Настоящее изобретение относится к мультивалентной и мультиспецифичной производной Fv-антитела, в частности к молекуле Fv-антитела, содержащей единицу диатела.
Биспецифичные антитела применяют для взаимодействия с двумя различными терапевтическими мишенями или выполнения двух различных функций. Такие антитела можно применять, например, для рекрутинга иммунных эффекторных клеток, например, Т- или NK-клеток к конкретной клетке-мишени. Известны и исследуются различные молекулы на основе фрагментов антител, например, при лечении рака.
Биспецифичные антитела можно конструировать с использованием только вариабельных доменов антител. Например, линкерную последовательность между VH- и VL-доменами можно укоротить до такой степени, чтобы они не могли укладываться друг на друга и попарно ассоциировать, вступая во внутримолекулярное взаимодействие. Такие короткие линкеры, например, из 2-12 остатков, предотвращают указанное образование мономерной молекулы одноцепочечного вариабельного фрагмента (scFv) и благоприятствуют межмолекулярному сопряжению VH/VL между комплементарными вариабельными доменами различных полипептидных цепей с образованием димерного "диатела" (Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90, 6444-6448). Такое диатело можно использовать для конструирования биспецифичных антител, получаемых за счет нековалентной ассоциации двух одноцепочечных полипептидных гибридных продуктов, каждый из которых состоит из VH-домена одного антитела, соединенного коротким линкером с VL-доменом другого антитела (или наоборот).
В WO 03/025018 описана биспецифичная антигенсвязывающая молекула, структура которой образована идентичными одноцепочечными полипептидами с четырьмя связывающими доменами. VH- и VL-домен в концевой части каждой полипептидной цепи связаны коротким линкером и межмолекулярно ассоциированы с соответствующими VH- и VL-доменами другой полипептидной цепи, а другие VH- и VL-домены каждой полипептидной цепи внутримолекулярно связаны друг с другом в пределах той же цепи с образованием антигенсвязывающей scFv-единицы. Такие конструкты являются гомодимерами, т.е. состоят из идентичных одноцепочечных полипептидов, которые попарно ассоциируют друг с другом.
Кроме того, желательны триспецифичные антитела, мишенью которых являются два опухолевых антигена, что обеспечивает повышенную селективность по отношению к раковым клеткам, сохранение здоровой ткани и приводит к расширению диапазона доз и терапевтической применимости, при которых указанное лекарственное вещество может эффективно уничтожать раковые клетки. Например, одно триспецифичное антитело можно применять для нацеленного воздействия на два различных опухолевых антигена, причем третья специфичность активирует цитотоксическое действие Т-клеток или NK-клеток.
В WO 2009/007124 описан триспецифичный гибрид одноцепочечного Fv (scFv), названный «тройным телом» и содержащий три scFv-фрагмента, расположенные в виде тандема, имеющие три различных специфичности (CD123 и CD33 (опухолевые маркеры) и одну к CD16 на NK-клетках). Эту молекула дополнительно стабилизирована дисульфидными связями в центральном scFv. Такие молекулы обеспечивают двойное нацеливание дважды положительные опухолевые клетки и одновалентное связывание с CD16 на NK-клетках.
Настоящее изобретение относится к мультивалентному Fv-антителу, содержащему единицу диатела, состоящую из двух пар вариабельных доменов, ассоциированных с двумя антигенсвязывающими сайтами. Термин «антигенсвязывающий сайт» обозначает антигенсвязывающий сайт Fv-фрагмента, состоящего из пары доменов VH/VL, т.е. антигенсвязывающий сайт VH/VL, или однодоменный антигенсвязывающий сайт. Каждая пара вариабельных доменов связана один за другим друг за другом в полипептиде. Единица диатела состоит из одного полипептида (единица одноцепочечного диатела) или двух полипептидов (единица диатела). По меньшей мере одна пара вариабельных доменов соединена в полипептиде с еще одним вариабельным доменом, находящимся со стороны N-конца от этой пары вариабельных доменов, и с еще одним вариабельным доменом - со стороны С-конца. Соответственно, такой полипептид содержит по меньшей мере четыре вариабельных домена, соединенных друг за другом, причем два расположенных рядом друг с другом вариабельных домена, соединенных друг за другом, представляют собой одну пару вариабельных доменов единицы диатела, а еще один вариабельный домен расположен со стороны N-конца от этой пары вариабельных доменов единицы диатела, и другие дополнительные вариабельные домены расположены со стороны С-конца от этой пары вариабельных доменов единицы диатела. Вариабельный домен, соединенный с N-концом этой пары вариабельных доменов единицы диатела, может представлять собой вариабельный домен легкой цепи (VL) или вариабельный домен тяжелой цепи (VH), также, как и домен, соединенный с С-концом этой пары вариабельных доменов единицы диатела, может представлять собой вариабельный домен легкой цепи (VL) или вариабельный домен тяжелой цепи (VH). Такой полипептид представляет собой Fv-полипептид, содержащий по меньшей мере четыре соединенных друг за другом вариабельных домена, т.е. вариабельный домен, соединенный к N-концом пары вариабельных доменов единицы диатела, присоединенной по С-концу к еще одному вариабельному домену. В конкретных вариантах реализации шесть, восемь или десять вариабельных доменов соединены друг за другом в составе такого Fv-полипептида.
Мультивалентное Fv-антитело является по меньшей мере четырехвалентным и содержит по меньшей мере четыре антигенсвязывающих сайта. Соответственно, Fv-полипептид Fv-антитела, содержащего по меньшей мере четыре вариабельных домена, где два из по меньшей мере четырех вариабельных доменов представляют собой пару из двух расположенных рядом друг с другом вариабельных доменов единицы диатела, ассоциирующих с другой парой вариабельных доменов единицы диатела с образованием двух (первого и второго) антигенсвязывающих сайтов, содержит по меньшей мере дополнительный (третий) вариабельный домен по N-концу Fv-полипептида, ассоциирующий с соответствующим вариабельным доменом с образованием дополнительного (третьего) антигенсвязывающего сайта, и по меньшей мере дополнительный (четвертый) вариабельный домен по С-концу Fv-полипептида, ассоциирующий с соответствующим вариабельным доменом с образованием дополнительного (четвертого) антигенсвязывающего сайта. В некоторых вариантах реализации по меньшей мере четыре антигенсвязывающих сайта мультивалентного Fv-антитела образованы двумя Fv-полипептидами, а в других вариантах реализации эти по меньшей мере четыре антигенсвязывающих сайта образованы в результате внутримолекулярного фолдинга одиночного Fv-полипептида.
Таким образом, в некоторых вариантах реализации каждый из других вариабельных доменов, связанных по N-концу и С-концу с парой вариабельных доменов единицы диатела в одном и том же полипептиде, является частью дополнительного антигенсвязывающего сайта. Соответственно, одна пара вариабельных доменов единицы диатела (первая пара) ассоциирована с другой парой вариабельных доменов единицы диатела (второй парой) с образованием двух антигенсвязывающих сайтов (первого и второго антигенсвязывающих сайтов), а дополнительный вариабельный домен, расположенный с N-конца, ассоциирует с соответствующим вариабельным доменом с образованием третьего антигенсвязывающего сайта, и дополнительный вариабельный домен, расположенный с С-конца, ассоциирует с соответствующим вариабельным доменом с образованием четвертого антигенсвязывающего сайта. Таким образом, такое мультивалентное Fv-антитело является по меньшей мере четырехвалентным. В некоторых вариантах реализации этот антигенсвязывающий сайт содержит дополнительный вариабельный домен, связанный по N-концу и/или С-концу с парой вариабельных доменов единицы диатела в scFv-единице или единице одноцепочечного диатела. В других вариантах реализации дополнительные вариабельные домены, соединенные по N-концу и С-концу с парой вариабельных доменов единицы диатела в составе одного и того же полипептида (первого полипептида), ассоциированы с соответствующими VH- или VL-доменами еще одного полипептида (второго полипептида), содержащего вторую (другую) пару вариабельных доменов единицы диатела, тем самым образуя еще два (третий и четвертый) VH/VL антигенсвязывающих сайта между вариабельными доменами двух (первого и второго) полипептидов.
В некоторых вариантах реализации две пары вариабельных доменов единицы диатела представляют собой пару вариабельных доменов легкой цепи (VL-VL), соединенных друг за другом в полипептиде, и пару вариабельных доменов тяжелой цепи (VH-VH), соединенных друг за другом в полипептиде, причем пара VL-VL и пара VH-VH ассоциируют в два (первый и второй) VL/VH-антигенсвязывающих сайта.
В некоторых вариантах реализации молекула мультивалентного антитела содержит единицу диатела, т.е. пару вариабельных доменов единицы диатела, встроенные в полипептидную цепь, содержащую по меньшей мере шесть, например, шесть, восемь или десять вариабельных доменов, соединенных друг за другом
Единица диатела состоит из пары двух вариабельных доменов, соединенных друг за другом так, что эти домены не могут образовывать внутримолекулярную структуру функциональной Fv-единицы, т.е. VH/VL-антигенсвязывающий сайт, а вместо этого ассоциируют с еще одной парой двух вариабельных доменов, соединенных друг за другом, с образованием двухвалентного димера, т.е. диатела, обеспечивая два антигенсвязывающих сайта. Соединение пары вариабельных доменов легкой цепи (VL-VL) друг за другом и пары вариабельных доменов тяжелой цепи (VH-VH) друг за другом предотвращает внутримолекулярное спаривание доменов в пределах каждой пары за счет доменов одного типа, т.е. VH-VH или VL-VL. Жесткая и компактная структура единицы диатела облегчает получение, правильный фолдинг (сворачивание) мультивалентного антитела, и повышает стабильность антитела. Такая единица диатела образует два VH/VL-антигенсвязывающих сайта в пределах молекулы антитела за счет двух ковалентно связанных VH- и VL-доменов, что выгодно для стабильности молекулы антитела, поскольку приводит к образованию более компактной молекулы. В некоторых вариантах реализации пара из двух вариабельных доменов соединена коротким линкером.
В некоторых случаях два одноцепочечных Fv-фрагмента (scFv-фрагмента) дистально соединены с единицей диатела, образуя два дополнительных VH/VL-антигенсвязывающих сайта (фиг. 1 и 2). Таким образом, такая молекула мультивалентного антитела является по меньшей мере четырехвалентной, поскольку содержит по меньшей мере четыре антигенсвязывающих сайта - два за счет единицы диатела и два за счет двух scFv-единиц. Каждый из дистально расположенных scFv-фрагментов можно расположить в полипептиде в порядке VH-VL или VL-VH.
Такие четырехвалентные молекулы антител выгодны для образования четырехвалентных, пятивалентных или шестивалентных триспецифичных антител. Эти новые триспецифичные антитела можно применять, например, для рекрутинга эффекторных клеток иммунной системы для уничтожения клеток-мишеней, например, опухолевых клеток или клеток, инфицированных вирусом. Поскольку такие триспецифичные антитела согласно настоящему изобретению являются по меньшей мере четырехвалентными, они обладают усиленной функциональной активностью по сравнению с трехвалентными и триспецифичными одноцепочечными фрагментами. Триспецифичные и четырехвалентные антитела согласно настоящему изобретению двухвалентно связываются посредством двух из четырех связывающих сайтов как с клеткой-мишенью, так и с эффекторными клетками иммунной системы. Например, двухвалентное связывание с клеткой-мишенью повышает не только авидность, но и специфичность адресного воздействия, если две из трех антигенных специфичностей направлены на два различных антигена, например, два различных опухолевых антигена на поверхности клетки-мишени. С другой стороны, можно модулировать, в частности, увеличивать цитотоксическую эффективность рекрутированной эффекторной клетки иммунной системы при двухвалентном связывании антитела с эффекторной клеткой иммунной системы. В других случаях такое триспецифичное антитело может иметь две специфичности к двум различным антигенам на эффекторных клетках, и третью специфичность для антигену на опухолевой клетке, нейроне или клетке, инфицированной вирусом, или связываться с и в конечном итоге нейтрализовать растворимый белок, например, фактор роста, цитокины или другие лиганды, не связанные с клетками.
В некоторых случаях вариабельные домены единицы диатела, соединенные коротким линкером, предотвращающим внутримолекулярное спаривание, представляют собой вариабельные домены легкой цепи (VL-VL) или вариабельные домены тяжелой цепи (VH-VH) (фиг. 1 и 2). Обнаружено, что данное конкретное расположение доменов облегчает правильный фолдинг молекул мультиспецифичных и мультивалентных антител согласно настоящему изобретению. В частности, для молекул четырехвалентных триспецифичных или тетраспецифичных Fv-антител можно принять эти меры для правильной ассоциации молекулы Fv-антитела, содержащей вариабельные домены для трех или более различных специфичностей, и для предотвращения неправильной ассоциации в пределах одиночного полипептида (мономера) или гомодимеризации между двумя идентичными Fv-полипептидами вместо правильной гетеродимеризации первого и второго полипептида (пример 2). Авторы настоящего изобретения получили правильную ассоциацию между вариабельными доменами для получения трех специфичностей и двух различных полипептидов, содержащих вариабельные домены, путем объединения в молекулу триспецифичного антитела, например, единицы диатела, образованной первой парой двух вариабельных доменов тяжелой цепи, связанных коротким линкером, ассоциированной со второй парой двух соответствующих вариабельных доменов легкой цепи, связанных коротким линкером. Таким образом, такая аранжировка вариабельных доменов в единице диатела, т.е. VL-VL в первом полипептиде вариабельных доменов и VH-VH во втором полипептиде вариабельных доменов, обеспечивает правильную ассоциацию и фолдинг четырехвалентного и триспецифичного или тетраспецифичного Fv-антитела. Авторы настоящего изобретения обнаружили, что такое расположение VL-VL/ VH-VH в составе единицы диатела или единицы одноцепочечного диатела стимулирует правильный фолдинг длинного полипептида, содержащего более шести вариабельных доменов, соединенных друг за другом с образованием молекулы Fv-антитела (например, фиг. 1), или правильный фолдинг и гетеродимеризацию двух полипептидов различной длины с образованием функциональной молекулы димерного Fv-антитела (например, фиг. 2, 5, 6а, 6b).
В дополнительных вариантах реализации по меньшей мере одна единица одноцепочечного диатела (scDb) дистально соединена с единицей диатела, образуя по меньшей мере один дополнительный антигенсвязывающий сайт. Таким образом, по меньшей мере один полипептид единицы диатела содержит одну пару вариабельных доменов единицы диатела, связанную с по меньшей мере одним scDb в полипептиде. Например, две scDb-единицы дистально соединены с единицей диатела, содержащей дополнительные два антигенсвязывающих сайта на каждую единицу одноцепочечного диатела (фиг. 4 и 5). Таким образом, такая мультивалентная молекула антитела является по меньшей мере четырехвалентной, поскольку она содержит по меньшей мере четыре антигенсвязывающих сайта. Такое мультивалентное антитело является по меньшей мере шестивалентным в вариантах реализации, где по меньшей мере две scDb-единицы соединены с единицей диатела, поскольку две дистально ориентированные scDb-единицы содержат четыре антигенсвязывающих сайта, а два антигенсвязывающих сайта находятся в единице диатела. Вариабельные домены в каждой из дистальных scDb-единиц можно расположить в порядке VH-VL-VH-VL, VL-VH-VL-VH, VL-VL-VH-VH или VH-VH-VL-VL в полипептиде.
В некоторых вариантах реализации единица диатела состоит из двух пар вариабельных доменов, расположенных на двух полипептидах, ассоциирующих друг с другом с образованием двух антигенсвязывающих сайтов, причем каждый из этих двух полипептидов содержит по меньшей мере один другой одноцепочечный фрагмент и/или фрагмент одноцепочечного диатела, расположенный дистально по отношению к каждой из двух пар вариабельных доменов единицы диатела. (Фиг. 6А, 6b, 7, 8а и 8b). Таким образом, такая молекула мультивалентного антитела является по меньшей мере семивалентной, поскольку она содержит по меньшей мере пять антигенсвязывающих сайтов - два связывающих сайта в единице диатела, по меньшей мере два связывающих сайта в двух scFv-единицах или scDb-единицах со стороны N-конца и С-конца от первой пары вариабельных доменов единицы диатела и по меньшей мере один связывающий сайт со стороны N-конца и С-конца от другой пары вариабельных доменов единицы диатела другого полипептида единицы диатела.
В некоторых случаях настоящее изобретение относится к молекуле мультивалентного антитела, содержащей полипептид, содержащий по меньшей мере шесть вариабельных доменов, соединенных друг за другом, причем единица диатела, содержащая первые два вариабельных домена из указанных по меньшей мере шести вариабельных доменов, встроена в указанный полипептид, и указанные первые два вариабельных домена единицы диатела ассоциированы с другими двумя вариабельными доменами единицы диатела, т.е. вторыми двумя вариабельными доменами, с образованием двух антигенсвязывающих сайтов. Указанные вторые два вариабельных домена могут быть расположены в том же полипептиде, что и первые два вариабельных домена, или в отдельном втором полипептиде, ассоциированном с первым полипептидом. Два антигенсвязывающих сайта, содержащиеся в единице диатела, образованы между первым и вторым вариабельными доменами, причем каждый вариабельный домен первой пары вариабельных доменов образует антигенсвязывающий сайт с другим вариабельным доменом второй пары вариабельных доменов. Соответственно, единица диатела образована указанным первым и вторым двумя вариабельными доменами, причем эти два вариабельных домена соединены коротким пептидным линкером в первом, а также втором двух вариабельных доменах для предотвращения внутримолекулярного спаривания.
Так, в некоторых вариантах реализации настоящее изобретение относится к молекуле мультивалентного антитела, содержащей полипептид, содержащий, т.е. включающий по меньшей мере шесть вариабельных доменов, соединенных друг за другом, причем два вариабельных домена данного полипептида связаны пептидным линкером, предотвращающим внутримолекулярное спаривание, и указанные два вариабельных домена ассоциированы с другими двумя соответствующими вариабельными доменами, связанными пептидным линкером, предотвращающим внутримолекулярное спаривание, причем указанные четыре вариабельных домена образуют два антигенсвязывающих сайта между указанными четырьмя вариабельными доменами. Например, такое антитело представляет собой Fv-антитело, в частности, четырехвалентное и триспецифичное Fv-антитело.
Эта молекула антитела является мультивалентной, т.е. содержит более одного антигенсвязывающего сайта. Она является четырехвалентной, если содержит четыре антигенсвязывающих сайта; пятивалентной, если содержит пять антигенсвязывающих сайтов, и шестивалентной, если содержит шесть антигенсвязывающих сайтов. Термин «четырехвалентное» относится к молекуле антитела, содержащей, в частности, состоящей из четырех антигенсвязывающих Fv-сайтов, где каждый из антигенсвязывающих Fv-сайтов содержит пару VH/VL, содержащую вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL) одной и той же антигенной специфичности, ассоциированные друг с другом. Таким образом, такая четырехвалентная молекула антитела содержит по меньшей мере восемь вариабельных доменов антитела, а именно четыре вариабельных домена тяжелой цепи (VH) и четыре вариабельных домена легкой цепи (VL). Поскольку четырехвалентная антигенсвязывающая молекула содержит по меньшей мере восемь вариабельных доменов антитела, ее молекулярная масса составляет более 100 Да, что приводит к увеличению периода полужизни такой молекулы по сравнению с трехвалентными и триспецифичными одноцепочечными Fv-молекулами.
В некоторых случаях молекула антитела является мультиспецифичной, т.е. обладает специфичностью в отношении различных эпитопов антигенов. В некоторых случаях молекула антитела является триспецифичной.
Триспецифичная и четырехвалентная молекула антитела содержит антигенсвязывающий сайт, обладающий специфичностью по отношению к первому эпитопу антигена, антигенсвязывающий сайт, обладающий специфичностью по отношению ко второму эпитопу антигена, и два антигенсвязывающих сайта, обладающих специфичностью по отношению к третьему эпитопу антигена. Таким образом, такая триспецифичная и четырехвалентная молекула антитела имеет, т.е. включает различные специфичности в отношении трех различных эпитопов антигенов. Например, такая антигенсвязывающая молекула содержит первый антигенсвязывающий сайт, обладающий специфичностью к первому эпитопу антигена, второй антигенсвязывающий сайт, обладающий специфичностью ко второму эпитопу антигена, третий и четвертый антигенсвязывающие сайты, обладающие специфичностью к третьему эпитопу антигена. Пятивалентная и триспецифичная молекула антитела содержит два антигенсвязывающих сайта, обладающих специфичностью к первому эпитопу антигена, два антигенсвязывающих сайта, обладающих специфичностью ко второму эпитопу антигена, и один антигенсвязывающий сайт, обладающий специфичностью к третьему эпитопу антигена. В качестве альтернативы, пятивалентная и триспецифичная молекула антитела может содержать три антигенсвязывающих сайта против первого эпитопа антигена и по одному антигенсвязывающему сайту для второго и третьего эпитопов антигенов. Шестивалентная и триспецифичная молекула антитела в некоторых вариантах реализации содержит по два антигенсвязывающих сайта для каждого из трех эпитопов антигенов, или, в качестве альтернативы, от одного до трех антигенсвязывающих сайтов для каждого из трех эпитопов антигенов.
В некоторых случаях молекула антитела является молекулой Fv-антитела. "Fv-антитело" относится к Fv-производному иммуноглобулина, содержащему только вариабельные (V) домены антитела, но не содержащему константных областей антитела или их фрагментов. Каждый вариабельный домен легкой цепи (VL) ассоциирован с соответствующим вариабельным доменом тяжелой цепи (VH), образуя антигенсвязывающий Fv-сайт (VH/VL-антигенсвязывающий сайт). Вариабельные домены антитела соединены друг за другом в гибридном полипептиде посредством пептидного линкера или пептидной связи. Fv-антитело, т.е. антигенсвязывающая молекула согласно настоящему изобретению может являться мономером одиночного полипептида или мультимерным полипептидом. Мультимерная антигенсвязывающая молекула, т.е. Fv-антитело, в частности, мультивалентное Fv-антитело, может представлять собой, например, димер, содержащий два полипептида, тример, содержащий три полипептида, или тетрамер, содержащий четыре полипептида. Димер является гетеродимерным, если он состоит из двух полипептидов различного аминокислотного состава, или гомодимерным, если он состоит из двух идентичных полипептидов.
Термин "полипептид" относится к полимеру из аминокислотных остатков, соединенных амидными связями. Предпочтительно полипептид является одноцепочечным гибридным неразветвленным белком. Вариабельные (Fv) домены антитела в полипептиде соединены друг за другом. Термин «Fv-полипептид» обозначает гибридный полипептид, в котором вариабельные (Fv) домены антитела соединены друг за другом. Полипептид может содержать смежные аминокислотные остатки в дополнение к N-концу и/или С-концу. Например, полипептид может содержать маркерную последовательность, предпочтительно на С-конце, которую можно использовать при очистке, а также обнаружении полипептида. Примером маркерной последовательности являются маркер His, например, маркер His, состоящий из шести остатков His, маркер FLAG, например, октапептид DYKDDDDK (SEQ ID NO:38), или маркер STREP® II, например, октапептид WSHPQFEK (SEQ ID NO:39), или С-маркер, например, тетрапептид EPEA (SEQ ID NO:40). В мультимерных антигенсвязывающих молекулах для различных полипептидов можно использовать различные маркерные последовательности, например, His-маркер для первого полипептида и FLAG-маркер для второго полипептида димерной молекулы. В некоторых вариантах реализации полипептид может содержать вариабельные домены, обеспечивающие получение антигенсвязывающих сайтов, а также константные домены антитела, например, CL, CH и/или Fc-домены. Например, такие варианты реализации могут включать Fv-полипептид или Fv-антитело, объединенное с по меньшей мере одним константным доменом антитела, например, Fc-доменом. В дополнительных вариантах реализации полипептид, содержащий вариабельные домены, можно соединить с другим агентом, например, токсином, иммуномодулятором или агентом, генерирующим сигнал.
Термин «линкер» относится к пептиду, соединяющему два расположенных рядом вариабельных домена в полипептиде между С-концом одного домена и N-концом другого расположенного рядом домена или наоборот. С точки зрения аминокислотного состава выбирают пептид, не мешающий образованию Fv-, т.е. VH/VL-антигенсвязывающих сайтов, а также не мешающий мультимеризации, например, димеризации мультиспецифичных, например, триспецифичных молекул. Например, линкер, содержащий остатки глицина и серина, обычно обеспечивает устойчивость к протеазам. В некоторых вариантах реализации используют пептидные линкеры (G2S)x, в которых, например, х=1-20, например, используют пептидные линкеры (G2S), (G2S)2, (G2S)3, (G2S)4, (G2S)5, (G2S)6, (G2S)7 или (G2S)8 или (G3S)x, в которых, например, х=1-15, или используют пептидные линкеры (G4S)x, в которых, например, х=1-10, предпочтительно 1-6. Аминокислотную последовательность линкера можно оптимизировать с целью улучшения связывания антигена и выхода продукции полипептида, например, с помощью способов фагового дисплея.
Длина линкеров влияет на гибкость антигенсвязывающего полипептидного димера. Желательная гибкость антигенсвязывающего полипептидного димера зависит от плотности антигена-мишени и доступности антигена-мишени, т.е. эпитопов антигена-мишени. Более длинные линкеры позволяют получать более гибкие антигенсвязывающие полипептиды с более гибкими антигенсвязывающими сайтами. Влияние длины линкеров на образование димерных антигенсвязывающих полипептидов описан, например, в Todorovska et al., 2001 Journal of Immunological Methods 248:47-66; Perisic et al., 1994 Structure 2:1217-1226; Le Gall et al., 2004, Protein Engineering 17:357-366 и WO 94/13804.
Единицу диатела встраивают в полипептид молекулы антитела. Термин "единица диатела" обозначает двухвалентную Fv-молекулу, состоящую из двух пар вариабельных доменов - первой пары и второй пары, которые ассоциируют с образованием двух VL/VH-антигенсвязывающих сайтов. Каждая пара вариабельных доменов соединена друг за другом в полипептиде. В некоторых вариантах реализации двухвалентный Fv-модуль состоит из первой и второй пар двух вариабельных доменов, расположенных рядом друг с другом, причем в каждой паре два вариабельных домена соединены коротким пептидным линкером, предотвращающим внутримолекулярную ассоциацию между вариабельными доменами, соединенными коротким линкером. Первая пара вариабельных доменов вынужденно ассоциировать со второй парой вариабельных доменов, образуя два антигенсвязывающих Fv-сайта с двумя парами вариабельных доменов. Соответственно, каждый из двух антигенсвязывающих Fv-сайтов образован одним вариабельным доменом первой пары вариабельных доменов и одним вариабельным доменом второй пары вариабельных доменов. Таким образом, такая единица диатела содержит по меньшей мере один антигенсвязывающий сайт из двух вариабельных доменов, непрямым образом соединенных коротким пептидным линкером 3, 3а (фиг. 1 и 2). Две пары вариабельных доменов, расположенных рядом друг с другом, располагаются на двух отдельных полипептидах с образованием димерной единицы диатела (фиг. (2, 3, 5, 8), или две пары вариабельных доменов, расположенных рядом друг с другом, располагаются на одном и том же полипептиде с образованием единицы одноцепочечного диатела (фиг.1, 4). В каждой паре вариабельных доменов короткий линкер 3, 3а соединяет С-конец одного вариабельного домена и N-концом другого вариабельного домена или наоборот. В каждой паре вариабельные домены могут быть ориентированы от N- к С-концу, например, VL-VH, VH-VL, VH-VH или VL-VL, причем два вариабельных домена пары имеют специфичности против различных эпитопов антигенов или специфичности против одного и того же эпитопа антигена. В некоторых случаях два вариабельных домена непосредственно связаны пептидной связью между N-концом одного вариабельного домена и С-концом другого вариабельного домена пары. Длина короткого пептидного линкера, соединяющего два вариабельных домена в каждой из первой и второй пары вариабельных доменов единицы диатела, является такой, которая позволяет предотвратить внутримолекулярную ассоциацию между вариабельными доменами, соединенными линкером. Такой линкер является «коротким», т.е. состоит из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или приблизительно 12 аминокислотных остатков. В случае 0 аминокислотных остатков линкер представляет собой пептидную связь. Такой короткий линкер благоприятствует правильной димеризации между двумя парами вариабельных доменов и образованию двух антигенсвязывающих Fv-сайтов. Укорачивание линкера до приблизительно 12 или менее аминокислотных остатков обычно предотвращает взаимодействие смежных областей одной и той же полипептидной цепи друг с другом. В одном варианте реализации настоящего изобретения эти линкеры состоят из от приблизительно 3 до приблизительно 12, например, от 5 до 10, в частности, от 7 до 9 смежных аминокислотных остатков. Длину линкера можно корректировать с целью получения конкретной ориентации доменов в единице диатела. Например, линкер (G2S)2 можно использовать для пары VH-VL или для пары VL-VH, а линкер (G2S)3 можно использовать для пары VH-VH или для пары VL-VL, или линкер (G2S)2 можно использовать для пары VH-VH и линкер (G2S)3 можно использовать для пары VL-VL единицы диатела (или наоборот). Между прочим, в принципе возможно, что два полипептида, содержащие линкер из более чем 12 аминокислотных остатков между вариабельными доменами антитела пары, правильно димеризуются друг с другом (см., например, Le Gall et al., 2004, Protein Engineering 17:357-366).
В некоторых вариантах реализации единица диатела представляет собой единицу одноцепочечного диатела (фиг. 1). «Единица одноцепочечного диатела» состоит из первой пары вариабельных доменов, соединенной со второй парой вариабельных доменов длинным линкером, допускающим внутримолекулярную ассоциацию первой и второй пар вариабельных доменов, в соответствии с «длинным линкером», используемым в scFv-единицах и описанным в следующем абзаце. Например, такой длинный линкер может состоять из более чем 12, в частности, от приблизительно 15 до приблизительно 50, предпочтительно от приблизительно 15 до приблизительно 35, в частности, от приблизительно 15 до приблизительно 25 смежных (последовательных) аминокислотных остатков.
В некоторых вариантах реализации домены одноцепочечного диатела в полипептиде, соединенные друг за другом, можно расположить в порядке VL-VH-VL-VH, VL-VL-VH-VH, VH-VH-VL-VL или VH-VL-VH-VL от N-конца к С-концу единицы одноцепочечного диатела.
В некоторых вариантах реализации молекула антитела согласно настоящему изобретению состоит из одиночного полипептида, содержащего единицу одноцепочечного диатела (фиг. 1, 4). В конкретных вариантах реализации молекула антитела содержит по меньшей мере три единицы одноцепочечного диатела, соединенные друг за другом в полипептиде (фиг. 4). В других вариантах реализации молекула антитела согласно настоящему изобретению содержит по меньшей мере одну единицу одноцепочечного диатела, дистально соединенную с единицей диатела (фиг. 5).
«Единица одноцепочечного Fv (scFv)» обозначает антигенсвязывающий Fv-сайт, образованный фрагментом одноцепочечного полипептида, состоящим из вариабельного домена легкой цепи (VL) и вариабельного домена тяжелой цепи (VH). Вариабельные домены можно ориентировать в виде VL-VH или VH-VL от N-конца к С-концу единицы scFv. Вариабельные домены соединены пептидным линкером между С-концом одного вариабельного домена и N-концом другого вариабельного домена или наоборот. Пептидный линкер является длинным и гибким (в общем случае состоит из приблизительно 12 или более аминокислотных остатков) для внутримолекулярного фолдинга и образования антигенсвязывающего Fv-сайта. Дополнительные аминокислотные остатки обеспечивают дополнительную гибкость. Например, такой длинный линкер может состоять из более чем 12, в частности, от приблизительно 15 до приблизительно 50, предпочтительно от приблизительно 15 до приблизительно 35, в частности, от приблизительно 15 до приблизительно 25 смежных аминокислотных остатков. Длину линкера можно корректировать для конкретной ориентации доменов от N-конца к С-концу единицы scFv. Например, линкер (G2S)6 можно использовать для единицы scFv VH-VL, а линкер (G2S)7 можно использовать для единицы scFv VL-VH.
