Изобретение относится к способам получения дсРНК, впРНК, бета-глюканов и маннанов при комплексной переработке биомассы дрожжей Saccharomyces cerevisiae и может быть использовано в биотехнологии, фармацевтической промышленности и ветеринарии для производства противовирусных и иммуномодулирующих препаратов, в том числе адъювантов для вакцин.
Разработка новых адъювантных систем как способ повышения эффективности вакцин нового поколения является весьма актуальной задачей в связи с рециркуляцией старых патогенов и возникновением новых. Адъюванты способствуют формированию более выраженного адаптивного иммунитета путем активации разных звеньев системы врожденного иммунитета и/или стимуляции процессов транспорта, процессинга и презентации антигенов, прямо или косвенно воздействуя на антиген-презентирующие клетки.
В настоящее время по-прежнему самыми распространенными и широко используемыми адъювантами являются адъюванты на основе соединений алюминия, вошедшие в клиническую практику еще в 20-годах прошлого века [Landsteiner K. The Specificity of serological reactions / rev. ed. - Dover: New York, 1945. - 310 р.]. Помимо солей алюминия, в разных странах мира в составе лицензированных вакцин содержатся: масляно-водные наноэмульсии (AS04, AS03) (США), виросомы; Montanide™ ISA51 (водно-масляная эмульсия); аналоги липида А бактерий (страны Европы), полиоксидоний (Россия). В последние годы назрела необходимость в разработке нового поколения адъювантов, чему есть несколько причин. Во-первых, известно, что соли алюминия, эффективные для бактериальных анатоксинов, не являются оптимальными для вакцин нового поколения. Во-вторых, существует проблема токсичности адьювантов. Показано, что все вышеперечисленные адъюванты, наряду со стимуляцией специфических иммунных реакций, вызывают ряд токсических, воспалительных реакций. Соли алюминия и масляные эмульсии, в частности, приводят к развитию грануломатозных реакций, хроническому воспалению. Соединения алюминия способны вызывать развитие аллергических реакций [Исаенко Е.Ю., Бабич Е.М., Елисеева И.В., Ждамарова Л.А., Белозерский В.И., Колпак С.А. Адъюванты в современной вакцинологии // Annals of Mechnikov Institute. -2013. - №. 4.-С. 5-21].
Двуспиральные РНК (дсРНК) относятся к классу иммуномодуляторов, индукторов интерферона с широким спектром биологического действия. Индукторы интерферона на основе дсРНК оказывают защитный эффект в отношении значительного спектра вирусов [Ершов Ф.И., Наровлянский А.Н. Использование индукторов интерферона при вирусных инфекциях // Вопр. вирусол. - 2015. - Т. 60, №2. - С. 5-10; Wong J.P., Nagata L.P., Christopher М.Е., Salazar A.M., Dale R.M. Prophylaxis of acute respiratory virus infections using nucleic acid-based drugs // Vaccine. - 2005. - Vol. 23, No. 17-18. - P. 2266-2268. doi: 10.1016/j.vaccine.2005.01.037]. Описана способность дсРНК усиливать иммуногенность антигенов [Scallan CD., Tingley D.W., Lindbloom J.D., Toomey J.S., Tucker S.N. An adenovirus-based vaccine with a double-stranded RNA adjuvant protects mice and ferrets against H5N1 avian influenza in oral delivery models // Clin. Vaccine Immunol. - 2013. - Vol.20, No. 1. - P. 85-94. doi: 10.1128/CVI.00552-12; Uppu D.SSM., Turvey M.E., Sharif A.R.M., Bidet K., He Y., Но V., Tambe A.D., Lescar J., Tan E.Y., Fink K., Chen J., Hammond P.T. Temporal release of a three-component protein subunit vaccine from polymer multilayers // J. Control Release. - 2020. - Vol.317. - P. 130-141. doi: 10.1016/j.jconrel.2019.11; Wolferstatter M, Schweneker M., Spath M, Lukassen S., Klingenberg M., Brinkmann K., Wielert U., Lauterbach H., Hochrein H., Chaplin P., Suter M, Hausmann J. Recombinant modified vaccinia virus Ankara generating excess early double-stranded RNA transiently activates protein kinase R and triggers enhanced innate immune responses // J. Virol. - 2014. Vol. 88, No. 24. - P. 14396-14411. doi: 10.1128/JVI.02082-14; Каплина О.H., Гамалей С.Г., Иванова О.С., Даниленко Е.Д. Двуспиральные РНК - перспективные адъюванты для повышения иммуногенности вакцин // Журнал микробиологии, эпидемиологии и иммунологии. - 2022. - №6. - С. 661-668. doi: 10.36233/0372-9311-342], что свидетельствует о перспективности их использования для повышения иммуногенности вакцин нового поколения.
Высокополимерные одноцепочечные РНК (впРНК) дрожжей обладают выраженными иммуномодулирующими свойствами. Продемонстрирована способность впРНК повышать пролиферативную активность клеток селезенки, продукцию спленоцитами ИФН-гамма, усиливать активность натуральных киллеров [Игнатьев Г.М., Кашенцева Е.А. Использование иммуномодулятора полирибоната совместно с вакциной желтой лихорадки в эксперименте. // Тез. докл. 1 Съезда иммунологов России, Новосибирск, 1993. - Новосибирск, 1992. - С. 184]. Следует отметить вклад впРНК в рост количества CD86+ спленоцитов in vitro и CD86+ клеток в популяции дендритных клеток после внутривенного введения препарата дсРНК [Цыпленкова Е.С., Вязовая Е.А., Даниленко Е.Д. Изучение влияния двуспиральных РНК на активность мышиных спленоцитов методом проточной цитометрии // Мед. иммунол. - 2022. - Т. 25, №. 2. - С. 387-394. doi: 10.15789/1563-0625-SOT-2572].
Многие природные полисахариды, такие как альфа-, бета-глюканы, бета-фруктаны, маннан и хитозан, обладают адъювантной активностью [Petrovsky N. Comparative safety of vaccine adjuvants: a summary of current evidence and future needs // Drug Saf. - 2015. - V. 38, No. 11. - P. 1059-1074. doi: 10.1007/s40264-015-0350-4; Fesel P. H., Zuccaro A. (3-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants // Fungal Genet. Biol. - 2015. - V. 38, No. 11. - P. 1059-1074. doi: 10.1016/j.fgb.2015.12.004]. Бета-глюканы характеризуются минимальным риском образования токсических метаболитов или накопления в тканях организма, сочетаются с разными типами вакцин, а их производство не отличается трудоемкостью. Благодаря этим преимуществам их применение в сочетании с любым типом вакцин, в том числе, векторными или ДНК-вакцинами, может быть целесообразным и перспективным [Курашова С.С, Дзагурова Т.К., Ишмухаметов А.А., Егорова М.С., Баловнева М.В., Соцкова С.Е., Ткаченко Е.А. Адъюванты на основе углеводов для производства вакцин // БИОпрепараты. Профилактика, диагностика, лечение. - 2018. - Т. 18, №2. -С. 81-91. doi: 10.30895/2221-996Х-2018-18-2-81-91].
Маннаны и их производные широко применяют для доставки антигенов в дендритные клетки, особенно в составе вакцин против опухолей человека. Связывание маннанов с помощью маннан-связывающего лектина и других лектинов С-типа семейства маннозных рецепторов приводит к активации комплемента, опсонизации, фагоцитозу, активации инфламмосом, каспазы 1 и выработке провоспалительных цитокинов (в том числе IL-1β, IL-6, TNF-α) [Lamkanfi М., Malireddi R.K., Kanneganti T.D. Fungal zymosan and mannan activate the cryopyrin inflammasome // J. Biol. Chem. - 2009. - Vol. 284, No. 31. -P. 20574-20581. doi: 10.1074/jbc. M109.023689; Takahara K., Yashima Y., Omatsu Y., Yoshida H., Kimura Y., Kang Y.S., Steinman R.M., Park C.G., Inaba K. Functional comparison of the mouse DC-SIGN, SIGNR1, SIGNR3 and Langerin, C-type lectins // Int. Immunol. - 2004. - Vol. 16, No. 6. - P. 819-829. doi: 10.1093/intimm/dxh084]. Известно, что маннаны смещают развитие иммунного ответа в сторону Тп2-реакции [Katsara М., Yuriev Е., Ramsland Р.А., Tselios Т., Deraos G., Lourbopoulos A, Grigoriadis N., Matsoukas J., Apostolopoulos V. Altered peptide ligands of myelin basic protein (MBP87-99) conjugated to reduced mannan modulate immune responses in mice. -Immunology. - 2009. - Vol. 128, No. 4. - P. 521-533. doi: 10.1111/j. 1365-2567.2009.03137.x], приводят к повышению активности В-клеток и усилению продукции антител [Ярилин А.А. Иммунология: учебник для студентов [Текст]. - Москва: ГЭОТАР-Медиа, 2010. - С.416-423], что повышает эффективность иммунизации против ряда патогенов [Vassilaros S., Tsibanis A., Tsikkinis A., Pietersz G.A., McKenzie I.F., Apostolopoulos V. Up to 15-year clinical follow-up of a pilot phase III immunotherapy study in stage II breast cancer patients using oxidized mannan-MUC 1. - Immunotherapy. - 2013. - Vol. 5, No. 11. - P. 1177-1182. doi: 10.2217/imt.l3.126; Katsara M., Yuriev E., Ramsland P.A., Tselios Т., Deraos G., Lourbopoulos A., Grigoriadis N., Matsoukas J., Apostolopoulos V. Altered peptide ligands of myelin basic protein (MBP87-99) conjugated to reduced mannan modulate immune responses in mice. -Immunology. - 2009. - Vol. 128, No. 4. - P. 521-533. doi: 10.1111/j.1365-2567.2009.03137.x; Petrushina I., Ghochikyan A., Mkrtichyan M., Mamikonyan G., Movsesyan N., Ajdari R., Vasilevko V., Karapetyan A., Lees A., Agadjanyan M.G., Cribbs D.H. Mannan-Abeta28 conjugate prevents Abeta-plaque deposition, but increases microhemorrhages in the brains of vaccinated Tg2576 (APPsw) mice // J. Neuroinflammation. - 2008. - Vol. 5. - Article: 42. doi: 10.1186/1742-2094-5-42].
