Изобретение касается усовершенствования способа для определения иммунологически связываемого поливалентного вещества в присутствии по меньшей мере двух специфических для иммунологически связываемого вещества реактивов и в присутствии ингибитора для компенсации вредных факторов в пробах сыворотки крови человека, также пригодного для этого реактива.
Чувстительное определение иммунологически связываемых веществ, как, например, поливалентных антигенов (пептидов, протеинов, полисахаридов, вирусов, бактерий, специфических клеток) при использовании двух или при необходимости большего количества антител, которые направлены против пространственно различных антигенных детерминантов известно в виде иммунорадиометрического или иммуноэнзимометрического слоистого анализа (иммуноанализ с двумя участками). Наиболее употребительской является реализация этого способа определения, причем определяемый антиген (проба) инкубируется с помощью первого антитела, которое или в твердой фазе соединено с пригодным материалом-носителем, как например, сефарозой, агарозой, трубочками из синтетического полимера и тому подобное, или имеется гомогенно, как например, битинилированно в растворе и с помощью определенного количества второго или других маркированных антител в жидкой фазе. Специфичность второго и при необходимости других антител выбирается предпочтительно таким образом, чтобы она была направлена против других детерминантов определяемого антигена как первое антитело, чтобы исключить конкурентную борьбу между антителами за те же самые места присоединения к определяемому антигену, так как этим была бы снижена чувствительность пробы. Как первое, соединенное в твердой фазе или биотинилированное, так и второе или другое имеющееся в растворе маркированное антитело добавляются в избытке. Соответствующий антиген может определяться или через зафиксированную на первом антителе или оставшуюся в растворе, иммунологически не связанную активность. В последнем случае необходимо добавлять определенное количество маркированных вторых антител.
По причине необходимой специфичности для иммунологических слоистых анализов используются, в частности, выведенные из моноклональных антител (МАК) совершенные иммуноглобулины IgG или их иммунологически реактивные фрагменты (Fab, F(a,bl)2). Однако кроме того в эти пробы можно вводить иммунореактивные составные части, которые выводятся из поликлональных антител (РАК).
Хотя в описанных иммуноанализах с двумя участками для определяемых аналитов используются специфические антитела, со значительной частотой появляются пробы сыворотки крови человека, которые вызывают неспецифические реакции. Последние приводят к ошибочным результатам испытаний с соответственно отягчающими последствиями для терапевтических мероприятий. Появление неспецифических реакций можно объяснить имеющимися в пробе веществами, которые точно также, как и определяемый аналит (поливалентный антиген), связывают специфические иммуноглобулиновые реагенты (вредные факторы), однако в других точках воздействия.
Поэтому обычно к таким иммуноанализам превентивно в избытке добавляют неспецифические иммуноглобулины или фрагменты иммуноглобулинов (как правило, речь при этом идет об иммуноглобулине С, 1 С) тех же видов животных, от которых происходят специфические антитела (М.М. Addison in Radio immunoassay and Related Procedures in Medicine Vol. 1, 131 147 (1974); европейский патент N 0174026). С момента применения специфических моноклональных антител, которые, как правило, происходят от мыши, указанное мероприятие по устранению помех в иммуноанализах требует большого количества неспецифического мышиного иммуноглобулина IgG в форме мышиной сыворотки, мышиного асцита или изолированного мышиного иммуноглобулина (примерно 300-500 мкг/мл; CLin, Chem. 32, 1491 1495 1986). Например, в соответствии с европейским патентом N 0174026 благодаря добавке примерно 30 мкг/мл важного неспецифического мышиного или крысиного иммуноглобулина IgG к иммунопробам описанного типа только в определенных сыворотках и в особых случаях (отрицательных пробах) достигается полная компенсация нарушений. Однако приготовление мышиного иммуноглобулина в масштабе порядка килограммов невозможно при существующих сегодня методах в экономически интересных условиях и к тому же вызывает критику с этических точек зрения.
Важным мероприятием для избежания помех в иммунопробах указанного типа является применение Fab или F(abl)2 фрагментов по меньшей мере для одного из примененных в иммунопробе специфических антител. Тем самым все вредные факторы в пробе, которые направлены на Fc-части иммуноглобулина IgG (ревматические факторы, анти-Fc-иммуноглобулины, как например, IgM), устраняются в точке его воздействия на одном из специфических реагентов и таким образом не нуждаются в компенсации.
Однако несмотря на использованные специфические реагенты антител, свободные от фрагментов Fc, в некоторых сыворотках крови человека в иммунопробах появляются нарушения. Эти нарушения можно объяснить наличием в сыворотке субстанций, которые направлены на фрагменты Fab или F(a,bl)2. В соответствии с европейским патентом N 0083869 такие вредные факторы зоны Fab распознаются только тогда, когда они отделены от Fc-части. В соответствующих иммунопробах эти факторы могут быт устранены путем добавки нативных или сетчатых, неспецифических или определяемого антигена фрагментов Fab или F(abl)2 (европейский патент N 0083869). При этом агрегированые фрагменты Fab или F(abl)2 обнаруживают в 2-3 раза более высокое защитное действие по сравнению с нативными составными частями. Напротив, полные неспецифические иммуноглобулины IgG в соответствии с европейским патентом N 0083869 не способствуют защите от вредных факторов в указанных иммунопробах как в нативной, так и в агрегированной форме. Кроме того, способ имеет существенный недостаток, заключающийся в том, что при этом наряду со специфическими для определяемого антигена иммуноглобулиновыми реагентами требуется значительное количество высококачественных неспецифических иммуноглобулиновых реагентов, что привносит с собой существенные экономические, а также этические проблемы. В соответствии с этим для защиты от вредных факторов используются, например, по меньшей мере в количестве 100 мкг/мл, оказавшиеся эффективными агрегированные неспецифические фрагменты иммуноглобулина, свободные от фрагмента Fc.
Поэтому в основе изобретения лежит задача предоставления в распоряжение новых возможностей для компенсации неспецифических реакций при иммуноанализах с помощью содержащих Fc-фрагменты и свободных от Fc-фрагментов специфических реагентов антител в качестве специфических реактантов, благодаря чему в сыворотках крови человека надежно устраняются направленные на полные иммуноглобулины IgG и Fab- или F(abl)2-фрагменты вредные факторы, и которые с учетом этических точек зрения докускают экономическое использование усовершенствованных тестовых систем.
Эта задача решается с помощью способа для определения иммунологически связываемого вещества в биологической жидкости первого вида животных или людей в присутствии по меньшей мере двух специфических для иммунологически связываемого вещества реактантов второго вида животных и в присутствии ингибитора для компенсации вредных факторов, отличающегося тем, что для компенсации вредных факторов перемешивают реакционную смесь с 0,1-50 мкг/мл неспецифических для иммунологически связываемого вещества, выведенных из моноклональных или поликлональных антител агрегатов второго и/или третьего вида животных.
В качестве иммунологически связываемого вещества, в частности, используют такие поливалентные антигены, как пептиды, протеины, полисахариды, вирусы, бактерии и другие специфические клетки или их составные части. В качестве специфических реактантов дя соответственно определяемой иммунологически связываемого вещества могут использоваться как поликлональные, так и моноклональные антитела, при этом предпочтительно комбинированно используются иммуноглобулины IgG и такие их иммунореактивные составные части, как Fab или F(abl)2-фрагменты. Особенно предпочтительным является использование двух или при необходимости нескольких специфических реактантов, из которых по меньшей мере один является Fab или F(abl)2-фрагментом и остальные являются специфическими реактантами иммуноглобулинов IgG.
