Данная заявка содержит "Перечень последовательностей", который был подан в электронном виде в формате ASCII и настоящим полностью включен в данную заявку посредством ссылки. Указанная копия ASCII, созданная 1 декабря 2015 года, имеет название 0966216, и ее размер составляет 20,0 байт.
Уровень техники
Биспецифические антитела способны связываться с двумя различными эпитопами и часто изготовляются на основе тяжелых и легких цепей иммуноглобулина двух разных моноспецифических исходных антител. Способность связываться с двумя различными эпитопами или антигенами делает биспецифические антитела привлекательным инструментом для терапевтических применений, в случае если имеется преимущество в нацеливании на более чем один антиген или эпитоп при лечении болезни. Однако эффективное получение биспецифических антител в формате, сходном с антителами естественного происхождения, может быть затруднено, поскольку в процессе эволюции тяжелые цепи антител приобрели свойство связывать легкие цепи антител относительно неспецифическим способом. В результате такого неспецифического спаривания одновременная экспрессия двух различных тяжелых цепей и двух различных легких цепей биспецифического антитела естественным образом приводит к случайной перетасовке спариваний "тяжелая цепь - легкая цепь". Такая перетасовка остается серьезной проблемой для создания биспецифических терапевтических средств, где однородное спаривание является необходимым условием хорошей технологичности и биологической эффективности.
Некоторые подходы были описаны для получения биспецифических антител в формате, сходном с антителами естественного происхождения. Однако эти подходы были разработаны и приведены в качестве примеров для случаев, когда оба исходных антитела, используемых для получения биспецифического антитела, имеют легкие цепи семейства генов каппа.
Несмотря на то, что большинство известных лекарственных средств на основе моноклональных антител (и, следовательно, потенциальных исходных антител) имеют легкие цепи каппа, присутствуют некоторые, которые имеют легкие цепи лямбда. Легкие цепи каппа и лямбда отличаются друг от друга как по структуре, так и по последовательности.
Обзор различных подходов для получения биспецифических антител в формате, сходном с антителами естественного происхождения из двух исходных антител может быть найден в Klein и соавт., (2012 год) мАт 4:6, 1-11. Международная заявка на патент № PCT/EP2011/056388 (WO 2011/131746) описывает способ получения in vitro гетеродимерного белка, в котором асимметрические мутации вводятся в области СН3 двух моноспецифических исходных белков для того, чтобы управлять направленным обменом "Fab-фрагментами" или "полумолекулами" между двумя моноспецифическими IgG4- или IgG4-подобными антителами, при инкубации в восстановительных условиях.
Публикация патента США №2009/0182127 (Novo Nordisk, Ink.) описывает синтез биспецифических антител путем модификации аминокислотных остатков в области контакта Fc и области контакта CH1:CL легких-тяжелых пар цепей, которые уменьшают способность легкой цепи одной пары взаимодействовать с тяжелой цепью другой пары. Международные патентные публикации № WO 2014/081955 (Amgen) и WO2014/150973 (Eli Lilly) описывают аминокислотные остатки в легкой цепи лямбда, которые потенциально могут быть модифицированы для обеспечения требуемой специфичности спаривания. Ни в одной из этих публикаций не описаны комплементарные аминокислотные модификации, которые могут быть использованы для получения биспецифического антитела из одного исходного антитела с легкой цепью каппа и другого исходного антитела с легкой цепью лямбда. Международная патентная публикация № WO2012/131555 (Glenmark) описывает замену области контакта между тяжелой цепью и легкой цепью лямбда антитела на область контакта домена РТК (рецептор Т-клетки).
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В данном описании предложены мультиспецифические антигенсвязывающие полипептиды, содержащие легкую цепь иммуноглобулина лямбда и легкую цепь иммуноглобулина лямбда. В одном аспекте антигенсвязывающий полипептид представляет собой конструкцию, содержащую первый гетеродимер и второй гетеродимер. В одном варианте реализации изобретения первый гетеродимер (H1L1) содержит полипептидную последовательность тяжелой цепи первого иммуноглобулина (H1) и полипептидную последовательность легкой цепи иммуноглобулина лямбда (L1), которые образуют первую область Fab, которая специфически связывается с первым антигеном; и второй гетеродимер (H2L2) содержит полипептидную последовательность тяжелой цепи второго иммуноглобулина (H2) и полипептидную последовательность легкой цепи иммуноглобулина каппа (L2), которые образуют вторую область Fab, которая специфически связывается со вторым антигеном. В некоторых вариантах реализации изобретения Н1 отличается от Н2. В некоторых вариантах реализации изобретения H1 и H2 содержат вариабельный домен тяжелой цепи (домен VH) и константный домен 1 тяжелой цепи (домен CH1). В одном варианте реализации изобретения L1 содержит вариабельный домен легкой цепи лямбда (VL-лямбда) и константный домен легкой цепи лямбда (CL-лямбда). В одном варианте реализации изобретения L2 содержит вариабельный домен легкой цепи каппа (VL-каппа) и константный домен легкой цепи каппа (CL-каппа). В некоторых вариантах реализации изобретения одна или более H1, H2, L1 и L2 содержат аминокислотные модификации по сравнению с соответствующими полипептидными последовательностями H1, H2, L1 и L2 дикого типа, причем аминокислотные модификации способствуют преимущественному спариванию H1 с L1 по сравнению с L2, и/или способствуют преимущественному спариванию H2 с L2 по сравнению с L1. В некоторых вариантах реализации изобретения аминокислотные модификации не вносят новый остаток цистеина. В некоторых вариантах реализации изобретения аминокислотные модификации не удаляют остаток цистеина, который встречается в природе.
В некоторых вариантах реализации изобретения конструкция содержит аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и/или которые способствуют преимущественному спариванию H2 с L2 по сравнению с L1, в случае если H1, H2, L1 и L2 коэкспрессируются в клетке или клетке млекопитающего, или в случае если H1, H2, L1 и L2 коэкспрессируются в системе бесклеточной экспрессии, или в случае если H1 и L1 продуцируются в (первой) клетке, а H2 и L2 продуцируются во второй (например, другой) клетке и продукты двух клеток смешиваются по окислительно-восстановительному способу производства, или в случае если H1 и L1 продуцируются в первой системе бесклеточной экспрессии, а H2 и L2 продуцируются во второй (например, другой) системе бесклеточной экспрессии и продукты двух систем бесклеточной экспрессии смешиваются.
В некоторых вариантах реализации конструкции, каждый гетеродимер содержит один Fab.
В некоторых вариантах реализации антигенсвязывающих полипептидных конструкций, описанных в данном документе:
a. H2 содержит аминокислотную замену в положении 143; L2 содержит аминокислотную замену в положении 124; а также
i. H1 содержит аминокислотную замену в положении 186 или 179, а L1 содержит аминокислотную замену в положении 180;
ii. H1 содержит аминокислотную замену в положении 186; а L1 содержит аминокислотные замены в положении 133;
iii. H1 содержит аминокислотную замену в положении 143; а L1 содержит аминокислотные замены в положении 133; или
iv. H1 содержит аминокислотную замену в положении 188; а L1 содержит аминокислотные замены в положении 178;
b. H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 131; а также
i. H2 содержит аминокислотную замену в положениях 186 или 124 и 186, а L2 содержит аминокислотную замену в положениях 133 или 133 и 160, или 124 и 133, или 176 и 180; или
ii. H2 содержит аминокислотную замену в положении 188; а L2 содержит аминокислотные замены в положении 131;
iii. H2 содержит аминокислотную замену в положении 143; а L2 содержит аминокислотные замены в положениях 124 и 133, или 124 и 133 и 180;
c. H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 131; а также
i. H2 содержит аминокислотную замену в положениях 124 или 186, или 124 и 179, или 188, а L2 содержит аминокислотную замену в положениях 176 и 178, или 176 и 180, или 131; или
ii. H2 содержит аминокислотную замену в положениях 143 и 188, или 143, или 124 и 143; а L2 содержит аминокислотные замены в положениях 124 и 176 и 178, или 124 и 178, или 124 и 180, или 124 и 176 и 180, или 124, или 124 и 176;
d. H1 содержит аминокислотную замену в положении 179, 186, 143 и/или 188; L1 содержит аминокислотную замену в положении 180, 133 и/или 176 и 178; H2 содержит аминокислотную замену в положении 143, а L2 содержит аминокислотную замену в положении 131 и/или 124;
e. H1 содержит аминокислотную замену в положении 39 или не содержит аминокислотных замен, которые способствуют преимущественному спариванию; L1 содержит аминокислотную замену в положении 38 или не содержит аминокислотных замен, которые способствуют преимущественному спариванию; Н2 содержит аминокислотную замену в положении 39 и L2 содержит аминокислотную замену в положении 38;
f. H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 131; а также
i. H2 содержит аминокислотную замену в положениях 188 или 124 и 186, и L2 содержит аминокислотную замену в положениях 176 и 178, или 176 и 180, или 131; или
ii. H2 содержит аминокислотную замену в положении 143 или 186; а L2 содержит аминокислотные замены в положениях 124 и 133, или 124 и 133 и 180;
g. H1 содержит аминокислотную замену в положении 188; L1 содержит аминокислотную замену в положениях 176 и 178, или 178; и
i. H2 содержит аминокислотную замену в положениях 177 и 188, и L2 содержит аминокислотную замену в положениях 176 и 178; или
ii. H2 содержит аминокислотную замену в положении 186 или 124 или 124 и 179; и L2 содержит аминокислотные замены в положениях 176, или 131 и 176;
h. H1 содержит аминокислотную замену в положении 186; L1 содержит аминокислотную замену в положении 133; а также
i. H2 содержит аминокислотную замену в положении 188, а L2 содержит аминокислотную замену в положении 131; или
ii. H2 содержит аминокислотную замену в положениях 177 и 188; и L2 содержит аминокислотные замены в положениях 176 и 178; или
H1 содержит аминокислотную замену в положениях 124 и 190; L1 содержит аминокислотную замену в положении 135; Н2 содержит аминокислотную замену в положениях 124 или 188, и L2 содержит аминокислотную замену в положениях 176, или 176 и 178;
i. H1 содержит аминокислотную замену в положениях 177 и 188; L1 содержит аминокислотную замену в положении 176 и 178; и
a. Н2 содержит аминокислотную замену в положении 188, и L2 содержит аминокислотную замену в положениях 176 и 178, или 131;
b. Н2 содержит аминокислотную замену в положении 186; и L2 содержит аминокислотные замены в положениях 133, или 124 и 160, и 180;
c. Н2 содержит аминокислотную замену в положении 124, или 124 и 179, или 124 и 186; и L2 содержит аминокислотные замены в положениях 176, или 176 и 178, или 176 и 180; или
d. H2 содержит аминокислотную замену в положении 143; а L2 содержит аминокислотные замены в положениях 133, или 124 и 133;
j. H1 содержит аминокислотную замену в положении 188; L1 содержит аминокислотную замену в положении 178; Н2 содержит аминокислотную замену в положении 124 или 188, и L2 содержит аминокислотную замену в положениях 176 и 178, или 176 и 180, или 176;
k. или H1 содержит аминокислотную замену в положениях 145 и 188; L1 содержит аминокислотную замену в положении 178; и Н2 содержит аминокислотную замену в положении 124 и/или 188, и L2 содержит аминокислотную замену в одном или более положениях 124, 133 и 178;
l. H1 содержит аминокислотную замену в положении 174, 179 или 186; L1 содержит аминокислотную замену в положении 176 или 180; Н2 содержит аминокислотную замену в положении 143 или 190, и L2 содержит аминокислотную замену в положении 131, 135 или 124; или
m. H1 содержит аминокислотную замену в положении 174; L1 содержит аминокислотную замену в положении 176; Н2 содержит аминокислотную замену в положении 190; и L2 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положении 135;
n. H1 содержит аминокислотную замену в положениях 143 и 190; L1 содержит аминокислотную замену в положении 133; Н2 содержит аминокислотную замену в положении 124, и L2 содержит аминокислотную замену в положениях 131 и 135;
o. H1 содержит аминокислотную замену в положении 143 и/или 186; L1 содержит аминокислотную замену в положении 133; Н2 содержит аминокислотную замену в положении 124, и L2 содержит аминокислотную замену в положении 131;
p. H1 содержит аминокислотную замену в положениях 143 и 179; L1 содержит аминокислотную замену в положениях 124 и 178; Н2 содержит аминокислотную замену в положении 186, и L2 содержит аминокислотную замену в положениях 178 и 180, или 160 и 180;
q. H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 124; Н2 содержит аминокислотную замену в положении 179 или 186, и L2 содержит аминокислотную замену в положении 124 и 160 и 180;
r. H1 содержит аминокислотную замену в положении 186; L1 содержит аминокислотную замену в положениях 180 или 178 и 180; Н2 содержит аминокислотную замену в положении 143 и/или 179, и L2 содержит аминокислотную замену в положениях 124 и 178, или 131;
s. H1 содержит аминокислотную замену в положении 179; L1 содержит аминокислотную замену в положении 180; Н2 содержит аминокислотную замену в положении 143, и L2 содержит аминокислотную замену в положении 124;
t. H1 содержит аминокислотную замену в положении 143 или 186; L1 содержит аминокислотную замену в положении 180 или не содержит аминокислотных замен, которые способствуют преимущественному спариванию; Н2 содержит аминокислотную замену в положениях 143 и 145, и L2 содержит аминокислотную замену в положении 124;
u. H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию; L1 содержит аминокислотную замену в положении 135; Н2 содержит аминокислотную замену в положении 139, и L2 содержит аминокислотную замену в положении 116;
v. H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положении 45; L1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию; Н2 содержит аминокислотную замену в положении 45, и L2 содержит аминокислотную замену в положении 44;
w. H1 содержит аминокислотную замену в положении 139; L1 содержит аминокислотную замену в положении 116; Н2 не содержит аминокислотных замен, которые способствуют преимущественному спариванию, и L2 содержит аминокислотную замену в положении 135; или
x. H1 содержит аминокислотную замену в положении 124; L1 содержит аминокислотную замену в положении 176; Н2 содержит аминокислотную замену в положении 124, и L2 содержит аминокислотную замену в положении 176.
В некоторых вариантах реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 25, 30, 35, 40, 45, 50 или 100-кратной-кратной аффинности области Fab, образованной соответствующими полипептидными последовательностями дикого типа H1 и L1 к первому антигену, и/или аффинность второй области Fab ко второму антигену находится в пределах около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 25, 30, 35, 40, 45, 50 или 100-кратной аффинности области Fab, образованной соответствующими полипептидными последовательностями дикого типа H2 и L2 ко второму антигену.
В некоторых вариантах реализации изобретения температура плавления (Tm) первой области Fab находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 20°C Tm области Fab, образованной соответствующими полипептидными последовательностями дикого типа H1 и L1 для первого антигена, и/или температура плавления (Tm) второй области Fab находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 20°C Tm области Fab, образованной соответствующими полипептидными последовательностями дикого типа H2 и L2 для второго антигена.
В некоторых вариантах реализации антигенсвязывающей полипептидной конструкции:
a. H1 и L1 представляют собой полипептидные последовательности дикого типа, а Н2 и L2 каждая содержат, по меньшей мере, одну аминокислотную модификацию;
b. одна или более из H1, L1, и H2 содержат, по меньшей мере, одну аминокислотную модификацию, а L2 представляет собой полипептидную последовательность дикого типа;
c. одна или более из H1, L1, и L2 содержат, по меньшей мере, одну аминокислотную модификацию, а H2 представляет собой полипептидную последовательность дикого типа;
d. одна или более из H1, H2, и L2 содержат, по меньшей мере, одну аминокислотную модификацию, а L1 представляет собой полипептидную последовательность дикого типа;
e. одна или более из L1, H2, и L2 содержат, по меньшей мере, одну аминокислотную модификацию, а H1 представляет собой полипептидную последовательность дикого типа; или
f. H1, L1, H2 и L2 каждая содержат, по меньшей мере, одну аминокислотную модификацию.
В некоторых вариантах реализации изобретения аминокислотные модификации находятся в:
a. доменах CH1 H1 и H2, домене CL-лямбда L1 и домене CL-каппа L2; или
b. доменах CH1 и VH H1 и H2, домене CL-лямбда и домене VL-лямбда L1, а также домене CL-каппа и домене VL-каппа L2.
В некоторых вариантах реализации изобретения аминокислотные модификации находятся в:
a. по меньшей мере двух из домена CH1 H1, домена CH1 H2, домена CL-лямбда L1 и домена CL-каппа L2;
b. по меньшей мере двух из доменов CH1 и VH H1 и H2, домена CL-лямбда и домена VL-лямбда L1, а также домена CL-каппа и домена VL-каппа L2, или
c. по меньшей мере двух из домена VH H1, домена VH H2, домена VL-лямбда L1 и домена VL-каппа L2.
В некоторых вариантах реализации изобретения H1, L1, H2 и/или L2 содержат, по меньшей мере, 1, 2, 3, 4, 5, 6, 7 или 8 аминокислотных мутаций в области Fab. В некоторых вариантах реализации изобретения, по меньшей мере, одна из H1, H2, L1 и L2 содержит, по меньшей мере, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных модификаций, по меньшей мере, одного константного домена и/или, по меньшей мере, одного вариабельного домена.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1, или H2 с L2 по сравнению с L1 с образованием H2L2, так что относительное спаривание, по меньшей мере, одного из H1L1 или H2L2 является, по меньшей мере, на 10% большим по сравнению с диким типом, а относительное спаривание другого находится в пределах около 10% от дикого типа или, по меньшей мере, на около 10% больше по сравнению с диким типом.
В некоторых вариантах реализации изобретения аминокислотные модификации способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1, или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, 40:60, и соотношение H2L2:H2L1 составляет, по меньшей мере, 60:40; или аминокислотные модификации способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1, или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, 40:60, и соотношение H1L1:H1L2 составляет, по меньшей мере, 60:40.
В некоторых вариантах реализации изобретения термическая стабильность первой области Fab находится в пределах около 0, 1, 2 или 3°C Tm области Fab, образованной соответствующими полипептидными последовательностями H1 и L1 дикого типа. В некоторых вариантах реализации изобретения термическая стабильность второй области Fab находится в пределах около 0, 1, 2 или 3°C Tm области Fab, образованной соответствующими полипептидными последовательностями H2 и L2 дикого типа.
В некоторых вариантах реализации изобретения аминокислотные модификации выбраны из группы, состоящей из уникальных идентификаторов наборов конструкций мАт, представленных в таблице 4A или 4B. В некоторых вариантах реализации изобретения аминокислотные модификации выбраны из группы, состоящей из уникальных идентификаторов наборов конструкций мАт, представленных в одной или более таблицах с 10-A1 по 10-A12. В некоторых вариантах реализации изобретения аминокислотные модификации выбраны из группы, состоящей из уникальных идентификаторов наборов конструкций мАт, представленных в любой из таблиц с 10-B1 по 10-B10.
В некоторых вариантах реализации изобретения конструкция дополнительно содержит димерный Fc, имеющий два полипептида Fc, каждый из которых содержит последовательность домена CH3 и связан с или без помощи линкеров с одной из первой области Fab и второй области Fab. В некоторых вариантах реализации изобретения Fc представляет собой Fc человека, Fc IgG1 человека, Fc IgA человека, Fc IgG человека, Fc IgD человека, Fc IgE человека, Fc IgM человека, Fc IgG2 человека, Fc IgG3 человека или Fc IgG4 человека. В одном варианте реализации изобретения Fc содержит одну или более модификаций по сравнению с диким типом, в, по меньшей мере, одной из последовательностей домена CH3, которые способствуют образованию гетеродимерного Fc.
В некоторых вариантах реализации изобретения Fc содержит:
i) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392M_T394W во втором полипептиде Fc;
ii) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392L_T394W во втором полипептиде Fc;
iii) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392L_T394W во втором полипептиде Fc;
iv) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392M_T394W во втором полипептиде Fc; или
v) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_S400E_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_N390R_K392M_T394W во втором полипептиде Fc.
В некоторых вариантах реализации изобретения Fc дополнительно содержит, по меньшей мере, одну последовательность домена CH2. В одном варианте реализации изобретения Fc содержит одну или более модификаций для способствования селективному связыванию рецепторов Fc-гамма, для уменьшения или устранения связывания с рецепторами Fc-гамма или для способствования связыванию с FcRn.
В некоторых вариантах реализации изобретения, в случае если H1, L1, H2 и L2 коэкспрессируются, изменение в количестве полного правильного спаривания, измеренное суммой спаривания H1L1 и H2L2, составляет более, чем около 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40% или 45%, по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию; или изменение в количестве полного правильного спаривания, измеренное количеством биспецифического антитела, полученного в процентах от видов, отличных от полученных полуантител, составляет более, чем около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% или 80%, по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию, или изменение количества полного правильного спаривания, измеренное количеством биспецифического антитела, полученного в процентах от всех видов, составляет более, чем около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% или 80%, по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию.
В некоторых вариантах реализации изобретения линкеры содержат один или более полипептидных линкеров, одну или более шарнирных областей антитела, или одну или более шарнирных областей IgG1. В одном варианте реализации изобретения один или более полипептидных линкеров содержат одну или более модификаций по сравнению с полипептидным линкером дикого типа.
В некоторых вариантах реализации изобретения аминокислотные модификации включают аминокислотные замены.
В некоторых вариантах реализации изобретения последовательности одной или более из H1, H2, L1 и L2 получают из человеческих последовательностей или гуманизированных последовательностей.
В некоторых вариантах реализации изобретения, описанные в данном документе конструкции конъюгированы с терапевтическим агентом или лекарственным средством.
В другом аспекте описания предложен выделенный рекомбинантный полинуклеотид или набор выделенных рекомбинантных полинуклеотидов, который кодирует(-ют) конструкции, описанные в данном документе. В некоторых вариантах реализации изобретения предложен вектор или набор векторов, содержащих один или более полинуклеотидов или наборов полинуклеотидов, описанных в данном документе. В некоторых вариантах реализации изобретения вектор или, по меньшей мере, один вектор из набора векторов является мультицистронным.
В другом аспекте описания предложена выделенная клетка, содержащая полинуклеотид или набор полинуклеотидов, или вектор или набор векторов, описанных в данном документе. В некоторых вариантах реализации изобретения клетка представляет собой дрожжевую клетку, бактериальную клетку, клетку насекомого или клетку млекопитающего. В некоторых вариантах реализации изобретения выделенная клетка стабильно трансфицируется или транзиентно трансфицируется вектором или набором векторов, описанными в данном документе.
В другом аспекте предложена фармацевтическая композиция, содержащая антигенсвязывающие полипептидные конструкции, описанные в данном документе. В некоторых вариантах реализации изобретения фармацевтическая композиция содержит фармацевтически приемлемый носитель. В некоторых вариантах реализации изобретения фармацевтическая композиция дополнительно содержит одно или более веществ, выбранных из группы, состоящей из буфера, антиоксиданта, молекулы с низкой молекулярной массой, лекарственного средства, белка, аминокислоты, углевода, липида, хелатирующего агента, стабилизатора и наполнителя.
В другом аспекте описан способ получения конструкций, описанных в данном документе. В некоторых вариантах реализации изобретения способ включает в себя этапы:
(a) получение клетки-хозяина, содержащей полинуклеотид или набор полинуклеотидов, кодирующих антигенсвязывающую полипептидную конструкцию;
(b) культивирование клетки-хозяина в культуре клеток-хозяев в условиях, которые позволяют экспрессировать антигенсвязывающую полипептидную конструкцию, и
(c) сбор антигенсвязывающей полипептидной конструкции из культуры клеток-хозяев.
В некоторых вариантах реализации изобретения клетка-хозяин транзиентно трансфицируется или стабильно трансфицируется полинуклеотидом или набором полинуклеотидов, описанными в данном документе.
В другом аспекте предлагается машиночитаемый носитель данных. В некоторых вариантах реализации изобретения машиночитаемый носитель данных хранит набор данных, содержащий данные, представляющие комплементарные аминокислотные модификации в первом гетеродимере, содержащем полипептидную последовательность тяжелой цепи (H1) первого иммуноглобулина и полипептидную последовательность легкой цепи лямбда (L1) иммуноглобулина; и/или втором гетеродимере, содержащем полипептидную последовательность тяжелой цепи (Н2) второго иммуноглобулина и полипептидную последовательность легкой цепи каппа (L2) второго иммуноглобулина. В некоторых вариантах реализации изобретения полипептидные последовательности H1 и H2, хранящиеся в наборе данных, содержат, по меньшей мере, вариабельный домен тяжелой цепи (домен VH) и константный домен тяжелой цепи (домен CH1) и отличаются друг от друга. В некоторых вариантах реализации изобретения полипептидные последовательности L1 и L2, хранящиеся в наборе данных, содержат, по меньшей мере, вариабельный домен легкой цепи (домен VL) и константный домен легкой цепи (домен CL). В некоторых вариантах реализации изобретения комплементарные аминокислотные модификации, хранящиеся в наборе данных, способствуют преимущественному спариванию H1 с L1 по сравнению с L2, и H2 с L2 по сравнению с L1. В некоторых вариантах реализации изобретения набор данных содержит данные, представляющие те модификации, перечисленные в таблице 4A или таблице 4B, или подмножество этих модификаций. В некоторых вариантах реализации изобретения набор данных содержит данные, представляющие те модификации, перечисленные в одной или более таблицах с 10-A1 по 10-A12 или таблицах с 10-B1 по 10-B10, или подмножество этих модификаций.
В другом аспекте описан способ получения биспецифической антигенсвязывающей полипептидной конструкции. В некоторых вариантах реализации изобретения биспецифическая антигенсвязывающая полипептидная конструкция, полученная способом, содержит:
a. первый гетеродимер, содержащий полипептидную последовательность тяжелой цепи (H1) первого иммуноглобулина и полипептидную последовательность легкой цепи лямбда (L1) первого иммуноглобулина; и
b. второй гетеродимер, содержащий полипептидную последовательность тяжелой цепи (H2) второго иммуноглобулина и полипептидную последовательность легкой цепи каппа (L2) второго иммуноглобулина.
В некоторых вариантах реализации изобретения полипептидные последовательности H1 и H2, полученные способом, содержат, по меньшей мере, вариабельный домен тяжелой цепи (домен VH) и константный домен тяжелой цепи (домен CH1) и отличаются друг от друга. В некоторых вариантах реализации изобретения полипептидные последовательности L1 и L2, полученные способом, содержат вариабельный домен легкой цепи (домен VL) и константный домен легкой цепи (домен CL). В некоторых вариантах реализации изобретения одна или более полипептидных последовательностей H1, L1, H2 и L2, полученные способом, содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и H2 с L2 по сравнению с L1.
В некоторых вариантах реализации изобретения способ получения биспецифической антигенсвязывающей полипептидной конструкции содержит:
a. введение одной или более комплементарных аминокислотных модификаций из набора данных, описанного в данном документе, в H1, L1, H2 и/или L2; а также
b. коэкспрессия H1, L1, H2 и L2 в клетке-хозяине для получения продукта экспрессии, содержащего биспецифическую антигенсвязывающую полипептидную конструкцию.
В некоторых вариантах реализации изобретения способ дополнительно включает в себя определение количества биспецифической антигенсвязывающей полипептидной конструкции в продукте экспрессии относительно других полипептидных продуктов для отбора предпочтительного подмножества комплементарных аминокислотных модификаций, которые обеспечивают повышенное количество указанной биспецифической антигенсвязывающей полипептидной конструкции по сравнению с количеством биспецифической антигенсвязывающей полипептидной конструкции в продукте экспрессии, полученном в результате коэкспрессии H1, L1, H2 и L2 дикого типа. В некоторых вариантах реализации изобретения биспецифическая антигенсвязывающая полипептидная конструкция продуцируется с чистотой более 70% по сравнению с другими полипетидными продуктами. В некоторых вариантах реализации изобретения конструкция, полученная способом, содержит Fc, содержащий, по меньшей мере, две последовательности домена CH3, и Fc спарен с помощью или без помощи одного или более линкеров, с первым гетеродимером и вторым гетеродимером. В некоторых вариантах реализации изобретения Fc представляет собой гетеродимерный Fc, содержащий одну или более аминокислотных модификаций, которые способствуют образованию гетеродимерного Fc относительно гомодимерного Fc.
В некоторых вариантах реализации способа получения биспецифической антигенсвязывающей полипептидной конструкции, в случае если H1, L1, H2 и L2 коэкспрессируются, изменение в количестве полного правильного спаривания, измеренное суммой полученных % H1L1 и % H2L2, составляет более, чем около 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40% или 45%, по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию; или изменение в количестве полного правильного спаривания, измеренное количеством биспецифического антитела, полученного в процентах от видов, отличных от полученных полуантител, составляет более, чем около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% или 80%, по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию, или изменение количества полного правильного спаривания, измеренное количеством биспецифического антитела, полученного в процентах от всех видов, составляет более, чем около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% или 80%, по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 изображены аминокислотные последовательности тяжелой цепи и легкой цепи D3H44, пертузумаба и CAT-2200, выровненные против последовательностей зародышевой линии человека для вариабельных и константных доменов. Транслируемые последовательности белка для каждого домена, зародышевой линии и аллеля были получены путем прямого запроса к IMGT/GENE-DB (http://www.imgt.org/genedb/query). Для определения ближайшего гена/аллеля использовался IMGT/DomainGapAlign (http://www.imgt.org/3Dstructure-DB/cgi/DomainGapAlign.cgi). Идентификация консенсусных последовательностей была выполнена с помощью BoxShade (http://www.ch.embnet.org/software/BOX_form.html) с параметром исключения 0,8. Аминокислотные остатки, затушеванные в черный цвет, представляют собой идентичность аминокислотной последовательности, тогда как те, которые затушеваны серым цветом, представляют собой сходство аминокислотной последовательности. Отнесение аминокислот к каждому домену на Фиг. 1 было выполнено согласно определениям IMGT, как описано в Lefranc M.-P. и соавт. "IMGT unique numbering for immunoglobulin and T cell receptor constant domains and Ig superfamily C-like domains" Dev. Comp. Immunol., 2005, 29, 185-203, и Lefranc, M.-P., Pommié, C., Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. и Lefranc, G. "IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains" Dev. Comp. Immunol., 27, 55-77 (2003 год). На Фиг. 1A изображены вариабельные тяжелые (VH) домены пертузумаба и D3H44, выровненные по подгруппам IGHV и IGHJ зародышевой линии человека (для каждого гена и аллели отображается одна представительная последовательность). Наиболее близкими последовательностями гена и аллеля к IGHV и IGHJ пертузумаба являются X92218|IGHV3-66*01 и J00256|IGHJ4*01, соответственно. Наиболее близкими последовательностями гена и аллеля к IGHV и IGHJ D3H44 являются X92218|IGHV3-66*01 и J00256|IGHJ4*01, соответственно. На Фиг. 1B изображены вариабельные легкие (VL) домены пертузумаба и D3H44, выровненные по подгруппам каппа IGKV и IGKJ зародышевой линии человека (от каждого гена и аллеля отображается одна представительная последовательность). Наиболее близкими последовательностями гена и аллеля к IGKV и IGKJ пертузумаба являются Y14865|IGKV1-NL1*01 и J00242|IGKJ2*01, соответственно. Наиболее близкими последовательностями гена и аллеля к IGKV и IGKJ D3H44 являются X59315|IGKV1-39*01 и J00242|IGKJ1*01, соответственно. На Фиг. 1C изображены константные тяжелые 1 (CH1) домены Пертузумаба и D3H44, выровненные по подгруппам CH1 IGHG зародышевой линии человека. Наиболее близкой последовательностью гена и аллеля к IGHG пертузумаба и D3H44 является J00228|IGHG1*01. На Фиг. 1D изображены константные легкие (CL) домены Пертузумаба и D3H44, выровненные по подгруппам каппа IGKC зародышевой линии человека. Наиболее близкой последовательностью гена и аллеля к IGKC Пертузумаба и D3H44 является J00241|IGKC*01. На Фиг. 1E изображен домен VH CAT-2200, выровненный по подгруппам IGHV и IGHJ зародышевой линии человека (для каждого гена и аллели отображается одна репрезентативная последовательность). Наиболее близкими последовательностями гена и аллеля к IGHV и IGHJ CAT-2200 являются M99660|IGHV3-23*01 и J00256|IGHJ4*01, соответственно. На Фиг. 1F изображен домен VL CAT-2200, выровненный по подгруппам лямбда IGLV и IGLJ зародышевой линии человека (от каждого гена и аллеля отображается одна представительная последовательность). Наиболее близкими последовательностями гена и аллеля к IGLV и IGLJ CAT-2200 являются Z73673|IGLV6-57*01 и M15641|IGLJ2*01, соответственно. На Фиг. 1G изображен домен CH1 CAT-2200, выровненный по подгруппам CH1 IGHG зародышевой линии человека. Наиболее близкой последовательностью гена и аллеля к IGHG CAT-2200 является J00228|IGHG1*01. На Фиг. 1H изображен домен CL CAT-2200, выровненный по подгруппам лямбда IGLC зародышевой линии человека. Наиболее близкой последовательностью гена и аллеля к IGLC CAT-2200 является J00253|IGLC2*01.
На Фиг. 2 изображена блок-схема для идентификации остатков области контакта и для компьютерного моделирования конструкций с преимущественным спариванием тяжелой-легкой цепи.
На Фиг. 3 изображено трехмерное структурное выравнивание между константными доменами D3H44 (PDB ID 1JPT) и CAT-2200 (PDB-ID 2VXS). На Фиг. 3A проиллюстрированы типичные конформационные различия, наблюдаемые между легкой цепью каппа и лямбда, когда они выровнены по их соответствующей тяжелой цепи. На Фиг. 3B изображен вид модели области контакта легкой цепи (без тяжелой цепи), представленной на Фиг. 3A, в качестве дальнейшего примера конформационных различий. Пунктирные стрелки указывают на конформационную перестановку элементов вторичной структуры в области контакта тяжелых и легких цепей.
На Фиг. 4 изображен схематический обобщенный обзор технических требований для образования биспецифического антитела, а также требований для анализа, необходимых для количественной оценки пар легкой цепи и тяжелой цепи (H-L). Цель разработки биспецифического антитела с высокой степенью чистоты (то есть, с малым количеством или без каких-либо, неправильно спаренных H-L взаимодействий) может быть достигнута за счет рациональной разработки (с помощью введения специфических аминокислотных мутаций) преимущественного спаривания двух уникальных тяжелых цепей с их уникальными когнатными легкими цепями. Этот процесс показан схематично; здесь H1 была разработана для преимущественного спаривания с L1 (обозначается галочкой), а не с L2 (обозначается буквой "X"). Аналогичным образом, H2 была спроектирована для преимущественного спаривания с L2, а не L1. Стрелки на гетеродимерах H1L1 и H2L2 представляют собой упрощенное спаривание между этими парами H-L, в то время как стрелки на гетеродимерах H1L2 и H2L1 представляют собой нарушение спаривания между последними парами H-L. Экспериментальный скрининг конструкций, способствующих преимущественному спариванию требует анализа, способного одновременно количественно определять H1L1:H1L2 и H2L2:H2L1. Эти требования анализа можно упростить, если предположить, что каждое биспецифическое плечо Fab может быть разработано независимо от другого. В этом случае анализ необходим только для количественной оценки H1L1:H1L2 или H2L2:H2L1, а не обоих одновременно.
На Фиг. 5 представлена схема, изображающая, как тяжелые цепи и легкие цепи могут быть помечены, и как определяется преимущественное спаривание. В этой схеме круговая граница представляет собой клетку, в которой трансфицируют 3 конструкции (одна тяжелая цепь и две уникальные легкие цепи). Продукты экспрессии секретируются из клетки, и супернатант (СПНТ) пропускают через устройство обнаружения, в данном случае чип ППР. На основании обнаружения двух различных меток, слитых с двумя легкими цепями, конкурирующими для спаривания с тяжелой цепью, может быть получена количественная оценка преимущественного спаривания тяжелой цепи с двумя легкими цепями.
На Фиг. 6 изображены критерии фильтрации эффективности, основанные на двух результатах LCCA для каждой конструкции. Эти критерии фильтрации эффективности были использованы для идентификации библиотеки конструкций K-L и библиотеки конструкций K-L, полученных из K-K. Для того, чтобы быть включенной, конструкция должна содержать положительный результат LCCA над диапазоном нейтральных значений данных (диапазон нейтральных значений данных определяется как область между соотношениями спаренный:неправильно спаренный 40:60 и 60:40), а другая LCCA должна быть выше нижнего предела диапазона нейтральных значений данных (соотношение спаренный:неправильно спаренный 40:60). Сценарии A и B представляют конструкции, которые проходят критерии фильтрации. Следует отметить, что сценарий B включен, поскольку LCCA H2L2:H2L1 находится выше диапазона нейтральных значений данных, а LCCA H1L1:H1L2 в пределах диапазона нейтральных значений данных (не ниже его). Сценарий C представляет конструкцию, которая отфильтровывается, и где оба результата LCCA являются положительными, но не находятся выше диапазона нейтральных значений данных. Сценарии D и E представляют конструкции, которые отсеиваются, поскольку, по меньшей мере, один из результатов LCCA находится ниже диапазона нейтральных значений данных, даже если другой LCCA находится выше диапазона нейтральных значений данных (см. сценарий D). Также включены конструкции, в которых поменяны обозначения H1L1 и H2L2. В описании этой фигуры предполагается, что соотношение спаривания дикого типа составляет 50:50.
На Фиг. 7 изображена эффективность выбранных конструкций K-L, а также конструкций K-L, полученных из K-K (основанная на данных LCCA для набора конструкции мАт в таблицах 4A и 4B), как определено "прочностью" конструкции =ΔH1:L1:L2_скаляр+ΔH2:L2:L1_скаляр. Этот показатель является показателем общего успеха спаривания на уровне разработки.
На Фиг. 8 изображены возможные продукты, связанные с тяжелой цепью, которые ожидаются, в случае если в клетке две различные легкие цепи коэкспрессируются с двумя различными тяжелыми цепями.
На Фиг. 9 изображен общий способ получения биспецифической антигенсвязывающей полипептидной конструкции с использованием библиотеки набора конструкций мАт, представленной в данном документе.
На Фиг. 10 изображены блочные диаграммы мин-макс, суммирующие эффективность всех конструкций K-L, протестированных в SMCA в трех биспецифических системах, с помощью кластера конструкции. На Фиг. 10A изображена эффективность, измеренная с помощью общего биспецифического вычисления (ΔБиспецифический%). На Фиг. 10B изображена эффективность, измеренная с помощью общего вычисления спаривания (ΔСпаривание%).
На Фиг. 11 изображен график блочной диаграммы мин-макс, суммирующий эффективность конструкций K-L и конструкций K-L, полученных из K-K, на биспецифическую систему, группой переносимости; "kl 3/3" указывает на конструкции K-L, переносимые в 3/3 биспецифических систем, "kl 3/3+2/3" указывает на конструкции K-L, переносимые, по меньшей мере, в 2 биспецифических системах, а "все kl" указывает на все протестированные конструкции K-L. Аналогичным образом, "kk 3/3" указывает на конструкции K-L, полученные из K-K, переносимые в 3/3 биспецифических систем, "kk 3/3+2/3" указывает на конструкции K-L, полученные из K-K, переносимые, по меньшей мере, в 2 биспецифических системах, а "все kk" указывает на все протестированные конструкции K-L, полученные из K-K; результаты представлены на основе общего биспецифического вычисления (ΔБиспецифический%).
На Фиг. 12 изображены сенсограммы ДСК биспецифических антител, полученных с использованием набора конструкций мАт 3972 (идентификатор конструкции SMCA) (в каждой из трех биспецифических систем) и исходных антител дикого типа для каждой системы. На Фиг. 12A изображено мАт CAT-2200 дикого типа (темно-серый), мАт Пертузумаба дикого типа (средне-серый) и конструкция 3972 SMCA CAT-2200/Пертузумаба (светло-серый); на Фиг. 12B изображено мАт CAT-2200 дикого типа (темно-серый), мАт SGN-CD19a дикого типа (средне-серый) и конструкция 3972 SMCA CAT-2200/SGN-CD19a (светло-серый); на Фиг. 12C изображено мАт SGN-CD19a дикого типа (темно-серый), мАт CR8071 дикого типа (средне-серый) и конструкция 3972 SMCA CR8071/SGN-CD19a (светло-серый).
На Фиг. 13 изображены блочные диаграммы мин-макс, суммирующие влияние аминокислотных замен конструкций мАт на Tm тестируемых Fab. Результаты представлены как изменение в Tm Fab по сравнению с ДТ и показаны для всех конструкций, для которых измеряется Tm ("Все"), и разделены антигенсвязывающим центром.
На Фиг. 14 изображены блочные диаграммы мин-макс, суммирующие влияние аминокислотных замен конструкций мАт на аффинность тестируемых Fab к его антигену. Результаты представлены как разница в log(KD) соответствующего Fab биспецифического из ДТ (-(log(KD_вариант)-log(KD_дт)). Результаты показаны для всех конструкций, для которых измеряется аффинность ("Все"), и разделены антигенсвязывающим центром.
На Фиг. 15 показаны профили СВЭЖХ-ЭХ биспецифических антител, очищенных с помощью белка A и преп-ЭХ, и исходных антител. На Фиг. 15A изображено исходное мАт CAT-2200 дикого типа; на Фиг. 15B изображено исходное мАт CR8071 дикого типа; на Фиг. 15C изображено исходное мАт SGN-CD19a дикого типа; на Фиг. 15D изображено исходное мАт Пертузумаб дикого типа; на Фиг. 15E изображено биспецифическое антитело, полученное с использованием конструкции 3972 SMCA CAT-2200/Пертузумаба; на Фиг. 15F изображено биспецифическое антитело, полученное с использованием конструкции 3972 SMCA CAT-2200/SGN-CD19a; и на Фиг. 15G изображено биспецифическое антитело, полученное с использованием конструкции 3972 SMCA CR8071/SGN-CD19a.
На Фиг. 16 изображен процесс отбора референтных значений дикого типа для вычисления "изменения общего спаривания по отношению к ДТ" и "изменения общего биспецифического по отношению к ДТ", для конструкций, тестируемых в SMCA, в случаях, когда соответствующая биспецифическая конструкция ДТ не была оценена с помощью SMCA. На Фиг. 16A изображен процесс отбора референтных значений дикого типа для "общего спаривания" ("% спаривания H1L1 и % спаривания H2L2") для каждой из трех биспецифических систем; на Фиг. 16B изображен процесс отбора референтных значений дикого типа для "общего биспецифического" ("H1L1_H2L2 и H1L2_H2L1**") для каждой из трех биспецифических систем.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В данном документе предложены сконструированные антитела (также обозначенные в данном документе как мультиспецифические антигенсвязывающие полипептидные конструкции), которые могут содержать первый гетеродимер (H1L1), имеющий первую тяжелую цепь (H1) иммуноглобулина и легкую цепь лямбда (L1) иммуноглобулина, которые спариваются с образованием первой области Fab, и второй гетеродимер (H2L2), имеющий тяжелую цепь (H2) иммуноглобулина и легкую цепь каппа (L2) иммуноглобулина, которые спариваются с образованием второй области Fab. Первая область Fab обычно связывается с первым антигеном, а вторая область Fab обычно связывается со вторым антигеном. В некоторых вариантах реализации изобретения первый и второй антигены отличаются друг от друга. Н1 отличается от Н2. Одна или более тяжелых цепей и легких цепей иммуноглобулина сконструированы таким образом, что могут содержать аминокислотные модификации, которые способствуют преимущественному спариванию правильно спаренных тяжелых и легких цепей (H1L1 или H2L2) при коэкспрессии или совместном продуцировании. Более конкретно, аминокислотные модификации способствуют преимущественному спариванию между каждой тяжелой цепью и правильной легкой цепью, таким образом, что тяжелая цепь первого гетеродимера (H1) может преимущественно спариваться с L1, а не L2, и тяжелая цепь второго гетеродимера (H2) может преимущественно спариваться с L2, а не с L1. Вследствие этого коэкспрессия полипептидов H1, L1, H2 и L2 может обеспечивать получение правильно спаренного биспецифического антитела с уменьшенным или ограниченным неправильным спариванием, таким образом уменьшая число и количество полученных неправильно спаренных видов, потенциально улучшая возможности производства. В одном варианте реализации изобретения аминокислотные модификации в областях Fab спариваются с аминокислотными модификациями в области Fc, что способствует образованию гетеродимерной области Fc для дальнейшего сокращения количества неправильно спаренных тяжелых цепей. Аминокислотные модификации не оказывают существенного влияния на термическую стабильность правильно спаренных гетеродимеров или аффинность связывания каждого правильно спаренного гетеродимера с антигеном по сравнению с гетеродимерами, которые образованы из полипептидов H1 и L1 или H2 и L2 дикого типа.
Также в данном документе предложены способы получения описанных выше мультиспецифических антигенсвязывающих полипептидных конструкций.
Определения
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается специалистом в области техники, к которой принадлежит заявленный объект изобретения. В случае если имеется множество определений терминов в данном документе, то имеют преимущество те, которые упоминаются в данном разделе. Если сделана ссылка на URL или другой такой идентификатор или адрес, следует понимать, что такие идентификаторы могут изменяться и конкретная информация в интернете может появляться и исчезать, но эквивалентную информацию можно найти с помощью функции поиска в интернете. Ссылка на них свидетельствует о доступности и открытом распространении такой информации.
Следует понимать, что общее изложенное выше описание и последующее подробное описание являются только иллюстративными и пояснительными и не ограничивают какой-либо заявленный объект изобретения. В данной заявке использование единственного числа включает множественное число, если специально не указано иное.
В данном документе, любой диапазон концентраций, процентный диапазон, коэффициент амплитуды или область целочисленных значений следует понимать, как включающие значение любого целого числа в пределах перечисленного диапазона, и при необходимости и его частей (например, одна десятая и одна сотая целого числа), если не указано иное. Используемый в данном документе термин "около" означает ± 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9% или 10% от указанного диапазона, значения, последовательности или структуры, если не указано иное. Следует понимать, что используемые в данном документе формы терминов в единственном числе включают ссылки на множественное число перечисленных конструкций, если из контекста явно не следует иное. Использование альтернативы (например, "или") следует понимать, как одно, другое или любую их комбинацию из альтернатив. Используемые в данном документе термины "включает" и "содержит" используются в качестве синонимов. Кроме того, следует понимать, что отдельные одноцепочечные полипептиды или конструкции иммуноглобулина, полученные из различных комбинаций структур и заменителей, описанных в данном документе, описаны в настоящей заявке в той же степени, как если бы каждый одноцепочечный полипептид или гетеродимер был изложен в индивидуальном порядке. Таким образом, выбор конкретных компонентов для образования отдельных одноцепочечных полипептидов или гетеродимеров находится в пределах объема данного изобретения.
Заголовки разделов, используемые в данном документе, предназначены только организационных целей и не могут быть истолкованы как ограничивающие описанный объект. Все документы или части документов, указанные в данной заявке, включая, но без ограничений, патенты, заявки на патенты, статьи, книги, учебные пособия и монографии, в явной форме включены в данный документ посредством ссылки в полном объеме для всех целей.
Надо понимать, что описанные в данном документе способы и композиции не ограничиваются конкретной методологией, протоколами, клеточными линиями, конструкциями и реагентами, описанными в данном документе и, по сути могут различаться. Также следует понимать, что употребляемая в данном документе терминология предназначена исключительно для описания конкретных вариантов реализации изобретения, и не предназначена для ограничения объема описанных в данном документе способов и композиций, которые будут ограничены только прилагаемой формулой изобретения.
Все публикации и патенты, упомянутые в данном документе, включены в данный документ в полном объеме посредством ссылки в целях описания и раскрытия, например, конструкций и методологий, которые описаны в публикациях, которые могут быть использованы в связи со способами, композициями и соединениями, описанными в данном документе. Публикации, рассмотренные в данном документе, приведены исключительно для их раскрытия до даты подачи настоящей заявки. Ничего в данном документе не следует трактовать как допущение того, что авторы данного изобретения не имеют права датировать более ранним числом такое описание вследствие предшествующего изобретения или любой другой причины.
В данной заявке названия аминокислот и названия атомов (например, N, O, C и т. д.) используются, как определено Protein DataBank (PDB) (www.pdb.org), который основан на номенклатуре ИЮПАК ("Номенклатура и символика для аминокислот и пептидов ИЮПАК" (названия остатков, названия атомов и т. д.)), Eur. J. Biochem., 138, 9-37 (1984 год) вместе с их поправками в Eur. J. Biochem., 152, 1 (1985 год). Термин "аминокислотный остаток" в первую очередь предназначен для обозначения аминокислотного остатка, содержащегося в группе, состоящей из 20 аминокислот, встречающихся в природе, то есть остатков аланина (Ala или А), цистеина (Cys, или С), аспарагиновой кислоты (Asp или D), глутаминовой кислоты (Glu или Е), фенилаланина (Phe или F), глицина (Gly или G), гистидина (His или Н), изолейцина (Ile или I), лизина (Lys или K), лейцина (Leu или L), метионина (Met или M), аспарагина (Asn или N), пролина (Pro или P), глутамина (Gln или Q), аргинина (Arg или R), серина (Ser или S), треонина (Thr или Т), валина (Val или V), триптофана (Trp или W) и тирозина (Tyr или Y).
Термины "полипептид", "пептид" и "белок" используются равноправно в данном документе для обозначения полимерных аминокислотных остатков. То есть, описание, направленное на полипептид, в равной степени относится к описанию пептида и описанию белка, и наоборот. Эти термины относятся как к встречающимся в природе аминокислотным полимерам, так и к аминокислотным полимерам, в которых один или более аминокислотных остатков представляют собой не встречающуюся в природе аминокислоту. Используемые в данном документе термины охватывают аминокислотные цепи любой длины, включая полноразмерные белки, отличающиеся тем, что аминокислотные остатки соединены ковалентной пептидной связью.
Термин "нуклеотидная последовательность" или "последовательность нуклеиновой кислоты" подразумевает обозначение последовательного отрезка из двух или более нуклеотидных молекул. Нуклеотидная последовательность может быть геномного происхождения, кДНК, РНК, полусинтетического или синтетического происхождения, или представлять собой любую их комбинацию.
"Клетка", "клетка-хозяин", "линия клеток" и "культура клеток" используются в данном документе взаимозаменяемо, и, как следует понимать, все такие термины включают потомство, получаемое при выращивании или культивировании клеток. Термины "трансформация" и "трансфекция" используются взаимозаменяемо для обозначения процесса введения ДНК в клетку.
Термин "аминокислота" относится к встречающимся в природе и не встречающимся в природе аминокислотам, а также аминокислотным аналогам и аминокислотным миметикам, которые функционируют аналогично встречающимся в природе аминокислотам. Природными аминокислотами являются 20 распространенных аминокислот (аланин, аргинин, аспарагин, аспарагиновая кислота, цистеин, глутамин, глутаминовая кислота, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин) и пиролизин и селеноцистеин. Аминокислотные аналоги относятся к соединениям, которые имеют такую же основную химическую структуру, идентичную с аминокислотой, встречающейся в природе, то есть, углерод, который связан с водородом, карбоксильную группу, амино группу и R группу, например, гомосерин, норлейцин, метионилсульфоксид, метионинметилсульфоний. Такие аналоги могут иметь модифицированные R группы (например, норлейцин) или модифицированные пептидные остовы, но сохраняют такую же основную химическую структуру, что и аминокислота, встречающаяся в природе. Термин аминокислота включает, например, встречающиеся в природе протеогенные L-аминокислоты; D-аминокислоты, химически модифицированные аминокислоты, такие как варианты аминокислот и производные; встречающиеся в природе непротеогенные аминокислоты, такие как аланин, орнитин и т.д.; и химически синтезированные соединения, имеющие свойства, известных в данной области техники, характерные для аминокислот. Примеры не встречающихся в природе аминокислот включают, без ограничений, N-метил-аминокислоты (например, метилаланин), D-аминокислоты, гистидин-подобные аминокислоты (например,2-амино-гистидин, гидроксигистидин, гомогистидин), аминокислоты, имеющие дополнительный метилен в боковой цепи ("гомо" аминокислоты) и аминокислоты, в которых функциональная группа карбоновой кислоты в боковой цепи заменяется на группу сульфоновой кислоты (например, цистеиновая кислота). Включение неприродных аминокислот, включая синтетические ненативные аминокислоты, замещенные аминокислоты или одну, или более D-аминокислот в белки антигенсвязывающих полипептидных конструкций по данному изобретению может быть эффективно в ряде различных способов. Пептиды, содержащие D-аминокислоты, демонстрируют повышенную стабильность in vitro или in vivo по сравнению с аналогами, содержащими L-аминокислоты. Таким образом, конструирование пептидов и т.д., включение D-аминокислот может быть особенно полезным, в случае если более высокая внутриклеточная стабильность является желаемой или требуемой. Конкретнее, D-пептиды и т.д., резистентны к эндогенным пептидазам и протеазам, в связи с этим обеспечивается улучшенная биодоступность молекулы и увеличенное время жизни in vivo, в случае если такие свойства являются желаемыми. Дополнительно, D-пептиды и т.д., не могут эффективно подвергаться рестриктированному представлению по главному комплексу гистосовместимости класса II клеткам T-хелперам, и являются, таким образом, менее подходящими для индуцирования гуморального иммунного ответа в целом организме.
Аминокислоты обозначаются в данном документе либо с помощью общеизвестных трехбуквенных символов, либо с помощью однобуквенных символов, рекомендованных Комиссией по биохимической номенклатуре ИЮПАК-МБС. Нуклеотиды аналогичным образом могут обозначаться общепринятым однобуквенным кодом.
Термин "консервативно модифицированные варианты” применяют как к аминокислотным, так и нуклеотидным последовательностям. Принимая во внимание конкретные последовательности нуклеиновых кислот, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или, в случае если нуклеиновая кислота не кодирует аминокислотную последовательность, по существу идентичные последовательности. Из-за вырожденности генетического кода любой заданный белок кодируется значительным количеством функционально идентичных нуклеиновых кислот. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором аланин определяется кодоном, этот кодон может быть изменен на любой из соответствующих описанных кодонов без изменения этого кодированного полипептида. Такие вариации нуклеиновых кислот являются "молчащими вариациями", которые представляют собой один вид консервативно модифицированных вариаций. В данном документе каждая последовательность нуклеиновой кислоты, которая кодирует полипептид, также описывает любую возможную молчащую вариацию нуклеиновой кислоты. Среднему специалисту в данной области техники будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно представляет собой единственный кодон для метионина, и TGG, который обычно представляет собой единственный кодон для триптофана), может быть модифицирован с получением функционально идентичной молекулы. Таким образом, в каждой описанной последовательности подразумевают каждую молчащую вариацию нуклеиновой кислоты, которая кодирует полипептид.
Что касается аминокислотных последовательностей, среднему специалисту в данной области техники будет понятно, что отдельные замены, делеции или вставки в последовательность нуклеиновой кислоты, пептида, полипептида или белка, которые приводят к изменению, вставке или делеции одной аминокислоты или небольшого количества в процентах аминокислот в кодируемой последовательности, относятся к "консервативно модифицированному варианту", причем указанное изменение приводит к делеции аминокислоты, вставке аминокислоты или замене аминокислоты на химически подобную аминокислоту. Таблицы консервативных замен, в которых приведены аминокислоты с подобными функциональными свойствами, известны среднему специалисту в данной области техники. Такие консервативно модифицированные варианты являются дополнительными и не исключают полиморфные варианты, межвидовые гомологи и аллели последовательностей данного изобретения.
Таблицы консервативных замен, в которых приведены аминокислоты с подобными функциональными свойствами, известны среднему специалисту в данной области техники. Далее представлены восемь групп каждая из которой содержит аминокислоты, которые могут представлять собой консервативные замены друг для друга:
Аланин (A), Глицин (G);
Аспарагиновая кислота (D), Глутаминовая кислота (Е);
Аспарагин (N), Глутамин (Q);
Аргинин (R), Лизин (К);
Изолейцин (I), Лейцин (L), Метионин (M), Валин (V);
Фенилаланин (F), Тирозин (Y), Триптофан (W); и
Серин (S), Треонин (T);
(см., например, Creighton, Proteins: Structures and Molecular Properties (W H Freeman & Co. ;2-е издание (декабрь 1993 год).
Термины "идентичный" или процентная "идентичность" в контексте двух или более последовательностей нуклеиновых кислот или полипептидных последовательностей, относятся к двум или более последовательностям, или подпоследовательностям, которые являются одинаковыми. Последовательности являются "по существу идентичными" или "по существу подобными", если они содержат определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (то есть, по меньшей мере, около 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности в пределах конкретной области) при сравнении и выравнивании для максимального соответствия в окне сравнения или определенной области, что измеряется с использованием одного из следующих алгоритмов сравнения последовательностей (или других алгоритмов, доступных средним специалистам в данной области техники), или путем выравнивания вручную и визуального наблюдения. Это определение также относится к последовательности, комплементарной исследуемой последовательности. Идентичность может распространяться на область, длина которой составляет, по меньшей мере, около 50 аминокислот или нуклеотидов, или на область, длина которой составляет 75-100 аминокислот или нуклеотидов, или, если не указано иное, на всю последовательность полинуклеотида или полипептида. Полинуклеотид, кодирующий полипептид антигенсвязывающих полипептидных конструкций, описанных в данном документе, включая гомологи из видов, отличных от человека, может быть получен способом, включающим этапы скрининга библиотеки в жестких условиях гибридизации с меченым зондом, содержащим полинуклеотидную последовательность антигенсвязывающих полипептидных конструкций, описанных в данном документе, или их фрагмент, и выделения полноразмерной кДНК и геномных клонов, содержащих указанную полинуклеотидную последовательность. Такие технологии гибридизации хорошо известны специалисту в данной области техники.
Примерами алгоритма, который пригоден для определения процента идентичности последовательностей и сходства последовательностей являются алгоритмы BLAST™ и BLAST™ 2,0, которые описаны в работах Altschul и соавт. (Nuc. Acids Res. 25:3389-402, 1977) и Altschul и соавт. (J.Mol.Biol.215:403-10, 1990), соответственно. Программное обеспечение для проведения анализов BLAST™ находится в открытом доступе в Национальном центре биотехнологической информации (the National Center for Biotechnology Information) (см. веб-сайт www.ncbi.nlm.nih.gov). Совокупные баллы рассчитываются с использованием, для нуклеотидных последовательностей, параметров M (оценка вознаграждения для пары комплементарных остатков, всегда >0) и N (оценка штрафа за некомплементарные остатки, всегда <0). Для аминокислотных последовательностей для вычисления совокупных баллов используется матрица замен. Расширение соседних слов в каждом направлении приостанавливается, в случае если: совокупный балл выравнивания уменьшается на величину X от ее максимального достигнутого значения; совокупный балл доходит до нуля или ниже, из-за накопления одного или более выравниваний остатков с отрицательным счетом; или достигается конец любой последовательности. Параметры алгоритма BLAST W, T и X определяют чувствительность и скорость выравнивания. Примерами параметров алгоритма для программы BLASTN (для нуклеотидных последовательностей) являются длина слова (W) 11, ожидаемое значение (E) 10, M=5, N=-4 и сравнение обеих цепей. Для аминокислотных последовательностей примерами параметров алгоритма для программы BLASTP являются длина слова 3, ожидаемое значение (E) 10 и матрица замен BLOSUM62 (см., Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89:10915, 1989).
Считается, что производное или вариант полипептида характеризуется "гомологичностью" или является "гомологичным" пептиду, если аминокислотные последовательности производного или варианта, по меньшей мере, на 50% идентичны в последовательности, которая составляет 100 аминокислот в длину от исходного пептида. В некоторых вариантах реализации изобретения производное или вариант характеризуется сходством, по меньшей мере, на 75% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения производное или вариант характеризуется сходством, по меньшей мере, на 85% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения аминокислотная последовательность производного характеризуется сходством, по меньшей мере, на 90% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения аминокислотная последовательность производного характеризуется сходством, по меньшей мере, на 95% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное. В некоторых вариантах реализации изобретения производное или вариант характеризуется сходством, по меньшей мере, на 99% с пептидом или фрагментом пептида, имеющим такое же число аминокислотных остатков, что и производное.
В данном контексте "выделенный" полипептид или конструкция означает конструкцию или полипептид, который был идентифицирован и отделен, и/или извлечен из компонента его природной среды клеточной культуры. Контаминирующие компоненты его природной среды представляют собой вещества, которые, как правило, мешают диагностическим или терапевтическим применениям гетеромультимера, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества.
В определенных вариантах реализации изобретения, в данном контексте, "выделенные" антигенсвязывающие полипептидные конструкции описывают антигенсвязывающие полипептидные конструкции, которые были идентифицированы и отделены и/или извлечены из компонента его природной среды клеточной культуры. Например, выделенная биспецифическая антигенсвязывающая полипептидная конструкция, описанная в данном документе, содержит гетеродимерные пары или "выделенные" гетеродимерные пары, которые содержат гетеродимер или гетеродимерную пару, которая была идентифицирована и отделена, и/или извлечена из компонента его природной среды клеточной культуры. Контаминирующие компоненты его природной среды представляют собой вещества, которые мешают диагностическим или терапевтическим применениям гетеродимера или антигенсвязывающих полипептидных конструкций, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества.
Гетеродимеры и антигенсвязывающие полипептидные конструкции и могут быть очищены до существенной гомогенности. Фразы "по существу гомогенный", "по существу гомогенная форма" и "существенная гомогенность" используются для указания того, что правильно спаренный продукт по существу лишен побочных продуктов, происходящих от нежелательных полипептидных комбинаций (например, гомодимеров или неправильно спаренных гетеродимеров). В контексте набора конструкции LCCA (H1L1L2) правильно спаренным продуктом является гетеродимер, содержащий H1 и L1 (H1L1). В контексте набора конструкции LCCA (H2L1L2) правильно спаренным продуктом является гетеродимер, содержащий H2 и L2 (H2L2). В одном варианте реализации изобретения в контексте биспецифической антигенсвязывающей полипептидной конструкции, где экспрессируются H1, L1, H2 и L2, правильно спаренный продукт представляет собой гетеродимерную пару, содержащую правильно спаренные H1L1 и H2L2 (H1L1H2L2). В некоторых вариантах реализации изобретения в контексте биспецифической антигенсвязывающей полипептидной конструкции, где экспрессируются H1, L1, H2 и L2, правильно спаренный продукт может содержать дополнительные продукты, которые демонстрируют правильное спаривание, по меньшей мере, в одной области Fab, в такой как, например, H1L1H2L1 или H1L2H2L2, или где вырабатываются "полуантитела", H1L1 или H2L2. Выраженная в терминах чистоты, в одном варианте реализации изобретения, существенная гомогенность означает, что количество полностью неправильно спаренных побочных продуктов не превышает 20%, например, составляет менее 10%, менее 5%, менее 1% или менее 0,5% от общей интенсивности ЖХ-МС из всех видов, присутствующих в смеси, причем процентное содержание отражает результаты масс-спектрометрического анализа.
Термины, понятные специалистам в области техники антител, имеют значение, приобретенное в данной области техники, если в данном документе прямо не указанно иное. Как известно, антитела имеют вариабельные области, шарнирную область и константные домены. Структура и функции иммуноглобулина рассматриваются, например, в Harlow и соавт., ред., Antibodies: A Laboratory Manual, глава 14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, 1988).
В данном контексте термины "антитело", "иммуноглобулин" или "антигенсвязывающая полипептидная конструкция" используются как взаимозаменяемые. "Антигенсвязывающая полипептидная конструкция" относится к полипептиду, который по существу кодируется геном иммуноглобулина или генами иммуноглобулина, или к одному, или более его фрагментам, которые специфически связывают анализируемое вещество (антиген). Все распознаваемые гены иммуноглобулинов включают каппа, лямбда, альфа, гамма, дельта, эпсилон и мю константные области генов, а также множество генов вариабельной области иммуноглобулина. Легкие цепи классифицируют как каппа или лямбда. Тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон, которые в свою очередь определяют изотипы иммуноглобулинов, IgG, IgM, IgA, IgD и IgE, соответственно. Дополнительно антитело может принадлежать к одному из нескольких подтипов, например, IgG может принадлежать к подклассам IgG1, IgG2, IgG3 или IgG4.
Структурная единица типового иммуноглобулина (антитела) состоит из двух пар полипептидных цепей, причем каждая пара имеет одну "легкую" (около 25 кДа) цепь иммуноглобулина и одну "тяжелую" цепь иммуноглобулина (около 50-70 кДа). Этот тип структурной единицы иммуноглобулина или антитела считается "естественного происхождения". Термин "легкая цепь" включает полноразмерную легкую цепь и ее фрагменты, содержащие последовательность вариабельного домена, достаточную, чтобы обеспечивать специфичность связывания. Полноразмерная легкая цепь содержит вариабельный домен, VL, и константный домен, CL. Вариабельный домен легкой цепи представляет собой амино-конец полипептида. Легкие цепи включают каппа цепи и лямбда цепи. Термин "тяжелая цепь" включает полноразмерную тяжелую цепь и ее фрагменты, содержащие последовательность вариабельной области, достаточную, чтобы обеспечивать специфичность связывания. Полноразмерная тяжелая цепь содержит вариабельный домен, VH, и три константных домена, CH1, CH2 и CH3. Домен VH представляет собой амино-конец полипептида, а домены CH находятся в карбоксильном конце, при этом CH3 находится наиболее близко к карбокси-концу полипептида. Тяжелые цепи могут принадлежать любому изотипу, включая IgG (включая подклассы IgG1, IgG2, IgG3 и IgG4), IgA (включая подклассы IgA1 и IgA2), IgM, IgD и IgE. Термин "вариабельная область" или "вариабельный домен" относится к части легких и/или тяжелых цепей антитела, как правило, ответственной за распознавание антигена, как правило, включающей приблизительно от 120 до 130 аминоконцевых аминокислот в тяжелой цепи (VH) и приблизительно от 100 до 110 аминоконцевых аминокислот в легкой цепи (VL).
