельной области легкой цепи вариабельную область легкой цепи с аминокислотной последовательностью, которая на 98%, 99% или 100% идентична аминокислотной последовательности, выбранной из группы SEQ ID NO:31 или SEQ ID NO:35, SEQ ID NO:59 или SEQ ID NO:63; и второй связывающий фрагмент, специфически связывающий CD3, содержит в качестве вариабельной области тяжелой цепи вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:1, и в качестве вариабельной области легкой цепи вариабельную область легкой цепи с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:5, SEQ ID NO:9, SEQ ID NO:13, SEQ ID NO:17 и SEQ ID NO:21.
В одном варианте осуществления биспецифическое антитело по изобретению отличается содержанием первого связывающего фрагмента, специфического для CEA, содержащего вариабельный домен легкой цепи каппа и константный домен легкой цепи каппа, и второго связывающего фрагмента, специфического для CD3ε, содержащего вариабельный домен легкой цепи лямбда и константный домен легкой цепи лямбда.
В одном варианте осуществления биспецифическое антитело по изобретению отличается содержанием первого связывающего фрагмента, специфического для CEA, содержащего вариабельный домен легкой цепи лямбда и константный домен легкой цепи каппа, и второго связывающего фрагмента, специфического для CD3ε, содержащего вариабельный домен легкой цепи лямбда и константный домен легкой цепи лямбда.
В одном варианте осуществления биспецифическое антитело по изобретению отличается содержанием первого связывающего фрагмента, специфического для CEA, содержащего вариабельный домен легкой цепи лямбда и константный домен легкой цепи лямбда, и второго связывающего фрагмента, специфического для CD3ε, содержащего вариабельный домен легкой цепи лямбда и константный домен легкой цепи каппа.
В конкретном варианте осуществления Fc-домен проявляет сниженную аффинность связывания с Fc-рецептором и/или сниженную эффекторную функцию в сравнении с естественным/дикого типа Fc-доменом IgG1. В конкретных вариантах осуществления Fc-домен модифицирован для снижения аффинности связывания с Fc-рецептором и/или снижения эффекторной функции в сравнении с не модифицированным Fc-доменом. В одном варианте осуществления Fc-домен имеет одну или более аминокислотных замен, которые приводят к уменьшению связывания с одним или более Fc-рецепторами и/или снижению эффекторных функций. В одном варианте осуществления такие одна или более замен выбраны из группы, состоящей из Pro238, Asp265, Asp270, Asn297 (потеря углевода Fc), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435 (Shields, R. L., et al., J. Biol. Chem. 276 (2001) 6591-6604; Lund, J., et al., FASEB J. 9 (1995) 115-119; Morgan, A., et al., Immunology 86 (1995) 319-324; EP 0 307 434). В одном варианте осуществления антитело, в отношении связывания с FcR, относится к подклассу IgG4, либо подклассу IgG1 или IgG2 с мутацией в S228, L234, L235 и/или D265, и/или имеет мутацию PVA236. В одном варианте осуществления мутации в Fc-домене представляют собой S228P, L234A, L235A, L235E и/или PVA236. В другом варианте осуществления мутации в Fc-домене представляют собой в IgG4 S228P, и в IgG1 L234A и L235A. В одном варианте осуществления одна или более аминокислотных замен в Fc-домене, которые уменьшают связывание с одним или более Fc-рецепторами и/или снижают эффекторные функции, имеют место в одном или более положениях, выбранных из группы, состоящей из L234, L235 и P329 (нумерация Kabat EU-индекс). В конкретных вариантах осуществления каждая субъединица Fc-домена имеет две аминокислотные замены, которые уменьшают связывание с Fc-рецептором и/или снижают эффекторную функцию, где указанные аминокислотные замены представляют собой L234A и L235A (нумерация Kabat EU-индекс). В конкретных вариантах осуществления каждая субъединица Fc-домена имеет три аминокислотные замены, которые уменьшают связывание с Fc-рецептором и/или снижают эффекторную функцию, где указанные аминокислотные замены представляют собой L234A, L235A и P329A (нумерация Kabat EU-индекс). В одном таком варианте осуществления Fc-домен представляет собой Fc-домен IgG1, в частности Fc-домен IgG1 человека (нумерация Kabat EU-индекс). В одном варианте осуществления Fc-домен относится к подклассу IgG4, и в одном варианте осуществления к подклассу IgG4 с мутацией S228P.
В одном варианте осуществления Fc-рецептор представляет собой Fcγ-рецептор. В одном варианте осуществления Fc-рецептор представляет собой Fc-рецептор человека. В одном варианте осуществления Fc-рецептор представляет собой активирующий Fc-рецептор. В конкретном варианте осуществления Fc-рецептор представляет собой FcγRIIIA, FcγRI и/или FcγRIIIA человека. В одном варианте осуществления эффекторная функция представляет собой антителозависимую опосредуемую клетками цитотоксичность (ADCC), но не ограничена только ADCC.
Один из вариантов осуществления изобретения относится к области CL каппа с SEQ ID NO:58 для использования во втором связывающем фрагменте в качестве области CL при создании биспецифического антитела, содержащего общую тяжелую цепь с SEQ ID NO:43, 44 или 45, и к области CL лямбда с SEQ ID NO:41 в качестве области CL в первом связывающем фрагменте.
Один из вариантов осуществления изобретения относится к области CL каппа с SEQ ID NO:58 для использования во втором связывающем фрагменте, специфически связывающем CD3, в качестве области CL при создании биспецифического антитела по изобретению, содержащего общую тяжелую цепь с SEQ ID NO:43, 44 или 45, и к области CL лямбда с SEQ ID NO:41 в качестве области CL в первом связывающем фрагменте, специфически связывающем CEACAM5.
Один из вариантов осуществления изобретения относится к области CL каппа с SEQ ID NO:58 для использования во втором связывающем фрагменте, специфически связывающем CD3, в качестве области CL при создании биспецифического антитела по изобретению, содержащего общую тяжелую цепь с SEQ ID NO:43, 44 или 45, вариабельную область легкой цепи с SEQ ID NO:5, 9, 13, 17 или 21 и область CL лямбда с SEQ ID NO:41 в качестве области CL в первом связывающем фрагменте, специфически связывающем CEACAM5.
Следующий вариант осуществления изобретения относится к олигонуклеотиду, выбранному из группы, состоящей из SEQ ID NO:76, 77, 78 и 79, для использования в созревании аффинности антитела путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов соответствующей вариабельной области легкой цепи с SEQ ID NO:31, 35, 59 и 63.
Другой аспект относится к способу получения биспецифического антитела по изобретению, включающему этапы a) культивирования клетки-хозяина по изобретению в условиях, подходящих для экспрессии биспецифического антитела, и b) извлечения биспецифического антитела. Изобретение также относится к биспецифическому антителу, полученному способом по изобретению.
Изобретение также относится к фармацевтической композиции, содержащей биспецифическое антитело по изобретению и фармацевтически приемлемый носитель. Изобретение также относится к способам применения биспецифического антитела и фармацевтической композиции по изобретению. В одном аспекте изобретение относится к биспецифическому антителу или фармацевтической композиции по изобретению для применения в качестве лекарственного препарата. Один аспект относится к биспецифическому антителу или фармацевтической композиции по изобретению для применения в лечении заболевания у индивидуума, который нуждается в этом. В конкретном варианте осуществления заболевание представляет собой рак.
Также предложено биспецифическое антитело по изобретению для применения в производстве лекарственного препарата для лечения заболевания у индивидуума, который нуждается в этом; а также способ лечения заболевания у индивидуума, включающий введение указанному индивидууму терапевтически эффективного количества композиции, содержащей биспецифическое антитело по изобретению в фармацевтически приемлемой форме. В конкретном варианте осуществления заболевание представляет собой рак. В любом из вышеуказанных вариантов осуществления индивидуум предпочтительно является млекопитающим, в частности, человеком.
Изобретение также относится к способу индукции лизиса клетки-мишени, в частности, опухолевой клетки, включающему создание контакта клетки-мишени с биспецифическим антителом по изобретению в присутствии T-клетки, в частности, цитотоксической T-клетки.
Следующий вариант осуществления изобретения относится к биспецифическому антителу по изобретению для применения в производстве лекарственного препарата для лечения субъекта, имеющего рак, экспрессирующий CEA.
Следующий вариант осуществления изобретения относится к биспецифическому антителу по изобретению для применения в производстве лекарственного препарата по изобретению, при этом рак выбран из группы, состоящей из колоректального рака, немелкоклеточного рака легких (НМРЛ), рака пищевода, рака пищеводно-желудочного перехода, рака поджелудочной железы и рака молочной железы.
Следующий вариант осуществления изобретения относится к биспецифическому антителу по изобретению для применения в одновременном, раздельном или последовательном комбинированном введении с анти-CD47 антителом. В одном варианте осуществления анти-CD47 антитело представляет собой магролимаб, ALX148 или TTI-621 и/или TTI-622.
Следующий вариант осуществления изобретения относится к биспецифическому антителу по изобретению для применения в одновременном, раздельном или последовательном комбинированном введении со вторым биспецифическим антителом, содержащим третий связывающий фрагмент, специфически связывающий CEACAM5 человека, и четвертый связывающий фрагмент, специфически связывающий CD47 человека, для лечения субъекта, имеющего рак, экспрессирующий CEA.
Такие вторые биспецифические анти-CEAxCD47 антитела описаны в PCT/IB2019/054559 и US16/428359.
Следующий вариант осуществления изобретения относится к биспецифическому антителу по изобретению для применения по изобретению, отличающегося тем, что биспецифическое антитело по изобретению и второе биспецифическое анти-CEAxCD47 антитело вводят указанному субъекту попеременно с интервалом от 6 до 15 дней, но не ограничиваясь такими интервалами.
Следующий вариант осуществления изобретения относится к первому биспецифическому антителу по изобретению, содержащему первый связывающий фрагмент, специфически связывающий CEACAM5 человека, и второй связывающий фрагмент, специфически связывающий CD3 человека, по изобретению и второму биспецифическому антителу анти-CEAxCD47 для применения в лечении рака по изобретению, отличающегося тем, что указанный рак представляет собой колоректальный рак, немелкоклеточный рак легких (НМРЛ), рак желудка, рак пищевода, рак поджелудочной железы и рак молочной железы.
Следующий вариант осуществления изобретения относится к композиции, содержащей биспецифическое антитело по изобретению, отличающееся отсутствием конкуренции с указанным вторым анти-CEAxCD47 биспецифическим антителом, описанным выше, для применения в лечении субъекта, имеющего рак, экспрессирующий CEA.
Следующий вариант осуществления изобретения относится к способу лечения пациента-человека, у которого диагностирована опухоль (рак), в частности, солидная опухоль, в частности, солидный рак, экспрессирующий CEA, в частности, колоректальный рак, немелкоклеточный рак легких (НМРЛ), рак желудка, рак пищевода, рак поджелудочной железы и рак молочной железы, включающему введение эффективного количества биспецифического антитела по изобретению и второго биспецифического антитела, описанного в PCT/IB2019/054559 и US16/428359, против CEA и CD47, пациенту-человеку, где способ последовательно включает:
введение пациенту дозы 0,1-30 мг/кг, в следующем варианте осуществления 0,5-10 мг/кг, в следующем варианте осуществления 1-10 мг/кг указанного второго анти-CEAxCD47 антитела, например, еженедельно в течение 4-12 недель,
введение пациенту указанного второго антитела q1, q2w, q3w или, необязательно, q4w,
введение пациенту после этих 4-12 недель и после дополнительных 2 или 3, или 4 периодов полувыведения указанного анти-CEAxCD47 антитела дозы 0,1-10 мг/кг антитела по изобретению,
введение пациенту указанного антитела по изобретению q1, q2w, q3w или, необязательно, q4w,
ожидание в течение 2 или 3, или 4 периодов полувыведения указанного антитела по изобретению, а затем, необязательно, повторение указанного цикла введения анти-CEAxCD47 биспецифического антитела, с последующим введением анти-CEAxCD3 биспецифического антитела и, необязательно, вновь повторение этого цикла.
Такой способ «попеременного» введения применяют, если антитело по изобретению и второе биспецифическое антитело конкурируют за связывание с CEA.
В случае, если указанное анти-CEAxCD47 биспецифическое антитело и анти-CEAxCD3 биспецифическое антитело по настоящему изобретению не конкурируют, эти два биспецифических антитела также можно вводить таким образом («одновременно»), что у пациента в плазме и в тканях присутствуют параллельно терапевтически эффективные концентрации обоих биспецифических антител, например, за счет введения пациенту примерно в одно и то же время дозы 0,1-30 мг/кг, в следующем варианте осуществления 0,5-10 мг/кг, в следующем варианте осуществления 1-10 мг/кг анти-CEAxCD47 биспецифического антитела и 0,1-10 мг/кг анти-CEAxCD3 биспецифического антитела по настоящему изобретению, с последующими одним или более такими комбинированными введениями с частотой q1w или q2w, или q3w, или, необязательно, q4w.
Термин «q1w» означает введение один раз в неделю; q2w означает введение один раз в две недели и так далее.
Следующий вариант осуществления изобретения относится к фармацевтической композиции, содержащей антитело по изобретению и фармацевтически приемлемый эксципиент или носитель.
Следующий предпочтительный вариант осуществления изобретения относится к фармацевтической композиции, содержащей антитело по изобретению для применения в качестве лекарственного препарата.
Следующий предпочтительный вариант осуществления изобретения относится к фармацевтической композиции, содержащей антитело по изобретению для применения в качестве лекарственного препарата для лечения солидных опухолей, экспрессирующих CEA.
Следующий предпочтительный вариант осуществления изобретения относится к фармацевтической композиции, содержащей антитело по изобретению для применения в качестве лекарственного препарата для лечения колоректального рака, НМРЛ (немелкоклеточного рака легких), рака желудка, рака пищевода, рака поджелудочной железы или рака молочной железы.
Следующий вариант осуществления изобретения относится к композиции по изобретению, при этом рак представляет собой колоректальный рак, немелкоклеточный рак легких (НМРЛ), рак желудка, рак пищевода, рак поджелудочной железы или рак молочной железы.
Следующий вариант осуществления изобретения относится к антителу по изобретению для применения в производстве фармацевтической композиции.
Следующий вариант осуществления изобретения относится к антителу по изобретению и фармацевтически приемлемому эксципиенту или носителю для применения в производстве фармацевтической композиции.
Следующий вариант осуществления изобретения относится к антителу по изобретению для применения в производстве лекарственного препарата для лечения солидных опухолей.
Следующий вариант осуществления изобретения относится к антителу по изобретению для применения в лечении колоректального рака, НМРЛ (немелкоклеточного рака легких), рака желудка, рака пищевода, рака поджелудочной железы или рака молочной железы.
Другой аспект изобретения относится к способу индукции клеточного лизиса опухолевой клетки, включающему создание контакта опухолевой клетки с биспецифическим антителом по любому из вышеописанных вариантов осуществления. В некоторых вариантах осуществления опухолевая клетка представляет собой клетку колоректального рака, клетку НМРЛ (немелкоклеточного рака легких), клетку рака желудка, клетку рака пищевода, клетку рака поджелудочной железы или клетку рака молочной железы.
В одном варианте осуществления клеточный лизис индуцируется за счет направляемой T-клетками клеточной цитотоксичности (TDCC).
Другой аспект изобретения относится к способу лечения субъекта, имеющего рак, экспрессирующий CEA, включающему введение субъекту терапевтически эффективного количества биспецифического антитела по любому из вышеописанных вариантов осуществления.
Другой аспект изобретения относится к способу лечения субъекта, имеющего рак, экспрессирующий CEA, включающему введение субъекту терапевтически эффективного количества биспецифического антитела по любому из вышеописанных вариантов осуществления в сочетании с биспецифическим антителом, связывающим CEA человека и CD47 человека. Если анти-CEAxCD47 антитело и анти-CEAxCD3 антитело являются конкурирующими, они будут конкурировать за CEA-рецепторы на поверхности опухолевой клетки, и занятость рецепторов и эффективность каждого партнера в сочетании зависят от их аффинности связывания и их концентраций в плазме, и, таким образом, являются труднопредсказуемыми и варьирующими с течением времени, если два лекарственных средства имеют разный период полувыведения и, соответственно, клиренс из организма. Таким образом, конкурирующие анти-CEAxCD3 и анти-CEAxCD47 биспецифические антитела следует вводить последовательно (попеременно). Если анти-CEAxCD3 и анти-CEAxCD47 биспецифические антитела являются не конкурирующими или лишь минимально конкурирующими, их можно вводить не только последовательно, но также и параллельно (одновременно), что вполне может быть преимуществом, поскольку ожидается, что уничтожение опухолевых клеток посредством вовлечения T-клеток за счет анти-CEAxCD3 биспецифического антитела, и в то же время, посредством вовлечения макрофагов за счет анти-CEAxCD47 биспецифического антитела, будет аддитивным, или даже может быть синергетическим, это означает, что эффективность возрастает при параллельном введении обоих лекарственных средств.
Другой аспект изобретения относится к способу увеличения периода выживаемости без прогрессирования и/или общей выживаемости у субъекта, имеющего рак, экспрессирующий CEA, включающему введение указанному субъекту терапевтически эффективного количества биспецифического антитела по любому из вышеописанных вариантов осуществления. В одном варианте осуществления рак представляет собой колоректальный рак, немелкоклеточный рак легких (НМРЛ), рак желудка, рак пищевода, рак поджелудочной железы, рак молочной железы, карциному головы и шеи, рак тела матки, рак мочевого пузыря или другой рак, экспрессирующий CEA.
В конкретных вариантах осуществления этих способов биспецифическое антитело вводят в сочетании с химиотерапией или радиотерапией. В одном варианте осуществления субъект является пациентом, страдающим от колоректального рака или рака легких, или рака желудка, рака пищевода или рака поджелудочной железы, или рака молочной железы или другого рака, экспрессирующего CEA.
В конкретных вариантах осуществления этих способов биспецифическое антитело по изобретению вводят пациенту в диапазоне доз от 0,1 до 100 мг/кг массы тела в день или в неделю в одной или в разделенных дозах, или путем непрерывной инфузии. В конкретных вариантах осуществления биспецифическое антитело по изобретению вводят пациенту в диапазоне доз от 1 до 20 мг/кг.
Другой аспект изобретения относится к способу лечения субъекта, имеющего рак, экспрессирующий CEA, включающему введение субъекту терапевтически эффективного количества биспецифического антитела по любому из вышеописанных вариантов осуществления в сочетании с биспецифическим антителом против CEA человека и CD47 человека. В конкретных вариантах осуществления этих способов биспецифическое антитело вводят в сочетании с биспецифическим анти-CEAxCD47 антителом одновременно, раздельно или последовательно. В конкретных вариантах осуществления этих способов биспецифическое анти-CEAxCD47 антитело вводят попеременно с антителом по изобретению, с интервалами 6-15 дней между введением антитела по изобретению и биспецифического анти-CEAxCD47 антитела. В конкретных вариантах осуществления анти-CEAxCD47 антитело вводят пациенту в диапазоне доз от 0,1 до 100 мг/кг массы тела в день или в неделю в одной или в разделенных дозах, или путем непрерывной инфузии.
В конкретных вариантах осуществления этих способов биспецифическое антитело вводят в сочетании с антагонистом оси PD-1 одновременно, раздельно или последовательно. В конкретных вариантах осуществления этих способов биспецифическое антитело вводят в сочетании с биспецифическим анти-CEAxCD47 антителом и антагонистом оси PD-1 одновременно, раздельно или последовательно. В конкретных вариантах осуществления антагонист оси PD-1 вводят пациенту в диапазоне доз от 0,1 до 100 мг/кг массы тела в день или в неделю в одной или в разделенных дозах, или путем непрерывной инфузии.
Другой аспект изобретения относится к способу увеличения периода выживаемости без прогрессирования и/или периода общей выживаемости у субъекта, имеющего рак, аномально экспрессирующий CEA, включающему введение указанному субъекту терапевтически эффективного количества биспецифического антитела по любому из вышеописанных вариантов осуществления. В одном варианте осуществления рак представляет собой колоректальный рак, немелкоклеточный рак легких (НМРЛ), рак желудка, рак пищевода, рак поджелудочной железы или рак молочной железы.
В конкретных вариантах осуществления этих способов биспецифическое антитело вводят в сочетании с химиотерапией или радиотерапией. В одном варианте осуществления субъект является онкологическим пациентом с колоректальным раком или раком легких, или раком желудка, раком пищевода или раком поджелудочной железы, или раком молочной железы, или другим CEA-экспрессирующим раком.
Другой вариант осуществления изобретения относится к биспецифическому антителу по изобретению для применения в любом из вышеописанных способов лечения. В одном варианте осуществления рак выбран из группы, состоящей из колоректального рака, немелкоклеточного рака легких (НМРЛ), рака желудка, рака пищевода, рака поджелудочной железы и рака молочной железы.
Другой вариант осуществления изобретения относится к полинуклеотидам, кодирующим биспецифические антитела, раскрытые в настоящем документе, или их домен (например, вариабельную область легкой цепи и/или вариабельную область тяжелой цепи), который иммуноспецифически связывает CEACAM5 или CD3ε. В некоторых аспектах настоящее изобретение относится к полинуклеотидам, содержащим нуклеотидную последовательность, кодирующую легкую цепь или тяжелую цепь антитела, описанного в настоящем документе. Полинуклеотиды могут содержать нуклеотидные последовательности, кодирующие тяжелую цепь, содержащую области VH, или области CDR тяжелой цепи антител, описанных в настоящем документе. Полинуклеотиды могут содержать нуклеотидные последовательности, кодирующие легкую цепь, содержащую области VL, или области CDR легкой цепи антител, описанных в настоящем документе.
Некоторые варианты осуществления относятся к векторам, содержащим выделенные полинуклеотиды, раскрытые в настоящем документе. Некоторые другие варианты осуществления относятся к клеткам, содержащим выделенные полинуклеотиды или векторы, кодирующие биспецифические антитела, раскрытые в настоящем документе. В некоторых вариантах осуществления клетка выбрана из группы, состоящей из клетки Streptomyces, дрожжей, CHO, YB/20, NS0, PER-C6, HEK-293T, NIH-3T3, HeLa, BHK, Hep G2, SP2/0, R1.1, B-W, L-M, COS 1, COS 7, BSC1, BSC40, BMT10, клетки растения, клетки насекомого и человеческой клетки в культуре ткани.
Некоторые варианты осуществления относятся к способам получения антител, раскрытых в настоящем документе. В некоторых вариантах осуществления способ получения антитела включает экспрессию антитела с использованием клеток, содержащих выделенные полинуклеотиды или векторы, кодирующие биспецифические антитела, раскрытые в настоящем документе. В некоторых вариантах осуществления способ получения антитела включает культивирование клетки, содержащей выделенный полинуклеотид или вектор, кодирующий биспецифические антитела, раскрытые в настоящем документе, и выделение экспрессируемого в ней антитела.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1: Эпитоп-специфическая сортировка новых CEA-связывающих компонентов, используемых в каппа/лямбда (KL) анти-CEAxCD3 биспецифических антителах (подробности в примере 5c)
Для определения характеристик новых CEA-связывающих антител по настоящему изобретению используют конкурентный иммуноферментный анализ. Профиль конкурентного блокирования определяют против антител, все из которых связывают CEACAM5 (CEA), и для которых уже определен и опубликован связывающий эпитоп. Такими антителами являются SM3E, MEDI (=MEDI-565), SAR, T84.66, лабетузумаб и CH1A1A, подробное описание данных антител и анализа смотри в примере 5c.
Фиг. 2 Agilent профиль очищенных биспецифических антител
В примере 6 описаны экспрессия, очистка и анализы новых каппа-лямбда биспецифических анти-CEAxCD3 антител. Очищенные биспецифические антитела анализировали методом электрофореза в денатурирующих и восстанавливающих условиях. Использовали биоанализатор Agilent 2100, на Фигуре представлен типичный результат, полученный для каппа-лямбда антител по настоящему изобретению AB1 L3-1/D (KL анти-CEAxCD3 биспецифическое антитело с лямбда CD3 LC SEQ ID NO:28 и каппа CEA LC SEQ ID NO:40, и общей HC SEQ ID NO 44) и L3-1AB8 H-CK5/D (гибридное KL анти-CEAxCD3 биспецифическое антитело с гибридной каппа CD3 LC SEQ ID NO:70 и лямбда CEA LC SEQ ID NO:42, и общей HC SEQ ID NO:44). Y4 L3-1/D представляет собой биспецифическое антитело с тем же плечом CD3, что и AB1 L3-1/D и L3-1AB8 H-CK5/D, но со вторым плечом, не связывающим CEA, это антитело часто используют в фармакологических анализах в качестве контрольного антитела.
Фиг. 3 Связывание CD3пол клеток Jurkat (и CD3отр клеток TIB-153)
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB1 L3-1/D и L3-1AB8 H-CK5/D с CD3-экспрессирующими клетками Jurkat. Оба KL биспецифических антитела имеют одно и то же анти-CD3 плечо. Y4L3-1/D представляет собой KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA. Справа на Фигуре показано отсутствие связывания CD3-отрицательной линии клеток TIB-153 даже при концентрации 100 нМ биспецифических антител.
Фиг. 4 Связывание CEAпол клеток MKN-45 (и CEAотр клеток MKN-45_hCEAKO)
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB1 L3-1/D и L3-1AB8 H-CK5/D с CEA-экспрессирующими клетками MKN-45. Оба KL биспецифических антитела имеют одно и то же анти-CD3 плечо. Y4L3-1/D, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, не связывает клетки MKN-45. Справа на Фигуре показано отсутствие связывания клеток линии MKN-45 после нокаута CEACAM5 даже при концентрации 100 нМ.
Фиг. 5 Связывание CEAПОЛ клеток LS-174T
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB1 L3-1/D и L3-1AB8 H-CK5/D с CEA-экспрессирующими клетками LS-174T. Оба KL биспецифических антитела имеют одно и то же анти-CD3 плечо. Y4L3-1/D, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, не связывает клетки LS-174T.
Фиг. 6 Уничтожение CEAПОЛ клеток MKN-45 (и CEAОТР клеток MKN-45_hCEAKO)
Зависимое от концентрации T-клеточное перенаправленное уничтожение/лизис клеток MKN-45 KL биспецифическими антителами по изобретению AB1 L3-1/D и L3-1AB8 H-CK5/D (анализ описан в примере 8a). Оба KL биспецифических антитела имеют одно и то же анти-CD3 плечо. Оба KL биспецифических антитела демонстрируют одинаковую эффективность уничтожения клеток. Это было удивительно/неожиданно, поскольку связывание AB1 L3-1/D с MKN-45 является намного более слабым, чем связывание L3-1AB8 H-CK5/D (смотри Фигуру 4).
Y4L3-1/D, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, демонстрирует лишь очень низкую эффективность уничтожения/лизиса при примерно в 100 раз более высоких концентрациях.
Справа на Фигуре: лишь очень незначительное неспецифическое уничтожение/лизис клеток MKN-45 с нокаутом CEA всеми 3 биспецифическими антителами.
Фиг. 7 Уничтожение CEAПОЛ LS-174T
Зависимое от концентрации T-клеточное перенаправленное уничтожение/лизис клеток LS-174T KL биспецифическими антителами по изобретению AB1 L3-1/D и L3-1AB8 H-CK5/D (анализ описан в примере 8a). Оба KL биспецифических антитела имеют одно и то же анти-CD3 плечо. L3-1AB8 H-CK5/D демонстрирует более высокую эффективность уничтожения клеток, чем AB1 L3-1/D. Связывание L3-1AB8 H-CK5/D с клетками LS-174T является более сильным, чем связывание AB1 L3-1/D (смотри Фигуру 5).
Y4L3-1/D, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, демонстрирует лишь очень низкую эффективность уничтожения/лизиса при более чем в 100 раз более высоких концентрациях.
Фиг. 8 Связывание моноклональных анти-CEA мАт 1B4 с разными рекомбинантными белками (ELISA).
