ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет по заявке на патент Китая № 202010309238.8, поданной 17 апреля 2020 г.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области антител, в частности к антителу к рецептору интерлейкина и способу его получения и его применению.
УРОВЕНЬ ТЕХНИКИ
При стимуляции антигеном антигенспецифические лимфоциты в организме человека идентифицируют антиген, отвечают активацией, пролиферацией, дифференцировкой и т. п. и, наконец, устраняют проникший антиген. Т-клетки и В-клетки являются первичными эффекторными клетками. Что касается различных типов антигенов, Т-клетки могут размножаться и усиливать иммунные ответы, непосредственно убивая клетки-мишени и секретируя различные типы цитокинов, тем самым достигая иммунологического эффекта. Исследования показали, что цитокины Th2, такие как интерлейкин (IL)-4, IL-5, IL-9 и IL-13, опосредуют основное патологическое развитие при аллергических заболеваниях, таких как аллергическая астма.
Астма представляет собой распространенное респираторное заболевание, которое обычно характеризуется воспалением дыхательных путей, гиперреактивностью бронхов и структурными изменениями стенок бронхов (ремоделированием дыхательных путей). Генетический фон и стимуляция факторов окружающей среды, включая аллергены и респираторные вирусы, вызывают возникновение астмы с основными патологическими проявлениями в виде рецидивирующих свистящих хрипов, одышки, стеснения в груди и кашля. Во всем мире насчитывается 2,3 миллиона пациентов с астмой, и ожидается, что в ближайшие десятилетия ее распространенность будет продолжать расти. В настоящее время астму лечат последовательно для облегчения симптомов и контроля рисков. Ингаляционные кортикостероиды являются стандартной терапией при астме средней степени тяжести; пациентов, страдающих тяжелой астмой, лечат в комбинации с бета-антагонистом длительного действия; а для пациентов, страдающих наиболее тяжелой астмой, может потребоваться дополнительное лекарственное средство. Эти виды лечения могут вызвать нарушения иммунной системы и другие серьезные побочные эффекты. Тем не менее, для 5-10% пациентов лекарственное средство до сих пор недоступно.
При аллергической астме была обнаружена аномально высокая экспрессия цитокинов Th2 в бронхах, и вместе с тем было подтверждено, что цитокины Th2 опосредуют возникновение и развитие воспалительных реакций и способствуют патологическим изменениям и т. п. в дыхательных путях. Эти цитокины способствуют активации эозинофилов, тучных клеток и других воспалительных клеток, а также хемотаксису в очаги воспаления. IL-4 и IL-13 нацелены на В-клетки, трансформируя секретируемые ими антитела из IgM в IgE, и вместе с тем вызывают ремоделирование бронхов посредством индукции гиперплазии бокаловидных клеток, превращения бронхиальных фибробластов в миофибробласты, отложения коллагена и пролиферации гладкомышечных клеток дыхательных путей.
Как IL-4, так и IL-13 могут активировать соответствующий сигнальный путь посредством связывания с альфа-рецептором интерлейкина-4 (IL-4R-альфа), поэтому антагонист IL-4R-альфа может блокировать патологические реакции IL-4 и IL-13, и ожидается, что он будет использоваться для лечения заболеваний, связанных с IL-4R-альфа, включая астму.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения, проведя большое количество экспериментов, получили группу моноклональных антител, которые могут блокировать передачу сигналов IL-4 и IL-13 посредством специфического блокирования связывания IL-4 и IL-13 с альфа-рецептором IL-4 (IL-4R-альфа) на поверхности клеток. Моноклональные антитела могут блокировать биологическую активность, опосредованную IL-4/IL-13.
В соответствии с первым аспектом в данной заявке предусмотрено антитело, специфически связывающееся с альфа-рецептором IL-4, или его антигенсвязывающий фрагмент, включая вариабельную область тяжелой цепи, где вариабельная область тяжелой цепи включает последовательность HCDR3, и необязательно дополнительно включает последовательность HCDR1 и/или HCDR2. В некоторых вариантах осуществления последовательность HCDR3 включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 107, 113, 119, 125, 131, 137, 143, 149, 155, 161, 167, 173, 179, 185, 191, 197 и 203. В некоторых вариантах осуществления последовательность HCDR1 включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 105, 111, 117, 123, 129, 135, 141, 147, 153, 159, 165, 171, 177, 183, 189, 195 и 201. В некоторых вариантах осуществления последовательность HCDR2 включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 106, 112, 118, 124, 130, 136, 142, 148, 154, 160, 166, 172, 178, 184, 190, 196 и 202. В необязательном варианте осуществления антигенсвязывающий фрагмент выбран из группы, состоящей из Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv-фрагмента, Fd-фрагмента и однодоменного антитела.
В некоторых вариантах осуществления вариабельная область тяжелой цепи включает аминокислотную последовательность, характеризующуюся по меньшей мере 80% гомологией с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 2, 6, 10, 14, 18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 76, 82, 88, 94 и 100, или вариабельная область тяжелой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 6, 10, 14, 18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 76, 82, 88, 94 и 100.
В некоторых вариантах осуществления антитело, специфически связывающееся с альфа-рецептором IL-4, или его антигенсвязывающий фрагмент дополнительно включают вариабельную область легкой цепи, где вариабельная область легкой цепи включает последовательность LCDR1, LCDR2 и/или LCDR3. В некоторых вариантах осуществления последовательность LCDR1 включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 108, 114, 120, 126, 132, 138, 144, 150, 156, 162, 168, 174, 180, 186, 192, 198 и 204. В некоторых вариантах осуществления последовательность LCDR2 включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 109, 115, 121, 127, 133, 139, 145, 151, 157, 163, 169, 175, 181, 187, 193, 199 и 205. В некоторых вариантах осуществления последовательность LCDR3 включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 110, 116, 122, 128, 134, 140, 146, 152, 158, 164, 170, 176, 182, 188, 194, 200 и 206.
В некоторых вариантах осуществления вариабельная область легкой цепи включает аминокислотную последовательность, характеризующуюся по меньшей мере 80% гомологией с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 56, 60, 64, 68, 73, 79, 85, 91, 97 и 103, или вариабельная область легкой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 56, 60, 64, 68, 73, 79, 85, 91, 97 и 103.
В некоторых вариантах осуществления антитело, специфически связывающееся с альфа-рецептором IL-4, или его антигенсвязывающий фрагмент включают тяжелую цепь, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 71, 77, 83, 89, 95 и 101, или аминокислотную последовательность, характеризующуюся по меньшей мере 80% гомологией с последовательностью. Необязательно, антитело или его антигенсвязывающий фрагмент включают легкую цепь, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 74, 80, 86, 92, 98 и 104, или аминокислотную последовательность, характеризующуюся по меньшей мере 80% гомологией с последовательностью.
В некоторых вариантах осуществления антитело, специфически связывающееся с альфа-рецептором IL-4 в соответствии с первым аспектом, представляет собой моноклональное антитело.
В некоторых вариантах осуществления антитело, специфически связывающееся с альфа-рецептором IL-4 в соответствии с первым аспектом, представляет собой гуманизированное антитело.
В некоторых вариантах осуществления антитело, специфически связывающееся с альфа-рецептором IL-4, или его антигенсвязывающий фрагмент, описанные в данном документе, связываются с тем же эпитопом на альфа-рецепторе IL-4, что и антитело 131-Hu, 136-Hu или 236-Hu, или конкурируют с антителом 131-Hu, 136-Hu или 236-Hu за связывание с альфа-рецептором IL-4, где антитело 131-Hu имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 83, и последовательность легкой цепи, представленную под SEQ ID NO: 86; антитело 136-Hu имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 89, и последовательность легкой цепи, представленную под SEQ ID NO: 92; и антитело 236-Hu имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 101, и последовательность легкой цепи, представленную под SEQ ID NO: 104.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, способны ингибировать секрецию IgE В-клетками. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с IL-4R-альфа при KD менее 600 пМ, предпочтительно менее 350 пМ. В некоторых других вариантах осуществления антитело или его антигенсвязывающий фрагмент способны ингибировать IL-4-индуцированную пролиферацию клеток TF-1.
В соответствии со вторым аспектом в данной заявке предусмотрена Нуклеотидная молекула, кодирующая антитело, специфически связывающееся с альфа-рецептором IL-4, или его антигенсвязывающий фрагмент.
В соответствии с третьим аспектом, в данной заявке предусмотрен вектор экспрессии, включающий нуклеотидную молекулу, которая уже упоминалась.
В некоторых вариантах осуществления вектор экспрессии представляет собой pTT5, pUC57, pDR1, pcDNA3.1(+), pDHFF или pCHO 1.0 и т. п.
В соответствии с четвертым аспектом в данной заявке предусмотрена клетка-хозяин, включающая вектор экспрессии, который уже упоминался. В некоторых вариантах осуществления клетка-хозяин представляет собой HEK293, COS, CHO, NS0, sf9, sf21, DH5α, BL21(DE3) или TG1 и т. п.
В соответствии с пятым аспектом в данной заявке предусмотрен способ получения антитела, специфически связывающегося с альфа-рецептором IL-4, или его антигенсвязывающего фрагмента в соответствии с первым аспектом, при этом способ включает следующие стадии:
а) культивирование клетки-хозяина в соответствии с четвертым аспектом в условиях экспрессии, которые позволяют клетке-хозяину продуцировать антитело или его антигенсвязывающий фрагмент, за счет чего обеспечивается экспрессия антитела или его антигенсвязывающего фрагмента; и
b) отделение и очистка антитела или его антигенсвязывающего фрагмента, экспрессированных на стадии а).
В соответствии с шестым аспектом в данной заявке предусмотрена фармацевтическая композиция, включающая антитело к альфа-рецептору IL-4 или его антигенсвязывающий фрагмент в соответствии с первым аспектом и фармацевтически приемлемые носители.
В некоторых вариантах осуществления композицию применяют для лечения заболеваний, связанных с IL-4R-альфа.
В соответствии с седьмым аспектом в данной заявке предусмотрено применение антитела к альфа-рецептору IL-4 или его антигенсвязывающего фрагмента в соответствии с первым аспектом, или композиции в соответствии с шестым аспектом в получении лекарственных средств для предупреждения или лечения заболеваний, связанных с IL-4R-альфа, таких как иммуноопосредованные воспалительные реакции или воспалительные заболевания.
В некоторых вариантах осуществления иммуноопосредованные воспалительные реакции или воспалительные заболевания включают: астму, аллергию, атопический дерматит, хронический синусит, эозинофильный эзофагит, полипы носа, псориаз, ревматоидный артрит, псориатический артрит, анкилозирующий спондилоартрит, рассеянный склероз, увеит, увеит Бехчета, ксерофтальмию и хроническую спонтанную крапивницу.
В соответствии с другими аспектами, в данной заявке предусмотрен способ предупреждения или лечения заболеваний, связанных с IL-4R-альфа, при этом способ включает введение антитела или его антигенсвязывающего фрагмента в соответствии с первым аспектом или фармацевтической композиции в соответствии с шестым аспектом нуждающемуся в этом индивиду.
Антитело к IL-4R-альфа или его антигенсвязывающий фрагмент по настоящему изобретению могут специфически связываться с IL-4R-альфа и обладают одним или несколькими из следующих эффектов: блокирование связывания IL-4 или IL-13 с IL-4R-альфа; ингибирование IL-4- или IL-13-индуцированной пролиферации линии клеток (таких как клетки TF-1); и/или ингибирование секреции IgE В-клетками. Фармакодинамические эксперименты in vivo полностью демонстрируют, что антитело по настоящему изобретению может противодействовать возникновению ответов Th2 ниже по сигнальному каскаду путем ингибирования сигнальных путей IL-4 и IL-13, обладает сильной функцией ингибирования астмы и быстро начинает действовать. Антитело к IL-4R-альфа или его антигенсвязывающий фрагмент по настоящему изобретению можно применять для предупреждения или лечения заболеваний, связанных с IL-4R-альфа, таких как иммуноопосредованные воспалительные заболевания.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг. 1 показаны результаты определения аффинности мышиных моноклональных антител к hIL-4R-альфа в отношении IL-4R-альфа человека.
На фиг. 2 показаны результаты ингибирования индуцированной посредством IL-4 человека пролиферации клеток TF-1 с помощью мышиных моноклональных антител к hIL-4R-альфа.
На фиг. 3 показаны результаты ингибирования индуцированной посредством IL-4 человека пролиферации клеток TF-1 с помощью гуманизированного моноклонального антитела к hIL-4R-альфа.
На фиг. 4 показаны результаты ингибирования секреции IgE В-клетками из свежей периферической крови с помощью гуманизированного моноклонального антитела к hIL-4R-альфа.
На фиг. 5 показаны результаты фармакокинетических экспериментов на гуманизированных моноклональных антителах к hIL-4R-альфа
На фиг. 6 показано ингибирование посредством 305-В сывороточного IgE в мышиных моделях.
На фиг. 7 показано соотношение эозинофилов к лейкоцитам в жидкости бронхоальвеолярного лаважа животных в каждой модельной группе.
На фиг. 8 показана эозинофильная инфильтрация в тканях легких животных в каждой модельной группе.
На фиг. 9 показана секреция слизи в тканях легких животных в каждой модельной группе.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
В данной заявке предусмотрено новое антитело к IL-4R-альфа, специфически связывающееся с IL-4R-альфа, или его антигенсвязывающий фрагмент. В предпочтительном варианте осуществления антитело или его антигенсвязывающий фрагмент в соответствии с данной заявкой связываются с IL-4R-альфа человека с высокой аффинностью и ингибируют активность IL-4R-альфа. В данной заявке также предусмотрены полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент, вектор, содержащий полинуклеотид, клетка-хозяин, содержащая полинуклеотид или вектор, способ получения и очистки антитела и медицинское и биологическое применение антитела или его антигенсвязывающего фрагмента, такое как предупреждение или лечение заболеваний или нарушений, связанных с IL-4R-альфа. Данная заявка также охватывает способ обнаружения IL-4R-альфа и регулирования активности IL-4R-альфа с использованием антитела или его антигенсвязывающего фрагмента.
Чтобы упростить понимание данной заявки, сначала будут определены некоторые используемые в данном документе термины.
Используемый в данном документе термин "антитело" относится к молекуле иммуноглобулина, содержащей четыре полипептидные цепи, а именно две тяжелые цепи (Н) и две легкие цепи (L), соединенные между собой дисульфидными связями, и ее мультимеру (например, IgM). Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (VH) и константную область тяжелой цепи (СН). Константная область тяжелой цепи содержит три домена: CH1, CH2 и CH3. Каждая легкая цепь содержит вариабельную область легкой цепи (VL) и константную область легкой цепи (CL). Константная область легкой цепи содержит домен (CL1). Области VH и VL можно дополнительно подразделить на гипервариабельные области, называемые определяющими комплементарность областями (CDR), перемежающиеся с консервативными областями, называемыми каркасными областями (FR).
Используемый в данном документе термин "антигенсвязывающий фрагмент" антитела относится к части или сегменту полной молекулы антитела, ответственной за связывание антигена. Антигенсвязывающий домен может включать вариабельную область тяжелой цепи (VH), вариабельную область легкой цепи (VL) или и то, и другое. Антигенсвязывающий фрагмент антитела может быть получен из полной молекулы антитела с использованием любой подходящей стандартной технологии, включая технологию протеолитического расщепления или рекомбинантной генной инженерии и т. п. Неограничивающие примеры антигенсвязывающего фрагмента включают: Fab-фрагмент; F(ab')2-фрагмент; Fd-фрагмент; Fv-фрагмент; молекулу одноцепочечного Fv (scFv); монодоменное антитело; dAb-фрагмент и минимальную распознающую единицу (например, отделенную CDR), состоящую из аминокислотных остатков, имитирующих гипервариабельную область антитела. Термин "антигенсвязывающий фрагмент" также включает другие сконструированные молекулы, такие как биантитело, триантитело, тетраантитело и микроантитело.
Используемые в данном документе термины "вариабельная область тяжелой цепи (VH)" и "вариабельная область легкой цепи (VL)" относятся к вариабельным областям тяжелой цепи и легкой цепи одного антитела соответственно, включая FR1, 2, 3 и 4 и CDR 1, 2 и 3.