Единица scFv связана с единицей диатела пептидным линкером между вариабельным доменом единицы scFv и вариабельным доменом единицы диатела. Выбирают длину пептидного линкера, позволяющую избегать стерических помех между расположенными рядом друг с другом вариабельными доменами и поддерживать стабильность молекулы; эта длина может составлять, например, от 5 до 50, в частности, от 5 до 35, предпочтительно по меньшей мере 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 смежных аминокислотных остатков.
В некоторых случаях единица диатела расположена в центре молекулы антитела для облегчения фолдинга и улучшения стабильности молекулы антитела. В таких случаях единица диатела соединена по N-концу и С-концу с дополнительным дистальным вариабельным доменом. В некоторых вариантах реализации единица диатела соединена с двумя (фиг. 1, 2, 8а, 8b), тремя (фиг. 6а, 6b) или четырьмя (фиг. 7) единицами scFv. В некоторых случаях молекула антитела, в частности, молекула триспецифичного Fv-антитела, содержит полипептид, содержащий по меньшей мере шесть вариабельных доменов, причем указанный полипептид содержит единицу scFv на N-конце, единицу scFv на С-конце и первую пару двух вариабельных доменов единицы диатела между двумя единицами scFv (фиг. 1 и 2). Эта первая пара двух вариабельных доменов единицы диатела не ассоциирует с образованием антигенсвязывающего Fv-сайта. Например, если единица диатела является димерной единицей диатела, каждый из двух вариабельных доменов первой пары расположенных рядом друг с другом вариабельных доменов соединен с дополнительным вариабельным доменом. В частности, каждый из двух вариабельных доменов первой пары расположенных рядом друг с другом вариабельных доменов соединен с дополнительным вариабельным доменом единицы scFv (фиг. 2). В других случаях один или оба вариабельных домена второй пары расположенных рядом друг с другом вариабельных доменов можно дополнительно соединить с дополнительным вариабельным доменом (фиг. 6а, 6b, 7). Если единица диатела представляет собой единицу одноцепочечного диатела, первая пара вариабельных доменов, расположенных рядом друг с другом, соединена по N-концу с дополнительным вариабельным доменом, а вторая пара вариабельных доменов, расположенных рядом друг с другом, соединена по С-концу с дополнительным вариабельным доменом. В частности, первая пара расположенных рядом друг с другом вариабельных доменов соединена по N-концу с дополнительным вариабельным доменом единицы scFv, а вторая пара расположенных рядом друге другом вариабельных доменов соединена по С-концу с дополнительным вариабельным доменом еще одной единицы scFv (фиг. 1). В других вариантах реализации первая пара вариабельных доменов первой единицы одноцепочечного диатела соединена по N-концу с дополнительным вариабельным доменом второй единицы одноцепочечного диатела, а вторая пара вариабельных доменов первого одноцепочечного диатела соединена по С-концу с дополнительным вариабельным доменом третьей единицы одноцепочечного диатела в полипептиде, содержащем три единицы одноцепочечного диатела, соединенные друг за другом (фиг. 4) В других случаях между первой и второй парами вариабельных доменов, расположенных рядом друг с другом, может располагаться по меньшей мере один дополнительный вариабельный домен.
В некоторых случаях, если единица диатела представляет собой единицу одноцепочечного диатела, мультивалентное антитело состоит из одиночного полипептида, содержащего два вариабельных домена, расположенных рядом друг с другом, т.е. первую пару вариабельных доменов, расположенных рядом друг с другом, ассоциированных с двумя другими вариабельными доменами этого полипептида, расположенными рядом друг с другом, т.е. второй парой вариабельных доменов (фиг. 1). Такая структура антитела благоприятствует получению мультиспецифичных, в частности, би-, три- или тетраспецифичных антител. В некоторых вариантах реализации такое мультивалентное, в частности, триспецифичное Fv-антитело содержит по меньшей мере восемь вариабельных доменов, соединенных друг за другом от N-конца к С-концу, причем первый и второй вариабельные домены образуют единицу scFv на N-конце, и указанная единица scFv на N-конце соединена по С-концу с третьиму вариабельным доменом первой пары вариабельных доменов единицы диатела, содержащей третий вариабельный домен и четвертый вариабельный домен, причем указанный четвертый вариабельный домен соединен по С-концу с пятым вариабельным доменом второй пары вариабельных доменов единицы диатела, содержащей пятый и шестой вариабельные домены, причем указанный шестой вариабельный домен соединен по С-концу с седьмым вариабельным доменом единицы scFv на С-конце, и указанная единица scFv образована седьмым и восьмым вариабельными доменами (фиг. 1). Вариабельные домены могут быть расположены, например, в одной из следующих ориентации от N-конца к С-концу полипептида: VH-VL-VL-VH-VL-VH-VL-VH, VH-VL-VH-VH-VL-VL-VL-VH, VL-VH-VL-VH-VL-VH-VH-VL или VH-VL-VL-VL-VH-VH-VL-VH. В конкретном варианте реализации ориентация одной пары вариабельных доменов единицы диатела представляет собой VH-VH, а ориентация другой пары вариабельных доменов единицы диатела - VL-VL (фиг. 1).
Четырехвалентные варианты реализации молекулы антитела, состоящей из одиночного полипептида, содержащего восемь вариабельных доменов, описанных выше, благоприятствуют получению триспецифичных антител. Например, такие триспецифичные антитела могут иметь первую и вторую специфичности в отношении антигена на клетке-мишени, например, опухолевой клетке, а третью специфичность - в отношении эффекторной клетки иммунной системы, например, Т- или NK-клетки. В других случаях такое триспецифичное антитело может иметь две специфичности для различных антигенов на эффекторных клетках, и третью специфичность в отношении антигена на опухолевой клетке. В некоторых вариантах реализации две дистальные единицы scFv могут иметь первую и вторую специфичности в отношении клетки-мишени, а единица диатела, расположенная между этими двумя единицами scFv, может иметь специфичность в отношении эффекторной клетки иммунной системы; т.е., от N-конца к С-концу, первый и второй вариабельные домены имеют первую специфичность в отношении клетки-мишени; третий и пятый вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы, четвертый и шестой вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы, а седьмой и восьмой вариабельные домены имеют вторую специфичность в отношении клетки-мишени (фиг. 1). В других вариантах реализации две дистальные единицы scFv могут иметь специфичность в отношении эффекторной клетки иммунной системы, а два антигенсвязывающих сайта, образованные единицей диатела, могут иметь две специфичности для клетки-мишени; т.е., от N-конца к С-концу, первый и второй, а также седьмой и восьмой вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы; третий и пятый вариабельные домены имеют первую специфичность в отношении клетки-мишени, а четвертый и шестой вариабельные домены имеют вторую специфичность в отношении клетки-мишени. В дополнительных альтернативных вариантах реализации одиночный полипептид может содержать более восьми, например, 10, 12 или более вариабельных доменов, и содержать более двух единиц scFv и/или более одной единицы диатела.
В дополнительных вариантах реализации мультивалентная молекула антитела, состоящая из одного полипептида, содержит три единицы одноцепочечного диатела, соединенные друг за другом (фиг. 4). Такая молекула антитела содержит по меньшей мере 12 вариабельных доменов, соединенных друг за другом от N-конца к С-концу. В конкретном варианте реализации ориентация одной пары вариабельных доменов второй единицы одноцепочечного диатела представляет собой VH-VH, а ориентация другой пары вариабельных доменов второй единицы одноцепочечного диатела - VL-VL (Fig. 4). Такая молекула антитела является шестивалентной и может содержать антигенсвязывающие сайты для от одной до шести различных антигенных специфичностей, в частности, двух или трех различных антигенных специфичностей.
В некоторых случаях молекула антитела, в частности, Fv-антитела содержит единицу диатела в формате димерной единицы диатела. В таких случаях молекула антитела представляет собой димер двух полипептидов, причем первая пара двух вариабельных доменов единицы диатела, расположенных рядом друг с другом, встроена в первый полипептид, содержащий по меньшей мере шесть вариабельных доменов, соединенных друг за другом, и указанная первая пара вариабельных доменов, расположенных рядом друг с другом, ассоциирована с еще одной второй парой двух вариабельных доменов, расположенных рядом друг с другом во втором полипептиде. Первый и второй полипептиды предпочтительно нековалентно связаны (фиг. 2, 3, 5, 6а, 6b, 7, 8а и 8b8b). Однако в некоторых случаях первый и второй полипептиды могут быть ковалентно связаны, например, дисульфидной связью или химическим линкером.
В некоторых вариантах реализации первый полипептид содержит по меньшей мере шесть вариабельных доменов, а второй полипептид содержит по меньшей мере два вариабельных домена (фиг. 2). В таких вариантах реализации второй полипептид входит в состав единицы диатела и, предпочтительно нековалентно, ассоциирован с другой парой двух вариабельных доменов, расположенных рядом друг с другом, встроенной в первый полипептид. В вариантах реализации, где первая полипептидная цепь состоит из шести вариабельных доменов, а второй полипептид состоит из двух вариабельных доменов, эти вариабельные домены можно расположить от N-конца к С-концу полипептида, например, в следующей ориентации: VH-VL-VH-VH-VL-VH (первый полипептид) и VL-VL (второй полипептид); VL-VH-VH-VH-VH-VL (первый полипептид) и VL-VL (второй полипептид); VH-VL-VL-VL-VH-VL (первый полипептид) и VH-VH (второй полипептид); VL-VH-VL-VL-VH-VL (первый полипептид) и VH-VH (второй полипептид) или VH-VL-VL-VL-VL-VH (первый полипептид) и VH-VH (второй полипептид). Единицы диател, содержащие одну пару двух вариабельных доменов в ориентации VH-VH и другую пару двух вариабельных доменов в ориентации VL-VL, благоприятствуют правильному фолдингу, в частности, мультиспецифичных, например, триспецифичных молекул антител.
Четырехвалентные варианты реализации молекулы антитела, содержащей первый полипептид, содержащий по меньшей мере шесть вариабельных доменов, и второй полипептид, содержащий по меньшей мере двух вариабельных домена, как описано выше, благоприятствуют получению триспецифичных антител. Из-за различного размера первого и второго полипептидов эти полипептиды легко выделить из надосадочной жидкости по отдельности. Например, такие триспецифичные антитела могут иметь первую и вторую специфичности в отношении клетки-мишени, например, опухолевой клетки, а третью специфичность в отношении эффекторной клетки иммунной системы, например, Т- или NK-клетки. В других вариантах реализации молекулы триспецифичных антител имеют первую и вторую специфичности в отношении первого и второго вирусного антигена или эпитопа вирусного антигена, а третью специфичность в отношении эффекторной клетки, например, Т- или NK-клетки. В дополнительных вариантах реализации молекула триспецифичного антитела имеет первую специфичность в отношении вирусного антигена, вторую специфичность к антигену на клетке-мишени, а третью специфичность в отношении эффекторной клетки, например, Т- или NK-клетки. В других случаях такое триспецифичное антитело может содержать первую и вторую специфичность для эффекторных клеток, например, NK-клеток или Т-клеток, и третью специфичность в отношении клетки-мишени, например, опухолевого антигена на опухолевой клетке или вирусного антигена. Первая и вторая специфичности для эффекторной клетки могут соответствовать различным антигенам или эпитопам одного и того же антигена на эффекторных клетках одного и того же типа.
В некоторых вариантах реализации две дистальные единицы scFv, образованные в первом полипептиде, могут иметь первую и вторую специфичности в отношении клетки-мишени, а единица диатела, расположенная между этими двумя единицами scFv и образованная первым и вторым полипептидами, может иметь специфичность в отношении эффекторной клетки иммунной системы; т.е., от N-конца к С-концу в первом полипептиде, первый и второй вариабельные домены имеют первую специфичность в отношении клетки-мишени; третий и четвертый вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы, пятый и шестой вариабельные домены имеют вторую специфичность в отношении клетки-мишени, а во втором полипептиде первый и второй вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы (фиг. 2). В других вариантах реализации две дистальные единицы scFv в первом полипептиде, содержащем шесть вариабельных доменов, могут иметь специфичность в отношении эффекторной клетки иммунной системы, а два антигенсвязывающих сайта, образованные единицей диатела первого полипептида, а также два вариабельных домена второго полипептида могут иметь разные специфичности в отношении клетки-мишени; т.е. от N-конца к С-концу первого полипептида, первый и второй, а также пятый и шестой вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы; третий вариабельный домен имеет первую специфичность в отношении клетки-мишени, четвертый вариабельный домен имеет вторую специфичность в отношении клетки-мишени, а во втором полипептиде первый вариабельный домен имеет вторую специфичность в отношении клетки-мишени, а второй вариабельный домен имеет первую специфичность в отношении клетки-мишени.
В дополнительных вариантах реализации первый полипептид содержит по меньшей мере шесть вариабельных доменов, а второй полипептид содержит по меньшей мере четыре (фиг. 6а, 6b) или шесть (фиг. 7) вариабельных доменов. В таких вариантах реализации первый полипептид содержит первую пару вариабельных доменов единицы диатела, а второй полипептид содержит вторую пару вариабельных доменов единицы диатела, которые, предпочтительно нековалентно, ассоциированы друг с другом, образуя два антигенсвязывающих сайта между первым и вторым полипептидами. В вариантах реализации, где первая полипептидная цепь состоит из шести вариабельных доменов, а второй полипептид состоит из четырех вариабельных доменов, пара вариабельных доменов, расположенных рядом друг с другом в единице диатела во втором полипептиде, соединена с единицей scFv, образуя дополнительный антигенсвязывающий сайт со стороны N-конца или С-конца второго полипептида (фиг. 6а, 6b). Такие варианты реализации являются пятивалентными и могут содержать антигенсвязывающие сайты для от одной до пяти различных антигенных специфичностей, в частности, двух или трех различных антигенных специфичностей. Единицы диател, содержащие одну пару двух вариабельных доменов в ориентации VH-VH в первом полипептиде и другую пару двух вариабельных доменов в ориентации VL-VL во втором полипептиде, благоприятствуют правильному фолдингу, в частности, мультиспецифичных, например, триспецифичных молекул антител.
В других вариантах реализации, где первая полипептидная цепь состоит из шести вариабельных доменов, а второй полипептид состоит из четырех вариабельных доменов, пара вариабельных доменов единицы диатела, расположенных рядом друг с другом во втором полипептиде, соединены по N-концу и С-концу с единицами scFv с образованием двух дополнительных антигенсвязывающих сайтов (фиг. 7). Такие варианты реализации являются шестивалентными и могут содержать антигенсвязывающие сайты для от одной до шести различных антигенных специфичностей, в частности, двух или трех различных антигенных специфичностей. В конкретных вариантах реализации единица диатела содержит одну пару вариабельных доменов в ориентации VH-VH в первом полипептиде и другую пару вариабельных доменов в ориентации VL-VL во втором полипептиде. В дополнительных альтернативных вариантах реализации мультивалентная, например, триспецифичная молекула Fv-антитела может содержать более одной димерной единицы диатела. В таких альтернативных вариантах реализации первый полипептид может содержать по меньшей мере шесть вариабельных доменов, а второй полипептид может содержать по меньшей мере четыре (фиг. 3) или шесть (фиг. 8а, 8b) вариабельных доменов. В вариантах реализации, где первый полипептид содержит по меньшей мере шесть вариабельных доменов, а второй полипептид содержит четыре вариабельных домена, четыре вариабельных домена второго полипептида, расположенные рядом друг с другом, ассоциированы с соответствующими четырьмя вариабельными доменами, расположенными рядом друге другом и встроенными в первый полипептид, тем самым образуя тандем из двух единиц диател, расположенных рядом друг с другом. Остальные по меньшей мере два дополнительных вариабельных домена первого полипептида расположены дистально по отношению к тандемным единицам диатела и образуют единицу scFv (фиг. 8а, 9b). В других вариантах реализации, где первый полипептид содержит шесть вариабельных доменов, а второй полипептид содержит шесть вариабельных доменов, шесть вариабельных доменов первого полипептида ассоциированы с соответствующими шестью вариабельными доменами второго полипептида, тем самым образуя триплет из единиц диател, расположенных рядом друге другом (фиг. 3). Последний вариант реализации, состоящий из первого и второго полипептида, где каждый полипептид содержит шесть вариабельных доменов, обладает преимуществом в том смысле, что он может позволить получить триспецифичное антитело в формате гомодимера, т.е. двух идентичных полипептидов, ассоциированных друг с другом и обеспечивающих двухвалентное связывание для каждой из трех антигенных специфичностей.
В некоторых случаях четырехвалентное триспецифичное Fv-антитело представлено в виде тандемного диатела. Такая молекула триспецифичного Fv-антитела состоит из первого и второго полипептида, причем каждый полипептид содержит четыре вариабельных домена, соединенных друг с другом. В такой молекуле Fv-антитела длина линкера предотвращает внутримолекулярное спаривание вариабельных доменов, так что молекула не может укладываться сама на себя, но принудительно сопрягается, т.е. ассоциирует с комплементарными доменами другого полипептида. Домены располагаются таким образом, что соответствующие VH- и VL-домены ассоциируют друг с другом во время этой димеризации. Несмотря на отсутствие межмолекулярных ковалентных связей, этот димер отличается высокой стабильностью после образования, остается интактным и не переходит обратно в мономерную форму. В некоторых вариантах реализации молекула триспецифичного Fv-антитела содержит димерную единицу диатела, причем ориентация одной пары вариабельных доменов, расположенных рядом друг с другом, представляет собой VH-VH, а ориентация другой пары вариабельных доменов, расположенных рядом друг с другом, представляет собой VL-VL. Такая ориентация вариабельных доменов в единице диатела облегчает правильную ассоциацию двух триспецифичных полипептидов. В частности, такая ориентация позволяет получить молекулы триспецифичных Fv-антител в формате тандемного диатела, поскольку оно является гетеродимером двух различных полипептидов. Соответственно, такая ориентация благоприятствует правильной гетеродимеризации триспецифичного тандемного диатела. Таким образом, в некоторых вариантах реализации молекула триспецифичного Fv-антитела представляет собой тандемное диатело (фиг. 3). В таких триспецифичных тандемных диателах вариабельные домены первого и второго полипептида можно расположить, например, в следующих ориентациях от N-конца к С-концу полипептида: VL-VH-VH-VL (первый полипептид) и VH-VL-VL-VH (второй полипептид) или наоборот (фиг. 3). Такие триспецифичные антитела в формате тандемного диатела могут иметь первую и вторую специфичности в отношении клетки-мишени, например, опухолевой клетки, и третью специфичность в отношении эффекторной клетки иммунной системы, например, Т- или NK-клетки. В других случаях такое триспецифичное антитело может иметь две специфичности для различных антигенов на эффекторных клетках, и третью специфичность в отношении антигена на опухолевой клетке. В некоторых вариантах реализации вариабельные домены, расположенные на внешних участках полипептидов, могут иметь первую и вторую специфичности в отношении клетки-мишени, а два вариабельных домена, расположенных в центральной части полипептида между вариабельными доменами, расположенными на внешних участках полипептидов, имеют третью специфичность в отношении эффекторной клетки иммунной системы; т.е., от N-конца к С-концу в первом полипептиде, первый вариабельный домен имеет первую специфичность в отношении клетки-мишени; второй и третий вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы, четвертый вариабельный домен имеет вторую специфичность в отношении клетки-мишени, а во втором полипептиде первый вариабельный домен имеет вторую специфичность в отношении клетки-мишени, второй и третий вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы, а четвертый вариабельный домен имеет первую специфичность в отношении клетки-мишени (фиг. 3). В других вариантах реализации вариабельные домены, расположенные на внешних участках полипептидов, могут содержать третью специфичность в отношении эффекторной клетки иммунной системы, а два вариабельных домена, расположенных в центральной части полипептида между вариабельными доменами, расположенными на внешних участках полипептидов, иметь первую и вторую специфичности для клетки-мишени; т.е. от N-конца к С-концу первого полипептида, первый вариабельный домен имеет третью эпитопную специфичность для эффекторной клетки иммунной системы; второй вариабельный домен имеет первую специфичность в отношении клетки-мишени, третий вариабельный домен имеет вторую специфичность в отношении клетки-мишени, четвертый вариабельный домен содержит третью специфичность в отношении эффекторной клетки иммунной системы, а во втором полипептиде первый вариабельный домен содержит третью специфичность в отношении эффекторной клетки иммунной системы, второй вариабельный домен имеет вторую специфичность в отношении клетки-мишени, третий вариабельный домен имеет первую специфичность в отношении клетки-мишени, а четвертый вариабельный домен содержит третью специфичность в отношении эффекторной клетки иммунной системы. В других вариантах реализации вариабельные домены, содержащие третью специфичность в отношении эффекторной клетки иммунной системы, расположены в молекуле антитела латерально; т.е., от N-конца к С-концу первого полипептида, первый и второй вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы; третий вариабельный домен имеет первую специфичность в отношении клетки-мишени, четвертый вариабельный домен имеет вторую специфичность в отношении клетки-мишени, а во втором полипептиде первый вариабельный домен имеет вторую специфичность в отношении клетки-мишени, второй вариабельный домен имеет первую специфичность в отношении клетки-мишени, а третий и четвертый вариабельные домены имеют третью специфичность в отношении эффекторной клетки иммунной системы.
В дополнительных вариантах реализации тандемное диатело конъюгировано с по меньшей мере одним дополнительным антигенсвязывающим доменом, в частности, с по меньшей мере одной единицей scFv со стороны N-конца или С-конца полипептида. В предпочтительных вариантах реализации тандемное диатело конъюгировано с по меньшей мере одной единицей scFv посредством пептидного линкера. В частности, такая молекула антитела состоит из первого и второго полипептида, причем оба полипептида состоят из шести вариабельных доменов, и каждый из первого полипептида и второго полипептида содержат единицу scFv со стороны N-конца каждого полипептида (фиг. 8b) или С-конца каждого полипептида (фиг. 8а). Такие молекулы антитела являются шестивалентными и могут содержать антигенсвязывающие сайты для от одной до шести различных антигенных специфичностей, в частности, двух или трех различных антигенных специфичностей. В конкретных вариантах реализации единица диатела, расположенная в центральной части молекулы антитела, содержит одну пару вариабельных доменов в ориентации VH-VH в первом полипептиде и другую пару вариабельных доменов в ориентации VL-VL во втором полипептиде. В некоторых случаях молекула мультивалентного антитела, в частности, молекула мультивалентного Fv-антитела, описанная в настоящем документе, является по меньшей мере триспецифичной и по меньшей мере четырехвалентной молекулой антитела. Такая молекула антитела содержит по меньшей мере два антигенсвязывающих сайта, обладающих специфичностью по отношению к одному и тому же эпитопу антигена. За счет этого повышается авидность, т.е. сила взаимодействия между эпитопом антигена и антигенсвязывающей молекулой. Авидность молекул триспецифичных антител можно дополнительно повысить за счет пятивалентных и шестивалентных вариантов реализации настоящего изобретения. Пятивалентные молекулы могут содержать по меньшей мере два связывающих сайта для двух или трех эпитопных специфичностей триспецифичного антитела, а шестивалентные варианты реализации могут содержать два антигенсвязывающих сайта для каждой из трех эпитопных специфичностей. В качестве альтернативы, полиспецифичность, т.е. количество специфичностей, можно увеличить за счет четырехвалентных, пятивалентных и шестивалентных вариантов реализации. Например, молекула антитела может являться тетраспецифичной. Преимущества повышенной авидности представляют собой повышенную стабильность взаимодействия и удерживания на мишени. Например, если мишенью является цитотоксическая эффекторная клетка иммунной системы, например, Т-клетка или NK-клетка, повышенная авидность может привести к повышенному цитотоксическому потенциалу молекулы антитела. В еще одном примере, если мишень является опухолевой клеткой, повышенная авидность улучшает время удерживания на мишени и снижает скорость диссоциации с мишени. В определенном варианте реализации настоящего изобретения молекула триспецифичного и четырехвалентного Fv-антитела содержит первый и второй антигенсвязывающие сайты, специфичные по отношению к двум различным эпитопам антигенов опухолевых клеток одного и того же типа, и третий и четвертый антигенсвязывающие сайты, специфичные по отношению к эпитопу антигена эффекторной клетки иммунной системы, например, Т-клетки или NK-клетки. Такая молекула антитела дает повышенную специфичность, а также авидность по отношению к опухолевой клетке определенного типа, а также повышенную авидность для активации или ингибирования рецептора на эффекторной клетке иммунной системы, что приводит к выгодно увеличенному специфичному цитотоксическому потенциалу антигенсвязывающей молекулы Связывание с двумя различными эпитопами опухолевого антигена приводит к увеличению специфичности адресного воздействия и расширению терапевтического окна путем снижения неспецифичной токсичности. Таким образом, в настоящем изобретении предложены молекулы мультиспецифичных антител, благоприятным образом повышающие авидность и/или биологическую активность вследствие наличия по меньшей мере двух антигенсвязывающих сайтов для по меньшей мере одной эпитопной специфичности, например, двух сайтов связывания для одной, двух или трех эпитопных специфичностей, другими словами, мишеней.
Важно отметить, что, несмотря на структурную сложность, такие мультиспецифичные, например, триспецифичные и мультивалентные, например, четырехвалентные молекулы антитела согласно настоящему изобретению являются стабильными.
В других случаях такое мультиспецифичное, например, триспецифичное антитело может иметь две специфичности для различных антигенов на эффекторной клетке, например, NK-клетке или Т-клетке, и третью специфичность в отношении антигена на опухолевой клетке.
Таким образом, молекулу антитела согласно настоящему изобретению можно применять различными способами для изменения мишеней цитотоксического потенциала эффекторных клеток иммунной системы для уничтожения опухолевых клеток или инфекционных агентов, например, клеток, инфицированных вирусами. В некоторых вариантах реализации мультиспецифичная, например, триспецифичная молекула антитела может связываться с двумя различными эпитопами антигена на мишени. Например, эти два различных эпитопа могут находиться на одном и том же антигене для предотвращения появления «ускользающих» мутантов или повышения эффективности, или эти два эпитопа могут находиться на двух различных антигенах мишени. В некоторых вариантах реализации триспецифичная молекула антитела может связываться с двумя различными эпитопами антигена на эффекторных клетках иммунной системы. Например, первый антигенсвязывающий сайт обладает специфичностью к активирующему рецептору, например, CD16, CD16A или CD3, а второй антигенсвязывающий сайт обладает специфичностью к костимулирующему антигену, например, CD137, ОХ-40 или CD28. В другом примере первый антигенсвязывающий сайт обладает специфичностью к CD16 или CD16A, а второй антигенсвязывающий сайт специфичен к другому активирующему рецептору на NK-клетках, например, NKG2D, DNAM, NCR.
В еще одном варианте реализации молекула триспецифичного, в частности, Fv-антитела содержит первый антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена на опухолевой клетке, второй антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена на эффекторной клетке иммунной системы, и третий антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена на растворимом белке, выбранном из группы факторов роста, цитокинов, хемокинов, митогенов и альбуминов. Примеры такого растворимого белка представляют собой ИЛ-6, BAFF, APRIL, ТФР-бета, ИЛ-10, VEGF-A, ТФР-альфа, ЭФР, HB-EGF, херегулины, ангиопоэтин-2 и человеческий сывороточный альбумин (ЧСА).
В альтернативном варианте реализации молекула антитела содержит один антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена, присутствующего на клетках одного типа, и три антигенсвязывающих сайта, обладающие специфичностью к эпитопам антигенов на клетках одного или более типов.
"Эффекторные клетки» представляют собой клетки иммунной системы, способные стимулировать или активировать цитотоксичность, фагоцитоз, презентацию антигена, высвобождение цитокинов. Такие эффекторные клетки являются, например, Т-клетками, естественными киллерами (NK-клетками), гамма-дельта (gd) Т-клетками, естественными киллерными Т-клетками (NKT), гранулоцитами, моноцитами, макрофагами, дендритными клетками, врожденными лимфоидными клетками (ILC) и антиген-презентирующими клетками, но не ограничиваются ими. Примеры специфичностей для эффекторных клеток включают CD2, CD3 и субъединицы CD3, например, CD3s, CD5, CD28 и другие компоненты Т-клеточного рецептора (TCR) или CD134 (ОХ40) для Т-клеток; CD16A, CD25, CD38, CD44, CD56, CD69, CD94, CD335 (NKp46), CD336 (NKp44), CD337 (NKp30), NKp80, NKG2A, NKG2C и NKG2D, DNAM, NCR для NK-клеток; CD18, CD64 и CD89 для гранулоцитов; CD18, CD32, CD47, CD64, CD89 и рецептор маннозы для моноцитов и макрофагов; CD64 и рецептор маннозы для дендритных клеток; а также CD35, но не ограничиваются ими. В некоторых вариантах реализации настоящего изобретения указанные специфичности, т.е. молекулы клеточной поверхности эффекторных клеток, подходят для опосредования уничтожения клеток после связывания молекулы мультиспецифичного, например, триспецифичного антитела с такой молекулой клеточной поверхности, и, тем самым, индукции цитолиза или апоптоза.
Антиген CD3 представляет собой компонент комплекса Т-клеточного рецептора на Т-клетках. В случае, когда специфичность для эффекторной клетки представляет собой CD3, связывание антигенсвязывающей молекулы согласно настоящему изобретению с CD3 вызывает цитотоксическую активность Т-клеток. Связывание молекулы антитела с CD3 и с клеткой-мишенью, например, опухолевой клеткой, может индуцировать лизис клетки-мишени.
Антиген CD16A (FcγRIMA) является рецептором, экспрессируемым на поверхности NK-клеток. NK-клетки обладают характерной цитолитической активностью, и связывание молекулы антитела согласно настоящему изобретению с CD16 или CD16A может запустить цитотоксическую активность NK-клетки по отношению к мишени.
