ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к фармацевтической композиции для лечения злокачественной опухоли, содержащей в качестве активных ингредиентов димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля.
Уровень техники
Интерлейкин 2 (IL-2), также называемый T-клеточным фактором роста (TCGF), представляет собой глобулярный гликопротеин, который играет центральную роль в продуцировании, выживании и гомеостазе лимфоцитов. IL-2 имеет размер белка от 15,5 кДа до 16 кДа и состоит из 133 аминокислот. IL-2 опосредует различные иммунные действия путем связывания с рецептором IL-2, состоящим из трех различных субъединиц.
Кроме того, IL-2 синтезируется в основном активированными T-клетками, в частности, CD4+ хелперными T-клетками. IL-2 стимулирует пролиферацию и дифференцировку T клеток и индуцирует продуцирование цитотоксических T-лимфоцитов (CTL) и дифференцировку лимфоцитов периферической крови человека в цитотоксические клетки и лимфокин-активируемые киллеры (LAK-клетки).
Между тем, CD80, также известный как B7-1, является представителем семейства B7 мембраносвязанных белков, которые вовлечены в иммунную регуляцию путем связывания с лигандом посредством обеспечения костимулирующих ответов и коингибирующих ответов. CD80 представляет собой трансмембранный белок, экспрессируемый на поверхности T-клеток, B-клеток, дендритных клеток и моноцитов. Известно, что CD80 связывает CD28, CTLA4 (CD152) и PD-L1. CD80, CD86, CTLA4 и CD28 вовлечены в костимулирующую-коингибирующую систему. Например, они регулируют активность T-клеток и вовлечены в их пролиферацию, дифференцировку и выживаемость.
Кроме того, недавно привлекли внимание ингибиторы иммунной точки контроля, такие как Китруда®. Ингибиторы иммунной точки контроля представляют собой противораковые средства, которые помогают атаковать злокачественные клетки путем активации иммунной системы организма. На сегодняшний день лечение злокачественной опухоли сфокусировано на уничтожении быстроделящихся клеток, что является характеристикой злокачественных клеток, так что существуют побочные эффекты в виде действия не только на злокачественные клетки, но также на быстроделящиеся клетки, присутствующие среди нормальных клеток. Однако известно, что в иммунных противораковых средствах используется иммунная система пациента со злокачественной опухолью для воздействия на злокачественные клетки, так что у существующих противораковых средств мало типичных побочных эффектов. Антитело против PD-1, такое как Китруда, связывается со специфическим рецептором (PD-1) T-клеток и блокирует каскад злокачественных клеток для избегания системы надзора активными T-клетками, тем самым проявляя противораковый эффект через иммунную реактивацию, которая позволяет T-клеткам в организме человека атаковать злокачественные клетки (выложенная публикация патента Кореи № 2018-0030580A).
Подробное описание изобретения
Техническая проблема
Таким образом, в результате исследования для разработки безопасного и эффективного противоракового средства, авторы настоящего изобретения обнаружили, что новый димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент в одной молекуле, и ингибитор иммунной точки контроля, демонстрирует превосходный противораковый эффект, и тем самым осуществили настоящее изобретение.
Решение проблемы
Для достижения указанной выше задачи один из аспектов настоящего изобретения относится к фармацевтической композиции для лечения злокачественной опухоли, содержащей в качестве активных ингредиентов димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля.
Эффект изобретения
Димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, может активировать иммунные клетки посредством IL-2. Кроме того, было подтверждено, что димерный слитый белок проявляет синергические эффекты при введении в комбинации с ингибитором иммунной точки контроля. Таким образом, фармацевтическая композиция для лечения злокачественной опухоли, содержащая в качестве активных ингредиентов димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля, может быть полезной для предупреждения и лечения злокачественной опухоли.
Краткое описание чертежей
На фиг. 1 проиллюстрирован схематический вариант осуществления димерного слитого белка.
На фиг. 2 проиллюстрировано схематическое изображение механизма, посредством которого димерный слитый белок действует в лимфатическом узле.
На фиг. 3 проиллюстрировано схематическое изображение механизма, посредством которого димерный слитый белок действует в опухолевом микроокружении.
На фиг. 4 проиллюстрировано схематическое изображение структуры слитого белка. Здесь, каждый из GI101 и mGI101 является вариантом осуществления слитого белка, и GI101C1, GI101C2 и mGI101C1 являются сравнительными примерами для сравнения с активностью слитого белка.
На фиг. 5 проиллюстрированы различные варианты осуществления слитого белка. Для получения слитого белка можно комбинировать белки человека и мыши. Белок CD80 и белок IL-2 могут быть связаны через различные линкеры, отличные от Fc.
На фиг. 6 проиллюстрирован результат, полученный путем идентификации полученного димерного слитого белка (GI101) посредством SDS-PAGE.
На фиг. 7 проиллюстрированы количества слитого белка (GI101) в зависимости от поглощения.
На фиг. 8 проиллюстрирован результат, полученный путем анализа полученного димерного слитого белка (GI101) посредством эксклюзионной хроматографии (SEC).
На фиг. 9 проиллюстрирован результат, полученный путем идентификации полученного димерного слитого белка mGI101 посредством SDS-PAGE.
На фиг. 10 проиллюстрированы результаты, полученные путем идентификации полученного димерного слитого белка GI101C1 посредством SDS-PAGE.
На фиг. 11 проиллюстрированы результаты, полученные путем идентификации полученного димерного слитого белка GI101C2 посредством SDS-PAGE.
На фиг. 12 проиллюстрирован результат, полученный путем идентификации полученного димерного слитого белка mGI101C1 посредством SDS-PAGE.
На фиг. 13 проиллюстрированы результаты, полученные путем идентификации полученного димерного слитого белка GI102-M45 посредством SDS-PAGE.
На фиг. 14 проиллюстрированы результаты, полученные путем идентификации полученного димерного слитого белка GI102-M61 посредством SDS-PAGE.
На фиг. 15 проиллюстрированы результаты, полученные путем идентификации полученного димерного слитого белка GI102-M72 посредством SDS-PAGE.
На фиг. 16 проиллюстрирована аффинность связывания между hCTLA4 и GI101.
На фиг. 17 проиллюстрирована аффинность связывания между hPD-L1 и GI101.
На фиг. 18 проиллюстрирована аффинность связывания между hPD-L1 и hPD-1.
На фиг. 19 проиллюстрирована аффинность связывания между mCTLA4 и mGI101.
На фиг. 20 проиллюстрирована аффинность связывания между mPD-L1 и mGI101.
На фиг. 21 проиллюстрированы результаты, полученные путем идентификации способности к связыванию GI-101 (hCD80-Fc-hIL-2v) с CTLA-4. Было идентифицировано, что GI-101 (hCD80-Fc-hIL-2v) обладает высокой способностью к связыванию CTLA-4.
На фиг. 22 проиллюстрированы результаты, полученные путем идентификации аффинности связывания GI101 с IL-2Rα или IL-2Rβ.
На фиг. 23 проиллюстрированы результаты, полученные путем идентификации аффинности связывания GI101 с IL-2Rα.
На фиг. 24 проиллюстрированы результаты, полученные путем идентификации аффинности связывания GI101 с IL-2Rβ.
На фиг. 25 проиллюстрированы результаты, полученные путем идентификации аффинности связывания IL-2Rα с GI102-M45.
На фиг. 26 проиллюстрированы результаты, полученные путем идентификации аффинности связывания IL-2Rα с GI102-M61.
На фиг. 27 проиллюстрированы результаты, полученные путем идентификации аффинности связывания IL-2Rα с GI102-M72.
На фиг. 28 проиллюстрированы результаты, полученные путем идентификации аффинности связывания IL-2Rβ с GI102-M45.
На фиг. 29 проиллюстрированы результаты, полученные путем идентификации аффинности связывания IL-2Rβ с GI102-M61.
На фиг. 30 проиллюстрированы результаты, полученные путем идентификации аффинности связывания IL-2Rβ с GI102-M72.
На фиг. 31 и 32 проиллюстрированы результаты, полученные путем определения количеств IFN-γ, секретируемого из клеток, когда клетки подвергали обработке GI101, GI101C1, GI101C2 или IL-2 в соответствующих концентрациях и проводили инкубацию.
На фиг. 33 проиллюстрированы результаты, полученные путем идентификации эффектов GI101, GI101C1, GI101C2 и IL-2 (пролейкин) на пролиферацию CD8+ T-клеток.
На фиг. 34 представлено схематическое изображение механизма, посредством которого GI101 действует на эффекторные T-клетки.
На фиг. 35 проиллюстрированы результаты, полученные путем идентификации эффектов GI101 и GI102 на пролиферацию CD8+ T-клеток и CD4+ T-клеток. Здесь, на (A) проиллюстрированы доли CD8+ T-клеток и CD4+ T-клеток, на (B) проиллюстрирована способность CD8+ T-клеток к пролиферации, и на (C) проиллюстрирована доля CD4+/FoxP3+ Treg клеток.
На фиг. 36 и 37 проиллюстрированы результаты, полученные путем идентификации эффектов GI101 и GI101w на пролиферацию CD8+ T-клеток и NK-клеток.
На фиг. 38 и 39 проиллюстрированы результаты, полученные путем идентификации эффекта GI101 на эффекторные T-клетки.
На фиг. 40 проиллюстрированы результаты, полученные путем идентификации эффектов mGI101 и mGI102-M61 на иммунные клетки мыши.
На фиг. 41 и 42 проиллюстрированы результаты, полученные путем идентификации эффекта GI101 на ингибирование активности T-клеток злокачественными клетками, экспрессирующими PD-L1 и CTLA-4.
На фиг. 43 проиллюстрированы результаты, полученные путем идентификации ингибирующего опухоль эффекта mGI101, в зависимости от его дозы, у мышей с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки.
На фиг. 44 проиллюстрированы результаты, полученные путем идентификации выживаемости мышей с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки, которым вводили mGI101.
На фиг. 45 проиллюстрированы результаты, полученные путем идентификации ингибирующего опухоль эффекта GI101 у мышей, трансплантированных происходящими из мыши клетками рака ободочной и прямой кишки.
На фиг. 46 проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа CD8+ T-клеток, IFN-γ T-клеток, CD4+ T-клеток и Treg-клеток в тканях злокачественной опухоли.
На фиг. 47 графически проиллюстрированы результаты, полученные путем лечения мышей с трансплантированными происходящим из мыши клетками рака ободочной и прямой кишки посредством hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа CD8+ T-клеток, IFN-γ T-клеток, CD4+ T-клеток и Treg-клеток в злокачественных тканях.
На фиг. 48 проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа макрофагов в злокачественных тканях.
На фиг. 49 графически проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа макрофагов в злокачественных тканях.
На фиг. 50 проиллюстрированы результаты, путем введения мышам с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа дендритных клеток в злокачественных тканях.
На фиг. 51 графически проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа дендритных клеток в злокачественных тканях.
На фиг. 52 проиллюстрированы результаты, полученные путем идентификации ингибирующего опухоль эффекта GI101 у мышей с трансплантированными происходящими из мыши клетками рака легкого.
На фиг. 53 графически проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящим из мыши клетками рака легкого hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа CD8+ T-клеток, IFN-γ T-клеток, CD4+ T-клеток и Treg-клеток в злокачественных тканях.
На фиг. 54 графически проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящими из мыши клетками рака легкого hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа макрофагов в злокачественных тканях.
На фиг. 55 графически проиллюстрированы результаты, полученные путем введения мышам с трансплантированными происходящими из мыши клетками рака легкого hIgG4, антитела против PD-1 или GI101, а затем FACS-анализа дендритных клеток в злокачественных тканях.
На фиг. 56 проиллюстрированы результаты, полученные путем идентификации ингибирующего опухоль эффекта mGI102-M61 у мышей с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки.
На фиг. 57 проиллюстрированы результаты, полученные путем анализа выживаемости мышей с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки, которым вводили mGI102-M61.
На фиг. 58 проиллюстрированы результаты, полученные путем идентификации ингибирующего опухоль эффекта mGI101 у мышей с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки.
На фиг. 59 проиллюстрирована степень ингибирования опухоли посредством mGI101 у мышей с трансплантированными происходящими из мыши клетками рака ободочной и прямой кишки.
На фиг. 60 представлен график роста опухоли, когда GI101 и Китруду использовали в комбинации у мышей с трансплантированными происходящими из человека клетками рака молочной железы. Рост опухоли ингибировался в группах введения GI101 или Китруды отдельно по сравнению с контрольной группой (hIgG4). Рост опухоли ингибировался в группе комбинированного введения GI101 и Китруды по сравнению с контрольной группой. Рост опухоли ингибировался в группе комбинированного введения GI101 и Китруды по сравнению с группами введения GI101 или Китруды отдельно.
На фиг. 61 представлена степень ингибирования роста опухоли, когда GI-101 и Китруду использовали в комбинации у мышей с трансплантированными происходящими из человека клетками рака молочной железы. Группа введения IgG4 включала 2 мыши, имевших степень ингибирования роста опухоли 30% или более, 1 мышь, имевшую степень ингибирования роста опухоли 50% или более и 1 мышь, имевшую степень ингибирования роста опухоли 80%. Группа введения GI101 включала 5 мышей, имевших степень ингибирования роста опухоли 30% или более, 5 мышей, имевших степень ингибирования роста опухоли 50% или более, и 2 мыши, имевших степень ингибирования роста опухоли 80%. Группа введения Китруды включала 7 мышей, имевших степень ингибирования роста опухоли 30% или более, 5 мышей, имевших степень ингибирования роста опухоли 50% или более, и 3 мыши, имевших степень ингибирования роста опухоли 80%. Группа комбинированного введения GI101 и Китруды включала 8 мышей, имевших степень ингибирования роста опухоли 30% или более, 8 мышей, имевших степень ингибирования роста опухоли 50% или более, и 6 мышей, имевших степень ингибирования роста опухоли 80%.
На фиг. 62 представлена степень роста опухоли у индивидуальных экспериментальных животных в каждой из групп лечения, когда GI101 и Китруду использовали в комбинации у мышей с трансплантированными происходящими из человека клетками рака молочной железы.
На фиг. 63 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе введения hIgG4 мышам с трансплантированными происходящими из человека клетками рака молочной железы.
На фиг. 64 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе введения GI101 мышам с трансплантированными происходящими из человека клетками рака молочной железы.
На фиг. 65 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе введения Китруды мышам с трансплантированными происходящими из человека клетками рака молочной железы.
На фиг. 66 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе комбинированного введения GI101 и Китруды мышам с трансплантированными происходящими из человека клетками рака молочной железы.
На фиг. 67 представлен график роста опухоли, когда mGI101 и антитело против PD-1 вводили в комбинации мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 68 представлена степень ингибирования роста опухоли, когда mGI101 и антитело против PD-1 вводили в комбинации мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 69 представлена степень роста опухоли у индивидуальных экспериментальных животных в каждой группе введения, когда mGI101 и антитело против PD-1 вводили в комбинации мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 70 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе введения hIgG4 мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 71 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе введения mGI101 мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 72 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе введения антитела против PD-1 мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 73 представлена степень роста опухоли у индивидуальных экспериментальных животных в группе комбинированного введения mGI101 и антитела против PD-1 мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 74 представлена степень роста опухоли у индивидуальных экспериментальных животных, когда происходящие из грызуна клетки рака ободочной и прямой кишки повторно инъецировали экспериментальным животным, которые продемонстрировали полную ремиссию в группе комбинированного введения mGI101 и антитела против PD-1 мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 75 представлен график роста опухоли, когда mGI101 и антитело против PD-L1 вводили в комбинации мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
На фиг. 76 представлен график роста опухоли, когда mGI101 и антитело против TIGIT вводили в комбинации мышам с трансплантированными происходящими из грызунов клетками рака ободочной и прямой кишки.
Наилучшие способы осуществления изобретения
Один из аспектов настоящего изобретения относится к фармацевтической композиции для лечения злокачественной опухоли, содержащей в качестве активных ингредиентов димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля.
Ингибитор иммунной точки контроля
Как используют в рамках изобретения, термин "иммунная точка контроля" относится к внутриклеточной сигнальной системе, которая поддерживает толерантность к своему и защищает ткани от чрезмерных иммунных ответов, которые вызывают повреждение. Белки иммунной точки контроля представляют собой белки клеточной мембраны, которые регулируют иммунную точку контроля и могут ингибировать дифференцировку, пролиферацию и активность иммунных клеток. В частности, белок иммунной точки контроля экспрессируется на активированных T-клетках, функционируя путем снижения пролиферации T-клеток, секреции цитокинов и цитотоксичности, и ингибирования чрезмерной активности T-клеток. Известно, что некоторые иммунные точки контроля являются одним из основных механизмов опухолевых клеток, приводящих к ускользанию от иммунного надзора. Таким образом, "ингибитор иммунной точки контроля" нацелен на белки иммунной точки контроля для ингибирования или блокирования иммунных точек контроля и повышения активации T-клеток, тем самым повышая противоопухолевый иммунитет и, таким образом, демонстрируя противораковый эффект. В дополнение к преимуществам наличия меньших побочных эффектов, таких как рвота и потеря волос, чем у общепринятых цитотоксических противораковых средств, и более высокого терапевтического эффекта, известно, что ингибиторы иммунной точки контроля имеют длительный терапевтический эффект даже после прекращения введения лекарственного средства вследствие использования системы иммунного ответа с превосходной способностью к памяти.
В частности, ингибитор иммунной точки контроля может быть нацелен на CTLA-4, PD-1, PD-L1, PD-L2, B7-H4, HVEM (медиатор проникновения вируса герпеса), BTLA, TIM3, GAL9, LAG3, VISTA, KIR или TIGIT.
В частности, ингибитор иммунной точки контроля может включать, но не ограничиваться ими, антитело против CTLA-4, антитело против PD-1, антитело против PD-L1, антитело против PD-L2, антитело против B7-H4, антитело против HVEM, антитело против BTLA, антитело против TIM3, антитело против GAL9, антитело против LAG3, антитело против VISTA, антитело против KIR и антитело против TIGIT.
Как используют в рамках изобретения, термин "ассоциированный с цитотоксическими T-лимфоцитами антиген 4 (CTLA-4)" обозначают как CD152, и он экспрессируется на поверхности мембраны активированных T-клеток. CTLA-4 связывается с CD80 (B7-1) и CD86 (B7-2) антигенпредставляющих клеток и ингибирует активность T-клеток. Ингибиторы CTLA-4 могут представлять собой ипилимумаб (ЕРВОЙ®) и тремелимумаб.
Как используют в рамках изобретения, термин "белок запрограммированной клеточной смерти 1 (PD-1)" обозначается как CD279, и он экспрессируется на поверхности активированных T-клеток. PD-1 реагирует с белком на поверхности злокачественных клеток, PD-L1 (B7-H1) и PD-L2 (B7-DC), и ингибирует активность T-клеток, продуцирование факторов роста и продуцирование цитокинов, которые опосредуются TCR (T-клеточный рецептор) и CD28, индуцируя передачу негативного сигнала. Ингибитор PD-1 может представлять собой, например, пембролизумаб (Китруда®), MK-3475, ниволумаб (Опдиво®), цемиплимаб (Либтайо®), JTX-4014, спартализумаб, камрелизумаб, синтилимаб, тислелизумаб, торипалимаб, достарлимаб, INCMGA00012, AMP-224 и AMP-514.
Как используют в рамках изобретения, термин "лиганд белка запрограммированной смерти 1 (PD-L1)" обозначается как CD274 или B7-H1, и он представляет собой белок, презентируемый на поверхности злокачественных клеток и на кроветворных клетках. PD-L1, презентируемый на поверхности злокачественных клеток, может связываться с PD-1, презентируемым на поверхности T-клеток. Ингибитор PD-L1 может представлять собой, например, атезолизумаб, авелумаб (Бавенсио®), дурвалумаб (Имфинзи®), KN035, CK-301, AUNP12, CA-170 и BMS-986189.
Как используют в рамках изобретения, термин "B7-H4" обозначается как содержащий домен V-set ингибитор активации T-клеток 1 (VTCN1), и он экспрессируется на поверхности мембраны антигенпредставляющих клеток. B7-H4 связывается с белком CD28 T-клеток и ингибирует активность, рост и продуцирование цитокинов T-клетками, негативно регулируя опосредуемый T-клетками иммунный ответ.
Как используют в рамках изобретения, термин "медиатор проникновения вируса герпеса (HVEM)" обозначается как CD270, и он также известен как представитель 14 суперсемейства рецепторов фактора некроза опухоли (TNFRSF14). HVEM экспрессируется на поверхности мембраны различных иммунных клеток, включая T-клетки, и связывается с различными белками-партнерами, регулируя воспаление и иммунный ответ. Когда HVEM связывается с аттенуатором B- и T-лимфоцитов (BTLA, CD272) или CD160 T-клеток, происходит ингибирование иммунной активности T-клеток. С другой стороны, когда HVEM связывается с TNFSF14 (LIGHT), HVEM индуцирует созревание дендритных клеток, пролиферацию T-клеток и продуцирование цитокинов, активируя воспаление и иммунный ответ.
Как используют в рамках изобретения, термин "T-клеточный мембранный белок 3 (TIM3)" также называется клеточным рецептором 2 вируса гепатита A (HAVCR2), и он экспрессируется в различных иммунных клетках. Когда TIM3 активируется посредством связывания с растворимым в воде белком GAL9 (галектин 9), внутриклеточный приток кальция возрастает, индуцируя апоптоз T-клеток, что в свою очередь вызывает иммунную толерантность. Кроме того, вместе с GAL9 TIM3 связывается с молекулой клеточной адгезии 1 (CEACAM1), которая представляет собой белок клеточной поверхности, ингибирующий иммунную активность T-клеток, и связывается с белком 1 группы с высокой подвижностью (HMGB1) или фосфатидилсерином (PTdSer), который является растворимым в воде белком, ингибируя иммунную активность. Ингибиторы TIM3 могут представлять собой LY3321367, MBG453 и TSR-022.
Как используют в рамках изобретения, термин "ген активации лимфоцитов 3 (LAG3)" обозначается как CD223, и он связывается с основным комплексом гистосовместимости (MHC) класса II, ингибируя пролиферацию и активность T-клеток. Ингибиторы LAG3 могут представлять собой IMP321, релатлимаб и GSK2831781.
Как используют в рамках изобретения, термин "Ig-супрессор активации T-клеток с V-доменом (VISTA)" принадлежит семейству B7 (B7-H5) и экспрессируется в различных иммунных клетках, ингибируя пролиферацию, активность и продуцирование цитокинов T-клетками. Ингибиторы VISTA могут представлять собой JNJ-63723283.
Как используют в рамках изобретения, термин "иммуноглобулин-подобный рецептор киллерных клеток (KIR)" представляет собой мембраносвязанный белок, экспрессируемый в NK-клетках и T-клетках, и представляет собой белок семейства, имеющего генетическое разнообразие и гомологию. Среди этих белков, KIR2DL1, KIR2DL2/L3, KIR3DL1 и KIR3DL2 могут связываться с MHC класса I, ингибируя иммунную активность NK-клеток.
Как используют в рамках изобретения, термин "T-клеточный иммуноглобулин и ITIM-домен (TIGIT)" представляет собой мембраносвязанный белок, экспрессируемый на поверхности NK-клеток и T-клеток, и он может связываться с CD155, CD112 и CD113, ингибируя иммунную активность.
Слитый белок, содержащий белок IL-2 и белок CD80, и его димер
Как используют в рамках изобретения, термин "IL-2" или "интерлейкин-2", если нет иных указаний, относится к любому IL-2 дикого типа, получаемому из любого источника, являющегося позвоночным, включая млекопитающих, например, приматов (таких как люди) и грызунов (таких как мыши и крысы). IL-2 может быть получен из клеток животных, и он также включает IL-2, полученный из рекомбинантных клеток, способных продуцировать IL-2. Кроме того, IL-2 может представлять собой IL-2 дикого типа или его вариант.
В настоящем описании, IL-2 или его вариант могут в совокупности обозначаться термином "белок IL-2" или "полипептид IL-2". IL-2, белок IL-2, полипептид IL-2 и вариант IL-2 специфически связывается, например, с рецептором IL-2. Это специфическое связывание может быть идентифицировано способами, известными специалистам в данной области.
Один из вариантов осуществления IL-2 может иметь аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36. Здесь, IL-2 также может быть в зрелой форме. В частности, зрелый IL-2 может не содержать сигнальную последовательность и может иметь аминокислотную последовательность SEQ ID NO: 10. Здесь, IL-2 может использоваться в рамках концепции, охватывающей фрагмент IL-2 дикого типа, где IL-2 дикого типа укорочен с N-конца или C-конца.
Кроме того, фрагмент IL-2 может иметь форму, где N-конец белка, имеющего аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36, укорочен на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 последовательно расположенных аминокислот. Кроме того, фрагмент IL-2 может иметь форму, где C-конец белка, имеющего аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36, укорочен на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 последовательно расположенных аминокислот.
Как используют в рамках изобретения, термин "вариант IL-2" относится к форме, где часть аминокислот в полноразмерном IL-2 или описанном выше фрагменте IL-2 заменена. Таким образом, вариант IL-2 может иметь аминокислотную последовательность, отличную от IL-2 дикого типа или его фрагмента. Однако вариант IL-2 может иметь активность, эквивалентную или сходную с IL-2 дикого типа. Здесь, "активность IL-2" может относиться, например, к специфическому связыванию с рецептором IL-2, которое можно определять способами, известными специалистам в данной области.
В частности, вариант IL-2 можно получать путем замены части аминокислот в IL-2 дикого типа. Вариант осуществления варианта IL-2, полученного путем аминокислотной замены, может быть получен путем замены по меньшей мере одной из 38-й, 42-й, 45-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может быть получен путем замены по меньшей мере одной из 38-й, 42-й, 45-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10 другой аминокислотой. Кроме того, когда IL-2 имеет форму, где часть N-конца в аминокислотной последовательности SEQ ID NO: 35 укорочена, аминокислота в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10, может быть заменена другой аминокислотой. Например, когда IL-2 имеет аминокислотную последовательность SEQ ID NO: 35, его вариант IL-2 может быть получен путем замены по меньшей мере одной из 58-й, 62-й, 65-й, 81-й или 92-й аминокислот в аминокислотной последовательности SEQ ID NO: 35 другой аминокислотой. Эти аминокислотные остатки соответствуют 38-му, 42-му, 45-му, 61-му и 72-му аминокислотным остаткам в аминокислотной последовательности SEQ ID NO: 10, соответственно. Согласно одному из вариантов осуществления, одна, две, три, четыре, пять, шесть, семь, восемь, девять или десять аминокислот могут быть заменены при условии, что такой вариант IL-2 сохраняет активность IL-2. Согласно другому варианту осуществления могут быть заменены от одной до пяти аминокислот.
В одном из вариантов осуществления вариант IL-2 может иметь форму, где две аминокислоты заменены. В частности, вариант IL-2 может быть получен путем замены 38-й и 42-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й и 45-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 42-й и 45-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 42-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 42-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 45-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 45-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Более того, вариант IL-2 может иметь форму, где три аминокислоты заменены. В частности, вариант IL-2 может быть получен путем замены 38-й, 42-й и 45-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 42-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 42-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 45-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 45-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 42-й, 45-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 42-й, 45-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 45-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может иметь форму, где четыре аминокислоты заменены. В частности, вариант IL-2 может быть получен путем замены 38-й, 42-й, 45-й и 61-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 42-й, 45-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 45-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 38-й, 42-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замены 42-й, 45-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Более того, вариант IL-2 может иметь форму, где пять аминокислот заменены. В частности, вариант IL-2 может быть получен путем замены каждой из 38-й, 42-й, 45-й, 61-й и 72-й аминокислот в аминокислотной последовательности SEQ ID NO: 10 другой аминокислотой.
Здесь, "другая аминокислота", внесенная путем замены, может представлять собой любую аминокислоту, выбранную из группы, состоящей из аланина, аргинина, аспарагина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глутамина, гистидина, изолейцина, лейцина, лизина, метионина, фенилаланина, пролина, серина, треонина, триптофана, тирозина и валина. Однако, что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 38-я аминокислота не может быть заменена на аргинин, 42-я аминокислота не может быть заменена на фенилаланин, 45-я аминокислота не может быть заменена на тирозин, 61-я аминокислота не может быть заменена на глутаминовую кислоту и 72-я аминокислота не может быть заменена на лейцин.
Что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 38-я аминокислота, аргинин, может быть заменена аминокислотой, отличной от аргинина. Предпочтительно, что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 38-я аминокислота, аргинин, может быть заменена на аланин (R38A).
Что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 42-я аминокислота, фенилаланин, может быть заменена аминокислотой, отличной от фенилаланина. Предпочтительно, что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 42-я аминокислота, фенилаланин, может быть заменена на аланин (F42A).
Что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 45-я аминокислота, тирозин, может быть заменена на аминокислоту, отличную от тирозина. Предпочтительно, что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 45-я аминокислота, тирозин, может быть заменена на аланин (Y45A).
Что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10, 61-я аминокислота, глутаминовая кислота, может быть заменена на аминокислоту, отличную от глутаминовой кислоты. Предпочтительно, что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 61-я аминокислота, глутаминовая кислота, может быть заменена на аргинин (E61R).
Что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10 72-я аминокислота, лейцин, может быть заменена на аминокислоту, отличную от лейцина. Предпочтительно, что касается аминокислотной замены для варианта IL-2, в аминокислотной последовательности SEQ ID NO: 10, 72-я аминокислота, лейцин, может быть заменена на глицин (L72G).
В частности, вариант IL-2 может быть получен путем по меньшей мере одной замены, выбранной из группы, состоящей из R38A, F42A, Y45A, E61R и L72G, в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может быть получен путем аминокислотных замен в двух, трех, четырех или пяти положениях из положений, выбранных из группы, состоящей из R38A, F42A, Y45A, E61R и L72G.
Кроме того, вариант IL-2 может иметь форму, в которой заменены две аминокислоты. В частности, вариант IL-2 может быть получен путем замен R38A и F42A. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A и Y45A. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A и E61R. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A и Y45A. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A и E61R. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен E61R и L72G.
Более того, вариант IL-2 может иметь форму, где три аминокислоты заменены. В частности, вариант IL-2 может быть получен путем замен R38A, F42A, и Y45A. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, F42A и E61R. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, F42A и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, Y45A и E61R. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, Y45A и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A, Y45A и E61R. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A, Y45A и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A, E61R и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен Y45A, E61R и L72G.
Кроме того, вариант IL-2 может иметь форму, где четыре аминокислоты заменены. В частности, вариант IL-2 может быть получен путем замен R38A, F42A, Y45A и E61R. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, F42A, Y45A и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, F42A, E61R и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен R38A, Y45A, E61R и L72G. Кроме того, в одном из вариантов осуществления вариант IL-2 может быть получен путем замен F42A, Y45A, E61R и L72G.
Более того, вариант IL-2 может быть получен путем замен R38A, F42A, Y45A, E61R и L72G.
Предпочтительно, вариант осуществления варианта IL-2 может содержать любую из следующих выбранных комбинаций замен (a)-(d) в аминокислотной последовательности SEQ ID NO: 10:
(a) R38A/F42A
(b) R38A/F42A/Y45A
(c) R38A/F42A/E61R
(d) R38A/F42A/L72G.
Здесь, когда IL-2 имеет аминокислотную последовательность SEQ ID NO: 35, аминокислотная замена может присутствовать в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10. Кроме того, даже когда IL-2 представляет собой фрагмент аминокислотной последовательности SEQ ID NO: 35, аминокислотная замена может присутствовать в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может иметь аминокислотную последовательность SEQ ID NO: 6, 22, 23 или 24.
Кроме того, вариант IL-2 может характеризоваться наличием низкой токсичности in vivo. Здесь, низкая токсичность in vivo может представлять собой побочный эффект, вызванный связыванием IL-2 с альфа-цепью рецептора IL-2 (IL-2Rα). Были разработаны различные варианты IL-2 для облегчения побочного эффекта, вызванного связыванием IL-2 с IL-2Rα, и такие варианты IL-2 могут представлять собой варианты, описанные в патенте США № 5229109 и патенте Кореи № 1667096. В частности, варианты IL-2, описанные в настоящей заявке, имеют низкую способность связываться с альфа-цепью рецептора IL-2 (IL-2Rα) и, таким образом, имеют более низкую токсичность in vivo, чем IL-2 дикого типа.
Как используют в рамках изобретения, термин "CD80", также называемый "B7-1", представляет собой мембранный белок, присутствующий в дендритных клетках, активированных B-клетках и моноцитах. CD80 обеспечивает костимулирующие сигналы, необходимые для активации и выживания T-клеток. CD80 известен в качестве лиганда для двух различных белков, CD28 и CTLA-4, присутствующих на поверхности T-клеток. CD80 состоит из 288 аминокислот, и, в частности, он может иметь аминокислотную последовательность SEQ ID NO: 11. Кроме того, как используют в рамках изобретения, термин "белок CD80" относится к полноразмерному CD80 или фрагменту CD80.
Как используют в рамках изобретения, термин "фрагмент CD80" относится к расщепленной форме CD80. Кроме того, фрагмент CD80 может представлять собой внеклеточный домен CD80. Один из вариантов осуществления фрагмента CD80 может быть получен путем удаления с N-конца аминокислот с 1-й по 34-ю, которые представляют собой сигнальную последовательность CD80. В частности, один из вариантов осуществления фрагмента CD80 может представлять собой белок, состоящий из аминокислот с 35-й по 288-ю в SEQ ID NO: 11. Кроме того, один из вариантов осуществления фрагмента CD80 может представлять собой белок, состоящий из аминокислот с 35-й по 242-ю в SEQ ID NO: 11. Кроме того, один из вариантов осуществления фрагмента CD80 может представлять собой белок, состоящий из аминокислот с 35-й по 232-ю в SEQ ID NO: 11. Кроме того, один из вариантов осуществления фрагмента CD80 может представлять собой белок, состоящий из аминокислот с 35-й по 139-ю в SEQ ID NO: 11. Кроме того, один из вариантов осуществления фрагмента CD80 может представлять собой белок, состоящий из аминокислот с 142-й по 242-ю в SEQ ID NO: 11. В одном из вариантов осуществления фрагмент CD80 может иметь аминокислотную последовательность SEQ ID NO: 2.
