Предлагаемое изобретение относится к биотехнологии, в частности к генной и белковой инженерии. Оно включает сконструированную in vitro рекомбинантную плазмидную ДНК pteilox4, обусловливающую биосинтез гибридного белка ILOX4, аминокислотная последовательность которого включает 63 N-концевых аминокислоты человеческого интерлейкина-3 и тетрамер окситоциноил-лизина, штамм E.coli BL21(DE3)/pteilox4 - продуцент гибридного белка ILOX4 и способ получения окситоцина на основе вышеуказанной рекомбинантной ДНК и штамма-продуцента.
Пептидный гормон окситоцин 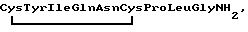 вырабатывается в человеческом организме нейросекреторными клетками гипоталамуса и накапливается в задней доле гипофиза [1, 2]. Он стимулирует сокращение гладкой мускулатуры матки и секрецию молока молочными железами [3,4]. Благодаря этому окситоцин находит широкое применение в медицине и ветеринарии [5].
вырабатывается в человеческом организме нейросекреторными клетками гипоталамуса и накапливается в задней доле гипофиза [1, 2]. Он стимулирует сокращение гладкой мускулатуры матки и секрецию молока молочными железами [3,4]. Благодаря этому окситоцин находит широкое применение в медицине и ветеринарии [5].
Трудности выделения и очистки природного гормона делают его малодоступным. Практически используется окситоцин, полученный химическим синтезом. Недостатком химического метода является трудоемкость и необходимость очистки препарата от сопутствующих близких по химической природе примесей, присутствующих в препарате вследствие ограничений, присущих синтетическому процессу (неколичественные выходы пептидного синтеза и реакций удаления защитных групп и др.).
В связи с этим более перспективным является способ получения гормона микробиологическим синтезом, который обеспечивает возможность получения целевого продукта с высоким выходом из сравнительно недорогого исходного сырья. Известно, однако, что короткие пептиды в бактериальной клетке быстро деградируют. Поэтому, чтобы избежать деградации пептида, его необходимо получать в составе гибрида с белком-носителем и выделять после химического или ферментативного расщепления. В литературе описаны многие такого рода гибриды, в которых роль белка-носителя выполняет β -галактозидаза, фактор некроза опухолей (TNF), α -интерферон и др. [6, 7, 8], а присоединенными пептидами являются энкефалин, соматостатин и др. Недостатком известных конструкций является небольшая доля пептида в составе гибридного белка [6, 7, 8]. Окситоцин в составе гибридного белка ILOX3 описан нами в 1993 г. и на способ его получения в 1997 г. выдан патент [9]. Этот способ является наиболее близким предлагаемому в настоящем изобретении и включает культивирование штамма-продуцента, полученного трансформацией клеток Escherichia coli TG1 плазмидной ДНК рTOTEilox3 с последующим разрушением клеток в буферном растворе, выделением гибридного белка ILOX3, расщеплением его ферментативным путем с последующим или одновременным восстановлением дисульфидных связей и заменой в образовавшемся декапептиде C-концевого остатка аминокислоты на амидогруппу. Однако, как показали наши дальнейшие исследования, гибридный белок ILOX3 и его высшие гомологи оказались неустойчивыми и деградировали в процессе биосинтеза, так что целевой гибридный белок содержал в среднем менее трех мономерных единиц окситоцина [10].
Изобретение решает задачу повышения уровня биосинтеза и устойчивости нового рекомбинантного белка ILOX4, и упрощения технологии получения рекомбинантного окситоцина.
Поставленная задача решается за счет того, что в способе получения окситоцина, включающем культивирование штамма-продуцента, полученного трансформацией клеток Escherichia coli плазмидной ДНК с последующим разрушением клеток в буферном растворе, выделением гибридного белка, расщеплением его ферментативным путем с последующим или одновременным восстановлением дисульфидных связей и заменой в образовавшемся декапептиде C-концевого остатка аминокислоты на амидогруппу, и отличающимся тем, что в качестве плазмидной ДНК используют специально сконструированную ДНК pteilox4, в качестве штамма-продуцента используют штамм Escherichia coli BL21(DE3)/pteilox4, который культивируют в богатой среде и после разрушения клеток выделяют гибридный белок ILOX4, который превращают в целевой продукт.
