.Культуру, аидуцироваиную митомицином С, центрифугируют с 2400 кратным ускорением в течение 30 мин для удаления клеточных остатков и нодучающийся в результате отстаивания верхний слой обрабатывают хлороформом (5 об. %). Эта всплыв(пая наверх фракция и есть сырой ниоцин.
Сырой пиоцин далее очищают путем лрибавлеНИЯ i.M MnCb (60 мл на 1 л лизата), примешивая при этом смесь к сырому ниоцину. После достижения рН 7,5 с по:мощью 1М NaOH полученный осадок удаляют цен1)рифугированием. Всплывшая наверх фаза является очищенным пиоцином.
Дальнейшую очистку проводят добавлением (NH4)2SO4 до 70%-ного насыщения и культивированием в течение ночи нри
,1° с
-I .
После центрифугирования в течение 30 мин при 4° С осадок от центрифугирования, содерл ащий компонент с пиоциновой активностью, растворяют в 50 мл O.OIM трис-(оксиметил)-аминометан-(трис)хлоргидрата рН 7,5, содержащем 0,0IM MgCls и 0,01М MgSO, и подвергают диализу в течение ночи при 4° С нротив 2 л того же буфера. При необходимости препарат осветляют центрифугированием в течение 90 мин. Л елатиноподобный осадок от центрифугирования осторожно растворяют в 20 мл буфера и хроматографируют на целлюлозе ДЕАЕ, предвар.ительно шромытой 1И доведенной до равновесного состояния с помощью того же буфера. В колонку (размером 1,5X28 см- вносят 8 мл образца ниоцина и адсорбируют в течение ч. Колонку промывают 200 мл буфера для удаления веществ, неадсорбированных на целлюлозе типа ДЕАЕ. Затем пиоцин элюируют с помощью 800 мм в 0,01М буфере. Собирают фракции по 5 мл и анализ груют на цоглощение при 280 нм и на теоциновую активность.
Фракции, проявляющие пиодиновую активность, днализируют против 0,01М трисбуфера, рН 7,5 для удале.ния NaCl, а затем концентрируют с помощью ультрацентрифугирования в течение 90 мин.
Типизацию пиоцина осуществляют, используя метод жидкой среды, описанный Джоном.
Штаммы Neisseria gonorrhoeae, которые испытывают на восприимчивость к пиоцину, выращивают в течение ночи в чашках с агаровой средой. Суспензию этих микроорганизмов приготавливают в разбавителе, состоящем ,из 0,85% NaCl и 0,1% НС1 (рН 6,4), и доводят до по1каза.н1ий Клетта, paiBiKbix 50-60. Чаш.ки с агаровой средой заражают с помощью тампона, погруженного в клеточную суспензию. Неразбавленные или серийно разбавленные препараты пиоцина (5 мкл вносят на поверхность чашек с агаровой средой. Все чащ-ки культи ., ... ;,
, -, ,.-. , :, «,
.....
731904
вируют в течение ночи при 37° С с повышенным содержанием COs.
Пиоцины были приготовлены для изучения под электронным микроскопом с помощью метода негативного пятнистого окрашивания. Препараты диоцина центрифугируют с ускорением в 100000 q в течение 1 ч, .и осадок от центриф.угирования повторно суспендируют в малом объеме
1М NH4C2H302 (рН 7,0). Медные сеточки, покрытые формваром, вносят в каплю образца в течение I-2 мин, а затем промокают досуха с помощью фильтровальной бумаги. Эти сеточ-ки вносят затем в каплю 1,5%-ного раст13Ора фосфовольфрамата натрия (рН 7,0) в течение 30 с. Избыток жидкости удаляют фильтровальной бумагой.
