Область техники, к которой относится изобретение
Настоящее изобретение относится к растворимому АСЕ2 (ангиотензинпревращающий фермент 2) и его слитым белкам, а также к их применению.
Предпосылки создания изобретения
В декабре 2019 года произошла вспышка заболевания пневмонией, вызванная инфицированием новым коронавирусом, - коронавирусом тяжелого острого респираторного синдрома 2 (SARS-CoV2) -, и коронавирусная инфекция быстро распространилась по всему миру. По состоянию на февраль 2021 года число инфицированных в мире достигло более 100 миллионов человек, а смертность составила приблизительно 2,2%. Эпидемия не только вызвала глобальный кризис в области здравоохранения, но и повлекла за собой далеко идущие последствия для общества, экономики и иных сфер деятельности.
Коронавирусы относятся к порядку Torovirus, семейству Coronaviridae и роду Coronaviridae. Коронавирусы относятся к типу вирусов с оболочкой и линейной одноцепочечной плюс-нитью РНК и представляют собой большую группу вирусов, широко распространенных в природе. У пациентов они вызывают заболевания, которые имеют различные клинические симптомы - от обычной простуды до тяжелой легочной инфекции. За последние два десятилетия коронавирусы вызвали две масштабные эпидемии: тяжелый острый респираторный синдром (SARS) в 2002/2003 годах и ближневосточный респираторный синдром (MERS) в 2012 году. С конца 2019 года новая эпидемия коронавируса (SARS-CoV2) значительно превзошла эпидемии SARS и MERS не только по масштабам эпидемии и совокупному числу инфицированных людей, но и по количеству летальных исходов.
Вирусный геном SARS-CoV2 очень похож на штамм RaTG13, выделенный из летучей мыши (китайской подковоносой (хризантемовой) летучей мыши), обнаруженный в 2013 году в Юньнане, Китай, с идентичностью последовательности до 96,2% (Zhou et al., 2020). Таким образом, можно сделать вывод, что происхождение данного нового коронавируса соответствует происхождению коронавирусов, вызвавших SARS и MERS, то есть, все они произошли от летучих мышей. Путь, ведущий к эпидемии этого нового коронавируса, SARS-CoV2, у людей, вероятно, соответствует SARS и MERS. Вирус произошел от летучих мышей, развивался и усиливался в промежуточных организмах-хозяевах (например, в организмах животных, имеющих более близкие контакты с человеком) и, в конечном счете, инфицировал человека. Вирус продолжал эволюционировать в организме человека и быстро распространился, что привело к вспышке вирусной инфекции. В природе коронавирусы, сходные с SARS, уже давно существуют в организме летучих мышей, обитающих во многих частях света, и большинство из них не способны инфицировать человека. Тем не менее, человек может случайно заражаться некоторыми природно-очаговыми болезнями через другого промежуточного хозяина. Эпидемия тяжелого острого респираторного синдрома в марте 2002 года и вспышка инфекции SARS-CoV2 через 17 лет показывают, что до тех пор, пока существует естественный хозяин, существует вероятность того, что в будущем могут возникнуть другие патогенные коронавирусные инфекции.
В ноябре 2020 года Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США разрешило экстренное применение двух моноклональных антител - бамланивимаба компании «Eli Lilly» и комбинации касиривимаба и имдевимаба компании «Regeneron» - для лечения инфекции SARS-CoV2. Оба антитела были одобрены для негоспитализированных взрослых и детей старше 12 лет с легкими и умеренными симптомами SARS-CoV2 и при риске обострения заболевания. Обычно при использовании моноклональных антител трудно найти баланс между высокой эффективностью и активностью широкого спектра действия. В процессе развития глобальной эпидемии, вызванной SARS-CoV2, появлялись варианты вируса, которые и далее будут появляться под давлением отбора, например, для дальнейшей адаптации к организму человека-хозяина и иммунитету человека. Варианты могут иметь изменения в иммунодоминантных сайтах, которые делают существующие нейтрализующие антитела неэффективными.
Как коронавирус SARS-CoV, так и родственные ему вирусы животных используют ангиотензинпревращающий фермент 2 (АСЕ2) в качестве рецептора для вторжения в клетки-мишени и их заражения. На поверхности коронавируса имеется множество S-белков в виде тримеров, обладающих высокой аффинностью к АСЕ2. Таким образом, для достижения эффективной нейтрализации вирусов необходимо одновременно блокировать многовалентное высокоаффинное связывание между S-белками и АСЕ2. Растворимый рецептор, который образуется путем слияния внеклеточной области АСЕ2 с константной областью антитела, имеет сходный механизм действия с нейтрализующими антителами и способен блокировать инфекцию вариантами, в которых произошли мутации, но которые по-прежнему используют АСЕ2 в качестве рецептора. Растворимые слитые белки АСЕ2 могут быть разработаны в качестве терапевтических препаратов, обладающих нейтрализующей способностью широкого спектра действия, и не будут ограничены мутациями вируса. Такие слитые белки не только могут быть использованы в качестве терапевтических препаратов для лечения инфекций SARS-CoV-2, но и позволят купировать подобные эпидемии, которые могут возникнуть в будущем.
Краткое изложение сущности изобретения
Растворимый АСЕ2, растворимый АСЕ2 с мутациями в активном центре (NN) и слитый белок ACE2-Fc, содержащий растворимый АСЕ2 или растворимый АСЕ2 с мутациями в активном центре (NN) и Fc-сегмент человеческого IgG1, способны эффективно нейтрализовать SARS-CoV2 и SARS-CoV и блокировать образование многоядерных синцитиев, которые могут быть индуцированы путем связывания спайкового белка (S) (содержащего сайт расщепления протеазой фурин) SARS-CoV2 с его человеческим рецептором АСЕ2.
В соответствии с первым аспектом в настоящем изобретении предлагается растворимый АСЕ2 или его процессированная форма. Растворимый АСЕ2 или его процессированная форма могут включать или содержать внеклеточный домен АСЕ2 или его фрагмент, который сохраняет способность связываться с коронавирусом.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать домен металлопротеазы (19-615аа) во внеклеточной области человеческого АСЕ2. В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать домен металлопротеазы (1-740аа) во внеклеточной области человеческого АСЕ2. В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать Q24, Т27, F28, D30, K31, Н34, Е37, D38, Y41, Q42, L45, М82, Y83, Q325, Е329, N330, K353, G354, D355, R357 и R393 человеческого АСЕ2, в частности, K31 и K353.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать человеческий АСЕ2, или любой его гомолог или ортолог, или его фрагмент, который сохраняет способность связываться с коронавирусом.
Растворимый АСЕ2 или его процессированная форма способны эффективно нейтрализовать вирус, использующий АСЕ2 в качестве рецептора, связывающегося с клеткой-хозяином. Вирус может включать SARS-CoV, HCoV-NL63 или SARS-CoV2.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать АСЕ2, содержащий мутацию в активном центре, или его процессированную форму. Предпочтительно, чтобы АСЕ2, содержащий мутацию в активном центре, или его процессированную форму, представлял собой человеческий растворимый АСЕ2 или его процессированную форму, имеющие мутацию (-и) H374N и/или H378N в положении (-ях) 374 и/или 378 (ACE2-NN).
Растворимый АСЕ2 или его процессированная форма могут представлять собой растворимый АСЕ2 или его процессированную форму, обладающую ферментативной активностью АСЕ2.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут быть гликозилированы, которые, предпочтительно, могут быть гликозилированы в положении (-ях) 53, 90, 103, 322, 432, 546 и/или 690 на N-конце человеческого АСЕ2.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут включать аминокислотную последовательность, представленную SEQ ID NO: 1.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут включать аминокислотную последовательность, представленную SEQ ID NO: 2.
В соответствии со вторым аспектом в настоящем изобретении предлагается слитый белок ACE2-Fc, который получают путем слияния растворимого АСЕ2 или его процессированной формы с Fc-доменом антитела.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать или содержать внеклеточный домен АСЕ2 или его фрагмент, который сохраняет способность связываться с коронавирусом.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать домен металлопротеазы (19-615аа) во внеклеточной области человеческого АСЕ2. Растворимый АСЕ2 или его процессированная форма могут включать домен металлопротеазы (1-740аа) во внеклеточной области человеческого АСЕ2. Растворимый АСЕ2 или его процессированная форма могут включать Q24, Т27, F28, D30, K31, Н34, Е37, D38, Y41, Q42, L45, М82, Y83, Q325, Е329, N330, K353, G354, D355, R357 и R393 человеческого АСЕ2, в частности, K31 и K353.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать человеческий АСЕ2, или любой его гомолог или ортолог, или его фрагмент, обладающий способностью связываться с коронавирусом.
Растворимый АСЕ2 или его процессированная форма способны эффективно нейтрализовать вирус, который использует АСЕ2 в качестве рецептора, связывающегося с клеткой-хозяином. Вирус может включать SARS-CoV, HCoV-NL63 или SARS-CoV2.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать АСЕ2, содержащий мутацию в активном центре, или в его процессированной форме. Предпочтительно, АСЕ2, содержащий мутацию в активном центре или в его процессированной форме, может включать человеческий растворимый АСЕ2 или его процессированную форму (ACE2-NN), включающие мутацию (-и) H374N и/или H378N в положении (-ях) 374 и/или 378.
Растворимый АСЕ2 или его процессированная форма могут включать растворимый АСЕ2 или его процессированную форму, обладающую ферментативной активностью АСЕ2.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут быть гликозилированы, которые, предпочтительно, могут быть гликозилированы в положении (-ях) 53, 90, 103, 322, 432, 546 и/или 690 на N-конце человеческого АСЕ2.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут включать аминокислотную последовательность, представленную SEQ ID NO: 1.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут включать аминокислотную последовательность, представленную SEQ ID NO: 2.
Растворимый АСЕ2 или его процессированная форма могут эффективно нейтрализовать вирус, который использует АСЕ2 в качестве рецептора, связывающегося с клеткой-хозяином. Вирус может включать SARS-CoV, HCoV-NL63 или SARS-CoV2.
В некоторых вариантах осуществления настоящего изобретения антитело может являться антителом IgG. Антитело может быть человеческим IgG, таким как IgG1, IgG2, IgG3 или IgG4, предпочтительно IgG1. Fc-домен антитела может представлять собой Fc-домен антитела, содержащий два Fc-домена тяжелых цепей антитела. Предпочтительно, каждый из Fc-доменов тяжелых цепей имеет шарнирную область на своем N-конце. Предпочтительно, каждый из Fc-доменов тяжелых цепей может включать СН3-домен, полученный из IgG1, IgG2, IgG3 или IgG4. Предпочтительно, каждый из Fc-доменов тяжелых цепей может включать СН2 и СН3 домены, полученные из IgG1, IgG2, IgG3 или IgG4. Fc-домены могут способствовать димеризации двух доменов АСЕ2.
Растворимый АСЕ2 или его процессированная форма могут быть связаны с С-концевой областью Fc-домена тяжелых цепей или с N-концевой областью Fc-домена тяжелых цепей.
В некоторых вариантах осуществления настоящего изобретения 2n (n равно 1, 2 или 3) растворимых АСЕ2 или их процессированных форм могут быть связаны с С- и/или N-концевой областью (-ями) двух Fc-доменов тяжелых цепей.
В некоторых вариантах осуществления настоящего изобретения два растворимых АСЕ2 или их процессированные формы могут быть связаны соответственно с N-концевой областью двух Fc-доменов тяжелых цепей с образованием димера. Альтернативно, два растворимых АСЕ2 или их процессированные формы могут быть связаны с С-концевой областью двух Fc-доменов тяжелых цепей с образованием димера.
В некоторых вариантах осуществления настоящего изобретения два растворимых АСЕ2 или их процессированные формы могут быть связаны соответственно с N-концевой областью двух Fc-доменов тяжелых цепей, в то время как два других растворимых АСЕ2 или их процессированные формы могут быть связаны соответственно с С-концевыми областями двух Fc-доменов тяжелых цепей, тем самым образуя тетрамерный слитый белок ACE2-Fc. Далее, каждый из двух растворимых АСЕ или их процессированные формы на N-конце тетрамерного слитого белка ACE2-Fc далее, на своем N-конце, связываются с растворимым АСЕ или его процессированной формы в тандем, тем самым образуя гексамерный литый белок ACE2-Fc. Растворимые АСЕ2 или их процессированные формы могут быть связаны в тандем с помощью линкера. Линкер может представлять собой линкер цистеина AAA. Альтернативно, каждый из двух растворимых АСЕ или их процессированные формы на С-конце тетрамерного слитого белка ACE2-Fc далее на своем С-конце связывается с растворимым АСЕ или его процессированной формой в тандем, тем самым образуя гексамерный слитый белок ACE2-Fc. Растворимые АСЕ2 или их процессированные формы связываются в тандем с помощью линкера. Линкер может представлять собой линкер цистеина AAA.
В некоторых вариантах осуществления настоящего изобретения каждый из двух Fc-доменов тяжелых цепей может на своем N-конце быть связан с двумя растворимыми АСЕ2 или их процессированными формами, которые связаны в тандем, тем самым образуя тетрамерный слитый белок ACE2-Fc. Два растворимых АСЕ2 или их процессированные формы связаны в тандем с помощью линкера. Линкер может представлять собой линкер цистеина AAA. Далее, растворимый АСЕ или его процессированная форма могут быть дополнительно связаны на каждой из N-концевых областей тетрамерного слитого белка ACE2-Fc с растворимым АСЕ или его процессированной формой в тандем, тем самым образуя гексамерный слитый белок ACE2-Fc. Растворимые АСЕ2 или их процессированные формы связаны в тандем с помощью линкера. Линкер может представлять собой линкер цистеина AAA. Альтернативно, каждый из двух Fc-доменов тяжелых цепей тетрамерного слитого белка ACE2-Fc может быть связан в своей С-концевой области с растворимым АСЕ или его процессированной формой, тем самым образуя гексамерный слитый белок ACE2-Fc.
Альтернативно каждый из двух Fc-доменов тяжелых цепей может быть связан в своей С-концевой области с двумя растворимыми АСЕ2 или их процессированными формами, которые связаны в тандем, тем самым образуя тетрамерный слитый белок ACE2-Fc. Два растворимых АСЕ2 или их процессированные формы могут быть связаны в тандем с помощью линкера. Линкер может представлять собой линкер цистеина AAA. Далее, каждый из двух Fc-доменов тяжелых цепей тетрамерного слитого белка ACE2-Fc может быть связан в своей N-концевой области с растворимым АСЕ или его процессированной формой, тем самым образуя гексамерный слитый белок ACE2-Fc.
Предпочтительно, слитый белок ACE2-Fc может фактически представлять собой димерный ACE2-Fc слитый белок, при этом одна процессированная форма АСЕ2 и один Fc-домен тяжелых цепей могут включать аминокислотную последовательность, представленную SEQ ID NO: 3.
Предпочтительно, одна процессированная форма АСЕ2 и один Fc-домен тяжелых цепей в слитом белке ACE2-Fc могут включать аминокислотную последовательность, представленную SEQ ID NO: 4.
Предпочтительно, слитый белок ACE2-Fc может дополнительно включать сигнальный пептид, предпочтительно сигнальный пептид CD33.
Предпочтительно, одна процессированная форма АСЕ2 и один Fc-домен тяжелых цепей в слитом белке ACE2-Fc могут включать аминокислотную последовательность, представленную SEQ ID NO: 5.
Предпочтительно, одна процессированная форма АСЕ2 и один Fc-домен тяжелых цепей в слитом белке ACE2-Fc могут включать аминокислотную последовательность, представленную SEQ ID NO: 6.
Предпочтительно, в тетрамерном слитом белке ACE2-Fc одна процессированная форма АСЕ2 может быть связана с одним Fc-доменом тяжелых цепей, который дополнительно связан с одним АСЕ2 (ACE2-Fc-ACE2), в результате чего появляется аминокислотная последовательность, представленная SEQ ID NO: 13.
Предпочтительно, в тетрамерном слитом белке ACE2-Fc два АСЕ2 или их процессированные формы могут быть связаны с одним Fc-доменом тяжелых цепей (ACE2-ACE2-Fc), в результате чего появляется аминокислотная последовательность, представленная SEQ ID NO: 14.
Слитый белок ACE2-Fc способен эффективно нейтрализовать вирус, который использует АСЕ2 в качестве рецептора, связывающегося с клеткой-хозяином. Вирус может включать SARS-CoV, HCoV-NL63 или SARS-CoV2.
Слитый белок ACE2-Fc по настоящему изобретению способен увеличить период полужизни и выход растворимого АСЕ2, и в максимальной степени удовлетворять потребности в оперативном развитии процессов и использовании в экстренных ситуациях.
В соответствии с третьим аспектом в настоящем изобретении предлагается мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы, которая может включать n полипептидных мономерных единиц, при этом каждая из полипептидных мономерных единиц может быть димером, в котором два растворимых АСЕ2 или их процессированные формы связаны с N-концевыми областями двух Fc-доменов тяжелых цепей соответственно, и n полипептидных мономерных единиц собраны в мультимер посредством хвоста, расположенного на каждом из С-концов Fc-доменов антитела.
В некоторых вариантах осуществления настоящего изобретения каждый из Fc-доменов тяжелых цепей в каждой полипептидной мономерной единице может быть на своем С-конце связан с хвостом. Таким образом, каждый из двух Fc-доменов тяжелых цепей полипептидной мономерной единицы на своем С-конце связан с одним хвостом, и n полипептидных мономерных единиц имеют в совокупности 2n хвостов, которые соединены друг с другом с образованием замкнутого кольцевого мультимера.
Хвост может включать любые подходящие аминокислотные последовательности и может быть хвостом, присутствующим в антителах, образующихся естественным путем. Альтернативно, указанный хвост может быть модифицированным хвостом, отличающимся от нативного хвоста по длине и/или композиции. Альтернативно, хвост может быть искусственно синтезированным хвостом, подходящим для мультимеризации, например, хвост, состоящий из гибкой Cys-последовательности подходящей длины. Альтернативно, хвост может включать вариант или фрагмент из естественной последовательности, такой как, хвост IgM PTLYNVSLVMSDTAGTCY (SEQ ID NO: 15) или хвост IgA PTHVNVSVVVMAEVDGTCY (SEQ ID NO: 16). Альтернативно, вариант из хвоста IgM или IgA, как правило, может включать аминокислотную последовательность, содержащую 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 или 18 аминокислот из хвоста IgM PTLYNVSLVMSDTAGTCY (SEQ ID NO: 15) или хвоста IgA PTHVNVSVVMAEVDGTCY (SEQ ID NO: 16). Хвост также может быть гибридным IgM/IgA хвостом. Предпочтительно, хвост может включать аминокислотную последовательность TGKPTLYNVSLVMSDTAGTCY (SEQ ID NO: 17).
