ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к биотехнологии, в частности к биспецифическим моноклональным антителам, которые специфически связываются с шиповидным белком коронавируса (SARS-COV-2), а также к способу получения таких антител.
УРОВЕНЬ ТЕХНИКИ
В декабре 2019 г. в г. Ухань, провинция Хубэй, было выявлено заболевание, вызванное новым коронавирусом (SARS-COV-2), которое поставило перед специалистами в области здравоохранения и врачами трудные задачи, связанные с быстрой диагностикой и клиническим ведением больных. Указанный вирус быстро распространился по всему миру, вызвав беспрецедентную по своим масштабам пандемию. Только в России согласно данным Росстата, в декабре прошлого года умерли 54 630 зараженных Covid-19. При этом основной причиной смерти коронавирус стал в 49 122 случаях (в 44 390 случаях коронавирус установлен как основная причина, в 4 732 - нужно провести дополнительные исследования). В качестве сопутствующего заболевания коронавирус был диагностирован у 5 508 умерших (у 1 000 человек он способствовал появлению осложнений, ускоривших смерть, а еще в 4 508 случаях - не повлиял на наступление смерти). В настоящее время сведения об эпидемиологии, клинических особенностях, профилактике и лечении этого заболевания ограничены. Известно, что наиболее распространенным клиническим проявлением нового варианта коронавирусной инфекции является пневмония, у значительного числа пациентов зарегистрировано развитие острого респираторного дистресс-синдрома (ОРДС). Вирус отнесен ко II группе патогенности, как и некоторые другие представители этого семейства (вирус SARS-COV, MERS-COV). В отношении нового коронавирусного заболевания отсутствуют средства как специфической профилактики, так и этиотропного лечения. Высокий процент смертности, быстрое географическое распространение SARS-COV-2 и нечетко определенная этиология заболевания создали острую необходимость в создании эффективных средств профилактики заболеваний, вызываемых данным вирусом.
Направления исследований по разработке новых моноклональных антител против SARS-COV-2 зачастую ориентированы на оптимизацию последовательностей антител к домену RBD шиловидного (spike) белка вируса SARS-COV-2. Так например, в публикации WO 2021196268 А1 предложены антитело, специфически связывающееся с белком S коронавируса, мультиспецифическое антитело и композиция антител. Кроме того, предложены нуклеиновая кислота, кодирующая антитело или антигенсвязывающий фрагмент, и мультиспецифическое антитело, клетка-хозяин, содержащая нуклеиновую кислоту, и способ получения антитела, а также мультиспецифическое антитело. Также, дополнительно предусмотрены профилактические, лечебные и/или диагностические применения антитела или антигенсвязывающего фрагмента, мультиспецифического антитела и композиции антитела.
Также в уровне техники описаны рекомбинантные моноклональные антитела (мАт) и их фрагменты, которые специфически связываются с шиповидным белком коронавируса (патент CN 111620946 В). В данном исследовании моноклональные антитела были рекомбинантно получены из выделенных В-клеток от людей, инфицированных коронавирусом. Такие антитела связывают различные эпитопы шиловидного белка коронавируса и нейтрализуют его. Изобретение обеспечивает способы применения антител по изобретению в профилактических и/или терапевтических способах предотвращения или лечения новой коронавирусной инфекции.
В публикации WO 2022054068 A1 раскрыто моноклональное антитело к новому коронавирусу или его антигенсвязывающая часть, которая специфически связывается с S-белком нового коронавируса. Описанные моноклональные антитела можно с успехом применять для обнаружения присутствия новых коронавирусов. Кроме того, указанные моноклональные антитела обладают нейтрализующей активностью и, таким образом, могут применяться для приготовления лекарственного средства для профилактики или лечения новой коронавирусной инфекции.
В публикации WO 2022/093745 А1 предложена аминокислотная последовательность человеческого антитела, содержащего антигенсвязывающий домен, который связывает антигенную детерминанту коронавируса, для применения в профилактике, лечении или выявлении коронавирусной инфекции у субъекта, нуждающегося в этом.
В публикации WO 2021/195385 А1 описаны полипептиды, которые специфически связываются с шиповидным белком (S) коронавируса человека, выбранные из шести библиотек фагового дисплея однодоменных антител VHH верблюда. Полипептиды, специфичные для S-белка, нарушают связывание S-белка SARS-COV-2 и/или SARS-COV с клеточным рецептором АСЕ2, что важно для нейтрализации вируса. Описано использование полипептидов, специфичных для белка S, для диагностики и лечения SARS-COV-2 и/или SARS-COV.
Человеческие моноклональные антитела к тяжелому острому респираторному синдрому коронавируса 2 (SARS-COV-2) также известны из публикации WO 2022/006562 А1. Также предложен способ выявления инфекции COVID-19, вызванной SARS-COV-2, у субъекта, включающий (а) приведение в контакт образца от указанного субъекта с антителом или фрагментом антитела, имеющим клон-спаренные тяжелые и последовательности CDR легкой цепи из таблиц 3 и 4 соответственно; и (b) обнаружение SARS-COV-2 в указанном образце путем связывания указанного антитела или фрагмента антитела с антигеном SARS-COV-2 в указанном образце. Образец может представлять собой биологическую жидкость, такую как кровь, мокрота, слезы, слюна, слизь или сыворотка, сперма, цервикальный или вагинальный секрет, амниотическая жидкость, ткани плаценты, моча, экссудат, транссудат, соскобы тканей или фекалии. Обнаружение может включать ELISA, RIA, анализ в латеральном потоке или вестерн-блоттинг.
Также опубликованы исследования по разработке биспецифических моноклональных антител (мАт) к SARS-COV-2.
Основной гипотезой в разработке биспецифических мАт к SARS-COV-2 является гипотеза о эффективности нейтрализующих антител, нацеленных на рецептор-связывающий домен (RBD) шиловидного белка SARS-COV-2, в борьбе с COVID-19. Так, например, в работе De Gasparo et al. была разработана биспецифическая IgG1-подобная молекула (COV-X2) на основе С121 и С135, двух антител, полученных от доноров, выздоровевших от COVID-19. В работе показано, что COV-X2 одновременно связывает два независимых сайта на RBD и, в отличие от его родительских антител, предотвращает обнаруживаемое пиковое связывание с клеточным рецептором вируса, ангиотензинпревращающим ферментом 2 (АСЕ2). Кроме того, COV-X2 нейтрализует SARS-COV-2 дикого типа и его варианты, вызывающие озабоченность, а также ускользающие мутанты, генерируемые родительскими моноклональными антителами. В исследовании также обнаружили, что в мышиной модели инфекции SARS-COV-2 с воспалением легких COV-X2 защищает мышей от болезни и подавляет утечку вируса. Таким образом, одновременное нацеливание на неперекрывающиеся эпитопы RBD с помощью IgG-подобных биспецифических антител возможно и эффективно и сочетает в себе преимущества коктейлей антител с преимуществами одномолекулярных подходов [De Gasparo, R., Pedotti, M., Simonelli, L. et al. Bispecific IgG neutralizes SARS-COV-2 variants and prevents escape in mice. Nature 593, 424-428 (2021)].
Также описана неконкурирующая пара человеческих нейтрализующих антител (В38 и Н4), блокирующих связывание SARS-COV-2 с его рецептором АСЕ2. Yanwu et al. разработали биспецифическое моноклональное антитело (bsAb15) на основе В38 и Н4. bsAb15 обладает большей нейтрализующей эффективностью, чем эти исходные антитела, приводит к меньшему селективному давлению и сохраняет нейтрализующую способность в отношении большинства вызывающих озабоченность вариантов SARS-COV-2 (с более сильной нейтрализующей активностью в отношении дельта-варианта). Также в исследовании отобрали в качестве ускользающих мутантов двух родительских мАт смесь мАт и bsAb15, демонстрируя, что bsAb15 может эффективно нейтрализовать все ускользающие мутанты с одним мАт. Кроме того, профилактическое и терапевтическое применение bsAb15 снижало титр вируса у инфицированных приматов, не являющихся человеком, и мышей, трансгенных по АСЕ2 человека. Таким образом, согласно исследованию, bsAb является осуществимой и эффективной стратегией лечения и профилактики тяжелой формы COVID-19 [Li, Z., Li, S., Zhang, G. et al. An engineered bispecific human monoclonal antibody against SARS-COV-2. Nat Immunol 23, 423-430 (2022)].
