Предполагаемое изобретение относится к биотехнологии, в частности к генной и белковой инженерии. Оно включает сконструированную in vitro рекомбинантную плазмидную ДНК p3PTEIL3, обусловливающую биосинтез рекомбинантного человеческого интерлейкина-3 (rhIL3), штамм E. coli/p3PTEIL3 - продуцент rhIL3 и способ получения рекомбинантного интерлейкина-3 на основе вышеуказанной рекомбинантной ДНК и штамма-продуцента.
Человеческий интерлейкин-3 (hIL3) гликопротеин, продуцируемый Т-лимфоцитами, является медиатором роста и дифференции клеток различных линий. Он играет важную роль в регуляции гемопоэза и иммунного ответа [1, 2] hIL3 представляет собой неспецифический гемопоэтин, действующий на предшественники всех клеток крови и даже на зрелые клетки [3] Эти свойства hIL3 определяют возможность использования его в качестве медицинского препарата регулятора кроветворения. В связи с малой доступностью природного hIL3 большое значение приобретает использование рекомбинантного hIL3 (rhIL3), который, несмотря на отсутствие в структуре гликозилирующих групп, обладает той же биологической активностью, что и природный hIL3 [4, 5]
Ранее для получения rhIL3 микробиологическим синтезом в E.coli уже была сконструирована экспрессионная плазмида pTE21L3 [6] с которой был достигнут высокий уровень биосинтеза rhIL3 в некоторых штаммах E.coli. Однако высокопродуктивный штамм при продолжительном культивировании или при многих пересевах терял свою продуктивность вследствие сегрегации плазмиды, обусловленной чрезмерным уровнем конститутивного биосинтеза рекомбинантного белка.
Более стабильная система, сохраняющая высокий выход rhIL3, была получена в конструкции экспрессионной плазмиды pTOTE2IL3, использующей схему биосинтеза с индукцией транскрипции на поздних стадиях роста культуры продуцента [7] В этой плазмиде инициация транскрипции осуществлялась с тандемом двух промоторов P3 фага fd и Ptac, контролируемым тандемом двух lac-операторов (7). При этом в штаммах E.coli JM109 и E.coli TGI был достигнут высокий уровень биосинтеза rhIL3, однако продуктивность их была недостаточной вследствие либо медленного накопления клеточной массы (JM 109), либо рекомбинационной неустойчивости плазмиды (в гесA + штамме TGI) [7]
Предлагаемая группа изобретений позволяет получать рекомбинантный человеческий интерлейкин-3 в стабильном штамме-продуценте с высоким выходом, позволяющим эффективно производить очистку по простой технологии.
Для этой цели использована новая конструкция экспрессионной плазмиды, содержащая дополнительно промотор РНК-полимеразы фага T7 (Pt7), в штамме E. coli BL21(DE3), в хромосоме которого содержится ген Т7-РНК-полимеразы под контролем индуцируемого промотора Ptac [8]
Предлагаемый способ получения интерлейкина-3 заключается в том, что рекомбинантной плазмидной ДНК p3PTYIL3 трансформируют клетки штамма Escherichia coli BL21 (DE3), полученный штамм Escherichia coli BL21(DE3)/p3PTEIL3 культивируют в богатой среде, выращенные клетки разрушают в буферном растворе и выделяют белок rhIL3.
Предлагаемая рекомбинантная плазмидная ДНК p3PTEIL3 характеризуется следующими признаками:
кодирует аминокислотную последовательность белка rhIL3;
имеет мол. м. 2,34 МДа;
содержит:
тандем промоторов P8 фага fd и Ptac, два lac-оператора, промотор P7, усилитель трансляции гена 10 фага T7, ген интерлейкина-3 и терминатор транскрипции фага fd;
в качестве генетического маркера ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой p3PTEIL3 клеток E.coli к пенициллиновым антибиотикам;
уникальные сайты узнавания рестрикционных эндонуклеаз, расположенные на следующем расстоянии вправо от сайта BamHI:
XbaI 325bp, EcoRV 361bp, Ndel 365 и 1292 bp, HpaI 411 bp, Hind III-545 bp, EcoRI 774 bp, Bg III 1128 bp, PstI 2527 bp.
