Изобретение относится к биотехнологии и касается вьщеления оксидазы D-аминокислот, проявляющих активность против цефалоспорина С.
Цель изобретения - повышение чистоты целевого продукта.
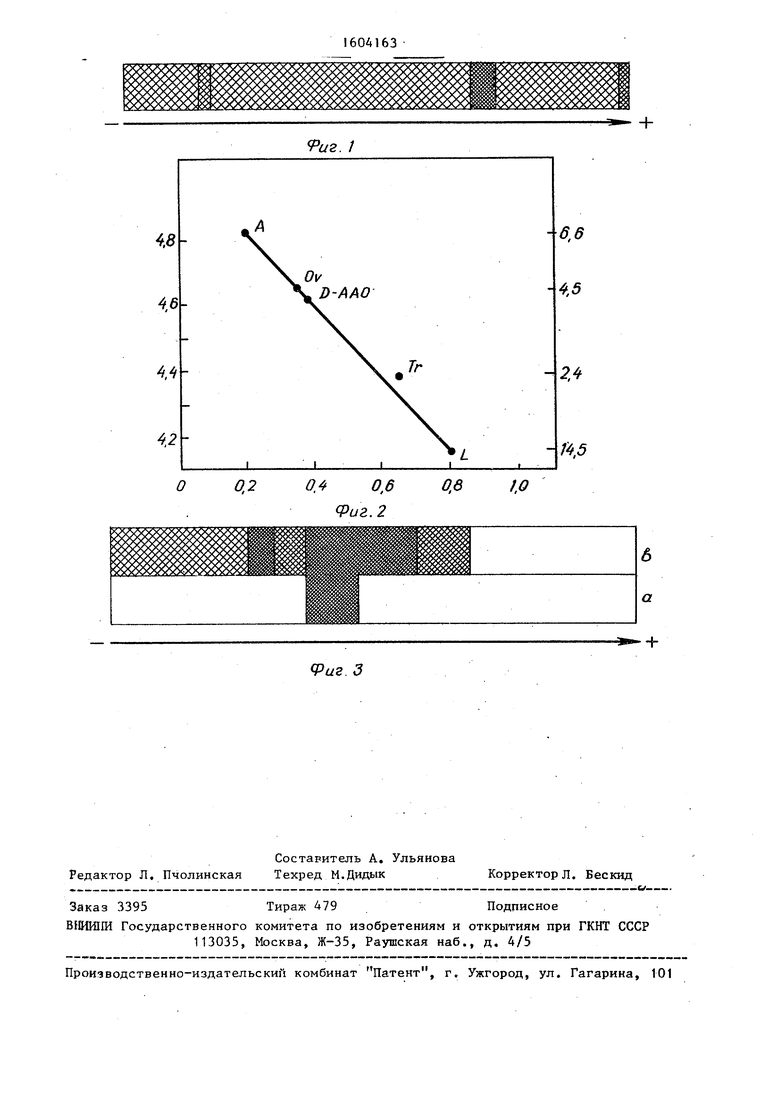
На фиг.1 приведены результаты электрофореза очищенной оксидазы D-аминокислот в полиакриламидном геле. Электрофорез в г чистого белка в додецилсульфате натрия показывает только одну полосу белка (темная меньшая полоса, видимая слева указывает границу двух оконденсиро- ванных гелей). ДoбaвляeмyкJ к гелям
метку отрезают перед подкрашиванием белка.
На фиг.2 приведены результаты определения молекулярной массы оксидазы D-аминокислот из Trigonopsis variabilis. Электрофорез в геле с додецилсульфатом натрия (ДСН) проводят в 12%-ном пOJIиaкpилaмидe согласно Laeimnli(Nature, 1970, 227, 680- 685). Использованы следующие белковые метки: А - альбумин (66000); OV - овальбумин (45000); Тг - трипси- ноген (24000); L- лизозим (14500).
На фиг.З показана активность окрашивания гелей по отношению к ок- ,
О
О 4
О5
оо
СМ
сидазе D-аминокислот, Показанные фракции взяты со стадии 111. Гели инкубируют: а) с цефалоспорином С; б) с D-лейцином.
Материалы.
