Изобретение касается стереоселективного синтеза хиральных аминов.
Известно, что биологическая активность химических продуктов, таких как фармацевтические и агрохимические препараты, обладающие хиральным центром, часто присуща в основном одной из возможных хиральных форм. Поскольку большинство химических синтезов по существу не являются стереоселективными, то возникает серьезная химическая технологическая проблема.
Многие из хиральных соединений этого типа являются аминами. Кроме того, из-за своей синтетической универсальности амины также являются хорошими объектами для разделения, после которого может быть эффективно осуществлено стереоселективное превращение в хиральное соединение. Химическое получение хирального амина, свободного от его энантиомера, до сих пор было главным образом основано на разделении смеси двух хиральных форм посредством образования диастереоизомерных производных, таких как соль хиральной кислоты, стереоселективных синтезах или использовании хиральных хроматографических колонок (патент США N 3944608 и EP-A 36265).
Некоторые структурные типы аминов поддаются ферментативному разделению. Ферментативные реакции, включающие альфа-аминокислоты, хорошо известны, и их предполагалось использовать для получения стереорегулярных образцов. В патенте США N 3871958, например, описано ферментативное получение альфа-аминокислоты-серина - взаимодействием альдегида с глицином в присутствии треонинальдолазы, полученной из образцов E. coli, а также родственный синтез серинола с использованием этаноламина.
Относительно мало сообщалось о ферментативных реакциях для аминокислот, в которых аминогруппа не соседствует с карбоксильной группой. Йонаха и др. (Agric. Biol. Chem., 42, (12), 2363-2367 (1978)) описывают омега-аминокислотную пируват-трансаминазу, обнаруженную в образцах Pseudomonas, для которой пируват был единственным аминным акцептором. Этот фермент, который ранее был кристаллизован и охарактеризован (Йонаха и др., Agric. Biol. Chem., 41 (9), 1701-1706 (1977)), имел низкую специфичность по субстрату для омега-аминокислот, таких как гипотаурин, 3-аминопропансульфонат, бета-аланин, 4-аминобутират и 8-аминооктаноат, и катализировал переаминирование между первичными аминоалканами и пируватом.
Накано и др. (J. Biochem., 81, 1375-1381 (1977)) идентифицировали две омега-аминокислотных трансаминазы в B. cereus: бета-аланин-трансаминазу, которая соответствует омега-аминокислота: пируват-трансаминазе, описанной Йонахой и др., и -аминобутират-трансаминазе. Они могут быть различны по своим резко отличным активностям по отношению к бета-аланину (100 против 3) и к -аминобутирату (43 против 100) соответственно, а также по своим различным требованиям к аминному акцептору.
Бернетт и др. (J.C.S. Chem. Comm., 1979, 826-828) предположили, что омега-аминокислотная пируват-трансаминаза и -аминобутират-трансаминаза характеризуются различной предпочтительностью по отношению к двум концевым атомам в -аминобутирате, меченном тритием.
Танизава и др. (Biochem., 21, 1104-1108 (1982)) исследовали бактериальную L-лизин-ε-аминотрансферазу и L-орнитин-δ-аминотрансферазу и отметили, что, хотя обе являются специфичными по отношению к L-аминокислотам, они действуют на расстоянии от центра и с такой же стереоспецифичностью, как -аминобутират-трансаминаза, изученная ранее Бернеттом и др.
Йонаха и др. (Agric. Biol. Chem., 47 (10), 2257-2265 (1983)) в дальнейшем охарактеризовали омега-аминокислотную пируват-трансаминазу и -аминобутират-трансаминазу (EC 2.6.1.18 и EC 2.6.1.19) и подтвердили документально их распределение в целом ряде организмов.
Уотерс и др. (FEMS Micro. Lett., 34 (1986), 279-282), сообщая о полном катаболизме бета-аланина и бета-аминоизобутирата с помощью P. aeruginosa, отметили, что первая стадия включала трансаминирование с бета-аланин : пируват-аминотрасферазой.
В наиболее широком смысле изобретение предусматривает использование омега-аминокислотной трансаминазы в присутствии аминного акцептора для обеспечения стереоселективного синтеза хиральных аминов, у которых аминогруппа связана с неконцевым хирально-замещенным атомом углерода. Таким образом, изобретение основано на открытии того, что омега-аминокислотные трансаминазы стереоселективно воздействуют на аминогруппы, которые не находятся в омега-положении, и что это действие может быть использовано для стереоселективного синтеза хирального амина только одной конфигурации.
