Изобретение относится к биотехнологии, в частности, к генной и белковой инженерии, и может быть использовано для получения рекомбинантного интерферона α-2b человека.
Интерферон α-2b человека (ИФН α-2b) относится к белковому семейству интерферонов α (синонимы: интерферон В-клеток, лейкоцитарный интерферон, лимфобластный интерферон), состоящему из более чем 23 форм с общим консервативным участком. ИФН α-2b секретируется активированными моноцитами, макрофагами, лимфобластами и фибробластами. ИФН α-2b человека представляет собой гликопротеин с молекулярной массой около 20-25 кДа. Пептидная часть ИФН α-2b представлена 165 аминокислотными остатками и содержит 4 цистеиновых аминокислотных остатка и два внутримолекулярных дисульфидных мостика. ИФН α-2b обладает иммуномодулирующей, антипролиферативной, антивирусной и антимикробной активностью.
[Streuli M., Nagata S., Weissmann С. // At least three human type alpha interferons: structure of alpha 2. Science, 1980, v. 209(4463), p 1343-1347].
ИФН α-2b осуществляет свое действие через связывание со специфическим рецептором [Stark G.R., Kerr I.M., Williams B.R., Sirverman R.H., Schreiber R.D. // How cells respond to interferons. Annual Review of Biochemistry, 1998, v.67, p.227-264].
Лекарственные формы ИФН α-2b (синонимы - Интрон (Шеринг-Плау), Виферон (Ферон) и Реальдирон (Биофа)) широко используются в медицине для терапии вирусных гепатитов В, С и D и различных онкологических заболеваний.
Известен способ получения рекомбинантного ИФН α-2b в дрожжевых клетках Pichia Pastoris [заявка PCT/IB 00/00339, МКИ C 12 N 15/00, 2000]. Недостатком этого способа является низкий выход - 100-150 мг с 1 л культуры клеток.
Известен способ получения ИФН α-2b, основанный на химическом синтезе фрагментов белковой молекулы ИФН α-2b с последующей их сборкой в целую молекулу [заявка РСТ/ЕР 03/01745, МКИ С 07 К 14/555, 2003]. Недостатками этого способа являются его высокая себестоимость и трудоемкость.
Известен наиболее близкий к заявленному способ получения ИФН α-2b, заключающийся в биосинтезе клетками бактерий Escherichia coli рекомбинантного ИФН α-2b [пат. РФ №2165455, МКИ C 12 N 15/21, 2001]. Получение ИФН α-2b в этом изобретении достигается посредством конструирования плазмиды pSS5 и продуцента SS5, полученного трансформацией штамма Escherichia coli BL21(DE3) этой плазмидой. Плазмида pSS5 несет сразу три последовательных промотора различной природы: лактозный промотор, промотор фага Т7 и триптофановый промотор; полусинтетический ген ИФН α-2b и терминатор транскрипции rrsBT1T2. Недостатками плазмиды pSS5 и продуцента на ее основе являются полусинтетический ген ИФН α-2b и триптофановый промотор. При получении полусинтетических генов сложно устранить все проблемы, связанные с переносом эукариотического гена в прокариотический организм, так как их получают путем мутагенеза природного гена. К таким проблемам относятся наличие редких кодонов и образующих "шпильки" последовательностей. Расположение триптофанового промотора в конце триады промоторов в плазмиде pSS5 ограничивает использование питательных сред для ферментации только средами, не содержащими триптофан. Это приводит к снижению выхода и значительному увеличению времени ферментации до 15-25 часов.
Изобретение решает задачу конструирования плазмиды, детерминирующей синтез рекомбинантного ИФН α-2b, и создания на ее основе высокопродуктивного бактериального штамма-продуцента, позволяющего получать рекомбинантный ИФН α-2b с высоким выходом (от 1 г с 1 л культуры) и по упрощенной технологии за счет уменьшения общего времени ферментации до 6 часов.
