Группа изобретений относится к области биотехнологии, иммунологии и вирусологии, а именно к однодоменному антителу ламы Н5, и к однодоменному антителу ламы Н5, слитому с Fc-фрагментом IgG1 человека (H5-Fc), специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, нейтрализующие вирус SARS-CoV-2, к фрагментам ДНК, кодирующим указанные антитела.
В декабре 2019 года в г. Ухань (Китай) наблюдался резкий рост пациентов с пневмонией неизвестного происхождения с тяжелым протеканием заболевания. Вскоре был установлен возбудитель - РНК вирус, относящийся к роду Бетакоронавирусов, а заболевание, вызываемое им, было названо COVID-19. Оказалось, что новый коронавирус, названный SARS-CoV-2, является родственным двум другим вирусам, SARS-CoV и MERS-CoV, вызвавших вспышки пневмонии в 2003 году в Азии и 2012 году на Ближнем Востоке, соответственно (Tomlinson В., Cockram С.Lancet. 2003;361(9368):1486-7. doi: 10.1016/S0140-6736(03) 13218-7; Geng Н., Tan W. Sci China Life Sci. 2013;56(8):683-7. doi: 10.1007/s 11427-013-4519-8). SARS-CoV распространился no 20 странам, всего было 8098 случаев, умерло 774 человека, таким образом летальность составила 9,6%. В 2004 году вспышка SARS-CoV закончилась, новых случаев больше зарегистрировано не было. MERS-CoV распространился по 23 странам, зафиксировано 2506 случаев, 912 смертей, с уровнем летальности в 36%. Все три вируса, SARS-CoV, MERS-CoV и SARS-CoV-2, передаются воздушно-капельным путем и имеют зоонозное происхождение, однако только последний смог распространиться по всему миру и вызвать пандемию, которая была объявлена Всемирной организацией здравоохранения (ВОЗ) 11 марта 2020 года. На 11 октября 2022 года, по данным ВОЗ, всего в мире зафиксировано 618 521 620 случаев заражения SARS-CoV-2, из них погибло 6 534 725 человек, тем самым уровень летальности составил 1,06%. В Российской Федерации, по данным ВОЗ, зафиксировано 21 218 993 случаев заражения, из них 388 295 смертей, летальность составила 1,83% (https://covidl9.who.int/table). Таким образом, хотя смертность от SARS-CoV-2 в 10-30 раз ниже, чем от SARS-CoV и MERS-CoV, ввиду его широкого распространения, принявшего характер пандемии, общее число смертей от SARS-CoV-2 в тысячи раз выше, что не может не вызывать глубокую озабоченность.
Как известно, на сегодняшний момент, вирус SARS-CoV-2 передается воздушно-капельным, воздушно-пылевым и контактным путями, может вызывать как легкую форму ОРВИ, так и тяжелую пневмонию, приводящую к острой дыхательной недостаточности, а также серьезное поражение других органов и тканей, в том числе и нервной.
Молекулярный механизм заражения SARS-CoV-2 заключается главным образом в распознавании тримером шиповидного Spike (S) белка на поверхности вириона рецептора АСЕ2 на поверхности клетки человека. Распознавание АСЕ2 происходит особым доменом на поверхности S-белка, а именно рецептор-связывающим доменом (RBD). При контакте S-белка и АСЕ2-рецептора запускается каскад изменений, приводящих, в конечном счете, к слиянию мембран вириона и клетки и к проникновению в нее генетического материала вируса (Bian J., Li Z. Acta Pharm Sin B. 2021;11(1):1-12. doi: 10.1016/j.apsb.2020.10.006; Chambers J. P., Yu J., Valdes J. J. et al. J Pathog. 2020;2020:9238696. doi: 10.1155/2020/9238696).
На первых этапах пандемии для ее сдерживания, ввиду отсутствия иных средств, применялись только ограничительные методы, включающие самоизоляцию, ношение масок и перчаток в общественных местах, запрет на проведение публичных мероприятий и т.п.Далее, в кратчайшие сроки, были разработаны вакцины против коронавируса, причем одной из первых была разработана вакцина Sputnik V в НИИЦЭМ им. Н. Ф. Гамалеи (Logunov D. Y., Dolzhikova I. V., Shcheblyakov D. V. Lancet. 2021;397(10275):671-681. doi: 10.1016/S0140-6736(21)00234-8). На сегодняшний момент в мире насчитывается более 15 одобренных вакцин против SARS-CoV-2 и более 63% людей на Земле прошли полную вакцинацию по данным ВОЗ (https://covidl 9.who.int/table).
Несмотря на высокий процент вакцинированных, до сих пор остается необходимость в дополнительных средствах профилактики и терапии COVID-19. Это обусловлено, во-первых, появлением новых штаммов SARS-CoV-2, против которых вакцины на основе Уханьского штамма Wu-1 имеют значительно меньшую эффективность. Во-вторых, существуют группы людей, для которых вакцинация противопоказана по состоянию здоровья. В-третьих, существуют группы риска, которые ввиду наличия хронических заболеваний или скомпрометированного иммунитета, даже не смотря на вакцинацию, подвержены тяжелому течению заболевания и высокой смертности от него. Для снижения смертности среди упомянутых категорий людей, а также для терапии заболевания при его тяжелом протекании могут быть использованы моноклональные антитела, нейтрализующие SARS-CoV-2.
Моноклональные терапевтические антитела хорошо зарекомендовали себя в мировой практике для лечения онкологических, аутоиммунных и инфекционных заболеваний у человека. В настоящее время одобрено или находятся на одобрении в США и Европе 173 моноклональных антитела (www.antibodysociety.org/antibody-therapeutics-product-data).
За неполные три года пандемии учеными со всего мира были получены сотни распознающих SARS-CoV-2 моноклональных антител человека, многие из них являются вирус-нейтрализующими (согласно базе данных CoV-AbDab (http://opig.stats.ox.ac.uk/webapps/covabdab/) и были запущены в доклинические и клинические испытания. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (US FDA) одобрило для экстренного использования моноклональное антитело человека Бебтеловимаб и коктейль (Тиксагевимаб+Цилгавимаб). В то же время, в связи с появлением новых штаммов SARS-CoV-2, было приостановлено применение ранее одобренных Сотровимаба, а также коктейлей (Касиривимаб+Имдевимаб) и (Бамланивимаб+Этесивимаб) (https://www.fda.gov/dmgs/emergency-preparedness-dmgs/coronavirus-covid-19-drugs).
Введение Бебтеловимаба или Сотровимаба в моно-формате, как было показано в результате проведения клинических испытаний, резко снижает риск тяжелого протекания болезни у пациентов из группы риска (Razonable R.R., Tulledge-Scheitel S.M., Hanson S.N. Open Forum Infect Dis. 2022;9(10):ofac411. doi: 10.1093/ofid/ofac411). При профилактическом введении коктейля (Тиксагевимаб+Цилгавимаб) иммунноскомпрометированным пациентам наблюдался низкий уровень заражения и тяжелых последствий от COVID-19 (Nguyen Y., Flahault A., Chavarot N. et al. Clin Microbiol Infect. 2022;S1198-743X(22)00383-4. doi: 10.1016/j.cmi.2022.07.015). Также было показано, что введение коктейля (Касиривимаб+Имдевимаб) снижает вирусную нагрузку, а также значительно снижает частоту обращения к врачу, особенно у серанегативных пациентов (Weinreich D.M., Sivapalasingam S., Norton Т., et al. N Engl J Med. 2021;384(3):238-251. doi: 10.1056/NEJMoa2035002). Наконец, введение коктейля (Бамланивимаб+Этесивимаб) приводило к значительному снижению вирусной нагрузки у пациентов с легким и среднетяжелым протеканием болезни (Gottlieb R.L., Nirula А., Chen P. et al. JAMA. 2021;325(7):632-644. doi: 10.100l/jama.2021.0202). Таким образом, эффективность действия моноклональных антител, нейтрализующих SARS-CoV-2, была показана во многих экспериментах и клинических исследованиях.
