Изобретение относится к pGH(A), полученному путем стадий:
а) культивирования одноклеточного организма, содержащего рекомбинатный вектор, включающий фрагмент ДНК, кодирующий указанный гормон, и
б) выделения указанного гормона из культуры.
Экспрессия генов эукариотами и прокариотами, хотя и использует одни и те же основные стадии транскрипции генов в информационную PHK (иPHK) и последующей трансляции этой иРНК в протеины, использует при этом различные совокупности внутриклеточных регуляторов для этих стадий.
Кроме того, у эукариотов многие зрелые протеины сначала транслируются как предпротеины, то есть как полипептиды, содержащие последовательность зрелого протеина, слитую с сигнальной последовательностью. Эукариотная иPHK кодирует полный предпротеин, в котором после трансляции удаляется предпоследовательность и получается "зрелый" протеин. Несмотря на то, что эукариотные клетки снабжены всем необходимым для того, чтобы специальной обработкой превратить такие предпротеины в зрелые протеины, прокариотические клетки в общем случае не способны распознавать сигналы обработки, содержащиеся в эукариотических протеинах. Таким образом, если полные транскрипты комплементарной ДНК (кДНК) эукариотической иРНК используются в качестве ДНК-последовательностей для экспрессии в прокариотах, то получают в результате предпротеин, а не зрелый протеин. Имеется возможность превратить предпротеин в зрелые протеины в лабораторных условиях, но эта стадия требует значительных затрат.
В том случае, когда для экспрессии зрелого протеина в прокариотах используют ДНК-последовательность, кодирующую зрелый протеин, эта последовательность не содержит эукариотических сигналов послетрансляционной обработки. Следовательно, для экспрессии клонированных эукариотических генов или других последовательностей гетерологичной ДНК в прокариотических системах, как было установлено, желательно использовать прокариотические контрольные сигналы ввиду их эффективности и вследствие того, что эукариотические сигналы не могут распознаваться прокариотической клеткой-хозяином.
Термин "гетерологичная ДНК", как он используется в соответствии с настоящим изобретением, означает ДНК, по крайней мере часть которой не содержится в общем случае внутри генома клетки-хозяина. Примеры гетерологичной ДНК включают, но ими не исчерпывается полный список, вирусные и эукариотические гены, фрагменты генов, аллели и синтетические последовательности ДНК. Термин "гетерологичный протеин" или "гетерологичный полипептид" означает здесь протеин или полипептид, по крайней мере часть которого в общем случае не кодируется геном клетки-хозяина.
Прокариотические регуляторные сигналы содержат промотор, который способствует инициации транскрипции, регулирующие трансляцию сигналы, содержащие сайт связывания рибосомы, сигнал начала трансляции и сигнал завершения трансляции. Все эти сигналы, за исключением сигнала завершения трансляции, должны быть расположены перед геном или другой ДНК, которые должны быть экспрессированы.
Известны несколько подходов с целью экспрессии гетерологичной ДНК (например, эукариотических генов) в прокариотах. В соответствии с одним из подходов сегмент ДНК, кодирующий искомый протеин, подвергают лигированию с ДНК, кодирующей полный бактериальный протеин или его некоторую часть под контролем бактериального промото ра. Эндогенная прокариотическая ДНК обязательно содержит также сайт связывания с рибосомой и сигнал начала трансляции. Экспрессия такой лигированной ДНК дает то, что называют протеином слияния, содержащим эукариотический полипептид, связанный с полным или частью бактериального протеина. Выделение эукариотического протеина затем можно осуществлять при помощи ориентированного на определенный сайт ферментного или химического расщепления в сайте слияния.
Примерами опубликованных работ, касающихся получения в бактериях эукариотических протеинов слияния, являются: Европейская патентная заявка N 47600 (опубликованная 17 марта 1982г.), которая относится к протеинам слияния и "однородным" протеином, содержащим бычий предгормон роста или бычий гормон роста ("бГР") на карбокси (С-) конце с порцией прокариотического протеина на амино (N)-конце; патентная заявка Великобритании N 2073245А (опубликованная 14 октября 1981г. ), касающаяся протеинов слияния бГР и β-лактамазы Е. соIi; (Кешент и др. Nucleic Acid Research, 9:19 30 (1981), касающаяся протеина слияния бГР и b-лактамазы Е.coli; Европейская патентная заявка N 95361 (опубликованная 30 ноября 1983г.), касающаяся протеина слияния, содержащего последовательно эндогенный протеин на N-конце, аминокислоту сигнала начала трансляции, сайт расщепления энтерокиназой и экзогенный протеин (например, гормон роста) на C-конце.
Однако преимущества этого подхода оказываются недостаточными ввиду необходимости в последующем расщеплять гетерологичный протеин из эндогенного полипептида в соответствии с описанием, приведенным выше.
В соответствии с другим подходом сигнал начала трансляции, ATG, под контролем бактериального промотора расположен непосредственно перед ДНК-последовательностью, кодирующей гетерологичный (например, эукариотический) протеин. Хотя протеины, полученные при помощи таких генных конструкций, не требуют последующего расщепления для образования искомого протеина, они в общем случае включают метионин (в некоторых случаях формилметионин) на N-конце, так как стартовый сигнал ATG также является кодоном метионина. Таким образом, если целевой зрелый протеин не начинается с метионина, то такой протеин будет иметь N-конец, измененный включением метионинового остатка.
К примерам таких генных конструкций относятся: работа Гарента и др. Cell, (1980), т. 20, с. 543-553, в соответствии с которой ген b-глобина кролика, который обладает N-концевым валином, экспрессируется в Е.coli с использованием только что описанной генной конструкции. В результате исследований ими было установлено, что, в то время, как в b-глобине кролика отсутствует амино-концевой метионин, а лейцины содержатся в позициях 3, 14, 28, 31, 32, в меченом протеине лейцины были обнаружены в позициях 4, 15, 29, 32 и 33, а метионин был найден в позиции I. Этот результат показывает, что этот протеин является b-глобином кролика плюс амино-концевой метионин, которые не удаляются в Е.coli (см. там же, с.546-547).
Другой пример относится к получению гормонов роста в бактериях с использованием описанной выше генной конструкции. Номер и др. Proc. Nat'L Acad. Sci. USA, (1984) т.81, с. 5403-5407 описывают систему с высокой степенью экспрессии в бактерии для получения бГР, которая приводит к получению N-Met бГР, то есть соединения, содержащего аминокислотную последовательность, подобную аминокислотной последовательности одного из встречающихся в природе видов бГР плюс метионин на его N-конце. Присоединение N-концевого метионина к различным видам гормона роста, производимого бактериями, уже обсуждалось в Европейской патентной заявке N 103395, опубликованной 21 марта 1984г.) и Европейской патентной заявке N 74444 (опубликованной 30 марта 1983г.) для бГР Сибургом и др. DNA (1983) т.2, с.37-45, для бГР и свиного гормона роста ("сГР").
Присоединение N-концевого метионина к природному N-концу может быть нежелательным по нескольким причинам. Во-первых, вполне возможно (хотя в настоящее время представляется мало правдоподобным), что метионин имеет тенденцию превращать протеин в антигенный в организме, в котором протеин без N-метионина является эндогенным. Во-вторых, присоединение метионина к N-терминальной части протеина может оказать нежелательный эффект на его биологическую активность или его физические свойства. В-третьих, эта измененная форма протеина может служить препятствием для научных исследований, направленных на определение связи между функцией природного протеина и его структурой.
Способность таких прокариотов, как бактерии удалять N-концевой метионин из протеинов либо во время их получения, либо уже после их получения, привлекает в последнее время большое внимание. Например, Уоллер, J. Mol. Biol. (1963), т. 7, с. 483-496 исследовал состав N-терминальных аминокислот "растворимых" и рибосомальных протеинов их экстракта Е.coli, не содержащего клеток, а в Европейской заявке N 103395 (опубликованной 21 марта 1984г.) предложен способ удаления N-концевого метионина из эукариотического протеина, синтезированного культурой Е.coli. В частности, метионин удаляется из одного из двух упомянутых, производимых бактериями протеинов бГР, причем оба они содержат остаток серина сразу же после содержащегося с самого начала N-концевого метионина. Генная конструкция, которая была использована в этих исследованиях, однако содержала синтетическую стартовую последовательность, кодирующую 5'-метионин-серин-лейцин-3', восстановленную непосредственно рядом с 5'-концом бГР-кодирующей последовательности, в которой предварительно были удалены основания, кодирующие первые 4 или 9 встречающихся в естественных условиях аминокислот. Таким образом, полученный в результате протеин, синтезированный в культуре Е.coli отличался от природного. В патентной заявке Великобритании N 2073245А (опубликованной 14 октября 1981г.) указано, что если met pro заменить ala в зрелом бГР-протеине, то "Met" может быть переработан бактерией таким образом, что в результате получается модифицированный бГР, начинающийся в аминокислотной последовательности Pro Phe Ala Pro.
Таким образом, имеется необходимость в экономическом и предсказуемом средстве для получения в таких микроорганизмах, как бактерии гетерологичных (например, эукариотических) протеинов, которые не содержат N-концевого метионина. Особенно желательно разработать такой способ, в соответствии с которым такие протеины, полученные в бактериях, не требовали бы в лабораторных условиях обработки после ферментации и не содержали бы дополнительного, неприродного N-концевого метионина.