Пероксидаза (тип N, из хрена обыкновенного), все аминокислоты, цефалоспорин С, динитрофенилгидра- зин и о-дианизин поставлены фирмой Sigma (Сан-Луи, США), акриламид и N,N - метиленбисакриламид - фирмой Merk-Schuchardt (Мюнхен, ФРГ), N,N, N ,N -тетраметилендиамин, перосуль- фатаммония и пластинки, покрытые 0,25 мл силикагеля Фирмой Merk (ФРГ), питательные среды - фирмой Difco (Дейтройт, США); кислород- ньш электрод - фирмой Rank Broth ers Bottisham (Кембридж, Англия).
Методики,
Аналитическое изоэлектрическое фокусирование проводят, используя системы 1KB 1804-101, Используемые несущие амфолиты имеют рН в интервале 3,5-9,5. Изоэлектрическое фокусирование проводят в соответствии с опытами фирмы 1KB на амфолиновых пластинках с полиакриламидным гелем (ПА1) (1KB - Produkter АВ, Бромма, Швеция, 1979). Железо определено с помощью атомной абсорбционной спектроскопии.
Состояние культур.
В исследованиях использованы дрожжи Trigonopsis variabilis.
Загружают большой объем культур в ферментатор емкостью 8 л. Питательная среда состоит из дрожжевого экстракта (1%), солодового экстракта (1,5%), в которые добавлено 0,2% D, L-метионина. Среду инкубируют 44 ч при 28°С, Исходную культуру помещают в косую среду, изготовленную из той же среды с добавкой 2% агара. Клетки отделяют центрифугированием при 4000g в течение 30 мин, промывают и хранят в замороженном состоянии до использования. Средний выход клеток при различных фермектациях состаляет 45-50 г клеточной пасты.
Электрофорез.
С аналитической целью проводят дисковый электрофорез в ПАГ так же, как в гелях с ДСН, при 8-10°С. Первоначально на трубку подают низкий ток в 2 мА до момента миграции красителя в разделительный гель, после чего ток повьпнают до постоянного
5
0
5
0
5
0
5
0
5
значения 4 мА на трубку. Диаметр и длина трубок соответственно 0,8 и 9,5 см. Для локализации белка гели окрашивают красителем Coomassie Brilliant Bluek.
Ферментативную активность на гелях обнаруживают согласно Hedrick и Smith, причем гели окрашивают детектированием образовавшейся перекиси водорода с использованием о-диани- зидина. Гели инкубируют в 0,1 М натри- фосфатном буфере (рН 7,2), содержащем 5 мМ цефалоспорин С (или других аминокислот), пероксидазу (0,025%) и о-дианизидин (0,025%). Инкубирование проводят 30-60 мин ДО образования красновато-коричневого красителя.
Для препаративного вьщеления фермента разработано специальное оборудование. Диаметр стеклянной трубки 2 см, ее длина 14 см. Гель (8% поли- акриламида) получают согласно Hedrick и Smith, используя в качестве буфера 0,19 М трисглицин (рН 8,3). Верхний гель полимеризован в присутствии персульфата аммония и тетраме- тиленэтилендиамина, а вместо рибофлавина ( количество катализатора взято согласно Laemmle. Электрофорез ведут 3-4 ч при 10°С, используя в nep-j вые 30 мин ток в 20 мА, а затем ток в 40 мА.
Определение молекулярной массы.
Молекулярную массу определяют по методике Hedrick и Smith, а электрофорез в геле с ДСН проводят согласно Laemmli. В первом случае используют метки для молекулярной массы при электрофорезе в гель неденатурированного полиакриламида (Sigma), во втором случае - метки молекулярной массы в геле с ДСН, Определение белка, проводят согласно Lowry.
Анализ активности оксидазы D-аминокислот.
Путем определения скорости потребления кислорода с помощью кислородного электрода фирмы Rank проводят испытания оксидазы D-амиНокислот при 50°С в течение 5-10 мин, в течение которых не происходит денатурирование фермента. Конечный объем испытуемой смеси составляет 2 мл, из которых 1,8 мл приходится на пирофосфат- ный буфер (20 , рН 8), 0,1 мл - на цефалоспорин С (100 мМ) и соответствующее количество фермента, необходимое для получения правильной начальной скорости потребления кислорода; I единица (Е) соответствует поглощению 1 ммоль кислорода в 1 мин в данных условиях. Время.от времени прове-j ряют активность, измеряя колометри- - ческим методом количество образовавшейся кетокислоты с использованием 2,4-динитрофенилгидразина, а также о-диаиизидинпероксидазы при 30 С.
