ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящее изобретение испрашивает приоритет по китайской патентной заявке № 202010397572.3, поданной 12 мая 2020 года, содержание которой включено в настоящий документ посредством ссылки в полном объеме и для всех целей.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к выделенному антителу. В частности, в настоящем изобретении предложены мышиные, химерные и гуманизированные моноклональные антитела или их антигенсвязывающие фрагменты, которые специфически связываются с ST2, а также предложены нуклеиновые кислоты, векторы экспрессии и клетки-хозяева для получения антител или их антигенсвязывающих фрагментов. В настоящем изобретении дополнительно предложены слитые полипептиды, мультиспецифические молекулы, вирусные векторы и фармацевтические композиции, которые содержат антитела или их антигенсвязывающие фрагменты, а также предложен способ применения антител или их антигенсвязывающих фрагментов в диагностике и лечении.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Интерлейкин 33 (IL33) является членом семейства цитокинов IL-1. Он экспрессируется эндотелиальными клетками или эпителиальными клетками тканей. Когда клетки подвергаются воздействию стимулов, таких как стресс, инфекция и повреждение, IL33 экспрессируется как «алармин» в поврежденных клетках и передает внешние сигналы стимуляции антигена в путь Th2. Рецептор IL33 состоит из двух белков: белка, ассоциированного с рецептором IL-1 (IL-1RL1, ST2), и вспомогательного белка рецептора IL-1 (IL-1RAP). Связывание IL33 с ST2 может индуцировать передачу сигнала только в присутствии IL-1RAP, но IL-1RAP не может напрямую связываться с IL33 или ST2 (Chackerian et al., (2007) J Immunol. 179:2551-2555).
ST2 принадлежит к семейству рецепторов Toll/IL1. Он имеет три подтипа: трансмембранная форма ST2L, растворимая форма sST2 и вариантная форма ST2V. В основном ST2L опосредует внутриклеточный сигнальный ответ IL33. Главным образом ST2L экспрессируется на поверхностях Th2-клеток, ILC2-клеток, тучных клеток и Treg-клеток, а также может экспрессироваться на поверхностях NK-клеток (естественные киллерные клетки), NKT-клеток, макрофагов, эозинофилов и базофилов.
IL33 связывается с ST2L на поверхности клетки и рекрутирует IL-1RAcP, тем самым активируя путь клеток MyD88/NFκB и индуцируя тучные клетки, Th2-клетки, Treg-клетки, ILC2-клетки и т.д., чтобы секретировать провоспалительные факторы, такие как IL-5, IL-6, IL-13, TNF и INF-γ, и хемокины, такие как CCL17, CCL22 и CXCL8, чтобы индуцировать воспаление. Экспрессия IL-33 аномально повышена при заболеваниях воспаления слизистой ткани, воспалении суставов и хроническом воспалении кожи, таких как аллергический ринит (Kamekura R et al., (2012) Clin Exp Allergy. 42:218-28), ревматоидный артрит, анкилозирующий спондилоартрит, атопический дерматит (Savinko T et al., (2012) J Invest Dermatol. 132:1392-1400) и псориаз; кроме того, дисрегуляция сигнального пути IL33/ST2 тесно связана с иммуноопосредованными заболеваниями, такими как астма, хроническое обструктивное заболевание легких (Hacker, Lambers et al., (2009) J Clin Lab Anal. 23:372-9), бронхит, воспалительное заболевание кишечника (Beltran CJ et al., (2010) Inflamm Whel Dis. 16:1097-107), системная красная волчанка, фиброз печени и системный склероз. Анализ на уровне генов показывает, что гены IL33 и ST2 имеют некоторые сайты однонуклеотидного полиморфизма (SNP), которые связаны с количеством базофилов, а также тесно связаны с возникновением атопического дерматита и астматических заболеваний (Shimizu M et al., (2005) Hum Mol Genet. 14:2919-27; Gudbjartsson DF et al., (2009) Nat Genet. 41:342-7). Таким образом, блокирование связывания IL33 с ST2 и ингибирование сигнального пути IL33/ST2 и индуцированного им воспаления стали важными темами в лечении воспалительных заболеваний, связанных с иммунной системой.
КРАТКОЕ ОПИСАНИЕ
Настоящее изобретение относится к антителу к ST2 с высокой аффинностью, высокой специфичностью и высокой биоактивностью, пригодному для применения при лечении заболеваний, опосредованных IL33/ST2.
Настоящее изобретение относится к выделенным антителам, например, мышиным, человеческим, химерным или гуманизированным моноклональным антителам или их антигенсвязывающим фрагментам, которые связываются с ST2 (например, ST2 человека и ST2 обезьяны).
Антитела или их антигенсвязывающие фрагменты, раскрытые в настоящем документе, имеют различные применения, включая обнаружение белков ST2 и лечение и предотвращение заболеваний и состояний, опосредованных IL33/ST2, причем указанные заболевания и состояния, опосредованные IL33/ST2, включают астму, аллергический ринит, хроническое обструктивное заболевание легких, эозинофильный бронхит, эозинофильный эзофагит, атопический дерматит, псориаз, системную красную волчанку, пемфигоид, ревматоидный артрит, анкилозирующий спондилоартрит, воспалительное заболевание кишечника, фиброз легких, фиброз печени, системный склероз, саркоидоз, болезнь «трансплантат против хозяина» (GVHD), диабет, сердечно-сосудистые заболевания или их комбинации.
В одном аспекте настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, содержащему CDR тяжелой цепи, причем указанные CDR тяжелой цепи включают CDR1 тяжелой цепи, CDR2 тяжелой цепи и CDR3 тяжелой цепи, где (1) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 15, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 20, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (2) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 16, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 21, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (3) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (4) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 13, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 23, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (5) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 14, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 19, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; или (6) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 24, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N или A, X2 представляет собой Q, E или K, и X3 представляет собой K или Q.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N или A, X2 представляет собой Q, и X3 представляет собой K или Q; X1 представляет собой N или A, X2 представляет собой E, и X3 представляет собой K или Q; или X1 представляет собой N или A, X2 представляет собой K, и X3 представляет собой K или Q.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N, X2 представляет собой Q, и X3 представляет собой K или Q; X1 представляет собой A, X2 представляет собой Q, и X3 представляет собой K или Q; X1 представляет собой N, X2 представляет собой E, и X3 представляет собой K или Q; X1 представляет собой A, X2 представляет собой E, и X3 представляет собой K или Q; X1 представляет собой N, X2 представляет собой K, и X3 представляет собой K или Q; или X1 представляет собой A, X2 представляет собой K, и X3 представляет собой K или Q.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N, X2 представляет собой Q, и X3 представляет собой K; X1 представляет собой A, X2 представляет собой E, и X3 представляет собой Q; или X1 представляет собой A, X2 представляет собой K, и X3 представляет собой K.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи, и аминокислотная последовательность вариабельной области тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 35, 37, 39, 41, 43, 45, 71 или 73, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с последовательностью, представленной в SEQ ID NO: 35, 37, 39, 41, 43, 45, 71 или 73.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых конкретных вариантах осуществления аминокислоты, представленные в SEQ ID NO: 71, могут кодироваться нуклеиновой кислотой, представленной в SEQ ID NO: 72; аминокислоты, представленные в SEQ ID NO: 73, могут кодироваться нуклеиновой кислотой, представленной в SEQ ID NO: 74.
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR легкой цепи, причем указанные CDR легкой цепи включают CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, где (1) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 25, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 29, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 32, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (2) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 26, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (3) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; или (4) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 28, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 31, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 33, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, X4 представляет собой M или L.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, и аминокислотная последовательность вариабельной области легкой цепи содержит последовательность, представленную в SEQ ID NO: 36, 38, 40, 42, 44, 46 или 79, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с последовательностью, представленной в SEQ ID NO: 36, 38, 40, 42, 44, 46 или 79.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых конкретных вариантах осуществления аминокислоты, представленные в SEQ ID NO: 79, могут кодироваться нуклеиновой кислотой, представленной в SEQ ID NO: 80.
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит CDR1 тяжелой цепи, CDR2 тяжелой цепи и CDR3 тяжелой цепи и CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, где (1) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 15, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 20, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 25, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 29, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 32, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (2) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 16, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 21, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 26, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (3) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (4) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 13, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 23, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (5) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 14, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 19, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; или (6) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 24, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 28, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 31, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 33, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N или A, X2 представляет собой Q, E или K, и X3 представляет собой K или Q; и аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, где X4 представляет собой M или L.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N или A, X2 представляет собой Q, и X3 представляет собой K или Q; X1 представляет собой N или A, X2 представляет собой E, и X3 представляет собой K или Q; X1 представляет собой N или A, X2 представляет собой K, и X3 представляет собой K или Q, и аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, где X4 представляет собой M или L.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N, X2 представляет собой Q, и X3 представляет собой K или Q; X1 представляет собой A, X2 представляет собой Q, и X3 представляет собой K или Q; X1 представляет собой N, X2 представляет собой E, и X3 представляет собой K или Q; X1 представляет собой A, X2 представляет собой E, и X3 представляет собой K или Q; X1 представляет собой N, X2 представляет собой K, и X3 представляет собой K или Q; или X1 представляет собой A, X2 представляет собой K, и X3 представляет собой K или Q; и аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, где X4 представляет собой M или L.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2, где аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где X1 представляет собой N, X2 представляет собой Q, и X3 представляет собой K; X1 представляет собой A, X2 представляет собой E, и X3 представляет собой Q; или X1 представляет собой A, X2 представляет собой K, и X3 представляет собой K; и аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, где X4 представляет собой M или L.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где (1) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 35, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 36, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (2) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 37, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 38, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (3) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 39, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 40, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (4) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 41, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 42, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (5) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 43, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 44, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (6) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 45, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 46, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (7) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 71, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 79, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; или (8) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 73, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 79, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, и аминокислотная последовательность тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 47, 51, 55, 59, 63, 67, 75 или 77, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с последовательностью, представленной в SEQ ID NO: 47, 51, 55, 59, 63, 67, 75 или 77.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых конкретных вариантах осуществления аминокислоты, представленные в вышеупомянутых SEQ ID NO: 47, 51, 55, 59, 63, 67, 75 и 77, могут кодироваться нуклеиновыми кислотами, представленными в SEQ ID NO: 48, 52, 56, 60, 64, 68, 76 и 78, соответственно.
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит легкую цепь, и аминокислотная последовательность легкой цепи содержит последовательность, представленную в SEQ ID NO: 49, 53, 57, 61, 65, 69 или 81, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с последовательностью, представленной в SEQ ID NO: 49, 53, 57, 61, 65, 69 или 81.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых конкретных вариантах осуществления аминокислоты, представленные в вышеупомянутых SEQ ID NO: 49, 53, 57, 61, 65, 69 и 81, могут кодироваться нуклеиновыми кислотами, представленными в SEQ ID NO: 50, 54, 58, 62, 66, 70 и 82, соответственно.
В некоторых вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь и легкую цепь, где (1) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 47, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 49, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (2) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 51, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 53, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (3) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 55, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 57, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (4) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 59, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 61, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (5) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 63, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 65, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (6) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 67, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ним, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 69, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; (7) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 75, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 81, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней; или (8) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 77, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 81, или аминокислотную последовательность, имеющую по меньшей мере 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность с ней.
В некоторых конкретных вариантах осуществления выделенное антитело или его антигенсвязывающий фрагмент связывается с ST2 и представляет собой моноклональное антитело (например, мышиное, химерное или гуманизированное антитело).
В некоторых вариантах осуществления выделенное антитело (например, мышиное, химерное или гуманизированное антитело) или его антигенсвязывающий фрагмент содержит тяжелую цепь и легкую цепь, причем тяжелая цепь содержит вариабельную область тяжелой цепи и константную область тяжелой цепи, и легкая цепь содержит вариабельную область легкой цепи и константную область легкой цепи, где вариабельная область тяжелой цепи и вариабельная область легкой цепи содержат аминокислотные последовательности, описанные выше; константная область тяжелой цепи содержит константную область IgG1, IgG2 или IgG4 человека, предпочтительно константную область IgG2 человека; константная область легкой цепи содержит константную область κ человека или константную область λ человека, где антитело или его антигенсвязывающий фрагмент связывается с ST2.
В некоторых вариантах осуществления антитело, описанное в данном документе, содержит или состоит из двух тяжелых цепей и двух легких цепей, причем тяжелые цепи и легкие цепи связаны друг с другом дисульфидными связями, где каждая тяжелая цепь содержит константную область тяжелой цепи, вариабельную область тяжелой цепи или последовательности CDR, описанные выше, и каждая легкая цепь содержит константную область легкой цепи, вариабельную область легкой цепи или последовательности CDR, описанные выше, где С-конец вариабельной области тяжелой цепи связан с N-концом константной области тяжелой цепи, а С-конец вариабельной области легкой цепи связан с N-концом константной области легкой цепи. Антитело, описанное в настоящем документе, может представлять собой, например, полноразмерное антитело изотипа IgG1, IgG2 или IgG4. В другом варианте осуществления антитело, описанное в настоящем документе, может представлять собой одноцепочечное антитело (scFv) или фрагмент антитела, такой как Fab, фрагмент F(ab')2, фрагмент Fd, фрагмент Fv, dAb или выделенная CDR.
В одном аспекте настоящее изобретение также относится к слитому полипептиду, содержащему антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, и другие функциональные молекулы, где указанные другие функциональные молекулы могут представлять собой пептиды, белки или небелки. Слитый полипептид обладает функцией связывания с ST2 и одной или более другими функциями. В одном аспекте настоящее изобретение также относится к мультиспецифической молекуле, содержащей антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, и по меньшей мере одну другую функциональную часть со специфичностью, которая отличается от специфичности антитела или антигенсвязывающего фрагмента, где указанная другая функциональная часть может представлять собой другой пептид или белок. Мультиспецифическая молекула способна связываться с ST2 и по меньшей мере с одним другим ассоциированным с заболеванием белком, например, IgE. В другом аспекте антитело к ST2 или его антигенсвязывающий фрагмент, описанные в настоящем документе, также могут кодироваться или переноситься вирусным вектором.
В одном аспекте настоящее изобретение также относится к фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, или содержащей мультиспецифическую молекулу, слитый полипептид или вирусный вектор, описанные в настоящем документе, и фармацевтически приемлемый эксципиент, разбавитель или носитель.
В одном аспекте настоящее изобретение также относится к выделенной молекуле нуклеиновой кислоты, кодирующей антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, вектору экспрессии, содержащему такую молекулу нуклеиновой кислоты, и клетке-хозяину, содержащей такой вектор экспрессии.
В одном аспекте настоящее изобретение также относится к способу получения антитела к ST2 или его антигенсвязывающего фрагмента, включающему следующие стадии: (i) экспрессия антитела или его антигенсвязывающего фрагмента в клетке-хозяине и (ii) выделение антитела или его антигенсвязывающего фрагмента из указанной клетки-хозяина или ее клеточной культуры.
В другом аспекте настоящее изобретение относится к способу определения уровня экспрессии ST2 в образце, полученном от субъекта, включающему стадию приведения образца в контакт с антителом или его антигенсвязывающим фрагментом, описанным в настоящем документе, в таких условиях, что антителу или его антигенсвязывающему фрагменту дают возможность связаться с ST2 (или образовать комплекс с ST2).
В другом аспекте настоящее изобретение относится к способу лечения или облегчения заболеваний и состояний, опосредованных IL33/ST2, у субъекта, нуждающегося в этом, причем указанные заболевания и состояния, опосредованные IL33/ST2, включают, но не ограничиваются, астму, аллергический ринит, хроническую обструктивную болезнь легких, эозинофильный бронхит, эозинофильный эзофагит, атопический дерматит, псориаз, системную красную волчанку, пемфигоид, ревматоидный артрит, анкилозирующий спондилоартрит, воспалительное заболевание кишечника, фиброз легких, фиброз печени, системный склероз, саркоидоз, болезнь «трансплантат против хозяина» (GVHD), диабет, сердечно-сосудистые заболевания или их комбинации, причем способ включает введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента или фармацевтической композиции, описанной в настоящем документе. В некоторых вариантах осуществления способ включает введение субъекту терапевтически эффективного количества молекулы нуклеиновой кислоты, кодирующей антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе. В некоторых вариантах осуществления способ включает введение субъекту терапевтически эффективного количества слитого полипептида, мультиспецифической молекулы, вирусного вектора или их фармацевтической композиции, описанной в настоящем документе. В некоторых вариантах осуществления мультиспецифическая молекула способна связываться с ST2 и по меньшей мере с другим ассоциированным с заболеванием белком, например, TSLP, IgE, IL4, IL13 или IL-5. В некоторых вариантах осуществления по меньшей мере одно другое антитело, например, антитело к TSLP, антитело к TSLPR, антитело к IL4, антитело к IL4R или антитело к IgE, можно вводить вместе с антителом или его антигенсвязывающим фрагментом, описанными в настоящем документе. В некоторых вариантах осуществления по меньшей мере одно дополнительное лекарственное средство, например, лекарственное средство против астмы, лекарственное средство против хронической обструктивной болезни легких, лекарственное средство против язвенного колита, лекарственное средство против атопического дерматита или лекарственное средство против псориаза, можно вводить вместе с антителом или его антигенсвязывающим фрагментом, описанными в настоящем документе.
Другие признаки и преимущества настоящего изобретения будут очевидны из следующего подробного описания и примеров, которые не следует рассматривать как ограничивающие. Содержание всех документов, данных из Genbank, патентов и опубликованных патентных заявок, цитируемых в настоящем документе, непосредственно включено в настоящий документ посредством ссылки.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: связывание химерных антител к ST2 с hST2, обнаруженное с помощью ELISA (иммуноферментный анализ), в диапазоне концентраций. На фиг. 1 показано связывание химерных антител xi17E2, xi8F4, xi26A1, xi31C8, xi3C6 и xi7D1 с hST2, причем связывание RG6149 показано в качестве контроля.
Фиг. 2: связывание химерных антител к ST2 с cyno-ST2 (ST2 яванского макака), обнаруженное с помощью ELISA, в диапазоне концентраций. На фиг. 2 показано связывание химерных антител xi17E2, xi8F4, xi26A1, xi31C8, xi3C6 и xi7D1 с cyno-ST2, причем связывание RG6149 показано в качестве контроля.
Фиг. 3: блокирование связывания IL33/ST2 химерными антителами к ST2, обнаруженное с помощью ELISA, в диапазоне концентраций антител. На фиг. 3 показано блокирование связывания IL33/ST2 химерными антителами xi26A1, xi7D1, xi17E2, xi31C8, xi3C6 и xi8F4, причем блокирование RG6149 показано в качестве контроля.
Фиг. 4: связывание гуманизированных антител к ST2 с hST2, обнаруженное с помощью ELISA, в диапазоне концентраций. На фиг. 4 показано связывание гуманизированных антител hz31C8-1.1 и hz31C8-1.2 с hST2, причем связывание химерных антител xi31C8, RG6149 и GSK3772847 показано в качестве контролей, и IgG2 показано в качестве отрицательного контроля.
Фиг. 5: связывание гуманизированных антител к ST2 с cyno-ST2, обнаруженное с помощью ELISA, в диапазоне концентраций. На фиг. 5 показано связывание гуманизированных антител hz31C8-1.1 и hz31C8-1.2 с cyno-ST2, причем связывание химерных антител xi31C8, RG6149 и GSK3772847 показано в качестве контролей, и IgG2 показано в качестве отрицательного контроля.
Фиг. 6: блокирование связывания IL33/ST2 гуманизированными антителами к ST2, обнаруженное с помощью ELISA, в диапазоне концентраций антител. На фиг. 6 показано блокирование связывания IL33/ST2 гуманизированными антителами hz31C8-1.1 и hz31C8-1.2, причем блокирование химерными антителами xi31C8, RG6149 и GSK3772847 показано в качестве контролей.
Фиг. 7: связывание химерных антител к ST2 с клетками, сверхэкспрессирующими hST2, обнаруженное с помощью FACS (метод анализа сортировки клеток с активированной флуоресценцией), в диапазоне концентраций антител. На фиг. 7 показано связывание химерных антител xi31C8, xi26A1, xi7D1, xi3C6, xi8F4 и xi17E2 с клетками HEK293T-hST2-NFκB-люцифераза, сверхэкспрессирующими hST2, причем связывание RG6149 показано в качестве контроля, и IgG4 показано в качестве отрицательного контроля.
Фиг. 8: связывание химерных антител к ST2 с клетками, сверхэкспрессирующими cyno-ST2, обнаруженное с помощью FACS, в диапазоне концентраций антител. На фиг. 8 показано связывание химерных антител xi31C8, xi26A1, xi7D1, xi3C6, xi8F4 и xi17E2 с клетками HEK293T-Cyno-ST2-NFκB-люцифераза, сверхэкспрессирующими cyno-ST2, причем связывание RG6149 показано в качестве контроля, и IgG4 показано в качестве отрицательного контроля.
Фиг. 9: блокирование связывания IL33/ST2 химерными антителами к ST2, обнаруженное с помощью анализов жизнеспособности клеток, в диапазоне концентраций антител. На фиг. 9 показано блокирование взаимодействия между белком IL33 и клетками HEK293T-hST2-NFκB-люцифераза, экспрессирующими hST2, химерными антителами xi31C8, xi26A1, xi7D1 и xi3C6, причем блокирование с помощью RG6149 показано в качестве контроля.
Фиг. 10: связывание гуманизированных антител к ST2 с клетками, сверхэкспрессирующими hST2, обнаруженное с помощью FACS, в диапазоне концентраций антител. На фиг. 10 показано связывание гуманизированных антител hz31C8-1.1 и hz31C8-1.2 с клетками HEK293T-hST2-NFκB-люцифераза, сверхэкспрессирующими hST2, причем связывание химерных антител xi31C8, RG6149 и GSK3772847 показано в качестве контролей, и IgG2 показано в качестве отрицательного контроля.
Фиг. 11: блокирование связывания IL33/ST2 гуманизированными антителами к ST2, обнаруженное с помощью анализов жизнеспособности клеток, в диапазоне концентраций антител. На фиг. 11 показано блокирование взаимодействия между белком IL33 и клетками HEK293T-hST2-NFκB-люцифераза, экспрессирующими hST2, гуманизированными антителами hz31C8-1.1 и hz31C8-1.2, причем блокирование химерными антителами xi31C8, RG6149 и GSK3772847 показано в качестве контролей, и IgG2 показано в качестве отрицательного контроля.
Фиг. 12: блокирование связывания IL33/ST2 гуманизированными антителами к ST2, обнаруженное с помощью анализов жизнеспособности клеток, в диапазоне концентраций антител. На фиг. 12 показано блокирование взаимодействия между белком IL33 и клетками KU812-NFκB-люцифераза, экспрессирующими hST2, гуманизированными антителами hz31C8-1.1 и hz31C8-1.2, причем блокирование химерными антителами xi31C8, RG6149 и GSK3772847 показано в качестве контролей, и блокирование IgG2 показано в качестве отрицательного контроля.
Фиг. 13: блокирование секреции IL5, индуцированной IL33/ST2, на CD4+ Т-клетках гуманизированными антителами к ST2, обнаруженное с помощью анализов жизнеспособности клеток, в диапазоне концентраций антител. На фиг. 13 показано блокирование секреции IL5, индуцированной IL33/ST2, на CD4+ Т-клетках гуманизированными антителами hz31C8-1.1 и hz31C8-1.2, причем блокирование химерными антителами xi31C8, RG6149 и GSK3772847 показано в качестве контролей.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следует понимать, что терминология, используемая в настоящем описании, предназначена только для описания конкретных вариантов осуществления изобретения и не является ограничивающей. Если не указано иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, которое обычно подразумевается специалистом в области техники, к которой относится настоящее изобретение.
Термин «ST2» включает варианты, гомологи, ортологи и паралоги ST2. Например, в некоторых вариантах осуществления антитело, специфичное к белку ST2 человека, может перекрестно реагировать с белком ST2 других видов (например, обезьяны) в определенных обстоятельствах. В других вариантах осуществления антитело, специфичное к белку ST2 человека, может быть полностью специфичным к белку ST2 человека и не вступать в перекрестную реакцию с белками других видов или других типов, или может перекрестно реагировать с белками ST2 некоторых других видов, но не всех других видов.
Термины «ST2 человека» и «hST2» и тому подобное являются взаимозаменяемыми в настоящем описании и относятся к белку с аминокислотной последовательностью ST2 человека, например, аминокислотной последовательностью ST2 человека, представленной в SEQ ID NO: 1, и белок состоит из нескольких доменов: лидерной последовательности, соответствующей аминокислотам 1-18, внеклеточного домена, соответствующего аминокислотам 19-331, трансмембранного домена, соответствующего аминокислотам 332-350, и внутриклеточного домена, соответствующего аминокислотам 351-556. Термины «ST2 обезьяны» и «cyno-ST2» и тому подобное являются взаимозаменяемыми в настоящем описании и относятся к белку с аминокислотной последовательностью ST2 обезьяны, например, аминокислотной последовательностью ST2 обезьяны, представленной в SEQ ID NO: 10.
«Антитело», описанное в настоящем документе, включает полноразмерное антитело и его любые антигенсвязывающие фрагменты (т.е. «антигенсвязывающую часть») или одиночные цепи. Полноразмерное антитело представляет собой гликопротеин, содержащий две тяжелые (H) цепи и две легкие (L) цепи, причем тяжелые и легкие цепи связаны дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, то есть CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. Области VH и VL могут быть дополнительно разделены на гипервариабельные области, то есть определяющие комплементарность области (CDR), и каркасные области (FR) с более консервативными последовательностями. Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца к карбоксиконцу следующим образом: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых цепей и легких цепей содержат связывающие домены, которые взаимодействуют с антигенами. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
«Антигенсвязывающий фрагмент» или «антигенсвязывающая часть» антитела относится к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигенами (например, белками ST2). Было показано, что антигенсвязывающая функция антитела может быть выполнена фрагментами полноразмерного антитела. Примеры, охватываемые термином «антигенсвязывающая часть/фрагмент» антитела, включают: (i) фрагмент Fab: одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, двухвалентный фрагмент, содержащий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; (v) фрагмент dAb, состоящий из домена VH (Ward et al., Nature. 341:544-546 (1989)); (vi) выделенная определяющая комплементарность область (CDR) и (vii) нанотело, вариабельные области тяжелой цепи, содержащие один вариабельный домен и два константных домена. Кроме того, хотя два домена фрагмента Fv, VL и VH, кодируются различными генами, они могут быть соединены с использованием рекомбинантных способов с помощью синтетического линкера с образованием одной белковой цепи, в которой VL и VH спариваются с образованием одновалентной молекулы (называемой одноцепочечным Fv (scFv), см., например, Bird et al., Science. 242:423-426 (1988); Huston et al., Proc. Natl. Acad. Sci. 85:5879-5883 (1988)). Такие одноцепочечные антитела также охватываются термином антигенсвязывающая часть/фрагмент. Эти фрагменты антител могут быть получены с использованием обычных методик, известных специалистам в данной области техники, и фрагменты могут быть подвергнуты функциональному скринингу с использованием того же способа, что и полноразмерные антитела.