Конструкция рекомбинантной плазмидной ДНК p3PTEIL3 обеспечивает высокий уровень экспрессии гена интерлейкина-3.
Предлагаемый штамм-продуцент Escherichia coli BL21 (DE3)/p3PTEIL3 характеризуется следующими признаками.
Морфологические признаки. Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или YT-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки. Клетки растут при температуре от 4oC до 40oC при оптимуме pH от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к пенициллиновым антибиотикам (до 500 мкг/мл).
Штамм-продуцент E. coli BL21(DE3)/p3PTEIL3 отличается от штамма-реципиента E. coli BL21(DE3) только наличием рекомбинантной плазмидной ДНК p3PTEIL3, которая и придает ему устойчивость к пенициллиновым антибиотикам.
Штамм-продуцент получают путем трансформации компетентных клеток E.coli BL21 (DE3) соответствующей рекомбинантной плазмидной ДНК.
Клетки E. coli BL21(DE3)/p3PTEIL3 являются продуцентом белка rhIL3. При индукции изопропилтио-b-D-галактозидом происходит эффективный биосинтез белка, который накапливается в клетках в виде телец включения, и его выход составляет более 30% суммарного белка бактерий.
Изобретение осуществляют следующим образом. Конструируют рекомбинантную плазмидную ДНК p3PTEIL3. Для этого сначала получают векторный фрагмент плазмиды pTE2IL3 [6] путем ее расщепления эндонуклеазами BamHI и XbaI. После очистки электрофорезом в 1% агарозном геле векторный фрагмент лигируют с BamHI/EcoRI-фрагментом плазмиды pKK223-3, содержащим промотор Ptac, синтетическим lac-оператором [7] и синтетическим фрагментом, содержащим промотор Pt7. Лигазной смесью трансформируют компетентные клетки E.coli BL21(DE3) и высевают на YT-агар, содержащий 50 мкг/мл ампициллина или другого пенициллинового антибиотика. Полученные клоны анализируют гибридизацией с 32P-мечеными олигонуклеотидами, составляющими синтетические фрагменты, и из гибридизующихся с зондами клонов выделяют плазмидные ДНК, которые подвергают рестриктному анализу с помощью рестриктаз HindIII, EcoRI, BamHI и BglII, а также определением нуклеотидной последовательности между сайтами BamHI и EcoRI. Штамм-продуцент E/coli BL21(DE3)/p3PTEIL3 выращивают в богатой среде (YT-, LB-бульон и др. ), индуцируют изопропилтио-b-D-галактозидом и снова выращивают до достижения максимальной плотности культуры.
Выделение белка rhIL3 из клеток продуцента включает следующие стадии:
разрушение выращенных клеток одним из обычно применяемых способов;
отмывку буферными растворами телец включения от растворимых компонентов клетки;
солюбилизацию денатурированного целевого белка из телец включения либо в 5 8 М растворе мочевины, либо в 5 М растворе гидрохлорида гуанидина, либо в другом подходящем растворителе;
очистку целевого белка в результате ступенчатого разбавления растворов в мочевине или гидрохлориде гуанидина, либо другими методами;
ренатурацию rhIL3 путем обработки денатурированного белка смесью восстановленной и окисленной форм глутатиона;
заключительную очистку rhIL3 с помощью ВЭЖХ.
На рис. 1 изображена частичная структура плазмиды p3PTEIL3 в промоторной области, гена rhIL3 и соответствующая гену аминокислотная последовательность rhIL3.
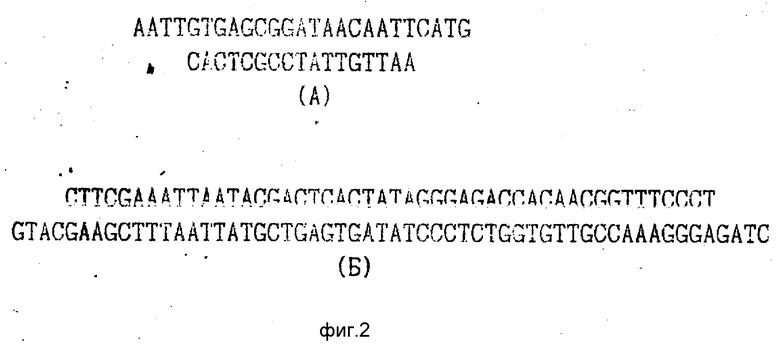
На рис. 2 приведены структуры синтетических фрагментов с lac-оператором (А) и промотором Pt7 (Б).