Гормоны роста (называемые также самототропинами) представляют собой полипептиды, продуцируемые и секретируемые клетками гипофиза и в значительной степени видоспецифичные. Наряду с их ролью в ускорении роста скелета, гормоны роста оказывают влияние на многочисленные метаболические процессы, включая стимулирование выделения молока, увеличение высвобождения инсулина из поджелудочной железы и выделения глюкогона, кроме того, они оказывают эффект мобилизации липидов. Экзогенное применение бГР к крупному рогатому скоту, например, приводило к увеличению надоев молока, эффективности корма и/или скорости роста, снижению времени откармливания и увеличению отношения: постная часть мяса/жир. Однако до сих пор не полностью ясно, как этот гормон вызывает столь многочисленные эффекты.
Известно, что бГР (бычий гормон роста) имеет несколько форм. В частности, синтезируется четыре вида бГР, которые отличаются в двух позициях протеина N-концевая аминокислота может варьировать из-за вероятной неоднозначности при удалении сигнальной пептидной последовательности таким образом, что зрелый протеин начинается с NH2-phe-pro, либо с NH2-ala-phe-pro. Кроме того, имеется некоторая неоднородность в аминокислоте 126, которая является либо лейцином, либо валином.
Четыре молекулярные формы (вида) гипофизарного бГР здесь обозначаются и эти обозначения сокращаются следующим образом:
Сокращения Структура
бГР (Л) NH2-phe (I)-pro (2).Leu(126).COOH
бГР (А,Л) NH2-ala (-I)-phe (I) -pro(2). Leu(126).COOH
бГР (В) NH2-phe(I)-pro(2).Val(126).COOH
бГР (A,В) NH2-ala(-I)-phe(I)-pro(2).Val(126).COOH
Миллс и др. (1970) J. Biol. Chem, 245, с.3407-3415, в точности так же идентифицировал два N-концевых фрагмента свиного гормона роста (сГР). В частности, один фрагмент содержал N-концевой фенилаланин, а другой - дополнительный N-концевой аланин. Эти молекулярные формы сГР в дальнейшем коротко обозначаются через сГР(Ф) и сГР(А) соответственно.
Полные ДНК-кодирующие последовательности и соответствующие аминокислотные последовательности для бГР(Л) и сГР(Ф) были опубликованы Сибургом и др. DNA(1983), 2, с.37-45 и эта работа используется здесь в качестве ссылки.
Целью настоящего изобретения является получение в бактериях аланиновой формы свиного гормона роста pGH(A), не содержащей дополнительного N-концевого метионина и свободной от формы с phe на N-конце. При этом осуществляют следующие стадии:
а) культивирования одноклеточного организма, содержащего рекомбинантный вектор, включающий фрагмент ДНК, кодирующий указанный гормон, и
б) выделения указанного гормона из культуры.
На приведенных фигурах штриховым контуром изображена кодирующая последовательность ДНК бактериального промотора, зачерненный контур представляет кодирующую последовательность гетерологичной ДНК, черточками изображены дополнительные кодирующие последовательности ДНК (они конкретно указаны на рисунках), а направленная стрелка указывает на ориентацию от 5' и 3' кодирующих последовательностей ДНК. Указаны также соответствующие сайты рестрикции эндонуклеаз. Отмеченные области ДНК приведены только с целью наглядного представления и они не связаны с реальными размерами этих областей.
На фиг.1 представлена конструкция M13mp8/Xbal, содержащая вектор M13mp8, имеющий вставленный в сайт рестрикции Sma I сайт рестрикции Xbal.
На фиг. 2 представлена конструкция M13mp 8/BG Hex-I, содержащая M13mp8/Xbal, несущую кодирующую бГР(Л) последовательность ДНК.
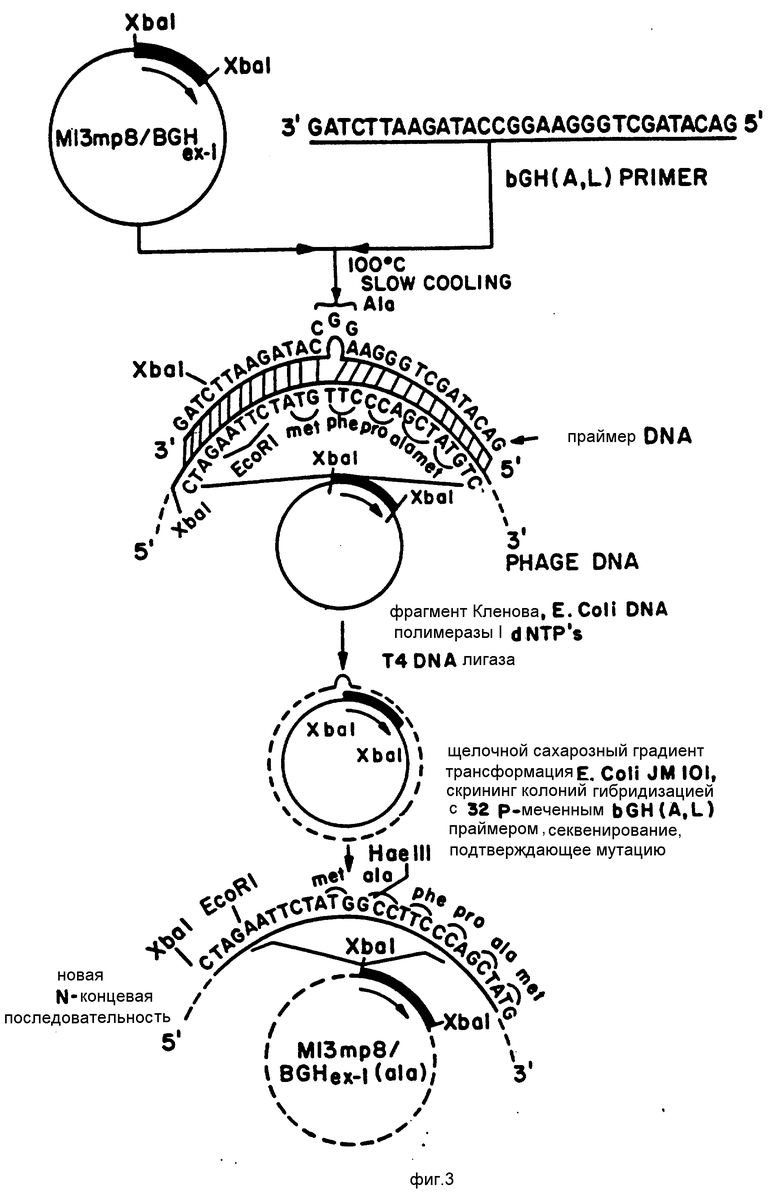
На фиг.3 представлена конструкция кодирующей бГР(А,Л) последовательности ДНК, полученная при помощи сайт-направленного мутагенеза.
На фиг. 4 представлена конструкция кодирующей бГР(А,B) последовательности ДНК, полученной при помощи сайт-направленного мутагенеза.
На фиг. 5 представлена конструкция M13mp9/PG Hex-I, содержащая M13mp 9 и кодирующую сГР(Ф) последовательность ДНК.
На фиг. 6 представлена конструкция кодирующей сГР(А) последовательности ДНК, полученной при помощи сайт-направленного мутагенеза.