«Мишень» является сайтом, на котором расположен эпитоп антигена, с которым должна связываться молекула антитела. Примерами мишеней являются растворимые агенты, антигены или клетки, инфекционные агенты, например, антигены вирусов или бактерий, например, происходящие от вируса денге, вируса простого герпеса, вируса гриппа, ВИЧ, ВГС, ЦМВ, или антигены на клетках, облегчающие проникновение вирусов и бактерий, или клетки, например, нейроны, экспонирующие антигены, или клетки, несущие аутоиммунные мишени, например, ИЛ-2/IL2R, аутоиммунный маркер или аутоиммунный антиген, или опухолевые клетки. В вариантах реализации, где по меньшей мере один из антигенсвязывающих сайтов обладает специфичностью к эффекторной клетке, мишень может представлять собой опухолевую клетку, по отношению к которой можно настроить активность эффекторной клетки с целью индукции или активации соответствующего биологического, например, иммунного ответа.
Подходящие специфичности в случае опухолевых клеток могут быть представлены опухолевыми антигенами и антигенами клеточной поверхности на соответствующей опухолевой клетке, Например, определенными опухолевыми маркерами. Термин "опухолевый антиген", используемый в настоящем документе, включает опухоль-ассоциированный антиген (ТАА) и опухольспецифичный антиген (TSA). Термин "опухоль-ассоциированный антиген" (ТАА), используемый в настоящем документе, относится к белку, присутствующему на опухолевых клетках, а также на нормальных клетках в течение эмбрионального периода (эмбриональные онкоантигены), и после родов в отдельных органах, однако в значительно более низкой концентрации, чем на опухолевых клетках. ТАА могут также присутствовать в строме в непосредственной близости от опухолевой клетки, однако в строме других частей тела они экспрессируются в низких количествах. В противоположность этому, термин "опухольспецифичный антиген" (TSA) относится к белку, экспрессируемому опухолевыми клетками. Термин "антиген клеточной поверхности" относится к молекуле любого антигена или его фрагмента, которая может распознаваться антителом на поверхности клетки.
Примеры специфичностей для опухолевых клеток включают CD19, CD20, CD26, CD29, CD30, CD33, CD52, CD200, CD267, EGFR, EGFR2, EGFR3, EGFRvIII, HER2, HER3, IGFR, IGF-1R, Ep-CAM, PLAP, антиген Томсона-Фриденрайха (TF), TNFRSF17, gpA33, MUC-1 (муцин), IGFR, CD5, ИЛ-4-R альфа, ИЛ13-R, FcεRI, комплексы МНС I класса/пептид и IgE, но не ограничиваются ими.
Молекулы антител согласно настоящему изобретению, в которых опухолевая специфичность направлена к антигену CD19, можно применять для иммунотерапии В-клеточных злокачественных новообразований, поскольку антиген CD19 экспрессируется практически на всех злокачественных новообразованиях В-ряда от лимфобластного лейкоза (ALL, ОЛЛ) до неходжкинской лимфомы (NHL).
Молекулы антител согласно настоящему изобретению, в которых опухолевая специфичность направлена против CD30, могут быть особенно полезны при лечении болезни Ходжкина и Т-клеточных лимфом.
Молекулы антитела согласно настоящему изобретению, в которых опухолевая специфичность направлена против рецептора эпидермального фактора роста (EGFR) или мутантной формы EGFRvIII, можно применять при лечении глиом, опухолей молочной железы, яичников, предстательной железы, легких, головы и шеи; заболеваниях печени, например, гепатоцеллюлярном раке, циррозе печени или хроническом гепатите.
Для увеличения времени полужизни молекул антител согласно настоящему изобретению в организме молекулу антитела при желании можно объединить с альбумином, например, человеческим сывороточным альбумином (ЧСА), или пэгилировать, сиалировать, пасилировать или гликозилировать (см., например, Stork et al., 2008, J. Biol. Chem., 283:7804-7812). В некоторых вариантах реализации молекула антитела является по меньшей мере триспецифичной и содержит по меньшей мере один, например, один или два антигенсвязывающих сайта, обладающих специфичностью по отношению к альбумину, например, ЧСА. Такая по меньшей мере триспецифичная молекула антител может являться, например, четырехвалентной, пятивалентной или шестивалентной молекулой антитела.
Вариабельные домены, обладающие специфичностью по отношению к эпитопам на клетках-мишенях или эффекторных клетках, можно получить путем отбора вариабельных фрагментов (Fv), специфичных по отношению к нужным антигенам. Эти процедуры можно выполнить, например, путем скрининга библиотек фагового дисплея одноцепочечных Fv или с использованием гибридомной технологии. Например, библиотеки фагового дисплея последовательностей scFv человека на основе IgM можно подвергать нескольким циклам отбора in vitro с целью обогащения связывающими агентами, специфичными по отношению к желательному антигену. Затем можно повысить сродство отобранных scFv путем созревания аффинности.
В некоторых вариантах реализации настоящего изобретения по меньшей мере один, предпочтительно все вариабельные домены антитела являются полностью человеческими, гуманизированными или химерными доменами. Гуманизированные антитела можно получить с помощью общепринятых способов, например, пересадки CDR (см., например, Antibody engineering: methods and protocols / edited by Benny K.C. Lo; Benny K.C. II Series: Methods in molecular biology (Totowa, NJ.). Таким образом, специалист в данной области техники легко может получить гуманизированный или полностью человеческий вариант антигенсвязывающей молекулы и вариабельных доменов из нечеловеческих источников, например, мыши или животных, не являющихся приматами, с помощью стандартных молекулярно-биологических методик, известных в данной области техники с целью снижения иммуногенности и улучшения эффективности указанной антигенсвязывающей молекулы в иммунной системе человека. В предпочтительном варианте реализации настоящего изобретения все вариабельные домены антитела являются гуманизированными или полностью человеческими; наиболее предпочтительно, молекула антитела согласно настоящему изобретению является гуманизированной или полностью человеческой. Термин "полностью человеческий" в настоящем документе означает, что аминокислотные последовательности вариабельных доменов и пептидов, связывающих вариабельные домены в полипептиде, происходят или могут обнаруживаться в организме человека. В некоторых вариантах реализации настоящего изобретения вариабельные домены, но не пептиды, связывающие вариабельные домены, могут являться доменами человека или гуманизированными доменами антитела.
Специалист легко может сконструировать и получить молекулу антитела, описанную в настоящем документе, используя установленные методики и стандартные способы, известные в данной области техники, см., например, Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y.; The Protein Protocols Handbook, edited by John M. Walker, Humana Press Inc. (2002); или Antibody engineering: methods and protocols / edited by Benny K.C. Lo; Benny K.C. II Series: Methods in molecular biology (Totowa, N.J.); Antibody Engineering / edited by Roland E. Kontermann and Stefan Dubel, Springer Verlag Berlin Heidelberg (2010)).
Молекулу антитела по любому из вариантов реализации, описанных в настоящем документе, можно получить путем экспрессии полинуклеотидов, кодирующих отдельные полипептидные цепи, образующие молекулу антитела. Таким образом, дополнительными вариантами реализации настоящего изобретения являются полинуклеотиды, например, ДНК или РНК, кодирующие полипептиды молекулы антитела, описанной выше в настоящем документе.
Указанные полинуклеотиды можно сконструировать с применением способов, известных специалистам, например, путем комбинирования генов, кодирующих вариабельные домены антитела, разделенные пептидными линкерами или непосредственно связанные между собой пептидными связями полипептидов, в генетическом конструкте, функционально связанном с подходящим промотором и, необязательно, подходящим терминатором транскрипции, и их экспрессии в бактерии или другой соответствующей системе экспрессии, например, клетках СНО. В зависимости от выбранных векторной системы и хозяина, можно использовать любой набор подходящих элементов транскрипции и трансляции, включая конститутивные и индуцируемые промоторы. Промотор выбирают так, чтобы он управлял экспрессией полинуклеотидов в соответствующей клетке-хозяине.
Полинуклеотиды можно внедрить в векторы, предпочтительно экспрессирующие векторы, которые представляют собой дополнительный вариант реализации настоящего изобретения. Указанные рекомбинантные векторы можно сконструировать в соответствии со способами, хорошо известными специалистам в данной области техники.
Для встраивания и экспрессии полинуклеотидов, кодирующих полипептидные цепи по настоящему изобретению, можно использовать множество систем экспрессирующих векторов/хозяев. Примеры экспрессирующих векторов для экспрессии в E.coli представляют собой pSKK (LeGalI et al., J Immunol Methods. (2004) 285(1):111-27) или pcDNA5 (Invitrogen) для экспрессии в клетках млекопитающих.
Таким образом, молекулу антитела, описанную в настоящем документе, можно получить путем внедрения вектора, кодирующего полипептиды, описанные выше, в клетку-хозяина и культивирования указанной клетки-хозяина в условиях, при которых указанные полипептиды экспрессируются, их можно выделить и, необязательно, дополнительно очистить.
В дополнительном варианте реализации настоящего изобретения предложены композиции, например, фармацевтические композиции, содержащие молекулу антитела, описанную выше, и по меньшей мере один дополнительный компонент.
В дополнительных вариантах реализации молекулы антител предназначены для применения в качестве медикамента или диагностического средства. В частности, молекулы антител предназначены для применения в иммунотерапии. Например, молекул антител предназначены для применения при лечении опухолевого, вирусного или нейродегенеративного заболевания. Таким образом, настоящее изобретение дополнительно включает способ лечения индивида посредством иммунотерапии, в частности, индивида, страдающего заболеваниями, выбранными из опухолевых, вирусных или нейродегенеративных заболеваний, включающий этап введения молекулы антитела в соответствии с настоящим изобретением.
В настоящем изобретении дополнительно предложен способ, в котором молекулу антитела, в частности, композицию, содержащую молекулу антитела, описанную выше в настоящем документе, и по меньшей мере один дополнительный компонент, вводят в эффективной дозе субъекту, например, пациенту, для лечения рака (например, неходжкинской лимфомы; хронического лимфоцитарного лейкоза). Антигенсвязывающая молекула согласно настоящему изобретению может быть предназначена для применения в качестве медикамента.
Краткое описание чертежей:
На фигуре 1 показана четырехвалентная триспецифичная антигенсвязывающая Fv-молекула, состоящая из одиночного полипептида, содержащего восемь вариабельных доменов, соединенных друг за другом в ориентации VH-VL-VH-VH-VL-VL-VL-VH от N-конца 7 к С-концу 8 полипептида. Такая ориентация доменов приведена лишь в качестве примера, возможны и другие аранжировки, описанные выше. Две одноцепочечные Fv (scFv) единицы 2 дистально соединены с единицей одноцепочечного диатела 1 пептидными линкерами 5, причем вариабельный домен легкой цепи (VL) одной единицы scFv 2 соединен по С-концу пептидным линкером 5 с N-концом вариабельного домена тяжелой цепи (VH) первой пары вариабельных доменов единицы диатела 1. VL единицы scFv 2 соединен по N-концу с VH единицей scFv длинным пептидным линкером 4. VH первой пары вариабельных доменов соединен коротким пептидным линкером 3 с еще одним доменом VH первой пары вариабельных доменов. Первая пара вариабельных доменов соединена по С-концу длинным пептидным линкером 4 с N-концом второй пары вариабельных доменов диатела, оба из которых являются VL-доменами. Два VL-домена второй пары вариабельных доменов соединены коротким пептидным линкером 3. Вторая пара вариабельных доменов диатела присоединена по С-концу к N-концу VL-домена еще одной единицы scFv 2, расположенной на С-конце полипептида. Единица scFv 2 на С-конце полипептида состоит из VL-домена, соединенного длинным линкером 4 с VH-доменом. Две единицы scFv 2 содержат антигенсвязывающие сайты с различными специфичностями, а единица диатела 1 содержит два антигенсвязывающих сайта, обладающих одинаковой специфичностью.
На фигуре 2 показана четырехвалентная триспецифичная антигенсвязывающая Fv-молекула, состоящая из первого полипептида, содержащего шесть вариабельных доменов, соединенных друг за другом в ориентации VH-VL-VH-VH-VL-VH от N-конца 7 к С-концу 8 первого полипептида, и второго полипептида, содержащего два вариабельных домена, соединенных друг за другом в ориентации VL-VL от N-конца 7а к С-концу 8а второго полипептида. Такая ориентация доменов приведена лишь в качестве примера, возможны и другие варианты расположения, описанные выше. Антигенсвязывающая Fv-молекула содержит димерную единицу диатела 1, образованную первой парой двух вариабельных доменов, встроенных в первый полипептид, ассоциированных со второй парой двух вариабельных доменов во втором полипептиде. Каждый из двух вариабельных доменов первой пары двух вариабельных доменов, встроенных в первый полипептид, соединен с единицей scFv 2. В каждой из единиц scFv 2 вариабельные домены соединены длинным пептидным линкером 4. N-концевая единица scFv 2 соединена по С-концу пептидным линкером 5 с N-концом первой пары вариабельных доменов единицы диатела 1. Первая пара вариабельных доменов состоит из двух VH-доменов, соединенных коротким пептидным линкером 3. Первая пара вариабельных доменов единицы диатела 1 соединена по С-концу пептидным линкером 5 с N-концом единицы scFv 2, расположенной на С-конце первого полипептида. Две единицы scFv 2 содержат антигенсвязывающие сайты с различными специфичностями, а единица диатела 1 содержит два антигенсвязывающих сайта, обладающих одинаковой специфичностью.
На фигуре 3 показана четырехвалентная триспецифичная антигенсвязывающая Fv-молекула в формате триспецифичного тандемного диатела, состоящего из первого и второго полипептида, образующих нековалентно ассоциированный гетеродимер. Тандемное диатело содержит димерную единицу диатела 1, образованную парой доменов VH-VH, расположенных в центральной части первого полипептида, с парой доменов VL-VL, расположенных в центральной части второго полипептида. Вариабельные домены в первом и втором полипептиде соединены друг за другом короткими пептидными линкерами 3, 3а от N-конца 7 к С-концу 8 в ориентации VL-VH-VH-VL в первом полипептиде и от N-конца 7а к С-концу 8а в ориентации VH-VL-VL-VH во втором полипептиде. Вариабельные домены, расположенные на N-конце и С-конце, содержат антигенсвязывающие сайты с различными специфичностями по N-концу и С-концу, а единица диатела 1, расположенная в центральной части, содержит два антигенсвязывающих сайта, обладающих одинаковой специфичностью.
На фигуре 4 показана шестивалентная антигенсвязывающая Fv-молекула, состоящая из одиночного полипептида, содержащего двенадцать вариабельных доменов, соединенных друг за другом от N-конца 7 к С-концу 8 полипептида. Ориентация домена приведена только для примера, возможны и другие аранжировки, указанные для каждой пары вариабельных доменов, расположенных рядом друг с другом. Две единицы одноцепочечного диатела 1а, 1b дистально соединены с единицей одноцепочечного диатела 1с пептидными линкерами 5, причем вариабельный домен одной единицы одноцепочечного диатела 1а соединен по С-концу пептидным линкером 5 с N-концом вариабельного домена первой пары вариабельных доменов единицы диатела 1с, а вариабельный домен единицы одноцепочечного диатела 1b соединен по N-концу линкером 5 с С-концом второй пары вариабельных доменов единицы одноцепочечного диатела 1с. Первая и вторая пары вариабельных доменов каждой из трех единиц одноцепочечного диатела 1а, 1b, 1с соединены друг с другом длинным пептидным линкером 4. В каждой из единиц одноцепочечного диатела 1а, 1b, 1с один из вариабельных доменов первой пары вариабельных доменов соединен коротким пептидным линкером 3 с другим вариабельным доменом первой пары вариабельных доменов, а два вариабельных домена второй пары вариабельных доменов соединены коротким пептидным линкером 3.
На фигуре 5 показана шестивалентная антигенсвязывающая Fv-молекула, состоящая из первого полипептида, содержащего 10 вариабельных доменов, соединенных друг за другом от N-конца 7 к С-концу 8 первого полипептида, и второго полипептида, содержащего два вариабельных домена, соединенных друг за другом от N-конца 7а к С-концу 8а второго полипептида. Конкретная ориентация домена на данной фигуре приведена только для примера, возможны и другие аранжировки, указанные для каждой пары вариабельных доменов, расположенных рядом друг с другом. Антигенсвязывающая Fv-молекула содержит димерную единицу диатела 10, образованную первой парой двух вариабельных доменов первого полипептида, ассоциированных со второй парой двух вариабельных доменов во втором полипептиде. Каждый из двух вариабельных доменов первой пары двух вариабельных доменов первого полипептида соединен к единицей одноцепочечного диатела 1а, 1b.
В каждой из единиц одноцепочечного диатела 1а, 1b две пары вариабельных доменов, расположенных рядом друг с другом, соединены длинным пептидным линкером 4. N-концевая единица одноцепочечного диатела 1а соединена по С-концу пептидным линкером 5 с N-концом первой пары вариабельных доменов единицы диатела 10. Первая пара вариабельных доменов состоит из двух VH-доменов, соединенных коротким пептидным линкером 3. Первая пара вариабельных доменов единицы диатела 10 соединена по С-концу пептидным линкером 5 с N-концом единицы одноцепочечного диатела 1b, расположенной на С-конце первого полипептида.
На фигуре 6 показана пятивалентная антигенсвязывающая Fv-молекула, состоящая из первого полипептида, содержащего шесть вариабельных доменов, соединенных друг за другом от N-конца 7 к С-концу 8 первого полипептида, и второго полипептида, содержащего четыре вариабельных домена, соединенных друг за другом от N-конца 7а к С-концу 8а второго полипептида. Ориентация домена приведена только для примера, для каждой пары вариабельных доменов, расположенных рядом друг с другом, возможны и другие аранжировки, указанные на фигуре. Антигенсвязывающая Fv-молекула содержит димерную единицу диатела 10, образованную первой парой двух вариабельных доменов первого полипептида, ассоциированных со второй парой двух вариабельных доменов во втором полипептиде. Каждый из двух вариабельных доменов первой пары двух вариабельных доменов первого полипептида соединен с единицей scFv 2. В каждой из единиц scFv 2 вариабельные домены соединены длинным пептидным линкером 4. N-концевая единица scFv 2 присоединена по С-концу пептидным линкером 5 с N-концом первой пары вариабельных доменов единицы диатела 10. Первая пара вариабельных доменов состоит из двух вариабельных доменов, соединенных коротким пептидным линкером 3. Первая пара вариабельных доменов единицы диатела 10 соединена о С-концу пептидным линкером 5 с N-концом единицы scFv 2, расположенной на С-конце первого полипептида. А) второй полипептид, содержащий четыре вариабельных домена, состоит из второй пары вариабельных доменов единицы диатела 10, соединенных с единицей scFv 2 по С-концу, и В) второй полипептид, содержащий четыре вариабельных домена, состоит из второй пары вариабельных доменов единицы диатела 10, соединенных с единицей scFv 2 по N-концу.
На фигуре 7 показана шестивалентная антигенсвязывающая Fv-молекула, состоящая из первого полипептида, содержащего шесть вариабельных доменов, соединенных друг за другом от N-конца 7 к С-концу 8 первого полипептида, и второго полипептида, содержащего шесть вариабельных домена, соединенных друг за другом от N-конца 7а к С-концу 8а второго полипептида. Ориентация домена приведена только для примера, для каждой пары вариабельных доменов, расположенных рядом друг с другом, возможны и другие аранжировки, указанные на фигуре. Антигенсвязывающая Fv-молекула содержит димерную единицу диатела 10, образованную первой парой двух вариабельных доменов первого полипептида, ассоциированных со второй парой двух вариабельных доменов во втором полипептиде. Каждый из вариабельных доменов единицы диатела 10 соединен с единицей scFv 2 в первом и втором полипептиде. В каждой из единиц scFv 2 вариабельные домены соединены длинным пептидным линкером 4. Каждая из четырех единиц scFv 2 соединена пептидным линкером 5 с вариабельным доменом единицы диатела 10. Эта первая пара вариабельных доменов состоит из двух вариабельных доменов, соединенных коротким пептидным линкером 3, а вторая пара вариабельных доменов единицы диатела состоит из двух вариабельных доменов, соединенных коротким пептидным линкером 3а.
На фигуре 8 показана шестивалентная антигенсвязывающая Fv-молекула, содержащая единицу тандемного диатела, состоящего из первого и второго полипептида, образующих нековалентно ассоциированный димер. Тандемное диатело содержит димерную единицу диатела 10а, образованную парой доменов, расположенных в центральной части первого полипептида, с парой вариабельных доменов, расположенных в центральной части второго полипептида. Вариабельные домены в первом и втором полипептиде соединены друг за другом короткими пептидными линкерами 3, 3а от N-конца 7 к С-концу 8 в каждом полипептиде. Ориентация домена приведена только для примера, для каждой пары вариабельных доменов, расположенных рядом друг с другом, возможны и другие аранжировки, указанные на фигуре. Каждый из первого и второго полипептида содержит единицу scFv 2, соединенную линкером 5 с единицей тандемного диатела. А) единицы scFv 2 присоединены по С-концу первого и второго полипептида. В) единицы scFv 2 присоединены по N-концу первого и второго полипептида.
На фигуре 9 показан анализ продуктов aTriFlex посредством электрофореза в ДСН-ПААГ. Длинные и короткие полипептиды хорошо экспрессируются (стрелки), а) aTriFlex_140 и aTriFlex_142, b) aTriFlex_138 и с) aTriFlex_139.
На фигуре 10 показана IMAC-очистка продуктов aTriFlex: a) aTriFlex_140, b) aTriFlex_138 и с) и aTriFlex_142 и d) aTriFlex_139.
На фигуре 11 показан анализ продуктов aTriFlex посредством электрофореза в ДСН-ПААГ. Длинные и короткие полипептиды хорошо экспрессируются. A) aTriFlex_101, В) aTriFlex_102, С) aTriFlex_103 и D) aTriFlex_104.
На фигуре 12 показано связывание молекул триспецифичного антитела с однократно или дважды положительными линиями клеток. Однократно положительные клетки CD19+/CD30-Raji и CD19-/CD30+ KARPAS-299, а также дважды положительные клетки CD19+/CD30+ МЕС-1 окрашивали последовательными разведениями молекул триспецифичного антитела; антитела, связавшиеся с поверхностью клеток, обнаруживали мАт против His с последующей обработкой конъюгированными с FITC IgG козы против антител мыши. Средние значения интенсивности флуоресценции, определяемые проточной цитометрией, использовали для расчета значений KD посредством нелинейной регрессии. Значения KD для молекул триспецифичного антитела определяли в двух независимых экспериментах.
На фигуре 13 показана цитотоксическая активность молекул триспецифичного антитела по отношению к однократно или дважды положительным линиям клеток. Анализ цитотоксичности с высвобождением кальцеина в течение 4 ч выполняли для однократно положительных клеток CD19+/CD30- Raji и CD19-/CD30+ KARPAS-299, а также дважды положительных клеток-мишеней CD19+/CD30+ МЕС-1 с использованием обогащенных NK-клеток в качестве эффекторных клеток в присутствии последовательных разведений указанных молекул триспецифичных антител. Флуоресцирующий кальцеин, высвободившийся из лизированных клеток-мишеней, использовали для расчета антитело-опосредованного лизиса клеток-мишеней и моделирования сигмоидных кривых зависимости реакции от дозы с целью определения значений EC50 посредством нелинейной регрессии. Значения EC50 для конструктов aTriFlex определяли в двух независимых экспериментах и показывали на графике.
Примеры, приведенные ниже, дополнительно иллюстрируют настоящее изобретение без ограничения рамок настоящего изобретения.
ПРИМЕР 1
Клонирование экспрессирующих ДНК-конструктов, кодирующих триспецифичние антигенсвязывающие Fv-молекулы
Для экспрессии триспецифичных конструктов в клетках СНО кодирующие последовательности всех молекул клонировали в виде конструктов из одного или двух генов в системе экспрессии в клетках млекопитающих, модифицированной соответствующим образом и происходящей от вектора pcDNA5/FRT. Вектор с двумя промоторами использовали для клонирования и экспрессии конструктов из двух генов (гетеродимерных триспецифичных конструктов). Для триспецифичных конструктов из одного гена использовали обычный экспрессирующий вектор pcDNA5/FRT. Вкратце, последовательности генов, кодирующих домены VH и VL, разделенные различными линкерами (длинными и короткими линкерами) синтезировали с помощью Life Technologies GeneArt (Регенсбург, Германия) и субклонировали. Полученные конструкты расщепляли различными ферментами рестрикции (BamHI, HindIII, XhoI, EcoRV, PacI), или различные домены VH/VL, связывающиеся с эффекторами и мишенями, амплифицировали посредством ПЦР с соответствующими праймерами. После этого различные перекрывающиеся фрагменты ДНК и 2-промоторный вектор с линеаризованным каркасом или вектор pcDNA5/FRT соединяли друг с другом с помощью Gibson Assembly в одной изотермической реакции. Конструкция всех экспрессирующих конструктов содержала кодирующие последовательности N-концевого сигнального пептида и С-концевого гексагистидинового (6xHis) маркера для первой кассеты генов или С-концевой FLAG-маркер для второй кассеты генов в целях облегчения секреции и очистки антитела, соответственно. Последовательности всех конструктов подтверждали секвенированием ДНК в GATC (Констанц, Германия) с использованием пары праймеров 5'-AGAGCTCGTTTAGTGAACCG-3' и 5'-TCATGTCTGGATCCGGCCTTG-3' для 1 кассеты генов или 5'-GCCTGGAGACGCCATCC-3' и 5'-GCAGAATTCCACCACACTGG-3' для 2 кассеты генов.
Культура клеток-хозяев
Клетки Flp-In СНО (Life Technologies), производные клеток яичника китайского хомячка CHO-KI (АТСС, CCL-61) (Kao and Puck, 1968), культивировали в питательной среде Хэма F-12 с добавлением L-глутамина, 10% ЭТС и 100 мкг/мл зеоцина. Адгезивные клетки отделяли с использованием 0,25% трипсина-ЭДТА и субкультивировали в соответствии со стандартными протоколами культивирования клеток, предоставленными Life Technologies.
Для адаптации к росту в суспензии клетки отделяли от флаконов для тканевых культур и помещали в бессывороточную среду HyClone CDM4 СНО для последующего инкубации в качалочных колбах при 37°С, 5% СО2 и 120 об/мин. Стандартная среда для культивирования адаптированных к суспензии клеток-хозяев Flp-In СНО представляла собой HyClone CDM4 СНО с добавлением L-глутамина, добавки НТ, пенициллина/стрептомицина и 100 мкг/мл зеоцина. Адаптированные к суспензии клетки консервировали с замораживанием в среде с 10% ДМСО и проверяли на предмет отрицательного результата анализа Mycoplasma с использованием набора MycoAlert для обнаружения Mycoplasma (Lonza).
Получение пулов стабильно трансфицированных клеток
Линии рекомбинантных клеток Flp-In СНО, стабильно экспрессирующих секретируемые триспецифичные антитела из одного или двух генов, получали путем трансфекции клеток-хозяев, адаптированных к суспензии. Для этого клетки помещали в стандартную среду без зеоцина за один день до совместной трансфекции экспрессирующими плазмидами (2,5 мкг), кодирующими нужный белок (pcDNA5/FRT), и рекомбиназу Flp (pOG44, Life Technologies) с использованием полиэтиленимина (ПЭИ). Вкратце, векторную ДНК и реагент для трансфекции смешивали в соотношении ДНК : ПЭИ 1:3 (мкг/мкг) в общем объеме среды OptiMEM I 100 мкл и инкубировали в течение 10 минут, а затем добавляли 2⋅106 клеток Flp-In СНО, суспендированных в 1 мл среды CHO-S-SFMII (Life Technologies). Через 24 ч инкубации начинали отдор стабильно трансфицированных клеток путем добавления 500 мкг/мл гигромицина В после разбавления культур до плотности 0,1⋅106 жизнеспособных клеток/мл в среде CHO-S-SFMII и посева в флаконы для культивирования Т75. Рекомбиназа Flp опосредует инсерцию экспрессирующих конструктов в геном клеток Flp-In СНО по встроенному сайту FRT за счет сайт-специфичной рекомбинации ДНК (O'Gorman et al., 1991). При отборе измеряли плотность жизнеспособных клеток дваджы в неделю, клетки центрифугировали и ресуспендировали в свежей селективной среде при максимальной плотности 0,1⋅106 жизнеспособных клеток/мл. Пулы клеток, стабильно экспрессирующих рекомбинантные белковые продукты, выделяли после 2-3 недель отбора; в этот момент клетки переносили на стандартную культуральную среду в качалочные колбы. Экспрессию рекомбинантных секретируемых белков подтверждали гель-электрофорезом белков в надосадочной жидкости культур клеток с использованием технологии Criterion Stain-Free (Bio-Rad). Стабильные пулы клеток криоконсервировали в среде, содержащей 50% ProFreeze (Lonza) и 7,5% ДМСО.
Продукция рекомбинантного белка при периодическом культивировании клеток СНО в суспензии с добавлением субстрата
Рекомбинантные белки продуцировали при 10-дневном периодическом культивировании стабильно трансфицированных линий клеток СНО с добавлением субстрата путем секреции в надосадочную жидкость культуры клеток. Для этого пулы клеток, стабильно экспрессирующих триспецифичные антитела, высевали при начальной плотности 6⋅105 клеток/мл в стандартную культуральную среду в поликарбонатные колбы Эрленмейера с газопроницаемыми крышками (Corning) и инкубировали при 37°С и 5% СО2 с перемешиванием при 140 об/мин. При периодическом культивировании с добавлением субстрата в среду добавляли 40 мл/л ActiCHO Feed А и 4 мл/л ActiCHO Feed В (GE Healthcare) в 0 день (день начала культивирования) и двойное количество этих добавок в 3, 5 и 7 дни. Надосадочную жидкость культур клеток собирали через 10 суток культивирования при жизнеспособности культуры >75%. Отбор образцов продуцирующих культур выполняли через день до подпитки и оценивали плотность и жизнеспособность клеток. В день сбора надосадочную жидкость культур клеток просветляли центрифугированием и вакуумной фильтрацией (0,22 мкм) через мембранные фильтры Millipore Express PLUS (Millipore) перед дальнейшим использованием.
Очистка триспецифичных Fv-антител
Триспецифичные антитела выделяли из просветленной надосадочной жидкости культур клеток СНО в ходе двустадийной процедуры, включавшей IMAC и препаративную эксклюзионную хроматоргафию. При необходимости для дополнительной тонкой очистки использовали третий хроматографический этап (FLAG-аффинную хроматографию). Для IMAC просветленную непосадочную жидкость загружали на колонку HisTrap с сефарозой. После промывки трис/NaCl-буфером (рН 7,5) белок элюировали в трехстадийном градиенте с использованием 15 мМ, 125 мМ и 500 мМ имидазола, соответственно. Чистоту фракций анализировали с помощью эксклюзионной ВЭЖХ и электрофореза в ДСН-ПААГ. Фракции, демонстрировавшие приемлемую чистоту, объединяли и подвергали препаративной гель-фильтрации с использованием препаративной колонки Superdex 200. В некоторых случаях для дополнительной тонкой очистки использовали FLAG-аффинную хроматографию. Так, белок загружали на FLAG-аффинную колонку, промывали PBS и элюировали с использованием глицин/HCl-буфера при рН 3,5. Фракции элюата, содержавшие очищенные триспецифичные антитела, объединяли, диализовали против 10 мМ ацетата натрия (рН 5,0) и концентрировали ультрафильтрацией до типичной концентрации приблизительно 1 мг/мл. Чистоту и однородность (прибл. 90%) конечных образцов оценивали посредством электрофореза в ДСН-ПААГ в восстанавливающих и невосстанавливающих условиях с последующим иммуноблоттингом с использованием антител, специфичных по отношению к His-маркеру и FLAG-маркеру, а также аналитической эксклюзионной ВЭЖХ. Очищенные белки хранили в виде аликвот при -80°С до использования.
Примеры биофизических данных для молекул триспецифичных Fv-антител описаны в примере 3; aTriFlex_101 (пример 3а)), aTriFlex_102 (пример 3b)), aTriFlex_103 (пример 3с)) и aTriFlex_104 (пример 3d) показаны в таблице 1. Результаты электрофореза фракций в ДСН-ПААГ показаны на Фигуре 11: Все молекулы триспецифичных четырехвалентных Fv-антител экспрессировали с приемлемыми титрами, независимо от порядка доменов, например, VH-VL-VH-VH-VL-VH (aTriFlex_101) or VL-VH-VH-VH-VH-VL (aTriFlex_102). Термическая стабильность и стабильность молекул триспецифичного четырехвалентного Fv-антитела были очень хорошими и сопоставимыми.
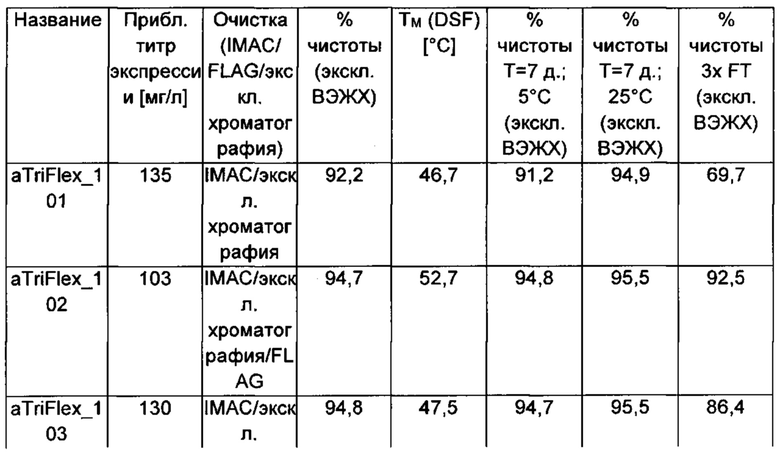
ПРИМЕР 2
Производство молекул триспецифичных антител с различным порядком доменов в единице диатела
Четыре различных молекулы Fv-антител в формате, соответствующем фигуре 2, сконструировали согласно описанию в примере 1; они характеризуются различным порядком доменов в двух парах вариабельных доменов в единице диатела (от N-конца к С-концу). Вариабельные домены единицы антитела показаны ниже в таблицах полужирным шрифтом:
aTriFlex_138: Единица антитела VH-VL/VH-VL:

