Изобретение относится к биотехнологии, в частности к генной и белковой инженерии. Оно включает сконструированную in vitro рекомбинантную плазмидную ДНК pERPIns1, обусловливающую биосинтез рекомбинантного человеческого проинсулина, и штамм Е. coli BL2l(DE3)/pERPInsl - суперпродуцент указанного белка. Изобретение может быть использовано для получения рекомбинантного человеческого инсулина.
Гормон поджелудочной железы инсулин синтезируется в организме человека специализированными клетками (островки Лангерганса) в виде предшественника - препроинсулина. В процессе биосинтеза происходит отщепление сигнального пептида из 23 аминокислот, в результате чего образуется проинсулин. В дальнейшем проинсулин претерпевает процессинг, который приводит к образованию самого пептидного гормона в результате специфического выщепления С-пептида, состоящего из 35 аминокислотных остатков. Зрелый биологически активный инсулин состоит из двух цепей, А и В, соединенных двумя дисульфидными связями. Цепь А содержит 21, а цепь В - 30 аминокислотных остатков.
Инсулин широко используется в медицине в качестве лекарственного препарата для лечения инсулинзависимого сахарного диабета. До недавнего времени с этой целью в промышленности получали свиной инсулин как наиболее близкий по структуре к человеческому, что зачастую приводило к нежелательным побочным эффектам при применении препарата. В настоящее время все больше находит применение сам человеческий инсулин, для получения которого наиболее перспективными являются способы, основанные на технологии рекомбинантных ДНК.
Ген, кодирующий проинсулин человека, получен как синтезом кДНК на матрице мРНК, выделенной из поджелудочной железы [1, 2], так и химико-ферментативным синтезом [3, 4]. Однако долгое время не удавалось достичь высокого уровня экспрессии гена в бактериях Escherichia coli, наиболее удобных для промышленного получения рекомбинантных белков. Наибольшее распространение получили методы, в которых использовались измененные структуры рекомбинантного проинсулина, включающие лидерные последовательности, которые, как предполагалось, должны защищать рекомбинантный белок от протеолитической деградации. Для этой цели использовали разнообразные лидерные последовательности [5-8], для отщепления которых чаще всего использовали обработку бромцианом [9, 10]. Наилучшим вариантом удаления лидерной последовательности оказалось расщепление гибридного белка, состоящего из двух усредненных иммуноглобулинсвязывающих (IgG) доменов белка А из S. aureus и проинсулина человека, соединенных через остатки Arg, LysArg или Lys после выделения из "тел включения" и ренатурации, в результате гидролиза смесью трипсина и карбоксипептидазы В [11].
Известен способ получения проинсулина [12], в котором используется структура гена проинсулина, полученного химико-ферментативным синтезом. Однако в предложенной структуре рекомбинантной плазмиды pPINS07 ген проинсулина связан с лидерным геном белка А из S. aureus, в результате содержание структуры проинсулина в рекомбинантном белковом продукте составляет лишь около 50%. Кроме того, регуляторный участок транскрипции плазмиды pPTNS07 предполагает обязательное использование индуцированного ИГПТ биосинтеза целевого белка; это удорожает процесс и в то же время не позволяет достигнуть столь высокого, как в настоящей заявке, выхода рекомбинантного белка (25-30% вместо 50-60%).
Известен наиболее близкий к заявленному способ получения проинсулина [13], в котором используется структура гена проинсулина, лишенного лидерного пептида (его заменяет последовательность двух аминокислот MetArg), однако в самой структуре гена проинсулина используются кодоны, не являющиеся оптимальными для продукции белка в Escherichia coli, что снижает выход целевого белка. Кроме того, регуляторный участок транскрипции использованной векторной плазмиды предполагает обязательное использование индуцированного ИПТГ биосинтеза целевого белка; это удорожает процесс.
Изобретение решает задачу получения высокопродуктивного рекомбинантного бактериального штамма-продуцента, позволяющего получать рекомбинантный человеческий проинсулин с высоким выходом.
Поставленная задача решается за счет того, что штамм-продуцент, полученный трансформацией клеток Escherichia coli плазмидной ДНК, культивируют до накопления рекомбинантного человеческого лроинсулина в количестве 40-50% от суммарного белка клеток, разрушают клетки в буферном растворе и выделяют тельца включения.
Используют штамм-продуцент Escherichia coli BL21(DE3), содержащий плазмидную ДНК pERPInsl - суперпродуцент рекомбинантного человеческого проинсулина.