Взаимодействие пиоцинов с клетками
Neisseria gonorrhoeae обнаруживают по подобной методике. Спустя 30 мин после добавления пиоцина к Ж1идкой к ультуре N. gonorrhoeae 72Н870, образец удаляют и обрабатывают так же, как это было описано. Препарат с негативно окрашенными пятнами был изучен с помощью электронного микроскопа RCA (ори 50 кВ)
В табл, 1 даны результаты производства гонококкового фактора .ингибирования, синтезированного во время выращивания Pseudomona.s aeruginosa АТСС 29260 в указанной среде. Добавление митомицина С (1 мкг1мл) вызывает широкий лизис культуры в пределах 3 ч и приводит к 16-кратному увеличению концентрации фактора инпибирования, определенного путем титрования иа Psendomonas aernginosa Рр7 и 1947. В таблице также показан титр факштаммах N. gonorrhoeae тила JW-31 и DGI тора ингибированля, изменяющийся с изменением различных штаммов Neisseria gonorrhoeae, причем не наблюдается различий с колониальными вариантами 1единичного штамма.
Таблица I
Примечание: а) Лнтолпщин С (I мкг на 1 мл среды) прк пос.тедовательном разбавлеиии не был способен ингибировать рост указанных организмов при разбавлении 1 :2 (0,5 мкг/мл);
б) N,D - не определялось.
Фактор ингибирования был частично очищен от всплывающих наверх жидвих фаз культур Pseudomonas aeruginosa штамма АТСС 29260, индуцированных митомицином С, по методике, описанной выше. Фактор ингибирования элюируют из целлюлозы ДЕАЕ с помошд ю NaCl градиента от О до 1,ОМ. Обнаруживают два пика, содержащие фактор ингибирования. Главный пик (а) злюируется при концентрации NaCl 0,06М .и обладает более, чем 90%-ной ингибиторной активностью. Слабый пик (6) элюируется при концентрации NaCl 0,91М и обладает менее, чем 10%-ной активностью. Фракции, составляющие пик а, объединяют, диализируют против 0,01 трисхлоргидратного буфера (рН 7,5) для удаления NaCl и концентрируют ультрацентрифугированием (100000-кратное ускорение в течение 90 мин}. Этот препарат используют в описываемых ниже испытаниях.
Изучение с помощью электронного микроскопа препарата с негативно окрашенными пятнами из очищенного фактора ингибирования показывает, что частицы напоминают пиоцины R-типа как в несокращенном, так и в сокращенном состояниях. В несокращенном состоянии эти частицы имеют длину 11 5 нж и ширину 15,3 нм. В сокращенном состоянии частицы состоят из внутреннего ядра (длиной 105 нж и щириной 6,5 «лг), окруженного сокращенной оболочкой (длиной 44,4 нм и шириной 18,6 нм.}. В со,кращенном состоянии находилось 20- 30% частиц, наблюдаемых в этих препаратах.
Тип пиоцина как в частично очищениых, так II в очищенных препаратах определяют по вышеописанной методике, причем результяты показывают, что модель ie изменяете;; зс время очистки. Пиоцин относится к модели 611131.
Действие пиоцина 611131 на рост клинического изолята Neisseria gonorrhoeae (штамм 72Н870).
Действие шиодина на рост клеток микроорганизма было определено добавлением различных концентраций очищенного препарата к экспоненциально растущим культурам.
При высоких концентрациях пиоцина полное ингибирование .роста, происходящее в пределах 1 ч, сопровождается широким ЛИЗИСОМ культуры. Изучение с помощью электронного микроскопа показывает, что между пиоцином и чувствительными клетками Neisseria gonorrhoeae происходит непосредстзепное взаимодействие. Взаимодействие пиоцина с клетками приводит к сокращению пиоцина.
Спектр ингиб)Ирования пиоцина типа 611131.
Аликвотные части очищенного препарата пиоцина наносят в виде пятен на посевы, полученные из клинических изолятов N. gonorrhoeae. Зона ингибирования отчетливо Внд.на во всех изученных штаммах. Не 1-аблюдается различий между колониями типа Т-1 и Т-4 из одного и того же штамма. Включен также отрицательный контрольный тест продуцирующего штамл а Р. aeruginosa АТСС 29260.
