Изобретение относится к генной инженерии, более конкретно к трансфекционной композиции для высших эукариотных клеток, комплексу нуклеиновой кислоты и эндосомолитическому пептиду в качестве компонентов композиции, конъюгату в качестве компонента комплекса.
В особенности в терапии генами требуется эффективная система для введения нуклеиновой кислоты в живые клетки. Гены переводят в клетки с тем, чтобы достигнуть синтеза терапевтических активных генетических продуктов в живом организме, например, с целью замены дефектного гена в случае генетического дефекта. "Стандартные" терапии генами основаны на принципе достижения длительного лечения путем осуществления одного единственного приема. Однако также существует потребность в методах лечения, в которых терапевтически активную ДНК (или мРНК) дают в качестве лекарства ("терапевтического генного агента") раз или же в случае необходимости повторно. Примерами генетически вызываемых заболеваний, при которых терапия генами представляет многообещающий подход, являются гемофилия, β-талласемия и "серозная комбинированная иммунная слабость", синдром, вызываемый генетически индуцированным отсутствием энзима деаминазы аденозина. Другой областью применения является иммунная регуляция, при которой гуморальный или внутриклеточный иммунитет обеспечивается дачей функциональной нуклеиновой кислоты, кодирующей выделяемый протеиновый антиген или не выделяемый протеиновый антиген, который можно рассматривать как вакцинацию. Другими примерами генетических дефектов, при которых можно давать нуклеиновую кислоту, кодирующую дефектный ген, например, в согласованной в каждом конкретном случае форме, включают мышечную дистрофию (ген дистрофина), фиброзно-кистозную дегенерацию (регуляторный ген трансмембранной проводимости висцидозы), гиперолестиролемию (ген рецептора липопротеина низкой плотности). Терапия генами также включает метод, согласно которому гормоны, факторы роста или белки цитотоксичной или проявляющей модуляцию иммунной системы активностью должны синтезироваться в теле.
Терапия генами также рассматривается как многообещающий метод лечения рака путем дачи так называемых "раковых вакцин". Для повышения иммуногенности опухолевых клеток раковым вакцинам сообщают либо большую антигенность либо способность к производству определенных веществ, способных к модуляции иммуносистемы, например, цитокинов, с тем, чтобы вызвать иммунную реакцию. Этот метод осуществляется путем трансфекции клеток ДНК, кодирующей цитокин, например, IL-2, IL-4, γ-интерферон, α-фактор опухолевого некроза. Вплоть до настоящего времени введение гена в аутогенные опухолевые клетки осуществлялось через ретровирусные векторы.
Известна трансфекционная композиция для гепатоцитов, представляющая собой конъюгат поликатиона, обладающего способностью к модифицированию, например, полилисина, и гликопротеина, обладающего способностью к модифицированию, причем конъюгат связывается рецепторами на поверхности гепатоцитов (см. Г.И. Ву, С.Н. Ву, J. Biol. Chem. 263, 29, стр. 14621 - 14624, 1988).
Недостаток известной композиции заключается в том, что она может использоваться только для переноса нуклеиновых кислот в гепатоциты. Поэтому эксплуатационные возможности известной композиции ограничены.
Задачей изобретения является расширение эксплуатационных возможностей трансфекционных систем с помощью композиции, позволяющей перенос нуклеиновых кислот в высшие эукариотные клетки.
Под термином "перенос" понимается в рамках данного изобретения не только введение комплексов нуклеиновой кислоты в клетку через клеточную мембрану, но и транспорт выделяющихся из них комплексов или нуклеиновой кислоты внутри клетки до достижения подлежащего экспрессии подходящего сайта. Высшие эукариотные клетки общеизвестны. Они не включают дрожжей.
Множество вирусов входят в эукариотный хозяин на основании механизмов, которые в принципе соответствуют механизму эндоцитоза, осуществляемого с участием рецептора. Основанная на этом механизме вирусная инфекция обычно начинается с того, что частицы вируса связываются с рецепторами на клеточной мембране, после чего вирус входит вовнутрь клетки. Процесс захода вовнутрь клетки осуществляется тем же образом, что и заход физиологических лигандов или макромолекул в клетку. Сначала рецепторы на поверхности клетки классифицируют себя по группам и мембрана инвертируется во внутрь и образует везикулу, снабженную сетчатым покрытием. После того, как везикула освободилась от сетчатого покрытия, в ней имеет место окисление при помощи пропонного насоса в мембране. Это приводит к отделению вируса от ядрышка. В зависимости от того, снабжен ли вирус липидным покрытием или нет, возможны два вида отделения вируса от ядрышка. В случае так называемых "голых" вирусов (таких, как, например, аденовирус, полиовирус, риновирус) полагалось, что низкое значение pH вызывает изменение конфигурации белков вируса, что приводит к разделению гидрофобных доменов, которые не доступны при физиологическом значении pH. Таким образом эти домены приобретают способность к взаимодействию с мембраной ядрышка и тем самым обуславливают отделение генома вируса от ядрышка и его заход в цитоплазму. Для вирусов с липидным покрытием (таких, как, например, вирусы везикулярного стоматита, вирус Semliki Forest, вирус гриппа) предполагают, что низкая величина pH модифицирует структуру или конфигурацию некоторых белков вируса, в результате чего имеет место слияние мембраны вируса с мембраной ядрышка. Вирусы, которые проникают в клетку с помощью этого механизма, имеют определенные молекулярные способности, которые сообщают им способность к разрушению мембраны ядрышка с тем, чтобы зайти в цитоплазму.
Другие вирусы, как, например, покрытые вирусы Sendai, вирус СПИДа и некоторые штаммы вируса лейкоза Молонея, или непокрытые вирусы SV40 и полиомы, не требуют низкого значения pH для проникновения в клетку. Либо они могут вызывать слияние с мембраной непосредственно на поверхности клетки (вирус Sendai, возможно вирус СПИДа), либо они способны к осуществлению механизмов по разложению мембраны клетки или прохода через нее. Предполагают, что вирусы, которые не зависят от конкретной величины pH, могут также использовать механизм эндоцитоза. Исходной точкой решения проблемы изобретения являлось использование механизма, по которому некоторые вирусы проникают в эукариотные клетки с тем, чтобы достичь эффективной передачи комплексов нуклеиновой кислоты в клетку и тем самым повышения экспрессии.
В рамках данного изобретения неожиданно было найдено, что определенные агенты (например, вирусы, компоненты вирусов или другие соединения), которые проявляют способность определенных вирусов к заходу в эукариотные клети, обеспечивают существенное повышение степени экспрессии нуклеиновой кислоты, вводимой в клетку в качестве части комплекса. Решение по изобретению является неожиданным из-за того, что ассортимент комплексов нуклеиновой кислоты, способных к проникновению в клетку, является очень широким.
Таким образом, данное изобретение относится к трансфекционной композиции для высших эукариотных клеток, включающей комплекс нуклеиновой кислоты и вещество со сродством для нуклеиновой кислоты, при этом она дополнительно содержит эндосомолитический агент в эффективном количестве.
Эндосомолитический агент представляет собой вещество, способное к поглощению клеткой и к переносу в цитоплазму содержащего ядрышек, в которых они находятся после захода в клетку. Такая способность эндосомолитического агента далее обозначается как "поглотительная функция". Эта поглотительная функция включает способность к активному включению клеткой, то есть при помощи зависящего от рецептора механизма эндоцитоза, либо к пассивному включению клеткой, то есть при помощи жидкой фазы или в качестве компонента комплекса нуклеиновой кислоты, и способность к разрушению ядрышек, которая обычно обозначается как эндосомолиз ядрышка.
Согласно одному варианту изобретения в качестве эндосомолитического агента используют вирус или компонент вируса. Далее вирус или компонент вируса обозначается как "свободный" вирус (компонент).
В рамках настоящего изобретения исследовалось действие повышающихся доз свободного аденовируса на степень передачи постоянного количества конъюгата-трансферина и полилизина в клетках HeLa с использованием в качестве репортерного гена люциферазы. Повышение степени передачи гена путем добавления свободного аденовируса достигло максимума 1 • 104 частиц вируса на клетку, то есть числа, которое соответствует примерному числу рецепторов аденовируса на клетку HeLa. Повышение экспрессии люциферазы, которая составляет до 2000 раз по сравнению с экспрессией, достигаемой одним конъюгатом трансферина и полилизина, обусловлено большей дозой вируса. Другая серия опытов была направлена на выявление действия ограничения количеств комплексов конъюгата и ДНК в присутствии постоянной дозы свободного аденовируса. Было найдено, что поглощение клеткой аденовирусов привело к повышению степени передачи гена с помощью трансферина/полилизина в широких пределах количества ДНК. Максимальная интенсивность экспрессии гена, достигаемая при помощи комплекса коньюгата и ДНК, соответствовала интенсивности, достигаемой при 100 раз меньшем количестве ДНК в случае использования аденовируса для повышения эффективности трансфекции.
Действие эндовирусной инфекции на передачу гена исследовалось с применением как некомплексированной ДНК, так и комплексов ДНК и полилизина или конъюгата трансферина и полилизина. В этих экспериментах установлено, что аденовирусная инфекция не привела к существенному увеличению передачи некомплексированной ДНК во время трансфекции. В противоположность этому, передача ДНК в виде комплекса с полилизином или в виде комплекса с конъюгатом трансферина и полилизина в значительной степени улучшалась аденовирусной инфекции. Максимальное действие достигается в случае применения конъюгата трансферина и полилизина. Так как поликатионная часть конъюгата не только служит для связывания трансферина с ДНК, но и вызывает существенные структурные изменения внутри ДНК, эти эксперименты сначала не позволяли установить, являлось ли обнаруженное действие результатом улучшенного жидкостного транспорта конденсированной поликатионом ДНК или улучшенной вирусом доставки связанного с рецептором комплекса конъюгата и ДНК. Поэтому проводились опыты по связыванию и определению последовательности с тем, чтобы выявить причину обнаруженного действия. Связывание комплекса конъюгата трансферина и полилизина и ДНК или комплекса полилизина и ДНК при низкой температуре позволяла удаление избыточного комплекса в жидкой фазе перед инфекцией аденовирусом. Когда поступали указанным образом, отдача связанных с рецептором комплексов конъюгата трансферина и полилизина и ДНК значительно увеличивалась в результате добавления аденовируса. В случае применения комплексом полилизина и ДНК такого эффекта не наблюдалось. Таким образом то, что существенно улучшается, это заход ДНК с помощью способствующего рецептором механизма эндоцитоза.
Кроме того, проводился анализ специфичной функции аденовируса, который вызывает улучшенный перенос гена при помощи рецептора. Слабая термообработка вирионов не меняет их способность к связыванию с соответствующими клеточными мембранами, но влияет на их свойство разрушать ядрышка после поглощения клеткой.
Таким образом различные эффекты по связыванию вируса и заходу вируса в клетку можно было выявлять в отдельных экспериментах. При этом было также установлено, что термическая инактивация аденовирусов привела к полному уничтожению их способности к улучшению переноса гена при помощи рецептора в качестве посредника. Это позволяет предполагать, что вызываемое аденовирусами разрушение ядрышек является частью их механизма захода в клетку, который специфично обуславливает улучшение отдачи гена при помощи конъюгата трансферина или полилизина. Тот факт, что штамм дефектного в отношении репликации вируса мог обеспечивать улучшение экспрессии гена, подтверждает предположение о том, что этот феномен обусловлен не функцией репликацией, а поглотительной функцией вириона.
Для выяснения того, что улучшение экспрессии гена можно приписывать к возможной трансактивации введенного гена вирусом, проводились опыты с применением клеточной линии, которая по своей структуре способна к экспрессии гена люциферазы RSV-LTR. В этой клеточной линии аденовирусы не проявляют действия, тогда как в родительской клеточной линии, в которой ген был введен при помощи конъюгата трансферина и полилизина, наблюдалось существенное повышение экспрессии гена. Результаты этих опытов показывают, что аденовирус влияет на событие, которое имеет место до транскрипции, и что его улучшающее перенос гена действие проявляется не на уровне экспрессии гена, а на уровне переноса гена.
В рамках данного изобретения также проводились исследования, направленные на выявление действия свободного аденовируса на перенос гена при помощи конъюгатов трансферина и полилизина в подобранных клеточных линиях. Было найдено, что присутствие рецепторов трансферина на целевой клетке является необходимым, но недостаточным в каждом случае с тем, чтобы обеспечить перенос гена при помощи конъюгата трансферина и полилизина. Предполагают, что специфичные у клетки факторы, относящиеся к результату находящихся в ядрышке комплексов конъюгата и ДНК, представляют собой крайне важный определяющий фактор для уровня передачи гена, достигаемого указанным путем. В этом отношении проводились опыты на подобранных клеточных линиях, направленные на исследование как переноса гена при помощи конъюгатов трансферина и полилизина, так и улучшение переноса гена при помощи аденовирусов. Клетки клеточной линии мукофисцидоза (CFT1) показывали небольшой уровень экспрессии гена люциферазы после обработки комплексами ДНК и конъюгата трансферина и полилизина. Этот уровень экспрессии существенно повысился в результате обработки адновирусом d1312. В противоположность этому клетки KB, обработанные комплексами ДНК и конъюгата трансферина и полилизина, показывали уровень экспрессии гена люциферазы лишь немного выше уровня фона, несмотря на присутствие рецептора трансферина. Однако в результате обработки аденовирусом d1312 сразу же обнаруживалась активность люциферазы в этих клетках. Обработка аденовирусами клеток HeLa имела подобный эффект, хотя этот эффект был значительно сильнее в этих клетках. Так как клетки HeLa и KB обладают примерно тем же числом рецепторов для аденовируса, различие в улучшении переноса гена может быть обусловлено числом рецепторов для трансферина, характерным для каждого типа клеток. В явную противоположность этим результатам клеточные линии WI-38 и MRC5), которые, как известно, лишь слабо поддерживают аденовирусную инфекцию, показывали в присутствии d1312 лишь незначительное улучшение экспрессии гена после обработки комплексом ДНК и конъюгата. Поэтому обработка свободным вирусом, например, аденовирусом, очевидно приводит к улучшению переноса гена при помощи комплексов ДНК и конъюгата в таких случаях, где перенос гена возможен за счет осуществляемого в присутствии рецептора в качестве посредника эндоцитоза, как, например, в случае клеток CFT1, а также в некоторых случаях, где перенос гена этим путем оказывается неэффективным, как, например, в случае клеток HeLa и КВ. Уровень достигаемого улучшения значительно меняется среди различных целевых клеток, так что можно предполагать, что этот эффект является функцией как числа рецепторов вируса, например, рецепторов аденовируса, у определенной клетки, так и числа рецепторов трансферина.
В случае применения свободного вируса вещество со сродством для нуклеиновой кислоты, предпочтительно органический поликатион, предпочтительно связано с фактором, содействующим поглощению клеткой. Однако в рамках данного изобретения было также найдено, что ДНК в виде комплекса с одним веществом со сродством для нуклеиновой кислоты, то есть без содействующего поглощению клеткой фактора, в определенных условиях может вводиться в клетку в присутствии свободного вируса. Кроме того, было найдено, что в случае некоторых клеточных линий комплексы, состоящие из нуклеиновой кислоты и вещества со сродством для нуклеиновой кислоты, могут вводиться через жидкую фазу при условии достаточно высокой концентрации комплекса. Опыты, которые проводились в рамках данного изобретения, показывали, что существенным признаком для поглотительной способности комплексов нуклеиновой кислоты является их компактность, которую можно приписывать к конденсации нуклеиновой кислоты веществом со сродством для нуклеиновой кислоты. Если вещество со сродством нуклеиновой кислоты имеет достаточную способность к связыванию с поверхностью клетки с тем, чтобы проникнуть в клетку вместе с вирусом, а также к сообщению комплексу в основном больной электронейтральности и конденсации нуклеиновой кислоты до компактной структуры, то может отпадать необходимость в улучшении способности к проникновению в клетку за счет ковалентного связывания содействующего поглощению клеткой фактора с веществом со сродством для нуклеиновой кислоты с тем, чтобы обеспечить перенос комплекса в клетку при помощи эндоцитоза, осуществляемого в присутствии рецептора в качестве посредника. Много клеток проявляет относительно большое сродство для определенных веществ со сродством для нуклеиновой кислоты, так что конъюгаты нуклеиновой кислоты и факторы связывания переносятся в клетку без потребности в наличии содействующего поглощению клеткой фактора. Это является действительным, например, для гепатоцитов, которые поглощают комплексы ДНК и полилизина.
Согласно предпочтительному варианту изобретения в качестве эндосомолитического агента используют вирус, связанный с веществом со сродством для нуклеиновой кислоты и имеющий способность к заходу в клетку в качестве части комплекса конъюгата и нуклеиновой кислоты и к переносу в цитоплазму содержимого ядрышек, в котором комплекс находится после захода в клетку.
Согласно другому предпочтительному варианту изобретения в качестве эндосомолитического агента используют компонент вируса, связанный с веществом со сродством для нуклеиновой кислоты и имеющий способность к заходу в клетку в качестве части комплекса конъюгата и нуклеиновой кислоты и к переносу в цитоплазму содержимого ядрышек, в которых комплекс находится после захода в клетку. Вирусы и компоненты вирусов, которые связаны с доменом связывания нуклеиновой кислоты, далее обозначаются как "вирусные конъюгаты" (несмотря на тип связывания).
Вирусные конъюгаты, которые также являются объектом данного изобретения, содержат вирус или компоненты вируса в качестве интегральной части функционального конструкта. Они объединяют преимущества векторных систем на основе конъюгатов содействующего поглощению клеткой фактора с преимуществами, которые вирусы вносят в эти системы.
Кроме того, вирусные конъюгаты согласно изобретению имеют то преимущество, что в отличие от известных конъюгатов, которые можно применять для переноса генов при помощи эндоцитоза в присутствии рецепторов в качестве посредника, они обладают специфичным механизмом, обеспечивающим их выделение из системы везикул клетки.
Векторная система, использующая вирусные конъюгаты, представляет собой качественно новый подход по сравнению с рекомбинантными вирусными векторами, который заключается в том, что транспортируемая чужеродная ДНК находится на наружной стороне вириона. Следовательно, вирусные конъюгаты согласно изобретению могут транспортировать в клетку очень большие генные конструкты без каких-либо ограничений в части последовательности.
Пригодность вируса, который можно применять в качестве свободного или связанного вируса или части вируса в качестве вирусного компонента в рамках данного изобретения, определена вышеупомянутой поглотительной функцией. Подходящими вирусами являются, например, такие вирусы, которые способны к заходу в клетку при помощи эндоцитоза в присутствии рецептора в качестве посредника во время трансфекции клеток комплексом нуклеиновой кислоты и к осуществлению своего выделения и тем самым выделения нуклеиновой кислоты из ядрышка в цитоплазму. Такими вирусами являются те вирусы, которые упоминаются в рамках данной заявки, а также другие вирусы, способные к поглощению определенной клеткой, и те вирусы, которые могут осуществлять выделение содержимого ядрышка в цитоплазму.
Восприимчивость клеточной линии к трансформации вирусом в качестве средства содействия заходу конъюгата в качестве "свободного вируса" зависит от присутствия и числа рецепторов на поверхности целевой клетки.
В число подходящих вирусов входят такие вирусы, которые способны к проникновению в клетку при помощи эндоцитоза в присутствии рецептора в качестве посредника во время трансфекции клеток комплексом нуклеиновой кислоты и к осуществлению своего выделения и тем самым выделения нуклеиновой кислоты из ядрышка в цитоплазму. Без претензий в части этой теории можно сказать, что в случае применения свободного вируса эта активность эндосомолиза могла бы оказать благотворное влияние на переносимый в клетку комплекс нуклеиновой кислоты, заключающееся в том, что эти комплексы транспортируются вместе с вирусами из ядрышка в цитоплазму, при этом предполагают, что они заходят в эти ядрышка, что и вирусы. Если комплексы содержат вирус в связанном виде, то они получают пользу от эндосомолитической активности вируса и транспортируются из ядрышек в цитоплазму. Этим достигается слияние между ядрышками и лизосомами и таким образом энзиматическая деградация, которая обычно имеет место в этих органеллах клетки.
В частности вирусами, которые пригодны для осуществления изобретения и поглотительная функция которых в начале инфекции осуществляется за счет эндоцитоза в присутствии рецептора в качестве посредника, являются вирусы без липидного покрытия, такие, как, например, аденовирус, полиовирус, риновирус, а также вирусы с оболочкой, такие как, например, вирус везикулярного стоматита, вирус Semliki Forest, вирус гриппа. Подходящими являются также зависящие от pH штаммы вируса Молонея. В частности, предпочитаются вирусы из группы, включающей аденовирус, подгруппу С, тип 5, вирус Semliki Forest, вирус везикулярного стоматита, полиовирус, риновирусы и вирус лейкоза Молонея.
Применение в рамках данного изобретения вирусов РНК, которые не имеют обратную транскриптазу, имеет то преимущество, что трансфекция в присутствии такого вируса не приводит к образованию вирусной ДНК в трансфектированной клетке. В результате опытов по изобретению было выявлено, что риновирус HRV2, представитель групп пикорнавируса, проявляет повышенную экспрессию репортерного гена. Эффективность риновируса проявлялась как в свободной форме, так и в виде конъюгатов.
В рамках данного изобретения под термином "вирус "(при условии, что он поглощается клеткой и высвобождает содержимое ядрышек, в которые он заходит) понимаются в дополнение к диким типам мутанты, которые в результате одной или больше мутаций потеряли определенные функции дикого типа (отличные от поглотительной функции), в частности способность к репликации.
Мутанты образуются в процессах обычного мутагенеза за счет мутаций в белковых областях, которые ответственны за репликационные функции и которые могут дополняться упаковочной линией. В случае аденовируса такими мутантами являются ts-мутанты (термочувствительные мутанты), мутанты Е1А и Е1В, мутанты, которые показывают мутации в стимулированных MLP генах, и мутанты, которые проявляют мутации в областях определенных капсидных белков. Также подходящими являются вирусные штаммы, которые имеют соответствующие естественные мутации. Способность вирусов к репликации можно исследовать известными из литературы методами. Так, например, на культуры клеток подают суспензии с различными концентрациями вируса и число растворенных клеток, которые можно видеть в виде пятен, регистрируют.
Другими вирусами, которые можно применять в рамках данного изобретения, являются так называемые дефектные вирусы, то есть вирусы, которые не обладают функцией, требуемой для автономной репликации вируса в одном или больше генах, для которых они требуют наличие вируса-помощника. Примерами такого типа вируса являются дефектные интерферирующие частицы, которые являются производными инфекционного стандартного вируса, имеют те же структурные белки, что и стандартный вирус, имеют мутации и нуждаются в стандартном вирусе в качестве вируса-помощника для осуществления репликации. Примерами такой группы вируса также являются сателлитные вирусы. Другой группой является класс парвовирусов, которые обозначаются как аденоассоциированные вирусы.
Так как циклы проникновения в клетки многих вирусов полностью не характеризованы, по-видимому, есть и другие вирусы, которые будут проявлять эндосомолитическую активность, требуемую для их пригодности в рамках данного изобретения. В рамках данного изобретения также пригодны живые вакцины или вакцинальные штаммы.
Под термином "вирус" понимаются в рамках данного изобретения и инактивированные вирусы, например, вирусы, инактивированные химической обработкой, такой, как, например, обработка формальдегидом, облучение УФ-лучами, химическая обработка в сочетании с облучением УФ-лучами, например, обработка псораленом и облучение УФ-лучами, облучение γ-лучами, бомбардировка нейтронами. Инактивированные вирусы, например, такие, которые также применяются для вакцин, можно получать известными из литературы методами, после чего их можно исследовать на пригодность повышения переноса комплексов ДНК. В рамках данного изобретения проводились опыты, в которых препараты аденовируса инактивировались при помощи стандартной УФ-стерилизационной лампы или обработкой формальдегидом. При этом неожиданно было найдено, что степень инактивации вирусов намного повышает степень снижения переноса гена, который достигается при добавлении аденовируса к среде трансфекции. В этих опытах применялись препараты инактивированного обработкой псораленом и облучением УФ-лучами биотинированного аденовируса, связанного с полилизином, связанным с стрептавидином. Эти опыты показывали, что инактивация привела к снижению титра вируса, который значительно превышает снижение способности к переносу гена. Это является четким указанием на то, что механизмы, связанные с нормальной репликацией в активном вирусе, можно разрушать без отрицательного влияния на эффект, необходимый для переноса гена.
Под термином "компоненты вируса" подразумеваются части вирусов, например, белковая часть свободного от нуклеиновой кислоты (пустой капсид вируса, который можно получать рекомбинантными способами), белки, получаемые фракционированием, или пептиды, которые имеют эндосомолитические функции интактного вируса. Эти компоненты вируса можно получать синтетическим путем, либо пептидным синтезом, либо рекомбинантными способами в зависимости от их размеров. В результате опытов в рамках данного изобретения было установлено, что белки аденовируса, связанные через биотин/стрептавидин с полилизином, позволяют улучшение переноса гена. Примером фрагментов или белков, отличных от аденовируса вирусов, которые являются существенными для обеспечения включения клеткой, является гемагглютинин (HA) вируса гриппа. N-концевая последовательность подединицы НА2 гемагглютинина вируса гриппа является ответственной для выделения вируса из ядрышка. Было установлено, что пептиды, состоящие из 20 аминокислот этой последовательности, способны к слиянию липидных мембран и к их частичному разрушению. В рамках данного изобретения с успехом применялись аутентичные и модифицированные пептиды вируса гриппа. Другим примером являются белки оболочки ретровирусов, например, gp41 HIV или части таких белков.
Применение вирусов, которые как таковые способны к проникновению в клетки и таким образом действуют в качестве содействующего поглощению клеткой фактора, является дальнейшим аспектом данного изобретения.
Вирусы или компоненты вирусов, которые сами по себе не могут связаться с клеткой и проникать в нее, предпочтительно используются в качестве вирусных конъюгатов в соответствии с вышеприведенным определением. В результате связывания с доменом, связывающим ДНК, например, с поликатионом, вирусу или компоненту вируса сообщается большое сродство к молекулам ДНК, так что он образует комплекс с ними и переносится в клетку в качестве компонента комплекса нуклеиновой кислоты, которая также содержит конъюгат содействующего поглощению клеткой фактора и связывающий ДНК домен. Кроме достигаемого при этом эффекта переноса связывание вируса или компонента вируса со связывающим нуклеиновую кислоту доменом может также привести к улучшению его эндосомолитических свойств.
Путем подбора другого содействующего поглощению клеткой фактора практически любая высшая эукариотная клетка может трансфектироваться предлагаемой системой.
В результате простого скрининга можно определять, проявляет ли данный вирус или компонент вируса поглотительную функцию и можно ли ее применять для осуществления данного изобретения. Такой анализ, например, для исследования вируса на его пригодность в качестве свободного вируса, целевые клетки приводят в контакт с комплексом ДНК в присутствии или отсутствии вируса. Количество комплекса ДНК, которое переносится в цитоплазму, можно затем простым образом определять путем выявления маркерного генного продукта, например, люциферазы. Если присутствие вируса приводит к тому, что комплекс ДНК поглощается и вводится в цитоплазму на большем уровне чем без вируса, то это можно приписывать поглотительной функции вируса. Также возможно сравнивать уровень поглощения комплекса ДНК в присутствии исследуемого вируса с другим вирусом, о котором известно подходящая поглотительная функция, например, с подгруппой С, типом 5, аденовируса. Таким опытам можно также подвергать вирусные конъюгаты. В этих опытах можно также исследовать дополнительные параметры, такие, как, например, конъюгаты с различным содействующим поглощению клеткой фактором в различных количествах. Кроме того, специалист в данной области может без труда проводить такого вида опыты, в случае необходимости в сочетании с другими опытами, например, с опытами по определению утечки липосома, с тем, чтобы исследовать компоненты вируса и другие компоненты с потенциальной эндосомолитической активностью на их способность к улучшению экспрессии гена.
Если применяют интактные вирусы, то проводят опыты, направленные на выявление способности вируса к репликации. Эти опыты предпочтительно проводят параллельно с предварительными опытами, направленными на выявление способности вируса к улучшению переноса гена. Исследование на способность к репликации проводят известными методами, такими как, например, метод с образованием пятен, метод по определению цитопатогенного действия или метод по определению поздней экспрессии гена в случае цитопатогенных вирусов или вирусов, которые в значительной степени содействуют росту клеток-хозяев. Для других вирусов применяют специфичные для данного вируса методы, например, метод по определению гемагглютинации, или химико-физические методы, например с применением электронного микроскопа.
В рамках данного изобретения в качестве предпочтительных вирусов, в частности таких, которые применяются в качестве свободных вирусов, применяют вирусы, которые можно получать с высоким титром, которые устойчивы, имеют низкую патогенность в нативном состоянии и, в случае необходимости позволяют упразднение способности к репликации. В частности предпочитаются аденовирусы. Если трансфекции подлежит специфичная клеточная популяция, то можно применять вирусы, которые проявляют специфичную способность к этой популяции. Если трансфекции подлежат различного типа клетки, то можно применять вирусы, которые являются инфекционными для широкого круга клеток.
Требования к композициям свободного вируса в основном заключаются в том, что вирус должен иметь наибольшую чистоту и что следует применять стабилизирующий буфер, выбираемый с учетом данного вируса.
В любом случае согласно изобретению можно применять только такие вирусы или компоненты вирусов, у которых риск безопасности является как можно минимальным, в частности риск репликации вируса в целевой клетке и рекомбинации вирусной ДНК с ДНК хозяина.
Преимущественно использовать механизм проникновения в клетки вирусов, инфицирующих животных по-другому, чем людей, для улучшения поглощения и переноса ДНК в высшие эукариотные клетки, в частности людей, пока вирус проявляет активность по разрушению ядрышка в высших эукариотных клетках. Члены семейства аденовирусов были выделены из птиц, амфибий и ряда других животных. Аденовирусы из птиц, амфибий, крупного рогатого скота, собак, мышей, овец, свиней и обезьян, а также аденовирусы человека можно получать от американской коллекции типовых культур в городе Роквиль в штате Мариленд.
Одно возможное преимущество применения вируса, например, аденовируса, из редкого вида может заключаться в уменьшенной токсичности в целевых клетках (например, от аденовируса куриц или лягушек нельзя ожидать репликации или раннего инициирования экспрессии гена в клетках млекопитающих), уменьшенной опасности для лица, занимающегося получением редкого аденовируса, по сравнению с аденовирусом человека, и уменьшенной интерференции в целевом организме от антител против аденовируса человека или мышей. Отсутствие интерференции антителами человека или мышей является особенно важным при применении вирусов в генной терапии на людях и мышах.
Крупный аденовирус CELO не проявляет реактивности в отношении антител, которые распознают основную группу эпитопов аденовирусов, инфицирующих клетки млекопитающих. Кроме того, аденовирус CELO можно выращивать в оплодотворенных яйцах с получением высокого выхода (0,5 мг/яйцо). В экспериментальной части показывается, что конъюгаты CELO и полилизина улучшают перенос ДНК в клетки HeLa, на уровне, который можно сравнивать с уровнем, достигаемым при применении аденовируса d1312 человека. Таким образом, применение конъюгатов CELO для улучшения транспорта ДНК можно рассматривать как многообещающий подход в экспериментах по генной терапии на людях. Вирусы редких видов предпочтительно используются в качестве компонентов вирусных конъюгатов в комбинированных комплексах согласно изобретению.
Связывание вируса со связывающим нуклеиновую кислоту доменом в вируссодержащих конъюгатах согласно изобретению может быть ковалентным или же нековалентным, например, при помощи биотино- стрептавидинового мостика. Возможно также ионное связывание в случае вируса с кислыми участками в поверхностных белках, что позволяет связь с поликатионом.
В рамках данного изобретения образовались комплексы в условиях, позволяющих ионное взаимодействие между аденовирусом и полилизином до образования комплекса с ДНК. При этом также проводились контрольные опыты в условиях, в которых полилизин сначала нейтрализуют ДНК и поэтому он не может связывать аденовирус. Комплексы с ионным связыванием аденовируса показывали лучшие результаты.
Примерами вирусных компонентов в эндосомолитических конъюгатах согласно изобретению являются пустые вирусные капсиды и вирусные пептиды. Связывание вирусного компонента со связывающим нуклеиновую кислоту доменом может быть ковалентным, например, путем химического связывания вирусного пептида с полилизином, или же нековалентным, например, ионным в случае вирусного компонента с кислотными остатками, которые связываются с поликатионом.
Соотношение вируса или компонента вируса и вещества со сродством для нуклеиновой кислоты можно варьировать в широких пределах. В случае конъюгата пептида гемагглютинина гриппа и полилизина перенос гена можно улучшать в большем размере при высоком содержании в конъюгатах вирусного пептида.
Конъюгаты вируса или вирусного компонента и вещества со сродством нуклеиновой кислоты можно получать (также как и конъюгаты содействующего поглощению клеткой фактора и поликатиона) путем связывания компонентов или же при помощи рекомбинантного метода, если вирусные компоненты и связывающий нуклеиновую кислоту домен представляют собой полипептиды.
Связывание вируса или вирусных белков или пептидов с полиаминами химическими методами можно осуществлять известным для связывания пептидов способом, причем, если требуется, отдельные компоненты перед реакцией связывания снабжают связывающими веществами (это мероприятие необходимо тогда, когда не имеется подходящей для связывания функциональной группы, например, меркаптогруппы или спиртовой группы). Связывающие вещества являются бифункциональными соединениями, которые сначала вступают в реакцию с функциональными группами отдельных компонентов, после чего осуществляют связывание модифицированных отдельных компонентов.
Связывание можно осуществлять с помощью
а) дисульфидных мостиков, которые в восстанавливающих условиях могут снова расщепляться (например, при применении сукцинимидил- пиридилдитиопропионата);
б) в основном стойких в биологических условиях соединений (например, простых тиоэфиров, реакцией малеимидо-связывающих веществ с сульфгидрильными группами связанного со вторым компонентом связывающего вещества);
в) неустойчивых в биологических условиях мостиков, например, сложноэфирных связей, или неустойчивых в слабокислых условиях ацетальных или кетальных связей.
В рамках данного изобретения эндосомолитические пептиды гемагглютинина НА2 гриппа связывались с полилизином химическим путем с применением сукцинимидил-пиридилдитиопропионата. Было установлено, что модификация пептида полилизином приводит к повышению эндосомолитической активности. Опыты по трансфекции показывали, что эффективность переноса гена в присутствии конъюгата трансферина и полилизина в качестве посредника значительно повышается, если комплекс ДНК кроме трансферина/полилизина содержит еще конъюгаты пептида вируса гриппа и полилизина.
В рамках данного изобретения аденовирус связывался с полилизином самыми различными методами. Один метод образования конъюгата вируса и полилизина осуществлялся аналогично образованию конъюгатов трансферина и полилизина (см. Вагнер и др., Proc. Natl. Acad. Sci., США 87, 1990, стр. 3410 - 3414) после модификации дефектного аденовируса d1312 с применением гетеробифункционального реагента. Несвязанный полилизин удалялся путем центрифугирования. Связывающая способность ДНК исследовалась с применением радиоактивно меченой ДНК (при применении клеток К562 было установлено, что в отсутствие хлорокрина значительно больший перенос гена достигается в присутствии комплексов, состоящих из ДНК, аденовируса, связанного с полилизином, и трансферина, связанного с полилизином, чем в присутствии немодифицированного аденовируса, который не связан с ДНК. Кроме того, было найдено, что значительная экспрессия гена имела место уже в присутствии 0,0003 мкг/ДНК в 5•105 клеток HeLa с применением модифицированного полилизином аденовируса).
Если вирус или вирусный компонент (или дополнительный, содействующий поглощению клеткой фактор, как, например, в случае трансферина) содержит подходящие углеводные цепи, их можно связывать с веществом, имеющим сродство для нуклеиновой кислоты, через одну или больше углеводных цепей гликопротеина.
Другой предпочтительный метод получения вирусных конъюгатов согласно изобретению заключается в энзиматическом связывании вируса или вирусного компонента с веществом со сродством для нуклеиновой кислоты, в особенности полиамина, при помощи трансглютаминазы.
Категория трансглютаминаз включает ряд различных ферментов, которые имеются, например, в эпидермисе (эпидермальная трансглютаминаза), крови (фактор XIII) и в клетках различных тканей (тканевая трансглютаминаза, например, трансглютаминаз печени).
Трансглютаминазы катализуют образование связей ε-(γ-глютамил)лизина в присутствии кальциевых ионов с расщеплением аммиака. Предпосылкой является наличие глютаминов и лизинов в белках, способных к взаимодействию с ферментом. Кроме ε-аминогрупп лизина в качестве субстрата можно также применять (поли)амины, такие, как, например, этаноламин, путресцин, спермин и спермидин.
До сих пор еще не ясно, какие факторы являются ответственными за то, что глютамин или лизин белка или полиамин вступает в реакцию с ферментом. Известно лишь то, что полиамины можно связывать с многочисленными клеточными белками, такими, как, например, цитокератины, тубулин, белками мембраны клеток, поверхностными белками вирусов гриппа при помощи трансглютаминазы.
В рамках данного изобретения было установлено, что полилизин может связываться с аденовирусами при помощи трансглютаминазы. При этом процесс связывания можно проводить в присутствии глицерина, что имеет то преимущество, что препарат вируса, например аденовируса, можно подавать непосредственно на связывание, так как содержащийся в нем буфер включает глицерин. Применение конъюгатов аденовируса и полилизина, имеющихся в виде комплекса с плазмидной ДНК и конъюгатами трансферина и полилизина, позволяет экспрессию гена, которая во много раз превышает действие, достигаемое при применении конъюгатов трансферина и полилизина в присутствии не связанного с полилизином аденовируса.
Другой предпочтительный метод получения конъюгатов согласно изобретению заключается в связывании вируса или его компонента с поликатионом через биотиново-белковый мостик, предпочтительно через биотиново-стрептавидиновый мостик.
Известная сильная связь биотина со стрептавидином или авидином используется для связывания адиновируса с полилизином путем модификации аденовируса биотином и химического связывания стрептавидина с полилизином.
Комплексы, состоящие из ДНК и конъюгата стрептавидина и полилизина, с которыми связан модифицированный биотином вирус, и, в случае необходимости не ковалентно связанного полилизина, проявляют очень высокую эффективность трансфекции, даже при низких концентрациях ДНК. Особенно эффективные комплексы образуются за счет того, что модифицированный биотином вирус сначала связывают с конъюгатом стрептавидина и полилизина, после чего осуществляют связывание ДНК. Связывание с биотином можно также осуществлять при помощи авидина.
Связь между вирусом или его компонентом и полилизином можно также осуществлять следующим образом. Вирус биотинилируют, антитела против биотина связывают с полилизином и связь между биотином и антителом используют для связывания вируса и полилизина. При этом используют общедоступные стандартные поликлональные или моноклональные антитела против биотина.
Связывание вируса с полилизином можно также осуществлять за счет связывания полилизина с лектином, имеющим сродство для поверхностного гликопротеина вируса, при этом процесс связывания в таком конъюгате осуществляют при помощи связи между лектином и гликопротеином. Если вирус не имеет подходящих боковых углеводных цепей, то его можно модифицировать соответствующим образом.
Вирус можно также связывать с веществом со сродством для нуклеиновой кислоты за счет того, что поверхность вируса модифицируют антигеном, чужеродным для вируса (например дигоксигенином, продуктом инофирмы Берингер Маннхайм, DE, или биотином) и связь между модифицированным вирусом и веществом со сродством для нуклеиновой кислоты устанавливают через антитело, связывающееся с антигеном. Конкретное осуществление процесса образования конъюгатов согласно изобретению зависит от различных критериев. Так, например, связывание при помощи биотина представляет собой меньше всего специфичный и поэтому широко применяемый метод, при этом связывание при помощи биотина приводит к образованию очень сильной нековалентной связи. Энзиматическая реакция с трансглютаминазой имеет то преимущество, что его можно также проводить в очень малом масштабе. К химическому связыванию обычно прибегают тогда, когда необходимо синтезировать большие количества конъюгата. Этот метод обычно является и наилучшим для связывания протеинов или пептидов вируса. Если применять инактивированные вирусы, то инактивацию обычно осуществляют до связывания при условии, что инактивация отрицательно не сказывается на связывании.
Если вирус, например, аденовирус, или его компонент с эндосомолитической активностью, имеет доступные связывающие домены, например кислые домены для связывания с поликатионом, то связывание вируса или его компонента с поликатионом можно также осуществлять ионным путем. В этом случае положительные заряды поликатиона, который может иметься в виде конъюгата с содействующим поглощению клеткой фактором, частично нейтрализуются кислым доменом вируса или его компонента, а остальные положительные заряды нейтрализуются нуклеиновой кислотой.
Если вещество со сродством для нуклеиновой кислоты представляет собой вещество интеркаляции, то его модифицируют связывающим веществом, подходящим в процессе связывания конкретного вируса или его компонента. Так, например, в случае связывания при помощи трансглютаминазы его модифицируют спермином или бифункциональной группой, пригодной для осуществления химического связывания, например активной сложноэфирной группой.
Соотношение вируса или его компонента и связывающих нуклеиновую кислоту веществ может колебаться в широких пределах. Обычно его определяют эмпирическим путем, например, связывания определенного количества вируса или его компонента с различным количеством полилизина. При этом оптимальный конъюгат используют для трансфекции.
Согласно другому варианту изобретения компонент вируса, например вирусный пептид с эндосомолитической активностью, можно модифицировать с таким расчетом, что его можно непосредственно связывать с ДНК. В этом случае сам пептид может содержать связывающий ДНК домен, получаемый за счет синтеза пептида и включения в него, предпочтительно за счет удлинения, положительно зараженных аминокислот, предпочтительно на C-конце.
Согласно другому варианту изобретения эндосомолитический агент представляет собой невирусный, в случае необходимости синтетический пептид. Если предлагаемая система содержит пептид такого типа, то он предпочтительно ионным путем связан с веществом со сродством для нуклеиновой кислоты, например с полилизином в случае наличия комплексов ДНК, содействующего поглощению клеткой фактора и полилизина. При этом включение эндосомолитического пептида в комплекс нуклеиновой кислоты осуществляют за счет связывания с положительно заряженным связывающим нуклеиновую кислоту доменом предпочтительно полилизином, через его кислые аминокислотные остатки.
В зависимости от химической структуры пептида, в частности его концевой группы, связывание с полилизином можно также осуществлять методами, используемыми в рамках данного изобретения для связывания пептидов с полилизином. Если применять естественный пептид, то его можно модифицировать подходящей концевой аминогруппой с тем, чтобы сделать его пригодным для осуществления связывания.
Другой метод включения невирусных эндосомолитических пептидов в комплексы нуклеиновой кислоты заключается в том, что их снабжают последовательностями, способными к связыванию с ДНК. Такая последовательность должна находиться в том месте, где она отрицательно не влияет на эндосомолитическую активность пептида. Поэтому, например, пептиды, N-конец которых ответствен за эту активность, удлиняют связывающими ДНК последовательностями на C-конце. Такими последовательностями могут быть гомологовые или гетерогенные катионные олигопептиды, например, олиголизиновый хвост, или естественный связывающий ДНК домен, например пептид, производимый от гистона. Эти связывающие ДНК последовательности в качестве интегральной части эндосомолитического пептида предпочтительно содержат примерно 10 - 40 аминокислот. Этот вариант изобретения позволяет повысить соотношение эндосомолитиеской последовательности и связывающей ДНК последовательности по сравнению с конъюгатами пептида с большим содержанием поликатиона с тем, чтобы достичь большей эффективности комплексов.
Невирусные эндосомолитические пептиды должны отвечать следующим требованиям. В отношении эндосомолитической активности утечка липидных мембран, достигаемая пептидом, предпочтительно должна быть больше при низкой величине pH (5 - 6), чем при величине pH 7. Кроме того, разрушенные зоны мембраны должны иметь размер, достаточный для обеспечения прохода комплексов ДНК (небольшие поры являются недостаточными). Для определения того, отвечает ли конкретный пептид этим требованиям, можно проводить опыты в пробирке за счет того, что пептиды применяют в свободном или связанном виде и/или в включенном в комплексе ДНК виде. В таких опытах с применением липосом, эритроцитов или клеточных культур определяют увеличением экспрессии гена. Такие опыты являются объектом примеров осуществления изобретения. Оптимальное количество пептида можно определять в рамках предварительных титрований путем анализа эффективности получаемого переноса гена. Следует учесть, что эффективность различных пептидов и оптимальный состав комплекса зависят от типа клетки.
Разрушающие мембрану пептиды обычно содержат амфипатические последовательности, а именно гидрофобный участок, который может взаимодействовать с липидной мембраной, и гидрофильный участок стабилизирует водную фазу в сайте разрушения мембраны.