Специалистам в области биологии хорошо известно, что определяющие комплементарность области (CDR, обычно относящиеся к CDR1, CDR2 и CDR3), представляют собой вариабельные области, оказывающие наибольшее влияние на аффинность и специфичность антител. Два общих определения доступны для последовательностей CDR VH или VL, а именно определения по Kabat и Chothia, например см. Kabat et al, “Sequences of Proteins of Immunological Interest”, National Institutes of Health, Bethesda, Md. (1991); A1-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); и Martin et al., Proc. Natl. Acad. Sci.USA86:9268-9272 (1989). Для последовательности вариабельной области данного антитела последовательности области CDR в последовательностях VH и VL могут быть определены на основе определений по Kabat или Chothia. В вариантах осуществления данной заявки последовательность CDR определяется по Kabat. В данной заявке CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи называются HCDR1, HCDR2 и HCDR3 соответственно; а CDR1, CDR2 и CDR3 вариабельной области легкой цепи называются LCDR1, LCDR2 и LCDR3 соответственно.
Для последовательности вариабельной области данного антитела последовательность области CDR в последовательности вариабельной области можно анализировать несколькими путями, например, последовательность области CDR можно определить с помощью онлайн-программного обеспечения Abysis (http://www.abysis.org/).
Используемый в данном документе термин "специфическое связывание" относится к реакции неслучайного связывания между двумя молекулами, такой как связывание антитела с эпитопом антигена, например к способности антитела связываться со специфическим антигеном с аффинностью, которая по меньшей мере в два раза превышает его аффинность в отношении неспецифического антигена. Однако следует понимать, что антитело может специфически связываться с двумя или более антигенами, родственными последовательностям антитела. Например, антитело по настоящему изобретению может специфически связываться с человеческим и отличным от человеческого (например, полученным от мышей или отличных от человека приматов) IL-4R-альфа.
Используемый в данном документе термин "моноклональное антитело" относится к антителу, полученному из в основном гомогенной популяции антител, то есть антитела, составляющие популяцию, идентичны, за исключением возможных встречающихся в природе мутаций у небольшого числа индивидов. Описанное в данном документе моноклональное антитело в частности включает "химерное" антитело, в котором часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующей последовательности в антителе определенного вида или принадлежащем к определенному классу или подклассу антител, в то время как остальная часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующей последовательности в антителе другого вида или принадлежащем к другому классу или подклассу антител; и моноклональное антитело также включает фрагменты такого антитела, если они проявляют требуемую биологическую активность (см. патент США № 4816567; и Morrison et al, Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)).
Используемый в данном документе термин "гомология" определяется как процент идентичных остатков в вариантах аминокислотной или нуклеотидной последовательности после выравнивания последовательностей и введения гэпов, который при необходимости может достигать максимального процента. Способы и компьютерные программы для выравнивания хорошо известны в данной области техники. Используемый в данном документе термин "по меньшей мере 80% гомология" относится к любому значению от 80% до 100% гомологии, например 85%, 90%, 95% и 99%.
Используемый в данном документе термин "заболевания, связанные с IL-4R-альфа" включает заболевания и/или симптомы, связанные с активацией сигнальных путей IL-4R-альфа. Иллюстративные заболевания или нарушения, связанные с IL-4R-альфа, включают иммуноопосредованные воспалительные реакции, такие как аллергические заболевания и астма.
Используемые в данном документе термины "период полужизни" и "период полужизни в сыворотке" относятся к времени, затраченному на снижение концентрации в сыворотке антигенсвязывающего белка согласно настоящему изобретению на 50% in vivo.
В соответствии с первым аспектом в данной заявке предусмотрено антитело, специфически связывающееся с IL-4R-альфа, или его антигенсвязывающий фрагмент, включая вариабельную область тяжелой цепи и/или вариабельную область легкой цепи. Аминокислотные последовательности CDR, VH, VL, тяжелой и легкой цепей и соответствующие нуклеотидные последовательности, применимые к антителу, раскрытому в данной заявке, перечислены в следующих таблицах 1-5. В некоторых вариантах осуществления антитело к IL-4R-альфа или его антигенсвязывающий фрагмент включают последовательности HCDR3, HCDR2 и/или HCDR1, которые независимо выбраны из любой из последовательностей HCDR3, HCDR2 или HCDR1, показанных в таблице 1. В некоторых вариантах осуществления антитело к IL-4R-альфа в соответствии с данной заявкой может дополнительно включать CDR легкой цепи, которая независимо выбрана из любой из последовательностей CDR1, CDR2 или CDR3 легкой цепи, показанных в таблице 2. Например, антитело к IL-4R-альфа в соответствии с данной заявкой может включать любой из вариабельных доменов тяжелой цепи, показанных в таблицах 3 и 4, необязательно в паре с любым из вариабельных доменов легкой цепи, показанных в таблицах 3 и 4.
(SEQ ID NO.)
(SEQ ID NO.)
В некоторых вариантах осуществления HCDR3 антитела или его антигенсвязывающего фрагмента, раскрытых в данном документе, выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 107, 113, 119, 125, 131, 137, 143, 149, 155, 161, 167, 173, 179, 185, 191, 197 и 203. В некоторых вариантах осуществления HCDR2 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 106, 112, 118, 124, 130, 136, 142, 148, 154, 160, 166, 172, 178, 184, 190, 196 и 202; и/или HCDR1 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 105, 111, 117, 123, 129, 135, 141, 147, 153, 159, 165, 171, 177, 183, 189, 195 и 201.
В конкретном варианте осуществления HCDR3 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 107, 113, 119, 125 и 131. В другом конкретном варианте осуществления HCDR3 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 137, 143, 149, 155, 161, 167, 173, 179, 185, 191, 197 и 203. В предпочтительном варианте осуществления HCDR3 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 125, 131 и 203.
В конкретном варианте осуществления HCDR2 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 106, 112, 118, 124 и 130. В другом конкретном варианте осуществления HCDR2 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 136, 142, 148, 154, 160, 166, 172, 178, 184, 190, 196 и 202. В предпочтительном варианте осуществления HCDR2 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 124, 130 и 202.
В конкретном варианте осуществления HCDR1 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 105, 111, 117, 123 и 129. В другом конкретном варианте осуществления HCDR1 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 135, 141, 147, 153, 159, 165, 171, 177, 183, 189, 195 и 201. В предпочтительном варианте осуществления HCDR1 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 123, 129 и 201.
В некоторых вариантах осуществления вариабельная область тяжелой цепи антитела, раскрытого в данном документе, включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 6, 10, 14, 18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 76, 82, 88, 94 и 100. В конкретном варианте осуществления вариабельная область тяжелой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, 6, 10, 14, 18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 76, 82, 88, 94 и 100.
В некоторых вариантах осуществления вариабельная область тяжелой цепи антитела, раскрытого в данном документе, включает аминокислотную последовательность, характеризующуюся по меньшей мере 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомологией с последовательностью, представленной под SEQ ID NO: 2, 6, 10, 14, 18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 76, 82, 88, 94 или 100. В предпочтительном варианте осуществления вариабельная область тяжелой цепи характеризуется более чем 99% гомологией с аминокислотной последовательностью, представленной под SEQ ID NO: 82, 88 или 100.
Антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, могут дополнительно включать вариабельную область легкой цепи в дополнение к вариабельной области тяжелой цепи.
В некоторых вариантах осуществления вариабельная область легкой цепи включает CDR3 (LCDR3), выбранную из аминокислотной последовательности, представленной под SEQ ID NO: 110, 116, 122, 128 и 134, или выбранную из аминокислотной последовательности, представленной под SEQ ID NO: 140, 146, 152, 158, 164, 170, 176, 182, 188, 194, 200 и 206. В предпочтительном варианте осуществления LCDR3 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 128, 134 и 206.
В некоторых вариантах осуществления LCDR2 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 109, 115, 121, 127 и 133, или выбранную из аминокислотной последовательности, представленной под SEQ ID NO: 139, 145, 151, 157, 163, 169, 175, 181, 187, 193, 199 и 205. В предпочтительном варианте осуществления LCDR2 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 127, 133 и 205.
В некоторых вариантах осуществления LCDR1 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 108, 114, 120, 126, 132, 138, 144, 150, 156, 162, 168, 174, 180, 186, 192, 198 и 204, или выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 108, 114, 120, 126, 132, 138, 144, 150, 156, 162, 168, 174, 180, 186, 192, 198 и 204. В предпочтительном варианте осуществления LCDR1 выбрана из аминокислотных последовательностей, представленных под SEQ ID NO: 126, 132 и 204.
В некоторых вариантах осуществления вариабельная область легкой цепи антитела, раскрытого в данном документе, включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 56, 60, 64, 68, 73, 79, 85, 91, 97 и 103. В конкретном варианте осуществления вариабельная область легкой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 56, 60, 64, 68, 73, 79, 85, 91, 97 и 103.
В некоторых вариантах осуществления вариабельная область легкой цепи антитела, раскрытого в данном документе, включает аминокислотную последовательность, характеризующуюся по меньшей мере 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% гомологией с последовательностью, представленной под SEQ ID NO: 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 56, 60, 64, 68, 73, 79, 85, 91, 97 или 103. В предпочтительном варианте осуществления вариабельная область тяжелой цепи характеризуется более чем 99% гомологией с аминокислотной последовательностью, представленной под SEQ ID NO: 85, 91 или 103.
В конкретном варианте осуществления раскрытые в данном документе антитело или его антигенсвязывающий фрагмент содержат тяжелую цепь, характеризующуюся по меньшей мере 80% гомологией с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 71, 77, 83, 89, 95 и 101, например 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99% гомологией. В более конкретном варианте осуществления тяжелая цепь антитела включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 71, 77, 83, 89, 95 и 101. В предпочтительном варианте осуществления тяжелая цепь антитела имеет аминокислотную последовательность, представленную под SEQ ID NO: 83, 89 или 101.
В конкретном варианте осуществления раскрытое в данном документе антитело включает легкую цепь, характеризующуюся по меньшей мере 80% гомологией с последовательностью, выбранной из группы, состоящей из SEQ ID NO: 74, 80, 86, 92, 98 и 104, например 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% и 99% гомологией. В более конкретном варианте осуществления легкая цепь антитела включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 74, 80, 86, 92, 98 и 104. В предпочтительном варианте осуществления легкая цепь антитела имеет аминокислотную последовательность, представленную под SEQ ID NO: 86, 92 или 104.
В некоторых вариантах осуществления по меньшей мере одна аминокислота может быть подвергнута замене, делеции или добавлению в соответствующие конкретные аминокислотные последовательности, перечисленные выше, в тяжелой цепи или вариабельной области тяжелой цепи, легкой цепи или вариабельной области легкой цепи антитела, раскрытого в данном документе, и при этом полученный вариант все еще сохраняет активность связывания IL-4R-альфа.
В некоторых вариантах осуществления количество аминокислотных замен, делеций или добавлений составляет 1-30, предпочтительно 1-20 и более предпочтительно 1-10. В предпочтительном варианте осуществления вариант последовательности отличается от исходной аминокислотной последовательности заменой, делецией или добавлением приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот. В более предпочтительном варианте осуществления вариант последовательности отличается от исходной аминокислотной последовательности заменой, делецией или добавлением приблизительно 1, 2, 3, 4 или 5 аминокислот. В конкретном варианте осуществления аминокислотная замена является консервативной.
В предпочтительном варианте осуществления раскрытое в данном документе антитело представляет собой антитело 131-Hu, 136-Hu или 236-Hu, где антитело 131-Hu имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 83, и последовательность легкой цепи, представленную под SEQ ID NO: 86, с такой же последовательностью CDR, как у антитела 131; антитело 136-Hu имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 89, и последовательность легкой цепи, представленную под SEQ ID NO: 92, с такой же последовательностью CDR, как у антитела 136; и антитело 236-Hu имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 101, и последовательность легкой цепи, представленную под SEQ ID NO: 104, с такой же последовательностью CDR, как у антитела 236.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, связываются с тем же эпитопом на альфа-рецепторе IL-4, что и антитело 131-Hu, 136-Hu или 236-Hu, или конкурируют с антителом 131-Hu, 136-Hu или 236-Hu за связывание с альфа-рецептором IL-4.
В некоторых вариантах осуществления антитело, раскрытое в данном документе, представляет собой моноклональное антитело. В конкретном варианте осуществления антитело, раскрытое в данном документе, представляет собой гуманизированное антитело.
Антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, могут специфически связываться с IL-4R-альфа. В конкретном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связывается с IL-4R альфа человека или IL-4R альфа мыши.
В предпочтительном варианте осуществления антитело или его антигенсвязывающий фрагмент специфически связываются с IL-4R-альфа человека.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связываются с IL-4R-альфа при KD менее 600 пМ. В предпочтительном варианте осуществления антитело или его антигенсвязывающий фрагмент связываются с IL-4R-альфа (например, IL-4R-альфа человека) при KD менее 350 пМ.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, способны ингибировать IL-4-индуцированную пролиферацию клеток TF-1. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, способны ингибировать секрецию IgE В-клетками.
Например, авторы данной заявки провели биологические эксперименты in vitro и in vivo с моноклональным антителом к hIL-4R-альфа, раскрытым в данном документе, и результаты показали, что антитело может хорошо связываться с IL-4R-альфа.
В частности, авторы данной заявки провели тестирование аффинности, экспериментальный анализ по блокированию связывания IL-4/IL-13 с IL-4R-альфа, тестирование клеточной функции in vitro и другие эксперименты с моноклональным антителом к hIL-4R-альфа. Результаты показывают, что моноклональное антитело к hIL-4R-альфа, раскрытое в данном документе может связывать IL-4R-альфа на клеточных поверхностях, блокировать передачу сигнала между IL-4/IL-13 и IL-4R-альфа и ингибировать возникновение воспалительных реакций.
В данной заявке дополнительно предусмотрена нуклеотидная молекула, кодирующая антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, вектор, содержащий полинуклеотид, клетка-хозяин, содержащая полинуклеотид или вектор, и способ получения и очистки антитела.
В некоторых вариантах осуществления нуклеотидная молекула, кодирующая антитело или его антигенсвязывающий фрагмент, функционально связана с регуляторной последовательностью, которая может распознаваться клеткой-хозяином, трансфицированной вектором.
В некоторых вариантах осуществления в данной заявке можно использовать любой подходящий вектор экспрессии. Например, вектор экспрессии может представлять собой один из следующих: pTT5, pUC57, pDR1, pcDNA3.1(+), pDHFF и pCHO 1.0. Вектор экспрессии может включать слитую последовательность ДНК, связанную с соответствующими последовательностями, регулирующими транскрипцию и трансляцию.
В некоторых вариантах доступная клетка-хозяин представляет собой клетку, содержащую вектор экспрессии, которая может представлять собой эукариотическую клетку. Например, можно использовать систему культур клеток-хозяев млекопитающего или насекомого для экспрессии антитела или его антигенсвязывающего фрагмента в соответствии с данной заявкой. Например, клетки HEK293, COS, CHO, NS0, sf9 и sf21 применимы к настоящему изобретению. Клетка-хозяин также может представлять собой прокариотическую клетку, содержащую вектор экспрессии, такой как DH5-альфа, BL21 (DE3) или TG1.
В некоторых вариантах осуществления способ получения моноклонального антитела к hIL-4R-альфа, раскрытого в данном документе, включает следующие стадии: культивирование клетки-хозяина в условиях экспрессии для экспрессии моноклонального антитела к hIL-4R-альфа; и выделение и очистку экспрессированного моноклонального антитела к hIL-4R-альфа. С помощью этого способа рекомбинантный белок может быть очищен до по существу гомогенного вещества, такого как вещество, демонстрирующее одну полосу на SDS-PAGE.
В некоторых вариантах осуществления антитело к IL-4R-альфа, раскрытое в данном документе, может быть выделено и очищено с помощью аффинной хроматографии. Основываясь на свойствах используемой колонки для аффинной хроматографии, антитело к IL-4R-альфа, связывающееся с колонкой для аффинной хроматографии, может быть элюировано традиционным способом, таким как использование высокосолевого буфера и изменение pH.
В некоторых вариантах осуществления гуманизированное моноклональное антитело к hIL-4R-альфа, раскрытое в данном документе, получают следующим способом: иммунизация мышей Balb/c посредством антигена IL-4R-альфа, полученного в лаборатории, степень слияния клеток селезенки мыши с клетками гибридомы после титрования выше при повторной иммунизации, и отбор штаммов клеток гибридомы, ингибирующих функциональную активность IL-4. Более конкретно, авторы данной заявки экспрессировали антиген внеклеточного домена IL-4R-альфа, IL-4 и IL-13 соответственно, посредством большого количества экспериментов. Исходя из этого, различные адъюванты смешивают с антигеном IL-4R-альфа для иммунизации мышей, а затем клетки селезенки мыши дополнительно сливают со штаммом sp2/0 клеток гибридомы. Положительный клеточный штамм отбирают из слитой гибридомы с использованием антигена внеклеточного домена IL-4R-альфа. После подтверждения того, что положительный клеточный штамм блокирует связывание IL-4/IL-13 с IL-4R-альфа и действительно ингибирует функцию IL-4/IL-13, получают целевой клеточный штамм. После гуманизации целевой молекулы гены как легкой цепи, так и тяжелой цепи клонируют в эукариотический вектор экспрессии pCHO1.0. Вектор экспрессии трансфицируют в клетку СНО путем липофекции, положительные клеточные клоны подвергают скринингу с пуромицином и метотрексатом, и прошедшие скрининг клоны с высокой экспрессией размножают в бессывороточной среде для выделения или очистки гуманизированного моноклонального антитела к hIL-4R-альфа с помощью колонки для аффинной хроматографии с белком A.
