(21)4073635/28-13
(22)02,06.86
(46) 30.04.88. БЮЛ. № 16
(71)Всесоюзньй научно-исследовательский институт молекулярной биологии
и Институт цитологии и генетики СО АН СССР
(72)В.В.Кравченко, Л.А.Петренко, С.Х.Дегтярев, Н.А.Нетесова, М.И.Ривкин и К.Д.Кузнеделов
(53)575.224.2:577.2/048/(088.8)
(56)PNAS, 1983, V. 80, р. 1830-1834.
(54)РЕКОМБИНАНТНАЯ ПЛАЗМВДНАЯ ДНК РСЕК10, КОДИРУЮЩАЯ СИНТЕЗ ФРАГМЕНТА Ю1ЕНОВА ДНК-ПОЛИМЕРАЗЫ I E.COLI И СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ
(57)Изобретение относится к биотехнологии и генетической инженерии. Цель изобретения - повьпиение уровня синтеза фрагмента Кленова ДНК-поли- меразы I E.coli. На основе векторной части плазмиды pCEZ12 и фрагмента BamHI-PstI плазмиды рС155, содержащего- последовательность нуклеотидов.
кодирующую синтез фрагмента Кленова ДНК-полимеразы I E.coli (ФК), сконструирована рекомбинантная гшазмида рСЕК 10 размером 7,21 т.п.о, Плазмида позволяет получить штаммы-продуценты, с повышенным уровнем синтеза ФК (не менее 375000-470000 ед. акт. на
1г сырого веса биомассы клеток). Последовательность, кодирующая синтез ФК, находится под контролем промотора Рр, регулируемого встроенным в плаз- Mi-еду геном термочувствительного ре- прессора с 1857. Двойной селективный контроль обеспечивается генайи устойчивости к ампициллину (bla) и тетрациклину (tet). Плазмида не содержит дополнительных фаговык генов, понижающих ФК, и не требует специального штамма Е. coli. Правильность ориентации последовательности, кодирующей ФК, относительно промотора PR и восг становление целостности гена Ыа при сборке обеспечивается направленным клонированием фрагмента в векторе.
2с.п. ф-лы, 2 ил., 1 табл.
б
СО
со
ГчЭ
О со
4;
Изобретение относится к биотехноогии, в частности к генетической нженерии, и представляет собой ре- омбинантную плазмидную ДНК, обеспеивающую синтез фрагмента Кпенова НК-полимеразы I E.coli, и способ онструирования данной рекомбинантной лазмидной ДНК.
Цель изобретения - повьппение уров-ю ня синтеза фрагмента Кпенова ДНК- олимеразы I E.coli.
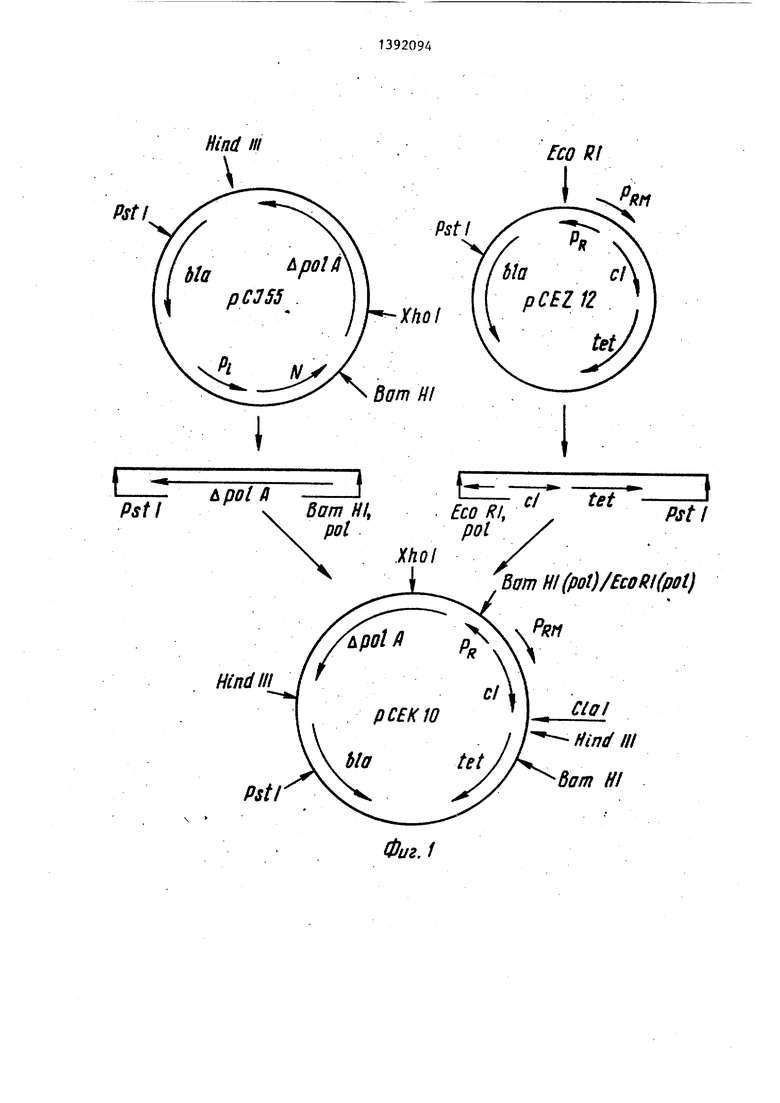