Тем не менее, у классических моноклональных антител имеется ряд следующих недостатков:
1) трудность и дороговизна их получения, так как из-за наличия у таких антител пары цепей (тяжелой и легкой), необходимо клонировать их из единичной В-клетки (низкопроизводительный метод) или использовать высокопроизводительные, но дорогостоящие методы микрофлюидики или анализа транскриптомов единичных клеток (Ehlers А.М,. den Hartog Jager С.F., Kardol-Hoefnagel Т. et al. Front Immunol. 2021;12:660037. doi: 10.3389/fimmu.2021.660037; Busse С.E., Czogiel I., Braun P. Eur J Immunol. 2014; 44(2):597-603. doi: 10.1002/eji.201343917);
2) сложности в обнаружении некоторых труднодоступных или скрытых эпитопов;
3) дороговизна лечения из-за необходимости введения нескольких граммов таких антител внутривенно, а также из-за сложностей в их биотехнологическом производстве (большой размер белка, его сложная четвертичная структура).
Для преодоления перечисленных недостатков наиболее перспективным является применение однодоменных антител верблюдовых. Однодоменные антитела получают из особых антител, состоящих из тяжелых цепей (hcAb), которые в природе встречаются у представителей семейства верблюдовых (например, двугорбый верблюд, лама, альпака). hcAb состоят всего из двух одинаковых цепей, каждая из которых состоит из двух константных (СН) и одного вариабельного домена (VHH). VHH является независимой антиген-распознающей единицей, формирующей функциональный паратоп, и этот домен можно использовать в качестве самостоятельной молекулы, распознающей антиген. Такие молекулы и называют однодоменными антителами, наноантителами или нанотелами.
Однодоменные антитела обладают рядом преимуществ, по сравнению с классическими антителами: 1) низкая или отсутствующая иммуногенность, ввиду небольшого размера однодоменных антител (15 кДа); 2) высокая термостабильность; 3) особая структура паратопа VHH, позволяющая ему распозновать эпитопы, недоступные для классических антител, например вогнутые эпитопы, или эпитопы, доступ к которым стерически ограничен; 4) дешевизна в наработке, в том числе возможность использовать прокариотические экспрессирующие системы; 5) малый размер антител, что позволяет доставлять однодоменные антитела напрямую в дыхательные пути в виде аэрозоля; 6) возможность использования в виде гомо- или гетеродимеров и тримеров, то есть молекул, обладающих или повышенной авидностью, или мультипаратопностью, устойчивых к мутационному ускользанию вируса (Walter J. D., Scherer M., Hutter С.A. J. et al. EMBO Rep.2022;23(4):e54199. doi: 10.15252/embr.202154199).
У однодоменных антител имеются также и недостатки: короткое время выведения из организма из-за сверхмалого размера и отсутствие у молекулы эффекторной части, запускающей иммунный ответ в организме. Эти проблемы легко решаются путем создания молекул, по структуре напоминающих hcAb, в которых Fc-фрагмент верблюдовых заменен на Fc-фрагмент IgG1 человека (Esmagambetov I. В., Shcheblyakov D. V., Egorova D. A. Et al. Acta Naturae. 2021;13(4):53-63. doi: 10.32607/actanaturae. 11487). Такие химерные антитела действуют аналогично классическим антителам человека, но являются более стабильными и дешевыми в производстве в эукариотической системе, так как имеют меньший размер и более простую структуру. Также Fc-химерные антитела можно производить и в прокариотической системе, что может еще в большей степени удешевить их производство (Ye G., Gallant J., Zheng J. et al. Elife. 2021;10:e64815. doi: 10.7554/eLife.64815).
В настоящее время препаратов на основе однодоменных антител и их производных для профилатики и лечения COVID-19 не имеется.
Известно мышиное моноклональное антитело, специфически связывающее RBD фрагмент в составе S белка вируса SARS-CoV-2, обладающее способностью нейтрализовать живой SARS-CoV-2 in vitro (патент RU 2744274 С1, опубл. 04.03.2021).
Недостатком данного антитела является его высокая иммуногенность по отношению к человеку, что делает критически опасным его внутривенное введение. Кроме того, отсутствуют данные по эффективности этого антитела in vivo на животной модели.
Известно однодоменное антитело (VHH), специфически связывающееся с RBD-доменом S-белка SARS-CoV-2 и нейтрализующее лентивирус, псевдотипированный S-белком SARS-CoV-2 (заявка WO 2022020668A1, опубл. 14.04.2016).
Известно семейство (большая библиотека) однодоменных антител к S-белку SARS-CoV-2, в том числе к его RBD части, в том числе нейтрализующих in vitro живой вирус SARS-CoV-2, а также лентивирус, псевдотипированный S-белком SARS-CoV-2. В этом же решении, наравне с некоторыми однодоменными антителами, были испытаны также и их димеры и тримеры (заявка WO 2022040603A2, опубл. 24.02.2022).
Недостатками упомянутых выше однодоменных антител является их низкий молекулярный вес, что способствует их быстрой элиминации из кровотока и требует их многократного введения. Также отсутствуют данные по эффективности этих антител in vivo на животной модели.
Наиболее близкими к заявляемым однодоменным антителам - прототипом, являются однодоменные антитела, связывающие RBD-домен S-белка SARS-CoV-2, а также их производные в виде простых димеров, а также мономеров и димеров, слитых с Fc-фрагментом IgG1 человека, самодимеризующихся при наработке в эукариотической системе. В данном решении показаны нейтрализующие свойства нескольких однодоменных антител, а также способность одного из этих антител (р2 с5) в виде химерного белка, слитого с Fc-фрагментом IgG1 человека (p2c5-fc), нейтрализовать SARS-CoV-2 in vivo в модели АСЕ2-трансгенных мышей и в модели сирийского хомячка (патент RU 2763001С1, опубл. 24.12.2021).
Недостатками прототипа являются:
1) Низкая вирус-нейтрализующая активность известных однодоменных антител. Лучшее однодоменное антитело р2 с5, выбранное авторами для дальнейшей модификации и использования в in vivo экспериментах, обладает очень низкой вирус-нейтрализующей активностью (664 нг/мл), что препятствует его использованию в профилактике и терапии COVID-19 у человека, так как потребует высоких доз введения препарата, а, следовательно, больших затрат на его производство, а также приведет к высокой нагрузке на организм пациента;
2) Константа диссоциации для взаимодействия р2 с5 с RBD-доменом S-белка SARS-CoV-2 является средней в сравнении с константами для классических антител (Ко=2,8 пМ), но слабой для однодоменных антител, для которых не представляет сложности получить Ко в пикомолярном диапазоне.