Инлибирование различных видов Neisseria с помощью этого пиоцина показано з табл. 2. Все изоляты N. gonorrhoeae кал
Таблица 2





| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВЫДЕЛЕНИЯ БАКТЕРИОЦИНОВ | 2011 |
|
RU2492231C2 |
| Способ очистки мундтицина Р436 (КС) | 2021 |
|
RU2794803C1 |
| БАКТЕРИОЦИН, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ШТАММ MICROCOCCUS VARIANS, НУКЛЕОТИДНЫЙ ФРАГМЕНТ, СИГНАЛЬНЫЙ ПЕПТИД | 1996 |
|
RU2172324C2 |
| ШТАММ ENTEROCOCCUS FAECIUM LVP1073, ПРОДУЦЕНТ БАКТЕРИОЦИНА ПРОТИВ БАКТЕРИАЛЬНЫХ ПАТОГЕНОВ, БАКТЕРИОЦИН E1073 ПРОТИВ БАКТЕРИАЛЬНЫХ ПАТОГЕНОВ, ШТАММ LACTOBACILLUS PLANTARUM 1 LVP7 - ИНДУКТОР СИНТЕЗА БАКТЕРИОЦИНА E1073, СИГНАЛЬНЫЙ ПЕПТИД СП1073 - РЕГУЛЯТОР СИНТЕЗА БАКТЕРИОЦИНА E1073, СПОСОБ ПОЛУЧЕНИЯ БАКТЕРИОЦИНА E1073 | 2009 |
|
RU2409661C2 |
| ГИБРИДНАЯ ЭКСПРЕССИЯ БЕЛКОВ NEISSERIA | 2001 |
|
RU2304617C2 |
| ГЕТЕРОЛОГИЧНАЯ ЭКСПРЕССИЯ БЕЛКОВ NEISSERIA | 2001 |
|
RU2299906C2 |
| ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ МУТАНТНУЮ РЕКОМБИНАНТНУЮ IgA1 ПРОТЕАЗУ Neisseria meningitidis СЕРОГРУППЫ В, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ УКАЗАННЫЙ ПОЛИНУКЛЕОТИД, КЛЕТКА-ХОЗЯИН, СОДЕРЖАЩАЯ УКАЗАННУЮ ПЛАЗМИДНУЮ ДНК, РЕКОМБИНАНТНАЯ IgA1 ПРОТЕАЗА Neisseria memingitidis СЕРОГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОЙ ФОРМЫ IgA1 ПРОТЕАЗЫ | 2011 |
|
RU2486243C1 |
| ШТАММ LACTOCOCCUS LACTIS SUBSP.LACTIS F-119 - ПРОДУЦЕНТ АНТИБИОТИЧЕСКОГО КОМПЛЕКСА LGS И СПОСОБ ПОЛУЧЕНИЯ АНТИБИОТИЧЕСКОГО КОМПЛЕКСА LGS С ЕГО ИСПОЛЬЗОВАНИЕМ | 2007 |
|
RU2374320C2 |
| СПОСОБ ЭКСПРЕСС-ОБНАРУЖЕНИЯ ВОЗБУДИТЕЛЯ ВНУТРИБОЛЬНИЧНЫХ ИНФЕКЦИЙ Burkholderia cepacia И ДИФФЕРЕНЦИАЦИИ ЕГО ОТ ДРУГИХ ВИДОВ БУРКХОЛЬДЕРИЙ И ПСЕВДОМОНАД | 2004 |
|
RU2281501C1 |
| НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНУЮ РЕКОМБИНАНТНУЮ IgA1 ПРОТЕАЗУ NEISSERIA MENINGITIDIS СЕРОГРУППЫ B, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ НУКЛЕОТИДНУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩУЮ АКТИВНУЮ IgA1 ПРОТЕАЗУ, ШТАММ-ПРОДУЦЕНТ, СОДЕРЖАЩИЙ ПЛАЗМИДНУЮ ДНК, ПРОДУЦИРУЮЩИЙ ЗРЕЛУЮ ФОРМУ IgA1 ПРОТЕАЗЫ, РЕКОМБИНАНТНАЯ Ig ПРОТЕАЗА NEISSERIA MENINGITIDIS СЕРОГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОЙ ФОРМЫ IgA1 ПРОТЕАЗЫ, ОБЛАДАЮЩЕЙ ИММУНОГЕННЫМИ И ПРОТЕКТИВНЫМИ СВОЙСТВАМИ | 2011 |
|
RU2453599C1 |
ИЗ рассеянных, так и нерассеянных заражеПИЙ ингибируются. Однако ингибируются40 только 3 из 20 штаммов N. meningitidis и 5 из 16 щтаммов N. lactamica. Не наблюдается какой-либо корреляции между серологической группой и ингибированием N. meningitidis. Ни один ;-:з других пяти испытанных видов не пнгнбируется этим пиоцином. Быстрая идентификация Neisseria gonorrhoeae.