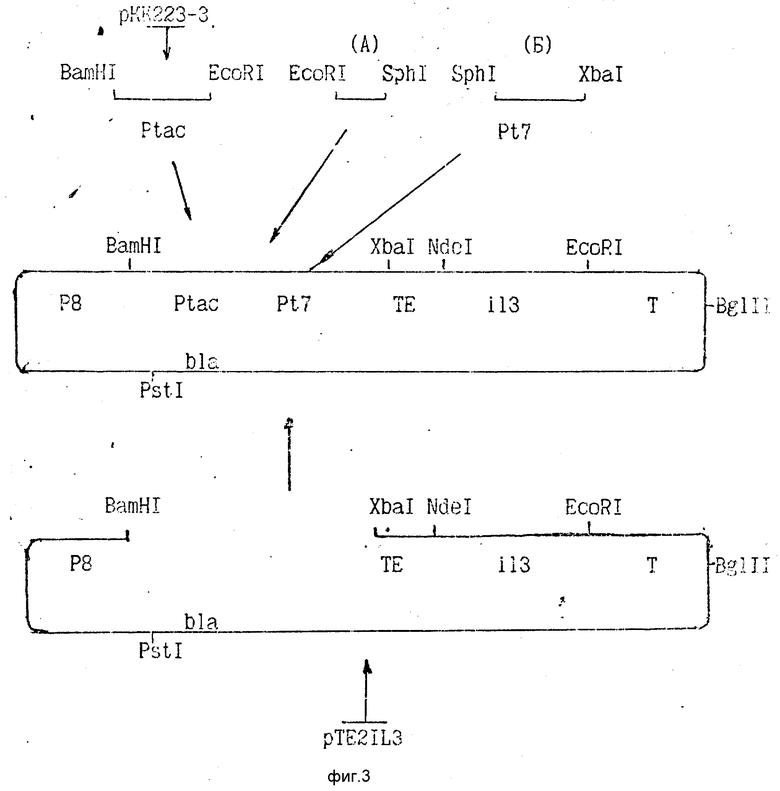
На рис. 3 приведена схема конструирования плазмиды p3PTEIL3.
Пример 1. Химический синтез олигонуклеотидов.
Синтез олигонуклеотидов выполняют твердофазным фосфоамидитным методом на ДНК-синтезаторе ASM-102U (НПО "Биосан", Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов 5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O-(b- цианэтилдиизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5 0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500 А), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20 30 мкмоль/г). Используя синтетический цикл, описанный в работе [9]
Пример 2. Конструирование рекомбинантной плазмидной ДНК p3PTEIL3.
Для приготовления вектора ДНК плазмиды pTE2IL3 (3 мкг) обрабатывают в 40 мкл буфера KGB (100 мМ K-глутамат, pH 8,8, 25 мМ трис-ацетат, pH 7,6, 10 мМ Mg-ацетат, 1 мМ меркаптоэтанол) рестриктазами BamHI (10 ед. акт.) и XbaI (10 ед. акт. ) в течение 1 ч при 37oC. Векторный фрагмент величиной 3,34 т.п.о. после электрофореза в 1% агарозном геле электрофоретически перемещают в слой геля 0,5% LGT-агарозы и затем элюируют методом вымораживания агарозы и последующего осаждения ДНК из раствора этанолом.
В тех же условиях из 3 мкг плазмиды pKK223-3 с помощью рестриктаз BamHI (10 ед. акт.) и EcoRI (10 ед. акт.) получают фрагмент, содержащий промотор Ptac, длиной 0,26 т.п.о.
Синтетические фрагменты (А) и (Б) получают отжигом соответствующих комплементарных 5'-фосфорилированных олигонуклеотидов.