В природе существуют некоторые примеры разрушающих мембрану пептидов, обычно небольших пептидов или пептидных доменов больших полипептидов. Такие пептиды можно классифицировать в соответствии с их функцией в природном контексте, а именно либо на разрушающие мембрану пептиды (например, пептиды обнаженных вирусов) и/или на вызывающие слияние мембран пептиды (например, вирусы с оболочкой). В случае синтетических пептидов для осуществления разрушения ядрышек пригодны оба класса пептидных последовательностей. Большинство естественных пептидов способно к образованию амфипатических α-спиралей.
Специфичность в отношении величины pH можно получать путем включения кислых остатков в гидрофильный участок предполагаемой амфипатической альфа-спирали с таким расчетом, что спираль может образоваться только при кислой величине pH, но не при нейтральной величине pH, при которой отталкивание между отрицательно зараженными кислыми остатками предотвращает образование спирали. Это свойство также обнаружено в случае имеющихся в природе последовательностей (например у N-конца вируса гриппа НА-2).
Полностью синтетический амфипатический пептид со способностью к разрушению мембраны в зависимости от величины pH известный. Этот пептид (в свободном виде) обеспечивает образование только небольших пор в мембранах, которые позволяют только высвобождение соединений небольшой величины. В рамках варианта изобретения, согласно которому используют невирусные, в случае необходимости синтетические, пептиды, обычно осуществляют следующие операции. Амфипатическую пептидную последовательность выбирают из группы имеющихся естественных или искусственных пептидов, известных специалисту. В случае необходимости включают кислые остатки (Gin, Asp) с тем, чтобы сделать разрушающую мембрану активность пептида более специфичной в отношении величины pH (двойной кислый мутант гемагглютининового пептида гриппа согласно примеру 35, обозначенный р50). В случае необходимости кислые остатки можно также включать с тем, чтобы улучшить способность пептида к связыванию с полилизином. Такой связывающий с поликатионом домен можно получать за счет включения C-концевых кислых удлинений, например, олиго-Glu-хвоста.
Пригодные для осуществления данного изобретения эндосомолитические пептиды можно также получать за счет слияния имеющихся естественных и искусственных последовательностей. В рамках данного изобретения проводились опыты с применением различных пептидов, которые производились от синтетического пептида GALA. Некоторые производные, которые применялись в этих опытах, получались за счет сочетания пептида GALA или его модификаций с последовательностями пептида гриппа или его модификаций, например с пептидами, обозначенными EALA-Inf и EALA-Р50 согласно примеру 35.
Длина последовательности пептида может быть критической в отношении стабильности амфипатической спирали. Повышение стабильности коротких доменов, которые производятся от естественных белков, которым не хватает стабилизирующих белковых участков, можно обеспечивать за счет удлинения спирали.
Эндосомолитическую активность пептидов можно повышать за счет образования гомодимеров, гетеродимеров или олигомеров. В результате проведения опытов по изобретению выявилось, что димер Р50 проявляет намного большую активность чем мономер.
Кроме того, опыты также показывали действие синтетических пептидов на поглощение ДНК при помощи конъюгатов трансферина и полилизина. При этом синтезировались различные пептиды, которые исследовались на способность к утечке липосом и эритроцитов, а также на их действие на экспрессию люциферазы в клетках TIB 73 и NIH 3Т3.
В качестве эндосомолитического агента можно также применять непептидное амфипатическое вещество. Требования к пригодности такого вещества для применения в рамках данного изобретения являются в основном теми же самыми, что и в случае амфипатических пептидов, а именно способность включаться в комплекс нуклеиновой кислоты, специфичность в отношении pH и так далее.
Другой вариант изобретения относится к комплексам, содержащим нуклеиновую кислоту и конъюгат, способный к образованию комплекса с нуклеиновой кислотой, которые служат для переноса нуклеиновой кислоты в высшие эукариотные клетки. Комплексы характеризуются тем, что конъюгат состоит из вещества со сродством для нуклеиновой кислоты и эндосомолитического агента, связанного с веществом со сродством для нуклеиновой кислоты, причем эндосомолитический агент способен к включению в клетку в качестве части комплекса конъюгата и нуклеиновой кислоты и к высвобождению содержимого ядрышек, в которых комплекс находится после захода в клетку, в цитоплазму.
В рамках данного изобретения предпочтительно применяют такие комплексы, в которых нуклеиновая кислота связана с веществом со сродством для нуклеиновой кислоты так, что комплексы являются в основном электронейтральными. Согласно предпочтительному варианту изобретения в качестве эндосомолитического агента используют вирус или его компонент, ковалентно связанный с поликатионом.
Используемые в рамках данного изобретения эндосомолитические конъюгаты также охватывают (дополнительно к конъюгатам, в которых эндосомолитические агенты ионным путем связаны со связывающим ДНК доменом) эндосомолитические агенты, которые непосредственно связываются с ДНК, например через их основное удлинение, хотя такого рода "конъюгат", строго говоря, не получается за счет сопряжения связи, то есть за счет связывания двух соединений друг с другом. Функция такого типа эндосомолитических агентов в качестве компонента предлагаемой системы является независимой от того, синтезированы ли они путем сопряжения эндосомолитического агента и связывающего ДНК домена или содержится ли первоначально в нем связывающий ДНК домен.
Согласно другому предпочтительному варианту изобретения кроме эндосомолитического конъюгата комплексы содержат другой конъюгат, в котором вещество со сродством для нуклеиновой кислоты, в случае эндосомолитического конъюгата поликатиона обычно тот же поликатион, что и в конъюгате, сопрягают с содействующим поглощению клеткой фактором, проявляющим сродство для целевой клетки. Этот вариант изобретения используют, в частности, тогда, когда целевая клетка не имеет или же имеет лишь немногие рецепторы для вируса, применяемого в качестве части эндосомолитического конъюгата. Этот вариант изобретения употребляется и тогда, когда используют компонент вируса, например имеющийся в природе, возможно модифицированный пептид, невирусный, в случае необходимости синтетический эндосомолитический пептид или вирус несвойственного вида, которые не обладают способностью к проникновению в клетки, которые подлежат трансфекции. Присутствие дополнительного конъюгата содействующего поглощению клеткой фактора и связывающего фактора приводит к улучшению характеристики эндосомолитических конъюгатов за счет того, что они образуют комплексы с нуклеиновой кислотой вместе со вторым конъюгатом и переносятся в клетку в качестве части результирующего комплекса, который в нижеследующем обозначается как "комбинационный комплекс" или "тройной комплекс". Предполагается, что комбинационные комплексы поглощаются клетками либо за счет связывания с поверхностным рецептором, специфичным для дополнительного содействующего поглощению клеткой фактора или, например, в случае применения вируса или его компонента, за счет связывания с рецептором вируса или же за счет связывания с обоими рецепторами, причем поглощение имеет место в результате эндоцитоза, вызываемого рецептором. Когда эндосомолитические агенты высвобождаются из ядрышек, ДНК, которая содержится в комплексах, также выделяется в цитоплазму, при этом она избегает лизосомальной деградации.
В опытах, которые проводились в соответствии с данным изобретением, почти все клетки HeLa поддались трансфекции свободным аденовирусом. В случае гепатоцитов эффективность можно было еще далее улучшать за счет использования тройных комплексов ДНК, в которых репортерная ДНК имеется в виде комплекса с конъюгатами полилизина и трансферина и связана с аденовирусом. В этом случае обеспечивается наличие эндосомолитического вируса и комплекса лиганда и рецептора в ядрышке. При этом почти во всех клетках указанного типа, например BNL. -CL2 и HepG2, имеет место трансфекция. В этом случае имеющиеся на поверхности клетки рецепторы как для вируса, так и для трансферина, могут захватывать тройные комплексы ДНК. Однако также возможно, что тройные комплексы ДНК могут проникать в клетку исключительно за счет действия совокупности клеточного лиганда и рецептора. Такая возможность исследовалась в опытах, в которых содержащие трансферин тройные комплексы ДНК проникали в клетки К562 в основном через рецептор трансферина, нежели через рецептор аденовируса.
Неожиданным образом было выявлено, что тройные комплексы переносили ДНК в клетку даже в случае очень низких концентраций ДНК. Так, например, в случае 30 пг ДНК/3 • 105 клеток получается 1,8 • 104 световых единиц (получаемых в результате экспрессии плазмиды, кодирующей люциферазу). Такая концентрация означает 60 молекул ДНК и 1 бляшкообразующую единицу вируса на клетку. Следовательно, данное изобретение обеспечивает эффективную трансформацию высших эукариотных клеток очень малыми количествами ДНК.
Присутствие вирусов, их компонентов или невирусных эндосомолитических агентов в комплексах ДНК в качестве компонентов эндосомолитических конъюгатов имеет следующие преимущества.
1. Широкая применимость технологии переноса генов с применением комплексов нуклеиновой кислоты, так как сам эндосомолитический агент, в частности в случае использования вируса или его компонента, может представлять собой содействующий поглощению клеткой фактор или же может также переводиться в комплекс с ДНК и с другим содействующим поглощению клеткой фактором, например, трансферином, асиалофетуином. Таким путем возможно использование положительного эффекта вирусов даже в случае таких клеток, которые не имеют рецептора данного вируса.
2. Улучшение эффективности переноса генов, так как связывание эндосомолитических конъюгатов с ДНК обеспечивает то, что они вместе проникают в клетки. Согласованный процесс поглощения и высвобождения вирусов и ДНК также позволяет снизить количества ДНК и вирусов, необходимые для эффективного переноса генов, что является крайне важным при применении предлагаемой системы в живом организме.
Под термином "содействующий поглощению клеткой фактор" в рамках данного изобретения понимаются лиганды или их фрагменты, которые после связывания с клеткой проникают в нее в результате эндоцитоза, предпочтительно вызываемого рецептором, или же факторы, связывание и проникновение которых осуществляют путем слияния с элементами клеточной мембраны.
Подходящими содействующими поглощению клеткой факторами являются лиганды трансферин, кональбумин, асиалогликопротеины (такие, как, например, асиалотрансферин, асиалорозомуцоид и асиалофетуин), лектины, вещества, которые содержат галактозу и проникают в клетки при помощи рецептора асиалогликопротеина, маннозилированные гликопротеины, лизосомальные энзимы, LDL, модифицированный LDL, липопротеины, которые проникают в клетки при помощи рецепторов (apo B100/LDL), вирусные протеины, такие, как, например протеин gp120 СПИДа, антитела или их фрагменты против поверхностных антигенов клетки, как, например, анти-CD4, анти-CD7, цитокины, такие, как, например интерлейкин-1, интерлейкин-2, фактор опухолевого некроза, интерферон, стимулирующий колонии фактор, факторы роста, такие, как, например инсулин, фактор роста эпидермиса, выделяющийся из тромбоцитов фактор роста, фактор β трансформирующего роста, фактор роста нервов, подобный инсулину фактор I роста, пролан Б, пролан А, гормон роста, пролактин, глюкагон, тироидные гормоны, α-2-макроглобулиновая протеаза и "бесплечевые" токсины. Другими примерами являются иммуноглобулины или их фрагменты в качестве лигандов для рецептора Fc или же антитела против иммуноглобулинов, которые связываются с поверхностными иммуноглобулинами. Лиганды могут быть естественного или синтетического происхождения.
В нижеследующем приводятся основные требования, предъявляемые к пригодности вышеприведенных факторов в рамках данного изобретения.
а) Они могут поглощаться клеткой, в которую следует перенести нуклеиновую кислоту, причем их способность к проникновению не ухудшается или же лишь в незначительной степени ухудшается в случае сопряжения со связывающим фактором.
б) Они способны к переносу нуклеиновой кислоты в клетку путем метода "ношения на спине" по любому пути, который они выбирают.
Нижеследующая экспериментальная часть иллюстрирует широкие возможности применения содействующего поглощению клеткой фактора или дополнительного содействующего поглощения клеткой фактора в рамках осуществления данного изобретения на примере конъюгатов трансферина и полилизина, комплексов асиалофетуина или галактозы и конъюгатов трансферина и полилизина, конъюгатов агглютинина пшеничного зачатка, комплексов конъюгата трансферина и полилизина и специфичного в отношении Т-клеток белка gp120, комплексов конъюгатов трансферина и полилизина и анти-СD7, также на примере комплексов полилизина и ДНК, которые не содержат содействующего поглощению клеткой фактора. Кроме того, возможность осуществления изобретения при применении вирусных конъюгатов иллюстрируется на примере комплексов ДНК и сопряженного с полилизином вируса или его компонента, которые не содержат конъюгата дополнительного содействующего поглощению клеткой фактора и связывающего фактора.
Возможность или необходимость применения содействующего поглощению клеткой фактора в случае применения в качестве эндосомолитического агента свободного вируса или же дополнительного содействующего поглощению клеткой фактора в случае применения в качестве эндосомолитического агента вируса или компонента вируса или невирусного пептида, являющегося частью эндосомолитического конъюгата, для обеспечения или улучшения процесса поглощения клеткой комплексов нуклеиновой кислоты можно выявлять за счет проведения соответствующих предварительных опытов. В этих опытах осуществляют параллельные трансфекции комплексами нуклеиновой кислоты, в одном случае без (дополнительного) содействующего поглощению клеткой фактора, например в случае комплексов нуклеиновой кислоты и вирусного конъюгата, а в другом случае применяют комплексы нуклеиновой кислоты с другим конъюгатом, состоящим из дополнительного содействующего поглощению клеткой фактора, для которого целевые клетки имеют соответствующий рецептор, и вещества со сродством для нуклеиновой кислоты.
Применение содействующего поглощению клеткой фактора или дополнительного содействующего поглощению клеткой фактора (в случае применения комбинационного комплекса) определяется, в частности, целевой клеткой, например, специфичными для данной клетки поверхностными антигенами или рецепторами, которые таким образом позволяют желаемый перенос нуклеиновой кислоты в данную клетку.
Пригодными в рамках данного изобретения веществами со сродством для нуклеиновой кислоты являются, например, гомологовые органические поликатионы, такие, как, например полилизин, полиаргинин, полиорнитин, гетерогенные поликатионы, имеющие по меньшей мере две аминокислоты с различным положительным зарядом, при этом они могут также иметь различную длину цепи, а также непептидные синтетические поликатионы, такие, как, например полиэтиленимин. Другими пригодными веществами со сродством для нуклеиновой кислоты являются естественные связывающие ДНК белки поликатионной природы, такие, как, например гистоны, протамины или их аналоги или фрагменты, а также спермин или спермидин.
Длина поликатиона не является критической при условии, что комплексы являются в основном электронейтральными. Полилизин имеет предпочтительную длину цепи, составляющую примерно 20 - 1000 лизиновых мономеров. Однако для данной длины ДНК не существует критической длины поликатиона. Так, например, если ДНК состоит из 6000 пар оснований (далее "п.о.") и 12000 отрицательных зарядов, то на моль ДНК применяют следующие количества поликатиона:
60 моль полилизина с молярным весом 200
30 моль полилизина с молярным весом 400
120 моль полилизина с молярным весом 100, и так далее.
Любой специалист может подобрать поликатион с подходящей длиной и в подходящем мольном количестве за счет проведения стандартных опытов с учетом вышесказанного.
Другими подходящими веществами со сродством для нуклеиновой кислоты в качестве компонента конъюгатов являются проявляющие интеркаляцию вещества, такие, как, например этидиевые димеры, акридин, или же проявляющие интерколяцию пептиды, содержащие триптофан и/или тирозин и/или фенилаланин.
Для составления качественного состава комплексов нуклеиновой кислоты обычно сначала определяют нуклеиновую кислоту, которая подлежит переносу в клетку. В первую очередь нуклеиновую кислоту выбирают с учетом биологического действия, которое должно проявляться в клетке, а в случае использования изобретения для генной терапии, - с учетом гена или участка гена, который подлежит экспрессии, например для замены дефектного гена, или же с учетом последовательности гена, которая подлежит торможению. Нуклеиновые кислоты, которые должны переноситься в клетку, могут быть ДНК или РНК, так как при этом не существует ограничение в отношении нуклеотидной последовательности.
Если применять опухолевые клетки в качестве раковой вакцины, то подлежащая введению в клетку ДНК предпочтительно кодирует модулирующее иммунную систему вещество, например, цитокин, как интерлейкин (далее: "ИЛ")-2, ИЛ-4, γ-интерферон (далее: "ИФ") α-фактор опухолевого некроза (далее: "ФОН"). Возможно также применять комбинации кодирующих цитокин ДНК, например ИЛ-2 и γ-ИФ. Другим полезным геном, который можно вводить в опухолевые клетки, может являться устойчивый ко многим лекарственным средствам ген (далее: "mdr").
Также возможно вводить в клетку две или больше различные последовательности нуклеиновой кислоты, например плазмидсодержащие кДНК, кодирующие два различных белка с управлением подходящими регуляторными последовательностями, или два различных плазмидных конструкта, которые содержат различные кДНК.
Терапевтически активные ингибирующие нуклеиновые кислоты, которые переносятся в клетки с тем, чтобы тормозить специфичные генные последовательности, могут представлять собой генные конструкты, которые служат для транскрипции анти-мРНК или рибозимов. Кроме того, возможно также введение в клетку олигонуклеотидов, например анти-м-олигонуклеотидов. Анти-м-олигонуклеотиды включают предпочтительно по меньшей мере 15 нуклеотадов. Олигонуклеотиды могут также представлять собой мультимеры. Если включению в клетку подлежат рибозимы, то они предпочтительно применяются в качестве компонента генного конструкта, включающего стабилизирующие генные элементы, например элементы гена т-РНК.
Кроме молекул нуклеиновой кислоты, которые тормозят гены, например вирусные гены, можно также применять гены, проявляющие различный образ тормозящего действия. В качестве примеров можно назвать гены, кодирующие вирусные белки, которые имеют так называемые транс-доминантные мутации.
В результате экспрессии генов в клетке получаются белки, которые доминируют соответствующий белок такого типа и, таким образом, защищают клетки, которые приобретают "клеточную иммунность" путем торможения вирусной репликации.
Подходящими являются транс-доминантные мутации вирусных белков, которые требуются для репликации и экспрессии, например Gag-, Tat- и Rev-мутанты. Последние показывали торможение репликации ретровируса СПИДа. Другой механизм достижения внутриклеточной иммунности заключается в экспрессии молекул РНК, включающих сайт связывания существенного вирусного белка, например, так называемые приманки TAR.
Примерами генов, которые можно применять при соматической терапии и которые можно переносить в клетки в качестве компонента генных конструктов, являются фактор VIII (гемофилия А), фактор IX (гемофилия Б), деаминаза аденозина, α-1 антитрипсин (эмфиземии легких) или регуляторный ген трансмембранной проводимости муковисцидоза.
Величина нуклеиновых кислот может колебаться в широких пределах. Так, например, генные конструкты размером примерно 0,15 кило пар оснований (далее: "к.п.о.") (в случае гена т-РНК, включающего рибозим) до примерно 50 к. п.о. или больше можно с успехом переносить в клетки при помощи предлагаемой системы. Небольшие молекулы нуклеиновой кислоты можно применять в качестве олигонуклеотидов. В связи с этим следует отметить, что применение предлагаемой системы не ограничено ни в отношении последовательности генов, ни в отношении их размера.
После выбора нуклеиновой кислоты подбирают вещество со сродством для нуклеиновой кислоты, предпочтительно органическое поликатионное вещество, с тем, чтобы обеспечить образование комплекса нуклеиновой кислоты, при этом получаемый комплекс должен быть предпочтительно в основном электронейтральным. Если комплексы включают конъюгат дополнительного содействующего поглощению клеткой фактора и вещества со сродством для нуклеиновой кислоты, то поликатионный компонент обоих конъюгатов подбирают с учетом вопроса электронейтральности.
Известно, что оптимальный перенос нуклеиновой кислоты в клетку может достигаться тогда, когда соотношение конъюгата и нуклеиновой кислоты выбирают с таким расчетом, что комплексы содействующего поглощению клеткой фактора и поликатиона/нуклеиновой кислоты являются в основном электронейтральными. Было найдено, что количество поглощаемой клеткой нуклеиновой кислоты не снижается, если некоторое количество конъюгата трансферина и поликатиона заменяют на не-ковалентно связанный поликатион. Наоборот, в некоторых случаях можно даже существенно повысить поглощение ДНК. Было обнаружено, что ДНК комплексов имеется в виде тороидальных структур диаметром 80-100 нм. Таким образом количество поликатиона подбирают с учетом электронейтральности и достижения компактной структуры. При этом количество поликатиона, получаемое в результате осуществления зарядки нуклеиновой кислоты с учетом обеспечения электронейтральности, обычно также обеспечивают компактную структуру ДНК.
Таким образом согласно другому варианту изобретения комплексы также включают связывающие нуклеиновую кислоту вещества в не-ковалентно связанном виде, которые могут быть идентичны или отличны от связывающего фактора. Если эндосомолитический агент представляет собой свободный вирус, то комплексы включают конъюгат нуклеиновой кислоты и содействующего поглощению клеткой фактора. В случае применения эндосомолитического конъюгата, например, вирусного конъюгата, нуклеиновая кислота переводится в комплекс с этим конъюгатом, в случае необходимости в сочетании с конъюгатом дополнительного содействующего поглощению клеткой фактора. Вид и количество не-ковалентно связанного "свободного" вещества со сродством для нуклеиновой кислоты также определяется конъюгатом или конъюгатами, в частности с учетом связывающего фактора в конъюгате. Так, например, если связывающий фактор представляет собой вещество, которое не обладает или же обладает лишь ограниченной способностью к конденсации ДНК, то в целях достижения высокой степени поглощения клеткой комплексов целесообразно применять вещества со сродством для ДНК, которое в значительной степени обладает этим свойством. Но если же сам связывающий фактор представляет собой конденсирующее нуклеиновую кислоту вещество и позволяет получить компактную структуру нуклеиновой кислоты, достаточную для обеспечения эффективного поглощения клеткой, то целесообразно применять вещество со сродством для нуклеиновой кислоты, которое обеспечивает усиление экспрессии благодаря другим механизмам.
Подходящими "свободными" веществами со сродством для нуклеиновой кислоты согласно изобретению являются, например, соединения, способные к конденсации нуклеиновой кислоты и/или защите их от нежелательной деградации в клетках, в частности вышеупомянутые вещества поликатионного типа. Другими подходящими веществами являются такие вещества, которые в результате связывания с нуклеиновой кислотой улучшают ее доступность для экспрессионной системы клетки, благодаря чему улучшается ее транскрипция/экспрессия. Примером такого вещества является хромозомальный не-гистонный белок HMG1, который обладает способностью к сообщению ДНК компактной структуры и к содействию экспрессии в клетках.
При определении молярного соотношения эндосомолитического агента и/или содействующего поглощению клеткой фактора и вещества со сродством для нуклеиновой кислоты или нуклеиновых кислот следует учесть, что имеет место комплексообразование нуклеиновой кислоты, образующийся комплекс может связаться с клеткой и поглощаться ею и что он выделяется из ядрышек либо с помощью эндосомолитического агента либо самим собой.
Соотношение содействующего поглощению клеткой фактора, связывающего фактора и нуклеиновой кислоты зависит, в частности, от размера молекул поликатиона и числа распределения положительно зараженных групп. Поэтому эти критерии должны согласоваться с размером и структурой переносимой нуклеиновой кислотой. Молярное соотношение содействующего поглощению клеткой фактора и вещества со сродством нуклеиновой кислоты предпочтительно составляет примерно 10 : 1 - 1 : 10.
После конструкции и синтеза конъюгатов и определения оптимального в отношении эффективной трансфекции соотношения конъюгата и ДНК количество конъюгата, которое можно заменять свободным веществом со сродством нуклеиновой кислоты, можно определять путем титрования. Если поликатионы применяют как в качестве связывающего фактора, так и в качестве свободного вещества со сродством для нуклеиновой кислоты, то поликатионы могут быть идентичными или же различными.
При получении предлагаемой системы, включающей конъюгаты вируса, целесообразно поступать следующим образом. Сначала подбирают генный конструкт, который подлежит переносу в клетку, и вирус или его компонент, пригодный для данной трансфекции. Затем вирус или его компонент связывают с поликатионом и переводят в комплекс с генным конструктом. Исходя из определенного количества конъюгата вируса можно осуществлять титрование путем обработки целевых клеток этим (постоянным) количеством конъюгата и уменьшения концентрации ДНК, или наоборот. Таким образом определяют оптимальное соотношение ДНК и конъюгата вируса. Если применять дополнительный содействующий поглощению клеткой фактор, то оптимальное соотношение конъюгата вируса и конъюгата содействующего поглощению клеткой фактора можно определить путем титрования с применением определенного количества ДНК.
Комплексы можно получать путем смешивания нуклеиновой кислоты, конъюгата вируса и, в случае необходимости, конъюгата содействующего поглощению клеткой фактора и связывающего фактора, а также не-ковалентно связанного вещества со сродством для нуклеиновой кислоты. При этом все компоненты могут иметься в виде разбавленных растворов. Если поликатионы применяют в качестве связывающего фактора и одновременно в качестве "свободных" поликатионов, то целесообразно сначала получать смесь конъюгатов со "свободными" поликатионами и затем объединять эту смесь с ДНК. Оптимальное соотношение ДНК и конъюгата или конъюгатов и поликатионов определяют путем многократного титрования, при этом осуществляют серию процессов трансфекции с применением постоянного количества ДНК и повышающихся количеств смеси конъюгата или конъюгатов и поликатиона. Оптимальное соотношение конъюгата или конъюгатов и поликатионов в смеси можно определять, например, путем сравнивания оптимальных составов смесей, применяемых в процессах многократного титрования.
Комплексы ДНК можно получать при физиологических концентрациях солей. Другая возможность заключается в применении высоких концентраций солей (примерно 2М NaCl) и последующем доведении до физиологических условий путем медленного разбавления или диализа.
Наиболее целесообразна последовательность смешения компонентов нуклеиновой кислоты, конъюгата или конъюгатов и, в случае необходимости, не-ковалентно связанного вещества со сродством для нуклеиновой кислоты предварительно определяют в рамках серии соответствующих опытов. В некоторых случаях может быть целесообразным предварительное образование комплекса нуклеиновой кислоты и конъюгата или конъюгатов с последующим добавлением "свободного" вещества со сродством для нуклеиновой кислоты, например, поликатиона, например, в случае конъюгатов трансферина и димерного этидия и полилизина.
Согласно предпочтительному варианту данного изобретения в качестве содействующего поглощению клеткой фактора и дополнительного содействующего поглощению клеткой фактора применяют трансферин, а в качестве связывающего фактора - поликатион. Под термином "трансферин" понимаются естественный и синтетический трансферины, а также модификации трансферина, которые связываются с рецептором и переносятся в клетку.
Нуклеиновая кислота поглощается в виде комплексов, в которых конъюгаты содействующего поглощению клеткой фактора и поликатионы имеются в виде комплекса с нуклеиновой кислотой. Если комплексы еще включают не-ковалентно связанное вещество со сродством для нуклеиновой кислоты, то этим веществом является предпочтительно поликатион. Этот второй поликатион идентичен или же отличен от поликатиона, содержащегося в конъюгате или в обоих конъюгатах.
В случае "комбинационных комплексов " нуклеиновая кислота поглощается клеткой в виде комплекса, в котором конъюгаты содействующего поглощению клеткой фактора и эндосомолитические конъюгаты комплексированы с нуклеиновой кислотой. Конъюгаты содействующего поглощению клеткой фактора и поликатиона, которые применяют вместе со свободным вирусом или же вместе с вирусными конъюгатами в комбинационных комплексах, можно получать химическим методом или же рекомбинантным методом в случае применения полипептида в качестве поликатиона.
В рамках данного изобретения, предпочтительно применяют конъюгаты, в которых гликопротеин, например, трансферин, и связывающий фактор, связанный друг с другом по меньшей мере через одну углеводную цепь гликопротеина. В отличие от получаемых традиционными методами связывания конъюгатов получаемые данным методом конъюгаты свободны от модификаций, происходящих от применяемого связывающего агента. В случае применения гликопротеинов, по меньшей мере с одной углеводной группой, пригодной для связывания, например, трансферина, получаемые конъюгаты также имеют то преимущество, что их места связывания с гликопротеинами и связывающим фактором точно определены.
Количество применяемого эндосомолитического агента и его концентрация зависят от целевой трансфекции. Целесообразно применять минимальное количество вируса или вирусного конъюгата, которое необходимо для обеспечения поглощения клеткой вируса или вирусного конъюгата и комплекса нуклеиновой кислоты и высвобождения из ядрышек. Количество вируса или вирусного конъюгата следует согласовать с данным типом клетки, причем в первую очередь следует учесть эффективность вируса для данного типа клетки. Другим критерием является применяемый конъюгат содействующего поглощению клеткой фактора и связывающего фактора, в частности в отношении содействующего поглощению клеткой фактора, для которого целевая клетка имеет определенное число рецепторов. Кроме того, количество вируса или вирусного конъюгата зависит от количества переносимой ДНК. Обычно небольшое количество вируса достаточно для обеспечения устойчивой трансфекции, для которой требуется лишь небольшое количество ДНК. В противоположность этому в случае осуществления временной трансфекции, требующей большего количества ДНК, вирус должен применяться в большем количестве. В определенных случаях целесообразно проводить предварительные эксперименты с применением предназначенных для трансфекции целевых клеток, в случае необходимости в виде смешанной популяции, и предназначенной для трансфекции векторной системы с тем, чтобы определить оптимальную концентрацию вируса путем титрования. При этом в качестве ДНК целесообразно применять генный конструкт, который по размеру в основном совпадает с тем веществом, предназначенным для данного конкретного случая, и содержит репортерный ген для более простого определения эффективности переноса гена. Подходящими репортерными генами для осуществления таких опытов являются люцифераза и β-галактосидаза.
При осуществлении данного изобретения предпочитается одновременное применение комплекса нуклеиновой кислоты и эндосомолитического агента. Но вполне возможно и их последовательное применение. В случае раздельного применения последовательность не является критической при условии, что мероприятия проводят друг за другом с соблюдением только короткого промежутка времени, чтобы обеспечить эффективный одновременный контакт компонентов.
Если применять свободный вирус в виде отдельного препарата, то одновременная дача препарата свободного вируса и комплексов может обеспечиваться за счет того, что препараты вируса являются частью среды трансфекции, содержащей комплекс нуклеиновой кислоты. В случае одновременной аппликации свободного вируса комплексы нуклеиновой кислоты и препарат вируса предварительно смешивают друг с другом. Согласно предпочтительному варианту данного изобретения эндосомолитический агент является компонентом комбинационного комплекса.
В определенных случаях может быть целесообразным применение лизосоматропного вещества дополнительно к эндосомолитическому агенту, например, если эндосомолитический агент представляет собой эндосомолитический конъюгат пептидов или ретровирус, эндосомолитическая активность которых не зависит от кислой величины pH. Известно, что лизосомотропные вещества тормозят активность протеаз и нуклеаз и поэтому могут тормозить деграцию нуклеиновых кислот. Такими веществами являются, например, хлорохин, моненсин, нигерицин и метиламин. Было найдено, что моненсин позволяет повысить экспрессию репортерного гена в случае применения ретровируса Молонея. Присутствие хлорохина приводит к экспрессии репортерного гена, завезенного переносом ДНК в присутствии трансферина, почти 100%-но в клетках К562. Клетки BNL.CL2 или гепатоциты HepG2 не реагируют на воздействие хлорохина, они могут трансфецироваться до уровня 5 -10% в случае использования эндосомолитических свойств дефектного повторной репликацией или химически инактивированного свободного аденовируса. Данное изобретение позволяет усилить преимущества биологических векторов. В результате распределения рецепторов получается тропизм для содействующего поглощению клеткой фактора и вируса. Согласование этих двух компонентов с применяемой популяцией клеток позволяет достичь более высокой селективности, что, в частности, имеет важное значение при применении изобретения для терапевтических целей.
Другим объектом данного изобретения являются фармацевтические композиции, которые содержат в качестве активного вещества комплекс терапевтически активной нуклеиновой кислоты, предпочтительно в качестве частей генного конструкта, эндосомолитический агент (в случае необходимости в виде конъюгата) и, в случае необходимости, конъюгат содействующего поглощению клеткой фактора. При этом можно применять любой инертный фармацевтический приемлемый носитель, такой как, например, солевой раствор, который может содержать фосфатный буфер, или же любой носитель, в котором комплексы ДНК являются растворимыми в требуемой степени. Способ приготовления фармацевтической композиции широко известен.
Данное изобретение обеспечивает максимальную гибкость при его осуществлении, например, при его применении в виде фармацевтической композиции. Предлагаемая система может иметься в виде лиофилизата или же содержаться в подходящем буфере в глубоко замороженном состоянии. Ее можно также применять в качестве готового к применению реагента в растворе, предпочтительно в охлажденном виде. Необходимые для трансфекции компоненты, то есть ДНК, эндосомолитический агент, в случае необходимости в виде конъюгата или в готовом к образованию конъюгата с соответствующим партнером, связывающее ДНК вещество, в случае необходимости в виде конъюгата с содействующим поглощению клеткой фактором, в случае необходимости свободный поликатион, могут содержаться в подходящей буферной системе или же могут представлять собой градиенты трансфекционного комплекта. Если трансфекционный комплект применяют в качестве модульной системы, то он может содержать различные ДНК, например, ДНК, кодирующие различные антигены, и/или различные конъюгаты содействующего поглощению клеткой фактора. Решение вопроса применения компонентов в виде готового к применению препарата или же их смешение непосредственно перед применением зависит, между прочим, от устойчивости комплексов, которая может заранее определяться известными методами. Согласно предпочтительному варианту изобретения в качестве ингредиента комплекта может применяться полученный с помощью трансглютаминазы конъюгат аденовируса и полилизина, который является устойчивым при хранении. Согласно другому предпочтительному варианту изобретения биотинилированный аденовирус и стрептавидин/полилизин смешивают друг с другом перед применением.
Предлагаемую систему можно апплицировать системно, предпочтительно внутривенным путем, в качестве частей фармацевтической композиции. Целевыми органами, на которые направлена данная аппликация, могут быть печень, селезенка, легкие, костный мозг и опухоли. Примером местной аппликации является легочная ткань (применение предлагаемой системы в качестве частей фармацевтической композиции, представляющей собой жидкость для инстиляции или аэрозоль для ингаляции). Кроме того фармацевтические композиции можно также апплицировать путем непосредственного впрыскивания в печень, мышечную ткань, опухоль или же путем местной дачи в желудочно-кишечный тракт. Другим методом дачи фармацевтической композиции является введение через дренажную систему желчи. Этот метод обеспечивает непосредственный доступ к мембранам гепатоцитов на канальцах желчи, чем избегается взаимодействия композиции с компонентами крови.
Известно, что миобласты (незрелые мышечные клетки) способны к транспорту генов в мышечные волокна мышей. Так как миобласты могут выделять генный продукт в кровь, этот подход может получать более широкое применение, чем обработка генетических дефектов мышечных клеток, как, например, дефекта, выраженного мышечной дистрофией. Следовательно, соответственно подготовленные миобласты можно применять для выделения генных продуктов, которые либо действуют в крови, либо транспортируются кровью. Эксперименты в соответствии с данным изобретением показывали, что культуры миобластов и мышечных трубочек, даже первичные, можно подвергать высокоэффективной трансфекции. Наиболее пригодная среда трансфекции содержала комбинационные комплексы биотинилированного аденовируса, трансферина/полилизина и стрептавидина/полилизина. Кроме репортерных генов люциферазы и β-галактозидазы в мышечных клетках экспрессировался фактор VIII. Кроме того, куриный аденовирус CELO применялся в комбинационных комплексах, содержащих агглютинин пшеничного зачатка в качестве дополнительного содействующего поглощению клеткой фактора.
Терапевтическое применение может также осуществляться ex vivo, причем обработанные клетки, например, клетки костного мозга, гепатоциты или миобласты, возвращаются в тело. Другое применение ex vivo предлагаемой системы сводится к аппликации так называемых "раковых вакцин". Принцип этого терапевтического подхода заключается в выделении опухолевых клеток из пациента и трансфекции клеток ДНК, кодирующей цитокин. Следующий прием может заключаться в инактивации клеток, например, путем облучения, которое проводится с таким расчетом, что клетки больше не обладают способностью к репликации, но все еще способны к экспрессии цитокина. Затем генетически модифицированные клетки в качестве вакцины дают пациенту, из которого клетки были изолированы. В зоне сайта вакцинации выделяемые цитокины активируют иммунную систему, например, за счет активации цитотоксичных Т-клеток. Эти активированные клетки способны к проявлению своего действия в других частях тела и к воздействию на необработанные опухолевые клетки. Таким образом риск повторного образования опухоли и образования метастазов снижается.
В экспериментах согласно данному изобретению первичные меланомные клетки успешно подвергались трансфекции репортерным геном, который содержался в комбинационных комплексах связанного с полилизином аденовируса и трансферина/полилизина.
Данное изобретение может также использоваться для определения иммунореакции хозяина на заданный антиген. Такие испытания основаны на переносе гена в экспрессирующие антиген клетки. Специфичные в отношении антигена цитотоксичные Т-лимфоциты (далее: "ЦТЛ"), которые разрушают инфицированные клетки, играют важную роль в защите хозяина от вирусных инфекций или опухолей. Взаимодействие между Т-клеткой и представляющей антиген клеткой (далее "ПАК") ограничено лимфоцитными антигенами человека (далее: "ЛАЧ"), которые являются молекулами большой тканевой совместимости. Для осуществления исследования степени разрушения ЦТЛ экспрессирующих антиген клеток в рамках опыта in vitro необходимо представлять антиген ЦТЛ в правильном контексте ЛАЧ. Такое исследование можно осуществлять следующим образом. ПАК подвергают трансфекции конструктом ДНК, содержащим последовательность, кодирующую антиген. Эпитропы антигена будут связываться с МНС класса I и будут иметься на поверхности клетки в качестве мишени для специфичной в отношении ЦТЛ реакции. После инкубации вместе с пробой сыворотки пациента, зависящей от присутствия специфичных ЦТЛ, ПАК будет подвергаться лизису. При этом лизис измеряют за счет определения выделения, например, радиоактивного хрома, который включился в ПАК перед добавлением к сыворотке.
Индуцированные Б-лимфопластоидные клеточные линии можно применять для экспрессии генов антигена путем трансфекции рекомбинантными вирусами вакцинии. Однако клетки, которые эффективно экспрессируют антиген в течение примерно одного дня, умирают в результате литического эффекта вакцинии. Эти трудности можно преодолеть при применении предлагаемой системы для введения кодирующих антиген конструктов ДНК, например, конструктов, кодирующих антигены СПИДа или опухолевые антигены, в фибробласты с тем, чтобы сообщить им способность к экспрессии антигена. Первичные фибробласты можно легко получать путем биопсии, легко выращивать и поддаются высокоэффективной трансфекции (со степенью примерно 50 - 70%) предлагаемой системой. Такие анализы полезны для идентификации распознаваемых клетками-килерами эпитопов в отношении вакцин. Кроме того, они могут преимущественно применяться для определения ограниченной ЛАЧ иммунной реакции индивидуума на заданный антиген.
Изобретение может использоваться для производства рекомбинантных белков, так как почти во всех клетках достигается высокая степень экспрессии переносимых генов. При этом в отношении последовательностей и молекулярного веса переносимой ДНК существуют лишь некоторые ограничения или же ограничений вообще не существует. Имеется широкий спектр различного рода клеток, которые поддаются трансфекции предлагаемыми конструктами ДНК. Так, например, клетки почти любого типа можно применять для производства рекомбинантных белков с высокой биологической активностью.
Перенос гена в клетки можно осуществлять тем же образом, что и показано для люциферазы и α-интерферона, то есть практически любой генный конструкт, который обеспечивает получение желаемого белкового продукта, может транспортироваться указанным образом. Желаемый белковый продукт можно выделять из трансфецированной клеточной культуры (либо надосадочной жидкости культуры либо подходящего продукта ее гомогенизации) через 24 часа или же через одну неделю после трансфекции. Выделение возможно также по истечении более продолжительного периода после трансфекции.
Использование предлагаемой системы для производства рекомбинантных белков имеет следующие преимущества.
1. Благодаря высокой эффективности трансфекции (более 90% трансфецированных клеток способны к экспрессии соответствующего гена на высоком уровне), предварительный отбор положительно трансфецированных клеток не требуется и, кроме того, нет необходимости в подборе устойчивых клеточных линий. Небольшая клеточная культура может быть достаточной для производства белка в необходимом количестве.
2. Большие генные конструкты могут транспортироваться. До сих пор с успехом переносились в клетки конструкты размером до 48 к.п.о.
3. Экспрессия гена может осуществляться в клетках, которые обеспечивают подходящие посттрансляционные переработку и модификацию (например, зависящее от витамина А карбоксилирование факторов свертывания крови или специфичного для данного типа клетки гликозилирования).
4. Возможен более широкий подбор типов целевых клеток для экспрессии гена.
Изобретение далее описывается с помощью нижеследующих примеров, которые представлены в целях иллюстрации, а не какого-то ни было ограничения его объема. В этих примерах используют следующие материалы и методы, если иного не указано.
Получение комплексов ДНК и конъюгата трансферина и полилизина
а) Конъюгаты трансферина и полилизина человека
Раствор 280 мг (3,5 мкмоль) человеческого, свободного от железа трансферина в 6 мл 30 мМ буфера ацетата натрия с pH 5 охлаждают до температуры 0oC и добавляют 750 мкл 30 мМ буфера ацетата натрия с pH 5, содержащего 11 мг (51 мкмоль) периодата натрия. Смесь оставляют стоять в темноте на ледяной бане в течение 90 минут. В целях удаления низкомолекулярных продуктов осуществляют гель-фильтрацию на йоните Sephadex G-25. Получают раствор, содержащий примерно 250 мг окисленного трансферина (определение при помощи нингидрина). (Для доказательства окисленной формы, содержащей альдегиды и дающей цветную реакцию при окрашивании анисовым альдегидом, смесь каплями подают на тонкую слоистую пластину силикагеля, сушат, пластины погружают в смесь п-анисового альдегида, серной кислоты и этанола в соотношении 1 : 1 : 18, сушат и нагревают). Раствор модифицированного трансферина быстро добавляют (в течение 10 - 15 минут) к раствору, содержащему 1,5 мкмоль меченого флуоресцеином поли(L)лизина со средней длиной 190 лизиновых мономеров, в 4,5 мл 100 мМ ацетата натрия с pH 5. pH раствора доводят до 7,5 путем добавления 1 М буфера бикарбоната натрия. С промежутком одного часа к смеси добавляют 4 раза 28,5 мг (450 мкмоль) цианоборгидрида натрия.
Через 17 часов добавляют 2 мл 5 М хлористого натрия с тем, чтобы довести раствор до общей концентрации 0,75 М. Реакционную смесь подают на катионообменную колонку марки Mono S HR 10/10 фирмы Фармация, Швеция, после чего элюируют солевым градиентом 0,75 М-2,5 М хлористого натрия, имеющим постоянное содержание 25 мМ HEPES, pH 7,3. Высокая солевая концентрация при подаче на колонку и в начале осуществления градиентной элюации является существенным для получения конъюгатов. Некоторые количества трансферина (примерно 30%) со слабой флуоресценцией элюируют в начале процесса. Основное количество меченого флуоресценцией конъюгата элюируют при солевой концентрации 1,35 -1,9 М и его собирают в виде 3 фракций. После двух серий диализа с применением 2 л 25 мМ буфера HEPES, pH 7,3, эти фракции (в порядке их элюации) дают фракцию А, содержащую 45 мг (0,56 мкмоль) трансферина, модифицированного 366 нмоль полилизина (далее этот конъюгат обозначается "TfpL190A"), фракцию Б, содержащую 72 мг (0,90 мкмоль) трансферина, модифицированного 557 нмоль полилизина (далее этот конъюгат обозначается "TfpL190B) и фракцию В, содержащую 7 мг (85 нмоль) трансферина, модифицированного 225 нмоль полилизина (далее этот конъюгат обозначается "TfpL190C"). Если конъюгаты сразу же не подаются на дальнейшее применение, то их замораживают в атмосфере жидкого азота и хранят при температуре -20oC в свободной от железа форме. Перед введением железа пробы (0,5 - 1 мг) доводят до физиологической солевой концентрации (150 мМ) добавлением хлористого натрия. Включение железа осуществляется за счет добавления 4 мкл 10 мМ цитратного буфера железа (III) (содержащего 200 мМ цитрата; pH буфера доведен до 7,8 добавлением бикарбоната натрия) на мг содержащегося в конъюгате трансферина. Железосодержащие конъюгаты подразделяют на небольшие аликвоты перед их применением для образования комплекса с ДНК, мгновенно замораживают в атмосфере жидкого азота или в сухой атмосфере льда и этанола и хранят при температуре -20oC (этот подход оправдал себя, так как найдено было, что повторяющиеся процессы оттаивания и замораживания приводят к потере активности конъюгатов).