В качестве неспецифических для иммунологически связываемого вещества агрегатов антител дя компенсации нарушений предпочтительно используются агрегаты не специфические для иммунологически связываемого вещества иммуноглобулины IgB, особенно предпочтительным является агрегат иммуноглобулина IgG, состоящий из неспецифического иммуноглобулина IgG и другой макромолекулы. При этом предпочтительным являются неспецифические иммуноглобулины, которые происходят от того же самого вида животных, что и по меньшей мере один из специфических реактантов. В данном случае в качестве макромолекул пригодны, например, Fab или F(abl)2-фрагменты, т.е. свободные от Fc фрагменты иммуноглобулина IgG, которые могут происходить от получаемых от мыши или другого вида моноклональных или поликлональных антител, а также протеины (например, альбумин), полисахариды (например, декстран) или другие растворимые в воде полимеры. Кроме того, для компенсации нарушений могут использоваться гетерополимеры, которые образуются, например, из мышиного иммуноглобулина IgG, с помощью образующего мостиковые связи иммуноглобулина IgG крупного рогатого скота. Особенно предпочтительно используются агрегаты, состоящие из молекулы иммуноглобулина IgG и свободного от FC-фракции фрагмента иммуноглобулина IgG, причем как иммуноглобулины IgG, так и свободные от Fc-фракции фрагменты иммуноглобулина IgG происходят от того же вида животных, что и один из специфических реактантов. Такие IgG/Fab или F(abl)2-полимеры обнаруживают даже в три раза лучшее действие защиты от вредных факторов, чем чистые агрегаты иммуноглобулина. В зависимости от индивидуальной пробы сыворотки для успешной защиты от мешающих факторов достаточно 0,1-50 мкг/мл смешанного полимера иммуноглобулина IgG, однако предпочтительно работают с концетрациями от 5 до 25 мкг/мл и особенно предпочтительно с концентрациями от 5 максимум до 10 мкг/мл, так как в большинсте случаев полностью эффективным является уже значительно меньшее добавление неспецифических агрегатов иммуноглобулина.
В соответствии с предпочтительным вариантом осуществления изобретения для компенсации вредных факторов в сыворотках крови человека используются неспецифические гомополимерные или гетерополимерные агрегаты иммуноглобулина IgG или IgG/Fab F(abl)2-агрегаты с молекулярным весом 320000 дальтонов и более. При этом агрегаты иммуноглобулина IgG содержат иммуноглобулин IgG вида, который происходит по меньшей мере из одного из специфических реактантов и относятся к тем же подклассам, что и по меньшей мере один из специфических реактантов. При этом предпочтительно используются полимерные препараты иммуноглобулина IgG с молекулярными весами от 30 тысяч до 10 миллионов дальтонов, причем компенсирующее воздействие относительно нарушающих сыворотку факторов возрастает с увеличением молекулярного веса. Особенно предпочтительными вариантами осуществления изобретения являются иммуноанализы с двумя участками с двумя или несколькими специфическими реактантами мышиных или крысиных моноклональных антител, из которых по меньшей мере один реактант является Fab или F(abl)2-фрагментом и по меньшей мере один из других реактантов является содержащим Fc-фракцию иммуноглобулином IgG, в которых для компенсации вредных факторов добавляются гомополимерные или гетерополимерные неспецифические агрегаты иммуноглобулина IgG или IgG/Fab или F(abl)2-агрегаты с молекулярными весами более 320000 дальтонов, причем содержащиеся в неспецифических агрегатах иммуноглобулины IgG и Fab или F(abl)2-фрагменты являются моноклональными и происходят из вида или подкласса, которые являются по меньшей мере одним из специфических реактантов. При этом безразлично, получается ли неспецифический иммуноглобулин IgG или Fab-фрагмент с помощью асцита, ферментации или генетической инженерии путем экспрессии в микроорганизмах.
Пригодные по способу в соответствии с изобретением для компенсации нарушений полимеризованные неспецифические молекулы иммуноглобулина или неспецифические IgG/Fab или F(abl)2-агрегаты должны выводиться из другого вида, чем те, из которых происходит анализируемая проба. Как правило, иммунологические способы определения используются для анализа сывороток крови человека с помощью специфических выводимых от мыши моноклональных антител или иммунореактивных фрагментов. Однако специфические реагенты антител могут также происходить от другого вида животных. Добавленные для защиты от вредных факторов неспецифические агрегаты антител могут соответствовенно происходить от мыши или в комбинации с иммуноглобулинами мыши ии их Fab-фрагментов другого, отличного от первого вида животных. Например, для защиты от нарушающих факторов в иммуноанализах могут использоватсья гетерополимеры иммуноглобулина мыши, иммуноглобулина крупного рогатого скота (смотри выше), которые применяются в качестве специфического реактива моноклональных антител мыши или иммуноглобулинов мыши.
Благодаря добавке неспецифических агрегатов иммуноглобулина IgG или IgG/Fab или F(abl)2-агрегатов по сравнению с нативными иммуноглобулинами IgG достигается увеличенная на коэффициент 20-1000 и по сравнению с нативными ии агрегированными свободными от Fc-фракции фрагментами иммуноглобулина IgG на коэффициент 20-1500 или в три раза эффективность компенсации неспецифических реакций в иммуноанализах по меньшей мере с двумя специфическими для определяемого иммуногена реактантами, из которых по меньшей мере один выводится из Fab или F(abl)2-фрагмента. Это удивительно, так как нельзя было предусмотреть, что содержащие иммуноглобулин IgG неспецифические реактанты в иммунопробах, в которых участвуют свободные от Fc-фракции специфические иммуноглобулиновые реагенты моноклональных или поликлональных антител, вообще способствуют компенсации направленных на Fab или F(abl)2-фрагменты нарушений. Так как в этом патенте недвусмысленно подчеркивается, что как нативные, так и агрегированные иммуноглобулины IgG не прояввляют никакого устраняющего нарушения эффекта при указанном там иммунологическом тесте агглютинации и добавленные для защиты от нарушений, как и для специфической агглютинации агенты должны быт свободны от иммуноглобулина IgG.
