Изобретение относится к генетическим исследованиям микроорганизмов и касается идентификации ауксотрофов. Ауксотрофные мутанты микроорганизмов находят широкое применение в научных исследованиях при картировании генов и создании генетических карт микроорганизмов, для выяснения путей биосинтеза тех или иных метаболитов, крайне необходимы в генетической инженерии.
При этом особенно большой интерес представляют множественные ауксотрофные мутанты. Получение таких мутантов - длительный и многоступенчатый процесс в силу сложности их идентификации.
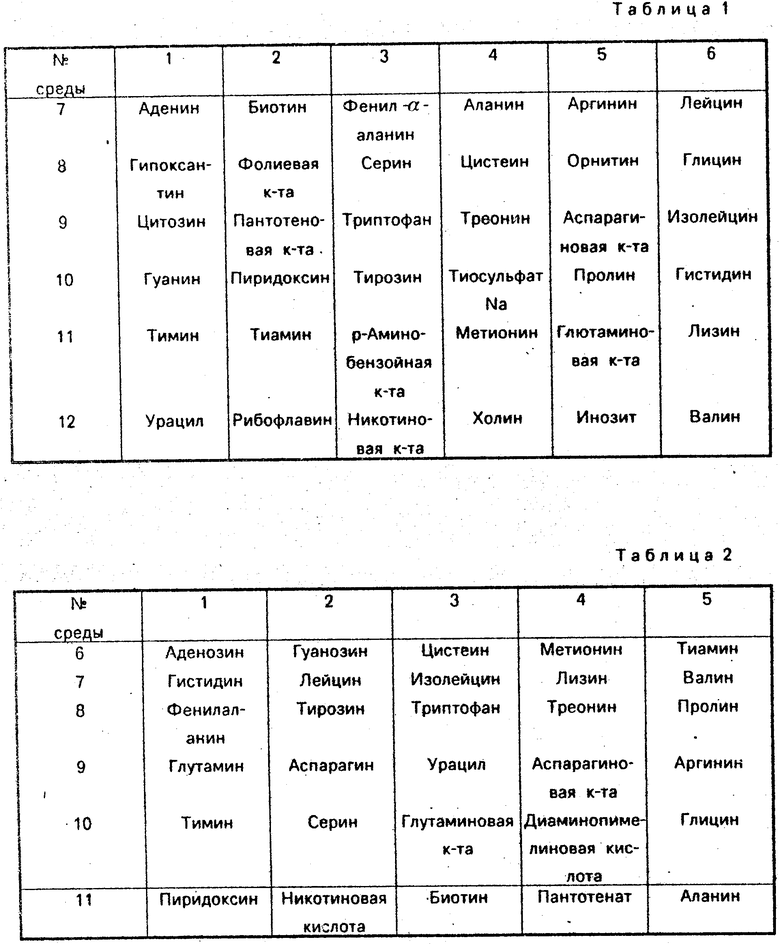
Известен и широко используется способ идентификации биохимических мутантов, в соответствии с которым анализируемые мутанты высеваются на 12 питательных сред, содержащих помимо минимального агара группы определенных аминокислот, витаминов и оснований (табл. 1). При этом каждый из 36 факторов роста содержится в двух средах. Этим способом могут быть определены ауксотрофные мутанты 1) по какому-либо одному метаболиту; 2) мутанты с нарушением общих путей биосинтеза, например val, leu или trp, tyr; 3) мутанты, требующие одного из двух факторов (т. е. имеющие альтернативные потребности в факторах роста). Мутант первого типа вырастет на каких-либо двух средах, если он нуждается в факторе, общем для этих сред. Мутант второго типа вырастет на одной среде, если необходимые ему для роста факторы входят в эту среду. Точная идентификация мутанта требует дополнительных экспериментов. Мутант третьего типа вырастает больше, чем на двух средах.