Известен патент RU 2216595 CI, опубл. 20.11.2003 г. на способ получения бета-глюканов клеточной стенки дрожжей. Клетки дрожжей, выращенных традиционным образом, разрушают механическим способом, предпочтительно в мельнице с бусами баллотини. Бета-глюканы отделяются от других полисахаридов, составляющих клеточную стенку дрожжей, в результате обработки изолированных клеточных стенок дрожжей ультразвуком с частотой 18-30 кГц/с, предпочтительно 18-22 кГц/с.
Суспензию отмытых клеточных стенок подвергают воздействию ультразвуком с частотой 18-30 кГц/с в течение 20-40 мин. Оптимальная концентрация клеточных стенок в суспензии составляет от 1 до 5%. Мощность ультразвукового воздействия зависит от объема суспензии и концентрации в ней клеточных стенок. В результате ультразвуковой обработки в данном режиме внутренний слой клеточной стенки, состоящий преимущественно из бета-глюканов, физически отделяется от внешнего слоя клеточной стенки, образуемого другими полисахаридами. Фактически дрожжевая клеточная стенка расслаивается на всем протяжении, и это четко видно при микроскопическом исследовании. Для удаления других полисахаридов, загрязняющих препарат β-1,3-глюканов, их разрушают щелочным или ферментативным гидролизом с помощью ферментов, деградирующих эти полисахариды. Ферментативный гидролиз может быть проведен обработкой суспензии, полученной в результате ультразвукового воздействия, ферментным препаратом эндоманнаназой в концентрации 2-3 Е/мл при температуре 45-48°С в течение 90 мин. В результате гидролиза маннаны и другие полисахариды, кроме бета-глюканов, превращаются в водорастворимые соединения и могут быть отделены от нерастворимых в воде бета-глюканов известными приемами, в частности, центрифугированием. В процессе такого рода обработки бета-глюканы не расщепляются и остаются нерастворимыми в воде. Осажденные бета-глюканы промываются и высушиваются. Полученные предложенным способом β-1,3-глюканы или β-1,6-глюканы содержат 80-85% β-глюкана. Маннаны при этом не рассматриваются в качестве целевого продукта и разрушаются в процессе очистки.
В Евразийском патенте ЕА 09677, опубл. 28.02.2008 г. «Средство для лечения гипергликемии и его применение» описан совмещенный способ получения бета-глюканов и маннанов из клеточных стенок дрожжей. Под термином «клеточные стенки дрожжей» здесь понимается нерастворимая фракция клеток дрожжей, не обогащенных хромом, полученная после автолиза или ферментативного гидролиза, главным образом, с помощью протеаз, приводящего к солюбилизации ≥50 мас. %, предпочтительно ≥60 мас.% сухого вещества от общей массы клеток и сохраняющего структурные полисахариды клеточной стенки, т.е. бета-глюканы и маннаны, причем маннаны находятся в форме маннопротеидов. Автолиз или ферментативный гидролиз проводят таким образом, чтобы солюбилизировать основную часть запасных сахаров клеток дрожжей, которыми являются гликоген и трегалоза. Клеточные стенки дрожжей получают разделением фракции, солюбилизированной путем автолиза или ферментативного гидролиза, имеющего продолжительность ≥18 ч. Предпочтительные способы автолиза водной суспензии дрожжевых клеток описаны в работе [Reed G., Nogodawithana T.W. Yeast Technology. - second ed., 1991. - New York. - P. 370-377]. Полученные таким образом клеточные стенки дрожжей обычно высушивают традиционными способами сушки, такими как распыление или сушка на нагреваемых валках. Согласно одному из вариантов изобретения для полного или практически полного удаления гликогена из клеточных стенок дрожжей клеточные стенки суспендируют в водной среде, нагревают в щелочной среде при температуре от 70 до 100°С в течение ≤3 ч и удаляют фракцию, солюбилизированную в результате данной обработки. Оставшуюся несолюбилизированной фракцию собирают и высушивают обычным образом. Такая обработка позволяет получить клеточные стенки дрожжей, имеющие общее содержание глюканов и маннанов 55-70 мас. % от массы сухого вещества, предпочтительно 60-70 мас. % и еще более предпочтительно 60-65 мас. % от массы сухого вещества.
Однако данный патент не содержит описания способа разделения маннанов и глюканов, кроме того, существует возможность потери в процессе выделения значительного количества маннопротеидов.
Варианты получения неразделенных продуктов маннанов и бета-глюканов описаны также в патенте России RU 2463065 C2, опубл. 10.10.2012 г. на применение дрожжевых клеточных стенок для лечения и/или предупреждения гиперинсулинемии. Для выделения и очистки полисахаридов используются ферментативный и щелочной гидролиз.
Патент России RU 2355190 С1, опубл. 20.05.2009 г. на способ получения биологически активной добавки не содержит этапов разделения маннанов и глюканов, выделение выполняется при достаточно низких рН, чтобы избежать потери маннопротеидов. Однако очевидно, что в данном виде технология получения приводит к существенным потерям β-глюканов. Кроме того, она предусматривает температурный режим, несовместимый с возможностью выделения дрожжевых РНК - не менее ценных биологически активных компонентов.
Известным аналогом в части технологии совмещенного получения маннанов и β-глюканов является патент RU 2504384 C2, опубл. 20.01.2014 г. на способ получения водорастворимых фракций маннопротеинов и Р-глюканов. В нем биомассу дрожжей сначала подвергают механической гомогенизации с ускорением мелющих тел 60-600 м/с2 в течение 0,5-15 мин, к полученному механокомпозиту добавляют раствор ферментативного комплекса с β-глюканазной или протеазной активностью. Ферментативный гидролиз проводят при рН 4-7, концентрации ферментного комплекса 0,5-10 мас. %, концентрации полученного механокомпозита 5-20 мас. %, при температуре 50-65°С в течение 10-30 ч, полученный гидролизат разделяют на маннопротеидную и β-глюкановую фракции, которые подвергают последующей очистке. Работа с материалом в кислых рН позволяет сохранить О-маннанолигосахариды в составе маннопротеидной фракции в отличие от многих других технологий их получения. В результате получают препараты маннопротеидов с содержанием основного вещества около 90% и бета-глюканов с содержанием 80%. Недостатком такой технологии является то, что маннаны выделяются не в чистом виде, а конъюгированном с протеинами; кроме того, при выделении β-глюканов и маннопротеидов не предусмотрено выделение из гомогената клеток других важных компонентов, таких как РНК.
Температурный режим на ферментативной стадии неприемлем для выделения дсРНК.
Известен аналог, касающийся получения суммарной РНК из клеток дрожжей (патент RU 2781832 С1, «Способ получения суммарной РНК из биомассы клеток дрожжей Saccharomyces cerevisiae», опубл. 18.10.2022 г.). В нем биомассу дрожжей инкубируют с буферным раствором (предпочтительный состав: натрий сернокислый - 100-150 мМ; ЭДТА - 5-10 мМ; калий фосфорнокислый однозамещенный - 20-80 мМ; калия хлорид -250-600 мМ; SDS - 3-8 мМ) и ферментативным препаратом зимолиазы при температуре 28-35°С до получения однородной суспензии с последующим охлаждением. Суспензию механически гомогенизируют под высоким давлением и центрифугируют.В полученный гомогенат добавляют хлорид натрия до концентрации 1,5-2,5 М, инкубируют 30-60 мин и центрифугируют. К супернатанту добавляют одноатомный спирт до концентрации 40-60 об.%. Полученную смесь перемешивают 4-6 ч, инкубируют ≥12 ч при 0-8°С, после чего центрифугируют для отделения осадка суммарной РНК.
Недостатком технологии, описанной в данном патенте, является использование зимолиазы, разрушающей β-глюканы.
Известен способ фракционирования нуклеиновых кислот (авторское свидетельство SU 1692986 A1, опубл. 23.11.1991 г.), в котором для выделения дсРНК к раствору нуклеиновых кислот добавляют уксуснокислый калий до концентрации 1,4-2,2 М, взятый в качестве замены хлористого лития, вносимого до концентрации 2,0 М, и выдерживают при 2-4°С. Осадок, содержащий впРНК, собирают центрифугированием. К надосадочной жидкости, содержащей дсРНК, ДНК и тРНК, добавляют уксуснокислый калий до полной концентрации 2,5-3,0 М, взятый в качестве замены хлористого лития, вносимого до концентрации 4,0 М, и выдерживают при 2-4°С. Осадок, содержащий дсРНК, собирают центрифугированием. ДНК и тРНК в надосадочной жидкости осаждают, добавляя 1-2 объема этанола, осадок собирают центрифугированием. Замена хлористого лития на уксуснокислый калий была проведена с целью удешевления технологии. Однако в данном патенте не описан способ выделения нуклеотидного материала из самих дрожжевых клеток.
Известен способ получения двуспиральной рибонуклеиновой кислоты (дсРНК) из клеток дрожжей (патент RU 2558256 C1, опубл. 27.07.2015 г.). В данном изобретении клетки дрожжей разрушают в растворе, содержащем 10 мМ Трис, 20 мМ ЭДТА и 0,5 М NaCl, рН 7,4, обработкой додецилсульфатом натрия в концентрации 0,5-1,0% в течение 20-25 мин при 20°С и хлороформом в концентрации до 25% в течение 20-25 мин при 20°С. Полученную смесь центрифугируют при 6000 об/мин в течение 20 мин, отделяют надосадочную жидкость. Концентрирование дсРНК из надосадочной жидкости проводят в 7-8%-ном растворе ПЭГ 6000 в течение ≥5 ч при 6°С с последующим центрифугированием при 6000 об/мин в течение 20 мин и растворением полученного осажденного концентрата в воде. Отделение одноцепочечных РНК от дсРНК осуществляют в 2 моль/л растворе LiCl в течение 5 ч при 6°С с последующим центрифугированием смеси при 6000 об/мин в течение 20 мин и сбором водной фазы. Затем проводят осаждение дсРНК из водной фазы в 3,5 моль/л растворе LiCl в течение 5 ч при 6°С с последующим центрифугированием при 6000 об/мин в течение 20 мин и растворением осадка в воде. Окончательную очистку и осаждение дсРНК из раствора осуществляют 55%-ным раствором этанола. Однако в данном патенте описывается выделение только дсРНК. При этом клеточные стенки, источник маннанов и глюканов, могут быть использованы для их выделения, а в процессе получения дсРНК возможно выделение и впРНК.