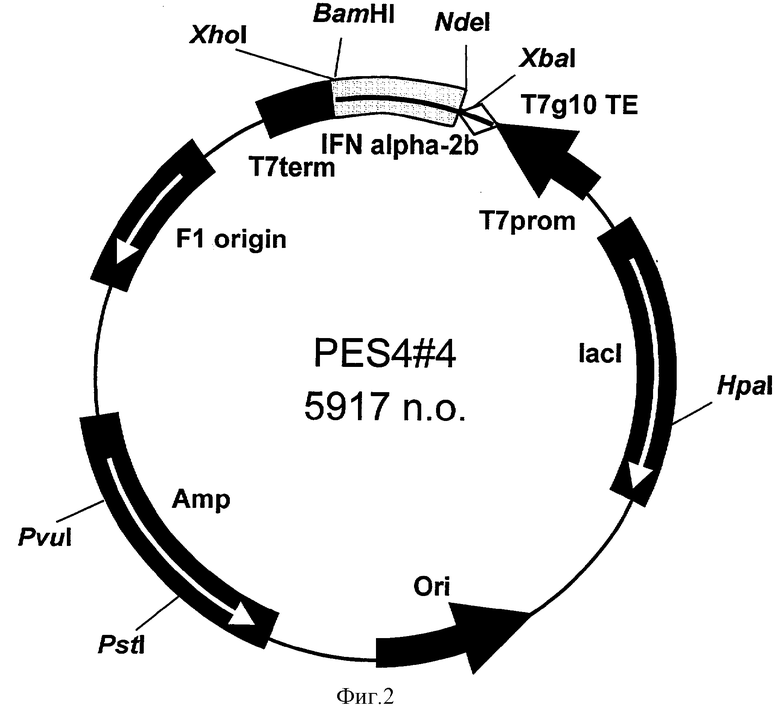
Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pES4-4, кодирующей полипептид с последовательностью ИФН α-2b человека, имеющей молекулярную массу 3,64 МДа (5917 п.о.), состоящей из NdeI/BamHI-фрагмента ДНК плазмиды рЕТ22b(+), содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы и усилитель трансляции гена 10 фага Т7, и NdeI/BamHI-фрагмента ДНК, содержащего последовательность искусственного гена рекомбинантного интерферона α-2b человека; содержащей в качестве генетического маркера ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pES4-4 клеток Е.coli к ампициллину; содержащей уникальные сайты узнавания эндонуклеаз рестрикции, расположенные на следующем расстоянии вправо от сайта Ndel: XbaI - 38 п.о., HpaI - 1332 п.о., PstI - 4065 п.о., PvuI - 4190 п.о., XhoI - 5363 п.о., а также за счет штамма Escherichia coli BDEES4, содержащего рекомбинантную плазмидную ДНК pES4-4, - продуцента рекомбинантного ИФН α-2b человека.
Предлагаемый штамм обеспечивает синтез рекомбинантного ИФН α-2b в количестве 20-30% от суммарного содержания белка клеток.
Преимущества предлагаемого изобретения заключаются, во-первых, в использовании полностью синтентетического, в отличие от плазмиды pSS5, гена ИФН α-2b с максимально широким набором кодонов, являющихся оптимальными для продукции белка в Escherichia coli; расположение которых устраняет возможность образования на синтезируемой мРНК протяженных "шпилек", потенциально ингибирующих трансляцию. На фиг.1 приведена аминокислотная последовательность ИФН α-2b и соответствующая ей нуклеотидная последовательность гена: верхняя строка - природная кДНК ИФН α-2b, нижняя строка - синтезированный искусственный ген. Во-вторых, применение для биосинтеза рекомбинантного белка оптимальных регуляторных элементов, контролирующих его экспрессию: Т7-lac промотора для предотвращения базальной экспрессии гена до момента начала индукции и высокого уровня транскрипции, соответствующей мРНК при индукции, высокоэффективного терминатора транскрипции Т7, и блока стоп-кодонов, исключающих биосинтез удлиненных вариантов рекомбинантного ИФН α-2b. Преимущества предлагаемого штамма Е. coli BL21(DE3) заключаются в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого de novo рекомбинантного ИФН α-2b и загрязнения выделяемого белка наиболее активными протеазами Е.coli. Также BL21(DE3) несет ген Т7-РНК полимеразы, что при использовании Т7-lac промотора и Т7 терминатора в плазмиде pES4-4 приводит к быстрой и эффективной продукции белка клетками Е.coli и, в отличие от плазмиды pSS5, существенно расширяет выбор питательных сред для ферментации.