б) Конъюгаты мышиного трансферина и полилизина
Применяют подобный метод, что и в случае трансферина человека, при этом связывание осуществляют при помощи боковых углеводных цепей. Применяют 4,1 мг (51 нмоль) мышиного трансферина и 2,1 мг (34 нмоль) полилизина с молекулярным весом 290 (далее: "pL 290"). Получают конъюгаты, состоящие из 15,5 нмоль мышиного трансферина и 13 нмоль pL 290.
Плазмидная ДНК
а) ДНК pRSVL.
Получение плазмидной ДНК pRSVL (содержащей ген люциферазы Photinus pyralis под контролем вируса Rous Sarcoma LTR) осуществляют известными приемами с применением тритона Х. Далее используют равновесное центрифугирование в градиенте плотности с применением хлорида цезия и бромида этидия, обесцвечивание буганолом-1 и диализ с применением 10 мМ буфера Tris/HCl (pH 7,5, 1 мМ ЭДТК). Образование комплекса осуществляют за счет смешения 6 мкг плазмидной ДНК в 350 мкл буфера HBS (150 мМ хлористого натрия, 20 мМ HEPES, pH 7,3) с 12 мкг конъюгата трансферина и полилизина в 150 мкл буфера HBS. Комплексообразование осуществляют за 30 минут до добавления комплекса к клеткам.
б) ДНК pCMVL
Плазмиду pCMVL (репортерный генный конструкт, содержащий ген люциферазы Photinus pyralis под контролем промотора цитомегаловируса) получают за счет удаления вставки BamHI из плазмиды pSTCX556, обработки плазмиды фрагментом Кленова и введения обработанного HindIII/Ssp1 и фрагментом Кленова фрагмента из плазмиды pRSVL, содержащего последовательность, кодирующую люциферазу. ДНК получают аналогично получению ДНК pRSVL.
Получение препарата вируса
а) Препараты аденовируса
Используют штамм d1312 аденовируса, имеющий делецию в зоне Ela. Репликацию вируса осуществляют в Ela-транс-комплементирующей клеточной линии 293 с последующей очисткой известным приемом. Очищенный вирус подают в предназначенный для хранения буфер (100 мМ Tris, pH 8,0, 100 мМ хлористого натрия, 0,1% альбумина сыворотки крупного рогатого скота (далее: "АСС"), 50% глицерина) или в буфер HBS с содержанием 40% глицерина и аликвоты хранят при температуре -70oC. Концентрацию вирионов определяют УФ-спектрофотометрическим анализом экстрагированной геномной вирусной ДНК (формула: одна единица оптической плотности соответствует 1012 вирусным частицам на мл).
б) Препарат ретровируса
Ретровирус Молонея N2 мышиного лейкоза упаковывают в экотропную упаковочную линию. Надосадочные жидкости, получаемые от экспрессирующих вирус клеток, собирают, мгновенно замораживают в атмосфере жидкого азота и хранят при температуре -20oC. Применяемый в примерах надосадочные жидкости имеют титр примерно 106 колониеобразующих единиц на мл (далее: "кое/мл"), который определялся путем образования устойчивых к неомицину колоний с применением клеток NIH3T3. Надосадочные жидкости пропускают через мембрану марки FILTRON, находящуюся в перемешиваемом аппарате для концентрации клеток под давлением азота. При этом 10-30 мл надосадочной жидкости обычно сгущают в 10 раз.
Клетки и среда
Клетки HeLa культивируют в среде DMEM, дополненной 5% инактивированной термообработкой телячьей эмбриональной сыворотки (далее: "ТЭС"), 100 единицами/мл пенициллина, 100 мкг на мл стрептомицина и 2 мМ глутамина. Клетки WI-38, MRC-5 и KB культивируют в среде ЕМЕМ (модифицированной Иглем основной среде), дополненной 10% инактивированной термообработкой ТЭС, антибиотиками, содержащимися в среде DMEM, 10 мМ заменимых аминокислот и 2 мМ глутамина. Эпительную клеточную линию дыхательного муковисцитоза CFT1 культивируют в среде F12-7Х. Для осуществления переноса генов клетки культивируют в снабженных ячейками диаметром 6 см чашках до достижения степени выращивания 5 • 105 клеток. Среду удаляют и добавляют 1 мл среды DMEM или ЕМЕМ/2% ТЭС. Затем добавляют комплексы конъюгата и ДНК с последующей немедленной подачей аденовируса d1312 (0,05 - 3,2 • 104 частиц в клетку) или сравнимого объема буфера (1 - 80 мкл). Чашки возвращают на инкубацию при температуре 37oC в присутствии 5% двуокиси углерода в течение одного часа, после чего добавляют 3 мл полной среды. После дополнительной 24-часовой инкубации осуществляют сбор клеток с тем, чтобы определить экспрессию гена люциферазы. В случае клеток CFT1 осуществляют культивацию в среде F12-7Х в течение 4 часов без трансферина человека перед осуществлением переноса гена.
Полученные от американской коллекции АТСС клетки имеют следующие регистрационные номера: клетки HeLa: CCL 2; клетки К562: CCL 243; клетки HepG2: НВ 8065; клетки TIB-73: TIB 73 (BNL CL.2); клетки NIH3T3: CRL 1658; клетки 293: CRL 1573; клетки KB: CCL 17; клетки WI-38: CCL 75; клетки MRC 5: CCL 171. Клетки Н9 с регистрационным номером 87 получены от американской организации AIDS Research and Reference Reagent Program, U.S. Department of Health and Human Services.
Первичные лимфоциты получают путем подачи 25 мл пробы пуповинной крови в подопытной трубке, содержащей ЭДТК. Под аликвотами размещают нижний слой 4,5 мл продукта Фиколл-гипак фирмы Фармация, Швеция, и центрифугируют при 2500 об/мин в течение 15 минут. Коричневатый слой между верхним слоем плазмы и прозрачным слоем Фиколла удаляют (примерно 10 мл). Добавляют 40 мл среды IMDM плюс 10% ТЭС, пробу центрифугируют при 1200 об/мин в течение 15 минут и клеточный материал суспендируют в 50 мл свежей среды IMDM плюс 10% ТЭС (плотность клеток составляет примерно 2 • 106 клеток/мл). Добавляют 250 мкл аликвота фитогемагглютинина, культуру инкубируют при температуре 37oC в течение 48 часов в присутствии 5% двуокиси углерода и добавляют рекомбинантный интерлейкин-2 (концентрация: 20 единиц на мл). Клетки разбавляют в отношении 1 : 3 средой IMDM плюс 20% ТЭС, содержащей 2 единицы интерлейкина-2 на мл. Аликвоты клеток подвергают глубокому замораживанию в атмосфере жидкого азота в присутствии 5% метилсульфоксида. Перед применением клетки выращивают в среде IMDM, содержащей 20% ТЭС и 2 единицы интерлейкина-2 на мл.
Для осуществления последующего процесса связывания клетки HeLa уравновешивают при температуре 4oC в 1 мл среды DMEM, дополненной 2% ТЭС. Комплексы конъюгата и ДНК добавляют тем же образом, что и в других случаях, и чашки инкубируют при температуре 4oC в течение 2 часов. Затем чашки интенсивно промывают холодной как лед средой DMEM, содержащей 2% ТЭС, после чего добавляют 2 мл этой среды. Добавляют аденовирус d1312 или буфер, клеткам дают медленно нагреваться, после чего их подают на 24-часовую инкубацию. После инкубации осуществляют сбор клеток и исследование на экспрессию гена люциферазы.
Определение активности люциферазы
Приготовление клеточных экстрактов, стандартизацию содержания белка и определение активности люциферазы осуществляют широкоизвестными методами.
Пример 1. Определение действия аденовируса на перенос гена с применением конъюгатов трансферина и полилизина. Сначала исследуют действие повышающихся доз вируса на способность определенного количества комплекса конъюгата и ДНК к осуществлению переноса гена. В целях образования комплекса 6 мкг плазмиды pRSVL смешивают с 12 мкг конъюгата человеческого трансферина и полилизина (далее: "hTfpL 190В"). Комплекс конъюгата и ДНК, а также различные количества аденовируса d1312 (0,05 - 3,2 • 104 вирусных частиц на клетку) добавляют к клеткам HeLa. Результаты данного опыта представлены на фиг. 1. Активность люциферазы выражается в световых единицах 50 мкг общего клеточного белка. Согласно данному анализу повышающиеся количества добавляемого аденовируса приводят к соответствующему улучшению переноса гена. На фиг. представлены средние значения 2 - 4 отдельных опытов. Балки указывают стандартное отклонение.
Пример 2. Действие, достигаемое в зависимости от количества комплекса конъюгата и ДНК. Комплексы конъюгата ДНК в виде полученных тем же образом, что и в примере 1 логарифмических разбавленных препаратов добавляют к клеткам HeLa вместе с постоянным количеством аденовируса d1312 (1 • 104 вирусных частиц на клетку) или же без него. Активность люциферазы определяют тем же образом, что и в примере 1. Результаты опыта представлены на фиг. 2.
Пример 3. Улучшение переноса гена, достигаемое конъюгатом трансферина и полилизина в присутствии аденовируса, имеет место через вызываемый рецептором эндоцитоз.
а) Действие аденовируса на перенос имеющейся в виде комплекса ДНК
Для осуществления трансфекции используют следующие компоненты. 6 мкг ДНК pRSVL без конъюгата трансферина и полилизина (DNA); 6 мкг ДНК pRSVL и 6 мкг несопряженного полилизина с молекулярным весом 270 (DNA + pL); 6 мкг ДНК pRSVL и 12 мкг конъюгата трансферина и полилизина с молекулярным весом 190, применяемого в предыдущих примерах (DNA + hTfpL190B). Указанные материалы добавляют к клеткам HeLa вместе с аденовирусом d1312 в концентрации 1 • 104 вирусных частиц на клетку (d1312) или без него. Получение клеточных экстрактов, стандартизация общего клеточного белка и определение активности люциферазы осуществляют вышеуказанным образом. Результаты опыта представлены на фиг. 3а.
б) Действие аденовируса на перенос связанной с рецептором ДНК
Комплексы конъюгата и ДНК (DNA + hTfpll90B) или комплексы полилизина и ДНК (DNA + pL) связывают с клетками HeLa за счет инкубации при температуре 4oC. Несвязанный комплекс удаляют перед добавлением аденовируса d1312 в концентрации 1 • 104 вирусных частиц на клетку (d1312) или сравнимого количества буфера. Затем осуществляют инкубацию при температуре З7oC, направленную на обеспечение включения клеткой связанного комплекса ДНК и аденовируса. Активность люциферазы определяют вышеуказанным образом. Результаты представлены на фиг. 3b.
в) Действие аденовируса на перенос гена с применением конъюгатов трансферина и полилизина
Комплексы конъюгата и ДНК, содержащие 6 мкг ДНК pRSVL и 12 мкг конъюгата трансферина и полилизина (DNA + hTfpL190B) добавляют к клеткам HeLa вместе с аденовирусом в концентрации 1 • 104 вирусных частиц на клетку (d1312) или инактивированным термообработкой аденовирусом д1312 в сравнимой концентрации (д1312 h. i.). Инактивацию термообработкой осуществляют путем инкубации при температуре 45oC в течение 30 минут. Активность люциферазы определяют вышеуказанным образом. Результаты опыта представлены на фиг. 3с.
Пример 4. Действие аденовируса на перенос гена при применении конъюгатов трансферина и полилизина в выбранных клеточных линиях, Комплексы конъюгата и ДНК (6 мкг pRSVL + 12 мкг hTfpL190B) добавляют к клеткам клеточных линий CFT1, KB, HeLa, W138 и MRC5 вместе с аденовирусом d1312 в концентрации 1 • 104 вирусных частиц на клетку или же без него. Эффективность переноса гена для каждой клеточной линии определяют описанным выше образом за счет определения активности люциферазы. Результаты опыта представлены на фиг. 4.
Пример 5. Улучшение экспрессии гена люциферазы на уровне переноса гена (а не на уровне трансактивации). Клеточную линию, обозначенную К562 10/6, которая конституционально экспрессирует люциферазу, приготовляют за счет трансфекции плазмидой, содержащей фрагмент гена люциферазы RSV (Apa1/Pvu1-фрагмент плазмиды pRSVL), который клонирован в сайт CIaI локуса pUC μ. Эту плазмиду переводят в комплекс с конъюгатом трансферина и полилизина и полученным комплексом трансфецируют клетки К562. Так как плазмида локуса pUC μ содержит устойчивый к неомицину ген, возможно подобрать экспрессирующие люциферазу клоны с учетом устойчивости к неомицину. Для осуществления дальнейших опытов подбирают клон К562 10/6.
Аликвоты родительской клеточной линии К562 (в 200 мкл среды RPMI 1640, содержащей 2% ТЭС; 500000 клеток на пробу) обрабатывают либо 13 мкг TfpL и 6 мкг pRSVL или же 4 мкг pL 90 и 6 мкг pRSVL, которые применяют в среде 500 мкл буфера HBS. После добавления аденовируса d1312 в указанной на фиг. 5 концентрации клетки выдерживают при температуре 37oC в течение 90 минут, после чего добавляют 2 мл среды RPMI, содержащей 10% ТЭС. Инкубацию продолжают еще при температуре 37oC в течение дальнейших 24 часов, после чего клетки подготовляют к определению активности люциферазы. Было обнаружено, что инкубация в присутствии аденовируса приводит к значительному повышению активности люциферазы (см. фиг. 5А). Это относится как к комплексам TfpL (2000 световых единиц против 25000 световых единиц), так и к комплексам pL90 (0 против 1,9 • 106 световых единиц). Это свидетельствует о том, что клеточная линия К562 обладает способностью к включению комплексов pRSVL и полилизина и что данный эффект в значительной степени усиливается присутствием аденовируса, что видно по экспрессии люциферазы.
Аналогичные опыты проводят с применением клеток К561 10/6, которые конституционально экспрессируют ген люциферазы RSVL. При этом применяют подобные количества аденовируса d1312. Аликвоты 500000 клеток (в 200 мкл среды RPMI, содержащей 2% ТЭС) инкубируют при температуре 37oC в течение 90 минут в присутствии аденовируса d1312 в указанной на фиг. 5В концентрации. Потом, также как и в случае родительской клеточной линии, добавляют среду RPMI, содержащую 10% ТЭС, инкубацию продолжают в течение дальнейших 24 часов, после чего определяют активность люциферазы. Как видно на фиг. 5В, обработка этих клеток аденовирусом не приводит к заметному действию на активность люциферазы, т. е. результаты контрольных опытов того же порядка, что и результаты опытов, которые проводят в присутствии вируса.
Пример 6. Трансфекция клеток печени конъюгатами асиалофетуина и полилизина (далее: "AfpL") или конъюгатами тетра-галактозного пептида и полилизина (далее: gal 4pL) в присутствии аденовируса. а) Получение лактосилированного пептида. 3,5 мг (1,82 мкмоль) разветвленного пептида Lys-(N∈-Lys)Lys- Gly-Ser-Gly-Gly-Ser-Gly-Gly-Ser-Gly-Gly-Cys, содержащего дитиопиридиновую группу для Сув, обрабатывают раствором 7,85 мг лактозы в 40 мкл 10 мМ водного раствора ацетата натрия с pH 5 при температуре 37oC. К раствору добавляют 4 аликвота 0,6 мг (10 мкмоль) цианоборгидрида натрия с соблюдением промежутка времени примерно 10 часов. По истечении всего 64 часов при температуре 37oC добавляют 0,5 мл буфера HEPES с pH 7,3 и 15 мг дитиотреитола. В результате фракционирования гель-фильтрацией (Сефадекс G-10, 12 х 130 мм, элюент: 20 мМ хлористого натрия) в атмосфере аргона получают 3,6 мл раствора лактозилированного пептида в свободной меркапто-форме (1,74 мкмоль согласно анализу Эльманна; выход: 84%). Пробы модифицированного пептида показывают цветную реакцию с анисовым альдегидом, но с нингидридом цветной реакции не имеет место. Это явление согласуется с предположением о том, что все 4 N-концевые аминогруппы лактозилированы. Конъюгат тетра-галактозного пептида и полилизина представлен на фиг. 6.
б) Получение модифицированного 3-дитиопиридинпропионатом полилизина
Интенсивно размешивая, 400 мкл 15 мМ этанольного раствора сукцинимидилгиопиридинпропионата (6,0 мкмоль) добавляют к прошедшему гель-фильтрацию раствору 0,60 мкмоль поли-L-лизина в виде гидробромида со средней длиной 290 лизиновых мономеров (далее: "pL290") в 1,2 мл 100 мМ буфера HEPES с pH 7,9. Через час добавляют 500 мкл 1 М ацетата натрия с pH 5 после гель- фильтрации на Сефадексе Г-25 с применением 100 мМ ацетата натрия. Раствор содержит 0,56 мкмоль pL2990 с 5,77 мкмоль связывающего вещества на основе дитиопиридина.
в) Сопряжение пептида с полилизином
Конъюгаты получают путем смешивания 1,5 мкмоль полученного на стадии а) лактолизированного пептида в 3 мл 20 мМ хлористого натрия с 0,146 мкл полученного на стадии б) модифицированного pL290 в 620 мкл 100 мМ буфера ацетата натрия в атмосфере аргона. После добавления 100 мкл 2 М буфера HEPES с pH 7,9 реакционную смесь оставляют стоять при комнатной температуре в течение 18 часов. Добавлением хлористого натрия концентрацию соли доводят до 0,66 М и конъюгаты выделяют путем катионообменной хроматографии на колонке HR 5/5 фирмы Фармация, Швеция, на ионите марки Mono S. Осуществляют градиентную элюацию с применением буфера А: 50 мМ HEPES с pH 7,3 и буфера В: 50 мМ HEPES плюс 3 М хлористого натрия. Элюируемые при концентрациях соли примерно 1,2 М - 1,8 М фракции собирают и объединяют в две конъюгатные фракции, обозначаемые ga14pL1 и ga14pL2. В результате диализа с применением 25 мМ буфера HEPES с pH 7,3 получают конъюгат gal4pLI, содержащий 24 нмоль модифицированного pL290, и конъюгат ga14pL2, содержащий 24,5 нмоль модифицированного pL290.
г) Получение конъюгатов асиалофетуина
Конъюгаты получают практически тем же образом, что и конъюгаты трансферина. Связывание асиалофетуина с полилизином осуществляют через дисульфидные мостики после модификации сукцинимидилпиридил-дитиопропионатом. Раствор 100 мг (2,2 мкмоль) асиалофетуина в 2 мл 100 мМ буфера HEPES с pH 7,9 подвергают гель-фильтрации на колонке Сефадекс Г-25. К получаемому раствору в количестве 4 мл, интенсивно размешивая, добавляют 330 мкл 15 мМ метанольного раствора сукцинимидилпиридил-дитиопропионата (5,0 мкмоль). Через час при комнатной температуре осуществляют очистку за счет гель-фильтрации на Сефадексе Г-25. При этом получают 5 мл раствора 1,4 мкмоль асиалофетуина, модифицированного 2,5 мкмоль связывающего вещества на основе дитиопиридина.
Конъюгаты получают путем смешивания 1,4 мкмоль асиалофетуина в 5 мл 100 мМ буфера HEPES с pH 7,9 с 0,33 мкмоль модифицированного pL190 (содержащего 1,07 мкмоль меркаптопропионатных групп; поступают тем же образом, что и для получения конъюгатов трансферина) в 6,5 мл 200 мМ буфера HEPES с pH 7,6 в атмосфере аргона. Реакционную смесь оставляют стоять при комнатной температуре в течение 24 часов. Конъюгаты выделяют из реакционной смеси путем катионообменной хроматографии на колонке марки HR 10/10 с применением ионита Mono S. Осуществляют градиентную элювию с применением буфера А: 50 мМ HEPES с pH 7,9 и буфера В: 50 мМ HEPES плюс 3 М хлорида натрия. До нагрузки колонки добавляют еще хлористый натрий до достижения конечной концентрации 0,6 М. Целевую фракцию элюируют при концентрации соли примерно 1,5 М. В результате диализа с применением буфера HBS получают конъюгаты, содержащие 0,52 мкмоль асиалофетуина, модифицированного 0,24 мкмоль pL190.
д) Трансфекция клеток HepG2 комплексами ДНК pRSVL
Клетки HepG2 выращивают в среде DMEM, содержащей 10% ТЭС, 100 единиц пенициллина/мл, 100 мкг/мл стрептомицина и 2 мМ глутамина в колбах марки Т25. Трансфекции осуществляют при плотности 400000 клеток на колбу. Перед трансфекции клетки промывают 4 мл свежей среды, содержащей 10% ТЭС. Непосредственно перед трансфекцией добавляют 7-хлор-4-[(4-дитиламино-1-метилбутил)-амино] -хинолина (далее: "хлорохин") в количестве, обеспечивающем конечную концентрацию в клеточной суспензии (плюс раствор ДНК), равную 100 мкМ. 10 мкг ДНК pRSVL в 330 мкл буфера HBS смешивают с указанными на фиг. 7 количеством конъюгата трансферина и полилизина с молекулярным весом 190 (TfpL), конъюгата асиалофетуина и полилизина с молекулярным весом 90 (AfpL), полилизина с молекулярным весом 290 (pL) и конъюгата тетра-галактозного пептида и полилизина (ga14pL), которые применяют в 170 мкл буфера HBS. В рамках конкурентных опытов применяют 240 мкг асиалофетуина [(gal)4pL + Af] или 30 мкг лактозилированного пептида [(gal)4pL + (gal)4], которые добавляют по истечении 30 минут. Смесь добавляют к клеткам, которые инкубируют при температуре 37oC в течение 4 часов, после чего среду трансфекции заменяют на 4 мл свежей среды DMEM, содержащей 10% ТЭС. По истечении 24 часов клетки собирают для осуществления анализа активности люциферазы. На фиг. 7 представлена общая активность люциферазы в трансфецированных клетках. По результатам опытов видно, что pL и TfpL показывают незначительную активность люциферазы, (gal)4pL дает почти ту же активность, что и AfpL, а (gal)4 и Af, соответственно, выступают в качестве конкурента в процессе связывания с асиалогликопротеином и поэтому их эффект незначителен.
е) Трансфекция клеток HepG2 комплексами ДНК pCMVL
Клетки HepG2 выращивают в чашках описанным на стадии г) образом до плотности 300000 клеток на чашку. Перед трансфекцией клетки промывают 1 мл свежей среды, содержащей 2% ТЭС. 6 мкг ДНК pCMVL в буфере HBS смешивают с указанными на фиг. 8 количествами конъюгата трансферина и полилизина с молекулярным весом 190 (TfpL), конъюгата асиалофетуина и полилизина (AfpL), полилизина с молекулярным весом 290 (pLys290), конъюгата (gal)4pL1 и (gal)4pL2, которые содержатся в 170 мкл HBS. По истечении 30 минут к каждому комплексу конъюгата и ДНК добавляют 1 мл среды DMEM, содержащей 2% ТЭС, и 50 мкл раствора аденовируса d1312C. В рамках конкурентных экспериментов добавляют 30 мкг лактозилированного пептида (gal)4pL [(gal)4pL1 + (gal)4] или [(gal)4pL2 + (gal)4]. Смесь добавляют к клеткам, которые подвергают инкубации при температуре 37oC в течение 2 часов, после чего добавляют 1,5 мл среды, содержащей 10% ТЭС. По истечении 24 часов клетки собирают для осуществления анализа активности люциферазы. Представленные на фиг. 8 значения представляют собой общую активность люциферазы в трансфецированных клетках. Видно, что pLys290 проявляют определенный эффект, (gal)4pL обеспечивают получение усиленного эффекта, а добавление (gal)4, который выступает в качестве конкурента в процессе связывания с рецептором асиалогликопротеина, приводит к уменьшению эффекта до величины, достигаемой полилизином.
ж) Трансфекция клеток TIB 73 комплексами ДНК pCMVL
Клетки, выделенные из эмбриональной мышиной печени клеточной линии АТСС Т1В73 (BNL CL.2) выращивают при температуре 37oC в содержащей 0,4% глюкозы среде DMEM, дополненной 10% инактивированной термообработкой ТЭС, содержащей 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ глутамина, в присутствии 5% двуокиси углерода. Трансфекции осуществляют при плотности 300000 клеток на чашку. Перед трансфекцией клетки промывают 1 мл свежей среды, содержащей 2% ТЭС.
6 мкг ДНК pCMVL в 300 мкл HBS смешивают с указанными на фиг. 9 количествами конъюгата мышиного трансферина и полилизина с молекулярным весом 290 (mTfpL), конъюгата асиалофетуина и полилизина (AfpL), полилизина с молекулярным весом 290 (pLys290), конъюгата (gal)4pL1 и конъюгата (gal)4pL2, которые применяют в среде 170 мл HBS. По истечении 30 минут к каждому комплексу конъюгата и ДНК добавляют 1 мл среды DMEM, содержащей 2% ТЭС, и 50 мкл раствора аденовируса d1312. Смесь добавляют к клеткам, которые подвергают инкубации при температуре 37oC в течение 2 часов, после чего добавляют 1,5 мл среды, содержащей 10% ТЭС. Через два часа среду трансфекции заменяют на 4 мл свежей среды. По истечении 24 часов клетки собирают для осуществления анализа активности люциферазы. На фиг. 9В представлена общая активность люциферазы в трансфецированных клетках.
Проводят сравнительные опыты, в которых трансфекцию осуществляют без аденовируса в присутствии хлорохина. При этом процесс тоже осуществляют при плотности 300000 клеток на чашку. Перед трансфекцией клетки промывают 1 мл свежей среды, содержащей 2% ТЭС. Непосредственно перед трансфекцией хлорохин добавляют в количестве, обеспечивающем конечную концентрацию в клеточной суспензии (плюс раствор ДНК), равную 100 мкмоль. 6 мкг ДНК pCMVL в 330 мкл HBS смешивают с указанным на фиг. 9В количеством mTfpL, AfpL, pLys290, (gal)4pL1 и (gal)4pL2, которые применяют в среде 170 мкл HBS. По истечении 30 минут комплексы ДНК добавляют к клеткам, которые подвергают инкубации при температуре 37oC в течение 2 часов, после чего добавляют 1,5 мл среды, содержащей 10% ТЭС и 100 мкмоль хлорохина. Через два часа среду трансфекции заменяют на 4 мл свежей среды. По истечении 24 часов клетки собирают для осуществления анализа активности люциферазы. Результаты опытов представлены на фиг. 9А.
Пример 7. Введение ДНК в клетки Т
а) Получение конъюгатов антиCD7 и полилизина с молекулярным весом 190
Раствор 1,3 мг антитела антиCD7 в 50 мМ буфера HEPES с pH 7,9 смешивают с 49 мкл 1 мМ этанольного раствора сукцинимидилпиридил- дитиопропионата. Через час при комнатной температуре смесь подвергают гель-фильтрации на колонке сефадекс G-25 с применением в качестве элюента 50 мМ буфера HEPES с pH 7,9.
При этом получают 1,19 мг (7,5 нмоль) антиCD7, модифицированного 33 нмоль пиридилдитиопропионатных групп. Поли-(L)-лизин с молекулярным весом 190, меченый флуоресцирующим агентом, аналогично модифицируют сукцинимидилпиридил- дитиопропионатом и переводят в форму, модифицированную свободными меркапто-группами, за счет обработки дитиотреитолом с последующей гель-фильтрацией. Раствор 11 нмоль полилизина с молекулярным весом 190, модифицированного 35 нмоль меркапто-групп, в 0,2 мл 30 мМ буфера ацетата натрия смешивают с модифицированным антиCD7 в 0,5 мл 300 мМ буфера HEPES с pH 7,9 в условиях исключения кислорода и смесь оставляют стоять в течение ночи при комнатной температуре. Реакционную смесь доводят до концентрации примерно 0,6 М путем добавления 5 М хлористого натрия. Выделение конъюгатов осуществляют путем ионообменной хроматографии на йоните Mono S (элюент: 50 мМ буфера HEPES с pH 7,3, солевой градиент: 0,6-3 М хлористого натрия). После диализа с применением 10 мМ буфера HEPES с pH 7,3 получают конъюгат, состоящий из 0,51 мг (3,2 нмоль) антитела антиCD7, модифицированного 6,2 нмоль полилизина с молекулярным весом 190.
б) Получение конъюгатов gp120 и полилизина с молекулярным весом 190.
Связывание осуществляют через тиоэфирные группы после модификации N-окси-сукцинимидным эфиром 6-малеимидокапроновой кислоты. Процесс осуществляют следующим образом.
Раствор 2 мг рекомбинантного gp120 в 0,45 мл 100 мМ буфера HEPES с pH 7,9 смешивают с 17 мкл 10 мМ раствора N-оксисукцинимидного эфира 6-малеимидокапроновой кислоты в диметилформамиде. Через час при комнатной температуре осуществляют гель-фильтрацию на колонке Сефадекс Г-25 с применением в качестве элюента 100 мМ буфера HEPES с pH 7,9. Получают 1,2 мл целевого раствора, который в условиях исключения кислорода немедленно подвергают реакции с раствором 9,3 нмоль полилизина с молекулярным весом 190, меченого флуоресцирующим агентом и модифицированного 30 нмоль меркапто- групп в 90 мкл 30 мМ ацетата натрия с pH 5,0. Реакционную смесь оставляют стоять в течение ночи при комнатной температуре, после чего ее концентрацию доводят примерно до 0,6 М путем добавления 5 М хлористого натрия. Конъюгаты выделяют путем ионообменной хроматографии на колонке Mono S с применением в качестве элюента 50 мМ HEPES с pH 7,3 (солевой градиент: 0,6 - 3 М хлористого натрия). После фракционирования и диализа с применением 25 мМ буфера HEPES с pH 7,3 получают конъюгаты А, Б и В следующего состава. Конъюгат А состоит из 0,40 мг gp120, модифицированного 1,9 нмоль полилизина 190, конъюгат Б - 0,25 мг gp120, модифицированного 2,5 нмоль полилизина 190, а конъюгат В - 0,1 мг gp120, модифицированного 1,6 нмоль полилизина 190.
ДНК pCMVL в количестве 6 мкг на пробу переводят в комплекс с полилизном 190 или его конъюгатами в 500 мкл HBS. Полученные комплексы полилизина и ДНК добавляют к клеткам Н9 (106 клеток в 5 мл среды RPMI, содержащей 2% ДЕС) или к первичным лимфоцитам человека (3 • 106 клеток в среде IMDM, содержащей 2% ДЕС). Через 5 минут добавляют аденовирус d1312 в указанном на фиг.10 количестве. Клетки инкубируют при температуре 37oC в течение 90 минут, после чего 15 мл среды RPMI (в случае клеток Н9) или IMDM (в случае первичных лимфоцитов) и 20% ДЕС добавляют к каждой пробе. Клетки инкубируют при температуре 37oC в течение лишь 24 часов, после чего собирают для осуществления анализа активности люциферазы. Результаты опытов представлены на фиг. 10 A (в случае клеток Н9) и фиг. 10 В (в случае первичных лимфоцитов). На фиг. 10 А видно, что конъюгат анти-CD7 (следы 7-9) и конъюгат gp120 (следы 10-12) показывают наилучшие результаты в отношении переноса гена, достигаемого аденовирусом, тогда как конъюгат gp120 проявляет явную экспрессию гена люциферазы даже в отсутствие аденовируса. Следует еще отметить, что только конъюгат gp120 проявляет способность к введению ДНК в первичные лимфоциты при условии наличия аденовируса (см. фиг. 10 Б, следы 7,8).
Пример 8. Инактивация аденовирусов
а) Инактивация УФ-облучением.
Полученный выше указанным образом препарат аденовируса d1312 подают в снабженную ячейками диаметром 2 см чашку (300 мкл на ячейку). На расстоянии 8 см от чашки устанавливают две УФ лампы марки G15 Т8 инофирмы Филипс. Вирус подвергают облучению ультрафиолетовыми лучами в течение указанного на рис. 11А времени. Аликвоты каждого препарата исследуют на титр вируса и определяют способность к улучшению переноса гена в клетки гена при применении конъюгатов полилизина и трансферина. Культивацию клеток и трансфекцию осуществляют в основном описанным выше образом. Применяемые для трансфекции компоненты представлены на фиг.11А. В 500 мкл буфера HBS получают комплексы ДНК pCMVL и 12 мкг TfpL, которые добавляют к 3 • 105 клеток HeLa в 1 мл среды DMEM, содержащей 2% ТЭС. Через 5 минут к каждой культуре добавляют 54 мкл каждого препарата вируса и культуру инкубируют при температуре 37oC в течение 60-120 часов. Потом к каждой культуре добавляют 5 мл аликвота среды DMEM, содержащей 10% ТЭС, инкубацию продолжают при температуре 37oC в течение 24 часов, после чего осуществляют сбор культур на исследование активности люциферазы. Количество 54 мкл необлученного вируса не входит в пределы насыщения, то есть опыт чувствителен к количеству вируса, которое превышает указанное по крайней мере в 3 раза. Результаты, полученные для экспрессии люциферазы, представлены на фиг. 11 Б (темные прямоугольники). Титр вируса каждого препарата определяют с применением Ela-комплиментирующей клеточной линии 293. В среде DMEM, содержащей 2% ТЭС, приготовляют ряд разбавленных групп облученного и необлученного вируса. Параллельно в 200 мкл среды DMEM, содержащей 2% ТЭС, приготовляют пробы 5 • 104 клеток 293 (с применением чашки с ячейками диаметром 2 см). Аликвот 5 мкл каждой разбавленной пробы подают в каждую ячейку. С целью достижения связывания вируса с клетками осуществляют инкубацию при температуре 37oC в течение 90 минут, после чего в каждую ячейку подают 2 мл среды DMEM, содержащей 10% ТЭС. Через 48 часов культуры исследуют на степень заболевания клеток. Разбавленная проба вируса с менее 50% клеток в культуре, показывающая значительную степень заболевания через 48 часов, указывает на относительное количество инфекционного вируса в каждом препарате. Полученные результаты представлены на фиг. 11 Б (открытые прямоугольники). Результаты опыта данного примера показывают, что вызываемое УФ-облучением снижение титра вируса на 4 log связано только с 20-кратным уменьшением степени переноса гена люциферазы. Таким образом решающие для инфективности вируса механизмы можно упразднять без существенного отрицательного влияния на способность вируса к улучшению переноса гена. Было обнаружено, что в малых дозах вируса обеспечиваемое вирусом улучшение переноса гена немного падает (см. фиг. 11А, следы 3-6), и что этот эффект проявляется ярче при высоких концентрациях (см. следы 7 -10).
б) Инактивация аденовирусов формальдегидом.
2 мл препарата аденовируса пропускают через колонку сефадекс Г-25 емкостью 10 мл, которую предварительно уравновешивают при помощи 150 мМ хлористого натрия, 25 мМ буфера HEPES с pH 7,9 и 10% глицерина. Аликвоты прошедшего гель-фильтрацию препарата вируса инкубируют на льду в течение 20 часов в присутствии 0,01%, 0,1% и 1% формальдегида. Кроме того, проводят контрольный опыт без применения формальдегида. Добавляют буфер трис с pH 7,4 до установления концентрации 100 мМ, после чего пробы подвергают двухстадийному диализу, причем на первой стадии работают с 1 л 150 мМ хлористого натрия, 50 мМ трис с pH 7,4 и 50% глицерина в течение двух часов, а на второй стадии диализ осуществляют в течение ночи с применением 2 • 1 л 150 мМ хлористого натрия, 20 мМ HEPES с pH 7,9 и 50% глицерина. Затем аликвоты вируса исследуют на титр с применением клеток 293. Затем определяют действие обработанного формальдегидом вируса на перенос гена в клетки HeLa (300000) за счет измерения активности люциферазы указанным выше образом. Обработка вируса формальдегидом в количестве 0,01% и 0,1% приводит к небольшому снижению активности по переносу гена (примерно 10-кратное уменьшение при 0,1%). Хотя обработка 1% формальдегида вызывает существенную потерю активности по переносу гена, 90 мкл вируса все еще проявляют способность к экспрессии гена, соответствующей 104 световым единицам. Наблюдаемое при обработке 0,1% формальдегида снижение титра вируса до 105 бляшкообразующих единиц связано с уменьшением активности люциферазы только на 10%. Результаты данного опыта представлены на фиг.12 А.
в) Инактивация аденовирусов длинноволновым УФ-облучением и обработкой 8-метокси-псораленом (псорален = 7Н-фуро[3,3- g][1]бензопиран-7-он).
Аликвоты очищенного вируса добавляют к 0,33 мкг/мкл 8-метокси-псоралена в диметилсульфоксиде и на льду подвергают воздействию источника УФ света длиной 365 нм, размещенного на расстоянии 4 см от проб. Облучение осуществляют в течение 15 - 30 минут. Затем пробы вируса пропускают через колонку сефадекс Г-25, которую предварительно уравновешивают при помощи буфера HBS и 40% глицерина. Затем вирусные препараты исследуют на их способность к улучшению переноса комплекса pCMVL и конъюгата hTfpL в клетки HeLa путем определения световых единиц активности люциферазы, а также на их способность к репликации в клетках 293 за счет определения титра. Результаты опыта представлены на фиг.12 В.
В нижеследующих примерах, которые иллюстрируют улучшение поглощения клеткой комплексов ДНК и конъюгата трансферина и полилизина в присутствии ретровирусов, используют следующие материалы и методы.
Конъюгаты трансферина и полилизина с молекулярным весом 190 и комплексы конъюгата и ДНК получают описанным выше образом. Клетки NIH3T3 выращивают в среде DMEM с добавкой 10% ТЭС, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ глутамина. Клетки выращивают в количестве 5 - 7 • 105 на каждую пробу за 18-24 часов до трансфекции. Непосредственно перед трансфекцией клетки подают в свежую среду, после чего последовательно добавляют: 100 мкмоль (если ничего другого не указано) хлорохина, комплекс ДНК и конъюгата полилизина и трансферина и препарат ретровируса. Затем клетки инкубируют при температуре 37oC в течение 4 часов, после чего среду заменяют и через 24 часа клетки собирают. Получение экстрактов осуществляют тремя циклами замораживания и оттаивания. После стандартизации в отношении содержания белка аликвоты экстрактов исследуют на активность люциферазы указанным выше образом.
Пример 9. Трансфекция клеток NIH3T3 вирусом Молонеа
106 клеток NIH3T3 подвергают трансфекции комплексом ДНК и конъюгата TfpL в присутствии 100 мкмоль хлорохина или же без применения последнего. Результаты опыта представлены на рис.13. Было найдено, что без применения хлорохина величины по активности люциферазы достигают только фонового уровня (след 1), тогда как в присутствии хлорохина можно определять высокую экспрессию репортерного гена pRSVL (см. след 2). Повышающиеся количества вируса лейкоза Молонеа (обозначенного RVS на рис.13), которые добавляют к клеткам одновременно с указанным комплексом, приводят к дальнейшему улучшению экспрессии гена люциферазы.
Пример 10. Влияние вируса Молонеа на перенос гена в случае трансфекции клеток NIH3Т3 комплексом ДНК и конъюгата трансферина и полилизина. Используемые в примере 9 препараты вируса представляют собой сырую, не фракционированную надосадочную жидкость экспрессирующих ретровирус клеток. Для подтверждения того, что достигаемое в присутствии препарата вируса улучшение переноса ДНК можно действительно приписывать вирусу, надосадочную жидкость подвергают очистке диализом и концентрацией на фактор 10. Если ретровирус ответствен за улучшение, то удерживаемая мембраной активность (несмотря на любую инактивацию крайне неустойчивого ретровируса во время концентрации) должна превышать активность первоначальной надосадочной жидкости примерно в 10 раз. Как и в предыдущем примере клетки NIH3T3 в количестве 106 подвергают трансфекции в условиях, представленных на фиг. 14. На фиг. 14 видно, что улучшающее перенос гена действие свойственно продукту удержания мембраной (применяют 20 - 600 мкл, следы 3 - 6). Кроме того, было найдено, что 200 и 600 мкл 10-кратно концентрированного препарата проявляют примерно половину активности 2 и 6 мл первоначального, то есть неконцентрированного препарата ретровируса (следы 7, 8). Осуществляют еще параллельные опыты с применением клеток К 562 человека, которые не имеют рецептора для экотропного мышиного ретровируса. Как и следовало ожидать, при этом улучшение экспрессии гена не наблюдается.
Пример 11. Взаимодействие между трансферином и его рецептором играет роль в переносе гена, которому содействует вирус Молонеа
Ретровирус исследуют на способность к переносу в клетку плазмидной ДНК в виде комплекса с одним полилизином с тем, чтобы выяснить вопрос о приписании переноса комплекса pRSVL и конъюгата TfpL в клетки к неспецифичной связи полилизина с ретровирусом, а также дальше изучать механизм захода в клетку. Количество применяемого полилизина соответствует оптимальному количеству, которое обеспечивает полную конденсацию плазмидной ДНК. Опыты, результаты которых представлены на фиг. 15, показывают, что в отсутствие хлорохина репортерный ген не экспрессируется ни при применении комплексов pRSVL и конъюгата TfpL, ни при применении комплекса pRSVL и полилизина (см. следы 1, 2). Однако в присутствии ретровируса репортерная ДНК, которую применяют в виде комплекса с конъюгатом TfpL, экспрессируется, тогда как при применении комплекса ДНК и полилизина экспрессии не наблюдается (см. следы 3, 4 по сравнению со следами 5, 6). Кроме того, данные опыты показывают, что присутствие избыточного свободного трансферина приводит к снижению переноса ДНК, достигаемого ретровирусом (см. следы 7, 8). Эти результаты подкрепляют предположение о том, что взаимодействие между трансферином и его рецептором играет существенную роль в улучшении поглощения ДНК, обеспечиваемого ретровирусом.
Пример 12. Влияние pH на обеспечиваемый ретровирусами перенос гена. Опыты данного примера направлены на исследование влияния pH на способность ретровирусов к улучшению переноса гена. В этих опытах трансфекцию осуществляют вышеуказанным образом. Применяют два общеизвестных ингибитора снижения pH ядрышка, то есть монензин и хлористый аммоний. Результаты опытов представлены на фиг. 16. Исследование действия обоих веществ на перенос комплекса ДНК и конъюгата TfpL показывает, что в функциональном отношении ни одно из обоих веществ не может заменить хлорохин. Однако при более высоких концентрациях хлористого аммония наблюдается небольшое улучшение экспрессии гена люциферазы (см. следы 1 - 5). Один ретровирус показывает небольшое улучшение переноса ДНК, что и наблюдается в предыдущих примерах (см. след 6). Резкое улучшение наблюдается при применении ретровируса в присутствии 1 мкмоль монензина (см. след 7). Менее сильное действие наблюдается при более высокой концентрации монензина (см. след 8), а также в присутствии хлористого аммония (см. следы 9, 10).
Пример 13. Улучшение переноса гена, достигаемое конъюгатами трансферина в присутствии N-концевого эндосомолитического пептида гемагглютинина НА2 гриппа
а) Синтез пептида
Пептид с последовательностью:
Gly-Leu-Phe-Glu-Ala-Ile-Ala-Gly-Phe-Ile-Glu-Asn-Gly-Trp-Glu- Gly-Met-Ile-Asp-Gly-Gly-Gly-Cys синтезируют известными приемами с применением флуоренилметоксикарбонила. В качестве защитных групп боковой цепи применяют третичный бутил (для Cys, Glu и Asp) и тритил для Asn. После реакции осуществляют анализ степени связывания с применением нингидрина. При этом устанавливают степень > 98% на каждой стадии. Начиная с аминокислоты 19 осуществляют двойное сочетание. N-концевые флуоренилметоксикарбонильные группы удаляют с части пептидной смолы путем обработки 20%-ным пиперидином в N-метилпирролидине. Затем защищенные флуоренилметоксикарбонилом фракции и не защищенные фракции промывают дихлорметаном и сушат в высоком вакууме. Получают 294 мг свободной от флуоренилметоксикарбонильных групп пептидной смолы и 367 мг защищенной флуоренилметоксикарбонильными группами пептидной смолы. 111 мг свободной от флуоренилметоксикарбонилльных групп пептидной смолы расщепляют путем обработки смесью, состоящей из 10 мл трифторуксусной кислоты, 0,75 г фенола, 300 мкл этандитиола, 250 мкл этилметилсульфида и 500 мкл воды, в течение 90 минут. Пептид отделяют от смолы путем фильтрации. Смолу промывают дихлорметаном и добавляют к фильтрату, который сгущают до объема примерно 2 мл, после чего, размешивая, его каплями добавляют к 40 мл простого диэтилового эфира. Осадок пептида удаляют центрифугированием и эфирную надосадочную жидкость удаляют. Осадок промывают три раза 40 мл простого диэтилового эфира и сушат в высоком вакууме. Получают 58 мг сырого продукта, который растворяют в 3,5 мл 20 мМ бикарбоната аммония, содержащего 300 мкл 25%-го аммиака/л. Раствор подвергают гель-фильтрации на колонке сефадекс Г-25 с применением того же буфера. Затем элюат подают на колонку марки Mono Q размером 100 х 14 мм, в которой процесс осуществляют в следующих условиях. Градиент: 0 - 10 минут: 100% А, 10-100 минут: 0-100% Б, при этом А = 20 мМ бикарбоната аммония + 300 мкл 25% аммиака/л, аБ=А+3М хлористого натрия. Измеряют при 280 нм, а детекция флуоресценции Тrp при 354 нм. Скорость подачи: 1 мл/мин. Продукт элюируют 1 М хлористым натрием. Основную фракцию колонки далее очищают путем обратнофазной жидкостной хроматографии, которую осуществляют под давлением в колонке размером 250 х 10 мм марки BIORAD-Hi-Pore RP 304. При этом соблюдают следующие условия. Градиент: 50-100% буфера А в течение 12,5 мин, 12,5 - 25 мин: 100% Б; при этом А = 20 мМ бикарбоната аммония + 300 мкл 25% аммиака/л, а Б = А в 98%-ном метаноле. Скорость подачи: 3 мл/мин. Измеряют при 237 нм. Продукт элюируют при 100% Б.