CEA ECD=внеклеточный домен CEA; CEA A3B3 представляет собой A3B3 домен CEA. Рекомбинантный мезотелин MSLN использован в качестве контроля для определения неспецифического связывания (связывания, не специфического для ECD CEA, соответственно, A3B3 домена CEA).
Фиг. 9 Связывание моноклональных анти-CEA мАт C11 с разными рекомбинантными белками (ELISA).
CEA ECD=внеклеточный домен CEA; CEA A3B3 представляет собой A3B3 домен CEA. Рекомбинантный мезотелин MSLN использован в качестве контроля для определения неспецифического связывания (связывания, не специфического для ECD CEA, соответственно, A3B3 домена CEA).
Фиг. 10 Связывание антител 1 цикла оптимизации направленности с CD3ПОЛ клетками Jurkat (и CD3ОТР клетками TIB-153)
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB13L3-1/N, AB14L3-1/N, AB15L3-1/N, AB17L3-1/N и AB20L3-1/N с CD3-экспрессирующими клетками HUT-78. Все KL биспецифические антитела имеют одно и то же анти-CD3 плечо. AB1L3-1/N является исходным KL биспецифическим антителом, из которого вышеуказанные бсАт были получены путем оптимизации направленности. Y4L3-1/N представляет собой KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA. TCB2014 представляет собой другое анти-CEAxCD3 T-клеточное биспецифическое антитело, описанное в US20140242079, которое было использовано в качестве эталонного антитела. чIgG1 представляет собой не связывающее человеческое IgG1 антитело, использованное в качестве контроля по изотипу. Справа на Фигуре показано отсутствие связывания с CD3-отрицательной линией клеток JKTβ-дел даже при концентрации 100 нМ биспецифических антител. Способ описан в примере 7b.
Фиг. 11 Связывание антител 1 цикла оптимизации направленности с CEAПОЛ клетками MKN-45, клетками HPAF-II и клетками LS-174T (ATCC® CL-188™) (и CEAОТР клетками MKN-45_hCEAKO)
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB13L3-1/N, AB14L3-1/N, AB15L3-1/N, AB17L3-1/N и AB20L3-1/N с CEA-положительными клетками MKN-45 (A); HPAF-II (C) и LS-174T (D) или CEA-отрицательными клетками MKN-45_hCEAKO (B). Все KL биспецифические антитела имеют одно и то же анти-CD3 плечо. AB1L3-1/N является исходным KL биспецифическим антителом, из которого вышеуказанные бсАт были получены путем созревания аффинности. Y4L3-1/N представляет собой KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA. TCB2014 представляет собой другое анти-CEAxCD3 T-клеточное биспецифическое антитело, описанное в US20140242079, которое было использовано в качестве эталонного антитела. чIgG1 представляет собой не связывающее человеческое IgG1 антитело, использованное в качестве контроля по изотипу. На Фигуре 10B показано отсутствие связывания с CEA-отрицательной линией клеток MKN-45_hCEAKO даже при концентрации 100 нМ биспецифических антител. Связывание антител по настоящему изобретению с клетками MKN-45 и/или HPAF-II при концентрации 100 нМ на 40% или более выше, чем связывание TCB2014. Способ описан в примере 7a.
Фиг. 12 Уничтожение CEAПОЛ клеток MKN-45, HPAF-II и LS-174T (и CEAОТР клеток MKN-45_hCEAKO) антителами 1 цикла оптимизации направленности
Зависимое от концентрации T-клеточное перенаправленное уничтожение/лизис CEA-положительных клеток MKN-45 (A); HPAF-II (C) и LS-174T (D) или CEA-отрицательных клеток MKN-45_hCEAKO (B) KL биспецифическими антителами по изобретению AB13L3-1/N, AB14L3-1/N, AB15L3-1/N, AB17L3-1/N и AB20L3-1/N (анализ описан в примере 8a). Все KL биспецифические антитела имеют одно и то же анти-CD3 плечо. Все KL биспецифические антитела демонстрируют улучшенное уничтожение клеток в сравнении с исходным антителом AB1L3-1/N, из которого они были получены путем оптимизации направленности, при использовании CEA-положительных клеток (A, C, D), но лишь очень незначительное неспецифическое уничтожение/лизис клеток MKN-45 с нокаутом CEA (B).
Y4L3-1/N, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, демонстрирует лишь очень низкую эффективность уничтожения/лизиса при максимальных протестированных концентрациях. TCB2014 представляет собой другое анти-CEAxCD3 T-клеточное биспецифическое антитело, описанное в US20140242079, которое было использовано в качестве эталонного антитела. Значения EC50 для биспецифических антител по настоящему изобретению ниже, чем EC50 для TCB2014, это указывает на повышенную эффективность уничтожения опухолевых клеток. Способ описан в примере 8a.
Фиг. 13 Связывание антител 2 цикла оптимизации направленности с CD3ПОЛ первичными T-клетками (CD4+ и CD8+) и CD3ОТР B-клетками и моноцитами
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB54L3-1/N, AB60L3-1/N, AB66L3-1/N, AB71L3-1/N, AB72L3-1/N и AB73L3-1/N с CD3-положительными первичными CD4+ T-клетками (A) и первичными CD8+ T-клетками (B) или с CD3-отрицательными B-клетками (C) и моноцитами (D). Все KL биспецифические антитела имеют одно и то же анти-CD3 плечо и связываются аналогичным образом с популяциями CD3-положительных T-клеток. Y4L3-1/N, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, связывается одинаково хорошо с CD3-положительными T-клетками. TCB2014 представляет собой другое анти-CEAxCD3 T-клеточное биспецифическое антитело, описанное в US20140242079, которое было использовано в качестве эталонного антитела. Антитело против CD47 человека B6H12 (а-CD47 мАт) было использовано в качестве положительного контроля (T-клетки, B-клетки и моноциты экспрессируют CD47). Вторичное Ат использовано только в варианте, в котором лишь обнаруживающее антитело добавляли к клеткам, и служит для определения фонового сигнала (отрицательный контроль). Все протестированные антитела продемонстрировали отсутствие связывания с популяциями CD3-отрицательных клеток даже при концентрации 200 нМ.
Фиг. 14 Связывание антител 2 цикла оптимизации направленности с CEAПОЛ клетками MKN-45, клетками HPAF-II и клетками LS-174T (и CEAОТР клетками MKN-45_hCEAKO)
Зависимое от концентрации связывание KL биспецифических антител по изобретению AB54L3-1/N, AB60L3-1/N, AB66L3-1/N, AB71L3-1/N, AB72L3-1/N и AB73L3-1/N с CEA-положительными клетками MKN-45 (A); HPAF-II (C) и LS-174T (D) или CEA-отрицательными клетками MKN-45_hCEAKO (B). Все KL биспецифические антитела имеют одно и то же анти-CD3 плечо. Y4L3-1/N представляет собой KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA. TCB2014 представляет собой другое анти-CEAxCD3 T-клеточное биспецифическое антитело, описанное в US20140242079, которое было использовано в качестве эталонного антитела. На Фигуре 14B показано отсутствие связывания с CEA-отрицательной линией клеток MKN-45_hCEAKO даже при концентрации 200 нМ биспецифических антител. Для Фигуры 14A значения EC50 приведены в таблице 4. Связывание биспецифических антител по изобретению при концентрации 200 нМ на 40% или более выше, чем связывание TCB2014.
Фиг. 15 Уничтожение CEAПОЛ клеток MKN-45, HPAF-II и LS-174T (и CEAОТР клеток MKN-45_hCEAKO) антителами 2 цикла оптимизации направленности
Зависимое от концентрации T-клеточное перенаправленное уничтожение/лизис CEA-положительных клеток MKN-45 (A); HPAF-II (C) и LS-174T (D) или CEA-отрицательных клеток MKN-45_hCEAKO (B) KL биспецифическими антителами по изобретению AB54L3-1/N, AB60L3-1/N, AB66L3-1/N, AB71L3-1/N, AB72L3-1/N и AB73L3-1/N (анализ описан в примере 8a). Все KL биспецифические антитела имеют одно и то же анти-CD3 плечо. Y4L3-1/N, KL биспецифическое антитело с тем же анти-CD3 плечом, но со вторым плечом, не связывающим CEA, демонстрирует лишь очень низкую эффективность уничтожения/лизиса при максимальных протестированных концентрациях. TCB2014 представляет собой другое анти-CEAxCD3 T-клеточное биспецифическое антитело, описанное в US20140242079, которое было использовано в качестве эталонного антитела. В сравнении с этим эталонным антителом TCB2014, все из KL биспецифических антител по изобретению AB54L3-1/N, AB60L3-1/N, AB66L3-1/N, AB71L3-1/N, AB72L3-1/N и AB73L3-1/N демонстрировали более низкие значения EC50, соответственно, более эффективное уничтожение CEA-положительных клеток-мишеней (A, C, D), но эквивалентно низкое неспецифическое уничтожение/лизис клеток MKN-45 с нокаутом CEA (B). Для Фигуры 15A, C и D значения EC50 приведены в таблице 5.
Фиг. 16 Секреция цитокинов после опосредованного T-клетками уничтожения опухолевых клеток MKN45
Секреция перфорина (B); гранзима B (C); IFNγ (D); TNFα (E); IL-2 (F); IL-6 (G); IL-10 (H), опосредованная KL биспецифическими антителами по изобретению AB17L3-1/N, AB72L3-1/N и AB73L3-1/N после опосредованного T-клетками уничтожения опухолевых клеток MKN45 (Э:М (человеческие МКПК: опухолевые клетки)=10:1, 48-ч инкубация, специфический лизис показан в (A). Способ описан в примере 8e.
Фиг. 17 T-клеточная активация после опосредованного T-клетками уничтожения опухолевых клеток MKN45
Повышение регуляции CD25 (A и B) и CD69 (C и D) на человеческих CD4+ и CD8+ T-клетках, опосредованное KL биспецифическими антителами по изобретению AB17L3-1/N, AB72L3-1/N и AB73L3-1/N, через 2 дня после опосредованного T-клетками уничтожения CEA-положительных опухолевых клеток MKN45 (уничтожение/лизис опухолевых клеток показано на Фиг. 16A).
Несмотря на отсутствие статистически значимых отличий лизиса опухолевых клеток при 100 нМ, меньшая T-клеточная активация антителами по изобретению в сравнении с TCB2014 предполагает более низкие побочные эффекты при одинаковом лизисе опухолей, способ описан в примере 8c.
Фиг. 18 In vivo противоопухолевая эффективность в модели HPAF-II на мышах NOG с переносом чМКПК
In vivo противоопухолевая эффективность KL биспецифических антител по изобретению AB17L3-1/N, AB72L3-1/N и AB73L3-1/N, а также TCB2014 (все вводили инъекцией в дозе 10 мг/кг) в модели с линией опухолевых клеток HPAF-II на мышах NOG с переносом чМКПК. Мыши были рандомизированы в день 11, когда средний объем опухоли был близок к 150 мм3. Результаты представлены в виде средних значений и SEM для 8 мышей объема опухоли, измеренного штангенциркулем, в разных группах исследования. Не было обнаружено никакой статистически значимой разницы между AB73L3-1/N и TCB2014. Способ описан в примере 9a.
Определения
Термины, используемые в настоящем документе, имеют общепринятое в данной области значение, если иное не определено, как описано далее.
Используемый в настоящем документе термин «антигенсвязывающий фрагмент», или «связывающий фрагмент», означает в самом широком смысле фрагмент антитела, который специфически связывает антигенную детерминанту, такую как CEA, CD47 и CD3.
Более конкретно, при использовании в настоящем документе «связывающий фрагмент» означает фрагмент, который связывает мембраносвязанный карциноэмбриональный антиген человека (CEA, то же, что и CEACAM5) или CD3 человека, специфически связывает CEA или CD3, более конкретно, клеточный поверхностный или мембраносвязанный CEA или CD3. Выражения «специфически связывающий», «специфический для», «связывающийся с» означают, что связывание является избирательным для антигена, и может быть отличимо от нежелательных или неспецифических взаимодействий. В некоторых вариантах осуществления степень связывания направленного на мишень антитела с посторонним, нецелевым белком примерно в 10 раз, предпочтительно более чем в 100 раз, меньше, чем степень связывания антитела с указанной мишенью, при измерении, например, методом поверхностного плазмонного резонанса (ППР), например, Biacore®, методом иммуноферментного анализа (ELISA) или проточной цитометрии (FACS). Мишенями являются белки, описанные в настоящем документе, например, CEA, CD47 и CD3ε.
Термины «специфически связывает CEA, CD3», «связывает CEA, CD3» в одном варианте осуществления относятся к антителу, которое способно связывать мишени CEA и CD3, соответственно, с аффинностью, достаточной для того, чтобы антитело могло быть использовано в качестве лекарственного средства для перенаправления T-клеток на опухолевые клетки за счет связывания CD3 и CEA, соответственно.
Предпочтительно, биспецифическое антитело по изобретению связывает эпитоп CD3, который является консервативным у разных биологических видов, предпочтительно, у человека и яванского макака.
Используемый в настоящем документе термин «антитело» относится к антителу, содержащему две тяжелые цепи и две легкие цепи. В одном варианте осуществления антитело представляет собой полноразмерное антитело. Используемый в настоящем документе термин «тяжелая цепь антитела» относится к тяжелой цепи антитела, состоящей из вариабельной области (вариабельный домен) и константной области (константный домен), как определено для полноразмерного антитела. Используемый в настоящем документе термин «легкая цепь антитела» относится к легкой цепи антитела, состоящей из вариабельной области и константной области, как определено для полноразмерного антитела. Константные области легких цепей, используемые по настоящему изобретению, содержатся в легких цепях, описанных в настоящем документе.
Термин «полноразмерное антитело» означает антитело, состоящее из двух «полноразмерных тяжелых цепей антитела» и двух «полноразмерных легких цепей антитела». «Полноразмерная тяжелая цепь антитела» представляет собой полипептид, состоящий из расположенных в направлении от N-конца к C-концу тяжелой цепи антитела: вариабельного домена (VH), домена 1 константной области тяжелой цепи антитела (CH1), шарнирной области антитела (HR), домена 2 константной области тяжелой цепи антитела (CH2) и домена 3 константной области тяжелой цепи антитела (CH3), сокращенно обозначаемых VH-CH1-HR-CH2-CH3. «Полноразмерная легкая цепь антитела» представляет собой полипептид, состоящий из расположенных в направлении от N-конца к C-концу легкой цепи антитела: вариабельного домена (VL) и константного домена легкой цепи антитела (CL), сокращенно обозначаемых VL-CL. Константный домен легкой цепи антитела (CL) может быть типа κ (каппа) или λ (лямбда). Два домена полноразмерного антитела связаны между собой межполипептидными дисульфидными связями между доменом CL и доменом CH1, а также между шарнирными областями тяжелых цепей полноразмерного антитела. Примерами типичных полноразмерных антител являются природные антитела, такие как IgG (например, IgG1 и IgG2), IgM, IgA, IgD и IgE. В одном варианте осуществления полноразмерное антитело по изобретению представляет собой человеческое антитело IgG1 типа, в другом варианте осуществления оно имеет одну или более аминокислотных замен в Fc-фрагменте, как описано ниже. Полноразмерное антитело по изобретению содержит два связывающих фрагмента, каждый из которых образован парой VH и VL, один из них связывает CEA, и другой связывает CD3.
Используемый в настоящем документе термин «Fc-область», или «Fc-домен», относится к C-концевой области тяжелой цепи IgG; в случае IgG1 антитела C-концевая область содержит -CH2-CH3 (смотри выше). Хотя границы Fc-области тяжелой цепи IgG могут слегка варьироваться, Fc-область тяжелой цепи человеческого IgG, как правило, определяют как отрезок от аминокислотного остатка в положении Cys226 до карбоксильного конца.
Константные области хорошо известны в данной области и описаны, например, Kabat E.A., (смотри, например, Johnson, G., and Wu, T.T., Nucleic Acids Res. 28 (2000) 214-218; Kabat, E.A., et al., Proc. Natl. Acad. Sci. USA 72 (1975) 2785- 2788).
Термин «эпитоп» охватывает любую полипептидную детерминанту, способную к специфическому связыванию с антителом. В конкретных вариантах осуществления детерминанты эпитопов включают химически активные поверхностные группировки молекул, такие как аминокислоты, сахарные боковые цепи, фосфорил или сульфонил и, в конкретных вариантах осуществления, они могут иметь специфические трехмерные структурные характеристики и/или специфические характеристики заряда. Эпитоп представляет собой область мишени, которую связывает антитело.
Используемый в настоящем документе термин «общая тяжелая цепь (cHC)» относится к полипептиду, содержащему в направлении от N-конца к C-концу тяжелой цепи антитела: вариабельный домен (VH), константный домен 1 тяжелой цепи антитела (CH1), шарнирную область антитела (HR), константный домен 2 тяжелой цепи антитела (CH2) и константный домен 3 тяжелой цепи антитела (CH3), сокращенно обозначаемые VH-CH1-HR-CH2-CH3. Общими тяжелыми цепями, подходящими для биспецифических антител по изобретению, являются тяжелые цепи, описанные в WO2012023053, WO2013088259, WO2014087248, WO2019175658 и WO2016156537 (полное содержание всех из которых включено в настоящий документ посредством ссылки). В одном варианте осуществления cHC биспецифического антитела по изобретению содержит в качестве областей CDR тяжелой цепи CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4. В одном варианте осуществления cHC биспецифического антитела по изобретению содержит в качестве вариабельной области тяжелой цепи область VH с SEQ ID NO:1. В одном варианте осуществления cHC биспецифического антитела по изобретению содержит SEQ ID NO:43, 44 или 45.
Формат биспецифических антител по изобретению и содержание общей тяжелой цепи позволяет производить аффинную очистку биспецифических антител, которые неотличимы от стандартной молекулы IgG и имеют характеристики, неотличимые от характеристик стандартного моноклонального антитела (смотри, например, WO2013088259, WO2012023053), что предполагает низкий потенциал иммуногенности, или его отсутствие, у пациентов.
При использовании в настоящем документе обозначения AB1L3-1, AB17L3-1, AB54L3-1, AB60L3-1, AB66L3-1, AB71L3-1, AB72L3-1, AB73L3-1, и тому подобные, относятся к биспецифическим анти-CEAxCD3 антителам по изобретению, содержащим общую тяжелую цепь, содержащую в качестве областей CDR тяжелой цепи области CDR с SEQ ID NO:2, 3 и 4, и во втором связывающем фрагменте легкую цепь, содержащую в качестве областей CDR легкой цепи области CDR с SEQ ID NO:18, 19 и 20. Таким образом, AB1, и так далее, обозначает первый связывающий фрагмент (анти-CEACAM5 связывающий фрагмент) и L3-1 обозначает второй связывающий фрагмент (анти-CD3-связывающий фрагмент).
В одном варианте осуществления AB1L3-1, AB17L3-1, AB54L3-1, AB60L3-1, AB66L3-1, AB71L3-1, AB72L3-1, AB73L3-1, и тому подобные, содержат общую тяжелую цепь с SEQ ID NO:43 (чIgG1 ДТ), и во втором связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:28. Такие биспецифические антитела по изобретению в примерах также обозначены как AB1L3-1, AB17L3-1, AB71L3-1, AB72L3-1, AB73L3-1.
В одном варианте осуществления AB1L3-1, AB17L3-1, AB54L3-1, AB60L3-1, AB66L3-1, AB71L3-1, AB72L3-1, AB73L3-1, и тому подобные, содержат общую тяжелую цепь с SEQ ID NO:44 (чIgG1 с мутациями L234A+L235A), и во втором связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:28. Такие биспецифические антитела по изобретению обозначены в примерах как AB1L3-1/D, AB17L3-1/D, AB71L3-1/D, AB72L3-1/D, AB73L3-1/D и тому подобные.
В одном варианте осуществления AB1L3-1, AB17L3-1, AB54L3-1, AB60L3-1, AB66L3-1, AB71L3-1, AB72L3-1, AB73L3-1, и тому подобные, содержат общую тяжелую цепь с SEQ ID NO:45 (IgG1 с мутациями L234A+L235A+P329A), и во втором связывающем фрагменте в качестве легкой цепи легкую цепь с SEQ ID NO:28. Такие биспецифические антитела по изобретению обозначены в примерах как AB1L3-1/N, AB17L3-1/N, AB54L3-1/N, AB60L3-1/N, AB66L3-1/N, AB71L3-1/N, AB72L3-1/N, AB73L3-1/N и тому подобные.
Биспецифические антитела по изобретению, содержащие общую тяжелую цепь, могут быть получены, например, как описано в WO2012023053. Способами, описанными в WO2012023053, получают биспецифические антитела, которые идентичны по структуре человеческому иммуноглобулину. Молекула такого типа состоит из двух копий уникального полипептида тяжелой цепи, первой вариабельной области легкой цепи, слитой с каппа-константным доменом, и второй вариабельной области легкой цепи, слитой с лямбда-константным доменом.
В биспецифических антителах по изобретению один связывающий сайт проявляет специфичность для CEA, и другой сайт проявляет специфичность для CD3, в каждую из которых вносят вклад тяжелая и соответствующая легкая цепь. Вариабельные области легкой цепи могут относиться к семейству лямбда или каппа, и предпочтительно слиты с константными доменами лямбда и каппа, соответственно. Это предпочтительно во избежание создания неестественных полипептидных соединений. Однако можно также получать плечо антитела, подходящее для создания биспецифических антител по изобретению, путем слияния вариабельного домена легкой цепи каппа с константным доменом лямбда для любой из двух специфичностей или путем слияния вариабельного домена легкой цепи лямбда с константным доменом каппа, также для любой из двух специфичностей. Биспецифические антитела, описанные в WO 2012023053, представляют собой «κλ-антитела». Этот формат κλ-антител позволяет производить аффинную очистку биспецифического антитела, которое неотличимо от стандартной молекулы IgG с характеристиками, неотличимыми от стандартных характеристик моноклонального антитела, и, таким образом, обладает преимуществом в сравнении с предыдущими форматами, включая, например, аминокислотные мосты или другие неестественные элементы.
Важнейшим этапом способа является идентификация двух Fv-областей антитела (каждая из которых образована вариабельным доменом легкой цепи и вариабельным доменом тяжелой цепи), обладающих разной антигенной специфичностью, которые имеют одинаковый вариабельный домен тяжелой цепи. Описано множество способов получения моноклональных антител и их фрагментов. (Смотри, например, Antibodies: A Laboratory Manual, Harlow E, and Lane D, 1988, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY). Полностью человеческие антитела представляют собой молекулы антитела, в которых последовательность как легкой, так и тяжелой, цепи, включая области CDR1 и 2, закодированы генами человека. Область CDR3 может иметь человеческое происхождение или быть создана синтетическим путем. Такие антитела называют «человеческими антителами» или «полностью человеческими антителами». Человеческие моноклональные антитела могут быть получены с использованием технологии триомы; технологии человеческой B-клеточной гибридомы (смотри Kozbor, et al., 1983 Immunol Today 4: 72) и технологии EBV гибридомы для получения человеческих моноклональных антител (смотри Cole, et al., 1985 В: Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96). Человеческие моноклональные антитела могут быть использованы и могут быть получены с использованием человеческих гибридом (смотри Cote, et al., 1983. Proc Natl Acad Sci USA 80: 2026-2030) или путем трансформации человеческих B-клеток вирусом Эпштейна-Барр in vitro (смотри Cole, et al., выше).
Используемый в настоящем документе термин «CD3ε или CD3» относится к CD3ε человека, имеющей идентификатор UniProt P07766 (CD3E_HUMAN). Термин «антитело против CD3», или анти-CD3 антитело», относится к антителу, связывающему CD3ε.
Используемые в настоящем документе термины «CEA» и «CEACAM5» означают карциноэмбриональный антиген (CEA, CEACAM-5 или CD66e; UniProtKB - P06731) человека, который представляет собой клеточный поверхностный гликопротеин и опухоль-ассоциированный антиген (Gold and Freedman, J Exp. Med., 121:439-462, 1965; Berinstein NL, J Clin Oncol., 20:2197-2207, 2002). Используемый в настоящем документе термин «CEACAM6» означает CEACAM6 человека (CD66c; UniProtKB - P40199), который также является членом семейства родственных карциноэмбриональным антигенам молекул клеточной адгезии (CEACAM). Используемый в настоящем документе термин «CEACAM1» означает CEACAM1 человека (UniProtKB - P13688 (CEAM1_HUMAN), который также является членом семейства родственных карциноэмбриональным антигенам молекул клеточной адгезии (CEACAM). Используемый в настоящем документе термин «CEACAM8» означает CEACAM8 человека (UniProtKB - P31997 (CEAM8_HUMAN), который также является членом семейства родственных карциноэмбриональным антигенам молекул клеточной адгезии (CEACAM). Дополнительную информацию и информацию по другим членам семейства CEA можно получить по сетевому адресу http://www.uniprot.org.
Используемые в настоящем документе термины «специфически связывающий CEA фрагмент», «связывающий CEA фрагмент», «CEA-связывающий фрагмент» в контексте биспецифических антител по изобретению относятся к специфичности для CEACAM5 на поверхности клетки. Связывание CEA на клетках может быть измерено с использованием клеток MKN-45 аденокарциномы желудка, содержащих 100000-400000 копий CEA на клетку. Концентрация антитела по изобретению варьируется в соответствующем диапазоне в отношении полученного значения EC50 для связывания клеток MKN-45, упомянутых выше. Биспецифические антитела по изобретению специфически связывают такой мембраносвязанный CEACAM5.
Используемый в настоящем документе термин «мембраносвязанный CEA человека» относится к карциноэмбриональному антигену (CEA) человека, связанному с мембраной клетки или с поверхностью клетки, в частности, поверхностью опухолевой клетки. В некоторых обстоятельствах термин «мембраносвязанный CEA человека» может означать CEA, который не связан с мембраной клетки, но который был спроектирован для сохранения мембраносвязанного эпитопа CEA, который связывает антитело по изобретению.
Используемый в настоящем документе термин «без перекрестной реактивности в отношении CEACAM8» в контексте биспецифических антител по изобретению означает, что связывание биспецифического антитела по изобретению тестируют на клетках PEAK, экспрессирующих CEACAM8, в сравнении со связыванием с клетками PEAK ДТ (для подробностей смотри примеры 1 и 5), и отсутствие перекрестной реактивности означает, что значение СИФ, измеренное для клеток PEAK, экспрессирующих CEACAM8, не более чем в два раза превышает значение СИФ, измеренное для клеток PEAK ДТ. Используемый в настоящем документе термин «без перекрестной реактивности в отношении определенных CEACAM» в контексте биспецифических антител по изобретению означает отсутствие указанной перекрестной реактивности в той же экспериментальной процедуре и в соответствии с теми же определениями, что и в случае CEACAM8.
Используемые в настоящем документе термины «биспецифическое антитело, связывающее CEA человека и CD3 человека» или «анти-CEAxCD3 бсАт» означают биспецифическое антитело, связывающее CEACAM5 и CD3ε человека.
Используемый в настоящем документе термин «определяющая комплементарность область» («CDR») означает несмежные участки связывания антигена (также известные как антигенсвязывающие области), присутствующие в полипептидах вариабельной области как тяжелой, так и легкой, цепи. Области CDR также называют «гипервариабельными областями», и в настоящем документе этот термин используется взаимозаменяемо с термином «CDR» применительно к участкам вариабельной области, которые образуют антигенсвязывающие области. Эта конкретная область описана в публикации Kabat et al., U.S. Dept. of Health and Human Services, «Sequences of Proteins of Immunological Interest» (1983) и публикации Chothia et al., J. Mol. Biol. 196:901-917 (1987). В публикации Kabat et al. также определена система нумерации для последовательностей вариабельного домена, применимая к любому антителу. Специалист в данной области может однозначно применять эту систему «нумерации Kabat» к любой последовательности вариабельного домена, не опираясь на какие-либо экспериментальные данные по самой последовательности. При использовании в настоящем документе «нумерация Kabat» означает систему нумерации, приведенную в публикации Kabat et al., U.S. Dept. of Health and Human Services, «Sequence of Proteins of Immunological Interest» (1983). Если не указано иначе, ссылка на нумерацию положений определенных аминокислотных остатков в биспецифическом антителе по изобретению (например, последовательностей CDR) соответствует системе нумерации Kabat.
