Область техники
Настоящая заявка относится к новому варианту малой субъединицы синтазы ацетогидроксикислот (ilvN), полинуклеотиду, кодирующему этот вариант, вектору экспрессии, включающему полинуклеотид, микроорганизму, продуцирующему L-валин, включающему вариант малой субъединицы синтазы ацетогидроксикислоты (ilvN), и способу получения L-валина с использованием микроорганизма.
Предшествующий уровень техники
L-аминокислоты представляют собой основные структурные единицы белков и используются в качестве важных веществ для фармацевтических препаратов, пищевых добавок, кормов для животных, питательных веществ, пестицидов, бактерицидов и тому подобного. Конкретно, аминокислота с разветвленной цепью (ВСАА) представляет собой общий термин для L-валина, L-лейцина и L-изолейцина, которые представляют собой незаменимые аминокислоты, и известно, что аминокислоты с разветвленной цепью обладают антиоксидантным эффектом и эффектом непосредственно стимулировать синтез белка в мышечных клетках.
При этом, аминокислоты с разветвленной цепью продуцируются в основном микроорганизмами рода Escherichia или Corynebacterium и, как известно, биосинтезируются из 2-кетоизокапроата в качестве предшественника после прохождения нескольких стадий из пировиноградной кислоты (публикации патентов Кореи №10-0220018 и 10-0438146). Однако продуцирование L-аминокислот с разветвленной цепью посредством микроорганизмов является проблематичным для достижения крупномасштабного промышленного производства.
При данных обстоятельствах авторы настоящего изобретения подтвердили, что введение варианта с усиленной активностью гена ilvN (малой субъединицы синтазы ацетогидроксикислоты), кодирующего фермент, вовлеченный в микробный биосинтез L-валина, с целью улучшения способности продуцировать L-валин путем использования микроорганизмов, значительно увеличивало способность продуцирования L-валина.
Техническая задача Авторы настоящего изобретения разработали новый вариант малой субъединицы синтазы ацетогидроксикислоты, увеличивающий продуцирование L-валина, полинуклеотид, кодирующий этот вариант, вектор экспрессии, включающий полинуклеотид, микроорганизм, продуцирующий L-валин, включающий вариант малой субъединицы синтазы ацетогидроксикислоты, и способ получения L-валина с использованием микроорганизма, и, таким образом, завершили настоящую заявку.
Техническое решение
Согласно одному аспекту настоящей заявки предложен вариант малой субъединицы синтазы ацетогидроксикислоты (ilvN), в котором аминокислота, соответствующая положению 44 в аминокислотной последовательности SEQ ID NO: 1, заменена другой аминокислотой.
Согласно другому аспекту настоящей заявки, предложен полинуклеотид, кодирующий вариант по настоящей заявке, и вектор экспрессии, содержащий полинуклеотид.
Согласно еще одному аспекту настоящей заявки предложен микроорганизм, включающий вариант по настоящей заявке, или полинуклеотид, кодирующий этот вариант.
Согласно еще одному аспекту настоящей заявки предложен способ получения L-валина, где способ включает культивирование микроорганизма в среде.
Согласно еще одному аспекту настоящей заявки предложена композиция для получения L-валина, где композиция содержит микроорганизм, включающий вариант по настоящей заявке, полинуклеотид, кодирующий вариант, вектор, содержащий полинуклеотид, или полинуклеотид по настоящей заявке; среда, полученная путем культивирования микроорганизма; или комбинация двух или более из них.
Согласно еще одному аспекту настоящей заявки предложено применение варианта малой субъединицы синтазы ацетогидроксикислоты для получения L-валина, где вариант содержит замену аминокислоты в положении 44 в аминокислотной последовательности SEQ ID NO: 1 другой аминокислотой.
Полезные эффекты
Культура микроорганизмов, включающих вариант малой субъединицы синтазы ацетогидроксикислот по настоящей заявке, дает возможность получать L-валин с высоким выходом по сравнению с микроорганизмами, имеющими известный немодифицированный полипептид.
Лучшее воплощение изобретения
Настоящая заявка будет конкретно описана следующим образом. Каждое описание и воплощение, раскрытые в настоящей заявке, также можно применять к другим описаниям и воплощениям. То есть все комбинации различных элементов, раскрытых в настоящей заявке, попадают в объем настоящей заявки. Кроме того, объем настоящей заявки не ограничивается приведенным ниже конкретным описанием. Кроме того, по всему описанию изобретения в целом предлагаются ссылки на многие статьи и патентные документы и цитаты из них. Раскрытия цитируемых статей и патентных документов включены посредством ссылки в настоящее описание во всей полноте, и уровень области техники, к которой относится настоящее изобретение, и подробности настоящего изобретения объясняются более четко.
Согласно одному аспекту настоящей заявки предложен вариант малой субъединицы синтазы ацетогидроксикислоты (ilvN), в котором аминокислота, соответствующая положению 44 в аминокислотной последовательности SEQ ID NO: 1, заменена другой аминокислотой.
При использовании здесь термин «L-валин» относится к L-аминокислоте, соответствующей одной из незаменимых аминокислот, которая имеет химическую формулу (CH3)2CHCH(NH2)COOH, относящейся по структуре, вместе с L-лейцином и L-изолейцином, к аминокислотам с разветвленной цепью.
При использовании здесь термин «синтаза ацетогидроксикислоты» относится к первому ферменту в биосинтезе L-валина и также называется ацетолактатсинтазой. Синтаза ацетогидроксикислоты может катализировать декарбоксилирование пирувата и реакцию конденсации с другой молекулой пирувата с образованием ацетолактата, который представляет собой предшественник валина, или катализировать декарбоксилирование пирувата и реакцию конденсации с 2-кетобутиратом с образованием ацетогидроксибутирата, который представляет собой предшественник изолейцина.
Синтаза ацетогидроксикислот кодируется двумя генами ilvB и ilvN, где ген ilvB кодирует большую субъединицу синтазы ацетогидроксикислоты, и ген ilvN кодирует малую субъединицу синтазы ацетогидроксикислоты. Среди них, считается, что малая субъединица, кодируемая геном ilvN, в значительной степени вовлечена в ингибирование конечным продуктом.
Синтаза ацетогидроксикислоты, кодируемая геном ilvN, может иметь аминокислотную последовательность SEQ ID NO: 1, но без ограничения этим.
Вариант по настоящей заявке может иметь усиленную активность вследствие замены аминокислоты в конкретном положении в существующей аминокислотной последовательности малой субъединицы синтазы ацетогидроксикислоты (ilvN), но без ограничения этим.
В одном воплощении вариант малой субъединицы синтазы ацетогидроксикислоты (ilvN) может представлять собой вариант малой субъединицы синтазы ацетогидроксикислоты (ilvN), включающий, по меньшей мере, одну аминокислотную замену в аминокислотной последовательности SEQ ID NO: 1, но без ограничения этим.
Конкретно, в варианте по настоящей заявке аминокислоты, соответствующие положению 44 и/или 42, могут быть заменены другими аминокислотами, но без ограничения этим. Более конкретно, в этом варианте аминокислоты в любом одном или обоих из приведенных выше положений или в соответствующих им положениях могут быть заменены другими аминокислотами, но без ограничения этим.
«Другая аминокислота» или «другие аминокислоты» без ограничения до тех пор, пока аминокислоты отличаются от аминокислот до замен. Например, формулировка «аминокислоту, соответствующую положению 44 в SEQ ID NO: 1, заменяют другой аминокислотой» может означать, что аминокислота заменяется аланином, фенилаланином, глицином, аргинином, аспартатом, цистеином, глутаматом, аспарагином, глутамином, гистидином, пролином, серином, тирозином, изолейцином, лизином, триптофаном, валином, метионином или лейцином, за исключением треонина, и формулировка «аминокислоту, соответствующую положению 42 в SEQ ID NO: 1, заменяют другой аминокислотой» означает, что аминокислоту заменяют валином, аспарагином, глицином, аргинином, аспартатом, цистеином, глутаминовой кислотой, глутамином, гистидином, пролином, серином, тирозином, изолейцином, лейцином, лизином, фенилаланином, триптофаном, метионином или треонином, за исключением аланина, но без ограничения этим.
При этом, специалист в данной области техники может узнать аминокислоты, соответствующие положениям 44 и 42 в SEQ ID NO: 1 по настоящей заявке, в любой аминокислотной последовательности посредством выравнивания последовательностей, известного в данной области техники, и даже если это не описывается здесь отдельно, было бы очевидно, что формулировка «аминокислота в конкретном положении в конкретном номере последовательности» означает включение даже «аминокислоты в положении, соответствующем» ему в любой аминокислотной последовательности. Таким образом, аминокислотная последовательность, в которой по меньшей мере одна аминокислота, выбранная из группы, состоящей из аминокислот, соответствующих положениям 44 и 42 в SEQ ID NO: 1, заменена другой аминокислотой, также включается в диапазон настоящей заявки.
Например, замена по меньшей мере одной аминокислоты из аминокислот, соответствующих положениям 44 и 42 в SEQ ID NO: 1, другой аминокислотой может порождать вариант, имеющий более высокую активность, чем незамененная (немодифицированная) аминокислотная последовательность.
Конкретно, в варианте по настоящей заявке аминокислоты, соответствующие положению 44 и/или положению 42, могут быть заменены другими аминокислотами, но без ограничения этим.
В качестве конкретного примера, в варианте по настоящей заявке треонин, аминокислота, соответствующая положению 44 в SEQ ID NO: 1, может быть заменена аланином и/или аланин, аминокислота, соответствующая положению 42 в SEQ ID NO: 1, может быть заменена валином, но без ограничения этим.
В качестве более конкретного примера, вариант по настоящей заявке может иметь аминокислотную последовательность, указанную в SEQ ID NO: 3 или 5, или может по существу состоять из аминокислотной последовательности.
Кроме того, вариант по настоящей заявке может включать не только вариант, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 3 или 5, но также вариант, имеющий по меньшей мере 80%, конкретно по меньшей мере 90%, более конкретно по меньшей мере 95% и еще более конкретно по меньшей мере 99% гомологии с аминокислотной последовательностью, где аминокислоты в положениях, соответствующих аминокислоте в положении 44 и/или аминокислоте в положении 42 с N-конца SEQ ID NO: 1, могут быть заменены другими аминокислотами. Было бы очевидно, что любой вариант, который имеет аминокислотную последовательность с делецией, модификацией, заменой, консервативной заменой или добавлением ее части, также может быть включен в диапазон настоящей заявки при условии, что аминокислотная последовательность имеет такую гомологию или идентичность и проявляет эффективность, соответствующую варианту по настоящей заявке.
Их примеры могут включать варианты, имеющие добавление или делецию, встречающуюся в природе мутацию, молчащую мутацию или консервативную замену, которая не изменяет функции варианта по настоящей заявке, на N-конце, С-конце и/или внутри аминокислотной последовательности.