В исследовании Dogan et al. сконструировали первичные Т-клетки CD8 для экспрессии CAR, специфичных для белка SARS-COV-2 Spike, с использованием внеклеточной области АСЕ2, и продемонстрировали их высокоспецифичную и мощную цитотоксичность по отношению к клеткам-мишеням, экспрессирующим шиловидный белок. В качестве потенциального терапевтического средства, исследователями был разработан биспецифический активатор Т-клеток, сочетающий АСЕ2 с анти-CD3 scFv (ACE2-Bite) для нацеливания на инфицированные клетки и вирус. Подобно подходу CAR-Т-клеток, ACE2-Bite обеспечивал способность цитотоксических клеток избирательно убивать мишени, экспрессирующие шиловидный белок. Кроме того, ACE2-Bite нейтрализовал псевдовирусы SARS-COV, SARS-COV-2 дикого типа и варианты, включая Delta и Omicron, в качестве белка-приманки. Примечательно, что молекула ACE2-Bite продемонстрировала более высокую аффинность к связыванию и нейтрализации вариантов Delta и Omicron по сравнению с шиловидными белками дикого типа SARS-COV-2, что указывает на потенциал этого подхода в качестве защищенной от вариантов терапевтической стратегии для будущего SARS-COV-2 варианты, использующие как гуморальное, так и клеточное звено адаптивного иммунного ответа [Dogan, Mikail; Kozhaya, Lina; Placek, Lindsey; Karabacak, Fatih; Yigit, Mesut; Unutmaz, Derya, Targeting SARS-COV-2 Infection Through CAR-T Like Bispecific T Cell Engagers Incorporating АСЕ2].
Моноклональные антитела (мАт) стали важным компонентом лечения коронавирусной инфекции 2019 (COVID-19). Большинство мАт вводятся в виде коктейля из нескольких антител, которые нацелены на различные области вириона коронавируса тяжелого острого респираторного синдрома 2 (SARS-COV-2). Так, например, было продемонстрировано, что усовершенствование этой стратегии путем выявления мАт, которые в сочетании с биспецифическими антителами нейтрализуют SARS-COV-2 лучше, чем смеси исходных мАт. Такие биспецифические антитела связывались с отдельными областями шиповидного белка SARS-COV-2, нейтрализовали вызывающие озабоченность варианты, включая вариант Delta, и обеспечивали защиту при введении хомякам до заражения SARS-COV-2. В совокупности эти данные свидетельствуют о том, что биспецифические антитела обладают большим потенциалом в качестве терапевтических средств против SARS-COV-2, устойчивых к вариантам [Hyeseon Cho, et al. Bispecific antibodies targeting distinct regions of the spike protein potently neutralize SARS-CoV-2 variants of concern//Sci Transl Med. 2021 Oct 20;13(616)].
Однако, несмотря на успехи в лечении антителами и разработке вакцин, заболевание COVID-19, вызванное инфекцией SARS-COV-2, остается серьезной проблемой для здоровья, приводящей к чрезмерной заболеваемости и смертности, а появление новых вариантов снижает эффективность существующих вакцин. Таким образом, все еще существует потребность в разработке дополнительных антител против SARS-COV-2.
Авторы настоящего изобретения разработали биспецифические антитела, нейтрализующие SARS-COV-2. Описанные здесь антитела можно использовать для лечения инфекций SARS-COV-2 и их симптомов.
РАСКРЫТИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном аспекте настоящего изобретения предложено биспецифическое антитело или его антиген-связывающий фрагмент, специфически связывающееся по меньшей мере с одним эпитопом в рецептор-связывающем домене (RBD) шиповидного белка (S) коронавируса тяжелого острого респираторного синдрома (SARS-COV-2).
Согласно одному варианту реализации настоящего изобретения указанный эпитоп является нелинейным.
Согласно дополнительному варианту реализации настоящего изобретения указанный эпитоп содержит область в пределах аминокислот 319-541 шиповидного белка, предпочтительно, указанный эпитоп содержит область в пределах аминокислот 319-490 шиповидного белка.
Согласно дополнительному варианту реализации настоящего изобретения указанное антитело ингибирует слияние вирусной и клеточной мембраны.
Согласно еще одному дополнительному варианту реализации настоящего изобретения антитело блокирует связывание шиповидного белка SARS-COV2 с рецептором ангиотензинпревращающего фермента 2 (АСЕ2) на клеточной поверхности.
Согласно еще одному варианту реализации настоящего изобретения антитело содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-7.
Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 1. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO:2. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 3. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 4. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 5. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 6. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 7.
Согласно предпочтительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 1.
Согласно дополнительному варианту реализации настоящего изобретения указанное антитело представляет собой полностью человеческое антитело.
В другом аспекте настоящего изобретения предложен способ получения антитела, включающий следующие этапы: а) конструирование рекомбинантной экспрессионной плазмидной ДНК, кодирующей антитело по любому из пп. 1-9, б) трансфекцию полученной ДНК клеток СНО, в) культивирование полученной линии клеток, и г) выделение указанного антитела.
Согласно одному из вариантов реализации изобретения конструирование рекомбинантной экспрессионной плазмидной ДНК осуществляется способами, широко известными специалисту в области техники.
Согласно дополнительному варианту реализации настоящего изобретения после трансфекции клетки культивировали на среде BalanCD с добавлением 500000 ЕД пенициллина, 100 мкг/см3 стрептомицина и 4 мМ L-глютамина при температуре 37°С, содержании CO2-5%, влажности не более 75% в течении 7 дней до точки снижения жизнеспособности ниже 80%.
Необходимо понимать, что настоящее изобретение не ограничивается конкретными описанными способами и условиями экспериментов, поскольку такие способы и условия могут варьировать. Также необходимо понимать, что терминология, используемая в настоящей заявке, предназначена лишь для целей описания конкретных вариантов осуществления и не предполагает ограничительный характер, поскольку объем настоящего изобретения будет ограничиваться лишь прилагаемой формулой изобретения. Если не определено иное, все технические и научные термины, используемые в настоящей заявке, имеют такое же значение, которое обычно понимается специалистом в области, к которой принадлежит настоящее изобретение. Хотя при практическом осуществлении или тестировании настоящего изобретения можно применять любые способы и материалы, сходные с описанными в данном документе или эквивалентные им, в данном документе описаны только предпочтительные способы и материалы. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки в полном объеме.
Используемые в настоящей заявке термины «биспецифическое моноклональное антитело», «биспецифическое антитело» «бАт», «мАт», «антитело» являются взаимозаменяемыми. Указанный термин предназначен для обозначения молекул иммуноглобулинов, состоящих из четырех полипептидных цепей, двух тяжелых (Н) цепей и двух легких (L) цепей, соединенных между собой дисульфидными связями (т.е. «молекулы полного антитела»), а также их мультимеров или их антигенсвязывающих фрагментов. Каждая тяжелая цепь состоит из вариабельного участка тяжелой цепи («HCVR» или «VH») и константного участка тяжелой цепи (состоящего из доменов СН1, СН2 и СН3). Каждая легкая цепь состоит из вариабельного участка легкой цепи («LCVR» или «VL») и константного участка легкой цепи (CL). Участки VH и VL могут быть дополнительно подразделены на участки гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с более консервативными участками, называемыми каркасными участками (FR). Каждый VH и VL состоит из трех CDR и четырех FR, расположенных от амино- до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В определенных вариантах осуществления настоящего изобретения FR антитела (или его антигенсвязывающий фрагмент) могут быть идентичными последовательностям зародышевой линии человека или могут быть естественно или искусственно изменены. Аминокислотная консенсусная последовательность может быть определена на основании анализа «бок о бок» двух или более CDR.
Используемый в настоящем изобретении термин «SARS-COV-2» относится к вирусу, который является возбудителем коронавирусной инфекции COVID-19 и представляет собой оболочечный одноцепочный РНК-вирус, относящийся к семейству Coronaviridae и роду Betacoronavirus. Геном коронавирусов кодирует 4 главных структурных белка: шиловидный (S), оболочечный (Е), мембранный (М) и нуклеокапсидный (N). S-белок (шиловидный белок) представляет собой очень крупный трансмембранный белок, который собирается в тримеры и образует характерные «шипы» на поверхности коронавирусов. Каждый S-мономер состоит из N-концевой субъединицы S1 и мембранно-проксимальной субъединицы S2. Вирус проникает в клетку-хозяина путем связывания S-белка с ангиотензин-превращающим ферментом 2-го типа (АСЕ2), который находится на поверхности клеток разных типов. АСЕ2 работает как рецептор вируса и взаимодействует с рецептор-связывающим доменом (RBD) субъединицы S1.
Используемый в настоящем изобретении термин «эпитоп» относится к антигенной детерминанте, которая взаимодействует со специфическим антигенсвязывающим сайтом в вариабельном участке молекулы антитела, известным как паратоп. Один антиген может иметь более одного эпитопа. Таким образом, различные антитела могут связываться с различными областями на антигене и могут иметь различные биологические эффекты. Термин «эпитоп» также относится к сайту на антигене, с которым вступают в реакцию В- и/или Т-клетки. Он также относится к участку антигена, с которым связывается антителом. Эпитопы можно определить как структурные или функциональные. Функциональные эпитопы в общем представляют собой подгруппу структурных эпитопов и содержат те остатки, которые непосредственно обусловливают свойство аффинности взаимодействия. Также эпитопы могут быть конформационными, т.е. состоять из нелинейных аминокислот. В определенных вариантах осуществления эпитопы могут включать детерминанты, которые представляют собой химически активные поверхностные группы молекул, таких как аминокислоты, боковые цепи Сахаров, фосфорильные группы или сульфонильные группы, и в определенных вариантах осуществления могут иметь специфические характеристики трехмерных структур и/или специфические характеристики заряда.