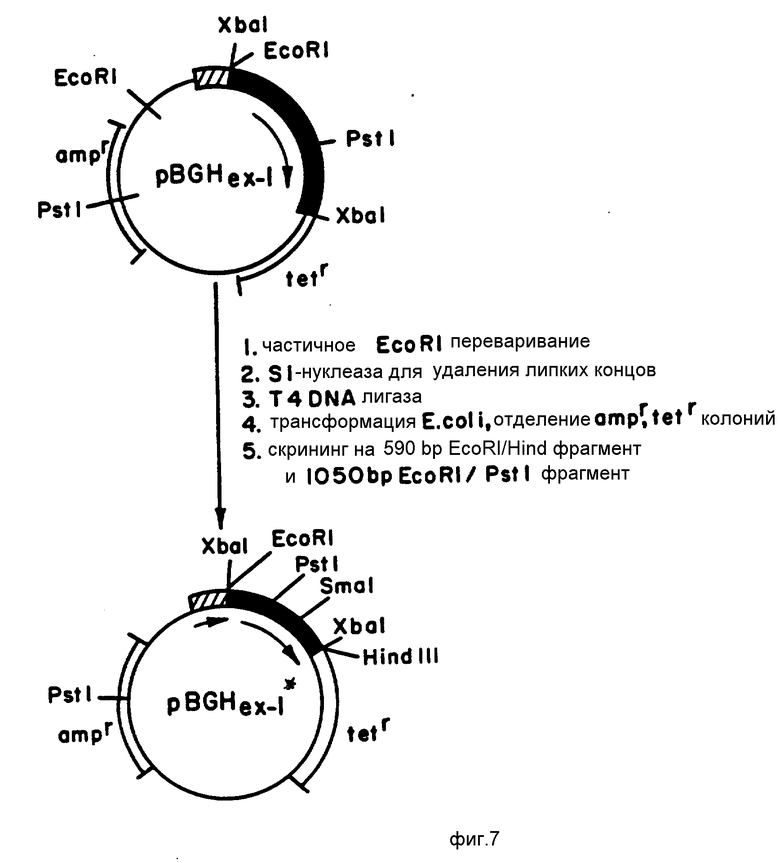
На фиг. 7 представлена конструкция pBGH
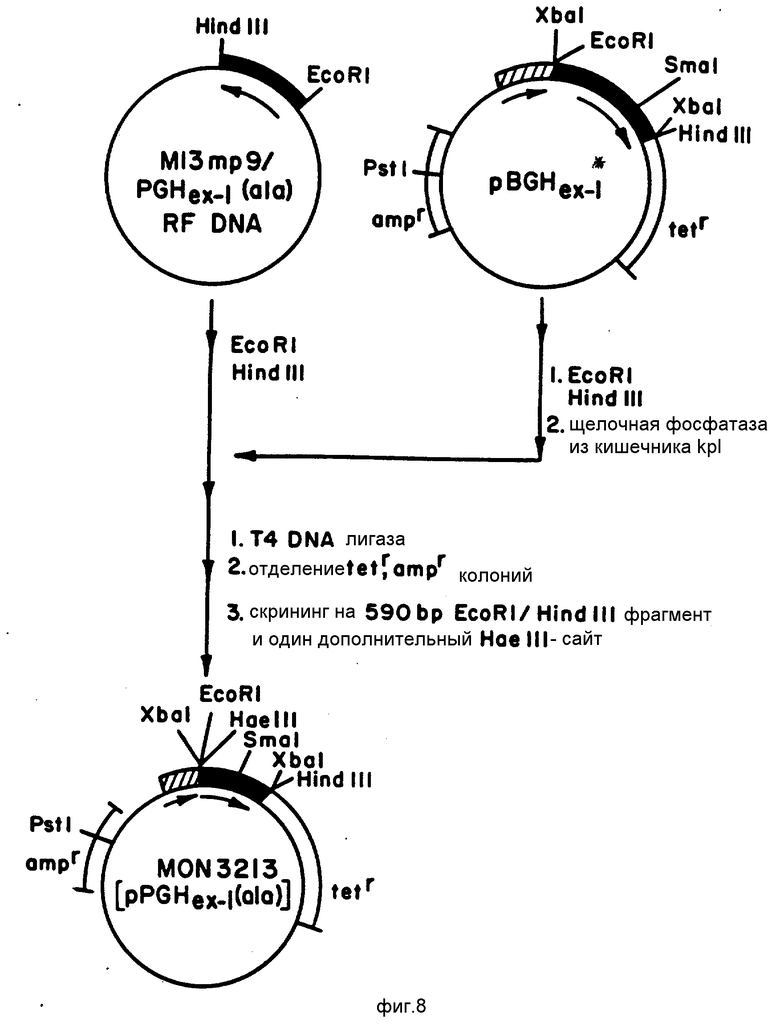
На фиг. 8 представлена конструкция вектора экспрессии pMON 3213, содержащего вектор pBGHex-I, несущий кодирующую сГР(А) последовательность ДНК вместо кодирующей бГР(Л) последовательности ДНК.
Настоящее изобретение представляет важный способ получения в прокариотах гетерологичного полипептида такого, как эукариотический (например, млекопитающего или птиц) протеин, который содержит N-концевой аланин. Таким образом, непосредственно получают полипептид без N-концевого метионина, что исключает необходимость в обработке предшественника в лабораторных условиях. Устойчивое получение такого полипептида без N-концевого метионина, не содержащегося в исходно кодирующей последовательности гена, является новым и совершенно неожиданным результатом.
Настоящее изобретение представляет весьма эффективный способ получения существенно чистых протеинов, которые имеют N-концевой аланин. К таким протеинам относятся без каких-либо ограничений упомянутые виды бычьего и свиного самототропина и его варианты, растительные протеины, а именно, малая субъединица рибулозо-1,5-бисфосфаткарбоксилазы, S-трансфераза глутатиона и протеин 70 теплового шока. Кроме того, настоящее изобретение можно эффективно использовать для получения других полипептидов, когда желательно иметь на N-конце аланин, а не метионин.
До сих пор во всех работах, посвященных экспрессии бГР и сГР в бактериальных клетках, в которых N-конец содержит N-концевую аминокислотную последовательность, гомологичную встречающимся в природе бГР, описано присутствие N-концевого метионина.
В статье Сибурга и др. DNA (1983) 2, с.37-45, на с.44 указано, что последовательность гена для N-концевых фенилаланиновых видов бГР, например бГР(Л), умышленно выбирается для экспрессии в культуре E.coli частично, чтобы избежать ожидаемого присоединения второй гидрофобной аминокислоты (метионина) к гидрофобному N-концевому аланину. Таким образом, доступные до настоящего времени исследования указывают на наличие N-концевого метионина в синтезированных бактериями видах ГР.
Как это более подробно описано в примерах реализации настоящего изобретения, подход заявителя с целью получения с ГР(А) или pGH(A) в бактериях, если очень коротко, заключается в следующем. Кодирующие вышеупомянутые протеины последовательности ДНК получают при помощи сайт-направленного мутагенеза последовательностей ДНК, кодирующих бычьи и свиные виды самототропина, содержащие N-концевой фенилаланин, как это показано на фиг. 3, 4 и 6. Затем полученные последовательности вставляют в векторы экспрессии таким образом, что окончательная последовательность содержит последовательно промотор, сайт связывания с рибосомами, стартовый кодон ATG (метионин), непосредственно примыкающий к последовательности ДНК, кодирующей pGH(A), и, наконец, стоп-кодон трансляции. Культуру E.coli затем подвергают трансфекции полученным вектором экспрессии, несущим искомую последовательность гена, и культивируют при условиях, которые позволили бы осуществить экспрессию искомой гетерологичной ДНК, и получить тем самым искомый гетерологичный протеин. Затем полученные таким образом протеины анализируют с точки зрения анализа их последовательности и соответствующей биологической активности.
Таким образом, в соответствии с настоящим изобретением устанавливается что, когда ДНК-последовательность, содержащая стартовый кодон (метионин), а затем сразу же кодоны гетерологичного N-аланилового полипептида, подвергаемого экспрессии, то протеин, выделенный из прокариотического организма, действительно содержит аланин на N-конце, а не метионин. Представляется правдоподобным, что аналогичный результат может быть получен, если этот аланиновый кодон расположен вслед за примерно тремя соседними метиониновыми кодонами, которые включают сигнал начала трансляции иРНК, кодирующей целевой полипептидный продукт. Например, ДНК, включающая этот сигнал начала трансляции и кодоны целевого полипептидного продукта, может включать любую последовательность, которая соответствующим образом кодирует met ala, met met ala, met met met ala или любой его функциональный эквивалент.
В предпочтительном варианте воплощения настоящего изобретения три различных штамма E. coli K12, каждый из которых был депонирован в Американской коллекции Типов Культур, Роквил, Мерилэнд, под номерами ATCC 39936, 53010 и 53009 соответственно обладают способностью удалять N-концевые метионины в том случае, когда после вышеупомянутых N-концевых метионинов следует сразу же аланин.
Это открытие представляется значительным, так как оно дает способ получения в прокариотах гетерологичных полипептидов, которые содержат N-концевые аланины.
В одном из предпочтительных вариантов осуществления способа, являющегося предметом настоящего изобретения он используется для получения двух видов бГР, бГР(А,Л) и бГР(А,В), и особенно одного вида сГР(А), которые не содержат других бычьих и свиных протеинов и/или сГР видов соответственно. В частности, этот способ обеспечивает получение pGH(A), как одного единственного вида.
В соответствии с одной из своих самых широких реализаций настоящее изобретение представляет собой углубление в использовании технологии рекомбинантной ДНК с целью непосредственного получения в прокариотах гетерологичных полипептидов.
Таким образом, описание настоящего изобретения предполагает знакомство с основными приемами, которые используются в технологии рекомбинантной ДНК с целью изоляции и клонирования последовательностей ДНК, кодирующих полипептид, с перераспределением или изменением клонированных последовательностей ДНК и экспрессией клонированных или модифицированных последовательной ДНК в трансформированных микроорганизмах. С приемами знаком любой специалист в этой области техники (например, Молекулярное клонирование. Лабораторное Руководство, под ред. Маниатиса и др. 1982г.).
Выделение или/и конструирование гетерологичной ДНК
В соответствии с одним из воплощений настоящего изобретения ДНК-последовательность, кодирующую искомый гетерологичный полипептид, подлежащий продуцированию в прокариоте, выбирают и выделяют, или ДНК-последовательность, которая его кодирует, конструируют или синтезируют химически. Во многих важных вариантах воплощения таким полипептидом является эукариотический протеин. Если полипептид является небольшим и известна полная аминокислотная последовательность, то можно построить синтетическую ДНК-молекулу или последовательность, кодирующую этот полипептид. Если аминокислотная последовательность полипептида неизвестна или ее размеры слишком велики для того, чтобы практически реализовать синтез соответствующей ДНК-последовательности, можно получить последовательность кДНК при помощи обратной транскрипции из соответствующей иРНК, полученной из тканей или клеток, экспрессирующих этот полипептид. Например, в одной из реализаций настоящего изобретения такая последовательность для бГР может быть получена из бычьих гипофизов при помощи известных в настоящее время приемов, описанных Гудмэном и др. в Методах Ферментологии, т.68, с.75-90 (1979). В качестве альтернативы кДНК-последовательность может быть получена из иРНК, выделенной из клеток, трансформированных геномной ДНК, выделенной из нативного генного белка при помощи соответствующего зонда. Геномная ДНК может быть также модифицирована в различных векторных системах таким образом, что ДНК может быть экспрессирована в прокариотах. Эти приемы известны каждому специалисту в этой области техники.