Кроме того, белок IL-2 и белок CD80 могут быть соединены друг с другом через линкер или носитель. В частности, IL-2 или его вариант и CD80 (B7-1) или его фрагмент могут быть связаны друг с другом через линкер или носитель. В настоящем описании "линкер" и "носитель" могут использоваться взаимозаменяемо.
Линкеры соединяют два белка. Один из вариантов осуществления линкера может включать 1-50 аминокислот, альбумин или его фрагмент, Fc-домен иммуноглобулина и т.п. Здесь, Fc-домен иммуноглобулина относится к белку, который содержит константную область тяжелой цепи 2 (CH2) и константную область тяжелой цепи 3 (CH3) иммуноглобулина и не содержит вариабельные области тяжелой и легкой цепей и константную область легкой цепи 1 (CH1) иммуноглобулина. Иммуноглобулин может представлять собой IgG, IgA, IgE, IgD или IgM, и предпочтительно может представлять собой IgG4. Здесь, Fc-домен иммуноглобулина G4 дикого типа может иметь аминокислотную последовательность SEQ ID NO: 4.
Кроме того, Fc-домен иммуноглобулина может представлять собой вариант Fc-домена, а также Fc-домен дикого типа. Кроме того, как используют в рамках изобретения, термин "вариант Fc-домена" может относиться к форме, которая отличается от Fc-домена дикого типа паттерном гликозилирования, имеет высокое гликозилирование по сравнению с Fc-доменом дикого типа или имеет низкое гликозилирование по сравнению с Fc-доменом дикого типа или дегликозилированную форму. Кроме того, он включает агликозилированный Fc-домен. Fc-домен или его вариант может быть адаптирован для наличия скорректированного количества сиаловых кислот, фукозилирований или гликозилирований, посредством условий культивирования или генетического манипулирования с хозяином.
Кроме того, гликозилирование Fc-домена иммуноглобулина может быть модифицировано общепринятыми способами, такими как химические способы, ферментативные способы и способы генной инженерии с использованием микроорганизмов. Кроме того, вариант Fc-домена может иметь смешанную форму из соответствующих Fc-областей иммуноглобулинов: IgG, IgA, IgE, IgD и IgM. Кроме того, Вариант Fc-домена может иметь форму, в которой некоторые аминокислоты Fc-домена заменены другими аминокислотами. Один из вариантов осуществления варианта Fc-домена может иметь аминокислотную последовательность SEQ ID NO: 12.
Слитый белок может иметь структуру, в которой с использованием Fc-домена в качестве линкера (или носителя) белок CD80 и белок IL-2 или белок IL-2 и белок CD80 связаны с N-концом и C-концом линкера или носителя, соответственно. Связывание между N-концом или C-концом Fc-домена и CD-80 или IL-2 необязательно может быть осуществлено посредством линкерного пептида.
В частности, слитый белок может иметь следующую структурную формулу (I) или (II):
N'-X-[линкер (1)]n-Fc-домен-[линкер (2)]m-Y-C' (I)
N'-Y-[линкер (1)]n-Fc-домен-[линкер (2)]m-X-C' (II)
Здесь, в структурных формулах (I) и (II),
N′ представляет собой N-конец слитого белка,
C′ представляет собой C-конец слитого белка,
X представляет собой белок CD80,
Y представляет собой белок IL-2,
линкеры (1) и (2) представляют собой пептидные линкеры, и
каждый из n и m независимо равен 0 или 1.
Предпочтительно, слитый может иметь структурную формулу (I). Белок IL-2 является таким, как описано выше. Кроме того, белок CD80 является таким, как описано выше. Согласно одному из вариантов осуществления белок IL-2 может представлять собой вариант IL-2 с от одной до пяти аминокислотных замен по сравнению с IL-2 дикого типа. Белок CD80 может представлять собой фрагмент, полученный путем укорочения на вплоть до приблизительно 34 последовательно расположенных аминокислотных остатков с N-конца или C-конца CD80 дикого типа. Альтернативно белок CD может представлять собой внеклеточный иммуноглобулин-подобный домен, обладающий активностью связывания с рецепторами T-клеточной поверхности CTLA-4 и CD28.
В частности, слитый белок может иметь аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30. Согласно другому варианту осуществления, слитый белок включает полипептид, обладающий идентичностью последовательности 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% с аминокислотной последовательностью SEQ ID NO: 9, 26, 28 или 30. Здесь, идентичность представляет собой, например, процентную гомологию, и она может быть определена посредством программного обеспечения для сравнения гомологии, такого как программное обеспечение BlastN Национального центра биотехнологической информации (NCBI).
Между белком CD80 и Fc-доменом может быть включен пептидный линкер (1). Пептидный линкер (1) может состоять из 5-80 последовательно расположенных аминокислот, 20-60 последовательно расположенных аминокислот, 25-50 последовательно расположенных аминокислот или 30-40 последовательно расположенных аминокислот. В одном из вариантов осуществления пептидный линкер (1) может состоять из 30 аминокислот. Кроме того, пептидный линкер (1) может содержать по меньшей мере один остаток цистеина. В частности, пептидный линкер (1) может содержать один, два или три остатка цистеина. Кроме того, пептидный линкер (1) может происходить из шарнирной области иммуноглобулина. В одном из вариантов осуществления пептидный линкер (1) может представлять собой пептидный линкер, состоящий из аминокислотной последовательности SEQ ID NO: 3.
Пептидный линкер (2) может состоять из 1-50 последовательно расположенных аминокислот, 3-30 последовательно расположенных аминокислот, или 5-15 последовательно расположенных аминокислот. В одном из вариантов осуществления пептидный линкер (2) может представлять собой (G4S)n (где n представляет собой целое число от 1 до 10). Здесь, в (G4S)n n может составлять 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. В одном из вариантов осуществления пептидный линкер (2) может представлять собой пептидный линкер, состоящий из аминокислотной последовательности SEQ ID NO: 5.
В другом аспекте настоящего изобретения предусматривается димер, полученный путем связывания двух слитых белков, каждый из которых содержит белок IL-2 и белок CD80. Слитый белок, содержащий IL-2 или его вариант и CD80 или его фрагмент, является таким, как описано выше.
Здесь, связывание между слитыми белками, составляющими димер, может быть достигнуто посредством, но не ограничиваясь ими, дисульфидной связи, образованной остатками цистеина, присутствующими в линкере. Слитые белки, составляющие димер, могут представлять собой одинаковые или отличающиеся друг от друга слитые белки. Предпочтительно, димер может представлять собой гомодимер. Вариантом осуществления слитого белка, составляющего димер, может быть белок, имеющий аминокислотную последовательность SEQ ID NO: 9.
Фармацевтическая композиция по настоящему изобретению, содержащая в качестве активных ингредиентов димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля, демонстрирует эффективность в отношении предупреждения или лечения злокачественной опухоли.
Злокачественная опухоль может быть выбрана из группы, состоящей из рака желудка, рака печени, рака легкого, рака ободочной и прямой кишки, рака молочной железы, рака предстательной железы, рака яичника, рака поджелудочной железы, рака шейки матки, рака щитовидной железы, рака гортани, острого миелоидного лейкоза, опухоли головного мозга, нейробластомы, ретинобластомы, рака головы и шеи, рака слюнных желез и лимфомы.
Предпочтительная доза фармацевтической композиции варьируется в зависимости от состояния и массы тела пациента, тяжести заболевания, формы лекарственного средства, пути и длительности введения, и она может быть надлежащим образом выбрана специалистами в данной области. В фармацевтической композиции для лечения или предупреждения злокачественной опухоли по настоящему изобретению активный ингредиент может содержаться в любом количестве (эффективное количество) в зависимости от применения, дозированной формы, цели смешивания и т.п. при условии, что активный ингредиент может демонстрировать противораковую активность. Его обычное эффективное количество будет определяться диапазоном от 0,001% до 20,0% по массе в расчете на общую массу композиции. Здесь, термин "эффективное количество" относится к количеству активного ингредиента, способному индуцировать противораковый эффект. Такое эффективное количество может быть экспериментально определено на основе общей информации, известной специалистам в данной области.
Как используют в рамках изобретения, термин "лечение" может использоваться для обозначения как терапевтического, так и профилактического лечения. Здесь, "профилактика" может использоваться для указания на то, что патологическое состояние или заболевание индивидуума облегчается или смягчается. В одном из вариантов осуществления термин "лечение" включает как применение, так и любую форму введения, для лечения заболевания у млекопитающего, в том числе человека. Кроме того, термин включает ингибирование или замедление заболевания или прогрессирования заболевания; и включает значения восстановления или возобновления сниженной или утраченной функции, так чтобы происходило частичное или полное облегчение заболевания; стимуляции неэффективных процессов или облегчения серьезного заболевания.
Как используют в рамках изобретения, термин "эффективность" относится к способности, которая может быть определена посредством одного или нескольких параметров, например, выживаемости или выживаемости без прогрессирования на протяжении определенного периода времени, такого как один год, пять лет или десять лет. Кроме того, параметр может включать ингибирование увеличения размера по меньшей мере одной опухоли у индивидуума.
Также на эффективность могут влиять фармакокинетические параметры, такие как биодоступность и лежащие в ее основе параметры, такие как скорость выведения. Таким образом, "улучшенная эффективность" (например, повышение эффективности) может быть следствием улучшения фармакокинетических параметров и повышения эффективности, которые могут быть измерены путем сравнения скорости выведения и роста опухоли у тестируемых животных или людей, или путем сравнения параметров, таких как выживаемость, рецидив или выживаемость без прогрессирования.
Как используют в рамках изобретения, термин "терапевтически эффективное количество" или "фармацевтически эффективное количество" относится к количеству соединения или композиции, эффективному для предупреждения или лечения рассматриваемого заболевания, которое является достаточным для лечения заболевания с приемлемым соотношением польза/риск, применимым для медицинского лечения, и не вызывает побочных эффектов. Уровень эффективного количества может быть определен в зависимости от факторов, включающих состояние здоровья пациента, тип и тяжесть заболевания, активность лекарственного средства, чувствительность пациента к лекарственному средству, способ введения, время введения, путь введения и скорость экскреции, длительность лечения, состав или одновременно применяемые лекарственные средства, и другие факторы, хорошо известные в области медицины. В одном из вариантов осуществления терапевтически эффективное количество означает количество лекарственного средства, эффективное для лечения злокачественной опухоли.
Здесь, фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый носитель. Фармацевтически приемлемый носитель может представлять собой любой носитель при условии, что носитель является нетоксичным веществом, пригодным для доставки пациенту. В качестве носителя могут присутствовать дистиллированная вода, спирт, жир, воск и инертное твердое вещество. Также в фармацевтической композиции может содержаться фармацевтически приемлемый адъювант (буфер, диспергирующий агент).
В частности, путем включения фармацевтически приемлемого носителя в дополнение к активному ингредиенту, фармацевтическая композиция может быть получена в виде парентерального состава в зависимости от ее пути введения с использованием общепринятых способов, известных в данной области. Здесь, термин "фармацевтически приемлемый" означает, что носитель не обладает более высокой токсичностью, чем может выдержать индивидуум, которому проводят введение (назначение), при этом не ингибируя активность активного ингредиента.
Когда фармацевтическую композицию получают в виде парентерального состава, его можно изготавливать в виде препаратов в форме инъекций, трансдермальных пластырей, назальных средств для ингаляции или суппозиториев с подходящими носителями в соответствии со способами, известными в данной области. В случае изготовления в форме инъекций, в качестве подходящего носителя можно использовать стерильную воду, этанол, многоатомный спирт, такой как глицерин или пропиленгликоль, или их смесь; и предпочтительно можно использовать изотонический раствор, такой как раствор Рингера, фосфатно-солевой буфер (PBS), содержащий триэтаноламин или стерильную воду для инъекций, и 5% декстроза, и т.п. Состав фармацевтических композиций известен в данной области, и, в частности, может быть упомянут Remington's Pharmaceutical Sciences (19th ed., 1995) и т. п. Этот документ считается частью настоящего описания.
Предпочтительная доза фармацевтической композиции может находиться в диапазоне от 0,01 мкг/кг до 10 г/кг, или от 0,01 мг/кг до 1 г/кг в сутки в зависимости от состояния пациента, массы тела, пола, возраста, тяжести состояния пациента и пути введения. Дозу можно вводить один раз в сутки или можно разделять на несколько раз в сутки. Такая доза не должна считаться ограничивающей объем настоящего изобретения ни в каком из аспектов.
Индивидуумами, у которых фармацевтическую композицию можно применять (назначать), являются млекопитающие и люди, особенно предпочтительно люди. В дополнение к активному ингредиенту, фармацевтическая композиция, описанная в настоящей заявке, может дополнительно содержать любое соединение или природный экстракт, безопасность которого уже подтверждена и о котором известно, что он обладает противораковой активностью, чтобы таким образом усилить или закрепить противораковую активность.
В другом аспекте настоящего изобретения предусматривается набор для лечения злокачественной опухоли, содержащий димерный слитый белок, содержащий белок IL-2 или вариант его и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля.
В другом аспекте настоящего изобретения предусматривается применение композиции для комбинированного введения, содержащая димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля, для предупреждения или лечения злокачественной опухоли.
В другом аспекте настоящего изобретения предусматривается применение композиции для комбинированного введения, содержащей слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля для усиления терапевтического эффекта на злокачественную опухоль.
В другом аспекте настоящего изобретения предусматривается применение композиции для комбинированного введения, содержащей слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, и ингибитор иммунной точки контроля для производства лекарственного средства для лечения злокачественной опухоли.
В другом аспекте настоящего изобретения предусматривается способ предупреждения или лечения злокачественной опухоли и/или способ повышения терапевтического эффекта в отношении злокачественной опухоли, включающий введение индивидууму композиции для комбинированного введения, содержащей димерный слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, или димерный слитый белок, где эти два слитых белка связаны, и ингибитор иммунной точки контроля.
Индивидуум может представлять собой индивидуума, страдающего от злокачественной опухоли. Кроме того, индивидуум может представлять собой млекопитающее, предпочтительно человека. Слитый белок, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, или димерный слитый белок, где эти два слитых белка связаны, является таким, как описано выше.
Путь введения, дозу и частоту введения слитого белка или димерного слитого белка и NK-клеток можно варьировать в зависимости от состояния пациента и наличия или отсутствия побочных эффектов, и, таким образом, слитый белок или димерный слитый белок можно вводить индивидууму различными путями и в различных количествах. Оптимальный способ введения, доза и частота введения могут быть выбраны из соответствующего диапазона специалистами в данной области.
Вследствие активности IL-2, слитый белок в одном из вариантов осуществления настоящего изобретения может активировать иммунные клетки, такие как натуральные киллеры. Таким образом, слитый белок может эффективно использоваться против злокачественной опухоли. В частности, было идентифицировано, что, по сравнению с диким типом, вариант IL-2 с от двух до пяти аминокислотных замен, в частности, вариант IL-2, который содержит аминокислотные замены в двух, трех, четырех или пяти положениях среди положений, выбранных из группы, состоящей из R38A, F42A, Y45A, E61R и L72G, обладает низкой связывающей способностью в отношении альфа-цепи рецептора IL-2 и, таким образом, демонстрирует улучшенные характеристики в отношении фармакологических побочных эффектов общепринятого IL-2. Таким образом, такой вариант IL-2, когда он используется отдельно или в форме слитого белка, может снижать встречаемость синдрома сосудистой (или капиллярной) (VLS), что является общеизвестной проблемой для IL-2.
Способ осуществления изобретения
Далее в настоящем описании, настоящее изобретение описано более подробно с помощью следующих примеров. Однако следующие примеры приведены только для иллюстрации настоящего изобретения и объем настоящего изобретения не ограничивается ими.
I. Получение слитого белка
Пример получения 1. Получение варианта hCD80-Fc-IL-2 (2M): GI101
Для получения слитого белка, содержащего фрагмент CD80 человека, Fc-домен и вариант IL-2, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 8), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2M) (R38A, F42A) (SEQ ID NO: 6), имеющий две аминокислотные замены, в указанном порядке, с N-конца. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 9. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI101".
Очистку проводили с использованием хроматографии, включающей смолу с белком A MabSelect SuRe. Слитый белок связывался с ней в условиях 25 мМ Tris, 25 мМ NaCl, pH 7,4. Затем проводили элюирование посредством 100 мМ NaCl, 100 мМ уксусной кислоты, pH 3. 20% 1 M Tris-HCl при pH 9 помещали в пробирку для сбора, а затем собирали слитый белок. Для собранного слитого белка буфер заменяли в ходе диализа на буфер PBS на 16 часов.
После этого определяли поглощение при длине волны 280 нм с течением времени посредством эксклюзионной хроматографии с использованием колонки TSKgel G3000SWXL (TOSOH Bioscience) до получения высококонцентрированного слитого белка. Здесь, выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR), и окрашивали кумасси синим для проверки его чистоты (фиг. 6). Было идентифицировано, что слитый белок содержался в концентрации 2,78 мг/мл, когда детекцию проводили посредством NanoDrop (фиг. 7). Кроме того, результаты, полученные посредством анализа с использованием эксклюзионной хроматографии, приведены на фиг. 8.
Пример получения 2. Получение варианта mCD80-Fc-IL-2 (2M): mGI101
Для получения слитого белка, содержащего CD80 мыши, Fc-домен и вариант IL-2, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 14), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), mCD80 (SEQ ID NO: 13), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2M) (R38A, F42A) (SEQ ID NO: 6) с двумя аминокислотными заменами, в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 15. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "mGI101".
Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 9). Было обнаружено, что слитый белок содержался в концентрации 1,95 мг/мл при детекции по поглощению при 280 нм с использованием NanoDrop.
Пример получения 3. Получение hCD80-Fc: GI101C1
Для получения слитого белка, содержащего фрагмент CD80 человека и Fc-домен, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 16), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнирную область Ig (SEQ ID NO: 3) и Fc-домен (SEQ ID NO: 4). Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 17. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI101C1".
Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 10). Было выявлено, что слитый белок содержался в концентрации 3,61 мг/мл при детекции по поглощению при 280 нм с использованием NanoDrop.
Пример получения 4. Получение варианта Fc-IL-2 (2M): GI101C2
Для получения слитого белка, содержащего Fc-домен и вариант IL-2, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 18), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2M) (R38A, F42A) (SEQ ID NO: 6) с двумя аминокислотными заменами, в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 19. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI101C2".
Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 11). Было обнаружено, что слитый белок содержался в концентрации 4,79 мг/мл при детекции по поглощению при 280 нм с использованием NanoDrop.
Пример получения 5. Получение mCD80-Fc: mGI101C1
Для получения слитого белка, содержащего CD80 мыши и Fc-домен, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 20) которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), mCD80 (SEQ ID NO: 13), шарнирную область Ig (SEQ ID NO: 3) и Fc-домен (SEQ ID NO: 4), в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHOTM) для экспрессии слитого белка SEQ ID NO: 21. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "mGI101C1".
Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 12). Было выявлено, что слитый белок содержался в концентрации 2,49 мг/мл при детекции по поглощению при 280 нм с использованием NanoDrop.
Слитые белки, полученные согласно примерам получения 1-5, обобщенно представлены в таблице 1 ниже.
[Таблица 1]
Пример получения 6. Получение CD80-Fc-IL-2: GI101w
Для получения слитого белка, содержащего фрагмент CD80 человека, Fc-домен и IL-2 человека, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 31), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и зрелый IL-2 человека (SEQ ID NO: 10), в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 32. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI101w". Очистку и сбор слитого белка проводили аналогично примеру получения 1.
Пример получения 7. Получение варианта hCD80-Fc-IL-2 (3M): GI102-M45
Для получения слитого белка, содержащего фрагмент CD80 человека, Fc-домен и вариант IL-2 (3M) (R38A, F42A, Y45A) (GI102-M45) с тремя аминокислотными заменами, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 25), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (SEQ ID NO: 22), в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 26. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI102-M45". Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 13).
Пример получения 8. Получение варианта hCD80-Fc-IL-2 (3M): GI102-M61
Для получения слитого белка, содержащего фрагмент CD80 человека, Fc-домен и вариант IL-2 (3M) (R38A, F42A, E61R) (GI102-M61) с тремя аминокислотными заменами, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 27), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (SEQ ID NO: 23), в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 28. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI102-M61".
Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 14).
Пример получения 9. Получение hCD80-Fc-IL-3M: GI102-M72
Для получения слитого белка, содержащего фрагмент CD80 человека, Fc-домен и вариант IL-2 (3M) (R38A, F42A, L72G) (GI102-M72) с тремя аминокислотными заменами, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 29), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (SEQ ID NO: 24), в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 30. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "GI102-M72".
Очистку и сбор слитого белка проводили аналогично примеру получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE в условиях с восстановлением (R) или без восстановления (NR) и окрашивали кумасси синим для проверки его чистоты (фиг. 15).
Пример получения 10. Получение mCD80-Fc-IL-3M: mGI102-M61
Для получения слитого белка, содержащего фрагмент CD80 мыши, Fc-домен и вариант IL-2 (3M) (R38A, F42A, E61R) (GI102-M61) с тремя аминокислотными заменами, синтезировали полинуклеотид с использованием услуги Invitrogen GeneArt Gene Synthesis от ThermoFisher Scientific. В частности, полинуклеотид содержал нуклеотидную последовательность (SEQ ID NO: 33), которая кодирует слитый белок, который содержит сигнальный пептид (SEQ ID NO: 1), фрагмент mCD80 (SEQ ID NO: 13), шарнирную область Ig (SEQ ID NO: 3), Fc-домен (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (SEQ ID NO: 23), в указанном порядке, на N-конце. Полинуклеотид встраивали в вектор pcDNA3_4. Затем вектор вводили в клетки CHO (Expi-CHO™) для экспрессии слитого белка SEQ ID NO: 34. После введения вектора проводили культивирование в течение 7 суток в условиях 37°C, 125 об/мин и 8% концентрации CO2. Затем культуру собирали и из нее очищали слитый белок. Очищенный слитый белок был обозначен как "mGI102-M61".
Очистку и сбор слитого белка проводили аналогично примеру получения 1.
II. Идентификация аффинности связывания между слитым белком и его лигандом
Для идентификации аффинности связывания между слитым белком и его лигандом измеряли аффинность связывания с использованием Octet RED 384.
Экспериментальный пример 1. Идентификация аффинности связывания между hCTLA-4 и GI101
Биосенсор AR2G (Amine Reactive 2nd gen, ForteBio, каталожный номер: 18-5092) предварительно гидратировали посредством 200 мкл дистиллированной воды 96-луночном микропланшете (GreinerBio-one, каталожный номер: 655209). Лиганд (CTLA-4, CTLA-4/CD152 человека, His-метка, Sino Biological, каталожный номер: 11159-H08H), подлежащий присоединению к биосенсору AR2G, разбавляли 10 мМ ацетатным буфером (pH 5, AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) до концентрации 5 мкг/мл. Кроме того, GI101, подлежащий связыванию с лигандом, разбавляли 1X буфером для кинетических исследований AR2G (AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) до концентрации 1000 нМ, 500 нМ, 250 нМ, 125 нМ или 62,5 нМ. Активирующий буфер получали путем смешения 20 мМ EDC и 10 мМ s-NHS (AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) в дистиллированной воде. 80 мкл каждого реагента помещали в 384-луночный микропланшет (Greiner Bio-one, каталожный номер: 781209) и запускали программу.
В результате, была измерена аффинность связывания между hCTLA-4 и GI101, как проиллюстрировано на фиг. 16.
Экспериментальный пример 2. Идентификация аффинности связывания между hPD-L1/GI101 и hPD-L1/PD-1
Ni-NTA (Nickel charged Tris-NTA, Ni-NTA Biosensors, ForteBio, 18-5101) предварительно гидратировали посредством 200 мкл 1X буфера для кинетических исследований Ni-NTA (10X Kinetics buffer, ForteBio, 18-1042) в 96-луночном микроплашнете (GreinerBio-one, каталожный номер: 655209). Лиганд (белок PD-L1/B7-H1 человека, His-метка, Sino biological, каталожный номер: 10084-H08H), подлежащий связыванию с биосенсорами Ni-NTA, разбавляли 1X буфером для кинетических исследований Ni-NTA до концентрации 5 мкг/мл. GI101, подлежащий присоединению к лиганду, разбавляли 1X буфером для кинетических исследований Ni-NTA в концентрации 1000 нМ, 500 нМ, 250 нМ, 125 нМ или 62,5 нМ. Кроме того, PD-1/PDCD1 человека (PD-1/PDCD1 человека, Fc-метка, Sino Biological, каталожный номер: 10377-H02H), подлежащий присоединению к лиганду, разбавляли 1X буфером для кинетических исследований Ni-NTA до концентрации 2000 нМ, 1000 нМ, 500 нМ, 250 нМ или 125 нМ. Затем 80 мкл каждого реагента помещали в 384-луночный микропланшет и запускали программу.
В результате, была измерена аффинность связывания между hPD-L1 и GI101, как проиллюстрировано на фиг. 17. Кроме того, была измерена аффинность связывания между hPD-L1 и hPD-1, как проиллюстрировано на фиг. 18.
Экспериментальный пример 3. Идентификация аффинности связывания между mCTLA-4 и mGI101
Аффинность связывания между mCTLA-4 и mGI101 исследовали аналогично экспериментальному примеру 1. Здесь, использованное оборудование является следующим: биосенсор: AR2G, лиганд: mCTLA-4 (рекомбинантная химера CTLA-4 Fc мыши, R&D Systems, каталожный номер: 434-CT-200), анализируемое соединение: mGI101 (500 нМ, 250 нМ, 125 нМ, 62,5 нМ, 31,3 нМ).
В результате была измерена аффинность связывания между mCTLA-4 и mGI101, как проиллюстрировано на фиг. 19.
Экспериментальный пример 4. Идентификация аффинности связывания между mPD-L1 и mGI101
Аффинность связывания между mPD-L1 и mGI101 идентифицировали аналогично тому, как в экспериментальном примере 1. Здесь, использованное оборудование является следующим. Биосенсор: AR2G, лиганд: mPD-L1 (рекомбинантная химера B7-H1/PD-L1 Fc мыши mGI101, R&D Systems, каталожный номер: 434-CT-200), анализируемое соединение: mGI101 (500 нМ, 250 нМ, 125 нМ, 62,5 нМ, 31,3 нМ).
В результате была измерена аффинность связывания между mPD-L1 и mGI101, как проиллюстрировано на фиг. 20.
Экспериментальный пример 5. Идентификация аффинности связывания GI-101 (hCD80-Fc-hIL-2v) с CTLA-4
Измерение кинетики связывания проводили с использованием устройства Octet RED 384 (ForteBio, Pall Life Science) при встряхивании при 30°C и 1000 об/мин. Способность к связыванию CTLA-4 измеряли с использованием биосенсорного чипа Amine Reactive 2 generation (AR2G), и способность к связыванию PD-L1 измеряли с использованием биосенсорного чипа Nickel charged Tris-NTA (Ni-NTA). Биосенсорный чип AR2G активировали посредством комбинации 400 мМ EDC и 100 мМ сульфо-NHS. Затем CTLA-4 человека-His-метку (Sino Biological, каталожный номер: 11159-H08H) разбавляли 10 мМ ацетатным буфером (pH 5) до 5 мкг/мл, и наносили на биосенсорный чип AR2G в течение 300 секунд и фиксировали.
Затем измеряли связывание CTLA-4 с GI-101 (hCD80-Fc-hIL-2v), GI-101C1 (hCD80-Fc), ипилимумабом (Bristol-Myers Squibb) и GI-101C2 (Fc-hIL-2v) в различных концентрациях в течение 300 секунд, а также измеряли их диссоциацию в течение 300 секунд. Анализ кинетики связывания проводили с использованием программного обеспечения Octet Data Analysis HT software ver. 10, предоставленного Pall Corporation. Результаты проиллюстрированы на фиг. 21.
Экспериментальный пример 6. Идентификация аффинности связывания между IL-2Rα или IL-2Rβ и GI101
Способность к связыванию IL-2Rα измеряли с использованием биосенсора AR2G, и способность к связыванию IL-2Rβ измеряли с использованием биосенсоров Ni-NTA (Nickel charged Tris-NTA, Ni-NTA Biosensors, ForteBio, 18-5101).
Лиганд (IL-2Rα-His-метка, Acro, каталожный номер: ILA-H52H9), подлежащий присоединению к биосенсору AR2G, разбавляли 10 мМ ацетатным буфером (pH 5, AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) до концентрации 5 мкг/мл. Биосенсор AR2G активировали буфером, полученном путем смешения 400 мМ EDC и 100 мМ сульфо-NHS, а затем разбавленный лиганд наносили на биосенсор AR2G на 300 секунд и фиксировали.
Между тем, лиганд (IL-2Rβ-His Tag, Acro, каталожный номер: CD2-H5221), подлежащий связыванию с биосенсором Ni-NTA, разбавляли 1X буфером для кинетических исследований Ni-NTA до концентрации 5 мкг/мл. Разбавленный лиганд наносили на биосенсор Ni-NTA на 600 секунд и фиксировали.
После этого наносили GI101, GI101w или Пролейкин (Novartis, hIL-2) в различных концентрациях для связывания с лигандом на 300 секунд. Затем определяли их связывание, а также определяли их диссоциацию в течение 300 секунд. Анализ кинетики связывания проводили с использованием программного обеспечения Octet Data Analysis HT software ver. 10, предоставленного Pall Corporation. Результаты проиллюстрированы на фиг. 22-24.
В результате было идентифицировано, что GI101 обладает низкой способностью к связыванию с альфа-цепью рецептора IL-2, IL-2Rα, и высокой способностью к связыванию IL-2Rβ, по сравнению с GI101w и Пролейкином.
Экспериментальный пример 7. Измерение аффинности связывания между слитым белком и лигандом
Для идентификации аффинности связывания между слитым белком и его лигандом проводили измерение аффинности связывания с использованием Octet RED 384.
Экспериментальный пример 7.1. Идентификация аффинности связывания между альфа-рецептором IL-2 и GI101-M45, GI101-M61 или GI101-M72
Биосенсор AR2G (Amine Reactive 2nd gen, ForteBio, каталожный номер: 18-5092) предварительно гидратировали посредством 200 мкл дистиллированной воды (DW) в 96-луночном микропланшете (GreinerBio-one, каталожный номер: 655209). Лиганд (белок IL-2R альфа человека, His-метка, Acro, ILA-H52H9), подлежащий связыванию с биосенсором, разбавляли 10 мМ ацетатным буфером (pH 5) (AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) до концентрации 5 мкг/мл. Анализируемое соединение (GI101-M45, GI101-M61, GI101-M72), подлежащее присоединению к лиганду, разбавляли 1X буфером для кинетических исследований AR2G (AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) до 500 нМ, 250 нМ, 125 нМ и 62,5 нМ, соответственно. Активирующий буфер получали путем смешения 20 мМ EDC и 10 мМ s-NHS (AR2G Reagent Kit, ForteBio, каталожный номер: 18-5095) в DW. 80 мкл каждого реагента помещали в 384-луночный микропланшет (Greiner Bio-one, каталожный номер: 781209) и запускали программу.
В результате, аффинность связывания между рецептором IL-2 альфа и GI101-M45 проиллюстрирована на фиг. 25. Кроме того, аффинность связывания между рецептором IL-2 альфа и GI101-M61 проиллюстрирована на фиг. 26, и аффинность связывания между рецептором IL-2 альфа и GI101-M72 проиллюстрирована на фиг. 27.
Экспериментальный пример 7.2. Идентификация аффинности связывания GI102-M45, GI102-M61 и GI102-M72 с IL-2Rβ
Биосенсоры Ni-NTA предварительно гидратировали посредством 200 мкл 1X буфера для кинетических исследований Ni-NTA (10X буфер для кинетических исследований, ForteBio, 18-1042) в 96-луночном микропланшете. Лиганд (белок IL-2R бета человека, His-метка, Acro, CD2-H5221), подлежащий присоединению к биосенсору, разбавляли 1X буфером для кинетических исследований Ni-NTA до концентрации 2 мкг/мл. GI102-M45, GI102-M61 или GI102-M72, подлежащие присоединению к лиганду, разбавляли 1X буфером для кинетических исследований Ni-NTA до концентрации 500 нМ, 250 нМ, 125 нМ или 62,5 нМ. 80 мкл каждого реагента помещали в 384-луночный микропланшет и запускали программу.
В результате, была измерена аффинность связывания между IL-2Rβ и GI102-M45, как проиллюстрировано на фиг. 28, и аффинность связывания между IL-2Rβ и GI102-M61 была измерена, как проиллюстрировано на фиг. 29. Кроме того, была измерена аффинность связывания между IL-2Rβ и GI102-M72, как проиллюстрировано на фиг. 30.
III. Идентификация иммунной активности слитого белка
Экспериментальный пример 8. Идентификация продукции IFN-γ, вызванной слитым белком
Экспериментальный пример 8.1. Культивирование PBMC, меченных CFSE
Мононуклеарные клетки периферической крови (PBMC), полученные от человека, метили сукцинимидильным сложным эфиром карбоксифлуоресцеина (CFSE) путем реакции с 1 мкМ красителем CellTrace CFSE при 37°C в течение 20 минут. CFSE, не связанный с клетками, удаляли путем реакции в течение 5 минут с культуральной средой, имеющей 5-кратный объем реакционного раствора для окрашивания, а затем центрифугирования при 1300 об/мин в течение 5 минут. CFB-меченные PBMC ресуспендировали в культуральной среде (среда RPMI1640, содержавшая 10% эмбриональную телячью сыворотку (FBS), 10 мМ HEPES, 100 Е/мл пенициллина/стрептомицина, 1 мМ пируват натрия, 55 мкМ 2-меркаптоэтанол, 1 мМ неосновные аминокислоты и 2 мМ L-глутамин), а затем добавляли в 96-луночный микропланшет в количестве 1×105 клеток на лунку. Проводили обработку посредством 5 мкг/мл PHA (лектин из Phaseolus Vulgaris, красная фасоль, Sigma-Aldrich, St. Louis, MO, США, каталожный номер № L1668-5MG), и GI101, GI101C1, GI101C2 или IL-2 (альдеслейкин; рекомбинантный IL-2 человека, Novartis) и проводили инкубацию в инкубаторе с 5% CO2 при 37°C в течение 6 суток.
