Молекулы, активные в отношении кальциевого рецептора
Родственная заявка
Настоящая заявка является продолжающей заявкой, сделанной Nemeth и др., озаглавленной "Молекулы, активные в отношении кальциевого рецептора", зарегистрированной 11 февраля 1992 г., которая, в свою очередь, является продолжающей по отношению к заявке Nemeth и др., озаглавленной "Активаторы кальциевых рецепторов", U.S.Serial N 07/749,451, зарегистрированной 23 августа 1991; обе последние заявки, включая рисунки, целиком включены в настоящий документ в качестве ссылки.
Область изобретения
Настоящее изобретение относится к созданию, разработке, композиции и применению новых кальцимиметических молекул, способных действовать тем же образом, что и внеклеточные ионы кальция действуют на клетки, к кальцилитическим молекулам, которые блокируют активность внеклеточных ионов кальция по отношению к клеткам, и к способам их применения и идентификации.
Предпосылки к созданию изобретения
Следующее описание суммируют информацию, относящуюся к настоящему изобретению. Оно не является признанием того, что какая-либо информация, приведенная здесь, является прототипом настоящего изобретения, а также того, что какие-либо публикации, конкретно или косвенным образом упоминаемые здесь, являются его прототипом.
Определенные клетки тела отвечают не только на химические сигналы, но также и на ионы, такие как внеклеточные ионы кальция (Ca2+). Изменения концентрации внеклеточного кальция (упоминаемого в настоящем документе как "[Ca2+]" изменяет функциональные реакции этих клеток. Одной из таких специализированных клеток является клетка паращитовидной железы, секретирует паратироидный гормон (ПТГ). ПТГ является главным эндокринным фактором, регулирующим гомеостаз Ca2+ в крови и внеклеточных жидкостях.
ПТГ, действуя на костные и почечные клетки, повышает уровень Ca2+ в крови. Это повышение уровня Ca2+ затем действует как сигнал отрицательной обратной связи, подавляя секрецию ПТГ. Эта взаимная связь между уровнем Ca2+ и секрецией ПТГ формирует главный механизм, поддерживающий гомеостаз Ca2+ в организме.
Внеклеточный Ca2+ действует непосредственно на клетки паращитовидной железы, регулируя секрецию ПТГ. Предполагают, что существует поверхностный белок клетки паращитовидной железы, который улавливает изменения концентрации Ca2+. Этот белок действует как рецептор для внеклеточного Ca2+ (кальциевый рецептор), и предполагают, что он улавливает изменения концентрации Ca2+ и инициирует функциональную реакцию клетки, секрецию ПТГ. Роль рецепторов кальция и внеклеточного Ca2+ в регуляции внутриклеточного Ca2+ и клеточной функции обсуждается в Netneth и др., II Cell Caicium 319,1990; роль рецепторов Ca2+ в парафолликулярных и паращитовидных клетках обсуждается в Nemeth, II Cell Calcium 323, 1990; и роль рецепторов Ca2+ для костных остеокластов обсуждается у Zaidi, 10 Biscience Reports 493, 1990.
Остальные клетки тела, а именно остеокласты кости, юкстагломерулярные почечные клетки и клетки проксимальных почечных канальцев, кератиноциты эпидермиса, парафолликулярные клетки щитовидной железы и трофобласты плаценты, обладают способностью воспринимать изменения в концентрации Ca2+. Предполагают, что рецепторы Ca2+ клеточной поверхности также могут присутствовать на этих клетках, сообщая им способность улавливать изменения Ca2+ и инициировать или облегчать реакцию на эти изменения.
В клетках паращитовидной железы, парафолликулярных клетках (C-клетках), кератиноцитах, юкстагломерулярных клетках и трофобластах увеличение Ca2+ вызывает увеличение внутриклеточной концентрации свободного Ca2+ . Такое увеличение может быть вызвано притоком в клетку внеклеточного Ca2+ или мобилизацией Ca2+ из внутриклеточных органелл. Изменения в концентрации Ca2+ без труда можно отследить и определить количественно, используя флюориметрические индикаторы, такие как fura-2 или indo-1 (Molecular Probes, Eugene, OP). Изменение уровня Ca2+ обеспечивает анализ для определения способности молекул действовать на рецепторы Ca2+ как агонисты или как антагонисты.
В клетках паращитовидной железы повышение концентрации Ca2+ снаружи вызывает быстрое и скоротечное повышение концентрации Ca2+, за которым следует более низкое и длительное повышение концентрации Ca2+. Скоротечное повышение концентрации Ca2+ происходит за счет мобилизации внутриклеточного кальция, в то время как более низкое и продолжительное повышение является результатом притока внеклеточного кальция. Мобилизация внутриклеточного кальция сопровождается увеличением образования инозит-1,4,5-трифосфата (ИФ3) и диацилглицерола, двух биохимических индикаторов, которые связаны с рецепторзависимой мобилизацией внутриклеточного кальция в различных других клетках.
В добавление к Ca2+, различные другие двух- и трехвалентные катионы, такие как Mg2+, Sr2+, Ba2+, La3+ и Gd3+ также вызывают мобилизацию внутриклеточного Ca2+, в клетках паращитовидной железы. Mg2+ и La3+ также увеличивают образование ИФ3; все эти неорганические катионы подавляют секрецию ПТГ. Следовательно, эти рецепторы, постулированные как кальциевые, являются смешанными, поскольку они улавливают целый ряд внеклеточных двух- и трехвалентных катионов.
Способность ряда соединений имитировать внеклеточный кальций in vitro обсуждается в Nemeth и др. , (спермин и спермидин). ''Белки, связывающие кальций, в норме и патологии", 1987, Academic Press, Inc., стр. 33-35; Brown и др. , (например, неомицин) 128 Endocrinology 3047, 1991; Chen и др., (дилтиазем и его аналог, ТА-3090) 5 J. Bone and Mineral Res., 581, 1990; и Zaidi и др., (верапамил) 167 Biochem, Biophys. Res. Comm. 807, 1990.
Brown и др. , 6 J.Bone and Mineral Res.II, 1991 обсуждают существующие теории, касающиеся действия ионов Ca2+ на клетки паращитовидной железы, и предполагают, что результаты могут быть объяснены как рецепторподобным механизмом, так и рецепторнезависимым механизмом:
Поливалентные катионы (например, двух- и трехвалентные катионы) оказывают многостороннее влияние на функцию паращитовидной железы, такое как подавление секреции паратироидного гормона (ПТГ) и аккумуляции цАМФ, стимуляция накопления инозитфосфатов и подъем цитозольной концентрации кальция. Полагают, что эти действия опосредуются рецепторподобным механизмом. Подавление аккумуляции цАМФ, стимулированной агонистами, двух- и трехвалентными катионами, например, блокируется вслед за преинкубацией с коклюшным токсином. Так, предполагаемый рецептор поливалентного катиона может быть связан с ингибированием аденилциклазы ингибирующим гуаниловым нуклеотидом регуляторного (G) белка, Gi.
Недавно мы показали, что поликатионный антибиотик, неомицин, имитирует действие двух- и трехвалентных катионов на некоторые стороны функции паращитовидной железы. Для того, чтобы определить, было ли это действие специфичным для данного агента или представляло собой более общее действие поликатионов, мы испытывали действие высокоосновных пептидов, полиаргинина и полилизина, а также протамина, на те же параметры диспергированных клеток паращитовидной железы крупного рогатого скота. Результаты показали, что клетка паращитовидной железы реагирует на ряд поликатионов, а также поливалентных катионов, возможно, посредством сходных биохимических путей. Эти результаты обсуждены с точки зрения недавно постулированной "рецепторнезависимой" модуляции G белков поликатионами в других системах.
Считалось доказанным, что рецепторы Ca2+ аналогичны другим связанным с G белками рецепторам (например, гликопротеин), однако недавние исследования с клетками других видов подняли вопрос о возможности того, что поликатионы могут модулировать функцию клетки с помощью альтернативного или дополнительного механизма. В маст-клетках (mast), например, ряд амфипатических катионов, включая мастопаран, пептид из яда осы, 48/80, синтетический поликатион, полилизин усиливают секрецию посредством механизма, чувствительного к коклюшному токсину, наводя на мысль о причастности G белка. Не было установлено ни одного классического рецептора поверхности клетки, который мог бы опосредовать действие этих разнообразных агентов. Более того, было показано, что те же соединения непосредственно активировали очищенные G белки в растворе или в искусственных фосфолипидных пузырьках. На основе этих наблюдений предполагают, что амфипатические катионы активируют G белки и, в свою очередь, секрецию маст-клеток посредством "рецепторнезависимого" механизма.
Было также показано, что поликатионы мощно взаимодействуют с кислыми фосфолипидами. Полилизины различной длины цепи (20-1000 аминокислот) связывают искусственные фосфолипидные пузырьки с константой диссоциации в пределах 0,5 нМ-1,5 мкМ. Сродство к связыванию прямо относится к длине цепи полилизина; полимеры в 1000 аминокислот имеют Kd=0,5 нМ, более короткие полимеры имеют более высокие значения Kd, и лизин не взаимодействует в сколько-нибудь значительной степени. Эта связь между активностью и длиной цепи подобна той, которая наблюдается при действии полилизина10200 полилизина3800 и лизина на функцию паращитовидной железы.
Возможно, что присоединение поликатионов к биологическим мембранам производит некоторые из этих биологических эффектов. Проницаемость плазматической мембраны, индуцированная в некоторых видах клеток разными порообразующими агентами, включая поликатионы, как предполагают, опосредуется их взаимодействием с фосфатидилсеринподобными структурами. Вдобавок "рецепторнезависимая" активация очищенных G белков амфипатическими катионами усиливается, если эти белки инкорпорированы в фосфолипидные пузырьки.
Ионы кальция в миллимолярных концентрациях также вызывают выраженные изменения в строении мембраны. В некоторых случаях кальций может как противодействовать, так и усиливать действие поликатионов на липиды мембраны. Эти заключения поднимают вопрос о возможности того, что действие как поливалентных катионов, так и поликатионов на клетки паращитовидной железы может вовлекать рецепторнезависимый механизм, не требующий присутствия классического рецептора поверхности клеток, связывающегося с G белком. Дальнейшие исследования, однако, призваны пролить свет на молекулярную основу чувствительности к Ca2+ этих и других клеточных типов. Snoback и Chen (6 Приложение 1), J. Bone and Mineral. Res. 1991, 135) и Racke и др., (6 Приложение 1) J.Bone and Mineral Res. 1991, 118) описывают эксперименты, о которых говорят, как об устанавливающих, что в клетках паращитовидной железы присутствуют рецепторы Ca2+ или сенсоры Ca2+. Транспортную РНК, выделенную из этих клеток, можно экспрессировать в ооцитах, и оказалось, что она снабжает эти ооциты таким фенотипом, который можно объяснить присутствием Ca2+ рецепторного белка.
Резюме
Заявитель продемонстрировал, что Ca2+ рецепторные белки дают возможность определенным специализированным клеткам, вовлеченным в метаболизм Ca2+ в организме, улавливать и реагировать на изменения концентрации внеклеточного Ca2+. Хотя эти рецепторы обладают определенными общими для них характеристиками, на них можно избирательно воздействовать различными фармакологическими агентами. Как уточнено ниже, определенные молекулы идентифицируются с избирательной активностью рецепторами Ca2+ на клетках паращитовидной железы, остеокластах и C-клетках.
Рецепторы Ca2+ представляют дискретные молекулярные мишени для нового класса молекул, которые имитируют ("кальцимиметики") или являются антагонистами ("кальцилитики") действие внеклеточного кальция. Такие рецепторы присутствуют на клеточных поверхностях и обладают малым сродством к внеклеточному Ca2+ (кажущийся Kd в основном выше чем приблизительно 0,5 мМ). Такие рецепторы могут включать свободный или связанный эффекторный механизм), как определяет Cooper, Bloom и Roth, "The Biochemical Basis of Neuropharmacology", Гл. 4. Следовательно, такие рецепторы отличаются от внутриклеточных рецепторов Ca2+, например, калмодулина и тропонинов. Кальцимиметики, например, действуют на рецепторы Ca2+ избирательно, прямо или косвенно подавляя функцию клеток паращитовидной железы или остеокластов, или стимулируя функцию C-клеток. Кальцимиметики и кальцилитики настоящего изобретения создают новые лекарственные средства для лечения гиперфункции паращитовидной железы, остеопороза и других связанных с метаболизмом кальция заболеваний. Настоящая заявка касается воздействия на рецепторы Ca2+ каждого из этих видов клеток и других видов клеток, которые улавливают и отвечают на изменения концентрации Ca2+.
Заявитель впервые продемонстрировал Ca2+ рецепторный белок в клетках паращитовидной железы, и дифференцировал фармокологически такие рецепторы Ca2+ в других клетках, таких как C-клетки и остеокласты. Заявитель также впервые описал способы, позволяющие идентифицировать молекулы, активные по отношению к этим рецепторам Ca2+, и использовать их в качестве основных молекул для открытия, разработки, оформления, модификации и/или конструирования кальцимиметиков или кальцилитиков, активных в отношении рецепторов Ca2+. Эти кальцимиметики или кальцилитики полезны для лечения различных болезненных состояний, которые характеризуются аномальными уровнями одного или более компонентов, например, полипептидов, таких как гормоны, энзимы, или факторы роста, экспрессия и/или секреция которых регулируется или подвержена воздействию со стороны активности одного или более рецепторов Ca2+. Далее, идентификация различных рецепторов Ca2+ в различных видах клеток и специфическая реакция этих рецепторов на различные основные молекулы позволяет синтезировать специфические молекулы, активные при лечении специфических заболеваний, на которые можно влиять путем воздействия на такие специфические рецепторы Ca2+. Например, на аномальные уровни секреции паратироидного гормона можно воздействовать такими специфическими молекулами без влияния на уровень секреции других регулируемых Ca2+ гормонов и подобных веществ.
Идентификация таких основных молекул затруднялась вследствие отсутствия скрининговой системы с большой пропускной способностью для обнаружения активных молекул, и отсутствия данных о структуре, базируясь на которых можно было обозначать эффективные лекарства-кандидаты. Эти препятствия в настоящее время устранены клонированием Ca2+ рецепторов клеток паращитовидной железы и функционально родственных рецепторов, и систематическим изучением строения определенных основных молекул, которые активируют такие клонированные рецепторы Ca2+ и функционально родственные рецепторы. Клонирование рецептора Ca2+ также облегчает разработку клеточных линий, подвергшихся трансфекции, подходящих для высокоэффективного скрининга натуральных продуктов или синтетических молекул. Это, вместе с изучением строения, обсуждаемым ниже, создает технологию, необходимую для разработки новых кальцимиметиков и кальцилитиков.
Заявитель в настоящей заявке дает возможность для таких методик. Например, кДНК рецептора Ca2+ клетки паращитовидной железы может быть клонирована скринингом на функциональную экспрессию в Xenopus ооцитах, а особенности строения органических молекул, необходимых для деятельности рецепторов Ca2+, можно определить посредством испытания натуральных продуктов или других молекул и последующего изучения их активности в зависимости от строения.
Так, в первом аспекте, настоящее изобретение обрисовывает фармацевтическую композицию, включающую молекулу, которая как имитирует действие внеклеточного Ca2+, вызывая повышение концентрации [Ca2+], внутри клетки, так и блокирует повышение концентрации [Ca2+], вызванное внеклеточным Ca2+. Эта молекула имеет EC50 меньшее или равное 5 мкМ и не является протамином.
Под "имитированием" подразумевается, что молекула обладает одним или более специфических воздействий, свойственным внеклеточному Ca2+, на клетки, реагирующие на внеклеточный Ca2+. Термин этот не означает, что все биологические функции внеклеточного Ca2+ имитируются, а то, что имитируется по крайней мере одна из них. Вдобавок не требуется, чтобы молекула связывалась с тем же участком рецептора, что и внеклеточный Ca2+ (см., например, новое соединение NPS 467 и его действие в примере 20 ниже). Под термином "блокирование" подразумевается, что одно такое воздействие Ca2+ уменьшается или предотвращается этой молекулой. Величина EC50 может быть определена в исследованиях, как описано ниже, в которых измерялась имитированная активность, а концентрация молекул, при которой имитировалась половина максимального имитирующего эффекта, равнялась EC50. Наоборот, IC50 кальцилитика - это то количество его, которое блокирует половину максимальной активности. Предпочтительно, такие исследования измеряют повышения [Ca2+], и являются специфическими для рецепторов Ca2+, что подтверждают способы, описанные ниже, или их эквивалент.
В предпочтительных вариантах осуществления изобретения, биологически исследования, описанные в настоящем документе, продемонстрировали, что повышение [Ca2+] , в клетке является скоротечным, его продолжительность была менее минуты, а скорость проявления этого повышения была высокой, повышение [Ca2+] i наблюдалось через 30 секунд и менее; и молекула также (а) вызывала более продолжительное повышение (больше тридцати секунд) [Ca2+]i; (b) вызывала повышение уровней инозит-1,4,5-трифосфата и/или диацилглицерола, например, менее чем за 60 секунд, и (c) подавляла образование циклического АМФ, стимулированное допамином или изопротеренолом. В добавление, кратковременное повышение [Ca2+]i устраняется предварительной обработкой клеток в течение 10 минут 10 мМ фторидом натрия и уменьшается в результате краткой предварительной обработки (не более 10 минут) клеток активатором протеинкиназы C, например, форбол миристатацетатом (ФМА), мезереином или (-)-индолактамом V.
В случае клеток паращитовидной железы те молекулы, которые оказывались активными по результатам всех анализов, описанных выше, особенно полезны в настоящем изобретении, поскольку они специфичны по своему действию на рецепторы Ca2+ этих клеток. Это в особенности относится к эффекту предварительной обработки ФМА, описанному выше.
В более предпочтительном варианте осуществления изобретения, клетка является клеткой паращитовидной железы, и молекула подавляет секрецию клеткой паратироидного гормона; и молекула вызывает повышение проводимости Cl- в ооцитах Xanopus, с внедренной мДНК клетки паращитовидной железы, костного остеокласта, юкстагломерулярной почечной клетки, клетки проксимального почечного канальца, кератиноцита, парафолликулярной клетки щитовидной железы или плацентарного трофобласта.
В других предпочтительных вариантах осуществления изобретения, молекула вызывает мобилизацию внутриклеточного Ca2+, что является причиной повышения [Ca2+] i; клетка является C-клеткой или остеокластом, а молекула ингибирует резорбцию кости in vivo; клетка является остеокластом, а молекула ингибирует резорбцию кости in vitro; или клетка является C-клеткой, а молекула стимулирует секрецию кальцитонина in vitro или in vivo; наиболее предпочтительно, если молекула является как кальцимиметиком, так и кальцилитиком, и имеет величины EC50 или IC50 воздействия на рецепторы Ca2+ меньше чем или равные 5 мкМ и даже более предпочтительно менее чем или равные 1 мкМ, 100 нмоляр, 10 нмоляр или 1 нмоляр. Такие более низкие величины EC50 и IC50 более выгодны, поскольку позволяют использовать более низкие концентрации молекул при использовании in vivo или in vitro для лечения или диагностики. Открытие молекул с такими низкими значениями EC50 и IC50 облегчает замысел и синтез столь же патентных и эффективных молекул.
Под молекулой- "кальцимиметиком" подразумевается любая молекула, обладающая одним или более воздействием внеклеточного Ca2+ и, предпочтительно, имитирующая действие Ca2+ на рецепторы Ca2+. Например, при использовании применительно к клеткам паращитовидной железы, эта молекула, которая при испытании на клетках паращитовидной железы in vitro обладала одной или более, а предпочтительно, всеми следующими характеристиками, измеренными с помощью методик, хорошо известных специалистам.
1. Молекула вызывает быстрое (время до пика < 5 сек) и кратковременное повышение [Ca2+] i, которое невосприимчиво к ингибированию 1 мк М La3+ или Gd3+. Повышение [Ca2+]i сохраняется в отсутствии внеклеточного Ca2+, но устраняется предварительной обработкой иономицином (в отсутствии внеклеточного Ca2+.
2. Повышение [Ca2+]i, вызванное внеклеточным Ca2+, не ингибируется дигидропиридинами.
3. Кратковременное повышение [Ca2+]i, вызванное молекулой, устраняется предварительной обработкой в течение 10 минут 10 мМ фторидом натрия.
4. Кратковременное повышение [Ca2+]i, вызванное молекулой, уменьшается в результате предварительной обработки активатором протеинкиназы C (ПКС), таким как форбол миристатацетат (ФМА), мезереин или (-)-индолактам. Общий эффект активатора протеинкиназы C состоит в том, чтобы сдвинуть кривую ответа на концентрацию молекулы вправо без влияния на максимальный ответ.
5. Молекула вызывает быстрое (< 30 с) повышение образования инозит-1,4,5-трифосфата и/или диацилглицерола;
6. Молекула ингибирует образование циклического АМФ, стимулированное допамином или протеренолом.
7. Молекула ингибирует секрецию ПТГ.
8. Предварительная обработка коклюшным токсином (100 нг/мл в течение 4 ч) блокирует ингибирующее действие молекулы на образование циклического АМФ, но не вызывает повышения [Ca2+]i, инозит-1,4,5-трифосфата или диацилглицерола, и не уменьшает секрецию ПТГ.
9. Молекула вызывает повышение проводимости Cl- в ооцитах Xenopus, в которые внедрена обогащенная поли(A)+ мРНК из клеток паращитовидной железы человека и крупного рогатого скота, но не действует на ооциты Xenopus, в которые вводились вода или мРНК из мозга или печени крысы.
10. Подобно предыдущему пункту, при использовании клонированного рецептора из клеток паращитовидной железы, молекула будет вызывать реакцию ооцитов Xehopus, в которые введены специфические кДНК и мРНК, кодирующие рецептор.
Под молекулой-"кальцилитиком" подразумевается любая молекула, которая блокирует одно или более воздействий внеклеточного Ca2+ на клетки, чувствительные к воздействию внеклеточного Ca2+, предпочтительно, действуя как антагонист рецептора Ca2+. Например, при использовании применительно к клеткам паращитовидной железы, это молекула, которая при испытании на клетках паращитовидной железы in vitro, обладала одной или более, а предпочтительно, всеми следующими характеристиками, измеренными с помощью методик, хорошо известных специалистам:
1. Молекула блокирует, как частично, так и полностью, способность возросших концентраций внеклеточного Ca2+:
а) повышать [Ca2+]i;
б) мобилизовать внутриклеточный Ca2+;
в) увеличивать образование инозит-1,4,5-трифосфата;
г) уменьшать образование циклического АМФ, стимулированное допамином или изопротеренолом; и
д) подавлять секрецию ПТГ;
2. При низких концентрациях Ca2+, например, 0,5 мМ, сама молекула не изменяет [Ca2+]i;
3. Молекула блокирует проводимость Cl- в ооцитах Xenopus, в которые введена обогащенная поли(A)+ мРНК из клеток паращитовидной железы человека или крупного рогатого скота, вызванную внеклеточным Ca2+ или кальцимиметическими соединениями; но не в ооцитах Xenopus, в которые введены вода или мРНК из мозга или печени крысы;
4. Подобно предыдущему пункту, при использовании клонированного рецептора из клеток паращитовидной железы, молекула будет блокировать реакцию ооцитов Xenopus, в которые введены специфические кДНК или мРНК, кодирующие рецептор Ca2+ , вызванную внеклеточным Ca2+ или кальцимиметическим соединением.
Параллельные определения подходящих кальцимиметиков и кальцилитиков по отношению к рецепторам Ca2+ клеток других видов очевидны из примеров, приведенных ниже.
Рецепторы Ca2+ способны улавливать и реагировать на определенные неорганические поликатионы или поликатионные органические молекулы. Например, клетка паращитовидной железы не способна различать повышения концентрации внеклеточного Ca2+ от добавления этих органических поликатионов, предположительно, потому что эти органические молекулы действуют на рецептор Ca2+ точно так же, как внеклеточный Ca2+. Кальцимиметические молекулы настоящего изобретения обладают особенно хорошим сродством к рецепторам Ca2+ и могут применяться в качестве лекарств, изменяющих отдельные клеточные функции, например, секрецию ПТГ клетками паращитовидной железы. В отличие от Ca2+, большинство из этих молекул действуют только на один или больше, но не на все рецепторы Ca2+, и, таким образом, обеспечивают способность специфически нацеливаться на один рецептор Ca2+.
Эти молекулы также обеспечивают основные структуры для разработки дополнительных новых лекарственных средств, эффективных при лечении различных заболеваний, в которых играют роль Ca2+ и [Ca2+], таких как гиперфункция паращитовидной железы, остеопороз, деформирующая остеодистрофия, гипертония, почечные болезни и рак.
Кальцимиметики и кальцилитики можно оформить в виде формацевтических композиций, полезных для регулирования уровня внеклеточного свободного Ca2+ у пациента и имитирования воздействия внеклеточного Ca2+ на клетки из описанной выше группы, путем введения пациенту такой фармацевтической композиции. До момента настоящего изобретения заявителю не были известны подобные молекулы, действующие на рецепторы Ca2+ и полезные при лечении заболеваний, вызванных нерегулярностью действия или регуляции рецепторов Ca2+ или заболеваний животных, имеющих нормальные рецепторы Ca2+, но которые можно лечить активацией или инактивацией таких рецепторов.
В еще одном предпочтительном варианте осуществления изобретения, молекула имеет величину EC50 меньше или равную 5 мкМ по отношению к одному или более, но не ко всем клеткам из следующей группы: клетки паращитовидной железы, остеокласты кости, юкстагломерулярные почечные клетки, клетки проксимальных почечных канальцев, кератиноциты, парафолликулярные клетки щитовидной железы (C-клетки) и плацентарные трофобласты.
Особенное преимущество настоящего изобретения заключается в специфичности действия таких молекул, поскольку это допускает специфическую in vivo и in vitro терапию и диагностику и открытие дополнительных кальцимиметических и кальцилитических молекул.
В специфических предпочтительных вариантах осуществления изобретения, молекула заряжена положительно при физиологическом уровне pH и избирается из следующей группы: разветвленные или циклические амины, позитивно заряженные полиаминокислоты, и арилалкиламины, например, полиамин с разветвленной цепью имеет формулу H2N-(CH2)j -(NRi-(CH2)j)k-NH2, где k - целое число от 1 до 10, каждое j - одно или то же или разное и является целым числом от 2 до 20, каждое Ri - одно и то же или разное и избирается из следующей группы; водород и -(CH2)j-NH2, где j равно описанному выше, и по крайней мере один Ri не является водородом.
В альтернативном варианте осуществления изобретения, молекула имеет формулу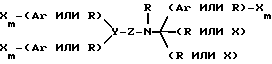
где X каждый независимо избирается из следующей группы: H, CH3, CH3O, CH3CH2O, Br, Cl, F, CF3, CHF2, CH2F, CF3O, CH3S, OH, CH2OH, CONH2, CN, NO2 и CH3CH2; Ar - гидрофобная форма; каждое R независимо избирается из следующей группы: водород, метил, этил, пропил, изопропил, бутил, изобутил, циклопентил, циклогексил, циклогептил, циклооктил, инденил, инданил, дигидроиндолил, тиодигидроиндолил, 2-, 3-, или 4-пиперид(ин)ил; Y избирается из следующей группы; CH, азот и ненасыщенный углеводород; Z избирается из следующей группы: кислород, азот, сера,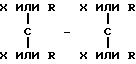