Под омега-аминокислотными трансаминазами подразумевают любой фермент, обладающий свойством превращать концевую -CH2-NH2-группу омега-аминокислоты в -CH=O-группу.
Равновесная ферментативная реакция, рассматриваемая в изобретении, может быть изображена следующим образом: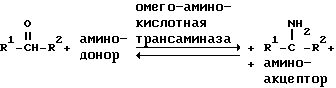
где каждый из R1 и R2, когда их выбирают независимо, представляет собой алкильную или арильную группу, которая является незамещенной или замещенной одной или более ферментативно неингибирующими группами, и R1 отличается от R2 по структуре или хиральности или R1 и R2, взятые вместе, представляют собой углеводородную цепь из 4 или более атомов углерода, содержащую центр хиральности.
Здесь под "амино-акцептором" подразумевают различные карбонильные соединения, более полно обсуждаемые ниже, которые способны акцептировать аминогруппу от рассматриваемого амина под влиянием омега-аминокислотной трансаминазы. Под "амино-донором" подразумевают различные аминосодержащие соединения, более полно обсуждаемые ниже, которые способны отдавать аминогруппы рассматриваемому кетону, тем самым превращаясь в карбонильные соединения, также под влиянием аналогичной омега-аминокислотной трансаминазы.
Описанная выше ферментативная реакция характеризуется, во-первых, тем, что омега-аминокислотная трансаминаза воздействует на первичный амин, в котором аминогруппа не находится в омега- (или концевом) положении. Во-вторых, трансаминаза воздействует на амин, который не должен быть аминокислотой. В-третьих, расходуемое в процессе ферментативного превращения аминное соединение не является необратимо метаболируемым, а может быть стереоселективно превращено в исходный амин одинаковой хиральности.
Изобретение обеспечивает способ стереоселективного синтеза одной хиральной формы амина формулы IA или IB в количестве, существенно превосходящем другую форму путем воздействия на кетон формулы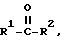
где R1 и R2 - такие, как определено выше, с омега-аминокислотной трансаминазой в присутствии донора аминогруппы.
Изобретение основано на открытии того, что действие омега-аминокислотной трансаминазы не ограничено омега-аминогруппами и, кроме того, является весьма исключительно стереоселективным по отношению к аминам определенного класса, превращая только одну хиральную форму амина в соответствующий кетон, который больше не является хиральным (по крайней мере по отношению к карбонильному атому углерода), и в свою очередь превращая этот кетон в одну единственную хиральную форму амина.
Термин "существенно выше", используемый здесь при описании стереоселективного синтеза одной хиральной формы по сравнению с другой, относится к соотношению, составляющему по крайней мере около 3 : 1, когда величина "ее" равна по крайней мере около 50%.
Поскольку реакция является равновесной, то или прямой, или обратной реакциям можно благоприятствовать добавлением дополнительных исходных соединений или удалением продуктов реакции.
Когда стереоселективно синтезируют одну хиральную форму амина, то следует либо добавить дополнительное количество кетона (вплоть до насыщения) и/или удалить образующийся амин. Это может быть реализовано на практике. Следовательно, стереоселективный синтез одной хиральной формы амина формулы IA или IB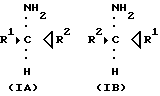
в количестве, существенно более высоком, чем количество другой формы, может быть достигнут путем воздействия на кетон формулы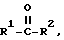
где R1 и R2 являются, как определено выше, омега-аминокислота трансаминазы в присутствии донора аминогруппы до тех пор, пока не образуется существенного количества одного из хиральных аминов. В примере, представленном выше, ацетофенон, например, подвергают воздействию трансаминазы в присутствии донора аминогруппы, чтобы получить исключительно S-1-амино-1-фенилэтан или получить это соединение с существенно более высоким процентным выходом, чем R-1-амино-1-фенилэтан.
Донорами аминогруппы являются амины, включающие нехиральную аминокислоту, глицин, и хиральные аминокислоты, имеющие S-конфигурацию, такие как L-аланин или L-аспарагиновая кислота. Амины, как хиральные, так и нехиральные, которые не являются аминокислотами, такие как S-2-аминобутан, пропиламин, бензиламин и т.д., также могут использоваться.