По 5 мкл растворов синтетических фрагментов (А) и (Б), содержащих по 20 пмоль, прибавляют к раствору 1 мкг описанного выше векторного фрагмента и 0,1 мкг фрагмента, содержащего промотор Ptac, в 10 мкл буфера L (20 мМ трис-HCl, pH 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреит) и лигируют с помощью 20 ед. акт. Т4-ДНК-лигазы в течение 6 ч при 10oC.
Аликвоту реакционной смеси используют для трансформации компетентных клеток E.coli BL21(DE3). Трансформанты высевают на чашку с YT-агаром, содержащим 50 мкг/мл ампициллина. Скрининг рекомбинантов проводят с помощью гибридизации колоний in situ с 32P-мечеными олигонуклеотидами, составляющими фрагменты (А) и (Б). Из гибридизующихся клонов выделяют ДНК и анализируют с помощью эндонуклеаз HindIII, BamHI, EcoRI и BglII и определением нуклеотидной последовательности регуляторного участка и структурного гена.
Пример 3. Получение штамма E.coli BL21(DE3)/p3PTEIL3 (ВКПМ В6511) - продуцента белка rhIL3 и определение его продуктивности.
Клетки E. coli BL21(DE3), несущие плазмиду p3PTEIL3, структура которой подтверждена данными анализа (см. пример 2), являются продуцентом белка rhIL3.
Штамм продуцента E.coli BL21(DE3)/p3PTEIL3 выращивают при 37oC в 100 мл YT-бульона (pH 7,0) с 50 мкг/мл ампициллина в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности A550 0,7 0,8, прибавляют изопропилтио-b-D-галактозид до концентрации 0,2 мМ и продолжают процесс еще 6 ч. Каждый час отбирают пробу по 2 мл, определяют A550 и количество культуры, соответствующее 1 мл с A550 1,0, центрифугируют 5 мин при 6000 об/мин. Осажденные клетки в 100 мкл лизирующего буфера с красителем бромфеноловым синим обрабатывают 20 с ультразвуком, нагревают 3 мин при 100oC и пробы по 1 мкл используют для электрофореза в 15% SDS-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют для определения относительного количества белка в полосе, соответствующей белку rhIL3.
Пример 4. Выделение и первичная очистка белка rhIL3. Влажные клетки (100 г) суспендируют в 200 мл буфера (50 мМ Na-фосфат, 1 мМ EDTA, pH 7,5), добавляют лизоцим (100 мкг/мл), инкубируют 30 мин при 20oC, добавляют раствор MgCl2 (до 10 мМ) и ДНКазу (10 мкг/мл), инкубируют при 20oC до потери вязкости (30 мин), разбавляют 2 л буфера (50 мМ Na-фосфат, 10 мМ EDTA, 4 М мочевина, 1% тритон X-100, pH 7,5) и центрифугируют 20 мин при 10000 g. Осадок суспендируют в 2 л буфера (50 мМ Na-фосфат, 10 мМ EDTA, 4 М мочевина, 1% тритон X-100, pH 7,5) с помощью ультразвукового дезинтегратора Sonifier 240 (Branson) и центрифугируют 20 мин при 10000 g. Отмывку буфером повторяют еще раз, затем аналогично промывают буфером (50 мМ Na-фосфат, 1 мМ EDTA, pH 7,5).
Пример 5. Ренатурация и заключительная очистка рекомбинантного интерлейкина-3 (rhIL3).
Осадок денатурированного rhIL3 после первичной очистки, описанной в примере 4, ресуспендируют в 2 л буфера (50 мМ Na-фосфат, 1 мМ EDTA, 5 М гуанидингидрохлорид, pH 7,5) и инкубируют при 20oC. Полученный раствор разбавляют при перемешивании 8 л буфера (50 мМ Na-фосфат, 1 мМ EDTA, pH 7,5), доводят pH суспензии до 8,5 и добавляют смесь окисленного и восстановленного глутатиона до концентрации 1,5 мМ. Смесь перемешивают при 20oC, контролируя ход окисления тиоловых групп в rhIL3 с помощью метода Эллмана [10] и ренатурацию считают завершенной при отрицательной реакции. По окончании окисления раствор концентрируют в ячейке для ультрафильтрации (Amicon) на мембране PM-10, добавляют трифторуксусную кислоту до 0,1% и центрифугируют 20 мин при 10000 g. Осветленный раствор хроматографируют на препаративной колонке с Nucleosil 300-7C4 (Masherey-Nagel) в линейном градиенте 0 60% ацетонитрила в 0,1% трифторуксусной кислоте. Фракцию очищенного rhIL3 (выходит при 40 43% ацетонитрила) лиофилизуют. Чистота препарата составляет более 98% (по данным аналитической ВЭЖХ). Выход чистого rhIL3 составляет 20 25% белка, определяемого во влажных клетках продуцента.