3) Отсутствует доказательство связывания химерной p2c5-fc молекулы с RBD доменом S-белка SARS-CoV-2 и существования ее вирус-нейтрализующей активности за счет р2 с5 части, а не fc-фрагмента. Эти факты ставят под сомнение наличие специфической терапевтической и профилактической активности химерной p2c5-fc на моделях in vivo;
4) недостаточные функциональные возможности известных однодоменных антител, поскольку вирус-нейтрализующая активность р2 с5 показана только на примере Уханьского штамма SARS-CoV-2, но не для других его вариантов. Появление новых штаммов SARS-CoV-2 требует тестирования кандидатных терапевтических антител против разных вариантов SARS-CoV-2 для поиска антител с широким спектром действия.
Таким образом, существует необходимость в создании антител, специфически связывающихся с RBD-доменом S-белка SARS-CoV-2, а также нейтрализующих различные варианты SARS-CoV-2, которые могут быть использованы для диагностики, профилактики и терапии заболевания человека, вызываемого вирусом SARS-CoV-2, лишенных перечисленных выше недостатков.
Задачей группы изобретений является создание однодоменного антитела и его производного, которые могут специфически и эффективно связывать RBD-домен S-белка SARS-CoV-2 различных вариантов вируса, а также получение ДНК фрагментов, кодирующих однодоменное антитело и его производное.
Техническим результатом группы изобретений является создание нового однодоменного антитела ламы Н5, а также его производного - химерного антитела Н5-Fc, представляющего однодоменное антитело Н5, слитое с Fc-фрагментом IgG1 человека, способных к связыванию с RBD-доменом S-белка SARS-CoV-2, отличающиеся по аминокислотной последовательности вариабельных VHH доменов от известных RBD-специфических однодоменных антител.
Также техническим результатом является повышение аффинности созданных антител, что позволяет снизить их дозировку при применении, что в значительной степени снижает их стоимость при производстве, а значит - повышает их привлекательность для коммерциализации.
Также техническим результатом является способность созданных антител высокоэффективно нейтрализовать вирус SARS-CoV-2 как в in vitro схемах, так и in vivo в профилактических и терапевтических схемах, что позволяет использовать их для профилактики и терапии заболевания, вызываемого вирусом SARS-CoV-2.
Также техническим результатом является способность созданных антител нейтрализовать вирус SARS-CoV-2 разных вариантов, что позволяет использовать их для профилактики и терапии заболеваний, вызываемых вирусом SARS-CoV-2 разных штаммов.
Поставленная задача решается созданием однодоменного антитела ламы Н5, специфически связывающего RBD-домен S-белка SARS-CoV-2, обладающего вируснейтрализующей активностью, имеющего аминокислотную последовательность SEQ ID NO: 1, а также созданием кодирующего его фрагмента ДНК, имеющего нуклеотидную последовательность SEQ ID NO: 3.
Поставленная задача решается также созданием химерного антитела H5-Fc, представляющего собой однодоменное антитело, слитое с Fc-фрагментом IgG1 человека, специфически связывающего RBD-домен S-белка SARS-CoV-2, обладающего вируснейтрализующей активностью, имеющего аминокислотную последовательность SEQ ID NO: 2, а также созданием кодирующего его фрагмента ДНК, имеющего нуклеотидную последовательность SEQ ID NO: 4.
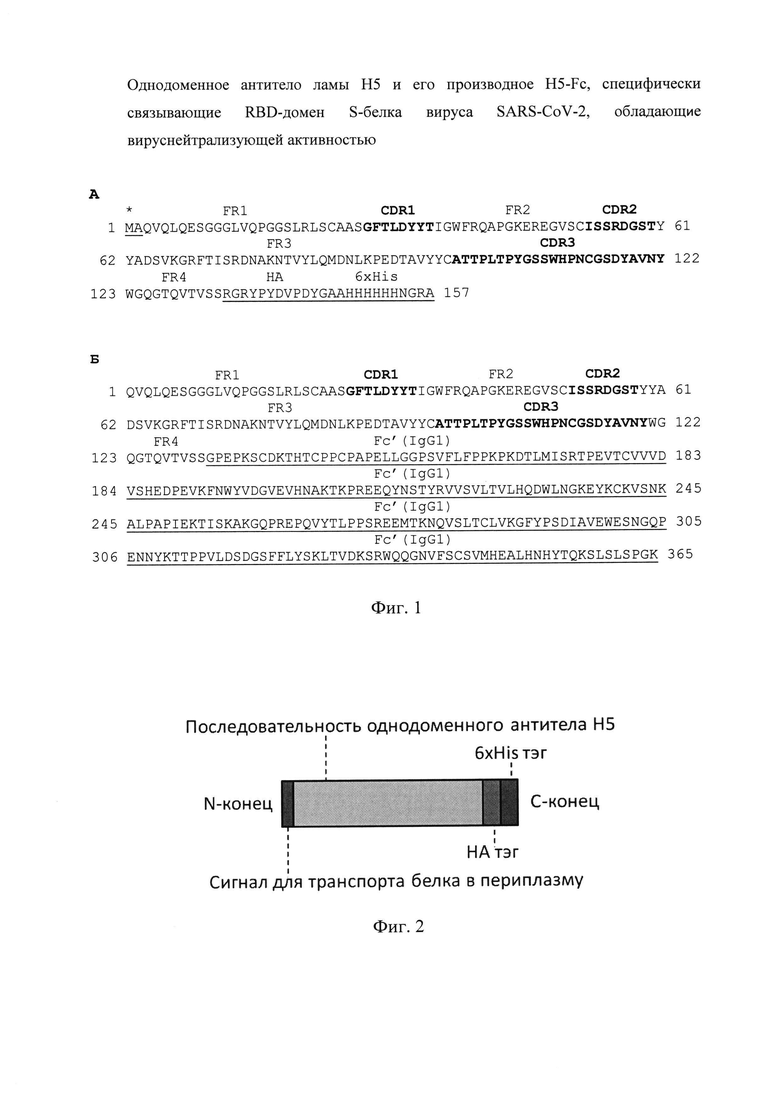
Аминокислотная последовательность однодоменного антитела ламы Н5 (SEQ ID NO: 1), а также нуклеотидная последовательность кодирующего его фрагмента ДНК (SEQ ID NO: 3) и аминокислотная последовательность химерного антитела H5-Fc (SEQ ID NO: 2), а также нуклеотидная последовательность кодирующего его фрагмента ДНК (SEQ ID NO: 4) представлены в перечне последовательностей.
Специалистам в данной области техники известно, что области любого однодоменного антитела могут подразделяться на гипервариабельные районы, которые называются определяющими комплементарность областями (CDR), перемежаемые областями с более высоким уровнем консервативности, называемыми рамочными областями (FR). Каждое однодоменное антитело образовано тремя CDR и четырьмя FR, расположенными от амино-терминального конца к карбокси-терминальному концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Участки CDR и FR антител могут быть определены по номенклатуре Международной информационной системы по иммуногенетике (International Immunogenetics Information System, www.imgt.org). У антител, созданных в рамках заявляемого технического решения, CDR участки являются идентичными, то есть CDR1 участок однодоменного антитела ламы Н5 совпадает с CDR1 участком химерного антитела H5-Fc, то же самое верно и для CDR2, и для CDR3 участков. CDR участки H5/H5-Fc имеют следующие аминокислотные последовательности: CDR1 - SEQ NO: 5, CDR2 - SEQ NO: 6 и CDR3 -SEQ NO: 7. Аминокислотные последовательности CDR участков SEQ NO: 5-7 представлены в перечне последовательностей.