При использовании здесь термин «консервативная замена» относится к замене одной аминокислоты другой аминокислотой, имеющей сходные структурные и/или химические свойства. Такая аминокислотная замена, как правило, может быть выполнена на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; отрицательно заряженные (кислые) аминокислоты включают глутаминовую кислоту и аспарагиновую кислоту; ароматические аминокислоты включают фенилаланин, триптофан и тирозин; и гидрофобные аминокислоты включают аланин, валин, изолейцин, лейцин, метионин, фенилаланин, тирозин и триптофан. Кроме того, аминокислоты могут быть классифицированы на аминокислоты с электрически заряженными боковыми цепями и аминокислоты с не заряженными электрически боковыми цепями. Аминокислоты с электрически заряженными боковыми цепями включают аспарагиновую кислоту, глутаминовую кислоту, лизин, аргинин и гистидин, и аминокислоты с не заряженными электрически боковыми цепями можно дополнительно классифицировать на неполярные аминокислоты или полярные аминокислоты. Неполярные аминокислоты могут включать глицин, аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан и пролин, и полярные аминокислоты включают серии, треонин, цистеин, тирозин, аспарагин и глутамин. Обычно, консервативные замены имеют незначительное влияние или не имеют никакого влияния на активность продуцируемого полипептида. Обычно консервативные замены могут иметь незначительное влияние или не иметь никакого влияния на активность белка или полипептида.
Кроме того, вариант может включать делеции или добавления аминокислот, которые имеют минимальное влияние на свойства и вторичные структуры полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью N-конца белка, вовлеченного в перенос белка ко-трансляционно или посттрансляционно. Полипептид также может быть конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
При использовании здесь термин «вариант» относится к полипептиду, который имеет другую последовательность, но сохраняет функции или свойства по сравнению с аминокислотной последовательностью до мутации в результате консервативной замены и/или модификации по меньшей мере одной аминокислоты. Такой вариант, как правило, может быть идентифицирован посредством модификации по меньшей мере одной аминокислоты в аминокислотной последовательности полипептида и оценки свойств модифицированного полипептида. То есть способность варианта может быть увеличена, не изменена или понижена по сравнению с полипептидом до мутации. Некоторые варианты могут включать вариант, в котором удалена по меньшей мере одна часть, такая как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать вариант, в котором удаляется часть N-конца и/или С-конца зрелого белка.
Термин «вариант» также можно использовать взаимозаменяемо с «модификацией», «модифицированным полипептидом», «модифицированным белком», «мутантом», «мутеином», «дивергентом» или подобным, и любой термин, который используется в смысле выполнения мутации, можно использовать без ограничения этим. Для целей настоящей заявки вариант может представлять собой полипептид, включающий: аминокислотную последовательность, указанную в SEQ ID NO: 3, в которой треонин, аминокислота, соответствующая положению 44 в SEQ ID NO: 1, заменен аланином; или аминокислотную последовательность, указанную в SEQ ID NO: 5, в которой аланин, аминокислота, соответствующая позиции 42 в SEQ ID NO: 1, заменен на валин.
При использовании здесь термин «гомология» или «идентичность» относится к степени сходства между двумя заданными аминокислотными последовательностями или нуклеотидными последовательностями и может быть выражен в процентах. Термины «гомология» и «идентичность» часто могут использоваться взаимозаменяемо друг с другом.
Гомология последовательностей или идентичность консервативных полинуклеотидов или полипептидов может быть определена посредством стандартных алгоритмов выравнивания, и совместно можно использовать штрафы за пробелы по умолчанию, установленные используемой программой. По существу, гомологичные или идентичные последовательности, как правило, могут гибридизоваться друг с другом в целом или частично при умеренных или очень строгих условиях. Было бы очевидно, что гибридизация также включает гибридизацию с полинуклеотидом, содержащим обычные кодоны, или кодоны, учитывающие вырождение кодонов в полинуклеотиде.
Гомология, сходство или идентичность между любыми двумя полинуклеотидными или полипептидными последовательностями могут быть определены с использованием известного компьютерного алгоритма, такого как программа "FASTA", с использованием параметров по умолчанию, таких как в Pearson et al., (1988) Proc. Natl. Acad. Sci. USA 85:2444. В качестве альтернативы они могут быть определены с помощью алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453), который выполняют в программе Needleman в европейском открытом программном пакете для молекулярной биологии (EMBOSS) (Rice et al., 2000, Trends Genet. 16:276-277) (версия 5.0.0 или более поздняя) (включая пакет программ GCG (Devereux, J., et al,Nucleic Acids Research 12:387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S. F., et al, J MOLEC BIOL 215:403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994 и CARILLO et al. (1988) SIAM J Applied Math 48:1073. Например, гомология, сходство или идентичность могут быть определены, используя BLAST информационной базы данных Национального центра биотехнологии или ClustalW.
Гомология, сходство или идентичность между полинуклеотидами или полипептидами может быть определена путем сравнения информации о последовательности с использованием компьютерной программы GAP, например, Needleman et al., (1970), J Mol Biol. 48:443, которая известна в Smith and Waterman, Adv. Appl. Math (1981) 2:482. Вкратце, программа GAP определяет сходство как количество выровненных символов (т.е. нуклеотидов или аминокислот), которые являются сходными, деленное на общее количество символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) двоичную матрицу сравнения (содержащую значение 1 для идентичности и 0 для неидентичности) и взвешенную матрицу сравнения Gribskov et al. (1986) Nucl. Acids Res. 14:6745 которые раскрыты в Schwartz and DayhofF, eds., Atlas of Protein Sequence And Structure, National Biomedical Research Foundation, pp.353-358 (1979) (или матрица замещения EDNAFULL (версия EMBOSS NCBI NUC4.4)); (2) штраф в размере 3,0 за каждый пробел и дополнительный штраф в размере 0,10 за каждый символ в каждом пробеле (или штраф за открытие пробела в размере 10 и штраф за расширение пробела в размере 0,5); и (3) отсутствие штрафа за конечные пробелы.
В одном воплощении настоящей заявки вариант по настоящей заявке может быть получен посредством мутации микроорганизма, продуцирующего L-валин. Мутацию микроорганизма можно выполнять различными способами, известными в данной области техники, и можно использовать один из физических или химических мутагенных факторов. Примеры химического мутагенного фактора, подходящего для настоящего изобретения, включают N-метил-N'-нитро-N-нитрозогуанидин (NTG), диэпоксибутан, этилметансульфонат, соединения иприта, гидразин и азотистую кислоту, но без ограничения этим. Примеры физического мутагенного фактора могут включать ультрафиолетовое и гамма-излучение, но без ограничения этим.
В настоящей заявке экспрессия малой субъединицы синтазы ацетогидроксикислоты (ilvN) может быть усилена, и усиление экспрессии может увеличить способность продуцирования L-валина.
При использовании здесь термин «соответствующий» относится к аминокислотным остаткам в положениях, указанных в полипептиде, или аминокислотным остаткам, которые подобны, идентичны или гомологичны тем, которые указаны в полипептиде. Идентификация аминокислоты в соответствующем положении может представлять собой определение конкретной аминокислоты в последовательности, которая относится к конкретной последовательности. При использовании здесь термин «соответствующий участок» обычно относится к аналогичному или соответствующему положению в родственном белке или эталонном белке.
Например, любую аминокислотную последовательность выравнивают по SEQ ID NO: 1, и на основании этого каждый аминокислотный остаток аминокислотной последовательности может быть нумерован со ссылкой на числовое положение аминокислотного остатка, соответствующего аминокислотному остатку SEQ ID NO: 1. Например, алгоритм выравнивания последовательностей, который описан в настоящей заявке, может идентифицировать положение аминокислоты или положение возникновения модификации, такой как замена, вставка или делеция, по сравнению с запрашиваемой последовательностью (также называемой «эталонной последовательностью»).
Например, можно использовать соответствующим образом алгоритм Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48:443-453) или программа Needle пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al, 2000, Trends Genet. 16:276-277) можно использовать для такого выравнивания, но без ограничения этим, программа выравнивания последовательностей, алгоритм парного сравнения последовательностей, или подобные, которые известны в данной области техники.
В соответствии с другим аспектом настоящей заявки предложен полинуклеотид, кодирующий вариант по настоящей заявке.
При использовании здесь термин «полинуклеотид» относится к полимеру из нуклеотидных мономеров, связанных ковалентной связью в цепь нуклеотидов, вытянутую в длину, и, в общем, к цепи ДНК или РНК определенной длины, и, более конкретно, может обозначать полинуклеотидный фрагмент, кодирующий вариант.
Полинуклеотид, кодирующий вариант по настоящей заявке, может включать нуклеотидную последовательность, кодирующую аминокислотную последовательность, указанную в SEQ ID NO: 3 или 5. В качестве примера, полинуклеотид по настоящей заявке может иметь или включать последовательность SEQ ID NO: 4 или 6. Альтернативно, полинуклеотид по настоящей заявке может состоять или по существу состоять из последовательности SEQ ID NO: 4 или 6.
В кодирующей области полинуклеотида по настоящей заявке можно выполнять различные модификации до тех пор, пока аминокислотная последовательность варианта по настоящей заявке изменится с учетом вырождения кодонов или кодонов, предпочтительных в организме, который предназначен для экспрессии варианта по настоящей заявке. Конкретно, полинуклеотид по настоящей заявке может иметь или включать нуклеотидную последовательность, имеющую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97% или по меньшей мере 98%, но менее 100% гомологию или идентичность с последовательностью SEQ ID NO: 4 или 6, или может состоять или состоять по существу из нуклеотидной последовательности, имеющей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97% или по меньшей мере 98%, но менее 100% гомологию или идентичность с последовательностью SEQ ID NO: 4 или 6, но без ограничения этим. Конкретно, в последовательностях, имеющих гомологию или идентичность, кодон, кодирующий аминокислоту, соответствующую положению 44 в SEQ ID NO: 3 или 5, может представлять собой один из кодонов, кодирующих аланин, и кодон, кодирующий аминокислоту, соответствующую положению 42 в SEQ ID NO: 5, может представлять собой один из кодонов, кодирующих валин.
Кроме того, полинуклеотид по настоящей заявке может включать, без ограничения, последовательность, которая при строгих условиях может гибридизоваться с зондом, который может быть получен из известной последовательности гена, например, последовательности, комплементарной части или всей полинуклеотидной последовательности по настоящей заявке. Термин «строгие условия» относится к условиям, которые обеспечивают специфическую гибридизацию между полинуклеотидами. Эти условия конкретно описаны в литературе (смотри J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; и F. M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York, 9.50 9.51, 11.7 11.8). Например, строгие условия могут включать в себя состояние, при котором полинуклеотиды, имеющие более высокую гомологию или идентичность, например, полинуклеотиды, имеющие по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% гомологии или идентичности, гибридизуются друг с другом, но полинуклеотиды, имеющие более низкую гомологию или идентичность, не гибридизуются друг с другом; или условия, при которых промывку выполняют один раз, конкретно два или три раза, при концентрации соли и температуре, эквивалентных 60°С, 1×SSC, 0,1% SDS, конкретно 60°C, 0,1×SSC, 0,1% SDS; и более конкретно 68°C, 0,1×SSC, 0,1% SDS, которые представляют собой условия для обычной Саузерн-гибридизации.
