Изобретение относится к кандидатной рекомбинантной вакцине против COVID-19, представлящей собой комплекс иммуногенов, включающих рекомбинантную плазмидную ДНК pVAX-RBD, обеспечивающую экспрессию гена рецепторсвязывающего домена белка S вируса SARS-CoV-2 в клетках эукариот, покрытую конъюгатом спермидин-полиглюкин с рекомбинантным белком RBD и может быть использована в области биотехнологии, в частности к медицине и иммунологии. На модели лабораторных животных вакцина обеспечивает высокий титр SARS-CoV-2-специфических антител, обладающих вируснейтрализующей активностью, а также стимулирует вирус специфический цитотоксический иммунный ответ.
Во время пандемии COVID-19 за последние два года заболеванию подверглось более 500 миллионов человек [1]. Несмотря на введение строгих мер по увеличению социальной дистанции для предотвращения массового инфицирования, а также на массовую вакцинацию, заболеваемость по всему миру неуклонно остается высокой. К тому же ВОЗ регулярно сообщает о появлении новых штаммов SARS-CoV-2, что не исключает усиления пандемической угрозы. Главной мерой, способной решить проблему защиты населения от вирусного инфекционного заболевания, приводящего к большому количеству долгосрочных негативных последствий, является вакцинация.
В результате поиска эффективных вакцин были реализованы различные подходы: классические инактивированные вакцины [2, 3], рекомбинантные векторные вакцины на основе аденовирусов [4, 5, 7], рекомбинантные субъединичные вакцины [7, 8], вакцины на основе мРНК [9, 10] и ДНК [11].
Данные подходы с разной степенью эффективности позволили ограничить распространение COVID-19, однако победить пандемию до сих пор не удалось. Перспективным подходом к созданию вакцин является комбинирование различных платформ, в частности платформ, основанных на ДНК и рекомбинантных белках.
Известны схемы совместного введения субъединичной и ДНК-вакцины в режиме прайм-буст [12, 13]. Такое введение подразумевает разделение по времени введение разных иммуногенов, таких как ДНК и белок. То есть сначала иммунная система «праймируется», проходит первичную подготовку ДНК-вакциной, а затем «бустируется», то есть подвергается более сильному воздействию с помощью субъединичной вакцины, чаще всего с адъювантами. Такой режим позволяет достичь уровней нейтрализующих антител гораздо более высоких, чем при иммунизации по отдельности.
Известны подходы, при которых введение разных типов иммуногенов происходит единовременно в разные участки тела или в одно и то же место [14, 15]. При реализации такого подхода на обезьянах было показано, что именно совместное введение белкового и ДНК иммуногенов приводит к индукции более сильного гуморального и клеточного вирус-специфичного ответа. Эти данные свидетельствуют о важности одновременного распознавания, процессинга и презентации иммуногенов в одних и тех же дренирующих лимфоузлах для развития защитного иммунитета.
Известна вакцина против ВИЧ-1 «КомбиВИЧвак» (патент РФ №2317107, МПК А61К 39/21, опубл. 20.02.2008 г.) [20], которая представляет собой комбинацию иммуногенов, состоящую из рекомбинантной плазмидной ДНК pcDNA-TCI с геном, кодирующим искусственный белок-иммуноген TCI, содержащий набор большого числа цитотоксических эпитопов ВИЧ-1 и обеспечивающая экспрессию искусственного гена в клетках млекопитающих, покрытую конъюгатом спермидин-полиглюкин с рекомбинантным полиэпитопным белком TBI, содержащим консервативные вируснейтрализующие В- и Т- клеточные эпитопы ВИЧ-1 [16, 17]. При иммунизации животных созданной конструкцией индуцируется выработка специфических антител, обладающих вирус-нейтрализующей активностью, а также специфический цитотоксический (ЦТЛ) иммунный ответ.При этом в отношении гуморального ответа наблюдается эффект синергизма при объединении в одной конструкции двух В- и Т-клеточных иммуногенов (TBI и TCI). Уровень антител при иммунизации отдельно взятыми конструкциями только с TBI или только с pcDNA-TCI был достоверно ниже в эксперименте, чем при использовании "КомбиВИЧвак". Для получения оболочки PGS-TBI использовался полиглюкин 60000 Да, который после активации раствором периодата натрия в течение 20-30 мин отделяли гель-фильтрацией на колонке с сефадексом G-50 в 20 мМ фосфатном буфере (рН 7,6). После этого в раствор активированного полиглюкина вносили 5,5 мг белка TBI (0,25 мкМ) и 0,4 мг спермидина (2,6 мкМ). Смесь инкубировали в течение 5 ч, затем добавляли боргидрид натрия, перемешивали еще 2 ч и образовавшийся конъюгат отделяли гель-фильтрацией на сефадексе G-50, используя 100 мМ Трис-HCl (рН 8,3). Так как исходные компоненты (TBI и спермидин) вносились в соотношении на 0,25 мкМ полиглюкина 0,25 мкМ TBI и 2,6 мкМ спермидина, то можно предположить, что и полученный конъюгат содержит их в соотношении на одну молекулу полиглюкина - одна молекула TBI и десять молекул спермидина. Данный аналог отличается от заявляемого изобретения не только иммуногеном, но и технологией создания конъюгата полиглюкин-спермидина и белка.
Наиболее близким аналогом по назначению является комплекс мРНК-RBD и полиглюкин-спермидина, индуцирующий SARS-CoV-2-специфичные антитела [18]. Данная мРНК получена с использованием pVAX-RBD в качестве матрицы и кодирует белок рецепторсвязывающего домена вируса SARS-CoV-2. Каждая наночастица этого комплекса имеет размер в диапазоне до 900 нм и содержит ядро, состоящее из молекул мРНК-RBD и покрытое слоем конъюгата полиглюкин-спермидин, удерживаемого за счет ионного взаимодействия между отрицательно заряженной нуклеиновой кислотой и положительно заряженным спермидином (патент РФ №2754230, МПК C12N15/30, опубл. 30.08.2022 г.)[21]. Недостаток вакцины на основе мРНК заключается в сложной и ресурсоемкой технологии получения препарата мРНК, низкой стабильности и необходимости соблюдения холодовой цепи при хранении и транспортировке.
Техническим результатом заявляемого изобретения является создание такого вакцинного иммунного комплекса в виде наночастиц, который имеет более простой состав и технологию изготовления, а при внутримышечном введении способен индуцировать высокий уровень антител, обладающих вируснейтрализующей активностью, и обеспечивать вирус-специфический Т-клеточный иммунный ответ.
Указанный технический результат достигается тем, что создан конъюгат белка рецепторсвязывающего домена (RBD) поверхностного гликопротеина S вируса SARS-CoV-2, 308V-542N, наработанного в клетках CHO, имеющего длину 244 а.о., молекулярный вес 35 кДа, аминокислотную последовательность SEQ ID N 2 и связанного с полимером полиглюкин-спермидин (PGS) при расчетном соотношении молекул белка RBD и молекул PGS - 1:1.
Указанный технический результат достигается также тем, что получен вакцинный комплекс против коронавирусной инфекции COVID-19 в виде наночастиц, индуцирующих формирование Т-клеточного иммунитета и SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, и состоящих из молекул конъюгата полиглюкин- спермидин-белок RBD и молекул плазмидной ДНК pVAX-RBD, причем каждая наночастица имеет размер не менее 50 нанометров, содержит ядро, состоящее из молекул плазмидной ДНК pVAX-RBD и расположенного вокруг ядра слоя конъюгата полиглюкин-спермидин-белок RBD, удерживаемого за счет ионного взаимодействия между отрицательно заряженным ядром и положительно заряженным спермидином.
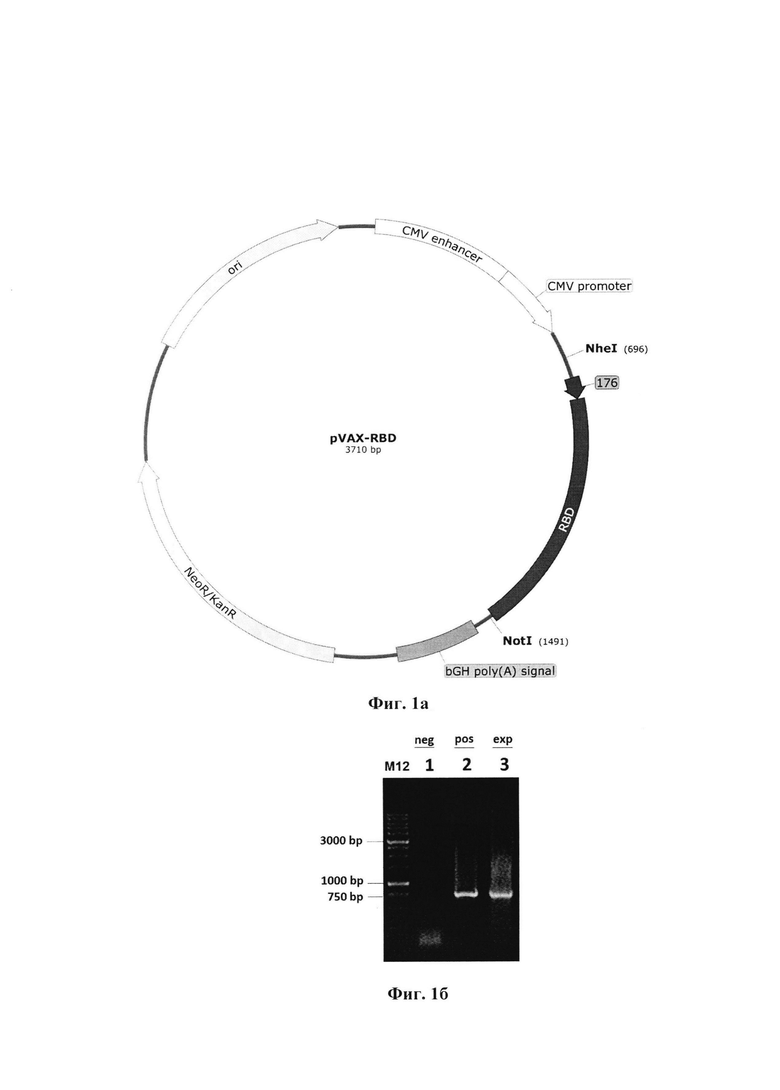
Причем плазмидная ДНК pVAX-RBD имеет нуклеотидную последовательность SEQ ID N 1 размером 3710 п.н., молекулярный вес 2,3 МДа, содержащая целевой ген, кодирующий химерный белок 176-RBD, перед которым находится промотор РНК-полимеразы фага Т7, обеспечивающий транскрипцию гена, и состоящей из следующих фрагментов:
- векторный фрагмент 1484-754 ДНК-плазмиды pVAX размером 2981 п.н., содержащий промотор РНК-полимеразы фага Т7 (664-682, размером 19 п.н.);
- ген устойчивости к неомицину/канамицину (NeoR/KanR) (1944-2738, 795 п.н.) и точка начала репликации ColE1 ori (3064-3652, 589 п.н.), обеспечивающие селекцию и амплификацию целевой плазмиды в клетках бактерий Escherichia coli;
- фрагмент 755-1483 размером 729 п.н., содержащий искусственный ген 176-RBD с инициирующим кодоном ATG.