«Выделенное антитело» относится к антителу, которое по существу не содержит других антител с отличающейся антигенной специфичностью (например, выделенное антитело, которое специфически связывается с белком ST2, по существу не содержит антител, которые специфически связываются с антигенами, отличными от белка ST2). Однако выделенное антитело, которое специфически связывается с белком ST2 человека, может перекрестно связываться с другими антигенами (например, белками ST2 других видов). Кроме того, выделенное антитело по существу не содержит других клеточных компонентов и/или химических веществ.
Термин «мышиное антитело» относится к антителу, в котором каркасная область и CDR в вариабельной области получены из последовательностей иммуноглобулина зародышевой линии мыши. Кроме того, если указанное антитело содержит константную область, указанная константная область также происходит из последовательностей иммуноглобулинов зародышевой линии мыши. Раскрытое в данном документе мышиное антитело может содержать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов зародышевой линии мыши (например, мутации, введенные случайной мутацией in vitro или точечной мутацией или соматической мутацией in vivo), но «мышиное антитело» не включает антитела, в которых последовательности CDR, полученные от других видов млекопитающих, были привиты на каркасную последовательность мыши.
«Химерное антитело» относится к антителу, образованному путем объединения генетических материалов человеческого и нечеловеческого происхождения. В более общем смысле, химерное антитело относится к антителу, содержащему генетические материалы одного вида и генетические материалы другого вида. В настоящем описании химерные антитела также обозначены как «Xi».
«Гуманизированное антитело» относится к антителу нечеловеческого вида, белковая последовательность которого была модифицирована для повышения его сходства с встречающимся в природе антителом человека. В настоящем описании гуманизированные антитела также обозначены как «hz».
«Изотип» относится к классу антител, который кодируется генами константной области тяжелой цепи (например, IgM или IgG1).
Термины «антигенраспознающее антитело» и «антигенспецифическое антитело» используются взаимозаменяемо в настоящем описании с термином «антитело, которое специфически связывается с антигеном».
Антитело, которое «специфически связывается с ST2 человека», относится к антителу, которое связывается с белком ST2 человека (и возможно другими белками ST2 из видов, не являющихся человеческими), но по существу не связывается с белками, не относящимися к ST2. Предпочтительно, антитело связывается с ST2 человека с «высокой аффинностью», то есть KD составляет 5,0×10-8 M или менее, 1,0×10-8 M или менее, предпочтительно 5,0×10-9 M или менее, 1,0×10-9 M или менее, и более предпочтительно 5,0×10-10 M или менее, 1,0×10-10 M или менее.
Термин «по существу не связывающийся с» белком или клеткой относится к несвязыванию с белком или клеткой или несвязыванию с ним с высокой аффинностью, то есть связыванию с белком или клеткой с KD 1,0×10-6 М или выше, предпочтительно 1,0×10-5 М или выше, 1,0×10-4 М или выше и более предпочтительно 1,0×10-3 М или выше, 1,0×10-2 М или выше.
Для IgG термин «высокая аффинность» относится к KD 1,0×10-6 M или менее, 1,0×10-7 M или менее, предпочтительно 1,0×10-8 M или менее, 5,0×10-9 M или менее и более предпочтительно 1,0×10-9 M или менее. Однако для других изотипов антител «высокоаффинное» связывание может отличаться. Например, «высокоаффинное» связывание изотипа IgM относится к KD 10-6 М или менее, предпочтительно 10-7 М или менее и более предпочтительно 10-8 М или менее.
«Идентичность» относится к сходству между двумя последовательностями нуклеиновых кислот или между двумя полипептидами. Идентичность последовательностей, описанных в настоящем документе, составляет по меньшей мере 80 %, 85 %, 90 % или 95 %, предпочтительно по меньшей мере 95 %. Неограничивающие примеры включают: 80 %, 81 %, 82 %, 83 %, 84 %, 85 %, 86 %, 87 %, 88 %, 89 %, 90 %, 91 %, 92 %, 93 %, 94 %, 95 %, 96 %, 97 %, 98 %, 99 % или 100 %. Сравнение последовательностей и определение процента идентичности между двумя последовательностями могут быть выполнены с использованием алгоритма BLASTN/BLASTP Национального центра биотехнологического института с настройками по умолчанию.
Термин «EC50», также известный как полумаксимальная эффективная концентрация, относится к концентрации антитела, которая может вызывать 50 % максимального эффекта после определенного времени воздействия.
Термин «IC50», также известный как полумаксимальная ингибирующая концентрация, относится к концентрации антитела, которая ингибирует специфическую биологическую или биохимическую функцию на 50 % по сравнению с отсутствием антитела.
Термины «ингибировать» и «блокировать» используются взаимозаменяемо и включают частичное и полное ингибирование/блокирование. В некоторых вариантах осуществления антитело к ST2 ингибирует связывание IL33/ST2 на по меньшей мере около 50 %, например, по меньшей мере около 60 %, 70 %, 80 %, 90 %, 95 %, 99 % или 100 %.
Термин «субъект» включает любого человека или животное, не являющееся человеком. Термин «животное, не являющееся человеком» включает всех позвоночных, например, млекопитающих и не млекопитающих, предпочтительно млекопитающих, например, приматов, не являющихся человеком, овец, собак, кошек, коров и лошадей.
Термин «терапевтически эффективное количество» относится к количеству, достаточному для предотвращения или облегчения заболевания или состояния и/или уменьшения тяжести заболевания или состояния, предпочтительно к количеству, которое вызывает уменьшение тяжести симптомов заболевания или увеличение частоты и продолжительности бессимптомных фаз или способно предотвратить повреждение или недееспособность, вызванные заболеванием. Терапевтически эффективное количество связано с заболеванием, подлежащим лечению, причем фактическое эффективное количество может быть легко определено специалистами в данной области техники.
Использование форм единственного числа включает формы множественного числа, если не указано иное. Формы единственного числа относятся к «по меньшей мере одному», если не указано иное. Фраза «по меньшей мере один» означает то же, что и «один или более», и «и/или» используется для обозначения «и» или «или», если не указано иное.
Аспекты настоящего изобретения более подробно описаны ниже.
Антитело к ST2 специфически связывается с ST2 и блокирует взаимодействие IL33/ST2, среди других полезных функциональных характеристик.
Антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, специфически связывается с ST2 человека с высокой аффинностью. Антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, специфически связывается с ST2 (например, ST2 человека и ST2 обезьяны) и блокирует связывание IL33/ST2 и передачу сигналов. Антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, блокирует секрецию IL5, индуцированную IL33/ST2, на CD4+ Т-клетках. Антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, по существу не связывается с другими рецепторами семейства IL-1R, например, IL1R1, IL1R2, IL1R3, IL1R7, IL1R8 и IL1R9. Антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, обладает хорошей физической стабильностью (например, термостабильностью). Антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, обладает длительным периодом полужизни in vivo.
Предпочтительно антитело к ST2, описанное в настоящем документе, представляет собой моноклональное антитело. Кроме того, антитело может представлять собой, например, мышиное, химерное или гуманизированное моноклональное антитело.
Моноклональное антитело к ST2
Предпочтительно антитело к ST2 или его антигенсвязывающий фрагмент, описанное в настоящем документе, представляет собой антитело, имеющее структурные и химические свойства, описанные ниже. Антитело к ST2 или его антигенсвязывающий фрагмент содержит CDR тяжелой цепи и CDR легкой цепи, где примеры последовательностей CDR тяжелой цепи и последовательностей CDR легкой цепи представлены ниже в Таблице 1 и Таблице 2; антитело к ST2 или его антигенсвязывающий фрагмент содержит вариабельные области тяжелой цепи и вариабельные области легкой цепи, где примеры вариабельных областей тяжелой цепи и вариабельных областей легкой цепи представлены ниже в Таблице 3. Некоторые антитела имеют одинаковые CDR, и некоторые антитела имеют одинаковые VH или VL. Константная область тяжелой цепи антитела может представлять собой константную область тяжелой цепи IgG2 человека, и константная область легкой цепи антитела может представлять собой константную область легкой цепи κ человека. Эти антитела также могут содержать константную область тяжелой цепи IgG1 или IgG4 мыши и/или константную область легкой цепи κ мыши.
CDR тяжелой цепи в таблице 1 и CDR легкой цепи в таблице 2 определены с использованием системы нумерации по Кабату. Однако, как хорошо известно в данной области техники, CDR также могут быть определены с помощью других систем нумерации, таких как система/способ нумерации Чотиа, IMGT, AbM или Contact, на основе последовательности вариабельной области тяжелой/легкой цепи.
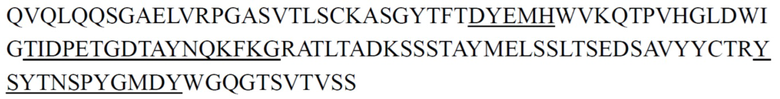

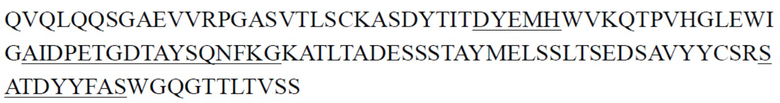
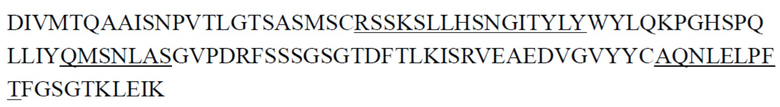
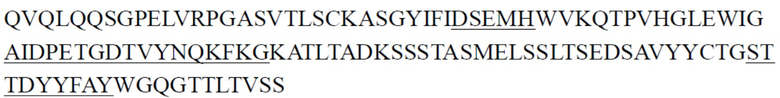
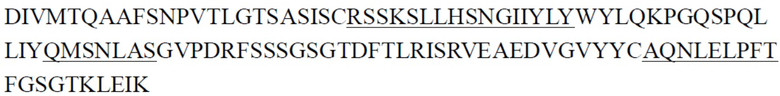
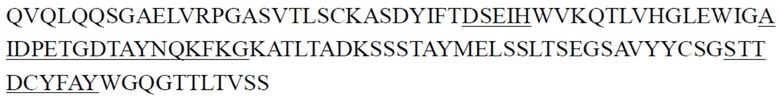
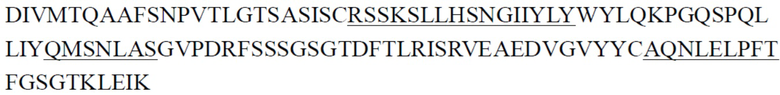
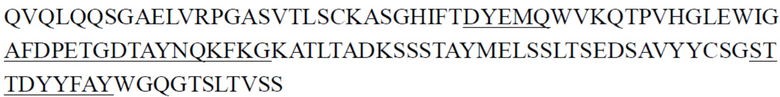
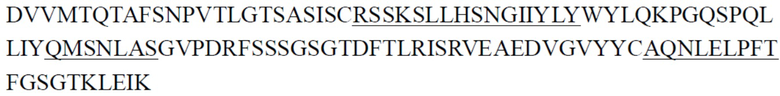
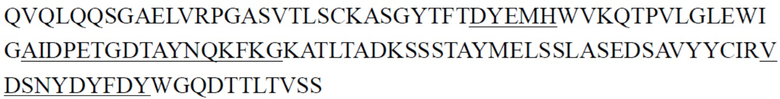
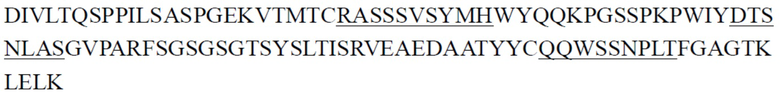
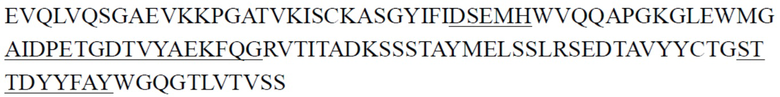


















Последовательности VH и/или VL (или последовательности CDR) других антител к ST2, которые связываются с ST2 человека, могут быть «смешаны и составлены» с последовательностями VH и/или VL (или последовательностями CDR) антитела, описанного в настоящем документе. Предпочтительно, когда цепи VH и VL (или CDR в таких цепях) смешивают и составляют, последовательность VH из конкретного спаривания VH/VL может быть заменена структурно подобной последовательностью VH. Аналогично, предпочтительно последовательность VL из конкретного спаривания VH/VL может быть заменена структурно подобной последовательностью VL.
Следовательно, в одном варианте осуществления антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, содержат:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в Таблице 3; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность, представленную в Таблице 3, или VL другого антитела к ST2, где указанное антитело специфически связывается с ST2 человека.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, содержат:
(a) CDR1, CDR2 и CDR3 тяжелой цепи, перечисленные в Таблице 1; и
(b) CDR1, CDR2 и CDR3 легкой цепи, перечисленные в Таблице 2, или CDR другого антитела к ST2, где указанное антитело специфически связывается с ST2 человека.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, описанное в настоящем документе, содержит CDR2 тяжелой цепи антитела к ST2, описанного в настоящем документе, и CDR других антител, которые связываются с ST2 человека, например, CDR1 и/или CDR3 тяжелой цепи и/или CDR1, CDR2 и/или CDR3 легкой цепи другого антитела к ST2.
Кроме того, в данной области техники хорошо известно, что домен CDR3, независимо от домена(ов) CDR1 и/или CDR2, сам по себе может определять специфичность связывания антитела с идентичными антигенами, и что на основании последовательности CDR3 можно прогнозировать множественные антитела с одинаковой специфичностью связывания. См., например, Klimka et al., British J. of Cancer. 83(2):252-260 (2000); Beiboer et al., J. Mol.Biol. 296:833-849 (2000); Rader et al., Proc. Natl. Acad. Sci. U.S.A. 95:8910-8915 (1998); Barbas et al., J. Am. Chem. Soc. 116:2161-2162 (1994); Barbas et al., Proc. Natl. Acad. Sci. U.S.A. 92:2529-2533 (1995); Ditzel et al., J. Immunol. 157:739-749 (1996); Berezov et al., BIAjournal 8: Scientific Review 8 (2001); Igarashi et al., J. Biochem (Tokyo). 117:452-7 (1995); Bourgeois et al., J. Virol. 72:807-10 (1998); Levi et al., Proc. Natl. Acad. Sci. U.S.A. 90:4374-8 (1993); Polymenis and Stoller, J. Immunol.152:5218-5329 (1994) и Xu and Davis, Immunity. 13:37-45 (2000); и патенты США № 6,951,646; 6,914,128; 6,090,382; 6,818,216; 6,156,313; 6,827,925; 5,833,943; 5,762,905 и 5,760,185. Все эти ссылки включены в настоящий документ посредством ссылки в полном объеме.
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, описанный в данном документе, содержит CDR2 тяжелой цепи антитела к ST2, описанного в данном документе, и по меньшей мере CDR3 тяжелой цепи и/или легкой цепи антитела к ST2, описанного в данном документе, или CDR3 тяжелой цепи и/или легкой цепи другого антитела к ST2, где указанное антитело специфически связывается с ST2 человека. Предпочтительно, эти антитела (a) конкурируют за связывание с ST2; (b) сохраняют функциональные характеристики; (c) связываются с тем же эпитопом и/или (d) имеют такую же аффинность связывания, что и антитело к ST2, описанное в настоящем документе. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, описанное в данном документе, может дополнительно содержать CDR2 легкой цепи антитела к ST2, описанного в данном документе, или CDR2 легкой цепи другого антитела к ST2, где указанное антитело специфически связывается с ST2 человека. В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, описанное в данном документе, может дополнительно содержать CDR1 тяжелой и/или легкой цепи антитела к ST2, описанного в данном документе, или CDR1 тяжелой и/или легкой цепи другого антитела к ST2, где указанное антитело специфически связывается с ST2 человека.
Консервативная модификация
В другом варианте осуществления антитело или его антигенсвязывающий фрагмент, описанное в настоящем документе, содержит последовательности CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи и/или вариабельной области легкой цепи, содержащие одну или более консервативных модификаций по сравнению с антителом к ST2, описанным в настоящем документе. В данной области техники известно, что некоторые консервативные модификации последовательности не устраняют антигенсвязывающую способность. См., например, Brummell et al., Biochem 32:1180-8 (1993); de Wildt et al., Prot. Eng. 10:835-41 (1997); Komissarov et al., J. Biol. Chem. 272:26864-26870 (1997); Hall et al., J. Immunol. 149:1605-12 (1992); Kelley and O’Connell Biochem. 32:6862-35 (1993); Adib-Conquy et al., Int. Immunol. 10: 341-6 (1998); и Beers et al., Clin. Can. Res. 6:2835-43 (2000).
Следовательно, в одном варианте осуществления антитело содержит вариабельную область тяжелой цепи и/или вариабельную область легкой цепи, каждая из которых содержит CDR1, CDR2 и CDR3, где:
(a) последовательность CDR1 вариабельной области тяжелой цепи содержит последовательности, перечисленные в Таблице 1, и/или их консервативные модификации; и/или
(a) последовательность CDR2 вариабельной области тяжелой цепи содержит последовательности, перечисленные в Таблице 1, и/или их консервативные модификации; и/или
(с) последовательность CDR3 вариабельной области тяжелой цепи содержит последовательности, перечисленные в Таблице 1, и/или их консервативные модификации; и/или
(d) последовательности CDR1, и/или CDR2, и/или CDR3 вариабельной области легкой цепи содержат последовательности, перечисленные в Таблице 2; и/или их консервативные модификации; и
(e) антитело специфически связывается с ST2 человека.
Антитело, описанное в настоящем документе, имеет одно или более из следующих функциональных свойств, описанных выше, например, обладает высокой аффинностью к ST2 человека и блокирует связывание IL33/ST2 и передачу сигналов.
В нескольких вариантах осуществления изобретения антитело может представлять собой мышиное, химерное или гуманизированное антитело или его антигенсвязывающий фрагмент.
Термин «консервативная модификация последовательности», при использовании в данном документе, относится к аминокислотной модификации, которая существенно не влияет на или не изменяет связывающее свойство антитела. Такие консервативные модификации включают аминокислотные замены, добавления и делеции. Модификации могут быть введены в антитело, описанное в настоящем документе, с использованием стандартных методик, известных в данной области техники, таких как точечная мутация и ПЦР-опосредованная мутация. Консервативные аминокислотные замены представляют собой замены, в которых аминокислотный остаток заменен аминокислотным остатком, имеющим схожую боковую цепь. Семейства аминокислотных остатков, имеющих схожие боковые цепи, известны в данной области техники. Эти семейства аминокислотных остатков включают аминокислоты с основными боковыми цепями (например, лизин, аргинин и гистидин), аминокислоты с кислыми боковыми цепями (например, аспарагиновая кислота и глутаминовая кислота), аминокислоты с незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин и триптофан), аминокислоты с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин и метионин), аминокислоты с β-разветвленными боковыми цепями (например, треонин, валин и изолейцин) и аминокислоты с ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан и гистидин). Следовательно, один или более аминокислотных остатков в CDR антитела, описанного в настоящем документе, могут быть заменены другими аминокислотными остатками того же семейства боковых цепей, и полученное антитело может быть протестировано на сохранение функций (т.е. функций, описанных выше) с использованием функциональных тестов, описанных в настоящем описании.
Сконструированное и модифицированное антитело
Антитело, описанное в настоящем документе, может быть сконструировано для получения модифицированных антител с использованием антител, имеющих одну или более последовательностей VH/VL антитела к ST2, описанного в настоящем документе, в качестве исходных материалов. Один или более остатков в пределах одной или обеих вариабельных областей (т.е. VH и/или VL) (например, в пределах одной или более CDR и/или одной или более каркасных областей) антитела могут быть генетически модифицированы. Дополнительно, или в качестве альтернативы, остатки в константной области антитела могут быть сконструированы, например, для изменения эффекторной функции антитела.
В некоторых вариантах осуществления трансплантация CDR может быть использована для генетической модификации вариабельных областей антитела. Антитело взаимодействует с антигеном-мишенью главным образом через аминокислотные остатки в шести определяющих комплементарность областях (CDR) тяжелой цепи и легкой цепи. Следовательно, аминокислотные последовательности в пределах CDR каждого антитела являются более разнообразными, чем последовательности вне CDR. Поскольку последовательности CDR отвечают за основные взаимодействия антитело-антиген, векторы экспрессии, в которых последовательности CDR конкретных встречающихся в природе антител привиты в каркасные последовательности других антител с другими свойствами, могут быть сконструированы для экспрессии рекомбинантных антител, моделирующих свойства конкретных встречающихся в природе антител (Riechmann et al., Nature. 332:323-327 (1998); Jones et al., Nature. 321:522-525 (1986); Queen et al., Proc. Natl. Acad. U.S.A. 86:10029-10033 (1989); и патенты США № 5,225,539; 5,530,101; 5,585,089; 5,693,762 и 6,180,370).
Соответственно, другой вариант осуществления настоящего изобретения относится к выделенному моноклональному антителу или его антигенсвязывающему фрагменту, которое содержит вариабельную область тяжелой цепи, содержащую CDR1, CDR2 и CDR3 с последовательностями, описанными выше в настоящем описании, и/или вариабельную область легкой цепи, содержащую CDR1, CDR2 и CDR3 с последовательностями, описанными выше в настоящем описании. Хотя эти антитела содержат последовательности CDR VH и VL моноклонального антитела, описанного в настоящем документе, они могут содержать различные каркасные последовательности.
Такие каркасные последовательности можно найти в общедоступных базах данных ДНК или общедоступных ссылках, включающие последовательности генов антител зародышевой линии. Например, последовательности ДНК зародышевой линии для генов вариабельной области тяжелой цепи и вариабельной области легкой цепи человека можно найти в базе данных последовательностей зародышевой линии человека Vbase (www.mrc-cpe.cam.ac.uk/Vbase) и Kabat et al, (1991), выше; Tomlinson et al., J. Mol. Biol. 227:776-798 (1992); и Cox et al., Eur. J. Immunol. 24:827-836 (1994). В другом варианте осуществления последовательности ДНК зародышевой линии для генов вариабельной области тяжелой цепи и вариабельной области легкой цепи человека можно найти в базе данных Genbank.
Последовательности белка антитела сравнивали с базами данных последовательностей белка с использованием одного из способов поиска сходства последовательностей Gapped BLAST (Altschul et al, (1997), выше), которые хорошо известны специалистам в данной области техники.
Каркасные последовательности антител, описанных в настоящем документе, предпочтительно являются такими, которые структурно схожи с каркасными последовательностям, используемыми для антител, описанных в настоящем документе. Последовательности CDR1, CDR2 и CDR3 VH могут быть привиты в каркасную область, которая содержит ту же последовательность, что и ген иммуноглобулина зародышевой линии, из которого были получены каркасные последовательности, или последовательности CDR могут быть привиты в каркасную область, которая содержит одну или более мутаций по сравнению с последовательностью зародышевой линии. Например, в некоторых случаях может быть полезно мутировать остатки в каркасной области; такие мутации могут поддерживать или усиливать антигенсвязывающую способность антитела (см., например, патенты США № 5,530,101; 5,585,089; 5,693,762 и 6,180,370).
Другой тип модификации вариабельной области представляет собой мутацию аминокислотных остатков в пределах CDR1, CDR2 и/или CDR3 VH и/или VL для улучшения одного или более свойств (например, аффинности и физико-химических свойств) целевого антитела. Мутации могут быть введены с помощью сайт-направленного мутагенеза или ПЦР-опосредованного мутагенеза, и влияние мутаций на связывание антитела или другие функциональные свойства может быть оценено с помощью анализов in vitro или in vivo, известных в данной области техники. Предпочтительно вводят консервативные модификации, известные в данной области техники. Консервативные модификации могут представлять собой аминокислотные замены, добавления или делеции, предпочтительно замены. Кроме того, как правило, изменяют не более одного, двух, трех, четырех или пяти остатков в пределах каждой CDR.
Кроме того, в другом варианте осуществления предложено выделенное моноклональное антитело к ST2 или его антигенсвязывающий фрагмент, которое содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, содержащие: (a) VH CDR1, содержащую последовательность по настоящему изобретению, или аминокислотную последовательность, содержащую 1, 2, 3, 4 или 5 аминокислотных замен, делеций или добавлений; (b) VH CDR2, содержащую последовательность по настоящему изобретению, или аминокислотную последовательность, содержащую 1, 2, 3, 4 или 5 аминокислотных замен, делеций или добавлений; (c) VH CDR3, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, содержащую 1, 2, 3, 4 или 5 аминокислотных замен, делеций или добавлений; (d) VL CDR1, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, содержащую 1, 2, 3, 4 или 5 аминокислотных замен, делеций или добавлений; (e) VL CDR2, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, содержащую 1, 2, 3, 4 или 5 аминокислотных замен, делеций или добавлений; и (f) VL CDR3, содержащую последовательность по настоящему изобретению или аминокислотную последовательность, содержащую 1, 2, 3, 4 или 5 аминокислотных замен, делеций или добавлений.
Генетически сконструированные антитела, описанные в настоящем документе, включают те антитела, в которых остатки каркасной области VH и/или VL были модифицированы для улучшения свойств антител. В целом, такие модификации каркасной области могут снижать иммуногенность антитела. Например, один или более каркасных остатков являются «обратно мутированными» в соответствующую последовательность зародышевой линии. Более конкретно, антитела, подвергающиеся соматической мутации, могут содержать остатки каркасной области, которые отличаются от полученной последовательности зародышевой линии антитела. Эти остатки могут быть идентифицированы путем сравнения каркасной последовательности антитела с последовательностью зародышевой линии полученного антитела.