В некоторых других вариантах осуществления для дальнейшего изменения мышиного исходного антитела с получением химерных или гуманизированных форм или других вариантных форм антитела можно использовать традиционную методику, такую как мутагенез с применением ПЦР. Исходное антитело в данной заявке может быть подвергнуто мутации в таком домене, как определяющая комплементарность антигену область (CDR), для получения варианта антитела, который можно подвергать скринингу в отношении наличия целевых свойств, таких как аффинность связывания (более низкая KD), IC50, специфичность и приоритетность связывания. Предпочтительно целевые свойства варианта антитела представляют собой улучшения свойств исходного антитела. Предпочтительно в вариант антитела вводят замены аминокислот, и по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных остатков в молекулах исходного антитела удаляют и вставляют другие остатки в соответствующие положения. Наиболее интересным сайтом для альтернативного мутагенеза является одна или несколько CDR, но также рассматриваются изменения в каркасной области (FR). Предпочтительными являются консервативные замены аминокислот, также могут быть введены неконсервативные замены аминокислот, и полученный вариант антитела можно использовать для скрининга целевых свойств.
В некоторых вариантах осуществления период полужизни антитела в сыворотке продлевается посредством трансформации Fc-области антитела. Идентифицированные сайты мутаций, которые могут улучшать способность связывания FcRn человека с антителами, в основном включают T250Q, M252Y, S254T, T256E, V308P, M428L, N434A и N434S. В этом варианте осуществления период полужизни антитела в сыворотке может быть продлен посредством мутации аминокислот в этих сайтах.
В данной заявке предусмотрена фармацевтическая композиция, включающая антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе, и фармацевтически приемлемый носитель. Антитело к IL-4R-альфа, раскрытое в данном документе, такое как моноклональное антитело к hIL-4R-альфа, может быть составлено в фармакологические средства вместе с фармацевтически приемлемым носителем для достижения более стабильной эффективности. В некоторых вариантах осуществления эти средства могут обеспечивать конформационную целостность аминокислотной основной последовательности антитела к IL-4R-альфа, раскрытого в данном документе, такого как моноклональное антитело к hIL-4R-альфа, при этом защищая многофункциональные группы белков от деградации (включая без ограничения агрегацию, дезамидирование или окисление). В некоторых вариантах осуществления жидкий состав обычно можно сохранять стабильным при 2-8°C в течение по меньшей мере одного года. В некоторых вариантах осуществления лиофилизированный состав сохраняется стабильным при 30°C в течение по меньшей мере шести месяцев.
В некоторых вариантах осуществления состав антитела к IL-4R-альфа, такого как моноклональное антитело к hIL-4R-альфа, может представлять собой суспензии, водные инъекции, лиофилизированный состав и т. п., обычно используемые в области фармацевтики, предпочтительно водные инъекции или лиофилизированный состав. Для водных инъекций или лиофилизированного состава антитела к IL-4R-альфа, раскрытого в данном документе, такого как моноклональное антитело к hIL-4R-альфа, фармацевтически приемлемые вспомогательные вещества включают без ограничения: поверхностно-активные вещества, стабилизаторы раствора, регуляторы изотоничности и буферы или их комбинации. В некоторых вариантах осуществления поверхностно-активные вещества включают без ограничения: неионогенные поверхностно-активные вещества, такие как сложный эфир жирной кислоты и полиоксиэтиленсорбитана (Tween 20 или 80), полоксамер (например, полоксамер 188), Triton, додецилсульфат натрия (SDS), лаурилсульфат натрия, тетрадецил, линолевую кислоту или октадецилсаркозин, Pluronics и MONAQUATTM, которые следует добавлять в таком количестве, которое может свести к минимуму тенденцию к грануляции моноклонального антитела к hIL-4R-альфа. В некоторых вариантах осуществления стабилизаторы раствора включают без ограничения одно или комбинацию из следующего: сахариды, такие как восстанавливающие сахариды и невосстанавливающие сахариды; аминокислоты, такие как глутамат натрия или гистидин; спирты, такие как триолы, спирты высших сахаров, пропиленгликоль и полиэтиленгликоль. Количество добавляемых стабилизаторов раствора должно быть таким, чтобы полученный конечный препарат оставался стабильным в течение времени, которое специалисты в данной области техники считают стабильным. Регуляторы изотоничности включают без ограничения: хлорид натрия, маннит или их комбинацию. Буферы включают без ограничения: Трис, гистидиновый буфер, фосфатный буфер или их комбинацию.
В данной заявке также предусмотрен способ предупреждения или лечения заболеваний, связанных с IL-4R-альфа, включающий: введение индивиду антитела к IL-4R-альфа или композиции, содержащей антитело к IL-4R-альфа, такое как моноклональное антитело к hIL-4R-альфа. В некоторых вариантах осуществления эффекты на иммуноопосредованные воспалительные реакции очевидны после введения животным, включая человека. В частности, антитело к IL-4R-альфа, раскрытое в данном документе может эффективно предупреждать и/или лечить иммуноопосредованные воспалительные реакции и может применяться в качестве противовоспалительного лекарственного средства.
В данной заявке дополнительно предусмотрено применение антитела к IL-4R-альфа или композиции, содержащей антитело к IL-4R-альфа, в получении лекарственных средств для предупреждения или лечения заболеваний или симптомов, связанных с IL-4R-альфа. В некоторых вариантах осуществления заболевания или симптомы, связанные с IL-4R-альфа, представляют собой иммуноопосредованные воспалительные реакции или иммуноопосредованные воспалительные заболевания.
В некоторых вариантах осуществления иммуноопосредованные воспалительные реакции или иммуноопосредованные воспалительные заболевания включают без ограничения: астму, аллергию, атопический дерматит, хронический синусит, эозинофильный эзофагит, полипы носа, псориаз, ревматоидный артрит, псориатический артрит, анкилозирующий спондилоартрит, рассеянный склероз, увеит, увеит Бехчета, ксерофтальмию и хроническую спонтанную крапивницу. В дополнение к заболеваниям, связанным с воспалением, антитело к IL-4R-альфа, раскрытое в данном документе, также может быть использовано для предупреждения или лечения рассеянного склероза, болезни Крона, колита, язвенного колита, системной красной волчанки, реакции "трансплантат против хозяина" и т. п.
В некоторых вариантах осуществления антитело к IL-4R-альфа, раскрытое в данном документе, можно использовать в качестве лекарственного средства для иммуноопосредованных воспалительных реакций. Лекарственное средство для иммуноопосредованных воспалительных реакций, как заявлено в данной заявке, относится к лекарственному средству, которое может ингибировать и/или лечить иммуноопосредованные воспалительные реакции. Например, лекарственное средство может задерживать развитие симптомов, связанных с иммуноопосредованными воспалительными реакциями, и/или уменьшать тяжесть этих симптомов. В некоторых вариантах осуществления лекарственное средство может облегчать сопутствующие симптомы существующей воспалительной реакции и предупреждать появление других симптомов. В некоторых вариантах осуществления лекарственное средство также может уменьшать или предупреждать метастазирование воспалительных реакций.
Когда моноклональное антитело к hIL-4R-альфа и его композицию, раскрытые в данном документе, вводят животным, включая людей, применяемая доза варьирует в зависимости от возраста и веса пациентов, характеристик и серьезности заболеваний и путей введения. Ссылаясь на результаты и всесторонние ситуации в экспериментах на животных, общая применяемая доза не должна превышать определенного диапазона. В конкретном варианте осуществления доза для внутривенной инъекции составляет 1-1800 мг/день.
Доза и частота применения антитела или его композиции могут варьировать в зависимости от применения предупреждения или лечения заболеваний. При превентивном применении композицию, содержащую антитело в соответствии с данной заявкой, или ее смесь вводят пациенту, который еще не находился в болезненном состоянии, для повышения резистентности пациента, и это количество определяется как "профилактически эффективная доза". При таком применении конкретная доза зависит от состояния здоровья и системного иммунитета пациента. Относительно низкую дозу обычно вводят через относительно длинные промежутки времени в течение длительного периода времени. При терапевтическом применении иногда требуются относительно высокие дозы через относительно короткие промежутки времени до тех пор, пока прогрессирование заболевания не замедлится или не остановится, и предпочтительно до тех пор, пока у пациента не проявится частичное или полное улучшение симптомов заболевания. После этого пациенту может быть назначена превентивная схема. Специалист в данной области техники может легко определить конкретную дозу и частоту, исходя из фактических потребностей.
В описании и формуле изобретения слова "включающий", "предусматривающий" и "содержащий" означают "включающий без ограничения", и не предназначены для исключения других частей, добавок, компонентов или стадий.
Следует понимать, что признаки, характеристики, компоненты или стадии, описанные в конкретных аспектах, вариантах осуществления или примерах данной заявки, могут быть применены к любым другим аспектам, вариантам осуществления или примерам, описанным в данном документе, если не имеет место противоречие.
Приведенное выше раскрытие в целом описывает настоящее изобретение. Следующие конкретные примеры представляют собой дополнительное описание настоящего изобретения и не должны рассматриваться как ограничивающие настоящее изобретение. Примеры не включают подробное описание традиционных способов. Такие способы хорошо известны специалистам в области биологии и описаны во многих публикациях, таких как Molecular Cloning Manual и Antibody Technology Laboratory Manual, опубликованных Cold Spring Harbor Laboratory. Реагенты без указания источников представляют собой традиционные реагенты.
Примеры
Пример 1. Получение антигенов растворимого внеклеточного домена IL-4R-альфа с Fc-меткой или Flag-меткой, а также эталонного антитела дупилумаба и IL-4
Последовательность антигена внеклеточного домена IL-4R-альфа человека получали из UniProt (UniProtKB-P24394). Оптимизацию кодонов проводили на основе предпочтения использования кодонов Cricetulus griseus и проводили генный синтез N-концевого аминокислотного фрагмента в сайтах 26-232. Последовательность подвергали субклонированию в вектор pUC57 для получения pUC57-hIL4Rα-ECD. Последовательность константной области IgG1 человека синтезировали на основе последовательности секукинумаба (см. WHO Drug Information Vol. 23, No. 4, 2009, P342). Последовательность подвергали субклонированию в вектор pUC57 для получения pUC57-IgG1-CH. hFc-фрагмент или Flag-метку (DYKDDDDK) встраивали в С-конец фрагмента hIL4R-альфа-ECD с помощью ПЦР и конструировали на векторе экспрессии pTT5 (который хранится в лаборатории) для получения pTT5 (hIL4R-альфа-ECD-hFc) и pTT5 (hIL4R-альфа-ECD-Flag) для секвенирования, и клоны с правильными последовательностями отбирали для трансфекции.
Аминокислотную последовательность дупилумаба (IgG4, κ) брали из who.int (см. WHO Drug Information Vol.26, No. 4, 2012. P412). После оптимизации кодонов синтезировали нуклеотидную последовательность и клонировали в вектор экспрессии pTT5. Валидация секвенирования подтвердила, что был получен правильный помеченный клонирующим вектором pTT5 (дупилумаб). Вектор pTT5 (дупилумаб) временно трансфицировали в линию клеток HEK293E. Клетки культивировали в среде Freestyle293, содержащей 3 мМ вальпроевой кислоты, в течение 5 дней. Затем антитело к дупилумабу очищали из супернатанта клеточной культуры с помощью колонки для аффинной хроматографии с протеином А (приобретенной у Pharmacia).
Последовательность IL-4 человека получали из UniProt (UniProtKB-P05112). Оптимизацию кодонов проводили на основе предпочтения использования кодонов Cricetulus griseus и синтезировали N-концевой аминокислотный фрагмент в сайтах 25-153. Последовательность подвергали субклонированию в вектор pUC57. Flag-метку (DYKDDDDK) встраивали в С-конец фрагмента hIL4 с помощью ПЦР и конструировали на векторе экспрессии pTT5 для получения pTT5 (hIL4-Flag) для секвенирования, и клоны с правильными последовательностями отбирали для трансфекции.
Плазмиду трансфицировали в линию клеток HEK293E (хранящуюся в лаборатории) посредством PEI. После культивирования клеток в среде Freestyle293, содержащей 3 мМ вальпроевой кислоты (приобретенной у Gibco) в течение 5 дней, целевой белок очищали из супернатанта клеточной культуры с помощью колонки для аффинной хроматографии с белком A (приобретенной у Pharmacia) или гранул для аффинной хроматографии Flag (приобретенных у Sigma). Белки количественно определяли с помощью бицинхониновой кислоты (BCA), и очищенные белки использовали для дальнейшего анализа и исследований. Очищенные белки использовали для последующей иммунизации мышей и дальнейшего анализа и исследований.
Пример 2. Иммунизация hIL-4R-альфа-ECD-Fc
Антиген hIL-4R-альфа-ECD-Fc в количестве 100 мкг/мышь разводили физиологическим солевым раствором до 75 мкл и смешивали с таким же объемом полного адъюванта Фрейнда. После полной ультразвуковой эмульгации мышам Balb/c в возрасте 4-5 недель (приобретенные у Shanghai Lingchang Biotechnology Co., Ltd., номер лицензии на животноводство: SCXK (Shanghai) 2013-0018) проводили подкожные инъекции в несколько мест. Через три недели белок в количестве 50 мкг/мышь разбавляли до 75 мкл и смешивали с таким же объемом неполного адъюванта Фрейнда. После завершения ультразвуковой эмульгации мышам проводили подкожные инъекции в несколько мест, а через две недели иммунизацию повторяли. Через одну неделю после третьей иммунизации всем мышам отрезали хвосты и собирали кровь для отделения сыворотки, а титр сывороточных антител к hIL-4R тестировали с помощью ELISA, с покрытием антигеном hIL-4R-альфа-Fc. Мышам с титром антител в сыворотке выше 10000 через неделю после взятия крови проводили повторную иммунизацию: инъекция в хвостовую вену 10 мкг антигена/100 мкл физиологического солевого раствора на мышь.
Титр определяли с помощью ELISA: планшет для ELISA покрывали антигеном hIL-4R-альфа-ECD-Fc в концентрации 1 мкг/мл, 100 мкл на лунку, и покрывали в течение ночи при 4°C. Планшеты дважды промывали посредством PBST (PBS, содержащего 0,5% Tween-20) и затем высушивали промакиванием. Каждую лунку блокировали посредством 200 мкл покрывающего раствора, содержащего 1% BSA, блокировали при комнатной температуре в течение 4 ч., высушивали промакиванием и хранили в холодильнике при -20°C для последующего использования. Во время тестирования в каждую лунку планшета для ELISA добавляли по 100 мкл мышиной сыворотки в разных концентрациях с двумя повторяющимися лунками для инкубации при комнатной температуре в течение 1,5 ч. Лунки промывали посредством PBST 3 раза и затем высушивали промакиванием. Затем добавляли 100 мкл кроличьих антител к IgG мыши, меченных посредством HRP, разбавленных посредством PBST в соотношении 1:10000 (приобретенного у Sigma), и планшеты инкубировали при комнатной температуре в течение 1 ч. Лунки промывали посредством PBST 3 раза и затем высушивали промакиванием. Затем в каждую лунку добавляли по 100 мкл хромогенного раствора (хромогенный раствор А для ELISA смешивали с хромогенным раствором B в объемном соотношении 1: 1 перед использованием) для хромогенной реакции, а затем в каждую лунку добавляли по 100 мкл 2М останавливающего раствора H2SO4 для прекращения реакции. Значение OD каждой лунки сразу же измеряли при 450 нм с помощью микропланшет-ридера (Molecular Device).