Описана также аналогичная схема сред (табл. 2), в которой количество факторов роста сокращено до 30 наиболее необходимых, кроме того, витамины вынесены в отдельную среду. При этом, если имеется потребность в каком-либо определенном витамине, необходима дополнительная проверка.
Эти схемы позволяют определить одиночные мутации, возникающие в результате точечного воздействия мутагена в каком-либо определенном участке хромосомы. В то же время известно, что встречаются множественные ауксотрофные мутации - двойные, тройные в результате действия мутагенов на разные участки хромосомы, а также в результате неоднократной обработки мутагенами. Ауксотрофные потребности таких мутантов не могут быть идентифицированы одномоментно с использованием вышеупомянутых схем и требуют трудоемкого последовательного многостадийного анализа, например, на базе метода реплик.
Цель изобретения - одномоментное определение множественных, прежде всего двойных и тройных, ауксотрофных потребностей мутантных штаммов.
Поставленная цель достигается тем, что предлагаемый набор питательных сред в отличие от известных содержит среды со всеми возможными парами и практически всеми возможными тройками исследуемых факторов роста.
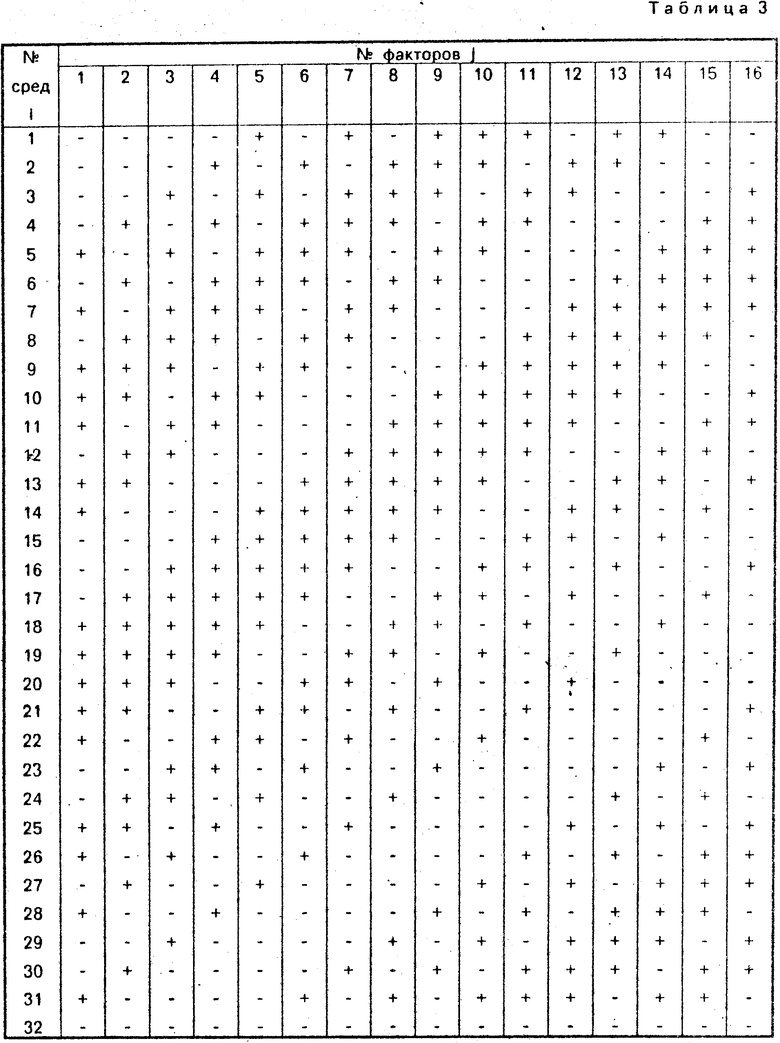
Сущность изобретения заключается в том, что исследуемые штаммы переносятся на 32 питательные среды, построенные специальным образом. Подробная схема, в соответствии с которой среды 1-31 получаются из минимальной питательной среды 32, приведены в табл. 3. В первом столбце представлены номера сред i, в первой строке - условные номера факторов роста j. Включение вещества j в среду i отмечено знаком + на пересечении j-го столбца и i-й строки. Максимально допустимое число факторов - 31. Если эксперимент проводится с меньшим числом факторов, то последние столбцы при составлении сред не учитываются. При необходимости одновременного исследования большего числа факторов можно ввести общественные факторы, представляющие совокупности каких-либо ростовых веществ, объединенных в группы (например, группа витаминов).