Гибридизация требует, чтобы две нуклеиновые кислоты имели комплементарные последовательности, хотя, в зависимости от строгости гибридизации, возможны ошибочные спаривания между основаниями. Термин «комплементарный» используется для описания взаимосвязи между нуклеотидными основаниями, которые способны гибридизоваться друг с другом. Например, для ДНК аденин комплементарен тимину и цитозин комплементарен гуанину. Соответственно, полинуклеотид по настоящей заявке также включает выделенные фрагменты нуклеиновых кислот, которые комплементарны полной последовательности, а также по существу подобные последовательности нуклеиновых кислот.
Конкретно, полинуклеотиды, имеющие гомологию или идентичность с полинуклеотидом по настоящей заявке, могут быть обнаружены при использовании условий гибридизации, включающих стадию гибридизации при Tm 55°С и используя вышеописанные условия. Кроме того, значение Tm может представлять собой 60°С, 63°С или 65°С, но без ограничения этим, и может быть соответствующим образом отрегулировано специалистом в данной области техники согласно цели.
Соответствующая степень жесткости для гибридизации полинуклеотидов зависит от длины полинуклеотидов и степени их комплементарности, и эти их показатели хорошо известны в данной области техники (например, Sambrook et al., выше).
В соответствии с еще одним аспектом настоящей заявки предложен вектор, включающий полинуклеотид по настоящей заявке. Вектор может представлять собой экспрессионный вектор для экспрессии полинуклеотида в микроорганизме, но без ограничения этим.
При использовании здесь термин «вектор» относится к конструкции ДНК, которая включает нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, и функционально связана с подходящей контрольной последовательностью так, что целевой белок может экспрессироваться в соответствующем хозяине. Область контроля экспрессии может включать промотор, способный инициировать транскрипцию, любую операторную последовательность для контроля такой транскрипции, последовательность для кодирования подходящего сайта связывания мРНК с рибосомой и последовательность для контроля терминации транскрипции и трансляции. Вектор после трансформации в подходящий микроорганизм может реплицироваться или функционировать независимо от генома хозяина или может быть интегрирован в его геном.
Вектор, используемый в настоящей заявке, не ограничивают конкретно и можно использовать любой вектор, известный в данной области техники. Пример вектора, который обычно используют, может включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A можно использовать в качестве фаговых или космидных векторов и векторы на основе pDZ, pBR, pUC, pBluescriptll, pGEM, pTZ, на основе pCL и pET можно использовать в качестве плазмидных векторов. Конкретно, можно использовать векторы pDZ, pDC, pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 и рСС1ВАС.
Например, полинуклеотид, кодирующий целевой полипептид, может быть вставлен в хромосому посредством вектора для хромосомной вставки в клетку. Вставку полинуклеотида в хромосому можно выполнять с использованием любого способа, известного в данной области техники, например, гомологичной рекомбинации, но не ограничения этим. Вектор может дополнительно включать селективный маркер для исследования вставки или невставки хромосомы. Селективный маркер предназначен для отбора клеток, трансформированных с помощью вектора, то есть для идентификации вставки или невставки целевой молекулы нуклеиновой кислоты, и можно использовать маркеры, придающие селективный фенотип, такой как устойчивость к лекарственным препаратам, ауксотрофия, устойчивость к цитотоксическим лекарственным средствам или поверхностная экспрессия полипептида. В условиях обработки селективным агентом могут выживать или проявлять другие фенотипические признаки только клетки, экспрессирующие селективные маркеры, поэтому могут быть отобраны трансформированные клетки.
При использовании здесь термин «трансформация» относится к введению вектора, содержащего полинуклеотид, кодирующий целевой полипептид, в микроорганизм таким образом, чтобы полипептид, кодируемый полинуклеотидом, мог экспрессироваться в микроорганизме. Примеры трансформированного полинуклеотида могут включать любой полипептид при условии, что он может экспрессироваться в микроорганизме, независимо от того, вставлен ли он и локализован в хромосоме микроорганизма или расположен вне хромосомы микроорганизма. Кроме того, полинуклеотид включает ДНК и/или РНК, кодирующие целевой полипептид. Полинуклеотид может быть введен в любой форме при условии, что он может быть введен в микроорганизм и экспрессироваться в нем. Например, полинуклеотид может быть введен в микроорганизм в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую все элементы, необходимые для его само-экспрессии. Обычно экспрессионная кассета может включать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, сайт связывания рибосом и сигнал терминации трансляции. Экспрессионная кассета может быть в форме самореплицирующегося вектора экспрессии. Кроме того, полинуклеотид может быть введен в микроорганизм в том виде, в каком он есть, и функционально связан с последовательностью, необходимой для экспрессии в микроорганизме, но без ограничения этим.
Термин «функционально связанный», как указано выше, относится к функциональной связи между полипептидной последовательностью и промоторной последовательностью для инициации и опосредования транскрипции полинуклеотида, кодирующего целевой вариант по настоящей заявке.
Согласно еще одному аспекту настоящей заявки предложен микроорганизм, включающий вариант по настоящей заявке или полинуклеотид по настоящей заявке.
Микроорганизм по настоящей заявке может включать модифицированный полипептид по настоящей заявке, полинуклеотид, кодирующий полипептид, или вектор, содержащий полинуклеотид по настоящей заявке.
При использовании здесь термин «микроорганизм (или штамм)» охватывает все микроорганизмы дикого типа или естественно или искусственно генетически модифицированные микроорганизмы, и он может представлять собой микроорганизм, в котором конкретный механизм ослабляют или усиливают вследствие вставки экзогенного гена или усиления или инактивации активности эндогенного гена и может представлять собой микроорганизм, включающий генетическую модификацию для получения желаемого полипептида, белка или продукта. В настоящей заявке термины «микроорганизм» и «штамм» имеют одинаковое значение и могут использоваться взаимозаменяемо без ограничений.
Конкретно, микроорганизм может представлять собой микроорганизмом рода Corynebacterium и, более конкретно, Corynebacterium glutamicum, но без ограничения этим.
Штамм по настоящей заявке может представлять собой: штамм, включающий по меньшей мере любой из вариантов по настоящей заявке, полинуклеотид по настоящей заявке и вектор, содержащий полинуклеотид по настоящей заявке; штамм, модифицированный для экспрессии варианта по настоящей заявке или полинуклеотида по настоящей заявке; штамм (например рекомбинантный штамм), экспрессирующий вариант по настоящей заявке, или полинуклеотид по настоящей заявке; или штамм (например рекомбинантный штамм), имеющий активность варианта по настоящей заявке, без ограничения этим.
Штамм по настоящей заявке может быть микроорганизмом, продуцирующим L-валин.
При использовании здесь термин «микроорганизм, продуцирующий L-валин» относится к штамму прокариотических или эукариотических микроорганизмов, способному продуцировать L-валин в организме. Для целей настоящего изобретения микроорганизм может представлять собой прокариотическую или эукариотическую клетку при условии, что она может продуцировать L-валин посредством включения малой субъединицы синтазы ацетогидроксикислоты, и его примеры могут включать штамм микроорганизма, относящийегося к роду Corynebacterium, например, Corynebacterium glutamicum.
Микроорганизм, продуцирующий L-валин, включающий вариант синтазы ацетогидроксикислоты, может включать как микроорганизм, который включает последовательность варианта малой субъединицы синтазы ацетогидроксикислоты, вызванного мутацией гена, кодирующего синтазу ацетогидроксикислоты на хромосоме, так и/или микроорганизм, который включает вариант малой субъединицы синтазы ацетогидроксикислоты, вызванный введением вектора, содержащего полинуклеотид, кодирующий вариант малой субъединицы синтазы ацетогидроксикислот, но без ограничения этим.
Кроме того, микроорганизм, продуцирующий L-валин, включающий вариант малой субъединицы синтазы ацетогидроксикислоты, может представлять собой микроорганизм, у которого активность варианта малой субъединицы синтазы ацетогидроксикислоты усиливают по сравнению с родительским штаммом.
Например, штамм по настоящей заявке представляет собой клетку или микроорганизм, который трансформируют посредством вектора, включающего полинуклеотид по настоящей заявке, или полинуклеотид, кодирующий вариант по настоящей заявке, для экспрессии варианта по настоящей заявке, и для цели настоящей заявки, применение штамма по настоящей заявке может охватывать все микроорганизмы, способные продуцировать L-валин, благодаря включению варианта по настоящей заявке. Например, штамм по настоящей заявке может представлять собой рекомбинантный штамм, имеющий усиленную способность продуцировать L-валин посредством экспрессии варианта малой субъединицы синтазы ацетогидроксикислоты (ilvN) вследствие введения полинуклеотида, кодирующего вариант по настоящей заявке, в нативный микроорганизм дикого типа или в микроорганизм, продуцирующий L-валин. Рекомбинантный штамм с усиленной способностью продуцировать L-валин может представлять собой микроорганизм с усиленной способностью продуцировать L-валин по сравнению с природным микроорганизмом дикого типа или микроорганизмом с немодифицированной синтазой ацетогидроксикислот (т.е. микроорганизм, экспрессирующий синтазу ацетогидроксикислоты дикого типа (SEQ ID NO: 1), или микроорганизм, не экспрессирующий мутантный белок (SEQ ID NO: 3 или 5)), но без ограничения этим.