aTriFlex_139: Единица диатела VL-VH/VL-VH:
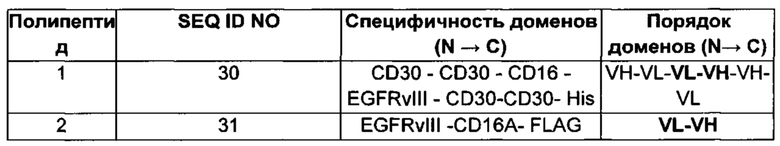
aTriFlex_140: Единица диатела VL-VL/VH-VH:

aTriFlex_142: Единица диатела VH-VH/VL-VL:

Молекулы Fv-антитела экспрессировали в клетках СНО; анализ надосадочной жидкости культур продуцирующих клеток посредством электрофореза в ДСН-ПААГ показал, что длинные полипептиды 1 и короткие полипептиды 2 хорошо экспрессировались и позволяли получить отдельные полосы при электрофорезе в денатурирующих условиях (фиг. 9а, 9b, 9с).
После одноэтапной (IMAC) очистки, описанной в примере 1, достигалась 76% чистота для aTriFlex_140 (фиг. 10а) и 78% чистота для aTriFlex_142 (фиг. 10с). Соответственно, молекулы мультиспецифичных Fv-антител с единицей диатела VL-VL/VH-VH (aTriFlex_140) или единицей диатела VH-VH/VL-VL (aTriFlex_142) демонстрировали значимо высокую долю молекул Fv-антител, подвергшихся правильному фолдингу. Вместе с тем получили несколько перекрывающихся разновидностей молекул, и из aTriFlex_138 и aTriFlex_139, содержащих домены в единицах диател в чередующемся порядке VH-VL/VH-VL и VL-VH/VL-VH, соответственно, не было выделено гетеродимерных молекул антител (фиг. 10b, 10d). Соответственно, ориентация вариабельных доменов в единице диатела VL-VL/VH-VH стимулирует ассоциацию двух различных полипептидов 1 и 2 друг с другом и их гетеродимеризацию в четырехвалентную триспецифичную молекулу антитела.
ПРИМЕР 3
Получили следующие молекулы мультивалентных антител, содержащих вариабельные домены антител против CD16A, CD19 и CD30:
Линкеры:
В примерах использовали следующие пептидные линкеры:
Линкер (4) в единице scFv: (G2S)6for VH-VL и (G2S)7for VL-VH
Линкер (3, 3а) в единице диатела: (G2S)3
Линкер (5), соединяющий единицу scFv с единицей диатела: (G2S)3
а) Молекула четырехвалентного триспецифичного Fv-антитела (aTriFlex_101, показанная на фиг. 2), имеющая специфичности для CD30, CD19 и CD16A, состоящая из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего два вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VH-VH в первом полипептиде и парой доменов VL-VL во втором полипептиде, что обеспечивает правильную гетеродимеризацию полипептида 1 и полипептида 2.

Аминокислотные последовательности:
Полипептид 1:

Полипептид 2:

b) Четырехвалентное триспецифичное Fv-антитело (aTriFlex_102), имеющее специфичности для CD30, CD19 и CD16A и состоящее из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего два вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VH-VH в первом полипептиде и парой доменов VL-VL во втором полипептиде.
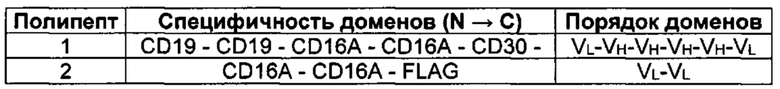
Аминокислотные последовательности:
Полипептид 1:

Полипептид 2:

с) Четырехвалентное триспецифичное Fv-антитело (aTriFlex_103), имеющее специфичности к CD30, CD19 и CD16A, состоящее из первого полипептида, содержащего шесть вариабельных доменов и С-концевой FLAG-маркер; и второго полипептида, содержащего два вариабельных домена и С-концевой His-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VH-VH в первом полипептиде и парой доменов VL-VL во втором полипептиде.
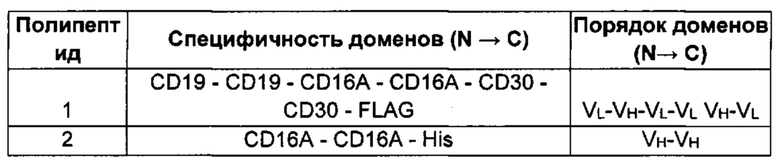
Аминокислотные последовательности:
Полипептид 1:

Полипептид 2:

d) Четырехвалентное триспецифичное Fv-антитело (aTriFlex_104), имеющее специфичности к CD30, CD19 и CD16A и состоящее из первого полипептида, содержащего шесть вариабельных доменов и С-концевой FLAG-маркер; и второго полипептида, содержащего два вариабельных домена и С-концевой His-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VH-VH в первом полипептиде и парой доменов VL-VL во втором полипептиде.

Аминокислотные последовательности:
Полипептид 1:

Полипептид 2:

Несмотря на различный размер полипептида 1 (шесть вариабельных доменов) и полипептида 2 (два вариабельных домена), молекул антител а), b), с) и d) могли удовлетворительно экспрессироваться. Термическая стабильность и стабильность молекул очищенных антител a)-d) были очень хорошими. Все конструкты a)-d) связывались с линиями первичных NK-клеток Raji, Karpas-299 и МЕС-1 при проточной цитометрии.
е) Четырехвалентное триспецифичное Fv-антитело (TeTrisAb_I), показанное на фиг. 3, имеющее специфичности к CD30, CD19 и CD16A и состоящее из первого полипептида, содержащего четыре вариабельных домена и С-концевой His-маркер; и второго полипептида, содержащего четыре вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VH-VH в первом полипептиде и парой доменов VL-VL во втором полипептиде; эта ориентация обеспечивает правильную гетеродимеризацию первого и второго полипептида.
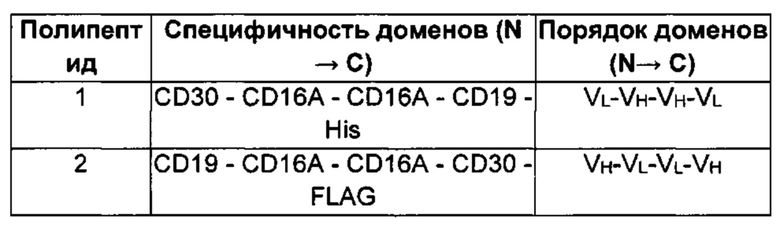
Аминокислотные последовательности
Полипептид 1:

Полипептид 2:


f) Четырехвалентное триспецифичное Fv-антитело (scTriFlex_2), показанное на фиг. 1, имеющее специфичности к CD30, CD19 и CD16A и состоящее из одного полипептида, содержащего восемь вариабельных доменов и His-маркер на С-конце. Димерная единица диатела, расположенная в центральной части молекулы, образована первой парой доменов VH-VH, ассоциированной со второй парой доменов VL-VL, расположенной рядом с ними в том же полипептиде. Пара VH-VH соединена с парой VL-VL линкером (G2S)6.

Аминокислотная последовательность:
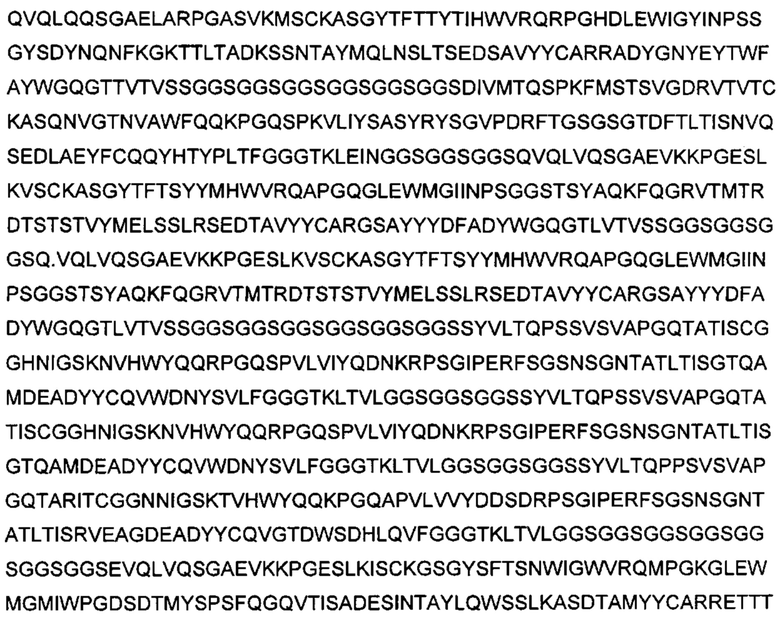

g) Молекула четырехвалентного триспецифичного Fv-антитела (показанная на фиг. 2), имеющая специфичности κCD30, CD19 и CD16A, состоящая из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего два вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VL-VL в первом полипептиде и парой доменов VH-VH во втором полипептиде, что обеспечивает правильную гетеродимеризацию полипептида 1 и полипептида 2.