Метод для быстрой идентификации Neisseria goni rrhoeae может быть осуществлен по любой из следующих методик: пиоцин 611131 наносят в виде пятна в чашку с агаровой средой, содержащую биологический образец, который испытывается, или диск, пропитанный пиоцином, вносится в чашку, зараженн1ую образцом, или же пиоцин, внесенный в одну половину чашки с расщепленной агаровой средой, заражается образцом. Вслед за культивированием осуществляют идентификацию микроорганизма в виде N. gonorrhoeae на основе зоны инпибирования, окружающей пятно, куда нанесен пиоцин, или ингибврования рометод типизации особенно полезен при эпидемиологических исследованиях, когда важно преобладание или появление новых штаммов, а также при определении обусловлена ли неудача обработк1И устойчивостью микроорганизма или повторным заражением новым типом микроорганизма. Дополнительные варианты изобретения описапы ниже. Идентификация N. gonorrhoeae, используя пиоцины, меченые флюоресцеином. По этому методу пиоцины, производимые и очищенные так, как это описано вытпе, размечают флюоресцеином согласно методу Джонсона. Меченые таким образом пиоцины реагируют о подозреваемой N. gonorrhoeae на предметном стекле, приготовленном из клинического материала или из изолированных колоний из чашки с агаровой средой. Избыток циоцинов удаляют промывкой предметного стекла 0,02 молярным физиологическим (рН 7,2) буфером, буферированным фосфатом, и предметное стекло рассматривают под уф-микроскопом. Клетки, имеющие типичную морфологию N. gonorrhoeae и показывающие флюоресценцию, рассматриваются как положительные для N. gonorrhoeae.
ста на той части чащки, в которую был внесен пиоцин.
Этот метод может быть применен для идентификации других бактерий.
Типизация Neisseria gonorrhoeae.
Типизация N. gonorrhoeae и других видов Neisseria включает использование нескольких типов пиоцина и основывается на ингибировании или наингибировании испытываемых микроорганизмов. Наблюдаемые результаты используются для идентификации типа изолята .путем соотнесения их с результатами, полученными с помощью целого ряда известных типов (табл. 3). Этот
Таблица 3 Идентификация бактерий с помощью пиоцинов, меченых радиоактивным изотопом. Радиоактивные пиоцины получают путем йодирования их изотопом j с помощью метода с применением хлорамина Т. Их можно получить мечеными с И или С путем ввода конкретных меченых аминокислот в культивируемую среду до начала синтеза пиоцина в Pseudomonas aeruginosa с по.мощью митомицина С. Пиоцины, производимые по этому методу, являются радиоактивными. Специфические бактерии легко идентифицируются путем взаимодействия радиоактивных пиоцинов с суспензией бактерий. Затем эту суспензию подвергают мембранной фильтрации или центрифугированию для отделения несвязанных пиоцинов от пиоцинов, которые связаны специфически с поверхностью бактериальных клеток. Фильтр или промытый клеточный осадок в пробирке после центрифугирования принимают затем во внимание, чтобы определить количество связанного пиоцина, который был меченым. Данные сравнивают со степенью неспецифического связывания, любое увеличение над уровнем неспецифического связывания показывает наличие неизвестного микроорганизма.
