Изобретение относится к биотехнологии и касается получения человеческого инсулина или его производных. 1 Цель изобретения - повышение выхода целевого продукта за счет избе
см
жания предварительного отщепления В-30 Ala.
Изобретение заключается в том, что осуществляют процесс ферментативного замещения В-30 аминокислоты в
свином инсулине формулы Ins - Ala - ОН с помощью сериновой карбоксипепти- дазы микробного происхождения, специфической для субстратов, имеющих С-концевую L-аминокислоту, водном растворе или дисперсии, содержащей амид треопина или сложный эфир треопина формулы Н - Thr - OR, где R 5 С - С6 -алкил, и представляет со бой, например, метильный, этильный, пропильный, изопропильный, бутильный изобутильный, трет-бутильный, амиль- ный, гексильный и им подобные радикалы, в которых для получения человеческого инсулина в качестве целевого продукта отщепляют одну из групп производных инсулина -NH4, -Thr - NH и OR3, используя ту же самую или иную L - специфическую се- риновую карбоксипептидазу.
Указанные группы могут быть замещены заместителями, которые являются инертными по отношению к энзиму, например атомами галогена (фтора, хло- ра, брома, иода), нитрогруппой, ал- коксильной группой (метоксильной, этоксильной и т.д.) или алкильной группой (метильной, этильной и т.д.)
Так, в случае типов сложных эфи- ров группа ORj предпочтительно принадлежит к ряду, состоящему из ал- коксильных групп, например метоксильной, этоксильной или трет-бутоксиль- ной, фенилоксильной группы к бензил- оксильной группы. Указанные группы могут быть замещены инертными растворителями, например нитрогруппами (п-нитробензилоксигруппа).
Первоначально свиной инсулин вводят в реакционный водный раствор или дисперсии, содержащей амид треопина или сложный эфир треопина указанной формулы, после чего добавляют энзим сериновой карбоксипептидазы. Предпочтительным энзимом является CPD-Y из пекарских дрожжей, но может также использоваться пеницилло- карбоксипептидаза или сериновая карбоксипептидаза из Aspergillus sp. прорастающего ячменя или других источников.
Предлагаемый способ осуществляют при значениях рН 7,0-10,5, предпочтительно 7,5-10,5„ Предпочтительное значение рН, которое часто лежит в очень узком диапазоне значений, зависит от оптимального и минимального значений рН соответственно для раз
0
Q
5
5
5
0
0
5
0
личных видов энзиматической активности используемого энзима. Значение рН следует выбирать таким образом, чтобы активности были уравновешены.
В том случае, когда в качестве энзима используют карбоксипептидаэу У, предпочтительные значения рН 7,5-10,5, более предпочтительные 9,0-10,5. Наиболее предпочтительным является нижнее значение рН из указанного предпочтительного диапазона в тех случаях, когда требуется проводить выделение промежуточных инсу- линамидов.
Выбранное предпочтительное значение рН следует поддерживать в ходе реакции сочетания, затем его можно изменить при осаждении продукта реакции, отщеплении защитных групп и т.д. Значение рН может быть выбрано таким образом, чтобы энзим проявлял амидаэную активность, в результате чего предотвращается осаждение образующегося инсулинамида и создается возможность для образования желаемого инсулина в одну стадию. Можно также выбрать значение рН, при котором энзим проявляет преимущественно пептидазную активность, в результате чего осуществляется режим, способствующий образованию стабильных промежуточных продуктов - инсу- линамидов.
Контроль над рН осуществляют введением стабильного буферного раствора для соответствующего диапазона рН в реакционную среду (например для этой цели можно использовать би- карбонатный буфер).
Значение рН поддерживают добавлением кислоты, например соляной, или основания, например гидрата окиси натрия, в процессе реакции. Это может быть осуществлено с использованием рН-стата.
Реакцию проводят в водной среде, которая может содержать до 50 об.% органического растворителя алкано- лы, например метанол и этанол, гли- коли, например этиленгликоль или пропиленгликоль, диметилформамид, диметилсульфоксид, тетрагидрофуран, диоксан и диметоксиэтан.
Выбор состава реакционной среды зависит, в частности, от растворимости, температуры и значения рН для компонентов реакционной среды и
получающегося инсулина, а также от стабильности энзима.
Реакционная среда может также включать компоненты, которые делают энзим нерастворимым, но сохраняют при этом значительную долю его энзи- матической активности, например ионообменные смолы.
В альтернативном варианте энзим может быть иммобилизован, например, в результате связывания в матрице, например в сшитом декстране или агарозе, или в двуокиси кремния, полиамиде или целлюлозе, или же путе капсулирования в полиакриламиде, альгинатах или волокнистых материалах. Кроме того, возможна модификация энзима химическими методами, предпочтительно солями ртути, с цель увеличения его стабильности или улучшения его энзиматических свойств
В том случае, когда желательно подавить какое-либо осаждение промежуточно образующихся инсулиамидов, реакционная среда содержит мочевину или хлоргидрат гуацидина в концентрации до 3 моль/л. Это может быть также предпочтительным при рН в среде, в который используемый в качестве субстрата инсулин имеет ограниченную растворимость.
Предпочтительная исходная концентрация для используемого в качестве субстрата инсулина лежит в пределах 0,002-0,5 моль/л, а аминного компонента- в пределах 0,05-3 ммоль/л.
Энзиматическая активность может меняться сама по себе, но концентрация энзима предпочтительно должна составлять величину 10 -10 моль/л наиболее предпочтительно - моль/л Наиболее пригодное значение активности зависит от концентрации субстрата, от концентрации амина и времени проведения реакции.
Согласно изобретению температура, при которой проводится реакция, предпочтительно лежит в пределах 20- 40°С. Наиболее удобная температура в условиях данного синтеза может быть определена экспериментально. В частности, она зависит от используемого в качестве аминного компонента вещества и от концентрации энзима. Пригодные температуры лежат обычно в пределах 20-30 С, предпочтительно 25°С. При температурах ниже 20 С время проведения реакции обычно слишком
1554766
велико,
а использование температур
0
5
выше 40 С часто вызывает проблемы, связанные со стабильностью энзимов и/или реагентов или продуктов реакции.
Аналогичные варианты возможны и для времени проведения реакции, которое очень сильно зависит от прочих параметров реакции, в особенности от концентрации энзима. Стандартное время проведения реакции согласно изобретению составляет примерно 2-6 ч.