Полученные наночастицы были названы КомбиКоронаВак-RBD (сокращенное наименование: CCV-RBD).
При иммунизации животных созданной конструкцией индуцируется выработка специфических антител, обладающих вируснейтрализующей активностью, а также специфический Т-клеточный иммунный ответ.При этом в отношении гуморального ответа наблюдается эффект синергизма при объединении в одной конструкции двух иммуногенов (ДНК и белок). Уровень антител при иммунизации отдельно взятыми конструкциями только белком RBD или только с pVAX-RBD был достоверно ниже в эксперименте, чем при использовании КомбиКоронаВак.
Изобретение иллюстрируется следующими графическими материалами.
Фиг. 1а. Карта плазмиды pVAX-RBD
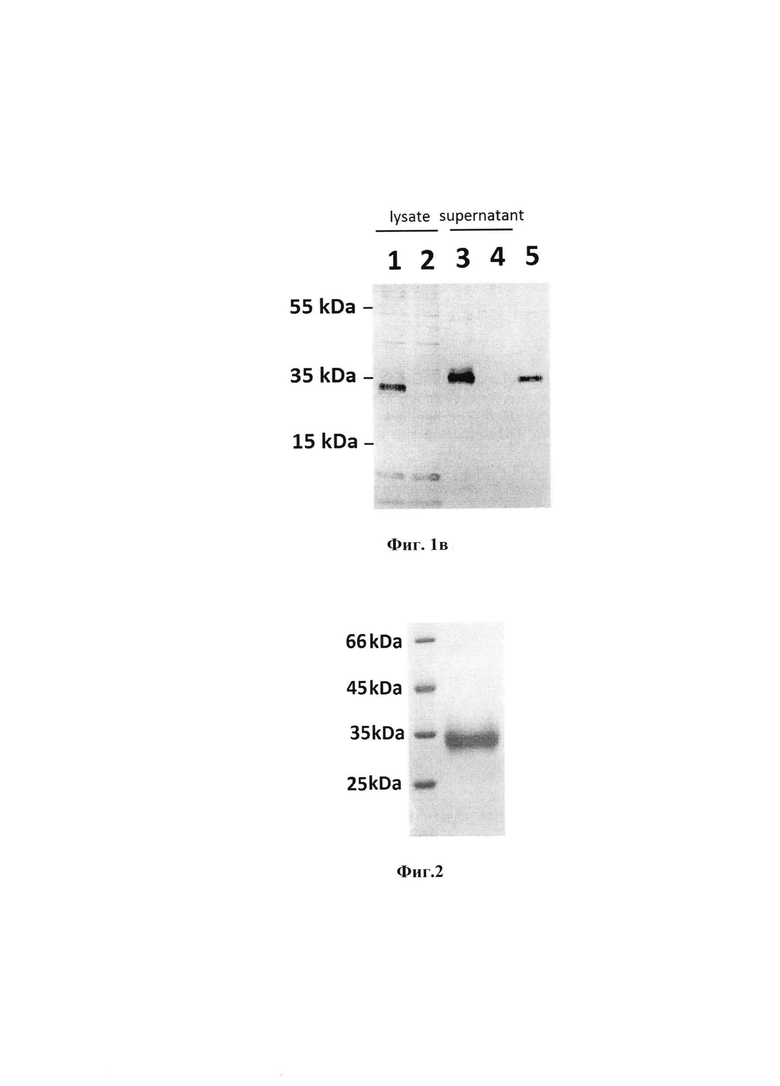
Фиг. 1б. Электрофоретический анализ экспрессии RBD в трансфицированных клетках в 1% агарозном геле:
Дорожка М12 - маркеры молекулярной массы;
Дорожки 1 и 3 - продукты, полученные с помощью ОТ-ПЦР из тотальной РНК клеток HEK293T, трансфицированных pVAX и pVAXrbd соответственно;
Дорожка 2 - продукт, полученный с помощью ПЦР плазмиды pVAXrbd.
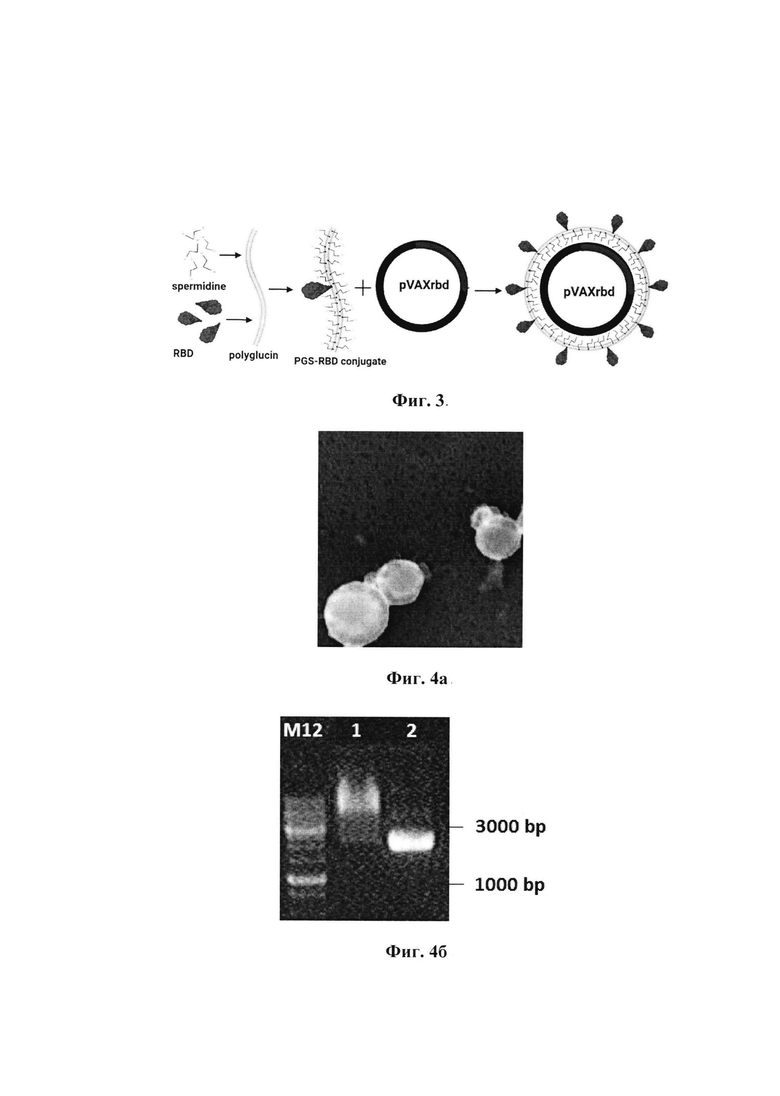
Фиг. 1в. Анализ продукции белка RBD в клетках HEK293T с помощью вестерн-блоттинга:
Дорожки 1 и 2 - лизаты HEK293T, трансфицированных pVAXrbd и pVAX, соответственно;
Дорожки 3 и 4 - культуральная среда из клеток HEK293T, трансфицированных pVAX и pVAXrbd, соответственно;
Дорожка 5 - очищенный рекомбинантный RBD.
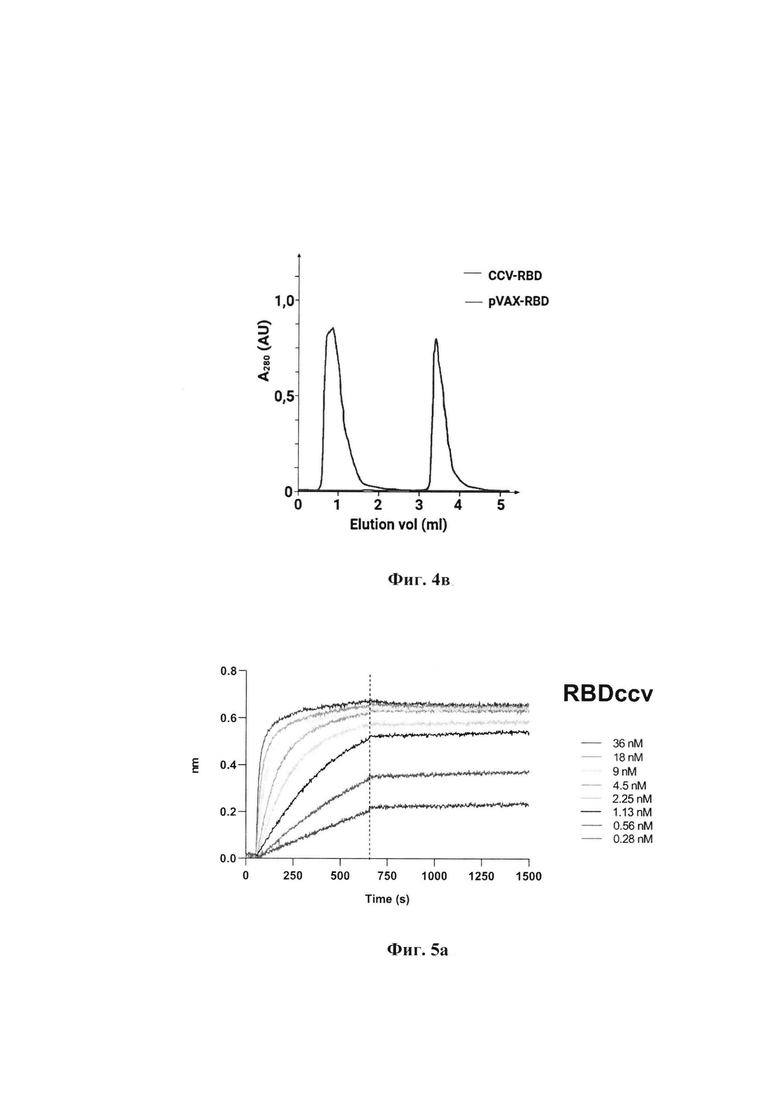
Фиг. 2. Электрофоретический анализ чистоты белка RBD в 12% ПААГ:
Дорожка 1 - маркеры молекулярной массы;
Дорожка 2 - очищенный белок RBD.