Например, рекомбинантный штамм с усиленной способностью может иметь способность продуцировать L-валин, которая усилена по меньшей мере примерно на 1%, конкретно, по меньшей мере примерно на 1%, по меньшей мере примерно на 2,5%, по меньшей мере примерно на 5%, по меньшей мере примерно на 6% по меньшей мере, примерно на 7%, по меньшей мере примерно на 8%, по меньшей мере примерно на 9%, по меньшей мере примерно на 10%, по меньшей мере, примерно на 10,5%, по меньшей мере примерно на 11%, по меньшей мере примерно на 11,5%, по меньшей мере примерно на 12%, по меньшей мере примерно на 12,5%, по меньшей мере примерно на 13%, по меньшей мере примерно на 13,5%, по меньшей мере примерно на 14%, по меньшей мере, примерно 14,5%, по меньшей мере, примерно на 15%, по меньшей мере примерно на 15,5%, по меньшей мере примерно на 16%, по меньшей мере примерно на 16,5%, по меньшей мере примерно на 17%, по меньшей мере примерно на 17,5%, по меньшей мере примерно на 18%, по меньшей мере примерно на 18,1%, по меньшей мере примерно на 18,2%, по меньшей мере примерно на 18,3%, по меньшей мере примерно на 18,4%, по меньшей мере примерно на 18,5% (верхняя граница конкретно не ограничена, и верхняя граница может представлять собой, например, самое большее примерно 200%, самое большее примерно 150%, самое большее примерно 100%, самое большее примерно 50%, самое большее примерно 45%, самое большее примерно 40%, самое большее примерно 35%, самое большее примерно 30%, самое большее примерно 25%, самое большее примерно 20%, или самое большее примерно 15%), по сравнению с родительским штаммом до модификации или немодифицированным микроорганизмом, но способность продуцировать L-валин этим не ограничивается до тех пор, пока она увеличивается на величину плюс (+) по сравнению с продуцирующей способностью родительского штамма до модификации или немодифицированного микроорганизма. В качестве другого примера, рекомбинантный штамм с усиленной продуцирующей способностью может иметь способность продуцировать L-валин, которая усилена по меньшей мере примерно в 1,1 раза, по меньшей мере примерно в 1,12 раза, по меньшей мере примерно в 1,13 раза, по меньшей мере примерно в 1,14 раза, по меньшей мере примерно в 1,15 раза, по меньшей мере примерно в 1,16 раза, по меньшей мере, примерно в 1,17 раза или, по меньшей мере, примерно в 1,18 раза (верхняя граница конкретно не ограничена, и верхняя граница может представлять собой, например, самое большее примерно в 10 раз, самое большее примерно в 5 раз, самое большее примерно в 3 раза, самое большее примерно в 2 раза, самое большее примерно в 1,5 раза, или самое большее примерно в 1,2 раза), по сравнению с таковой у родительского штамма до модификации или немодифицированного микроорганизма, но способность продуцировать L-валин этим не ограничивается. При использовании здесь термин «примерно» относится к диапазону, включающему ±0,5, ±0,4, ±0,3, ±0,2, ±0,1 и тому подобные, и таким образом включает все значения в диапазоне, эквивалентные или аналогичные тем, которые указаны после этого термина, но без ограничения ими.
При использовании здесь термин «немодифицированный микроорганизм» может относиться к штамму дикого типа или нативному штамму как таковому, или к штамму до трансформации вследствие генетической мутации, вызванной естественными или искусственными факторами, не исключая штаммы, включающие мутации, которые могут естественным образом возникать у микроорганизмов. Например, немодифицированный микроорганизм может относиться к штамму, в который не вводят описанный здесь вариант малой субъединицы синтазы ацетогидроксикислоты (ilvN), или к штамму до его введения. «Немодифицированный микроорганизм» можно использовать взаимозаменяемо со «штаммом до модификации», «микроорганизмом до модификации», «немутированным штаммом», «немодифицированным штаммом», «немутированным микроорганизмом» или «эталонным микроорганизмом».
В другом воплощении настоящей заявки микроорганизм по настоящей заявке может представлять собой Corynebacterium glutamicum, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium stationis, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium ammoniagenes, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens.
При использовании здесь термин «усиление» активности полипептида относится к увеличению активности полипептида по сравнению с его собственной активностью. Усиление может использоваться взаимозаменяемо с такими терминами, как активация, повышающая регуляция, сверхэкспрессия и увеличение. Конкретно, активация, усиление, повышающая регуляция, сверхэкспрессия и увеличение могут включать проявление активности, имевшейся первоначально, или проявление активности, которая была улучшена по сравнению с собственной активностью или активностью до модификации.
Термин «собственная активность» относится к активности конкретного полипептида, имевшейся первоначально у родительского штамма до трансформации, или немодифицированного микроорганизма, когда микроорганизм трансформируют посредством генетической мутации, вызванной естественными или искусственными факторами. Этот термин можно использовать взаимозаменяемо с «активностью до модификации». «Усиление», «повышающая регуляция», «сверхэкспрессия» или «увеличение» активности полипептида по сравнению с его собственной активностью означает, что активность и/или концентрация (уровень экспрессии) конкретного полипептида, которую первоначально имел родительский штамм до трансформации или немодифицированный штамм микроорганизма, улучшается.
Усиление может быть достигнуто посредством введения экзогенного полипептида или увеличения собственной активности и/или концентрации (уровня экспрессии) полипептида. Усиление активности полипептида можно идентифицировать по увеличению степени активности или уровня экспрессии соответствующего полипептида или увеличению количества продукта, высвобождаемого из соответствующего полипептида.
Для усиления активности полипептида можно использовать различные способы, хорошо известные в данной области техники, и способ не ограничивают до тех пор, пока он может усиливать активность целевого полипептида по сравнению с активностью микроорганизма до модификации. Конкретно, можно использовать генную инженерию и/или белковую инженерию, хорошо известные специалистам в данной области техники, которые представляют собой обычные способы молекулярной биологии, но способ этим не ограничивают (e.g., Sitnicka et al, Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol.2. 1-16; and Sambrook et al, Molecular Cloning 2012).
Конкретно, усиление активности полипептида по настоящей заявке может быть достигнуто посредством:
1) увеличения числа внутриклеточных копий полинуклеотида, кодирующего полипептид;
2) замены области контроля экспрессии гена, кодирующего полипептид на хромосоме, последовательностью с более высокой активностью;
3) модификации нуклеотидной последовательности, кодирующей инициирующий кодон, или 5-UTR транскрипта гена, кодирующего полипептид;
4) модификации аминокислотной последовательности полипептида таким образом, чтобы усилить активность полипептида;
5) модификации полинуклеотидной последовательности, кодирующей полипептид, таким образом, чтобы активность полипептида усиливалась (например, модификации полинуклеотидной последовательности гена полипептида таким образом, чтобы кодировать полипептид, который был модифицирован для усиления активности полипептида);
6) введения экзогенного полипептида, проявляющего активность полипептида, или экзогенного полинуклеотида, кодирующего то же самое;
7) оптимизации кодона полинуклеотида, кодирующего полипептид;
8) модификации или химического модифицирования экспонированного участка, выбранного путем анализа третичной структуры полипептида; или
9) комбинации двух или более, выбранных из пп. (1)-(8) выше, но конкретно не ограничивается этим.
Более конкретное описание приводится ниже.
Увеличение числа внутриклеточных копий полинуклеотида, кодирующего полипептид, в пункте (1) выше, может быть достигнуто посредством введения в микроорганизм вектора, функционально связанного с полинуклеотидом, кодирующим соответствующий полипептид, и способного реплицироваться и функционировать независимо от хозяина. Альтернативно, увеличение может быть достигнуто посредством введения одной или более копий или более полинуклеотидов, кодирующих соответствующий полипептид, в хромосому микроорганизма. Введение в хромосому может быть выполнено посредством введения вектора, который способен вставлять полинуклеотид в хромосому микроорганизма, в сам микроорганизм, но без ограничения этим. Вектор представляет собой такой, как описано выше.
Замена области контроля экспрессии (или последовательности контроля экспрессии) гена, кодирующего полипептид на хромосоме, на последовательность с более высокой активностью по п. 2 выше может представлять собой, например, индуцирование модификации последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации для дальнейшего усиления активности области контроля экспрессии, или замену последовательности последовательностью, обладающей более сильной активностью. Область контроля экспрессии может включать, но не ограничивается конкретно этим, промотор, операторную последовательность, последовательность, кодирующую сайт связывания рибосомы, последовательность, контролирующую терминацию транскрипции и трансляции, и тому подобное. Например, замена может представлять собой замену исходного промотора сильным промотором, но не без ограничения этим.
Примеры известных более сильных промоторов могут включать промоторы CJ1-CJ7 (US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага lamda, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (US 10584338 В2), промотор 02 (US 10273491 В2), промотор tkt, промотор уссА и тому подобное, но без ограничения этим.
Модификация нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR транскрипт гена, кодирующего полипептид по п. 3 выше, может представлять собой, например, замену нуклеотидной последовательности на нуклеотидную последовательность, кодирующую другой инициирующий кодон, имеющий более высокую степень экспрессии полипептида по сравнению с эндогенным инициирующим кодоном, но без ограничения этим.
Модификация аминокислотной последовательности или полинуклеотидной последовательности по пп. 4 и 5 выше может представлять собой индуцирование модификации последовательности посредством делеции, вставки, неконсервативной или консервативной замены или их комбинации в аминокислотной последовательности полипептида или полинуклеотидной последовательности, кодирующей полипептид, для дальнейшего усиления активности полипептида, или замену последовательности аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной для придания более сильной активности, или аминокислотной последовательностью или полинуклеотидной последовательностью, модифицированной для усиления ее активности, но без ограничения этим. Конкретно, замену можно выполнять посредством вставки полинуклеотида в хромосому путем гомологичной рекомбинации, но без ограничения этим. Вектор, используемый здесь, может дополнительно включать селективный маркер для идентификации вставки хромосомы. Селективный маркер представляет собой такой, как описано выше.
Введение экзогенного полипептида, проявляющего активность полипептида по п. 6 выше, может представлять собой введение в микроорганизм экзогенного полинуклеотида, кодирующего полипептид, проявляющий такую же/сходную активность, как у этого полипептида. Экзогенный полинуклеотид не ограничивают его происхождением или последовательностью до тех пор, пока экзогенный полинуклеотид проявляет такую же же/сходную активность, как у этого полинуклеотида. Введение можно выполнять посредством любого известного способа трансформации, соответствующим образом выбранного специалистом в данной области техники, и введенный полинуклеотид экспрессируется в клетке-хозяине, и, таким образом, полипептид продуцируется и его активность может быть усилена.
При использовании здесь термин «введение» относится к способу доставки полинуклеотида, кодирующего вариант малой субъединицы синтазы ацетогидроксикислоты, или вектора, включающего его, в микроорганизм. Такое введение можно легко выполнять посредством обычного в данной области техники способа. В общем, примеры способа включают способ осаждения CaCl2, способ Ханахана (a Hanahan method) с улучшенной эффективностью с использованием диметилсульфоксида (ДМСО) в качестве восстанавливающего агента в способе осаждения CaCl2, способ электропорации, способ осаждения фосфатом кальция, способ слияния протопластов, способ перемешивания с использованием карбид кремниевых волокон, способ трансформации с использованием ПЭГ, декстрансульфата, липофектамина и способ трансформации, опосредованный высушиванием/подавлением, и тому подобное. Способ трансформации вектора не ограничивается приведенными выше примерами способов, и любой способ трансформации или трансфекции, обычно используемый в данной области техники, можно использовать без ограничения. Доставленный полинуклеотид можно быть вставлен в хромосому микроорганизма и локализован в ней или может находиться вне хромосомы, при условии, что полинуклеотид может экспрессироваться в клетке-хозяине. Альтернативно, полинуклеотид может быть введен в любой форме при условии, что он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид можно вводить в микроорганизм в форме экспрессионной кассеты, которая представляет собой полинуклеотидную конструкцию, включающую все значимые элементы, необходимые для его самоэкспрессии, но без ограничения этим. Экспрессионная кассета обычно включает промотор, функционально связанный с открытой рамкой считывания (далее «ОРС») гена, сигнал терминации транскрипции, связывающий рибосому домен и сигнал терминации трансляции. Экспрессионная кассета может быть в форме самореплицирующегося вектора экспрессии. Полинуклеотид может быть введен в микроорганизм в том виде, в каком он есть, и функционально связан с последовательностью, необходимой для его экспрессии в микроорганизме, но без конкретного ограничения.