При использовании в качестве аминного компонента амида или замещенного амина желательно специальное разложение амидной группы в получаемом инсулинамиде. Б этом отношении карбоксипептидазы, в особенности карбоксипептидаза Y, являются весьма пригодными, поскольку указанная карбоксипептидаза Y проявляет ами- дазную активность при рН выше 9, в 5 то время как ее карбоксипептидазная активность становится незначительной
Указанная карбоксипептидаза может быть вообще использована для разложения сложноэфирных групп
0
0
0
5
OR
,, в промежуточно образующихся
сложных эфирах инсулина с получением готового инсулина, не содержа- щего С-конечной защищенной группы.
Исходные материалы.
Свиной инсулин был представлен 5 лабораторией инсулина Нордиск, Копенгаген. Карбоксипептидазу Y из пекарских дрожжей (промышленного производства Карлсбергских пекарен) выделяли модифицированным методом афинной хроматографии, получая в результате лиофилнзированный порошок, содержащий 10% энзима в цитрате натрия. Перед употреблением энзим подвергают обессоливанию на колонке 1,5-25 см с Сефадексом Г-25, применяя в качестве растворителя и элюен- та воду. Концентрацию энзима определяют спектрофотометр.ически, исполь0
5
зуя 14,8. Используемый энзим не содержит протеазу А.
L - Треонинамид поставляла фирма Вега-4 окс,Аризона, США, метиловый эфир L-треонина - фирма Фулка, Швейцария, а L-треонин, дансилхлорид, карбоксипептидазу А и трипсин - фирма Сигма, США. Хроматографические материалы поставлялись фирмой 1 Фармация, Швеция. Все прочие peaгенты и растворители марки Аналитически чистый поставлялись фирмой Мерк, ФРГ.
Анализ аминокислот. Образцы для анализа аминокислот получали в результате гидролиза 6 М НС1 при в вакууме в течение 24 ч. Анализ проводился на анализаторе аминокислот Дюррум Д-500. Состав аминокислот определялся из расчета на известное количество ас- парагиновой кислоты и глицина. Единственными аминокислотами, на которы оказывают влияние проводимые реакции, являются треонин, лизин и ала- нин. Соответствующие значения для указанных аминокислот, содержащихся в свином инсулине (инсулине человека), составляют: треонин 1,93 (2,87 лизин 0,97 (0,98) и аланин 2,00 (1,05). Количество непревращенного свиного инсулина в реакционной смеси определялось анализом аланина в инсулине хроматографическим методом на Сефадексе Г-50. Выход реакции сочетания определялся как количеств получаемого - продукта, поделенное на все количество инсулина, израсходованного в ходе реакции.
Приготовление карбоксипептидазы (пример 2 и 3).
К 100 мкл раствора инсулина или производных инсулина (0,7 мг/мл) в 0,1 М трис-HCl (рН 7,5) добавляют 10 мкг карбоксипептидаэы А После выдерживания при комнатной температре в течение 6 ч реакцию останавливают добавлением эквивалентного объ ема 0,5 М НС1, и количество освобожденных аминокислот определяют в результате проведения анализа аминокислот.
Энзиматическая обработка производных инсулина (пример 4).
Обработка различных производных инсулина карбоксипептидазами А и Y проводилась в 0,05 М трис-буфере (рН 7,5) при комнатной температуре с использованием инсулина в количестве, составляющем примерно 1,5 мМ, и карбоксипептидазы - 5 мкМ Время реакции 3 ч в случае карбоксипептидазы Аи 1,5 часа в случае карбоксипептидазы Y. При указанных условиях достигалось наибольшее количество освобожденных С-конечньгх аминокислот. После подкисления НС1 аликвоты брались непосредственно
для проведения анализа аминокислот на анализаторе.
Последовательность С-конечной части в различных производных инсулина определялась после обработки трипсином и реакции продукта этой обработки с дансилхлоридом с последующей идентификацией дансилпептиQ дов. Обработка производных инсулина трипсином производилась в 0,1 М кислого карбоната натрия при рН 8,2 с использованием 1 мМ инсулина, 40 мкМ ДРСС-трипсина и при времени
5 инкубирования 1 ч. Предварительные эксперименты со свиным инсулином показали, что эти условия пригодны для полного освобождения С-конеч- ного аланильного остатка из
Q В-цепи. Освобождаемые аминокислоты или дипептиды подвергались данси- лированию согласно следующему способу. Образец в 100 мкл продукта обработки трипсином гасят добавлени5 ем 100 мкл 0,5 М соляной кислоты, аликвоты разгоняют досуха и снова растворяют в 100 мкл 0,1 М кислого карбоната натрия (рН 3,2), после чего добавляют 10 мкл дансилхлорио |Да (5 мг/мл) в ацетоне и инкубируют реакционную смесь в течение 2 ч при 37°С. Затем реакционную смесь анализируют методом жидкостной хроматографии под высоким давлением, используя систему для жидкостной хроматографии фирмы Уотерс, состоящую из инжектора (Мод.бЮ, двух насосов (Мод.бОООА), программного устройства для раствора (Мод.660),
детектора-УФ (Мод 450), устройства для обработки данных Уотерс) и устройства радиального сжатия (Уотерс, РСМ-100), оборудованного колонкой радиального наполнения Уотерс -А (обратная фаза С-18). Были синтезированы следующие стандартные соединении: Днс-Ала-ОН, Днс-тре-ОН, Днс-Ала-Тре-ОН, Днс-Тре- Тре-ОН, Днс-Тре-Ш2, Днс-Tpe-Tpe-NHi |И Днс-Ала-Тре-МН. Все 7 дансильных производных могут быть легко разделены методом жидкостной хроматографии под высоким давлением с использованием двух различных программ.