После того, как получена гетерологичная ДНК-последовательность, содержащая кодоны искомого полипептида, может оказаться желательным осуществить некоторые модификации в нуклеотидной последовательности этой молекулы. Например, если эта молекула была получена при помощи обратной транскрипции из матрицы иРНК, то она будет часто содержать по крайней мере часть ДНК, кодирующую главную последовательность предпротеина. Таким образом, необходимо удалить всю ДНК главной последовательности перед первым кодоном искомого протеина. В некоторых случаях может оказаться необходимым добавить или включить кодон аланина взамен другого кодона в начале последовательности, кодирующей искомый протеин, если он еще имеет кодона N-концевого аланина. Затем вводят сигнал начала трансляции (который также является кодоном метионина) перед и непосредственно рядом с кодоном аланина. Несмотря на то, что комплекс стартовый сигнал/кодон метионина будет в общем случае и в предпочтительном варианте нуклеотидной последовательности ATG, последовательность GTC может также иногда служить в качестве комплекса стартовый сигнал/кодон метионина. Кроме того, присутствие более одного кодона метионина, например, двух, трех, и может быть еще большего количества соседних кодонов метионина, необходимо рассматривать как незначительную модификацию в рамках способа, предлагаемого в соответствии с настоящим изобретением.
По крайней мере один сигнал прекращения трансляции должен быть введен после кодона-C-концевой аминокислоты, если его там не было до этого. Примерами сигналов прекращения трансляции являются дезоксинуклеотидный триплет TAA, TGA, TAG. Таким образом, по существу, применяются методы рекомбинантной ДНК с целью конструирования последовательности рекомбинантной ДНК, содержащей последовательно сигнал начала трансляции/кодон метионина, кодоны для искомого полипептида с кодоном N-концевого аналина, примыкающим к стартовому сигналу, и по крайней мере один сигнал прекращения трансляции, примыкающий к кодону C-концевой аминокислоты.
Было установлено, что эффективной экспрессии иРНК могут препятствовать вторичные структуры, образованные водородом, связывающим две комплементарные серии нуклеотидов внутри иРНК. Исключение этих комплементарных последовательностей, в частности, в той части молекулы, которая кодирует N-конец, упрощает связывание рибосом с иРНК и, следовательно, увеличивает степень экспрессии. Может оказаться, следовательно, желательным заменить кодоны, которые участвуют в образовании таких вторичных структур, кодонами той же аминокислоты, но состоящими из другого нуклеотидного триплета (см. ЕП N 75444, опубликованный 30 марта 1983, Сибург и др. 1983, DNA 2, с.37-45, и Шонер и др. 1984, Proc. Nat'l. Acad Sci.USA, 81, с.5403-5407).
Другие подходы к построению гетерологичных последовательностей ДНК будут очевидны каждому специалисту в этой области техники. Например, если имеется в распоряжении молекула ДНК, которая кодирует полипептид, который подвергается экспрессии N-концевой структурой типа NH2-met-X-y. где x является аминокислотой, отличной от аланина, то кодон аланина может быть вставлен между комплексом сигнал начала трансляции/кодон метионина и кодоном для x. В качестве альтернативы, кодон для x может быть изъят, а кодон аланина вставлен вместо него. Таким образом, при помощи способа, являющегося предметом настоящего изобретения, получили бы протеин, имеющий N-концевую структуру NH2-ala-x-y, или NH2-ala-y, соответственно.
В точности так же можно осуществить удаления, добавления и/или замены в любом из кодонов аминокислоты внутри данной последовательности гена таким образом, что при помощи способа, являющегося предметом настоящего изобретения, можно осуществить экспрессию вариантного полипептида. "Вариантный" полипептид, содержит один или несколько аминокислотных пробелов, замен и/или добавлений по сравнению с встречающейся в природе аминокислотной последовательностью данного полипептида. Примеры таких вариантов включают (но ими не ограничивается весь список) е бГР/Л и е бГР(В), в которых аминокислотная последовательность этих вариантных видов бГР идентична бГР(Л) и бГР(В) соответственно полученным при помощи бычьих гипофизарных клеток, за исключением присутствия дополнительного метионина на N-конце. Эти вариантные полипептиды получают как имеющие аминокислотную последовательность, существенно такую же, что и аминокислотная последовательность встречающегося в природе полипептида, если только биологическая активность не снижается до недопустимого предела. Конструирование и экспрессия вариантных полипептидов может оказаться желательной для того, чтобы добиться более значительного накопления, более высокой стабильности протеина, чтобы упростить стадию очистки полипептида и/или оптимизировать его биологическую активность.
Описанные выше модификации молекулы ДНК, кодирующей целевой полипептид, могут быть осуществлены с использованием ферментов рестрикции, экзонуклеаз, эндонуклеаз и т. д. при помощи приемов, известных в этой области техники. Можно также использовать общие приемы сайт-направленного мутагенеза с тем, чтобы осуществить вышеуказанные модификации в структуре или последовательности молекулы ДНК, эти приемы также известны каждому специалисту в этой области техники. См. например, Золлер Смит (1982) Nac. Acids. Res. т.10, с. 6487-6500; Золлер и Смит (1983) Meth.Enzymol. 100, с.468-500, Норрис и др. (1983) Nuc. Acids. Res. т.11, с.5103-5112.
В соответствии с известными приемами технологии рекомбинантных ДНК после того, как получена искомая последовательность гетерологичной ДНК, далее эту последовательность вставляют в соответствующий клонирующий вектор, который обеспечивает средство для репликации ДНК-последовательности. При этом можно использовать любой подходящий клонирующий вектор в предпочтительном варианте, содержащий маркерную функцию, например, плазмидные векторы E.coli, которые содержат Col EI Херифилд и др. Proc. Nat'l. Acad. Sci. USA, (1974), т. 71, с.3455, pEP322; Боливар и др. Gene, (1977), т.2, с.95, pBP325; Себерон и др. Gene, (1978), т.4, с.122, и pkc7; Рао и др. Gene (1979), т.7 с.79; и векторы бактериофага E.coli, которые включают λL 47.1, Шарон, Лоенен и др. Gene, (1980), т.10, с.249; и M13mp8 и M13m p9, Мессинг и др. Gene (1982), т. 19, с. 269. Общие приемы введения вышеупомянутой последовательности ДНК в клонирующий вектор с целью создания рекомбинантного вектора известны каждому специалисту в этой области техники. См. например, книгу "Молекулярное клонирование Лабораторное Руководство", под ред. Маниатиса, и др. 1982 г.
После того, как получены несколько копий искомой последовательности гетерологичной ДНК, эти последовательности можно выделить из рекомбинантных векторов и вставить в систему экспрессии с целью получения и выделения целевого гетерологичного протеина в соответствии с описанием, которое более детально дано ниже. Перед вставкой этих последовательной ДНК в вектор экспрессии можно осуществить различные модификации последовательности гетерологичной ДНК при помощи приемов, известных каждому специалисту в этой области техники, при помощи известных приемов модификации можно также осуществить уже после вышеупомянутой вставки.
В примерах реализации настоящего изобретения наряду с M13mp9, описанным Мессингом и др. Gene (1982), т.19, с.269, а качестве клонирующего вектора был выбран M13mp8, описанный там же, модифицированный таким образом, чтобы он содержал сайт рестрикции Xbal, как это указано на фиг. 1. Векторы M13mp8 и M13mp9, которые вместе именуются как "векторы M13", позволяют изолировать рекомбинантные векторы как в двухнитевой или репликативной форме (РФ), так и однонитевой форме ДНК. Изоляция рекомбинантных векторов РФ ДНК упрощает последующее введение искомых последовательностей ДНК после репликации в векторы экспрессии, как это показано например, на фиг. 8. В качестве альтернативы изоляция однонитевой формы этих рекомбинантных векторов упрощает как изоляцию рекомбинантных векторов, которые содержат искомую последовательность ДНК с правильной 5' --> 3' ориентацией для экспрессии, так и построение любой модификации последовательности ДНК при помощи таких приемов, как сайт-направленный мутагенез, как это показано на черт. 2, 4 и 6. Кроме того, эти векторы M13 могут "согласовывать" фрагменты ДНК или гены длиной до 4 килооснований (кб), которые обеспечивают клонирование типичной, полной, эукариотической генетической последовательности.
Маркерная функция, используемая в векторах M13, как это описано Мессингом и др. Gene, (1982), т.19, с.269, включает фермент β галактозидазу. В частности, искомую последовательность гетерологичной ДНК вставляют во фрагмент lac z гена, который содержится на векторе M13, что нарушает нормальную комплектацию фрагмента lac z гена, содержащегося на векторе M13, частичным фрагментом lac z гена, содержащегося в хромосомной ДНК клетки-хозяина (например, jMIOI E. coli), так, что вышеупомянутый хозяин уже не способен включать в обмен веществ лактозу, содержащуюся в среде для роста бактерий E.coli после инфицирования векторами M13, которые не содержат инородной генетической последовательности, вставленной во фрагмент lac z гена, способны включать в обмен веществ лактозу, содержащуюся в среде для выращивания бактерий, и вырабатывают характерные голубые бляшки, если эти бактерии выращиваются на агаре, включающем среду I x YT, содержащую 0,8% (в/о) триптона, 0,5% (в/о) экстракта дрожжей, 0,5% (в/во) NaCl и цветовой индикатор для b галактозидазы. Когда бактерии растут на вышеупомянутой среде, бляшки E. coli после инфицирования рекомбинантными векторами, несущими вставленную последовательность гетерологичной ДНК в генном фрагменте lac z вектора M13, прозрачны или бесцветны. Следовательно, положительная вставка последовательности гетерологичной ДНК в эти клонирующие векторы устанавливается в результате образования бесцветных бляшек инфицирования клетки-хозяина E.coli рекомбинантным вектором. Вставка последовательностей ДНК, кодирующих бГР(Л) и сГР(Ф), в векторы М13 представлена на фиг. 2 и 7, соответственно.