Здесь, обработку GI101, GI101C1, GI101C2 и IL-2 проводили в концентрации 1 нМ, 10 нМ или 100 нМ. Клетки анализировали посредством FACS и определяли IFN-γ человека, присутствующий в культуральной среде, с использованием набора для ELISA (Biolegend, San Diego, CA, США, каталожный номер № 430103).
Экспериментальный пример 8.2. Анализ FACS
Клеточные осадки, полученные путем удаления супернатанта, промывали буфером FACS (3% эмбриональная телячья сыворотка (FBS), 10 мМ EDTA, 1M HEPES, 100 единиц/мл пенициллина, стрептомицин, 10 мкг/мл, 1 мМ пируват натрия), а затем подвергали реакции с блокатором Fc (Biolegend, каталожный номер № 422302) при 4°C в течение 5 минут. Затем проводили обработку Ab против CD3 с APC (Biolegend, каталожный номер № 300412) и Ab против CD8a с PE (Biolegend, каталожный номер № 300908) и реакции позволяли протекать при 4°C в течение 20 минут. Затем полученный материал промывали буфером для FACS. Клеточные осадки ресуспендировали в буфере для FACS, а затем анализировали с использованием BD LSR Fortessa (BD Biosciences, San Diego, CA, США) и программного обеспечения FlowJo.
Экспериментальный пример 8.3. ELISA для IFN-γ человека
Количество IFN-γ, секретируемое в супернатант каждого образца, в котором проводили культивирование клеток, определяли с использованием набора для ELISA для IFN-γ человека (Biolegend, каталожный номер № 430103). В кратком изложении, в планшет для ELISA добавляли антитела против IFN-γ человека и реакции позволяли протекать в течение ночи при 4°C, так чтобы эти антитела покрывали его. Затем проводили блокирование при комнатной температуре в течение 1 часа посредством раствора PBS, к которому был добавлен 1% BSA. Проводили промывание промывочным буфером (0,05% Tween-20 в PBS), а затем стандартный раствор и каждый образец надлежащим образом разбавляли и добавляли к нему. Затем реакции позволяли протекать при комнатной температуре в течение 2 часов.
После завершения реакции планшет промывали и в него добавляли вторичные антитела (антитела для детекции). Реакции позволяли протекать при комнатной температуре в течение 1 часа. Проводили промывание буфером для промывания, а затем добавляли раствор авидин-HRP. Реакции позволяли протекать при комнатной температуре в течение 30 минут. В него добавляли раствор субстрата и индуцировали реакцию развития окраски в темноте при комнатной температуре в течение 20 минут. Наконец, добавляли H2SO4 для остановки реакции развития окраски, и измеряли поглощение при 450 нм посредством спектрофотометра для микропланшетов Epoch (BioTek Instruments, Inc., Winooski, VT, США).
В результате, было обнаружено, что клетки, обработанные GI101, демонстрировали заметное повышение секреции IFN-γ по сравнению с клетками, обработанными GI101C1, GI101C2 или IL-2 (фиг. 31 и 32).
Экспериментальный пример 9. Идентификация эффекта GI101 на пролиферацию CD8+ T-клеток
Мононуклеарные клетки периферической крови (PBMC), полученные от человека, метили CFSE путем реакции с 1 мкМ красителем CellTrace CFSE при 37°C в течение 20 минут. CFSE, не связанный с клетками, удаляли путем реакции в течение 5 минут с культуральной средой, имеющей 5-кратный объем реакционного раствора для окрашивания, а затем центрифугирования при 1300 об/мин в течение 5 минут. CFB-меченные PBMC ресуспендировали в культуральной среде (среда RPMI1640, содержавшая 10% эмбриональную телячью сыворотку (FBS), 10 мМ HEPES, 100 Е/мл пенициллина/стрептомицина, 1 мМ пируват натрия, 55 мкМ 2-меркаптоэтанол, 1 мМ неосновные аминокислоты и 2 мМ L-глутамин), а затем добавляли в 96-луночный микропланшет в количестве 1×105 клеток на лунку.
После этого проводили обработку 1 мкг/мл антитела против CD3ε (Biolegend, каталожный номер № L1668-5MG) и GI101, GI101C1, GI101C2, или Пролейкина (Novartis), и проводили инкубацию в инкубаторе с 5% CO2 при 37°C в течение 6 суток. Здесь, клетки обрабатывали GI101, GI101C1, GI101C2 и IL-2 в концентрации 100 нМ. Инкубированные клетки исследовали в отношении их степени пролиферации путем измерения посредством FACS-анализа с использованием антител APC-TCRαβ и PE-CD8α доли CD8+ T-клеток, которые не были меченными посредством CFSE.
В результате, было обнаружено, что GI101 активировал пролиферацию CD8+ T-клеток in vitro аналогично IL-2 дикого типа Пролейкину (фиг. 33 и 34).
Экспериментальный пример 10. Идентификация эффекта GI101 и GI102 на пролиферацию CD8+ T-клеток
PBMC человека приобретали от Allcells (партия № 3014928, CIF). Использовали 1 M краситель CellTrace CFSE, который подвергали реакции с PBMC человека в условиях блокирования светом при комнатной температуре в течение 20 минут. Клетки метили CFSE путем реакции с 1 мкМ красителем CellTrace CFSE при 37°C в течение 20 минут. CFSE, не связанный с клетками, удаляли путем реакции в течение 5 минут с культуральной средой, имевшей 5-кратный объем реакционного раствора для окрашивания, а затем центрифугирования при 1300 об/мин в течение 5 минут. CFB-меченные PBMC ресуспендировали в культуральной среде (среда RPMI1640, содержавшая 10% эмбриональную телячью сыворотку (FBS), 10 мМ HEPES, 100 Е/мл пенициллина/стрептомицина, 1 мМ пируват натрия, 55 мкМ 2-меркаптоэтанол, 1 мМ неосновные аминокислоты и 2 мМ L-глутамин), а затем добавляли в 96-луночный микропланшет в количестве 1×105 клеток на лунку.
После этого CFB-меченные PBMC подвергали обработке посредством 1 мкг/мл антитела против CD3ε (OKT3, eBioscience, США) и GI101, GI101C1, GI101C2 или Пролейкина (Novartis), и проводили инкубацию в инкубаторе с 5% CO2 при 37°C в течение 7 суток. Здесь, клетки подвергали обработке GI101, GI101C1, GI101C2 и IL-2 в концентрации 10 мкМ.
Инкубированные клетки исследовали в отношении их степени пролиферации путем измерения посредством FACS-анализа с использованием антитела против CD4 человека с PE (BioLegend, США), антитела против CD8 человека с PE/Cy7 (BioLegend, США), и антитела против FoxP3 человека с APC (BioLegend, США), доли CD8+ T-клеток, которые не были мечены посредством CFSE.
В результате, группы обработки GI101, GI102_M61, GI101C2 и Пролейкином демонстрировали значительное повышение доли CD8+ T-клеток по сравнению с контрольной группой (без стимула), группой обработки антителом против CD3 отдельно и группой обработки GI101C1. Кроме того, по сравнению с группой отрицательного контроля (без стимула) и группой обработки CD3 отдельно, группы обработки GI101, GI101C2 и Пролейкином продемонстрировали значительное повышение пролиферации CD4+/FoxP3+ Treg клеток, в то время как группы обработки GI102 и GI101C1 не продемонстрировали значительного повышения пролиферации CD4+/FoxP3+ Treg-клеток (фиг. 35).
Экспериментальный пример 11. Идентификация эффекта GI101 или GI101w на пролиферацию CD8+ T-клеток и NK-клеток
Мышей C57BL/6 в возрасте 7 недель, приобретенных в Orient Bio (Корея), распределяли на 3 группы, причем каждая из групп включала 3 мыши, и им внутрибрюшинно инъецировали PBS, GI101 или GI101w. Здесь, GI101 и GI101w соответственно получали в количестве 40,5 мкг в 200 мкл PBS, и инъецировали внутрибрюшинно. Через пять суток после инъекции у мышей из каждой группы извлекали селезенку. Из нее выделяли клетки, и определяли общее количество клеток с использованием гемоцитометра. Спленоциты исследовали в отношении соотношений CD8+ T-клеток и NK-клеток в них посредством FACS-анализа с использованием антитела против CD3ε с APC (Biolegend; 145-2C11), антитела против NK1.1 с PE (Biolegend; PK136) и антитела против CD8α с Pacific blue (BD; 53-6.7). Иными словами, вычисляли количества CD8+ T-клеток и NK-клеток, присутствующих в селезенке.
В результате, было идентифицировано, что GI101 активировал пролиферацию CD8+ T-клеток и NK-клеток in vivo по сравнению с GI101w (фиг. 36 и 37).
Экспериментальный пример 12. Идентификация эффекта GI101 на функцию T-клеток
Проводили эксперимент с использованием набора для биоанализа блокады CTLA-4 (Promega, каталожный номер № JA4005). В кратком изложении, эксперимент является следующим. Эффекторные клетки с CTLA-4, поддерживаемые в жидком азоте, размораживали на водяной бане при постоянной температуре 37°C в течение 3 минут и 0,8 мл эффекторных клеток с CTLA-4 хорошо перемешивали с 3,2 мл предварительно нагретого буфера для анализа (90% RPMI+10% эмбриональная телячья сыворотка). Затем смесь добавляли в 96-луночный белый планшет для культивирования клеток (SPL, каталожный номер № 30196) в количестве 25 мкл на лунку. Затем добавляли 25 мкл GI101 в различных концентрациях. В качестве отрицательного контроля добавляли 25 мкл буфера для анализа. Затем 96-луночный белый планшет для культивирования клеток накрывали и помещали в условия комнатной температуры до получения клеток aAPC/Raji.
Клетки aAPC/Raji, поддерживаемые в жидком азоте, размораживали на водяной бане при постоянной температуре 37°C в течение 3 минут, и 0,8 мл клеток aAPC/Raji хорошо перемешивали с 3,2 мл предварительно нагретого буфера для анализа. Затем 25 мкл смеси добавляли в каждую лунку планшета и реакции позволяли протекать в инкубаторе с 5% CO2 при 37°C в течение 16 часов. После завершения реакции полученному материалу позволяли стоять при комнатной температуре в течение 15 минут, а затем к нему добавляли реагент Bio-Glo, следя за тем, чтобы избегать пузырей. В три наружных лунки добавляли реагент Bio-Glo, и эти лунки использовали в качестве пустых образцов для внесения поправки на фоновый сигнал. Реакции позволяли протекать при комнатной температуре в течение 10 минут, а затем определяли люминесцению посредством Cytation 3 (BioTek Instruments, Inc., Winooski, VT, США). Конечный анализ данных проводили путем вычисления RLU (GI101-фоновый уровень)/RLU (без обработки-фоновый уровень).
В результате, было обнаружено, что GI101, связанный с CTLA-4, экспрессировался на эффекторных T-клетках и активировал функцию T-клеток, а не ингибировал ее (фиг. 38 и 39).
Экспериментальный пример 13. Идентификация эффекта mGI101 и mGI102 на иммунные клетки
Мышей C57BL/6 в возрасте 7 недель, приобретенных в Orient Bio (Корея), распределяли на 3 группы, причем каждая из групп включала 3 мыши, и им внутривенно вводили PBS, 3 мг/кг, 6 мг/кг или 12 мг/кг GI101, или 3 мг/кг, 6 мг/кг или 12 мг/кг mGI102 (mGI102-M61). На 1, 3, 5, 7 и 14 сутки после инъекции у мышей каждой группы извлекали селезенки. После этого для ткани селезенки вычисляли количества эффекторных CD8+ T-клеток, NK-клеток и Treg-клеток посредством FACS-анализа с использованием соответствующих антител, и вычисляли соотношения эффекторных CD8+ T-клеток и NK-клеток относительно Treg-клеток. Информация, касающаяся антител, использованных в каждом клеточном анализе, является следующей:
Эффекторные CD8+ T-клетки: антитело против CD3ε мыши с PB (Biolegend, # 155612; KT3.1.1), антитело против CD8α мыши с FITC (BD, # 553031, 53-6.7), антитело против CD44 мыши с PE/Cy7 (Biolegend, # 103030; IM7), антитело против CD122 мыши с APC (Biolegend, # 123214; TM-β1).
NK-клетки: антитело против CD3ε мыши с PB (Biolegend, # 155612; KT3.1.1), антитело против NK-1.1 мыши с PE (Biolegend, # 108708; PK136).
Treg-клетки: антитело против CD3 мыши с FITC (Biolegend, # 100204; 17A2), антитело против CD4 мыши с PB (Biolegend, # 100531; RM4-5), антитело против CD25 мыши с PE (Biolegend, # 102008; PC61), антитело против Foxp3 мыши с APC (Invitrogen, # FJK-16s, 17-5773-82).
В результате, группа, в которой вводили mGI101 или mGI102 (mGI102-M61), продемонстрировала значительное увеличение количеств CD8+ T-клеток и NK-клеток в моменты времени от 3 суток до 14 суток после введения по сравнению с группой введения PBS. Кроме того, было обнаружено, что группа, в которой вводили mGI102, продемонстрировала значительное увеличение долей активированных CD8+ T-клеток/Treg-клеток и NK-клеток/Treg-клеток в моменты времени от 3 суток до 14 суток после введения по сравнению с группой введения PBS (фиг. 40).
IV. Идентификация противоракового эффекта слитого белка
Экспериментальный пример 14. Идентификация эффекта GI101 на ингибирование активности T-клеток злокачественными клетками, экспрессирующими PD-L1 и CTLA-4
Линию злокачественных клеток NCl-H292, экспрессирующих PD-L1 и CTLA-4, культивировали в течение 3 часов в культуральной среде, содержавшей 10 мкг/мл митомицина C (Sigma), а затем митомицин C удаляли путем промывания культуральной средой. После этого, 5×104 клеток обработанной митомицином C линии злокачественных клеток NCl-H292 инкубировали с 1×105 PBMC человека в 96-луночном микропланшете. Здесь, проводили обработку 5 мкг/мл PHA (Sigma) для активации T-клеток. Кроме того, GI101C1 и GI101 в концентрации 50 нМ подвергали реакции с IgG1-Fc (Biolegend) или абатацептом (= Оренсия; Bristol-Myers Squibb) в концентрации 50 нМ в течение 30 минут при 4°C, а затем полученный материал использовали для обработки злокачественных клеток NCl-H292. Через 3 суток супернатант инкубированных клеток собирали и количество IFN-γ определяли с использованием набора для ELISA (Biolegend).
В качестве группы положительного контроля использовали PBMC человека, стимулированные PHA в отсутствии обработанной митомицином C линии злокачественных клеток NCl-H292; и в качестве группы отрицательного контроля использовали PBMC человека, стимулированные PHA в присутствии обработанной митомицином C линии злокачественных клеток NCl-H292. Экспериментальный способ с использованием набора для ELISA для IFN-γ проводили аналогично экспериментальному примеру 9.3.
В результате, GI101 эффективно активировал иммунный ответ, который ингибировался линией злокачественных клеток, сверхэкспрессирующих PD-L1. Кроме того, было обнаружено, что GI101 ингибировал передачу сигнала CTLA-4, экспрессированного на эффекторных T-клетках (фиг.41 и 42).
Экспериментальный пример 15. Идентификация противоракового эффекта mGI101 у мышей, которым были трансплантированы происходящие из мыши клетки рака ободочной и прямой кишки
Мышей BALB/c (самки, возраст 7 недель), приобретенных в Orient Bio, подвергали акклиматизации в течение периода 7 суток. Затем, 5×106 клеток линии злокачественных клеток CT-26 (ATCC, США) смешивали с 0,05 мл матрикса на основе матригеля, свободного от фенолового красного (BD), и проводили аллотрансплантацию смеси посредством подкожного введения в количестве 0,1 мл в правую область спины мышей. Через определенный период времени после трансплантации злокачественных клеток определяли объем опухоли и отбирали особей, которые достигали приблизительно 28 мм3, а затем отобранных мышей равномерно распределяли на группы, исходя из размера опухоли и массы тела, причем каждая группа содержала 10 мышей. После этого с использованием одноразового шприца (31G, 1 мл), hIgG4 вводили в дозе 6 мг/кг в группе отрицательного контроля. Для экспериментальных групп проводили внутривенное введение mGI101 в дозе 3 мг/кг, 6 мг/кг или 12 мг/кг. После первого введения проводили всего три введения один раз каждые трое суток. Размер опухоли измеряли каждые сутки.
В результате, в группах, в которых вводили mGI101 в дозе 6 мг/кг и 12 мг/кг, соответственно, увеличение размера опухоли значительно ингибировалось по сравнению с группой отрицательного контроля в те же самые моменты времени измерения и в конце теста (фиг. 43). Кроме того, в результате измерения выживаемости, в группе, в которой вводили mGI101 в дозе 6 мг/кг, наблюдали значительное улучшение по сравнению с группой отрицательного контроля в те же самые моменты времени измерения и в конце теста (фиг. 44).
Экспериментальный пример 16. Идентификация противоракового эффекта GI101 у мышей, которым трансплантировали происходящие из мыши клетки рака ободочной и прямой кишки
Экспериментальный пример 16.1. Идентификация эффекта ингибирования опухоли
Мышей BALB/c (самки, возраст 7 недель), приобретенных в Orient Bio, подвергали акклиматизации в течение периода 7 суток. Затем 5×106 клеток линии злокачественных клеток CT-26 (ATCC, США) суспендировали в 0,1 мл PBS, и проводили аллотрансплантацию суспензию посредством подкожного введения в количестве 0,1 мл в правую область спины мышей. Через определенный период времени после трансплантации злокачественных клеток определяли объем опухоли и отбирали особей, которые достигали от 50 мм3 до 200 мм3, а затем отобранных мышей равномерно распределяли на группы, исходя из размера опухоли и массы тела, причем каждая группа содержала 10 мышей. После этого в группе отрицательного контроля лекарственное средство не вводили, и в группах положительного контроля с использованием одноразового шприца (31G, 1 мл), внутривенно вводили антитело против PD-1 в дозе 5 мг/кг или антитело против PD-1 в дозе 5 мг/кг и антитело против CTLA-4 в дозе 5 мг/кг. Для экспериментальных групп внутривенно вводили GI101 в дозе 0,1 мг/кг или 1 мг/кг. После первого введения проводили всего три введения один раз каждые трое суток. Размер опухоли измеряли каждые сутки.
В результате, у мышей с трансплантированной клеточной линией злокачественной опухоли CT-26 все группы, которым вводили антитело против PD-1, антитело против PD-1 и антитело против CTLA-4 или GI101 в дозе 0,1 мг/кг или 1 мг/кг, демонстрировали значительное ингибирование роста опухоли по сравнению с группой отрицательного контроля. В частности, экспериментальная группа, в которой вводили GI101 в дозе 0,1 мг/кг, продемонстрировала значительный эффект ингибирования опухоли по сравнению с группой введения антитела против PD-1 (*p<0,05) (фиг. 45).
Экспериментальный пример 16.2. Анализ иммунных клеток в тканях злокачественной опухоли
Мышей в каждой группе экспериментального примера 16.1 умерщвляли, когда объем опухоли достигал в среднем 200 мм3, и собирали ткани злокачественной опухоли. После этого ткани злокачественной опухоли разделяли до уровня единичных клеток для анализа иммунных клеток в них, а затем проводили FACS-анализ иммунных клеток в тканях злокачественной опухоли с использованием приведенных ниже антител. В частности, в качестве антител использовали антитело против CD3 мыши (Biolegend, каталожный номер № 100320), антитело против CD4 мыши (Biolegend, каталожный номер № 100526), антитело против CD8 мыши (Biolegend, каталожный номер № 100750), антитело против FoxP3 мыши (eBioscience, каталожный номер 12-5773-82), антитело против CD25 мыши (Biolegend, каталожный номер № 102049), антитело против CD44 мыши (eBioscience, каталожный номер № 61-0441-82), антитело против PD-1 мыши (Biolegend, каталожный номер № 135218), антитело против IFN мыши (Biolegend, каталожный номер № 505832), антитело против CD49b мыши (Biolegend, каталожный номер № 108906), антитело против H2 мыши (Invitrogen, каталожный номер № A15443), антитело против CD11c мыши (Biolegend, каталожный номер № 117343), антитело против CD80 мыши (eBioscience, каталожный номер № 47-4801-82), антитело против CD86 мыши (Biolegend, каталожный номер № 104729), антитело против F4/80 мыши (eBioscience, каталожный номер № 47-4801-82) и антитело против CD206 мыши (eBioscience, каталожный номер № 17-2061-80).
В результате, экспериментальная группа, в которой вводили GI101 в дозе 0,1 мг/кг, продемонстрировала значимое повышение уровня CD8+ T-клеток по сравнению с группой положительного контроля, в которой вводили только антитело против PD-1 в дозе 5 мг/кг (*p<0,05, фиг. 46 и 47). Более того, все экспериментальные группы, в которых вводили GI101, продемонстрировали значимо повышенный уровень экспрессии IFN-γ в T-клетках по сравнению с группой отрицательного контроля (*p<0,05, фиг. 46 и 47). Кроме того, экспериментальная группа, в которой вводили GI101 в дозе 0,1 мг/кг, продемонстрировала повышение уровня макрофагов M1 по сравнению с группой отрицательного контроля и группой положительного контроля, в которой вводили только антитело против PD-1 (фиг. 48 и 49). Кроме того, все экспериментальные группы, в которых вводили GI101, продемонстрировали повышенный уровень экспрессии CD86 в макрофагах и дендритных клетках (* p < 0,05, фиг. 48-51).
Экспериментальный пример 17. Идентификация противоракового эффекта GI101 у мышей, которым трансплантировали происходящие из мыши клетки рака легкого
Экспериментальный пример 17.1. Идентификация эффекта ингибирования опухоли
Мышей BALB/c (самки, возраст 7 недель), приобретенных в Orient Bio, подвергали акклиматизации в течение периода 7 суток. Затем 5×106 клеток линии злокачественных клеток LLC2 (ATCC, США) суспендировали в 0,1 мл PBS, и проводили аллотрансплантацию суспензии посредством подкожного введения в количестве 0,1 мл в правую область спины мышей. Через определенный период времени после трансплантации злокачественных клеток определяли объем опухоли и отбирали особей, которые достигали от 50 мм3 до 200 мм3, а затем отобранных мышей равномерно распределяли на группы, исходя из размера опухоли и массы тела, причем каждая группа содержала 10 мышей. После этого в группе отрицательного контроля лекарственное средство не вводили, и в группах положительного контроля с использованием одноразового шприца (31G, 1 мл) внутривенно вводили антитело против PD-1 в дозе 5 мг/кг или антитело против PD-1 в дозе 5 мг/кг и антитело против CTLA-4 в дозе 5 мг/кг. Для экспериментальных групп внутривенно вводили GI101 в дозе 0,1 мг/кг или 1 мг/кг. После первого введения проводили всего три введения один раз каждые трое суток. Размер опухоли измеряли каждые сутки.
В результате, все экспериментальные группы продемонстрировали значимый эффект ингибирования опухоли по сравнению с группой отрицательного контроля (* p<0,05) (фиг. 52).
Экспериментальный пример 17.2. Анализ иммунных клеток в тканях злокачественной опухоли
Мышей в каждой группе экспериментального примера 17.1 умерщвляли, когда объем опухоли достигал в среднем 200 мм3, и ткани злокачественной опухоли собирали. После этого проводили FACS-анализ аналогично экспериментальному примеру 16.2 для анализа иммунных клеток в тканях злокачественной опухоли.
В результате, экспериментальная группа, в которой вводили GI101 в дозе 0,1 мг/кг, продемонстрировала значимое повышение уровня CD8+ T-клеток по сравнению с группой положительного контроля, в которой вводили только антитело против PD-1 (*p<0,05, фиг.59). Более того, все экспериментальные группы, в которых вводили GI101, продемонстрировали значимо повышенный уровень экспрессии IFN-γ по сравнению с группой отрицательного контроля (*p<0,05, фиг. 59). Кроме того, все экспериментальные группы, в которых вводили GI101, продемонстрировали повышенный уровень экспрессии CD86 в макрофагах и дендритных клетках (* p<0,05, фиг. 53-55).
Экспериментальный пример 18. Идентификация противоракового эффекта mGI102-M61 у мышей, которым трансплантировали происходящие из мыши клетки рака ободочной и прямой кишки
Мышей BALB/c (самки, возраст 7 недель), приобретенных в Orient Bio, подвергали акклиматизации в течение периода 7 суток. Затем 5×106 клеток линии злокачественных клеток CT-26 (ATCC, США) смешивали с 0,05 мл матрикса на основе матригеля, свободного от фенолового красного, (BD), и проводили аллотрансплантацию смеси посредством подкожного введения в количестве 0,1 мл в правую область спины мышей. Через определенный период времени после трансплантации злокачественных клеток определяли объем опухоли и отбирали особей, которые достигали примерно 28 мм3, а затем отобранных мышей равномерно распределяли на группы, исходя из размера опухоли и массы тела, причем каждая группа содержала 10 мышей. После этого с использованием одноразового шприца (31G, 1 мл), hIgG4 вводили в дозе 6 мг/кг в группе отрицательного контроля. Для экспериментальных групп внутривенно вводили mGI102-M61 в дозе 3 мг/кг, 6 мг/кг или 12 мг/кг. После первого введения проводили всего три введения один раз каждые трое суток. Размер опухоли измеряли каждые сутки.
В результате, было идентифицировано, что экспериментальная группа, в которой вводили mGI102-M61 в дозе 12 мг/кг, продемонстрировала значительное ингибирование роста опухоли в те же самые моменты времени измерения и в конце теста, по сравнению с группой отрицательного контроля (фиг. 56). Кроме того, в результате определения выживаемости было идентифицировано, что экспериментальная группа, в которой вводили mGI102-M61 в дозе 12 мг/кг, продемонстрировала значимое улучшение в те же самые моменты времени измерения и в конце теста, по сравнению с группой отрицательного контроля (фиг. 57).
Экспериментальный пример 19. Идентификация противоракового эффекта mGI101 у мышей, которым трансплантировали происходящие из мыши клетки рака ободочной и прямой кишки
Мышей BALB/c (самки, возраст 7 недель), приобретенных от Orient Bio, подвергали акклиматизации в течение периода 7 суток. Затем, 5×106 клеток линии злокачественных клеток CT-26 (ATCC, USA) смешивали с 0,05 мл матрикса на основе матригеля, свободного от фенолового красного, (BD), и проводили аллотрансплантацию смеси посредством подкожного введения в количестве 0,1 мл в правую область спины мышей. Через определенный период времени после трансплантации злокачественных клеток определяли объем опухоли и отбирали особей, которые достигали приблизительно от 200 мм3 до 250 мм3, а затем отобранных мышей равномерно распределяли на группы, исходя из размера опухоли и массы тела, причем каждая группа содержала 10 мышей.
После этого с использованием одноразового шприца (31G, 1 мл) в группе отрицательного контроля вводили hIgG4 в дозе 4 мг/кг. Для экспериментальных групп mGI101 вводили внутривенно в дозе 1 мг/кг, 4 мг/кг или 6 мг/кг. Кроме того, в качестве контрольных групп были выбраны группы, в которых вводили mCD80 в дозе 4,9 мг/кг или Fc-IL-2v (GI101C2) в дозе 2,8 мг/кг. Кроме того, в качестве контрольной группы была выбрана группа, в которой одновременно вводили mCD80 в дозе 4,9 мг/кг и Fc-IL-2v (GI101C2) в дозе 2,8 мг/кг.
При измерении объема опухоли было идентифицировано, что группа, в которой вводили mGI101 в дозе 6 мг/кг, продемонстрировала значительное ингибирование в те же самые моменты времени измерения и в конце теста по сравнению с отрицательным контролем. Наблюдали превосходное ингибирование роста опухоли по сравнению с группой, в которой вводили комбинацию mCD80 и Fc-IL-2v (GI101C2) (фиг.58 и 59).
В заключение, в тесте ингибирования роста опухоли на мышах BALB/c, которым проводили аллотрансплантацию CT-26, происходящей из мышей BALB/c клеточной линии рака ободочной и прямой кишки, было продемонстрировано, что тестируемое вещество mGI101 имело эффект ингибирования опухоли в этих условиях тестирования по сравнению с препаратами mCD80 и IL-2v отдельно; и было идентифицировано, что mGI101 продемонстрировал превосходную противораковую эффективность по сравнению с группой, в которой вводили комбинацию mCD80 и IL-2v (фиг.58 и 59). В частности, группа, в которой вводили mGI101 в дозе 6 мг/кг, продемонстрировала значительное ингибирование увеличения размера опухоли по сравнению с группой отрицательного контроля и группой, в которой вводили комбинацию mCD80 и Fc-IL2v (GI101C2).
V. Определение противоракового эффекта комбинированного введения димерного слитого белка и ингибитора иммунной точки контроля
Экспериментальный пример 20. Определение противоракового эффекта посредством комбинированного введения GI101 и антитела против PD-1 у мышей с трансплантированными происходящими из человека клетками рака молочной железы
Этот тест оценивал эффект ингибирования роста опухоли в модели опухоли с трансплантированными ксеногенными клетками MDA-MB-231, которые представляют собой происходящие из человека клетки рака молочной железы, с использованием модели на гуманизированных мышах, полученных посредством ксеногенной трансплантации PBMC человека мышам NSGb2m после внутрибрюшинного введения GI101 в качестве тестируемого материала, и Китруды (Пембролизумаб, MSD), антитела против PD-1, в качестве положительного контроля, отдельно или в комбинации.
Исходные растворы тестируемого материала, отрицательного контроля и положительного контроля, описанных в таблице 2, разбавляли добавлением эксципиента в соответствии с каждой дозой.
[Таблица 2]
Происходящие из человека клетки рака молочной железы MDA-MB-231 (Homo sapiens, молочная железа/грудь человека; происходящие из метастаза: плевральный выпот) приобретали в банке Korea cell line bank (Корея) и использовали в тесте. Среду для культивирования клеток использовали путем смешения эмбриональной телячьей сыворотки (FBS, 16000-044, Thermofisher scientific, США), пенициллина-стрептомицина; 10000 единиц/мл пенициллина и 10000 мкг/мл стрептомицина (15140122, Thermofisher scientific, США) и RPMI1640 (A1049101, Thermofisher scientific, США) на 100 мл в качестве композиции, представленной в таблице ниже.
[Таблица 3]
Клетки для применения в тесте размораживали, помещали во флакон для культивирования клеток и культивировали при 37°C в инкубаторе с 5% CO2 (MCO-170M, Panasonic, Япония). Культивированные клетки суспендировали с использованием трипсин-EDTA (каталожный номер № 25200-072, Thermofisher scientific, США). Суспендированные клетки собирали центрифугированием (125×g, 5 минут) с использованием центрифуги, переносили в новую среду и новый флакон, и субкультивировали. В день трансплантации клеточной линии культивированные клетки помещали в центрифужную пробирку, выделяли, а затем центрифугировали (125×g, 5 минут) для удаления супернатанта. Затем, получали суспензию клеток (5×106 клеток/0,05 мл) PBS (каталожный номер LB 001-04, Welgene Inc., Корей) и хранили на льду до инокуляции.
Для теста самок мышей NSGb2m (NOD.Cg-B2mtm1UncPrkdcscidIl2rgtm1Wjl/SzJ) в возрасте 8 недель приобретали в JoongABio (Корея) и использовали. Массу тела определяли на следующие сутки после окончания карантина и периода акклиматизации, а затем суспензией клеток PBMC человека (5×106 клеток/0,2 мл), полученной для здоровых животных, заполняли одноразовый шприц, и ее вводили в хвостовую вену животных. Наблюдение за основными симптомами после трансплантации клеток проводили один раз в сутки.
Раствором, полученным добавлением свободного от фенолового красного матрикса на основе матригеля (0,05 мл, 356237, BD, США) к полученной суспензии клеток MDA-MB-231 (5×106 клеток/0,05 мл), заполняли одноразовый шприц, и его трансплантировали путем подкожного введения (0,1 мл/животное) в правый бок животного, которому трансплантировали PBMC человека. Наблюдение за основными симптомами проводили один раз в сутки в ходе приживления и периода роста после трансплантации клеточной линии.
Через определенный период времени после трансплантации клеток определяли объем опухоли для мышей, не демонстрирующих ненормального состояния здоровья, и отбирали 32 особи, так чтобы среднее значение в каждой группе достигало от 40 до 80 мм3. Отобранных животных распределяли на 4 группы (8 животных на группу) настолько равномерно, насколько возможно, исходя из объема опухоли и массы тела.
Тестируемые группы были организованы, как показано в таблице 4. Тестируемый материал вводили животным с использованием одноразового шприца (31G, 1 мл), и частота введения составляла два раза в неделю всего с 4 введениями.
[Таблица 4]
(мл/кг)
В ходе периода наблюдения идентифицировали основные симптомы, такие как внешний вид, подведение, экскременты и т.п., один раз в сутки, и определяли умерших животных. Массу тела определяли в день трансплантации клеточной линии, два раза в неделю и в день умерщвления животного.
В ходе периода наблюдения проводили измерение максимальной длины (L) и перпендикулярной ширины (W) опухоли три раза в неделю с использованием цифрового толщиномера (mitutoyo, Япония) и применяли в следующем уравнении для вычисления объема опухоли (TV).
<Уравнение 1>
TV (мм3)=(W2XL)/2
<Уравнение 2>
%TGI(ингибирование роста опухоли)=(1-(Ti-T0)/(Vi-V0))X100
В качестве объема опухоли у каждой особи до введения брали величину, измеренную в момент распределения на группы.
На 21 сутки, 25 сутки, 28 сутки и 31 сутки после трансплантации опухоли, вводили лекарственные средства, представленные в таблице 4, соответственно, и в результате в группах введения происходило ингибирование роста опухоли GI101 или Китруды отдельно по сравнению с контрольной группой (hIgG4). В группах комбинированного введения GI101 и Китруды по сравнению с контрольной группой происходило ингибирование роста опухоли. В группе комбинированного введения GI101 и Китруды по сравнению с группами введения GI101 или Китруды отдельно происходило ингибирование роста опухоли (фиг. 60).