Ключевой особенностью схемы табл. 3 является то, что каждая пара исследуемых факторов входит в 8 питательных сред, что позволяет надежно определить парную ауксотрофность штаммов. Далее практически каждая (96,55% всего возможного многообразия троек) входит в 4 питательных среды, что позволяет определить ауксотрофные потребности мутантов, нуждающихся в трех факторах роста. Наконец, каждый из факторов входит в 16 сред, что позволяет с большой надежностью идентифицировать одиночные ауксотрофные потребности.
В качестве минимальной питательной среды можно использовать какой-либо вариант "голодного агара", например, среду Фогеля-Боннера, среду Спицайзена, а также другие среды по усмотрению экспериментатора в зависимости от исследуемого объекта.
В примерах использована минимальная агаризованная среда Спицайзена, содержащая на 1 л: (NH4)2SO4 - 2 г, КН2РО4 - 6 г, К2НРО4 ˙ 3Н2О - 18,34 г, цитрат Na˙ 5H2O - 1,34 г, MgSO4 ˙ 7H2O - 0,2 г, глюкоза - 10 г, агар - 20 г. В соответствии с табл. 3 этой среде присваивается N 32, а каждая из сред NN 1-31 дополнительно содержит до 16 из 31 фактора роста. Принятое далее соответствие исследованных факторов роста (аминокислот, азотистых оснований, витаминов) номерам j табл. 3 приведено в табл. 4, но подчеркнем, что оно может быть произвольным.
Целесообразно заранее приготовить стерильные водные растворы каждого фактора, скомбинировать их для образования пулов по средам и хранить на холоду. Исследуемые штаммы переносятся на 32 среды схемы, при этом результаты роста по всем штаммам анализируются одновременно.
Для получения необходимых в дальнейшем анализе количественных оценок использована дискретная двухбалльная модель: визуально определяемое наличие роста оценивается единицей, отсутствие роста - нулем. Соответственно для каждого штамма k записывается совокупность значений Yi(k), так что Yi(k) = 1, если штамм k на среде i вырос и Yi(k) = 0, если роста не наблюдалось.
Математический этап анализа заключается в подсчете коэффициентов аj(k) по формуле где Xij = +1, если фактор j входит в среду i (+ в клетке i, j таблицы 3) и Xij = -1, если не входит. При проведении расчетов целесообразно воспользоваться программируемым далее, выписывая только ненулевые произведения,
а2 = 1 - 1 + 1 - 1 - 1 + 1 - 1 + 1 = 0,
a3 = -1 + 1 + 1 - 1 + 1 + 1 - 1 - 1 = 0,
a4 = 1 + 1 + 1 + 1 + 1 + 1 + 1 + 1 = 8, далее, опуская все выкладки,
а5 = а6 = 0, а7 = 8,
а8 = 0, а9 = -8,
аj = 0, j ≥ 10.
Итак, ненулевых коэффициента три: а4 = = а7 = 8 и а9 = -8. Рассматривая совместно столбцы j = 4 и j = 7 табл. 3 видим, что факторы N 4 и N 7 одновременно присутствуют в средах с номерами i = 4, 7, 8, 15, 16, 19, 22, 25. Именно на этих восьми средах получен рост штамма и шло подсуммирование единиц при вычислении коэффициентов а4 и а7, в результате чего получились восьмерки. Что касается коэффициента а9 = -8, легко понять, что он представляет не отрицательный линейный эффект фактора N 9, а связан с коэффициентом b47 при произведении Х4Х7 в модели процесса Y= Σ bjXj+ +Σ bjmXjXm (используя теорию планирования эксперимента, можно показать, что b47 = -a47/16). Переходя к новым переменным Xj = (xj + 1)/2, получим модель роста штамма Bac. subtilis 17-15: Y = X4X7, которая показывает, что рост (Y = 1) имеет место лишь при одновременном присутствии в среде факторов j = 4 и j = 7, так как в этом случае Х4 = Х7 = 1 и Х4˙Х7 = 1 (при отсутствии в среде одного из факторов, например, Х4 = 0, получаем Y = 0, т. е. отсутствие роста).