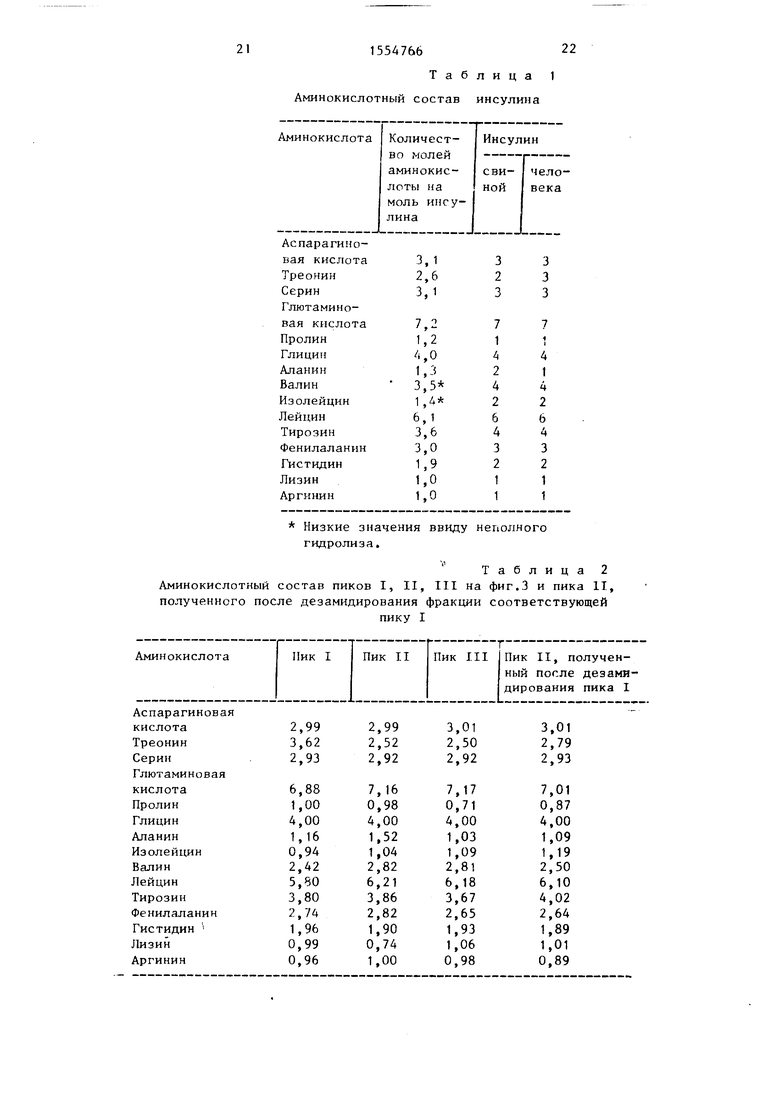
На фиг.1 изображено протекание реакции между свиным инсулином и L-треонинамидом с использованием в качестве катализатора карбоксипептидазы Y (результаты анализа
5
5
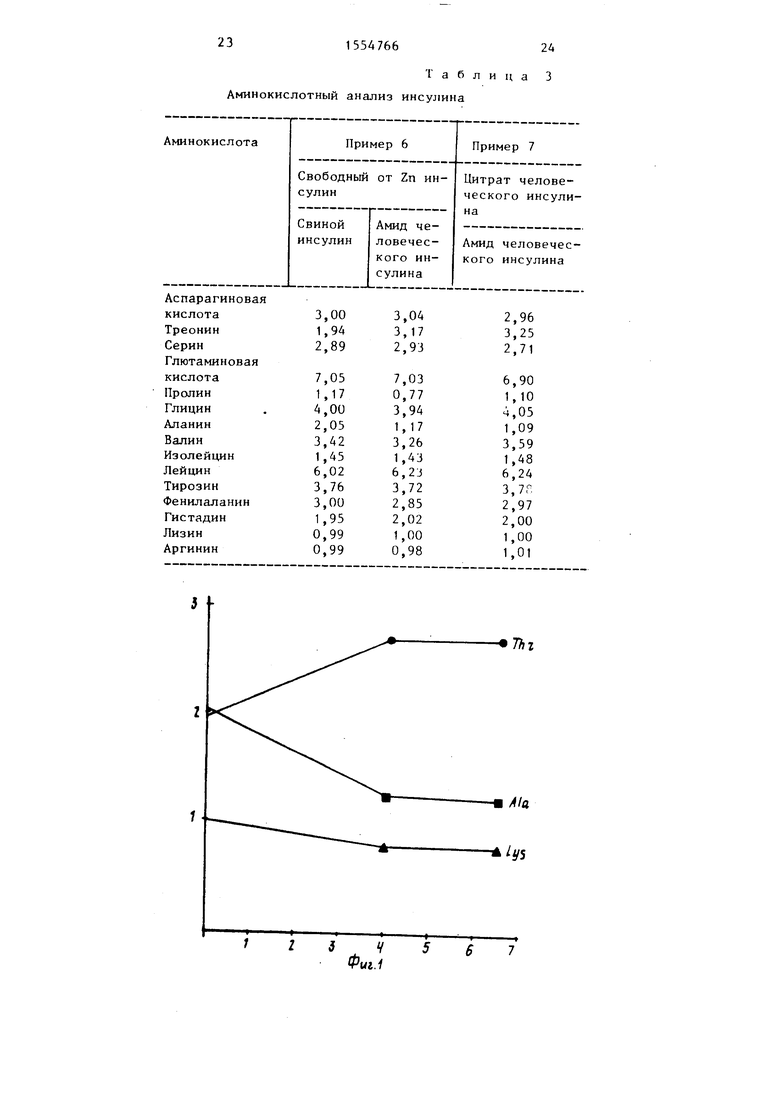
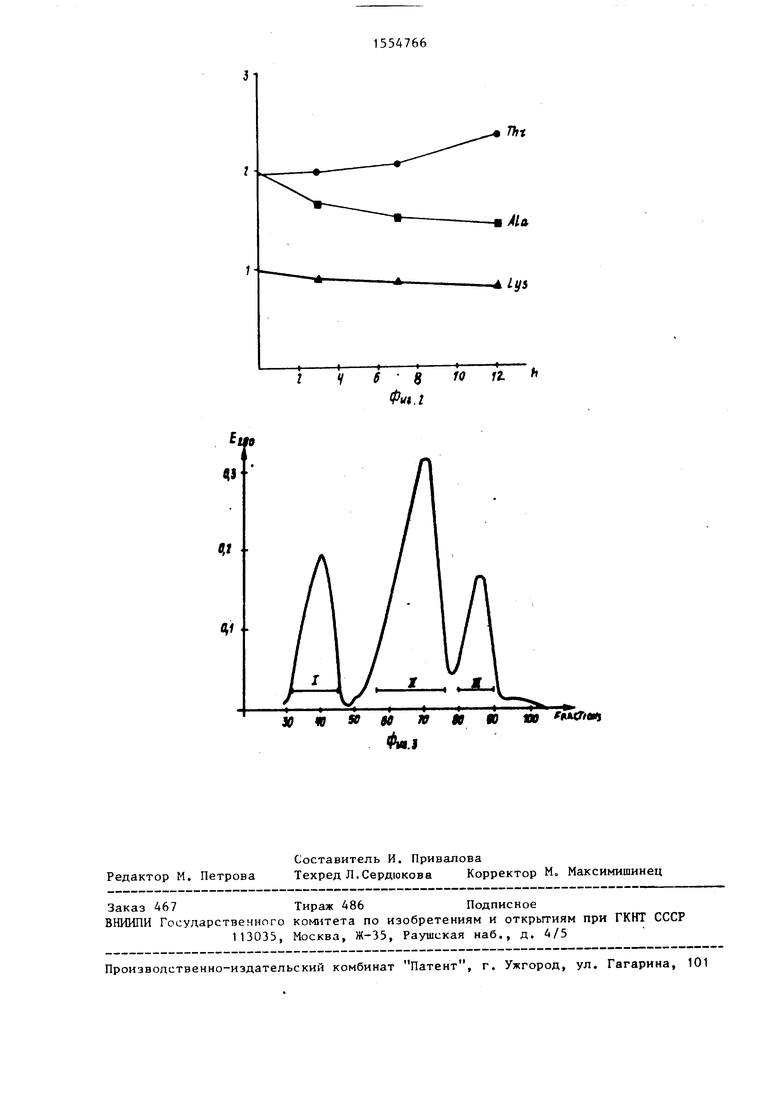
аминокислот приведены в зависимости от времени проведения реакции.) ; на фиг.2 - протекание аналогичной реакции, в которой в качестве аминного компонента используется метиловый эфир L-треонина} на фиг.З - профиль элюиронания для ионообменной хроматографии продукта реакции между свиным инсулином и треонинамидом.