Используемый в настоящем изобретении термин «человеческое антитело» предназначен для включения антител, имеющих вариабельные и константные участки, полученные из последовательностей иммуноглобулинов зародышевой линии человека.
Используемый в настоящем изобретении термин «специфически связывает», или «специфически связывается с», или подобный означает, что антитело или его антигенсвязывающий фрагмент образуют комплекс с антигеном, который является относительно устойчивым в физиологических условиях. Специфическое связывание можно характеризовать при помощи равновесной константы диссоциации, составляющей по меньшей мере приблизительно 1×10-8 М или менее (например, значение меньше KD означает более сильное связывание). Способы определения наличия специфического связывания двух молекул хорошо известны в данной области и включают, например, равновесный диализ, поверхностный плазмонный резонанс и им подобные.
Используемый в настоящем изобретении термин «выделенное антитело», предназначен для обозначения антитела, которое, по сути, не содержит антител, специфичных в отношении другого антигена.
В одном аспекте настоящего изобретения предложено биспецифическое антитело или его антиген-связывающий фрагмент, специфически связывающееся по меньшей мере с одним эпитопом в рецептор-связывающем домене (RBD) шиповидного белка (S) коронавируса тяжелого острого респираторного синдрома (SARS-COV-2).
Разработанная молекула биспецифического моноклонального антитела Bite anti-Spike/CD3 представляет собой рекомбинантное моноклональное химерное антитело сконструированное генно-инженерным способом, селективно связывающееся с поверхностным шиповидным (Spike) белком капсида вируса SARS-COV-2, и Т-клеточным ко-рецептором (CD3) в составе протеинового комплекса, экспрессируемым на поверхности Т-клеток. Антитело способно связываться с рецептор связывающим доменом шиповидного белка вируса SARS-COV-2, тем самым обладая нейтрализующим потенциалом в отношении вируса SARS-COV-2. Антитело Bite anti-Spike/CD3 активирует эндогенные Т-клетки, соединяя CD3 в комплексе Т-клеточного рецептора (ТКР) с Spike SARS-COV-2 на поверхности зараженных клетках. Биологическая активность антитела Bite anti-Spike/CD3 проявляется через опосредованное образование цитолитического синапса между Т-клеткой и зараженной клеткой, что приводит к гибели клетки-мишени. Доказано, что антитело Bite anti-Spike/CD3 способно связываться с CD3-рецептором на поверхности CD4-/ CD8-положительных клеток. За счет биспецифической активности антитело Bite anti-Spike/CD3 может иметь терапевтический потенциал для лечения COVID-19 и профилактическое применение для предупреждения инфекционных заболеваний вызванных вариантами SARS-COV-2.
Согласно одному варианту реализации настоящего изобретения указанный эпитоп является нелинейным.
Согласно дополнительному варианту реализации настоящего изобретения указанный эпитоп содержит область в пределах аминокислот 319-541 шиповидного белка.
Согласно предпочтителному варианту реализации настоящего изобретения, указанный эпитоп содержит область в пределах аминокислот 319-490 шиповидного белка.
Согласно дополнительному варианту реализации настоящего изобретения указанное антитело ингибирует слияние вирусной и клеточной мембраны.
Согласно еще одному дополнительному варианту реализации настоящего изобретения антитело блокирует связывание шиповидного белка SARS-COV-2 с рецептором ангиотензинпревращающего фермента 2 (АСЕ2) на клеточной поверхности.
Согласно еще одному варианту реализации настоящего изобретения антитело содержит аминокислотную последовательность, выбранную из SEQ ID NO: 1-7.
Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 1. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 2. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 3. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 4. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 5. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 6. Согласно дополнительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 7.
Согласно предпочтительному варианту реализации настоящего изобретения указанное антитело содержит аминокислотную последовательность, представленную в SEQ ID NO: 1.
Согласно дополнительному варианту реализации настоящего изобретения указанное антитело представляет собой полностью человеческое антитело.
В другом аспекте настоящего изобретения предложен способ получения антитела, включающий следующие этапы: а) конструирование рекомбинантной экспрессионной плазмидной ДНК, кодирующей антитело по любому из пл. 1-9, б) трансфекцию полученной ДНК клеток СНО, в) культивирование полученной линии клеток, и г) выделение указанного антитела.
Согласно одному из вариантов реализации изобретения конструирование рекомбинантной экспрессионной плазмидной ДНК осуществляется способами, широко известными специалисту в области техники.
Согласно дополнительному варианту реализации настоящего изобретения после трансфекции клетки культивировали на среде BalanCD с добавлением 500000 ЕД пенициллина, 100 мкг/см3 стрептомицина и 4 мМ L-глютамина при температуре 37°С, содержании CO2-5%, влажности не более 75% в течении 7 дней до точки снижения жизнеспособности ниже 80%.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
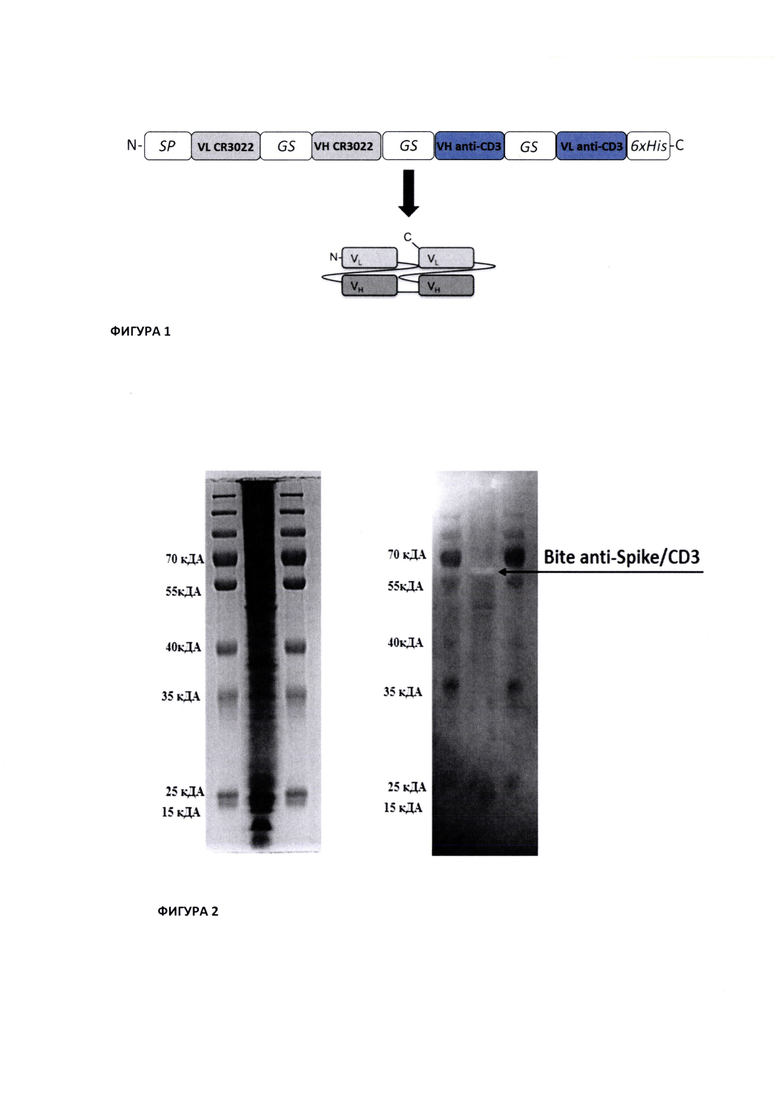
На Фиг. 1 показана доменная организация моноклонального биспецифического антитела (Bite) anti-Spike/CD3.
На Фиг. 2 показаны результаты Western Blot анализа антитела Bite anti-Spike/CD3 в культуральной жидкости транзиента.