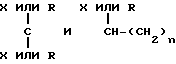
где каждое n независимо между 1 и 4 включением, а каждое m независимо между 0 и 5 включением. Наиболее предпочтительна молекула, которая является и кальцимиметиком и кальцилитиком.
В предпочтительных вариантах осуществления изобретения, гидрофобная форма избирается из следующей группы: фенил, 2-, 3-, или 4-пиридил, 1- или 2-нафтил, 1- или 2-хинолинил, 2- или 3-индолил, бензил, и фенокси; молекула является производным от R-дифенилпропил- α- этоксифениламина молекула имеет формулу: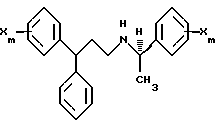
с каждым X, предпочтительно, независимо избираемым из следующей группы: Cl, F, CF3, CH3 и CH3O.
Согласно предпочтительному варианту осуществления изобретения, новые аналоги фенил- α- этоксифениламина и их производные имеют следующую формулу: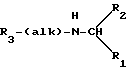
где alk является алкиленом с прямой или разветвленной цепью из от 1 до 6 атомов углерода; R1 - низший алкил от 1 до 3 атомов углерода или низший галоалкил из от 1 до 3 атомов углерода, замещенных от 1 до 7 атомами галогенов; R2 и R3 - независимо избираемые карбоциклический арил- или циклоалкильные группы, как моноциклические, так и бициклические, C 5 или 6 кольцами, факультативно замещенными 1 - 5 заместителями, независимо избираемыми из следующей группы: низший алкил из от 1 до 3 атомов углерода, замещенных от 1 до 7 атомами галогенов; низший алкокси из от 1 до 3 атомов углерода; галоген; нитрогруппа; аминогруппа; алкиламиногруппа; амидогруппа; низший алкиламиногруппа из от 1 до 3 атомов углерода; цианогруппа; гидроксильная группа; ацил из от 2 до 4 атомов углерода; низший гидроксиалкил из от 1 до 3 атомов углерода или низший тиоалкил из от 1 до 3 атомов углерода. Подходящими карбоциклическими арильными группами являются группы с 1 или 2 кольцами, из которых по крайней мере одно имеет ароматический характер, и включают карбоциклические арильные группы, такие как фенил и бициклические карбоциклические арильные группы, такие как нафтил. Из приведенной выше формулы очевидно, что заключенные в ней соединения могут существовать в качестве рацемических смесей и в виде индивидуальных стереоизомеров. Особенно предпочтительными являются производные R-фенилпропил- α- этоксифениламина, которые, как полагают, проявляют усиленную активность в понижении содержания ионизированного кальция в сыворотке крови.
Предпочтительными соединениями являются те, которые включают в качестве "alk" n-пропилен. Также предпочтительны соединения, в которых R1 является метилом. Также предпочтительны такие соединения, в которых R2 и R3 являются факультативно замещенным фенилом.
Особенно предпочтительными являются соединения, в которых R2 является однозамещенным фенилом, более предпочтительно - метазамещенным. Особенно предпочтительные R3 группы включают незамещенный или однозамещенный фенил, особенно орто-замещенный. Предпочтительные заместители для R2 включают галоген, галоалкил, предпочтительно, тригалометил, и алкокси, предпочтительно, метокси. Предпочтительные заместители для R3 включают галоген.
В другом аспекте, настоящее изобретение представляет способ лечения пациентов с заболеванием или состоянием, которые характеризуются аномальной [Ca2+] или [Ca2+] i в одном или более видах клеток или в крови, плазме или внеклеточных жидкостях. Способ включает этап введения пациенту терапевтически эффективного количества молекул, которые как имитируют активность внеклеточного Ca2+, вызывая повышение [Ca2+]i в клетке, так и блокируют повышение [Ca2+]i, вызванное внеклеточным Ca2+.
Под "аномальным" подразумевается пациент, имеющий, по сравнению с основной популяцией, отличающийся метаболизм Ca2+, на который оказывают воздействие один или более белков (например, гормоны) крови или внеклеточных жидкостей организма, или другие молекулы, которые влияют на уровень внеклеточного и/или внутриклеточного Ca2+. Так, эти заболевания включают гиперфункцию паращитовидной железы, остеопороз, другие костные и связанные с неорганическими веществами нарушения (как описано, например, в стандартных медицинских учебниках, таких как "Harrison's Principles of Internal Medicine"). Согласно настоящему изобретению, подобные заболевания лечат с помощью молекул, которые имитируют или блокируют одно или более воздействий Ca2+ и, таким образом, непосредственно или косвенно воздействуют на уровне белков или других молекул в организме пациента.
Под "терапевтически эффективным количеством" подразумевается количество, которое до некоторой степени облегчает один или более симптомов заболевания или состояния пациента. Дополнительно под "терапевтически эффективным количеством" подразумевается количество, которое возвращает к норме, как частично, так и полностью, физиологические или биохимические параметры, связанные или вызвавшие заболевание или состояние. В общем, количество составляет между приблизительно 1 нмоль и 1 мкмоль молекул, в зависимости от ее EC50, самого заболевания, его тяжести и возраста пациента,
В предпочтительных вариантах осуществления изобретения, молекулы имеют EC50 менее, чем или равное 5 мкМ и не являются протамином; и, наиболее предпочтительно, взаимодействуют с рецептором Ca2+ как кальцилитик или кальцимиметик. Наиболее предпочтительно, если молекула избирается из описанных выше.
В других предпочтительных вариантах осуществления изобретения, у пациента наблюдается заболевание, которое характеризуется аномальным уровнем одного или более компонентов, чей уровень регулируется или подвержен влиянию активности одного или более рецепторов Ca2+, и молекула является активной по отношению к рецептору Ca2+ клетки из следующей группы: клетки паращитовидной железы, остеокласты кости, юкстагломерулярные почечные клетки, клетки проксимального почечного канальца, кератиноциты, парафолликулярные клетки щитовидной железы и трофобласты плаценты.
В еще некоторых предпочтительных вариантах осуществления изобретения, молекула снижает уровень паратироидного гормона в сыворотке крови пациента, например, до уровня, существующего у здоровых людей, или до степени, достаточной, чтобы вызвать снижение Ca2+ в плазме; и молекула вводится в количествах, достаточных для оказания уместного терапевтического эффекта на пациента.
В третьем аспекте, настоящее изобретение представляет способ диагностики заболевания или состояния пациента, путем идентификации количества и/или локализации (и/или функциональной целостности) одного или более рецепторов Ca2+ у пациента и сравнения этого количества и/или локализации (и/или функциональной целостности) с таковыми здоровых людей, в качестве показателя наличия заболевания или состояния.
В предпочтительных вариантах осуществления изобретения, способ является иммунологическим анализом, в котором используются антитела к рецепторам Ca2+ для установления количества и/или локализации и/или функциональной целостности рецепторов Ca2+, или анализом, в котором используются меченые кальцимиметические или кальцилитические молекулы, которые связывают рецепторы Ca2+; а диагностируемое заболевание является раком, например, эктопической опухолью паращитовидной железы, или состоянием, которое характеризуется повышенным уровнем остеокластов в кости или повышенным уровнем их активности.
В четвертом аспекте, настоящее изобретение представляет способ определения молекул, полезных в качестве терапевтических. Способ включает скрининг потенциально полезных молекул по способности имитировать активность внеклеточного Ca2+ в клетке, или по способности блокировать повышение [Ca2+]i, вызванное внеклеточным Ca2+, и определение имеет ли выбранная молекула EC50 или IC50 меньше или равную 5 мкМ.
В других аспектах, изобретение представляет рекомбинантный рецептор Ca2+, клетку, включающую рекомбинантный рецептор Ca2+, выделенную нуклеиновую кислоту, кодирующую рецептор Ca2+, биологическую активность и применение молекул NPS 019, новые соединения или композиции на основе NPS 459, NPS 467, NPS 551, NPS 568 (см. фиг. 36), и способ идентификации полезных кальцимиметических или кальцилитических молекул, путем определения молекул, которые имитируют или блокируют одно или более воздействий Ca2+ на первый рецептор Ca2+, но не на второй рецептор Ca2+, например, посредством рекомбинантного рецептора Ca2+.
Под "рекомбинантным", подразумевается любой рецептор Ca2+, полученный методиками рекомбинантной ДНК, отличающийся от естественных рецепторов Ca2+ по локализации, чистоте или строению. В основном, такой рецептор будет присутствовать в клетке в количестве, отличном от естественно наблюдающегося у здорового человека.
Под "выделенным" подразумевается антитело или нуклеиновая кислота, которые отличаются от естественно наблюдающихся антитела или нуклеиновой кислоты и которые были отделены от антитела или нуклеиновой кислоты, с которыми они встречаются в естественных условиях, например, в векторной системе, и могут быть использованы для экспрессии рекомбинантного рецептора Ca2+. Предпочтительно, чтобы антитело или нуклеиновая кислота были в виде гомогенного препарата, полученного с помощью стандартных методик.
Такие клонированные рецепторы можно экспрессировать в желательной клетке, изолировать и кристаллизовать для определения структуры. Подобная структура поможет в разработке полезных молекул настоящего изобретения, которые могут присоединяться к рецепторам Ca2+. В дополнение, эквивалент подобных рецепторов может быть клонирован, используя первый клон в качестве пробы для клонов в других клетках, кДНК или геномных коллекций.
Антитела к клонированному рецептору можно изолировать и использовать в качестве лекарственных препаратов настоящего изобретения, или в качестве диагностического инструмента для определения количества и/или локализации и/или функциональной целостности рецепторов Ca2+, для диагностики связанных с Ca2+ заболеваний или состояний. Такие антитела можно также применять путем введения внутривенно в качестве кальцимиметиков или кальцилитиков.
Таким образом, в основном, настоящее изобретение представляет кальцимиметические или кальцилитические молекулы, способные действовать в качестве как избирательных агонистов, так и в качестве антагонистов соответственно, на рецепторы Ca2+ одного или более, но не на все клетки из следующей группы: клетки паращитовидной железы, остеокласты кости, юкстагломерулярные почечные клетки, клетки проксимальных почечных канальцев, кератиноциты, парафолликулярные клетки щитовидной железы и трофобласты плаценты. Такая композиция может включать любой фармацевтически приемлемый носитель, известный специалистам.
Настоящее изобретение также представляет модулирование количества рецепторов Ca2+ у пациента, посредством стандартных методик, например, десенсибилизации и родственных методик (напр., рибозимы), в качестве терапевтического средства для лечения болезненного состояния.
Настоящее изобретение создает способы идентификации молекул, которые действуют на активность рецепторов Ca2+, применяющие методики, описанные ниже, для определения кальцимиметиков и/или кальцилитиков. Далее, молекулы, оказавшиеся эффективными в плане ослабления или усиления экспрессии рецептора Ca2+ на уровне транскрипции или трансляции, подвергаются анализу либо антителами, либо с помощью других методик, описанных ниже, чтобы определить их пригодность для терапевтических целей.
Другие особенности и преимущества настоящего изобретения будут очевидны из следующего описания его предпочтительных вариантов осуществления и их формулы изобретения.
Описание предпочтительных вариантов осуществления изобретения
Фиг. 1a -1f изображает характерные молекулы, полезные для настоящего изобретения.
Фиг. 2 является графическим изображением, показывающим повышения [Ca2+] i, индуцированные внеклеточным Ca2+, в клетках паращитовидной железы крупного рогатого скота, нагруженных хин-2 или фура-2. Начальная концентрация Ca2+ была 0,5 мМ (применяли CaCl2), и на каждой стрелке увеличивалась на 0,5 мМ.
Фиг. 3 является графическим изображением, показывающим мобилизацию [Ca2+] i в клетках паращитовидной железы крупного рогатого скота. Начальная концентрация Ca2+ была 0,5 мМ и уменьшалась до < 1 мкМ после добавления этиленгликольтетрауксусной кислоты (EGTA) (ЭГТУ), как обозначено. (a) Внеклеточный Mg2+ (8 мМ, конечная) вызывает повышение [Ca2+]i в отсутствии внеклеточного Ca2+. (b) Предварительная обработка иономицином (1 мкМ) блокирует ответ на Mg2+. (c) Предварительная обработка 5 мкМ молекулой 1799 (митохондриальный разобщитель) не оказывает влияния на ответ на Mg2+.
Фиг. 4 является графическим изображением, показывающим предпочтительные ингибирующие эффекты низкой концентрации Gd3+ на стационарные повышения [Ca2+] i, а также то, что высокая концентрация Gd3+ вызывает кратковременные повышения [Ca2+]i. (a) Верхнее изображение: контроль. Исходная концентрация внеклеточного Ca2+ составляет 0,5 мМ и повышалась на 0,5 мМ на каждой стрелке. (b) Среднее изображение Gd3+ (5 мкМ) блокирует стационарное, но не кратковременное повышение [Ca2+]i, вызванное внеклеточным Ca2+. (c) Нижнее изображение: Gd3+ (50 мкМ) вызывает кратковременное повышение [Ca2+]i и устраняет как кратковременный, так и поддерживаемый ответы на внеклеточный Ca2+. На среднем и нижнем изображениях было добавлено достаточно ЭГТУ, чтобы связать избирательно Gd3+: блок притока Ca2+ снят и [Ca2+]i быстро повышается.
Фиг. 5a- 5c является графическим изображением, показывающим, что воздействия ФМА на [Ca2+]i, образование ИФ3 и секрецию ПТГ преодолеваются повышением концентраций внеклеточного Ca2+ Для каждого варианта существует сдвиг вправо в кривой ответа концентрации на внеклеточный Ca2+. Обратите также внимание на то, что в то время, как [Ca2+] изменяется линейно, реакция концентрации изменяется нелинейно.
Фиг. 6 является графическим изображением, показывающим, что повышения [Ca2+] i, вызванные спермином, прогрессивно подавляются повышением [Ca2+]. Спермин (200 мкМ) добавляли в моменты времени, показанные стрелками. На этом и на всех последующих рисунках числа, сопровождающие графики, являются [Ca2+]i в нМ.
Фиг. 7 является графическим изображением, показывающим, что спермин мобилизует внутриклеточный Ca2+ в клетках паращитовидной железы крупного рогатого скота. EGTA добавляли для уменьшения [Ca2+] до <1 мкМ до того, как добавляли спермин (200 мкМ), как обозначено (левый график). Предварительная обработка иономицином (1 мкМ) блокирует ответ на спермин (правый график).
Фиг. 8a и 8b являются графическими изображениями, показывающими, что спермин повышает [Ca2+]i и ингибирует секрецию ПТГ в клетках паращитовидной железы крупного рогатого скота, подобно внекелеточному Ca2+. Данные указывают, что кривые ответа на концентрацию спермина являются средними по результатам двух экспериментов.
Фиг. 9a-9c является графическим изображением, показывающим контрастирующие эффекты ФМА на ответы на внеклеточный Ca2+ и на ответы на АТФ γ S в клетках паращитовидной железы крупного рогатого скота. Изображение слева: Кривая реакции концентрации на ингибирование образования циклического АМФ, индуцированное внеклеточным Ca2+, сдвигается вправо добавлением ФМА (100 нМ). Изображение среды: ФМА не влияет на способность АТФ γ S повышать [Ca2+] i. Обратите также внимание, что кривая реакции концентрации на АТФ γ S демонстрирует классическое сигмоидальное поведение в качестве функции от логарифма концентрации, в противоположность кривой для внеклеточных двухвалентных катионов.
Фиг. 10 графически представляет мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы человека, вызванную внекелточным Mg2+. Клетки были получены из аденомы и содержались в буфере, содержащем 0,5 мМ внеклеточного Ca2+. Кратковременное и сохраняющееся повышения [Ca2+]i, вызванные внеклеточным Mg2+ (10 мМ, конечная), показывают, что на сохраняющееся повышение не оказывает влияния нимодипин (1 мкМ), но оно подавляется La3+ (1 мкМ) и быстро возвращается, когда La3+ избирательно связывается низкой концентрацией ЭГТУ. (b) La3+ (1 мкМ) блокирует сохраняющееся, но не кратковременное повышение [Ca2+]i, вызванное внеклеточным Mg2+. (c) Кратковременное повышение концентрации щитозольного Ca2+, вызванное внеклеточным Mg2+, сохраняется в отсутствии внеклеточного Ca2+.
Фиг. 11 графически изображает мобилизацию внутриклеточного Ca2+, вызванную неомицином или протамином, в клетках паращитовидной железы крупного рогатого скота. На всех графиках начальные [Ca2+] и [Mg2+] были 0,5 и 1 мМ, соответственно. На графиках (a) и (b) концентрации Ca2+ и Mg2+ повысились до 2 и 8 мМ, с 0,5 и 1 мМ соответственно. На других графиках от (c) до (i), неомицин B (30 мкМ) или протамин (1 мг/мл) добавляли, как обозначено. La3+ (1 мкМ), ЭГТУ (1 мМ) или иономицин (100 нМ) добавляли, как обозначено. Каждый график представляет образец, прошедший 5 или более испытаний с использованием по крайней мере трех различных клеточных препаратов. Интервал времени = 1 мин.
Фиг. 12 графическое изображение, показывающее, что неомицин B блокирует кратковременное, но не блокирует стационарное повышение [Ca2+]i, вызванное внеклеточным Ca2+. Слева контроль: [Ca2+] была вначале 0,5 мМ и повышалась на 0,5 мМ на каждой открытой стрелке, до добавления неомицина B (30 мкМ), Справа: неомицин B (30 мкМ) добавили до кальция. Интервал = 1 мин.
Фиг. 13 является графическим изображением, показывающим, что неомицин B (a) или протамин (b) ингибирует секрецию ПТГ в концентрациях, которые вызывают повышение [Ca2+]. Клетки инкубировали с указанными концентрациями органического поликатиона в течение 30 мин в присутствии 0,5 мМ внеклеточного Ca2+. Открытые символы: контрольные ответы для ПТГ секреции в присутствии 0,5 (кружки) или 2 мМ (бриллиант) внеклеточного Ca2+. Величины [Ca2+]i обозначены символом "бриллиант". В экспериментах с протамином использовали клетки паращитовидной железы крупного рогатого скота, а в экспериментах с неомицином B использовали клетки аденомы паращитовидной железы человека. Каждая точка является средней ± SEM от трех экспериментов.
Фиг. 14 графически изображает предпочтительные ингибирующие воздействия ФМА на кратковременное повышение цитозольной [Ca2+], вызванной спермином. Начальная [Ca2+] i составляла 0,5 мМ; спермин (200 мкМ) или АТФ (50 мкМ) добавляли, как обозначено. Интервал - 1 мин.
Фиг. 15 графически показывает, что ФМА сдвигает вправо кривые реакции концентрации на внеклеточный Ca2+ и неомицин-B-индуцированные повышения [Ca2+] i(b). Клетки предварительно обрабатывали ФМА в течение 1 мин до повышения [Ca2+] или до прибавления неомицина B, как обозначено. Каждая точка является средней ± SEM от 3 до 5 экспериментов.
Фиг. 16 графически показывает, что ФМА сдвигает вправо кривые реакции концентрации на внеклеточный Ca2+(a) и спермин-индуцированное подавление секреции ПТГ(b). Клетку инкубировали с обозначенными [Ca2+] и спермином в течение 30 мин в присутствии (закрашенные кружки) или в отсутствии (незакрашенные кружки) 100 нМ ФМА. Каждая точка является средней ± SEM от 3 экспериментов.
Фиг. 17 графически показывает, что протамин увеличивает образование инозитфосфатов. Клетки паращитовидной железы инкубировали в течение ночи в культуральной среде, содержащей 4 ед Ci /мл 3H-мио-инозита. отмывали и инкубировали с обозначенной концентрацией протамина при 37oC. Через 30 с с реакцию прекращали добавлением CHCl3: MeOH:HCl и ИФ1 (кружки) и ИФ3 (треугольники) отделяли с помощью анионообменной хроматографии. Каждая точка является средней от 2 экспериментов, выполненных с тремя повторностями.
Фиг. 18 графически показывает, что ФМА подавляет образование ИФ1, вызванное внеклеточным Ca2+ (a) или спермином (b). Клетки, меченные 3H-мио-инозитом, выдерживали с обозначенной [Ca2+] или спермином в течение 30 с перед прекращением реакции и определением ИФ1 с помощью анионообменной хрмоатографии. Заштрихованные колонки: Клетки предварительно обрабатывали ФМА (100 нМ) в течение 5 мин до увеличения [Ca2+] или прибавления спермина. Каждая величина является средней от 2 экспериментов, выполненных с тремя повторностями.
Фиг. 19 графически изображает кратковременное и сохраняющееся повышение [Ca2+]i, вызванное неомицином B в клетках аденомы паращитовидной железы человека. [Ca2+] была 0,5 мМ. (a) Сохраняющееся повышение [Ca2+]i, вызванное неомицином B (10 мкМ) подавлялось La3+. (b) Кратковременное повышение [Ca2+] i, вызванное неомицином B, не реагировало на La3+. (c) Кратковременные повышения [Ca2+]i сохранялись в отсутствии внеклеточного Ca2+.
Фиг. 20 графически показывает, что неомицин B вызывает колеблющиеся повышения проводимости Cl- в ооцитах Xenopus, экспрессирующих рецептор Ca2+. (a) Верхняя запись от ооцита, спустя три дня, после введения поли(A)+-мРНК из гиперпластических клеток паращитовидной железы человека. (b) Нижняя запись от ооцита с введенной водой. Неомицин B не смог вызвать ответ в пяти ооцитах с введенной водой, а карбохол вызвал ответ в одном, что и показано. В обеих записях сохраняющийся потенциал был - 76 mV.
Фиг. 21 графически показывает, что неомицин B не оказывал влияния на основное или вызванное повышение в C-клетках. Контроль, левая запись: Нагруженные фура-2 клетки гМТС 6-23 сначала орошались буфером, содержащим 1 мМ Ca2+, до того, как концентрацию повышали до 3 мМ. Правая запись: Предварительная обработка 5 мМ неомицином B.
Фиг. 22 графически показывает, что внеклеточный Ca2+ вызывает повышения [Ca2+] i в остеобластях крысы. Микрофлюориметрическая запись в одиночном остеокласте крысы, нагруженном индо-1 и орошаемом в обозначенное время (интервале) буферным раствором, содержащим обозначенную [Ca2+]. Нормальный буфер, которым орошали в промежутках, содержал 1 мМ Ca2+.
Фиг. 23 графически показывает, что спермин или неомин B не вызывали повышений [Ca2+]i в остеокластах крысы. Нагруженный индо-2 остеоклатс орошали обозначенными концентрациями спермина или неомицина B отдельно (плоские интервалы) или вместе с 20 мМ Ca2+ (пики).
Фиг. 24 графически представляет различные эффекты аргиотоксина (показанного на чертеже как аргиопин, строение приведено на фиг. 1) 659 и аргиотоксина 636 на [Ca2+]i на клетки паращитовидной железы крупного рогатого скота. Начальная [Ca2+] составляла 0,5 мМ и повышалась до 1,5 мМ, где обозначено (правая запись). Где обозначено, добавляли аргиотоксин 659 (300 М) или аргиотоксин 636 (400 М).
Фиг. 25 графически показывает, что внеклеточный Mg2+ или Gd3+ вызывают колеблющиеся повышения проводимости Cl- в ооцитах Xenopus, содержащих внедренную поли(A)+-мРНК из клеток паращитовидной железы крупного рогатого скота, B записи (a) концентрация внеклеточного Ca2+ была < 1 мкМ, а в записи (b) - 0,7 мМ. Запись (c) показывает, что внеклеточный Mg2+ не мог вызвать ответа в ооците, с введенной мРНК только для вещества K рецептора, хотя орошение веществом K вызывало ответ. Сохраняющийся потенциал составлял от -70 до -80 mV.
Фиг. 26 графически показывает, что внеклеточный Ca2+ вызывает колеблющиеся повышения проводимости Cl- в ооцитах Xenopus, содержащих внедренную поли(A)+-мРНК из гиперпластической ткани паращитовидной железы человека. Ооцит испытывали на чувствительность к внеклеточному Ca2+ спустя три дня после введения 50 нг поли(a)+-мРНК. Сохраняющийся потенциал составлял -80 mV.
Фиг. 27 графически представляет мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы крупного рогатого скота, вызванную будмуншиамином (budmunchiamine). Будмуншиамин (300 мкМ, строение также указано), добавляли, где обозначено.
Фиг. 28 графически показывает, что способность клеток паращитовидной железы мобилизовать внутриклеточный Ca2+ является стереоспецифичной. Клетки паращитовидной железы крупного рогатого скота, нагруженные фура-2, сначала суспендировали в буфере, содержащем 0,5 мМ внутриклеточного Ca2+, перед добавлением обозначенной концентрации каждой молекулы.
Фиг. 29 графически показывает эффект La3+ на [Ca2+]i в остеокластах. Показана запись из одиночного остеокласта крысы, нагруженного индо-1. При низких концентрациях La3+ частично блокирует повышения [Ca2+]i, вызванные внеклеточным Ca2+.
Фиг. 30a и 30b графически показывают мобилизацию внутриклеточного Ca2+, вызванную внеклеточным Mn2+, в остеокластах крысы. Внеклеточный M2+ вызывает зависящие от концентрации повышения [Ca2+]i (фиг. 30a), которые сохраняются в отсутствии внеклеточного Ca2+ (фиг. 30b).
Фиг. 31a и 31b графически показывают мобилизацию внутриклеточного Ca2+ в остеокластах крысы, вызванную молекулой NPS 449 (см. фиг. 38). Изолированные остеокласты крысы, нагруженные индо-1, орошали обозначенными концентрациями NPS 449 в присутствии (фиг. 31a) или отсутствии (фиг. 31b) 1 мМ внеклеточного CaCl2.
Фиг. 32 графически показывает мобилизацию внутриклеточного Ca2+ и C-клетках вызванную NPS 019 (см. фиг. 1). Клетки rМТС 6-23 были нагружены фура-2 и орошались буфером, содержащим 0,5 мМ [Ca2+]. Где обозначено, добавляли NPS 019 до конечной концентрации 10 мкМ. Записи показывают, что кратковременное повышение [Ca2+], вызванное NPS 019, невосприимчиво к ингибированию La3+ (средняя запись) и сохраняется в отсутствии внеклеточного Ca2+ (правая запись).
Фиг. 33 графически показывает, что NPS 456 (см. фиг. 36) вызывает колеблющиеся повышения тока Cl- в ооцитах Xenopus, в которые была введена поли(A)+-мРНК клеток паращитовидной железы крупного рогатого скота.
Фиг. 34 графически показывает, что внеклеточный Ca2+ вызывает колеблющиеся повышения тока Cl- в ооцитах Xenopus, в которые была введена мРНК остеокласта человека. Ооциты испытывали на чувствительность к внеклеточному Ca2+ спустя три дня после введения 50 нг поли(A)+-мРНК.
Фиг. 35 графически показывает, что рецептор Ca2+ клетки паращитовидной железы кодируется мРНК в размерах 2,5 - 3,5 т.п.н. Поли-(A)+-мРНК была фракционирована по размеру в денатурирующем градиенте глицерола и разделена на десять фракций. Каждую фракцию отдельно (50 нг) вводили в ооциты Xenopus. Спустя три дня ооциты испытывали на способность отвечать на внеклеточный Ca2+ повышением проводимости Cl-.
Фиг. 36a - 36t изображает химическое строение молекул, производных от дифенилпропил- α- этоксифениламина, представляющих семейство молекул, которые приготавливали и отбирали, чтобы об наружить подходящие для настоящего изобретения.
Фиг. 37a и 37b графически изображает, что NPS 021 является кальцилитиче ским соединением, которое блокирует воздействие внеклеточного Ca2+ на [Ca2+] i в клетках паращитовидной железы крупного рогатого скота. Вначале клетки находились в буфере, содержащем 0,5 мМ CaCl2, и, когда обозначено, [Ca2+] повышали до конечной концентрации 2 мМ (левая запись). Добавление NPS 0,21 (200 мкМ) не вызывало изменений [Ca2+]i, но ингибировало повышение [Ca2+]i, вызванное внеклеточным Ca2+ (правая запись).
Фиг. 38 графически показывает ответ Ca2+ in vivo на NPS 467.
Фиг. 39 графически показывает ответ ПТГ in vivo на NPS 467.
Фиг. 40 графически показывает ответ Ca2+ in vivo на 25 мг/кг NPS 467.
Фиг. 41 и 42 графически показывает ответы Ca2+ in vivo на различные энантиомеры NPS 467.
Фиг. 43а изображает схему реакции для приготовления фендилина или аналогов фендилина или производных, изображенных на фиг. 36.
Фиг. 43b изображает схему реакции для синтеза NPS 467.
Фиг. 44 изображает кривую реакции на дозу, показывающую, что NPS 467 снижает содержание ионов кальция в сыворотку крови при оральном введении.
Кальцимиметические и кальцилитические молекулы
Кальцилитические и кальцимиметические молекулы, подходящие для настоящего изобретения, были, в основном, описаны выше. Эти молекулы можно без труда идентифицировать, используя скрининговые процедуры для определения молекул, которые имитируют или противодействуют активности Ca2+ на рецепторы Ca2+. Примеры таких процедур приводятся ниже. Эти примеры не являются ограничивающими в настоящем изобретении, а просто иллюстрируют способы, которые могут без труда использовать или адаптировать специалисты в данной области.
В общем случае, кальцимиметические и кальцилитические молекулы идентифицируют путем скрининга молекул, которые смоделированы на основе описанных ниже (названных основными молекулами). Как будет показано ниже, существуют несколько специфических кальцимиметиков и кальцилитиков для различных рецепторов Ca2+. Производные молекулы могут быть синтезированы и испытаны с помощью стандартных процедур специалистами. Множество молекул можно без труда подвергнуть скринингу с целью поиска наиболее подходящих для настоящего изобретения.
Органические катионные молекулы, которые имитируют или противостоят действию Ca2+ в других системах, содержат необходимые структуры или воздействия на рецепторы Ca2+. Рациональный дизайн других подходящих молекул включает изучение молекулы, являющейся кальцимиметиком или кальцилитиков, и затем модификацию структуры известной молекулы. Например, полиамины являются потенциальными кальцимиметиками, поскольку спермин имитирует действие Ca2+ в нескольких системах in vitro. Результаты показывают, что спермин действительно вызывает изменения [Ca2+]i и секреции ПТГ, напоминающие изменения, вызванные внеклеточными двух- и трехвалентными катионами (см. ниже). Эксперименты, приведенные ниже, имеют своей целью продемонстрировать, что эти явления, полученные со спермином, задействуют те же механизмы, что и внеклеточный Ca2+. Для того, чтобы осуществить это, определяли целый ряд физиологических и биохимических параметров, характеризующих активацию рецептора Ca2+. Те молекулы, которые имеют сходный эффект подходит для настоящего изобретения и могут быть открыты отбором или созданием молекул, имеющих строение, сходное со строением спермина. При нахождении другой подходящей молекулы всю процедуру отбора можно без труда повторить.
Для ясности, ниже приведен ряд специфических скрининговых процедур для определения таких подходящих молекул, которые активны в отношении рецептора Ca2+ клетки паращитовидной железы или которые действуют как агонисты или антагонисты клеточного ответа на изменения [Ca2+]. Эквивалентные исследования можно применять для молекул, активных в отношении других рецепторов Ca2+, или для тех, которые иным образом имитируют или противодействуют клеточным функциям, которые регулируются [Ca2+]. Эти анализы служат примерами процедур, полезных для поиска кальцимиметических молекул настоящего изобретения. Эквивалентные процедуры можно применять для поиска кальцилитических молекул путем скрининга молекул, наиболее антагонистических действию внеклеточного Ca2+. Исследования in vitro можно также применять, чтобы установить избирательность, насыщаемость и обратимость этих кальцимиметиков и кальцилитиков с помощью стандартных методик.
Процедура скрининга
В общих чертах, клетки паращитовидной железы крупного рогатого скота, нагруженные фура-2, сначала суспендировали в буфере, содержащем 0,5 мМ CaCl2. Испытуемое вещество добавляли в кювету в малом объеме (5-15 мкл) и отмечали любое изменение в сигнале флюоресценции. В кювете производили кумулятивные повышения концентрации испытуемого вещества до тех пор, пока не достигали некоторой определенной заранее концентрации или не замечали изменений флюоресценции. Если изменений флюоресценции не наблюдалось, молекулу признавали неактивной и не подвергали ее дальнейшим испытаниям. При начальных исследованиях, например, молекул полиаминного типа, молекулы испытывали при концентрациях 5 или 10 мМ. С тех пор как стало известно больше сильных молекул (см. ниже), предельные концентрации были снижены. Например, более новые молекулы испытывались при концентрациях до 500 мкМ или менее. Если при этих концентрациях не происходило изменений флюоресценции, молекулу считали неактивной.
Молекулы, вызывавшие повышения [Ca2+]i, подвергали дополнительным испытаниям. Двумя существенно важными характеристиками, позволяющими отнести молекулу к кальцимиметикам, являются мобилизация внутриклеточного Ca2+ и чувствительность к активаторам ПКС. Молекулы, вызывающие мобилизацию внутриклеточного Ca2+ ФМА-чувствительным образом, все без исключения являлись кальцимиметиками и ингибировали секрецию ПТГ. Если необходимо, можно выполнить дополнительные исследования, чтобы укрепить эти выводы. В типичном случае, все различные тесты на кальцимиметическую или кальцилитическую активность (см. выше) не выполнялись. Более того, если молекула вызывала мобилизацию внутриклеточного Ca2+ ФМА-чувствительным образом, ее сразу подвергали скринингу на клетках паращитовидной железы человека. Например, для определения EC50 выполняли изменения [Ca2+]i, а также для измерения способности молекулы ингибировать секрецию ПТГ в клетках паращитовидной железы человека, которые были получены от пациентов, подвергавшихся оперативному вмешательству, по поводу первичной или вторичной гиперфункции паращитовидной железы. Чем ниже EC50 или IC50, тем более сильной является молекула, как кальцимиметик или кальцилитик.
Измерения [Ca2+] i с фура-2 обеспечивает очень быстрый способ скрининга новых органических молекул на активность. В течение половины рабочего дня можно испытать 10 - 15 молекул и оценить их способность мобилизировать внутриклеточный Ca2+ (или неспособность). Можно оценить также чувствительность любого повышения [Ca2+]i к депрессии ФМА. Более того, один клеточный препарат может предоставить данные о [Ca2+]i, уровнях циклического АМФ, ИФ3 и секреции ПТГ. Типичная процедура заключается в том, чтобы нагрузить клетки фура-2 и затем разделить клеточную суспензию на две части; большинство клеток используют для измерения [Ca2+]i, а оставшиеся инкубируют с молекулами для оценки их воздействия на секрецию ПТГ и циклического АМФ. Благодаря чувствительности радиоиммунологического анализа для циклического АМФ и ПТГ, оба параметра можно определить в одной инкубационной пробирке, содержащей 0,3 мл суспензии (около 500 000 клеток). Измерения количества инозитфосфатов являются трудоемким этапом скрининга. Однако, ионообменные колонки с хлоридом (лучше, чем с формиатом) обеспечивают очень быстрый способ для скрининга на ИФ3 образованием, поскольку ротационное выпаривание (которое занимает около 30 ч) не требуется. Этот метод может пропустить около ста образцов за половину рабочего дня. Те молекулы, которые вызвали к себе интерес, исходя из измерений [Ca2+] i, циклического АМФ, ИФ3 и ПТГ, затем подвергают более тщательным испытания путем исследования образования различных инозитфосфатов и определения их изомерической формы посредством жидкостной хроматографии высокого разрешения.
Молекулы, представляющие интерес, найденные с помощью этих методик, затем оценивают на специфичность, например, посредством изучения их воздействия на [Ca2+] i в секретирующих кальцитонин C-клетках при использовании, например, линии МТС - 6-23 клеток крысы.
Последующее является иллюстрацией способов, подходящих для этих скрининговых процедур. Примеры типичных результатов для различных испытуемых кальцимиметических или кальцилитических молекул приведены на фиг. 2 - 34.
Подготовка клеток паращитовидной железы.
Паращитовидые железы получали от телят (12 - 15-недельного возраста) непосредственно после убоя на местной скотобойне и перевозили в лабораторию в ледяном буфере для клеток паращитовидной железы (БКП), содержащем (мМ): NaCl, 126; KCl, 4; MgCl2, 1: N-2-гидроксиэтилпиперазин-N'-2-этансульфонат натрия, 20; pH 7,4; глюкоза, 5,6 и различные количества CaCl2, например, 1,25 мМ. Паращитовидные железы человека получали от пациентов, подвергавшихся хирургическому удалению ткани паращитовидной железы по поводу первичной или уремической гиперфункции паращитовидной железы (ГП), далее с ними поступали так же, как с тканями телят. С желез обрезали лишний жир и соединительную ткань, затем измельчали ножницами на маленькие кусочки кубовидной формы со стороной приблизительно 2 - 3 мм. Разрозненные клетки паращитовидной железы получали путем обработки коллагеназой. Разрозненные клетки затем очищали центрифугированием в буфере Percoll. Получавшийся в результате препарат клеток паращитовидной железы был в значительной степени очищен от красных кровяных клеток, адипоцитов и капиллярной ткани, что определялось фазовоконтрастной микроскопией и окрашиванием Суданом черным B. Разрозненные и очищенные клетки присутствовали в виде маленьких групп, состоящих из 5 - 20 клеток. Жизнеспособность клеток, по результатам вытеснения трипанового синего или этидиум бромида, была обычно 95%.
Хотя клетки можно использовать для экспериментальных целей уже на этой стадии, их физиологические реакции (подавляемость секреции ПТГ и остаточные уровни [Ca2+]i) лучше после культивирования клеток в течение ночи. Первичная культура имеет также то преимущество, что клетки можно метить изотопами до почти изотопного равновесия, как это необходимо для исследования, включающих измерения метаболизма инозитфосфата (см. ниже). После очистки в градиентах Percoll клетки отмывали несколько раз в смеси 1:1. Ham's F12 модифицированной по способу Дульбекко среде Игла (GIBCO) с добавленными 50 мкг/мл стрептомицина, 100 ед/мл пенициллина, 5 мкг/мл гентамицина и ИТС+. ИТС+ - это заранее смешанный раствор, содержащий инсулин, трансферрин, селен и бычий сывороточный альбумин (БСА)-линоленовую кислоту (Collaborative Research, Bedford, MA). Клетки затем переносили в пластиковые флаконы (75 или 150 см2; Falcon) и инкубировали при 37oC во влажной атмосфере с содержанием 5% CO2. К этим ночным культурам не добавляли сыворотку, поскольку ее присутствие позволяет клеткам прикрепиться к пластику, претерпевать пролиферацию и дедифференцироваться. Клетки, культивируемые при таких условиях, легко удалялись из флаконов сцеживанием и демонстрировали ту же жизнеспособность, что и свежеприготовленные клетки.
Измерение цитозольного Ca2+
Очищенные клетки паращитовидной железы ресуспендировали в 1,25 мМ CaCl2 - 2% БСА-БКП, содержащей 1 мкМ фура-2-ацетоксиметилового сложного эфира и инкубировали при 37oC в течение 20 мин. Полученный после центрифугирования осадок с клетками ресуспендировали в том же буфере, но без сложного эфира, и инкубировали еще 15 мин при 37oC. Клетки затем дважды отмывали БКП, содержащим 0,5 мМ CaCl2 и 0,5% БСА и содержали при комнатной температуре (около 20oC). Немедленно перед использованием клетки разводили в пять раз предварительно подогретым 0,5 мМ CaCl2-БКП для достижения конечной концентрации БСА 0,1%. Концентрация клеток в кювете для исследования флуоресценции была 1 - 2 • 106мл.
Флюоресценцию нагруженных индикатором клеток измеряли при 37oC на спектрофлюориметре (Biomedical Instrumentation Group, University of Pennsylvania, Philadelphia, PA), оборудованным обогреваемым держателем кюветы, и магнитной мешалкой, использующей длины волн намагничивания и излучения 340 и 510 нм соответственно. Эта флюоресценция показывает уровень цитозольного Ca2+. Сигналы флюоресценции калибровали дигитонином (50 мкг/мл, конечная концентрация) для получения максимума флюоресценции и ЭГТУ (10 мМ, pH 8,3, конечная концентрация) для получения минимальной флюоресценции (Фмакс, и Фмин) и константа диссоциации составляла 224 нМ. Утечка красителя зависела от температуры и большая ее часть наблюдалась в первые две минуты после подогревания в кювете; затем утечка красителя возрастала очень медленно. Чтобы откорректировать калибрование по утечке красителя, клетки помещали в кювету и перемешивали при 37oC 2 - 3 мин. Потом клеточную суспензию центрифугировали, а надосадочную жидкость возвращали в чистую кювету. Надосадочную жидкость затем обрабатывали дигитонином и ЭГТУ, как описано выше, чтобы получить стандарт для оценки утечки красителя, который обычно составлял 10 - 15% от общего Ca2+-зависимого флюоресцентного сигнала. Эту оценку затем вычитали из полученных значений Фмин.
Измерение секреции ПТГ
В большей части экспериментов, клетки, нагруженные фура-2, использовали для изучения секреции ПТГ. Нагруживание клеток паращитовидной железы фура-2 не изменяет их секреторного ответа на внеклеточный Ca2+. Клетки суспендировали в БКП, содержащем 0,5 мМ CaCl2 и 0,1% БСА. Инкубировали клетки в пластиковых пробирках (Falcon 2058), содержащих 0,3 мл клеточной суспензии с малыми объемами CaCl2 или органических поликатионов или без них. После инкубации при 37oC в течение различных отрезков времени (обычно 30 мин) пробирки помещали в лед и клетки собирали в осадок центрифугированием при 2oC. Образцы надосадочной жидкости доводили до pH 4,5 уксусной кислотой и хранили при -70oC. Эту процедуру использовали для клеток паращитовидной железы как человека, так и крупного рогатого скота.
Для клеток крупного рогатого скота количество ПТГ в образце надосадочной жидкости определяли гомологичным радиоиммунологическим исследованием с использованием GW-1 антитела или его эквивалента при финальном разведении 1/45 000. 1251-ПТГ (65-84, INC-STAR, Stillwater, MN) использовали в качестве радиоизотопного индикатора, а фракции разделяли с помощью нагруженной декстраном угольной пыли. Счет образцов выполняли на Packard Cobra 5005 счетчике гамма-излучения.
Для клеток человека использовали коммерчески доступный набор для радиоиммуноанализа (INS-РТН; Nichols Institute, Los Angeles, CA), который распознает интактный и N-терминальный ПТГ человека, поскольку антитела GW-1 плохо распознают ПТГ человека.
Измерение циклического АМФ
Клетки инкубировали, как описано выше для исследований секреции ПТГ и в конце инкубации брали 0,15 мл образца и переносили в 0,85 мл горячей (70oC) воды и поддерживали при этой температуре 5 - 10 мин. Затем пробирки замораживали и оттаивали несколько раз и клеточные остатки осаждали центрифугированием. Порции надосадочной жидкости ацетилировали и концентрации циклического АМФ определяли радиоиммуноанализом.
Измерение образования инозитфосфата
Фосфолипиды мембраны метили путем инкубации клеток паращитовидной железы 4 мкCi/мл 3H-мио-инозитом в течение 20 - 24 ч. Затем клетки отмывали и ресуспендировали в БКП, содержащем 0,5 мМ CaCl2 и 0,10 БСА. Инкубирование производили в микроцентрифужных пробирках в присутствии или отсутствии различных концентраций органического поликатиона в течение различных отрезков времени. Реакции прекращали добавлением 1 мл смеси хлороформ/метанол/12 NHCl (200510051; по объему). Затем добавляли кислый гидролизат фитина (200 мкл; 25 мкг фосфата на пробирку). Пробирки центрифугировали и 600 мкл водной фазы разводили в 10 мл воды.
Инозитфосфаты отделяли путем ионообменной хроматографии с использованием AG1-X8 как с хлоридом, так и с формиатом. Когда нужно было определить только уровни ИФ3, использовали хлорид, тогда как формиат использовали для сепарации общего количества инозитфосфатов (ИФ3, ИФ2 и ИФ1). Для определения ИФ3 разведенный образец помещали на колонку с хлоридом и отмывали 10 мл 30 мМ HCl, затем 6 мл 90 мМ HCl и ИФ3 элюировали 3 мл 500 мМ HCl. Последний элюат разводили и подсчитывали. Для определения общего количества инозитфосфатов разведенный образец помещали на колонку с формиатом и ИФ1, ИФ2 и ИФ3 элюировали последовательно путем повышения концентрации формиатного буфера. Элюированные образцы их формиатных колонок подвергали ротационному выпариванию, остатки смешивали и подсчитывали.