Пример 3. Слияние и скрининг гибридомы
Клетки гибридомы sp2/0 (из банка клеток Комитета по коллекции типовых культур Китайской академии наук, под коллекционным номером TCM-18) культивировали в инкубаторе при 5% CO2 при 37 °C, среду меняли за сутки до слияния. Через три дня после повторной иммунизации клетки селезенки собирали у мышей для слияния. Способ слияния и скрининга описан следующим образом: селезенку мыши измельчали и промывали. Затем клетки селезенки подсчитывали и смешивали с клетками sp2/0 в соотношении 10:1, полученную смесь центрифугировали при 1500 об/мин в течение 7 мин. Супернатант смывали. Затем в течение 1 мин.добавляли 1 мл PEG (1450) и осторожно встряхивали в течение 90 с., в течение 2,5 мин.добавляли 5 мл бессывороточной среды DMEM (приобретенной у Gibco), затем добавляли еще 5 мл бессывороточной среды за один раз для остановки реакции, полученную смесь выдерживали 5 мин. и центрифугировали при 1280 об/мин. в течение 8 мин. Имея 2 миллиона клеток sp2/0 в 96-луночном планшете, клетки равномерно инокулировали в 96-луночный планшет по 200 мкл на лунку. Для скрининга использовали среду HAT, содержащую гипоксантин (H), аминоптерин (A) и тимидин (T), затем половину среды меняли каждые 3-4 дня, а в день 10 использовали среду HT. Через десять дней, когда клетки гибридомы покрывали более 10% дна 96-луночного планшета, супернатант тестировали с помощью ELISA с 96-луночным планшетом, покрытым антигеном hIL-4R-альфа-ECD-Fc. Способ ELISA был таким же, как описанный в примере 2. Положительные гибридомы отбирали и подвергали клонированию в 24-луночный планшет для размножения, а размноженные гибридомы подвергали субклонированию путем предельного разведения для получения штаммов гибридом, стабильно экспрессирующих целевое антитело, а затем создавали банк клеток из сохраненных штаммов гибридом.
Пример 4. Блокирование связывания IL-4 с hIL-4R-альфа-ECD-Fc мышиными моноклональными антителами к hIL-4R-альфа
Блокирование связывания IL-4 с hIL-4R-альфа-ECD-Fc мышиными моноклональными антителами к hIL-4R-альфа исследовали с помощью ELISA. Планшеты для ELISA покрывали белком hIL-4, и после блокирования добавляли hIL-4R-альфа-ECD-Fc и супернатанты культуры клеток гибридомы, состоящие из мышиных моноклональных антител к hIL-4R-альфа, из 300 мкл субклонов, и, наконец, добавляли HRP-меченные овечьи антитела к IgG человека для хромогенного тестирования. Штаммы клеток, которые могли блокировать связывание IL-4 с IL-4R-альфа-ECD-Fc, сохраняли для следующего цикла субклонирования.
Пример 5. EC50 мышиных моноклональных антител к hIL-4R-альфа, связывающихся с IL-4R-альфа человека
Оптимизированные мышиные моноклональные антитела к hIL-4R-альфа очищали на колонке для аффинной хроматографии с белком G и количественно определяли с помощью BCA. EC50 моноклональных антител к hIL-4R-альфа, связывающихся с hIL-4R-альфа, тестировали с помощью ELISA. Способ тестирования был таким же, как описанный в примере 3. Планшет для ELISA покрывали посредством 1 мкг/мл антигена hIL-4R-альфа-ECD-Fc, и затем для тестирования добавляли мышиные моноклональные антитела к hIL-4R-альфа в различных концентрациях.
Авторы настоящего изобретения проанализировали 266 антител из сохраненных гибридом, и репрезентативные экспериментальные результаты показаны на фиг. 1. Данные ЕС50 для некоторых оптимизированных антител приведены в таблице 6. Эти антитела обладали высокой аффинностью к IL-4R-альфа человека, и значение ЕС50 составляло приблизительно 10 нг/мл.
Пример 6. Ингибирование мышиными моноклональными антителами к hIL-4R-альфа hIL-4- или hIL-13-индуцированной пролиферации клеток TF-1
Штаммы клеток гибридомы, подвергнутые третьему циклу субклонирования, размножали в бессывороточной среде, и затем собирали клеточные супернатанты для очистки антител с помощью колонок для аффинной хроматографии с белком G (приобретенных у GE). Очищенные антитела определяли количественно и подтверждали их функциональную активность.
Клетки TF-1 представляют собой цитокинзависимые клеточные штаммы, выделенные из костного мозга пациентов с эритроидным лейкозом человека. Исследования показали, что клетки TF-1 хорошо растут при стимуляции посредством hIL-4 или hIL-13, и поэтому клетки TF-1 могут стать лучшей моделью для подтверждения функций сигнального пути IL-4.
Клетки TF-1 (от ATCC, с коллекционным номером CRL-2003) в состоянии хорошего роста подсчитывали и ресуспендировали с рекомбинантным hIL-4 или hIL-13 (приобретенным у R&D Systems) в конечной концентрации 20 нг/мл, затем формировали клеточную суспензию при 2x105/100 мкл. Среда представляет собой среду RPMI1640 (приобретенную у Gibco), содержащую 10% фетальную бычью сыворотку (приобретенную у Sigma), 100 ЕД/мл пенициллина (приобретенного у Gibco) и 100 мг/мл стрептомицина (приобретенную у Gibco), которая называется полная среда RPMI-1640. Мышиные моноклональные антитела к hIL-4R-альфа (20 мкг/мл-3 нг/мл, серийные разведения при коэффициенте разведения 3, 9 разных концентраций) в разных концентрациях разводили раствором среды по 100 мкл на лунку и добавляли в 96-луночные плоские планшеты для культивирования клеток (приобретенные у Corning), и затем в каждую лунку добавляли по 100 мкл клеточной суспензии. В каждой группе устанавливали по две повторяющиеся лунки для инкубации при 5% CO2 при 37°C в течение 72 ч. Затем в каждую лунку добавляли по 20 мкл раствора CCK-8 (приобретенного у Dojindo) для продолжения культивирования в течение 8 ч. После того, как раствор хорошо перемешали, значение OD каждой лунки измеряли при 450 нм с помощью микропланшет-ридера и рассчитывали коэффициент пролиферации клеток.
Авторы настоящего изобретения проанализировали функциональную активность вышеуказанных 266 мышиных моноклональных антител к hIL-4R-альфа в отношении ингибирования IL-4-индуцированной пролиферации клеток TF-1 (фиг. 2), и данные в отношении 17 оптимизированных антител представлены в таблице 7.
Пример 7. Секвенирование мышиных моноклональных антител к hIL-4R-альфа
Общую РНК каждого штамма клеток гибридомы экстрагировали с помощью Trizol (приобретенного у Sangon Biotech (Шанхай)), и мРНК подвергали обратной транскрипции в кДНК с помощью набора для обратной транскрипции (приобретенного у Takara). Гены вариабельной области легкой цепи и вариабельной области тяжелой цепи мышиных моноклональных антител к hIL-4R-альфа амплифицировали с помощью ПЦР с праймерами, описанными в литературе, и затем продукты ПЦР клонировали в вектор pGEM-T, и последовательности генов вариабельной области секвенировали и анализировали. Основываясь на результатах различных функциональных экспериментов и анализа раннего развития, авторы настоящего изобретения наконец выбрали 17 антител, перечисленных в таблице 2, в качестве ведущих антител и секвенировали их для получения нуклеотидных последовательностей вариабельных областей легкой и тяжелой цепи. Трансформированные аминокислотные последовательности подвергали алгоритму BLAST в GenBank, и все последовательности соответствовали характеристикам генов вариабельной области IgG мыши. Дальнейший анализ последовательностей показал, что последовательности CDR легкой и тяжелой цепей антител 54, 55, 57, 64, 75, 81, 83, 84, 88, 100, 228 и 236 были очень подобными, с различиями только в нескольких аминокислотах, и последовательности CDR легкой и тяжелой цепей антител 29, 59, 120, 131 и 136 были очень подобными. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 29 представлена под SEQ ID NO: 1, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 2; нуклеотидная последовательность вариабельной области легкой цепи антитела 29 представлена под SEQ ID NO: 3, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 4. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 59 представлена под SEQ ID NO: 5, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 6; нуклеотидная последовательность вариабельной области легкой цепи антитела 59 представлена под SEQ ID NO: 7, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 8. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 120 представлена под SEQ ID NO: 9, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 10; нуклеотидная последовательность вариабельной области легкой цепи антитела 120 представлена под SEQ ID NO: 11, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 12. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 131 представлена под SEQ ID NO: 13, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 14; нуклеотидная последовательность вариабельной области легкой цепи антитела 131 представлена под SEQ ID NO: 15, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 16. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 136 представлена под SEQ ID NO: 17, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 18; нуклеотидная последовательность вариабельной области легкой цепи антитела 136 представлена под SEQ ID NO: 19, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 20. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 54 представлена под SEQ ID NO: 21, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 22; нуклеотидная последовательность вариабельной области легкой цепи антитела 54 представлена под SEQ ID NO: 23, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 24. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 55 представлена под SEQ ID NO: 25, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 26; нуклеотидная последовательность вариабельной области легкой цепи антитела 55 представлена под SEQ ID NO: 27, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 28. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 57 представлена под SEQ ID NO: 29, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 30; нуклеотидная последовательность вариабельной области легкой цепи антитела 57 представлена под SEQ ID NO: 31, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 32. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 64 представлена под SEQ ID NO: 33, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 34; нуклеотидная последовательность вариабельной области легкой цепи антитела 64 представлена под SEQ ID NO: 35, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 36. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 75 представлена под SEQ ID NO: 37, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 38; нуклеотидная последовательность вариабельной области легкой цепи антитела 75 представлена под SEQ ID NO: 39, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 40. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 81 представлена под SEQ ID NO: 41, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 42; нуклеотидная последовательность вариабельной области легкой цепи антитела 81 представлена под SEQ ID NO: 43, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 44. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 83 представлена под SEQ ID NO: 45, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 46; нуклеотидная последовательность вариабельной области легкой цепи антитела 83 представлена под SEQ ID NO: 47, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 48. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 84 представлена под SEQ ID NO: 49, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 50; нуклеотидная последовательность вариабельной области легкой цепи антитела 84 представлена под SEQ ID NO: 51, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 52. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 88 представлена под SEQ ID NO: 53, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 54; нуклеотидная последовательность вариабельной области легкой цепи антитела 88 представлена под SEQ ID NO: 55, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 56. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 100 представлена под SEQ ID NO: 57, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 58; нуклеотидная последовательность вариабельной области легкой цепи антитела 100 представлена под SEQ ID NO: 59, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 60. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 228 представлена под SEQ ID NO: 61, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 62; нуклеотидная последовательность вариабельной области легкой цепи антитела 228 представлена под SEQ ID NO: 63, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 64. Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 236 представлена под SEQ ID NO: 65, и аминокислотная последовательность вариабельной области тяжелой цепи представлена под SEQ ID NO: 66; нуклеотидная последовательность вариабельной области легкой цепи антитела 236 представлена под SEQ ID NO: 67, и аминокислотная последовательность вариабельной области легкой цепи представлена под SEQ ID NO: 68.
Пример 8. Гуманизация моноклональных антител к hIL-4R-альфа
На основании результатов анализа последовательностей антитела 29, 59, 131, 136, 228 и 236 выбирали для конструирования химерных антител и гуманизированных антител. Химерные антитела конструировали путем перехвата вариабельных областей тяжелой цепи и вариабельных областей легкой цепи мышиных антител и соединения этих областей с константными областями легкой и тяжелой цепи IgG4 человека (из антитела дупилумаба) с помощью ПЦР с перекрывающимися праймерами.
Аминокислотные последовательности вариабельных областей легкой цепи и вариабельных областей тяжелой цепи мышиных моноклональных антител к hIL-4R-альфа анализировали на основе правила Kabat и определяли три CDR и четыре FR. Взяв в качестве примера антитело 136, аминокислотные последовательности определяющих комплементарность областей тяжелой цепи антитела представляли собой HCDR1: DYGMH (SEQ ID NO: 129), HCDR2: YISSGSTTIYYADTVKG (SEQ ID NO: 130) и HCDR3: ISTVVAKRYAMDY (SEQ ID NO: 131), и аминокислотные последовательности определяющих комплементарность областей легкой цепи представляли собой LCDR1: RASQDISNYLN (SEQ ID NO: 132), LCDR2: YTSRLHS (SEQ ID NO: 133) и LCDR3: QQINALPLT (SEQ ID NO: 134).
На основании сравнения гомологии IgBlast NCBI и последовательностей зародышевой линии IgG человека в качестве матрицы для прививки CDR тяжелой цепи выбрали IGHV3-48*01, и область CDR тяжелой цепи мышиного моноклонального антитела 136 к hIL-4R-альфа привили в каркасную область IGHV3-48*01 для конструирования антитела с привитым CDR тяжелой цепи. Аналогичным образом, на основании сравнения гомологии с последовательностями зародышевой линии IgG человека в качестве матрицы для прививки CDR легкой цепи выбрали IGKV1-33*01, и область CDR легкой цепи мышиного моноклонального антитела 136 к hIL-4R-альфа привили в каркасную область IGKV1-33*01 для конструирования антитела с привитым CDR легкой цепи, и полученное антитело определяли как 136-Gr (136-прививка). На этом основании некоторые аминокислотные сайты в каркасной области подвергали обратной мутации Во время обратной мутации аминокислотные последовательности кодировали по Kabat, и расположение сайтов обозначали кодами по Kabat. Предпочтительно, чтобы для последовательности вариабельной области легкой цепи F в сайте 73 кода по Kabat заменили обратно на мышиный L, а L в сайте 96 CDR3 подвергли мутации в F. В вариабельной области тяжелой цепи не имели место обратные мутации. Последовательности генов вариабельной области оптимизировали и синтезировали Sangon Biotech на основе предпочтения использования кодонов Cricetulus griseus. Синтезированную гуманизированную последовательность вариабельной области связывали с константной областью IgG4 человека, и это антитело определяли как гуманизированное антитело (136-гуманизация, 136-Hu) антитела 136.
По тому же принципу гуманизировали остальные пять антител. Временно экспрессируемые векторы гуманизированных тяжелых цепей и легких цепей конструировали с помощью вектора pTT5 (приобретенного у Научно-исследовательского института по проблемам биотехнологий NRC), затем плазмиды временно трансфицировали системой HEK293 (приобретенной у Научно-исследовательского института по проблемам биотехнологий NRC) и экспрессировали антитела. Клетки HEK293 культивировали в экспрессионной среде FreeStyle 293 (приобретенной у Gibco). Плазмиды трансфицировали в клетки путем трансфекции PEI. Через пять дней супернатант клеток собирали и очищали с помощью белка А для получения антитела.
Наконец, гуманизированная вариабельная область тяжелой цепи антитела 29 имеет генную последовательность общей длиной 366 п. н., кодирующую 122 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 69, и аминокислотной последовательностью, представленной под SEQ ID NO: 70; и гуманизированная вариабельная область легкой цепи имеет генную последовательность общей длиной 321 п. н., кодирующую 107 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 72, и аминокислотной последовательностью, представленной под SEQ ID NO: 73. После того, как вариабельные области тяжелой и легкой цепи связывали с константной областью IgG4 человека, наконец получали гуманизированную тяжелую цепь 29-Hu, состоящую из 448 аминокислот (с последовательностью, представленной под SEQ ID NO: 71) и гуманизированную легкую цепь 29-Hu, состоящую из 214 аминокислот (с последовательностью, представленной под SEQ ID NO: 74).
Гуманизированная вариабельная область тяжелой цепи антитела 59 имеет генную последовательность общей длиной 354 п. н., кодирующую 118 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 75, и аминокислотной последовательностью, представленной под SEQ ID NO: 76; и гуманизированная вариабельная область легкой цепи имеет генную последовательность общей длиной 318 п. н., кодирующую 106 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 78, и аминокислотной последовательностью, представленной под SEQ ID NO: 79. После того, как вариабельные области тяжелой и легкой цепи связывали с константной областью IgG4 человека, наконец получали гуманизированную тяжелую цепь 59-Hu, состоящую из 444 аминокислот (с последовательностью, представленной под SEQ ID NO: 77) и гуманизированную легкую цепь 59-Hu, состоящую из 213 аминокислот (с последовательностью, представленной под SEQ ID NO: 80).
Гуманизированная вариабельная область тяжелой цепи антитела 131 имеет генную последовательность общей длиной 366 п. н., кодирующую 122 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 81, и аминокислотной последовательностью, представленной под SEQ ID NO: 82; и гуманизированная вариабельная область легкой цепи имеет генную последовательность общей длиной 321 п. н., кодирующую 107 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 84, и аминокислотной последовательностью, представленной под SEQ ID NO: 85. После того, как вариабельные области тяжелой и легкой цепи связывали с константной областью IgG4 человека, наконец получали гуманизированную тяжелую цепь 131-Hu, состоящую из 448 аминокислот (с последовательностью, представленной под SEQ ID NO: 83) и гуманизированную легкую цепь 131-Hu, состоящую из 214 аминокислот (с последовательностью, представленной под SEQ ID NO: 86).