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать или содержать внеклеточный домен АСЕ2 или его фрагмент, который сохраняет способность связываться с коронавирусом.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать домен металлопротеазы (19-615аа) во внеклеточной области человеческого АСЕ2. Растворимый АСЕ2 или его процессированная форма могут включать домен металлопротеазы (1-740аа) во внеклеточной области человеческого АСЕ2. Растворимый АСЕ2 или его процессированная форма могут включать Q24, Т27, F28, D30, K31, Н34, Е37, D38, Y41, Q42, L45, М82, Y83, Q325, Е329, N330, K353, G354, D355, R357 и R393 человеческого АСЕ2, в частности, K31 и K353.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать человеческий АСЕ2 или любой его гомолог или ортолог, или его фрагмент, сохраняющий способность связываться с коронавирусом.
Растворимый АСЕ2 или его процессированная форма могут эффективно нейтрализовать вирус, который использует АСЕ2 в качестве рецептора, связывающегося с клеткой-хозяином. Вирус может включать SARS-CoV, HCoV-NL63 или SARS-CoV2.
В некоторых вариантах осуществления настоящего изобретения растворимый АСЕ2 или его процессированная форма могут включать АСЕ2, содержащий мутацию в активном центре, или в его процессированной форме. Предпочтительно, АСЕ2, содержащий мутацию в активном центре, или в его процессированной форме может включать человеческий растворимый АСЕ2 или его процессированную форму (ACE2-NN), имеющие мутацию (-и) H374N и/или H378N в положении 374 и/или в положении 378.
Растворимый АСЕ2 или его процессированная форма могут включать растворимый АСЕ2 или его процессированную форму, обладающую ферментативной активностью АСЕ2.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут быть гликозилированы в положении (-ях) 53, 90, 103, 322, 432, 546 и/или 690 в N-концевой части человеческого АСЕ2.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут включать аминокислотную последовательность, представленную SEQ ID NO: 1.
Предпочтительно, растворимый АСЕ2 или его процессированная форма могут включать аминокислотную последовательность, представленную SEQ ID NO: 2.
Растворимый АСЕ2 или его процессированная форма могут эффективно нейтрализовать вирус, который использует АСЕ2 в качестве рецептора, связывающегося с клеткой-хозяином. Вирус может включать SARS-CoV, HCoV-NL63 или SARS-CoV2.
В некоторых вариантах осуществления настоящего изобретения антитело может являться антителом IgG. Антитело может быть человеческим IgG, таким как IgG1, IgG2, IgG3 или IgG4, предпочтительно IgG1. Fc-домен антитела может относиться к Fc-домену антитела, содержащему два Fc-домена тяжелых цепей антитела. Предпочтительно, каждый из Fc-доменов тяжелых цепей может иметь шарнирную область на своем N-конце.
Предпочтительно, каждый из Fc-доменов тяжелых цепей может включать домен СН3, полученный из IgG1, IgG2, IgG3 или IgG4. Предпочтительно, каждый из Fc-доменов тяжелых цепей может включать СН2 и СН3 домены, полученные из IgG1, IgG2, IgG3 или IgG4. Fc-домены могут способствовать димеризации двух доменов АСЕ2.
В некоторых вариантах осуществления настоящего изобретения Fc-домен тяжелых цепей может включать Fc-домен тяжелых цепей, имеющий мутацию L309C в положении 309.
Растворимый АСЕ2 или его процессированная форма могут быть связаны с С-концевой областью Fc-домена тяжелых цепей или с N-концевой областью Fc-домена тяжелых цепей.
Предпочтительно, в каждой полипептидной мономерной единице одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом могут включать аминокислотную последовательность, представленную SEQ ID NO: 7.
Предпочтительно, в каждой полипептидной мономерной единице одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом могут включать аминокислотную последовательность, представленную SEQ ID NO: 8.
Предпочтительно, в каждой полипептидной мономерной единице одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом могут включать аминокислотную последовательность, представленную SEQ ID NO: 18.
В некоторых вариантах осуществления настоящего изобретения мультимер ACE2-hFc(n) слитого белка может включать ACE2-hFc5, ACE2-NN-hFc5 или ACE2-NN-hFc5 L309C, при этом:
ACE2-hFc5 обозначает пентамер, собранный из 5 полипептидных мономерных единиц посредством 10 хвостов, расположенных на С-концах 5 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc5 обозначает пентамер, собранный из 5 полипептидных мономерных единиц посредством 10 хвостов, расположенных на С-концах 5 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc5-L309C обозначает пентамер, собранный из 5 полипептидных мономерных единиц посредством 10 хвостов на С-концах 5 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18, и при этом Fc-домен тяжелых цепей включает мутацию L309C в положении 309.
В некоторых вариантах осуществления настоящего изобретения мультимер ACE2-hFc(n) слитого белка может включать ACE2-hFc6, ACE2-NN-hFc6 или ACE2-NN-hFc6 L309C, при этом:
ACE2-hFc6 обозначает гексамер, собранный из 6 полипептидных мономерных единиц посредством 12 хвостов на С-концах 6 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc6 обозначает гексамер, собранный из 6 полипептидных мономерных единиц посредством 12 хвостов на С-концах 6 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc6-L309C обозначает гексамер, собранный из 6 полипептидных мономерных единиц посредством 12 хвостов на С-концах 6 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18.
В некоторых вариантах осуществления настоящего изобретения мультимер слитого белка может являться одним или несколькими мультимер ами, выбранными из нижеприведенных мультимеров слитого белка:
ACE2-hFc4, представляющий собой тетрамер, собранный из 4 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc4, представляющий собой пентамер, собранный из 4 полипептидных мономерных единиц с хвостами на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc4-L309C, представляющий собой тетрамер, собранный из 4 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18, и при этом Fc-домен тяжелых цепей включает мутацию L309C в положении 309;
ACE2-hFc5, представляющий собой пентамер, собранный из 5 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc5, представляющий собой пентамер, собранный из 5 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc5-L309C, представляющий собой пентамер, собранный из 5 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18, и при этом Fc-домен тяжелых цепей включают мутацию L309C в положении 309;
ACE2-hFc6, представляющий собой гексамер, собранный из 6 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACESARS-CoV22 и двух Fc-доменов тяжелых цепей, при этом одна АСЕ2 процессированная форма, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc6, представляющий собой гексамер, собранный из 6 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc6-L309C, представляющий собой гексамер, собранный из 6 полипептидных мономерных единиц посредством хвостов на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18.
Мультимеры слитого белка могут значительно повысить аффинность к вирусному S-белку и при этом усилить эффекторную функцию Fc-молекулы.
В соответствии с четвертым аспектом в настоящем изобретении предлагается экспрессирующий вектор, включающий ген, кодирующий растворимый АСЕ2 или его процессированную форму по первому аспекту, слитый белок ACE2-Fc по второму аспекту, или мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту.
В соответствии с пятым аспектом в настоящем изобретении предлагается клеточная линия млекопитающих, включающий ген, кодирующий растворимый АСЕ2 по первому аспекту, слитый белок ACE2-Fc по второму аспекту или мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту. Клеточная линия может включать, в частности, клеточную линию СНО, клеточную линию 293, клеточную линию Vero и полученную из них клеточную линию, например, клетки Vero Е6 или клетки HEK293T.
В соответствии с шестым аспектом в настоящем изобретении предлагается способ получения растворимого АСЕ2 по первому аспекту, слитого белка ACE2-Fc по второму аспекту или мультимера Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту, включающий следующие этапы:
(1) трансфицирование клеточной линии млекопитающих экспрессирующим вектором по четвертому аспекту для получения клеточной линии млекопитающих, экспрессирующего растворимый АСЕ2 по первому аспекту, слитый белок ACE2-Fc по второму аспекту или мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту;
(2) культивирование клеточной линии млекопитающих, полученного на этапе (1), в условиях культивирования для получения растворимого АСЕ2 по первому аспекту, слитого белка ACE2-Fc по второму аспекту или мультимера Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту таким образом, чтобы получить рекомбинантный белок; и
(3) очистка рекомбинантного белка, полученного на этапе (2).
В соответствии с седьмым аспектом в настоящем изобретении предлагается фармацевтическая композиция, включающая: растворимый АСЕ2 по первому аспекту, слитый белок ACE2-Fc по второму аспекту, или мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту; и фармацевтически приемлемый носитель.
В соответствии с восьмым аспектом в настоящем изобретении предлагается применение растворимого АСЕ2 по первому аспекту, слитого белка ACE2-Fc по второму аспекту или ACE2-hFc(n) мультимера Fc-слитого белка растворимого АСЕ2 или его процессированной формы по третьему аспекту при получении лекарственного средства для лечения или профилактики заболевания, связанного с АСЕ2.
Заболевание может являться заболеванием, выбранным из группы любых других заболеваний, в результате инфицирования вирусом, использующим АСЕ2 в качестве рецептора. Вирус может включать коронавирус, например, SARS-CoV, HCoV-NL63 или SARS-CoV2. Заболевание может быть выбрано из группы заболеваний, включающих пневмонию, тяжелую острую респираторную инфекцию, почечную недостаточность, сердечную недостаточность, респираторный дистресс-синдром взрослых (ARDS), поражение печени, желудочно-кишечное заболевание или тяжелый острый респираторный синдром.
Растворимый АСЕ2 по первому аспекту, слитый белок ACE2-Fc по второму аспекту или мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированная форма по третьему аспекту в настоящем изобретении могут быть использованы для введения в неотложных ситуациях, тем самым позволяя избежать высоких уровней заболеваемости и смертности в результате инфекций, вызванных вирусами, использующими АСЕ2 в качестве рецептора, в частности, коронавирусных инфекций.
Растворимый АСЕ2 по первому аспекту, слитый белок ACE2-Fc по второму аспекту или мультимер Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированная форма по третьему аспекту в настоящем изобретении также могут быть применены для пассивной иммунизации медицинских работников и лиц, подверженных риску заражения вирусом, в частности, коронавирусом, использующим АСЕ2 в качестве рецептора.
Синцитии представляют собой многоядерные гигантские клетки, которые в конечном счете образуются в результате слияния клеток после инфицирования вирусами клеток-хозяев. У тяжелых пациентов, инфицированных SARS-CoV2, могут наблюдаться диффузные повреждения альвеолярного эпителия, что приводит к образованию слившихся многоядерных клеток (синцитий). Синцитии вызваны связыванием вирусного S-белка с АСЕ2, что является основной причиной проявления цитопатического эффекта. Между тем, цитокиновый шторм также может вызвать повреждение альвеол. Мультимерный Fc-слитый белок (ACE2-NN-hFcn) растворимого АСЕ2 способен эффективно предотвратить связывание вирусного S-белка с АСЕ2, тем самым позволяя избежать образование слитых многоядерных клеток (синцития), а также последующего цитопатического эффекта.
Предпочтительно, лекарственное средство может быть введено путем ингаляции, интраназально или путем инсталляции в дыхательные пути, глазной инъекции и инъекции в среднее ухо, закапывания в уши, топической, трансдермальной, парентеральной, подкожной и внутривенной инъекции, внутрикожной инъекции, внутримышечной инъекции, внутриплевральной инсталляции, внутрибрюшинной инъекции, путем интрализионального введения, нанесения на слизистую или трансплантации носителя с пролонгированным высвобождением. Предпочтительно, лекарственное средство вводят путем ингаляции с помощью небулайзера.
В соответствии с девятым аспектом в настоящем изобретении предлагается способ скрининга лекарственного средства против инфекции, вызванной вирусом, в частности, коронавирусом, использующим АСЕ2 в качестве рецептора, при этом способ включает: скрининг лекарственного средства с использованием растворимого АСЕ2 по первому аспекту, слитого белка ACE2-Fc по второму аспекту, мультимера Fc-слитого белка (ACE2-hFc(n)) растворимого АСЕ2 или его процессированной формы по третьему аспекту, вектора по четвертому аспекту или клеточной линии млекопитающих по пятому аспекту.
В соответствии с десятым аспектом в настоящем изобретении предлагается способ скрининга лекарственного средства против инфекции, вызванной вирусом, в частности, коронавирусом, использующим АСЕ2 в качестве рецептора, при этом способ включает: использование протеазы фурин или участка расщепления фурином в S-белке коронавируса в качестве мишени для скрининга лекарственного средства.
Лекарственное средство может являться ингибитором протеазы фурин.
Лекарственное средство способно блокировать образование синцития посредством S-белка, содержащего сайт расщепления фурином, при инфицировании вирусом, в частности, коронавирусом, использующим АСЕ2 в качестве рецептора.
В соответствии с одиннадцатым аспектом в настоящем изобретении предлагается применение реагента, нацеленного на протеазу фурин или на сайт расщепления фурином в S-белке коронавируса, при приготовлении лекарственного средства против инфекции, вызванной вирусом, в частности, коронавирусом, использующим АСЕ2 в качестве рецептора.
В частности, настоящее изобретение относится к использованию реагента, блокирующего образование синцития посредством S-белка, содержащего сайт расщепления фурином, во время инфекции, вызванной вирусом, в частности, коронавирусом, использующим АСЕ2 в качестве рецептора, в приготовлении лекарственного средства для лечения инфекции, вызванной вирусом, использующим АСЕ2 в качестве рецептора, в частности, коронавирусом.
Предпочтительно, реагент может являться ингибитором протеазы фурин.
Коронавирус может представлять собой SARS-CoV2.
В соответствии с двенадцатым аспектом в настоящем изобретении предлагается мутантный S-белок, представляющий собой процессированную форму и/или форму, в которой мутирован сайт расщепления фурином.
Процессированная форма может включать S-белок, содержащий только внеклеточный домен (S1+S2), т.е. делетирующий трансмембранную и внутриклеточную области.
Сайт расщепления фурином может быть мутирован путем делеции, замены или добавления одной или нескольких аминокислот таким образом, чтобы сайт расщепления фурином более не являлся активным как сайт расщепления фурином.
Предпочтительно, форма, в которой сайт расщепления фурином мутирован, может включать мутацию от RRAR к SRAS в сайте расщепления фурином S-белка.
Предпочтительно, в дополнение к мутации в сайте расщепления фурином, мутированный S-белок может содержать только внеклеточный домен (S1+S2), т.е. делетирующий трансмембранную и внутриклеточную области.
Мутированный S-белок может иметь аминокислотную последовательность, представленную SEQ ID NO: 10, SEQ ID NO: 11 или SEQ ID NO: 12.
SEQ ID NO: 10 представляет собой аминокислотную последовательность, в которой сайт расщепления фурином S-белка мутировал от RRAR к SRAS. SEQ ID NO: 11 представляет собой аминокислотную последовательность S-белка в процессированной форме с делетированными трансмембранной и внутриклеточной областями. SEQ ID NO: 12 представляет собой аминокислотную последовательность, в которой сайт расщепления фурином мутировал от RRAR к SRAS, и S-белок имеет процессированную форму с делетированием трансмембранной и внутриклеточной областей.
В соответствии с тринадцатым аспектом настоящее изобретение относится к использованию мутантного S-белка по двенадцатому аспекту при приготовлении лекарственного средства для лечения или профилактики заболевания, связанного с АСЕ2.
Заболевание представляет собой инфекционное заболевание, вызванное вирусом, использующим АСЕ2 в качестве рецептора, и, предпочтительно, вирус может являться коронавирусом, предпочтительно SARS-CoV, HCoV-NL63 или SARS-CoV2.
Заболевание может быть выбрано из группы заболеваний, включающих пневмонию, тяжелую острую респираторную инфекцию, почечную недостаточность, сердечную недостаточность, респираторный дистресс-синдром взрослых (ARDS), поражение печени, желудочно-кишечное заболевание или тяжелый острый респираторный синдром.
Лекарственное средство может быть применено для пассивной иммунизации медицинских работников и лиц, подверженных риску инфицирования вирусами, использующими АСЕ2 в качестве рецептора, в частности, коронавирусами.
В соответствии с четырнадцатым аспектом в настоящем изобретении предлагается рекомбинантная вакцина для профилактики инфекции, вызванной SARS-CoV2, которая включает S-белок по двенадцатому аспекту.
Сайт расщепления фурином S-белка SARS-CoV2 и протеаза фурин могут быть использованы в качестве мишеней для скрининга лекарственных препаратов для лечения инфекционного заболевания, вызванного вирусом, использующим АСЕ2 в качестве рецептора.
Настоящее изобретение подтверждает, что сайт расщепления фурином в S-белке SARS-CoV2 необходим для формирования синцития. В частности, спайковый (S) белок ARS-CoV 2 имеет мотив RRAR около остатков 681-685 (около участка соединения S1/S2), который может быть расщеплен протеазой, такой как фурин. Выравнивание последовательностей показывает, что указанный мотив присутствует во всех известных штаммах SARS-CoV2, но не в штамме летучей мыши RaTG13, который наиболее близок к SARS-CoV2. При выравнивании последовательностей вирусов летучих мышей в базе данных подобная последовательность (RRAT) имеется только в вирусе SARS-HKU5 летучей мыши. Мы обнаружили, что клетки 293Т, трансфицированные плазмидами, экспрессирующими S-белок дикого типа ARS-CoV2 или S-белок SARS-CoV2 с мутированным сайтом расщепления протеазой фурин, могут связываться с Fc ACE2-IgG1. При этом контрольные клетки, трансфицированные пустым вектором, не связывались с ACE2-IgG1 Fc. S-белок дикого типа ARS-CoV2 и S-белок ARS-CoV2 с мутированным сайтом расщепления протеазой фурин могли нормально экспрессироваться на поверхности клеток 293Т. Кроме того, S-белок ARS-CoV2 с мутированным сайтом расщепления протеазой фурин (RRAR мутировал в SRAS) мог связываться прочнее с Fc ACE2-IgG1.