Таким образом, основным недостатком представленных выше аналогов является отсутствие комплексной технологии получения из дрожжей Saccharomyces cerevisiae биологически активных веществ полисахаридной и нуклеиновой природы, что увеличивает расход исходного сырья и затраты на их получение.
Из уровня техники неизвестны технологии комплексной переработки дрожжей Saccharomyces cerevisiae для одновременного получения дсРНК, впРНК и полисахаридов в виде глюканов и маннанов.
Техническим результатом заявляемого изобретения является создание такой технологии комплексной переработки биомассы Saccharomyces cerevisiae, которая позволяет в одном производственном цикле получать препараты дсРНК, впРНК, бета-глюканов и маннанов, обладающих адъювантными свойствами, при котором обеспечивается снижение отходов сырья и уменьшаются затраты на получение препаратов по сравнению с затратами на получение указанных препаратов из дрожжевых клеток в отдельности.
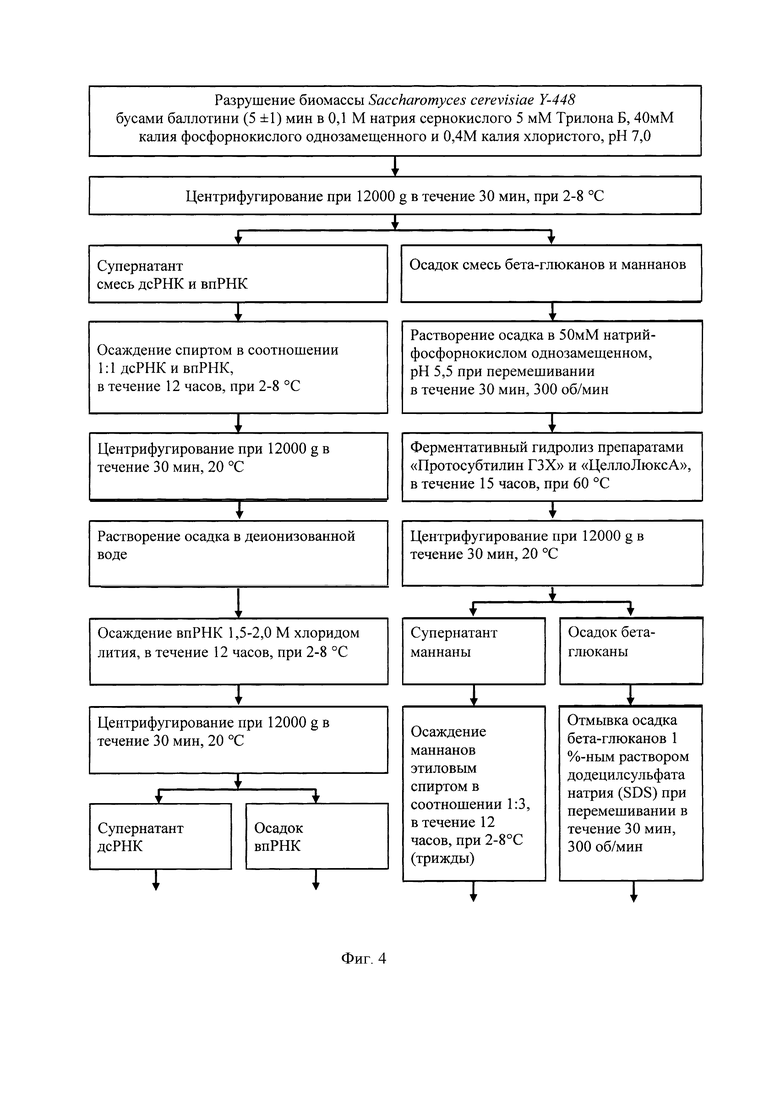
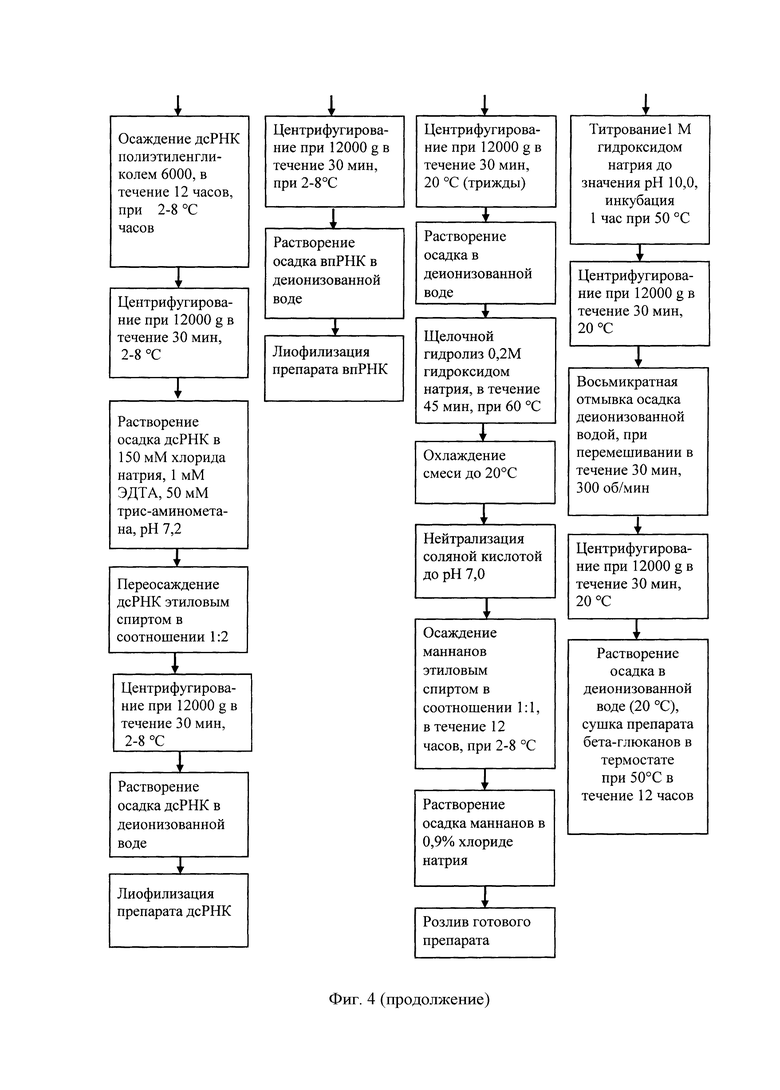
Указанный технический результат достигается тем, что способ получения препаратов дсРНК, впРНК, бета-глюканов и маннанов из биомассы Saccharomyces cerevisiae, согласно изобретения, включает суспендирование биомассы дрожжей в растворе, содержащем 0,1 М натрия сернокислого, 5 мМ Трилона Б, 40 мМ калия фосфорнокислого однозамещенного и 0,4 М калия хлористого при рН 7,0; помещение указанной суспензии в сосуд для разрушения, содержащий стеклянные бусы баллотини, в пропорции 2:1 и гомогенизацию смеси до получения лизата клеток; лизат клеток отделяют от бус баллотини и центрифугируют при оборотах и времени, обеспечивающих формирование плотного осадка, содержащего смесь бета-глюканов и маннанов, и супернатанта, содержащего смесь дсРНК и впРНК; из супернатанта выделяют смесь дсРНК и впРНК добавлением этилового спирта в соотношении 1:1 с последующей инкубацией при температуре и времени, обеспечивающих выпадение осадка дсРНК и впРНК; полученную смесь дсРНК и впРНК центрифугируют при скорости и времени, обеспечивающих образование плотного осадка, который растворяют в деионизованной воде до достижения раствором оптической плотности 200 о.е. при длине волны 260 нм; осаждение впРНК проводят хлоридом лития, для чего сухой хлорид лития добавляют к раствору до достижения концентрации 1,5-2,0 М, смесь инкубируют с последующим центрифугированием до образования плотного осадка; для дробного выделения дсРНК супернатант смешивают с этиловым спиртом в соотношении 1:4 с последующей инкубацией при температуре и времени, обеспечивающих выпадение осадка с дсРНК, который центрифугируют при скорости и времени до образования плотного осадка, который растворяют в растворе, содержащем 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2 до достижения раствором оптической плотности 150 о.е. при длине волны 260 нм; далее для освобождения от примесных ДНК и РНК дсРНК осаждают хлоридом лития, добавляя его к раствору до концентрации 3,5-4 М с инкубацией до получения осадка и последующим центрифугированием этого осадка до образования плотного осадка дсРНК, который растворяют в растворе, содержащем 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2, до достижения раствором оптической плотности 150 о.е. при длине волны 260 нм; для депротеинизации к раствору дсРНК добавляют полиэтиленгликоль 6000 до концентрации 8% с последующей инкубацией и центрифугированием до получения осадка дсРНК, который растворяют в 40 мл раствора, содержащего 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис-аминометана, рН 7,2, переосаждают этиловым спиртом в соотношении 1:2 и центрифугируют при скорости и времени до получения осадка, содержащего дсРНК, который растворяют в деионизованной воде и лиофильно высушивают с получением сухого препарата дсРНК, количество которого с цикла составляет до 0,125% от исходной биомассы дрожжей; для дробного выделения впРНК осадок впРНК растворяют в деионизованной воде и инкубируют при температуре 85-90°С, далее смесь охлаждают и центрифугируют при скорости и времени, достаточном для получения и отделения осадка, а впРНК из супернатаната осаждают 1,5 М хлоридом лития и осадок центрифугируют при скорости и времени до образования плотного осадка, содержащего впРНК, который растворяют в деионизованной воде до достижения раствором оптической плотности 200 о.е. при длине волны 260 нм, полученный раствор очищают гель-фильтрацией на колонке с Сефарозой 2 В, фракции которого с оптической плотностью, превышающей 200 о.е. при длине волны 260 нм, объединяют и осаждают впРНК из раствора этиловым спиртом в соотношении 1:3 с последующим центрифугированием при скорости и времени, обеспечивающих образование осадка, содержащего впРНК, который растворяют в деионизованной воде и лиофильно высушивают с получением сухого препарата впРНК, количество которого с цикла составляет до 0,25% от биомассы дрожжей; для выделения бета-глюканов и маннанов осадок после разрушения дрожжевой биомассы и центрифугирования растворяют в растворе, содержащем 50 мМ натрий-фосфорнокислый однозамещенный, рН 5,5, и перемешивают, к полученному раствору добавляют ферментные препараты «Протосубтилин ГЗХ» и «ЦеллоЛюксА» в соотношении 13,3:1, инкубируют в течение времени и температуре, достаточных для проведения полного ферментативного гидролиза, смесь компонентов которого центрифугируют при скорости, времени и температуре до получения осадка, содержащего бета-глюканы и супернатанта, содержащего маннаны; к раствору супернатанта добавляют 3 части этилового спирта для осаждения маннанов и центрифугируют при скорости, времени и температуре, достаточных для образования осадка, который подвергают водно-спиртовой отмывке трижды до осветления промывных вод, проводят щелочной гидролиз 0,2 М гидроксидом натрия, после чего рН полученной смеси доводят до 7,0 соляной кислотой; полученный полупродукт осаждают добавлением этилового спирта в соотношении 1:1 и растворяют осадок 0,9%-ным раствором хлорида натрия с получением препарата маннанов, составляющим 0,0725%) от биомассы дрожжей; для выделения бета-глюканов осадок, полученный после разрушения биомассы дрожжей, отмывают 1%-ным раствором додецилсульфата натрия (SDS) и центрифугируют при скорости, времени и температуре, достаточных для образования осадка, полученный осадок пятикратно отмывают деионизованной водой, растворяют в деионизованной воде и подвергают воздействию ультразвука с частотой 22 кГц/с, тремя циклами с получением суспензии, которую титруют 1 М гидроксидом натрия до значения рН 10.0 и центрифугируют с получением осадка, который отмывают 8 раз деионизованной водой, суспендируют в деионизованной воде и сушат в термостате при 50°С 12 часов с получением препарата бета-глюканов в сухой форме, составляющего 2% от биомассы дрожжей Saccharomyces cerevisiae.