Используемый в настоящем документе термин «олигонуклеотид-направленный мутагенез» означает способ с использованием вырожденных олигонуклеотидов. Для мутагенеза каждой области CDR используют сочетание различных вырожденных олигонуклеотидов. Они содержат (но не ограничиваются ими) вырожденные кодоны NNS, HMT, DMT, NHT.
Используемый в настоящем документе термин «экспрессионный вектор» относится к одному или более векторам, которые содержат последовательности для тяжелой и легкой цепей антитела по изобретению соответствующим образом, известным на данном уровне техники. Используемый в настоящем документе термин «клетка-хозяин» охватывает клеточную систему любого вида, которая может быть генетически модифицирована для получения биспецифических антител по настоящему изобретению. В одном варианте осуществления клетка-хозяин генетически модифицирована для продуцирования антигенсвязывающей молекулы.
Используемый в настоящем документе термин «замена аминокислоты» означает замену одной аминокислоты другой аминокислотой из группы 20 стандартных протеиногенных аминокислот.
Терапевтические варианты применения и способы применения анти-CEAxCD3 антител по изобретению
Анти-CEACAMxCD3 биспецифические антитела по изобретению оптимизированы для лечения солидных опухолей, либо в монотерапии, либо в комбинированной терапии, в частности, в сочетании с анти-CD47 антителом, анти-CEAxCD47 антителом и/или антагонистом оси PD-1. Антитело по изобретению и анти-CD47 антитело или анти-CEAxCD47 антитело можно вводить как описано ниже.
В конкретном варианте осуществления заболевание, соответственно, солидная опухоль, представляет собой рак, клетки которого экспрессируют или избыточно экспрессируют CEA, включая, но без ограничения, группу, состоящую из колоректальной опухоли, немелкоклеточной опухоли легких, опухоли желудка, опухоли пищевода, опухоли поджелудочной железы и опухоли молочной железы. В конкретном варианте осуществления опухоль представляет собой колоректальную опухоль. Все терапевтические варианты применения, способы применения, сочетания и так далее, описанные в настоящем документе, в частности, являются вариантами осуществления для лечения этих опухолей/заболеваний.
Авторы изобретения признают, что антитела по изобретению обладают низким, или не обладают, потенциалом образования ADA и, соответственно, потери воздействия вследствие нейтрализующих ADA, и, соответственно, потери эффективности.
В одном варианте осуществления изобретение относится к способу лечения карцином (рака, опухолей, например, карцином человека), в частности, CEA-экспрессирующих опухолей, in vivo. Данный способ включает введение субъекту фармацевтически эффективного количества композиции, содержащей биспецифическое антитело по изобретению. Термин «субъект» относится к субъекту-человеку, в одном варианте осуществления - к пациенту, страдающему от рака/опухоли/карциномы.
Экспрессия CEA в различных опухолях, как правило, является очень высокой, в частности, при колоректальной карциноме, раке пищевода, аденокарциноме поджелудочной железы, раке желудка, немелкоклеточном раке легких, раке молочной железы, карциноме головы и шеи, раке тела матки и раке мочевого пузыря, в числе прочих. В здоровом нормальном железистом эпителии в желудочно-кишечном тракте CEA в основном экспрессируется поляризованным образом на апикальной поверхности клеток. Этот поляризованный паттерн экспрессии ограничивает доступность для анти-CEA моно- или биспецифических антител, которые вводят системно, следовательно, с потенциальной токсичностью. Этот поляризованный паттерн экспрессии теряется в клетках злокачественных опухолей желудочно-кишечного тракта. CEA экспрессируется в равной степени по всей клеточной поверхности раковых клеток, это означает, что раковые клетки намного более доступны для антитела по изобретению, чем нормальные здоровые клетки, и могут быть избирательно уничтожены анти-CEAxCD3 биспецифическими антителами по изобретению и, соответственно, сочетаниями, указанными выше.
В одном варианте осуществления биспецифические антитела по настоящему изобретению могут быть использованы в монотерапии для лечения прогрессирующих солидных опухолей, в одном варианте осуществления - CEA-экспрессирующих опухолей. В одном варианте осуществления биспецифическое антитело по изобретению используют в сочетании с анти-CEAxCD47 бсАт одновременно, раздельно или последовательно. В одном варианте осуществления биспецифическое антитело по изобретению используют в сочетании с анти-CEAxCD47 бсАт и/или антагонистом оси PD-1 одновременно, раздельно или последовательно. В одном варианте осуществления биспецифическое антитело по изобретению используют в сочетании с антагонистом оси PD-1 одновременно, раздельно или последовательно. Такие антагонисты оси PD-1 описаны, например, в WO2017118675. Такие сочетания обеспечивают атаку на солидные опухоли макрофагов и T-клеток. Анти-CD47 антитела описаны, например, в WO2009091601, WO2009091547, WO2011143624, WO2009131453, WO2013119714, WO2015105995, WO2017181033, WO2018026600, WO2019157432 и WO2013032948, и биспецифические антитела против CEA и CD47 описаны в PCT/IB2019/054559 и US16/428539.
Используемые в настоящем документе термины «сочетание», «одновременное, раздельное или последовательное введение сочетания» антитела по изобретению и второго антитела, связывающего CD47 человека или CEA человека и CD47 человека, относятся к любому введению двух антител (или трех антител в случае сочетания антитела по изобретению, анти-CD47 мАт или анти-CEAxCD47 бсАт и антагониста оси PD-1), либо раздельно, либо совместно, когда два или три антитела вводят в рамках соответствующего режима дозирования, разработанного для получения пользы от комбинированной терапии, например, путем раздельного, последовательного, одновременного, параллельного, хронологически смещенного или попеременного введения. Таким образом, два или три антитела можно вводить либо в виде части одной и той же фармацевтической композиции, либо в отдельных фармацевтических композициях. Антитело по изобретению можно вводить до, в процессе или после, введения второго биспецифического антитела, или в определенной комбинации этих режимов. Если антитело по изобретению вводят пациенту через определенные интервалы времени, например, в течение стандартного курса лечения, второе биспецифическое антитело можно вводить до, в процессе или после, каждого введения антитела по изобретению, или в определенной комбинации этих режимов, или через разные интервалы времени относительно введения антитела по изобретению, или в одной дозе до, в любой момент в процессе, или после курса лечения антителом по изобретению. В одном варианте осуществления антитело по изобретению и второе биспецифическое антитело вводят попеременным введением, в одном варианте осуществления с интервалами от 6 до 15 дней между введением антитела по изобретению и второго антитела. При таком попеременном введении введение первая доза может представлять собой антитело по изобретению или второе антитело.
Термин «антагонист оси PD-1» относится к анти-PD-1 антителу или анти-PD-L1 антителу. Анти-PD-1 антителами являются, например, пембролизумаб (кейтруда®, MK-3475), ниволумаб, пидилизумаб, ламбролизумаб, MEDI-0680, PDR001 и REGN2810. Анти-PD-1 антитела описаны, например, в WO200815671, WO2013173223, WO2015026634, US7521051, US8008449, US8354509, WO20091/14335, WO2015026634, WO2008156712, WO2015026634, WO2003099196, WO2009101611, WO2010/027423, WO2010/027827, WO2010/027828, WO2008/156712 и WO2008/156712. Анти-PD-L1 антителами являются, например, атезолизумаб, MDX-1105, дурвалумаб и авелумаб. Анти-PD-L1 антитела описаны, например, в WO2015026634, WO2013/019906, W02010077634, US8383796, WO2010077634, WO2007005874 и WO2016007235.
Что касается комбинированного введения антитела по изобретению и второго биспецифического антитела, оба соединения могут присутствовать в одной лекарственной форме или в отдельных лекарственных формах, например, в двух разных или идентичных лекарственных формах.
Если антитело по изобретению и второе антитело не конкурируют за связывание с CEACAM5, в одном варианте осуществления оба антитела вводят одновременно. Если антитело по изобретению и второе антитело конкурируют за связывание с CEACAM5, в одном варианте осуществления антитела вводят попеременно.
Антитело по изобретению, как правило, будут вводить пациенту в режиме дозирования, обеспечивающем наиболее эффективное лечение рака (как с точки зрения эффективности, так и с точки зрения безопасности), в связи с которым пациент получает лечение, как известно в данной области. Предпочтительно, опухолевые клетки атакуются одновременно T-клетками и макрофагами для достижения полного терапевтического потенциала этого подхода, анти-CEAxCD3 и анти-CEAxCD47 биспецифические антитела не должны конкурировать за связывание с CEA на клеточной поверхности.
Как описано выше, введенное количество антитела и время введения антитела по изобретению может зависеть от типа (например, пола, возраста, массы тела) и состояния здоровья пациента, получающего лечение, степени тяжести заболевания или состояния, подвергаемого лечению, и от пути введения. Например, антитело по изобретению и второе антитело можно вводить пациенту в диапазоне доз от 0,1 до 100 мг/кг массы тела в день или в неделю в одной или в разделенных дозах, или путем непрерывной инфузии. В одном варианте осуществления каждое из антитела по изобретению и второго антитела вводят пациенту в диапазоне доз от 1 до 20 мг/кг. В некоторых случаях уровни доз ниже нижнего предела вышеуказанного диапазона могут быть достаточными, в то время как в других случаях могут быть использованы даже большие дозы без вызывания какого-либо вредного побочного эффекта.
Используемый в настоящем документе термин «период полувыведения антитела» означает период полувыведения указанного антитела при измерении в обычном фармакокинетическом анализе. Антитело по изобретению и второе биспецифическое антитело против CEA и CD47 имеют период полувыведения, составляющий 3-14 дней.
В другом аспекте изобретение также относится к применению биспецифического антитела по изобретению в лечении заболевания, в частности, нарушений клеточной пролиферации, при которых экспрессируется CEA, в частности, при которых CEA экспрессируется аномально (например, избыточно экспрессируется или экспрессируется в ином паттерне на клеточной поверхности) в сравнении с нормальной тканью того же клеточного типа. Такие нарушения включают, но без ограничения, колоректальный рак, НМРЛ (немелкоклеточный рак легких), рак желудка, рак пищевода, рак поджелудочной железы и рак молочной железы. Уровни экспрессии CEA могут быть определены способами, известными в данной области (например, с использованием иммуногистохимического анализа, иммунофлуоресцентного анализа, иммуноферментного анализа, ELISA, проточной цитометрии, радиоиммунного анализа и так далее).
В одном аспекте биспецифические антитела по настоящему изобретению могут быть использованы для направления in vivo или in vitro на клетки, которые экспрессируют CEA. Биспецифические антитела по изобретению особенно полезны для предотвращения образования опухолей, уничтожения опухолей и ингибирования роста или метастазирования опухолей за счет индукции TDCC опухолевых клеток. Биспецифические антитела по изобретению могут быть использованы для лечения любой опухоли, экспрессирующей CEA. Конкретные злокачественные новообразования, которые можно лечить биспецифическими антителами по изобретению, включают, но без ограничения, колоректальный рак, немелкоклеточный рак легких, рак желудка, рак пищевода, рак поджелудочной железы и рак молочной железы.
Биспецифические антитела по изобретению вводят млекопитающему, предпочтительно человеку, в фармацевтически приемлемой лекарственной форме, такой как те, которые описаны ниже, включая те, которые можно вводить человеку внутривенно в виде болюса или путем непрерывной инфузии в течение определенного периода времени, внутримышечным, внутрибрюшинным, интрацереброспинальным, подкожным, внутрисуставным, интрасиновиальным, интратекальным, пероральным, топическим или ингаляционным путем введения. Биспецифические антитела по изобретению также можно вводить внутриопухолевым, перитуморальным, интралезиональным или перилезиональным путем введения для оказания как местного, так и системного терапевтического воздействия.
Соответствующая доза биспецифических антител по изобретению для лечения заболевания будет зависеть от типа заболевания, подвергаемого лечению, степени тяжести и течения заболевания, предшествующей терапии, истории болезни пациента и его ответа на антитело, а также от решения лечащего врача. Биспецифическое антитело по изобретению можно вводить пациенту один раз или в течение серии процедур. Настоящее изобретение относится к способу избирательного уничтожения опухолевых клеток, экспрессирующих CEA.
Данный способ включает взаимодействие биспецифических антител по изобретению с указанными опухолевыми клетками. Эти опухолевые клетки могут быть из карциномы человека, включая колоректальную карциному, немелкоклеточную карциному легких (НМРЛ), карциному желудка, карциному пищевода, карциному поджелудочной железы и карциному молочной железы.
В другом аспекте изобретение относится к биспецифическим антителам по изобретению для применения в производстве лекарственного препарата для лечения заболевания, связанного с аномальной экспрессией CEA. В конкретном варианте осуществления заболевание представляет собой рак, клетки которого экспрессируют или даже избыточно экспрессируют CEA, включая, но без ограничения, колоректальную опухоль, немелкоклеточную опухоль легких, опухоль желудка, опухоль пищевода, опухоль поджелудочной железы и опухоль молочной железы. В конкретном варианте осуществления опухоль представляет собой колоректальную опухоль.
Композиции, препараты, дозы и пути введения
В одном аспекте настоящее изобретение относится к фармацевтическим композициям, содержащим биспецифические антитела по настоящему изобретению и фармацевтически приемлемый носитель. Настоящее изобретение также относится к таким фармацевтическим композициям, используемым в способе лечения заболевания, такого как рак, или в производстве лекарственного препарата для лечения заболевания, такого как рак. В частности, настоящее изобретение относится к способу лечения заболевания, и более конкретно, лечения рака, включающему введение терапевтически эффективного количества фармацевтической композиции по изобретению.
В одном аспекте настоящее изобретение относится к фармацевтическим композициям, сочетаниям и способам лечения карцином, опухолей человека, описанных выше. Например, изобретение относится к фармацевтическим композициям, содержащим фармацевтически эффективное количество антитела по настоящему изобретению и фармацевтически приемлемый носитель, для применения в лечении карцином человека.
Композиции биспецифического антитела по изобретению можно вводить с использованием общепринятых способов введения, включая, но без ограничения, внутривенный, внутрибрюшинный, пероральный, интралимфатический или прямой внутриопухолевый путь введения. Внутривенное введение или подкожное введение являются предпочтительными.
В одном аспекте изобретения терапевтические препараты, содержащие биспецифические антитела по изобретению, готовят для хранения путем смешивания антитела, имеющего достаточную степень чистоты, с необязательными фармацевтически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16-е издание, Osol, A. Ed. (1980)), в виде лиофилизированных препаратов или жидких препаратов. Приемлемые носители, эксципиенты или стабилизаторы нетоксичны для реципиентов в используемых дозах и концентрациях. Препараты, используемые для in vivo введения, должны быть стерильными. Это легко достигается путем фильтрования через стерильные фильтрационные мембраны. Наиболее эффективный способ введения и режим дозирования для фармацевтических композиций по настоящему изобретению зависит от степени тяжести и течения заболевания, состояния здоровья пациента и его ответа на лечение, а также от мнения лечащего врача. Соответственно, дозы композиций могут быть фиксированными дозами или могут быть адаптированы для отдельного пациента, например, зависеть от массы тела. Тем не менее, эффективная доза композиций по настоящему изобретению, как правило, будет находиться в диапазоне от 0,1 до 20 мг/кг.
Биспецифические антитела по настоящему изобретению имеют молекулярную массу в размере 150 кДа на моль. В одном варианте осуществления они имеют Fc-фрагмент. Период полувыведения в организме пациентов находится в диапазоне 3-14 дней. Этот период полувыведения допускает, но без ограничения, введение один раз в день, один раз в неделю или один раз в две недели.
Биспецифические антитела по настоящему изобретению, и их соответствующие композиции, могут находиться в различных лекарственных формах, которые включают, но без ограничения, жидкие растворы или суспензии, таблетки, пилюли, порошки, суппозитории, полимерные микрокапсулы или микровезикулы, липосомы, а также инъекционные или инфузионные растворы. Предпочтительная форма зависит от способа введения и варианта терапевтического применения.
Композиция, содержащая биспецифическое антитело по настоящему изобретению, будет сформулирована, разделена на дозы и введена в соответствии с надлежащей медицинской практикой. Факторы для рассмотрения в данном контексте включают конкретное заболевание или нарушение, подвергаемое лечению, конкретное млекопитающее, получающее лечение, клиническое состояние отдельного пациента, причину заболевания или нарушения, участок доставки средства, способ введения, расписание введения и другие факторы, известные практикующим врачам.
Изделия
Другой аспект изобретения относится к изделию, включающему материалы, полезные для лечения, предотвращения и/или диагностирования заболеваний, описанных выше. Изделие включает контейнер и этикетку или вкладыш в упаковку на контейнере, или в связке с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для в/в растворов и так далее. Контейнеры могут быть выполнены из различных материалов, таких как стекло или пластик. Контейнер заключает в себе композицию, которая может быть сама по себе, или в сочетании с другой композицией, эффективной для лечения, предотвращения и/или диагностирования состояния, и может иметь порт стерильного доступа (например, контейнер может представлять собой мешок для внутривенного раствора или флакон, имеющий пробку, прокалываемую иглой для подкожных инъекций). Одно активное средство в композиции представляет собой биспецифическое антитело по изобретению. На этикетке или вкладыше в упаковку указано, что композиция используется для лечения конкретного заболевания. Кроме того, изделие может включать (a) первый контейнер с композицией, содержащейся в нем, где композиция содержит биспецифическое антитело по изобретению; и (b) второй контейнер с композицией, содержащейся в нем, где композиция содержит дополнительное цитотоксическое или иное лекарственное средство. В данном варианте осуществления изобретения изделие может также включать вкладыш в упаковку, указывающий, что композиции могут быть использованы для лечения определенного заболевания. Альтернативно или дополнительно, изделие может также включать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-солевой буфер, раствор Рингера и раствор декстрозы. Оно также может включать другие материалы, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Таблица 1
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
ПРИМЕРЫ
Пример 1. Клонирование, экспрессия и очистка представителей семейства CEACAM человека
Клонирование
Последовательности, соответствующие целому внеклеточному домену (ECD) и A3-B3 доменам CEACAM5, синтезировали и субклонировали в экспрессионный вектор млекопитающих pEAK8 (Edge Biosystems, Gaithersburg, Md.). Векторы были модифицированы для введения Avitag™ (Avidity, Denver Colo.) и либо гексагистидиновой метки, человеческой Fc-области, либо мышиной Fc-области на C-конце. Конструкции подтверждали методом секвенирования ДНК. Очистку рекомбинантного растворимого белка проводили методом IMAC (аффинная хроматография с иммобилизованными ионами металла) с аффинной матрицей FcXL или CaptureSelect™ IgG-Fc (ms) (Thermo Fisher Scientific).
Также были получены векторы, кодирующие полноразмерный вариант CEACAM 1, 3, 4, 5, 6, 7, 8, 18, 19, 20, 21 человека, а также CEACAM5 и CEACAM6 яванского макака, для экспрессии на клеточной поверхности клеток PEAK и/или CHO. Растворимый полноразмерный CEACAM16 человека клонировали аналогичным образом.
Кроме того, также были получены векторы, кодирующие следующие укороченные варианты CEACAM5 человека, для экспрессии на клеточной поверхности клеток PEAK и/или CHO: A1-B1-A2-B2-A3-B3; B1-A2-B2-A3-B3; A2-B2-A3-B3; B2-A3-B3; A3-B3. Субдомен B3 экспрессируется в виде слитого белка с первыми 140 ак человеческого белка CD86.
Экспрессия
Плазмиды, указанные выше, трансфицировали в клетки млекопитающих с использованием реагента для трансфекции на основе липосом, такого как липофектамин 2000 (Thermo Fisher Scientific). Для этапа трансфекции необходимы лишь небольшие количества ДНК и клеток, как правило, 4×105 клеток и 2 мкг плазмидной ДНК на лунку, трансфекцию проводили в 6-луночном планшете. Хотя можно использовать разные линии клеток млекопитающих, в примерах, приведенных ниже, трансфицировали монослой трансформированных эпителиальных клеток зародышевой почки человека (клетки PEAK). Эти клетки стабильно экспрессируют ген EBNA-1, дополнительно поддерживая процесс эписомной репликации, являются полуприкрепляющимися и могут расти в стандартных условиях культивирования клеток (5% CO2; 37°C в среде DMEM с добавлением 10% эмбриональной телячьей сыворотки). Через 24 ч клетки помещали в условия селекции путем добавления среды, содержащей 0,5-2 мкг/мл пуромицина: клетки, содержащие эписомный вектор, являются устойчивыми к этому антибиотику.
Через две-три недели после трансфекции размноженные и отобранные клетки инъецировали в биореакторы CELLine™ одноразового использования (Sigma Aldrich) для этапа продуцирования. CELLine™ представляет собой двухкамерный биореактор, который можно использовать в стандартном инкубаторе для клеточных культур. Меньший отсек (15 мл) содержит клетки и отделен от большего (один литр) отсека, содержащего среду, полупроницаемой мембраной с порогом отсечения 10 кДа (Bruce et al. 2002, McDonald et al. 2005). Эта система допускает диффузию питательных веществ, газов и продуктов метаболизма, при этом удерживая клетки и секретируемые белки в меньшем отсеке. Культуру поддерживали в течение 7-10 дней до сбора супернатанта. Поскольку среда содержит сыворотку, клетки сохраняют хорошую жизнеспособность, и может быть получено несколько производственных партий с использованием одних и тех же клеток и контейнеров.
Очистка
После сбора супернатанты клеточных культур осветляли центрифугированием. Затем в супернатант добавляли 100 мМ имидазол и наносили на смолу Ni-NTA для аффинной хроматографии (Qiagen). Относительно высокая концентрация имидазола сводит к минимуму связывание примесей со смолой. После промывания колонки белки элюировали при скорости потока 2 мл/мин с использованием 30 мл градиента имидазола (20-400 мМ имидазол) на хроматографической системе AKTA Prime (Cytiva). Градиентное элюирование еще больше повышает чистоту рекомбинантного белка, но может быть заменено на ступенчатое элюирование, если хроматографическая система не доступна. Элюированные фракции могут быть проанализированы методом SDS-ПААГ или ELISA для определения содержания в них рекомбинантного белка. Интересующие фракции объединяли и обессоливали на колонках 10-кДа Amicon® (Millipore), уравновешенных фосфатно-солевым буфером или другим подходящим буфером. Обессоленные белки затем можно количественно определять различными методами, и их чистоту анализировать методом SDS-ПААГ. Рекомбинантные белки биотинилировали in vitro с использованием биотин-лигазы (Avidity, Denver Colo.) согласно инструкциям производителя. После обессоливания уровень биотинилирования оценивали в анализах с осаждением, используя магнитные гранулы со стрептавидином и метод SDS-ПААГ.
Пример 2. Селекция методом фагового дисплея CEACAM5 Fv с использованием библиотек scFv человека, содержащих фиксированный вариабельный домен тяжелой цепи
Общие методы создания и использования библиотек scFv человека, экспонированных на бактериофаге M13, описаны в публикации Vaughan et al., (Nat. Biotech. 1996, 14:309-314), полное содержание которой включено в настоящий документ посредством ссылки. Библиотеки для селекции и скрининга кодируют scFv, все из которых имеют общий одинаковый для всех VH-домен и отличаются только VL-доменом. Способы создания библиотек с фиксированным VH-доменом, и их использования для идентификации и сборки биспецифических антител, описаны в US 2012/0184716 и WO 2012/023053, полное содержание всех из которых включено в настоящий документ посредством ссылки. Методы определения связывания scFv с CEACAM5 человека описаны ниже.
Селекция белков
Аликвоты библиотек scFv в фаге (1012 БОЕ) блокировали PBS, содержащим 3% (масс/об) обезжиренного молока, в течение одного часа при комнатной температуре на роторной мешалке. Проводили селекцию блокированных фагов на магнитных гранулах со стрептавидином (Dynabeads™ M-280) в течение одного часа при комнатной температуре на роторной мешалке. Подвергнутые селекции фаги инкубировали с 100 нМ биотинилированным либо CEACAM5, либо A3-B3 доменом, человека, удерживаемыми на магнитных гранулах со стрептавидином, в течение двух часов при комнатной температуре на роторной мешалке. Гранулы собирали с использованием магнитной подставки, с последующим пятикратным промыванием раствором PBS/0,1% Tween® 20 и двукратным промыванием PBS. Фаги элюировали 100 нМ TEA в течение 30 минут при комнатной температуре на роторной мешалке. Элюированные фаги и гранулы нейтрализовали 1M Tris-HCl, pH 7,4, непосредственно добавляли к 10 мл клеток TG1 в экспоненциальной фазе роста (штаммы E. coli, обычно используемые в фаговом дисплее) и инкубировали в течение одного часа при 37°C с медленным встряхиванием (90 об/мин). Аликвоту инфицированных TG1 серийно разбавляли для титрования результатов селекции. Оставшиеся инфицированные TG1 центрифугировали при 3800 об/мин в течение 10 минут, ресуспендировали в 2 мл 2xTY и распределяли на чашках с агаром для биоанализа 2xTYAG (2xTY среда, содержащая 100 мкг/мл ампициллина и 2% глюкозы). После инкубации в течение ночи при 30°C в чашки добавляли 10 мл 2xTY, клетки соскребали с поверхности и переносили в 50-мл полипропиленовую пробирку. К клеточной суспензии добавляли 50% раствор глицерина для получения конечной концентрации 17% глицерина. Аликвоты после раундов селекции хранили при -80°C.
Извлечение фага
50 мкл клеточной суспензии, полученной в предыдущих раундах селекции, добавляли к 50 мл 2xTYAG и выращивали при 37°C с перемешиванием (240 об/мин) до достижения значения OD600, составляющего 0,3-0,5. Затем культуру суперинфицировали 1,2×1011 M13K07 фага-помощника и инкубировали в течение одного часа при 37°C (90 об/мин). Среду заменяли путем центрифугирования клеток при 3800 об/мин в течение 10 минут, удаления среды и ресуспендирования осадка в 50 мл 2xTYAK (среда 2xTY, содержащая 100 мкг/мл ампициллина; 50 мкг/мл канамицина). Затем культуру выращивали в течение ночи при 30°C (240 об/мин). На следующий день супернатант, содержащий фаг, использовали для следующего раунда селекции.
Селекция на клеточной поверхности
Супернатанты, содержащие фаги, блокировали PBS, содержащим 3% (масс/об) обезжиренного молока, в течение одного часа при комнатной температуре на роторной мешалке. Затем проводили отсев блокированных фагов в течение одного часа на клетках MKN-45 CEACAM5KO, которые не экспрессируют CEACAM5 человека. Фаги после отсева инкубировали с 2×107 клеток MKN-45, экспрессирующих CEACAM5 (блокированных в PBS, 3% БСА, 0,1% NaN3), в течение двух часов при комнатной температуре с осторожным встряхиванием. Клетки осаждали и промывали шесть раз PBS. Связанные фаги элюировали 76 мМ лимонной кислотой при встряхивании в течение 10 минут. После нейтрализации при помощи 1M Tris-HCl, pH 8, клетки добавляли непосредственно к 10 мл TG1 в экспоненциальной фазе роста и инкубировали в течение одного часа при 37°C с медленным встряхиванием. Аликвоту инфицированных TG1 серийно разбавляли для титрования результатов селекции. Инфицированные TG1 центрифугировали при 3800 об/мин в течение 10 минут, ресуспендировали в 2 мл среды 2xTY и распределяли на чашках с агаром для биоанализа 2xTYAG. После инкубации в течение ночи при 30°C в чашки добавляли 10 мл 2xTY, клетки соскребали с поверхности и переносили в 50-мл полипропиленовую пробирку. К клеточной суспензии добавляли 50% раствор глицерина для получения конечной концентрации 17% глицерина. Аликвоты после раундов селекции хранили при -80°C.