Целевые фракции упаривают, повторно растворяют в буфере А и затем лиофилизуют. Получают 8,4 мг очищенного жидкостной хроматографией под давлением продукта в цистеин-защищенной форме. Этот пептид обозначается Р16. В целях получения Р16 в свободной меркапто-форме, защищенное третичным бутилом вещество подвергают обработке смесью тиоанизола, этандиэтиола, трифторуксусной кислоты и трифторметансульфокислоты в соотношении 2 : 1 : 40 : 3 (трифторметансульфокислоту добавляют в качестве последнего компонента) при комнатной температуре в течение 30 минут. Пептид выделяют путем осаждения простым диэтиловым эфиром и последующей гель-фильтрации на Сефадексе Г-25 с применением вышеупомянутого буфера А в атмосфере аргона.
б) Связывание пептида гриппа с полилизином
б1) Непосредственное связывание при помощи сукцинимидилпиридилдитиопропионата
19,8 мг гидробромида полилизина с молекулярным весом 300 подвергают гель-фильтрации на колонке марки Сефадекс Г-25 с применением раствора ацетата натрия с pH 5, направленной на удаление низкомолекулярных фракций. Согласно данным анализа с применением нингидрина после гель-фильтрации концентрация полилизина составляет 3,16 мг/мл. Добавлением 1 М гидроокиси натрия pH раствора доводят до 7-8. К 2,5 мл раствора полилизина (содержание полилизина 7,9 мг = 0,13 мкмоль) добавляют 0,64 мкмоль сукцинимидилпиридилдитиопропионата в виде 40 мМ раствора в абсолютном этаноле. Таким образом мольное соотношение сукцинимидилпиридилдитиопропионата и полилизина составляет 5:1. Смеси дают реагироваться в течение ночи, после чего ее подвергают гель- фильтрации на колонке марки Сефадекс G-25 с применением 20 мМ бикарбоната аммония с pH 8,2. После восстановления аликвота фильтрата дитиотреитолом определяют содержание тиопиридона. Согласно данным этого анализа реакция протекала полностью. 0,3 мкмоль модифицированного пиридилдитиопропионатом полилизина (в расчете на мкмоль пиридилдитиопропионата) подвергают взаимодействию с 0,35 мкмоль пептида в тиольной форме. Белый осадок, который получается при смешивании пептида и полилизина, растворяют в 2 М гидрохлорида гуанидиния. Реакционную смесь оставляют стоять в течение ночи, после чего осуществляют фотометрическое определение содержания тиопиридона в реакционной смеси. Данные этого анализа подтверждают полноту реакции. Затем смесь два раза диализуют с применением 2 л 20 мМ буфера HEPES, содержащего 0,5 М гидрохлорида гуанидиния. Получаемый раствор подают на колонку марки Mono S размером 0,7 х 6 см. При этом соблюдают следующие условия. Градиент: 0 - 20 мин: 100% А, 20-140 мин: 0-100% Б, причем А означает 20 мМ HEPES с pH 7,3/0,5 М гидрохлорида гуанидиния, а В - 20 мМ HEPES с pH 7,3 плюс 3 М гидрохлорида гуанидиния. Скорость подачи: 0,3 мл/мин. Детекция при 280 нм, детекция флуоресценции при 354 нм, а возбуждение при 280 нм. Целевую фракцию элюируют 1,5 М гидрохлорида гуанидиния с последующим диализом с применением 2 л HBS. Согласно последующему определению концентрации полилизина с применением нингидрина его концентрация составляет примерно 1,14 мг/мл. Количество пептида в растворе конъюгата рассчитывают с учетом его абсорбции при 280 нм. По этим данным молярное соотношение пептида и полилизина составляет 4 : 1.
б2) Связывание при помощи связывающего вещества на основе полиэтиленгликоля
14,6 мг гидробромида полилизина с молекулярным весом 300 подвергают гель-фильтрации аналогично стадии б1). Согласно данным опыта с применением нингидрина концентрация полилизина после гель- фильтрации составляет 4,93 мг/мл. Добавлением 1 М гидроокиси натрия pH раствора доводят до 7-8. К 2,7 мл раствора полилизина (содержание полилизина 13,3 мг = 0,22 мкмоль) добавляют 4,33 мкмоль пиридилдитиопропионата в виде 30 мМ раствора в абсолютном этаноле. Таким образом молярное соотношение пиридилдитиопропионата и полилизина составляет 20:1. Через 90 минут реакционную смесь подвергают гель-фильтрации на колонке Сефадекс Г-25 с применением 0,1 М ацетата натрия и 3М гидрохлорида гуанидиния. После восстановления аликвота фильтрата дитиотреитолом определяют содержание тиопиридона. Согласно данному анализу целевая фракция содержит 3,62 мкмоль пиридилдитиопропионата. Модифицированный пиридилдитиопропионатом полилизин подвергают восстановлению применяемым в количестве 79 мг дитиотреитолом в течение 2 часов. Раствор снова фильтруют на колонке Г-25 в указанных условиях. Согласно данным метода Эльмана концентрация тиола составляет 3,15 мкмоль в 2,224 мл.
17,6 мг (5 мкмоль) полиоксиэтилен-бис(6-аминогексила) растворяют в 500 мкл 20 мМ бикарбоната натрия и 3 М гидрохлорида гуанидиния с pH 7-8, после чего подвергают взаимодействию с 13,8 мг N-оксисукцинимидного эфира ε -малеимидо-капроновой кислоты (= 44,6 мкмоль), растворенного в 300 мкл диметилформамида. По истечении 30 минут раствор подвергают гель-фильтрации на колонке Г-25 с применением 20 мМ бикарбоната натрия и 3 М гидрохлорида гуанидиния. Согласно данным фотометрического определения малеимидо-группы при 300 нм концентрация прореагировавшего полиоксиэтилен-бис(6- аминогексила) составляет 6,36 мкмоль в 2 мл раствора. К 1,05 мл этого раствора (соответствует 3,34 мкмоль указанного эфира) каплями добавляют 1,39 мкмоль пептида в тиольной форме в 2,5 мл 20 мМ бикарбоната натрия и 3 М гидрохлорида гуанидиния. При этом смесь интенсивно размешивают в атмосфере аргона. По истечении 15 минут свободные тиольные группы не могут больше обнаруживаться.
Раствор восстановленного меркапто-модифицированного полилизина доводят до pH 7-8 путем добавления 1 М гидроокиси натрия. 1,37 мл этого раствора, интенсивно размешивая, добавляют к вышеуказанной реакционной смеси. Таким образом молярное соотношение пептида-SH, N-оксисукцинимидного эфира ε-малеимидо-капроновой кислоты и полилизина-SH составляет 1 : 2,4 : 1,4 (из расчета на указанный эфир и SH). Свободные тиольные группы не могут больше обнаруживаться через 150 минут. Реакционную смесь подвергают диализу в течение ночи с применением 2 л 20 мМ HEPES с pH 7,3 плюс 0,6 М хлористого натрия, после чего подают на колонку марки Mono S. Процесс осуществляют в следующих условиях. Градиент: 1 мл водной фазы (10 мМ HEPES с pH 7,3; 100 мМ натриевой соли флуоресцеин-2,7-бис-метилениминодиуксусной кислоты (далее: кальцеин); 150 мМ хлористого натрия) тщательно промывают эфирной фазой и обрабатывают ультразвуком при температуре 0oC в течение 5 минут. Через 30 минут на льду материал еще раз обрабатывают ультразвуком в течение дальнейших 10 минут. Получаемую устойчивую эмульсию медленно упаривают. После удаления диэтилового эфира при 100 мбар добавляют 0,75 мл вышеупомянутой водной фазы. Остаток диэтилового эфира удаляют путем дополнительного упаривания при давлении 50 мбар в течение 30 минут. Получают 1,7 мл остатка, который подвергают центрифугированию при 500 об/мин. 1,0 мл продукта центрифугирования пропускают через микропористую поликарбонатную мембрану величиной пор 0,1 мкм с получением 0,7 мл раствора липосом. Липосомы отделяют от невключенного материала путем гель-фильтрации (Сефадекс Г-50 медиум фирмы Фармация, Швеция; 23 мл объема геля, 10 мМ HEPES с pH 7,3/150 мМ хлористого натрия). Собирают 6 фракций объемом 500 мкл. Липидный фосфор определяют при 2 мМ.
г) Определение выделения из липосом содержимого
Выделение из липосом содержимого определяют за счет того, что измеряют высвобождение заключенного кальцеина и получаемого разбавления, которое прекращает гашение флуоресценции. Флуоресценцию кальцеина определяют при помощи спектрального флуорометра типа Kontron SMF 25 (возбуждение при 490 нм, эмиссия при 515 нм). Для этой цели 100 мкл аликвотов вышеуказанного раствора липосом разбавляют 100 раз 0,1 М ацетата натрия/50 мМ хлористого натрия или 10 мМ НЕPES/150 мМ хлористого натрия с соответствующей величиной pH (4,3, 4,5, 5,0, 6,0, 7,3). К получаемому раствору объемом 1 мл добавляют 2,5 мкл пептида в защищенной третичным бутилом форме (1 мкг/мкл раствора в HBS) при одновременном смешивании в атмосфере аргона (конечная концентрация: 400 нМ пептида). Флуоресценцию кальцеина определяют в разные моменты после добавления пептида. Значения для обеспечения 100%-ного выделения определяют путем добавления 2 мкл тритона X-100. Такие же приемы применяют для определения флуоресценции кальцеина после добавления к раствору липосом конъюгатов пептида и полилизина. При этом к 1 мл раствора липосом добавляют 2,5 мкг конъюгата (1 мкг/мкл концентрации в пересчете на количество одного полилизина). При этом конечная концентрация составляет 20 нм модифицированного пептида. Аналогичным образом анализу подвергают 2,5 мкг конъюгата пептида и полилизина после инкубации вместе с 5 мкг ДНК в течение 15 минут. Было найдено, что пептид вызывает выделение из липосом содержимого только в кислой области pH (см. фиг. 17). Конъюгат пептида является активным при значительно более низкой величине pH, хотя даже при нейтральной величине pH наблюдают значительную активность, которая далее увеличивается по мере снижения величины pH. Комплекс конъюгата и ДНК вызывает упразднение активности при нейтральной величине pH, тогда, как при кислой величине pH наблюдается заметная активность. На фиг. 17 пептид обозначен Influ.
д) Трансфекция клеток К562
Клетки К562 выращивают в виде суспензии в среде RPMI 1640 (плюс 2 г бикарбоната натрия/л, 10% ТЭС, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ глутамина) до достижения плотности 500000 клеток/мл. За 12 до 20 часов до трансфекции клетки подают в свежую среду, содержащую 50 мкМ десферриоксамина, 30-амино-3.14.25-триокси-3.9.14.20.25- пентаазатриаконтан-2.10.13.21.24-пентаона (в целях повышения числа рецепторов трансферина). В день трансфекции клетки собирают, суспендируют в свежей среде, содержащей 10% ТЭС и 50 мкм десферриоксамина (250000 клеток на мл) и порциями по 2 мл подают в чашку, снабженную 24 ячейками. 6 мкг ДНК pCMVL в 160 мкл HBS смешивают с указанным на фиг. 18 количеством конъюгата TfpL или с pL300 в 160 мкл HBS, через 15 минут добавляют указанные количества конъюгата пептида гриппа и полилизина (P16pL) и через дальнейшие 15 минут смесь добавляют к клеткам К562, которые инкубируют при температуре 37oC в течение 24 часов, после чего осуществляют их сбор для определения активности люциферазы описанным в предыдущих примерах образом. Представленные на фиг. 18 значения представляют собой общую активность люциферазы трансфецированных клеток.
е) Трансфекция клеток HeLa
Клетки HeLa культивируют в чашках диаметром 6 см описанным выше образом. Трансфекцию осуществляют при плотности 300000 клеток на чашку. Перед трансфекцией клетки инкубируют в 1 мл свежей среды, содержащей 2% ТЭС. 6 мкг ДНК pCMVL в 160 мкл HBS смешивают с указанным на фиг. 19 количеством конъюгата TfpL или полилизином с молекулярным весом 300 (pL300) или же смесью конъюгата pL300 в 160 мкл HBS. Через 15 минут добавляют указанные количества конъюгата пептида гриппа и pL (P16pL) и через дальнейшие 15 минут смесь добавляют к клеткам, которые инкубируют при температуре 37oC в течение 2 часов, после чего добавляют 2,5 мл свежей среды, содержащей 10% ТЭС. Клетки инкубируют при температуре 37oC в течение 24 часов, после чего осуществляют их сбор для определения активности люциферазы описанным в предыдущих примерах образом. Представленные на фиг. 19 значения представляют собой общую активность люциферазы трансфецированных клеток.
Пример 14. Улучшение переноса гена, достигаемое конъюгатами трансферина с помощью второго N-концевого эндосомолитического пептида гемаглютинина НА2 гриппа
а) Синтез конъюгата пептида гриппа и полилизина
Пептид последовательностью Gly-Leu-Phe-Gly-Ala-Ile-Ala-Gly- Phe-Ile-Glu-Asn-Gly-Trp-Glu-Gly-Met-Ile-Asp-Gly-Gly-Gly-Cys (обозначенный P41) синтезируют известным методом. Связывание пептида гриппа с полилизином (pL300) осуществляют тем же образом, что и в примере 13, b1), т.е. при помощи сукцинимидил- пиридилдитиопропионата. При этом получают конъюгаты (P41pL), в которых мольное соотношение пептида и полилизина составляет 4 : 1.
б) Трансфекция клеток HeLa
Клетки HeLa выращивают в среде DMEM, содержащей 5% ТЭС, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ глутамина, что осуществляют в чашках диаметром 6 см. Трансфекцию осуществляют при плотности 300000 клеток на чашку. Перед трансфекцией клетки инкубируют в 165 мл свежей среды, содержащей 2% ТЭС. 6 мкг ДНК pCMVL в 160 мкл HBS (150 мМ хлористого натрия, 20 мМ HEPES с pH 7,3) смешивают с 6 мкг конъюгата TfpL190B в 160 мкл HBS, через 15 минут добавляют 10 мкг конъюгата P41pL или, в качестве сравнения, 18 мкг конъюгата P16pL (см. пример 13). Указанные количества обоих пептидсодержащих конъюгатов являются оптимальными для достижения улучшения переноса гена. Через дальнейшие 15 минут смесь добавляют к клеткам, которые инкубируют при температуре 37oC в течение 4 часов, после чего добавляют 2 мл среды, содержащей 18% ТЭС. По истечении 24 часов клетки собирают для определения активности люциферазы. Представленные на фиг. 20 значения представляют собой общую активность люциферазы трансфецированных клеток. Сравнение результатов, получаемых при применении обоих пептидсодержащих конъюгатов, показывает, что конъюгат P41pL обеспечивает улучшение переноса гена, которое в 3,5 раз превышает результат, достигаемый другим пептидсодержащим конъюгатом.
в) Трансфекция клеток BNL CL.2 конъюгатами пептида вируса гриппа и полилизина
Клетки BNL CL.2 выращивают аналогично примеру 6. Пептид Р41 подвергают сопряжению с полилизином с молекулярным весом 300 при молярном соотношении 1 : 1, 3 : 1 и 8: 1, соответственно. Комплексы 6 мкг ДНК pCMVL и 20 мкг конъюгатов добавляют к клеткам. В качестве сравнения используют 20 мкг полилизина с молекулярным весом 300 (pL300) или 20 мкг конъюгата Р16 по примеру 13. Клетки инкубируют при температуре 37oC в течение 4 часов, после чего добавляют 2 мл среды, содержащей 18 % ТЭС. По истечении 24 часов клетки собирают для определения активности люциферазы. Результаты опыта представлены на фиг. 20В. Содержание пептида в конъюгатах коррелирует с улучшением экспрессии гена. Согласно результатам анализа по выделению содержимого из липосом (см. фиг. 20С), который осуществляют описанным в примере 13 образом, активность конъюгатов (при pH 5, что эквивалентно 2,5 мкг полилизина) увеличивается по мере повышения содержания пептида. На фиг. 20 C пептид Р41 обозначен "influ2".
Пример 15. Трансфекция клеток HeLa β-галактозидазным репортерным ген-конструктом и доказательство экспрессии β галактозидазы in situ
а) Выращивание и трансфекция клеток
Клетки HeLa аналогично предыдущим примерам выращивают в среде DMEM, содержащей 5% ТЭС, пенициллин, стрептомицин и глутамин, причем процесс осуществляют в чашках диаметром 3 см. Плотность: 3 • 104 клеток на чашку.
6 мкг β-галактозидазного репортерного ген-конструкта (далее: pCMV-β-gal) в 160 мкл HBS переводят в комплекс с 12 мкг TfpL190B в 160 мкл HBS путем инкубации при комнатной температуре в течение 30 минут. Кроме того, 6 мкг pCMV- β -gal в 160 мкл HBS инкубируют с 6 мкг TfpL190B в 80 мкл HBS при комнатной температуре в течение 15 минут. Затем добавляют 12 мкг конъюгата пептида гриппа (P16pL) по примеру 13 в 80 мкл HBS и смесь инкубируют в течение 15 минут. Полученные комплексы ДНК и поликатиона смешивают с 1 мл среды DMEM, содержащей 2% ТЭС, антибиотики и глутамин, описанным выше образом. С целью выявления действия хлорохина и аденовируса на успех трансфекции, проводят дополнительные опыты с применением хлорохина в конечной концентрации 100 мкМ и раствора штамма аденовируса d1312 в конечной концентрации 50 мкл, который добавляют к среде, содержащей комплексы ДНК и поликатиона.
Перед трансфекцией первоначальную питательную среду удаляют и к клеткам добавляют 1 мл среды, содержащей комплексы ДНК и хлорохин или аденовирус, или же 1 мл среды, которая содержит только комплексы ДНК. После двухчасовой инкубации при температуре 37oC к клеткам добавляют 1 мл среды DMEM, содержащей 10% ТЭС, антибиотики и глутамин, и процесс инкубации продолжают в течение дальнейших 2 часов. Затем всю среду удаляют и клетки культивируют в 3 мл свежей среды DMEM, содержащей 10% ТЭС, антибиотики и глутамин.
б) Определение экспресс β-галактозидазы
Через 48 часов после трансфекции среду удаляют, клетки промывают фосфатсодержащим солевым раствором (далее: буфер PBS) и подвергают взаимодействию с 0,5% глутарьдиальдегида в буфере PBS при комнатной температуре в течение 5 минут. Затем непрореагировавший диальдегид удаляют и клетки промывают буфером PBS, после чего осуществляют инкубацию вместе с окрашивающим раствором (10 мМ фосфатного буфера с pH 7,0, 150 мМ хлористого натрия, 1 мМ хлористого магния, 3,3 мМ K4Fe(CN)6 • 3H2O, 3,3 мМ K3Fe(CN)6 и 0,2% 5-бром-4-хлор-3-индолил-β-галактопиранозида) при температуре 37oC в течение 20 минут до 3 часов. Затем крышки чашек промывают буфером PBS, водой и 96%-ным этанолом. После сушки осуществляют анализ с применением микроскопа марки Аксиофот фирмы Цайсс. На фиг. 21 представлены снимки микроскопических увеличений (112 раз). А: клетки HeLa, трансфецированные комплексом 6 мкг pCMV-β gal и 12 мкг TfpL190B. Реакцию окрашивания осуществляют в течение 3 часов. На фиг. видно, что только небольшое число клеток (55 клеток; группа окрашенных клеток указана стрелкой) экспрессирует ген β-галактозидазы. В: клетки HeLa, трансфецированные комплексом 6 мкг pCMV-β-gal, 6 мкг TfpL190B и 12 мкг P16pL. Реакцию окрашивания осуществляют также в течение 3 часов. Небольшое число клеток (250) экспрессирует ген β-галактозидазы. Однако реакция клеток сильнее чем в случае А. С: клетки HeLa, трансфецированные комплексом 6 мкг pCMV- β- gal, 6 мкг TfpL190B и 12 мкг P16pL в присутствии 100 мкМ хлорохина. При этом реакцию окрашивания также осуществляют в течение 3 часов. Большое число клеток проявляет значительную положительную реакцию (более 1000 клеток). D: Клетки HeLa, трансфецированные комплексом 6 мкг pCMV-β-gal и 12 мкг TfpL190B в присутствии аденовируса d1312. При этом реакцию окрашивания осуществляют в течение 20 минут. Почти все клетки (более 90%) проявляют положительную реакцию. E: Не-трансфецированные клетки HeLa (контрольный опыт для определения специфичности реакции β-галактозидазы). При этом реакцию окрашивания осуществляют в течение 3 часов.
Пример 16. Трансфекция клеток HeLa козмидой размером 48 к.п.о. в присутствии аденовируса
а) Получение козмиды, содержащей последовательность, кодирующую люциферазу
Фрагмент SalI размером 3,0 к.п.о., содержащий кодирующую люциферазу P. pyralis последовательность под контролем промотора RSV, выделяют из плазмиды p220RSVLuc α и легируют в сайт SAlI козмидного клона С1-7аА1 с тем, чтобы образовать конкатамеры. Козмидный клон С1-7аА1 включает фрагмент Sau3A геномной ДНК человека с размером 37 к.п.о. (частичное переваривание), который не кодирует определенные гены и клонирован в сайт BamHI козмидного вектора pWE15. Продукты легирования упаковывают in vitro, аликвот получаемых фаговых частиц подают на инфекцию в Е. coli NM544 и подают на пластинки марки LB amp. Рекомбинантные продукты подвергают скринингу путем гибридизации колоний с применением в качестве пробы гибридизации фрагмента SalI размером 3,0 к.п. о. , меченого 32P. Ряд положительных продуктов обнаруживают путем рестрикционного картирования. Козмидный конструкт (CosLuc), включающий одиночную копию вставки SalI, выращивают и очищают на градиенте хлористого цезия (общий размер: 48 к.п.о.).
Небольшую контрольную козмиду pWELuc размером 12 к.п.о. получают путем переваривания CosLuc с помощью NotI, повторного легирования, трансформации бактерий и выделения клона, содержащего целевую плазмиду. Получают молекулу ДНК размером 12 к.п.о., которая недостаточна вставкой ДНК человека и частью полилинкера CosLuc. Плазмида pSPNeoLuc размером 8 к.п.о. представляет собой описанную в примере 5 плазмиду, которая содержит фрагмент гена люциферазы RSV (Apa1/Pvu1-фрагмент pRSVL, клонированный в сайт Cla1 локуса pUC μ).
б) Введение козмиды в клетки HeLa
На клетки HeLa (3 • 104 клеток на чашку диаметром 6 см) подают 1 мл среды DMEM + 2% ТЭС и их инкубируют вместе с полученными вышеописанным образом комплексами ДНК и конъюгата трансферина и полилизина (TfpL), содержащими указанные на фиг. 22А количества hTfpL, свободного полилизина (pL) и ДНК. Среда инкубации дополнительно еще включает либо 100 мкМ хлорохина (см. следы 1, 2), либо 10 мкл аденовируса d1312, содержащего 5 • 1011 частиц на мл (следы 3-12). После двухчасовой инкубации при температуре 37oC добавляют 4 мл среды DMEM + 10% ТЭС. Через 24 часа клетки собирают для определения активности люциферазы. Результаты опыта представлены на фиг. 22А.
в) Введение козмиды в нейробластомные клетки
На клетки нейробластомной клеточной линии Gl-ME-N (1 • 106 клеток на чашку диаметром 6 см) подают 1 мл среды DMEM + 2% ТЭС, после чего их инкубируют вместе с комплексами ДНК и конъюгата TfpL, содержащими указанные на рис. 22В количество hTfpL, свободного полилизина pL и ДНК. Кроме того, среда инкубации дополнительно еще содержит либо 100 мкМ хлорохина (следы 3, 4), либо 10 мкл аденовируса d1312, содержащего 5 • 1011 частиц на мл (следы 5, 6). После двухчасовой инкубации при температуре 37oC добавляют 4 мл среды DMEM, содержащей 10% ТЭС. По истечении 24 часов клетки собирают для определения активности люциферазы. Результаты опыта представлены на рис. 22В.
Пример 17. Перенос гена с помощью конъюгата химически связанных аденовируса и полилизина
а) Получение конъюгата аденовируса и полилизина путем химического связывания.
2,35 мл прошедшего гель-фильтрацию (с применением колонки сефадекс Г-25) раствора аденовируса d1312 (примерно 1011 частиц) в 150 мМ хлористого натрия /25 мМ HEPES, pH 7,9+10% глицерина смешивают с 10 мкл (10 нмоль) 1 мМ раствора сукцинимидилпиридилдитиопропионата. Через 3,5 часа при комнатной температуре модифицированный вирус отделяют от избыточного реагента путем гель-фильтрации, осуществляемой вышеуказанным образом. Через 2,5 мл раствора пропускают аргон и в условиях исключения кислорода в атмосфере аргона подвергают взаимодействию с 42 мкл раствора 1 нмоль меченого изотиоцианатом флуоресциина полилизина, модифицированного 2,3 нмоль меркаптопропионатных групп. Через 18 часов при комнатной температуре половину раствора переводят в пробирку, на него подают 1 мл раствора хлористого цезия (плотность 1,33 г/мл) и центрифугируют при 35000 об./мин в течение двух часов при комнатной температуре. Вирус собирают в качестве 200 мкл фракции хлористого цезия и разбавляют до 1 мл буфером HBS, содержащим 50% глицерина. 300 мкл раствора модифицированного вируса исследуют на способность к связыванию ДНК следующим образом. Раствор вируса разбавляют 1 мл буфера HBS и смешивают с 100 мкл раствора 15 нг ДНК pRSVL, меченой 35S. В качестве контрольного опыта применяют немодифицированный вирус d1312 в том же количестве. Через 30 минут пробы переводят в пробирки, на них подают 1мл раствора хлористого цезия (плотность 1,33/мл) и центрифугируют при 35000 об./мин в течение двух часов при комнатной температуре. Градиент подразделяют на 5 фракций: фракция 1 = 1 мл; фракция 2 = 0,6 мл; фракции 3-5 = по 200 мкл. Определяют радиоактивность фракции объемом 200 мл. Полученные данные представлены на фиг. 23. Вирус, содержащий фракции 3-5, в частности фракция 3, проявляет значительно более высокую радиоактивность, чем контрольная проба. Это можно приписывать специфичной связи модифицированного полилизином аденовируса с меченой ДНК, а в присутствии хлористого цезия, может быть, вызывает частичную диссоциацию комплекса.
б) Трансфекция клеток К562
Клетки К562 (АТСС CCL 243) выращивают в виде суспензии в среде RPMI 1640 (содержащей 2 г/л бикарбоната натрия, 10% ТЭС, 100 единиц/мл пенициллина, 100 мкл/мкл стрептомицина и 2 мМ глутамина) до плотности 500000 клеток/мл. За 12 до 20 часов до трансфекции клетки подают в свежую среду, содержащую 50 мкМ десферриоксима (служащего для повышения числа рецепторов трансферина). В день трансфекции клетки собирают, суспендируют в свежей среде, содержащей 10% ТЭС и 50 мкМ десферриоксима (250000 клеток/мл) и порции по 2 мл подают в снабженную 24 ячейками чашку. 6, 0,6 и 0,06 мкг, соответственно, ДНК pCMVL в 100 мкл HBS смешивают с 50 мкл модифицированного полилизином аденовируса (на фиг. 24: "pLadeno") или с соответствующим количеством (37 мкл) контрольного аденовируса d1312. По истечении 20 минут добавляют 12, 1,2 и 0,12 мкг, соответственно, конъюгата TfpL190B в 150 мкл HBS. По истечении дальнейших 20 минут смесь добавляют к клеткам К562, которые инкубируют при температуре 37oC в течение 24 часов, после чего их собирают для определения активности люциферазы описанным в предыдущих примерах образом. На фиг. 24 представлена общая активность люциферазы трансфецированных клеток.
в) Трансфекция клеток HeLa
Один из методов исследования активности конъюгата вируса и полилизина заключается в определении способности конъюгата к переносу минимальных количеств ДНК (менее 0,1 мкг). Повышенная способность к переносу ДНК ожидается в том случае, если аденовирус непосредственно связан с конденсируемой полилизином ДНК, так как содействующие поглощению клеткой факторы (трансферин и аденовирус (или его белок) непосредственно связываются с переносимой ДНК. Для проверки этого предположения поступают следующим образом. Постоянное количество конъюгата полилизина и аденовируса (2,5 мкл, примерно 5 • 107 вирусных частиц) переводят в комплекс с различными количествами (3 мкг - 0,0003 мкг) репортерной плазмиды в 475 мкл HBS. После 15-минутной инкубации при комнатной температуре к каждой пробе добавляют конъюгат трансферина и полилизина в количестве, соответствующем массе ДНК (такое количество конъюгата TfpL обеспечивает полную "упаковку" (электронейтральность) 50% плазмидной ДНК и одновременно предоставляет еще место связывания конъюгату вируса и полилизина. После добавления конъюгата TfpL смеси инкубируют в течение 15 минут, после чего каждую смесь подают в чашку диаметром 6 см, содержащую 300000 клеток HeLa в 1 мл среды DMEM/2% ТЭС. Клетки инкубируют при температуре 37oC в течение 90 минут, после чего добавляют 4 мл среды DMEM/10% ТЭС. В параллельных опытах эквивалентное количество ДНК переводят в комплекс с двукратным массовым избытком конъюгата TfpL (это количество обеспечивает полную конденсацию ДНК) и применяют для осуществления переноса гена в клетках HeLa (в одном случае в присутствии 25 мл несвязанного с полилизином аденовируса d1312, в другом случае - без последнего). По истечении 24 часов клетки собирают, приготовляют экстракты и аликвоты исследуют на активность люциферазы. Результаты опытов представлены на фиг. 25, где видно, что в отсутствие аденовируса активность люциферазы на может определяться в количестве ДНК менее 0,3 мкг. Как связанный с полилизином аденовирус, так и несвязанный с ним аденовирус проявляют хорошее действие в случае больших количеств ДНК (3 мкг и 0,3 мкг). Однако в случае несвязанного с полилизином аденовируса наблюдается примерно 100-кратное снижение активности при 0,03 мкг ДНК, а в случае применения ДНК в количестве менее 0,03 мкг активность ничтожна. В противоположность этому связанный с полилизином аденовирус сохраняет свою способность к переносу гена как при 0,003, так и при 0,0003 мкг ДНК. Это количество ДНК соответствует примерно 150 молекул ДНК на клетку и примерно одной вирусной частице на клетку.
Пример 18. Перенос гена с помощью энзиматически связанных с полилизином аденовирусов
а) Реакция с ферментом
2 мл препарата аденовируса (штамм d1312; 5 • 1010 бляшкообразующих единиц/мл) подвергают гель-фильтрации в колонке Сефадекс Г-25, которую предварительно уравновешивают с помощью 25 мл реакционного буфера (0,1 М Tris-НС1; pH 8,0, 2 мМ дитиотреитола, 30% глицерина). Элюацию осуществляют 3,5 мл реакционного буфера. Подвергаемая энзиматическому связыванию реакционная смесь состоит из 1150 мкл вируссодержащего элюата, 0,5 нмоль выделенной из печени морской свинки трансглутаминазы (далее: "TG"), 2 нмоль или 20 нмоль полилизина с молекулярным весом 290, 10 мМ хлористого кальция и реакционного буфера до конечного объема 1500 мкл. Реакцию осуществляют при температуре 37oC в течение 1 часа, после чего ее прекращают путем добавления 30 мкл 0,5 М ЭДТУК. В целях определения специфичности связывания приготовляют те же реакционные смеси, но без трансглутаминазы. Несвязанный полилизин отделяют от вирусов путем центрифугирования в градиенте хлористого цезия (плотностью 1,33 г/мл; 170000 • g, 2 часа). Вируссодержащую фракцию собирают, смешивают с одинаковым объемом глицерина, замораживают в среде жидкого азота и хранят при температуре -70oC.
б) Анализ связывания полилизина с аденовирусом
Вышеупомянутую реакцию осуществляют с применением полилизина, меченого 125J. После центрифугирования в градиенте хлористого цезия вируссодержащую фракцию отводят и разделяют с помощью другого градиента хлористого цезия. Градиент фракционируют и в каждой фракции определяют радиоактивность с применением сцинтилляционного счетчика. Результаты опыта представлены на фиг. 26, где видно, что в случае содержащей TG реакционной смеси (d1312/TG-pL) радиоактивный полилизин накапливается в вируссодержащей фракции. В случае контрольной смеси, т. е. реакционной смеси без содержания TG (d1312/pL), накопления радиоактивного полилизина в вируссодержащей фракции не наблюдается.
в) Исследование фракций, содержащих модифицированный полилизином аденовирус, их влияние на эффективность трансфекции
1) Клетки и среда
5 • 105 клеток (мышиные гепатоциты; АТСС N: TIB 73) в среде DMEM, содержащей 10% инактивированной термообработкой ТЭС, 2 мМ глутамина, 100 единиц/мл пенициллина и 100 мкг/мл стрептомицина, выращивают в чашках диаметром 6 см.
2) Образование состоящего из вируса, ДНК и трансферина комплекса
50 мкл модифицированной полилизином вируссодержащей фракции смешивают с 6 мкг плазмидной ДНК pCMVL в 10 мкл HBS и смесь инкубируют при комнатной температуре в течение 20 минут. Затем к смеси добавляют 8 мкг конъюгата мышиного трансферина и полилизина с молекулярным весом 290 (mTflpL 290В), после чего инкубацию продолжают в течение дальнейших 20 минут.
3) Трансфекция мышиных гепатоцитов
Состоящий из модифицированного вируса, ДНК и трансферина комплекс смешивают с 1,5 мл среды DMEM, содержащей 2% ТЭС, 2 мМ глутамина и антибиотики, и добавляют к клеткам после предварительной замены на свежую среду. После двухчасовой инкубации при температуре 37oC к клеткам добавляют 2 мл среды DMEM, содержащей 10% ТЭС, глутамин и антибиотики. После культивации в течение дальнейших 2 часов всю среду удаляют и к клеткам добавляют 4 мл свежей среды DMEM, содержащей 10% ТЭС, глутамин и антибиотики.
4) Определение экспрессии люциферазы
Через 24 часа после трансфекции клетки собирают и определяют активность люциферазы указанным выше образом. Как видно на фиг. 27 вирусный препарат, содержащий обработанный TG и 20 нмоль полилизина (d1312/TG-20 нмоль pL) аденовирус, проявляет наибольшую экспрессию (153 540 000 световых единиц). Вирусный препарат, содержащий аденовирус, обработанный TG и 2 нмоль полилизина (d1312 /TG-2 нмоль pL), проявляет несколько меньшую активность (57 880 000 световых единиц). Контрольная фракция, которая содержит только обработанный 20 нмоль полилизина аденовируса, проявляет примерно на фактор 500 меньшую активность. В качестве сравнения применяют комплексы, которые содержат аденовирус, который не обработан ни TG, ни полилизином. При этом активность люциферазы составляет 4 403 000 световых единиц.
5) Эффективность трансфекции, достигаемая с помощью модифицированного полилизином аденовируса по сравнению с немодифицированным аденовирусом, можно повышать в частности небольшими количествами ДНК
Трансфекцию осуществляют описанным на стадии 3) образом с применением 50 мкл фракции аденовируса d1312/TG-20 нмоль pL и 6 мкг pCMVL/8 мкг TfpL, 0,6 мкг pCMVL/0,8 мкг mTfpL или 0,06 мкг pCMVL/0,08 мкг mTfpL для образования комплекса. В качестве сравнения осуществляют трансфекцию с применением 6 мкг, 0,6 мкг и 0,06 мкг, соответственно, комплекса pCMVL/mTfpL и немодифицированного аденовируса d1312. Результаты опыта представлены на фиг. 28. Видно, что содержащие модифицированный полилизином аденовирус комплексы проявляют высокую экспрессию даже в случае небольших количеств ДНК, тогда как экспрессия резко снижается при применении комплекса, содержащего немодифицированный аденовирус.
Пример 19. Перенос гена при помощи конъюгатов, в которых связь между аденовирусом и полилизином осуществлена при помощи биотиново- стрептавидинового мостика
а) Биотинилирование аденовируса d1312
2,4 мл прошедшего гель-фильтрацию в колонке Сефадекс Г-25 раствора аденовируса d1312 (примерно 1011 частиц) в 150 мМ хлористого натрия/5 мМ HEPES, pH 7,9 /10% глицерина смешивают с 10 мкл (10 нмоль) 1 мМ раствора биотина марки NHS-LC. По истечении 3 часов при комнатной температуре модифицированный биотином вирус отделяют от избыточного реагента при помощи гель-фильтрации, осуществляемой описанным выше образом. К раствору добавляют глицерин в количестве, обеспечивающем его концентрацию в растворе 40% и раствор общим объемом 3,2 мл хранят при -25oC. Факт биотинилирования вируса исследуют путем накапывания раствора различной степени разбавления на целлюлозно-нитратную мембрану, после чего осуществляют сушку при температуре 80oC в течение 2 часов в вакуумной сушилке, блокировку альбумином сыворотки крупного рогатого скота, инкубацию с сопряженной со стрептавидином щелочной фосфатазой, промывку и одночасовую инкубацию вместе с проявляющим раствором тетразолевой соли нитросинего и толуидиновой соли 5-бром-4-хлор-3- индолилфосфата. При этом имеет место положительная цветная реакция, что свидетельствует о состоявшемся биотинилировании.
б) Получение конъюгатов стрептавидина и полилизина
79 нмоль (4,7 мг) стрептавидина в 1 мл 200 мМ буфера HEPES с pH 7,9 и 300 мМ хлористого натрия обрабатывают 15 мМ этанольным раствором сукцинимидилпиридилдитиопропионата (236 нмоль). По истечении 90 минут при комнатной температуре модифицированный белок подвергают гель-фильтрации в колонке Сефадекс Г-25. При этом получают 75 нмоль стрептавидина, модифицированного 196 нмоль указанного дитиопиридина. Модифицированный белок подвергается взаимодействию с 75 нмоль модифицированного 190 нмоль 3-меркаптопропионата полилизина со средней длиной лизиновых мономеров 290 в 2,6 мл 100 мМ буфера HEPES с pH 7,9, содержащего 150 мМ хлористого натрия. Конъюгаты выделяют путем катионообменной хроматографии в колонке Mono S HR5 (градиент: 20-100% буфера, состоящего из 50 мМ HEPES pH 7,9 плюс 3 М хлористого натрия). Целевую фракцию элюируют при концентрации соли между 1,2 М и 1,7 М. В результате диализа с применением буфера HBS (20 mM HEPES с pH 7,3, 150 мМ хлористого натрия) получают конъюгат, состоящий из 45 нмоль стрептавидина и 53 нмоль полилизина.
в) Трансфекция клеток HeLa
Клетки HeLa выращивают в чашках диаметром 6 см указанным в примере 13 образом. Трансфекцию осуществляют при плотности 300000 клеток на чашку. Перед трансфекцией клетки инкубируют в 1 мл свежей среды, содержащей 2% ТЭС.
6 мкг ДНК pCMVL в 100 мкл HBS смешивают с 0,8 мкг модифицированного стрептавидином полилизина в 170 мкл HBS. По истечении 20 минут добавляют 3 мкг полилизина с молекулярным весом 300 в 170 мкл HBS. По истечении дальнейших 20 минут добавляют 65 мкл биотинилированного аденовируса или, в качестве контроля, соответствующее количество аденовируса d1312 (30 мкл, т.е. количество, в котором вирус применяют для осуществления модификации). Получаемые при этом смеси ("биотин.AdV/комплекс А" и "AdV (контроль)"; см. фиг. 29) оставляют стоять в течение дальнейших 20 минут.
Кроме того, 50 мкл биотинилированного аденовируса смешивают с 0,8 мкг модифицированного стрептавидином полилизина в 50 мкл HBS, через 20 минут добавляют 6 мкг ДНК pCMVL в 170 мкл HBS и через дальнейшие 20 минут добавляют еще 3 мкг полилизина с молекулярным весом 300 в 200 мкл HBS. Получаемый комплекс обозначается "биотинAdV/комплекс Б".
0,6 мкг ДНК pCMVL в 67 мкл HBS смешивают с 0,3 мкг модифицированного стрептавидином полилизина в 33 мкл HBS. По истечении 20 минут добавляют 65 мкл биотинилированного аденовируса или, в качестве контроля, соответствующее количество аденовируса d1312 (30 мкл, т.е. количество, в котором вирус применяют для осуществления модификации). Получаемые комплексы ("биотин. AdV/комплекс А" или "AdV (контроль)"; см. фиг. 29) оставляют стоять в течение дальнейших 20 минут, после чего комплексы разбавляют HBS до 500 мкл. Кроме того, получают еще другой комплекс за счет смешения 50 мкл биотинилированного аденовируса с 0,3 мкг модифицированного стрептавидином полилизина в 50 мкл HBS и последующего добавления через 20 минут 0,6 мкг ДНК pCMVL в 50 мкл HBS. Получаемый комплекс ("Биотин.AdV/комплекс Б") оставляют стоять в течение дальнейших 20 минут, после чего его разбавляют HBS до 500 мкл.
Указанные комплексы добавляют к клеткам, которые инкубируют при температуре 37oC в течение 2 часов, после чего добавляют 2,5 мкл свежей среды, содержащей 10% ТЭС. Клетки инкубируют еще в течение 24 часов при 37oC, после чего их собирают на определение активности люциферазы описанным выше образом. Результаты опыта представлены на рис. 29, показывающем общую активность люциферазы трансфецированных клеток.
Параллельно осуществляют трансфекцию клеток HeLa с применением в качестве вирусного компонента конъюгата биотинилированного вируса, инактивированного обработкой УФ-лучами и псораленом. Инактивацию осуществляют следующим образом. Пробы по 200 мкл биотинилированного вируса подают в снабженную двумя ячейками пластинку диаметром 1,6 см. К каждой пробе добавляют 2 мкл (33 мг/мл) 8-метоксипсоралена в диметилсульфоксиде, пластинки подают на лед и подвергают облучению УФ-светом с помощью лампы UVP TL-33, размещенной на расстоянии 4 см, в течение 10 минут при 365 нм. После облучения обе пробы объединяют и подвергают гель-фильтрации в колонке Г50, которую предварительно уравновешивают 40%-ным раствором глицерина в HBS. Аликвоты 75 мкл переводят в комплекс с 0,8 мкг модифицированного стрептавидином полилизина и полученные таким образом комплексы применяют для трансфекции клеток HeLa описанным выше образом.
В результате цитопатического анализа устанавливают, что инактивация приводит к снижению титра вируса на фактор более 104, тогда как способность к переносу гена уменьшается на менее 50% при высоких концентрациях и на фактор 5 при низких концентрациях.
г) Трансфекция клеток К562
Клетки К562 в виде суспензии выращивают в среде RPMI 1640, содержащей 2 г/л бикарбоната натрия, 10% ТЭС, 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина и 2 мМ глутамина, до достижения плотности 500000 клеток/мл. За 16 часов до трансфекции клетки подают в свежую среду, содержащую 50 мкмоль десферриоксима. В утро трансфекции клетки собирают, повторно суспендируют в свежей среде, содержащей 10% ТЭС (плюс 50 мкМ десферриоксима) при плотности 250000 клеток/мл и подают в снабженную 24 ячейками чашку (2 мл на ячейку).