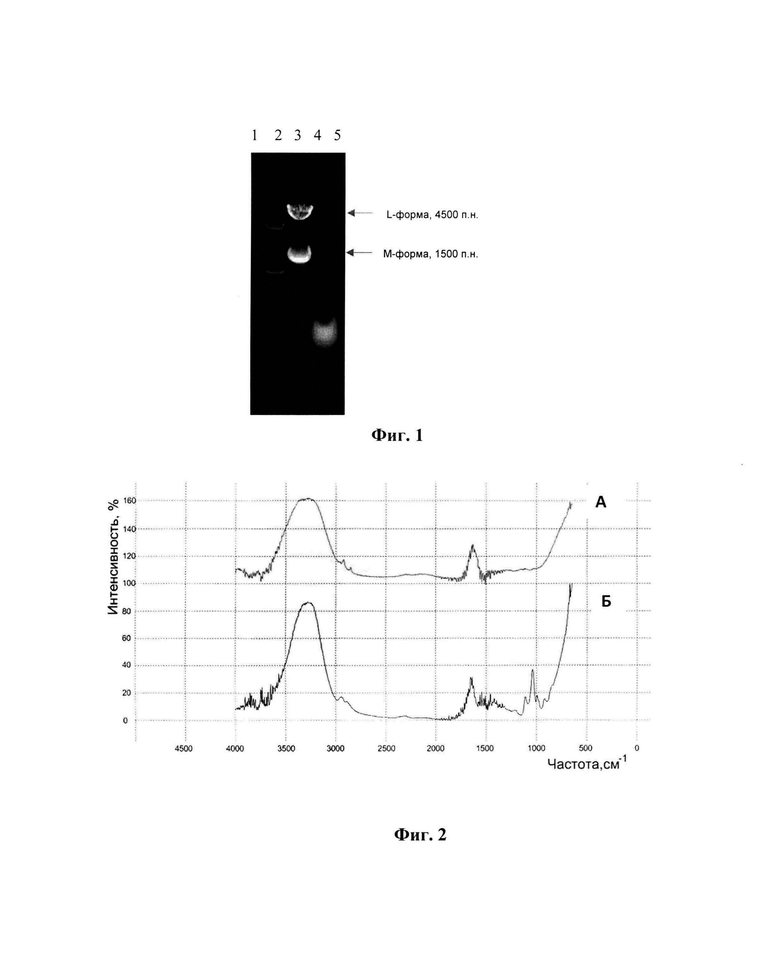
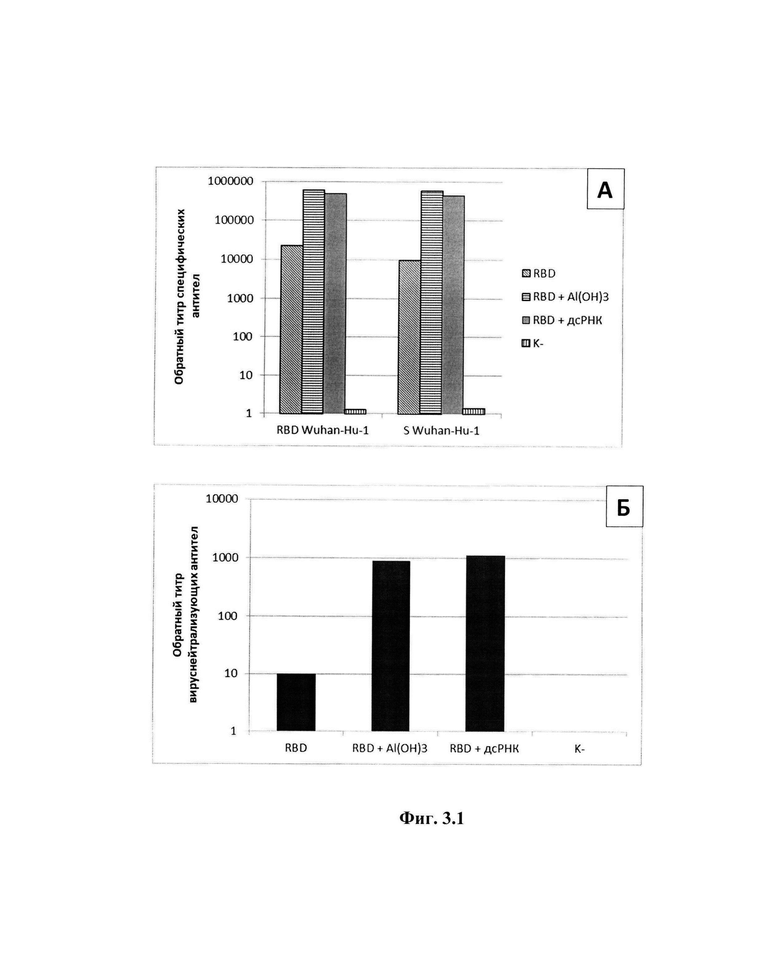
Изобретение поясняется следующими графическими материалами, представленными на фиг.1-4. На фиг.1 представлена электрофореграмма препаратов дсРНК и впРНК после электрофореза в 1%-ном геле агарозы и окрашивания этидием бромидом. Дорожки: 1 - маркеры молекулярных масс 250 - 10000 п. н. (СибЭнзим, Россия); 2 - препарат дсРНК; 3 - препарат впРНК. На фиг.2 представлены ИК-спектры препаратов бета-глюканов (А), в качестве препарата сравнения используется Зимозан (Sigma, США), (Б).
