(5) СПОСОБ ИНГИБИРОВАНИЯ СИНТЕЗА РИБОСОМНЫХ РНК


| название | год | авторы | номер документа |
|---|---|---|---|
| Ингибитор активности фермента | 1977 |
|
SU627162A1 |
| Способ получения синтетической двунитчатой РНК, обладающей интерферониндуцирующей активностью | 1979 |
|
SU933001A3 |
| Способ определения величины синтеза ДНК в лейкоцитах периферической крови | 1989 |
|
SU1718122A1 |
| БЕЗОПАСНЫЙ ЭКСТРАКЦИОННО-ФЛУОРИМЕТРИЧЕСКИЙ СПОСОБ ОПРЕДЕЛЕНИЯ СЕЛЕНА В ВОДЕ | 2015 |
|
RU2597769C1 |
| СПОСОБ ОЦЕНКИ ЦИТОТОКСИЧЕСКОЙ АКТИВНОСТИ ЛИМФОЦИТОВ НАТУРАЛЬНЫХ КИЛЛЕРОВ | 2012 |
|
RU2514019C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ПТИЦ С ИСПОЛЬЗОВАНИЕМ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК | 2007 |
|
RU2473688C2 |
| СПОСОБ ПОВЫШЕНИЯ СПЕЦИФИЧНОСТИ ОБРАТНОЙ ТРАНСКРИПЦИИ | 2008 |
|
RU2459872C2 |
| Способ получения функционально активных мембранных рецепторных белков | 1990 |
|
SU1723126A1 |
| Способ диагностики ювенильного ревматоидного артрита | 1989 |
|
SU1725132A1 |
| АЗИДОНАФТАЛИН-1-СУЛЬФОНИЛГИДРАЗИНЫ В КАЧЕСТВЕ ПОЛУПРОДУКТОВ ДЛЯ ПОЛУЧЕНИЯ ТРИЛИТИЕВЫХ СОЛЕЙ (1`R, 5`R) -3`-АЗА-1` -(2-АМИНО -1,6-ДИГИДРО-6-ОКСОПУРИНИЛ-9) -3`-ДЕЗОКСИ-3`- (АЗИДОНАФТАЛИН-1-СУЛЬФАМИДО) -ГЕКСОПИРАНОЗИЛ -6`- ТРИФОСФАТОВ - СПЕЦИФИЧЕСКИХ ФОТОАКТИВИРУЕМЫХ НЕОБРАТИМЫХ ИНГИБИТОРОВ РНК-ПОЛИМЕРАЗЫ | 1991 |
|
RU2036903C1 |
I
Изобретение относится к молекулярной биологии и генетике, касается техники ингибирования синтеза рибосомных РНК и может быть использовано в качестве тест-метода для изучений синтеза рибонуклеиновых кислот в клетках высших организмов.
Известен способ ингибирования синтеза рибосомных РНК путем инкубирования биологического материала с раствором селенита натрия I}.
Однако известный способ применим лишь для ингибирования активности А-формы (фермента только в системе бесклеточного синтеза.
Для опытов in vivo в литературе способы ингибирования синтеза рибосомных РНК неизвестны.
Цель изобретения - ингибирование синтеза рибосомных РНК в нативных клетках.
Указанная цель достигается тем, что в способе ингибирования синтеза рибосомных РНК путем инкубирования
биологического материала с раствором селенита натрия интактные бластодермы -отделяют от желтка, инкубируют с селенитом натрия, затем добавляют в инкубационную смесь Н -уридин в концентрации 0-50 мк Ки/мл, отмывают от невключившейся метки, добавляют РНК-носитель, уридин и контролируют степень ингибирования определением фракции РНК-ДНК.
to
Способ осуществляют следующим образом.
Оплодотворяют зрелую икру, определяют стадии развития по Нейфаху, отделяют интактную бластодерму от
15 желтка, инкубируют с lO-jO селенита натрия , охлаждают, добавляют трихлоруксусную кислоту (ТХУ) и измеряют радиоактивность.
Инкубацию проводят после отделе20ния бластодермы от желтка в присутствии антибиотиков. В инкубационную смесь добавляют смесь Н -уридин в концентрации 0-50 мк Ки/мл, отмывают от невключившейся метки, добавляют РНКгноситель 0,1 мл (5 мг/мл намеченный уридин мл (10 мг/мл) и контролируют степень ингибирования определением фракции РНК-ДНК методом Шмидта-Таннгаузера.