Степень ингибирования роста опухоли вычисляли в конце эксперимента (42 сутки после трансплантации опухоли) по сравнению с сутками 1 введения лекарственного средства (17 сутки после трансплантации опухоли). В результате, в группе введения hIgG4 было 2 мыши со степенью ингибирования роста опухоли 30% или более, 1 мышь со степенью ингибирования роста опухоли 50% или более и 1 мышь со степенью ингибирования роста опухоли 80% или более; в группе введения GI101 было 5 мышей со степенью ингибирования роста опухоли 30% или более, 5 мышей со степенью ингибирования роста опухоли 50% или более и 2 мыши со степенью ингибирования роста опухоли 80% или более; в группе введения Китруды было 7 мышей, имевших степень ингибирования роста опухоли 30% или более, 5 мышей со степенью ингибирования роста опухоли 50% или более, и 3 мыши со степенью ингибирования роста опухоли 80% или более; и в группе комбинированного введения GI101 и Китруды было 8 мышей со степенью ингибирования роста опухоли 30% или более, 8 мышей со степенью ингибирования роста опухоли 50% или более, и 6 мышей со степенью ингибирования роста опухоли 80% или более (фиг. 61).
Кроме того, степень роста опухоли у индивидуальных экспериментальных животных в каждой группе введения, когда GI101 и Китруду использовали в комбинации у мышей, которым трансплантировали происходящие из человека клетки рак молочной железы, представлены на фиг. 62-66.
Экспериментальный пример 21. Определение противоракового эффекта посредством комбинированного введения mGI101 и антитела против PD-1 у мышей, которым трансплантировали происходящие из мыши клетки рака ободочной и прямой кишки
В этом тесте оценивали эффект ингибирования роста опухоли в модели опухоли, в которой аллогенные клетки аденокарциномы толстой кишки мыши (MC38) трансплантировали мышам C57BL/6 после внутрибрюшинного введения mGI101 в качестве тестируемого материала и антитела против PD-1 в качестве положительного контроля, отдельно или в комбинации.
Клетки аденокарциномы толстой кишки мыши (MC38), которые представляют собой происходящие из грызуна клетки рака ободочной и прямой кишки, приобретали от Kerafast Inc. (США) и использовали в тесте. Клетки MC38 культивировали в среде RPMI1640 (Gibco), содержавшей 10% эмбриональную телячью сыворотку (Gibco) и 1% антибиотик/противогрибковое средство (Gibco). Культивируемые клетки собирали с использованием трипсина и суспендировали в PBS. 1×106 клеток MC38 инъецировали п/к в правый бок самок мышей C57BL/6 (в возрасте 7 недель) для получения модели аллотрансплантата опухоли.
Мышей случайным образом распределяли (5 на группу) на основе объема опухоли (30 мм3). Трансплантаты опухолей идентифицировали приблизительно через 2 суток после инокуляции клеток. Тестируемые группы организовывали, как показано в таблице 5, и вводили тестируемые материалы.
[Таблица 5]
(cloneRMP1-14, InVivoMab)
В ходе периода тестирования идентифицировали клинические симптомы, такие как изменения заболевания и поведения и т.п., один раз в сутки, и выявляли умерших животных. Животных умерщвляли после периода тестирования. Размер солидной злокачественной опухоли MC38 измеряли с использованием 3D-сканера опухоли (TM900, Peria, Бельгия). Для каждой экспериментальной группы вычисляли среднее снижение массы тела и процентное изменение, и среднее ингибирование роста опухоли. Противоопухолевую эффективность вычисляли по сравнению с контрольной группой носителя.
Все статистические вычисления проводили с использованием Prism 8.0 (Graph Pad Software Inc., США). Результаты измерения объема опухоли сравнивали посредством одностороннего ANOVA (конечный момент времени), а затем критерия множественных сравнения Бонферрони. Значимым считали значение "p"<0,05.
Все протестированные животные оставались здоровыми, не демонстрируя признаков патологических нарушений после введения mGI101 и совместного введения mGI101 и антитела против PD-1. Результаты комбинированной терапии с использованием mGI101 и/или антитела против PD-1 в отношении опухолей MC38 представлены на фиг. 67-73. Противораковый эффект наблюдали в группе лечения лекарственным средством по сравнению с контрольной группой, и было заметным отличие размера опухоли в ходе периода тестирования в течение 16 суток. Опухоль MC38 известна в качестве модели, отвечающей на антитела против PD-1, из предшествующих источников литературы, и противораковый эффект также наблюдали в группе введения антитела против PD-1 этого теста (p>0,01). Противораковый эффект также был обнаружен в группе введения mGI101 (6 мг/кг) отдельно, также как и в группе введения антитела против PD-1 (p>0,01). Группа комбинированного введения mGI101 (0,6 мг/кг) + антитело против PD-1 (5 мг/кг) продемонстрировала превосходный противораковый эффект (p>0,0001).
Индивидуальные размеры опухолей для каждой тестируемой группы представлены на фиг. 69-73. В соответствии с результатами индивидуальных размеров опухолей наблюдали небольшую регрессию опухоли у некоторых животных в группе введения антитела против PD-1. Группа введения mGI101 (6 мг/кг) отдельно продемонстрировала лучший эффект ингибирования роста опухоли, чем группа введения антитела против PD-1. Размер опухоли оставался тем же до 5-7 суток, однако она вновь начинала расти после 7 суток. Группа комбинированного введения (GI101 (0,6 мг/кг) + антитела против PD-1 (5 мг/кг)) продемонстрировала примечательное превосходное ингибирование роста опухоли. В частности, двое в группе комбинированного введения продемонстрировали полный ответ (отсутствие опухоли).
Клетки MC38 вновь инъецировали в левый бок (в противоположность первоначальной области инъекции злокачественных клеток) двух мышей группы комбинированного введения, которые продемонстрировали полную ремиссию. Этим мышам продолжали вводить антитело против PD-1 (5 мг/кг, BIW) до 32 суток (фиг. 74). Небольшая опухоль (>30 мм3) была обнаружена у одной из двух мышей, однако размер опухоли не увеличивался до 35 суток (фиг. 69). У другой мыши опухоль после повторной инъекции не была обнаружена (фиг. 69 и 74).
В заключение, проводили тестирование противоопухолевой эффективности mGI101 отдельно и в комбинации с антителом против PD-1 в модели с аллогенной опухолью MC38, и в результате группа комбинированного введения (GI101 (0,6 мг/кг) + антитело против PD-1 (5 мг/кг)) продемонстрировала наилучшую противоопухолевую эффективность. Двое экспериментальных животных в группе комбинированного введения продемонстрировали полный ответ, и мыши, продемонстрировавшие полный ответ, продемонстрировали противораковый эффект, когда MC38 реинъецировали (таблица 6).
[Таблица 6]
Объем опухоли (мм3)
Объем опухоли (мм3)
Экспериментальный пример 22. Определение противоракового эффекта посредством комбинированного введения mGI101 и антитела против PD-L1 у мышей, которым трансплантировали происходящие из мыши клетки рака ободочной и прямой кишки
В этом тесте оценивали эффект ингибирования роста опухоли в модели опухоли, в которой аллогенные клетки карциномы толстой кишки мыши (CT26) трансплантировали мышам BALB/c, после внутрибрюшинного введения mGI101 в качестве тестируемого материала и антитела против PD-1 (BioXcell, каталожный номер № BE0101) в качестве положительного контроля, отдельно или в комбинации.
Клетки CT26 культивировали в среде RPMI1640 (Gibco), содержавшей 10% эмбриональную телячью сыворотку (Gibco) и 1% антибиотик/противогрибковое средство (Gibco). Культивируемые клетки собирали с использованием трипсина и суспендировали в PBS. 5×105 клеток CT26 инъецировали п/к в правый бок самок мышей C57BL/6 (в возрасте 7 недель) для получения модели аллотрансплантата опухоли.
5×105 клеток CT26 подкожно инъецировали в правый бок самок мышей BALB/c (возраст 7 недель) для получения модели с трансплантатом опухоли.
Мышей случайным образом распределяли (4 на группу) на основе объема опухоли (50~120 мм3). Трансплантаты опухолей идентифицировали приблизительно через 2 суток после инокуляции клеток. Тестируемые группы организовывали, как показано в таблице 7, и вводили тестируемые материалы.
[Таблица 7]
В ходе периода тестирования идентифицировали клинические симптомы, такие как изменения заболевания и поведения и т.п., один раз в сутки, и выявляли умерших животных. Животных умерщвляли после периода тестирования. Размер солидной злокачественной опухоли CT26 измеряли с использованием 3D-сканера опухоли (TM900, Peria, Бельгия). Для каждой экспериментальной группы вычисляли среднее снижение массы тела и процентное изменение, и среднее ингибирование роста опухоли. Противоопухолевую эффективность вычисляли по сравнению с контрольной группой носителя.
Все статистические вычисления проводили с использованием Prism 8.0 (Graph Pad Software Inc., США). Результаты измерения объема опухоли сравнивали посредством одностороннего ANOVA (конечный момент времени), а затем критерия множественных сравнения Бонферрони. Значимым считали значение "p"<0,05.
Противоопухолевую эффективность mGI101 отдельно и в комбинации с антителом против PD-L1 тестировали в модели с аллогенной опухолью CT26, и в результате группа комбинированного введения (mGI101 (3 мг/кг) + антитело против PD-L1 (10 мг/кг)) продемонстрировала наилучшую противоопухолевую эффективность (фиг. 75).