Специалистам в данной области техники известно, что делеция, добавление, замена одной или даже нескольких аминокислот могут приводить к получению аллельной формы исходного белка, которая будет сохранять свою активность. Также известен способ «перевивки» CDR-петель одного антитела в структуру рамочных последовательностей другого антитела с сохранением исходных свойств, что в частности позволяет производить гуманизацию антител, полученных в других млекопитающих. Таким образом, антитела предлагаемого решения включают те варианты, аминокислотная последовательность которых вне CDR петель отличается от приведенных в настоящей заявке последовательностей, но которые сохранили ту же антигенсвязывающую активность.
Как известно специалисту в данной области технологии, консервативные Fc-фрагменты классических антител необходимы для их функционирования, в частности они обеспечивают связь между гуморальным и клеточным ответом и могут усилить действие терапевтических моноклональных антител (Gogesch P., Dudek S., van Zandbergen G. et al. Int J Mol Sci. 2021;22(16):8947. doi: 10.3390/ijms22168947). Fc-фрагмент IgG1 человека в составе антитела, имеющего последовательность SEQ ID NO: 2, согласно заявляемому решению может быть заменен на Fc-фрагмент другого изотипа (IgG2, IgG3, IgG4) или класса (IgM, IgD, IgE, IgA) человека.
Специалистам в данной области техники также известно, что ввиду вырожденности генетического кода могут быть различия в последовательности ДНК. Поэтому последовательности нуклеиновых кислот, согласно заявляемому решению могут отличаться, при условии, что они кодируют участки цепей антитела, содержащего последовательность нуклеотидов, не менее чем на 90% гомологичную SEQ ID NO: 2 и SEQ ID NO: 4, и последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 1-2 и SEQ ID NO: 5-7.
Антитела заявляемого решения могут быть человеческим антителом, гуманизованным антителом, мышиным антителом, химерным антителом и другими генно-инженерными антителами.
Краткое описание чертежей, иллюстрирующих изобретение
На фиг. 1 представлены аминокислотные последовательности однодоменного антитела ламы Н5 (А) и химерного антитела H5-Fc, представляющего собой однодоменное антитело ламы Н5, слитое с Fc-фрагментом IgG1 человека (Б). CDR районы отмечены жирным шрифтом. Введенные генно-инженерным способом последовательности отмечены подчеркиванием (сигнал транспорта в периплазму (*), НА и 6×His тэги, Fc'-фрагмент IgG1 человека).
На фиг. 2 представлена схематическая структура однодоменного антитела ламы Н5, полученного в прокариотической экспрессионной системе (Е. coli). Сигнал для секреции в периплазму бактерии необходим для правильного фолдинга белка, 6xHis тэг - для металл-хелатной очистки белка.
На фиг. 3 представлена схематическая структура химерного антитела H5-Fc, представляющего собой однодоменное антитело ламы Н5, слитое с Fc-фрагментом IgG1 человека, полученное в эукариотической экспрессионной системе (НЕК 293Т фибробласты). Данный белок представляет собой самособирающийся димер из двух одинаковых цепей, связанных двумя дисульфидными мостиками.
На фиг. 4 представлен анализ чистоты однодоменного антитела ламы Н5 (дорожка 1) и химерного антитела H5-Fc (дорожка 2) с помощью электрофореза по Лэммли в разделяющем 15% полиакриламидном геле с додецил сульфатом натрия и меркаптоэтанолом. На дорожке М нанесен маркер молекулярного веса. Однодоменное антитело ламы Н5 было наработано в периплазме бактерий Е. coli и затем очищено с помощью аффинной металл-хелатной хроматографии на Ni-NTA агарозе. Антитело Н5-Fc было наработано в эукариотических клетках НЕК293Т и очищено с помощью аффинной хроматографии на белке А.
На фиг. 5 представлен конкурентный иммуноферментный анализ взаимодействия рекомбинантного RBD-домена SARS-CoV-2 с Н5 однодоменным антителом ламы или рекомбинантным АСЕ2 растворимым рецептором. Однодоменное антитело ламы Н5 брали в последовательных разведениях, смешивали с биотинилированным RBD и добавляли в лунки с иммобилизованным АСЕ2. Детекцию RBD проводили стрептавидин-HRP конъюгатом. В качестве отрицательного контроля использовали нерелевантное однодоменное антитело А10.
На фиг. 6 представлена динамика веса сирийских хомячков, зараженных вирусом SARS-CoV-2. Хомячкам вводили по 10 мг антител/1 кг веса за 24 часа до (профилактика) или за 6 часов после заражения (терапия). В качестве положительного контроля использовали iB14 моноклональное антитело человека, нейтрализующее SARS-CoV-2. В качестве отрицательного контроля - суммарные IgG человека. Химерное антитело H5-Fc являлось тестируемым.
На фиг. 7 представлена вирусная нагрузка SARS-CoV-2 в легких сирийских хомячков, определенная методом RT-qPCR. В качестве положительного контроля использовали iB14 моноклональное антитело человека, нейтрализующее SARS-CoV-2. В качестве отрицательного контроля - суммарные IgG человека. Химерное антитело H5-Fc являлось тестируемым.
Последующие примеры приведены для целей объяснения и не ограничивают каким-либо образом рамки заявленного изобретения.
Пример 1. Получение иммунной библиотеки кДНК однодоменных антител ламы с использованием рекомбинантного RBD-домена S-белка SARS-CoV-2 вируса дикого типа.
Иммунизация ламы (Llama glama) была проведена согласно стандартному полному протоколу с использованием в качестве адьюванта GERBU LQ (Baral T.N., MacKenzie R., Arbabi Ghahroudi M. Curr Protoc Immunol. 2013;103:2.17.1-2.17.57. doi: 10.1002/0471142735.im0217sl03). Необходимое количество рекомбинантного RBD (от 600 до 900 мкг на одну инъекцию) смешивали с адьювантом и вводили ламе в две точки в нижней части шеи подкожно. Цикл иммунизаций включал три инъекции с перерывом в три недели. Через неделю после третьей иммунизации из яремной вены ламы брали 25 мл крови. Для выделения лейкоцитов из крови иммунизированной ламы использовали центрифугирование в градиенте плотности фиколла (ПанЭко, Россия). Всего было выделено 1,5×10 лейкоцитов.
Выделение суммарной РНК В-лимфоцитов проводили согласно одностадийному методу с использованием гуанидин изотиоцианата (Chomczynski P., Sacchi N. Nat Protocol. 2006;1(2):581-5. doi: 10.1038/nprot.2006.83). Все работы по выделению проводились со стандартными предосторожностями, направленными на сохранение интактности РНК. Количество суммарной РНК полученной из периферических В-лимфоцитов, составило 200 мкг.
Синтез кДНК проводили с использованием набора RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific) по протоколу производителя. Параллельно проводили 2 реакции с использованием 10 мкг суммарной РНК в каждой и двух разных праймеров - Олиго (dT)ig и статистического гексамерного праймера. Амплификацию VHH кДНК-фрагментов проводили в 2 этапа методом ПЦР с использованием Phusion ДНК-полимеразы (Thermo Fischer Scientific) (Pardon E., Laeremans Т., Triest S. et al Nat Protoc. 2014;9(3):674-93. doi: 10.1038/nprot.2014.039).
Для создания кДНК-библиотеки, позволяющей проводить скрининг методом фагового дисплея был выбран вектор pHEN4 (Arbabi Ghahroudi М., Desmyter A., Wyns L., Hamers R., Muyldermans S. FEBS Lett. 1997; 414(3):521-6. doi: 10.1016/s0014-5793(97)01062-4). Данный вектор не позволяет проводить выделение и очистку однодоменных антител после завершения процедуры скрининга. В связи с этим вектор pHEN4 был модифицирован генно-инженерным путем. Полученный в итоге вектор pHEN4-HH содержал в своей последовательности 6xHis-tag, что позволяет проводить очистку продуктов экспрессии методом металл-хелатной аффинной хроматографии.