Оптимизация кодона полинуклеотида, кодирующего полипептид по п. 7, может представлять собой оптимизацию кодона эндогенного полинуклеотида с целью увеличения его транскрипции или трансляции в микроорганизме, или оптимизацию кодона экзогенного полинуклеотида с целью выполнения оптимизированной транскрипции или трансляции в микроорганизме.
Модификация или химическая модификация экспонированного участка, выбранного путем анализа третичной структуры полипептида по п. 8 выше, может представлять собой, например, модификацию или химическую модификацию экспонированного участка, подлежащего модификации или химической модификации посредством сравнения информации о последовательности полипептида, подлежащего анализу, с помощью базы данных, в которой хранится информация о последовательностях существующих белков для определения матричного белка-кандидата по данным о сходстве последовательностей и идентификации структуры на основе определенного кандидата.
Такое усиление активности полипептида может означать, что активность, концентрация, уровень экспрессии соответствующего полипептида повышается относительно активности или концентрации полипептида, экспрессируемого в микробном штамме дикого типа или микробном штамме до модификации, или что количество продукта, получаемого из соответствующего полипептида, увеличивается, но без ограничения этим.
В микроорганизме по настоящей заявке модификация полинуклеотида в целом или частично может быть индуцирована посредством (а) гомологичной рекомбинации с использованием вектора для хромосомной вставки в микроорганизм или редактирование генома с использованием генетически сконструированной нуклеазы (например CRISPR-Cas9) и/или (б) обработки светом, таким как ультрафиолетовые лучи и радиоактивные лучи и/или химическими веществами, не ограничиваясь этим. Способ модификации гена в целом или частично может включать в себя способ с использованием технологии рекомбинантной ДНК. Например, часть или весь ген может быть удален посредством инъецирования нуклеотидной последовательности или вектора, содержащего нуклеотидную последовательность, гомологичную гену-мишени, в микроорганизм, чтобы вызвать гомологичную рекомбинацию. Инъецированная нуклеотидная последовательность или вектор может содержать доминантный селективный маркер, но без ограничения этим.
В микроорганизме по настоящей заявке вариант, полинуклеотид, L-валин и тому подобное представляют собой такие, как описано в других аспектах.
Согласно еще одному аспекту настоящей заявки предложен способ получения L-валина, где способ включает культивирование микроорганизма в среде.
Конкретно, способ получения L-валина по настоящей заявке может включать культивирование в среде штамма Corynebacterium glutamicum, включающего вариант по настоящей заявке, полинуклеотид по настоящей заявке или вектор по настоящей заявке, но без ограничения этим.
При использовании здесь термин «культивирование» относится к выращиванию микроорганизма по настоящей заявке в надлежащим образом отрегулированных условиях окружающей среды. Культивирование по настоящей заявке можно выполнять согласно подходящим средам или условиям культивирования, известным в данной области техники. Такое культивирование может легко регулироваться специалистом в данной области техники согласно выбранному штамму. Конкретно, культивирование можно выполнять периодическим образом, непрерывным образом и/или периодическим образом с подпиткой, но без ограничения этим.
При использовании здесь термин «среда» относится к смеси, содержащей в качестве основных ингредиентов питательные вещества, необходимые для культивирования микроорганизма по настоящей заявке, где среда поставляет питательные вещества, факторы роста и тому подобное, включая воду, которая необходима для выживания и роста. Конкретно, среда и другие условия культивирования для культивирования микроорганизма по настоящей заявке могут представлять собой любую среду, которую используют для обычного культивирования микроорганизмов без конкретного ограничения. Однако микроорганизм по настоящей заявке можно культивировать в обычной среде, содержащей подходящие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины в аэробных условиях, с одновременным регулированием температуры, рН и тому подобного.
Например, питательные среды для штаммов рода Corynebacterium можно найти в литературе ("Manual of Methods for General Bacteriology" Американского общества бактериологии (Washington D.C., USA, 1981)).
В настоящей заявке примеры источников углерода могут включать: углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, сахароза и мальтоза; сахарные спирты, такие как маннит и сорбит; органические кислоты, такие как пировиноградная кислота, молочная кислота и лимонная кислота; и аминокислоты, такие как глутаминовая кислота, метионин и лизин. Кроме того, можно использовать натуральные органические источники питательных веществ, такие как гидролизаты крахмала, меласса, сырая меласса, рисовые отруби, маниок, жмых и жидкий кукурузный отвар, и, конкретно, могут быть использованы углеводы, такие как глюкоза и стерильная предварительно обработанная меласса (т.е. меласса, превращенная в восстановленные сахара), и подходящие количества других источников углерода можно использовать без ограничения. Эти источники углерода можно использовать по отдельности или в комбинации двух или более, но без ограничения этим.
Примеры источников азота могут включать: неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; аминокислоты, такие как глутаминовая кислота, метионин и глутамин; и органические источники азота, такие как пептон, NZ-амин, мясные экстракты, дрожжевые экстракты, солодовые экстракты, жидкий кукурузный экстракт, гидролизаты казеина, рыба или продукты ее разложения, обезжиренный соевый жмых или продукты его разложения. Эти источники азота можно использовать по отдельности или в комбинации двух или более из них, но без ограничения этим.
Примеры источников фосфора могут включать одноосновный фосфат калия, двухосновный фосфат калия и соответствующие им натрийсодержащие соли. Примеры неорганических соединений могут включать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и подобные, и, кроме того, могут включать аминокислоты, витамины и/или подходящие предшественники. Эти составляющие ингредиенты или предшественники можно добавлять в среду периодическим или непрерывным способом. Однако примеры источников фосфора этим не ограничиваются.
рН среды можно регулировать путем добавления соединений, таких как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, в среду надлежащим образом во время культивирования микроорганизма по настоящей заявке. Кроме того, для подавления образования пены во время культивирования можно добавлять пеногаситель, такой как полигликоле вый сложный эфир жирных кислот. Дополнительно, для поддержания аэробного состояния среды можно вводить в среду кислород или кислородсодержащий газ, или можно вводить газообразный азот, водород или диоксид углерода без введения газа для поддержания анаэробного или неаэробного состояния среды, но без ограничения этим.
В настоящей заявке температуру культивирования можно поддерживать на уровне от 20°С до 45°С, конкретно от 25°С до 40°С, и культивирование можно выполнять в течение примерно от 10 часов до 160 часов, но без ограничения этим.
L-валин, полученный путем культивирования по настоящей заявке, может высвобождаться в среду или может оставаться в клетках.
Способ получения L-валина по настоящей заявке может дополнительно включать получение микроорганизма по настоящей заявке, получение среды для культивирования штамма или комбинацию этих стадий (независимо от порядка, в любом порядке), например, перед стадией культивирования.
Способ получения L-валина по настоящей заявке может дополнительно включать извлечение L-валина из среды, полученной в результате культивирования (среды, где было выполнено культивирование), или из микроорганизма по настоящей заявке. Способ по настоящей заявке может дополнительно включать стадию извлечения после стадии культивирования.
Извлечение может представлять собой сбор желаемого L-валина с использованием соответствующего способа, известного в данной области техники в соответствии со способом культивирования микроорганизма по настоящей заявке, например, периодического, непрерывного типа культивирования или культивирования с подпиткой. Примеры способа могут включать центрифугирование, фильтрацию, обработку агентом, осаждающим кристаллизованный белок (высаливание), экстракцию, разрушение ультразвуком, ультрафильтрацию, диализ, различные типы хроматографии, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, ВЭЖХ (высокоэффективная жидкостная хроматография) и комбинацию этих способов, и желаемый L-валин может быть выделен из среды или микроорганизма посредством использования соответствующего способа, известного в данной области техники.
Способ получения L-валина по настоящей заявке может дополнительно включать стадию очистки. Очистку можно выполнять, используя подходящий способ, известный в данной области техники. В качестве примера, когда способ получения L-валина по настоящей заявке включает как стадию извлечения, так и стадию очистки, стадию извлечения и стадию очистки можно выполнять непрерывно или с перерывами, независимо от порядка, или можно выполнять одновременно или объединять в одну стадию, но без ограничения этим.
В способе по настоящей заявке вариант, полинуклеотид, вектор, штамм и тому подобные представляют собой такие, как описано в других аспектах.
В еще одном аспекте настоящей заявки предложена композиция для получения L-валина, где композиция содержит микроорганизм, включающий вариант по настоящей заявке, полинуклеотид, кодирующий вариант, вектор, содержащий полинуклеотид, или полинуклеотид по настоящей заявке; среду, полученную путем культивирования микроорганизма; или комбинацию двух или более из них.
Композиция по настоящей заявке может дополнительно содержать любой подходящий эксципиент, который обычно используют в композициях для получения аминокислот, и примеры эксципиента могут включать консервант, смачивающий агент, диспергирующий агент, суспендирующий агент, буфер, стабилизирующий агент или изотонический агент, но без ограничения этим.
В композиции по настоящей заявке вариант, полинуклеотид, вектор, штамм, среда, L-валин и подобные представляют собой такие, как описано в других аспектах.
Согласно другому аспекту настоящей заявки предложено применение варианта малой субъединицы синтазы ацетогидроксикислоты для получения L-валина, варианта, имеющего замену аминокислоты в положении 44 в аминокислотной последовательности SEQ ID NO: 1 другой аминокислотой.
При использовании настоящей заявки вариант, L-валин и тому подобные представляют собой такие, как описано в других аспектах.
Способ осуществления изобретения Далее в настоящая заявка будет подробно описана со ссылкой на Примеры. Однако эти примеры представляют собой только предпочтительные воплощения для иллюстрации настоящей заявки и, следовательно, не предназначены для ограничения объема прав настоящей заявки. Между тем, технический материал, не описанный здесь, может быть достаточно понятным и легко выполняемым для специалиста в данной области техники или аналогичных технических областях настоящей заявки.