"Определяющая комплементарность область" или "CDR" представляет собой аминокислотную последовательность, которая определяет антигенсвязывающую специфичность и аффинность. "Каркасные" области (FR) могут способствовать поддержанию правильной конформации CDR для содействия связывания между антигенсвязывающей областью и антигеном. В структурном плане каркасные области могут быть расположены в антителах между CDR. Вариабельные области, как правило, обладают аналогичной общей структурой относительно консервативных каркасных областей (FR), соединенных тремя гипервариабельными областями, CDR. CDR из двух цепей каждой пары, как правило, выравниваются по каркасным областям, которые могут обеспечивать связывание со специфическим эпитопом. От N-конца к C-концу вариабельные области, как легкой, так и тяжелой цепи, как правило, содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Как правило, аминокислоты присваивают к каждому домену в соответствии с определениями по Kabat Sequences of Proteins of Immunological Interest (National Institutes of Health, Бетесда, Мэриленд (1987 год и 1991 год)), если не указано иное.
"Мультиспецифическая антигенсвязывающая полипептидная конструкция" или "мультиспецифическое антитело" нацелено или связывается с более чем одним отдельным антигеном или эпитопом. "Биспецифическая", "обладающая двойной специфичностью" или "бифункциональная" антигенсвязывающая полипептидная конструкция или антитело представляет собой разновидность мультиспецифической антигенсвязывающей полипептидной конструкции, которая нацеливается или связывается с двумя различными антигенами или эпитопами. В общем, биспецифическая антигенсвязывающая полипептидная конструкция может иметь два разных антигенсвязывающих домена. Два антигенсвязывающих домена биспецифической антигенсвязывающей полипептидной конструкции или антитела связывают два разных эпитопа, которые могут находиться на одной и той же или на разных молекулярных мишенях. В одном варианте реализации изобретения биспецифическая антигенсвязывающая полипептидная конструкция находится в естественном формате. Другими словами, биспецифическая антигенсвязывающая полипептидная конструкция имеет тот же формат, что и встречающееся в природе антитело IgG, IgA, IgM, IgD или IgE.
Тяжелые цепи антител спариваются с легкими цепями антител и встречаются или контактируют друг с другом в одной или более "областей контакта". "Область контакта" включает в себя один или более "контактных" аминокислотных остатков в первом полипептиде, которые взаимодействуют с одним или более "контактными" аминокислотами второго полипептида. Например, область контакта существует между двумя доменами СН3 димеризованной области Fc, между доменом CH1 тяжелой цепи и доменом CL легкой цепи, и между доменом VH тяжелой цепи, и доменом VL легкой цепи. "Область контакта" может быть получена из антитела IgG и, например, из человеческого антитела IgG1.
Термин "аминокислотные модификации" в данном контексте включает, но без ограничений, аминокислотные вставки, делеции, замены, химические модификации, физические модификации и перестройки.
Аминокислотные остатки для тяжелых и легких цепей иммуноглобулина могут быть пронумерованы в соответствии с несколькими конвенциями, включая Кабат (как описано в Kabat and Wu, 1991 год; Kabat и соавт, Sequences of proteins of immunological interest. 5-е издание - US Department of Health and Human Services, публикация NIH №91-3242, стр. 647 (1991 год)), IMGT (как изложено в Lefranc, M.-P., и соавт. IMGT®, international ImMunoGeneTics information system® Nucl. Acids Res, 37, D1006-D1012 (2009 год), и Lefranc, M.-P., IMGT, International ImMunoGeneTics Information System, Cold Spring Harb Protoc. 1-е июня 2011 года; 2011 год (6)), 1JPT (как описано в Katja Faelber, Daniel Kirchhofer, Leonard Presta, Robert F Kelley, Yves A Muller, The 1. 85 Å resolution crystal structures of tissue factor in complex with humanized fab d3h44 and of free humanized fab d3h44:revisiting the solvation of antigen combining sites1, Journal of Molecular Biology, том 313, выпуск 1, стр. 83-97,) и ЕС (согласно индексу ЕС, как в Kabat, ссылаясь на нумерацию антитела ЕС (Edelman и соавт., 1969 год, Proc Natl Acad Sci USA 63:78-85)). Нумерация Кабат используется в данном документе для доменов VH, CH1, CL и VL, если не указано иное. Нумерация ЕС используется в данном документе для доменов CH3 и CH2 и шарнирной области, если не указано иное. В таблице 22А представлена таблица соответствия, показывающая нумерацию аминокислот для выбранных положений в полипептиде тяжелой цепи IgG1 с использованием систем нумерации IMGT, Кабат, 1JPT и ЕС. В таблице 22B представлена таблица соответствия, показывающая нумерацию аминокислот для выбранных положений в полипептиде легкой цепи лямбда с использованием систем нумерации IMGT и Кабат. В таблице 22C представлена таблица соответствия, показывающая нумерацию аминокислот для выбранных положений в полипептиде легкой цепи каппа с использованием систем нумерации IMGT, 1JPT и Кабат.
Антигенсвязывающие полипептидные конструкции
Антигенсвязывающие полипептидные конструкции (то есть антитела), описанные в данном документе, могут быть мультиспецифическими или биспецифическими. Мультиспецифическая антигенсвязывающая полипептидная конструкция может содержать, по меньшей мере, первый гетеродимер (H1L1), имеющий полипептидную последовательность тяжелой цепи (H1) первого иммуноглобулина и полипептидную последовательность легкой цепи лямбда (L1) первого иммуноглобулина, которые образуют первую область Fab и, по меньшей мере, второй гетеродимер (H2L2), имеющий полипептидную последовательность тяжелой цепи (H2) иммуноглобулина и полипептидную последовательность легкой цепи каппа (L2) иммуноглобулина, которые образуют вторую область Fab, где H1 и H2 отличаются друг от друга. В одном варианте реализации изобретения биспецифическая антигенсвязывающая полипептидная конструкция содержит первый гетеродимер (H1L1), имеющий полипептидную последовательность тяжелой цепи (H1) первого иммуноглобулина и полипептидную последовательность легкой цепи лямбда (L1) первого иммуноглобулина, которые образуют первую область Fab и второй гетеродимер (H2L2), имеющий полипептидную последовательность тяжелой цепи (H2) иммуноглобулина и полипептидную последовательность легкой цепи каппа (L2) иммуноглобулина, которые образуют вторую область Fab, где H1 и H2 отличаются друг от друга. В одном варианте реализации изобретения каждый гетеродимер содержит одну область Fab. В данном контексте термин "область Fab" относится к области, образующейся в результате спаривания одной полипептидной последовательности легкой цепи иммуноглобулина с одной полипептидной последовательностью тяжелой цепи иммуноглобулина, и состоит из доменов VH и CH1 полипептидной последовательности тяжелой цепи иммуноглобулина, и доменов VL и CL полипептидной последовательности легкой цепи иммуноглобулина. В некоторых вариантах реализации изобретения первая область Fab связывается с первым антигеном, а вторая область Fab связывается со вторым антигеном. Первый и второй антигены могут быть одинаковыми или разными. Одна или более тяжелых цепей и легких цепей иммуноглобулина могут содержать аминокислотные модификации, которые способствуют преимущественному спариванию правильно спаренных тяжелых и легких цепей при коэкспрессии или совместном продуцировании.
Когда антигенсвязывающая полипептидная конструкция представляет собой биспецифическую антигенсвязывающую полипептидную конструкцию (то есть биспецифическое антитело), она также может упоминаться как "гетеродимерная пара".
С целью иллюстрации, первый гетеродимер антигенсвязывающей полипептидной конструкции упоминается как H1L1 и содержит полипептидную последовательность тяжелой цепи (H1) первого иммуноглобулина, спаренную с полипептидной последовательностью легкой цепи лямбда (L1) первого иммуноглобулина, а второй гетеродимер упоминается как H2L2 и содержит полипептидную последовательность тяжелой цепи (Н2) второго иммуноглобулина, спаренную с полипептидной последовательностью легкой цепи каппа (L2) второго иммуноглобулина. Следует, однако, понимать, что это обозначение является произвольным и предназначено только для указания того, что один гетеродимер содержит легкую цепь каппа иммуноглобулина, а другой содержит легкую цепь лямбда иммуноглобулина. Область Fab первого гетеродимера H1L1 также может упоминаться в данном документе как "лямбда Fab"; в то время как область Fab второго гетеродимера H2L2 также может упоминаться данном документе как "каппа Fab".
Исходные антитела
Полипептидные последовательности тяжелой цепи иммуноглобулина, также называемые "тяжелые цепи", и полипептидные последовательности легкой цепи иммуноглобулина, также называемые "легкие цепи", каждого гетеродимера могут быть получены из одного или более исходных антител, где, по меньшей мере, одно исходное антитело содержит легкую цепь каппа, и, по меньшей мере, одно другое исходное антитело содержит легкую цепь лямбда, а аминокислотные модификации, которые способствуют преимущественному спариванию, спроектированы в этих тяжелых и легких цепях. Исходные последовательности легкой цепи иммуноглобулина и тяжелой цепи иммуноглобулина, не обладающие аминокислотными модификациями, которые способствуют преимущественному спариванию, называются полипептидными последовательностями тяжелой цепи иммуноглобулина дикого типа, полипептидными последовательностями легкой цепи каппа иммуноглобулина дикого типа и полипептидными последовательностями легкой цепи лямбда иммуноглобулина дикого типа. В одном варианте реализации изобретения тяжелые и легкие цепи гетеродимеров антигенсвязывающей полипептидной конструкции получают из двух исходных антител. Как правило, два исходных антитела отличаются друг от друга; однако это не всегда так. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция представляет собой биспецифическую антигенсвязывающую полипептидную конструкцию, где каждый гетеродимер получают из другого исходного антитела. В другом варианте реализации изобретения антигенсвязывающая полипептидная конструкция представляет собой биспецифическую антигенсвязывающую полипептидную конструкцию, в которой оба исходных антитела связываются с одним и тем же антигеном, но нацелены на разные эпитопы на одном и том же антигене. В одном варианте реализации изобретения, по меньшей мере, одно исходное антитело является моноспецифическим, то есть может связываться только с одним эпитопом. В другом варианте реализации изобретения, по меньшей мере, одно исходное антитело может связываться с более чем одним эпитопом.
Тяжелая цепь и легкая цепь каждого гетеродимера антигенсвязывающей полипептидной конструкции спариваются с целью образования области Fab, которая специфически связывается с тем же антигеном, что и исходное антитело, из которого они были получены. Например, если биспецифическую антигенсвязывающую полипептидную конструкцию получали на основе исходных антител CAT-2200 (содержащих легкую цепь лямбда и связывание с IL-17A) и D3H44 (содержащую легкую цепь каппа и связывание с тканевым фактором) тяжелая цепь и легкая цепь одного гетеродимера будут спариваться с образованием области Fab, которая связывается с IL-17A, а тяжелая и легкая цепь второго гетеродимера будет спариваться с образованием области Fab, которая связывается с тканевым фактором.
Исходные антитела могут быть получены из видов, включая, но без ограничений, людей, мышей, крыс, кроликов, овец, коров, коз или верблюдов. В одном варианте реализации изобретения исходные антитела могут быть получены от людей или мышей.
Исходные антитела могут также включать те, которые получены из гибридом с использованием стандартных протоколов получения моноклональных антител, таких как описанные Kohler and Milstein (Nature, 256: 495-497, 1975 год).
Антитела, связывающиеся с конкретной мишенью, также могут быть идентифицированы с помощью ряда различных стратегий, включая фаговый дисплей, in vitro дисплей и другие способы. Эти стратегии приводят к образованию антител в формате scFv, формате Fab или полноразмерном формате IgG. Обзор этих стратегий содержится в главе 4 Therapeutic Antibody Engineering, William R. Strohl и Lila M. Strohl, Woodhead Publishing series in Biomedicine № 11, ISBN 1 907568 37 9, октябрь 2012 года. В одном варианте реализации изобретения исходные антитела включают антитела, идентифицированные с помощью фагового дисплея или in vitro дисплея. Антитела, идентифицированные в форматах, отличных от Fab или полноразмерных форматов IgG, могут быть преобразованы в те же, что известны в данной области техники. Способы преобразования scFvs в Fab хорошо известны в данной области техники (см., например, Steinwand и соавт. Mabs 6:204-218 или Zuberbuhler и соавт. Protein Engineering, Design & Selection 22:169-174). В одном варианте реализации изобретения антитела, первоначально идентифицированные как scFvs, но где scFv был преобразован в формат Fab и сконструирован в виде стандартного или встречающегося в природе антитела, также могут быть использованы в качестве исходных антител.
В одном варианте реализации изобретения тяжелые цепи и легкие цепи каждого гетеродимера антигенсвязывающей полипептидной конструкции могут быть получены из исходного антитела, которое представляет собой гуманизированное антитело. Гуманизированные антитела могут быть получены путем замещения определяющей комплементарность области (CDR) антитела человека на CDR антитела, полученного из млекопитающего, отличного от человека, например, мыши. Способы идентификации CDR известны в данной области техники (Kabat и соавт., Sequence of Proteins of Immunological Interest (1987 год), National Institute of Health, Бетесда, Мэриленд; Чотиа и соавт., Nature (1989 год) 342:877). Известны также общие методы генетической рекомбинации, подходящие для этой цели (см. Публикацию европейской патентной заявки № EP 125023; и WO 96/02576). Например, CDR антитела мыши может быть определена известными способами, а ДНК может быть получена таким образом, чтобы кодировать антитело, в котором CDR лигируется с каркасной областью (FR) антитела человека. Затем гуманизированное антитело может быть получено с использованием системы, которая использует стандартные экспрессионные векторы. Такие ДНК могут быть синтезированы с помощью ПЦР, используя в качестве праймеров несколько олигонуклеотидов, разработанных таким образом, чтобы включать части, которые перекрывают концы как областей CDR, так и FR (см. способ, описанный в WO 98/13388). FR антитела человека, связанные через CDR, выбраны таким образом, что CDR образуют подходящий антигенсвязывающий участок. Если необходимо, аминокислоты в FR из вариабельной области антитела могут быть модифицированы таким образом, что CDR реконструированного человеческого антитела могут образовывать подходящий антигенсвязывающий домен (Sato, K. и соавт., Cancer Res. (1993 год) 53:851-856). Модифицируемые аминокислотные остатки в FR включают части, которые непосредственно связываются с антигеном через нековалентные связи (Amit и соавт., Science (1986 год) 233:747-53), части, которые оказывают определенное влияние или воздействие на структуру CDR (Чотиа и соавт., J. Mol. Biol. (1987 год) 196:901-17) и части, участвующие в взаимодействии между VH и VL (EP 239400).
В одном варианте реализации изобретения тяжелые цепи и легкие цепи каждого гетеродимера антигенсвязывающей полипептидной конструкции могут быть получены из исходного антитела, которое представляет собой химерное антитело. Химерные антитела представляют собой антитела, полученные путем совмещения последовательностей, полученных от разных животных. Например, химерное антитело может быть получено путем совмещения вариабельных доменов тяжелой цепи и легкой цепи из мышиного антитела с константными доменами тяжелой цепи и легкой цепи из человеческого антитела. Химерные антитела могут быть получены известными способами. С целью получения таких химерных антител, например, ДНК, кодирующая вариабельный домен антитела, может быть лигирована с нуклеиновой кислотой, кодирующей константный домен антитела человека; полученный продукт лигирования можно вставить в экспрессионный вектор; и конструкция может быть введена в клетку-хозяина для получения химерного антитела.
Тяжелая и легкая цепи гетеродимеров могут быть получены из многочисленных исходных антител, известных в данной области техники. Большинство антител могут служить в качестве исходного антитела при условии, что они содержат полипептидную последовательность тяжелой цепи иммуноглобулина, которая спаривается с полипептидной последовательностью легкой цепи иммуноглобулина с образованием области Fab, которая связывается с антигеном. В одном варианте реализации изобретения, по меньшей мере, одно исходное антитело представляет собой терапевтическое антитело, то есть антитело, которое используется для лечения заболевания. Неограничивающие примеры подходящих терапевтических антител, содержащих легкую цепь каппа и антигены, которые они связывают, указаны в таблице А ниже:
Неограничивающие примеры подходящих терапевтических антител, содержащих легкую цепь лямбда и антигены, которые они связывают, указаны в таблице B ниже:
Подклассы иммуноглобулина
Иммуноглобулиновые тяжелые цепи исходных антител относятся к следующим классам: IgA1, IgA2, IgM, IgD, IgE, IgG1, IgG2, IgG3 и IgG4. В одном варианте реализации изобретения первый и второй гетеродимер антигенсвязывающей полипептидной конструкции содержит тяжелую цепь IgG. В одном варианте реализации изобретения первый и второй гетеродимер антигенсвязывающей полипептидной конструкции содержит тяжелую цепь IgG1. Иммуноглобулиновые легкие цепи исходных антител представляют собой либо легкие цепи каппа, либо легкие цепи лямбда.
Антигенсвязывающие полипептидные конструкции, описанные в данном документе, содержат, по меньшей мере, один гетеродимер, имеющий полипептидную последовательность тяжелой цепи иммуноглобулина и полипептидную последовательность легкой цепи каппа иммуноглобулина, и, по меньшей мере, другой гетеродимер, имеющий полипептидную последовательность тяжелой цепи иммуноглобулина и полипептидную последовательность легкой цепи лямбда иммуноглобулина. В одном варианте реализации изобретения антигенсвязывающие полипептидные конструкции содержат один гетеродимер, имеющий полипептидную последовательность тяжелой цепи IgG и полипептидную последовательность легкой цепи каппа иммуноглобулина, и другой гетеродимер, имеющий полипептидную последовательность тяжелой цепи IgG и полипептидную последовательность легкой цепи лямбда иммуноглобулина.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с доменом VH, выбранным из домена VH групп зародышевой линии IGHV1, IGHV2, IGHV3, IGHV4, IGHV5, IGHV6 или IGHV7. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с доменом VH из подгруппы зародышевой линии IGHV3. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с сегментом J, выбранным из сегмента J генов зародышевой линии IGHJ1, IGHJ2, IGHJ3, IGHJ4, IGHJ5 или IGHJ6. В другом варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с сегментом J из подгруппы зародышевой линии IGHJ4. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с доменом CH1, выбранным из домена CH1 подгрупп зародышевой линии IGHG1, IGHG2, IGHG3 или IGHG4. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с доменом CH1 из подгруппы зародышевой линии IGHG1.
Для гетеродимеров, содержащих полипептидную последовательность легкой цепи лямбда, легкие цепи лямбда могут содержать домен CL-лямбда, выбранный из генов зародышевой линии IGLC1, IGLC2, IGLC3, IGLC6 или IGLC7. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий легкую цепь лямбда, содержащую домен CL-лямбда из подгруппы зародышевой линии IGLC2. Легкие цепи лямбда могут содержать домен VL-лямбда, выбранный из подгрупп зародышевой линии IGLV1, IGLV2, IGLV3, IGLV4, IGLV5, IGLV6, IGLV7, IGLV8, IGLV9, IGLV10 или IGLV11. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер с легкой цепью лямбда, имеющий домен VL-лямбда из подгруппы зародышевой линии IGLV6. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий легкую цепь лямбда с сегментом лямбда J, выбранным сегмента J генов зародышевой линии IGLJ1, IGLJ2, IGLJ3, IGLJ6 или IGLJ7. В другом варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий тяжелую цепь с сегментом лямбда J из подгруппы зародышевой линии IGLJ2.
Для гетеродимеров, содержащих полипептидную последовательность легкой цепи каппа, легкие цепи каппа могут содержать домен CL-каппа, выбранный из аллелей CL зародышевой линии IGKC*01, IGKC*02, IGKC*03, IGKC*04 или IGKC*05. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий легкую цепь каппа, содержащую домен CL-каппа из подгруппы зародышевой линии IGKC*01. Легкие цепи каппа могут содержать домен VL-каппа, выбранный из подгрупп зародышевой линии IGKV1, IGKV1D, IGKV2, IGKV3, IGKV4, IGKV5 или IGKV6. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер с легкой цепью каппа, имеющий домен VL-каппа из подгруппы зародышевой линии IGKV1. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий легкую цепь каппа с сегментом J, выбранным сегмента J генов зародышевой линии IGKJ1, IGKJ2, IGKJ3, IGKJ4 или IGKJ5. В другом варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит гетеродимер, имеющий легкую цепь каппа с сегментом J из подгруппы зародышевой линии IGKJ1 или IGKJ2.
Тяжелые цепи иммуноглобулина обычно содержат, по меньшей мере, один вариабельный домен (VH) и три константных домена, CH1, CH2 и CH3. В одном варианте реализации изобретения каждая тяжелая цепь первого гетеродимера и второго гетеродимера антигенсвязывающей полипептидной конструкции содержит домен VH, домен CH1, домен CH2 и домен CH3. В одном варианте реализации изобретения каждая тяжелая цепь первого гетеродимера и второго гетеродимера содержит домен VH, домен CH1 и домен CH3. В еще одном варианте реализации изобретения каждая тяжелая цепь первого гетеродимера и второго гетеродимера содержит домен VH и домен CH1. Легкие цепи иммуноглобулина обычно содержат один вариабельный домен (VL) и один константный домен (CL). В одном варианте реализации изобретения легкая цепь каждого гетеродимера содержит домен VL и домен CL.
Как было указано выше, в некоторых вариантах реализации изобретения полипептидная последовательность тяжелой цепи иммуноглобулина и полипептидная последовательность легкой цепи иммуноглобулина каждого гетеродимера могут быть получены из известного терапевтического антитела, или из антитела, которое связывается с различными молекулами-мишенями или раковыми антигенами. Аминокислотные и нуклеотидные последовательности многочисленных таких молекул легко доступны (см., например, GenBank Accession №: AJ308087.1 (домен CL и вариабельная область легкой цепи гуманизированного антитела D3H44 к человеческому тканевому фактору); GenBank Accession №: AJ308086.1 (домен CH1 и вариабельная область тяжелой цепи гуманизированного антитела D3H44 к человеческому тканевому фактору); GenBank Accession №: HC359025.1 (генетический модуль легкой цепи Fab Пертузумаба); GenBank Accession №: HC359024.1 (генетический модуль тяжелой цепи Fab Пертузумаба); GenBank Accession №:GM685465.1 (антитело Трастузумаб (= Герцептин) - дикий тип; легкая цепь); GenBank Accession №:GM685463.1 (антитело Трастузумаб (= Герцептин) - дикий тип; тяжелая цепь); GenBank Accession №:GM685466.1 (антитело Трастузумаб (= Герцептин) - ГЦ-оптимизированная легкая цепь); и GenBank Accession №:GM685464. 1 (антитело Трастузумаб (= Герцептин) - ГЦ-оптимизированная тяжелая цепь. Последовательности каждого из описанных выше полипептидов доступны на веб-сайте NCBI по состоянию на 28 ноября 2012 года и содержание которых включено в данный документ посредством ссылки в полном объеме для любых целей. Аминокислотные и нуклеотидные последовательности для цетуксимаба также известны в данной области техники, см., например, веб-сайт Drug Bank при поддержке Канадского института исследований в области здравоохранения, Alberta Innovates - Health Solutions и Инновационного центра метаболомики (TMIC - The Metabolomics Innovation Centre), Accession № DB00002.
Аминокислотные модификации, которые способствуют преимущественному спариванию
Одна или более тяжелых и легких цепей H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями, которые сконструированы в тяжелые и легкие цепи исходных антител. В одном варианте реализации изобретения две из тяжелых и легких цепей H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями. В одном варианте реализации изобретения три из тяжелых и легких цепей H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями.
В некоторых вариантах реализации изобретения аминокислотные модификации могут быть асимметричными, таким образом, что модифицированные аминокислотные положения отличаются между H1 и H2, и между L1 и L2.
В одном варианте реализации изобретения H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями, тогда как H1 и L1 не содержат таких аминокислотных модификаций. В одном варианте реализации изобретения H1, L1 и H2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями, тогда как L2 не содержит таких аминокислотных модификаций. В одном варианте реализации изобретения H1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями, тогда как L1 не содержит таких аминокислотных модификаций. В одном варианте реализации изобретения L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию между тяжелыми и легкими цепями, тогда как H1 не содержит таких аминокислотных модификаций.
В одном варианте реализации изобретения одна или более аминокислотных модификаций содержат одну или более аминокислотных замен. Аминокислотные модификации способствуют преимущественному спариванию H1 с L1 и H2 с L2, в случае если H1 или H2 коэкспрессируются с L1 и L2, или в случае если коэкспрессируются H1, L1, H2 и L2. Как указано выше, с целью иллюстрации, гетеродимеры антигенсвязывающей полипептидной конструкции будут определены следующим образом:гетеродимер H1L1 содержит легкую цепь лямбда L1, а гетеродимер H2L2 содержит легкую цепь каппа L2.
В данном контексте "конструкция мАт" или "набор конструкций мАт" относится к определенному набору аминокислотных модификаций, которые способствуют преимущественному спариванию, которые присутствуют в одном наборе H1, L1, H2 и L2, а также определяются как H1L1H2L2. Аминокислотные модификации в одной или более из H1, L1, H2 и L2, которые способствуют преимущественному спариванию, упоминаются и представлены в виде конструкций мАт или наборов мАт (то есть H1L1H2L2). Наборы конструкций мАт изначально тестируются как наборы LCCA (то есть H1L1L2 или H2L1L2) для определения силы специфичности спаривания, где H1 и H2 отдельно коэкспрессируются с L1 и L2.
В одном варианте реализации изобретения аминокислотные модификации могут быть сделаны с одной или более аминокислотами, которые являются частью области контакта между легкой цепью и тяжелой цепью. В одном варианте реализации изобретения аминокислотные модификации, введенные в последовательности полипептида тяжелой цепи иммуноглобулина и последовательности полипептида легкой цепи иммуноглобулина, являются комплементарными друг к другу. Комплементарность в области контакта тяжелой и легкой цепи может быть достигнута на основе стерических и гидрофобных контактов, электростатических/зарядовых взаимодействий или их комбинации и множества других взаимодействий. Комплементарность между белковыми поверхностями широко описана в литературе в терминах соответствия замка и ключа, выступа-во-впадину, выступа и полости, донора и акцептора и т.д., всего, что подразумевает характер структурной и химической совместимости между двумя взаимодействующими поверхностями. В одном варианте реализации изобретения, по меньшей мере, один из гетеродимеров содержит аминокислотную модификацию, введенную в тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина, которая вносит новую водородную связь в область контакта между легкой и тяжелой цепью. В одном варианте реализации изобретения, по меньшей мере, один из гетеродимеров содержит аминокислотную модификацию, введенную в тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина, которая вносит новый соляной мостик в область контакта между легкой и тяжелой цепью.
В одном варианте реализации изобретения аминокислотные модификации набора конструкции мАт способствуют преимущественному спариванию в основном посредством электростатического притяжения и отталкивания. В одном варианте реализации изобретения аминокислотные модификации набора конструкции мАт способствуют преимущественному спариванию через преимущественно пространственные механизмы. Такие конструкции мАт включены в таблицы 4A, 4B, 7A и 7B, с выбранными примерами, включая те, которые имеют уникальные идентификаторы 10771-11335, 10771-11360, 10780-11417. В одном варианте реализации изобретения аминокислотные модификации набора конструкции мАт способствуют преимущественному спариванию с использованием как пространственных, так и электростатических механизмов.
В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, причем H1 и L1 не включают аминокислотных модификаций, которые способствуют преимущественному спариванию, а H2 и L2 каждая содержат, по меньшей мере, одну аминокислотную модификацию, которая способствует преимущественному спариванию. В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, причем одна или более из H1, L1 и H2 содержат, по меньшей мере, одну аминокислотную модификацию, которая способствует преимущественному спариванию, а L2 не включает аминокислотные модификации, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, причем одна или более из H1, L1 и L2 содержат, по меньшей мере, одну аминокислотную модификацию, которая способствует преимущественному спариванию, а H2 не включает аминокислотные модификации, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, причем одна или более из H1, H2 и L2 содержат, по меньшей мере, одну аминокислотную модификацию, которая способствует преимущественному спариванию, а L1 не включает аминокислотные модификации, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, причем одна или более из L1, H2 и L2 содержат, по меньшей мере, одну аминокислотную модификацию, которая способствует преимущественному спариванию, а H1 не включает аминокислотные модификации, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, причем каждая из L1, H2 и L2 содержит, по меньшей мере, одну аминокислотную модификацию, которая способствует преимущественному спариванию.
Аминокислотные модификации могут встречаться в константных доменах и/или в вариабельных доменах одной или более из H1, L1, H2 и L2. В одном варианте реализации изобретения аминокислотные модификации могут встречаться в доменах CH1 H1 и H2, домене CL-лямбда L1 и домене CL-каппа L2. В другом варианте реализации изобретения аминокислотные модификации могут встречаться в доменах CH1 и VH H1 и H2, доменах CL-лямбда и VL-лямбда L1 и доменах CL-каппа и VL-каппа L2. В другом варианте реализации изобретения аминокислотные модификации могут встречаться в доменах VH H1 и H2, домене VL-лямбда L1 и домене VL-каппа L2.
В одном варианте реализации изобретения аминокислотные модификации могут находиться в каркасных областях одной или более из H1, L1, H2 и L2. В одном варианте реализации изобретения аминокислотные модификации ограничиваются консервативными остатками каркаса вариабельных (VH, VL) и константных (CH1, CL) доменов, как обозначено в нумерации остатков по Кабату. Например, Almagro [Frontiers In Bioscience (2008 год) 13:1619-1633] предоставляет определение остатков каркасной области на основе схем нумерации Кабат, Чотиа и IMGT.
Количество аминокислотных модификаций в каждой конструкции мАт или наборе конструкции мАт может варьироваться. В одном варианте реализации изобретения H1 содержит от 0 до 8 аминокислотных модификаций, от 0 до 7 аминокислотных модификаций, от 0 до 6 аминокислотных модификаций, от 0 до 5 аминокислотных модификаций, от 0 до 4 аминокислотных модификаций, от 0 до 3 аминокислотных модификаций, от 0 до 2 аминокислотных модификаций, одну аминокислотную модификацию или не содержит аминокислотных модификаций. В одном варианте реализации изобретения L1 содержит от 0 до 8 аминокислотных модификаций, от 0 до 7 аминокислотных модификаций, от 0 до 6 аминокислотных модификаций, от 0 до 5 аминокислотных модификаций, от 0 до 4 аминокислотных модификаций, от 0 до 3 аминокислотных модификаций, от 0 до 2 аминокислотных модификаций, одну аминокислотную модификацию или не содержит аминокислотных модификаций. В одном варианте реализации изобретения H2 содержит от 0 до 8 аминокислотных модификаций, от 0 до 7 аминокислотных модификаций, от 0 до 6 аминокислотных модификаций, от 0 до 5 аминокислотных модификаций, от 0 до 4 аминокислотных модификаций, от 0 до 3 аминокислотных модификаций, от 0 до 2 аминокислотных модификаций, одну аминокислотную модификацию или не содержит аминокислотных модификаций. В одном варианте реализации изобретения L2 содержит от 0 до 8 аминокислотных модификаций, от 0 до 7 аминокислотных модификаций, от 0 до 6 аминокислотных модификаций, от 0 до 5 аминокислотных модификаций, от 0 до 4 аминокислотных модификаций, от 0 до 3 аминокислотных модификаций, от 0 до 2 аминокислотных модификаций, одну аминокислотную модификацию или не содержит аминокислотных модификаций.
В одном варианте реализации изобретения общее количество аминокислотных модификаций в H1, L1, H2 и L2 составляет менее чем 20, менее чем 15, менее чем 12, менее чем 11, менее чем 10, менее чем 9, менее чем 8, менее чем 7, менее чем 6, менее чем 5, менее чем 4 или менее чем 3. В одном варианте реализации изобретения общее количество аминокислотных модификаций в H1, L1, H2 и L2 равно 2.
В одном варианте реализации изобретения аминокислотные модификации могут быть сконструированы специально для системы каппа-лямбда, где одно исходное антитело содержит полипептидную последовательность легкой цепи каппа и одно исходное антитело содержит полипептидную последовательность легкой цепи лямбда. Такие аминокислотные модификации или конструкции упоминаются в данном документе как конструкции K-L. Примеры таких аминокислотных модификаций или конструкций K-L представлены в таблице 4A, таблице 7A и таблицах с 10-A1 по 10-A12.
В другом варианте реализации изобретения аминокислотные модификации могут быть первоначально идентифицированы по отношению к системе каппа-каппа (конструкции K-K), где оба исходных антитела содержат полипептидную последовательность легкой цепи каппа, и впоследствии перенесены в систему каппа-лямбда. Для специалиста в данной области техники будет понятно, как эти конструкции можно перенести в систему каппа-лямбда. Например, тяжелые и легкие цепи исходных антител каппа и лямбда могут быть выровнены для определения эквивалентных положений легкой цепи лямбда, соответствующих конструкциям K-K. Эквивалентные положения легкой цепи лямбда могут быть затем изменены в соответствии с конструкцией K-K. Такие аминокислотные модификации или конструкции упоминаются в данном документе как конструкции K-L, полученные из K-K и могут относиться к следующим группам: а) те, в которых изменения не требуются для конструкций, а аминокислотные остатки, модифицированные в системе каппа-каппа, идентичны аминокислотам, модифицированным в системе каппа-лямбда; b) те, которые содержат молчащие модификации, где, по меньшей мере, одна модификация, сделанная в системе каппа-каппа, является не нужной в системе каппа-лямбда, поскольку эта модификация существует естественным образом в полипептидной последовательности легкой цепи лямбда; c) те, которые содержат аминокислотные модификации, по меньшей мере, одного аминокислотного остатка в том же относительном положении в полипептидной последовательности легкой цепи каппа и полипептидной последовательности легкой цепи лямбда, но где исходный аминокислотный остаток в этих положениях отличается между полипептидными последовательностями легкой цепи каппа и лямбда, что приводит к той же аминокислотной модификации в положении, и d) те, которые содержат, по меньшей мере, одну дополнительную аминокислотную модификацию в системе каппа-лямбда по сравнению с системой каппа-каппа. Примеры таких конструкций K-L, полученных из K-K, приведены в таблице 4B, таблице 7B и в таблицах с 10-B1 по 10-B10. Конкретные примеры группы a) отмечены звездочкой в таблице 4B. Конкретный пример группы b) демонстрируется набором конструкции мАт с уникальным идентификатором 10689-10707. Молчащая модификация находится в L1 (Q160E отсутствует в ДТ в лямбда, так как остаток в положении 160 является E, а не Q). Конкретный пример группы c) демонстрируется набором конструкции мАт с уникальным идентификатором 10652-10734, где в L1 аминокислотный остаток 124 представляет собой E в ДТ лямбда и Q в ДТ каппа. Конкретный пример группы b) демонстрируется набором конструкции мАт с уникальным идентификатором 10684-10706, который включает модификацию аминокислоты K129T.
В одном варианте реализации изобретения одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и H2 с L2 по сравнению с L1, где аминокислотные модификации содержат консервативные аминокислотные замены наборов конструкции мАт, представленных в таблице 4A, таблице 4B, таблице 7A, таблице 7B, таблицах с 10-A1 по 10-A12 и таблицах с 10-B1 по 10-B10.
В одном варианте реализации изобретения аминокислотные модификации не вносят новый остаток цистеина и не удаляют встречающийся в природе остаток цистеина в тяжелых цепях иммуноглобулина или легких цепях иммуноглобулина в пределах одной и той же конструкции.
Комбинация аминокислотных модификаций в H1, L1, H2 и L2, которые способствуют преимущественному спариванию, в общем относится к конструкциям. Конструкции могут быть более конкретно обозначены как "конструкции LCCA" (в контексте H1, L1, L2 или H2, L1, L2) или "конструкции мАт" (в контексте H1, L1, H2, L2). Как правило, конструкции LCCA сконструированы с одной или более конкретными комплементарными конструкциями LCCA на основе каждой тяжелой цепи желаемого биспецифического антитела и соответственно обычно представлены в формате, в котором идентифицированы модификации аминокислот во всех четырех полипептидных цепях биспецифического антитела (см., например, таблицы 4А и 4В). Хотя конкретные аминокислотные замены могут быть идентифицированы повсюду, следует понимать, что консервативная замена в каждом положении аминокислоты также может быть рассмотрена. Кроме того, для иллюстрации, гетеродимер H1L1 представляет собой гетеродимер, содержащий легкую цепь лямбда, а гетеродимер H2L2 представляет собой гетеродимер, содержащий легкую цепь каппа, если не указано иное. Наконец, все аминокислотные остатки или положения нумеруются в соответствии с системой нумерации Кабата, если не указано иное.
Конструкции содержат наборы драйверов комплементарных аминокислотных замен, которые способствуют преимущественному спариванию, и могут также содержать вторичные замены. Вторичные замены могут действовать с целью оптимизации эффективности наборов драйверов.
Один или более наборов драйверов могут использоваться для способствования преимущественному спариванию. Эти наборы драйверов могут использоваться индивидуально или в комбинации для способствования преимущественному спариванию. В одном варианте реализации изобретения набор драйверов представляет собой набор электростатических драйверов, в котором, как ожидается, электростатическое притяжение и отталкивание являются преобладающими факторами, способствующими преимущественному спариванию. Например, конструкция, в которой H1 содержит аминокислотную замену 186K, L1 содержит аминокислотную замену 133D, H2 содержит аминокислотную замену 188D, а L2 содержит аминокислотную замену 131K, может способствовать преимущественному спариванию с помощью электростатического механизма. Во всех примерах можно найти множество других примеров электростатических драйверов. В одном варианте реализации изобретения один или более наборов электростатических драйверов могут быть выбраны из набора, указанного в таблице C:
Таблица C: Типовые наборы электростатических драйверов
L143D_Q179E
L124E/L124E_Q 179E
L143K_V190K
или
L124K_L143K
В одном варианте реализации изобретения набор драйверов представляет собой набор дисульфидных направляющих драйверов, который может оказать неблагоприятное воздействие на формирование дисульфидной связи в неправильно спаренных гетеродимерах. Пример этого типа набора драйверов будет включать 125R в H1, 122D в L1, 228D в H2 и 121K в L2.
В одном варианте реализации изобретения набор драйверов может представлять собой набор стерических драйверов, который может способствовать стерически комплементарным взаимодействиям между правильно спаренными гетеродимерами и стерической несовместимости между неправильно спаренными гетеродимерами. Неограничивающие примеры наборов стерических драйверов представлены в таблице D, где "-" указывает на отсутствие аминокислотных замен, которые способствует преимущественному спариванию.
Таблица D: Типовые наборы стерических драйверов
В одном варианте реализации изобретения набор драйверов представляет собой набор драйверов переменной конструкции. Такие наборы драйверов с переменной конструкцией содержат одну или более аминокислотных модификаций в вариабельных доменах каппа и/или лямбда Fab, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения набор драйверов переменной конструкции способствует преимущественному спариванию на основе стерических механизмов. В одном варианте реализации изобретения набор драйверов переменной конструкции способствует преимущественному спариванию на основе электростатических механизмов. Неограничивающие примеры наборов драйверов переменной конструкции представлены в таблице E, где "-" указывает на отсутствие аминокислотных модификаций в этом полипептиде, что способствует преимущественному спариванию.
В одном варианте реализации изобретения одна или более не встречающихся в природе дисульфидных связей могут быть сконструированы в один или оба гетеродимера антигенсвязывающей полипептидной конструкции. Примером такого типа аминокислотной модификации является тот, в котором тяжелая цепь содержит замену 122C, спаренную с заменой 124C в легкой цепи каппа.
Вторичные замены могут быть включены в конструкцию с целью оптимизации эффективности спаривания конструкции. Например, вторичные замены могут действовать с целью A) оптимизации количества контактов между тяжелой и правильно спаренной легкой цепью, B) обеспечения благоприятных условий для драйверов, C) оптимизации сети водородной связи для наборов драйверов, или D) обеспечения стерического размещения драйверов. Неограничивающие примеры этих типов вторичных замен приведены в таблице F, где "Lk" обозначает специфическую замену легкой цепи каппа, "L1" обозначает специфическую замену легкой цепи лямбда. "L" обозначает специфическую замену легкой цепи как в легкой цепи каппа так и в лямбда, а "H" обозначает специфическую замену тяжелой цепи.
Таблица F: Типовые вторичные замены
Антигенсвязывающие полипептидные конструкции могут быть сконструированы с различными комбинациями аминокислотных модификаций, соответствующих наборам драйверов и вторичным заменам, описанным выше. Неограничивающие конкретные примеры таких комбинаций, сгруппированных в кластеры на основе общих признаков, описаны ниже. Как описано в данном документе, комбинации аминокислотных модификаций в нескольких положениях в пределах одной цепи идентифицируются с использованием "_" между каждым измененным положением. Например, "124_186" указывает, что оба положения 124 и 186 модифицированы в упомянутой полипептидной цепи. Аналогичным образом, "124_133_180" указывает, что все положения 124, 133 и 180 модифицированы в упомянутой полипептидной цепи.
Кластер K-L 1:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 1, при этом:
H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 131; а также
a) H2 содержит аминокислотную замену в положениях 188 или 124_186, а L2 содержит аминокислотную замену в положениях 176_178 или 176_180, или 131; или
b) H2 содержит аминокислотную замену в положении 143 или 186; а L2 содержит аминокислотные замены в положениях 124_133 или 124_133_180.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 125, 145 и 179, L1 дополнительно содержит аминокислотные замены в одном или более положениях 122, 124 и 133, H2 дополнительно содержит аминокислотную замену в положении 228, и/или L2 дополнительно содержит аминокислотные замены в положении 121.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 1, при этом: H1 содержит аминокислотную замену в положениях 125_143_145; L1 содержит аминокислотную замену в положениях 122_124_131; и H2 содержит аминокислотную замену в положении 228, а L2 содержит аминокислотную замену в положениях 121_133. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 179, L1 дополнительно содержит аминокислотную замену в положении 133, H2 дополнительно содержит аминокислотную замену в одном или более положениях 124, 143, 186 и 188, а L2 дополнительно содержит аминокислотные замены в одном или более положениях 124, 131, 176, 178 и 180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 1, при этом H1 содержит аминокислотные замены в положениях 125_143_145 или 125_143_145_179; L1 содержит аминокислотные замены в положениях 122_124_131 или 122_124_131_133; H2 содержит аминокислотные замены в положениях 124_186_228, 143_228, 143_186_228, 186_228 или 188_228, а L2 содержит аминокислотные замены в положениях 121 _ 124_ 133, 121_124_133_180, 121_131_133_178, 121_133_176_178 или 121_133_176_180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 1, при этом аминокислотные замены в H1 выбраны из 125R, 145T, 143D, 143E, 179E и их консервативных замен; аминокислотные замены в L1 выбраны из 124Q, 122D, 131K, 131R, 133S и их консервативных замен; аминокислотные замены в H2 выбраны из 228D, 124R, 143I, 143R, 186K, 186R, 188K и их консервативных замен; аминокислотные замены в L2 выбраны из 124E, 121K, 131D, 176D, 178E, 178F, 180D, 180E, 133D, 133G, 133I и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 1, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В одном варианте реализации изобретения H1 содержит 125R_143E_145T_179E, а L1 содержит 122D_124Q_131R. В другом варианте реализации изобретения H1 содержит 125R_143E_145T, а L1 содержит 122D_124Q_131R. В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 1, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения H1 содержит 125R_143E_145T_179E, L1 содержит 122D_124Q_131R, H2 содержит 188K_228D, а L2 содержит 121K_133I_176D_178E. В другом варианте реализации изобретения H1 содержит 125R_143D_145T, L1 содержит 122D_124Q_131R, H2 содержит 143R_228D, а L2 содержит 121K_124E_133D.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 1, содержащую набор дисульфидных направляющих драйверов.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 1 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 1, как указано в одной или более конструкциях в таблице 10-A1.
Кластер K-L 2:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 2, при этом:
H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 131; а также
c) H2 содержит аминокислотную замену в положениях 186 или 124_186, а L2 содержит аминокислотную замену в положениях 133 или 133_160, или 124_133, или 176_180; или
d) H2 содержит аминокислотную замену в положении 188; а L2 содержит аминокислотные замены в положении 131;
e) H2 содержит аминокислотную замену в положении 143; а L2 содержит аминокислотные замены в положениях 124_133 или 124_133_180;
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 139, 145, 174 и 179, L1 дополнительно содержит аминокислотные замены в одном или более положениях 116, 124, 133 и 176, H2 дополнительно содержит аминокислотную замену в одном или более положениях 190, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 135, 178, 38 и 44.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 2, причем H1 содержит аминокислотную замену в положениях 143_145; L1 содержит аминокислотную замену в положении 131; и H2 содержит аминокислотные замены в положении 143 или 186, или 188, а L2 содержит аминокислотную замену в положении 133. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 139, 174 и 179, L1 дополнительно содержит аминокислотную замену в одном или более положениях 116, 124, 133 и 176, H2 дополнительно содержит аминокислотную замену в одном или более положениях 124, 190, 39 и 45, и L2 дополнительно содержит аминокислотные замены в одном или более положениях 124, 131, 135, 160, 176, 178, 180, 38, 44.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 2, причем H1 содержит аминокислотные замены в положениях 139_143_145, 143_145, 143_145_174 или 143_145_179; L1 содержит аминокислотные замены в положениях 116_ 124_ 131_176, 124_131, 124_131_133, 124_131_133_176, 131 или 131_133; H2 содержит аминокислотные замены в положениях 124_186_190, 143, 143_186, 143_186_190, 143_190, 186, 186_190, 188, 39_143 или 45_143, и L2 содержит аминокислотные замены в положениях 124_133, 124_133_135, 124_133_135_180, 124_133_180, 131_133_135_178, 131_133_178, 133, 133_135_176_180, 133_160, 38_124_133 или 44_124_133.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 2, при этом аминокислотные замены в H1 выбраны из 139W, 143D, 145T, 174G, 179E и их консервативных замен; аминокислотные замены в L1 выбраны из 124Q, 176F, 116F, 131K, 131R, 133S и их консервативных замен; аминокислотные замены в H2 выбраны из 124R, 143I, 143K, 143R, 186K, 188K, 190F, 39E, 45P и их консервативных замен; аминокислотные замены в L2 выбраны из 124E, 131D, 131E, 133D, 133G, 135A, 135W, 160E, 176D, 178F, 180D, 180E, 38R, 44F и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 2, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 2, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 2, имеющую один или более наборов стерических 1, стерических 2, стерических или электростатических драйверов вариабельного домена.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 2 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 2, как указано в одной или более конструкциях в таблице 10-A2.
Кластер K-L 3:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 3, при этом:
H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 131; а также
a) H2 содержит аминокислотную замену в положениях 124_186 или 124_179, или 188, а L2 содержит аминокислотную замену в положениях 176_178 или 176_180, или 131; или
b) H2 содержит аминокислотную замену в положениях 143_188 или 143, или 124_143; а L2 содержит аминокислотные замены в положениях 124_176_178 или 124_178, или 124_180, или 124_176_180, или 124, или 124_176.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 139, 145, 174 и 179, L1 дополнительно содержит аминокислотную замену в одном или более положениях 116, 124 и 176, H2 дополнительно содержит аминокислотную замену в одном или более положениях 177, 190, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 133, 135, 178, 38 и 44.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 3, при этом: H1 содержит аминокислотную замену в положениях 143_145; L1 содержит аминокислотную замену в положениях 124_131; и H2 содержит аминокислотную замену в одном или более положениях 143, 124 и 188, а L2 содержит аминокислотную замену в положениях 133 или 176_178. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 139, 174 и 179, L1 дополнительно содержит аминокислотную замену в положении 116 и 176, H2 дополнительно содержит аминокислотную замену в одном или более положениях 177, 179, 186, 190, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 124, 131, 135, 180, 38 и 44.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 3, причем H1 содержит аминокислотные замены в положениях 139_143_145, 139_143_145_179, 143_145, 143_145_174_179 или 143_145_179; L1 содержит аминокислотные замены в положениях 116_124_131_176 или 124_131; H2 содержит аминокислотные замены в положениях 124_143, 124_179, 124_186, 143, 143_188, 177_188, 188, 188_190, 39_124_179 или 45_124_179, и L2 содержит аминокислотные замены в положениях 124_133, 124_133_176, 124_ 133_ 176_ 178, 124_ 133_ 176_ 180, 124_133_178, 124_133_180, 131_133_178, 133_135_176_178, 133_135_176_180, 133_176_178, 133_176_180, 176_178, 38_133_176_180 или 44_133_176_180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 3, при этом аминокислотные замены в H1 выбраны из 139W, 174G, 145T, 143D, 143E, 179E и их консервативных замен; аминокислотные замены в L1 выбраны из 116F, 124Q, 131K, 131R, 176F и их консервативных замен; аминокислотные замены в H2 выбраны из 124R, 143K, 143R, 177I, 179K, 186K, 186R, 188K, 190F, 39E, 45P и их консервативных замен; аминокислотные замены в L2 выбраны из 135A, 135W, 44F, 124E, 38R, 131D, 131E, 176D, 176E, 178D, 178E, 178F, 180D, 180E, 133D, 133G, 133I, 133L и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 3, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 3, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 3, имеющую один или более наборов стерических 1, стерических 2, стерических или электростатических драйверов вариабельного домена.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 3 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 3, как указано в одной или более конструкциях в таблице 10-A3.
Кластер K-L 4:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, при этом: H1 содержит аминокислотную замену в положении 188; L1 содержит аминокислотную замену в положениях 176_178 или 178; и
a) H2 содержит аминокислотную замену в положениях 177_188, а L2 содержит аминокислотную замену в положениях 176_178; или
b) H2 содержит аминокислотную замену в положении 186 или 124, или 124_179; а L2 содержит аминокислотные замены в положениях 176 или 131_176.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 125, 139 и 177, L1 дополнительно содержит аминокислотную замену в одном или более положениях 122, 129 и 133, H2 дополнительно содержит аминокислотную замену в одном или более положениях 145, 228, 45 и 39, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 135, 44, 38, 121 и 133.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, причем: H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положении 188; L1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положениях 176_178; H2 содержит аминокислотную замену в положениях 188 или 186_188, и L2 содержит аминокислотную замену в положениях 176_178.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, причем: H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положении 188; L1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положениях 178; H2 содержит аминокислотную замену в одном или более положениях 124, 186 и 188, а L2 содержит аминокислотную замену в положении 176. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 125, 139 и 177, L1 дополнительно содержит аминокислотную замену в одном или более положениях 122, 129, 133 и 176, H2 дополнительно содержит аминокислотную замену в одном или более положениях 145, 228, 45, 177, 179 и 39, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 135, 44, 38, 121, 131, 178 и 133.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, причем H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотные замены в положениях 125_188, 139_188, 188 или 177_188; L1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотные замены в положениях 129_176_178, 129_ 178, 122_129_176_178, 176_178 или 133_176_178; H2 содержит аминокислотные замены в положениях 145_186, 145_186_228, 145_177_188, 124, 124_145_179, 124_145_179_186_188, 124_145_179_188, 124_186_188, 124_188, 45_124_145_179, 39_124_145_179, или 186_188 и L2 содержит аминокислотные замены в положениях 44_131_133_176, 38_131_133_176, 121_131_176, 131_135_176, 131_176, 131_133_176, 131_133_176_178, 176, 176_178, 133_176 или 133_176_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, при этом аминокислотные замены в H1 выбраны из 125R, 139W, 188A, 188K и 177I и их консервативных замен; аминокислотные замены в L1 выбраны из 129T, 122D, 176A, 176D, 176E, 133I, 133L, 178D, 178E, 178T и 178W и их консервативных замен; аминокислотные замены в H2 выбраны из 124E, 145T, 177D, 179E, 186E, 186I, 186L, 188D, 188W, 228D, 39E и 45P и их консервативных замен; аминокислотные замены в L2 выбраны из 121K, 131K, 131R, 133A, 133G, 135W, 176A, 176K, 176R, 176V, 178A, 178K, 178L, 178R, 38R и 44F и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 4, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 4, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, имеющую один или более наборов электростатических, дисульфидных направляющих, стерических 3, стерических или электростатических драйверов вариабельного домена.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 4 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 4, как указано в одной или более конструкциях в таблице 10-A4.
Кластер K-L 5:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 5, при этом:
H1 содержит аминокислотную замену в положении 186; L1 содержит аминокислотную замену в положении 133; а также
a) H2 содержит аминокислотную замену в положении 188, а L2 содержит аминокислотную замену в положении 131; или
b) H2 содержит аминокислотную замену в положениях 177_188; а L2 содержит аминокислотные замены в положениях 176_178; или
при этом H1 содержит аминокислотную замену в положениях 124_190; L1 содержит аминокислотную замену в положении 135; Н2 содержит аминокислотную замену в положениях 124 или 188, и L2 содержит аминокислотную замену в положениях 176 или 176_178.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 143 и/или 188, L1 дополнительно содержит аминокислотную замену в положении 131 и/или 178, H2 дополнительно содержит аминокислотную замену в положениях 143 и/или 145, и/или L2 дополнительно содержит аминокислотные замены в положении 133 и/или 178.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 5, причем H1 содержит аминокислотную замену в положении 186 или 124; L1 содержит аминокислотную замену в положении 133 и/или 135; и H2 содержит аминокислотные замены в одном или более положениях 188, 177 и 124, а L2 содержит аминокислотную замену в положении 176 и/или 131.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 143, 188 и 190, L1 дополнительно содержит аминокислотную замену в положениях 131 и/или 178, H2 дополнительно содержит аминокислотную замену в положениях 143 и/или 145, и/или L2 дополнительно содержит аминокислотные замены в положениях 133 и/или 178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 5, причем H1 содержит аминокислотные замены в положениях 124_190, 143_186_188 или 186_188; L1 содержит аминокислотные замены в положениях 131_133_178, 133_135, 133_135_178, 133_178 или 135_178; H2 содержит аминокислотные замены в положениях 124, 143_188, 145_177_188 или 177_188, а L2 содержит аминокислотные замены в положениях 131_176_178, 131_178, 133_176, 133_176_178 или 176_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 5, при этом аминокислотные замены в H1 выбраны из 124E, 143S, 186K, 188T, 190D, 190E и их консервативных замен; аминокислотные замены в L1 выбраны из 131S, 133D, 133I, 135K, 135R, 178F, 178T и их консервативных замен; аминокислотные замены в H2 выбраны из 124R, 143T, 145T, 177D, 177I, 188D, 188K и их консервативных замен; аминокислотные замены в L2 выбраны из 131K, 133G, 133L, 176A, 176D, 176K, 178E, 178K, 178R, 178S и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 5, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 5, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 является таковой, где H1 содержит 186K_188T, L1 содержит 133D_178T, H2 содержит 145T_177D_188D, а L2 содержит 176K_178K.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 5 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 5, как указано в одной или более конструкциях в таблице 10-A5.
Кластер K-L 6:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 6, при этом:
H1 содержит аминокислотную замену в положениях 177_188; L1 содержит аминокислотную замену в положении 176_178; и
a) Н2 содержит аминокислотную замену в положении 188, и L2 содержит аминокислотную замену в положениях 176_178 или 131;
b) H2 содержит аминокислотную замену в положении 186; а L2 содержит аминокислотные замены в положениях 133 или 124_160_180;
c) Н2 содержит аминокислотную замену в положении 124 или 124_179, или 124_186; и L2 содержит аминокислотные замены в положениях 176 или 176_178, или 176_180; или
d) H2 содержит аминокислотную замену в положении 143; а L2 содержит аминокислотные замены в положениях 133 или 124_133.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в 145 и/или 146, и/или H2 дополнительно содержит аминокислотную замену в одном или более положениях 143 и/или 177.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 6, при этом:
H1 содержит аминокислотную замену в положениях 177_188; L1 содержит аминокислотную замену в положениях 176_178; и H2 содержит аминокислотную замену в одном или более положениях 124, 143, 179, 186 и 188, а L2 содержит аминокислотную замену в одном или более положениях 133, 176 и 178.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положениях 145 и/или 146, H2 дополнительно содержит аминокислотную замену в положении 177, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 124, 131, 160 и 180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 6, причем H1 содержит аминокислотные замены в положениях 145_177_188 или 146_177_188; L1 содержит аминокислотные замены в положениях 176_178; H2 содержит аминокислотные замены в положениях 124, 124_179, 124_186, 143, 143_186_188, 177_188, 179, 186, 186_188 или 188, а L2 содержит аминокислотные замены в положениях 124_133, 124_160_176_178_180, 124_160_180, 131_133_178, 133, 133_176, 133_176_178, 133_176_180 или 176_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 6, при этом аминокислотные замены в H1 выбраны из 145T, 146T, 177D, 188D и их консервативных замен; аминокислотные замены в L1 выбраны из 176K, 178K, 178L, 178R и их консервативных замен; аминокислотные замены в H2 выбраны из 124R, 143K, 143R, 143S, 177I, 179K, 186K, 186R, 188K, 188T, 188W и их консервативных замен; аминокислотные замены в L2 выбраны из 124E, 131D, 131E, 133D, 133G, 133I, 133L, 160E, 176A, 176D, 176E, 178A, 178D, 178E, 178F, 180E и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 6, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 6, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 6, имеющую набор стерических драйверов.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 6 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 6, как указано в одной или более конструкциях в таблице 10-A6.
Кластер K-L 7:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 7, при этом: H1 содержит аминокислотную замену в положении 188; L1 содержит аминокислотную замену в положении 178; H2 содержит аминокислотную замену в положении 124 или 188, а L2 содержит аминокислотную замену в положениях 176_ 178 или 176-180, или 176. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 125, 139, 145 и 177, L1 дополнительно содержит аминокислотную замену в положении 122, H2 дополнительно содержит аминокислотную замену в одном или более положениях 143, 177, 179, 186, 228, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 121, 124, 133, 135, 160, 38 и 44.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 7, при этом: H1 содержит аминокислотную замену в положениях 145_188; L1 содержит аминокислотную замену в положении 178; и H2 содержит аминокислотную замену в положении 124 и/или 188, а L2 содержит аминокислотную замену в одном или более положениях 124, 133 и 178.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 125, 139 и 177, L1 дополнительно содержит аминокислотную замену в положении 122, H2 дополнительно содержит аминокислотную замену в одном или более положениях 143, 177, 179, 186, 228, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 121, 135, 160, 176, 180, 38 и 44.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 7, причем H1 содержит аминокислотные замены в положениях 125_145_188, 139_145_188, 145_177_188 или 145_188; L1 содержит аминокислотные замены в положениях 122_178 или 178; H2 содержит аминокислотные замены в положениях 124, 124_143, 124_179, 124_186, 124_186_228, 124_188, 124_228, 143_188, 177_188, 179_188, 186_188, 188, 188_228, 39_124_179 или 45_124_179, а L2 содержит аминокислотные замены в положениях 121_133_176, 121_133_176_180, 121_176_178, 124_133_176, 124_133_176_178, 124_133_176_178_180, 124_133_176_180, 124_133_178, 124_160_176_178, 124_160_176_178_180, 124_176_178_180, 124_176_180, 133_135_176, 133_135_176_180, 133_176, 133_176_178, 133_176_180, 135_176_178, 176_178, 38_133_176_180 или 44_133_176_180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 7, при этом аминокислотные замены в H1 выбраны из 125R, 139W, 145T, 177T, 188E и их консервативных замен; аминокислотные замены в L1 выбраны из 122D, 178K и их консервативных замен; аминокислотные замены в H2 выбраны из 124K, 124R, 143K, 143R, 177I, 179K, 186R, 188K, 228D, 39E, 45P и их консервативных замен; аминокислотные замены в L2 выбраны из 121K, 124E, 133D, 133G, 133L, 135W, 160E, 176D, 176E, 178D, 178E, 180E, 38R, 44F и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 7, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 7, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция кластера K-L 7 содержит комбинацию аминокислотных замен, имеющую один или более наборов драйверов, выбранных из набора дисульфидного направляющего драйвера, набора стерического 2 драйвера и набора драйвера вариабельного домена.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 7 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 7, как указано в одной или более конструкциях, изложенных в таблице 10-A7.
Кластер K-L 8:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, при этом:
H2 содержит аминокислотную замену в положении 143; L2 содержит аминокислотную замену в положении 124; и
a) H1 содержит аминокислотную замену в положении 186 или 179, а L1 содержит аминокислотную замену в положении 180;
b) H1 содержит аминокислотную замену в положении 186; а L1 содержит аминокислотные замены в положении 133;
c) H1 содержит аминокислотную замену в положении 143; а L1 содержит аминокислотные замены в положении 133; или
d) H1 содержит аминокислотную замену в положении 188; а L1 содержит аминокислотные замены в положении 178.
В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 124, 139, 177 и 190, L1 дополнительно содержит аминокислотную замену в одном или более положениях 129, 131, 135 и 176, H2 дополнительно содержит аминокислотную замену в одном или более положениях 122, 124, 145, 179, 186, 188, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 129, 133, 135, 160, 176, 178, 38 и 44.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, при этом: H1 содержит аминокислотную замену в положениях 143, 186, 179 и/или 188; L1 содержит аминокислотную замену в положениях 129 и/или 178; и H2 содержит аминокислотную замену в положении 143, а L2 содержит аминокислотную замену в положении 124. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 124, 139, 177 и 190, L1 дополнительно содержит аминокислотную замену в одном или более положениях 131, 133, 135, 176 и 180, H2 дополнительно содержит аминокислотную замену в одном или более положениях 122, 124, 145, 179, 186, 188, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 129, 133, 135, 160, 176, 178, 38 и 44.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, при этом H1 содержит аминокислотные замены в положениях 124_143, 139_143, 139_143_186, 139_186, 139_188, 143, 143_179, 143_186, 143_186_188, 143_190, 177_188, 179, 179_190, 186 или 186_188, 188; L1 содержит аминокислотные замены в положениях 129_ 131_133, 129_133, 129_133_135, 129_133_135_180, 129_133_178, 129_133_180, 129_176_178, 129_176_178_180, 129_178, 129_178_180, 129_180, 133_176_178, 133_178 или 176_178; H2 содержит аминокислотные замены в положениях 122_143_145, 122_143_145_179, 124_143_145, 124_143_145_179, 143_145, 143_145_179, 143_145_179_186_188, 143_145_179_188, 143_145_188, 39_143_145_179 или 45_143_145_179, и L2 содержит аминокислотные замены в положениях 124_129_160_178, 124_129_178, 124_133_178, 124_135_160_178, 124_135_178, 124_160_176_178, 124_160_178, 124_176_178, 124_178, 38_124_178 или 44_124_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, при этом аминокислотные замены в H1 выбраны из 124K, 139W, 143A, 143I, 143K, 143S, 177I, 179K, 186K, 186R, 188K, 188T, 190K и их консервативных замен; аминокислотные замены в L1 выбраны из 129T, 131D, 131E, 133D, 133L, 133W, 135S, 176A, 176D, 176E, 178D, 178E, 178T, 178W, 180E и их консервативных замен; аминокислотные замены в H2 выбраны из 122C, 124W, 143E, 145T, 179E, 186I, 188L, 188W, 39E, 45P и их консервативных замен; аминокислотные замены в L2 выбраны из 124C, 124K, 124R, 129K, 133A, 135W, 160K, 160R, 176A, 178R, 38R, 44F и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 8, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 8, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, имеющую один или более наборов стерических 2, стерических 3, стерических 4 драйверов и драйверов вариабельного домена. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, которая вводит не встречающуюся в природе дисульфидную связь.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 8 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 8, как указано в одной или более конструкциях в таблице 10-A8.
Кластер K-L 9:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 9, при этом:
H1 содержит аминокислотную замену в положении 179, 186, 143 и/или 188; L1 содержит аминокислотную замену в положении 180, 133 и/или 176_178; H2 содержит аминокислотную замену в положении 143, а L2 содержит аминокислотную замену в положении 131 и/или 124. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 125, L1 дополнительно содержит аминокислотную замену в положении 122 или 129, H2 дополнительно содержит аминокислотную замену в одном или более положениях 145, 179 и 228, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 121, 129, 135, 160 и 178.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 9, при этом: H1 содержит аминокислотную замену в положении 125; L1 содержит аминокислотную замену в положениях 122_129; а H2 содержит аминокислотную замену в положении 145, и L2 содержит аминокислотную замену в положении 121 и/или 124. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 143, 179, 186 и 188, L1 дополнительно содержит аминокислотную замену в одном или более положениях 133, 176, 178 и 180, H2 дополнительно содержит аминокислотную замену в одном или более положениях 143, 179 и 228, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 129, 131, 135, 160 и 178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 9, причем H1 содержит аминокислотные замены в положениях 125_143, 125_179, 125_186 или 125_188; L1 содержит аминокислотные замены в положениях 122_ 129_ 133, 122_ 129_ 176_ 178 или 122_129_180; H2 содержит аминокислотные замены в положениях 143_145, 143_145_179, 143_145_179_228, 143_145_228 или 145_179_228, и L2 содержит аминокислотные замены в положениях 121_124_160_178, 121_124_178, 121_129_131, 121_131, 124_135_160_178 или 124_135_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 9, при этом аминокислотные замены в H1 выбраны из 125R, 143K, 179K, 186R, 188K и их консервативных замен; аминокислотные замены в L1 выбраны из 122D, 129T, 133D, 176D, 176E, 178E, 178T, 180E и их консервативных замен; аминокислотные замены в H2 выбраны из 143E, 145T, 179E, 228D и их консервативных замен; аминокислотные замены в L2 выбраны из 135W, 124R, 160K, 121K, 131K, 129K, 178R и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 9, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 9, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 9, имеющую набор дисульфидных направляющих драйверов.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 9 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 9, как указано в одной или более конструкциях в таблице 10-A9.