Неспецифический иммуноглобулин IgG сам по себе или с фрагментами антител, а также с протеинами, полисахаридами или другими растворимыми в воде макромолекулами благодаря воздействию высокой температуры может образовывать мостиковые связи химически или нековалентно через биородственное взаимодействие в соответствии с известными из литературы методами. В любом случае в водных буферных растворах появляются растворимые агрегаты. Химическое образование сетчатых молекул может осуществляться, например, с помощью гомобифункциональных или гетеробифункциональных химических соединительных ветвей (линкеров), с помощью протеинов или активированного декстрана или путем самообразования сетчатых молекул иммуноглобулина IgG или их и/или других свободных от Fc-фракции фрагментов с помощью карбодиимида. Кроме того, можно связыват мономеры антител с помощью восстановления дисульфидом и повторного окисления или с помощью самоокисления их углеводной фракции или подходящей макромолекулой. В качестве химических соединителей в расчет принимаются, например, сложный эфир бис(малеинимидо)-метила, димети-суберимидат, димидил-суберат янтарной кислоты, глутардиальдегид, N-суцинимидидил-3-(2-пиридилдитио)пропионат, N-5-азидо-2-нитробензоил-суци- нимид, N-суцинимидил(4-иодацетил)-аминобензоат или комбинация малеинимидогексаноилсуцинимидата и S-ацетилангидрид меркаптоянтарной кислоты или аналогичные соединения. Получение активированного декстрана может осуществляться, например, из аминодекстрана определеного молекулярного веса с помощью малеинимидогексаноисуцинимида, который затем связывается с полученным производным меркаптоацетила неспецифических молекул иммуноглобулина IgG. С помощью биородственного взаимодействия подразумевались в общем и в целом такие связи в том виде, в каком они имеются, например, в парах биотин/авидин (или стрептавидин), гептен/антигаптен-антитела, антиген/антиантиген-антите- ла, лиганд/связующий протеин (например, тироксин/тироксиносвязующий протеин), гормон/рецептор. Надлежащий вариант исполнения получается путем по меньшей мере двойного биотинилирования неспецифического иммуноглобулина IgG и добавления стрептавидина или путем образования производных иммуноглобулина IgG, по меньшей мере с двумя молекулами дигоксигенина и образования мостиковых связей с моноклональным или поликлональным антителом антидигоксина. Точно также пригодными являются агрегаты, которые появляются в результате реакции обмена полиальбумина человека с моноклональным антителом антиальбумина человека.
Соответственно полученные сырые смеси агрегатов могут непосредствено использоваться после диализа или разделяться, например, с помощью гельфильтрации на фракции с увеличивающимся молекулярным весом и затем использоваться непосредственно для компенсации нарушающих факторов в иммунопробах.
Один из добавленных в испытательную смесь специфических реактантов, предпочтительно содержащее Fc-фракцию антитело иммуноглобулина IgG, связывается известным способом в твердой фазе с материалом-носителем, например агарозой или синтетическими трубочками и тому подобное, или имеется, например, в связанном с биотином виде, в жидкой фазе. Если используется биотинилированное первое антитело, вначале образуется комплекс из антигена и маркированного второго специфического реактанта, который затем в нужном месте связывается с материалом-носителем, покрытым связывающим биотин протеином, например, авидином или стрептавидином. Однако наряду с этим возможны также другие варианты проведения испытаний с первым битинилированным антителом: антиген вступает в реакцию с биотинилированным моноклональным антителом и в нужном месте или после определенного прединкубационного периода связывается с соединяющей биотин твердой фазой и затем лишь связывается с другим специфическим реактантом. Или возможен случай, когда вначале антиген вступает в реакцию со вторым специфическим реактантом и затем с биотинилированным антителом. Маркировка второго и при необходимости других специфических реактантов, в которых речь предпочтительно идет о свободных от Fc-фракции фрагментах иммуноглобулина IgG, осуществляется путем соединения с ферментом, с флуоресцирующей, хемилюминесцирующей или радиоактивной субстанцией. Способы маркирования подобного рода производных антител известны.
Изобретение пригодно для определения всех антигенов по меньшей мере с двумя антигенными детерминантами. Примерами этого являются тиреотропин (ТSH), карциноэмбриональный антиген (СЕА), вирусы гепатита (поверхностный антиген гипатита В, НВS), 1-фетопротеин (AFР), человеческий хорионгонадотропин (HCG), лютеинизированный гормон (LH), фоликулярностимулирующий гормон (FSH), β2-микроглобулин, кислая простатафосфатаза, пролактин, ферритин и инсулин.
Далее изобретение поясняют нижеследующие примеры.
П р и м е р. Получение моноклонального агрегата мышиного иммуноглобулина IgG путем "смешивания" с гомобиофункциональным реактивом
Моноклональный иммуноглобулин МАКЗЗ-IgG (более 95% чистый); подклассовый состав К, γ I: специфичность антикреатинкиназа) изолируется из асцетической жидкости с помощью осаждения сульфата аммония и хроматографии на ионнообменнике DEAE (см. A. Johnstone und R. Thorpe, Immunocheuistry in practice, Blachwell Scientific Publications 1982, с. 44-45).
50 мг иммуноглобулина IgG растворяются в 3 мл, 0,025 молярного бикарбонаткарбонатного буффера с водородным показателем 9,5. В этот раствор выпускают пипеткой 17 мкл 12,5%-ного раствора глутардиальдегида и инкубируют в течение 2 ч при 25оС. Затем раствор охлаждается в ледяной ванне и с помощью 50 ммолей тританоламинового буфера устанавливается водородный показатель 8,2. В этот растор добавляют 0,6 мл свежеприготовленного раствора боргидрида натрия (8 мг боргидрида натрия) 1 мл бидестиллированной воды), и инкубируют в течение последующих 2,5 ч при 0оС. Через 16 ч диализа при 0-4оС около 10 ммолей фосфатного буфера с водородным показателем 75, содержащего 0,2 М NaCl, удаляются избыточные реактивы. Диализат путем улльтрафильтрации концентрируется до объема 1,25 мл. Часть этого концентрата (сырой смеси) используется непосредственно для устранения нарушающих факторов, 1,0 м хроматографируется с помощью гельфилльтрационной колонки с наполнителем АсА22 (LKB). Размер колонки 2 х 40 см, рабочий буфер 10 ммолей фосфата/100 ммолей NaCl c водородным показателем 7,5. Элюат колонки исследуется с помощью ультрафиолетового монитора при 280 нм на содержание протеина и фракционируется. Фракции всего содержащего протеин элюционного участка объединяются в 4 группы с одинаковым объемом. Путем калибровки гельхроматографической колонки с помощью протеинов известного молекулярного веса с группами могут быть сопряжены участки молекулярного веса 160000 400000, 400000 1000000, 1 2 млн и 2 10 млн дальтонов. После концентрирования групп до концентрации протеина около 2 мг/мл агрегатные растворы иммуноглобулина IgG различных участков молекулярного веса могут использоваться для устранения нарушающих факторов.
П р и м е р 2. Получение моноклонального агрегата мышиного иммуноглобулина путем "сшивания" с гетеробифункционалльным реактивом
а) Получение МАКЗЗ-IgG МН (малеинимидогексаноил-МАКЗЗ-IgG).
10 мг МАКЗЗ-I С растворяются в 4 мл 30 милимолярного фосфатного буфера с рН7,1. К этому раствору пипеткой добавляют 20 мкл (4 мкмоля), 0,2 молярного раствора малеинимидогексаноилсуциннимидата в диметилсульфооксиде. Реакционная смесь инкубируется в течение 1 ч при 25оС, затем в течение 16 ч около 10 ммолей фосфатного буфера с рН 6,1, содержащего 50 ммолей NaCl, диализируются при 0-4оС. Получают 4,45 мл растора с 22 мг МАКЗЗ-1gG-МН/мл.
б) Получение МАКЗЗ-IgG-SAMS (S-ацетилмеркаптосуцинил-МАКЗЗ-IgG).
100 мг МАКЗЗ-IgG растворяются в 4 м 0,1 молярного фосфатного буфера с водородным показателем 8,0. К этому раствору пипеткой добавляют 40 мкл 0,25 молярного раствора S-ацетилмеркаптоангидрида янтарной кислоты в диметилсульфоксиде. Реакционная смесь инкубируется в течение 1 ч при 25оС, затем в течение 16 ч около 10 ммолей фосфатного буфера с рН 6,1, содержащего 50 ммолей NaCl, диализируется при 0-4оС. Получают 4,45 мл с 21,8 мг МАКЗЗ-IgG-SAMS/мл.