Полипептид 1:

Полипептид 2:


h) Молекула пятивалентного тетраспецифичного Fv-антитела (показанная на фиг. 6а), имеющая специфичности κCD30, CD19, EGFRvIII и CD16A, состоящая из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего четыре вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, содержащая два антигенсвязывающих сайта для CD16A, образована парой доменов VL-VL в первом полипептиде и парой доменов VH-VH во втором полипептиде, что обеспечивает правильную гетеродимеризацию полипептида 1 и полипептида 2.

Полипептид 1:

Полипептид 2:


i) Молекула шестивалентного триспецифичного Fv-антитела (показанная на фиг. 5), имеющая специфичности κCD30, CD19 и CD16A, состоящая из первого полипептида, содержащего десять вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего два вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, расположенная в центральной части молекулы и содержащая два антигенсвязывающих сайта для CD16A, образована парой доменов VL-VL в первом полипептиде и парой доменов VH-VH во втором полипептиде, что обеспечивает правильную гетеродимеризацию полипептида 1 и полипептида 2.
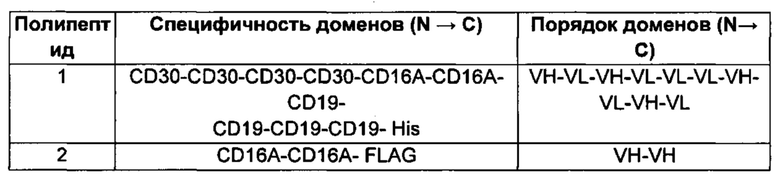
Полипептид 1:


Полипептид 2:

j) Молекула шестивалентного тетраспецифичного Fv-антитела (показанная на фиг. 7), имеющая специфичности κCD30, CD19, EGFRvIII и CD16A, состоящая из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего шесть вариабельных доменов и С-концевой FLAG-маркер.

Полипептид 1:


Полипептид 2:

k) Молекула пятивалентного триспецифичного Fv-антитела (показанная на фиг. 6b), имеющая специфичности κCD30, CD19 и CD16A, состоящая из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего четыре вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, содержащая два антигенсвязывающих сайта для CD16A, образована парой доменов VL-VL в первом полипептиде и парой доменов VH-VH во втором полипептиде, что обеспечивает правильную гетеродимеризацию полипептида 1 и полипептида 2.
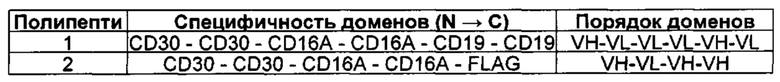
Полипептид 1:


Полипептид 2:

I) Четырехвалентное триспецифичное Fv-антитело, показанное на фиг. 3, имеющее специфичности κCD30, CD19 и CD16A и состоящее из первого полипептида, содержащего четыре вариабельных домена и С-концевой His-маркер; и второго полипептида, содержащего четыре вариабельных домена и С-концевой FLAG-маркер. Димерная единица диатела, расположенная в центральной части молекулы, образована парой доменов VL-VL в первом полипептиде и парой доменов VH-VH во втором полипептиде; эта ориентация обеспечивает правильную гетеродимеризацию первого и второго полипептида.

Полипептид 1:

Полипептид 2:

m) Шестивалентное тетраспецифичное Fv-антитело, показанное на фиг. 8а, имеющее специфичности κCD30, CD19, EGFRvIII и CD16A и состоящее из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего шесть вариабельных доменов и С-концевой FLAG-маркер. Каждый из первого и второго полипептида содержит единицу scFv на С-конце.

Полипептид 1:


Полипептид 2:

n) Шестивалентное тетраспецифичное Fv-антитело, показанное на фиг. 8b, имеющее специфичности κCD30, CD19, EGFRvIII и CD16A и состоящее из первого полипептида, содержащего шесть вариабельных доменов и С-концевой His-маркер; и второго полипептида, содержащего шесть вариабельных доменов и С-концевой FLAG-маркер. Каждый из первого и второго полипептида содержит единицу scFv на N-конце.
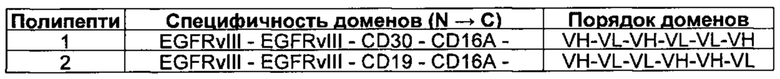
Полипептид 1:

Полипептид 2:

о) Четырехвалентное триспецифичное Fv-антитело, показанное на фиг. 1, имеющее специфичности κCD30, CD19 и CD16A и состоящее из одного полипептида, содержащего восемь вариабельных доменов и His-маркер на С-конце. Единица одноцепочечного диатела, расположенная в центральной части молекулы, образована первой парой доменов VH-VH, ассоциированной со второй парой доменов VL-VL, расположенной рядом с ними в том же полипептиде. Пара VH-VH соединена с парой VL-VL линкером (G2S)6.
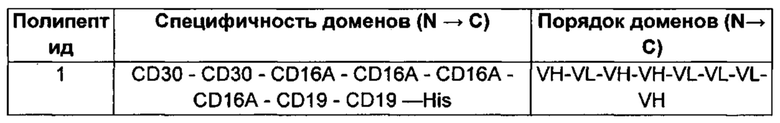
Полипептид 1
р) Шестивалентное триспецифичное Fv-антитело, показанное на фиг. 4, имеющее специфичности κCD30, CD19 и CD16A и состоящее из одного полипептида, содержащего 12 вариабельных доменов и His-маркер на С-конце. Полипептид содержит три единицы одноцепочечного диатела, соединенные друг за другом.
Полипептид 1