Пример 1. (исследования проводили согласно имеющемуся уровню техники)„
Свиной инсулин инокулируют с карбоксипептидазон Y при 25 С и рН 5-7 (в диапазоне рН, в котором указанный энзим проявляет максимальную пептидазную активность). В результате обработки из С-конца В-цепи освобоадаются следующие аминокислоты: аланин 1,0 лизин 1,0; пролин 1,0; треонин 1,0 тирозин 1,0 и фенилаланин 2,0. В положении В-23 (глицин) карбоксипептидаза Y прекращает свое воздействие, следовательно, можно освободить первые 7 аминокислот в В-цепи инсулина. Неожиданным является тот факт, что С-конечный аспарагин (A-2U цепи А вообще не освобождается. При рН 9,5 С-конечный аланин в цепи В освобождается значительно быстрее последующих аминокислот. Важным является тот факт, что связь лейцин - тира- зин (В-15-16) не подвергалась гидролизу вообще, когда использовалась очищенная карбоксипептидаза Y, не содержащая протеазы А (эндопептида- зы), в противоположность препаратам поставляемым промышленностью из других источников.
Пример 2. Превращение свиного инсулина в инсулин человека с использованием треонинамида в качесве аминного компонента без выделения промежуточного инсулинамида.
К 2 мМ раствору свиного инсулина не содержащему цинк, в 10 мМ ЭДТА добавляют раствор 0,1 М КС1, содержащий 0,5 М L-треонинамида, а при рН 9,5 и температуре 25 С вводят 50 мкМ карбоксипептидазы Y. рН реакционной смеси поддерживают на постоянном уровне в результате введения 0,5 М раствора гидрата окиси натрия, используя для этой цели рН-стат. От реакционной смеси в разное время отбирают аликвоты, а
10
15
20
25
0
5
0
5
0
5
в конце реакцию останавливают в результате введения 6 М раствора соляной кислоты, в результате чего рН доходит до 1-2. Образцы подвергают хроматографированию на колонне 1чЗО см, наполненной Сефадексом Г-50 с использованием в качестве растворителя 1 М уксусной кислоты для отделения инсулина от энзима и свободных аминокислот. Фракции, содержащие инсулин, лиофилизируют и определяют состав аминокислот согласно описанному способу.
Протекание реакции показано на фиг.1. Из анализа аминокислот рассчитано, что содержание треонина увеличивается до 0,7 моль на 1 моль инсупина, в то время как 0,8 моль аланина и 0,2 моль лизина исчезают (фиг.1). Эти результаты подтверждают тот факт, что спустя 6,5 ч проведения реакции 20% свиного инсулина остается в непрореагировавшем состоянии, в то время как 20% инсулина наряду с аланином теряет также аминокислоту лизин Потери 0,8 моль аланина сопровождаются введением 0,7 моль треонина.
Продукты реакции подвергают дальнейшему анализу при обработке кар- боксипептидазой А, которая освобождает только аминокислоты, содержащие свободные L-карбоксильные группы, и не освобождает лизин. В результате обработки карбоксипепти- даэой А свиного инсулина освобождаются только аланин и аспарагин из С-концов цепей В и А соответственно. В результате инкубирования образцов инсулина, полученных после проведения реакции в течение 6,5 ч (фиг.U, следует ожидать освобождения наряду с аспарагином еще только ана- лина в количестве, соответствующем доле непрореагировавшегося свиного инсулина. Однако неожиданно помимо аланина освобождается также треонин в количествах, эквивалентных количеству треонинамида, вошедшего с образованием промежуточного инсулинамида человека. Это должно быть обусловлено тем, что в дополнение к пептидаэной и экстераэной активности карбоксипептидаза Y обладает также гидролаз- ной активностью по отношению к пепти- дамидам. Очевидно в процессе реакции свиного инсулина в присутствии карбоксипептидазы Y С-конечный аланин первоначально обменивается с треонинамидом в реакции транспепти- дирования с образованием инсулинамид человека, который затем гидролизует- ся карбоксипептидазой Y с получением инсулина человека с общим выходом около 60%,а 20% непрореагировавшего свиного инсулина и 20% других продуктов составляют остальное. Другие опыты подтверждают указанную последовательность реакций. Используя меньшие количества энзима или более короткое время реакции, после обработки карбоксипептидазой А продуктов реакции можно наблюдать, что количество освободившегося треонина меньше количества вошедшего треонина, т.е. в основном образуется инсулинамид человека.
Для отделения продуктов реакции образец, полученный после проведения реакции в течение 6,5 ч (фиг.1), подвергают жидкостной хроматографии с использованием колонки с обратной фазой Ликросорб РР-18 (5 мкм, 0, см), насоса Уотерс (мод. 6000А) и детектора (Мод. 750-УФ), функционирующего при 220 нм. В качестве элюента используют смесь, содержащую 28,75% CHNC и 5 мМ тартрат- ного буфера (рН 3), содержащего 5 мМ н-бут S03Na и 50 мМ сульфата натрия. Скорость потока 1,0 мл/мин. С использованием этой системы нельзя разделить инсулин человека и свиной инсулин, однако все остальные побочные продукты удаляются. Результат анализа аминокислот материала, полученного после хроматографирования, приведены в табл.1. Результаты анализа хорошо согласуются с теоретическими предпосылками. Наличие 2,6 моль треонина и 1,3 моль аланина предполагает наличие в образце примерно 70% инсулина человека и 30% свиного инсулина.