Кластер K-L 10:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 10, при этом: H1 содержит аминокислотную замену в положении 174, 179 или 186; L1 содержит аминокислотную замену в положении 176 или 180; H2 содержит аминокислотную замену в положении 143 или 190, а L2 содержит аминокислотную замену в положении 131, 135 или 124; или H1 содержит аминокислотную замену в положении 174; L1 содержит аминокислотную замену в положении 176; H2 содержит аминокислотную замену в положении 190; и L2 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит аминокислотную замену в положении 135. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 143, L1 дополнительно содержит аминокислотную замену в одном или более положениях 116, 129 и 133, H2 дополнительно содержит аминокислотную замену в одном или более положениях 145, 179 и 188, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 133, 160 и 178.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 10, при этом: H1 содержит аминокислотную замену в положениях 174 и/или 186; L1 содержит аминокислотную замену в положениях 176 и/или 180; и H2 содержит аминокислотную замену в положении 145, 190 и/или 188, а L2 содержит аминокислотную замену в положениях 135, 131, 178 или 133, или не содержит аминокислотных замен, которые способствуют преимущественному спариванию. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 143 и/или 179, L1 дополнительно содержит аминокислотную замену в одном или более положениях 116, 129 и 133, H2 дополнительно содержит аминокислотную замену в положениях 143 и/или 179, и/или L2 дополнительно содержит аминокислотные замены в положениях 124 и/или 160.
В некоторых вариантах осуществления антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 10, при этом H1 содержит аминокислотные замены в положениях 143_174, 174, 174_179, 174_186 или 186; L1 содержит аминокислотные замены в положениях 116_129_133_176, 116_129_176_180, 116_176, 129_180 или 176; H2 содержит аминокислотные замены в положениях 143_145_179_188_190, 143_145_179_190, 143_145_190, 143_190, 145_179, 145_179_188_190, 188 или 190, а L2 содержит аминокислотные замены в положениях 124_135_160_178, 124_135_178, 131, 131_135, 133, 135, 135_178, 178, или не содержит аминокислотных замен, которые способствуют преимущественному спариванию.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 10, при этом аминокислотные замены в H1 выбраны из 143K, 174G, 179K, 186R и их консервативных замен; аминокислотные замены в L1 выбраны из 116F, 129T, 133D, 176F, 180E и их консервативных замен; аминокислотные замены в H2 выбраны из 143E, 143I, 145T, 179E, 188F, 190F и их консервативных замен; аминокислотные замены в L2 выбраны из 124K, 124R, 131K, 133A, 135A, 160K, 178F, 178R и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 10, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 10, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 10, имеющую набор стерических 1 драйверов.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 10 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 10, как указано в одной или более конструкциях в таблице 10-A10.
Кластер K-L 11:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 11, при этом: H1 содержит аминокислотную замену в положениях 143_190; L1 содержит аминокислотную замену в положении 133; H2 содержит аминокислотную замену в положении 124, а L2 содержит аминокислотную замену в положениях 131_135. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 125, L1 дополнительно содержит аминокислотную замену в одном или более положениях 122, 129 и 135, H2 дополнительно содержит аминокислотную замену в одном или более положениях 139, 145, 190 и 228, и/или L2 дополнительно содержит аминокислотные замены в положении 121.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 11, при этом:
H1 содержит аминокислотную замену в положениях 143_190; L1 содержит аминокислотную замену в положениях 129_133_135; Н2 содержит аминокислотную замену в положении 124_145, и L2 содержит аминокислотную замену в положениях 131_135. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 125, L1 дополнительно содержит аминокислотную замену в одном или более положении 122, H2 дополнительно содержит аминокислотную замену в одном или более положениях 139, 190 и 228, и/или L2 дополнительно содержит аминокислотные замены в положении 121.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 11, при этом H1 содержит аминокислотные замены в положениях 125_143_190 или 143_190; L1 содержит аминокислотные замены в положениях 122_129_133_135 или 129_133_135; H2 содержит аминокислотные замены в положениях 124_139_145_190, 124_139_145_190_228 или 124_145 и L2 содержит аминокислотные замены в положениях 121_131_135 или 131_135.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 11, при этом аминокислотные замены в H1 выбраны из 125R, 143K, 190K и их консервативных замен; аминокислотные замены в L1 выбраны из 122D, 129T, 133D, 135S и их консервативных замен; аминокислотные замены в H2 выбраны из 124E, 139I, 145T, 190I, 228D и их консервативных замен; аминокислотные замены в L2 выбраны из 121K, 131K, 135K и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 11, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 11, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 является таковой, в которой H1 содержит 143K_190K, L1 содержит 129T_133D_135S, H2 содержит 124E_145T и L2 содержит 131K_135K.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 11, имеющую набор дисульфидных направляющих драйверов.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 11 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 11, как указано в одной или более конструкциях в таблице 10-A11.
Кластер K-L 12:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 12, при этом: H1 содержит аминокислотную замену в положениях 143 и/или 186; L1 содержит аминокислотную замену в положении 133; H2 содержит аминокислотную замену в положении 124, а L2 содержит аминокислотную замену в положениях 131. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в одном или более положениях 124, 125, 139 и 188, L1 дополнительно содержит аминокислотную замену в одном или более положениях 122, 129, 131 и 178, H2 дополнительно содержит аминокислотную замену в одном или более положениях 143, 145, 179, 186, 188, 228, 39 и 45, и/или L2 дополнительно содержит аминокислотные замены в одном или более положениях 121, 133, 135, 176, 178, 38 и 44.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 12, при этом H1 содержит аминокислотные замены в положениях 124_143, 125_143, 125_143_186, 125_186, 125_186_188, 139_143, 139_143_186, 139_186, 139_186_188, 143 или 186_188; L1 содержит аминокислотные замены в положениях 122_129_133, 122_129_133_178, 122_133_178, 129_131_133, 129_133, 129_133_178 или 133_178; H2 содержит аминокислотные замены в положениях 124_143_145, 124_145_179, 124_145_179_186_188, 124_145_179_188, 124_145_179_228, 124_145_186, 39_124_145_179 или 45_124_145_179, и L2 содержит аминокислотные замены в положениях 121_131_133_176, 131_133_135, 131_133_135_176, 131_133_135_178, 131_133_176, 131_133_176_178, 38_131_133_176 или 44_131_133_176.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 12, при этом аминокислотные замены в H1 выбраны из 124K, 125R, 139W, 143I, 143K, 186K, 188T и их консервативных замен; аминокислотные замены в L1 выбраны из 122D, 129T, 131D, 131E, 133D, 178T и их консервативных замен; аминокислотные замены в H2 выбраны из 124E, 143E, 145T, 179E, 186E, 186I, 188W, 228D, 39E, 45P и их консервативных замен; аминокислотные замены в L2 выбраны из 121K, 131K, 131R, 133G, 133S, 133T, 135K, 135W, 176R, 178A, 178S, 38R, 44F и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 12, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-L 12, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 представляет собой одно из следующего:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 12, имеющую один или более наборов дисульфидных направляющих, стерических 2, стерических 3, электростатических или стерических драйверов вариабельного домена.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 12 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-L 12, как указано в одной или более конструкциях в таблице 10-A12.
Кластер K-K 1:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 1, при этом: H1 содержит аминокислотную замену в положениях 143_179; L1 содержит аминокислотную замену в положениях 124_178; H2 содержит аминокислотную замену в положении 186, а L2 содержит аминокислотную замену в положениях 178_180 или 160_180. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотную замену в положении 145.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 1, при этом: H1 содержит аминокислотную замену в положениях 143_145_179; L1 содержит аминокислотную замену в положениях 124_178; H2 содержит аминокислотную замену в положении 186, а L2 содержит аминокислотную замену в положении 180. В некоторых вариантах реализации изобретения L2 дополнительно содержит аминокислотные замены в положении 160 и/или 178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 1, при этом: H1 содержит аминокислотные замены в положении 143_145_179; L1 содержит аминокислотные замены в положениях 124_178; H2 содержит аминокислотные замены в положении 186, а L2 содержит аминокислотные замены в положениях 178_180 или 160_180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 1, при этом аминокислотные замены в H1 выбраны из 143E, 145T, 179E и их консервативных замен; аминокислотные замены в L1 выбраны из 124K, 178R и их консервативных замен; аминокислотная замена в H2 представляет собой 186R или ее консервативную замену; аминокислотные замены в L2 выбраны из 160E, 178E, 180E и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 1, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 содержит 143E_145T_179E, а L1 содержит 124K_178R. В дополнительном варианте реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 1, при этом гетеродимер H2L2 содержит следующие наборы аминокислотных замен: H2 содержит 124K_178R, а L2 содержит 178E_180E или 160E_180E. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию этих гетеродимеров H1L1 и H2L2.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 1, как указано в таблице 10-B1.
Кластер K-K 2:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 2, при этом: H1 содержит аминокислотную замену в положении 143; L1 содержит аминокислотную замену в положении 124; H2 содержит аминокислотную замену в положении 179 или 186, а L2 содержит аминокислотную замену в положении 124_160_180. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотную замену в положении 145, и/или H2 дополнительно содержит аминокислотную замену в положении 146.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 2, при этом: H1 содержит аминокислотную замену в положениях 143_145; L1 содержит аминокислотную замену в положении 124; H2 содержит аминокислотную замену в положениях 179 или 186, а L2 содержит аминокислотную замену в положениях 124_160_180. В некоторых вариантах реализации изобретения H2 дополнительно содержит аминокислотную замену в положении 146.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 2, при этом H1 содержит аминокислотные замены в положениях 143_145; L1 содержит аминокислотные замены в положении 124; H2 содержит аминокислотные замены в положении 186, 179 или 146_179, а L2 содержит аминокислотные замены в положениях 124_160_180.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 2, при этом аминокислотные замены в H1 выбраны из 143E, 145T и их консервативных замен; аминокислотные замены в L1 представляют собой 124R или ее консервативные замены; аминокислотные замены в H2 выбраны из 186R, 179K, 146G и их консервативных замен; аминокислотные замены в L2 выбраны из 124E, 160E, 180E и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 2, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 содержит 143E_145T, а L1 содержит 124R. В дополнительном варианте реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 2, при этом гетеродимер H2L2 содержит следующие наборы аминокислотных замен: H2 содержит 186R, 179K или 146G_179K, а L2 содержит 124E_160E_180E. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 является таковой, в которой H1 содержит 143E_145T, L1 содержит 124R, H2 содержит 179K, а L2 содержит 124E_160E_180E.
В одном варианте реализации изобретения аминокислотная комбинация кластера K-K 2 содержит одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 2, как указано в одной или более конструкциях в таблице 10-B2.
Кластер K-K 3:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 3, при этом:
H1 содержит аминокислотную замену в положении 186; L1 содержит аминокислотную замену в положениях 180 или 178_180; Н2 содержит аминокислотную замену в положении 143 и/или 179, и L2 содержит аминокислотную замену в положениях 124_178 или 131. В некоторых вариантах реализации изобретения H2 дополнительно содержит аминокислотную замену в положении 145.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 3, при этом: H1 содержит аминокислотную замену в положении 186; L1 содержит аминокислотную замену в положении 180; H2 содержит аминокислотную замену в положении 145, а L2 содержит аминокислотную замену в положении 124 или 131. В некоторых вариантах реализации изобретения L1 дополнительно содержит аминокислотную замену в положении 178, H2 дополнительно содержит аминокислотную замену в положениях 143 и/или 179, и/или L2 дополнительно содержит аминокислотные замены в положении 178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 3, при этом H1 содержит аминокислотные замены в положении 186; L1 содержит аминокислотные замены в положении 180 или 178_180; H2 содержит аминокислотные замены в положениях 143_145, 143_145_179 или 145_179, а L2 содержит аминокислотные замены в положениях 131 или 124_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 3, при этом аминокислотные замены в H1 выбраны из 186R и ее консервативных замен; аминокислотные замены в L1 выбраны из 178E, 180E и их консервативных замен; аминокислотные замены в H2 выбраны из 143E, 145T, 179E и их консервативных замен; аминокислотные замены в L2 выбраны из 124K, 131K, 178R и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 3, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 содержит 186R, а L1 содержит 178E_180E или 180E. В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 3, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 является таковой, где H1 содержит 186R, L1 содержит 180E, H2 содержит 143E_145T_179E, а L2 содержит 124K_178R.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-K 3 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 3, как указано в одной или более конструкциях в таблице 10-B3.
Кластер K-K 4:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 4, при этом: H1 содержит аминокислотную замену в положении 179; L1 содержит аминокислотную замену в положении 180; H2 содержит аминокислотную замену в положении 143, а L2 содержит аминокислотную замену в положении 124. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 146, H2 дополнительно содержит аминокислотную замену в положении 145, и/или L2 дополнительно содержит аминокислотные замены в положении 160 и/или 178.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 4, при этом: H1 содержит аминокислотную замену в положении 179; L1 содержит аминокислотную замену в положении 180; H2 содержит аминокислотную замену в положениях 143_145, а L2 содержит аминокислотную замену в положении 124. В некоторых вариантах реализации изобретения H1 дополнительно содержит аминокислотные замены в положении 146, и/или L2 дополнительно содержит аминокислотные замены в положении 160 и/или 178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 4, при этом H1 содержит аминокислотные замены в положениях 146_179 или 179; L1 содержит аминокислотные замены в положении 180; H2 содержит аминокислотные замены в положениях 143_145 и L2 содержит аминокислотные замены в положениях 124 или 124_160_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 4, при этом аминокислотные замены в H1 выбраны из 146G, 179K и их консервативных замен; аминокислотные замены в L1 представляют собой 180E или ее консервативные замены; аминокислотные замены в H2 выбраны из 143E, 145T и их консервативных замен; аминокислотные замены в L2 выбраны из 124R, 160K, 178R и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 4, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 содержит 179K или 146G_179K, а L1 содержит 180E. В других вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 4, при этом гетеродимер H2L2 содержит следующие наборы аминокислотных замен: H2 содержит 143E_145T, а L2 содержит Q124R_Q160K_T178R или Q124R. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 является таковой, в которой H1 содержит 179K, L1 содержит 180E, H2 содержит 143E_145T и L2 содержит 124R_160K_178R.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-K 4 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 4, как указано в одной или более конструкциях в таблице 10-B4.
Кластер K-K 5:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 5, при этом: H1 содержит аминокислотную замену в положении 143 или 186; L1 содержит аминокислотную замену в положении 180 или не содержит аминокислотных замен, которые способствуют преимущественному спариванию; H2 содержит аминокислотную замену в положениях 143_145, а L2 содержит аминокислотную замену в положении 124. В некоторых вариантах реализации изобретения L2 дополнительно содержит аминокислотные замены в одном или более положениях 160 и/или 178. В дополнительном варианте реализации изобретения L2 содержит аминокислотные замены в положениях 124, 124_178 или 124_ 160_178.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 5, при этом аминокислотные замены в H1 выбраны из 186R, 143R, 143K и их консервативных замен; аминокислотная замена в L1 представляет собой 180E и ее консервативные замены; аминокислотные замены в H2 выбраны из 143E, 145T и их консервативных замен; аминокислотные замены в L2 выбраны из 124R, 160K, 178R и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 5, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 5, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-K 5 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 5, как указано в одной или более конструкциях в таблице 10-B5.
Кластер K-K 6:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 6, при этом: H1 содержит аминокислотную замену в положении 39 или не содержит аминокислотных замен, которые способствуют преимущественному спариванию; L1 содержит аминокислотную замену в положении 38 или не содержит аминокислотных замен, которые способствуют преимущественному спариванию; Н2 содержит аминокислотную замену в положении 39 и L2 содержит аминокислотную замену в положении 38.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 6, при этом аминокислотные замены в H1 выбраны из 39D, 39E, 39K, 39R и их консервативных замен; аминокислотные замены в L1 выбраны из 38D, 38E, 38K, 38R и их консервативных замен; аминокислотные замены в H2 выбраны из 39D, 39E, 39K, 39R и их консервативных замен; аминокислотные замены в L2 выбраны из 38D, 38E, 38K, 38R и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 6, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен:
В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 6, при этом гетеродимер H2L2 содержит один из следующих наборов аминокислотных замен:
В некоторых вариантах реализации изобретения антигенсвязывающая конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2. В одном варианте реализации изобретения комбинация гетеродимера H1L1 и гетеродимера H2L2 является таковой, где H1 содержит 39R, L1 содержит 38E, H2 содержит 39D, и L2 содержит 38R.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 6, как указано в одной или более конструкциях в таблице 10-B6. В одном варианте реализации изобретения конструкция кластера K-K 6 не является конструкцией, соответствующей уникальному идентификатору LCCA 10674-10749 или 10679-10744.
Кластер K-K 7:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 7, при этом: H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию; L1 содержит аминокислотную замену в положении 135; H2 содержит аминокислотную замену в положении 139, а L2 содержит аминокислотную замену в положении 116. В некоторых вариантах реализации изобретения L2 дополнительно содержит аминокислотную замену в положении 135.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 7, при этом аминокислотная замена в L1 представляет собой 135W или ее консервативные замены; аминокислотная замена в H2 представляет собой 139W или ее консервативные замены; и аминокислотные замены в L2 выбраны из 116A, 135V и их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 7, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию, а L1 содержит 135W. В дополнительном варианте реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 7, при этом гетеродимер H2L2 содержит следующие наборы аминокислотных замен: H2 содержит 139W, а L2 содержит 116A или 116A_1335V. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию одного гетеродимера H1L1 с одним гетеродимером H2L2.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 7, как указано в одной или в другой конструкциях в таблице 10-B7.
Кластер K-K 8:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 8, при этом: H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержат аминокислотную замену в положении 45; L1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию; H2 содержит аминокислотную замену в положении 45 и L2 содержит аминокислотную замену в положении 44.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 8, при этом аминокислотная замена в H1 представляет собой 45F или ее консервативные замены; аминокислотная замена в H2 представляет собой 45P, 45A или их консервативные замены; аминокислотная замена в L2 представляет собой 44F или ее консервативные замены.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 8, при этом гетеродимер H1L1 содержит один из следующих наборов аминокислотных замен: H1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию или содержит 45F, а L1 не содержит аминокислотных замен, которые способствуют преимущественному спариванию. В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 8, при этом гетеродимер H2L2 содержит следующий набор аминокислотных замен: H2 содержит 45A или 45P, а L2 содержит 44F. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию гетеродимеров H1L1 и H2L2, где H1 и L1 не содержат аминокислотных замен, которые способствуют преимущественному спариванию, H2 содержит 45A и L2 содержит 44F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 8, как указано в одной или более конструкциях в таблице 10-B8.
Кластер K-K 9:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 9, при этом: H1 содержит аминокислотную замену в положении 139; L1 содержит аминокислотную замену в положении 116; Н2 не содержит аминокислотных замен, которые способствуют преимущественному спариванию, а L2 содержит аминокислотную замену в положении 135.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 9, при этом аминокислотная замена в H1 представляет собой 139W или ее консервативные замены; аминокислотная замена в L1 представляет собой 116A или ее консервативные замены; и аминокислотная замена в L2 представляет собой 135W или ее консервативные замены.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 9, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 содержит 139W, а L1 содержит 116A. В дополнительном варианте реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 9, при этом гетеродимер H2L2 содержит следующий набор аминокислотных замен: H2 не содержит аминокислотных замен, которые способствуют преимущественному спариванию, а L2 содержит 135W.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 9, как указано в таблице 10-B9.
Кластер K-K 10:
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 10, при этом: H1 содержит аминокислотную замену в положении 124; L1 содержит аминокислотную замену в положении 176; H2 содержит аминокислотную замену в положении 124, а L2 содержит аминокислотную замену в положении 176. В некоторых вариантах реализации изобретения L1 и/или L2 дополнительно содержат аминокислотную замену в положении 133.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 10, при этом: H1 содержит аминокислотную замену в положении 124; L1 содержит аминокислотную замену в положениях 133_176; H2 содержит аминокислотную замену в положении 124, а L2 содержит аминокислотную замену в положениях 133_176.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 10, при этом аминокислотная замена в H1 представляет собой 124E или ее консервативные замены; аминокислотные замены в L1 выбраны из 133G, 176R или их консервативных замен; аминокислотная замена в H2 представляет собой 124R или ее консервативные замены; аминокислотные замены в L2 выбраны из 133G, 176D или их консервативных замен.
В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H1L1, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 10, при этом гетеродимер H1L1 содержит следующий набор аминокислотных замен: H1 содержит 124E, а L1 содержит 133G_176R. В дополнительных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция имеет гетеродимер H2L2, содержащий комбинацию аминокислотных замен в соответствии с кластером K-K 10, при этом гетеродимер H2L2 содержит следующий набор аминокислотных замен: H2 содержит 124R, а L2 содержит 133G_176D. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию этих гетеродимеров H1L1 и H2L2.
В одном варианте реализации изобретения аминокислотные комбинации кластера K-L 9 содержат одну или более вторичных замен, выбранных из таблицы F.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит комбинацию аминокислотных замен в соответствии с кластером K-K 10, как указано в таблице 10-B10.
Преимущественное спаривание в наборах конструкции LCCA
Одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и H2 с L2 по сравнению с L1. В общем, при отсутствии аминокислотных модификаций и любого встречающегося в природе смещения, последовательность тяжелой цепи (H1) иммуноглобулина дикого типа, при коэкспрессии с двумя различными последовательностями легкой цепи (L1 и L2) иммуноглобулина дикого типа, будет достоверно спариваться в равной степени с обеими легкими цепями, что приводит к приблизительной смеси 50:50 H1 спаренной с L1 (H1L1, правильно спаренная) и H1 спаренной с L2 (H1L2, неправильно спаренная). Аналогичным образом, если H2 дикого типа коэкспрессируется с L1 и L2 дикого типа, тяжелая цепь будет достоверно спариваться в равной степени с обеими легкими цепями, что приводит к приблизительной смеси 50:50 H2 спаренной с L1 (H2L1, неправильно спаренная) и H2 спаренной с L2 (H2L2, правильно спаренная). Термин "преимущественное спаривание" используется в данном документе для описания специфичности спаривания или предпочтения спаривания полипептидной последовательности тяжелой цепи иммуноглобулина с одной полипептидной последовательностью легкой цепи иммуноглобулина по сравнению с другой полипептидной последовательностью легкой цепи иммуноглобулина. В данном контексте, преимущественное спаривание будет происходить между, например, H1 и L1, если количество гетеродимера H1L1 является большим, чем количество гетеродимера H1L2, в случае если Н1 коэкспрессируется и с L1, и с L2. Подобным образом, преимущественное спаривание будет происходить между, например, H2 и L2, если количество гетеродимера H2L2 является большим, чем количество гетеродимера H2L1, в случае если Н2 коэкспрессируется и с L1, и с L2.
Однако в некоторых случаях существует унаследованное смещение спаривания, наблюдаемое в полипептидных последовательностях тяжелой и легкой цепей дикого типа, полученных из исходных антител. Это унаследованное смещение спаривания можно наблюдать в контексте набора конструкции LCCA, в котором исходные H1 или H2 дикого типа коэкспрессируются с исходными L1 и L2 дикого типа, где одна из легких цепей преимущественно спаривается с тяжелыми цепями обоих исходных антитела. В одном варианте реализации изобретения преимущественное спаривание происходит, когда аминокислотные модификации в одной или более из H1, L1, H2 и L2 способствуют преимущественному спариванию, которое является большим, чем преимущественное спаривание, которое происходит в соответствующей системе дикого типа.
Степень преимущественного спаривания или "прочности" конструкции является мерой способности аминокислотных модификаций способствовать преимущественному спариванию. Степень преимущественного спаривания может быть оценена, как описано в данном документе в другом месте, и в примерах, и основана на измерении правильно спаренных гетеродимеров (то есть, H1L1 и H2L2) по сравнению с неправильно спаренными гетеродимерами (то есть, H1L2 и H2L1). Степень преимущественного спаривания может быть оценена в контексте наборов конструкции LCCA (H1L1L2 или H2L1L2), где одна тяжелая цепь коэкспрессируется с двумя уникальными легкими цепями, или набора конструкции мАт (H1L1H2L2), где коэкспрессируется тяжелая и легкая цепи исходного антитела.
Следующие варианты реализации изобретения относятся к контексту наборов конструкции LCCA. Во всех вариантах реализации изобретения этого раздела термин "около" означает ±5% от указанного отношения, а преимущественное спаривание сравнивается по отношению к дикому типу, если не указано иное. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 60:40. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 60:40.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 65:35. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 65:35.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 70:30. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 70:30.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 75:25. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 75:25.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 80:20. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 80:20.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 85:15. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 85:15.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 90:10. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 90:10.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 95:5. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 95:5.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H1L1:H1L2 составляет, по меньшей мере, около 40:60, а соотношение H2L2:H2L1 составляет, по меньшей мере, около 99:1. В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, где соотношение H2L2:H2L1 составляет, по меньшей мере, около 40:60, а соотношение H1L1:H1L2 составляет, по меньшей мере, около 99:1.
В других вариантах реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1 или H2 с L2 по сравнению с L1 с образованием H2L2, так что количество H1L1 или H2L2 превышает около 40, 45, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 около 99%.
В одном варианте реализации изобретения преимущественное спаривание измеряется LCCA, как описано в примерах. Результаты LCCA обычно прогнозируют результаты в контексте преимущественного спаривания в наборах конструкции мАт (описанных ниже), в которых коэкспрессируются H1, L1, H2 и L2.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1, или H2 с L2 по сравнению с L1 с образованием H2L2, так что относительное спаривание, по меньшей мере, одного из H1L1 или H2L2 является, по меньшей мере, на 10% большим по сравнению с диким типом, а относительное спаривание другого находится в пределах около 10% от дикого типа или, по меньшей мере, на около 10% больше по сравнению с диким типом.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1, или H2 с L2 по сравнению с L1 с образованием H2L2, так что относительное спаривание, по меньшей мере, одного из H1L1 или H2L2 является, по меньшей мере, на 20% большим по сравнению с диким типом, а относительное спаривание другого составляет около 10% от дикого типа или, по меньшей мере, на около 10% больше по сравнению с диким типом.
В одном варианте реализации изобретения одна или более из H1, H2, L1 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 с образованием H1L1, или H2 с L2 по сравнению с L1 с образованием H2L2, так что относительное спаривание, по меньшей мере, одного из H1L1 или H2L2 является, по меньшей мере, на 30% большим по сравнению с диким типом, а относительное спаривание другого составляет около 10% от дикого типа или, по меньшей мере, на около 10% больше по сравнению с диким типом.
Преимущественное спаривание в наборах конструкции мАт
Преимущественное спаривание также может быть оценено в контексте наборов конструкции мАт, где H1, L1, H2 и L2 коэкспрессируются, и одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 и H2 с L2 с образованием биспецифической антигенсвязывающей полипептидной конструкции, которая содержит правильно спаренный первый гетеродимер (H1L1) и правильно спаренный второй гетеродимер (H2L2). В этом варианте реализации изобретения, как показано на Фиг. 8, когда две различные полипептидные последовательности тяжелой цепи иммуноглобулина коэкспрессируются с двумя различными полипептидными последовательностями легкой цепи иммуноглобулина, может появиться ряд потенциальных продуктов, четырнадцать из которых показаны на Фиг. 8, только одним из которых является желаемое или правильно спаренное биспецифическое антитело H1L1H2L2 (виды антител A на Фиг. 8). Однако в контексте оценки правильного спаривания между тяжелыми цепями и легкой цепью на основе наборов конструкции мАт, некоторые из дополнительных продуктов также могут рассматриваться как демонстрирующие правильное спаривание в контексте областей Fab, поскольку они содержат правильно спаренные гетеродимеры на уровне Fab (см., например, виды антител E, H, K и M на Фиг. 8). В некоторых вариантах реализации изобретения Fc-часть антигенсвязывающей полипептидной конструкции содержит асимметричные аминокислотные модификации, которые способствуют образованию гетеродимерного Fc. В этих вариантах реализации изобретения ожидается, что число и количество видов E-J будут уменьшаться.
Что касается наборов конструкции LCCA, то в контексте набора конструкции мАт, в котором коэкспрессируются все четыре полипептидные последовательности иммуноглобулина H1, L1, H2 и L2, в некоторых случаях может иметь место унаследованное смещение при спаривании, приводящее к тому, что одна из легких цепей (либо L1, либо L2), преимущественно спаривается как с H1, так и с H2. Таким образом, при определении прочности конструкции мАт в контексте биспецифической антигенсвязывающей полипептидной конструкции, может потребоваться оценка степени спаривания с аминокислотными модификациями конструкции мАт по сравнению с количеством правильного спаривания, наблюдаемым в соответствующей исходной системе дикого типа (полипептидные последовательности H1, L1, H2, L2 без аминокислотных модификаций конструкции мАт). Таким образом, в одном варианте реализации изобретения предполагается, что конструкция мАт проявляет преимущественное спаривание, если количество правильно спаренной биспецифической антигенсвязывающей полипептидной конструкции больше, чем количество правильно спаренного биспецифического антитела, полученного в соответствующей исходной системе дикого типа. В альтернативном варианте предполагается, что конструкция мАт проявляет преимущественное спаривание, если процент правильно спаренной биспецифической антигенсвязывающей полипептидной конструкции в общем продукте экспрессии больше, чем процент правильно спаренного биспецифического антитела, полученного в общем продукте экспрессии в соответствующей исходной системе дикого типа. В одном варианте реализации изобретения общий продукт экспрессии может содержать виды антител А-N на Фиг. 8. В одном варианте реализации изобретения общий продукт экспрессии может быть только теми видами антител, которые имеют две тяжелые цепи и две легкие цепи (виды антител А-J на Фиг. 8). В последнем варианте реализации изобретения преимущественное спаривание измеряется в процентах от общего биспецифического антитела, за исключением полуантител (таких как виды K-N на Фиг. 8).
В другом варианте реализации изобретения предполагается, что конструкция мАт проявляет преимущественное спаривание, если количество правильного спаривания увеличивается в гетеродимере биспецифической антигенсвязывающей полипептидной конструкции, которая проявляет высокую степень ошибочного спаривания в соответствующей исходной системе дикого типа. В другом варианте реализации изобретения предполагается, что конструкция мАт проявляет преимущественное спаривание, если общее количество правильного спаривания между H1 и L1 и между H2 и L2 является большим, чем то, которое наблюдается в соответствующей исходной системе дикого типа. Например, со ссылкой на Фиг. 8, виды A, B, H, I и M будут считаться правильно спаренными по отношению к H1L1, а виды A, C, E, F и K будут считаться правильно спаренными по отношению к H2L2. В этом варианте реализации изобретения преимущественное спаривание измеряется как процент от общего спаривания.
В одном варианте реализации изобретения преимущественное спаривание измеряется SMCA, как описано в данном документе.
В некоторых вариантах реализации изобретения предполагается, что конструкция мАт считается способствующей преимущественному спариванию, в случае если изменение в количестве полного правильного спаривания, измеренное суммой спаривания H1L1 и H2L2, составляет более, чем около 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40% или 45% по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab, которые способствуют преимущественному спариванию.
В некотором варианте реализации изобретения предполагается, что конструкция мАт считается способствующей преимущественному спариванию, в случае если изменение количества полного правильного спаривания, измеренное количеством биспецифического антитела, полученного в процентах от видов, отличных от полученных полуантител, составляет более, чем около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% или 80% по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab которые способствуют преимущественному спариванию.
В одном варианте реализации изобретения предполагается, что конструкция мАт считается способствующей преимущественному спариванию, в случае если изменение количества полного правильного спаривания, измеренное количеством биспецифического антитела, полученного в процентах от всех полученных видов, составляет более, чем около 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70% или 80% по сравнению со спариванием соответствующих полипептидных цепей H1, L1, H2 и L2 без аминокислотных замен в области Fab которые способствуют преимущественному спариванию.
Термическая стабильность областей Fab
Аминокислотные модификации в одной или более полипептидных последовательностях H1, L1, H2 и L2 способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и H2 с L2 по сравнению с L1 и минимально влияют на термическую стабильность каждого гетеродимера антигенсвязывающей полипептидной конструкции. Влияние аминокислотных модификаций на каждый гетеродимер определяется путем измерения термической стабильности областей Fab, образованных H1 и L1 или H2 и L2, и ее сравнения с термической стабильностью области Fab, образованной соответствующей полипептидной последовательностью H1 и L1 дикого типа (первая область Fab дт) или соответствующими полипептидными последовательностями H2 и L2 дикого типа (вторая область Fab дт). Термины "соответствующие полипептидные последовательности H1 и L1 дикого типа" и "соответствующие полипептидные последовательности H2 и L2 дикого типа" предназначены для описания соответствующих полипептидных последовательностей H1, L1, H2 и L2, которые не имеют аминокислотных модификаций, способствующих преимущественному спариванию, как описано в данном документе.
Термическую стабильность можно измерить различными способами, известными в данной области техники и описанными в данном документе, включая дифференциальную сканирующую калориметрию (ДСК) или дифференциальную сканирующую флуориметрию (ДСФ). Последние способы обеспечивают измерение термической стабильности в терминах "температуры плавления" или Tm.
В контексте следующих вариантов реализации изобретения термин "около" означает ±10% от указанной температуры. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 20°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 15°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 10°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 9°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 8°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 7°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 6°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 5°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 4°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 3°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 2°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm в пределах около 1°C Tm соответствующей дт первой области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первую область Fab, которая имеет Tm, которая примерно такая же, как у соответствующей дт первой области Fab.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 20°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 15°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 10°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 9°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 8°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 7°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 6°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 5°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 4°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 3°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 2°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm в пределах около 1°C Tm соответствующей дт второй области Fab. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит вторую область Fab, которая имеет Tm, которая примерно такая же, как у соответствующей дт второй области Fab.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит первый гетеродимер и второй гетеродимер, причем температура плавления (Tm) первой области Fab находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 20°C Tm области Fab, образованной соответствующими полипептидными последовательностями дикого типа H1 и L1 для первого антигена (первая область Fab дт), и/или температура плавления (Tm) второй области Fab находится в пределах около 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или 20°C Tm области Fab, образованной соответствующими полипептидными последовательностями дикого типа H2 и L2 для второго антигена (вторая область Fab дт).
Кроме того, в некоторых вариантах реализации изобретения Tm первой или второй области Fab больше, чем у соответствующей первой Fab дт или соответствующей второй Fab дт. Таким образом, в одном варианте реализации изобретения Tm первой или второй области Fab увеличивается на около 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 3,0, 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,5, 5,0°C или больше по сравнению с соответствующей первой областью Fab дт или соответствующей второй областью Fab дт.
В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит аминокислотные замены, соответствующие номерам конструкции K-L 2979, 3018, 3041, 3102, 3898 и/или 3947. В одном варианте реализации изобретения антигенсвязывающая полипептидная конструкция содержит аминокислотные замены, соответствующие номерам конструкции K-L 3025, 3109, 3113, 3878, 3890, 3910, 3931, 3954, 3967, 4010 и/или 4040.
Способность областей Fab связывать антиген
Аминокислотные модификации в одной или более полипептидных последовательностях H1, L1, H2 и L2 способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и H2 с L2 по сравнению с L1 и минимально влияют на способность каждого гетеродимера антигенсвязывающей полипептидной конструкции связываться с его антигеном. Влияние аминокислотных модификаций на каждый гетеродимер определяется путем измерения способности областей Fab, образованных H1 и L1 или H2 и L2, связываться с их соответствующими антигенами и ее сравнения со способностью соответствующей первой области Fab дт или соответствующей второй области Fab дт связываться с их соответствующими антигенами.
Способность областей Fab связываться с их соответствующими антигенами может быть измерена с помощью ряда способов, известных в данной области техники, некоторые из которых описаны в данном документе в другом месте. Например, поверхностный плазмонный резонанс (ППР) или анализы связывания целых клеток могут быть использованы для оценки способности первой области Fab связывать первый антиген и второй области Fab связывать второй антиген. Последние два способа измеряют способность областей Fab связываться с их соответствующими антигенами путем определения аффинности области Fab к его антигену.
В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 100-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 50-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 40-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 30-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 20-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 10-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 9-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 8-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 7-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 6-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 5-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах 4-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 3-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену находится в пределах около 2-кратной аффинности первой области Fab дт к первому антигену. В одном варианте реализации изобретения аффинность первой области Fab к первому антигену примерно такая же, как аффинность первой области Fab дт к первому антигену.
В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 100-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 50-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 40-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 30-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 20-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 10-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 9-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 8-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 7-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 6-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 5-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 4-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 3-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену находится в пределах около 2-кратной аффинности второй области Fab дт ко второму антигену. В одном варианте реализации изобретения аффинность второй области Fab ко второму антигену примерно такая же, как аффинность второй области Fab дт ко второму антигену.
Переносимость аминокислотных модификаций или наборов конструкций
Аминокислотные модификации или наборы конструкций, описанные в данном документе, могут быть применены для получения биспецифической антигенсвязывающей полипептидной конструкции, в которой полипептидные последовательности тяжелой цепи иммуноглобулина и полипептидные последовательности легкой цепи иммуноглобулина каждого гетеродимера могут быть получены из одного или более исходных антител, где, по меньшей мере, одно исходное антитело содержит легкую цепь каппа, и, по меньшей мере, одно другое исходное антитело содержит легкую цепь лямбда. Основываясь на следующем обсуждении, наборы конструкций мАт могут быть применены к большинству таких биспецифических антигенсвязывающих полипептидных конструкций.
Остатки области контакта VH:VL и CH1:CL в области контакта между тяжелыми и легкими цепями иммуноглобулина являются консервативными в достаточной степени (Padlan и соавт., 1986 год, Mol. Immunol. 23(9):951-960). Данная консервативность последовательности, результат эволюционных ограничений, увеличивает вероятность того, что функционально активные антигенсвязывающие домены будут формироваться во время комбинаторного спаривания легких и тяжелых цепей. В результате данной консервативности последовательности следует, что описанные в данном документе наборы конструкций мАт, основанные на моделировании структур D3H44 Fab каппа и CAT-2200 Fab лямбда, которые приводят к преимущественному спариванию, могут быть перенесены в Fab каппа и Fab лямбда других исходных антител для стимулирования преимущественного спаривания, поскольку эта область отображает высокую консервативность последовательности по антителам. Кроме того, в случае если возникают отличия в последовательности, они обычно находятся дистально от области контакта CH1:CL. Это особенно относится к доменам CH1 и CL. В одном варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab каппа содержит одну или более аминокислотных модификаций в доменах CL и/или CH1, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab лямбда содержит одну или более аминокислотных модификаций в доменах CL и/или CH1, которые способствуют преимущественному спариванию.
Существует, однако, некоторая вариабельность последовательности антигенсвязывающего сайта по отношению к остаткам петли (и длине) CDR (определяющим комплементарность областям), в частности, для CDR-H3. Таким образом, в одном варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab каппа содержит одну или более аминокислотных модификаций в доменах VH и/или VL, которые находятся дистально по отношению к петлям CDR, в случае если аминокислотная последовательность антигенсвязывающего сайта значительно отличается от таковой в антителе D3H44. В другом варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab лямбда содержит одну или более аминокислотных модификаций в доменах VH и/или VL, которые способствуют преимущественному спариванию и находятся дистально по отношению к петлям CDR, в случае если аминокислотная последовательность антигенсвязывающего сайта значительно отличается от таковой в антителе CAT-2200. В другом варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab каппа содержит одну или более аминокислотных модификаций, которые способствуют преимущественному спариванию в доменах VH и/или VL, которые находятся проксимально или дистально по отношению к петлям CDR, в случае если аминокислотная последовательность антигенсвязывающего сайта по существу аналогична аминокислотной последовательности антитела D3H44. В другом варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab лямбда содержит одну или более аминокислотных модификаций в доменах VH и/или VL, которые способствуют преимущественному спариванию и находятся проксимально или дистально по отношению к петлям CDR, в случае если аминокислотная последовательность антигенсвязывающего сайта по существу аналогична аминокислотной последовательности антитела CAT-2200. В одном варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab каппа содержит одну или более аминокислотных модификаций в доменах CL и/или CH1, а также модификации в доменах VH и/или VL, которые способствуют преимущественному спариванию. В одном варианте реализации изобретения, описанные в данном документе антигенсвязывающие полипептидные конструкции содержат гетеродимеры, где Fab лямбда содержит одну или более аминокислотных модификаций в доменах CL и/или CH1, а также модификации в доменах VH и/или VL, которые способствуют преимущественному спариванию.
В одном варианте реализации изобретения аминокислотные модификации в одной или более из H1, L1, H2 и L2 антигенсвязывающей полипептидной конструкции могут способствовать преимущественному спариванию в антигенсвязывающей полипептидной конструкции, где Fab каппа одного исходного антитела является человеческим или гуманизированным IgG1/κ. Неограничивающие примеры таких исходных антител включают Офатумумаб (человеческий) или Трастузумаб, или Бевацизумаб (гуманизированный). В одном варианте реализации изобретения аминокислотные модификации в одной или более из H1, L1, H2 и L2 антигенсвязывающей полипептидной конструкции могут способствовать преимущественному спариванию в антигенсвязывающей полипептидной конструкции, где Fab лямбда одного исходного антитела является человеческим или гуманизированным IgG1/лямбда. Неограничивающие примеры таких человеческих антител включают Бриакинумаб или Сифалимумаб, а примером гуманизированного антитела является Бронтиктузумаб.
В другом варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи иммуноглобулина антител с использованием широко используемых подгрупп VH и VL.
В одном варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи антител, имеющих каркас близкий к каркасу зародышевой линии. Примеры таких антител включают Обинутузумаб.
В одном варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи антител, имеющих междоменный угол VH:VL близкий к среднему значению, наблюдаемому для пар тяжелой и легкой цепи. Пример такого типа антител включает, но без ограничений, Пертузумаб. В другом варианте реализации изобретения аминокислотные модификации, описанные в данном документе, являются переносимыми на тяжелые и легкие цепи иммуноглобулина антител, имеющих канонические домены CL и СН1. Подходящие примеры таких антител включают, но без ограничений, Трастузумаб.
Примеры, Фигуры и Таблицы демонстрируют, что аминокислотные модификации (например, в пределах одного или более фрагментов Fab, содержащих вариабельную область и константную область), которые способствуют преимущественному спариванию являются переносимыми на другие тяжелые и легкие цепи иммуноглобулина, что приводит к аналогичным наборам преимущественного спаривания одной тяжелой цепи иммуноглобулина с одной из двух легких цепей иммуноглобулина.
Каркасные структуры
Гетеродимеры антигенсвязывающей полипептидной конструкции могут быть связанны с каркасной структурой. Каркасной структурой может быть пептид, полипептид, полимер, наночастица или другая химическая структурная единица. Гетеродимеры антигенсвязывающей полипептидной конструкции могут быть связаны с N- или С-концом каркасной структуры, где каркасная структура представляет собой полипептид. В одном варианте реализации изобретения каркасная структура представляет собой полипептид альбумина.
В другом варианте реализации изобретения каркасная структура представляет собой Fc иммуноглобулина (Fc) или его часть. В некоторых вариантах реализации изобретения Fc содержит, по меньшей мере, одну или две последовательности домена CH3. В некоторых вариантах реализации изобретения Fc дополнительно содержит, по меньшей мере, одну или две последовательности домена CH2. В некоторых вариантах реализации изобретения антигенсвязывающая полипептидная конструкция содержит Fc, спаренный, с помощью одного или более линкера, или без линкеров, с первым гетеродимером и/или вторым гетеродимером. В некоторых вариантах реализации изобретения Fc представляет собой Fc человека. В некоторых вариантах реализации изобретения Fc представляет собой Fc IgG или IgG1 человека. В некоторых вариантах реализации изобретения Fc представляет собой гетеродимерный Fc. В некоторых вариантах реализации изобретения Fc состоит из одного полипептида. В некоторых вариантах реализации изобретения Fc состоит из множества пептидов, например, двух полипептидов.
В некоторых вариантах реализации изобретения Fc содержит одну или более аминокислотных модификаций в, по меньшей мере, одной из последовательностей домена CH3. Аминокислотные модификации могут быть сделаны в Fc иммуноглобулина с целью стимулирования преимущественного спаривания между гетеродимерными последовательностями домена CH3 по отношению к гомодимерным последовательностям домена CH3. Такие аминокислотные модификации известны в данной области техники и включают, например, те, которые описаны в патентной публикации США №2012/0149876. Альтернативные стратегии для стимулирования преимущественного спаривания между последовательностями гетеродимерного домена CH3, по отношению к последовательностям гомодимерного CH3, включают в себя, например, "выступы-во-впадины", заряженные остатки с ионными взаимодействиями и разработанный домен обмена цепями (SEED). Последние стратегии были описаны в данной области техники и рассмотрены выше в Klein и соавт.дальнейшее обсуждение доменов Fc следует ниже.
В некоторых аспектах Fc представляет собой Fc, описанный в патентных заявках PCT/CA2011/001238, поданной 4 ноября 2011 года, или PCT/CA2012/050780, поданной 2 ноября 2012 года, причем полное описание каждой из них включено в данный документ в полном объеме посредством ссылки для всех целей.
В некоторых аспектах антигенсвязывающая полипептидная конструкция, описанная в данном документе, содержит гетеродимерный Fc, содержащий модифицированный домен CH3, который был асимметрично модифицирован. Гетеродимерный Fc может содержать два полипептида константных доменов тяжелой цепи:первый полипептид тяжелой цепи и второй полипептид тяжелой цепи, которые могут быть использованы взаимозаменяемо при условии, что Fc содержит один первый полипептид тяжелой цепи и один второй полипептид тяжелой цепи. Как правило, первый полипептид тяжелой цепи содержит первую последовательность CH3, а второй полипептид тяжелой цепи содержит вторую последовательность CH3.
В результате введения одной или более асимметричных аминокислотных модификаций в две последовательности CH3, как правило, образуется гетеродимерный Fc, а не гомодимер, при этом происходит димеризация двух последовательностей CH3. Используемый в данном документе термин "асимметричные аминокислотные модификации" относится к любой модификации, при которой аминокислота в определенном положении в первой последовательности CH3 отличается от аминокислоты, находящейся во второй последовательности CH3 в том же самом положении, и первая, и вторая последовательность CH3 преимущественно спариваются с образованием гетеродимера, а не гомодимера. Эта гетеродимеризация может происходить в результате модификации только одной из двух аминокислот в одном и том же положении соответствующей аминокислоты в каждой последовательности; или модификации обеих аминокислот в каждой последовательности в одном и том же соответствующем положении в каждой первой и второй последовательности CH3. Первая и вторая последовательность CH3 гетеродимерного Fc могут содержать одну или более, чем одну асимметричную аминокислотную модификацию.
В таблице X представлена аминокислотная последовательность домена Fc IgG1 человека, соответствующая аминокислотам с 231 по 447 полноразмерной тяжелой цепи IgG1 человека. Последовательность CH3 содержит аминокислоты 341-447 полноразмерной тяжелой цепи IgG1 человека.
Как правило, Fc может включать две непрерывные последовательности тяжелой цепи (A и B), которые способны к димеризации. В некоторых аспектах, одна или обе последовательности Fc включают одну или более мутаций или модификаций в следующих местах: L351, F405, Y407, T366, K392, T394, T350, S400 и/или N390 согласно нумерации EU. В некоторых аспектах Fc включает мутантную последовательность, проиллюстрированную в таблице X. В некоторых аспектах Fc содержит мутации Варианта 1 A-B. В некоторых аспектах Fc содержит мутации Варианта 2 A-B. В некоторых аспектах Fc содержит мутации Варианта 3 A-B. В некоторых аспектах Fc содержит мутации Варианта 4 A-B. В некоторых аспектах Fc содержит мутации Варианта 5 A-B.
В некоторых вариантах реализации изобретения Fc может содержать одну или более аминокислотных модификаций в, по меньшей мере, одной из последовательностей домена CH2. Ряд мутаций в последовательности тяжелой цепи Fc является известным в данной области техники для выборочного изменения аффинности антитела Fc к различным Fc-гамма рецепторам. В некоторых вариантах изобретения Fc содержит одну или более модификаций для изменения связывания рецепторов Fc-гамма с антигенсвязывающей полипептидной конструкцией.
Домен CH2 соответствует аминокислотам 231-340 последовательности, показанной в таблице X. Типовые, неограничивающие аминокислотные модификации, которые изменяют способность Fc антигенсвязывающей полипептидной конструкции связываться с рецепторами Fc-гамма, перечислены ниже:
S298A/E333A/K334A, S298A/E333A/K334A/K326A (Lu Y, Vernes JM, Chiang N, и соавт. J Immunol Methods. 28 февраля 2011 года; 365(1-2):132-41); F243L/R292P/Y300L/V305I/P396L, F243L/R292P/Y300L/L235V/P396L (Stavenhagen JB, Gorlatov S, Tuaillon N, и соавт. Cancer Res. 15 сентября 2007 года; 67(18):8882-90; Nordstrom JL, Gorlatov S, Zhang W, и соавт. Breast Cancer Res. 30 ноября 2011 года; 13(6):R123); F243L (Stewart R, Thom G, Levens M, и соавт. Protein Eng Des Sel. сентябрь 2011 года; 24(9):671-8. ), S298A/E333A/K334A (Shields RL, Namenuk AK, Hong K, и соавт. J Biol Chem. 2 марта 2001 года;276(9):6591-604); S239D/I332E/A330L, S239D/I332E (Lazar GA, Dang W, Karki S, и соавт. Proc Natl Acad Sci U S A. 14 марта 2006 года; 103(11):4005-10); S239D/S267E, S267E/L328F (Chu SY, Vostiar I, Karki S, и соавт. Mol Immunol. сентябрь 2008 года; 45(15):3926-33); S239D/D265S/S298A/I332E, S239E/S298A/K326A/A327H, G237F/S298A/A330L/ I332E, S239D/I332E/S298A, S239D/K326E/A330L/I332E/S298A, G236A/S239D/D270L/I332E, S239E/S267E/H268D, L234F/S267E/N325L, G237F/V266L/S267D и другие мутации, перечисленные в WO2011/120134 и WO2011/120135 включены в данный документ посредством ссылки. Therapeutic Antibody Engineering (по William R. Strohl и Lila M. Strohl, Woodhead Publishing series in Biomedicine №11, ISBN 1 907568 37 9, октябрь 2012 года) описывает дополнительные модификации Fc, которые влияют на связывание Fc с рецепторами Fc-гамма на стр. 283.
Дополнительные модификации для улучшения эффекторной функции.
В некоторых вариантах реализации изобретения Fc антигенсвязывающей полипептидной конструкции, описанной в данном документе, может быть модифицирован с целью улучшения его эффекторной функции. Такие модификации известны в данной области техники и включают в себя абнормальное фукозилирование или разработку аффинности части Fc антител к активирующему рецептору, главным образом FCGR3a для АЗКЦ и к C1q для КЗЦ. В приведенной ниже таблице Y обобщены различные конструкции, описанные в литературе для разработки эффекторной функции.
Таким образом, в одном варианте реализации изобретения, описанная в данном документе антигенсвязывающая полипептидная конструкция может включать в себя димерный Fc, который содержит одну или более аминокислотных модификаций, как указано в приведенной выше таблице, придающих улучшенную эффекторную функцию. В других вариантах реализации изобретения антигенсвязывающая полипептидная конструкция может быть афукозилированна с целью улучшения эффекторной функции.
Связывание FcRn и параметры ФК
Как известно специалистам в данной области техники, связывание с FcRn приводит к рециркуляции подвергшегося эндоцитозу антитела из эндосомы назад в кровоток (Raghavan и соавт., 1996 год, Annu Rev Cell Dev Biol 12:181-220; Ghetie и соавт., 2000 год, Annu Rev Immunol 18:739-766). Этот процесс в сочетании с уменьшением почечной фильтрации, обусловленным большим размером полноразмерной молекулы, приводит к более продолжительному периоду полувыведения антитела из сыворотки крови в пределах от одной до трех недель. Связывание Fc с FcRn также играет ключевую роль в транспорте антитела. Таким образом, в одном варианте реализации изобретения, Fc содержит одну или более аминокислотных модификаций, которые изменяют или способствуют способности Fc связывать FcRn.
Линкеры
Конструкции, описанные в данном документе, могут содержать один или более гетеродимеров, описанных в данном документе, функционально спаренных с Fc, описанным в данном документе. В некоторых аспектах Fc спарен с одним или более гетеродимерами с помощью или без одного, или более линкеров. В некоторых аспектах Fc непосредственно спарен с одним или более гетеродимерами. В некоторых аспектах Fc спарен с одним или более гетеродимерами с помощью одного или более линкеров. В некоторых аспектах Fc спарен с тяжелой цепью каждого гетеродимера с помощью линкера.
В некоторых аспектах один или более линкеров представляют собой один или более полипептидных линкеров. В некоторых аспектах один или более линкеров содержат одну или более шарнирных областей антитела. В некоторых аспектах один или более линкеров содержат одну или более шарнирных областей IgG1.
Дополнительные необязательные модификации
В одном варианте реализации изобретения тяжелые и легкие цепи иммуноглобулина антигенсвязывающей полипептидной конструкции, описанные в данном документе, могут быть дополнительно модифицированы (то есть, путем ковалентного присоединения различных типов молекул) таким образом, что ковалентное присоединение не препятствует преимущественному спариванию между тяжелой цепью и легкими цепями или влиянию на способность гетеродимера связываться с его антигеном или влиянию на его стабильность. Такая модификация включает, например, но не в качестве исчерпывающего перечня, гликозилирование, ацетилирование, пегилирование, фосфорилирование, амидирование, дериватизацию известных защитных/блокирующих групп, протеолитическое расщепление, связывание с клеточным лигандом или другим белком и т.д. Любая из многочисленных химических модификаций может быть осуществлена известными методами, в том числе, но без ограничений, специфическим химическим расщеплением, ацетилированием, формилированием, метаболическим синтезом туникамицина и т.д.
В другом варианте реализации изобретения тяжелые и легкие цепи иммуноглобулина антигенсвязывающей полипептидной конструкции, описанные в данном документе, могут быть конъюгированы (прямо или опосредованно) с лекарственным средством или молекулой лекарственного средства, модифицирующего данную биологическую реакцию. В определенных вариантах реализации изобретения антигенсвязывающая полипептидная конструкция конъюгирована с лекарственным средством, например, токсином, химиотерапевтическим средством, иммуномодулятором или радиоизотопом. Несколько способов получения ADC (конъюгат антитела с лекарственным средством или конъюгат антигенсвязывающей полипептидной конструкции с лекарственным средством) известны в данной области техники и описаны, например, в патентах США 8,624,003 (способ реакции, проводящейся в одном реакционном сосуде без выделения промежуточных соединений), 8,163,888 (одноэтапный) и 5,208,020 (двухэтапный способ), например. В некоторых вариантах реализации лекарственный препарат выбран из майтанзина, ауристатина, калихимицина или их производных. В других вариантах реализации изобретения лекарственный препарат представляет собой майтанзин, выбранный из DM1 и DM4.
В некоторых вариантах реализации антигенсвязывающая полипептидная конструкция конъюгирована с цитотоксическим средством. Используемый в данном документе термин "цитотоксическое средство" относится к веществу, которое ингибирует или подавляет функциональную активность клеток и/или вызывает разрушение клеток. Подразумевается, что термин охватывает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32 и Lu177), химиотерапевтические средства и токсины, такие как, например, низкомолекулярные токсины, или обладающие ферментативной активностью токсины бактериального, грибного, растительного или животного происхождения, включая их фрагменты и/или варианты.
Лекарственные средства или молекулы лекарственного средства не должны быть истолкованы как ограниченные лишь классическими химическими лекарственными средствами. Например, молекула лекарственного средства может быть белком или полипептидом, обладающим желаемой биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин A, Онконазу (или другую цитотоксическую РНКазу), экзотоксин синегнойной палочки, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, альфа-интерферон, бета-интерферон, нейрональный ростовой фактор, тромбоцитарный фактор роста, тканевый активатор плазминогена, апоптотический агент, например, ФНО-альфа, ФНО-бета, AIM I (см., международная публикация № WO 97/33899), AIM II (см., международная публикация № WO 97/34911), лиганд Fas (Takahashi и соавт., 1994 год, J. Immunol., 6:1567) и VEGI (см., международная публикация № WO 99/23105), тромботический препарат или анти-ангиогенный препарат, например, ангиостатин или эндостатин; или модификатор биологической реакции, такой как, например, лимфокин (например, интерлейкин-1 ("ИЛ-1"), интерлейкин-2 ("ИЛ-2"), интерлейкин-6 ("ИЛ-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("ГМКСФ"), и гранулоцитарный колониестимулирующий фактор ("ГКСФ")), или фактор роста (например, гормон роста ("ГР")).
Более того, в альтернативном варианте реализации изобретения антигенсвязывающая полипептидная конструкция может быть конъюгирована с терапевтическими молекулами, такими как радиоактивные вещества или макроциклические хелатирующие агенты, используемые для конъюгации ионов радиоактивного металла (см. выше примеры радиоактивных материалов). В определенных вариантах реализации изобретения макроциклическим хелатирующим агентом является 1,4,7,10-тетраазациклододекан-N, N', N'', N''-тетрауксусная кислота (DOTA), которая может быть присоединена к антителу с помощью линкерной молекулы. Такие линкерные молекулы, как правило, известны в данной области техники и описаны в Denardo и соавт., 1998 год, Clin Cancer Res. 4:2483; Peterson и соавт., 1999 год, Bioconjug. Chem. 10:553; и Zimmerman и соавт., 1999 год, Nucl. Med. Biol. 26:943.
В некоторых вариантах реализации изобретения тяжелые и легкие цепи иммуноглобулинов антигенсвязывающей полипептидной конструкции экспрессируются в виде слитых белков, содержащих метку для облегчения очистки и/или тестирования и т.д. Согласно изложенному в данном документе "метка" представляет собой любую дополнительную последовательность аминокислот, которая содержится в белке либо на С-конце, на N-конце или внутри последовательности, что способствует идентификации или очистке данного белка. Подходящие метки включают, но без ограничений, метки известные специалистам в данной области техники, которые являются полезным в очистке и/или тестировании, такие метки как альбумин-связывающий домен (АСД), гистидиновую метку, FLAG-метку, глутатион-S-трансферазу, гемагглютинин (HA) и связывающий мальтозу белок. Такие меченые белки также могут быть сконструированы таким образом, чтобы содержать сайт расщепления, такой как сайт расщепления тромбином, энтерокиназой или фактором X, для простоты удаления метки до, во время или после очистки.
Способы получения антигенсвязывающих полипептидных конструкций
Как описано выше, описанные в данном документе антигенсвязывающие полипептидные конструкции могут содержать первый гетеродимер и второй гетеродимер, первый гетеродимер, содержащий тяжелую цепь иммуноглобулина или ее фрагмент, имеющую, по меньшей мере, домен VH и CH1, и легкую цепь лямбда иммуноглобулина, имеющую VL и домен CL, и второй гетеродимер, содержащий тяжелую цепь иммуноглобулина или ее фрагмент, имеющую, по меньшей мере, домен VH и CH1, и легкую цепь каппа иммуноглобулина, имеющую домен VL и домен CL. Полипептидные последовательности иммуноглобулина сконструированы для включения аминокислотных модификаций, которые способствуют преимущественному спариванию, как описано в данном документе. Соответственно, в случае биспецифической антигенсвязывающей полипептидной конструкции обычно имеются четыре различные полипептидные последовательности, две полипептидные последовательности тяжелой цепи иммуноглобулина или их фрагменты и две полипептидные последовательности легкой цепи иммуноглобулина, которые составляют антигенсвязывающую полипептидную конструкцию. Полипептидные последовательности тяжелой цепи иммуноглобулина и полипептидные последовательности легкой цепи иммуноглобулина антигенсвязывающей полипептидной конструкции могут быть легко получены с использованием технологии рекомбинантной ДНК, известной в данной области техники. Стандартные методы, такие как, например, те, которые описаны в Sambrook и Russell, Molecular Cloning:A Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y., 3-е изд., 2001 год); Sambrook и соавт., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y., 2-е изд., 1989 год); Short Protocols in Molecular Biology (Ausubel и соавт., John Wiley and Sons, New York, 4-е изд., 1999 год); и Glick and Pasternak, Molecular Biotechnology:Principles and Applications of Recombinant DNA (ASM Press, Washington, D. C., 2-е изд., 1998 год) могут быть использованы для методов рекомбинантной нуклеиновой кислоты, синтеза нуклеиновых кислот, культивирования клеток, инкорпорации трансгена и экспрессии рекомбинантных белков.
Полинуклеотидные и аминокислотные последовательности тяжелой и легкой цепей иммуноглобулина исходных антител, которые составляют антигенсвязывающую полипептидную конструкцию, являются либо известными в данной области техники, либо могут быть легко определены с использованием способов секвенирования нуклеиновой кислоты и/или белка.
Соответственно, также предложены полинуклеотиды или набор полинуклеотидов, кодирующих тяжелые и легкие цепи иммуноглобулина антигенсвязывающей полипептидной конструкции. Такие полинуклеотиды включают ДНК и РНК как в одноцепочечной, так и в двухцепочечной форме, а также соответствующие комплементарные последовательности. ДНК включает, например, кДНК, геномную ДНК, химически синтезированную ДНК, ДНК, амплифицированную с помощью ПЦР, и их комбинации. Полинуклеотиды включают полноразмерные гены или молекулы кДНК, а также комбинацию их фрагментов.
Полинуклеотиды, кодирующие сконструированные полипептиды тяжелой и легкой цепи иммуноглобулина, описанные в данном документе, могут быть получены с помощью сайт-направленного мутагенеза нуклеотидов в ДНК, кодирующей полипептид, с использованием кассетного или ПЦР-мутагенеза или других методов, хорошо известных в данной области техники, для получения ДНК, кодирующей сконструированные полипептиды тяжелой и легкой цепей иммуноглобулина, и последующей экспрессии рекомбинантной ДНК в клеточной культуре, как описано в данном документе. Однако полинуклеотиды, кодирующие сконструированные полипептиды тяжелой и легкой цепи иммуноглобулина, также могут быть получены синтезом гена in vitro с использованием установленных методов.
Специалисту в данной области техники будет понятно, что из-за вырожденности генетического кода может быть сделано чрезвычайно большое количество полинуклеотидов, все из которых кодируют сконструированные полипептиды тяжелой и легкой цепи иммуноглобулина, описанные в данном документе. Таким образом, определив конкретную аминокислотную последовательность, специалисты в данной области техники могут сделать любое количество различных полинуклеотидов путем простого изменения последовательности одного или более кодонов способом, который не изменяет аминокислотную последовательность кодируемого белка.
Также предложены экспрессирующие системы и конструкции в виде плазмид, экспрессионных векторов, транскрипционных или экспрессионных кассет, которые содержат, по меньшей мере, один полинуклеотид, как указано выше. Также предложены клетки-хозяева, содержащие такие экспрессирующие системы или конструкции.
Обычно экспрессионные векторы, используемые в клетках-хозяевах, будут содержать последовательности для обеспечения функционирования плазмиды, а также для клонирования и экспрессии экзогенных нуклеотидных последовательностей. Такие последовательности, которые в совокупности называются "фланкирующими последовательностями", в некоторых вариантах реализации изобретения обычно включают одну или более из следующих нуклеотидных последовательностей: промотор, одну или более последовательностей энхансера, точку начала репликации, последовательность терминации транскрипции, полную последовательность интрона, содержащую донорные и акцепторные сайты сплайсинга, последовательность, кодирующую лидерную последовательность для секреции полипептидов, сайт связывания рибосом, последовательность полиаденилирования, полилинкерную область для встраивания полинуклеотида, кодирующего полипептид, который необходимо экспрессировать, и селектируемый маркерный элемент. Вектор может быть мультицистронным, то есть экспрессирующим два или более полинуклеотида, кодирующих тяжелую и легкую цепи иммуноглобулина антигенсвязывающей полипептидной конструкции, или антигенсвязывающая полипептидная конструкция может быть экспрессирована с помощью набора векторов, где каждый вектор экспрессирует один или более полинуклеотидов. Антигенсвязывающая полипептидная конструкция также может быть экспрессирована с использованием набора векторов, содержащих комбинацию мультицистронных векторов и векторов, которые содержат один полинуклеотид, кодирующий одну из тяжелой и легкой цепей иммуноглобулина.
В некоторых вариантах реализации изобретения вектор может содержать последовательность, кодирующую "метку", то есть молекулу олигонуклеотида, расположенную в 5' или 3' конце кодирующей последовательности полипептида; олигонуклеотидная последовательность кодирует поли-His (такую как гекса-His) или другую "метку", такую как FLAG, HA (гемагглютинин вируса гриппа) или myc, для которой существуют коммерчески доступные антитела. После экспрессии полипептида эта метка обычно слита с полипептидом и может служить средством для аффинной очистки или выявления полипептида из клетки-хозяина. Аффинная очистка может быть осуществлена, например, с помощью колоночной хроматографии с использованием антител к метке в качестве аффинной матрицы. Необязательно, метка может быть впоследствии удалена из очищенного полипептида различными способами, такими как расщепление пептидазой.
Векторы обычно содержат промотор, который распознается организмом-хозяином и функционально связан с полинуклеотидом, кодирующим полипептид. Промоторы представляют собой нетранскрибированные последовательности, расположенные в 3'-5' направлениипо отношению к стартовому кодону структурного гена (обычно в пределах от 100 до 1000 пар оснований), которые контролируют транскрипцию структурного гена. Промоторы обычно классифицируют в один из двух классов: индуцируемые промоторы и конститутивные промоторы. Индуцируемые промоторы инициируют повышенные уровни транскрипции из ДНК под их контролем в ответ на некоторое изменение условий в культуре, такое как наличие или отсутствие питательного вещества или изменение температуры. С другой стороны, конститутивные промоторы равномерно транскрибируют ген, к которому они функционально связаны, то есть без или с минимальным контролем над экспрессией гена. Известно большое количество промоторов, распознаваемых различными потенциальными клетками-хозяевами.