Используют рекомбинантную плазмидную ДНК pteilox4,
- кодирующую аминокислотную последовательность гибридного белка ILOX4, включающую 63 N-концевых аминокислоты человеческого интерлейкина-3 и тетрамер окситоцина, фланкированного остатками лизина,
- имеющую молекулярную массу 2,19 МДа,
- состоящую из:
HindIII/EcoRI-фрагмента I ДНК плазмиды pTE2IL3, содержащего промотор Р8 гена 8 фага fd, промотор и усилитель трансляции гена 10 фага Т7, терминатор транскрипции фага fd, ген β -лактамазы и N-концевую часть гена интерлейкина-3,
HindIII/EcoRI-фрагмента II, представляющего собой синтетический ген тетрамера окситоциноил-лизина,
- содержащую:
в качестве генетического маркера ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pteilox4 клеток E.coli к пенициллиновым антибиотикам,
уникальные сайты узнавания рестрикционных эндонуклеаз, расположенные на следующем расстоянии вправо от сайта NdeI: HpaI - 46 п.о., EcoRI - 251 п.о., BglII - 674 п.о., PstI - 2073 п.о., BamHI - 3215 п.о., XbaI - 3273 п.о., EcoRV - 3309 п.о.
Используют штамм-продуцент Escherichia coli BL21(DE3), содержащий рекомбинантную плазмидную ДНК pteilox4, - продуцент гибридного белка ILOX4.
Предлагаемое изобретение позволяет получать рекомбинантный окситоцин по простой технологии и с высоким выходом.
Конструкция рекомбинантной плазмидной ДНК pteilox4 обеспечивает высокий уровень экспрессии гибридного гена, содержащего ген тетрамера окситоциноил-лизина.
Для конструирования плазмиды использован химический подход, позволяющий создавать оптимальные для бактериальной экспрессии варианты структурного гена, а также регуляторных элементов, контролирующих его экспрессию.
Ген тетрамера окситоцина, фланкированного остатками лизина, получают из соответствующих синтетических олигонуклеотидов путем ферментативной сшивки с помощью ДНК-лигазы с векторными фрагментами в три этапа.
Источником первого векторного фрагмента служит известная рекомбинантная плазмидная ДНК pTE21L3, кодирующая человеческий интерлейкин-3 [11]. Экспрессия гена интерлейкина-3 в этой плазмиде контролируется промотором Р8 фага fd, а также промотором и усилителем трансляции гена 10 фага T7. Штамм E.coli BL21(DE3), содержащий плазмиду pTE2IL3, обеспечивает высокий уровень биосинтеза интерлейкина-3.
Источником второго векторного фрагмента служит известная рекомбинантная плазмидная ДНК pte2il3g, модифицированная путем вставки в сайт рестриктазы EcoRI полилинкерного фрагмента (плазмида pte2il3ghll) [12]. В конце гена il3 этой плазмиды отсутствует терминирующий кодон и после сайта рестриктазы EcoRI содержится сайт рестриктазы SalI.
Рекомбинантную плазмидную ДНК pTE21L3 расщепляют эндонуклеазами HindIII и EcoRI, полученный фрагмент лигируют с первым синтетическим HindIII/EcoRI-фрагментом, содержащим ген димера окситоцина (дуплекс A, фиг. 2), и получают рекомбинантную плазмидную ДНК pteilox2-1.
Рекомбинантную плазмидную ДНК pte2il3gh11 расщепляют эндонуклеазами EcoRI и SalI, полученный фрагмент лигируют с вторым синтетическим EcoRI/SalI-фрагментом, содержащим ген димера окситоцина (дуплекс B, фиг. 2), и получают рекомбинантную плазмидную ДНК pteilox2-2.
Обе рекомбинантные плазмидные ДНК, pteilox2-1 и pteilox2-2, расщепляют рестриктазами EcoRI и EcoRV и выделенные после электрофореза в агарозном геле фрагменты (из плазмиды pteilox2-1- малый, 255 п.о., из плазмиды pteilox2-2 - большой, 3058 п.о.) лигируют и получают рекомбинантную плазмидную ДНК pteilox4.
Предлагаемый штамм-продуцент Escherichia coli BL21(DE3)/pteilox4 характеризуется следующими признаками:
Морфологические признаки. Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" - колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или YT-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки. Клетки растут при температуре от 4 до 40oC при оптимуме pH от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к пенициллиновым антибиотикам (до 500 мкг/мл).