Конструирование нового гена, кодирующего ИФН α-2b человека, осуществляют на основе плазмиды рЕТ22b(+). Искусственный ген, кодирующий ИФН α-2b, фланкированный сайтами рестриктаз NdeI и BamaHI, получают химико-ферментативным синтезом набора олигонуклеотидных фрагментов с последующей их сборкой и амплификацией при помощи полимеразной цепной реакции (ПЦР). Перед лигированием для генерации липких концов амплификат и векторную плазмиду обрабатывают рестриктазами NdeI и BamHI. Лигазную смесь используют для трансформации компетентных клеток Е.coli DH5α. Отбор положительных клонов проводят при помощи ПЦР с использованием специфических праймеров, с последующим рестриктным анализом выделенной плазмидной ДНК рестриктазами NdeI и BamHI. Структуру гена, кодирующего рекомбинантный ИФН α-2b, определяют секвенированием по методу Сенгера. Она должна полностью соответствовать нуклеотидной последовательности исходного искусственного гена ИФН α-2b (фиг.1).
Рекомбинантная плазмидная ДНК pES4-4, кодирующая полипептид ИФН α-2b, характеризуется следующими признаками:
- имеет молекулярную массу 3,64 МДа;
- кодирует полипептид ИФН α-2b;
- состоит из: Ndel/BamHI-фрагмента ДНК плазмиды рЕТ22b(+), содержащей промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фагаТ7, ген β-лактамазы, и Ndel/BamHI - фрагмента ДНК, включающего искусственный ген ИФН α-2b;
- имеет уникальную совокупность признаков: промотор и терминатор транскрипции РНК-полимеразы бактериофага Т7, усилитель трансляции гена 10 бактериофага Т7; искусственный ген, кодирующий ИФН α-2b; ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pES4-4 клеток Е. coli к ампициллину; уникальные сайты узнавания рестрикционных эндонуклеаз, расположенные на следующем расстоянии вправо от сайта NdeI: XbaI - 38 п.о., HpaI - 1332 п.о., PstI - 4065 п.о., PvuI - 4190 п.о., XhoI - 5363 п.о.
Для получения штамма-продуцента рекомбинантного ИФН α-2b плазмидную ДНК pES4-4 используют для трансформации компетентных клеток Escherichia coli BL21(DE3) и проводят отбор клонов, сохраняющих уровень биосинтеза рекомбинантного полипептида не ниже 20-30% от суммарного клеточного белка в течение по крайней мере шести последовательных пассирований. Для этого клоны трансформированных плазмидой pES4-4 клеток Е.coli BL21(DE3) выращивают в богатой среде (YT-, LB-бульон и др.) с добавлением ампициллина до 100 мкг/мл и раствора глюкозы до 1% в течение 12-14 часов, инокулируют новую порцию питательной среды в соотношении 1:100, растят культуру до достижения оптической плотности 1 O.Е., индуцируют изопропилтио-β-D-галактозидом, и растят еще 3-6 часов. Получение из клеток продуцента рекомбинантного ИФН α-2b включает следующие стадии: отделение бактерий от культуральной среды, их разрушение одним из обычно применяемых способов; отмывку буферными растворами телец включения от водорастворимых компонентов клетки; солюбилизацию и восстановление целевого белка, его рефолдинг и окончательную очистку.