Sal. kadack
Sal. miarembe
Sal. onderspoort
Sal. thompson
Sal. spp. Group C
Serratia mercescen
S. marcescens
Shigella flexneri
Sh. spp. Group D
Brucella abortus
B. bronchisepticus
B. suis
Neisserla gonorrho
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. gonorrhoeae
N. meningitidis
N. meningitidis
N. meningitidis
N. meningitidis
N. meningitidis
N. flava
N. flavescens
N. laciamia
N. lactamica
N. mucosa
N. ovis
N. peiflava
N. subfiava
Acinetobacler anitra
A. anitratus
A. calcoaceiicus
Aeromonrjs hydioph
Alcaligenes viscosib
Azotomanas iiisoliid
EnteroDactcr aeroge
E. cloaceae
E. cloaceae
E. cloaceae
E. cloaceae
Exhericliia coli
E. cnii
E. coli
E. ooH
E. coli
E coli
E. coli B.
E cnli iiidole CP
E, Cdli Se rs
E. coli 1. С 411
E. coli K. 1
E. coli K. 12
E. cuii
Haffni.i Spp
Herrt le;i Vaginocoi
H VagiII с .la
H. Vaguii4-,lH V;igirioi()bi
Hk-bbiclla oz I II.:l
K. pneu ioi;i;k
K. pneumonia
K. pneuinuniae
Mima polymorpha Proteus rnorganic Providencia s tuartie
Таблица 4
+
+
+
+ + + + + + + + + + 4- + +
+ +
+
/- +
+/+
+
+
+/+7+/+/+ /+/I +
+/+
+
+
+
+
+
+//+
+ -
+/+/+
+ +/+
+
+ I -
+
+
+
j
-- i +/- i +
II
Орга:.изн
Pse i: d от on ; s .i er ug i;: os л
P. dertiijir:c-T
PS. aerugin s;i non p;f.T
F alkuligc, ;ifs
PS. irniolo.MC.
PS. inaltopliiha
PS. п;а1Ор)
PS saccliarcphili.i
S;:ln;onell;i ariz .
Sa. bergen
Sal. bornura
. cholerssuis
Sal. dahlem
Sal. deversoir
Sal. djakarta
Sal. diigbe
Sal. enieriditis
Sal. florida
Bacteriocjn I Bacteriocin 2 3-acleriocin 3 Bacteriocin 4 Bacterjocin 5 Baceriocin 6 BfiCteriocin 7
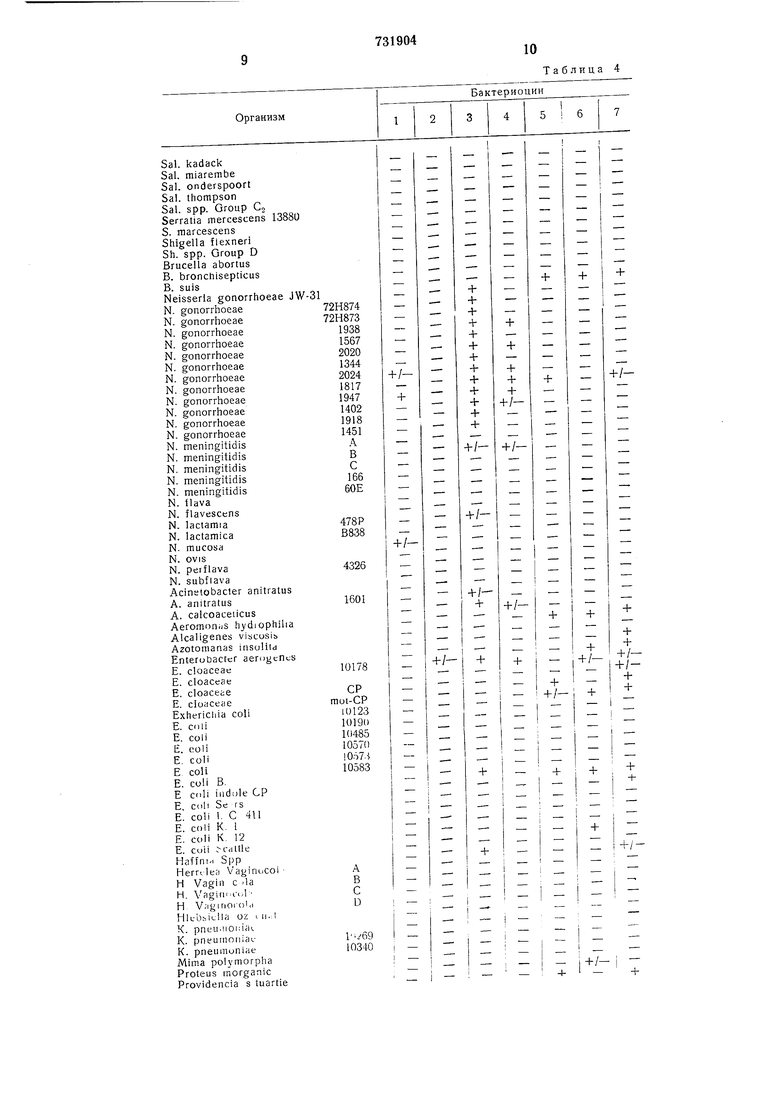
Изобретение может быть использовано при идентификации антигенов, которые являются свойственными для бактерий таксономически несвязанных родов. Как известно, бактериоцины связываются с специфическими рецепторными участками на внешней поверхности бактерий. Эти рецепторные участки подвергаются также действию механизМов защиты хозяина и могут таким образом стимулировать