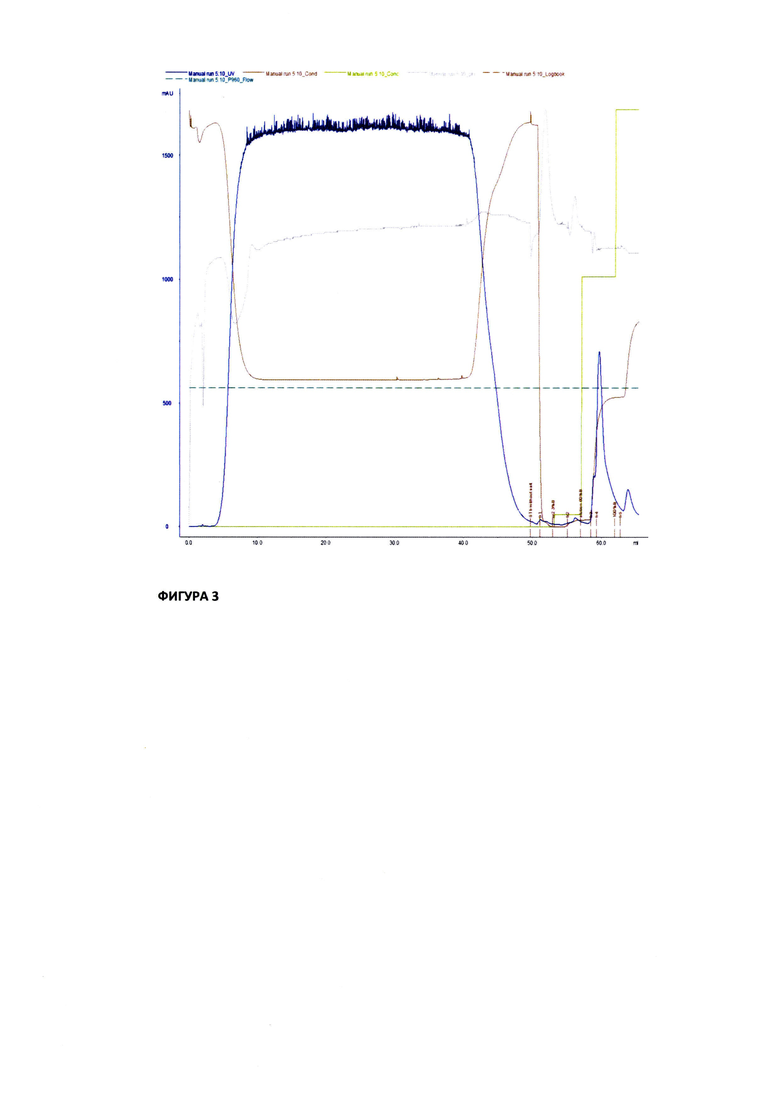
На Фиг. 3 показаны результаты хроматографической очистки и выделения антитела Bite anti-Spike/CD3 из культуральной жидкости на Hi Trap IMAC HP 5 мл (GE)
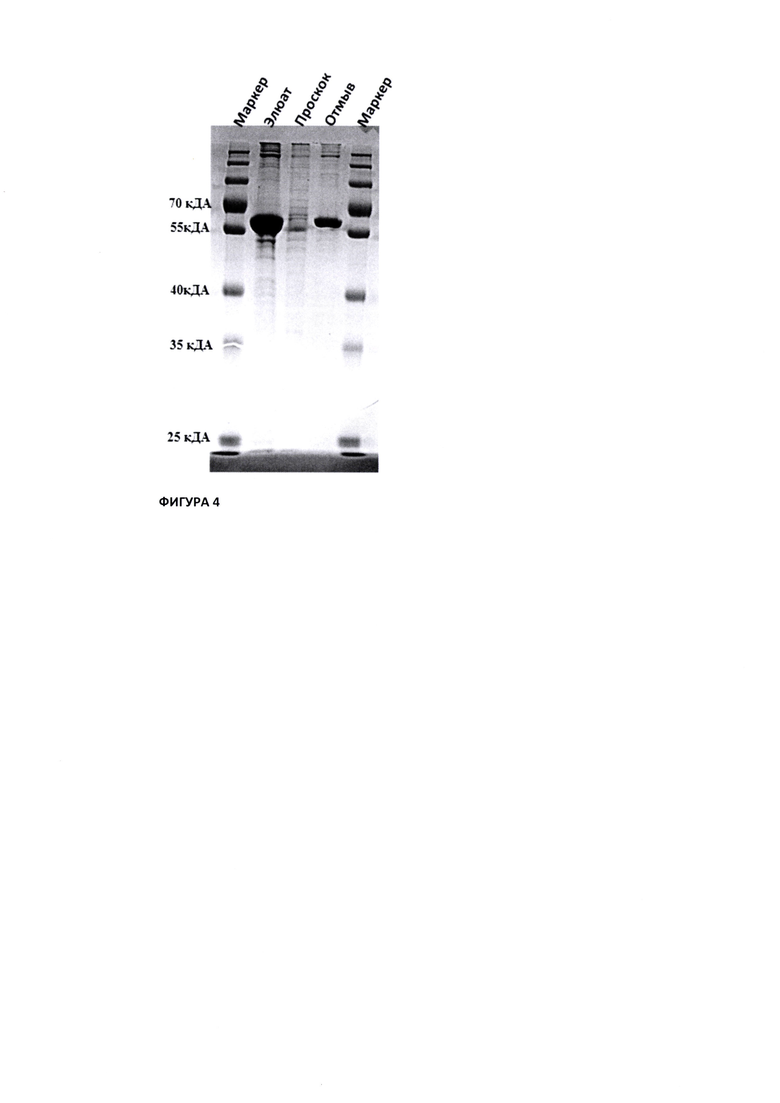
На Фиг. 4 показана электрофореграмма аналитического SDS Page на наличие антитела Bite anti-Spike/CD3 после стадии Hi Trap IMAC HP 5 мл (GE)
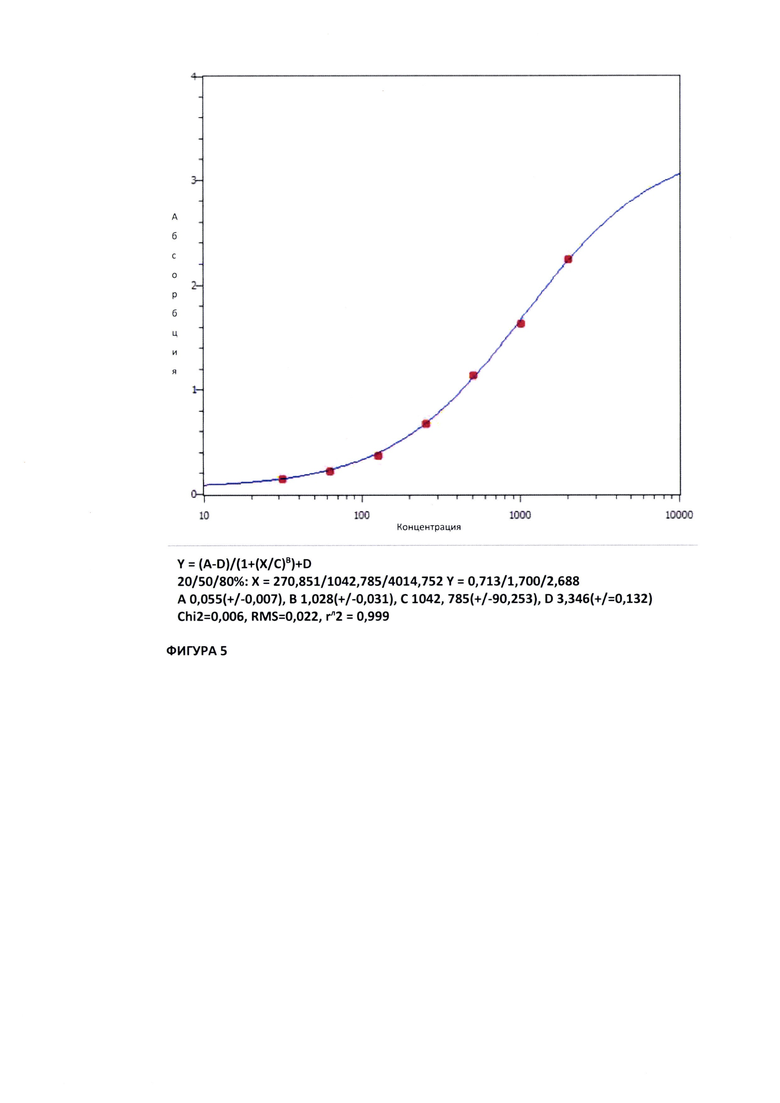
На Фиг. 5 показана кривая взаимодействия белка SARS-COV-2 (2019-nCOV) Spike S1+S2 (Sino Biological, кат. №40589-V08B1) с антителом Bite anti-Spike/CD3 в дозозависимом диапазоне.