Полученный штамм-продуцент Escherichia coli BDEES4 характеризуется следующими признаками.
Морфологические признаки: клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки: при росте на агаризованной среде LB колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или YT-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки: клетки растут при температуре от 4°С до 40°С при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам: клетки проявляют устойчивость к ампициллину (до 500 мкг/мл), обусловленную наличием в плазмиде гена β-лактамазы (bla).
Преимущества предлагаемого штамма-продуцента заключаются в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого de novo рекомбинантного ИФН α-2b и загрязнения выделяемого белка наиболее активными протеазами Е.coli, а также в использовании богатых сред, что позволяет проводить ферментацию за 6 часов.
Клетки Е.coli BDEES4 являются суперпродуцентом. При индукции изопропилтио-β-D-галактозидом происходит эффективный биосинтез рекомбинантного ИФН α-2b, который накапливается в клетках в количестве более 20% суммарного белка бактерий.
На фиг.1 представлена нуклеотидная последовательность и кодируемая ею аминокислотная последовательность Ndel/BamHI-фрагмента плазмиды pES4-4; на фиг.2 - физическая карта плазмиды pES4-4.
Изобретение иллюстрируют примеры.
Пример 1.
Конструирование рекомбинантной плазмидной ДНК pES4-4.
Нуклеотидную последовательность, соответствующую гену ИФН α-2b, получают химико-ферментативным синтезом. Для этого теоретически расчитанную последовательность ДНК разбивают на перекрывающиеся фрагменты размером 45-55 п.о. Химический синтез олигонуклеотидов, соответствующих этим фрагментам, выполняют твердофазным фосфоамидитным методом при помощи, например, ДНК-синтезатора ASM-102U (БИОССЕТ, Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов -5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O-(β-цианэтил-диизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500 Å), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Полученные олигонуклеотиды подвергают 5'-концевому фосфорилированию с использованием Т4 полинуклеотидкиназы (Fermentas, Литва). Для этого олигонуклеотиды в количестве 20 пмоль смешивают с ферментом в количестве 10 ед. в буферном растворе, содержащем 50 мМ Tris-HCl (pH 7.6 при 25°С), 10 мМ MgCl2, 5 мМ дитиотреита, 1 мМ спермидина, 0,1 мМ АТФ и 0,1 мМ ЭДТА. Реакцию ведут 30 минут, полинуклеотидкиназу инактивируют нагреванием до 65°С в течение 10 мин.
Фосфорилированные олигонуклеотиды смешивают в эквимолярном соотношении в 50 мкл буфера, содержащего 20 мМ трис-HCl, pH 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреита, прогревают до 65°С, медленно охлаждают до 37°С в течение часа и добавляют 10 ед. ак. Т4-ДНК-лигазы. Реакцию лигирования ДНК проводят 4 ч при 37°С, 0,1 мкл полученного раствора используют в качестве матрицы в полимеразной цепной реакции (ПЦР) в присутствии термостабильной ДНК-полимеразы Pfu и специфических праймеров:
5' CTTCCTTCATATGTGTGATCTGCCTCAAACCCACAGCCTTGGTAGCCGTCGT3' и
5' AGAAGAGGATCCAGAAGCTTATTATTCCTTGGAACGTAAAGATTCTTGCAAGT 3'.