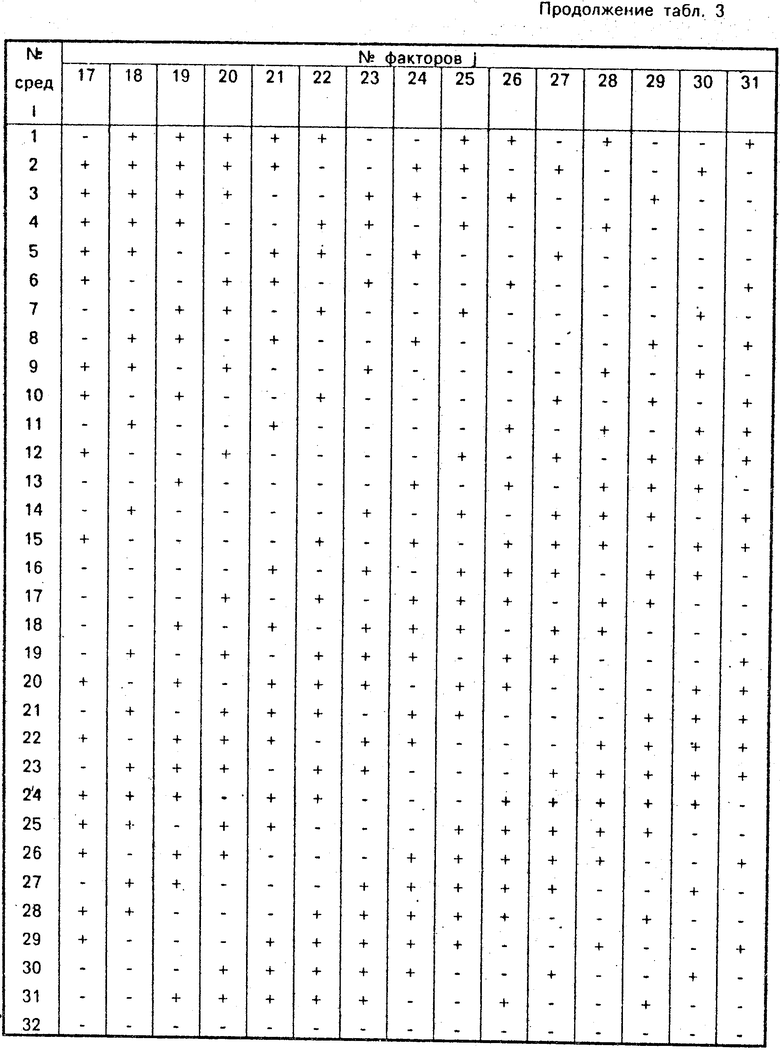
Таким образом, мы имеем здесь дело с парной ауксотрофностью штамма Bac. subtilis 17-15 по лейцину и метионину (см. соответствие факторов номерам в табл. 4). Определить же ауксотрофные потребности данного штамма по способу-прототипу оказалось невозможно: как видно из табл. 6 (второй столбец) ни на одной диагностических сред роста этого штамма не наблюдается, ведь ни одна из сред не содержит лейцина и метионина одновременно. Точности ради отметим, что схемы-прототипы, содержат около 30% всех возможных парных сочетаний исследуемых факторов и в этих случаях определить парную ауксотрофность в принципе можно, но требуется дополнительный эксперимент, по трудоемкости не уступающий первому.