Пример. Очистка оксидазы D-аминокислот из Trigon opsivaria- bilis. Все операции проводят при .
Стадия I, Получение сырого экстракта.
32 ч. замороженной клеточной пасты (-20°С) оттаивают и суспендируют в 1,5 ч. буфера, содержащего 20 мМ пирофосфат натрия (рН 8,3). Клеточную суспензию смешивают с равным объемом сухого льда и размельчают в миксере. Оксидаза D-аминокислот извлекается вместе с другими растворимыми белками. Разрушенные клетки центрифугируют 30 мин при 12000g
Стадия II. Тепловое осаждение.
К подкисленному всплывшему на поверхность продукту со стадии I (730 мл) для защиты фермента добавляют 25 мМ Р, L-метионина.Продукт нагревают до 50°С и оставляют при
этой температуре на водяной бане на 10 мино Образовавшийся осадок отделяют центрифугированием 30 мин при 1200g, после чего отбрасывают. Всплывший на поверхность продукт диа- лизуют в течение 1 сут против буфера, содержащего 20 мМ пирофосфат натрия (рН 8,3).
Стадия III. Осаждение сульфатом аммония.
Образец со стадии II концентрируют продуванием теплового воздуха над мешком для диализа (содержащим образец) с целью увеличения количества белка на 1%. Добавлением 2 М уксусной кислоты значение рН образца снижают до 6,3, после чего перемешивают 1,5 ч с твердым сульфатом аммония. Фракцию, подсоленную 30% сульфата аммония, оставляют для образования осадка, после чего центрифугируют 30 мин при 12000g, Осадок отбрасывают, а всплывший на поверхность продукт (после доведения рН до 6)смешивают с твердым сульфатом аммония (55%). Образовавшийся спустя 2 ч осадок центри- фугируют 30 мин при 12000g и диализу55
0
5
0
5
0
5
0
5
0
5
ют против буфера, содержащего 20 мМ пирофосфат натрия (рН 8,3).
Стадия IV. Изоэлектрическое осаждение.
Образец со стадии III (10,3 мл) диализуют 12 ч против буфера, содержащего 25 мМ ацетат натрия (рН 5,1). Образовавшийся осадок удаляют центрифугированием 30 мин при 12000 g. J В осадке обнаруживается следы ферментативной активности. Всплывший на поверхность продукт снова диализуют 12 ч против буфера, содержащего 0,1 М ацетат натрия (рН 4-6). В образовавшемся осадке обнаруживается большая часть активности оксидазы D-аминокислот. Осадок растворяют в буфере, содержащем 20 мМ пирофосфат натрия (рН 8,3), и диализуют сутки против того же буфера. х
Препаративный дисковый электрофорез в геле.
Образец со стадии IV используют в препаративном электрофорезе в геле. Для электрофоретического испытания используют 5 мг белка со сл-адии IV. Полосу, показывающую активность оксидазы аминокислот относительно це- фалоспорина С, отрезают и гомогенизируют 5 мин в гемогенизаторе марки Potter (1000 об/мин) в буфере, содержащем 20 мМ пирофосфат натрия (рН 8,3), затем полученную суспензию центрифугируют 20 мин при lOOOg Гель промывают 3 раза равным коли-- чеством буфера и всплывший на поверхность продукт концентрируют с помощью теплого, воздуха до конечного объема в 3 мл, которые используют для дальнейших исследований.
Степень очистки оксидазы D-амино- . кислот из Trigonopsxsvariabilis, активность, измеренная относительно цефалоспорина С, содержание белка, определенное по методике Lovjry, приведены в таблице.
. Предлагаемый способ упрощает очистку оксидазы D-аминокислот до однородного состояния. Препаративным электрофорезом в геле после изоэлект- рического осаждения удаляют две дополнительные слабые полосы, наблюдаемые при электрофорезе в геле. При этом удельная активность уменьшается с 5,8 Е/мг (см.таблицу) до 3,8Е/мг. Это можно объяснить частичным денатурированием, происходяп;им в ходе экстракции, или потерей еще неидентифицй- рованных софакторов.