Изомерические формы ИФ3 оценивали посредством жидкостной хроматографии высокого разрешения. Реакции прекращали добавлением 1 мл 0,45 М хлорной кислоты и содержали во льду 10 мин. После центрифугирования надосадочной жидкости доводили до pH 7 - 8 с помощью NaHCO3. Экстракт затем помещали на анионообменную колонку Partisil SAX и элюировали линейным градиентом формиата аммония. Различные фракции затем обессоливали с помощью Dowex и подвергали ротационному выпариванию перед сцинтилляционным подсчетом на Packard Tricarb 1500 LSC.
Для всех методов сепарации инозитфсофатов применяли соответствующие контроли с использованием аутентичных стандартов, чтобы определить, не создают ли органические поликатионы помехи для сепарации. На тот случай, если это действительно так, образцы обрабатывали катионообменной смолой для удаления мешающих молекул до того, как приступить к отделению инозитфосфатов.
Измерения цитозольного Ca2+ в C-клетках
Неопластические C-клетки, выделенные из карциномы мозгового вещества щитовидной железы крысы (rМТС 6-23), получили из Американской коллекции культур тканей (АТСС N 1607) и культивировали монослоем в модифицированной по способу Дульбекко среде Игла (МДСИ) с добавлением 15% лошадиной сыворотки в отсутствии антибиотиков. Для измерения [Ca2+]i клетки собирали с помощью этилендиаминтетрауксусной кислоты (ЭДТК) с 0,05% трипсина, дважды отмывали БКП, содержащим 1,25 мМ CaCl2 и 0,5% БСА, и нагружали фура-2, как описано выше для клеток паращитовидной железы. Измерения [Ca2+]i выполняли, как описано выше, с соответствующей корректировкой на утечку красителя.
Измерения [Ca2+]i в остеокластах крысы
Остеокласты получали от 1 - 2-дневных крысят Sprague-Dawley в асептических условиях. Крысят умерщвляли декапитацией, задние лапы удаляли, бедренные кости быстро очищали от мягких тканей и помещали в предварительно подогретую Ф-12/МДСИ (МДСИ, содержащую 10% сыворотки теленка и антибиотики (пенициллин-стрептомицин-гентамицин; 100 ед/мл - 100 мкг/мл - 100 мкг/мл)). Кости от двух крысят разрезали вдоль и помещали в 1 мл культуральной среды. Костные клетки получали посредством осторожного измельчения фрагментов костей пластиковой пипеткой и разводили культуральной средой. Костные фрагменты оседали, а равные порции (около 1 мл) среды переносили в 6-луночный лоток для культивирования, содержащий 25 мм покровные стекла. Клеткам давали осесть в течение 1 ч при 37oC в увлаженной атмосфере с 5% CO2. Покровные стекла затем трижды отмывали свежей средой для удаления незакрепившихся клеток. Измерения [Ca2+]i в остеокластах производили в течение 6 - 8 ч после удаления незакрепившихся клеток.
Клетки, закрепившиеся на покровных стеклах, нагружали индо-1 посредством инкубации с 5 мкМ индо-1 ацетоксиметиловым сложным эфиром/0,01% Pluronic F 28 в течение 30 мин, при 37oC в Ф-12/МДСИ, не содержащей сыворотки, а взамен содержащей 0,5% БСА. Покровные стекла затем отмывали и инкубировали дополнительно 15 мин при 37oC в Ф-12/МДСИ, не содержащей сложного эфира, перед помещением в орошающую камеру, смонтированную на столике Nikon Diaphot инвертированного микроскопа, оборудованного для микрофлюориметрии. Остеокласты легко идентифицировать по их большим размерам и присутствию множества ядер. Клетки орошали буфером (обычно БКП, содержащим 0,1% БСА и 1 мМ Ca2+) со скоростью 1 мл/мин, с испытуемым веществом или без него. Флюоресценция, испускаемая на 340 нм, направлялась через обзорное отверстие микроскопа на 440 нм дихроическое зеркало и интенсивность флюоресценции на 495 и 405 нм собиралась в трубки фотоэлектрического умножителя на выходе усиливали, переводили в цифровую форму и хранили в 80386 РС. Соотношения интенсивности флюоресценции применяли для оценки Ca2+.
Экспрессия в ооцитах
В дополнительных исследованиях в ооциты Xenopus вводили мРНК из клеток паращитовидной железы крупного рогатого скота или человека и использовали в скрининговых процедурах, а проводимость Cl-1 измеряли как способ косвенной оценки повышенной [Ca2+]i. Ниже в качестве примера проводится исследование эффекта неомицина. Ооциты получали мРНК, обогащенную поли(A+), из ткани паращитовидной железы человека (гиперпластические железы от случая вторичной ГП). Спустя 3 дня ооциты испытывали на ответ неомицину. Неомицин B вызывал колеблющиеся повышения проводимости Cl-1, которые исчезали при орошении не содержащим лекарства физиологическим раствором (см. фиг. 20). Ответные реакции на неомицин B наблюдались при концентрациях между 100 мкМ и 10 мМ. Чтобы убедиться в том, что ответная реакция, вызванная неомицином B, зависела от введения мРНК паращитовидных клеток, было изучено воздействие неомицина B на токи в ооцитах с введенной водой. В каждом из испытанных пяти ооцитов, неомицин B (10 мМ) не смог вызвать каких-либо изменений тока. Известно, что приблизительно 40% ооцитов реагирует на карбахол, благодаря эффекту, опосредованному эндогенным мускариновым рецептором. В пяти испытаниях ооцитах, один показал внутренние токи в ответ на карбахол, и это показано на нижней записи фиг. 20. Таким образом, в клетках, экспрессирующих мускариновый рецептор, связанный с повышениями [Ca2+]i и проводимостью Cl-, неомицин B не смог вызвать ответной реакции. Это свидетельствует о том, что ответная реакция на неомицин B зависит от экспрессии специфичного белка, который кодирует мРНК клетки паращитовидной железы. Это обстоятельство настойчиво наводит на мысль, что в интактных клетках неомицин B действует непосредственно на рецепторы Ca2+ и измеряет функцию клеток паращитовидной железы.
Приготовление лекарства из основных молекул.
В настоящем документе демонстрируются определенные органические молекулы, которые имитируют или противодействуют воздействиям внеклеточного Ca2+ путем воздействия на рецепторы Ca2+. Испытанные молекулы, однако, вовсе необязательно подходят в качестве потенциальных лекарств, однако они служат примером того, что гипотеза лекарственных средств, действующих на рецепторы Ca2+, является правильной. Эти молекулы можно использовать для определения особенностей строения, которые дают им возможность действовать на рецепторы Ca2+, и таким образом, отбирать молекулы, подходящие для целей настоящего изобретения.
Приводим следующий пример такой аналитической процедуры:
Этот пример подробно описан ниже и используется здесь только для демонстрации логического обоснования, которое можно применять для разработки подходящих молекул настоящего изобретения на основе основных молекул, обсуждаемых в настоящем документе. Специалисты в данной области легко поймут этапы, обозначенные в примере, и аналогичный анализ можно проводить в отношении других ведущих молекул, пока не будут разработаны желательные кальцимиметики или кальцилитики.
Остальные примеры также приводятся ниже. Вместе представленные данные показывают, что подходящие основные молекулы будут иметь ароматические группы, предпочтительно замещенные в одном или более положений, и могут иметь неразветвленные или разветвленные замещенные или незамещенные алкильные группы. В добавление можно сказать, что важно выбирать молекулы с правильной стереоспецифичностью, чтобы обеспечить большее сродство к рецептору Ca2+. Таким образом, эти сведения указывают на подходящие молекулы, от которых можно получать производные, чтобы найти оптимальные молекулы настоящего изобретения, во многом, как описано ниже.
Хотя испытуемые молекулы имеют структурные различия, они обладают также общими чертежами строения, которые могут быть изучены. В этом примере, коррекция между суммарным положительным зарядом и потенцией в мобилизации внутриклеточного Ca2+ были тщательно изучена. Протамин (+21; EC50=40 нМ) был более эффективным, чем неомицин B (+6; EC50 = 20 мкМ в клетках паращитовидной железы человека и 40 мкМ в клетках паращитовидной железы крупного рогатого скота), который, в свою очередь, был эффективнее, чем спермин (+4; EC50 = 150 мкМ) в вызывании мобилизации [Ca2+]i в клетках паращитовидной железы. Эти результаты ставят вопрос о том, определяет ли только положительный заряд потенцию молекулы или осуществляют другие особенности строения, вносящие свою лепту в активность в отношении рецептора Ca2+. Это важно определить в самом начале, поскольку это предположение серьезно сталкивается с точкой зрения, что рецептор Ca2+ может служить мишенью для эффективных и специфичных терапевтических молекул. Таким образом, целый ряд других органических поликатионов, родственных неомицину B и спермину, можно изучать, чтобы определить взаимосвязь между суммарным положительным зарядом молекул и ее способностью вызывать мобилизацию внутриклеточного Ca2+.
Первым рядом изучения молекул были аминогликозиды. Молекулы испытывали на клетках пароащитовидной железы крупного рогатого скота и определяли их EC50 для мобилизации внутриклеточного Ca2+. Для аминогликозидов порядок способности вызывать кратковременные повышения цитозольного Ca2+ был следующим: неомицин B (EC50 = 20 или 40 мкМ) > гентамицин (150 мкМ) > бекамицин (200 мкМ) > стрептомицин (600 мкМ). Канамицин и линкомицин не оказывали действия при концентрации 500 мкМ. Суммарный положительный заряд этих аминогликозидов при pH 7,3 был следующий: неомицин B (+6) > гентамицин (+5) = беканамицин (+5) > канамицин (средний +4,5) > стрептомицин (+3) > линкомицин (+1). Так, внутри аминогликозидного ряда существует некоторая коррекция между суммарными положительными зарядами, но она не является абсолютной, и канамицин, в отношении которого можно предсказать, что он будет активнее стрептомицина, вообще не обладал активностью.
Испытывая различные полиамиды, мы выявили дополнительные и более выраженные расхождения между суммарным положительным зарядом и потенцией. Были испытаны три структурных класса полиаминов: (1) с прямой цепью, (2) с разветвленной цепью и (3) циклические. Строение испытанных полиамидов приведено на фиг. 1. Среди полиамидов с прямой цепью спермин (+4; EC50 = 150 мкМ) оказался более активным, чем пентаэтиленгексамин (+6; EC50 = 500 мкМ) и тетраэтиленпентамин (%5; EC50 = 2,5 мкМ), несмотря на то, что две последние молекулы имели больший положительный заряд.
Мы синтезировали некоторые разветвленные полиамины, которые имели различное количество вторичных и первичных аминогрупп и, таким образом, различались по суммарному положительному заряду. Две из эти молекул, NPS, 381 и NPS 382, испытывали на действие на [Ca2+]i в клетках паращитовидной железы крупного рогатого скота. NPS 382 (+8; EC50 = 50 мкМ) была в два раза активнее, чем NPS 381 (+10; EC50 = 100 мкМ), несмотря на то, что у нее на два положительных заряда меньше.
Сходное расхождение между величиной положительного заряда и активностью было отмечено в экспериментах, с циклическими полиаминами. Например, гексациклен (+6; EC50 = 20 мкМ) был активнее, чем NPS 383 (+8; EC50 = 150 мкМ). Результаты, полученные с этими полиаминами, показывают, что положительный заряд не является единственным фактором, обуславливающим активность.
Дополнительные исследования проникают в суть структурных особенностей молекул, которые сообщают им активность по отношению к рецепторам Ca2+ клеток паращитовидной железы. Одной из важных особенностей строения является внутримолекулярное расстояние между атомами азота (которые несут положительный заряд). Так, спермин в 50 раз активнее, чем триэтилентетрамин (EC50 = 8 мМ), вызывает повышения [Ca2+]i в клетках паращитовидной железы крупного рогатого скота, хотя обе молекулы несут положительный заряд, равный +4. Единственным различием в структуре этих двух полиаминов является количество метиленовых групп, разделяющих атомы азота: в спермине 3-4-3, в то время как к триэтилентетрамине 2-2-2. Это, кажущееся небольшим, изменение расстояния между атомами азота в очень большой степени причастно к активности и предполагает, что эти структурные взаимоотношения между атомами азота в молекуле являются решающими. Этот вывод подтверждают данные, полученные в экспериментах с гексацикленом и пентаэтиленгексамином. Первая молекула является просто циклическим аналогом последней и содержит то же количество метиленовых групп между всеми атомами азота, однако присутствие кольцевой структуры увеличивает активность в 25 раз. Эти результаты указывают на то, что положительный заряд по себе не является решающим фактором, определяющим активность органической молекулы по отношению к рецептору Ca2+.
Другая серия экспериментов обнаружила, что ароматические группы важны для активности по отношению к рецепторам Ca2+. Эти результаты были получены с двумя арилалкиламинами, выделенными из яда паука Argiope lobata. Эти молекулы, аргиотоксин 636 и аргиотоксин 659, имели идентичные поликатионные части, связанные с различными ароматическими группами (фиг. 24). Агриотоксин 659 вызывал кратковременные повышения [Ca2+]i в клетках паращитовидной железы крупного рогатого скота при испытаниях с концентрациями 100 - 300 мкМ. Напротив, арготиоксин 636 был неэффективным при этих концентрациях (фиг. 24). Единственным различием в строении между этими двумя арилалкиламинами является ароматическая часть молекул: аргиотоксин 659 содержит 4-гидроксииндол, в то время как аргиотоксин 363 содержит 2,4-дигидроксифенильную группу. Суммарный положительный заряд у этих двух арилалкиламинов одинаковый (+4), поэтому их разная активность может быть результатом разных ароматических групп. Это свидетельствует о то, что суммарный положительный заряд один не определяет активность. Однако реальная важность этих данных состоит в открытии того факта, что ароматические группы вносят существенный вклад в способность молекул активировать рецептор Ca2+.
Агатоксин 489 (NPS 017) и Агатоксин 505 (NPS 015) оба вызывают мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы с EC50 6 и 22 мкМ соответственно. Единственным различием в строении этих молекул гидроксильная группа в индольной части (фиг. 1). Это показывает, что замещения в ароматической части молекул может повилять на активность. Это означает, что дополнительные основные молекулы, которые нужно изучить, будут включать молекулы с замещенными ароматическими долями.
Структурные черты, которые систематически разнятся у основных молекул, описанных здесь, включают (1) суммарный положительный заряд, (2) количество метиленовых групп, разделяющих атомы азота, и (3) циклические варианты, например, полиаминов, имеющих или не имеющих изменений в шаге метиленовых групп и суммарном положительном заряде. Дополнительно можно исследовать систематические вариации в структуре и локализации ароматических групп, например, в ряду арилалкиламинов, выделенных из яда ос и пауков; и можно приготовить синтетические молекулы, соединяя коммерчески доступные ароматические доли и полиамидную долю аргиотоксина. Полиаминную долю аргиотоксина можно без труда присоединить к любой ароматической доле, содержащей карбоновую кислоту. Таким образом, нетрудно систематически отбирать гидрокси- и метоксипроизводные фенилуксусной кислоты и бензойной кислоты, а также гидроксииндолуксусной кислоты. Аналоги, включающие гетероароматические соединения с функциональными группами, также можно изготовить и оценить на предмет активности.
Сравнения возможностей и эффективности подобных молекул откроют оптимальную структуру и локализацию ароматической группы при постоянном положительном заряде.
Одной из структурных вариаций полиаминов, которая, как представляется, способна повысить активность, является присутствие циклической версии родительской молекулы с прямой цепью. Будмуншиамин A, изолированный из растения Apbizia amara, является циклическим производным спермина (фиг. 1). Добавление будмуншиамина A к клеткам паращитовидной железы крупного рогатого скота вызывало быстрое и кратковременное повышение [Ca2+]i , которое сохранялось в отсутствии внеклеточного Ca2+ и притуплялось предварительной обработкой ФМА. Следовательно, он вызывает мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы, возможно, воздействуя на рецепторы Ca2+. Он обладает почти равной активностью со спермином (EC50 около 200 мкМ), хотя его положительный заряд на единицу меньше (+3), чем у спермина.
Результаты, полученные с будмуншиамином A, демонстрируют предсказывающие возможности исследования структура-активность, и необходимо получать новую информацию о структуре путем испытаний натуральных продуктов. Так, при скрининге натуральных продуктов, рационально подобранных на основе уже полученной информации о структуре, можно, например, отобрать молекулы на основании тщательно установленных хемотаксономических принципов, используя подходящие базы данных, такие как Napralert. Например, макроциклические полиамидные алкалоиды, выделенные из мотыльковоподобных (papilionoid) бобовых растений, родственных Albiria, таких как Pithecdobium и другие молекулы, полученные из растений, можно подвергнуть скринингу.
Фиг. 36 приводит второй пример серии молекул, подвергавшихся скриннингу, для определения подходящих для целей настоящего изобретения молекул. Это молекулы были, в основном, производными фендилина и испытывались для определения их EC50. Кроме того, тестирование родственных молекул, таких как NPS 447 и NPS 448, открыло стереоспецифичные эффекты молекулярной структуры. Наиболее активными соединениями, испытанными к настоящему моменту, являются новые соединения, обозначенные NPS 467 и NPS 568, которые имеют величины EC50 менее 5 мкМ. Специалисты в данной области, просматривая эти серии молекул, могут установить другие подходящие производные, которые можно испытывать в рамках настоящего изобретения.
Эти примеры демонстрируют общий вид и процесс скрининга, полезные для настоящего изобретения, и указывают, что другие дополнительные хранилища соединений и натуральных продуктов можно сделать источником для скрининга, чтобы определить другие подходящие основные молекулы или новые молекулы настоящего изобретения.
Как обсуждалось выше, примерами молекул, подходящих в качестве кальцимиметиков, могут служить разветвленные или циклические полиамины, положительно заряженные полиаминокислоты и арилалкиламины. Вдобавок другие положительно заряженные органические молекулы, включающие естественно встречающиеся молекулы и их аналоги, являются подходящими кальцимиметиками. Эти естественно встречающиеся молекулы и их аналоги предпочтительно имеют положительные соотношения заряд:масса, которые коррелируют с такими соотношениями для молекул, приведенных в качестве примеров в настоящем документе. (Примеры включают материалы, выделенные от морских животных, из ядов членистоногих, наземных растений и культуральных сред, на которых росли бактерии и грибы). Предполагается, что одна группа предпочтительных естественно встречающихся молекул и аналогов, подходящих в качестве кальцимиметиков, будет иметь соотношение положительный заряд:молекулярный вес (в дальтонах) от приблизительно 1: 40 до 1:200, предпочтительно от приблизительно 1:40 до 1:100. Более специфические примеры таких молекул приводятся ниже.
Полиамины.
Полиамины, подходящие в качестве кальцимиметиков настоящего изобретения, могут быть циклическими, а также нециклическими с разветвленной цепью. Разветвленные или циклические полиамины потенциально обладают более высокой кальцимиметической активностью, чем их аналоги с неразветвленными цепями. То есть, разветвленные или циклические полиамины имеют тенденцию показывать более низкие величины EC50, чем их соответствующие линейные полиамины с тем же эффективным зарядом при физиологическом уровне PH (см. таблицу 1).
Термин "разветвленные полиамины" относятся к молекуле, построенной в виде цепи, состоящей из коротких алкильных мостиков или алкильных групп, соединенных друг с другом аминными связями, а также содержащей пункты, в которых цепь ветвится. Такие "пункты ветвления" могут быть расположены как на атоме углерода, так и на атоме азота, предпочтительно, на атоме азота. Атом азота пункта ветвления, в типичном случае, третичный амин, но также может быть и четвертичным. Разветвленный полиамин может иметь от 1 до 20 пунктов ветвления, предпочтительно от 1 до 10 пунктов ветвления.
В основном, алкильные мостики и алкильные ветви в разветвленном полиамине имеют в длину от 1 до 50 атомов углерода, предпочтительно от 2 до 6 атомов углерода. Алкильные ветви могут также прерываться одним или более гетероатомов (азот, кислород или сера) или замещаться функциональными группами, как-то: гало, включая фторо, хлоро, бромо, иодо; гидрокси; нитро; ацилокси (R'COO-), ациламидо (R'CONH-), алкокси (-OR'), где R' может состоять из 1 до 4 атомов углерода. Алкильные ветви также могут замещаться группами, которые заряжена положительно при физиологическом уровне pH, такими как амино или гуанидо. Эти функциональные заместители могут усиливать или изменять физические свойства, такие как растворимость, повышая активность, доставку или биологическую доступность молекул.
Разветвленные полиамины, могут иметь три или более цепей и точки окончания ветвления. Эти конечные точки могут быть метильными группами или аминогруппами, предпочтительно аминогруппами.
Одной предпочтительной группой молекул является группа разветвленных полиаминов с формулой:
H2N-(CH2)j-(NRi- (CH2)j)k-NH2,
где k целое число от 1 до 10, каждое j - то же или отличается и является целым от 2 до 20, каждое Ri - то же или отличается и избирается из группы, состоящей из водорода и -(CH2)j-NH2, где j определено выше, и по крайней мере одно Ri не является водородом.
Особенно предпочтительными разветвленными полиаминами настоящего изобретения являются молекулы N1, N1, N5, N10, N14, N14 - гексакис-(3-аминопропил)спермин и N1, N1, N5, N14, N14 - тетракис-(3-аминопропил)спермин, обозначенные как NPS 381 и NPS 382, соответственно, на фиг. 1.
"Циклические полиамины" относятся к гетероциклам, содержащим два или более гетероатомов (азот, кислород или сера), по крайней мере два из которых являются атомами азота. В основном, гетероциклы состоят из 6 - 20 атомов в кольцах, предпочтительно от 10 до 18. Гетероатомы азота разделены 2 - 10 атомами углерода. Гетероциклы также могут быть замещены по азоту аминоалкильными или аминоарильными группами (NR2R-), в которых R - аминоарил или низший алкил, состоящий из 2 - 6 атомов углерода.
Особенно предпочтительны циклические полиамины настоящего изобретения, показанные на фиг. 1, такие как гексациклен (1,4,7,10,13,16-гексааза-циклооктадекан) и NPS 383.
Полиаминокислоты.
Полиаминокислоты, подходящие для настоящего изобретения, могут содержать два или более положительно заряженных аминокислотных остатков при физиологическом уровне pH. Эти положительно заряженные аминокислоты включают гистидин, лизин и аргинин. Эти полипептиды варьируют по длине от 2 до 800 аминокислот, более предпочтительно от 20 до 300 аминокислот. Эти полипептиды могут состоять из одного повторяющегося аминокислотного остатка, или обладать разнообразием естественно встречающегося белка.
Аминокислотные остатки, составляющие полиаминокислоты, могут быть любыми из двадцати естественно встречающихся аминокислот, или других альтернативных остатков. Альтернативные остатки включают, например, ω- аминокислоты формулы H2N(CH2)nCOOH, где n равно числу от 2 до 6. Это нейтральные неполярные аминокислоты, такие как саркозин, t-бутилаланин, t-бутилглицин, t-метилизолейцин, норлейкин, фенилглицин, цитруллин, метионин сульфоксид, циклогексилаланин и гидроксипролин. Орнитин является альтернативным положительно заряженным аминокислотным остатком. От полиаминокислот настоящего изобретения также можно получать химические производные с помощью известных специалистам способов.
Особенно предпочтительные полиаминокислоты настоящего изобретения включают полиаргинин, полилизин, и поли(аргинил-тирозин), состоящие из 20 - 300 остатков. Другой предпочтительной полиаминокислотой является протамин или аналог протамина.
Арилалкиламины.
"Арилалкиламины" в настоящем документе относятся к классу положительно заряженных токсинов, выделенных из яда членистоногих. Предпочтительные арилалкиламины настоящего изобретения включают филантотоксин-433, аргиотоксин-636, и аргиотоксин-659, агатоксин-505, агатоксин 489, строение которых показано на фиг. 1, и другие синтетические молекулы, смоделированные на основе этих натуральных продуктов, такие как NPS 019.
Композиция настоящего изобретения включают не только по крайней мере один кальцимиметик или кальцилитик, но также могут включать определенные дополнительные компоненты. Эти дополнительные компоненты включают нацеливающие компоненты, метки и другие функциональные вещества, которые могут быть полезными в настоящей заявке, например, для скрининга агонистов или антагонистов внеклеточного Ca2+.
Например, иммуноглобулин или его часть или лиганд, специфичный для клеток паращитовидной железы или рецепторов Ca2+, можно использовать как специфичный нацеливающий компонент. Иммуноглобулин может быть поликлональным или моноклональным антителом и может включать целые антитела или иммунологически реактивные фрагменты этих антител, такие как Fab, Fав′ , или ( Fав′ )2. Также можно использовать рецептороспецифичные лиганды.
Композиции настоящего изобретения могут также содержать компоненты, преобразованные с помощью молекулы или иона, которые действуют как метки. Можно использовать целый ряд меток, включая радиоизотопы, хромофоры и флюоресцентные метки. В частности, мечение радиоизотопами без труда может улавливаться in vivo. Радиоизотопы можно присоединять координационной связью в качестве катионов в порфириновой системе. Подходящие катионы включают технеций, галлий и индий. В композициях положительно заряженная молекула может быть связана или ассоциирована с меткой.
Способы синтеза.
Стратегия для синтезов и модификации полиаминов включает использование целого ряда аминоблокирующих групп (фталимидо, БОК, CBZ, бензил и нитрил), которые можно выборочно удалять для построения конструктивной молекулы. Методы синтеза, применяемые в настоящем изобретении, смоделированы на основе тех, которые использовали для построения аргиопинов 636 и 659 и других арилалкиламинов, полученных из ядов пауков.
Цепи длиной 2 - 4 метиленовые группы обычно выполнялись алкилированием с соответствующим N-(бромоалкил)фталимидом. Смесь амина и бромоалкилфталимида 1: 1,2 нагревался с вертикальным холодильником в ацетонитриле в присутствии 50% KF на Celite. Цепи выполнялись также алкилированием данного амина акрилонитрилом или этилакрилатом. Ход реакции прослеживали с помощью тонкослойной хроматографии и промежуточных соединений, очищенных в силикагеле с применением комбинаций дихлорметана, метанола и изопропиламина. Конечные продукты очищали катионным обменом (HEMA-SB) и жидкостной хроматографией высокого разрешения с обращенной фазой (Vydac C-18). Проверку чистоты и структуры выполняли с помощью 1H- и 13C-ядерного магнитного резонанса и масс-спектрометрией высокого разрешения (EI, Cl и/или FAB).
БОК-блокирующие группы добавляли воздействием на амин (1o или 2o) ди-трет-бутил-дикарбонатом в дихлорметане в присутствии каталитического количества диметиламинопиридина. Бензил-блокирующие группы получали одним из двух путей: (1) конденсация 1o амина с бензальдегидом с последующим восстановлением борогидридом натрия или (2) алкилирование 2o амина бензилбромидом в присутствии KF. Амидные связи и циклизации обычно выполняли реакцией амина (1o или 2o) с N-гидроксисукцинимидэфиром данной кислоты. Это выполнялось непосредственно (в случае циклизаций) воздействием на "аминокислоту" дициклогексилкарбодиимидом в разбавленном виде.
Разблокировка фталимидо функциональности выполнялась восстановлением гидразином в обратном токе метанола. Разблокировки БОК функциональности выполняли в безводной трифторуксусной кислоте (TFA в тексте). Разблокировка бензил-, нитрил- и CBZ блокирующих функциональностей выполняли восстановлением в ледяной уксусной
кислоте при 55psi водорода в присутствии каталитического количества гидроксида палладия на угле. Нитрильные функциональности (в присутствии бензила и CBZ-групп) выборочно восстанавливали под водородом в присутствии пористого Raney никеля.
Конкретно, разветвленные полиамины в типичном случае приготавливали из простых диаминоалканов формулы NH2-(CH2)n-NH2 или простых полиаминов, таких как спермидин или спермин. Один из двух первичных (терминальных) аминов защищен или "замаскирован" блокирующей группой, такой как БОК (t-бутилоксикарбонил), фталимидо, бензил, 2-этилнитрил (продукт из амина и акрилонитрила Michael condensation production) или амид. Типичная реакция - добавление БОК блокирующей группы воздействием ди-t-бутил-дикарбоната (БОК ангидрид):
Моноблокированный продукт отделяется от неблокированного и ди-блокированного продуктов простыми методиками хроматографии или дистилляции.
Оставшийся свободный амин в моноблокированном продукте затем выборочно алкилируют (или ацилируют) алкилирующим (или ацилирующим) агентом. Чтобы гарантировать моноалкилирование, свободный амин частично защищен конденсацией с бензальдегидом с последующим восстановлением борогидратом натрия для образования N-бензил производного:
N-бензил производное затем реагирует с алкилирующим агентом. Типичным алкилирующим агентом является N-(бромоалкил)фталимид, который реагирует следующим образом:
Например, N-(бромбутил)фталимид применяется, чтобы удлинить или разветвить цепь с четырьмя единицами метилена. Напротив, реакция с акрилонитрилом с последующим восстановлением цианогруппы удлинит цепь тремя метиленами и аминогруппой.
Блокирующие группы получившейся молекулы с удлиненной цепью затем можно выборочно удалить для получения нового свободного амина. Например, трифторуксусная кислота используется для удаления БОК группы; каталитическое гидрирование используют для восстановления нитрильной функциональности и удаления бензила; гидразин используют для удаления фталимидо групп следующим образом:
Новый свободный амин можно алкилировать (или ацилировать) далее, как описано выше, для увеличения длины полиамина. Этот процесс повторяют, пока не будет достигнута желаемая длина цепи и количество ветвей. На заключительном этапе при разблокировании продукта получается желаемый полиамин. Однако до разблокирования, можно произвести дополнительные модификации на блокированном конце, следующим образом.
Например, до БОК-разблокирования, полиамин ацилируют N-гидроксисукцинимидэфиром, 3,4-диметоксифенилуксусной кислоты для получения диблокированного полиамина: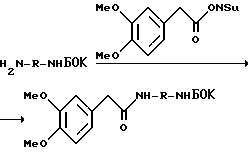
Это в конечном счете приведен к образованию арилполиамина. БОК группа затем может быть выборочно удалена трифторуксусной кислотой для освобождения другого аминоконца, который можно наращивать, как описано выше.
Некоторые разветвленные полиамины могут быть получены одновременным алкилированием или ацилированием свободных первичного и вторичного аминов в полиамине, полученном, как описано выше. Например, в результате действия на спермин избытка акрилонитрила с последующим каталитическим восстановлением, получается следующее: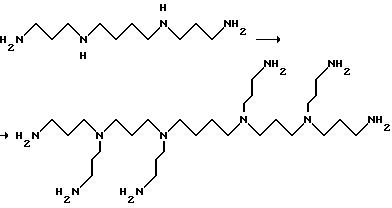
Циклические полиамины можно приготовить, как описано выше, начиная с исходных материалов, таких как гексациклен (Aldrich Chem).
Полиаминокислоты в рамках настоящего изобретения можно приготовить с помощью рекомбинантных методик, хорошо известных специалистам, или могут быть синтезированы с помощью стандартных твердофазных методик. Твердофазный синтез начинается с C-конца пептида с использованием α- аминозащищенной аминокислоты. БОК-блокирующие группы можно применять для всех аминогрупп, даже если подходят другие блокирующие группы. Например, БОК-лиз-OH можно эстерифицировать на носителях - хлорметилированных полистирольных смолах. Полистирольный носитель предпочтительно является сополимером стирола и приблизительно от 0,5 до 2% дивинилбензола, осуществляющего связь между цепями и придающего полистирольному полимеру абсолютную нерастворимость в определенных органических растворителях. См. Steward и др., Solid-Phase Peptide Synthesis (1969), W.H. Freeman Co., San Francisco; and Merrifield, J. Am. Chem. Soc. (1963) 85: 2149 - 2154. Эти и другие способы синтеза пептидов также иллюстрируются примерами в U.S. Patent Nos. 3862925; 3842067; 3972859; 4105602.
Полипептидный синтез можно осуществлять вручную или автоматически, применяя, например, синтезатор белка Applied Biosystems 403A (Foster City, California) или автоматический синтезатор белка Biosearch SAM, II (Biosearch, Inc. , San Rafael, California), следуя инструкциям, приложенным производителем.
Арилалкиламины настоящего изобретения являются натуральными продуктами, выделенными с помощью известных методик, или синтезированными как описано в Jasys и др., Tetrahedron Lett. (1988) 29: 6223 - 6226 и Nason и др., Tetrahedron Lett. (1989) 30: 2337 - 2340.
Одна основная методика для приготовления фендилина (или аналогов фендилина, показанных на фиг. 36) следующая. В 10 мл флакон с круглым дном, снабженный магнитной мешалкой и каучуковой перегородкой, помещают 1,0 ммоль 3,3'-бисфенилпропиламин (или первичный алкиламин) в 2 мл этанола и воздействуют на него 1,1 ммоль фенола и 1,0 ммоль ацетофенона (или замещенного ацетофенона). Туда же добавляют 2,0 ммоль MgSO4 и 1,0 ммоль NaCNBH3. Смесь перемешивают под атмосферой азота при комнатной температуре (около 20oC) в течение 24 ч. Реакционную смесь затем вливают в 50 мл эфира и отмывают 3 раза 1N NaOH и один раз солевым раствором. Эфирный слой высушивали безводным K2CO3 и восстанавливали в вакууме. Продукт затем очищали посредством колоночной хроматографии или жидкостной хроматографии высокого разрешения, инкорпорирующей в кремниевую стационарную фазу комбинации CH2Cl2-метанолизопропиламина (обычно 3% метанола и 0,1% изопропиламина в хлористом метилене).
Предпочтительная методика приготовления фендилина или аналогов фендилина (таких, которые изображены на фиг. 36) применяет изопропилат титана (IV) и является модификацией методик, описанных в J. Org. Chem. 55: 2552 (1990). Для синтеза NPS 544 тетрахлорид титана (способ, описанный в Tetrahedron Letters 31: 5547 (1990) использовали вместе изопропилата титана (IV). Схема реакции изображена на фиг. 43a. На этом чертеж R, R' и R'' изображают гидрокарбонильные группы. Согласно одному варианту осуществления изобретения, в 4 мл флаконе смешивали 1 ммоль амина (1) (обычно первичный амин) и 1 ммоль кетона или альдегида (2) (обычно ацетофенон), затем воздействовали 1,25 ммоль изопропилата титана (1) (3) и оставляли стоять при комнатной температуре на примерно 30 мин при периодическом перемешивании. В другом варианте вместо (1) можно использовать вторичный амин. Необходимо отметить, что некоторые реакции дадут обильный осадок или твердые вещества, которые нагревают до точки их плавления, чтобы можно было перемешать реакционную смесь несколько раз за время хода реакции. На реакционную смесь действует 1 мл этанола, содержащего 1 ммоль цианоборогидрида натрия (4), а получившуюся смесь оставляют стоять при комнатной температуре на приблизительно 16 ч при периодическом перемешивании. По истечении этого времени реакцию охлаждают заливанием приблизительно 500 л воды. Реакционную смесь затем разводят этиловым эфиром до общего объема приблизительно 4 мл и центрифугируют. Верхнюю органическую фазу удаляют и выпаривают на ротационном выпаривателе. Получившийся продукт (6) частично очищают хроматографией на короткой кремниевой колонке (или тонкослойной хроматографией (препаративной) на кремнии), применяя комбинацию дихлорметан : метанол : изопропиламин (обычно 95:5:0,1), перед очисткой посредством жидкостной хроматографии высокого разрешения (нормальная фаза с применением кремния с дихлорметан : метанол : изопропиламин или обращенная фаза с применением C-18 с 10,1% ТФУ с ацетонитрилом или метанолом).
Если необходимо или желательно, можно выполнять хиральное разделение методами, описанными в примере 21.
Рецептура и применение
Как было показано в настоящем документе, молекулы настоящего изобретения можно применять для того, чтобы: (а) имитировать или противодействовать одному или более воздействий внеклеточного Ca2+; (б) воздействовать на уровень внеклеточного свободного Ca2+ в организме; и (в) лечить заболевания, такие как гиперфункция паращитовидной железы, остеопороз и гипертония. В то время как было показано, что эти молекулы, в основном, воздействуют на клетки паращитовидной железы, они также могут воздействовать на рецепторы Ca2+ других клеток, включая остеокласты кости, юкстагломерулярные почечные клетки, клетки проксимальных почечных канальцев, кератиноциты, парафолликулярные клетки щитовидной железы и трофобласты плаценты.
Несмотря на то, что эти молекулы, в типичном случае, будут применять для лечения больных людей, их также можно применять и для лечения сходных заболеваний других теплокровных животных, таких как остальные приматы, скота, например свиней и коров, домашней птицы, спортивных животных и домашних, таких как лошади, собаки и кошки.
Для терапевтического и/или диагностического применения, рецептуры, включающие молекулы настоящего изобретения, можно приготавливать для разных способов введения пациенту, включая местное, наружное и системное применение. Методики и рецептуры в общем виде можно найти в Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, PA.
Для системного применения предпочтительно оральное введение. С другой стороны, можно применять и инъекции, например, внутримышечные, внутривенные, интраперитонеальные и подкожные. Для инъекций молекулы настоящего изобретения входят в рецептуры в форме жидких растворов, предпочтительно, в физиологически совместимых буферах, таких как раствор Хенка или раствор Рингера. В добавление, молекулы могут входить в состав твердой рецептуры, которую растворяют или суспендируют непосредственно перед употреблением. Возможны также лиофилизированные формы.
Системное введение также может осуществляться с помощью чрезслизистых или чрезкожных средств или орально. Для чрезкожного или чрезслизистого введения в рецептуре применяют пенетранты, подходящие для того барьера, через который необходимо проникнуть лекарственному веществу. Такие пенетранты хорошо известны специалистам и включают, например, для чрезслизистого применения соли желчных кислот. Для облегчения проникновения можно также применять детергенты. Чрезслизистое введение может осуществляться, например, с помощью назальных аэрозольных баллонов или с помощью суппозиториев. Для орального введения для молекул формируют общепринятые формы, такие как капсулы и таблетки.
Для наружного применения молекулы помещают в мази, гели, линименты и кремы, как принято и известно в данной области.
Как показано в нижеприведенных примерах, количества различных соединений настоящего изобретения, которые должны вводиться пациенту, можно определить с помощью стандартных методик. В основном, это количество составляет от приблизительно 1 до 50 мг/кг веса организма.
Рекомбинантные рецепторы Ca2+.
Для обнаружения основных структур для разработки различных лекарственных молекул традиционно применяют скрининг натуральных продуктов. Однако процедура скрининга с высокой пропускной способностью для коллекций натуральных продуктов и других молекул для обнаружения активности в отношении рецепторов Ca2+, ранее не представлялось возможной. Чтобы преодолеть это затруднение, лучше всего клонировать кДНК, кодирующую рецептор Ca2+, и затем создать трансформированные клеточные линии, удобные для осуществления скрининга с высокой пропускной способностью. Структуру рецептора дополнительно можно использовать также для того, чтобы лучше понять молекулярную геометрию лиганд-связывающего сайта (сайтов), и использовать такую информацию для расширения программы рациональной разработки лекарств, как обсуждалось выше. Ограниченные исследования зависимости активности от структуры и испытания избранных молекул из натуральных продуктов обеспечат начальную базу данных о структуре, необходимую для руководства рациональным скринингом натуральных продуктов и разработки лекарств.
ДНК рецептора Ca2+ клетки паращитовидной железы человека и крупного рогатого скота могут быть клонированы путем функциональной экспрессии в ооцитах Xenopus. За увеличением внутриклеточного Ca2+ в ооцитах Xenopus возможно наблюдать косвенно, посредством измерения тока через канал Cl-, активированный эндогенным Ca2+. Усиление ответа, которое приносит этот путь трансдукции, облегчает обнаружение рецепторных белков, кодируемых мРНК, при очень низких уровнях. Это позволяет обнаружение клонов рецептор-специфичной кДНК без потребности в лигандах высокого сродства, специфической антисыворотке или в информации о последовательности в белке или нуклеиновой кислоте. Примером такой процедуры является следующее:
Взрослых самок Xenopus laevis получали из Xenopus (Ann Arbor, MI) и содержали в стандартных условиях. У амфибий при гипотермической анестезии иссекали доли яичника. Группы ооцитов переносили в модифицированный физраствор Барта (МФБ). Отдельные ооциты получали инкубацией в МФБ, содержащем 2 мг/мл коллагеназы (Sigma, Type IA) в течение 2 ч при 21oC и для дальнейших манипуляций отбирали ооциты V - VI.
Стеклянные капиллярные пипетки (1 мм диаметр) вытягивали в длину и вручную обламывали, чтобы получить диаметр кончика приблизительно 15 мкМ. Каплю мРНК (1 нг/нл в воде, обработанной диэтилпирокарбонатом (ДЭПК)) помещали на парафильм и набирали в капиллярную пипетку путем всасывания. Капиллярную пипетку затем соединяли с микрошприцем (picospritzer WPI Instruments) и объем выталкиваемых воздухом капель регулировали таким образом, чтобы они содержали 50 нг мРНК (обычно 50 нл). 35 мм культуральные чашки с кусочками нейлона, прикрепленном ко дну, применяли, чтобы закрепить ооциты во время инъекции мРНК в вегетативный полюс. Инъецированные ооциты помещали в 35 мм чашки для культивирования, содержащие МФБ, 100 мкг/мл пенициллина и 100 ед/мл стрептомицина, и инкубировали при 18oC в течение 3 дней.
После инкубации ооцит помещали в 100 мкл пластиковую камеру и орошали МФБ со скоростью потока 0,5 мл/мин, применяя перистальтический насос. Испытуемые молекулы или неорганические поликатионы добавляли путем быстрого помещения трубопровода в различные буферы. Записывающие и проводящие электроды были изготовлены из тонкостенных капиллярных трубок с сопротивлением 1 - 3 Мом и наполненных 3М KCl. Ооциты прокалывали (на анимальном полюсе) обоими электродами под контролем микроскопа и соединяли с усилителем напряжения Axon Instruments Axoclamp 2A, который использовался для поддержания разности потенциалов (-70 до -80 mV) и измерения токов, протекающих для поддержания разности потенциалов. Токи записывались непосредственно ленточным самописцем.