Подходящие промоторы для применения с дрожжевыми хозяевами, бактериальными хозяевами и клетками насекомых хорошо известны в данной области техники. Дрожжевые энхансеры преимущественно используются с промоторами дрожжей. Подходящие промоторы для применения с клетками-хозяевами млекопитающих хорошо известны и включают в себя, но не без ограничений, те, которые получены из геномов вирусов, таких как вирус полиомы, вирус оспы кур, аденовирус (например, аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и наиболее предпочтительно вирус обезьян 40 (SV40). Другие подходящие промоторы млекопитающих включают гетерологичные промоторы млекопитающих, например, промоторы теплового шока и промотор актина.
Вектор может содержать один или более элементов, которые содействуют экспрессии, когда вектор интегрирован в геном клетки-хозяина. Примеры включают элемент EASE (Aldrich и соавт. 2003 Biotechnol Prog. 19:1433-38) и участок прикрепления к матриксу (MAR). MAR опосредуют структурную организацию хроматина и могут изолировать интегрированный вектор от эффектов "позиционирования". Таким образом, MAR особенно полезны, когда вектор используется для создания стабильных трансфектантов. Ряд природных и синтетических нуклеиновых кислот, содержащих MAR, известен в данной области техники, например, в патентах США №6,239,328; 7,326,567; 6,177,612; 6,388,066; 6,245,974; 7,259,010; 6,037,525; 7,422,874; 7,129,062.
После конструирования вектора и вставки полинуклеотида в надлежащий сайт вектора, готовый вектор может быть вставлен в подходящую клетку-хозяина для амплификации и/или экспрессии полипептида. Трансформация экспрессионного вектора в выбранную клетку-хозяина может быть осуществлена с помощью известных способов, включая трансфекцию, инфекцию, совместным осаждением фосфатом кальция, электропорацию, микроинъекцию, липофекцию, опосредованную DEAE-декстраном трансфекцию или другими известными методиками. Выбранный способ частично будет зависеть от типа используемой клетки-хозяина. Клетки-хозяева могут быть транзиентно трансфицированными или клетки-хозяева могут быть стабильно трансфицированными. Эти способы и других подходящие способы хорошо известны специалисту в данной области техники и изложены, например, в Sambrook и соавт., 2001 год, выше.
Для долгосрочного, высокопроизводительного продуцирования рекомбинантных белков предпочтительной является стабильная экспрессия. Например, могут быть получены клеточные линии, которые стабильно экспрессируют сконструированные тяжелые и легкие цепи антигенсвязывающей полипептидной конструкции. Вместо того, чтобы использовать экспрессионные векторы, которые содержат вирусные сайты инициации репликации, клетки-хозяева могут быть трансформированы с помощью ДНК, контролируемой подходящими регуляторными элементами экспрессии (например, промоторной, энхансерной последовательностями, терминаторами транскрипции, сайтами полиаденилирования и т.д.) и селектируемым маркером. После введения чужеродной ДНК или полипептида, сконструированные клетки могут расти в течение 1-2 дней в обогащенной среде, а затем их перемещают на селективную среду. Селектируемый маркер в рекомбинантной плазмиде придает устойчивость к отбору и позволяет клеткам стабильно интегрировать эту плазмиду в их хромосомы и расти с образованием очагов, которые, в свою очередь, могут быть клонированы и размножены в клеточные линии.
Может быть использован ряд систем селекции, в том числе, но без ограничений, ген тимидинкиназы вируса простого герпеса (Wigler и соавт., 1977 год, Cell 11:223), ген гипоксантингуанин-фосфорибозилтрансферазы (Szybalska & Szybalski, 1962 год, Proc. Natl. Acad. Sci. USA 48:2026) и ген аденин-фосфорибозилтрансферазы (Lowy и соавт., 1980 год, Cell 22:817) могут быть использованы в tk-, hgprt- или aprt-клетках, соответственно. Кроме того, устойчивость к антиметаболитам может быть использована в качестве основы селекции на ген dhfr, который придает устойчивость к метотрексату (Wigler и соавт., 1980 год, Natl. Acad. Sci. USA 77:3567; O'Hare и соавт., 1981 год, Proc. Natl. Acad. Sci. USA 78:1527); ген gpt, который придает устойчивость к микофеноловой кислоте (Mulligan & Berg, 1981 год, Proc. Natl. Acad. Sci. USA 78:2072); ген neo, который придает устойчивость к аминогликозиду G-418 (Colberre-Garapin и соавт., 1981 год, J. Mol. Biol. 150:1); и ген hygro, который придает устойчивость к гигромицину (Santerre и соавт., 1984 год, Gene 30:147).
Клетка-хозяин, при культивировании в соответствующих условиях, продуцирует антигенсвязывающую полипептидную конструкцию, которая может быть выделена из культуральной среды (если клетка-хозяин секретирует ее в среду) или непосредственно из клетки-хозяина, продуцирующего ее (если она не секретируется). Выбор соответствующей клетки-хозяина будет зависеть от различных факторов, таких как требуемые уровни экспрессии, полипептидные модификации, которые желательны или необходимы для проявления активности (такие как гликозилирования или фосфорилирование) и облегчают фолдинг в биологически активную молекулу. Клетка-хозяин может быть эукариотической или прокариотической. Например, экспрессия в бактериальной системе приведет к созданию негликозилированного продукта, а экспрессия в дрожжах приведет к созданию гликозилированного продукта. Могут быть использованы эукариотические клетки-хозяева, которые обладают клеточным механизмом для правильного процессинга первичного транскрипта (например, гликозилирование и фосфорилирование) продукта гена.
Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, хорошо известны в данной области техники и включают, но без ограничений, иммортализованные клеточные линии, доступные из Американской коллекции типовых культур (АТСС), и любые клеточные линии, используемые в экспрессирующей системе, известной в данной области техники, могут применяться для создания рекомбинантных полипептидов, описанных в данном документе. В общем, клетки-хозяева трансформируют рекомбинантным экспрессионным вектором, который содержит ДНК, кодирующую антигенсвязывающую полипептидную конструкцию. К клеткам-хозяевам, которые могут быть использованы, относятся прокариоты, дрожжи или клетки высших эукариот. Прокариоты включают грамотрицательные или грамположительные организмы, например, E. coli или бациллы. Клетки высших эукариот включают клетки насекомых и стабильные клеточные линии млекопитающих. Примеры подходящих линий клеток-хозяев млекопитающих включают линию клеток почек обезьян COS-7 (ATCC CRL 1651) (Gluzman и соавт., 1981 г., Cell 23:175), клетки L, клетки C127, клетки 3T3 (ATCC CCL 163), клетки яичника китайского хомячка (CHO) или их производные, такие как Veggie CHO и подобные клеточные линии, которые растут в бессывороточной среде (Rasmussen и соавт., 1998 год, Cytotechnology 28:31), клетки HeLa, клеточные линии BHK (ATCC CRL 10) и клеточная линия CV1/EBNA, полученная из клеточной линии почек африканской зеленой обезьяны CV1 (ATCC CCL 70), как описано в McMahan и соавт., 1991 год, EMBO J. 10:2821, человеческие эмбриональные клетки почек, такие как 293, 293 EBNA или MSR 293, человеческие эпидермальные клетки A431, клетки человека Colo205, другие трансформированные клеточные линии приматов, нормальные диплоидные клетки, клеточные штаммы, полученные из in vitro культуры первичной ткани, первичные эксплантаты, клетки HL-60, U937, HaK или Jurkat. В альтернативном варианте возможно получить полипептид с помощью клеток низших эукариот, таких как дрожжи или клеток прокариот, таких как бактерии. Подходящие штаммы дрожжей включают в себя Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces, Candida или любой штамм дрожжей, способный экспрессировать гетерологичные полипептиды. Подходящие бактериальные штаммы включают в себя Escherichia coli, Bacillus subtilis, Salmonella typhimurium или любой бактериальный штамм, способный экспрессировать гетерологичные полипептиды.
Если антигенсвязывающая полипептидная конструкция продуцируется клеткой дрожжей или бактерий, может быть желательным модифицировать продукт, полученный таким образом, например, путем фосфорилирования или гликозилирования соответствующих сайтов, чтобы получить функциональный продукт. Такое ковалентное присоединение может быть достигнуто с использованием известных химических или ферментативных способов. Антигенсвязывающая полипептидная конструкция может также быть получена путем оперативного связывания набора полинуклеотидов с подходящими контрольными последовательностями в одном или более экспрессионных векторах насекомых, и с использованием экспрессирующей системы насекомых. Материалы и способы для экспрессирующих систем бакуловируса/клеток насекомых коммерчески доступны в виде набора в, например,Invitrogen, Сан-Диего, Калифорния, США (набор MaxBac™), и такие способы хорошо известны в данной области техники, как описано в Summers and Smith, Texas Agricultural Experiment Station Bulletin №1555 (1987 год) и Luckow and Summers, Bio/Technology 6:47 (1988 год). Соответствующие клонирующие и экспрессионные векторы для применения с бактериальными, грибковыми, дрожжевыми клетками-хозяевами и клетками-хозяевами млекопитающих описаны Pouwels и соавт. (Cloning Vectors:A Laboratory Manual, Elsevier, New York, 1985 год).
В некоторых вариантах реализации изобретения внеклеточные системы экспрессии белков могут использоваться для коэкспрессии полипептидов (например, полипептидов тяжелой и легкой цепи) из набора полинуклеотидов, без использования живых клеток. Вместо этого все компоненты, необходимые для транскрипции ДНК в РНК и трансляции РНК в белок (например, рибосомы, тРНК, ферменты, кофакторы, аминокислоты), предоставляются в растворе для применения in vitro. В некоторых вариантах реализации изобретения экспрессия in vitro требует (1) генетическую матрицу (мРНК или ДНК), кодирующую тяжелую и легкую цепь полипептидов и (2) реакционный раствор, содержащий необходимую для транскрипции и трансляции молекулярную машинерию. В некоторых вариантах реализации изобретения клеточные экстракты, по существу, поставляют компоненты реакционного раствора, например: РНК-полимеразы для транскрипции мРНК, рибосомы для трансляции полипептида, тРНК, аминокислоты, ферментные кофакторы, источник энергии и клеточные компоненты, необходимые для правильного фолдинга белков. Внеклеточные системы экспрессии белка могут быть приготовлены с использованием лизатов, полученных из бактериальных клеток, дрожжевых клеток, клеток насекомых, клетки растений, клеток млекопитающих, человеческих клеток или их комбинации. Такие клеточные лизаты могут обеспечить правильный состав и пропорции ферментов, и строительных блоков, необходимых для трансляции. В некоторых вариантах реализации изобретения клеточные мембраны удаляются, чтобы оставить только цитозольные компоненты и органеллы клетки.
Несколько внеклеточных систем экспрессии белков известны в данной области техники, как рассмотрено у Carlson и соавт. (2012 год) Biotechnol. Adv. 30:1185-1194. Например, внеклеточные системы экспрессии белков доступны на основе прокариотических или эукариотических клеток. Примеры внеклеточных прокариотических систем экспрессии включают те, которые из E. coli. Внеклеточные системы экспрессии белков доступны на основе экстрактов из, например, ретикулоцитов кролика, зародышей пшеницы и клеток насекомых. Такие внеклеточные прокариотические и эукариотические системы экспрессии белков коммерчески доступны от таких компаний, как Roche, Invitrogen, Qiagen и Novagen. Специалист в данной области техники легко сможет выбрать подходящие внеклеточные системы экспрессии белков, которые будут производить полипептиды (например, полипептиды тяжелой цепи и легкой цепи), которые способны к спариванию друг с другом. Кроме того, внеклеточная система экспрессии белков также может быть дополнена шаперонами (например, BiP) и изомеразами (например, дисульфидизомераза), чтобы повысить эффективность фолдинга IgG.
Коэкспрессия тяжелых цепей и легких цепей
Сконструированные тяжелые цепи и легкие цепи иммуноглобулина антигенсвязывающей полипептидной конструкции, описанные в данном документе, могут коэкспрессироваться в клетках млекопитающих, как указано выше. В одном варианте реализации изобретения тяжелые цепи иммуноглобулина и легкие цепи иммуноглобулина антигенсвязывающей полипептидной конструкции коэкспрессируются в клетке-хозяине. Таким образом, в случае биспецифической антигенсвязывающей полипептидной конструкции две тяжелые цепи иммуноглобулина и две легкие цепи иммуноглобулина коэкспрессируются в клетке-хозяине. Однако, альтернативные способы получения биспецифических антигенсвязывающих полипептидных конструкций, которые не полагаются на использовании одной клональной или стабильной клеточной линии, экспрессирующей все четыре цепи, также известны в данной области техники (Gramer и соавт. (2013 год) mAbs 5, 962; Strop и соавт. (2012 год) J Mol Biol 420, 204). Эти способы основаны на обмене плечами двух пар тяжелой и легкой цепи после изготовления, участвующих в образовании биспецифического антитела в окислительно-восстановительных условиях (окислительно-восстановительное производство). При таком подходе гетеродимеры H1L1 и H2L2 могут быть экспрессированы в двух разных клеточных линиях для независимого производства двух гетеродимеров. Впоследствии эти два гетеродимера смешивают в выбранных окислительно-восстановительных условиях для достижения реассоциации двух уникальных тяжелых цепей H1 и H2 с образованием биспецифической антигенсвязывающей полипептидной конструкции, содержащей H1L1H2L2.
Хотя преимущественное спаривание обусловлено главным образом включением аминокислотных модификаций набора конструкции мАт в полипептиды тяжелой и легкой цепи иммуноглобулина, количество правильно спаренных гетеродимеров может быть дополнительно оптимизировано путем изменения соотношения полинуклеотидов, кодирующих каждый полипептид друг к другу, как показано в примерах.
Тестирование антигенсвязывающих полипептидных конструкций
Как описано выше, антигенсвязывающие полипептидные конструкции содержат первый гетеродимер, содержащий H1 и L1, а также второй гетеродимер, содержащий H2 и L2, где L1 представляет собой легкую цепь лямбда и L2 представляет собой легкую цепь каппа, а H1 и H2 отличаются друг от друга. Одна или более из H1, L1, H2 и L2 содержат аминокислотные модификации, которые способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и H2 с L2 по сравнению с L1. Первая область Fab гетеродимера H1L1, и вторая область Fab гетеродимера H2L2 способны связываться с антигеном с аффинностью, которая аналогична соответствующей первой области Fab дт или второй области Fab дт. Первая область Fab гетеродимера H1L1 и вторая область Fab гетеродимера H2L2 также обладают термической стабильностью, которая сопоставима с термической стабильностью соответствующей первой области Fab дт или второй области Fab дт.
Аффинность каждого гетеродимера пары гетеродимеров к его соответствующему антигену может быть испытана, как описано ниже. Термическая стабильность каждого гетеродимера пары гетеродимеров также может быть испытана, как описано ниже.
В одном варианте реализации изобретения одна тяжелая цепь коэкспрессируется с двумя различными легкими цепями в наборе конструкции LCCA, как описано выше, где тяжелая цепь преимущественно спаривается с одной из двух легких цепей. В другом варианте реализации изобретения две уникальные тяжелые цепи коэкспрессируются с двумя уникальными легкими цепями, где каждая тяжелая цепь преимущественно спаривается с одной из легких цепей.
Способы измерения преимущественного спаривания
Степень преимущественного спаривания может быть оценена, например, с использованием способов, описанных ниже и в примерах. Преимущественное спаривание может быть оценено в контексте наборов конструкции LCCA (H1L1L2 или H2L1L2) или наборов конструкции мАт (H1L1H2L2).
В одном варианте реализации изобретения конкурентный анализ легкой цепи (LCCA - Light Chain Competition Assay) может использоваться для оценки преимущественного спаривания в контексте наборов конструкции LCCA. Совместная патентная заявка PCT/US2013/063306, поданная 3-го октября 2013 года, описывает различные варианты реализации LCCA и включена в данный документ посредством ссылки в полном объеме для любых целей. Способ позволяет провести количественный анализ спаривания тяжелых цепей с определенными легкими цепями внутри смеси коэкспрессируемых белков, и может быть использован для определения, может ли одна конкретная тяжелая цепь иммуноглобулина селективно связываться с одной из двух легких цепей иммуноглобулина, в случае если тяжелая цепь и легкие цепи коэкспрессируются. Способ кратко описан следующим образом: по меньшей мере одна тяжелая цепь и две различные легкие цепи коэкспрессируются в клетке в таких соотношениях, что тяжелая цепь является ограничивающим спаривание реактантом. Тяжелые цепи и легкие цепи могут быть помечены для облегчения обнаружения. Секретируемые белки могут быть отделены от клетки, а полипептиды легкой цепи иммуноглобулина, связанные с тяжелой цепью, выделены из других секретируемых белков, с целью получить выделенную спаренную фракцию тяжелой цепи. Затем определяют количество каждой другой легкой цепи в выделенной фракции тяжелой цепи и анализируют относительное количество каждой другой легкой цепи в выделенной фракции тяжелой цепи, чтобы определить способность тяжелой цепи избирательно спариваться с одной из легких цепей. Дополнительные подробности, касающиеся вариантов реализации изобретения по этому способу, приведены в примерах.
В другом варианте реализации изобретения преимущественное спаривание оценивается в контексте наборов конструкции мАт, где H1, L1, H2 и L2 коэкспрессируются. В этом варианте реализации изобретения одна или более из H1, L2, H2 и L2 могут быть помечены для облегчения обнаружения и анализа. Полученные виды спаренных тяжелых цепей и легких цепей оценивают с использованием ЖХ-МС (жидкостной хромато-масс-спектрометрии) на основе различий в молекулярной массе каждого вида. Анализ на антигенную активность также может быть использован для количественного определения относительных совокупностей гетеродимеров, содержащих каждую легкую цепь, в результате чего измеренная степень связывания (по отношению к контролю) будет использоваться для оценки каждой соответствующей совокупности гетеродимеров.
Термическая стабильность
Термическая стабильность гетеродимеров может быть определена с помощью любого способа, известного в данной области техники. Температура плавления каждого гетеродимера свидетельствует о его термической стабильности. Температура плавления гетеродимера может быть измерена с помощью таких методов, как дифференциальная сканирующая калориметрия (Chen и соавт. (2003 год) Pharm Res 20:1952-60; Ghirlando и соавт. (1999 год) Immunol Lett 68:47-52). В альтернативном варианте термическая стабильность гетеродимеров может быть измерена с помощью кругового дихроизма (Murray и соавт. (2002 год) J. Chromatogr Sci 40:343-9).
Аффинность к антигену
Аффинность связывания гетеродимеров с их соответствующими антигенами и скорость диссоциации взаимодействия может быть определена с помощью конкурентно-связывающих анализов в соответствии со способами, хорошо известными в данной области техники. Один из примеров конкурентно-связывающего анализа представляет собой радиоиммунологический анализ, включающий инкубацию меченого антигена (например, 3Н или 125I с молекулой интереса (например, гетеродимерами, описанными в данном документе) в присутствии возрастающего количества немеченого антигена и обнаружения молекулы, связанной с меченым лигандом. Аффинность гетеродимера к антигену и связывающая скорость диссоциации может быть определена по данным насыщения с помощью анализа Скатчарда.
Кинетические параметры гетеродимеров, описанные в данном документе, также могут быть определены с помощью анализов на основе поверхностного плазмонного резонанса (ППР), известных в данной области техники (например, кинетического анализа BIAcore). Для обзора технологии на основе ППР см. Mullet и соавт., 2000 год, Methods 22:77-91; Dong и соавт., 2002 год, Review in Mol. Biotech. 82303-23; Fivash и соавт., 1998 год, Current Opinion in Biotechnology 9:97-101; Rich и соавт., 2000 год, Current Opinion in Biotechnology 11:54-61. Кроме того, любой из приборов ППР и способов на основе ППР для измерения белок-белковых взаимодействий, описанных в патентах США №6,373,577; 6,289,286; 5,322,798; 5,341,215; 6,268,125 рассматриваются в способах по данному изобретению. FACS (сортировка клеток с активированной флуоресценцией) также может быть использован для измерения аффинности, как известно в данной области техники.
Фармацевтические композиции
Также в настоящем изобретении предложены фармацевтические композиции, содержащие антигенсвязывающие полипептидные конструкции, описанные в данном документе. Такие композиции содержат терапевтически эффективное количество антигенсвязывающей полипептидной конструкции и фармацевтически приемлемый носитель. В конкретном варианте реализации изобретения термин "фармацевтически приемлемый" одобренный регуляторным ведомством Федерального правительства или правительства штата, или приведенный в Фармакопее США или другой общепризнанной фармакопее с целью использования на животных и, более конкретно, на людях. Термин "носитель" относится к разбавителю, адъюванту, вспомогательному веществу или носителю, с которым терапевтическое средство вводится. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включая нефтяные масла, масла животного, растительного или синтетического происхождения, такие как ореховое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. Вода является предпочтительным носителем, в случае если фармацевтическая композиция вводится внутривенно. Солевые растворы и водные растворы декстрозы и глицерина также могут быть использованы в качестве жидких носителей, особенно для инъекционных препаратов. Подходящие фармацевтические вспомогательные вещества включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикатный гель, стеарат натрия, глицеролмоностеарат, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропилен, гликоль, воду, этанол и тому подобное. При желании композиция может также содержать незначительные количества увлажнителей или эмульгаторов, или pH-буферных средств. Эти композиции могут быть в виде растворов, суспензий, эмульсии, таблеток, драже, капсул, порошков, препаратов длительного высвобождения и тому подобного. Композиция может быть приготовлена в виде суппозитория с традиционным связующими веществами и носителями, такими как триглецириды. Состав для перорального применения может содержать стандартные носители, такие как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, целлюлозу, карбонат магния и т.д., фармацевтической степени чистоты. Примеры подходящих фармацевтических носителей описаны в "Remington's Pharmaceutical Sciences" E. W. Martin. Такие композиции содержат терапевтически эффективное количество композиции, преимущественно в очищенном виде, вместе с подходящим количеством носителя для получения требуемой формы для введения пациенту. Препарат должен соответствовать способу введения.
В определенных вариантах реализации изобретения композицию, содержащую антигенсвязывающую полипептидную конструкцию, получают по стандартным методикам в виде фармацевтической композиции, адаптированной для внутривенного введения человеку. Как правило, композиции для внутривенного введения представляют собой растворы в стерильном изотоническом буфере на водной основе. При необходимости композиция также может содержать солюбилизирующее средство и местный анестетик, такой как лидокаин, для облегчения боли в участке инъекции. Как правило, ингредиенты поставляются либо по отдельности, либо смешанные друг с другом в виде единичной лекарственной формы, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметичной емкости, такой как ампула или пакет-саше с указанием количества действующего вещества. Если композицию вводят путем инфузии ее можно распределить с использованием инфузионного флакона, содержащего стерильную воду или солевой раствор фармацевтической степени чистоты. Если композицию вводят путем инъекции, может предоставляться ампула со стерильной водой для инъекций или физиологическим раствором, для того, чтобы можно было смешать ингредиенты до введения.
В некоторых вариантах реализации изобретения композиции, описанные в данном документе, получают в форме нейтральных солей. Фармацевтически приемлемые соли включают соли, образующиеся с анионами, как, например, соли производные соляной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и соли, образующиеся с катионами, как, например, соли производные натрия, калия, аммония, кальция, гидроксида железа, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и т.д.
Количество композиции, описанное в данном документе, которое будет эффективно в лечении, ингибировании и профилактике заболевания или нарушения, связанного с неправильной экспрессией и/или активностью терапевтического белка, можно определить стандартными клиническими методами. В дополнение, анализы in vitro могут быть необязательно использованы для определения оптимального диапазона доз. Точная доза используется в препарате также в зависимости от способа введения и серьезности заболевания или расстройства, и решения в соответствии с оценкой практикующего врача и обстоятельств каждого пациента. Эффективные дозы экстраполированы из кривых зависимости доза-эффект, полученные in vitro или из тестовых систем экспериментальной модели на животном.
Применение антигенсвязывающих полипептидных конструкций
Как описано выше, описанные в данном документе антигенсвязывающие полипептидные конструкции получены из исходных антител, где каждый гетеродимер антигенсвязывающей полипептидной конструкции соответствует одному из исходных антител и был сконструирован для включения аминокислотных модификаций, которые способствуют преимущественному спариванию тяжелых и легких цепей иммуноглобулина, которые составляют гетеродимеры. Соответственно, описанные в данном документе антигенсвязывающие полипептидные конструкции могут быть использованы в лечении или профилактике одного и того же заболевания, расстройства или инфекции, для которых используется исходное антитело или комбинация исходных антител.
В другом варианте реализации изобретения антигенсвязывающие полипептидные конструкции, описанные в данном документе, также могут быть использованы в комбинации с другими лекарственными средствами, известными в данной области техники, для лечения или профилактики рака, аутоиммунных заболеваний, воспалительных заболеваний или инфекционных заболеваний. В конкретном варианте реализации изобретения антигенсвязывающие полипептидные конструкции, описанные в данном документе, могут быть использованы в комбинации с моноклональными или химерными антителами, лимфокинами или гемопоэтическими факторами роста (такими как, например, ИЛ-2, ИЛ-3 и ИЛ-7), которые, например, служат для увеличения количества или активности эффекторных клеток, которые взаимодействуют с молекулами и увеличивать иммунный ответ. Антигенсвязывающие полипептидные конструкции, описанные в данном документе, также могут быть использованы в комбинации с одним или более лекарственными средствами, используемыми для лечения заболевания, нарушения или инфекции, такими как, например, противораковые агенты, противовоспалительные агенты или противовирусные агенты.
Создание биспецифических антител с использованием библиотеки набора конструкций мАт
В одном варианте реализации изобретения наборы конструкций мАт, описанные в данном документе, могут быть использованы для получения биспецифических антигенсвязывающих полипептидных конструкций. Наборы конструкций мАт, описанные в данном документе, могут быть использованы в виде библиотеки набора конструкций мАт, где библиотека набора конструкций мАт включает в себя наборы конструкций мАт, которые являются полезными в способствовании преимущественному спариванию для образования биспецифических антигенсвязывающих полипептидных конструкций. В одном варианте реализации изобретения библиотеки набора конструкций мАт представлены наборами конструкций мАт, включенными в таблицу 4A и таблицу 4B. В одном варианте реализации изобретения библиотека набора конструкций мАт представлена наборами конструкций мАт в одной или более таблицах с 10-A1 по 10-A12 и с 10-B1 по 10-B10. В другом варианте реализации изобретения библиотека набора конструкций мАт представлена в одной или более таблицах с 10-A1 по 10-A12. В одном варианте реализации изобретения библиотека набора конструкций мАт представлена в одной или более таблицах 10-B1, 10-B2, 10-B3, 10-B4, 10-B6, 10-B8 и 10-B10. Библиотека набора конструкций мАт может использоваться для создания антигенсвязывающей полипептидной конструкции, начиная с двух исходных антител (то есть, мАт1 и мАт2) следующим образом. С целью иллюстрации, мАт1 содержит Fab лямбда и содержит полипептид тяжелой цепи H1 иммуноглобулина и полипептид легкой цепи L1 иммуноглобулина, тогда как мАт2 содержит Fab каппа и содержит полипептид тяжелой цепи H2 иммуноглобулина и полипептид легкой цепи L2 иммуноглобулина.
Аминокислотные модификации набора конструкций мАт (H1L1H2L2) библиотеки набора конструкций мАт вводятся в тяжелую и легкую цепь иммуноглобулина мАт1 (H1 и L2) и тяжелую и легкую цепь иммуноглобулина мАт2 (H2 и L2). Затем H1, L1, H2 и L2 коэкспрессируются и определяется количество правильно спаренной биспецифической антигенсвязывающей полипептидной конструкции. Один или более наборов конструкций мАт библиотеки набора конструкций мАт могут быть отдельно испытаны или проскринированы для определения того, который обеспечивает требуемое количество биспецифической антигенсвязывающей полипептидной конструкции. Каждый гетеродимер биспецифической антигенсвязывающей полипептидной конструкции может быть дополнительно испытан для оценки способности каждого гетеродимера связываться с антигеном или оценки его термической стабильности, как описано в данном документе. Дополнительные свойства, которые могут быть оценены, включают растворимость, агрегацию, скорости константы ассоциации и константы диссоциации, способность выдерживать воздействие кислот, щелочи, окисления, циклов замораживания/оттаивания, перемешивания, давления и т.д. биспецифической антигенсвязывающей полипептидной конструкции по сравнению с исходными антителами или областями Fab исходных антител. Последние свойства могут оказать влияние на определяющие комплементарность области (CDR) антитела интереса, и, таким образом, могут быть испытаны для каждой полученной биспецифической антигенсвязывающей полипептидной конструкции.
В некоторых вариантах реализации изобретения количество правильно спаренной биспецифической антигенсвязывающей полипептидной конструкции оценивается с помощью ЖХ-МС. В некоторых вариантах реализации изобретения количество правильно спаренной биспецифической антигенсвязывающей полипептидной конструкции оценивается с помощью методов разделения на основе заряда, таких как метод капиллярного изоэлектрического фокусирования (кИЭФ) или хроматографический метод. Процедура получения биспецифической антигенсвязывающей полипептидной конструкции из мАт1 и мАт2 с использованием библиотеки наборов конструкции мАт схематически изображена на фигуре 9.
В одном варианте реализации изобретения библиотека набора конструкций мАт хранится на машиночитаемом носителе данных, чтобы облегчить использование библиотеки набора конструкций мАт для разработки биспецифических антигенсвязывающих полипептидных конструкций.
Реализация с помощью компьютера
В одном варианте реализации изобретения компьютер содержит, по меньшей мере, один процессор, соединенный с чипсетом. Также соединенными с чипсетом являются запоминающее устройство, устройство для хранения данных, клавиатура, графический адаптер, указательное устройство и сетевой адаптер. Дисплей соединен с графическим адаптером. В одном варианте реализации изобретения функциональность чипсета обеспечивается с помощью контроллера концентратора памяти и контроллера концентратора ввода/вывода. В другом варианте реализации изобретения запоминающее устройство соединено непосредственно с процессором, а не с чипсетом.
Устройство для хранения данных представляет собой любое устройство, способное хранить данные, например, жесткий диск, компакт-диск "только для чтения" (CD-ROM), DVD или твердотельное запоминающее устройство. Запоминающее устройство хранит команды и данные, используемые процессором. Указывающее устройство может быть мышью, трекболом или другим типом указывающего устройства и используется в комбинации с клавиатурой для ввода данных в компьютерную систему. Графический адаптер отображает изображения и другую информацию на дисплее. Сетевой адаптер соединяет компьютерную систему с локальной или глобальной сетью.
Как известно специалистам в данной области техники, компьютер может иметь различные и/или другие компоненты, чем описанные ранее. Кроме того, в компьютере могут отсутствовать определенные компоненты. Кроме того, устройство для хранения данных быть локальным и/или удаленным от компьютера (например, встроенным в сетевую систему хранения данных (SAN)).
Как известно специалистам в данной области техники, компьютер применяется для выполнения компьютерных программных модулей для обеспечения функциональных возможностей, описанных в данном документе. В данном контексте термин "модуль" относится к логике компьютерной программы, используемой для обеспечения установленной функциональности. Таким образом, модуль может быть реализован в аппаратном обеспечении, встроенном программном обеспечении и/или программном обеспечении. В одном варианте реализации изобретения программные модули хранятся на устройстве хранения данных, загружаются в запоминающее устройство и выполняется процессором.
Понятно, что примеры и варианты реализации, описанные в данном документе, приводятся только в качестве иллюстрации и что различные модификации или изменения в их свете будут предложены специалистам в данной области техники и должны быть включены в сущность и сферу действия настоящей заявки и объема прилагаемой формулы изобретения.
ПРИМЕРЫ
Ниже приведены примеры конкретных вариантов реализации изобретения для изготовления и использования антигенсвязывающих полипептидных конструкций, описанных в данном документе. Примеры приводятся исключительно в иллюстративных целях и никоим образом не направлены на ограничение объема настоящего описания. Были предприняты усилия для того, чтобы обеспечить точность в отношении использованных чисел (например, количеств, температур и т.д.), однако, безусловно, необходимо делать поправку на некоторые экспериментальные ошибки и отклонения.
Конструкции и способы, описанные в данном документе, могут быть получены и осуществлены с использованием, если не указано иное, обычных способов химии белков, биохимии, технологий рекомбинантных ДНК и фармакологии, которые соответствуют данной области техники. Такие технологии подробно описаны в литературе. См., например, T. E. Creighton, Proteins: Structures and Molecular Properties (W. H. Freeman and Company, 1993 год); A. L. Lehninger, Biochemistry (Worth Publishers, Inc., текущее дополнение); Sambrook и соавт., Molecular Cloning:A Laboratory Manual (2-е издание, 1989 год); Methods In Enzymology (S. Colowick and N. Kaplan ред., Academic Press, Inc.); Remington's Pharmaceutical Sciences, 18-е издание (Easton, Pennsylvania:Mack Publishing Company, 1990 год); Carey and Sundberg Advanced Organic Chemistry 3-е изд. (Plenum Press) том A и B (1992 год).
ПРИМЕРЫ
Пример 1: Молекулярное моделирование и компьютерная инженерия области контакта Fab
Структурно-вычислительный подход, основанный на молекулярном моделировании, был использован для создания библиотеки конструкций каппа-лямбда (K-L) для получения биспецифических антител, где один Fab имеет легкую цепь каппа, а другой Fab имеет легкую цепь лямбда (то есть, система каппа-лямбда или система K-L). Библиотека конструкций K-L содержит конструкции с аминокислотными модификациями в тяжелых и легких цепях Fab, которые способствуют преимущественному образованию желаемого биспецифического антитела, когда эти тяжелые и легкие цепи коэкспрессируются. Библиотека конструкций K-L использует преимущества унаследованных различий между легкими цепями каппа и лямбда в биспецифических антителах и, следовательно, оптимизирована для систем каппа-лямбда. Библиотека конструкций K-L была создана путем изучения структуры репрезентативных Fab с D3H44 (антитело к тканевому фактору) в качестве репрезентативного Fab, содержащего легкую цепь каппа (Fab каппа) и CAT-2200 (антитело к ИЛ-17A), в качестве репрезентативного Fab, содержащего легкую цепь лямбда (Fab лямбда), но библиотека может использоваться в контексте других биспецифических антител системы K-L или их фрагментов для идентификации конструкций, которые проявляют желательную специфичность спаривания в антителах интереса.
Репрезентативные Fab были выбраны на основе критериев, указанных в таблице 1. Согласно этим критериям Fab должны быть человеческими или гуманизированными, обычно использоваться подгруппы VH и VL, и содержать минимальное количество мутаций каркасной области. Кроме того, была выполнена парная трехмерная суперпозиция с доступным неизбыточными (порог идентичности последовательности - 90%) структурами каппа и лямбда (структуры, полученные из RSCB PDB, базы данных, поддерживаемой Университетом Ратджерса (Камден, штат Нью-Джерси) и Калифорнийским университетом в Сан-Диего (Сан-Диего, штат Калифорния), см. в интернете по адресу www.rcsb.org. Вместе с другими параметрами, описанными в таблице 1, для определения репрезентативной структуры для каждой из систем каппа и лямбда использовались низкие междоменные RMSD (среднеквадратичные отклонения) H-L для VH-VL или CH1-CL. После отбора D3H44 (PDB ID 1JPT) и CAT-2200 (PDB ID 2VXS) в качестве репрезентативных Fab был проведен структурный анализ этих областей контакта Fab in silico, с целью выявления и понимания остатков, важных для взаимодействия между тяжелыми и легкими цепями, с использованием двухстороннего подхода.
Во-первых, общий анализ консервативности последовательностей в вариабельных и константных областях контакта Fab, выполнялся путем выравнивания последовательностей и структур известных антител. На фи. 1 показано выравнивание последовательностей константного и вариабельного доменов из различных подгрупп антител по сравнению с последовательностями D3H44 и антителом к HER2 пертузумабом (оба содержат легкие цепи каппа), и CAT-2200 (содержит легкую цепь лямбда). На Фиг. 1А и Фиг. 1Е проиллюстрировано выравнивание, сравнивающее репрезентативные подгруппы человеческих зародышевых линий VH с такими D3H44/Пертузумаба и CAT-2200, соответственно. На Фиг. 1B проиллюстрировано выравнивание, сравнивающее D3H44/Пертузумаб с репрезентативными подгруппами каппа VL зародышевой линии человека. На Фиг. 1C и Фиг. 1G проиллюстрировано выравнивание, сравнивающее последовательности человеческих аллелей CH1 с последовательностями D3H44/Пертузумаба и CAT-2200, соответственно. На Фиг. 1D проиллюстрировано выравнивание, сравнивающее последовательности D3H44/Пертузумаба с последовательностями аллеля каппа человека. На Фиг. 1F проиллюстрировано выравнивание, сравнивающее CAT-2200 с репрезентативными подгруппами лямбда VL зародышевой линии человека. На Фиг. 1H проиллюстрировано выравнивание, сравнивающее последовательности CAT-2200 с последовательностями аллеля лямбда человека. Этот анализ показал, что пертузумаб и D3H44 демонстрируют высокую степень консервативности последовательностей с константными и вариабельными доменами каппа зародышевых линий, тогда как CAT-2200 демонстрирует высокую степень консервативности последовательностей с константными и вариабельными доменами лямбда зародышевых линий. Эти выравнивания также иллюстрируют более высокую степень консервативности в константных доменах как тяжелых, так и легких цепей, что делает более вероятной переносимости на другие антитела конструкций в области контакта константного домена.
Второе направление предусматривало анализ кристаллической структуры областей контакта D3H44 и CAT-2200 с использованием ряда инструментов молекулярного моделирования, показанных на фигуре 2 (например, ResidueContacts™ и AffinityDecomposition™). С целью улучшения переносимости на другие антитела или фрагменты, анализ фокусировался на константном домене, где консервативность последовательности выше (см. Фиг. 1). Эти анализы были использованы для определения различий в положениях "горячих" точек (ключевых остатков области контакта) между репрезентативными структурами Fab каппа (D3H44) и Fab лямбда (CAT-2200), как показано в таблице 2. Присутствуют заметные конформационные различия в константном домене между структурами CH1-CL (каппа) и CH1-CL (лямбда). Наложение доступных структур Fab на домен CH1 показало, что структуры CL (каппа) принимают очень сходную конформацию и образуют жесткий кластер. Конформации CL (лямбда), однако, имеют тенденцию находиться в двух разных кластерах: одна ближе к ориентации каппа (каппа-подобный кластер) и еще одна несоответствующая ориентация (кластер лямбда). В результате этого анализа D3H44 представляет собой типичную структуру каппа, а CAT-2200 представляет собой типичную структуру лямбда (кластер лямбда). На Фиг. 3 показано различие в конформации типичного константного домена каппа-лямбда, использующее D3H44 и CAT-2200 в качестве репрезентативных структур. Эти различия, по-видимому, обусловлены главным образом движением жесткого тела константного домена легкой цепи относительно домена CH1, что изменяет характер области контакта H-L в константном домене каппа по сравнению с системой лямбда. Выявленные несоответствия "горячих" точек (таблица 2), а также вышеупомянутые конформационные различия (Фиг. 3) служили отправной точкой для разработки конструкций спаривания H-L для биспецифической системы, содержащей Fab каппа и Fab лямбда. Нумерация аминокислот в последовательностях исходных D3H44 и CAT-2200 по Кабату приведена в таблице 3A и таблице 3B.
Затем путем мутагенеза in silico и упаковки/моделирования с использованием программного обеспечения ZymepackM были сымитированы и определены потенциальные мутации в положениях "горячих" точек, а также в положениях по соседству с положениями "горячих" точек интереса в трехмерной кристаллической структуре. Zymepack™ - это пакет программного обеспечения, который, при наличии вводной структуры и набора мутаций, изменяет типы остатков во вводной структуре согласно заданным мутациям и создает новую структуру, представляющую собой аппроксимацию физической структуры мутантного белка. Кроме того, Zymepack™ позволяет оценить свойства мутантного белка, вычисляя ряд количественных показателей. Эти показатели включают измерения стерической и электростатической комплементарности, которые могут коррелировать со стабильностью, аффинностью связывания или гетеродимерной специфичностью мутантного белка.
Путем использования несоответствий в области контакта были введены мутации для обеспечения селективного спаривания желаемых или предпочтительных полипептидных цепей или гетеродимеров, избегая образования некорректно спаренных или неправильно спаренных полипептидных цепей или гетеродимеров. Например, для получения биспецифического антитела с одной Fab каппа и одной Fab лямбда требуется четыре полипептидных цепи. Fab каппа содержит тяжелую цепь 1 (H1) и легкую цепь каппа 1 (L1), в то время как Fab лямбда содержит тяжелую цепь 2 (H2) и легкую цепь лямбда 2 (L2). В этом случае желательными парами H-L будут H1L1 и H2L2, тогда как нежелательными парами будут H1L2 и H2L1. Следует отметить, что обозначение/нумерация полипептидных цепей здесь произвольна. Таким образом, для получения целенаправленных конструкций Fab каппа - Fab лямбда в константном домене, были идентифицированы мутации, которые благоприятствуют спаренной области контакта CH1-CL (каппа) (H1L1) над неправильно спаренной области контакта CH1-CL (лямбда) (H1L2) и мутации, которые благоприятствуют спаренной области контакта CH1-CL (лямбда) (H2L2) над неправильно спаренной области контакта CH1-CL (каппа) (H2L1). С использованием компьютерных методов, включая программное обеспечение Zymepack™, была смоделирована стерическая комплементарность, которая также рассчитывалась на основании энергетических факторов, таких как упаковка с учетом ван-дер-ваальсовых взаимодействий, эффекты кавитации и близкие контакты гидрофобных групп. Аналогичным образом, были смоделированы энергии электростатического взаимодействия и оценены на основании кулоновских взаимодействий между зарядами, водородных связей и эффектов десольватации. Для расчета относительных стерических и электростатических оценок были имитированы модели предпочтительных пар как H1L1, так и H2L2 тяжелой и легкой цепей, а также модели неправильных пар H1L2 и H2L1, полученные путем введения мутаций интереса. Это позволило определить, приводил ли конкретный набор мутаций к благоприятным энергиям, то есть большей стерической и/или электростатической комплементарности для предпочтительных пар тяжелой-легкой цепей относительно неправильных пар. Рассчитанные стерические и электростатические энергии являются компонентами свободной энергии, ассоциированной со спариванием легкой и тяжелой цепей. Следовательно, большая стерическая и электростатическая комплементарность свидетельствует о большем изменении свободной энергии, ассоциированном со спариванием предпочтительной пары, относительно спаривания неправильной пары. Большая стерическая и/или электростатическая комплементарность приводит к преимущественному (селективному) спариванию желаемых тяжелых и легких цепей относительно стерического штрафа и/или электростатического отталкивания для неправильной пары.
Пример 2: Выбор и описание конструкций
Подход, описанный в примере 1, использовался для разработки гетеродимерных пар тяжелой цепи - легкой цепи (то есть, H1L1 и H2L2) которые проявляют селективное или преимущественное спаривание, в случае если одна из гетеродимерных пар H-L содержит легкую цепь каппа, а другая содержит легкую цепь лямбда. Гетеродимеры сконструированы парами, называемыми "конструкция мАт" или "набор конструкции мАт", и включают набор аминокислотных замен в цепях H1, L1, H2 и L2, которые способствуют преимущественному спариванию. Наборы конструкций мАт были первоначально протестированы как "конструкции LCCA", где одна тяжелая цепь набора конструкции мАт коэкспрессируется с двумя легкими цепями набора конструкции мАт, одной каппа и одной лямбда, чтобы оценить относительное спаривание. Аминокислотные замены, описанные во всех примерах, определены со ссылкой на таблицу 3A и таблицу 3B (для Fab пертузумаба и CAT-2200), используя систему нумерации Кабат, как описано в Kabat and Wu, 1991 год; Kabat и соавт, Sequences of proteins of immunological interest. 5-е издание - US Department of Health and Human Services, публикация NIH №91-3242, стр 647 (1991 год), если не указано иное. Однако, для ссылок, таблицы 22А, 22В и 22С обеспечивают идентификацию выбранных положений аминокислот в легкой цепи, легкой цепи каппа и легкой цепи лямбда с использованием, если это может быть применимо, систем нумерации IMGT, 1JPT и EU.
Конструкции мАт были наложены на молекулярную модель D3H44 и CAT-2200 и количественные показатели были рассчитаны, как описано в примере 1. Затем лучшие конструкции выбирались на основе риска (минимизируя влияние на стабильность, а также иммуногенность) и воздействия (что учитывает предполагаемую силу специфичности стимулирования спаривания). Затем лучшие конструкции тестировали методом конкурентного анализа легкой цепи (LCCA), чтобы экспериментально определить их специфичность спаривания (см. пример 4). Хотя конструкции мАт были идентифицированы с использованием D3H44 и CAT-2200 в качестве репрезентативных Fab, они были протестированы в системе K-L с использованием Пертузумаба в качестве Fab каппа и CAT-2200 в качестве Fab лямбда (система K-L пертузумаб-CAT-2200). Как изображено на Фиг. 1С и Фиг. 1D, D3H44 и Пертузумаб имеют идентичные последовательности в константном домене, что обеспечивает беспрепятственное взаимопревращение между этими двумя системами. Эти конструкции мАт упоминаются как конструкции K-L.
Второй набор конструкций был также протестирован в системе K-L пертузумаб/CAT-2200. Эти конструкции были предложены с использованием репрезентативных конструкций, которые отражали разнообразие конструкций из библиотеки конструкций каппа-каппа (K-K), описанной в таблице 30 Международной патентной заявки номер PCT/CA2013/050914 (Международная патентная публикация № WO2014/082179) в качестве отправной точки. Эти репрезентативные, полученные из K-K конструкции включали подмножество конструкций, которые по возможности переносились без модификации в систему K-L или были адаптированы к системе каппа-лямбда путем модификации аминокислотных остатков по мере необходимости для учета различий в последовательности и структуре между легкими цепями каппа и лямбда. Оба типа репрезентативных конструкций, полученных из K-K, упоминаются как конструкции K-L, полученные из K-K.
Специфичность спаривания для конструкций K-L и конструкций K-L, полученных из K-K, была экспериментально оценена как конструкции LCCA с помощью LCCA в системе каппа-лямбда, как описано в примере 4.
Пример 3: Получение конструкций Fab, кодирующих тяжелые цепи IgG пертузумаба, легкие цепи IgG пертузумаба, тяжелые цепи IgG CAT-2200 и легкие цепи IgG CAT-2200.
Fab тяжелых и легких цепей дикого типа и антитело Пертузумаб к HER2, и для антитела CAT-2200 к IL17 были получены следующим образом. Белковые последовательности Fab легкой цепи пертузумаба (GenBank Accession № HC359025.1, SEQ ID NO: 2) и тяжелой цепи пертузумаба (GenBank Accession № HC359024.1, SEQ ID NO: 1) были обратно переведены в ДНК, кодон-оптимизированы для экспрессии в клетках млекопитающих и синтезированы в ген (SEQ ID NO: 8 и 7, соответственно). Белковые последовательности легкой цепи Fab CAT-2200 (цепь L 2VXS, SEQ ID NO: 4) и тяжелой цепи (цепь H 2VXS, SEQ ID NO: 3) были взяты из ввода PDB 2VXS, обратно переведены в ДНК, кодон-оптимизированы для экспрессии в клетках млекопитающих и синтезированы в ген (SEQ ID NO: 10 и 9, соответственно). Полипептидные последовательности и последовательности ДНК этих тяжелых и легких цепей антител представлены в таблице 3C.
Вставки вектора легкой цепи, состоящие из сайта расщепления 5'-EcoRI - сигнального пептида HLA-A - метки HA или FLAG - клона легкой цепи Ig - 'стоп TGA или TAA' - сайта расщепления 3'-BamH1, были лигированы в вектор pTT5 (Durocher Y и соавт., Nucl. Acids Res. 2002 год; 30, №2, e9) для получения экспрессионных векторов легкой цепи. Полученные экспрессионные векторы легкой цепи секвенировали для подтверждения правильной рамки считывания и последовательности, кодирующей ДНК. Аналогично, вставки векторов тяжелой цепи, состоящие из сайта рестрикции 5'-EcoR1 - сигнального пептида HLA-A - клона тяжелой цепи (заканчивающегося в T238, см. таблицу 3A) -ABD2-метки His6 - стоп TGA - сайта расщепления 3'-BamH1, были лигированы в вектор pTT5 (ABD2= две копии белка связывающего домена альбумина, тандемно) для получения экспрессионных векторов тяжелой цепи. Полученные экспрессионные векторы тяжелой цепи также были секвенированы для подтверждения правильной рамки считывания и последовательности, кодирующей ДНК. Различные конструкции Fab Пертузумаба или CAT-2200, содержащие аминокислотные замены для наборов конструкции мАт, были получены либо путем синтеза генов, либо путем сайт-направленного мутагенеза (Braman J, Papworth C & Greener A., Methods Mol. Biol. (1996 год) 57:31-44).
Тяжелые и легкие цепи были помечены соответственно на C- и N-концах, для облегчения оценки преимущественного спаривания путем теста конкурентного анализа-ППР (LCCA). Метка тяжелой цепи ABD2-His6 позволила захватить комплексы H-L на поверхности чипа ППР с меткой к His, в то время как метки легкой цепи FLAG и HA позволяют количественно определять относительные популяции L1 и L2.
Пример 4: Оценка преимущественного спаривания сконструированных гетеродимеров Fab с помощью конкурентного анализа легкой цепи (LCCA)
Конструкции, кодирующие тяжелые и легкие цепи IgG Пертузумаба и CAT-2200 в формате Fab, содержащие аминокислотные модификации, были получены, как описано в примере 3. Способность конструкций преимущественно спариваться с образованием желаемого гетеродимера H-L в контексте набора конструкций LCCA (H1, L1, L2 или H2, L2, L1) определяли с использованием конкурентного анализа легкой цепи (LCCA).
LCCA количественно определяет относительное спаривание одной тяжелой цепи для, по меньшей мере, для двух уникальных легких цепей, одной каппа и одной лямбда и может быть охарактеризован следующим образом. Чтобы определить специфичность спаривания для Fab лямбда, одна конструкция Fab тяжелой цепи CAT-2200 была коэкспрессирована с одной конструкцией Fab легкой цепи CAT-2200 (для спаренного Fab продукта H1L1) и одной конструкцией Fab легкой цепи Пертузумаба (для неправильно спаренного Fab продукта H1L2) и относительную специфичность спаривания легкой цепи (H1L1:H1L2) определяли путем теста конкурентного анализа-ППР, который проводили дважды. И наоборот, чтобы определить специфичность спаривания для Fab каппа, одна конструкция Fab тяжелой цепи Пертузумаба была коэкспрессирована с одной конструкцией Fab легкой цепи Пертузумаба (для спаренного Fab продукта H2L2) и одной конструкцией Fab легкой цепи CAT-2200 (для неправильно спаренного Fab продукта H2L1) и относительную специфичность спаривания легкой цепи (H2L2:H2L1) определяли путем теста конкурентного анализа-ППР, который проводили дважды.
Анализ LCCA проводили путем сопутствующей экспрессии тяжелых и легких цепей при соотношениях 1:1:1 (по массе) для H1:L1:L2 или H2:L2:L1, причем тяжелая цепь находилась в предельных количествах. Количество каждого образованного гетеродимера (то есть, H1L1 и H1L2 или H2L2 и H2L1) определяли путем связывания тяжелых цепей с чипом ППР посредством аффинной адсорбции гистидиновой метки с последующим определением количества каждой метки легкой цепи (HA или FLAG) с использованием антител, специфичных к этим меткам. Впоследствии выбранные совпадения гетеродимеров были проверены с помощью конкурентного анализа легкой цепи, в соответствии с которым соотношения ДНК L1:L2 снова проверялись на соотношение 1:1, сохраняя при этом тяжелую цепь в предельных количествах. Для этих конкретных комбинаций системы каппа-лямбда метки легкой цепи могут влиять на спаривание LCCA дикого типа, что приводит к отклонениям от ожидаемого нейтрального соотношения 50%: 50%. По этой причине расположение с наименьшим количеством отклонения от нейтрального было выбрано путем слияния легкой цепи Пертузумаба с меткой HA и легкой цепи CAT-2200 с меткой FLAG. Схематическое изображение, представляющее конструкцию анализа изображено на Фиг. 4. На Фиг. 5 изображено как метятся тяжелые цепи и легкие цепи, и как оценивается преимущественное спаривание. Ниже приведены экспериментальные подробности LCCA.
Способ трансфекции
Конструкции LCCA, содержащие одну конструкцию тяжелой цепи и две конструкции легкой цепи, полученные, как описано в примере 3, трансфицировали в клетки CHO-3E7 следующим образом. Клетки CHO-3E7 с плотностью 1,7-2×106 клеток/мл, культивировали при 37°C в среде FreeStyle™ F17 (Invitrogen cat# A-1383501), дополненной 4 мМ глутамином и 0,1% KoliphorP188 (Sigma #K4894). Общий объем 2 мл трансфицировали в общей сложности с 2 μг ДНК, используя PEI-pro (Polyplus transfection # 115-375) при соотношении ДНК:PEI 1:2,5. Все трансфекции проводили с использованием 333 нг каждой тяжелой цепи и каждой легкой цепи (то есть, H1:L1:L2 или H2:L2:L1=соотношение 1:1:1), 300 нг AKTdd pTT22 (вектор, содержащий конститутивно активный мутант протеинкиназы B) и 700 нг оцДНК (ДНК спермы лосося). Спустя 24 часа после добавления смеси ДНК-PEI к клеткам добавляли 0,5 мМ вальпроевой кислоты (конечная концентрация) и 1% масса/объем Триптона (конечная концентрация) и клетки затем переносили в 32°C. Супернатанты тестировали на экспрессию на 7-й день с помощью анализа электрофореза ДСН-ПААГ в невосстанавливающих условиях с последующим синим окрашиванием Кумасси для визуализации полос.
Способ конкурентного анализа ППР
Степень преимущественного спаривания тяжелых цепей с каждой из двух легких цепей в конструкциях LCCA оценивали с использованием считывания на основе ППР уникальных меток эпитопа, расположенных на N-конце каждой легкой цепи. Конструкция LC CAT-2200 содержит метку FLAG, а конструкция LC Пертузумаба содержит метку HA.
Материалы для поверхностного плазмонного резонанса (ППР). Сенсорные чипы GLC, набор связывания амина Biorad ProteOn (1-этил-3-(3-диметиламинопропил) карбодиимид гидрохлорид (EDC), N-гидроксисульфосукцинимид (sNHS) и этаноламин) и 10 мМ натрий-ацетатных буферов были приобретены у Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио). 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота (HEPES) буфер, этилендиаминтетрауксусная кислота (ЭДТА) и NaCl были приобретены у Sigma-Aldrich (Оквилл, штат Онтарио). 10% раствор Твин 20 был приобретен у Teknova (Холлистер, штат Калифорния).
Биосенсорные анализы ППР. Все анализы поверхностного плазмонного резонанса проводились с использованием прибора BioRad ProteOn XPR36 (Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио)) с электродным буфером ФСБТ (ФСБ Teknova Inc с 0,05% Твин 20) при температуре 25°C. Поверхность захвата anti-penta His™ была получена с использованием сенсорного чипа GLC, активированного разбавлением 1:5 стандартных растворов sNHS/EDC BioRad, введенных в течение 140 с при 100 мкл/мин в (горизонтальном) направлении анализируемого вещества. Сразу же после активации, 25 мкг/мл раствора антитела anti-penta His™ (Qiagen Inc.) в 10 мМ NaOAc pH 4,5 вводили в (горизонтальном) направлении анализируемого вещества при скорости потока 25 мкл/мин до иммобилизации приблизительно 3000 резонансных единиц (РЕ). Оставшиеся активные группы блокировали 140-секундной инъекцией 1 М этаноламина при 100 мкл/мин в направлении анализируемого вещества, и это также обеспечивало создание активированных имитатором точек для сравнения.
Тестирование гетеродимеров для связывания с моноклональными антителами к FLAG (Sigma Inc.) и к HA (Roche Inc.) происходило в два этапа:непрямой захват гетеродимеров на поверхности anti-penta His™ в направлении лиганда, с последующим введением антитела к FLAG и к НА в направлении анализируемого вещества. Во-первых, для стабилизации исходной линии использовали одно введение ФСБТ в течение 30 с при 100 мкл/мин в направлении лиганда. Для каждого захвата гетеродимера неочищенные гетеродимеры в клеточных культурах разбавляли до 4% в ФСБТ. От одного до пяти гетеродимеров или контролей (то есть контролей, содержащих либо 100% меченную HA легкую цепь, либо 100% меченную FLAG легкую цепь) одновременно вводили в отдельные каналы лигандов в течение 240 с при потоке 25 мкл/мин, в результате чего насыщающий гетеродимер захватывал приблизительно от 300 до 400 РЕ на поверхности anti-penta His. Первый канал лиганда оставался пустым для использования в качестве холостой пробы, если это было необходимо. Этот этап захвата гетеродимера немедленно сопровождался двумя введениями буфера в направлении анализируемого вещества для стабилизации исходной линии, а затем 5 нМ анти-FLAG и 5 нМ анти-НА каждый вводили в двух повторностях при 50 мкл/мин в течение 120 секунд с 180 секундной фазой диссоциации, что приводило к набору сенсограмм связывания со сравнительным буфером для каждого захваченного гетеродимера. Гетеродимеры восстанавливали с помощью выброса 0,85% фосфорной кислоты в течение 18 с при 100 мкл/мин с целью подготовки поверхности anti-penta His™ для следующего цикла введения. Сенсограммы были выровнены и дважды проверены с использованием контрольного введения буфера и точек, и полученные сенсограммы были проанализированы с использованием программного обеспечения ProteOn Manager v3. 0.
Анализ данных и описание показателей LCCA
Оценка преимущественного спаривания для каждой конструкции, представленной в формате H1L1H2L2, была произведена двумя комплементарными наборами конструкций LCCA, один для комбинации H1L1L2, а другой для комбинации H2L1L2. Каждый набор конструкции LCCA обозначается "уникальным идентификатором" (например, 10629 или 10695). Каждый набор конструкции мАт (H1L1H2L2), следовательно, идентифицируется с помощью номеров набора конструкций LCCA для двух составляющих LCCA (например, 10629-10695).
Наборы конструкций LCCA (H1L1L2 или H2L1L2) каждого набора конструкций мАт были протестированы в LCCA, а наборы конструкций интереса были определены на основе следующих критериев включения. Для того, чтобы считаться конструкцией интереса, конструкция должна содержать, по меньшей мере, один положительный результат LCCA над диапазоном нейтральных значений данных (соотношение > 60:40 правильно:неправильно спаренных), в то время как другой комплементарный LCCA должен был быть выше нижнего предела диапазона нейтральных значений данных (соотношение 40:60 правильно:неправильно спаренных), как проиллюстрировано на Фиг. 6. Диапазон нейтральных значений был определен как область между 40%: 60% и 60%: 40% соотношениями правильно:неправильно спаренных с использованием срединного нормализованного значения для H1-L1:H1-L2 и H2-L2:H2-L1. Например, сценарии A и B на Фиг. 6 изображают конструкции, которые проходят критерии включения. Сценарий B включен, поскольку LCCA H2L2:H2L1 находится выше диапазона нейтральных значений данных, а LCCA H1L1:H1L2 в пределах диапазона нейтральных значений данных (не ниже его). Сценарий C представляет собой конструкцию, в которой оба результата LCCA являются положительными, но не находятся выше диапазона нейтральных значений данных и, следовательно, не включены. Сценарии D и E представляют конструкции, которые отсеиваются, поскольку, по меньшей мере, один из результатов LCCA находится ниже диапазона нейтральных значений данных, даже если другой LCCA находится выше диапазона нейтральных значений данных (см. сценарий D). В таблицах 4A (конструкции K-L) и 4B (конструкции K-L, полученные из K-K) перечислены наборы конструкций мАт, которые соответствуют этим критериям, а в таблицах 5A и 5B показаны результаты LCCA для этих конструкций. В таблицах 4A и 4B и всех последующих таблицах, в которых перечислены аминокислотные замены наборов конструкций LCCA или наборов конструкций мАт, отсутствие замещения в таблице или "-" указывает на то, что полипептидная цепь не включает аминокислотную замену, которая способствует преимущественному спариванию.
Эффективность каждого набора конструкций LCCA описывается двумя скалярными значениями: ln(r1/f1) или ln(r2/f2), где r1 и r2 соответствуют срединным значениям соотношений H1L1:H1L2 и H2L2:H2L1, соответственно, и f1 и f2 соответствуют соответствующим срединным значениям соотношений H1L1:H1L2 и H2L2:H2L1. Нормализованные скалярные значения генерируются путем вычитания вклада системы ДТ (дикого типа) из конструкции; ln(r1/f1)конструкция - ln(r1/f1)ДТ или ln(r2/f2)конструкция - ln(r2/f2)ДТ.Для (HC) CAT-2200 дикого типа, конкурирующего с (LC-FLAG) CAT-2200 плюс (LC-HA) Пертузумаба ln(r1/f1)ДТ=-0,48. Для (HC) Пертузумаба дикого типа, конкурирующего с (LC-HA) Пертузумаба плюс (LC-FLAG) CAT-2200 ln(r2/f2)ДТ=0,66. Эта процедура эффективно удаляет любые существующие смещения спаривания ДТ и нормализует специфичность по сравнению с нейтральной 50%: 50% системой L1:L2 ДТ. Эти значения представлены в столбцах 2 и 5 в таблицах 5А и 5В.
Кроме того, нормализованные скалярные значения также были преобразованы обратно в чистое нормализованное срединное соотношение H1L1:H1L2 и H2L2:H2L1. Это преобразование эффективно нормализовало соотношения LCCA до 100%, как это было отмечено для некоторых сконструированных Fab, таким образом, что общее количество H1L1 и H1L2, или H2L2 и H2L1 значительно отличалось от 100%. Это расхождение в общем процентном отношении легкой цепи, как полагается, частично объясняется появлением вариабельного неспецифического связывания во время первоначального захвата гетеродимера на чипе ППР. Эти значения представлены в столбцах 4 и 7 в таблицах 5А и 5В.
Кроме того, показан скалярный диапазон для каждой нормализованной конструкции LCCA (эксперименты H1L1:H1L2 и H2L2:H2L1), а также диапазон нормализованного соотношения правильно спаренных к неправильно спаренным гетеродимерам Fab (диапазон представляет собой один и тот же процент от L1 и процент L2 в нормализованном соотношении). Для результатов, полученных из единичного измерения, значения диапазона отмечены как NA (не доступно). Эти значения представлены в столбцах 3 и 6 в таблицах 5А и 5В.
Для сведения, в таблице 23 представлено соответствие между % спаренных:неправильно спаренных гетеродимеров и соответствующими скалярными значениями LCCA.
Результаты
Результаты LCCA показаны в таблицах 5A и 5B в контексте наборов конструкций мАт. Таблица 5A содержит данные LCCA, относящиеся к конструкциям K-L, описанным в примере 2, тогда как в таблице 5B приведены данные LCCA, относящиеся к конструкциям K-L, полученным из K-K, также описанным в примере 2. Следует отметить, что все эксперименты LCCA выполнялись на конструкциях, содержащих межцепочечную Fab дисульфидную связь, расположенную в константном домене (H/C233-L/C214, нумерация Кабат).
В таблице 4A перечислена 231 конструкция K-L, которые соответствовали критериям включения. Данные LCCA, относящиеся к этим конструкциям, показаны в таблице 5A. В таблице 4В перечислены 44 конструкции K-L, полученные из K-K, которые соответствовали критериям включения, а в таблице 5B приведены данные LCCA, относящиеся к этим конструкциям. Конструкции, которые поддерживают 100% идентичности между исходной библиотекой конструкций K-K из таблицы 30 в вышеупомянутой заявке PCT Application №PCT/CA2013/050914 и новая система каппа-лямбда (то есть были перенесены без модификаций) обозначены звездочкой (*) в таблице 4B. Большинство конструкций в таблице 4B имеют аминокислотные модификации только в константной области, однако некоторые из конструкций также включают аминокислотные модификации в вариабельной области. Эти конструкции были предложены для дальнейшего обусловливания специфичности спаривания, в то же время способствуя переносимости в другие системы антител.
"Прочность" конструкции может быть определена как сумма двух скалярных значений LCCA для конкретной конструкции. Сравнивая "прочность" конструкций K-L, полученных из K-K, с конструкциями K-L, было очевидно, что те конструкции, специально предназначенные для системы K-L, как правило, демонстрируют лучшую специфичность спаривания, чем конструкции K-K, перенесенные в систему K-L, как показано на Фиг. 7. Средняя "прочность" конструкции для конструкций K-L, полученных из K-K, составляла 1,86 с максимальной "прочностью" конструкции 3,53. Средняя "прочность" конструкции для конструкций K-L составляла 2,69 с максимальной "прочностью" конструкции 5,23.
Пример 5: Получение сконструированных гетеродимеров Fab для биофизической характеризации в большом количестве
Правильно спаренные гетеродимеры H1L1 или H2L2, как указано в наборах уникальных идентификаторов (таблицы 4A и 4B), были получены в большом количестве (обычно до 20 мл) и очищены следующим образом, чтобы проверить термическую стабильность и связывание антигена. Для этих экспериментов тяжелые цепи Fab были экспрессированы без метки ABD2, тогда как легкие цепи были экспрессированы с помощью той же метки HA или FLAG, что и в LCCA. Тяжелая и легкая цепи каждого гетеродимера экспрессировались в 20 мл культуры клеток CHO-3E7. Клетки CHO-3E7 с плотностью 1,7-2×106 клеток/мл, культивировали при 37°C в среде FreeStyle™ F17 (Invitrogen cat# A-1383501), дополненной 4 мМ глутамином и 0,1% Koliphor P188 (Sigma #K4894). Общий объем 20 мл трансфицировали в общей сложности с 20 мкг ДНК, используя PEI-pro (Polyplus cat# 115-375) при соотношении ДНК:PEI 1:2,5. Спустя 24 часа после добавления смеси ДНК-PEI к клеткам добавляли 0,5 мМ вальпроевой кислоты (конечная концентрация) и 1% масса/объем Триптона (конечная концентрация) и клетки затем переносили в 32°C.