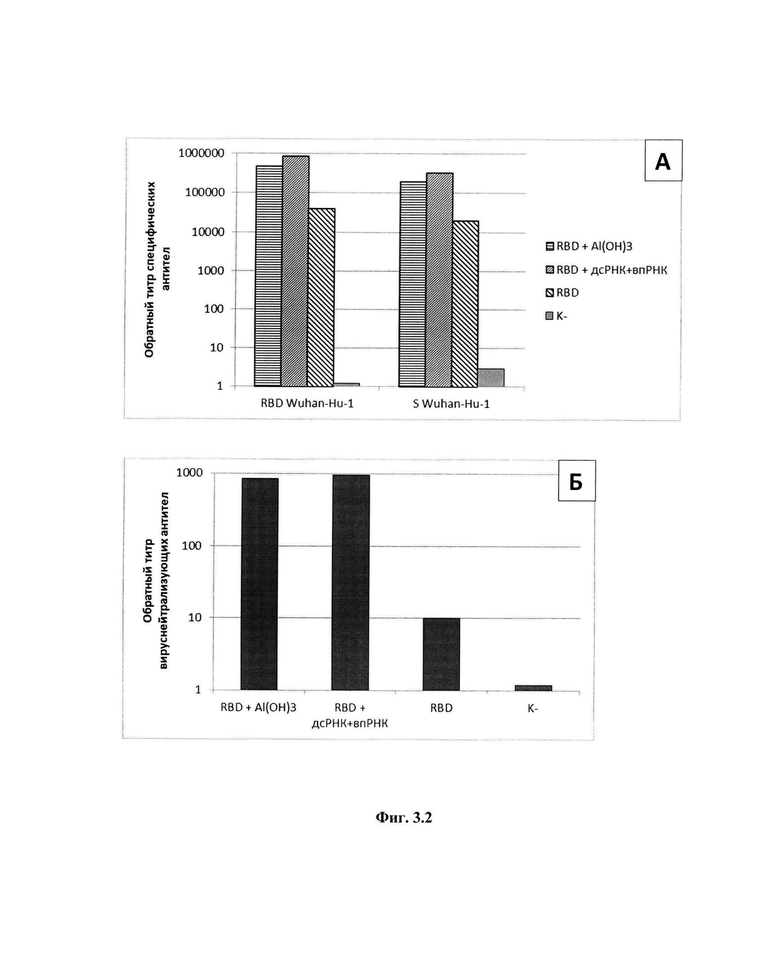
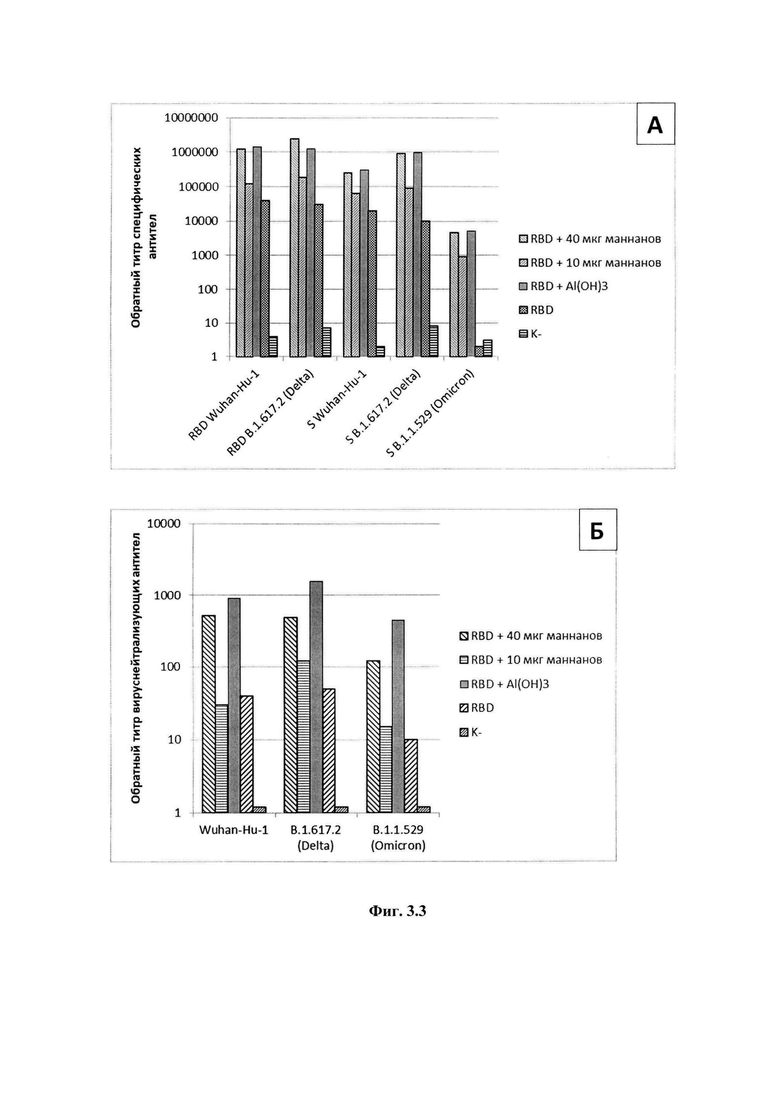
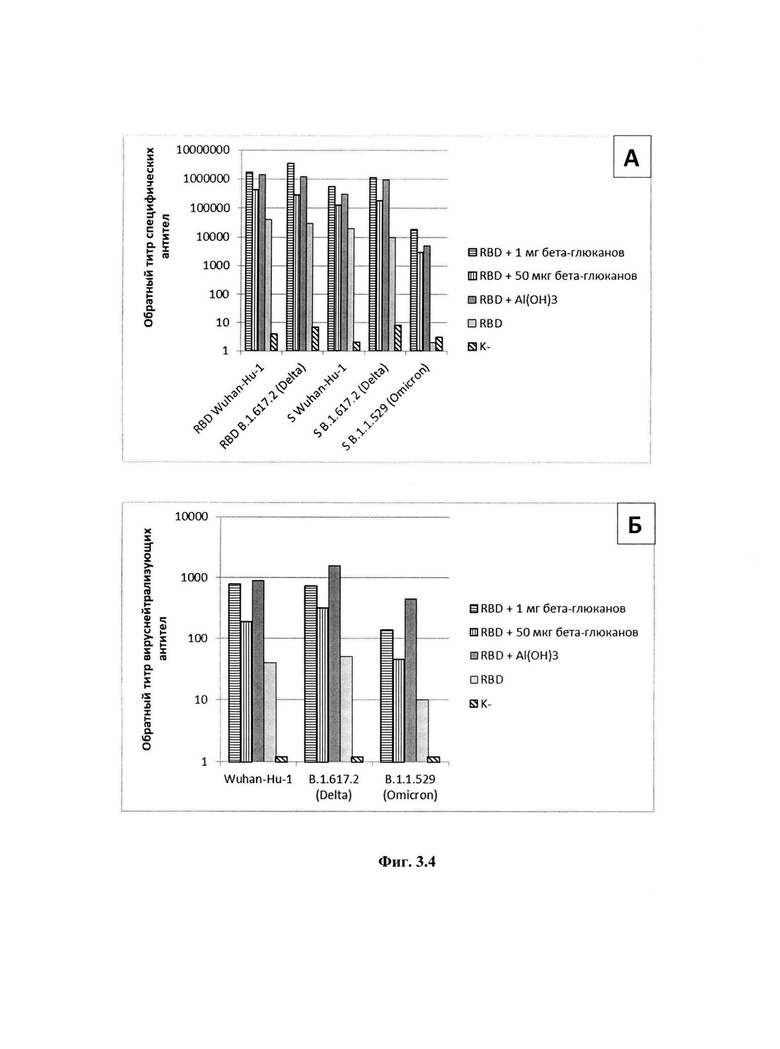
На фиг 3.1-3.4 представлены экспериментальные данные, полученные в ходе изучения адъювантных свойств препаратов: фиг.3.1, А - Титры специфических антител в крови мышей после иммунизации белком RBD в сочетании с дсРНК; Б - Титры вируснейтрализующих антител крови мышей после иммунизации белком RBD в сочетании с дсРНК; фиг.3.2, А - Титры специфических антител в крови мышей после иммунизации белком RBD в сочетании с дсРНК и впРНК; Б - Титры вируснейтрализующих антител крови мышей после иммунизации белком RBD в сочетании с дсРНК и впРНК; фиг.3.3, А - Титры специфических антител крови мышей после иммунизации белком RBD в сочетании с препаратом маннанов; Б - Титры вируснейтрализующих антител крови мышей после иммунизации белком RBD в сочетании с препаратом маннанов; фиг.3.4, А - Титры специфических антител крови мышей после иммунизации белком RBD в сочетании с препаратом глюканов; Б - Титры вируснейтрализующих антител крови мышей после иммунизации белком RBD в сочетании с препаратом бета-глюканов. На фиг.4 приведена технологическая схема получения препаратов дсРНК, впРНК, бета-глюканов и маннанов при комплексной переработке киллерных дрожжей Saccharomyces cerevisiae.
Сущность изобретения поясняется, но не ограничивается, следующими примерами 1-4. Подробная технологическая схема получения препаратов дсРНК, впРНК, бета-глюканов и маннанов при комплексной переработке киллерных дрожжей Saccharomyces cerevisiae представлена на фиг.4.
Пример 1. Разрушение биомассы клеток Saccharomyces cerevisiae Y-448 механическим способом с помощью бус баллотини
200 г биомассы суспендировали в 2 л раствора, содержащего 0,1 М натрия сернокислого, 5 мМ Трилона Б, 40 мМ калия фосфорнокислого однозамещенного и 0,4 М калия хлористого, рН 7,0. Суспензию помещали в сосуд для разрушения, содержащий 100 г стеклянных бус баллотини, и гомогенизировали в течение 5±1 мин. Контроль разрушения клеток вели микроскопически. Лизат клеток отделяли от бус баллотини центрифугированием при 12000 g в течение 30 мин, 2-8°С. Супернатант объемом 2 л использовали для выделения дсРНК и впРНК, а осадок - для получения бета-глюканов и маннанов.
Пример 2. Выделение дсРНК и впРНК
Супернатант из примера 1 объемом 2 л осаждали этиловым спиртом в соотношении 1:1 и инкубировали при 2-8°С 12 часов. Смесь центрифугировали при 12000 g в течение 30 мин. Осадок растворяли в 200 мл воды деионизованной до достижения раствором оптической плотности 200 о.е. при длине волны 260 нм. Осаждение впРНК проводили хлоридом лития, для чего сухой хлорид лития добавляли к раствору до достижения концентрации 1,5-2,0 М. Смесь инкубировали при 2-8°С 12 часов и центрифугировали при 12000 g в течение 30 мин.