Пример 3. Превращение свиного инсулина в инсулин человека с использованием в качестве аминного компонента метилового эфира треонина без выделения промежуточных продуктов.
К раствору не содержащего цинк свиного инсулина (8 мМ) в 10 мл ЭДТА 0,1 М КС1, содержащему 0,5 М метилового эфира L-треонина, при рН 9,5 и при 25°С, добавляют 60 мкМ карбок- сипептидазы Y. Значения рН в ходе реакции поддерживают в результате
0
5
0
5
0
5
0
5
0
5
введения 0,5 М раствора гидрата окиси натрия с использованием рН-стата. В различное время отбирают аликвоты реакционной смеси, а саму реакцию останавливают в результате введения 6 М раствора соляной кислоты, в результате чего рН становится равным 1-2. Затем образец хроматографируют на колонке см, заполненной Сефадексом Г-50, с использованием в качестве растворителя 1 М раствор уксусной кислоты для отделения инсулина от инзима и свободных аминокислот. Фракции, содержащие инсулин, лиофилизируют и определяют состав аминокислот согласно описанному способу.
Ход реакции показан на фиг.2„ Результаты, полученные при проведении этой реакции, аналогичны описанным при использовании треонинамида: треонин вводится одновременно с освобождением аланина и небольшого количества лизина. В результате обработки карбоксипептидаэой А образца, полученного после проведения реакции в течение 17 ч, освобождаются как ала- нин, так и треонин, что приводит к предположению, что С-конечный аланин цепи В первоначально обменивается с метиловым эфиром треонина с образованием монометилового эфира инсулина человека, который затем гидролизуется карбоксипептидазой У с получением инсулина человека. Хотя количественные соотношения в реакции те же, что и в реакции с участием треонинамида, конечная реакционная смесь содержит только 40% инсулина человека, 40% свиного инсулина, не подвергшегося превращению, и 20% других продуктов гидролиза.
Пример 4. Превращение свиного инсулина в инсулин человека с использованием в качестве аминного компонента треонинамина и с изоляцией промежуточных инсулинамидов.
К раствору не содержащего цинк свиного инсулина (2 мМ) в 2 мМ ЭДТА, 0,1 М КС1 и 1,5 М хлоргидрата гуани- дина, содержащего 0,5 М треонинамида (Ь-тре-МНг) при рН 7,5 и температуре 25 С добавляют карбоксипептидаэу У (15 мкМ). Значение рН в ходе реакции поддерживают в результате добавления 0,5 М гидрата окиси натрия с использованием рН-стата. В ходе реакции отбирают аликвоты реакционной смеси в
чества ннсулиновой пробы. В процессе реакции превращается примерно 75% исходного свиного инсулина. Однако хотя содержание треонина в пике I (3,65) больше 3,0 (соответствует инсулину человека), очевидно, что пер- воначальньй продукт транспептиднрова- ння (-I1PO-J H3-TPK-NH2) не является достаточно стабильным для того, чтобы не происходила олигомеризация с образованием -ПРО-ЛИЗ-ТРЕ-ЫН4.
На первый взгляд образование смеси промежуточных амидов говорит о го состава лиофнлизированного инсулн--| ; том, что в результате описанной старлзное время, а саму реакцию оста- навминают в результате гашения спустя 2 ч доведением pll до 1,0-2,0 добавлением 1 М расгвора соляной кислоты. Фракцию, содержащую инсулин, отделяют от энзима и низкомслекулчрных компонентов в результате хроматографнро- вания на колонке 1 t30 см, заполненной Сефадексом Г-50 мепким с использованием в качестве растворителя 1 М раствора уксусной кислоты, и лнофилн- знруют ее. Проведенный согласно описанному способу анализ аминокислотно10
на показывает, что 78/ свиного инсулина расходуется в ходе реакции.
Для дальнейшего анализа реагентов, содержащихся в инсулнновон пробе проводят хроматографнрованис на ионообменном Сефадексе-К -ДЕЛИ Л-25 согласно способу, описанному Морихара и др. Образец лиофилизированного инсулина (75 мг) растворяют в 0,01 М ТРИС, 2,5 М мочевине, 0,05 М NaCl при рН 7,5 и хроматографируют на колонке, заполненной Сефадексом
R-ДЕЛЕ Л-25 (2,5x25 см) с использованием в качестве растворителя того же самого буферного раствора. Инсулин злюируют растворами хлористого натрия с градиентом концентраций от 0,05 до 0,30 М в том же буфере, собирают фракции по 8 мл на колонке, заполненной Сефадексом Г-25 и лиофили- зируют их.
Профиль элюирования изображен на фиг.З, на котором наблюдаются три пика. Кроме того, определяются составы аминокислот и состав веществ пиков в результате обработки карбокси- пептидазами Y и Л согласно описанному способу. Поскольку в указанных реакциях участвует только С-конец В-цепи инсулина, в качестве модели свиного инсулина используется последовательность -ПРО-ЛИЗ-Л. 1А-ОН. Аналогичным образом сокращения для других производных инсулина будут представлять собой: -ПРО-ЛИЗ-ТРК-ОН - для инсулина человека, -ПРО-ЛИЗ-ТРЕ- -МНг - для инсулинамида человека и т.д. При рН 7,5, используемом в ходе реакции, при котором амидазная активность карбоксипептидазы У обычно ниже ее пептидазной активности, образующейся пептидамид в значительной мере стабилен, поскольку пик I соответствует примерно 20% всего коли20
25
30
35
40
45
50
55
дни дезамидирования посредством карбоксипептидазы Y не образуется чистый инсчпин человека, поскольку в результате дезлмидиоованил очевидно следует ожидать получение смеси -НРО-ЛИЗ-ТРК-ОН и -ПРО-ЛИХ-ТРЕ-ТРЕ-ОН Однако при обработке вещества пика I с целью дезамидирования 10 мкМ карбоксипептидлзы У при рН 10,0 в 0,1 М НС1, 2 мМ ЭДТА в течение 20 мин получают неожиданно практически чистьп инсулин человека. Этот опыт протекал следующим образом.
После отделения прореагировавшего инсулина от знзима и низкомолекулярных продуктов хроматографированием на Сефадексе Г-50 полученный продукт хроматографируют на ДЕАЕ-Сефадексе А-25 с использованием способа, аналогичного описанному на фиг.З. Продукт реакции элюируется в пике II (что ожидалось), тогда как непроре- агировавшнй продукт (менее 10%) элюируется в пике I. Пик III вообще отсутствует, т.е. производные инсулина, не содержание лиэин;не образуются в ходе реакции. Как следует из результатов, приведенных в табл.2, полученных при обработке карбоксипептида- зами А и У и трипсином, указанная реакция влияет только на содержание треонина в сколько-нибудь значительной степени, что находится в соответствии с ожидаемым отсутствием пепти- даэной активности карбоксипептидазы У при данном значении рН.