В предпочтительном варианте воплощения настоящего изобретения последовательности ДНК, кодирующие бГР(Л) и сГР(Ф), содержащиеся на бактериальных плазмидах pBGHex-I и pPGHex-I соответственно, как это описано у Сибурга и др. DNA (1983), 2(1), с. 37-45, выделяют из этих плазмид при помощи рестриктазы в специальном сайте. Необходимо отметить, что бактерии после трансфекции бактериальными плазмидами pBGHex-I или pPGHex-I соответственно и последующего культивирования при условиях, которые позволяют получить экспрессию последовательностей, кодирующих бГР(Л) и сГР(Ф) соответственно продуцируют соматотропин с N-концевым метионином (например, met-бГР(Л)met-сГР(Ф) соответственно). Соответствующие последовательности затем вставляют в РФ ДНК модифицированного вектора М13mp8 (M13mp8/Xbal) и вектора М13mp9, как это показано на фиг. 2 и 5, соответственно. Вставка искомых последовательностей ДНК бГР(Л) и сГР(Ф) и РФ ДНК M13mp8/Xbal и М13mp9 подтверждалась при помощи рестрикционного анализа, как это снова показано на фиг. 2 и 5 соответственно jMIOI E.coli затем подвергают трансфекции одним из этих рекомбинантных векторов, как это описано Мессингом и др. Methods in Enzymology (1983), т.101, с. 20, и однонитевую ДНК рекомбинатных векторов изолируют в соответствии с описанием, данным Мессингом и др. Gene (1982), т.19, с.269. Соответствующие абзацы этих работ Мессинга и др. используются здесь в качестве ссылок.
После выделения различные однонитевые ДНК этих рекомбинантных векторов модифицируют при помощи сайт-направленного мутагенеза с тем, чтобы получить кодирующие последовательности ДНК для бГР(А,В), бГР(А,Л), бГР(В) и сГР(А). В частности, бГР(Л) модифицируют добавлением кодона аланина, например, GCC, на 5' конце кодирующей бГР(Л) последовательности, как это показано на фиг. 3. Следует ожидать, что таким образом можно добавлять любой из четырех кодонов аланина. Предпочтительным кодоном аланина для оптимального выхода соматотропина в системе экспрессии, используемой в соответствии с настоящим изобретением, является GCC. Подтверждение присоединения кодона аланина при конструировании кодирующей бГР(А, Л) последовательности достигается в результате анализа полной последовательности ДНК бГР(А,Л) или ее 5'-конца с использованием метода Сэнгера и др. Proc. Nat'l. Acad. Sci. USA, (1077), т.74, с.5468.
Кодирующая бГР(А,В) последовательность конструируется при помощи сайт-направленного метагенеза кодирующей бГР(А, Л) последовательности, как это показано на фиг. 4, при помощи превращения кодона лейцина в аминокислотной позиции 127 [в бГР(А,Л)] в кодон валина, например, GTG. Снова можно ожидать, что при таком превращении можно использовать любой кодон для валина. Правильность конструкции кодирующей бГР(А,В) последовательности снова подтверждается при помощи анализа результирующей кодирующей бГР(А,В) последовательности.
Кодирующая бГР(В) последовательность, которая проявляется в синтезе протеинов met-бГР(В) бактериями после трансфекции векторами экспрессии, содержащими кодирующую бГР(В) последовательность, конструируется аналогично при помощи сайт-направленного мутагенеза кодирующей бГР(Л) последовательности при помощи превращения кодона лейцина в аминокислотной позиции 126[бГР(Л)] в кодон валина, например, GTG.
Конструирование кодирующей сГР(А) последовательности при помощи сайт-направленного мутагенеза кодирующей сГР(В) последовательности осуществляют в точности так же, как это показано на фиг. 6 и описано более полно ниже, а правильность конструкции подтверждается анализом последовательности ДНК.
После выделения и конструирования искомых последовательностей гетерологичной ДНК по аналогии с примерами для бГР(А,Л), бГР(А,В) и сГР(А), эти последовательности могут быть подвернуты репликации и получены многочисленные копии в результате амплификации соответствующих рекомбинантных векторов с использованием приемов, известных каждому специалисту в этой области техники, и которые уже были указаны выше. Такие последовательности гетерологичной ДНК могут быть вставлены в любые подходящие векторы экспрессии с целью продуцирования в прокариотах искомых гетерологичных полипептидов.
Получение полипептидов с N-концевым аланином.
Как уже было указано выше, подходящий вектор экспрессии должен содержать необходимые сигналы транскрипции и трансляции с целью получения гетерологичного протеина в выбранной клетке-хозяине вместе с маркерной функцией для идентификации тех векторов экспрессии, в которых были вставлены искомые последовательности гетерологичной ДНК. При помощи использования прокариотического вектора экспрессии последовательности рекомбинантной ДНК могут быть добавлены к генетическому комплементу прокариотического организма через трансдукцию, трансформацию или трансфекцию (все эти приемы в дальнейшем именуются одним термином "трансфекция") и затем вышеупомянутый организм можно культивировать при условиях (которые в общем случае диктуются используемыми промоторами и хозяином), которые стимулируют продуцирование целевого полипептида. Таким образом, "геномные" ДНК организмов, используемых в соответствии с настоящим изобретением, содержат как хромосомную ДНК, так и эписомную ДНК.
Известно несколько векторов экспрессии для экспрессии гетерологичного гена и продуцирования гетерологичного протеина в прокариотической клетке-хозяине, они известны теперь каждому специалисту в этой области техники.
В одном из предпочтительных вариантов воплощения настоящего изобретения использовали экспрессии pBGHex-I, см. Сибург и др, DNA (1983), 2 (1), с. 37-45, и модифицированный вектор pBGHex-I.
Вектором экспрессии pBGHex-I является бактериальная плазмида pBR322, несущая ген для бГР(Л). Этот ген содержит последовательно триптофановый промотор (ptrp), последовательность Шайн-Дольгарно, комплекс стартовый сигнал трансляции ATG /кодон метионина, примыкающий непосредственно к кодирующей последовательности N-концевого фенилаланина, первую аминокислоту полипептида бГР(Л), кодирующую бГР(Л) последовательность и стоп-кодон трансляции. Маркерной функцией на векторе экспрессии pBGHex-1 является устойчивость к антибиотику. В частности, pBGHex-1 несет два гена устойчивости к антибиотику, один ген устойчивости к ампициллину (ampr и второй устойчивости в тетрациклину (tetr), которые переносят специфическую устойчивость к антибиотику чувствительной к другим антибиотикам клетке-хозяину, стабильно трансформированной вектором экспрессии. Таким образом, стабильные трансформированные клетки могут быть выбраны по росту в среде, содержащие либо тетрациклин, либо ампициллин, либо оба этих антибиотика.
В примере реализации настоящего изобретения при получении вектора экспрессии pMON3213, несущего кодирующую сГР(А) последовательность вместо кодирующей бГР(Л) последовательности, использовали pBGH
Продуцирование бГР(А,Л), бГР(А,В) или сГР(А) в культуре E.coli осуществляли в результате трансформации либо E.coli W3110, либо E.coli E392, E.coli штамма 294, которые депонированы под номерами ATCC 39936, 53010 и 53009, соответствующим вектором экспрессии, используя приемы, которые более подробно описаны ниже. Трансформированные клетки E.coli W3110 затем культивируют при условиях, которые позволяют осуществить экспрессию генов соматотропина и продуцирование искомых гетерологичных полипептидов.
Очистка полученного гетерологичного пептида будет зависеть как от типа протеина, так от клетки-хозяина. Было, например, замечено, что гетерологичные протеины, полученные в такой бактерии, как E.coli очень часто осаждаются внутри клетки в форме "преломляющих" тел. Здесь используют термин "преломляющий" ввиду того, что эти тела можно действительно наблюдать с использованием фазово-контрастного микроскопа. Способ, который используют для выделения гетерологичных протеинов в биологически активной форме, описан в Европейской патентной заявке N 114506 (опубликованной 1 августа 1984г.), которая здесь используется в качестве ссылки. Очень коротко, этот способ очистки содержит концентрирование клеток-хозяев, лизирование этих клеток с целью получения экстракта или гомогената клеток и, наконец, выделение преломляющих тел при помощи дифференциального центрифугирования: все эти стадии хорошо известны каждому специалисту в этой области техники. Выделенные преломляющие тела растворяют в сильном денатурирующем средстве таком, как хлоргидрат гуанидина и солюбилизированный протеин, затем подвергают реакции обмена в подходящем растворителе (например, мочевине), подвергают очистке при помощи хроматографии и, наконец, биологической активности, то есть заставляют принять их активную конфигурацию, а затем окисляют так, чтобы сохранить эту конфигурацию, поддерживая дисульфидные связи между их цистеиновыми остатками, как описано в Европейской патентной заявке N 114506. Более детальное описание такой очистки гетерологичных протеинов дано в двух одновременно находящихся в стадии рассмотрения патентных заявках США, одной, поданной С.Б. Сторрсом и озаглавленной "Способ солюбилизации соматотропинов", и второй, поданной Л. Э. Бентлом, С.Б.Сторрсом и Дж. Митчелом и озаглавленной "Способ натурализации соматотропина", которые здесь используются а качестве ссылок. Две вышеупомянутые одновременно находящиеся на стадии рассмотрения патентные заявки США и настоящая заявка поданы от имени фирмы Монсанто Компани. Последующая очистка гетерологичного полипептида с целью его освобождения от загрязняющих бактериальных протеинов может быть осуществлена с использованием известных методов хроматографии таких, как гелевая фильтрация или ионообменная хроматография. В общем случае полученная в результате очистки композиция будет содержать по массе от примерно 90% до примерно 99,5% N-аланинового полипептида и от примерно 0,5% до примерно 10% протеина, родственного бактерии или другому прокариотическому хозяину, в котором полипептид был синтезирован.