Электрофорез в геле полиакрилами- да в додецилсульфате натрия дает единственную полосу (фиг,1) для молекулярной массы примерно 43000 (фиг,2). Молекулярная масса белка составляет примерно 86000,
Таким образом, оксидаза D-амино- кислот в своей активной форме существует в виде димера молекулярной мас-i сы примерно 86000 и с двумя субъеди-л ницами.
Ферментное окрашивание гелей после инкубирования соответственно с D-лейцином и цефалоспорином показывает только одну полосу на образце, полученном препаративным электрофорезом в геле, в то время как менее чистый препарат (стадия III) дает три основные полосы после инкубирования с D-лейцином и только одну полосу с цефалоспорином С (фиг.З), Это указывает на присутствие нескольких оксидаз D-аминокислот в Trigonopsis но только один из них проявляет ак-j тивность против цефалоспорина С.
Выделенная оксидаза D-аминокислот с активностью относительно цефалоспорина С обладает следующими свойствами
Изоэлектрическая точка фермента - примерно 4,6, Атомный абсорбционный анализ доказывает, что фермент содержит 2 моль железа, что соответствует 1 моль на субъединицу,-После диализа против 10 мМ этилендиаминтетрауксус- ной кислоты и 1 мМ о-фенантролина соответственно активность остается неизменной. Обработка арсенитом снижает активность до 46%, а цианидом - до 34% при их концентрации 0,5 мм.
Значения Km для цефалоспорина С, фенил- аланина,аланина,метионина и лейцина составляет соответственно 13, 10, 76,
0,76 и 0,12, Образец, полученный после препаративного электрофореза в геле при концентрации 5 мг/мл, не дает спектра, аналогичного спектрам зависимых от флавина ферментов, поскольку оксидаза аминокислот, извлеченная из почек свиньи, проявляющая активность относительно цефалоспорина С, зависи: ма от FAD (Р, Mazzeo, А, Romeo, I,C,S, Perkin, 1972 I (РЗ), 2532), Добавление различных софакторов флавйна, таких как FMN или FAD, не увеличивает удельную активностьi Попыт
0
S
5
0
5
0
5
j ки выделить из фермента фпавины успе- ха не имели. Так, экстенсивный диа- jЛИЗ против KB в кислотном растворе в щелочной среде результата не дал. Так же как ни трипсинизацией, ни кипячением или экстрагированием 85%- Iным раствором фенола не были получены флавины,
Чистый фермент в 20 мМ пирофосфат- ном буфере при рН 8,3 устойчив в замороженном состоянии. Оттаивание и замораживание не ведет к потере активности. При 8°С активность падает очень медленно, но при комнатной температуре устойчивость относительно низка. Для увеличения тепловой устойчивости фермент иммобилизуют различны ми способами. Добавление 17,5 Е/мл фермента, полученного после стадии III, при 30°С в течение 30 мин к се- фарозе В, активированной CNBr, в 0,1 боратном буфере (рН 8,3) приводит к постоянной активности в 7 Е/мл сефарозы. Формула йзоб-ретения
1,Способ выделения оксидазы D-ами- нокислоты из Frigonopsis, variabilis, предусматривающий получение сырого бесклеточного экстракта, фракционирование сульфатом аммония до концентра- . ции примерно 30%, отделение образовавшегося осадка, последующее фракционирование сульфатом аммония до достижения концентрации примерно 50%, отделение второго образовавшегося осадка, содержащего целевой продукт, перевод его в растворимое состояние суспендированием в буфере, содержащем пирофосфат натрия, отличаю- и.
щ и и с я тем, что с целью, повьштения чистоты целевого продукта, фракционированию сульфатом аммония подвер- - гают супернатантнзто фракцию, полученную путем подкисления бесклеточного экстракта до рН 4-6, нагревания и удаления осадка, а после переведения второго осадка в растворимое состояние оксидазу D-аминокислот собирают изозлектрическим осаждением,
2,Способ поп,I, отличающийся тем, что подкисление ведут уксусной кислотой.
3,Способ ПОП.1, отличающийся тем, что перед нагреванием добавляют D,L-мeтиoнин до конечной концентрации 25 мМ.
4.Способ по п.1, отличающий с я тем, что перед фракционированием сульфатом аммония осуществляют диализ супернатанта против буфе- ра с рН 8,3, содержащего 20 мМ пиро- фосфат натрия.