Далее проводили гидролиз и лигирование полученных VHH кДНК фрагментов в фагмидный вектор для фагового дисплея, pHEN4-HH, по сайтам для эндонуклеаз рестрикции Pst I и Not I. Полученными плазмидными конструкциями трансформировали штамм Е. coli TG1. В данном амбер-супрессорном штамме VHH при экспрессии сливаются с поверхностным белком III фага. Представительность созданной иммунной библиотеки VHH-последовательностей составила 10 независимых клонов.
Пример 2. Получение библиотеки бактериофагов.
Процедура скрининга кДНК библиотеки методом фагового дисплея требует ее перевода в фаговую форму, поскольку сам принцип метода заключается в том, что белок-мишень, иммобилизованный на твердой подложке, связывается, в данном случае, с однодоменными антителами, которые фаг экспонирует на своей оболочке. Это достигается путем слияния интересующих нас генов с геном фага, отвечающего за синтез белка капсида, что и было достигнуто в результате получения библиотеки VHH-фрагментов в векторе pHEN4-HH. Перевод библиотеки в фаговую форму осуществлялся при ее культивировании совместно с фагом-помощником VCSM13. Титр полученного препарата фага, несущего VHH, составил 8x10, что вполне достаточно для проведения минимум 10 процедур скрининга.
Пример 3. Селекция однодоменных антител, специфически узнающих RBD-домен S-белка SARS-CoV-2 вируса, определение их последовательности.
Аффинное обогащение фаговой библиотеки проводили согласно базовому протоколу по генерации наноантител с целью повышения содержания в ней фаговых клонов, экспонирующих VHH против RBD (Esparza T.J., Martin N.P., Anderson G.P., Goldman E.R., Brody D.L. Sci Rep.2020; 10(1):22370. doi: 10.1038/s41598-020-79036-0). Отбор RBD-связывающих фагов проводили на SS-биотинилированном рекомбинантном RBD, связанным с магнитным сорбентом со стрептавидином (MagnaBind Streptavidin, Thermo Fisher Scientific). После интенсивных отмывок неспецифически связавшихся фагов, дисульфидные связи SS-биотина восстанавливали дитиотритолом и высвобождали фаговые частицы. Затем фаг в количестве 105-106 частиц добавляли к клеткам Е. coli штамма TG, в результате чего геном фага попадал в цитоплазму бактерии-хозяина с последующей репликация фагмиды через ori Е. coli., таким образом получали суббиблиотеку с преобладающим количеством клонов, способных связывать RBD.
Иммуноферментный анализ проводили с целью поиска отдельных фаговых клонов, экспонирующих VHH против RBD. Для этого рекомбинантый RBD иммобилизовали на дне лунок планшета и добавляли в них отдельные клоны суббиблиотеки, переведенные с помощью фага помощника в форму фага, экспонирующего индивидуальные VHH. Не связавшиеся с RBD фаги тщательно отмывали и добавляли в лунки мышиные антитела против фага М13, после чего вносили RAM-HRP конъюгат.При добавление субстратного буфера в лунках, в которых фаги связались с иммобилизованным RBD, происходило проявление цветовой индикации в ходе ферментативной реакции. Результаты реакции оценивали визуально, а в числовом виде получали с помощью микропланшетного спектрофотометра Multiskan GO (Thermo Scientific) при длине волны 491 нм. В результате проведенных процедур был отобран Н5 клон, взаимодействующий с RBD SARS-CoV-2.
В дальнейшем проводили культивацию бактерий, содержащих вектор pHEN4-Н5, кодирующий последовательность Н5 однодоменного антитела ламы, и выделяли из них плазмидную ДНК с помощью набора GeneJET Plasmid Miniprep Kit (Thermo Fischer Scientific) согласно рекомендациям производителя. Нуклеотидную последовательность однодоменного антитела определяли методом секвенирования по Сэнгеру с использованием набора реактивов BigDye Terminator v 3.1 Cycle Sequencing Kit (Thermo Fischer Scientific) на приборе Gene Analyzer 3500 (Applied Biosystems). В результате была определена нуклеотидная последовательность (SEQ ID NO: 3), кодирующая однодоменное антитело ламы Н5 (SEQ ID NO: 1), для которого были найдены границы CDR и FR участков и определены аминокислотные последовательности CDR: CDR1 - SEQ NO: 5, CDR2 - SEQ NO: 6 и CDR3 - SEQ NO: 7 (Фиг. 1 А).
Пример 4. Получение и очистка однодоменного антитела ламы Н5 Наработку однодоменного антитела ламы Н5, имеющего аминокислотную последовательность SEQ ID NO: 1 и структуру, указанную на Фиг. 2, проводили в периплазматическом пространстве Е. coli штамма НВ2151. Очистку полученных VHH проводили с помощью аффинной хроматографии на Ni-NTA агарозном сорбенте (Novagen). После тщательных промывок, VHH элюировали с сорбента с помощью буфера элюции (25 мМ Трис-HCl рН 8,0, 300 мМ NaCl, 250 мМ имидазол) и измеряли оптическую плотность (А280). Фракции, содержащие белки, объединяли и проводили диализ против PBS с двукратной сменой буфера.
Пример 5. Получение, продукция и очистка химерного антитела H5-Fc, представляющего собой однодоменное антитело ламы Н5, слитое с Fc-фрагментом IgG1 человека.
Для получения химерного антитела H5-Fc на первом этапе была проведена амплификация кДНК последовательности, кодирующей однодоменное антитело ламы Н5, с использованием метода ПЦР. Затем эта последовательность была клонирована по сайтам Agel-Apal в эукариотический экспрессирующий вектор pCDH3, содержащий последовательность Fc-фрагмента IgG1 человека. В результате был получен вектор pCDH3-H5-Fc' содержащий нуклеотидную последовательность (SEQ ID NO: 4), кодирующую химерные H5-Fc' цепи (SEQ ID NO: 2), которые при экспрессии в эукариотической системе самостоятельно объединяются по двум S-S мостиками и формируют димер (H5-Fc')2, который мы называем химерным антителом H5-Fc (Фиг. 1Б, Фиг. 3).
Экспрессирующие конструкции pCDH3-H5-Fc'' доставляли в клетки НЕК293Т с использованием кальций-фосфатной трансфекции (Kutner R. Н., Zhang X. Y., Reiser J. Nat Protoc. 2009; 4(4):495-505. doi: 10.1038/nprot.2009.22). Через 8 ч среду IMDM с добавлением 10% FBS для выращивания клеток заменяли бессывороточной средой EX-CELL (Сигма-Олдрич, США) для НЕК293 клеток. Трансфицированные клетки выдерживали в течение 6 дней в СО2-инкубаторе при 37°С, 5% CO2. Клеточные супернатанты, очищенные от клеточного дебриса кратким центрифугированием, фильтровали через 0,22 мкм PES-фильтр (ТРР, Швейцария), а белковые продукты очищали на колонке с агарозой с белком A (McLab, США, #РРА-503) в соответствии с инструкциями производителя. Затем элюат с химерным антителом H5-Fc диализовали в течение ночи против PBS и концентрировали с использованием концентраторов Ultra-15 Ultracel-IOOK (Amicon) до конечной концентрации 2 мг/мл.