Омега-аминокислотные трансаминазы, пригодные для использования в настоящем способе, представляют собой известные пиридоксалфосфат-зависимые ферменты, обнаруженные в различных микроорганизмах, включающих Pseudomonas, Escherichia, Bacillus, Saccharomyces, Hansenula, Candida, Streptomyces, Aspergillus и Neurospora. Две омега-аминокислотные трансаминазы, которые особенно подходят для использования в изобретении, EC 2.6.1.18 и EC 2.6.1.19, были кристаллизованы и охарактеризованы Йонахой и др. (Agric. Biol. Chem., 47 (10), 2257 - 2265 (1983)).
Микроорганизмы, имеющие требуемую активность, легко могут быть выделены с помощью хемостатной культуры, т.е. выращиванием в постоянной, но ограниченной химической среде с акцептором аминогруппы и амином в качестве одного-единственного источника азота. Амин может быть, но не должен быть хиральным, поскольку в нормальной среде омега-аминокислотные трансаминазы метаболизируют первичные амины. Нехиральные амины, которые успешно использовали для получения омега-аминокислотной трансаминазы, включают н-октиламин, циклогексиламин, 1,4-бутандиамин, 1,6-гександиамин, 6-аминогексановую кислоту, 4-аминомасляную кислоту, тирамин и бензиламин. Хиральные амины, такие как 2-аминобутан, альфа-фенэтиламин и 2-амино-4-фенилбутан, также успешно использовать, когда имеется аминокислота, такая как L-лизин, L-орнитин, бета-аланин и таурин.
С помощью таких процедур культура будет обогащена этими микроорганизмами, продуцирующими требуемые омега-аминокислотные трансаминазы. Например, в одном таком хемостате, проведенном со случайными образцами почвы, не обработанными конкретным амином, выдержку проводили приблизительно в течение одного месяца. Согласно этому доминирующие микроорганизмы независимо идентифицировали с помощью ATCC как Bacillus megaterium, которая незначительно отличается от известных штаммов и фенотипически подобна им.
Организмы, выделяемые таким образом, могут быть выращены с помощью целого ряда способов. Во-первых, можно использовать стандартную солевую среду, дополненную фосфатным буфером, ацетатом натрия как источником углерода, 2-кетоглутаратом как акцептором аминогруппы и азотсодержащим соединением, таким как н-пропиламин, н-октиламин, 2-аминобутан, 2-аминогептан, циклогексиламин, 1,6-гександиамин, путресцин, 6-аминогексановая кислота, 4-аминомасляная кислота, L-лизин, L-орнитин, бета-аланин, альфа-фенэтиламин, 1-фенил-3-аминобутан, бензиламин, тирамин, таурин и т.д.
Альтернативно микроорганизм может выращиваться с использованием амина в качестве одного-единственного источника углерода, тем самым ограничивая рост тех организмов, которые могут катаболизировать этот амин для получения углерода. В-третьих, микроорганизм может выращиваться с использованием сукцината натрия, ацетата натрия или любого другого источника углерода и аммониевой соли или белкового гидролизата как основного источника азота и с последующим добавлением или в начале, или в процессе роста амина, такого как 2-аминобутан, 1-фенил-3-аминобутан, альфа-фенэтиламин и т.д., для индуцирования получения необходимой трансаминазной активности.
Омега-аминокислотная трансаминаза может быть в свободной форме или в виде экстракта, свободного от клеток, или в виде цельного клеточного препарата, как отмечено выше, или иммобилизована на подходящей подложке или матрице, такой как сшитый декстран или агароза, кремнезем, полиамид или целлюлоза. Ее можно также инкапсулировать в полиакриламиде, альгинатах, волокнах и т. п. Методы такой иммобилизации описаны (Methos of Enzymology, 44, 1976). Последний вариант особенно полезен, поскольку к однажды иммобилизованному ферменту необходимо только подавать донор аминогруппы и кетон соответствующей формулы для реакции стереоселективного синтеза и затем удалять образовавшиеся хиральные амины.
Ниже описаны используемые методики и материалы, за которыми следуют типичные примеры.
Методики и материалы.