Для приготовления мРНК брали ткань от телят или пациентов с вторичной ГП, подлежащей хирургическому удалению паращитовидных желез. Нет необходимости приготавливать очищенные клетки; использовали целые кусочки железы, чтобы приготовить мРНК, управляющую экспрессией рецептора Ca2+ в ооцитах Xenopus. Общую клеточную РНК приготавливали путем гуанидин тиоцианат/фенольной экстракции из гомогенизированных желез. Олиго-dT целлюлозная хроматография применялась для отделения поли(A)+ мРНК с помощью стандартной методики. Для разделения по размеру мРНК применяли центрифугирование в градиентах глицерола. мРНК денатурировали 20 мМ ртуть-гидроксидметилом и помещали (50 - 100 мкг при концентрации 1 мг/мл) в линейный градиент 15 - 30% глицерола, приготовленный в пробирках Beckman TLS 55. после центрифугирования на 34 000 об/мин в течение 16 ч собирали по 0,3 мл градиентных фракций и разводили их равным объемом воды, содержащей 5 мМ бета-меркаптоэтанола. мРНК затем регенерировали двумя циклами этанольной преципитации. По желанию мРНК (50 - 100 мкг поли(A)+) можно отделять в 1,2% агароза/6,0М мочевина препаративном геле с рядом маркеров размером РНК. После визуального наблюдения мРНК путем окрашивания этидиум бромидом, срезы геля, содержащие РНК размером ≈ 1,5 - 2,0 т. п. н., иссекаются. мРНК извлекают из срезов с применением РНК связывающего матрикса (в соответствии с процедурой, описанной производителем; Stratagene, Inc. ) и извлеченные фракции мРНК вымывают в обработанной ДЭПК воде.
Количества восстановленной мРНК определяли путем измерения У/Ф абсорбции. Размеры мРНК, содержащейся в каждой фракции, определяли электрофорезом в формальдегид/агарозном геле, используя малое количество (0,5 мкг) каждого образца. Работоспособность мРНК оценивали трансляцией in vitro каждого образца. Лизаты ретикулоцитов (коммерчески доступные наборы, BRL) использовали для трансляции 0,05 - 0,5 мкг каждой фракции мРНК. Полученные 35S-меченные белки анализировали с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия. Интактная мРНК была способна управлять полномасштабным синтезом белков в соответствии с размерами отдельных фракций мРНК.
Коллекции кДНК была встроена в вектор λ ZAPII после модификаций по методике Gubler и Hoffman. РНК из фракции (фракций), которая давала наилучшие результаты в исследованиях на ооцитах, использовали в качестве исходного материала. Синтез первичной цепи кДНК запускался с помощью олиго-dT/NotI затравкой-линкером. Синтез второй цепи осуществляли с помощью РНКаза Н/ДНК-полимеразы 1 способом самозатравки. Двойная цепь кДНК дефосфорилировалась с помощью T4 ДНК-полимеразы и EcoRI адапторов, присоединяемых с концами кДНК посредством T4 лигазы. После воздействия NotI для отщепления линкера, кДНК полной длины отбиралась по размеру путем хроматографии вытеснения на Sephacryl 500HA. Первая цепь кДНК метилась радиоактивным α- 32P-dАТФ и все этапы синтеза и извлечения прослеживания по инкорпорации радиоактивности. Полномасштабная кДНК, извлеченная из калибровочной колонки, сшивалась EcoRI/NotI обработанными λ ZAPII плечами. Упаковку вектора лигированной смесью производили с помощью коммерчески доступного высокоэффективного упаковывающего экстракта (Stratagene, Inc.), и переносили на подходящий штамм-хозяин. (XL1-blue). Процент рекомбинантного фага определяли соотношением белых и голубых бляшек, когда библиотеку высевали на IPTG и X-gal.
Средний размер вставки определяли из десяти произвольно выбранных клонов. Фаговые ДНК "mini-preps" обрабатывали EcoRI и NotI для высвобождения вставки и размеры определяли электрофорезом в агарозе. Библиотека содержала > 90% рекомбинантного фага, а размер вставки варьировал от 1,5 до 4,2 т.п.н. Рекомбинантная сшивка была плотно упакована для генерации 800 000 первичных клонов. Упаковочная смесь титровалась и высеивалась с плотностью 50 000 бляшек на 15 см чашки. Каждый пул в 50 000 клонов вымывали в SM буфере и хранили индивидуально.
Лизатные линии каждого из клонированных пулов использовали для приготовления малого количества ДНК. Фаговые частицы концентрировали преципитацией в полиэтиленгликоле, а ДНК фага очищали воздействием протеиназы K с последующей экстракцией фенол : хлороформом. Двадцать микрограммов ДНК подвергали воздействию NotI и использовали как матрицу in vitro для транскрипции РНК. Транскрипция in vitro производилась по стандартной методике, с использованием T7 РНК-полимеразы. После воздействия ДНКазы 1/ протеиназы K и экстракции фенол/хлороформом, РНК концентрировали этанольной преципитацией и использовали для инъекции ооцитам.
В ооциты инъецировали синтетическую мРНК (кРНК) от каждого из 16 субпулов библиотеки, содержащих 50 000 независимых клонов каждый. После инкубации в течение 3 - 4 дней ооциты оценивали на способность 10 мМ неомицина вызывать в них Ca2+ - зависимый ток Cl. Пул, обозначенный 6, показал положительный ответ и, следовательно, содержит клон кДНК, кодирующий функциональный кальциевый рецептор. Чтобы снизить сложность пула 6 и, таким образом, достигнуть прогресса в очищении клона рецептора кальция, содержащегося в этом пуле, фаг пула 6 вновь высеяли ≈ 20 000 бляшек на чашку и собрали его с 12 чашек. Из каждого из этих субпулов приготовили ДНК и синтезировали кРНК. Ооциты вновь инъецировали этой кРНК и 3 - 4 днями позже оценили на способность неомицина вызывать в них Ca2+ -зависимый ток Cl. Субпул 6 - 3 был положительным и стал объектом для нового раунда посевов, снижающих сложность пула до приблизительно 5 000 клонов. Пул вновь оценивали путем приготовления кРНК и инъекции в ооциты. Субпул 6-3.4 был положительным. Чтобы продвинуть дальнейшую очистку позитивного клона в пуле 6-3.4, фаговая ДНК из этого пула была спасена как ДНК плазмида путем суперинфекции с фагом-помощником, ExAssist. Трансфекция спасенных плазмид в бактериальный штамм DH5a lph aF' привела к появлению трансформированных бактериальных колоний на чашках с ампициллином. Они были собраны в пул из 900 клонов каждый. Затем из каждого субпула была приготовлена ДНК плазмида и синтезирована кРНК, которая оценивалась обычным путем. Субпул 6-3.4.4 был положительным. Бактерия, содержавшая плазмиду субпула 6-3.4.4, была впоследствии высеяна в субпулы по ≈ 50 клонов каждый. Продолжение этого процесса, как ожидается, приведет к выделению единственного клона, кодирующего функциональный рецептор Ca2+.
Начальные эксперименты использовали ооциты Xenopus, инъецированные водой или поли(A)+-обогащенной мРНК (50 нг) из клеток паращитовидной железы крупного рогатого скота. Спустя три дня, ооциты исследовали на их способность повышать внутриклеточный Ca2+ в ответ на повышения концентрации внеклеточных двух- и трехвалентных катионов. Ооциты, прокалывали записывающими и проводящими электродами и [Ca2+]i оценивали косвенно путем измерения токов через Cl- канал, активированный эндогенным Ca2+. В ооцитах, инъецированных поли(A)+-обогащенной мРНК из клеток паращитовидной железы крупного рогатого скота (или человека, фиг. 26), повышение концентрации внеклеточного Ca2+ с 0,7 до 3,5 или 10 мМ вызывало быстрое и кратковременное повышение проводимости Cl-, которая затем колебалась около высшего значения основной проводимости. Повышение концентрации внеклеточного Mg2+ от 1 до 10 мМ также вызывало колеблющиеся повышения проводимости Cl-. Ответ проводимости Cl- на внеклеточный Mg2+ сохранялся, когда концентрация внеклеточного Ca2+ снижалась до > 1 мкМ (фиг. 25).
Непроникающий трехвалентный катион Gd3+ (600 мкМ) также вызывал колеблющиеся повышения проводимости Cl- (фиг. 25). Такие повышения проводимости Cl-, которые осциллируют и сохраняются при номинальном отсутствии внеклеточного Ca2+, были замечены, когда ооцитам позволяли экспрессировать другие Ca2+-мобилизующие рецепторы и стимулировали их соответствующим лигандом (например, веществом K, фиг. 25). В этих случаях повышение проводимости Cl- отражает мобилизацию внутриклеточного Ca2+. Эти начальные исследования также показывают, что внеклеточные поликатионы мобилизуют внутриклеточный Ca2+ в ооцитах, инъецированных мРНК клетки паращитовидной железы.
Ооциты, инъецированные водой, не показали каких-либо изменений в токе Cl-, будучи подвергнуты воздействию внеклеточного Ca2+ (10 мМ) или Mg2+ (20 или 30 мМ). В одной серии экспериментов, ооциты инъецировали мРНК, кодирующей рецептор вещества K. В этих ооцитах внеклеточный Mg2+ (20 мМ) не вызывал никакого тока, но клетки бурно реагировали на добавление вещества K (фиг. 25). Эти эксперименты, показывают, что ооцит не обладает эндогенной чувствительностью к внеклеточному Ca2+ или Mg2+.
Сходные эксперименты были выполнены на ооцитах, инъецированных поли(A)+-обогащенной мРНК, приготовленной из паращитовидных желез человека (гиперпластической ткани при случае вторичной ГП). В этих ооцитах повышение концентрации внеклеточного Ca2+ вызывало обратимое повышение проводимости Cl-, которое осциллировало (фиг. 26). Добавление 300 мкМ La3+ также вызывало колеблющиеся повышения проводимости Cl-. Повышение концентрации внеклеточного Mg2+ от 1 до 10 мМ вызывало повышение проводимости Cl-, которое сохранялось в отсутствии внеклеточного Ca2+. Дополнительные эксперименты наводят на мысль о том, что ответ на внеклеточный Ca2+ зависит от концентрации. Таким образом, в трех мРНК-инъецированных ооцитах проводимость Cl- повышалась до максимума в 111 ± 22 нА при 3 мМ и 233 ± 101 нА при 10 мМ внеклеточного Ca2+.
Результаты, полученные в ооцитах Xenopus, демонстрируют присутствие в клетках паращитовидной железы мРНК, кодирующей (кодирующих) белок (белки), который может сообщать в норме нереагирующей клетке чувствительность к внеклеточному Ca2+. Более того, способность внеклеточного Mg2+ вызывать осциллирующие повышения тока Cl- в отсутствии внеклеточного Ca2+ показывает, что ток Cl- зависит от мобилизации внутриклеточного Ca2+ в большей степени, чем от притока внеклеточного Ca2+. Результаты, полученные с La3+ также показывают, что экспрессируемый белок (белки) связан с мобилизацией внутриклеточного Ca2+. Все вместе, эти данные говорят о том, что экспрессируемый белок (белки) действует скорее как поверхностный рецептор, чем как канал. Эти исследования обеспечивают неотразимые доказательства существования Ca2+ рецепторного белка на поверхности клеток паращитовидной железы и демонстрируют возможность применения системы ооцитов Xenopus для молекулярного клонирования кДНК, кодирующей рецептор Ca2+.
В другой серии экспериментов, мРНК клетки паращитовидной железы, денатурированная гидроксидом метил-ртути, была фракционирована по размеру центрифугированием в градиенте глицерола, и были собраны фракции. Каждая группа была инъецирована в ооциты Xenopus и спустя три дня инкубационного периода ооциты оценивали на экспрессию Ca2+ рецептора. Те ооциты, которые были инъецированы фракциями 4 - 6, показали самые большие и наиболее стойкие повышения проводимости Cl- в ответ на внеклеточный Ca2+ (фиг. 35). Эти результаты показывают, что рецептор Ca2+ кодируется мРНК размером 2,5 - 3,5 т.п.н. Это показывает, что стратегия использования прямой экспрессии РНК, синтезированной с вектора транскрипции кДНК библиотеки, является осуществимой. Эксперименты такого рода с фракционированием по размеру были выполнены, и были получены аналогичные результаты в каждом из трех экспериментах с различными фракциями.
Фракции мРНК, полученные и охарактеризованные в предыдущих экспериментах, можно исследовать инъекцией в ооциты. Для каждой фракции мРНК по 10 - 20 ооцитов инъецировали 50 нг РНК при концентрации 1 нг/мл в воде. Инъецированные ооциты поддерживали при 18oC в течение 48 - 72 ч, после чего их оценивали на экспрессию рецептора Ca2+, применяя измерения тока Cl-. Для каждой группы инъецированных ооцитов, количество положительных по экспрессии рецептора, а также величина Ca2+-зависимого тока Cl-, были определены. В качестве отрицательного контроля ооциты инъецировали поли(A)+-обогащенной мРНК, РНК дрожжей или водой.
Ожидается, что мРНК размером 2,5 - 3,5 т.п.н., будет кодировать рецептор. мРНК большего размера может сделать необходимым клонирующие процедуры, основанные на гибридном истощении мРНК перед инъекцией ооцитам. Эта стратегия не зависит от генерации клонов полномасштабной кДНК для своего успешного осуществления. Если экспрессия рецептора с единственной фракцией мРНК не получена, ооциты инъецируют смесью фракций для определения комбинации, которая даст такую экспрессию. Если все же окажется, что множественные субъединицы необходимы для образования функционального рецептора, применяют стратегию гибридного истощения. При таком подходе, клоны выбирают на основании их способности истощать виды специфичной мРНК из общей популяции мРНК. Клон, кодирующий единичную субъединицу, идентифицируют по его способности предотвращать образование активного многоединичного комплекса. Путем скрининга истощения возможно определить клоны, кодирующие все необходимые субъединицы.
Этот подход позволяет изолировать клоны, кодирующие индивидуальные субъединицы, которые требуются для образования функционального рецепторного комплекса. Синтетические РНК из пулов клонов исследуют на их способность индуцировать экспрессию Ca2+ рецептора в ооцитах Xenopus теми же методиками, что и для анализа оригинальных фракция мРНК. Первоначально исследовали 10 пулов, представляющих 100 000 первичных клонов. Пулы клонов, показывающие положительный ответ, подвергали скринингу на меньшую (обычно в 4 - 10 раз) сложность, и опять положительные пулы дополнительно подразделяли и подвергали скринингу. Этот процесс субфракционирования библиотеки продолжали, пока не были идентифицированы индивидуальные положительные клоны. В качестве отрицательного контроля для анализа экспрессии в ооцитах, вырабатывали античувствительные транскрипты путем T7 транскрипции тех ДНК-матриц, которые индуцировали положительный ответ. Античувствительные транскрипты не способны к повышению экспрессии аутентичного рецептора, и это будет контролировать любой неспецифичный положительный сигнал, являющийся результатом инъекции синтетической РНК. Другое соображение то, что является фактом, что синтетическая РНК может случайно "отравить" трансляцию в инъецированных ооцитах посредством неизвестного механизма. Чтобы контролировать такую возможность, совместно с мРНК клетки паращитовидной железы инъецируют синтетические РНК, дающие отрицательный ответ, в разных разведениях, чтобы определить, не создают ли они неспецифические помехи для экспрессии рецептора Ca2+.
Когда индивидуальный клон, кодирующий рецептор Ca2+, идентифицирован, кДНК вставка будет вырезана из вектора и использована для широкомасштабной продукции синтетической РНК. Инъекция в ооциты этого единственного вида РНК позволяет строго оценить экспрессированный рецептор.
Если размер мРНК, кодирующей рецептор Ca2+, слишком велик, для клонирования прямой транскрипцией и экспрессией, или если вовлечены множественные субъединицы, применяют методику гибридного истощения пулов, подвергающихся скринингу. Приготавливают ДНК со вставкой кДНК из пулов клонов из библиотеки, подобранных по размеру кДНК клетки паращитовидной железы. Эту ДНК гибридизируют с мРНК клетки паращитовидной железы при условиях, позволяющих образованию ДНК/РНК дуплексов. Негибридизированная, истощенная гибридизацией РНК восстанавливается и используется для инъекции ооцитам. ДНК из пулов клонов, содержащих последовательности, представляющие мРНК рецептора Ca2+, истощается из этой мРНК из тотальной популяции мРНК клеток паращитовидной железы, и экспрессия рецептора уменьшается или отсутствует при инъекции ооцитам. Процесс субфракционирования продолжают на пулах клонов снижающейся сложности, на каждом этапе анализируя клонированную ДНК, которая истощает мРНК, кодирующую рецептор Ca2+, из тотальной популяции мРНК клеток паращитовидной железы. Применение внутреннего контроля во время гибридного истощения гарантирует, что гибридно-истощенная РНК является интактной и способной к трансляции в ооците.
Клетки паращитовидной железы человека экспрессируют бета-адренергический рецептор, связанный с аденилатциклазой. Этот рецептор можно экспрессировать в ооцитах, где он способен к агонист-индуцированной активации эндогенной аденилатциклазы. Во время скрининга истощением гибридизацией клонов рецептора Ca2+, исследуют ооциты, инъецированные истощенной гибридизацией мРНК, на предмет изопротеренол-индуцированной активности аденилатциклазы. Положительный ответ в этом анализе служит показателем того, что любое угнетение ответа рецепторов Ca2+ является специфичным, и не имеет отношения к общему угнетению тотальной популяции мРНК.
Стратегия скрининга гибридным истощением может привести к изоляции клонов, которые не содержат полного участия, кодирующего белок. Положительные клоны, изолированные этой стратегией скрининга, секвенируют для определения их способности кодировать белок. Анализ норзерн блоттинга РНК паращитовидной железы человека позволяет определить размер полных мРНК, соответствующих специфичным клонам. Если оказывеатся, что положительные клоны не имеют полной длины, используют клонированную ДНК в качестве зонда гибридизации для скрининга кДНК библиотеки паращитовидной железы на полные кДНК.
Множество клеточных линий обладают способностью связывания экзогенно экспрессированных рецепторов с эндогенными функциональными ответами. Ряд клеточных линий (например, NIH-3T3, HeLa, CHO, NG115, HEK, 293 и COS7) можно испытать для подтверждения того, что они не обладают эндогенным рецептором Ca2+. Эти линии, не обладающие ответом на внешний Ca2+, можно использовать для создания стабильных трансформированных клеточных линий, экспрессирующих клонированный рецептор Ca2+.
Анализ последовательностей клонов кДНК рецептора Ca2+, идентифицированных экспрессирующим клонированием, сможет трансдифференцировать открытую рамку считывания, кодирующую рецепторный белок. Кодирующий участок кДНК будет субклонирован в множественный клонирующий сайт эукариотического экспрессирующего вектора pMSG. Этот вектор обеспечивает высокий уровень транскрипции, управляемой промотором вируса опухоли молочной железы мышей (ВОММ), и является активным в отношении множества видов клеток млекопитающих. Этот вектор также содержит ген gpt резистентности к микофенолокислоте, которая находится под контролем начального промотора SV40, и последовательности, необходимые для селекции и роста бактерии. Большие количества экспрессирующего вектора-плазмиды рецепторной кДНК можно вырастить и выделить от E.coli.
Наиболее эффективные способы трансфекции эукариотической клетки ДНК-плазмидой различаются в зависимости от данного типа клеток. Экспрессирующая Ca2+ рецептор конструкция будет введена в культивируемые клетки с помощью соответствующих методик, таких как преципитация фосфатом Ca2+, трансфекция диэтиламиноэтил (DEAE)-декстраном, липофекция или электропорация. После процедуры трансфекции клетки выращивают в присутствии антибиотика G418 для отбора клеток, экспрессирующих ген гезистентности к неомицину. Колонии G418 устойчивых трансфектантов будут субклонированы и оформлены как самостоятельные клеточные линии. Экспрессия Ca2+ рецепторного белка в G418 устойчивых клетках будет оцениваться несколькими способами. Саузерн блот и слот блот анализы подтвердят присутствие и количество копий последовательностей кДНК рецептора. Анализ нозерн блоттинга будут применять для демонстрации того, что рецепторная мРНК транскрибируется с плазмидной конструкцией. Функциональная экспрессия рецепторного белка будет определена измерением внутриклеточного Ca2+ в ответ на поступившие извне вещества, обладающие сродством к рецептору Ca2+.
Клонирование рецептора Ca2+ облегчает как структурные, так и функциональные исследования этого нового рецептора. Рецептор, полученный рекомбинантным путем, может быть кристаллизован для структурных исследований. Стабильные трансфектированные клеточные линии, экспрессирующие рецептор, можно использовать для скрининга с высокой пропускной способностью для библиотек натуральных продуктов или других соединений. Молекулы требуемой активности и специфичности можно метить (радиоизотопами или флюоресцентами). Способность испытуемых молекул или экстрактов вытеснять такую меченую молекулу образует основу для скрининговой процедуры с высокой пропускной способностью.
Приданные соответствующим клеткам или тканям, экспрессирующим другие рецепторы Ca2+, эти рецепторы могут быть клонированы теми же способами, которые описаны выше для кальциевого рецептора клетки паращитовидной железы. Например, мРНК из ткани остеокластомы человека кодирует кальциевый рецептор остеокласта (фиг. 34). Таким образом, чтобы изолировать клон для рецептора остеокласта человека, нужно только изолировать мРНК из ткани остеокластомы, изготовить библиотеку кДНК и исследовать/фракционировать субпулы, как описано выше. Более того, предпочтительными рецепторами для скрининга лекарств являются рецепторы человека. Клон, кодирующий кальциевый рецептор, из одного вида можно использовать для получения соответствующего клона кДНК человека путем перекрестной гибридизации, как хорошо известно специалистам. Вдобавок клон Ca2+ рецептора клеток паращитовидной железы или других клеток позволяет изолировать гены, кодирующие сходные улавливающие Ca2+ белки, в других клетках и экспрессировать эти белки. Это достигается множеством различных подходов. Анализ саузерн блоттинга геномной ДНК человека, непользующий кДНК Ca2+ рецептора в качестве зонда гибридизации, даст показатель числа родственных последовательностей, закодированных в геноме; гибридизация на разных генонемах даст показатель степени дивергенции родственных последовательностей. Это обеспечит информацию о потенциальном количестве генов, кодирующих родственные рецепторные белки. Анализ норзерн блоттинга с кДНК рецептора Ca2+ в качестве зонда, определит, присутствуют ли в различных тканях одни и те же или родственные транскрипции. Если будут определены родственные транскрипты, гомологично рецептору Ca2+ клеток паращитовидной железы, становится относительно простой задачей получение клонов этих мРНК, как скринингом соответствующих библиотек кДНК, так и реакцией цепочки с полимеразой (РЦП). Полученные этим способом клоны нового рецептора можно оценивать функционально путем экспрессии, а также в ооцитах или в трансфектированных клеточных линиях. Трансфектированные клеточные линии, экспрессирующие клеточноспецифичный Ca2+ рецептор, затем могут служить средством скрининга с высокой пропускной способностью для молекул, которые специфично действуют на Ca2+ - чувствительный механизм, например, остеокластов или юкстагломерулярных клеток.
В альтернативном способе кальциевый рецептор можно клонировать экспрессией в эукариотических клетках. Например, библиотеку кДНК можно приготовить из мРНК клеток паращитовидной железы и клонировать в эукариотический экспрессирующий вектор, pCДНК1. Субпулы из этой библиотеки можно трансфектировать в эукариотические клетки, такие как COS7 или HEK293, в результате чего проявится относительно высокий уровень кратковременной экспрессии кодируемых кДНК последовательностей. Клетки, трансфектированные клоном функционального кальциевого рецептора, будут экспрессировать кальциевый рецептор, который затем можно активировать кальцием, неомицином или другими кальцимиметическими соединениями. Если клетки предварительно нагружают флюориметрическим индикатором [Ca2+] i, активация кальциевого рецептора приводит к усилению флюоресценции. Таким образом, библиотечные субпулы, содержащие кальциевый рецептор, идентифицируются по их способности, после трансфекции в эукариотические клетки, индуцировать кальцимиметически специфичное повышение флюоресценции. Эту флюоресценцию можно уловить, применяя как флюориметр, так и клеточный сортер с активацией флюоресценции (КСАФ).
В альтернативном способе, рецептор Ca2+ можно клонировать применением моноклонального антитела, выработанного против рецептора. Моноклональные антитела обеспечивают мощный инструмент для выделения специфических белков по иммунологическому средству. Для выделенного белка, представляющего интерес, получают сведения об аминокислотной последовательности и используют их для конструирования олигонуклеотидных зондов последовательности с целью скрининга клонов с полной последовательностью кДНК.
Для приготовления гибридом в качестве иммуногена используют целые клетки паращитовидной железы крупного рогатого скота. Получают очищенные разъединенные клетки, и живые клеточные препараты вводят интраперитонеально мышам подходящей линии согласно стандартной процедуре. Стандартной процедуры придерживаются и для режима иммунизации и приготовления гибридом. Применяют двухступенчатую процедуру скрининга для идентификации гибридом, секретирующих моноклональные антитела, узнающие Ca2+ рецептор. Начальный скрининг будет идентифицировать те моноклональные антитела, которые узнают поверхностные антигены клетки паращитовидной железы. Затем применяют иммуногистохимические методики для скрининга гибридомных надосадочных жидкостей на присутствие мышиных антител, которые связываются с поверхностью клеток паращитовидной железы. Этот скрининг можно выполнять на фиксированных кусочках ткани парщитовидной железы или на разрозненных клетках первичной культуры. Методики этого исследования хорошо представлены в литературе.
Этот скрининг выявит гибридомы, производящие моноклональные антитела к множеству детерминант клеточной поверхности, а моноклональные антитела, специфичные по отношению к Ca2+ рецептору, как ожидается, будут содержать только их малую субпопуляцию. Чтобы идентифицировать моноклональные антитела, которые связываются с Ca2+ рецептором, гибридомные надосадочные жидкости, которые оказались положительными при начальном скрининге, исследуют на их способность блокировать ответ культивируемых клеток паращитовидной железы на вещества, обладающие сродством к Ca2+ рецептору. Некоторые антитела, которые связываются с внеклеточным домейном рецептора, как ожидается, будут ингибировать или активировать лиганд, связывающий или каким-либо другим образом мешающий или влияющий на активацию рецептора.
Моноклональные антитела, положительные по результатам обоих скринингов, характеризуют посредством вестерн блоттинга, иммунопреципитации и иммуногистохимического метода. Это позволяет определить размеры антигена, который узнается, и его распределение в тканях. Подходящие моноклональные антитела затем используют для выделения Ca2+ рецепторного белка путем иммуноаффинной хроматографии согласно стандартной методике. Достаточные количества белка получают, чтобы определить ограниченную аминокислотную последовательность. Затем конструируются дегенеративные олигонуклеотидные зонды на основании информации о пептидной последовательности. Эти зонды затем применяют для скрининга библиотек кДНК паращитовидной железы на клоны Ca2+ полной длины. Полученные клоны характеризуют секвенированием ДНК и функциональной экспрессией в ооцитах и в культурах клеток млекопитающих.
С другой стороны, антитела можно применять для скрининга библиотек экспрессии, например библиотек кДНК в λ gt II или в их эквивалентах, для определения тех клонов, которые экспрессируют антигенно реактивный белок. Такие клоны можно секвенировать для определения, кодируют ли они белок, который может быть Ca2+ рецептором, или нет.
Специалистам в данной области понятно, что библиотеки фагов также могут быть использованы для клонирования и анализа кальциевых рецепторов вместо моноклональных антител. В этих библиотеках, различные участки антител или произвольные пептиды дозированно клонируют в фаговые экспрессирующие векторы, таким образом, что участки антител или пептиды распределяются по поверхности фаговой частицы. Фаг, несущий участки антител или пептиды, способные с высокой специфичностью связывать кальциевые рецепторы, будет присоединяться к клеткам, которые несут эти рецепторы. (например, клетки паращитовидной железы, C-клетки, остеокласты и т.д.). Миллионы таких фагов можно просеять через эти клеточные типы, которые будут выбирать только тот фаг, который может присоединяться к этим клеткам (включая сюда фаги, присоединяющиеся к кальциевым рецепторам). Этим способом сложность библиотеки может быть значительно снижена. Впоследствии процедуры скрининга, описанные выше для моноклональных антител, можно применить для изоляции фага, который несет связывающиеся с Ca2+ рецептором участки антитела или пептида, и этот фаг можно использовать для выделения кальциевого рецептора с целью структурной идентификации и клонирования. Наборы для приготовления таких библиотек фагов являются коммерчески доступными (например, Stratacyte или Cambridge Antibody Technology Limited). Рекомбинантный фаг, наделенный такими свойствами связываться с кальциевыми рецепторами, можно также использовать вместо моноклональных антител в различных исследованиях кальциевых рецепторов. Такой фаг также можно использовать в скрининге конкурентным связыванием с высокой пропускной способностью для идентификации органических соединений, способных к функциональному связыванию кальциевых рецепторов, которые могут служить основой для разработки лекарственных средств для человека, действующих на кальциевые рецепторы.
В альтернативном варианте, можно применять перекрестное сшивание радиолигандов к их рецепторам для выделения рецепторного белка, как описано в Pilch и Czech, 1 Receptor Biochem, Methodol. 161, 1984. Ковалентное присоединение радиолиганда допускает продолжительное отмывание для удаления неспецифичного связывания. Например, молекула с высоким сродством, например, произвольный сополимер аргинина и тирозина (MW = 22K; соотношение аргинина и тирозина = 4: 1), которая мобилизует внутриклеточный Ca2+ с величиной EC50 около 100 нМ или менее, иодируется 125I и перекрестно сшивается. Протамины, благодаря их намного меньшим размерам, могут быть предпочтительными для исследований с перекрестным сшиванием и могут быть восстановительно алкилированы, как описано у Dottavio-Martin и Ravel, 87 Analyt, Biochem, 562, 1978.
Неспецифичное мечение сводится к минимуму перекрестным сшиванием в присутствии немеченных поликатионов и двух- и трехвалентных катионов. При высоких концентрациях этих молекул неспецифические взаимодействия метки с клеточной поверхностью могут быть уменьшены.
Первичная гиперфункция паращитовиной железы (ГП) характеризуется гиперкальциемией и повышенными уровнями циркулирующего ПТГ. Одним из основных дефектов при ГП является пониженная чувствительность клеток паращитовидной железы к регуляции внеклеточным Ca2+ по принципу отрицательной обратной связи. Так, в ткани пациентов с первичной ГП "точка отсчета" для внеклеточного Ca2+ сдвинута вправо так, что более высокие, чем в норме, концентрации внеклеточного Ca2+ требуются, чтобы подавить секрецию ПТГ. Более того, при первичной ГП даже высокие концентрации внеклеточного Ca2+ часто подавляют секрецию ПТГ только частично. При вторичной (уремической) ГП наблюдается сходное повышение точки отсчета для внеклеточного Ca2+, несмотря на то, что степень, до которой Ca2+ подавляет секрецию ПТГ, является нормальной. Изменения секреции ПТГ идут параллельно изменениям [Ca2+]i, точка отсчета для повышений [Ca2+] i, индуцированных внеклеточным Ca2+, смещена вправо, а величина таких повышений уменьшена. Кроме того, окрашивание ткани моноклональным антителом, которое узнает рецептор Ca2+, ослаблено в аденоматозных и гиперпластических клеток паращитовидной железы.
Рецептор Ca2+ представляет дискретную молекулярную организацию для фармакологического воздействия. Молекулы, которые имитируют или противодействуют воздействию внеклеточного Ca2+, благоприятны при длительном применении как в случае первичной, так и вторичной ГП. Такие молекулы обеспечивают дополнительный толчок, который необходим для подавления секреции ПТГ, которой не может достичь только одно состояние гтперкальциемии. Такие молекулы с большей эффективностью, чем внеклеточный Ca2+, могут преодолеть кажущийся неподавляемым компонент секреции ПТГ, которая особенно создает проблемы в аденоматозной ткани. Альтернативно или дополнительно такие молекулы могут подавлять синтез ПТГ, тогда как длительная гиперкальциемия подавляет уровни препроПТГ мРНК в клетках ткани паращитовидной железы крупного рогатого скота и аденоматозной ткани паращитовидной железы человека. Длительная гиперкальциемия также подавляет пролиферацию клеток паращитовидной железы in vitro, так что кальцимиметики также могут быть эффективны в ограничении гиперплазии клеток паращитовидной железы, характерной для вторичной ГП.
Другие клетки тела могут непосредственно реагировать на физиологические изменения концентрации внеклеточного Ca2+. Секреция кальцитонина парафолликулярными клетками щитовидной железы (C-клетки) регулируется изменениями концентрации внеклеточного Ca2+. Секреция ренина юкстагломерулярными клетками почек, подобно секреции ПТГ, подавляется увеличенными концентрациями внеклеточного Ca2+. Внеклеточный Ca2+ вызывает мобилизацию внутриклеточного Ca2+ в этих клетках. Изолированные остеокласты отвечают на повышения концентрации внеклеточного Ca2+ соответствующими повышениями [Ca2+]i, которые происходят частично вследствие мобилизации внутриклеточного Ca2+. Повышения [Ca2+] i в остеокластах связаны с подавлением функциональных ответов (резорбция кости), аналогично секреции ПТГ клетками паращитовидной железы. Таким образом, имеются существенные предпосылки для предположения, что Ca2+, в дополнение к его повсеместной роли внутриклеточного сигнала, функционирует так же как внеклеточный сигнал, регулирующий реакции определенных специализированных клеток. Молекулы настоящего изобретения можно применять в лечении заболеваний, связанных с нарушенными реакциями на Ca2+ в этих клетках.
Клонирование рецептора Ca2+ в клетках паращитовидной железы, а также в других клетках позволит непосредственно оценить присутствие гомологичных белков в других клетках. Таким образом можно получить семейство структурно гомологичных Ca2+ рецепторных белков. Такие рецепторы позволят понять, каким образом эти клетки улавливают внеклеточный Ca2+ и облегчить оценку механизма (или механизмов) как мишени для лекарственных средств, описанных здесь, которые являются эффективными при лечении ГП, остеопороза и гипертонии, и новых лекарственных средств для лечения других костных и минералзависимых болезней.
Иные области применения описаны выше. Например, рекомбинантные Ca2+ рецепторные белки могут применяться в терапии и вводиться стандартными способами, например, трансфекцией нуклеиновой кислоты, кодирующей этот белок. Вдобавок такой белок полезен для исследований кальцимиметических молекул настоящего изобретения.
Следующие примеры иллюстрируют настоящее изобретение, но не ограничивают его рамок.
Примеры
В исследованиях, описанных в настоящем документе, было обнаружено, что множество органических молекул мобилизуют внутриклеточный Ca2+ и подавляют секрецию ПТГ в клетках паращитовидной железы. Эти молекулы различаются по строению, но большинство имеет суммарный положительный заряд при физиологическом pH. Катионная
природа органических молекул играет важную роль, но не является единственным фактором, определяющим их активность.
Пример 1.
Скрининг кальцимиметических молекул на клетках паращитовидной железы крупного рогатого скота.
Разъединенные клетки паращитовидной железы крупного рогатого скота очищали в градиентах Percoll и культивировали в течение ночи в не содержащей сыворотки среде. Затем клетки нагружали фура-2 и флюориметрически измеряли концентрацию свободного внутриклеточного Ca2+. Изменения [Ca2+]i применяли для скрининга молекул, активных в отношении рецептора Ca2+. Чтобы считаться кальцимиметиком, молекула должна была продемонстрировать нормальные эффекты, вызываемые повышением внеклеточного Ca2+ и запускаемые активацией рецептора Ca2+. То есть:
1) молекула должна вызывать повышение [Ca2+]i, которое сохраняется в отсутствии внеклеточного Ca2+ (демонстрируя мобилизацию внутриклеточного Ca2+);
2) молекула должна вызывать понижение изопротеренол-стимулированного образования циклического АМФ, блокированного токсином коклюша;
3) молекула должна ингибировать секрецию ПТГ при тех же концентрациях, которые вызывали повышение [Ca2+]i; и
4) кривая ответа на концентрацию для мобилизации Ca2+ и секреции ПТГ, вызванного молекулой, должна быть сдвинута вправо активатором ПКС, таким как форбол миристатацетат (ФМА).
Было испытано несколько классов структурно различных классов молекул: полиамины, аминогликозиды, протамин и полимеры лизина или аргинина. Структура этих молекул изображена на фиг. 1. В фиг. 1 включены молекулы с суммарным положительным зарядом и значения их EC50 при вызывании мобилизации внутриклеточного Ca2+ в клетках паращитовидной железы крупного рогатого скота.
В целом, чем больше суммарный положительный заряд молекулы, тем больше ее способность вызывать мобилизацию внутриклеточного Ca2+. Однако было обнаружено несколько поразительных исключений из этого очевидного правила, как обсуждалось ниже.
Как можно видеть из чертежей, спермин, неомицин B и протамин вызывали быстрые и кратковременные повышения [Ca2+]i, в фура-2-нагруженных клетках паращитовидной железы крупного рогатого скота (фиг. 6, 7, 11). Они, однако, не вызывали стационарных, поддерживаемых повышений [Ca2+]i в клетках паращитовидной железы крупного рогатого скота (фиг. 6, 11), хотя они достигали этого в клетках паращитовидной железы человека (фиг. 19). С этой точки зрения, они напоминали ответ цитозольного Ca2+, вызванный внеклеточным Mg2+, который вызывает мобилизацию внутриклеточного Ca2+, не сопровождаемую притоком внеклеточного Ca2+, в клетках крупного рогатого скота (фиг. 11b). Кратковременные повышения [Ca2+]i, вызванные спермином, неомицином B или протамином, не блокировались низкими концентрациями (1 мкМ) La3+ или Gd3+ (фиг. 11). Кратковременные повышения цитозольного Ca2+, вызванные молекулярными поликатионами, сохранялись в отсутствии внеклеточного Ca2+, но блокировали, когда клеточные запасы Ca2+ истощали предварительной обработкой иономицином (фиг. 7, 11h, i). Следовательно, все эти молекулы вызывают мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы.
Дополнительно было показано, что молекулярные поликатионы мобилизуют тот же пул внутриклеточного Ca2+, что использует внеклеточный Ca2+. Так, повышение концентрации внеклеточного Ca2+ постепенно ингибировало кратковременные повышения [Ca2+]i, вызванные спермином (фиг. 6). Наоборот, максимально эффективные концентрации спермина или неомицина B (фиг. 12) блокировали кратковременные, но не стационарные повышения [Ca2+]i, вызванные внеклеточным Ca2+.
Важно, что спермин, неомицин B и протамин ингибировали секрецию ПТГ в той степени, что и внеклеточный Ca2+. Эти ингибирующие секрецию эффекты были получены при концентрациях, которые вызывали мобилизацию внутриклеточного Ca2+ (фиг. 8, 13). Эти результаты уместны для понимания механизмов, участвующих в регуляции секреции ПТГ внеклеточных Ca2+. Поскольку множество неорганических поликатионов ингибируют секрецию, и только внеклеточный Ca2+ вызывает поддерживаемые стационарные повышения [Ca2+] i, такие повышения [Ca2+] i, не могут сколько-нибудь значительным образом участвовать в регуляции секреции. Мобилизация внутриклеточного Ca2+, скорее, чем приток внеклеточного Ca2+, является главным механизмом, связанным с подавлением секреции ПТГ. Это важно, поскольку это определяет главный механизм, на который надо воздействовать, если молекула, должна подействовать на секрецию ПТГ; молекулы, стимулирующие избирательно приток внеклеточного Ca2+, будут относительно неэффективным в подавлении секреции ПТГ. Напротив, молекулы вызывающие только мобилизацию внутриклеточного Ca2+, будут столь же эффективными в подавлении секреции ПТГ, что и внеклеточный Ca2+.
Подобно мобилизации внутриклеточного Ca2+, вызванной внеклеточным Ca2+, мобилизация, вызванная молекулярными поликатионами, подавлялась ФМА. Показательный эксперимент, демонтирующий предпочтительные ингибирующие эффекты ФМА на кратковременные повышения цитозольного Ca2+, вызванные спермином, показан на рис. 14. Кратковременные повышения цитозольного Ca2+, вызванные АТФ, оставались без изменений, даже когда применялись субмаксимальные концентрации АТФ. Эффект ФМА на кратковременные повышения цитозольного Ca2+, вызванные молекулярными поликатионами, был аналогичен его эффекту на ответы на внеклеточный Ca2+; в обоих случаях присутствовал сдвиг вправо в кривой ответа концентрации (фиг. 15). Депрессивные эффекты ФМА на [Ca2+]i сопровождались потенцирующими эффектами на секрецию, которые преодолевались при более высоких концентрациях органических поликатионов (фиг. 16).
Мобилизация внутриклеточного Ca2+, вызванная молекулярными поликатионами, была связана с увеличением образования инозитфосфатов. Например, протамин вызывал быстрое (в течение 30 с) повышение образования ИФ3, которое сопровождалось повышением уровня ИФ1. Оба эти эффекта зависели от концентрации внеклеточного протамина (фиг. 17). Кроме того, предварительная обработка ФМА снижала образование инозитфосфатов, вызванное молекулярными поликатионами. Результаты, полученные со спермином, представлены на фиг. 18.
Спермин, неомицин B и протамин подавляли изопроперенол-индуцированные повышения циклического АМФ. Подобно ингибирующим эффектам внеклеточного Ca2+ на циклический АМФ, эффекты, вызванные молекулярными поликатионами, блокировались предварительной обработкой токсином коклюша (табл. 2).
Токсин коклюша (ТК) блокирует ингибирующие эффекты внеклеточного Ca2+ и молекулярных поликатионов на образование циклического АМФ. Клетки паращитовидной железы крупного рогатого скота культивировали 16 ч в присутствии или отсутствии 1000 нг/мл токсина коклюша. Затем клетки отмывали и инкубировали 15 мин с 10 мкМ изопротеренола при указанных концентрациях внеклеточного Ca2+ или молекулярных поликатионов или без них. Общее количество циклического АМФ (клетки + надосадочная жидкость) определяли посредством радиоиммуноанализа (РИА) и результаты выражали в процентах от уровней, полученных при 0,5 мМ Ca2+ (112 ± 17 пмоль/106 клеток). Каждая величина = среднее ± СКО среднего от трех экспериментов.
В клетках паращитовидной железы человека внеклеточный Mg2+ вызывал поддерживаемое стационарное повышение [Ca2+]i в добавление к быстрому кратковременному повышению (фиг. 10). Как в клетках паращитовидной железы крупного рогатого скота, отвечающих на внеклеточный Ca2+, стационарное повышение [Ca2+]i, вызванное Mg2+ в клетках паращитовидной железы человека, происходит вследствие притока Ca2+ через потенциалонечувствительные каналы (фиг. 10a). Этот эффект Mg2+ на стационарное состояние [Ca2+]i в клетках паращитовидной железы человека наблюдается как в аденоматозной, так и в гиперпластической ткани.