в) "Сшивание" МАКЗЗ-IgG-МН с МАКЗЗ-IgG-MS (меркаптосицинил-МАКЗЗ-IgG).
50 мг МАКЗЗ-IgG-SAMS разбавляют до концентрации 15 мг/мл в 25 ммолярном фосфатном буфере с рН 6,5, содержащем 2 ммоля этилендиаминтетраацетата (буфер А). К этому растору пипеткой добавляют 75 мкл 1 молярного раствора гидроксиламина и инкубируют смесь в течение 20 мин при 25оС.
3 мл этого раствора с 44 мг МАКЗЗ-IgG-MS разбавяют буфером А до 6,0 мл. В этот раствор добавляют 4 м раствора с 88 мг МАКЗЗ-IgG-МН. Смесь инкубируется в течение 40 мин при 25оС, затем для обрыва реакции "сшивания" разбаляется до 2 ммолей цистеином. После дальнейшей 30-минутной инкубации при 25оС добавляется до 5 ммолей N-метималеинимид и раствор выдерживается в течение 1 ч при 25оС. Реакционный раствор диализируется относительно 10 ммолей фосфатного буфера с рН 7,5, содержащего 0,2 моля NaCl. Диализат с помощью ультрафильтрации концентрируется до концентрации протеина 35 мг/мл и путем центрифугирования освобождается от небольшой мути. Агрегатная смесь может использоваться непосредственно или после разделения на фракции молекулярных весов (путем хроматографии АсА22, как в примере 1) для устранения нарушающих факторов.
П р и м е р 3. Получение агрегата МАКЗЗ-IgG путем "сшивания" с помощью аминодекстрана
а) Получение S-ацетилтиоацетил-аминодекстрана
100 мг аминодекстрана (декстан Т40, фармация; 28 аминогрупп/молекулярный вес 40000) растворяются в 4 мл 30 ммолярного фосфатного буфера с рН 7,1. К этому раствору пипеткой добавляют 0,25 мл 0,2 молярного раствора S-ацетилтиоацетилсуцинимидата в диметилсульфоксиде. Реакционная смесь инкубируется в течение 1 ч при 25оС, затем диализируется относительно 10 ммолей фосфатного буфера с рН 6,1, содержащего 50 молей NaCl. Выход составляет 90 мг S-ацетилтиоацетил-аминодекстрана в 4,5 мл раствора.
б) "Сшивание" МАКЗЗ-IgG-МН с тиоацетил-аминодекстраном
2 мл раствора с 20 мг/мл S-ацетилтиоацетил-аминодекстрана осторожно с помощью 0,1 молярного NaOH доводят до рН 6,5 и добавляют этилендиаминтетраацетат до 2 ммолей. К этому раствор пипеткой добавляют 40 мкл 1 молярного раствора гидроксиламина и инкубируют в течение 20 мин при 25оС. Затем раствор разбавляют 25 молярным фосфатным буфером с рН 6,5 до 6,8 м и добавляют 7,2 растора с 158,4 мг МАКЗЗ-IgG-MH (получение, как в примере 2а). После 30 мин инкубации при 25оС к раакционной смеси добавляют цистеин до 2 ммолей и после 30 мин дополнительно N-метилмалеинимид до 5 ммолей. После дальнейшей инкубации в течение 1 ч при 25оС полимеризат диализируется относительно 10 ммолей фосфатного буфера с рН 7,5/50 ммолей NaCl. После отделения центрифугированием слабой мути получают 19,5 мл раствора МАКЗЗ-IgG-декстран-агрегата с содержанием протеина 7,1 мг/мл.
П р и м е р 4. Получение агрегата МАКЗЗ-IgG путем "сшивания" с иммуноглобулином крупного рогатого скота IgG
а) Получение иммуноглобулина IgG-МН крупного рогатого скота
100 мг поликлонального иммуноглобулина крупного рогатого скота преобразовываются по аналогии с примером 2а с помощью 4 микромолей малеинимидогексаноилсуцинимидата. Выход составляет 4,45 мл раствора с 22 мг иммуноглобулина крупного рогатого скота IgG-MH/мл.
б) Получение МАКЗЗ-SATA (S-ацетилтиоацетил-МАКЗЗ-IgG)
100 мг МАКЗЗ-IgG растворяются в 4 м 30 ммолярного фосфатного буфера с рН 7,1. К этому раствору пипеткой добавляют 20 мкл (мкмоля) 0,1 молярного раствора S-ацетилтиоацетилсуцинимидата в диметисульфоксиде. Реакционная смесь инкубируется в течение 1 ч при 25оС, затем в течение 16 ч диализируется при 0-4оС относительно ммолярного фосфатного буфера с рН 6,1/50 ммолей NaCl. Получают 4,45 мол раствора с 22 мг МАКЗЗ-IgG-SATA/мл.
в) "Сшивание" тиоацетил-МАКЗЗ-IgG с иммуноглобулином крупного рогатого скота IgG-МН.
50 мг МАКЗЗ-IgG-SATA разбавляются 25 ммолярным фосфатным буфером с рН 6,5, содержащим 2 ммолярный этилендиаминтетраацетат (буфер А) до концентрации 15 мг/мл. К этому раствору пипеткой добавляют 75 мкл 1 молярного раствора гидроксиламина и инкубируют в течение 20 мин при 25оС.
К 3 мл этого раствора с 44 мг тиоацетил-МАКЗЗ-IgG добавляют 4 мл раствора с 88 мг иммуноглобулина крупного рогатого скота IgG-МН и инкубируют в течение 25 мин при 25оС. Для обрыва реакции "сшивания" добавляется затем цистеин до 5 ммолей и после 30 мин инкубации при 25оС иодацетомид до 5 ммолей. После дальнейшей инкубации в течение 1 ч при 25оС раствор диализируется в течение 16 ч при 0-4оС относительно 10 ммолярного фосфатного буфера с рН 7,5, содержащего 50 ммолей NaCl. После центрифугирования диализата получают 8,9 мл агрегата иммуноглобулина крупного рогатого скота МАКЗЗ-IgG с концентрацией протеина 13 мг/мл.
П р и м е р 5. Получение агрегата МАКЗЗ-IgG "сшитого" с помощью МАКЗЗ-Fab
а) Получение МАКЗЗ-Fab-SAТА
200 мг МАКЗЗ-IgG расщепляются с помощью папаина на Fab/Fc и МАКЗ-Fab изолируется из смеси путем хроматографии на DE52 целлюлозы (ватманская бумага) (методы смотри в источнике: A. Johnstone und R. Thorpe, Immunochemistry in Practice, Blackwelle Scientific Publications 1982, 52-53). Выход составляет 95 мг МАКЗЗ-Fab в виде специфического лиофилизата.
50 мг МАКЗЗ-Fab растворяются в 2 мл 30 ммолярного фосфатного буфера с рН 7,1. К этому раствору пипеткой добавляют 30 мкл (3 мкмоля) 0,1 молярного раствора S-ацетилтиоацетилсуцинимидата в диметилсульфоксиде. Реакционная смесь инкубируется в течение 1 ч при 25оС, затем в течение 16 ч при 0-4оС диализируется относительно 10 ммолярного фосфатного буфера с рН 6,1, содержащего 50 ммолей NaCl, 2 ммоля этилендиаминтетраацетата. Получают 2,6 мл раствора с 18,5 мг МАКЗЗ-Fab-SATA/мл.