Другой тип каркасной модификации включает мутацию одного или более остатков каркасной области или даже одной или более CDR для удаления Т-клеточных эпитопов, тем самым снижая иммуногенность, которую может продуцировать антитело. Этот способ также известен как «деиммунизация» и более подробно описан в публикации патента США № 20030153043.
Кроме того, в дополнение к каркасной области или модификациям CDR, для изменения одного или более функциональных свойств антитела, таких как период полужизни в сыворотке, фиксация комплемента, связывание Fc-рецептора и/или антиген-зависимая цитотоксичность, часто используют другой тип модификации, например, модификации Fc-области антитела, описанного в настоящем документе, с помощью генной инженерии. Кроме того, антитело, описанное в настоящем документе, также может быть химически модифицировано (например, быть связанным с одной или более химическими функциональными группами) или модифицировано для изменения его гликозилирования, для изменения одного или более функциональных свойств антитела.
В одном варианте осуществления CH1-шарнирная область модифицирована, например, путем увеличения или уменьшения количества остатков цистеина в шарнирной области. Этот способ подробно описан в патенте США № 5,677,425. Изменение количества остатков цистеина в CH1-шарнирной области может, например, облегчить сборку легкой цепи и тяжелой цепи или улучшить или понизить стабильность антитела.
В другом варианте осуществления Fc-шарнирную область антитела подвергают мутации для увеличения или уменьшения биологического периода полужизни антитела. Более конкретно, одну или более аминокислотных мутаций вводят в область CH2-CH3 Fc-шарнирной области таким образом, что антитело имеет сниженное связывание со стафилококковым белком A (SpA) по сравнению с антителом с природным Fc-шарнирным доменом. Этот способ более подробно описан в патенте США № 6,165,745.
В еще одном варианте осуществления модифицируют гликозилирование антитела. Например, могут быть получены дегликозилированные антитела (т.е. антитела не имеют гликозилирования). Такие модификации гликозилирования могут быть достигнуты, например, путем изменения одного или более сайтов гликозилирования в последовательности антитела. Например, одна или более аминокислотных замен могут быть сделаны для устранения сайтов гликозилирования в одном или более каркасах вариабельной области и, таким образом, гликозилирования в этих сайтах. Такое дегликозилирование может увеличивать аффинность антитела к антигену. См., например, патенты США № 5,714,350 и 6,350,861.
Кроме того, могут быть получены антитела с измененным гликозилированием, такие как низкофукозилированные антитела с уменьшенными остатками фукозы или антитела с увеличенными биссекционирующими структурами GlcNac. Было продемонстрировано, что измененное гликозилирование повышает активность ADCC (антителозависимая клеточная цитотоксичность) антитела. Такие модификации гликозилирования могут быть достигнуты, например, путем экспрессии антитела в клетке-хозяине, в которой система гликозилирования была изменена, что известно в данной области техники и может быть использовано в качестве клетки-хозяина для экспрессии рекомбинантных антител, описанных в настоящем документе, с получением антител с измененным гликозилированием. Например, поскольку в клеточных линиях Ms704, Ms705 и Ms709 отсутствует ген фукозилтрансферазы FUT8 (α(1,6)-фукозилтрансфераза), антитела, экспрессируемые в клеточных линиях Ms704, Ms705 и Ms709, не содержат фукозы. FUT8-/- клеточные линии Ms704, Ms705 и Ms709 получали путем целевого разрушения гена FUT8 в клетках CHO/DG44 с использованием двух альтернативных векторов (см. патент США № 20040110704 и Ohnuki et al., Biotechnol Bioeng. 87:614-22 (2004)). В качестве другого примера, EP1,176,195 описывает клеточную линию с нарушенной функцией гена FUT8. Ген кодирует фукозилтрансферазу, так что антитела, экспрессируемые в такой клеточной линии, демонстрируют низкое фукозилирование путем снижения или устранения ферментов, связанных с α-1,6-связью. EP1,176,195 также описывает клеточную линию с низкой или отсутствующей ферментативной активностью для добавления фукозы в N-ацетилглюкозамин, связанный с Fc-областью антитела, например, клеточную линию миеломы крыс YB2/0 (ATCC CRL 1662). WO03/035835 описывает клетки Lec13, вариантную клеточную линию CHO (яичник китайского хомячка), которая имеет сниженную способность к добавлению фукозы к Asn(297)-родственным сахарам, что приводит к низкому фукозилированию в антителах, экспрессируемых клеткой-хозяином (см. Shields et al., J. Biol. Chem. 277:26733-26740 (2002)). Антитела с измененными характеристиками гликозилирования также могут быть получены в яйцах, как описано в WO06/089231. Альтернативно, антитела с измененными характеристиками гликозилирования могут быть получены в растительных клетках, таких как Lemna (ряска). WO99/54342 раскрывает клеточную линию, генетически сконструированную для экспрессии гликозилтрансферазы, которая модифицирует гликопротеины (например, β(1,4)-N-ацетилглюкозаминтрансферазу III (GnTIII)) таким образом, что антитела, экспрессируемые в клеточной линии, демонстрируют увеличенные биссекционирующие структуры GlcNac и, таким образом, обладают повышенной активностью ADCC (см. также Umana et al., Nat. Biotech. 17:176-180, (1999)). Альтернативно, фукозидазу используют для отсечения остатков фукозы антитела; например, α-L-фукозидаза удаляет остатки фукозы из антитела (Tarentino et al., Biochem.14:5516-23 (1975)).
Другим типом модификации антитела, описанного в настоящем документе, является пегилирование. Антитело может быть пегилировано, например, для увеличения биологического (например, сывороточного) периода полужизни антитела. Для получения пегилированного антитела указанное антитело или его фрагмент подвергают взаимодействию с полиэтиленгликолем (ПЭГ), например, реакционноспособным сложным эфиром или альдегидным производным ПЭГ, в таких условиях, что одна или более групп ПЭГ присоединяется к антителу или фрагменту антитела. Предпочтительно, пегилирование осуществляют ацилированием или алкилированием с реакционноспособной молекулой ПЭГ (или аналогичного реакционноспособного водорастворимого полимера). Термин «полиэтиленгликоль» в данном контексте включает любую форму ПЭГ для получения других производных белка, такую как моно(C1-C10)алкокси- или арилокси-полиэтиленгликоль или полиэтиленгликоль-малеимид. В некоторых вариантах осуществления антитело, подлежащее пегилированию, представляет собой дегликозилированное антитело. Способы пегилирования известны в данной области техники и могут быть применены к антителу, описанному в настоящем документе. См., например, EP0154316 и EP0401384.
Физические свойства антитела
Антитела, описанные в настоящем документе, могут быть охарактеризованы их различными физическими свойствами, и, таким образом, определяются и/или различаются категории, к которым они принадлежат. Например, антитело может содержать один или более сайтов гликозилирования в вариабельной области легкой цепи или тяжелой цепи. Эти сайты гликозилирования могут приводить к повышению иммуногенности антитела или изменению значения pK антитела, вызванному измененным связыванием антигена (Marshall et al., Annu Rev Biochem. 41:673-702 (1972); Gala and Morrison. J Immunol. 172: 5489-94 (2004); Wallick et al., J Exp Med. 168: 1099-109 (1988); Spiro, Glycobiology. 12: 43R-56R (2002); Parekh et al., Nature 316:452-7 (1985); и Mimura et al., Mol Immunol 37:697-706 (2000)). Известно, что гликозилирование происходит в мотивах, содержащих последовательности N-X-S/T. В некоторых случаях предпочтительно, чтобы антитело к ST2 не содержало гликозилирования вариабельной области. Это может быть достигнуто либо путем отбора антител, которые не содержат мотив гликозилирования в вариабельных областях, либо путем мутации остатков в области гликозилирования.
В одном предпочтительном варианте осуществления антитело не содержит сайт изомеризации аспарагина. Дезаминирование аспарагина может происходить в последовательностях N-G или D-G и приводить к образованию остатков изоаспарагиновой кислоты, снижая стабильность.
Каждое антитело имеет уникальную изоэлектрическую точку (pI), обычно в диапазоне рН от 6 до 9,5. Значение pI антител IgG1 обычно находится в диапазоне рН от 7 до 9,5, тогда как рН антител IgG4 обычно находится в диапазоне рН от 6 до 8. Предполагается, что антитела со значениями pI, выходящими за пределы нормального диапазона, могут подвергаться некоторому разворачиванию и быть нестабильными в условиях in vivo. Следовательно, антитела к ST2, имеющие значение pI в пределах нормального диапазона, являются предпочтительными. Это может быть достигнуто либо путем отбора антител с pI в пределах нормального диапазона, либо путем мутации поверхностных остатков.
Молекула нуклеиновой кислоты, кодирующая антитело, описанное в настоящем документе
В другом аспекте настоящее изобретение относится к молекуле нуклеиновой кислоты, кодирующей вариабельные области или CDR тяжелой цепи и/или легкой цепи антитела, описанного в настоящем документе. Нуклеиновая кислота может присутствовать в интактных клетках, в клеточных лизатах или в частично очищенной или по существу чистой форме. Нуклеиновая кислота является «выделенной» или «по существу чистой» при очистке от других клеточных компонентов или других загрязнений, например, других клеточных нуклеиновых кислот или белков, с помощью стандартных методик. Нуклеиновая кислота, описанная в настоящем документе, может представлять собой, например, ДНК или РНК, и может содержать или не содержать интронные последовательности. В предпочтительном варианте осуществления нуклеиновая кислота является кДНК молекулой.
Нуклеиновая кислота, описанная в настоящем документе, может быть получена с использованием стандартных методов молекулярной биологии. Для антител, экспрессируемых гибридомами (например, гибридомами, полученными от трансгенных мышей, несущих гены иммуноглобулина человека), кДНК легкой цепи и тяжелой цепи, кодирующие антитела, полученные из гибридом, могут быть получены с помощью стандартной ПЦР-амплификации или с использованием методов клонирования кДНК. Для антител, полученных из библиотек генов иммуноглобулинов (например, с использованием технологии фагового дисплея), нуклеиновые кислоты, кодирующие такие антитела, могут быть выделены из библиотек генов.
Предпочтительно молекула нуклеиновой кислоты, раскрытая в настоящем документе, включает такие, которые кодируют последовательности VH и VL или CDR моноклонального антитела к ST2, раскрытого в настоящем документе. После получения фрагментов ДНК, кодирующих фрагменты VH и VL, могут быть дополнительно проведены такие операции, как преобразование генов вариабельной области в гены цепи полноразмерного антитела, гены Fab-фрагментов или гены scFv, с использованием стандартных методов рекомбинантной ДНК. В этих операциях фрагмент ДНК, кодирующий VL или VH, функционально связан с другим фрагментом ДНК, кодирующим другой белок, например, константной областью антитела или гибким линкером. Термин «функционально связанный», используемый в данном контексте, означает, что два фрагмента ДНК объединены так, что аминокислотные последовательности, кодируемые этими двумя фрагментами ДНК, остаются в рамке считывания.
Выделенная ДНК, кодирующая область VH, может быть преобразована в ген полноразмерной тяжелой цепи путем функционального связывания ДНК, кодирующей VH, с другой молекулой ДНК, кодирующей константную область тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константной области тяжелой цепи человека известны в данной области техники, и эти фрагменты ДНК могут быть получены с помощью стандартной ПЦР-амплификации. Константная область тяжелой цепи может представлять собой константную область IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно представляет собой константную область IgG2. Для гена Fab-фрагмента тяжелой цепи ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей только константную область CH1 тяжелой цепи.
Выделенная ДНК, кодирующая область VL, может быть преобразована в ген полноразмерной легкой цепи (а также в ген Fab легкой цепи) путем функционального связывания ДНК, кодирующей VL, с другой молекулой ДНК, кодирующей константную область легкой цепи (CL). Последовательности генов константной области легкой цепи человека известны в данной области техники, и эти фрагменты ДНК могут быть получены с помощью стандартной ПЦР-амплификации. В предпочтительных вариантах осуществления константная область легкой цепи может представлять собой константную область легкой цепи κ или λ.
Для получения гена scFv фрагмент ДНК, кодирующий VH и VL, функционально связан с другим фрагментом, кодирующим гибкий линкер, например, другим фрагментом, кодирующим аминокислотную последовательность (Gly4-Ser)3, так что последовательности VH и VL могут быть экспрессированы в виде непрерывного одноцепочечного белка, в котором области VL и VH связаны гибким линкером (см., например, Bird et al., Science 242:423-426 (1988); Huston et al., Proc Nat. Acad. Sci. USA 85:5879-5883 (1988); и McCafferty et al., Nature 348:552-554 (1990)).
Получение моноклонального антитела, описанного в настоящем документе
Моноклональное антитело (mAb), описанное в настоящем документе, может быть получено с использованием методики соматической гибридизации (гибридомы), описанной в Kohler and Milstein Nature. 256:495 (1975). Другие варианты осуществления изобретения для получения моноклонального антитела включают вирусную или онкогенную трансформацию В-лимфоцитов и методики фагового дисплея. Химерные или гуманизированные антитела также хорошо известны в данной области техники. См., например, патенты США № 4,816,567; 5,225,539; 5,530,101; 5,585,089; 5,693,762 и 6,180,370.
Трансфектома для получения моноклонального антитела
Антитело, описанное в настоящем документе, также может быть получено в трансфектомах клеток-хозяев с использованием, например, методов рекомбинантной ДНК в комбинации со способами трансфекции генов (например, Morrison, S. Science 229:12021985). В одном варианте осуществления ДНК, кодирующая частичные или полноразмерные легкие и тяжелые цепи, полученная стандартными методами молекулярной биологии, встраивается в один или более векторов экспрессии таким образом, что гены функционально связаны с транскрипционными и трансляционными регуляторными последовательностями. В этом случае термин «функционально связанный» относится к связыванию гена антитела с вектором таким образом, что транскрипционные и трансляционные регуляторные последовательности в векторах выполняют свои предполагаемые функции регулирования транскрипции и трансляции гена антитела.
Термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования), которые контролируют транскрипцию или трансляцию гена антитела. Такие регуляторные последовательности описаны в Goeddel (Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, CA (1990)). Предпочтительно регуляторные последовательности для экспрессии в клетке-хозяине млекопитающего включают вирусные элементы, которые направляют экспрессию белка высокого уровня в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовирусов, например, главный поздний промотор аденовируса (AdMLP). Альтернативно используют невирусные регуляторные последовательности, такие как промоторы убиквитина или промоторы β-глобина. Кроме того, регуляторные элементы состоят из последовательностей различного происхождения, такие как система промотора SRα, которая содержит последовательность раннего промотора SV40 и последовательность длинного концевого повтора вируса Т-клеточного лейкоза человека I типа (Takebe et al., Mol Cell. Biol. 8:466-472 (1988)). Вектор экспрессии и экспрессионные регуляторные последовательности совместимы с используемой экспрессирующей клеткой-хозяин.
Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть вставлены в один и тот же вектор экспрессии или различные векторы экспрессии. В предпочтительных вариантах осуществления вариабельные области вставляют в вектор экспрессии, который кодирует константную область тяжелой цепи и константную область легкой цепи требуемого подтипа, для конструирования полноразмерного гена антитела, так что VH функционально связан с CH в векторе, а VL функционально связан с CL в векторе. Альтернативно рекомбинантный вектор экспрессии может кодировать сигнальный пептид, который облегчает секрецию цепей антител из клетки-хозяина. Ген цепи антитела может быть клонирован в векторе таким образом, что сигнальный пептид связан в рамке считывания с амино-концом гена цепи антитела. Сигнальный пептид может быть сигнальным пептидом иммуноглобулина или гетерологичным сигнальным пептидом (то есть сигнальным пептидом белка, не являющегося иммуноглобулином).
В дополнение к генам цепей антитела и регуляторным последовательностям, рекомбинантный вектор экспрессии, описанный в настоящем документе, может нести другие последовательности, такие как последовательность, которая регулирует репликацию вектора в клетке-хозяине (например, начало репликации), и селектируемый маркерный ген. Селектируемый маркерный ген может быть использован для отбора клеток-хозяев, в которые был введен вектор (см., например, патенты США № 4,399,216; 4,634,665 и 5,179,017). Например, селектируемый маркерный ген обычно придает устойчивость к лекарственному средству, такому как G418, гигромицин или метотрексат, клетке-хозяину, в которую был введен вектор. Предпочтительные селектируемые маркерные гены включают ген дигидрофолатредуктазы (DHFR) (для селекции/амплификации метотрексата в DHFR клетках-хозяевах) и ген neo (для селекции G418).
Для экспрессии легкой цепи и тяжелой цепи клетки-хозяева трансфицируют векторами экспрессии, кодирующими тяжелую цепь и легкую цепь, с использованием стандартных методик. Термин «трансфект» охватывает различные способы введения экзогенной ДНК в прокариотические или эукариотические клетки-хозяева, такие как электропорация, осаждение фосфатом кальция и трансфекция ДЭАЭ-декстраном (диэтиламиноэтилдекстран). Хотя экспрессия антитела, описанного в настоящем документе, в прокариотических или эукариотических клетках-хозяевах теоретически возможна, экспрессия антитела в эукариотических клетках является предпочтительной, и экспрессия антитела в клетках-хозяевах млекопитающих является наиболее предпочтительной. Это связано с тем, что эукариотические клетки, особенно клетки млекопитающих, чаще собираются и секретируют правильно сложенные и иммунологически активные антитела, чем прокариотические клетки.
Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантного антитела, описанного в настоящем документе, включают клетки яичника китайского хомячка (клетки CHO) (включая клетки dhfr-CHO, вводимые селектируемым маркером DHFR, как описано в Urlaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220 (1980); селектируемый маркер DHFR является таким, как описано в RJ Kaufman and PA Sharp. J. Mol. Biol. 159: 601-621 (1982)), клетки миеломы NSO, клетки COS и клетки SP2. Другой предпочтительной системой экспрессии, в частности, при использовании клеток миеломы NSO, является система экспрессии гена GS, раскрытая в WO87/04462, WO89/01036 и EP338,841. Когда рекомбинантный вектор экспрессии, кодирующий ген антитела, вводят в клетку-хозяина млекопитающего, антитело получают путем культивирования клетки-хозяина в течение периода времени, достаточного для обеспечения экспрессии антитела в клетке-хозяине или, предпочтительно, достаточного для обеспечения секреции антитела в среде, в которой растет клетка-хозяин. Антитело может быть выделено из культуральной среды с использованием способов очистки белка.
Слитый полипептид
В другом аспекте настоящее изобретение относится к слитому полипептиду, содержащему одно или более антител или их антигенсвязывающих фрагментов, описанных в настоящем документе, где антитела или их антигенсвязывающие фрагменты, описанные в настоящем документе, связаны с по меньшей мере одной другой функциональной молекулой, которая может представлять собой пептид, или белок, или небелок. В одном варианте осуществления слитый полипептид, описанный в данном документе, включает иммуноконъюгаты, содержащие одно или более антител или их антигенсвязывающих фрагментов, описанных в данном документе, и по меньшей мере один терапевтический агент, такой как стероид, связанный с антителами или их антигенсвязывающими фрагментами, описанными в данном документе. В одном варианте осуществления слитый полипептид, описанный в данном документе, включает бифункциональные молекулы, содержащие одно или более антител или их антигенсвязывающих фрагментов, описанных в данном документе, и по меньшей мере один иммунный цитокин или лиганд рецептора, связанный с антителом или его антигенсвязывающим фрагментом, описанным в данном документе, причем иммунный цитокин или лиганд рецептора может быть для IgE, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-9, IL13, IL13R, IL-17, IL-23, IL-33, OX40 лиганда (OX40L), GM-CSF, или TSLP, или TSLPR/IL7R. В одном варианте осуществления бифункциональные молекулы имеют третью функцию в дополнение к функции связывания Fc-рецептора и функции связывания ST2. Третья функция может быть для IgE, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-9, IL13, IL13R, IL-17, IL-23, IL-33, OX40 лиганда (OX40L), GM-CSF, или TSLP, или TSLPR/IL7R. В данном контексте термин «бифункциональная молекула» охватывает молекулы, имеющие три или более функций. В других вариантах осуществления слитый полипептид, описанный в настоящем документе, также включает другие формы. Эти и другие формы слитого полипептида могут быть получены с помощью генной инженерии, с помощью химических способов и т.д.
Мультиспецифическая молекула
В другом аспекте настоящее изобретение относится к мультиспецифической молекуле, содержащей одно или более антител или их антигенсвязывающих фрагментов, описанных в настоящем документе, где антитела или их антигенсвязывающие фрагменты, описанные в настоящем документе, связаны с по меньшей мере одной другой функциональной частью со специфичностью, отличной от специфичности антител или антигенсвязывающих фрагментов, описанных в настоящем документе, причем указанная другая функциональная часть содержит другой пептид или белок (например, другое антитело или его антигенсвязывающий фрагмент) для получения мультиспецифической молекулы, которая связывается с по меньшей мере двумя различными сайтами связывания или мишенями. Таким образом, «мультиспецифическая молекула», как описано в данном документе, охватывает молекулы с двумя типами специфичности (т.е. биспецифические молекулы), тремя типами специфичности (т.е. триспецифические молекулы), четырьмя типами специфичности (т.е. тетраспецифические молекулы) или более специфичностями.
В одном конкретном варианте осуществления мультиспецифическая молекула, описанная в настоящем документе, может иметь один или более других типов специфичности в дополнение к специфичности связывания к Fc и специфичности связывания ST2. В качестве примера, другие типы специфичности могут быть для IgE, IL4, IL4R, IL5, IL5R, IL6, IL9, IL13, IL13R, IL17, IL23, IL33, или TSLP, или TSLPR/IL7R.
В одном конкретном варианте осуществления мультиспецифическая молекула может присутствовать в различных формах и размерах. В качестве примера, с точки зрения биспецифической молекулы, на одном конце спектра размеров биспецифическая молекула остается в форме обычного антитела, за исключением того, что она имеет два связывающих плеча с различными типами специфичности, а не два связывающих плеча с одинаковой специфичностью. На другом конце биспецифическая молекула состоит из двух одноцепочечных фрагментов антитела (scFv), связанных пептидной цепью, известная как конструкция Bs(scFv)2. Биспецифические молекулы среднего размера содержат два различных фрагмента F(ab), связанных пептидным линкером. Эти и другие формы биспецифической молекулы могут быть получены с помощью генной инженерии или соматической гибридизации или с помощью химических методов. См., например, Kufer et al., процитирован выше; Cao and Suresh, Bioconjugate Chemistry. 9(6).635-644 (1998); и van Spriel et al., Immunology Today. 21(8):391-397 (2000).
Вирусный вектор
В другом аспекте антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, также могут кодироваться или переноситься с помощью вирусного вектора. Кроме того, антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, также могут быть использованы с вирусным вектором, или вирусный вектор, кодирующий или несущий антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем документе, вводят людям.
Настоящее изобретение также относится к фармацевтической композиции, содержащей слитый полипептид, мультиспецифическую молекулу и вирусный вектор, описанные в настоящем документе, и фармацевтически приемлемый эксципиент, разбавитель или носитель.
Фармацевтическая композиция
В другом аспекте настоящее изобретение относится к фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, составленные вместе с фармацевтически приемлемым эксципиентом, разбавителем или носителем. Фармацевтическая композиция может необязательно содержать один или более других фармацевтически активных ингредиентов, таких как другое антитело или лекарственное средство, например, другое антитело к ST2, антитело к IgE, другое противовоспалительное лекарственное средство, лекарственное средство против астмы, лекарственное средство против хронической обструктивной болезни легких, лекарственное средство против язвенного колита, лекарственное средство против атопического дерматита или лекарственное средство против псориаза.
Предпочтительно, фармцевтическая композиция является пригодной для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, инъекцией или инфузией). В зависимости от способа введения, активный ингредиент может быть энкапсулирован в материал, чтобы защитить его от действия кислот и других природных условий, которые могут инактивировать его. «Парентеральное введение» относится к режиму, который отличается от энтерального введения и местного введения и который обычно осуществляют путем инъекции, включая, но не ограничиваясь, внутривенную, внутримышечную, внутриартериальную, внутрипозвоночную, внутрикапсульную, внутриорбитальную, внутрисердечную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, субкапсулярную, субарахноидальную, внутриспинальную, эпидуральную и внутригрудинную инъекцию и болюсную инъекцию. Альтернативно фармацевтическая композиция, описанная в настоящем документе, может быть введена непарентеральным путем, например, местное, эпидермальное или мукозальное введение, например, интраназальное, пероральное, вагинальное, ректальное, сублингвальное или местное введение.
Фармацевтические композиции могут быть в форме стерильных водных растворов или дисперсий. Они также могут быть составлены в виде микроэмульсий, липосом или других упорядоченных структур, подходящих для высоких концентраций лекарственных средств.
Схему введения корректируют для обеспечения наилучшего желаемого ответа (например, терапевтического ответа). Например, может быть введена одна большая доза, могут быть введены несколько разделенных доз в течение времени или доза может быть пропорционально уменьшена или увеличена, на что указывает острота терапевтической ситуации. Особенно предпочтительно составление парентеральных композиций в виде единиц дозирования для обеспечения простоты введения и однородности дозы. Единицы дозирования относятся к физически отдельным единицам, которые подходят для однократного введения субъекту, получающему лечение; каждая единица содержит заранее определенное количество активного ингредиента, рассчитанное для получения желаемого терапевтического эффекта, в сочетании с фармацевтическим носителем. Альтернативно антитело можно вводить в составе с замедленным высвобождением, и в этом случае частота введения уменьшается.
Для введения композиции доза может составлять от около 0,0001 до 100 мг/кг массы тела хозяина, чаще от 0,01 до 5 мг/кг. Например, доза может составлять 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела или находиться в диапазоне от 1 до 10 мг/кг массы тела. Примеры схем лечения требуют введения два раза в неделю, один раз в неделю, один раз в две недели, один раз в три недели, один раз в четыре недели, один раз в месяц, один раз в два месяца, один раз в три месяца или один раз в три-шесть месяцев.
Фармацевтическая композиция может представлять собой агент с замедленным высвобождением, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биоразлагаемые и биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота.
В некоторых вариантах осуществления антитело, описанное в настоящем документе, может быть составлено для обеспечения надлежащего распределения in vivo. Например, для обеспечения того, чтобы терапевтическое антитело, описанное в настоящем документе, пересекало гематоэнцефалический барьер, антитело может быть составлено в липосомы, которые могут дополнительно содержать нацеливающие функциональные группы для усиления селективной доставки в определенные клетки или органы.