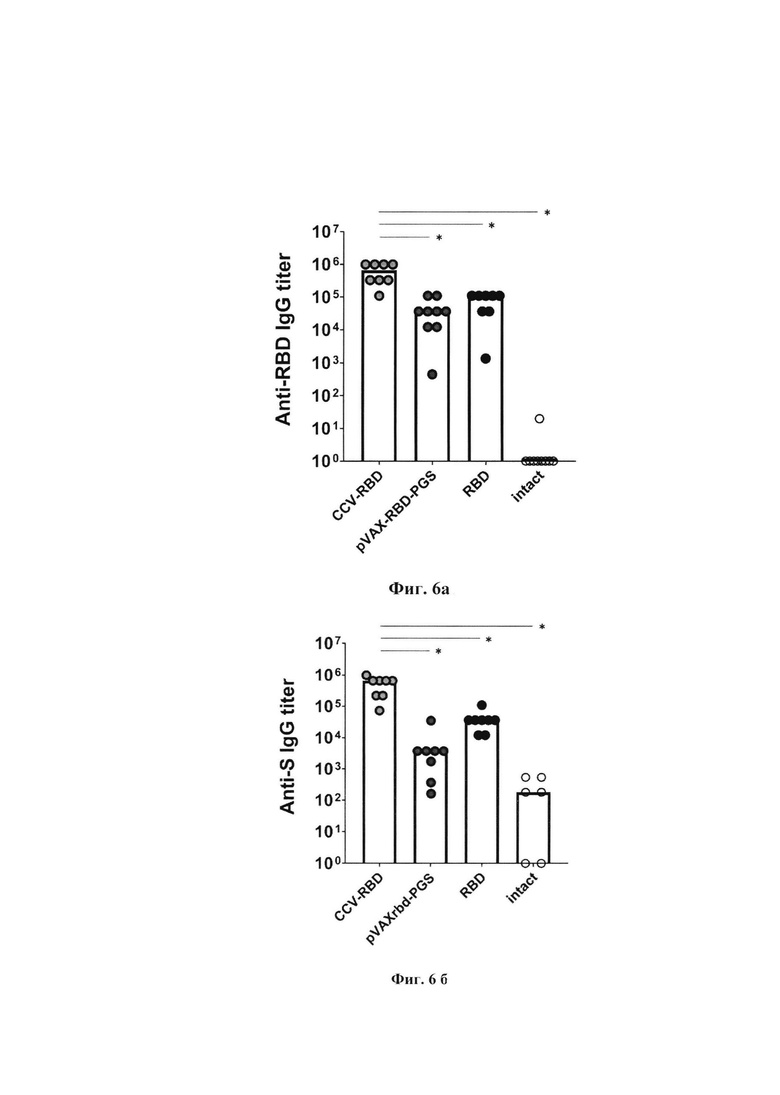
Фиг. 3. Схематическое изображение сборки частиц комбинированной вакцины КомбиКоронаВак (CCV-RBD): внутри плазмида pVAX-RBD, а на поверхности - белок RBD.
Фиг. 4а. Электронная микрофотография частиц CCV-RBD.
Фиг. 4б. Подтверждение инкапсуляции ДНК в оболочке PGS и PGS-RBD с помощью электрофореза в 1% агарозном геле:
Дорожка М12 - маркеры молекулярной массы;
Дорожка 1 - CCV-RBD;
Дорожка 2 - голая плазмида pVAXrbd.
Фиг. 4в. Гель-фильтрация на колонке Sepharose CL-6B (хроматографический профиль).
Левая линия - CCV-RBD;
Правая линия - плазмида pVAXrbd.
Фиг. 5а. Связывание CCV-RBD с iB14, nAb SARS-CoV-2, с помощью прибора Octet RED96.
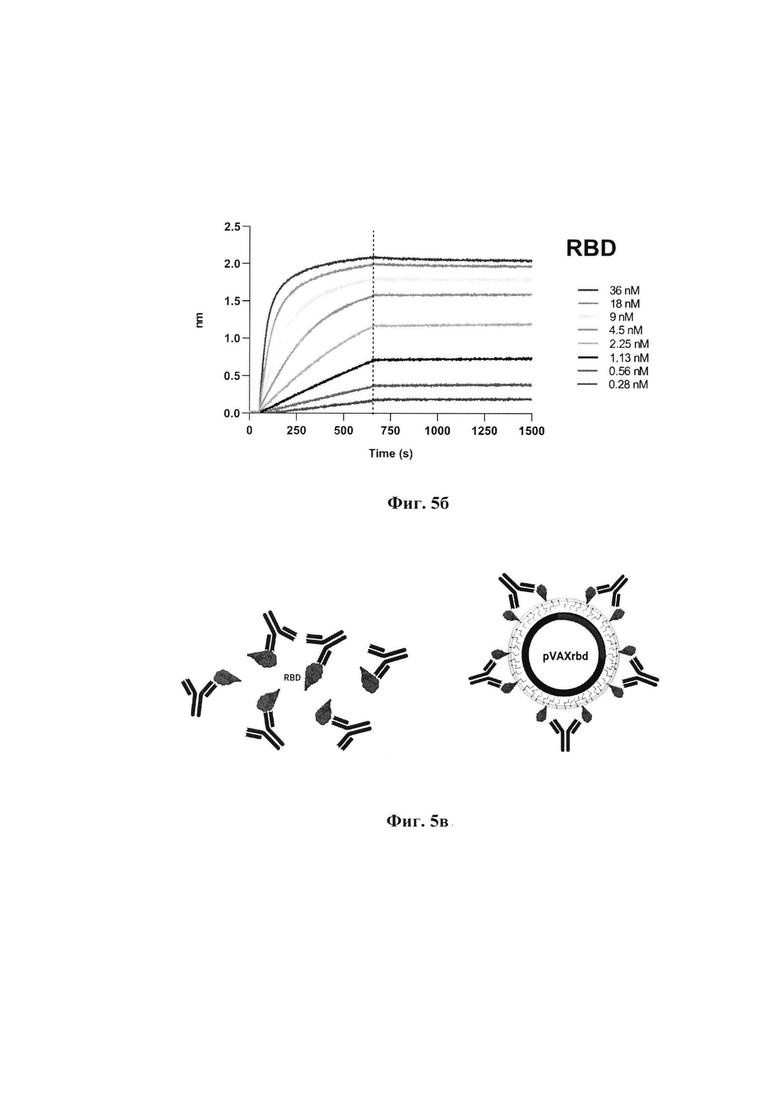
Фиг. 5б. Связывание RBD с iB14, nAb SARS-CoV-2, с помощью прибора Octet RED96.
Фиг. 5в. Схематическое представление взаимодействия белка RBD и частиц CCV-RBD и нейтрализующего антитела iB14.
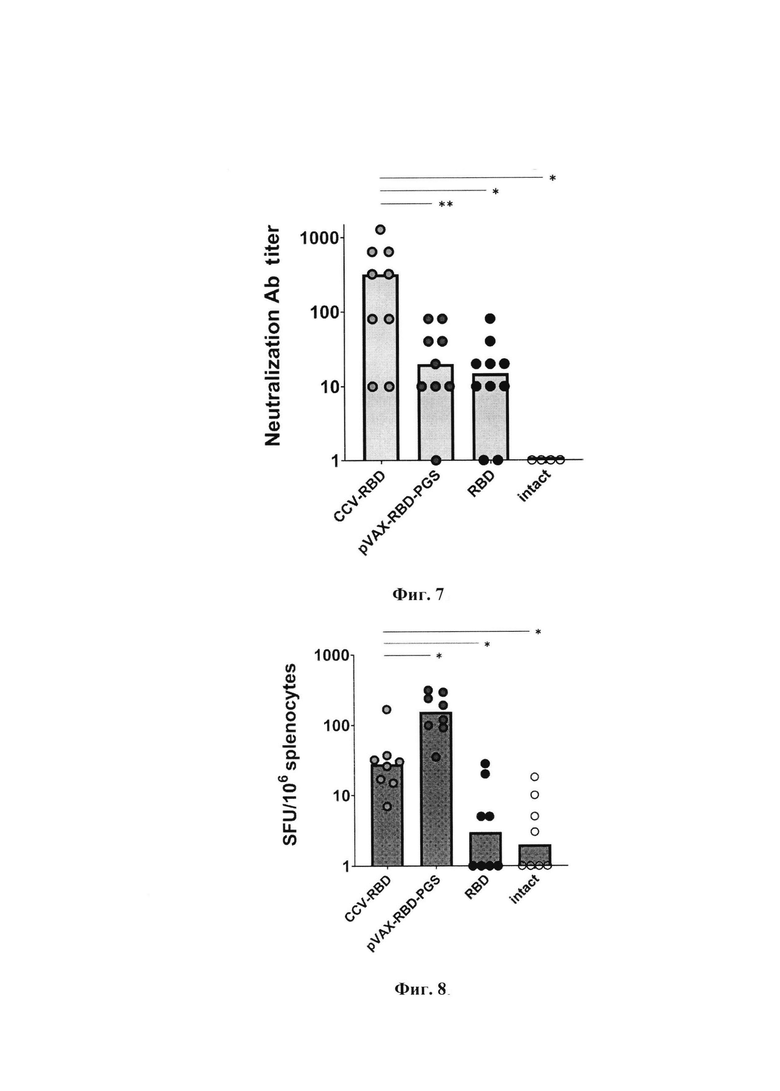
Фиг. 6а. Титры антител в сыворотке иммунизированных животных. В качестве антигена в ИФА был использован рекомбинантный белок RBD.