б) "Сшивание" МАКЗЗ-IgG-МН с тиоацетил-МАКЗЗ-Fab
30 мг МАКЗЗ-Fab-SAТА разбавляются 25 ммолярным фосфатным буфером с рН6,5 до концентрации 15 мг/мл. К этому раствору пипеткой добавляют 50 мкл 1 молярного раствора гидроксиламина и инкубируют в течение 20 мин при 25оС.
В 1,5 мл этого раствора с 22 мг тиоацетил- МАКЗЗ-Fab добавляется 55 мг МАКЗЗ-IgG-МН (получение, как в примере 2а) и эта смесь разбавляется бидистиллированной водой до общего объема 10 мл. Исходная смесь инкубируется в течение 35 мин при 25оС и затем для обрыва реакции "сшивания" выводится с помощью цистеина до 2 ммолей. После 30 мин при 25оС добавляется L-метилмалеинимид до 5 ммолей и вновь смесь инкубируется в течение 1 ч при 25оС. Затем реакционный раствор диализируется в течение 16 ч при 0-4оС относительно 10 ммолярного фосфатного буфера с рН 7,5, содержащего 0,1 молярный NaCl. После центрифугирования получают 12,8 мл светлого раствора с 5,3 мг агрегата МАКЗЗ-IgG-Fab/мл.
П р и м е р 6. Получение агрегата МАКЗЗ-IgG путем "сшивания" с помощью биотин/стрептавидина
а) Получение биотинилированного МАКЗЗ-IgG
50 мг МАКЗЗ-IgG в 2,5 мл 0,1 молярного калийфосфатного буфера с рН 8,5 смешивают с 1,13 мг биотинил- ε -аминокрапновой кислоты-N-гидроксисуцинимида (фирмы "Берингер Маннхайм ГмбХ"), растворенного в 50 мл диметилсульфоксида (молярное соотношение иммуноглобулина IgG к биотину 1 7,5) и после перемешивания инкубируют в течение 90 мин при 25оС. Биотинилированный иммуноглобулин IgG диализирован в течение ночи при 4оС относительно 2 ммолярного калийфосфатного буфера с рН 7,0.
Выход: 45 мг МАКЗЗ-IgG-биотина в 6,4 мл.
б) "Сшивание" со стрептавидином
4 мл раствора МАКЗЗ-IgG-биотина с промежутками 5 мин смешивались с 3 частями по 1,5 мг стрептавидина (фирмы "Берингер Маннхайм ГмбХ") в 50 ммолярном калийфосфатном буфере/0,1 молярный хлорид натрия и инкубировались в течение 20 мин при 25оС. Молярное отношение иммуноглобулина 1 С к стрептавидину 2,5 1. Исходная смесь концентрирована с помощью ультрафильтрации до 2 мл и, как в примере 1, хроматографирована на колонке АсА22. Все фракции с молекулярным весом более 320000 объединены.
Выход: 21 мг агрегата МАКЗЗ-IgG-стрептавидина в 12 мл раствора с 17 мг содержания иммуноглобулина IgG.
П р и м е р 7. "Сшивание" моноклонального антитела антиальбумина человека с альбумином человека
40 мг альбумина человека (фирма "Берингверке", Марбург) агрегируют в 1 мл 50 ммолярного калийфосфатного буфера с рН 7,0 путем нагревания в течение 3 ч до 70оС. После охлаждения до 25оС часть с 25 мг агрегата альбумина человека разбавляют 50 ммолярным калийфосфатным буфером, содержащим 0,1 молярный NaCl, с рН 7,0 до концентрации 2,5 мг/мл, и через промежутки в 5 мин примешиваются 4 части по 2,5 мг МАК МI-антиальбумина человека (гамма I, каппа), растворенного в 0,2 мл 50 ммолярного калийфосфатного буфера с рН 7,0. После 20 мин инкубации при 25оС смесь концентрируют с помощью ультрафильтрации до 2 мл и хроматографирована, как и выше, на гелевой колонке АсА. Содержание протеин фракции с молекулярным весом более 320000 были объединены.
Выход: 12,4 мг агрегата МАК МI-IgG-альбумина в 7,8 мл с содержанием 8,86 мг иммуноглобулина IgG.
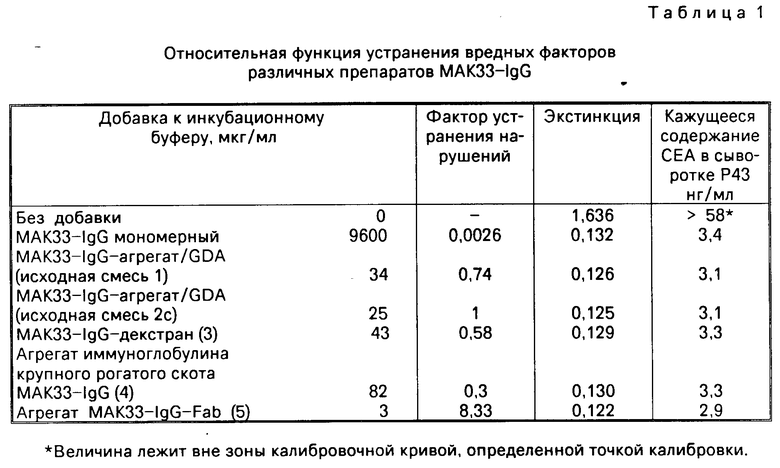
П р и м е р 8. Сравнение управнения мешающих факторов нативного МАКЗЗ-IgG и различных агрегатов МАКЗЗ-IgG в СЕА энзим-иммунопробе с двумя специфическими реактантами моноклональных антител
Используют реактивы из тест-упаковки энзимин R СЕА (фирма "Берингер Маннхайм ГмбХ"). Содержавшиеся в этом тесте пробирки покрываются моноклональным СЕА-специфическим мышиным иммуноглобулином (IgG I, K). Энзиммаркированный реактант является моноклональным, СЕА-специфическим конъюгатом мышиной Fab-пероксидазы; подклассовым составом Fab является К, γ I).
Различные препараты МАКЗЗ-IgG добавляют во все возрастающей концентрации соответственно к инкубационному буферу и испытание проводят с сывороткой человека (Р43), в которой были обнаружены необычайно эффективные нарушающие факторы. Табл. 1 показывает концентрацию препарата МАКЗЗ-IgG (в мкг протеина/мл) в инкубационном буфере, которая подавляла нарушение настолько, что было обнаружено правильное содержание аналита в нормальном диапазоне (1 3,5 нг/мл).