Повышение синтеза обеспечивается за счет того-, что сконструирована екомбинантная плазмидная ДНК рСЕК tO 5 размером 7, 21 т.п.о., состоящая из векторной части рекомбинантной плазидной ДНК pCEZ 12 длиной 3,50 т.п.о. и чужеродного фрагмента рекомбинантной плазмидной ДНК рС1 55 размером 20 3,71 т.п.о., имеющая уникальные сайты рестрикдии Xhol, PstI, Sal GI, BaraHI, Clal, ген с 1857 фага A для автономного, штаммонезависимого синтеза термочувствительного pefipeccopa, 25 последовательность нуклеотидов, кодирующую фрагмент Кпенова ДНК-полимеразы I E.coli, промоторно-операторную область Рр/Ррц, ДНК фага Л , обеспечивающую регулируемую репрессором jQ транскрипцию гена с1857 и последовательности, кодирующей фрагмент ДНК- полимеразы I, а также гены устойчивости к антибиотикам тетрациклину и ампициллину с собственными промотор- ными участками, обеспечивающие возможность двойного селективного контроля .
Способ конструирования рекомбинантной ДНК рСЕК 10 заключается в Q том, что плазмидную ДНК pCEZ 12 обрабатывают последовательно эндонук- леазами рестрикции Е coRI, ДНК-поли- меразой 1 E.coli в присутствии дез- оксирибонуклеотидтрифосфатов для до- ., стройки липких концов ДНК до тупых, и после удаления этих ферментов из смеси фенольной экстракцией, эндонук- леазой рестрикции Pstl. Из полученной смеси вьщеляют методом электрофореза в полиакриламидном геле и электроэлюции векторную часть плазмидной ДНК pCEZ 12 размером 3,50 т.п.о. Рекомбинантную плазмидную ДНК рС1 55 обрабатывают последовательно эндонуклеазой рестрикции Ват HI, ДНК-55 полимеразой 1 E.coli в присутствии дезоксирибонуклеотид трифосфатов, чем достигается достройка образовав35
50
5 0 5 Q
Q , 5
5
0
шихся липких концов до тупых, и после удаления ферментов из смеси фенольной экстракцией, эндонуклеазой рестрикции Pstl, и из полученной смеси вьщеляют методом электрофореза и электроэлюции фрагмент рекомбинантной плазмидной ДНК рС155 размером 3,71 т.п.о. Затем полученные векторную часть плазмидной ДНК pCEZ12 и фрагмент плазмидной ДНК рС155 сшивают ДНК-лигазой фага ТА в присутствии АТР, при этом ковалентная сшивка фрагмента и векторной части происходит по тупым концам ДНК с одной стороны и липким концам, образуемым рестрик- ционной эндонуклеазой Pstl, с другой стороны, чем достигается направленная встройка фрагмента в векторную часть, обеспечивающая правильную ориентацию последовательности, кодирующей фрагмент ДНК-полимеразы I, относительно промоторно-операторной области Рр/Рр, и восстановление устойчивости к ампициллину, служащей селективным маркером при отборе целевой рекомбинантной ДНК рСЕК 10. Полученной смесью трансформируют CaCl-j-обработанные клетки и из клойов, -устойчивых к ампициллину и тетрациклину, вьщеляют целевую рекомбинантную гшазмиду рСЕК 10.