Штамм-продуцент E. coli BL21(DE3)/pteilox4 отличается от штамма-реципиента Е. coli BL21 (DЕ3) только наличием рекомбинантной плазмидной ДНК pteilox4, которая и придает ему устойчивость к пенициллиновым антибиотикам.
Штаммы-продуценты получают путем трансформации компетентных клеток E. coli BL21(DE3) соответствующей рекомбинантной плазмидной ДНК.
Клетки E. coli BL21(DE3)/pteilox4 являются продуцентом гибридного белка ILOX4. При индукции изопропилтио- β -D-галактозидом, а также и без индукции при конститутивном синтезе, происходит эффективный биосинтез гибридного белка, который накапливается в клетках в виде телец включения, и его выход составляет более 30% суммарного белка бактерий.
Штамм-продуцент депонирован во Всероссийской коллекции промышленных микроорганизмов, N ВКПМ В-7757 от 16 марта 1999 г.
Изобретение осуществляют следующим образом. Конструируют рекомбинантную плазмидную ДНК pteilox4 в три этапа, как описано выше. На третьем этапе лигируют EcoRI/EcoRV-фрагменты плазмид pteilox2-1 и pteilox2-2.
Лигазной смесью трансформируют компетентные клетки E.coli BL21(DE3) и высевают на YT-агар, содержащий 50 мкг/мл ампициллина или другого пенициллинового антибиотика. Полученные клоны анализируют гибридизацией 32P-мечеными олигонуклеотидами C и D (фиг.2), и из гибридизующихся клонов выделяют плазмидную ДНК, которую подвергают рестриктному анализу с помощью рестриктаз HindIII, EcoRI и BglII, а также определением нуклеотидной последовательности между сайтами EcoRV и EcoRI. Штамм-продуцент E.coli BL21(DE3)/pteilox4 выращивают в богатой среде (YT-, LB-бульон и др.) (или индуцируют изопропилтио β -D-галактозидом, и снова выращивают) до достижения максимальной плотности культуры.
Выделение гибридного белка ILOX4 из клеток продуцента включает следующие стадии:
- разрушение выращенных клеток одним из обычно применяемых способов;
- отмывку буферными растворами телец включения от растворимых компонентов клетки;
- солюбилизацию денатурированного целевого белка из телец включения либо в 5-8 М растворе мочевины, либо в 5 М растворе гидрохлорида гуанидина, либо в другом подходящем растворителе;
- очистку гибридного белка в результате ступенчатого разбавления растворов в мочевине или гидрохлориде гуанидина, либо другими методами.
Расщепление гибридного белка проводят ферментативным путем с последующим или одновременным восстановлением дисульфидных связей для разъединения образующихся при расщеплении пептидных фрагментов. При ферментативном расщеплении гибридного белка, в котором мономерные единицы окситоцина фланкированы остатками лизина, используют трипсин, а затем восстанавливают дисульфидные связи с помощью меркаптоэтанола.
Удаление C-концевого аминокислотного остатка из окситоциноил-лизина, образующегося при расщеплении гибридного белка, проводят либо с помощью карбоксипептидазы Y (с одновременным амидированием концевого остатка глицина в окситоцине), либо с помощью карбоксипептидазы В с последующим химическим амидированием концевой аминокислоты (глицина) и получают окситоцин.
На фиг. 1 изображена частичная структура плазмиды pteilox4 в области гена гибридного белка ILOX4 и соответствующая ей аминокислотная последовательность гибридного белка.
На фиг. 2 приведена структура синтетических фрагментов (дуплексы A и B), содержащих димеры гена окситоцина, и указаны полусайты рестриктаз, по которым эти дуплексы встраивались в векторные плазмиды pte2i13 (дуплекс A) и pte2il3gh11 (дуплекс B).
Изобретение иллюстрируется нижеследующими примерами.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pteilox4
Химический синтез олигонуклеотидов выполняют твердофазным фосфоамидитным методом на ДНК-синтезаторе ASM-102U (БИОССЕТ, Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов-5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O- ( β-цианэтилдиизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500 A), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Используют синтетический цикл, описанный в работе [13].