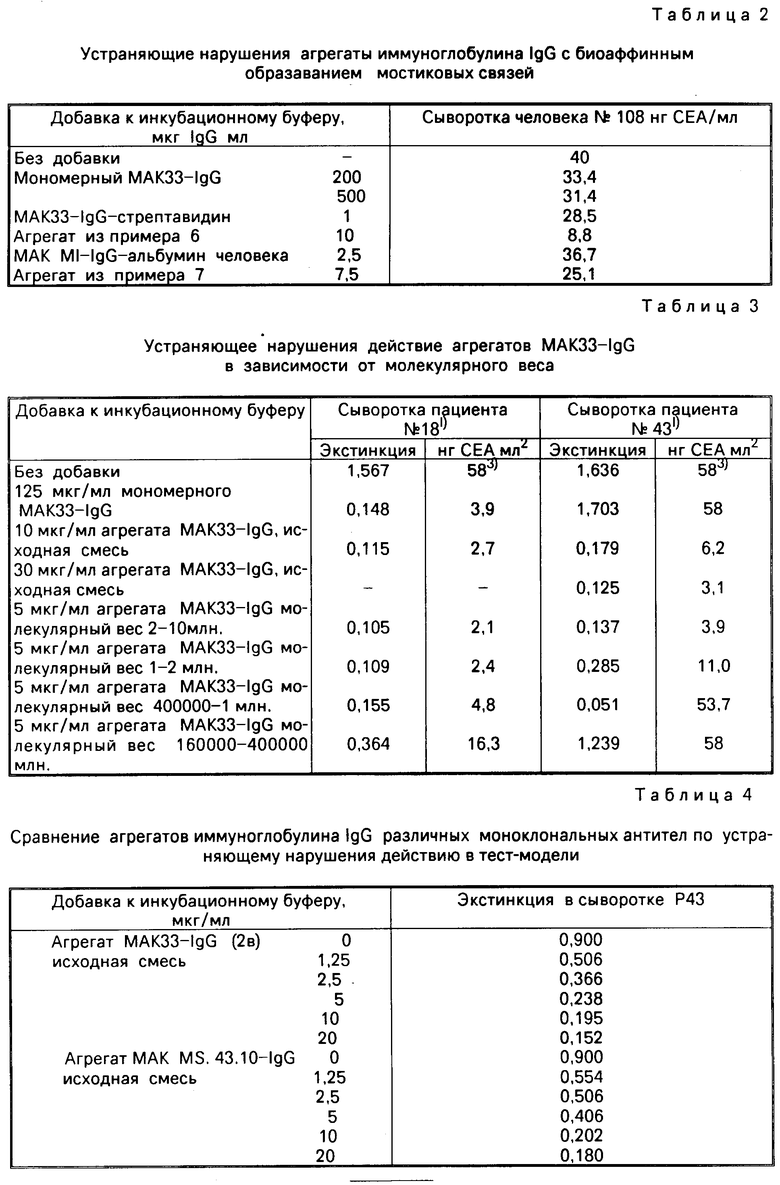
П р и м е р 9. Устраняющее нарушения действия агрегатов МАК-IgG, которые были "сшиты" с помощью нековалентных биоаффинных связей, в СЕА энзимин-тесте
Реактивы и проведение испытания см. в примере 8.
Фактическое содержание карциноэмбрионального антигена (СЕА) использованной пробы сыворотки человека, определенное с помощью базового метода, составляет 2,8-4,2 нг/мл. Табл. 2 показывает считанные на эталонной кривой кажущиеся концентрации СЕА для пробы сыворотки человека (N 108), которая содержит нарушающие факторы, при различных добавках агрегата иммуноглобулина к инкубационному буферу.
Из табл. 2 видно, что биоаффинно-связанные агрегаты иммуноглобулина IgG, отнесенные к весовому количеству иммуноглобулина IgG, значительно эффективнее устраняют вредные факторы, чем несвязанный иммуноглобулин IgG: коэффициент более 500 или более 60. Используемое количество выбрано таким образом, что получилось только частичное устранение вредных факторов, именно при таких условиях наилучшим образом оценивается относительное устраняющее нарушения действие различных препаратов.
П р и м е р 10. Зависимость устраняющей нарушения функции от молекулярного веса агрегата МАКЗЗ-IgG
Реактивы и способ проведения этого опыта такие же, как и в примере 8. Препараты МАКЗЗ-IgG получены в соответствии с примером 2.
1) Определенное с помощью базового метода действительное содержание в СЕА находится в диапазоне 1 3,5 нг/мл.
2) Кажущееся содержание СЕА определяется по экстинции и эталонной кривой с помощью стандартных проб СЕА в соответствии с инструкцией по работе для энзимуна СЕА.
3) Кроме определенного с помощью эталонной точки участка эталонной кривой.
П р и м е р 11. Сравнение устраняющего нарушения действия агрегатов МАКЗЗ-IgG полученных из различных моноклональных антител
Используют пробирки из упаковки энзимин RTSH (фирма "Берингер Маннхайм ГмбХ"), которые покрыты моноклональным антитиреотропином мышиного иммуноглобулина (IgG I, K). Остальные реагенты, как и в примере 8, заимствованы из энзимин СЕА-упаковки. Испытание проводят в соответствии с инструкцией по работе с этой упаковкой. При этом проведении испытания не может быть подан никакой сигнал с помощью содержания аналита пробы сыворотки, потому что оба специфических реактанта имеют различную специфичность. Таким образом, речь идет только о модели теста, который с помощью пробы сыворотки выдает только тогда сигнал по величине гашения, когда проявляются вредные факторы. Привлеченные для сравнения агрегаты моноклонального антитела иммуноглобулина МАК-IgG получены по аналогии с примером 2b из МАКЗЗ.
Экстинция для пробы сыворотки без СЕА и без нарушающих факторов: 0,162.
Результат: устраняющее нарушение действия различных моноклональных агрегатов МАК-IgG одинаково в рамках границ ошибок.
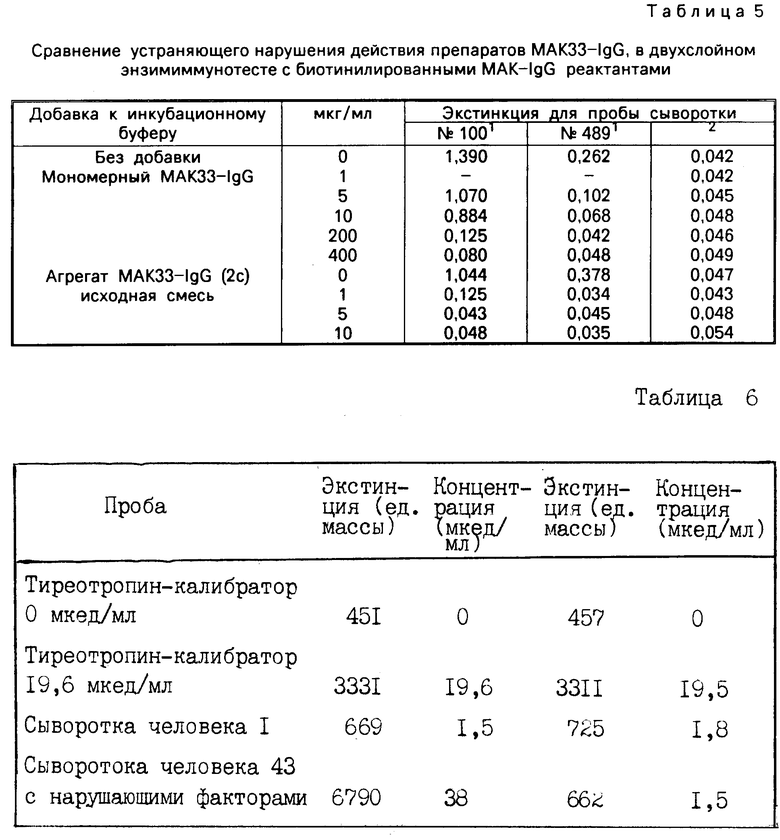
П р и м е р 12. Устраняющее нарушения действия агрегата МАКЗЗ-IgG и мономерного иммуноглобулина МАКЗЗ-IgG в тесте с биотинилированными реактантами МАК-IgG
В этом тесте используются два моноклональных гептатито-поверхностных (НВА)-специфических иммуноглобулина моноклональных антител МАК-I C в биотинилированной форме (биотинилирование проведено в соответствии с Т. V. Updyke und G. L. Nicolson in Methods in Enzy mology, 121 (1986) 717 725). Один из них, МАК6Е7-IgG является подтипом IgG 1/K, другой, МАК5А10-IgG, является подтипом I C2a, К. В качестве энзиммаркированного реактанта используют МАК510-Fab-POD-коньюгат.