Применение и способ согласно настоящему изобретению
Антитело или его антигенсвязывающий фрагмент (кодирующая молекула нуклеиновой кислоты, фармацевтическая композиция, слитый полипептид, мультиспецифическая молекула или вирусный вектор), описанные в данном документе, имеют множество применений in vitro и in vivo, относящихся к диагностике, лечению и/или предотвращению заболеваний и состояний, опосредованных IL33/ST2. Заболевания и состояния, опосредованные IL33/ST2, включают, но не ограничиваются, астму, аллергический ринит, хроническую обструктивную болезнь легких, эозинофильный бронхит, эозинофильный эзофагит, атопический дерматит, псориаз, системную красную волчанку, пемфигоид, ревматоидный артрит, анкилозирующий спондилоартрит, воспалительное заболевание кишечника, фиброз легких, фиброз печени, системный склероз, саркоидоз, болезнь «трансплантат против хозяина» (GVHD), диабет, сердечно-сосудистые заболевания или их комбинации. Антитело или его антигенсвязывающий фрагмент, описанные в настоящем документе, можно вводить субъектам-людям для облегчения, уменьшения или лечения заболеваний или состояний. В процессе диагностики антитело, описанное в настоящем документе, может быть приведено в контакт с образцом субъекта в условиях, которые позволяют антителу или его антигенсвязывающему фрагменту, описанному в настоящем документе, связываться с ST2, для обнаружения количества белка ST2 в образце субъекта.
Эти и другие способы согласно настоящему изобретению более подробно обсуждаются ниже.
Комбинированная терапия
Настоящее изобретение относится к совместному введению антитела к ST2 или его антигенсвязывающего фрагмента (кодирующей молекулы нуклеиновой кислоты, фармацевтической композиции, слитого полипептида, мультиспецифической молекулы или вирусного вектора), описанных в настоящем документе, с одним или более другими антителами или лекарственными средствами, которые могут уменьшать, облегчать или лечить заболевания и состояния, опосредованные IL33/ST2, у субъекта, причем указанные заболевания и состояния, опосредованные IL33/ST2, включают, но не ограничиваются, астму, аллергический ринит, хроническую обструктивную болезнь легких, эозинофильный бронхит, эозинофильный эзофагит, атопический дерматит, псориаз, системную красную волчанку, пемфигоид, ревматоидный артрит, анкилозирующий спондилоартрит, воспалительное заболевание кишечника, фиброз легких, фиброз печени, системный склероз, саркоидоз, болезнь «трансплантат против хозяина» (GVHD), диабет, сердечно-сосудистые заболевания или их комбинации. В одном варианте осуществления предложен способ лечения астмы, хронического обструктивного заболевания легких или атопического дерматита у субъекта, где антитело к ST2 или его антигенсвязывающий фрагмент, описанные в данном документе, вводят вместе с одним или более другими антителами, например, антителом к TSLP, антителом к TSLPR, антителом к IL4, антителом к IL4R, антителом к IL13, антителом к IL13R, антителом к IL5, антителом к IL5R и/или антителом к IgE. В другом варианте осуществления предложен способ лечения астмы, хронического обструктивного заболевания легких или атопического дерматита у субъекта, где антитело к ST2 или его антигенсвязывающий фрагмент, описанные в данном документе, вводят вместе с по меньшей мере одним дополнительным лекарственным средством, например, лекарственным средством против астмы, лекарственным средством против хронического обструктивного заболевания легких или лекарственным средством против атопического дерматита. В некоторых вариантах осуществления субъектом является человек. Другие виды терапии, которые можно применять в комбинации с антителом к ST2 или его антигенсвязывающим фрагментом, описанным в настоящем документе, также включают: уменьшение или предотвращение воздействия аллергена, гормональную терапию, хирургическое вмешательство и т. д.
Комбинацию терапевтических агентов, описанных в настоящем документе, можно вводить одновременно в виде одной композиции в фармацевтически приемлемом носителе или вводить одновременно в виде отдельных композиций, где каждый агент находится в фармацевтически приемлемом носителе. В другом варианте осуществления комбинацию терапевтических агентов можно вводить последовательно.
Кроме того, если комбинированную терапию вводят несколько раз, а агенты вводят последовательно, то последовательность в последовательном введении в каждый момент времени может быть изменена или сохранена, и последовательное введение может быть скомбинировано с одновременным введением или любой их комбинацией.
Хотя вышеуказанное изобретение было подробно описано с помощью иллюстраций и примеров с целью четкого понимания, специалистам в данной области техники будет очевидно в свете идей настоящего изобретения, что определенные изменения и модификации могут быть внесены в настоящее изобретение без отступления от сущности или объема прилагаемой формулы изобретения. Настоящее изобретение дополнительно поясняется описанием следующих примеров, которые не предназначены для ограничения. Специалисты в данной области техники легко идентифицируют множество несущественных параметров, которые могут быть изменены или модифицированы для получения по существу аналогичных результатов.
Если не указано иное, при практическом осуществлении настоящего изобретения будут использоваться обычные способы в области химии белка, биохимии, технологии рекомбинантной ДНК и фармакологии в данной области техники.
Примеры
Пример 1. Получение антигена ST2 и тестового белка
Полноразмерный ген изоформы A (SEQ ID NO: 1) рецептора, подобного интерлейкину-1 (ST2), человека (hST2), по базе UniProt, использовали в качестве матрицы ST2 по настоящему изобретению для получения последовательности гена, кодирующей антиген и белок для обнаружения по настоящему изобретению, которую можно рекомбинировать с Fc-фрагментом тяжелой цепи мышиного антитела (таким как IgG2a мыши) с образованием hST2-mFc или рекомбинировать с Fc-фрагментом тяжелой цепи человеческого антитела с образованием hST2-hFc для иммунизации мышей или более поздних скрининговых анализов. кДНК, кодирующие рекомбинантный белок, содержащий внеклеточный домен hST2 с Fc-меткой тяжелой цепи мышиного антитела (hST2-ECD-mFc, SEQ ID NO: 2), и рекомбинантный белок, содержащий внеклеточный домен hST2 с Fc-меткой тяжелой цепи человеческого антитела (hST2-ECD-hFc, SEQ ID NO: 4), (SEQ ID NO: 3 и 5, соответственно) получали путем синтеза гена и субклонировали, соответственно, в векторы экспрессии pDNA3.1 (Invitrogen, V-790), и клетки Expi293 (Thermo, A14527) трансфицировали вектором, сконструированным выше, для транзиентной экспрессии. Затем рекомбинантные белки hST2-ECD-mFc и hST2-ECD-hFc очищали с помощью колонки с белком A (GE Healthcare).
кДНК, кодирующую рекомбинантный белок, содержащий IL33 человека с Avitag (SEQ ID NO: 7), получали путем синтеза гена и клонировали в вектор экспрессии, меченный GST (глутатион-S-трансфераза), и вектор, сконструированный выше, трансформировали в BL21-компетентные клетки для индуцированной экспрессии. Белок GST-IL33 Avitag очищали с помощью колонки для очистки GST и расщепляли ферментом тромбин (Sigma, T4648-1KU), а затем GST удаляли с получением hIL33 Avitag (SEQ ID NO: 6).
кДНК, кодирующую рекомбинантный белок IL33 человека (SEQ ID NO: 9), получали путем синтеза гена и клонировали в вектор экспрессии, меченный GST, и вектор, сконструированный выше, трансформировали в BL21-компетентные клетки для индуцированной экспрессии. Белок GST-IL33 очищали с помощью колонки для очистки GST и расщепляли ферментом тромбин (Sigma, T4648-1KU), а затем GST удаляли с получением белка hIL33 (SEQ ID NO: 8).
Антитело к ST2 RG6149 в примерах соответствует антителу Ab2 в CN104334582, которое было получено в лаборатории, и аминокислотные последовательности тяжелой цепи и легкой цепи которого представлены в SEQ ID NO: 83 и SEQ ID NO: 84 по настоящему изобретению. Антитело к ST2 GSK3772847 соответствует антителу STLM208 в CN104411333, которое было получено в лаборатории, и аминокислотные последовательности тяжелой цепи и легкой цепи которого представлены в SEQ ID NO: 85 и SEQ ID NO: 86 по настоящему изобретению.
Пример 2. Получение гибридомных моноклональных антител к ST2
Очищенный рекомбинантный белок hST2-ECD-mFc (100 мкг/мышь) хорошо смешивали с равным объемом полного адъюванта Фрейнда (Sigma, F5881-10X10ML) (первичная иммунизация) или неполного адъюванта Фрейнда (Sigma, F5506-10X10ML) (бустерная иммунизация) и эмульгировали, и мышей BALB/c иммунизировали подкожно полученной эмульсией каждые 2 недели в течение 8 недель. За три дня до слияния мышей иммунизировали антигеном hST2-ECD-mFc без адъюванта (50 мкг/мышь) в качестве бустера путем внутрибрюшинной инъекции. Клетки селезенки иммунизированных мышей (1×108) сливали с клетками миеломы SP2/0 (2×107), следуя процедуре слияния, опосредованного ПЭГ, для получения клеток гибридомы. После слияния клетки ресуспендировали в полной среде HAT (Gibco, 21060017), распределяли в 96-луночный планшет при 0,1 мл/лунку и культивировали в инкубаторе с 5% CO2 при 37 °C. В общем, через около 10-15 дней после слияния супернатанты клеточной культуры тестировали на связывающую активность в отношении ST2 с помощью ELISA (см. Пример 5), и супернатанты клеточной культуры в лунках, определенных как положительные, отбирали и тестировали на блокирующую активность в отношении связывания IL33/ST2 с помощью ELISA (см. Пример 6).
Отбирали лунки, в которых ST2 может быть специфически связан и связывание IL33/ST2 может быть заблокировано, и содержимое в лунках расширяли в 24-луночные планшеты во времени в зависимости от плотности клеток. После повторного тестирования клеточных штаммов, перенесенных в 24-луночный планшет, проводили консервацию и первое субклонирование. Клеточные штаммы, определенные как положительные после первого субклонирования, отбирали для консервации, и проводили второе субклонирование. Клеточные штаммы, определенные как положительные после второго субклонирования, подвергали консервации и экспрессии белка.
Пример 3. Получение кДНК антитела к ST2 и конструирование химерных антител
Тотальную РНК выделяли в виде матрицы из вышеуказанных клеток гибридомы с функциями связывания и блокирования с использованием набора для выделения тотальной РНК, и первую цепь кДНК синтезировали с использованием обратной транскриптазы superscript III (Thermo, 18080051) в соответствии с инструкциями. Затем последовательности вариабельных областей антител амплифицировали с помощью ПЦР-реакций с использованием вырожденных праймеров IgG мыши.
Смесь для ПЦР электрофоретически разделяли в 1% геле агароза/Tris-борат, содержащем 0,5 мкг/мл этидия бромида. Фрагмент ДНК ожидаемого размера (около 500 п.о. (пара оснований) для тяжелых и легких цепей) вырезали из геля и очищали. Очищенный продукт ПЦР клонировали в вектор pMD-19T (Takara, 6013) и трансформировали в DH5α-компетентную Escherichia coli (Takara, 9057). Пять колоний отбирали из планшета с твердой культурой LB и подвергали секвенированию ДНК. Получали последовательности вариабельной области тяжелой цепи и последовательности вариабельной области легкой цепи антител: 7D1 (SEQ ID NO: 35, 36), 3C6 (SEQ ID NO: 37, 38), 31C8 (SEQ ID NO: 39, 40), 26A1 (SEQ ID NO: 41, 42), 8F4 (SEQ ID NO: 43, 44) и 17E2 (SEQ ID NO: 45, 46).
Конструирование и экспрессия химерных антител: синтетический фрагмент гена области VL мыши лигировали с константной областью κ-цепи человека путем двойного расщепления для конструирования химерной легкой цепи. Синтетический фрагмент гена области VH мыши лигировали с константной областью IgG2 человека путем двойного расщепления для конструирования химерной тяжелой цепи.
Клетки Expi CHO (система 50 мл, клетки 6×106/мл, 1 мкг/мл ДНК) совместно трансфицировали вектором ДНК, содержащим химерную легкую цепь, описанную выше, и вектором ДНК, содержащим химерную тяжелую цепь, описанную выше, для транзиентной экспрессии и культивировали в течение 7 дней. Затем химерные антитела в супернатанте клеточной культуры очищали с помощью колонки с белком A (GE Healthcare).
Пример 4. Определение аффинности антител к ST2 к антигену hST2
Антитело к IgG мыши (для захвата антигена ST2 человека, Cytiva, 29215281) или антитело к his (для захвата антигена ST2 яванского макака, Cytiva, 29234602) присоединяли к биочипу CM5 (Cytiva, BR-1000-12) с использованием способа, описанного в инструкциях к продукту, и позволяли антителу к ST2 с диапазоном концентраций (32,8 нМ, 16,4 нМ, 8,2 нМ, 4,1 нМ, 2,05 нМ, 1,0259 нМ и 0,51297 нМ) течь по поверхности чипа. Сигналы реакции детектировали в реальном времени с помощью прибора Biacore (Cytiva, BiacoreT200), а затем получали кривую ассоциация-диссоциация. После завершения диссоциации в каждом цикле биочип регенерировали c помощью регенерационного раствора для следующего захвата; процедуру повторяли до определения аффинности различных антител к ST2. Полученные данные окончательно анализировали с использованием программного обеспечения GE BIAevaluation с моделью связывания 1:1 (Ленгмюр) для определения констант скорости ассоциации ka (kon) и констант скорости диссоциации kd (koff), и константы диссоциации KD определяют как KD = kd/ka. Определенная аффиннность антитела к ST2 к антигену ST2 человека (hST2-ECD-mFc), антигену ST2 яванского макака (Cyno-ST2-his, Sino biologica1, 90915-C08H) и определенная аффинность химерных антител к ST2 в отношении ST2 показаны в таблице 5.
Пример 5. Анализы связывания антител к ST2 на основе ELISA
Скрининг связывания ST2 и анализ антител проводили с помощью ELISA с использованием hST2-ECD-mFc (для анализов химерных и гуманизированных антител), белка hST2-ECD-hFc (для анализов гибридомных антител) и Cyno-ST2-his (Sino Biological, 90915-C08H) в качестве антигенов. 96-луночный планшет с высокой адсорбцией (Costar, 9018) покрывали 2 мкг/мл hST2-ECD-mFc или антигена Cyno-ST2-his при 100 мкл/лунку и инкубировали в течение ночи при 4 °C. После того, как неадсорбированный антиген хорошо вымывали, неспецифические сайты связывания блокировали с помощью блокирующего буфера (PBS, содержащий 2% бычий сывороточный альбумин). После того, как планшет трижды промывали с помощью промывочного буфера (PBS, содержащий 0,05 об.% Tween 20), добавляли антитела к ST2 (серийно разбавленные 3-кратно от исходной концентрации 10 нМ до 8 концентраций: 3,333 нМ, 1,111 нМ, 0,370 нМ, 0,123 нМ, 0,041 нМ, 0,014 нМ, 0,005 нМ и 0,002 нМ) при 100 мкл/лунку, и планшет инкубировали при комнатной температуре в течение 1 часа. После промывания планшета с помощью промывочного буфера добавляли вторичное антитело, конъюгированное с пероксидазой хрена (HRP), и планшет инкубировали в течение еще 60 минут. После промывания планшета с помощью промывочного буфера добавляли раствор тетраметилбензидинового (ТМБ) субстрата (Thermo, 00-4201-56) при 100 мкл/лунку и планшет инкубировали при комнатной температуре в течение 2 мин. Останавливающий раствор (2 Н H2SO4) добавляли при 100 мкл/лунку для прекращения реакций. Колориметрические сигналы генерировали и считывали при 450 нм с помощью считывающего устройства для микропланшетов (PE, Envision). Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения EC50. Значения EC50 для связывания химерных антител к ST2 с hST2 и Cyno-ST2 показаны в таблице 6 и на фиг. 1-2.
Пример 6. Анализы блокирования антител к ST2 на основе ELISA
На основе ELISA определяли способны ли антитела к ST2 блокировать связывание hIL33 с hST2. 96-луночный планшет с высокой адсорбцией покрывали 2 мкг/мл hST2-ECD-hFc при 100 мкл/лунку и инкубировали в течение ночи при 4 °C. После того, как неадсорбированный антиген хорошо вымывали, неспецифические сайты связывания блокировали с помощью блокирующего буфера (PBS, содержащий 1% бычий сывороточный альбумин). После того, как планшет трижды промывали с помощью промывочного буфера (PBST, PBS, содержащий 0,05 об.% Tween 20), добавляли антитела к ST2 (серийно разбавленные 2-кратно от исходной концентрации 66,67 нМ до 8 концентраций) при 100 мкл/лунку и планшет инкубировали при 37 °C в течение 1 часа. После того, как планшет промывали 3 раза промывочным буфером, в каждую лунку добавляли 100 мкл меченного биотином hIL33 Avitag (с концентрацией 0,1 мкг/мл) и планшет инкубировали при 37 °C в течение 1 часа. После того, как планшет промывали 3 раза промывочным буфером, добавляли вторичное антитело Avidin HRP, разбавленное 1:1000 (Jackson immunoresearch, 016-030-084), при 100 мкл/лунку, и планшет инкубировали при комнатной температуре в течение 1 часа. После того, как планшет промывали с помощью промывочного буфера, добавляли раствор ТМБ субстрата (Thermo, 00-4201-56) при 100 мкл/лунку и планшет инкубировали при комнатной температуре в течение 3 мин. Добавляли 50 мкл останавливающего раствора (2 Н H2SO4) для прекращения реакций. Колориметрические сигналы генерировали и считывали при 450 нм с помощью считывающего устройства для планшетов (PE, Envision). Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения IC50. Значения IC50 для блокирования связывания IL33/ST2 химерными антителами к ST2 показаны в таблице 7 и на фиг. 3.
Пример 7. Клеточные анализы связывания антител к ST2
Связывающую способность антител к ST2 с линиями клеток, сверхэкспрессирующих hST2, и линиями клеток, сверхэкспрессирующих cyno-ST2, анализировали с использованием метода на основе FACS путем анализов связывания с линией клеток HEK293T, сверхэкспрессирующих hST2 (SEQ ID NO: 1), (HEK293T-hST2-NFκB-люцифераза) и линией клеток HEK293T, сверхэкспрессирующих ST2 яванского макака (SEQ ID NO: 10), (HEK293T-Cyno-ST2-NFκB-люцифераза). Клетки HEK293 трансфицировали плазмидой pLenti6.3-hST2 и плазмидой pLenti6.3-NFκB-люцифераза с использованием лентивирусной системы трансфекции для конструирования клеточной линии HEK293T-hST2-NFκB-люцифераза. Клетки HEK293T трансфицировали плазмидой pLenti6.3-cynoST2 и плазмидой pLenti6.3-NFκB-люцифераза для конструирования клеточной линии HEK293T-Cyno-ST2-NFκB-люцифераза.
Добавляли 3×105 клеток HEK293T-hST2-NFκB-люцифераза или HEK293T-Cyno-ST2-NFκB-люцифераза в 96-луночный культуральный планшет. К клеточным суспензиям добавляли серийно разбавленные антитела к ST2 (серийно разбавленные 4-кратно от исходной концентрации 667 нМ до 8 концентраций в анализах связывания с клетками HEK293T-hST2-NFκB-люцифераза; серийно разбавленные 4-кратно от исходной концентрации 400 нМ до 8 концентраций в анализах связывания с клетками HEK293T-Cyno-ST2-NFκB-люцифераза). После того, как планшет инкубировали при комнатной температуре в течение 60 минут, клетки промывали 3 раза PBS. Вторичное PE антитело козы к IgG человека, разбавленное 1:200 (Jackson immunoresearch, 109-116-170), добавляли при 100 мкл/лунку, и планшет инкубировали при комнатной температуре в течение 30 минут. Клетки промывали 3 раза PBS и ресуспендировали в 100 мкл PBS, а затем анализировали и детектировали сигналы флуоресценции с помощью проточного цитометра (BD, Accuri C6). Связывающую способность антител к ST2 в отношении hST2 и cyno-ST2 на поверхностях клеточной линии измеряли с точки зрения средней интенсивности флуоресценции (MFI) окрашивания. Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения EC50. Значения EC50 для связывания химерных антител к ST2 с hST2 и Cyno-ST2 на клеточном уровне показаны в таблице 8 и на фиг. 7-8.
н.д.: предел обнаружения не достигнут.
Пример 8. Анализы антител к ST2, блокирующих взаимодействие между белком IL33 и линией клеток, сверхэкспрессирующих hST2 (HEK293T-hST2-NFκB-люцифераза)
Клетки HEK293T естественным образом экспрессируют белок IL1RAcP на высоком уровне. На этом основании клетки HEK293 трансфицировали плазмидой pLenti6.3-hST2 и плазмидой pLenti6.3-NFκB-люцифераза с использованием лентивирусной системы трансфекции для конструирования стабильно трансфицированной клеточной линии HEK293T-hST2-NFκB-люцифераза. IL33 связывается с ST2 и рекрутирует IL-1RAcP. Таким способом нижележащий сигнальный путь NFκB может быть активирован, и, таким образом, начинается экспрессия люциферазы. Когда в системе присутствует антитело к ST2, связывание IL33 с ST2 и, следовательно, экспрессия люциферазы могут быть заблокированы. Блокирующую активность антител к ST2 оценивали путем обнаружения изменений интенсивности флуоресценции реакционного субстрата.
Клетки HEK293T-hST2-NFκB-люцифераза в логарифмической фазе в хороших условиях отбирали и разбавляли до 2,5×105 клеток/мл. Разбавление клеток добавляли к 384-луночному планшету (Thermo, 262360) при 20 мкл/лунку. Затем серийно разбавленные антитела к ST2 (серийно разбавленные 4-кратно от начальной концентрации 667 нМ) добавляли при 15 мкл/лунку. Планшет инкубировали при 37 °С в течение 30 мин. После инкубации в каждую лунку добавляли 15 мкл белка hIL33 (с конечной концентрацией 50 пМ). Смеси хорошо перемешивали и инкубировали при 37 °С в течение 5 часов. Затем в каждую лунку добавляли 50 мкл ONE-Glo™ для анализа люциферазы (Promega, E6120), и смеси реагировали в темноте в течение 2-5 минут. Хемилюминесцентные сигналы считывали с помощью считывающего устройства для микропланшетов (PE, Envision). Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения IC50. Значения IC50 для блокирования химерными антителами к ST2 сигнального пути IL33/ST2 на клеточном уровне показаны в таблице 9 и на фиг. 9.
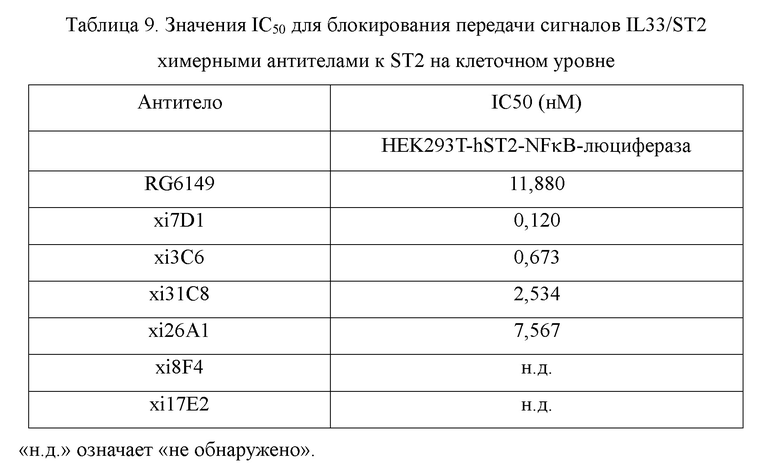
Пример 9. Анализы антител к ST2, блокирующих взаимодействие между белком IL33 и клетками, естественно экспрессирующими hST2 (KU812-NFκB-люцифераза)
Клетки KU812 естественным образом экспрессируют белок ST2 и IL1RAcP. Клетки KU812 трансфицировали pLenti6.3-NFκB-люциферазой с использованием лентивирусной системы трансфекции для конструирования стабильно трансфицированной клеточной линии KU812-NFκB-люцифераза. Экспериментальный принцип согласно примеру 8. Клетки KU812-NFκB-люцфераза в логарифмической фазе в хороших условиях отбирали и разбавляли до 2,5×105 клеток/мл. Разбавление клеток добавляли к 384-луночному планшету (Thermo, 262360) при 20 мкл/лунку. Затем серийно разбавленные антитела к ST2 (серийно разбавленные 4-кратно от начальной концентрации 667 нМ) добавляли при 15 мкл/лунку. Планшет инкубировали при 37 °С в течение 30 мин. После инкубации в каждую лунку добавляли 15 мкл белка hIL33 (с конечной концентрацией 200 пМ). Смеси хорошо перемешивали и инкубировали при 37 °С в течение 5 часов. Затем в каждую лунку добавляли 50 мкл ONE-Glo™ для анализа люциферазы (Promega, E6120), и смеси реагировали в темноте в течение 2-5 минут. Хемилюминесцентные сигналы считывали с помощью считывающего устройства для микропланшетов (PE, Envision). Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения IC50.
Пример 10. Анализы антител к ST2, блокирующих костимулирующее взаимодействие между IL33 и IL2 и CD4+ Т-клетками
При костимуляции IL33 и IL2, IL33 связывается с ST2 на поверхности CD4+ T-клеток, активируя нижележащий сигнальный путь для индуцирования секреции цитокина IL5. Когда в системе присутствует антитело к ST2, оно может блокировать связывание IL33 с ST2 на CD4+ Т-клетках, ингибируя секрецию IL5. Способность антител к ST2 блокировать костимулирующее взаимодействие между IL33 и IL2 и CD4+ Т-клетками анализировали путем измерения содержания IL5.