Для приготовления вектора ДНК плазмиды pte2i13 (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера КGВ (100 мМ К-глутамат, pH 8,8, 25 мМ трис-ацетат, pH 7,6, 10 мМ Mg-ацетат, 1 мМ меркаптоэтанол) рестриктазами HindIII (10 ед.акт. ) и EcoRI (10 ед.акт.) в течение 1 ч при 37oC. Векторный фрагмент величиной 3,19 т.п.о. после электрофореза в 1% агарозном геле электрофоретически перемещают в слой DEAE-бумаги, затем элюируют 1M NaCl и осаждают ДНК из раствора этанолом.
Синтетический фрагмент с геном димера окситоцина (A) (фиг. 2) в количестве 2 пмоль, прибавляют к раствору 1 мкг описанного выше векторного фрагмента в 10 мкл буфера L (20 мМ трис-HCl, pH 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреит) и лигируют с помощью 10 ед.акт. Т4-ДНК-лигазы в течение 6 ч при 10oC.
Аликвоту реакционной смеси используют для трансформации компетентных клеток E.coli BL21(DE3). Трансформанты высевают на чашки с YT-агаром, содержащим 50 мкг/мл ампициллина. Скрининг рекомбинантов проводят с помощью гибридизации колоний in situ с 32P-меченым олигонуклеотидом D (фиг.2). Из гибридизующихся клонов выделяют ДНК плазмиды pteilox2-1 и анализируют с помощью эндонуклеаз HindIII и EcoRI, и определением нуклеотидной последовательности регуляторного участка, начала структурного гена и клонированного синтетического фрагмента.
Аналогично, из ДНК плазмиды pte2i13gh11 (3 мкг, 1 пмоль) с помощью рестриктаз EcoRI (10 ед.акт.) и SalI (10 ед.акт.) приготавливают EcoRI/SalI-вектор величиной 3,3 т.п.о., 1 мкг которого лигируют с 2 пмоль синтетического фрагмента с геном димера окситоцина (B) (фиг.2), и после трансформации компетентных клеток E.coli BL21(DE3) получают клоны рекомбинантов, из которых выделяют ДНК плазмиды pteilox2-2.
Обе рекомбинантные плазмидные ДНК, pteilox2-1 и pteilox2-2, расщепляют рестриктазами EcoRI и EcoRV и выделенные после электрофореза в агарозном геле фрагменты (из плазмиды pteilox2-1 - малый, 255 п.о., из плазмиды pteilox2-2 - большой, 3058 п.о.) лигируют и получают и после трансформации компетентных клеток E. coli BL21(DE3) получают клоны рекомбинантов, скрининг которых проводят с помощью гибридизации колоний in situ с 32P-мечеными олигонуклеотидами C и D (фиг.2). Из гибридизующихся с двумя олигонуклеотидами клонов выделяют плазмидную ДНК pteilox4, которую анализируют с помощью эндонуклеаз XbaI, HindIII, EcoRI и SalI, и определением нуклеотидной последовательности регуляторного участка, начала структурного гена и клонированного синтетического фрагмента.
Пример 2. Получение штамма E.coli BL21(DE3)/pteilox4 (ВКПМ 7757) - продуцента гибридного белка ILOX4 и определение его продуктивности.
Клетки E. coli BL21(DE3), несущие плазмиду pteilox4, структура которой подтверждена данными анализа (см. пример 2), являются продуцентом гибридного белка ILOX4.
Штамм продуцента E.coli BL21(DE3)ЮЯpteilox4 выращивают при 37oC в 100 мл YT-бульона (pH 7,0) с 50 мкг/мл ампициллина в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности A550 0,7-0,8, прибавляют изопропилтио- β -D-галактозид до концентрации 0,2 мМ и продолжают процесс еще 6 ч. Каждый час отбирают пробу по 2 мл, определяют A550 и количество культуры, соответствующее 1 мл с A550 1,0, центрифугируют 5 мин при 6000 об/мин. Осажденные клетки в 100 мкл лизирующего буфера с красителем бромфеноловым синим обрабатывают 20 сек ультразвуком, нагревают 3 мин при 100oC и пробы по 1 мкл используют для электрофореза в 15% SDS-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют для определения относительного количества белка в полосе гибридного белка.
Пример 3. Получение окситоцина.
1) Выделение гибридного белка ILOX4.