2.1. Выделение дсРНК
ДсРНК из супернатанта из примера 2 осаждали этиловым спиртом в соотношении 1:4 и инкубировали при 2-8°С 12 часов. Смесь центрифугировали при 12000 g в течение 30 мин. Осадок растворяли в 200 мл раствора, содержащего 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2, до достижения раствором оптической плотности 150 о.е. при длине волны 260 нм. Для освобождения от примесных ДНК и РНК дсРНК осаждали хлоридом лития, добавляя его к раствору до концентрации 3,5-4 М с инкубацией в течение 12 часов при 2-8°С и центрифугировали при 12000 g в течение 30 мин, 2-8°С. Осадок растворяли в 170 мл раствора, содержащего 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2, до достижения раствором оптической плотности 150 о.е. при длине волны 260 нм. С целью депротеинизации к раствору дсРНК добавляли 13,6 мл полиэтиленгликоля 6000 до концентрации 8% с инкубацией 12 часов при 2-8°С и центрифугировали при 12000 g в течение 30 мин, 2-8°С. Осажденную дсРНК растворяли в 40 мл раствора, содержащего 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис-аминометана, рН 7,2, переосаждали этиловым спиртом в соотношении 1:2 (80 мл) и центрифугировали при 12000 g в течение 30 мин, 2-8°С. Осадок, содержащий дсРНК, растворяли в 50 мл воды деионизованной и лиофильно высушивали. Количество выделенной дсРНК с цикла составляло 250 мг. Электрофореграмма полученного препарата дсРНК представлена на фиг.1.
2.2. Выделение впРНК
Осадок из примера 2 растворяли в 50 мл воды деионизованной и инкубировали 45 мин при температуре 85-90°С. Смесь охлаждали и центрифугировали при 12000 g в течение 20 мин при 4°С. ВпРНК из супернатанта осаждали 1,5 М хлоридом лития в течение 12 часов при 2 - 8°С и центрифугировали при 12000 g в течение 30 мин, 2-8°С. Осадок, содержащий впРНК, растворяли в 60 мл воды деионизованной до достижения раствором оптической плотности 200 о.е. при длине волны 260 нм. Полученный раствор очищали гель-фильтрацией на колонке с Сефарозой 2 В объемом 130 мл. Фракции с оптической плотностью, превышающей 200 о.е. при длине волны 260 нм, объединяли (объем 110 мл). Осаждение впРНК из раствора проводили этиловым спиртом в соотношении 1:3 и центрифугировали при 12000 g в течение 30 мин. Осадок, содержащий впРНК, растворяли в 50 мл воды деионизованной и лиофильно высушивали. Количество выделенной впРНК с цикла составляло 500 мг. Электрофореграмма полученного препарата впРНК представлена на фиг.1.
Пример 3. Выделение бета-глюканов и маннанов
Осадок из примера 1 массой 100 г растворяли в 200 мл раствора, содержащего 50 мМ натрий-фосфорнокислого однозамещенного, рН 5,5, и перемешивали на магнитной мешалке в течение 30 мин при 300 об/мин. К полученному раствору добавляли 40 г «Протосубтилин ГЗХ» и 3 мл «ЦеллоЛюксА» (ООО ПО «Сиббиофарм»), ферментативный гидролиз вели 15 часов при температуре 60°С. Смесь охлаждали до температуры 20°С и центрифугировали при 12000 g в течение 20 мин при 20°С. Супернатант использовали для выделения маннанов, а осадок для получения бета-глюканов.
3.1. Выделение маннанов
К раствору супернатанта из примера 3 объемом 200 мл добавляли 3 части спирта этилового для осаждения маннанов и центрифугировали при 12000 g в течение 20 мин при 20°С. Осадок подвергали водно-спиртовой отмывке трижды, до осветления промывных вод. Проводили щелочной гидролиз 0,2 М гидроксидом натрия в течение 45 мин при температуре 60°С. После гидролиза смесь охлаждали до 20°С, рН доводили до 7,0 соляной кислотой. Полученный полупродукт осаждали 100 мл этилового спирта, добавленного в объеме 1:1, и растворяли в 75 мл 0,9% хлорида натрия. В результате получали препарат маннанов объемом 75 мл, с концентрацией маннанов 1,9 мг/мл (142,5 мг).
3.2. Выделение бета-глюканов
Осадок из примера 3 отмывали 1%-ным раствором додецилсульфата натрия (SDS), вызывающего денатурацию примесных белков. Для этого к осадку, содержащему бета-глюканы, добавляли 300 мл раствора SDS, емкость с суспензией помещали на магнитную мешалку при 300 об/мин на 30 мин и центрифугировали при 12000 g в течение 20 мин при 20°С. Полученный осадок пятикратно отмывали деионизованной водой объемом 300 мл. После отмывки осадок растворяли в 100 мл деионизованной воды (20°С) и подвергали воздействию ультразвука с частотой 22 кГц/с, тремя циклами по 5 мин с интервалами 2 мин для охлаждения. Полученную суспензию титровали 1 М гидроксидом натрия до значения рН 10,0 и инкубировали в термостате 1 час при 50°С. Полученный полупродукт центрифугировали при описанных ранее условиях и далее осадок отмывали 8 раз деионизованной водой, объемом 300 мл, с последующим центрифугированием. Конечный осадок суспендировали в 50 мл деионизованной воды, разливали на чашки Петри и сушили в термостате при 50°С 12 часов. Количество препарата бета-глюканов, полученного с одного цикла, составляет 4 г. ИК-спектры препарата бета-глюканов в сравнении с препаратом - аналогом зимозаном (Sigma, США) представлены на фиг.2. Из представленных данных видно, что основные пики поглощения препарата бета-глюканов и препарата сравнения совпадают.
Пример 4. Оценка адъювантных свойств дсРНК, впРНК, бета-глюканов и маннанов
4.1. Изучение адъювантных свойств дсРНК
Изучение адъювантных свойств препарата дсРНК проводили на мышах линии BALB/c, самцах, с массой тела 16-18 г. Две группы животных по 10 голов в каждой иммунизировали рекомбинантным белком-аналогом рецептор-связывающего домена (RBD) спайкового белка SARS-CoV-2 (Delta (В. 1.617.2)) в дозе 50 мкг в объеме 100 мкл двукратно с интервалом 14 суток, внутримышечно. Первая группа получала только RBD, второй, помимо RBD, вводили препарат дсРНК в дозе 100 мкг на мышь. В качестве положительного контроля использовали мышей, которым внутримышечно двукратно с интервалом 14 суток вводили RBD и гидрооксид алюминия Al(ОН)3 в дозе 250 мкг на мышь. В качестве отрицательного контроля использовали мышей, которым по той же схеме вводили физиологический раствор в эквивалентном объеме. Через 10 суток после второй иммунизации из ретроорбитального синуса брали образец крови для определения титров специфических антител и вируснейтрализующих антител. Для получения сыворотки образцы крови инкубировали сначала при 37°С в течение 1 часа, затем при +4°С в течение 20 часов, после чего сгусток крови в пробирке обводили стерильной иглой и центрифугировали в течение 10 мин при 5000g. Препараты сывороток хранили при температуре - 20°С.
Титры специфичных антител в сыворотках крови определяли методом иммуноферментного анализа (ИФА). В качестве антигена использовали рекомбинантный RBD поверхностного белка вируса SARS-CoV-2 штамм Wuhan-Hu-1 и рекомбинантный спайковый белок вируса SARS-CoV-2 штамм Wuhan-Hu-1. Сорбцию антигенов проводили в буфере 0,1 М NaHCO3, внося по 200 нг очищенного белка в лунке. После 16-часовой инкубации при +4°С и удаления растворов в лунки добавляли по 150 мкл блокирующего буфера (фосфатно-солевой буфер, содержащий 1,06 мМ KH2PO4, 155,17 мМ NaCl, 2,97 мМ Na2HPO4, рН 7,4 с 1% бычьим сывороточным альбумином), продолжали инкубацию в течение 1 часа при 37°С. После удаления блокирующего буфера проводили трехкратную промывку раствором фосфатно-солевого буфера, содержащего 0,5% Tween-20. Далее в лунки добавляли сыворотки крови иммунизированных животных в объеме 50 мкл/лунка в блокирующем буфере и инкубировали в течение 1 часа при 37°С. После трехкратной отмывки лунок раствором фосфатно-солевого буфера, содержащего 0,5% Tween-20, в них добавляли по 100 мкл конъюгатов антимышиных антител с пероксидазой хрена в рабочем разведении 1:2000, инкубировали в течение 1 ч при 37°С. После трехкратной отмывки лунок раствором фосфатно-солевого буфера с 0,5% Tween-20 добавляли 50 мкл жидкого субстрата на основе ТМБ (3,3',5,5'-Тетраметилбензидин) и ацетатного буфера (100 мМ Na ацетат, рН 4,5), содержащего Н2О2 в рабочей концентрации 1%, инкубировали в течение 15 минут при комнатной температуре в защищенном от света месте. Реакцию блокировали добавлением 50 мкл 1М HCl в каждую лунку. Оптическую плотность измеряли на мультимодальном ридере Thermo Scientific Varioskan LUX при длине волны 450 нм. Титр определяли по значению максимального разведения, при котором сигнал оптической плотности превышал значение оптической плотности лунок с отрицательным контролем.
Титр вируснейтрализующих антител определяли при помощи реакции вируснейтрализации. С этой целью были использован штамм вируса Wuhan-hCoV-19/Australia/VICO 1/2020 (Wuhan-Hu-1), SARS-CoV-2, полученный из Государственной коллекции возбудителей вирусных инфекций и риккетсиозов ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора. Вируссодержащий материал хранился при температуре -70°С. Концентрация вируса в наработанном для анализа препарате составляла 5×106 ТЦД50/мл (тканевая цитопатическая доза на миллилитр).
Определение нейтрализующих титров сывороток проводили путем учета неповрежденного монослоя культуры клеток Vero-Е6 в лунках 96-луночного планшета (Corning, США). Культуру клеток выращивали в 96-луночных культуральных планшетах в ростовой питательной среде DMEM с добавлением 10% бычьей фетальной сыворотки и антибиотиков. Заражение клеток вирусами проводили в дозе 100 ТЦД50/лунку. Исследуемые образцы сыворотки крови разводили последовательно в среде DMEM (ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора, Россия) с глутамином и антибиотиками с 2-кратным шагом, начиная с разведения 1:10 до 1:2560. К сывороткам в разных разведениях добавляли вирус в равной пропорции 1:1 и инкубировали в течение 1 ч при 37°С. Затем смесь вируса и сыворотки наносили в дублях на монослой культуры клеток в объеме 100 мкл/лунку. Планшеты инкубировали в течение 4 сут при 37°С во влажной атмосфере 5% СО2. Для окрашивания в каждую лунку планшета вносили 100 мкл 0,2%-го раствора генцианвиолета. Через 4 сут жидкость из лунок удаляли и промывали лунки водой. Учет результатов проводили визуально. Любое специфическое поражение культуры клеток в лунке учитывали как цитопатическое действие (ЦПД).