ПРИМЕР 4
Оценка связывания клеток и цитотоксической активности, опосредованных молекулами триспецифичных антител
Процедуры исследования:
Клетки и культура клеток
Клетки CD19+/CD30+ МЕС-1 (DSMZ, № по каталогу: АСС 497), CD19+/CD30- Raji (DSMZ, №по каталогу: АСС 319) и CD19-/CD30+ KARPAS-299 (DSMZ, №по каталогу: АСС 31) культивировали при стандартных условиях в среде RPMI 1640 (№по каталогу: 21875-034) или IMDM (№ по каталогу: 12440-053) с добавлением 10% термически инактивированной эмбриональной телячьей сыворотки (ЭТС) (№ по каталогу: 10270-106), 100 ед/мл пенициллина G, 100 мкг/мл стрептомицина (№ по каталогу: 1540-122) и 2 мМ L-глутамина (№ по каталогу: 25030-024; все реагенты приобретены в Invitrogen, Карлсруэ, Германия), в настоящем документе называемой полной средой RPMI, в соответствии с рекомендациями поставщика линий клеток.
МПК выделяли из лейкотромбоцитарного слоя крови здоровых добровольцев (German Red Cross, Мангейм, Германия) путем центрифугирования в градиенте плотности. Образцы лейкотромбоцитарного слоя разбавляли двух-трехкратным объемом PBS (Invitrogen, №по каталогу: 14190-169), наслаивали на Histopaque-1077 (Stemcell Technologies, №по каталогу: 07861) и центрифугировали при 800×g в течение 25 мин при комнатной температуре без использования торможения. МПК, расположенные на границе слоев, собирали и 3 раза промывали PBS перед использованием для обогащения NK-клеток. NK-клетки обогащали популяцией МПК с использованием набора для обогащения NK-клеток человека EasySep™ Human NK Cell Enrichment Kit Stemcell Technologies, № по каталогу: 19055) для иммуномагнитного выделения интактных NK-клеток человека и Big Easy EasySep™ Magnet в соответствии с инструкциями производителя.
Анализ связывания клеток и проточно-цитометрический анализ
Аликвоты указанных линий клеток инкубировали со 100 мкл последовательных разведений His-меченых антител в буфере для FACS (PBS, Invitrogen, № по каталогу: 14190-169), содержащем 2% термически активированную ЭТС (Invitrogen, №по каталогу: 10270-106), 0,1% азид натрия (Roth, Карлсруэ, Германия, №по каталогу: А1430.0100) в течение 45 мин на льду. Окрашивание NK-клеток выполняли в буфере для FACS с добавлением 1 мг/мл поликлональных антител человека (Gammanorm, Octapharma, Лангенфелд, Германия, № по каталогу: PZN-2451445) для блокирования связывания антител с Fc-рецепторами.
После повторной промывки буфером для FACS антитела, связанные с клетками, обнаруживали с использованием 10 мкг/мл мАт 13/45/31-2 против His (Dianova, Гамбург, Германия, № по каталогу: DIA910-1MG), а затем 15 мкг/мл конъюгированного с FITC антитела козы против антител мыши (Dianova, № по каталогу: 115-095-062). Затем клетки повторно промывали и ресуспендировали в 0,2 мл буфера для FACS, содержащего 2 мкг/мл иодида пропидия (PI) (Sigma, № по каталогу: Р4170) с целью исключения мертвых клеток. Флуоресценцию 2-5×103 живых клеток измеряли с использованием проточного цитометра Beckman-Coulter FC500 MPL и программного обеспечения МХР (Beckman-Coulter, Крефельд, Германия) или проточного цитометра Millipore Guava EasyCyte (Merck Millipore, Швальбах, Германия). Среднюю интенсивность флуоресценции образцов клеток рассчитывали с использованием программного обеспечения СХР (Beckman-Coulter) или Incyte (Merck Millipore, Швальбах, Германия). После вычитания значений интенсивности флуоресценции клеток, окрашенных только вторичными и третичными реагентами, значения использовали для анализа с использованием GraphPad Prism (GraphPad Prism версии 6.00 for Windows, GraphPad Software, Ла-Хойя, Калифорния, США). Для расчета KD использовали уравнение для одного сайта связывания (гиперболическое).
Анализ цитотоксичности
Для анализа цитотоксичности с высвобождением кальцеина клетки-мишени метили 10 мкМ кальцеина AM (Invitrogen, № по каталогу: С3100МР) в течение 30 мин в среде RPMI 1640 при 37°С, промывали и высевали 1×104 клеток в отдельные лунки 96-луночного микропланшета вместе с эффекторными клетками в общем объеме 200 мкл при соотношении эффекторных клеток и клеток-мишеней (Е : Т) 5:1 в присутствии возрастающих концентраций антитела. После инкубации в течение указанных периодов времени при 37°С в увлажненной атмосфере 5% СО2 измеряли флуоресценцию (F) кальцеина, высвобожденного в надосадочную жидкость, с помощью планшет-ридера при 520 нм (Victor 3, Perkin Elmer, Турку, Финляндия, № по каталогу: 1420-012). Удельный лизис клеток рассчитывали по формуле: [F(образец)-F(спонтанная)]/[F(максимальная)-F(спонтанная)]×100%. F(спонтанная) представляет собой флуоресценцию, высвобожденную из клеток-мишеней в отсутствие эффекторных клеток и антител, а F (максимальная) представляет собой полный лизис клеток, вызванный добавлением 1% тритона X 100 (Roth, Карлсруэ, Германия, № по каталогу: 3051.2). Кривые регрессии аппроксимировали для расчета ЕС50 (GraphPad Prism версии 6.00 for Windows, GraphPad Software, Ла-Хойя, Калифорния, США).
Активность молекул триспецифичных антител
Связывание молекул триспецифичных антител aTriFlex_101, aTriFlex_102, aTriFlex_103 и aTriFlex_104, описанных в примере 3, представляющих молекулу триспецифичного антитела, показанную на фигуре 2, содержащую вариабельные домены того же антитела, расположенные в другой ориентации (см. пример 3а), 3b), 3с) и 3d)), продемонстрировали на однократно положительных клетках CD19+/CD30- Raji и CD19-/CD30+ KARPAS-299, а также дважды положительных клетках CD19+/CD30+ МЕС-1 с помощью проточной цитометрии. Константы диссоциации для CD19 находились в диапазоне от 6 до 64 нМ, а для CD30 - от 16 до 45 нМ (фигура 12). На дважды положительных клетках МЕС-1 наблюдали константы диссоциации в диапазоне от 9 до 47 нМ, что в целом подтверждало данные, полученные для однократно положительных клеток Raji и KARPAS-299, соответственно.
При анализах цитотоксичности наблюдали значения EC50 в диапазоне 89-580 пМ на однократно положительных клетках CD30+/CD19- Raji и 155-952 пМ на клетках CD19-/CD30+ KARPAS-299. На дважды положительных клетках CD19+/CD30+ МЕС-1 наблюдали значения EC50 в диапазоне 67-235 мкМ, что означало 11-кратное увеличение активности (фигура 13).
В целом молекулы триспецифичных антител демонстрировали аналогичную активность по отношению к линиям клеток, протестированным в данном исследовании; однако при использовании других пар антигенов плотность и геометрия антигена может оказывать более сильное влияние на активность конструктов, что приведет к более выраженному увеличению кажущегося сродства к дважды положительным клеткам по сравнению с однократно положительными клетками.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Affimed GmbH
<120> МУЛЬТИВАЛЕНТНЫЕ FV-АНТИТЕЛА
<130> A 3305PCT
<150> EP 15189665.1
<151> 13.10.2015
<160> 40
<170> Версия PatentIn 3.5
<210> 1
<211> 780
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 1
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
260 265 270
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
275 280 285
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
290 295 300
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
305 310 315 320
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
325 330 335
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
340 345 350
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
355 360 365
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
385 390 395 400
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
405 410 415
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
420 425 430
Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln
435 440 445
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
450 455 460
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
465 470 475 480
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
485 490 495
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser
500 505 510
Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala
515 520 525
Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser
530 535 540
Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu
545 550 555 560
Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe
565 570 575
Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val
580 585 590
Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp
595 600 605
Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
610 615 620
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val
645 650 655
Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr
660 665 670
Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys
675 680 685
Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met
690 695 700
Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser
705 710 715 720
Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
725 730 735
Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr
740 745 750
Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
755 760 765
Ser Ala Ala Ala Gly Ser His His His His His His
770 775 780
<210> 2
<211> 234
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 2
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser
100 105 110
Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala
115 120 125
Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser
130 135 140
Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu
145 150 155 160
Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe
165 170 175
Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr
180 185 190
Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr
195 200 205
Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ala Ala
210 215 220
Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
225 230
<210> 3
<211> 780
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 3
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp His
85 90 95
Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser
100 105 110
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
130 135 140
Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr
145 150 155 160
Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu
165 170 175
Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro
180 185 190
Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr
195 200 205
Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr
210 215 220
Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala
225 230 235 240
Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
245 250 255
Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala
260 265 270
Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser
275 280 285
Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro
290 295 300
Gly Gln Gly Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser
305 310 315 320
Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp
325 330 335
Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu
340 345 350
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp
355 360 365
Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly
370 375 380
Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly
385 390 395 400
Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala
405 410 415
Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala
420 425 430
Pro Gly Gln Gly Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly
435 440 445
Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg
450 455 460
Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser
465 470 475 480
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr
485 490 495
Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
500 505 510
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
515 520 525
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
530 535 540
Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His Trp Val Arg Gln
545 550 555 560
Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Ser
565 570 575
Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys Thr Thr Leu Thr
580 585 590
Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu Asn Ser Leu Thr
595 600 605
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg Ala Asp Tyr Gly
610 615 620
Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val
625 630 635 640
Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
645 650 655
Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe
660 665 670
Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser
675 680 685
Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln
690 695 700
Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val
705 710 715 720
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
725 730 735
Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln
740 745 750
Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
755 760 765
Asn Ala Ala Ala Gly Ser His His His His His His
770 775 780
<210> 4
<211> 234
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 4
Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu
85 90 95
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser
100 105 110
Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala
115 120 125
Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser
130 135 140
Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu
145 150 155 160
Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe
165 170 175
Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr
180 185 190
Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr
195 200 205
Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ala Ala
210 215 220
Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
225 230
<210> 5
<211> 752
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 5
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp His
85 90 95
Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser
100 105 110
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
130 135 140
Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr
145 150 155 160
Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu
165 170 175
Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro
180 185 190
Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr
195 200 205
Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr
210 215 220
Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala
225 230 235 240
Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
245 250 255
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser
260 265 270
Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His
275 280 285
Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln
290 295 300
Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile
305 310 315 320
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr
325 330 335
Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val
340 345 350
Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val
355 360 365
Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
370 375 380
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys
385 390 395 400
Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg
405 410 415
Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro
420 425 430
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
435 440 445
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
450 455 460
Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys
465 470 475 480
Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln
485 490 495
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
500 505 510
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His
515 520 525
Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile
530 535 540
Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys
545 550 555 560
Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu
565 570 575
Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg
580 585 590
Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln
595 600 605
Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
610 615 620
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln
625 630 635 640
Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr
645 650 655
Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln
660 665 670
Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg
675 680 685
Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp
690 695 700
Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr
705 710 715 720
Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr
725 730 735
Lys Leu Glu Ile Asn Ala Ala Ala Gly Ser His His His His His His
740 745 750
<210> 6
<211> 262
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 6
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
130 135 140
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
145 150 155 160
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
165 170 175
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
180 185 190
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
195 200 205
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
210 215 220
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp Tyr
245 250 255
Lys Asp Asp Asp Asp Lys
260
<210> 7
<211> 754
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 7
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly
260 265 270
Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn
275 280 285
Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile
290 295 300
Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
305 310 315 320
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala
325 330 335
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val
340 345 350
Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly
355 360 365
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
370 375 380
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
385 390 395 400
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
405 410 415
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
420 425 430
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
435 440 445
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
450 455 460
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
465 470 475 480
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser
485 490 495
Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn
500 505 510
Asn Ile Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln
515 520 525
Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile
530 535 540
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr
545 550 555 560
Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val
565 570 575
Gly Thr Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys
580 585 590
Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
595 600 605
Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser
610 615 620
Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys
625 630 635 640
Gly Ser Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln
645 650 655
Met Pro Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp
660 665 670
Ser Asp Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser
675 680 685
Ala Asp Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys
690 695 700
Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr
705 710 715 720
Val Gly Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
725 730 735
Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp Tyr Lys Asp Asp Asp
740 745 750
Asp Lys
<210> 8
<211> 260
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 8
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
130 135 140
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
145 150 155 160
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
165 170 175
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
180 185 190
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
195 200 205
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
210 215 220
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser His His
245 250 255
His His His His
260
<210> 9
<211> 494
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 9
Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
20 25 30
Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly
100 105 110
Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
115 120 125
Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr
130 135 140
Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly
145 150 155 160
Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr
165 170 175
Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr
180 185 190
Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
195 200 205
Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp
210 215 220
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
225 230 235 240
Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val
245 250 255
Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr
260 265 270
Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln
275 280 285
Gly Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser
290 295 300
Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser
305 310 315 320
Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
325 330 335
Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala
340 345 350
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser
355 360 365
Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val
370 375 380
Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn
385 390 395 400
Ile Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
405 410 415
Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro
420 425 430
Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile
435 440 445
Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly
450 455 460
Thr Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu
465 470 475 480
Thr Val Leu Ala Ala Ala Gly Ser His His His His His His
485 490
<210> 10
<211> 499
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 10
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Asn
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala Met Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser
130 135 140
Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile
145 150 155 160
Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro
165 170 175
Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu
180 185 190
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
195 200 205
Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
210 215 220
Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
225 230 235 240
Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser
245 250 255
Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly
260 265 270
His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly
275 280 285
Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly
290 295 300
Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu
305 310 315 320
Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln
325 330 335
Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr
340 345 350
Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln
355 360 365
Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser
370 375 380
Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His Trp Val
385 390 395 400
Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile Asn Pro
405 410 415
Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys Thr Thr
420 425 430
Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu Asn Ser
435 440 445
Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg Ala Asp
450 455 460
Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln Gly Thr
465 470 475 480
Thr Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp Tyr Lys Asp Asp
485 490 495
Asp Asp Lys
<210> 11
<211> 1019
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 11
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
260 265 270
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
275 280 285
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
290 295 300
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
305 310 315 320
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
325 330 335
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
340 345 350
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
355 360 365
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
385 390 395 400
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
405 410 415
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
420 425 430
Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln
435 440 445
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
450 455 460
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
465 470 475 480
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
485 490 495
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser
500 505 510
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu
515 520 525
Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile
530 535 540
Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln
545 550 555 560
Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys
565 570 575
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn
580 585 590
Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp
595 600 605
Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly
610 615 620
Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser
625 630 635 640
Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr
645 650 655
Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His
660 665 670
Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln
675 680 685
Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn
690 695 700
Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp
705 710 715 720
Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe
725 730 735
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly
740 745 750
Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro
755 760 765
Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys
770 775 780
Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val
785 790 795 800
Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser
805 810 815
Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu
820 825 830
Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser
835 840 845
Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
850 855 860
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
865 870 875 880
Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
885 890 895
Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser
900 905 910
Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly
915 920 925
Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr
930 935 940
Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile
945 950 955 960
Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala
965 970 975
Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr
980 985 990
Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
995 1000 1005
Ala Ala Ala Gly Ser His His His His His His
1010 1015
<210> 12
<211> 749
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 12
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly
260 265 270
Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn
275 280 285
Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile
290 295 300
Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
305 310 315 320
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala
325 330 335
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val
340 345 350
Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly
355 360 365
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
370 375 380
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
385 390 395 400
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
405 410 415
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
420 425 430
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
435 440 445
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
450 455 460
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
465 470 475 480
Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala
485 490 495
Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser
500 505 510
Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro
515 520 525
Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp
530 535 540
Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp
545 550 555 560
Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser
565 570 575
Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly
580 585 590
Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
595 600 605
Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
610 615 620
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
625 630 635 640
Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile
645 650 655
Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
660 665 670
Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu
675 680 685
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
690 695 700
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr
705 710 715 720
Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr
725 730 735
Val Leu Ala Ala Ala Gly Ser His His His His His His
740 745
<210> 13
<211> 262
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 13
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
130 135 140
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
145 150 155 160
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
165 170 175
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
180 185 190
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
195 200 205
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
210 215 220
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp Tyr
245 250 255
Lys Asp Asp Asp Asp Lys
260
<210> 14
<211> 749
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 14
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly
260 265 270
Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn
275 280 285
Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile
290 295 300
Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
305 310 315 320
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala
325 330 335
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val
340 345 350
Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly
355 360 365
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
370 375 380
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
385 390 395 400
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
405 410 415
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
420 425 430
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
435 440 445
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
450 455 460
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
465 470 475 480
Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala
485 490 495
Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser
500 505 510
Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro
515 520 525
Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp
530 535 540
Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp
545 550 555 560
Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser
565 570 575
Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly
580 585 590
Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
595 600 605
Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
610 615 620
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
625 630 635 640
Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile
645 650 655
Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
660 665 670
Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu
675 680 685
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
690 695 700
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr
705 710 715 720
Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr
725 730 735
Val Leu Ala Ala Ala Gly Ser His His His His His His
740 745
<210> 15
<211> 522
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
130 135 140
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
145 150 155 160
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
165 170 175
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
180 185 190
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
195 200 205
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
210 215 220
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
245 250 255
Gly Ser Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro
260 265 270
Gly Gln Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln
275 280 285
Tyr Ala Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val
290 295 300
Ile Tyr Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser
305 310 315 320
Gly Ser Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln
325 330 335
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly
340 345 350
Thr Pro Leu Ile Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gly
355 360 365
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
370 375 380
Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
385 390 395 400
Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser
405 410 415
Phe Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly
420 425 430
Leu Glu Trp Met Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr
435 440 445
Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile
450 455 460
Ser Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala
465 470 475 480
Met Tyr Tyr Cys Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala Phe
485 490 495
Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ala Ala
500 505 510
Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
515 520
<210> 16
<211> 1264
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 16
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser
130 135 140
Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn
145 150 155 160
Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro
165 170 175
Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp
180 185 190
Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
195 200 205
Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His
210 215 220
Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly
225 230 235 240
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
260 265 270
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr
275 280 285
Phe Thr Thr Tyr Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp
290 295 300
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr
305 310 315 320
Asn Gln Asn Phe Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser
325 330 335
Asn Thr Ala Tyr Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala
340 345 350
Val Tyr Tyr Cys Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr
355 360 365
Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
370 375 380
Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro
385 390 395 400
Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys
405 410 415
Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro
420 425 430
Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser
435 440 445
Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr
450 455 460
Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys
465 470 475 480
Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu
485 490 495
Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu
500 505 510
Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile
515 520 525
Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln
530 535 540
Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys
545 550 555 560
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn
565 570 575
Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp
580 585 590
Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly
595 600 605
Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser
610 615 620
Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr
625 630 635 640
Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His
645 650 655
Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln
660 665 670
Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn
675 680 685
Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp
690 695 700
Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe
705 710 715 720
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly
725 730 735
Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
740 745 750
Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr
755 760 765
Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu
770 775 780
Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro
785 790 795 800
Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr
805 810 815
Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr
820 825 830
Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala
835 840 845
Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
850 855 860
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser
865 870 875 880
Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn
885 890 895
Asn Ile Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln
900 905 910
Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile
915 920 925
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr
930 935 940
Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val
945 950 955 960
Gly Thr Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys
965 970 975
Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
980 985 990
Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser
995 1000 1005
Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys
1010 1015 1020
Lys Gly Ser Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val
1025 1030 1035
Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp
1040 1045 1050
Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln
1055 1060 1065
Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr Ala Tyr Leu Gln
1070 1075 1080
Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala
1085 1090 1095
Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala Met Asp
1100 1105 1110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser
1115 1120 1125
Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser
1130 1135 1140
Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly
1145 1150 1155
Asn Asn Ile Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro
1160 1165 1170
Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro
1175 1180 1185
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr
1190 1195 1200
Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp
1205 1210 1215
Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp His Leu Gln Val
1220 1225 1230
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ala Ala Gly Ser
1235 1240 1245
His His His His His His Ala Arg Thr Ile Phe Ile Cys Ile Ala
1250 1255 1260
Leu
<210> 17
<211> 262
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
130 135 140
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
145 150 155 160
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
165 170 175
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
180 185 190
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
195 200 205
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
210 215 220
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp Tyr
245 250 255
Lys Asp Asp Asp Asp Lys
260
<210> 18
<211> 749
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 18
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly
260 265 270
Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn
275 280 285
Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile
290 295 300
Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
305 310 315 320
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala
325 330 335
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val
340 345 350
Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly
355 360 365
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
370 375 380
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
385 390 395 400
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
405 410 415
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
420 425 430
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
435 440 445
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
450 455 460
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
465 470 475 480
Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala
485 490 495
Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser
500 505 510
Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro
515 520 525
Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp
530 535 540
Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp
545 550 555 560
Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser
565 570 575
Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly
580 585 590
Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
595 600 605
Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
610 615 620
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
625 630 635 640
Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile
645 650 655
Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
660 665 670
Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu
675 680 685
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
690 695 700
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr
705 710 715 720
Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr
725 730 735
Val Leu Ala Ala Ala Gly Ser His His His His His His
740 745
<210> 19
<211> 778
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 19
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
260 265 270
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
275 280 285
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
290 295 300
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
305 310 315 320
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
325 330 335
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
340 345 350
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
355 360 365
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
385 390 395 400
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
405 410 415
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
420 425 430
Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln
435 440 445
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
450 455 460
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
465 470 475 480
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
485 490 495
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser
500 505 510
Gly Gly Ser Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser
515 520 525
Pro Gly Gln Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys
530 535 540
Gln Tyr Ala Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu
545 550 555 560
Val Ile Tyr Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe
565 570 575
Ser Gly Ser Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val
580 585 590
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser
595 600 605
Gly Thr Pro Leu Ile Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu
610 615 620
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val
645 650 655
Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr
660 665 670
Ser Phe Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys
675 680 685
Gly Leu Glu Trp Met Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg
690 695 700
Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser
705 710 715 720
Ile Ser Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
725 730 735
Ala Met Tyr Tyr Cys Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala
740 745 750
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ala
755 760 765
Ala Gly Ser Asp Tyr Lys Asp Asp Asp Asp
770 775
<210> 20
<211> 749
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 20
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly
260 265 270
Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn
275 280 285
Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile
290 295 300
Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
305 310 315 320
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala
325 330 335
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val
340 345 350
Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly
355 360 365
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
370 375 380
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
385 390 395 400