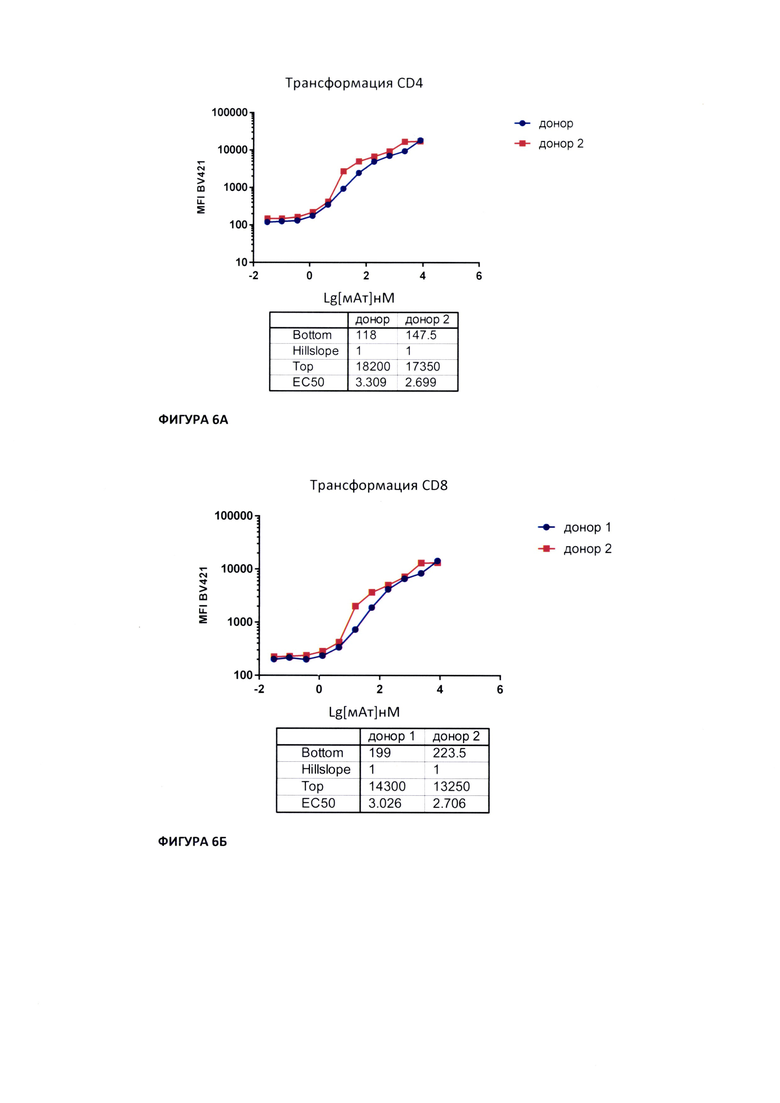
На Фиг. 6А показано определение полумаксимальной эффективной дозы (ЕС50) мАт для популяций CD4-положительных клеток.
На Фиг. 6Б показано определение полумаксимальной эффективной дозы (ЕС50) мАт для популяций CD4-положительных клеток.
Следующие примеры предлагаются с тем, чтобы обеспечить специалистам в данной области техники полное раскрытие и описание того, как получать и применять изобретение, но они не предназначены для ограничения объема того, что изобретатели рассматривают в качестве своего изобретения.
ПРИМЕРЫ
Пример 1. Способ получения антитела Bite anti-Spike/CD3
1.1 Дизайн антитела Bite anti-Spike/CD3.
Для сборки Bite формата антитела использовали вариабельные области двух антител: антитело формата IgG1, связывающий шиловидный домен вируса SARS-COV-2-CR3022; антитело против CD3 Т-клеток - блинатумомаб. Аминокислотные последовательности антител CR3022 и anti-CD3 взяты из научных публикаций и открытых источников информации. Сигнальный пептид белковой молекулы описан в уровне техники [Correnti, С.Е., Laszlo, G.S., de van der Schueren, W.J. et al. Simultaneous multiple interaction T-cell engaging (SMITE) bispecific antibodies overcome bispecific T-cell engager (BiTE) resistance via CD28 co-stimulation. Leukemia 32, 1239-1243 (2018)].
Между вариабельной областью легкой цепи CR3022 и вариабельной областью тяжелой цепи anti-CD3 (блинатумомаб) использовали длинный линкер GGGGGGGGGGGGS; между вариабельной областью тяжелой цепи anti-CD3 (блинатумомаб) и вариабельной областью тяжелой цепи CR3022 использовали короткий линкер GGGGS; между вариабельной областью тяжелой цепи CR3022 и вариабельной области легкой цепи anti-CD3 (блинатумомаб) использовали длинный линкер -GGSGGSGGSGGSGG. С- концевой конец молекулы тагировали 6 аминокислотами гистидина - 6xHis-tag (см. SEQ ID NO: 1). Доменная организация полученного антитела Bite anti-Spike/CD3 представлена на Фиг. 1.
1.2. Создание конструкции pCMV_anti-Spike/CD3.
Нуклеотидную последовательность антитела Bite anti-Spike/CD3 синтезировали с оптимизацией кодонового состава под экспрессию в клетках млекопитающих: клетки HEK293 и СНО. Ген Bite anti-Spike/CD3 и вектор pCMV3-untagged (Sino Biological) подвергали рестрикционной обработке двумя эндонуклеазами HindIII и XbaI для получения липких концов и затем очищали от эндонуклеаз и продуктов рестрикционного расщепления с помощью набора («Qiagene», Германия). Полученные фрагменты лигировали по липким концам, формирующимся в результате рестрикционного расщепления. Реакцию лигирования вектора и вставки осуществляли с помощью ДНК-лигазы фага Т4 («Neb», США), в эквимолярном соотношении (вектор/вставка - 1/3). Отобранные единичные колонии пересевали в жидкую среду 2YT с содержанием селективного антибиотика ампициллина 100 мкг/см3 и культивировали на шейкере при 37°С в течение 16 часов при 220 об/мин. Затем из полученной ночной культуры с помощью набора «Plasmide MiniPrep» («Qiagene», Германия) выделяли плазмидную ДНК, которую далее использовали в аналитической рестрикции. Полученные клоны на этапе скрининга анализировали на наличие специфической вставки с помощью аналитической рестрикции по фланкирующим эндонуклеазам (HindIII и XbaI).
1.3. Транзиторная экспрессия антитела Bite anti-Spike/CD3 в клетках СНО.
Для получения рекомбинантных белков была выбрана эукариотическая система экспрессии с использованием клеточной линии яичников китайского хомячка (Chinese hamster ovary - СНО) «FreeStyle CHO-S» (ThermoFisher, США).
Трансфекцию клеток СНО, находящиеся в фазе логарифмического роста, осуществляли с помощью электропорации приборе Gene Pulse Xcell («Bio-Rad», США). После трансфекции клетки культивировали на среде BalanCD с добавлением 500000 ЕД пенициллина, 100 мкг/см3 стрептомицина и 4 мМ L-глютамина при температуре 37°С, содержании CO2-5%, влажности не более 75% в течении 7 дней до точки снижения жизнеспособности ниже 80%. Образцы культуральной жидкости отбирали для оценки экспрессии антитела. Оценку наличия антитела Bite anti-Spike/CD3 в культуральной жидкости проводили с помощью анализа Western Blot.
1. 4. Анализ Western Blot антитела Bite anti-Spike/CD3 в культуральной жидкости транзиента.
В качестве образцов для анализа использовали образцы культуральной жидкости транзиента. Для проведения анализа WB на 12,5% полиакриламидный гель наносили по 10 мкл образца, подготовленных для белкового фореза. Перенос белковых образцов проводили полусухим способом с использованием PVDF мембраны. Перенос проводили при силе тока в 150 мА, в течение 50 мин. Неспецифическое блокирование мембраны PVDF после переноса проводили буфером PBST с добавлением сухого молока в концентрации 2%.
Мембрану инкубировали в растворе антител Rb pAb to 6х His tag HRP-conjugate (разведение раствора 1:5000), по окончании инкубации мембрану промывали однократным буфером PBST, затем на мембрану наносили раствор для появления хемилюминесценции. Результат реакции фиксировался системой документирования гелей.
Для сопоставления анализа уровня экспрессии также наносили тот же образец культуральной жидкости транзиента на 2,5% полиакриламидный гель (SDS Page) с дальнейшим окрашиванием раствора Кумасси (см. Фиг. 2).
Анализ Western Blot образца в культуральной жидкости транзиента показал наличие специфичной полосы молекулярным весом от 55 до 70 кДа. Наличие единственной полосы в данном области указывает на характерную электрофоретическую подвижность антитела Bite anti-Spike/CD3 имеющую схожую теоретическую молекулярную массу, рассчитанной по аминокислотной последовательности. При анализе 12,5% SDS Page четкой полосы невозможно определить из-за большой нагрузки по белку на лунку.
1.5. Хроматографическая очистка и выделение антитела Bite anti-Spike/CD3 из культуральной жидкости.