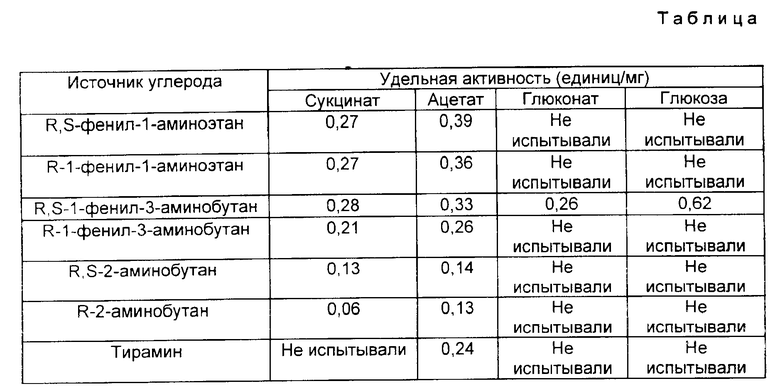
Ферментативная активность. Ферментативная активность выражена здесь в единицах/мг. Единица активности фермента определяется как активность, которая продуцирует 1 мкмоль кетона в 1 мин. Для однозначности ее измеряют в виде микромолей 1-фенилбутан-3-она, образующегося из R,S-1-фенил-3-аминобутана. Для измерения активности омега-аминокислота трансаминазы применяли следующие стандартные тесты, изложенные в примерах, данных ниже.
Известный объем испытуемого препарата фермента инкубируют при 37oC и pH 7 в растворе, имеющем следующий состав, ммоль/л:
Пируват натрия - 100
R,S-Фенил-3-аминобутан - 30
Пиродоксальфосфат - 0,5
Образец удаляют и добавляют 20 об.% водного раствора трихлоруксусной кислоты с концентрацией 12%. Выпавший в осадок белок удаляют центрифугированием и определяют концентрацию 1-фенилбутан-3-она в надсадочной жидкости с помощью жидкостной хроматографии на 4-мкм колонке типа Novopak phenyl, 100 x 8 мм, элюируя 40% изопропанола и 0,09% фосфорной кислоты в воде. При этих условиях 1-фенилбутан-3-он элюируется за 5,3 мин.
Чистота аминов. Чистоту продуцируемых аминов определяли с помощью газовой хроматографии на колонке типа Chrom Q размером 6 фут х 2 мм с 10% жидкой фазы марки E-30 на носителе с размером частиц 100/120 меш при 210oC и скорости несущего газового потока 10 мл/мин.
Определение энантиомерного обогащения.
Величину "ее" данного продукта определяли по реакции с (-)-альфа-(трифторметилфенил)метоксиацетилхлоридом (Гэл, J. Pharm. Sci. , 66, 169 (1977); Мошер и др., J. Org. Chem., 34, 25430 (1969)) с последующей капиллярной газовой хроматографией получаемого в результате соединения на колонке типа Chrompack из плавленой двуокиси кремния.
Стандартная солевая среда. Подходящая солевая среда для микробиологических превращений, описанная в следующих примерах, имеет следующий состав:
MgSO4 - 1,00 г/л
CaCl2 - 0,021 г/л
ZnSO4 • 7H2O - 0,20 мг/л
MnSO4 • 4H2O - 0,10 мг/л
H3BO3 - 0,02 мг/л
CuSO4 • 5H2O - 0,10 мг/л
CoCl2 • 6H2O - 0,05 мг/л
NiCl2 • 6H2O - 0,01 мг/л
FeSO4 - 1,50 мг/л
NaMoO4 - 2,00 мг/л
FebD ТА - 5,00 мг/л
KH2PO4 - 20,00 ммоль/л
NaOH - До pH 7
Данный состав не является критическим, но был стандартизирован для всех методик, чтобы исключить их непостоянство.
Микроорганизмы. Культуры или получали из указанного источника, или выделяли, как описано, и затем независимо идентифицировали.
Обогащение микроорганизмов, продуцирующих омега-аминокислотную трансаминазу. Хемостат поддерживают с помощью 0,5% (вес/объем) R,S-2-аминобутана и 10 ммоль/л 2-кетоглутарата при скорости разбавления 0,03/ч в стандартной солевой среде. Хемостат инокулируют и ингибируют приблизительно в течение одного месяца при pH 6,8 - 7,0. Штаммы, которые развиваются, выделяют и выращивают на минимальном агаре, содержащем стандартную солевую среду, дополненную 10 мМ 2-кетоглутарата и 5 мМ R,S-1-фенил-3-аминобутана.