Клетки центрифугировали через 7 дней после трансфекции, а гетеродимеры очищали от супернатанта путем аффинного захвата с использованием IgG-CH1 CaptureSelect™ (Life Technologies™, Catalog №:194-320-005) при 4°C следующим образом. Супернатанты разбавляли до 20-25% супернатанта клеточной культуры в уравновешивающем буфере (фосфатно-солевой буфер Дульбекко (DPBS) без кальция, магния и фенолового красного (HyClone™# SH30028.02)), а затем инкубировали с перемешиванием в течение 16 часов с IgG-CH1 CaptureSelect™, также предварительно уравновешенным уравновешивающим буфером. Затем смолу собирали с помощью центрифугирования, переносили на 96-луночный планшет, три раза промывали уравновешивающим буфером. Связанные образцы элюировали объемом от 1 до 4 слоев буфера для элюирования:100 мМ глицина pH 2,6. Элюированные образцы собирали центрифугированием в 96-луночный УФ-прозрачный приемный планшет, каждая лунка содержала нейтрализующий буфер:1M Трис pH 9,0 при 10% объема элюирования.
После очистки экспрессию гетеродимера оценивали с помощью анализа High Throughput Protein Express в невосстанавливающих условиях с использованием Caliper LabChip GXII (Perkin Elmer #760499). Процедуры выполнялись в соответствии с руководством пользователя HT Protein Express LabChip версии 2 LabChip GXII User Manual, со следующими изменениями. Образцы гетеродимера при 2 мкл или 5 мкл (диапазон концентраций 5-2000 нг/мкл), добавляли к отдельным лункам в 96-луночные планшеты (BioRad # HSP9601) вместе с 7 мкл буфера для образцов HT Protein Express (Perkin Elmer # 760328). Затем образцы гетеродимера денатурировали при 70°C в течение 15 минут. Прибором LabChip пользовались с использованием HT Protein Express Chip (Perkin Elmer # 760499) и настройкой анализа Ab-200. После использования чип был очищен водой MilliQ и хранился при 4°C.
Включение модификаций набора конструкций мАт не оказало существенного влияния на экспрессию белка. Уровни экспрессии, полученные для Fab CAT-2200, содержащей модификации конструкции, составляют 0,45 мг/20 мкл экспрессии (SD 0,16 мг) по сравнению с 0,64 мг/20 мл (SD 0,1 мг) для дикого типа Fab. Fab Пертузумаба с модификациями конструкции экспрессируется при 0,39 мг/20 мл экспрессии (SD 0,14 мг) по сравнению с 0,4 мл/20 мкл экспрессии (SD 0,03 мг) для Fab дикого типа. Анализ с помощью калипера показал уровни чистоты, сравнимые с Fab ДТ.
Пример 6: Измерения аффинности сконструированных гетеродимеров Fab к антигену.
Была оценена способность гетеродимеров Fab CAT-2200 связываться с ИЛ-17A и Fab Пертузумаба связываться с HER2-ECD, с целью определения, повлияла ли замена аминокислот на способность гетеродимера связываться с антигеном. Гетеродимеры Fab получали, как описано в примере 5. Аффинность каждого гетеродимера Fab к его антигену определяли с помощью ППР следующим образом.
Материалы для SPR. Сенсорные чипы GLC, набор связывания амина Biorad ProteOn (EDC, sNHS и этаноламин) и 10 мМ натрий-ацетатные буферы были приобретены у Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио). Электронный буфер ФСБ с 0,05% Твин 20 (ФСБТ) был приобретен у компании Teknoca Inc. (Холлистер, штат Калифорния). Рекомбинантный белок HER2 был приобретен у компании eBioscience (Сан Диего, штат Калифорния). Козьи поликлональные антитела к человеческому Fc были приобретены у компании Jackson Immuno Research Laboratories Inc. (Вест Гроув, штат Пенсильвания). Рекомбинантный человеческий ИЛ-17A был приобретен у R&D Systems (Миннеаполис, штат Миннесота).
Определение аффинности CAT-2200:ИЛ-17A. Все анализы поверхностного плазмонного резонанса проводились с использованием прибора BioRad ProteOn XPR36 (Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио)) с электродным буфером ФСБТ при температуре 25°C. Поверхность ИЛ-17A была получена с использованием сенсорного чипа GLC, активированного разбавлением 1:10 стандартных растворов sNHS/EDC BioRad, введенных в течение 140 с при 100 мкл/мин в (вертикальном) направлении лиганда. Сразу же после активации, 2 мкг/мл раствора ИЛ-17A в 10 мМ NaOAc pH 4,5 вводили в (вертикальном) направлении лиганда при скорости потока 25 мкл/мин до иммобилизации приблизительно 50 резонансных единиц (РЕ). Оставшиеся активные группы блокировали 140-секундной инъекцией 1 М этаноламина при 100 мкл/мин также в направлении лиганда.
Скрининг сконструированных Fab для связывания с ИЛ-17A проводили путем инъекции очищенных Fab CAT-2200 в (горизонтальном) направлении анализируемого вещества. Во-первых, для стабилизации исходной линии использовали два введения буфера в течение 30 с при 100 мкл/мин в (горизонтальном) направлении анализируемого вещества. Пять концентраций трехкратной серии разбавлений каждого из сконструированных Fab CAT-2200 (60 нм, 20 нм, 6,7 нм, 2,2 нм, 0,74 нм) с контролем пустого буфера одновременно вводили в при 50 мкл/мин в течение 120 секунд с 15-минутной фазой диссоциации, что приводило к набору сенсограмм связывания со сравнительным буфером. Комплексы CAT-2200:ИЛ-17A на поверхности ППР диссоциировали и поверхность ИЛ-17A восстанавливали для получения следующего цикла введения двумя выбросами 0,85% фосфорной кислоты в течение 18 с при 100 мкл/мин. Сенсограммы были выровнены и дважды проверены с использованием контрольного введения буфера и точек, и полученные сенсограммы были проанализированы с использованием программного обеспечения ProteOn Manager v3. 1. Дважды проверенные сенсограммы соответствовали модели связывания Ленгмюра.
Определение аффинности Пертузумаб:HER2 Все анализы поверхностного плазмонного резонанса проводились с использованием прибора BioRad ProteOn XPR36 (Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио)) с электродным буфером ФСБТ при температуре 25°C. Поверхность захвата антитела anti-penta-His™ (Qiagen) была получена с использованием сенсорного чипа GLC, активированного разбавлением 1:10 стандартных растворов sNHS/EDC BioRad, введенных в течение 140 с при 100 мкл/мин в (горизонтальном) направлении анализируемого вещества. Сразу же после активации, 10 мкг/мл раствора антитела anti-penta-His в 10 мМ NaOAc pH 4,5 вводили в (вертикальном) направлении лиганда при скорости потока 25 мкл/мин до иммобилизации приблизительно 3000 резонансных единиц (РЕ). Оставшиеся активные группы блокировали 140-секундной инъекцией 1 М этаноламина при 100 мкл/мин в направлении анализируемого вещества, с целью гарантии создания активированных имитатором точек для сравнения.
Скрининг сконструированных Fab на наличие связывания с HER2 происходил в два этапа:непрямой захват сконструированных Fab на поверхности антитела anti-penta His™ в направлении лиганда, с последующим одновременным введением 5 концентраций очищенного антигена HER2 и одного буферного контроля для двойной проверки в направлении анализируемого вещества. Во-первых, для стабилизации исходной линии использовали одно введение буфера в течение 30 с при 100 мкл/мин в направлении лиганда. Для каждого захвата, сконструированного Fab, сконструированные Fab разбавляли до 2 мкг/мл в ФСБТ. От одного до пяти сконструированных Fab или контролей одновременно вводили в отдельные каналы лигандов в течение 240 с при потоке 25 мкл/мин. Это привело к захвату приблизительно от 400 до 600 РЕ на поверхности антитела к Fc человека. Первый канал лиганда оставался пустым для использования в качестве холостой пробы, если это было необходимо. Этот этап захвата немедленно сопровождался двумя введениями буфера в направлении анализируемого вещества для стабилизации исходной линии, а затем 100 нМ, 33,3 нМ, 11,1 нМ, 3,7 нМ и 0,41 нМ Her2 вместе с буферным контролем одновременно вводили при 50 мкл/мин в течение 120 секунд с 1200-секундной фазой диссоциации. Захваченные поверхности антитела восстанавливали с помощью выброса 0,85% фосфорной кислоты в течение 18 с при 100 мкл/мин с целью подготовки для следующего цикла введения. Сенсограммы были выровнены и дважды проверены с использованием контрольного введения буфера и точек, и полученные сенсограммы были проанализированы с использованием программного обеспечения ProteOn Manager v3. 1. Дважды проверенные сенсограммы соответствовали модели связывания 1:1.
Результаты Значения антигенной аффинности (KD) для сконструированных образцов гетеродимера Fab приведены в таблице 6A (конструкции K-L) и в таблице 6B (конструкции K-L, полученные из K-K) в контексте спаривания конструкции. Значения KD H1L1 и H2L2 Fab включены в столбцы 2 и 4 (KD гетеродимера H1L1 Fab CAT-2200 и гетеродимера H2L2 Fab Пертузумаба, соответственно), и 3 и 5 (изменение KD гетеродимера H1L1 или H2L2 Fab по сравнению с его соответствующим диким типом). Значения KD определяли только для образцов гетеродимера Fab, которые показали захват гетеродимера Fab по меньшей мере в 100 РЕ. Контрольное значение KD для CAT-2200 дикого типа, связанного с ИЛ-17A, используемое для сравнения со сконструированными гетеродимерами, составляло 0,273 нМ. Это значение является срединным значениям, основанным на многочисленных измерениях гетеродимера Fab CAT-2200 дикого типа, где легкая цепь содержит метку FLAG. Аналогичным образом, контрольное значение KD для пертузумаба дикого типа, связанного с HER2, составляло 8,055 нМ. Это значение является срединным значениям, основанным на многочисленных измерениях гетеродимера Fab пертузумаба дикого типа, где легкая цепь содержит метку HA. В таблице 6 разница в KD по отношению к аффинности связывания дикого типа антигена показана с использованием расчета -(log(KD_конструкции) - log(KD_дт)), так что положительные значения указывают на более низкие значения KD, тогда как отрицательные значения указывают на увеличенные значения KD гетеродимера Fab по сравнению с аффинностью связывания дикого типа для антигена. НС указывает на то, что связывание не обнаружено. Следует отметить, что некоторые гетеродимеры Fab не имеют измеренных значений KD и поэтому обозначены как не определенные (НО). В некоторых из этих случаев были оценены гетеродимеры Fab, но технические трудности (такие как слабый захват гетеродимера Fab в эксперименте ППР, то есть менее 100 РЕ), не позволяли точно определять значения KD.
Для большинства испытанных сконструированных гетеродимеров Fab, аминокислотные замены не влияли или практически не влияли на значения KD. Три конструкции (уникальные идентификаторы 10881-11412, 10883-11236 и 10888-11415) в библиотеке конструкций K-L показали> 5-кратное снижение KD на Fab CAT-2200. В библиотеке конструкций K-L, полученных из K-K, один из гетеродимеров Fab Пертузумаба (конструкции, содержащие код 10724 в уникальном идентификаторе) показал> 5-кратное снижение KD на стороне Пертузумаба.
Пример 7: Измерения термической стабильности сконструированных гетеродимеров Fab с помощью ДСФ.
Дифференциальная Сканирующая Флуориметрия (ДСФ) использовалась для измерения термической стабильности правильно спаренных гетеродимеров по сравнению с термической стабильностью немодифицированной пары тяжелая цепь-легкая цепь дикого типа. Хотя известно, что ДСФ является менее точным методом измерения температуры плавления, он использовался здесь, поскольку ДСФ является высокопроизводительным методом измерения термической стабильности и, таким образом, подходит для некоторого измерения термической стабильности для большого количества гетеродимеров Fab. Гетеродимеры Fab получали, как описано в примере 5.
Измерение термической стабильности с помощью ДСФ
Термическая стабильность сконструированных пар гетеродимеров Fab была измерена с помощью ДСФ следующим образом. Каждый гетеродимер очищали, как описано в примере 5, и разбавляли до 0,5 мг/мл в DPBS (фосфатно-солевом буфере Дульбекко, HyClone Cat # SH30028.02). Для большинства образцов рабочий состав краски геля Sypro Orange (Life Technologies Cat # S-6650) готовили разбавлением 4 мкл краски геля Sypro Orange до 2 мл DPBS. Образцы ДСФ были приготовлены путем добавления 14 мкл белка 0,5 мг/мл к 60 мкл разбавленного рабочего состава краски геля Sypro Orange. Однако для белков, которые имели менее 0,5 мг/мл, каждый образец ДСФ получали путем добавления 14 мкл неразбавленного белка к 60 мкл рабочего состава красителя Sypro Orange (который разбавляли до 1:1500 в DPBS). Затем анализ ДСФ проводили в двух экземплярах по 20 мкл аликвот с использованием прибора кПЦР Rotor-Gene 6000 (QiaGen Inc). Каждый образец сканировали с 30°C по 94°C с интервалом 1°C с 10-секундным балансом между каждым этапом и 30-секундным временем ожидания в начале. Использовали фильтр возбуждения 470 нМ и эмиссионный фильтр 610 нМ с усилением 9. Данные анализировали с помощью программного обеспечения Rotor-Gene 6000, используя максимальное значение от первой производной кривой денатурации как Tm. Остальные образцы ДСФ были подготовлены и проанализированы аналогичным образом со следующими модификациями протокола, которые не изменяют измеренные значения Tm:1) рабочий состав приготовили путем разбавления 1 мкл краски геля Sypro Orange до 2 мл DPBS, 2) анализировали аликвоты по 30 мкл и 3) использовали усиление 10.
Результаты ДСФ для сконструированных образцов гетеродимера Fab приведены в таблицах 6A (конструкции K-L) и 6B (конструкции K-L, полученные из K-K) в контексте наборов конструкции мАт. Термическая стабильность H1L1 и H2L2 Fab включена в столбцы 6 и 8 (H1L1 относится к гетеродимеру Fab CAT-2200 и H2L2 к гетеродимеру Fab Пертузумаба) таблиц 6A и 6B. Для гетеродимеров Fab, где проводились повторы, указанное значение Tm представляет собой срединное значение. Сравнение значений Tm сконструированного гетеродимера Fab по отношению к значению Tm гетеродимера Fab типа дикого типа приведено в столбце 7 для H1L1 CAT-2200 и в столбце 9 для H2L2 Пертузумаба. Было определено, что средняя Tm для конструкции Fab CAT-2200 дикого типа, содержащей метку FLAG, составляет 76,9°C. Было определено, что средняя Tm для гетеродимера Fab Пертузумаба дикого типа, содержащего метку HA, составляет 82,4°C. Учитывая ограничения пропускной способности в масштабной процедуре для большого количества гетеродимеров Fab, полученных с помощью наборов конструкции мАт, измерения ДСФ выполнялись путем выборочного исследования подмножества сконструированных гетеродимеров Fab. По этой причине некоторые гетеродимеры Fab не имеют значений Tm, измеренных с помощью ДСФ, и поэтому обозначены как не определенные (НО). Большинство тестируемых конструкций показали хорошую термическую стабильность в диапазоне ожидаемых значений Tm, обычно наблюдаемых для Fab дикого типа. Сконструированные гетеродимеры Fab, содержащие мутации K145T_V177D_S188D в HC и S176K_Y178L в LC лямбда или S176K_T178L в LC каппа, отрицательно влияли на значения Tm на> 10°C в гетеродимере Fab, содержащем эти мутации.
Пример 8: Измерения термической стабильности сконструированных гетеродимеров Fab с помощью ДСК.
В дополнение к измерению термической стабильности с помощью ДСФ, более точный метод дифференциальной сканирующей калориметрии (ДСК) также использовался для измерения термической стабильности правильно спаренных, сконструированных гетеродимеров Fab по сравнению с термической стабильностью немодифицированной пары тяжелая цепь-легкая цепь дикого типа. Гетеродимеры Fab получали, как описано в примере 5.
Измерение термической стабильности с помощью ДСК
Термическая стабильность сконструированных пар гетеродимеров Fab была измерена с помощью ДСК следующим образом. 400 мкл очищенных образцов в основном с концентрациями либо 0,2 мг/мл или 0,4 мг/мл в ФСБ использовались для анализа ДСК с VP-капиллярным ДСК (GE Healthcare). В начале каждого прогона ДСК 5 холостых введений буфера были выполнены для стабилизации исходного уровня, и введение буфера наносили перед каждым вводом пробы для сравнения. Каждый образец сканировали от 20 до 100°С со скоростью 60°С/ч, с ограниченной обратной связью, 8-секундным фильтром, 5-минутным preTstat и давлением азота 4,9 килограмм на квадратный сантиметр. Полученные термограммы проверялись и анализировались с использованием программного обеспечения Origin 7.
Термическая стабильность H1L1 и H2L2 Fab включена в столбцы 10 и 12 (H1L1 относится к гетеродимеру Fab CAT-2200 и H2L2 к гетеродимеру Fab Пертузумаба, соответственно) таблицы 6. Для каждого гетеродимера Fab, где проводились повторы, указанное значение Tm представляет собой срединное значение. Сравнение значений Tm сконструированного гетеродимера Fab по отношению к значению Tm гетеродимера Fab типа дикого типа приведено в столбце 11 для CAT-2200 H1L1 (конструкция Fab CAT-2200 дикого типа, содержащая метку FLAG, со средней Tm 70,7°C) и столбце 13 для H2L2 Пертузумаба (конструкция Fab Пертузумаба дикого типа, содержащая метку HA, со средней Tm 78,7°C). Учитывая высокие требования к количествам проб и низкую пропускную способность ДСК, измерения выполнялись путем выборочного исследования подмножества конструкций. По этой причине некоторые гетеродимеры Fab не имеют значений Tm, измеренных с помощью ДСК, и поэтому обозначены как не определенные (НО). Все сконструированные гетеродимеры Fab, анализируемые с помощью ДСК, показали хорошую термическую стабильность. Значения Tm, полученные с помощью ДСК, как правило были ниже, чем у ДСФ (пример 7).
Пример 9: Оптимизация конструкций
Эффективность исследований конструкций K-L и конструкций K-L, полученных из K-K, в системе пертузумаб/CAT-2200, как показано в таблицах 5A и 5B, соответственно, были рассмотрены с целью лучшего понимания способности конструкций управлять селективным спариванием. Этот анализ сделал возможным следующий цикл оптимизации конструкции, чтобы, по возможности, улучшить специфичность спаривания и/или повысить стабильность гетеродимеров. Оптимизация конструкции включала модификацию конструкций K-L и конструкций K-L, полученных из K-K, на конкретном замещенном аминокислотном остатке для оценки влияния замены этого остатка на альтернативную аминокислоту, добавления аминокислотных замен в дополнительных аминокислотных остатках и/или удаления аминокислотных замен на конкретном остатке (то есть, возвращая его обратно в остаток дикого типа). Процесс конструирования и выбор конструкций выполнялся, как описано в примерах 1 и 2, а полученные конструкции испытывались в системе K-L пертузумаба/CAT-2200.
Дополнительный набор конструкций K-L был предложен с использованием альтернативного способа увеличения разнообразия конструкций с улучшенной специфичностью спаривания, в котором выбранные конструкции константного домена из таблиц 4A и 4B, а также выбранные кандидаты на оптимизацию конструкции константного домена, описанные в предыдущем абзаце, были объединены с двумя конструкциями вариабельного домена K-K. Две конструкции вариабельных доменов K-K включены в таблицу 4B и имеют уникальные идентификаторы 10621-10733 и 10623-10745. Эти конструкции не способствовали сильной специфичности спаривания сами по себе и не влияли на связывание антигена, но, как было показано, улучшали специфичность спаривания в сочетании с определенными конструкциями константного домена Fab в системе антител каппа-каппа. Этот дополнительный набор конструкций K-L также включал в себя только комбинации конструкций константного домена, где конструкции константного домена выбраны из таблиц 4A, 4B и кандидатов на оптимизацию конструкции, описанных в предыдущем абзаце.
Наконец, был предложен еще один дополнительный набор конструкций K-L, полученных из K-K, с использованием репрезентативных конструкций из библиотеки конструкций K-K, описанных в таблице 5 заявки PCT Application №PCT/IB2015/054107 в качестве отправной точки и испытан в системе K-L пертузумаба/CAT-2200. Как описано в примере 2, репрезентативные конструкции, полученные из K-K, были адаптированы к системе каппа-лямбда путем модификации аминокислотных остатков по мере необходимости для учета различий в последовательности и структуре между легкими цепями каппа и лямбда.
Специфичность спаривания дополнительных конструкций K-L и конструкций K-L полученных из К-К, описанных в этом примере, была экспериментально оценена с помощью LCCA в системе K-L пертузумаб-CAT-2200, как описано в примере 10.
Пример 10: Получение и оценка преимущественного спаривания сконструированных гетеродимеров Fab с помощью LCCA
Конструкции Fab пертузумаба и CAT-2200, содержащие аминокислотные замены, соответствующие наборам конструкции мАт, были получены, как описано в примере 3. Как описано в примере 4, все эксперименты LCCA выполнялись на конструкциях, содержащих межцепочечную Fab дисульфидную связь, расположенную в константном домене (H/C233-L/C214, нумерация Кабат). Для оценки преимущественного спаривания, показанного наборами конструкции мАт, преимущественное спаривание, показанное соответствующими наборами конструкции LCCA, было измерено, как описано в примере 4, со следующими исключениями.
Оценка преимущественного спаривания в наборах конструкции LCCA проводилась с помощью LCCA, как описано в примере 4, за исключением того, что трансфекцию проводили с использованием тяжелых и легких цепей в соотношении 1:1:3 (по массе) для H1:L1:L2 или H2:L2:L1 вместо 1:1:1, причем тяжелая цепь присутствует в ограничивающих количествах, а неправильно спаренная легкая цепь присутствует в избытке. Поскольку конструкции, описанные в примере 9, были оптимизированными версиями тех, которые описаны в таблицах 4А и 4В, или представляли собой комбинации двух конструкций, некоторые из которых уже показывают относительно высокую специфичность спаривания, возможно, что дальнейшее улучшение такой специфичности (если оно может быть достигнуто) может не выявляться в LCCA при H1:L1:L2 или H2:L2:L1 1:1:1 из-за ограничений анализа. Таким образом, для определения улучшения специфичности использовалось соотношение ДНК 1:1:3, а не 1:1:1. Все трансфекции проводили с использованием 300 нг H1, 300 нг L1 и 900 нг L2 (то есть H1:L1:L2 или H2:L2:L1=соотношение 1:1:3), 100 нг AKTdd pTT22 (вектор, содержащий конститутивно активный мутант протеинкиназы B) и 400 нг оцДНК (ДНК спермы лосося).
Анализ данных и описание показателей LCCA
Анализ данных проводили с использованием модифицированной версии анализа, описанной в примере 4, как изложено ниже.
Все наборы конструкций LCCA были протестированы в LCCA, и конструкции интереса были определены на основе следующих критериев включения. Для того, чтобы считаться конструкцией интереса, конструкция должна содержать, по меньшей мере, один положительный результат LCCA выше верхнего предела 95% ДТ скалярного доверительного интервала, тогда как другой комплементарный LCCA должен быть выше нижнего предела 95% ДТ скалярного доверительного интервала. Все конструкции, которые прошли вышеприведенные критерии, были включены во вторую библиотеку конструкций K-L, как указано в таблице 7A, или вторую библиотеку конструкций K-L, полученных из K-K, как указано в таблице 7B.
Таблица 8A и таблица 8B описывают данные LCCA, полученные для второй библиотеки конструкций K-L, и второй библиотеки конструкций K-L, полученных из K-K, соответственно, в контексте конструкций мАт. Эффективность каждого набора конструкций LCCA в этом примере описывалась двумя скалярными значениями, которые соответствовали натуральному логарифму соотношения H1L1:H1L2 (срединное значение) и соотношения H2L2:H2L1 (срединное значение), соответственно. Следует отметить, что эти скалярные значения немного отличаются от скалярных значений, рассчитанных в примере 4, но аналогично используются для оценки преимущественного спаривания. Эти скалярные значения представлены в столбцах 2 и 5 в таблице 8A для второй библиотеки конструкций K-L и в таблице 8B для второй библиотеки конструкций K-L, полученных из K-K. Для (HC) CAT-2200 дикого типа, конкурирующего с (LC-FLAG) CAT-2200 плюс (LC-HA) Пертузумаба, 95% скалярный доверительный интервал составлял от -3,24 (нижний предел) до -2,94 (верхний предел). Для (HC) Пертузумаба дикого типа, конкурирующего с (LC-HA) Пертузумаба плюс (LC-FLAG) CAT-2200, 95% скалярный доверительный интервал составлял от -0,9 (нижний предел) до -0,6 (верхний предел).
Скалярный диапазон для каждой конструкции LCCA (эксперименты H1L1L2 и H2L1L2) показан в столбцах 3 и 6 в таблицах 8A и 8B как показатель воспроизводимости результатов и представляет собой разницу между максимальными и минимальными скалярными значениями, измеренными для конструкции. Для результатов, полученных из единичного измерения, значения диапазона отмечены как NA (не доступно). Кроме того, скалярные значения также были преобразованы обратно в чистое срединное соотношение для H1L1:H1L2 и H2L2:H2L1. Это преобразование эффективно нормализовало соотношения LCCA до 100%, поскольку в некоторых случаях наблюдалось, что общее количество H1L1 и H1L2, или H2L2 и H2L1 значительно отличалось от 100%. Эти значения представлены в столбцах 4 и 7 в таблицах 8А и 8В.
Результаты
Как отмечено выше, результаты LCCA показаны в таблицах 8A и 8B в контексте наборов конструкций мАт. В таблице 8A приведены данные LCCA, относящиеся к конструкциям K-L, описанным в примере 9. 151 конструкция K-L соответствовала критериям включения, и большинство конструкций имеют аминокислотные модификации только в константной области, причем некоторые из конструкций также включают аминокислотные модификации в вариабельной области (см. таблицу 7A). Эти конструкции были предложены для дальнейшего обусловливания специфичности и стабильности спаривания, в то же время способствуя переносимости в другие системы антител.
В таблице 8B приведены данные LCCA, относящиеся к конструкциям K-L, полученным из K-K, также описанным в примере 9. 21 конструкция K-L, полученные из K-K, отвечали критериям включения. Эти конструкции включают только конструкции с константным доменом.
Пример 11: Получение сконструированных гетеродимеров Fab для биофизической характеризации в большом количестве
Для изучения биофизических характеристик правильно спаренных гетеродимеров, получение выбранных правильно спаренных гетеродимеров H1L1 или H2L2 (сконструированных Fab), идентифицированных в наборах уникальных идентификаторов, показанных в таблицах 7A и 7B, было увеличено, как описано в примере 5, за исключением того, что вместо 20 мл было выполнено увеличение на 50 мл. Экспрессированные Fab очищали, как описано в примере 5, для проверки термической стабильности и связывания антигена. Протокол очистки модифицировали для инкубации супернатантов с помощью смолы IgG-CH1 CaptureSelect™ без какого-либо начального разбавления в уравновешивающем буфере, а элюирование связанных образцов проводили с 4 объемами слоя элюирующего буфера.
После очистки экспрессию гетеродимера оценивали с помощью анализа High Throughput Protein Express в невосстанавливающих и восстанавливающих условиях с использованием Caliper LabChip GXII, как указано ранее в примере 5.
Данные показали, что включение модификаций набора конструкций мАт в сконструированные Fab не оказало существенного влияния на экспрессию белка, судя по количеству продукта после очистки. Количество продукта после очистки, полученное для сконструированных Fab CAT-2200, составляло 1,3 мг/50 мл экспрессии (SD 0,6) по сравнению с 1,9 мг/50 мл экспрессии (SD 0,5) для Fab дикого типа. Сконструированные Fab пертузумаба получали в количестве 0,9 мг/50 мл экспрессии (SD 0,4) по сравнению с 1,1 мг/50 мл экспрессии (SD 0,2) для Fab дикого типа. Анализ с помощью калипера показал степень чистоты, сравнимую с Fab ДТ (данные не показаны). Ряд сконструированных Fab имел осадок, связанный с ними (отмечены с помощью "*" в таблицах 9A и 9B) после очистки. Указанную выше информацию о количестве продукта измеряли после удаления осадка.
Пример 12: Измерения аффинности сконструированных гетеродимеров Fab к антигену
Была оценена способность сконструированных гетеродимеров Fab CAT-2200 связываться с ИЛ-17A и сконструированных гетеродимеров Fab Пертузумаба связываться с HER2-ECD, с целью определения, повлияла ли замена аминокислот на способность гетеродимера связываться с антигеном. Связывание с антигеном оценивали, как описано в примере 6, за исключением того, что сконструированные Fab CAT-2200 вводили при 50 мкл/мин в течение 120 секунд с фазой диссоциации 600-800 секунд вместо 900 секунд фазы диссоциации. Кроме того, скрининг сконструированных Fab пертузумаба для связывания с HER2 проводили, как описано в примере 6, со следующими модификациями: для каждого захвата Fab, Fab разбавляли до 2,5 мкг/мл вместо 2 мкг/мл с временем введения 120 с вместо 240 с. Самая низкая концентрация введенного антигена HER2, составляла 1,23 нМ вместо 0,41 нМ. Фаза диссоциации для серии концентраций составляла 600 с вместо 1200 с.
Результаты Значения антигенной аффинности (KD) для сконструированных образцов гетеродимера Fab приведены в таблице 9A (конструкции K-L) и в таблице 9B (конструкции K-L, полученные из K-K) в контексте спаривания конструкции мАт. Значения KD H1L1 и H2L2 Fab включены в столбцы 2 и 4 (KD гетеродимера H1L1 Fab CAT-2200 и гетеродимера H2L2 Fab Пертузумаба, соответственно), и 3 и 5 (изменение KD гетеродимера H1L1 или H2L2 Fab по сравнению с его соответствующим диким типом). Значения KD определяли только для образцов гетеродимера Fab, которые показали захват гетеродимера Fab по меньшей мере в 100 РЕ. Контрольное значение KD для Fab CAT-2200 дикого типа, связанного с ИЛ-17A, используемое для сравнения со сконструированными гетеродимерами, составляло 0,209 нМ. Это значение является срединным значениям, основанным на многочисленных измерениях гетеродимера Fab CAT-2200 дикого типа, где легкая цепь содержит метку FLAG и не отличается существенно от значения, указанного в примере 6. Аналогичным образом, контрольное значение KD для пертузумаба дикого типа, связанного с HER2, составляло 7,8 нМ. Это значение также является срединным значениям, основанным на многочисленных измерениях гетеродимера Fab пертузумаба дикого типа, где легкая цепь содержит метку HA и не отличается существенно от значения, указанного в примере 6. В таблицах 9A и 9B разница в KD по отношению к аффинности связывания дикого типа антигена показана с использованием расчета -(log(KD_конструкции) - log(KD_дт)), так что положительные значения указывают на более низкие значения KD, тогда как отрицательные значения указывают на увеличенные значения KD гетеродимера Fab по сравнению с аффинностью связывания дикого типа для антигена. Как отмечалось ранее, из-за ограничений пропускной способности в масштабной процедуре для большого количества гетеродимеров Fab, полученных с помощью наборов конструкции мАт, измерения ППР выполнялись путем выборочного исследования подмножества сконструированных гетеродимеров Fab. По этой причине некоторые гетеродимеры Fab не имеют значений KD, измеренных с помощью ППР, и поэтому обозначены как не определенные (НО). Для испытанных сконструированных гетеродимеров Fab, аминокислотные замены не влияли или практически не влияли на значения KD (в пределах 2-кратного ДТ).
Пример 13: Измерения термической стабильности сконструированных гетеродимеров Fab с помощью ДСФ
Дифференциальная Сканирующая Флуориметрия (ДСФ) использовалась для измерения термической стабильности правильно спаренных сконструированных гетеродимеров Fab по сравнению с термической стабильностью Fab дикого типа. Гетеродимеры Fab получали, как описано в примере 11.
Измерение термической стабильности с помощью ДСФ
Термическая стабильность сконструированного гетеродимера Fab была измерена с помощью ДСФ следующим образом. Каждый очищенный гетеродимер разбавляли до 0,67 мг/мл в DPBS (фосфатно-солевом буфере Дульбекко, HyClone Cat # SH30028.02). Рабочий состав краски геля Sypro Orange (Life Technologies Cat # S-6650) был приготовлен разведением 1:1000 в DPBS. Образцы ДСФ были приготовлены путем добавления 15 мкл белка 0,67 мг/мл к 15 мкл рабочего состава краски геля Sypro Orange. Однако для белков, которые имели концентрацию менее 0,67 мг/мл, каждый образец ДСФ получали путем добавления 15 мкл неразбавленного белка к 15 мкл рабочего состава красителя Sypro Orange. Затем анализ ДСФ проводили на 30 мкл аликвоты с использованием прибора кПЦР Rotor-Gene 6000 (QiaGen Inc). Каждый образец сканировали с 30°C по 94°C с интервалом 1°C с 10-секундным балансом между каждым этапом и 30-секундным временем ожидания в начале. Использовали фильтр возбуждения 470 нМ и эмиссионный фильтр 610 нМ с усилением 8. Данные анализировали с помощью программного обеспечения Rotor-Gene 6000, используя максимальное значение от первой производной кривой денатурации как Tm.
Результаты ДСФ для сконструированных образцов гетеродимера Fab приведены в таблицах 9A (конструкции K-L) и 9B (конструкции K-L, полученные из K-K) в контексте наборов конструкции мАт. Термическая стабильность H1L1 и H2L2 Fab включена в столбцы 6 и 8 (H1L1 относится к гетеродимеру Fab CAT-2200 и H2L2 к гетеродимеру Fab Пертузумаба) таблиц 9A и 9B. Для гетеродимеров Fab, где проводились повторы, указанное значение Tm представляет собой срединное значение. Сравнение значений Tm сконструированного гетеродимера Fab по отношению к значению Tm гетеродимера Fab типа дикого типа приведено в столбце 7 для H1L1 CAT-2200 и в столбце 9 для H2L2 Пертузумаба. Было определено, что средняя Tm для конструкции Fab CAT-2200 дикого типа, содержащей метку FLAG, составляет 77,0°C. Было определено, что средняя Tm для гетеродимера Fab Пертузумаба дикого типа, содержащего метку HA, составляет 81,7°C. Как упоминалось ранее, измерения ДСФ выполнялись путем выборочного исследования подмножества сконструированных гетеродимеров Fab, поэтому некоторые гетеродимеры Fab не имеют значений Tm, измеренных ДСФ, и обозначены как не определенные (НО). Большинство тестируемых конструкций показали хорошую термическую стабильность (демонстрируя уменьшение Tm по сравнению с ДТ до 4-5°C) в диапазоне ожидаемых значений Tm, обычно наблюдаемых для Fab дикого типа, со следующим исключением. Наличие мутаций S131K_L135K в легкой цепи каппа в сочетании с мутациями L124E_K145T в тяжелой цепи каппа в двух гетеродимерах Fab отрицательно влияло на их значения Tm на> 10°C. Также следует отметить, что для некоторых из сконструированных Fab CAT-2200 в спектрах ДСК наблюдалось плечо к основному пику Fab (эти Fabs отмечены "#" в таблицах 9A и 9B). Такой профиль также наблюдался для Fab CAT-2200 дикого типа.
Пример 14: Измерения термической стабильности сконструированных гетеродимеров Fab с помощью ДСК
В дополнение к измерению термической стабильности с помощью ДСФ, более точный метод дифференциальной сканирующей калориметрии (ДСК) также использовался для измерения термической стабильности правильно спаренных, сконструированных гетеродимеров Fab по сравнению с термической стабильностью немодифицированной пары тяжелая цепь-легкая цепь дикого типа. Гетеродимеры Fab получали, как описано в примере 11, и измеряли термическую стабильность с помощью ДСК, как описано в примере 8. Однако для гетеродимеров Fab, которые имели концентрацию образцов ниже, чем указано в протоколе, ДСК проводили при имеющейся концентрации.
Измерения термической стабильности для H1L1 и H2L2 Fabs показаны в столбцах 10 и 12 (H1L1 относится к гетеродимеру Fab CAT-2200 и H2L2 к гетеродимеру Fab Пертузумаба, соответственно) таблицы 9A и 9B. Для каждого гетеродимера Fab, где проводились повторы, указанное значение Tm представляет собой срединное значение. Сравнение значений Tm сконструированного гетеродимера Fab по отношению к значению Tm гетеродимера Fab типа дикого типа приведено в столбце 11 для CAT-2200 H1L1 (конструкция Fab CAT-2200 дикого типа, содержащая метку FLAG, со средней Tm 70,7°C) и столбце 13 для H2L2 Пертузумаба (конструкция Fab Пертузумаба дикого типа, содержащая метку HA, со средней Tm 78,1°C). Учитывая высокие требования к количествам проб и низкую пропускную способность ДСК, измерения выполнялись путем выборочного исследования подмножества конструкций, которые характеризовались с помощью ДСФ. По этой причине некоторые гетеродимеры Fab не имеют значений Tm, измеренных с помощью ДСК, и поэтому обозначены как не определенные (НО). Все сконструированные гетеродимеры Fab, анализируемые с помощью ДСК, показали хорошую термическую стабильность (уменьшение Tm по сравнению с ДТ до 2-3°C). Значения Tm, полученные с помощью ДСК, как правило были ниже, чем у ДСФ (пример 13). Несмотря на то, что абсолютная разница в Tm сконструированных Fab по сравнению с ДТ, как было замечено, зависит от используемого метода, будь то ДСФ или ДСК, относительный рейтинг сконструированных Fab относительно Fab типа дикого типа был таким же независимо от используемого метода.
Пример 15: Группирование конструкций и отбор репрезентативных конструкций
В примерах 2-14 описана конструкция и тестирование конструкций K-L и конструкций K-L, полученных из K-K. Способность этих конструкций способствовать спариванию была протестирована в LCCA в формате Fab, где была определена способность одной тяжелой цепи каждого набора конструкций LCCA спариваться с соответствующей легкой цепью. Также оценивали влияние аминокислотных замен на стабильность и связывание с антигеном полученных правильно спаренных гетеродимеров (одна тяжелая цепь и одна легкая цепь).
Чтобы проверить, могут ли быть использованы результаты преимущественного спаривания для наборов конструкций мАт (H1L1H2L2), полученных в формате LCCA (коэкспрессия H1, L1 и L2 или H2, L1 и L2) для конструкций K-L и конструкций K-L, полученных из K-K, в случае если полипептидные цепи набора конструкций мАт коэкспрессируются (то есть, когда коэкспрессируются H1, L1, H2 и L2), было отобрано репрезентативное число из более чем 400 конструкций, описанных в таблицах 4А, 4В, 7А и 7В, поскольку было невозможно проверить все конструкции таким образом. Репрезентативные конструкции тестировали в анализе, обозначенном как "SMCA" (подробно описано в примере 16).
Для выбора репрезентативных конструкций, конструкции K-L, идентифицированные в таблицах 4А и 7А (n=404), объединяли и группировали в соответствии с идентичностью в замещенных положениях аминокислотных остатков. В некоторых случаях, когда конструкция K-L, полученная из K-K, из таблицы 7B была значительно модифицирована из оригинальной конструкции K-K, она также была включена в эту группу конструкций K-L для группирования. В таблицах с 10-A1 по 10-A12 показаны полученные кластеры с 1 по 12 для конструкций K-L. Аналогичным образом, конструкции K-L, полученные из K-K, представленные в таблицах 4B и 7B (n=43), за исключением тех, которые были перемещены в группу конструкций K-L, также были объединены и сгруппированы. В таблицах с 10-B1 по 10-B10 показаны полученные кластеры с 1 по 10 для конструкций K-L, полученных из K-K. Затем были выбраны репрезентативные конструкции из каждого кластера, как описано ниже. Окончательный набор выбранных репрезентативных конструкций также включен в таблицу 11A (конструкции K-L) и 11B (конструкции K-L, полученные из K-K), включая обозначение ID конструкций SMCA.
Репрезентативные конструкции для SMCA
Репрезентативные конструкции были отобраны на основе первичных критериев демонстрации специфичности спаривания и минимального влияния или отсутствия влияния на связывание антигена, а также с учетом вторичных критериев, таких как: стабильность конструкции, минимизация числа замен, экспрессия, сопоставимая с ДТ. Максимальная специфичность спаривания оценивалась с учетом двух Fab и каждого Fab индивидуально, где один Fab спаривался менее хорошо, чем другой Fab в гетеродимерной паре, но все же показывал спаривание лучше, чем у дикого типа (сценарии A и B, как показано на Фиг. 6). Кроме того, разнообразие в кластере и количество конструкций в кластере дополнительно определяли тип и количество выбранных представителей кластера.
В общей сложности 33 репрезентативные конструкции K-L были выбраны из кластеров с 1 по 12 для тестирования в формате SMCA. Аналогичным образом, 7 конструкций K-L, полученных из K-K, со значениями эффективности LCCA от высоких до средних, были выбраны из 7 кластеров 10 идентифицированных кластеров. Три из кластеров конструкций K-L, полученных из K-K, не включали конструкции, которые соответствовали критериям репрезентативной конструкции по основным критериям, таким как критерии эффективности LCCA или отсутствие влияния на связывание антигена.
По меньшей мере одна репрезентативная конструкция была выбрана из каждого кластера. Кластеры, которые были представлены только одной репрезентативной конструкцией, включали кластеры 5 и 11 конструкций K-L, а также все 7 кластеров конструкций K-L, полученных из K-K. Остальные кластеры имели более одной репрезентативной конструкции, поскольку кластеры были либо большими (то есть кластеры 2, 3, 4, 6, 7, 8) и/или имели большее различие в конструкциях (небольшие кластеры) (то есть кластеры 1, 9, 10, 12). Несмотря на то, что конструкции в каждом кластере разделяли сходства последовательностей, небольшие кластеры внутри кластера отличались, по меньшей мере, одним драйвером (заменой аминокислоты). Репрезентативные конструкции для каждого кластера выделены жирным шрифтом в таблицах с 10-A1 по 10-A12 и с 10-B1 по 10-B10. Как уже отмечалось в данном документе в другом месте, "-" в этих таблицах указывает на то, что аминокислотные замены, которые способствуют преимущественному спариванию, не присутствуют в этой конкретной полипептидной цепи.
Драйверы и вторичные замены
Здесь описываются кластеры, для которых были выбраны репрезентативные конструкции, с акцентом на определение "драйверов" или "наборов драйверов" (положений аминокислот и/или аминокислотных замен), которые способствовали желаемой специфичности спаривания в каждом кластере, как оценивается с помощью LCCA. Термин "набор драйверов" относится к набору положений аминокислот и аминокислотных замещений, которые способствуют преимущественному спариванию, тогда как термин "драйвер" относится к одной позиции/замене в наборе драйверов. Однако другие остатки и замены иногда ассоциировались с конструкцией или набором конструкций в кластере. Эти остатки и замены называются "вторичными" заменами и иногда включаются в набор конструкций мАт с целью улучшения эффективности используемого набора драйверов или наборов драйверов. Механически эти вторичные замены могут действовать так, чтобы i) способствовать стерическому размещению драйверов, ii) увеличивать количество контактов или заменять контакты, потерянные при заменах в положениях набора драйвера, в области контакта между тяжелыми и легкими цепями; iii) оптимизировать сеть водородных связей на основе наборов драйверов и/или iv) создавать условия, способствующие повышению эффективности наборов драйверов. Один или более из этих механизмов могут использоваться/предназначены для повышения эффективности драйверов в каждом кластере.
Некоторые из этих вторичных замен были общими для нескольких кластеров. Например, замена K145T в тяжелой цепи использовалась как в H1L1, так и в H2L2 для удаления положительного заряда, в случае если поблизости были спроектированы отрицательно заряженные драйверы или драйвер. В ряде кластеров замена K129T в легкой цепи лямбда иногда использовалась для удаления положительного заряда, в случае если в окрестностях были спроектированы отрицательно заряженные драйверы или драйвер (например, кластер 4 конструкций K-L и кластеры 8-12). Кроме того, замена E124Q в легкой цепи лямбда использовалось в некоторых кластерах для удаления отрицательного заряда вблизи положительно заряженного драйвера или драйверов (например, кластеры 1-3 конструкций K-L). Замена V133G применялась преимущественно в легкой цепи каппа в некоторых конструкциях для стерического размещения конкретных наборов драйверов (например, преимущественно в кластерах 1, 3, 4, 6, 7, 10, 12 конструкций K-L и кластере 10 конструкций K-L, полученных из K-K). Y178T в легкой цепи лямбда вводили в конструкции для стерического размещения конкретных наборов драйверов в соответствующем H1L1 (например, кластеры K-L 4, 5, 8 и 12). V133S в легкой цепи применяли еще в одном наборе драйверов (например, кластеры K-L 1, 2 и 12), нацеленных на улучшения сети водородных связей.
Описание кластеров конструкций K-L
Для представления кластера 1 были выбраны две конструкции (13175-13486 и 13180-13515), поскольку в этих конструкциях использовались аналогичные электростатические драйверы, занимающие аналогичное пространство (см. таблицу 10-A1). Для всех членов этого кластера была использована комбинация из двух наборов драйверов. Первый набор электростатических драйверов был сконструирован для обеспечения отрицательно заряженной замены (преимущественно L143D) в H1 с образованием соляных мостиков с положительно заряженной заменой (T131K/R) в L1, тогда как H2 была сконструирована так, чтобы обеспечить a) положительно заряженные замены S188K или L124R_S186R, для образования соляных мостиков с отрицательно заряженными заменами в L2 (S176D_T178E или S176D_T180E) или b) образования соляных мостиков между L143R или S186K в H2 и Q124E_V133D в L2. Второй набор электростатических драйверов, называемый "набор дисульфидных направляющих драйверов", был добавлен для дисбаланса образования дисульфидной связи H-L или гетеродимера в неправильно спаренных гетеродимерах на ранних этапах процесса сборки H-L. Набор дисульфидных направляющих драйверов использовал положительно заряженную замену в H1 (A125R), отрицательно заряженную замену в L1 (S122D), отрицательно заряженную замену в H2 (K228D) и положительно заряженную замену в L2 (S121K). Эти аминокислотные замены расположены вблизи дисульфидной связи, вдали от углубленной (основной) области контакта H-L, где расположено большинство аминокислотных замен в конструкциях K-L. Два набора драйверов были сконструированы таким образом, чтобы образовывать соляные мостики в преимущественно спаренных гетеродимерах, в то время как неправильные пары были бы нежелательны в основном из-за электростатического отталкивания.
Для кластера 2 были выбраны четыре репрезентативные конструкции (13282-13484, 13287-13494, 13218-13482 и 10945-11377), отражающие степень разнообразия в этом кластере (см. таблицу 10-А2). Все члены этого кластера основаны на H1, сконструированном аналогично первому набору электростатических драйверов кластера 1, чтобы позволить отрицательно заряженным заменам (L143D или L143D_Q179E) образовывать соляные мостики с положительно заряженной заменой (T131K/R) в L1, тогда как H2 была сконструирована таким образом, чтобы позволить a) положительно заряженной замене (S186K) образовывать соляные мостики с отрицательно заряженными заменами в L2, прежде всего V133D или V133D_Q160E; b) S188K спариваться с S131D/E L2; или c) L143R/K спариваться с Q124E_V133D L2. Некоторые члены кластера 2 также содержали набор стерических драйверов в дополнение к набору электростатических драйверов. Использовались два набора стерических драйверов, называемых "стерический 1" и "стерический 2". Набор стерических драйверов 1 включал F174G в H1, предназначенный для того, чтобы быть стерически комплементарным к S176F или T116F_S176F L1, и V190F H2, стерически комплементарный к L135A L2. Набор стерических драйверов 2 включал A139W в H1, стерически совместимый с остатками L1 ДТ и H2 ДТ, стерически совместимый с L135W L2. Следовательно, в этом кластере неправильное спаривание H1L2 и H2L1 было бы нежелательным в первую очередь из-за электростатического отталкивания или электростатического отталкивания в сочетании со стерической несовместимостью. Существуют также два члена кластеров, которые помимо набора электростатических драйверов содержат компонент вариабельной конструкции ("набор драйверов вариабельных доменов") в цепи H2 (Q39E или L45P) и цепи L2 (соответственно Q38R или P44F), для которых не были выбраны репрезентативные конструкции, но, как ожидается, улучшат спаривание.
Для кластера 3 были выбраны пять репрезентативных конструкций (10931-11263, 13221-13411, 10987-11257, 10983-11306 и 10947-11392) (см. таблицу 10-A3). Члены этого кластера использовали подобные наборы электростатических драйверов в H1 (L143D или L143E_Q179E) и L1 (T131R/K), как в кластере 2, но отличались по отношению к конструкции H2L2. Конструкция H2L2 основывалась главным образом на a) L124R_S186R/K или L124R_Q179K или S188K H2 спаривания с S176D_T178E или S176D_T180E L2; или b) L143R_S188K H2 спаривания с Q124E_V133D_S176D_T178D/E L2, что приводит к нежелательному неправильному спариванию в основном из-за электростатического отталкивания. Как и в кластере 2, некоторые члены кластера также использовали набор стерических драйверов 1 или стерических драйверов 2 в дополнение к набору электростатических драйверов. Представителем такой комбинации набора драйверов является 13221-13411. Этот кластер также включал в себя две конструкции, содержащие "набор драйверов вариабельных доменов" в дополнение к набору электростатических драйверов.
Для кластера 4 были выбраны три репрезентативные конструкции (13335-13361, 13209-13364 и 11100-11205) (см. таблицу 10-А4). Большинство членов этого кластера использовали подобные электростатические драйверы на H1 (S188K) и L1 (S176E/D_Y178E) с несколькими вариациями в паре H2L2:a) V177D_S188D из H2 спаривались с S176K_T178R/K из L2; или b) S186E или L124E или L124E_Q179E из H2 спаривались с S176K/R или S131K/R_S176R/K из L2, что позволило бы сформировать соляные мостики в преимущественно спаренных гетеродимерах, в то время как неправильно спаренные пары были бы нежелательны в основном из-за электростатического отталкивания. Некоторые члены этого кластера использовали только набор стерических драйверов, то есть набор стерических драйверов 3 или некоторые компоненты набора стерических драйверов 3, в дополнение к наборам электростатических драйверов. Набор стерических драйверов 3 основывался на остатках ДТ в H1L1 или S188A в H1, и S176A_Y178W в L1, и S188W или S186I/L_S188W в H2, сконструированных стерически комплементарными S176V или S176A_T178A в L2. Кроме того, некоторые члены этого кластера также содержали набор дисульфидных направляющих драйверов в дополнение к наборам электростатических драйверов. Кроме того, несколько членов кластера содержали набор(ы) драйверов вариабельных доменов в дополнение к электростатическому драйверу.
Кластер 5 был представлен уникальным идентификатором 11066-11181 (см. таблицу 10-A5). Все члены этого кластера использовали наборы электростатических драйверов. Первый набор электростатических драйверов основывался на спаривании S186K в H1 с V133D в L1 и спаривании S188D или V177D_S188D из H2 с S131K или S176K_T178R/K, соответственно, в L2. Второй набор драйверов имел обратную ориентацию заряда, в первую очередь спаривание L124E_V190D в H1 с L135K в L1 и спаривание L124R или S188K из H2 с S176D или S176D_T178E из L2.
Для кластера 6 были выбраны две репрезентативные конструкции (10808-11308 и 10814-11233) (см. таблицу 10-A6). Все члены этого кластера использовали аналогичные наборы электростатических драйверов в паре H1L1:V177D_S188D из H1 с S176K_Y178K/R из L1 и более разнообразным набором драйверов в паре H2L2, а именно a) спаривании S188K из H2 с S176E/D_T178E или S131D/E из L2; b) спаривании S186K/R из H2 с V133D или Q124E_Q160E_T180E из L2; c) спаривании L124R или L124R_Q179K из H2 с S176D или S176D_T178D или S176D_T180E из L2; или d) спаривании L143K/R из H2 с V133D или Q124E_V133D из L2. Эти замены позволили образовывать соляные мостики в преимущественно спаренных гетеродимерах, в то время как неправильные пары были бы нежелательны в основном из-за электростатического отталкивания.
Для представления кластера 7 были выбраны три конструкции (13167-13514, 13263-13493 и 12878-11240) (см. таблицу 10-A7). Все члены кластера использовали наборы электростатических драйверов, на основе общего набора драйверов в H1L1:S188E в H1 и Y178K в L1 с некоторым разнообразием набора драйверов в паре H2L2: главным образом спаривание L124R или S188K (также в комбинации с L143K или S186R, или Q179K) из H2 с S176D/E_T178D/E или S176D/E_T180E или в сочетании с Q124E из L2. Некоторые из членов кластера также содержали набор дисульфидных направляющих драйверов, набор стерических драйверов 2 или набор драйверов вариабельных доменов в дополнение к набору электростатических драйверов.
Для представления кластера 8 были выбраны пять конструкций (13320-13370, 12938-13469, 13226-13434, 11026-11270 и 13316-13451) (см. таблицу 10-A7). Члены этого кластера обладали большим разнообразием в электростатической паре H1L1, в то время как пара H2L2 была основана на спаривании драйверов L143E или L143E_Q179E в H2 в основном с драйверами Q124R/K_T178R или Q124R_Q160R/K_T178R в L2. Разнообразие драйверов H1L1 было следующим: a) S186R или Q179K, используемые в качестве драйверов в H1, в основном спариваются с драйвером S180E из L1 (вариация S186K в H1 спаренная с драйвером V133D в L1); b) драйвер L143K из H1 спаренный с V133D или T131E/D_V133D из L1; или c) S188K из H1 спаренная в основном с S176D/E_Y178E из L1. Некоторые из членов кластера содержали стерический набор драйверов 2 или стерический набор драйверов 3, или стерический набор драйверов 4, основанный на L124W в H2, сконструированный таким образом, чтобы быть стерически комплементарным V133A в L2 (некоторые члены кластера, как часть последней конструкции, также использовали замену L143A в H1 и V133W в L1) в дополнение к набору электростатических драйверов. Дополнительно, несколько членов кластера содержали набор драйверов вариабельных доменов в дополнение к электростатическому набору, для которого была выбрана одна репрезентативная конструкция, или содержали сконструированную дисульфидную связь между F122C из H2 и Q124C из L2.
Для кластера 9 были выбраны две репрезентативные конструкции (13208-13455 и 13194-13427) (см. таблицу 10-A9). Почти все члены этого кластера использовали комбинацию из двух наборов электростатических драйверов. Первый набор драйверов в основном состоял из Q179K или S186R в H1 и S180E в L1 с L143E или L143E_Q179E в H2, и S131K или Q124R_T178R, или Q124R_Q160K_T178R в L2, а второй набор драйверов был набором дисульфидных направляющих драйверов.
Для кластера 10 были выбраны две репрезентативные конструкции (13238-13463 и 13304-13466) (см. таблицу 10-A10). Члены этого кластера основывались только на стерических драйверах 1 или в сочетании с наборами электростатических драйверов, используемых в кластере 9.
Для кластера 11 был выбран одна репрезентативная конструкция (13306-13375) (см. таблицу 10-A11). Все члены этого кластера использовали электростатические замены для обеспечения преимущественного спаривания гетеродимеров на основе следующих драйверов: L143K_V190K в H1 и V133D в L1 и L124E в H2 и S131K_L135K в L2.
Для кластера 12 были выбраны три репрезентативные конструкции (13227-13383, 11065-11206 и 11034-11204) (см. таблицу 10-A12). Члены этого кластера в основном использовали электростатические замены для обеспечения преимущественного спаривания гетеродимеров. К ним относятся L143K или S186K в H1 и V133D или T131D/E_V133D в L1 и преимущественно L124E_Q179E в H2 и S131K/R_S176R в L2. Некоторые члены кластера дополнительно включали в себя наборы дисульфидных направляющих драйверов, стерических драйверов 2, стерических драйверов 3 или наборы драйверов вариабельных доменов.
Описание кластеров конструкций K-L, полученных из K-K.
В LCCA был протестирован значительно меньший набор конструкций K-L, полученных из K-K, (n=43) по сравнению с конструкциями K-L. Следовательно, каждый из десяти перенесенных кластеров каппа-каппа представлял собой единую конструкцию. Большинство кластеров использовали наборы электростатических драйверов, за исключением двух, которые использовали наборы стерических драйверов. Кроме того, два кластера состояли из конструкций только в вариабельном домене, в то время как остальные кластеры содержали конструкций константных доменов.
Кластер 1 включал электростатические конструкции на основе следующего набора драйверов: L143E_Q179E в H1, E124K_Y178R в L1, S186R в H2, и T178E_T180E или Q160E_T180E в L2 и представлен конструкцией 10657-10760.
Кластер 2 включал в себя комплекты электростатических драйверов с L143E в H1, E124R в L1 и S186R или Q179K в H2, Q124E_Q160E_T180E в L2 (репрезентативная конструкция 10652-10734).
Кластер 3 содержал положительно заряженный драйвер S186R в H1, комплементарный отрицательно заряженному драйверу S180E в L1, и отрицательно заряженный драйвер L143E или Q179E, или их комбинацию в H2, преимущественно комплементарный положительно заряженному драйверу Q124K_T178R в L2 (репрезентативная конструкция 10685-10726).
В кластере 4 использовались следующие драйверы: Q179K в H1, S180E в L1 и L143E в H2, Q124R или Q124R_Q160K_T178R в L2 (репрезентативная конструкция 10665-10724).
Члены кластера 6 содержали конструкции вариабельных доменов на основе электростатической пары Q39K/R и Q38E/D в одном плече и Q39E/D с Q38K/R в другом плече (обе ориентации пар зарядов использовались в плечах H1L1 и H2L2) (репрезентативная конструкция 10681-10741).
Члены кластера 8 включали второй набор драйверов вариабельных доменов, стерический по своей природе: ДТ или L45F в H1 комплементарный ДТ в L1, и L45A/P в H2 комплементарный P44F из L2 (репрезентативная конструкция 10621-10733).
Кластер 10 включал в себя единую конструкцию на основе L124E в H1, S176R в L1 и L124R в H2 и S176D в L2 (конструкция 10640-10713).
Репрезентативные конструкции кластеров 5, 7 и 9 не были выбраны, поскольку они не соответствовали критериям эффективности, таким как стимулирование (от умеренного до сильного) или отсутствие воздействия на связывание антигена относительно исходного Fab.
Пример 16: Оценка преимущественного спаривания сконструированных гетеродимеров в наборах мАт в формате биспецифического антитела
В примерах с 2 по 4 и 10 показана способность сконструированных Fabs преимущественно спариваться в контексте конструкции LCCA (то есть, H1, L1, L2 или H2, L1, L2, где одна тяжелая цепь коэкспрессируется с двумя легкими цепями в формате Fab). В этом примере оценивались сконструированные гетеродимеры в наборах конструкций мАт (H1, L1, H2, L2), чтобы определить, способствовали ли они преимущественному спариванию в формате биспецифического антитела (то есть, где H1, L1, H2 и L2 коэкспрессируются). В дополнение к аминокислотным заменам набора конструкций мАт, область Fc полноразмерной тяжелой цепи каждого гетеродимера была асимметрично модифицирована, так что одна тяжелая цепь содержала аминокислотные замены T350V, L351Y, F405A и Y407V (цепь A) а другая тяжелая цепь содержала аминокислотные замены T350V, T366L, K392L и T394W (цепь B), где нумерация аминокислот Fc соответствует системе нумерации ЕС. Эти асимметричные модификации Fc были включены для содействия гетеродимеризации уникальных тяжелых цепей.
Способность сконструированных гетеродимеров в наборах мАт способствовать преимущественному спариванию в контексте биспецифического антитела оценивалась в 3 биспецифических системах с использованием комбинаций антител CAT-2200 и пертузумаба, описанных в предыдущих примерах, и CR8071 (антитело к гемагглютининовому белку гриппа B) и SGN-CD19a (антитело к CD19). Были испытаны 3 биспецифические системы: A) CAT-2200 (имеющее легкую цепь лямбда)/пертузумаб (имеющее легкую цепь каппа), B) CAT-2200 (имеющее легкую цепь лямбда)/SGN-CD19a (имеющее легкую цепь каппа) и C) CR8071 (имеющее легкую цепь лямбда)/SGN-CD19a (имеющее легкую цепь каппа). CAT-2200 и CR8071 являются человеческими антителами, тогда как пертузумаб и SGN-CD19a являются гуманизированными антителами.
Формат анализа (SMCA)
Способность гетеродимеров, содержащих аминокислотные замены конструкции мАт, преимущественно спариваться с образованием правильно спаренного биспецифического антитела, оценивалась, как описано ниже. Анализ основан на коэкспрессии четырех цепей (цепей H1 и L1 из одного антитела и цепей H2 и L2 из другого антитела) и выявления присутствия правильно сформированного биспецифического антитела с использованием масс-спектрометрии (ЖХ-МС). На Фиг. 8 изображена схема, иллюстрирующая четыре исходные полипептидные цепи и потенциальные продукты, полученные в результате коэкспрессии этих исходных полипептидных цепей при отсутствии преимущественного спаривания между тяжелыми и легкими цепями (в областях Fab и Fc) гетеродимерных пар. Две уникальные конструкции полноразмерных тяжелых цепей коэкспрессировались с двумя уникальными конструкциями легких цепей, что дало десять возможных видов антител (также называемых видами мАт):H1L1_H1L1, H1L2_H1L2, H1L1_H1L2, H2L1_H2L1, H2L2_H2L2, H2L1_H2L2, H1L1_H2L1, H1L2_H2L2, H1L2_H2L1 и H1L1_H2L2. Вид H1L1_H2L2 представляет собой корректно спаренное биспецифическое антитело (см. Фиг. 8). Возможны также четыре вида полуантител (полу-Ат), как показано на Фиг. 8. Когда модификации вводятся в область Fc для содействия гетеродимеризации уникальных тяжелых цепей, число потенциальных видов мАт уменьшается (то есть наблюдается меньше видов E-J). Относительную специфичность спаривания по количеству правильно спаренных видов биспецифических антител H1L1_H2L2 по сравнению с другими видами определяли с использованием ЖХ-МС после очистки белка A (pA) и дегликозилирования. Когда это возможно, цепи оставались немечеными, при условии, что все виды мАт и полу-Аб отличаются друг от друга по меньшей мере на 50 Да. В случае если различия в массе препятствовали этой возможности, к легким цепям добавляли N-концевые метки (HA или FLAG) для обеспечения достаточной массовой дифференциации между видами, с упором на использование метки FLAG, где это возможно, так как иногда наблюдается расщепление метки HA (в случае конструкций, содержащих CR8071, использовалась только метка FLAG).
Этот анализ, включающий экспрессию H1, L1, H2 и L2 биспецифического антитела и последующий анализ спаренных продуктов, называется SMCA.
Получение конструкций
Конструкции, кодирующие тяжелые и легкие цепи IgG CAT-2200, пертузумаба, CR8071 и SGN-CD19a, содержащие аминокислотные модификации в соответствии с конструкциями, были получены следующим образом. Последовательности легкой цепи CAT-2200 и пертузумаба были получены, как описано в примере 3, тогда как полноразмерные последовательности тяжелой цепи для этих антител были созданы путем добавления последовательности ДНК IgG1*01, кодирующей частичные шарнирные домены CH2-CH3 [SEQ ID NO: 6] и модифицированной для содействия гетеродимеризации, на С-конец домена CH1 тяжелых цепей Fab (содержащий часть шарнира и исключающий расширение ABD2, используемое для конструкций в LCCA), получение которых описано в примере 3. Конструкции, содержащие аминокислотные замены для наборов конструкций мАт, были получены либо путем синтеза гена, либо путем сайт-направленного мутагенеза, как отмечено в примере 3. Следует отметить, что канонический C-концевой остаток лизина тяжелой цепи был удален для предотвращения гетерогенности сигнала ЖХ-МС из-за удаления C-концевого лизина (Lawrence W. Dick Jr. и соавт., Biotechnol. Bioeng. (2008 год) 100:1132-43).
Нуклеотидные последовательности ДНК для части Fab тяжелой цепи (SEQ ID NO: 18) и последовательности ДНК легкой цепи (SEQ ID NO: 19) CR8071 показаны в таблице 3C. Аминокислотные последовательности Fab CR8071 для тяжелой цепи (SEQ ID NO: 14) и легкой цепи (SEQ ID NO: 15) соответствуют аминокислотным последовательностям ввода PDB 4FQJ (комплекс Fab CR8071 и гемагглютинина), где замена R222K в тяжелой цепи была сделана для преобразования в наиболее распространенную последовательность IGHG1*01.
Нуклеотидные последовательности ДНК для части Fab тяжелой цепи (SEQ ID NO: 16) и последовательности ДНК легкой цепи (SEQ ID NO: 17) SGN-CD19a также показаны в таблице 3C. Последовательности SGN-CD19a соответствуют SEQ ID NO:7 и 17 описанным в патенте США №8,242,252 и включены здесь как SEQ ID NO: 12 (тяжелая цепь) и SEQ ID NO: 13 (легкая цепь).
Как отмечалось выше, последовательности легкой цепи CAT-2200, CR8071, пертузумаба и SGN-CD19a в некоторых из сконструированных гетеродимеров в наборах мАт были приготовлены с использованием меток FLAG или HA, или без них для обеспечения достаточной массовой дифференциации между видами. Легкие цепи с метками были получены, как описано в примере 3.