Выделение рекомбинантного антитела Bite anti-Spike/CD3 проводили с помощью металл-хелатной хроматографии с применением IMAC Sepharose FF. Н первом этапе очистки использовали колонку Hi Trap IMAC HP 5 мл (GE). Хроматографию проводили при 4°С с использованием системы  pure. Колонку предварительно иммобилизировали раствором 0, 1 М NiCl2, затем уравновешивали 5 объемами колонки (CV) раствора уравновешивающего буфера 20 mM Трис-HCl, 0.3 М NaCl, рН 7.8. К осветленному транзиенту обьемом 50 мл добавляли раствор 5М NaCl до конечной концентрации 0,3 М, доводили рН до 7,8 раствором 5М NaOH и наносили (3 мл/мин) на колонку с Ni2+ IMAC HP (5 мл). После нанесения колонку последовательно промывали 3 CV уравновешивающего буфера. 3 CV буферного раствора 20 mM Трис-HCl, рН 7.8 и 3 CV буферного раствора 20 mM Трис-HCl, 0,015 М имидазол, рН 7.8. Целевой белок элюировали 0,3 М имидазола в буфере 20 mM Трис-HCl, рН 7.8 (Фиг. 3.). Аликвоты по 10 мкл из собранных фракций элюата, проскока и промывок анализировали на присутствие белка в SDS-PAGE (см. Фиг. 4).
pure. Колонку предварительно иммобилизировали раствором 0, 1 М NiCl2, затем уравновешивали 5 объемами колонки (CV) раствора уравновешивающего буфера 20 mM Трис-HCl, 0.3 М NaCl, рН 7.8. К осветленному транзиенту обьемом 50 мл добавляли раствор 5М NaCl до конечной концентрации 0,3 М, доводили рН до 7,8 раствором 5М NaOH и наносили (3 мл/мин) на колонку с Ni2+ IMAC HP (5 мл). После нанесения колонку последовательно промывали 3 CV уравновешивающего буфера. 3 CV буферного раствора 20 mM Трис-HCl, рН 7.8 и 3 CV буферного раствора 20 mM Трис-HCl, 0,015 М имидазол, рН 7.8. Целевой белок элюировали 0,3 М имидазола в буфере 20 mM Трис-HCl, рН 7.8 (Фиг. 3.). Аликвоты по 10 мкл из собранных фракций элюата, проскока и промывок анализировали на присутствие белка в SDS-PAGE (см. Фиг. 4).
Анализ наличия антитела Bite anti-Spike/CD3 с помощью SDS Page (см. Фиг. 4) показал, что в основной фракции элюата был обнаружен целевой белок с мажорной полосой в диапазоне молекулярного веса от 55 до 70 кДа. Молекулярный вес целевого белка совпадает с молекулярным весом идентифицированного белка при анализе культуральнйо жидкости транзиента на этапе анализа наличия в культуральной жидкости транзиента антитела Bite anti-Spike/CD3 с помощью анализа Western Blot (Фиг. 2). Таким образом, можно отметить, что авторами настоящего изобретения была выделена целевая фракция, характерная для антитела Bite anti-Spike/CD3. После подтверждения подлинности антитела Bite anti-Spike/CD3 основную фракцию элюата диализовали в буфер готовой формы - PBS.
1.6. Измерение концентрации антитела Bite anti-Spike/CD3.
Определение концентрации проводили спектрофотометрическим методом при длине волны 280 нм в кюветах с длиной оптического пути 1 см по сравнению с буфером готовой формы (БГФ) - PBS. Рассчитанный коэффициент экстинкции для антитела Bite anti-Spike/CD3 в программе PROTEIN CALCULATOR v3.4 (http://protcalc.sourceforge.net/) составил - ε'X=280nm=2.1483 (мг/мл)-1 см-1. Концентрацию выражали в мг/мл и рассчитывали по формуле:
ODcp - среднее значение оптической плотности,
R - значение разведения,
Кэ (Коэффициент экстинкции) = 2,1483
Полученные результаты:

Пример 2. Подтверждение активности антитела Bite anti-Spike/CD3. Связывание Spike белком SARS-COV-2.
Метод определения основан на твердофазном иммуноферментном анализе - ИФА с применением рекомбинантных антигена S1 коронавируса SARS-COV-2 и моноклонального Bite anti-Spike/CD3. Метод основан на взаимодействии S-белка с одной вариабельной областей (CR3022) антитела Bite anti-Spike/CD3. Образующийся комплекс после инкубации и отмывки обнаруживали с помощью мышиных антител к 6xHis-tag (Ab1187, Abeam), конъюгированных с пероксидазой хрена.
Белок SARS-COV-2 (2019-nCoV) Spike S1+S2 (Sino Biological, кат. №40589-V08B1) с концентрацией 100 мкг/мл наносили на планшеты по 100 мкл в лунку от концентрации 2000 нг на лунку 0 до нг на лунку разведенные в фосфатно-солевом буферном растворе для иммуноферментного анализа рН 7,2-7,6 (PBSt). Планшет заклеивали пленкой и инкубировали в течение 60±10 минут при температуре (37±2)°С при 250 об/мин. После инкубации лунки планшетов промывали 4 раза раствором PBSt по 800 мкл/лунку.
Антитело Bite anti-Spike/CD3 разводили в PBSta в 1000 раз и наносили по 100 мкл/лунку раствора антител. Планшеты заклеивали пленкой и инкубировали в течение 60±10 минут при температуре (37±2)°С при 250 об/мин. После инкубации лунки планшетов промывали 4 раза по 800 мкл/лунку раствором PBSt. Далее вносили мышиные антитела к 6xHis-tag (Ab1187, Abeam), конъюгированные с пероксидазой хрена, разведенные в 2500 раз, также в PBSt. И инкубировали в течение 60±10 минут при температуре (37±2)°С при 250 об/мин. После инкубации лунки планшетов промывали 4 раза раствором PBSt по 800 мкл/лунку.
После промывки в лунки планшетов вносили по 100 мкл раствора субстрата 3,3',5,5'-тетраметилбензидина («БиоТест Системы», кат. №01016-1). Инкубировали планшеты при комнатной температуре, до появления интенсивного синего окрашивания (в течение 5 мин). Реакцию останавливали внесением 100 мкл/лунку стоп-раствора (0,5 М раствор серной кислоты). Измеряли оптическую плотность на спектрофотометре при длине волны 450 нм.
В результате было показано специфиячное образование комплекса антиген-антитело. Белок SARS-COV-2 (2019-nCOV) Spike S1+S2 (Sino Biological, кат. №40589-V08B1) связывался с антителом Bite anti-Spike/CD3 в дозозависимом диапазоне. На Фиг. 5 показана кривая взаимодействия белка SARS-COV-2 (2019-nCOV) Spike S1+S2 (Sino Biological, кат. №40589-V08B1) с антителом Bite anti-Spike/CD3 в дозозависимом диапазоне.
Пример 3. Связывание антитела Bite anti-Spike/CD3 с CD3, экспрессированными на поверхности Т- клеток
Подготовка клеток и инкубация с исследуемым мАт. Ампулу с мононуклеарными клетками периферической крови (далее - МКПК) размораживали на водяной бане при температуре (37±2)°С в течение 2-3 мин. Содержимое ампулы переносили в центрифужную пробирку вместимостью 15 мл, содержащую 14,0 мл теплой питательной среды DMEM/F12 с 10% фетальной бычьей сыворотки. Центрифугировали в течение 5 мин при скорости 400g в условиях комнатной температуры (22±2)°С. Надосадочную жидкость удаляли, осадок клеток ресуспендировали в 1,0 мл стерильного фосфатно-солевого буферного раствора, доводили объем до 15,0 мл ФСБ и центрифугировали в течение 5 мин при скорости 450g при температуре (22±2)°С. Ресуспендировали в 1 мл FACS и считали концентрацию клеток. В эксперименте использовали клетки с жизнеспособностью не менее 95%. Готовили суспензию клеток с концентрацией 1 млн/мл в нагретом до 20 (±5°С) FACS буфере.
Переносили суспензию клеток в лунки V-образного планшета в объеме 100 мкл. В лунки планшета вносили 100 мкл разведений исследуемого мАт и осторожно пипетировали. Инкубировали в течение (30±5) мин при 5 (±3°С). По окончанию инкубации дважды отмывали клетки охлажденным до 5 (±3°С) FACS буфером, осаждая при 450 g 5 минут при 5 (±3°С).
Цитофенотипирование клеток. Готовили раствор для блокирования неспецифического связывания с Fc-рецепторами клеток (человеческий Fc-блок) в разведении 1:200 в охлажденном FACS буфере. В растворе Fc-блока готовили смесь красок: anti-Human CD4-PerCP Су5.5 (PerCP-Cy™5.5 Mouse Anti-Human CD4 Clone L200 cat. №552838, 1:150), Anti-Human CD8-BB515 (BB515 Mouse Anti-Human CD8, Clone RPA-T8, cat. №1:200), Streptavidin BV421 (BV421 Streptavidin6 cat. №563259, разведение 1:200).
Инкубировали в течение 20 (±5) мин во льду. По окончанию инкубации, клетки однократно отмывали 2,0 мл охлажденного FACS-буфера и осаждали центрифугированием 5 минут при 500g при температуре (5±3)°С. Фиксировали клетки в 100 мкл FIX буфера.
Гейтирование целевых популяций. Интенсивность флуоресценции клеток измеряли на проточном цитофлуориметре BD LSR Fortessa. В координатах прямого и бокового светорассеяния выделяли гейт синглетных событий. В гейте синглетных событий выделяли клеточный гейт. Из клеточного гейта выделяли CD4- и CD8-положительные клетки. Для определения процента клеток, связанных с исследуемым мАт в популяциях CD4- и CD8-положительных клеток выделяли популяцию клеток, положтельную по BV421. Клетки, положительные по BV421 определяли как клетки, связанные с исследуемым мАт. Определения значений ЕС50 проводили по медиане флуоресцентного сигнала по BV421 (далее - MFI).