Фиг. 6б. Титры антител в сыворотке иммунизированных животных. В качестве антигена в ИФА рекомбинантный белок S.
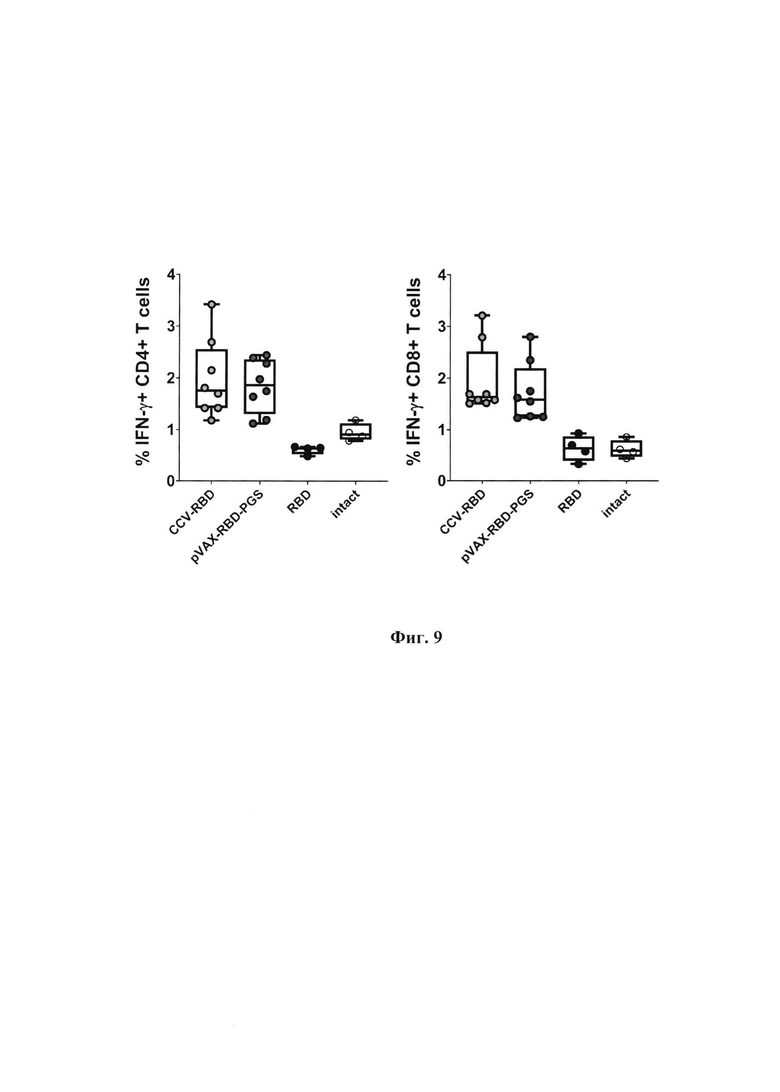
Фиг. 7. Вируснейтрализующая активность сывороток мышей, иммунизированных CCV-RBD, pVAXrbd-PGS и белком RBD, определялась с использованием штамма SARS-CoV-2 nCoV/Victoria/1/2020 (100 TCID50).
Фиг. 8. Результаты оценки вирус-специфических Т-лимфоцитов у иммунизированных животных на 31 сутки после начала иммунизации с помощью метода ELISpot.
Фиг. 9. Результаты оценки специфических цитотоксических и хэлперных Т-лимфоцитов, продуцирующих IFN-γ, с помощью метода внутриклеточного окрашивания цитокинов и проточной цитометрии.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Дизайн плазмиды pVAX-RBD
Для конструирования иммуногенов использовали последовательность гена, кодирующего полноразмерный белок S SARS-CoV-2 (GenBank Acc. No. MN908947). Для оптимизации состава кодонов, а также вторичной структуры РНК для эффективной экспрессии в клетках млекопитающих использовали программу GeneOptimizer (https://www.thermofisher.com/ru/en/home/life-science/cloning/gene-synthesis/geneart-gene-synthesis/geneoptimizer.html). Полученную нуклеотидную последовательность синтезировали в ООО “ДНК-синтез” (Россия) и клонировали в составе вектора pGH. Для амплификации фрагмента, кодирующего RBD белка S (320V-542N) и сигнальную последовательность 176 (MMRTLILAVLLVYFCATVHC) на 5'-конце, использовали праймеры: 5'-TAATACGACTCACTATAGGCTAGCCT-3' (прямой) и 5'-AAAAAAGCGGCCGCTCATTAGTTGAAGTTCACGCATTTGTTCTTC-3' (обратный), - и матрицу, содержащую указанный фрагмент. Продукт амплификации встраивали в вектор pVAX (“Thermo Fisher Scientific”, США) по сайтам NheI и NotI под немедленный ранний промотор цитомегаловируса человека (CMV). Структуру полученной конструкции подтверждали секвенированием по методу Сэнгера в ЦКП “Геномика” (Россия). В результате была получена ДНК-конструкция pVAX-RBD, кодирующая RBD с сигнальной последовательностью 176, обеспечивающей секрецию нарабатываемого белка из клетки.
Пример 2. Наработка и очистка рекомбинантной плазмиды pVAX-RBD
Клетки Escherichia coli Stbl3 трансформировали плазмидами pVAX и pVAX-RBD с помощью хлористого кальция. Наработку плазмидной ДНК для иммунизации проводили в 2.7 л питательной среды LB с добавлением канамицина (25 мкг/мл). Плазмидную ДНК выделяли и очищали с помощью набора EndoFree Plasmid Giga Kit (“QIAGEN”, Германия) согласно рекомендациям производителя. Данный набор позволяет очень эффективно очистить препарат плазмидной ДНК от бактериальных эндотоксинов. После очистки структуру плазмиды проверяли рестрикционным анализом и спектрофотометрией.
Пример 3. Получение и очистка рекомбинантного белка RBD
Для создания клеточной линии, продуцирующей белок RBD (SEQ ID N 2), клетки CHO-K1 трансфицировали плазмидой pVEAL2-S-RBD (обеспечивает экспрессию и секрецию белка RBD коронавируса SARS-CoV-2 в клетках млекопитающих, патент РФ №2752858) с использованием липофектамина 3000 (Thermo Fisher Scientific) согласно рекомендациям производителя. Для интеграции экспрессионной кассеты вектора в клеточный геном вместе с целевой плазмидой добавляли плазмиду pCMV (CAT) T7-SB100, кодирующую транспозазу SB100. Через 3 дня в культуральную среду добавляли селективный антибиотик пуромицин (InvivoGen, Сан-Диего, Калифорния, США) в конечной концентрации 10 мкг/мл (вектор pVEAL2 содержит ген устойчивости к пуромицину). Отбор устойчивых клонов продолжался в течение 3 дней. Клоны, показавшие наибольшую продуктивность, выращивали на вальцовых установках и собирали питательную среду. Рекомбинантный RBD был выделен из культуральной среды клеток CHO-K1. Его центрифугировали, фильтровали (0,22 М) и подвергали двухстадийной хроматографической очистке. Первый этап включал металлохелатную хроматографию на колонке Ni-NTA (Qiagen, Hilden, Düsseldorf, Germany) согласно инструкции производителя. Последующую очистку проводили методом ионообменной хроматографии на последовательно соединенных колонках с катионообменным (СП-Сефароза) и анионообменным (Q-Сефароза) сорбентами в 20 мМ Трис-HCl (рН 8,2). Полученный белок диализовали против фосфатно-солевого буфера (PBS) и подвергали стерилизующей фильтрации через фильтры 0,22 мкм. Концентрацию белка определяли по методу Лоури. Препарат белка RBD анализировали с помощью денатурирующего электрофореза в 15% PAGE.
Пример 4. Получение и характеризация конъюгата белка RBD с полиглюкин-спермидином
На первом этапе декстран активировали следующим образом: 1 моль декстрана 40000 (MP Biomedicals, Irvine, CA, USA) обрабатывали 40 моль перйодата натрия в течение 60 мин, после чего остатки окислителя удаляли из активированного декстран гель-фильтрацией на колонке с сефадексом G-25, уравновешенным 50 мМ карбонатным буфером (рН 8,6). Далее к раствору активированного декстрана добавляли белок RBD - 1 моль декстрана и 1 моль белка - и смесь инкубировали в течение 2 ч. Затем к смеси добавляли боргидрид натрия в соотношении 80 моль боргидрида на 1 моль декстрана. После инкубации в течение 2 ч полученные конъюгаты очищали от непрореагировавших компонентов гель-фильтрацией на Sephadex G-25, уравновешенном PBS. Препарат конъюгата стерильно фильтровали через фильтры 0,22 мкм и обозначили как PGS-RBD.
Присоединение белка к полиглюкин-спермидину наблюдали по изменению выхода фракции из гель-фильтрующей колонки и «размытию» бэнда белка в 15% PAGE электрофорезе в денатурирующих условиях. Наличие белка в структуре также было подтверждено с помощью ультрафиолетовой (УФ) спектроскопии: белок RBD и конъюгат PGS-RBD имели характерный для белков пик поглощения при 280 нм.
Пример 5. Сборка вакцинной конструкции и ее характеризация
Схематичное изображение получения комплексов ДНК и PGS представлено на фиг. 3. Для формирования комплексов CCV-RBD плазмиду pVAX-RBD смешивали с конъюгатом PGS-RBD в соотношении 100 мкг ДНК на каждые 100 мкг белка, инкубировали 5 мин, затем добавляли избыток PGS. Добавляли так, чтобы положительного заряда молекул спермидина было достаточно для электростатического взаимодействия с отрицательно заряженным конъюгатом нуклеиновой кислоты и для полного оборачивания плазмиды (инкубация 1 ч при 2-8°С).