Пример 3. Скрининг на связывание/не связывание scFv с растворимыми CEACAM5, CEACAM6 и CEACAM1
Периплазматический препарат scFv для тестов на связывание и функциональных тестов
Отдельные трансформированные клоны TG1 из полученных продуктов селекции инокулировали в микропланшет для титрования с глубокими лунками, содержащими 0,9 мл на лунку среды 2xTYAG (среда 2xTY, содержащая 100 мкг/мл ампициллина 0,1% глюкозы) и выращивали при 37°C в течение 5-6 часов (240 об/мин). Затем добавляли 100 мкл на лунку 0,2 мМ IPTG в среде 2xTY для получения конечной концентрации 0,02 мМ IPTG. Планшет инкубировали в течение ночи при 30°C со встряхиванием при 240 об/мин. Планшет с глубокими лунками центрифугировали при 3200 об/мин в течение 10 минут при 4°C, и супернатант осторожно удаляли. Осадки ресуспендировали в 150 мкл буфера TES (50 мМ Tris-HCl (pH 8), 1 мМ ЭДТА (pH 8), 20% сахароза, с добавлением полного ингибитора протеаз, Roche). Создавали гипотонический шок путем добавления 150 мкл разбавленного буфера TES (разбавление 1:5 TES:вода) и инкубации на льду в течение 30 минут. Планшет центрифугировали при 4000 об/мин в течение 10 минут при 4°C для осаждения клеток и детрита. Супернатанты осторожно переносили в другой микропланшет для титрования и хранили на льду для немедленного тестирования в функциональных анализах или анализах связывания.
Связывание
Скрининг scFv на связывание с CEACAM5 проводили в гомогенном анализе с использованием технологии CellInsight™. Следующие реагенты смешивали в каждой лунке 384-луночного планшета с прозрачными лунками (Corning): 30 мкл суспензии полистирольных гранул со стрептавидином (Polysciences; 3000 гранул/лунку), покрытых или биотинилированным CEACAM5, биотинилированным доменом A3-B3, или биотинилированным NusA в качестве контрольного белка; 60 мкл блокированного периплазматического препарата scFv; 10 мкл обнаруживающего буфера (PBS, содержащий мышиное анти-c-myc антитело в концентрации 5 мкг/мл; конъюгат антител против Fc мыши с AlexaFluor® 647, разбавленный 1:200). После смешивания при 600 об/мин в течение 5 минут 384-луночный планшет инкубировали при комнатной температуре, и через 2 часа снимали показания на платформе для одновременного многопараметрического анализа CellInsight™ CX5 (ThermoFisher Scientific). Клоны, экспрессирующие scFv со специфическим сигналом для CEACAM5, но не NusA, выбирали для дальнейшего анализа или секвенирования.
Связывание с CEACAM1, CEACAM6 и другими CEACAM можно определять таким же образом.
Секвенирование клонов
Отдельные клоны инокулировали в 96-луночный микропланшет для титрования с глубокими лунками, содержащими 1 мл среды LBAG (среда LB с 100 мкг/мл ампициллина и 2% глюкозы) на лунку, и выращивали в течение ночи при 37°C, 300 об/мин. ДНК экстрагировали с использованием набора Zyppy-96 Plasmid Miniprep kit (Zymo Research) и секвенировали.
Пример 4. Перевод кандидатов с фиксированной VH в формат IgG и временная экспрессия в клетках млекопитающих
После скрининга и секвенирования scFv-кандидаты с желаемыми свойствами связывания переводили в формат IgG и осуществляли экспрессию путем временной трансфекции в клетки PEAK. Последовательности VH и VL выбранных scFv амплифицировали со специфическими олигонуклеотидами, клонировали в экспрессионный вектор, содержащий последовательности для константных областей тяжелой и легкой цепей, и конструкции подтверждали секвенированием. Экспрессионные векторы трансфицировали в клетки млекопитающих при помощи липофектамина 2000 (Thermo Fisher Scientific) согласно инструкциям производителя. Вкратце, 4×106 клеток PEAK культивировали во флаконах T75 в 25 мл среды для культивирования, содержащей эмбриональную бычью сыворотку. Трансфицированные клетки культивировали в течение 5-6 дней при 37°C, и продуцирование IgG количественно определяли с использованием прибора Octet RED96. Супернатант собирали для очистки IgG на аффинной смоле FcXL (Thermo Fisher Scientific) согласно инструкциям производителя. Вкратце, супернатанты от трансфицированных клеток инкубировали в течение ночи при 4°C с соответствующим количеством смолы FcXL. После промывания смолы PBS образцы наносили на колонку Amicon Pro и затем IgG элюировали в 50 мМ глициновом буфере, pH 3,5. Затем проводили диализ элюированной фракции IgG с использованием 50 кДа Amicon против буфера гистидин-NaCl, pH 6,0, и количественное содержание IgG определяли по поглощению при 280 нм. Чистоту и целостность и IgG подтверждали методом электрофореза с использованием Agilent Bioanalyzer 2100 в соответствии с инструкциями производителя (Agilent Technologies, Santa Clara, Calif., USA).
Пример 5. Определение характеристик анти-CEACAM5 моноклональных антител
a) Связывание анти-CEACAM5 плеч с клетками, трансфицированными разными членами семейства CEACAM
Специфичность анти-CEACAM5 плеч антител (протестированных в виде двухвалентных мАт или одновалентных бсАт) показана методом проточной цитометрии с использованием клеток PEAK и/или CHO, трансфицированных разными членами семейства CEACAM.
Векторы, кодирующие полноразмерный вариант CEACAM 1, 3, 4, 5, 6, 7, 8, 18, 19, 20 и 21 и 20 человека, использовали для экспрессии этих белков на поверхности клеток PEAK и/или CHO, как описано в примере 1. Аналогично, векторы, кодирующие полноразмерный вариант CEACAM 5 и 6 яванского макака, также использовали для экспрессии этих белков на поверхности клеток PEAK и/или CHO. Не трансфицированные клетки PEAK и/или CHO использовали в качестве отрицательного контроля. Клетки собирали, подсчитывали, проверяли на жизнеспособность и ресуспендировали при плотности 3×106 клеток/мл в FACS-буфере (PBS 2% БСА, 0,1% NaN3). 100 мкл клеточной суспензии вносили в лунки 96-луночных планшетов с V-образным дном (3×105 клеток/лунку). Супернатант удаляли центрифугированием в течение 3 минут при 4°C, 1300 об/мин, и клетки инкубировали в течение 15 минут при 4°C с антителом по изобретению в возрастающих концентрациях. Антитела с анти-CEACAM5 плечами для тестирования разбавляли в FACS-буфере, диапазон концентраций составлял 30 пМ-500 нМ. Клетки промывали дважды холодным FACS-буфером и повторно инкубировали еще в течение 15 минут при 4°C с совместимым вторичным антителом против IgG человека. Клетки промывали дважды холодным FACS-буфером и ресуспендировали в 300 мкл FACS-буфера с разведенным 1:1500 TOPRO-3 (Invitrogen). Флуоресценцию измеряли с использованием FACSCalibur™ (BD Biosciences) или платформы Cytoflex (Beckman Coulter). Строили кривые зависимости ответа от дозы для связывания с использованием программы GraphPad Prism8. Таким же образом можно определять характеристики для CEACAM1, CEACAM6 и других CEACAM.
Результаты, полученные с использованием экспериментальных методов, описанных в примерах 1 и 5a, представлены в таблицах 2 и 3 (протестированы полноразмерные антитела в концентрации 10 мкг/мл; BiTE MEDI-565 тестировали в эквимолярной концентрации). Показано, что в случае биспецифических антител AB17L3-1/N, AB71L3-1/N, AB72L3-1/N, AB73L3-1/N значение СИФ, измеренное для связывания трансфицированных CEACAM5 клеток, составляло от 29000 до 41000 (Таблица 2). Напротив, значение СИФ, определенное с использованием клеток PEAK, трансфицированных CEACAM 1,3,4,6,8, было ниже 1000, с единственным исключением в виде сильного сигнала для AB72L3-1/N на трансфицированных CEACAM8 клетках. Если значения СИФ, полученные на трансфицированных клетках, экспрессирующих любой конкретный CEACAM, разделить на значения, полученные на клетках PEAK ДТ, можно рассчитать «коэффициенты относительно PEAK ДТ» (Таблица 3). За исключением AB72L3-1/N, все антитела по изобретению являются специфическими для CEACAM5, поскольку все значения «коэффициента относительно PEAK ДТ» были ниже 2,0. Напротив, для MEDI-565 BiTE был получен «коэффициент относительно PEAK ДТ», превышающий 2, в случае CEACAM8, это указывало на перекрестную реактивность с этим членом семейства CEACAM. Это может приводить к уничтожению, например, нейтрофилов, поскольку, как описано выше, человеческие нейтрофилы экспрессируют CEACAM8 на поверхности.
ТАБЛИЦА 2. Связывание с CEACAMx, временно экспрессированным на клетках PEAK [СИФ]
ТАБЛИЦА 3. Связывание с CEACAMx, временно экспрессированным на клетках PEAK [коэффициент относительно PEAK ДТ]
b) Связывание анти-CEACAM5 моноклональных антител с рекомбинантными белками в твердофазном иммуноферментном анализе (ELISA)
Биотинилированный рекомбинантный белок CEACAM5 человека в концентрации 0,5 мкг/мл был захвачен в лунках покрытого стрептавидином 96-луночного микропланшета. Планшет промывали, моноклональные анти-ОАА двухвалентные антитела по настоящему изобретению добавляли в широком диапазоне концентраций (например, от 5×10-4 до 1 мкг/мл) и инкубировали в течение 1 часа. Планшет промывали, и связанные антитела обнаруживали при помощи конъюгата антител против человеческого IgG (Fc)-HRP (Jackson ImmunoResearch). После промывания в лунки планшета вносили реагент Amplex Red® (Molecular Probes). Сигнал флуоресценции измеряли на планшетном ридере Synergy HT (Biotek).
Связывание с другими рекомбинантными членами семейства CEACAM, такими как CEACAM1 и CEACAM6, можно оценивать аналогичным образом. Результаты связывания представлены на Фигурах 8 и 9, для мАт 1B4 и мАт C11, соответственно.
c) Эпитоп-специфическая сортировка антител по изобретению путем конкуренции с эталонными антителами
Эпитоп-специфическая сортировка представляет собой конкурентный иммуноанализ, используемый, например, для определения характеристик связывания новых моноклональных антител с целевым белком. Профиль конкурентного блокирования при связывании нового антитела с целевым белком анализировали в сравнении с антителами, также связывающими этот целевой белок, для которых связываемый эпитоп уже был установлен/опубликован. Конкуренция с одним из этих эталонных антител указывает на то, что новое антитело имеет тот же самый, или близко расположенный, эпитоп, и они «сгруппированы» вместе.
Способность анти-CEACAM5 мАт по настоящему изобретению конкурировать с анти-CEACAM5 эталонными антителами тестировали методом ELISA на рекомбинантном CEACAM5 человека со следующими эталонными антителами, имеющими мышиную Fc-область: SM3E, мАт, полученное из sm3E, описанного в патенте US20050147614A1; MEDI, мАт, полученное из MEDI-565, описанного в патенте WO2016036678A1; SAR, мАт, полученное из Mab2_VLg5VHg2, описанного в патенте EP3199552A1; CH1A1A, мАт, полученное из CH1A1A-2F1, описанного в патенте US20120251529 и в публикации Klein et al. Oncoimmunology, 2017 Jan 11;6(3); гуманизированное T84.66, мАт, полученное из варианта 1, описанного в патенте WO2017055389; LAB, мАт, полученное из hMN14, описанного в патенте US 2002/0165360 A1.
SM3E связывается, например, больше с N-концом, дистальной по отношению к клеточной мембране частью CEA, MEDI со средней частью, и CH1A1A связывается вблизи мембраны.
Биотинилированное CEACAM5 человека наносили в концентрации 0,5 мкг/мл в покрытые стрептавидином лунки 96-луночного планшета и инкубировали с эталонным мАт в концентрации 10 мкг/мл или с посторонним мАт, имеющим мышиную Fc-область, в течение 1 часа. Анти-CEACAM5 мАт по настоящему изобретению (то есть, двухвалентные моноклональные анти-CEA антитела) добавляли в концентрации 0,2 мкг/мл на 1 час при комнатной температуре. Планшет промывали, и связанные анти-CEACAM5 мАт обнаруживали при помощи конъюгата антител против человеческих IgG (Fc)-HRP (Jackson ImmunoResearch). После промывания в лунки планшета вносили реагент Amplex Red®. Сигнал флуоресценции измеряли на планшетном ридере Synergy HT (Biotek).
Исходя из результатов, полученных с анти-CEACAM5 мАт, производное анти-CEAxCD3 бсАт по изобретению считали конкурирующим с эталонным антителом в случае, если связывание с CEACAM5 уменьшалось более чем на 80% при сравнении результатов с добавлением и без добавления инструментального антитела. Анти-CEAxCD3 антитело считали не конкурирующим с инструментальным антителом в случае, если связывание с CEACAM5 уменьшалось на менее чем 20% при сравнении результатов с добавлением и без добавления инструментального антитела. На Фигуре 1 схематически показаны области связывания эталонных антител, используемых в примере 5c.
d) Определение домена CEACAM5, связываемого антителами по изобретению, с использованием укороченной формы CEACAM5
Используя укороченные формы CEACAM5, лишенные одного или более из его внеклеточных субдоменов, можно определять субдомен, который связывают антитела по изобретению.
Векторы, кодирующие полноразмерный вариант CEACAM5 человека (содержащий все его внеклеточные домены, а именно, N-A1-B1-A2-B2-A3-B3), а также векторы, кодирующие только подгруппу внеклеточных доменов CEACAM5 (A1-B1-A2-B2-A3-B3; B1-A2-B2-A3-B3; A2-B2-A3-B3; B2-A3-B3; A3-B3 и B3), использовали для экспрессии этих белков на поверхности клеток PEAK и/или CHO, как описано в примере 1. Не трансфицированные клетки PEAK и/или CHO использовали в качестве отрицательного контроля. Окрашивание методом проточной цитометрии и получение результатов производили, как описано в примере 5, подраздел a).
Антитело по изобретению считали связывающим конкретный укороченный белок CEACAM5, если связанное антитело было обнаружено при помощи PE-конъюгированного вторичного антитела против Fc IgG человека.
Пример 6. Экспрессия и очистка биспецифических антител, имеющих лямбда и каппа легкую цепь
Одновременная экспрессия одной тяжелой цепи и двух легких цепей в одной и той же клетке может приводить к сборке трех разных антител. Одновременная экспрессия может быть достигнута разными путями, такими как трансфекция несколькими векторами, экспрессирующими одну из цепей, для их совместной экспрессии или использование векторов, которые управляют экспрессией нескольких генов. Вектор, кодирующий разные анти-CEACAM5 антитела, трансфицировали совместно с другим вектором, экспрессирующим тяжелую и легкую цепи анти-CD3 антитела. Альтернативно, две легкие цепи клонировали в вектор pNovi κHλ, который получали ранее, для возможности совместной экспрессии одной тяжелой цепи, одной легкой цепи каппа и одной легкой цепи лямбда, как описано в US 2012/0184716 и WO 2012/023053, которые включены в настоящий документ посредством ссылки в полном объеме. Экспрессия трех генов управляется промоторами человеческого цитомегаловируса (hCMV), и вектор также содержит ген глутамин-синтетазы (GS), который позволяет проводить селекцию и получать стабильные линии клеток. Общие гены VH и VL анти-CEACAM5 IgG и анти-CD3 IgG клонировали в вектор pNovi κHλ для временной экспрессии в клетках млекопитающих. Клетки Expi293 культивировали в суспензии в соответствующей колбе Эрленмейера, используя подходящее количество клеток и объем среды для культивирования. Плазмидную ДНК в итоге трансфицировали в клетки Expi293 с использованием PEI. Концентрацию антитела в супернатанте трансфицированных клеток определяли в процессе продуцирования с использованием Octet RED96. В зависимости от концентрации антитела супернатанты собирали через 5-7 дней после трансфекции и осветляли центрифугированием при 1300 g в течение 10 мин. Процесс очистки включал три этапа. Во-первых, аффинную матрицу CaptureSelect™ FcXL (Thermo Fisher Scientific) промывали PBS, а затем добавляли в осветленный супернатант. После инкубации в течение ночи при +4°C и 20 об/мин супернатанты центрифугировали при 2000 g в течение 10 мин, фильтрат сохраняли, и смолу промывали дважды PBS. Затем смолу переносили на колонки Amicon Pro, и для элюции использовали раствор, содержащий 50 мМ глицин с pH 3,0. Получали несколько элюированных фракций, их нейтрализовали Tris-HCl pH 7,4 и объединяли. Пул, содержащий суммарные человеческие IgG, (биспецифическое и два моноспецифических антитела) количественно анализировали с использованием спектрофотометра Nanodrop (NanoDrop Technologies, Wilmington, Del.), а затем инкубировали в течение 30 мин при RT и 20 об/мин с соответствующим объемом аффинной матрицы CaptureSelect™ KappaXL (Thermo Fisher Scientific).GE Healthcare). Этапы извлечения и промывания смолы, элюирования и нейтрализации выполняли как описано ранее. Последний этап аффинной очистки выполняли с использованием аффинной матрицы CaptureSelect™ для лямбда-Fab (Thermo Fisher Scientific), используя тот же метод, что и для этапа очистки каппа. Все элюированные фракции объединяли и обессоливали против буфера для препарата His-NaCl, pH 6, с использованием 50-кДа ячеек для фильтрования центрифугированием Amicon Ultra (Merck Millipore). Конечный продукт количественно оценивали с использованием Nanodrop.
Очищенные биспецифические антитела анализировали методом электрофореза в денатурирующих и восстанавливающих условиях с использованием биоанализатора Agilent 2100 с набором Protein 80 в соответствии с инструкциями производителя (Agilent Technologies, Santa Clara, Calif., USA). 4 мкл очищенных образцов смешивали с буфером для образцов, содержащим дитиотреитол (DTT; Sigma Aldrich, St. Louis, Mo.). Образцы нагревали при 95°C в течение 5 мин, а затем наносили на чип. Все образцы тестировали на примеси эндотоксина с использованием реакции с лизатом амебоцитов мечехвоста (LAL; Charles River Laboratories, Wilmington, Mass.).
Пример 7. In vitro характеристики одновалентных и биспецифических антител
a) Связывание одновалентных и биспецифических антител с клетками, экспрессирующими CEACAM5, и клетками, не экспрессирующими CEACAM5
Для демонстрации связывания анти-CD3xCEACAM5 κλ-антител с клетками-мишенями можно проводить серию экспериментов методом проточной цитометрии, сравнивая связывание анти-CD3xCEACAM5 κλ-антител и их одновалентных аналогов. Примеры клеток, которые могут быть использованы, включают линии CEACAM5-положительных клеток, такие как линия клеток MKN45 аденокарциномы желудка (экспрессирующих 155000 молекул CEACAM5 на клетку) или линия клеток HPAF-II аденокарциномы поджелудочной железы (экспрессирующих 108000 молекул CEACAM5 на клетку), или линия клеток LS-174T колоректальной аденокарциномы (экспрессирующих 26000 молекул CEACAM5 на клетку), и линии CEACAM5-отрицательных клеток, такие как линия клеток A549 карциномы легких и линия клеток MKN45 с нокаутом CEACAM5, полученная при помощи методологии CRISPR-CAS9. Окрашивание клеток и оценку связывания можно проводить, как описано выше. Полученные кривые связывания представлены на Фигурах 4, 5, 11 и 14. Значения EC50 могут быть рассчитаны с использованием GraphPad Prism8 для связывания с клетками MKN45; данные представлены в Таблице 4. Связывание бсАт по изобретению в концентрациях 200 нМ, 1000 нМ и 5000 нМ составляет 40% и более в сравнении со связыванием TCB2014 (смотри Таблицу 4, а также Фигуры 11 и 14).
ТАБЛИЦА 4: Значения EC50 для связывания с клетками MKN-45. Н/П: не применимо
b) Связывание одновалентных и биспецифических антител с клетками, экспрессирующими CD3, и клетками, не экспрессирующими CD3
Для демонстрации связывания анти-CD3xCEACAM5 κλ-антител с эффекторными T-клетками можно проводить серию экспериментов методом проточной цитометрии, сравнивая связывание анти-CD3xCEACAM5 κλ-антител и их одновалентных аналогов. Примеры клеток, которые могут быть использованы, включают человеческие первичные T-клетки, а также линии CD3-положительных (Jurkat и/или HuT 78) или CD3-отрицательных (TIB-153 и/или JKT-бета-дел) клеток. Окрашивание клеток и оценку связывания можно проводить, как описано выше. Результаты представлены на Фигурах 3 и 10.
c) Эпитоп-специфическая сортировка анти-CEACAM5 антител путем конкуренции с эталонными антителами
Эпитоп-специфическая сортировка представляет собой конкурентный иммуноанализ, используемый для определения характеристик связывания антител по изобретению или, например, связывания соответствующих анти-CEA (целевой белок) антител первого связывающего фрагмента. Профиль конкурентного блокирования при связывании антитела с целевым белком анализировали в сравнении с антителами, также связывающими этот целевой белок, для которых связываемый эпитоп уже был установлен/опубликован. Конкуренция с одним из этих эталонных антител указывает на то, что антитело имеет тот же самый, или близко расположенный, эпитоп, и они «сгруппированы» вместе. Способность анти-CEACAM5 плеч, которые являются частью биспецифических антител по настоящему изобретению, конкурировать с эталонными анти-CEACAM5 антителами тестировали методом ELISA с рекомбинантным CEACAM5 человека и следующими эталонными антителами, имеющими мышиную Fc-область: SM3E, последовательности мАт получены из SM3E, описанного в патенте US20050147614A1, мАт получено стандартными методами; MEDI, мАт получено из MEDI-565, описанного в патенте WO2016036678A1; CH1A1A, мАт получено из CH1A1A-2F1, описанного в патенте US20120251529 и в публикации Klein et al. Oncoimmunology, 2017 Jan 11;6(3). SM3E связывается больше с N-концом, дистальной по отношению к клеточной мембране частью CEA, MEDI со средней частью, и CH1A1A связывается вблизи мембраны.
κλ-антитела, используемые в концентрации 1 мкг/мл, были захвачены антителами козы против IgG (Fcγ) человека (Jackson ImmunoResearch), нанесенными в концентрации 10 мкг/мл в черные лунки 96-луночного планшета, после блокирования блокирующим буфером (PBS, 2% БСА, 0,05% Tween 20). Конкурирующие IgG (0,03-20 мкг/мл) преинкубировали в течение 1 часа с 0,1 мкг/мл биотинилированного CEACAM5 человека в блокирующем буфере. Планшет с κλ-антителами промывали и инкубировали с преинкубированной смесью конкурирующего IgG/CEACAM5 в течение 1 часа. После промывания CEACAM5 обнаруживали при помощи конъюгата стрептавидин-HRP (Jackson ImmunoResearch). В лунки планшета вносили реагент Amplex Red® (Molecular Probes). Сигнал флуоресценции измеряли на планшетном ридере Synergy HT (Biotek).
Если связывание CEACAM5 с κλ-антителами уменьшалось за счет соответствующего инструментального антитела на 80% или более, можно было сделать вывод, что анти-CEAxCD3 биспецифическое антитело конкурирует за связывание с инструментальным антителом. Следовательно, анти-CEAxCD3 антитело считали не конкурирующим с инструментальным антителом в том случае, если связывание CEACAM5 с соответствующим κλ-антителом уменьшалось на 20% или менее при сравнении результатов с добавлением и без добавления инструментального антитела.
d) Связывание биспецифических антител с первичными человеческими клетками крови
Для демонстрации связывания анти-CD3xCEACAM5 κλ-антител с первичными T-клетками и отсутствия связывания с первичными B-клетками и моноцитами (CEA-отрицательные популяции), можно проводить серию экспериментов методом проточной цитометрии. Окрашивание клеток и оценку связывания можно проводить, как описано в примере 7a. Данные представлены на Фигуре 13.
Пример 8. Зависимая от T-клеток клеточная цитотоксичность (TDCC), опосредованная биспецифическими антителами
a) TDCC в отношении линий CEACAM5-положительных и CEACAM5-отрицательных клеток
Зависимую от T-клеток клеточную цитотоксичность (TDCC) в отношении разных линий CEACAM5-положительных и CEACAM5-отрицательных опухолевых клеток, индуцируемую анти-CEAxCD3 биспецифическими антителами по настоящему изобретению, оценивали с использованием либо человеческих МКПК, либо очищенных первичных T-клеток в качестве эффекторных клеток.
Клетки-мишени отделяли от подложки обработкой трипсином или раствором для диссоциации клеток после двух промываний PBS. После этапа центрифугирования клетки ресуспендировали в аналитической среде, доводили до нужной концентрации и помещали в 96-луночные планшеты.
Эффекторными клетками могли быть либо человеческие мононуклеарные клетки периферической крови (МКПК), либо очищенные T-клетки. МКПК выделяли из лейкоцитарных пленок, полученных от здоровых людей-доноров, с использованием пробирок SepMate™ (Stemcell Technologies) с буфером Lymphoprep™ (Stemcell Technologies). Если в качестве эффекторных клеток использовали очищенные T-клетки, выполняли дополнительный этап очистки, в котором T-клетки выделяли методом отрицательной селекции из МКПК с использованием набора для отрицательной иммуномагнитной селекции T-клеток (STEMCELL Technologies).
При проведении анализа TDCC, если в качестве эффекторных клеток использовали МКПК, их добавляли к клеткам-мишеням при конечном соотношении Э:М, составляющем 10:1; в случае использования очищенных T-клеток конечное соотношение Э:М составляло 5:1. Затем добавляли анти-CEAxCD3 антитела по изобретению и соответствующие контрольные антитела в диапазоне концентраций (до 100 нМ в двойных повторах) к предварительно внесенным в лунки клеткам-мишеням и эффекторным клеткам. Уничтожение клеток-мишеней оценивали через 24 ч, 48 ч или 72 ч инкубации в условиях 37°C и 5% CO2 путем количественного определения высвобождения LDH в среду апоптотическими/некротическими клетками (Cytotoxicity Detection Kit PLUS (LDH), Roche). Максимального высвобождения LDH (= 100% лизис) добивались путем инкубации клеток-мишеней с 1% Triton X-100. Спонтанное высвобождение LDH (= 0% лизис) определяли при совместной инкубации клеток-мишеней с эффекторными клетками без добавления какого-либо антитела. Кривые TDCC (Фигуры 6, 7, 12 и 15) и значения EC50 (Таблица 5) можно рассчитывать с использованием GraphPad Prism8. В случае линий клеток MKN-45 и LS-174T значения EC50, определенные с бсАт по изобретению и приведенные в Таблице 5, были значительно ниже, чем значения EC50, измеренные с TCB2014, это указывало на более высокую способность к уничтожению in vitro опухолевых клеток этими бсАт по изобретению.
ТАБЛИЦА 5. Значения EC50 при уничтожении трех линий CEA+ клеток
b) Анализ уничтожения клеток сочетанием анти-CEAxCD3 и анти-CEAxCD47 биспецифических антител
Сочетания биспецифического антитела по настоящему изобретению с анти-CD47 мАт (таким как описанное в US20140140989 и в WO2017196793) или с анти-CEAxCD47 биспецифическим антителом (описанным в PCT/IB2019/054559, содержание которого включено в настоящий документ посредством ссылки) можно тестировать в моделях, описанных выше. В дизайн эксперимента добавляли дополнительный вариант тестирования, когда такое направленное на CD47 антитело (моно- или биспецифическое) использовали либо отдельно, либо в сочетании с анти-CEAxCD3 антителом по настоящему изобретению, в разных дозах.
c) Повышение регуляции маркеров активации T-клеток при уничтожении CEA-экспрессирующих опухолевых клеток, индуцированном анти-CEAxCD3 бсАт
Для уничтожения CEA-положительных опухолевых клеток, индуцированного анти-CEAxCD3 бсАт, необходима активация T-клеток, которую можно количественно определять методом проточной цитометрии с использованием антител, узнающих специфические маркеры активации T-клеток, такие как CD69 (ранний маркер активации) или CD25 (поздний маркер активации).