Обработка результатов. Обработку результатов проводили в программе FlowJo версии 7.6.5. Средние значения, стандартное отклонение (SD) и коэффициент вариации (CV) рассчитывали для каждой экспериментальной точки в программе Microsoft Excel. Расчет критерия приемлемости R2 (коэффициент достоверности аппроксимации) и обработку результатов проводили в программе GraphPadPrism 8.0, аппроксимируя зависимость MFI по BV421 от десятичного логарифма разведения мАт 4-х параметрической логистической функцией «доза-ответ». Определяли полумаксимальную эффективную дозу (ЕС50) мАт для популяций CD4- и CD8-положительных клеток (Фиг. 6А и 6Б).
1. При инкубации МКПК условно здорового донора с 500 нМ исследуемого мАт, 97,8% CD4-положительных и 61,05% CD8-положительных клеток были связаны с исследуемым мАт.
2. Значение ЕС50 связывания исследуемого мАт с CD3-рецептором на поверхности CD4- положительных клеток условно-здоровых доноров составило 2,699 и 3,309 нМ для доноров 1 и 2 соответственно.
3. Значение . ЕС50 связывания исследуемого мАт с CD3-рецептором на поверхности CD8- положительных клеток условно здоровых доноров составило 2,706 и 3,026 нМ для доноров 1 и 2 соответственно.
Таким образом, исследуемое мАт способно связываться с CD3-рецептором на поверхности CD4-/CD8-положительных клеток. Значения полумаксимальной эффективной концентрации связывания с CD3-рецептором на поверхности CD4-/ CD8-положительных клеток различается для разных доноров.
Следует понимать, что объем настоящего изобретения не ограничен конкретными вариантами осуществления, описанными в данном документе. Действительно, различные модификации настоящего изобретения, помимо описанных в настоящей заявке, будут очевидны специалистам в данной области техники исходя из изложенного выше описания и прилагаемых фигур. Такие модификации находятся в пределах объема прилагаемой формулы изобретения.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>
<120> БИСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ SARS-CoV-2
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 504
<212> Антитело
<213> Искусственная последовательность
<400> 1
EIVLT QSPGT LSLSP GERAT LSCRA SQSVS SSYLA WYQQK PGQAP RLLIY GASSR
ATGIP DRFSG SGSGT DFTLA ISRLE PEDFA VYYCQ QYGSS PTFGQ GTKVE IKGGG
GSGGG GSGGG GS2QV QLQQS GAELV RPGSS VKISC KASGY AFSSY WMNWV KQRPG
QGLEW IGQIW PGDGD TNYNG KFKGK ATLTA DESSS TAYMQ LSSLA SEDSA VYFCA
RRETT TVGRY YYAMD YWGQG TTVTV SSGGG GS3EV QLLES GGGLV QPGGS LRLSC AASGF
TFSWY AMSWV RQAPG KGLEW VSYIR HQAGS TEYAD SVKGR FTISR DNSKN TLYLQ MNSLR
AEDTA VYYCA RGDYK RRAYS AIDYW GQGTL VTVSS GGSGG SGGSG GSGG4 VDDIQ LTQSP
AIMSA SPGEK VTMTC RASSS VSYMN WYQQK SGTSP KRWIY DTSKV ASGVP YRFSG SGSGT
SYSLT ISSME AEDAA TYYCQ QWSSN PLTFG AGTKL ELKHH HHHH
<210> 2
<211> 504
<212> Антитело
<213> Искусственная последовательность
<400> 2
EIVLT QSPGT LSLSP GERAT LSCRA SQSVS SSYLA WYQQK PGQAP RLLIY GASSR
ATGIP DRFSG SGSGT DFTLA ISRLE PEDFA VYYCQ QYGSS PTFGQ GTKVE IKGGG
GSGGG GSGGG GSDIQ LTQTP ASLAV SLGQR ATISC KASQS VDYDG DSYLN WYQQI
PGQPP KLLIY DASNL VSGIP PRFSG SGSGT DFTLN IHPVE KVDAA TYHCQ QSTED
PWTFG GGTKL EIKGG GGSEV QLLES GGGLV QPGGS LRLSC AASGF TFSWY AMSWV
RQAPG KGLEW VSYIR HQAGS TEYAD SVKGR FTISR DNSKN TLYLQ MNSLR AEDTA
VYYCA RGDYK RRAYS AIDYW GQGTL VTVSS GGSGG SGGSG GSGGQ VQLQQ SGAEL
VRPGS SVKIS CKASG YAFSS YWMNW VKQRP GQGLE WIGQI WPGDG DTNYN GKFKG
KATLT ADESS STAYM QLSSL ASEDS AVYFC ARRET TTVGR YYYAM DYWGQ GTSVT
VSSHH HHHH
<210> 3
<211> 497
<212> Антитело
<213> Искусственная последовательность
<400> 3
EIVLT QSPGT LSLSP GERAT LSCRA SQSVS SSYLA WYQQK PGQAP RLLIY GASSR ATGIP
DRFSG SGSGT DFTLA ISRLE PEDFA VYYCQ QYGSS PTFGQ GTKVE IKGGG GSGGG GSGGG
GSDVV MTQSH RFMST SVGDR VSITC RASQD VNTAV SWYQQ KPGQS PKLLI FSASY RYTGV
PDRFT GSGSG ADFTL TISSV QAEDL AVYYC QQHYS TPWTF GGGTK LDIKG GGGSE VQLLE
SGGGL VQPGG SLRLS CAASG FTFSW YAMSW VRQAP GKGLE WVSYI RHQAG STEYA DSVKG
RFTIS RDNSK NTLYL QMNSL RAEDT AVYYC ARGDY KRRAY SAIDY WGQGT LVTVS SGGSG
GSGGS GGSGG QIQLV QSGPD LKKPG ETVKL SCKAS GYTFT NFGMN WVKQA PGKGF KWMAW
INTYT GESYF ADDFK GRFAF SVETS ATTAY LQINN LKTED TATYF CARGE IYYGY DGGFA
YWGQG TLVTV SHHHH HH
<210> 4
<211> 497
<212> Антитело
<213> Искусственная последовательность
<400> 4
EIVLT QSPGT LSLSP GERAT LSCRA SQSVS SSYLA WYQQK PGQAP RLLIY GASSR ATGIP
DRFSG SGSGT DFTLA ISRLE PEDFA VYYCQ QYGSS PTFGQ GTKVE IKGGG GSGGG GSGGG
GSAIQ MTQSP SSLSA SVGDR VTITC RASED IYNGL AWYQQ KPGKA PKLLI YGASS LQDGV
PSRFS GSGSG TEFTL TISSL QPEDE ATYYC AGPHK YPLTF GGGTK VEIKG GGGSE VQLLE
SGGGL VQPGG SLRLS CAASG FTFSW YAMSW VRQAP GKGLE WVSYI RHQAG STEYA DSVKG
RFTIS RDNSK NTLYL QMNSL RAEDT AVYYC ARGDY KRRAY SAIDY WGQGT LVTVS SGGSG
GSGGS GGSGG EVQLL ESGGG LVQPG GSLRL SCAAS GFTFS NYDMA WVRQA PGKGL EWVSS
ISTRG DITSY RDSVK GRFTI SRDNS KNTLY LQMNS LRAED TAVYY CARQD YYTDY MGFAY
WGQGT LVTVS SHHHH HH
<210> 5
<211> 502
<212> Белок
<213> Искусственная последовательность
<400> 5
DIQLT QSPDS LAVSL GERAT INCKS SQSVL YSSIN KNYLA WYQQK PGQPP KLLIY WASTR ESGVP
DRFSG SGSGT DFTLT ISSLQ AEDVA VYYCQ QYYST PYTFG QGTKV EIKGG GGSGG GGSGG GGSQV
QLQQS GAELV RPGSS VKISC KASGY AFSSY WMNWV KQRPG QGLEW IGQIW PGDGD TNYNG KFKGK
ATLTA DESSS TAYMQ LSSLA SEDSA VYFCA RRETT TVGRY YYAMD YWGQG TTVTV SSGGG GSQMQ
LVQSG TEVKK PGESL KISCK GSGYG FITYW IGWVR QMPGK GLEWM GIIYP GDSET RYSPS FQGQV
TISAD KSINT AYLQW SSLKA SDTAI YYCAG GSGIS TPMDV WGQGT TVTVG GSGGS GGSGG SGGVD
DIQLT QSPAI MSASP GEKVT MTCRA SSSVS YMNWY QQKSG TSPKR WIYDT SKVAS GVPYR FSGSG
SGTSY SLTIS SMEAE DAATY YCQQW SSNPL TFGAG TKLEL KHHHH HH
<210> 6
<211> 463
<212> Антитело
<213> Искусственная последовательность
<400> 6
DIQLT QSPDS LAVSL GERAT INCKS SQSVL YSSIN KNYLA WYQQK PGQPP KLLIY WASTR ESGVP
DRFSG SGSGT DFTLT ISSLQ AEDVA VYYCQ QYYST PYTFG QGTKV EIKGG GGSGG GGSGG GGSAI
QMTQS PSSLS ASVGD RVTIT CRASE DIYNG LAWYQ QKPGK APKLL IYGAS SLQDG VPSRF SGSGS
GTEFT LTISS LQPED EATYY CAGPH KYPLT FGGGT KVEIK GGGGS QMQLV QSGTE VKKPG ESLKI