Эффективность образования комплексов CCV-RBD оценивали по изменению электрофоретической подвижности ДНК в 1%-ном агарозном геле. Инкапсулированная плазмида теряла подвижность в электрическом поле при покрытии конъюгатом PGS-RBD (фиг. 4б).
Для оценки размера и формы полученных частиц их суспензии наносили на медные сетки для электронной микроскопии, покрытые пленкой стабилизированного углеродом формвара. Препараты окрашивали 2% водным раствором фосфорно-вольфрамовой кислоты и исследовали с помощью электронного микроскопа JEM-1400 (JEOL, Akishima, Tokyo, Japan). Для получения изображений использовалась цифровая камера Veleta (SIS, Schwentinental, Германия), а для анализа и обработки изображений использовался программный пакет iTEM (SIS, Schwentinental, Германия). Изображения частиц представлены на фиг. 4а.
Гель-фильтрацию CCV-RBD и pVAXrbd проводили на колонке Sepharose CL-6B объемом 10 мл в PBS (pH 7,4). Образцы исследуемых препаратов наносили в эквимолярных количествах на нуклеотидный материал. Как показано на фиг. 4в, пик, соответствующий фракции CCV-RBD, появляется в свободном объеме перед плазмидной ДНК.
УФ-спектры поглощения CCV-RBD и их отдельных компонентов (PGS, RBD, PGS-RBD и плазмиды pVAXrbd) исследовали на спектрофотометре Nanodrop One в диапазоне длин волн 220-300 нм (Thermo Fisher Scientific). Сохранение структуры плазмидной ДНК как в отдельности, так и в комплексах CCV-RBD было подтверждено наличием характерного для ДНК пика поглощения при 260 нм.
Кинетику вакцинного антигена (CCV-RBD), рекомбинантного фрагмента спайкового белка SARS-CoV-2 (RBD) по сравнению со специфичным для SARS-CoV-2 RBD антителом iB14, измеряли с помощью прибора Octet RED96 (ForteBio, Pall Life Sciences, Нью-Йорк, США). Все анализы проводили при перемешивании со скоростью 1000 об/мин в кинетическом буфере ForteBio 1 (PBS, 0,05% Tween, 0,05% NaN3). Конечный объем для всех растворов составил 200 мкл на лунку. Анализы проводили при 30°С в сплошных черных 96-луночных планшетах (Greiner Bio-One, Кремсмюнстер, Австрия). Кинетический буфер лигандов (CCV-RBD и RBD) 1 загружали на поверхность Ni-NTA биосенсоров (NTA) на 300 с.Перед анализом ассоциации антигена на биосенсоре с антителом в растворе в течение 600 с применяли 60-секундный базовый этап биосенсора. Двукратный градиент концентрации антитела iB14, начиная с 144 нМ для RBD и CCV-RBD, использовали в серии титрования из восьми. За диссоциацией взаимодействия следили в течение 900 с.Коррекцию дрейфа базовой линии проводили путем вычитания среднего значения сдвигов, зарегистрированных для сенсора, нагруженного антигеном, но не инкубированного с антителом, и сенсора без антигена, но инкубированного с антителом. Данные октета обрабатывались программным обеспечением для сбора данных ForteBio v.11.1.1.19. Экспериментальные данные были сопоставлены с уравнениями связывания, описывающими взаимодействие 2:1 (глобальная адаптация, бивалентный аналит), для достижения оптимальной аппроксимации. Связывание nAb iB14 с эпитопом, который частично перекрывается с местом взаимодействия RBD-ACE2 [], достаточно эффективно распознает RBD в CCV-RBD с константой диссоциации 6,6*10-11 по сравнению с 1,47*10-10 для RBD белка.
Пример 6. Проверка иммуногенных свойств вакцинного комплекса CCV-RBD. Иммунизация лабораторных животных, забор и подготовка образцов
Работа с животными проводилась в соответствии с «Руководством по уходу и использованию лабораторных животных». Протоколы были одобрены Комитетом по уходу и использованию животных (IACUC) при Государственном научном центре вирусологии и биотехнологии «Вектор» (номер разрешения: НИЦ ВБ «Вектор» / 10.09.2020).
Для оценки иммуногенности созданных конструкций использовали самок мышей BALB/c массой 16-18 г.Мышей разделили на четыре группы по 8-9 животных в каждой и иммунизировали следующим образом: группа CCV-RBD - комбинированная вакцина, содержащая 100 мкг ДНК и 100 мкг белка; группа pVAXrbd-PGS, 100 мкг плазмиды pVAXrbd, инкапсулированной в оболочку PGS без белка; группа RBD, 100 г рекомбинантного белка RBD; интактная группа, без иммунизации. Животные первых трех групп получали препараты в объеме 200 мкл внутримышечно в верхнюю часть задних конечностей (по 100 мкл на конечность). Мышей иммунизировали двукратно с интервалом между иммунизациями 3 недели.
Через десять дней после второй иммунизации у животных брали кровь для анализа гуморального иммунного ответа и собирали селезенку для анализа клеточного ответа. Сыворотку отделяли от клеточных элементов центрифугированием (9000 g, 15 мин), нагревали в течение 30 мин при 56°С и исследовали на наличие антител, специфически связывающихся с белками S и RBD, в ИФА. Образцы также были проанализированы на их нейтрализующую активность. Селезенки последовательно измельчали на нейлоновых фильтрах для клеток с диаметром пор 70 и 40 мкм (BD Falcon, Franklin Lakes, NJ, USA). После лизиса эритроцитов лизирующим буфером (Sigma, Burlington, MA, USA) спленоциты дважды промывали полной средой RPMI и помещали в 1 мл среды RPMI с 2 мМ L-глутамина, 50 мкг/мл гентамицина и 10% FBS (Thermo Fisher Scientific, США). Клетки подсчитывали с использованием автоматического счетчика клеток TC20 (Bio-Rad, Hercules, CA, USA). Среди спленоцитов количество IFN-γ-продуцирующих клеток определяли методами ELISpot и ICS.
Пример 6а. Иммуноферментный анализ сывороток, исследование их вируснейтрализующих свойств
В качестве антигенов для ИФА использовали рекомбинантные эукариотические белки RBD и S, сорбированные на 96-луночные планшеты (Corning, NY, USA) в концентрации 1 мкг/мл в 2 М мочевине в течение ночи при 4°С. Планшеты трижды промывали и инкубировали в течение 1,5 ч при комнатной температуре с 1% казеина в PBS с 0,05% Tween 20 для блокирования неспецифического связывания белка. Затем планшеты инкубировали с трехкратными серийными разведениями мышиной сыворотки в течение 1 ч при комнатной температуре, после промывки PBST добавляли кроличьи антимышиные IgG, конъюгированные с пероксидазой хрена (Sigma), разведенные 1:3000, и инкубируют 1 ч при комнатной температуре. После окончательной промывки планшеты проявляли с использованием субстрата TMB (Amresco, Radnor, PA, USA). Реакцию останавливали 1N HCl и анализировали при 450 нм на многорежимном ридере для микропланшетов Varioskan LUX (Thermo Fisher Scientific, Waltham, MA, USA).
По результатам RBD-специфического ИФА в конечной точке эксперимента отмечено>32-кратное увеличение медианы титра специфических антител у животных, иммунизированных комбинированной ДНК/белковой вакциной CCV-RBD, по сравнению с группой, инъецировали pVAXrbd-PGS, и наблюдалось 8-кратное увеличение по сравнению с группой, получавшей белок RBD без адъювантов фиг. 6а, p<0,01). S-специфический ИФА для обнаружения белка S показал 75-кратное увеличение в группе, иммунизированной CCV-RBD, по сравнению с группой, которой вводили pVAXrbd-PGS, и 13-кратное увеличение по сравнению с группой, иммунизированной RBD (фиг.6б), р<0,01).
Нейтрализующие свойства антител сыворотки крови определяли по ингибированию цитопатического действия (ЦПД) вируса в культуре клеток in vitro. Вкратце, штамм SARS-CoV-2 nCoV/Victoria/1/2020 (100 TCID50), инкубированный с серийными разведениями исследуемых сывороток, высевали на 100% конфлюэнтный монослой клеток Vero E6. Планшеты инкубировали 4 дня при 37°С в присутствии 5% СО2, затем клетки окрашивали 0,2% раствором генцианвиолета. Результаты фиксировались визуально. Нейтрализующую активность сывороток иммунизированных животных оценивали по титру (разведению) сывороток, при котором в 50% лунок регистрировали защиту клеток от ЦПД вируса.
Результаты анализа коррелируют с результатами ИФА: сыворотки животных, иммунизированных CCV-RBD, показали более высокую активность в отношении ингибирования цитопатического действия вируса (фиг. 7, р<0,05). Таким образом, иммунизация комбинированной ДНК/белковой вакциной CCV-RBD индуцировала более сильный гуморальный иммунитет (IgG и нейтрализующие антитела), чем иммунизация только ДНК или белком RBD.
Пример 6б. Исследование поствакцинального Т-клеточного ответа с помощью ELISpot и ICS. Анализ Т-клеточного иммунного ответа проводили с использованием набора Mouse IFNgamma ELISpot (R&D Systems, Миннеаполис, Миннесота, США) в соответствии с инструкциями производителя. Спленоциты высевали по 5*105 клеток/лунку и добавляли среду RPMI с 10% FBS (отрицательный контроль), или смесью пептидов (каждого в концентрации 20 мкг/мл), или конканавалином А (положительный контроль). Клетки инкубировали в течение 20 ч при 37°С в присутствии 5% СО2. Затем планшеты промывали и инкубировали с первичными антителами против IFN-γ. Планшеты снова промывали и затем инкубировали со вторичным антителом, конъюгированным со щелочной фосфатазой. Наконец, планшеты снова промывали, а затем инкубировали с BCIP/NBT. Количество клеток, продуцирующих ИФН, подсчитывали с использованием считывающего устройства ELISpot (Carl Zeiss, Оберкохен, Германия). Количество пятнообразующих единиц (SFU) на миллион клеток рассчитывали путем вычитания среднего значения из лунок отрицательного контроля. Спленоциты, выделенные от иммунизированных мышей BALB/c, стимулировали пулом пептидов из последовательности S-белка SARS-CoV-2, рестриктированных главным комплексом гистосовместимости (MHC) класса I (H2-Dd, H-2-Kd и H- 2-Ld) и молекулы MHC класса II (H2-IAd и H2-IEd) мышей BALB/c (таблица 1). Пептиды были отобраны с использованием инструментов IEDB Analysis Resource и синтезированы AtaGenix Laboratories (Ухань, Китай); чистота пептидов>80%.