Экспериментальный пример 23. Определение противоракового эффекта посредством комбинированного введения mGI101 и антитела против TIGIT у мышей, которым трансплантировали происходящие из мыши клетки рака ободочной и прямой кишки
В этом тесте оценивали эффект ингибирования роста опухоли в модели опухоли, в которой аллогенные клетки карциномы толстой кишки мыши (CT26) трансплантировали мышам BALB/c, после внутрибрюшинного введения mGI101 в качестве тестируемого материала и антитела против TIGIT, специфически связывающегося с внеклеточным доменом (ECD) TIGIT, имеющего аминокислотную последовательность SEQ ID NO: 39, в качестве положительного контроля, отдельно или в комбинации.
Клетки CT26 культивировали в среде RPMI1640 (Gibco), содержавшей 10% эмбриональную телячью сыворотку (Gibco) и 1% антибиотик/противогрибковое средство (Gibco). Культивируемые клетки собирали с использованием трипсина и суспендировали в PBS. 5×105 клеток CT26 инъецировали п/к в правый бок самок мышей C57BL/6 (в возрасте 7 недель) для получения модели аллотрансплантата опухоли.
Мышей случайным образом распределяли (5 на группу) на основе объема опухоли (50~120 мм3). Трансплантаты опухолей идентифицировали приблизительно через 2 суток после инокуляции клеток. Тестируемые группы организовывали, как показано в таблице 8, и вводили тестируемые материалы.
[Таблица 8]
(Merck, MK-7684)
В ходе периода тестирования идентифицировали клинические симптомы, такие как изменения заболевания и поведения и т.п., один раз в сутки, и выявляли умерших животных. Животных умерщвляли после периода тестирования. Размер солидной злокачественной опухоли CT26 измеряли с использованием 3D-сканера опухоли (TM900, Peria, Бельгия). Для каждой экспериментальной группы вычисляли среднее снижение массы тела и процентное изменение, и среднее ингибирование роста опухоли. Противоопухолевую эффективность вычисляли по сравнению с контрольной группой носителя.
Все статистические вычисления проводили с использованием Prism 8.0 (Graph Pad Software Inc., США). Результаты измерения объема опухоли сравнивали посредством одностороннего ANOVA (конечный момент времени), а затем критерия множественных сравнения Бонферрони. Значимым считали значение "p"<0,05.
Противоопухолевую эффективность mGI101 отдельно и в комбинации с антителом против TIGIT тестировали в модели с аллогенной опухолью CT26, и в результате группа комбинированного введения (mGI101 (3 мг/кг) + антитело против TIGIT (20 мг/кг)) продемонстрировала наилучшую противоопухолевую эффективность (фиг. 76). Противоопухолевый эффект не наблюдался в группе введения антитела против TIGIT отдельно по сравнению с контрольной группой. Однако комбинированное введение антитела против TIGIT и mGI101 продемонстрировала значительно лучший противоопухолевый эффект по сравнению с группой введения mGI101 отдельно.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GI Innovation,Inc.
<120> ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ, СОДЕРЖАЩАЯ
СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ БЕЛОК IL-2 И БЕЛОК CD80, И ИНГИБИТОР ИММУННОЙ
ТОЧКИ КОНТРОЛЯ
<130> POPB214529PCT
<150> KR 10-2019-0154632
<151> 2019-11-27
<160> 39
<170> KoPatentIn 3.0
<210> 1
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> сигнальный пептид (TPA)
<400> 1
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala
20 25
<210> 2
<211> 208
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hB7-1:35-242
<400> 2
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
<210> 3
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> шарнирная область
<400> 3
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10 15
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
20 25 30
<210> 4
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> fc иммуноглобулина
<400> 4
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly
210 215
<210> 5
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер
<400> 5
Gly Gly Gly Gly Ser
1 5
<210> 6
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hIL-2M
<400> 6
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 7
<211> 617
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок, содержащий варианты IL-2 и фрагменты CD80
<400> 7
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Ile His Val Thr Lys Glu
20 25 30
Val Lys Glu Val Ala Thr Leu Ser Cys Gly His Asn Val Ser Val Glu
35 40 45
Glu Leu Ala Gln Thr Arg Ile Tyr Trp Gln Lys Glu Lys Lys Met Val
50 55 60
Leu Thr Met Met Ser Gly Asp Met Asn Ile Trp Pro Glu Tyr Lys Asn
65 70 75 80
Arg Thr Ile Phe Asp Ile Thr Asn Asn Leu Ser Ile Val Ile Leu Ala
85 90 95
Leu Arg Pro Ser Asp Glu Gly Thr Tyr Glu Cys Val Val Leu Lys Tyr
100 105 110
Glu Lys Asp Ala Phe Lys Arg Glu His Leu Ala Glu Val Thr Leu Ser
115 120 125
Val Lys Ala Asp Phe Pro Thr Pro Ser Ile Ser Asp Phe Glu Ile Pro
130 135 140
Thr Ser Asn Ile Arg Arg Ile Ile Cys Ser Thr Ser Gly Gly Phe Pro
145 150 155 160
Glu Pro His Leu Ser Trp Leu Glu Asn Gly Glu Glu Leu Asn Ala Ile
165 170 175
Asn Thr Thr Val Ser Gln Asp Pro Glu Thr Glu Leu Tyr Ala Val Ser
180 185 190
Ser Lys Leu Asp Phe Asn Met Thr Thr Asn His Ser Phe Met Cys Leu
195 200 205
Ile Lys Tyr Gly His Leu Arg Val Asn Gln Thr Phe Asn Trp Asn Thr
210 215 220
Thr Lys Gln Glu His Phe Pro Asp Asn Gly Ser Gly Gly Gly Gly Ser
225 230 235 240
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly
245 250 255
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
260 265 270
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg
275 280 285
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
290 295 300
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
305 310 315 320
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
325 330 335
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
340 345 350
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
355 360 365
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
370 375 380
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
385 390 395 400
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
405 410 415
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
420 425 430
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
435 440 445
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala
450 455 460
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly
465 470 475 480
Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
485 490 495
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
500 505 510
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe
515 520 525
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
530 535 540
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
545 550 555 560
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
565 570 575
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
580 585 590
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
595 600 605
Cys Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 8
<211> 1857
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101)
<400> 8
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac ctgatga 1857
<210> 9
<211> 592
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101)
<400> 9
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 10
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hIL-2
<400> 10
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 11
<211> 288
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CD80
<400> 11
Met Gly His Thr Arg Arg Gln Gly Thr Ser Pro Ser Lys Cys Pro Tyr
1 5 10 15
Leu Asn Phe Phe Gln Leu Leu Val Leu Ala Gly Leu Ser His Phe Cys
20 25 30
Ser Gly Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu
35 40 45
Ser Cys Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile
50 55 60
Tyr Trp Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp
65 70 75 80
Met Asn Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr
85 90 95
Asn Asn Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly
100 105 110
Thr Tyr Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg
115 120 125
Glu His Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr
130 135 140
Pro Ser Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile
145 150 155 160
Ile Cys Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu
165 170 175
Glu Asn Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp
180 185 190
Pro Glu Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met
195 200 205
Thr Thr Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg
210 215 220
Val Asn Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro
225 230 235 240
Asp Asn Leu Leu Pro Ser Trp Ala Ile Thr Leu Ile Ser Val Asn Gly
245 250 255
Ile Phe Val Ile Cys Cys Leu Thr Tyr Cys Phe Ala Pro Arg Cys Arg
260 265 270
Glu Arg Arg Arg Asn Glu Arg Leu Arg Arg Glu Ser Val Arg Pro Val
275 280 285
<210> 12
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> модифицированный Fc
<400> 12
Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys Pro Lys
1 5 10 15
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
20 25 30
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
35 40 45
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
50 55 60
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
65 70 75 80
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
85 90 95
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
100 105 110
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