CD4+ Т-клетки сортировали из МКПК (мононуклеарные клетки периферической крови) человека с использованием набора для сортировки CD4+ Т-клеток человека (stemcell, 17952) и высевали в 96-луночный планшет для культивирования клеток с U-образным дном при 2,5×105 клеток/лунку, 60 мкл/лунку. Затем разбавленные антитела к ST2 (серийно разбавленные 3-кратно от начальной концентрации 53 нМ) добавляли при 15 мкл/лунку. Планшет инкубировали при 37 °С в течение 30 мин. После инкубации в каждую лунку добавляли 15 мкл белка hIL33 (с конечной концентрацией 4 нг/мл) и 15 мкл IL2 (с конечной концентрацией 10 нг/мл) (PrimeGene, GMP-101-02). Смеси хорошо перемешивали и инкубировали при 37 °С в течение 48 часов. После инкубации супернатанты собирали и анализировали на содержание IL5, следуя инструкциям в наборе для анализа IL5 (R&D, DY205). Значения OD450 (оптическая плотность) считывали с помощью считывающего устройства для микропланшетов (PE, Envision). Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения IC50. Результаты IC50 для блокирования химерными антителами к ST2 секреции IL5, индуцированной IL33/ST2, на CD4+ Т-клетках показаны в таблице 10.
«н.д.» означает «не обнаружено».
Пример 11. Гуманизация гибридомных моноклональных антител к ST2 человека
31C8 (аминокислотная последовательность вариабельной области тяжелой цепи, представленная в SEQ ID NO: 39, и аминокислотная последовательность вариабельной области легкой цепи, представленная в SEQ ID NO: 40) выбирали для гуманизации. Использовали утвержденный способ трансплантации CDR, и выбирали каркасную область человеческого антитела, которая может быть использована для гуманизации мышиного антитела. Выбирали антитело зародышевой линии человека или его подтип с наивысшей идентичностью последовательности (т.е. сходством последовательностей) с аминокислотами в вариабельной области мышиного антитела. CDR вариабельной области тяжелой цепи и CDR вариабельной области легкой цепи мышиного антитела вставляли в выбранную каркасную область, и остатки в каркасной области мутировали. Некоторые CDR дополнительно мутировали для улучшения свойств антитела, таких как физико-химические свойства или лекарственная способность. Получали гуманизированные антитела hz31C8-1.1 и hz31C8-1.2. Аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи hz31C8-1.1 представляли собой SEQ ID NO: 71 и SEQ ID NO: 79, соответственно. Последовательности ДНК, кодирующие вариабельную область тяжелой цепи и вариабельную область легкой цепи гуманизированного антитела hz31C8-1.1, представляли собой SEQ ID NO: 72 и SEQ ID NO: 80, соответственно. Аминокислотные последовательности вариабельной области тяжелой цепи и вариабельной области легкой цепи hz31C8-1.2 представляли собой SEQ ID NO: 73 и SEQ ID NO: 79, соответственно. Последовательности ДНК, кодирующие вариабельную область тяжелой цепи и вариабельную область легкой цепи гуманизированного антитела hz31C8-1.2, представляли собой SEQ ID NO: 74 и SEQ ID NO: 80, соответственно.
Синтетический фрагмент гена гуманизированный области VL лигировали с константной областью κ-цепи человека путем двойного расщепления для конструирования гуманизированной легкой цепи, и синтетический фрагмент гена гуманизированной области VH лигировали с константной областью IgG2 человека путем двойного расщепления для конструирования гуманизированной тяжелой цепи. Векторы экспрессии трансфицировали гуманизированной тяжелой цепью и легкой цепью ДНК, соответствующей каждому антителу. Экспрессию белка проводили с использованием системы экспрессии ExpiCHO. Затем гуманизированные антитела в супернатантах клеточной культуры очищали с помощью колонки с белком А.
Пример 12. Анализы биологической функции гуманизированных антител к ST2
Гуманизированные антитела hz31C8-1.1 (аминокислотная последовательность тяжелой цепи, представленная в SEQ ID NO: 75, и аминокислотная последовательность легкой цепи, представленная в SEQ ID NO: 81) и hz31C8-1.2 (аминокислотная последовательность тяжелой цепи, представленная в SEQ ID NO: 77, и аминокислотная последовательность легкой цепи, представленная в SEQ ID NO: 81) подвергали анализам аффинности, связывания, блокирования и клеточной функции с использованием способов, описанных в примере 4, примере 5, примере 6, примере 7, примере 8, примере 9 и примере 10. В настоящем описании «hz» и «xi» обозначают гуманизированные антитела и химерные антитела, соответственно; например, «hz31C8-1.1» и «hz31C8-1.2» представляют собой гуманизированные антитела 31C8-1.1 и 31C8-1.2, а «xi31C8» представляет собой химерное антитело 31C8.
Аффинность гуманизированных антител к ST2 в отношении ST2 показана в таблице 11. Значения EC50 для связывания гуманизированных антител к ST2 с hST2 и Cyno-ST2 в анализах ELISA показаны в таблице 12 и на фиг. 4-5. Значения IC50 для блокирования связывания IL33/ST2 гуманизированными антителами к ST2 в анализах ELISA показаны в таблице 13 и на фиг. 6. Значения EC50 для связывания гуманизированных антител к ST2 с hST2 на клеточном уровне показаны в таблице 14 и на фиг. 10. Значения IC50 для блокирования гуманизированными антителами к ST2 передачи сигналов IL33/ST2 на клеточном уровне показаны в таблице 15 и на фиг. 11-12. Значения IC50 для блокирования гуманизированными антителами к ST2 секреции IL5, индуцированной IL33/ST2, на CD4+ T-клетках показаны в таблице 16 и на фиг. 13.
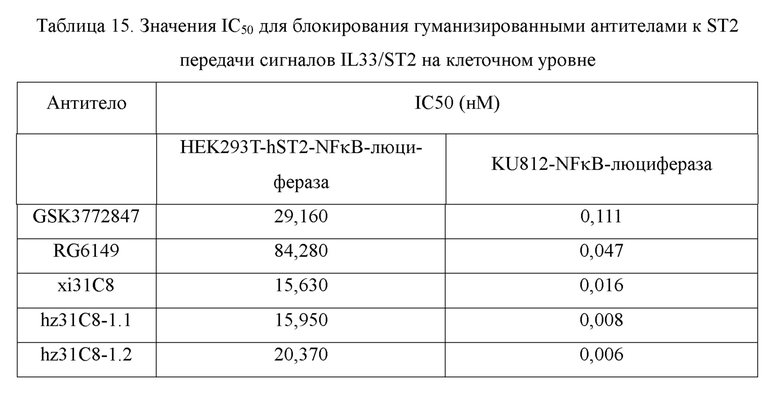
Пример 13. Анализы стабильности гуманизированных антител к ST2
Термостабильность различных антител определяли с помощью ДСК (дифференциальная сканирующая калориметрия). Образцы растворяли в буфере PBS (pH 7,4) и проводили определение с использованием MicroCal VP-Capillary DSC (Malvern Panalytical). Как показано в Таблице 17, гуманизированные антитела к ST2 hz31C8-1.1 и hz31C8-1.2 по настоящему изобретению демонстрируют хорошую термостабильность и превосходят контрольные антитела к ST2 RG6149 и GSK3772847.
Периодическую стабильность антител определяли с помощью SEC-HPLC (эксклюзионная ВЭЖХ (высокоэффективная жидкостная хроматография)) при определенных концентрациях, например, концентрацию антител контролировали на уровне около 1 мг/мл и хранили в буфере PBS (рН 7,2), например, при 40 °C в течение 2 недель. Определение проводили с использованием LC-20ADXR/DGU (Shimadzu). Как показано ниже в Таблице 18, гуманизированные антитела к ST2 hz31C8-1.1 и hz31C8-1.2 по настоящему изобретению продемонстрировали хорошую стабильность.
Антитела к ST2 разбавляли буфером PBS до концентрации 1,5 мг/мл. После обработки при 72 °С в течение 5 минут образцы анализировали с помощью ELISA на предмет их связывающей активности в отношении антигена. Процедура ELISA следующая: 96-луночный планшет покрывали 2 мкг/мл hST2-ECD-mFc в качестве антигена при 100 мкл/лунку и инкубировали при 4 °C в течение ночи. После того, как неадсорбированный антиген хорошо вымывали, неспецифические сайты связывания блокировали с помощью блокирующего буфера (PBST, содержащий 1% бычий сывороточный альбумин). После того, как планшет трижды промывали с помощью промывочного буфера (PBST, PBS, содержащий 0,05 об.% Tween 20), добавляли антитела к ST2 (серийно разбавленные 3-кратно от исходной концентрации 1,5 мкг/мл до 8 концентраций) при 100 мкл/лунку и планшет инкубировали при 37 °C в течение 1 часа. После того, как планшет промывали с помощью промывочного буфера, добавляли вторичное антитело к Fcγ IgG человека (Jackson ImmunoResearch, 109-035-008) и планшет инкубировали при 37 °C в течение 1 часа. После того, как планшет промывали с помощью промывочного буфера, добавляли раствор ТМБ субстрата (Thermo, 00-4201-56) при 100 мкл/лунку для проявления цвета. После того, как планшет инкубировали при комнатной температуре в течение 5 минут, добавляли останваливающий раствор (2 Н H2SO4) для остановки реакций. Сигналы считывали при 450 нм с помощью считывающего устройства для планшетов (PE, Envision). Данные анализировали с использованием GraphPad Prism5 и рассчитывали значения EC50. Как показано ниже в Таблице 19, гуманизированные антитела к ST2 hz31C8-1.1 и hz31C8-1.2 по настоящему изобретению продемонстрировали хорошую термостабильность.
Пример 14. Перекрестная реактивность гуманизированных антител к ST2 с рецепторами семейства IL1R
ST2 принадлежит к семейству рецепторов IL1R. Перекрестная реактивность антител к ST2 с рецепторами семейства IL1R, определенная с помощью ELISA. Рецепторы семейства IL1R включают IL1R1/CD121a (Sino biological, 10126-H08H), IL1R2/CD121b (Sino biological, 10111-H08H), IL1R3/IL1RAP (Sino biological, 10121-H08H), IL1R7/IL-18RAcP (Sino biological, 10176-H08H), IL1R8/IL1RAPL1 (Sino biological, 10177-H08H) и IL1R9/IL1RAPL2 (Sino biological, 10156-H08H). Анализы ELISA проводили с использованием белков, описанных выше в качестве антигенов (2 мкг/мл). См. Пример 13 для получения информации о конкретной процедуре ELISA. Отсутствие связывания означает, что антитело остается несвязанным с антигеном при самой высокой концентрации. Слабое связывание означает, что антитело слабо связывается с антигеном при самой высокой концентрации и не связывается при низких концентрациях. Сильное связывание означает, что очень хорошая кривая связывания может быть получена на основании данных связывания.
(-: нет связывания; +: слабое связывание; +++++: сильное связывание)
Пример 15. Фармакокинетическая оценка гуманизированных антител к ST2
Восемнадцать лабораторных мышей Balb/c, разделенных на группы по 6, при 12/12-часовом визуальном контроле, которые имели свободный доступ к пище и воде (ad libitum), приобретали в Модельном центре исследований животных Нанкинского университета. В день эксперимента каждой мыши вводили образец 1 мг/мл путем внутривенной инъекции в дозе 10 мг/кг. Орбитальную кровь собирали перед введением дозы (0 мин или один день или больше перед введением дозы) и через 30 мин, 8 ч, 24 ч, 2 д (48 ч), 4 д (96 ч), 7 д и 14 д после введения дозы, а затем собирали сыворотку и анализировали концентрацию антител с помощью ELISA. Процедура ELISA следующая: 96-луночный планшет покрывали 2 мкг/мл ST2-His (sinobiological, 10105-H08H) в качестве антигена при 100 мкл/лунку и инкубировали при 4 °C в течение ночи. После того, как неадсорбированный антиген хорошо вымывали, неспецифические сайты связывания блокировали с помощью блокирующего буфера (PBST, содержащий 1% бычий сывороточный альбумин). После того, как планшет трижды промывали с помощью промывочного буфера (PBST, PBS, содержащий 0,05 об.% Tween 20), добавляли 100 мкл тестируемой сыворотки и планшет инкубировали при 37 °C в течение 1 часа. После того, как планшет промывали с помощью промывочного буфера, добавляли вторичное антитело к Fcγ IgG мыши и планшет инкубировали при 37 °C в течение 1 часа. После того, как планшет промывали с помощью промывочного буфера, добавляли раствор ТМБ субстрата (Thermo, 00-4201-56) при 100 мкл/лунку для проявления цвета. После того, как планшет инкубировали при комнатной температуре в течение 5 минут, добавляли останваливающий раствор (2 Н H2SO4) для остановки реакций. Сигналы считывали при 450 нм с помощью считывающего устройства для планшетов (PE, Envision). Данные анализировали с помощью GraphPad Prism5. Ссылаясь на Таблицу 21, фармакокинетический (PK) анализ показывает, что гуманизированные антитела к ST2 hz31C8-1.1 и hz31C8-1.2 по настоящему изобретению имеют периоды полужизни около 9,7 дней и 7,33 дней, превосходящие контрольное антитело к ST2 RG6149.
Несмотря на то, что настоящее изобретение было подробно описано в данном документе выше с помощью общих пояснений и конкретных вариантов осуществления, специалистам в данной области техники будет очевидно, что на основе настоящего изобретения могут быть сделаны модификации или улучшения. Соответственно, все такие модификации и усовершенствования, выполненные без отступления от сущности настоящего изобретения, подпадают под объем защиты настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Chia Tai Tianqing Pharmaceutical Group Co., Ltd.
<120> БЕЛОК, СВЯЗЫВАЮЩИЙ АНТИГЕН ST2
<130> 6291-2143480IB
<150> CN202010397572.3
<151> 2020-05-12
<160> 86
<170> SIPOSequenceListing 1.0
<210> 1
<211> 556
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> НЕОПРЕДЕЛЕННЫЙ
<222> (78)
<223> X представляет собой E или A
<400> 1
Met Gly Phe Trp Ile Leu Ala Ile Leu Thr Ile Leu Met Tyr Ser Thr
1 5 10 15
Ala Ala Lys Phe Ser Lys Gln Ser Trp Gly Leu Glu Asn Glu Ala Leu
20 25 30
Ile Val Arg Cys Pro Arg Gln Gly Lys Pro Ser Tyr Thr Val Asp Trp
35 40 45
Tyr Tyr Ser Gln Thr Asn Lys Ser Ile Pro Thr Gln Glu Arg Asn Arg
50 55 60
Val Phe Ala Ser Gly Gln Leu Leu Lys Phe Leu Pro Ala Xaa Val Ala
65 70 75 80
Asp Ser Gly Ile Tyr Thr Cys Ile Val Arg Ser Pro Thr Phe Asn Arg
85 90 95
Thr Gly Tyr Ala Asn Val Thr Ile Tyr Lys Lys Gln Ser Asp Cys Asn
100 105 110
Val Pro Asp Tyr Leu Met Tyr Ser Thr Val Ser Gly Ser Glu Lys Asn
115 120 125
Ser Lys Ile Tyr Cys Pro Thr Ile Asp Leu Tyr Asn Trp Thr Ala Pro
130 135 140
Leu Glu Trp Phe Lys Asn Cys Gln Ala Leu Gln Gly Ser Arg Tyr Arg
145 150 155 160
Ala His Lys Ser Phe Leu Val Ile Asp Asn Val Met Thr Glu Asp Ala
165 170 175
Gly Asp Tyr Thr Cys Lys Phe Ile His Asn Glu Asn Gly Ala Asn Tyr
180 185 190
Ser Val Thr Ala Thr Arg Ser Phe Thr Val Lys Asp Glu Gln Gly Phe
195 200 205
Ser Leu Phe Pro Val Ile Gly Ala Pro Ala Gln Asn Glu Ile Lys Glu
210 215 220
Val Glu Ile Gly Lys Asn Ala Asn Leu Thr Cys Ser Ala Cys Phe Gly
225 230 235 240
Lys Gly Thr Gln Phe Leu Ala Ala Val Leu Trp Gln Leu Asn Gly Thr
245 250 255
Lys Ile Thr Asp Phe Gly Glu Pro Arg Ile Gln Gln Glu Glu Gly Gln
260 265 270
Asn Gln Ser Phe Ser Asn Gly Leu Ala Cys Leu Asp Met Val Leu Arg
275 280 285
Ile Ala Asp Val Lys Glu Glu Asp Leu Leu Leu Gln Tyr Asp Cys Leu
290 295 300
Ala Leu Asn Leu His Gly Leu Arg Arg His Thr Val Arg Leu Ser Arg
305 310 315 320
Lys Asn Pro Ile Asp His His Ser Ile Tyr Cys Ile Ile Ala Val Cys
325 330 335
Ser Val Phe Leu Met Leu Ile Asn Val Leu Val Ile Ile Leu Lys Met
340 345 350
Phe Trp Ile Glu Ala Thr Leu Leu Trp Arg Asp Ile Ala Lys Pro Tyr
355 360 365
Lys Thr Arg Asn Asp Gly Lys Leu Tyr Asp Ala Tyr Val Val Tyr Pro
370 375 380
Arg Asn Tyr Lys Ser Ser Thr Asp Gly Ala Ser Arg Val Glu His Phe
385 390 395 400
Val His Gln Ile Leu Pro Asp Val Leu Glu Asn Lys Cys Gly Tyr Thr
405 410 415
Leu Cys Ile Tyr Gly Arg Asp Met Leu Pro Gly Glu Asp Val Val Thr
420 425 430
Ala Val Glu Thr Asn Ile Arg Lys Ser Arg Arg His Ile Phe Ile Leu
435 440 445
Thr Pro Gln Ile Thr His Asn Lys Glu Phe Ala Tyr Glu Gln Glu Val
450 455 460
Ala Leu His Cys Ala Leu Ile Gln Asn Asp Ala Lys Val Ile Leu Ile
465 470 475 480
Glu Met Glu Ala Leu Ser Glu Leu Asp Met Leu Gln Ala Glu Ala Leu
485 490 495
Gln Asp Ser Leu Gln His Leu Met Lys Val Gln Gly Thr Ile Lys Trp
500 505 510
Arg Glu Asp His Ile Ala Asn Lys Arg Ser Leu Asn Ser Lys Phe Trp
515 520 525
Lys His Val Arg Tyr Gln Met Pro Val Pro Ser Lys Ile Pro Arg Lys
530 535 540
Ala Ser Ser Leu Thr Pro Leu Ala Ala Gln Lys Gln
545 550 555
<210> 2
<211> 566
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hST2-ECD-mFc
<400> 2
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Gly Val His
1 5 10 15
Ser Lys Phe Ser Lys Gln Ser Trp Gly Leu Glu Asn Glu Ala Leu Ile
20 25 30
Val Arg Cys Pro Arg Gln Gly Lys Pro Ser Tyr Thr Val Asp Trp Tyr
35 40 45
Tyr Ser Gln Thr Asn Lys Ser Ile Pro Thr Gln Glu Arg Asn Arg Val
50 55 60
Phe Ala Ser Gly Gln Leu Leu Lys Phe Leu Pro Ala Ala Val Ala Asp
65 70 75 80
Ser Gly Ile Tyr Thr Cys Ile Val Arg Ser Pro Thr Phe Asn Arg Thr
85 90 95
Gly Tyr Ala Asn Val Thr Ile Tyr Lys Lys Gln Ser Asp Cys Asn Val
100 105 110
Pro Asp Tyr Leu Met Tyr Ser Thr Val Ser Gly Ser Glu Lys Asn Ser
115 120 125
Lys Ile Tyr Cys Pro Thr Ile Asp Leu Tyr Asn Trp Thr Ala Pro Leu
130 135 140
Glu Trp Phe Lys Asn Cys Gln Ala Leu Gln Gly Ser Arg Tyr Arg Ala
145 150 155 160
His Lys Ser Phe Leu Val Ile Asp Asn Val Met Thr Glu Asp Ala Gly
165 170 175
Asp Tyr Thr Cys Lys Phe Ile His Asn Glu Asn Gly Ala Asn Tyr Ser
180 185 190
Val Thr Ala Thr Arg Ser Phe Thr Val Lys Asp Glu Gln Gly Phe Ser
195 200 205
Leu Phe Pro Val Ile Gly Ala Pro Ala Gln Asn Glu Ile Lys Glu Val
210 215 220
Glu Ile Gly Lys Asn Ala Asn Leu Thr Cys Ser Ala Cys Phe Gly Lys
225 230 235 240
Gly Thr Gln Phe Leu Ala Ala Val Leu Trp Gln Leu Asn Gly Thr Lys
245 250 255
Ile Thr Asp Phe Gly Glu Pro Arg Ile Gln Gln Glu Glu Gly Gln Asn
260 265 270
Gln Ser Phe Ser Asn Gly Leu Ala Cys Leu Asp Met Val Leu Arg Ile
275 280 285
Ala Asp Val Lys Glu Glu Asp Leu Leu Leu Gln Tyr Asp Cys Leu Ala
290 295 300
Leu Asn Leu His Gly Leu Arg Arg His Thr Val Arg Leu Ser Arg Lys
305 310 315 320
Asn Pro Ile Asp His His Ser Ile Glu Gly Arg Met Asp Glu Pro Arg
325 330 335
Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala Pro Asn
340 345 350
Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile Lys Asp
355 360 365
Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val Val Asp
370 375 380
Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val Asn Asn
385 390 395 400
Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp Tyr Asn
405 410 415
Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln Asp Trp
420 425 430
Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn Lys Asp Leu Pro
435 440 445
Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val Arg Ala
450 455 460
Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr Lys Lys
465 470 475 480
Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu Asp Ile
485 490 495
Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr Lys Asn
500 505 510
Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr Ser Lys
515 520 525
Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr Ser Cys
530 535 540
Ser Val Val His Glu Gly Leu His Asn His His Thr Thr Lys Ser Phe
545 550 555 560
Ser Arg Thr Pro Gly Lys
565
<210> 3
<211> 1650
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hST2-ECD-mFc
<400> 3
aagttttcta agcagtcctg gggcctggag aacgaggctc tgatcgtgag atgtccccgc 60
cagggcaagc ctagctacac agtggactgg tactatagcc agaccaacaa gtctatccca 120
acacaggaga gaaatcgcgt gttcgcctct ggccagctgc tgaagtttct gcccgccgct 180
gtggctgata gcggcatcta tacctgcatc gtgagatctc ctaccttcaa ccgcacaggc 240
tacgccaatg tgacaatcta taagaagcag tccgactgta acgtgcccga ttacctgatg 300
tattccaccg tgtccggcag cgagaagaat agcaagatct actgccctac catcgacctg 360
tataactgga cagccccact ggagtggttc aagaattgtc aggccctgca gggctctagg 420
taccgggctc acaagtcctt cctggtcatc gataacgtga tgaccgagga cgctggcgat 480
tacacatgca agttcatcca taacgagaat ggcgccaatt attccgtgac cgctacaagg 540
agctttacag tgaaggacga gcagggcttc tccctgtttc cagtgatcgg cgcccccgct 600
cagaacgaga tcaaggaggt ggagatcggc aagaacgcca atctgacctg ctccgcctgt 660
ttcggcaagg gcacacagtt tctggccgct gtgctgtggc agctgaatgg caccaagatc 720
acagatttcg gcgagcctag gatccagcag gaggagggcc agaaccagtc tttttccaat 780
ggactggctt gcctggacat ggtgctgagg atcgctgacg tgaaggagga ggatctgctg 840
ctgcagtatg attgtctggc tctgaacctg cacggcctga ggcggcatac cgtgaggctg 900
tctcggaaga atccaatcga ccaccattcc atcgagggaa ggatggatga gccaaggggc 960
ccaaccatca agccatgccc cccttgcaag tgtcctgctc caaacctgct gggcggccct 1020
tccgtgttca tctttccacc caagatcaag gacgtgctga tgatctctct gtcccctatc 1080
gtgacatgcg tggtggtgga cgtgtctgag gacgatccag atgtgcagat ctcctggttc 1140
gtgaacaatg tggaggtgca caccgcccag acccagacac atagggagga ttacaattcc 1200
acactgaggg tggtgtccgc cctgccaatc cagcatcagg actggatgtc cggcaaggag 1260
tttaagtgca aggtgaacaa taaggatctg cccgccccta tcgagaggac catcagcaag 1320
cccaagggct ctgtgcgggc tcctcaggtg tatgtgctgc ctccacccga ggaggagatg 1380
accaagaagc aggtgaccct gacatgtatg gtgacagact tcatgccaga ggatatctac 1440
gtggagtgga ccaacaatgg caagacagag ctgaactata agaataccga gcccgtgctg 1500
gactccgatg gcagctactt tatgtatagc aagctgagag tggagaagaa gaactgggtg 1560
gagcgcaata gctactcttg ttccgtggtg cacgagggcc tgcataacca ccataccaca 1620
aagagctttt ctcggacacc tggcaagtga 1650
<210> 4
<211> 565
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hST2-ECD-hFc
<400> 4
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Gly Val His
1 5 10 15
Ser Lys Phe Ser Lys Gln Ser Trp Gly Leu Glu Asn Glu Ala Leu Ile
20 25 30
Val Arg Cys Pro Arg Gln Gly Lys Pro Ser Tyr Thr Val Asp Trp Tyr
35 40 45
Tyr Ser Gln Thr Asn Lys Ser Ile Pro Thr Gln Glu Arg Asn Arg Val
50 55 60
Phe Ala Ser Gly Gln Leu Leu Lys Phe Leu Pro Ala Ala Val Ala Asp
65 70 75 80
Ser Gly Ile Tyr Thr Cys Ile Val Arg Ser Pro Thr Phe Asn Arg Thr
85 90 95