Получают 3 различных комплекса ДНК: а) Раствор 6 мкг ДНК pCMVL в 160 мкл HBS (150 mM хлористого натрия, 20 мМ HEPES с pH 7,3) смешивают с 12 мкг конъюгата TfpL190B в 160 мкл HBS, через 30 минут добавляют 20 мкл аденовируса d1312 и получаемую смесь добавляют к клеткам К562. б) Раствор 800 нг модифицированного стрептавидином полилизина (StpL) в 160 мкл HBS смешивают с 20 мкл биотинилированного аденовируса, полученного на вышеописанной стадии а), через 30 минут добавляют раствор 6 мкг ДНК pCMVL в 160 мкл HBS и через дальнейшие 30 минут раствор смешивают с 10 мкг конъюгата TfpL190B в 160 мкл HBS. По истечении 30 минут смесь добавляют к клеткам. в) Этот комплекс получают аналогично пункту б) с той разницей, что вместо конъюгата TfpL190B добавляют раствор 3,5 мкг поли(L)лизина с молекулярным весом 290 (pL290). Клетки инкубируют при температуре 37oC в течение 24 часов, после чего их собирают на определение активности люциферазы. Результаты опытов представлены на фиг. 30, показывающем общую активность люциферазы трансфецированных клеток.
Пример 20. Перенос гена в первичные клетки костного мозга
а) Выделение первичных клеток костного мозга
Первичные клетки костного мозга отбирают из мышей при помощи снабженного инъекционной иглой (диаметром 0,4 -0,5 мм) шприца емкостью 1 мл, вводимого в изолированные бедренную и большеберцовую кость. Отбор осуществляют с применением среды IMDM, содержащей 10% ТЭС, 5 • 10-5 М β-меркаптоэтанола, 1% интерлейкина 3 и антибиотики. Отобранные клетки промывают указанной средой за счет центрифугирования при 100 • g в течение 8 минут. Затем клетки повторно суспендируют в указанной среде при концентрации 107/мл и культивируют в пробирках марки Т25. По истечении 4 часов непроросшие клетки переводят в другие пробирки марки Т25, в которых культивируют в течение ночи в присутствии 50 мкМ десферриоксима.
б) Образование комплекса, состоящего из аденовируса, ДНК и трансферина
50 мкл биотинилированного аденовируса инкубируют вместе с 400 нг модифицированного стрептавидином полилизина в 20 мкл HBS в течение 20 минут. Затем добавляют 20 мкл буфера HBS, содержащего 6 мкг ДНК pCMVL. После 20-минутной инкубации добавляют 7 мкг конъюгата мышиного трансферина и полилизина (mTfpL) в 160 мкл HBS, после чего смесь инкубируют в течение дальнейших 20 минут.
в) Трансфекция
Клетки костного мозга отделяют от питательной среды путем центрифугирования при 100 • g в течение 8 минут. Полученные клетки повторно суспендируют в 3 мл указанной среды, содержащей 2% ТЭС и 250 мкл состоящего из аденовируса, ДНК и трансферина комплекса, после чего смесь культивируют в пробирках марки Т25 при температуре 37oC в течение 3 часов. Потом добавляют 3 мл указанной среды, содержащей 10% ТЭС. По истечении дальнейших 2 часов добавляют еще 6 мл указанной среды.
г) Определение экспрессии люциферазы
После 48-часовой трансфекции клетки костного мозга собирают и подвергают анализу экспрессии люциферазы описанным выше образом. При этом устанавливают, что трансфекция приводит к экспрессии активности люциферазы, соответствующей 310 • 103 световых единиц/100 мкг общего клеточного белка.
Пример 21. Трансфекция нейробластомных клеток козмидой размером 48 к.п. о. в присутствии аденовируса
а) Получение козмиды, включающей последовательность, кодирующую люциферазу
Фрагмент Sal I размером 3,0 к.п.о., включающий кодирующую люциферазу последовательность из P. pyralis под контролем промотора RSV, легируют в единственный сайт Sal I козмидного клона С1-7аА1 с получением конкатамера (С1-7аА1 включает Sau 3А-фрагмент размером 37 к.п.о. человеческой геномной ДНК (частичное переваривание), не кодирующий определенных генов, клонированный в сайт Bam HI козмидного вектора pWE15). Продукт легирования упаковывают in vitro и аликвот получаемых фаговых частиц инфицируют штамм Е. coli NM544, который подают на пластинки марки LB amp. Получаемые рекомбинантные продукты подвергают скринингу путем гибридизации колоний с применением в качестве зонда фрагмента Sal I размером 3,0 к.п.о., меченого 32P. Ряд положительных продуктов подвергают анализу путем рестрикционного картирования. Козмидный конструкт (CosLuc), включающий одну единственную копию вставки Sal I, выращивают и очищают на градиенте хлористого цезия (общий размер: 48 к.п.о.).
Получают небольшую контрольную плазмиду pWELuc размером 12 к.п.о. путем переваривания указанного козмидного конструкта с помощью Not I, повторного легирования, трансформации бактерий и выделения клона, включающего целевую плазмиду. Получают молекулу ДНК размером 12 к.п.о., в которой отсутствуют вставка человеческой ДНК и часть полилинкера козмидного конструкта CosLuc.
б) Введение козмиды в нейробластомные клетки
На клетки нейробластомной линии Gl-ME-N (1 • 106 клеток на чашку диаметром 6 см) подают слой 1 мл среды DMEM, содержащей 2% ТЭС, после чего клетки инкубируют вместе с описанными выше комплексами ДНК и TfpL комплексами, содержащими указанные количества hTfpL, свободного полилизина и ДНК. Подвергаемые инкубации смеси содержат еще либо 100 мкМ хлорохина (см. следы 3, 4) либо 10 мкл аденовируса d1312, включающего 5 • 1011 частиц/мл (см. следы 5, 6). Обе последние пробы (обозначенные как StpL/биотин.), включающие 15 мкл биотинилированного аденовируса d1312 (1 • 1011 частиц), инкубируют вместе с 0,8 мкг полученного в примере 19, модифицированного стрептавидином полилизина в 150 мкл HBS в течение 30 минут. Затем добавляют 6 мкг ДНК в 150 мкл HBS, инкубируют при комнатной температуре в течение 30 минут и добавляют 150 мкл буфера HBS, содержащего 6 мкг hTfpL и 1 мкг свободного полилизина. После дополнительной инкубации при комнатной температуре в течение 30 минут смесь добавляют к клеткам, которые инкубируют при температуре 37oC в течение 2 часов, после чего к каждой пробе добавляют 4 мл среды DMEM, содержащей 10% ТЭС. Через 24 часа клетки собирают для определения активности люциферазы. Результаты опыта представлены на фиг. 31.
Пример 22. Перенос гена в первичные клетки эпителиальных дыхательных путей с применением конъюгатов согласно изобретению. Направленные на выявление пригодности употребления переноса генов с применением конъюгатов согласно изобретению для генетической поправки муковисцидоза исследования показали, что иммортализованные клеточные линии из эпителия дыхательных путей человека проявляют чувствительность к такому методу осуществления переноса генов. Для исключения того, что этот дефект является результатом вызываемых иммортализацией изменений эпителия дыхательных путей, первичные клетки эпителия дыхательных путей (далее: клетки 1o АЕ) подвергают воздействию конъюгатов трансферина и полилизина. Клетки 1o AE выделяют из назального полипа человека известными приемами. При этом ткань последовательно промывают стерильным солевым раствором и средой MEM, содержащей 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 40 мкг/мл гентамицина и переводят в лабораторию при 4oC. Хрящ и избыточную подслизистую ткань очищают и эпителиальные слои инкубируют в содержащем протеазу (тип 14 фирмы Зигма, ДЕ; 0,1 мг/дл) растворе в среде MEM при температуре 4oC в течение 16 до 48 часов. Добавляют 10% ТЭС для нейтрализации протеазы, после чего смесь слабо размешивают. Получаемую суспензию фильтруют через найлоновую сетку величиной ячеек 10 мкм с тем, чтобы удалить инородные вещества, и фильтрат подвергают центрифугированию при 150 • g в течение 5 минут, после чего промывают в среде F12, содержащей 10% ТЭС. Затем клетки 1oAE обрабатывают конъюгатами трансферина и полилизина (hTfpL), содержащими в качестве репортерного гена кодирующую люциферазу плазмиду (pRSVL). В этом опыте первичные клетки не проявляют ту чувствительность к действию этого вектора, которая проявляется соответствующими иммортализованными клеточными линиями (1o АЕ: фон = 429 ± 41; + hTfpL = 543 ± ед. люциферазы), что, вероятно, свидетельствует об относительно небольшом числе рецепторов трансферина на клетках 1o AE.
В целях исследования альтернативного целевого рецептора на клетках 1o AE конструируют конъюгаты, которые включают в качестве лиганда некомпетентный к репликации аденовирус (bAdpL; см. пример 19 а) и 19 б) для получения биотинилированного аденовируса и конъюгатов стрептавидина и полилизина; кодирующую люциферазу плазмиду применяют в качестве репортерного гена). Клетки 1o AE человека, обработанные этим конъюгатом, проявляют степень экспрессии репортерного гена, которая значительно превышает фон (+ bAdpL = 2 585 753 ± 453 585 ед. люциферазы). Кроме того, клетки 1o AE другого происхождения также проявляют высокую степень чувствительности к переносу гена этим методом (из мышей = 3 230 244 ± 343 153; из обезьян = 53 498 880 ± 869 481 ед. люциферазы). Таким образом способность к осуществлению переноса гена в клетки 1o AE подтверждает потенциальную пригодность этого подхода по осуществлению непосредственной доставки генов in vivo.
Пример 23. Перенос генов в гепатоциты с применением предлагаемых конъюгатов
а) Трансфекция клеток
Клетки мышиной эмбриональной гепатоцитной клеточной линии BNL CL.2 (АТСС TIB 73) выращивают описанным в примере 6 образом. Клетки HeLa и гепатоциты выращивают в пластмассовых чашках Петри диаметром 6 см. Трансфекцию осуществляют при плотности клеток примерно 3 • 105/чашку. Перед трансфекцией стандартную питательную среду заменяют на 1 мл свежей среды, содержащей 2% ТЭС.
б) Образование двойного комплекса
Биотинилированный аденовирус (примерно 109 бляшкообразующих единиц; см. пример 19 а) и 19 б)) подвергают взаимодействию с 800 нг модифицированного стрептавидином полилизина в 50 мкл HBS. По истечении 30 минут при комнатной температуре добавляют 6 мкг ДНК pCMVL в 170 мкл HBS, инкубируют в течение 30 минут и добавляют 3 мкг полилизина с молекулярным весом 300 в 200 мкл HBS. По истечении дальнейших 30 минут раствор применяют для осуществления трансфекции.
в) Образование тройного комплекса
Тройной комплекс ДНК, содержащий аденовирус и трансферин, получают следующим образом. Биотинилированный аденовирус (примерно 109 вирусных частиц) смешивают с 800 нг модифицированного стрептавидином полилизина. По истечении 30 минут при комнатной температуре раствор смешивают с 6 мкг плазмидной ДНК в 170 мл HBS, инкубируют в течение 30 минут, после чего добавляют 10 мкг конъюгата полилизина и трансферина TfpL 190В в 200 мкл HBS. По истечении дальнейших 30 минут раствор применяют для осуществления трансфекции.
г) Определение β-галактозидазы
Клетки BNL CL. 2 засеивают на покровных стеклах и через 24 часа клетки трансфецируют репортерным геном pCMV-β-gal. Через 48 часов определяют содержание β-галактозидазы.
Применяемые для переноса двойные комплексы. Биотинилированный аденовирус объединяют с модифицированным стрептавидином полилизином. Альтернативно аденовирус ковалентно связывают с полилизином при помощи трансглутаминазы. Конъюгат аденовируса и полилизина добавляют к ДНК в условиях, обеспечивающих образование комплекса между ДНК и полилизином. При этом нейтрализуется определенная часть (примерно 1/4) отрицательных зарядов ДНК. Затем рассчитанное количество полилизина добавляют с тем, чтобы нейтрализовать остаток отрицательных зарядов. Комплексы, состоящие из ДНК, связанные с конъюгатом аденовируса и полилизина и с полилизином, далее обозначаются как двойные комплексы переноса.
В основном существуют два пути получения двойных комплексов переноса. ДНК можно сначала связывать со стрептавидинилированным полилизином и затем с биотинилированным аденовирусом, или же аденовирус сначала связывают с полилизином и затем переводят в комплекс ДНК. Последний метод дает намного лучшие результаты, в частности при низкой концентрации ДНК. Поэтому этот подход представляет собой предпочтительный метод получения двойного и тройного комплексов.
Тройные комплексы переноса, содержащие трансферин. Конъюгаты аденовируса и полилизина добавляют к ДНК в условиях, обеспечивающих образование комплекса и нейтрализацию части (примерно 1/4) отрицательных зарядов ДНК. Затем к комплексу добавляют рассчитанное количество конъюгата трансферина и полилизина с тем, чтобы нейтрализовать остаток отрицательных зарядов ДНК. Комплексы, состоящие из ДНК, конъюгатов аденовируса и полилизина и трансферина и полилизина, далее обозначаются как тройные комплексы. В принципе такой тройной комплекс должен обладать способностью к включению клеткой в результате связывания с клеточными рецепторами аденовируса или рецепторами трансферина.
Связь между конденсатами ДНК и аденовирусом существенно улучшает экспрессию репортерного гена люциферазы. Физическую связь между аденовирусным штаммом d1312 и полилизином можно осуществлять либо за счет инкубации обоих компонентов вместе с трансглутаминазой, либо за счет биотинилирования аденовируса и стрептавидинилирования полилизина. Эффект такого связывания на эффективность трансфекции четко показан на фиг. 32, где представлены результаты инкубации гепатоцитов вместе с комплексами ДНК и конъюгата трансферина и полилизина (TfpL) в присутствии хлорохина или в присутствии аденовируса (AdV + TfpL). Трансфериновая инфекция, т.е. способствуемая трансферином трансфекция, в присутствии свободного аденовируса улучшается так, что приводит к типичному усилению высвобождения комплексов ДНК и конъюгата трансферина и полилизина в клетки. В случае комплексов pLAdV/TpfL аденовирус сопряжают с полилизином при помощи трансглутаминазы и затем подвергают взаимодействию с ДНК, что приводит к нейтрализации части отрицательных зарядов ДНК. Потом добавляют конъюгат трансферина и полилизина для нейтрализации остатка отрицательных зарядов. Таким путем синтезируют тройной комплекс ДНК и конъюгатов аденовируса и полилизина и трансферина и полилизина. Видно, что при этом можно достигать чрезвычайно высокой степени экспрессии люциферазы, т.е. 1,5 • 109 световых единиц люциферазы (примерно 5000 световых единиц на клетку). В случае комплексов AdV + pL + TfpL аденовирус смешивают с полилизином тем же образом, что и в случае обработки трансглутаминазой. С целью выяснения специфичности способствуемого трансглутаминазой связывания полилизина с вирусом, процесс осуществляют в отсутствие фермента. Затем препарат вируса переводят в комплекс с тем же количеством ДНК и TfpL, что и в комплексах pLAdV/TfpL. В этом случае трансфекция является умеренной также как и в случае содержащих AdV и TfpL комплексов, так как в обоих экспериментах взаимное размещение вируса, трансферина и ДНК представляет собой случайный процесс, тогда, как в случае содержащих pLAdV и TfpL комплексов включение клеткой обеспечивается физическим связыванием вируса с ДНК в тройном комплексе. При этом достигается высокая степень трансфериновой инфекции.
Трансфекция клеток К562 показывает эндосомолитические свойства аденовируса. Человеческая эритролейкозная клеточная линия К562 содержит примерно 150000 рецепторов трансферина. Как уже указывалось выше, в присутствии хлорохина эти клетки можно подвергать высокоэффективной трансфериновой инфекции в результате обработки комплексами репортерной ДНК и конъюгата полилизина и трансферина даже в отсутствие аденовируса (TfpL, фиг. 33). Те же самые комплексы с добавленным свободным аденовирусом (AdV/TfpL), но в отсутствие хлорохина, обеспечивают лишь относительно небольшую степень экспрессии репортерного гена, предположительно из-за того, что клетки К562, также как и другие клетки крови, имеют низкое число рецепторов аденовируса. Если же аденовирус связывают с полилизином через биотиново-стрептавидиновый мостик и репортерную ДНК полностью конденсируют добавлением соответствующего количества полилизина для получения тройного комплекса (pLAdV/pL), то степень поддерживаемой аденовирусом трансфекции является средней. Предполагается, что небольшое число рецепторов аденовируса на клетках К562 используется эффективным образом. Однако если связанную с аденовирусом и полилизином репортерную ДНК полностью конденсируют и нейтрализуют добавлением конъюгата трансферина и полилизина для получения тройного комплекса pLAdV/TfpL и имеются многочисленные клеточные рецепторы трансферина, то благодаря эффективному связыванию трансферина и эндосомолитическим свойствам вируса эффективность трансфекции увеличивается по меньшей мере на 2 порядка (см. фиг. 33).
Тройные комплексы ДНК обеспечивают экспрессию репортерного гена почти во всех гепатоцитах. Эффективность предлагаемых комплексов ДНК также исследуют на мышиных гепатоцитах (BNL CL.2) за счет определения процента их трансфекции. При этом в качестве репортерного гена применяют ген β-галактозидазы под контролем промотора CMV. После фиксации клеток определяют активность β-галактозидазы.
На фиг. 34 представлены результаты анализа активности β-галактозидазы в случае мышиных гепатоцитов после А) трансфериновой инфекции в присутствии хлорохина, В) трансфериновой инфекции в присутствии свободного аденовируса d1312 и С) трансфекции тройным комплексом, состоящим из аденовируса d1312, полилизина, трансферина и репортерной ДНК. После обычной трансфериновой инфекции репортерного ДНК в отсутствие аденовируса только несколько клеток проявляют экспрессию репортерного гена. Процентная трансфекция составляет менее 0,1%. Если добавлять хлорохин, то процент трансфекции повышается примерно до 0,2% (см. фиг. 34А). В случае присутствия свободного аденовируса 5-10% клеток экспримируют репортерный ген (см. фиг. 34В), тогда как тройные комплексы, включающие модифицированные трансглутаминазой вирусы, обеспечивают экспрессию почти во всех клетках (см. фиг. 34С). Так как тройной комплекс можно применять в высоко-разбавленном виде, токсичный эффект, который наблюдается в случае высоких доз свободного (инактивированного) аденовируса, обычно не имеет место. Но следует отметить, что в случае применения тройного комплекса в высокой концентрации, обеспечивающей вовлечение в процесс 100% клеток, подобный токсичный эффект начинает проявляться. Токсичные эффекты могут являться результатом остаточной активности вирусного гена, эндосомолитических свойств добавляемого вируса или же могут просто являться причиной очень высокого уровня экспрессии трансфецированного гена.
Экспрессия трансфецированного репортерного гена является временным явлением, но продолжается неделями в неделящихся гепатоцитах. Получают тройные комплексы (pLAdV/TfpL), включающие модифицированный полилизином аденовирус d1312 и модифицированный аденовирус d1312, инактивированный обработкой псораленом. Аналогично опыту по фиг. 34В 2/3 сливающейся клеточной культуры гепатоцитов трансфецируют репортерным геном люциферазы CMVL и активность люциферазы определяют в разные моменты. Как видно на фиг. 35, активность люциферазы является максимальной через три дня, то есть в момент слияния клеточной культуры и прекращения деления клеток. Экспрессия репортерного гена продолжается в неделящихся клетках по меньшей мере в течение 6 недель без необходимости отбора для обеспечения жизнедеятельности гена, в частности тогда, когда инактивированный обработкой псораленом аденовирус применяется для образования тройного комплекса.
Пример 24. Применение куриного аденовируса CELO для улучшения переноса ДНК в клетки человека.
2 мл куриного аденовируса CELO (штамм Phelps, серотип FAV-1,7) подвергают гель-фильтрации в колонке PD-10, которую предварительно уравновешивают буфером 20 мМ HEPES с pH 7,3 и буфером 150 мМ хлористого натрия, содержащим 10% глицерина. 2 мл получаемого элюата подвергают взаимодействию с 20 мкл 1 мМ биотина марки NHS-LC при комнатной температуре в течение 3 часов. Получаемый биотинилированный вирус подвергают диализу с применением 3 х 300 мл буфера HBS, содержащего 40 % глицерина, при температуре 4oC, после чего аликвоты вируса хранят при температуре -70oC. Клетки HeLa (5 • 105 клеток на чашку диаметром 6 см) в 2 мл среды DMEM, содержащей 2% ТЭС, инкубируют с 6 мкг плазмиды pCMVL, имеющейся в виде комплекса с полилизином (pL) или конъюгатом трансферина и полилизина (TfpL) в 500 мкл буфера HBS (комплекс предварительно инкубируют при комнатной температуре в течение 30 минут). Затем пробы добавляют к клеткам при температуре 37oC в присутствии вируса, взятого в количестве, указанном на рис. 36. В случае проб, содержащих биотинилированный вирус CELO, указанное количество вируса предварительно инкубируют вместе с указанным количеством модифицированного стрептавидином полилизина (StpL) в 200 мкл буфера HBS при комнатной температуре в течение 30 минут, после чего добавляют 6 мкг плазмиды pCLus в 100 мкл HBS. После 30-минутной инкубации при комнатной температуре указанное количество содержащего TfpL комплекса добавляют к клеткам при температуре 37oC. Через 2 часа к клеткам добавляют 5 мл среды DMEM, содержащей 10% ТЭС, а по истечении 24 часов клетки собирают и подают на определение активности люциферазы. Результаты опыта представлены на фиг. 36.
Как видно на фиг. 36, вирус CELO в свободном виде улучшает доставку ДНК в клетки HeLa (см. следы 1 - 6). Однако если вирус CELO модифицируют биотином и включают в комплекс вместе с стрептавидином, либо вместе с конъюгатом трансферина и полилизина, либо без него, то вирус обеспечивает улучшение доставки ДНК в такой степени, которую можно сравнивать с тем случаем, когда применяют аденовирус d1312 человека. Данная линия клеток HeLa имеет большую емкость связывания включающих полилизин и ДНК комплексов в отсутствие трансферина (ср. активность люциферазы проб 1 и 4 на фиг. 36). Таким образом включение вируса CELO в комплекс полилизина и ДНК является достаточным для "пуска" поглощения вируса.
Пример 25. Трансфекция миобластов
а) Трансфекция миобластов и миотрубочек включающими ДНК, трансферин и полилизин комплексами в присутствии свободного аденовируса и в присутствии биотинилированного и стрептавидинилированного аденовируса.
Миобласты C2Cl2 (N АТСС: CRL 1772) и миобласты G8 (No АТСС: CRL 1456) выращивают в имеющей высокую концентрацию глюкозы среде DMEM, содержащей 10% ТЭС. Миобластовые культуры подвергают трансфекции с подачей примерно 5 • 105 клеток на чашку диаметром 6 см. Культуры миотрубочек получают за счет подачи миобластов в чашки диаметром 6 см (примерно 5 x105 клеток на чашку) и замены среды на имеющую высокую концентрацию глюкозы среду DMEM, содержащую 2% лошадиной сыворотки, при достижении слияния клеток. Трансфекцию миотрубочек осуществляют через 5 - 7 дней. Используемые для трансфекции комплексы получают описанным в примере 19 образом с применением указанных на фиг. 37 количеств TfpL, StpL и биотинилированного аденовируса d1312. Через 20 часов после трансфекции клетки собирают для определения активности люциферазы. Результаты опыта представлены на фиг. 37, где активность люциферазы для всей пробы указана в световых единицах. Видно, что как миобласты, так и миотрубочки можно трансфецировать с высокой эффективностью. После дифференциации в миотрубочки наблюдается снижение эффективности трансфекции на менее 1 log (в случае C2Cl2) или же значительного уменьшения не наблюдается (в случае G8). В случае линии G8 участие в образовании миотрубочек имеет место с более низкой частотой, за что частично может быть ответственным отсутствие определяемого уменьшения эффективности в дифференцированной культуре. В случае данной клетки взаимодействие между трансферином и его рецептором в переносе ДНК не является решающим. Во всех 4 клеточных препаратах наблюдается лишь слабая доставка ДНК при применении комплексов TfpL и ДНК в присутствии свободного аденовируса d1312 (см. следы 1, 4, 7, 10). Эффективность трансфекции повышается в присутствии биотинилированного аденовируса, модифицированного стрептавидином (см. следы 2, 3, 5, 6, 8, 9, 11, 12). Эффективность доставки, достигаемая при применении комбинационного комплекса, включающего только вирус, pL и StpL, повышается менее чем на 1 log по сравнению с комплексами, которые включают только конъюгат трансферина и полилизина (TfpL) (ср., например, след 2 (без трансферина) со следом 3 (в присутствии трансферина)). По поводу незначительной трансфекции в случае применения свободного вируса и высокой трансфекции в случае применения включающих связанный вирус комплексов либо в присутствии, либо в отсутствие конъюгата трансферина и полилизина предполагается, что аденовирус служит в качестве лиганда в этих клетках, а в случае отсутствия связывания свободный вирус может проникать в клетки, однако комплекс ДНК и конъюгата трансферина и полилизина не проникает эффективно (в данном примере в качестве ДНК применяют pCMVL).
б) Гистохимический анализ частоты трансфекции в миотрубочках
Аналогично стадии а) подготовляют культуры миотрубочек С2Сl2 (5 • 105 клеток высеивают в чашки диаметром 6 см в качестве миобластов, которые потом дифференцируют в миотрубочки). 6 мкг ДНК pCMV β-gal переводят в комплекс с 8 мкг TfpL в среде 500 мкл буфера HBS и добавляют к клеткам в присутствии 18 мкл аденовируса d1312 (1 • 1012 вирусов на мл) в 2 мл среды DMEM, содержащей 2% ТЭС. Кроме того, 6 мкг ДНК pCMVLacZ переводят в комплекс с 7 мкг TfpL и 800 нг StpL, включающего 18 мкл биотинилированного аденовируса d1312 (1 • 1012 вирусов на мл) в 500 мкл буфера HBS и добавляют к клеткам в 2 мл среды DMEM, содержащей 2% ТЭС. После 24-часовой инкубации клетки окрашивают описанным в примере 15 образом с тем, чтобы определить активность β-галактозидазы. Результаты окрашенных проб согласуются с результатами трансфекции с применением в качестве репортерного гена люциферазы (см. пункт а). В случае применения свободного вируса наблюдается лишь очень низкая экспрессия гена в культурах миотрубочек, тогда как связывание вируса с ДНК приводит к высокому уровню экспрессии гена. Присутствие окрашенных в синий цвет канальцев указывает на успешный перенос гена в эти дифференцированные клетки в присутствии свободного аденовируса.
в) Перенос ДНК в первичные мышиные миобласты и миотрубочки
Основные скелетные мышцы обеих задних ног мыши-самца породы С57В1/6 в возрасте 4 недель стерильно изолируют в буфере PBS и разрезают на куски примерно 5 мм. Ткань суспендируют в 20 мл буфера PBS, оставляют стоять в течение примерно 2 минут и надосадочную жидкость аспирируют. Указанные операции повторяют 3 раза. Затем ткань смешивается с 3,5 мл буфера PBS, содержащего 0,5 мл трипсина /ЭДТУК, 0,5 мл 1%-ной по весу коллагеназы и 0,5 мл 1%-ного АСС (фракция V, в 4 мМ хлористого кальция) и смесь подвергают инкубации при температуре 37oC в течение 30 минут с частым незначительным размешиванием. После 30-минутной инкубации оставшиеся ткани дают осаждаться, надосадочную жидкость удаляют и смешивают с 5 мл среды DMEM, включающей 20% ТЭС. Инкубацию с протеазой повторяют 3 - 4 раза до полной дисперсии ткани. Получаемую суспензию клеток фильтруют и центрифугируют при 500 g в течение 15 минут. Продукт центрифугирования повторно суспендируют в 10 мл среды DMEM, содержащей 20% ТЭС, и фибропласты удаляют за счет высеивания клеток на непокрытые чашки диаметром 15 см в течение 60 минут. Несвязанные клетки тщательно удаляют и высеивают на 5 покрытых ламинином чашек Петри диаметром 10 см, каждая из которых содержит 15 мл среды DMEM, содержащей 20% ТЭС. После достижения слияния (примерно через одну неделю) клетки обрабатывают трипсином и снова высеивают на покрытые ламинином чашки Петри диаметром 6 см в количестве примерно 1 • 106 клеток на чашку. Для получения культур миотрубочек примерно через 5 дней (когда достигнуто слияние клеток) среду заменяют на среду DMEM, содержащую 2% лошадиной сыворотки, и через одну неделю осуществляют процесс трансфекции. Культуры миобластов подвергают трансфекции в чашках диаметром 6 см примерно при 80%-ном слиянии.
Покрытые ламинином чашки приготовляют следующим образом. В чашки подают 0,025 мг/мл полилизина с молекулярным весом 30000 - 70000 в стерильной воде при комнатной температуре в течение 30 минут. Затем чашки промывают 3 раза стерильной водой и сушат воздухом. Затем чашки покрывают ламинином, подаваемым в воде в количестве 8 мкг/мл, при комнатной температуре в течение ночи, после чего чашки промывают 3 раза стерильной водой перед высеиванием клеток.
Применяемые для трансфекции комплексы ДНК получают за счет разбавления указанного на фиг. 38 количества биотинилированного аденовируса d1312, инактивированного обработкой псораленом и ультрафиолетовыми лучами (полученным описанным в примере 19 образом; на фиг.: "инакт. AdV") в 150 мкл буфера HBS и добавления 1 мкг StpL в 150 мкл буфера HBS с последующей инкубацией при комнатной температуре в течение 30 минут. Затем к каждой пробе добавляют 100 мкл буфера HBS, содержащего 6 мкг pCMVL, с последующей инкубацией при комнатной температуре в течение 30 минут. В конце концов к каждой пробе добавляют 7 мкг TfpL в 100 мкл буфера HBS, инкубируют при комнатной температуре в течение 30 минут и добавляют к культурам миобластов и миотрубочек в чашках диаметром 6 см, содержащих 2 мл среды DMEM, содержащей 2% ТЭС. После одночасовой инкубации среду заменяют на 5 мл среды DMEM, содержащей 20% ТЭС (в случае миобластов), или на 5 мл среды DMEM, содержащей 2% лошадиной сыворотки (в случае миотрубочек). Через 48 часов осуществляют сбор клеток для определения активности люциферазы. Результаты опыта представлены на фиг. 38.
Пример 26. Улучшение переноса вируса CELO в миобласты с применением пектина в качестве лиганда
а) Сопоставительный анализ аденовируса d1312 и вируса CELO в клетках HeLa и миобластах C2Cl2
Состоящие либо из клеток HeLa либо из миобластов C2Cl2 пробы (5 • 105 клеток на чашку диаметром 6 см) подвергают трансфекции 6 мкг ДНК pCMVL в виде комплекса с 1 мкг StpL/7 мкг TfpL и 5 мкл биотинилированного аденовируса d1312 (см. пример 19; 1 • 1012 частиц/мл) или 18 мкл биотинилированного вируса CELO (см. пример 24; 0,3 • 1012 частиц на мл). После 20-часовой инкубации клетки собирают для определения активности люциферазы. Результаты опыта представлены на фиг. 39.
Трансфекция в клетках HeLa может осуществляться со сравнимой эффективностью как при применении состоящих из аденовируса d1312 человека, StpL, TfpL и ДНК комплексов, которые проникают в клетки либо при помощи рецептора аденовируса, либо при помощи рецептора трансферина, так и состоящих из вируса CELO, StpL, TfpL и ДНК комплексов, которые могут проникать в клетки при помощи рецептора трансферина. Перенос ДНК в миобласты C2Cl2 может эффективно осуществляться при применении включающих аденовирус d1312 комплексов, тогда как включающие вирус CELO комплексы являются менее эффективными в этих клетках. Результаты опытов по предыдущим примерам свидетельствуют о том, что рецептор трансферина играет лишь незначительную роль в процессе переноса комбинационного комплекса в эти клетки. Предполагается, что рецептор аденовируса представляет собой основной сайт проникновения. Незначительная активность вируса CELO в миобластах может представлять собой результат плохого связывания как вируса CELO, так и трансферина с миобластами C2Cl2.
б) Улучшение трансфекции миобластов C2Cl2 вирусом CELO при применении агглютинина пшеничного зачатка в качестве лиганда
Ввиду малоэффективного переноса в опыте по пункту а) трансферин заменяют на новый лиганд, получаемый следующим образом.
Биотинилированный вирус CELO (3 • 109 клеток) и 1 мкг биотинилированного агглютинина пшеничного зачатка (далее: "АПЗ") в 150 мкл буфера HBS смешивают с раствором 0,8 мкг StpL в 150 мкл буфера HBS и смесь инкубируют при комнатной температуре в течение 30 минут. 6 мкг ДНК pCMVL в 100 мкл буфера HBS добавляют к раствору вируса, АПЗ и StpL в 300 мкл HBS с последующей инкубацией при комнатной температуре в течение 30 минут. Затем к смеси добавляют 7 мкг TfpL в 100 мкл буфера HBS, после чего снова инкубируют при комнатной температуре в течение 30 минут. Получаемые комплексы добавляют к миобластам C2Cl2 (5 • 105 клеток на чашку диаметром 6 см) в 2 мл среды DMEM, содержащей 2% ТЭС. Через час к клеткам добавляют 5 мл среды DMEM, содержащей 10% ТЭС, и по истечении 20 часов осуществляют сбор клеток для определения активности люциферазы (в световых единицах). Результаты опыта представлены на фиг. 40. На этой фиг. видно, что в отсутствие вируса перенос ДНК является очень незначительным как в присутствии АПЗ, так и в его отсутствие (см. следы 1, 6). Перенос незначительной эффективности достигается при применении связанного вируса CELO (см. след 2). Но 16-кратное повышение эффективности переноса достигается в случае включения в комплекс АПЗ. Увеличение содержания АПЗ в комплексы (от 1 мкг до 5 мкг) приводит к незначительному снижению эффективности переноса (ср. следы 3 и 4), тогда как в результате увеличения содержания StpL в комплексе (от 1 мкг до 2 мкг) наблюдается незначительное повышение эффективности переноса (ср. следы 3 и 5). Таким образом, результаты данного опыта свидетельствуют о том, что АПЗ в качестве лиганда обеспечивает повышение эффективности переноса ДНК в клетки C2Cl2 при помощи вируса CELO.
г) Экспрессия гена полного фактора VIII в миобластах и миотрубочках C2Cl2
Получение миобластов и миотрубочек C2Cl2 осуществляют описанным выше образом. Трансфекцию осуществляют с применением 6 мкг плазмиды, кодирующей кДНК полного фактора VIII человека, в виде комплекса с 5 или 15 мкл биотинилированного аденовируса d1312, 0,5 или 1 мкг StpL и 7 или 6 мкг TfpL. Комплексы вируса и ДНК добавляют к клеткам в среде DMEM, содержащей 2% ТЭС. После 4-часовой инкубации при температуре 37oC к каждой чашке добавляют 3 мл свежей среды DMEM, содержащей 10% ТЭС. По истечении 18 часов осуществляют сбор клеток для определения присутствия фактора VIII. Представленные на фиг. 41 результаты указаны как милли-единицы за 24 часа на 1 • 106 клеток.
Применение белка аденовируса для переноса ДНК. Аденовирус wt300 выращивают в клетках HeLa, очищают и биотинилируют описанным выше для аденовируса d1312 образом. 1,2 мл вируса подвергают диализу с применением 3 х 300 мл 5мМ среды MES, содержащей 1 мМ ЭДТУК, pH 6,25, при температуре 4oC в течение 18 часов. Затем осуществляют центрифугирование при 27 g в течение 30 минут с применением ротора типа SW60. Надосадочную жидкость осторожно удаляют и остаток повторно суспендируют в буфере HBS, содержащем 40% глицерина. К надосадочной жидкости добавляют 20 мМ буфера HEPES с pH 7,4 и 150 мМ хлористого натрия и как осадок (содержащий сердцевину вируса и основную массу гексонового капсида, на фиг. 42 обозначены как "серд."), так и фракции надосадочной жидкости (содержащие оболочки, обозначенные на фиг. 42 как "обол.") применяют для определения активности переноса ДНК в мышиных фибропластах Mov13 и клетках HeLa.
Образование включающего ДНК комплекса осуществляют следующим образом. Указанные на фиг. 42 количества каждой фракции, то есть разрушенный вирус перед центрифугированием и интактный вирус (выраженные как мкг белка согласно методу Брэдфорда), разбавляют 300 мкл буфера HBS. Затем добавляют 3 мкг модифицированного стрептавидином полилизина в 50 мкл буфера HBS с последующей инкубацией при комнатной температуре в течение 30 минут. 6 мкг ДНК pCMVL разбавляют 100 мкл буфера HBS и добавляют к первому раствору с последующей инкубацией в течение 30 минут. В заключение добавляют 2 мкг TfpL в 100 мкл HBS с последующей инкубацией в течение дальнейших 30 минут. Для получения проб, содержащих только TfpL, 8 мкг TfpL в 170 мкл HBS смешивают с 6 мкг pCMVL в 330 мкл HBS при комнатной температуре в течение 30 минут. Указанные количества белка вируса разбавляют 300 мкл HBS, после чего добавляют к включающим TfpL и ДНК комплексам. Все пробы добавляют к 5 • 105 клеток HeLa или фибропластов Mov13, находящихся в чашках диаметром 6 см, содержащих 2 мл среды DMEM, содержащей 10% ТЭС, после чего пробы оставляют стоять в течение одного часа. Затем добавляют 5 мл свежей среды, содержащей 10% ТЭС, и через 20 часов осуществляют сбор клеток для определения активности люциферазы в световых единицах. Результаты опыта представлены на фиг. 42, где график А относится к опыту с применением клеток HeLa, а график В - к опыту с применением фибропластов Mov13.
Как видно на фиг. 42, при обоих типах клеток наблюдается зависящее от дозы повышение активности переноса ДНК в случае содержащей оболочку фракции (см. пробы 4-6 на обоих графиках). Если же такое количество белка биотинилированного вируса включено в комплексы из TfpL и ДНК, которые не содержат модифицированного стрептавидином полилизина, то наблюдается степень переноса ДНК, близкая к фоновому уровню (см. пробу 3 на обоих графиках).
Пример 28. Улучшенный перенос гена при применении тройных комплексов ДНК, включающих конъюгат галактозного лиганда и полилизина
а) Тройные комплексы, включающие конъюгат пептида гриппа
Было обнаружено, что в присутствии сопряженного с полилизином пептида, включающего последовательности, производящиеся от N-конца подединицей НА-2 гемагглютинина вируса гриппа, в комплексах ДНК и конъюгата трансферина и полилизина приводит к значительному улучшению переноса гена, обеспечиваемого трансферином/полилизином (см. примеры 13, 14). Подобные комбинационные комплексы ДНК, включающие конъюгат полилизина и лиганда галактозы, а также модифицированного полилизином пептида гриппа (далее: "InflupL"; полученного описанным в примерах 6, 13 образом) получают путем добавления конъюгата лиганда и полилизина к плазмидной ДНК pCMVL с тем, чтобы нейтрализовать половину заряда ДНК. Остаток заряда используют для нагрузки комплексов конъюгатом полилизина и пептида гриппа. Добавление этих комплексов ДНК, включающих синтетический лиганд (gal)4, к гепатоцитам BNL CL.2 (трансфекцию осуществляют описанным в примере 6 ж) образом приводит к экспрессии гена люциферазы, которая значительно превышает экспрессию, получаемую с помощью трансферина в качестве лиганда (см. фиг. 43). Экспрессия превышает больше чем в 500 раз экспрессию, обнаруживаемую в контрольных опытах с применением комплексов ДНК, которые содержат то же самое количество полилизина, но не содержат пептида гриппа (см. также фиг. 43). Достигаемая комбинационными комплексами ДНК активность также примерно в 30 раз больше активности, достигаемой комплексами ДНК и (gal)4pL в присутствии хлорохина.
б) Тройные комплексы, включающие конъюгат аденовируса
Комплексы получают следующим образом. 2 мкл, 6 мкл и 18 мкл, соответственно, биотинилированного аденовируса d1312 (полученного как в примере 19) в количестве 1012 частиц на мл в 50 мкл HBS смешивают с 100 нг, 160 нг и 480 нг, соответственно, модифицированного стрептавидином полилизина в 100 мкл буфера HBS с последующей инкубацией в течение 30 минут. Затем добавляют раствор 6 мкг pCMVL в 200 мкл HBS и, через дальнейшие 30 минут, раствор 3,8 мкг (gal)4pL (полученного как в примере 6) или 7 мкг TfpL в 150 мкл HBS.
Растворы каждого комплекса ДНК добавляют к 300000 клеткам (АТСС ТIВ73, АТСС ТIВ74, АТСС ТIВ75, АТСС ТIВ76), которые выращивают в чашках диаметром 6 см, содержащих среду DMEM, имеющую высокое содержание глюкозы и 2% ТЭС. Далее работают описанным выше образом. Данные по экспрессии гена (через 24 часа) представлены на фиг. 44.
Пример 29. Перенос ДНК с помощью конъюгата трансферина и полилизина в присутствии свободного и сопряженного риновируса
а) Препараты риновируса HRV-2
400 мкл раствора риновируса (примерно 30 мкг) в буфере HBS (150 мМ хлористого натрия и 5 мМ HEPES, pH 7,9), содержащем 10% глицерина, обрабатывают 10 нмоль биотина марки NHS-LC. После инкубации при комнатной температуре в течение 3 часов вирус отделяют от невключенного биотина путем экстенсивного диализа с помощью буфера HBS, содержащего 40% глицерина, при температуре 4oC. Получаемый путем выращивания в присутствии акридинового оранжевого (на фиг. 45 обозначенного как "акрид.") светочувствительный риновирус подвергают инактивации известным образом.
б) Получение комплексов ДНК и проведение трансфекции
1) Комплексы ДНК и конъюгата трансферина и полилизина получают за счет смешивания раствора 6 мкг плазмидной ДНК pCMVL в 330 мкл буфера HBS (150 мМ хлористого натрия, 20 мМ HEPES, pH 7,3) с раствором 8 мкг TfpL290 в 170 мкл HBS. Комплексы ДНК смешивают с 1,5 мл среды DMEM, содержащей 2% ТЭС, и с 0,14 мкг - 3,5 мкг риновируса HRV-2 или инактивированного риновируса HRV-2. Смеси добавляют к клеткам NIH 3Т3 (300000 клеток на чашку диаметром 6 см). Через 4 часа среду заменяют на 4 мл свежей среды DMEM, содержащей 10% ТЭС. Через 24 часа клетки собирают для определения активности люциферазы описанным выше образом. Результаты опыта представлены на фиг. 45А.
2) Комбинационные комплексы ДНК, включающие конъюгат трансферина и полилизина и конъюгат риновируса и полилизина, получают следующим образом. Раствор 3,5 мкг биотинилированного риновируса HRV-2 в 100 мкл HBS смешивают с 1 мкг модифицированного стрептавидином полилизина в 100 мкл HBS. Через 30 минут при комнатной температуре раствор смешивают с 6 мкг плазмидной ДНК в 150 мкл HBS, инкубируют при комнатной температуре в течение дальнейших 30 минут и затем смешивают с 6 мкг TfpL290 в 150 мкл HBS. Аналогично получают другие комплексы, указанные на фиг. 45В. Комплексы ДНК смешивают с 1,5 мл среды DMEM, содержащей 2% ТЭС, и смесь добавляют к клеткам NIH 3Т3 (300000 клеток на чашку диаметром 6 см). Далее поступают аналогично пункту 1). Результаты опыта представлены на фиг. 45В.
Пример 30. Трансфекция клеток HeLa комбинационными комплексами, включающими связанный ионным путем аденовирус.
Сначала 30 мкл аденовируса d1312 (примерно 109 бляшкообразующих единиц) смешивают с 1 мкг полилизина с молекулярным весом 450, имеющего среднюю длину мономерных звеньев 450, в 170 мкл HBS. Через 30 минут при комнатной температуре к смеси добавляют 6 мкг ДНК pCMVL в 170 мкл HBS. После инкубации в течение дальнейших 30 минут комплексы смешивают с 9 мкг TfpL190 в 170 мкл HBS. Аликвот получаемого комплекса (10% = 50 мкл раствора 600 нг ДНК или 1% = 5 мкл раствора 60 нг ДНК) разбавляют 1,5 мл среды DMEM, содержащей 2% ТЭС, после чего добавляют к 300000 клеткам HeLa. Через 4 часа добавляют 2 мл среды DMEM, содержащей 20% ТЭС. Через 24 часа после трансфекции осуществляют сбор клеток для определения активности люциферазы описанным выше образом. Активность люциферазы общего экстракта составляет 29 115 000 световых единиц (в случае 600 нг ДНК) или же 1090 000 световых единиц (в случае 60 нг ДНК).