115 120 125
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
130 135 140
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
145 150 155 160
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
165 170 175
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
180 185 190
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
195 200 205
Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 13
<211> 306
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> mCD80
<400> 13
Met Ala Cys Asn Cys Gln Leu Met Gln Asp Thr Pro Leu Leu Lys Phe
1 5 10 15
Pro Cys Pro Arg Leu Ile Leu Leu Phe Val Leu Leu Ile Arg Leu Ser
20 25 30
Gln Val Ser Ser Asp Val Asp Glu Gln Leu Ser Lys Ser Val Lys Asp
35 40 45
Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His Glu Asp Glu Ser
50 55 60
Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val Val Leu Ser Val
65 70 75 80
Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys Asn Arg Thr Leu
85 90 95
Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly Leu Val Leu Ser
100 105 110
Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys Glu Arg Gly Thr
115 120 125
Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser Ile Lys Ala Asp
130 135 140
Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro Ser Ala Asp Thr
145 150 155 160
Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro Lys Pro Arg Phe
165 170 175
Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile Asn Thr Thr Ile
180 185 190
Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser Ser Gln Leu Asp
195 200 205
Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu Ile Lys Tyr Gly
210 215 220
Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys Pro Pro Glu Asp
225 230 235 240
Pro Pro Asp Ser Lys Asn Thr Leu Val Leu Phe Gly Ala Gly Phe Gly
245 250 255
Ala Val Ile Thr Val Val Val Ile Val Val Ile Ile Lys Cys Phe Cys
260 265 270
Lys His Arg Ser Cys Phe Arg Arg Asn Glu Ala Ser Arg Glu Thr Asn
275 280 285
Asn Ser Leu Thr Phe Gly Pro Glu Glu Ala Leu Ala Glu Gln Thr Val
290 295 300
Phe Leu
305
<210> 14
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI101)
<400> 14
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg 120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa 180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag 240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc 300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag 360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag 420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct 480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt 540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc 600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt 660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc 720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca 780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg 1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt 1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtctct tggaggtggt 1440
ggcggttctg cccctacctc cagctctacc aagaaaaccc agctccagtt ggagcatctg 1500
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc 1560
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacttgcag 1620
tgcctggaag aggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac 1680
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa 1740
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt 1800
ctgaaccggt ggatcacctt ctgccagagc atcatctcca cactgacc 1848
<210> 15
<211> 616
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI101)
<400> 15
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
465 470 475 480
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
485 490 495
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
500 505 510
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
515 520 525
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu
530 535 540
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
545 550 555 560
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
565 570 575
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
580 585 590
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
595 600 605
Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 16
<211> 1437
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101C1)
<400> 16
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctaggtgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc cctgggc 1437
<210> 17
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101C1)
<400> 17
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly
450
<210> 18
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101C2)
<400> 18
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccatctc acgccgctga gtctaagtac ggccctcctt gtcctccatg tcctgctcca 120
gaagctgctg gcggaccctc tgtgttcctg tttcctccaa agcctaagga ccagctcatg 180
atctctcgga cccctgaagt gacctgcgtg gtggtggatg tgtctcaaga ggaccctgag 240
gtgcagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga 300
gaggaacagt tcaactccac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat 360
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca agggcctgcc ttccagcatc 420
gaaaagacca tctccaaggc taagggccag cctagggaac cccaggttta caccctgcct 480
ccaagccaag aggaaatgac caagaaccag gtgtccctga cctgcctggt caagggcttc 540
tacccttccg acattgccgt ggaatgggag tccaatggcc agcctgagaa caactacaag 600
accacacctc ctgtgctgga ctccgacggc tccttctttc tgtactctcg cctgaccgtg 660
gacaagtcta ggtggcaaga gggcaacgtg ttctcctgct ctgtgctgca cgaggccctg 720
cacaatcact acacccagaa gtccctgtct ctgtctcttg gcggaggcgg aggatctgct 780
cctacctcca gctccaccaa gaaaacccag ctccagttgg agcatctgct gctggacctc 840
cagatgatcc tgaatggcat caacaattac aagaacccca agctgaccgc catgctgacc 900
gctaagttct acatgcccaa gaaggccacc gagctgaagc acctccagtg cctggaagag 960
gaactgaagc ccctggaaga agtgctgaat ctggcccagt ccaagaactt ccacctgagg 1020
cctagggacc tgatctccaa catcaacgtg atcgtgctgg aactgaaagg ctccgagaca 1080
accttcatgt gcgagtacgc cgacgagaca gccaccatcg tggaatttct gaaccggtgg 1140
atcaccttct gccagtccat catctccaca ctgacc 1176
<210> 19
<211> 367
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101C2)
<400> 19
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
1 5 10 15
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
20 25 30
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
35 40 45
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
50 55 60
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
85 90 95
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
100 105 110
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
115 120 125
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
130 135 140
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
145 150 155 160
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
165 170 175
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
180 185 190
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
195 200 205
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
210 215 220
Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser
225 230 235 240
Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln
245 250 255
Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala
260 265 270
Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys
275 280 285
His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu
290 295 300
Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile
305 310 315 320
Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr
325 330 335
Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu
340 345 350
Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
355 360 365
<210> 20
<211> 1434
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI101C1)
<400> 20
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg 120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa 180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag 240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc 300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag 360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag 420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct 480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt 540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc 600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt 660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc 720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca 780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg 1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt 1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtccct gggc 1434
<210> 21
<211> 478
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI101C1)
<400> 21
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
465 470 475
<210> 22
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M45)
<400> 22
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Ala Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 23
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M61)
<400> 23
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Arg Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 24
<211> 133
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M72)
<400> 24
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Gly Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 25