Гуманизированная вариабельная область тяжелой цепи антитела 136 имеет генную последовательность общей длиной 366 п. н., кодирующую 122 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 87, и аминокислотной последовательностью, представленной под SEQ ID NO: 88; и гуманизированная вариабельная область легкой цепи имеет генную последовательность общей длиной 321 п. н., кодирующую 107 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 90, и аминокислотной последовательностью, представленной под SEQ ID NO: 91. После того, как вариабельные области тяжелой и легкой цепи связывали с константной областью IgG4 человека, наконец получали гуманизированную тяжелую цепь 136-Hu, состоящую из 448 аминокислот (с последовательностью, представленной под SEQ ID NO: 89) и гуманизированную легкую цепь 136-Hu, состоящую из 214 аминокислот (с последовательностью, представленной под SEQ ID NO: 92).
Гуманизированная вариабельная область тяжелой цепи антитела 228 имеет генную последовательность общей длиной 360 п. н., кодирующую 120 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 93, и аминокислотной последовательностью, представленной под SEQ ID NO: 94; и гуманизированная вариабельная область легкой цепи имеет генную последовательность общей длиной 318 п. н., кодирующую 106 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 96, и аминокислотной последовательностью, представленной под SEQ ID NO: 97. После того, как вариабельные области тяжелой и легкой цепи связывали с константной областью IgG4 человека, наконец получали гуманизированную тяжелую цепь 228-Hu, состоящую из 446 аминокислот (с последовательностью, представленной под SEQ ID NO: 95) и гуманизированную легкую цепь 228-Hu, состоящую из 213 аминокислот (с последовательностью, представленной под SEQ ID NO: 98).
Гуманизированная вариабельная область тяжелой цепи антитела 236 имеет генную последовательность общей длиной 360 п. н., кодирующую 120 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 99, и аминокислотной последовательностью, представленной под SEQ ID NO: 100; и гуманизированная вариабельная область легкой цепи имеет генную последовательность общей длиной 318 п. н., кодирующую 106 аминокислотных остатка, с нуклеотидной последовательностью, представленной под SEQ ID NO: 102, и аминокислотной последовательностью, представленной под SEQ ID NO: 103. После того, как вариабельные области тяжелой и легкой цепи связывали с константной областью IgG4 человека, наконец получали гуманизированную тяжелую цепь 236-Hu, состоящую из 446 аминокислот (с последовательностью, представленной под SEQ ID NO: 101) и гуманизированную легкую цепь 236-Hu, состоящую из 213 аминокислот (с последовательностью, представленной под SEQ ID NO: 104).
Пример 9. Аффинность гуманизированных моноклональных антител к hIL-4R-альфа, связывающихся с IL-4R-альфа
Аффинность шести очищенных гуманизированных антител определяли с помощью Biacore T200 (GE Healthcare) с эталонным антителом дупилумабом в качестве контроля. Конкретный экспериментальный способ был следующим: использовали сенсорный чип CM5 с белком A (GE Healthcare), с FC1 (проточной кюветой 1) в качестве эталонного канала и FC2 (проточной кюветой 2) в качестве канала для образцов. Гуманизированные антитела или контрольное антитело захватывали в канале FC2, а затем вводили hIL-4R-альфа-ECD-Flag в различных концентрациях. Условия циклирования были следующими: во все каналы FC вводили аналит со скоростью 50 мкл/мин в течение 4 мин. при времени диссоциации 20 мин., а затем вводили 6 М гидрохлорида гуанидина (Sinopharm Chemical Reagent Co., Ltd.) со скоростью 10 мкл/мин. в течение 30 с. для регенерации поверхности Затем различия сигналов и аффинность для взаимодействия между захваченными антителами и незахваченными антителами рассчитывали с помощью программного обеспечения для оценки Biacore T200, версия 1.0. Как показано в таблице 8, аффинность гуманизированных антител 136-Hu и 236-Hu к hIL-4R-альфа-ECD-Flag была значительно выше, чем у эталонного антитела дупилумаба.
Пример 10. Ингибирование гуманизированными моноклональными антителами к hIL-4R-альфа пролиферации клеток TF-1
Эксперимент по ингибированию пролиферации клеток TF-1 гуманизированными моноклональными антителами к hIL-4R-альфа проводили со ссылкой на пример 6. Результаты представлены на фиг. 3. Как показано в таблице 9, ингибирующая способность гуманизированных антител 131-Hu, 136-Hu и 236-Hu в отношении IL-4-опосредованной пролиферации клеток TF-1 была значительно выше, чем у эталонного антитела дупилумаба.
Пример 11. Ингибирование гуманизированными моноклональными антителами к hIL-4R-альфа секреции IgE В-клетками, полученными из периферической крови
При астме IL-4 и IL-13 индуцировали превращение IgM В-клеток в IgE. Затем секретировалось большое количество IgE. После связывания с рецепторами на поверхности нейтрофилов, лимфоцитов и тучных клеток IgE активирует иммунные реакции, которые сужают дыхательные пути и запускают воспалительную реакцию, что приводит к усугублению симптомов астмы. Таким образом, одним из видов функциональной активности моноклональных антител к hIL-4R-альфа было ингибирование секреции IgE В-клетками. Авторы настоящего изобретения подтвердили этот вид активности гуманизированных моноклональных антител к hIL-4R-альфа.
Мононуклеарные клетки отделяли из свежей периферической крови человека (предоставленной больницей Changhai) с помощью Histopaque-1077 (приобретенного у Sigma). Клетки ресуспендировали с образованием клеточной суспензии при 5x105/мл в полной среде RPMI-1640, и 100 мкл клеточной суспензии добавляли в 96-луночный планшет для культивирования клеток. Вместе с тем добавляли 100 мкл 20 нг/мл рекомбинантных hIL-4 и антител к hIL-4R-альфа в различных концентрациях (20 мкг/мл - 3 нг/мл) для инкубации при 5% CO2 при 37°C в течение 14 дней. Затем пипетировали 100 мкл клеточного супернатанта для определения концентрации IgE с помощью ELISA.
Способ анализа ELISA описан следующим образом: планшет для ELISA (приобретенный у R&D Systems) покрывали мышиным антителом к IgE человека в концентрации 2,5 мкг/мл, 100 мкл на лунку, и покрывали в течение ночи при 4°C. Планшет дважды промывали посредством PBST и затем высушивали промакиванием. Каждую лунку блокировали при комнатной температуре в течение 4 ч. посредством 200 мкл покрывающего раствора, содержащего 1% BSA, затем высушивали промакиванием и хранили в холодильнике при -20°C для последующего использования. Во время тестирования стандартное соединение IgE человека (приобретенное у R&D Systems) в конечных концентрациях 100 нг/мл, 33 нг/мл, 11 нг/мл, 3,7 нг/мл, 1,2 нг/мл, 410 пг/мл, 130 пг/мл и 46 пг/мл соответственно получали со средой. Затем в планшет для ELISA с покрытием добавляли стандартное соединение IgE человека, по 100 мкл на лунку, с 2 повторяющимися лунками для каждой концентрации; и 100 мкл клеточного супернатанта добавляли в планшет для ELISA с покрытием для инкубации при комнатной температуре в течение 1,5 ч. Лунки промывали посредством PBST 3 раза и затем высушивали промакиванием. В каждую лунку добавляли 100 мкл биотинилированного крысиного антитела к IgE человека (приобретенного у R&D Systems и разбавленного при коэффициенте разбавления 1000 с помощью PBST, содержащего 1% BSA) для инкубации при комнатной температуре в течение 1 ч. Лунки промывали посредством PBST 3 раза и затем высушивали промакиванием. В каждую лунку добавляли 100 мкл стрептавидина-HRP (приобретенного у BD Pharmingen и разбавленного при коэффициенте разбавления 1000 с помощью PBST, содержащего 1% BSA) для инкубации при комнатной температуре в течение 1 ч. Затем в каждую лунку добавляли по 100 мкл хромогенного раствора (хромогенный раствор А для ELISA смешивали с хромогенным раствором B в объемном соотношении 1: 1 перед использованием) для хромогенной реакции, а затем в каждую лунку добавляли по 100 мкл 2М останавливающего раствора H2SO4 для прекращения реакции. Значение OD каждой лунки сразу же измеряли при 450 нм с помощью микропланшет-ридера. Результаты показали, что по сравнению с дупилумабом 131-Hu, 136-Hu и 236-Hu оказывали более сильный ингибирующий эффект на секрецию IgE В-клетками периферической крови, что указывает на то, что эти антитела обладали лучшей функциональной активностью, чем дупилумаб (фиг. 4).
Пример 12. Фармакокинетическое исследование гуманизированных моноклональных антител к hIL-4R-альфа на яванских макаках
На основе гуманизированных антител модифицировали Fc-области антител. Сайты мутаций в Fc-областях представляли собой в основном следующие: T250Q, M252Y, S254T, T256E, V308P, M428L, N434A и N434S. Время полужизни антитела можно изменить посредством изменения Fc, и более длительный период полужизни может уменьшить частоту введения.
Образцы крови собирали в 0 ч., 1 ч., 4 ч., 10 ч., 24 ч., 48 ч., 72 ч., 120 ч., 168 ч., 240 ч., 336 ч., 408 ч., 504 ч., 672 ч., 840 ч., 1008 ч., 1176 ч., 1344 ч., 1512 ч., 1680 ч. и 1848 ч. после проведения однократного введения яванским макакам для фармакокинетического анализа. Концентрации антител в сыворотке определяли посредством валидированного способа ELISA. Количественный диапазон для анализа в этом исследовании составлял 15,63-1000,00 нг/мл, и стандартные образцы во всех точках концентрации были получены из 100% сыворотки яванских макаков. Фармакокинетические параметры антитела в контрольной группе без мутации в Fc-области и антитела с модификацией в Fc-области после подкожного введения представлены в таблице 10 и таблице 11. В каждой группе было по два животных, один самец и одна самка, и способ введения представлял собой подкожную инъекцию (sc) в дозе 5 мг/кг.
По сравнению с контрольной группой клиренс антитела с модификацией в Fc-области был относительно мягким после подкожного введения, а период полужизни антитела в сыворотке был значительно продлен (фиг. 5), где 305-A-sc представляет собой контрольную группу, и 305-B-sc представляет собой антитело с модификацией в Fc-области. Этот эксперимент был поручен United-Power Pharma Tech Co., Ltd.
Пример 13. Создание модели астмы на мышах с двойной гуманизацией B-hIL-4/hIL-4RA и оценка эффективности лекарственного средства
Целевую молекулу получали в масштабе опытного производства, и для целевой молекулы проводили разработку состава. Полученный стабильный продукт с концентрацией 150 мг/мл назвали 305-В. Для фармакодинамических исследований in vivo использовали мышей с двойной гуманизацией IL-4/IL-4R-альфа (мыши B-hIL-4/hIL-4RA или B-hIL-4/IL4RA). У этих мышей с фоном C57BL/6 мышиные IL-4 и IL-4R-альфа подвергали нокауту и замене на гуманизированные гены IL-4 и IL-4R-альфа. Гуманизированные мыши вели себя так же, как нормальные мыши, без каких-либо отклонений. Конструирование гуманизированных мышей и исследование фармакодинамики 305-В на мышиной модели астмы было поручено компании Biocytogen Pharmaceuticals (Beijing) Co., Ltd.
В эксперименте использовали модель астмы на мышах, сенсибилизированных куриным овальбумином (OVA). Мышей, сенсибилизированных OVA, подвергали аллергизации после аэрозольной ингаляции OVA, что вызывало иммунный ответ Th2. В легких наблюдали усиление эозинофильной инфильтрации, а также значительное повышение концентрации сывороточного IgE. Эту модель использовали для оценки противоаллергической функции антитела при астме. Дупилумаб использовали в качестве положительного контроля, а 305-В использовали в качестве тестового образца с основным компонентом, представляющим собой гуманизированное моноклональное антитело к hIL-4R-альфа.
Всего 49 животных случайным образом разделяли на 7 групп по весу, по 7 животных в каждой группе. G1 представляет собой модельную группу PBS, G2 представляет собой группу положительного контроля дупилумаба (50 мг/кг), G3 представляет собой группу тестового образца 305-B (0,5 мг/кг), G4 представляет собой группу тестового образца 305-B (5 мг/кг), G5 представляет собой группу тестового образца 305-B (25 мг/кг), G6 представляет собой группу тестового образца 305-B (50 мг/кг), а G7 представляет собой немодельную наивную группу. Всех животных в группах G1-G6 подвергали сенсибилизации посредством OVA: мышей подвергали сенсибилизации посредством внутрибрюшинной инъекции OVA в дни 0, 7 и 14 и подвергали стимуляции посредством ингаляции аэрозоля OVA с дня 21 по день 25. Все животные в группах получали лекарственные средства в день 20 и день 23.
По сравнению с наивной группой, уровень IgE в сыворотке в группе модели PBS значительно увеличился, и процент эозинофилов среди лейкоцитов в жидкости бронхоальвеолярного лаважа значительно увеличился. Гистопатологические результаты показали, что инфильтрация смешанных воспалительных клеток и эозинофилов в тканях легких была значительно усилена, и секреция слизи значительно увеличена, что свидетельствует об успешности моделирования.
По сравнению с модельной группой PBS, IgE в сыворотке группы тестового образца 305-B (0,5 мг/кг) значительно снизился (p<0,05), и IgE в сыворотке группы положительного контроля дупилумаба (50 мг/кг) и групп тестового образца 305-B (5 мг/кг), (25 мг/кг) и (50 мг/кг) значительно уменьшился (p<0,0001), как показано на фиг. 6; за исключением группы 305-В (0,5 мг/кг), процент эозинофилов среди лейкоцитов (Eos/WBC%) в каждой обработанной группе значительно снизился (p<0,0001), как показано на фиг. 7; за исключением группы 305-В (0,5 мг/кг), в тканях легких каждой обработанной группы отсутствовала явная эозинофильная инфильтрация, и количество эозинофилов значительно уменьшилось (p<0,05), как показано на фиг. 8; и образование слизи в легких значительно улучшилось (р<0,05) в группах 305-В (25 мг/кг) и (50 мг/кг), как показано на фиг. 9. По вышеуказанным показателям не выявили значимой разницы между 305-В и дупилумабом в одинаковой дозе (50 мг/кг).
В заключение, при астмоподобном воспалении легких, характеризующемся главным образом иммунными реакциями Th2, две терапевтические дозы (25 мг/кг или 50 мг/кг) 305-B могут значительно снизить уровень IgE в сыворотке и Eos/WBC% в жидкости альвеолярного лаважа, уменьшить степень эозинофильной инфильтрации легких и уменьшить секрецию слизи. Это полностью продемонстрировало, что 305-B может противодействовать возникновению ответов Th2 ниже по сигнальному каскаду путем ингибирования сигнальных путей IL-4 и IL-13, обладает сильной функцией ингибирования астмы и быстро начинает действовать. Этот ингибирующий эффект был дозозависимым. По оценочным показателям не выявили значимой разницы между 305-В и дупилумабом в одинаковой дозе.
Заключение. При определении аффинности и анализе различных видов функциональной активности in vitro антитело по настоящему изобретению проявляет стабильную биологическую активность, более сильную, чем активность контрольного антитела дупилумаба. Результаты фармакодинамических исследований in vivo показывают, что нет значимой разницы между антителом по настоящему изобретению и контрольным антителом дупилумабом.
Можно понять, что хотя данная заявка описана в той или иной форме, данная заявка не ограничена тем, что показано и описано в описании. Для специалистов в области биологии очевидно, что в примеры и/или признак или параметр могут быть внесены различные изменения, не выходя за объем данной заявки. Эти изменения включены в объем, установленный данной заявкой.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Шанхай Мабгик Биотек Ко., Лтд.