Неомицин B и спермин испытывали на воздействие на [Ca2+]i в клетках паращитовидной железы человека, приготовленных из аденоматозной ткани. Результаты с неомицином B проиллюстрированы на фиг. 19. Неомицин B вызывал не только кратковременное, но и дополнительно стационарное повышение [Ca2+]i в клетках паращитовидной железы человека (фиг. 19a). Таким образом, изменения [Ca2+]i, вызванные внеклеточным Ca2+, Mg2+ или неомицином B весьма сходны.
Кратковременные повышения цитозольного [Ca2+]i, вызванные неомицином B, сохранялись в присутствии La3+ (1 мкМ) и в отсутствии внеклеточного Ca2+. Следовательно, неомицин B вызывает мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы человека. Неомицин B ингибировал секрецию ПТГ в клетках паращитовидной железы человека в концентрациях, которые вызывали мобилизацию внутриклеточного Ca2+ (фиг. 13). Существовали, однако, некоторые различия в ответах на неомицин B в клетках паращитовидной железы человека и крупного рогатого скота. EC50 неомицина B для мобилизации внутриклеточного Ca2+ составляла 40 мкМ в клетках крупного рогатого скота (КРС) и 20 мкМ в клетках человека (фиг. 13 и 15), в то время как активность спермина была сходной как в клетках КРС, так и в клетках человека (EC50 = 150 мкМ). Таким образом, хотя клетки КРС могут использоваться для начальных исследований по отбору молекул по активности, важно выполнить последующие исследования с использованием клеток паращитовидной железы человека.
Для того, чтобы оценить эффекты молекулярных поликатионов на C-клетки, использовали линию неопластических клеток, выделенных из карциномы мозгового вещества щитовидной железы крысы (МТС 6-23 клетки). Как спермин (10 мМ), так и неомицин B (5 мМ) не влияли на [Ca2+]i в этих клетках. Также не влияли на реакцию на последующее добавление внеклеточного Ca2+ молекулы настоящего изобретения. Результаты, представляющие отсутствие влияния неомицина B, показаны на фиг. 21. Неомицин B (1 мМ) и спермин (1 или 5 мМ) не смогли вызвать какого-либо повышения [Ca2+]i в остеокластах (фиг. 23). Как показывает график, имелось некоторое потенцирование ответа на последующее повышение концентрации внеклеточного Ca2+, хотя эти результаты не были стабильными. В двух остальных клетках спермин (5 мМ) снова не оказывал влияния на исходную [Ca2+]i и вызывал небольшое ингибирование (около 15%) повышения [Ca2+] i, индуцированного внеклеточным Ca2+. В третьей клетке неомицин B (5 мМ) не оказывал влияния на исходную [Ca2+]i и на повышения [Ca2+]i, вызванные внеклеточным Ca2+. Общая картина, которая получается из результатов этих исследований, такова: спермин и неомицин B не влияют на исходные или стимулированные уровни цитозольного Ca2+ в остеокластах.
Неспособность молекулярных поликатионов воздействовать на Ca2+ - чувствительные механизмы C-клеток или остеокластов демонстрирует возможность открытия или разработки новых основных молекул, которые специфично действуют на рецепторы Ca2+ клеток паращитовидной железы или каким-либо другим образом модулируют одну или более функций нормального ответа клеток паращитовидной железы на [Ca2+]i.
Скрининг различных других молекул описан в подробностях ниже, а результаты суммированы в табл. 1.
Пример 2. Скрининг полиаминов.
Полиамины с прямой цепью (спермин, спермидин, ТЭТА, ТЭПА и ПЭГА) и два их производных (NPS 381 и NPS 382) подвергались скринингу как в примере 1. Было обнаружено, что все эти молекулы мобилизуют внутриклеточный Ca2+ в клетках паращитовидной железы КРС. Порядок их активности следующий, с суммарным положительным зарядом в круглых скобках:
Молекула - EC50 (в мкМ)
NPS 382 (+8) - 50
NPS 381 (+10) - 100
спермин (+4) - 150
ПЭГА (+6) - 500
Спермидин (+3) - 2000
ТЭПА (+5) - 2500
ТЭТА (+4) - 8000
Путресцин (+2) и кадаверин (+2) были неактивны при концентрации 2 мМ.
Другой полиамин с прямой цепью, ДАДД, вел себя несколько иначе по сравнению с другими полиаминами и описан в примере 7.
Пример 3. Скрининг циклических полиаминов.
Два циклических полиамина, гексациклен и NPS 383, подвергали скринингу, как описано в примере 1. Гексациклен (+6, EC50 = 20 мкМ) оказался в 7 раз более активным, чем NPS 383 (+8, EC50 = 150 мкМ). Обратное положение, как ожидается, будет базироваться только на суммарном положительном заряде, как структурной характеристике активности в отношении рецептора Ca2+.
Пример 4. Скрининг аминогликозидных антибиотиков.
Шесть антибиотиков подвергали скринингу, как описано в примере 1. Получившиеся EC50 для мобилизации внутриклеточного Ca2+, в порядке их активности, были следующие:
Антибиотик - EC50 (в мкМ)
неомицин (+6) - 10
гентамицин (+5) - 150
беканамицин (+5) - 200
стрептомицин (+3) - 600
Канамицин (+4,5) и линкомицин (+1) не оказывали влияния при концентрации 500 мкМ. В серии с аминогликозидами существует корреляция между суммарным положительным зарядом и активностью. Однако неомицин значительно более активен, чем различные полиамины (NPS 381, NPS 382, NPS 383, ПЭГА), имеющие такой же или больший положительный заряд.
Пример 5. Скрининг пептидов и полиаминокислот.
Протамин и полимеры лизина или аргинина, различные по длине, подвергали скринингу на их способность мобилизовать внутриклеточный Ca2+, так описано в примере 1. Полученные величины EC50 для мобилизации внутриклеточного Ca2+, в порядке их активности:
Пептид (MB в кД) - EC50 (в мМ)
полиарг (100) - 4
полиарг (40) - 15
полилиз (27) - 30
протамин (4,8) - 75
полиаргтир (22) - 200
полилиз (14) - 1000
полилиз (3,8) - 3000
Суммарный положительный заряд этих полимеров возрастает с возрастанием молекулярного веса. Таким образом, как и для аминогликозидов, существует прямая корреляция между суммарным зарядом и активностью в этой серии полиаминокислот. Протамин, по существу, является полиарг с суммарным положительным зарядом +21.
Пример 6. Скрининг арилалкиламинов.
Молекулы, выбранные из класса арилалкиламиновых токсинов, выделенных из ядов ос и пауков, подвергали скринингу, как описано в примере 1.
Филантотоксин-433 (+3) не оказывал влияния при концентрации 500 мкМ. По строению он схож с аргиотоксинами, описанными ниже.
Аргиотоксин-636 (400 мкМ) не вызывал повышений [Ca2+]i, но потенцировал ответы цитозольного Ca2+ на последующее добавление внеклеточного Ca2+. Это является чертой, общей для всех молекул, которые активируют рецептор Ca2+ и наблюдается также для множества внеклеточных двухвалентных катионов. Более подробно это явление рассматривается в примере 7.
В противоположность аргиотоксину-636, аргиотоксин-659 вызывал повышения [Ca2+] i с EC50 = 300 мкМ. Аргиотоксин-659 отличается от аргиотоксина-636 тем, что имеет гидроксилированную индольную часть, скорее, чем дигидроксилфенильную группу. Это является единственным структурным различием этих двух молекул. Таким образом, различие в активности заключается в природе ароматической группы, что не касается полиаминной цепи, которая несет положительный заряд.
Пример 7. Скрининг веществ, блокирующих Ca2+ каналы.
Вещества, блокирующие Ca2+ каналы, то есть молекулы, которые блокируют приток внеклеточного Ca2+ через потенциалочувствительные Ca2+ каналы, подвергали скринингу, как описано в примере 1. Существует три структурных класса веществ, блокирующих Ca2+ каналы: (1) дигидропиридины, (2) фенилалкиламины и (3) бензотиазинины.
Ни один из испытанных дигидропиридинов (нифедипин, нитрендипин, ВАУ К 8544 и (-) 202-791 и (+) 202-791) не влиял на исходную [Ca2+]i и на повышения [Ca2+]i, вызванные внеклеточным [Ca2+]i, когда их испытывали при концентрации 1 мкМ. Предыдущие исследования показали, что у клеток паращитовидной железы нет потенциалочувствительных Ca2+ каналов, но есть потенциалонечувствительные Ca2+ каналы, которые регулируются рецептором Ca2+.
Фенилалкиламины, которые изучались, были следующие: верапамил, D-600 (метоксипроизводное верапамила), TMB-8 и аналог TMB-8, NPS 384. Первые три молекулы испытывали при концентрации 100 мкМ. Фенилалкиламины вели себя иначе, чем другие изученные молекулы. Они не вызывали изменений [Ca2+]i, когда добавлялись к клеткам, орошаемым буфером, содержащим низкую концентрацию внеклеточного Ca2+ (0,5 мМ). Однако верапамил, D-600 и TMB-8 потенцировали мобилизацию внутриклеточного Ca2+, вызванную внеклеточными двухвалентными катионами, и, вдобавок, блокировали приток внеклеточного Ca2+. При промежуточных уровнях внеклеточного Ca2+ (1-1,5 мМ) эти молекулы были способны вызвать маленькое, но устойчивое повышение [Ca2+] i, которое появлялось вследствие мобилизации внутриклеточного Ca2+.
Фенилалкиламины действовали иначе, чем органические поликатионы, такие как неомицин. Данные исследований наводят на мысль, что верапамил, D-600 и TMB-8 обладают лишь частичным сродством к Ca2+ рецептору, в противоположность остальным испытанным молекулам, которые обладают полным сродством к этим рецепторам.
Молекула NPS 384 в концентрации 300 мкМ не вызывала повышения [Ca2+]i, но блокировала приток внеклеточного Ca2+. Испытания при более высоких концентрациях могут выявить способность этой молекулы вызывать мобилизацию внутриклеточного Ca2+.
Несмотря на то, что способность этих молекул блокировать приток представляет интерес и не явилась полной неожиданностью, важна, тем не менее, способность этих молекул вызывать кратковременные повышения [Ca2+] i (вследствие мобилизации внутриклеточного Ca2+). Значительный опыт в измерениях [Ca2+]i в клетках паращитовидной железы показывает, что кратковременные повышения [Ca2+]i почти неизменно являются результатом мобилизации внутриклеточного Ca2+ и, следовательно, отражают активацию рецептора Ca2+.
Испытанный бензотиазипин, дилтиазем, был во всех отношениях сходен с верапамилом и D-600 и также был эффективным при 100 мкМ.
Следует заметить, что, за исключением фенилалкиламинов, все вышеназванные активные изученные молекулы вызывали повышение [Ca2+]i, которое по величине соответствовало повышению, которое вызывала максимально эффективная концентрация внеклеточного Ca2+. Это свидетельствует о том, что эти молекулы обладают равной эффективностью с внеклеточными двухвалентными катионами. Это контрастирует с активностью фенилалкиламинов, которые, как представляется, действуют как вещества, обладающие лишь частичным сродством.
Для фенилалкиламинов выясняются некоторые интересные взаимоотношения между структурой и активностью. Значительными являются различия в активности молекул, подобных TMB-8 и NPS 384. TMB-8 потенцировала кратковременные повышения [Ca2+] i при 100 мкМ, в то время как NPS 384 не могла этого сделать даже при 300 мкМ, несмотря на то, что эти молекулы обладают одинаковым суммарным положительным зарядом. Из этого следует, что некоторые другие особенности строения, не связанные с суммарным зарядом, сообщают молекуле TMB-8 более высокую активность.
Пример 8. Скрининг молекул на клетках паращитовидной железы человека.
Спермин и неомицин испытывали на влияние на [Ca2+]i на клетках паращитовидной железы человека, полученных из желез, удаленных хирургическим путем и приготовленных, как описано в примере 1. Было обнаружено, что в клетках паращитовидной железы человека спермин вызывает только малое повышение [Ca2+]i при испытаниях при концентрации 300 мкМ.
С другой стороны, неомицин вызывал большое повышение [Ca2+]i в клетках паращитовидной железы человека, когда испытывался при концентрации 20 мкМ. Величина ответа, вызванного неомицином, была равна величине, вызванной максимально эффективной концентрацией внеклеточного Ca2+.
Пример 9. Скрининг молекула на ооцитах Xenopus.
Ооциты, инъецированные мРНК из клеток паращитовидной железы человека, экспрессируют Ca2+ рецептор и мобилизуют внутриклеточный Ca2+ в ответ на целый ряда внеклеточных неорганических двух- и трехвалентных катионов. Использование этого скрининга позволяет производить испытания воздействия непосредственно на Ca2+ рецептор. Ооциты, экспрессирующие Ca2+ рецептор, также реагировали на несколько молекул, активных в отношении интактных клеток паращитовидной железы, при таком скрининге. Гексациклен вызывал мобилизацию внутриклеточного Ca2+ при концентрации 135 мкМ. Неомицины (100 мкМ) и NPS 382 (5 мМ) также были эффективными. Это является довольно убедительным свидетельством того, что эти молекулы действуют на рецептор Ca2+ или на какой-то другой белок, тесно связанный с его функцией.
Например, мы имели возможность определить экспрессию Ca2+ рецептора в ооцитах измерением мобилизации 45Ca2+. В этих экспериментах в ооциты инъецировали мРНК из паращитовидной железы КРС или воду и, спустя 72 ч, подвергали воздействию сыворотки или 10 мМ неомицина. Перед стимуляцией ооциты нагружали 45Ca2+. Стимуляция сывороткой в течение 20 мин приводила к высвобождению внутриклеточного 45Ca2+, составляющему 45% повышения по сравнению с результатом стимуляции буфером. Стимуляция 10 мМ неомицином в течение 20 мин приводила к 76%-ному повышению высвобождения 45Ca2+. Анализ достаточно чувствителен для применения в клонировании Ca2+ рецептора и имеет преимущество в виде более высокой пропускной способности, чем электрофизиологическое измерение тока Cl-, активированного Ca2+.
В другом примере получали ткань остеобластомы человека при биопсии костной ткани. Ооциты инъецировали мРНК, изолированной из этой ткани, и стимулировали 30 мМ Ca2+. Контроли не реагировали, в то время как 8 из 12 ооцитов, инъецированных мРНК, отвечали соответствующим образом (фиг. 34). Эти эксперименты представили первое доказательство того, что ответ Ca2+ в остеокластах на внеклеточный Ca2+ на самом деле кодируется генетически. Результаты также указывают на то, что Ca2+ рецептор остеокласта можно клонировать экспрессией в ооцитах Xenopus.
Пример 10. Скрининг молекул в остеокластах крысы.
Различная чувствительность к внеклеточному Ca2+ клеток паращитовидной железы и остеокластов наводит на мысль, что их Ca2+ рецепторы различаются. В то время как клетки паращитовидной железы реагируют на концентрации внеклеточного Ca2+ между 0,5 и 3 мМ, остеокласты реагируют, только когда уровень внеклеточного Ca2+ превышает 5 мМ. Эта довольно высокая концентрация Ca2+, тем не менее, физиологична для остеокластов; поскольку они разрушают костную ткань, локальная концентрация внеклеточного Ca2+ может достигать уровней около 30 мМ.
Скрининг молекул в остеокластах выполняли следующим образом. Остеокласты получали из длинных костей новорожденных крысят. [Ca2+]i измеряли в отдельных клетках, используя флюориметрический индикатор индо-1. Испытывали спермин, спермидин, неомицин и верапамил, и ни один из них не вызывал значительного повышения [Ca2+] i в остеокластах (хотя малые ответы определялись).
При концентрации 1 мМ спермидин вызывал малое повышение [Ca2+]i (около 10% от того, которое вызывает максимальная концентрация внеклеточного Ca2+). Ни неомицин, ни спермин (10 мМ и 10 или 20 мМ, соответственно) не вызвали повышений [Ca2+]i в остеокластах крысы. Неомицин (10 мМ) не блокировал повышения [Ca2+]i, вызванного последующим добавлением 25 мМ внеклеточного Ca2+. Предварительная обработка спермином (20 мМ), однако, подавляла ответ на внеклеточный Ca2+. Верапамил (100 мкМ) не вызывал уловимых повышений [Ca2+] i, но блокировал ответ на внеклеточный Ca2+.
Сравнения остеокластов и клеток паращитовидной железы показали, что молекулы, активные в отношении последних, относительно неэффективны в остеокластах. Это показывает, что лекарства, которые нацелены на специфичный Ca2+ рецептор, не воздействуя в то же время на другие типы рецепторов в других Ca2+ - чувствительных клетках, уже получены. Подобно этим, лекарства, активные в отношении двух или более таких Ca2+ рецепторов, также могут быть разработаны.
Примеры других Ca2+ рецепторов.
Следующие примеры показывают, что, коль скоро существуют подтипы рецепторов для молекулярных лигандов, существуют и подтипы Ca2+ рецепторов, на которые можно по-разному воздействовать лекарствами. Ca2+ рецептор клетки паращитовидной железы улавливает уровни внеклеточного Ca2+ около 1,5 мМ, в то время как Ca2+ рецептор остеокласта отвечает на уровни около 10 мМ (фиг. 22). Неомицин или спермин, которые активируют рецептор Ca2+ клеток паращитовидной железы, не могли повлиять на Ca2+ рецепторы C-клеток или остеокластов (фиг. 21 и 23). Эти данные составляют первое доказательство существования фармакологически различающихся подтипов Ca2+ рецепторов и применяются для разработки лекарств, которые избирательно действуют на определенный тип Ca2+ рецепторов. На самом деле, испытания основных молекул демонстрируют такие клеточноспецифичные эффекты. Например, NPS 449, которая повышает [Ca2+]i в остеокластах, не оказывает влияния на [Ca2+]i в клетках паращитовидной железы. И наоборот, NPS 447, которая активирует Ca2+ рецепторы клеток паращитовидной железы, эффективна для активации Ca2+ рецепторов остеокластов только при концентрации в 10 раз выше. И наконец, агатоксин 489, несмотря на то, что он не очень способен активировать Ca2+ рецепторы C-клеток (EC50 = 150 мкМ), является достаточно патентным активатором Ca2+ рецепторов клеток паращитовидной железы (EC50 = 3 мкМ). Основные молекулы, находящиеся в настоящее время в стадии разработки, будут избирательно действовать на активность Ca2+-чувствительных клеток специфичного типа in vivo.
Лекарства с меньшей специфичностью не обязательно станут нежелательными в терапевтическом смысле. Так, подавление активности остеокластов и стимулирование секреции кальцитонина представляют два различных подхода к ингибированию костной резорбции. Лекарства, мишенями для которых служат Ca2+ рецепторы в обоих этих типах клеток, могут быть очень эффективным лекарственным средством при остеопорозе. Поскольку ПТГ также участвует в костном метаболизме, лекарства, действующие на Ca2+ рецепторы клеток паращитовидной железы, также могут быть полезны для лечения и/или профилактики остеопороза.
Результаты испытаний некоторых молекул показаны ниже. В табл. 3 показана сравнительная активность кальцимиметических молекул. Клетки паращитовидной железы КРС и C-клетки (rМТ 6-23) нагружали фура-2, а остеокласты крысы - индо-1, и активность указанных молекул в мобилизации внутриклеточного Ca2+ определяли построением кривой кумуляции ответа концентрации. Молекулы, обозначенные как "неактивна", не повышали [Ca2+]i, когда испытывались при концентрации 1 мМ.
Пример 11. Основные молекулы для Ca2+ рецептора клетки паращитовидной железы.
Исследования структуры и активности полиаминов и арилалкиламинов привели к испытаниям молекул, структурно похожих на NPS 456. NPS 456 является мощным активатором Ca2+ рецептора клетки паращитовидной железы. Эта молекула примечательна тем, что она имеет только один положительный заряда и при этом значительно сильнее многих многоосновных молекул. Краткая (2 мин) предварительная обработка ФМА сдвигает кривую ответа концентрации для NPS 456 вправо. Это означает, что NPS 456 действует посредством того же механизма, что и внеклеточный Ca2+. NPS 456 вызывает мобилизацию внутриклеточного Ca2+ в ооцитах Xenopus, экспрессирующих Ca2+ рецептор клетки паращитовидной железы, что свидетельствует с непосредственном действии на Ca2+ рецептор (фиг. 33). Кроме того, NPS 456 содержит хиральный углерод и, следовательно, существует в двух изомерических формах. Оба изомера были синтезированы и испытаны на активность. R-изомер, NPS 447, был в 12 раз сильнее S-изомера, NPS 448 (фиг. 28). Это является первой демонстрацией того, что Ca2+ рецептор способен узнавать органическую молекулу стереоспецифическим образом.
Поскольку NPS 447 является структурно простой молекулой с избирательными и сильными воздействиями на клетку паращитовидной железы (ее Ca2+ рецепторы), исследования структуры и активности на основе этой основной молекулы просты. Цель таких исследований - создание целого ряда родственных молекул с различными характеристиками, из которых может быть выбран конечный кандидат. Такая попытка уже выявила некоторые структурные участки NPS 447, которые вносят свой вклад в активность и мощность. Например, новое соединение NPS 459 является аналогом NPS 447, меньшим по размеру (MB<240), хотя почти таким же сильным как родительская молекула, в то время как несколько других аналогов относительно неактивны. Наиболее интересные молекулы из такого аналогового поиска можно запустить в испытании in vivo для изучения влияния на секрецию ПТГ и уровень Ca2+ в сыворотке (см. примеры 15, 16, 17, 18 и 23).
Новое соединение NPS 467 является еще более маленькой молекулой, чем NPS 447, хотя первая приблизительно в 3 раза сильнее последней в том, что касается способности вызывать мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы. Подобно NPS 456, NPS 467 является рацемической смесью. Можно предвидеть, что разделение NPS 467 на ее энантиомеры приведет к появлению изомера с еще большей мощностью, чем у рацемической смеси (см. пример 16). NPS 551 - другое новое соединение, такое же модное, как NPS 467 в вызывании мобилизации внутриклеточного Ca2+ в клетках паращитовидной железы. NPS 551 является рацемической смесью, и можно предвидеть, что разделение NPS 551 на ее энантиомеры приведет к появлению изомера с еще большей мощностью, чем у рацемической смеси. Дальнейшие исследования структуры и активности молекул, родственных NPS 447, NPS 467, NPS 551 и NPS 568, как ожидается, выявят чистые изомеры с большей мощностью, чем у этих молекул в их рацематных формах.
Результаты, полученные с NPS 456 (фиг. 33) показывают, что она вызывает осциллирующие повышения тока Cl- при концентрации 100 мкМ. NPS 456 является наиболее мощной молекулой, активной в ооцитах Xenopus, экспрессирующих Ca2+ рецептор клетки паращитовидной железы. Результаты, полученные в этой экспрессирующей системе с неомицином и NPS 456, демонстрируют, что эти молекулы действуют непосредственно на Ca2+ рецептор.
Пример 12. Основные молекулы для Ca2+ рецептора остеокласта.
Стратегия, примененная для выяснения механизма действия внеклеточного Ca2+ на остеокласт, сходна с той, которая оказалась эффективной на клетках паращитовидной железы. Первые эксперименты исследовали воздействия La3+ на [Ca2+] i в единичных остеокластах крысы, нагруженных флюориметрическим индикатором индо-1. Как описано выше, трехвалентные катионы, такие как La3+, не проникают в клетку и блокируют приток Ca2+. Низкие микромолярные концентрации La3+ частично подавляли повышения [Ca2+]i, индуцированные внеклеточным Ca2+ (фиг. 29). Демонстрация La3+ - резистентного повышения [Ca2+]i является свидетельством мобилизации внутриклеточного Ca2+. Результаты этих экспериментов соответствуют тем, которые получены на клетках паращитовидной железы и предполагают, что внеклеточный Ca2+ использует в обоих клеточных типах для регуляции [Ca2+]i сходные механизмы.
Другая серия экспериментов показала, что внеклеточный Mn2+ вызывал кратковременные повышения [Ca2+]i (фиг. 30a), которые сохранялись в отсутствии внеклеточного Ca2+ (фиг. 30b). Хотя Mn2+ способен проникать в некоторые клетки, маловероятно, что это произошло, в остеопласте, поскольку Mn2+ гасит флюоресценцию индо-1. Поэтому, если Mn2+ проник бы в клетку, наблюдалось бы понижение, а не повышение флюоресцентного сигнала.
Результаты, полученные с различными двух- и трехвалентными катионами, имеют отношение к присутствию Ca2+ рецептора на поверхности остеокласта, который связан с мобилизацией внутриклеточного Ca2+ и притоком внеклеточного Ca2+ через потенциалонечувствительные каналы. Результаты свидетельствуют о генетическом материале в остеокластах человека, который кодирует Ca2+ - рецепторный белок (см. ниже). Кратковременные повышения [Ca2+]i, получающиеся в результате мобилизации внутриклеточного Ca2+, достаточны для ингибирования остеокластической костной резорбции in vitro. Таким образом, как и в случае клеток паращитовидной железы, активация Ca2+ рецептора оказывается жизнеспособным средством ингибирования активности остеокластов.
NPS 449 в настоящее время является основной молекулой для разработки кальцимиметических лекарств для этого рецептора. Это небольшая молекула (MB<425), и она мобилизует внутриклеточный Ca2+ в остеокластах крысы с EC50 = 200 мкМ (фиг. 31a и 31b). Несмотря на то, что сила NPS 449 относительно мала, она обладает простой структурой и только одним положительным зарядом и, как ожидается, будет обладать желательными фармакодинамическими и фармакокинетическими свойствами.
NPS449 испытывали на способность ингибировать костную резорбцию in vitro. Это осуществляли морфометрическим анализом образования ямки на тонких срезах кортикальной кости КРС с применением сканирующего электронного микроскопа. Остеокласты крысы инкубировали 24 ч на срезах кости в присутствии или отсутствии различных концентраций NPS 449. NPS 449 вызывала зависящее от концентрации подавление костной резорбции с IC50 10 мкМ. Предвиденные результаты впервые продемонстрировали, что молекулы, действующие на этом новом участке, могут подавить остеокластическую резорбцию кости. Можно получить более мощные аналоги NPS 449, используя химию синтеза, которые можно оценить методами, описанными в настоящем документе.
Пример 13. Основные молекулы для Ca2+ рецептора C-клетки.
Активация Ca2+ рецептора C-клетки стимулирует секрецию кальцитонина, который затем действует на остеокласты, ингибируя резорбцию кости. Кальцимиметические лекарства, избирательно действующие на C-клетки, являются полезными для лечения остеопороза.
Мобилизация внутриклеточного Ca2+ применяется в качестве функционального индекса активности Ca2+ рецептора. Попытки скрининга в C-клетках облегчаются доступностью линий клеток, экспрессирующих фенотип C-клетки (например, клетки карциномы мозгового вещества щитовидной железы крысы; rМТС 6-23 клетки). Для начальных исследований выбрали три молекулы арилалкиламинов. Две из них встречаются в естественных условиях (агатоксин 489 и агатоксин 505), а другая (NPS 019) является синтетическим аналогом агатоксина. Обнаружено, что агатоксин 505 блокирует повышение [Ca2+]i, индуцированные внеклеточным Ca2+, с IC50 = 3 мкМ. Ингибирующий эффект является результатом блокирования потенциалочувствительного Ca2+ канала типа в этих клетках. Напротив, обнаружено, что агатоксин 489 мобилизует внутриклеточный Ca2+ в клетках МТС с EC50 = 150 мкМ. Он является первой обнаруженной органической молекулой, которая активирует Ca2+ рецепторы C-клеток. Его синтетический аналог, NPS 019, оказался даже более сильным и мобилизовал внутриклеточный Ca2+ с EC50 = 5 мкМ (фиг. 32). Важное обстоятельство заключается в том, что единственным структурным различием между NPS 019 и агатоксином 489 является присутствие или отсутствие гидроксильной группы. Тот факт, что такая малая разница в строении значительно влияет на потенцию молекулы, указывает на структурно специфичный связывающий участок Ca2+ рецептора. Это, в свою очередь, внушает оптимизм в деле создания очень сильных и избирательных активаторов Ca2+ рецепторов.
NPS 019, которая является маленькой молекулой (MB<500), - основная молекула для разработки кальцимиметиков для Ca2+ рецепторов C-клеток, и может быть испытана на способность стимулировать секрецию кальцитонина in vitro. Последующее испытание in vivo определит способность этой молекулы стимулировать секрецию кальцитонина и ингибировать резорбцию кости. Эти исследования in vivo будут выполнены на крысах. Результаты, полученные в этих исследованиях, которые, как можно предвидеть, будут положительными, обеспечат первое свидетельство того, что маленькая органическая молекула, действующая на новый рецептор, может стимулировать секрецию кальцитонина и подавлять резорбцию кости.
Пример 14. Кальцилитическая активность NPS 021 по отношению к клеткам паращитовидной железы.
Для того, чтобы соединение считалось кальцилитиком, оно должно блокировать воздействия внеклеточного Ca2+ или кальцимиметического соединения на чувствительные к внеклеточному Ca2+ клетки. Примером кальцилитического соединения является NPS 021, чье строение приведено на фиг. 1. В клетках паращитовидной железы КРС, нагруженных фура-2, NPS 021 блокирует повышения [Ca2+] i, вызванные внеклеточным Ca2+. IC50 NPS 021 для блокирования этого ответа составляет приблизительно 200 мкМ, а при концентрациях около 500 мкМ устраняется повышение [Ca2+] i, вызванное внеклеточным Ca2+. Важно, что NPS 021 сам по себе не вызывает никакого изменения [Ca2+]i, когда испытывается при низкой [Ca2+]i.
Пример 15. NPS 467 понижает содержание ионов кальция в сыворотке крови.
Соединения, в отношении которых было показано активирование Ca2+ рецепторов клеток паращитовидной железы КРС in vitro, были испытаны на гипокальцемическую активность in vivo. Крыс-самцов Sprague-Dawley (200 г) содержались на бедной кальцием диете в течение одной недели до получения испытуемых веществ или наполнителя в качестве контроля. Кровь брали из хвостовой вены спустя три часа после интраперитонеального введения NPS 467. Количество ионов Ca2+ в цельной крови или сыворотке измеряли анализатором Ciba-Corning 634, согласно инструкции к прибору. Общий сывороточный кальций, альбумин и фосфаты измеряли с помощью методик, хорошо известных в данной области. NPS 467 вызывало дозозависимое сокращение Ca2+ в цельной крови или сыворотке (фиг. 38). Падение Ca2+ в крови в это время шло параллельно падению уровней тотального кальция крови. Ни при одной из испытанных доз не было никаких изменений в уровнях сывороточного альбумина или фосфатов. В предварительных исследованиях NPS 467, при дозах, эффективных в понижении Ca2+ крови, вызывал дозозависимое сокращение в циркулирующих уровнях ПТГ (фиг. 39). Гипокальцемический эффект NPS 467 был максимальным в течение 3 ч и возвращался к контрольному уровню спустя 24 ч (фиг. 40).
NPS467 (РЭ изомер, см. пример 16) был также эффективен в понижении содержания сывороточного Ca2+ у крыс, которых содержали на нормальной, полноценной по кальцию диете. Единственная доза NPS 467 (РЭ изомер, 10 мг/кг внутрибрюшинно) вызывала быстрое падение сывороточных уровней ионизированного Ca2+, которое было максимальным в течение 1 ч (22% понижения от контрольного уровня) и оставалось пониженным на этом уровне или около него еще до 6 ч.
Пример 16. NPS 467 понижает содержание ионов кальция в сыворотке крови стереоспецифическим путем.
NPS 467 является рацемической смесью. Разделение NPS 467 на два его энантиомера было достигнуто сепарацией на хиральной колонке. РЭ-изомер ("ранний элюат", см. пример 21) был почти в 100 раз более сильным, чем ПЭ-изомер ("поздний элюат") в активации Ca2+ рецептора клетки паращитовидной железы КРС in vitro, что оценивалось по способности энантиомеров вызывать повышения [Ca2+] i в клетках паращитовидной железы (фиг. 41). Подобно этому, сходное разделение нового соединения NPS 568 на его энантиомеры показало, что РЭ-изомер был в 40 раз более сильным, чем ПЭ-изомер, в способности вызывать мобилизацию внутриклеточного Ca2+ в клетках паращитовидной железы КРС (см. таблицу 3, supra).
Изомеры NPS 467 испытывали на эффекты в отношении сывороточного Ca2+, как в примере 15. В соответствии с результатом in vitro РЭ-изомер NPS 467 оказался более способным, чем ПЭ-изомер, понижать сывороточный Ca2+ in vivo (фиг. 42; каждое соединение испытывали при концентрации 5 мг/кг веса тела).
Пример 17. NPS 467 понижает содержание ионов кальция в сыворотке крови в модели вторичной гиперфункции паращитовидной железы in vivo.
Принята и широко используется модель вторичной гиперфункции паращитовидной железы на животном, которая происходит вследствие хронической почечной недостаточности у крыс с удаленными на 5/6 почками. После такого хирургического вмешательства у животных сначала наблюдается гипокальциемия, а затем, для поддержания уровней сывороточного Ca2+, происходит компенсаторная гиперплазия паращитовидных желез и повышается уровень циркулирующего ПТГ. Крысы-самцы Sprague-Dawley (250 г) подвергались резекции почек на 5/6 и выздоравливали в течение 2 недель. В это время у них наблюдалась нормокальциемия (благодаря повышенным уровням сывороточного ПТГ). Введение NPS 467 (РЭ-изомер; 10 мг/кг внутрибрюшинно) вызывало быстрое (в течение 2 ч) падение сывороточного Ca2+ до 83% от контрольного у животных-моделей вторичной гиперфункции паращитовидной железы. Это наводит на мысль, что соединения такого рода будут эффективно подавлять секрецию ПТГ у пациентов с вторичной гиперфункцией паращитовидных желез и с гиперпластическими паращитовидными железами.
Пример 18: NPS 467 не понижает содержание ионов кальция в сыворотке крови у животных с удаленными паращитовидными железами.
Для того, что определить первичную ткань-мишень для действия NPS 467, при котором она вызывает гипокальциемический ответ, у крыс удаляли хирургическим путем паращитовидные железы. Животные после тотальной паратироидэктомии обнаруживали гипокальциемию и сильно зависели от получаемого с пищей кальция для поддержания гомеостаза сывороточного Ca2+. У паратироидэктомированных животных уровни сывороточного Ca2+ были 0,92 мМ, которые постепенно падали до 0,76 мМ после 6 ч голодания. Введение единичной дозы NPS 467 РЭ (10 мг/кг внутрибрюшинно) не вызывало каких-либо изменений за период в 6 ч. Эти результаты показывают, что для гипокальциемических эффектов NPS 467 РЭ требуются интактные паращитовидные железы. Эти данные дополнительно показывают, что NPS 467 РЭ может нацеливаться на паращитовидные железы in vivo. Эти результаты согласовываются с точкой зрения, что NPS 467 РЭ действует на Ca2+ рецептор паратироидной клетки, угнетая секрецию ПТГ, и, таким образом, вызывая падение уровней сывороточного Ca2+.
Пример 19. NPS 467 повышает содержание внутриклеточного кальция в паращитовидных железах человека.
Разъединенные клетки паращитовидной железы приготавливали из ткани аденомы паращитовидной железы, полученной при операциях от пациентов с первичной гиперфункцией паращитовидных желез. Клетки нагружали фура-2 и [Ca2+] i измеряли, как описывалось выше. И NPS 467 РЭ, и NPS 568 РЭ вызывали зависящее от концентрации повышение [Ca2+]i. Величины EC50 для NPS 568 РЭ составляли 20 и 3 мкМ соответственно. Таким образом, оба эти соединения способны повышать [Ca2+]i в патологической ткани человека, и, поэтому можно ожидать, что они будут понижать сывороточные уровни ПТГ и Ca2+ у пациентов с первичной гиперфункцией паращитовидных желез.
Пример 20. Механизма действия NPS 467 на кальциевый рецептор клетки паращитовидной железы.
Разъединенные клетки паращитовидной железы КРС применяли для дальнейшего изучения механизма действия NPS 467 на рецепторном уровне. В присутствии 0,5 мМ внеклеточного Ca2+ NPS 467 РЭ вызывал быстрое и кратковременное повышение [Ca2+]i, которое сохранялось в присутствии 1 мкМ La3+ и частично подавлялось предварительной обработкой ФМА (100 мМ в течение 2 мин). Кроме того, NPS 467 (РЭ-изомер, 30 мкМ) вызывал быстрое повышение проводимости Cl- в ооцитах Xenopus, инъецированных мРНК клеток паращитовидной железы. Все эти результаты согласовываются с действием NPS 467 на Ca2+ рецептор. Однако ответ цитозольного Ca2+ на NPS 467 устранялся, когда клетки паращитовидной железы суспендировали в лишенном Ca2+ буфере. Это предполагает, что NPS 467 сам по себе не может вызвать мобилизацию внутриклеточного Ca2+. Однако он вызывает ответ в клетках паращитовидной железы и ооцитах в присутствии малого количества внеклеточного Ca2+. Это наводит на мысль, что необходима частичная занятость Ca2+ - связывающего участка, чтобы NPS 467 смог вызвать ответ. Чтобы проверить это предположение, клетки паращитовидной железы суспендировали в не содержащем Ca2+ буфере и подвергали воздействию субмаксимальных концентраций неомицина. Неомицин применяли, поскольку он имитирует, почти во всех отношениях, воздействие внеклеточного Ca2+ на клетки паращитовидной железы и ооциты Xenopus, экспрессирующие Ca2+ рецептор клеток паращитовидной железы. Добавление 10 мкМ неомицина само по себе не вызывало повышения [Ca2+]i при этих условиях. Однако последующее добавление NPS 467 РЭ (30 мкМ) вызывало кратковременное повышение [Ca2+]i, которое, поскольку внеклеточный Ca2+ отсутствовал, могло происходить вследствие мобилизации внутриклеточного Ca2+. Когда клетки, которые находились в не содержащем Ca2+ буфере, подвергали воздействию 30 мкМ NPS 467, повышения [Ca2+]i не происходило. Эта концентрация NPS 467 является максимально эффективной в повышении [Ca2+]i в присутствии 0,5 мМ внеклеточного Ca2+. Однако последующее добавление 10 мкМ неомицина вызывало кратковременное повышение [Ca2+]i. Предположительно, неомицин связывает тот же участок, что и внеклеточный Ca2+ и может функционально его заменять. Применяя субмаксимальную концентрацию, которая сама по себе не вызывает ответной реакции, достигают частичной занятостью Ca2+ - связывающего участка и делают возможной активацию Ca2+ рецептора NPS 467.
Были проведены также дополнительные исследования для дальнейшего выяснения механизма действия NPS 467. Клетки еще раз суспендировали в не содержащем Ca2+ буфере, чтобы гарантировать, что любое наблюдаемое повышение [Ca2+] i происходит вследствие мобилизации внутриклеточного Ca2+. В этих экспериментах, однако, использовали максимально эффективную концентрацию (100 мкМ) неомицина. В отсутствии внеклеточного Ca2+ 100 мкМ неомицин вызывал быстрое и кратковременное повышение [Ca2+]i. Последующее добавление 30 мкМ NPS 467 РЭ не вызывало повышения [Ca2+]i. В обратном эксперименте, 30 мкМ NPS 467 РЭ добавляли раньше 100 мкМ неомицина. Как и ожидалось, NPS 467 РЭ не вызывал какого-либо повышения [Ca2+] i. Он, однако, не влиял на повышение [Ca2+]i, вызванное последующим добавлением 10 мкМ неомицина. Эти результаты, полученные с максимально эффективными концентрациями NPS 467 и неомицина, предполагают, что эти два соединения не действуют на один и тот же участок. Скорее, эти результаты можно удовлетворительно объяснить, если предположить, что существуют два отдельных участка на Ca2+ рецепторе, одного, к которому присоединяется внеклеточный Ca2+ и неомицин, и другого, к которому присоединяется NPS 467 и структурно родственно ему соединения (такие как NPS 568). Связывание лигандом первого участка может привести к полной активации Ca2+ рецептора, в то время как связывание лигандом последнего участка может наблюдаться и/или быть функционально уместным, когда участок, связывающий внеклеточный Ca2+, уже занят до некоторой, пока не определенной, степени. Возможно, что лиганд, связывающий участок, к которому присоединяется внеклеточный Ca2+, делает доступным прежде закрытый участок присоединения для NPS 467. Представляется, что NPS 467-связывающий участок является аллостерическим участком, который усиливает активацию рецептора в ответ на связывание лигандом участка присоединения внеклеточного Ca2+.
Эти данные показывают, что Ca2+ рецептор клетки паращитовидной железы обладает по крайней мере двумя различными участками для органических лигандов. Один участок связывает физиологический лиганд, внеклеточный Ca2+ и определенные органические поликатионы, такие как неомицин. Связывание этого участка приводит к полной активации Ca2+ рецептора, повышению [Ca2+]i и подавлению секреции ПТГ. NPS 467 определяет ранее неизвестный связывающий участок на Ca2+ рецепторе. Связывание этого участка может наблюдаться только и/или приводить к полной активации Ca2+ рецептора, когда участок, связывающий внеклеточный Ca2+, занят. Лиганды, связывающие тот или другой участок, являются эффективными в подавлении уровней сывороточного Ca2+ in vivo.
Пример 21. Приготовление NPS 467.
В 250 мл флакона с круглым дном смешивали 10,0 (100 ммоль) 3'-метокси-ацетофенона и 13,5 г (100 ммоль) 3-фенилпропиламина и на смесь действовали 125 ммоль (35,5 г) изопропилата титана (IV). Реакционную смесь перемешивали 30 мин при комнатной температуре под азотной атмосферой. По истечении этого времени добавляли по капле в течение 2 мин 6,3 г (100 ммоль) цианоборогидрида натрия в 100 мл этанола. Реакционную смесь перемешивали при комнатной температуре под азотом в течение 16 ч. Спустя это время реакционную смесь переносили в 2 л делительную воронку с 1,5 л этилового эфира и 0,5 л воды. Фазы уравновешивали и эфирный слой удаляли. Оставшуюся водную фазу тщательно экстрагировали четырьмя порциями по 1 л эфира. Смывы объединяли, высушивали над безводным карбонатом калия, получая прозрачное светло-янтарное масло.
Тонкослойная хроматография этого продукта на окиси кремния с применением хлороформ:метанол:изопропиламина (100:5:1) представила продукт при Rf 0,65 в виде следов двух стартовых материалов при Rf 0,99 (3'-метокси-ацетофенон) и Rf 0,0 (3-фенилпропиламин).
Реакционную смесь подвергли хроматографии через окись кремния (48 х 4,6 см) с использованием градиента хлороформ:метанол:изопропиламина (99:1:0,1), после чего собрали 13,66 г очищенного NPS 467. Этот материал растворяли в гексан-изопропаноле (99: 1), содержащем 0,1% диэтиламина для получения раствора с концентрацией 50 мг/кг. Хиральное разделение выполняли путем хроматографии 4 мл этого раствора (200 мг, максимум для достижения разделения) через Chiral Cel OD (25 х 2 см), с применением 0,7% изопропиламина, 0,07% диэтиламина в гексане при 100 мл/мин, наблюдая за оптической плотностью при 260 нм. При этих условиях (с введением 100 мг материала) ранний элюированный изомер (NPS 467 РЭ) начинал выходить из колонки на 26 минуте, поздний элюированный изомер (NPS 467 ПЭ) начинал выходить на 34 минуте. Каждый оптический изомер (свободное основание) превращали в соответствующий гидрохлорид растворением 3 г свободного основания в 100 мл этанола и воздействием им на 100 мл воды, содержащей 10 молярных эквивалентов HCl. Лиофилизация этого раствора давала твердое вещество белого цвета.
Пример 22. Приготовление NPS 568.
NPS 568 приготавливали, применяя методы, описанные в примере 21, заменяя 3-фенилпропиламин на эквивалентное количество 3-(2-хлорфенил)пропиламина. Было обнаружено, что, если позволить смеси 3'-метоксиацетофенона, 3-(2-хлорфенил)пропиламина и изопропилата титана (IV) перемешиваться 5 ч до воздействия на нее NaCNBH3/EtOH, то выход продукта значительно увеличивается (98%).
Пример 23. NPS 467 понижает содержание ионов кальция в сыворотке крови при оральном применении.
Крысы (самцы, Sprague-Dawley 250-300 г) получали стандартный рацион и выдерживались без пищи в течение ночи перед экспериментом. NPS 467 РЭ суспендировали в кукурузном масле и вводили разовой оральной дозой через зонд. Спустя три часа брали образцы крови из хвостовой вены и оценивали уровни ионов Ca2+. Фиг. 44 показывает, что NPS 467 РЭ вызывал дозозависимое понижение уровней ионизированного Ca2+ в сыворотке крови, при оральном применении.