Проводят 25 циклов амплификации (95°С, 20 с; 62°С, 40 с; 72°С, 60 с) для синтеза полноразмерного фрагмента ДНК, содержащего последовательность гена ИФН α-2b, фланкированного сайтами узнавания рестриктаз NdeI и BamHI. Продукт амплификации гидролизуют рестриктазами NdeI и BamHI, очищают электрофорезом в 5%-ном акриламидном геле, полосу ДНК величиной около 500 п.о. выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
Для приготовления вектора ДНК плазмиды рЕТ-22b(+) (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера Y (33 мМ трис-ацетат, рН 7,9, 10 мМ Mg-ацетат, 66 мМ К-ацетат 1, 0,5 мМ DTT, 0,1 мг/мл BSA) рестриктазой Ndel (10 ед. акт.), осаждают ДНК этанолом, растворяют в 40 мкл буфура для рестриктазы BamHI (10 мМ трис-HCl, рН 8,0, 5 мМ MgCl2, 100 мМ KCl, 0,02% Triton X-100, 0,1 мг/мл BSA) и обрабатывают рестриктазой BamHl (10 ед. акт.) в течение 1 ч при 37°С. Полученный фрагмент ДНК величиной 5,4 т.п.о. после электрофоретического разделения в 1%-ном агарозном геле выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
1 мкг полученного векторного фрагмента лигируют с 2 пмоль Ndel/BamHI - фрагмента размером 500 п.о., содержащего синтетический ген рекомбинантного ИФН α-2b, в 10 мкл буфера (20 мМ трис-HCl, рН 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреита) с помощью 10 ед. акт. Т4-ДНК-лигазы в течение 12 ч при 10°С.
1 мкл полученной лигазной смеси используют для электротрансформации компетентных клеток Е.coli BL21(DE3), которую проводят, например, при помощи аппарата для электротрансформации ВТХ 600 (ВТХ, США) при зазоре между пластинами электропорационной кюветы 1 мм и напряжении разряда 1.4 кВ. После трансформации суспензию бактерий смешивают с питательной средой SOC, растят 1 ч на +37°С и высевают на чашки Петри с LB-агаром, содержащим 50 мкг/мл ампициллина.
Первичный отбор клонов, содержащих нужную плазмиду, проводят методом "ПЦР с клонов" с использованием специфических праймеров:
5' CTTCCTTCATATGTGTGATCTGCCTCAAACCCACAGCCTTGGTAGCCGTCGT 3' и
5' AGAAGAGGATCCAGAAGCTTATTATTCCTTGGAACGTAAAGATTCTTGCAAGT 3'.
Проводят 25 циклов амплификации (95°С, 20с; 62°С, 40с; 72°С, 60с), с последующим электрофоретическим анализом ПЦР продуктов в 1%-ном агарозном геле на наличие ПЦР-продукта длиной около 500 п.о. Отобранные клоны используют для подроста в жидкой среде и выделения плазмидной ДНК плазмиды, которую анализируют на наличие вставки с помощью эндонуклеаз рестрикции NdeI и BamHI с последующим разделением продуктов гидролиза в 5%-ном полиакриламидном геле. Окончательное строение плазмид, содержащих Ndel/BamHI-фрагмет около 500 п.о., подтверждают определением нуклеотидной последовательности методом секвенирования по Сенгеру. По данным секвенирования отбирают ту плазмиду, нуклеотидная и соответствующая ей аминокислотная последовательности Ndel/BamHI - фрагмента которой полностью идентичны первоначально запланированной (фиг.1). Проводят трансформацию клеток Е.coli BL21(DE3) выбранной плазмидой, как описано выше, петлей переносят 5-10 колоний в 5 мл жидкой среды 2xYT, содержащей 50 мкг/мл ампициллина, в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности А550 0,7-0,8, отбирают аликвоту культуры для последующего анализа, прибавляют индуктор - изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают рост еще 2 ч. Равные аликвоты суспензии клеток, отобранных до внесения индуктора и после завершения роста центрифугируют, отделяют супернатант и анализируют осадок клеток электрофорезом в ПААГ, как описано ниже в примере 2. Появление отчетливо видимой полосы белка в районе 20 кДа в образце пробы, отобранной после индукции, свидетельствует о способности штамма синтезировать рекомбинантный ИФН α-2b при индукции IPTG и полностью подтверждает корректность сборки плазмиды.