5.Способ поп.1 отличаю- щ и и с я тем, что перед диализом супернатант концентрируют испарением,
6.Способ non.I отличающийся тем, что проводят дополнительную стадию очистки оксидазы Dаминокислоты, полученной изоэлектри- ческим осаждением, элекрофорезом в геле.
7. Способ поп.1,отличаю- щ и и с я тем, что изоэлекрическое осаждение ведут диализом против буфера с рН 5,1, содержащего 25 мМ ацетат натрия, с последующим удалением осадка и диализом раствора, полученного в результате указанной стадии диализа против буфера с рН 4,6, содержащего 100 мМ ацетат натрия.



| название | год | авторы | номер документа |
|---|---|---|---|
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ | 2009 |
|
RU2412998C1 |
| МУТАНТНЫЕ ОКСИДАЗЫ D-АМИНОКИСЛОТ | 2007 |
|
RU2362806C2 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ (ВАРИАНТЫ) | 2012 |
|
RU2507262C1 |
| РЕКОМБИНАНТНАЯ ДНК, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНЫЙ ГИБРИДНЫЙ БЕЛОК ОКСИДАЗЫ D-АМИНОКИСЛОТ С ХИТИНСВЯЗЫВАЮЩИМ ДОМЕНОМ (DAOcbd) РЕКОМБИНАНТНАЯ ПЛАЗМИДА pVR1, ОБЕСПЕЧИВАЮЩАЯ ЕГО СИНТЕЗ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli С 41(DE3)/pVR1 - ПРОДУЦЕНТ DAOcbd | 2006 |
|
RU2310688C1 |
| СПОСОБ ПОЛУЧЕНИЯ СУБСТАНЦИИ L-ЛИЗИН-АЛЬФА-ОКСИДАЗЫ | 2011 |
|
RU2471866C1 |
| СПОСОБ ПОЛУЧЕНИЯ ХОЛЕСТЕРОЛЭСТЕРАЗЫ, ТРИПСИНА, ДЕЗОКСИРИБОНУКЛЕАЗЫ И РИБОНУКЛЕАЗЫ ИЗ ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ КРУПНОГО РОГАТОГО СКОТА | 2005 |
|
RU2311455C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pETTvDAO2, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ D-АМИНОКИСЛОТ (DAO) ДРОЖЖЕЙ Trigonopsis variabilis В КЛЕТКАХ Escherichia coli И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli C41(DE3)/pETTvDAO2 - ПРОДУЦЕНТ DAO | 2006 |
|
RU2310687C1 |
| Способ очистки оксидазы д-аминокислот | 1975 |
|
SU572205A3 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pNhaDAAO, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ ОБЫЧНЫХ И N-ЗАМЕЩЕННЫХ D-АМИНОКИСЛОТ (NhaDAAO) АРХЕЙ Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO) В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 - ПРОДУЦЕНТ NhaDAAO | 2023 |
|
RU2829362C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pNhyDAAO, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ D-АМИНОКИСЛОТ БАКТЕРИЙ Natronosporangium hydrolyticum ACPA39 (NhyDAAO) В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pNhyDAAO ВКПМ В-14337 - ПРОДУЦЕНТ NhyDAAO | 2022 |
|
RU2803295C1 |
Изобретение относится к биотехнологии и касается выделения оксидазы Д-аминокислот, проявляющих активность против цефалоспорина С. Цель изобретения - повышение чистоты целевого продукта. Фермент выделяют из GRIGONOPSIS VARIABILIS способом, который проводят в 3 стадии, а именно: (а) подкисление и нагревание сырого клеточного экстракта с получением осадка и супернатанта (всплывшей фракции)
(б) обработку его сульфатом аммония в количестве, достаточном для образования второго осадка, содержащего оксидазу Д-аминокислот и (в) повторное суспендирование полученного на стадии (б) осадка и выделение изоэлектрическим осаждением. 6 з.п. ф-лы, 3 ил., 1 табл.
III (NH4)4SO| (30-55%)
IV (рН 4,6)
6,0 4,9 2,8 5,8
33 39
78 40
иг. /
Ov
46
D-AAO
,4
.2
О
0.2
.З
6,6 .5
Тг
2А
145
O.Q
10
| Appl Biochem | |||
| Biotechnol, 1981, 6, 293-308 | |||
| Патент США № 3658678, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Контрольный висячий замок в разъемном футляре | 1922 |
|
SU1972A1 |