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M45)
<400> 25
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttcgccatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 26
<211> 592
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M45)
<400> 26
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Ala Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 27
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M61)
<400> 27
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aaagggaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 28
<211> 592
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M61)
<400> 28
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Arg Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 29
<211> 1857
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M72)
<400> 29
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatggggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac ctgatga 1857
<210> 30
<211> 592
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M72)
<400> 30
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Gly Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 31
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101w)
<400> 31
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc 120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa 180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac 240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct 300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag 360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac 420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct 480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg 540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc 600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc 660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct 720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct 780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct 840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct 900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc 960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc 1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc 1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag 1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc 1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct 1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac 1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg 1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt 1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat 1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg 1560
acccgcatgc tgacctttaa gttctacatg cccaagaagg ccaccgagct gaagcacctc 1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag 1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg 1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa 1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c 1851
<210> 32
<211> 592
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101w)
<400> 32
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 33
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI102-M61)
<400> 33
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg 120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa 180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag 240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc 300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag 360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag 420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct 480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt 540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc 600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt 660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc 720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca 780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag 840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa 900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag 960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg 1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg 1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt 1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg 1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag 1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct 1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg 1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtctct tggaggtggt 1440
ggcggttctg cccctacctc cagctctacc aagaaaaccc agctccagtt ggagcatctg 1500
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc 1560
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacttgcag 1620
tgcctggaaa gggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac 1680
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa 1740
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt 1800
ctgaaccggt ggatcacctt ctgccagagc atcatctcca cactgacc 1848
<210> 34
<211> 616
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI102-M61)
<400> 34
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
465 470 475 480
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
485 490 495
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
500 505 510
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
515 520 525
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Arg
530 535 540
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
545 550 555 560
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
565 570 575
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
580 585 590
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
595 600 605
Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 35
<211> 153
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hIL-2 дикого типа
<400> 35
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
20 25 30
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
35 40 45
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe
50 55 60
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
65 70 75 80
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
85 90 95
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
100 105 110
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
115 120 125
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
130 135 140
Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150
<210> 36
<211> 158
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> IL-2 с сигнальной последовательностью
<400> 36
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Ala Pro Thr Ser Ser Ser Thr
20 25 30
Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met
35 40 45
Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met
50 55 60
Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His
65 70 75 80
Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn
85 90 95
Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser
100 105 110
Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe
115 120 125
Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn
130 135 140
Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150 155
<210> 37
<211> 474
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотидная последовательность, кодирующая IL-2, с сигнальной
последовательностью
<400> 37
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg 60
tctccttctc acgctgcccc taccagctcc tctaccaaga aaacccagct ccagttggag 120
catctgctgc tggacctcca gatgattctg aacgggatca acaactataa gaaccccaag 180
ctgacccgca tgctgacctt taagttctac atgcccaaga aggccaccga gctgaagcac 240
ctccagtgcc tggaagaaga actgaagccc ctggaagagg tgctgaatct ggcccagtcc 300
aagaacttcc acctgaggcc acgggacctg atcagcaaca tcaacgtgat cgtgctggaa 360
ctgaagggct ccgagacaac ctttatgtgc gagtacgccg acgagacagc caccatcgtg 420
gaatttctga accggtggat caccttctgc cagagcatca tctccacact gacc 474
<210> 38
<211> 591
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> mGI-101
<400> 38
Val Asp Glu Gln Leu Ser Lys Ser Val Lys Asp Lys Val Leu Leu Pro
1 5 10 15
Cys Arg Tyr Asn Ser Pro His Glu Asp Glu Ser Glu Asp Arg Ile Tyr
20 25 30
Trp Gln Lys His Asp Lys Val Val Leu Ser Val Ile Ala Gly Lys Leu
35 40 45
Lys Val Trp Pro Glu Tyr Lys Asn Arg Thr Leu Tyr Asp Asn Thr Thr
50 55 60
Tyr Ser Leu Ile Ile Leu Gly Leu Val Leu Ser Asp Arg Gly Thr Tyr
65 70 75 80
Ser Cys Val Val Gln Lys Lys Glu Arg Gly Thr Tyr Glu Val Lys His
85 90 95
Leu Ala Leu Val Lys Leu Ser Ile Lys Ala Asp Phe Ser Thr Pro Asn
100 105 110
Ile Thr Glu Ser Gly Asn Pro Ser Ala Asp Thr Lys Arg Ile Thr Cys
115 120 125
Phe Ala Ser Gly Gly Phe Pro Lys Pro Arg Phe Ser Trp Leu Glu Asn
130 135 140
Gly Arg Glu Leu Pro Gly Ile Asn Thr Thr Ile Ser Gln Asp Pro Glu
145 150 155 160
Ser Glu Leu Tyr Thr Ile Ser Ser Gln Leu Asp Phe Asn Thr Thr Arg
165 170 175
Asn His Thr Ile Lys Cys Leu Ile Lys Tyr Gly Asp Ala His Val Ser
180 185 190
Glu Asp Phe Thr Trp Glu Lys Pro Pro Glu Asp Pro Pro Asp Ser Gly
195 200 205
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
325 330 335
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser
450 455 460
Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln
465 470 475 480
Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala
485 490 495
Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys
500 505 510
His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu
515 520 525
Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile
530 535 540
Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr
545 550 555 560
Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu
565 570 575
Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 39
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ECD TIGIT
<400> 39
Met Met Thr Gly Thr Ile Glu Thr Thr Gly Asn Ile Ser Ala Glu Lys
1 5 10 15
Gly Gly Ser Ile Ile Leu Gln Cys His Leu Ser Ser Thr Thr Ala Gln
20 25 30
Val Thr Gln Val Asn Trp Glu Gln Gln Asp Gln Leu Leu Ala Ile Cys
35 40 45
Asn Ala Asp Leu Gly Trp His Ile Ser Pro Ser Phe Lys Asp Arg Val
50 55 60
Ala Pro Gly Pro Gly Leu Gly Leu Thr Leu Gln Ser Leu Thr Val Asn
65 70 75 80
Asp Thr Gly Glu Tyr Phe Cys Ile Tyr His Thr Tyr Pro Asp Gly Thr
85 90 95
Tyr Thr Gly Arg Ile Phe Leu Glu Val Leu Glu Ser Ser Val Ala Glu
100 105 110
His Gly Ala Arg Phe Gln Ile Pro
115 120
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ, СОДЕРЖАЩАЯ СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ БЕЛОК IL-2 И БЕЛОК CD80, И ПРОТИВОРАКОВОЕ СРЕДСТВО | 2021 |
|
RU2839599C1 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ БЕЛОК IL-2 И БЕЛОК CD80, И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2811541C2 |
| КОМПОЗИЦИЯ ДЛЯ ПРОТИВОРАКОВОГО ЛЕЧЕНИЯ, СОДЕРЖАЩАЯ NK-КЛЕТКИ И СЛИТЫЙ БЕЛОК, КОТОРЫЙ СОДЕРЖИТ БЕЛОК IL-2 И БЕЛОК CD80 | 2020 |
|
RU2832136C1 |
| СЛИТЫЙ БЕЛОК GDF15 ДЛИТЕЛЬНОГО ДЕЙСТВИЯ И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2020 |
|
RU2836280C1 |
| БЕЛКИ С ДВОЙНОЙ ФУНКЦИЕЙ И СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2016 |
|
RU2741345C2 |
| СЛИТЫЕ С FGF21 БЕЛКИ ДЛИТЕЛЬНОГО ДЕЙСТВИЯ И СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2016 |
|
RU2741087C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| НОВЫЙ МОДИФИЦИРОВАННЫЙ СЛИТЫЙ БЕЛОК FC-ФРАГМЕНТА ИММУНОГЛОБУЛИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800919C2 |
| СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ РЕЦЕПТОР TGF-БЕТА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2776204C1 |
| ВНЕКЛЕТОЧНЫЙ ДОМЕН АЛЬФА-СУБЪЕДИНИЦЫ FC-РЕЦЕПТОРА IGE, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЕГО, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2019 |
|
RU2796162C2 |
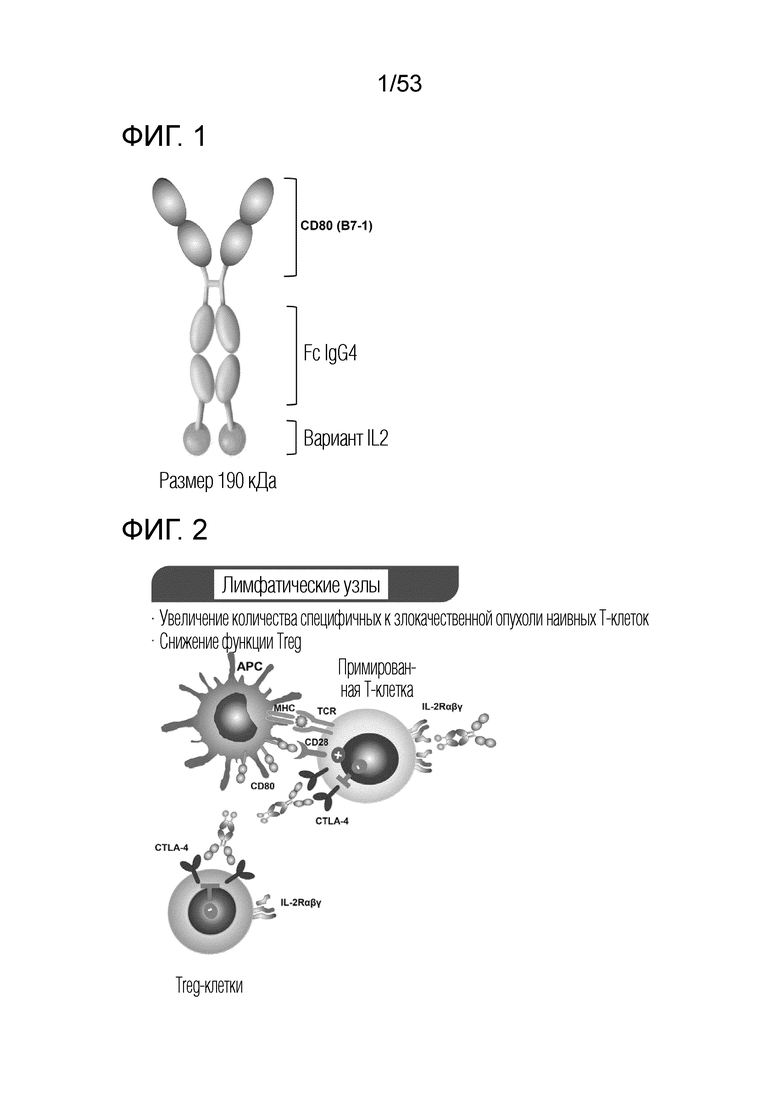
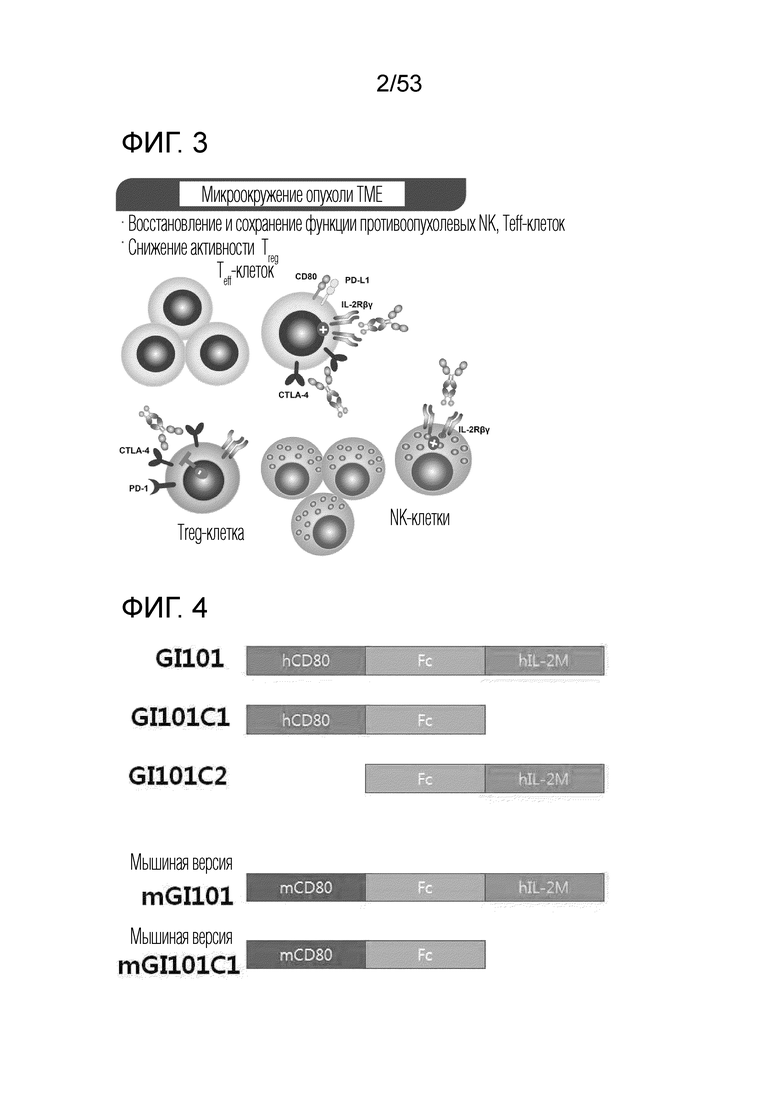
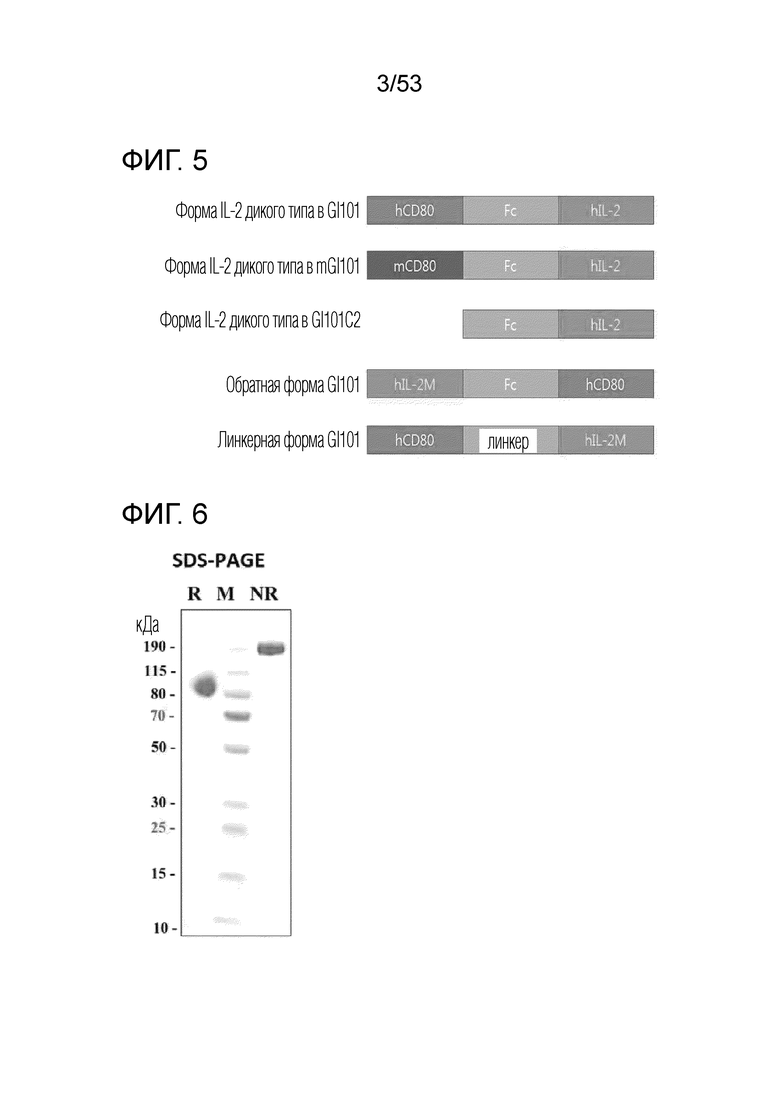
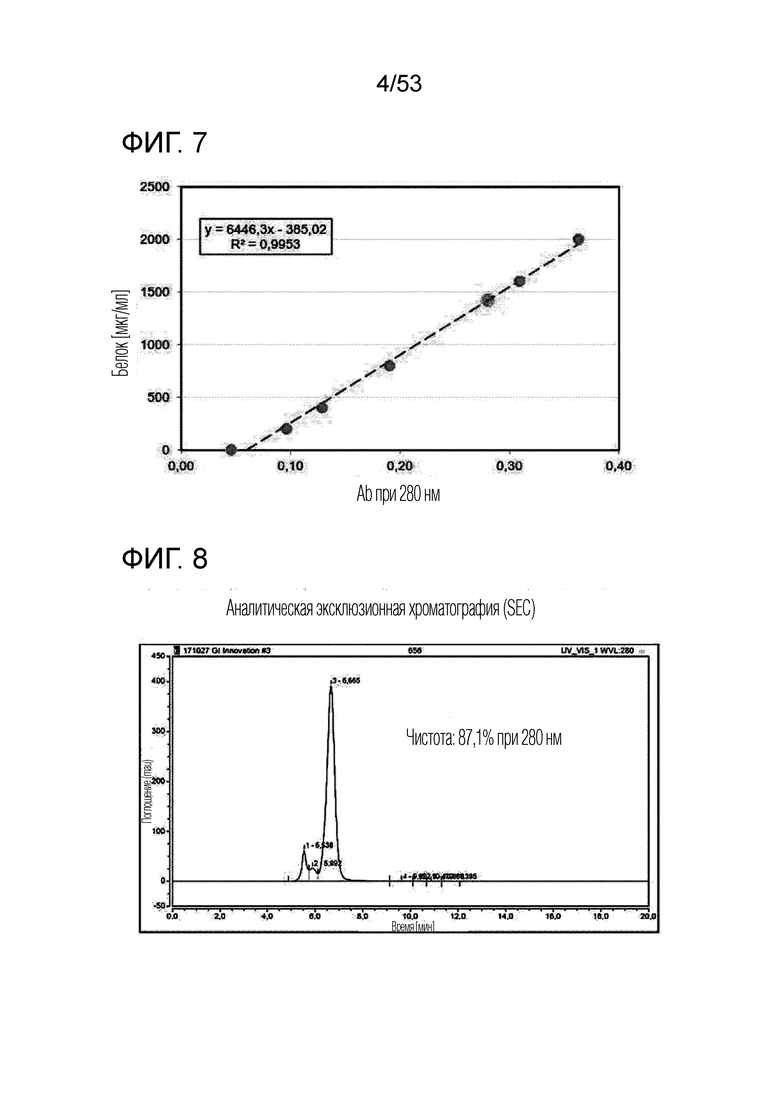
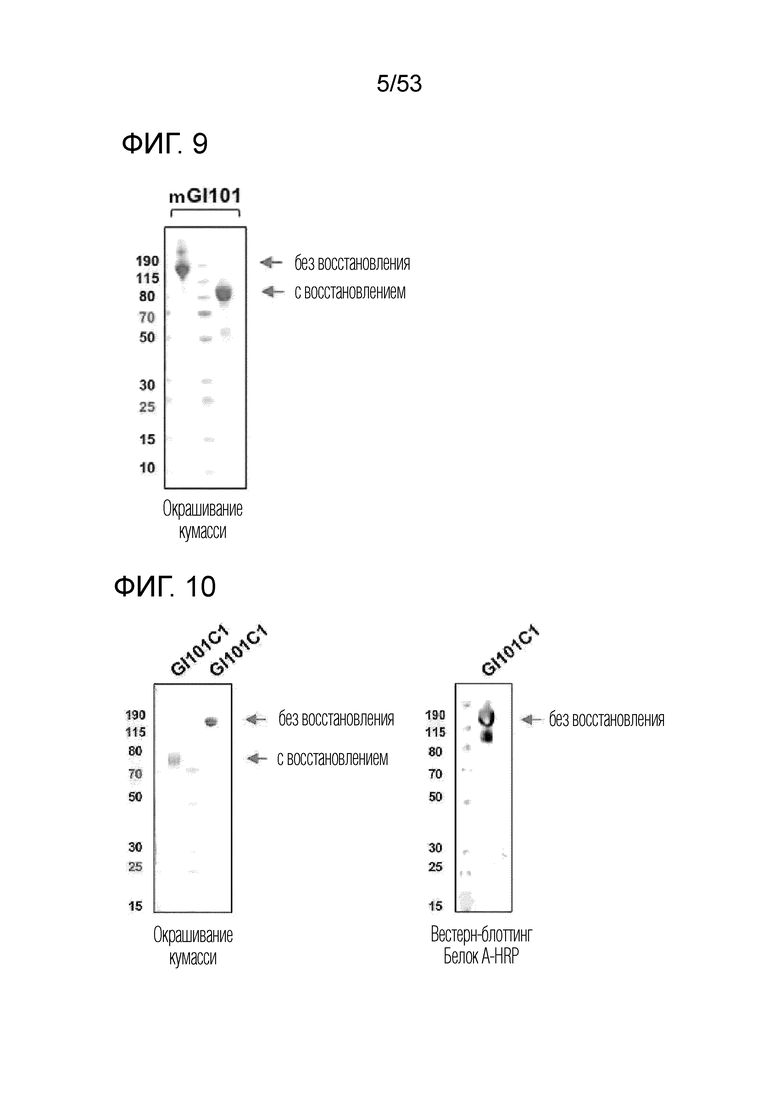
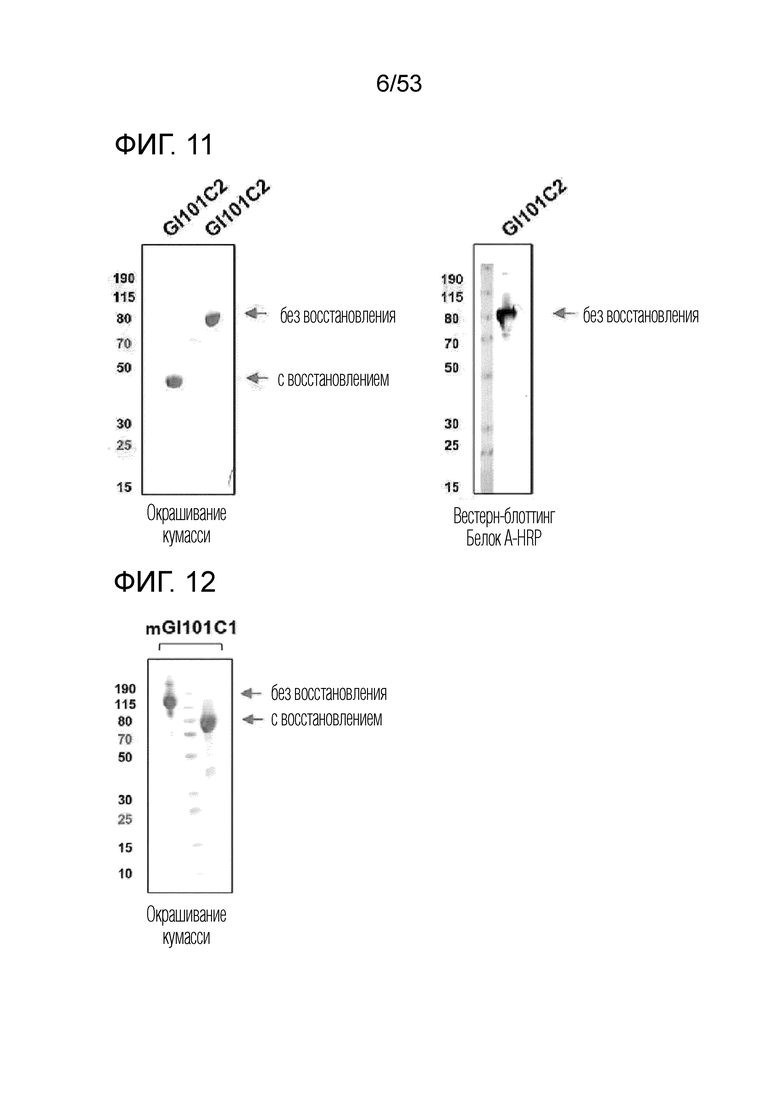
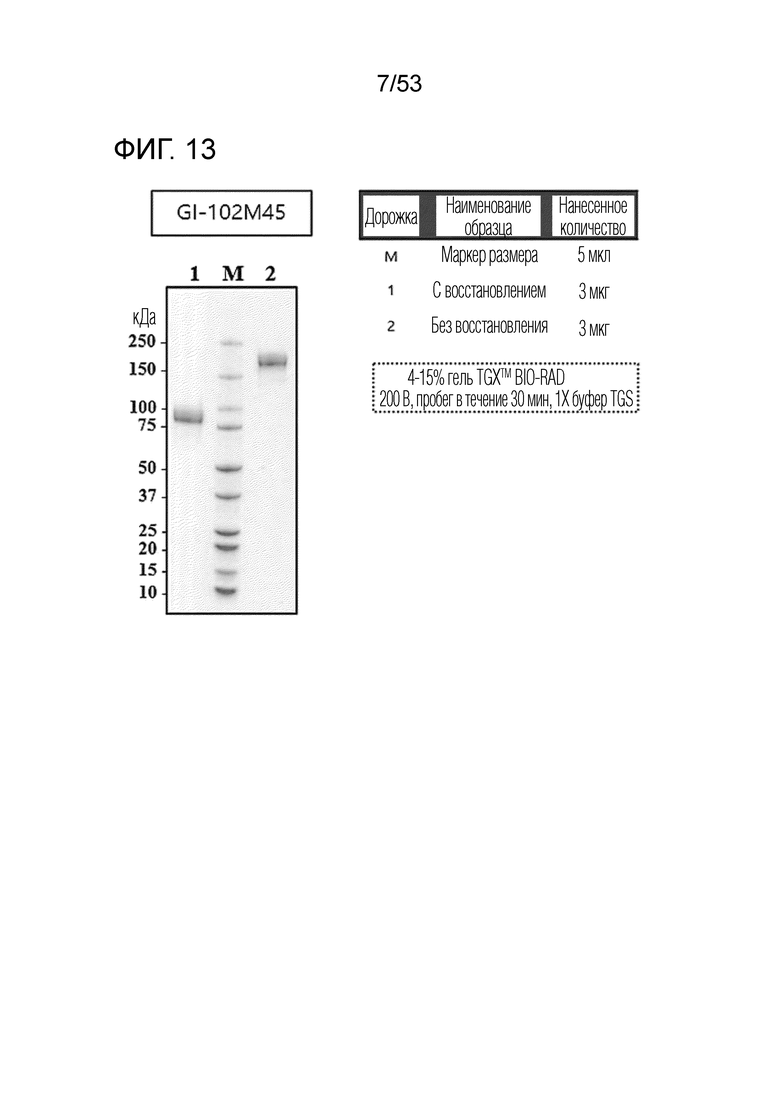

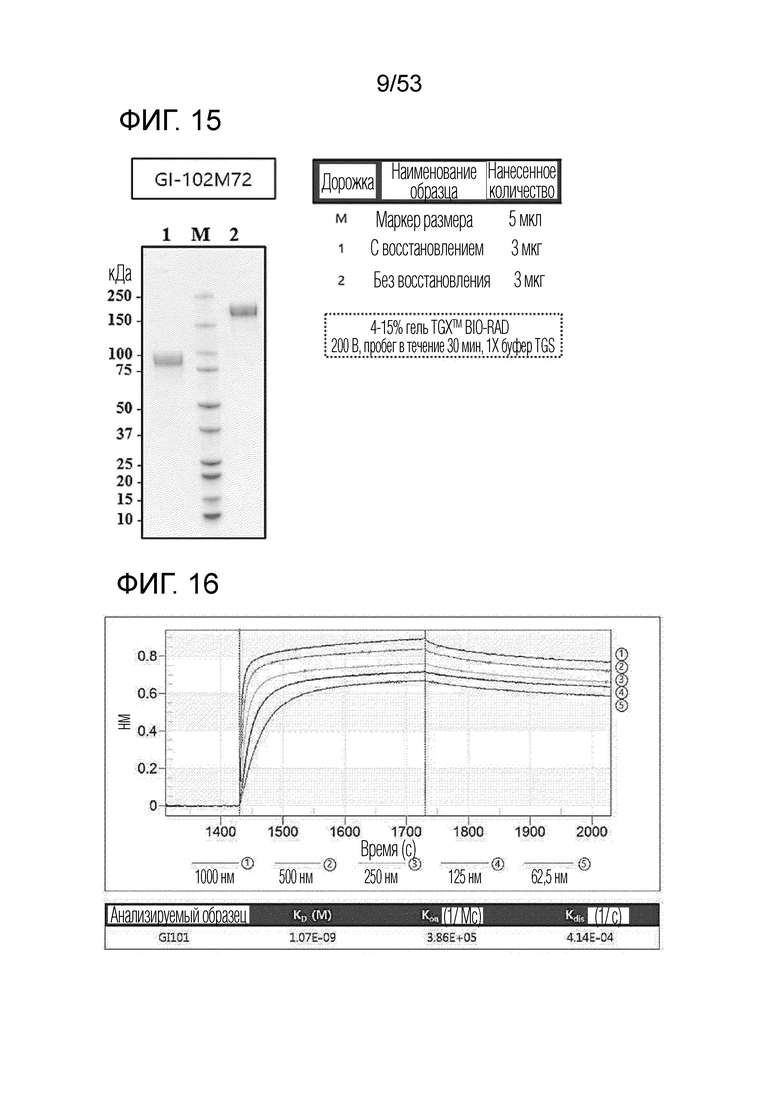
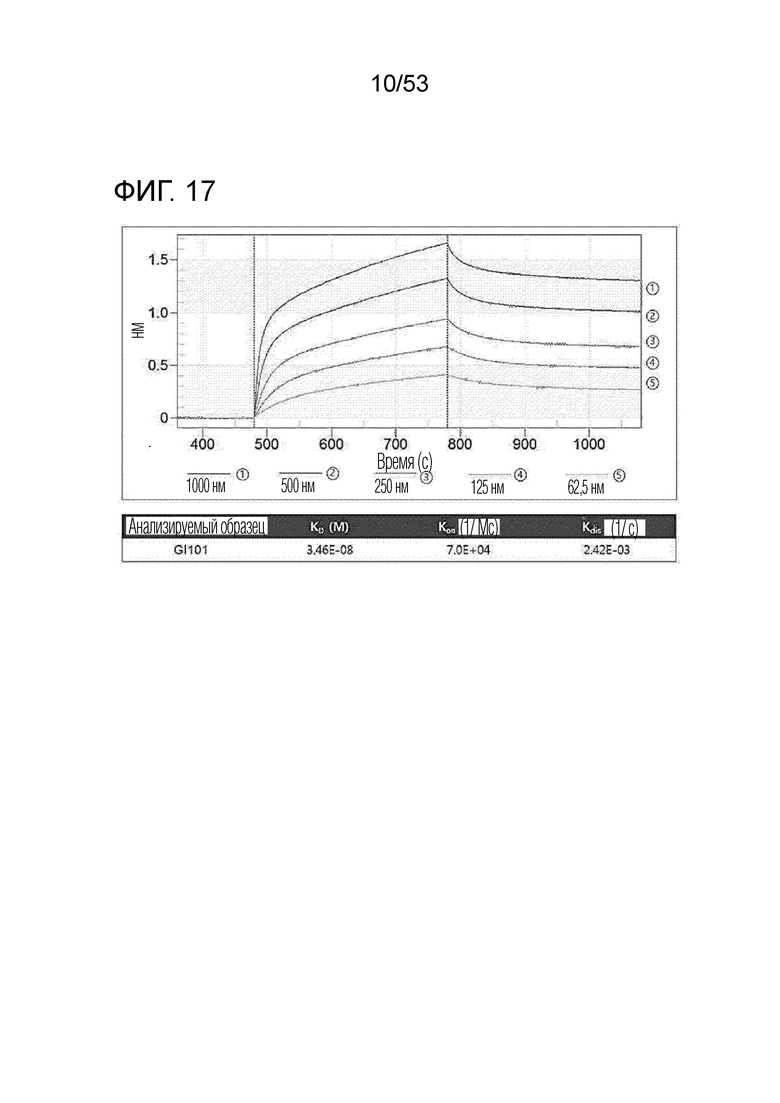
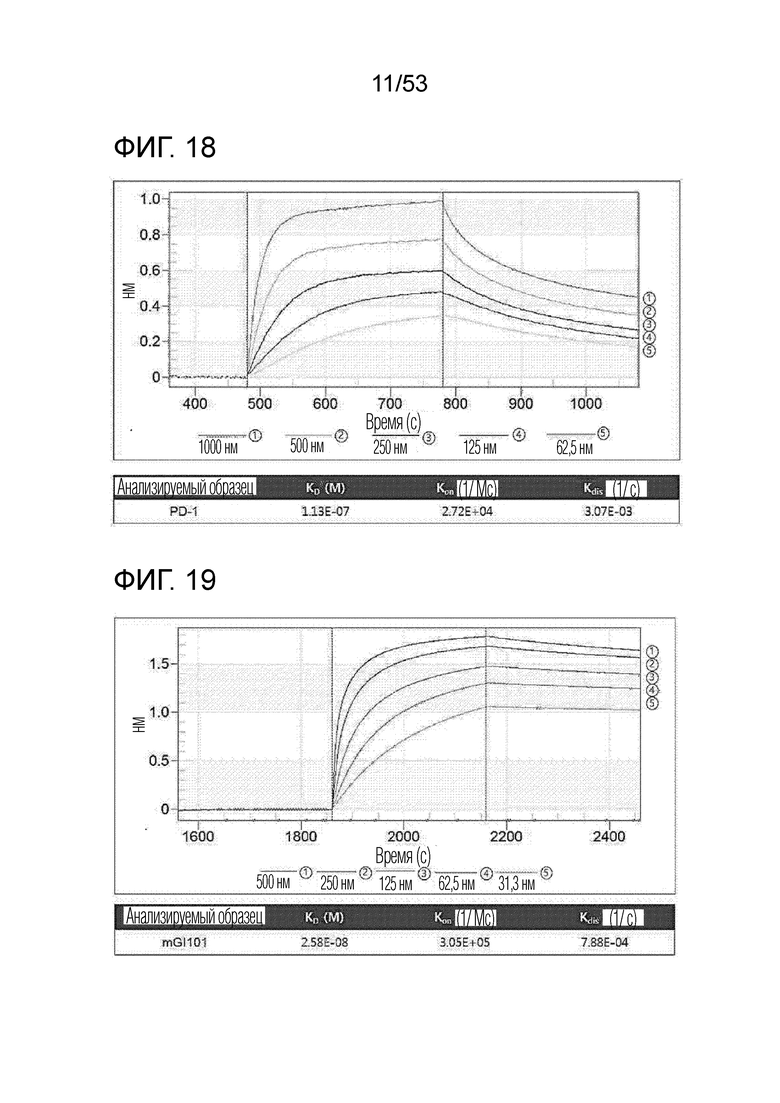
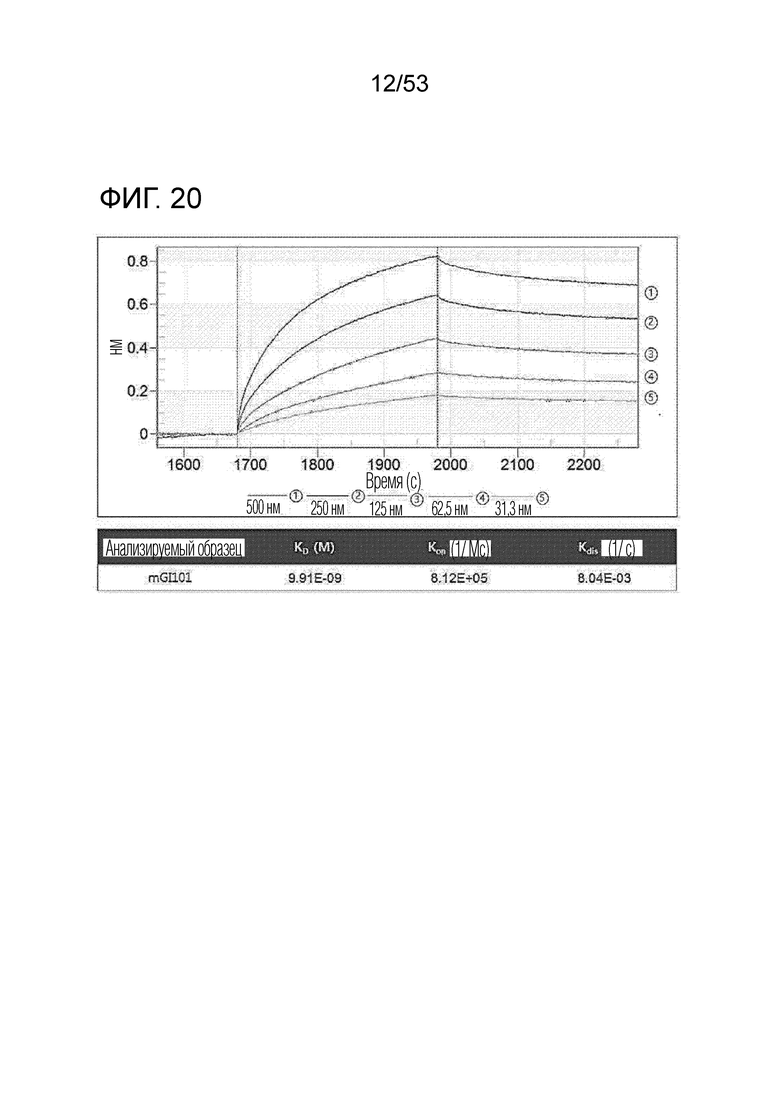
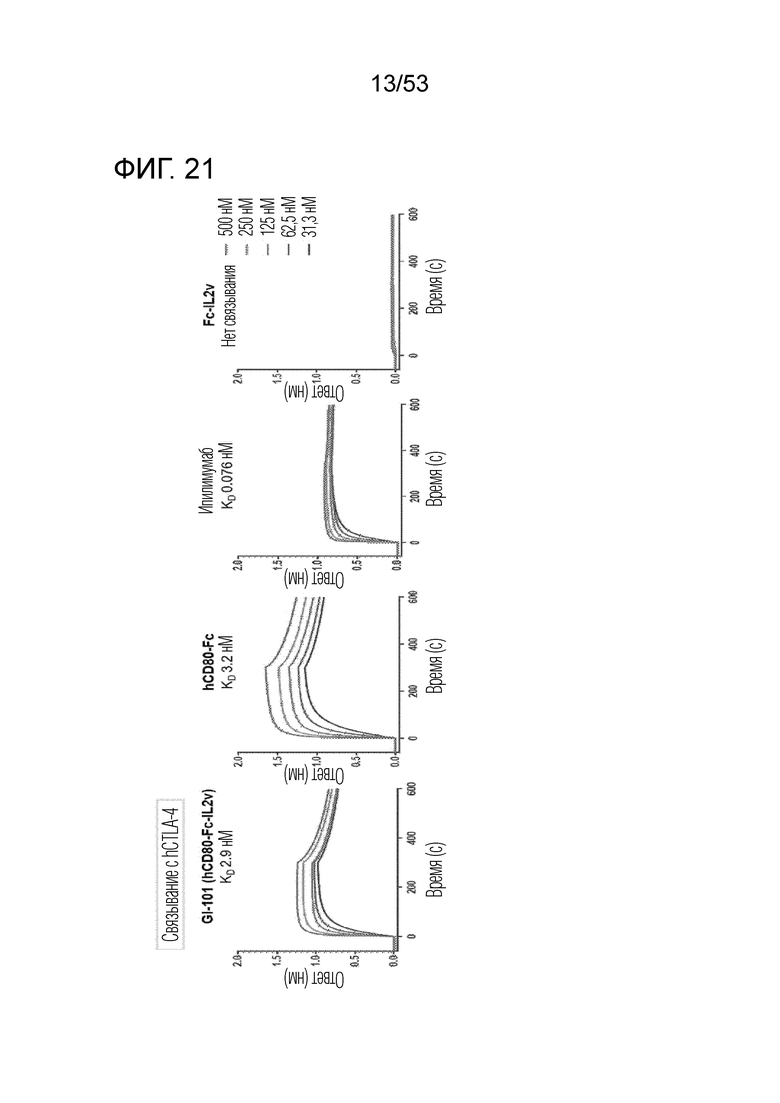
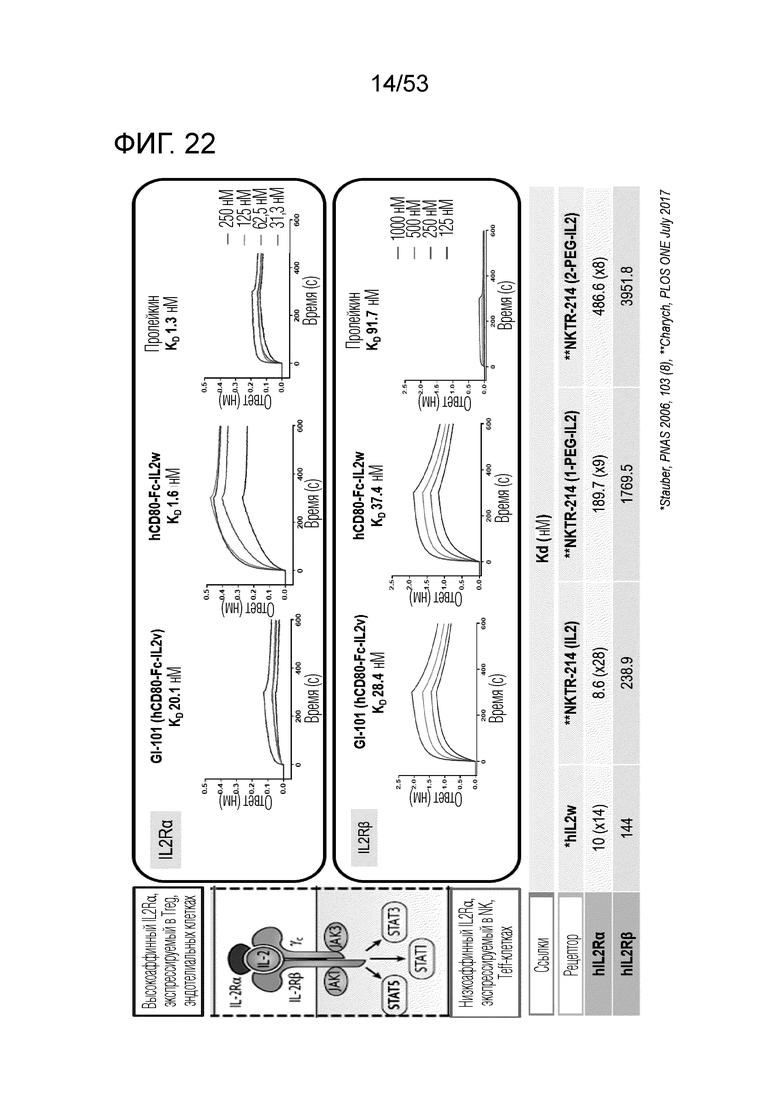
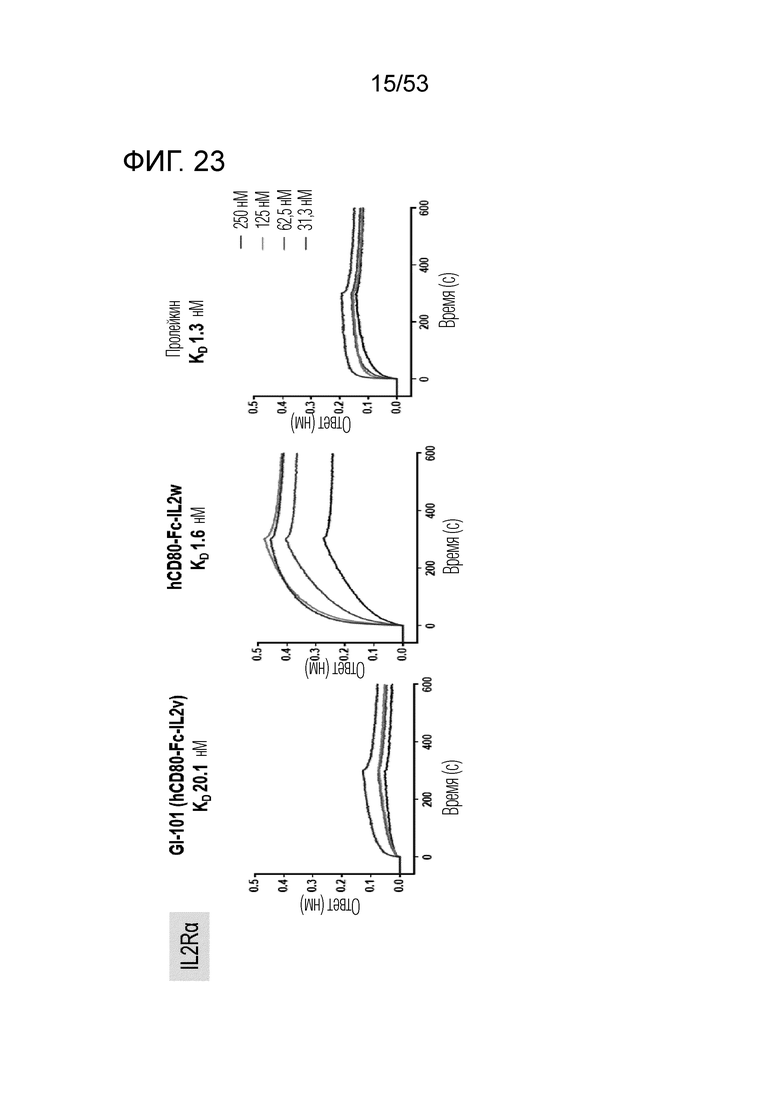
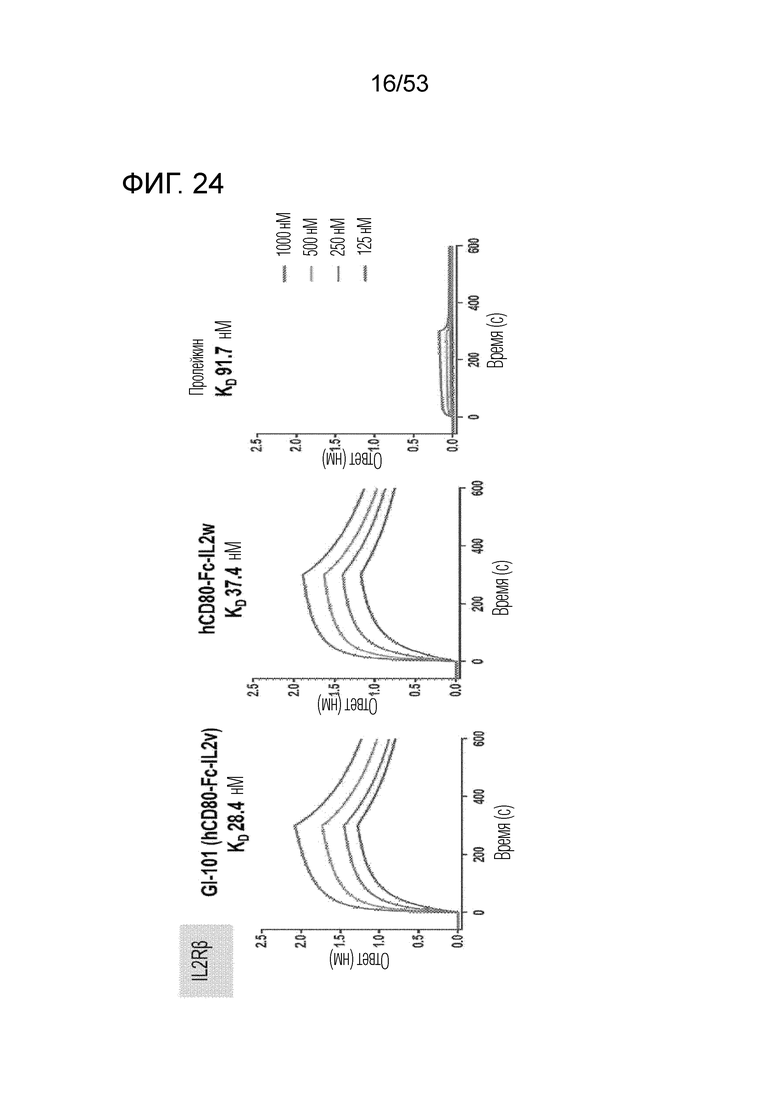
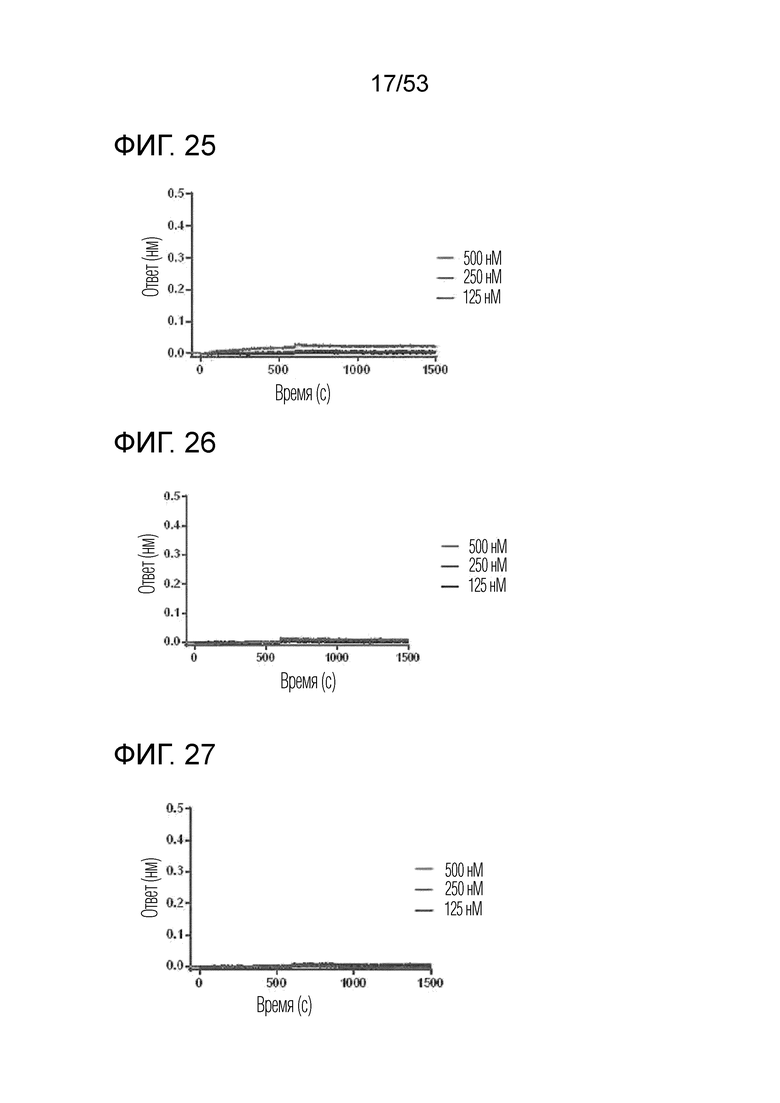
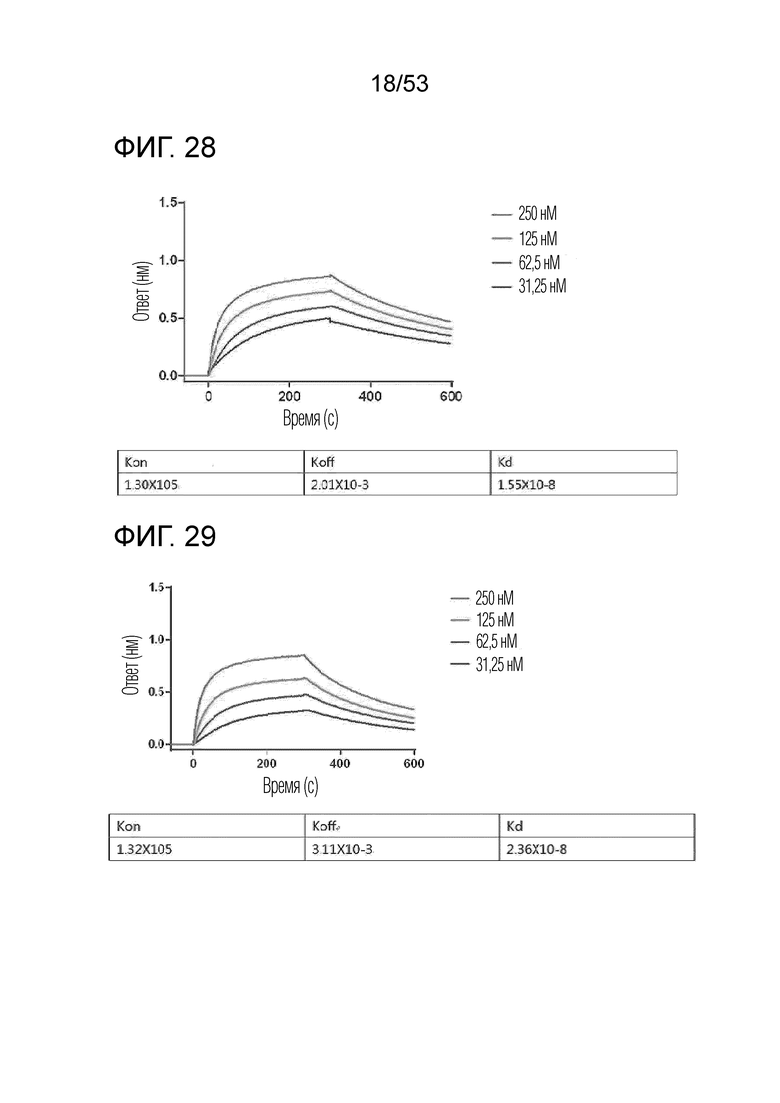
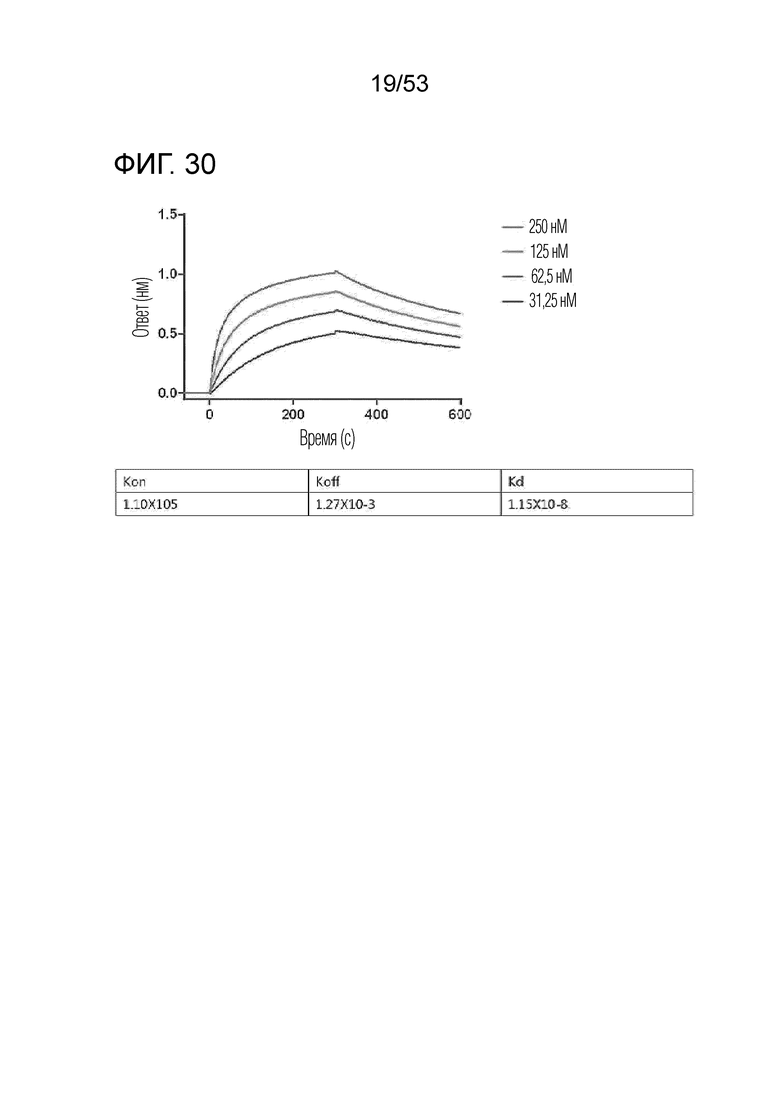
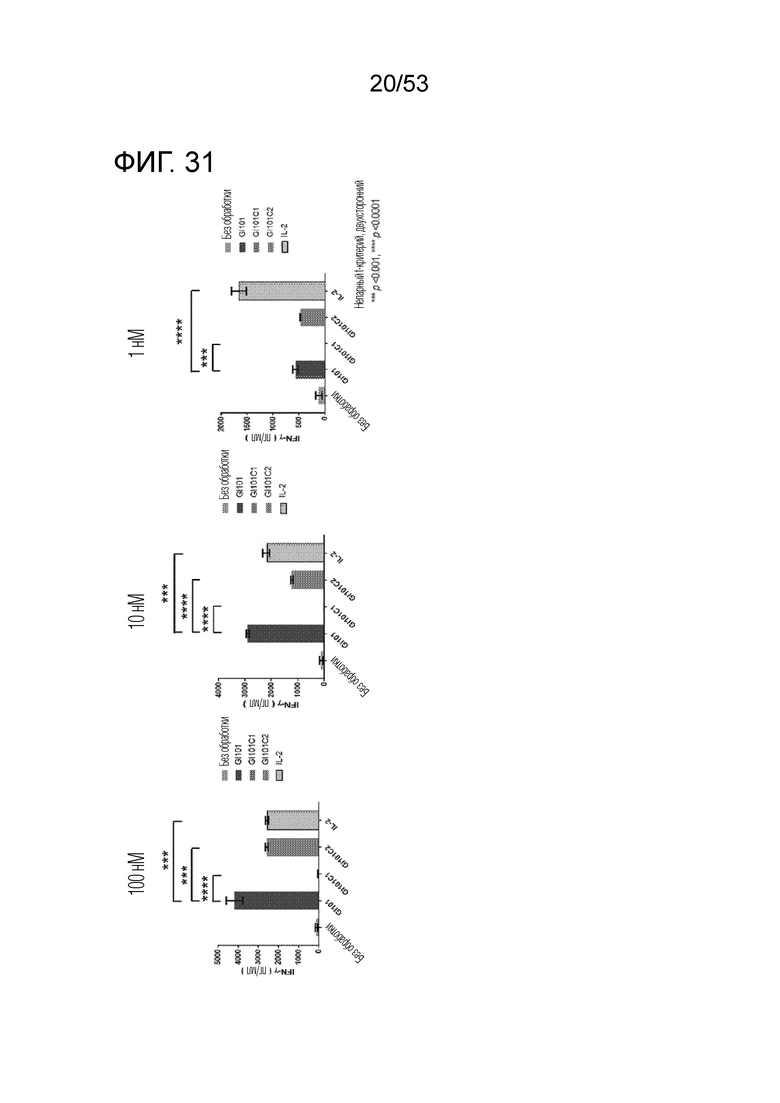
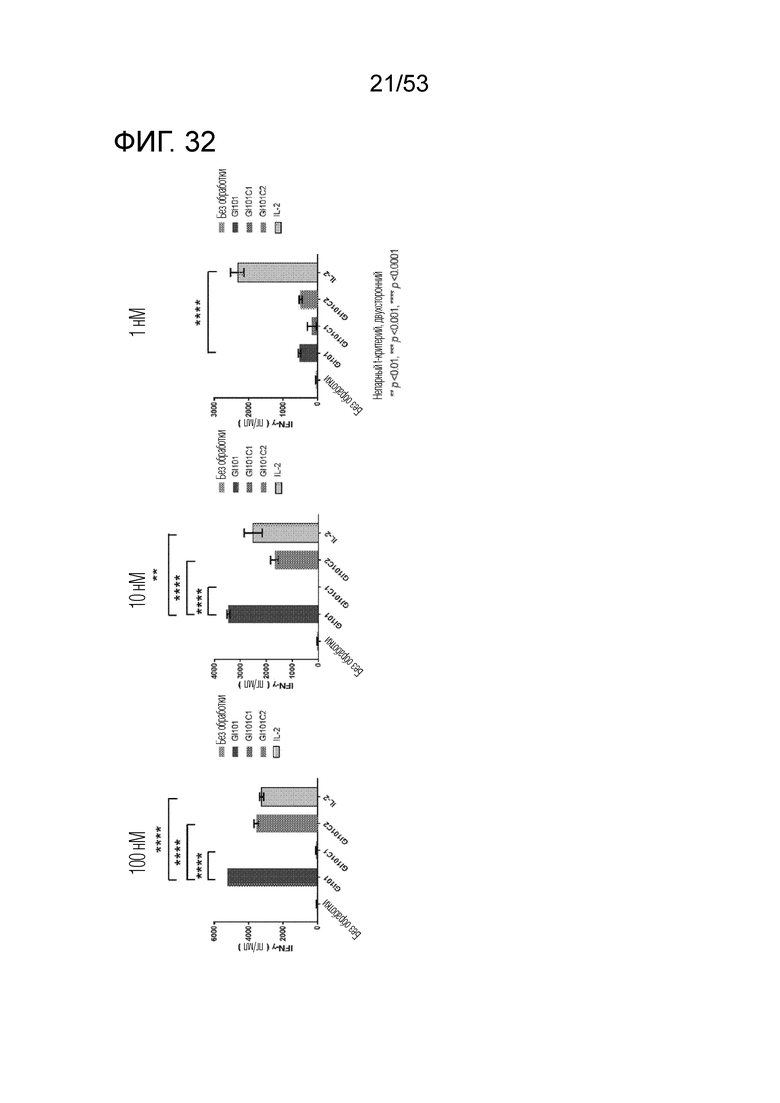
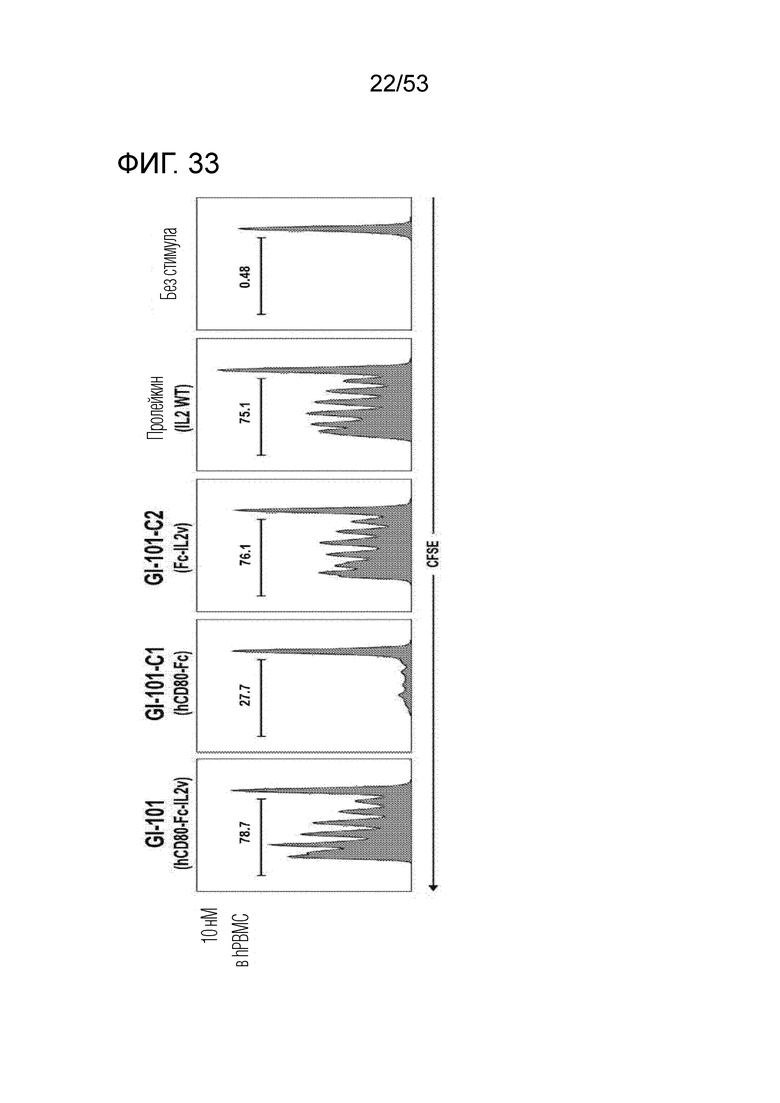
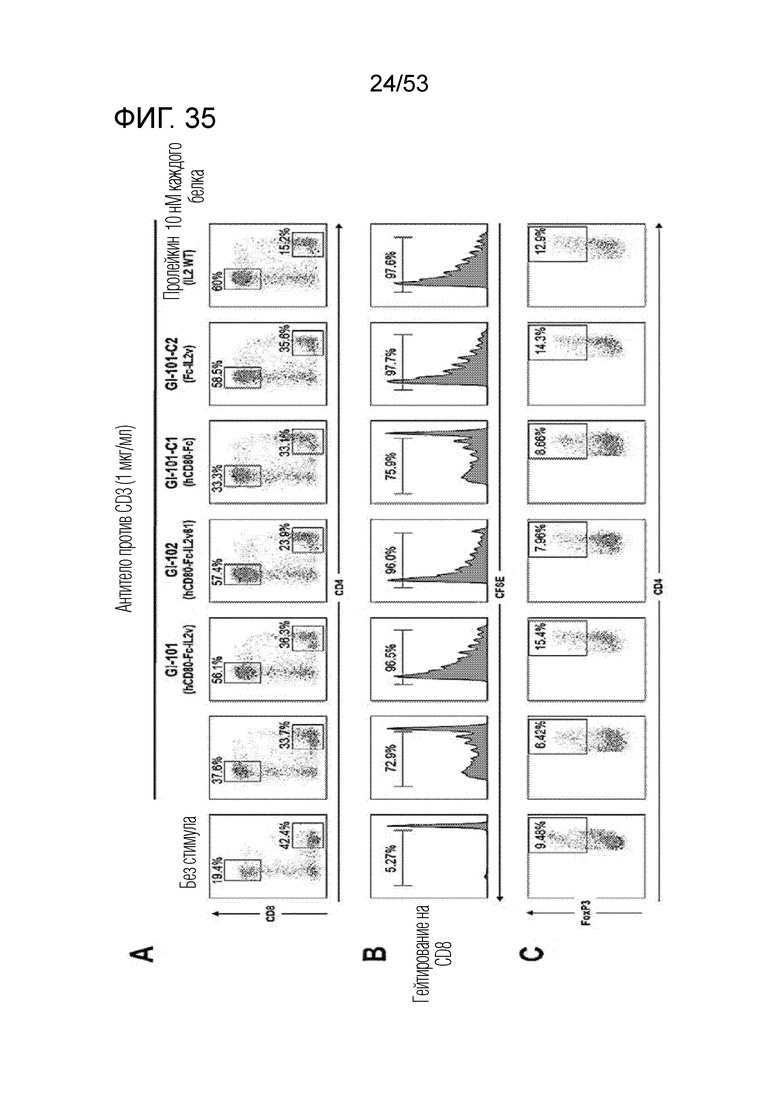
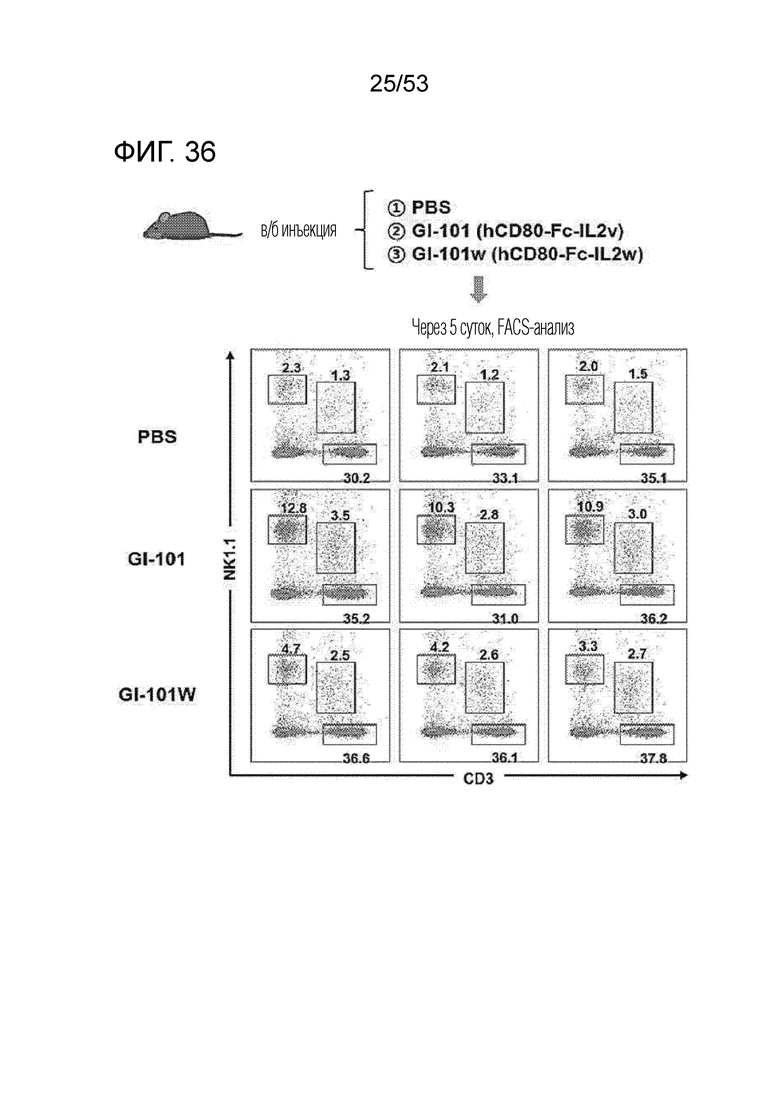
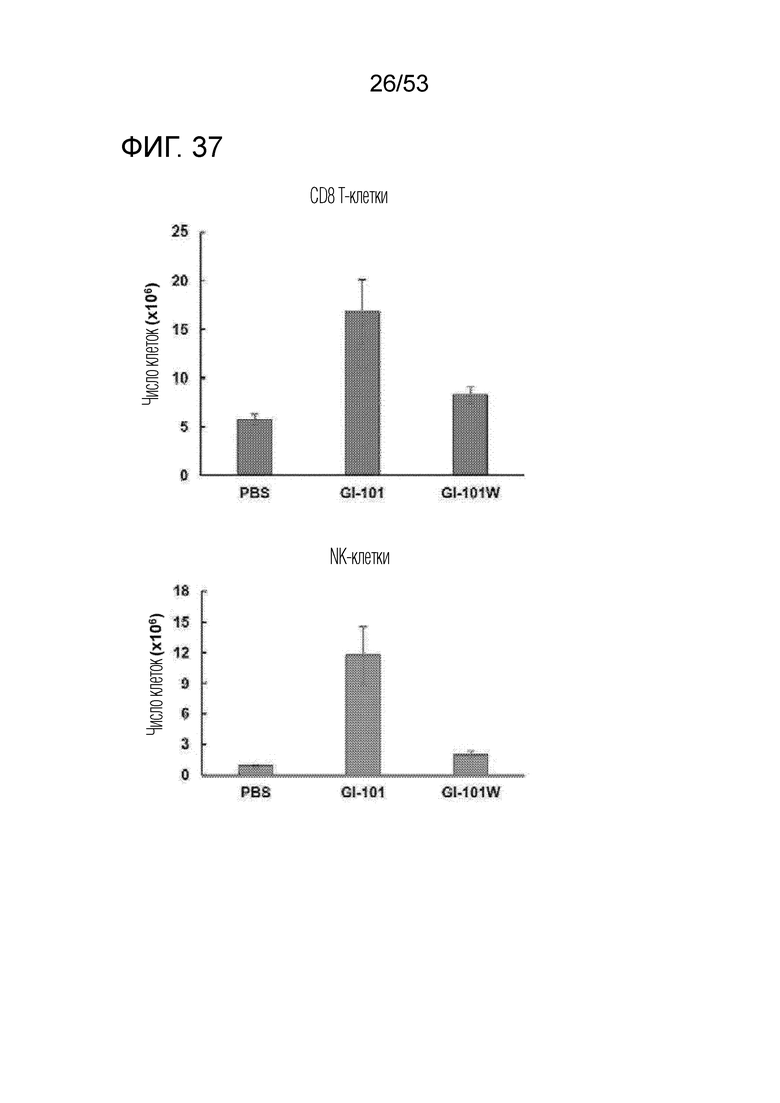
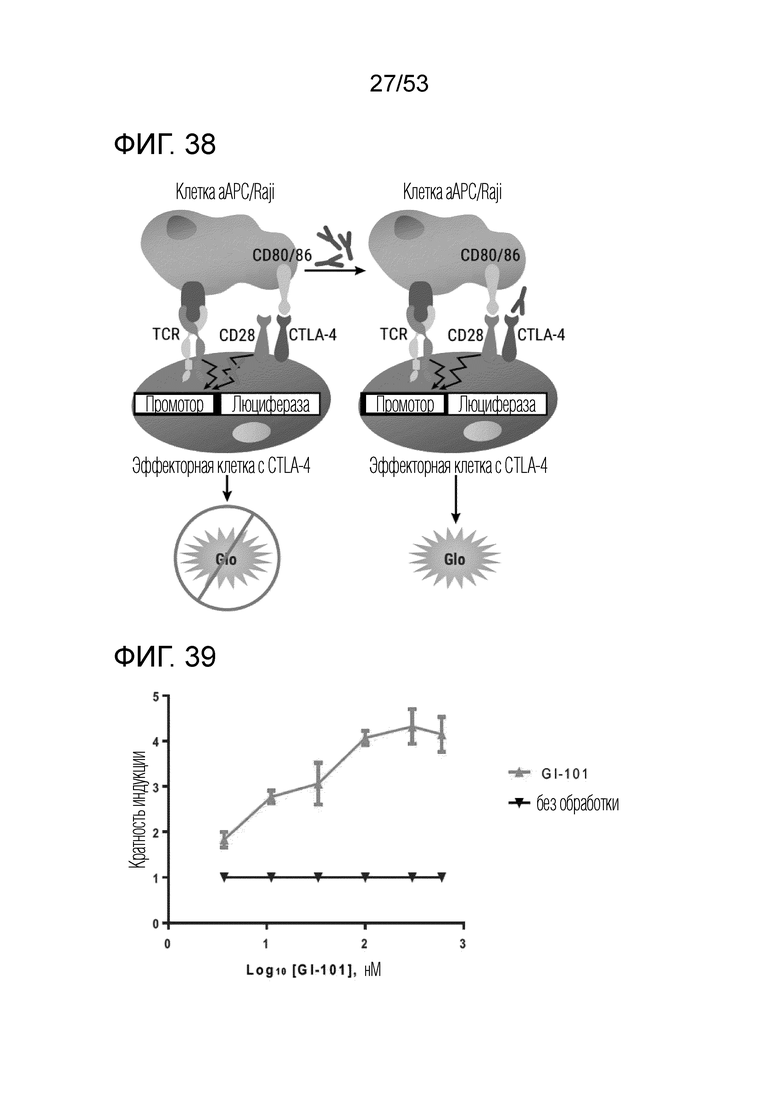
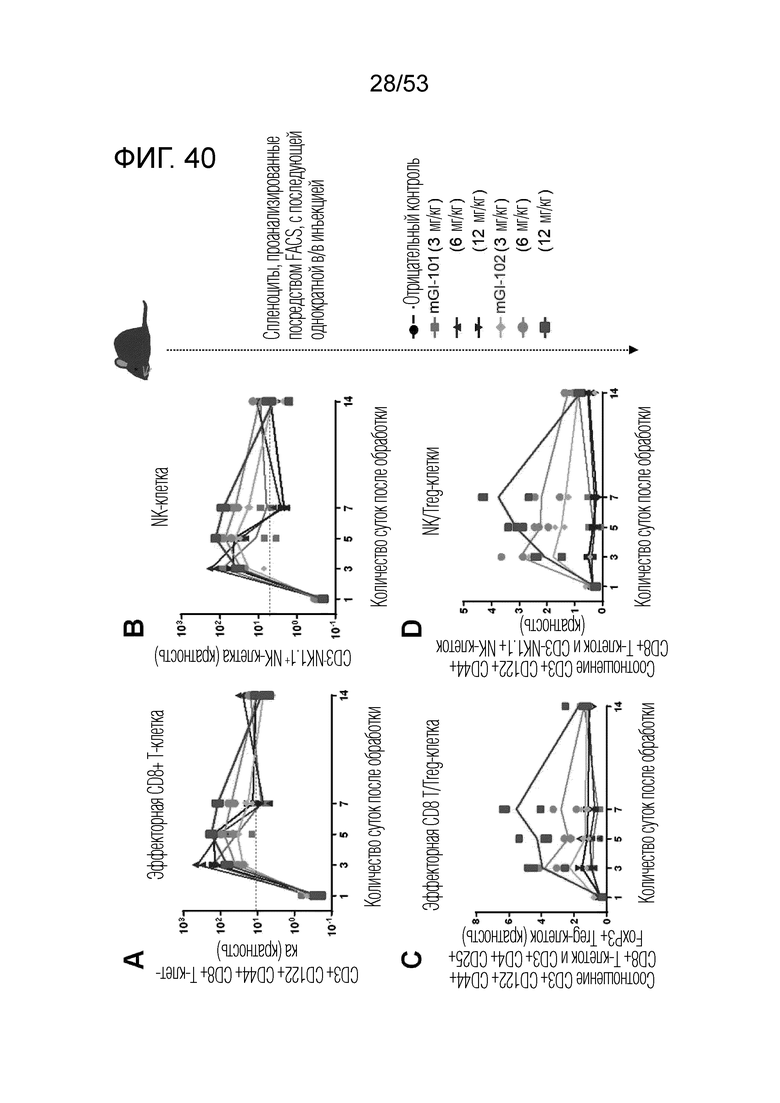
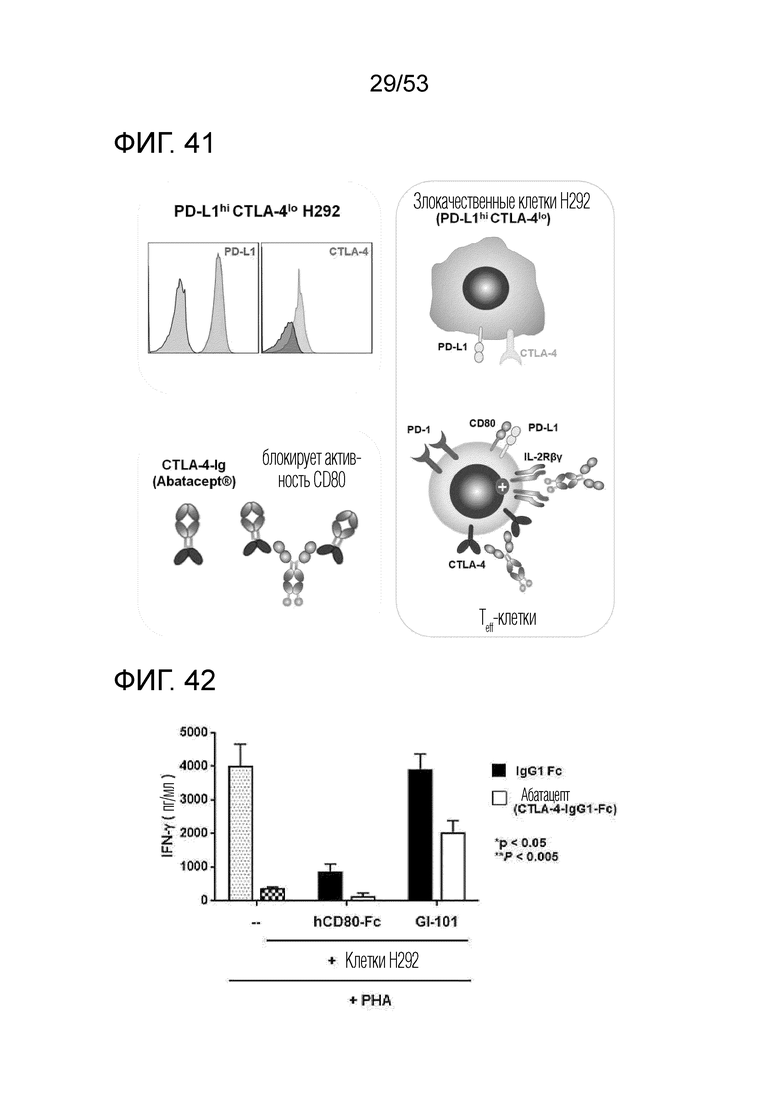
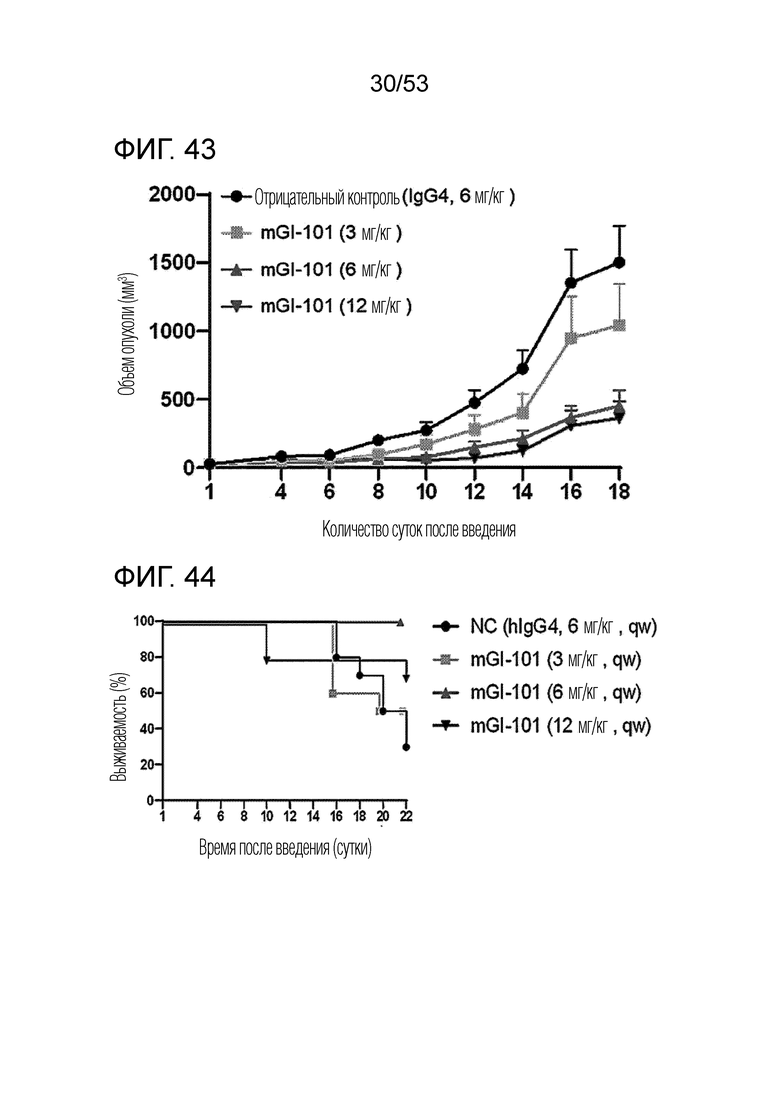
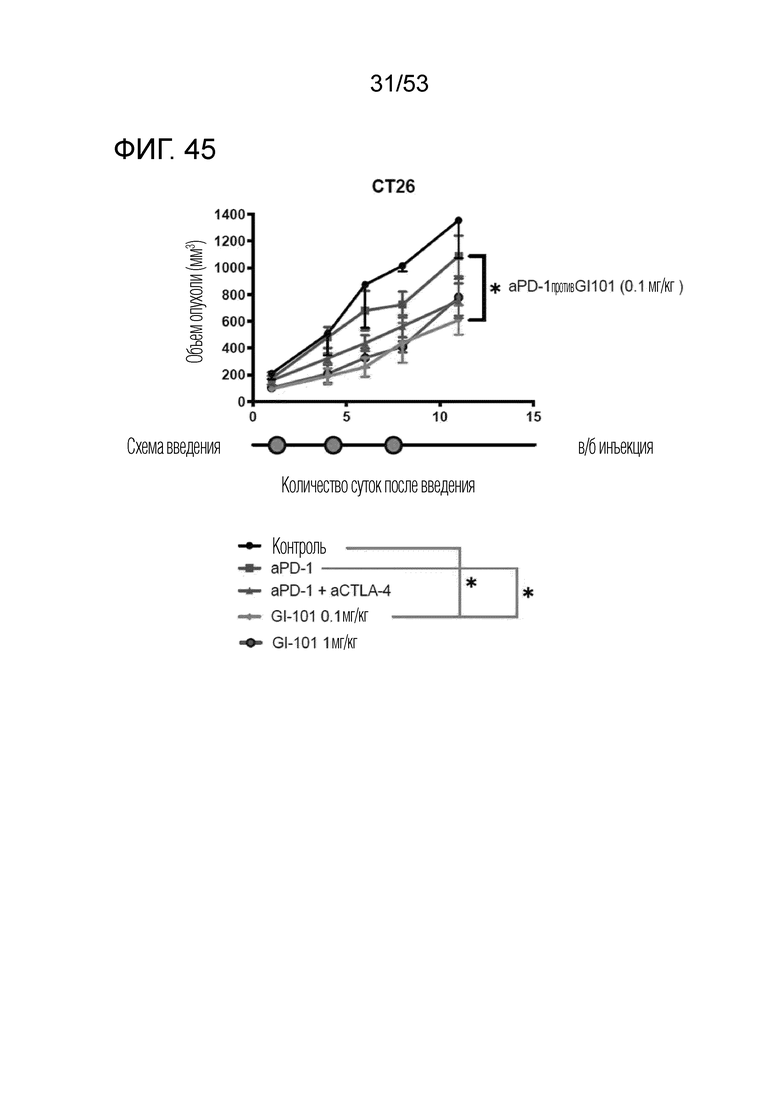
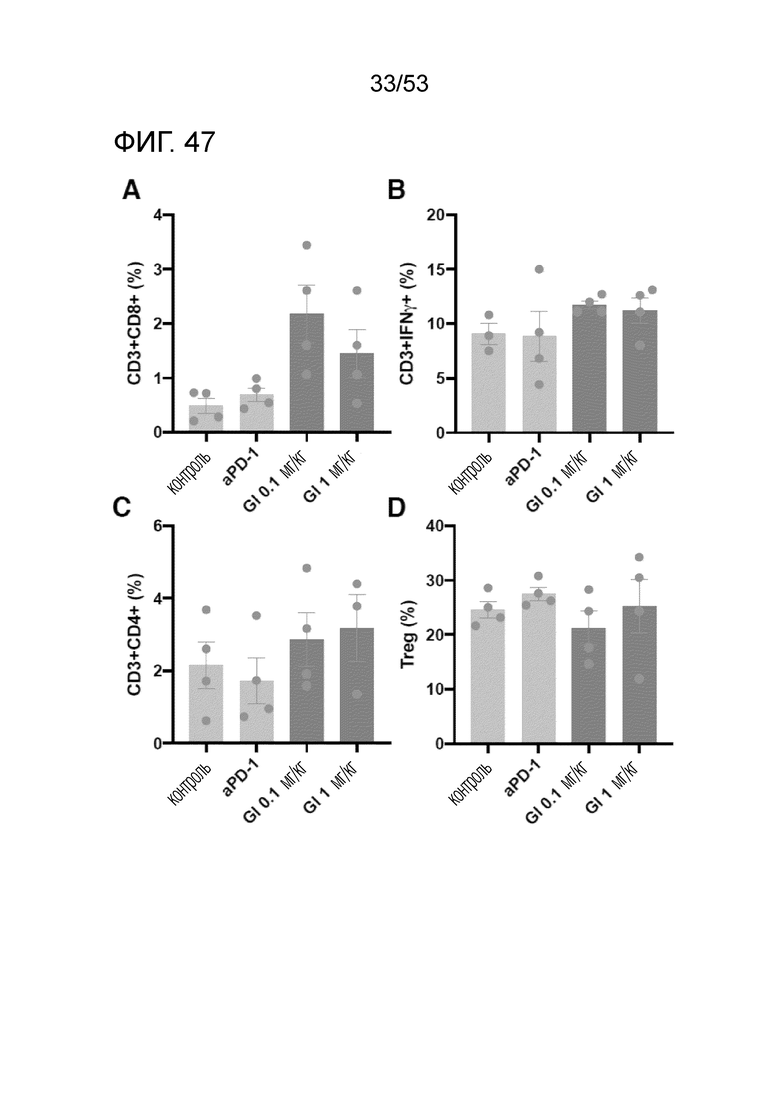
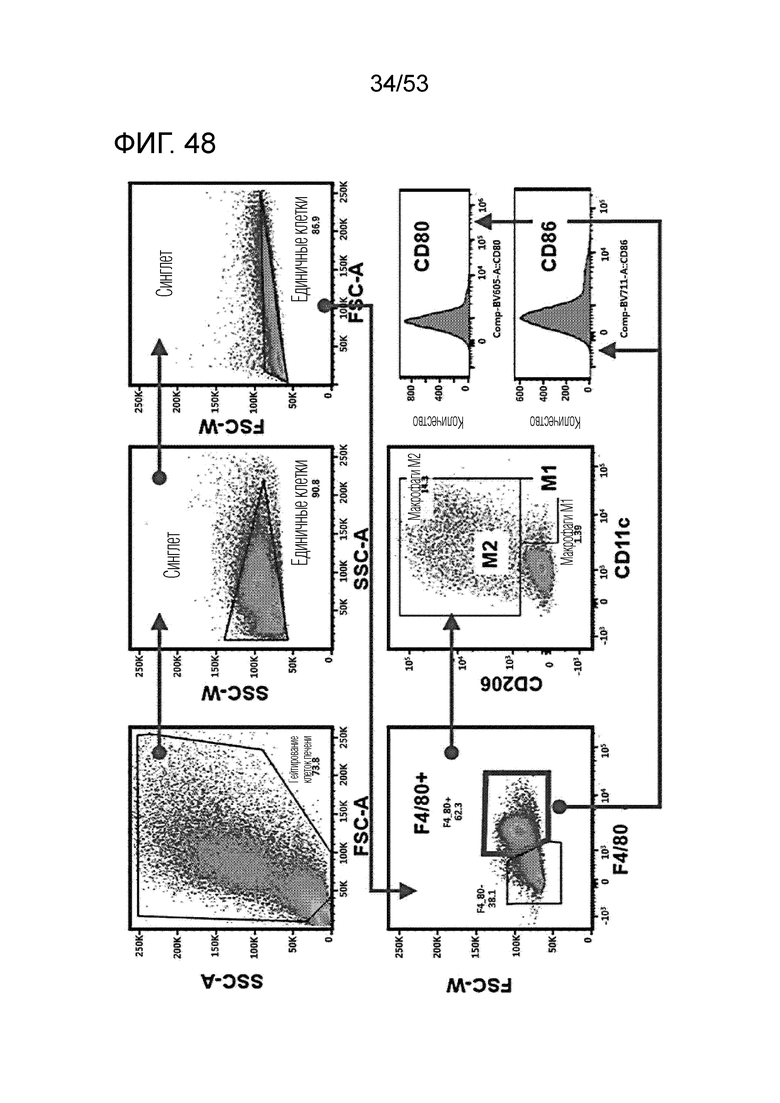
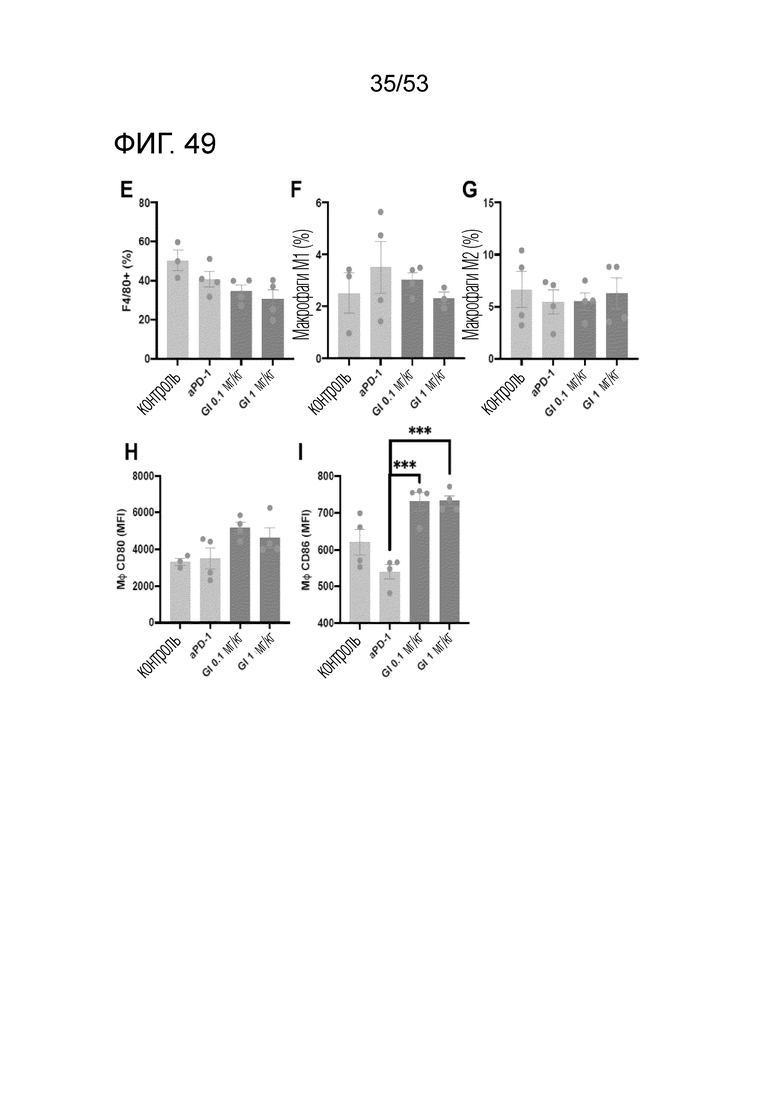
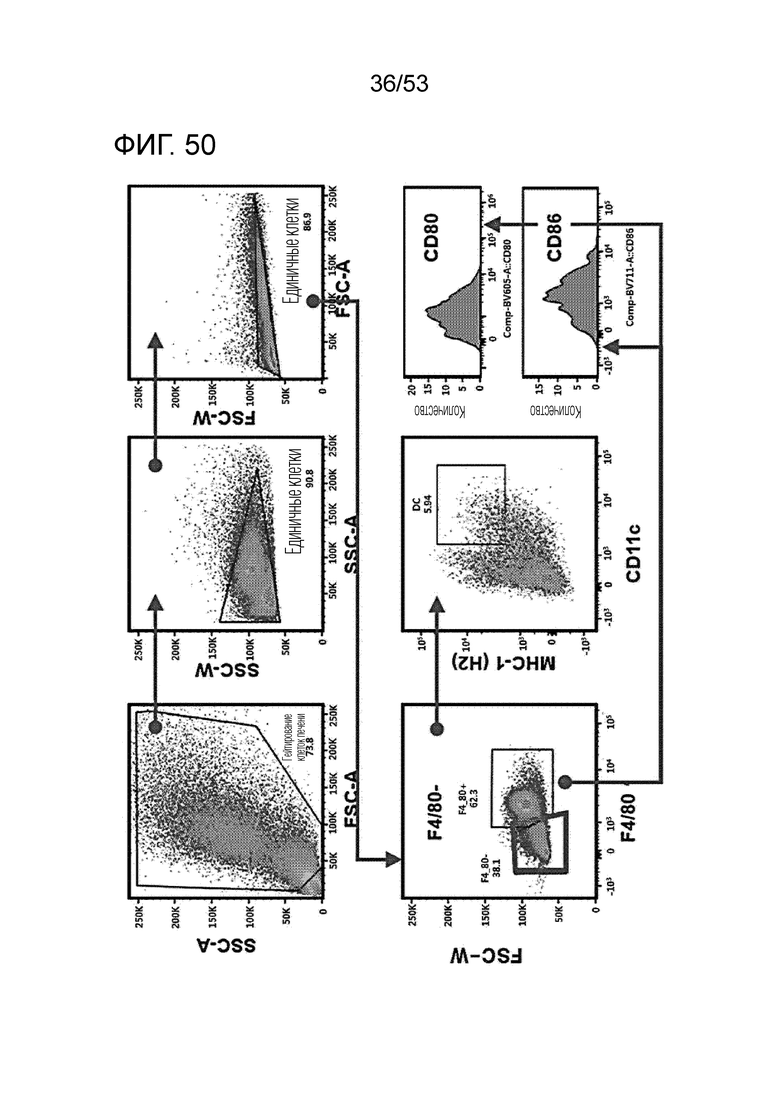
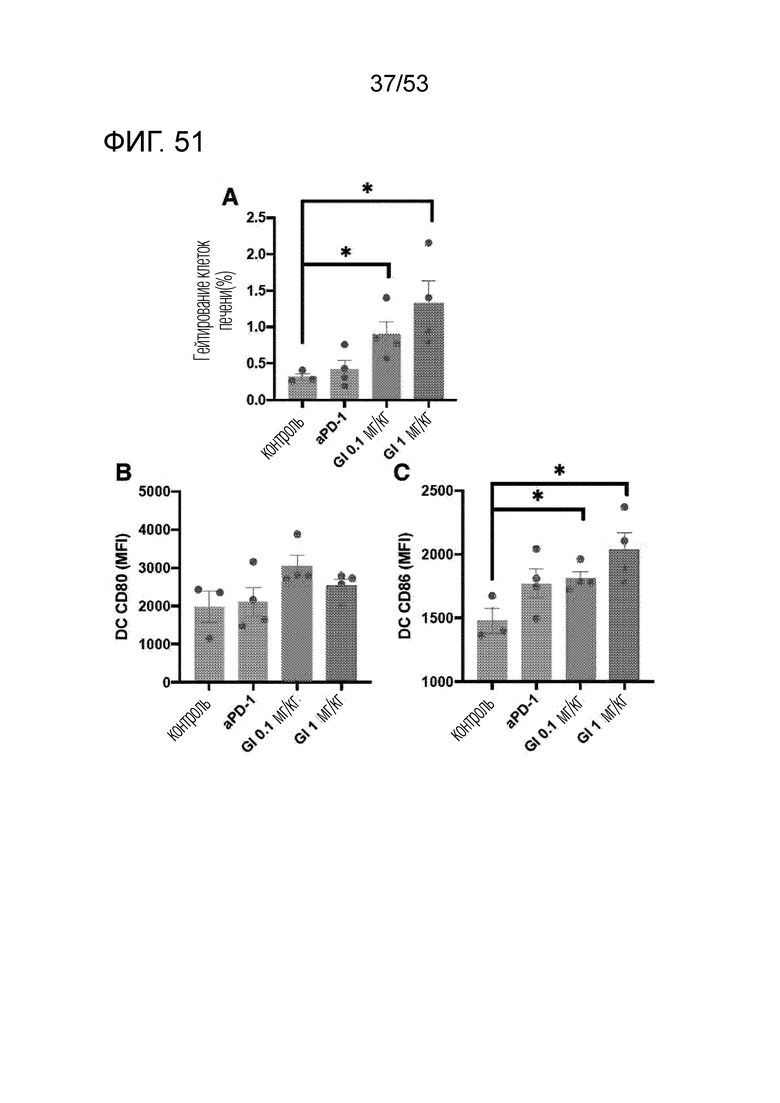
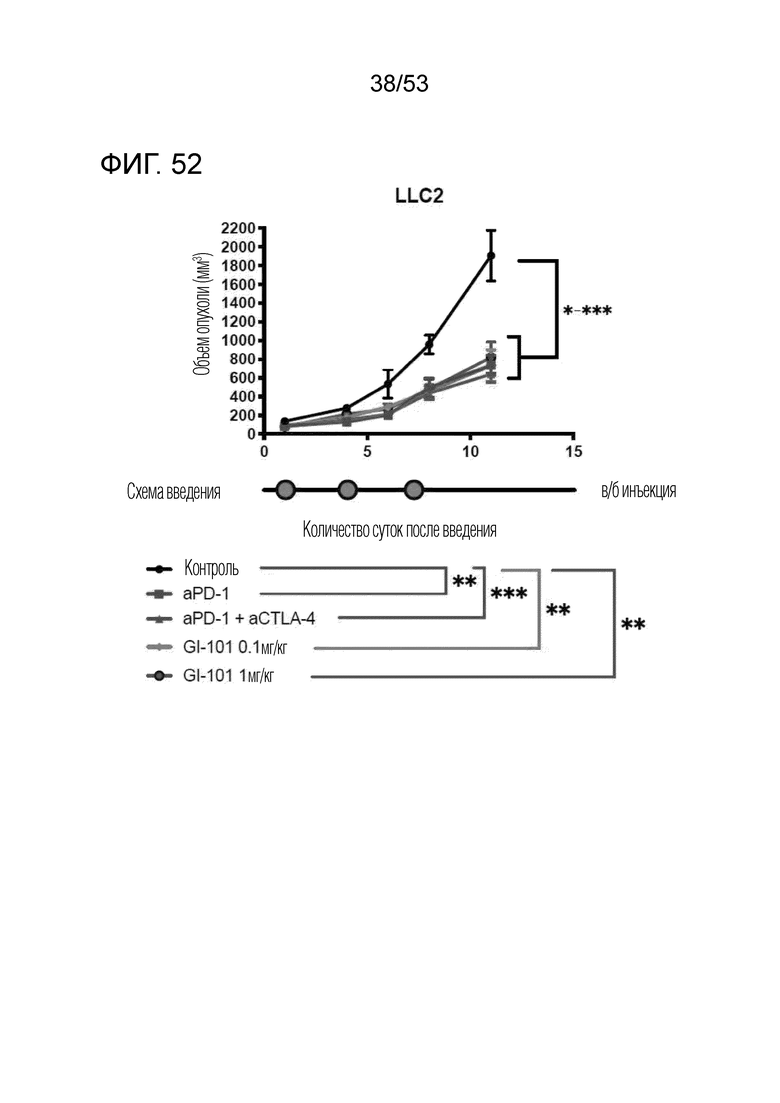
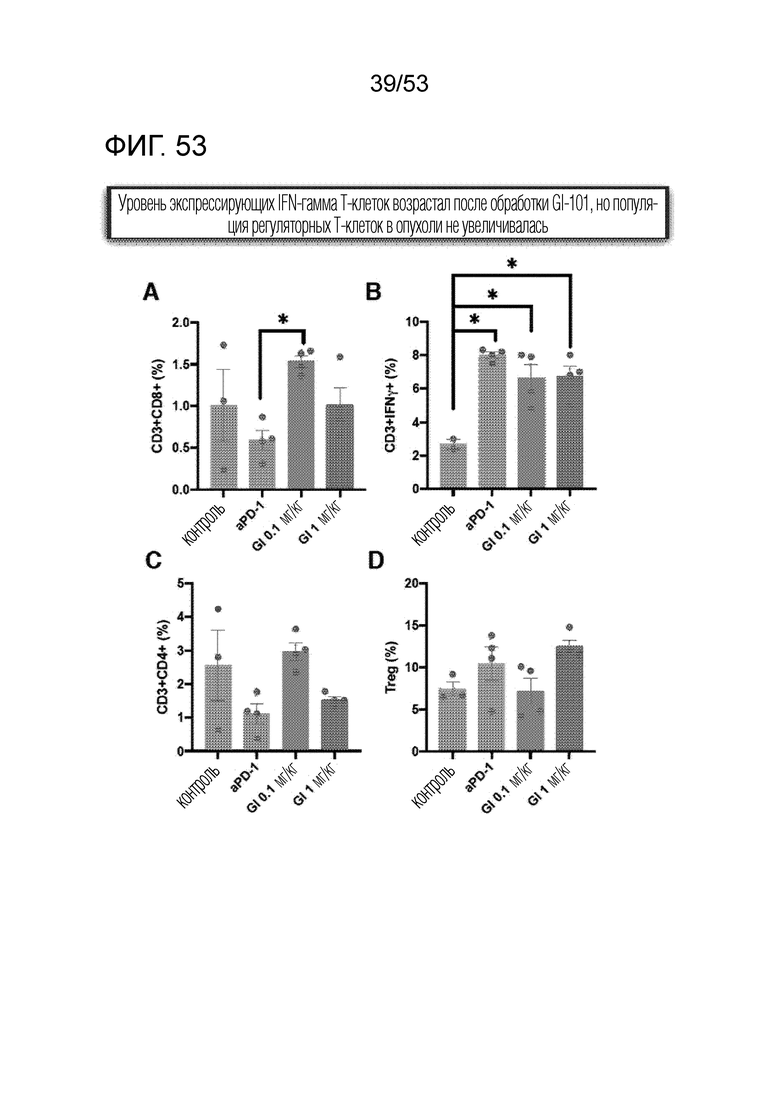
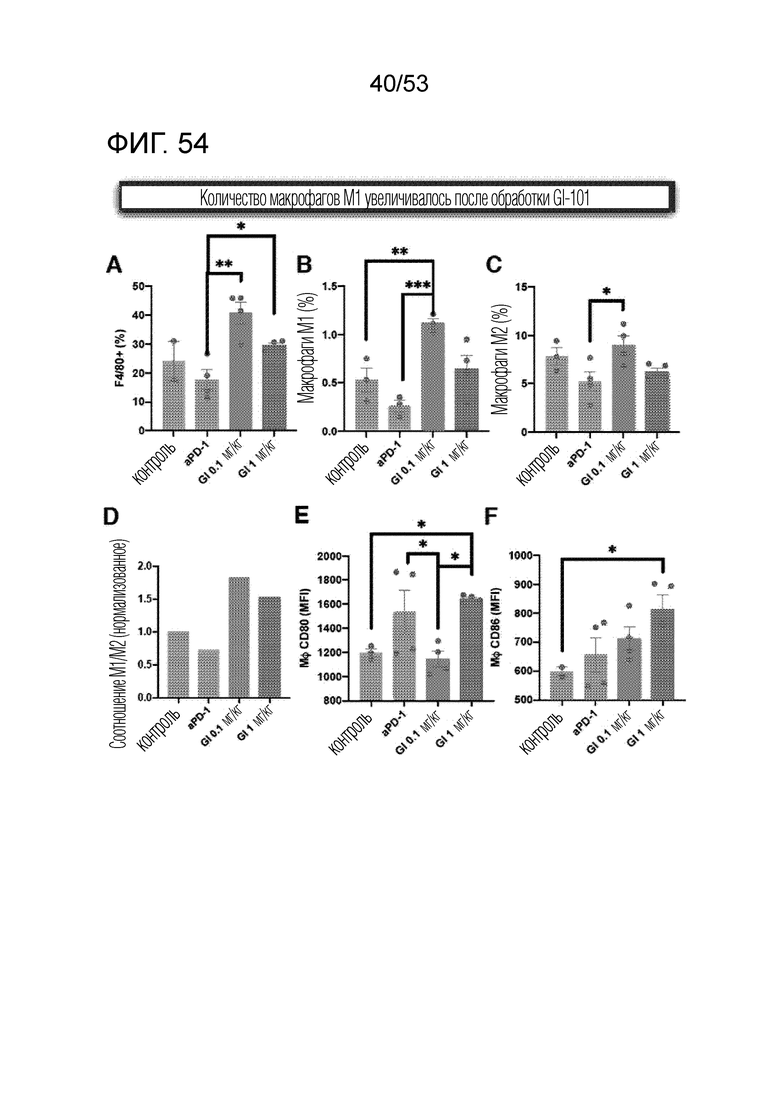
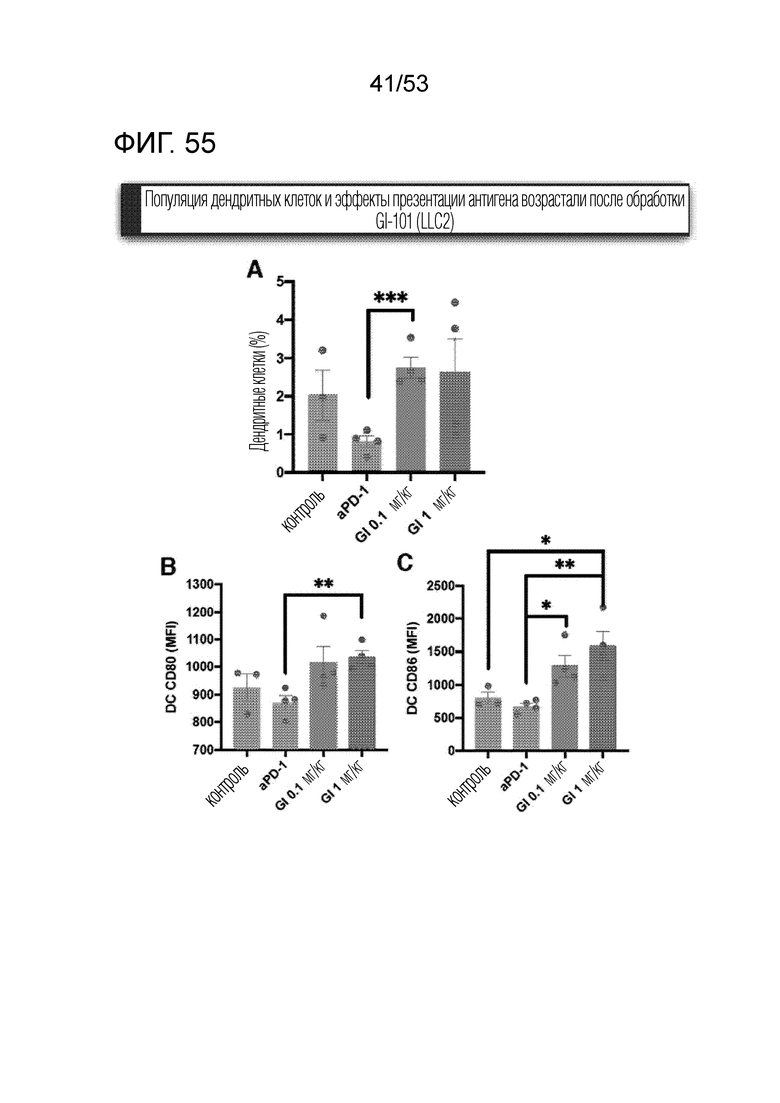
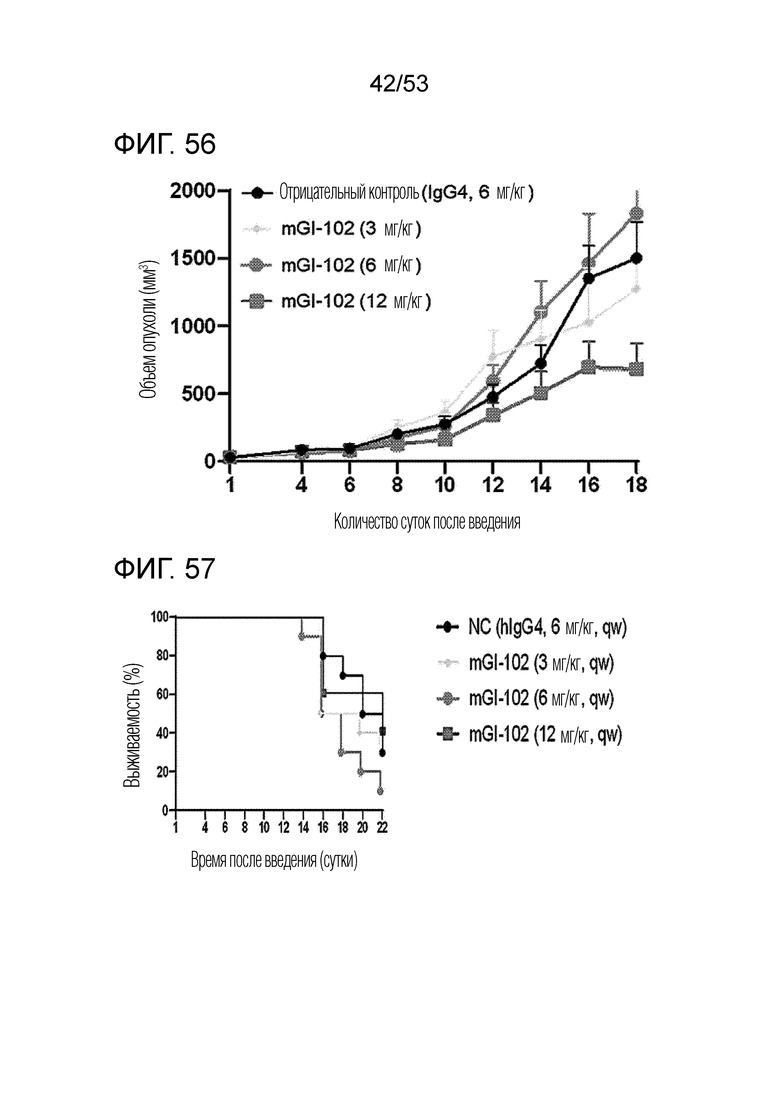
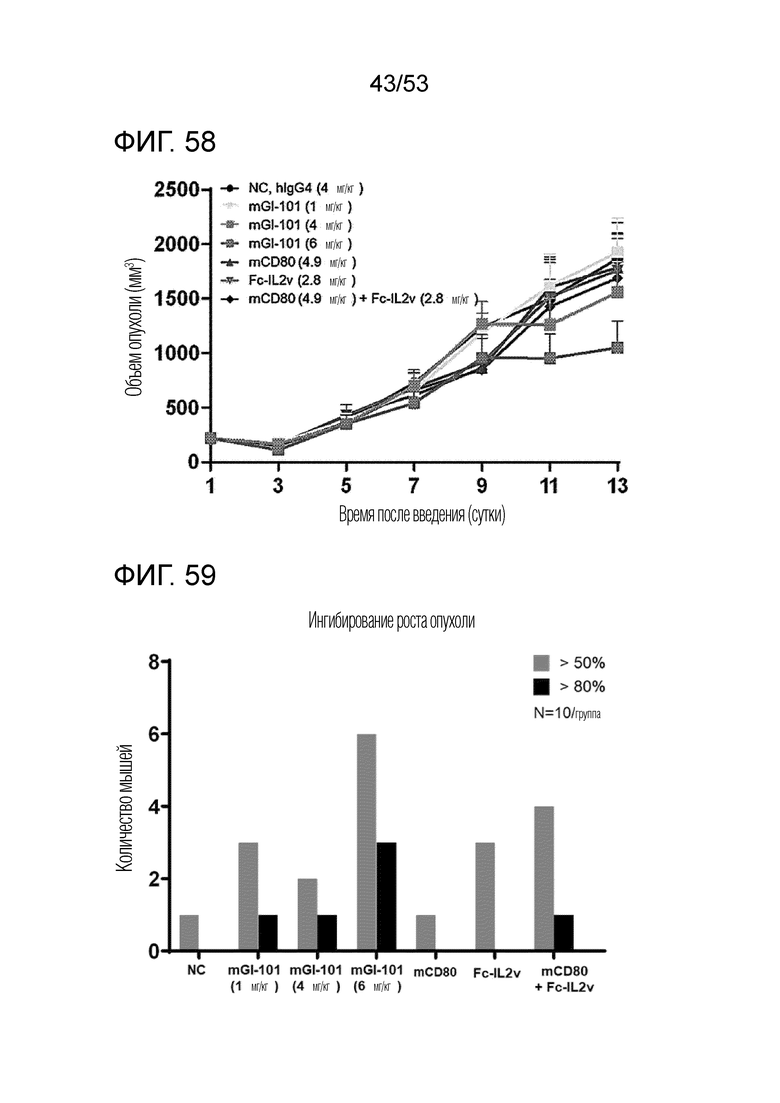
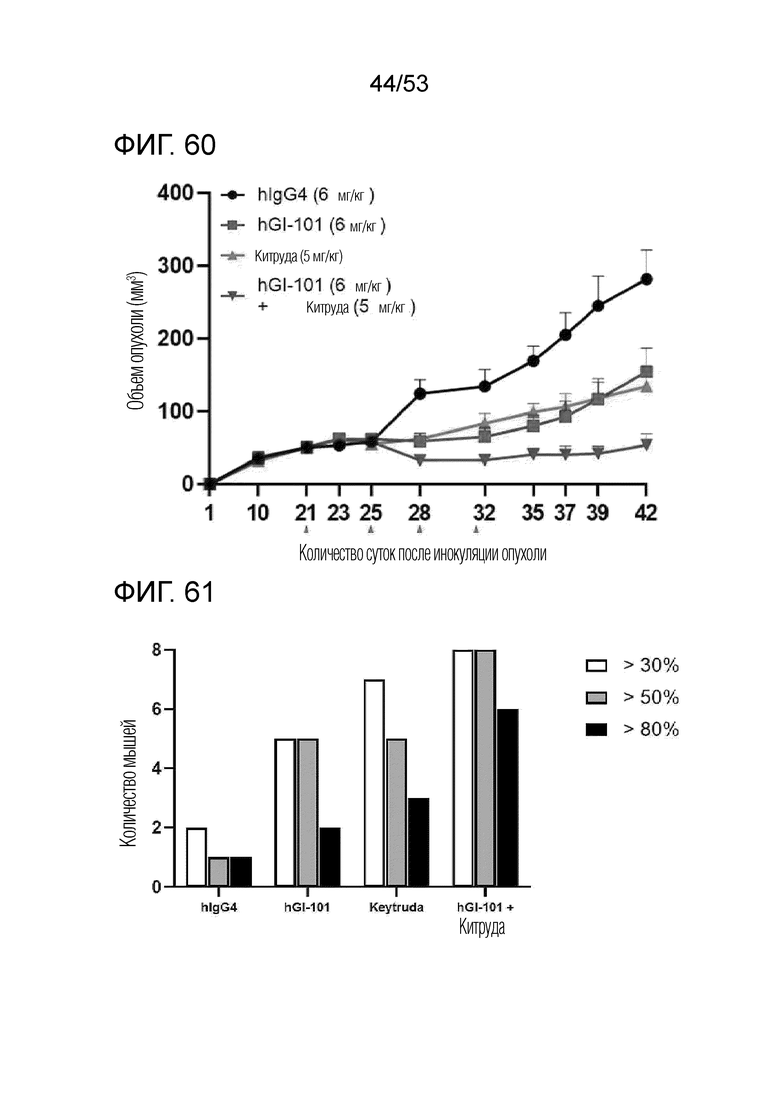
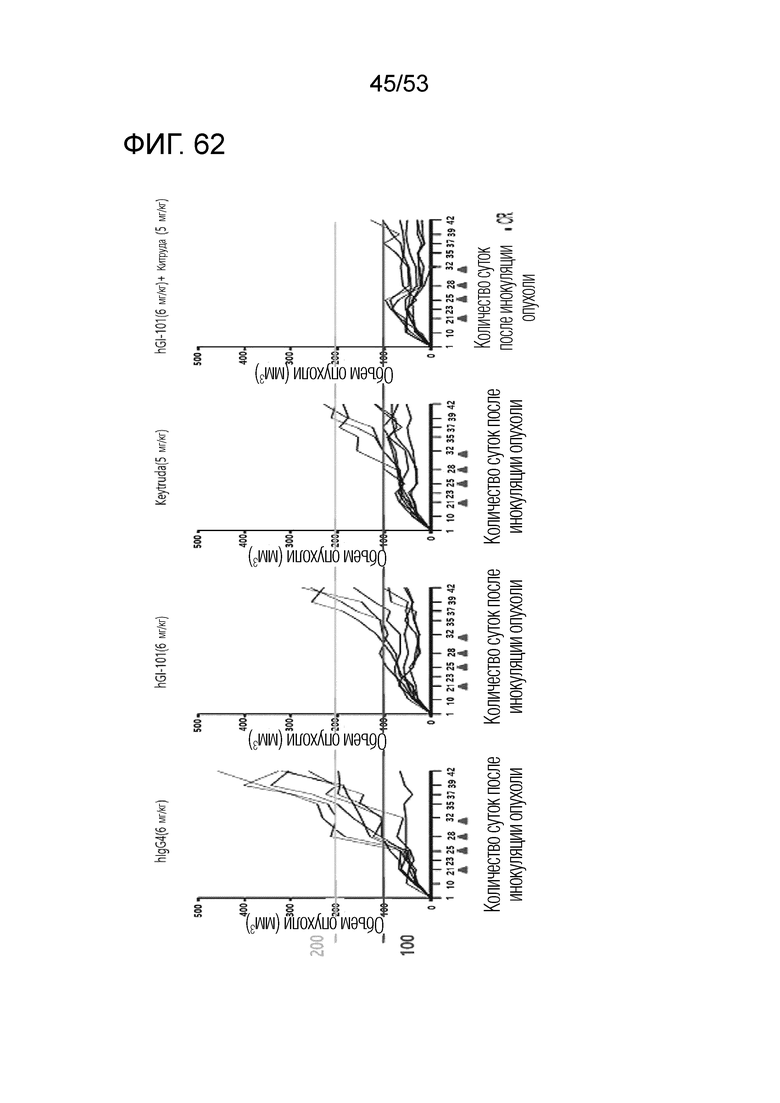
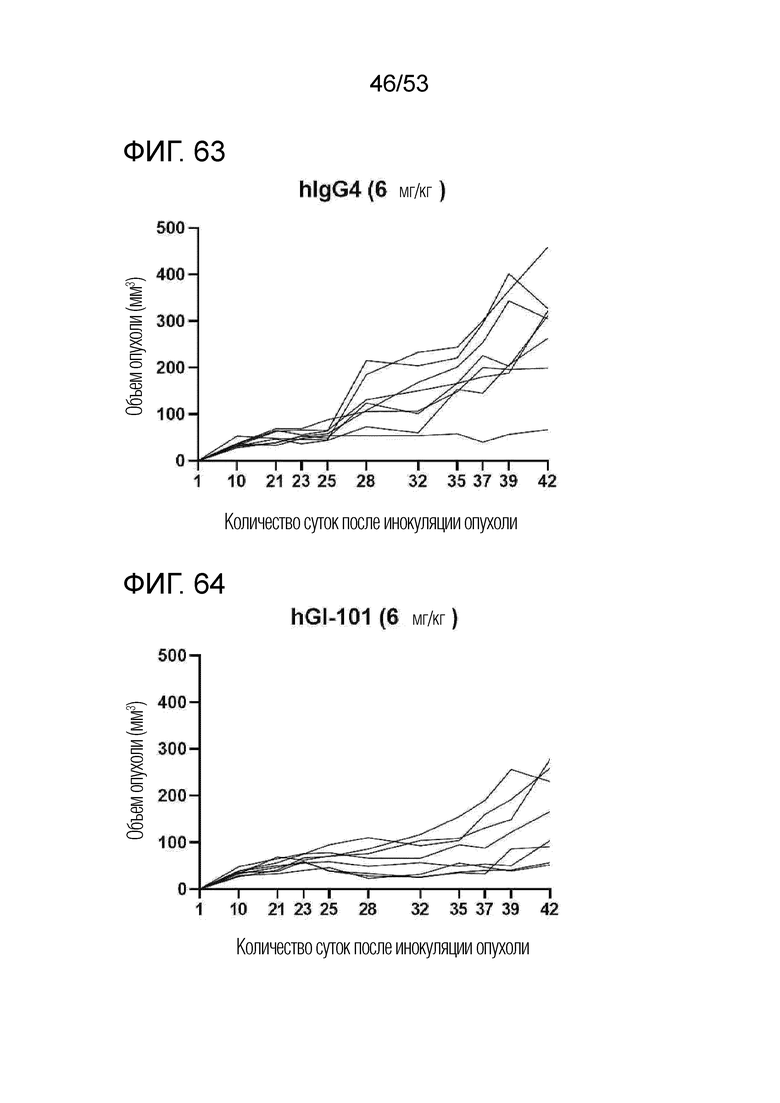
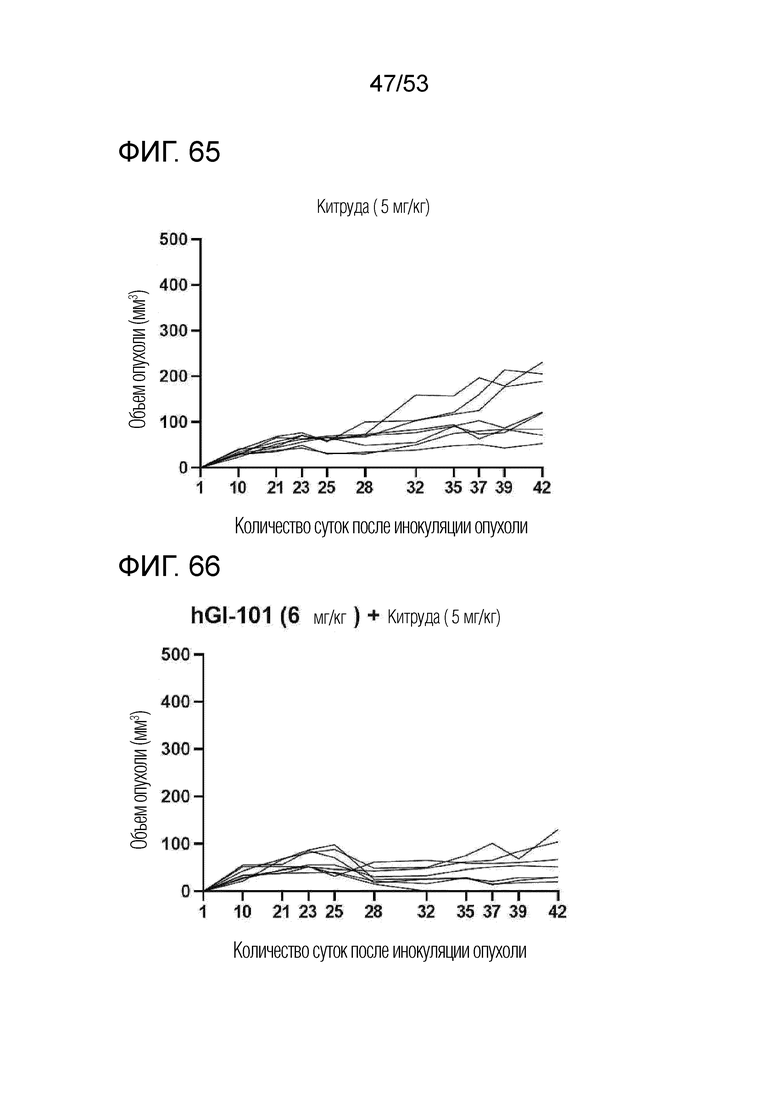
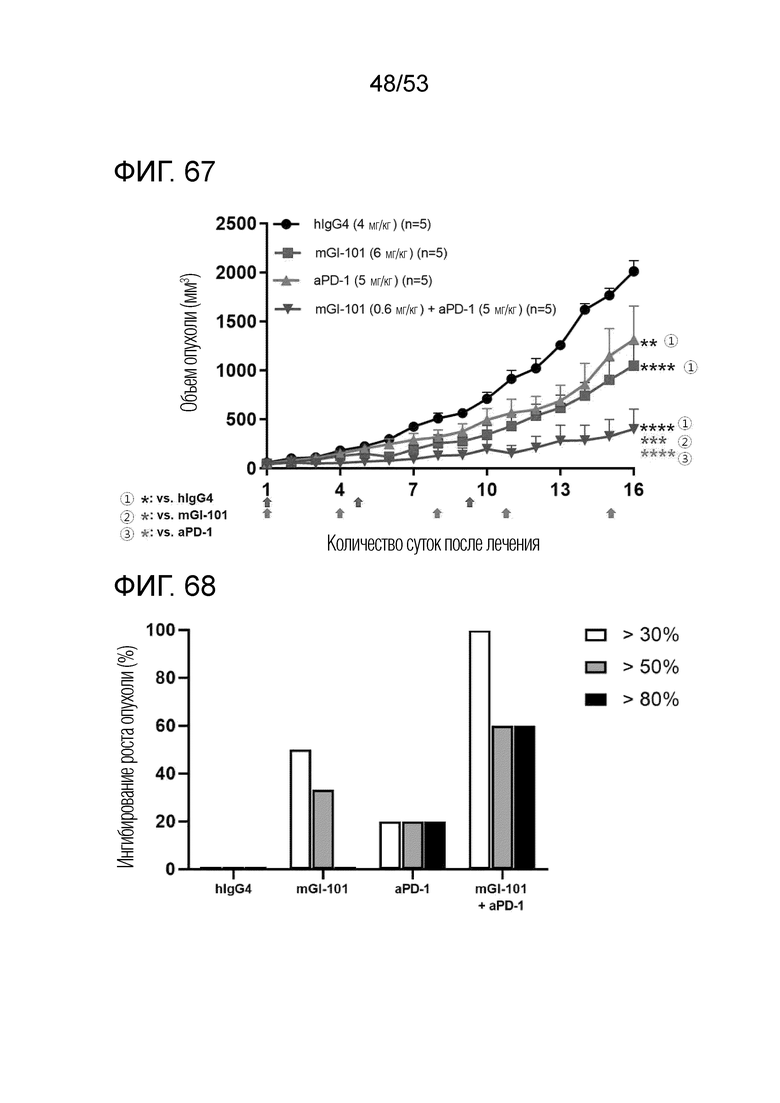
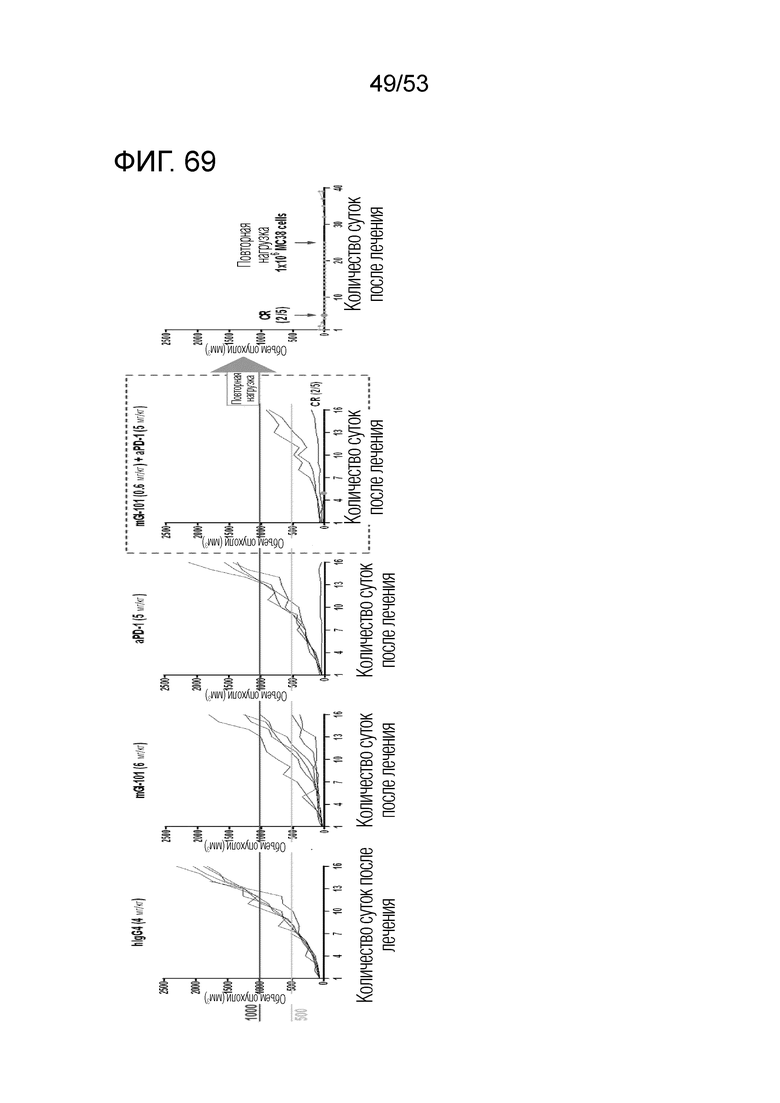
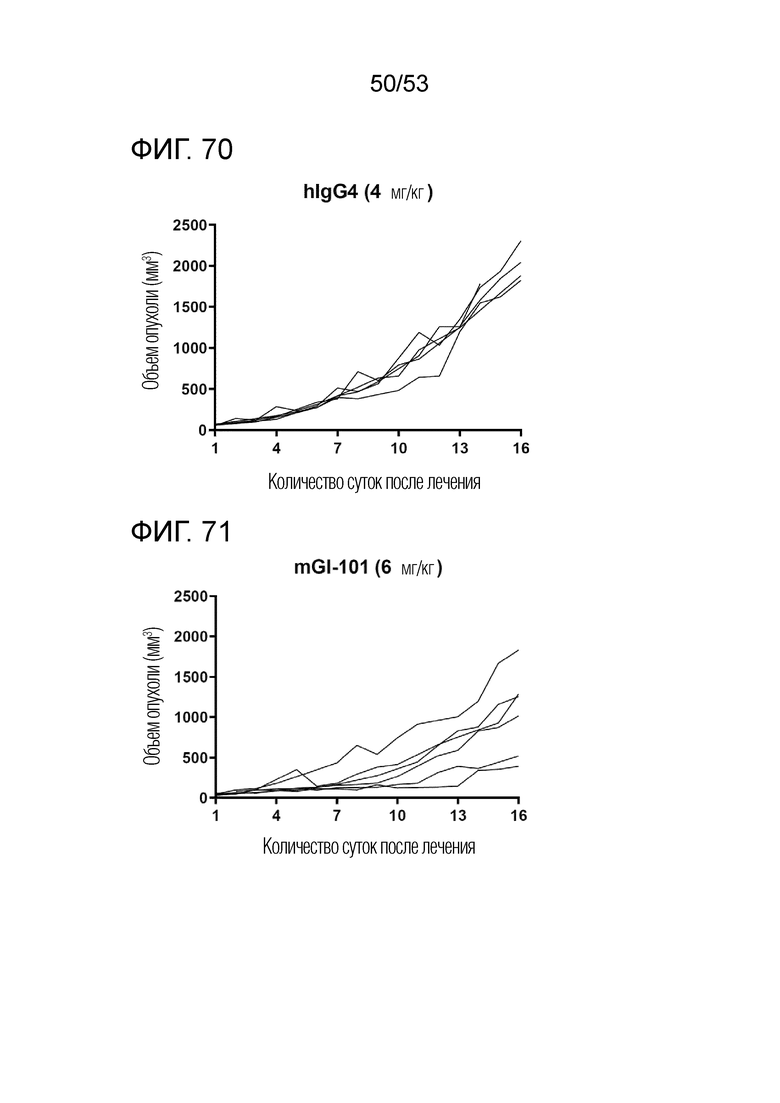
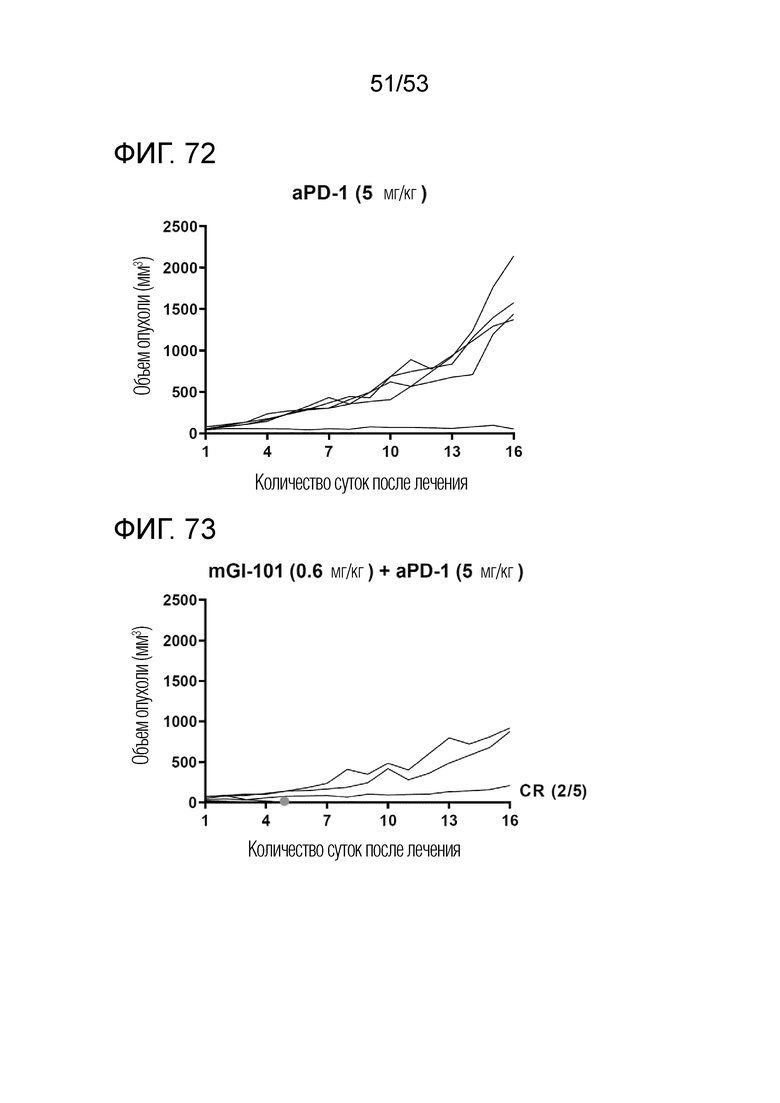
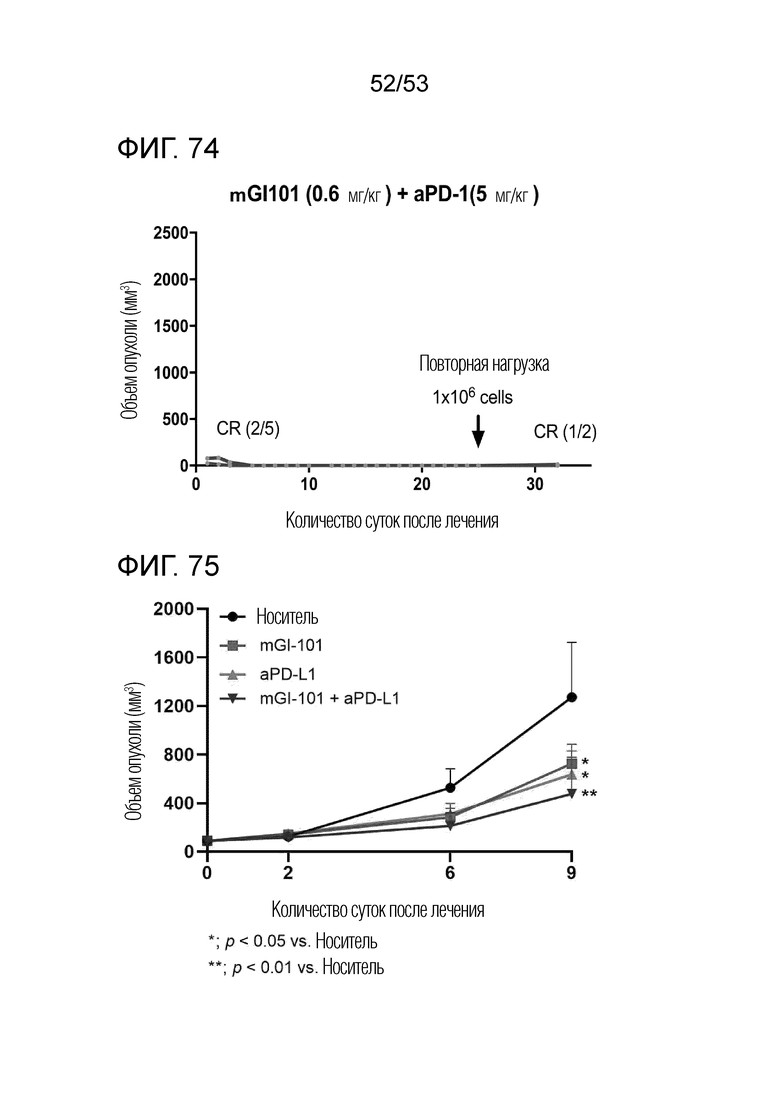
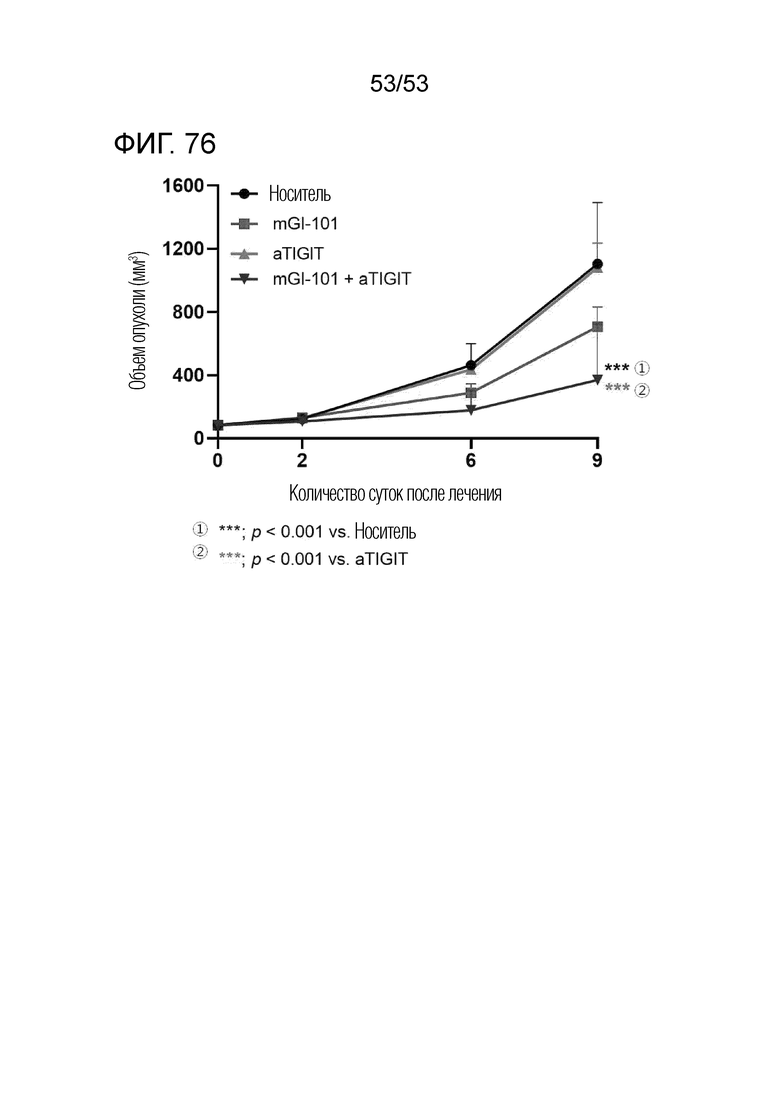
Группа изобретений относится к области фармацевтики и медицины. 1 объект представляет собой фармацевтическую композицию для предупреждения или лечения злокачественной опухоли, содержащую в качестве активных ингредиентов димерный слитый белок, содержащий вариант IL-2, содержащий аминокислотные замены R38A и F42A в SEQ ID NO: 10, и внеклеточный домен белка CD80, и ингибитор иммунной точки контроля, который выбирают из группы, состоящей из антитела против PD-1, антитела против PD-L1 и антитела против TIGIT. 2 объект – способ предупреждения или лечения злокачественной опухоли, включающий введение индивидууму, имеющему злокачественную опухоль, димерного слитого белка, содержащего вариант IL-2, содержащий аминокислотные замены R38A и F42A в SEQ ID NO: 10, и внеклеточный домен белка CD80, и ингибитора иммунной точки контроля, который выбирают из группы, состоящей из антитела против PD-1, антитела против PD-L1 и антитела против TIGIT. Технический результат заключается в синергетическом эффекте при введении димерного слитого белка в комбинации с ингибитором иммунной точки контроля. 2 н. и 4 з.п. ф-лы, 76 ил., 8 табл., 33 пр.
1. Фармацевтическая композиция для предупреждения или лечения злокачественной опухоли, содержащая в качестве активных ингредиентов димерный слитый белок, содержащий вариант IL-2 и внеклеточный домен белка CD80, и ингибитор иммунной точки контроля,
где слитый белок представлен следующей структурной формулой (I):
N'-X-[линкер (1)]n-Fc-домен-[линкер (2)]m-Y-C' (I), где
N' представляет собой N-конец слитого белка,
C' представляет собой C-конец слитого белка,
X представляет собой внеклеточный домен белка CD80,
Y представляет собой вариант IL-2, содержащий аминокислотные замены R38A и F42A в SEQ ID NO: 10,
линкер (1) представляет собой пептидный линкер, состоящий из 30-40 аминокислот,
линкер (2) представляет собой пептидный линкер, состоящий из 5-15 аминокислот,
n и m равны 1,
и где ингибитор иммунной точки контроля выбирают из группы, состоящей из антитела против PD-1, антитела против PD-L1 и антитела против TIGIT.
2. Фармацевтическая композиция для предупреждения или лечения злокачественной опухоли по п.1, где внеклеточный домен CD80 состоит из аминокислот с 35-й по 242-ю в SEQ ID NO: 11.
3. Фармацевтическая композиция для предупреждения или лечения злокачественной опухоли по п.1, где слитый белок имеет аминокислотную последовательность SEQ ID NO: 9.
4. Фармацевтическая композиция для предупреждения или лечения злокачественной опухоли по п.1, где
антитело против PD-1 представляет собой любое, выбранное из группы, состоящей из пембролизумаба, ниволумаба, цемиплимаба, JTX-4014, спартализумаба, камрелизумаба, синтилимаба, тислелизумаба, торипалимаба, достарлимаба, INCMGA00012, AMP-224 и AMP-514; или
антитело против PD-L1 представляет собой любое, выбранное из группы, состоящей из атезолизумаба, авелумаба, дурвалумаба, KN035, CK-301, AUNP12, CA-170 и BMS-986189.
5. Фармацевтическая композиция для предупреждения или лечения злокачественной опухоли по п.1, где злокачественная опухоль представляет собой любую, выбранную из группы, состоящей из рака желудка, рака печени, рака легкого, рака ободочной и прямой кишки, рака молочной железы, рака предстательной железы, рака яичника, рака поджелудочной железы, рака шейки матки, рака щитовидной железы, рака гортани, острого миелоидного лейкоза, опухоли головного мозга, нейробластомы, ретинобластомы, рака головы и шеи, рака слюнных желез и лимфомы.
6. Способ предупреждения или лечения злокачественной опухоли, включающий:
введение индивидууму, имеющему злокачественную опухоль, димерного слитого белка, содержащего вариант IL-2 и внеклеточный домен белка CD80, и ингибитора иммунной точки контроля,
где слитый белок представлен следующей структурной формулой (I):
N'-X-[линкер (1)]n-Fc-домен-[линкер (2)]m-Y-C' (I), где
N' представляет собой N-конец слитого белка,
C' представляет собой C-конец слитого белка,
X представляет собой внеклеточный домен белка CD80,
Y представляет собой вариант IL-2, содержащий аминокислотные замены R38A и F42A в SEQ ID NO: 10,
линкер (1) представляет собой пептидный линкер, состоящий из 30-40 аминокислот,
линкер (2) представляет собой пептидный линкер, состоящий из 5-15 аминокислот,
n и m равны 1,
и где ингибитор иммунной точки контроля выбирают из группы, состоящей из антитела против PD-1, антитела против PD-L1 и антитела против TIGIT.
| PARK J.C | |||
| et al | |||
| Приспособление для записи звуковых явлений на светочувствительной поверхности | 1919 |
|
SU101A1 |
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| - Vol | |||
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| - P | |||
| Способ обмыливания жиров и жирных масел | 1911 |
|
SU500A1 |
| US 2017216403 A1, 03.08.2017 | |||
| WO 2019191295 A1, 03.10.2019 | |||
| WO 2018201014 A1, 01.11.2018. | |||