Состав аминокислот для пика II, полученного при реакции дезамидирования, близок к составу инсулина человека: ТРЕ 2,87; АЛА 1,05; ЛИЗ 0,98. Обработка продукта деэамидирования карбоксипептидазами Y и А трипсином показывает, что образец содержит 90-95% чистого инсулина человека.Это
10
20
5
0
5
0
5
0
5
дни дезамидирования посредством карбоксипептидазы Y не образуется чистый инсчпин человека, поскольку в результате дезлмидиоованил очевидно следует ожидать получение смеси -НРО-ЛИЗ-ТРК-ОН и -ПРО-ЛИХ-ТРЕ-ТРЕ-ОН Однако при обработке вещества пика I с целью дезамидирования 10 мкМ карбоксипептидлзы У при рН 10,0 в 0,1 М НС1, 2 мМ ЭДТА в течение 20 мин получают неожиданно практически чистьп инсулин человека. Этот опыт протекал следующим образом.
После отделения прореагировавшего инсулина от знзима и низкомолекулярных продуктов хроматографированием на Сефадексе Г-50 полученный продукт хроматографируют на ДЕАЕ-Сефадексе А-25 с использованием способа, аналогичного описанному на фиг.З. Продукт реакции элюируется в пике II (что ожидалось), тогда как непроре- агировавшнй продукт (менее 10%) элюируется в пике I. Пик III вообще отсутствует, т.е. производные инсулина, не содержание лиэин;не образуются в ходе реакции. Как следует из результатов, приведенных в табл.2, полученных при обработке карбоксипептида- зами А и У и трипсином, указанная реакция влияет только на содержание треонина в сколько-нибудь значительной степени, что находится в соответствии с ожидаемым отсутствием пепти- даэной активности карбоксипептидазы У при данном значении рН.
Состав аминокислот для пика II, полученного при реакции дезамидирования, близок к составу инсулина человека: ТРЕ 2,87; АЛА 1,05; ЛИЗ 0,98. Обработка продукта деэамидирования карбоксипептидазами Y и А трипсином показывает, что образец содержит 90-95% чистого инсулина человека.Это
утверждение сделано в предположении, что производное инсулина -ПРО-ЛИЗ-ТРЕ -ТРЕ-ЫНг реагирует практически только при наличии гидролазной активности и пептидиламида аминокислоты, а -ПРО-ЛИЗ-ТРК-Ш2 реагирует главным образом при наличии амидазной активности. Общий выход для превращения свиного инсулина в инсулин человека составляет примерно 30% из расчета на превращенный инсулин.
Пример 5. К раствору свиного Zn-инсулина (7 мг/мл) в О,1 М растворе треонинамида при рН 6,9 добавля ют карбоксипептидазу Р из Penicillum jauthenellum (СРД-Р) до тех пор, пока концентрация не станет равной 0,42 мг/мл. После этого реакция идет в течение 2,5 ч при 25 С. 50% евино- го инсулина превращается в человеческий инсулин, что определяется при помощи тонкослойной хроматографии.
Примеры 6-14. Получение человеческого инсулина с помощью модифи- цированной СРД-V. Методы.
Аминокислотный анализ. Образцы для аминокислотного анализа гидролизуют в 5,7 М НС1 при 110°С в вакууме в течение 24 ч и анализируют на аминокислотном анализаторе Дуррум-Д-500.
Анализ высокоэффективной жидкостной хроматографией.
Превращение свиного инсулина в амид человеческого инсулина может быть прослежено с помощью жидкостной хроматографии при высоком давлении (HPLC) на обратимой фазе.
Используют радиально заполненную С8 колонку от Вотерс Асе., элюирова- ние достигается с помощью 0,1 М раствора сульфата аммония, содержащего 30% ацетонитрила. Образцы (30 мкл) из реакционной смеси разбавляют 1 М уксусной кислоты перед инжектированием в Дюррум Д-500.
Диск-ПЭГ электрофорез. Превращение свиного инсулина в амид человеческого инсулина также может быть прослежено с помощью Диск-ПЭГ электрофореза на 12,5%-ном полиакриламидном геле в буфере, состоящем из 0,005 М Трис - 0,04 М глицина при рН 8,3. Амид человеческого инсулина мигрирует медленнее свиного инсулина, соответствуя потере одного отрицательного заряда.
Q
5
,-
0
5
0
0
5
С-Конценой анализ.
СРД-У гидролиз используют для опрен1 лочия С-концевой последовательности И--Ц1Т1И инсулина. 500 мкг инсулина в 0,05 М ES буфере при рН 6,75 переваривают 30 мкг СРД-У при 37°С в течение 1 ч. Высвободившиеся аминокислоты определяют па аминокислотном анализаторе.
Препаративная хроматография на ДЕАЕ-Сефарозе.
Амид человеческого инсулина отделяют от непрореагировавшего свиного инсулина ионообменной хроматографией на ДКАК-Сефароэе. Лиофилизированный образец инсулина растворяют в 0,01 М Трис - 0,05 М NaCl - 2,5 М мочевине , при рН 7,5 и наносят на колонку, уравновешенную тем же самым буфером. Инсулины элюируют с градиентом NaCl от 0,05 до 0,30 М в том же самом буфере.
Получение модифицированной карбок- сипептидазы У (СРД-УМ).
К 1 мл водного раствора СРД-У (9,9 мг/мл, 150 мкмоль) прибавляют 100 мкл 0,5 М НЕРК S (рН 7,5) и последовательно 25 мкл водного раствора М HgCl.. Смесь оставляют при комнатной температуре на 10 мин, после чего она готова для использования. Обычно используют стехиометри- ческие количества фермента и ртути, но избыток ртути может быть легко удален гель-фильтрацией или диализом. Может быть приготовлена СРД-У, модифицированная метил-Нр, или этил-Hg, по аналогичной методике с использованием метил-HgCl или этил-Hgd соответственно .
Тест на возможное высвобождение ртути из СРД-УМ.
Поскольку специфическая экстераз- пая активность нативной СРД-У по отношению к субстрату В L - Lys - OMe практически равна нулю, тогда как СРД-УМ имеет очень высокую активность, это различие может быть использовано для определения возможного высвобождения ртути, которое будет приводить к снижению экстеразной активности. Однако ни добавление 10 мМ ЕДТА, ни гель-Фильтрация не приводят даже к малейшему снижению активности, показывая, что нет высвобождения ртути.
Результаты.