Состав инкубационного буфера:
40 ммолей фосфатного буфера с водородным показателем 7,0
0,2 молярный средний виннокислый натрий
0,5% (W/V) альбумина сыворотки крупного рогатого скота
0,1% (W/V) иммуноглобулина IgG крупного рогатого скота
0,5% (W/V) плуроника F 68
0,01 (W/V) фенола
200 нг/мл МАК6Е7-IgG (биотинилирован)
25 нг/мл МАК5А10-IgG (биотинилирован)
200 mu/мл МАК5А10-Fab-POD коньюгата
Устраняющая нарушение добавка протеина дана в табл. 5.
Исходые смеси агрегата МАКЗЗ-IgG были получены в соответствии с примером 2b.
Для теста пипеткой вводят по 0,2 мл пробы сыворотки и 1 мл инкубационного буфера в пробирку из полистирола, которая покрыта стрептавидином. Затем смесь инкубируют в течение 4 ч при комнатной температуре. Затем каждую пробирку промывают 3 х 1,5 мл водопроводной воды и добавляют по 1 мл раствора субстрата (АВТS/пероксид) из энзимун СЕА-тестовой упаковки. После инкубации в течение 1 ч при комнатной температуре измеряют экстинкция в преобразованном растворе субстрата при 405 нм.
Сравнение устраняющего нарушения действия препаратов МАКЗЗ-IgG, в двухслойном энзимиммунотесте с биотинилированными МАК-IgG реактанами
1) Пробы сыворотки N 100 и 489 получены из хранилища доноровской крови и свободно декларированы как НВSAg.
2) Проба здорового донора, которая не содержит ни НВsAg, ни нарушающего фактора.
Из данных следует, что для сыворотки N 100 достигается то же самое устранение нарушений по сигналу 0,125 с помощью 200 мкг/мл мономерного МАКЗЗ-IgG или 1 мкг/мл агрегата МАКЗЗ-IgG, и что для этого теста агрегат иммуноглобулина IgG в 200 раз активнее с точки зрения устранения нарушений.
П р и м е р 13. Устраняющее нарушения действие агрегата МАКЗЗ-IgG в тесте с сухими химическими носителями реактивов
Определение проводится с помощью сухих химических носителей реактивов в одностадийном тесте в соответствии с принципом слоистой твердой фазы двойного антитела в роторных вставных элементах с центробежным анализатором с помощю описанной в выложенной заявке ФРГ N 34250085 анализирующей аппаратуры.
1) Получение растворов реактивов
а) буфер 1
50 ммолей/л калийфосфатного буфера, рН 6,0, получается путем смешивания 50 ммолей/л раствора К2НРО4 и 50 ммолей раствора КН2РО4 до достижения рН 6,0.
б) буфер 11
Буфер 11 получают точно также, как и буфер 1, с той лишь разницей, что устанавливается рН 7,5 и что буфер дополнительно содержит 10 г/л альбумина сыворотки крупного рогатого скота и 150 ммолей/л NaCl.
в) Раствор реактанта R1, способного соединяться с тиреотропином (Т Н).
В качестве реактанта R1 используется моноклональное мышиное антитело тиреотропина. Содержащая это антитело асцитическая жидкость добавляется до 1,8 моля сульфатом аммония. Осадок растворяется в буфере из 15 ммолей фосфата натрия с рН 7,0 и 50 ммолей хлорида натрия. Полученный таким образом раствор подвергается пассажу с помощью DEAE-целлюлозы. Полученное таким образом антитело, способное соединяться с тиреотропином (TSH), биотинилируется (5 биотин/IgG) и разбавляется буфером 11 до концентрации протеина 1 мг/мл.
г) Энзиммаркированный раствор реактанта R2
В качестве реактанта R2 также используется моноклональное мышиное антитело тиреотропина, которое однако распознает иную антигенную детерминанту, чем реактант R1. Содержащая это антитело асцитическая жидкость очищается. Полное антитело расщепляется известным образом в соответствии с методом Р.Р. Портера, Биохимический журнал N 73, 1959, с. 119, на Fab-фрагмент. Полученные Fab-фрагменты в соответствии с методом, описанным в источнике Tshikawa etal. I. of Immunoassay 4 (1983), с. 209 327, соединяются в β -галоктозидазой. Раствор реактанта R2 разбавляется в буфере 11 до концентрации 500 mu/мл (соразмерно с о-нитрофенил- β-галоктозидазой при 37оС).
д) Устраняющий нарушения протеин
МАКЗЗ-IgG-полимер, полученный по аналогии с примером 2, раствоярется в буфере 11 до концентрации 1 мг/мл.
F) Раствор авидина
Авидин разбавляется в буфере 1 до концентрации протеина 50 мкг/мл.
е) Раствор субстрата
Красный хлорфенол-
β -галактозид 5 ммолей/л (3,05 г/л)
(полученный в соответствии
с выложенной заявкой
ФРГ N 3345748)
НЕРЕS 70 ммолей/л (16,7 г/л)
NaCl 154 ммоля/л (9 г/л)
Альбумин сыворотки
крупного рогатого скота 0,3% (3 г/л)
Твин 20 0,2% (2 г/л)
рН (с NaOH) 7,25
2. Получение носителей реактивов.
а) Носитель реактивов 1 (без устраняющего нарушения протеина)
40 мкл раствора, который на каждый литр содержит 100 ммолей фосфата натрия с рН 7,3 (37оС), 2 ммоля хлорида магния, 9 г хлорида натрия, 5 г альбумина сыворотки крупного рогатого скота, 0,5 мг биотинилированого антитиреотропинного моноклонального антитела мыши (реактант R1), 1000 единиц коньюгата мышиного антитиреотропинного антитела Fab-фрагмента- β-галактозидазы (раствор реактанта R2) (активность определяется с помощью орто-нитрофенил- β -D-галактозида при 37оС) наносится каплями на волокнистую массу, состоящую из коммерческой полиэфирной бумаги. Затем производится сушка при комнатной температуре. Эти волокнистые маты хранятся до их применения при 4оС и относительной влажности 20%
б) Носитель реактивов 1 (с устраняющим нарушением протеином)
Получение осуществляется также, как и носителя реактивов 1, с той лишь разницей, что дополнительно на каждый литр пропиточного раствора содержится 0,01 г агрегата МАКЗЗ-IgG из примера 2с (исходная смесь).
b) Носитель реактивов 2
На целлюлозном волокнистом мате в соответствии с бромциановым способом активизации (выложенная заявка ФРГ N 1768512) фиксируется авидин (раствор авидина), причем на каждый грамм волокнистого материала для фиксирования предлагается 10 мкг авидина. Путем промывки удаляется несоединившееся антитело и волокнистый мат осторожно высушивается при комнатной температуре. Хранение полученных таким образом волокнистых матов осуществляется по аналогии с хранением носителя реактивов 1.
3. Проведение определения.