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
405 410 415
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
420 425 430
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
435 440 445
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
450 455 460
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
465 470 475 480
Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala
485 490 495
Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser
500 505 510
Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro
515 520 525
Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp
530 535 540
Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp
545 550 555 560
Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser
565 570 575
Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly
580 585 590
Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
595 600 605
Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
610 615 620
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
625 630 635 640
Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile
645 650 655
Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
660 665 670
Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu
675 680 685
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
690 695 700
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr
705 710 715 720
Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr
725 730 735
Val Leu Ala Ala Ala Gly Ser His His His His His His
740 745
<210> 21
<211> 519
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 21
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
260 265 270
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
275 280 285
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
290 295 300
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
305 310 315 320
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
325 330 335
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
340 345 350
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
355 360 365
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
385 390 395 400
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
405 410 415
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
420 425 430
Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln
435 440 445
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
450 455 460
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
465 470 475 480
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
485 490 495
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp
500 505 510
Tyr Lys Asp Asp Asp Asp Lys
515
<210> 22
<211> 497
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 22
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
130 135 140
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
145 150 155 160
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
165 170 175
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
180 185 190
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
195 200 205
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
210 215 220
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
225 230 235 240
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser
245 250 255
Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His
260 265 270
Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln
275 280 285
Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile
290 295 300
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr
305 310 315 320
Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val
325 330 335
Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val
340 345 350
Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln
355 360 365
Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys
370 375 380
Lys Gly Ser Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg
385 390 395 400
Gln Met Pro Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly
405 410 415
Asp Ser Asp Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile
420 425 430
Ser Ala Asp Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu
435 440 445
Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr
450 455 460
Thr Val Gly Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
465 470 475 480
Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser His His His His His
485 490 495
His
<210> 23
<211> 493
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 23
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp His
85 90 95
Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser
100 105 110
Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala
165 170 175
Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser
180 185 190
Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
210 215 220
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly
225 230 235 240
Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
245 250 255
Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr
260 265 270
Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly
275 280 285
Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr
290 295 300
Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr
305 310 315 320
Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
325 330 335
Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp
340 345 350
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
355 360 365
Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met
370 375 380
Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln
385 390 395 400
Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser
405 410 415
Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro
420 425 430
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
435 440 445
Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr
450 455 460
His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn
465 470 475 480
Ala Ala Ala Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
485 490
<210> 24
<211> 757
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 24
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
130 135 140
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
145 150 155 160
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
165 170 175
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
180 185 190
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
195 200 205
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
210 215 220
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
225 230 235 240
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser
245 250 255
Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His
260 265 270
Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln
275 280 285
Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile
290 295 300
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr
305 310 315 320
Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val
325 330 335
Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val
340 345 350
Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln
355 360 365
Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys
370 375 380
Lys Gly Ser Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg
385 390 395 400
Gln Met Pro Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly
405 410 415
Asp Ser Asp Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile
420 425 430
Ser Ala Asp Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser Leu
435 440 445
Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr Thr
450 455 460
Thr Val Gly Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
465 470 475 480
Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser
485 490 495
Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln Thr
500 505 510
Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala Tyr
515 520 525
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Lys
530 535 540
Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser
545 550 555 560
Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu Asp
565 570 575
Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Pro Leu
580 585 590
Ile Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly
595 600 605
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
610 615 620
Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
625 630 635 640
Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser
645 650 655
Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp
660 665 670
Met Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser
675 680 685
Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala
690 695 700
Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr
705 710 715 720
Cys Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala Phe Asp Ile Trp
725 730 735
Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ala Ala Gly Ser His
740 745 750
His His His His His
755
<210> 25
<211> 753
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 25
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp His
85 90 95
Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser
100 105 110
Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
115 120 125
Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
130 135 140
Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
145 150 155 160
Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala
165 170 175
Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser
180 185 190
Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
195 200 205
Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr
210 215 220
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly
225 230 235 240
Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
245 250 255
Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr
260 265 270
Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly
275 280 285
Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr
290 295 300
Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr
305 310 315 320
Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
325 330 335
Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp
340 345 350
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
355 360 365
Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met
370 375 380
Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln
385 390 395 400
Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser
405 410 415
Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro
420 425 430
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
435 440 445
Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr
450 455 460
His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn
465 470 475 480
Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Glu Leu Thr Gln Pro
485 490 495
Pro Ser Val Ser Val Ser Pro Gly Gln Thr Ala Arg Ile Thr Cys Ser
500 505 510
Gly Asp Ala Leu Pro Lys Gln Tyr Ala Tyr Trp Tyr Gln Gln Lys Pro
515 520 525
Gly Gln Ala Pro Val Leu Val Ile Tyr Lys Asp Ser Glu Arg Pro Ser
530 535 540
Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Thr Thr Val Thr
545 550 555 560
Leu Thr Ile Ser Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys
565 570 575
Gln Ser Ala Asp Ser Ser Gly Thr Pro Leu Ile Val Phe Gly Thr Gly
580 585 590
Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
595 600 605
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val
610 615 620
Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser
625 630 635 640
Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr Trp Ile Gly Trp Val
645 650 655
Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met Gly Ile Ile Tyr Pro
660 665 670
Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr
675 680 685
Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr Leu Gln Trp Ser Ser
690 695 700
Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Arg Leu Gly Ser
705 710 715 720
Ser Trp Thr Asn Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val
725 730 735
Thr Val Ser Ser Ala Ala Ala Gly Ser Asp Tyr Lys Asp Asp Asp Asp
740 745 750
Lys
<210> 26
<211> 754
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 26
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Glu Leu Thr
130 135 140
Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln Thr Ala Arg Ile Thr
145 150 155 160
Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala Tyr Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Lys Asp Ser Glu Arg
180 185 190
Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Thr Thr
195 200 205
Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
210 215 220
Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Pro Leu Ile Val Phe Gly
225 230 235 240
Thr Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly
260 265 270
Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr
275 280 285
Tyr Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp
290 295 300
Ile Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn
305 310 315 320
Phe Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala
325 330 335
Tyr Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr
340 345 350
Cys Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala
355 360 365
Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser
385 390 395 400
Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile
405 410 415
Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro
420 425 430
Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu
435 440 445
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
450 455 460
Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
465 470 475 480
Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
485 490 495
Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser
500 505 510
Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly
515 520 525
His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly
530 535 540
Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly
545 550 555 560
Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu
565 570 575
Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln
580 585 590
Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr
595 600 605
Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val
610 615 620
Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser
625 630 635 640
Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val
645 650 655
Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro
660 665 670
Gly Asp Ser Asp Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr
675 680 685
Ile Ser Ala Asp Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser
690 695 700
Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu Thr
705 710 715 720
Thr Thr Val Gly Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly
725 730 735
Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser His His His His
740 745 750
His His
<210> 27
<211> 750
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 27
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Glu Leu Thr
130 135 140
Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln Thr Ala Arg Ile Thr
145 150 155 160
Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala Tyr Trp Tyr Gln Gln
165 170 175
Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Lys Asp Ser Glu Arg
180 185 190
Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Thr Thr
195 200 205
Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
210 215 220
Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Pro Leu Ile Val Phe Gly
225 230 235 240
Thr Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly
260 265 270
Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr
275 280 285
Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val
290 295 300
Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly
305 310 315 320
Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala
325 330 335
Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp
340 345 350
His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
355 360 365
Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
370 375 380
Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr
385 390 395 400
Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly
405 410 415
Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr
420 425 430
Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr
435 440 445
Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
450 455 460
Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp
465 470 475 480
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
485 490 495
Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val
500 505 510
Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr
515 520 525
Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln
530 535 540
Gly Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser
545 550 555 560
Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser
565 570 575
Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr
580 585 590
Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala
595 600 605
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser
610 615 620
Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe
625 630 635 640
Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser
645 650 655
Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln
660 665 670
Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val
675 680 685
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
690 695 700
Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln
705 710 715 720
Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
725 730 735
Asn Ala Ala Ala Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
740 745 750
<210> 28
<211> 1016
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 28
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
130 135 140
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
145 150 155 160
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
165 170 175
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
180 185 190
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
195 200 205
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
210 215 220
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
225 230 235 240
Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
260 265 270
Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser
275 280 285
Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
290 295 300
Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys
305 310 315 320
Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val
325 330 335
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
340 345 350
Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly
355 360 365
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
385 390 395 400
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
405 410 415
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
420 425 430
Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln
435 440 445
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
450 455 460
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
465 470 475 480
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
485 490 495
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser
500 505 510
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu
515 520 525
Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile
530 535 540
Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln
545 550 555 560
Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys
565 570 575
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn
580 585 590
Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp
595 600 605
Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly
610 615 620
Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser
625 630 635 640
Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr
645 650 655
Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys Asn Val His
660 665 670
Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln
675 680 685
Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn
690 695 700
Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp
705 710 715 720
Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe
725 730 735
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly
740 745 750
Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
755 760 765
Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr
770 775 780
Ser Asn Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu
785 790 795 800
Trp Met Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro
805 810 815
Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr
820 825 830
Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr
835 840 845
Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala
850 855 860
Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
865 870 875 880
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
885 890 895
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
900 905 910
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val
915 920 925
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
930 935 940
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
945 950 955 960
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
965 970 975
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp His
980 985 990
Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ala Ala
995 1000 1005
Gly Ser His His His His His His
1010 1015
<210> 29
<211> 1521
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 29
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser
130 135 140
Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn
145 150 155 160
Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro
165 170 175
Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp
180 185 190
Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
195 200 205
Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His
210 215 220
Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly
225 230 235 240
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
245 250 255
Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
260 265 270
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr
275 280 285
Phe Thr Thr Tyr Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp
290 295 300
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr
305 310 315 320
Asn Gln Asn Phe Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser
325 330 335
Asn Thr Ala Tyr Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala
340 345 350
Val Tyr Tyr Cys Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr
355 360 365
Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly
370 375 380
Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro
385 390 395 400
Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys
405 410 415
Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro
420 425 430
Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser
435 440 445
Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr
450 455 460
Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys
465 470 475 480
Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu
485 490 495
Glu Ile Asn Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu
500 505 510
Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Val
515 520 525
Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp
530 535 540
Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ile Ile Asn
545 550 555 560
Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val
565 570 575
Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu Ser
580 585 590
Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser
595 600 605
Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val
610 615 620
Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln
625 630 635 640
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu Lys
645 650 655
Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Tyr Met His
660 665 670
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Ile Ile
675 680 685
Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln Gly Arg
690 695 700
Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Glu Leu
705 710 715 720
Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly
725 730 735
Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp Gly Gln Gly Thr Leu
740 745 750
Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
755 760 765
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser
770 775 780
Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His
785 790 795 800
Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln
805 810 815
Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile
820 825 830
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr
835 840 845
Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val
850 855 860
Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val
865 870 875 880
Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln
885 890 895
Pro Ser Ser Val Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys
900 905 910
Gly Gly His Asn Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg
915 920 925
Pro Gly Gln Ser Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro
930 935 940
Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
945 950 955 960
Thr Leu Thr Ile Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr
965 970 975
Cys Gln Val Trp Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys
980 985 990
Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Glu Val Gln
995 1000 1005
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu Ser Leu
1010 1015 1020
Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Asn Trp
1025 1030 1035
Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
1040 1045 1050
Gly Met Ile Trp Pro Gly Asp Ser Asp Thr Met Tyr Ser Pro Ser
1055 1060 1065
Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Glu Ser Ile Asn Thr
1070 1075 1080
Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met
1085 1090 1095
Tyr Tyr Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr
1100 1105 1110
Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
1115 1120 1125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr
1130 1135 1140
Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg Ile
1145 1150 1155
Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val His Trp Tyr
1160 1165 1170
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr Asp Asp
1175 1180 1185
Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn
1190 1195 1200
Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
1205 1210 1215
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Gly Thr Asp Trp Ser Asp
1220 1225 1230
His Leu Gln Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
1235 1240 1245
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
1250 1255 1260
Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu
1265 1270 1275
Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser
1280 1285 1290
Gly Tyr Ser Phe Thr Ser Asn Trp Ile Gly Trp Val Arg Gln Met
1295 1300 1305
Pro Gly Lys Gly Leu Glu Trp Met Gly Met Ile Trp Pro Gly Asp
1310 1315 1320
Ser Asp Thr Met Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile
1325 1330 1335
Ser Ala Asp Glu Ser Ile Asn Thr Ala Tyr Leu Gln Trp Ser Ser
1340 1345 1350
Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys Ala Arg Arg Glu
1355 1360 1365
Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala Met Asp Tyr Trp Gly
1370 1375 1380
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser
1385 1390 1395
Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val
1400 1405 1410
Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile
1415 1420 1425
Gly Ser Lys Thr Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
1430 1435 1440
Pro Val Leu Val Val Tyr Asp Asp Ser Asp Arg Pro Ser Gly Ile
1445 1450 1455
Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu
1460 1465 1470
Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
1475 1480 1485
Gln Val Gly Thr Asp Trp Ser Asp His Leu Gln Val Phe Gly Gly
1490 1495 1500
Gly Thr Lys Leu Thr Val Leu Ala Ala Ala Gly Ser His His His
1505 1510 1515
His His His
1520
<210> 30
<211> 767
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 30
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
130 135 140
Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly
145 150 155 160
Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
165 170 175
Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile
180 185 190
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
210 215 220
Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly
245 250 255
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
260 265 270
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
275 280 285
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
290 295 300
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
305 310 315 320
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
325 330 335
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
340 345 350
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
355 360 365
Ser Gly Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala
370 375 380
Glu Val Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser
385 390 395 400
Gly Tyr Ser Phe Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Met Pro
405 410 415
Gly Lys Gly Leu Glu Trp Met Gly Ile Ile Tyr Pro Gly Asp Ser Asp
420 425 430
Thr Arg Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp
435 440 445
Lys Ser Ile Ser Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser
450 455 460
Asp Thr Ala Met Tyr Tyr Cys Ala Arg Leu Gly Ser Ser Trp Thr Asn
465 470 475 480
Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
485 490 495
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
500 505 510
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
515 520 525
Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His Trp Val Arg Gln
530 535 540
Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Ser
545 550 555 560
Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys Thr Thr Leu Thr
565 570 575
Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu Asn Ser Leu Thr
580 585 590
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg Ala Asp Tyr Gly
595 600 605
Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val
610 615 620
Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
625 630 635 640
Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser
645 650 655
Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys
660 665 670
Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys
675 680 685
Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr
690 695 700
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
705 710 715 720
Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe
725 730 735
Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys
740 745 750
Leu Glu Ile Asn Ala Ala Ala Gly Ser His His His His His His
755 760 765
<210> 31
<211> 251
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 31
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Pro
85 90 95
Leu Ile Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gly Gly Ser
100 105 110
Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu
115 120 125
Val Lys Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly
130 135 140
Tyr Thr Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Gln Gly Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr
165 170 175
Ser Tyr Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr
180 185 190
Ser Thr Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp
195 200 205
Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe
210 215 220
Ala Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala
225 230 235 240
Ala Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
245 250
<210> 32
<211> 769
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 32
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
130 135 140
Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly
145 150 155 160
Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
165 170 175
Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile
180 185 190
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
210 215 220
Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly
245 250 255
Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
260 265 270
Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr
275 280 285
Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly
290 295 300
Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr
305 310 315 320
Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr
325 330 335
Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
340 345 350
Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp
355 360 365
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gly Gly Ser Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser
385 390 395 400
Val Ser Pro Gly Gln Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu
405 410 415
Pro Lys Gln Tyr Ala Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
420 425 430
Val Leu Val Ile Tyr Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu
435 440 445
Arg Phe Ser Gly Ser Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser
450 455 460
Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp
465 470 475 480
Ser Ser Gly Thr Pro Leu Ile Val Phe Gly Thr Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln
500 505 510
Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser
515 520 525
Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His Trp Val
530 535 540
Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile Asn Pro
545 550 555 560
Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys Thr Thr
565 570 575
Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu Asn Ser
580 585 590
Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg Ala Asp
595 600 605
Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln Gly Thr
610 615 620
Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr
645 650 655
Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val
660 665 670
Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln
675 680 685
Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr
690 695 700
Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr
705 710 715 720
Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu
725 730 735
Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly
740 745 750
Thr Lys Leu Glu Ile Asn Ala Ala Ala Gly Ser His His His His His
755 760 765
His
<210> 33
<211> 249
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 33
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val Ala Pro
130 135 140
Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly Ser Lys
145 150 155 160
Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val Leu Val
165 170 175
Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser
180 185 190
Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln
195 200 205
Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn Tyr Ser
210 215 220
Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ala Ala Ala Gly
225 230 235 240
Ser Asp Tyr Lys Asp Asp Asp Asp Lys
245
<210> 34
<211> 781
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 34
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
130 135 140
Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly
145 150 155 160
Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
165 170 175
Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile
180 185 190
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
210 215 220
Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly
245 250 255
Ser Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
260 265 270
Lys Pro Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr
275 280 285
Phe Thr Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly
290 295 300
Leu Glu Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr
305 310 315 320
Ala Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr
325 330 335
Ser Thr Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala
340 345 350
Val Tyr Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp
355 360 365
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Ser Gly
370 375 380
Gly Ser Gly Gly Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val
385 390 395 400
Lys Lys Pro Gly Glu Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr
405 410 415
Ser Phe Thr Ser Tyr Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys
420 425 430
Gly Leu Glu Trp Met Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg
435 440 445
Tyr Ser Pro Ser Phe Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser
450 455 460
Ile Ser Thr Ala Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
465 470 475 480
Ala Met Tyr Tyr Cys Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala
485 490 495
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly
500 505 510
Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
515 520 525
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser
530 535 540
Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His Trp Val Arg Gln Arg Pro
545 550 555 560
Gly His Asp Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Ser Gly Tyr
565 570 575
Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys Thr Thr Leu Thr Ala Asp
580 585 590
Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu Asn Ser Leu Thr Ser Glu
595 600 605
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg Ala Asp Tyr Gly Asn Tyr
610 615 620
Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
625 630 635 640
Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
645 650 655
Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys
660 665 670
Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala
675 680 685
Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly
690 695 700
Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly
705 710 715 720
Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
725 730 735
Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln
740 745 750
Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu
755 760 765
Ile Asn Ala Ala Ala Gly Ser His His His His His His
770 775 780
<210> 35
<211> 237
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 35
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Lys Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Gly Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Thr Pro
85 90 95
Leu Ile Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu Gly Gly Ser
100 105 110
Gly Gly Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val
115 120 125
Ser Val Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn
130 135 140
Ile Gly Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser
145 150 155 160
Pro Val Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro
165 170 175
Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile
180 185 190
Ser Gly Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp
195 200 205
Asp Asn Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
210 215 220
Ala Ala Ala Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
225 230 235
<210> 36
<211> 755
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 36
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
130 135 140
Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly
145 150 155 160
Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
165 170 175
Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile
180 185 190
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly
195 200 205
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
210 215 220
Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly
245 250 255
Ser Gly Gly Ser Ser Tyr Val Leu Thr Gln Pro Ser Ser Val Ser Val
260 265 270
Ala Pro Gly Gln Thr Ala Thr Ile Ser Cys Gly Gly His Asn Ile Gly
275 280 285
Ser Lys Asn Val His Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Val
290 295 300
Leu Val Ile Tyr Gln Asp Asn Lys Arg Pro Ser Gly Ile Pro Glu Arg
305 310 315 320
Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly
325 330 335
Thr Gln Ala Met Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Asn
340 345 350
Tyr Ser Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
355 360 365
Ser Gly Gly Ser Gly Gly Ser Ser Tyr Glu Leu Thr Gln Pro Pro Ser
370 375 380
Val Ser Val Ser Pro Gly Gln Thr Ala Arg Ile Thr Cys Ser Gly Asp
385 390 395 400
Ala Leu Pro Lys Gln Tyr Ala Tyr Trp Tyr Gln Gln Lys Pro Gly Gln
405 410 415
Ala Pro Val Leu Val Ile Tyr Lys Asp Ser Glu Arg Pro Ser Gly Ile
420 425 430
Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Thr Thr Val Thr Leu Thr
435 440 445
Ile Ser Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser
450 455 460
Ala Asp Ser Ser Gly Thr Pro Leu Ile Val Phe Gly Thr Gly Thr Lys
465 470 475 480
Leu Thr Val Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln
485 490 495
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
500 505 510
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His
515 520 525
Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile
530 535 540
Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys
545 550 555 560
Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu
565 570 575
Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg
580 585 590
Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln
595 600 605
Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
610 615 620
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile Val
625 630 635 640
Met Thr Gln Ser Pro Lys Phe Met Ser Thr Ser Val Gly Asp Arg Val
645 650 655
Thr Val Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Asn Val Ala Trp
660 665 670
Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala
675 680 685
Ser Tyr Arg Tyr Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser
690 695 700
Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser Glu Asp Leu
705 710 715 720
Ala Glu Tyr Phe Cys Gln Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly
725 730 735
Gly Gly Thr Lys Leu Glu Ile Asn Ala Ala Ala Gly Ser His His His
740 745 750
His His His
755
<210> 37
<211> 263
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> полипептид Fv
<400> 37
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Ser Ser Trp Thr Asn Asp Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Met Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro
130 135 140
Gly Glu Ser Leu Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr
145 150 155 160
Ser Tyr Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu
165 170 175
Trp Met Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln
180 185 190
Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr
195 200 205
Val Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr
210 215 220
Tyr Cys Ala Arg Gly Ser Ala Tyr Tyr Tyr Asp Phe Ala Asp Tyr Trp
225 230 235 240
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ala Ala Gly Ser Asp
245 250 255
Tyr Lys Asp Asp Asp Asp Lys
260
<210> 38
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> FLAG-маркер
<400> 38
Asp Tyr Lys Asp Asp Asp Asp Lys
1 5
<210> 39
<211> 8
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> Маркер STREP II
<400> 39
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 40
<211> 4
<212> БЕЛОК
<213> искусственная последовательность
<220>
<223> C-маркер
<400> 40
Glu Pro Glu Ala
1
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| CD3-СВЯЗЫВАЮЩИЙ ДОМЕН | 2015 |
|
RU2742691C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| ТРИФУНКЦИОНАЛЬНАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2753882C2 |
| ВАРИАНТНЫЕ МОЛЕКУЛЫ IL-10 И СПОСОБ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2020 |
|
RU2836849C2 |
| КОМПОЗИЦИИ IL-12, НАЦЕЛЕННЫЕ НА EDB | 2019 |
|
RU2758143C1 |
| БИСПЕЦИФИЧЕСКИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК EGFR/CD16 | 2019 |
|
RU2792240C2 |
| БИСПЕЦИФИЧНЫЕ МОЛЕКУЛЫ, ИММУНОРЕАКТИВНЫЕ С ИММУННЫМИ ЭФФЕКТОРНЫМИ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ АКТИВИРУЮЩИЙ РЕЦЕПТОР | 2014 |
|
RU2721707C2 |
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
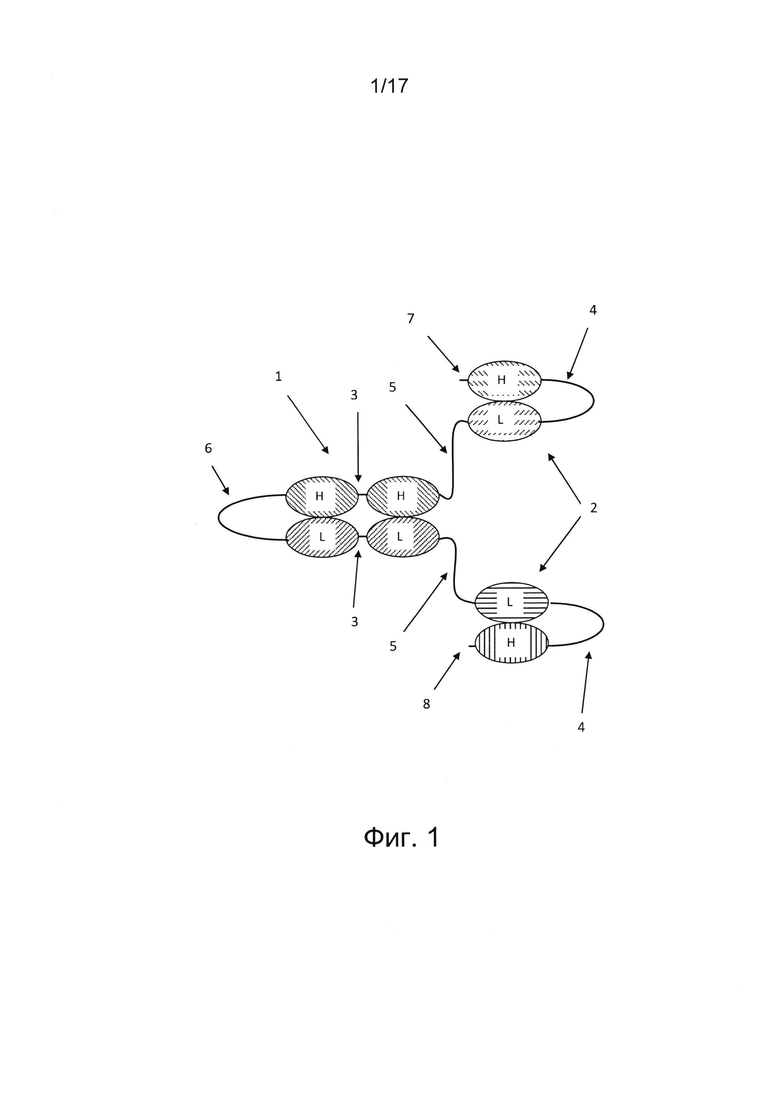
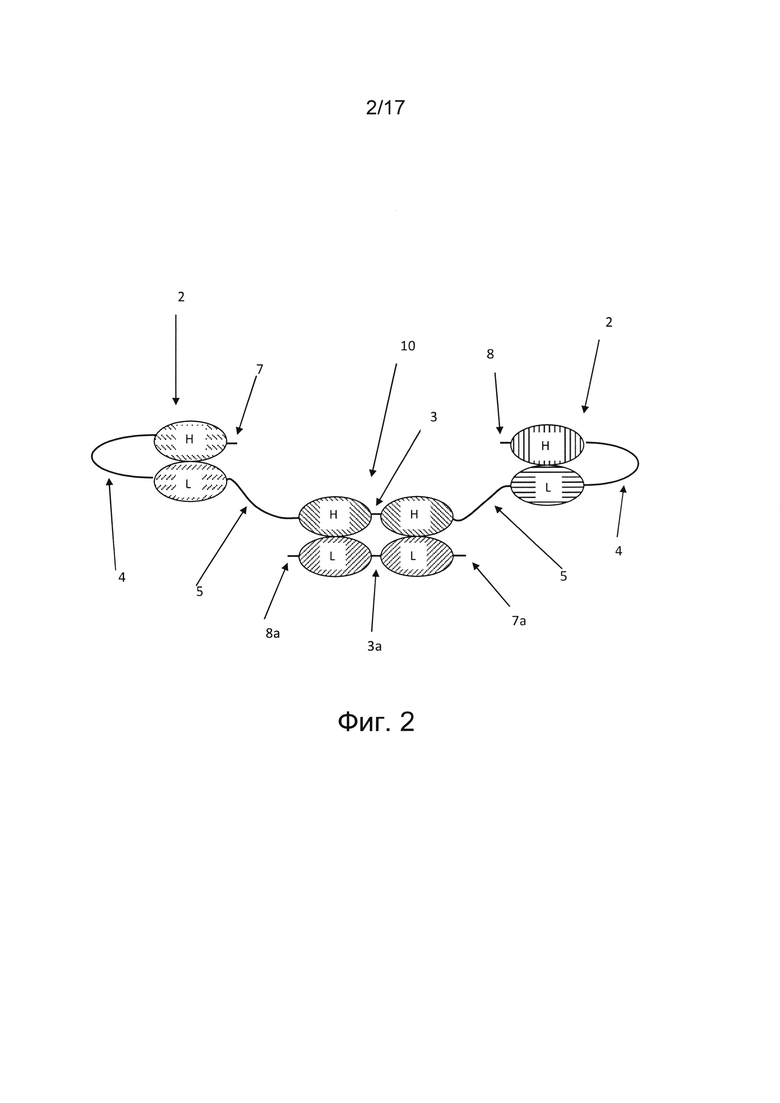
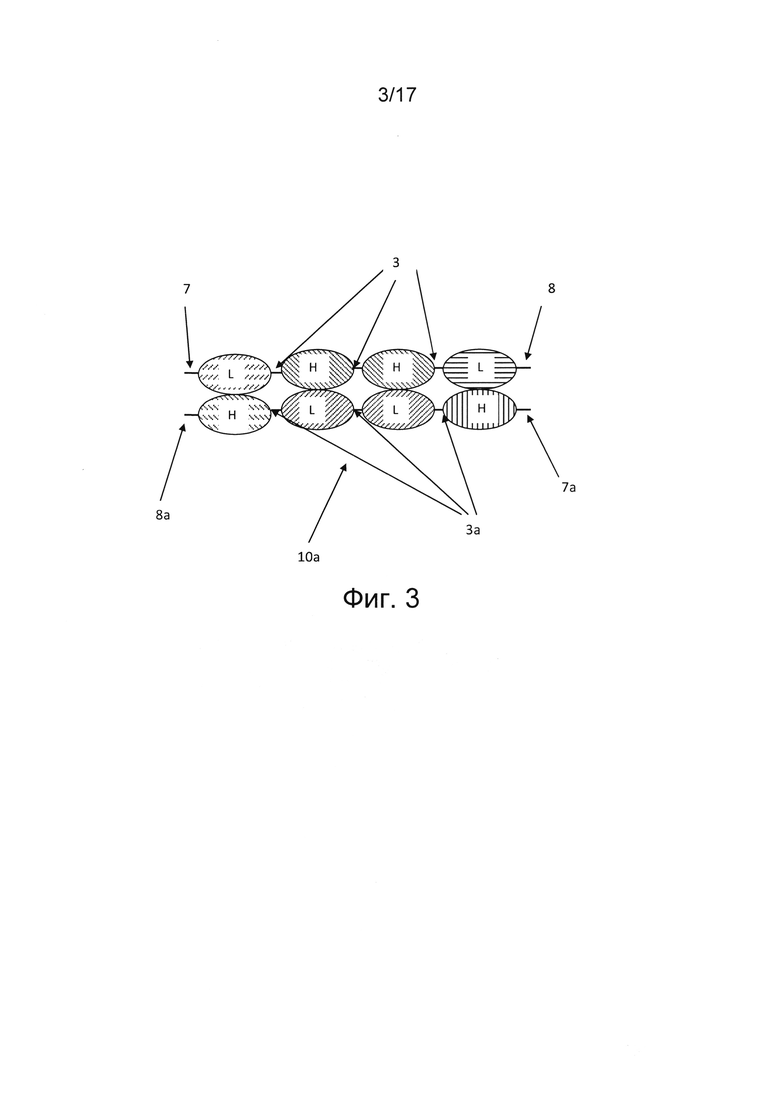
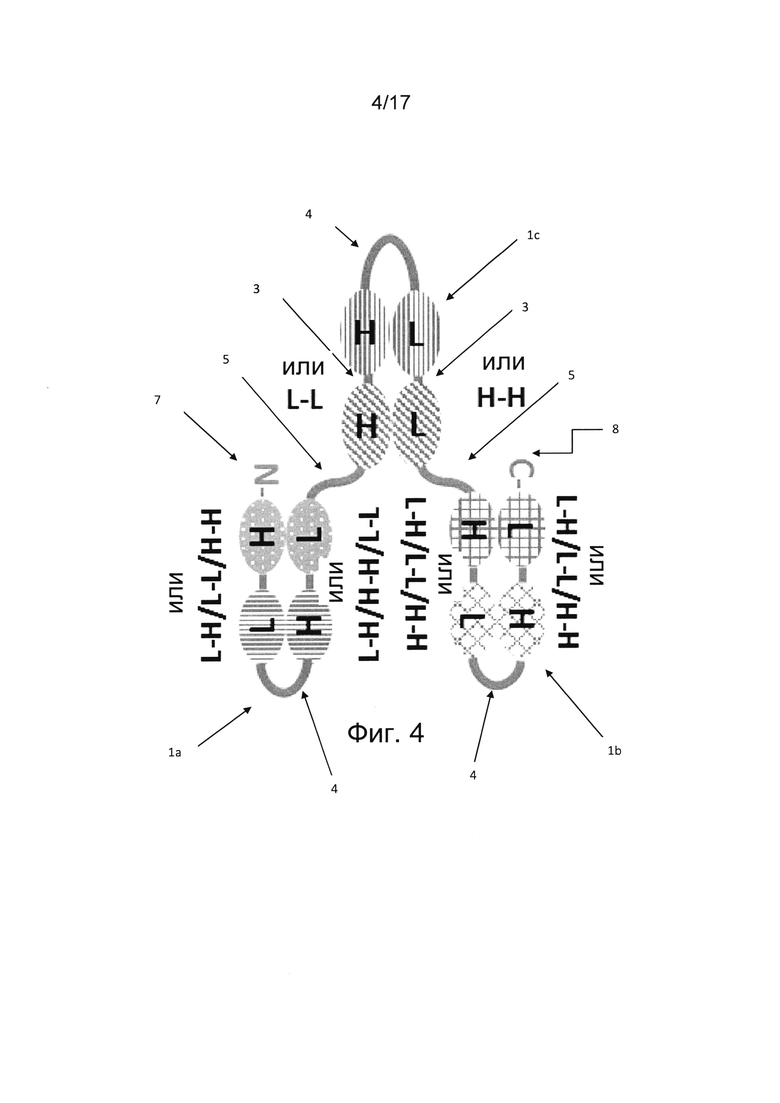
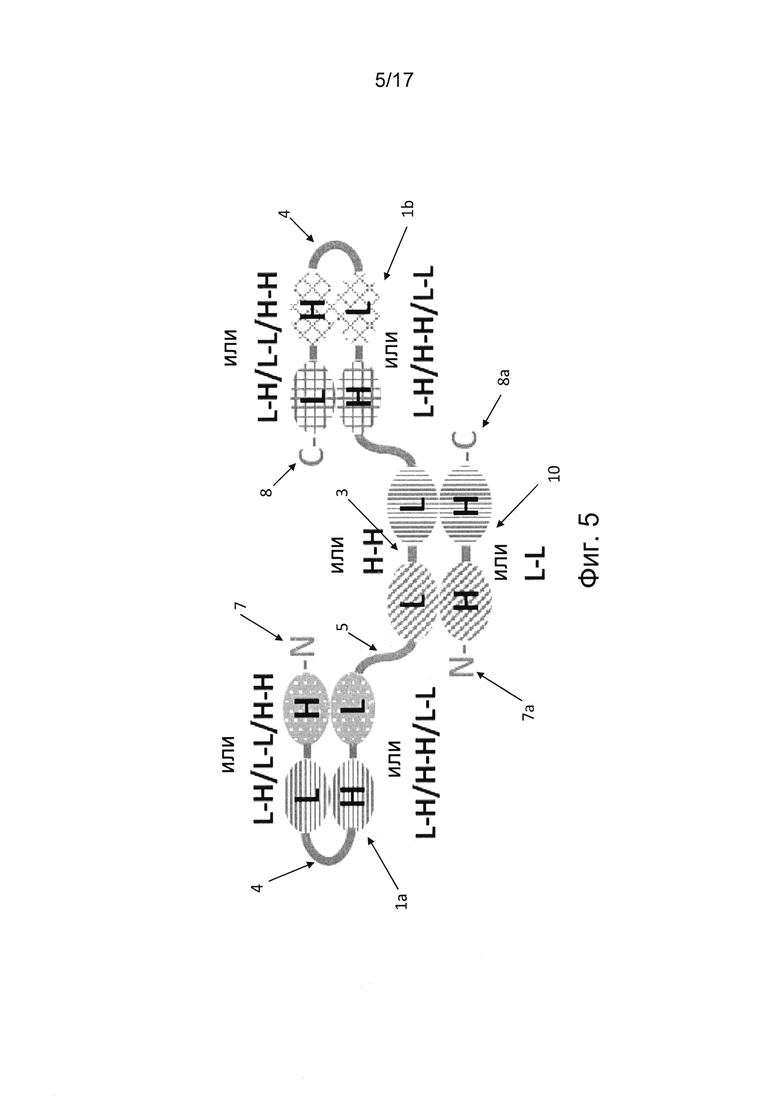
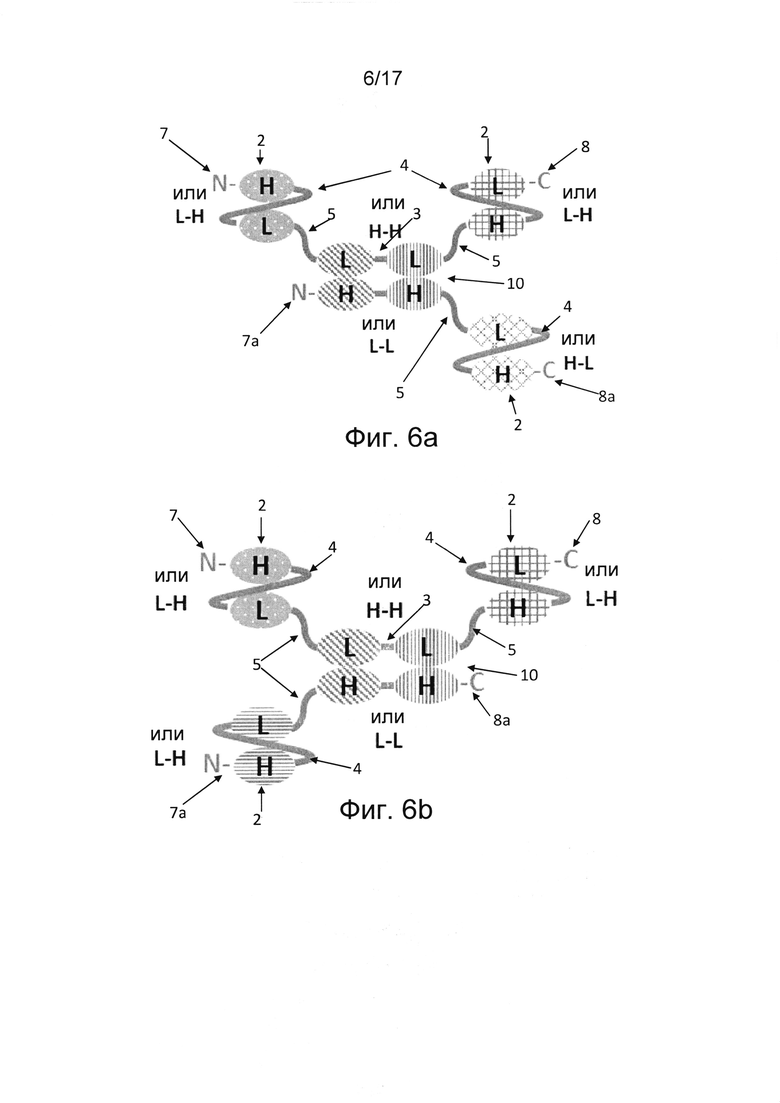
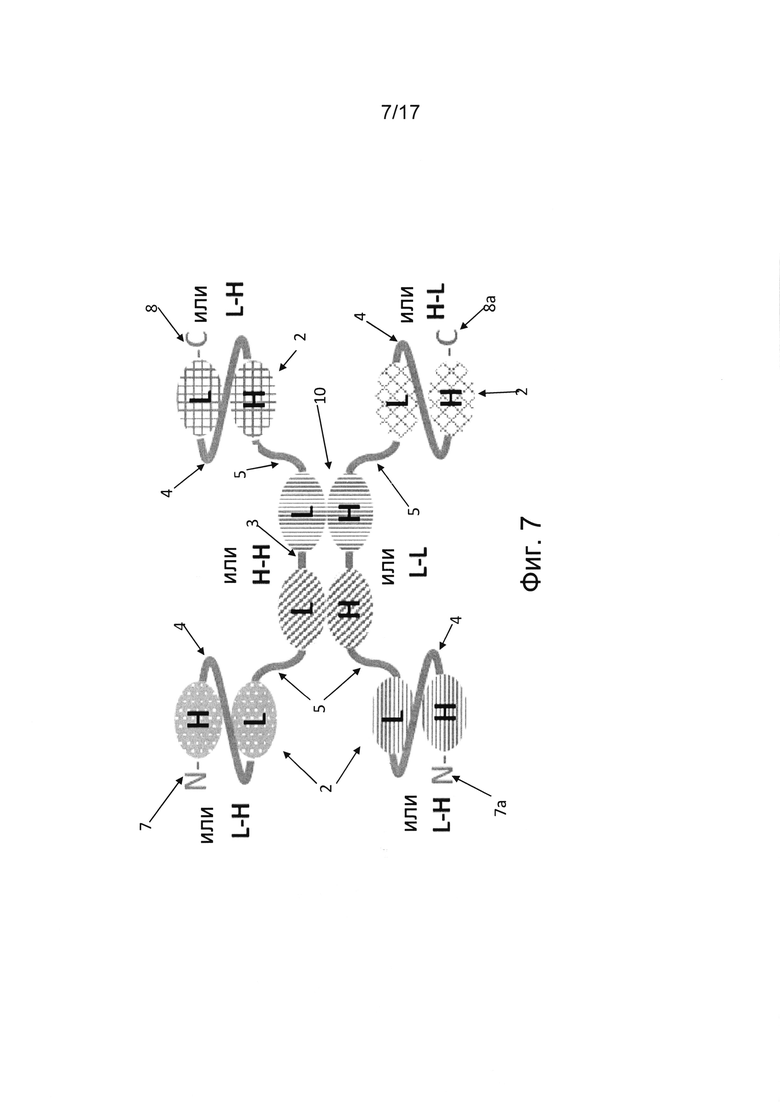
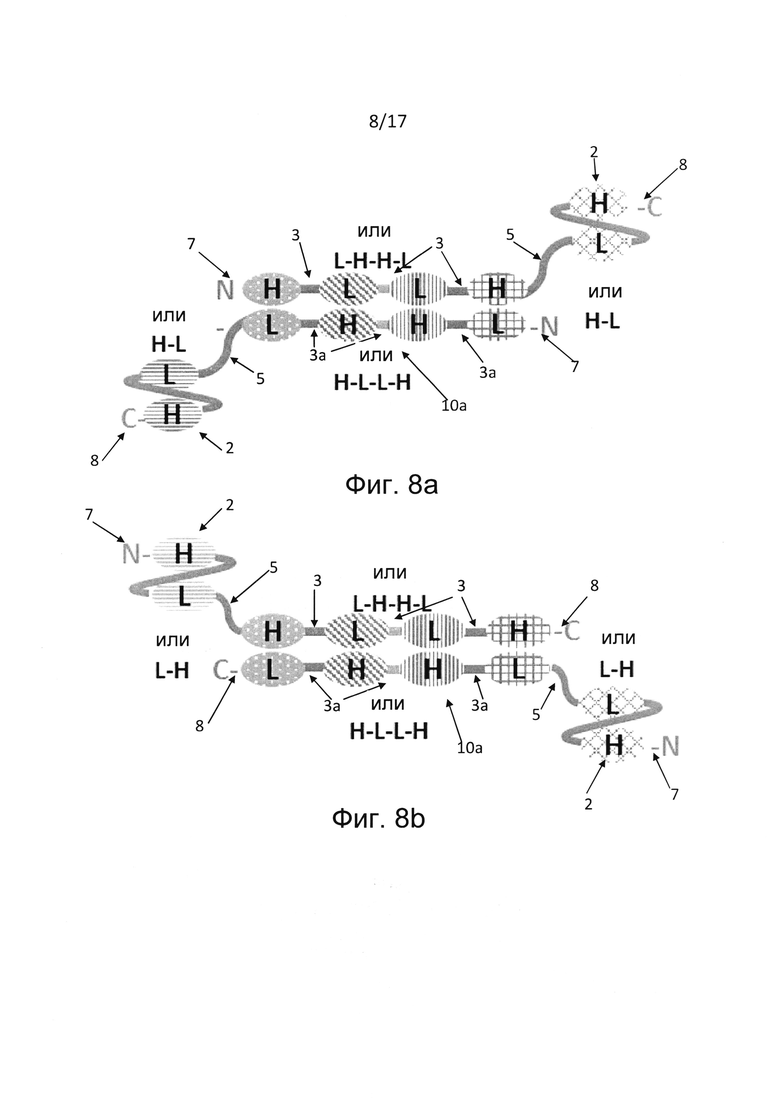
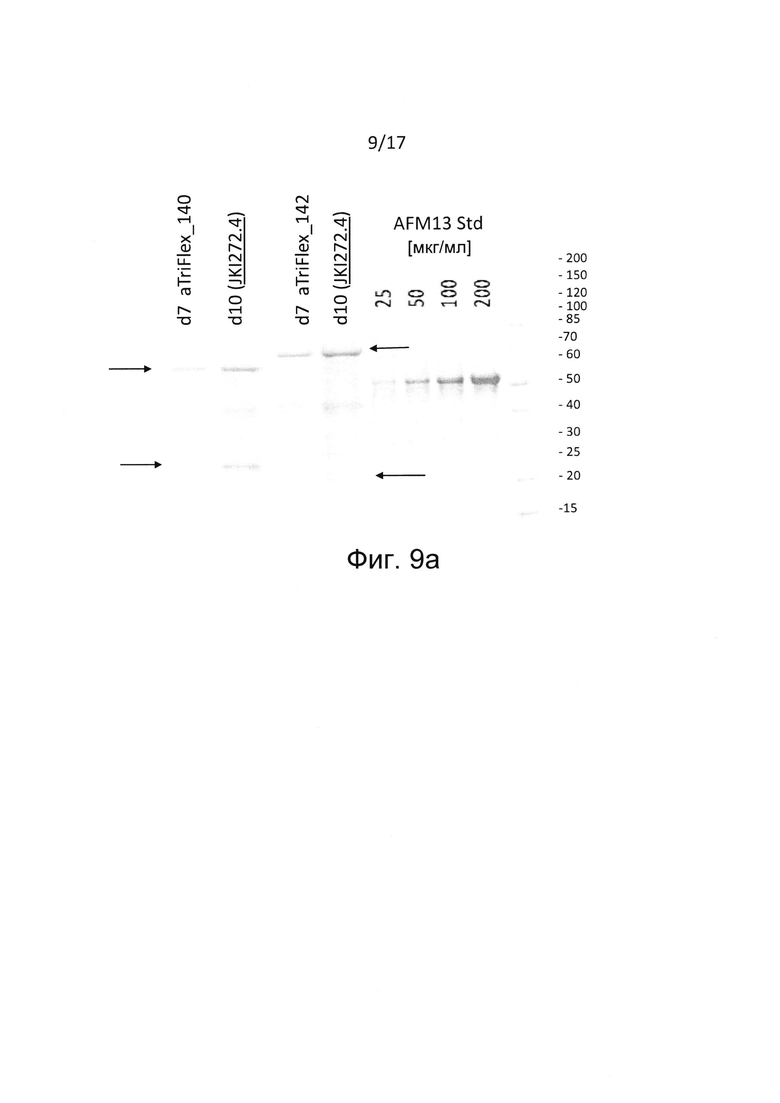