Пример 1. Селекция мутантного штамма с повышенной способностью продуцировать валин посредством искусственной мутации
Пример 1-1. Искусственный мутагенез посредством ультрафиолетового облучения
С целью отбора мутантного штамма с повышенной способностью продуцирования валина, Corynebacterium glutamicum КССМ1120 IP (Корейская патентная публикация №10-1117022), штамм, продуцирующий валин, высевали на чашки с питательной средой, содержащей агар, и культивировали при 30°С в течение 36 часов. Сотни полученных таким образом колоний облучали ультрафиолетовым излучением при комнатной температуре для индуцирования случайного мутагенеза в геноме штамма.
Пример 1-2. Оценка титров ферментации мутировавших штаммов и выбор штамма
С целью селекции мутантных штаммов, имеющих повышенную способность продуцировать L-валин по сравнению с Corynebacterium glutamicum КССМ11201Р, используемым в качестве родительского штамма, выполняли тест на титр ферментации на случайно выбранных мутировавших штаммах. После субкультивирования каждой колонии в питательной среде каждый штамм инокулировали в колбу 250 мл с угловыми перегородками, содержащую 25 мл среды для продуцирования, и культивировали при встряхивании при 200 об/мин в течение 72 часов при 30°С.После этого, используя ВЭЖХ, анализировали концентрацию L-валина и проанализированные концентрации L-валина сводили в Таблицу 1.
Питательная среда (рН 7,2)
глюкоза 10 г, мясной сок 5 г, полипептон 10 г, хлорид натрия 2,5 г, дрожжевой экстракт 5 г, агар 20 г и мочевина 2 г (из расчета на 1 л дистиллированной воды) Среда для продуцирования (рН 7,0)
глюкоза 100 г, сульфат аммония 40 г, соевый белок 2,5 г, кукурузный крахмал 5 г, мочевина 3 г, двухосновный фосфат калия 1 г, гептагидрат сульфата магния 0,5 г, биотин 100 мкг, тиамин-HCl 1 мг, пантотенат кальция 2 мг, никотинамид 3 мг, карбонат кальция 30 г (из расчета на 1 л дистиллированной воды)
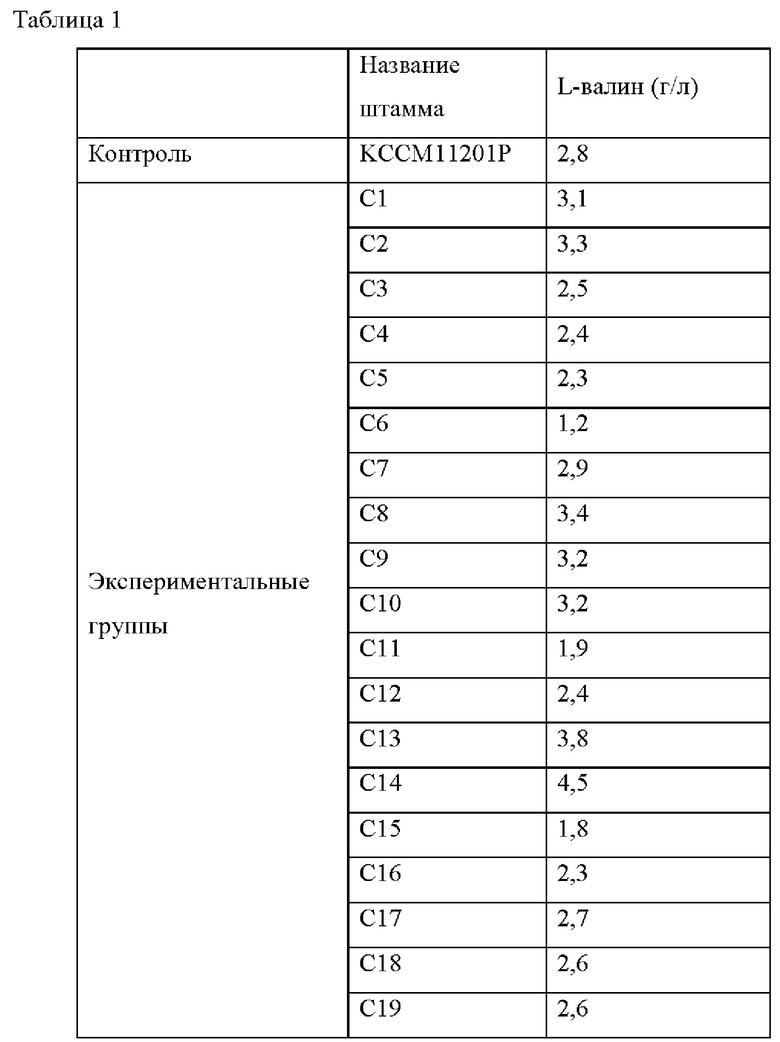

На основании результатов, приведенных в Таблице 1, выбрали штамм С14 с наибольшим увеличением количества продукции валина по сравнению со штаммом КССМ11201Р в качестве контроля.
Пример 2. Идентификация мутации посредством секвенирования генов
Основные гены штамма секвенировали и сравнивали с генами штамма КССМ11201Р и штамма Corynebacterium glutamicum АТСС14067 дикого типа. Результаты доказали, что штамм КССМ11201Р и штамм С14 с повышенной способностью продуцировать валин включали мутации нуклеотидной последовательности в конкретных положениях области открытой рамки считывания (ОРС) гена ilvN. Конкретно, КССМ11201Р имел форму, в которой аминокислота аланин в положении 42 была заменена на валин, что соответствует изменению существующего GCA на GTA посредством введения одной мутации в нуклеотид в положении 125 от инициирующего кодона гена ilvN.
Штамм С14 с наибольшим увеличением количества продуцирования валина включал мутацию A42V, включенную в родительский штамм КССМ11201Р, и имел форму, в которой аминокислота треонин в положении 44 была заменена аланином, что соответствует изменению существующего АСС на GCC посредством введения одной мутации в нуклеотид в положение 130 от инициирующего кодона гена ilvN.
В результате анализа областей мутации подтвердили, что области мутации влияют на домен, связывающий эффектор биосинтазы валина, предполагая усиление активности соответствующего белка. В следующих примерах предпринимались попытки исследования индивидуальных эффектов мутаций A42V и Т44А, вставленных в конкретные положения ОРС в гене ilvN, и влияние комбинированного применения на способность микроорганизма рода Corynebacterium продуцировать валин, аминокислоту с разветвленной цепью. Кроме того, предпринималась попытка исследования влияния замены аминокислотой, отличной от аланина, в отношении мутации треонина, аминокислоты в положении 44 на способность микроорганизма рода Corynebacterium продуцировать аминокислоты с разветвленной цепью - валин, изолейцин и лейцин.
Пример 3. Получение штаммов КССМ11201Р с введением мутаций ilvN и опеределение способности продуцировать валин
Пример 3-1. Получение штамма с введением мутации ilvN в штамм Corynebacterium glutamicum КССМ11201Р и оценка способности продуцировать L-валин
Для вставки мутанта ilvN(A42V+T44A), указанного в SEQ ID NO: 6, в Corynebacterium glutamicum КССМ11201Р конструировали вектор, содержащий целевую мутацию. Конкретно, геномную ДНК штамма С14 выделяли, используя мини-набор G-spin Total DNA Extraction Mini Kit (Intron, Cat. No. 17045) согласно протоколу, представленному в наборе, и выполняли ПЦР, используя геномную ДНК в качестве матрицы. Условия для ПЦР были следующими: денатурация при 94°С в течение 5 минут; 25 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 150 секунд; и затем полимеризация при 72°С в течение 7 минут. Продукт ПЦР (далее называемый «мутировавший фрагмент 1») длиной 1010 п. н.(пар нуклеотидов) получали с использованием SEQ ID NOs: 7 и 8.
Полученный мутировавший фрагмент 1 лигировали с вектором pDZ (публикация патента Кореи №10-0924065 и международная публикация патента №2008-033001), обработанным ферментом рестрикции XbaI (New England Biolabs, Beverly, MA) с использованием набора для инфузионного клонирования (Takara Bio Inc., Otsu, Japan), с последующей трансформацией в Е. coli DH5a. Полученный ген трансформировали в Е. coli DH5a и затем трансформированные штаммы отбирали в среде LB, содержащей канамицин, и получали из нее ДНК с помощью набора для очистки плазмидной ДНК «ДНК-spin» (iNtRON), таким образом конструируя вектор pDZ-ilvN(A42V+T44A), содержащий мутировавший фрагмент 1.

Конкретно, вектор pDZ-ilvN (A42V+T44A) трансформировали в Corynebacterium glutamicum KCCM1120IP посредством гомологичной рекомбинации на хромосоме (van der Rest et al., Appl. Microbiol Biotechnol 52: 541-545, 1999). Штаммы с вектором, вставленным в хромосому посредством гомологичной рекомбинации последовательности, отбирали в среде, содержащей канамицин (25 мг/л). Затем с использованием SEQ ID NO: 7 и 8 выполняли ПЦР на трансформантах Corynebacterium glutamicum, подвергнутых вторичной рекомбинации для идентификации штамма, в котором аланин был заменен валином в положении 42 и треонин был заменен аланином в положении 44 в аминокислотной последовательности SEQ ID NO: 1 в пределах ОРС гена ilvNnn хромосоме, соответственно. Рекомбинантный штамм назвали Corynebacterium glutamicum KCCM11201P::/7vJV(A42V+T44A).
Для сравнения способности продуцировать L-валин у штаммов Corynebacterium glutamicum КССМ11201Р и KCCM11201P::ilvN(A42V+T44A), продуцирующих валин, выполняли оценку в колбе. Каждый штамм субкультивировали в питательной среде, инокулировали в колбу 250 мл с угловыми перегородками, содержащую 25 мл питательной среды, и культивировали при встряхивании при 200 об/мин в течение 72 часов при 30°С.Затем с помощью ВЭЖХ анализировали концентрацию L-валина и проанализированные концентрации L-валина сводили в Таблицу 3.
Питательная среда (рН 7,2)
глюкоза 10 г, мясной сок 5 г, полипептон 10 г, хлорид натрия 2,5 г, дрожжевой экстракт 5 г, агар 20 г и мочевина 2 г (из расчета на 1 л дистиллированной воды) Среда для продуцирования (рН 7,0)
глюкоза 100 г, сульфат аммония 40 г, соевый белок 2,5 г, кукурузный крахмал 5 г, мочевина 3 г, двухосновный фосфат калия 1 г, гептагидрат сульфата магния 0,5 г, биотин 100 мкг, тиамин-HCl 1 мг, пантотенат кальция 2 мг, никотинамид 3 мг, карбонат кальция 30 г (из расчета на 1 л дистиллированной воды)
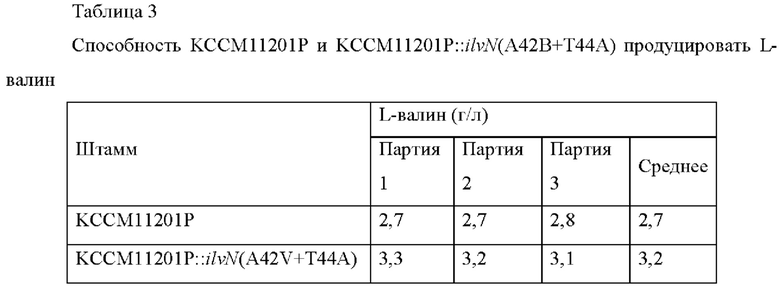
Результаты показали, что способность штамма KCCM11201P::ilvN(A42V+T44A) продуцировать L-валин увеличилась на 18,5% по сравнению с КССМ11201Р.