Для заявляемого способа математический анализ всех подобных ситуаций позволяет сформулировать следующее правило 1. Если в результате постановки эксперимента на средах, приготовленных в соответствии с табл. 3, подсчеты по формуле (1) дают два ненулевых коэффициента, равных +8 и третий, равный -8, то факторы, соответствующие первым двум, образуют пару, обеспечивающую рост (парная ауксотрофность).
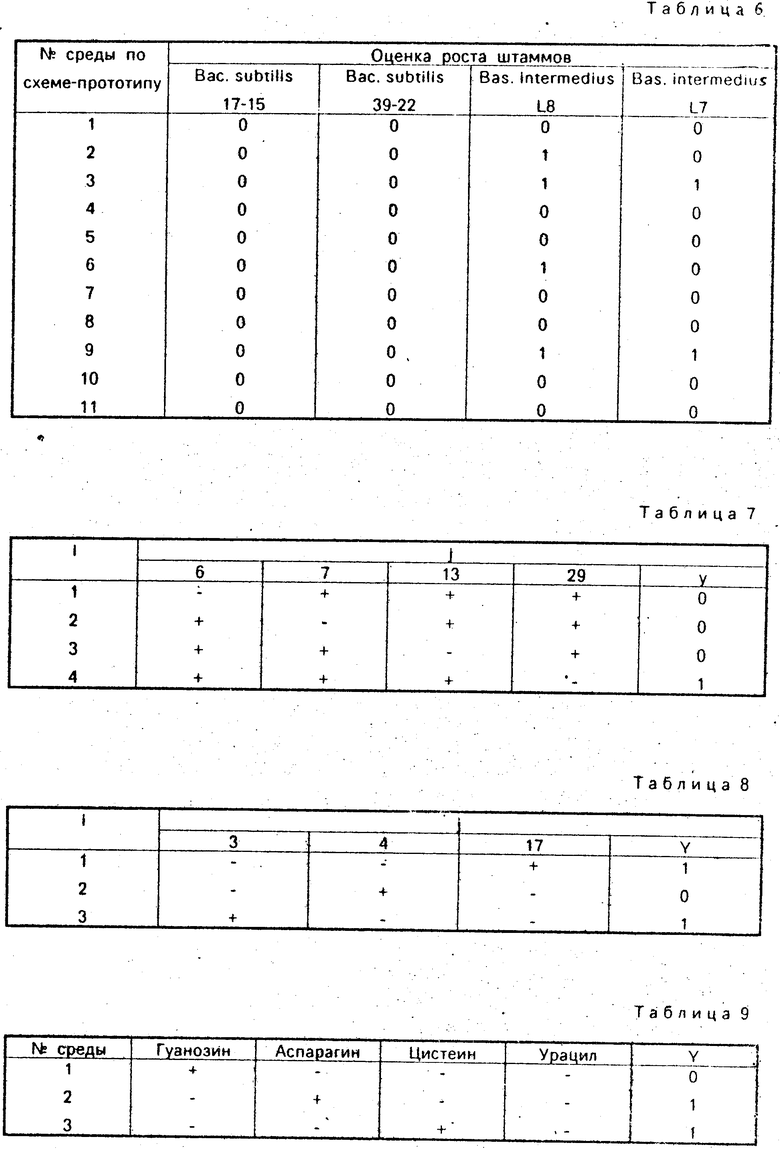
П р и м е р 2. Результаты роста штамма Bac. subtilis 39-22 приведены в третьем столбце табл. 5. Рост имеет место на четырех средах, а ненулевыми оказываются следующие коэффициенты а6 = а7 = а13 = а29 = 4, а17 = а20 = а22 = -4. Рассматривая совместно столбцы NN 6, 7, 13, 29, табл. 3, легко увидеть, что в тех средах, на которых наблюдался рост (т. е. в средах с номерами i = 8, 13, 14, 16) присутствуют одновременно все факторы с номерами j = 6, 7, 13, 29, в остальные же среды, на которых рост отсутствовал, факторы 6, 7, 13, 29 не входят вообще, или входят попарно в различных сочетаниях. Очевидно, здесь мы имеем дело со множественной ауксотрофностью. Нельзя утверждать, что необходимыми в данном случае являются все факторы NN 6, 7, 13, 29, но такую возможность следует допустить. Поставив дополнительный опыт с четырьмя средами, в каждой из которых отсутствует один из таких факторов, но присутствуют остальные, определим, есть ли среди рассматриваемых четырех факторов "лишний" или все они необходимы. По существу этот эксперимент заменяет обычную в таких случаях проверку. В данном случае рост наблюдался на среде, в которую не был включен фактор N 29 (см. табл. 7) откуда последовал вывод: штамм Bac. subtilis 39-22 - ауксотроф по сочетанию гистидин + лейцин + триптофан. (Математическая модель: Y = = X6X7X13).