Пример Ь. Растворяют 80 мг свободного от цинка инсулина в 6,5 мл
1,1 M Thr - NHa, 0,05 M HEPE S, 1,0 KC1 при рН 7,0 и прибавляют 12,7 мг СРД-УМ в 2,5 мл 0,05 M HEPES при рН 7,0. Реакцию ведут при 20°С, затем проводят Н L С и гель-электрофорез. HLC показывает превращение 78% после 21 ч реакции. Гель-электрофорез показывает подобную глубину конверсии.
После 21 ч реакции устанавливают рН 2 с помощью 1 M HC1 и обессоливают образец на Сефадексе R -G50 в 1 М уксусной кислоте, а затем лио- филизируют. Хроматография на ДЕАЕ-Се- фарозе показывает профиль элюции с ,двумя пиками. Гель-элек -рофорез показывает, что два пика являются очевидно чистыми. Пик I содержит 41,5 мг амида человеческого инсулина. Амино- кислотный анализ пика I (табл.3) показывает, что аланин обменивается на треонин. Пик II содержит 40% непрореагировавшего свиного инсулина, который может быть рециклизован.
Пример 7. Растворяют 70 мг кристаллизованного цитрата сырого инсулина в 2 мл 1,5 M Thr - NH2, 0,05 M HEPE S, 1,0 M HC1 при pll 7,0. Реакцию проводят при 20°С в присут- ствии 15 мкМ СРД-УМ. После 72 ч реакции HPLC показывает конверсию 77%. Гель-электрофорез показывает подобную глубину конверсии. Реакционную смесь анализируют, как описано в примере А. Хроматография на ДЕАЕ-Сефарозе показывает профиль элюции с двумя пиками в соотношении 2:1 для амида человеческого инсулина и свиного инсулина
соответственно. Аминокислотный анализ
пика I показан в табл.30 Пик II содержит 40% непрореагировавшего свиного инсулина.
Пример 8. Растворяют 100 мг свиного Zn-инсулина в 6,7 мл 1,0 M Thr - NH4, 1M мочевины при рН 7,5 (рН устанавливают НВг). Прибавляют 2,3 г СРД-УМ. Реакцию проводят при 32°С, затем проводят HPLC. После 2ч 10 ми 75% свиного инсулина превращается
в амид человеческого инсулина.
Пример 9. Растворяют 100 мг свиного Zn-инсулина в 6,7 t-л 1,0 М Thr-NHj, 1 М мочевины, рН 7,75 (рН устанавливают НВг). Добавляют 2,7 мг СРД-УМ. Реакцию проводят при 32°С, затем проводят HPLC. После 2 ч 10 мин 73% свиного инсулина поеQ
5 0 5
0
5
0
0
вращается в амид человеческого инсулина.
Пример 10. Растворяют 100 мг свиного Zn-инсулина в 6,7 мл 1,0 М Thr-NH, 1 М мочевины, рН 6,0 (рН устанавливают НВг). Прибавляют 3,6 мг СРД-УМ. Проводят реакцию при 32 С, затем проводят HPZC. После 1 ч 40 мин 75% свиного инсулина превращается в амид человеческого инсулина.
Пример 11. Растворяют 100 мг свиного Zn-инсулина в 6,7 мл 1,0 Н Thr-NHt, 1 М мочевины, рН 8,25 (рН устанавливают НВг). Прибавляют 3,6 мг СРД-УМ. Реакцию проводят при 32°С, а затем проводят HPLC. После 1 ч 20 мин 68% свиного инсулина.превращается в амид человеческого инсулина.
Пример 12. Растворяют 100 мл свиного Zn-инсулина в 6,7 мл 1,0 М Thr-NH2, 10 МКУ, 1 М мочевины, рН 8,0 -(рН устанавливают HNOj). Прибавляют 3,6 мг СРД-УМ. Реакцию проводят при 32 С, затем проводят HPLC. После 2 ч 5 мин 73% свиного инсулина превращается в амид человеческого инсулина.
Пример 13, Повторяют процедуру примера С, но используют 3 мг СРД-УМ, иммобилизованного на силнка- геле. После 2 ч получают 75% конверсии.
HPLC, Диск-ПЭГ и аминокислотный анализ продуктов, полученных в примерах, показывает, что они действительно являются амидом человеческого инсулина.
Последующее деамидирование человеческого инсулина может быть проведено по примеру 4.
Пример 14„ Деамидирование амида человеческого инсулина на немодифицированной СРД-У и модифицированной СРД-У.
А. Немодифицированная СРД-У.
К раствору 15 мг/мл человеческого инсулина в 1 мМ ЕДТА при рН 8,0 и температуре 25°С прибавляют СРД-У до конечной концентрации 7,2 мкМ. После 22 мин реакции 74% амида превращается, как определено HPLC. После разделения на ДЕАЕ-Сефарозе HPLC анализ и аминокислотный анализ показывают, что человеческий инсулин является чистым.
19
B.СРД-У, модифицированная ме- тил-Hg.
Аналогично примеру А, но с 4,9 мкМ метил-Не-СРД-У, рН 8,0. Пос- ле 39 мин реакции 61% амида инсулина превращается в человеческий инсулин.
C.СРД-У, модифицированная этил-Hg.
Аналогично примеру А, но с 7,2 мкМ этил-Н8-СРД-У, рН 8,0. После 36 мин реакции 74% конверсии наблюдается по данным HPLC.
Д. СРД-У, модифицированная ме- тил-Hg.
Аналогично примеру А, но с 19 мкМ метил-Нв-СРД-У, рН 7,5. После 2 ч реакции по данным HPLC наблюдается 90% конверсии.
Формула изобретения
1. Способ получения человеческого инсулина формулы Ins - Thr - ОН или его производных, имеющих формулы Ins - Thr - NHj,Ins - Thr - NH2 или Ins - Thr - OR3 ,
где Ins - DeS - Thr (.30) человеческого инсулина R 3 - С,-С6-алкил,
путем ферментативного замещения В-30 аминокислоты в свином инсулине формулы
Ins - Ala - ОН
с помощью карбоксипептидазы при оптимальной температуре процесса с последующим выделением целевого про- дукта, отличающийся тем, что, с целью повышения выхода целевого продукта за счет избежания предварительного отщепления В-30 Ala, свиной инсулин вводят в реак- цию с водным раствором или дисперсией, содержащей амид треонина или сложный эфир треонина общей формулы
1554766
20 Н - Thr - OR:
5
0
5
0
5
„ -
где RJ имеет указанные значения, при рН 7-9,5, причем исходные концентрации свиного инсулина и амида треонина или его сложного эфира устанавливают от 0,002 до 0,5 М И от 0,05 до 3 М соответственно, в качестве карбоксипептидазы используют сериновую карбоксипептидазу микробного происхождения, специфическую для субстратов, имеющих С-концевую L-аминокислоту с образованием при -этом одного из производных инсулина формул Ins-Thr-NH2, Ins - Thr - Thr - Ins - Thr - OR3, n которой для получения человеческого инсулина в качестве целевого продукта отщепляют одну из групп -NH2, -Thr-МЦ и -OR з путем взаимодействия той же самой или иной L-специ- фичнон серинопой карбоксипептидазы в водном растворе или дисперсии при рН 9 - 10,5.