Чистоту белковых проб контролировали, проводя 15% акриламидный гель-электрофорез (ДСН-ПААГ). Из Фиг. 4 видно, что полученные препараты однодоменного антитела ламы Н5 и химерного антитела H5-Fc имеют достаточную чистоту для их дальнейшего использования в in vitro и in vivo приложениях.
Пример 6. Демонстрация специфического связывания однодоменного антитела ламы Н5 и химерного антитела H5-Fc с RBD-доменом S-белка SARS-CoV-2 вируса дикого типа.
Конкурентный ИФА проводили для определения эффективности нейтрализации взаимодействия RBD-ACE2 одно доменным антителом ламы Н5. Для этого в лунках микропланшета иммобилизовали растворимый рекомбинантный АСЕ2, в которые затем добавляли смесь однодоменного антитела и биотинилированного рекомбинантного RBD. После тщательной промывки в лунки добавляли конъюгат Streptavidin-HRP (1:1000). Ферментативную реакцию проводили в субстратном буфере до проявления цветовой индикации. Результаты реакции оценивали визуально, а в числовом виде получали с помощью микропланшетного спектрофотометра Multiskan GO (Thermo Scientific) при длине волны 491 нм (Фиг. 5). Из Фиг. 5 видно, что однодоменное антитело ламы Н5 блокирует взаимодействие ACE2-RBD, в отличие от нерелевантного однодоменного антитела А10.
Кинетику связывания однодоменного антитела ламы Н5 и химерного антитела H5-Fc с рекомбинантным RBD SARS-CoV-2 дикого типа (Wuhan-1) анализировали на приборе Octet RED96 (ForteBio, Pall LifeSciences, NewYork, NY, США). Рекомбинантный RBD SARS-CoV-2 биотинилировали с помощью EZ-Link™ NHS-LC-LC-Biotin (#21343, ThermoFisher Scientific, США) и иммобилизовали на SA-сенсоре, далее инкубировали с EZ-Link™ Biocytin (#28022, ThermoFisher Scientific, США) для блокировки неспецифического связывания. Нагруженные сенсоры тестировали против панели из трех концентраций однодоменного антитела ламы Н5 (6 нМ, 12 нМ, 20 нМ). Параллельно рекомбинантный RBD SARS-CoV-2 иммобилизовали на NTA-сенсоры (18-5101, ForteBio), которые тестировали против панели концентраций химерного антитела H5-Fc (1,3 нМ, 2,5 нМ, 4,5 нМ, 9 нМ, 18 нМ, 36 нМ). Обработка данных проводилась с использованием программного обеспечения для сбора данных ForteBiov.l 1.1.1.19 (модель 1:1 с глобальным приближением), R2>0,9, Х2<3. Значения кинетических констант приведены в Таблице 1, где KD - константа диссоциации, kon - кинетическая константа ассоциации, koff- кинетическая константа диссоциации.
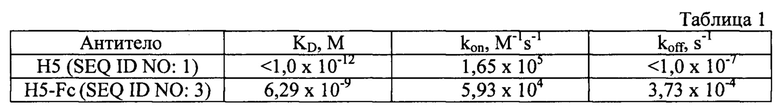
Полученные данные указывают на то, что химерное антитело H5-Fc, как и Н5, специфически распознает RBD SARS-CoV-2. KD взаимодействия однодоменного антитела ламы Н5 с RBD SARS-CoV-2 находится ниже пикомолярных значений и однодоменное антитело ламы Н5 может быть охарактеризовано как ультра-аффинное. В сравнении с прототипом (однодоменное антитело р2 с5, KD=2,8×10-9 М), аффинность связывания однодоменного антитела ламы Н5 с RBD-доменом S-белка SARS-CoV-2 не менее чем в 2800 выше. Химерное антитело H5-Fc сравнить с прототипом (p2c5-fc) не представляется возможным из-за отсутствия данных о константе равновесной диссоциации для взаимодействия p2c5-fc с RBD. Стоит отметить, что, строго говоря, наличие специфического взаимодействия с RBD-доменом S-белка SARS-CoV-2 для прототипа p2c5-fc не показано.
Пример 7. Демонстрация способности однодоменного антитела ламы Н5 и химерного антитела H5-Fc нейтрализовать лентивирусные частицы, псевдотипированные S-белком SARS-CoV-2 различных вариантов, in vitro в культуре клеток.
Лентивирусные частицы, псевдотипированные S-белком SARS-CoV-2, получали способом, описанным ниже. Клетки НЕК293Т трансфицировали в молярном отношении 4:6:3 смесью плазмид psPAX2, pLV-EGFP и pCAGGS-SpikeA19, кодирующей укороченный S-белок либо дикого типа, либо мутантного варианта. Через восемь часов после трансфекции культуральную среду заменяли средой Opti-Mem (Gibco, Thermo Fisher Scientific) с добавлением 2,5% термоинактивированной FBS (Gibco, Thermo Fisher Scientific). Через 48 часов супернатанты собирали и предварительно очищали низкоскоростным центрифугированием и фильтрацией через фильтр 0,45 мкм. Затем псевдотипированные лентивирусные частицы концентрировали центрифугированием.
Полученные лентивирусные частицы использовали для трансдукции клеток АСЕ2-НЕК293Т в присутствии различных концентраций нейтрализующих антител. Клетки АСЕ2-НЕК293Т, стабильно экспрессирующие АСЕ2 человека, высевали в культуральный планшет в день начала анализа нейтрализации. Антитела серийно разводили в Opti-MEM+2,5% FBS в два этапа до концентраций в диапазоне от 0,5 нг/мл до 1 мкг/мл и совместно инкубировали с 20 тыс.S-псевдотипированньгх лентивирусных частиц перед трансдукцией. Смесь антитело/8-псевдотипированный лентивирус добавляли к клеткам АСЕ2-НЕК293Т, и планшет возвращали в инкубатор с СО2. Через 72 часа после трансдукции процент трансдуцированных (GFP+) клеток измеряли с помощью проточной цитометрии. Полумаксимальную ингибирующую концентрацию (IC50) определяли с помощью нелинейной регрессии как концентрацию антитела, которая нейтрализовала 50% псевдотипированного лентивируса. Были использованы данные двух независимых экспериментов. В таблице 2 продемонстрирована способность антитела ламы Н5 и химерного антитела H5-Fc нейтрализовать лентивирусы, псевдотипированных S-белками SARS-CoV-2 вируса разных вариантов.

Из полученных результатов видно, что химерное антитело H5-Fc, несмотря на худшую константу диссоциации KD, чем у однодоменного антитела ламы Н5, нейтрализует лентивирусные частицы, псевдотипированные S-белком SARS-CoV-2 дикого типа, в чуть меньшей концентрации. Однако, стоит учесть, что молекулярная масса химерного антитела H5-Fc примерно в 6 раз больше, чем у антитела ламы Н5, таким образом в молярном выражении, H5-Fc действует значительно эффективнее, чем Н5 (числовые значения показаны в квадратных скобках). Большая эффективность нейтрализации у химерного антитела H5-Fc может объясняться наличием двух паратопов, а также большими физическими размерами. Кроме того, полученные данные свидетельствуют о том, что однодоменное антитело ламы Н5, а следовательно и химерное антитело H5-Fc, способны нейтрализовать не только вирусные частицы, несущие S-белок дикого типа (Wu-1), но и его мутантные варианты. Данные об псевдовирус-нейтрализующей активности у антител-прототипов (однодоменное антитело р2 с5 и фьюжн-антитело p2c5-fc) отсутствуют.Также для прототипов отсутствуют данные о нейтрализации вариантов SARS-CoV-2 вируса.