Для аналогичных случаев множественной ауксотрофности получено правило 2. Если среди коэффициентов aj четыре окажутся равными +4, то имеет место тройная или четверная ауксотрофность. Для полного разрешения вопроса надо поставить эксперимент, приготовив четыре среды, полученные из минимальной добавлением разных наборов троек из тех четырех факторов j, для которых получено aj= 4.
Результаты роста штамма Bac. subtilis 39-22 на средах схем-прототипа приведены в третьем столбце табл. 6. Ни на одной из сред роста не обнаружено и определить ауксотрофные потребности штамма оказывается невозможно. Как и в примере 1, отсутствие среды, содержащей одновременно необходимые данному штамму факторы роста, не является случайным. Совокупности отдельных троек факторов, входящих в тест-среды способа-прототипа, составляют ничтожно малую часть всех возможных их сочетаний.
Таким образом, примеры 1 и 2 иллюстрируют эффективность заявленного способа в наиболее часто встречающихся случаях полиауксотрофности в противоположность практически полному бессилию способа-прототипа.
П р и м е р 3. Результаты роста штамма Bac. intermedius L8 приведены в столбце 3 табл. 5. После расчетов по формуле (1) получаем следующие ненулевые коэффициенты а3 = а4 = а17 = 8. Рассматривая совместно столбцы 3, 4, 17 табл. 3, легко установить, что встречающиеся здесь комбинации факторов 3, 4, 17 образуют, по терминологии теории планирования эксперимента, восьмикратно повторенный дробный факторный план ДФЭ 23-1, что определяет смешение оценок для линейных эффектов и взаимодействий и не позволяет понять распределение "ролей" между факторами 3, 4, 17. Выход состоит в постановке небольшого дополнительного эксперимента, по существу заменяющего обычную проверку. Схема этого эксперимента приведена в табл. 8. Легко видеть, что фактор N 4 не является необходимым для роста. Что же касается факторов N 3 и N 17, ясно их альтернативное действие. Таким образом, для роста штамма Bac. intermedius L8 нужен либо цистеин, либо аспарагин. Математическим выражением этого факта является модель Y = = 0,5 X3 + 0,5 X17, означающая, если учесть дискретность величины Y, что возможность оказывать влияние на рост данного штамма присуща в равной мере (0,5 + 0,5 = 1) факторам j = 3 и j = 17.
Результаты роста штамма Bac. intermedius L8 на средах схемы-прототипа приведены в четвертом столбце табл. 6. Рост штамма на средах 2, 3, 6, 9 позволяет предположить возможными факторами роста следующие: гуанозин (пересечение 2-й и 6-й сред, табл. 2), аспарагин (пересечение 2-й и 9-й сред), цистеин (пересечение 3-й и 6-й сред), урацил (пересечение 3-й и 9-й сред). Для окончательного разрешения вопроса необходима постановка дополнительного эксперимента, что и было сделано (табл. 9). Анализ результата - рост на средах 2 и 3 - приводит к заключению об альтернативной ауксотрофности по цистеину и аспарагину.