Используют рекомбинантную плазмидную ДНК pERPIns1
- кодирующую аминокислотную последовательность рекомбинантного человеческого проинсулина;
- имеющую молекулярную массу 2,56 МДа;
- состоящую из:
NdeI/HindIII-фрагмента ДНК плазмиды рЕТ20b(+) [15], содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фага Т7, ген β-лактамазы, и NdеI/HindIII-фрагмента ДНК, содержащего адаптированную к этим сайтам последовательность гена рекомбинантного человеческого проинсулина.
- содержащую:
в качестве генетического маркера ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pERPIns1 клеток Е. coli к пенициллиновым антибиотикам;
уникальные сайты узнавания рестрикционных эндонуклеаз, расположенные на следующем расстоянии вправо от сайта NdeI: XbaI - 38 п.о., ВglII - 96 п.о., PvuII - 741 п.о., BglI - 2163 п.о, PvuI - 2413 п.о., HindIII - 4371 п.о.
Конструкция рекомбинантной плазмидной ДНК pERPIns1 обеспечивает высокий уровень экспрессии клонированного в ней гена рекомбинантного человеческого проинсулина.
Для конструирования плазмиды использован химический подход, позволяющий использовать для экспрессии клонированного структурного гена оптимальные регуляторные элементы, контролирующие его экспрессию.
Структурный ген проинсулина получают химико-ферментативным синтезом. Концевые его участки, содержащие начальные кодоны ATG-AAA (аминокислоты MetLys) и соответствующие вектору сайты рестриктаз, введены с помощью ПНР с синтетическими олигонуклеотидными праймерами А и В (см. схему) и затем ген клонируют в векторную плазмиду рЕТ20b(+) [15].
Предлагаемый штамм-продуцент Escherichia coli BL21(DE3)/pERPInsl характеризуется следующими признаками.
Морфологические признаки. Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" - колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или YT-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки. Клетки растут при температуре от 4 до 40oС при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к пенициллиновым антибиотикам (до 500 мкг/мл).
Штамм-продуцент Е. coli BL21(DE3)/ pERPIns1 отличается от штамма-реципиента Е. coli BL21(DE3) только наличием рекомбинантной плазмидной ДНК pERPIns1, которая и придает ему устойчивость к пенициллиновым антибиотикам. Штаммы-продуценты получают путем трансформации компетентных клеток Е. coli BL21(DE3) соответствующей рекомбинантной плазмидной ДНК.
Клетки Е. coli BL21(DE3)/pERPInsl являются суперпродуцентом. При индукции изопропилтио-β-D-галактозидом, а также и без индукции происходит эффективный биосинтез рекомбинантного человеческого проинсулина, который накапливается в клетках в количестве более 40% суммарного белка бактерий. Индентификация образующегося рекомбинантного человеческого проинсулина проведена с помощью масс-спектрометрии.
Штамм-продуцент депонирован во Всероссийской коллекции промышленных микроорганизмов, ВКПМ В-8018 от 27.09.2000 г.
Изобретение осуществляют следующим образом. Конструируют рекомбинантную плазмидную ДНК pERPIns1, для чего полученный химико-ферментативным синтезом ген рекомбинантного человеческого проинсулина амплифицируют с помощью ПЦР с синтетическими олигонуклеотидными праймерами (см. схему), содержащими сайты рестриктаз NdeI (N-конец гена, праймер А1) и HindIII (С - конец гена, праймер В1), полученную ДНК расщепляют соответствующими рестриктазами и затем лигируют с расщепленной по тем же сайтам векторной плазмидой рЕТ20b(+) [15].
Лигазной смесью трансформируют компетентные клетки Е. coli BL21(DE3) и высевают на YT-агар, содержащий 50 мкг/мл ампициллина или другого пенициллинового антибиотика. Полученные клоны анализируют гибридизацией 32Р-меченными олигонуклеотидами А1 и В1 (см. схему), и из гибридизующихся клонов выделяют плазмидную ДНК, которую подвергают рестриктному анализу с помощью рестриктаз NdeI и HindIII.
Штамм-продуцент Е. coli BL21(DE3)/pERPTnsl выращивают в богатой среде (YT-, LB-бульон и др.) (или индуцируют изопропилтио-β-D-галактозидом и снова выращивают) до достижения максимальной плотности культуры.
На схеме изображена структура гена рекомбинантного человеческого проинсулина и синтетических праймеров, использованных для выделения гена с помощью ПЦР и отбора клонов путем гибридизации.
Изобретение иллюстрируется нижеследующими примерами.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pERInsl.
Химический синтез олигонуклеотидов выполняют твердофазным фосфоамидитным методом на ДНК-синтезаторе ASM-102U (БИОССЕТ, Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов - 5'-димeтoкcитpитил-N-aцил-2'-дeзoкcинyклeoзид-3'-О-β-циaнэтил-диизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500  ), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Используют синтетический цикл, описанный в работе [14].
), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Используют синтетический цикл, описанный в работе [14].
Для приготовления вектора ДНК плазмиды рЕТ20b(+) (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера Y (33 мМ трис-ацетат, рН 7,9, 10 мМ Mg-ацетат, 66 мМ К-ацетат 1, 0,5 мМ DTT, 0,1 мг/мл BSA) рестриктазой NdeI (10 ед.акт.), а затем в 40 мкл буфeра R (10 мМ трис-НС1, рН 8,5, 10 мМ MgCl2, 100 мМ КС1, 0,1 мг/мл BSA) рестриктазой HindIII (10 ед.акт.) в течение 1 ч при 37oС. Векторный фрагмент величиной 3,6 т.п.о. после электрофореза в 1% агарозном геле электрофоретически перемещают в слой DEAE-бумаги, затем элюируют 1М NaCl и осаждают ДНК из раствора этанолом.
Для приготовления фрагмента гена проводят амплификацию с помощью ПЦР, используя в качестве матрицы плазмиду с искусственным геном проинсулина (0,01 мкг в образце), а в качестве праймеров - синтетические олигонуклеотиды А1 и В1 (по 60 пмоль каждого). ПЦР проводят в ДНК-амплификаторе, в буферном растворе, содержащем каждый из четырех dNTP в концентрации 0,5 мМ и 5 ед. акт, Taq-ДНК-полимеразы, в следующем режиме: денатурация 1 мин при 94oС, отжиг 30 с при 60oС, элонгация 40 с при 72oС, 30 циклов ПЦР. После этого реакционную смесь депротеинизируют хлороформом, упаривают досуха, остаток растворяют в 20 мкл воды, затем расщепляют теми же рестриктазами, которые использовались при приготовлении вектора, и выделяют целевой фрагмент из агарозного геля.
Полученный синтетический фрагмент с геном рекомбинантного человеческого проинсулина в количестве 2 пмоль прибавляют к раствору 1 мкг описанного выше векторного фрагмента в 10 мкл буфера (20 мМ трис-HCl, рН 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреит) и лигируют с помощью 10 ед.акт. Т4-ДНК-лигазы в течение 12 ч при 10oС.
Аликвоту реакционной смеси используют для трансформации компетентных клеток Е. coli BL21(DE3). Трансформанты высевают на чашки с YT-агаром, содержащим 50 мкг/мл ампициллина. Скрининг рекомбинантов проводят с помощью гибридизации колоний in situ с 32P-меченным олигонуклеотидом А1 (см. схему). Из гибридизующихся клонов выделяют ДНК плазмиды pERPInsl и анализируют с помощью эндонуклеаз NdeI и HindIII.
Пример 2. Получение штамма Е. coli BL21(DE3)/pERInsl (ВКПМ В-8018) - продуцента и определение его продуктивности.
Штамм-продуцент Е. coli BL21(DE3)/pERIns1 получают трансформацией компетентных клеток Е. coli BL21(DE3) плазмидой pERPIns1, как описано в примере 1. Клетки Е. coli BL21(DE3), несущие плазмиду pERIns1, структура которой подтверждена данными анализа (см. пример 1), являются суперпродуцентом.
Штамм продуцента Е. coli BL21(DE3)/ pERInsI выращивают при 37oС в 100 мл YT-бульона (рН 7,0) с 50 мкг/мл ампициллина в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности А550 0,7-0,8, прибавляют изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают процесс еще 6 ч или продолжают выращивание в отсутствие индуктора в течение 6 ч. Каждый час отбирают пробу по 2 мл, определяют А550 и количество культуры, соответствующее 1 мл с А5501,0, центрифугируют 5 мин при 6000 об/мин. Осажденные клетки в 100 мкл лизирующего буфера с красителем бромфеноловым синим обрабатывают 20 с ультразвуком, нагревают 3 мин при 100oС и пробы по 1 мкл используют для электрофореза в 15% SDS-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Shimadzu CS-930. По данным сканирования рекомбинантный проинсулин составляет 40-50% суммарного клеточного белка.
Идентификация образующегося рекомбинантного человеческого проинсулина проведена с помощью MALDI-масс-спектрометрии на масс-спектрометре Vision 2000. Полученный сигнал рекомбинантного проинсулина соответствует расчетному значению массы 9724 Да.