Титром считали последнее разведение, при котором регистрировали защиту монослоя культуры клеток в лунках от ЦПД вируса. В качестве положительного контроля использовали 20-кратное разведение образца сыворотки крови реконвалесцента COVID-19 с ранее установленным титром 1:80. В качестве отрицательного контроля использовали питательную среду.
Результаты представлены на фиг.3.1. Как видно, добавление дсРНК в качестве адъюванта в дозе 100 мкг усиливает гуморальный иммунный ответ иммунизированных мышей и наработку нейтрализующих антител. Среднее значение обратного титра антител в группе мышей, иммунизированных RBD в сочетании с дсРНК, сравнимо со значением параметра в группе мышей, иммунизированных RBD с Al(ОН)3, и значительно превышает показатели группы, иммунизированной только RBD (фиг.3.1, А). Аналогичные закономерности были обнаружены при определении обратных титров вируснейтрализующих антител в крови мышей этих групп (фиг.3.1, Б).
4.2. Изучение адъювантных свойств препарата, содержащего дсРНК и впРНК
Изучение адъювантных свойств препарата, содержащего смесь дсРНК и впРНК, проводили на мышах линии BALB/c, самцах, с массой тела 16-18 г. Две группы животных по 10 голов в каждой иммунизировали рекомбинантным белком RBD в дозе 50 мкг в объеме 100 мкл двукратно с интервалом 14 суток, внутримышечно. Первая группа получала только RBD, второй вводили RBD и препарат субстанции дсРНК, содержащей 21% дсРНК и 79% впРНК в дозе 100 мкг на мышь. В качестве положительного контроля использовали мышей, которым внутримышечно двукратно с интервалом 14 суток вводили RBD и гидрооксид алюминия Al(ОН)3 в дозе 250 мкг на мышь. В качестве отрицательного контроля использовали мышей, которым по той же схеме вводили физиологический раствор в эквивалентном объеме. Через 10 суток после второй иммунизации из ретроорбитального синуса брали образец крови для определения титров специфических антител и вируснейтрализующих антител.
Титры специфичных и вируснейтрализующих антител в сыворотках определяли как описано в примере 4.1.
Результаты представлены на фиг.3.2. Как видно, добавление дсРНК и впРНК (21% дсРНК и 79% впРНК) в качестве адъюванта в дозе 100 мкг обеспечивает усиление гуморального иммунного ответа. Среднее значение обратного титра вируснейтрализующих антител для группы мышей, иммунизированных RBD в сочетании с дсРНК и впРНК, составило 990, в то время как значение показателя группы, иммунизированной RBD с А1(ОН)3, составило 900, только RBD - 10 (фиг.3.2, А). Аналогичная картина наблюдается для значений обратного титра нейтрализующих антител (фиг.3.2, Б).
4.3. Изучение адъювантных свойств маннанов
Изучение адъювантных свойств маннанов проводили на самцах мышей линии BALB/c с массой тела16-18 г., иммунизированных рекомбинантным рецептор-связывающим доменом (RBD) спайкового белка SARS-CoV-2 (Delta (В. 1.617.2)) в дозе 50 мкг, в объеме 100 мкл. В каждой группе было по 10 голов. Иммунизацию проводили двукратно с интервалом 14 суток, внутримышечно. Первая группа получала только RBD, во второй и третьей использовали RBD с препаратом маннанов в дозе 40 мкг и 10 мкг на мышь, соответственно. В качестве отрицательного контроля использовали мышей линии BALB/c, которым внутримышечно вводили двукратно с интервалом 14 суток 100 мкл физиологического раствора. В качестве положительного контроля использовали мышей линии BALB/c, которым внутримышечно вводили двукратно с интервалом 14 суток RBD и гидроксид алюминия Al(ОН)3 в дозе 250 мкг. Через 10 суток после второй иммунизации из ретроорбитального синуса брали образец крови для определения гуморального иммунного ответа.
Титры специфичных антител в сыворотках определяли методом иммуноферментного анализа (ИФА) как описано в примере 4.1, однако в качестве антигенов использовали рекомбинантный RBD поверхностного белка SARS-CoV-2 (штаммы В. 1.1.7 (Alpha) и В. 1.617.2 (Delta)) и рекомбинантный спайковый белок (штаммы В. 1.1.7 (Alpha), В. 1.617.2 (Delta) и В. 1.1.529 (Omicron)).
Титры вируснейтрализующих антител в сыворотках определяли в тесте вируснейтрализации как описано в примере 4.1, однако при этом использовали штаммы вируса SARS-CoV-2 Wuhan-hCoV-19/Australia/VICO 1/2020 (Wuhan-Hu-1), Delta-hCoV-19/Russia/PSK-2804/2021 (Delta (B.1.617.2)) и Omicron 1-hCoV-19/Russia/Moscowl71619-031221/2021 (Omicron (B. 1.1.529)).
Результаты представлены на фиг.3.3. Можно заключить, что использование маннанов в качестве адъюванта обеспечивает усиление гуморального иммунного ответа и вызывает наработку нейтрализующих антител. Доза 40 мкг обладает большей адъювантной активностью, чем доза 10 мкг (фиг.3.3, А). Среднее значение обратного титра вируснейтрализующих антител для группы мышей, иммунизированных RBD с препаратом маннанов в дозе 10 мкг, почти на порядок меньше титров антител в группе, иммунизированной RBD с Al(ОН)3. В то же время обратный титр вируснейтрализующих антител для группы, иммунизированной RBD с препаратом маннанов в дозе 40 мкг, лишь незначительно ниже титров группы, иммунизированной RBD с Al(ОН)3 (фиг.3.3, Б).
4.4. Изучение адъювантных свойств бета-глюканов
Изучение адъювантных свойств бета-глюканов проводили на мышах, иммунизированных белком RBD - рекомбинантным аналогом рецептор-связывающего домена спайкового белка SARS-CoV-2 (штамм Delta (В.1.617.2). В экспериментах использовали самцов мышей линии BALB/c массой 16-18 г., группы по 10 голов.
Иммунизацию белком RBD проводили в дозе 50 мкг в объеме 100 мкл на мышь двукратно с интервалом 14 суток, внутримышечно. Первая группа получала только RBD, во второй и третьей использовали RBD с препаратом бета-глюканов в дозе 1 мг и 50 мкг на мышь. В качестве положительного контроля использовали мышей, которым внутримышечно двукратно с интервалом 14 суток вводили белок RBD и гидроокись алюминия Al(ОН)3 в дозе 250 мкг. В качестве отрицательного контроля использовали мышей, которым по той же схеме вводили 100 мкл физиологического раствора. Через 10 суток после второй иммунизации из ретроорбитального синуса брали образец крови для определения титров специфических антител и вируснейтрализующих антител.
Титры специфичных антител в сыворотках определяли методом иммуноферментного анализа как описано в примере 4.1, однако в качестве антигенов использовали рекомбинантный RBD поверхностного белка SARS-CoV-2 (штаммы В. 1.1.7 (Alpha) и В. 1.617.2 (Delta)) и рекомбинантный спайковый белок (штаммы В.1.1.7 (Alpha), В.1.617.2 (Delta) и В.1.1.529 (Omicron)).
Титры вируснейтрализующих антител в сыворотках определяли в тесте вируснейтрализации как описано в примере 4.1, однако при этом использовали штаммы вируса SARS-CoV-2 Wuhan-hCoV-19/Australia/VICO 1/2020 (Wuhan-Hu-1), Delta-hCoV-19/Russia/PSK-2804/2021 (Delta (B.l.617.2)) и Omicron 1-hCoV-19/Russia/Moscowl71619-031221/2021 (Omicron (B.l.1.529)).
Результаты представлены на фиг.3.4. Использование препарата бета-глюканов в качестве адъюванта в дозе 1 мг обеспечивает большую индукцию гуморального иммунитета в сравнении с дозой бета-глюканов 50 мкг. Полученные значения среднего обратного титра антител для группы мышей, иммунизированных RBD с препаратом бета-глюканов в дозе 1 мг на мышь, близки к значениям группы, иммунизированной RBD с Al(ОН)3 (фиг.3.4, А). Использование препарата бета-глюканов в дозе 1 мг обеспечивает наработку титров нейтрализующих антител, сравнимых с группой, иммунизированной RBD с Al(ОН)3 (фиг.3.4, Б).