Пример 3-2. Получение штамма с введением мутации ilvN в штамм Corynebacterium glutamicum КССМ11201Р и оценка способности продуцировать L-валин
Для того чтобы вставить мутант ilvN(T44A), указанный в SEQ ID NO: 3, в Corynebacterium glutamicum КССМ11201Р, конструировали вектор, содержащий целевую мутацию. В частности, геномная ДНК штамма АТСС 14067, Corynebacterium glutamicum дикого типа, была экстрагирована с использованием мини-набора для тотальной экстракции ДНК G-spin (Intron, Cat. №17045) в соответствии с протоколом, прилагаемым к набору, и была проведена ПЦР с использованием геномной ДНК в качестве матрицы. Условия для ПЦР были следующими: денатурация при 94°С в течение 5 минут; 25 циклов денатурации при 94°С в течение 30 секунд, отжиг при 55°С в течение 30 секунд и полимеризация при 72°С в течение 150 секунд; и затем полимеризация при 72°С в течение 7 минут. Продукт ПЦР (далее именуемый «мутировавший фрагмент 2») длиной 515 п. н. получали с использованием SEQ ID NO: 9 и 10, и продукт ПЦР (далее именуемый «мутировавший фрагмент 3») длиной 518 п. н. получали с использованием SEQ ID NO: 11 и 12.
Полученные мутировавшие фрагменты 2 и 3 лигировали с вектором pDZ (публикация патента Кореи №10-0924065 и международная публикация патента №2008-033001), обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MA) с использованием набора для инфузионного клонирования (Takara Bio Inc., Otsu, Japan) с последующей трансформацией в Е. coli DH5a. Полученный ген трансформировали в Е. coli DH5a и затем трансформированные штаммы отбирали в среде LB, содержащей канамицин, и получали из нее ДНК с помощью набора для очистки плазмидной ДНК «ДНК-spin» (iNtRON), тем самым конструируя вектор pDZ-ilvN(T44A), содержащий мутировавшие фрагменты 2 и 3.

Конкретно, вектор pDZ-ilvN(T44A) трансформировали в Corynebacterium glutamicum KCCM1120IP посредством гомологичной рекомбинации на хромосоме (van der Rest et al., Appl. Microbiol Biotechnol 52: 541-545, 1999). Гомологичная рекомбинация приводила к восстановлению от мутации A42V и введению мутации Т44А. Штаммы с вектором, вставленным в хромосому путем гомологичной рекомбинации последовательности, отбирали в среде, содержащей канамицин (25 мг/л). Затем, используя SEQ ID NOs: 9 и 12, выполняли ПЦР на трансформантах Corynebacterium glutamicum, подвергнутых вторичной рекомбинации для идентификации штамма, в котором треонин был заменен аланином в положении 44 в аминокислотной последовательности SEQ ID NO: 1 в ОРС гена ilvN на хромосоме. Рекомбинантный штамм назвали Corynebacterium glutamicum KCCM1120IP::ilvN(T44A).
Для сравнения способности полученных штаммов продуцировать L-валин штаммы культивировали и концентрацию L-валина анализировали посредством такого же способа, что и в Примере 3-1, и проанализированные концентрации L-валина сводили в Таблицу 5 ниже.
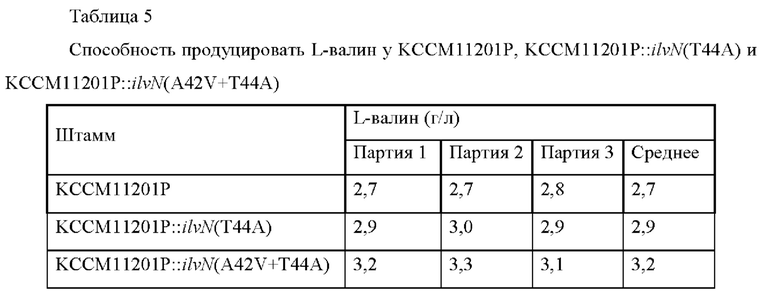
Результаты показали, что способность штаммов KCCM11201P::/7v7^(T44A) и KCCM11201P::ilvN(A42V+T44A) продуцировать L-валин увеличилась на 7,4% и 18,5% по сравнению с КССМ11201Р, соответственно.
Пример 3-3. Получение штамма с введением мутации ilvN в штамм Corynebacterium glutamicum CJ7V и оценка способности продуцировать L-валин
Чтобы определить, повлияла ли мутация на увеличение способности продуцировать L-валин и у других штаммов Corynebacterium glutamicum, продуцирующих L-валин, получали штамм с улучшенной способностью продуцировать L-валин посредством введения одного типа мутации (ilvN(A42V); (Biotechnology and Biopr ос ess Engineering, June 2014, Volume 19, Issue 3, 456-467) в Corynebacterium glutamicum ATCC14067 дикого типа.
Конкретно, геномную ДНК штамма АТСС 14067 Corynebacterium glutamicum дикого типа выделяли с использованием мини-набора для выделения тотальной ДНК G-spin (nitron, Cat. No 17045) согласно протоколу, представленному в наборе. ПЦР выполняли с использованием геномной ДНК в качестве матрицы. Для конструирования вектора для введения мутации A42V в ген ilvN получали фрагменты гена (А и В), используя пару праймеров SEQ ID NOs: 13 и 14 и пару праймеров SEQ ID NOs: 15 и 16, соответственно. Условия для ПЦР были следующими: денатурация при 94°С в течение 5 минут; 25 циклов денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд; а затем полимеризация при 72°С в течение 7 минут.
В результате для фрагментов А и В можно было получить полинуклеотиды длиной 528 п. н. и 509 п. н., соответственно. Перекрывающуюся ПЦР с использованием двух фрагментов в качестве матрицы с SEQ ID NOs: 13 и 16 выполняли с получением продукта ПЦР длиной 1010 п. н. (далее именуемого «мутировавший фрагмент 4»).
Полученный мутировавший фрагмент 4 обрабатывали ферментом рестрикции XbaI (New England Biolabs, Beverly, MA) и затем лигировали с вектором pDZ, обработанным тем же ферментом рестрикции лигазой Т4 (New England Biolabs, Беверли, Массачусетс). Полученный ген трансформировали в Е. coli DH5a, и затем трансформированные штаммы отбирали в среде LB, содержащей канамицин, и получали из них ДНК с помощью набора для очистки плазмидной ДНК «ДНК-spin» (iNtRON). Вектор, предназначенный для введения A42V в ген ilvN, назвали pDZ-ilvN(A42V).

Затем вектор pDZ-ilvN(A42V) трансформировали в Corynebacterium glutamicum АТСС14067 дикого типа посредством гомологичной рекомбинации на хромосоме (van der Rest et al., Appl. Microbiol Biotechnol 52: 541-545, 1999). Штаммы с вектором, вставленным в хромосому посредством гомологичной рекомбинации последовательности, отбирали в среде, содержащей канамицин (25 мг/л). Затем выполняли ПЦР с использованием SEQ ID NOs: 13 и 16 на трансформантах Corynebacterium glutamicum, подвергнутых вторичной рекомбинации для амплификации фрагмента гена, и затем штамм со вставленной мутацией идентифицировали посредством секвенирования генов. Рекомбинантный штамм назвали Corynebacterium glutamicum CJ7V.
Наконец, Corynebacterium glutamicum CJ7V трансформировали соответствующими векторами посредством таких же способов, как в Примере 3-1 и Примере 3-2, соответственно, и штаммы-трансформанты назвали Corynebacterium glutamicum CJ7V::ilvN(T44A) и CJ7V::ilvN (A42V+T44A), соответственно. Для сравнения способности полученных штаммов продуцировать L-валин штаммы культивировали и концентрацию L-валина анализировали посредством таких же способов, как в Примере 3-1, и проанализированные концентрации L-валина сводили в Таблицу 7 ниже.
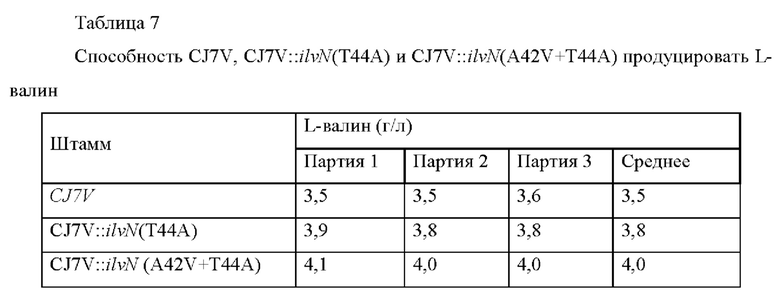
Результаты показали, что способность штаммов CJ7V::ilvN(T44A) и CJ7V::ilvN (A42V+T44A) продуцировать L-валин увеличилась на 8,5% и 12,3% по сравнению с CJ7V, соответственно.