В эксперименте по образованию синцития не наблюдалось слияния клеток между контрольными клетками, трансфицированными пустым вектором, и клетками, экспрессирующими полноразмерный человеческий АСЕ2 (естественный человеческий АСЕ2), и, таким образом, не наблюдалось образования многоядерного синцития. Напротив, наблюдалось образование большого количества многоядерных синцитиев после совместного культивирования клеток, экспрессирующих S-белок дикого типа SARS-CoV2, с клетками, экспрессирующими полноразмерный человеческий АСЕ2 в течение 3 часов. Образовавшиеся синцитии погибли через 24 часа непрерывного культивирования. Когда фуриновый сайт RRAR в S-белке SARS-CoV2 был мутирован в SRAS, слияние клеток было полностью ингибировано, и при этом не наблюдалось образование многоядерных синцитий, что является подтверждением того, что мутировавший S-белок потерял способность опосредовать слияние клеток, и что фуриновый сайт в S-белке играл важнейшую роль в SARS-CoV2-инфекции клеток.
Таким образом, препараты, нацеленные на сайт фурина S-белка (PRRAR) и протеазу фурин, способны подавлять инфекцию SARS-CoV2. Более того, противовирусные препараты для лечения инфекции SARS-Со V2 могут быть разработаны путем ингибирования слияния коронавируса в процессе проникновения в клетки.
Кроме того, когда фуриновый сайт RRAR в S-белке ARS-CoV2 был мутирован в SRAS, слияние клеток было полностью ингибировано, и не наблюдалось многоядерных синцитиев. Это указывает на то, что мутировавший S-белок утратил способность опосредовать слияние клеток. Таким образом, SARS-CoV2 S с мутированным сайтом расщепления протеазой фурина в настоящем изобретении, в частности, SARS-CoV2 S, имеющий мутацию (-и) в сайте расщепления протеазой фурин в виде делеции трансмембранной и внутриклеточной областей, может быть использован в качестве рекомбинантного белкового препарата для конкурентного блокирования связывания вируса, имеющего S-белок дикого типа, с клеточным рецептором, тем самым блокируя инфекцию. Кроме того, S-белок SARS-CoV2 с мутированным сайтом расщепления протеазой фурин также может быть использован в качестве молекулы-кандидата для рекомбинантной вакцины и является более стабильным, чем S-белок дикого типа.
SARS-CoV2 - β-коронавирус - имеет оболочку. Вирионы SARS-CoV2 представляют собой круглые или овальные частицы с полиморфизмом диаметром 60-140 нм. Спайковые гликопротеины (S-белок) на поверхности оболочки являются основными антигенными белками коронавируса и играют исключительно важную роль в возникновении инфекции и распространении вирусов. S-белок состоит из двух субъединиц, при этом субъединица S1 связывается с рецептором клеточной поверхности, в то время как субъединица S2 содержит основные мотивы, необходимые для процесса слияния мембран.
Аминокислотная последовательность спайкового (S) белка SARS-CoV2 имеет гомологию с S-белком SARS-CoV, составляющую приблизительно 76%. То есть, гомология является относительно низкой. Таким образом, большинство нейтрализующих антител вируса SARS-CoV не способны нейтрализовать SARS-CoV2. Однако SARS-CoV2 использует один и тот же рецептор клетки-хозяина, что и SARS-CoV, т.е. ангиотензинпревращающий фермент 2 (АСЕ2). Как и SARS-CoV, так и вызывающий инфекцию SARS-CoV2 должен использовать АСЕ2 в качестве рецептора для проникновения в клетки-мишени. Другими словами, несмотря на то, что между аминокислотной последовательностью S-белка SARS-CoV2 и S-белка SARS-CoV имеются многочисленные различия, оба вируса по-прежнему используют АСЕ2 в качестве рецептора для проникновения в клетки-хозяева. Это указывает на то, что белок АСЕ2 имеет высокую структурную совместимость на своей поверхности с S-белком таких коронавирусов. Следовательно, АСЕ2, как молекула белка-хозяина, может быть легко использована спайковыми белками, имеющими многочисленные различия в последовательностях. Таким образом, АСЕ2 все еще будет являться точкой проникновения для таких вирусов для инфицирования организма человека в будущем. Растворимый АСЕ2 и растворимый АСЕ2 (NN) с мутированным активным центром, полученные в соответствии с настоящим изобретением, способны блокировать связывание вируса, использующего АСЕ2 в качестве рецептора-хозяина, с рецептором АСЕ2, тем самым ингибируя вторжение вируса. Как растворимый АСЕ2, так и АСЕ2 (NN) имеют большое значение для профилактики и контроля над возможной будущей эпидемией.
Более того, в связи с эпидемией, вызванной SARS-CoV2 среди населения, многие варианты вирусов появляются под давлением отбора с целью дальнейшей адаптация к человеку-хозяину и иммунитету человека. Указанные варианты могут характеризоваться изменениями вирулентности и антигенных сайтов. Подобные изменения наблюдались и регистрировались несколько раз в отношении SARS-CoV. Последние данные секвенирования SARS-CoV2 указывают на то, что рецептор-связывающий домен (RBD) S-белка все еще находится в процессе мутации в последовательности. Преимущество растворимого АСЕ2 и растворимого АСЕ2 (NN) с мутированным активным центром, полученными по настоящему изобретению, также заключается в том, что SARS-CoV2 может быть эффективно нейтрализован, до тех пор, пока вирус использует АСЕ2 в качестве рецептора проникновения, при этом мутируют даже вирусные белки, в частности, S-белок. Для профилактики и лечения инфекции, вызванной вирусом, находящимся в процессе эволюции, рецептор растворимого АСЕ2 по настоящему изобретению, в отличие от моноклональных нейтрализующих антител к S-белку, может быть использован в качестве терапевтического препарата, обладающего нейтрализующей способностью широкого спектра действия и не ограниченного мутациями вируса, без необходимости скрининга антител. Таким образом, он вполне применим для решения неотложных задач по предотвращению вирусной эпидемии в настоящее время и в будущем. Результаты экспериментов показывают, что пентамерный ACE2-NN-hFc5 по настоящему изобретению обладает высокой нейтрализующей активностью против инфекций всех существующих основных вариантов псевдовирусов.
Кроме того, в отличие от лекарственных средств на основе антител, рецептор является собственным белком человеческого организма, и поэтому в отношении его не требуется анализ перекрестной реактивности с тканями. Слитые белки рецептора используют в качестве препарата неотложной помощи при острых инфекционных заболеваниях, они обладают длительным периодом полужизни и их обычно вводят 1-2 раза для проведения лечения. Таким образом, рецептор также позволяет избежать проведения исследований антител к лекарственному средству и длительной токсичности, а также сократить цикл разработки.
Слитый белок ACE2-Fc и мультимеры слитого белка (ACE2-NN-hFcn) растворимого ACE2-Fc по настоящему изобретению могут представлять собой терапевтические препараты, являющиеся специфичными для предотвращения новой эпидемии максимально быстрым способом. Слитые белки ACE2-Fc и их мультимеры по настоящему изобретению в основном обладают следующими преимуществами:
(1) предотвращение формирования устойчивости вируса в виде нейтрализующих антител;
(2) предотвращение образования синцитиев SARS-CoV2 и SARS-CoV;
(3) способность рекрутинга посредством соответствующих рецепторов комплементов, дендритных клеток, макрофагов и естественных клеток-киллеров против вирионов или инфицированных клеток благодаря сохранению эффекторной функции Fc-домена;
(4) продление периода полужизни в кровотоке молекул растворимого АСЕ2;
(5) ACE2-Fc, ACE2(NN)-Fc или ACE2-NN-hFcn по настоящему изобретению применимы для сострадательного использования в неотложных ситуациях (в последующем могут быть проведены формальные клинические испытания), так как была проведена оценка рекомбинантного человеческого АСЕ2 (rhACE2) в клиническом испытании II фазы, продемонстрировавшем хорошую переносимость и безопасность, несмотря на то, что у субъектов не наблюдалось значительного улучшения клинических симптомов;
(6) ACE2-Fc, ACE2(NN)-Fc или ACE2-NN-hFcn по настоящему изобретению могут найти широкое применение в ближайшие месяцы или годы для помощи инфицированным пациентам перед вакцинацией;
(7) растворимый АСЕ2, полученный в соответствии со способом настоящего изобретения, применимый для блокирования вирусной инфекции, не зависит от естественной ферментативной активности АСЕ2 и не влияет на активность естественного АСЕ2 у пациентов; при этом растворимый АСЕ2 (NN) с мутированным активным центром даже позволяет избежать потенциального побочного эффекта, вызванного ферментативной активностью АСЕ2 в организме, тем самым максимизируя безопасность использования в организме человека;
(8) по сравнению с антителами к анти-S-белку еще одно преимущество растворимого АСЕ2 заключается в том, что до тех пор, пока вирус использует АСЕ2 в качестве рецептора проникновения, мутация (-и) вируса не влияет (-ют) на эффективность растворимого АСЕ2, в том числе на повышенную аффинность к рецептору, вызванную мутацией (-ями) вируса.
АСЕ2 представляет собой металлопротеазу, катализирующую деградацию ангиотензина I до нонапептида ангиотензина (1-9) или ангиотензина II до гептапептида ангиотензина (1-7). Считается, что АСЕ2 участвует в регуляции функций сердечно-сосудистой системы и может играть защитную роль при острых повреждениях легких, например, в отношении вазодилатации, антипролиферативного и антиокислительного стресса. АСЕ2 экспрессируется в сосудистой системе, а также в большинстве органов, но в основном в легких, сердце, печени, почках и семенниках. Таким образом, потенциальные лекарственные препараты, подавляющие ферментативную активность АСЕ2, не являются идеальными лекарственными препаратами.
Человеческий природный АСЕ2 (называемый АСЕ2 мембранного типа) имеет полную длину 805 аминокислот, в которой область положений 1-740 является внеклеточным доменом, при этом остальные 65 аминокислот служат в качестве коротких трансмембранных и внутриклеточных областей. Ферментативная активность АСЕ2 осуществляется внеклеточным доменом. Растворимый АСЕ2 или его процессированная форма в настоящем изобретении не имеет трансмембранного домена.
Краткое описание рисунков
На Фиг. 1 приведены пики слитых белков ACE2-hFc-ACE2, ACE2-ACE2-hFc, ACE2-hFc5 и ACE2-hFc5-L309C, полученные после очистки методом эксклюзионной ВЭЖХ (SEC-HPLC).
На Фиг. 2 проиллюстрировано, что спайковый белок нового коронавируса может индуцировать образование синцитиев.
На Фиг. 3 проиллюстрировано, что слитый белок ACE2-hFc5 в значительной степени ингибирует слияние межклеточных мембран, опосредованное S-белком S RS in vitro.
На Фиг. 4 приведены аминокислотная последовательность (SEQ ID NO: 9) и фуриновый сайт спайкового белка (S-белок) SARS-CoV2.
На Фиг. 5 приведена аминокислотная последовательность (SEQ ID NO: 10) S-белка SARS-CoV2, имеющая мутации в сайте расщепления фурином.
На Фиг. 6 приведена экспрессия S-белка SARS-CoV2 на поверхности клетки и результат анализа клеточной поверхности S-белка SARS-CoV2, имеющего мутации в сайте расщепления фурином.
На Фиг. 7 проиллюстрировано, что между контрольными клетками, трансфицированными пустым вектором (отмечены красным) и клетками, экспрессирующими человеческий АСЕ2 (отмечены зеленым), не происходит слияния клеток без образования многоядерных синцитиев; в группе клеток, экспрессирующих S-белок дикого типа SARS-CoV2 (отмечены красным), наблюдается образование большого количества многоядерных синцитиев после совместного культивирования с клетками, экспрессирующими АСЕ2 (отмечены зеленым) в течение 3 часов, и образовавшиеся синцитии погибли после 24 часов непрерывного культивирования; и S-белок, имеющий мутации в сайте фурина, не приводит к образованию синцитиев после смешивания с клетками, экспрессирующими АСЕ.
На Фиг. 8 приведены внеклеточный домен (Ectdomain) (1-1208аа) (SEQ ID NO: 11) S-белка дикого типа SARS-CoV2 и внеклеточный домен (1-1208аа) (SEQ ID NO: 12) S-белка SARS-CoV2, имеющие мутации в сайте расщепления фурином.
На Фиг. 9 приведены аффинность и авидность слитых белков ACE2-hFc и ACE2-hFc5 к RBD-области спайкового S-белка SARS-CoV2.
На Фиг. 10 проиллюстрировано, что различные формы мультимеров слитого белка АСЕ2 подавляют инфекцию псевдовируса SARS-CoV2 (D614).
На Фиг. 11 проиллюстрировано, что слитый белок ACE2-hFc5 подавляет инфекции, вызванные псевдовирусами SARS-CoV2 D614 и G614, вирусом SARS, и псевдовирусом панголин-CoV.
На Фиг. 12 проиллюстрировано, что слитый белок ACE2-hFc5 подавляет инфекцию живого вируса SARS-CoV2.
На Фиг. 13 приведены физико-химические свойства ACE2-hFc5 до и после небулизации, проанализированные методом эксклюзионной ВЭЖХ (SEC-HPLC).
На Фиг. 14 приведены результаты оценки in vitro нейтрализующей активности молекулы ACE2-hFc5 против инфекции псевдовируса S ARS-CoV2 до и после небулизации.
На Фиг. 15 приведены результаты ОТ-кПЦР анализа содержания гРНК и сгРНК SARS-CoV2 в легких хомяка.
На Фиг. 16 схематически изображены молекулярные структуры различных форм слитых белков ACE2-IgG1.
Подробное описание вариантов осуществления настоящего изобретения
Нижеприведенные варианты осуществления предназначены для иллюстрации настоящего изобретения, но не для ограничения объема настоящего изобретения. Все изменения или замены, внесенные в способы, этапы или условия настоящего изобретения без отступления от характера и сути настоящего изобретения, входят в объем настоящего изобретения.
Если не указано иное, химические реагенты, используемые в вариантах осуществления настоящего изобретения, представляют собой обычные коммерчески доступные реагенты, при этом технические средства, используемые в вариантах осуществления настоящего изобретения, представляют собой обычные средства, хорошо известные специалистам в данной области техники. Fc во всех вариантах осуществления и на фигурах получен из IgG1. ACE2-hFc во всех вариантах осуществления настоящего изобретения и на фигурах представляет собой ACE2-NN-hFc, в котором одна процессированная форма АСЕ2 и один Fc-домен тяжелых цепей имеют аминокислотную последовательность, представленную SEQ ID NO: 4. Если не указано иное, различные формы ACE2-hFc относятся ко всем мультимерам и мутантам ACE2-hFc и ACE2-hFc. ACE2-hFc-ACE2, описанные в вариантах осуществления настоящего изобретения и на фигурах, представляют собой тетрамерные ACE2(NN)-hFc-ACE2(NN), в которых одна процессированная форма АСЕ2 связана с одним Fc-доменом тяжелых цепей и далее связана с одним АСЕ2 (ACE2-Fc-ACE2), что приводит к появлению аминокислотной последовательности, представленной SEQ ID NO: 13. ACE2-ACE2-hFc во всех вариантах осуществления и на фигурах представляют собой тетрамерные ACE2(NN)-ACE2(NN)-hFc, в которых один Fc-домен тяжелых цепей (ACE2-ACE2-Fc) связан с двумя АСЕ2 или их процессированными формами, которые связаны в тандем, что приводит к появлению аминокислотной последовательности, представленной SEQ ID NO: 14. ACE2-hFc5s во всех вариантах осуществления настоящего изобретения и на фигурах относится к ACE2(NN)-hFc(5), представляющий собой тетрамер, собранный из 5 полипептидных мономерных единиц посредством 10 хвостов, расположенных на С-концах 5 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8. ACE2-hFc5-L309C во всех вариантах осуществления настоящего изобретения и на фигурах относится к ACE2(NN)-hFc(5)-L309C, представляющий собой тетрамер, собранный из 5 полипептидных мономерных единиц посредством 10 хвостов, расположенных на С-концах 5 Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм АСЕ2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма АСЕ2, один Fc-домен тяжелых цепей вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18, и Fc-домен тяжелых цепей включает мутацию L309C в положении 309. См. Фиг. 16.
Пример 1: Конструирование плазмиды для экспрессии слитого белка ACE2-hFc, плазмиды для экспрессии S-белка SARS-CoV2 и плазмиды для экспрессии мутантного S-белка SARS-CoV2 сайта фурина
Используя праймеры, приведенные в таблице 1, последовательности ДНК, кодирующие сигнальный пептид CD33, внеклеточную область домена металлопротеазы АСЕ2 (содержащую мутации инактивации фермента H374N и H378N) и hlgGl Fc, были получены методом ПЦР с перекрывающимися праймерами (Overlap PCR). Конкретные этапы клонирования заключаются в следующем: для конструирования экспрессирующей плазмиды ACE2-hFc использовали pcDNA3.0-ACE2-NEMGE в качестве матрицы и использовали праймеры 2-FrontIn и 4-MidR для получения методом ПЦР фрагментов ДНК, кодирующих внеклеточную область АСЕ2 (19-615аа), содержащую мутации инактивации активности фермента H374N и H378N; использовали pCHOGS-HH009 в качестве матрицы и использовали праймеры 3-MidF и 5-IgG1Cterm для получения методом ПЦР фрагментов ДНК, кодирующих hlgGl Fc; затем, используя полученные продукты в качестве матриц, и используя праймеры 6-FrontOutXhoI и 5-IgG1Cterm, синтезировали посредством ПЦР с перекрывающимися праймерами полноразмерную ДНК, кодирующую CD33 сигнальный пептид-внеклеточную область-hFc АСЕ2, которую затем вставляли в экспрессирующий вектор pCHOGS между сайтами расщепления XhoI и Pad для получения экспрессирующей плазмиды pCHOGS-ACE2-NN-hIgG1.