Gly Tyr Ala Asn Val Thr Ile Tyr Lys Lys Gln Ser Asp Cys Asn Val
100 105 110
Pro Asp Tyr Leu Met Tyr Ser Thr Val Ser Gly Ser Glu Lys Asn Ser
115 120 125
Lys Ile Tyr Cys Pro Thr Ile Asp Leu Tyr Asn Trp Thr Ala Pro Leu
130 135 140
Glu Trp Phe Lys Asn Cys Gln Ala Leu Gln Gly Ser Arg Tyr Arg Ala
145 150 155 160
His Lys Ser Phe Leu Val Ile Asp Asn Val Met Thr Glu Asp Ala Gly
165 170 175
Asp Tyr Thr Cys Lys Phe Ile His Asn Glu Asn Gly Ala Asn Tyr Ser
180 185 190
Val Thr Ala Thr Arg Ser Phe Thr Val Lys Asp Glu Gln Gly Phe Ser
195 200 205
Leu Phe Pro Val Ile Gly Ala Pro Ala Gln Asn Glu Ile Lys Glu Val
210 215 220
Glu Ile Gly Lys Asn Ala Asn Leu Thr Cys Ser Ala Cys Phe Gly Lys
225 230 235 240
Gly Thr Gln Phe Leu Ala Ala Val Leu Trp Gln Leu Asn Gly Thr Lys
245 250 255
Ile Thr Asp Phe Gly Glu Pro Arg Ile Gln Gln Glu Glu Gly Gln Asn
260 265 270
Gln Ser Phe Ser Asn Gly Leu Ala Cys Leu Asp Met Val Leu Arg Ile
275 280 285
Ala Asp Val Lys Glu Glu Asp Leu Leu Leu Gln Tyr Asp Cys Leu Ala
290 295 300
Leu Asn Leu His Gly Leu Arg Arg His Thr Val Arg Leu Ser Arg Lys
305 310 315 320
Asn Pro Ile Asp His His Ser Ile Glu Gly Arg Met Asp Glu Pro Lys
325 330 335
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
340 345 350
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
355 360 365
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
370 375 380
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
385 390 395 400
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
405 410 415
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
420 425 430
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
435 440 445
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
450 455 460
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
465 470 475 480
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
485 490 495
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
500 505 510
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
515 520 525
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
530 535 540
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
545 550 555 560
Leu Ser Pro Gly Lys
565
<210> 5
<211> 1647
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hST2-ECD-hFc
<400> 5
aagttttcta agcagtcctg gggcctggag aacgaggctc tgatcgtgag atgtccccgc 60
cagggcaagc ctagctacac agtggactgg tactatagcc agaccaacaa gtctatccca 120
acacaggaga gaaatcgcgt gttcgcctct ggccagctgc tgaagtttct gcccgccgct 180
gtggctgata gcggcatcta tacctgcatc gtgagatctc ctaccttcaa ccgcacaggc 240
tacgccaatg tgacaatcta taagaagcag tccgactgta acgtgcccga ttacctgatg 300
tattccaccg tgtccggcag cgagaagaat agcaagatct actgccctac catcgacctg 360
tataactgga cagccccact ggagtggttc aagaattgtc aggccctgca gggctctagg 420
taccgggctc acaagtcctt cctggtcatc gataacgtga tgaccgagga cgctggcgat 480
tacacatgca agttcatcca taacgagaat ggcgccaatt attccgtgac cgctacaagg 540
agctttacag tgaaggacga gcagggcttc tccctgtttc cagtgatcgg cgcccccgct 600
cagaacgaga tcaaggaggt ggagatcggc aagaacgcca atctgacctg ctccgcctgt 660
ttcggcaagg gcacacagtt tctggccgct gtgctgtggc agctgaatgg caccaagatc 720
acagatttcg gcgagcctag gatccagcag gaggagggcc agaaccagtc tttttccaat 780
ggactggctt gcctggacat ggtgctgagg atcgctgacg tgaaggagga ggatctgctg 840
ctgcagtatg attgtctggc tctgaacctg cacggcctga ggcggcatac cgtgaggctg 900
tctcggaaga atccaatcga ccaccattcc atcgagggaa ggatggatga gccaaagtct 960
tgtgataaga cccatacatg ccccccttgt cctgctccag agctgctggg aggaccatcc 1020
gtgttcctgt ttccacccaa gcccaaggac accctgatga tctcccgcac accagaggtg 1080
acctgcgtgg tggtggacgt gagccacgag gatcccgagg tgaagtttaa ctggtacgtg 1140
gatggcgtgg aggtgcataa tgctaagacc aagccaagag aggagcagta taacagcaca 1200
taccgcgtgg tgtctgtgct gaccgtgctg caccaggact ggctgaacgg caaggagtac 1260
aagtgcaagg tgtctaataa ggccctgccc gctcctatcg agaagacaat ctccaaggcc 1320
aagggccagc ctagggagcc acaggtgtat accctgcctc catctcggga cgagctgaca 1380
aagaaccagg tgtccctgac ctgtctggtg aagggcttct accccagcga tatcgctgtg 1440
gagtgggagt ctaatggcca gcctgagaac aattataaga ccacaccccc tgtgctggac 1500
agcgatggct ctttctttct gtactctaag ctgacagtgg ataagtccag gtggcagcag 1560
ggcaacgtgt ttagctgctc tgtgatgcat gaggctctgc acaatcatta cacccagaag 1620
tccctgagcc tgtctcccgg caagtga 1647
<210> 6
<211> 174
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hIL33 Avitag
<400> 6
Ser Ile Thr Gly Ile Ser Pro Ile Thr Glu Tyr Leu Ala Ser Leu Ser
1 5 10 15
Thr Tyr Asn Asp Gln Ser Ile Thr Phe Ala Leu Glu Asp Glu Ser Tyr
20 25 30
Glu Ile Tyr Val Glu Asp Leu Lys Lys Asp Glu Lys Lys Asp Lys Val
35 40 45
Leu Leu Ser Tyr Tyr Glu Ser Gln His Pro Ser Asn Glu Ser Gly Asp
50 55 60
Gly Val Asp Gly Lys Met Leu Met Val Thr Leu Ser Pro Thr Lys Asp
65 70 75 80
Phe Trp Leu His Ala Asn Asn Lys Glu His Ser Val Glu Leu His Lys
85 90 95
Cys Glu Lys Pro Leu Pro Asp Gln Ala Phe Phe Val Leu His Asn Met
100 105 110
His Ser Asn Cys Val Ser Phe Glu Cys Lys Thr Asp Pro Gly Val Phe
115 120 125
Ile Gly Val Lys Asp Asn His Leu Ala Leu Ile Lys Val Asp Ser Ser
130 135 140
Glu Asn Leu Cys Thr Glu Asn Ile Leu Phe Lys Leu Ser Glu Thr Gly
145 150 155 160
Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
165 170
<210> 7
<211> 525
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIL33 Avitag
<400> 7
agcataacag gaatatcacc cataactgaa tatctagctt cattaagtac atataacgat 60
cagagcatta ccttcgcgct ggaggacgaa agctacgaga tctatgtgga agacctgaag 120
aaagatgaaa agaaagacaa ggttctgctg agctactatg agagccaaca cccgagcaac 180
gaaagcggtg acggcgtgga tggcaaaatg ctgatggtta ccctgagccc gaccaaggac 240
ttttggctgc acgcgaacaa caaagagcac agcgtggaac tgcacaagtg cgagaaaccg 300
ctgccggatc aggcgttctt tgtgctgcac aacatgcaca gcaactgcgt tagcttcgaa 360
tgcaagaccg atccgggtgt gtttattggc gttaaggaca accacctggc gctgatcaaa 420
gttgatagca gcgagaacct gtgcaccgaa aacattctgt tcaagctgag cgagaccggt 480
ctgaacgaca tttttgaagc gcaaaaaatc gagtggcacg aataa 525
<210> 8
<211> 159
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> hIL33 белок
<400> 8
Ser Ile Thr Gly Ile Ser Pro Ile Thr Glu Tyr Leu Ala Ser Leu Ser
1 5 10 15
Thr Tyr Asn Asp Gln Ser Ile Thr Phe Ala Leu Glu Asp Glu Ser Tyr
20 25 30
Glu Ile Tyr Val Glu Asp Leu Lys Lys Asp Glu Lys Lys Asp Lys Val
35 40 45
Leu Leu Ser Tyr Tyr Glu Ser Gln His Pro Ser Asn Glu Ser Gly Asp
50 55 60
Gly Val Asp Gly Lys Met Leu Met Val Thr Leu Ser Pro Thr Lys Asp
65 70 75 80
Phe Trp Leu His Ala Asn Asn Lys Glu His Ser Val Glu Leu His Lys
85 90 95
Cys Glu Lys Pro Leu Pro Asp Gln Ala Phe Phe Val Leu His Asn Met
100 105 110
His Ser Asn Cys Val Ser Phe Glu Cys Lys Thr Asp Pro Gly Val Phe
115 120 125
Ile Gly Val Lys Asp Asn His Leu Ala Leu Ile Lys Val Asp Ser Ser
130 135 140
Glu Asn Leu Cys Thr Glu Asn Ile Leu Phe Lys Leu Ser Glu Thr
145 150 155
<210> 9
<211> 477
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hIL33 белок
<400> 9
agcataacag gaatatcacc cataactgaa tatctagctt cattaagtac atataacgat 60
cagagcatta ccttcgcgct ggaggacgaa agctacgaga tctatgtgga agacctgaag 120
aaagatgaaa agaaagacaa ggttctgctg agctactatg agagccaaca cccgagcaac 180
gaaagcggtg acggcgtgga tggcaaaatg ctgatggtta ccctgagccc gaccaaggac 240
ttttggctgc acgcgaacaa caaagagcac agcgtggaac tgcacaagtg cgagaaaccg 300
ctgccggatc aggcgttctt tgtgctgcac aacatgcaca gcaactgcgt tagcttcgaa 360
tgcaagaccg atccgggtgt gtttattggc gttaaggaca accacctggc gctgatcaaa 420
gttgatagca gcgagaacct gtgcaccgaa aacattctgt tcaagctgag cgagacc 477
<210> 10
<211> 556
<212> БЕЛОК
<213> Яванский макак (Macaca fascicularis)
<400> 10
Met Gly Leu Trp Ile Leu Ala Ile Leu Thr Ile Leu Val Tyr Ser Thr
1 5 10 15
Ala Ala Lys Phe Ser Lys Gln Ser Trp Gly Leu Glu Asn Glu Ala Leu
20 25 30
Ile Val Arg Cys Pro Arg Gln Gly Lys Ser Ser Tyr Ile Val Asp Trp
35 40 45
Tyr Tyr Ser Gln Thr Asn Lys Ser Ile Pro Thr Gln Glu Arg Asn Arg
50 55 60
Val Phe Ala Ser Gly Gln Leu Leu Lys Phe Leu Pro Ala Glu Val Ala
65 70 75 80
Asp Ser Gly Ile Tyr Thr Cys Ile Val Arg Ser Pro Thr Phe Asn Arg
85 90 95
Thr Gly Tyr Ala Asn Val Thr Ile Tyr Lys Lys Gln Pro Asp Cys Asn
100 105 110
Val Pro Asp Tyr Leu Met Tyr Ser Thr Val Ser Gly Ser Glu Lys Asn
115 120 125
Ser Lys Ile Tyr Cys Pro Thr Ile Asp Leu Tyr Asn Trp Thr Ala Pro
130 135 140
Leu Glu Trp Phe Lys Asn Cys Gln Ala Leu Gln Gly Ser Arg Tyr Lys
145 150 155 160
Ala His Lys Ser Phe Leu Val Ile Asp Asn Val Met Thr Asp Asp Ala
165 170 175
Gly Asp Tyr Thr Cys Lys Phe Ile His Asn Glu Asn Gly Ala Asn Tyr
180 185 190
Ser Val Thr Ala Thr Arg Ser Phe Thr Val Lys Asp Glu Gln Gly Phe
195 200 205
Ser Arg Phe Pro Val Ile Arg Ala Pro Ala His Asn Glu Thr Lys Glu
210 215 220
Val Glu Ile Gly Glu Asn Thr Asn Leu Thr Cys Ser Ala Cys Phe Gly
225 230 235 240
Lys Gly Ala Gln Phe Leu Ala Ala Val Gln Trp Gln Leu Asn Gly Asn
245 250 255
Lys Ile Thr Asp Phe Ser Glu Pro Arg Ile Gln Gln Glu Glu Gly Gln
260 265 270
Asn Gln Ser Phe Ser Asp Gly Leu Ala Cys Val Asn Thr Val Leu Arg
275 280 285
Ile Ala Asp Val Lys Glu Glu Asp Leu Leu Leu Arg Tyr Asp Cys Leu
290 295 300
Ala Leu Asn Leu His Gly Leu Arg Arg His Thr Ile Arg Leu Ser Arg
305 310 315 320
Lys Asn Pro Ile Asp His Gln Ser Thr Tyr Cys Ile Ile Ala Val Cys
325 330 335
Ser Val Leu Leu Met Leu Ile Asn Val Leu Val Ile Ile Leu Lys Thr
340 345 350
Phe Trp Ile Glu Ala Thr Leu Leu Trp Arg Asp Ile Ala Lys Pro Tyr
355 360 365
Lys Thr Arg Asn Asp Gly Lys Leu Tyr Asp Ala Tyr Val Ile Tyr Pro
370 375 380
Arg Asn Tyr Thr Ser Ser Thr Asp Gly Ala Ser Arg Val Glu Tyr Phe
385 390 395 400
Val His Gln Ile Leu Pro Asp Val Leu Glu Asn Lys Cys Gly Tyr Thr
405 410 415
Leu Cys Ile Tyr Gly Arg Asp Met Leu Pro Gly Glu Asp Val Val Thr
420 425 430
Ala Val Glu Thr Asn Ile Arg Lys Ser Arg Arg His Ile Phe Ile Leu
435 440 445
Thr Pro Gln Ile Thr His Ser Glu Glu Phe Ala Tyr Glu Gln Glu Val
450 455 460
Ala Leu His Ser Ala Leu Ile Gln Asn Asp Ser Lys Val Ile Leu Ile
465 470 475 480
Glu Met Glu Ala Leu Ser Glu Leu Asp Met Leu Gln Ala Glu Ala Leu
485 490 495
Gln Asp Ser Leu Arg His Leu Met Glu Val Gln Gly Thr Ile Lys Trp
500 505 510
Arg Glu Asp His Val Ala Asn Lys Arg Ser Leu Asn Ser Lys Phe Trp
515 520 525
Lys His Val Arg Tyr Gln Met Pro Val Pro Ser Lys Met Pro Arg Lys
530 535 540
Ala Ser Ser Leu Thr Ser Leu Ala Ala Gln Lys Gln
545 550 555
<210> 11
<211> 5
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 11
Asp Tyr Glu Met His
1 5
<210> 12
<211> 5
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 12
Asp Ser Glu Met His
1 5
<210> 13
<211> 5
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 13
Asp Ser Glu Ile His
1 5
<210> 14
<211> 5
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 14
Asp Tyr Glu Met Gln
1 5
<210> 15
<211> 17
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 15
Thr Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 16
<211> 17
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 16
Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Ser Gln Asn Phe Lys
1 5 10 15
Gly
<210> 17
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> НЕОПРЕДЕЛЕННЫЙ
<222> (12)
<223> X представляет собой N или A
<220>
<221> НЕОПРЕДЕЛЕННЫЙ
<222> (13)
<223> X представляет собой Q или E или K
<220>
<221> НЕОПРЕДЕЛЕННЫЙ
<222> (16)
<223> X представляет собой K или Q
<220>
<223> HV-CDR2 антитела 31C8
<400> 17
Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Xaa Xaa Lys Phe Xaa
1 5 10 15
Gly
<210> 18
<211> 17
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 18
Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 19
<211> 17
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 19
Ala Phe Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 20
<211> 12
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 20
Tyr Ser Tyr Thr Asn Ser Pro Tyr Gly Met Asp Tyr
1 5 10
<210> 21
<211> 9
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 21
Ser Ala Thr Asp Tyr Tyr Phe Ala Ser
1 5
<210> 22
<211> 9
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 22
Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr
1 5
<210> 23
<211> 9
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 23
Ser Thr Thr Asp Cys Tyr Phe Ala Tyr
1 5
<210> 24
<211> 10
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 24
Val Asp Ser Asn Tyr Asp Tyr Phe Asp Tyr
1 5 10
<210> 25
<211> 11
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 25
Lys Ala Ser Gln Ser Val Ser Asn Asp Val Ala
1 5 10
<210> 26
<211> 16
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 26
Arg Ser Ser Lys Ser Leu Leu His Ser Asn Gly Ile Thr Tyr Leu Tyr
1 5 10 15
<210> 27
<211> 16
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 27
Arg Ser Ser Lys Ser Leu Leu His Ser Asn Gly Ile Ile Tyr Leu Tyr
1 5 10 15
<210> 28
<211> 10
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 28
Arg Ala Ser Ser Ser Val Ser Tyr Met His
1 5 10
<210> 29
<211> 7
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 29
Tyr Ala Ser Asn Arg Tyr Thr
1 5
<210> 30
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> НЕОПРЕДЕЛЕННЫЙ
<222> (2)
<223> X представляет собой M или L
<220>
<223> LV-CDR2 антител 3C6, 31C8, 26A1 и 8F4
<400> 30
Gln Xaa Ser Asn Leu Ala Ser
1 5
<210> 31
<211> 7
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 31
Asp Thr Ser Asn Leu Ala Ser
1 5
<210> 32
<211> 9
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 32
Gln Gln Asp Tyr Ser Ser Pro Tyr Thr
1 5
<210> 33
<211> 9
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 33
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1 5
<210> 34
<211> 9
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 34
Ala Gln Asn Leu Glu Leu Pro Phe Thr
1 5
<210> 35
<211> 121
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 35
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Asp Trp Ile
35 40 45
Gly Thr Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Tyr Ser Tyr Thr Asn Ser Pro Tyr Gly Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 36
<211> 107
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 36
Asp Val Val Met Thr Gln Thr Pro Lys Phe Leu Leu Val Ser Ala Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Ser Val Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Phe Thr Ile Asn Thr Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Phe Cys Gln Gln Asp Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 37
<211> 118
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 37
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Asp Tyr Thr Ile Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Ser Gln Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Ser Ala Thr Asp Tyr Tyr Phe Ala Ser Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 38
<211> 112
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 38
Asp Ile Val Met Thr Gln Ala Ala Ile Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Met Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly His Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 39
<211> 118
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 39
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Asp Ser
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Ser
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 40
<211> 112
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 40
Asp Ile Val Met Thr Gln Ala Ala Phe Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 41
<211> 118
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 41
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Asp Tyr Ile Phe Thr Asp Ser
20 25 30
Glu Ile His Trp Val Lys Gln Thr Leu Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Gly Ser Ala Val Tyr Tyr Cys
85 90 95
Ser Gly Ser Thr Thr Asp Cys Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 42
<211> 112
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 42
Asp Ile Val Met Thr Gln Ala Ala Phe Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 43
<211> 118
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 43
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly His Ile Phe Thr Asp Tyr
20 25 30
Glu Met Gln Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Phe Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ser Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Leu Thr Val Ser Ser
115
<210> 44
<211> 112
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 44
Asp Val Val Met Thr Gln Thr Ala Phe Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 45
<211> 119
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 45
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val Leu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ile Arg Val Asp Ser Asn Tyr Asp Tyr Phe Asp Tyr Trp Gly Gln Asp
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 46
<211> 106
<212> БЕЛОК
<213> Мышь (Mus sp.)
<400> 46
Asp Ile Val Leu Thr Gln Ser Pro Pro Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 47
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 7D1
<400> 47
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Asp Trp Ile
35 40 45
Gly Thr Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Tyr Ser Tyr Thr Asn Ser Pro Tyr Gly Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys
210 215 220
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
290 295 300
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 48
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 7D1
<400> 48
caggtgcagc tgcagcagtc tggagctgag ctggtgaggc caggagcttc cgtgaccctg 60
agctgcaagg cctctggcta caccttcaca gactatgaga tgcactgggt gaagcagacc 120
cccgtgcatg gcctggattg gatcggcaca atcgaccctg agaccggcga tacagcttac 180
aaccagaagt ttaagggcag ggccaccctg acagctgaca agtccagctc taccgcctat 240
atggagctgt ccagcctgac aagcgaggat tccgccgtgt actattgtac ccggtactcc 300
tatacaaata gcccctacgg catggactat tggggccagg gcacctccgt gacagtgtct 360
tccgctagca caaagggccc tagcgtgttc ccactggctc cctgctccag aagcacatct 420
gagtccaccg ccgctctggg ctgtctggtg aaggactact tccctgagcc agtgacagtg 480
tcctggaaca gcggcgccct gacatccgga gtgcacacct ttccagctgt gctgcagtcc 540
agcggcctgt acagcctgtc ttccgtggtg accgtgccca gctctaattt tggcacccag 600
acatatacct gcaacgtgga ccataagcct tctaatacaa aggtggataa gaccgtggag 660
aggaagtgct gcgtggagtg cccaccttgt ccagctccac cagtggctgg cccttccgtg 720
ttcctgtttc ctccaaagcc aaaggacaca ctgatgatct ctcggacacc cgaggtgacc 780
tgcgtggtgg tggacgtgtc ccacgaggat ccagaggtgc agttcaactg gtacgtggat 840
ggcgtggagg tgcataatgc caagaccaag cccagggagg agcagttcaa ctctacattt 900
cgggtggtgt ccgtgctgac cgtggtgcac caggattggc tgaacggcaa ggagtataag 960
tgcaaggtga gcaataaggg cctgcccgct cctatcgaga agacaatctc taagaccaag 1020
ggccagccta gagagccaca ggtgtacacc ctgccccctt cccgcgagga gatgacaaag 1080
aaccaggtga gcctgacctg tctggtgaag ggcttctatc cttctgacat ctccgtggag 1140
tgggagagca atggccagcc agagaacaat tacaagacca caccacccat gctggacagc 1200
gatggctctt tctttctgta tagcaagctg acagtggata agtctcgctg gcagcagggc 1260
aacgtgttta gctgctctgt gatgcatgag gccctgcaca atcattatac ccagaagtcc 1320
ctgagcctgt ctcccggcaa gtga 1344
<210> 49
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 7D1
<400> 49
Asp Val Val Met Thr Gln Thr Pro Lys Phe Leu Leu Val Ser Ala Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Ser Val Ser Asn Asp
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Tyr Gly Thr Asp Phe Thr Phe Thr Ile Asn Thr Val Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Phe Cys Gln Gln Asp Tyr Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 50
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 7D1
<400> 50
gacgtggtca tgacccagac acccaagttc ctgctggtga gcgccggcga cagggtgacc 60
atcacatgca aggcctctca gtccgtgagc aacgatgtgg cttggtacca gcagaagccc 120
ggccagtctc ctaagctgct gatctactat gcttccaata ggtacaccgg cgtgccagac 180
cggtttaccg gctctggcta tggcacagat ttcaccttta caatcaacac agtgcaggcc 240
gaggacctgg ccgtgtactt ctgtcagcag gattactcca gcccctatac ctttggcggc 300
ggcacaaagc tggagatcaa gcgtacggtg gccgctccca gcgtgttcat ctttccccct 360
tctgacgagc agctgaagtc tggcaccgct tccgtggtgt gcctgctgaa caatttctac 420
cccagagagg ccaaggtgca gtggaaggtg gataacgctc tgcagtccgg caatagccag 480
gagtctgtga ccgagcagga ctccaaggat agcacatatt ctctgtcttc caccctgaca 540
ctgtctaagg ccgactacga gaagcacaag gtgtatgctt gcgaggtgac ccatcagggc 600
ctgagctctc ctgtgacaaa gtcctttaat cgcggcgagt gttga 645
<210> 51
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 3C6
<400> 51
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Asp Tyr Thr Ile Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Ser Gln Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Ser Ala Thr Asp Tyr Tyr Phe Ala Ser Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 52
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 3C6
<400> 52