Таким образом, заявляемая группа изобретений позволяет получить рекомбинантный интерлейкин-3 путем микробиологического синтеза по простой технологии и с высоким выходом.
Источники информации
1. Kindler V. Thorens B. Kossodo S. Allet B. Eliason J.F. Thatcher D. Farber N. Vassalli P. Proc.Nat.Acad. Sci.USA. 1986. V. 83. No. 4, p. 1001 1005.
2. Ihle J.N. Keller J. Oroszlan S. Henderson L.E. Copeland D.T. J.Immunol. 1983. V. 131. No. 1, p. 282 287.
3. Райт А. Основы иммунологии. 1991. М. Мир, с. 130 134.
4. Kimmenade A. Bond M.W. Shumacher J.H. Laquoi C. Eur.J.Biochem. 1988. V. 173. No. 1, p. 109 114.
5. Луценко С. В. Гуревич А.И. Каневский В.Ю. Смирнов В.А. Назимов И.В. Ажикина Т.Л. Чернов И.П. Ростапшов В.М. Сонина Н.В. Ажаев А.В. Биоорган. химия. 1991. Т. 17. No. 12, с. 1649 1654.
6. Гуревич А.И. Скапцова Н.В. Луценко С.В. Смирнов В.А. Куркин А.Р. Ажаев А.В. Биоорган. химия. 1991. Т. 17. No. 5, с. 647 -652.
7. Луценко С. В. Гуревич А.И. Птицын Л.Р. Рязанова Л.А. Смирнов В.А. Биоорган. химия. 1992. Т. 19. No. 3, с. 391 397, прототип.
8. Studier F.W. Rosenberg A.H. Dunn J.J. Dubendorf J.W. Methods Enzymol. 1990. V. 185. p. 60 89.
9. Atkinson T. Smith M. In: Oligonucleotide synthesis; a practical approach. 1984. Ed. Gait M.J. p. 35 81. IRL Press, Oxford.
10. Ellman G.L. Arch.Biochem.Biophys. 1959. V. 82. No. 1, p. 70 77.
Использование: биотехнология, генная белковая инженерия. Сущность изобретения: способ получения человеческого интерлейкина-3 основан на использовании новой рекомбинантной плазмидной ДНК p3PTEIL3, в которой искусственный ген человеческого интерлейкина-3 находится под контролем тандема трех промоторов (P8, Ptac и Pt7), и его экспрессия происходит после индукции изопропилтиогалактозидом, и штамма-продуцента E.coli ВКПМ В-6968, обеспечивающего высокий уровень биосинтеза целевого белка (до 30% тотального клеточного белка). Рекомбинантная плазмида содержит тандем промоторов P8, PtaC и Pt7, два Lac-оператора, усилитель трансляции гена 10 фага T7, терминатор транскрипции фага fd, ген β-лактамазы и искусственный ген человеческого интерлейкина-3. Штамм Escherichia coli ВКПМ В-6568 получают путем трансформации компетентных клеток E. coli BL21 (DE3) ekfpvbljq p3PTEIL3. Штамм Escherichia coli ВКПМ В-6968 культивируют в богатой среде, выращенные клетки разрушают в буферном растворе и выделяют рекомбинантный человеческий интерлейкин-3 из телец включения, и затем ренатурируют его известным способом. 3 с.п. ф-лы, 3 ил.
| Биоорганическая химия | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| Ситценабивная машина | 1922 |
|
SU391A1 |