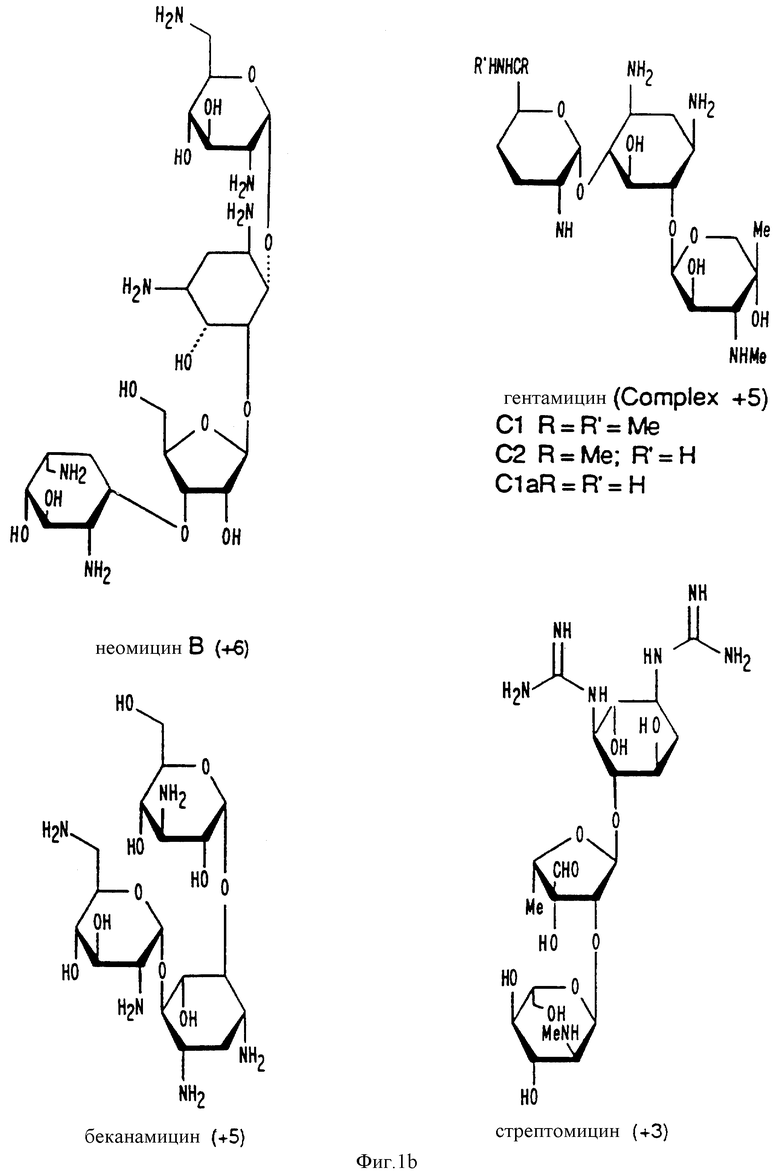
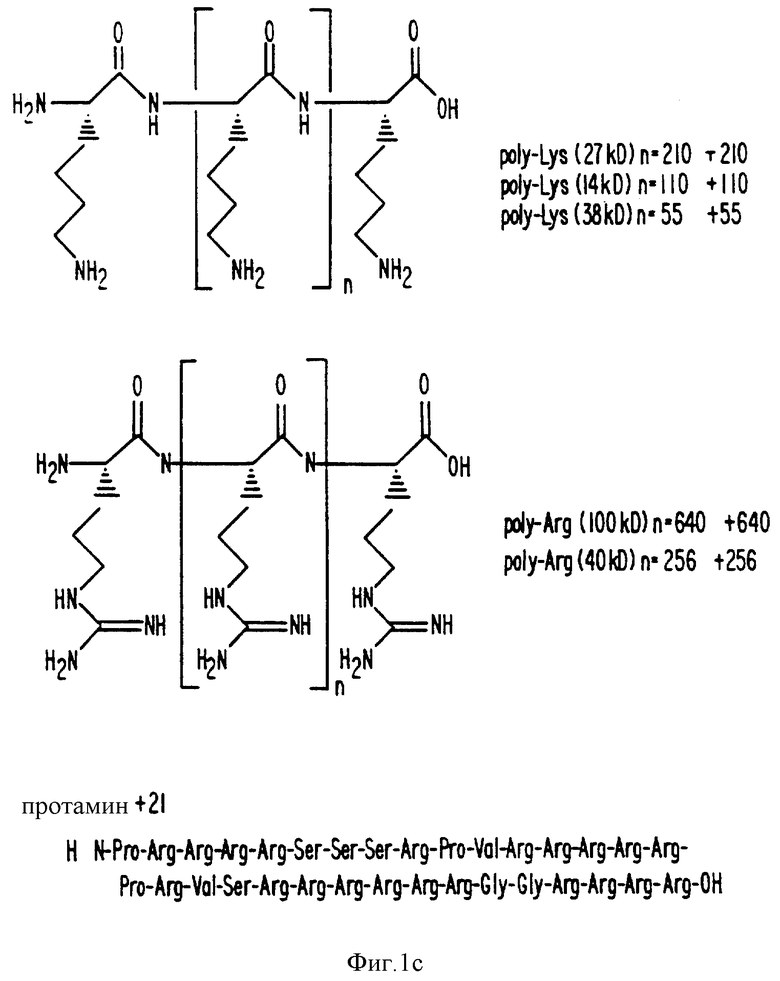
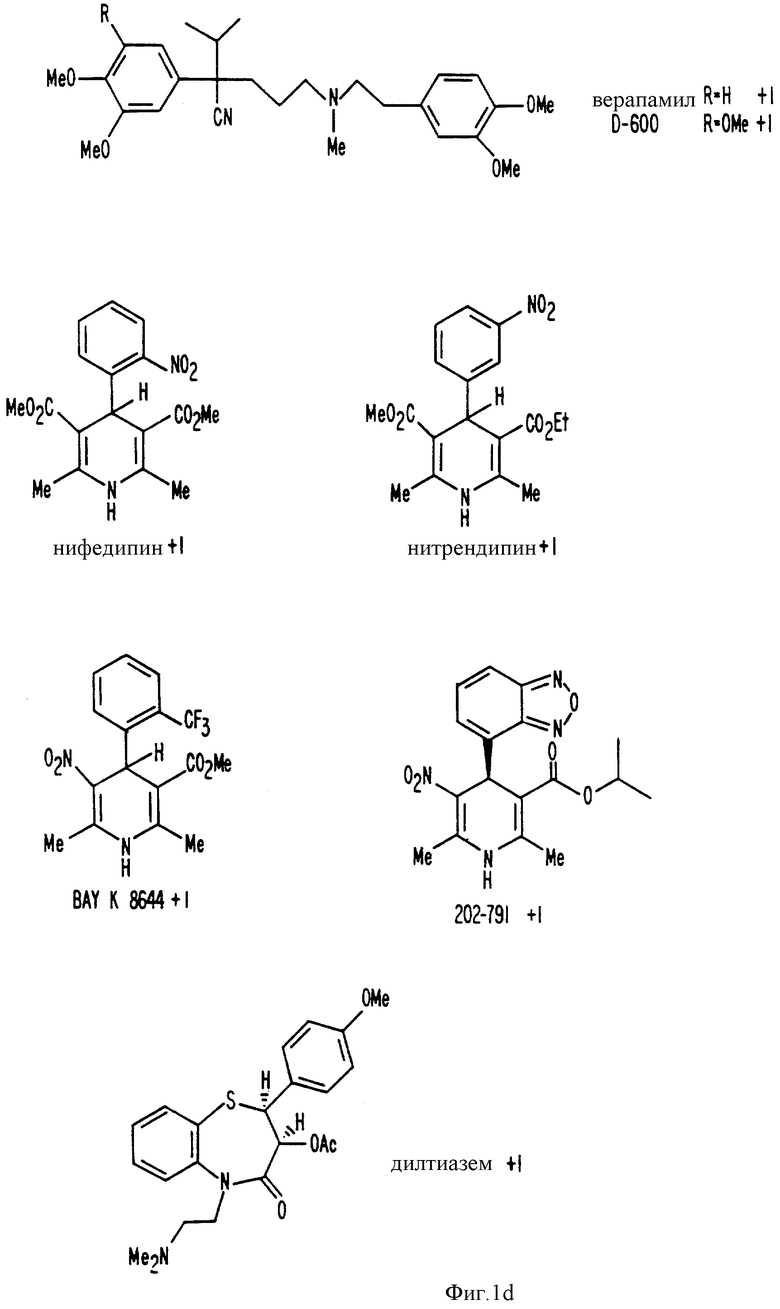
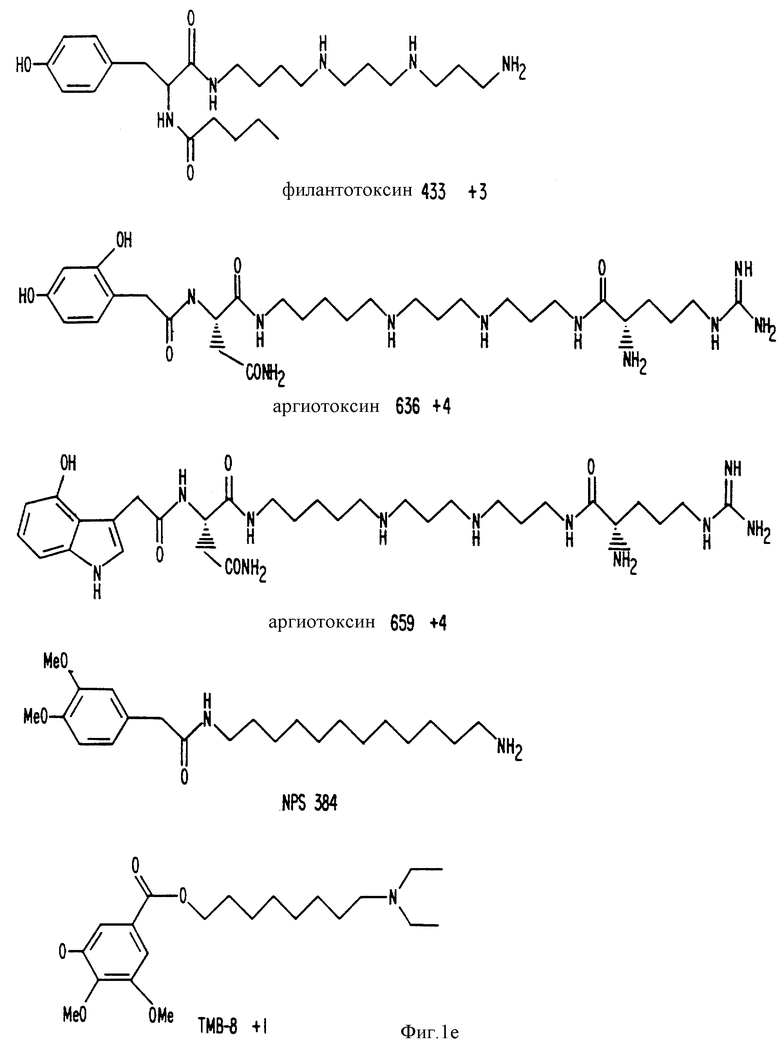
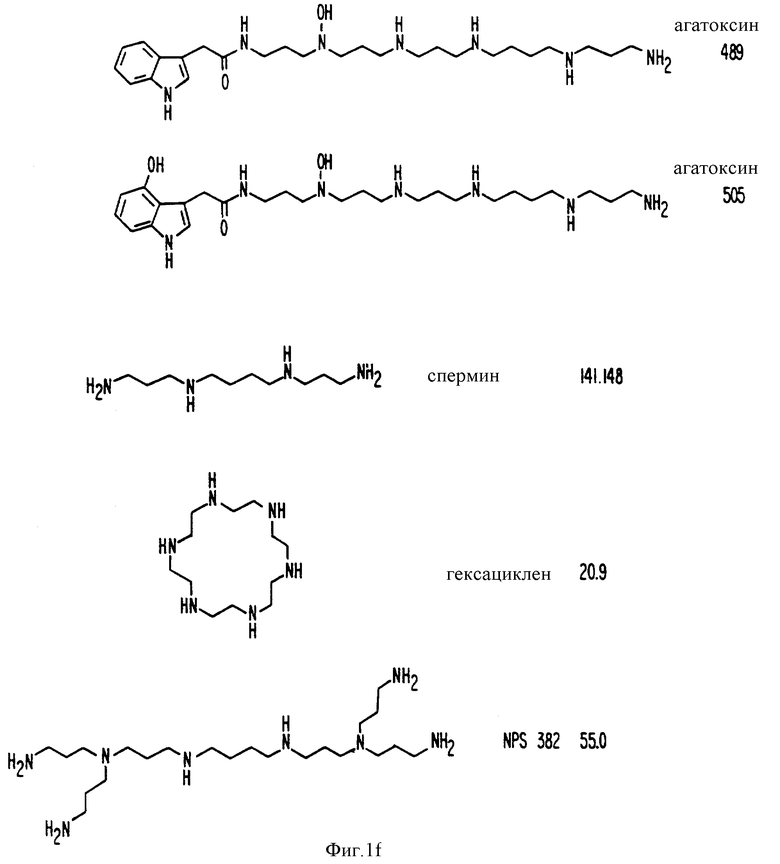