Было установлено, что в общем случае по крайней мере примерно 80% гетерологичного пептида, экспрессированного по способу настоящего изобретения, имеет N-концевую структуру типа NH2-ala. Оставшийся полипептид в общем случае имеет метиониловую форму и содержит N-концевую структуру типа NH2-met -ala. Однако варьируя условия роста и/или время индукции экспрессии гена можно увеличить долю полипептида с аланином на N-конце до по крайней мере 95% и даже больше.
В особенности предпочтительном варианте воплощения настоящего изобретения соматотропиновые полипептиды, продуцированные и выделенные в соответствии с описанием, приведенным выше, как было показано, проявляют биологическую активность, подобную активности соматотропина; этот факт был установлен при помощи анализа реакции рецептора печени кролика, осуществленного в соответствии с описанием, приведенным Пушима и Фризеном, J. Clin. Endocrinol. Metab. (1973), т.37, с. 334-337, и биоанализа на прирост веса крыс. В соответствии с последним анализом биоактивности соматотропинов, продуцированных клетками E.coli, определялись относительно известной партии соматотропина (например, бычьего или свиного гипофизарного соматотропина), связывая прирост веса крыс с удаленными гипофизами с изменением количества вводимого соединения. В частности, титрованные дозы (от 0 до 60 мкг) неизвестного или стандартного источника соматотропина вводились крысам с удаленными гипофизами (95 135 г) в форме инъекций на основе ежедневной дозы в течение 7 и более дней. Используя множественную регрессию, составляли регрессионное уравнение для прироста веса тела животных, получающих известные и неизвестные партии гормона, в зависимости от логарифмических доз. Наклоны контролировали с тем, чтобы гарантировать непараллелизм и обеспечить пересечение полученных множеств. Биоактивности представляли в виде отношения наклонов, умноженного на активность стандарта.
Приводимые ниже примеры служат иллюстрацией предпочтительных вариантов воплощения настоящего изобретения и их не следует рассматривать в качестве его ограничения. Несмотря на то, что настоящее изобретение было описано в связи с предпочтительными вариантами его воплощения, каждому специалисту в этой области техники ясно, что на основе приведенного выше патентного описания можно осуществить различные модификации, не выходя за область, охватываемую формулой изобретения.
Микроорганизмы и плазмиды.
Следующие микроорганизмы могут быть получены из Американского Собрания Типов Культур (ATCC), 12301 Парклаун Драйв, Роквилл, Мериленд, 20652, США:
ATCC 38835 E.coli W3110
ATCC 53010 E.coli LK392
ATCC 53009 E.coli W3110 штамм 294
ATCC 53024 E.coli W3110/pMO N 3209/
ATCC 53022 E.coli W3110/pMO N 3215/
ATCC 53023 E.coli W3110/pMO N 3213/
Эти депонированные организмы могут быть получены после выдачи патента США подателю настоящей заявки, фирме Монсанто Компани. Эти депонированные организмы будут доступны после выдачи любого патента США по патентной заявке, для которой будет установлен приоритет по дате подачи настоящей заявки. Однако следует иметь в виду, что доступность депонированного организма не дает право на лицензию с целью реализации целей настоящего изобретения, умаляющего патентные права заявителя, гарантированные патентными законодательствами. Кроме того, настоящее изобретение не ограничивается теми микроорганизмами, которые были депонированы, так как депонированные организмы только иллюстрируют предмет настоящего изобретения.
Пример 1. Все олигонуклеотиды синтезировали в отделе Биологических наук фирмы Монсанто, используя синтезаторы ДНК фирмы Апплайд Биосистемз, в соответствии с процедурой, преложенной производителем, фирмой Апплайд Биоситемз Инк, Фостер Сити, Калифорния, ферменты рестрикции и ферменты, модифицирующие ДНК, были получены от фирмы Нью Инглэнд Биолэбо (Беверли, Массачусетс), от фирмы Нью Ниглэнд Нюклса, (Бостон, Массачусетс) и от фирмы Бетесда Рисеч Лабораториз (БРЛ) (Гайтерсбург, Мэриленд), Линкер Xbal получали от фирмы Коллаборэтив Рисеч, Инк. (Лексингтон Массачутес), ДНК лигазу Т4 получали от фирмы БРЛ (Гайтерсбург, Мэрилэнд), Днк-лигазу Т4 получали от фирмы Нью-Ниглэнд Биолюбо (Беверли, Массачутес). Меченые 32P нуклеотиды получали от фирмы Амернэм (Арлингтон Хайтс, Иллинойз). Фрагмент Кленова ДНК-полимеразы I E. coli получали от фирмы Нью Инглэнд Нюклеа (Бостон, Массачутесе), vMIOI E. coli получали от доктора Джо Мессинга, Университет Миннесоты (Сент Паул, Миннесота).
Расщепление рестриктазами реакции ДНК-лигазы Т4 и фрагмента Кленова ДНК полимеразы I Ecoli могут быть осуществлены в соответствии с приемами, предложенными производителями этих препаратов. Предпочтительными буферами для описываемых ниже ферментов рестрикции являются следующие: для Xbal: 100 мМ NaCl, 50 мМ трис, pH 7,5, 10 мМ MgSO4, для PeoPl, Hind III и Sma 1:50 мМ NaCl, 10 мМ Трис, pH 7,5, 10 мМ MgSO4.
Реакцию ДНК-лигазы Т4 осуществляли в буферах, содержащих 25 мМ Трис, pH 8,0, 10мМ MgCl2, 10 мМ дитиотриола (ДТТ), 2 мМ спермидина и 0,2 мМ АТФ. Фрагмент Кленова ДНК полимеразы I E.coli использовали в буфере, содержащем 20 мМ Трис, pH 7,2 10 мМ MgCl2, 10 мМ (ДТТ), 1 мМ АТФ и 1 мМ каждого ДАТФ, дГТФ, дПТФ, дГТФ. Если необходимо было получить меченую, вновь синтезированную нить ДНК, то в реакции Кленова добавляли комплекс альфа 32P-ДАТФ (400 Ки/ммоль).
Меченые олинуклеотиды получали с использованием гамма-32P-АТФ (уд.акт. более 5000 Ки/ммоль) и ДНК-киназы Т4 в 100 мМ Трис, pH 8,0, 10 мМ MgCl2, 5 мМ ДТТ.
Плазмиды, несущие последовательности ДНК, кодирующие бГР(Л) и сГР (pBGHex-I и pPGHex-I соответственно), были получены от фирмы Генетек Инк, Сан-Франциско, Калифорния. Эти плазмиды могут быть получены в соответствии с описанием, приведенным в Европейской патентной заявке N 75444 (опубликованной 30 марта 1983 г.); Сибург и др. DNA (1983), 2(1), с. 37-45; Геддел и др. Nature (1979), т. 281, с. 544-548; Деббер и др. книга "Промоторы: Структура и функция (1982), редакторы: м. Дж. Чемберлин и Р. Родригес. Гл. 293; Миоццери и Янофски, J. Bacteriol (1978), т.133, с. 1457-1466; и Розенберг и Курт Annual Revia of Genetics т.13, с. 319-353.
M13mp8 и M13mp9 были получены от доктора Джо Мессинга из Университета Миннесоты (сент Паул, Миннесота).
Все компоненты среды для выращивания бактерий и антибиотики получали либо от фирмы Сигма (сент Луис, Мо), либо от фирмы Дифко Лабораториз (Дейтрот, Мичиган).
Пример 2. Приводимые ниже примеры иллюстрируют конструирование кодирующих последовательностей ДНК, которые в результате экспрессии обеспечивают прямое продуцирование в бактерии полипептидов, содержащих N-концевой аланин. В частности, кодирующие последовательности ДНК конструировали таким образом, что комплекс начало трансляции/кодон метионина (ATG) содержится непосредственно перед кодоном аланина (например, GCC). Эти три кодирующие последовательности, содержащие бГР(А, Л), бГР(А,В) и сГР(А), конструировали при помощи сайт-направленного мутагенеза в ранее выделенных последовательностях ДНК соматотропина.
а. Конструирование кодирующей бГР(А,Л) последовательности ДНК.