Влажные клетки (100 г) суспендируют в 200 мл буфера (50 мM Na-фосфат, 1 мМ EDTA, pH 7,5), добавляют лизоцим (100 мкг/мл), инкубируют 30 мин при 20oС, добавляют раствор MgCl2 (до 10 мМ) и ДНКазу (10 мкг/мл), инкубируют при 20oC до потери вязкости (30 мин), разбавляют 2 л буфера (50 мМ Na-фосфат, 10 мМ EDTA, 4 М мочевина, 1% тритон Х100, pH 7,5) и центрифугируют 20 мин при 10000 g. Осадок суспендируют в 2 л буфера (50 мМ Na-фосфат, 10 мМ EDTA, 4 М мочевина, 1% тритон X100, pH 7,5) с помощью ультразвукового дезинтегратора Sonifier 240 (Branson) и центрифугируют 20 мин при 10000 g. Отмывку буфером повторяют еще раз, затем аналогично промывают буфером (50 мМ Na-фосфат, 1 мМ EDTA, pH 7,5). Осадок ресуспендируют в 2 л буфера (50 мМ Na-фосфат, 1 мМ EDTA, 5 М гуанидингидрохлорид, pH 7,5) и инкубируют при 20oC. Полученный раствор разбавляют при перемешивании 8 л буфера (50 мМ Na-фосфат, 1 мМ EDTA, pH 7,5) и центрифугируют.
2) Ферментативный гидролиз гибридного белка ILOX4 и получение окситоциновой кислоты.
Суспензию 100 мг гибридного белка ILOX4 в 10 мл 0,1 М аммоний-карбонатного буфера, pH 8,5, обрабатывают 2 мг трипсина 6 ч при 37oC, до полного растворения осадка. Полноту гидролиза белка контролируют с помощью ВЭЖХ, используя смолу Sepharon C-18 в градиенте концентрации 20-80% ацетонитрила в 0,1% трифторуксусной кислоте.
К 10 мл трипсинового гидролизата ILOX4 прибавляют 1 мг карбоксипептидазы B и инкубируют 1 ч при 37oC. Раствор замораживают, лиофилизуют и выделяют дезамидоокситоцин обращенно-фазовой хроматографией на смоле Sepharon C-18 в градиенте концентрации 20-80% ацетонитрила в 0,1% трифторуксусной кислоте.
3) Превращение окситоциновой кислоты в окситоцин
50 мг окситоциновой кислоты растворяют в 2 мл 0,25 М HCl в метаноле и выдерживают 1.5 часа при комнатной температуре. К полученному раствору метилового эфира окситоциновой кислоты прибавляют 2 мл 10 М метанольного NH3, смесь выдерживают 18 ч при комнатной температуре и упаривают. Получают 40 мг окситоцина.
ЛИТЕРАТУРА
1. Lederis K.//Gen.Compar. Endocrinol. 1961. V. 1. N 1. p. 80-86.
2. Du Vigneaud V.//Science. 1956. V. 123. p.956.
3. Du Vigneaud V., Ressler Ch., Trippet S. // J.Biol.Chem. 1953. V. 205. p.949-957.
4. Dale H.H./Biochem.J. 1909. V.4. p.427-447.
5. Машковский М.Д.// "Лекарственные средства". М, Медицина. Т.2. с.72.
6. Черненькая Е.А., Гуревич А.И., Коробко В.Г., Биоорган, химия, т. 15, N 4, 1989, с. 508-513.
7. Авторское свидетельство СССР N 1321063, приоритет от 14.11.85, кл. C 12 N 15/00.
8. Авторское свидетельство СССР N 1724691, приоритет от 20.06.90, кл. C 12 N 15/42.
9. Патент РФ N 20085585, приоритет от 18.06.93, кл. C 12 N 15/16.
10. А.И.Гуревич, Т.А.Качалина, А.Л.Каюшин, М.Д.Коростелева, К.В.Мальцев, О. А. Миргородская, А. И.Мирошников. Биоорган, химия, т. 22, N 1, 1996, с. 14-19.
11. А.И.Гуревич, Н.В.Скапцова, С.В.Луценко, В.А.Смирнов, А.Н.Куркин, А. В.Ажаев. Биоорган, химия, т. 17, N 5, 1991, с. 647-653.
12. А.И.Гуревич, Р.С.Есипов, Т.А.Качалина, А.Л.Каюшин. Биоорган, химия, т.21, N 4, 1995, с. 282-288.