Итак, в случае альтернативной ауксотрофности оба способа - заявленный и прототип - дают ответ на вопрос об определении веществ, необходимых для роста штамма-ауксотрофа, в два этапа.
В рамках заявленного способа рассмотренная в примере 3 задача подпадает под правило 3. Если получено три ненулевых коэффициента aj, причем все они равны 8, следует поставить дополнительный эксперимент с тремя средами, в каждую из которых входит лишь один фактор из трех, для которых aj = 8. Отсутствие роста на среде, содержащей какой-либо из этих факторов, будет означать, что альтернативными факторами роста являются два других.
П р и м е р 4. В пятом столбце табл. 5 приведены результаты роста штамма Bac. intermedius L7. Подсчет по формуле (1) приводит к единственному ненулевому коэффициенту а18 = 16 (вклад в это число дают те 16 сред, в которые входит фактор N 18). Интерпретация этого результата проста и однозначна: единственным фактором ауксотрофности для штамма Bac. intermedius L7 является урацил. (Математическая модель: Y = X18).
Для всех аналогичных ситуаций действует правило 4. Если получен единственный ненулевой коэффициент aj (он будет равен 16), то исследуемый мутант будет ауксотрофом по единственному фактору с соответствующим номером.
Результаты роста штамма на средах способа-прототипа приведены в пятом столбце табл. 6. Анализируемый штамм вырос на средах 3 и 9. Общим фактором для этих сред является урацил, что говорит о соответствующей моноауксотрофности. Признавая, что новым способом тот же результат получен с помощью существенно большего числа сред, заметим, что 16-кратное дублирование вхождения соответствующего фактора в среды практически гарантирует его достоверность.
На расширенные возможности нового способа указывает следующее примечание. Описанные выше правила полностью справедливы, если в постановке эксперимента (составление сред, констатация роста) не допущено ни одной ошибки. Если допустить, что совершена одна ошибка (например, какое-либо вещество-фактор роста не добавлено в одну из сред или констатируется отсутствие роста ввиду сомнительности визуальной оценки), во всех рассмотренных случаях можно непосредственно сделать те же заключения, что и при отсутствии ошибки, учтя, что некоторые коэффициенты окажутся измененными на ± 1. Получение правильных выводов возможно и в некоторых из случаев, когда ошибок указанного рода совершено больше. При применении методик ( 1, 2, 3) аналогичные ошибки могут привести либо к невозможности выявления искомого эффекта, либо к неверному выводу.
Таким образом, разработан новый универсальный статистически обоснованный способ идентификации ауксотрофных мутантов, позволяющий определять как одиночную и альтернативную, так и, причем весьма эффективно, множественную ауксотрофность (по 2, 3 и даже четырем факторам).
Идентификация ауксотрофных мутантов предлагаемым способом может быть осуществлена в любой лаборатории, не требует дополнительного оборудования. Результаты могут быть обработаны вручную описанным выше способом, а при наличии микрокалькулятора типа МК-56 или компьютера обработка кардинально упрощается.