Для оценки состояния активации T-клеток после завершения анализа на уничтожение (описанного выше, пример 8a), использовали следующий способ: плавающие клетки (которые включают как CD4+, так и CD8+, T-клетки) переносили в новые 96-луночные планшеты с V-образным дном. Супернатант удаляли центрифугированием (3 минуты при 4°C, 1300 об/мин), и клетки дважды промывали холодным FACS-буфером (PBS, 2% БСА, 0,1% NaN3), с последующей инкубацией в течение 15 минут при 4°C с Fc-блокирующим реагентом (BD Biosciences). После двух промываний FACS-буфером клетки инкубировали в течение 15 минут при 4°C со следующими антителами (используемыми в соответствии с рекомендациями производителя): анти-CD45 (V500-конъюгированные, BD Biosciences), CD69 (FITC-конъюгированные, Biolegend), CD8 (PerCP-Cy5.5-конъюгированные, Biolegend), CD25 (PE-конъюгированные, Biolegend), CD4 (APC-конъюгированные, ThermoFisher), CD3 (APC-R700-конъюгированные, BD Biosciences).
Клетки дважды промывали холодным FACS-буфером и ресуспендировали в 200 мкл FACS-буфера. Флуоресценцию измеряли с использованием платформы Cytoflex (Beckman Coulter), и данные анализировали с использованием программы FlowJo™ v10 (BD Life Sciences). Результаты представлены на Фигуре 17.
d) Пролиферация T-клеток, индуцированная молекулами анти-CEAxCD3 бсАт
Анти-CEAxCD3 бсАт анализировали на их способность индуцировать пролиферацию T-клеток при перекрестном связывании в присутствии CEA-положительных опухолевых клеток-мишеней. В качестве отрицательного контроля также использовали CEA-отрицательные злокачественные клетки. Свежевыделенные человеческие МКПК доводили до концентрации 1 миллион клеток в мл теплым PBS и окрашивали 0,2 мкМ карбоксифлуоресцеин-диацетат-сукцинимидным эфиром (CFSE, ThermoFisher Scientific) в PBS в течение 15 мин при 37°C, промывали несколько раз полной средой RPMI (содержащей 10% ЭТС, 2 мМ L-глутамин, 1 мМ пируват натрия, 10 мМ HEPES, 50 мкМ 2-меркаптоэтанол и 25 мкг/мл гентамицин) и переносили в 96-луночные планшеты при концентрации 2×106 клеток в мл. 0,02×106 клеток-мишеней помещали в лунку плоскодонного 96-луночного планшета и добавляли разные анти-CEAxCD3 бсАт в указанных концентрациях. Добавляли CFSE-меченые МКПК для получения конечного соотношения Э:М, составляющего 10:1, и аналитические планшеты инкубировали в течение пяти дней в инкубаторе с увлажненной атмосферой при 37°C. На пятый день эффекторные клетки собирали и промывали дважды FACS-буфером (PBS, 2% БСА, 0,1% NaN3). Затем клетки окрашивали BD Horizon 620 (BD Biosciences, 564996) для исключения мертвых клеток, а также анти-CD45 (V500-конъюгированными, BD Biosciences), анти-CD4-APC (ThermoFischer, 17-0049-41) и анти-CD8-PerCP-Cy5.5 (Biolegend, 301032). Окрашивание CFSE анализировали на живых CD4+ или CD8+ клетках методом проточной цитометрии с использованием CytoFLEX (Beckman Coulter), и результаты оценивали при помощи программы FlowJo.
e) Цитокины, высвобождаемые в супернатант при уничтожении CEA-экспрессирующих опухолевых клеток, индуцированном анти-CEAxCD3 бсАт
Для уничтожения CEA-положительных опухолевых клеток, индуцированного анти-CEAxCD3 бсАт, необходима активация T-клеток. При активации T-клетки могут высвобождать множество цитокинов, которые в дальнейшем могут действовать как иммуномодулирующие средства. Способность биспецифических антител по настоящему изобретению индуцировать высвобождение цитокинов T-клетками при уничтожении CEA-экспрессирующих опухолевых клеток оценивали путем количественного определения конкретных цитокинов в супернатанте после завершения анализа TDCC, описанного в примере 8a. После 2 дней совместного культивирования CEA-положительных клеток-мишеней и CD3-положительных эффекторных T-клеток культуральные супернатанты собирали путем центрифугирования и хранили замороженными при -80° до дальнейшего анализа. Количества цитокинов/ферментов, таких как гранзим B, IL2, IL6, IL10, TNFα и IFNγ, определяли при помощи исследовательской платформы Mesoscale с использованием мультиплексных наборов, и результаты представлены на Фигуре 16.
f) TDCC CEACAM5-положительных клеток в присутствии растворимого CEA
Известно, что от CEA-положительных опухолей отделяется CEA. Такой растворимый CEA может отрицательно влиять на противоопухолевую эффективность направленных на CEA антител, которые не связывают предпочтительно мембраносвязанный CEA. Для оценки того, оказывает ли влияние на биспецифические антитела по настоящему изобретению растворимый CEA (sCEA), анализ зависимой от T-клеток клеточной цитотоксичности (TDCC), описанный в примере 8a, проводили в присутствии добавленного sCEA в различных концентрациях (BioRad №PHP282). Значения EC50 в присутствии sCEA затем сравнивали со значениями EC50, полученными в отсутствие sCEA (Таблица 6). Значение EC50, рассчитанное для конкретной концентрации sCEA (0,2, 1 или 1 мкг/мл), затем сравнивали с таковым, полученным в отсутствие sCEA (0 мкг/мл), и выражали в виде кратности изменения EC50 в сравнении с ситуацией без растворимого CEA. Такие значения приведены в Таблице 7.
ТАБЛИЦА 6. Значения EC50 для уничтожения клеток LS-174T в присутствии sCEA
ТАБЛИЦА 7. Кратность изменения EC50 в сравнении с вариантом без растворимого CEA (0 мкг/мл)
Значительно более высокие сдвиги EC50 для уничтожения опухолевых клеток были обнаружены, если sCEA добавляли в концентрации 1 и 5 мкг/мл sCEA, для TCB2014 и TCB2017, в сравнении с биспецифическими антителами по изобретению. У пациентов с CEA-положительными опухолями обнаруживают концентрации sCEA 1 мкг/мл и выше. Более низкий сдвиг кривых для уничтожения опухолевых клеток вследствие sCEA у бсАт по изобретению свидетельствует о меньшем ингибирующем влиянии высоких уровней sCEA на эффективность бсАт по изобретению в сравнении с TCB2014 и TCB2017.
g) TDCC CEACAM5-отрицательных популяций первичных клеток крови
Учитывая механизм действия анти-CEAxCD3 биспецифических антител, перекрестная реактивность с другими CEACAM могла бы приводить к истощению важных популяций циркулирующих здоровых клеток. Например, перекрестная реактивность с CEACAM8, который экспрессируется нейтрофилами, могла бы приводить к истощению таких клеточных популяций. Для подтверждения отсутствия связывания и, следовательно, уничтожения таких CEA-отрицательных популяций циркулирующих здоровых клеток, очищенные первичные клетки, такие как нейтрофилы, использовали в качестве «клеток-мишеней» вместо линий CEA-положительных клеток в экспериментальном методе, описанном в примере 8a.
Пример 9. Оценка противоопухолевой активности анти-CEAxCD3 перенаправляющей T-клетки молекулы в качестве единственного средства или в комбинированной терапии с направленными на CD47 антителами в гуманизированных моделях мышиных опухолей
a) Противоопухолевая активность молекул анти-CEAxCD3 в МКПК-гуманизированной модели мышиной опухоли
Мышам линии NOG (мыши NOD/Shi-scid/OL-2Rγnull, Taconic Biosciences) в возрасте 8-10 недель имплантировали подкожно (п/к) 1-5×106 CEA-положительных опухолевых клеток (из линии клеток или от пациента), и животных случайным образом распределяли в несколько групп лечения. Через четыре-семь дней всем мышам вводили в/б или в/в инъекцией 10 или 20×106 человеческих МКПК (мононуклеарные клетки периферической крови) для процесса гуманизации. Затем в/в вводили молекулы анти-CD3xCEA или контроль, начиная через 3-6 дней после инъекции МКПК, один или два раза в неделю в разных дозах. У мышей наблюдали развитие опухолей 3 раза в неделю, и опухоли измеряли цифровым штангенциркулем до конечной точки эксперимента (объем опухоли=1500 мм3 или начало симптомов РТПХ). Объем опухоли рассчитывали по формуле (длина x ширина2) x 0,5. Статистический анализ проводили с использованием одностороннего сравнительного анализа ANOVA по окончании исследования. Результаты эксперимента, в котором 1 миллион клеток HPAF-II подкожно вводили мышам NOG, с последующим введением 10 миллионов человеческих МКПК, представлены на Фигуре 19.
b) Противоопухолевая активность молекул анти-CEAxCD3 в CD34+-гуманизированной модели мышиной опухоли
Полностью гуманизированным мышам CD34+-huNOG (CD34+-привитые мыши NOD/Shi-scid/OL-2Rγnull, Taconic Biosciences) в возрасте 14 недель и с содержанием в крови >25% человеческих CD45+ клеток, имплантируют подкожно (п/к) 1-5×106 CEA-положительных опухолевых клеток (из линии клеток или от пациента), и животных случайным образом распределяют в несколько групп лечения. Когда средний объем опухоли достигает заданного значения (в диапазоне 100-200 мм3), в/в вводят молекулы анти-CD3xCEA или контроль один или два раза в неделю в разных дозах. У мышей наблюдают развитие опухолей 3 раза в неделю, и опухоли измеряют цифровым штангенциркулем до конечной точки эксперимента (объем опухоли=1500 мм3). Объем опухоли рассчитывают по формуле (длина x ширина2) x 0,5. Статистический анализ проводят с использованием одностороннего сравнительного анализа ANOVA по окончании исследования.
c) Противоопухолевая активность молекул анти-CEAxCD3 в сочетании с направленными на CD47 антителами (моноспецифическими или биспецифическими) в гуманизированной модели мышиной опухоли
Сочетания биспецифического антитела по настоящему изобретению с анти-CD47 мАт (такими как описанные в US20140140989 и в WO2017196793) или с анти-CEAxCD47 биспецифическим антителом (описанным в PCT/IB2019/054559, содержание которого включено в настоящий документ посредством ссылки) можно тестировать в моделях, описанных выше. Дополнительные группы добавляют в дизайн эксперимента, включая группы лечения, в которых направленное на CD47 антитело (моноспецифическое или биспецифическое) вводят в/в либо отдельно, либо в сочетании с анти-CEAxCD3 антителом по настоящему изобретению, один или два раза в неделю в разных дозах.
d) Противоопухолевая активность молекул анти-CEAxCD3 в сочетании с направленными на CD47 антителами (моноспецифическими или биспецифическими) в трансгенных моделях мышиной опухоли
Сочетания биспецифического антитела по настоящему изобретению с анти-CD47 мАт (такими как описанные в US20140140989 и в WO2017196793) или с анти-CEAxCD47 биспецифическим антителом (описанным в PCT/IB2019/054559, содержание которого включено в настоящий документ посредством ссылки) можно тестировать на трансгенных мышах, генетически модифицированных для экспрессии CD3 человека, CD47 человека и SIRPα человека, которым имплантированы подкожно (п/к) 0,5-5×106 мышиных опухолевых клеток, генетически модифицированных для экспрессии CEA человека и CD47 человека. Когда средний объем опухоли достигает заданного значения (в диапазоне 100-200 мм3), мышей случайным образом распределяют в группы лечения. Препараты вводят внутривенно, один или два раза в неделю в разных дозах. У мышей наблюдают развитие опухолей 3 раза в неделю, и опухоли измеряют цифровым штангенциркулем до конечной точки эксперимента (объем опухоли=1500 мм3). Объем опухоли рассчитывают по формуле (длина x ширина2) x 0,5. Статистический анализ проводят с использованием одностороннего сравнительного анализа ANOVA по окончании исследования.
Пример 10. Высвобождение цитокинов, протестированное в цельной крови и МКПК от здоровых людей - доноров крови
Анализ in vitro высвобождения цитокинов выполняют с использованием цельной крови (ЦК АВЦ) с минимальным разведением тестируемыми антителами (95% по объему крови) в водной среде. Считается, что этот формат анализа близко имитирует in vivo среду, содержащую факторы в физиологических концентрациях, которые могут влиять на механизм высвобождения цитокинов. Однако считается, что этот формат плохо позволяет прогнозировать опосредованное T-клетками высвобождение цитокинов (например, анти-CD28).
Альтернативно, можно выполнять анализ высвобождения цитокинов с использованием мононуклеарных клеток периферической крови (МКПК) от здоровых людей-доноров и с антителами в водной среде (водная фаза, ВФ) для оценки опосредованного T-клетками высвобождения цитокинов (МКПК АВЦ ВФ). В этом формате ограничено перекрестное связывание мАт во избежание высвобождения цитокинов в высокой степени, которое наблюдается с анти-CD3 антителами при перекрестном связывании.
Отрицательные контроли (анти-EGFR мАт и PBS), а также специфические положительные контроли (анти-CD52 мАт, анти-CEAxCD3 BiTE и/или анти-CD28 мАт) для каждого формата анализа тестируют параллельно с анти-CEAxCD3 биспецифическими антителами. Через 24 ч с случае ЦК АВЦ, и 48 ч в случае МКПК АВЦ ВФ, супернатанты тестируют на цитокины в мультиплексном анализе с использованием электрохемилюминесценции в качестве метода обнаружения (Mesoscale Discovery, Sector 600). IFNγ, TNFα и IL-6 измеряют в случае ЦК АВЦ, и IFNγ, IL-2, IL-10 и TNFα измеряют в случае МКПК АВЦ ВФ. По результатам строят графики для каждого цитокина, при это каждого донора представляют в виде одной точки данных.
Пример 11. Созревание аффинности анти-CEA антитела путем олигонуклеотид-направленного мутагенеза с использованием вырожденных олигонуклеотидов (оптимизация направленности; ОН)
Антитела, идентифицированные в процессе скрининга, описанного в примере 3, были выбраны для созревания аффинности с целью повышения их аффинности и эффективности. Все эти антитела имеют одну и ту же общую вариабельную область тяжелой цепи, но разные вариабельные области легких цепей. AB1 и C11 содержат легкую цепь каппа (IGKV3-11 и IGKV1-5, соответственно, в соответствии с номенклатурой IMGT), в то время как AB8 и 1B4 содержат легкую цепь лямбда (IGLV2-14 и IGLV3-21, соответственно). Было создано несколько фаговых библиотек, экспонирующих варианты scFv, за счет внесения разнообразия в CDR1, CDR2 и CDR3 вариабельной области легкой цепи, при этом вариабельная область тяжелой цепи была оставлена без изменения. Для создания библиотек для каждого кандидата использовали различные стратегии диверсификации, когда диверсифицировали CDRL1+CDRL2; или только CDRL3, или все три области CDRL (CDRL1+CDRL2+CDRL3) методом олигонуклеотид-направленного мутагенеза исходной последовательности с использованием вырожденных олигонуклеотидов. CDRL1 диверсифицировали по 1-5 аминокислотным положениям; CDRL2 по 1-4 аминокислотным положениям, в то время как CDRL3 по 1-5 аминокислотным положениям. Всего для каждого кандидата получали вплоть до 5×109 трансформантов, частично охватывающих теоретическое разнообразие вплоть до 1014.
Для кандидатов AB1 и 1B4 получали дополнительные библиотеки, диверсифицированные по вплоть до 17 аминокислотным положениям среди всех областей CDRL, с получением вплоть до 5×109 трансформантов, частично охватывающих теоретическое разнообразие вплоть до 1021.
Эти библиотеки использовали для отбора методом фагового дисплея, как описано в примере 2, за исключением того, что строгость отбора могла быть увеличена между раундами путем уменьшения концентрации рекомбинантного чCEACAM5 от 100 нМ постепенно до 0,01 нМ между разными раундами отбора или с использованием клеток, экспрессирующих более низкие уровни чCEACAM5, например, клеток линии SNUC-1. Отобранные варианты подвергали скринингу на способность связывать CEACAM5 с использованием анализа, описанного в примере 3. Положительные клоны переводили в формат IgG и характеризовали, как описано в примерах 4 и 5, соответственно.
Плечо анти-CEA AB1 (SEQ ID NO:31-34) было оптимизировано в двух последовательных циклах оптимизации направленности. Цикл 1 привел к получению плеч анти-CEA AB13, AB14, AB15, AB17 и AB20. Цикл 2 привел к получению плеч анти-CEA AB54, AB60, AB66, AB71, AB72 и AB73.
Пример 12: TDCC (зависимая от T-клеток клеточная цитотоксичность) и/или TDCC плюс ADCP органоидов, полученных из опухоли
Органоиды, полученные из клеток опухолей, являются передовой трансляционной моделью для тестирования перенаправляющих T-клеток соединений и/или перенаправляющих макрофаги и NK-клетки соединений.
Органоиды получают стандартными методами (Schütte et al., Nature Communications 2017; DOI:10.1038/ncomms14262) и инкубируют с соединениями в течение срока до 8 дней в совместной культуре с МКПК и in vitro полученными макрофагами. Среду меняют каждые 4 дня, заменяя ее свежей средой.
Органоиды собирают и ферментативно диссоциируют на отдельные клетки при 37°C в течение 5 мин с использованием Accutase. Клетки осаждают центрифугированием, ресуспендируют в FACS-буфере (PBS, 2% ЭБС, 2 мМ ЭДТА) и фильтруют через 400-мкм клеточное сито. Суспензии соответствующих групп клеток инкубируют с антителами против CD45, CD4, CD8, CEA и CD14 (все от компании Thermo Fisher Scientific, Dreieich, Germany) в течение 30 мин на льду. Для гейтирования живых клеток используют йодид пропидия, их количественное определение и анализ проводят с использованием программы FlowJo (FlowJo, LLC, Ashland, OR, USA).
Супернатант из каждой лунки замораживают при -80°C для анализа активности T-клеток методом ELISA.
Пример 13: TDCC и/или TDCC плюс ADCP фрагментов опухолевой ткани, полученной от пациента
Фрагменты свежей опухолевой ткани, полученной от пациента, являются еще одной передовой трансляционной моделью для тестирования соединений, перенаправляющих T-клетки и/или макрофаги, и/или NK-клетки.
Свежие образцы опухолевой ткани будут вырезаны стандартными методами, описанными ранее (Sönnichsen et al., Clinical Colorectal Cancer, 17 (2018) e189-e199). Вкратце, сразу после хирургической резекции и первой макроскопической патологоанатомической оценки образцы опухоли разрезают на срезы размером 350 мкм с использованием измельчителя тканей ((McIlwain TC752; Campden Instruments, Leicestershire, England). Затем диаметр среза ткани стандартизируют с использованием инструмента для отбора 3-мм образцов (kai Europe, Solingen, Germany). Три среза ткани случайным образом объединяют, помещают на мембранные вкладыши и культивируют в 6-луночных планшетах. Срезы инкубируют в стандартных условиях 37°C и 5% CO2. Среду заменяют через 2 часа и 24 часа после подготовки до начала воздействия.
После 24 часов предварительного культивирования в стандартной среде для культивирования клеток три среза ткани могут быть подвергнуты воздействию биспецифических антител по изобретению отдельно или в сочетании, соответственно, в течение срока вплоть до 120 часов. При необходимости время инкубации уменьшают до 72 часов. Среда будет заменена через 72 часа.
После воздействия соединениями срезы опухоли фиксируют в течение ночи с использованием 4% параформальдегида. Супернатант из каждой лунки будет заморожен при -80°C для анализа активности T-клеток методом ELISA.
Зафиксированные параформальдегидом срезы погружают в парафин и получают 5-мкм срезы. Выполняют окрашивание гематоксилином и эозином (HE) для оценки гистопатологических аспектов и относительного содержания опухолевых клеток. Общее количество клеток, количество опухолевых клеток и пролиферацию анализируют методом иммунофлуоресцентного окрашивания. Вкратце, парафиновые секции депарафинизируют. После извлечения антигена срезы промывают 0,3% PBS/Triton X и блокируют 5% нормальной козьей сывороткой (Jackson ImmunoResearch, Suffolk, UK) в течение 30 минут. Первичные антитела против цитокератинов (AE1þ3), Ki67 и расщепленного PARP, соответственно, разбавляют в 0,5% бычьем сывороточном альбумине и инкубируют при 4°C в течение ночи. Срезы промывают 0,3% фосфатно-солевым буфером/Triton X и метят вторичными антителами. Ядра окрашивают красителем Hoechst 33342 (Sigma-Aldrich, St. Louis, MO). Для дополнительного анализа используют антитела против CEA (опухолевые клетки), CD163 (макрофаги) и CD3, CD4, CD8, PD-L1, а также FoxP3 (все T-клетки) в зависимости от доступности срезов опухоли.
Область, содержащую клетки опухоли, анализируют на окрашенных HE срезах с использованием сканера срезов (Pannoramic SCAN и Pannoramic Viewer, 3D Histech, Budapest, Hungary) для изучения различных фракций опухолевых клеток. Срезы, содержащие больше доброкачественных эпителиальных клеток, чем неопластических эпителиальных клеток, исключают из анализа. Срезы, не содержащие опухолевые клетки, исключают из анализа фракции пролиферирующих опухолевых клеток, но включают в анализ опухолевых клеток для каждого варианта. Для дальнейшего анализа получают 5 фотографий (20x) каждого среза ткани из флуоресцентно окрашенных срезов с использованием флуоресцентного микроскопа Olympus BX51 (Olympus Deutschland, Hamburg, Germany). Положительное количество пикселей определяют для окрашивания Hoechst 33342, цитокератина, Ki67 и расщепленного PARP с Image J алгоритмами специфической для красителей сегментации. Площадь пролиферирующей/апоптотической опухоли рассчитывают путем анализа пикселей положительных по Ki67/расщепленному PARP ядер, окруженных пикселями цитокератин-положительной окраски.
Для каждой фотографии рассчитывают общее количество клеток (Hoechst-положительных), количество опухолевых клеток (Hoechst- и цитокератин-положительных) и количество пролиферирующих опухолевых клеток (Hoechst-, цитокератин- и Ki67-/расщепленный PARP-положительных). Количество опухолевых клеток нормализуют к общему количеству клеток, и количество пролиферирующих опухолевых клеток нормализуют к количеству опухолевых клеток для учета разных фракций опухолевых клеток на каждой фотографии. Затем рассчитывают средние значения для срезов из значений отдельных изображений. Средние значения для вариантов рассчитывают с использованием средних значений для срезов.