caggtgcagc tgcagcagtc tggagctgag ctggtgaggc caggagcttc cgtgaccctg 60
agctgcaagg cctctggcta caccttcaca gactatgaga tgcactgggt gaagcagacc 120
cccgtgcatg gcctggattg gatcggcaca atcgaccctg agaccggcga tacagcttac 180
aaccagaagt ttaagggcag ggccaccctg acagctgaca agtccagctc taccgcctat 240
atggagctgt ccagcctgac aagcgaggat tccgccgtgt actattgtac ccggtactcc 300
tatacaaata gcccctacgg catggactat tggggccagg gcacctccgt gacagtgtct 360
tccgctagca caaagggccc tagcgtgttc ccactggctc cctgctccag aagcacatct 420
gagtccaccg ccgctctggg ctgtctggtg aaggactact tccctgagcc agtgacagtg 480
tcctggaaca gcggcgccct gacatccgga gtgcacacct ttccagctgt gctgcagtcc 540
agcggcctgt acagcctgtc ttccgtggtg accgtgccca gctctaattt tggcacccag 600
acatatacct gcaacgtgga ccataagcct tctaatacaa aggtggataa gaccgtggag 660
aggaagtgct gcgtggagtg cccaccttgt ccagctccac cagtggctgg cccttccgtg 720
ttcctgtttc ctccaaagcc aaaggacaca ctgatgatct ctcggacacc cgaggtgacc 780
tgcgtggtgg tggacgtgtc ccacgaggat ccagaggtgc agttcaactg gtacgtggat 840
ggcgtggagg tgcataatgc caagaccaag cccagggagg agcagttcaa ctctacattt 900
cgggtggtgt ccgtgctgac cgtggtgcac caggattggc tgaacggcaa ggagtataag 960
tgcaaggtga gcaataaggg cctgcccgct cctatcgaga agacaatctc taagaccaag 1020
ggccagccta gagagccaca ggtgtacacc ctgccccctt cccgcgagga gatgacaaag 1080
aaccaggtga gcctgacctg tctggtgaag ggcttctatc cttctgacat ctccgtggag 1140
tgggagagca atggccagcc agagaacaat tacaagacca caccacccat gctggacagc 1200
gatggctctt tctttctgta tagcaagctg acagtggata agtctcgctg gcagcagggc 1260
aacgtgttta gctgctctgt gatgcatgag gccctgcaca atcattatac ccagaagtcc 1320
ctgagcctgt ctcccggcaa gtga 1344
<210> 53
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 3C6
<400> 53
Asp Ile Val Met Thr Gln Ala Ala Ile Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Met Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Thr Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly His Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 54
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 3C6
<400> 54
gacatcgtga tgacccaggc cgctatctct aaccccgtga ccctgggcac atctgcctcc 60
atgagctgca ggtccagcaa gtctctgctg cactccaatg gcatcacata cctgtattgg 120
tacctgcaga agcccggcca ttcccctcag ctgctgatct atcagatgag caacctggct 180
tctggcgtgc cagacaggtt ctcttccagc ggctccggca ccgacttcac cctgaagatc 240
agcagggtgg aggctgagga tgtgggcgtg tactattgtg ctcagaatct ggagctgccc 300
ttcacctttg gctccggcac aaagctggag atcaagcgta cggtggccgc tcccagcgtg 360
ttcatctttc ccccttctga cgagcagctg aagtctggca ccgcttccgt ggtgtgcctg 420
ctgaacaatt tctaccccag agaggccaag gtgcagtgga aggtggataa cgctctgcag 480
tccggcaata gccaggagtc tgtgaccgag caggactcca aggatagcac atattctctg 540
tcttccaccc tgacactgtc taaggccgac tacgagaagc acaaggtgta tgcttgcgag 600
gtgacccatc agggcctgag ctctcctgtg acaaagtcct ttaatcgcgg cgagtgttga 660
<210> 55
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 26A1
<400> 55
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Asp Tyr Ile Phe Thr Asp Ser
20 25 30
Glu Ile His Trp Val Lys Gln Thr Leu Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Gly Ser Ala Val Tyr Tyr Cys
85 90 95
Ser Gly Ser Thr Thr Asp Cys Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 56
<211> 1335
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 26A1
<400> 56
caggtgcagc tgcagcagtc tggagctgag ctggtgaggc caggagcttc tgtgaccctg 60
tcctgcaagg ccagcgacta catcttcacc gattccgaga tccactgggt gaagcagaca 120
ctggtgcatg gcctggagtg gatcggagct atcgaccctg agaccggcga tacagcttac 180
aaccagaagt ttaagggcaa ggccaccctg acagctgaca agtccagctc taccgcctat 240
atggagctgt ccagcctgac atccgagggc tccgccgtgt actattgcag cggctctacc 300
acagattgtt acttcgctta ttggggccag ggcaccacac tgacagtgtc ttccgctagc 360
acaaagggcc ctagcgtgtt cccactggct ccctgctcca gaagcacatc tgagtccacc 420
gccgctctgg gctgtctggt gaaggactac ttccctgagc cagtgacagt gtcctggaac 480
agcggcgccc tgacatccgg agtgcacacc tttccagctg tgctgcagtc cagcggcctg 540
tacagcctgt cttccgtggt gaccgtgccc agctctaatt ttggcaccca gacatatacc 600
tgcaacgtgg accataagcc ttctaataca aaggtggata agaccgtgga gaggaagtgc 660
tgcgtggagt gcccaccttg tccagctcca ccagtggctg gcccttccgt gttcctgttt 720
cctccaaagc caaaggacac actgatgatc tctcggacac ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccacgagga tccagaggtg cagttcaact ggtacgtgga tggcgtggag 840
gtgcataatg ccaagaccaa gcccagggag gagcagttca actctacatt tcgggtggtg 900
tccgtgctga ccgtggtgca ccaggattgg ctgaacggca aggagtataa gtgcaaggtg 960
agcaataagg gcctgcccgc tcctatcgag aagacaatct ctaagaccaa gggccagcct 1020
agagagccac aggtgtacac cctgccccct tcccgcgagg agatgacaaa gaaccaggtg 1080
agcctgacct gtctggtgaa gggcttctat ccttctgaca tctccgtgga gtgggagagc 1140
aatggccagc cagagaacaa ttacaagacc acaccaccca tgctggacag cgatggctct 1200
ttctttctgt atagcaagct gacagtggat aagtctcgct ggcagcaggg caacgtgttt 1260
agctgctctg tgatgcatga ggccctgcac aatcattata cccagaagtc cctgagcctg 1320
tctcccggca agtga 1335
<210> 57
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 26A1
<400> 57
Asp Ile Val Met Thr Gln Ala Ala Phe Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 58
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 26A1
<400> 58
gacatcgtga tgacccaggc cgctttctct aaccccgtga ccctgggcac atctgcctcc 60
atcagctgca gatccagcaa gtctctgctg cactccaatg gcatcatcta cctgtattgg 120
tacctgcaga agcccggcca gtcccctcag ctgctgatct atcagatgag caacctggct 180
tctggcgtgc cagaccgctt ctcttccagc ggctccggca ccgacttcac cctgaggatc 240
agcagggtgg aggctgagga tgtgggcgtg tactattgtg ctcagaatct ggagctgccc 300
ttcacctttg gctccggcac aaagctggag atcaagcgta cggtggccgc tcccagcgtg 360
ttcatctttc ccccttctga cgagcagctg aagtctggca ccgcttccgt ggtgtgcctg 420
ctgaacaatt tctaccccag agaggccaag gtgcagtgga aggtggataa cgctctgcag 480
tccggcaata gccaggagtc tgtgaccgag caggactcca aggatagcac atattctctg 540
tcttccaccc tgacactgtc taaggccgac tacgagaagc acaaggtgta tgcttgcgag 600
gtgacccatc agggcctgag ctctcctgtg acaaagtcct ttaatcgcgg cgagtgttga 660
<210> 59
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 8F4
<400> 59
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly His Ile Phe Thr Asp Tyr
20 25 30
Glu Met Gln Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Phe Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ser Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 60
<211> 1335
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 8F4
<400> 60
caggtgcagc tgcagcagtc tggagctgag ctggtgaggc caggagcttc tgtgaccctg 60
tcctgcaagg ccagcggcca catcttcacc gactacgaga tgcagtgggt gaagcagaca 120
ccagtgcatg gactggagtg gatcggagcc ttcgaccctg agaccggcga tacagcttac 180
aaccagaagt ttaagggcaa ggccaccctg acagctgata agtccagctc taccgcctat 240
atggagctgt ccagcctgac atccgaggac tccgccgtgt actattgtag cggctctacc 300
acagattact attttgctta ttggggccag ggcacctccc tgacagtgtc ttccgctagc 360
acaaagggcc ctagcgtgtt cccactggct ccctgctcca gaagcacatc tgagtccacc 420
gccgctctgg gctgtctggt gaaggactac ttccctgagc cagtgacagt gtcctggaac 480
agcggcgccc tgacatccgg agtgcacacc tttccagctg tgctgcagtc cagcggcctg 540
tacagcctgt cttccgtggt gaccgtgccc agctctaatt ttggcaccca gacatatacc 600
tgcaacgtgg accataagcc ttctaataca aaggtggata agaccgtgga gaggaagtgc 660
tgcgtggagt gcccaccttg tccagctcca ccagtggctg gcccttccgt gttcctgttt 720
cctccaaagc caaaggacac actgatgatc tctcggacac ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccacgagga tccagaggtg cagttcaact ggtacgtgga tggcgtggag 840
gtgcataatg ccaagaccaa gcccagggag gagcagttca actctacatt tcgggtggtg 900
tccgtgctga ccgtggtgca ccaggattgg ctgaacggca aggagtataa gtgcaaggtg 960
agcaataagg gcctgcccgc tcctatcgag aagacaatct ctaagaccaa gggccagcct 1020
agagagccac aggtgtacac cctgccccct tcccgcgagg agatgacaaa gaaccaggtg 1080
agcctgacct gtctggtgaa gggcttctat ccttctgaca tctccgtgga gtgggagagc 1140
aatggccagc cagagaacaa ttacaagacc acaccaccca tgctggacag cgatggctct 1200
ttctttctgt atagcaagct gacagtggat aagtctcgct ggcagcaggg caacgtgttt 1260
agctgctctg tgatgcatga ggccctgcac aatcattata cccagaagtc cctgagcctg 1320
tctcccggca agtga 1335
<210> 61
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 8F4
<400> 61
Asp Val Val Met Thr Gln Thr Ala Phe Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 62
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 8F4
<400> 62
gacgtggtca tgacccagac agccttctct aaccccgtga ccctgggcac atctgcttcc 60
atcagctgca gatccagcaa gtctctgctg cactccaatg gcatcatcta cctgtattgg 120
tacctgcaga agcccggcca gtcccctcag ctgctgatct atcagatgag caacctggcc 180
tctggcgtgc cagaccgctt ctcttccagc ggctccggca ccgacttcac cctgaggatc 240
agcagggtgg aggctgagga tgtgggcgtg tactattgtg ctcagaatct ggagctgccc 300
ttcacctttg gctccggcac aaagctggag atcaagcgta cggtggccgc tcccagcgtg 360
ttcatctttc ccccttctga cgagcagctg aagtctggca ccgcttccgt ggtgtgcctg 420
ctgaacaatt tctaccccag agaggccaag gtgcagtgga aggtggataa cgctctgcag 480
tccggcaata gccaggagtc tgtgaccgag caggactcca aggatagcac atattctctg 540
tcttccaccc tgacactgtc taaggccgac tacgagaagc acaaggtgta tgcttgcgag 600
gtgacccatc agggcctgag ctctcctgtg acaaagtcct ttaatcgcgg cgagtgttga 660
<210> 63
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 17E2
<400> 63
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val Leu Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ile Arg Val Asp Ser Asn Tyr Asp Tyr Phe Asp Tyr Trp Gly Gln Asp
100 105 110
Thr Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 64
<211> 1338
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 17E2
<400> 64
caggtgcagc tgcagcagtc cggagctgag ctggtgaggc caggagcttc cgtgaccctg 60
agctgcaagg cctctggcta caccttcaca gactatgaga tgcactgggt gaagcagaca 120
cccgtgctgg gactggagtg gatcggagct atcgaccctg agaccggcga tacagcttac 180
aaccagaagt ttaagggcaa ggccaccctg acagctgaca agtccagctc taccgcttat 240
atggagctgt ccagcctggc cagcgaggac tccgccgtgt actattgtat ccgggtggat 300
agcaattacg actatttcga ttactggggc caggatacca cactgacagt gtcttccgct 360
agcacaaagg gccctagcgt gttcccactg gctccctgct ccagaagcac atctgagtcc 420
accgccgctc tgggctgtct ggtgaaggac tacttccctg agccagtgac agtgtcctgg 480
aacagcggcg ccctgacatc cggagtgcac acctttccag ctgtgctgca gtccagcggc 540
ctgtacagcc tgtcttccgt ggtgaccgtg cccagctcta attttggcac ccagacatat 600
acctgcaacg tggaccataa gccttctaat acaaaggtgg ataagaccgt ggagaggaag 660
tgctgcgtgg agtgcccacc ttgtccagct ccaccagtgg ctggcccttc cgtgttcctg 720
tttcctccaa agccaaagga cacactgatg atctctcgga cacccgaggt gacctgcgtg 780
gtggtggacg tgtcccacga ggatccagag gtgcagttca actggtacgt ggatggcgtg 840
gaggtgcata atgccaagac caagcccagg gaggagcagt tcaactctac atttcgggtg 900
gtgtccgtgc tgaccgtggt gcaccaggat tggctgaacg gcaaggagta taagtgcaag 960
gtgagcaata agggcctgcc cgctcctatc gagaagacaa tctctaagac caagggccag 1020
cctagagagc cacaggtgta caccctgccc ccttcccgcg aggagatgac aaagaaccag 1080
gtgagcctga cctgtctggt gaagggcttc tatccttctg acatctccgt ggagtgggag 1140
agcaatggcc agccagagaa caattacaag accacaccac ccatgctgga cagcgatggc 1200
tctttctttc tgtatagcaa gctgacagtg gataagtctc gctggcagca gggcaacgtg 1260
tttagctgct ctgtgatgca tgaggccctg cacaatcatt atacccagaa gtccctgagc 1320
ctgtctcccg gcaagtga 1338
<210> 65
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 17E2
<400> 65
Asp Ile Val Leu Thr Gln Ser Pro Pro Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 66
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 17E2
<400> 66
gacatcgtgc tgacccagag cccacctatc ctgagcgcct ctccaggaga gaaggtgacc 60
atgacatgca gggcttccag ctccgtgtcc tacatgcact ggtatcagca gaagcctggc 120
tccagcccaa agccctggat ctacgacacc tctaacctgg cttccggagt gccagctagg 180
ttctccggca gcggctctgg cacctcctat agcctgacaa tctccagggt ggaggctgag 240
gatgctgcta catactattg tcagcagtgg tcttccaatc ctctgacctt tggcgctggc 300
acaaagctgg agctgaagcg tacggtggcc gctcccagcg tgttcatctt tcccccttct 360
gacgagcagc tgaagtctgg caccgcttcc gtggtgtgcc tgctgaacaa tttctacccc 420
agagaggcca aggtgcagtg gaaggtggat aacgctctgc agtccggcaa tagccaggag 480
tctgtgaccg agcaggactc caaggatagc acatattctc tgtcttccac cctgacactg 540
tctaaggccg actacgagaa gcacaaggtg tatgcttgcg aggtgaccca tcagggcctg 600
agctctcctg tgacaaagtc ctttaatcgc ggcgagtgtt ga 642
<210> 67
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 31C8
<400> 67
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Thr Leu Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Asp Ser
20 25 30
Glu Met His Trp Val Lys Gln Thr Pro Val His Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Ser
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 68
<211> 1335
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного антитела 31C8
<400> 68
caggtgcagc tgcagcagtc tggaccagag ctggtgaggc ctggagcctc cgtgaccctg 60
agctgcaagg cttctggcta catcttcatc gactccgaga tgcactgggt gaagcagaca 120
ccagtgcatg gactggagtg gatcggagct atcgacccag agaccggcga tacagtgtat 180
aaccagaagt ttaagggcaa ggccaccctg acagctgata agtccagctc taccgcctcc 240
atggagctgt ccagcctgac aagcgaggac tccgccgtgt actattgtac cggcagcacc 300
acagattact atttcgctta ctggggccag ggcaccacac tgacagtgtc ttccgctagc 360
acaaagggcc ctagcgtgtt cccactggct ccctgctcca gaagcacatc tgagtccacc 420
gccgctctgg gctgtctggt gaaggactac ttccctgagc cagtgacagt gtcctggaac 480
agcggcgccc tgacatccgg agtgcacacc tttccagctg tgctgcagtc cagcggcctg 540
tacagcctgt cttccgtggt gaccgtgccc agctctaatt ttggcaccca gacatatacc 600
tgcaacgtgg accataagcc ttctaataca aaggtggata agaccgtgga gaggaagtgc 660
tgcgtggagt gcccaccttg tccagctcca ccagtggctg gcccttccgt gttcctgttt 720
cctccaaagc caaaggacac actgatgatc tctcggacac ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccacgagga tccagaggtg cagttcaact ggtacgtgga tggcgtggag 840
gtgcataatg ccaagaccaa gcccagggag gagcagttca actctacatt tcgggtggtg 900
tccgtgctga ccgtggtgca ccaggattgg ctgaacggca aggagtataa gtgcaaggtg 960
agcaataagg gcctgcccgc tcctatcgag aagacaatct ctaagaccaa gggccagcct 1020
agagagccac aggtgtacac cctgccccct tcccgcgagg agatgacaaa gaaccaggtg 1080
agcctgacct gtctggtgaa gggcttctat ccttctgaca tctccgtgga gtgggagagc 1140
aatggccagc cagagaacaa ttacaagacc acaccaccca tgctggacag cgatggctct 1200
ttctttctgt atagcaagct gacagtggat aagtctcgct ggcagcaggg caacgtgttt 1260
agctgctctg tgatgcatga ggccctgcac aatcattata cccagaagtc cctgagcctg 1320
tctcccggca agtga 1335
<210> 69
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 31C8
<400> 69
Asp Ile Val Met Thr Gln Ala Ala Phe Ser Asn Pro Val Thr Leu Gly
1 5 10 15
Thr Ser Ala Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Gln Met Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 70
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного антитела 31C8
<400> 70
gacatcgtga tgacccaggc cgctttctct aaccccgtga ccctgggcac atctgcctcc 60
atcagctgca gatccagcaa gtctctgctg cactccaatg gcatcatcta cctgtattgg 120
tacctgcaga agcccggcca gtcccctcag ctgctgatct atcagatgag caacctggct 180
tctggcgtgc cagaccgctt ctcttccagc ggctccggca ccgacttcac cctgaggatc 240
agcagggtgg aggctgagga tgtgggcgtg tactattgtg ctcagaatct ggagctgccc 300
ttcacctttg gctccggcac aaagctggag atcaagcgta cggtggccgc tcccagcgtg 360
ttcatctttc ccccttctga cgagcagctg aagtctggca ccgcttccgt ggtgtgcctg 420
ctgaacaatt tctaccccag agaggccaag gtgcagtgga aggtggataa cgctctgcag 480
tccggcaata gccaggagtc tgtgaccgag caggactcca aggatagcac atattctctg 540
tcttccaccc tgacactgtc taaggccgac tacgagaagc acaaggtgta tgcttgcgag 600
gtgacccatc agggcctgag ctctcctgtg acaaagtcct ttaatcgcgg cgagtgttga 660
<210> 71
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HV гуманизированного антитела 31C8-1.1
<400> 71
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Asp Ser
20 25 30
Glu Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 72
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HV of гуманизированного антитела 31C8-1.1
<400> 72
gaggtgcagc tggtgcagtc tggcgccgag gtgaagaagc caggcgctac cgtgaagatc 60
agctgcaagg cctctggcta catcttcatc gactccgaga tgcactgggt gcagcaggct 120
ccaggcaagg gactggagtg gatgggcgct atcgaccctg agaccggcga tacagtgtac 180
gctgagaagt ttcagggcag ggtgaccatc acagccgata agtccagctc tacagcttat 240
atggagctgt ccagcctgcg gtccgaggac accgccgtgt actattgtac aggcagcacc 300
acagattact atttcgctta ttggggccag ggcaccctgg tgacagtgtc ttcc 354
<210> 73
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HV гуманизированного антитела 31C8-1.2
<400> 73
Gln Val Gln Leu Val Gln Ser Gly Ala Lys Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Asp Ser
20 25 30
Glu Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Ala Lys Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 74
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> HV гуманизированного антитела 31C8-1.2
<400> 74
caggtgcagc tggtgcagtc tggcgccaag gtgaagaagc caggcgctac cgtgaagatc 60
agctgcaagg cctctggcta catcttcatc gactccgaga tgcactgggt gcagcaggct 120
ccaggcaagg gactggagtg gatgggcgct atcgaccctg agaccggcga tacagtgtac 180
gctaagaagt ttaagggcag ggtgaccatc acagccgata agtccagctc tacagcttat 240
atggagctgt ccagcctgcg gtccgaggac accgccgtgt actattgtac aggcagcacc 300
acagattact atttcgctta ttggggccag ggcaccctgg tgacagtgtc ttcc 354
<210> 75
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь гуманизированного антитела 31C8-1.1
<400> 75
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Asp Ser
20 25 30
Glu Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Ala Glu Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 76
<211> 1335
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь гуманизированного антитела 31C8-1.1
<400> 76
gaggtgcagc tggtgcagtc tggcgccgag gtgaagaagc caggcgctac cgtgaagatc 60
agctgcaagg cctctggcta catcttcatc gactccgaga tgcactgggt gcagcaggct 120
ccaggcaagg gactggagtg gatgggcgct atcgaccctg agaccggcga tacagtgtac 180
gctgagaagt ttcagggcag ggtgaccatc acagccgata agtccagctc tacagcttat 240
atggagctgt ccagcctgcg gtccgaggac accgccgtgt actattgtac aggcagcacc 300
acagattact atttcgctta ttggggccag ggcaccctgg tgacagtgtc ttccgcctcc 360
accaagggcc ctagcgtgtt cccactggct ccctgctcca gaagcacatc tgagtccacc 420
gccgctctgg gctgtctggt gaaggactac ttccctgagc cagtgacagt gtcctggaac 480
agcggcgccc tgacatccgg agtgcacacc tttccagctg tgctgcagtc cagcggcctg 540
tacagcctgt cttccgtggt gaccgtgccc agctctaatt ttggcaccca gacatatacc 600
tgcaacgtgg accataagcc ttctaataca aaggtggata agaccgtgga gaggaagtgc 660
tgcgtggagt gcccaccttg tccagctcca ccagtggctg gcccttccgt gttcctgttt 720
cctccaaagc caaaggacac actgatgatc tctcggacac ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccacgagga tccagaggtg cagttcaact ggtacgtgga tggcgtggag 840
gtgcataatg ccaagaccaa gcccagggag gagcagttca actctacatt tcgggtggtg 900
tccgtgctga ccgtggtgca ccaggattgg ctgaacggca aggagtataa gtgcaaggtg 960
agcaataagg gcctgcccgc tcctatcgag aagacaatct ctaagaccaa gggccagcct 1020
agagagccac aggtgtacac cctgccccct tcccgcgagg agatgacaaa gaaccaggtg 1080
agcctgacct gtctggtgaa gggcttctat ccttctgaca tctccgtgga gtgggagagc 1140
aatggccagc cagagaacaa ttacaagacc acaccaccca tgctggacag cgatggctct 1200
ttctttctgt atagcaagct gacagtggat aagtctcgct ggcagcaggg caacgtgttt 1260
agctgctctg tgatgcatga ggccctgcac aatcattata cccagaagtc cctgagcctg 1320
tctcccggca agtga 1335
<210> 77
<211> 444
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь гуманизированного антитела 31C8-1.2
<400> 77
Gln Val Gln Leu Val Gln Ser Gly Ala Lys Val Lys Lys Pro Gly Ala
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ile Phe Ile Asp Ser
20 25 30
Glu Met His Trp Val Gln Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ala Ile Asp Pro Glu Thr Gly Asp Thr Val Tyr Ala Lys Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Gly Ser Thr Thr Asp Tyr Tyr Phe Ala Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys
210 215 220
Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr
290 295 300
Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 78
<211> 1335
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь гуманизированного антитела 31C8-1.2
<400> 78
caggtgcagc tggtgcagtc tggcgccaag gtgaagaagc caggcgctac cgtgaagatc 60
agctgcaagg cctctggcta catcttcatc gactccgaga tgcactgggt gcagcaggct 120
ccaggcaagg gactggagtg gatgggcgct atcgaccctg agaccggcga tacagtgtac 180
gctaagaagt ttaagggcag ggtgaccatc acagccgata agtccagctc tacagcttat 240
atggagctgt ccagcctgcg gtccgaggac accgccgtgt actattgtac aggcagcacc 300
acagattact atttcgctta ttggggccag ggcaccctgg tgacagtgtc ttccgcctcc 360
accaagggcc ctagcgtgtt cccactggct ccctgctcca gaagcacatc tgagtccacc 420
gccgctctgg gctgtctggt gaaggactac ttccctgagc cagtgacagt gtcctggaac 480
agcggcgccc tgacatccgg agtgcacacc tttccagctg tgctgcagtc cagcggcctg 540
tacagcctgt cttccgtggt gaccgtgccc agctctaatt ttggcaccca gacatatacc 600
tgcaacgtgg accataagcc ttctaataca aaggtggata agaccgtgga gaggaagtgc 660
tgcgtggagt gcccaccttg tccagctcca ccagtggctg gcccttccgt gttcctgttt 720
cctccaaagc caaaggacac actgatgatc tctcggacac ccgaggtgac ctgcgtggtg 780