Источники информации
1. Bell G.I., Swain W.F, Pictet R., Cordell В., Goodman H.M., Rutter W. J.//Nature, 1979, v. 282, p. 525-527.
2. Sures I., Goeddel D.V, Gray A., Ulrich A.// Science, 1980, v. 208, p. 57-59.
3. Williams D. C. , Van Frank R.M, Murth M.L., Burnett J.P.// Science, 1982, v. 215. p. 687-689.
4. Ovchinnikov Y.A., Efimov V.A., Ivanova I.N., Reverdatto S.V, Skiba N. P., Chakhmakhcheva O.G.//Gene, 1984, v. 31, p. 65-68.
5. Sung W. L. , Yao F.L., Zanab D.M., Narang S.A. // Proc. Natl. Acad. Sci. USA, 1986, v. 83, p. 561-565.
6. Tang J, Xue Y, Fan X., Fu Y.// Clin. J. Biotechnol., 1993, v. 9, p. 71-78.
7. Berg H., Walter M., Mauch L., Seissler J., Northemann W.J.// Immunol. Methods, 1993, v. 164, p. 221-231.
8. Wei G, Hu M.H., Tang L.G.// Biochem. Mol. Biol. Int., 1995, v. 35, p. 37-46.
9. McGregor W.C.//Ann. N.Y. Acad. Sci., 1983, v. 413. p. 231-237.
10. Cowley D.J, Mackin R.B. // FEBS Lett., 1997, v. 402, p. 124 - 130.
11. Johansson P., Nilsson L., Samuelsson E., Moks Т., Stahl S,, Uhlen M. // Eur. J. Biochem., 1996, v. 236, p. 656-661.
12. Пат. РФ 2144957, 27.01.2000, Бюл. 3.
13. Dabrowski S., Brillowska A., Kur J.//Prot. Express. Purif., 1999, v. 16, р. 315-323.
14. Atkinson Т., Smith M.// in: Oligonucleotide synthesis; a practical approach. 1984. Ed. Gait M.J. p. 35-81. IRL Press, Oxford.
15. Novagen Catalog 1996-1997.
Изобретение относится к биотехнологии, в частности к генной и белковой инженерии, и решает задачу получения высокопродуктивного рекомбинантного бактериального штамма-продуцента человеческого проинсулина. Для этого штамм-реципиент трансформируют рекомбинантной плазмидной ДНК рERPIns1, которая состоит из Nde I/Hind III - фрагмента ДНК плазмиды рЕТ 20b(+), содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фага Т7, ген β-лактамазы, и Nde I/Hind III - фрагмента ДНК, содержащего адаптированную к этим сайтам последовательность гена рекомбинантного человеческого проинсулина. Штамм-продуцент культивируют до накопления рекомбинантного человеческого проинсулина в количестве 60-70% от суммарного белка клеток, разрушают клетки в буферном растворе и выделяют тельца включения. Штамм-продуцент Е. coli BL21 (DE3)/pERPIns выращивают в богатой среде (YT-, LB-бульон др.) (или индуцируют изопропилтио-β-D-галактозидом и снова выращивают) до достижения максимальной плотности культуры. Изобретение позволяет получать рекомбинантный человеческий проинсулин по упрощенной технологии и с высоким выходом. 2 с.п. ф-лы, 1 ил.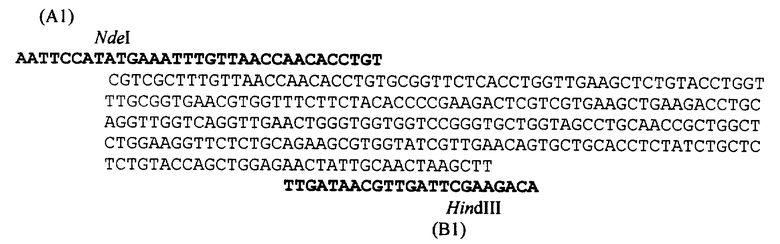
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PPINS16, КОДИРУЮЩАЯ ГИБРИДНЫЙ ПОЛИПЕПТИД, СОДЕРЖАЩИЙ ПРОИНСУЛИН ЧЕЛОВЕКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PPINS25, КОДИРУЮЩАЯ ГИБРИДНЫЙ ПОЛИПЕПТИД, СОДЕРЖАЩИЙ ПРОИНСУЛИН ЧЕЛОВЕКА, И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО ПОЛИПЕПТИДА, СОДЕРЖАЩЕГО ПРОИНСУЛИН ЧЕЛОВЕКА | 1999 |
|
RU2143493C1 |
| СПОСОБ ПОЛУЧЕНИЯ ШТАММА-ПРОДУЦЕНТА ГИБРИДНОГО БЕЛКА APRO INS ДЛЯ ПОЛУЧЕНИЯ ИНСУЛИНА ЧЕЛОВЕКА | 1998 |
|
RU2148641C1 |
| US 5840542, 24.11.1998 | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |