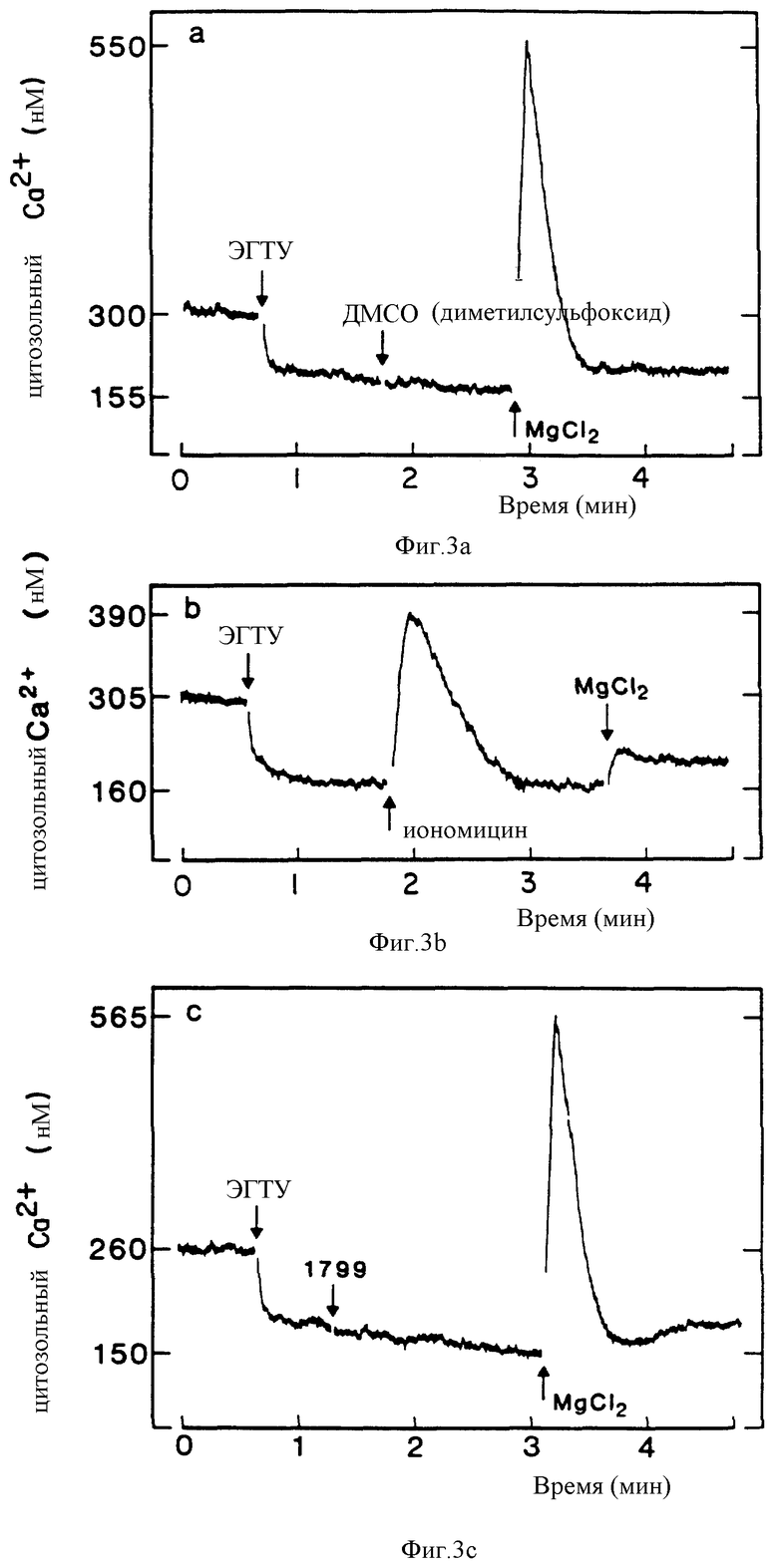

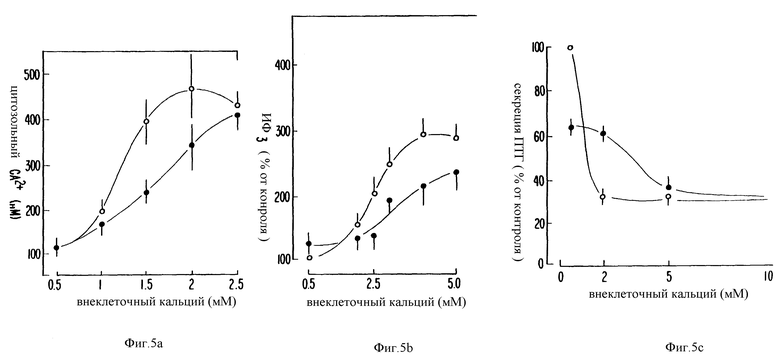
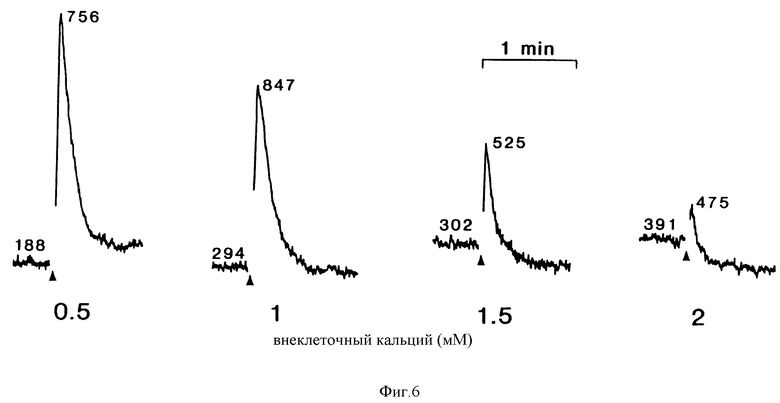
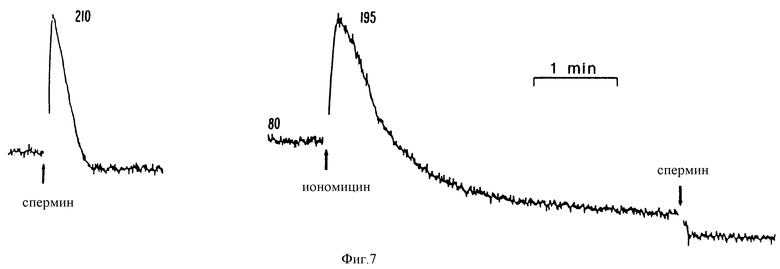

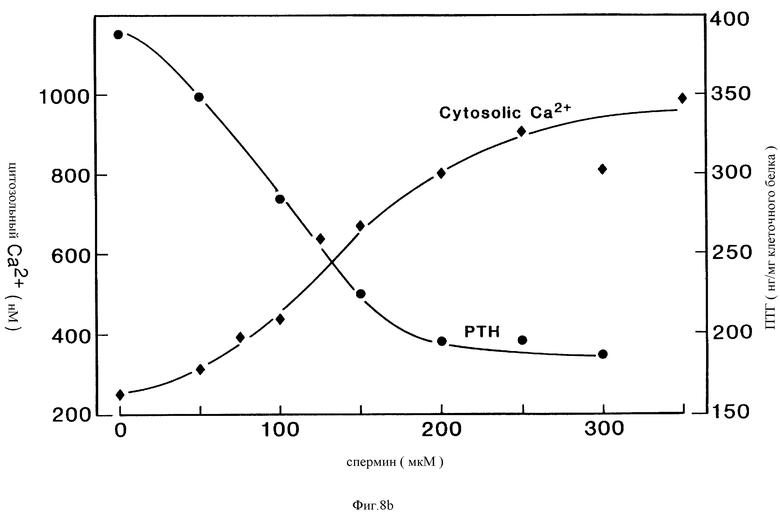

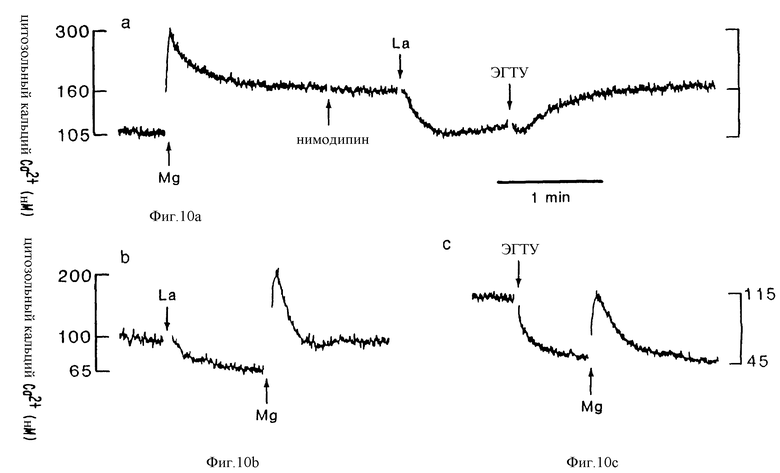
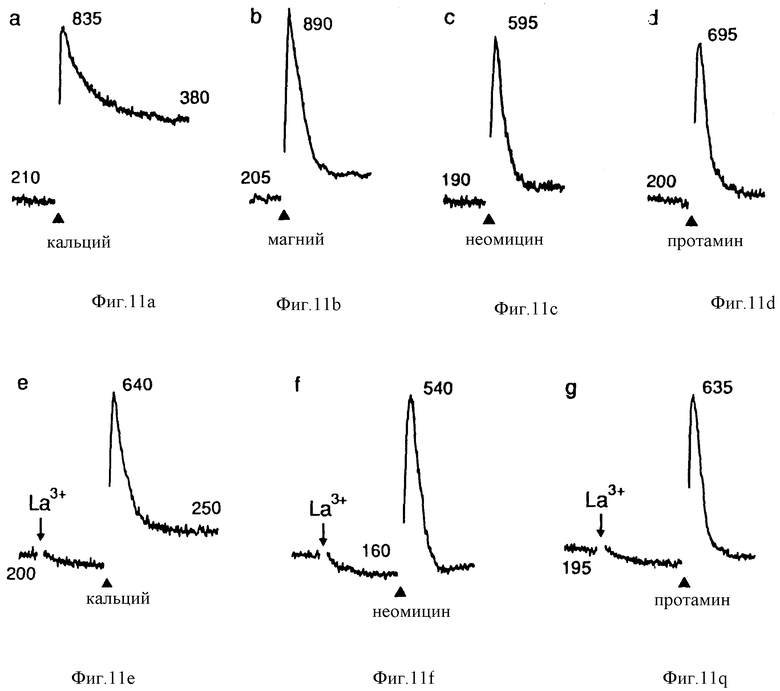
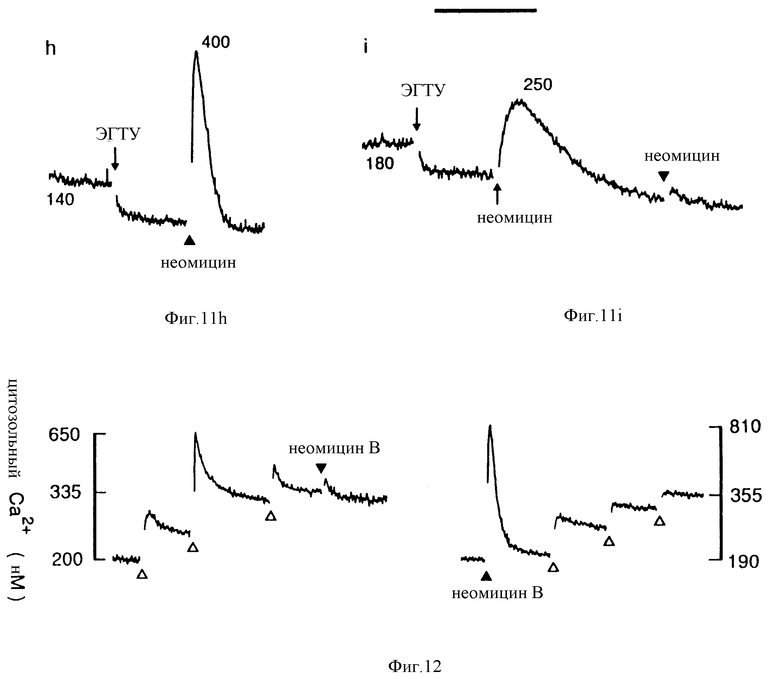

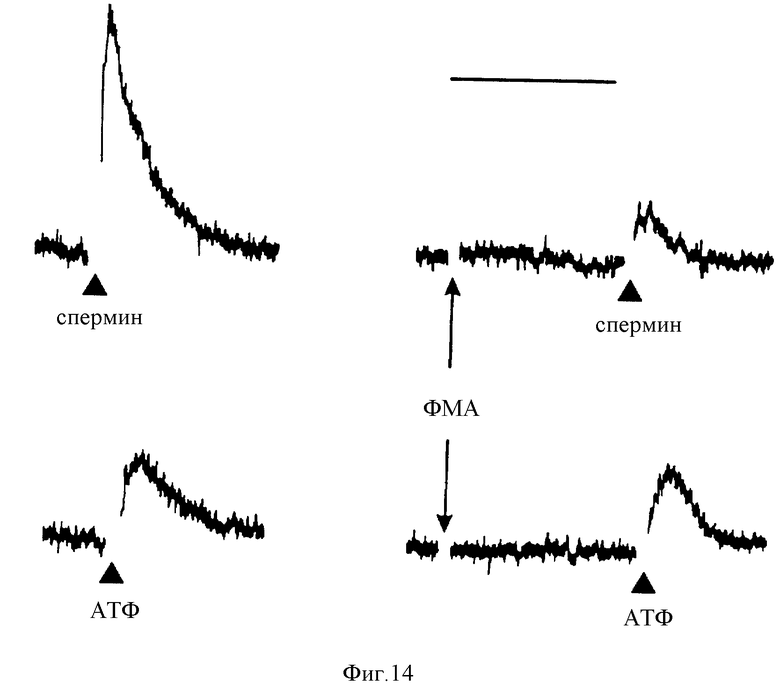
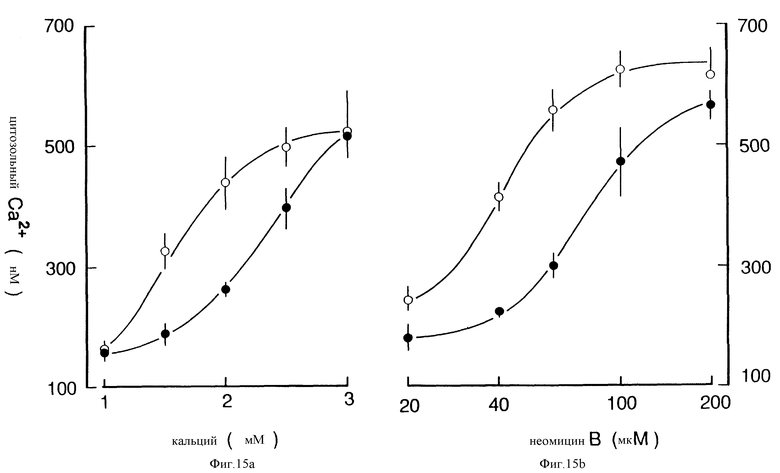

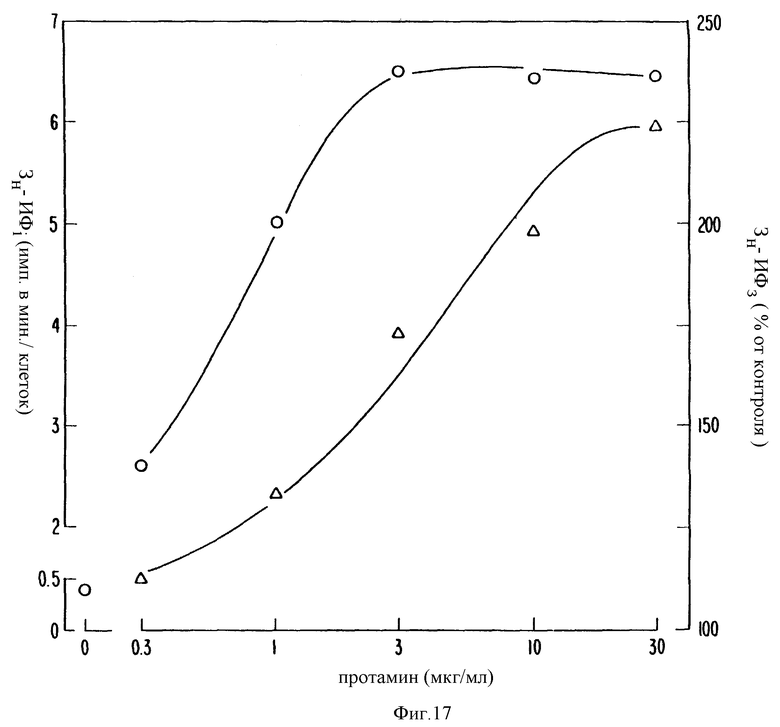


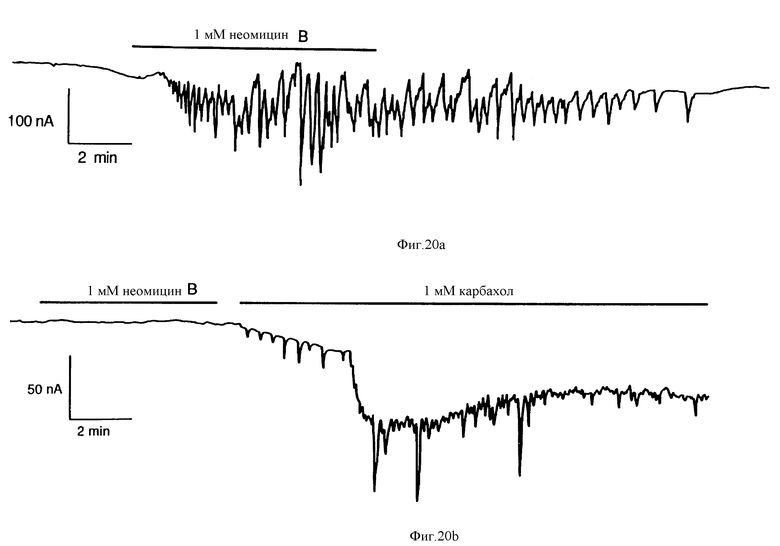
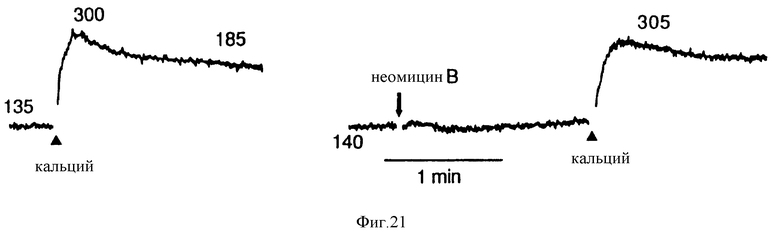

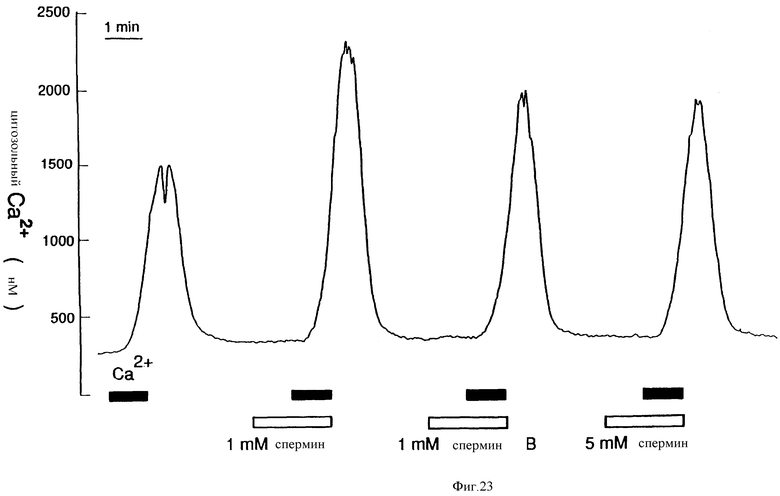


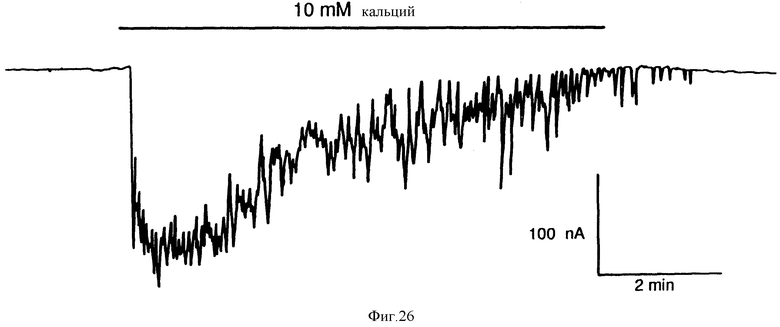
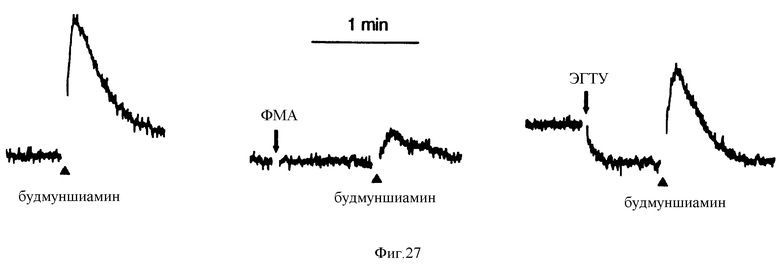
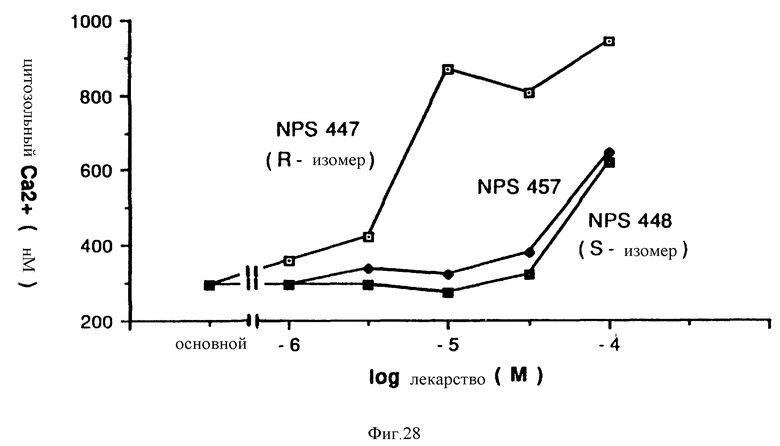
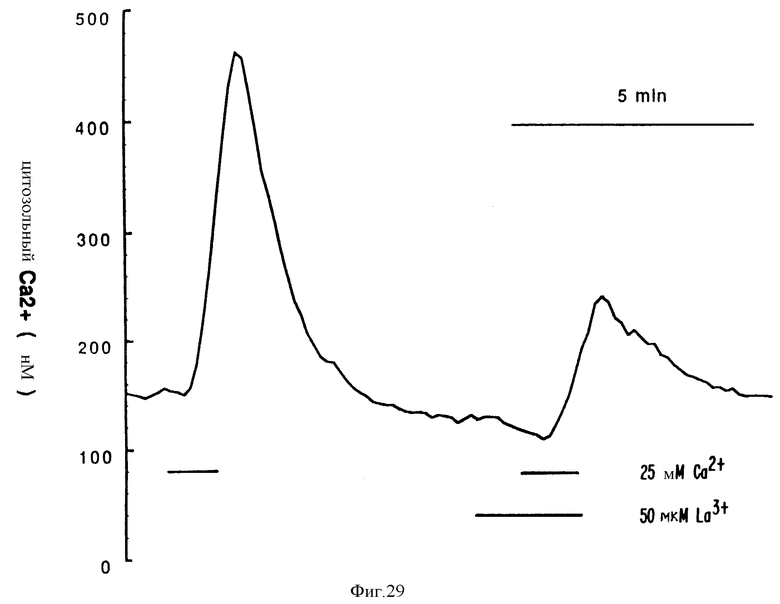
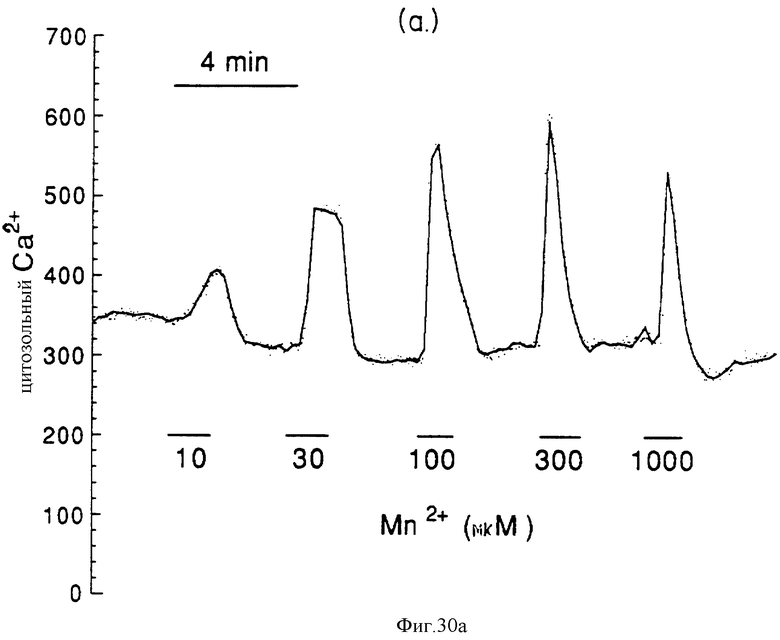