При использовании ELISpot группа, иммунизированная ДНК-вакциной, покрытой PGS, показала наибольший клеточный иммунитет (фиг. 8). В отличие от гуморального ответа мы не наблюдали повышения клеточного иммунитета в группе CCV-RBD. Однако следует отметить, что иммунизация одним белком RBD не вызывала индукцию клеточного иммунного ответа.
ICS проводили на спленоцитах, выделенных от иммунизированных мышей BALB/c. Для анализа 2*106 клеток высевали в лунки 24-луночных культуральных планшетов (ТРР, Трасадинген, Швейцария) и стимулировали смесью пептидов, указанной выше. Каждый пептид добавляли в концентрации 20 мкг/мл на лунку и клетки инкубировали в течение 4 ч при 37°С в присутствии 5% СО2 и еще 16 ч с брефельдином А (5 мкг/мл, GolgiPlug BD Biosciences). На следующий день клетки окрашивали анти-CD3, конъюгированным с Alexa Fluor 700 (BD), анти-CD4, конъюгированным с BV786 (BD), и анти-CD8, конъюгированным с FITC (BD); фиксировали 1% параформальдегидом в PBS; и пермеабилизировали 0,5% Tween 20 в PBS в соответствии с инструкциями производителя. Затем клетки окрашивали для выявления внутриклеточных цитокинов анти-IFN-APC (BD, США). Образцы анализировали с использованием проточного цитометра ZE5 (Bio-Rad) и программного обеспечения Everest.
Мы оценили процент IFN-γ-продуцирующих лимфоцитов среди всех CD4+ и CD8+ Т-клеток с помощью внутриклеточного окрашивания цитокинов (ICS) с использованием проточной цитометрии. Как показано на фиг. 9, после стимуляции пептидами CD4+ и CD8+ Т-лимфоциты реагировали более высоким высвобождением IFN-γ в группах, иммунизированных комбинированной и ДНК-вакциной. Индукция клеточного иммунитета, характеризующаяся продукцией IFN-γ, не происходила ни в группе, иммунизированной белком RBD, ни в интактной группе.
Таким образом, преимущество заявляемой вакцинной платформы по сравнению с другими аналогами заключается в том, что в качестве носителя используют декстран, который не является иммуногенным, а иммунный ответ формируется только против антигена-мишени. Включение плазмидной ДНК-вакцины в ядро частиц обеспечивает дополнительный адъювантный эффект на синтез антител. Вакцинный комплекс CCV-RBD является перспективной вакциной-кандидатом для использования в профилактике COVID-19, а платформа, используемая для создания комбинированных вакцин, будет полезна для разработки различных профилактических или терапевтических препаратов, способных индуцировать гуморальные и клеточные ответы для защиты [19].
Список источников патентной и научно-технической информации.
1. URL: https://covid19.who.int/
2. Xia S., Zhang Y., Wang Y., Wang H., Yang Y., Gao G.F., Tan W., Wu G., Xu M., Lou Z., Huang W., Xu W., Huang B., Wang H., Wang W., Zhang W., Li N., Xie Z., Ding L., You W., Zhao Y., Yang X., Liu Y., Wang Q., Huang L., Yang Y., Xu G., Luo B., Wang W., Liu P., Guo W., Yang X. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: a randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect Dis. 2021 Jan;21(1):39-51. doi: 10.1016/S1473-3099(20)30831-8.
3. Han B., Song Y., Li C., Yang W., Ma Q., Jiang Z., Li M., Lian X., Jiao W., Wang L., Shu Q., Wu Z., Zhao Y., Li Q., Gao Q. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine (CoronaVac) in healthy children and adolescents: a double-blind, randomised, controlled, phase 1/2 clinical trial. Lancet Infect Dis. 2021 Dec;21(12):1645-1653. doi: 10.1016/S1473-3099(21)00319-4.
4. Logunov D.Y., Dolzhikova I.V., Zubkova O.V., Tukhvatullin A.I., Shcheblyakov D.V., Dzharullaeva A.S., Grousova D.M., Erokhova A.S., Kovyrshina A.V., Botikov A.G., Izhaeva F.M., Popova O., Ozharovskaya T.A., Esmagambetov I.B., Favorskaya I.A., Zrelkin D.I., Voronina D.V., Shcherbinin D.N., Semikhin A.S., Simakova Y.V., Tokarskaya E.A., Lubenets N.L., Egorova D.A., Shmarov M.M., Nikitenko N.A., Morozova L.F., Smolyarchuk E.A., Kryukov E.V., Babira V.F., Borisevich S.V., Naroditsky B.S., Gintsburg A.L. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: two open, non-randomised phase 1/2 studies from Russia. Lancet. 2020 Sep 26;396(10255):887-897. doi: 10.1016/S0140-6736(20)31866-3.
5. Folegatti P.M., Ewer K.J., Aley P.K., Angus B., Becker S., Belij-Rammerstorfer S., Bellamy D., Bibi S., Bittaye M., Clutterbuck E.A., Dold C., Faust S.N., Finn A., Flaxman A.L., Hallis B., Heath P., Jenkin D., Lazarus R., Makinson R., Minassian A.M., Pollock K.M., Ramasamy M., Robinson H., Snape M., Tarrant R., Voysey M., Green C., Douglas A.D., Hill A.V.S., Lambe T., Gilbert S.C., Pollard A.J.; Oxford COVID Vaccine Trial Group.Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: a preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet. 2020 Aug 15;396(10249):467-478. doi: 10.1016/S0140-6736(20)31604-4.
6. Sadoff J., Le Gars M., Shukarev G., Heerwegh D., Truyers C., de Groot A.M., Stoop J., Tete S., Van Damme W., Leroux-Roels I., Berghmans P.J., Kimmel M., Van Damme P., de Hoon J., Smith W., Stephenson K.E., De Rosa S.C., Cohen K.W., McElrath M.J., Cormier E., Scheper G., Barouch D.H., Hendriks J., Struyf F., Douoguih M., Van Hoof J., Schuitemaker H. Interim Results of a Phase 1-2a Trial of Ad26.COV2.S Covid-19 Vaccine. N Engl J Med. 2021 May 13;384(19):1824-1835. doi: 10.1056/NEJMoa2034201.
7. Рыжиков А.Б., Рыжиков Е.А., Богрянцева М.П., Усова С. В., Даниленко Е.Д., Нечаева Е.А., Пьянков О.В., Пьянкова О.Г., Гудымо А.С., Боднев С.А., Онхонова Г.С., Слепцова Е.С., Кузубов В.И., Рындюк Н.Н., Гинько З.И., Петров В.Н., Моисеева А.А., Торжкова П.Ю., Пьянков С.А., Трегубчак Т.В., Антонец Д.В., Гаврилова Е.В., Максютов Р.А. Простое слепое плацебо-контролируемое рандомизированное исследование безопасности, реактогенности и иммуногенности вакцины «ЭпиВакКорона» для профилактики COVID-19 на добровольцах в возрасте 18-60 лет (фаза I-II) // Инфекция и иммунитет.- 2021. - Т. 11. - №2. - C. 283-296. doi: 10.15789/2220-7619-ASB-1699.
8. Keech C., Albert G., Cho I., Robertson A., Reed P., Neal S., Plested J.S., Zhu M., Cloney-Clark S., Zhou H., Smith G., Patel N., Frieman M.B., Haupt R.E., Logue J., McGrath M., Weston S., Piedra P.A., Desai C., Callahan K., Lewis M., Price-Abbott P., Formica N., Shinde V., Fries L., Lickliter J.D., Griffin P., Wilkinson B., Glenn G.M. Phase 1-2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N Engl J Med. 2020 Dec 10;383(24):2320-2332. doi: 10.1056/NEJMoa2026920.
9. Walsh E.E., Frenck R.W. Jr., Falsey A.R., Kitchin N., Absalon J., Gurtman A., Lockhart S., Neuzil K., Mulligan M.J., Bailey R., Swanson K.A., Li P., Koury K., Kalina W., Cooper D., Fontes-Garfias C., Shi P.Y., Türeci Ö., Tompkins K.R., Lyke K.E., Raabe V., Dormitzer P.R., Jansen K.U., Şahin U., Gruber W.C. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N Engl J Med. 2020 Dec 17;383(25):2439-2450. doi: 10.1056/NEJMoa2027906.
10. Baden L.R., El Sahly H.M., Essink B., Kotloff K., Frey S., Novak R., Diemert D., Spector S.A., Rouphael N., Creech C.B., McGettigan J., Khetan S., Segall N., Solis J., Brosz A., Fierro C., Schwartz H., Neuzil K., Corey L., Gilbert P., Janes H., Follmann D., Marovich M., Mascola J., Polakowski L., Ledgerwood J., Graham B.S., Bennett H., Pajon R., Knightly C., Leav B., Deng W., Zhou H., Han S., Ivarsson M., Miller J., Zaks T.; COVE Study Group.Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N Engl J Med. 2021 Feb 4;384(5):403-416. doi: 10.1056/NEJMoa2035389.
11. Khobragade A., Bhate S., Ramaiah V., Deshpande S., Giri K., Phophle H., Supe P., Godara I., Revanna R., Nagarkar R., Sanmukhani J., Dey A., Rajanathan T.M.C., Kansagra K., Koradia P.; ZyCoV-D phase 3 Study Investigator Group.Efficacy, safety, and immunogenicity of the DNA SARS-CoV-2 vaccine (ZyCoV-D): the interim efficacy results of a phase 3, randomised, double-blind, placebo-controlled study in India. Lancet. 2022 Apr 2;399(10332):1313-1321. doi: 10.1016/S0140-6736(22)00151-9.