Контрольный опыт.
6 мкг ДНК pCMVL в 170 мкл HBS смешивают с 1 мкг полилизина с молекулярным весом 450 (имеющего среднюю длину мономерных звеньев 450) в 170 мкл HBS и через 30 минут при комнатной температуре добавляют 9 мкг конъюгата TfpL190 в 170 мкл HBS. После 30-минутной инкубации комплексы смешивают с 30 мкл аденовируса d1312 (примерно 109 бляшкообразующих единиц). Аликвот получаемого комплекса (10 % = 50 мкл раствора 600 нг ДНК, или 1% = 5 мкл раствора 60 нг ДНК) разбавляют 1,5 мл среды DMEM, содержащей 2% ТЭС, и добавляют к 300000 клеткам HeLa. Через 4 часа добавляют 2 мл среды DMEM, содержащей 20% ТЭС. Сбор клеток осуществляют через 24 часа после трансфекции. Определение активности люциферазы осуществляют описанным выше образом. Общая активность люциферазы составляет 405 000 световых единиц (в случае 600 нг ДНК) и 200 световых единиц, соответственно, в случае 60 нг ДНК.
Пример 31. Местное введение включающего ДНК, аденовирус и конъюгат трансферина и полилизина комплекса в печень крысы.
а) Непосредственное впрыскивание
Комплексы получают описанным в примере 19 образом. Они содержат 200 мкл биотинилированного аденовируса d1312, 6,4 мкг модифицированного стрептавидином полилизина, 48 мкг ДНК pCMVL и 48 мкг конъюгата TfpL290. Комплекс содержится в 200000 мкл HBS. Крысу-самца породы "Sprague-Dawley" весом 240 г анестезируют авертином, после чего осуществляют гревосечение 4 см. Раствор комплекса впрыскивают в левую долю печени. Затем рану закрывают. Через 48 часов после впрыскивания комплекса крысу умерщвляют и определяют экспрессию люциферазы. В зоне впрыскивания определяют 5615 световых единиц/мг белка продукта гомогенизации печени. Общая активность люциферазы на сайте впрыскивания составляет 370 600 световых единиц.
б) Введение комплексов в печень через желчную дренажную систему
Комплексы получают следующим образом. 200 мкл биотинилированного аденовируса d1312 в 200 мкл HBS и 6,4 мкг модифицированного стрептавидином полилизина в 400 мкл HBS подвергают инкубации при комнатной температуре в течение 30 минут, после чего добавляют 48 мкг ДНК pCMVL в 800 мкл HBS. После 30-минутной инкубации добавляют еще 48 мкг конъюгата TfpL в 900 мкл HBS. Крысы-самцы породы "Spraque-Dawley" весом 250 г анестезируют авертином, после чего брюшную полость открывают путем срединого разреза. Кишку перемещают на левую сторону корпуса и в желчный проток вводят иглу 27 G, подключенную к трубе и шприцу объемом 1 мл. Впрыскивание комплексов осуществляют в течение 4 минут, после чего иглу выводят из желчного протока и место инъекции, а также рану закрывают. Через 30 часов крыс умерщвляют и из различных долей печени отбирают пробы на определение экспрессии гена люциферазы. Пиковая активность люциферазы составляет 19 000 световых единиц/мг белка, а рассчитанная общая экспрессия в печени составляет 2,7 • 106 световых единиц.
Пример 32. Местное введение включающего ДНК, аденовирус и конъюгат трансферина и полилизина комплекса в пережатую мышиную хвостовую вену. Комплексы получают описанным в примере 19 образом. Они включают 45 мкл биотинилированного аденовируса d1312, 0,8 мкг модифицированного стрептавидином полилизина, 6 мкг ДНК pCMVL и 24 мкг конъюгата TfpL290. Раствор комплексов в 150 мкл HBS впрыскивают в хвостовую вену мыши-самца породы С3Н/Не (в возрасте 2 месяцев) после предварительной анестезии авертином. Непосредственно после впрыскивания хвостовую вену пережимают на 20 минут на проксимальном и дистальном концах так, чтобы раствор комплекса находился в той части хвостовой вены, в которую он впрыскивался, и не мог промываться кровью. Через 48 часов после впрыскивания мышь умерщвляют и хвостовую вену приготовляют к определению экспрессии люциферазы. Она составляет 2600 световых единиц/3 см хвостовой вены.
Пример 33. Трансфекция первичных клеток меланомы человека, Первичные клетки меланомы выделяют из меланомы, которая хирургическим путем удалялась из пациента, за счет механического разрушения опухоли в среде RPM1 1640, содержащей 5% ТЭС, 2 мМ глютамина и антибиотики, и последующего прессования ткани через стальное сито. Опухолевые клетки промывают несколько раз путем центрифугирования и последующего повторного суспендирования и потом культивируют в колбах марки Т25. Через 24 часа после выделения опухолевые клетки трансфецируют комбинационными комплексами, включающими 3 мкл, 9 мкл или 27 мкл биотинилированного аденовируса d1312 (1 • 1012 вирусных частиц/мл), 0,5 мкг модифицированного стрептавидином полилизина, 6 мкг ДНК pCMVL и 7 мкг конъюгата TfpL290, применяемым в виде раствора в 500 мкл HBS. Через 36 часов после трансфекции клетки собирают и определяют экспрессию люциферазы. Результаты опыта представлены на фиг. 46.
Пример 34. Трансфекция первичных фибробластов человека. Кожную ткань подают в чашку петри диаметром 6 см, содержащую среду DMEM, 2 мМ глютамина, 20% ТЭС и антибиотики. Ткань тщательно режут на маленькие куски при помощи хирургического ножа и культивируют в присутствии 3 мл среды в течение 5 дней. Затем клетки промывают свежей средой DMEM, содержащей 2 мМ глютамина, 10% ТЭС и антибиотики, и культивируют в течение дальнейших 7 дней, после чего клетки обрабатывают трипсином и переводят на дальнейшее культивирование в новые чашки петри. Когда клетки являются почти сливающимися, они снова подвергаются обработке трипсином и хранят в замороженном состоянии. Перед трансфекцией клетки размораживают и высеивают в чашки петри диаметром 6 см, в которых их культивируют в среде DMEM, содержащей 2 мМ глютамина, 10% ТЭС и антибиотики. Служащие для трансфекции комплексы получают следующим образом. 3 мкл, 10 мкл, 20 мкл и 30 мкл, соответственно, биотинилированного аденовируса d1312 и 0,1 мкг, 0,3 мкг, 0,5 мкг и 0,8 мкг, соответственно, модифицированного стрептавидином полилизина в 150 мкл HBS инкубируют в течение 30 минут при комнатной температуре. Затем добавляют 6 мкг плазмиды pCMY-β-gal в 170 мкл HBS и смесь инкубируют в течение дальнейших 30 минут. Затем добавляют конъюгат TfpL в 170 мкг HBS в следующих количествах: 7,8 мкг в случае комплекса, включающего 3 мкл аденовируса d1312, 7 мкг в случае комплекса, включающего 10 мкл аденовируса d1312, и 6 мкг в случае комплексов, включающих 20 мкл и 30 мкл аденовируса d1312. После 30-минутной инкубации комплексы добавляют к клеткам в 2 мл среды DMEM, содержащей 2 мМ глютамина, 2% ТЭС и антибиотики, и клетки инкубируют при температуре 37oC в течение 4 часов, после чего среду удаляют и культивирование продолжают при температуре 37oC с применением среды DMEM, содержащей 2 мМ глютамина, 10% ТЭС и антибиотики. По истечении 48 часов вышеуказанным образом определяют экспрессию β-галактозидазы.
В случае трансфекции комплексом, включающим 3 мкл аденовируса d1312, 14% клеток продуцируют β-галактозидазу. В случае комплекса, включающего 10 мкл аденовируса d1312, получают 32% положительных клеток, в случае комплекса, включающего 20 мкл аденовируса d1312, - 39% положительных клеток, а в случае комплекса, включающего 30 мкл аденовируса d1312, - 64% положительных клеток.
Пример 35. Перенос гена с применением невирусных эндосомолитических агентов.
а) Синтез разрушающих мембрану пептидов
1) Синтез пептида
Синтез осуществляют при помощи автоматического аппарата типа АВI431А путем твердофазного метода с применением 0,97 ммоль/г пара- алкоксибензиловой смолы в качестве твердой основы и защищенных 9- флуоренметоксикарбонилом аминокислот. Аминокислоту с карбоксильной группой на конце связывают со смолой через симметричный ангидрид. Последующие аминокислоты связывают с помощью 1-оксибензотриазол- дициклогексилкарбодиимида. Применяют следующие защищающие боковую цепь группы: (Trt)Asn, (Trt)Cys [(треть.-бутил)Cys в случае пептидов EALA и GLF], (треть.-бутил)Glu, (Trt)His, (треть.- бутил)Ser.
EALA: Trp Glu Ala Ala Leu Ala Glu Ala Leu Ala
Glu Ala Leu Ala Glu His Leu Ala Glu Ala Leu Ala Glu Ala Leu Glu Ala Leu Ala Ala
Gly Gly Ser Cys
GLF Gly Leu Phe Gly Ala Leu Ala Glu Ala Leu
Ala Glu Ala Leu Ala Glu His Leu Ala Glu Ala Leu Ala Glu Ala Leu Glu Ala Leu Ala Ala Gly Gly Ser Cys
GLF-II Gly Leu Phe Gly Ala Leu Ala Glu Ala Leu
Ala Glu Ala Leu Ala Glu Ala Leu Ala Glu Ala Leu Ala Glu Ala Leu Glu Ala Leu Ala Ala Gly Gly Ser Cys
GLF-delta Gly Leu Phe Glu Leu Ala Glu Ala Leu Ala
Glu Ala Leu Ala Glu Ala Leu Ala Glu Ala Leu Ala Glu Ala Leu Glu Ala Leu Ala Ala
Gly Gly Ser Cys
EALA-lnf Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile Glu
Asn Gly Trp Glu Gly Leu Ala Glu Ala Leu Ala Glu Ala Leu Glu Ala Leu Ala Ala Gly
Gly Ser Cys
EALA-P50 Gly Leu Phe Glu Ala Ile Glu Gly Phe Ile Glu
Asn Gly Trp Glu Gly Leu Ala Glu Ala Leu Ala Glu Ala Leu Glu Ala Leu Ala Ala
Gly Gly Ser Cys
P50 Gly Leu Phe Glu Ala Ile Glu Gly Phe Ile Glu
Asn Gly Trp Glu Gly Met Ile Asp Gly Gly Gly Cys
Пептиды отделяют от резины и защищающие боковую цепь группы [за исключением (треть.-бутил) Cys] удаляют за счет обработки нагруженной 100 мг пептида основы, взятой в количестве 3 мл смесью фенола, этандитиола, тионизола, воды и трифтор-уксусной кислоты в объемном соотношении 0,75:0,25:0,5:0,5:10 при комнатной температуре в течение 90 минут. Сырые пептиды осаждают в простом диэтиловом эфире и промывают два раза. Защищенные трет.- бутилмеркаптогруппой пептиды EALA и GLF растворяют в небольшом объеме 1М бикарбоната триэтиламмония с pH 8, разбавляют до объема 100 мМ бикарбоната триэтиламмония и далее очищают путем обратнофазной жидкостной хроматографии под давлением на колонке Нуклеосил 500-5С4 с применением в качестве градиента 0,1% трифторуксусной кислоты и ацетонитрила. Оба пептида элюируют примерно при 50% ацетонитрила. Свободную форму Cys-SH пептидов получают за счет снятия защитных групп Trt аналогично вышеуказанному методу. Получают 5 мг сырых пептидов, которые растворяют в 100 мкл раствора 100 мМ бикарбоната триэтиламмония с pH 8, содержащего 1 мкл β-меркаптоэтанола, и очищают путем гель-фильтрации на колонке Сефадекс Г-25 (100 мМ бикарбоната триэтиламмония, 0,5 мМ ЭДТУК) и сушки замораживанием или же ионнообменной хроматографии на колонке Mono Q (20 мМ HEPES, pH 7,3, градиент: 0-3 М хлористого натрия; пептид элюируют при 1,5 М NaCl).
2) Модификация N-(оксиэтил)малеинимидом
После получения свободной меркаптоформы в результате гель- фильтрации на колонке Сефадекс Г-25 (20 мМ HEPES, pH 7,3, 0,5 мМ ЭДТУК) C-концевую меркаптогруппу пептидов GLF-delta, GLF-II, EALA- Inf, EALA-P50 и Р50 блокируют путем взаимодействия с взятым в 1,3-10-кратном молярном избытке N-(оксиэтил)-малеинимидом при комнатной температуре в течение одного часа. Избыточный малеинимид удаляют путем гель-фильтрации на колонке Сефадекс Г-25 (100 мМ бикарбоната триэтиламмония, pH 8) и после сушки замораживанием получают пептиды GLF-delta-mal, GLF-II-mal, EALA-Inf-mal, EALA-P50- mal, Р50-mal в виде триэтиламмониевой соли.
3) Модификация 2,2'-дитиобиспирид ином
Пептиды со свободной меркаптогруппой подвергают взаимодействию с 10 эквивалентами 2,2'-дитиобиспиридина в среде буфера 20 мМ HEPES с pH 7,9, содержащего 0,5 мМ ЭДТУК, при комнатной температуре в течение ночи. Избыточный реагент удаляют путем гель-фильтрации на колонке Сефадекс Г-25 (100 мМ бикарбоната триэтиламмония с pH 8) или ионообменной хроматографии на колонке Mono Q (20 мМ HEPES, pH 7,3, градиент: 0-3 М хлористого натрия; пептид элюируют при 1,5 М NaCl). Получают (2-пиридилтио)-Cys-пептиды GLF-delta-SSPy, GLF-H- SSPy, EALA-Inf-SSPy, EALA-P50-SSPy и P50-SSPy.
4) Димеризация пептидов
Гомодимерный Р50 (Р50 dim) получают путем взаимодействия эквимолярных количеств Р50-Cys-(2-пиридилтио) и P50-Cys-SH в среде 20 мМ HEPES, pH 7,3 при комнатной температуре в течение 3 дней. Реакционную смесь разделяют на колонке Mono Q HR-5/5 (20 мМ HEPES, pH 7,3, градиент: 0,09 - 3 М хлористого натрия; димерный Р50 элюируют при 1,1 М NaCl). Гетеродимерный GLF-SS- Р50 получают аналогичным образом путем взаимодействия пептида Р50 в свободной меркаптоформе с модифицированным пиридилтио-группой пептидом GLF.
б) Определение лизиса липосом
Способность синтетических пептидов к разрушению липосом исследуют за счет определения выделения флуоресцентного красителя из липосом, нагруженных самогасящей концентрацией кальцеина. Липосомы получают из яичного фосфатидилхолина путем обратнофазного упаривания из водной фазы, содержащей 100 мМ кальцеина, 375 мМ натриевых ионов, 50 мМ хлористого натрия, имеющего pH 7,3, и последующего пропускания через поликарбонатный фильтр с величиной пор 100 нм. Затем липосомы очищают путем гель-фильтрации на колонке Сефадекс Г-25 с применением изо-осмотического буфера (200 мМ хлористого натрия, 25 мМ HEPES, pH 7,3). Для определения лизиса при разных значениях pH основной раствор липосом (6 мкл/мл) разбавляют буфером, содержащим 400 мМ хлористого натрия и 40 мМ цитрата натрия. Аликвот 100 мкл добавляют к 80 мкл имеющего разную степень разведения водного раствора пептида в снабженной 96 ячейками пластинке микротитратора (конечная концентрация липида: 25 мкмоль) и после 30-минутной инкубации при комнатной температуре определяют флуоресценцию кальцеина при 600 нм (возбуждение при 490 нм). 100%-ный лизис получают за счет добавления 1 мкл 10%-ного раствора тритона X-100. Единицы лизиса рассчитывают как реципрокное значение концентрации пептида, при которой устанавливают 50% лизиса (то есть объем (мкл) раствора липосом, который подвергают 50%-ному лизису на мкг пептида). Значения ниже 20 единиц являются результатом экстраполяции. Результаты опыта представлены на фиг. 47, где видно, что пептиды GLF и EALA проявляют наивысшую специфичную в отношении pH активность.
в) Определение гемолиза
Свежие эритроциты человека промывают несколько раз буфером HBS и повторно суспендируют в содержащем 300 мМ хлористого натрия и 30 мМ цитрата натрия буфере при концентрации 6,6 • 107 /мл. Аликвот 75 мкл добавляют к 75 мкл имеющего разную степень разведения водного раствора пептида в снабженной 96 ячейками пластинке микротитратора и инкубируют при температуре 37oC в течение одного часа при постоянном взбалтывании. После удаления нерастворившихся эритроцитов путем центрифугирования при 1000 об./мин в течение 5 минут получают 100 мкл надосадочной жидкости, которую подают в новую пластинку микротитратора, после чего определяют абсорбцию гемоглобина при 450 нм (коррекция фона при 750 нм). 100%-ный лизис определяют путем добавления 1 мкл 10%-ного раствора тритона X-100 перед центрифугированием. Единицы гемолизиса рассчитывают как реципрокные значения концентрации пептида, при которой обнаруживают 50%-ный лизис (то есть объем (мкл) раствора эритроцитов, который подвергают 50%-ному лизису на мкг пептида). Значения ниже 3 единиц гемолиза являются результатом экстраполяции. Результаты опыта представлены на фиг. 48, где видно, что пептиды Р50 dim и EALA-Р50 mal проявляют наивысшую, специфичную в отношении pH активность по лизису клеток и/или высвобождению больших молекул, таких, как, например, гемоглобин. Мономерные Р50 mal и Р50 SS-Py проявляют низкую активность. Мелиттин проявляет наивысшую активность, которая, однако, не является специфичной в отношении кислых величин pH.
г) Получение включающих ДНК комбинационных комплексов
6 мкг ДНК pCMVL в 150 мкл HBS смешивают с 4 мкг конъюгата TfpL290 в 150 мкл HBS и через 30 минут при комнатной температуре добавляют 4 - 20 мкг поли(L)-лизина с молекулярным весом 290 в 100 мкл HBS. По истечении дальнейшей 30-минутной инкубации при комнатной температуре добавляют 0,3 - 30 мкг пептида в 100 мкл HBS, после чего инкубируют еще в течение 30 минут. Оптимальное количество эндосомолитического агента предварительно определяют с учетом результатов анализа эффективности переноса гена (см. таблицу, показывающую перенос гена в клетки BNL CL.2). Одновременное добавление полилизина (pL) и эндосомолитического агента, а также применение большего объема комбинационного комплекса (конечный объем: 1,5 мл) приводят к сравнимой или лучшей эффективности трансфекции. В данном опыте также установлено, что непептидные амфипатические вещества дезоксихолевая кислота и олеиновая кислота могут также улучшить перенос ДНК.
г) Трансфекция клеток
Прилипшие клеточные линии (гепатоциты BNL CL.2 и клетки NIH 3Т3, соответственно) выращивают в чашках Петри диаметром 6 см за 1-2 дней до трансфекции (среда: DMEM с 10 % ТЭС; 300000 клеток на чашку). Среду удаляют и добавляют 1,5 мл среды DMEM, содержащей 2% ТЭС, и 500 мкл комплекса. Альтернативно применяют 0,5 мл среды DMEM, содержащей 6% ТЭС, и 1,5 мл комплекса. После 4-часовой инкубации добавляют 2 мл среды DMEM, содержащей 18% ТЭС. Альтернативно среду трансфекции можно также заменять на 4 мл среды DMEM, содержащей 10% ТЭС. Через 24 часа после трансфекции осуществляют сбор клеток на определение активности люциферазы в световых единицах. Результаты трансфекции гепатоцитов BNL CL.2 представлены на фиг. 49, показывающей общую активность люциферазы в трансфецированных клетках. При этом применяемые в опыте согласно фиг. 49А. комплексы получают следующим образом. 6 мкг ДНК pCMVL в 250 мкл HBS смешивают с 4 мкг конъюгата TfpL290 в 250 мкл HBS и через 30 минут при комнатной температуре добавляют 20 мкг поли(L)лизина с молекулярным весом 290 в 750 мкл HBS. После дальнейшей инкубации в течение 30 минут при комнатной температуре добавляют указанные количества пептидов в 250 мкл HBS. После дальнейшей 30-минутной инкубации получаемые комплексы смешивают с 0,5 мл среды DMEM, содержащей 6% ТЭС, и добавляют к 450 000 клеткам.
Применяемые комплексы ДНК в опыте согласно фиг. 49В получают следующим образом. 6 мкг ДНК pCMVL в 500 мкл HBS смешивают с 4 мкг конъюгата TfpL290 в 250 мкл HBS и раствор оставляют стоять при комнатной температуре в течение 30 минут. Раствор 20 мкг поли(L)лизина с молекулярным весом 290 в 500 мкл HBS смешивают с указанными количествами пептидов в 250 мкл HBS и сразу же добавляют к смеси конъюгата TfpL и ДНК. После 30-минутной инкубации при комнатной температуре комплексы смешивают с 0,5 мл среды DMEM, содержащей 6% ТЭС, и добавляют к 450 000 клеткам. Через 24 часа после трансфекции осуществляют сбор клеток и определение активности люциферазы описанным выше образом.
Результаты опытов с применением клеток NIH3T3 представлены на фиг. 50. Применяемые в опытах согласно фиг. 50А. и В. комплексы получают тем же образом, что и комплексы для осуществления опытов по фиг. 49А. и В. На фиг. 49 и 50 видно, что комплексы, включающие Р50 dim, проявляют наибольшую активность.
Пример 36. Перенос гена с применением синтетического невирусного пептида с олигилизиновым удлинением на C-конце
Пептид последовательностью Met Ala Gln Asp Ile Ile Ser Thr Ile Gly Asp Leu Val Lys Tip Ile Ile Asp Thr Val Asn Lys Phe Thr Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys синтезируют и очищают известными приемами. Этот пептид производится от δ-токсина Staphylococcus aureus последовательностью Met Ala Gln Asp Ile Ile Ser Thr Ile Gly Asp Leu Val Lys Trp Ile Ile Asp Thr Val Asn Lys Phe Thr Lys Lys. Этот пептид обладает способностью к разрушению мембран при кислых значениях pH, если он удлинен на дополнительные 10 лизиновых остатков.
6 мкг ДНК pCMVL в 170 мкл HBS смешивают с 4 мкг конъюгата TfpL290 в 170 мкл HBS. Через 30 минут при комнатной температуре добавляют примерно 3 мкг пептида в 170 мкл HBS. После 30-минутной инкубации получаемый комплекс смешивают с 1,5 мл среды DMEM, содержащей 2% ТЭС, и добавляют к 450 000 гепатоцитам BNL CL.2. По истечении 2 часов добавляют 2 мл среды DMEM, содержащей 20% ТЭС. Через 24 часа после трансфекции осуществляют сбор клеток для определения активности люциферазы описанным выше образом. В общем экстракте она составляет 481000 световых единиц.
Пример 37. Трансфекция гепатоцитов в присутствии мелиттиновых пептидов, имеющих олиголизиновый хвост на C-конце.
Пептиды последовательностей (от N-конца до C-конца): Gly Ile Gly Ala Val Leu Lys Val Leu Thr Thr Gly Leu Pro Ala Leu Ile Ser Trp Ile Lys Arg Lys Arg Gln Gln Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys (данный пептид далее обозначается как mel 1) и Gly Ile Gly Ala Val Leu Glu Val Leu Glu Thr Gly Leu Pro Ala Leu Ile Ser Trp lle Lys Arg Lys AArg Gln Gln Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys (кислый мутант, который далее обозначается как mel 2) синтезируют и очищают известными приемами.
6 мкг ДНК pCMVL в 170 мкл HBS смешивают с 4 мкг конъюгата TfpL290 в 170 мкл HBS и оставляют стоять при комнатной температуре в течение 30 минут. Затем добавляют примерно 3 мкг пептида mel 1 или 5 мкг пептида mel 2 в 170 мкл HBS и после дальнейшей 30-минутной инкубации получаемый комплекс смешивают с 1,5 мл среды DMEM, содержащей 2% ТЭС, и добавляют к 450 000 клеткам BNL CL.2, которые предварительно выращивают описанным в примере 36 образом. По истечении 4 часов добавляют 2 мл среды DMEM, содержащей 20% ТЭС. Через 24 часа после трансфекции осуществляют сбор клеток и определение активности люциферазы описанным выше образом. В общем экстракте активность люциферазы составляет 9200 световых единиц (в случае пептида mel 1) и 9400 световых единиц (в случае пептида mel 2), соответственно.
Пример 38. Экспрессия альфа-интерферона в клетках HeLa.
5 • 105 клеток HeLa на чашку Петри диаметром 6 см подвергают трансфекции плазмидой pAD-CMV1-IFN, кодирующей интерферон альфа2с человека под контролем усилителя/промотора CMV (см. заявку ДЕ 40 21 917 A; pADCMVI- IFN получают путем клонирования вставки Hindlll-XbaI/IFN- α 2c в pAD-CMV1). Пробы по 6 мкг ДНК в 330 мкл HBS смешивают с 8 мкг конъюгата TfpL в 330 мкл HBS и смесь оставляют стоять при комнатной температуре в течение 30 минут. Пробы 6 - 10 содержат только 4 мкг конъюгата TfpL. После первой 30-минутной инкубации аликвот 20 мкг P16pL в 160 мкл HBS добавляют к пробам 6 и 7, а к пробам 8, 9 и 10 добавляют аликвот 20 мкг полилизина с молекулярным весом 290. После дополнительной 30-минутной инкубации добавляют аликвоты 160 мкл буфера HBS, содержащего 10 мкл (в случае пробы 8) или 50 мкл (в случае проб 9, 10) свободного Р16 (синтез Р16 и Р16р1 описан в примере 13). После дополнительной 30-минутной инкубации пробы добавляют к клеткам HeLa в 2 мл среды DMEM, содержащей 2% ТЭС в присутствии следующих дополнительных веществ. Пробы 2, 7 и 10 содержат 100 мкмоль хлорохина ("хл." на фиг. 51), пробы 3, 4 - соответственно 5 и 15 мкл аденовируса d1312 (1 • 1012 частиц/мл; "AdV" на фиг. 51), проба 5 - 15 мкл инактивированного псораленом аденовируса d1312 ("инак. ПС AdV" на фиг. 51). Пробы 11, 12 и 13, содержащие только аликвоты вируса по пробам 3, 4 и 5, служат в качестве контроля. Через два часа после трансфекции к клеткам добавляют 5 мл свежей среды DMEM, содержащей 10% ТЭС. Через 48 часов после трансфекции среду заменяют на 2 мл свежей среды DMEM, содержащей 10% ТЭС. Через 72 часа после трансфекции осуществляют сбор клеток для определения содержания интерферона с применением иммуноферментного твердофазного анализа. Результаты опытов, то есть содержание интерферона в нг/мл, представлены на фиг. 51, где видно, что конъюгат TfpL проявляет очень низкую активность по переносу интерферонового гена в эти клетки, что совпадает с результатами, достигнутыми при применении генов люциферазы и β-галактозидазы. Присутствие хлорохина приводит к получению обнаруживаемого сигнала (примерно 7 нг/мл; см. пробу 2). Аденовирус d1312 стимулирует перенос ДНК в зависимости от применяемой дозы (см. пробы 3 и 4). Обработка клеток сравнимыми количествами вируса в отсутствие включающего ДНК комплекса не приводит к получению обнаруживаемого сигнала интерферона (см. пробы 11, 12). В результате трансфекции эндосомолитическим пептидом Р16 по примеру 13 в виде конъюгата (см. пробы 6, 7) или же в качестве вещества, связанного ионным путем с поверхностью комплекса ДНК и конъюгата TfpL (см. пробы 8, 9 и 10), достигают обнаруживаемого уровня производства интерферона, который можно повышать при применении конъюгата этого пептида в присутствии хлорохина (см. пробу 7).
Пример 39. Перенос гена в В-лимфобластомные клетки.
Конъюгаты человеческого иммуноглобулина Г или анти- человеческого иммуноглобулина Г и полилизина получают нижеописанным методом, причем сопряжение осуществляют известными приемами путем введения дисульфидных мостиков после предварительной модификации сукцинимидилпиридилдитио-пропионатом.
а) Получение конъюгатов анти-человеческого иммуноглобулина Г и полилизина, содержащего 300 лизиновых остатков (далее: полилизин 300)
К раствору 2 мг анти-человеческого иммуноглобулина Г козы в буфере HBS, состоящего из 150 М хлористого натрия и 20 мМ буфера HEPES и имеющего значение pH, равное 7,8, добавляют 14 мкл 5 мМ этанольного раствора сукцинимидилпиридилтио-пропионата. Смесь оставляют стоять при комнатной температуре в течение 10 часов, после чего подвергают гель-фильтрации на ионите Сефадекс Г-25 с применением в качестве элюента 100 мМ буфера HEPES, имеющего значение pH, равное 7,3. Получают 1,3 мг анти-человеческого иммуноглобулина Г, модифицированного 30 нмоль пиридилдитиопропионатных остатков. Аналогичным методом осуществляют модификацию полилизина 300 сукцинимидилпиридилтио-пропионатом, и обработкой дитиотреитолом и последующей гель-фильтрацией переводят в форму, модифицированную свободными меркапто-группами. Раствор 12 нмоль полилизина 300, модифицированного 29 нмоль меркапто-групп, в 0,3 мл буфера HBS с исключением кислорода смешивают с вышеупомянутым модифицированным анти-человеческим иммуноглобулином Г, и полученную смесь оставляют стоять при комнатной температуре в течение ночи. Затем добавлением 5 М хлористого натрия содержание хлористого натрия в реакционной смеси доводят до 0,6 М. Выделение конъюгатов осуществляют путем ионообменной хроматографии на йоните марки Mono S HR 5/5 фирмы Pharmacia, Швеция. В результате диализа с применением 25 мМ буфера HEPES, pH 7,3, получают соответствующие конъюгаты, состоящие из 0,33 мг анти-человеческого иммуноглобулина Г, модифицированного 4 нмоль полилизина 300, причем мольное соотношение анти-человеческого иммуноглобулина Г и полилизина 300 составляет 1 : 2.
б) Получение конъюгатов человеческого иммуноглобулина Г и полилизина 300
К раствору 19,5 мг (122 нмоль) антител I-4506 фирмы Sigma, DE, в 2 мл буфера HBS добавляют 39 мкл 1 мМ этанольного раствора сукцинимидилпиридилтио-пропионата. Смесь оставляют стоять при комнатной температуре в течение 2,5 часов, после чего подвергают гель-фильтрации на йоните Сефадекс Г-25 с применением в качестве элюента 100 мМ буфера HEPES с pH 7,9, в результате чего получают 19 мг (119 нмоль) человеческого иммуноглобулина Г, модифицированного 252 нмоль пиридилдитиопропионатных остатков. Аналогичным методом осуществляют модификацию полилизина 300 сукцинимидилпиридилтио- пропионатом, и обработкой дитиотреитолом с последующей гель- фильтрацией переводят в форму, модифицированную свободными меркапто-группами. Раствор 119 нмоль полилизина 300, модифицированного 282 нмоль меркапто-групп, в 1 мл буфера HBS с исключением кислорода смешивают с вышеупомянутым модифицированным анти-человеческим иммуноглобулином Г, и получаемую смесь оставляют стоять при комнатной температуре в течение ночи. Затем добавлением 5 М хлористого натрия содержание хлористого натрия в реакционной смеси доводят примерно до 0,6 М. Выделение конъюгатов осуществляют путем ионообменной хроматографии на йоните марки Mono S фирмы Pharmacia, Швеция, причем в качестве элюента применяют 50 мМ буфера HEPES, pH 7,3 с солевым градиентом 0,6 М - 3 М хлористого натрия. В результате диализа с применением буфера HEPES, pH 7,3, получают соответствующие конъюгаты, состоящие из 9 мг (57 нмоль) антитела, модифицированного 90 нмоль полилизина 300. При этом мольное соотношение антитела и полилизина 300 составляет 1 : 1,6.
в) Образование комплексов и трансфекция
Комплексы получают следующим образом: 30 мкл биотинилированного аденовируса d1312 (1012 частиц на мл) в 50 мкл буфера HEPES смешивают с 800 нг модифицированного стрептавидином полилизина в 100 мкл буфера HBS. После инкубации в течение 30 минут добавляют раствор 9 мкг ДНК pCMVL в 200 мкл буфера HBS. Еще раз инкубируют в течение 30 минут, после чего добавляют раствор 5,1 мкг полилизина 450 (pL), 10,2 мкг конъюгата TfpL, 12 мкг конъюгата человеческого иммуноглобулина Г и полилизина или 10 мкг конъюгата анти-человеческого иммуноглобулина Г и полилизина в 150 мкл буфера HBS. Комплексы ДНК добавляют к 106 В-лимфобластомных клеток (клетки получают из периферийных моноядерных клеток крови человека в результате иммортализации вирусом Эпштейна и Барра, выращиваемых в снабженной 24 ячейками чашке в 1 мл среды RPMI 1640 с 2% ТЭС). Дальнейшую обработку клеток и опыты по определению активности люциферазы осуществляют описанным выше образом. Полученные данные по экспрессии гена (активность люциферазы в световых единицах) приведены на фиг. 52.
Пример 40. Трансфекция комбинированными комплексами конъюгата аденовируса и полилизина, и конъюгата липопротеина низкой плотности и полилизина.
а) Получение конъюгатов липопротеина низкой плотности и полилизина.
К раствору 10 мг (14,3 нмоль) липопротеина низкой плотности (условное сокращение на фиг. 53: LDL, продукт L-2139 с молекулярным весом 3500000 фирмы Sigma, DE, диаметр частиц примерно 26 нм) в 2 мл буфера HBS добавляют 143 мкл 10 мМ этанольного раствора сукцинимидилпиридилтио-пропионата (1,43 мкмоль), и получаемую смесь оставляют стоять при комнатной температуре в течение 2 часов. В результате гель-фильтрации на ионите Сефадекс Г-25 (колонка размером 14 х 140 мм) с использованием в качестве элюента буфера HBS получают 3,2 мл раствора примерно 10 мг липопротеина низкой плотности, модифицированного 0,70 мкмоль пиридилдитиопропионатных остатков. К данному раствору добавляют 1,2 мл 5 М хлористого натрия с тем, чтобы предотвратить образование осадка, которое иначе произошло бы при последующем добавлении полилизина. Полилизин 300 вышеописанным образом модифицируют сукцинимидилпиридилтио- пропионатом, и путем обработки дитиотреитолом и последующей гель- фильтрации переводят в форму, модифицированную меркапто-группами. Раствор модифицированного липопротеина низкой плотности в атмосфере аргона смешивают с раствором 0,33 мкмоль полилизина 300, модифицированного 0,76 мкмоль меркапто-групп, в 4 мл 2 М хлористого натрия и 0,5 М буфера HEPES, pH 7,9, затем реакционную смесь оставляют стоять при комнатной температуре в течение 48 часов. Получаемый реакционный раствор разбавляют добавлением стерильной воды до объема 32 мл (после чего концентрация хлористого натрия составляет примерно 0,5 М), и его очищают путем ионообменной хроматографии на колонке Biorad Macroprep S, 10 х 100 мм, с использованием в качестве элюента 20 мМ буфера HEPES с градиентом 0,2- 3 М хлористого натрия. Фракции продукта получают при концентрации соли, составляющей 1,8 - 2,2 М, и их собирают. В результате диализа с применением буфера HBS получают конъюгаты, состоящие из 2,35 мг (3,4 нмоль) липопротеина низкой плотности, модифицированного 190 нмоль полилизина, что соответствует 7,5 мг свободной основной формы полилизина. Таким образом липопротеин низкой плотности модифицирован 55 полилизиновых остатков.
б) Получение комплексов и трансфекция
18 мкл биотинилированного аденовируса d1312/16 разбавляют добавлением буфера HBS до объема 100 мкл. Кроме того, к 1,2 мкг полилизина, модифицированного стрептавидином, добавляют буфер HBS до объема 100 мкл, и смешивают с указанным аденовирусом. По истечении 30 минут добавляют 150 мкл буфера HBS, содержащего 6 мкг ДНК pCMVL. По истечении дальнейших 30 минут добавляют 300 мкл буфера HBS, содержащего 4 мкг конъюгата LDLpL (с содержанием полилизина, составляющим 20 мкг). Получаемый раствор размешивают с 1,5 мл DMEM (10 % ТЭС) и прибавляют к 3 • 105 первичных клеток меланома, получаемых по примеру 33 и обозначаемых НММ1 и НММ4, находящихся в инкубационной чашке диаметром 6 мм. Дальнейшие операции по выращиванию клеток и определение активности люциферазы осуществляют описанным в примере 33 образом. Данные по экспрессии гена приведены на фиг. 53; часть А) фиг. показывает результаты, получаемые в опытах с клетками меланома НММ1. Все опыты осуществляют с конъюгатами AdpL, не приведенными на данной фиг. В части Б) фиг. показаны результаты, получаемые в опытах с применением клеток меланома НММ4. И в этом случае все опыты осуществляют с конъюгатами AdpL, которые приведены, однако, лишь отдельно в последнем столбце ("d1312/16" относится к особенному препарату вируса). В опыте, результаты которого приведены во втором столбце, применяют липопротеин низкой плотности в 25-кратном избытке, что из-за конкуренции относительно рецептора липопротеина низкой плотности приводит к уменьшению эффективности переноса гена.
Пример 41. Перенос гена в эпителиальные клетки дыхательных путей крыс in vivo с помощью комбинированных комплексов
а) Получение комбинированных комплексов ДНК и конъюгатов TfpL и AdVpL
Путем сочетания 8 мкг конъюгата человеческого трансферина и полилизина (hTfpL) в 150 мкл буфера HBS, содержащего 150 Мм хлористого натрия и 20 мМ буфера HEPES, pH 7,3, с 6 мкг ДНК pCMVL в 350 мкл буфера HBS получают комплексы ДНК и указанного конъюгата, которые инкубируют при комнатной температуре в течение 30 минут. Аналогично описанному в примере 19в) методу получают конъюгаты аде- новируса и полилизина, причем используют аденовирус, инактивированный обработкой псораленом и УФ-лучами. Комбинированные комплексы ДНК и конъюгатов hTfpL и AdVpL получают согласно методу, описанному в примере 23в).
б) Введение комплексов по внутритрахеальному пути в живой организм
Для осуществления данного опыта используют крысы породы Sigmodon hispidus (т.н. "Cotton Rat"), которые представляют собой пригодные животные для исследования аденовирусных заболеваний легких у человека. Животные анестезируют метоксифлураном. На брюшной стороне горла выполняют вертикальный разрез, после чего трахею препарируют. Затем крысы укладывают на бок под углом 45o, и 250 -300 мкл комплексов, содержащих 3 мкг ДНК, впрыскивают непосредственно в трахею. По истечении после инъекции промежутков, указанных на фиг. 54, животных умерщвляют дачей двуокиси углерода, и трахею и связанные с ней легкие удаляют после предварительной промывки in situ холодным фосфатсодержащим раствором поваренной соли. Для определения активности люциферазы ткань легких гомогенизируют в среде буфера, получаемые экстракцией лизаты подвергают стандартизации относительно общего содержания протеина, и экспрессию гена люциферазы измеряют вышеописанным образом. Указанные световые единицы относятся к 1,250 мкг общего протеина, получаемого из лизатов легких. Опыты осуществляют по 3-4 раза, результаты представляют собой средние значения ± среднее отклонение.