Что касается конструирования плазмиды для экспрессии спайкового белка коронавируса pCAGGS-SARS-CoV2 S-C9 в качестве матрицы использовали PCMV3-2019-nCoV-Spike(Sl+S2)-long (Sino Biological, Cat#: VG40589-UT), и SB-S-NheI:
 и SB-S-C9-XhoI:
и SB-S-C9-XhoI:
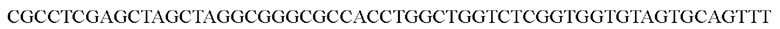
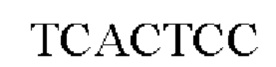 использовали в качестве праймеров для получения фрагментов ДНК, кодирующих спайковый белок SARS-CoV2, которые были вставлены в вектор pCAGGS человека между сайтами расщепления NheI и XhoI для получения плазмиды, обеспечивающей экспрессию S-белка SARS-CoV2. В плазмиде N-концевой сигнальный пептид представлял собой сигнальный пептид CD4, а на С-конце находилась метка С9.
использовали в качестве праймеров для получения фрагментов ДНК, кодирующих спайковый белок SARS-CoV2, которые были вставлены в вектор pCAGGS человека между сайтами расщепления NheI и XhoI для получения плазмиды, обеспечивающей экспрессию S-белка SARS-CoV2. В плазмиде N-концевой сигнальный пептид представлял собой сигнальный пептид CD4, а на С-конце находилась метка С9.
На основе указанной плазмиды были созданы праймеры SB-S-NheI, SB-S-C9-XhoI, SB-Drs-f: 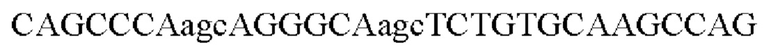 и SB-Drs-r:
и SB-Drs-r:
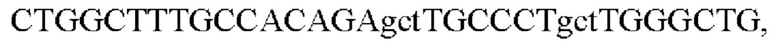 сайт расщепления протеазой фурин PRRAR в S-белке был мутирован к PSRAS методом ПЦР с перекрывающимися праймерами таким образом, чтобы обеспечивалось получение плазмиды для экспрессии S-белка S ARS-CoV2, содержащей мутантный сайт фурина.
сайт расщепления протеазой фурин PRRAR в S-белке был мутирован к PSRAS методом ПЦР с перекрывающимися праймерами таким образом, чтобы обеспечивалось получение плазмиды для экспрессии S-белка S ARS-CoV2, содержащей мутантный сайт фурина.
Пример 2: Конструирование плазмид для экспрессии слитого белка ACE2-hFc-ACE2, слитого белка ACE2-ACE2-hFc, слитого белка ACE2-hFc5 и слитого белка ACE2-hFc5-L309C
Конструирование плазмид для экспрессии слитого белка ACE2-hFc-ACE2, слитого белка ACE2-ACE2-hFc, слитого белка ACE2-hFc5 и слитого белка ACE2-hFc5-L309C было аналогичным конструированию плазмиды для экспрессии слитого белка ACE2-hFc и осуществлялось методом, аналогичным методу в Примере 1.
Пример 3: Экспрессия, очистка и анализ методом эксклюзионной ВЭЖХ (SEC-HPLC) слитого белка ACE2-hFc и слитого белка ACE2-hFc5
Плазмида экспрессии, экспрессирующая слитый белок ACE2-hFc, полученная в Примере 1, и плазмида экспрессии, экспрессирующая слитый белок ACE2-hFc5, полученная в Примере 2, были соответственно трансфицированы в клетки 293F с использованием ПЭИ. Супернатанты культуры собирали через 5 дней после трансфекции и очищали через колонку Protein А в один этап с целью получения очищенного слитого белка ACE2-hFc и слитого белка ACE2-hFc5, соответственно. После количественного определения белка с помощью спектрофотометра NanoDrop™ 2000 был проведен анализ чистоты методом эксклюзионной ВЭЖХ (Фиг. 1А и Фиг. 1С). Как видно из Фиг. 1А, анализ чистоты полученного слитого белка ACE2-hFc показывает только один пик, при этом основной пик составляет 99,34%. Как видно из Фиг. 1С, анализ чистоты полученного слитого белка ACE2-hFc5 показывает только один пик, при этом основной пик составляет 66,94%.
Пример 4: Экспрессия, очистка и анализ методом эксклюзионной ВЭЖХ (SEC-HPLC) слитого белка ACE2-hFc-ACE2, слитого белка ACE2-ACE2-hFc и слитого белка ACE2-hFc5-L309C
Экспрессионные плазмиды, экспрессирующие слитый белок ACE2-hFc-ACE2, слитый белок ACE2-ACE2-hFc и слитый белок ACE2-hFc5-L309C, полученные в Примере 2, соответственно трансфицировали в клетки 293F с помощью ПЭИ. Супернатанты культуры собирали через 5 дней после трансфекции и очищали через колонку Protein А и молекулярное сито в два этапа с целью получения очищенного слитого белка ACE2-ACE2-hFc, слитого белка ACE2-hFc-ACE2 и слитого белка ACE2-hFc5-L309C, соответственно. После количественного определения белков с помощью спектрофотометра NanoDrop™ 2000 для каждого из них был проведен анализ чистоты методом эксклюзионной ВЭЖХ (Фиг. IB, 1D и 1Е). Как видно из Фиг. 1 В, основной пик слитого белка ACE2-ACE2-hFc составляет 98,96%. Как видно из Фиг. 1D, основной пик слитого белка ACE2-hFc-ACE2 составляет 97,99%. Как видно из Фиг. 1Е, основной пик слитого белка ACE2-hFc5-L309C составляет 97,99%.
Пример 5: Экспрессия, очистка и анализ методом эксклюзионной ВЭЖХ слитого белка ACE2-hFc5
Экспрессионная плазмида, экспрессирующая слитый белок ACE2-hFc5, полученная в Примере 2, была трансфицирована в клетки 293F с помощью ПЭИ. Супернатант культуры собирали через 5 дней после трансфекции и очищали в несколько этапов с целью получения очищенного слитого белка ACE2-hFc5. После количественного определения белка с помощью спектрофотометра NanoDrop™ 2000 был проведен анализ чистоты методом эксклюзионной ВЭЖХ (Фиг. 1F). Как видно из Фиг. 1F, основной пик слитого белка ACE2-hFc5 составляет 99,2%.
Пример 6: Эксперимент по ингибированию слияния межклеточных мембран In Vitro
6.1 Способ
Эксперимент по использованию слитых белков АСЕ2, ингибирующих образование синцитиев коронавируса (SARS-CoV2)
Контрольный вектор pCAGGS и плазмиды, экспрессирующие S-белок SARS-CoV2 и S-белок SARS, котрансфицировали, соответственно, с pEGFP-Nl в клетки 293Т. Плазмида, экспрессирующая hACE2-C9 (сохраненная в нашей лаборатории) и вектор pmCherry-Cl котрансфицировали в клетки 293Т с помощью ПЭИ. Клетки переваривали через 24 часа после трансфекции, промывали один раз полной средой DMEM (минимальная эссенциальная среда Игла, модифицированная по способу Дульбекко) (10% FBS, 1×PS) и подсчитывали. Затем 10 мкг/мл контрольного белка и слитых белков ACE2-hFc5 коинкубировали, соответственно, с 2,5Е5/лунку клеток, трансфицированных контрольным вектором pCAGGS, трансфицированных SARS-CoV2-S и SARS-S клеток при 37°С в течение 30 мин. Затем добавляли трансфицированные hACE2-C9 клетки в количестве 2,5Е5/лунку. После совместного культивирования в клеточном инкубаторе в атмосфере 5% CO2 при 37°С в течение 3 часов ингибирующую активность слитых белков ACE2-hFc5 на предмет образования синцитиев коронавируса (SARS-CoV2) наблюдали под флуоресцентным микроскопом. Результаты эксперимента фотографировали и регистрировали.
6.2 Из Фиг. 2 видно, что большое количество синцитиев образовалось как в группе вируса SARS-CoV2, так и в группе вируса SARS. Наибольшее количество синцитиев образовалось в группе вируса SARS-CoV2. В противоположность этому в группе контрольного вектора слияния клеток не наблюдалось. Данный результат показывает, что S-белок вируса SARS-CoV2 похож на S-белок вируса SARS и обладает способностью индуцировать слияние клеток с образованием синцитиев. S-белок вируса SARS-CoV2 обладает более сильной индукционной способностью, чем S-белок вируса SARS (верхняя панель на Фиг. 3). Это также косвенно подтверждает, что S-белок вируса SARS-CoV2 имеет более сильную аффинность к рецептору hACE2, чем S-белок вируса SARS. При добавлении слитого белка ACE2-hFc5 было отмечено, что по сравнению с контрольным белком слитый белок ACE2-hFc5 мог значительно ингибировать индуцированное S-белком вируса SARS-CoV2 образование синцитиев со степенью ингибирования до 90% или более, и даже полностью ингибировать образование синцитиев, индуцированное вирусом SARS.
Кроме того, можно видеть (нижняя панель на Фиг. 3), что большое количество синцитиев образовывалось в том случае, если клетки, экспрессирующие S-белок коронавируса, были коинкубированы с клетками, экспрессирующими рецептор АСЕ2. Это указывает на то, что S-белок коронавируса может индуцировать процесс слияния мембран после связывания с рецептором АСЕ2. Многоядерные синцитии более явно проявлялись через 20 часов. При добавлении в экспериментальную систему 10 мкг/мл ACE2-hFc5 количество синцитиев значительно уменьшилось с коэффициентом ингибирования до 90% или более. В то время как ингибирующий эффект ACE2-hFc в аналогичных условиях был неочевиден. Кроме того, можно видеть, что ACE2-hFc5 ингибировал образование синцитиев дозозависимым образом. ACE2-hFc5 по-прежнему обладал значительной ингибирующей активностью в отношении образования синцитиев при низкой концентрации, составлявшей 0,3 мкг/мл.
Процесс слияния оболочечного вируса (включая коронавирусы) с клеточной (внутренней) мембраной является критически важным фактором для вирусной инфекции. Образование синцитиев представляет собой заметное патологическое изменение, возникающее в легких после заражения человека вирусом SARS-CoV2. Полученные нами экспериментальные результаты показывают, что слитый белок ACE2-hFc5 обладает высокой активностью ингибирования образования синцитиев и способен блокировать инфекцию, вызываемую коронавирусами, в частности, SARS-CoV2.
Пример 7: Анализы экспрессии на поверхности клеток и образования синцитиев для S-белка дикого типа SARS-CoV2 и мутированного S-белка, содержащего мутации в сайте расщепления протеазой фурин
Пустой вектор pCAGGS (контроль) и плазмиды, экспрессирующие S-белок SARS-CoV2 и мутированный S-белок SARS-CoV2 сайтом фурина (в котором PRRAR был мутирован в PSRAS), были соответственно котрансфицированы с помощью ПЭИ вектором pmCherry-C1 в клетки 293Т. Плазмида, экспрессирующая полноразмерный АСЕ2, имеющий внеклеточную, трансмембранную и внутриклеточную области, и вектор pEGFP-N1 были совместно трансфицированы в клетки 293Т с помощью ПЭИ. Клетки переваривали через 24 часа после трансфекции, промывали один раз полной средой DMEM (10% FBS, 1×PS) и подсчитывали. Часть контрольных клеток, трансфицированных пустым вектором, и клетки, экспрессирующие S-белок ARS-CoV2 S (SEQ ID NO: 9) (Фиг. 4) и мутировавший в сайте фурина S-белок ARS-CoV2 S (SEQ ID NO: 10) (Фиг. 5), затем исследовали на предмет экспрессии S-белков SARS-CoV2 на клеточной поверхности. Оставшуюся часть клеток использовали для исследования образования синцитиев.
В отношении исследования экспрессии на поверхности клеток S-белка SARS-CoV2 20 мкг/мл слитого белка ACE2-NN-Fc соответственно инкубировали с вышеуказанными клетками на льду в течение 45 минут, затем клетки трижды промывали буфером FACS (проточная цитофлуориметрия) (PBS, 0,5% BSA), после чего добавляли антитело ФИТЦ к Fc-вторичному антителу человека (F9512, Sigma), разбавленное в отношении 1:300, и инкубировали на льду в течение 30 минут. Поверхностные экспрессии S-белка SARS-CoV2 и мутировавшего S-белка SARS-CoV2 в сайте фурина анализировали проточным цитометром после трехкратной промывки клеток буфером FACS (PBS, 0,5% BSA). Результаты анализировали с помощью программного обеспечения FlowJo V10, и они приведены на Фиг. 6.
В отношении исследования по эксперименту образования синцитиев пустые трансфицированные вектором контрольные клетки и клетки, трансфицированные плазмидой SARS-CoV2 S и плазмидой SARS-CoV2 S, мутировавшей в сайте фурина, были отдельно высеяны в 48-луночный планшет для культивирования клеток в количестве 2,5Е5/лунку. Через 30 минут АСЕ2-трансфицированные клетки добавляли в вышеуказанные лунки с клетками при 2,5Е5/лунку. После непрерывного совместного культивирования в 5% CO2 клеточном инкубаторе при 37°С в течение 3 часов образование вирусных синцитиев наблюдали под флуоресцентным микроскопом. Результаты эксперимента фотографировали и регистрировали. Как показано на Фиг. 7, ни слияния клеток, ни образования многоядерных синцитиев не наблюдалось между контрольными клетками, трансфицированными пустым вектором (отмечены красным), и клетками, экспрессирующими полноразмерный АСЕ2 (отмечены зеленым). Напротив, большое количество многоядерных синцитиев образовывалось после совместного культивирования клеток, экспрессирующих S-белок дикого типа (отмечены красным) с клетками, экспрессирующими человеческий полноразмерный АСЕ2 (отмечены зеленым) в течение 3 часов. После того, как сайт расщепления фурином RRAR в S-белке SARS-CoV2 мутировал в SRAS, слияние клеточных мембран было полностью подавлено, и многоядерных синцитиев не наблюдалось. Это указывает на то, что мутировавший S-белок утратил способность опосредовать слияние клеточных мембран. Таким образом, сайт фурина в S-белке SARS-CoV2 необходим для опосредованного S-белком слияния клеток.
Более того, мутировавший в сайте расщепления протеазой фурин S-белок SARS-CoV2 оказывает значительное влияние на ингибирование слияния вирусов с клетками и образование многоядерных синцитиев (см. крайнее правое изображение на Фиг. 7).
Пример 8: Анализ аффинности и авидности различных форм слитых белков ACE2-hFc
8.1 Способы
Аффиность (BIAcore Т200) и авидность (Fortebio Octet RED384) слитых белков ACE2-NN-hFc и ACE2-NN-hFc5 к рецептор-связывающему домену (RBD) (аа331-527) S-белка коронавируса (SARS-CoV-2) определяли методом поверхностного плазмонного резонанса (SPR) и биослойной интерферометрии (BLI), соответственно.
При проведении анализа аффинности слитые белки ACE2-hFc и ACE2-hFc5 сначала фиксировали на поверхности биосенсорного чипа СМ5, покрытого античеловеческим Fc-антителом. Затем 2-кратные серийные разведения от 200 нМ до 6,25 нМ белка SARS-CoV-2 RBD, который имеет метку His6-Avi на С-конце, пропускали через чип со скоростью 30 мкл/мин для определения кинетики межмолекулярного связывания и диссоциации белков. Для расчета константы ассоциации (Ka), константы диссоциации (Kd) и равновесной константы диссоциации (KD) использовалась модель связывания Ленгмюра 1:1 (ΒΙΑ Evaluation Software). Что касается анализа на авидность, 20 мкг/мл белка SARS-CoV-2 RBD с С-концевой меткой His6-Avi сначала фиксировали на поверхности стрептавидинового биосенсора. Затем в качестве аналитов использовали различные концентрации слитых белков ACE2-hFc (0 нМ, 2-кратные серийные разведения в диапазоне I, 65-105,3 нМ) и ACE2-hFc5 (0 нМ, 2-кратные серийные разведения в диапазоне 8,22-526,3 нМ) для связывания с поверхностью сенсора, связанного с RBD (рецептор-связывающий домен) в течение 180 секунд с последующей диссоциацией в течение 300 секунд. Для расчета констант связывания Ka, Kd и KD использовали модель связывания 1:1 (программное обеспечение Fortebio data Analysis 11.1-knetics).
8.2 Результаты
Используя метод определения аффинности моновалентного связывания (BIAcore Т200) (Фиг. 9), было обнаружено, что слитые белки ACE2-hFc и ACE2-hFc5 имели одинаковую аффинность к RBD SARS-CoV-2, т.е. 26,4 нМ и 29,1 нМ, соответственно. Для анализа аффинности слитых белков ACE2-NN-hFc и ACE2-NN-hFc5 к RBD S-белка использовали ПО Fortebio. В экспериментах 20 мкг/мл белка RBD-His-avi фиксировали на поверхности стрептавидинового биосенсора, при этом в качестве подвижных фаз использовали различные концентрации слитых белков ACE2-NN-hFc и ACE2-NN-hFc5. Для анализа экспериментальных данных использовалось программное обеспечение Octet DataAnalysis 11.0, и в качестве фонового значения для вычитания использовали 0 нМ. Результаты показывают (Фиг. 9), что по сравнению со слитым белком ACE2-hFc мультимеризованный слитый белок ACE2-hFc5 значительно повышает аффинность к спайковому белку RBD-His-avi, при этом значения KD (<1,0 Е-12 М) выходят за пределы диапазона обнаружения прибора.
Пример 9: Эксперимент по нейтрализации псевдовируса коронавируса In Vitro
9.1 Способы
9.1.1 Упаковка псевдовируса коронавируса
Что касается упаковки псевдовируса штамма коронавируса (D614), клетки HEK293T инокулировали в 10-см блюдо для культуры клеток. При достижении клетками 80%-й конфлюентности плазмиду экспрессии полноразмерного S-белка коронавируса pSARS-CoV2 S-C9 (D614) котрансфицировали с упаковочной плазмидой psPAX2 и плазмидой pHFV-Luc, экспрессирующей флуоресцеин, в соотношении 1:3:4 с использованием реагента Lipofactamine 3000. Среду удаляли через 6 ч после трансфекции, добавляли свежую среду DMEM, содержащую 2% FBS и пенициллин, и проводили непрерывное культивирование в течение 48 часов. Затем супернатант культуры, содержащий частицы псевдовируса, собирали, центрифугировали и фильтровали для удаления клеточного остатка, и замораживали при -80°С для дальнейшего использования. Для упаковки других вариантов SARS-CoV-2, SARS и коронавирусов панголина условия приготовления были аналогичными, за исключением того, что pSARS-CoV2 S-C9 (D614) заменили на плазмиду, экспрессирующую S-белки вариантов. Псевдовирусы коронавируса, используемые в настоящем изобретении, включают: исходный штамм D614SARS-CoV2; исходный штамм D614 SARS-CoV2, имеющий мутированный сайт фурина; основной эпидемический штамм G614 SARS-CoV2; вариант D614 (L18F; A22V; V367F; N439K; Y453F; N501Y; T478I; P1263L); SARS-CoV2; SARS; и коронавирус панголина.