Определение с помощью этих обоих носителей реактивов 1 и 2 или 1l и 2 осуществляется с помощью описанного в выложенной заявке ФРГ N 3425008. 5 устройства для проведения аналитических определений.
Это устройство содержит вставной элемент ротора для центробежных анализирующих автоматов, состоящий из фасонного корпуса, который имеет камеру дя нанесения пробы, которая соединена с большим количеством полей реактивов, которые соответственно содержат пропитанный опредеенным реактивом гигроскопический материал-носитель, и имеет по меньшей мере одну смесительную клапанную камеру и одну измерительную камеру, которые совместно образуют путь транспортировки исследуемой жидкости, который ведет радиально изнутри радиально наружу, если вставной элемент закреплен на роторе, и кроме того, ведет по меньшей мере в другую камеру для проведения измерений и по меньшей мере частично идентичен с путем транспортировки исследуемой жидкости.
При этом путь для транспортировки исследуемой жидкости ведет от камеры для загрузки пробы через заполненную гигроскопичным материалом камеру (а), содержащую буфер, через камеру (b) и расположенную между камерами (а) и (b) первую клапанную камеру (КI) ко второй клапанной камере (VК2) и от нее через камеру (б) и приемную камеру (АК) к измерительной камере (К). Для загрузки другой жидкости предусмотрена выполненная в виде насосной камере субстратная камера (РК), которая через дозирующую камеру (DK) и состоящее из капилляров (Кар) дозирующее устройство и перепускную камеру соединена со второй клапанной камерой (VK2).
Для определения экстинции "а" (измерительный сигнал, включая сигнал помехи) используются носитель реактивов 1 и носитель реактивов 2, для определения экстинкции "b" (специфический измеренный сигнал без сигнала помехи) используются носители реактивов 1l и 2.
Носитель реактивов 1 или 1l размещается на поле "с" вставного элемента ротора и носитель реактивов 2 на поле "d". При этом 40 мкл концентрированной пробы вводится непосредственно пипеткой через отверстие на верхней кромке на пол "а". 270 мкл раствора субстрата вводится пипеткой в камеру РК. С помощью надлежащей программы центрифугирования, в которой высокая частота вращения чередуется с остановкой, проба и раствор субстрата транспортируется затем в направлении разделительной матрицы и кюветты.
При этом в процессе обработки программы реактанты R1 и R2 без устраняющего нарушения протеина или с ним вымытаются испытываемой жидкостью с поля "с" и затем гомогенная смесь подается для проведения реакции. На поле "d" образованные комплексы соединяются с помощью реактанта R1 с авидином. Перенос пробы с поля "с" на поле "d" осуществляется в течение очень короткого времени.
Раствор субстрата с помощью дозирующей камеры К разделяется на порции, из которых первая предназначена для вымывания избыточного некомплексированного конъюгата.
Нарушающие факторы, которые могли бы связывать реактант R2, нейтрализуются при использовании 10 мкг устраняющего нарушения протеина/мл исследуемого раствора и также вымываются (использованные носители реактивов 1).
Связанная через образование комплекса на поле "d" активность β -галактозидазы пропорциональна содержащемуся в пробе количеству тиреотропина или холостому значению пробы. Эта активность определяется с помощью другой порции субстрата, причем субстрат в ходе 5-минутной реакции преобразуется в цветные продукты. Образовавшиеся цвета и дальнейшее цветообразование/мл в жидкой фазе измеряются в кюветте при 576 нм.
При этих условиях получены следующие результаты.
Все измерения проведены при λ= 576 нм и толщине слоя 0,3 см и пересчитаны на толщину слоя d 1 см.
а) Определение тиреотропина при использовании носителя реактивов 1
б) Определение тиреотропина при использовании носителя реактивов 1l с полимером МАКЗЗ-IgG для нейтрализации специфических для М-Fab-фрагментов нарушающих факторов.
в) Стандартные тиреотропины калиброваны на 2. 1RP 80/588.
Содержание тиреотропина в сыворотке человека 43 было подтверждено с помощью энзимун ТSH теста фирмы "Берингер Маннхайм ГмбХ" для 1,4 мкед/мл. Этот тест в качестве специфических реактантов содержит нанесенный на тубу антитиреотропин моноклонального антитела и поликлональный амниотический анти-TSH-Fab-POD-конъюгат. Последний является инертным по отношению к нарушающим факторам в сыворотке человека 43.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения поливалентного антигена | 1984 |
|
SU1475492A3 |
| Способ получения дигиталис-антител | 1984 |
|
SU1455999A3 |
| АНТИТЕЛО VFF-18 И ЕГО АНАЛОГИ | 1995 |
|
RU2204606C2 |
| СПОСОБЫ ИЗМЕРЕНИЯ ЭЛЕКТРОХЕМИЛЮМИНЕСЦЕНТНЫХ ЯВЛЕНИЙ И ОПРЕДЕЛЕНИЯ АНАЛИЗИРУЕМОГО ВЕЩЕСТВА И РЕАКТИВ | 1994 |
|
RU2116647C1 |
| Способ определения креатинкиназы МВ в сыворотке крови | 1980 |
|
SU1336941A3 |
| СРЕДСТВО ДЛЯ КОЛОРИМЕТРИЧЕСКОГО ОПРЕДЕЛЕНИЯ АНАЛИТА | 1989 |
|
RU2015513C1 |
| Способ получения производных резоруфина | 1986 |
|
SU1621811A3 |
| Способ иммунологического определения одновалентных аналитов | 1990 |
|
SU1799471A3 |
| Способ определения @ -амилазы поджелудочной железы | 1986 |
|
SU1637670A3 |
| Способ получения гибридомы, продуцирующей моноклональные антитела к интерферону типа омега | 1987 |
|
SU1602393A3 |
Изобретение относится к медицине. Способ определения поливалентного антигена в биологической жидкости путем конкурентного связывания двух специфических тел или Fab-фрагментов Jg другого типа животных в присутствии ингибитора неспецифической реакции. Отличие способа заключается в том, что в качестве ингибитора неспецифической реакции используют неспецифические JgG-агрегаты второго и третьего типа животных, либо агрегаты, состоящие из JgG или Fab-фрагментов JgG и неспецифического JgG третьего вида животных или JgG свободного от FG-фракции или протеина, или полисахарида, или растворимого в воде полимера и имеющие молекулярный вес от 320 000 до 100 миллионов дальтон и добавляют их в количестве 0,1-50 мкт/мл.
СПОСОБ ОПРЕДЕЛЕНИЯ ПОЛИВАЛЕНТНОГО АНТИГЕНА В БИОЛОГИЧЕСКОЙ ЖИДКОСТИ путем конкурентного связывания двух специфических антител или Fab-фрагментов Jg другого типа животных в присутствии ингибитора неспецифической реакции, отличающийся тем, что в качестве ингибитора неспецифической реакции используют неспецифические JgG-агрегаты второго и третьего типов животных, причем агрегаты, состоящие из JgG-или Fab-фрагментов JgG и неспецифического Jg G третьего вида животных или Jg G, свободного от FG-фракции, или протеина, или полисахарида, или растворимого в воде полимера и имеющие мол.м. 320 000 100 000 000 дальтон, добавляют в количестве 0,1 50,0 мкт/мл.
| ПОГРУЗЧИК СИЛОСАГ''ТП"^""'1.'Пис -UU to S ' ""* " -•^-; лтснт-й! • V ••;-:;ч^^::дч •^i'A:;j'iii'2^ | 0 |
|
SU174026A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