Последовательность ДНК, кодирующую соматотропин бГР(Л), выделяли из pBGHex-I, как Xba I-фрагмент и клонировали в сайт Xbal модифицированного вектора M13mp8 (M13mP8/Xbal). Конструкция вектора M13mp8/Xbal, который содержит линкер Xbal в исходном сайте Smal, представлена на фиг.1. Как показано на фиг. 2 Xbal отщепляет на одном из концов кодирующую бГР(Л) последовательность ДНК, "вырезая" таким образом полную кодирующую бГР(Л) последовательность. Плазмиду pBGHex-I после обработки ферментом Xbal смешивали в присутствии ДНК-лигазы Т4 с РФ ДНК M13mP8/Xbal, линеаризованной при помощи расщепления ферментом рестрикции Xbal, и обрабатывали щелочной фосфатазой из кишечника крупного рогатого скота. Затем смесь инкубировали в течение ночи при температуре 14oC. Обработка щелочной фосфатазой из кишечника крупного рогатого скота предотвращает рециркуляризацию вектора M13mP8/Xbal. Вставку кодирующей бГР(Л) последовательности ДНК в вектор M13mP8/Xbal с целью образования рекомбинантного вектора M13mP8/BGHex-I первоначально контролировали образованием бесцветных бляшек на культуре бактерий, jMIOI E.coli, выращиваемой на среде I x УТ, используя процедуру покрытия мягким агаром, описанную в книге "Молекулярное клонирование: лабораторное руководство", ред. Маниатис и др. (1982), с. 64, который содержит 10 μл 100 мМ МПТТ (изопропил-β-D-тиогалактопиранозида) и 50 mл 2% (в/о) X-GAL (5-бром-4-хлор-3-индолио-β-D галактопиранозида) в 3 мл верхнего агара, и осуществляли трансфекцию вышеупомянутым рекомбинантным вектором, как это было описано выше. Вставку кодирующей бГР(Л) последовательности подтверждали при помощи рестрикции изолированной РФ-формы ДНК, "Молекулярное клонирование: лабораторное руководство", Маниатис и др. (ред.) (1982 г.), гл. 3, рекомбинантного вектора ферментом Xbal, в результате чего получали фрагмент в 590 пар оснований, содержащий вставленную последовательность. Фрагмент в 590 пар оснований идентифицировали при помощи электрофореза на агаровом геле в однопроцентном (в/о) агаре в соответствии с описанием, приведенным в книге "Молекулярное клонирование: лабораторное руководство", Маниатис и др. (ред.), (1982). Все последующие фрагменты после обработки ферментами идентифицировали именно при помощи этой процедуры. Ориентацию вставленных кодирующих бГР(Л) последовательностей контролировали при помощи рестрикции рекомбинантного вектора ферментами Smal и Hind III. Если кодирующие последовательности находятся в правильной 5'-->3' ориентации, в результате расщепления этими ферментами рестрикции должны получить фрагмент в 207 пар оснований. Выделение однонитевой ДНК фага проводили в соответствии с процедурой, предложенной Мессингом и др. Gene(1982), т. 19, с. 269. Затем вектор M13mp8/BGHex-I использовали в качестве матрицы в сайт-направленном мутагенезе по существу в соответствии с описанием, данным Золлером и Смитом, Nucl. Acids. Res. (1982), т. 10, с. 6487-6500; Золлером и Смитом Methods in Enzymol (1983), т. 100, с. 468-500; Норрисом и др. Nuc. Acids. Res. (1983), т. 11, с. 5103-5112, которые используются здесь в качестве ссылок.
На фиг. 3 приведена диаграмма процедуры мутагенеза при конструировании кодирующей бГР(А,Л) последовательности ДНК из кодирующего бГР(Л) последовательности. Если очень коротко, то олигонуклеотидную затравку (смотри табл. 1), содержащую последовательность для искомой мутации, используют для синтеза затравки копии замкнутой кольцевой ДНК матрицы M13mp8/BGHex-I однонитевой ДНК. Полученные таким образом замкнутые кольцевые молекулы двунитевой ДНК отделяли от неполных и однонитевых колец ДНК при помощи центрифугирования в градиенте щелочной сахарозы, как это описано Золлером и Смитом, Methods in Enzymol. (1983), т. 100, с. 468-500. Замкнутые кольцевые молекулы двунитевой ДНК затем использовали для трансформации VM101 E.co1i в соответствии с описанием, данным Мессингом и др. Gene (1982) т. 19, с. 269-276 и полученные в результате бесцветные бляшки переносили на фильтры Палл, полученные от фирмы Палл Ультрафайн Фильтреш Корп, и анализировали путем гибридизации с меченой 32P формой олигонуклеотидной затравки, использованной для осуществления мутагенеза в специальном сайте. Перенос вышеупомянутых бляшек осуществляли в соответствии с методами, описанными производителем фильтром Пэлл. Анализ путем гибридизации осуществляли с использованием нейлоновых фильтров Вилдайн, которые описаны фирмой Пэлл Ультрафайн Вильтрошн Корпорешн (Гленн Коув, Нью-Йорк), "Описательное Руководство для переноса ДНК на Нейлоновые фильтры Пэлл Виодайн ТМ" (1983). Фильтры промывали при увеличивающихся температурах до тех пор, пока не исчезнет радиоактивный сигнал с контрольного фильтра, который приготавливали с использованием фага M13mp8/BGHex-1. Общий протокол промывки фильтров включает промывку при комнатной температуре в 6хSS C/0,9H NaCl и 0,09 М цитрата натрия в течение десяти минут, затем промывку при температуре 50oC в 6хSSC в течение пяти минут и последующие промывки при увеличении температуры на 5oC. Бляшки, которые гибридизировались с меченой радиоактивным элементом олигонуклеотидной затравкой при температурах более высоких, чем контрольные фаги, рассматривали, как несущие новую созданную кодирующую бГР(Л) последовательность и их называли в дальнейшем потенциально положительными. В качестве альтернативы отдельные бесцветные бляшики отбирали из культуры трансформированных клеток jМФ01 E.coli и выращивали в 5 мл среды 2 х УТ (1,6%) (в/о) триптона, 1,0% (в/о) экстракта дрожжей, 0,5% (в/о) NaCl в течение ночи при температурах 37oC в условиях аэрации. ДНК фага, полученную в соответствии с описанием, данным Мессенгом и др. Gene (1982), т. 19, с. 289, затем наносили пятнами на нитроцеллюлозу, подвергали гибридизации с затравкой, меченой радиоактивным элементом, и промывали при увеличении температуры, как это было описано выше. ДНК фага, которые имели температуры гибридизации выше, чем контрольные бляшки M13mp8/BGHex-I, также называли потенциально положительными. Потенциально положительные бляшки из обеих процедур анализа выращивали в соответствии с описанием, приведенным выше, использовали для получения однонитевой ДНК фага, последовательность которой затем анализировали в соответствии с процедурой Сэнгера и др. Proc. Nat'l. Acad. Sci. USA, (1977), т. 74, с. 5463, с тем, чтобы подтвердить, что они действительно несут кодирующую сГР(А, Л) последовательность. РФ ДНК M13mp8/BGHex-I(ala) также анализировали при помощи ферментов рестрикции Hae II с тем, чтобы убедиться в присоединении кодона анализа после комплекса стартовый сигнал/кодон метионина ATG, так как при создании кодона аланина образуется дополнительный сайт рестрикции Hae III.
Вероятность присоединения кодона аланина к кодирующей бГР(Л) последовательности ДНК составляет примерно 2%
б. Конструирование кодирующего сГР(А) последовательности ДНК
Здесь также использовали сайт-направленный мутагенез для того, чтобы присоединить кодон аланина к кодирующей сГР(Ф) последовательности ДНК, описанной Сибургом и др. DNA (1983, 2(1), с. 37-45. Процедуру мутагенеза в соответствии с диаграммой, представленной на фиг. 8 осуществляли следующим образом.
Кодирующую сГР(Ф) последовательность ДНК в 590 пар оснований, содержащуюся на плазмиде pPGHex-I, выделяли на этой плазмиде при помощи расщепления ферментами рестрикции E. coRI и Hind III, которые расщепляют плазмиду на 5'-3'-концах кодирующей сГР(Ф) последовательности соответственно, как это показано на фиг. 7. Расщепленную плазмиду pPGHex-I затем смешивали с РФ ДНК M13mp9, также после рестрикции ферментами EcoRI и Hi nd III, но дополнительно обработанной щелочной фосфатазой из кишечника крупного рогатого скота, с тем, чтобы предотвратить повторное лигирование рестрикционных фрагментов M13mp9. Затем в вышеупомянутую смесь добавляли ДНК-лигазу Т4. Благодаря наличию липких концов на РФ ДНК фага и кодирующей сГР(Ф) последовательности ДНК, образованных при использовании двух различных ферментов рестрикции, ДНК сГР(Ф) может быть селективно вставлена в РФ ДНК фага и может быть вставлена в правильной 5'-->3' ориентации, как это показано на фиг. 7. Затем осуществляют трансформацию jMIOI E.coIi в соответствии с описанием, приведенным выше для M13mp8/BGHex-I, причем рекомбинантный вектор M13mp9/PGHex-I несет кодирующую сГР(Ф) последовательность ДНК. Затем трансформированные jMIOI E. coIi выращивали на среде I х УТ, содержащей колориметрические реагенты, и выбор осуществляли по образованию бесцветных бляшек, как это было уже описано выше. Вставку кодирующих сГР(Ф) последовательностей ДНК подтверждали следующим образом. Выбирали бесцветные бляшки, а РФ ДНК M13mp9/PGHex-I, выделенную в соответствии с описанием, приведенным выше, затем расщепляли ферментами EcoRI и Hind III и подвергали электрофорезу на агаровом геле, в результате чего получали фрагмент в 590 пар оснований, содержащий вставленную ДНК сГР(Ф). Затем фаг M13mp9/PGHex-I размножали в jMIOI E.coIi, а ДНК фага выделяли в соответствии с описанием, приведенным выше.