Все публикации, патенты, патентные заявки, сетевые адреса и регистрационные номера/базы данных последовательностей, включая как полинуклеотидные, так и полипептидные последовательности, упомянутые в настоящем документе, включены в настоящий документ посредством ссылки в полном объеме для всех целей в такой же степени, как если было бы специально и индивидуально указано, что каждая отдельная публикация, патент, патентная заявка, сетевой адрес или регистрационный номер/база данных последовательностей включены посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> ЛАМКАП БИО АЛЬФА АГ
<120> БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ CEACAM5 И CD3
<130> 4130.003PC02
<150> EP19198124
<151> 2019-09-18
<150> US62/902150
<151> 2019-09-18
<160> 138
<170> PatentIn version 3.5
<210> 1
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 VH
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 2
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 CDRH1
<400> 2
Thr Tyr Ala Met Asn
1. 5
<210> 3
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 CDRH2
<400> 3
Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1. 5 10 15
Val Lys Asp
<210> 4
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 CDRH3
<400> 4
His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe Ala Tyr
1. 5 10
<210> 5
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 VL 1B6
<400> 5
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ala Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 6
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1B6 CDRL1
<400> 6
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1. 5 10
<210> 7
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1B6 CDRL2
<400> 7
Gly Thr Asn Lys Arg Ala Pro
1. 5
<210> 8
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1B6 CDRL3
<400> 8
Ala Leu Trp Tyr Ala Asn Arg Trp Val
1. 5
<210> 9
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 VL 1A10
<400> 9
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Lys Gly
85 90 95
Tyr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 10
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A10 CDRL1
<400> 10
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1. 5 10
<210> 11
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A10 CDRL2
<400> 11
Gly Thr Asn Lys Arg Ala Pro
1. 5
<210> 12
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A10 CDRL3
<400> 12
Ala Leu Trp Tyr Lys Gly Tyr Trp Val
1. 5
<210> 13
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 VL 1F8
<400> 13
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Asp Gly
85 90 95
Lys Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 14
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1F8 CDRL1
<400> 14
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1. 5 10
<210> 15
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1F8 CDRL2
<400> 15
Gly Thr Asn Lys Arg Ala Pro
1. 5
<210> 16
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1F8 CDRL3
<400> 16
Ala Leu Trp Tyr Asp Gly Lys Trp Val
1. 5
<210> 17
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 VL 1A4
<400> 17
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Lys Gln
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 18
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A4 CDRL1
<400> 18
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1. 5 10
<210> 19
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A4 CDRL2
<400> 19
Gly Thr Asn Lys Arg Ala Pro
1. 5
<210> 20
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A4 CDRL3
<400> 20
Ala Leu Trp Tyr Lys Gln Arg Trp Val
1. 5
<210> 21
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 VL 1H4
<400> 21
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Asn Gln
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 22
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1H4 CDRL1
<400> 22
Arg Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1. 5 10
<210> 23
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1H4 CDRL2
<400> 23
Gly Thr Asn Lys Arg Ala Pro
1. 5
<210> 24
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1H4 CDRL3
<400> 24
Ala Leu Trp Tyr Asn Gln His Trp Val
1. 5
<210> 25
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1B6 LC
<400> 25
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ala Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 26
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A10 LC
<400> 26
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Lys Gly
85 90 95
Tyr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 27
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1F8 LC
<400> 27
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Asp Gly
85 90 95
Lys Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 28
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A4 LC
<400> 28
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Lys Gln
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 29
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1H4 LC
<400> 29
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Asn Gln
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 30
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константная область общей тяжелой цепи (WT IgG1)
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 31
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA VL AB1 (2F2)
<400> 31
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Leu Arg His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 32
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB1 CDRL1
<400> 32
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 33
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB1 CDRL2
<400> 33
His Gly Ser Asn Arg Ala Thr
1. 5
<210> 34
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB1 CDRL3
<400> 34
Gln Gln Phe Asp Leu Arg His Glu His Asn Thr
1. 5 10
<210> 35
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA VL AB8 (2A3)
<400> 35
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1. 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Ile Glu Asn
20 25 30
Ala Ile Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Pro
35 40 45
Met Ile Tyr Thr Leu Ser Asp Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Trp Asp Thr Phe
85 90 95
Ala Ile Gly Pro Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 36
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB8 CDRL1
<400> 36
Thr Gly Thr Ser Ser Asp Val Ile Glu Asn Ala Ile Val Ser
1. 5 10
<210> 37
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB8 CDRL2
<400> 37
Thr Leu Ser Asp Arg Pro Ser
1. 5
<210> 38
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB8 CDRL3
<400> 38
Ser Ser Trp Asp Thr Phe Ala Ile Gly Pro Ala Val
1. 5 10
<210> 39
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константная область каппа легкой цепи (CK)
<400> 39
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
1. 5 10 15
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
20 25 30
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
35 40 45
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
50 55 60
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
65 70 75 80
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
85 90 95
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
100 105 110
Asn Arg Gly Glu Cys
115
<210> 40
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь CEA AB1 (VKCK_2F2)
<400> 40
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Leu Arg His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 41
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Константная область лямбда легкой цеп (CL)
<400> 41
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala Ala
1. 5 10 15
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
20 25 30
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
35 40 45
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
50 55 60
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
65 70 75 80
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
85 90 95
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
100 105 110
Thr Glu Cys Ser
115
<210> 42
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь CEA AB8 (VLCL_2A3)
<400> 42
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1. 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Ile Glu Asn
20 25 30
Ala Ile Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Pro
35 40 45
Met Ile Tyr Thr Leu Ser Asp Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Trp Asp Thr Phe
85 90 95
Ala Ile Gly Pro Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
115 120 125
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
130 135 140
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
145 150 155 160
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
165 170 175
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
180 185 190
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
195 200 205
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 43
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Общая тяжелая цепь (дикий тип)
<400> 43
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly
450
<210> 44
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Общая тяжелая цепь (мутация LALA)
<400> 44
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly
450
<210> 45
<211> 454
<212> Белок
<213> Искусственная последовательность
<220>
<223> Общая тяжелая цепь (мутация LALA+P329A)
<400> 45
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly
450
<210> 46
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> VK_SM3E
<400> 46
Glu Asn Val Leu Thr Gln Ser Pro Ser Ser Met Ser Val Ser Val Gly
1. 5 10 15
Asp Arg Val Asn Ile Ala Cys Ser Ala Ser Ser Ser Val Pro Tyr Met
20 25 30
His Trp Leu Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile Tyr
35 40 45
Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Ser Val Gln Pro Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 47
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH_SM3E
<400> 47
Gln Val Lys Leu Glu Gln Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1. 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Ser
20 25 30
Tyr Met His Trp Leu Arg Gln Gly Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Asp Pro Glu Asn Gly Asp Thr Glu Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Phe Thr Thr Asp Thr Ser Ala Asn Thr Ala Tyr
65 70 75 80
Leu Gly Leu Ser Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Asn Glu Gly Thr Pro Thr Gly Pro Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 48
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> VL_MEDI
<400> 48
Gln Ala Val Leu Thr Gln Pro Ala Ser Leu Ser Ala Ser Pro Gly Ala
1. 5 10 15
Ser Ala Ser Leu Thr Cys Thr Leu Arg Arg Gly Ile Asn Val Gly Ala
20 25 30
Tyr Ser Ile Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Pro Pro Gln Tyr
35 40 45
Leu Leu Arg Tyr Lys Ser Asp Ser Asp Lys Gln Gln Gly Ser Gly Val
50 55 60
Ser Ser Arg Phe Ser Ala Ser Lys Asp Ala Ser Ala Asn Ala Gly Ile
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Met Ile Trp His Ser Gly Ala Ser Ala Val Phe Gly Gly Gly Thr Lys
100 105 110
Leu Thr Val Leu
115
<210> 49
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH_MEDI
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Gly Thr Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Asp Arg Gly Leu Arg Phe Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 50
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> VK_SAR
<400> 50
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Phe Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Thr Arg Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln His His Tyr Gly Thr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 51
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH_SAR
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Val Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Gly Gly Ile Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala His Tyr Phe Gly Ser Ser Gly Pro Phe Ala Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 52
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> VK_CH1A1A
<400> 52
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 53
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH_ CH1A1A
<400> 53
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 54
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> VK_T84.66
<400> 54
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Gly Glu Ser Val Asp Ile Phe
20 25 30
Gly Val Gly Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Arg Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Thr Asn
85 90 95
Glu Asp Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 55
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH_T84.66
<400> 55
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Asn Ser Lys Tyr Val Pro Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Pro Phe Gly Tyr Tyr Val Ser Asp Tyr Ala Met Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 56
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> VK_ЛАБЕТУЗУМАБ
<400> 56
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Gly Thr Ser
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Thr Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Leu Tyr Arg Ser
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 57
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH_ЛАБЕТУЗУМАБ
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Thr Thr Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile His Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Phe Gly Phe Pro Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser
115
<210> 58
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гибридная константная область каппа легкой цепи (H-CK 5)
<400> 58
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg Thr Val Ala Ala
1. 5 10 15
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