Восстановление фермента. Если не указано особо, клетки из культуры центрифугируют в течение 10 мин при 10000 g, повторно суспендируют в 10 мМ фосфатном буфере при pH 7 и 0,5 мМ пиридоксальфосфате и разрывают путем двух проходов через охлаждаемый французский пресс, работающий при давлении 15000 фунтов/дюйм2. Остатки клеток удаляют центрифугированием в течение 1 ч при 10000 g и собирают ферментсодержащую надосадочную жидкость.
Следующие примеры будут служить дальнейшей иллюстрацией существа изобретения, но они не должны истолковываться как ограничение его объема, который определяется только приведенной формулой изобретения.
Пример 1. Следующая методика дает пример выращивания микроорганизмов, продуцирующих омега-аминокислотную трансаминазу, при использовании в качестве единственного источника азота донора аминогруппы.
Bacillus megaterium выращивали в 3-литровой вибрирующей колбе (200 об/мин) в течение 17 ч при 30oC, куда было добавлено: 1 л вышеназванного солевого раствора, 60 мМ ацетата натрия, 30 мМ фосфатного буфера, 30 мМ динатрий 2-кетоглутарата и 100 мМ н-пропиламина как источника азота. Когда культура достигла плотности 0,6 г (сухого веса) на 1 л, клетки собирали и выделяли фермент, как описано выше. Удельная активность омега-аминокислотной трансаминазы, полученной таким образом, при испытании согласно представленной выше методике составила 0,49 ед./мг.
Используемый в вышеуказанной методике штамм Bacillus megaterium получали из образцов почвы без особой обработки аминами путем инокуляции ранее описанного хемостата и выделением доминантных организмов (тех, которые способны расти на R,S-1-фенил-3-аминобутане). Этот штамм независимо идентифицировали с помощью американской коллекции типов культур как Bacillus megaterium, который существенно не отличался от известного штамма АТСС 14581 и который по своему фенотипу был подобен штамму АТСС 49097В.
Пример 2. Следующая методика дает пример выращивания микроорганизмов, продуцирующих омега-аминокислотную трансаминазу, с использованием донора аминогруппы в качестве единственного источника углерода.
Pseudomonas aeruginosa АТСС 15692 выращивали на бета-аланине как единственном источнике углерода, как описали Уэй и др. (FEMS Micro Lett., 34, 279 (1986)), и затем получали клеточные экстракты, содержащие омега-аминокислотную трансаминазу, как здесь описано. При анализе по методике, описанной выше, удельная активность омега-аминокислотной трансаминазы составила 0,040 ед/мг.
Пример 3. Pseudomonas putida АТСС 39213 выращивали, как описано в примере 1, и затем получали ферментный экстракт, как здесь указано. Удельная активность омега-аминокислотной трансаминазы составила 0,045 ед/мг.
Пример 4. Следующая методика дает пример выращивания микроорганизмов с использованием аммония в качестве единственного источника азота и последующим индуцированием выделения омега-аминокислота трансаминазы путем добавления амина.
Bacillus megaterium выращивали в культурах объемом 1 л в стандартной солевой среде, дополненной 40 мМ указанного источника углерода, 5 мМ хлорида аммония, 80 мМ фосфатного буфера и 2 мМ аминного индуктора, указанного ниже. Спустя 30 - 40 ч фермент собирали и анализировали, как описано выше (таблица).
Пример 5. Следующая методика дает пример выращивания микроорганизмов с использованием обогащенного белком источника и последующим индуцированием выделения омега-аминокислотной трансаминазы путем добавления амина.
Bacillus megaterium выращивали в 121-литровом ферментере при pH 7 и 30oC с аэрацией и при перемешивании в вышеуказанной солевой среде, дополненной касаминовыми кислотами до концентрации 10 г/л. Постепенно добавляли ацетат натрия вплоть до достижения концентрации 120 мМ, когда образуются агрегаты. В этой точке плотность клеток составляла 3 г (сухого веса) на 1 л. 1-Фенил-3-аминобутан добавляли вплоть до концентрации 10 мМ, когда образуются агрегаты. Спустя 12 ч фермент собирали и анализировали, как описано выше. Удельная активность составляла 0,49 ед./мг.