12. Vaine M., Wang S., Hackett A., Arthos J., Lu S. Antibody responses elicited through homologous or heterologous prime-boost DNA and protein vaccinations differ in functional activity and avidity. Vaccine. 2010 Apr 9;28(17):2999-3007. doi: 10.1016/j.vaccine.2010.02.006.
13. Wang S., Arthos J., Lawrence J.M., Van Ryk D., Mboudjeka I., Shen S., Chou T.H., Montefiori D.C., Lu S. Enhanced immunogenicity of gp120 protein when combined with recombinant DNA priming to generate antibodies that neutralize the JR-FL primary isolate of human immunodeficiency virus type 1. J Virol. 2005 Jun;79(12):7933-7. doi: 10.1128/JVI.79.12.7933-7937.2005.
14. Li Y., Bi Y., Xiao H., Yao Y., Liu X., Hu Z., Duan J., Yang Y., Li Z., Li Y., Zhang H., Ding C., Yang J., Li H., He Z., Liu L., Hu G., Liu S., Che Y., Wang S., Li Q., Lu S., Cun W. A novel DNA and protein combination COVID-19 vaccine formulation provides full protection against SARS-CoV-2 in rhesus macaques. Emerg Microbes Infect. 2021 Dec;10(1):342-355. doi: 10.1080/22221751.2021.1887767.
15. Felber B.K., Lu Z., Hu X., Valentin A., Rosati M., Remmel C.A.L., Weiner J.A., Carpenter M.C., Faircloth K., Stanfield-Oakley S., Williams W.B., Shen X., Tomaras G.D., LaBranche C.C., Montefiori D., Trinh H.V., Rao M., Alam M.S., Vandergrift N.A., Saunders K.O., Wang Y., Rountree W., Das J., Alter G., Reed S.G., Aye P.P., Schiro F., Pahar B., Dufour J.P., Veazey R.S., Marx P.A., Venzon D.J., Shaw G.M., Ferrari G., Ackerman M.E., Haynes B.F., Pavlakis G.N. Co-immunization of DNA and Protein in the Same Anatomical Sites Induces Superior Protective Immune Responses against SHIV Challenge. Cell Rep.2020 May 12;31(6):107624. doi: 10.1016/j.celrep.2020.107624.
16. Karpenko L.I., Ilyichev A.A., Eroshkin A.M., Lebedev L.R., Uzhachenko R.V., Nekrasova N.A., Plyasunova O.A., Belavin P.A., Seregin S.V., Danilyuk N.K., Zaitsev B.N., Danilenko E.D., Masycheva V.I., Bazhan S.I. Combined virus-like particle-based polyepitope DNA/protein HIV-1 vaccine design, immunogenicity and toxicity studies. Vaccine. 2007 May 22;25(21):4312-23. doi: 10.1016/j.vaccine.2007.02.058.
17. Bazhan S.I., Karpenko L.I., Lebedev L.R., Uzhachenko R.V., Belavin P.A., Eroshkin A.M., Ilyichev A.A. A synergistic effect of a combined bivalent DNA-protein anti-HIV-1 vaccine containing multiple T- and B-cell epitopes of HIV-1 proteins. Mol Immunol. 2008 Feb;45(3):661-9. doi: 10.1016/j.molimm.2007.07.016.
18. Karpenko L.I., Rudometov A.P., Sharabrin S.V., Shcherbakov D.N., Borgoyakova M.B., Bazhan S.I., Volosnikova E.A., Rudometova N.B., Orlova L.A., Pyshnaya I.A., Zaitsev B.N., Volkova N.V., Azaev M.S., Zaykovskaya A.V., Pyankov O.V., Ilyichev A.A. Delivery of mRNA Vaccine against SARS-CoV-2 Using a Polyglucin:Spermidine Conjugate. Vaccines (Basel). 2021 Jan 21;9(2):76. doi: 10.3390/vaccines9020076.
19. Borgoyakova M.B., Karpenko L.I., Rudometov A.P., Volosnikova E.A., Merkuleva I.A., Starostina E.V., Zadorozhny A.M., Isaeva A.A., Nesmeyanova V.S., Shanshin D.V., Baranov K.O., Volkova N.V., Zaitsev B.N., Orlova L.A., Zaykovskaya A.V., Pyankov O.V., Danilenko E.D., Bazhan S.I., Shcherbakov D.N., Taranin A.V., Ilyichev A.A. Self-Assembled Particles Combining SARS-CoV-2 RBD Protein and RBD DNA Vaccine Induce Synergistic Enhancement of the Humoral Response in Mice. Int J Mol Sci. 2022 Feb 16;23(4):2188. doi: 10.3390/ijms23042188.
20. Патент РФ №2317107, МПК А61К 39/21, опубл. 20.02.2008 г.
21. Патент РФ №2754230, МПК C12N15/30, опубл. 30.08.2022 г.(прототип).--->
Приложение
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="pVAX-RBD.xml"
softwareName="WIPO Sequence" softwareVersion="2.0.0" productionDate=
"2022-06-09">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1</ApplicationNumberText>
<FilingDate>2022-06-09</FilingDate>
</ApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное бюджетное учреждение
науки «Государственный научный центр вирусологии и биотехнологии «Вектор»
Федеральной службы по надзору в сфере защиты прав потребителей и благополучия
человека (ФБУН ГНЦ ВБ «Вектор» Роспотребнадзора)</ApplicantName>
<ApplicantNameLatin>The State Research Center of Virology and
Biotechnology «VECTOR»</ApplicantNameLatin>
<InventionTitle languageCode="ru">Конъюгат белка рецепторсвязывающего
домена (RBD) поверхностного гликопротеина S вируса SARS-CoV-2 с полимером
полиглюкин-спермидин (PGS) и вакцинный комплекс против коронавирусной инфекции
COVID-19 на основе указанного конъюгата и плазмидной ДНК pVAX-RBD</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">(SEQ ID No1)
<INSDSeq>
<INSDSeq_length>3710</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..3710</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q1">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>gene</INSDFeature_key>
<INSDFeature_location>755..1483</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>gene</INSDQualifier_name>
<INSDQualifier_value>176-RBD</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>gene</INSDFeature_key>
<INSDFeature_location>1944..2738</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>gene</INSDQualifier_name>
<INSDQualifier_value>Neomycin/Canamycin resistance</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>unsure</INSDFeature_key>
<INSDFeature_location><754..>1484</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>map</INSDQualifier_name>
<INSDQualifier_value>pVAX sequence</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gctgcttcgcgatgtacgggccagatatacgcgttgacattgattattgact
Agttattaatagtaatcaattacggggtcattagttcatagcccatatatggagttccgcgttacataacttacgg
Taaatggcccgcctggctgaccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaac
Gccaatagggactttccattgacgtcaatgggtggactatttacggtaaactgcccacttggcagtacatcaagtg
Tatcatatgccaagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagtacatga
Ccttatgggactttcctacttggcagtacatctacgtattagtcatcgctattaccatggtgatgcggttttggca
Gtacatcaatgggcgtggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagt
Ttgttttggcaccaaaatcaacgggactttccaaaatgtcgtaacaactccgccccattgacgcaaatgggcggta
Ggcgtgtacggtgggaggtctatataagcagagctctctggctaactagagaacccactgcttactggcttatcga
Aattaatacgactcactatagggagacccaagctggctagcctcgagaattcgtcctgctgcgcacgtgggaagcc
Ctggccccggccgccaccatgatgcggacactgatcctggctgtgctgctggtgtacttctgtgccaccgtgcact
Gtgtgcagcccaccgaatccatcgtgcggttccccaatatcaccaatctgtgccccttcggcgaggtgttcaatgc
Caccagattcgcctctgtgtacgcctggaaccggaagcggatcagcaattgcgtggccgactactccgtgctgtac
Aactccgccagcttcagcaccttcaagtgctacggcgtgtcccctaccaagctgaacgacctgtgcttcacaaacg
Tgtacgccgacagcttcgtgatccggggagatgaagtgcggcagattgcccctggacagacaggcaagatcgccga
Ctacaactacaagctgcccgacgacttcaccggctgtgtgattgcctggaacagcaacaacctggactccaaagtc
Ggcggcaactacaattacctgtaccggctgttccggaagtccaatctgaagcccttcgagcgggacatctccaccg
Agatctatcaggccggcagcaccccttgtaacggcgtggaaggcttcaactgctacttcccactgcagtcctacgg
Ctttcagcccacaaatggcgtgggctatcagccctacagagtggtggtgctgagcttcgaactgctgcatgcccct
Gccacagtgtgcggccctaagaaaagcaccaatctcgtgaagaacaaatgcgtgaacttcaactaatgagcggccg
Ctcgagtctagagggcccgtttaaacccgctgatcagcctcgactgtgccttctagttgccagccatctgttgttt
Gcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgc
Atcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaa
Gacaatagcaggcatgctggggatgcggtgggctctatggcttctactgggcggttttatggacagcaagcgaacc
Ggaattgccagctggggcgccctctggtaaggttgggaagccctgcaaagtaaactggatggctttctcgccgcca
Aggatctgatggcgcaggggatcaagctctgatcaagagacaggatgaggatcgtttcgcatgattgaacaagatg
Gattgcacgcaggttctccggccgcttgggtggagaggctattcggctatgactgggcacaacagacaatcggctg
Ctctgatgccgccgtgttccggctgtcagcgcaggggcgcccggttctttttgtcaagaccgacctgtccggtgcc
Ctgaatgaactgcaagacgaggcagcgcggctatcgtggctggccacgacgggcgttccttgcgcagctgtgctcg
Acgttgtcactgaagcgggaagggactggctgctattgggcgaagtgccggggcaggatctcctgtcatctcacct
Tgctcctgccgagaaagtatccatcatggctgatgcaatgcggcggctgcatacgcttgatccggctacctgccca
Ttcgaccaccaagcgaaacatcgcatcgagcgagcacgtactcggatggaagccggtcttgtcgatcaggatgatc
Tggacgaagagcatcaggggctcgcgccagccgaactgttcgccaggctcaaggcgagcatgcccgacggcgagga
Tctcgtcgtgacccatggcgatgcctgcttgccgaatatcatggtggaaaatggccgcttttctggattcatcgac
Tgtggccggctgggtgtggcggaccgctatcaggacatagcgttggctacccgtgatattgctgaagagcttggcg
Gcgaatgggctgaccgcttcctcgtgctttacggtatcgccgctcccgattcgcagcgcatcgccttctatcgcct
Tcttgacgagttcttctgaattattaacgcttacaatttcctgatgcggtattttctccttacgcatctgtgcggt
Atttcacaccgcatacaggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaataca
Ttcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaataatagcacgtgctaaaacttcatttt
Taatttaaaaggatctaggtgaagatcctttttgataatctcatgaccaaaatcccttaacgtgagttttcgttcc
Actgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcgtaatctgctgctt
Gcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctaccaactctttttccgaaggta
Actggcttcagcagagcgcagataccaaatactgtccttctagtgtagccgtagttaggccaccacttcaagaact
Ctgtagcaccgcctacatacctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtct
Taccgggttggactcaagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacag
Cccagcttggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccg
Aagggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccaggggg
Aaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtca
Ggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctc
acatgttctt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">(SEQ ID No2)
<INSDSeq>
<INSDSeq_length>244</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..244</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>PEPTIDE</INSDFeature_key>
<INSDFeature_location>235..244</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q3">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>HIS-tag</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>VEKGIYQTSNFRVQPTESIVRFPNITNLCPFGEVFNATRFASVYAWNRKRIS
NCVADYSVLYNSASFSTFKCYGVSPTKLNDLCFTNVYADSFVIRGDEVRQIAPGQTGKIADYNYKLPDDFTGCVIA
WNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYRVV
VLSFELLHAPATVCGPKKSTNLVKNKCVNFHHHHHHHHHH</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Плазмидная ДНК-матрица pVAX-RBD, молекул мРНК-RBD, полученная с использованием ДНК-матрицы pVAX-RBD, обеспечивающая синтез и секрецию белка RBD SARS-CoV 2 в клетках организма млекопитающих и комплекс в виде наночастиц, содержащих молекулы мРНК-RBD, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью | 2021 |
|
RU2754230C1 |
| Плазмидная ДНК-кассета pVAX-C3-PolyA, плазмидная ДНК-матрица pVAX-C3-RBD-PolyA для синтеза мРНК и полинуклеотидная вакцина против SARS-CoV-2, представляющая собой молекулы мРНК-C3-RBD-PolyA, индуцирующие SARS-CoV-2-специфические антитела | 2024 |
|
RU2839841C1 |
| Плазмидная ДНК-матрица и молекула мРНК, используемая для создания вакцины против коронавируса, полученная с использованием ДНК-матрицы и обеспечивающая экспрессию гена искусственного полиэпитопного белка-иммуногена, содержащего консервативные эпитопы антигенов вируса SARS-Cov-2 и индуцирующего SARS-Cov-2-специфический иммунитет | 2024 |
|
RU2839372C1 |
| Искусственный ген, кодирующий белок-иммуноген BSI-COV-Ub, рекомбинантная плазмидная ДНК pBSI-COV-Ub, обеспечивающая экспрессию целевого гена, и искусственный полиэпитопный белок-иммуноген BSI-COV-Ub, содержащий убиквитин и эпитопы антигенов вируса SARS-CoV-2 и индуцирующий SARS-CoV-2-специфический Т-клеточный иммунитет | 2023 |
|
RU2806590C1 |
| Интегративный плазмидный вектор pVEAL3-RBDdel, обеспечивающий синтез и секрецию рекомбинантного белка рецепторсвязывающего домена RBDdelta коронавируса SARS-CoV-2 в клетках млекопитающих, рекомбинантный штамм клеточной линии CHO-K1- RBDdelta и рекомбинантный белок RBDdelta SARS-CoV-2, продуцируемый штаммом клеточной линии | 2023 |
|
RU2816175C1 |
| Искусственный ген, кодирующий белок-иммуноген BSI-COV, рекомбинантная плазмидная ДНК pBSI-COV, обеспечивающая экспрессию целевого гена, и искусственный полиэпитопный белок-иммуноген BSI-COV, содержащий эпитопы антигенов вируса SARS-CoV-2 и индуцирующий SARS-CoV-2-специфический T-клеточный иммунитет | 2023 |
|
RU2806556C1 |
| Интегративный плазмидный вектор pVEAL2-S-RBD, обеспечивающий экспрессию и секрецию рекомбинантного рецепторсвязывающего домена (RBD) коронавируса SARS-CoV-2 в клетках млекопитающих, рекомбинантный штамм клеточной линии CHO-K1-RBD и рекомбинантный белок RBD SARS-CoV-2, продуцируемый указанным штаммом клеточной линии CHO-K1-RBD | 2021 |
|
RU2752858C1 |
| РЕКОМБИНАНТНЫЙ ВИРУС ГРИППА, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПРОФИЛАКТИКИ COVID-19 И ГРИППА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2802058C1 |
| Вакцина на основе вирусоподобных частиц (VLP) для профилактики COVID-19 для парентерального применения | 2024 |
|
RU2834060C1 |
| Рекомбинантная плазмида pVEAL3-XBB.1.5, обеспечивающая синтез и секрецию рецептор-связывающего домена RBD S белка SARS-CoV-2 варианта XBB.1.5, и рекомбинантный штамм CHO-XBB.1.5 - продуцент рецептор-связывающего домена RBD S белка SARS-CoV-2 варианта XBB.1.5, предназначенного для создания иммунобиологических препаратов | 2024 |
|
RU2839373C1 |
Изобретение относится к биотехнологии. Описан конъюгат белка рецепторсвязывающего домена (RBD) поверхностного гликопротеина S вируса SARS-CoV-2, 308V-542N для получения вакцинного комплекса против коронавирусной инфекции COVID-19, наработанного в клетках CHO, имеющего длину 244 а.о., молекулярный вес 35 кДа, аминокислотную последовательность SEQ ID N 2, связанного с полимером полиглюкин-спермидин (PGS) при расчетном соотношении молекул белка RBD и молекул PGS - 1:1. Также описан вакцинный комплекс против коронавирусной инфекции COVID-19 в виде наночастиц, индуцирующих формирование Т-клеточного иммунитета и SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, и состоящих из молекул конъюгата полиглюкин-спермидин-белок RBD по п.1 и молекул плазмидной ДНК pVAX-RBD, причем каждая наночастица имеет размер не менее 50 нанометров, содержит ядро, состоящее из молекул плазмидной ДНК pVAX-RBD и расположенного вокруг ядра слоя конъюгата полиглюкин-спермидин-белок RBD, удерживаемого за счет ионного взаимодействия между отрицательно заряженным ядром и положительно заряженным спермидином. Техническим результатом заявляемого изобретения является создание более простой иммуногенной конструкции, которая при внутримышечном введении способна индуцировать высокий уровень антител, обладающих вируснейтрализующей активностью, а также обеспечивать вирус-специфический Т-клеточный ответ. 2 н. и 1 з.п. ф-лы, 16 ил.,1 табл., 6 пр.
1. Конъюгат белка рецепторсвязывающего домена (RBD) поверхностного гликопротеина S вируса SARS-CoV-2, 308V-542N для получения вакцинного комплекса против коронавирусной инфекции COVID-19, наработанного в клетках CHO, имеющего длину 244 а.о., молекулярный вес 35 кДа, аминокислотную последовательность SEQ ID N 2, связанного с полимером полиглюкин-спермидин (PGS) при расчетном соотношении молекул белка RBD и молекул PGS - 1:1.
2. Вакцинный комплекс против коронавирусной инфекции COVID-19 в виде наночастиц, индуцирующих формирование Т-клеточного иммунитета и SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью, и состоящих из молекул конъюгата полиглюкин-спермидин-белок RBD по п.1 и молекул плазмидной ДНК pVAX-RBD, причем каждая наночастица имеет размер не менее 50 нанометров, содержит ядро, состоящее из молекул плазмидной ДНК pVAX-RBD и расположенного вокруг ядра слоя конъюгата полиглюкин-спермидин-белок RBD, удерживаемого за счет ионного взаимодействия между отрицательно заряженным ядром и положительно заряженным спермидином.
3. Вакцинный комплекс по п.2, отличающийся тем, что плазмидная ДНК pVAX-RBD, имеющая нуклеотидную последовательность SEQ ID N 1 размером 3710 п.н., молекулярный вес 2,3 МДа, содержащая целевой ген, кодирующий химерный белок 176-RBD, перед которым находится промотор РНК-полимеразы фага Т7, обеспечивающий транскрипцию гена, состоит из следующих фрагментов:
- векторный фрагмент 1484-754 ДНК-плазмиды pVAX размером 2981 п.н., содержащий промотор РНК-полимеразы фага Т7 (664-682, размером 19 п.н.);
- ген устойчивости к неомицину/канамицину (NeoR/KanR) (1944-2738, 795 п.н.) и точка начала репликации ColE1 ori (3064-3652, 589 п.н.), обеспечивающие селекцию и амплификацию целевой плазмиды в клетках бактерий Escherichia coli;
- фрагмент 755-1483 размером 729 п.н., содержащий искусственный ген 176-RBD с инициирующим кодоном ATG.
| WO 2021239838 A2, 02.12.2021 | |||
| Плазмидная ДНК-матрица pVAX-RBD, молекул мРНК-RBD, полученная с использованием ДНК-матрицы pVAX-RBD, обеспечивающая синтез и секрецию белка RBD SARS-CoV 2 в клетках организма млекопитающих и комплекс в виде наночастиц, содержащих молекулы мРНК-RBD, индуцирующих SARS-CoV-специфические антитела, обладающие вируснейтрализующей активностью | 2021 |
|
RU2754230C1 |
| Тест-система и способ дифференцированного выявления антител к SARS-CoV-2 | 2021 |
|
RU2754340C1 |
| WO 2022031611 A1, 10.02.2022 | |||
| US 20090281031 A1, 12.11.2009 | |||
| LARISA I | |||
| KARPENKO et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Сборник тезисов Всероссийской конференции молодых ученых, | |||