<120> АНТИТЕЛО К AЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 ЧЕЛОВЕКА, СПОСОБ ЕГО
ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ
<130> P22416837RU
<160> 206
<170> PatentIn версия 3.5
<210> 1
<211> 366
<212> ДНК
<213> Mus musculus
<400> 1
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgaaactg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccggaaaaag gcctggaatg ggtggcgtat attaacagcg gcagcagcaa aatttatcat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgttt
240
ctgcagatga ccagcctgcg cagcgaagat accgcgatgt attattgcgc gcgctttccg
300
accgtggtgg cggcgcgcta tccgatggat tattggggcc agggcaccag cgtgaccgtg
360
agcagc
366
<210> 2
<211> 122
<212> БЕЛОК
<213> Mus musculus
<400> 2
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Asn Ser Gly Ser Ser Lys Ile Tyr His Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Phe Pro Thr Val Val Ala Ala Arg Tyr Pro Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 3
<211> 321
<212> ДНК
<213> Mus musculus
<400> 3
gatattcaga tgacccagac cccgagcagc ctgagcgcga gcctgggcga tcgcgtgacc
60
attagctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
gatggcaccg tgaaactgct gatttatttt accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcggcggcag cggcaccgat tatagcctga ccattagcaa cctggaacag
240
gaagatattg cgacctattt ttgccagcag ggcaacaccc tgccgtatac ctttggcggc
300
ggcaccaaac tggaaattaa a
321
<210> 4
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 4
Asp Ile Gln Met Thr Gln Thr Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Phe Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Gly Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 5
<211> 354
<212> ДНК
<213> Mus musculus
<400> 5
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgaaactg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca ttcattgggt gcgccaggcg
120
ccggataaag gcctggaatg ggtggcgtat attcgcagcg atagcagcat tattcattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgttt
240
ctgcagatga ccagcctgcg cagcgaagat accgcgatgt attattgcac ccgcggccgc
300
gatcgcggct attttgatta ttggggccag ggcaccaccc tgaccgtgag cagc
354
<210> 6
<211> 118
<212> БЕЛОК
<213> Mus musculus
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Ile His Trp Val Arg Gln Ala Pro Asp Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Arg Ser Asp Ser Ser Ile Ile His Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Thr Arg Gly Arg Asp Arg Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 7
<211> 318
<212> ДНК
<213> Mus musculus
<400> 7
cagattgtgc tgacccagag cccggcgctg atgagcgcga gcccgggcga aaaagtgacc
60
atgacctgca gcgcgagcag cagcgtgagc tatatgtatt ggtatcagca gaaaccgcgc
120
agcagcccga aaccgtggat ttatctgacc agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagcagcat ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg accagcattc cgtttacctt tggcagcggc
300
accaaactgg aaattaaa
318
<210> 8
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 8
Gln Ile Val Leu Thr Gln Ser Pro Ala Leu Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Arg Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Ile Pro Phe Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 9
<211> 366
<212> ДНК
<213> Mus musculus
<400> 9
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgaaactg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccggaaaaag gcctggaatg ggtggcgtat attagcagcg gcagcaccac catttattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acggcaaaaa caccctgttt
240
ctgcagatga ccagcctgcg cagcgaagat accgcgatgt attattgcgc gcgcattagc
300
accgtggtgg cgaaacgcta tgcgatggat tattggggcc agggcaccag cgtgaccgtg
360
agcagc
366
<210> 10
<211> 122
<212> БЕЛОК
<213> Mus musculus
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Thr Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Gly Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 11
<211> 321
<212> ДНК
<213> Mus musculus
<400> 11
gatattcaga tgacccagac cgcgagcagc ctgagcgcga gcctgggcga tcgcgtgacc
60
attagctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
gatggcaccg tgaaactgct gatttattat accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcagcggcag cggcaccgat tatagcctga ccattagcaa cctggaacag
240
gaagatattg cgacctattt ttgccagcag attaacgcgc tgccgctgac ctttggcgcg
300
ggcaccaaac tggaactgaa a
321
<210> 12
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 12
Asp Ile Gln Met Thr Gln Thr Ala Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Ile Asn Ala Leu Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 13
<211> 366
<212> ДНК
<213> Mus musculus
<400> 13
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgaaactg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccggaaaaag gcctggaatg ggtggcgtat attagcagcg gcagcagcac catttattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa caccctgttt
240
ctgcagatga ccagcctgcg cagcgaagat accgcgatgt attattgcgc gcgcattagc
300
accgtggtgg cgaaacgcta tgcgatggat tattggggcc agggcaccag cgtgaccgtg
360
agcagc
366
<210> 14
<211> 122
<212> БЕЛОК
<213> Mus musculus
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Ser Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 15
<211> 321
<212> ДНК
<213> Mus musculus
<400> 15
gatattcaga tgacccagac cgcgagcagc ctgagcgcga gcctgggcga tcgcgtgacc
60
attagctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
gatggcaccg tgaaactgct gatttattat accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcagcggcag cggcaccgat tatagcctga ccattagcaa cctggaacag
240
gaagatattg cgacctattt ttgccaggaa gtgaacatgc tgccgctgac ctttggcgcg
300
ggcaccaaac tggaactgaa a
321
<210> 16
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 16
Asp Ile Gln Met Thr Gln Thr Ala Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Glu Val Asn Met Leu Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 17
<211> 366
<212> ДНК
<213> Mus musculus
<400> 17
gaagtgcagc tggtggaaag cggcggcggc ctggtgaaac cgggcggcag cctgaaactg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccggaaaaag gcctggaatg ggtggcgtat attagcagcg gcagcaccac catttattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acggcaaaaa caccctgttt
240
ctgcagatga ccagcctgcg cagcgaagat accgcgatgt attattgcgc gcgcattagc
300
accgtggtgg cgaaacgcta tgcgatggat tattggggcc agggcaccag cgtgaccgtg
360
agcagc
366
<210> 18
<211> 122
<212> БЕЛОК
<213> Mus musculus
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Thr Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Gly Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Thr Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 19
<211> 321
<212> ДНК
<213> Mus musculus
<400> 19
gatattcaga tgacccagac cgcgagcagc ctgagcgcga gcctgggcga tcgcgtgacc
60
attagctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
gatggcaccg tgaaactgct gatttattat accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcagcggcag cggcaccgat tatagcctga ccattagcaa cctggaacag
240
gaagatattg cgacctattt ttgccagcag attaacgcgc tgccgctgac ctttggcgcg
300
ggcaccaaac tggaactgaa a
321
<210> 20
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 20
Asp Ile Gln Met Thr Gln Thr Ala Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Ile Asn Ala Leu Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 21
<211> 360
<212> ДНК
<213> Mus musculus
<400> 21
gaagtgcagc tgcagcagag cggcccggaa ctggtgaaac cgggcgcgag cgtgaaaatt
60
ccgtgcaaag cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa gcctggaatg gattggcgat attaacccga acaacggcaa cattctgttt
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
gtggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 22
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 22
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asn Gly Asn Ile Leu Phe Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Val Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 23
<211> 318
<212> ДНК
<213> Mus musculus
<400> 23
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgagc tatatgcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcagcaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 24
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 24
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 25
<211> 360
<212> ДНК
<213> Mus musculus
<400> 25
gatgtgcagc tgcagcagag cggcccggaa ctgctgaaac cgggcgcgag cgtgaaaatt
60
ccgtgccagg cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa acctggaatg gattggcaac attaacccga acaacggcgg caccttttat
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 26
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 26
Asp Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Leu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Gln Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 27
<211> 318
<212> ДНК
<213> Mus musculus
<400> 27
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggtgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 28
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 28
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 29
<211> 360
<212> ДНК
<213> Mus musculus
<400> 29
gaagtgcagc tgcagcagag cggcccggaa ctggtgaaac cgggcgcgag cgtgaaaatt
60
ccgtgcaaag cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa gcctggaatg gattggcgat attaacccga acaacggcaa cattctgttt
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
gtggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 30
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 30
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asn Gly Asn Ile Leu Phe Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Val Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 31
<211> 318
<212> ДНК
<213> Mus musculus
<400> 31
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgagc tatatgcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcagcaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 32
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 32
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 33
<211> 360
<212> ДНК
<213> Mus musculus
<400> 33
gaagtgcagc tgcagcagag cggcccggaa ctggtgaaac cgggcgcgag cgtgaaaatt
60
agctgcaaag cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa gcctggaatg gattggcacc accaacccga acaacggcgg caccctgtat
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgc gcgcggcggc
300
ctgcgccgcc gcggctttgt ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 34
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 34
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Thr Thr Asn Pro Asn Asn Gly Gly Thr Leu Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Leu Arg Arg Arg Gly Phe Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 35
<211> 318
<212> ДНК
<213> Mus musculus
<400> 35
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgaac tatctgcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcgacc agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcagcaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 36
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 36
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Asn Tyr Leu
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 37
<211> 360
<212> ДНК
<213> Mus musculus
<400> 37
gaagtgcagc tgcagcagag cggcccggaa ctggtgaaac cgggcgcgag cgtgaaaatt
60
ccgtgcaaag cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa gcctggaatg gattggcgat attaacccga acaacggcaa cattctgttt
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
gtggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 38
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 38
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Pro Asn Asn Gly Asn Ile Leu Phe Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Val Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 39
<211> 318
<212> ДНК
<213> Mus musculus
<400> 39
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgagc tatatgcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcagcaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 40
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 40
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 41
<211> 360
<212> ДНК
<213> Mus musculus
<400> 41
gatgtgcagc tgcagcagag cggcccggaa ctgctgaaac cgggcgcgag cgtgaaaatt
60
ccgtgccagg cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa acctggaatg gattggcaac attaacccga acaacggcgg caccttttat
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 42
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 42
Asp Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Leu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Gln Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 43
<211> 318
<212> ДНК
<213> Mus musculus
<400> 43
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 44
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 44
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 45
<211> 360
<212> ДНК
<213> Mus musculus
<400> 45
gaagtgcagc tgcagcagag cggcccggaa ctggtgaaac cgggcgcgag cgtgaaaatt
60
ccgtgcaaag cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
cgcggcaaaa gcctggaatg gattggcacc attaacccga acaacggcga taccatgtat
180
aaccagaaat ttaaagataa agcgaccctg accgtggata aaagcagcac caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccgcggca ccagcgtgac cgtgagcagc
360
<210> 46
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 46
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser Arg Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Thr Ile Asn Pro Asn Asn Gly Asp Thr Met Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Arg
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 47
<211> 318
<212> ДНК
<213> Mus musculus
<400> 47
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcgatt agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat tttctgacca ttagccgcgt ggaagtggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 48
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 48
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ile Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Phe Leu Thr Ile Ser Arg Val Glu Val Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 49
<211> 360
<212> ДНК
<213> Mus musculus
<400> 49
gatgtgcagc tgcagcagag cggccgcgaa ctgctgaaac gcggcgcgag cgtgaaaatt
60
ccgtgccagg cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa acctggaatg gattggcaac attaacccga acaacggcgg caccttttat
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 50
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 50
Asp Val Gln Leu Gln Gln Ser Gly Arg Glu Leu Leu Lys Arg Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Gln Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 51
<211> 318
<212> ДНК
<213> Mus musculus
<400> 51
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 52
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 52
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 53
<211> 360
<212> ДНК
<213> Mus musculus
<400> 53
gatgtgcagc tgcagcagag cggcccggaa ctgctgaaac cgggcgcgag cgtgaaaatt
60
ccgtgccagg cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa acctggaatg gattggcacc attaacccga acaacggcgg caccttttat
180
aaccagaact ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtgc
240
atggaactgc gcagcctgac cagcgatgat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 54
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 54
Asp Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Leu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Gln Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Thr Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Cys
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 55
<211> 318
<212> ДНК
<213> Mus musculus
<400> 55
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 56
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 56
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 57
<211> 360
<212> ДНК
<213> Mus musculus
<400> 57
gatgtgcagc tgcagcagag cggcccggaa ctgctgaaac cgggcgcgag cgtgaaaatt
60
ccgtgccagg cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa acctggaatg gattggcaac attaacccga acaacggcgg caccttttat
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttct ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 58
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 58
Asp Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Leu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Gln Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Leu Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 59