Пример 2.
Получение штамма-продуцента Е.coli BDEES4 с рекомбинантным ИФН α-2b и определение его продуктивности.
Штамм-продуцент BDEES4 получают трансформацией компетентных клеток Е.coli BL21(DE3) плазмидой pES4-4, как описано в примере 1.
Штамм продуцента Е.coli BDEES4 выращивают при 37°С в 100 мл YT бульона (рН 7,0) с 50 мкг/мл ампициллина в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности А 550 0,7-0,8, прибавляют изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают процесс еще 4 ч, или продолжают выращивание в отсутствие индуктора в течение 4 ч. Каждый час отбирают пробу по 2 мл, определяют А 550 и количество культуры, соответствующее 1 мл с А 550 1,0, центрифугируют 5 мин при 6000 об/мин. Осажденные клетки в 100 мкл лизирующего буфера с красителем бромфеноловым синим обрабатывают 20 с ультразвуком, нагревают 3 мин при 100°С и пробы по 1 мкл используют для электрофореза в 15% SDS-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют для определения относительного количества белка в полосе целевого белка. По данным сканирования содержание рекомбинантного ИФН α-2b составляет 20-30% от всех клеточных белков.

Изобретение относится к генной инженерии и может быть использовано для получения рекомбинантного интерферона α-2b человека. Конструируют рекомбинантную плазмидную ДНК pES4-4, которая имеет молекулярную массу 3,64 МДа (5917 п.о.) и состоит из Ndel/BamHI-фрагмента ДНК, содержащего последовательность искусственного гена рекомбинантного интерферона α-2b человека, гена β-лактамазы и Ndel/BamHI-фрагмента ДНК плазмиды рЕТ22b(+), содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фага Т7. Плазмида pES4-4 содержит в качестве генетического маркера ген β-лактамазы, детерминирующий устойчивость трансформированных этой плазмидой клеток Е. coli к ампициллину, и уникальные сайты узнавания эндонуклеаз рестрикции, расположенные на следующем расстоянии вправо от сайта Ndel: XbaI -38 п.о., HpaI-1332 п.о., PstI-4065 п.о., PvuI-4190 п.о., XhoI-5363 п.о. Полученной плазмидой трансформируют клетки Escherichia coli и получают штамм E. coli BDEES4 - суперпродуцент рекомбинантного интерферона α-2b человека. Изобретение позволяет получать рекомбинантный интерферон α-2b человека с высоким выходом и по упрощенной технологии. 2 н.п. ф-лы, 2 ил.
NdeI/BamHI-фрагмента ДНК плазмиды pET22b(+), содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы и усилитель трансляции гена 10 фага Т7;
NdeI/BamHI-фрагмента ДНК, содержащего последовательность искусственного гена рекомбинантного интерферона α-2b, приведенную на фиг.1, и содержащая
ген β-лактамазы, детерминирующий устойчивость трансформированных плазмидой pES4-4 клеток E. coli к ампициллину, в качестве генетического маркера,
уникальные сайты узнавания эндонуклеаз рестрикции, расположенные на следующем расстоянии вправо от сайта NdeI: XbaI - 38 п.о., HpaI - 1332 п.о., PstI - 4065 п.о., PvuI - 4190 п.о., XhoI - 5363 п.о.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pSS5, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА, ШТАММ ESCHERICHIA COLI SS5 - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ПОЛУЧЕНИЯ ИНТЕРФЕРОНА АЛЬФА-2b | 1999 |
|
RU2165455C1 |
| et al., Elimination of an HuIFN alpha 2b readthrough species, produced in Escherichia coli, by replacing its natural translational stop signal, J | |||
| Biotechnol., 1998, v.63, n.3, p.179-186.WO 03070764, 28.08.2003.RU 2118366 С1, 27.08.1998. | |||