Пример 8. Демонстрация способности однодоменного антитела ламы Н5 и химерного антитела H5-Fc нейтрализовать вирус SARS-CoV-2 дикого типа in vitro в культуре клеток.
Анализ нейтрализации вируса в микрокультуре проводили в 96-луночных планшетах (Costar) с использованием в качестве мишеней клеток VeroE6, выращенных в среде DMEM+5% FCS (Nurtop Е., Villarroel P. М. S., Pastorino В. et al. Virol J. 2018;15(1):192. doi: 10.1186/sl2985-018-1105-5). Равный объем серийно разбавленных антител смешивали с вирусным супернатантом (100 TCID50 (50% инфекционная доза культуры ткани)/100 мкл) и оставляли при 37°С на 1 час. Затем смесь вирус/антитело (100 мкл) переносили в лунки с монослоем клеток VeroE6. На 5-7-е сутки после инфицирования лунки визуально просматривали под инвертированным микроскопом. IC100 определяли как самую низкую концентрацию антитела, обеспечивающую полную защиту от 100 TCID50 в 100 мкл. В таблице 3 представлена вируснейтрализующая активность полученных антител в отношении SARS-COV-2 дикого типа

Для проводимых экспериментов использовали изолят hCoV-19/Russia/Moscow-PMVL-12/2020 (EPI_ISL_572398) (D614G, S686del, V687I). Полученные данные показывают, что однодоменное антитело ламы Н5 и химерное антитело H5-Fc нейтрализуют живой вирус примерно с одинаковой эффективностью (~0,1 нМ), что сравнимо с таковой у прототипа р2 с5 (5 нг/мл, ~0,06 нМ).
Пример 9. Демонстрация способа экстренной профилактики и терапии заболевания, вызванного вирусом SARS-CoV-2 дикого типа in vivo на модели золотистого хомячка с использованием химерного антитела H5-Fc
Для исследований защитных свойств химерного антитела H5-Fc на животных были использованы самки сирийских хомяков. Использовали две схемы, профилактическую (введение антитела за 24 часа до инфицирования) и терапевтическую (6 часов после), в каждой - по 2 группы животных (n=5), которым внутрибрюшинно вводили H5-Fc или общий IgG человека (отрицатательный контроль) в количестве 10 мг/кг веса. В качестве положительно контроля использовали профилактическую схему с использованием ранее полученного iB14 моноклонального антитела человека, нейтрализующего SARS-CoV-2 (Gorchakov A. A., Kulemzin S. V., Guselnikov S. V. et al. Cell Discov. 2021;7(1):96. doi: 10.1038/s41421-021-00340-8). В нулевой точке (инфицирование) вводили интраназально (50 мкл/ноздрю) SARS-CoV-2 в общей дозе 1,3×103 бляшкообразующих единиц, изолят SARS-CoV-2/human/RUS/Nsk-FRCFTM-1/2020 (EPIJSL 481284) (D614G). Животных ежедневно взвешивали (Фиг. 6). Через 5 дней после инфекции животных подвергли эвтаназии и извлекали из них легкие для количественного определения вирусной нагрузки (Фиг. 7).
Половину левой доли легкого хомячка взвешивали, помещали в раствор ЛИРА (Biolabmix) и хранили на холоде. Затем образцы тщательно гомогенизировали и выделяли РНК в соответствии с инструкциями производителя раствора. Набор для синтеза кДНК RevertAid (Thermo Scientific) использовали для превращения 1 мг суммарной РНК в кДНК с использованием случайных праймеров-гексамеров. RT-qPCR проводили с использованием кДНК в разведении 1:10 в качестве матрицы и двух наборов праймеров, специфичных для генов RdRp и Е SARS-CoV-2 (Corman V.M., Landt О., Kaiser М, et al. Euro Surveill. 2020;25(3):2000045. doi: 10.2807/1560-7917.ES.2020.25.3.2000045). Праймеры, обнаруживающие транскрипты RpU8 хомячка (Zivcec M., Safronetz, D., Haddock, E. et al. J. Immunol. Methods 2011;368, 24-35), использовали для нормализации. Каждый RT-qPCR анализ выполняли в трех повторениях с двумя техническими повторами, и каждый анализ включал отрицательные контроли без матрицы.
Вышеприведенные данные показывают, что однодоменное антитело ламы Н5 и химерное антитело H5-Fc в концентрациях 10 мг/1 кг веса животного эффективно предотвращают появление клинических признаков заражения SARS-CoV-2 (потеря веса) на модели сирийского хомячка в профилактической и терапевтической моделях (Фиг. 6). Такие же результаты демонстрирует прототип p2c5-fc, однако только в профилактической схеме, терапевтический же эффект не был исследован. Кроме того, стоит отметить, что в качестве отрицательного контроля прототипа (плацебо) был использован фосфатно-солевой буфер, что оставляет вопрос о том, какая конкретно часть p2c5-fc молекулы отвечает за отсутствие клинических проявлений SARS-CoV-2 инфекции у хомячков, т.е. является ли это р2 с5-специфическим ответом. В нашей схеме в качестве отрицательного контроля был использован суммарный IgG человека, обладающий Fc-частью, что разрешает эту проблему.
Для прототипа p2c5-fc также было показано протективное действие на модели АСЕ2-трансгенных мышей, заражаемых SARS-CoV-2 инфекцией. Эта модель хороша тем, что является летальной, то есть оказываемый эффект дискретен (выживание/смерть), но, в то же время, не лишена и недостатков. Так, она не соответствует тканевому распределению АСЕ2 в организме человека - у мыши АСЕ2 эксперссируется повсеместно и животные гибнут не только от поражения легких, но даже в большей степени головного мозга и почек. В заявляемом решении отказались от этой модели и изучали изменение вирусной нагрузки в легких тех же сирийских хомячках, для которых мы наблюдали динамику изменения веса. Данные количественного анализа содержания РНК вируса SARS-CoV-2 в легких хомячков на 5 день после заражения показали, что и в профилактической, и в терапевтической схеме введения химерного антитела H5-Fc вирусная нагрузка падает в 10000 раз по сравнению с контролем, в случае профилактики - практически до 0 (Фиг. 7). Таким образом, можно заключить, что химерное антитело H5-Fc обладает сходными протективными свойствами с прототипом р2 с5 в профилактической схеме, но дополнительно работает в терапевтической схеме на модели сирийского хомячка.
Таким образом, можно заключить, что созданные однодоменное антитело ламы Н5 и химерное антитело H5-Fc лишены недостатков прототипа и могут быть использованы для профилактики и терапии заболевания человека, вызываемого вирусом SARS-CoV-2.
Предлагаемые однодоменное антитело ламы Н5 и химерное антитело H5-Fc по сравнению с прототипом обладают следующими преимуществами: 1) более высокая вируснейтрализующая активность однодоменного антитела ламы Н5 по отношению к живому вирусу SARS-CoV-2; 2) более высокая аффинность по отношению к RBD SARS-CoV-2 дикого типа; 3) более широкие функциональные возможности за счет способности созданных антител нейтрализовать вирус SARS-CoV-2 разных вариантов, что позволяет использовать их для профилактики и терапии заболеваний, вызываемых вирусом SARS-CoV-2 разных штаммов.