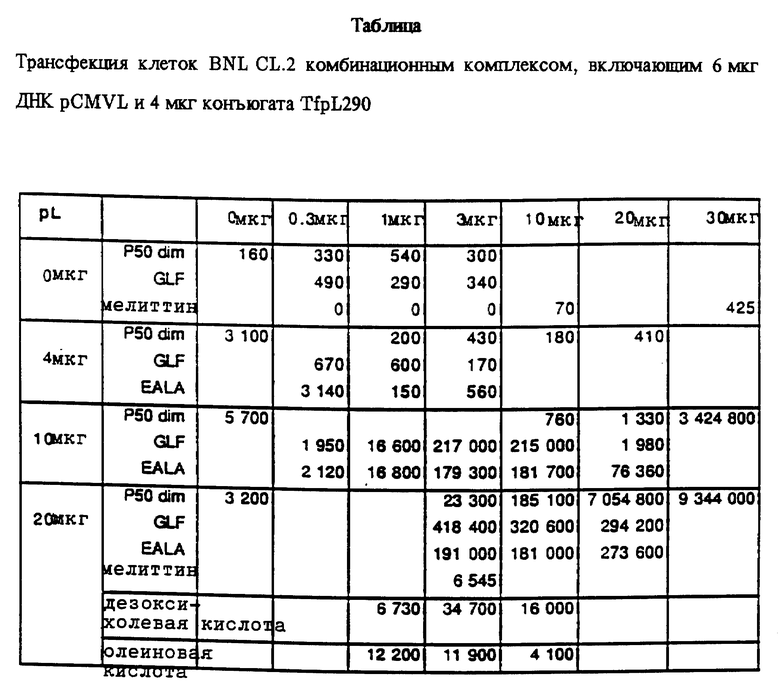
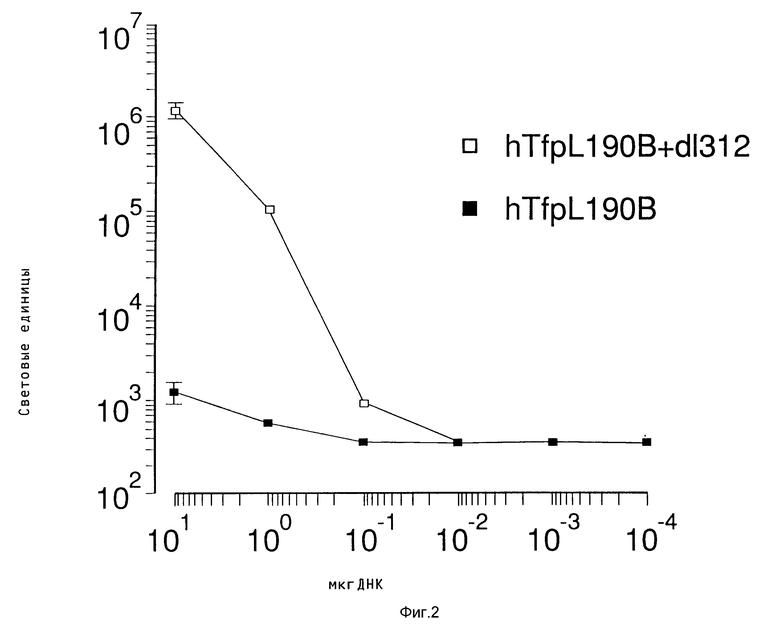
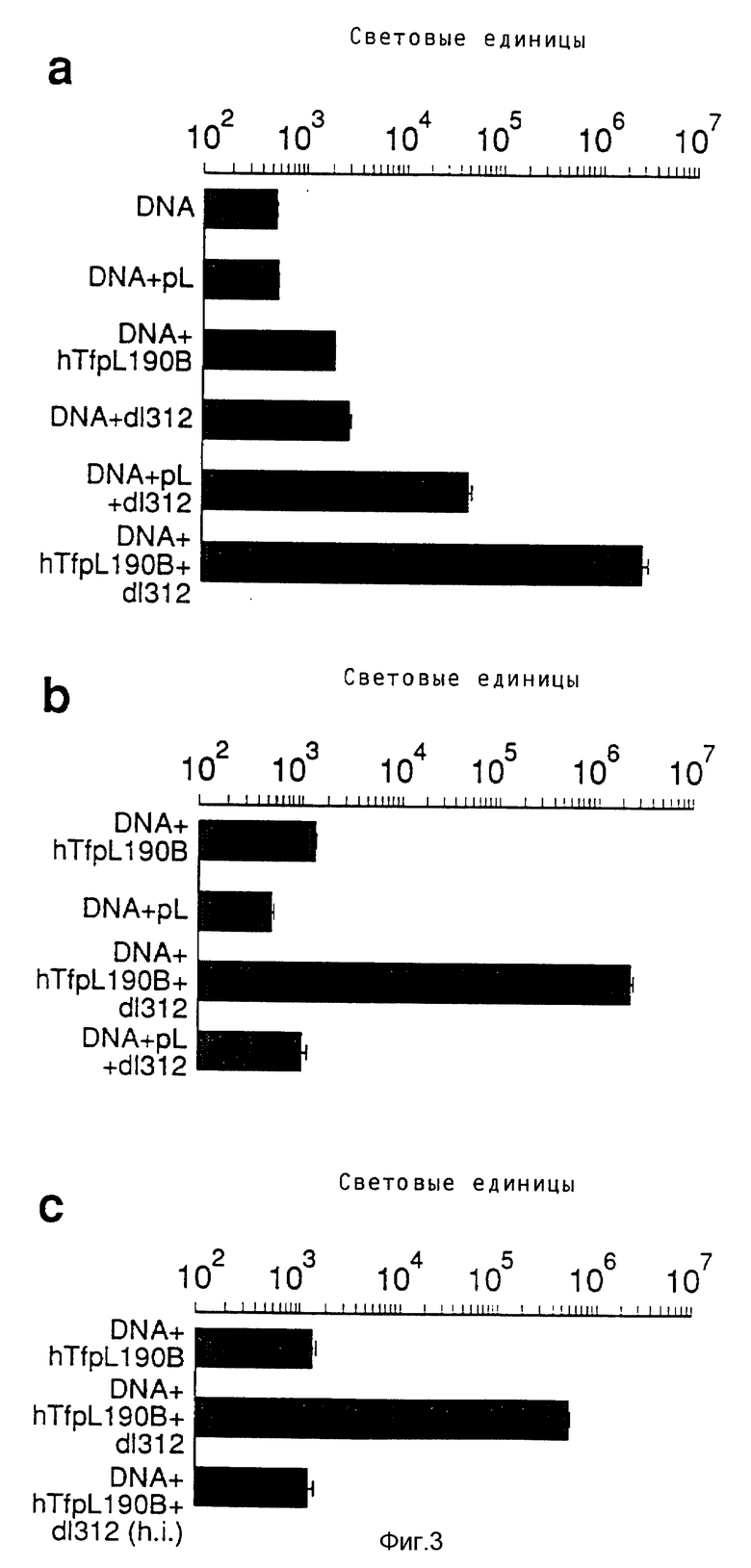
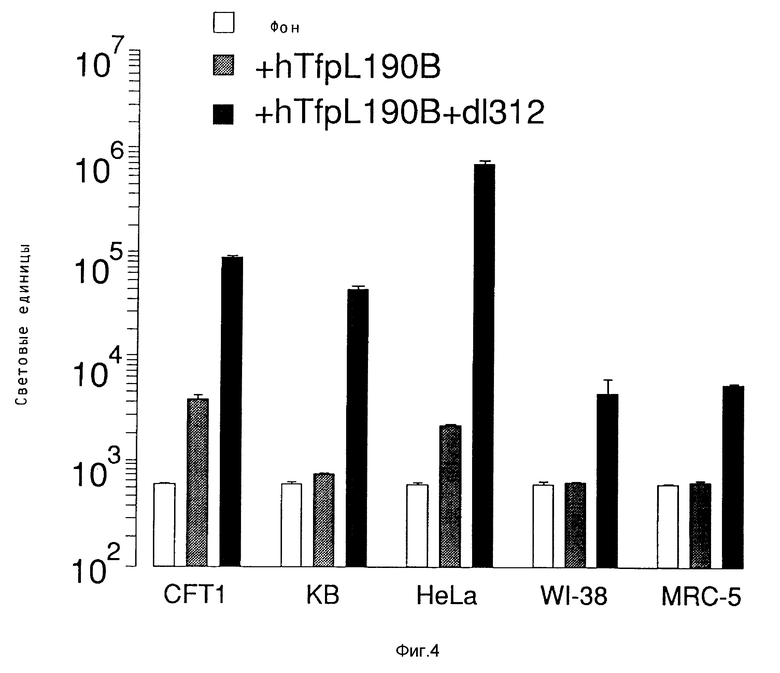
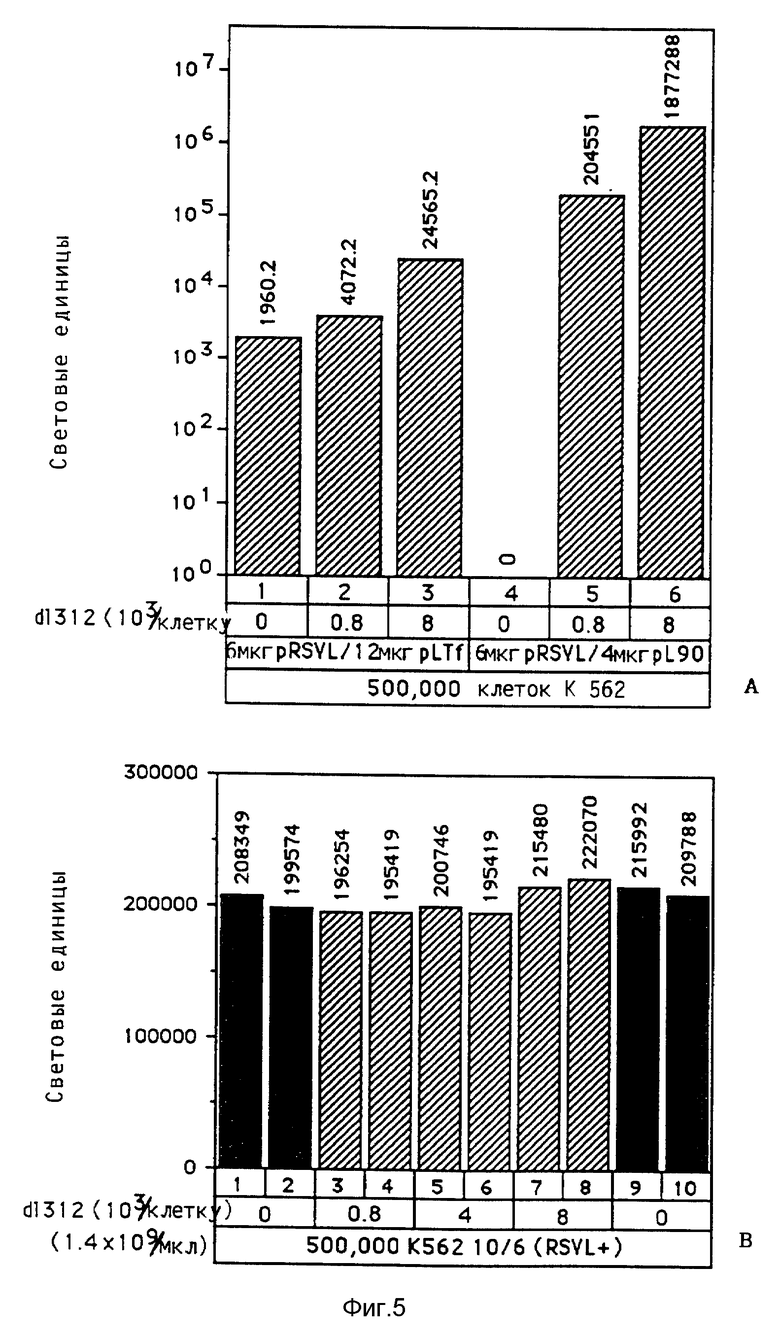
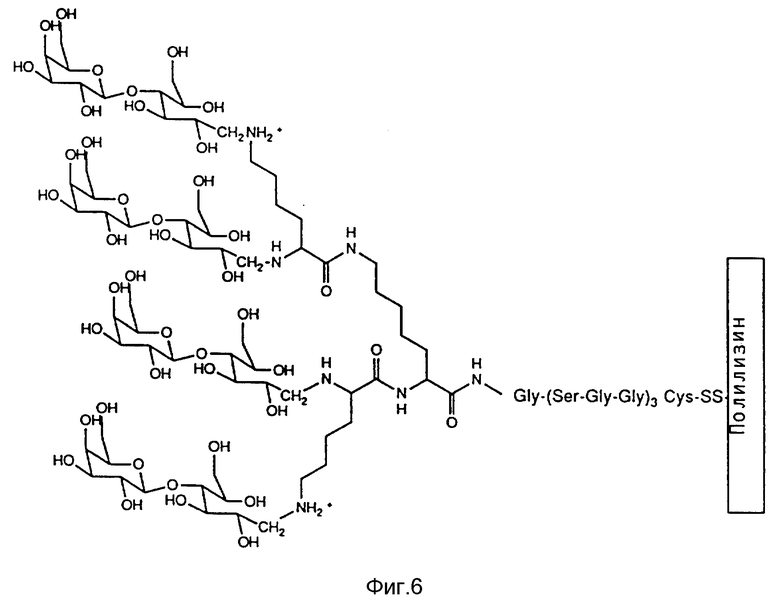
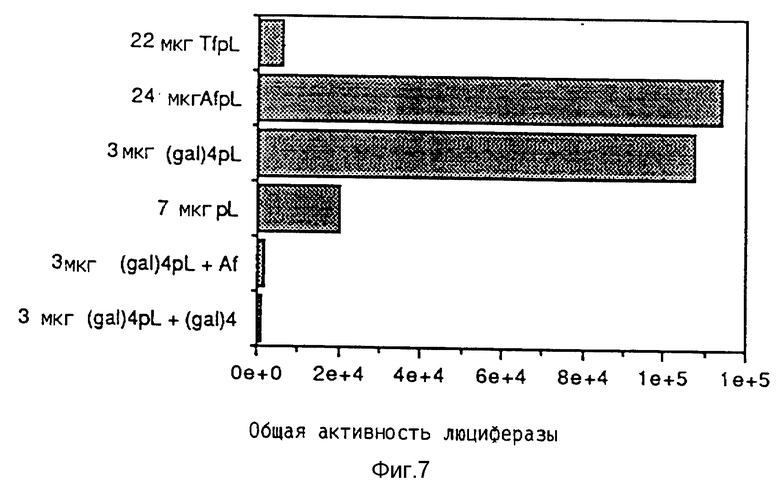
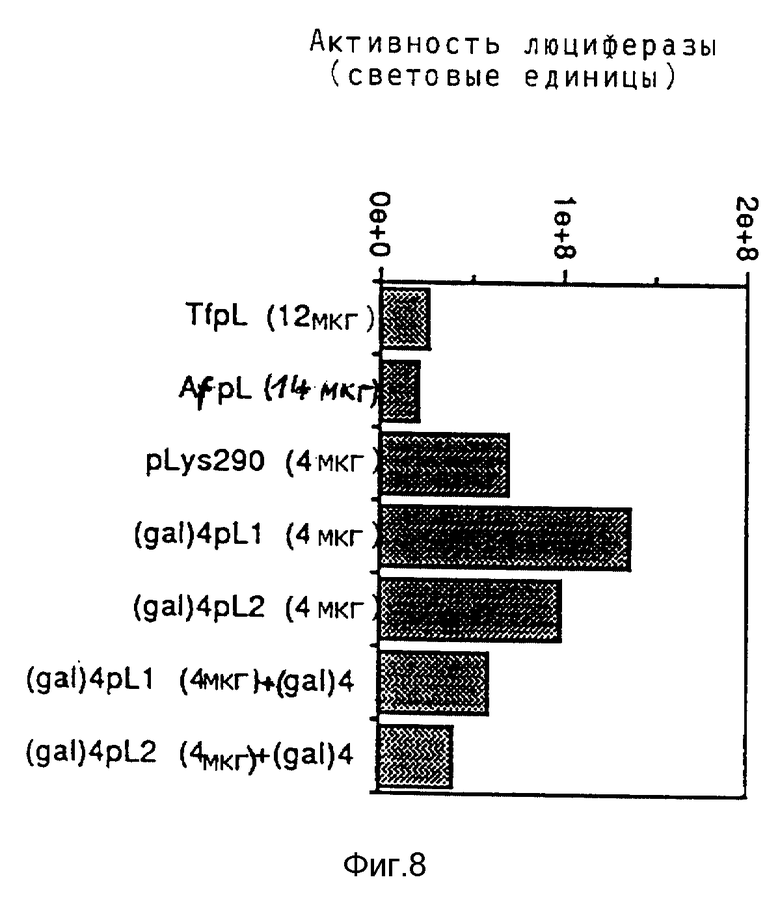
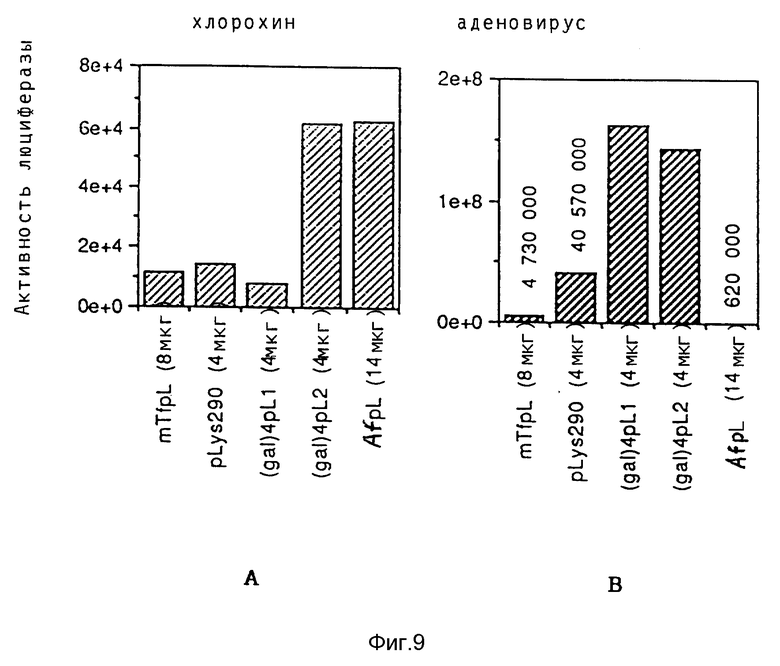
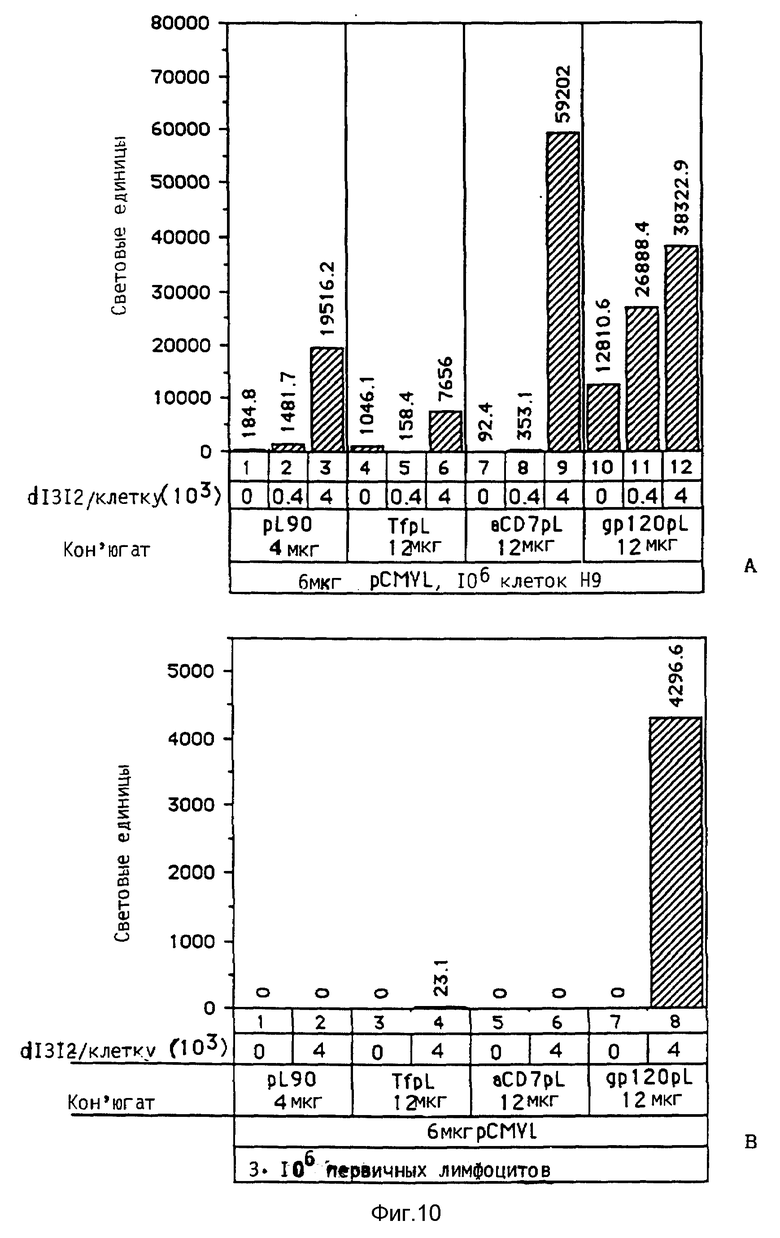
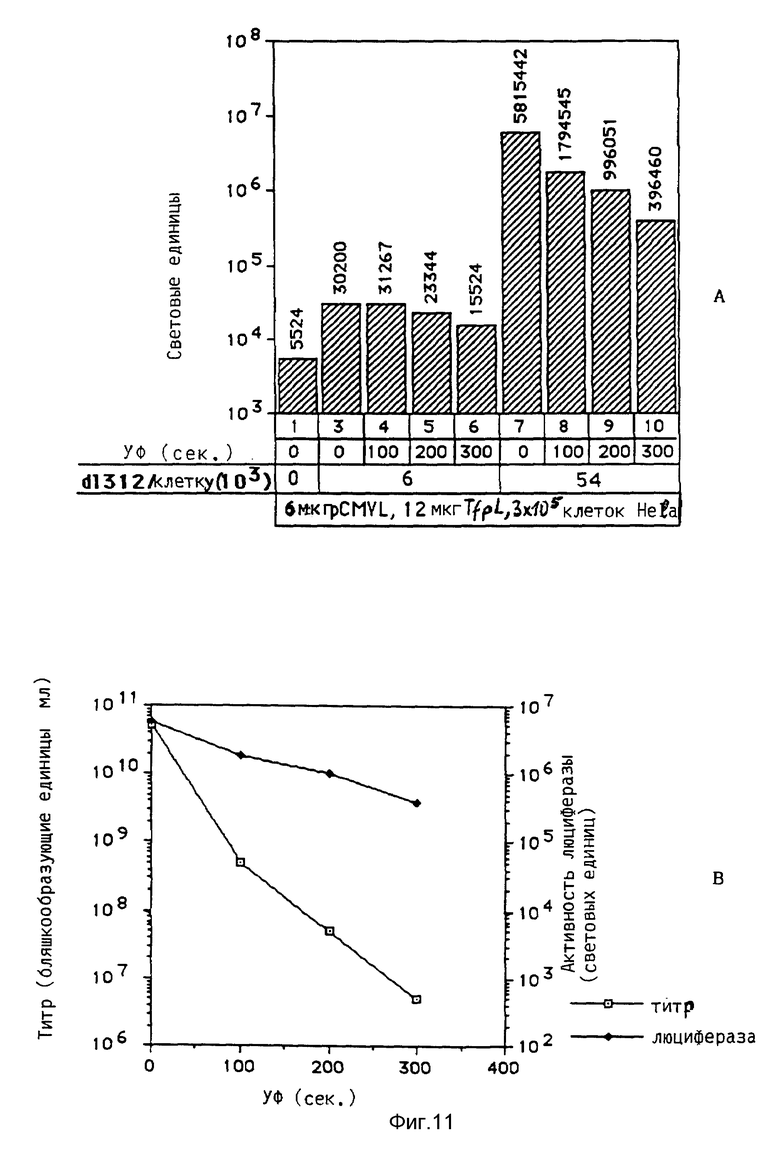
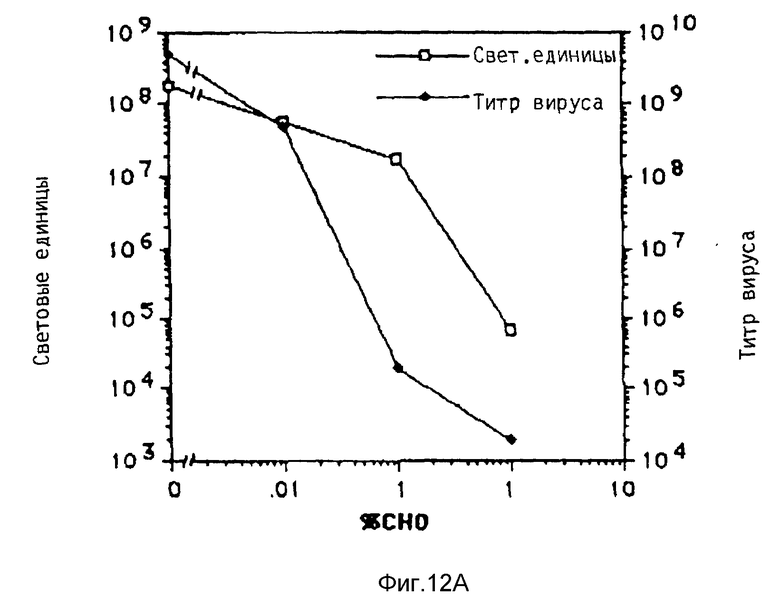
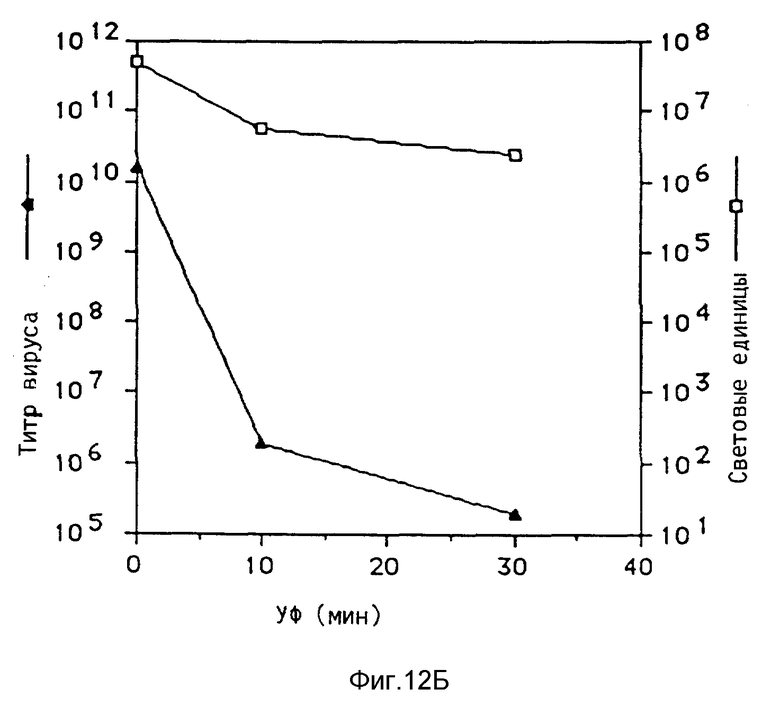
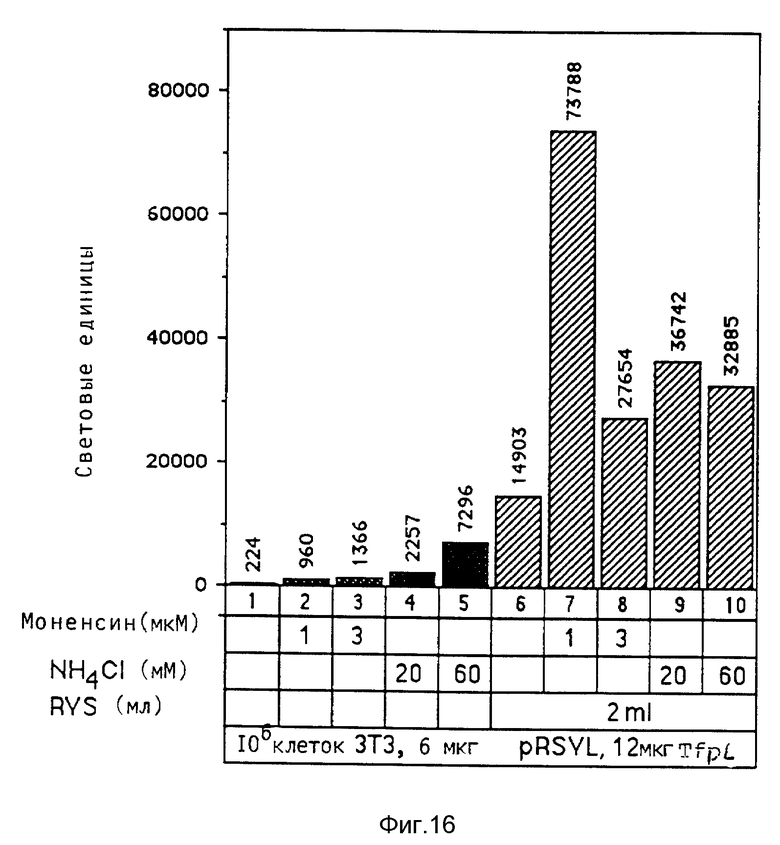
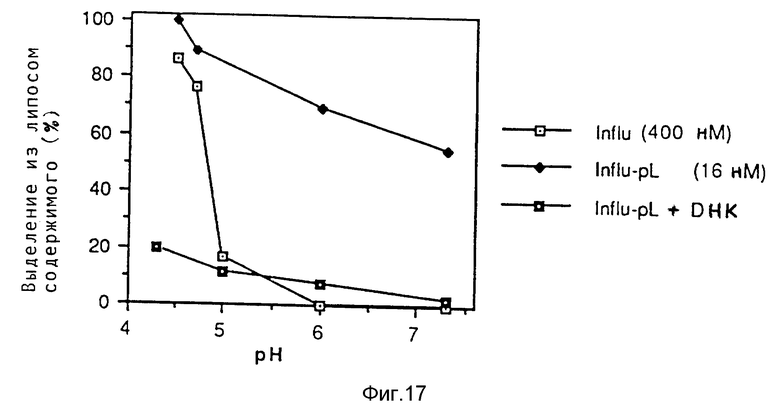
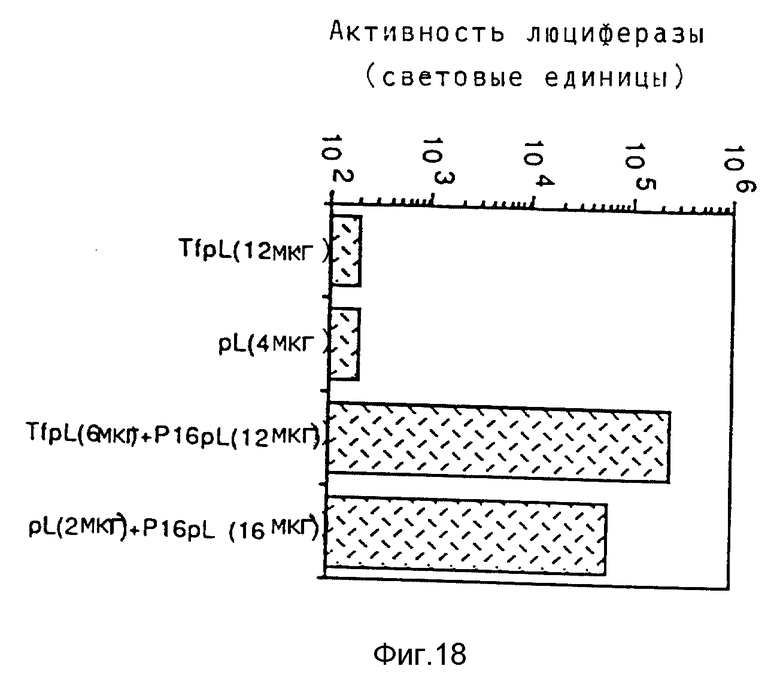
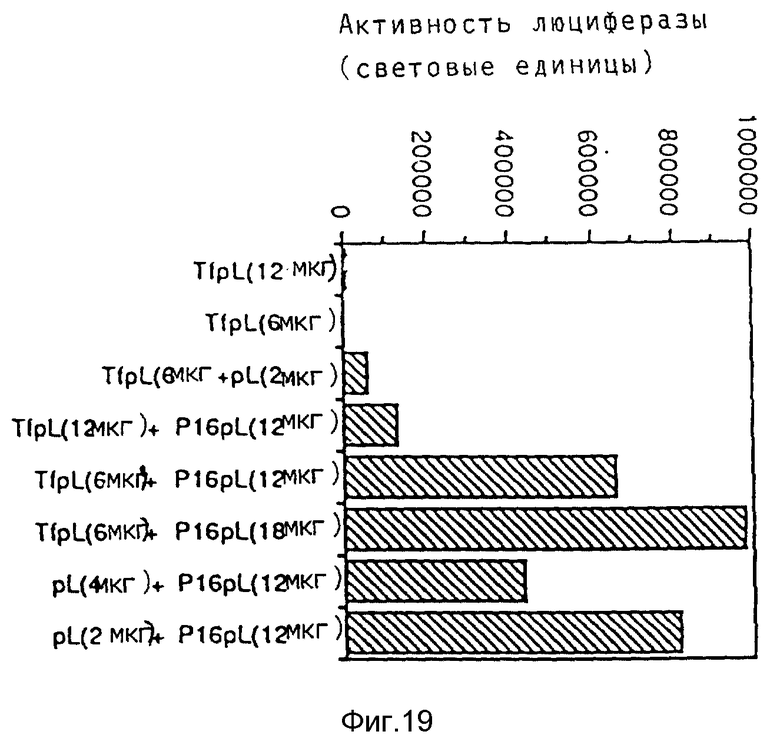
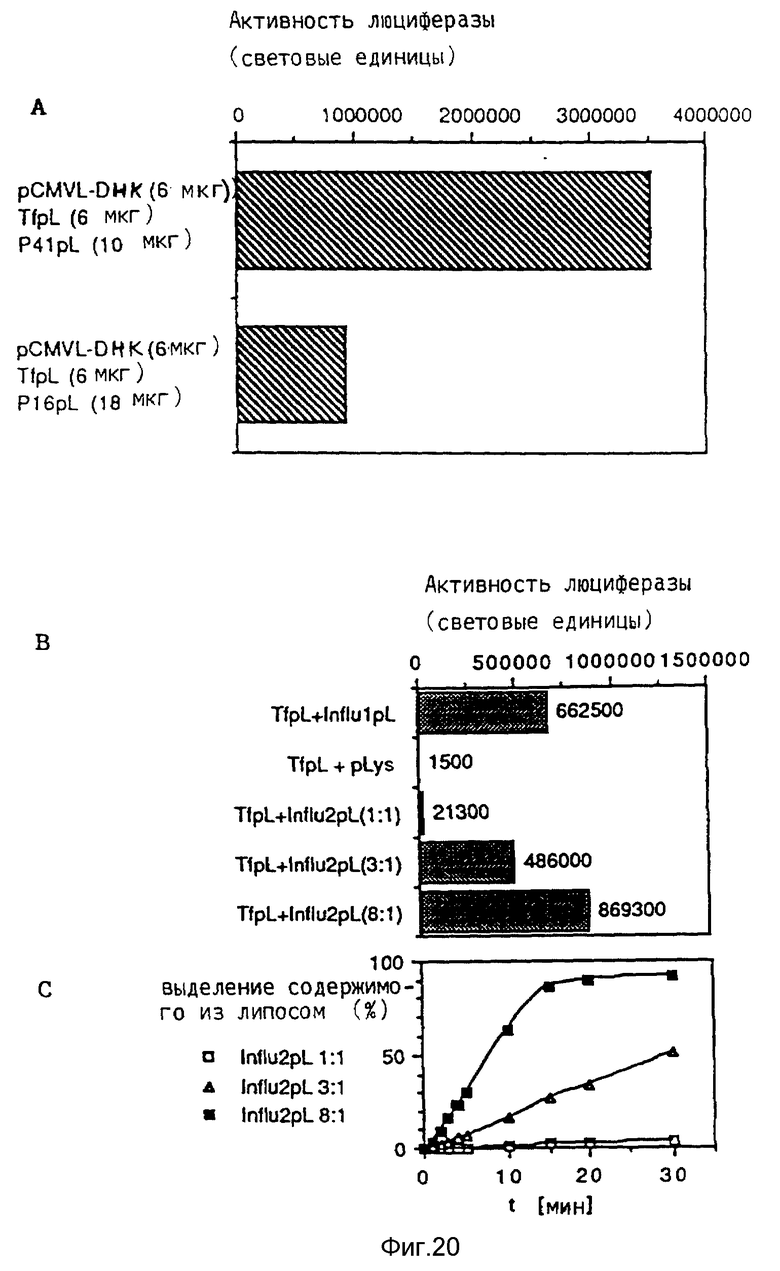
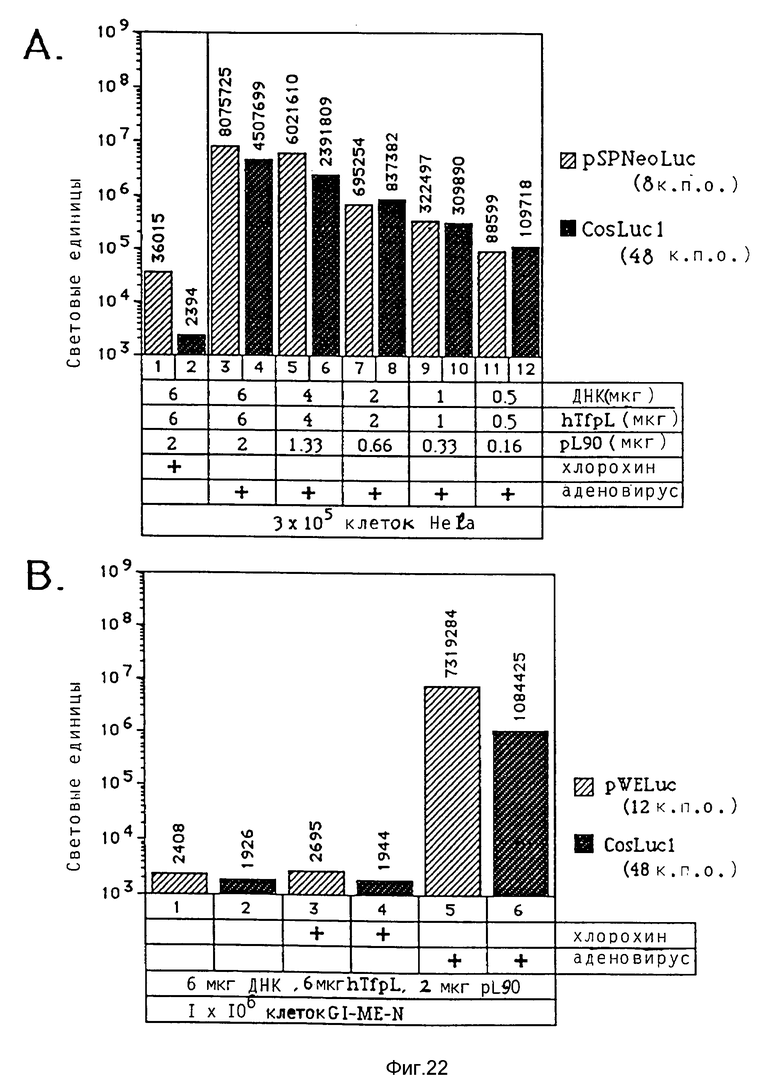
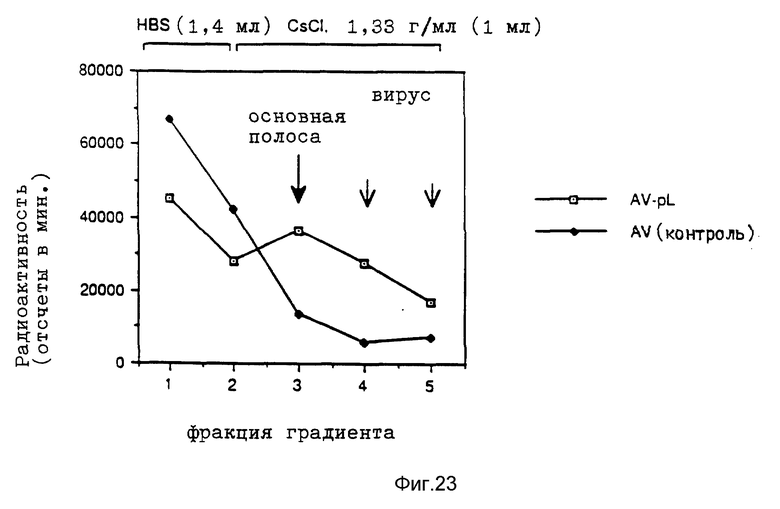
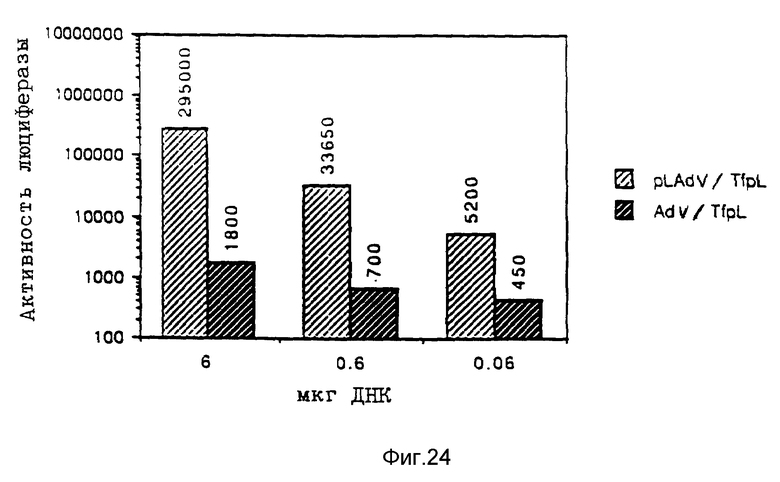
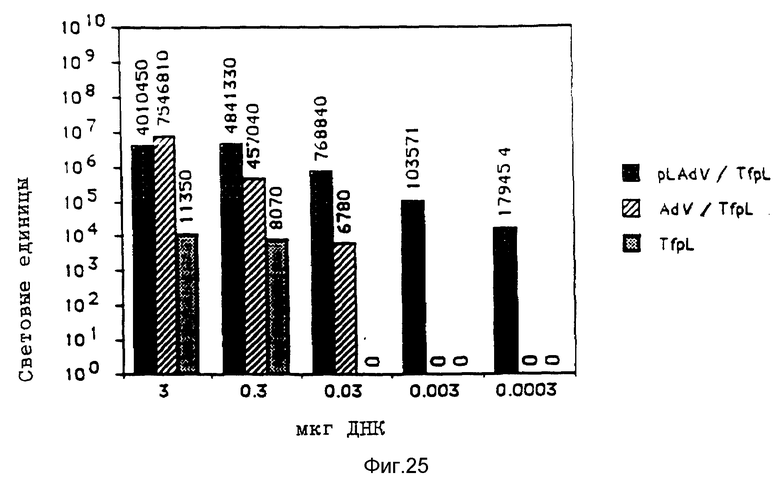
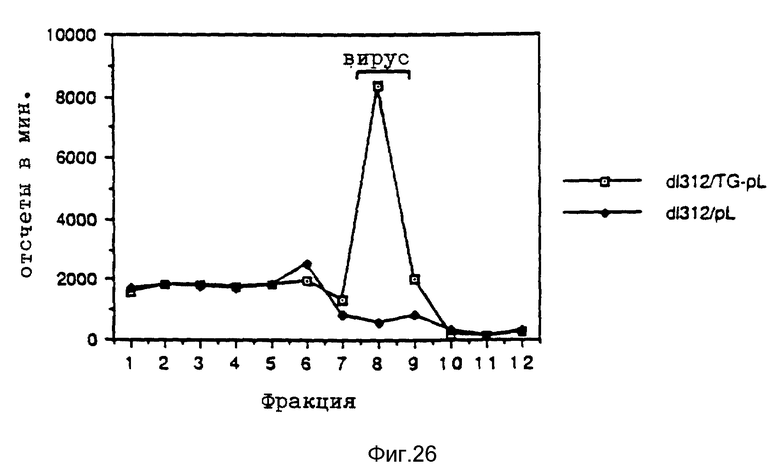
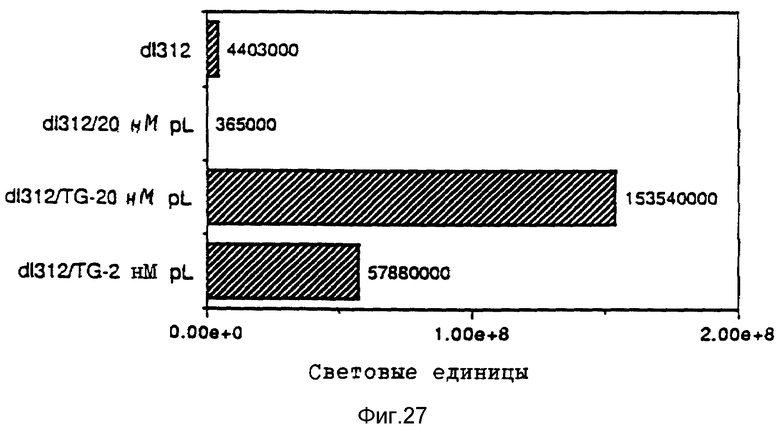
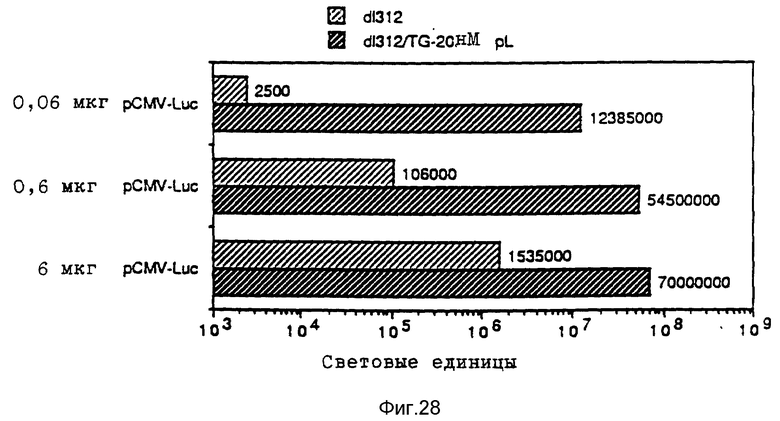
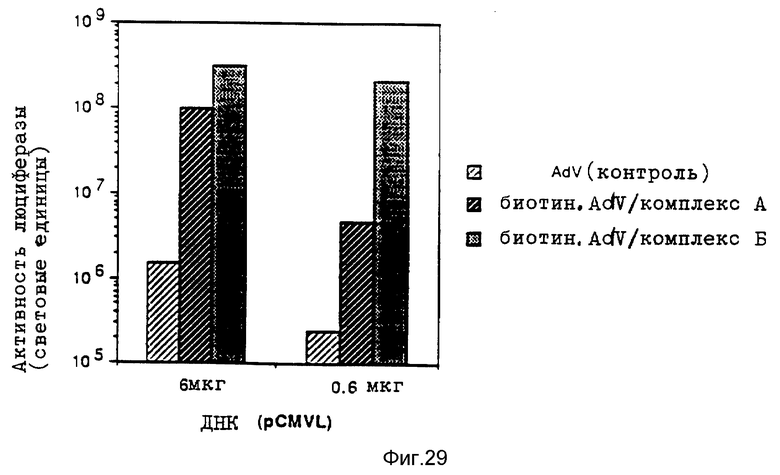
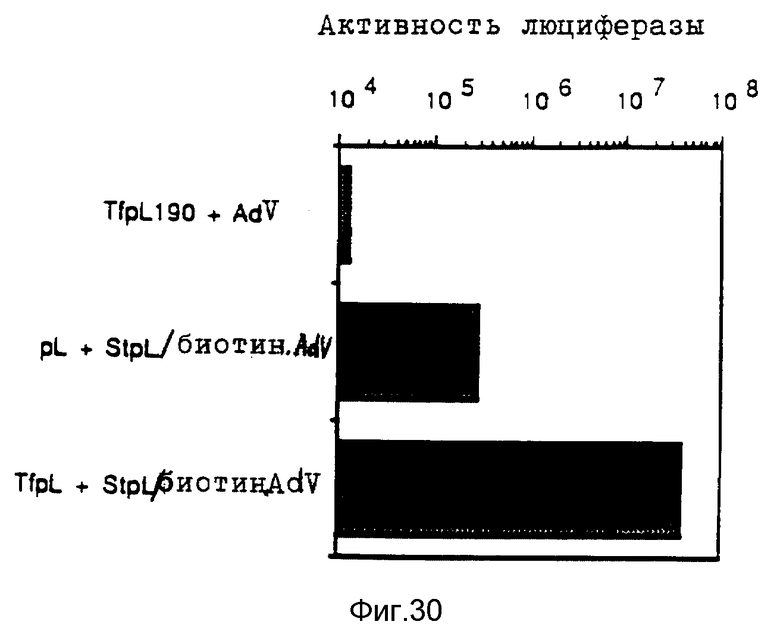
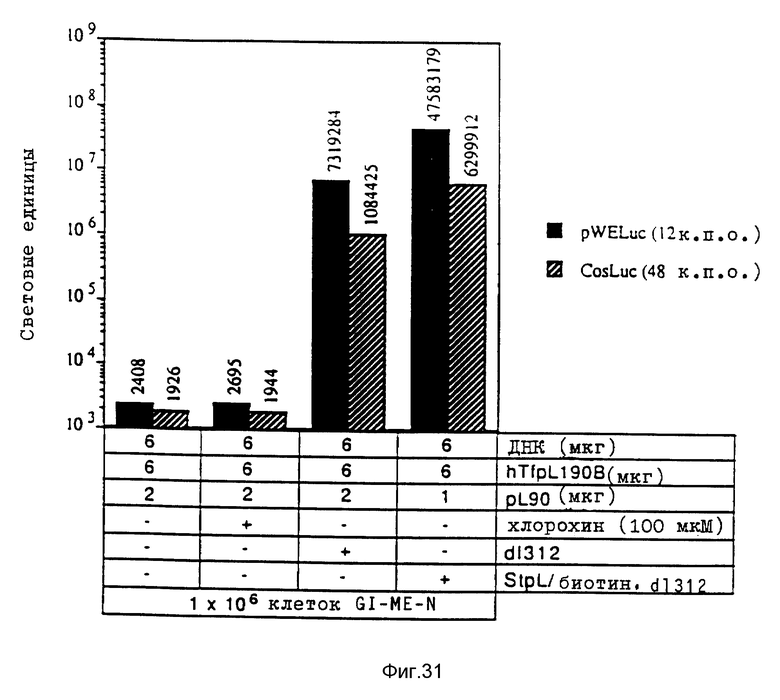
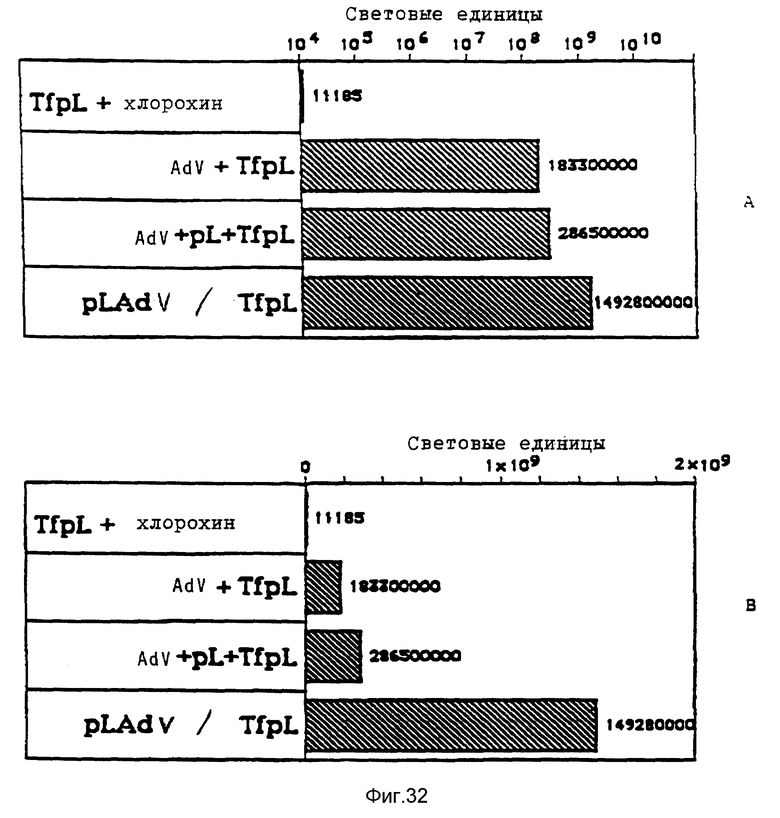
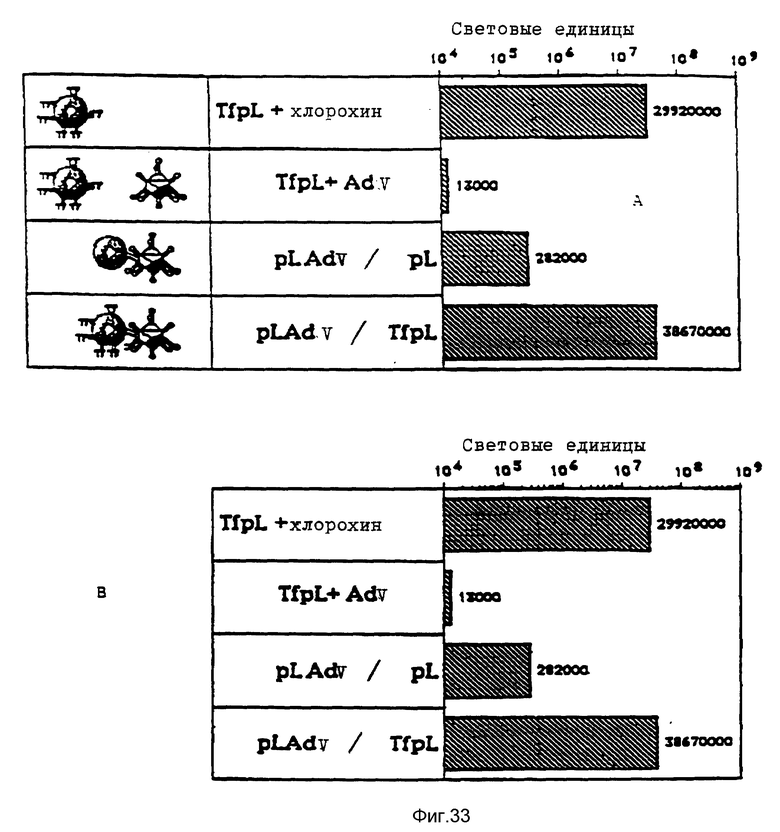
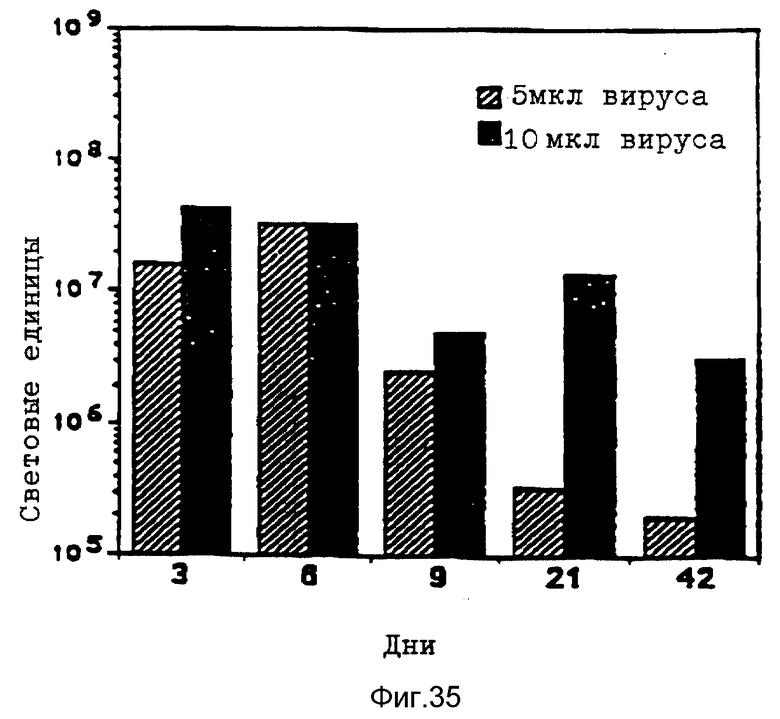
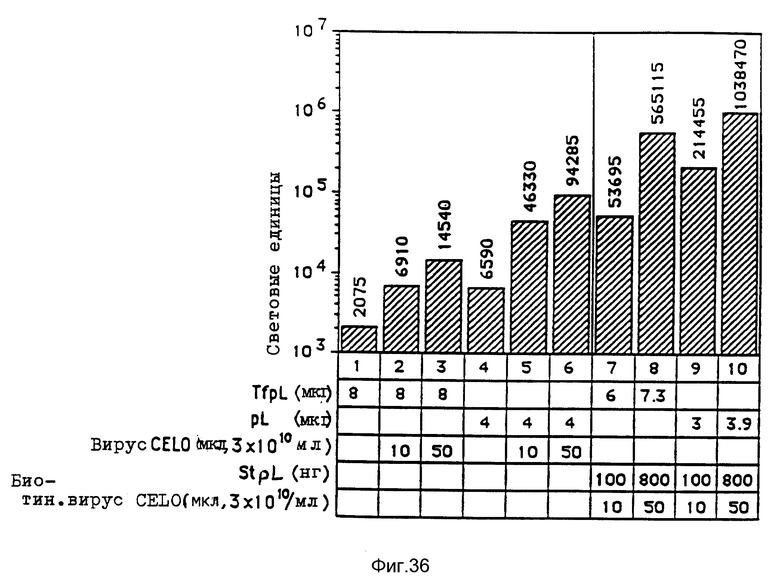
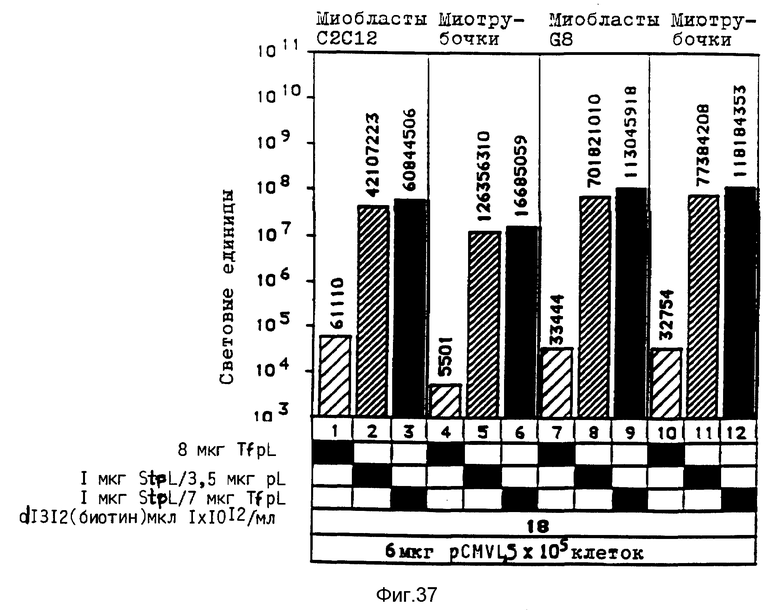
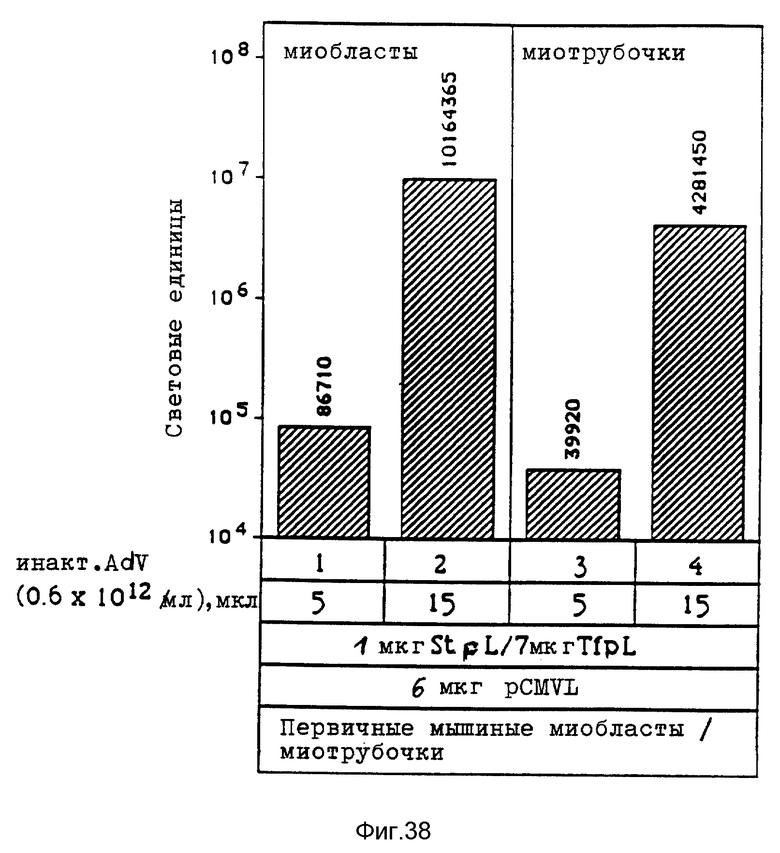
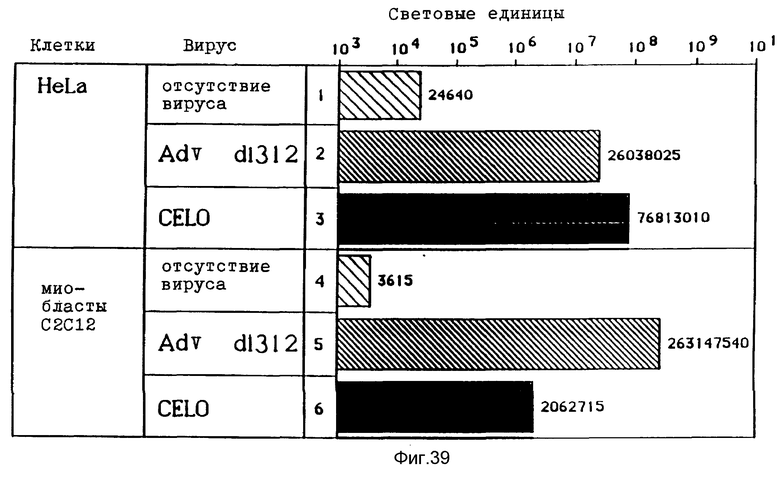
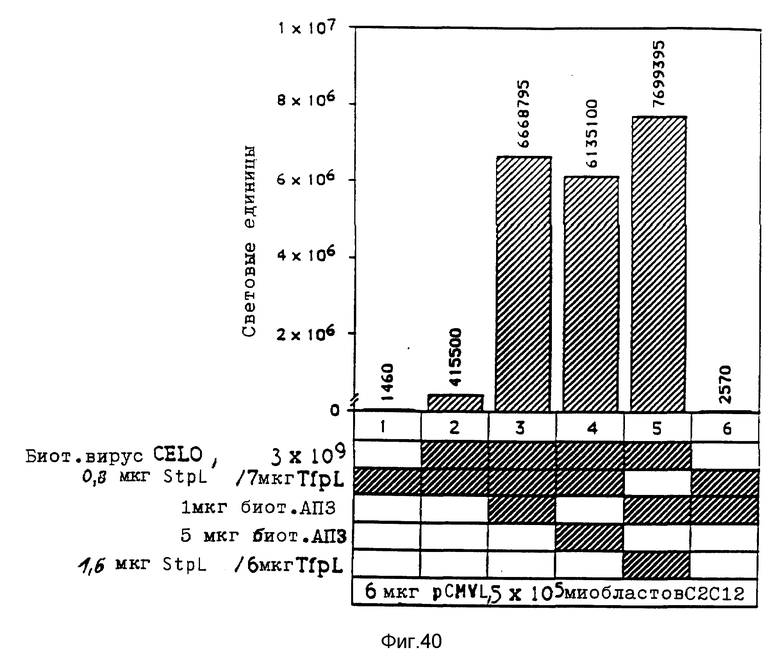

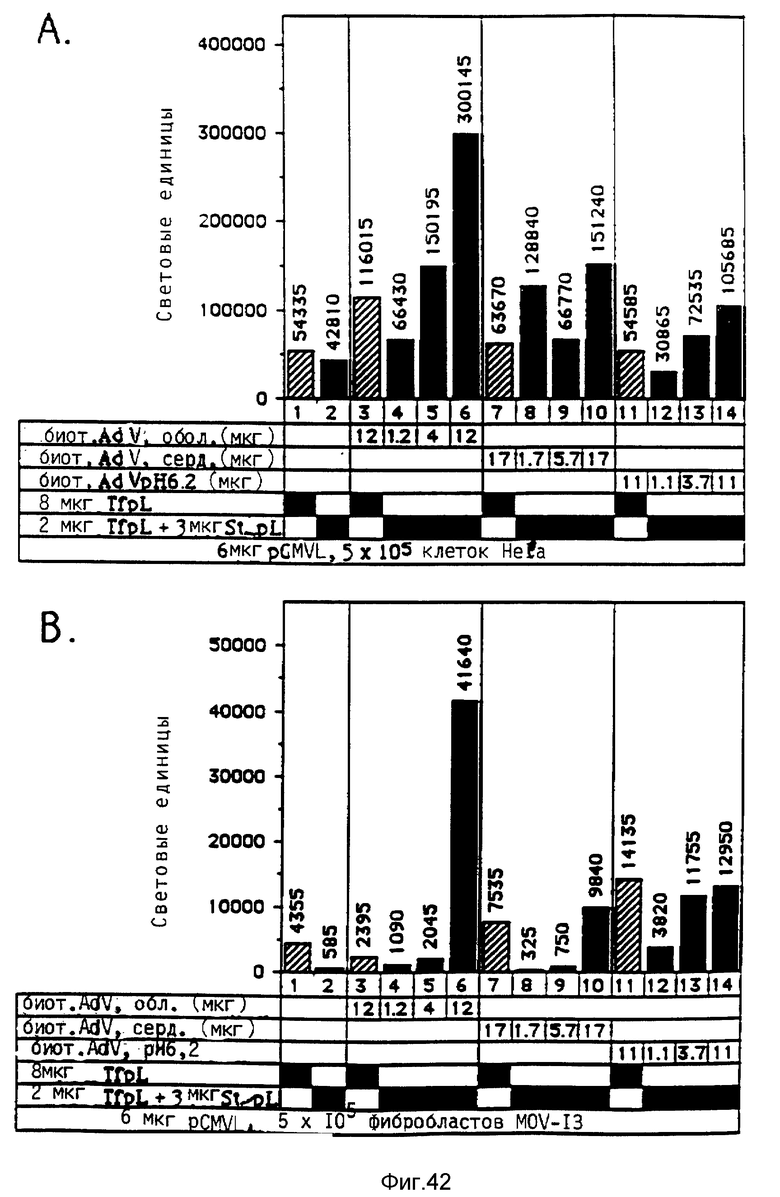
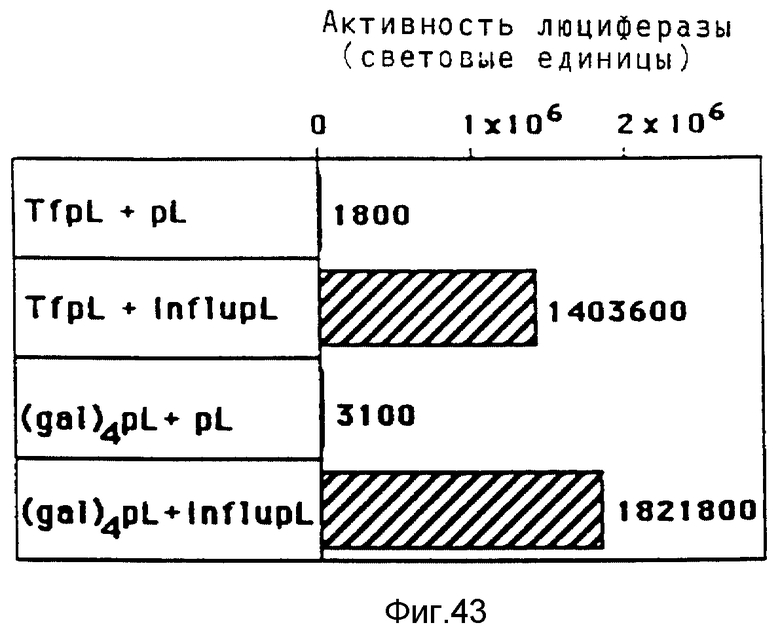