Версии этих конструкций дикого типа также были получены для оценки естественного смещения в каждой системе, выявления потенциального влияния меток HA или FLAG на спаривание и определения влияния сконструированных конструкций на преимущественное спаривание по дикому типу.
Трансфекция, экспрессия и очистка конструкций
Конструкции, кодирующие две тяжелые цепи и две легкие цепи каждой системы биспецифических антител, как дикого типа, так и с аминокислотными заменами, соответствующие каждому тестируемому набору конструкций мАт, были трансфицированы в клетки CHO-3E7 следующим образом. Клетки CHO-3E7 с плотностью 1,7-2×106 клеток/мл, культивировали при 37°C в среде FreeStyle(TM) F17 (Invitrogen cat# A-1383501), дополненной 4 мМ глутамина и 0,1% Koliphor P188 (Sigma #K4894). Общий объем 200 мл трансфицировали в общей сложности 200 мкг ДНК с различными соотношениями H1:H2:L1:L2 (который состоит из 100 мкг ДНК антитела и 100 мкг ДНК ЗФБ/AKT/"спейсерной ДНК") с использованием PEI-max (Polysciences Cat # 24765-2) при соотношении ДНК:PEI 1:4. Спустя 24 часа после добавления смеси ДНК-PEI к клеткам добавляли 0,5 мМ вальпроевой кислоты (конечная концентрация) и 1% масса/объем Триптона N1 (конечная концентрация) и клетки затем переносили в 32°C и инкубировали в течение 7 дней до сбора. Культуральную среду собирали центрифугированием и фильтровали в вакууме с использованием фильтра Stericup 0,22 мкл (Millipore Cat # SCGPU05RE). Отфильтрованную культуральную среду затем очищали, используя смолу MabSelect белка A SuRe (GE Healthcare #17-5438-02), которую предварительно уравновешивали ФСБ pH 7,4. Виды антител, связанные со смолой, затем промывали в ФСБ pH 7,4 и элюировали 100 мМ натрий-цитратным буфером с рН 3,6. Элюированные виды антител были сконцентрированы и извлечены из буфера ФСБ pH 7,4 путем центрифугирования с использованием центрифужного фильтра Amicon ultra 15 Ultracel 10K (Millipore # SCGP00525). Образовавшиеся в результате очистки белком A образцы SMCA, содержащие виды антител, оценивали с помощью Калипера перед дегликозилированием и ЖХ-МС.
Метод масс-спектрометрии
Степень преимущественного спаривания гетеродимеров, обусловленная наборами конструкций мАт в контексте биспецифического антитела, оценивали с использованием масс-спектрометрии после очистки белка А и неденатурирующего дегликозилирования. Поскольку гетеродимеры содержали только N-связанные гликаны Fc, образцы SMCA обрабатывали только одним ферментом, N-гликозидазой F (ПНГазой-F). Очищенные образцы дегликозилировали с помощью ПНГазы F следующим образом:0,2 ед. ПНГазы F/мкг антитела в 50 мМ Трис-HCl pH 7,0, инкубация в течение ночи при 37°C, конечная концентрация белка 0,5 мг/мл. После дегликозилирования образцы хранили при 4°C до анализа ЖХ-МС.
Образцы дегликозилированного белка анализировали с помощью интактной ЖХ-МС с использованием системы HPLC Agilent 1100, связанной с масс-спектрометром LTQ-Orbitrap XL (ThermoFisher Scientific) с помощью источника электрораспыления Ion Max (ThermoFisher). Образцы (5 мкг) инъецировали в колонку с обращенной фазой Poros R2 (Applied Biosystems) размером 2,1×30 мм и обрабатывали с использованием следующих условий градиента:0-3 мин:20% растворитель B; 3-6 мин:20-90% растворитель В; 6-7 мин:90-20% растворитель B; 7-9 мин:20% растворитель B. Растворитель А представлял собой дегазированный 0,1% водный раствор муравьиной кислоты, а растворитель В представлял собой дегазированный ацетонитрил. Скорость потока составляла 3 мл/мин. Поток был разделен после колонки, чтобы направить 100 мкл в электрораспылительный интерфейс. Колонку нагревали до температуры 82,5°C и растворители нагревались перед колонкой до 80°C для улучшения формы пика белка. LTQ-Orbitrap XL калибровали с использованием калибровочного раствора LTQ Positive Ion ESI фирмы ThermoFisher Scientific (кофеин, MRFA и Ultramark 1621) и настраивали с использованием растворов CsI 10 мг/мл. Напряжение на конусе (настройка фрагментации источника) составляло 48 В, разрешение FT составляло 7500, а диапазон развертки составлял 400-4,000 м/з. LTQ-Orbitrap XL был настроен для оптимального выявления больших белков (> 50 кДа).
Диапазоны, содержащие многозарядные ионы из полноразмерных антител (м/з 2000-3800) и полуантител (м/з 1400-2000) были отдельно деконволюционированы в профили молекулярного веса с использованием модуля MaxEnt 1 MassLynx, программного обеспечения для управления прибором и анализа данных (Waters). Вкратце, первичные данные ЖХ-МС белка были впервые открыты в QualBrower, модуле просмотра спектра Xcalibur (Thermo Scientific) и преобразованы для совместимости с MassLynx с использованием программы преобразования файлов Databridge, предоставляемой Waters. Преобразованные спектры белков были просмотрены в модуле Spectrum MassLynx и деконволюционированы с использованием MaxEnt 1. Количество каждого вида антител в каждом образце определяли непосредственно из полученных профилей молекулярного веса.
Анализ результатов ЖХ-МС
В целом, в большинстве случаев обработка дегликозилированием давала возможность установить все возможные различные виды антител, идентифицированные с помощью ЖХ-МС. Во многих случаях каждый вид антитела был представлен одним пиком ЖХ-МС. Исключения включают побочные пики, которые, вероятно, также соответствуют желаемым биспецифическим видам (возможно, аддукты или гетерогенность в расщеплении лидерных пептидов); однако, поскольку идентичность видов, приводящая к побочным пикам, была неясной, вклады этих побочных пиков по отношению к биспецифическим видам не учитывались. Кроме того, желаемые биспецифические виды, H1L1_H2L2, обычно не могут быть выделены экспериментально из неправильно спаренного типа H1L2_H2L1 на основе ЖХ-МС. Таким образом, когда содержание биспецифических антител предоставляется в таблицах, нельзя полностью исключить, что оно не содержит этот тип неправильно спаренных видов. Однако очень низкое содержание, наблюдаемое для таких видов, как H1L2_H1L2 и H2L1_H2L1, а также полуантител H1L2 и H2L1, свидетельствует о том, что произошло только незначительное, если таковое вообще имеет место, загрязнение биспецифических видов неправильно спаренными видами.
Оценка естественного смещения и влияния меток на спаривание в системах дикого типа
В качестве первого шага для каждой из трех тестируемых биспецифических систем (одна из легких цепей содержит метку, если это необходимо для достаточной массовой дифференциации между видами), немодифицированные тяжелые и легкие цепи дикого типа исходных антител для каждой системы были коэкспрессированы с использованием различных соотношений ДНК для H1:H2:L1:L2 и полученные виды антител были идентифицированы и количественно определены. Это позволило идентифицировать любое унаследованное (или введенное присутствием метки) смещение спаривания в каждой системе (например, если легкая цепь одного исходного антитела в биспецифической системе предпочтительно связывается с тяжелой цепью другого исходного антитела в биспецифической системе), а также позволило идентифицировать соотношение ДНК для H1:H2:L1:L2, которое обеспечивало наибольшее количество биспецифического антитела с наименьшим количеством полу-Ат. Соотношение ДНК было выбрано для компенсации естественных различий в уровнях экспрессии и/или внутренних смещений спаривания между тяжелыми и легкими цепями двух исходных антител и/или влияния метки для каждой системы. В тестируемых биспецифических системах A и B дикого типа L2 была помечена меткой FLAG. В биспецифической системе С ни одна из легких цепей не была помечена.
В таблице 12 показан результат этой оценки. Соотношение ДНК для H1:H2:L1:L2, которое обеспечивало наибольшее количество биспецифического антитела с наименьшим количеством полу-Ат, составляло 10:20:24:46 для системы A, 15:15:35:35 для системы B и 15:15:35:35 для системы C. Следует отметить, что в биспецифической системе Ат CAT-2200/пертузумаб (система A) наблюдалось смещение, где тяжелая цепь (H2) пертузумаба преимущественно спаривалась с легкой цепью (L1) CAT-2200. Это смещение было очевидным, когда рассматривались данные для эквивалентных соотношений тяжелой цепи 1 и тяжелой цепи 2, но при этом обратных соотношений легкой цепи 1 и легкой цепи 2 (например, рассматриваются обратные соотношения ДНК L1:L2 в H1:H2:L1:L2 при 8:22:17:53 и 8:22:53:17 в таблице 12). В условиях H1:H2:L1:L2=8:22:53:17, распространенные неправильно спаренные виды мАт H1L1_H2L1 являются намного более многочисленными (79,6%), чем в случае H1:H2:L1:L2=8:22:17:53, где распространенные неправильно спаренные виды H1L2_H2L2 составляли около 38,4%. Это указывало на то, что тяжелая цепь пертузумаба, по-видимому, преимущественно спаривалась с легкой цепью CAT-2200 в этой системе, когда легкая цепь пертузумаба имела метку FLAG.
Конструирование конструкций SMCA, содержащих N-концевые метки (HA или FLAG) на одной из легких цепей, способствовало получению аналогичных конструкций SMCA ДТ, которые исследовались на возможное влияние меток на способность легких цепей спариваться с тяжелыми цепями. В таблице 13 показаны результаты этого эксперимента и показано, что в некоторых случаях метка оказывала влияние на спаривание легких цепей с тяжелыми цепями. Например, в отношении случая системы А, отмеченного выше, где имело место преимущественное спаривание тяжелой цепи пертузумаба с легкой цепью CAT-2200, когда легкая цепь пертузумаба содержала метку FLAG, в случае если легкая цепь CAT-2200 содержала метку HA, а 4 полипептидные цепи экспрессировались при одинаковом соотношении ДНК, наблюдалось преимущественное спаривание тяжелой цепи CAT-2200 с легкой цепью пертузумаба (см. таблицу 13, "Система A, L1-HA" и сравните "Система A, L1-HA" с "Системой A, L2-FLAG" с тем же соотношением 10:20:24:46). Кроме того, для биспецифической системы SGN-CD19a/CAT-2200 (система B) степень неправильного спаривания тяжелой цепи SGN-CD19a с легкой цепью CAT-2200 в системе, которая содержит легкую цепь SGN-CD19a с меткой FLAG была больше, чем в двух других разновидностях одной и той же системы при одинаковом соотношении (таблица 13, "Система B, L1-FLAG" и противоположная "система B, L1-FLAG" с "системой B, L2-FLAG и L2-HA" при одинаковом соотношении 15:15:35:35). Следует отметить, что в таблице 13 отражены данные, собранные для всех выполненных повторений, поэтому процент видов может, в некоторых случаях, отличаться от данных на начальной стадии оценки, приведенных в таблице 12.
Для тестирования каждой из 40 репрезентативных конструкций в каждой биспецифической системе использовали соотношения ДНК H1:H2:L1:L2, соответствующие отношениям от соответствующих биспецифических систем дикого типа, которые давали наибольшее количество биспецифических видов Ат и наименьшее количество полу-Ат. Как указано выше, для системы CAT-2200/пертузумаб используемое соотношение составляло 10:20:24:46 (H1:H2:L1:L2), где H1 и L1 относятся к CAT-2200, а H2 и L2 относятся к пертузумабу. Для систем SGN-CD19a/CAT-2200 и SGN-CD19a/CR8071 использовалось соотношение 15:15:35:35 (H1:H2:L1:L2).
Результаты преимущественного спаривания в формате биспецифического антитела
Результаты SMCA с 40 репрезентативными конструкциями показаны в таблицах с 14 по 17. Таблицы с 14 по 17 относятся к наборам конструкций мАт как "конструкции" и идентифицированы 4-значным числом. Каждое 4-значное число соответствует уникальному идентификатору набора конструкций мАт (набор уникальных идентификаторов LCCA) в таблицах с 10-A1 по A12 и с 10-B1 по 10-B10. В таблице 24 представлена таблица соответствия между конструкциями, протестированными в SMCA и уникальные идентификаторы набора LCCA. Анализ преимущественного спаривания проводился на основе двух типов вычислений. Первый тип вычислений отмечен как "% спаривания H1L1 и % спаривания H2L2" в таблицах и представляет собой количество правильного спаривания между H1 и L1 и между H2 и L2 по всем наблюдаемым видам мАт и видам полу-Ат, включая виды мАт, которые могли иметь только одно правильно спаренное плече, в процентах от совокупного продукта (H1L1_H2L2_и_H1L2_H2L1+H1L1_H1L1+H2L2_H2L2+H1L1+H2L2+0,5 x (H1L1_H2L1+H1L1_H1L2+H1L2_H2L2+H2L1_H2L2)). Это вычисление называется вычислением "общего спаривания". Второй тип вычислений отмечен как "H1L1_H2L2 и H1L2_H2L1**" в таблицах и представляет собой количество правильного спаренного биспецифического антитела в процентах от всех наблюдаемых видов мАт (то есть, видов мАт с A по J на Фиг. 8) (H1L1_H2L2_и_H1L2_H2L1/(H1L1_H2L2_и_H1L2_H2L1+H1L1_H1L1+H2L2_H2L2+H1L1_H2L1+H1L2_H2L2+H1L1_H1L2+H2L1_H2L2+H1L2_H1L2+H2L1_H2L1)). Этот вычисление называется "общим биспецифическим" вычислением. В этом вычислении не учитывались полуантитела, поскольку, если они присутствуют, их можно удалить/свести к минимуму с помощью препаративной ЭХ (эксклюзионной хроматографии) или путем дальнейшего титрования ДНК H1:H2:L1:L2. В таблице 12 показано, что титрования ДНК эффективны для управления процентом экспрессированных видов полу-Ат. Эффективность очистки с помощью преп-ЭХ в снижении количества полу-Ат типа "Цепь A Fc" показана в примере 18.
Чтобы учесть смещения, которые наблюдались в некоторых биспецифических системах дикого типа, как описано выше, данные преимущественного спаривания были представлены как сравнение с соответствующими биспецифическими конструкциями дикого типа при одном и том же соотношении ДНК H1:H2:L1:L2. Это сравнение было представлено в столбцах "Изменение в % спаривания H1L1 и % спаривания H2L2 по отношению к дикому типу" (то есть изменение общего спаривания по отношению к ДТ) и "Изменение в H1L1_H2L2 и H1L2_H2L1** по отношению к дикому типу" (то есть изменение общего биспецифического (спаривания) по отношению к ДТ). Если соответствующая биспецифическая конструкция дикого типа не была оценена с помощью SMCA, проводилось сравнение с аналогичной конструкцией дикого типа. Эти случаи обозначаются как "‡" рядом с указанными значениями. Подобную конструкцию дикого типа для сравнения, выбирали следующим образом. Для каждой биспецифической системы и конструкции (с метками или без них) были взяты срединные значения для "H1L1_H2L2 и H1L2_H2L1**" и средние значения для "% спаривания H1L1 и % спаривания H2L2" из повторных экспериментов SMCA на соотношение ДНК. Были взяты наибольшие значения из любого соотношения в каждой конструкции, а среднее по всем конструкциям было использовано для сравнения с ДТ для конкретной системы (см. Фиг. 16A и 16B).
В таблицах 14 и 15 приводится краткое изложение результатов SMCA для тестируемых репрезентативных конструкций K-L, где конструкции были отсортированы в соответствии с "Изменением в % спаривания H1L1 и % спаривания H2L2 по отношению к дикому типу" (таблица 14) или "Изменением в H1L1_H2L2 и H1L2_H2L1** по отношению к дикому типу" (таблица 15). Аналогично, в таблицах 16 и 17 приводится краткое изложение результатов SMCA для тестируемых репрезентативных конструкций K-L, полученных из K-K, где конструкции были отсортированы в соответствии с изменением общего спаривания (таблица 16) или изменением общих биспецифических (таблица 17) вычислений.
Эффективность
Эффективность или способность способствовать преимущественному спариванию правильных тяжелых и легких цепей для каждой из репрезентативных конструкций оценивалась с использованием вычисления общего спаривания и использованием общего биспецифического вычисления. Во-первых, положительное значение в столбце "Изменение в % спаривания H1L1 и % спаривания H2L2 по отношению к дикому типу" (изменение общего спаривания по отношению к ДТ) показало, что набор конструкций мАт мог способствовать преимущественному спариванию. Во-вторых, положительное значение в столбце "Изменение в H1L1_H2L2 и H1L2_H2L1** по отношению к дикому типу" (изменение общего биспецифического по отношению к ДТ) также указывало на набор конструкций мАт, который мог способствовать преимущественному спариванию.
Все конструкции K-L и 18 из 21 конструкций K-L, полученных из K-K, показали сниженное количество первично неправильно спаренных видов (H1L2_H2L2 или H1L1_H2L1) по сравнению с диким типом, хотя в некоторых случаях количество различных неправильно спаренных видов было увеличено по сравнению с диким типом. В среднем, как конструкции K-L, так и конструкции K-L, полученные из K-K, вызывали наибольшее увеличение желаемого биспецифического по отношению к дикому типу (изменение общего биспецифического по отношению к ДТ) в системе A (48,8%), а затем в системе B (31,1%) и системе С (14,5%). В среднем, все тестируемые конструкции приводили к относительно незначительному увеличению общего количества полуантител по сравнению с диким типом, от 24,1% до 27,9%, но улучшали среднее соотношение спаренных полуантител к неправильно спаренным полуантителам от 1,5 до 6,9.
Переносимость
Способность репрезентативного набора конструкций мАт способствовать преимущественному спариванию во множестве систем была протестирована как показатель "переносимости" конструкций. Используя вычисление "изменения общего спаривания по отношению к ДТ", 29 из 33 конструкций K-L были переносимыми в 3 из 3 протестированных систем, а 3 конструкции были переносимыми в 2 из 3 протестированных систем (таблица 14). Другими словами, эти конструкции показали положительное значение "изменения спаривания по отношению к ДТ". Когда преимущественное спаривание измерялось с использованием вычисления "изменения общего биспецифического по отношению к ДТ", 24 из 33 конструкций K-L были идентифицированы как переносимые в 3 из 3 систем, а 8 конструкций были переносимыми в 2 из 3 систем (таблица 15). Для конструкций K-L, полученных из K-K, используя вычисление "изменения общего спаривания по отношению к ДТ" 2 из 7 конструкций были переносимыми в 3 из 3 системах, а 4 конструкции были переносимыми в 2 из 3 системах (таблица 16). Используя вычисление "изменения общего биспецифического по отношению к ДТ", 1 из 7 конструкций была переносимой в 3 из 3 системах, а 5 конструкций были переносимыми в 2 из 3 системах (таблица 17). 23 из 33 конструкций K-L и 1 из 7 конструкций K-L, полученных из K-K, были переносимыми в 3 из 3 протестированных систем при оценке с использованием обоих вычислений.
Что касается эффективности кластеров, все конструкции в 9 из 12 кластеров K-L были переносимыми в 3/3 системах в соответствии с вычислением "изменения общего спаривания по отношению к ДТ", а 4 из 12 в соответствии с вычислением "изменения общего биспецифического по отношению к ДТ". Все конструкции в кластерах kl-4, kl-9 и kl-12 были переносимыми в 3/3 системах с использованием обоих вычислений. На Фиг. 10 изображены блочные диаграммы, сравнивающие эффективность репрезентативных конструкций из каждого кластера на основе вычисления "изменения общего биспецифического по отношению к ДТ" (Фиг. 10А, изменение биспецифического%) или на основе вычисления "изменения общего спаривания по отношению к ДТ" (Фиг. 10B, изменение спаривания%). Для кластеров K-L, полученных из K-K, была протестирована только одна конструкция для каждого кластера, и поэтому переносимость кластера была эквивалентна рассмотренной выше переносимости конструкции.
В целом, конструкции K-L способствовали преимущественному спариванию более эффективно, чем конструкции K-L, полученные из K-K. На Фиг. 11 показаны блочные диаграммы, изображающие способность конструкций K-L и конструкций K-L, полученных из K-K, способствовать преимущественному спариванию в каждой тестируемой системе на основе вычисления "изменения общего биспецифического по отношению к ДТ". Например, на Фиг. 11 показано, что конструкции K-L обычно способствуют большему увеличению количества биспецифического антитела по сравнению с ДТ, чем конструкции K-L, полученные из K-K (сравнивая значение "все kl" с значением "все kk" для каждой системы). Это также верно при рассмотрении только конструкций, переносимых в 2 или более или в 3 три системы.
Пример 17: Препаративная эксклюзионная хроматография (ЭХ) выбранных биспецифических антител SMCA и исходных мАт для биофизического исследования.
Для оценки биофизических характеристик биспецифических антител, полученных в SMCA, описанной в примере 16, образцы SMCA, очищенные с помощью белка A, из подмножества тестируемых репрезентативных конструкций, были подвергнуты препаративной ЭХ для удаления видов полу-Ат.для этого этапа были выбраны образцы SMCA (36 в общей сложности), имеющие, по меньшей мере, 60% "H1L1_H2L2 и H1L2_H2L1**" (общий биспецифический). Некоторые образцы SMCA, не соответствующие этому ограничению, также были включены для полноты. Например, в тех случаях, когда образцы SMCA для данной конструкции соответствовали этому ограничению в двух из трех тестируемых биспецифических системах, и образец SMCA соответствовал этому ограничению, но не третьему, тогда для всех трех систем образцы SMCA были включены. Препаративную ЭХ проводили следующим образом. Виды антител в образцах SMCA были разделены с использованием колонки Superdex 200 10/300 Increase (GE Healthcare), установленной на системе GE Healthcare ÄKTA Avant 25, оборудованной автоматическим дозатором ALIAS Bio Cool (Spark-Holland), используемым для ввода образцов в колонку. Образцы SMCA (0,9 мл) в ФСБ, pH 7,4 (Hyclone DPBS/модифицированный, без кальция, без магния, кат. №SH-300028. 02) автоматически загружали в петлю объемом 2 мл, заполненную ФСБ. Затем образцы автоматически вводили в колонку и растворяли при 0,5 мл/мин с объемом элюирования 1 CV. Элюирование белками контролировали при ОП280 и собирали в 0,5 мл фракции. Для каждого образца SMCA фракции, которые составляли основной пик (фракции оценивали с помощью Калипера), объединяли и далее характеризовали биофизические параметры, как описано в примерах 19 и 20.
Пример 18: Удаление полу-антител с помощью препаративной эксклюзионной хроматографии
В примере 16 было отмечено, что полу-Ат исключаются из вычислений спаривания в соответствии с "общим биспецифическим" вычислением, так как процентным содержанием получаемых полу-Ат можно управлять либо путем титрования ДНК, либо, в случае полу-Ат типа "Цепь A Fc", полу-Ат могут быть удалены/сведены к минимуму с помощью очистки препаративной ЭХ образцов SMCA, очищенных с помощью белка A. Для демонстрации факта удаления "полу-Ат цепи A Fc" очищенные образцы SMCA из примера 17 для двух конструкций K-L во всех трех биспецифических системах были подвергнуты анализу ЖХ-МС, как описано в примере 16. Две выбранные конструкции K-L представляли собой 2901 и 3972 (как указано в таблице 11A). Результаты показаны в таблице 18.
Сравнение данных ЖХ-МС в таблице 18 с соответствующими данными ЖХ-МС для образцов SMCA, очищенных с помощью белка A, в таблице 14 показывает, что препаративная ЭХ может быть эффективной при удалении/минимизации видов полу-Ат типа "Цепь A Fc" (H1L1 в 2901 и 3972). Во всех тестируемых образцах наблюдалось увеличение процентного содержания желаемых видов биспецифических антител (H1L1_H2L2 и H1L2_H2L1), а также уменьшение процента видов полуантител типа Цепь А Fc (H1L1 в таблице 18).
Пример 19: Термическая стабильность биспецифических антител SMCA.
После препаративной ЭХ была измерена термическая стабильность очищенных с помощью преп-ЭХ биспецифических антител SMCA, и проведено сравнение с исходными моноклональными антителами CAT-2200, пертузумабом, CR8071 и SGN-CD19a с целью определения, повлияла ли замена аминокислот на термическую стабильность.
Измерение термической стабильности
Термическая стабильность выбранных биспецифических гетеродимерных антител и образцов контроля дикого типа была измерена с помощью дифференциальной сканирующей калориметрии (ДСК) следующим образом. После препаративной обработки ЭХ 400 мкл образцов в основном с концентрациями 0,4 мг/мл в ФСБ использовались для ДСК анализа с VP-капиллярным ДСК (GE Healthcare). В начале каждого прогона ДСК 5 холостых введений буфера были выполнены для стабилизации исходного уровня, и введение буфера наносили перед каждым вводом пробы для сравнения. Каждый образец сканировали от 20 до 100°С со скоростью 60°С/ч, с ограниченной обратной связью, 8-секундным фильтром, 3-минутным preTstat и давлением азота 4,9 килограмм на квадратный сантиметр. Полученные термограммы проверялись и анализировались с использованием программного обеспечения Origin 7.
Результаты показаны в таблице 20 и таблице 21. Значения Tm Fab дикого типа, используемые для расчета "Средней разницы в Tm Fab по сравнению с диким типом (°C)", как указано в таблицах, были получены из ДСК-термограмм исходных антител CAT-2200 (71,2°C), пертузумаба (76,7°C), CR8071 (66,9°C, соответствующей первичному пику) и SGN-CD19a (91.0°C). Исходное антитело CR8071 показало два температурных перехода для Fab. Для исходных антител, наблюдались дополнительные температурные переходы для доменов CH2 и CH3 Fc (при, приблизительно, 71°C и 82°C, соответственно), где они не перекрывались с переходом для Fab (Фиг. 12). Некоторые конструкции с большим количеством полуантител H2L2 "цепь B Fc" показывали дополнительный температурный переход при, приблизительно, 60°C, вероятно, из-за присутствия нековалентных гомодимеров. В некоторых случаях конструкции приводили к снижению Tm Fab пертузумаба, что приводило к перекрытию пиков Fab CAT-2200, Fab пертузумаба и CH2 в этих образцах (Фиг. 12A, репрезентативная конструкция 3972). Подобным образом, многие конструкции снизили Tm Fab SGN-CD19a до такой степени, что привели к перекрытию пиков CH3 в этих антителах (Фиг. 12B и 12C, репрезентативная конструкция 3972). Такие перекрытия привели к некоторой двойственности в определении Tm составляющих компонентов Fab, и, следовательно, значения Tm, указанные в таблицах 20 и 21, считаются приблизительными. Примечательно, что биспецифические антитела CR8071/SGN-CD19a, содержащие конструкции, показывали один переход для Fab CR8071 вместо двух переходов, наблюдаемых в биспецифических антителах дикого типа (например, Фиг. 12C).
Из всех проверенных конструкций K-L средняя разница в Tm Fab по сравнению с диким типом во всех системах составляла от 0°C до -1,5°C для 6 конструкций, от -1,5°C до -3,0°C для 11 конструкций и от -3,0°C до -4,5°C для 15 конструкций (таблица 20). Из всех проверенных конструкций K-L, полученных из K-K, средняя разница составляла от 0°C до -1,5°C для 2 конструкций, от -1,5°C до -3,0°C для 1 конструкции и от -3,0°C до -4,5°C для 1 конструкции (таблица 21). Средние разницы в Tm Fab по сравнению с диким типом среди проверенных конструкций составляли -0,9°C для CAT-2200 (STDEV=1,4), -4,6°C для пертузумаба (STDEV=1,6), -5,8°C для SGN-CD19a (STDEV=3,5) и+1,1°C для CR8071 (STDEV=0,8). На Фиг. 13 изображена блочная диаграмма изменения Tm Fab по сравнению с исходным антителом для тестируемых конструкций, для всех тестируемых конструкций ("Все") или с помощью антигенсвязывающего центра (CAT-2200, пертузумаба, CR8071 или SGN-CD19a).
Пример 20: Измерения аффинности к антигену биспецифических антител
Была оценена способность биспецифических антител связываться с соответствующими антигенами с целью определения, повлияла ли замена аминокислот на связывание антигена. Аффинность связывания антигена определяли с помощью ППР следующим образом.
Биосенсорные анализы ППР
Для исследований с Biacore T200:Сенсорный чип CM5 серии S, набор связывания амина Biacore (NHS, EDC и 1 М этаноламин) и 10 мМ натрий-ацетатные буферы были приобретены у GE Healthcare Life Science (Миссиссога, штат Онтарио, Канада). Для исследований с Biorad ProteOn:Сенсорные чипы GLC, набор связывания амина Biorad ProteOn (EDC, sNHS и этаноламин) и 10 мМ натрий-ацетатные буферы были приобретены у Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио). Электронный буфер ФСБ с 0,05% Твин 20 (ФСБТ) был приобретен у Teknova Inc. (Холлистер, штат Калифорния). Козьи поликлональные антитела к человеческому Fc были приобретены у компании Jackson Immuno Research Laboratories Inc. (Вест Гроув, штат Пенсильвания). Антигены:His-меченый рекомбинантный человеческий CD-19 был приобретен у Abcam (Кембридж, Великобритания) и His-меченый гемагглютинин вируса гриппа типа B (B/Brisbane/60/2008) был приобретен у Sino Biological (Пекин, Китай). Белок рекомбинантного внеклеточного домена (ECD) Her2 был приобретен у eBioscience (Сан Диего, штат Калифорния). Рекомбинантный человеческий ИЛ-17A был приобретен у R&D Systems (Миннеаполис, штат Миннесота).
Анализы поверхностного плазмонного резонанса (ППР) с антигенами Гемагглютинином и CD-19 проводились с использованием прибора Biacore T200 (GE Healthcare) с электродным буфером ФСБТ (с 0,5 М стоковым раствором ЭДТА, добавленным до конечной концентрации 3,4 мМ) при температуре 25°C. Анализы поверхностного плазмонного резонанса с антигенами HER2 ECD и ИЛ-17 проводились с использованием прибора BioRad ProteOn XPR36 (Bio-Rad Laboratories (Канада) Ltd. (Миссиссога, штат Онтарио)) с электродным буфером ФСБТ при температуре 25°C.
Определение аффинности SGN-CD19a:CD-19 проводилось следующим образом. Скрининг антител (образцов SMCA, очищенных с помощью препаративной ЭХ, и контролей OAA) на наличие связывания с антигеном CD-19 происходил в два этапа: непрямой захват His-меченого CD-19 на поверхности анти-His IgG с последующей инъекцией пяти концентраций очищенного антитела для кинетического анализа с использованием кинетического титрования. Поверхность захвата анти-His была приготовлена на сенсорном чипе CM5 серии S с помощью связывания амина при приблизительно 12000 РЕ для анти-His IgG (набор для захвата гистидина, GE Healthcare) в тестируемых и контрольной проточных ячейках в соответствии с инструкциями производителя. CD-19 вводили 10 мкг/мл через тестируемую проточную ячейку в течение 60 секунд при скорости потока 10 мкл/мин. В целом, это привело к захвату приблизительно от 250 РЕ CD-19 на поверхности анти-His IgG. CD-19 не был захвачен в контрольной (пустой) проточной ячейке. За этапом захвата следует введение пяти концентраций очищенного антитела (200 нМ и 2-кратные разведения, 80 нМ и 2-кратные разведения или 40 нМ и 2-кратные разведения; в зависимости от антитела), которые последовательно вводились как в тестируемые, так и в контрольную проточные ячейки по 40 мкл/мин в течение 180 секунд с фазой диссоциации 600 секунд. Захваченные поверхности антитела восстанавливали с помощью 10 мМ глицина с рН 1,5 в течение 120 секунд при 30 мкл/мин с целью подготовки поверхностей для следующего цикла введения. По меньшей мере, два введения буферного контроля выполнялись для каждого введения анализируемого вещества и использовались для сравнения. Полученные в результате кинетического титрования сенсограммы были проанализированы с помощью программного обеспечения Biacore T200 BiaEvaluation версии 3.0 и соответствовали модели связывания 1:1.
Определение аффинности CR8071:Гемагглютинин проводилось следующим образом. Гемагглютинин (НА) разбавляли в 10 мМ ацетатном буфере с рН 5,5 и непосредственно иммобилизировали с помощью связывания амина на сенсорном чипе CM5 серии S. Это привело к приблизительно 120-130 РЕ иммобилизованного НА. Контрольная проточная ячейка оставалась пустой (блокированная этаноламином) для использования в качестве холостой пробы. Антитела вводились на поверхность НА для кинетического анализа с использованием кинетического титрования. Пять концентраций (40 нМ и 2-кратные разведения) очищенного антитела (образцы SMCA, очищенные с помощью препаративной ЭХ, и контроли OAA) последовательно вводили как в тестируемые, так и в контрольную проточные ячейки по 50 мкл/мин в течение 300 секунд с фазой диссоциации 3600 секунд. Поверхности HA восстанавливали с помощью 2 циклов использования 10 мМ глицина с рН 1,5 в течение 120 секунд при 30 мкл/мин с целью подготовки поверхностей для следующего цикла введения. По меньшей мере, два введения буферного контроля выполнялись для каждого введения анализируемого вещества для использования в качестве контроля. Полученные в результате кинетического титрования сенсограммы были проанализированы с помощью программного обеспечения Biacore T200 BiaEvaluation версии 3.0 и соответствовали модели связывания 1:1.
Определение аффинности CAT-2200:ИЛ-17 проводилось, как описано ранее в примере 6, со следующими модификациями: тестируемые антитела представляли собой образцы SMCA, очищенные с помощью препаративной ЭХ, и контроли OAA вместо фрагментов Fab CAT-2200. Одно из контрольных антител 13612 в начале вводили при 10 нМ вместо 60 нМ.
Пертузумаб: Определение аффинности к HER2 проводилось следующим образом. Все линии были горизонтально иммобилизованы с 25 мкг/мл козьим антителом к IgG человека,γ специфическим к фрагменту Fc (к Fc человека). В среднем было зарегистрировано 4058 резонансных единиц (РЕ). Поверхность захвата антитела к Fc человека была получена с использованием сенсорного чипа GLC, активированного разбавлением 1:10 стандартных растворов sNHS/EDC BioRad, введенных в течение 140 секунд при 100 мкл/мин в (горизонтальном) направлении анализируемого вещества. Сразу же после активации, 25 мкг/мл раствора антитела к Fc человека в 10 мМ NaOAc с pH 4,5 вводили в (горизонтальном) направлении анализируемого вещества при скорости потока 25 мкл/мин в течении 240 секунд до иммобилизации приблизительно 4000 резонансных единиц (РЕ). Оставшиеся активные группы блокировали 140-секундной инъекцией 1 М этаноламина при 30 мкл/мин в течении 300 секунд также в направлении анализируемого вещества, с целью гарантии создания активированных имитатором точек для сравнения. Скрининг антител на наличие связывания с HER2 происходил в два этапа: непрямой захват антител (образцов SMCA, очищенных с помощью препаративной ЭХ, и контролей OAA) на поверхности антитела к IgG человека (специфического к фрагментуγ Fc) в направлении лиганда с последующим одновременным введением 5 концентраций очищенного ECD HER2 и одного буферного контроля для двойной проверки в направлении анализируемого вещества. Во-первых, для стабилизации исходной линии использовали одно введение буфера в течение 30 секунд при 100 мкл/мин в направлении лиганда. Для каждого захвата антитела, антитела разбавляли до 2 мкг/мл в ФСБТ. От одного до пяти антител или контролей одновременно вводили в отдельные каналы лигандов в течение 480 с при потоке 25 мкл/мин. Это привело к захвату приблизительно 800-1200 РЕ на поверхности антитела к Fc человека. Первый канал лиганда оставался пустым для использования в качестве холостой пробы, если это было необходимо. Этот этап захвата немедленно сопровождался одной инъекцией буфера в направлении анализируемого вещества для стабилизации исходной линии, а затем 100 нМ, 33,3 нМ, 11,1 нМ, 3,7 нМ и 0,41 нМ HER2 вместе с буферным контролем одновременно вводили при 50 мкл/мин в течение 120 секунд с 300-секундной фазой диссоциации. Захваченные поверхности антитела восстанавливали с помощью двух выбросов 0,85% фосфорной кислоты в течение 18 с при 100 мкл/мин с целью подготовки для следующего цикла введения. Сенсограммы были выровнены и дважды проверенны с использованием контрольного введения буфера и точек, и полученные сенсограммы были проанализированы с использованием программного обеспечения ProteOn Manager v3,1. Дважды проверенные сенсограммы соответствовали модели связывания Ленгмюра.
Антигенную аффинность гетеродимерных антител оценивали с учетом соответствующего контроля OAA (однорукого, one-armed) дикого типа: Аффинности к антигену также были получены для биспецифических антител дикого типа; однако, захват с помощью ППР биспецифических антител ДТ может быть гетерогенным (например, включать захват неправильно спаренных гетеродимеров), тем самым препятствуя определению KD. Кроме того, аффинности для дикого типа измеряли в форматах ОАА, содержащих соответствующие метки легкой цепи, присутствующие в антителах. Наблюдалось, что наличие меток понижало аффинность SGN-CD-19a к CD19 в целых 4-5 раза (таблица 19). Например, при добавлении метки FLAG к легкой цепи, измеренная аффинность связывания SGN-CD-19a с CD19 уменьшилась с 67,5 нМ до 224,5 нМ (таблица 19). Следовательно, все расчеты изменения аффинности по отношению к дикому типу были выполнены с использованием медианных значений меченных конструкций дикого типа, подходящих для спаривания.
При измерении связывания с CD-19, сенсограммы некоторых антител не соответствовали модели связывания 1:1. В таких случаях антитела содержали большое количество полуантител "цепь B Fc", которые склонны к гомодимеризации, что вызывало эффекты авидности и скрывало состояние связывания Fab. Для антител, поддавшихся связыванию, аффинности связывания были дополнительно оценены в формате ОАА.
Большинство протестированных конструкций (сделаны 36 конструкций, соответствующие 96 биспецифическим антителам) показали значения KD, сравнимые со значениями соответствующего дикого типа (таблица 20 и 21, Фиг. 14). Девять сконструированных Fab демонстрировали сниженную аффинность в 2-3 раза по сравнению с ДТ, и в одном сконструированном Fab наблюдалось снижение аффинности в 4 раза. Из этих 10 Fab только одна конструкция (#34) присутствовала дважды, и 8 из 10 затронутых Fab представляли CR8071. Средняя разница в log(KD) ДТ (-(log KD_антитела- log KD_дт)) для всех "наконечников", имеющих конструкции, составляла 0,003 со стандартным отклонением 0,187 (например, -0,3 соответствует 2-кратному снижению аффинности, -0,6 соответствует 4-кратному снижению аффинности). Эти результаты подытожены на Фиг. 14, где изображена блочная диаграмма изменения аффинности по сравнению с исходным антителом для тестируемых конструкций, для всех тестируемых конструкций ("Все") или с помощью антигенсвязывающего центра (CAT-2200, пертузумаба, CR8071 или SGN-CD19a).
Пример 21: Профили сверхэффективной жидкостной хроматографии эксклюзионной хроматографии (СВЭЖХ-ЭХ) для разработанных биспецифических антител сравнивались с исходными антителами.
Для оценки качества сконструированных биспецифических антител, антитела, очищенные с помощью препаративной ЭХ, как в Примере 17, подвергали СВЭЖХ-ЭХ. СВЭЖХ-ЭХ анализ осуществляли с использованием колонки ЭХ Waters BEH200 (2,5 мл, 4,6×150 мм, нержавеющая сталь, 1,7 мкм частицы) установленной на 30°C и монтировали на системе Waters Acquity UPLC с ФДМ детектором. Время разгона составляло 7 мин, и общий объем на инъекцию составлял 2,8 мл с электронным буфером ФСБ и 0,02% Твин 20 с pH 7,4 при 0,4 мл/мин. Элюирование контролировали с помощью УФ-поглощения в диапазоне 210-400 нм, и хроматограммы экстрагировали при 280 нм. Интегрирование пика осуществляли с помощью программного обеспечения Empower 3.
На Фиг. 15A-D изображены профили СВЭЖХ-ЭХ исходных антител, а на Фиг. 15E-G изображены профили, соответствующие антителам конструкции 3972 в трех биспецифических системах. Типовая конструкция 3972 использовалась в данном документе, чтобы продемонстрировать, что сконструированные/разработанные биспецифические антитела с высоким биспецифическим содержанием (>80%) имеют профили СВЭЖХ-ЭХ, которые сопоставимы с профилями исходных антител. Для таких сконструированных биспецифических антител средний процент мономерных видов составляет 95,2% (то есть, было обнаружено мало или вообще не обнаружено виды с высокой молекулярной массой).
Все ссылки, выданные патенты, патентные заявки и номера доступа последовательностей (например, номера доступа Genbank) приведенные в тексте данного описания изобретения, таким образом, включены посредством ссылки во всей своей полноте во всех отношениях.
Таблица 1: Критерии для модели Fab
Использует обычно подгруппы VH и VL
Каркас близкий к зародышевой линии
3D-структура представляет типичную конформацию каппа или лямбда (низкое, попарно вычисленное СКО)
Отсутствие серьезных структурных изменений, наблюдаемых при связывании антигена
Таблица 2: Положения "горячих" точек аминокислот (отмеченные звездочкой) в области контакта тяжелой и легкой цепей в D3H44 (1JPT, типичный Fab, содержащий легкую цепь каппа) и CAT-2200 (2VXS, типичный Fab, содержащий легкую цепь лямбда).
1JPT
Таблица 3А. Нумерация аминокислотных последовательностей тяжелой цепи Fab D3H44, Пертузумаба и CAT-2200 по Кабату.
(Seq ID No. 1)
(Seq ID No. 3)
Таблица 3В. Нумерация аминокислотных последовательностей легкой цепи D3H44, Пертузумаба и CAT-2200 по Кабату.
(Seq ID No. 2)
(Seq ID No. 4)
Таблица 3C. Последовательности аминокислот и ДНК
Таблица 4A: Библиотека конструкций K-L
Таблица 4B - Библиотека конструкций K-L, полученных из K-K
* Конструкции с точными соответствиями мутаций библиотеки конструкций каппа-каппа.
Таблица 5A: Результаты LCCA для конструкций K-L, которые прошли критерии исключения по эффективности
Таблица 5B: Результаты LCCA для конструкций K-L, полученных из K-K, которые прошли критерии исключения по эффективности
Таблица 6A: Оценка связывания антигена и термической стабильности конструкций K-L, которые прошли критерии исключения по эффективности LCCA
Таблица 6B: Оценка связывания антигена и термической стабильности конструкций K-L, полученных из K-K, которые прошли критерии исключения по эффективности LCCA
Таблица 7A: Вторая библиотека конструкций K-L
Таблица 7B - Вторая библиотека конструкций K-L, полученных из K-K
Таблица 8A: Результаты LCCA для конструкций K-L из второй библиотеки конструкций K-L, которые прошли критерии исключения по эффективности
значения
H2L2:H2L1
Таблица 8B: Результаты LCCA для конструкций K-L, полученных из K-K, из второй библиотеки конструкций K-L, полученных из K-K, которые прошли критерии исключения по эффективности
1
6
7
13064
13425
13424
13469
13471
13470
13472
13340
13093
13079
13079
13444
13444
13443
13451
13438
13106
13108
13461
13462
13341
Таблица 9A - Оценка связывания антигена и термической стабильности конструкций K-L, которые прошли критерии исключения по эффективности LCCA
H1L1 Fab представляет CAT-2200 и H2L2 Fab представляет Пертузумаб.
Ряд гетеродимеров имел связанный с ними осадок, отмеченный как "*" в таблицах 9А и 9В, если осадок был связан с Fab H1L1; отмеченный как ʺ†" в таблицах 9А и 9В, если осадок был связан с Fab H2L2; и отмеченный как "‡" в таблицах 9А и 9В, если осадок был связан как с Fab H1L1, так и с Fab H2L2. # - указывает на наличие плеча к основному пику Fab (переход) в термограмме ДСК
Таблица 9B - Оценка связывания антигена и термической стабильности конструкций K-L, полученных из K-K, которые прошли критерии исключения по эффективности LCCA
H1L1 Fab представляет CAT-2200 и H2L2 Fab представляет Пертузумаб.
Ряд гетеродимеров имел связанный с ними осадок, отмеченный как "*" в таблицах 9А и 9В, если осадок был связан с Fab H1L1; отмеченный как "†" в таблицах 9А и 9В, если осадок был связан с Fab H2L2; и отмеченный как "‡" в таблицах 9А и 9В, если осадок был связан как с Fab H1L1, так и с Fab H2L2. # - указывает на наличие плеча к основному пику Fab (переход) в термограмме ДСК
Таблица 10-A1 Кластер 1 конструкций K-L
Таблица 10-A2 Кластер 2 конструкций K-L
Таблица 10-A3 Кластер 3 конструкций K-L
Таблица 10-A4 Кластер 4 конструкций K-L
Таблица 10-A5 Кластер 5 конструкций K-L
Таблица 10-A6 Кластер 6 конструкций K-L
Таблица 10-A7 Кластер 7 конструкций K-L
Таблица 10-A8 Кластер 8 конструкций K-L
Таблица 10-A9 Кластер 9 конструкций K-L
Таблица 10-A10 Кластер 10 конструкций K-L
Таблица 10-A11 Кластер 11 конструкций K-L
Таблица 10-A12 Кластер 12 конструкций K-L
В таблицах с 10-A1 по 10-A12, Fab H1L1 представляет CAT-2200, а Fab H2L2 представляет Пертузумаб
Таблица 10-B1 Кластер 1 конструкций K-L, полученных из K-K
Таблица 10-B2 Кластер 2 конструкций K-L, полученных из K-K
Таблица 10-B3 Кластер 3 конструкций K-L, полученных из K-K
Таблица 10-B4 Кластер 4 конструкций K-L, полученных из K-K
Таблица 10-B5 Кластер 5 конструкций K-L, полученных из K-K
Таблица 10-B6 Кластер 6 конструкций K-L, полученных из K-K
Таблица 10-B7 Кластер 7 конструкций K-L, полученных из K-K
Таблица 10-B8 Кластер 8 конструкций K-L, полученных из K-K
Таблица 10-B9 Кластер 9 конструкций K-L, полученных из K-K
Таблица 10-B10 Кластер 10 конструкций K-L, полученных из K-K
В таблицах с 10-B1 по 10-B10, Fab H1L1 представляет собой CAT-2200, а Fab H2L2 представляет собой Пертузумаб.
Таблица 11A: Аминокислотные мутации в областях Fab H1, L1, H2 и L2 для конструкций K-L, протестированных в формате SMCA (нумерация по Кабату).
Таблица 11B: Аминокислотные мутации в областях Fab H1, L1, H2 и L2 для конструкций K-L, полученных из K-K, протестированных в формате SMCA (нумерация по Кабату).
Таблица 12: Влияние титрования соотношения ДНК на процент видов антител, как оценено с помощью ЖХ-МС, выбранных систем дикого типа
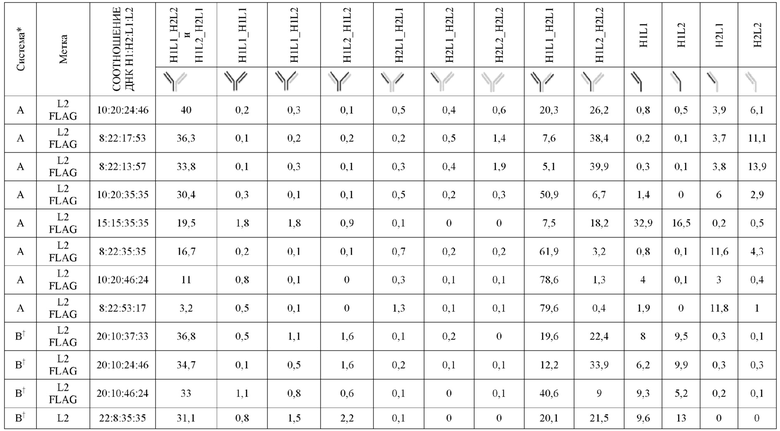
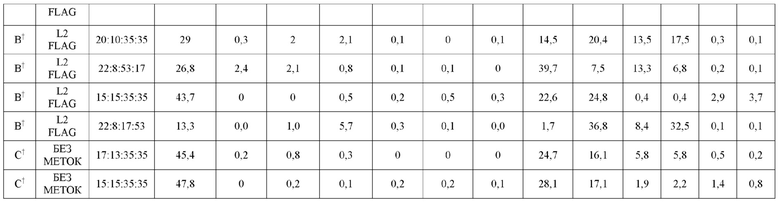
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба; Система B: H1/L1 SGN-CD19a и H2/L2 CAT-2200; Система C: H1/L1 SGN-CD19a и H2/L2 CR8071;
†Системы B† и C† для контроля диким типом отличаются от систем B и C конструкций в ориентации "наконечников" H1L1 против H2L2.
Таблица 13: Данные ЖХ-МС о спаривании и количестве продукта после очистки белком А (мг/л) для конструкций антител дикого типа и влияние меток на спаривание
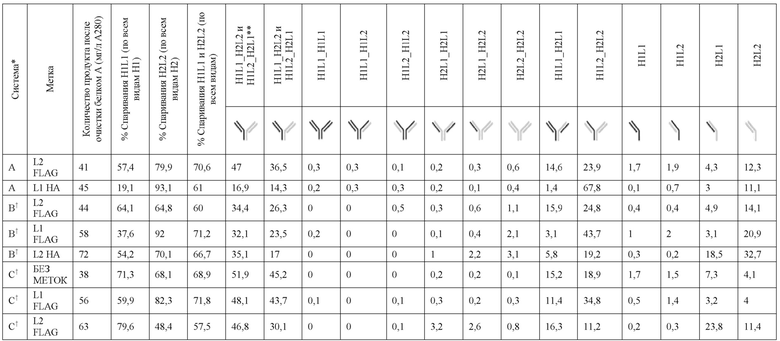
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 SGN-CD19a и H2/L2 CAT-2200 (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 SGN-CD19a и H2/L2 CR8071 (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2 );
Множественные эксперименты ЖХ-МС SMCA были выполнены для некоторых конструкций. В этих случаях сообщаются срединные значения спаривания и срединные количества каждого вида SMCA;
** с учетом только полных видов Ат;
†Системы B† и C† для контроля диким типом отличаются от систем B и C конструкций в ориентации "наконечников" H1L1 против H2L2.
Таблица 14: Данные ЖХ-МС о спаривании и количестве продукта после очистки белком А (мг/л) для конструкций K-L (общее спаривание)
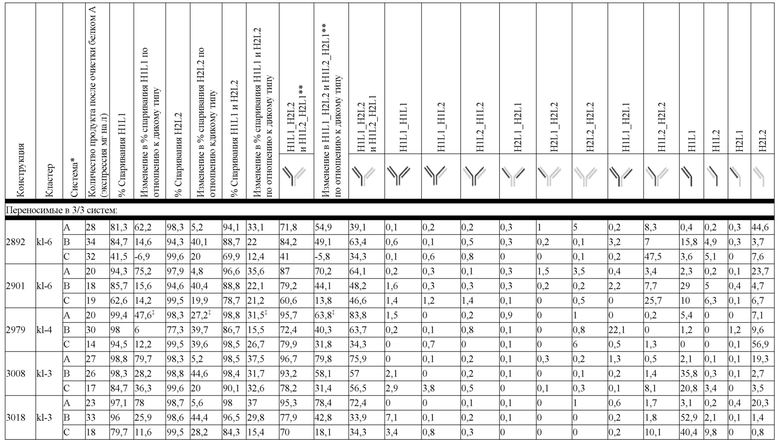
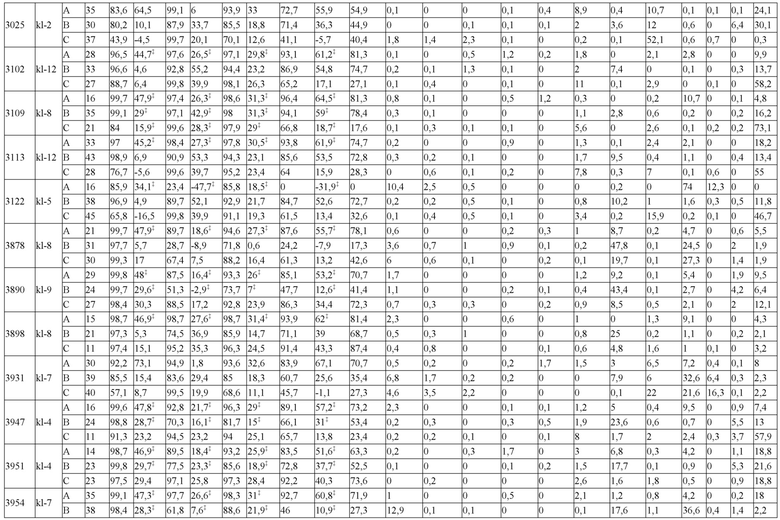
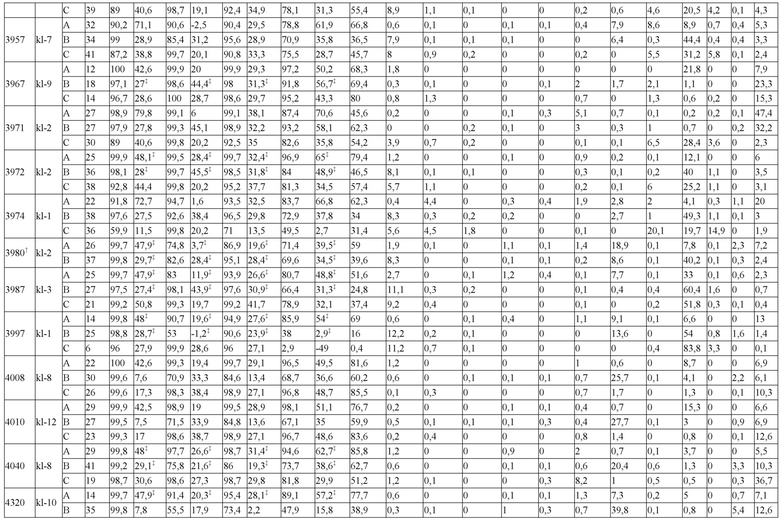
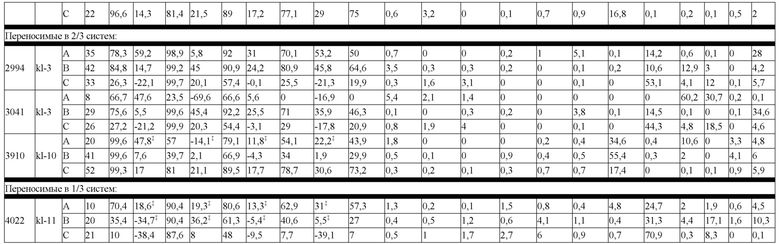
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2 );
** С учетом только полных видов Ат;
Конструкции заштрихованы с увеличением серого в соответствии с их переносимостью в 3/3 (не заштрихованы), 2/3 или 1/3 систем, основанной на положительном Изменении в % спаривания H1L1 и H2L2 по отношению к дикому типу;
‡ Оценочное изменение по отношению к дикому типу;
† Конструкцию 3980 оценивали только в 2-х системах.
Таблица 15: Данные ЖХ-МС о спаривании и количестве продукта после очистки белком А (мг/л) для конструкций K-L (общее биспецифическое)
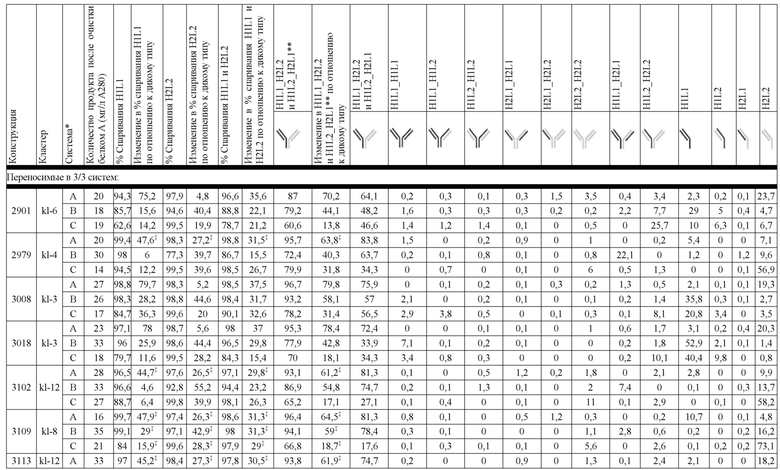
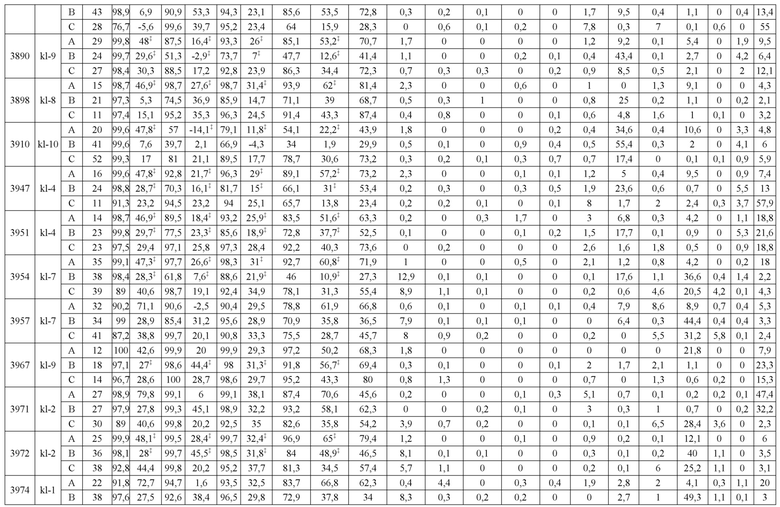
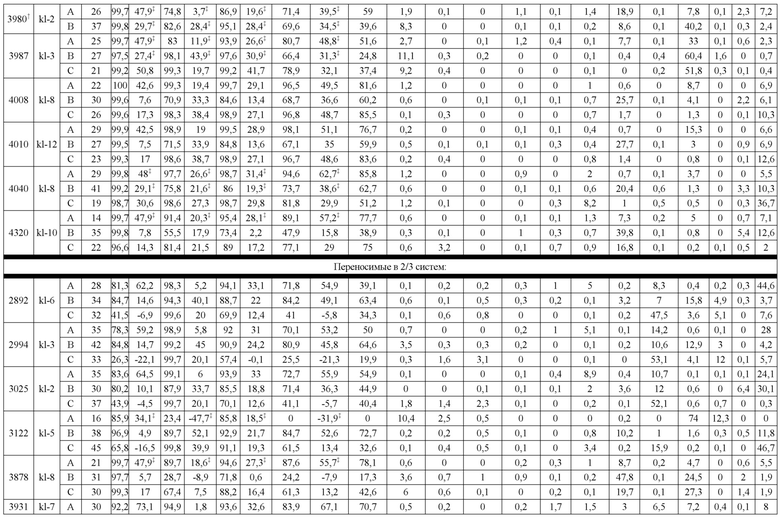
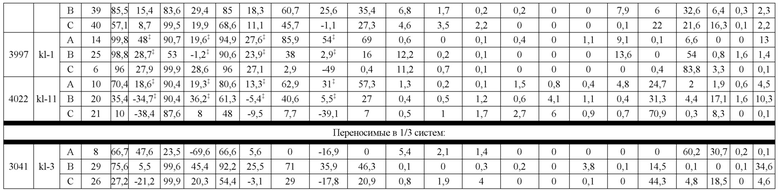
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2);
** С учетом только полных видов Ат;
Конструкции заштрихованы в соответствии с их переносимостью в 3/3 (не заштрихованы) или 2/3 (светло-серые) систем, основанной на положительном Изменении в H1L1_H2L2 и H1L2_H2L1 ** по отношению к дикому типу;
‡ Оценочное изменение по отношению к дикому типу;
† Конструкцию 3980 оценивали только в 2-х системах.
Таблица 16: Данные ЖХ-МС о спаривании и количестве продукта после очистки белком А (мг/л) для конструкций K-L, полученных из K-K (общее спаривание)
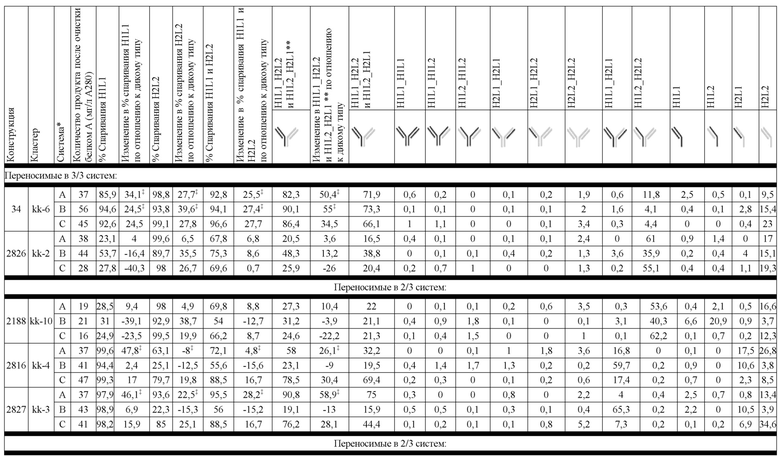
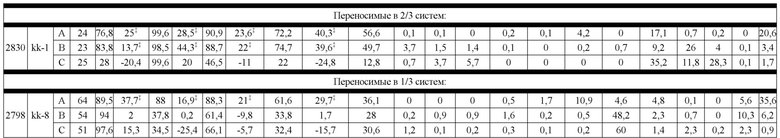
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2);
** С учетом только полных видов Ат.
Конструкции заштрихованы с увеличением серого в соответствии с их переносимостью в 3/3 (не заштрихованы), 2/3 или 1/3 систем, основанной на положительном Изменении в % спаривания H1L1 и H2L2 по отношению к дикому типу;
‡ Оценочное изменение по отношению к дикому типу.
Таблица 17: Данные ЖХ-МС о спаривании и количестве продукта после очистки белком А (мг/л) для конструкций K-L, полученных из K-K (общее биспецифическое)
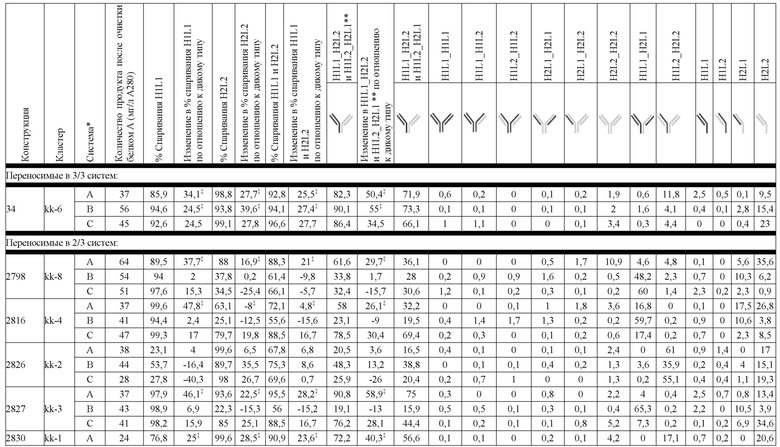
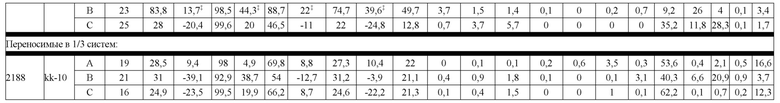
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2);
** С учетом только полных видов Ат;
Конструкции заштрихованы с увеличением серого в соответствии с их переносимостью в 3/3 (не заштрихованы), 2/3 или 1/3 систем, основанной на положительном Изменении в H1L1_H2L2 и H1L2_H2L1 ** по отношению к дикому типу;
‡ Оценочное изменение по отношению к дикому типу.
Таблица 18: Данные спаривания, полученные после преп-ЭХ ЖХ-МС для выбранных конструкций в трех системах
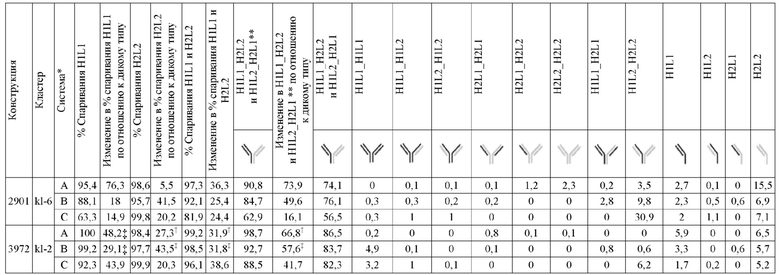
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2);
** С учетом только полных видов Ат;
‡ Оценочное изменение по отношению к дикому типу.
Таблица 19: Данные связывания антигена для антигенсвязывающих центров дикого типа в формате ОАА
* Обозначает другой способ захвата.
Таблица 20: Аффинность связывания антигена и оценка термической стабильности для выбранных конструкций K-L
(° C)††
(° C)††
(° C)‡‡
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2);
Конструкции сгруппированы и затенены тремя градиентами с "Средней разницей в Tm Fab (по системам) от дикого типа (° C)" от 0 до -1,5, от -1,5 до -3,0 и менее -3;
†НО= Не определено;
‡ MVB=Смешанное валентное связывание в формате SMCA;
** KD определена в формате OAA;
†† ДСК для антител проводили в формате SMCA и предоставлены температурные переходы, относящиеся к каждому плечу Fab;
‡‡ Tm Fab дикого типа получали из экспериментов ДСК в формате мАт.