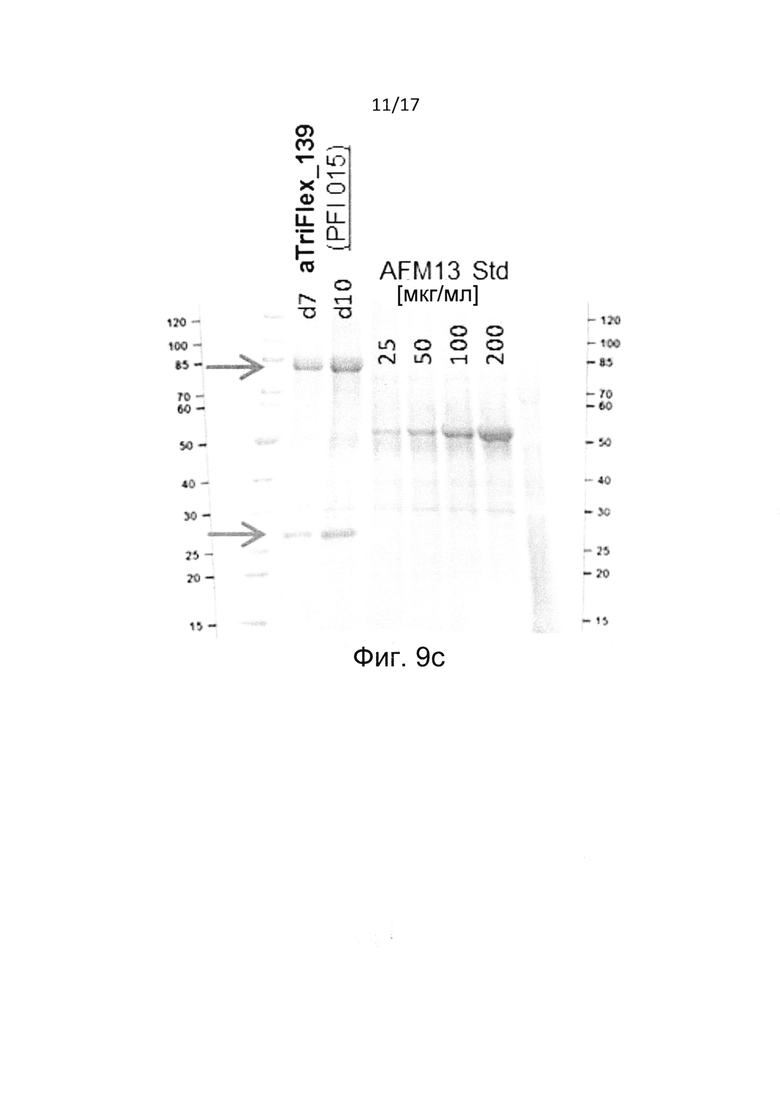
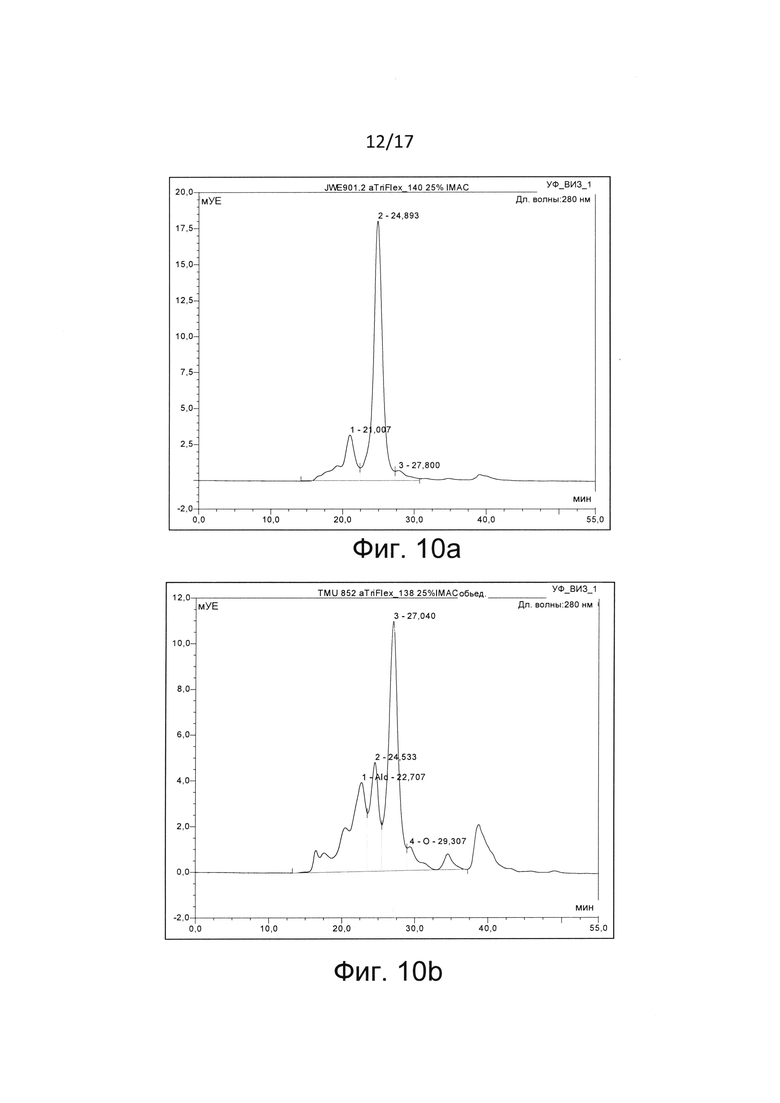
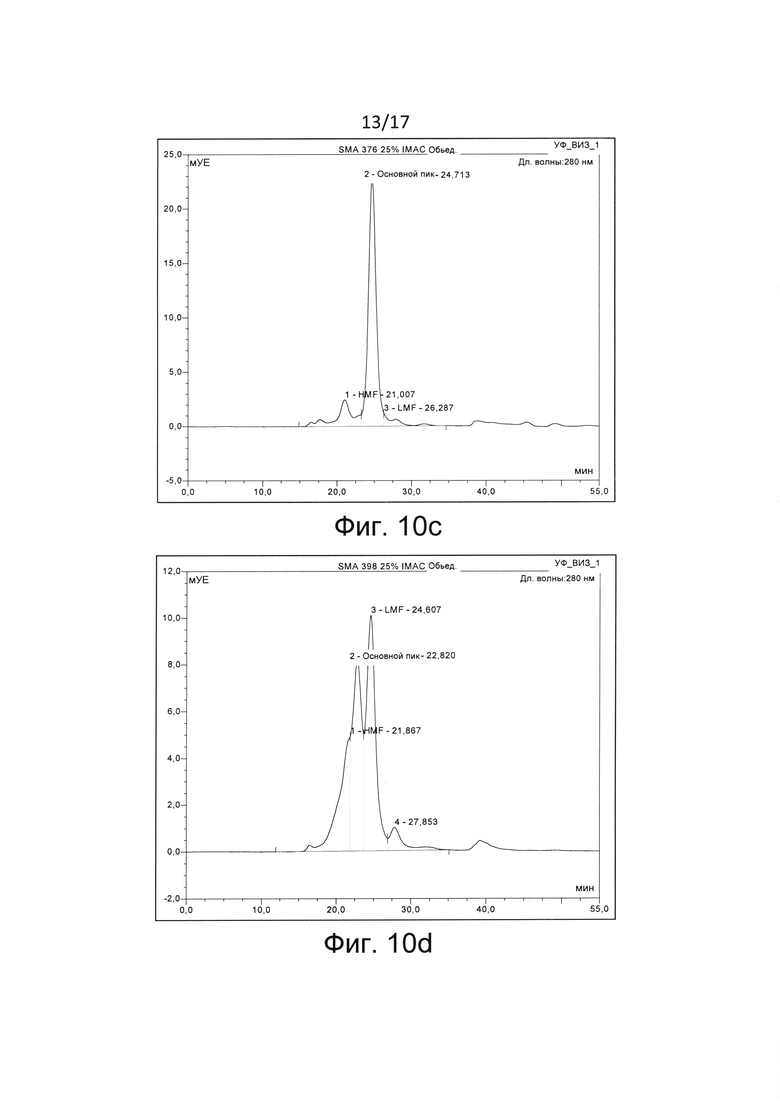
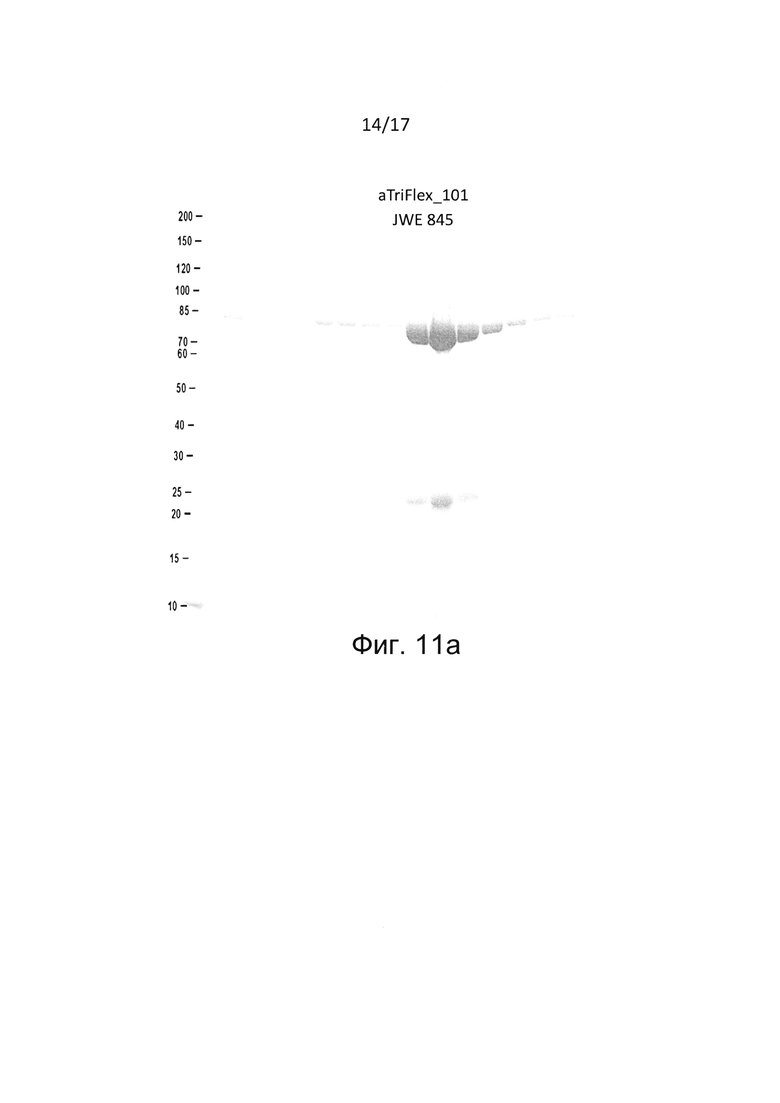
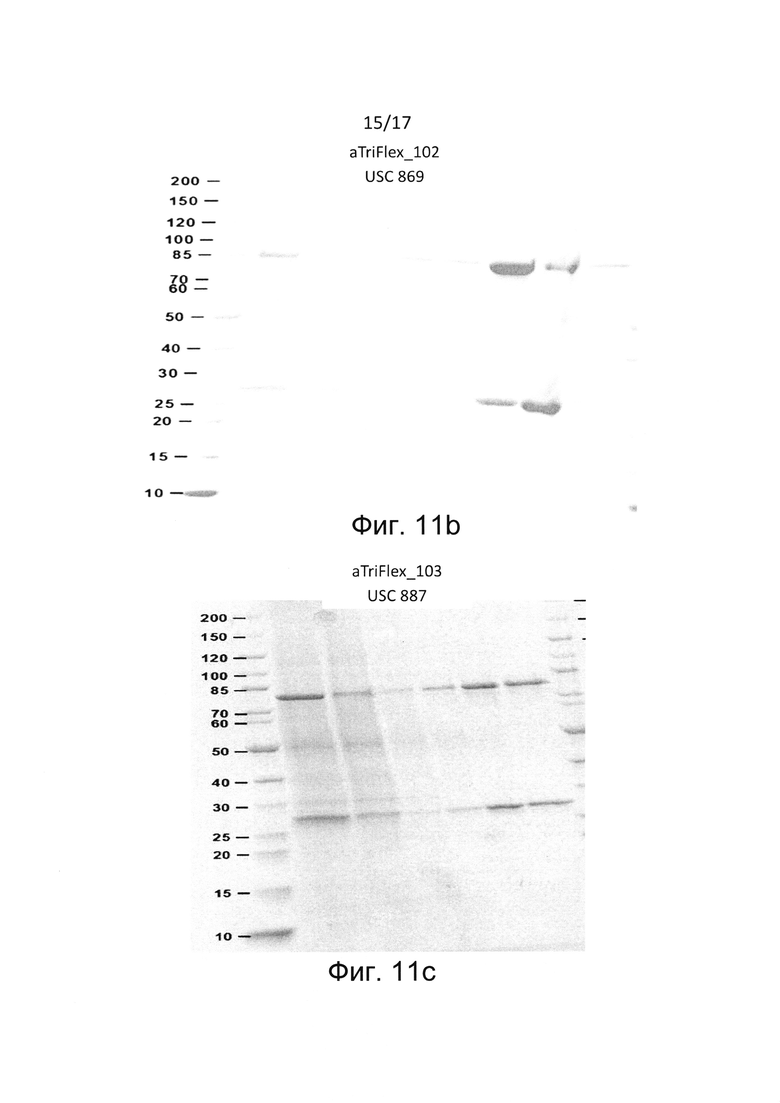
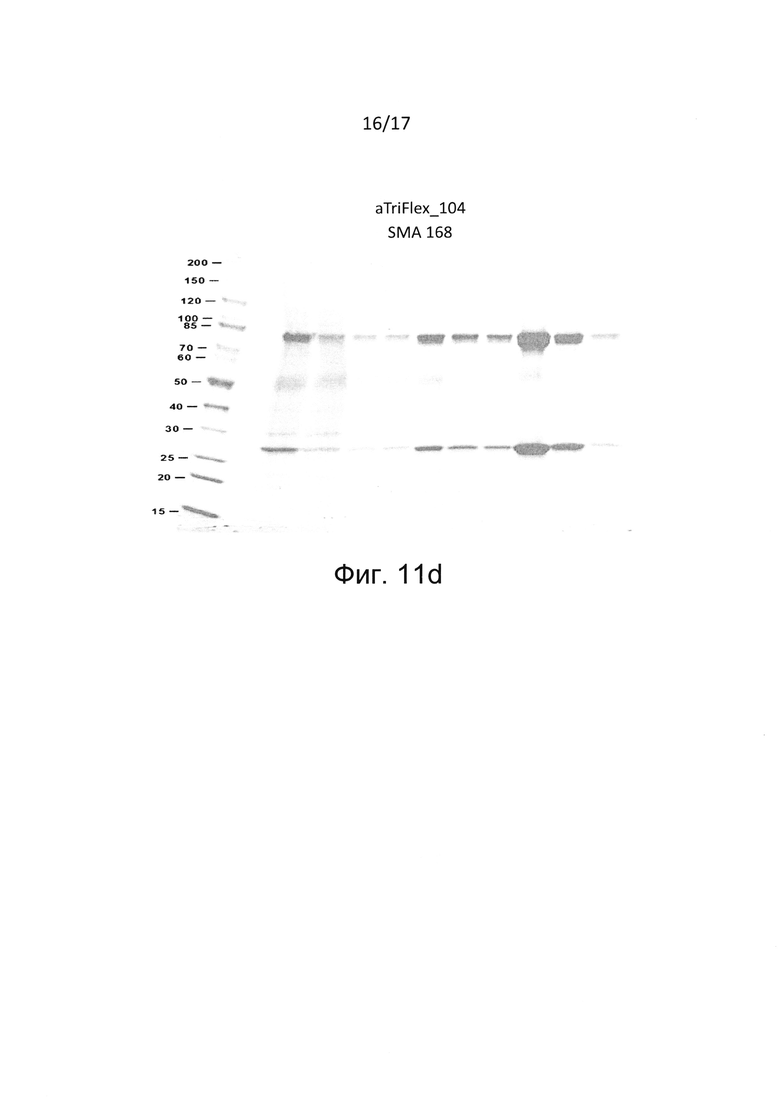
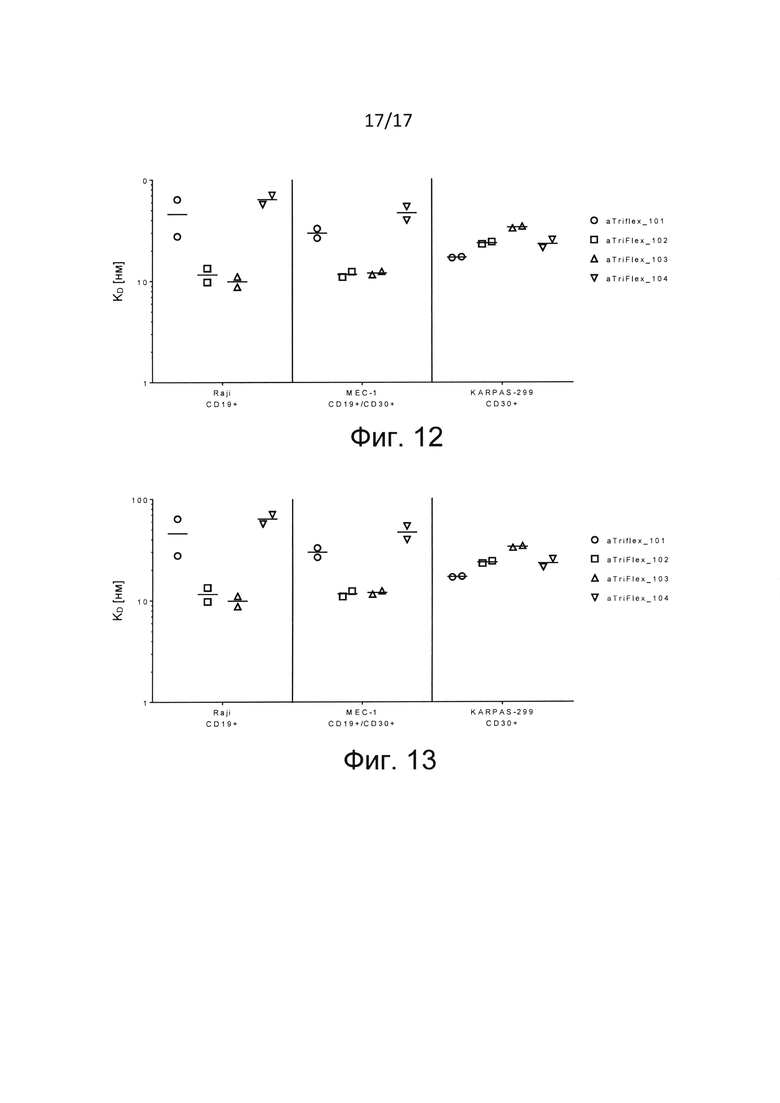
Настоящее изобретение относится к биохимии и иммунологии, в частности к молекуле триспецифичного антитела, содержащего единицу диатела, встроенную в полипептидную цепь, содержащую по меньшей мере шесть вариабельных доменов, соединенных друг за другом. В некоторых случаях два одноцепочечных Fv-фрагмента соединены дистально с единицей диатела с образованием двух дополнительных антигенсвязывающих сайтов. Антигенсвязывающая молекула может являться мономером одиночного полипептида или мультимерным полипептидом. Изобретение может применяться для нацеленного воздействия на два различных опухолевых антигена, причем третья специфичность активирует цитотоксическое действие эффекторных клеток, либо связывать две эффекторные клетки и один опухолевый антиген. Изобретение можно применять различными способами для изменения мишеней цитотоксического потенциала эффекторных клеток иммунной системы для уничтожения опухолевых клеток или инфекционных агентов. 16 з.п. ф-лы, 13 ил., 1 табл., 4 пр.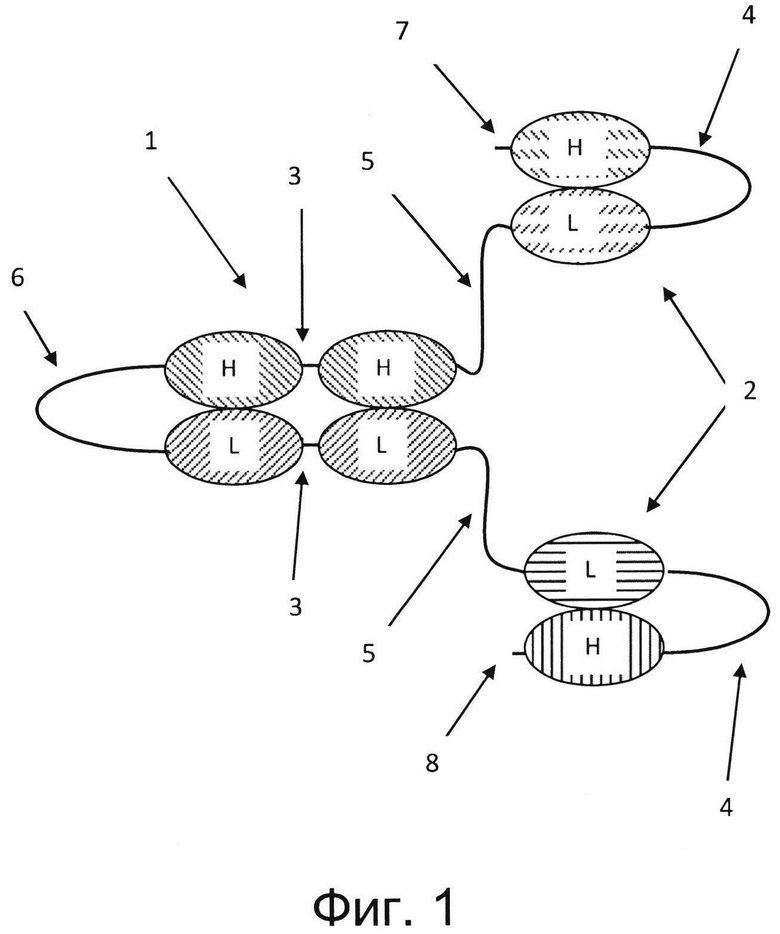
1. Мультивалентное Fv-антитело для обеспечения по меньшей мере четырех антигенсвязывающих сайтов для связывания антигенов, содержащее единицу диатела,
состоящую из двух пар вариабельных доменов, причем одна пара представляет собой пару вариабельных доменов легкой цепи (VL-VL), соединенных друг за другом в полипептид пептидным линкером, состоящим из 12 или менее аминокислотных остатков, а другая пара представляет собой пару вариабельных доменов тяжелой цепи (VH-VH), соединенных друг за другом в полипептид пептидным линкером, состоящим из 12 или менее аминокислотных остатков, причем пара VL-VL и пара VH-VH ассоциируют в два антигенсвязывающих сайта, и по меньшей мере одна пара вариабельных доменов встроена в полипептид, содержащий по меньшей мере шесть вариабельных доменов, соединенных друг за другом, причем
I)
(а) указанная одна пара вариабельных доменов встроена в первый полипептид, содержащий шесть вариабельных доменов, причем указанный первый полипептид содержит на N-конце scFv-единицу, соединенную с указанной одной парой вариабельных доменов, и на C-конце scFv-единицу, соединенную с указанной одной парой вариабельных доменов, и указанная одна пара вариабельных доменов ассоциирует с указанной другой парой вариабельных доменов во втором полипептиде, который состоит из двух вариабельных доменов;
(b) указанная одна пара вариабельных доменов встроена в первый полипептид, содержащий десять вариабельных доменов, причем указанный первый полипептид содержит на N-конце единицу одноцепочечного диатела, соединенную с указанной одной парой вариабельных доменов, и единицу одноцепочечного диатела на C-конце, соединенную с указанной одной парой вариабельных доменов, и указанная одна пара вариабельных доменов ассоциирует с другой парой вариабельных доменов во втором полипептиде, который состоит из указанных двух вариабельных доменов;
(c) в первом полипептиде, содержащем шесть вариабельных доменов, scFv-единица соединена по N-концу и scFv-единица соединена по C-концу с указанной парой вариабельных доменов легкой цепи (VL-VL), и во втором полипептиде scFv-единица соединена по N-концу и scFv-единица соединена по C-концу с указанной парой вариабельных тяжелых цепей (VH-VH); или
(d) указанное Fv-антитело состоит из единственного полипептида, содержащего восемь вариабельных доменов, и указанная одна пара из двух вариабельных доменов встроена в полипептид, причем указанный полипептид содержит на N-конце scFv-единицу, соединенную с указанной одной парой вариабельных доменов, и на C-конце scFv-единицу, соединенную с указанной другой парой вариабельных доменов, причем указанная одна пара вариабельных доменов соединена по С-концу с указанной другой парой вариабельных доменов линкером из более чем 12 аминокислотных остатков, и указанная одна пара вариабельных доменов ассоциирует с другой парой из двух вариабельных доменов указанного полипептида, и при этом
II) указанное одноцепочечное диатело состоит из первой пары вариабельных доменов, соединенных линкером из более чем 12, предпочтительно от 15 до 50 аминокислотных остатков, и каждая пара вариабельных доменов соединена линкером из 12 или менее аминокислотных остатков;
scFv-единица состоит из вариабельного домена легкой цепи, и вариабельного домена тяжелой цепи, соединенных линкером из 12 или более, предпочтительно, от 15 до 50 аминокислотных остатков.
2. Мультивалентное Fv-антитело по п. 1, характеризующееся тем, что Fv-антитело является четырехвалентным.
3. Мультивалентное Fv-антитело по любому из пп. 1 или 2, характеризующееся тем, что указанное антитело является триспецифичным.
4. Мультивалентное Fv-антитело по любому из пп. 1-3, характеризующееся тем, что указанные пептидные линкеры содержат остатки глицина и серина.
5. Мультивалентное Fv-антитело по п. 4, характеризующееся тем, что указанный пептидный линкер представляет собой линкер (G2S)X, где x=1-10.
6. Мультивалентное Fv-антитело по любому из пп. 1-5, характеризующееся тем, что в указанной единице одноцепочечного диатела вариабельные домены расположены в порядке VL-VH-VL-VH, VL-VL-VH-VH, VH-VH-VL-VL или VH-VL-VH-VL.
7. Мультивалентное Fv-антитело по любому из пп. 1-5, характеризующееся тем, что в scFv-единице вариабельные домены расположены как VL-VH или VH-VL.
8. Мультивалентное Fv-антитело по п. 1, характеризующееся тем, что указанный полипептид содержит шесть вариабельных доменов, соединенных друг за другом от N-конца до C-конца, причем первый и второй вариабельные домены образуют scFv-единицу на N-конце, где указанная scFv-единица на N-конце соединена по C-концу с третьим вариабельным доменом, указанный третий вариабельный домен соединен пептидным линкером, состоящим из 12 или менее аминокислотных остатков с четвертым вариабельным доменом, четвертый вариабельный домен соединен по C-концу с scFv-единицей на C-конце, и указанная scFv-единица образована пятым и шестым вариабельными доменами, причем вариабельные домены расположены от N-конца к С-концу в ориентации, выбранной из:
(a) VH-VL-VH-VH-VL-VH (первый полипептид) и VL-VL (второй полипептид);
(b) VL-VH-VH-VH-VH-VL (первый полипептид) и VL-VL (второй полипептид);
(c) VH-VL-VL-VL-VH-VL (первый полипептид) и VH-VH (второй полипептид);
(d) VL-VH-VL-VL-VH-VL (первый полипептид) и VH-VH (второй полипептид); или
(e) VH-VL-VL-VL-VL-VH (первый полипептид) и VH-VH (второй полипептид).
9. Мультивалентное Fv-антитело по любому из пп. 1-8, характеризующееся тем, что указанные вариабельные домены в указанной единице диатела имеют одну и ту же эпитопную специфичность или различные эпитопные специфичности.
10. Мультивалентное Fv-антитело по любому из пп. 1-9, характеризующееся тем, что указанное Fv-антитело обладает специфичностью к антигену, присутствующему на эффекторной клетке иммунной системы.
11. Мультивалентное Fv-антитело по п. 10, характеризующееся тем, что указанное Fv-антитело содержит два антигенсвязывающих сайта, обладающих специфичностью к одному и тому же типу эффекторной клетки иммунной системы.
12. Мультивалентное Fv-антитело по п. 11, характеризующееся тем, что указанное Fv-антитело является триспецифичным и двухвалентно связывается с эффекторной клеткой иммунной системы.
13. Мультивалентное Fv-антитело по п. любому из пп. 9-12, характеризующееся тем, что антиген выбран из CD3, CD16 и CD16A.
14. Мультивалентное Fv-антитело по любому из пп. 1-13, характеризующееся тем, что указанное Fv-антитело имеет по меньшей мере одну специфичность к опухолевому антигену.
15. Мультивалентное Fv-антитело по п. 14, характеризующееся тем, что указанное Fv-антитело имеет специфичности к двум опухолевым антигенам.
16. Мультивалентное Fv-антитело по любому из пп. 10-15, характеризующееся тем, что указанное Fv-антитело имеет по меньшей мере одну специфичность к вирусному антигену.
17. Мультивалентное Fv-антитело по любому из пп. 1-16 для применения в качестве медикамента.
| АДАПТИВНАЯ АНТЕННАЯ РЕШЕТКА | 1991 |
|
RU2014680C1 |
| KIPRIYANOV S | |||
| et al., Bispecific tandem diabody for tumor therapy with improved antigen binding and pharmacokinetics, Journal of molecular biology, 1999, vol | |||
| ПРИСПОСОБЛЕНИЕ ДЛЯ ПОДАЧИ УГЛЯ В ТЕНДЕР ПАРОВОЗА | 1920 |
|
SU293A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Топливник с глухим подом | 1918 |
|
SU141A1 |
| SCHUBERT I | |||
| et al., A single-chain triplebody with specificity for CD19 and CD33 mediates effective lysis of mixed lineage leukemia cells by | |||