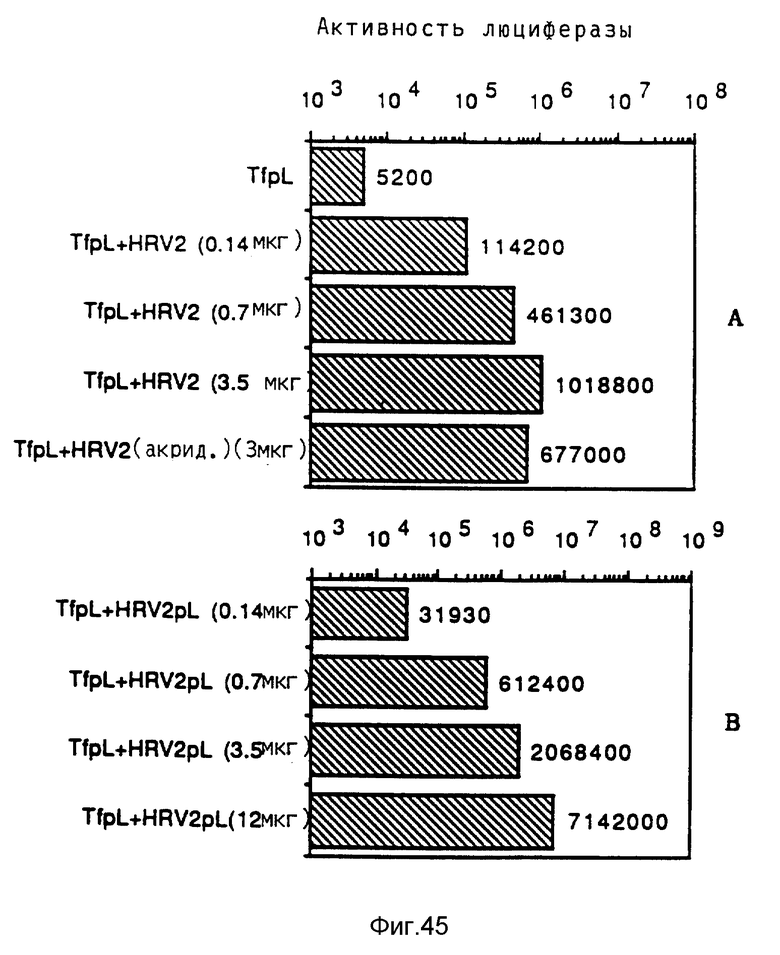
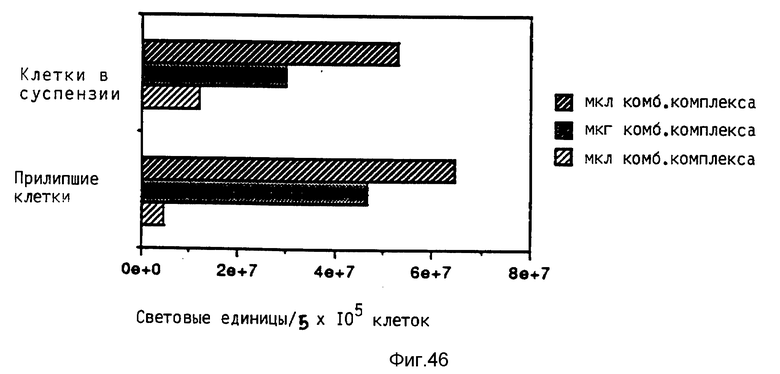
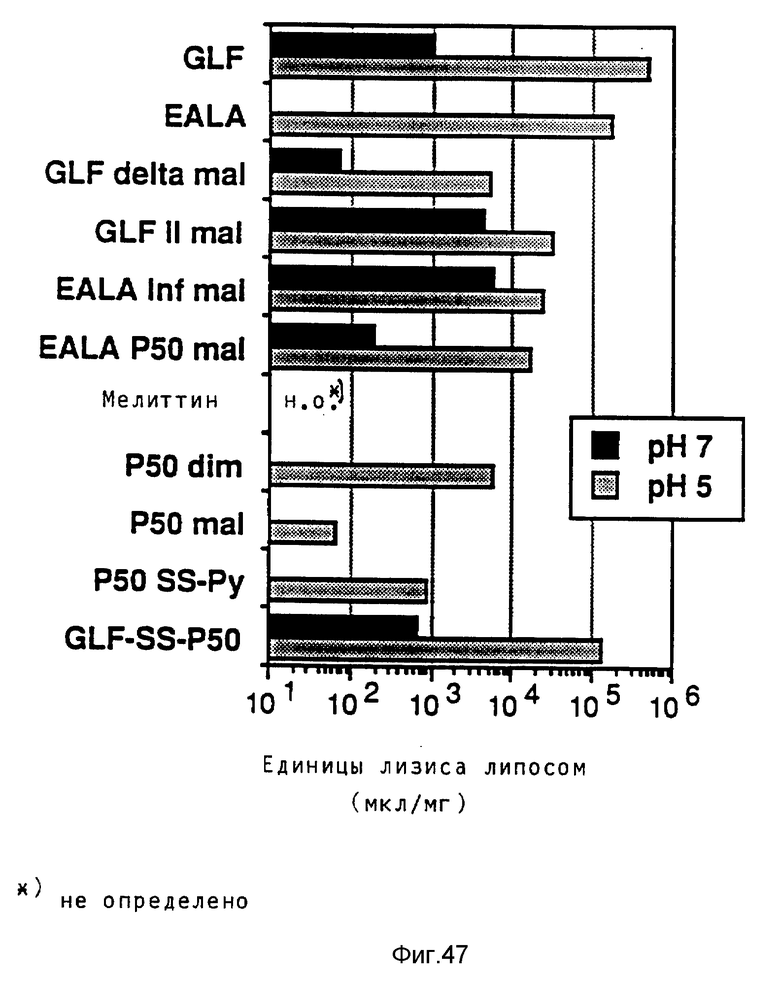
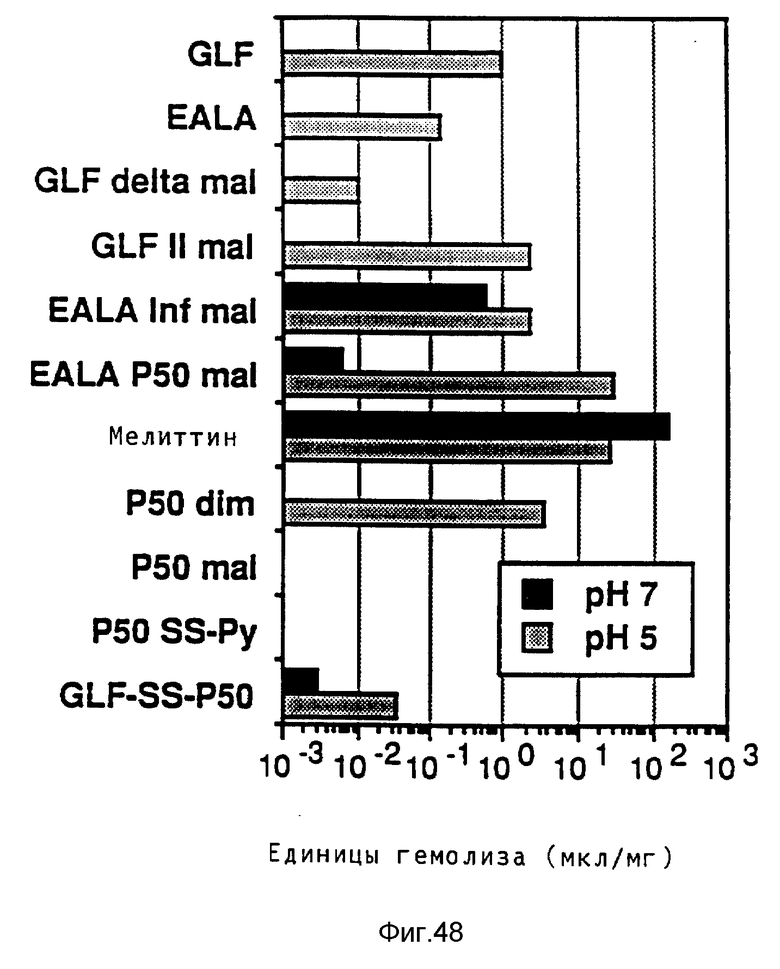
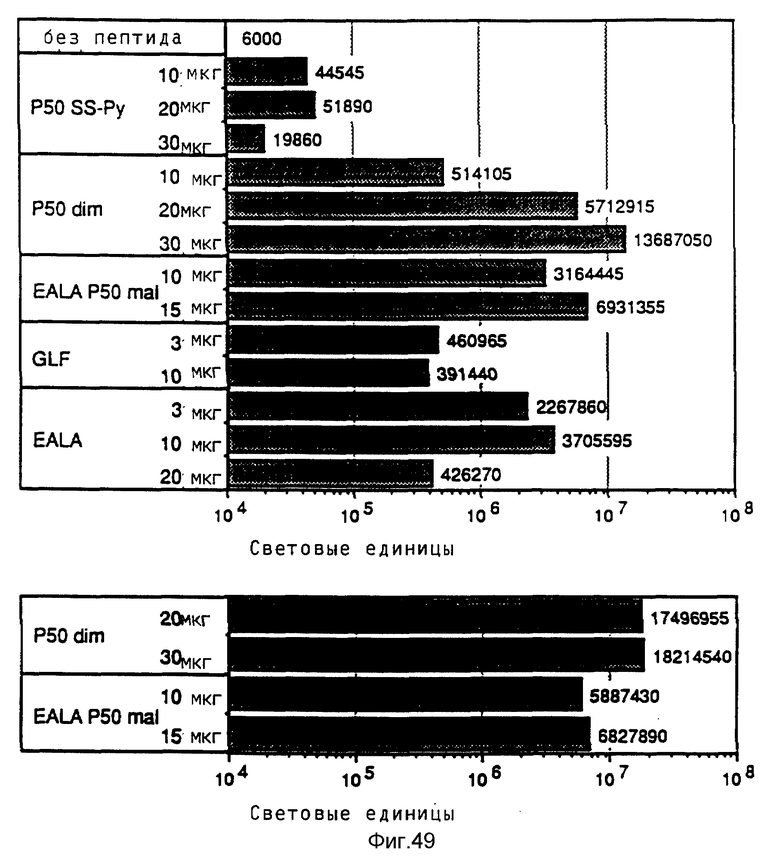
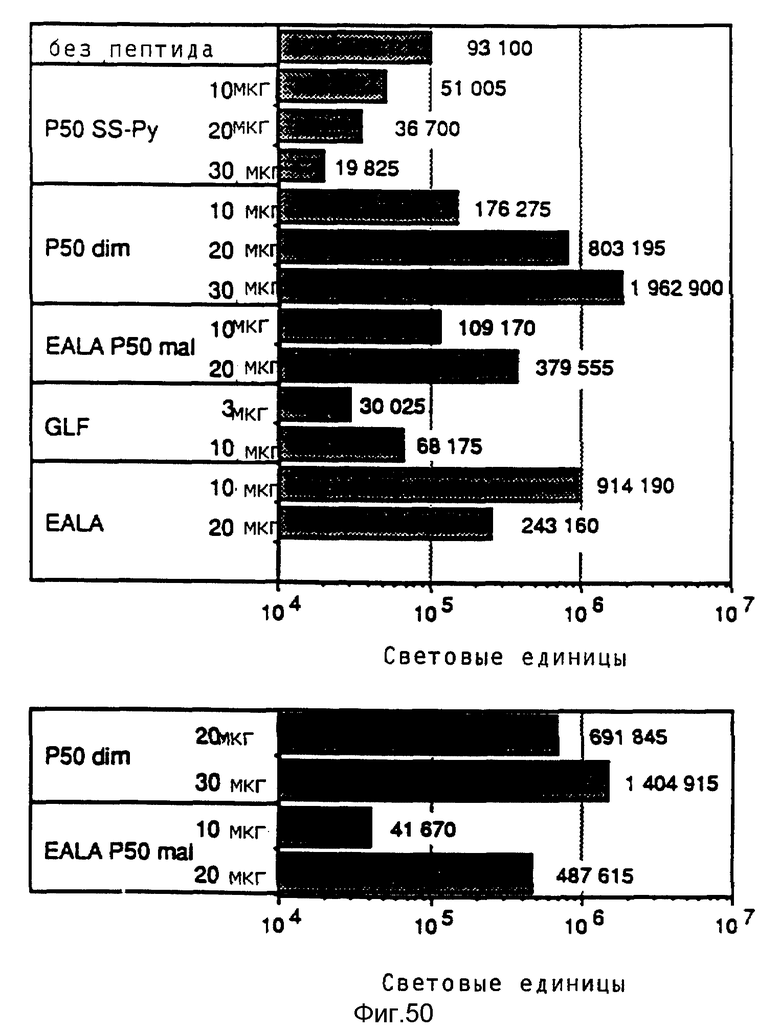
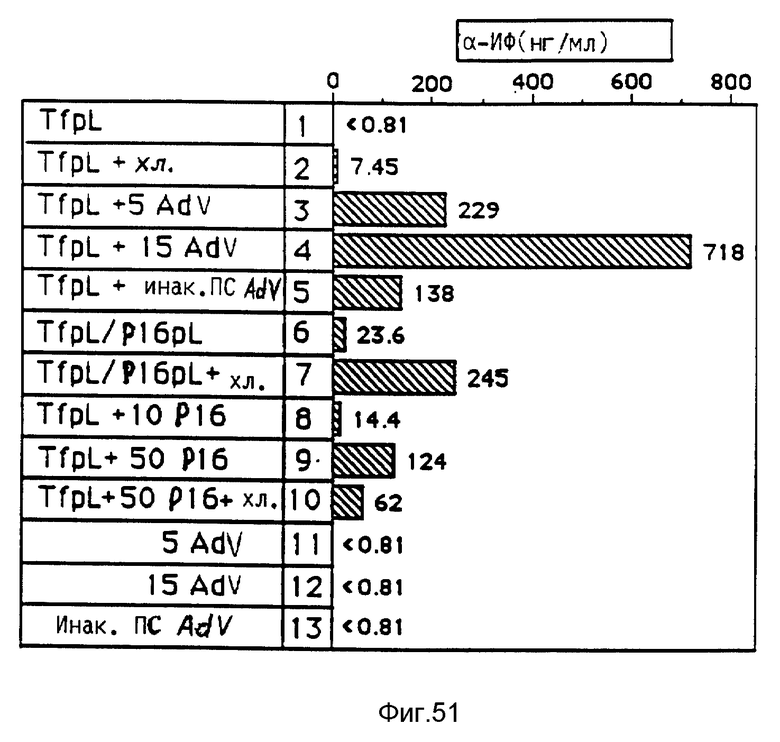
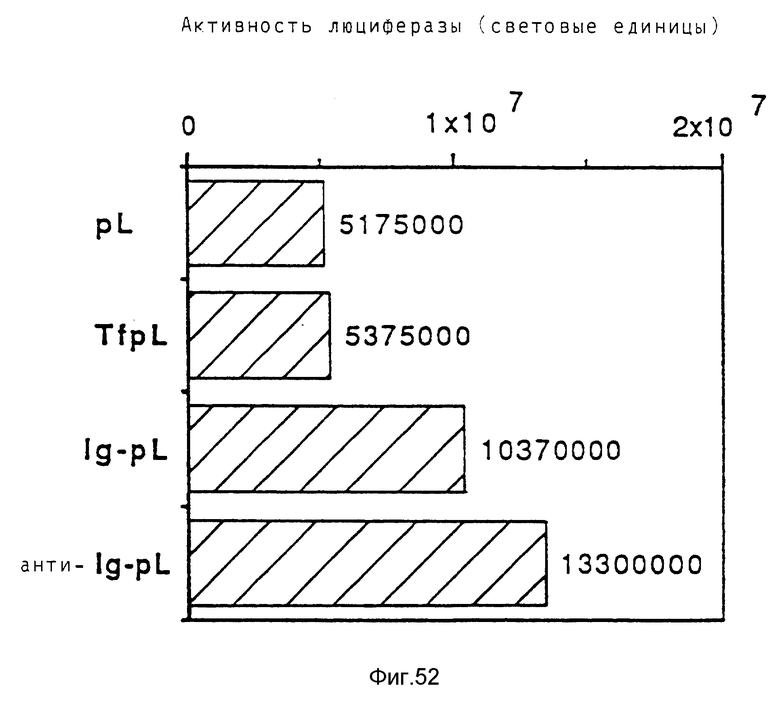
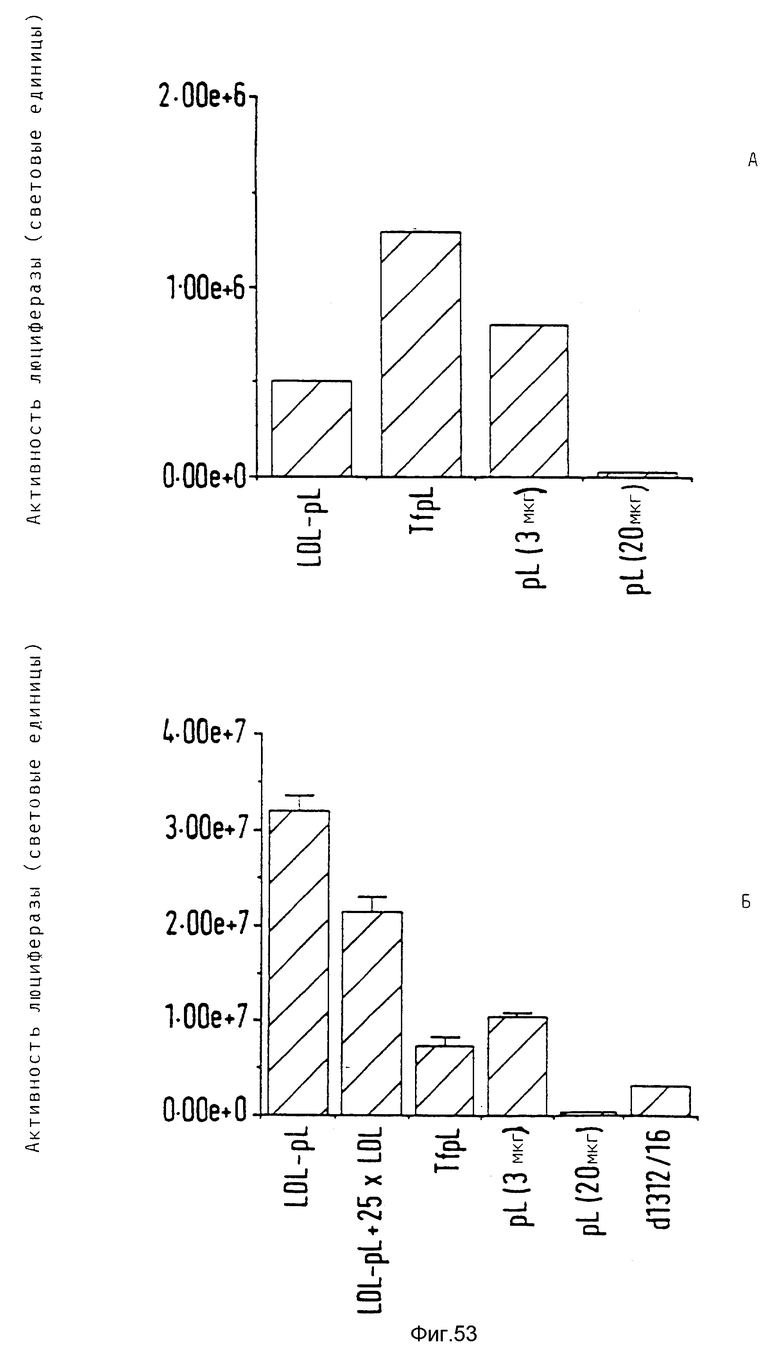
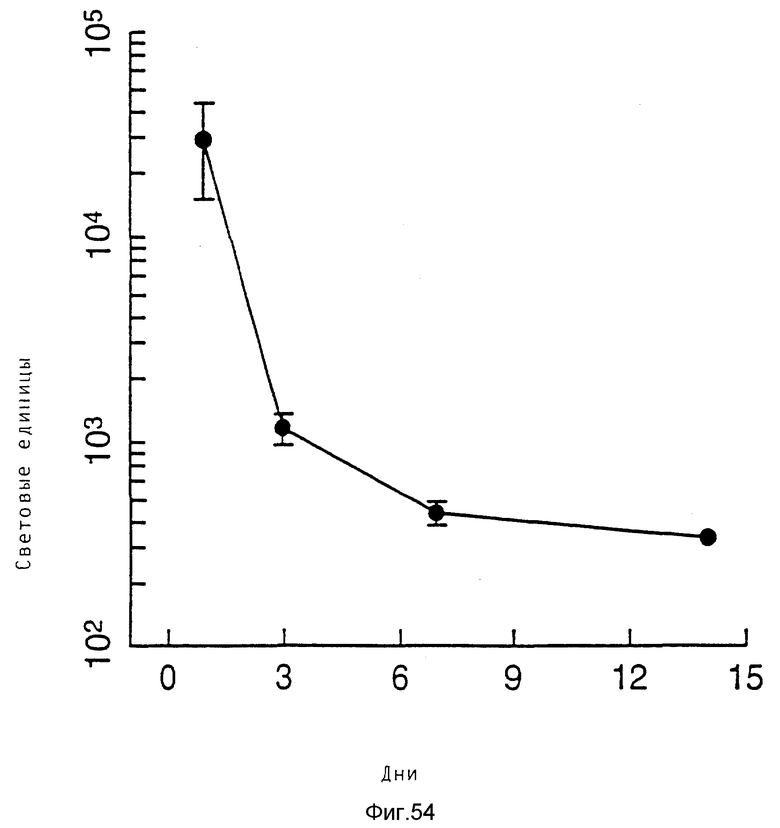
Изобретение относится к области биотехнологии и может быть применено для переноса нуклеиновых кислот в высшие эукариотные клетки. Охарактеризована трансфекционная композиция, содержащая комплекс нуклеиновой кислоты, вещества со сродством к нуклеиновой кислоте и эндосомолитический пептид. Использование изобретения позволит облегчить процесс введения нуклеиновых кислот в клетки. 4 с. и 71 з.п. ф-лы, 54 ил., 1 табл.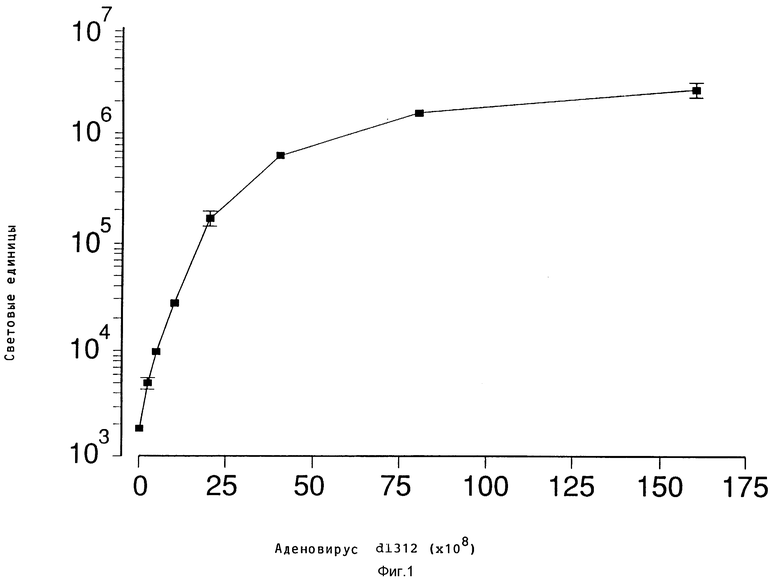
Приоритет по пунктам:
30.09.91 - по пп. 5; 8-10; 22, 39, 46-48, 50-52, 54, 56, 59-64, 69-71 (заявка 07/767788);
30.09.91 - по пп.2, 3, 6, 7, 14, 17-21, 23, 28-30, 34, 36, 40, 41, 43-45, 65, 66, 67 (заявка 07/768039);
07.04.92 - по пп.13, 15, 16, 37, 55, 57, 58, 72;
02.09.92 - по пп.1, 4, 11, 12, 24, 25-27, 31-33, 35, 38, 42, 49, 53, 68, 73-75.
| СПОСОБ ТРАНСПОРТИРОВКИ СЖИЖЕННОГО ПРИРОДНОГО ГАЗА | 1999 |
|
RU2228486C2 |
| СПОСОБ ПУТЕВОГО ОБСЛЕДОВАНИЯ И ДИАГНОСТИКИ ДЕЙСТВУЮЩИХ МАГИСТРАЛЬНЫХ ТРУБОПРОВОДОВ И СИСТЕМА ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2002 |
|
RU2228487C2 |
| СПОСОБ АВТОМАТИЧЕСКОГО УПРАВЛЕНИЯ РЕКТИФИКАЦИОННОЙ КОЛОННОЙ | 0 |
|
SU388758A1 |
| Огнетушитель | 0 |
|
SU91A1 |