Достижение технического результата заявляемого изобретения подтверждается примерами 1-4, в которых представлена новая технология комплексной переработки биомассы Saccharomyces cerevisiae, позволяющая в одном производственном цикле получать препараты дсРНК, впРНК, бета-глюканов и маннанов, обладающих адъювантными свойствами. При этом обеспечивается снижение отходов сырья и уменьшаются затраты на получение препаратов по сравнению с затратами на получение указанных препаратов из дрожжевых клеток в отдельности. Расчеты показывают, что использование предложенного способа получения препаратов позволяет получить одновременно с 1 кг биомассы до 1,25 г дсРНК, до 2,5 г впРНК, до 20 г бета-глюканов и до 712,5 мг маннанов. Для препарата дсРНК затраты на наработку биомассы составляют до 20% от общих затрат на получение указанной биомассы, для впРНК - до 30%, для бета-глюканов - до 50%, а для маннанов - до 35%. Таким образом, использование комплексной переработки позволяет достичь экономии до 50% от затрат на процедуру получения указанных препаратов из дрожжевых клеток в отдельности.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения биологически активных компонентов из клеток дрожжей Saccharomyces cerevisiae и лечебное средство на их основе | 2018 |
|
RU2722731C2 |
| СПОСОБ ПОЛУЧЕНИЯ СУММАРНОЙ РНК ИЗ БИОМАССЫ КЛЕТОК ДРОЖЖЕЙ SACCHAROMYCES CEREVISIAE | 2022 |
|
RU2781832C1 |
| СПОСОБ ПОЛУЧЕНИЯ ДВУСПИРАЛЬНОЙ РИБОНУКЛЕИНОВОЙ КИСЛОТЫ ИЗ КЛЕТОК ДРОЖЖЕЙ Saccharomyces cerevisiae | 2022 |
|
RU2781833C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА 1 ТИПА | 2006 |
|
RU2317107C2 |
| Рекомбинантный белок, связывающийся с RBD S-белка SARS-CoV-2 | 2022 |
|
RU2778942C1 |
| Конъюгат белка рецепторсвязывающего домена (RBD) поверхностного гликопротеина S вируса SARS-CoV-2 с полимером полиглюкин-спермидин (PGS) и вакцинный комплекс против коронавирусной инфекции COVID-19 на основе указанного конъюгата и плазмидной ДНК pVAX-RBD | 2022 |
|
RU2781294C1 |
| Плазмидная ДНК-матрица pVAX-RBD, молекул мРНК-RBD, полученная с использованием ДНК-матрицы pVAX-RBD, обеспечивающая синтез и секрецию белка RBD SARS-CoV 2 в клетках организма млекопитающих и комплекс в виде наночастиц, содержащих молекулы мРНК-RBD, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью | 2021 |
|
RU2754230C1 |
| СПОСОБ ПОЛУЧЕНИЯ ДВУСПИРАЛЬНОЙ РИБОНУКЛЕИНОВОЙ КИСЛОТЫ (дсРНК) ИЗ КЛЕТОК ДРОЖЖЕЙ Saccharomyces cerevisiae | 2014 |
|
RU2558256C1 |
| Штамм гибридных культивируемых клеток Homo sapiens/Mus musculus Hu-C6D7-RBD - продуцент человеческих моноклональных антител, специфичных к RBD домену S белка вируса SARS-CoV-2 | 2022 |
|
RU2788359C1 |
| ИСКУССТВЕННЫЕ МИКОБАКТЕРИАЛЬНЫЕ ЧАСТИЦЫ И ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИННАЯ КОМПОЗИЦИЯ НА ИХ ОСНОВЕ | 2003 |
|
RU2242245C2 |

Изобретение относится к биотехнологии. Предложен способ получения дсРНК, впРНК, бета-глюканов и маннанов из биомассы Saccharomyces cerevisiae, включающий суспендирование биомассы дрожжей в растворе, содержащем 0,1 М натрия сернокислого, 5 мМ Трилона Б, 40 мМ калия фосфорнокислого однозамещенного и 0,4 М калия хлористого при рН 7,0; помещение указанной суспензии в сосуд для разрушения, содержащий стеклянные бусы баллотини, в пропорции 2:1 и гомогенизацию смеси в течение 5±1 мин; лизат клеток отделяют от бус баллотини и центрифугируют при 12000 g в течение 30 мин, 2-8°С до формирования плотного осадка со смесью бета-глюканов и маннанов и супернатанта, содержащего смесь дсРНК и впРНК. Затем из супернатанта дробно выделяют дсРНК, впРНК, из плотного осадка выделяют бета-глюканы и маннаны. Изобретение обеспечивает комплексную переработку биомассы Saccharomyces cerevisiae в одном производственном цикле с получением препаратов дсРНК, впРНК, бета-глюканов и маннанов, обладающих адъювантными свойствами. 7 ил., 4 пр.
Способ получения дсРНК, впРНК, бета-глюканов и маннанов из биомассы Saccharomyces cerevisiae, включающий суспендирование биомассы дрожжей в растворе, содержащем 0,1 М натрия сернокислого, 5 мМ Трилона Б, 40 мМ калия фосфорнокислого однозамещенного и 0,4 М калия хлористого при рН 7,0; помещение указанной суспензии в сосуд для разрушения, содержащий стеклянные бусы баллотини, в пропорции 2:1 и гомогенизацию смеси в течение 5±1 мин; лизат клеток отделяют от бус баллотини и центрифугируют при 12000 g в течение 30 мин, 2-8°С до формирования плотного осадка со смесью бета-глюканов и маннанов и супернатанта, содержащего смесь дсРНК и впРНК; из супернатанта выделяют смесь дсРНК и впРНК добавлением этилового спирта в соотношении 1:1 с последующей инкубацией при 2-8°С 12 ч; полученную смесь дсРНК и впРНК центрифугируют при 12000 g в течение 30 мин, плотный осадок растворяют в деионизованной воде до достижения раствором оптической плотности 200 о.е. при длине волны 260 нм; осаждение впРНК проводят хлоридом лития, для чего сухой хлорид лития добавляют к раствору до достижения концентрации 1,5-2,0 М, смесь инкубируют при 2-8°С 12 ч и центрифугируют при 12000 g в течение 30 мин;
для дробного выделения дсРНК супернатант смешивают с этиловым спиртом в соотношении 1:4, инкубируют 12 ч при 2-8°С и центрифугируют при 2-8°С, 12000 g в течение 30 мин; образованный плотный осадок растворяют в растворе, содержащем 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2 до достижения раствором оптической плотности 150 о.е. при длине волны 260 нм; далее дсРНК осаждают хлоридом лития, добавляя его к раствору до концентрации 3,5-4 М с инкубацией в течение 12 ч при 2-8°С и центрифугировании при 12000 g и при 2-8°С в течение 30 мин, образованный плотный осадок дсРНК растворяют в растворе, содержащем 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2, до достижения раствором оптической плотности 150 о.е. при длине волны 260 нм; затем к раствору дсРНК добавляют полиэтиленгликоль 6000 до концентрации 8% с последующей инкубацией в течение 12 ч при 2-8°С и центрифугировании при 12000 g в течение 30 мин при 2-8°С; осадок дсРНК растворяют в 40 мл раствора, содержащего 150 мМ хлорида натрия, 1 мМ ЭДТА, 50 мМ трис аминометана, рН 7,2, переосаждают этиловым спиртом в соотношении 1:2 и центрифугируют при 12000 g в течение 30 мин при 2-8°С; осадок, содержащий дсРНК, растворяют в деионизованной воде и лиофильно высушивают;
для дробного выделения впРНК осадок впРНК растворяют в деионизованной воде и инкубируют 45 мин при температуре 85-90°С, далее смесь охлаждают и центрифугируют при 12000 g в течение 20 мин при 4°С; затем впРНК из супернатаната осаждают 1,5 М хлоридом лития, осадок центрифугируют при 12000 g в течение 30 мин при 2-8°С; плотный осадок, содержащий впРНК, растворяют в деионизованной воде до достижения раствором оптической плотности 200 о.е. при длине волны 260 нм, полученный раствор очищают гель-фильтрацией на колонке с Сефарозой 2В, фракции которого с оптической плотностью, превышающей 200 о.е. при длине волны 260 нм, объединяют и осаждают впРНК из раствора этиловым спиртом в соотношении 1:3 с последующим центрифугированием при 12000 g в течение 30 мин; осадок, содержащий впРНК, растворяют в деионизованной воде и лиофильно высушивают;
для выделения бета-глюканов и маннанов осадок после разрушения дрожжевой биомассы и центрифугирования растворяют в растворе, содержащем 50 мМ натрий-фосфорнокислый однозамещенный, рН 5,5, и перемешивают, к полученному раствору добавляют ферментные препараты «Протосубтилин ГЗХ» и «ЦеллоЛюксА» в соотношении 13,3:1, инкубируют 15 ч при температуре 60°С, смесь охлаждают до 20°С и центрифугируют при 12000 g в течение 20 мин при 20°С до получения осадка, содержащего бета-глюканы, и супернатанта, содержащего маннаны;
к раствору супернатанта добавляют 3 части этилового спирта и центрифугируют при 12000 g в течение 20 мин при 20°С, полученный осадок подвергают водно-спиртовой отмывке трижды до осветления промывных вод, проводят щелочной гидролиз 0,2 М гидроксидом натрия в течение 45 мин при температуре 60°С, после чего рН полученной смеси доводят до 7,0 соляной кислотой; полученный полупродукт осаждают добавлением этилового спирта в соотношении 1:1 и растворяют осадок 0,9%-ным раствором хлорида натрия с получением маннанов;
для выделения бета-глюканов осадок, полученный ферментативным гидролизом разрушенной дрожжевой биомассы, отмывают 1%-ным раствором додецилсульфата натрия (SDS) и центрифугируют при 12000 g в течение 20 мин при 20°С, полученный осадок пятикратно отмывают деионизованной водой, растворяют в деионизованной воде с получением суспензии, которую титруют 1 М гидроксидом натрия до значения рН 10,0; смесь инкубируют в термостате 1 ч при 50°С и центрифугируют при 12000 g в течение 20 мин при 20°С с получением осадка, который отмывают 8 раз деионизованной водой, суспендируют в деионизованной воде и сушат в термостате при 50°С 12 ч с получением бета-глюканов.
| СПОСОБ ПОЛУЧЕНИЯ ВОДОРАСТВОРИМЫХ ФРАКЦИЙ МАННОПРОТЕИНОВ И β-ГЛЮКАНА | 2011 |
|
RU2504384C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДВУСПИРАЛЬНОЙ РИБОНУКЛЕИНОВОЙ КИСЛОТЫ ИЗ КЛЕТОК ДРОЖЖЕЙ Saccharomyces cerevisiae | 2022 |
|
RU2781833C1 |
| СЕРБА Е.М | |||
| и др | |||
| Аппарат для сжигания нефти | 1920 |
|
SU985A1 |
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |
| Нивелир для отсчетов без перемещения наблюдателя при нивелировании из средины | 1921 |
|
SU34A1 |
| HARBAH R | |||
| et al | |||
| "Mannan: structure, biosynthesis and methods extraction from yeast Saccharomyces | |||