--->
Перечень последовательностей
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Перечень
последовательностей H5 H5-Fc CDR.xml" softwareName="WIPO Sequence"
softwareVersion="2.0.0" productionDate="2022-06-08">
<ApplicantFileReference>1</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение науки Институт молекулярной и клеточной биологии
Сибирского отделения Российской академии наук (ИМКБ СО
РАН)</ApplicantName>
<ApplicantNameLatin>Institute of Molecular and Cellular Biology,
Siberian Branch of Russian Academy of Sciences</ApplicantNameLatin>
<InventionTitle languageCode="ru">Однодоменное антитело ламы H5 и
его производное Н5-Fc, специфически связывающие RBD-домен S-белка
вируса SARS-CoV-2, обладающие вируснейтрализующей
активностью</InventionTitle>
<SequenceTotalQuantity>7</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>157</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..157</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
/INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MAQVQLQESGGGLVQPGGSLRLSCAASGFTLDYYTIGWFRQAPGKEREG
VSCISSRDGSTYYADSVKGRFTISRDNAKNTVYLQMDNLKPEDTAVYYCATTPLTPYGSSWHPNCGSDYA
VNYWGQGTQVTVSSRGRYPYDVPDYGAAHHHHHHNGRA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>365</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..365</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>QVQLQESGGGLVQPGGSLRLSCAASGFTLDYYTIGWFRQAPGKEREGVS
CISSRDGSTYYADSVKGRFTISRDNAKNTVYLQMDNLKPEDTAVYYCATTPLTPYGSSWHPNCGSDYAVN
YWGQGTQVTVSSGPEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDP
EVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKG
QPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLT
VDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>474</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..474</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
/INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ATGGCCCAGGTGCAGCTGCAGGAGTCTGGGGGAGGCTTGGTGCAGCCTG
GGGGGTCTCTGAGACTCTCCTGTGCAGCCTCTGGATTCACTTTGGATTATTATACCATAGGCTGGTTCCG
CCAGGCCCCAGGGAAGGAGCGCGAGGGGGTCTCATGTATTAGTAGTAGGGATGGTAGCACATACTATGCA
GACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACACGGTGTATCTGCAAATGGACA
ACCTGAAACCTGAGGACACGGCCGTTTATTACTGTGCGACGACCCCACTAACCCCGTACGGTAGTAGCTG
GCACCCAAACTGCGGTTCAGACTATGCGGTTAACTACTGGGGCCAGGGGACCCAGGTCACCGTCTCCTCA
CGCGGCCGCTACCCGTACGACGTTCCGGACTACGGTGCCGCACATCATCATCACCATCACAACGGCCGAG
CATAG</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>1098</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1098</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>CAGGTGCAGCTGCAGGAGTCTGGGGGAGGCTTGGTGCAGCCTGGGGGGT
CTCTGAGACTCTCCTGTGCAGCCTCTGGATTCACTTTGGATTATTATACCATAGGCTGGTTCCGCCAGGC
CCCAGGGAAGGAGCGCGAGGGGGTCTCATGTATTAGTAGTAGGGATGGTAGCACATACTATGCAGACTCC
GTGAAGGGCCGATTCACCATCTCCAGAGACAATGCCAAGAACACGGTGTATCTGCAAATGGACAACCTGA
AACCTGAGGACACGGCCGTTTATTACTGTGCGACGACCCCACTAACCCCGTACGGTAGTAGCTGGCACCC
AAACTGCGGTTCAGACTATGCGGTTAACTACTGGGGCCAGGGGACCCAGGTCACCGTCTCCTCGGGCCCA
GAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGT
CAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGT
GGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCAT
AATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCC
TGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCAT
CGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGG
GAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCG
TGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGG
CTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGC
TCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCCCCGGGTAAATGA<
/INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>8</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>GFTLDYYT</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>8</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ISSRDGST</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<SequenceData sequenceIDNumber="7">
<INSDSeq>
<INSDSeq_length>24</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..24</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ATTPLTPYGSSWHPNCGSDYAVNY</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
| Моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817697C1 |
| ПОЛИПЕПТИДНЫЙ МОДУЛЬ ДЛЯ СВЯЗЫВАНИЯ КОНСЕРВАТИВНОГО ЭПИТОПА РЕЦЕПТОР-СВЯЗЫВАЮЩЕГО ДОМЕНА БЕЛКА SPIKE КОРОНАВИРУСА SARS-COV-2 | 2022 |
|
RU2809183C1 |
| Экспрессионный вектор на основе аденоассоциированного вируса и способ его применения для экстренной профилактики и профилактики заболеваний, вызываемых вирусом SARS-CoV-2 (варианты) | 2022 |
|
RU2777404C1 |
| Экспрессионный вектор на основе аденоассоциированного вируса, несущий гены рекомбинантных антител, и его применение для профилактики заболеваний, вызываемых вирусом гриппа А и вирусом SARS-CoV-2 | 2023 |
|
RU2817792C1 |
| Плазмидная генетическая конструкция pET21a_SKP, обеспечивающая экспрессию в прокариотической системе E.coli рекомбинантного белка SKP, и рекомбинантный белок SKP, обладающий широконейтрализующими свойствами однодоменного наноантитела против SARS-CoV-2 | 2024 |
|
RU2839376C1 |
| Однодоменное антитело и его модификации, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, и способ их применения для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2763001C1 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
| Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2 | 2021 |
|
RU2794141C2 |
| БИСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ SARS-COV-2 | 2022 |
|
RU2791749C1 |


Изобретение относится к области биотехнологии. Описано однодоменное антитело ламы Н5, специфически связывающееся с RBD-доменом S-белка SARS-CoV-2 вируса, обладающее вируснейтрализующей активностью, и химерное антитело H5-Fc. В одном варианте реализации изобретения однодоменное антитело имеет аминокислотную последовательность SEQ ID NO: 1, кодируемую нуклеотидной последовательностью SEQ ID NO: 3, и имеет определяющие комплементарность области (CDR) с аминокислотными последовательностями SEQ ID NO: 5-7. Изобретение позволяет получать антитела с высокой аффинностью, распознающие RBD-домен SARS-CoV-2, и использовать их для диагностики, терапии и профилактики коронавирусной инфекции, вызванной SARS-CoV-2, у человека и других млекопитающих. 2 н.п. ф-лы, 7 ил., 3 табл., 9 пр.
1. Однодоменное антитело ламы Н5, специфически связывающееся с RBD-доменом S-белка SARS-CoV-2 вируса, обладающее вируснейтрализующей активностью, имеющее аминокислотную последовательность SEQ ID NO: 1, кодируемую нуклеотидной последовательностью SEQ ID NO: 3, имеющее определяющие комплементарность области (CDR) с аминокислотными последовательностями SEQ ID NO: 5-7.
2. Химерное антитело H5-Fc, представляющее собой однодоменное антитело по п. 1, слитое с Fc-фрагментом IgG1 человека, специфически связывающееся с RBD-доменом S-белка SARS-CoV-2 вируса, обладающее вируснейтрализующей активностью, имеющее аминокислотную последовательность SEQ ID NO: 2, кодируемую нуклеотидной последовательностью SEQ ID NO: 4, имеющее CDR с аминокислотными последовательностями SEQ ID NO: 5-7.
| Однодоменное антитело и его модификации, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, и способ их применения для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2763001C1 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
| Гуманизированное моноклональное антитело, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, средство и способ для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2765731C1 |