Этим способом можно пользоваться как основным при идентификации любых вновь получаемых ауксотрофов, или использовать его как дополнительный при идентификации тех штаммов, ауксотрофные потребности которых не удалось определить обычно употребляемыми способами. (56) Стент Г. и Камендар Р. Молекулярная генетика. М. : Мир, 1981, с. 117-130.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения ауксотрофных мутантов холерных вибрионов | 1980 |
|
SU982345A1 |
| ШТАММ BACILLUS SUBTILIS - ПРОДУЦЕНТ РИБОФЛАВИНА (ВАРИАНТЫ) | 1994 |
|
RU2081175C1 |
| ШТАММЫ БАКТЕРИЙ Bacillus subtilis И Bacillus amyloliquefaciens-ПРОДУЦЕНТЫ ИНОЗИНА И СПОСОБ ПРОДУКЦИИ ИНОЗИНА С ИХ ИСПОЛЬЗОВАНИЕМ | 2005 |
|
RU2333949C2 |
| Способ получения селективной питательной среды без аденина или урацила,или лейцина | 1983 |
|
SU1310425A1 |
| Штамм актиномицета Streptomyces parvulus ВКПМ Ас-2177, обладающий способностью к солюбилизации фосфора | 2023 |
|
RU2802956C1 |
| Штамм @ @ @ -тест-объект для определения продукции @ -триптофана | 1984 |
|
SU1191464A1 |
| СПОСОБ ПОЛУЧЕНИЯ РИБОФЛАВИНА, ШТАММ BACILLUS SUBTILIS - ПРОДУЦЕНТ РИБОФЛАВИНА (ВАРИАНТЫ) | 2002 |
|
RU2261273C2 |
| КОНСОРЦИУМ ПСИХРОТОЛЕРАНТНЫХ БАКТЕРИАЛЬНЫХ ШТАММОВ ДЛЯ БИОЛОГИЧЕСКОЙ ЗАЩИТЫ И СТИМУЛЯЦИИ РОСТА СЕЛЬСКОХОЗЯЙСТВЕННЫХ РАСТЕНИЙ | 2022 |
|
RU2795906C1 |
| Штамм бактерий SеRRатIа маRсеSсеNS - продуцент @ , @ -диаминопимелиновой кислоты | 1986 |
|
SU1377293A1 |
| ШТАММЫ БАКТЕРИЙ BACILLUS SUBTILIS И BACILLUS LICHENIFORMIS, ИСПОЛЬЗУЕМЫЕ В КАЧЕСТВЕ КОМПОНЕНТОВ ПРЕПАРАТА ПРОТИВ ВИРУСНЫХ И БАКТЕРИАЛЬНЫХ ИНФЕКЦИЙ, И ПРЕПАРАТ НА ОСНОВЕ ЭТИХ ШТАММОВ | 1997 |
|
RU2142287C1 |



Сущность изобретения: генетические исследования, идентификация ауксотрофов. Сущность изобретения: способ определения ауксотрофных потребителей мутантных штаммов предполагает составление набора питательных сред, содержащего все возможные пары и тройки исследуемых факторов роста. Проведено математическое обоснование и сформулированы правила вывода заключений о различных типах ауксотрофности. 9 табл.
СПОСОБ ОПРЕДЕЛЕНИЯ АУКСОТРОФНЫХ ПОТРЕБНОСТЕЙ МУТАНТНЫХ ШТАММОВ БАКТЕРИЙ, включающий выращивание бактерий на комплексе питательных сред, содержащих минимальный агар, а также факторы роста, отличающийся тем, что, с целью одновременного определения множественных ауксотрофных потребностей, бактерии выращивают на 32 средах, при этом в каждую из них, кроме контрольной 32-й, включают до 16 факторов из набора факторов роста общим числом до 31, который формируют с учетом ожидаемых потребностей данной культуры, составление сред осуществляют следующим образом: нумеруют факторы числом j от 1 до 31, среды - числом m, в первую среду добавляют факторы с номерами j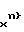 5, 7, 9, 10, 11, 13, 14, 18, 19, 20, 21, 22, 25, 26, 28, 31, а в последующие - с номерами jm,n, определяемыми по формуле jm,n = jm-1,n-1, m = 2 - 31, n = 1 - 16, если оказывается jm,n = 0, то полагают jm,n = 31, и по результатам роста на этих средах делают заключения об одиночной, альтернативной, двойной, тройной и четверной ауксотрофности.
5, 7, 9, 10, 11, 13, 14, 18, 19, 20, 21, 22, 25, 26, 28, 31, а в последующие - с номерами jm,n, определяемыми по формуле jm,n = jm-1,n-1, m = 2 - 31, n = 1 - 16, если оказывается jm,n = 0, то полагают jm,n = 31, и по результатам роста на этих средах делают заключения об одиночной, альтернативной, двойной, тройной и четверной ауксотрофности.