Пример .1. Каждой из 20 самок вьюна инъецируют 1 мл гонатропина с концентрацией 5000 ед., после Зб-часовой инкубации при икра созревает, ее выделяют у самок и у 10 самцов сперму. В полученную икру в чашках Петри добавляют суспензию выделенной у 10 самцов спермы, в течение 5 с перемешивают в присутствии 10 мл дехлорированной водопроводной воды и оставляют на 5 мин, Оплодо- , творенную икру промывают водой до . полного удаления полостной жидкости из спермы. Затем помещают в чашки Петри, заливают 200 мл воды, содержащей 100 ед,/мл пенициллина и стрептомицина 50 ед,/мл. Инкубацию опло-дотворенной икры проводят при , Под микроскопом по методу Нейфаха определяют 10-тичасовую стадию развития. Отделяют зародыши (бластодермы) от желтка, сначала промывая для разрушения оболочек зародышей в 200 мл 0, раствора трипсина в течение 7.мин, затем .неоднократно промывая зародыши раствором Гольфретера,
Затем отделенные зародыши помещаю в 9 пробирок., заливают буфером Гольф ретера двойной концентрации на 0,05 tris-HCl буфере рН 7,6, в присутствии 100 ед,/мл пенициллина, 50 ед,/мл стрептомицина до достижения уровня 1 мл. Переносят пробирки в ледяную баню на 5 мин, В каждую пробирку добавляют Н -уридин из расчета 40-50 мк Ки/мл, встряхивают несколько раз и оставляют в ледяной бане на 10 мин для отстаивания, В t-ю, 5-ю, 6-ю пробирки добавляют селенит натрия до конечной концентрации 4 ммоль, в , 8-ю, 9-ю пробирки добавляют селенит натрия до конечной концентрации kQ тмоль. Переносят все пробирки в. водяную баню при и инкубируют в течение 1 ч, После инкубации в каждую пробирку добавляют 3 мл раствора Гольфретера при оС для остёновки реакции, тщательно перемешивают содержимое пробирки и осторожно отсасывают раствор до уровня 1 мл. Процедуру повторяют
три раза для полного удаления невключающейся метки. В каждую пробир добавляют по 1 мл раствора Гольфретера и замораживают в бане (сухой лед+ацетон), дают возможность оттая в лебяной бане, затем добавляют в каждую пробирку 1 каплю РНК-носителя с концентрацией 5 мг/мл, затем в каждую пробирку добавляют по 20 мкл раствора немеченного уридина (10 мг/мл). Дальнейшую обработку полученной смеси для количественног определения РНК проводят по методу Шмидта-Таннгаузера, для чего в каждую-пробирку добавляют 50 ТХУ до конечной концентрации 10%, гомогенезируют стеклянным пестиком, затем центрифугируют при 4000 об/мин 10 мин, Надосадочную жидкость (кислоторастворимая фракция) переливают в отдельные пробирки, подвергают анализу по методу Ives и др.
Осадок высушивают и проводят гидролиз следующйм образом,
В каждую пробирку вливают по 2 мл 0,5 Н NaCH, осторожно размешивают, проводят инкубацию в течение 2 ч. Пробирки после гидролиза переносят в ледяную баню, добавляют 50% ТХУ до концентрации 10%, в осадок при этом выпадают ДНК-белки, а в растворе остаются рибонуклеотиды РНК, Растворы центрифугируют при чООО об/мин. Осадок выбрасывают, Надосадочную жидкость обрабатывают эфиром до рН 7 и измердют радиоактивность на сцинтилляционном счетчике (SL-30 Intertedmik, France), По включению метки в РНК судят об интенсивности синтеза РНК. Устанавливают, что ингибирование синтеза РНК на этой стадии не происходит. Далее об относительной интенсивност синтеза РНК в зародышах на различных стадиях развития судят по включению Н -уридина в тотальную РНК с учетом внедрения метки в суммарную кислоторастворимую фракцию и в фосфорилированные производные уридина.
Примеры 2-5. Процесс проводят, как описано в примере 1, но рассматривают 11,16,17,18-часовы стадии развития зародышей. Степень ингибирования соответственно составляет 27,2, 21,3, 53, и 72,5%, т,е, возрастает.