20 25 30
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
35 40 45
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
50 55 60
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
65 70 75 80
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
85 90 95
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
100 105 110
Phe Asn Arg Gly Glu Cys
115
<210> 59
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA VL 1B4
<400> 59
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Leu Ile Gly Lys Asn Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asn Tyr Gly Ile Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Leu Lys Leu Glu
85 90 95
Pro Asp Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 60
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA 1B4 CDRL1
<400> 60
Gly Gly Asn Leu Ile Gly Lys Asn Ala Val His
1. 5 10
<210> 61
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA 1B4 CDRL2
<400> 61
Asn Tyr Gly Ile Arg Pro Ser
1. 5
<210> 62
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA 1B4 CDRL3
<400> 62
Gln Val Trp Asp Thr Leu Lys Leu Glu Pro Asp Ala Val
1. 5 10
<210> 63
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA VL C11
<400> 63
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Leu Ile Ser Asn Gly
20 25 30
Leu Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ile Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Leu Ser Trp Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 64
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA C11 CDRL1
<400> 64
Arg Ala Ser Gln Leu Ile Ser Asn Gly Leu Tyr
1. 5 10
<210> 65
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA C11 CDRL2
<400> 65
Asp Ala Ser Ile Leu Glu Ser
1. 5
<210> 66
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA C11 CDRL3
<400> 66
Gln Gln Arg Leu Ser Trp Pro Leu Thr
1. 5
<210> 67
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1B6 LC-гибридная область каппа
<400> 67
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ala Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 68
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A10 LC-гибридная область каппа
<400> 68
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Lys Gly
85 90 95
Tyr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 69
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1F8 LC-гибридная область каппа
<400> 69
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Asp Gly
85 90 95
Lys Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 70
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1A4 LC-гибридная область каппа
<400> 70
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Lys Gln
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 71
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> huCD3 1H4 LC-гибридная область каппа
<400> 71
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1. 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Asn Gln
85 90 95
His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 72
<211> 220
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA 2A3 LC-гибридная область каппа
<400> 72
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1. 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Ile Glu Asn
20 25 30
Ala Ile Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Pro
35 40 45
Met Ile Tyr Thr Leu Ser Asp Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Trp Asp Thr Phe
85 90 95
Ala Ile Gly Pro Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 73
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA C11 LC (VKCK_C11)
<400> 73
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Leu Ile Ser Asn Gly
20 25 30
Leu Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ile Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Leu Ser Trp Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 74
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA 1B4 LCEA 1B4 LC (VLCL_1B4)
<400> 74
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Leu Ile Gly Lys Asn Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asn Tyr Gly Ile Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Leu Lys Leu Glu
85 90 95
Pro Asp Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 75
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA 1B4 LC-гибридная область каппа (VLCK_1B4)
<400> 75
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Leu Ile Gly Lys Asn Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asn Tyr Gly Ile Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Leu Lys Leu Glu
85 90 95
Pro Asp Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 76
<211> 645
<212> DNA
<213> Искусственная последовательность
<220>
<223> VKCK_C11 (ДНК)
<400> 76
gacatccaga tgacccagtc tccttccacc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gcttattagt aatggtttgt attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgat gcttccattt tggaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagag ttcactctca ccatcagcag cctgcagcct 240
gatgattttg caacttatta ctgtcagcag cgtctcagct ggcctctcac tttcggccaa 300
gggaccaagg tggaaatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttaa 645
<210> 77
<211> 651
<212> DNA
<213> Искусственная последовательность
<220>
<223> VLCL_1B4 (ДНК)
<400> 77
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gaaaccttat tggaaagaat gctgtgcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctataattat ggtattcggc cctcagggat tcctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactctta agcttgagcc tgatgctgtg 300
ttcggcggag ggaccaagct gaccgtccta ggtcagccca aggctgcccc ctcggtcact 360
ctgttcccgc cctcctctga ggagcttcaa gccaacaagg ccacactggt gtgtctcata 420
agtgacttct acccgggagc cgtgacagtg gcttggaaag cagatagcag ccccgtcaag 480
gcgggagtgg agaccaccac accctccaaa caaagcaaca acaagtacgc ggccagcagc 540
tatctgagcc tgacgcctga gcagtggaag tcccacagaa gctacagctg ccaggtcacg 600
catgaaggga gcaccgtgga gaagacagtg gcccctacag aatgttcata a 651
<210> 78
<211> 651
<212> DNA
<213> Искусственная последовательность
<220>
<223> VKCK_2F2 (AB1) (ДНК)
<400> 78
gaaattgtgt tgacacagtc tccagccacc ctgtctttgt ctccagggga aagagccacc 60
ctctcctgca gggccagtca gtctgttaat tctaatttaa attggtacca acagaaacct 120
ggccaggctc ccaggctcct catctatcat gggtccaata gggccactgg catcccagcc 180
aggttcagtg gcagtgggtc tgggacagac ttcactctca ccatcagcag cctagagcct 240
gaagattttg cagtttatta ctgtcagcag tttgatctta ggcatgagca taataccttc 300
ggccaaggga ccaaggtgga aatcaaacgt acggtggctg caccatctgt cttcatcttc 360
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac 420
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccctcca atcgggtaac 480
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc 540
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat 600
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgtta a 651
<210> 79
<211> 657
<212> DNA
<213> Искусственная последовательность
<220>
<223> VLCL_2A3 (AB8) (ДНК)
<400> 79
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttatt gagaatgcta ttgtctcctg gtaccaacag 120
cacccaggca aagcccccaa acccatgatt tatactctta gtgatcggcc ctcaggggtt 180
tctaatcgct tctctggctc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatggg atacttttgc gattggtcct 300
gctgtgttcg gcggagggac caagctgacc gtcctaggtc agcccaaggc tgccccctcg 360
gtcactctgt tcccgccctc ctctgaggag cttcaagcca acaaggccac actggtgtgt 420
ctcataagtg acttctaccc gggagccgtg acagtggctt ggaaagcaga tagcagcccc 480
gtcaaggcgg gagtggagac caccacaccc tccaaacaaa gcaacaacaa gtacgcggcc 540
agcagctatc tgagcctgac gcctgagcag tggaagtccc acagaagcta cagctgccag 600
gtcacgcatg aagggagcac cgtggagaag acagtggccc ctacagaatg ttcataa 657
<210> 80
<211> 1365
<212> DNA
<213> Искусственная последовательность
<220>
<223> Общая тяжелая цепь VHCH (дикий тип; ДНК)
<400> 80
gaggtgcagc tggtggagtc tgggggaggc ttggtccagc ctggggggtc cctgaaactc 60
tcctgtgcag cctctgggtt caccttcaac acctatgcta tgaactgggt ccgccaggct 120
cccgggaaag ggctggagtg ggttggccgt attagaagca aatataacaa ttacgcgaca 180
tactatgctg actcggtgaa agacaggttc accatctcca gagatgattc aaagaacacg 240
gcgtatctgc aaatgaacag cctgaaaacc gaggacacgg ccgtgtatta ctgtgtgaga 300
cacgggaatt tcggcaattc ttatgtctcg tggttcgctt actggggcca agggactctg 360
gtcacagtct cgagcgcctc caccaagggc ccatcggtct tccccctggc accctcctcc 420
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 480
ccggtgacag tctcgtggaa ctcaggagcc ctgaccagcg gcgtgcacac cttcccggct 540
gtcctacagt cctcaggact ctactccctc agcagcgtgg tgactgtgcc ctccagcagc 600
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtggac 660
aagagagttg agcccaaatc ttgtgacaaa actcacacat gcccaccgtg cccagcacct 720
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 780
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 840
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 900
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 960
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 1020
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta taccctgccc 1080
ccatctcggg aggagatgac caagaaccag gtcagcctga cttgcctggt caaaggcttc 1140
tatcccagcg acatcgccgt ggagtgggag agcaacgggc agccggagaa caactacaag 1200
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctatagcaa gctcaccgtg 1260
gacaagtcca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 1320
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gttaa 1365
<210> 81
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB13 CDRL1
<400> 81
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 82
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB13 CDRL2
<400> 82
His Gly Ser Asn Arg Ala Thr
1. 5
<210> 83
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB13 CDRL3
<400> 83
Gln Gln Phe Asp Tyr Phe Met Asn Lys Asn Thr
1. 5 10
<210> 84
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB14 CDRL1
<400> 84
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 85
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB14 CDRL2
<400> 85
His Gly Ser Asn Arg Ala Thr
1. 5
<210> 86
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB14 CDRL3
<400> 86
Gln Gln Phe Asp Tyr Phe Arg Glu Ser Asn Thr
1. 5 10
<210> 87
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB15 CDRL1
<400> 87
Arg Ala Ser Gln Thr Val Asn Asn Asn Leu Asn
1. 5 10
<210> 88
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB15 CDRL2
<400> 88
Tyr Ala Ser Asn Arg Ala Thr
1. 5
<210> 89
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB15 CDRL3
<400> 89
Gln Gln Phe Asn Tyr His His Glu His Asn Thr
1. 5 10
<210> 90
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB17 CDRL1
<400> 90
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 91
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB17 CDRL2
<400> 91
His Gly Ser Asn Arg Ala Thr
1. 5
<210> 92
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB17 CDRL3
<400> 92
Gln Gln Phe Asp Tyr Phe Lys Glu His Asn Thr
1. 5 10
<210> 93
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB20 CDRL1
<400> 93
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 94
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB20 CDRL2
<400> 94
His Gly Ser Asn Arg Ala Thr
1. 5
<210> 95
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB20 CDRL3
<400> 95
Gln Gln Phe Asp Tyr Phe Arg Glu Leu Asn Thr
1. 5 10
<210> 96
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB54 CDRL1
<400> 96
Arg Ala Ser Gln Glu Val His Lys Asn Leu Asn
1. 5 10
<210> 97
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB54 CDRL2
<400> 97
His Ser Ser Asn Arg Ala Thr
1. 5
<210> 98
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB54 CDRL3
<400> 98
Gln Gln Phe Asn Tyr His His Glu His Asn Thr
1. 5 10
<210> 99
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB60 CDRL1
<400> 99
Arg Ala Ser Gln Thr Val Asn Ala Asn Leu Asn
1. 5 10
<210> 100
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB60 CDRL2
<400> 100
Tyr Ala Ser Asn Arg Ala Thr
1. 5
<210> 101
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB60 CDRL3
<400> 101
Gln Gln Phe Asp Tyr His Asn Glu His Asn Thr
1. 5 10
<210> 102
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB66 CDRL1
<400> 102
His Ser Ser Gln Val Val Asn Lys Asn Leu Asn
1. 5 10
<210> 103
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB66 CDRL2
<400> 103
His Gly Ser Asn Arg Ala Thr
1. 5
<210> 104
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB66 CDRL3
<400> 104
Gln Gln Phe Asp Tyr Phe Arg Glu His Asn Thr
1. 5 10
<210> 105
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB71 CDRL1
<400> 105
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 106
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB71 CDRL2
<400> 106
His Ser Ser Asn Arg Pro His
1. 5
<210> 107
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB71 CDRL3
<400> 107
Gln Gln Phe Asp Tyr Phe Lys Glu His Asn Thr
1. 5 10
<210> 108
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB72 CDRL1
<400> 108
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 109
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB72 CDRL2
<400> 109
His Ser Thr Asn Arg Pro Arg
1. 5
<210> 110
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB72 CDRL3
<400> 110
Gln Gln Phe Asp Tyr Phe Lys Glu Tyr Asn Thr
1. 5 10
<210> 111
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB73 CDRL1
<400> 111
Arg Ala Ser Gln Ser Val Asn Ser Asn Leu Asn
1. 5 10
<210> 112
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB73 CDRL2
<400> 112
His Ser Asn Asn Arg Pro His
1. 5
<210> 113
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB73 CDRL3
<400> 113
Gln Gln Phe Asp Tyr Phe Arg Glu Tyr Asn Thr
1. 5 10
<210> 114
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB13 VL вариабельная область легкой цепи
<400> 114
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Met Asn
85 90 95
Lys Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 115
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB14 VL вариабельная область легкой цепи
<400> 115
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Thr Val Asn Asn Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asn Tyr His His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 116