Таблица 21: Аффинность связывания антигена и оценка термической стабильности для выбранных конструкций K-L, полученных из K-K
*Система A: H1/L1 CAT-2200 и H2/L2 Пертузумаба (с соотношением 10:20:24:46 ДНК H1:H2:L1:L2); Система B: H1/L1 CAT-2200 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2); Система C: H1/L1 CR8071 и H2/L2 SGN-CD19a (с соотношением 15:15:35:35 ДНК H1:H2:L1:L2);
Конструкции сгруппированы и затенены тремя градиентами с "Средней разницей в Tm Fab (по системам) от дикого типа (° C)" от 0 до -1,5, от -1,5 до -3,0 и менее -3;
†НО= Не определено;
†† ДСК для антител проводили в формате SMCA и предоставлены температурные переходы, относящиеся к каждому плечу Fab;
‡‡ Tm Fab дикого типа получали из экспериментов ДСК в формате мАт.
Таблица 22A: Нумерация аминокислотных остатков тяжелой цепи IgG1 в соответствии с системами нумерации IMGT, Кабат, 1JPT и ЕС
Таблица 22B: Нумерация аминокислотных остатков легкой цепи лямбда в соответствии с системами нумерации IMGT и Кабат
Таблица 22C: Нумерация аминокислотных остатков легкой цепи каппа в соответствии с системами нумерации IMGT, Кабат и 1JPT
Таблица 23: Таблица соответствия для %Спаренных/%Неправильно спаренных видов и скалярных значений LCCA
49
Таблица 24: Номера конструкции SMCA и соответствующие уникальные идентификаторы набора LCCA
* конструкция являлась идентичной конструкции LCCA 13304-13466, за исключением того, что положение Q124 в L2 было протестировано как Q124K.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЗАЙМВОРКС ИНК.
<120> АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ, СОДЕРЖАЩИЕ ЛЕГКИЕ
ЦЕПИ КАППА И ЛЯМБДА И ИХ ПРИМЕНЕНИЯ
<130> ZYME038US
<140> 15/765,574
<141> 2016-10-07
<150> US 62/239,206
<151> 2015-10-08
<150> US 62/261,769
<151> 2015-12-01
<160> 182
<170> PatentIn version 3.5
<210> 1
<211> 227
<212> Белок
<213> Искусственная последовательность
<220>
<223> синтетическая Fab тяжелой цепи пертузумаба (Границы доменов: VH; E1 -
S119, CH1; A120 - V217, шарнир (частичный); E218 - T227)
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr
225
<210> 2
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> синтетическая Fab легкой цепи (каппа) пертузумаба (Границы доменов: VL;
D1 - K107, CL; R108 - C214)
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 3
<211> 226
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи CAT-2200 (Границы доменов: VH; E1 - S118, CH1;
A119 - V216, Hinge(partial); E217 - T226)
<400> 3
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Ile His Gly Val Thr Arg Asn Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Gln Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr
225
<210> 4
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAT-2200 легкая цепь (лямбда) (Границы доменов: VL; N1 - L110,
CL; G111 - S216)
<400> 4
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Leu Ala Asn Tyr
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ser Pro Thr Ile Val
35 40 45
Ile Phe Ala Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Pro
85 90 95
Tyr Ser Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 5
<211> 222
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgG1 Fc (Границы доменов: шарнир (частичный); C1 - P5, CH2; A6 -
K115, CH3; G116 - K222)
<400> 5
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
1 5 10 15
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
20 25 30
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
35 40 45
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
50 55 60
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
65 70 75 80
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
85 90 95
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
100 105 110
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
115 120 125
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
130 135 140
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
145 150 155 160
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
165 170 175
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
180 185 190
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
195 200 205
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 6
<211> 666
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IgG1 Fc
<400> 6
tgccctccct gtccagctcc agaactgctg ggaggaccta gcgtgttcct gtttccccct 60
aagccaaaag acactctgat gatttccagg actcccgagg tgacctgcgt ggtggtggac 120
gtgtctcacg aggaccccga agtgaagttc aactggtacg tggatggcgt ggaagtgcat 180
aatgctaaga caaaaccaag agaggaacag tacaactcca cttatcgcgt cgtgagcgtg 240
ctgaccgtgc tgcaccagga ctggctgaac gggaaggagt ataagtgcaa agtcagtaat 300
aaggccctgc ctgctccaat cgaaaaaacc atctctaagg ccaaaggcca gccaagggag 360
ccccaggtgt acacactgcc acccagcaga gacgaactga ccaagaacca ggtgtccctg 420
acatgtctgg tgaaaggctt ctatcctagt gatattgctg tggagtggga atcaaatgga 480
cagccagaga acaattacaa gaccacacct ccagtgctgg acagcgatgg cagcttcttc 540
ctgtattcca agctgacagt ggataaatct cgatggcagc aggggaacgt gtttagttgt 600
tcagtgatgc atgaagccct gcacaatcat tacactcaga agagcctgtc cctgtctccc 660
ggcaaa 666
<210> 7
<211> 681
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи пертузумаба
<400> 7
gaagtgcagc tggtcgaatc tggaggagga ctggtgcagc caggagggtc cctgcgcctg 60
tcttgcgccg ctagtggctt cacttttacc gactacacca tggattgggt gcgacaggca 120
cctggaaagg gcctggagtg ggtcgccgat gtgaacccaa atagcggagg ctccatctac 180
aaccagcggt tcaagggccg gttcaccctg tcagtggacc ggagcaaaaa caccctgtat 240
ctgcagatga atagcctgcg agccgaagat actgctgtgt actattgcgc ccggaatctg 300
gggccctcct tctactttga ctattggggg cagggaactc tggtcaccgt gagctccgcc 360
tccaccaagg gaccttctgt gttcccactg gctccctcta gtaaatccac atctggggga 420
actgcagccc tgggctgtct ggtgaaggac tacttcccag agcccgtcac agtgtcttgg 480
aacagtggcg ctctgacttc tggggtccac acctttcctg cagtgctgca gtcaagcggg 540
ctgtacagcc tgtcctctgt ggtcaccgtg ccaagttcaa gcctgggaac acagacttat 600
atctgcaacg tgaatcacaa gccatccaat acaaaagtcg acaagaaagt ggaacccaag 660
tcttgtgata aaacccatac a 681
<210> 8
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь (каппа) пертузумаба
<400> 8
gatattcaga tgacccagtc cccaagctcc ctgagtgcct cagtgggcga ccgagtcacc 60
atcacatgca aggcttccca ggatgtgtct attggagtcg catggtacca gcagaagcca 120
ggcaaagcac ccaagctgct gatctatagc gcctcctacc ggtataccgg cgtgccctct 180
agattctctg gcagtgggtc aggaacagac tttactctga ccatctctag tctgcagcct 240
gaggatttcg ctacctacta ttgccagcag tactatatct acccatatac ctttggccag 300
gggacaaaag tggagatcaa gaggactgtg gccgctccct ccgtcttcat ttttccccct 360
tctgacgaac agctgaaaag tggcacagcc agcgtggtct gtctgctgaa caatttctac 420
cctcgcgaag ccaaagtgca gtggaaggtc gataacgctc tgcagagcgg caacagccag 480
gagtctgtga ctgaacagga cagtaaagat tcaacctata gcctgtcaag cacactgact 540
ctgagcaagg cagactacga gaagcacaaa gtgtatgcct gcgaagtcac acatcagggg 600
ctgtcctctc ctgtgactaa gagctttaac agaggagagt gt 642
<210> 9
<211> 678
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи CAT-2200
<400> 9
gaggtgcagc tgctggaatc tggggggggc ctggtgcagc ctggggggtc cctgagactg 60
tcatgtgctg ccagcgggtt tactttcagc tcctacgcta tgtcctgggt gcgacaggca 120
cccgggaagg gactggagtg ggtctctgca atcagtgggt caggcgggag tacttactat 180
gccgacagcg tgaagggacg gttcactatc tcaagagata acagcaagaa caccctgtat 240
ctgcagatga acagcctgag agcagaagac acagccgtgt actattgcgc cagggatctg 300
atccacggag tcactcgcaa ttggggccag gggactctgg tgaccgtctc tagtgctagc 360
acaaaggggc cctctgtgtt tccactggcc ccctcaagca aaagcacatc cggaggaact 420
gcagctctgg gatgtctggt gaaggactac ttcccccagc ctgtgaccgt ctcttggaac 480
agtggagccc tgaccagcgg cgtgcacaca tttcctgctg tcctgcagtc ctctggcctg 540
tactccctga gttcagtggt cacagtgcct agctcctctc tggggaccca gacatatatt 600
tgcaacgtga atcataaacc aagcaacact aaggtcgaca agaaagtgga gcccaagagc 660
tgtgataaaa ctcatacc 678
<210> 10
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> легкая цепь (лямбда) CAT-2200
<400> 10
aactttatgc tgactcagcc ccactccgtg tccgagagcc ctggcaaaac tgtgactatt 60
tcatgtaccc gatcatctgg aagcctggcc aactactatg tgcagtggta ccagcagagg 120
ccaggcagct cccccactat cgtgattttc gctaacaatc agcggccttc cggcgtccca 180
gacagatttt ccgggtctat cgattctagt tcaaatagtg catcactgac tatttccggg 240
ctgaagaccg aggacgaagc cgattactat tgccagacct acgaccccta ttctgtggtc 300
ttcggcgggg gaaccaagct gacagtgctg ggacagccaa aagcggcgcc cagtgtcaca 360
ctgtttcccc ctagctccga ggaactgcag gctaacaaag caacactggt gtgtctgatc 420
agcgacttct accctggagc tgtgactgtc gcctggaagg ctgattctag tccagtgaaa 480
gcaggcgtcg agaccacaac tccctctaag cagagtaaca acaagtacgc agcctcaagc 540
tatctgtcac tgaccccaga acagtggaag agccaccgga gctattcctg ccaggtcact 600
cacgaaggct ccactgtcga gaaaaccgtc gctcccaccg aatgttca 648
<210> 11
<211> 217
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fc последовательность 231-447 человеческого IgG1 (EU-нумерация), без шарнира
<400> 11
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 12
<211> 228
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи SGN-CD19a (Границы доменов: VH; Q1 - S120, CH1;
A121 - V218, шарнир (частичный); E219 - T228)
<400> 12
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Arg Tyr Asn Pro Ala
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Ala Tyr Tyr
85 90 95
Cys Ala Arg Met Glu Leu Trp Ser Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr
225
<210> 13
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> SGN-CD19a легкая цепь (каппа) (Границы доменов: VL; E1 - K106,
CL; R107 - C213)
<400> 13
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Phe Gln Gly Ser Val Tyr Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 14
<211> 233
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи CR8071 (Границы доменов: VH; Q1 - S125, CH1;
A126 - V223, шарнир (частичный); E224 - T233)
<400> 14
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Arg Val Ser Cys Arg Ala Ser Gly Tyr Ile Phe Thr Glu Ser
20 25 30
Gly Ile Thr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Gly Tyr Ser Gly Asp Thr Lys Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Lys Asp Thr Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Tyr Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Val Gln Tyr Ser Gly Ser Tyr Leu Gly Ala Tyr Tyr Phe
100 105 110
Asp Tyr Trp Ser Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr
225 230
<210> 15
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> CR8071 легкая цепь (лямбда) (Границы доменов: VL; Q1 - L110,
CL; R111 - S216)
<400> 15
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Thr Asn
20 25 30
Tyr Val Tyr Trp Tyr Gln Gln Phe Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Ser Tyr Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Ser Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asp Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 16
<211> 684
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи SGN-CD19a
<400> 16
caggtgacac tgagagaatc cggcccagcc ctggtgaagc ccactcagac cctgacactg 60
acttgcacct tctctgggtt ttccctgtct acaagtggga tgggagtggg atggatcagg 120
cagccacctg gaaaagccct ggagtggctg gctcacattt ggtgggacga tgacaagcgg 180
tacaacccag cactgaaaag cagactgaca atcagcaagg atacttccaa aaaccaggtg 240
gtcctgacaa tgactaatat ggaccccgtg gacacagccg cttactattg cgcccgcatg 300
gaactgtgga gctactattt cgactactgg gggcagggaa cactggtcac tgtgagctcc 360
gctagcacta aggggccttc cgtgtttcca ctggctccct ctagtaaatc cacctctgga 420
ggcacagctg cactgggatg tctggtgaag gattacttcc ctgaaccagt cacagtgagt 480
tggaactcag gggctctgac aagtggagtc catacttttc ccgcagtgct gcagtcaagc 540
ggactgtact ccctgtcctc tgtggtcacc gtgcctagtt caagcctggg cacccagaca 600
tatatctgca acgtgaatca caagccatca aatacaaaag tcgacaagaa agtggagccc 660
aagagctgtg ataaaactca tacc 684
<210> 17
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<223> SGN-CD19a легкая цепь (каппа)
<400> 17
gagatcgtgc tgacccagtc tccagccaca ctgtctctga gcccaggaga gagggccacc 60
ctgtcctgct ctgccagctc cagcgtgagc tacatgcact ggtatcagca gaagccagga 120
caggccccta ggctgctgat ctacgacacc agcaagctgg cctccggcat ccccgcaaga 180
ttcagcggct ccggctctgg cacagacttt accctgacaa tcagctccct ggagcctgag 240
gatttcgccg tgtactattg ttttcagggc agcgtgtatc cattcacctt tggccagggc 300
acaaagctgg agatcaagcg gacagtggcg gcgcccagtg tcttcatttt tccccctagc 360
gacgaacagc tgaagtctgg gacagccagt gtggtctgtc tgctgaacaa cttctaccct 420
agagaggcta aagtgcagtg gaaggtcgat aacgcactgc agtccggaaa ttctcaggag 480
agtgtgactg aacaggactc aaaagatagc acctattccc tgtcaagcac actgactctg 540
agcaaggccg actacgagaa gcataaagtg tatgcttgtg aagtcaccca ccaggggctg 600
agttcaccag tcacaaaatc attcaacaga ggggagtgc 639
<210> 18
<211> 699
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fab тяжелой цепи CR8071
<400> 18
caggtgcagc tggtccagtc cggggctgaa gtgaaaaaac ctggggcatc cgtgcgggtg 60
tcatgtcggg caagcgggta tatctttact gagtctggaa tcacctgggt gaggcaggct 120
cccggacagg gactggaatg gatgggatgg atttctggat acagtggcga cacaaagtat 180
gcacagaaac tgcagggccg cgtcaccatg acaaaggata cttcaaccac aactgcctac 240
atggagctgc ggagcctgag atatgacgat acagccgtgt actattgcgc ccgggacgtg 300
cagtacagcg ggtcctacct gggggcatac tacttcgatt actggtcacc tggaactctg 360
gtcaccgtct cttcagctag caccaagggc ccttctgtgt ttccactggc accctcaagc 420
aaaagcacct ccggaggaac agcagcactg ggatgtctgg tcaaggacta tttccccgag 480
cctgtgaccg tctcatggaa tagcggcgca ctgactagtg gggtgcacac ctttcccgcc 540
gtcctgcagt cctctgggct gtacagcctg agttcagtgg tcacagtgcc aagctcctct 600
ctgggaactc agacctatat ctgcaacgtc aatcataaac ccagcaacac aaaggtcgac 660
aagaaagtgg agcccaagag ctgtgataaa actcatacc 699
<210> 19
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CR8071 легкая цепь (лямбда)
<400> 19
cagagcgtcc tgactcagcc tccctccgcc tccggaacac ctgggcagag agtgactatc 60
tcctgtagcg gatcaagctc aaacattgga accaactacg tgtattggta ccagcagttc 120
cccggcacag ctcctaagct gctgatctat cggagctacc agagaccaag cggggtcccc 180
gacaggtttt ctggcagtaa atcagggagc tccgccagcc tggctatttc cggcctgcag 240
tctgaggacg aagcagatta ctattgcgcc acctgggacg attccctgga tggatgggtc 300
ttcggcggcg gcacaaaact gaccgtcctg aggcagccaa aggcggcgcc cagtgtcaca 360
ctgtttcccc ctagctccga ggaactgcag gctaacaaag caacactggt gtgtctgatc 420
agcgacttct accctggagc tgtgactgtc gcctggaagg ctgattctag tccagtgaaa 480
gcaggcgtcg agaccacaac tccctctaag cagagtaaca acaagtacgc agcctcaagc 540
tatctgtcac tgaccccaga acagtggaag agccaccgga gctattcctg ccaggtcact 600
cacgaaggct ccactgtcga gaaaaccgtc gctcccaccg aatgttca 648
<210> 20
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Пертузумаб
<400> 20
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Thr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asp Val Asn Pro Asn Ser Gly Gly Ser Ile Tyr Asn Gln Arg Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Arg Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Gly Pro Ser Phe Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 21
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> D3H44
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Glu Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Leu Ile Asp Pro Glu Gln Gly Asn Thr Ile Tyr Asp Pro Lys Phe
50 55 60
Gln Asp Arg Ala Thr Ile Ser Ala Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Thr Ala Ala Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 22
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92218|IGHV3-66*01
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg
<210> 23
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99641|IGHV1-18*01
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 24
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X62106|IGHV1-2*02
<400> 24
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 25
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99642|IGHV1-24*01
<400> 25
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Val Ser Gly Tyr Thr Leu Thr Glu Leu
20 25 30
Ser Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Gly Phe Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Glu Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr
<210> 26
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X62109|IGHV1-3*01
<400> 26
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Ala Gly Asn Gly Asn Thr Lys Tyr Ser Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Arg Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 27
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92209|IGHV1-45*01
<400> 27
Gln Met Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Thr Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Tyr Arg
20 25 30
Tyr Leu His Trp Val Arg Gln Ala Pro Gly Gln Ala Leu Glu Trp Met
35 40 45
Gly Trp Ile Thr Pro Phe Asn Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Asp Arg Val Thr Ile Thr Arg Asp Arg Ser Met Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg
<210> 28
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92343|IGHV1-46*01
<400> 28
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 29
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M29809|IGHV1-58*01
<400> 29
Gln Met Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Ser Ser
20 25 30
Ala Val Gln Trp Val Arg Gln Ala Arg Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Trp Ile Val Val Gly Ser Gly Asn Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Glu Arg Val Thr Ile Thr Arg Asp Met Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala
<210> 30
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> L22582|IGHV1-69*01, KC713934|IGHV1-69D*01
<400> 30
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 31
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> KF698734|IGHV1-69-2*01
<400> 31
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Val Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Leu Val Asp Pro Glu Asp Gly Glu Thr Ile Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Asp Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr
<210> 32
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99637|IGHV1-8*01
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asp Ile Asn Trp Val Arg Gln Ala Thr Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Met Asn Pro Asn Ser Gly Asn Thr Gly Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asn Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 33
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99648|IGHV2-26*01
<400> 33
Gln Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Asn Ala
20 25 30
Arg Met Gly Val Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Phe Ser Asn Asp Glu Lys Ser Tyr Ser Thr Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg
<210> 34
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> X62111|IGHV2-5*01
<400> 34
Gln Ile Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Val Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Tyr Trp Asn Asp Asp Lys Arg Tyr Ser Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala His
<210> 35
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> L21969|IGHV2-70*01
<400> 35
Gln Val Thr Leu Arg Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Cys Val Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Asp Trp Asp Asp Asp Lys Tyr Tyr Ser Thr Ser
50 55 60
Leu Lys Thr Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg
<210> 36
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> KC713935|IGHV2-70D*04
<400> 36
Gln Val Thr Leu Lys Glu Ser Gly Pro Ala Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Met Arg Val Ser Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Arg Ile Asp Trp Asp Asp Asp Lys Phe Tyr Ser Thr Ser
50 55 60
Leu Lys Thr Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg
<210> 37
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92287|IGHV3-11*03
<400> 37
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Ser Tyr Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 38
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92217|IGHV3-13*01
<400> 38
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met His Trp Val Arg Gln Ala Thr Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Gly Thr Ala Gly Asp Thr Tyr Tyr Pro Gly Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Glu Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Gly Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg
<210> 39
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92216|IGHV3-15*01
<400> 39
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Lys Thr Asp Gly Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Thr
100
<210> 40
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99657|IGHV3-20*01
<400> 40
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Asn Trp Asn Gly Gly Ser Thr Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr His Cys
85 90 95
Ala Arg
<210> 41
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AB019439|IGHV3-21*01
<400> 41
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 42
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99660|IGHV3-23*01
<400> 42
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys
<210> 43
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M83134|IGHV3-30*01, X92283|IGHV3-30-3*01
<400> 43
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 44
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AC244456|IGHV3-30-5*01
<400> 44
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys
<210> 45
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AB019439|IGHV3-33*01
<400> 45
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 46
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99672|IGHV3-43*01
<400> 46
Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys
<210> 47
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> KC713950|IGHV3-43D*01
<400> 47
Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys
<210> 48
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99675|IGHV3-48*01
<400> 48
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 49
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> AB019438|IGHV3-49*03
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Gly Asp Tyr
20 25 30
Ala Met Ser Trp Phe Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Ser Lys Ala Tyr Gly Gly Thr Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ile
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Arg
100
<210> 50
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99679|IGHV3-53*01
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg
<210> 51
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99682|IGHV3-64*01
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Ala Ile Ser Ser Asn Gly Gly Ser Thr Tyr Tyr Ala Asn Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 52
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99649|IGHV3-7*01
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 53
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92206|IGHV3-72*01
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp His
20 25 30
Tyr Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Thr Arg Asn Lys Ala Asn Ser Tyr Thr Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg
100
<210> 54
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X70197|IGHV3-73*01
<400> 54
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Ser Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Ser Lys Ala Asn Ser Tyr Ala Thr Ala Tyr Ala Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Arg
100
<210> 55
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> L33851|IGHV3-74*01
<400> 55
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Arg Ile Asn Ser Asp Gly Ser Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 56
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99651|IGHV3-9*01
<400> 56
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Trp Asn Ser Gly Ser Ile Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Lys
<210> 57
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> HM855939|IGHV3-NL1*01
<400> 57
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys
<210> 58
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X05714|IGHV4-28*01
<400> 58
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Asp
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Ser Ser
20 25 30
Asn Trp Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Val Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 59
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> L10089|IGHV4-30-2*01
<400> 59
Gln Leu Gln Leu Gln Glu Ser Gly Ser Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly Tyr Ser Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Arg Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg
<210> 60
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z14238|IGHV4-30-4*01
<400> 60
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Asp Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg
<210> 61
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> L10098|IGHV4-31*01
<400> 61
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly Tyr Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Leu Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg
<210> 62
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> AB019439|IGHV4-34*01
<400> 62
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg
<210> 63
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z12367|IGHV4-38-2*01
<400> 63
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Ser Gly
20 25 30
Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Ser Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 64
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> AB019439|IGHV4-39*01
<400> 64
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg
<210> 65
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X05713|IGHV4-4*01
<400> 65
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Pro Gly
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Asn Trp Trp Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Glu Ile Tyr His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Lys Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Cys Cys
85 90 95
Ala Arg
<210> 66
<211> 97
<212> Белок
<213> Искусственная последовательность
<220>
<223> AB019438|IGHV4-59*01
<400> 66
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg
<210> 67
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> M29811|IGHV4-61*01
<400> 67
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Ser Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg
<210> 68
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X92227|IGHV5-10-1*01
<400> 68
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Ser Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly His Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg
<210> 69
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> M99686|IGHV5-51*01
<400> 69
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg
<210> 70
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> J04097|IGHV6-1*01
<400> 70
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Asn
20 25 30
Ser Ala Ala Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Ser Lys Trp Tyr Asn Asp Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Asn
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg
100
<210> 71
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X62110|IGHV7-4-1*02
<400> 71
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Asn Thr Gly Asn Pro Thr Tyr Ala Gln Gly Phe
50 55 60
Thr Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg
<210> 72
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00256|IGHJ1*01
<400> 72
Ala Glu Tyr Phe Gln His Trp Gly Gln Gly Thr Leu Val Thr Val Ser
1 5 10 15
Ser
<210> 73
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00256|IGHJ2*01
<400> 73
Tyr Trp Tyr Phe Asp Leu Trp Gly Arg Gly Thr Leu Val Thr Val Ser
1 5 10 15
Ser
<210> 74
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00256|IGHJ3*01
<400> 74
Asp Ala Phe Asp Val Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
1 5 10 15
<210> 75
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00256|IGHJ5*01
<400> 75
Asn Trp Phe Asp Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10 15
<210> 76
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00256|IGHJ6*01
<400> 76
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 77
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00256|IGHJ4*01
<400> 77
Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10 15
<210> 78
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Pertuzumab
<400> 78
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Gly
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 79
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> D3H44
<400> 79
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Arg Asp Ile Lys Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Tyr Ala Thr Ser Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Gly Glu Ser Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 80
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> Y14865|IGKV1-NL1*01
<400> 80
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Leu
35 40 45
Tyr Ala Ala Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro
85 90 95
<210> 81
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X59315|IGKV1-39*01, X59312|IGKV1D-39*01
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro
85 90 95
<210> 82
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X17263|IGKV1D-12*01, V01577|IGKV1-12*01
<400> 82
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro
85 90 95
<210> 83
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z00006|IGKV1-13*02
<400> 83
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Ala
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn Ser Tyr Pro
85 90 95
<210> 84
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00248|IGKV1-16*01
<400> 84
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro
85 90 95
<210> 85
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X72808|IGKV1-17*01
<400> 85
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Asn Ser Tyr Pro
85 90 95
<210> 86
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X63398|IGKV1-27*01
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Tyr Asn Ser Ala Pro
85 90 95
<210> 87
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> M64856|IGKV1-33*01, M64855|IGKV1D-33*01
<400> 87
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Asn Leu Pro
85 90 95
<210> 88
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z00001|IGKV1-5*01
<400> 88
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser
85 90 95
<210> 89
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> M64858|IGKV1-6*01
<400> 89
Ala Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Tyr Asn Tyr Pro
85 90 95
<210> 90
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z00014|IGKV1-8*01
<400> 90
Ala Ile Arg Met Thr Gln Ser Pro Ser Ser Phe Ser Ala Ser Thr Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Cys Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Tyr Pro
85 90 95
<210> 91
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z00013|IGKV1-9*01
<400> 91
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Ser Tyr Pro
85 90 95
<210> 92
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X17262|IGKV1D-13*01
<400> 92
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Ala
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn Asn Tyr Pro
85 90 95
<210> 93
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> K01323|IGKV1D-16*01
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Glu Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro
85 90 95
<210> 94
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X63392|IGKV1D-17*01
<400> 94
Asn Ile Gln Met Thr Gln Ser Pro Ser Ala Met Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Arg Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Val Pro Lys His Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Asn Ser Tyr Pro
85 90 95
<210> 95
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X72817|IGKV1D-43*01
<400> 95
Ala Ile Arg Met Thr Gln Ser Pro Phe Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Trp Ala Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Ala Lys Ala Pro Lys Leu Phe Ile
35 40 45
Tyr Tyr Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro
85 90 95
<210> 96
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z00008|IGKV1D-8*01
<400> 96
Val Ile Trp Met Thr Gln Ser Pro Ser Leu Leu Ser Ala Ser Thr Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Met Ser Gln Gly Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Cys Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Phe Pro
85 90 95
<210> 97
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X12684|IGKV2-24*01
<400> 97
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Ser Trp Leu Gln Gln Arg Pro Gly Gln Pro
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Ile Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Thr Gln Phe Pro
100
<210> 98
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X12691|IGKV2D-28*01, X63397|IGKV2-28*01
<400> 98
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Gln Thr Pro
100
<210> 99
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> U41645|IGKV2-29*02
<400> 99
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Val Ser Ser Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Ile His Leu Pro
100
<210> 100
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X63403|IGKV2-30*01
<400> 100
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Tyr Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Lys Val Ser Asn Arg Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Thr His Trp Pro
100
<210> 101
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> X59311|IGKV2D-40*01, X59314|IGKV2-40*01
<400> 101
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu Asp Ser
20 25 30
Asp Asp Gly Asn Thr Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln
35 40 45
Ser Pro Gln Leu Leu Ile Tyr Thr Leu Ser Tyr Arg Ala Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys
65 70 75 80
Ile Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln
85 90 95
Arg Ile Glu Phe Pro
100
<210> 102
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> AP001216|IGKV2D-26*01
<400> 102
Glu Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Ile Thr Pro Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asp Gly Tyr Thr Tyr Leu Tyr Trp Phe Leu Gln Lys Ala Arg Pro Val
35 40 45
Ser Thr Leu Leu Ile Tyr Glu Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Phe Gly Val Tyr Tyr Cys Met Gln Asp
85 90 95
Ala Gln Asp Pro
100
<210> 103
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> M31952|IGKV2D-29*01
<400> 103
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Pro
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ser
85 90 95
Ile Gln Leu Pro
100
<210> 104
<211> 100
<212> Белок
<213> Искусственная последовательность
<220>
<223> X63402|IGKV2D-30*01
<400> 104
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val Tyr Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Asn Trp Phe Gln Gln Arg Pro Gly Gln Ser
35 40 45
Pro Arg Arg Leu Ile Tyr Lys Val Ser Asn Trp Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Thr His Trp Pro
100
<210> 105
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X01668|IGKV3-11*01
<400> 105
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro
85 90 95
<210> 106
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X72815|IGKV3D-15*01, M23090|IGKV3-15*01
<400> 106
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asn Asn Trp Pro
85 90 95
<210> 107
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X12686|IGKV3-20*01
<400> 107
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
<210> 108
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X17264|IGKV3D-11*01
<400> 108
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Gly Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Pro Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp His
85 90 95
<210> 109
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X12687|IGKV3D-20*01
<400> 109
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Gly Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
<210> 110
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X72820|IGKV3D-7*01
<400> 110
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asp Tyr Asn Leu Pro