Пример 6. Следующая методика дает типичный синтез хирального амина. Растворимый ферментный препарат получали из Bacillus megaterium способом, описанным в примере 1. Анализ, выполненный по описанной выше методике, показал, что удельная активность составляла 0,58 ед./мг. К 200 мл водного раствора 350 г этого препарата, 0,4 мМ пиридоксальфосфата и 40 мМ фосфата натрия добавляют 4,2 мМ 1-фенилбутан-3-она и 100 мл 2-аминобутана как донора аминогруппы. Смесь инкубировали при pH 7 и 30oC в течение 4 ч, когда концентрация R-1-фенил-3-аминобутана достигала 3,35 мМ, что соответствовало 80%-ному превращению. Продукт выделяли путем добавления 40 мл 10N раствора гидроксида натрия и экстрацией щелочного водного раствора 250 мл н-гептана. После упаривания гептановых экстрактов было получено 100,5 г продукта, который анализировали по его производному, как описано выше, и обнаружили, что он содержал 96,4% S-1-фенил-3-аминобутана.
По аналогии с этим из 1-фенилпропан-2-она был получен S-1-фенил-2-аминопропан с величиной "ее" 96,4 и выходом 94,8%. Из ацетофенона был получен S-1-амино-1-фенилэтан с величиной "ее" 100 и выходом 44%.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЭНАНТИОМЕРНОГО ОБОГАЩЕНИЯ СМЕСИ ДВУХ ЭНАНТИОМЕРНЫХ ХИРАЛЬНЫХ АМИНОВ | 1990 |
|
RU2087536C1 |
| СПОСОБ ПОЛУЧЕНИЯ ХИРАЛЬНОГО АМИНА | 1999 |
|
RU2213142C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ФОСФИНОТРИЦИНА ПУТЕМ ФЕРМЕНТАТИВНОГО ТРАНСАМИНИРОВАНИЯ С ПОМОЩЬЮ АСПАРТАТА | 2000 |
|
RU2275428C2 |
| СПОСОБ ЭНАНТИОМЕРНОГО ОБОГАЩЕНИЯ СМЕСИ D- И L-ТРЕО-2-АМИНО-3-ГИДРОКСИ-3-ФЕНИЛПРОПИОНОВЫХ КИСЛОТ, ИЛИ ИХ ПРОИЗВОДНЫХ, ИЛИ ИХ СОЛЕЙ | 1992 |
|
RU2112805C1 |
| ПОЛУЧЕНИЕ 1,4 ДИАМИНОБУТАНА | 2010 |
|
RU2577967C2 |
| ХИМИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ СПИРОИНДОЛОНОВ И ИХ ПРОМЕЖУТОЧНЫХ СОЕДИНЕНИЙ | 2013 |
|
RU2662815C2 |
| Фермент трансаминаза D-аминокислот из Haliscomenobacter hydrossis с расширенной субстратной специфичностью | 2023 |
|
RU2824015C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОИЗВОДНОГО ГЛЮТАМАТА (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МОНАТИНА | 2002 |
|
RU2280078C2 |
| Новые трансаминазы и способ дезаминирования аминосоединения с их применением | 2016 |
|
RU2718778C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНДОЛ-3-ПИРОВИНОГРАДНОЙ КИСЛОТЫ И ЕЕ ПРОИЗВОДНЫХ | 2006 |
|
RU2325442C2 |
Изобретение касается способа стереоселективного синтеза одной хиральной формы амина в количестве, существенно превосходящем другую форму, путем воздействия на кетон ферментом омега-аминокислотной трансаминазой в присутствии донора аминогруппы. Донор аминогруппы используют в значительном молярном избытке. Кетон и донор аминогруппы приводят во взаимодействие с целыми клетками микроорганизма, которые продуцируют омега-аминокислотную трансаминазу, или с водным раствором фермента, или с ферментом, иммобилизованным на носителе. Способ может использоваться в химических технологиях при производстве фармацевтических и агрохимических препаратов. 5 з.п. ф-лы, 1 табл.
в количестве, существенно более высоком, чем другой формы, где каждый из R1 и R2 представляет собой алкильную или группу, незамещенную или замещенную ферментативно неингибирующей группой, и R1 отличается от R2 по структуре или хиральности,
отличающийся тем, что осуществляют взаимодействие кетона формулы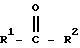
в которой R1 и R2 - такие, как определено выше,
с омега-аминокислотной трансаминазой в присутствии донора аминогруппы по крайней мере до тех пор, пока не образуется существенного количества одного из названных хиральных аминов.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| US, патент, 3944608, C 07 C 103/34, 1976 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| EP, заявка, 036265, C 07 B 19/00, 1981 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US, патент, 3871958, C 12 D 13/06, 1975. | |||