9.1.2 Эксперименты по нейтрализации псевдовируса коронавируса
В экспериментах по нейтрализации псевдовирусов коронавируса клетки 293Т-АСЕ2, стабильно экспрессирующие человеческий АСЕ2, сначала высевали на непрозрачный 96-луночный планшет для культур клеток в количестве 1Е5/лунку и культивировали в CO2-инкубаторе при 37°С в течение 20 часов для проведения эксперимента по нейтрализации. В день проведения эксперимента 75 мкл псевдовирусов коронавирусов равномерно смешивали с 25 мкл различных форм серийно разведенных растворимых слитых белков АСЕ2 с последующим инкубированием при комнатной температуре в течение 30 мин. Затем удаляли супернатант клеточной культуры в 96-луночном планшете для культур клеток. Далее предварительно перемешанные смеси псевдовирусов и слитых белков АСЕ2 добавляли в клетки 293Т-АСЕ2. После инкубирования в CO2-инкубаторе при 37°С в течение 24 часов добавляли свежую среду DMEM, содержащую 2% FBS (фетальную бычью сыворотку) вместо продолжения культивирования. Через 24 часа активность люциферазы измеряли с использованием набора реактивов для анализа гена люциферазы Bright-Glo™ и микропланшетного люминометра. В эксперименте было предусмотрено не менее двух дублирующих лунок и контрольная лунка с фетальной бычьей сывороткой.
9.2 Результаты
9.2.1 Нейтрализация инфекции, вызванной псевдовирусом коронавируса, различными формами мультимеров слитого белка ACE2-hFc
Что касается сравнения различных форм слитых белков ACE2-hFc для нейтрализации инфекции, вызванной псевдовирусом коронавируса, в качестве клеток-хозяев использовали клетки 293Т, стабильно экспрессирующие человеческий АСЕ2, при этом серийно разведенные слитые белки ACE2-NN-hFc смешивали с псевдовирусами SARS-CoV-2 для инфицирования 293Т-АСЕ2. Активность внутриклеточной люциферазы (RLU) определяли на второй день после инфицирования. Процентное ингибирование слитых белков ACE2-NN-hFc при различных концентрациях рассчитывали на основе активности внутриклеточной люциферазы инфицированной вирусом контрольной группы с фосфатно-солевым буферным раствором. Как показано в Фиг. 10, различные формы слитых белков ACE2-hFc обладают нейтрализующей активностью против инфекции псевдовируса D614 SARS-CoV-2. В частности, ACE2-hFc5 и ACE2-hFc5 L309C обладают самой высокой нейтрализующей активностью in vitro при IC50 9,86 нг/мл (концентрация полумаксимального ингибирования) и 12,4 нг/мл соответственно, за ними следуют тетрамер ACE2-hFc-ACE2 и тетрамер ACE2-ACE2-hFc при IC50 39,31 нг/мл и 260,8 нг/мл соответственно, в то время как ACE2-NN-hFc обладает наиболее низкой нейтрализующей активностью. Данный результат указывает на то, что активность слитых белков АСЕ2 для нейтрализации инфекции SARS-CoV-2 пропорциональна их аффинности к RBD S-белка, т.е. чем выше степень мультимеризации, тем выше нейтрализующий эффект. Таким образом, нейтрализующая активность слитого белка ACE2-hFc значительно усиливается за счет модификации мультимеризации.
9.2.2 Нейтрализующая активность ACE2-NN-hFc5 против коронавирусной инфекции широкого спектра действия
С целью оценки противоинфекционной активности широкого спектра действия ACE2-NN-hFc5 против вариантов SARS-CoV-2 и родственных коронавирусов было упаковано несколько одноточечных мутантных псевдовирусов SARS-CoV-2 на основе присутствующих в популяции превалирующих вариантов и проводили оценку нейтрализующей активности ACE2-NN-hFc5 с помощью модели инфекции стабильной клеточной линии 293Т-АСЕ2. Результаты показали, что ACE2-NN-hFc5 обладает высокой нейтрализующей активностью и противовирусной активностью широкого спектра действия против исходного штамма D614, основного эпидемического штамма G614 и других вариантов SARS-CoV-2, а также вируса SARS и псевдовируса коронавируса панголина. ACE2-NN-hFc5 обладает нейтрализующей активностью IC50 при 9,56 нг/мл для псевдовируса основного эпидемического штамма G614 (Фиг. 1 и Таблица 2 ниже). В том, что касается нейтрализующей активности в отношении псевдовирусной инфекции основных вариантов SARS-CoV-2 (см. Таблицу 3), результаты показывают, что мультимер ACE2-NN-hFc5 обладает высокой нейтрализующей способностью in vitro. В частности, ACE2-NN-hFc5 обладает более высокой нейтрализующей активностью при IC50 0,036 нг/мл для варианта N501Y. Данные результаты показывают, что ACE2-NN-hFc5 обладает высокоэффективной противокоронавирусной активностью широкого спектра действия in vitro.

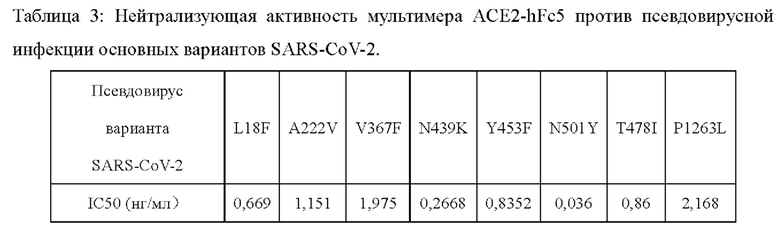
Пример 10: Анализ на нейтрализацию живых вирусов в отношении SARS-CoV-2 in vitro
10.1 Способ
Клетки Vero высевали в 96-луночный планшет при плотности приблизительно 2х104 клеток/лунку. На следующий день среду для культивирования клеток изменяли на 2% среду FBS-DMEM. Слитый белок ACE2-hFc разводили в 2% среде FBS-DMEM до рабочих концентраций 20 мкг/мл, 2 мкг/мл и 0,2 мкг/мл, по три реплицированные лунки. Вирус 2019-nCoV (штамм вируса: уханьский штамм 01 C-Tan-nCoV) разводили до 200 ЦПД50/100 мкл средой 2% FBS-DMEM. К 50 мкл разведенного образца добавляли равный объем вируса 200 ЦПД50 и инкубировали при 37°С в течение 1 часа. Затем 100 мкл комплекса антитела с вирусом добавляли в клетки и инкубировали при 37°С. Цитопатический эффект наблюдали после инкубации при 37°С в течение 48 часов. 100 мкл культурального супернатанта аспирировали через 48 ч для экстракции нуклеиновых кислот, и 80 мкл элюата использовали для окончательного элюирования. 5 мкл экстрактов нуклеиновых кислот брали для составления реакционной флуоресцентной смеси для проведения ОТ-ПЦР в реальном времени, которую анализировали с использованием системы флуоресцентной количественной ПЦР ABI Q5. Для определения ЦПД50 вируса в образцах на основе измеренных пороговых чисел циклов ПЦР образцов использовали стандартную кривую по следующей формуле: коэффициент ингибирования репликации вируса (%)=(контрольный ЦПД50-ЦПД50 слитого белка)/контрольный ЦПД50×100%. Вышеуказанный эксперимент был проведен в лаборатории 3-го уровня биологической безопасности.
10.2 Результаты
Оценку влияния нейтрализации с использованием ACE2-hFc5 на живой вирус 2019-nCoV (уханьский штамм C-Tan-nCoV 01) проводили в лаборатории биологической безопасности с использованием клеток Vero в качестве клеток-хозяев. Результаты эксперимента показали (Фиг. 12), что как ACE2-hFc5, так и ACE2-hFc обладают значительной нейтрализующей активностью против инфекции коронавируса при IC50 0,02-0,06 мкг/мл и 5,3-6,8 мкг/мл, соответственно. ACE2-NN-hFc5 позволяет достичь более высокого эффекта нейтрализации in vitro (в сотни раз превышающий эффект ACE2-NN-hFc), при этом 80% ингибирование вируса все еще наблюдается при концентрации 0,2 мкг/мл. Приведенные выше результаты полностью подтверждают, что ACE2-NN-hFc5 обладает эффективной противокоронавирусной активностью in vitro.
Пример 11: Оценка ингаляции небулайзером слитого белка ACE2-hFc5 у хомяков 11.1 Способы
11.1.1 Анализ осаждения и распределения ACE2-NN-hFc5 в дыхательных путях и легких после ингаляции небулайзером
Для доставки небулизированного ACE2-hFc5 нами была выбрана небулайзерная система доставки системной экспозиции для мелких животных, которая включала воздушный насос, массовый расходомер воздуха и бокс для определения системной экспозиции. Небулайзерная система доставки сопрягалась с небулайзером Aerogen® Solo, адаптером и устройством для сбора аэрозоля. Для анализа осаждения и распределения препарата в дыхательных путях жидкости назального лаважа хомяков собирали путем промывания нормальным физиологическим раствором через 0 ч, 6 ч и 24 ч после доставки препарата, соответственно. Собрали трахеи, главные бронхи и альвеолы, и часть каждой из них была лизирована буфером для лизиса тканей. Супернатант удаляли центрифугированием для выявления содержания мультимеров ACE2-hFc5. Для проведения анализа доз осаждения в легких у каждого хомяка промывали легкие 4 мл нормального физраствора для сбора жидкости бронхоальвеолярного лаважа (ЖБАЛ) через 15 мин, 30 мин и 60 мин после небулайзерной ингаляции. Затем легкие извлекали и гомогенизировали. Часть гомогената легких лизировали буфером для лизиса тканей, и затем супернатант отделяли центрифугированием для определения содержания мультимеров ACE2-hFc5.
11.1.2 ИФА на содержание ACE2-hFc5
Содержание мультимеров ACE2-hFc5 определяли с помощью иммуноферментного анализа (ELISA) для связывания SARS-CoV-2 RBD. В частности, наносили стрептавидин в концентрации 2 мкг/мл для фиксации 2 мкг/мл меченного биотином SARS-CoV-2 RBD. Добавляли тестируемые разведенные лаважную жидкость или супернатант лизиса тканей, в то время как очищенный ACE2-NN-hFc5 использовали в качестве стандарта. Для определения использовали анти-hFc вторичное антитело, меченое пероксидазой хрена (HRP). Значения OD450-OD630 считывали с помощью микропланшетного ридера.
11.1.3 Анализ мультимерных форм ACE2-hFc5 методом эксклюзионной ВЭЖХ до и после небулизации
Мультимеры ACE2-hFc5 до и после небулизации были подвергнуты анализу агрегации и деградации методом ВЭЖХ. Использовали систему высокоэффективной жидкостной хроматографии Agilent 1260, аналитическая колонка G4000 TSK G4000SWxl и предколонка TSK-GEL SWxl. Буфер включал 50 ммоль РВ (фосфатный буфер) и 300 ммоль NaCl при значении рН 6,7±0,1. Анализ проводили в течение 20 или 25 мин при расходе 0,8 мл/мин.
11.2 Результат: ACE2-NN-hFc5 может быть эффективно осажден в альвеолах хомяка путем небулайзерного введения.
Был изучен способ введения через дыхательные пути путем ингаляции с помощью небулайзера ввиду того, что SARS-CoV-2 в основном вызывает вирусную инфекцию дыхательных путей, и при этом очаг инфекции в основном находится в легких. ACE2-hFc5 распыляли с помощью небулайзера Aerogen и собирали распыленные капли с помощью подобранного комплекта стеклянных трубок на ледяной бане с коэффициентом извлечения 90% или более. Был проведен анализ физико-химических свойств ACE2-hFc5 методом эксклюзионной ВЭЖХ до и после небулизации, а также его нейтрализующей активности против инфекции псевдовируса SARS-CoV-2. Результаты показывают, что небулизация не вызывает агрегации и деградации ACE2-NN-hFc5 (Фиг. 13). Кроме того, показано, что ACE2-NN-hFc5 способен значительно подавлять инфекцию псевдовируса SARS-CoV-2, и его способность к противовирусному действию существенно не изменяется до и после распыления (Фиг. 14).
Систему небулайзерной доставки для мелких животных далее использовали для введения хомякам путем небулайзерной ингаляции 5 мг/мл ACE2-NN-hFc5 в течение 50 мин. Для анализа осаждения и распределения ACE2-NN-hFc5 в каждой части дыхательных путей брали жидкость назального лаважа, трахею, главные бронхи и альвеолы хомяков через 0 ч, 6 ч и 24 ч после небулизации. Было обнаружено, что ингалированный ACE2-hFc5 в основном распределялся в альвеолах (около 75%) через 0 ч, 6 ч и 24 ч после ингаляции. Распределение ACE2-hFc5 в жидкости назального лаважа быстро уменьшилось с 17,33% через 0 ч до 0,7% через 24 ч после ингаляции. Распределение ACE2-hFc5 в трахее было меньше и находилось в пределах от 0,35% до 3,4%. Распределение ACE2-hFc5 в бронхах постепенно увеличивалось с 5% до 19%. данные результаты показывают, что ингалируемый ACE2-NN-hFc5 может эффективно достигать альвеол и в основном распределяться в альвеолах в течение 24 часов после ингаляции.
Далее проводили исследование связи между ингаляционными дозами ACE2-hFc5 и отложением в легких хомяков, а также проанализировали нейтрализующую активность в отношении псевдовирусов в жидкости бронхоальвеолярного лаважа. Собирали жидкость бронхоальвеолярного лаважа и гомогенат легких хомяков после небулайзерной ингаляции ACE2-hFc5 в концентрации 5 мг/мл в течение 15 мин, 30 мин и 60 мин. Затем содержание ACE2-hFc5 определяли методом ИФА анализа. Количество ACE2-NN-hFc5 в жидкости бронхоальвеолярного лаважа и гомогенате легких рассчитывали как общее количество осаждения в легких. Результаты показывают, что после небулайзерной ингаляции ACE2-hFc5 количество отложений в легких хомяков увеличивается с увеличением дозы. Количество отложений соответствует 6,48 мкг, 18,69 мкг и 33,35 мкг при ингаляции в течение 15 мин, 30 мин и 60 мин, соответственно. Количество отложений ACE2-hFc5 в легких хомяков быстро уменьшалось после ингаляции. Через 12 часов после ингаляции количество отложений составляло 14,73%-46% от количества непосредственно после ингаляции. Тем не менее, при ингаляции в течение 15 мин и 30 мин жидкость бронхоальвеолярного лаважа, полученная от хомяков, органы дыхания каждого из которых были промыты 4 мл нормального физиологического раствора через 12 часов после ингаляции, сохраняла 90%-ю нейтрализующую активность против псевдовирусов SARS-CoV-2 или выше после 4-кратного разведения.
Пример 12: Оценка эффективности ACE2-NN-hFc5 в модели хомяка при инфицировании новым коронавирусом (SARS-CoV-2)
12.1 Способ
12.1.1 План эксперимента
Все эксперименты на животных были одобрены Комитетом по контролю этических норм обращения с животными Куныминского института биомедицинских наук Китайской академии медицинских наук и соответствовали лабораторной практике и рекомендациям Национальной Куныминской лаборатории биологической безопасности высокого уровня в Юньнани, Китай.
Взрослых свободных от специфических патогенов хомяков перевели в лабораторию биологической безопасности высокого уровня и выращивали по отдельности в соответствующих клетках. Хомяков разделили на три группы в соответствии с массой тела: контрольная группа без лечения; группа, подвергавшаяся небулайзерной терапии в течение 15 минут дважды в день; и группа, подвергавшаяся небулайзерной терапии в течение 30 минут дважды в день, при этом в каждой группе находилось по 7 хомяков. В эксперименте хомякам инокулировали 104 БОЕ вирусов SARS-CoV-2 (GD108#) через носовые полости. Через 2 часа проводили первую небулайзерную ингаляцию с небулизацией в течение 15 мин (25 мг ACE2-hFc5) и в течение 30 мин (50 мг ACE2-hFc5), соответственно. Небулайзерную ингаляцию проводили один раз каждые 12 часов, в общей сложности 6 раз. Хомяков взвешивали перед каждой небулайзерной ингаляцией. Эксперимент закончился через 62 часа после инфицирования вирусом. Ткани легких были взяты для анализа геномной нагрузки вируса SARS-CoV-2 (гРНК) и субвируса (сгРНК). На 100 мг легочной ткани (левое легкое) добавляли 1 мл фосфатно-солевого буферного раствора (PBS). 200 мкл брали для экстракции РНК после быстрой гомогенизации, затем проводили ОТ-кПЦР анализ для определения гРНК и сгРНК вирусной нагрузки SARS-CoV-2. Статистический анализ проводили с помощью программного обеспечения GraphPad Prism 8. Для анализа различий между двумя группами использовался двусторонний U-критерий Манна-Уитни.
12.2 Результаты: Небулайзерное введение ACE2-hFc5 позволяет эффективно снизить вирусную нагрузку в модели хомяка при инфекции, вызванной S ARS-CoV-2.
Основываясь на установленном способе ингаляционной доставки с помощью небулайзера, нами была проведена оценка противовирусной способности ACE2-NN-hFC5 in vivo с использованием модели хомяка в отношении инфекции, вызванной SARS-CoV-2. Для оценки эффективности ACE2-hFC5 в эксперименте использовали две группы доз: одна группа предусматривала небулайзерную ингаляцию в течение 15 минут, другая -небулайзерную ингаляцию в течение 30 минут.Титры вируса достигали максимума на третий день после инфицирования хомяков SARS-CoV-2, и с течением временем уровень виремии постепенно повысился. Таким образом, третий день после заражения вирусом был выбран в качестве конечной точки эксперимента. После лечения ACE2-NN-hFc5 наблюдалась некоторая тенденция к улучшению, заключающаяся в снижении массы тела хомяков. Согласно результатам количественной оценки гРНК и сгРНК в тканях легких небулайзерная ингаляция ACE2-hFc5 способна в значительной степени ингибировать репликацию вируса SARS-CoV-2 в тканях легких (Фиг. 15). На Фиг. 15 приведены результаты анализа кПЦР, проведенного по завершению эксперимента (через 62 часа после инфицирования) для определения изменений генома SARS-CoV-2 (гРНК) и субгенома (сгРНК) в легких хомяков.  обозначает контрольную группу без лечения;
обозначает контрольную группу без лечения;  обозначает группу ACE2-hFc5-15 мин-два раза в сутки;
обозначает группу ACE2-hFc5-15 мин-два раза в сутки;  обозначает группу ACE2-hFc5-30 мин-два раза в сутки; горизонтальная линия представляет медиану. Для статистического анализа различий между двумя группами использовался U-критерий Манна-Уитни. Таким образом, небулайзерная ингаляция ACE2-hFc5 позволяет эффективно снизить вирусную нагрузку в модели хомяка для инфекции, вызванной SARS-CoV-2, и обладает потенциалом для профилактики и лечения пневмонии, вызванной SARS-CoV-2.
обозначает группу ACE2-hFc5-30 мин-два раза в сутки; горизонтальная линия представляет медиану. Для статистического анализа различий между двумя группами использовался U-критерий Манна-Уитни. Таким образом, небулайзерная ингаляция ACE2-hFc5 позволяет эффективно снизить вирусную нагрузку в модели хомяка для инфекции, вызванной SARS-CoV-2, и обладает потенциалом для профилактики и лечения пневмонии, вызванной SARS-CoV-2.
Несмотря на то, что настоящее изобретение было подробно изложено выше на основе общего описания, специалистам в данной области техники должно быть очевидно, что могут быть выполнены конкретные варианты осуществления и проведены эксперименты, внесены некоторые изменения или усовершенствования. Таким образом, все указанные изменения или усовершенствования, внесенные без отступления от сути настоящего изобретения, входят в объем охраны настоящего изобретения.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing nonEnglishFreeTextLanguageCode="ru"
dtdVersion="V1_3" fileName="58381-Seq-ru.xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2024-10-02">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022125303</ApplicationNumberText>
<FilingDate>2022-09-27</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>WO21508HHRU</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>CN</IPOfficeCode>
<ApplicationNumberText>202010124368.4</ApplicationNumberText>
<FilingDate>2020-02-27</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">ХУАХУЭЙ ХЕЛС ЛТД.</ApplicantName>
<ApplicantNameLatin>HUAHUI HEALTH LTD.</ApplicantNameLatin>
<InventorName languageCode="ru">Ли Вэнхуэй</InventorName>
<InventorNameLatin>LI Wenhui</InventorNameLatin>
<InventionTitle languageCode="ru">РАСТВОРИМЫЙ АПФ2, ЕГО СЛИТЫЙ БЕЛОК
И СПОСОБЫ ИХ ПРИМЕНЕНИЯ</InventionTitle>
<SequenceTotalQuantity>18</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>596</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..596</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q1">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
<NonEnglishQualifier_value>Homo
sapiens</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNTNITEENVQNMNNAGDK
WSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNTMSTIYSTGKVCNPDNP
QECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMARANHYEDYGDYWRGDYE
VNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCLPAHLLGDMWGRFWTNL
YSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSMLTDPGNVQKAVCHPTA
WDLGKGDFRILMCTKVTMDDFLTAHHEMGHIQYDMAYAAQPFLLRNGANEGFHEAVGEIMSLSAATPKHL
KSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQWMKKWWEMKREIVGVV
EPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKCDISNSTEAGQKLFNML
RLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSPYAD</INSDSeq_seq
uence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>596</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..596</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q2">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..596</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q3">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNTNITEENVQNMNNAGDK
WSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNTMSTIYSTGKVCNPDNP
QECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMARANHYEDYGDYWRGDYE
VNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCLPAHLLGDMWGRFWTNL
YSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSMLTDPGNVQKAVCHPTA
WDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHEAVGEIMSLSAATPKHL
KSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQWMKKWWEMKREIVGVV
EPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKCDISNSTEAGQKLFNML
RLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSPYAD</INSDSeq_seq
uence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>828</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..828</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q4">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..828</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q5">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNTNITEENVQNMNNAGDK
WSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNTMSTIYSTGKVCNPDNP
QECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMARANHYEDYGDYWRGDYE
VNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCLPAHLLGDMWGRFWTNL
YSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSMLTDPGNVQKAVCHPTA
WDLGKGDFRILMCTKVTMDDFLTAHHEMGHIQYDMAYAAQPFLLRNGANEGFHEAVGEIMSLSAATPKHL
KSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQWMKKWWEMKREIVGVV
EPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKCDISNSTEAGQKLFNML
RLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSPYADEPKSSDKTHTCPP
CPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNS
TYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCL
VKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQ
KSLSLSPGK</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>828</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..828</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q6">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..828</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q7">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNTNITEENVQNMNNAGDK
WSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNTMSTIYSTGKVCNPDNP
QECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMARANHYEDYGDYWRGDYE
VNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCLPAHLLGDMWGRFWTNL
YSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSMLTDPGNVQKAVCHPTA
WDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHEAVGEIMSLSAATPKHL
KSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQWMKKWWEMKREIVGVV
EPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKCDISNSTEAGQKLFNML
RLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSPYADEPKSSDKTHTCPP
CPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNS
TYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCL
VKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQ
KSLSLSPGK</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>844</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..844</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q8">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..844</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q9">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHHEMGHIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGV
EVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNV
FSCSVMHEALHNHYTQKSLSLSPGK</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>844</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..844</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q10">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..844</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q11">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGV
EVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNV
FSCSVMHEALHNHYTQKSLSLSPGK</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="7">
<INSDSeq>
<INSDSeq_length>862</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..862</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q12">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..862</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q13">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHHEMGHIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGV
EVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNV
FSCSVMHEALHNHYTQKSLSLSTGKPTLYNVSLVMSDTAGTCY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="8">
<INSDSeq>
<INSDSeq_length>862</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..862</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q14">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..862</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q15">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGV
EVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNV
FSCSVMHEALHNHYTQKSLSLSTGKPTLYNVSLVMSDTAGTCY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="9">
<INSDSeq>
<INSDSeq_length>1273</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1273</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q16">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Coaxana purpurea</INSDQualifier_value>
<NonEnglishQualifier_value>Coaxana
purpurea</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MFVFLVLLPLVSSQCVNLTTRTQLPPAYTNSFTRGVYYPDKVFRSSVLH
STQDLFLPFFSNVTWFHAIHVSGTNGTKRFDNPVLPFNDGVYFASTEKSNIIRGWIFGTTLDSKTQSLLI
VNNATNVVIKVCEFQFCNDPFLGVYYHKNNKSWMESEFRVYSSANNCTFEYVSQPFLMDLEGKQGNFKNL
REFVFKNIDGYFKIYSKHTPINLVRDLPQGFSALEPLVDLPIGINITRFQTLLALHRSYLTPGDSSSGWT
AGAAAYYVGYLQPRTFLLKYNENGTITDAVDCALDPLSETKCTLKSFTVEKGIYQTSNFRVQPTESIVRF
PNITNLCPFGEVFNATRFASVYAWNRKRISNCVADYSVLYNSASFSTFKCYGVSPTKLNDLCFTNVYADS
FVIRGDEVRQIAPGQTGKIADYNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDIS
TEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKSTNLVKNKCV
NFNFNGLTGTGVLTESNKKFLPFQQFGRDIADTTDAVRDPQTLEILDITPCSFGGVSVITPGTNTSNQVA
VLYQDVNCTEVPVAIHADQLTPTWRVYSTGSNVFQTRAGCLIGAEHVNNSYECDIPIGAGICASYQTQTN
SPRRARSVASQSIIAYTMSLGAENSVAYSNNSIAIPTNFTISVTTEILPVSMTKTSVDCTMYICGDSTEC
SNLLLQYGSFCTQLNRALTGIAVEQDKNTQEVFAQVKQIYKTPPIKDFGGFNFSQILPDPSKPSKRSFIE
DLLFNKVTLADAGFIKQYGDCLGDIAARDLICAQKFNGLTVLPPLLTDEMIAQYTSALLAGTITSGWTFG
AGAALQIPFAMQMAYRFNGIGVTQNVLYENQKLIANQFNSAIGKIQDSLSSTASALGKLQDVVNQNAQAL
NTLVKQLSSNFGAISSVLNDILSRLDKVEAEVQIDRLITGRLQSLQTYVTQQLIRAAEIRASANLAATKM
SECVLGQSKRVDFCGKGYHLMSFPQSAPHGVVFLHVTYVPAQEKNFTTAPAICHDGKAHFPREGVFVSNG
THWFVTQRNFYEPQIITTDNTFVSGNCDVVIGIVNNTVYDPLQPELDSFKEELDKYFKNHTSPDVDLGDI
SGINASVVNIQKEIDRLNEVAKNLNESLIDLQELGKYEQYIKWPWYIWLGFIAGLIAIVMVTIMLCCMTS
CCSCLKGCCSCGSCCKFDEDDSEPVLKGVKLHYT</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="10">
<INSDSeq>
<INSDSeq_length>1273</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..1273</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q17">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1273</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q18">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MFVFLVLLPLVSSQCVNLTTRTQLPPAYTNSFTRGVYYPDKVFRSSVLH
STQDLFLPFFSNVTWFHAIHVSGTNGTKRFDNPVLPFNDGVYFASTEKSNIIRGWIFGTTLDSKTQSLLI
VNNATNVVIKVCEFQFCNDPFLGVYYHKNNKSWMESEFRVYSSANNCTFEYVSQPFLMDLEGKQGNFKNL
REFVFKNIDGYFKIYSKHTPINLVRDLPQGFSALEPLVDLPIGINITRFQTLLALHRSYLTPGDSSSGWT
AGAAAYYVGYLQPRTFLLKYNENGTITDAVDCALDPLSETKCTLKSFTVEKGIYQTSNFRVQPTESIVRF
PNITNLCPFGEVFNATRFASVYAWNRKRISNCVADYSVLYNSASFSTFKCYGVSPTKLNDLCFTNVYADS
FVIRGDEVRQIAPGQTGKIADYNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDIS
TEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKSTNLVKNKCV
NFNFNGLTGTGVLTESNKKFLPFQQFGRDIADTTDAVRDPQTLEILDITPCSFGGVSVITPGTNTSNQVA
VLYQDVNCTEVPVAIHADQLTPTWRVYSTGSNVFQTRAGCLIGAEHVNNSYECDIPIGAGICASYQTQTN
SPSRASSVASQSIIAYTMSLGAENSVAYSNNSIAIPTNFTISVTTEILPVSMTKTSVDCTMYICGDSTEC
SNLLLQYGSFCTQLNRALTGIAVEQDKNTQEVFAQVKQIYKTPPIKDFGGFNFSQILPDPSKPSKRSFIE
DLLFNKVTLADAGFIKQYGDCLGDIAARDLICAQKFNGLTVLPPLLTDEMIAQYTSALLAGTITSGWTFG
AGAALQIPFAMQMAYRFNGIGVTQNVLYENQKLIANQFNSAIGKIQDSLSSTASALGKLQDVVNQNAQAL
NTLVKQLSSNFGAISSVLNDILSRLDKVEAEVQIDRLITGRLQSLQTYVTQQLIRAAEIRASANLAATKM
SECVLGQSKRVDFCGKGYHLMSFPQSAPHGVVFLHVTYVPAQEKNFTTAPAICHDGKAHFPREGVFVSNG
THWFVTQRNFYEPQIITTDNTFVSGNCDVVIGIVNNTVYDPLQPELDSFKEELDKYFKNHTSPDVDLGDI
SGINASVVNIQKEIDRLNEVAKNLNESLIDLQELGKYEQYIKWPWYIWLGFIAGLIAIVMVTIMLCCMTS
CCSCLKGCCSCGSCCKFDEDDSEPVLKGVKLHYT</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="11">
<INSDSeq>
<INSDSeq_length>1208</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..1208</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q19">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1208</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q20">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MFVFLVLLPLVSSQCVNLTTRTQLPPAYTNSFTRGVYYPDKVFRSSVLH
STQDLFLPFFSNVTWFHAIHVSGTNGTKRFDNPVLPFNDGVYFASTEKSNIIRGWIFGTTLDSKTQSLLI
VNNATNVVIKVCEFQFCNDPFLGVYYHKNNKSWMESEFRVYSSANNCTFEYVSQPFLMDLEGKQGNFKNL
REFVFKNIDGYFKIYSKHTPINLVRDLPQGFSALEPLVDLPIGINITRFQTLLALHRSYLTPGDSSSGWT
AGAAAYYVGYLQPRTFLLKYNENGTITDAVDCALDPLSETKCTLKSFTVEKGIYQTSNFRVQPTESIVRF
PNITNLCPFGEVFNATRFASVYAWNRKRISNCVADYSVLYNSASFSTFKCYGVSPTKLNDLCFTNVYADS
FVIRGDEVRQIAPGQTGKIADYNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDIS
TEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKSTNLVKNKCV
NFNFNGLTGTGVLTESNKKFLPFQQFGRDIADTTDAVRDPQTLEILDITPCSFGGVSVITPGTNTSNQVA
VLYQDVNCTEVPVAIHADQLTPTWRVYSTGSNVFQTRAGCLIGAEHVNNSYECDIPIGAGICASYQTQTN
SPRRARSVASQSIIAYTMSLGAENSVAYSNNSIAIPTNFTISVTTEILPVSMTKTSVDCTMYICGDSTEC
SNLLLQYGSFCTQLNRALTGIAVEQDKNTQEVFAQVKQIYKTPPIKDFGGFNFSQILPDPSKPSKRSFIE
DLLFNKVTLADAGFIKQYGDCLGDIAARDLICAQKFNGLTVLPPLLTDEMIAQYTSALLAGTITSGWTFG
AGAALQIPFAMQMAYRFNGIGVTQNVLYENQKLIANQFNSAIGKIQDSLSSTASALGKLQDVVNQNAQAL
NTLVKQLSSNFGAISSVLNDILSRLDKVEAEVQIDRLITGRLQSLQTYVTQQLIRAAEIRASANLAATKM
SECVLGQSKRVDFCGKGYHLMSFPQSAPHGVVFLHVTYVPAQEKNFTTAPAICHDGKAHFPREGVFVSNG
THWFVTQRNFYEPQIITTDNTFVSGNCDVVIGIVNNTVYDPLQPELDSFKEELDKYFKNHTSPDVDLGDI
SGINASVVNIQKEIDRLNEVAKNLNESLIDLQELGKYEQ</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="12">
<INSDSeq>
<INSDSeq_length>1208</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..1208</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q21">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial Sequence</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная
последовательность</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1208</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q22">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MFVFLVLLPLVSSQCVNLTTRTQLPPAYTNSFTRGVYYPDKVFRSSVLH
STQDLFLPFFSNVTWFHAIHVSGTNGTKRFDNPVLPFNDGVYFASTEKSNIIRGWIFGTTLDSKTQSLLI
VNNATNVVIKVCEFQFCNDPFLGVYYHKNNKSWMESEFRVYSSANNCTFEYVSQPFLMDLEGKQGNFKNL
REFVFKNIDGYFKIYSKHTPINLVRDLPQGFSALEPLVDLPIGINITRFQTLLALHRSYLTPGDSSSGWT
AGAAAYYVGYLQPRTFLLKYNENGTITDAVDCALDPLSETKCTLKSFTVEKGIYQTSNFRVQPTESIVRF
PNITNLCPFGEVFNATRFASVYAWNRKRISNCVADYSVLYNSASFSTFKCYGVSPTKLNDLCFTNVYADS
FVIRGDEVRQIAPGQTGKIADYNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDIS
TEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKSTNLVKNKCV
NFNFNGLTGTGVLTESNKKFLPFQQFGRDIADTTDAVRDPQTLEILDITPCSFGGVSVITPGTNTSNQVA
VLYQDVNCTEVPVAIHADQLTPTWRVYSTGSNVFQTRAGCLIGAEHVNNSYECDIPIGAGICASYQTQTN
SPSRASSVASQSIIAYTMSLGAENSVAYSNNSIAIPTNFTISVTTEILPVSMTKTSVDCTMYICGDSTEC
SNLLLQYGSFCTQLNRALTGIAVEQDKNTQEVFAQVKQIYKTPPIKDFGGFNFSQILPDPSKPSKRSFIE
DLLFNKVTLADAGFIKQYGDCLGDIAARDLICAQKFNGLTVLPPLLTDEMIAQYTSALLAGTITSGWTFG
AGAALQIPFAMQMAYRFNGIGVTQNVLYENQKLIANQFNSAIGKIQDSLSSTASALGKLQDVVNQNAQAL
NTLVKQLSSNFGAISSVLNDILSRLDKVEAEVQIDRLITGRLQSLQTYVTQQLIRAAEIRASANLAATKM
SECVLGQSKRVDFCGKGYHLMSFPQSAPHGVVFLHVTYVPAQEKNFTTAPAICHDGKAHFPREGVFVSNG
THWFVTQRNFYEPQIITTDNTFVSGNCDVVIGIVNNTVYDPLQPELDSFKEELDKYFKNHTSPDVDLGDI
SGINASVVNIQKEIDRLNEVAKNLNESLIDLQELGKYEQ</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="13">
<INSDSeq>
<INSDSeq_length>1443</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..1443</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q23">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная</NonEnglishQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1443</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q24">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGV
EVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNV
FSCSVMHEALHNHYTQKSLSLSPGKAAATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNTNITEENVQN
MNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNTMSTIYSTGK
VCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMARANHYEDYGD
YWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCLPAHLLGDMW
GRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSMLTDPGNVQK
AVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHEAVGEIMSLS
AATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQWMKKWWEMK
REIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKCDISNSTEAG
QKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSPYAD</INSD
Seq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="14">
<INSDSeq>
<INSDSeq_length>1440</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..1440</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q25">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная</NonEnglishQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1440</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q26">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADTIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNTNITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQ
EIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNTMSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSL
DYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMARANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDV
EHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCLPAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDA
MVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSMLTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTM
DDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHEAVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEI
NFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQWMKKWWEMKREIVGVVEPVPHDETYCDPASLFHV
SNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKCDISNSTEAGQKLFNMLRLGKSEPWTLALENVVGA
KNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSPYADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPK
PKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNG
KEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQ
PENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK</INSDSeq
_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="15">
<INSDSeq>
<INSDSeq_length>18</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..18</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q27">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная</NonEnglishQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..18</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q28">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>PTLYNVSLVMSDTAGTCY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="16">
<INSDSeq>
<INSDSeq_length>18</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..18</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q29">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная</NonEnglishQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..18</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q30">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>PTHVNVSVVMAEVDGTCY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="17">
<INSDSeq>
<INSDSeq_length>21</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q31">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная</NonEnglishQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..21</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q32">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>TGKPTLYNVSLVMSDTAGTCY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="18">
<INSDSeq>
<INSDSeq_length>862</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>REGION</INSDFeature_key>
<INSDFeature_location>1..862</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier id="q33">
<INSDQualifier_name>note</INSDQualifier_name>
<INSDQualifier_value>Artificial</INSDQualifier_value>
<NonEnglishQualifier_value>Искусственная</NonEnglishQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..862</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q34">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
<NonEnglishQualifier_value>Синтетическая
конструкция</NonEnglishQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MPLLLLLPLLWAGALATIEEQAKTFLDKFNHEAEDLFYQSSLASWNYNT
NITEENVQNMNNAGDKWSAFLKEQSTLAQMYPLQEIQNLTVKLQLQALQQNGSSVLSEDKSKRLNTILNT
MSTIYSTGKVCNPDNPQECLLLEPGLNEIMANSLDYNERLWAWESWRSEVGKQLRPLYEEYVVLKNEMAR
ANHYEDYGDYWRGDYEVNGVDGYDYSRGQLIEDVEHTFEEIKPLYEHLHAYVRAKLMNAYPSYISPIGCL
PAHLLGDMWGRFWTNLYSLTVPFGQKPNIDVTDAMVDQAWDAQRIFKEAEKFFVSVGLPNMTQGFWENSM
LTDPGNVQKAVCHPTAWDLGKGDFRILMCTKVTMDDFLTAHNEMGNIQYDMAYAAQPFLLRNGANEGFHE
AVGEIMSLSAATPKHLKSIGLLSPDFQEDNETEINFLLKQALTIVGTLPFTYMLEKWRWMVFKGEIPKDQ
WMKKWWEMKREIVGVVEPVPHDETYCDPASLFHVSNDYSFIRYYTRTLYQFQFQEALCQAAKHEGPLHKC
DISNSTEAGQKLFNMLRLGKSEPWTLALENVVGAKNMNVRPLLNYFEPLFTWLKDQNKNSFVGWSTDWSP
YADEPKSSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGV
EVHNAKTKPREEQYNSTYRVVSVLTVCHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLP
PSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNV
FSCSVMHEALHNHYTQKSLSLSTGKPTLYNVSLVMSDTAGTCY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ SARS-COV-2 | 2022 |
|
RU2791749C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ НЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ К SARS-CoV-2 В СЫВОРОТКЕ ИЛИ ПЛАЗМЕ КРОВИ ЛЮДЕЙ, ПЕРЕНЕСШИХ COVID-19 ИЛИ ПРИВИТЫХ ВАКЦИНАМИ ДЛЯ ПРОФИЛАКТИКИ НОВОЙ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19, С ИСПОЛЬЗОВАНИЕМ НАБОРА РЕАГЕНТОВ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА, СОДЕРЖАЩЕГО РЕКОМБИНАНТНЫЙ РЕЦЕПТОР-СВЯЗЫВАЮЩИЙ ДОМЕН (RBD) ПОВЕРХНОСТНОГО ГЛИКОПРОТЕИНА S КОРОНАВИРУСА SARS-COV-2 И РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ РЕЦЕПТОР АСЕ2 | 2021 |
|
RU2784655C1 |
| Однодоменное антитело ламы Н5 и его производное H5-Fc, специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2022 |
|
RU2793967C1 |
| ПОЛИПЕПТИДНЫЙ МОДУЛЬ ДЛЯ СВЯЗЫВАНИЯ КОНСЕРВАТИВНОГО ЭПИТОПА РЕЦЕПТОР-СВЯЗЫВАЮЩЕГО ДОМЕНА БЕЛКА SPIKE КОРОНАВИРУСА SARS-COV-2 | 2022 |
|
RU2809183C1 |
| ПОЛИПЕПТИД С АНТИ-SARS-CoV-2 АКТИВНОСТЬЮ И ПРОЛОНГИРОВАННЫМ ВРЕМЕНЕМ ЦИРКУЛЯЦИИ В КРОВОТОКЕ | 2023 |
|
RU2822355C1 |
| Рекомбинантный белок, связывающийся с RBD S-белка SARS-CoV-2 | 2022 |
|
RU2778942C1 |
| Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2 | 2021 |
|
RU2794141C2 |
| ПОЛИВАЛЕНТНАЯ ВЕКТОРНАЯ ВАКЦИНА HVT | 2020 |
|
RU2832110C1 |
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
| ЛЕЧЕНИЕ И/ИЛИ ПРОФИЛАКТИКА ЗАБОЛЕВАНИЯ ИЛИ СИНДРОМА, СВЯЗАННОГО С ВИРУСНОЙ ИНФЕКЦИЕЙ | 2021 |
|
RU2823437C2 |
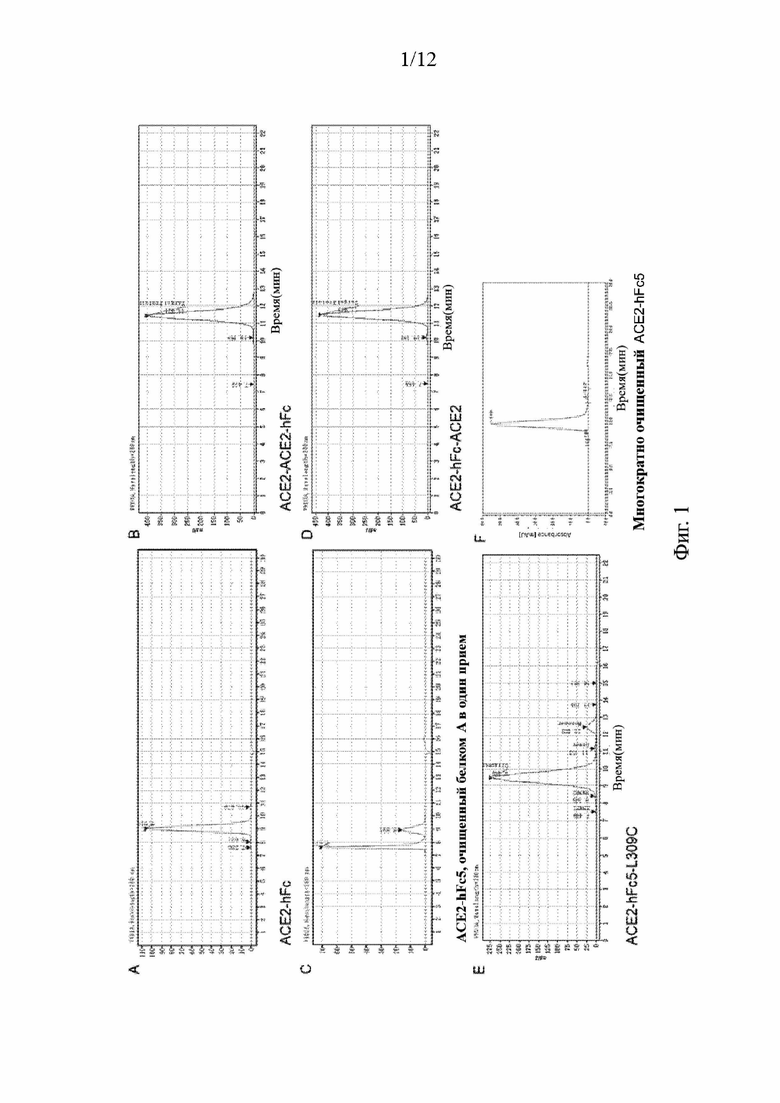
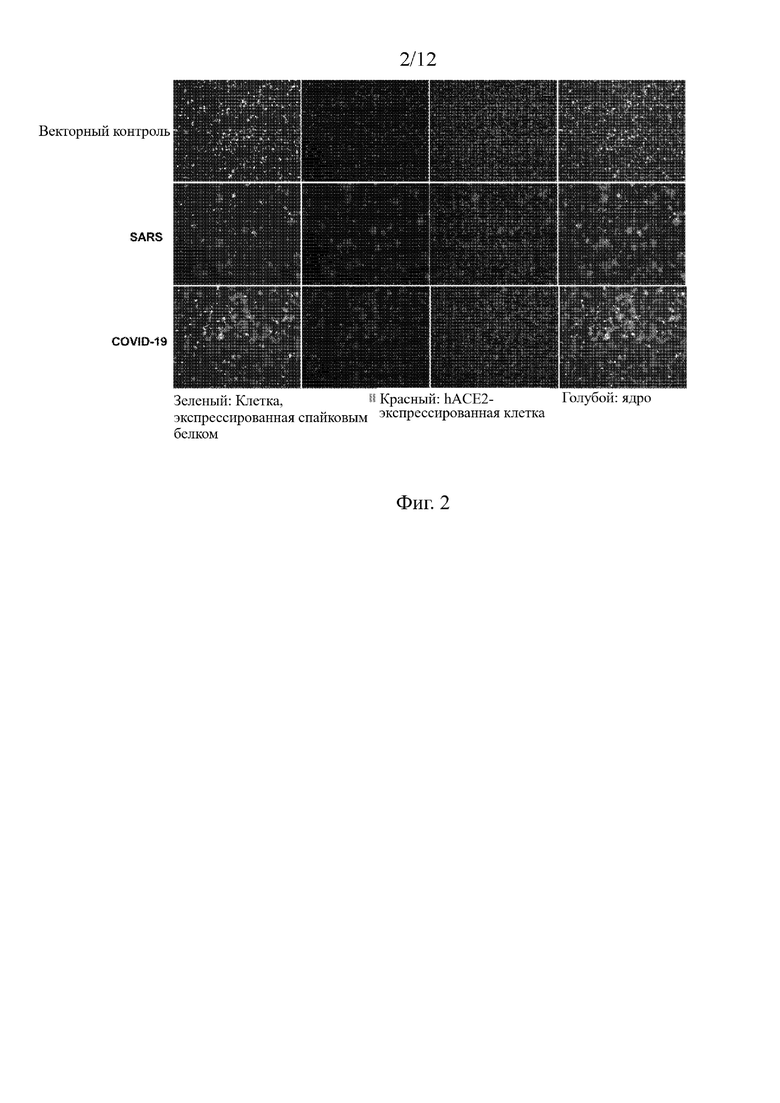


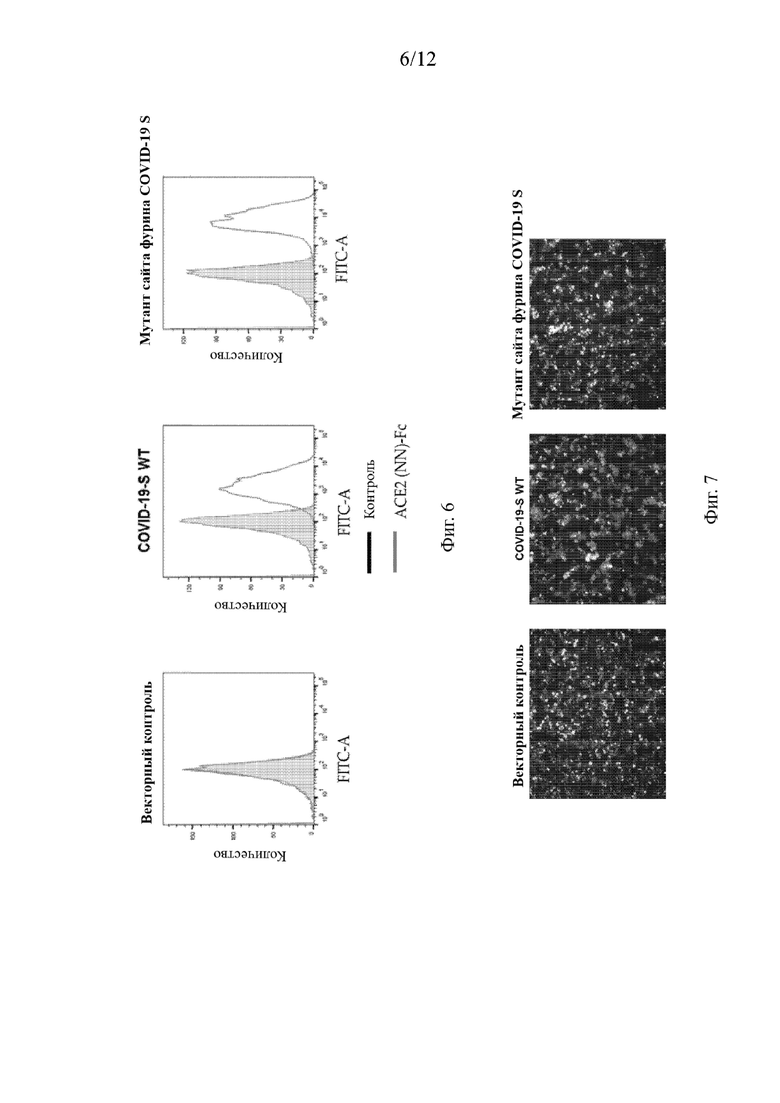

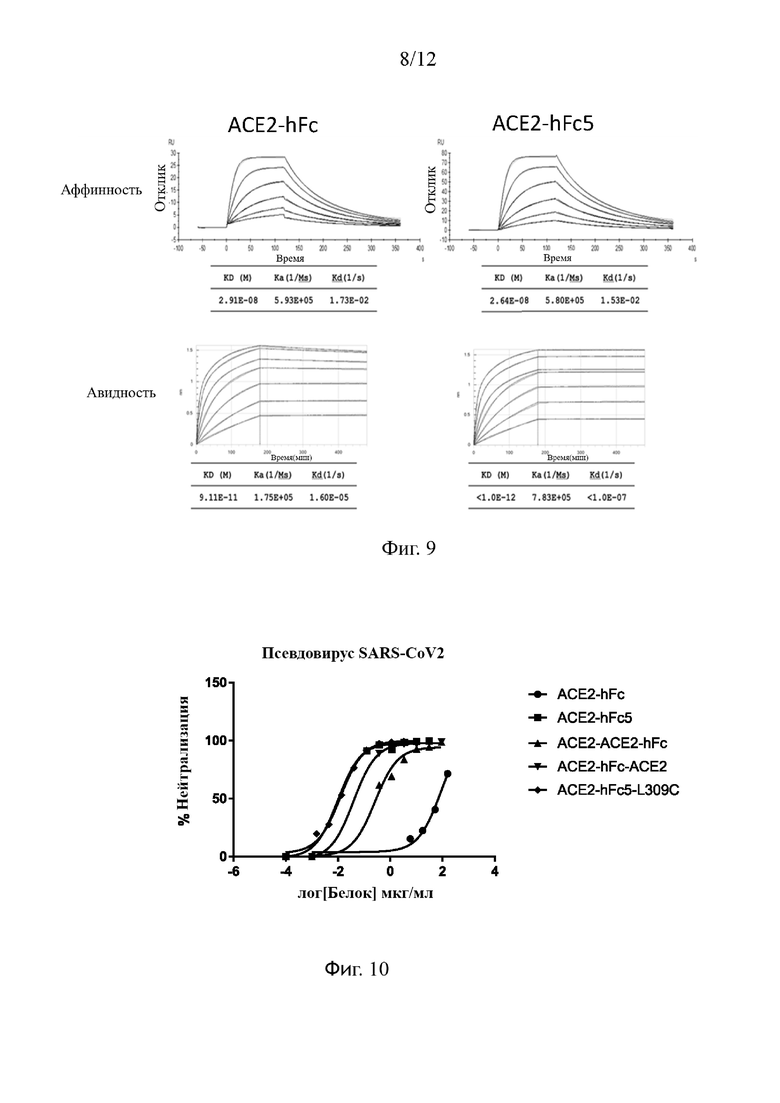
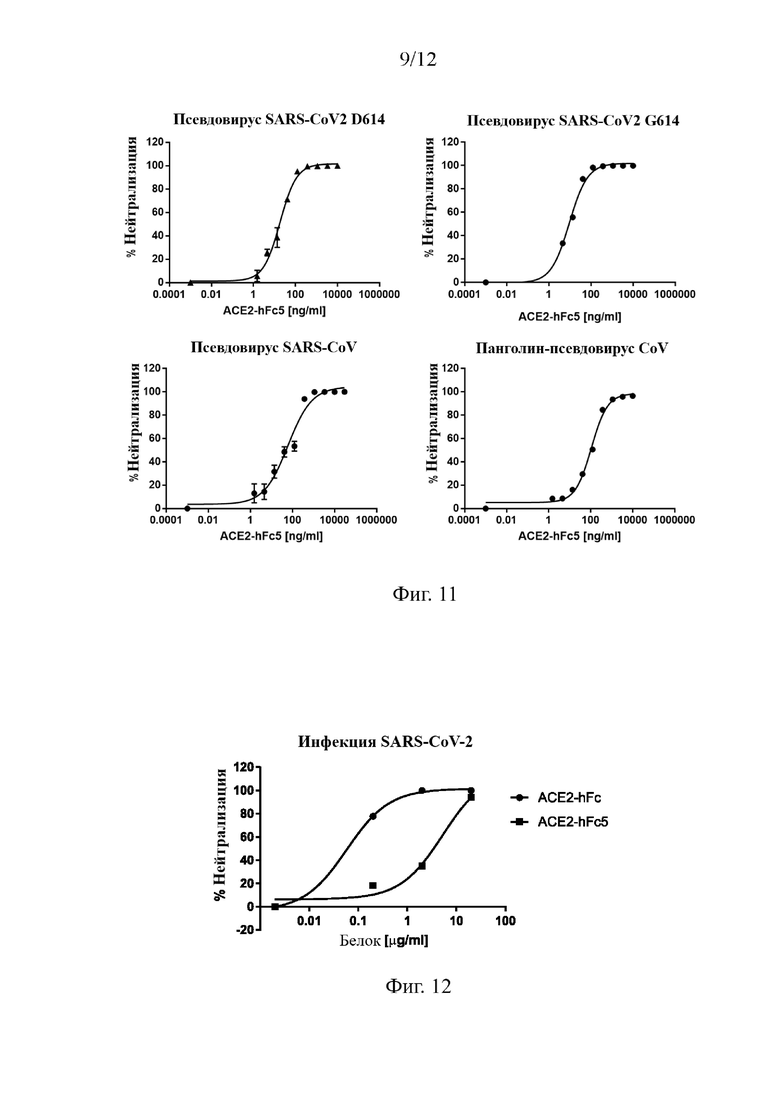
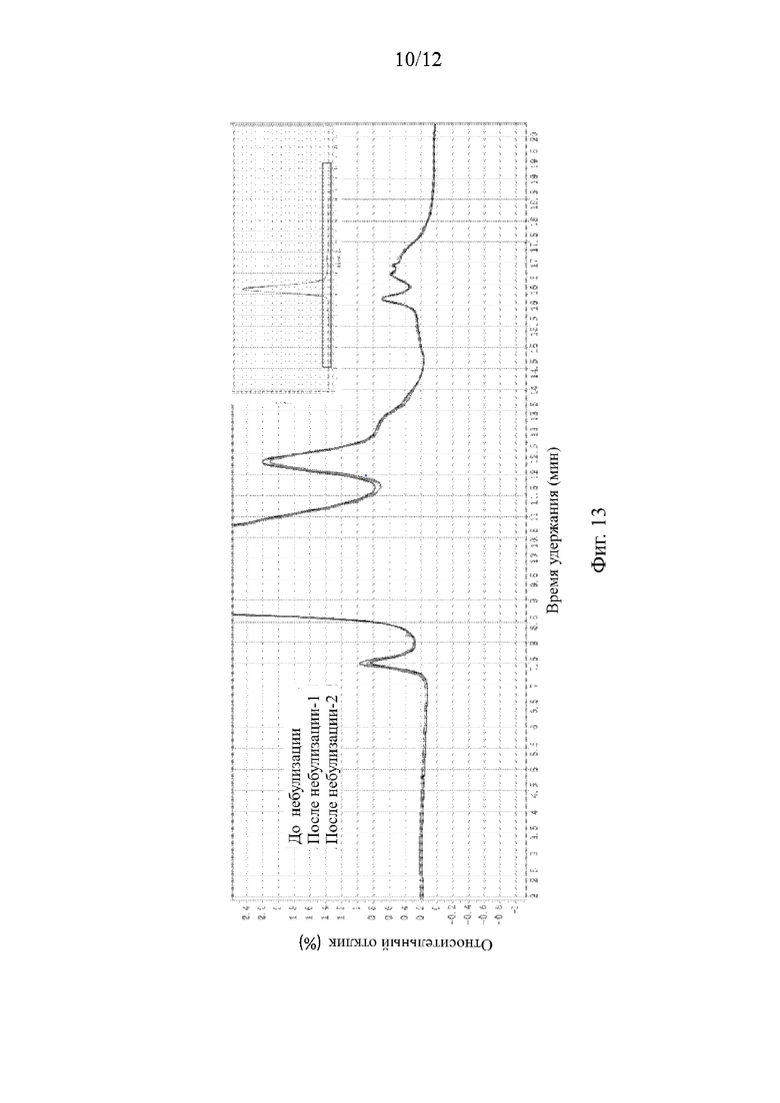
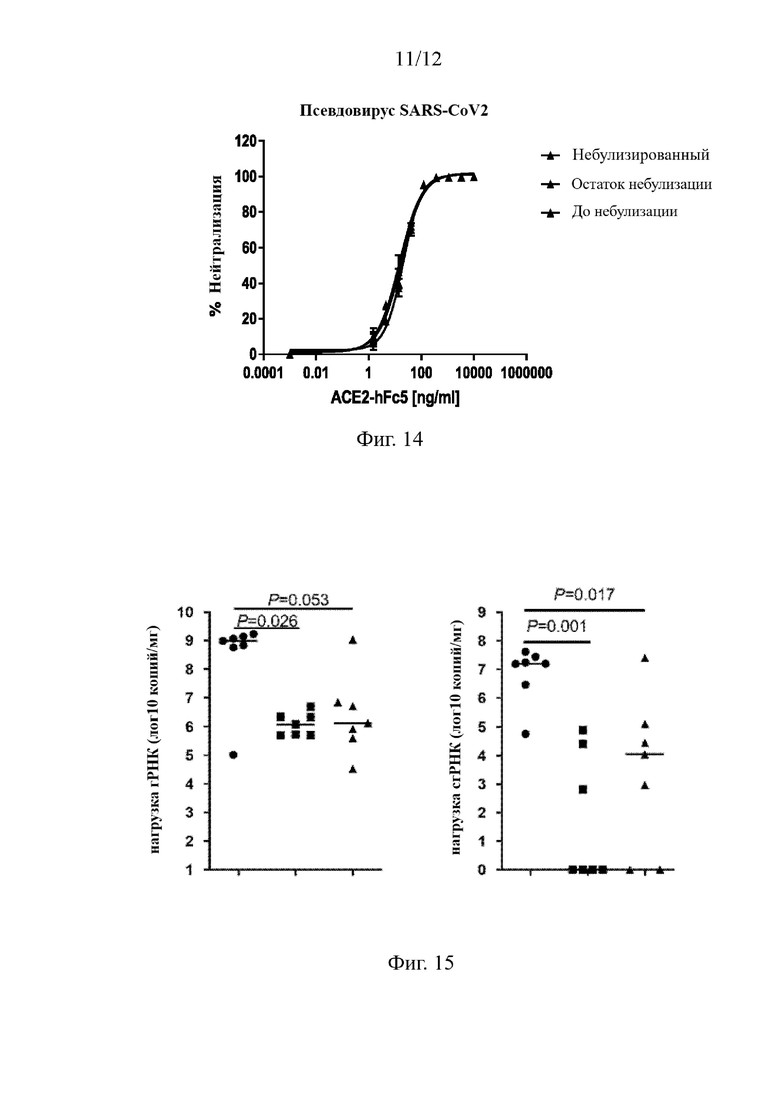
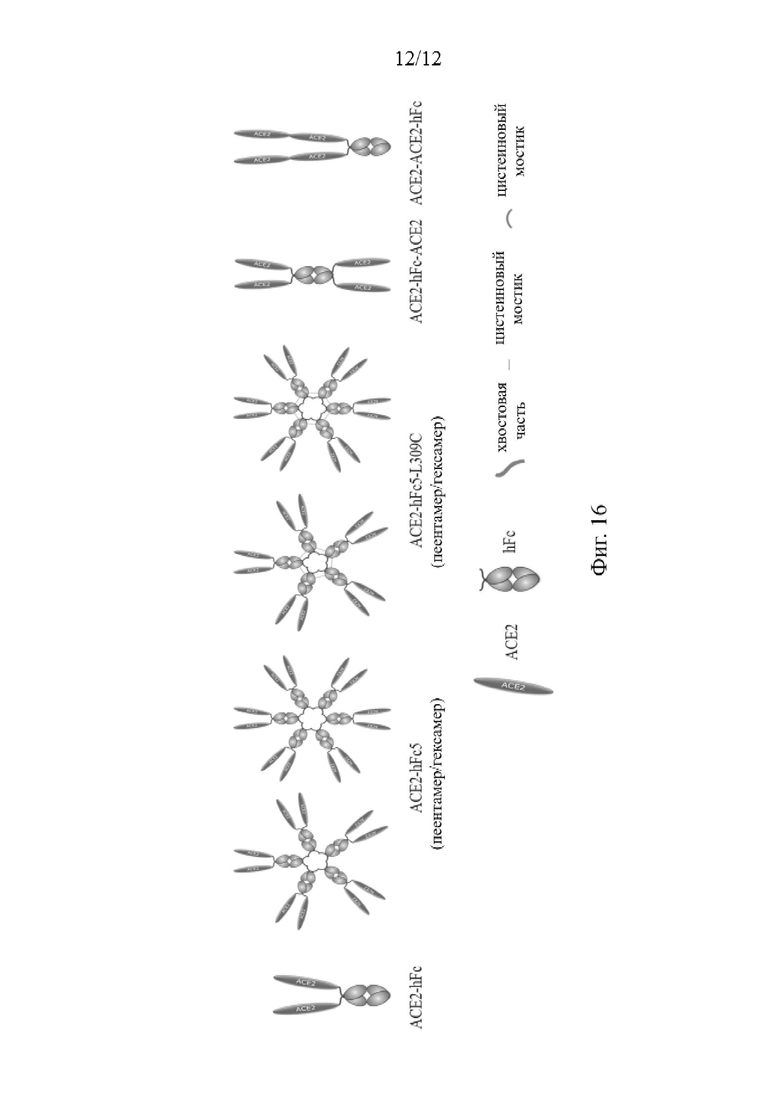
Изобретение относится к области биотехнологии, а именно к cлитому белку растворимого ACE2, который связывается с коронавирусом, включающему процессированную форму растворимого ACE2, Fc-домен антитела и хвост, причем процессированная форма растворимого ACE2 связана с N-концевой областью Fc-домена, а хвост расположен на C-концевой области Fc-домена, процессированная форма растворимого ACE2 содержит или состоит из внеклеточного домена ACE2 или фрагмента внеклеточного домена ACE2, сохраняющего способность связываться с коронавирусом, а также к мультимеру Fc-слитого белка, ACE2-hFc(n), который связывается с коронавирусом, включающему n полипептидных мономерных единиц, и который отличается тем, что каждая из полипептидных мономерных единиц представляет собой димер, в котором две процессированные формы растворимого ACE2 связаны с N-концевыми областями двух Fc-доменов тяжелых цепей, соответственно, и n полипептидных мономерных единиц собраны в мультимер посредством хвоста, расположенного на каждом из C-концов двух Fc-доменов тяжелых цепей, где n выбран из 4, 5 или 6. Также раскрыты вектор и клетка, содержащие ген, кодирующий вышеуказанный слитый белок, и клетка и вектор, содержащие ген, кодирующий вышеуказанный мультимер. Изобретение эффективно при приготовлении лекарственного средства для лечения или профилактики заболевания, связанного с ACE2. 7 н. и 10 з.п. ф-лы, 16 ил., 3 табл., 12 пр.
1. Слитый белок растворимого ACE2, который связывается с коронавирусом, включающий процессированную форму растворимого ACE2, Fc-домен антитела и хвост,
причем процессированная форма растворимого ACE2 связана с N-концевой областью Fc-домена, а хвост расположен на C-концевой области Fc-домена,
процессированная форма растворимого ACE2 содержит или состоит из внеклеточного домена ACE2 или фрагмента внеклеточного домена ACE2, сохраняющего способность связываться с коронавирусом, и
процессированная форма растворимого ACE2 включает аминокислотную последовательность, представленную SEQ ID NO: 1 или SEQ ID NO: 2, и
хвост представляет собой хвост, полученный из IgM и/или IgA и включающий последовательность, представленную SEQ ID NO: 17.
2. Слитый белок по п. 1, отличающийся тем, что антитело представляет собой антитело IgG, предпочтительно человеческое антитело IgG, такое как IgG1, IgG2, IgG3 или IgG4, предпочтительно IgG1,
предпочтительно, Fc-домен антитела включает в себя два Fc-домена тяжелых цепей антитела,
предпочтительно, каждый из Fc-доменов тяжелых цепей имеет шарнирную область на своем N-конце,
предпочтительно, каждый из Fc-доменов тяжелых цепей включает CH3-домен, полученный из IgG1, IgG2, IgG3 или IgG4,
предпочтительно, каждый из Fc-доменов тяжелых цепей включает CH2- и CH3-домены, полученные из IgG1, IgG2, IgG3 или IgG4, и
предпочтительно, каждый из Fc-доменов тяжелых цепей включает шарнирную область, CH2-домен и CH3-домен.
3. Слитый белок по п. 1, отличающийся тем, что 2n процессированных форм растворимого ACE2 связаны с N-концевой областью Fc-домена, где n выбран из 1, 2 или 3,
предпочтительно, две процессированные формы растворимого ACE2 связаны с N-концевыми областями двух Fc-доменов тяжелых цепей с образованием димера;
предпочтительно, процессированные формы растворимого ACE2 связаны в тандем линкером, предпочтительно представляющим собой цистеиновый ААА линкер.
4. Слитый белок по п. 1, отличающийся тем, что процессированная форма растворимого ACE2 гликозилирована, предпочтительно в положении(ях) 53, 90, 103, 322, 432, 546 и/или 690 человеческого ACE2.
5. Мультимер Fc-слитого белка, ACE2-hFc(n), который связывается с коронавирусом, включающий n полипептидных мономерных единиц, отличающийся тем, что каждая из полипептидных мономерных единиц представляет собой димер, в котором две процессированные формы растворимого ACE2 связаны с N-концевыми областями двух Fc-доменов тяжелых цепей, соответственно, и n полипептидных мономерных единиц собраны в мультимер посредством хвоста, расположенного на каждом из C-концов двух Fc-доменов тяжелых цепей,
причем процессированная форма растворимого ACE2 включает аминокислотную последовательность, представленную SEQ ID NO: 1 или SEQ ID NO: 2,
хвост представляет собой хвост, полученный из IgM и/или IgA и включающий последовательность, представленную SEQ ID NO: 17, и
n выбран из 4, 5 или 6.
6. Мультимер слитого белка по п. 5, отличающийся тем, что каждая из С-концевых областей двух Fc-доменов тяжелых цепей связана с хвостом в каждой полипептидной мономерной единице и n полипептидных мономерных единиц имеют в совокупности 2n хвостов, соединенных друг с другом и образующих мультимер.
7. Мультимер слитого белка по п. 5, отличающийся тем, что мультимер слитого белка включает один или несколько мультимеров, выбранных из нижеприведенной группы:
ACE2-hFc4, представляющий собой тетрамер, собранный из 4 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc4, представляющий собой тетрамер, собранный из 4 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc4-L309C, представляющий собой тетрамер, собранный из 4 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18, и Fc-домен тяжелой цепи включает мутацию L309C в положении 309;
ACE2-hFc5, представляющий собой пентамер, собранный из 5 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc5, представляющий собой пентамер, собранный из 5 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc5-L309C, представляющий собой пентамер, собранный из 5 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18, и при этом Fc-домен тяжелой цепи включает мутацию L309C в положении 309;
ACE2-hFc6, представляющий собой гексамер, собранный из 6 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 7;
ACE2-NN-hFc6, представляющий собой гексамер, собранный из 6 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелой цепи, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 8;
ACE2-NN-hFc6-L309C, представляющий собой гексамер, собранный из 6 полипептидных мономерных единиц посредством хвостов, расположенных на С-концах Fc-доменов, при этом каждая из полипептидных мономерных единиц включает димер, состоящий из двух процессированных форм ACE2 и двух Fc-доменов тяжелых цепей, при этом одна процессированная форма ACE2, один Fc-домен тяжелой цепи вместе с хвостом включают аминокислотную последовательность, представленную SEQ ID NO: 18.
8. Экспрессирующий вектор, включающий ген, кодирующий слитый белок по любому из пп. 1-4.
9. Экспрессирующий вектор, включающий ген, кодирующий мультимер слитого белка по любому из пп. 5-7.
10. Клетка млекопитающих для экспрессии слитого белка по любому из пп. 1-4, включающая ген, кодирующий слитый белок по любому из пп. 1-4.
11. Клетка млекопитающих по п. 10, отличающаяся тем, что клетка включает, в частности, клетку CHO, клетку 293, клетку Vero и клетку, полученную из них; предпочтительно включает клетку Vero E6 или клетку HEK293T.
12. Клетка млекопитающих для экспрессии мультимера слитого белка по любому из пп. 5-7, включающая ген, кодирующий мультимер слитого белка по любому из пп. 5-7.
13. Клетка млекопитающих по п. 12, отличающаяся тем, что клетка включает, в частности, клетку CHO, клетку 293, клетку Vero и клетку, полученную из них; предпочтительно включает клетку Vero E6 или клетку HEK293T.
14. Применение слитого белка по любому из пп. 1-4 или мультимера слитого белка по любому из пп. 5-7 при приготовлении лекарственного средства для лечения или профилактики заболевания, связанного с ACE2,
причем указанное заболевание выбрано из группы, состоящей из инфекции, вызванной вирусом, использующим ACE2 в качестве рецептора, пневмонии, тяжелой острой респираторной инфекции, почечной недостаточности, сердечной недостаточности, респираторного дистресс-синдрома взрослых (ARDS), поражения печени, желудочно-кишечного заболевания или тяжелого острого респираторного синдрома, и
при этом вирус представляет собой коронавирус.
15. Применение по п. 14, отличающееся тем, что коронавирус выбран из группы, включающей SARS-CoV, HCoV-NL63 или SARS-CoV2.
16. Применение по п. 14, отличающееся тем, что лекарственное средство используется для пассивной иммунизации медицинских работников и лиц, подверженных риску инфицирования вирусом, в частности, коронавирусом, использующим ACE2 в качестве рецептора.
17. Применение по любому из пп. 14-16, отличающееся тем, что лекарственное средство вводят путем ингаляции, интраназально или путем инстилляции в дыхательные пути, глазной инъекции и инъекции в среднее ухо, закапывания в уши, топической, трансдермальной, парентеральной, подкожной и внутривенной инъекции, внутрикожной инъекции, внутримышечной инъекции, внутриплевральной инстилляции, внутрибрюшинной инъекции, путем интрализионального введения, нанесения на слизистую оболочку или путем трансплантации носителя с пролонгированным высвобождением; предпочтительно лекарственное средство вводят путем ингаляции с помощью небулайзера.
| ROBERT L | |||
| KRUSE, Therapeutic strategies in an outbreak scenario to treat the novel coronavirus originating in Wuhan, China, F1000Research, 07 Feb 2020, 9:72 | |||
| HANNAH KLEINE-WEBER et al., Functional analysis of potential cleavage sites in the MERScoronavirus spike protein, SCientifiC REPOrtS, 2018, 8:16597, DOI:10.1038/s41598-018-34859-w | |||
| CN |