производство антител, которые могут быть либо бактерицидного, либо опсон1изирующего типов. Бактерии могут делить свойственные антигены. Эти свойственные антигены могут быть также бактериоциновыми рецепторными участками, поэтому взаимодействие бактериоцинов с разными видами бактерий может быть использовано в качестве индикатора разделяющихся антигенов (табл. 4). Это, в свою очередь, может быть использовано при выделеняи и очистке антигенов для производства вакцин. Таким образом, второстепенный антиген на одном микроорганизме может быть главным антигеном на другом н поэтому может облегчать очистку и может быть ползчен в больплих количествах.
Способ моЖет быть использован также для демонстрирования свойственных антигенов (или поверхностных компонентов) в клетках млекопитающих. Вследствие свойственной специфичности их рецепторов бактериоцины могут быть использованы вме12
Продолжение табл. 4
Бя КТСрИОЦКР
: PS aeruginosa FS-5 : PS aeruginosa FS-6 : PS aeruginosa PA 108 : PS aeruginosa PS-7 : PS Vibriocholera Ее Toz : PS Serratia marcesceus : Aeromonas hydro.philia
сто соединений, таких как лектины, к.л:. в методиках, включающих типизацию ткан.:;). Изобретение может быть использовано также при идентификации других бакгеэ зй. Таким образом, это показало в табл. 4, бактериоцины из Vibro cholerae, Aeroni s Hydrophila и Serratia marcescens обычно перекрестно реагируют с таксономичес.ки несвязанным микроорганизмом. Предлагаемый способ позволяет упростить определение наличия микроорган.чзма с использованием любого бактериазина.
Формула И310бретения
L Способ идентификации микрооргакизмов в биолопическо1М материале путан: госева на питательную среду с последующим контактированием выросщих колоний с бактериоцином, инкубированием и учетом наличия или отсутствия роста, о т л и ч а к.щ и и с я тем, что, с целью упрощения способа, выросщие колонии контактируют с бактериоцином R-типа из микроорганиз;у;оЕ таксономическ1и несвязанного рода.