13. Atkinson Т. , Smith M.//in: Oligonucleotide synthesis; apractical approach. 1984. Ed. Gait M.J. p. 35-81. IRL Press, Oxford.


Изобретение относится к биотехнологии, в частности к генной и белковой инженерии. Способ получения окситоцина основан на использовании новой рекомбинантной плазмидной ДНК pteilox4, кодирующей гибридный белок ILOX4, аминокислотная последовательность которого состоит из N-концевой (63 аминокислоты) последовательности человеческого интерлейкина-3 и олигомера окситоцина, содержащего мономерные единицы, фланкированные остатками лизина для специфического ферментативного расщепления, и штамма продуцента E.coli BL21(DE3)/pteilox4, обеспечивающего высокий уровень биосинтеза гибридного белка (до 30% тотального клеточного белка). Рекомбинантная плазмида pteilox4 состоит из HindIII/EcoR1-фрагмента ДНК плазмиды pTE21L3, содержащего промотор Р8 гена 8 фага fd, промотор и усилитель трансляции гена 10 фага Т7, терминатор транскрипции фага fd, ген β-лактамазы и N-концевую часть гена интерлейкина-3, и HindIII/EcoRI-фрагмента, представляющего собой синтетический ген тетрамера окситоциноил-лизина. Штамм Escherichia coli BL21 (DE3)/pteilox4 получают путем трансформации компетентных клеток E.coli BL21 (ВЕЗ) соответствующей плазмидой pteilox4. Штамм депонирован под номером ВКПМ В-7757. Штамм Escherichia coli BL21 (DE3)/pteilox4 культивируют в богатой среде, выращенные клетки разрушают в буферном растворе, выделяют гибридный белок ILOX4. Его расщепляют ферментативным путем с последующим или одновременным восстановлением дисульфидных связей. Удаление С-концевого аминокислотного остатка из окситоцинового декапептида, образующегося при расщеплении гибридного белка, проводят либо с помощью карбоксипептидазы Y (с одновременным амидированием концевого остатка глицина в окситоцине), либо с помощью карбоксипептидазы В с последующим химическим амидированием концевой аминокислоты (глицина). В результате получают устойчивый рекомбинантный белок ILOX4. Процесс является более простым, а целевой продукт более устойчив. 3 с.п. ф-лы, 2 ил.
| СПОСОБ ПОЛУЧЕНИЯ ОКСИТОЦИНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК РТОТЕILOX 3, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК ILOX3 ДЛЯ ПОЛУЧЕНИЯ ОКСИТОЦИНА, ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА ILOX3 | 1993 |
|
RU2085585C1 |
| Рекомбинантная плазмидная ДНК @ 10FMD, кодирующая гибридный белок Р204-AS @ Р @ С @ С @ - VPI/200-213/ Р @ Р @ S @ Р @ ( 131-160) и штамм бактерий ЕSснеRIснIа coLI -продуцент гибридного белка Р204- А @ Р @ С @ С @ V PI/200-213/-Р @ Р @ S @ Р @ - VPI /131-160/ | 1990 |
|
SU1724691A1 |
| Гуревич А.И., Качалина Т.А., Каюшин А.Л., Коростелева М.Д., Мальцев К.В., Миргородская О.А., Мирошников А.И., Биоорганическая химия, т.22, N1, 1996, с | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Гуревич А.И., Скапцова Н.В., Луценко С.В., Смирнов В.А., Куркин А.Н., Ажаев А.В., Биоорганическая химия, т.17, N 5, 1991, с | |||
| МАШИНА ДЛЯ ПРОКЛАДКИ ДРЕНАЖНЫХ ТРУБ | 1923 |
|
SU647A1 |
| Гуревич А.И., Есипов Р.С., Качалина Т.А., Каюшин А.Л., Биоорганическая химия, т.21, N 4, 1995, с | |||
| ПОРШНЕВОЙ ДВИГАТЕЛЬ | 1916 |
|
SU282A1 |
| Atkinson N., Smith M.// in: Oligonucleotide synthesis; apractical approach | |||
| Колосниковая решетка с чередующимися неподвижными и движущимися возвратно-поступательно колосниками | 1917 |
|
SU1984A1 |
| Ed | |||
| Gait M.J | |||
| p | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| IRL Press, Oxford. | |||