85 90 95
<210> 111
<211> 101
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z00023|IGKV4-1*01
<400> 111
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Thr Pro
100
<210> 112
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X02485|IGKV5-2*01
<400> 112
Glu Thr Thr Leu Thr Gln Ser Pro Ala Phe Met Ser Ala Thr Pro Gly
1 5 10 15
Asp Lys Val Asn Ile Ser Cys Lys Ala Ser Gln Asp Ile Asp Asp Asp
20 25 30
Met Asn Trp Tyr Gln Gln Lys Pro Gly Glu Ala Ala Ile Phe Ile Ile
35 40 45
Gln Glu Ala Thr Thr Leu Val Pro Gly Ile Pro Pro Arg Phe Ser Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn Ile Glu Ser
65 70 75 80
Glu Asp Ala Ala Tyr Tyr Phe Cys Leu Gln His Asp Asn Phe Pro
85 90 95
<210> 113
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X12683|IGKV6D-21*01, X63399|IGKV6-21*01
<400> 113
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Phe Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys His Gln Ser Ser Ser Leu Pro
85 90 95
<210> 114
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z70260|IGKJ2*02
<400> 114
Cys Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 115
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00242|IGKJ3*01
<400> 115
Phe Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
1 5 10
<210> 116
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00242|IGKJ4*01
<400> 116
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 117
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00242|IGKJ5*01
<400> 117
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
1 5 10
<210> 118
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00242|IGKJ2*01
<400> 118
Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 119
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00242|IGKJ1*01
<400> 119
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 120
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> JN582178|IGHG1*04, Z17370|IGHG1*02, Pertuzumab, D3H44,
AL122127|IGHG1*05, J00228|IGHG1*01
<400> 120
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 121
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Y14737|IGHG1*03
<400> 121
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 122
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00230|IGHG2*01, AF449616|IGHG2*03, AF449618|IGHG2*05,
AL928742|IGHG2*06
<400> 122
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val
<210> 123
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AJ250170|IGHG2*02
<400> 123
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Thr Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val
<210> 124
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AF449617|IGHG2*04
<400> 124
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val
<210> 125
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X99549|IGHG3*04,AJ390244|IGHG3*13,X16110|IGHG3*03,AJ390236|IGHG3*05,
AJ390242|IGHG3*09,AL122127|IGHG3*10,AJ390279|IGHG3*19,AJ390260|IGHG3*15,
AJ390237|IGHG3*06,AJ390238|IGHG3*07,AJ390247|IGHG3*11,AJ390252|IGHG3*12,
AJ390262|IGHG3*16,AJ390241|IGHG3*08,X03604|IGHG3*01,AJ390254|IGHG3*14
<400> 125
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 126
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AJ390272|IGHG3*17
<400> 126
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 127
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> AJ390276|IGHG3*18
<400> 127
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Tyr Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 128
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> K01316|IGHG4*01, AL928742|IGHG4*04
<400> 128
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val
<210> 129
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> D3H44, J00241|IGKC*01, Pertuzumab
<400> 129
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 130
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> M11736|IGKC*02
<400> 130
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Glu Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Gly Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 131
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> M11737|IGKC*03
<400> 131
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Arg Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Glu Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 132
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AF017732|IGKC*04
<400> 132
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Leu Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 133
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> AF113887|IGKC*05
<400> 133
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Asn Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 134
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAT-2200
<400> 134
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Ile His Gly Val Thr Arg Asn Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 135
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAT-2200
<400> 135
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Leu Ala Asn Tyr
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ser Pro Thr Ile Val
35 40 45
Ile Phe Ala Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Tyr Asp Pro
85 90 95
Tyr Ser Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 136
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73673|IGLV6-57*01
<400> 136
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Ala Ser Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ser Pro Thr Thr Val
35 40 45
Ile Tyr Glu Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser
85 90 95
Ser Asn
<210> 137
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73653|IGLV1-36*01
<400> 137
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Glu Ala Pro Arg Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asp Asp Leu Leu Pro Ser Gly Val Ser Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly
<210> 138
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> M94116|IGLV1-40*01
<400> 138
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Ser Gly
<210> 139
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73654|IGLV1-44*01
<400> 139
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly
<210> 140
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73663|IGLV1-47*01
<400> 140
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Tyr Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Gly
<210> 141
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73661|IGLV1-51*01
<400> 141
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Ser Ser Leu
85 90 95
Ser Ala
<210> 142
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73657|IGLV2-11*01
<400> 142
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Ser
85 90 95
Tyr Thr Phe
<210> 143
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73664|IGLV2-14*01
<400> 143
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Leu
<210> 144
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73642|IGLV2-18*01
<400> 144
Gln Ser Ala Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Ser Tyr
20 25 30
Asn Arg Val Ser Trp Tyr Gln Gln Pro Pro Gly Thr Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Thr Ser Ser
85 90 95
Ser Thr Phe
<210> 145
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> X14616|IGLV2-23*01
<400> 145
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Ser Tyr
20 25 30
Asn Leu Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Gly Ser Lys Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ala Gly Ser
85 90 95
Ser Thr Leu
<210> 146
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> X97462|IGLV2-8*01
<400> 146
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Gly Ser
85 90 95
Asn Asn Phe
<210> 147
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X57826|IGLV3-1*01
<400> 147
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Lys Leu Gly Asp Lys Tyr Ala
20 25 30
Cys Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser Thr Ala
85 90 95
<210> 148
<211> 95
<212> Белок
<213> Искусственная последовательность
<220>
<223> X97473|IGLV3-9*01
<400> 148
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Ala
85 90 95
<210> 149
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X97464|IGLV3-10*01
<400> 149
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Lys Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Ser Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Glu Asp Ser Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Met Ala Thr Leu Thr Ile Ser Gly Ala Gln Val Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Tyr Ser Thr Asp Ser Ser Gly Asn His
85 90 95
<210> 150
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73658|IGLV3-12*01
<400> 150
Ser Tyr Glu Leu Thr Gln Pro His Ser Val Ser Val Ala Thr Ala Gln
1 5 10 15
Met Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Asp Pro Val Leu Val Ile Tyr
35 40 45
Ser Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Pro Gly Asn Thr Thr Thr Leu Thr Ile Ser Arg Ile Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
<210> 151
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X97471|IGLV3-16*01
<400> 151
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Met Ala Arg Ile Thr Cys Ser Gly Glu Ala Leu Pro Lys Lys Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Phe Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Ile Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Leu Ser Ala Asp Ser Ser Gly Thr Tyr
85 90 95
<210> 152
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X56178|IGLV3-19*01
<400> 152
Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Val Arg Ile Thr Cys Gln Gly Asp Ser Leu Arg Ser Tyr Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser Ser Gly Asn His
85 90 95
<210> 153
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X71966|IGLV3-21*01
<400> 153
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
<210> 154
<211> 94
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73666|IGLV3-22*01
<400> 154
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Val Leu Gly Glu Asn Tyr Ala
20 25 30
Asp Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Glu Leu Val Ile Tyr
35 40 45
Glu Asp Ser Glu Arg Tyr Pro Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Asn Thr Thr Thr Leu Thr Ile Ser Arg Val Leu Thr Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Leu Ser Gly Asp Glu Asp Asn
85 90
<210> 155
<211> 96
<212> Белок
<213> Искусственная последовательность
<220>
<223> X97474|IGLV3-25*01
<400> 155
Ser Tyr Glu Leu Met Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Tyr
85 90 95
<210> 156
<211> 94
<212> Белок
<213> Искусственная последовательность
<220>
<223> D86994|IGLV3-27*01
<400> 156
Ser Tyr Glu Leu Thr Gln Pro Ser Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Val Leu Ala Lys Lys Tyr Ala
20 25 30
Arg Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Ala Gln Val Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Tyr Ser Ala Ala Asp Asn Asn
85 90
<210> 157
<211> 103
<212> Белок
<213> Искусственная последовательность
<220>
<223> X57828|IGLV4-3*01
<400> 157
Leu Pro Val Leu Thr Gln Pro Pro Ser Ala Ser Ala Leu Leu Gly Ala
1 5 10 15
Ser Ile Lys Leu Thr Cys Thr Leu Ser Ser Glu His Ser Thr Tyr Thr
20 25 30
Ile Glu Trp Tyr Gln Gln Arg Pro Gly Arg Ser Pro Gln Tyr Ile Met
35 40 45
Lys Val Lys Ser Asp Gly Ser His Ser Lys Gly Asp Gly Ile Pro Asp
50 55 60
Arg Phe Met Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Thr Phe Ser
65 70 75 80
Asn Leu Gln Ser Asp Asp Glu Ala Glu Tyr His Cys Gly Glu Ser His
85 90 95
Thr Ile Asp Gly Gln Val Gly
100
<210> 158
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73667|IGLV4-60*01
<400> 158
Gln Pro Val Leu Thr Gln Ser Ser Ser Ala Ser Ala Ser Leu Gly Ser
1 5 10 15
Ser Val Lys Leu Thr Cys Thr Leu Ser Ser Gly His Ser Ser Tyr Ile
20 25 30
Ile Ala Trp His Gln Gln Gln Pro Gly Lys Ala Pro Arg Tyr Leu Met
35 40 45
Lys Leu Glu Gly Ser Gly Ser Tyr Asn Lys Gly Ser Gly Val Pro Asp
50 55 60
Arg Phe Ser Gly Ser Ser Ser Gly Ala Asp Arg Tyr Leu Thr Ile Ser
65 70 75 80
Asn Leu Gln Leu Glu Asp Glu Ala Asp Tyr Tyr Cys Glu Thr Trp Asp
85 90 95
Ser Asn Thr
<210> 159
<211> 99
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73648|IGLV4-69*01
<400> 159
Gln Leu Val Leu Thr Gln Ser Pro Ser Ala Ser Ala Ser Leu Gly Ala
1 5 10 15
Ser Val Lys Leu Thr Cys Thr Leu Ser Ser Gly His Ser Ser Tyr Ala
20 25 30
Ile Ala Trp His Gln Gln Gln Pro Glu Lys Gly Pro Arg Tyr Leu Met
35 40 45
Lys Leu Asn Ser Asp Gly Ser His Ser Lys Gly Asp Gly Ile Pro Asp
50 55 60
Arg Phe Ser Gly Ser Ser Ser Gly Ala Glu Arg Tyr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Thr Trp Gly
85 90 95
Thr Gly Ile
<210> 160
<211> 104
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73672|IGLV5-37*01
<400> 160
Gln Pro Val Leu Thr Gln Pro Pro Ser Ser Ser Ala Ser Pro Gly Glu
1 5 10 15
Ser Ala Arg Leu Thr Cys Thr Leu Pro Ser Asp Ile Asn Val Gly Ser
20 25 30
Tyr Asn Ile Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Pro Pro Arg Tyr
35 40 45
Leu Leu Tyr Tyr Tyr Ser Asp Ser Asp Lys Gly Gln Gly Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Lys Asp Ala Ser Ala Asn Thr Gly Ile
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Met Ile Trp Pro Ser Asn Ala Ser
100
<210> 161
<211> 104
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73668|IGLV5-39*01
<400> 161
Gln Pro Val Leu Thr Gln Pro Thr Ser Leu Ser Ala Ser Pro Gly Ala
1 5 10 15
Ser Ala Arg Phe Thr Cys Thr Leu Arg Ser Gly Ile Asn Val Gly Thr
20 25 30
Tyr Arg Ile Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Leu Pro Arg Tyr
35 40 45
Leu Leu Arg Tyr Lys Ser Asp Ser Asp Lys Gln Gln Gly Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Lys Asp Ala Ser Thr Asn Ala Gly Leu
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Ala Ile Trp Tyr Ser Ser Thr Ser
100
<210> 162
<211> 104
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73670|IGLV5-45*01
<400> 162
Gln Ala Val Leu Thr Gln Pro Ala Ser Leu Ser Ala Ser Pro Gly Ala
1 5 10 15
Ser Ala Ser Leu Thr Cys Thr Leu Arg Ser Gly Ile Asn Val Gly Thr
20 25 30
Tyr Arg Ile Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Pro Pro Gln Tyr
35 40 45
Leu Leu Arg Tyr Lys Ser Asp Ser Asp Lys Gln Gln Gly Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Lys Asp Ala Ser Ala Asn Ala Gly Ile
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Met Ile Trp His Ser Ser Ala Ser
100
<210> 163
<211> 105
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73669|IGLV5-52*01
<400> 163
Gln Pro Val Leu Thr Gln Pro Ser Ser His Ser Ala Ser Ser Gly Ala
1 5 10 15
Ser Val Arg Leu Thr Cys Met Leu Ser Ser Gly Phe Ser Val Gly Asp
20 25 30
Phe Trp Ile Arg Trp Tyr Gln Gln Lys Pro Gly Asn Pro Pro Arg Tyr
35 40 45
Leu Leu Tyr Tyr His Ser Asp Ser Asn Lys Gly Gln Gly Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Asn Asp Ala Ser Ala Asn Ala Gly Ile
65 70 75 80
Leu Arg Ile Ser Gly Leu Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Gly Thr Trp His Ser Asn Ser Lys Thr
100 105
<210> 164
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> X14614|IGLV7-43*01
<400> 164
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Ala Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Tyr Tyr Pro Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Ala
35 40 45
Leu Ile Tyr Ser Thr Ser Asn Lys His Ser Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Leu Leu Tyr Tyr Gly Gly
85 90 95
Ala Gln
<210> 165
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73674|IGLV7-46*01
<400> 165
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
His Tyr Pro Tyr Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asp Thr Ser Asn Lys His Ser Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Leu Leu Ser Tyr Ser Gly
85 90 95
Ala Arg
<210> 166
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73650|IGLV8-61*01
<400> 166
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
Tyr Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Ser Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Val Leu Tyr Met Gly Ser
85 90 95
Gly Ile
<210> 167
<211> 104
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73675|IGLV9-49*01
<400> 167
Gln Pro Val Leu Thr Gln Pro Pro Ser Ala Ser Ala Ser Leu Gly Ala
1 5 10 15
Ser Val Thr Leu Thr Cys Thr Leu Ser Ser Gly Tyr Ser Asn Tyr Lys
20 25 30
Val Asp Trp Tyr Gln Gln Arg Pro Gly Lys Gly Pro Arg Phe Val Met
35 40 45
Arg Val Gly Thr Gly Gly Ile Val Gly Ser Lys Gly Asp Gly Ile Pro
50 55 60
Asp Arg Phe Ser Val Leu Gly Ser Gly Leu Asn Arg Tyr Leu Thr Ile
65 70 75 80
Lys Asn Ile Gln Glu Glu Asp Glu Ser Asp Tyr His Cys Gly Ala Asp
85 90 95
His Gly Ser Gly Ser Asn Phe Val
100
<210> 168
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> Z73676|IGLV10-54*01
<400> 168
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Lys Gly Leu Arg Gln
1 5 10 15
Thr Ala Thr Leu Thr Cys Thr Gly Asn Ser Asn Asn Val Gly Asn Gln
20 25 30
Gly Ala Ala Trp Leu Gln Gln His Gln Gly His Pro Pro Lys Leu Leu
35 40 45
Ser Tyr Arg Asn Asn Asn Arg Pro Ser Gly Ile Ser Glu Arg Leu Ser
50 55 60
Ala Ser Arg Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ala Trp Asp Ser Ser Leu
85 90 95
Ser Ala
<210> 169
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> X04457|IGLJ1*01
<400> 169
Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
1 5 10
<210> 170
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> M15641|IGLJ2*01, M15642|IGLJ3*01
<400> 170
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 171
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> M18338|IGLJ6*01
<400> 171
Asn Val Phe Gly Ser Gly Thr Lys Val Thr Val Leu
1 5 10
<210> 172
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> X51755|IGLJ7*01
<400> 172
Ala Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu
1 5 10
<210> 173
<211> 98
<212> Белок
<213> Искусственная последовательность
<220>
<223> CAT-2200
<400> 173
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Gln Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val
<210> 174
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> D87017|IGLC3*04, CAT-2200, X06875|IGLC2*02, J00253|IGLC2*01
<400> 174
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 175
<211> 104
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00252|IGLC1*01
<400> 175
Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
1 5 10 15
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
20 25 30
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys
35 40 45
Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr
50 55 60
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
65 70 75 80
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
85 90 95
Thr Val Ala Pro Thr Glu Cys Ser
100
<210> 176
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> X51755|IGLC1*02
<400> 176
Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 177
<211> 104
<212> Белок
<213> Искусственная последовательность
<220>
<223> J00254|IGLC3*01
<400> 177
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
1 5 10 15
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
20 25 30
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
35 40 45
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
50 55 60
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
65 70 75 80
Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
85 90 95
Thr Val Ala Pro Thr Glu Cys Ser
100
<210> 178
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> K01326|IGLC3*02
<400> 178
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Pro Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 179
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> X06876|IGLC3*03
<400> 179
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 180
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> J03011|IGLC6*01
<400> 180
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Lys Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Asn Thr Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Ala Glu Cys Ser
100 105
<210> 181
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> M61771|IGLC7*02, X51755|IGLC7*01
<400> 181
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Val Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Val Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Arg Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Ala Glu Cys Ser
100 105
<210> 182
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> KM455557|IGLC7*03
<400> 182
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Val Ser Asp
20 25 30
Phe Asn Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Val Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Arg Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Ala Glu Cys Ser
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СКОНСТРУИРОВАННЫЕ ПАРЫ ТЯЖЕЛАЯ-ЛЕГКАЯ ЦЕПИ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2826203C1 |
| СКОНСТРУИРОВАННЫЕ ПАРЫ ТЯЖЕЛАЯ-ЛЕГКАЯ ЦЕПИ ИММУНОГЛОБУЛИНА И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2720472C2 |
| ВАРИАНТЫ ПЕРТУЗУМАБА И ИХ АНАЛИТИЧЕСКАЯ ХАРАКТЕРИСТИКА | 2014 |
|
RU2737727C2 |
| МОДИФИЦИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДНЫЕ КОНСТРУКЦИИ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2832921C1 |
| АНТИТЕЛО-ПОДОБНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ОБЛАСТЯМИ, ИМЕЮЩИЕ ОРИЕНТАЦИЮ СВЯЗЫВАЮЩИХ ОБЛАСТЕЙ КРЕСТ-НАКРЕСТ | 2012 |
|
RU2823693C2 |
| КОНЪЮГАТЫ АНТИ-HER2 БИПАРАТОПНЫХ АНТИТЕЛ-ЛЕКАРСТВЕННЫХ ВЕЩЕСТВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2806049C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
| ТРИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ BCMA, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2829901C2 |
| АНТИТЕЛА К LY6G6D И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2818569C1 |
| АНТИТЕЛА ПРОТИВ PD-L1 И ИХ ВАРИАНТЫ | 2017 |
|
RU2770590C2 |
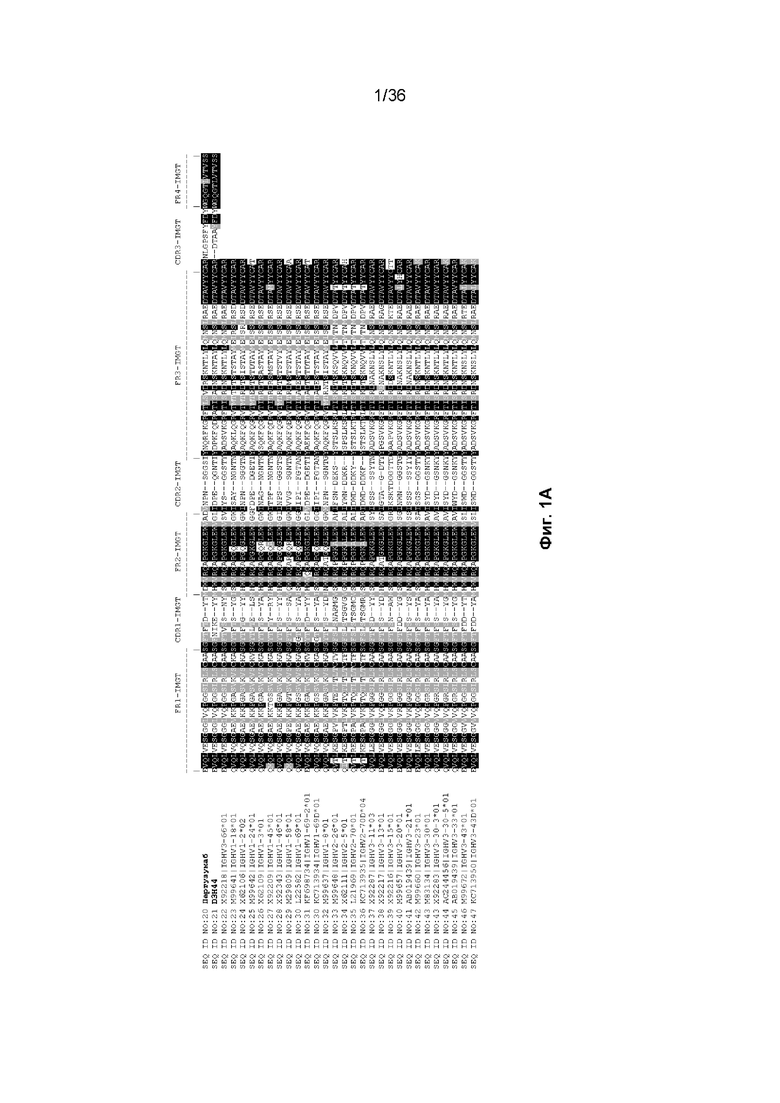





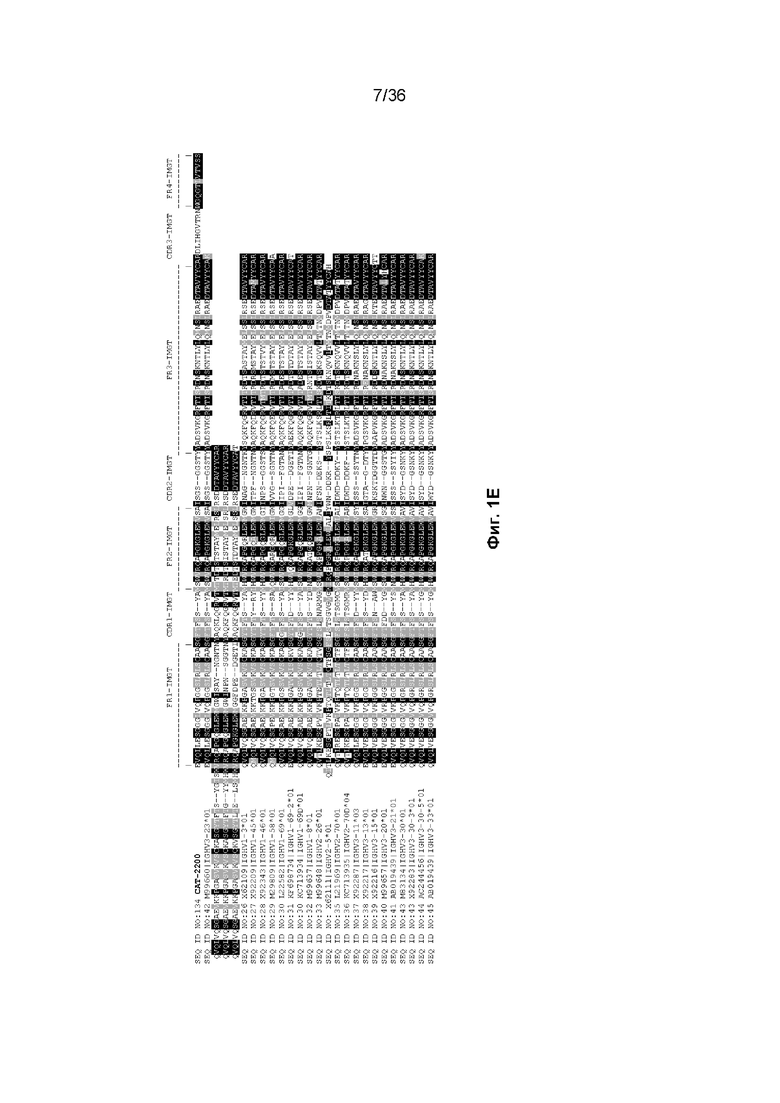

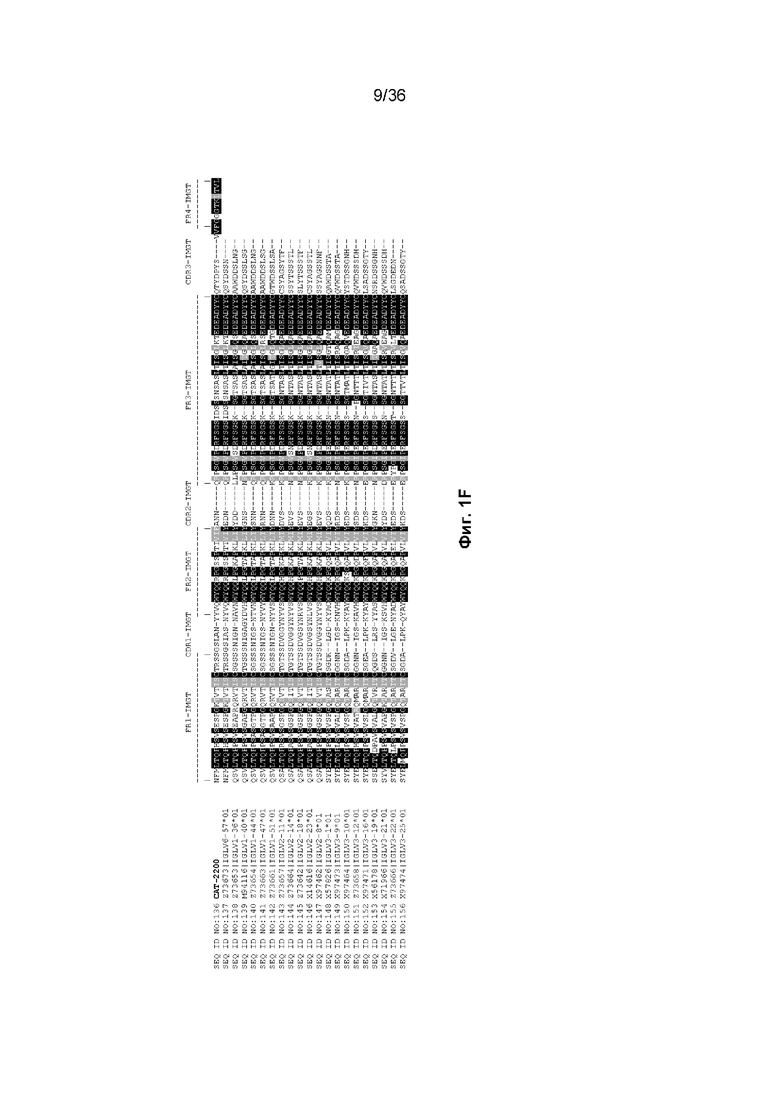


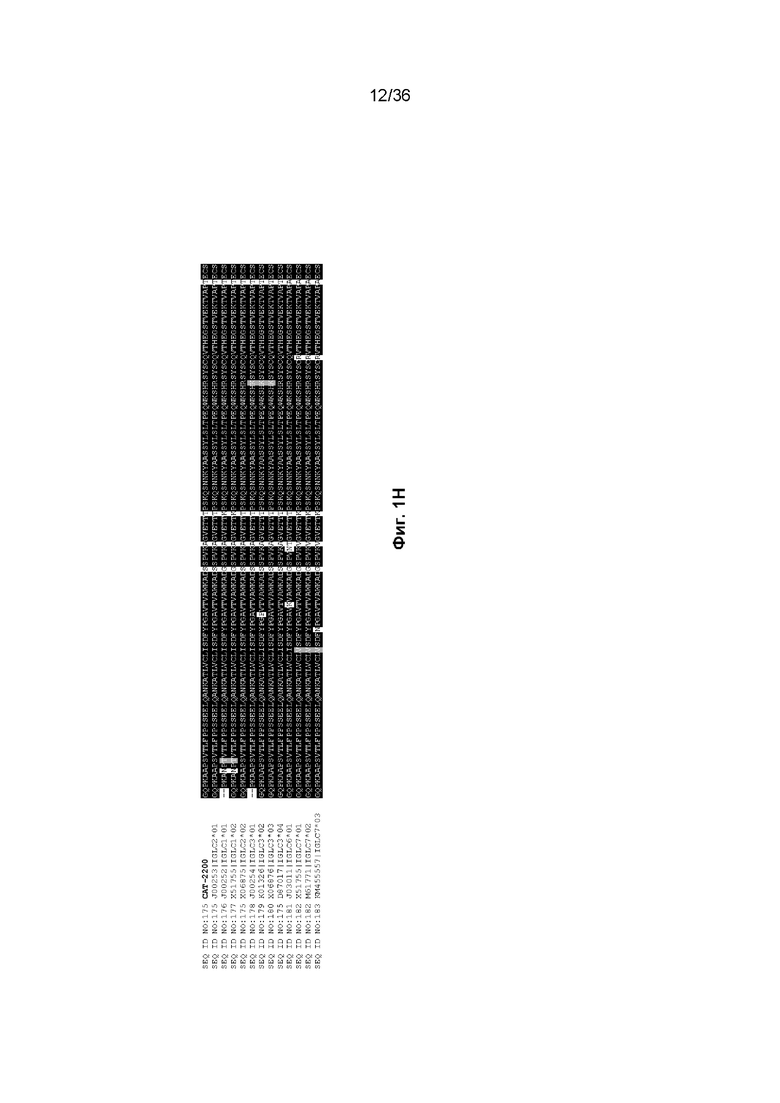
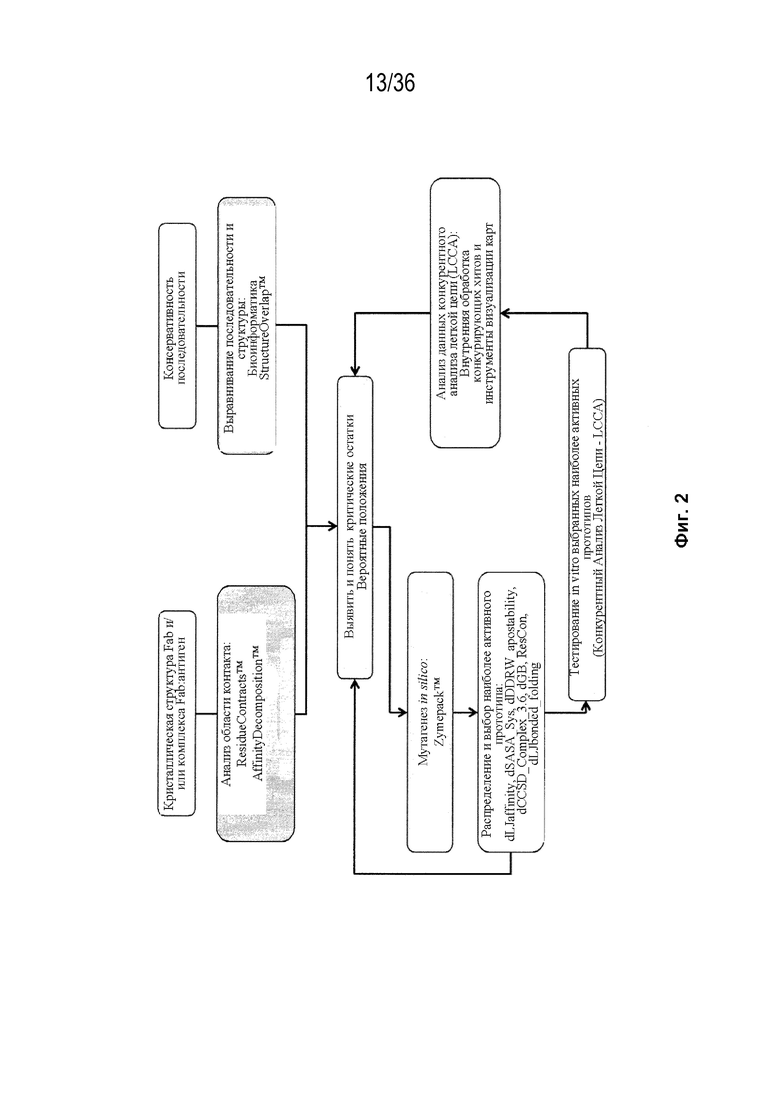
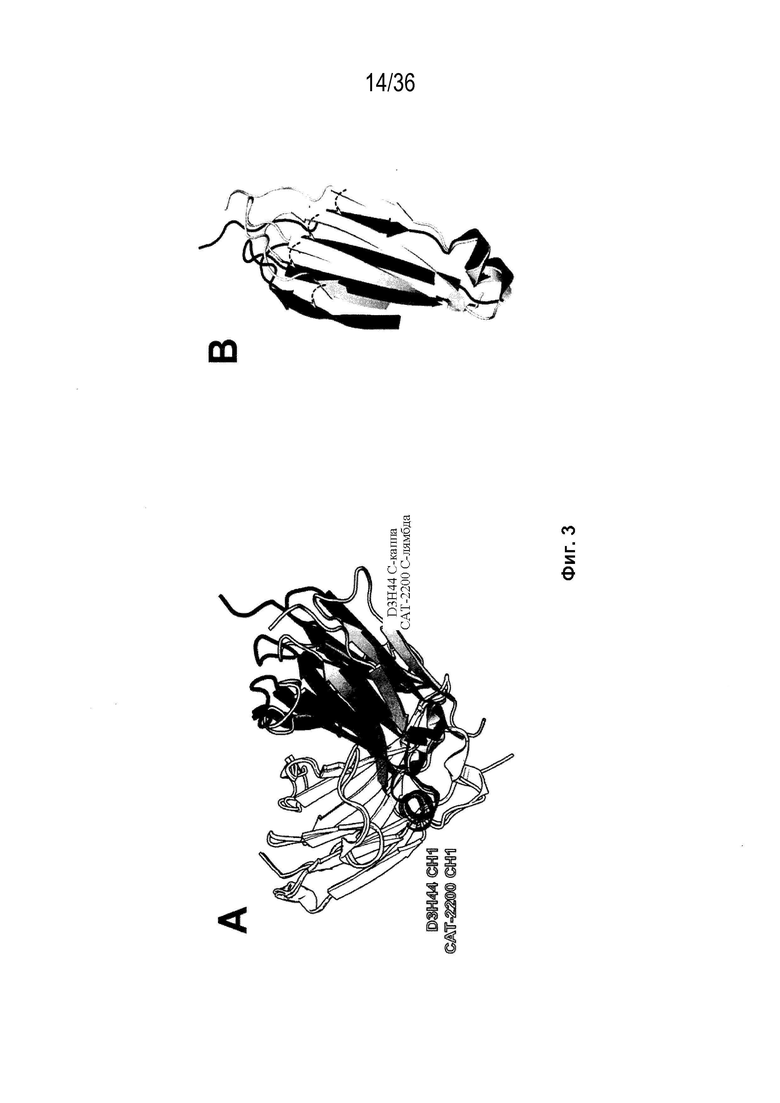
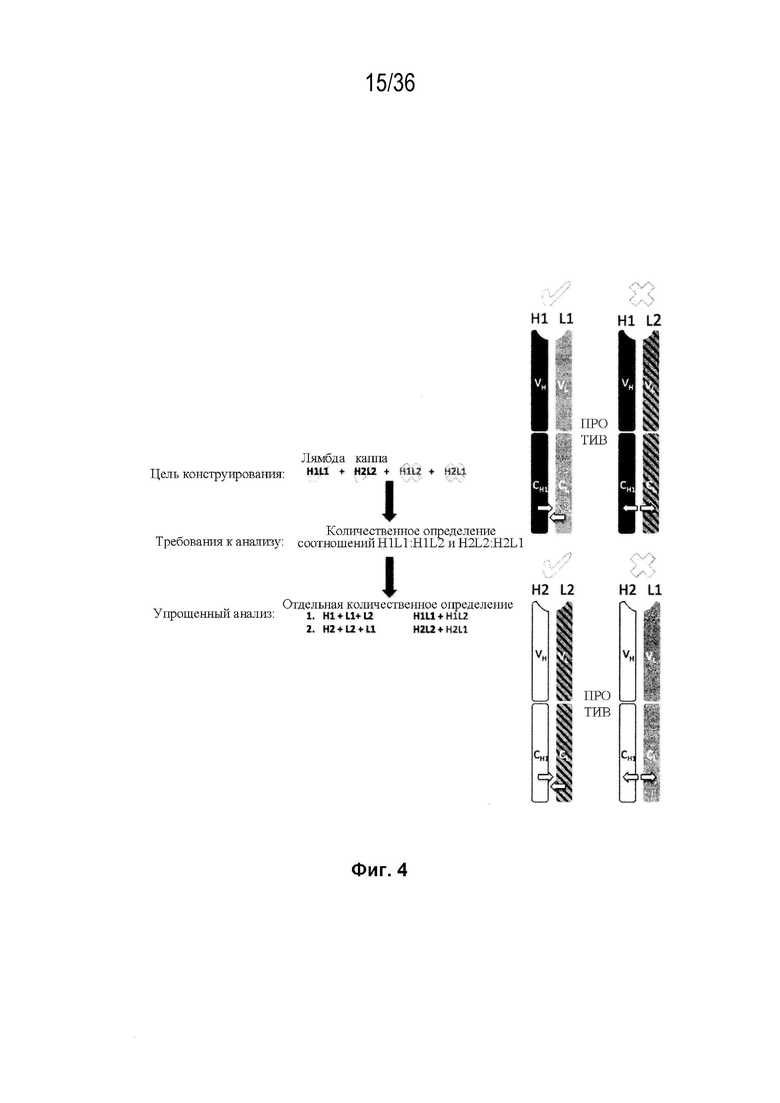
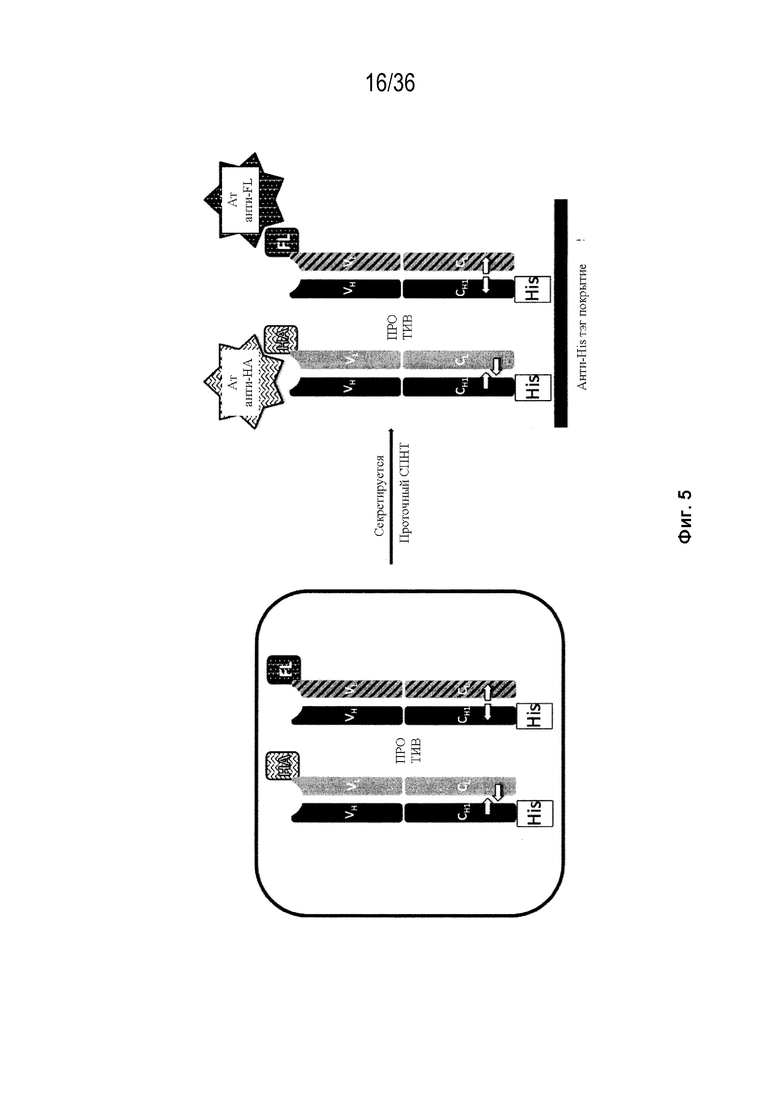
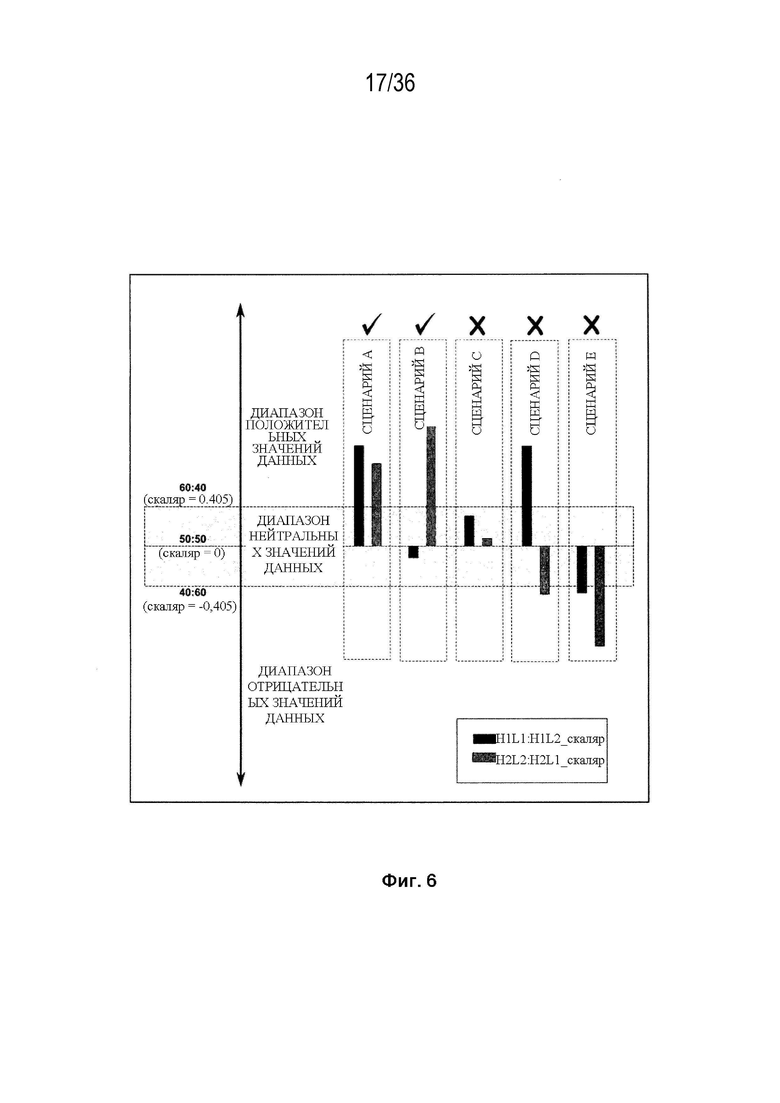
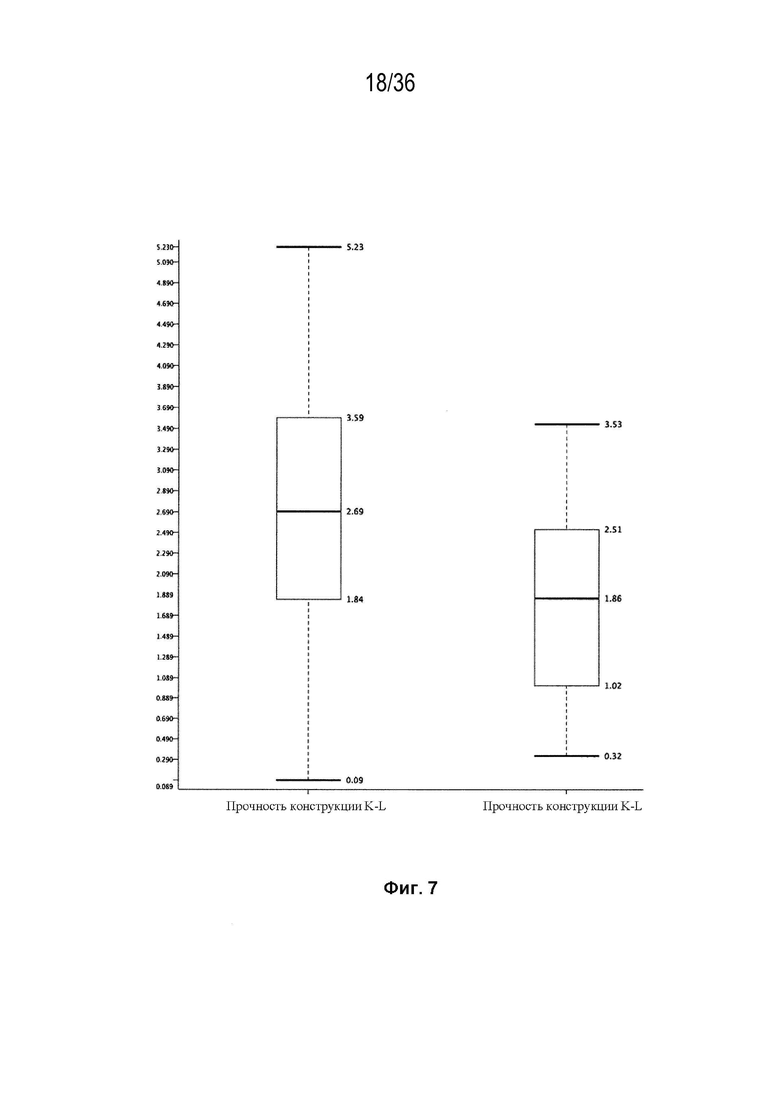
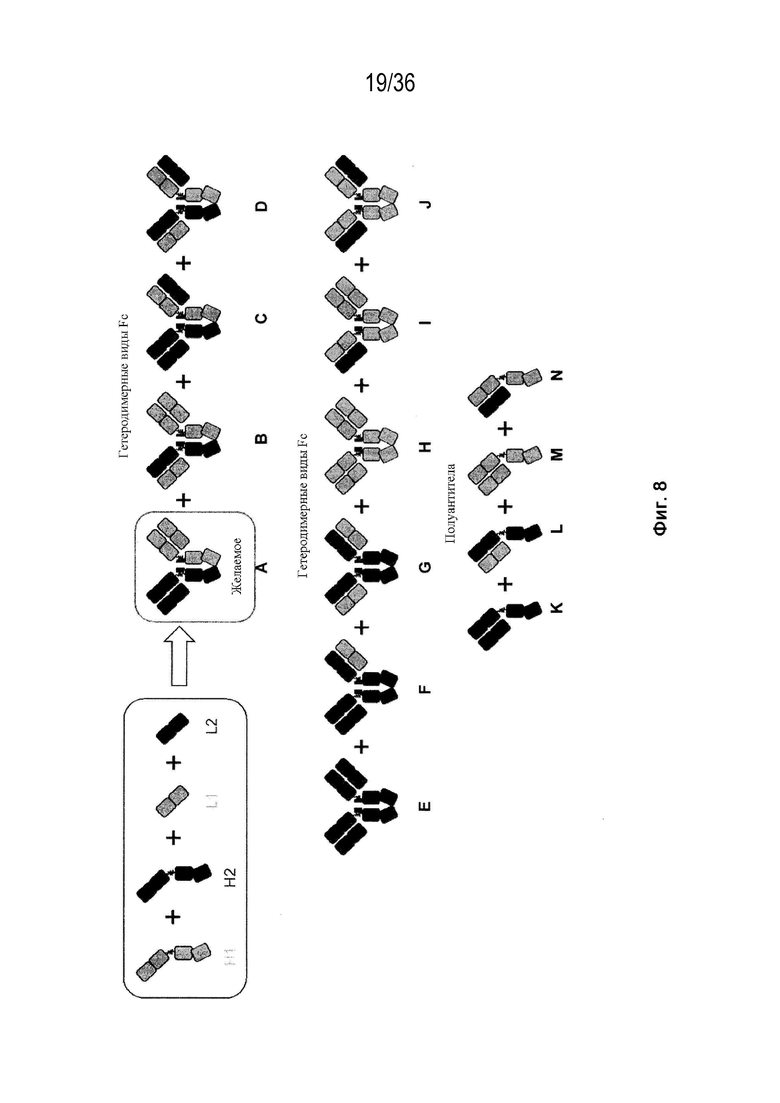
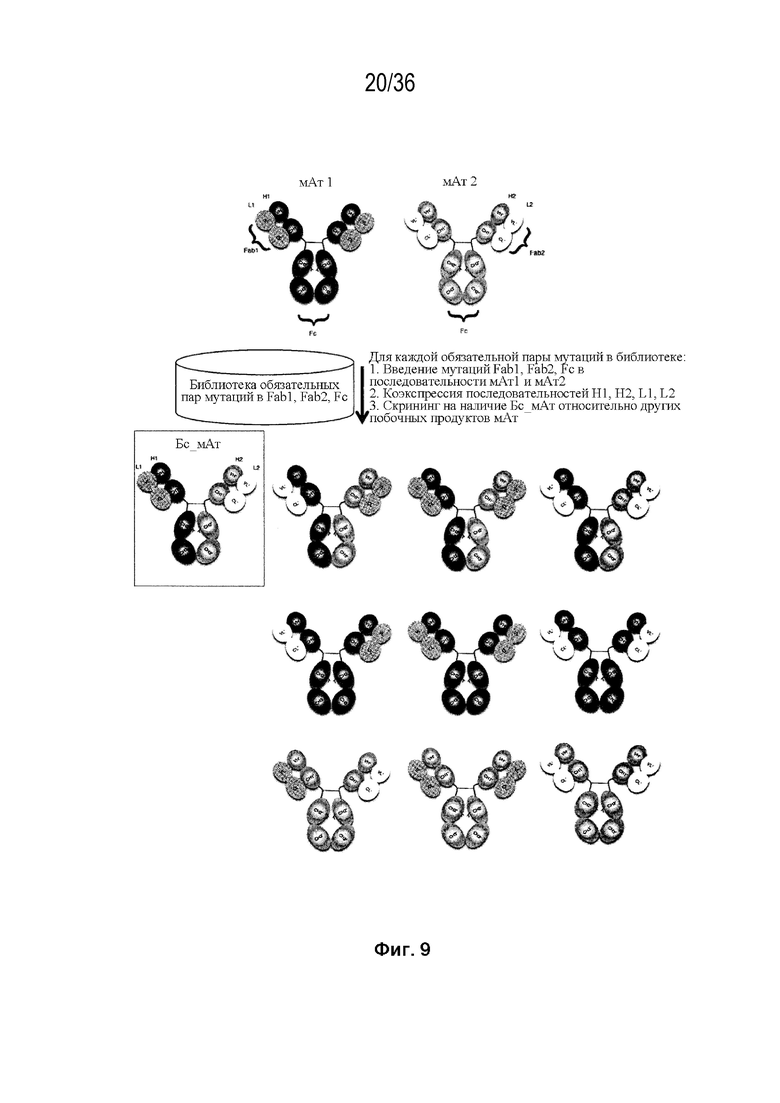
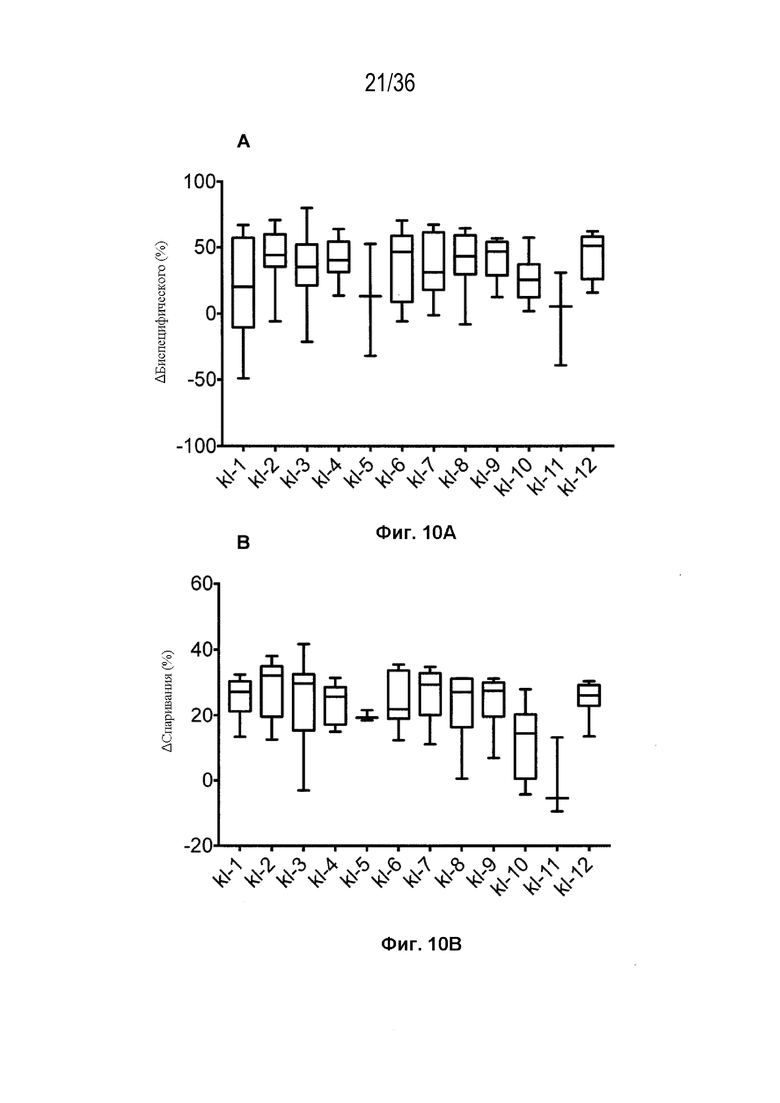

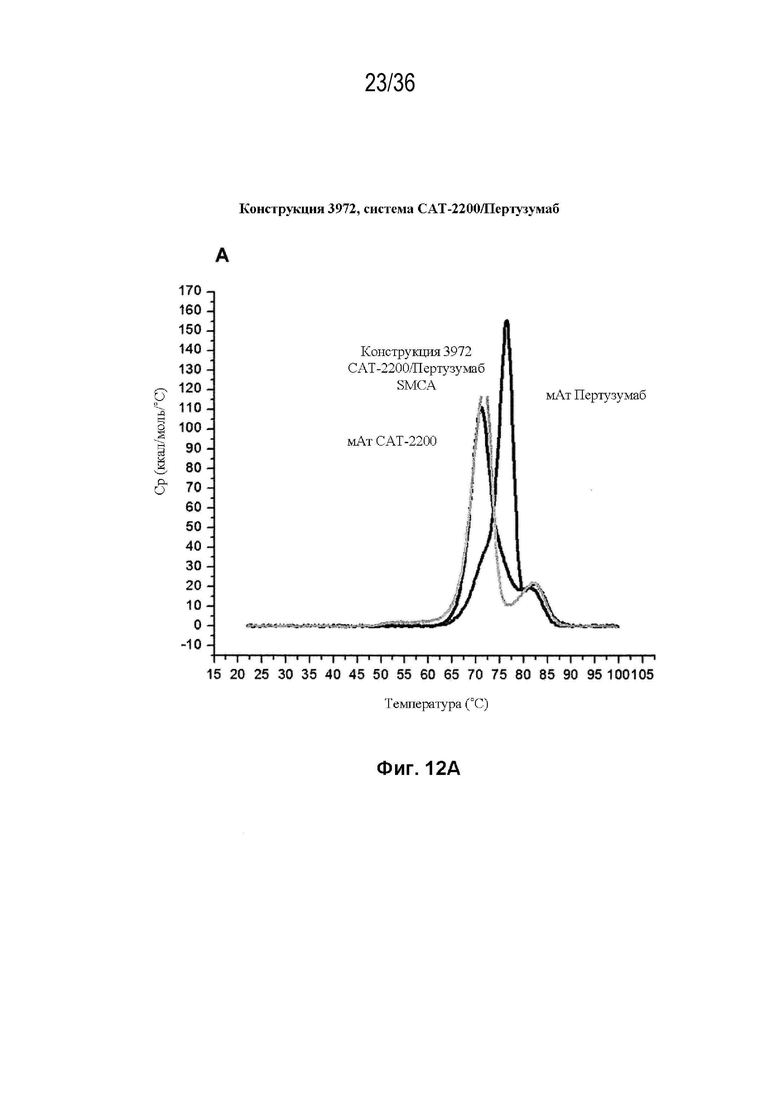
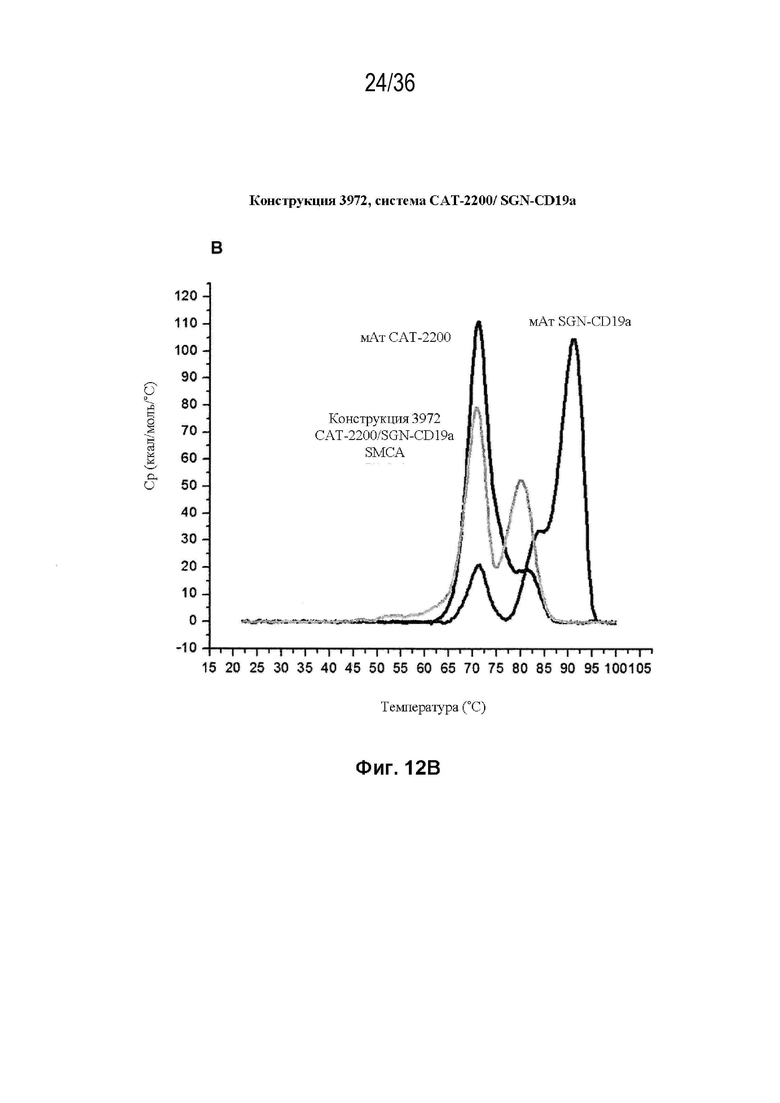
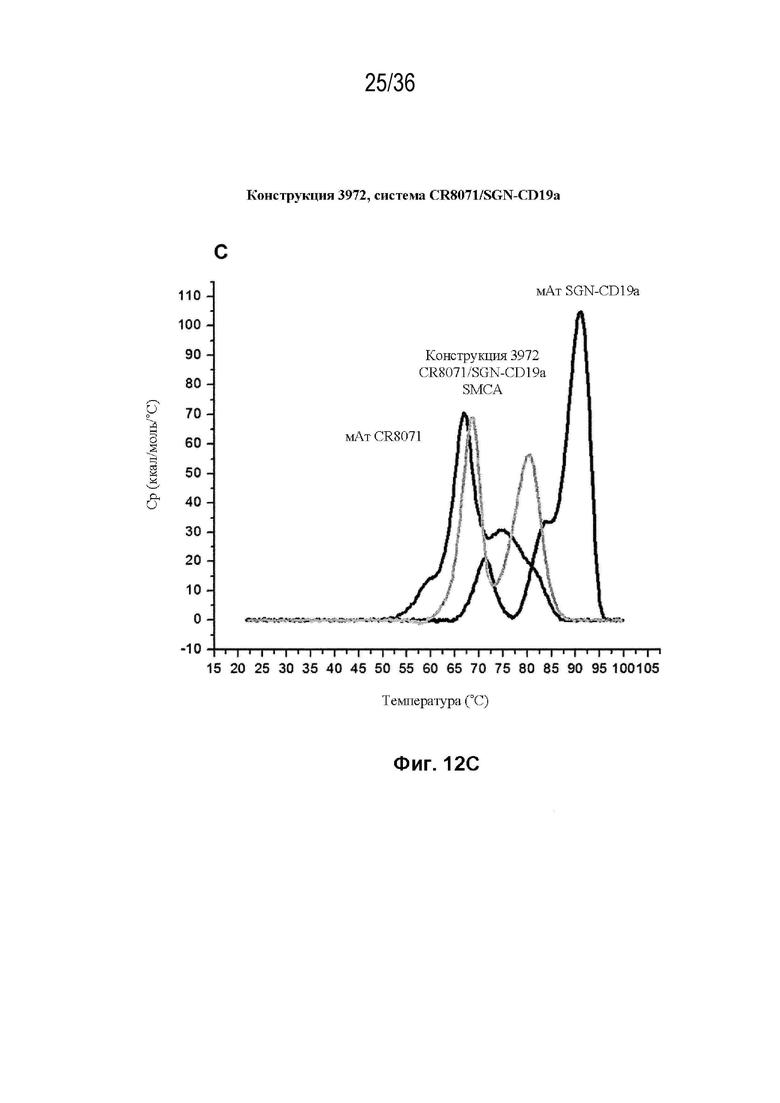
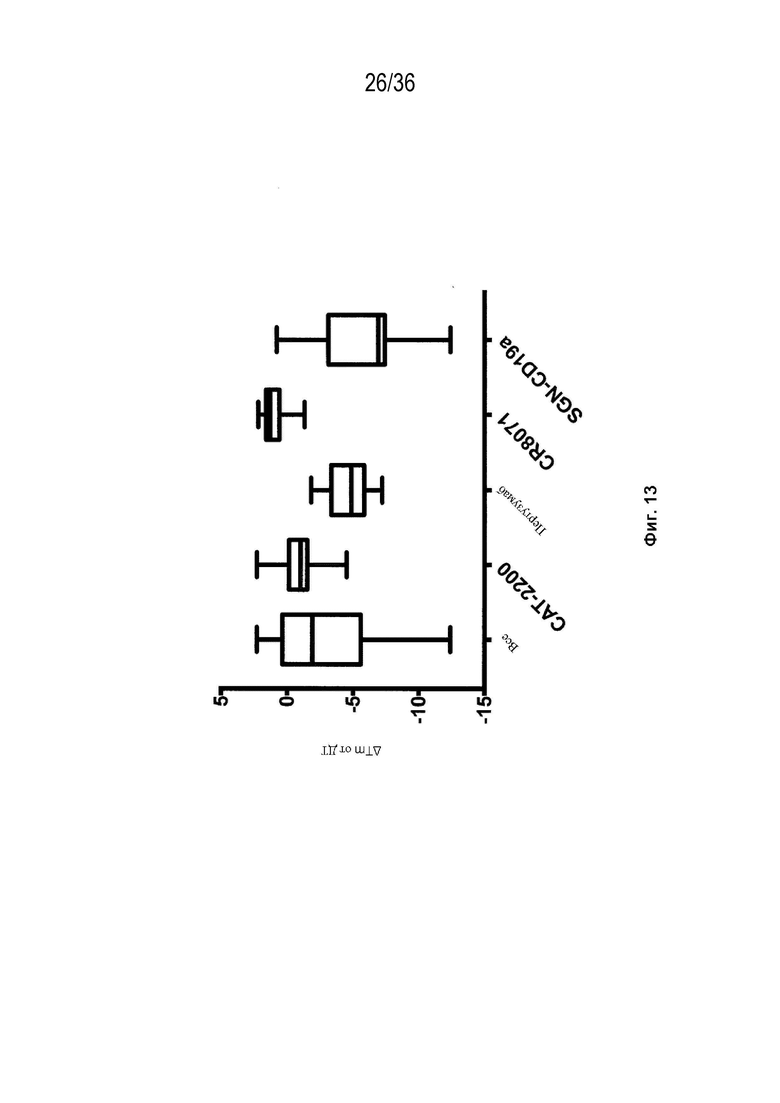
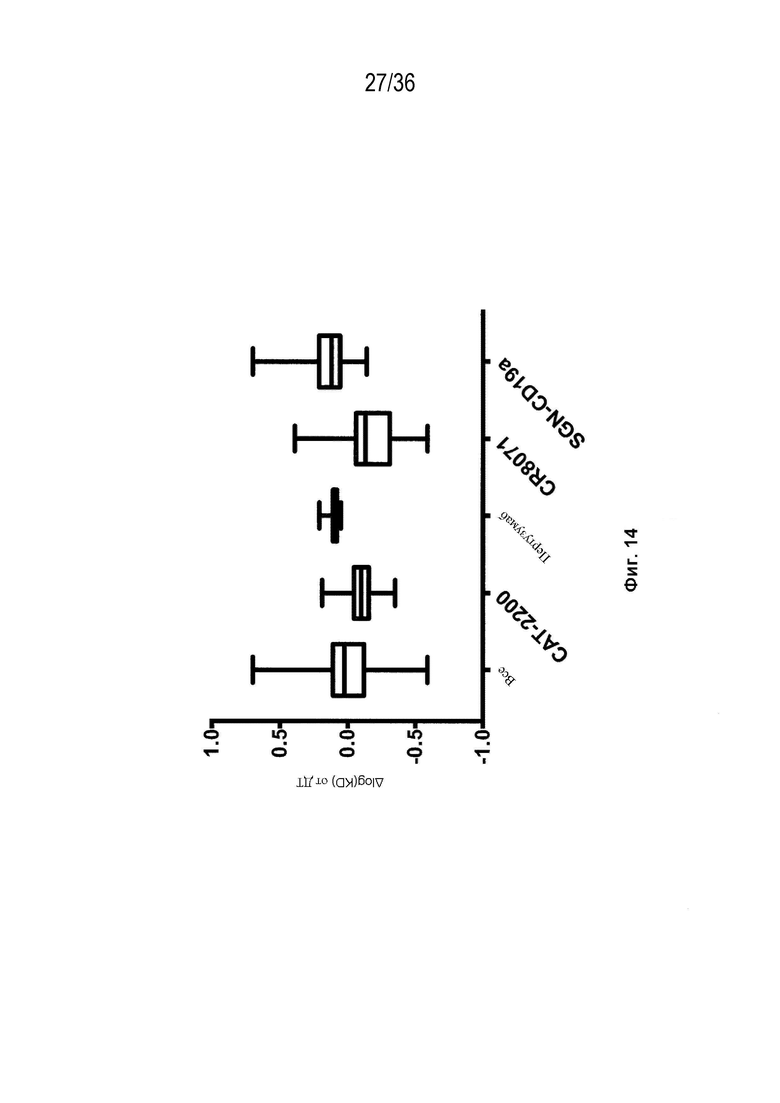
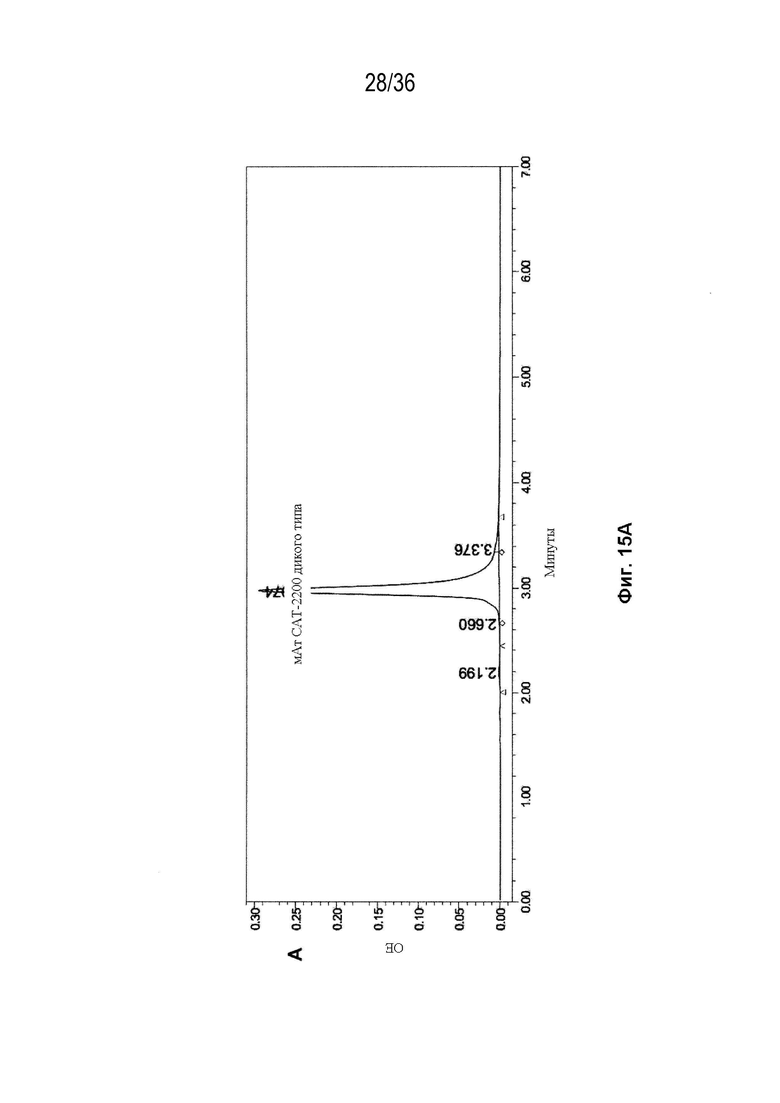
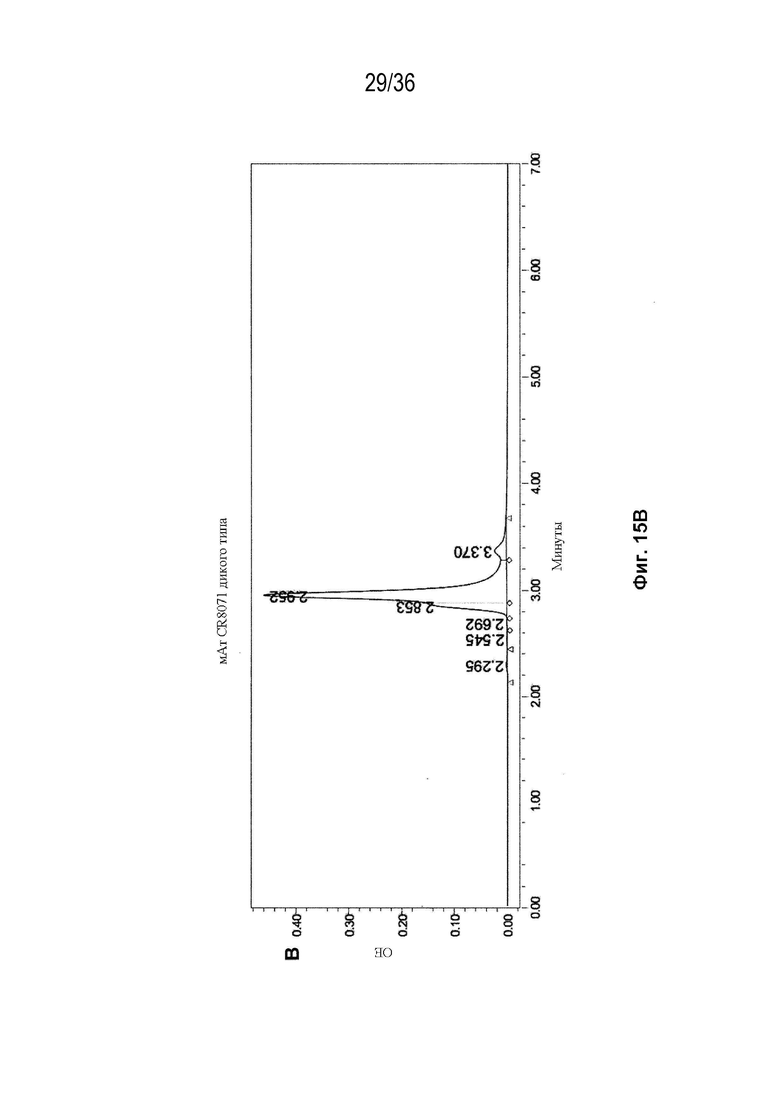
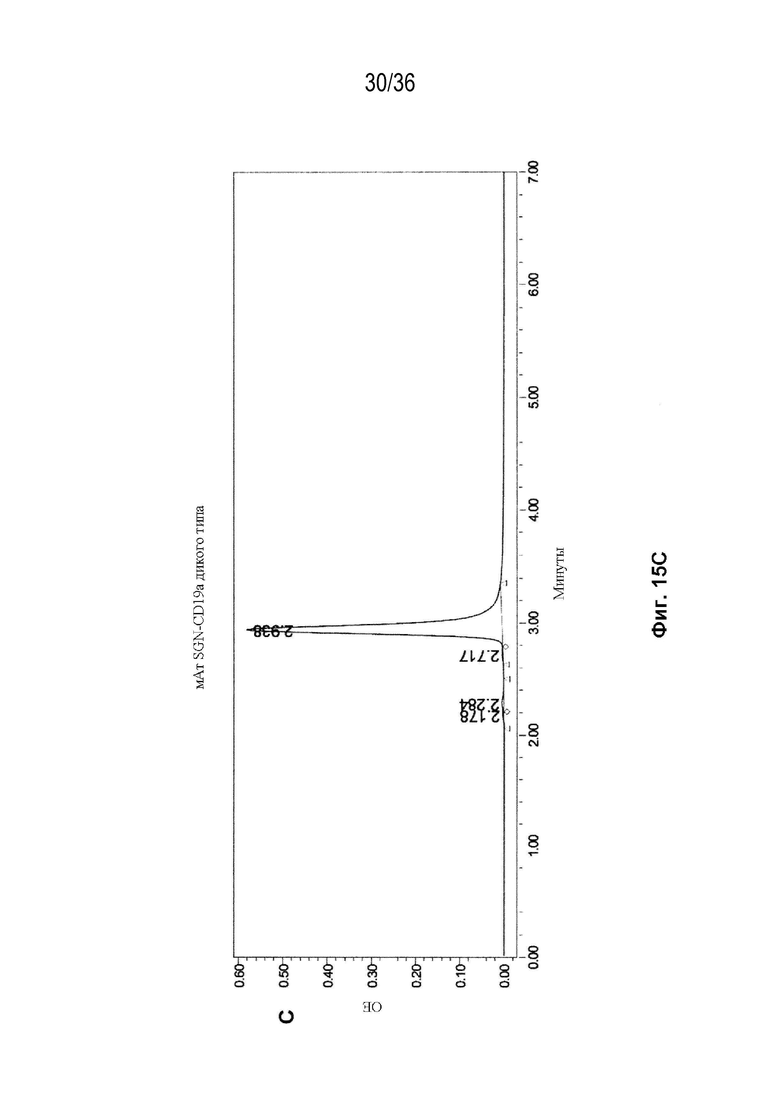
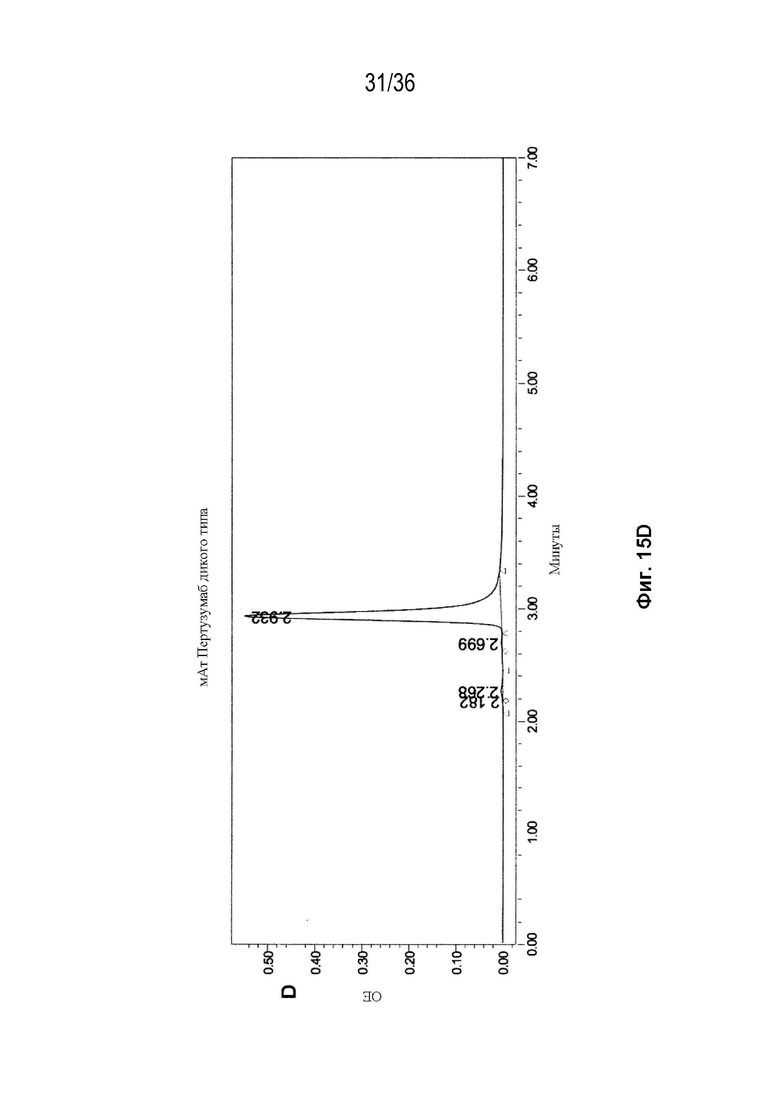
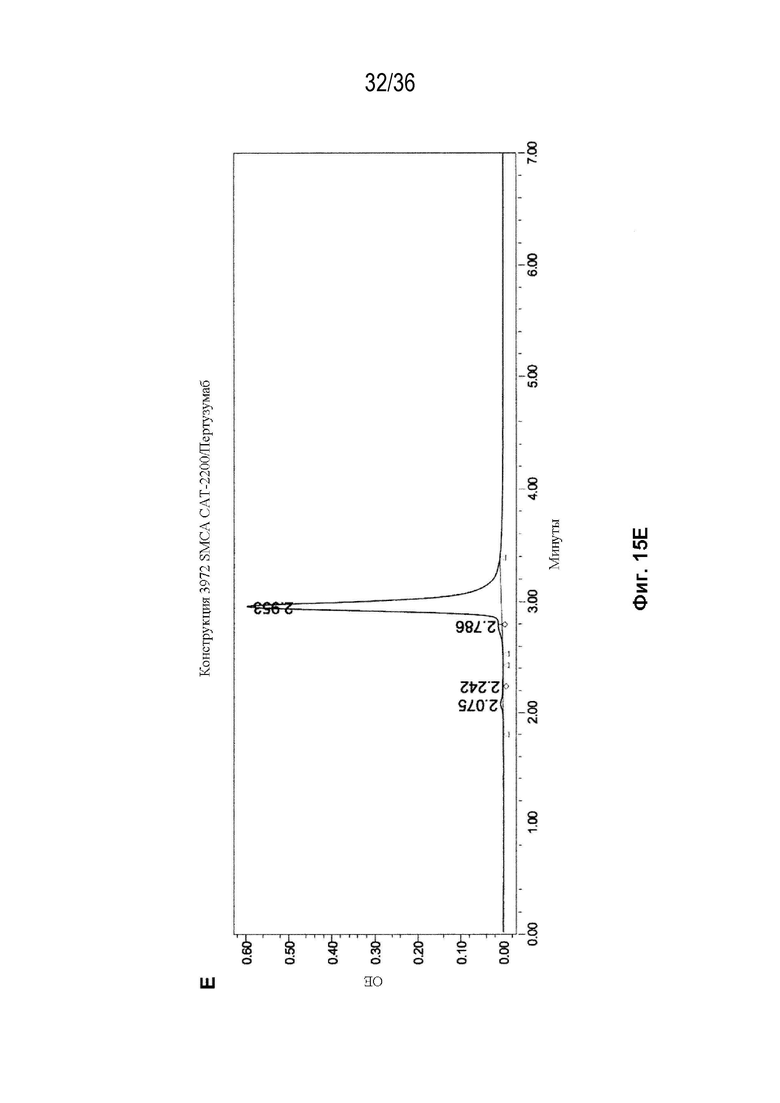
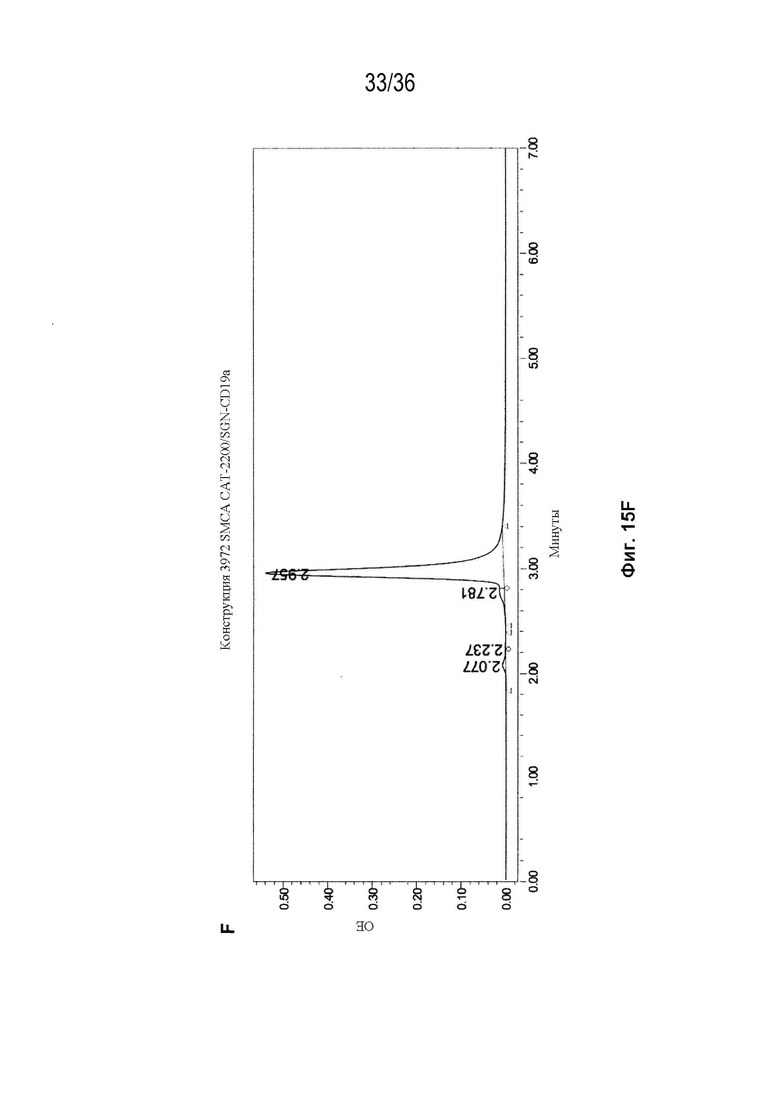
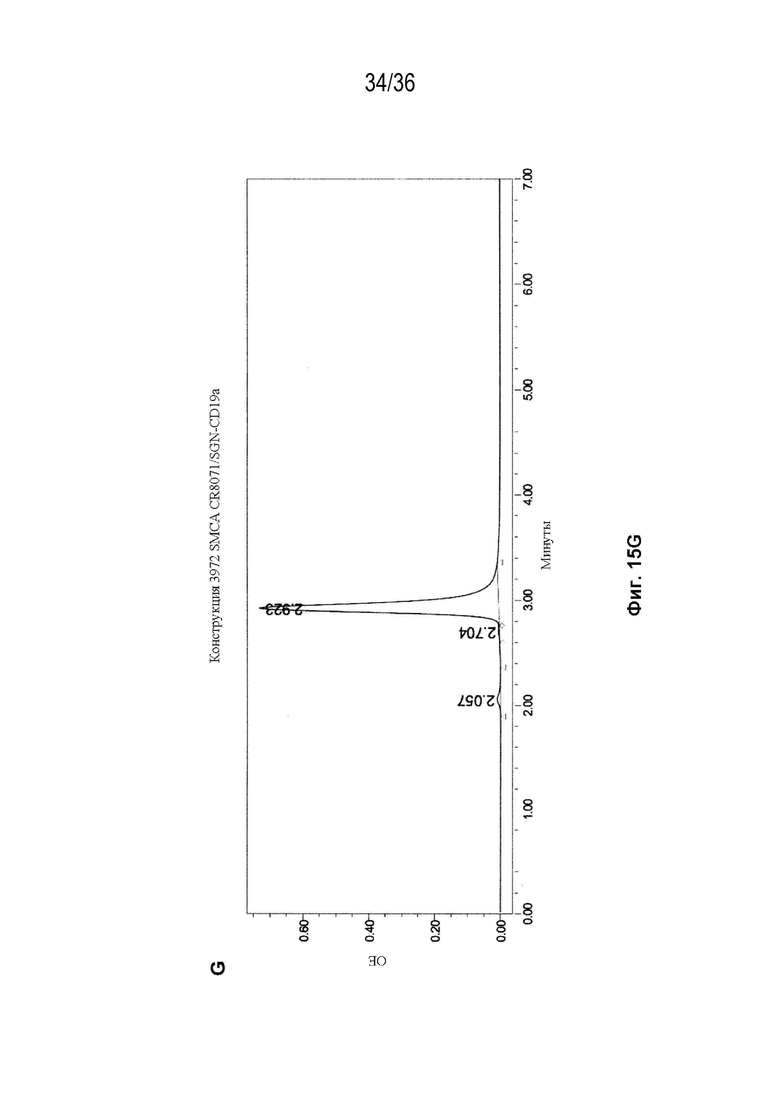
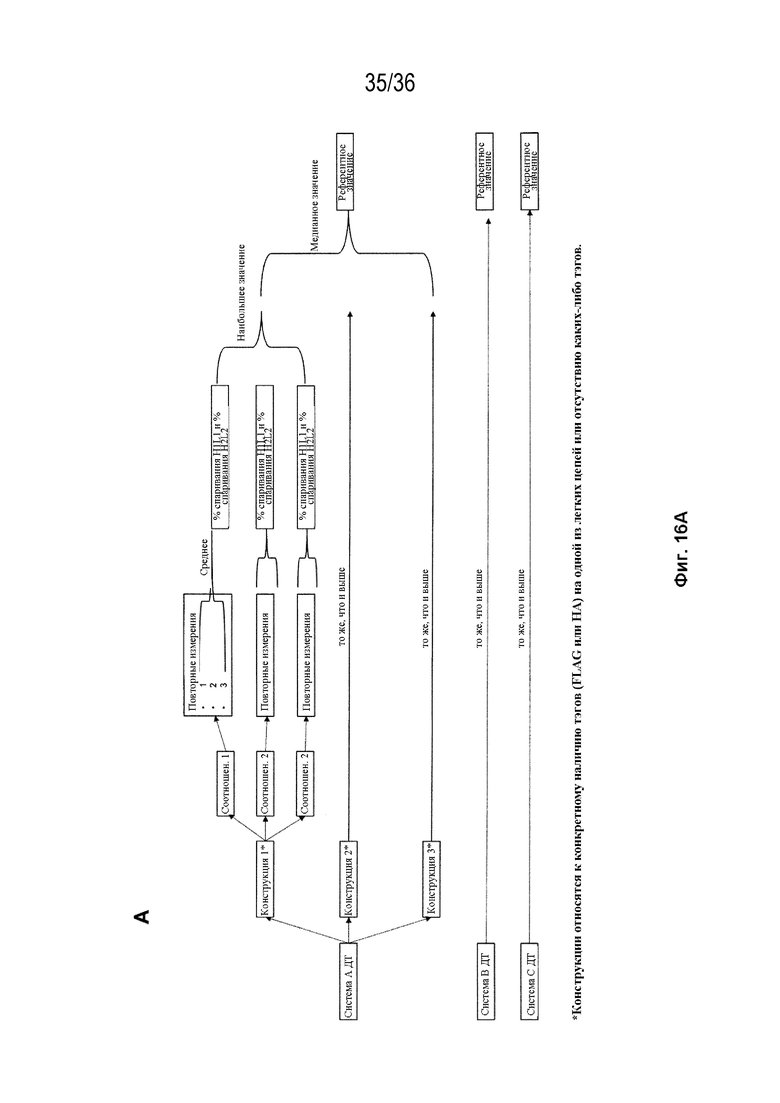
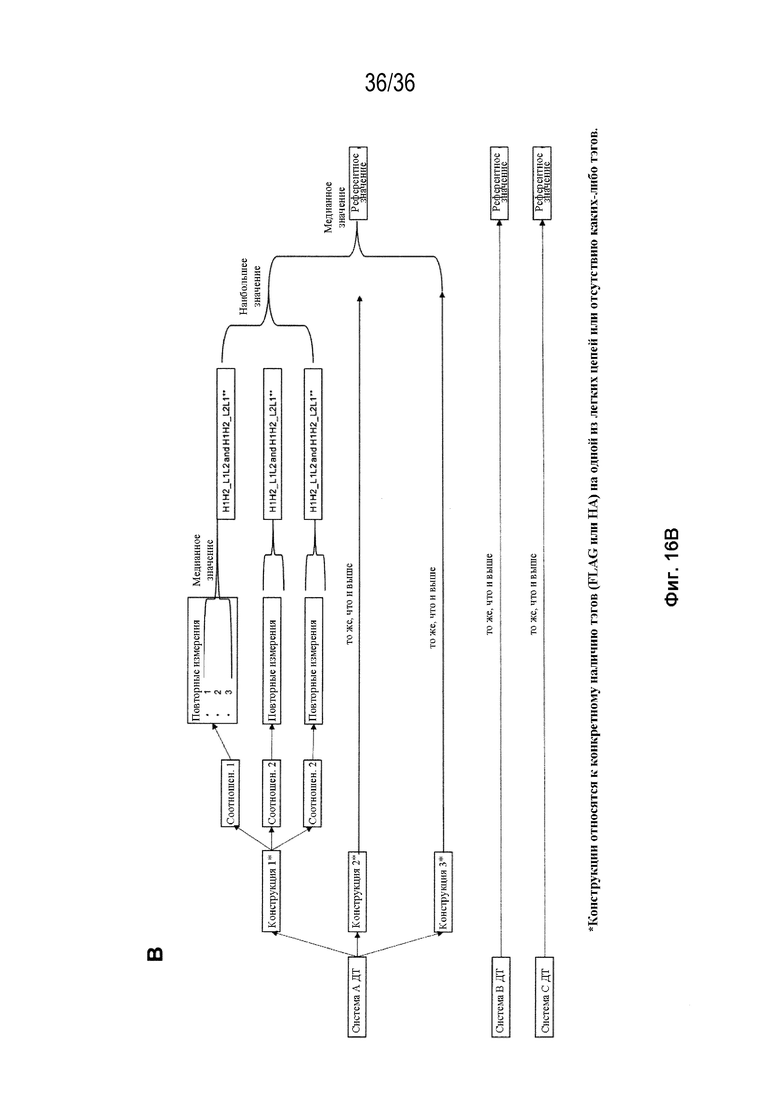
Изобретение относится к области биотехнологии, в частности к способу получения биспецифической антигенсвязывающей полипептидной конструкции. Указанная конструкция содержит первый (H1L1) и второй (H2L2) гетеродимеры, причем первый гетеродимер содержит первую область Fab, которая специфически связывается с первым антигеном; и второй гетеродимер (H2L2) содержит вторую область Fab, которая специфически связывается со вторым антигеном. Заявленный способ содержит следующие стадии: 1) введение одной или более замен в полинуклеотиды, кодирующие H1, H2, L1, L2; 2) введение указанных полинуклеотидов в клетку-хозяин и ее культивирование; 3) сбор биспецифической антигенсвязывающей полипептидной конструкции. Настоящее изобретение обеспечивает антигенсвязывающие полипептиды, содержащие легкую цепь иммуноглобулина лямбда и легкую цепь иммуноглобулина каппа, полученные с помощью комплементарных аминокислотных модификаций. 7 з.п. ф-лы, 16 ил., 25 табл., 21 пр.
1. Способ получения биспецифической антигенсвязывающей полипептидной конструкции, содержащей первый гетеродимер и второй гетеродимер,
причем первый гетеродимер (H1L1) содержит полипептид тяжелой цепи (H1) первого иммуноглобулина G (IgG) и полипептид легкой цепи (L1) иммуноглобулина лямбда, которые образуют первую область Fab, которая специфически связывается с первым антигеном; и второй гетеродимер (H2L2) содержит полипептид тяжелой цепи (H2) второго иммуноглобулина G (IgG) и полипептид легкой цепи (L2) иммуноглобулина каппа, которые образуют вторую область Fab, которая специфически связывается со вторым антигеном, и
при этом:
H1, L1, H2 и L2 являются человеческими или гуманизированными полипептидами, и причем H1 преимущественно спаривается с L1 по сравнению с L2, а H2 преимущественно спаривается с L2 по сравнению с L1;
причем способ включает стадии (1)-(3):
(1) введение одной или более аминокислотных замен в полинуклеотид, кодирующий константный домен 1 тяжелой цепи (CH1) H1 и H2, в полинуклеотид, кодирующий константный домен легкой цепи лямбда (CL-лямбда) L1 и в полинуклеотид, кодирующий константный домен легкой цепи каппа (CL-каппа) L2, что способствуют преимущественному спариванию H1 с L1 по сравнению с L2 и/или способствуют преимущественному спариванию H2 с L2 по сравнению с L1, причем одна или более аминокислотных замен выбраны из одного из следующих наборов аминокислотных замен, причем нумерация аминокислотных остатков приведена по Кабату:
a) H1 содержит аминокислотную замену 186K, L1 содержит аминокислотные замены 129T, 133D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
b) H1 содержит аминокислотные замены 186K и 188T, L1 содержит аминокислотные замены 133D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
c) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124R, 160K и 178R;
d) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T, 176A, 178W и 180E, H2 содержит аминокислотные замены 143E, 145T и 188W, и L2 содержит аминокислотные замены 124K, 176A и 178R;
e) H1 содержит аминокислотные замены 143A и 186R, L1 содержит аминокислотные замены 129T, 133W и 180E, H2 содержит аминокислотные замены 124W, 143E и 145T, и L2 содержит аминокислотные замены 124K, 133A и 178R;
f) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124R, 129K, 160K и 178R;
g) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 188L, и L2 содержит аминокислотные замены 124R и 178R;
h) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R и 178R;
i) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 188L, и L2 содержит аминокислотные замены 124R, 160K и 178R;
j) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R, 160K и 178R;
k) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124R, 160K и 178R;
l) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
m) H1 содержит аминокислотные замены 177I и 188K, L1 содержит аминокислотные замены 133L, 176D и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
n) H1 содержит аминокислотные замены 177I и 188K, L1 содержит аминокислотные замены 133L, 176D и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
o) H1 содержит аминокислотную замену 188K, L1 содержит аминокислотные замены 176E и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
p) H1 содержит аминокислотную замену 188K, L1 содержит аминокислотные замены 176E и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
q) H1 содержит аминокислотные замены 143S, 186K и 188T, L1 содержит аминокислотные замены 133D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
r) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T, 178E и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124R, 160K и 178R;
s) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R и 178R;
t) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
u) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T, 179E и 188W, и L2 содержит аминокислотные замены 124R, 176A и 178R;
v) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T, 179E, 186I и 188W, и L2 содержит аминокислотные замены 124R, 176A и 178R;
w) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160R и 178R;
x) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T, 179E и 188W, и L2 содержит аминокислотные замены 124R, 160R, 176A и 178R;
y) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T, 179E, 186I и 188W, и L2 содержит аминокислотные замены 124R, 160R, 176A и 178R;
z) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 124W, 143E, 145T и 179E, и L2 содержит аминокислотные замены 124K, 133A и 178R;
aa) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124K и 178R;
bb) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 122C, 143E, 145T и 179E, и L2 содержит аминокислотные замены 124C, 160K и 178R;
cc) H1 содержит аминокислотные замены 143A и 179K, L1 содержит аминокислотные замены 129T, 133W и 180E, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R, 160K и 178R;
dd) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R, 160K, и 178R;
ee) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T, 178E и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
ff) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
gg) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124R, 129K, 160K и 178R;
hh) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 188L, и L2 содержит аминокислотные замены 124R, 160K и 178R;
ii) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124K и 178R;
jj) H1 содержит аминокислотную замену 188K, L1 содержит аминокислотные замены 129T и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
kk) H1 содержит аминокислотные замены 139W, 143I и 186K, L1 содержит аминокислотные замены 129T, 133D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 135W, 160K и 178R;
ll) H1 содержит аминокислотные замены 139W и 186K, L1 содержит аминокислотные замены 129T, 133D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 135W, 160K и 178R;
mm) H1 содержит аминокислотные замены 139W и 143K, L1 содержит аминокислотные замены 129T и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 135W и 178R;
nn) H1 содержит аминокислотные замены 143K и 190K, L1 содержит аминокислотные замены 129T, 133D и 135S, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
oo) H1 содержит аминокислотные замены 124K и 143K, L1 содержит аминокислотные замены 129T, 131E и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160R и 178R;
pp) H1 содержит аминокислотные замены 124K и 143K, L1 содержит аминокислотные замены 129T, 131D и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160R и 178R;
qq) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T, 131E и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160R и 178R;
rr) H1 содержит аминокислотную замену 143K, L1 содержит аминокислотные замены 129T, 131D и 133D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160R и 178R;
ss) H1 содержит аминокислотные замены 139W и 188K, L1 содержит аминокислотные замены 129T, 176D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 135W и 178R;
tt) H1 содержит аминокислотные замены 139W и 188K, L1 содержит аминокислотные замены 129T, 176E и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 135W и 178R;
uu) H1 содержит аминокислотную замену 188K, L1 содержит аминокислотные замены 129T и 178D, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
vv) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 188L, и L2 содержит аминокислотные замены 124R и 178R;
ww) H1 содержит аминокислотную замену 188K, L1 содержит аминокислотные замены 129T, 176D и 178T, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
xx) H1 содержит аминокислотную замену 188K, L1 содержит аминокислотные замены 129T, 176E и 178E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
yy) H1 содержит аминокислотные замены 143A и 179K, L1 содержит аминокислотные замены 129T, 133W и 180E, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R и 178R;
zz) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T, 179E и 188L, и L2 содержит аминокислотные замены 124R и 178R;
aaa) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T, 178E и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
bbb) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 129K, 160K и 178R;
ccc) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 129K и 178R;
ddd) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 129K и 178R;
eee) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 160K и 178R;
fff) H1 содержит аминокислотную замену 186R, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R и 178R;
ggg) H1 содержит аминокислотные замены 179K и 190K, L1 содержит аминокислотные замены 129T, 133D, 135S и 180E, H2 содержит аминокислотные замены 143E и 145T, и L2 содержит аминокислотные замены 124R, 160K и 178R;
hhh) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 122C, 143E и 145T, и L2 содержит аминокислотные замены 124C, 160K и 178R;
iii) H1 содержит аминокислотные замены 143A и 179K, L1 содержит аминокислотные замены 129T, 133W и 180E, H2 содержит аминокислотные замены 143E, 145T и 188L, и L2 содержит аминокислотные замены 124R и 178R;
jjj) H1 содержит аминокислотные замены 143A и 179K, L1 содержит аминокислотные замены 129T и 133W и 180E, H2 содержит аминокислотные замены 143E, 145T и 188L, и L2 содержит аминокислотные замены 124R, 160K и 178R; или
kkk) H1 содержит аминокислотную замену 179K, L1 содержит аминокислотные замены 129T и 180E, H2 содержит аминокислотные замены 143E, 145T и 179E, и L2 содержит аминокислотные замены 124R, 129K, 160K и 178R;
(2) введение полинуклеотидов с одной или более аминокислотных замен в клетку-хозяин и культивирование клетки-хозяина в культуре клеток-хозяев в условиях, которые позволяют экспрессировать биспецифическую антигенсвязывающую полипептидную конструкцию, и
(3) сбор биспецифической антигенсвязывающей полипептидной конструкции из культуры клеток-хозяев.
2. Способ по п. 1, отличающийся тем, что конструкция дополнительно содержит димерный Fc, имеющий два полипептида Fc, каждый полипептид Fc из которых содержит последовательность домена CH2 и последовательность домена CH3 и связан с или без помощи линкеров с одной из первой области Fab и второй области Fab.
3. Способ по п. 2, отличающийся тем, что Fc представляет собой Fc человека, Fc IgG1 человека, Fc IgA человека, Fc IgG человека, Fc IgD человека, Fc IgE человека, Fc IgM человека, Fc IgG2 человека, Fc IgG3 человека или Fc IgG4 человека.
4. Способ по п. 2, отличающийся тем, что Fc содержит одну или более модификаций по сравнению с диким типом в, по меньшей мере, одной из последовательностей домена CH3, которые способствуют образованию гетеродимерного Fc.
5. Способ по п. 4, отличающийся тем, что Fc содержит:
i) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392M_T394W во втором полипептиде Fc;
ii) гетеродимерный Fc IgG1, имеющий модификации L351Y_F405A_Y407V в первом полипептиде Fc и модификации T366L_K392L_T394W во втором полипептиде Fc;
iii) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392L_T394W во втором полипептиде Fc;
iv) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_K392M_T394W во втором полипептиде Fc; или
v) гетеродимерный Fc IgG1, имеющий модификации T350V_L351Y_S400E_F405A_Y407V в первом полипептиде Fc и модификации T350V_T366L_N390R_K392M_T394W во втором полипептиде Fc,
причем нумерация аминокислотных остатков в полипептиде Fc приведена в соответствии с системой нумерации EU.
6. Способ по п. 2, отличающийся тем, что линкеры представляют собой один или более полипептидных линкеров.
7. Способ по п. 6, отличающийся тем, что линкеры содержат одну или более шарнирных областей антитела.
8. Способ по п. 7, отличающийся тем, что линкеры содержат одну или более шарнирных областей IgG1.
| FISCHER N | |||
| et al., Exploiting light chains for the scalable generation and platform purification of native human bispecific IgG, Nature communications, 2015, V | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| WO2014082179, 05.06.2014 | |||
| ДЕЕВ С.М | |||
| и др., Современные технологии создания неприродных антител для клинического применения, Acta Naturae, 2009, V | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ZHU Z | |||
| et | |||