<211> 318
<212> ДНК
<213> Mus musculus
<400> 59
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggtg agcaacctgg cgagcggcgt gccggtgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 60
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 60
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Val Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 61
<211> 360
<212> ДНК
<213> Mus musculus
<400> 61
gatgtgcagc tgcagcagag cggcccggaa ctgctgaaac cgggcgcgag cgtgaaaatt
60
ccgtgccagg cgagcggcta tacctttacc gattataaca tggattgggt gaaacagagc
120
catggcaaaa acctggaatg gattggcaac attaacccga acaacggcgg caccttttat
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata aaagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 62
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 62
Asp Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Leu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Gln Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 63
<211> 318
<212> ДНК
<213> Mus musculus
<400> 63
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcgtgaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 64
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 64
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 65
<211> 360
<212> ДНК
<213> Mus musculus
<400> 65
gaagtgcagc tgcagcagag cggcccggaa ctggtgaaac cgggcgcgag cgtgaaaatt
60
ccgtgcaaag cgagcggcta tacctttacc gattataacg tggattgggt gaaacagagc
120
catggccaga gcctggattg gattggcacc attaacccga acaacggcgg cattctgagc
180
aaccagaaat ttaaaggcaa agcgaccctg accgtggata ccagcagcag caccgcgtat
240
atggaactgc gcagcctgac cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccagcgtgac cgtgagcagc
360
<210> 66
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 66
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Pro Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Lys Gln Ser His Gly Gln Ser Leu Asp Trp Ile
35 40 45
Gly Thr Ile Asn Pro Asn Asn Gly Gly Ile Leu Ser Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 67
<211> 318
<212> ДНК
<213> Mus musculus
<400> 67
cagattgtgc tgagccagag cccggcgatt ctgagcgcga gcccgggcga aaaagtgacc
60
atgacctgcc gcgcgagcag cagcgtgatt tatattcatt ggtattatca gaaagcgggc
120
agcagcccga aaccgtggat ttatgcggcg agcaacctgc cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccagctat agcctgacca ttagccgcgt ggaagcggaa
240
gatgcggcga cctattattg ccagcagtgg agcagcaacc cgccgacctt tggcggcggc
300
accaaactgg aaattaaa
318
<210> 68
<211> 106
<212> БЕЛОК
<213> Mus musculus
<400> 68
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Tyr Gln Lys Ala Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Pro Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 69
<211> 366
<212> ДНК
<213> Синтетическая последовательность
<400> 69
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccgggcaaag gcctggaatg ggtggcgtat attaacagcg gcagcagcaa aatttatcat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gcgctttccg
300
accgtggtgg cggcgcgcta tccgatggat tattggggcc agggcaccct ggtgaccgtg
360
agcagc
366
<210> 70
<211> 122
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 70
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Asn Ser Gly Ser Ser Lys Ile Tyr His Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Pro Thr Val Val Ala Ala Arg Tyr Pro Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 71
<211> 448
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 71
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Asn Ser Gly Ser Ser Lys Ile Tyr His Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Pro Thr Val Val Ala Ala Arg Tyr Pro Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 72
<211> 321
<212> ДНК
<213> Синтетическая последовательность
<400> 72
gatattcaga tgacccagag cccgagcagc ctgagcgcga gcgtgggcga tcgcgtgacc
60
attacctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
ggcaaagcgc cgaaactgct gatttatttt accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcagcggcag cggcaccgat tataccctga ccattagcag cctgcagccg
240
gaagattttg cgacctatta ttgccagcag ggcaacaccc tgccgtatac ctttggccag
300
ggcaccaaac tggaaattaa a
321
<210> 73
<211> 107
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 73
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Phe Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 74
<211> 214
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 74
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Phe Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 75
<211> 354
<212> ДНК
<213> Синтетическая последовательность
<400> 75
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca ttcattgggt gcgccaggcg
120
ccgggcaaag gcctggaatg ggtgagctat attcgcagcg atagcagcat tattcattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gcgcggccgc
300
gatcgcggct attttgatta ttggggccag ggcaccctgg tgaccgtgag cagc
354
<210> 76
<211> 118
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 76
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Arg Ser Asp Ser Ser Ile Ile His Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Arg Asp Arg Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 77
<211> 444
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 77
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Arg Ser Asp Ser Ser Ile Ile His Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Arg Asp Arg Gly Tyr Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440
<210> 78
<211> 318
<212> ДНК
<213> Синтетическая последовательность
<400> 78
gaaattgtgc tgacccagag cccggcgacc ctgagcctga gcccgggcga acgcgcgacc
60
ctgagctgca gcgcgagcag cagcgtgagc tatatgtatt ggtatcagca gaaaccgggc
120
caggcgccgc gcccgtggat ttatctgacc agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccgatttt accctgacca ttagcagcct ggaaccggaa
240
gattttgcgg tgtattattg ccagcagtgg accagcattc cgtttacctt tggccagggc
300
accaaagtgg aaattaaa
318
<210> 79
<211> 106
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 79
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Trp Ile Tyr
35 40 45
Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Thr Ser Ile Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 80
<211> 213
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 80
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Trp Ile Tyr
35 40 45
Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Thr Ser Ile Pro Phe Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 81
<211> 366
<212> ДНК
<213> Синтетическая последовательность
<400> 81
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccgggcaaag gcctggaatg ggtgagctat attagcagcg gcagcagcac catttattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gcgcattagc
300
accgtggtgg cgaaacgcta tgcgatggat tattggggcc agggcaccct ggtgaccgtg
360
agcagc
366
<210> 82
<211> 122
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 82
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Ser Ser Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 83
<211> 448
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Ser Ser Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 84
<211> 321
<212> ДНК
<213> Синтетическая последовательность
<400> 84
gatattcaga tgacccagag cccgagcagc ctgagcgcga gcgtgggcga tcgcgtgacc
60
attacctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
ggcaaagcgc cgaaactgct gatttattat accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcagcggcag cggcaccgat tttaccctga ccattagcag cctgcagccg
240
gaagatattg cgacctatta ttgccaggaa gtgaacatgc tgccgtttac ctttggccag
300
ggcaccaaag tggaaattaa a
321
<210> 85
<211> 107
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 85
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Glu Val Asn Met Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 86
<211> 214
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Glu Val Asn Met Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 87
<211> 366
<212> ДНК
<213> Синтетическая последовательность
<400> 87
gaagtgcagc tggtggaaag cggcggcggc ctggtgcagc cgggcggcag cctgcgcctg
60
agctgcgcgg cgagcggctt tacctttagc gattatggca tgcattgggt gcgccaggcg
120
ccgggcaaag gcctggaatg ggtgagctat attagcagcg gcagcaccac catttattat
180
gcggataccg tgaaaggccg ctttaccatt agccgcgata acgcgaaaaa cagcctgtat
240
ctgcagatga acagcctgcg cgcggaagat accgcggtgt attattgcgc gcgcattagc
300
accgtggtgg cgaaacgcta tgcgatggat tattggggcc agggcaccct ggtgaccgtg
360
agcagc
366
<210> 88
<211> 122
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 88
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Ser Thr Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 89
<211> 448
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 89
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Gly Ser Thr Thr Ile Tyr Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu
420 425 430
Ala Leu His Ser His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 90
<211> 321
<212> ДНК
<213> Синтетическая последовательность
<400> 90
gatattcaga tgacccagag cccgagcagc ctgagcgcga gcgtgggcga tcgcgtgacc
60
attacctgcc gcgcgagcca ggatattagc aactatctga actggtatca gcagaaaccg
120
ggcaaagcgc cgaaactgct gatttattat accagccgcc tgcatagcgg cgtgccgagc
180
cgctttagcg gcagcggcag cggcaccgat tttaccctga ccattagcag cctgcagccg
240
gaagatattg cgacctatta ttgccagcag attaacgcgc tgccgtttac ctttggccag
300
ggcaccaaac tggaaattaa a
321
<210> 91
<211> 107
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 91
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Ile Asn Ala Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 92
<211> 214
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 92
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Ile Asn Ala Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 93
<211> 360
<212> ДНК
<213> Синтетическая последовательность
<400> 93
caggtgcagc tggtgcagag cggcgcggaa gtgaaaaaac cgggcgcgag cgtgaaagtg
60
agctgcaaag cgagcggcta tacctttacc gattataaca tggattgggt gcgccaggcg
120
ccgggccaga acctggaatg gattggcaac attaacccga acaacggcgg caccttttat
180
aaccagaaat ttaaaggccg cgcgaccctg accgtggata aaagcgcgag caccgcgtat
240
atggaactga gcagcctgcg cagcgaagat accgcggtgt attattgcgc gcgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccctggtgac cgtgagcagc
360
<210> 94
<211> 120
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 94
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 95
<211> 446
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 95
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Asn Leu Glu Trp Ile
35 40 45
Gly Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 96
<211> 318
<212> ДНК
<213> Синтетическая последовательность
<400> 96
gaaattgtgc tgacccagag cccggcgacc ctgagcctga gcccgggcga acgcgcgacc
60
ctgagctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
cagagcccgc gcccgtggat ttatgcggcg agcaacctgg cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccgattat accctgacca ttagccgcct ggaaccggaa
240
gattttgcgg tgtattattg ccagcagtgg agcgtgaacc cgccgacctt tggccagggc
300
accaaactgg aaattaaa
318
<210> 97
<211> 106
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 97
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 98
<211> 213
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 98
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Val Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 99
<211> 360
<212> ДНК
<213> Синтетическая последовательность
<400> 99
caggtgcagc tggtgcagag cggcgcggaa gtgaaaaaac cgggcgcgag cgtgaaagtg
60
agctgcaaag cgagcggcta tacctttacc gattataacg tggattgggt gcgccaggcg
120
ccgggccagg gcctggaatg gattggcacc attaacccga acaacggcgg cattctgagc
180
aaccagaaat ttaaaggccg cgtgaccatt accgtggata ccagcgcgag caccgcgtat
240
atggaactga gcagcctgcg cagcgaagat accgcggtgt attattgcgg ccgcggcggc
300
ctgcgccgcc gcggctttat ggattattgg ggccagggca ccctggtgac cgtgagcagc
360
<210> 100
<211> 120
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 100
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Asn Pro Asn Asn Gly Gly Ile Leu Ser Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Val Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 101
<211> 446
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 101
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Asn Val Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Thr Ile Asn Pro Asn Asn Gly Gly Ile Leu Ser Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Val Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
<210> 102
<211> 318
<212> ДНК
<213> Синтетическая последовательность
<400> 102
gaaattgtgc tgacccagag cccggcgacc ctgagcctga gcccgggcga acgcgcgacc
60
ctgagctgcc gcgcgagcag cagcgtgatt tatattcatt ggtatcagca gaaaccgggc
120
caggcgccgc gcccgtggat ttatgcggcg agcaacctgc cgagcggcgt gccggcgcgc
180
tttagcggca gcggcagcgg caccgatttt accctgacca ttagcagcct ggaaccggaa
240
gattttgcgg tgtattattg ccagcagtgg agcagcaacc cgccgacctt tggccagggc
300
accaaagtgg aaattaaa
318
<210> 103
<211> 106
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 103
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Pro Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 104
<211> 213
<212> БЕЛОК
<213> Синтетическая последовательность
<400> 104
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ile Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Ala Ser Asn Leu Pro Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 105
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 105
Asp Tyr Gly Met His
1 5
<210> 106
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 106
Tyr Ile Asn Ser Gly Ser Ser Lys Ile Tyr His Ala Asp Thr Val Lys
1 5 10 15
Gly
<210> 107
<211> 15
<212> БЕЛОК
<213> Mus musculus
<400> 107
Ala Arg Phe Pro Thr Val Val Ala Ala Arg Tyr Pro Met Asp Tyr
1 5 10 15
<210> 108
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 108
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 109
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 109
Phe Thr Ser Arg Leu His Ser
1 5
<210> 110
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 110
Gln Gln Gly Asn Thr Leu Pro Tyr Thr
1 5
<210> 111
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 111
Asp Tyr Gly Ile His
1 5
<210> 112
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 112
Tyr Ile Arg Ser Asp Ser Ser Ile Ile His Tyr Ala Asp Thr Val Lys
1 5 10 15
Gly
<210> 113
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 113
Thr Arg Gly Arg Asp Arg Gly Tyr Phe Asp Tyr
1 5 10
<210> 114
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 114
Ser Ala Ser Ser Ser Val Ser Tyr Met Tyr
1 5 10
<210> 115
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 115
Leu Thr Ser Asn Leu Ala Ser
1 5
<210> 116
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 116
Gln Gln Trp Thr Ser Ile Pro Phe Thr
1 5
<210> 117
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 117
Asp Tyr Gly Met His
1 5
<210> 118
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 118
Tyr Ile Ser Ser Gly Ser Thr Thr Ile Tyr Tyr Ala Asp Thr Val Lys
1 5 10 15
Gly
<210> 119
<211> 15
<212> БЕЛОК
<213> Mus musculus
<400> 119
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr
1 5 10 15
<210> 120
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 120
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 121
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 121
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 122
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 122
Gln Gln Ile Asn Ala Leu Pro Leu Thr
1 5
<210> 123
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 123
Asp Tyr Gly Met His
1 5
<210> 124
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 124
Tyr Ile Ser Ser Gly Ser Ser Thr Ile Tyr Tyr Ala Asp Thr Val Lys
1 5 10 15
Gly
<210> 125
<211> 15
<212> БЕЛОК
<213> Mus musculus
<400> 125
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr
1 5 10 15
<210> 126
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 126
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 127
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 127
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 128
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 128
Gln Glu Val Asn Met Leu Pro Leu Thr
1 5
<210> 129
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 129
Asp Tyr Gly Met His
1 5
<210> 130
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 130
Tyr Ile Ser Ser Gly Ser Thr Thr Ile Tyr Tyr Ala Asp Thr Val Lys
1 5 10 15
Gly
<210> 131
<211> 15
<212> БЕЛОК
<213> Mus musculus
<400> 131
Ala Arg Ile Ser Thr Val Val Ala Lys Arg Tyr Ala Met Asp Tyr
1 5 10 15
<210> 132
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 132
Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 133
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 133
Tyr Thr Ser Arg Leu His Ser
1 5
<210> 134
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 134
Gln Gln Ile Asn Ala Leu Pro Leu Thr
1 5
<210> 135
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 135
Asp Tyr Asn Met Asp
1 5
<210> 136
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 136
Asp Ile Asn Pro Asn Asn Gly Asn Ile Leu Phe Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 137
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 137
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 138
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 138
Arg Ala Ser Ser Ser Val Ser Tyr Met His
1 5 10
<210> 139
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 139
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 140
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 140
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<210> 141
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 141
Asp Tyr Asn Met Asp
1 5
<210> 142
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 142
Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 143
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 143
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 144
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 144
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 145
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 145
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 146
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 146
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 147
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 147
Asp Tyr Asn Met Asp
1 5
<210> 148
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 148
Asp Ile Asn Pro Asn Asn Gly Asn Ile Leu Phe Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 149
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 149
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 150
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 150
Arg Ala Ser Ser Ser Val Ser Tyr Met His
1 5 10
<210> 151
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 151
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 152
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 152
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<210> 153
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 153
Asp Tyr Asn Met Asp
1 5
<210> 154
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 154
Thr Thr Asn Pro Asn Asn Gly Gly Thr Leu Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 155
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 155
Ala Arg Gly Gly Leu Arg Arg Arg Gly Phe Val Asp Tyr
1 5 10
<210> 156
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 156
Arg Ala Ser Ser Ser Val Asn Tyr Leu His
1 5 10
<210> 157
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 157
Ala Thr Ser Asn Leu Ala Ser
1 5
<210> 158
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 158
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<210> 159
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 159
Asp Tyr Asn Met Asp
1 5
<210> 160
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 160
Asp Ile Asn Pro Asn Asn Gly Asn Ile Leu Phe Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 161
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 161
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 162
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 162
Arg Ala Ser Ser Ser Val Ser Tyr Met His
1 5 10
<210> 163
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 163
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 164
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 164
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<210> 165
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 165
Asp Tyr Asn Met Asp
1 5
<210> 166
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 166
Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 167
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 167
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 168
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 168
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 169
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 169
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 170
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 170
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 171
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 171
Asp Tyr Asn Met Asp
1 5
<210> 172
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 172
Thr Ile Asn Pro Asn Asn Gly Asp Thr Met Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 173
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 173
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 174
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 174
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 175
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 175
Ala Ile Ser Asn Leu Ala Ser
1 5
<210> 176
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 176
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 177
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 177
Asp Tyr Asn Met Asp
1 5
<210> 178
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 178
Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 179
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 179
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 180
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 180
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 181
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 181
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 182
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 182
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 183
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 183
Asp Tyr Asn Met Asp
1 5
<210> 184
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 184
Thr Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Asn Phe Lys
1 5 10 15
Gly
<210> 185
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 185
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 186
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 186
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 187
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 187
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 188
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 188
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 189
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 189
Asp Tyr Asn Met Asp
1 5
<210> 190
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 190
Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 191
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 191
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Leu Asp Tyr
1 5 10
<210> 192
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 192
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 193
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 193
Ala Val Ser Asn Leu Ala Ser
1 5
<210> 194
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 194
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 195
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 195
Asp Tyr Asn Met Asp
1 5
<210> 196
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 196
Asn Ile Asn Pro Asn Asn Gly Gly Thr Phe Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 197
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 197
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 198
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 198
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 199
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 199
Ala Ala Ser Asn Leu Ala Ser
1 5
<210> 200
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 200
Gln Gln Trp Ser Val Asn Pro Pro Thr
1 5
<210> 201
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 201
Asp Tyr Asn Val Asp
1 5
<210> 202
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 202
Thr Ile Asn Pro Asn Asn Gly Gly Ile Leu Ser Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 203
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 203
Gly Arg Gly Gly Leu Arg Arg Arg Gly Phe Met Asp Tyr
1 5 10
<210> 204
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 204
Arg Ala Ser Ser Ser Val Ile Tyr Ile His
1 5 10
<210> 205
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 205
Ala Ala Ser Asn Leu Pro Ser
1 5
<210> 206
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 206
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К ФАКТОРУ XI | 2017 |
|
RU2800719C2 |
| АНТИТЕЛО К CD40, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2779128C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2019 |
|
RU2781542C2 |
| СЛИТЫЙ БЕЛОК НА ОСНОВЕ АНТИТЕЛА К ЕТА И TGF-β TRAP, СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2832190C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДУПРЕЖДЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2017 |
|
RU2766586C2 |
| МОНОКЛОНАЛЬНЫЕ АГЕНТЫ, СВЯЗЫВАЮЩИЕ сМЕТ, ИХ КОНЪЮГАТЫ С ЛЕКАРСТВЕННЫМИ СРЕДСТВАМИ И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2744914C2 |
| АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО CTLA-4 | 2020 |
|
RU2818586C2 |
| АНТИТЕЛО К GIPR И ЕГО СЛИТЫЙ С GLP-1 БЕЛОК, А ТАКЖЕ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800370C2 |
| НАЦЕЛЕННЫЕ НА uPARAP КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2740311C2 |
| Антитело против PCSK9 и его применение | 2016 |
|
RU2756012C2 |
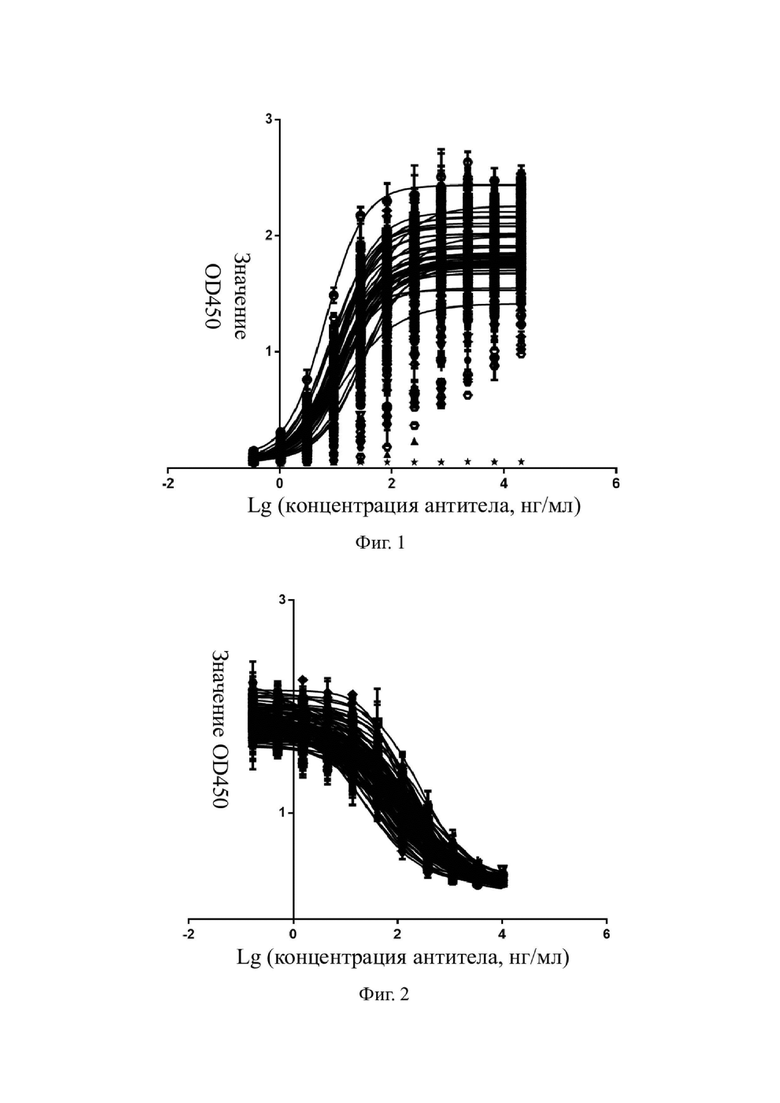
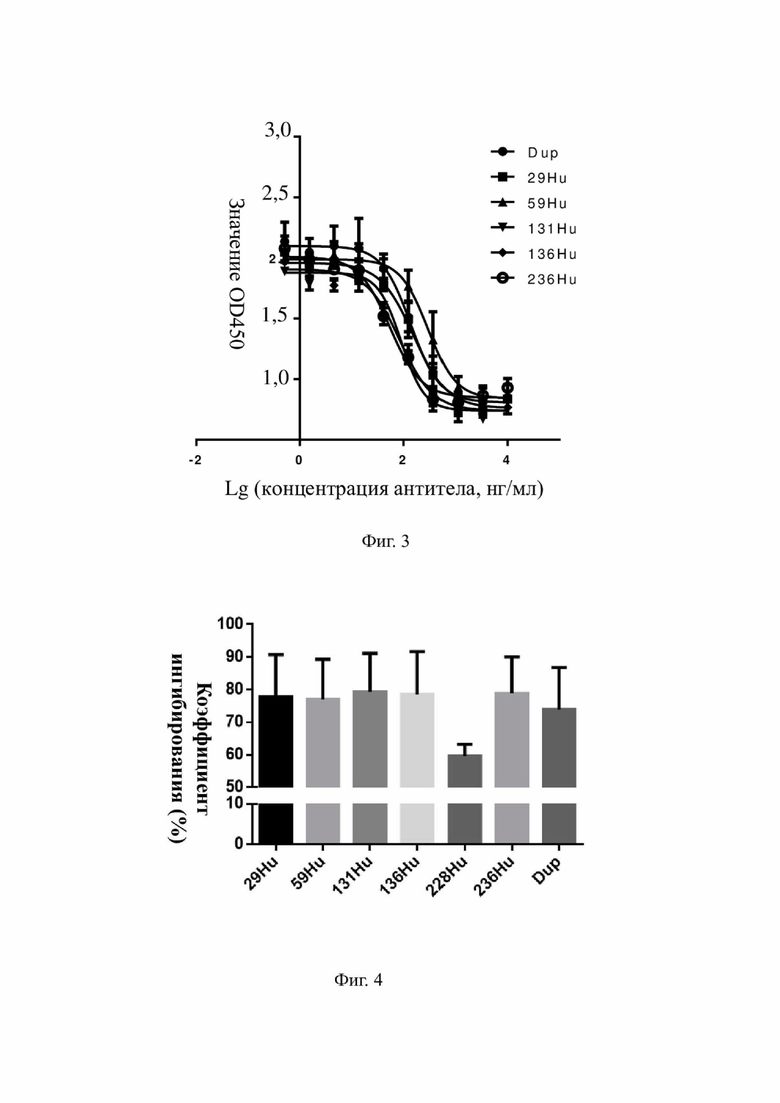
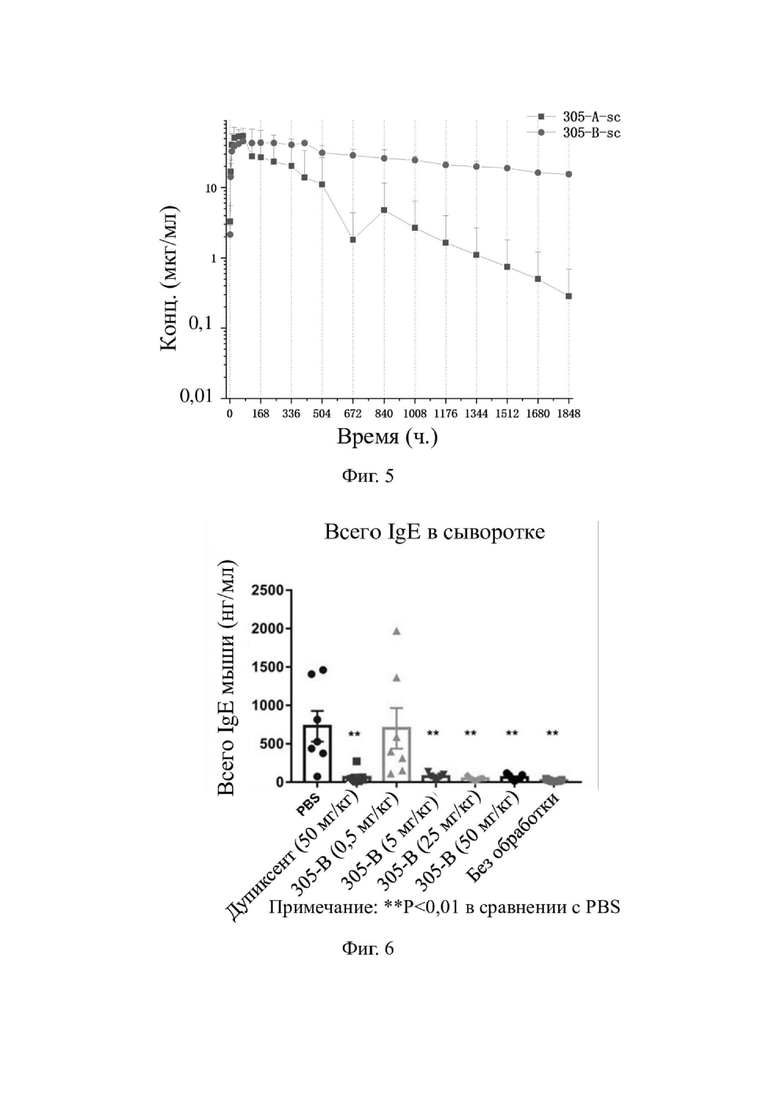
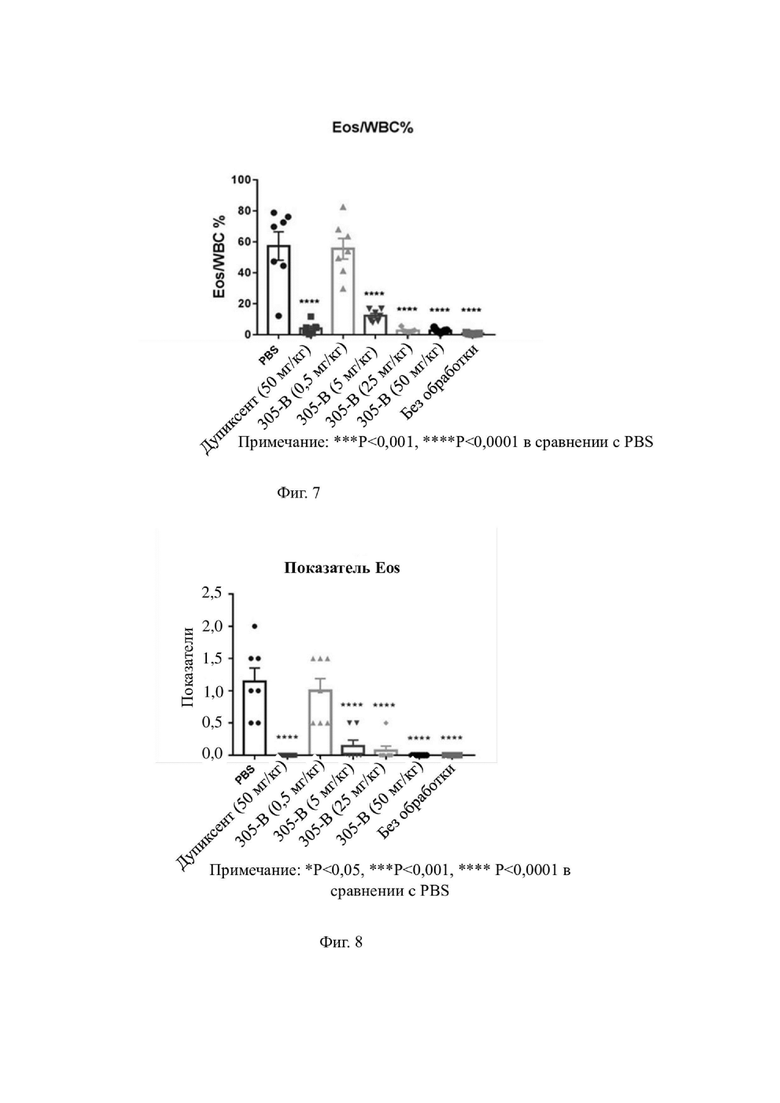
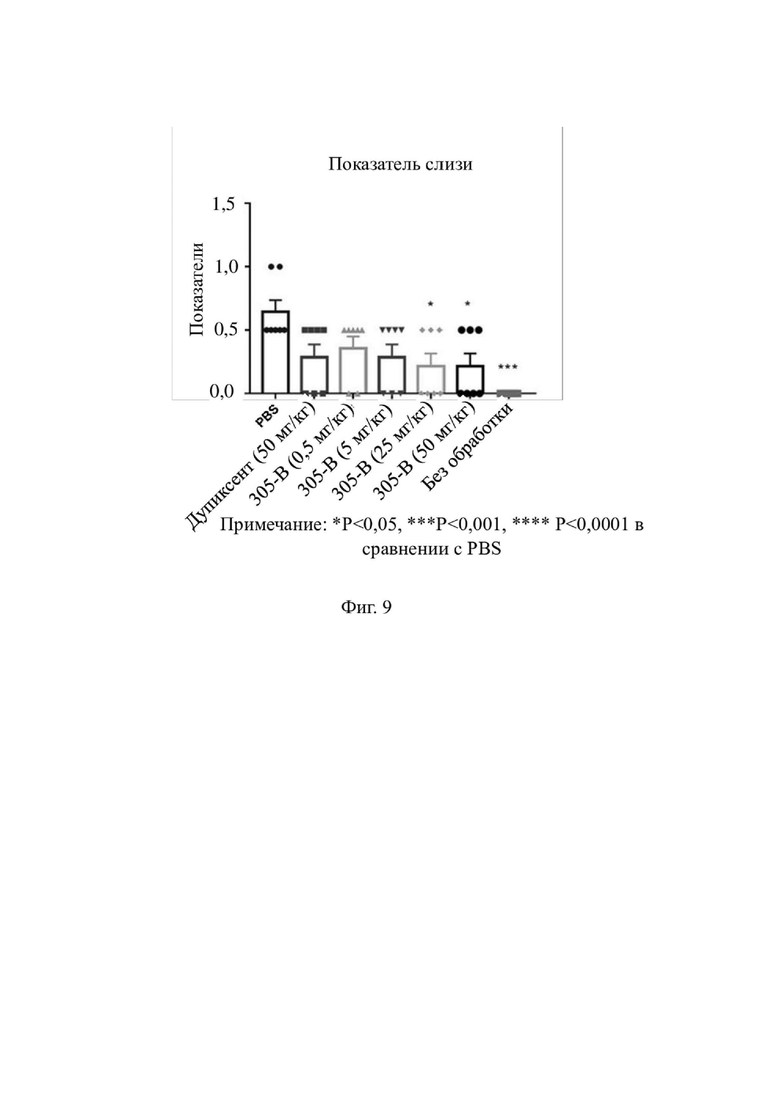
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая антитело, специфически связывающееся с альфа-рецептором интерлейкина-4, или его антигенсвязывающий фрагмент, фармацевтическую композицию для предупреждения или лечения заболеваний, связанных с IL-4R-альфа, нуклеотидную молекулу для получения вышеуказанного антитела, вектор экспрессии для получения вышеуказанного антитела, клетку-хозяин для получения вышеуказанного антитела, способ получения вышеуказанного антитела, применение антитела или его антигенсвязывающего фрагмента или композиции при получении лекарственных средств для предупреждения или лечения заболеваний, связанных с IL-4R-альфа. Изобретение расширяет арсенал средств, специфически связывающихся с IL-4R-альфа. 7 н. и 8 з.п. ф-лы, 9 ил., 11 табл., 13 пр.
1. Антитело, специфически связывающееся с альфа-рецептором интерлейкина-4, или его антигенсвязывающий фрагмент, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит последовательность HCDR3, последовательность HCDR1 и последовательность HCDR2, вариабельная область легкой цепи содержит последовательность LCDR1, последовательность LCDR2 и последовательность LCDR3, где
последовательность HCDR1, последовательность HCDR2, последовательность HCDR3, последовательность LCDR1, последовательность LCDR2 и последовательность LCDR3 соответственно содержат аминокислотные последовательности SEQ ID NO: 123-128; или
последовательность HCDR1, последовательность HCDR2, последовательность HCDR3, последовательность LCDR1, последовательность LCDR2 и последовательность LCDR3 соответственно содержат аминокислотные последовательности SEQ ID NO: 129-134; или
последовательность HCDR1, последовательность HCDR2, последовательность HCDR3, последовательность LCDR1, последовательность LCDR2 и последовательность LCDR3 соответственно содержат аминокислотные последовательности SEQ ID NO: 201-206.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 14 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 16; или
вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 82 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 85; или
вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 18 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 20; или
вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 88 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 91; или
вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 66 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 68; или
вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 100 и вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 103.
3. Антитело или его антигенсвязывающий фрагмент по п. 1 или 2, где антитело или его антигенсвязывающий фрагмент имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 83, и последовательность легкой цепи, представленную под SEQ ID NO: 86; или
антитело или его антигенсвязывающий фрагмент имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 89, и последовательность легкой цепи, представленную под SEQ ID NO: 92; или
антитело или его антигенсвязывающий фрагмент имеет последовательность тяжелой цепи, представленную под SEQ ID NO: 101, и последовательность легкой цепи, представленную под SEQ ID NO: 104.
4. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3, где антигенсвязывающий фрагмент выбран из группы, состоящей из Fab-фрагмента, Fab'-фрагмента, F(ab')2-фрагмента, Fv-фрагмента, scFv-фрагмента, Fd-фрагмента и однодоменного антитела.
5. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-4, где антитело или его антигенсвязывающий фрагмент связывается с альфа-рецептором интерлейкина-4 при KD менее 600 пМ.
6. Антитело или его антигенсвязывающий фрагмент по п. 5, где антитело или его антигенсвязывающий фрагмент связывается с альфа-рецептором интерлейкина-4 при KD менее 350 пМ.
7. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, где антитело или его антигенсвязывающий фрагмент специфически связывается с альфа-рецептором интерлейкина-4 человека или альфа-рецептором интерлейкина-4 мыши.
8. Антитело или его антигенсвязывающий фрагмент по п. 7, где антитело или его антигенсвязывающий фрагмент способен ингибировать IL-4-индуцированную пролиферацию клеток TF-1 и/или ингибировать секрецию IgE В-клетками.
9. Фармацевтическая композиция для предупреждения или лечения заболеваний, связанных с IL-4R-альфа, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-8 и фармацевтически приемлемый носитель.
10. Нуклеотидная молекула для получения антитела, специфически связывающегося с альфа-рецептором интерлейкина-4, или его антигенсвязывающего фрагмента, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-8.
11. Вектор экспрессии для получения антитела, специфически связывающегося с альфа-рецептором интерлейкина-4, или его антигенсвязывающего фрагмента, содержащий нуклеотидную молекулу по п. 10.
12. Клетка-хозяин для получения антитела, специфически связывающегося с альфа-рецептором интерлейкина-4, или его антигенсвязывающего фрагмента, содержащая нуклеотидную молекулу по п. 10 или вектор экспрессии по п. 11.
13. Способ получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-8, включающий следующие стадии:
а) культивирование клетки-хозяина по п. 12 в условиях экспрессии, которые позволяют клетке-хозяину продуцировать антитело или его антигенсвязывающий фрагмент, за счет чего обеспечивается экспрессия антитела или его антигенсвязывающего фрагмента; и
b) отделение и очистка антитела или его антигенсвязывающего фрагмента, экспрессированных на стадии а).
14. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-8 или композиции по п. 9 при получении лекарственных средств для предупреждения или лечения заболеваний, связанных с IL-4R-альфа.
15. Применение по п. 14, где иммуноопосредованные воспалительные заболевания выбраны из астмы, аллергии, атопического дерматита, хронического синусита, эозинофильного эзофагита, полипов носа, псориаза, ревматоидного артрита, псориатического артрита, анкилозирующего спондилоартрита, рассеянного склероза, увеита, увеита Бехчета, ксерофтальмии и хронической спонтанной крапивницы.
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| ЭЛЕМЕНТ, СВЯЗЫВАЮЩИЙСЯ С α-РЕЦЕПТОРОМ ИНТЕРЛЕЙКИНА-4 (IL-4Rα)-173 | 2008 |
|
RU2490278C2 |