2.Способ по п.1, отлича ю- щ и и с я тем, что используют не- модифицировэнную или модифицированную карбоксипептидаэу Y из пекарских дрожжей или карбоксипептидазу Р ич Penicilium jouthenellum.
3.Способ по п.1, отлича ю- щ и и с я тем, что используют карбоксипептидазу Y, модифицированную солями ртути или иммобилизованную.
4.Способ по п.1, отлича ю- щ и и с я тем, что до 50% органического растворителя используют этанол или метанол, или этнленгликоль, или пропиленгликоль, или диметил- формамид, или диметилсульфоксид, или тетрагидрофуран, или дноксан, или диметоксиэтан.
5.Способ по п.1, отлича ю- щ и и с я тем, что используют водный раствор или дисперсию, содержащую мочевину, или гидрохлорид гуани- дина в концентрации до 3 М.
21
155476622
Таблица 1 Аминокислотный состав инсулина









| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения инсулина человека | 1984 |
|
SU1611217A3 |
| Способ ферментативного получения пептидов | 1980 |
|
SU1378785A3 |
| СПОСОБ ПОЛУЧЕНИЯ ИНСУЛИНОВЫХ СОЕДИНЕНИЙ | 2002 |
|
RU2376379C2 |
| ХИМЕРНЫЙ БЕЛОК, ОБЕСПЕЧИВАЮЩИЙ ОБРАЗОВАНИЕ БИОАКТИВНОЙ КОНФОРМАЦИИ, СПОСОБ ПОЛУЧЕНИЯ БЕЛКА С ИНСУЛИНОВОЙ АКТИВНОСТЬЮ, СПОСОБ ИДЕНТИФИКАЦИИ ПЕПТИДИЛЬНОГО ФРАГМЕНТА | 1998 |
|
RU2238951C2 |
| Способ получения полипептидов | 1973 |
|
SU559653A3 |
| ОЧИСТКА ИНСУЛИНА | 2012 |
|
RU2603752C2 |
| ИНСУЛИН И ЕГО ПРОИЗВОДНЫЕ С ПОВЫШЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬ ЦИНК | 1997 |
|
RU2176646C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНСУЛИНА ИЛИ ПРОИЗВОДНЫХ ИНСУЛИНА С ПРАВИЛЬНО СОЕДИНЕННЫМИ ЦИСТИНОВЫМИ МОСТИКАМИ | 2002 |
|
RU2302882C2 |
| АНАЛОГИ ИНСУЛИНА, УСТОЙЧИВЫЕ К ПРОТЕАЗАМ | 2007 |
|
RU2524150C2 |
| Фармацевтическая композиция, содержащая инсулин и глюкагон | 2019 |
|
RU2823246C2 |
Изобретение относится к биохимии. Цель изобретения - повышение выхода целевого продукта за счет избежания предварительного отщепления аминокислоты В-30 в свином инсулине. Способ предусматривает введение свиного инсулина в реакцию ферментативного замещения с водным раствором или дисперсией, содержащей амид треопина или сложный эфир треопина формулы H-TR-OR3, где R3 является C1-C6-алкилом, при PH 7,0 - 9,5 при исходных концентрациях свиного инсулина и амида треопина или от 0,05 до 3 М соответственно. В качестве фермента используют сериновую карбоксипептидазу микробного происхождения специфическую для субстратов, имеющих C-концевую Z-аминокислоту. В результате образуется одно из производных инсулина. Для получения человеческого инсулина отщепляют одну из групп -NH2, - THR-NH2 и OR3 путем взаимодествия той же самой или иной 4 - специфической карбоксипептидазы в водном растворе или дисперсии при PH от 9 до 10,5. Выход инсулина повышается до 75%. 3 табл. 3 ил.
Низкие значения ввиду неполного гидролиза.
Таблица 2
Аминокислотный состав пиков I, II, III на фиг.З и пика II, полученного после дезамидирования фракции соответствующей
пику I
Аспарагиновая
кислота2,992,993,01
Треонин3,622,522,50
Серии2,932,922,92 Глютаминовая
кислота6,887,167,17
Пролин1,000,980,71
Глицин4,004,004,00
Алании1,161,521,03
Изолейцин0,941,041,09
Валин2,422,822,81
Лейцин5,806,216,18
Тирозин3,803,863,67
Фенилаланин2,742,822,65
Гистидин 1,961,901,93
Лизин0,990,741,06
Аргинин0,961,000,98
3,01 2,79 2,93
7,01 0,87 4,00 1,09 1,19 2,50 6,10 4,02 2,64 1,89 1,01 0,89
23155476624
Таблица 3 Аминокислотный анализ инсулина
Пг
1 ч 68 ю it Фиг. г
ячоявсюилт тн 0w.j
| Видмер Ф., Йохансен И.Т0 Энзи- матический пептидный синтез: катализируемое карбоксипептидазой-У образование пептидных связей о - Carls- berg Res Commun | |||
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |
| Пишущая машина | 1922 |
|
SU37A1 |
| Способ обработки медных солей нафтеновых кислот | 1923 |
|
SU30A1 |
| - Nature, т | |||
| СПОСОБ ПОЛУЧЕНИЯ ЧИСТОГО ГЛИНОЗЕМА И ЕГО СОЛЕЙ ИЗ СИЛИКАТОВ ГЛИНОЗЕМА, ПРОСТЫХ ГЛИН И. Т.П. | 1915 |
|
SU280A1 |
| Способ применения поваренной соли в нагревательной закалочной ванне при высоких температурах | 1923 |
|
SU412A1 |
| Морихара В0 и др„ Превращение свиного инсулина в инсулин человека, катализируемое Ахром-бактерпротеа- эой-1 | |||
| - Biochem | |||
| and Bioph, Res | |||
| Comm., т„ 92, 1980, № 2, c | |||
| Приспособление для подъема падающих гребней в машинах льнопрядильного, джутового и т.п. производств | 1913 |
|
SU396A1 |