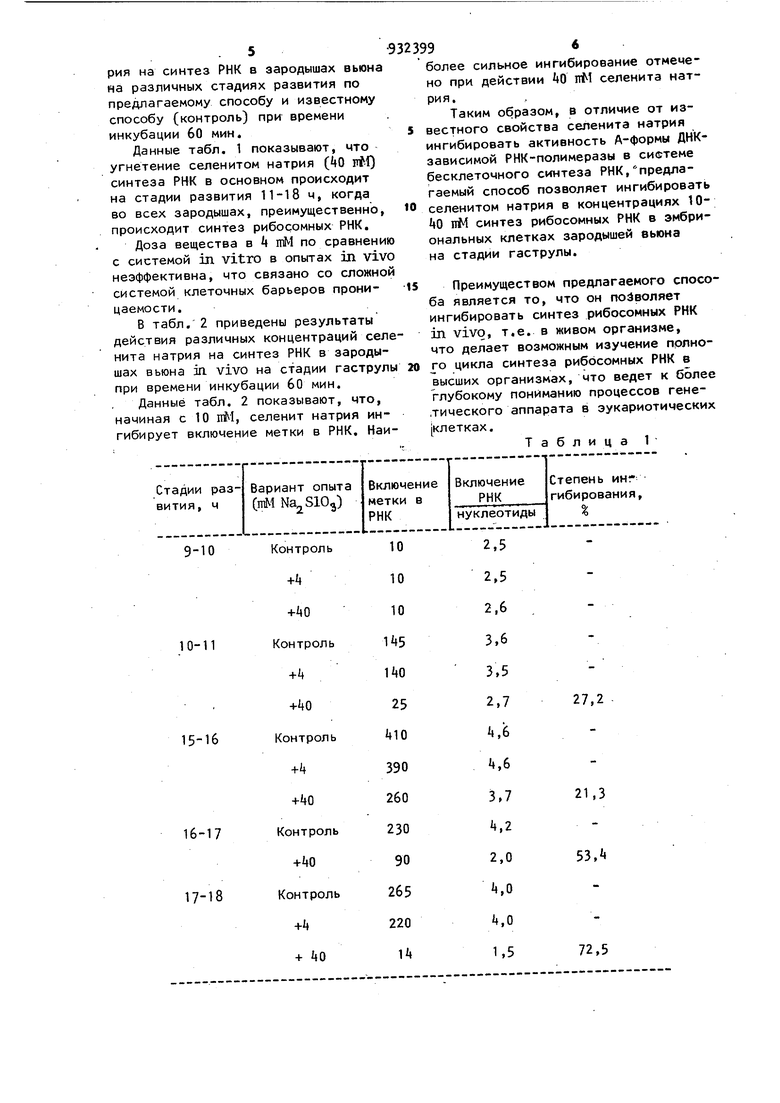
В табл,,1 приведены результаты исследований действия селенита натрия на синтез РНК в зародышах вьюна на различных стадиях развития по предлагаемому способу и известному способу (контроль) при времени инкубации 60 мин.
Данные табл. 1 показывают, что угнетение селенитом натрия (ijO rtl) синтеза РНК в основном происходит на стадии развития 11-18 ч, когда во всех зародышах, преимущественно, происходит синтез рибосомных РНК.
Доза вещества в 4 тМ по сравнению с системой in vitro в опытах in vivo неэффективна, что связано со сложной системой клеточных барьеров проницаемости.
В табл. 2 приведены результаты действия различных концентраций селенита натрия на синтез РНК в зародышах вьюна in. vivo на стадии гаструлы при времени инкубации 60 мин.
Данные табл. 2 показывают, что, начиная с 10 irM, селенит натрия ингибирует включение метки в РНК. Наи9-10
10-11
15-16
16-17
17-18
более силь-ное ингибирование отмечено при действии tO г селенита натрия.
Таким образом, в отличие от известного свойства селенита натрия ингибировать активность А-формы ДНКзависимой РНК-полимеразы в системе бесклеточного синтеза РНК,предлагаемый способ позволяет ингибировать селенитом натрия в концентрациях 10kQ nW синтез рибосомных РНК в эмбриональных клетках зародышей вьюна на стадии гаструлы.
Преимуществом предлагаемого способа является то, что он позволяет ингибировать синтез рибосомных РНК in vivo, т.е. в живом организме, что делает возможным изучение полного цикла синтеза рибосомных РНК в высших организмах, что ведет к более глубокому пониманию процессов генетического аппарата в эукариотических клетках.
Таблица 1
27,2
21,3
53,
72,5
Формула изобретения
Способ ингибирования синтеза рибосомных РНК путем инкубирования
,1 |ИолЬгического материала с раствором 1селенита натрия, о т л и чающийс я тем, дто, с целью ингибирования синтеза рибосомных РНК в нативных клетках, интактные бластодермы отделяют от желтка, инкубируют с селенитом натрия, затем добавляют в инкуТаблица 2
бационнуюсмесь Н -уридин в концентрации ijO-SO мк Ки/мл, .отмывают от невключившейся метки добавляют РНКноситель, уриди и контролируют степень ингибирования определением фракции РНК-ДНК. I
Источники информации, принятые во внимание при экспертизе 1. Авторское свидетельство СССР ff627t62, кл. С 12 К 1/02, 1978.