gtggacgtgt cccacgagga tccagaggtg cagttcaact ggtacgtgga tggcgtggag 840
gtgcataatg ccaagaccaa gcccagggag gagcagttca actctacatt tcgggtggtg 900
tccgtgctga ccgtggtgca ccaggattgg ctgaacggca aggagtataa gtgcaaggtg 960
agcaataagg gcctgcccgc tcctatcgag aagacaatct ctaagaccaa gggccagcct 1020
agagagccac aggtgtacac cctgccccct tcccgcgagg agatgacaaa gaaccaggtg 1080
agcctgacct gtctggtgaa gggcttctat ccttctgaca tctccgtgga gtgggagagc 1140
aatggccagc cagagaacaa ttacaagacc acaccaccca tgctggacag cgatggctct 1200
ttctttctgt atagcaagct gacagtggat aagtctcgct ggcagcaggg caacgtgttt 1260
agctgctctg tgatgcatga ggccctgcac aatcattata cccagaagtc cctgagcctg 1320
tctcccggca agtga 1335
<210> 79
<211> 112
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LV гуманизированных антител 31C8-1.1 и 31C8-1.2
<400> 79
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Gln Leu Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 80
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> LV гуманизированных антител 31C8-1.1 и 31C8-1.2
<400> 80
gacatccaga tgacccagag cccttccagc ctgtccgcca gcgtgggcga tagggtgacc 60
atcacatgcc ggtcttccaa gtctctgctg cactccaacg gcatcatcta cctgtattgg 120
taccagcaga agcccggcaa ggcccctaag ctgctgatct atcagctgag caatctggct 180
tctggcgtgc catccaggtt cagctcttcc ggctctggca ccgactttac cctgacaatc 240
agctctctgc agccagagga tttcgccaca tactattgtg ctcagaacct ggagctgccc 300
ttcacctttg gccagggcac aaaggtggag atcaag 336
<210> 81
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкие цепи гуманизированных антител 31C8-1.1 и 31C8-1.2
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Asn Gly Ile Ile Tyr Leu Tyr Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Gln Leu Ser Asn Leu Ala Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Ser Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Ala Gln Asn
85 90 95
Leu Glu Leu Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 82
<211> 660
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкие цепи гуманизированных антител 31C8-1.1 и 31C8-1.2
<400> 82
gacatccaga tgacccagag cccttccagc ctgtccgcca gcgtgggcga tagggtgacc 60
atcacatgcc ggtcttccaa gtctctgctg cactccaacg gcatcatcta cctgtattgg 120
taccagcaga agcccggcaa ggcccctaag ctgctgatct atcagctgag caatctggct 180
tctggcgtgc catccaggtt cagctcttcc ggctctggca ccgactttac cctgacaatc 240
agctctctgc agccagagga tttcgccaca tactattgtg ctcagaacct ggagctgccc 300
ttcacctttg gccagggcac aaaggtggag atcaagcgta cggtggccgc tcccagcgtg 360
ttcatctttc ccccttctga cgagcagctg aagtctggca ccgcttccgt ggtgtgcctg 420
ctgaacaatt tctaccccag agaggccaag gtgcagtgga aggtggataa cgctctgcag 480
tccggcaata gccaggagtc tgtgaccgag caggactcca aggatagcac atattctctg 540
tcttccaccc tgacactgtc taaggccgac tacgagaagc acaaggtgta tgcttgcgag 600
gtgacccatc agggcctgag ctctcctgtg acaaagtcct ttaatcgcgg cgagtgttga 660
<210> 83
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь RG6149
<400> 83
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Asn Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asn Ser Asp Thr Arg Phe Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Thr Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg His Gly Thr Ser Ser Asp Tyr Tyr Gly Leu Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys
210 215 220
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
290 295 300
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 84
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь RG6149
<400> 84
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Asp Asp Asn Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 85
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь GSK3772847
<400> 85
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ile Tyr
20 25 30
Asp Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Arg Gly Glu Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Pro Trp Ser Thr Glu Gly Ser Phe Phe Val Leu Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Thr Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val
195 200 205
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys
210 215 220
Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Ala Ala Ala Ser
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Ala Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 86
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь GSK3772847
<400> 86
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asp Asp Asp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Ile Thr Ala Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ДЛЯ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА 4 | 2017 |
|
RU2774446C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| КОНЪЮГАТ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО, ПРОМЕЖУТОЧНОЕ СОЕДИНЕНИЕ ДЛЯ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2800137C1 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2840044C1 |
| ПОЛИПЕПТИД ДЛЯ ЛЕЧЕНИЯ ПАТОЛОГИЧЕСКИХ ТРОМБОВ | 2019 |
|
RU2778566C1 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2801209C2 |
| АНТИТЕЛА К СОБАЧЬЕМУ АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 | 2020 |
|
RU2838943C1 |
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| АНТИТЕЛО К АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 ЧЕЛОВЕКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2807060C1 |
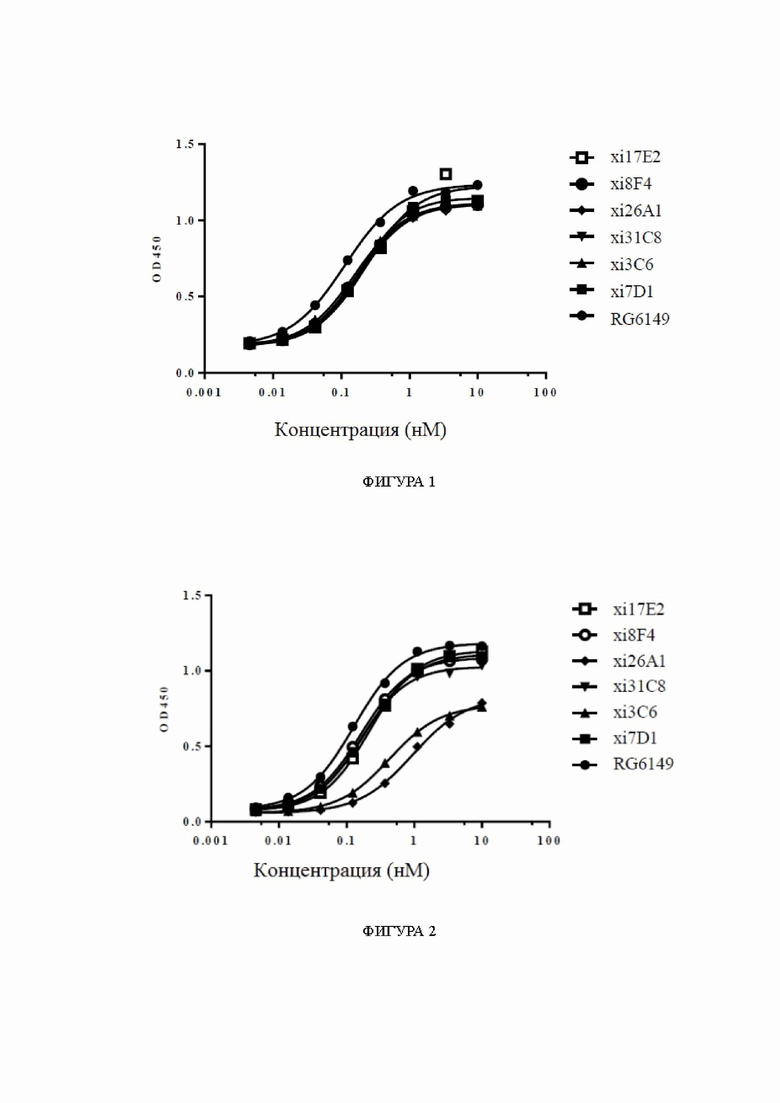
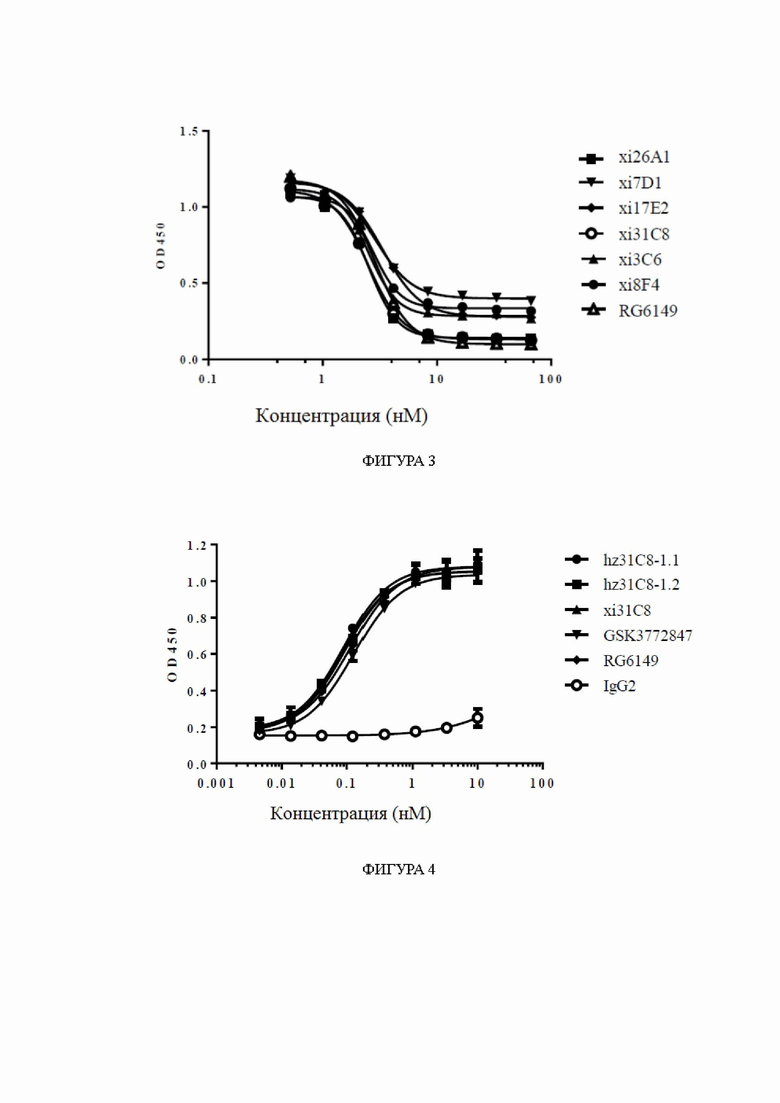
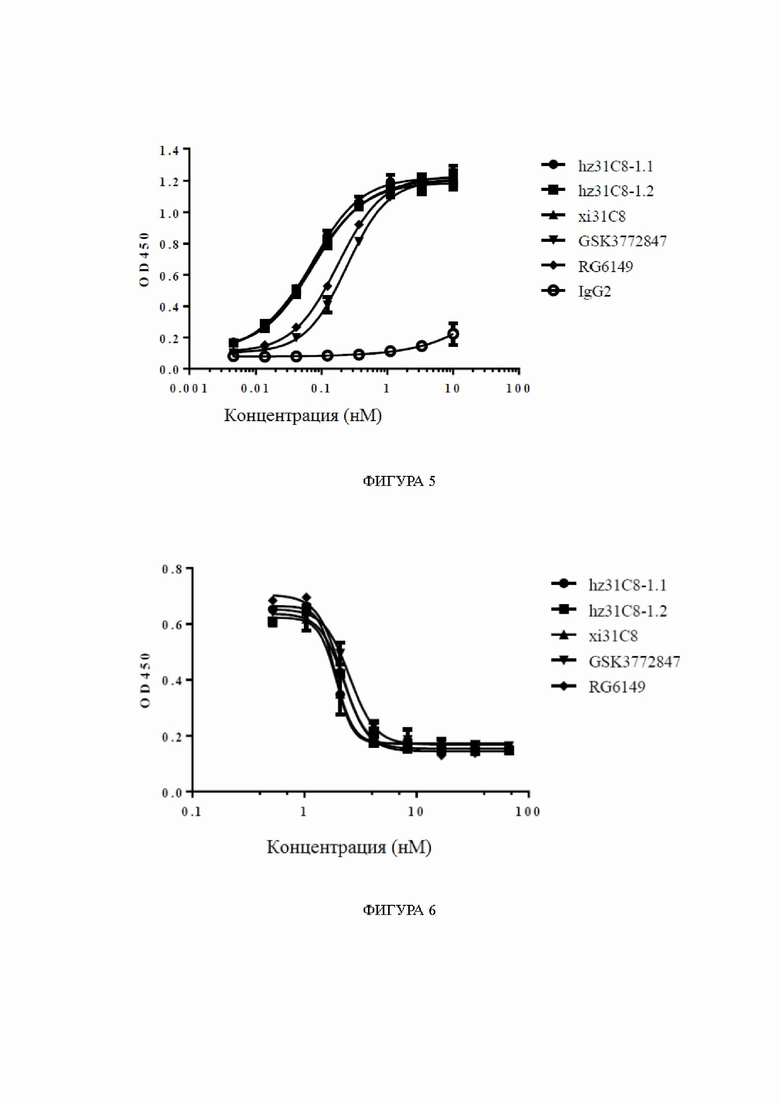
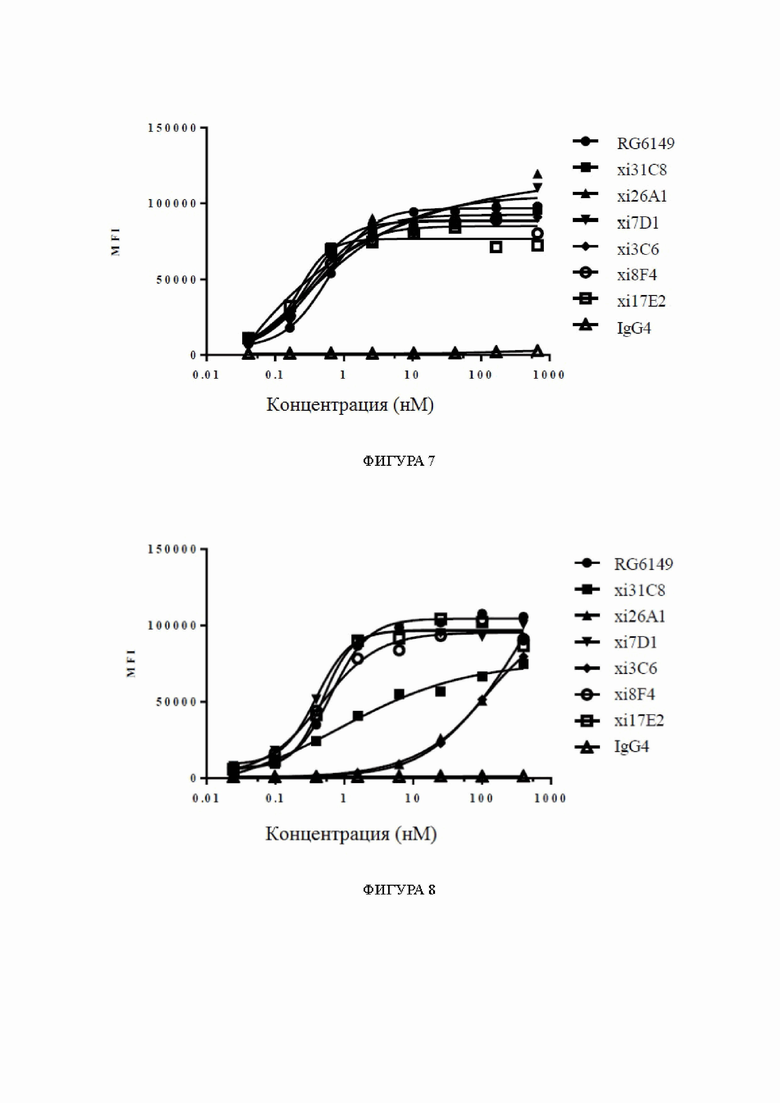
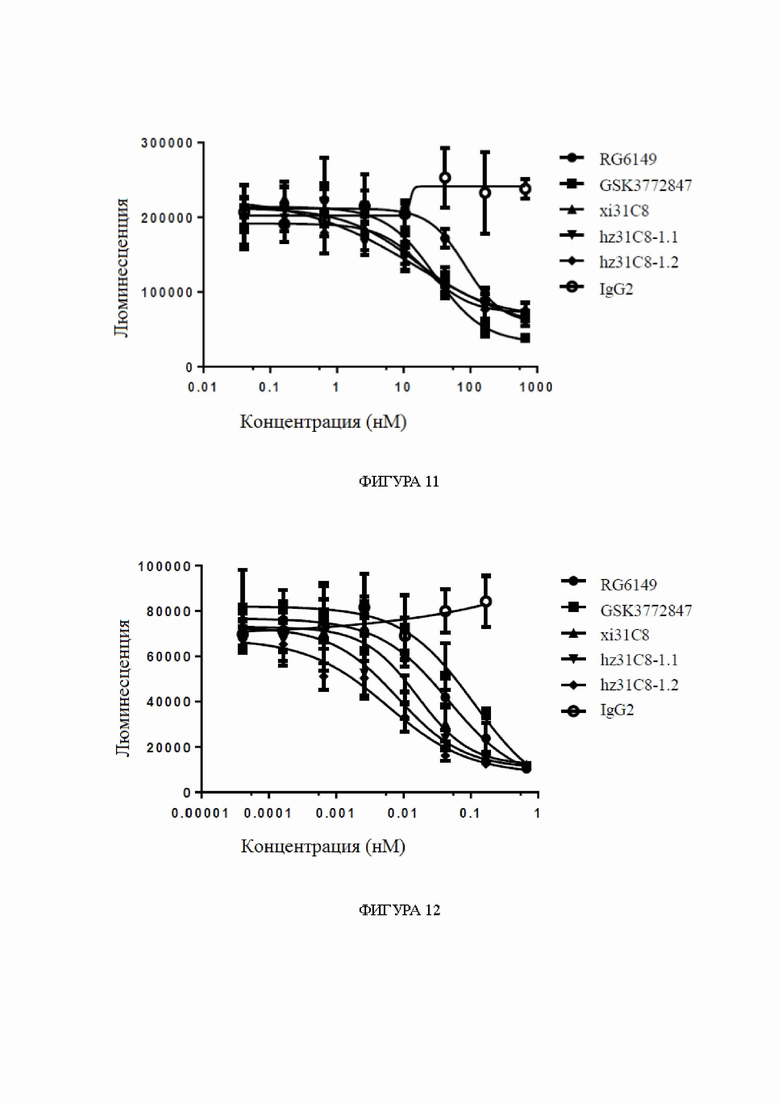
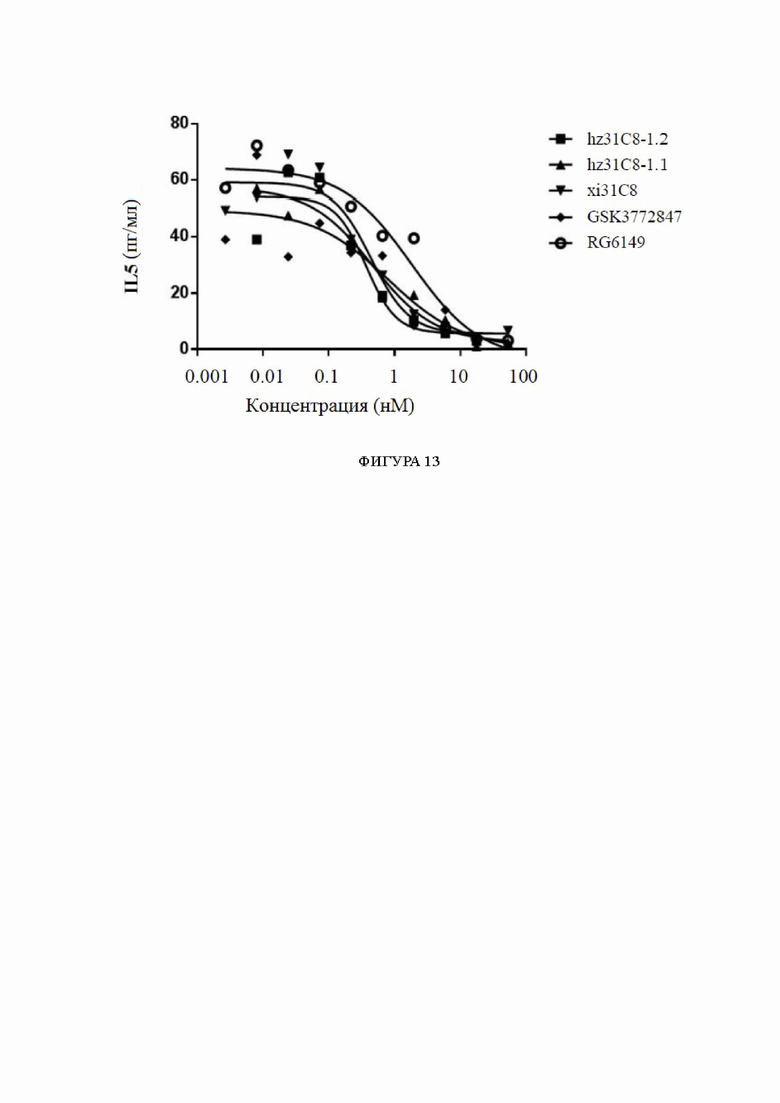
Изобретение относится к области биотехнологии, в частности к выделенному антителу к ST2 или его антигенсвязывающему фрагменту. Настоящее изобретение также относится к выделенной молекуле нуклеиновой кислоты, кодирующей указанное антитело, вектору экспрессии, клетке-хозяину, слитому полипептиду и мультиспецифической молекуле, содержащим указанное антитело, вирусному вектору, способу определения уровня экспрессии ST2 в образце, полученном от субъекта, а также к фармацевтической композиции для лечения или облегчения заболеваний и состояний, опосредованных IL33/ST2, и способу лечения таких заболеваний. Настоящее изобретение обеспечивает антитела или их антигенсвязывающие фрагменты, которые связываются с ST2 и способны влиять на этиопатогенез IL33/ST2-опосредованных заболеваний. 11 н. и 14 з.п. ф-лы, 13 ил., 21 табл., 15 пр.
1. Выделенное антитело к ST2 или его антигенсвязывающий фрагмент, содержащее:
(i) CDR1 тяжелой цепи, CDR2 тяжелой цепи и CDR3 тяжелой цепи, где:
(1) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 15, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 20;
(2) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 16, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 21;
(3) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22;
(4) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 13, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 23;
(5) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 14, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 19, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22; или
(6) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, и CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 24;
(ii) CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, где:
(1) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 25, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 29, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 32;
(2) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 26, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34;
(3) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34; или
(4) CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 28, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 31, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 33, где
аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где:
(i) X1 представляет собой N или A, X2 представляет собой Q, и X3 представляет собой K или Q;
(ii) X1 представляет собой N или A, X2 представляет собой E, и X3 представляет собой K или Q; или
(iii) X1 представляет собой N или A, X2 представляет собой K, X3 представляет собой K или Q;
дополнительно аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, где X4 представляет собой M или L.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащее CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи, CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, где:
(1) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 15, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 20, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 25, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 29, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 32;
(2) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 16, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 21, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 26, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34;
(3) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34;
(4) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 13, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 23, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34;
(5) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 14, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 19, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34; или
(6) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 24, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 28, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 31, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 33; где
аминокислотная последовательность SEQ ID NO: 17 представляет собой AIDPETGDTVYX1X2KFX3G, где:
(i) X1 представляет собой N или A, X2 представляет собой Q, и X3 представляет собой K или Q;
(ii) X1 представляет собой N или A, X2 представляет собой E, и X3 представляет собой K или Q; или
(iii) X1 представляет собой N или A, X2 представляет собой K, X3 представляет собой K или Q;
дополнительно аминокислотная последовательность SEQ ID NO: 30 представляет собой QX4SNLAS, где X4 представляет собой M или L.
3. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1 или 2, содержащее CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи, CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи, где:
(1) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 15, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 20, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 25, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 29, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 32;
(2) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 16, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 21, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 26, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, причем X4 = M;
(3) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, причем X1 = N, X2 = Q, X3 = K и X4 = M;
(4) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, причем X1 = A, X2 = E, X3 = Q и X4 = L;
(5) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 12, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 17, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, причем X1 = A, X2 = K, X3 = K и X4 = L;
(6) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 13, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 23, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, причем X4 = M;
(7) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 14, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 19, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 22, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 27, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 30, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 34, причем X4 = M; или
(8) CDR1 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 11, CDR2 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 18, CDR3 тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 24, CDR1 легкой цепи содержит последовательность, представленную в SEQ ID NO: 28, CDR2 легкой цепи содержит последовательность, представленную в SEQ ID NO: 31, и CDR3 легкой цепи содержит последовательность, представленную в SEQ ID NO: 33.
4. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3, которое является химерным или гуманизированным.
5. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-4, содержащее:
(i) вариабельную область тяжелой цепи, где аминокислотная последовательность вариабельной области тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 35, 37, 39, 41, 43, 45, 71 или 73, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с последовательностью, представленной в SEQ ID NO: 35, 37, 39, 41, 43, 45, 71 или 73;
(i) вариабельную область легкой цепи, где аминокислотная последовательность вариабельной области легкой цепи содержит последовательность, представленную в SEQ ID NO: 36, 38, 40, 42, 44, 46 или 79, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с последовательностью, представленной в SEQ ID NO: 36, 38, 40, 42, 44, 46 или 79; или
(iii) вариабельную область тяжелой цепи, представленную в (i), и вариабельную область легкой цепи, представленную в (ii).
6. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-5, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(1) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 35, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 36, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней;
(2) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 37, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 38, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней;
(3) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 39, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 40, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней;
(4) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 41, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 42, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней;
(5) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 43, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 44, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней;
(6) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 45, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 46, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней;
(7) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 71, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 79, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней; или
(8) вариабельная область тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 73, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней, и вариабельная область легкой цепи содержит последовательность, представленную в SEQ ID NO: 79, или аминокислотную последовательность, имеющую по меньшей мере 80% идентичность с ней.
7. Выделенное антитело к ST2 или его антигенсвязывающий фрагмент, содержащее:
(i) тяжелую цепь, где аминокислотная последовательность тяжелой цепи содержит последовательность, представленную в SEQ ID NO: 47, 51, 55, 59, 63, 67, 75 или 77; и
(ii) легкую цепь, где аминокислотная последовательность легкой цепи содержит последовательность, представленную в SEQ ID NO: 49, 53, 57, 61, 65, 69 или 81.
8. Антитело или его антигенсвязывающий фрагмент по п. 7, содержащее тяжелую цепь и легкую цепь, где:
(1) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 47, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 49;
(2) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 51, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 53;
(3) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 55, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 57;
(4) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 59, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 61;
(5) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 63, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 65;
(6) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 67, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 69;
(7) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 75, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 81; или
(8) тяжелая цепь содержит последовательность, представленную в SEQ ID NO: 77, и легкая цепь содержит последовательность, представленную в SEQ ID NO: 81.
9. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-8, выбранное из моноклонального антитела, фрагмента Fab, фрагмента F(ab')2, фрагмента Fd, фрагмента Fv, dAb, одноцепочечной молекулы Fv или их комбинаций.
10. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-9, специфически связывающееся с ST2 человека или обезьяны и блокирующее связывание IL33/ST2 и его передачу сигналов.
11. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-10, по существу не связывающееся с IL1R1, IL1R2, IL1R3, IL1R7, IL1R8 и IL1R9.
12. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-11, блокирующее секрецию IL5, индуцированную IL33/ST2, на CD4+ T-клетках.
13. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12.
14. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 13.
15. Клетка-хозяин, экспрессирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12, содержащая молекулу нуклеиновой кислоты по п. 13 или вектор экспрессии по п. 14.
16. Слитый полипептид, который связывается с ST2, содержащий антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12.
17. Mультиспецифическая молекула, которая связывается с ST2, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12.
18. Вирусный вектор, экспрессирующий антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12, содержащий молекулу нуклеиновой кислоты, кодирующую антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12, или молекулу нуклеиновой кислоты по п. 13.
19. Фармацевтическая композиция для лечения или облегчения заболеваний и состояний, опосредованных IL33/ST2, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12, и один или более фармацевтически приемлемых эксципиентов, разбавителей или носителей.
20. Способ определения уровня экспрессии ST2 в образце, полученном от субъекта, включающий приведение указанного образца в контакт с антителом или его антигенсвязывающим фрагментом по любому из пп. 1-12 в таких условиях, что антитело или его антигенсвязывающий фрагмент и ST2 образуют комплекс.
21. Способ лечения или облегчения заболеваний и состояний, опосредованных IL33/ST2, у субъекта, нуждающегося в этом, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-12 или молекулы нуклеиновой кислоты по п. 13.
22. Способ по п. 21, где заболевания и состояния, опосредованные IL33/ST2, включают астму, аллергический ринит, хроническое обструктивное заболевание легких, эозинофильный бронхит, эозинофильный эзофагит, атопический дерматит, псориаз, системную красную волчанку, пемфигоид, ревматоидный артрит, анкилозирующий спондилоартрит, воспалительное заболевание кишечника, фиброз лёгких, фиброз печени, системный склероз, саркоидоз, болезнь "трансплантат против хозяина" (GVHD), диабет, сердечно-сосудистые заболевания или их комбинации.
23. Способ по п. 21 или 22, где антитело или его антигенсвязывающий фрагмент или молекулу нуклеиновой кислоты вводят вместе с одним или несколькими другими антителами или лекарственными средствами.
24. Способ по п. 23, где указанные другие антитела включают антитело к TSLP, антитело к TSLPR, антитело к IL4, антитело к IL4R, антитело к IL13, антитело к IL13R, антитело к IL5, антитело к IL5R и/или антитело к IgE.
25. Способ по п. 23, где указанные другие лекарственные средства включают лекарственное средство против астмы, лекарственное средство против хронического обструктивного заболевания легких, лекарственное средство против язвенного колита, лекарственное средство против атопического дерматита или лекарственное средство против псориаза.
| AU 2018202097 A1, 31.05.2018 | |||
| CN 107446040 A, 08.12.2017 | |||
| АНТИТЕЛА ПРОТИВ РАСТВОРИМОГО ST-2 ЧЕЛОВЕКА И СПОСОБЫ АНАЛИЗА | 2011 |
|
RU2625014C2 |
| ДЫЛЕВА Ю.А | |||
| и др., Физиологическая и патофизиологическая роль стимулирующего фактора роста st2, Клиническая лабораторная диагностика, 2017, 62 (10), стр | |||
| Коллекторный альтернатор многофазного тока с переменным числом периодов при постоянном числе оборотов | 1921 |
|
SU599A1 |