SCKGS GYGFI TYWIG WVRQM PGKGL EWMGI IYPGD SETRY SPSFQ GQVTI SADKS INTAY LQWSS
LKASD TAIYY CAGGS GISTP MDVWG QGTTV TVGGS GGSGG SGGSG GEVQL LESGG GLVQP GGSLR
LSCAA SGFTF SNYDM AWVRQ APGKG LEWVS SISTR GDITS YRDSV KGRFT ISRDN SKNTL YLQMN
SLRAE DTAVY YCARQ DYYTD YMGFA YWGQG TLVTV SSHHH HHH
<210> 7
<211> 498
<212> Белок
<213> Искусственная последовательность
<400> 7
DIQLT QSPDS LAVSL GERAT INCKS SQSVL YSSIN KNYLA WYQQK PGQPP KLLIY WASTR ESGVP
DRFSG SGSGT DFTLT ISSLQ AEDVA VYYCQ QYYST PYTFG QGTKV EIKGG GGSGG GGSGG GGSDV
VMTQS HRFMS TSVGD RVSIT CRASQ DVNTA VSWYQ QKPGQ SPKLL IFSAS YRYTG VPDRF TGSGS
GADFT LTISS VQAED LAVYY CQQHY STPWT FGGGT KLDIK GGGGS QMQLV QSGTE VKKPG ESLKI
SCKGS GYGFI TYWIG WVRQM PGKGL EWMGI IYPGD SETRY SPSFQ GQVTI SADKS INTAY LQWSS
LKASD TAIYY CAGGS GISTP MDVWG QGTTV TVGGS GGSGG SGGSG GEVQL LESGG GLVQP GGSLR
LSCAA SGFTF SNYDM AWVRQ APGKG LEWVS SISTR GDITS YRDSV KGRFT ISRDN SKNTL YLQMN
SLRAE DTAVY YCARQ DYYTD YMGFA YWGQG TLVTV SSHHH HHH
| название | год | авторы | номер документа |
|---|---|---|---|
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
| Моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817697C1 |
| ПОЛИПЕПТИДНЫЙ МОДУЛЬ ДЛЯ СВЯЗЫВАНИЯ КОНСЕРВАТИВНОГО ЭПИТОПА РЕЦЕПТОР-СВЯЗЫВАЮЩЕГО ДОМЕНА БЕЛКА SPIKE КОРОНАВИРУСА SARS-COV-2 | 2022 |
|
RU2809183C1 |
| Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2 | 2021 |
|
RU2794141C2 |
| Однодоменное антитело ламы Н5 и его производное H5-Fc, специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2022 |
|
RU2793967C1 |
| Рекомбинантный RBD S-белок коронавируса SARS-CoV-2 и способ его получения | 2023 |
|
RU2833839C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ НЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ К SARS-CoV-2 В СЫВОРОТКЕ ИЛИ ПЛАЗМЕ КРОВИ ЛЮДЕЙ, ПЕРЕНЕСШИХ COVID-19 ИЛИ ПРИВИТЫХ ВАКЦИНАМИ ДЛЯ ПРОФИЛАКТИКИ НОВОЙ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19, С ИСПОЛЬЗОВАНИЕМ НАБОРА РЕАГЕНТОВ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА, СОДЕРЖАЩЕГО РЕКОМБИНАНТНЫЙ РЕЦЕПТОР-СВЯЗЫВАЮЩИЙ ДОМЕН (RBD) ПОВЕРХНОСТНОГО ГЛИКОПРОТЕИНА S КОРОНАВИРУСА SARS-COV-2 И РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ РЕЦЕПТОР АСЕ2 | 2021 |
|
RU2784655C1 |
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |
| 5-[4-(1,3-БЕНЗОТИАЗОЛ-2-ИЛ)ФЕНИЛ]-10,15,20-ТРИС(1-МЕТИЛПИРИДИНИЙ-3-ИЛ)ПОРФИРИН ТРИИОДИД, ПРОЯВЛЯЮЩИЙ СВОЙСТВО СВЯЗЫВАНИЯ СПАЙКОВОГО БЕЛКА ВИРУСА SARS-CoV-2 | 2021 |
|
RU2773397C1 |
| СПОСОБ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ КОНЦЕНТРАЦИИ S-АНТИГЕНА В ЦЕЛЬНОВИРИОННЫХ ИНАКТИВИРОВАННЫХ АДСОРБИРОВАННЫХ НА ГИДРООКИСИ АЛЮМИНИЯ, СУБЪЕДИНИЧНЫХ НА ОСНОВЕ S-БЕЛКА, РЕКОМБИНАНТНЫХ ИЛИ ПОЛИПЕПТИДНЫХ, СОДЕРЖАЩИХ ДОМЕН RBD SPIKE-БЕЛКА ВИРУСА SARS-COV ВАКЦИНАХ ДЛЯ ПРОФИЛАКТИКИ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19 И/ИЛИ ДРУГИХ КОРОНАВИРУСНЫХ ИНФЕКЦИЙ | 2023 |
|
RU2825291C1 |
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая биспецифическое антитело и способ его получения. Биспецифическое антитело содержит в себе первый антигенсвязывающий фрагмент, специфически связывающийся с эпитопом в рецептор-связывающем домене RBD спайкового белка S коронавируса тяжелого острого респираторного синдрома SARS-COV2, и второй антигенсвязывающий фрагмент, специфически связывающийся с CD3 человека, причем указанное антитело содержит аминокислотную последовательность SEQ ID NO: 1. Изобретение расширяет арсенал средств, специфически связывающихся с эпитопом в рецептор-связывающем домене RBD спайкового белка S коронавируса тяжелого острого респираторного синдрома SARS-COV2 и специфически связывающихся с CD3 человека. 2 н. и 6 з.п. ф-лы, 7 ил., 1 табл., 3 пр.
1. Биспецифическое антитело, специфически связывающееся с эпитопом в рецептор-связывающем домене RBD спайкового белка S коронавируса тяжелого острого респираторного синдрома SARS-COV2 и CD3 человека, содержащее первый антигенсвязывающий фрагмент, специфически связывающийся с эпитопом в рецептор-связывающем домене RBD спайкового белка S коронавируса тяжелого острого респираторного синдрома SARS-COV2, и второй антигенсвязывающий фрагмент, специфически связывающийся с CD3 человека, причем указанное антитело содержит аминокислотную последовательность SEQ ID NO: 1.
2. Антитело по п. 1, отличающееся тем, что эпитоп является нелинейным.
3. Антитело по п. 1, отличающееся тем, что эпитоп содержит область в пределах аминокислот 319-541 белка S.
4. Антитело по п. 3, отличающееся тем, что эпитоп содержит область в пределах аминокислот 319-490 белка S.
5. Антитело по п. 1, где антитело ингибирует слияние вирусной и клеточной мембраны.
6. Антитело по п. 1, где антитело блокирует связывание белка S SARS-COV2 с рецептором ангиотензинпревращающего фермента 2 (АСЕ2) на клеточной поверхности.
7. Антитело по п. 1, где антитело представляет собой полностью человеческое антитело.
8. Способ получения антитела по любому из пп. 1-7, включающий следующие этапы:
а) конструирование рекомбинантной экспрессионной плазмидной ДНК, кодирующей антитело по любому из пп. 1-7,
б) трансфекцию полученной ДНК клеток СНО,
в) культивирование полученной линии клеток и
г) выделение антитела.
| О.Н | |||
| Солопова, В.А | |||
| Мисюрин, Биспецифические антитела в клинике и клинических исследованиях (обзор литературы), Клиническая онкогематология | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Плуг с фрезерным барабаном для рыхления пласта | 1922 |
|
SU125A1 |
| М.Ю | |||
| Щелканов и др | |||
| ИСТОРИЯ ИЗУЧЕНИЯ И СОВРЕМЕННАЯ КЛАССИФИКАЦИЯ КОРОНАВИРУСОВ (NIDOVIRALES: CORONAVIRIDAE), Инфекция и иммунитет, 2020, т | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Способ изготовления замочных ключей с отверстием для замочного шпенька из одной болванки с помощью штамповки и протяжки | 1922 |
|
SU221A1 |
| Zhaohui Li et al | |||
| An | |||