Как указано выше, специалист в данной области техники, к которой относится настоящая заявка, сможет понять, что настоящая заявка может быть воплощена в других конкретных формах без отступления от ее технического духа или существенных характеристик. Таким образом, воплощения, описанные выше, следует рассматривать как иллюстрирующие и не ограничивающие настоящую заявку. Объем настоящей заявки следует понимать так, что все изменения или модификации, вытекающие из определений и объема формулы изобретения и их эквивалентов, попадают в объем заявки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>CJCheilJedangCorporation
<120>Новый вариант субъединицы синтазыацетогидроксикислоты и
способ получения L-валина c использованием этого варианта
<130> OPA22038
<150> KR 10-2021-0110890
<151> 2021-08-23
<160> 16
<170>KoPatentIn 3.0
<210> 1
<211> 172
<212> PRT
<213> Unknown
<220>
<223> Вариант малой субъединицы синтазыацетогидроксикислоты
<400> 1
Met Ala AsnSer Asp Val ThrArg His Ile LeuSer Val Leu Val Gln
1 5 10 15
Asp Val Asp Gly Ile IleSerArg Val SerGly Met PheThrArgArg
20 25 30
Ala PheAsnLeu Val SerLeu Val Ser Ala Lys ThrGluThrLeuGly
35 40 45
Ile AsnArg Ile Thr Val ValVal Asp Ala Asp GluLeuAsn Ile Glu
50 55 60
Gln Ile Thr Lys GlnLeuAsn Lys Leu Ile Pro Val Leu Lys Val Val
65 70 75 80
ArgLeu Asp GluGluThrThr Ile Ala Arg Ala Ile Met Leu Val Lys
85 90 95
Val Ser Ala Asp SerThrAsnArg Pro Gln Ile Val Asp Ala AlaAsn
100 105 110
Ile PheArg Ala Arg Val Val Asp Val Ala Pro Asp Ser Val Val Ile
115 120 125
GluSerThrGlyThr Pro Gly Lys LeuArg Ala LeuLeu Asp Val Met
130 135 140
Glu Pro PheGly Ile ArgGluLeu Ile GlnSerGlyGln Ile Ala Leu
145 150 155 160
AsnArgGly Pro Lys Thr Met Ala Pro Ala Lys Ile
165 170
<210> 2
<211> 519
<212> DNA
<213>Unknown
<220>
<223> малая субъединица синтазыацетогидроксикислоты
<400> 2
atggctaattctgacgtcacccgccacatcctgtccgtactcgttcaggacgtagacgga 60
atcatttcccgcgtatcaggtatgttcacccgacgcgcattcaacctcgtgtccctcgtg 120
tctgcaaagaccgaaacactcggcatcaaccgcatcacggttgttgtcgacgccgacgag 180
ctcaacattgagcagatcaccaagcagctcaacaagctgatccccgtgctcaaagtcgtg 240
cgacttgatgaagagaccaccatcgcccgcgcaatcatgctggttaaggtctctgcggat 300
agcaccaaccgtccgcagatcgtcgacgccgcgaacatcttccgcgcccgagtcgtcgac 360
gtggctccagactctgtggttattgaatccacaggcaccccaggcaagctccgcgcactg 420
cttgatgtgatggaaccattcggaatccgcgaactgatccaatccggacagattgcactc 480
aaccgcggtccgaagaccatggctccggccaagatctaa 519
<210> 3
<211> 172
<212> PRT
<213>Unknown
<220>
<223> вариант малой субъединицы синтазыацетогидроксикислоты (T44A)
<400> 3
Met Ala AsnSer Asp Val ThrArg His Ile LeuSer Val Leu Val Gln
1 5 10 15
Asp Val Asp Gly Ile IleSerArg Val SerGly Met PheThrArgArg
20 25 30
Ala PheAsnLeu Val SerLeu Val Ser Ala Lys Ala GluThrLeuGly
35 40 45
Ile AsnArg Ile Thr Val ValVal Asp Ala Asp GluLeuAsn Ile Glu
50 55 60
Gln Ile Thr Lys GlnLeuAsn Lys Leu Ile Pro Val Leu Lys Val Val
65 70 75 80
ArgLeu Asp GluGluThrThr Ile Ala Arg Ala Ile Met Leu Val Lys
85 90 95
Val Ser Ala Asp SerThrAsnArg Pro Gln Ile Val Asp Ala AlaAsn
100 105 110
Ile PheArg Ala Arg Val Val Asp Val Ala Pro Asp Ser Val Val Ile
115 120 125
GluSerThrGlyThr Pro Gly Lys LeuArg Ala LeuLeu Asp Val Met
130 135 140
Glu Pro PheGly Ile ArgGluLeu Ile GlnSerGlyGln Ile Ala Leu
145 150 155 160
AsnArgGly Pro Lys Thr Met Ala Pro Ala Lys Ile
165 170
<210> 4
<211> 519
<212> DNA
<213>Unknown
<220>
<223> вариант малой субъединицы синтазыацетогидроксикислоты
(T44A)
<400> 4
atggctaattctgacgtcacccgccacatcctgtccgtactcgttcaggacgtagacgga 60
atcatttcccgcgtatcaggtatgttcacccgacgcgcattcaacctcgtgtccctcgtg 120
tctgcaaaggccgaaacactcggcatcaaccgcatcacggttgttgtcgacgccgacgag 180
ctcaacattgagcagatcaccaagcagctcaacaagctgatccccgtgctcaaagtcgtg 240
cgacttgatgaagagaccaccatcgcccgcgcaatcatgctggttaaggtctctgcggat 300
agcaccaaccgtccgcagatcgtcgacgccgcgaacatcttccgcgcccgagtcgtcgac 360
gtggctccagactctgtggttattgaatccacaggcaccccaggcaagctccgcgcactg 420
cttgatgtgatggaaccattcggaatccgcgaactgatccaatccggacagattgcactc 480
aaccgcggtccgaagaccatggctccggccaagatctaa 519
<210> 5
<211> 172
<212> PRT
<213>Unknown
<220>
<223> вариант малой субъединицы синтазыацетогидроксикислоты (T44A+A42V)
<400> 5
Met Ala AsnSer Asp Val ThrArg His Ile LeuSer Val Leu Val Gln
1 5 10 15
Asp Val Asp Gly Ile IleSerArg Val SerGly Met PheThrArgArg
20 25 30
Ala PheAsnLeu Val SerLeu Val Ser Val Lys Ala GluThrLeuGly
35 40 45
Ile AsnArg Ile Thr Val ValVal Asp Ala Asp GluLeuAsn Ile Glu
50 55 60
Gln Ile Thr Lys GlnLeuAsn Lys Leu Ile Pro Val Leu Lys Val Val
65 70 75 80
ArgLeu Asp GluGluThrThr Ile Ala Arg Ala Ile Met Leu Val Lys
85 90 95
Val Ser Ala Asp SerThrAsnArg Pro Gln Ile Val Asp Ala AlaAsn
100 105 110
Ile PheArg Ala Arg Val Val Asp Val Ala Pro Asp Ser Val Val Ile
115 120 125
GluSerThrGlyThr Pro Gly Lys LeuArg Ala LeuLeu Asp Val Met
130 135 140
Glu Pro PheGly Ile ArgGluLeu Ile GlnSerGlyGln Ile Ala Leu
145 150 155 160
AsnArgGly Pro Lys Thr Met Ala Pro Ala Lys Ile
165 170
<210> 6
<211> 519
<212> DNA
<213>Unknown
<220>
<223> вариант малой субъединицы синтазыацетогидроксикислоты (T44A+A42V)
<400> 6
atggctaattctgacgtcacccgccacatcctgtccgtactcgttcaggacgtagacgga 60
atcatttcccgcgtatcaggtatgttcacccgacgcgcattcaacctcgtgtccctcgtg 120
tctgtaaaggccgaaacactcggcatcaaccgcatcacggttgttgtcgacgccgacgag 180
ctcaacattgagcagatcaccaagcagctcaacaagctgatccccgtgctcaaagtcgtg 240
cgacttgatgaagagaccaccatcgcccgcgcaatcatgctggttaaggtctctgcggat 300
agcaccaaccgtccgcagatcgtcgacgccgcgaacatcttccgcgcccgagtcgtcgac 360
gtggctccagactctgtggttattgaatccacaggcaccccaggcaagctccgcgcactg 420
cttgatgtgatggaaccattcggaatccgcgaactgatccaatccggacagattgcactc 480
aaccgcggtccgaagaccatggctccggccaagatctaa 519
<210> 7
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 7
cggggatcctctagaaggacggtactcaaatactaaactt c 41
<210> 8
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 8
cggggatcctctagagacaactacattattattataccac a 41
<210> 9
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 9
cggggatcctctagaaggacggtactcaaatactaaactt c 41
<210> 10
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 10
ggcctttgcagacacgagggacacgagg 28
<210> 11
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 11
tgtccctcgtgtctgcaaaggccgaaacactcggc 35
<210> 12
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 12
cggggatcctctagagacaactacattattattataccac a 41
<210> 13
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 13
cggggatcctctagaaggacggtactcaaatactaaactt c 41
<210> 14
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 14
tgccgagtgtttcggtctttacagacacgagggacacg 38
<210> 15
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер odhA_down_F
<400> 15
tgtctgtaaagaccgaaacactcggcatca a 31
<210> 16
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223>праймер
<400> 16
cggggatcctctagagacaactacattattattataccac a 41
<---
Изобретение относится к области биотехнологии, конкретно, к мутантным белкам, вовлеченным в продуцирование L-валина, и может быть использовано для получения L-валина в Corynebacterium glutamicum. Предложены мутантные белки модифицированной малой субъединицы синтазы ацетогидроксикислоты (ilvN) с аминокислотной последовательностью SEQ ID NO: 1, в которой аминокислота, соответствующая положению 44, заменена на аланин, а также дополнительно может быть произведена замена аминокислоты, соответствующей положению 42 в аминокислотной последовательности SEQ ID NO: 1, на валин. Изобретение обеспечивает получение модифицированных ilvN, увеличивающих продуцирование L-валина в Corynebacterium glutamicum. 6 н. и 3 з.п. ф-лы, 7 табл., 3 пр.
1. Малая субъединица синтазы ацетогидроксикислоты (ilvN), вовлеченная в увеличение продуцирования L-валина, содержащая аминокислотную последовательность SEQ ID NO: 1, в которой аминокислота, соответствующая положению 44 в аминокислотной последовательности SEQ ID NO: 1, заменена аланином.
2. Малая субъединица синтазы ацетогидроксикислоты (ilvN), вовлеченная в увеличение продуцирования L-валина, содержащая аминокислотную последовательность SEQ ID NO: 1, в которой аминокислота, соответствующая положению 44 в аминокислотной последовательности SEQ ID NO: 1, заменена аланином и аминокислота, соответствующая положению 42 в аминокислотной последовательности SEQ ID NO: 1, заменена валином.
3. Малая субъединица синтазы ацетогидроксикислоты (ilvN) по п. 1, состоящая из аминокислотной последовательности SEQ ID NO: 3.
4. Малая субъединица синтазы ацетогидроксикислоты (ilvN) по п. 2, состоящая из аминокислотной последовательности SEQ ID NO: 5.
5. Полинуклеотид, кодирующий малую субъединицу синтазы ацетогидроксикислоты (ilvN) по любому из пп. 1-4.
6. Вектор экспрессии, содержащий полинуклеотид по п. 5.
7. Микроорганизм Corynebacterium glutamicum для продуцирования L-валина, экспрессирующий малую субъединицу синтазы ацетогидроксикислоты (ilvN) по любому из пп. 1-4, или полинуклеотид, кодирующий эту малую субъединицу синтазы ацетогидроксикислоты (ilvN).
8. Микроорганизм по п. 7, имеющий повышенную способность продуцировать L-валин по сравнению с микроорганизмом, содержащим полипептид с SEQ ID NO: 1 или полинуклеотид, кодирующий этот полипептид.
9. Способ получения L-валина, включающий культивирование микроорганизма по п. 7 в среде и извлечение L-валина из среды.
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| et al., Genetic analysis of protein stability and function | |||
| Anna | |||
| Rev | |||
| Genet | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |