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование рекомбинантной плаз шдной ДНК рСЕК 10 (фиг. 1). 10 мкг плазмидной ДНК pCEZ 12 гидролизуют 20 ед, рестрикци- онной эндонуклеазы Е coRI в 10 мкл С буфета для гидролиза, содержащего 20 мМ трис-НС рН 7,8, 10 мМ MgCl, 50 мМ NaCl, 2 мМ 2-меркаптоэтанола, при 37°С 1 ч. В полученную рестрик- ционную смесь добавляют dATP, dGTR и TIP до 0,05 мМ каждого, 10 ед. ДНК- полимеразы I и инкубируют 30 мин при 12°С. Реакцию останавливают добавлением Na/j- ЭДТА до концентрации 20 мМ, белок удаляют экстракцией водным фенолом, рН 8,0, а ДНК трижды переосаждают 96%-ным этиловым спиртом из раствора 0,3 М ацетата натрия, рН 7,0. Полученный препарат ДНК растворяют в 100 мкл буфера для гидролиза, содержащего 20 мМ трис-НС1 рН 7,8,10 мМ MgCl,, 50 мМ NaCl, 2 мМ 2-меркаптоэтанола, -и гидролизуют 20 ед. рестрикционной эндонуклеазы Pstl при 37°С 1 ч. Реак1щю останавливают добавлением Na ЭДТА до концентрации 20 мМ, белок удаляют экстракцией водным фенолом, рН 8,0, ДНК трижды переосаждают 96%-ным этиловым спиртом из раствора 0,3 М ацетата натрия, рН 7,0. Полученный препарат ДНК растворяют в 50 мкл буфера, содержащего 20 мМ трис-НС1 рН 7,8,
pCEZ12 и и,5 мкг препарата фрагмента ДНК плазмиды рС155 растворяют в ... 30 мкл буфера для лигирования, содержащего 20 мМ трис-НС рН 7,5, 10 мМ MgCl, 10 мМ 2-меркаптоэтанола и 0,5 мМ АТР, и обрабатывают 2 ед, ДНК-лигазы фага Т4 при 12°С в течение 12 ч. Полученную реакционную



40 мМ Na2 ЭДТА, 0,02% красителя бром- п смес.ь используют для трансформации
фенолового синего, и разделяют рест- рикционные фрагменты ДНК электрофорезом в 4%-ном полиакриламидном геле. Гель окрашивают в растворе бромистого этидия 2 мкг/мл, вырезают из геля зо-|5 ну, содержащую векторную часть ДНК плазмиды pCEZ 12, и вьщеляют ДНК из геля методом электроэлюции. 10 мкг плазмидной ДНК рС155 гидролизуют 20 ед, рестрикционной эндонуклеазы 20 BamHI в 100 мкл буфера для гидролиза, содержащего 20 мМ трис-НС1 рН 7,8, 10 мМ MgCl,f, 50 мМ NaCl, 2 мМ 2-мер- каптоэтанола, при 37°С 1 ч. В полученную реакционную смесь добавляют. 25 dATP, dCTP, dGTP и TIP. до 0,05 мМ каждого, 10 ед. ДНК-полимеразы I и инкубируют 30 мин при . Реакцию останавливают добавлением до концентрации 20 мМ, белок удаляют 0 экстракцией водным фенолом, рН 8,0 и ДНК трижды переосаждают 96%-ным этиловым спиртом из раствора 0,3 М ацетата натрия, рН 7,0. Полученньм препарат ДНК растворяют в 100 мкл буфера для гидролиза, содержащего 20 мМ трис-НС рН 7,8, 10 мМ MgCl-, 50 мМ ЫаС1, 2мМ 2-меркаптоэтанола, и гидролизуют 20 ед. рестрикционной эндонуклеазы PstI при 37°С 1 ч. Реакцию останавливают добавлением Na j ЭДТА до концентрации 20 мМ, белок удаляют экстракцией водным фенолом, рН 8,0, ДНК трижды переосаждают 96%-ным этиловым спиртом из раствора 0,3 М ацетата натрия, рН 7,0. Полученный препарат ДНК растворяют в 50 мкл буфера, содержащего 20 мМ трис-НС рН 7,8, 40 мМ Каг-ЭДТА, 0,02% красителя бромфенолового синего, и разделяют рестрикционные фрагменты ДНК электрофорезом в 4%-ном полиакриламидном геле. Гель окрашивают в растворе бромида этидия 2 мкг/ /мл, вырезают из геля зону, содержащую фрагмент ДНК плазмиды рС155, и вьщеляют ДНК из геля методом электроэлюции. 0,5 мкг полученного препарата векторной части ДНК-плазмиды ;.
35
40
45
50
55
СаС - обработанных клеток E., которые затем высевают на 1,5% агар LB, содержащий 20 мкг/мл тетрацикли на и 50 мкг/мл ампициллина, и инкубируют при 30°С 16 ч. Из выросших индивидуальных колоний вьщеляют пла мидную ДНК известным методом.
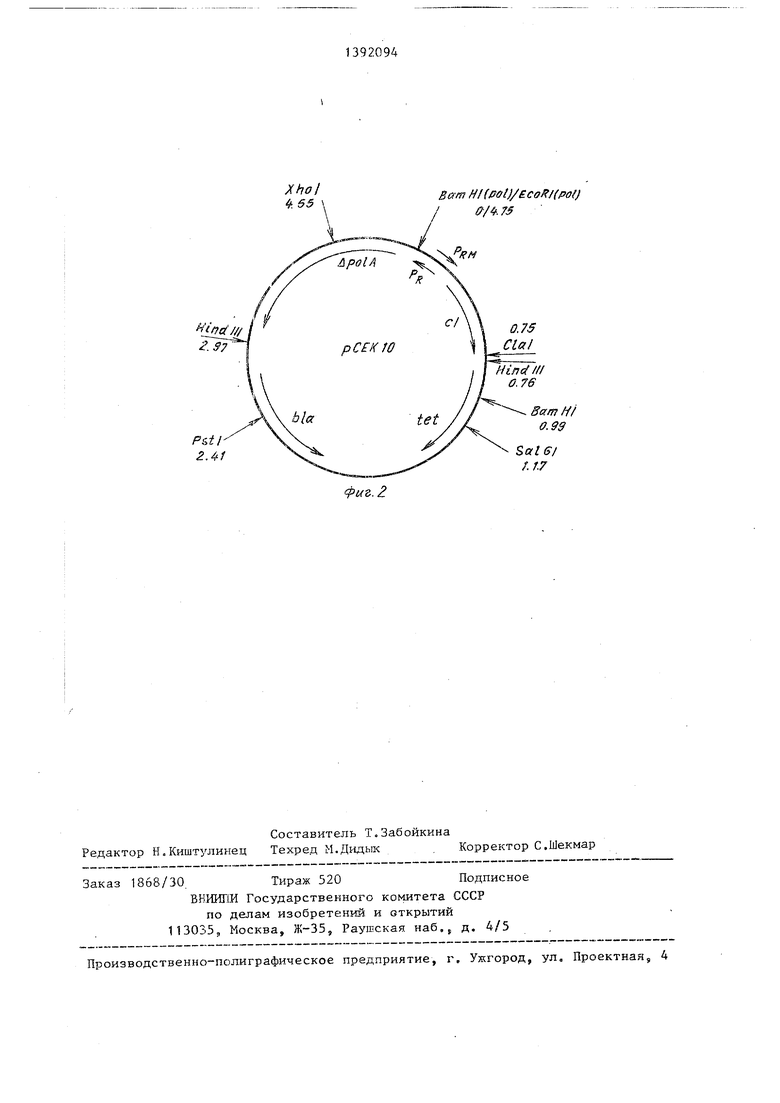
Полученную целевую рекомбинантну плазмидную ДНК рСЕК 10 анализируют помощью эндонуклеаз рестрикции с п следующим электрофорезом в 4%-ном полиакриламидном геле (фиг. 2).
П р и М е р 2. Использование ре- комбинантной плазмидной ДНК рСЕК 10 для создания штаммов E. - проду центов фрагмента Кленова ДНК-полиме разы E..
0,02 мкг рекомбинантной плазмидн ДНК рСЕК 10. берут для трансформации СаС, - обработанных клеток E. каждого из трех исследуемых штаммов Y 10Ь9, WR 3110 и N 4830, Индивидуальные колонии каждого из штаммов, выросшие при 30°С на чашках с 1,5% агаром LB, содержащим ампициллин - 50 мкг/мл и тетрациклин - 20 мкг/мл пересевают и выращивают в 50 мл сре ды LB с антибиотиками ампициллином 50 мкг/мл и тетрациклином 20 мкг/мл до плотности 0,8-1,0 о,е,/мл, затем растят еще 2 ч при , Клетки осаждают центрифугированием, промывают буфером, содержащим 50 мМ трис НС рН 7,5, замораживают в жидком азоте и хранят при -70°С.
Из 1 г биомассы клеток E, каждого штамма выделяют фермент. Фе мент характеризуют по ферментативно активности, гомогенность препарата доказывают анализом в полиакриламид ном геле с додецилсульфатом натрия,
Данные по определению количественного содержания ферментативно ак тивного фрагмента ДНК-полимеразы I в различных штаммах E. приведен в таблице.
Из данных таблицы видно, что пре лагаемая плазмида рСЕК 10 обеспечивает почти десятикратное по сравнени
смес.ь используют для трансформации
СаС - обработанных клеток E., которые затем высевают на 1,5% агар LB, содержащий 20 мкг/мл тетрациклина и 50 мкг/мл ампициллина, и инкубируют при 30°С 16 ч. Из выросших индивидуальных колоний вьщеляют плаз- мидную ДНК известным методом.
Полученную целевую рекомбинантную плазмидную ДНК рСЕК 10 анализируют с помощью эндонуклеаз рестрикции с последующим электрофорезом в 4%-ном полиакриламидном геле (фиг. 2).
П р и М е р 2. Использование ре- комбинантной плазмидной ДНК рСЕК 10 для создания штаммов E. - продуцентов фрагмента Кленова ДНК-полиме разы E..
0,02 мкг рекомбинантной плазмидной ДНК рСЕК 10. берут для трансформации СаС, - обработанных клеток E. каждого из трех исследуемых штаммов - Y 10Ь9, WR 3110 и N 4830, Индивидуальные колонии каждого из штаммов, выросшие при 30°С на чашках с 1,5% агаром LB, содержащим ампициллин - 50 мкг/мл и тетрациклин - 20 мкг/мл, пересевают и выращивают в 50 мл среды LB с антибиотиками ампициллином 50 мкг/мл и тетрациклином 20 мкг/мл до плотности 0,8-1,0 о,е,/мл, затем растят еще 2 ч при , Клетки осаждают центрифугированием, промывают буфером, содержащим 50 мМ трис- НС рН 7,5, замораживают в жидком азоте и хранят при -70°С.
Из 1 г биомассы клеток E, каждого штамма выделяют фермент. Фермент характеризуют по ферментативной активности, гомогенность препарата доказывают анализом в полиакриламидном геле с додецилсульфатом натрия,
Данные по определению количественного содержания ферментативно активного фрагмента ДНК-полимеразы I в различных штаммах E. приведены в таблице.
Из данных таблицы видно, что предлагаемая плазмида рСЕК 10 обеспечивает почти десятикратное по сравнению
с прототипом pCI 55 увеличение выхода не отличающегося по удельной активности от прототипа активного фермен та, при этом плазмида не требует специального штамма, содержащего профаг (либо его фрагмент) Pi с 1857,
Суммарное количестно фермента в , единицах активности.,
Изобретение позволяет получить штаммы-продуценты фрагмента Кленова
ДНК-полимеразы I E.coli. Уровень синтеза фермента в полученных штаммах
E.coli Y 1089, WR 3110 и N 4830, содержащих плазмиду рСЕК 10j не менее 3750GO-47QOOO ед. сырого веса биомассы клеток5 что в 7,5 - 9 раз выше уровня синтеза этого фермента в сравнении с лучшими из известных штаммов E.coli N 4830, содержащим плазмиду рС1 55, Полученный положительный эффект изобретения достигается за счет свойств сконструированной новым способом плазмйды рСЕК 10о
Формула изобретения
Предща Количество фермента, очищенного по методу, изложенному в прототипе, вьщеленного из 1 г клеток, Суммарное количество фермента в единицах активности.
„
5
5
0
т,п. уникальные сайты эндонуклеаз рестрикции Pst I, Hind III - два сайта Xho TO Cla I, Bam I, Sal GI, генетические маркеры гшазмиды: Ыа - ген, обеспечивающий синтез /э-лактамазы, tet - ген, обеспечиваы: щй устойчивость к тетрациклину, ро1А,- после довательность, кодирующую синтез фрагмента Кпенова ДНК-полимеразы I E.coli, cl ген репрёссора фага с 18575 и промоторы PRs Rw
Psti
a
3
Bam HI,ECO Ri,
potpol
.Khol
Hind III
PstI
PstI
Khu
Bom HI
ECO Ri,
pol
c/ tet
PstI
Bam HI (pol)/ECO Rl(pot)
«
Ciai
НЫ III Bam HI
Фиг.1
Ват Hf(Pot)/ecoRl(pol) &/fy.75
Sam Hi 0.99
фиг. Z