Далее ДНК M13mp9/PGHex-I использовали в качестве матрицы для сайт-направленного мутагенеза, как это показано на фиг. 8, в соответствии с описанием, приведенным выше при конструировании кодирующей бГР(А, Л) последовательности, используя специальную затравку (смотри табл. 1). Вероятность присоединения кодона аланина, в данном случае GCC, к кодирующей сГР(Ф) последовательности составила примерно 12% Полученную в результате кодирующую сГР(Ф) последовательность снова исследовали при помощи анализа ДНК-последовательности.
Пример 3. Служит иллюстрацией способа конструирования рекомбинантных векторов экспрессии, которые обеспечивают прямое продуцирование в бактериях полипептидов, содержащих N-концевой аланин. С помощью сконструированных векторов получают три полипептида соматотропина видов бГР(А,Л), бГР(А,В) и сГР(А). Для сравнения используют прямое получение met-бГР(В) и met-(бГР(А).
а) Экспрессия кодирующей сГР(А) последовательности ДНК.
Как показано на фиг. 10, кодирующую сГР(А) последовательность, содержащуюся на рекомбинантном векторе M13mp9/PGHex-I (ala) использовали для того, чтобы заменить кодирующую бГР(Л) последовательность ДНК, содержащуюся на модифицированном векторе экспрессии pBGHex-I. Модифицированный вектор экспрессии pBGH
Вектор pBGHex-I после рестрикции подвергали рециркуляризации (замыканию) с использованием ДНК-лигазы Т4 и затем использовали для трансформации культур E.coli M101; все это осуществляли в соответствии с описаниями, приведенными ранее. ДНК плазмиды из каждой из колоний затем анализировали на присутствие фрагмента EcoRI/Hind III в 590 пар оснований, несущего кодирующую сГР(А) последовательность, и фрагмента EcoRI/Pst 1 в 1050 пар оснований, несущего последовательность Ptrp (смотри фиг. 9). Удаление этого сайта рестрикции EcoRI упрощает вставку в специальный сайт последовательности, кодирующей сГР(А), в вектор экспрессии pBGHex-I в правильной ориентации, как это описано на фиг. 10. Рекомбинантный вектор экспрессии, полученный таким образом, в дальнейшем обозначается pMON3213. Смесь, содержащую pMON3213, далее использовали для трансформации культуры E.coli W 3110, затем трансформированные клетки выращивали и подвергали селекции. Культура E.coli W3110, содержащая pMON3213, депонирована под номером ATCC 53023. Замену кодирующей бГР(Л) последовательности кодирующей сГР(А) последовательностью в векторе экспрессии pBGH
Экспрессию кодирующей сГР(А) последовательности ДНК и продуцирование сГР(А) в культуре E.coli W3110 осуществляли при помощи традиционных приемов. Были получены высокие концентрации протеинов в 22000 Да, что устанавливали при помощи анализа ДСН/ПАГЭ.
Бактерии, несущие плазмиды экспрессии pMON3209, pMON3214, pBGHex-1 и pMON3215, хранили следующим образом. Каждую колонию культуры E.coli W3110, трансформированную при помощи только одной плазмиды (pMON3209 или pMON3215, pMON3214 или PBGHex-I), выращивали в течение ночи при температуре 37oC в 5 мл БЛ плюс 12,5 μл/мг тетрациклина при аэрации. Порцию в 1 мл каждой культуры после выращивания в течение ночи добавляли отдельно в индивидуальные колбы, содержащие 25 мл БЛ плюс 12,5 мг/мл тетрациклина и выращивали до достижения ОП600= 1,0. Затем клетки из каждой колбы собирали центрифугированием при ускорении 6000 x g при температуре 4oC в течение 5 мин. Осадки отдельно суспендировали снова в 12 мл БЛ плюс 7,5% (в/о) ДМСО и очень быстро замораживали на сухом льду порциями в 1 мл. Далее клетки хранили в холодильнике с жидким азотом. Кроме того, примерно 10 мкм очищенной ДНК плазмиды хранили при температуре -80oC.
Клетки E.coli 3110, несущие вектор экспрессии pMON3213, хранили и далее использовали для получения в больших количествах сГР(А) ферментацией в 10-100-литровых емкостях. Содержание протеина с ГР(А) в реакторе ферментации емкостью 100 л составило приблизительно 1г/л бульона, это факт устанавливали при помощи радиоиммуноанализа Рознера и др. J. Immunol. Methods (1982), т. 52, с. 175-181.
Пример 4. Этот пример осуществляли с тем, чтобы определить N-концевую аминокислотную последовательность гетерологичных протеинов бГР(А,Л), бГР(А, В), met-бГР(АБ), met-бГР(Л) и сГР(А), полученных при помощи бактерий.
Соматотропиновые полипептиды, синтезированные в культуре E.coli, подвергали очистке из неочищенных, солюбилизованных тел включения, содержащих один из бГР(А, Л), бГР(А,В), met-бГР(В), met-бГР(Л) или бГР(А) при помощи иммуносорбентной хроматографии в соответствии с описанием, данным Криви и Роулдом, Hybridoma (1984), т. 3, с. 151-161.
Все три вида соматотропина, очищенные при помощи иммуносорбентной хроматографии, как оказалось, имели чистоту, превышающую 95% этот факт был установлен при помощи анализа ДНС-ПАГЭ с использованием 1 мг очищенного протеина на геле с градиентом 7,5-15% осуществленного в соответствии с процедурой, описанной Лаеммли, Nature (1970), т. 227, с. 680-685.
Концентрации протеинов очищенных видов бГР определяли при помощи известного анализа с использованием высокоразрешающей жидкостной хроматографии.
Протеины, очищенные при помощи хроматографии, основанной на принципе иммунного средства, для использования в анализе на N-концевую последовательность подвергали диализу для полного удаления воды, а затем подвергали лиофилизации. Перед выполнением анализа N-концевой последовательности очищенные протеины снова суспендировали в буфере бикарбоаната аммония, содержащего 50 мМ бикарбоната аммония плюс 0,1% (в/о ДСН и подвергали диализу против того же буфера с тем, чтобы удалить остаточные трис/оксиметил/аминометил/трис/и глицерин. Затем использовали анализатор последовательностей Протеинов Модели 470А фирмы Эпплайд Биосистемз (фирма Эпплайд Тиосистема, Инж. Фостер Сити, Калифорния) для анализа всех N-концевых последовательностей, как это описано Ханкапиллером и др. (1983), Methods in Enzymol. т. 91, с. 486-493.
В табл.2 приведены результаты анализа последовательностей для нескольких препаратов полипептидов бГР(А,Л), бГР(А,В), met-бГР(В), met-бГР(Л) и сГР(А). Количество протеина с N-концевым метионином приведено в процентах от общего содержания соматотропина в пробе. Для определения количества метионина использовали два метода. При помощи непрямого метода количество NH2-met-ala-phe в преобладающей популяции NH2ala-phe. вычисляли, исходя из различий в lag-сигналах. Так как эта процедура зависит от некоторой оценки "нормальной" последовательности lag, которая изменяется от цикла к циклу, она является лишь весьма грубой оценкой для содержащейся последовательности NH2-ala-phe. Прямой метод, содержащий реакцию молекулярной леградации Эдмана, заключается в сравнении мощности сигналов от РТН-met до РТН-ala после отделения pTH-met от химического шума с использованием высокоразрешающей жидкостной хроматографии (ВРЖХ). "РТН" обозначен фенил-тиохидантоин. В частности, реакция последовательной деградации по Эдману заключается во взаимодействии N-концевой аминокислоты с реагентом, который отщепляет эту аминокислоту, и в ее последующем высвобождении в форме РНТ-производиного этой аминокислоты. Последняя процедура будет давать хорошие оценки в для met-ala-phe. в том случае, если загрязнение свободной аминокислоты является низким.
Как показывают результаты относительно N-концевой последовательности, представленные в табл. 2, нет никаких очевидных признаков отщепления метионина, если после N-концевого метионина следует фенилаланин. Однако 80% и более молекул, продуцированных генетическими конструкциями МВ (А,Л) и МВ(А,В), имеют аланин на N-конце, а не метионин. Степень отщепления N-концевого метионина варьирует в клетках от ферментации, но всегда составляет не менее 80% от всех молекул соматотропина. Кроме того, уровни продуцирования и приблизительно 10-15% от общего количества бактериального протеина были достигнуты для соматотропина, синтезированного в трансформированных микроорганизмах.






Использование: биотехнология, сельское хозяйство. Сущность изобретения: из штамма E.coli, предварительно трансформированного рекомбинатным вектором, в котором непосредственно после сайта инициации трансляции, представленного метиониновым кодоном, встроен фрагмент ДНК, кодирующий свиной гормон роста с N-концевым аланином pGH (А), получен без дополнительной обработки полипептид pGH(A), не содержащий на N-конце остатка метионина. 8 ил., 2 табл.
Рекомбинантный полипепдит pGH(A) с N-концевым аланином, непосредственно полученный в штамме бактерий Escherichia coli, трансформированном рекомбинантной плазмидной ДНК, содержащей нуклеотидную последовательность, кодирующую полипептид pGH(A) и расположенную сразу после сайта инициации трансляции, представленного кодоном для метионина, с мол.м. 22000 Дальтон, не содержащий дополнительного остатка метионина на N-конце.
| Seeburg et al., DNA, v | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Пишущая машина | 1922 |
|
SU37A1 |