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB15 VL вариабельная область легкой цепи
<400> 116
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Thr Val Asn Asn Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asn Tyr His His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 117
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB17 VL вариабельная область легкой цепи
<400> 117
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Lys Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 118
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB20 VL вариабельная область легкой цепи
<400> 118
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
Leu Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 119
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB54 VL вариабельная область легкой цепи
<400> 119
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Glu Val His Lys Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arиg Leu Leu Ile
35 40 45
Tyr His Ser Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asn Tyr His His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 120
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB60 VL вариабельная область легкой цепи
<400> 120
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Thr Val Asn Ala Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr His Asn Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 121
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB66 VL вариабельная область легкой цепи
<400> 121
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys His Ser Ser Gln Val Val Asn Lys Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 122
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB71 VL вариабельная область легкой цепи
<400> 122
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Ser Asn Arg Pro His Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Lys Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 123
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB72 VL вариабельная область легкой цепи
<400> 123
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Thr Asn Arg Pro Arg Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Lys Glu
85 90 95
Tyr Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 124
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB73 VL вариабельная область легкой цепи
<400> 124
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Asn Asn Arg Pro His Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
Tyr Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 125
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB13
<400> 125
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Met Asn
85 90 95
Lys Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 126
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB14
<400> 126
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
Ser Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 127
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB15
<400> 127
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Thr Val Asn Asn Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asn Tyr His His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 128
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB17 LC
<400> 128
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Lys Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 129
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB20
<400> 129
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
Leu Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 130
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепиь LC CEA AB54
<400> 130
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Glu Val His Lys Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asn Tyr His His Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 131
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB60
<400> 131
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Thr Val Asn Ala Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr His Asn Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 132
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB66 LC light chain
<400> 132
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys His Ser Ser Gln Val Val Asn Lys Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Gly Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 133
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> CEA AB71 LC light chain
<400> 133
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Ser Asn Arg Pro His Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Lys Glu
85 90 95
His Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 134
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB72
<400> 134
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Thr Asn Arg Pro Arg Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Lys Glu
85 90 95
Tyr Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 135
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Легкая цепь LC CEA AB73
<400> 135
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn Ser Asn
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr His Ser Asn Asn Arg Pro His Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Tyr Phe Arg Glu
85 90 95
Tyr Asn Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 136
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Мутация в CDRL1 CEA AB1
<220>
<221> прочий признак
<222> (1)..(2)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий признак
<222> (5)..(5)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий признак
<222> (7)..(8)
<223> Xaa может быть любой природной аминокислотой
<400> 136
Xaa Xaa Ser Gln Xaa Val Xaa Xaa Asn Leu Asn
1. 5 10
<210> 137
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутация в CDRL2 CEA AB1
<220>
<221> прочий признак
<222> (1)..(3)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий признак
<222> (6)..(7)
<223> Xaa может быть любой природной аминокислотой
<400> 137
Xaa Xaa Xaa Asn Arg Xaa Xaa
1. 5
<210> 138
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> мутация CDRL3 CEA AB1
<220>
<221> прочий признак
<222> (2)..(2)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий признак
<222> (4)..(7)
<223> Xaa может быть любой природной аминокислотой
<220>
<221> прочий признак
<222> (9)..(9)
<223> Xaa может быть любой природной аминокислотой
<400> 138
Gln Xaa Phe Xaa Xaa Xaa Xaa Glu Xaa Asn Thr
1. 5 10
22
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| КОМБИНАЦИИ АНТИТЕЛ ДЛЯ ЛЕЧЕНИЯ РАКА У КОНКРЕТНЫХ ПАЦИЕНТОВ | 2020 |
|
RU2816531C2 |
| ТЕРАПЕВТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЛИГАНДА CD40 | 2018 |
|
RU2770209C2 |
| АНТИТЕЛА, ИНДУЦИРУЮЩИЕ ИММУННУЮ ТОЛЕРАНТНОСТЬ, ИНДУЦИРОВАННЫЕ ЛИМФОЦИТЫ И ТЕРАПЕВТИЧЕСКИЙ АГЕНТ/СПОСОБ КЛЕТОЧНОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ИНДУЦИРОВАННЫХ ЛИМФОЦИТОВ | 2019 |
|
RU2816592C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
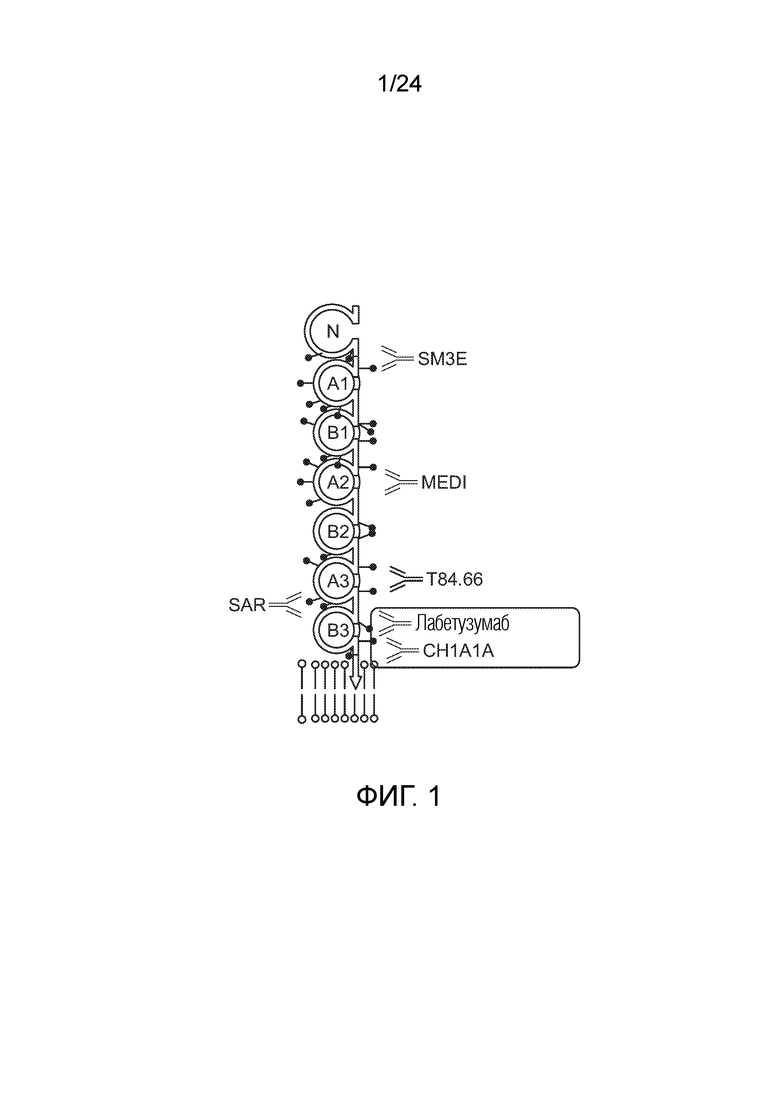
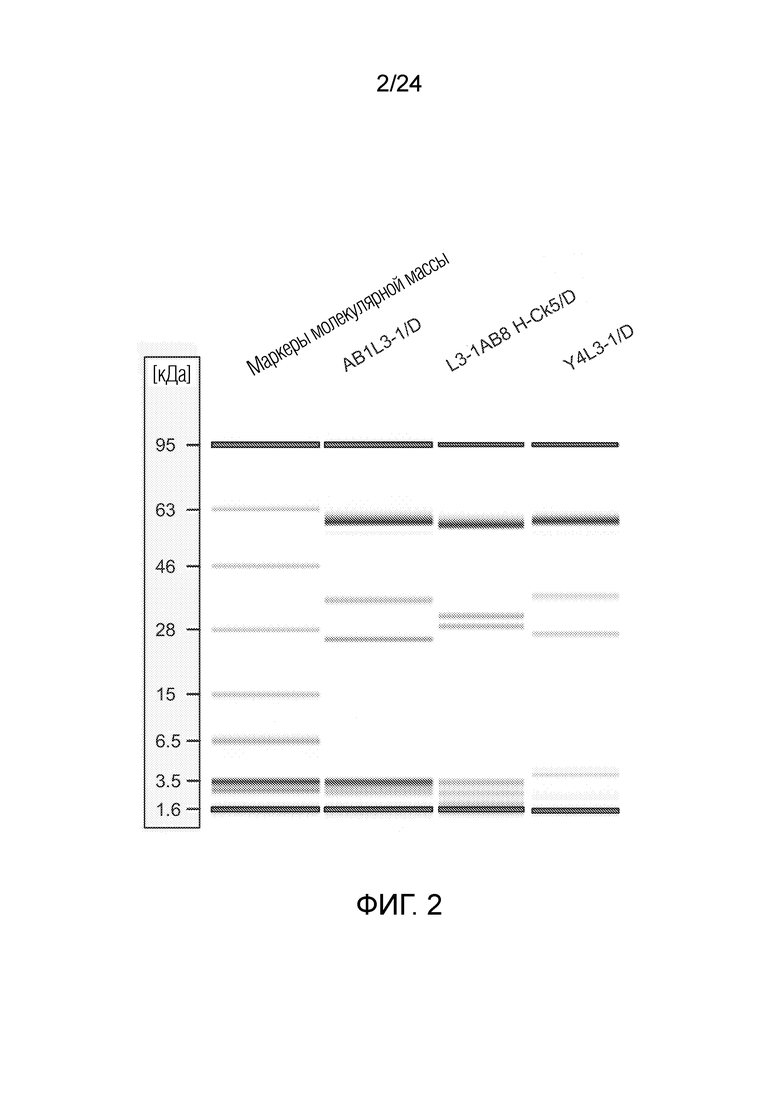
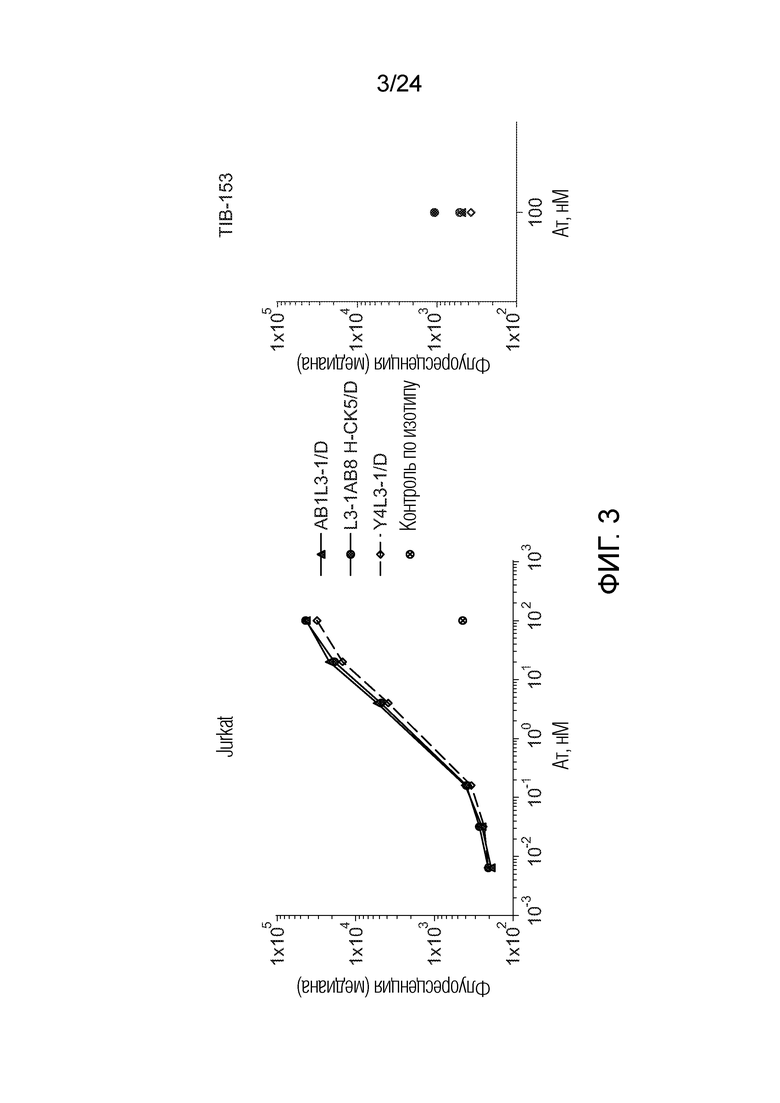
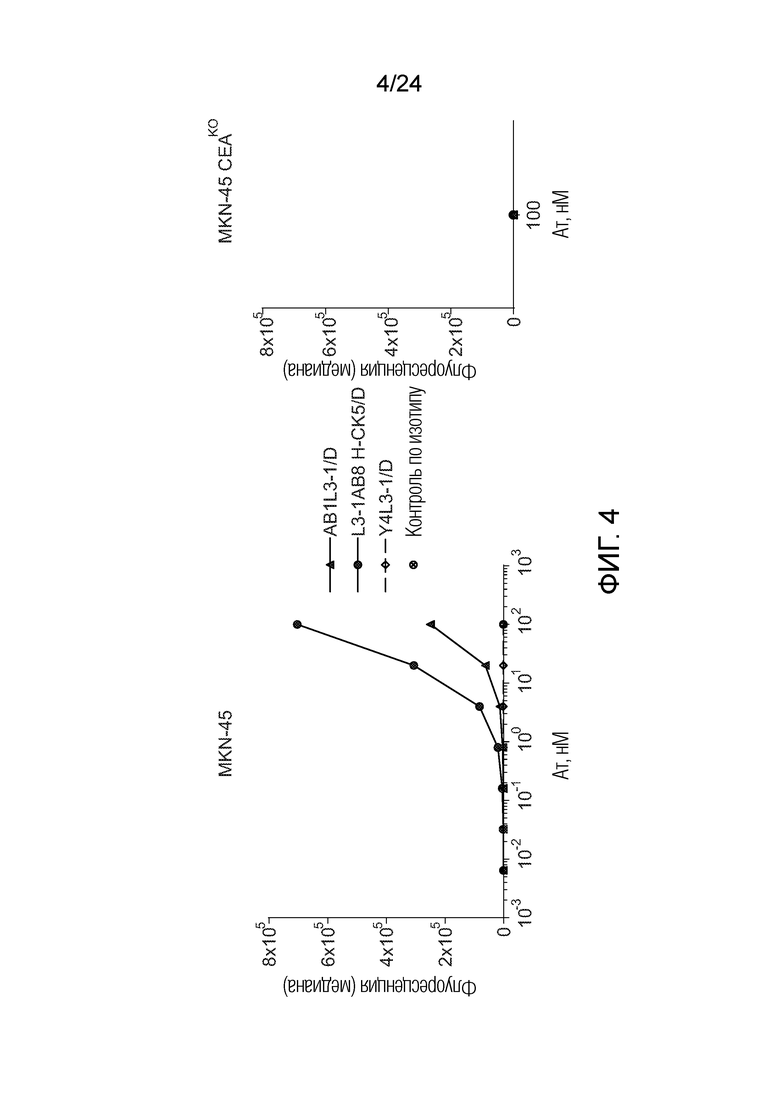
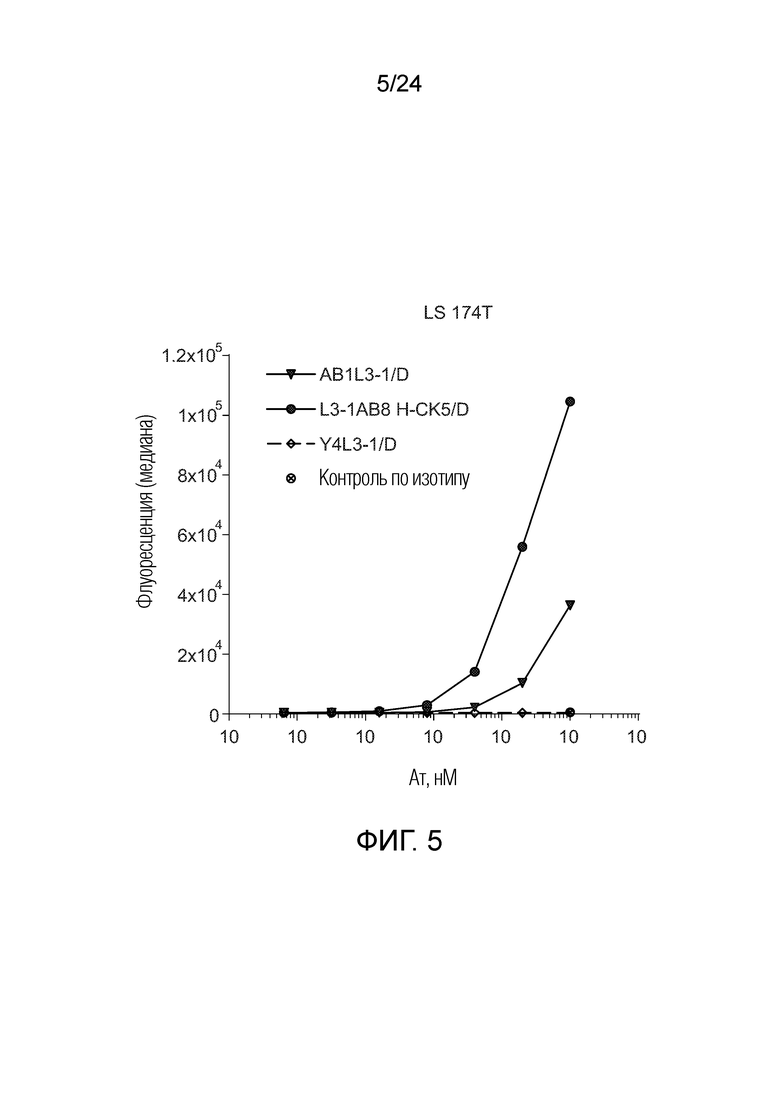

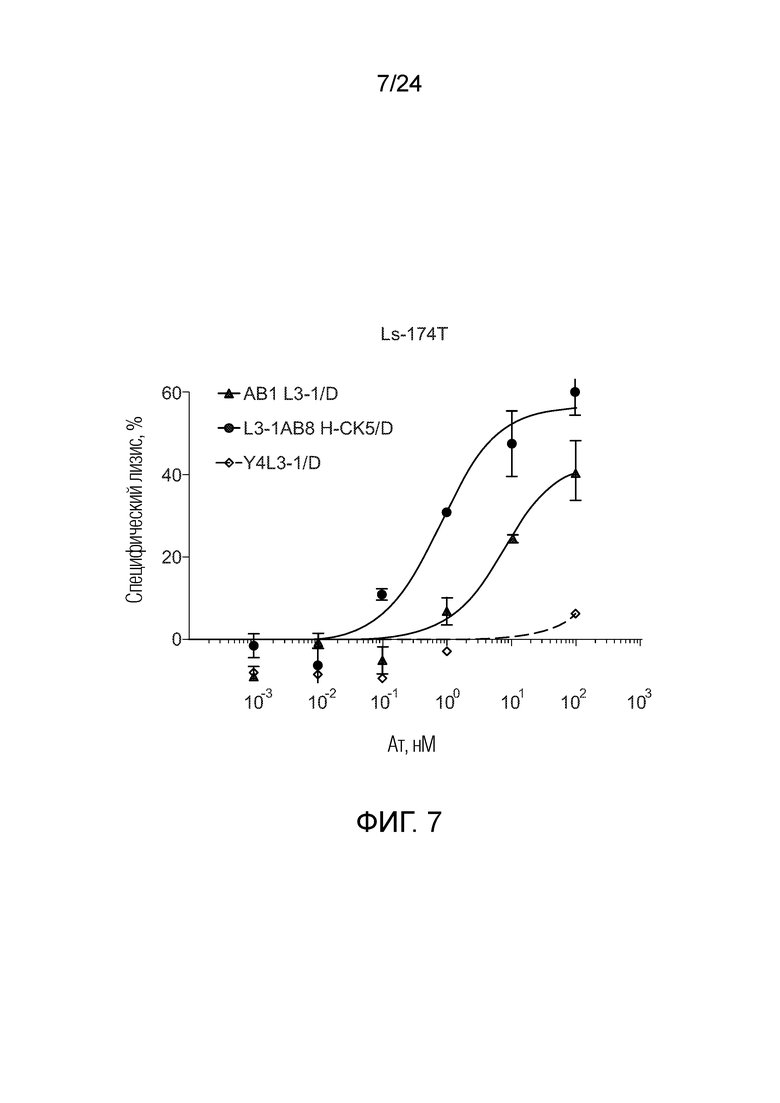
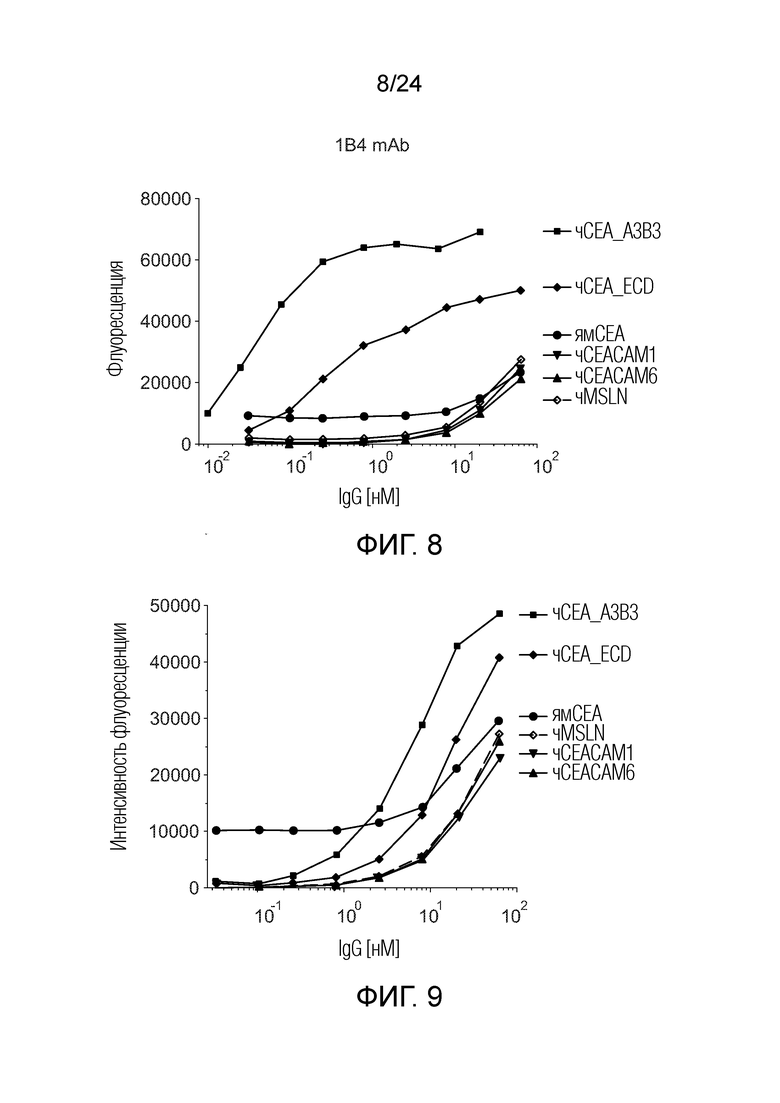
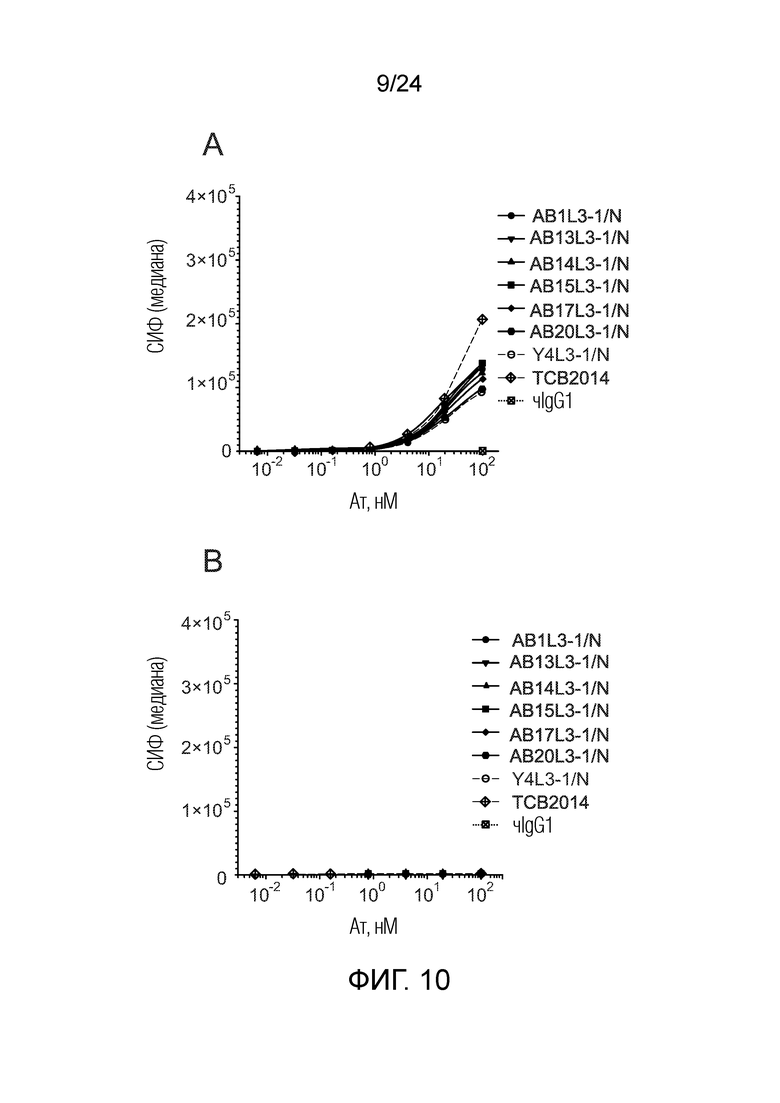
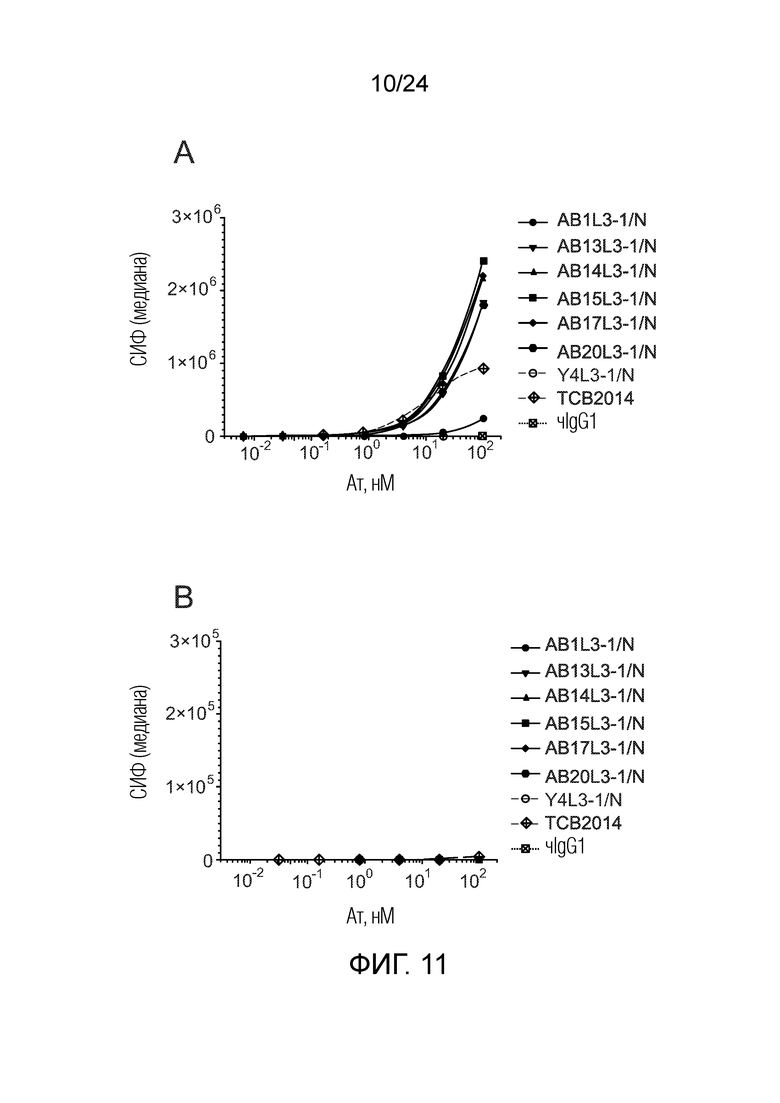
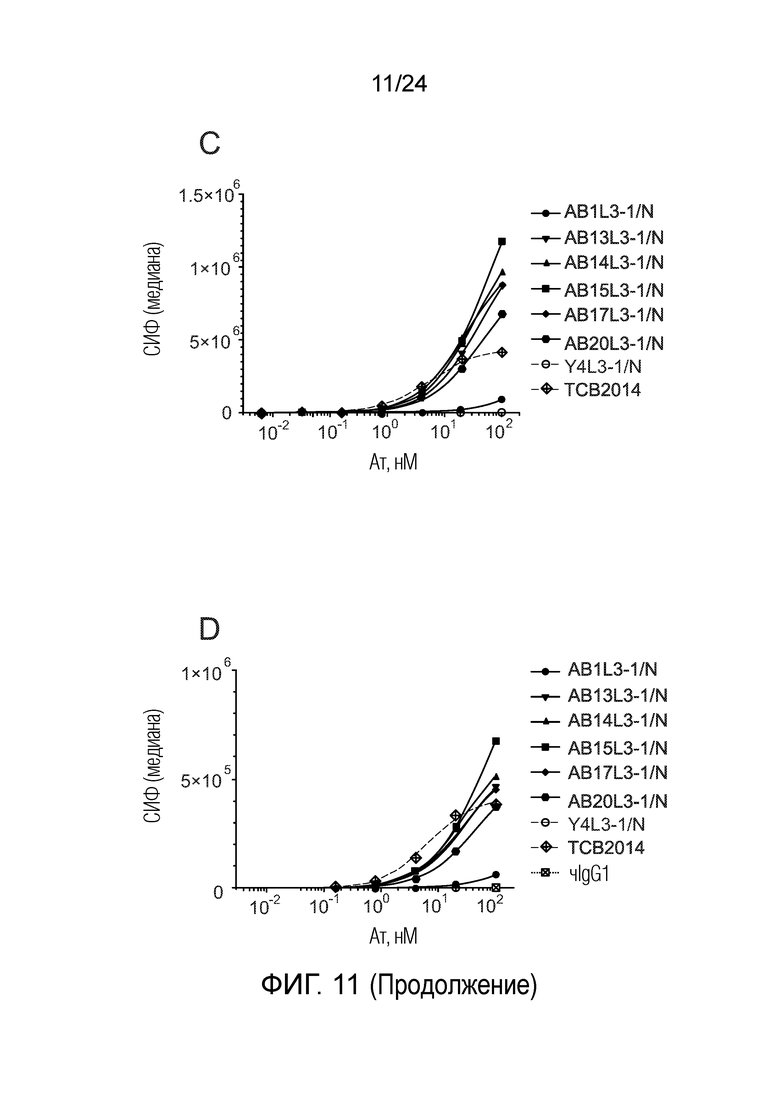
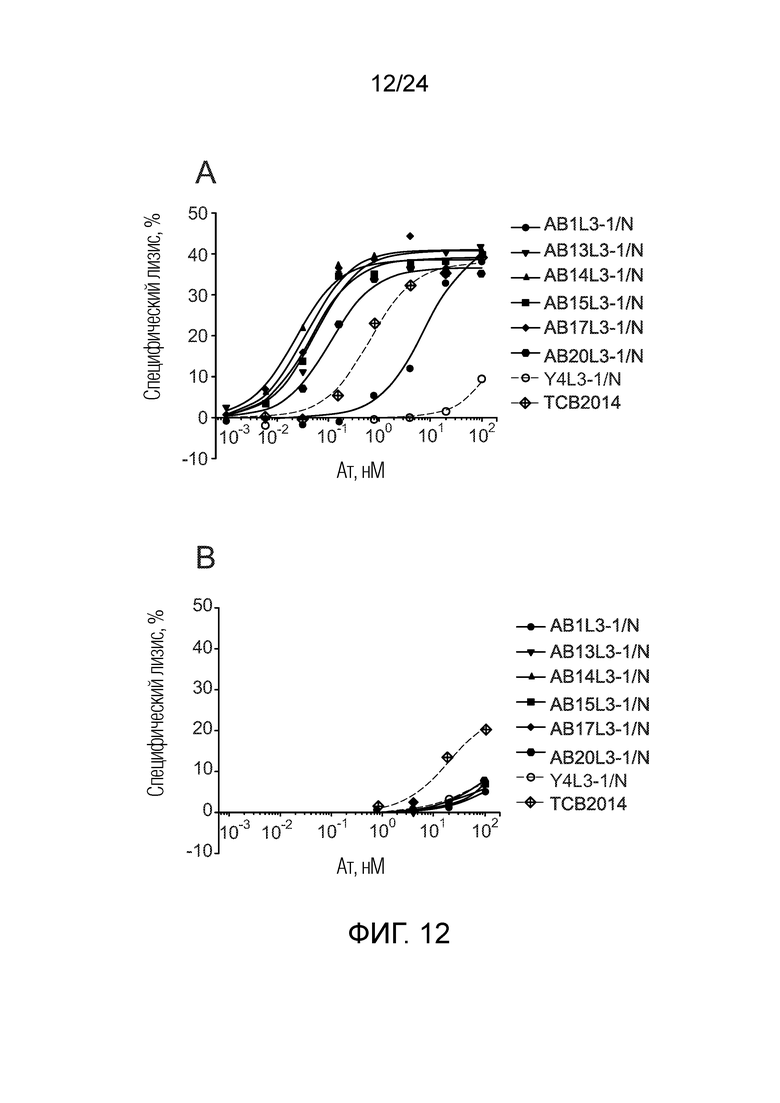
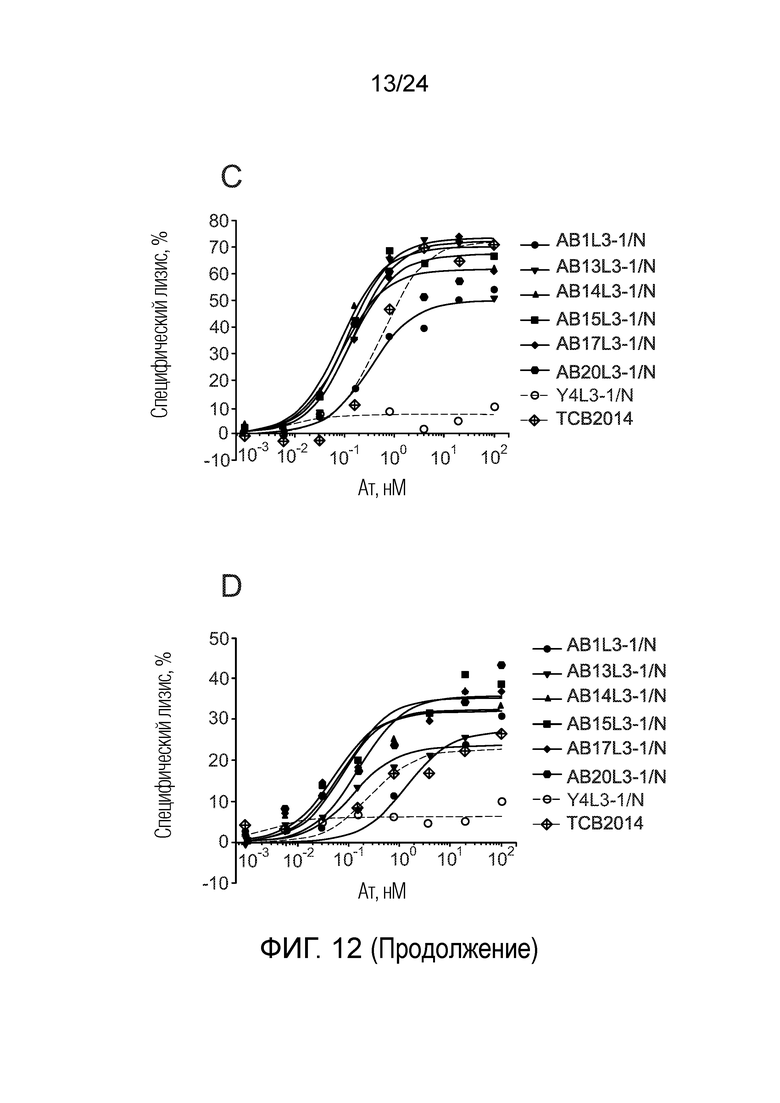
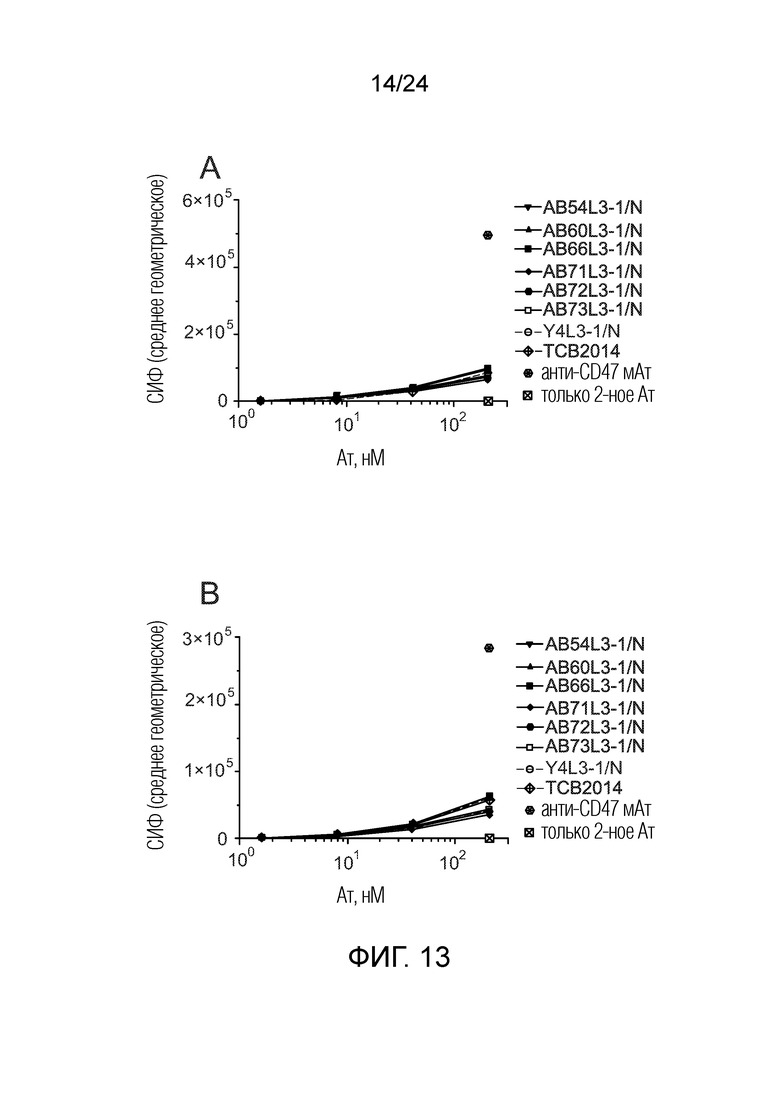
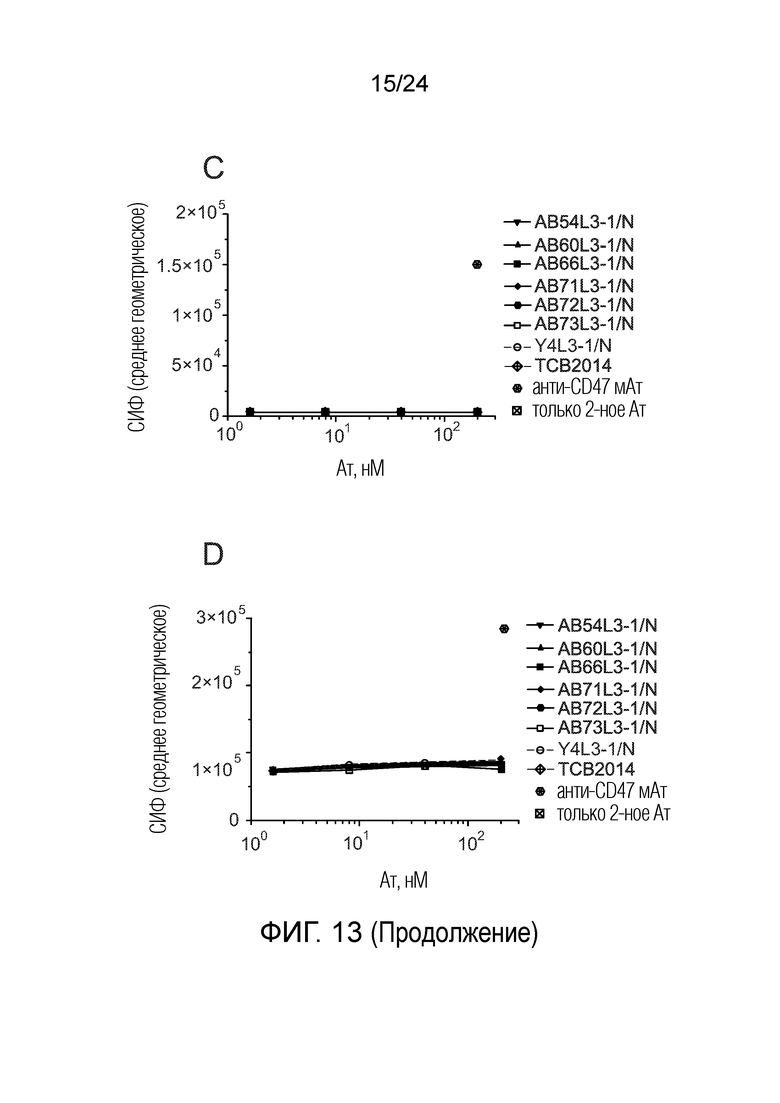
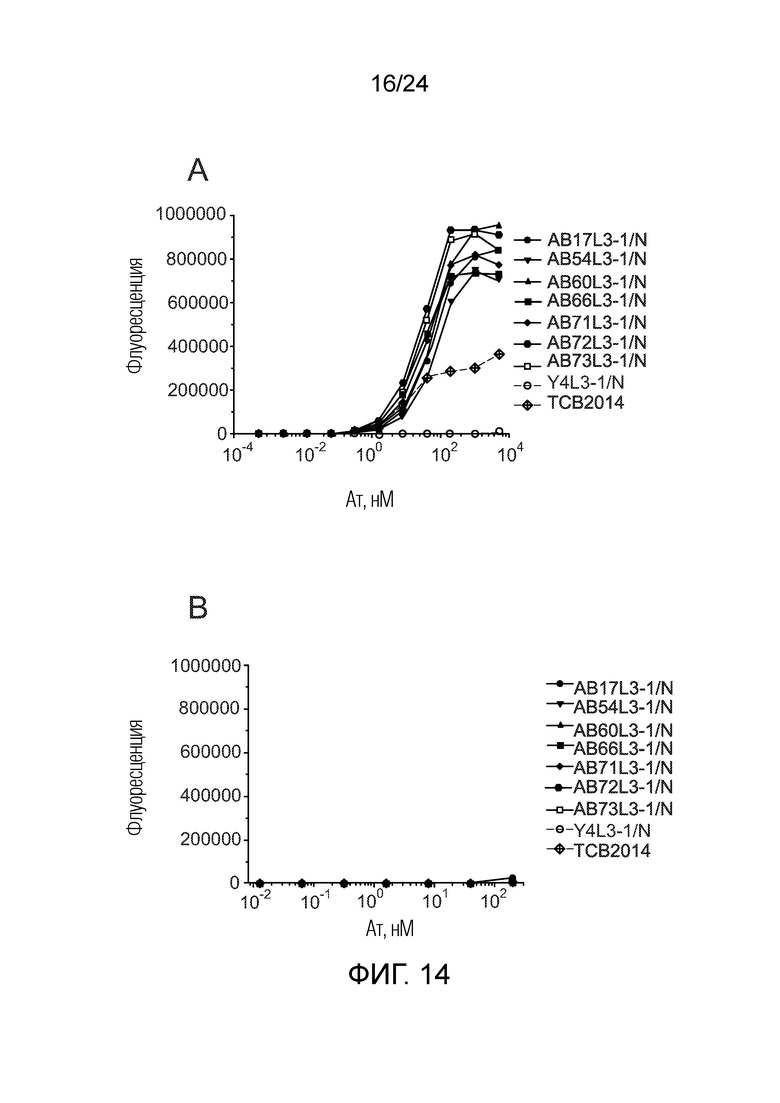
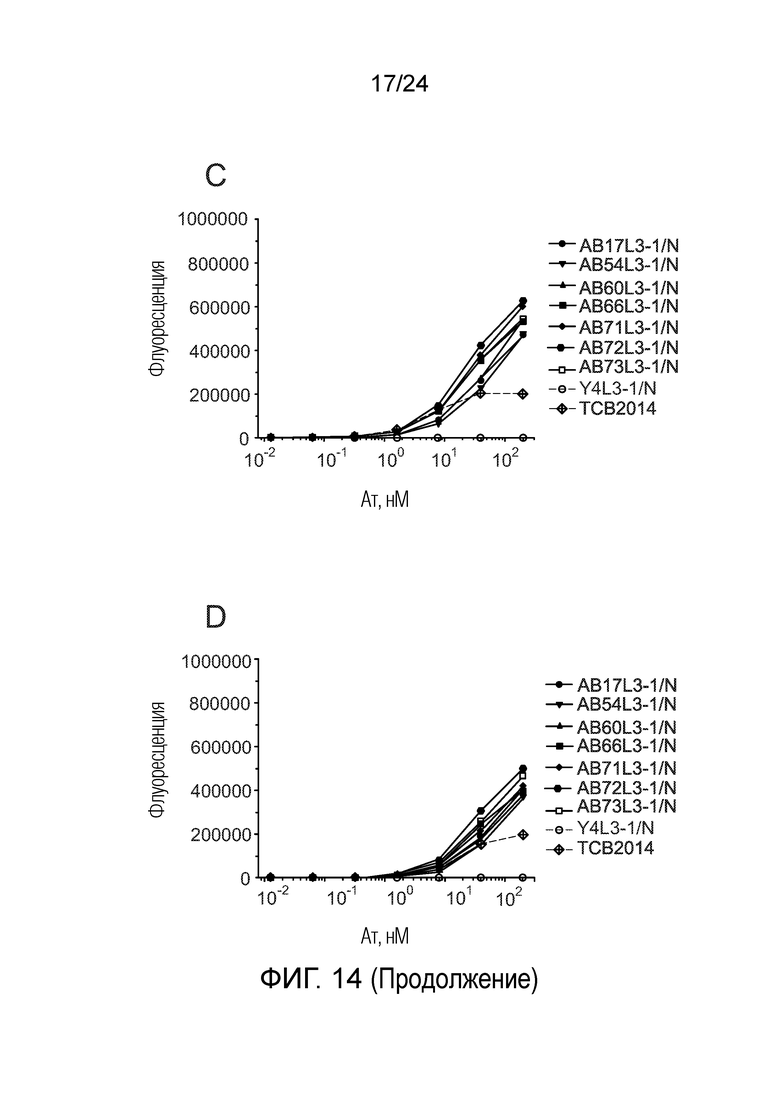

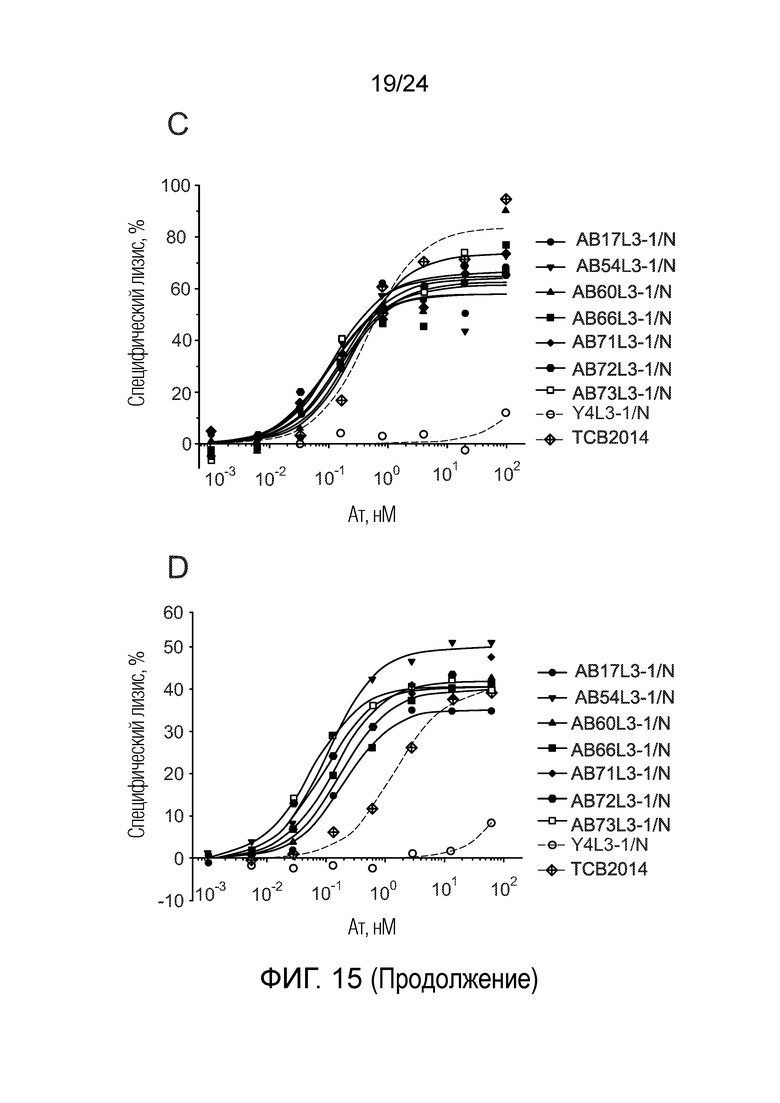
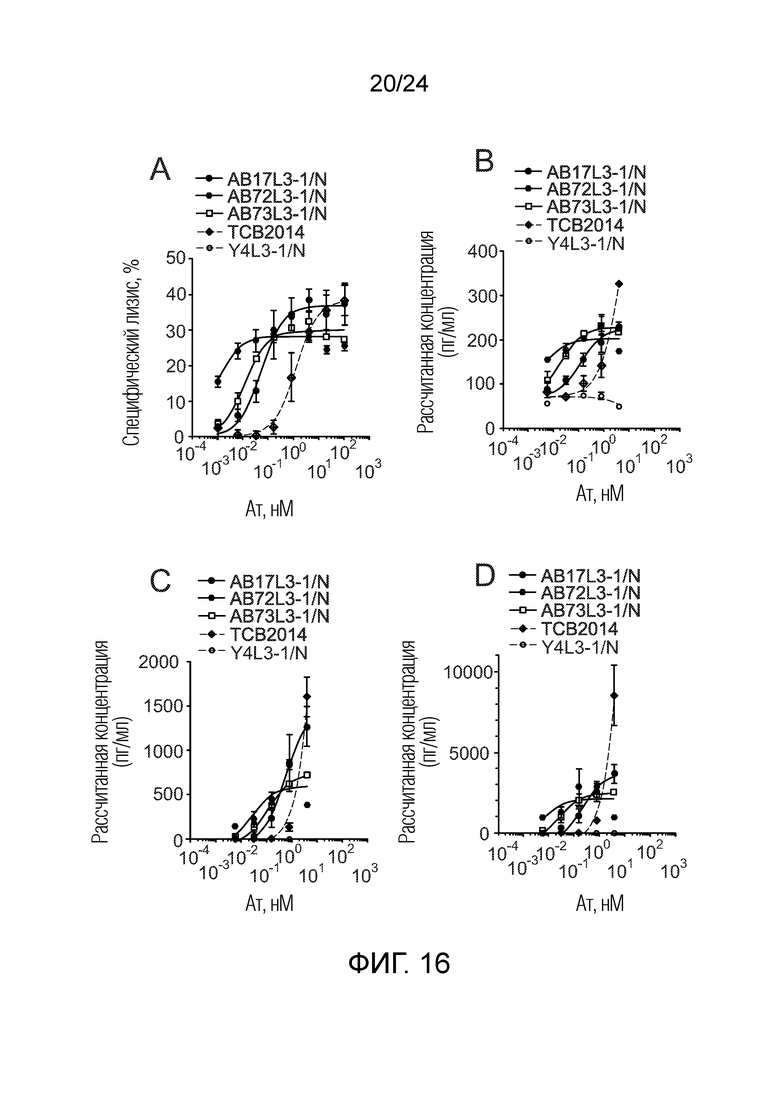
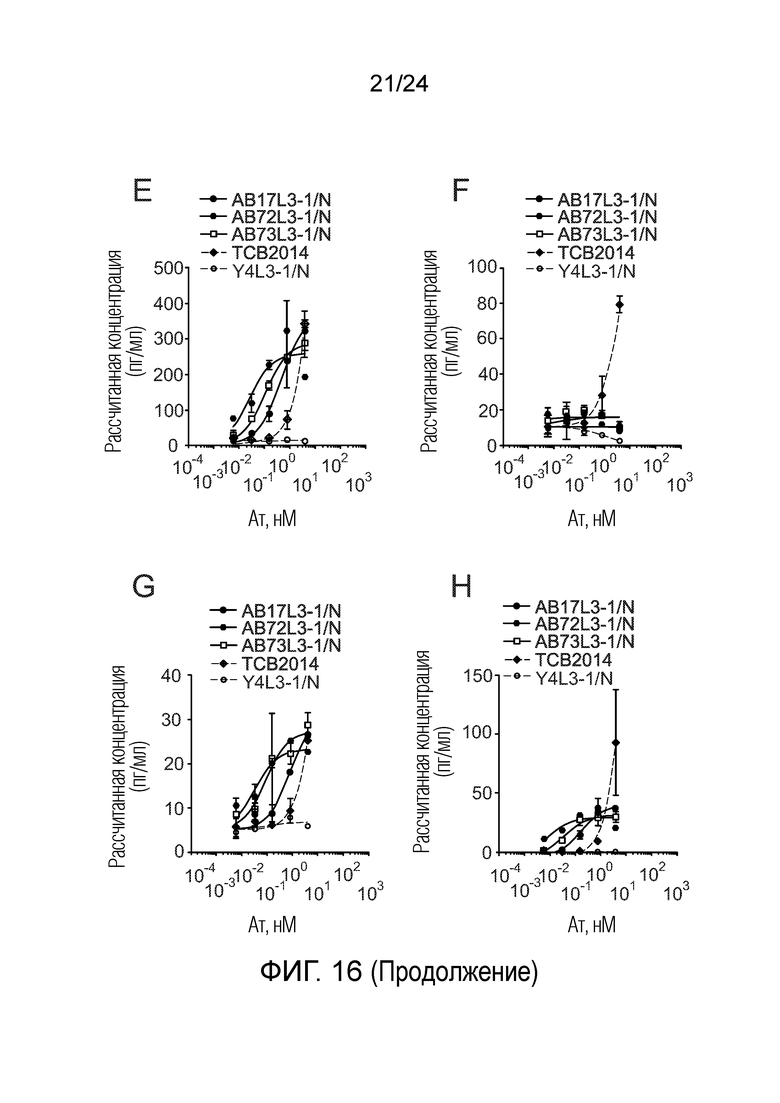

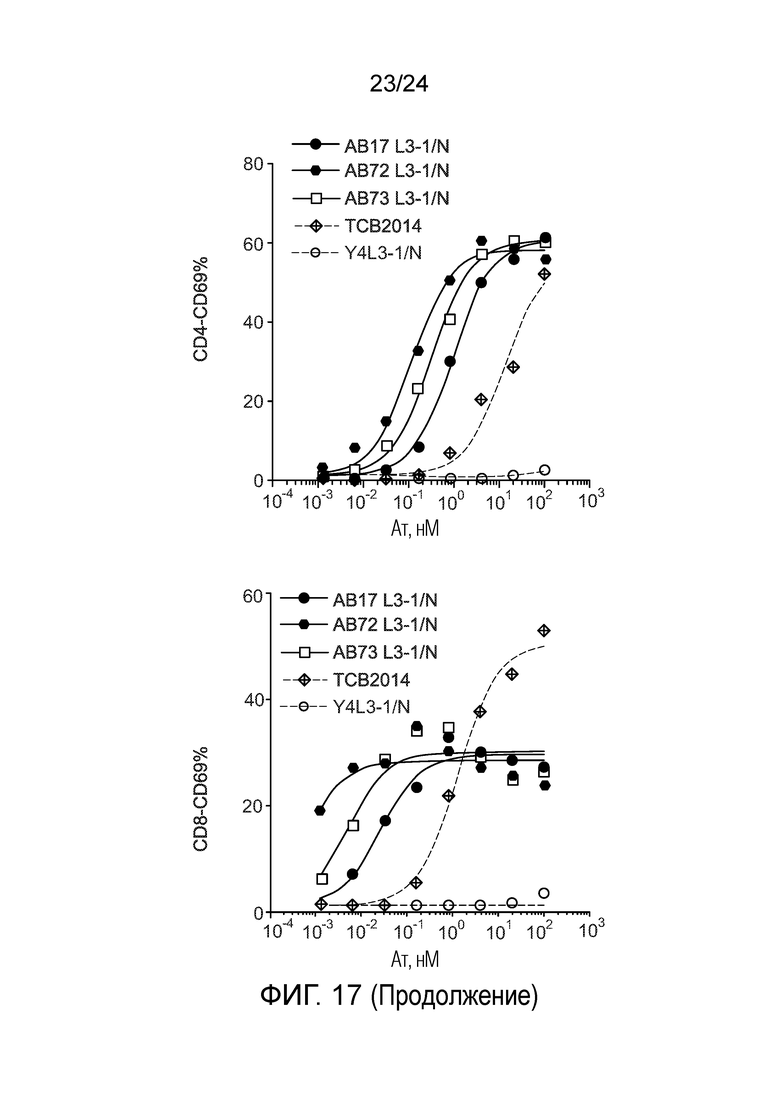
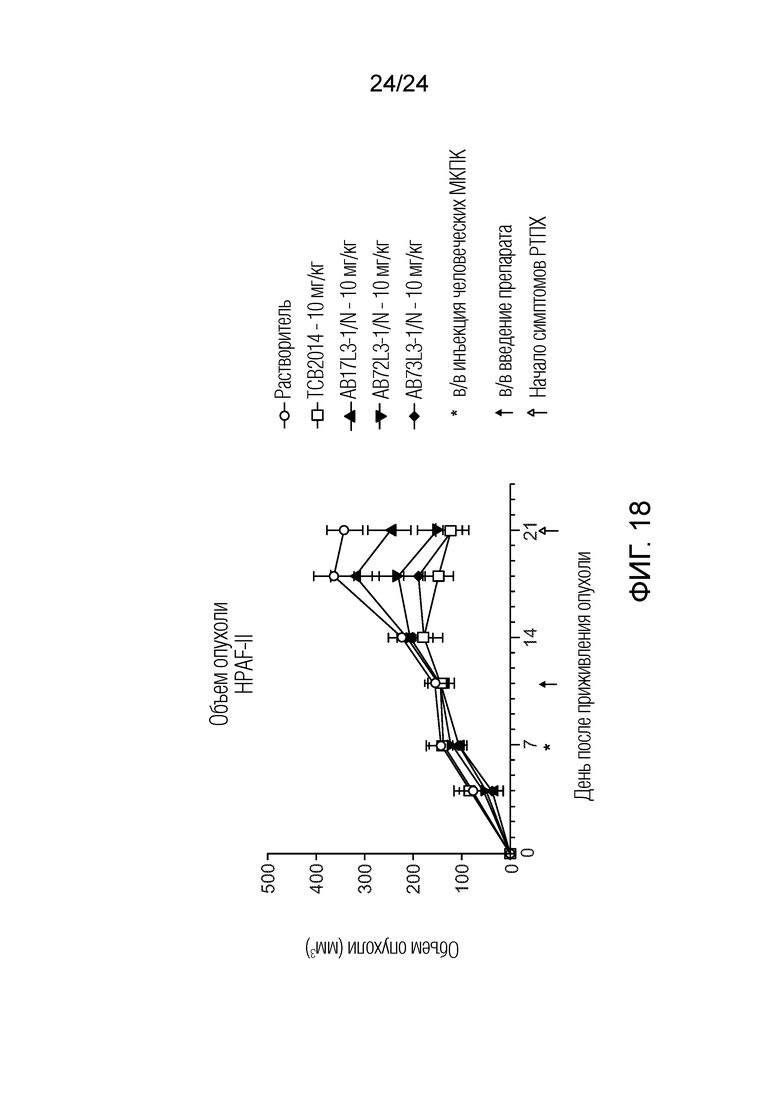
Изобретение относится к области биотехнологии, в частности к биспецифическому антителу, специфически связывающему CEACAM5 человека и CD3ε человека. Указанное антитело содержит связывающий фрагмент, специфически связывающий CEACAM5 человека, и второй связывающий фрагмент, специфически связывающий CD3ε человека. Настоящее изобретение также относится к применению указанного антитела для лечения солидных опухолей, экспрессирующих CEA, in vivo, фармацевтической композиции его содержащей, полинуклеотиду, вектору и клетке для получения указанного антитела. Изобретение обеспечивает получение новых эффективных биспецифических анти-CEAxCD3. 6 н. и 8 з.п. ф-лы, 18 ил., 7 табл., 13 пр.
1. Биспецифическое антитело, специфически связывающее CEACAM5 человека и CD3ε человека, отличающееся тем, что оно содержит первый связывающий фрагмент, специфически связывающий CEACAM5 человека, и второй связывающий фрагмент, специфически связывающий CD3ε человека, где:
a) первый связывающий фрагмент содержит вариабельную область тяжелой цепи (VH), содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4,
b) первый связывающий фрагмент содержит вариабельную область легкой цепи (VL), содержащую набор CDRL, выбранный из группы, состоящей из
b1) CDRL1 с SEQ ID NO:90, CDRL2 с SEQ ID NO:91 и CDRL3 с SEQ ID NO:92,
b2) CDRL1 с SEQ ID NO:96, CDRL2 с SEQ ID NO:97 и CDRL3 с SEQ ID NO:98,
b3) CDRL1 с SEQ ID NO:99, CDRL2 с SEQ ID NO:100 и CDRL3 с SEQ ID NO:101,
b4) CDRL1 с SEQ ID NO:102, CDRL2 с SEQ ID NO:103 и CDRL3 с SEQ ID NO:104,
b5) CDRL1 с SEQ ID NO:105, CDRL2 с SEQ ID NO:106 и CDRL3 с SEQ ID NO:107,
b6) CDRL1 с SEQ ID NO:108, CDRL2 с SEQ ID NO:109 и CDRL3 с SEQ ID NO:110, и
b7) CDRL1 с SEQ ID NO:111, CDRL2 с SEQ ID NO:112 и CDRL3 с SEQ ID NO:113,
c) второй связывающий фрагмент содержит VH, содержащую CDRH1 с SEQ ID NO:2, CDRH2 с SEQ ID NO:3 и CDRH3 с SEQ ID NO:4, и
d) второй связывающий фрагмент содержит VL, содержащую CDRL1 с SEQ ID NO:18, CDRL2 с SEQ ID NO:19 и CDRL3 с SEQ ID NO:20.
2. Биспецифическое антитело по п.1, отличающееся тем, что оно содержит
a) в первом связывающем фрагменте вариабельную область тяжелой цепи VH с SEQ ID NO:1,
b) в первом связывающем фрагменте вариабельную область легкой цепи VL, выбранную из группы, состоящей из
b1) вариабельной области легкой цепи VL с SEQ ID NO:117,
b2) вариабельной области легкой цепи VL с SEQ ID NO:119,
b3) вариабельной области легкой цепи VL с SEQ ID NO:120,
b4) вариабельной области легкой цепи VL с SEQ ID NO:121,
b5) вариабельной области легкой цепи VL с SEQ ID NO:122,
b6) вариабельной области легкой цепи VL с SEQ ID NO:123,
b7) вариабельной области легкой цепи VL с SEQ ID NO:124, и
c) во втором связывающем фрагменте вариабельную область тяжелой цепи VH с SEQ ID NO:1 и вариабельную область легкой цепи VL с SEQ ID NO:17.
3. Биспецифическое антитело по п.1, отличающееся тем, что оно содержит
a) в первом связывающем фрагменте вариабельную область тяжелой цепи VH с SEQ ID NO:1,
b) в первом связывающем фрагменте легкую цепь, выбранную из группы, состоящей из
b1) легкой цепи с SEQ ID NO:128,
b2) легкой цепи с SEQ ID NO:130,
b3) легкой цепи с SEQ ID NO:131
b4) легкой цепи с SEQ ID NO:132,
b5) легкой цепи с SEQ ID NO:133,
b6) легкой цепи с SEQ ID NO:134,
b7) легкой цепи с SEQ ID NO:135, и
c) во втором связывающем фрагменте вариабельную область тяжелой цепи VH с SEQ ID NO:1 и легкой цепи с SEQ ID NO:28.
4. Биспецифическое антитело по п.1, отличающееся тем, что оно содержит общую тяжелую цепь с SEQ ID NO:44 или 45 и во втором связывающем фрагменте легкую цепь с SEQ ID NO:28, и в первом связывающем фрагменте легкую цепь с SEQ ID NO:128.
5. Биспецифическое антитело по п.1, отличающееся тем, что оно содержит общую тяжелую цепь с SEQ ID NO:44 или SEQ ID NO:45 и во втором связывающем фрагменте легкую цепь с SEQ ID NO:28, и в первом связывающем фрагменте легкую цепь с SEQ ID NO:133.
6. Биспецифическое антитело по п.1, отличающееся тем, что оно содержит общую тяжелую цепь с SEQ ID NO:44 или SEQ ID NO:45 и во втором связывающем фрагменте легкую цепь с SEQ ID NO:28, и в первом связывающем фрагменте легкую цепь с SEQ ID NO:135.
7. Применение биспецифического антитела по любому из пп.1-6 для получения лекарственного средства для лечения солидных опухолей, экспрессирующих CEA, in vivo.
8. Биспецифическое антитело по любому из пп.1-6 для применения для лечения солидных опухолей, экспрессирующих CEA.
9. Биспецифическое антитело по любому из пп.1-6 для применения в лечении солидных опухолей, экспрессирующих CEA, в сочетании с биспецифическим анти-CEAxCD47 антителом одновременно, раздельно или последовательно.
10. Биспецифическое антитело по любому из пп.1-6 для лечения солидных опухолей, экспрессирующих CEA, в сочетании с антагонистом оси PD-1 одновременно, раздельно или последовательно.
11. Фармацевтическая композиция, содержащая биспецифическое антитело по любому из пп.1-6 и фармацевтически приемлемый носитель для лечения солидных опухолей, экспрессирующих CEA.
12. Выделенный полинуклеотид, кодирующий антитело по любому из пп.1-6, или его связывающий фрагмент.
13. Вектор для экспресссии антитела по любому из пп.1-6, содержащий выделенный полинуклеотид, кодирующий антитело по любому из пп.1-6.
14. Клетка для получения антитела по любому из пп.1-6, содержащая выделенный полинуклеотид, кодирующий антитело по любому из пп.1-6, или его связывающий фрагмент.
| EP 2961771 A1, 06.01.2016 | |||
| CA 3079039 A1, 20.06.2019 | |||
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АНТИГЕНАМ, АКТИВИРУЮЩИМ Т-КЛЕТКИ, И ОПУХОЛЕВОМУ АНТИГЕНУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2605390C2 |
| MIZUTANI K., et al., Response to nivolumab in metastatic collecting duct carcinoma expressing PD-L1: A case report, Molecular and Clinical Oncology 7, 2017, no | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Паробензиновая турбина | 1921 |
|
SU988A1 |
| BAYLOT V | |||
| et al., TCTP Has a Crucial Role in the Different Stages of Prostate Cancer | |||