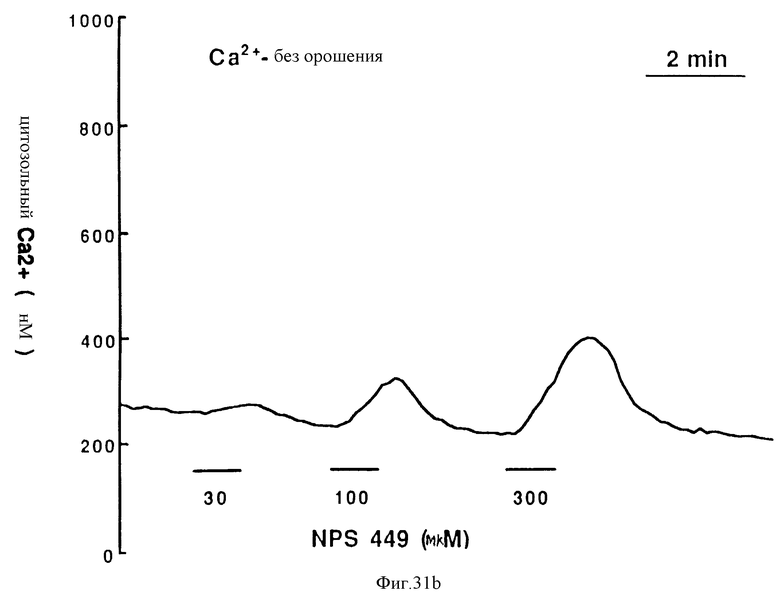


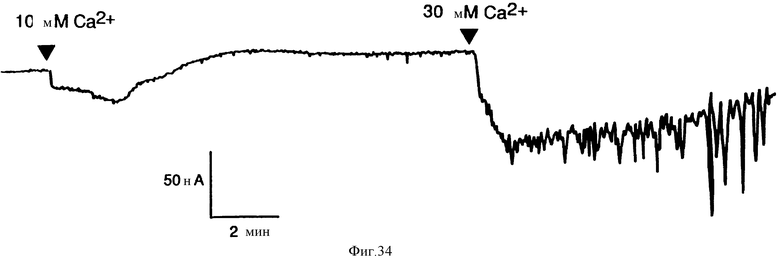

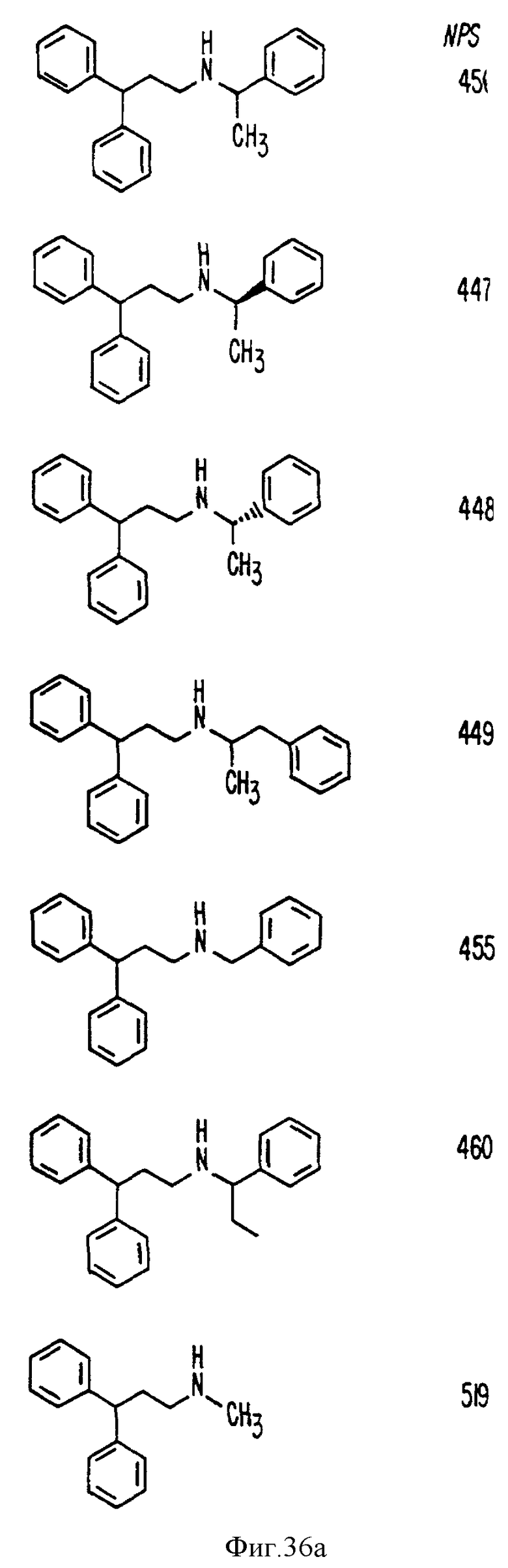

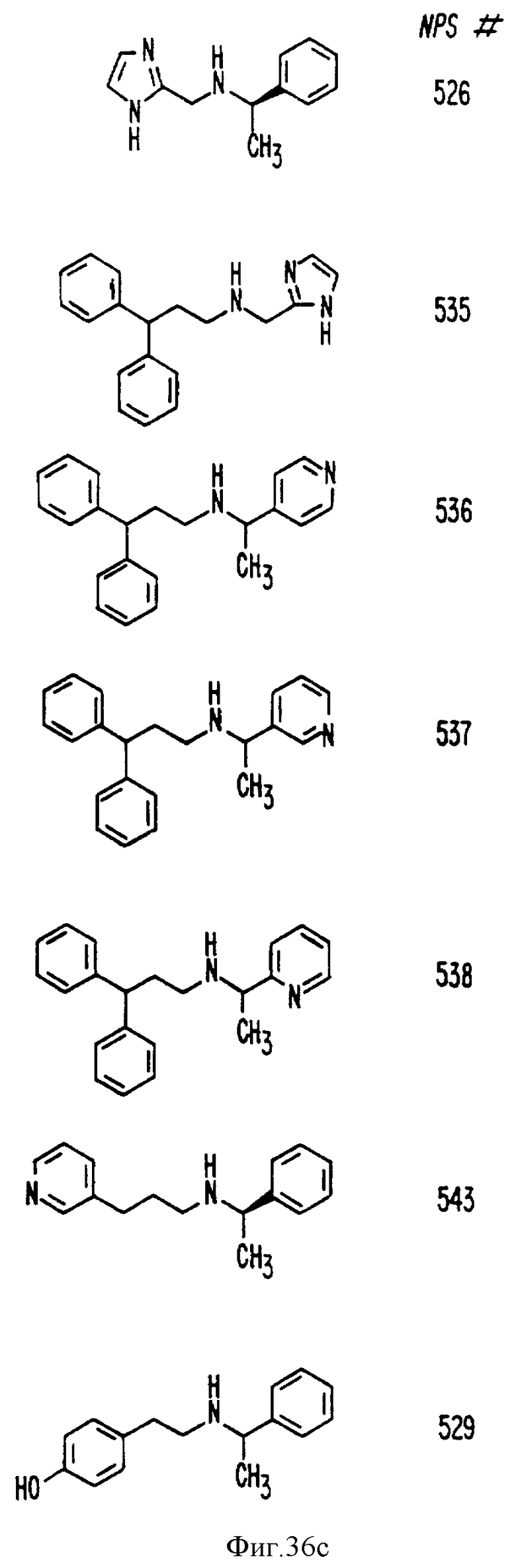

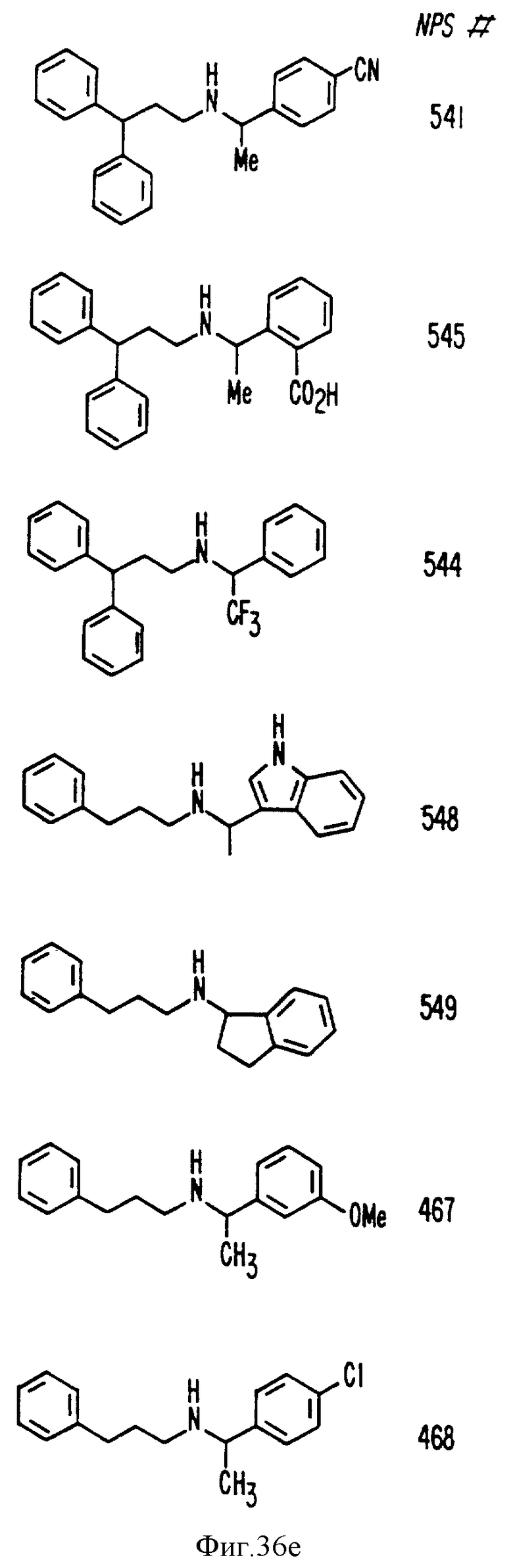

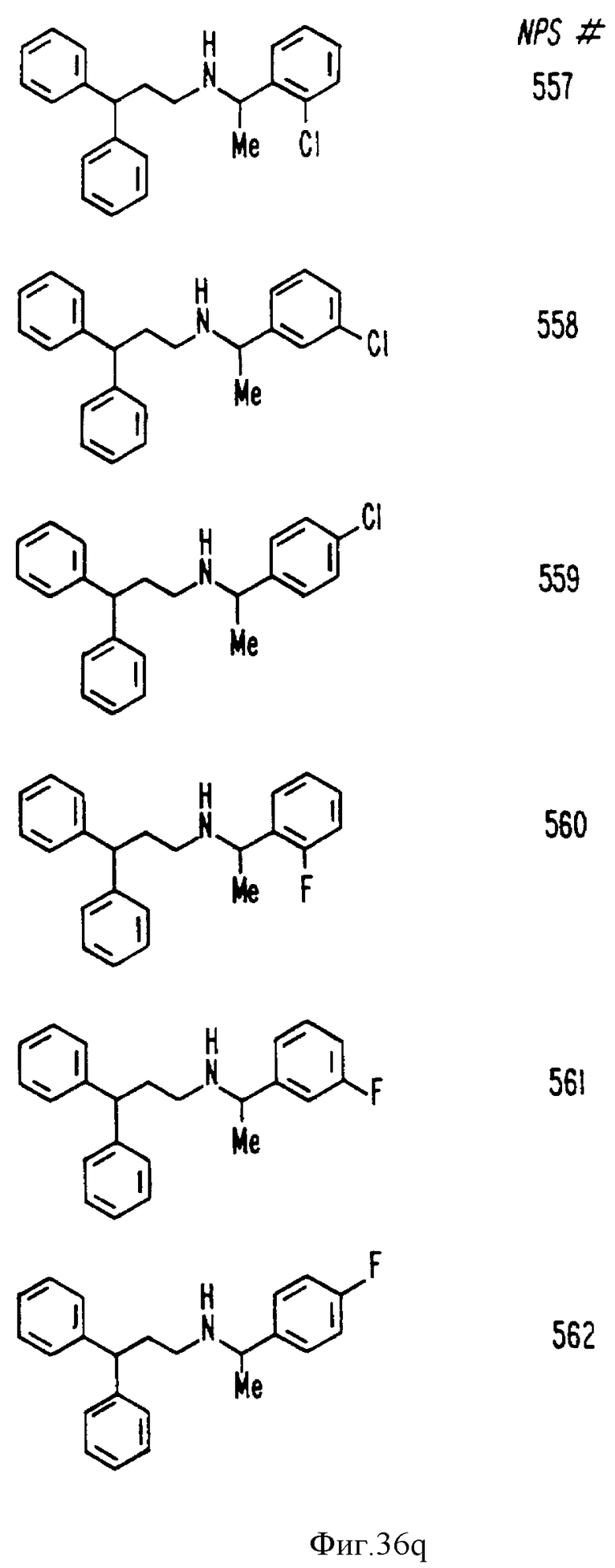
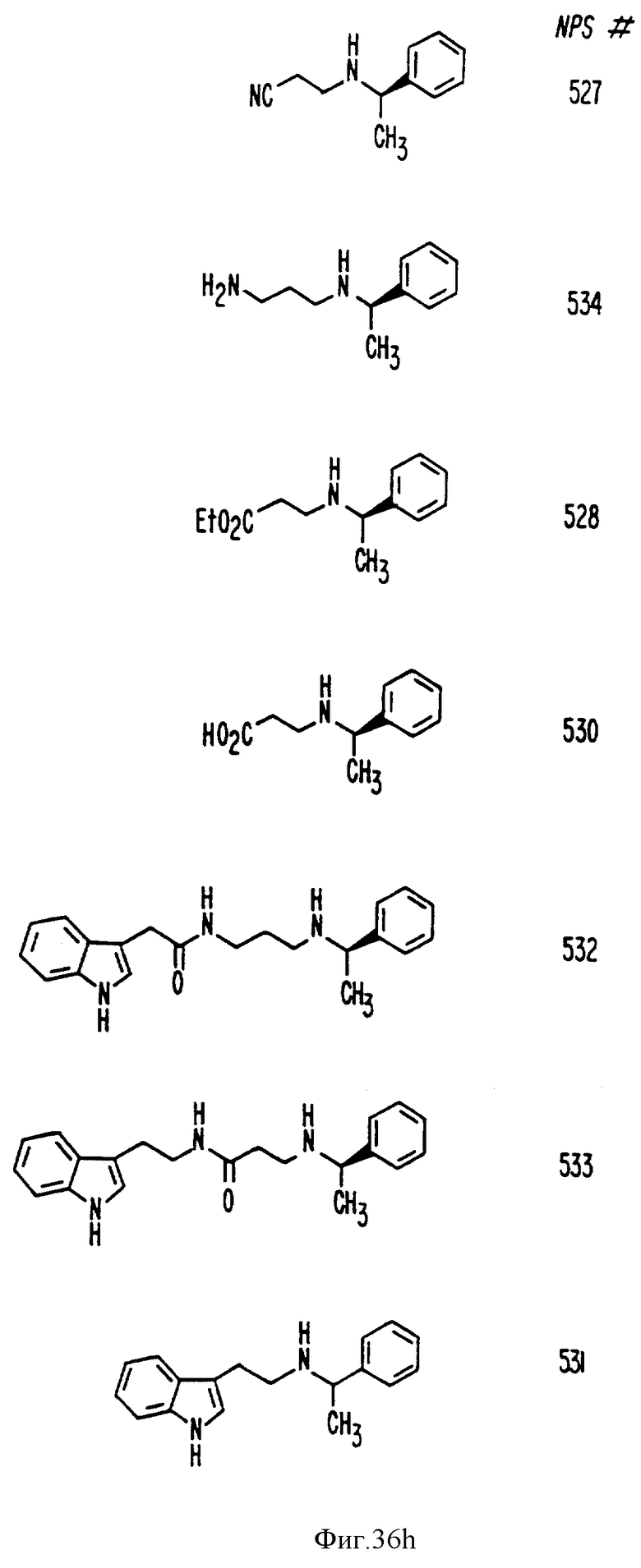
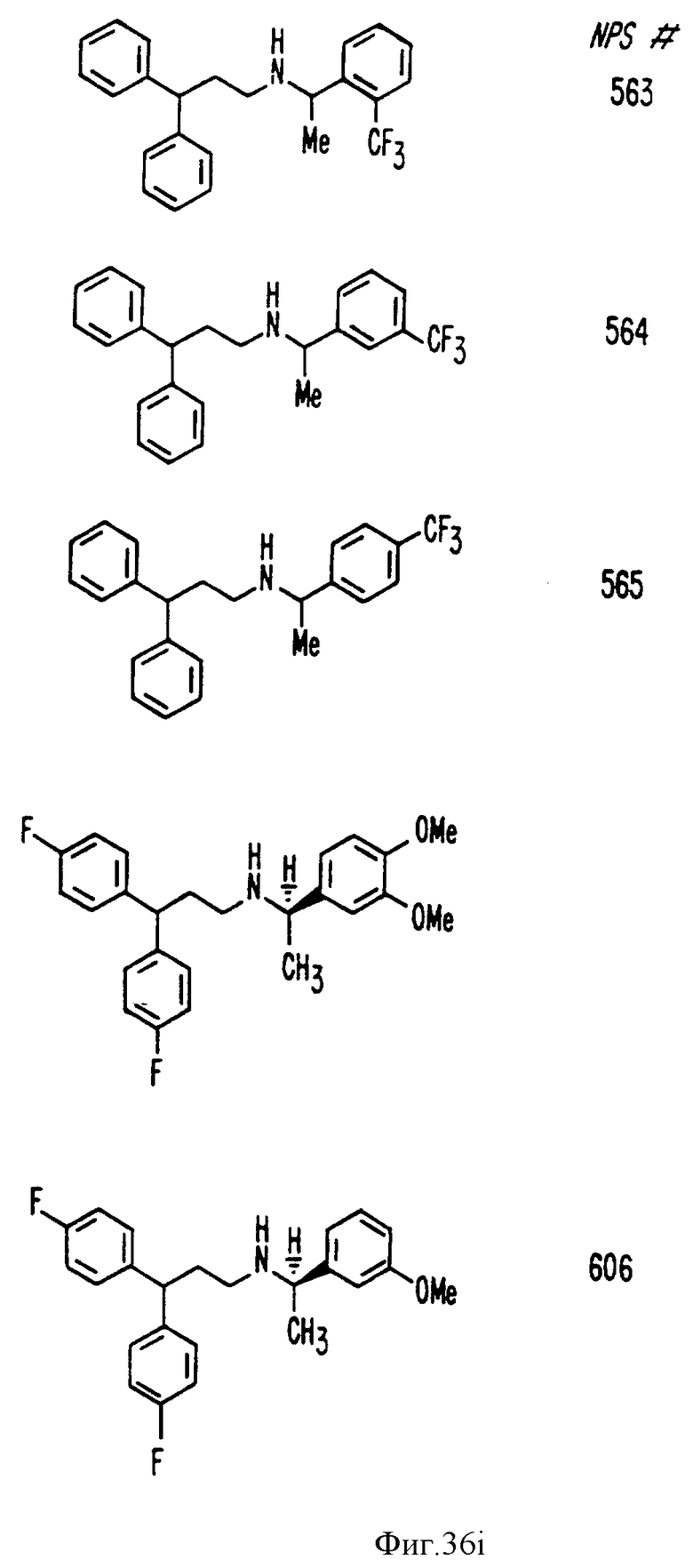
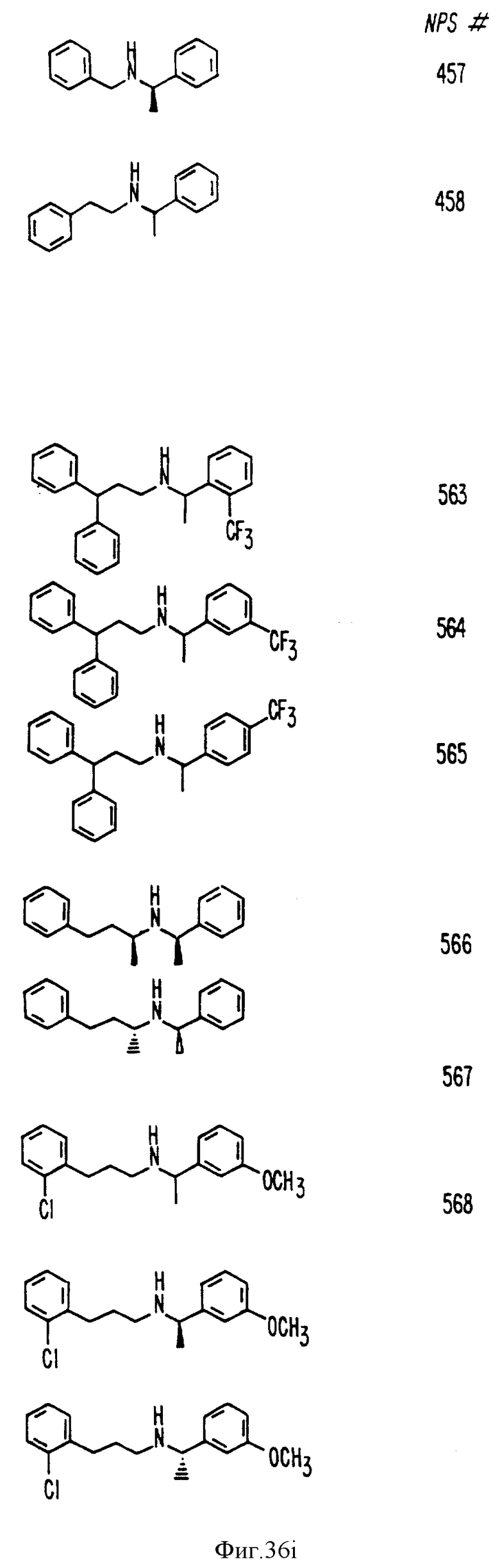
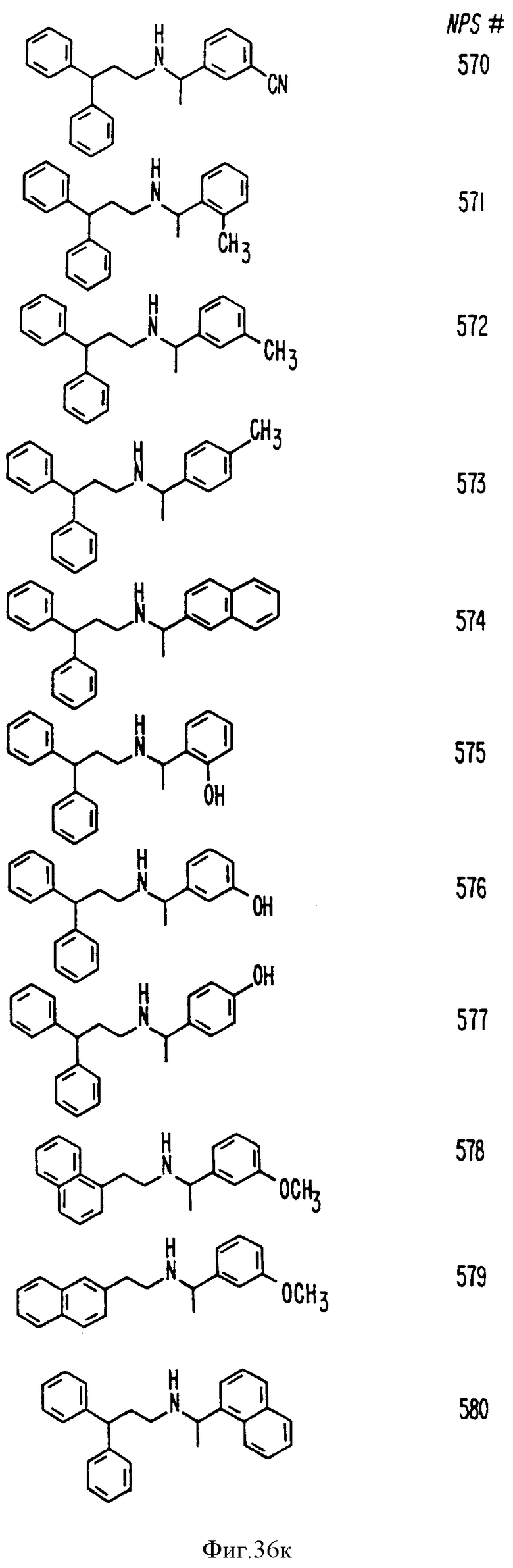


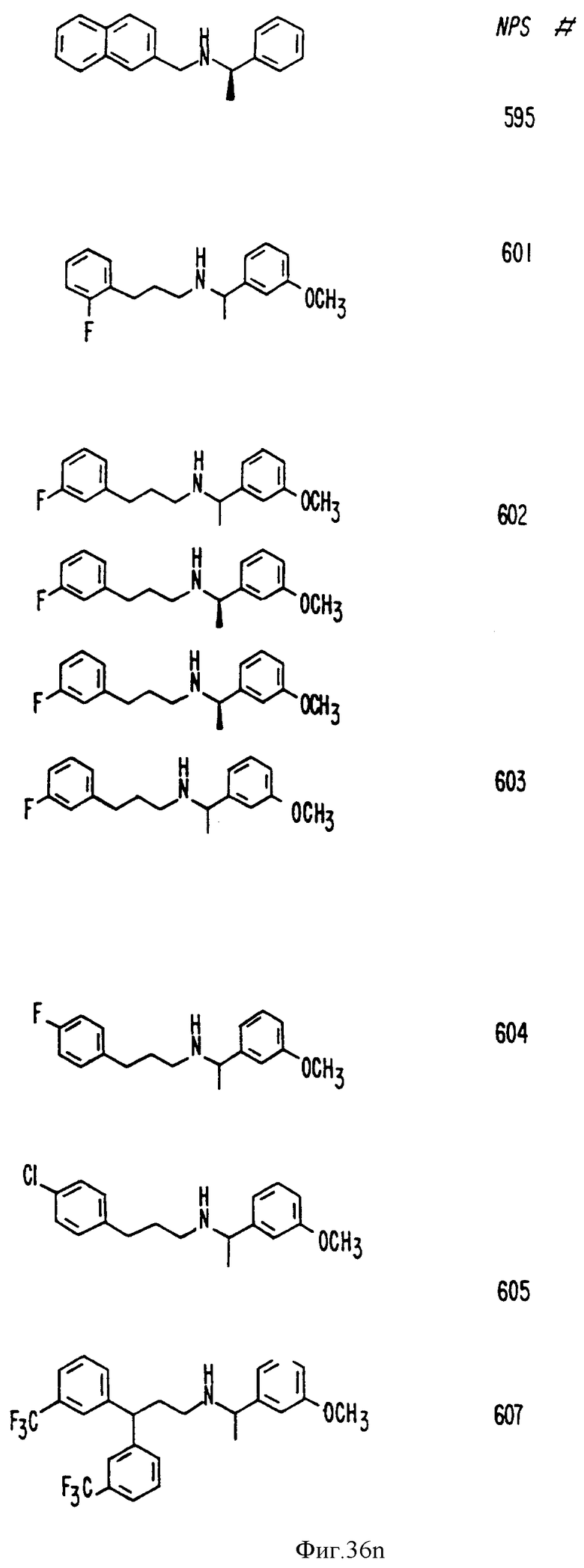
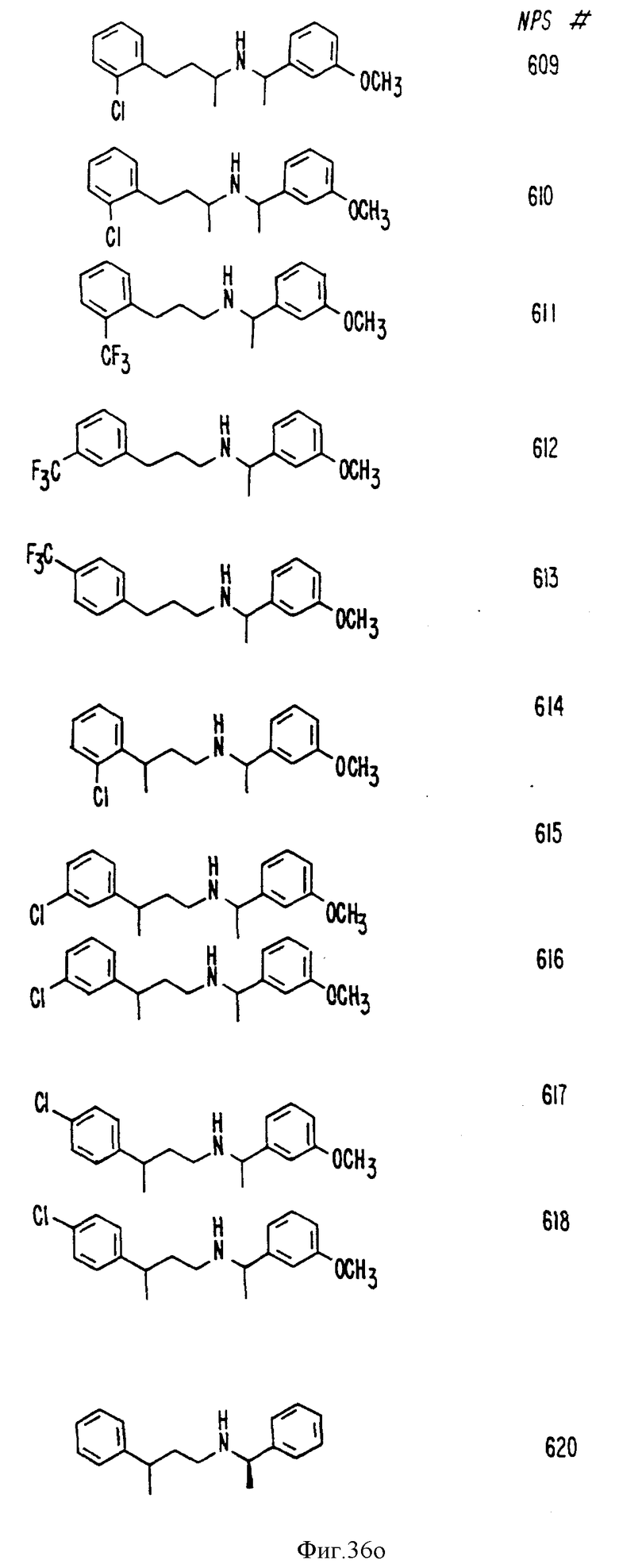
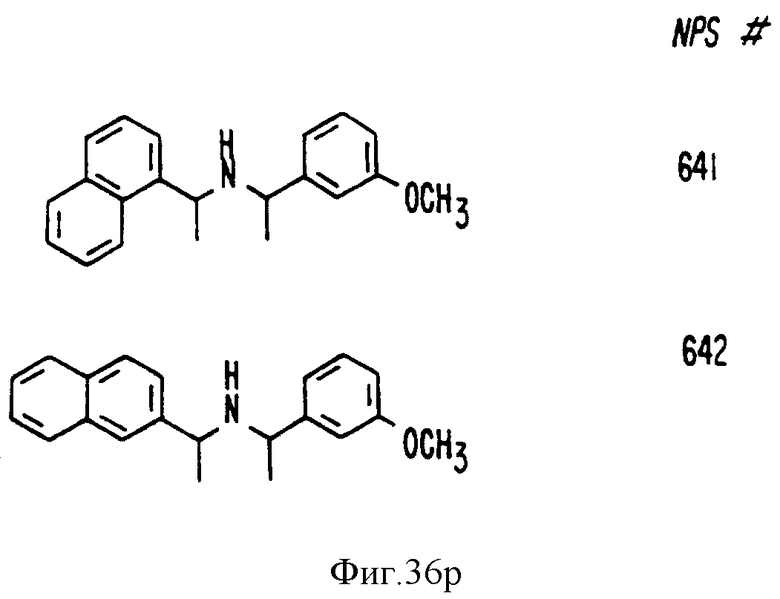
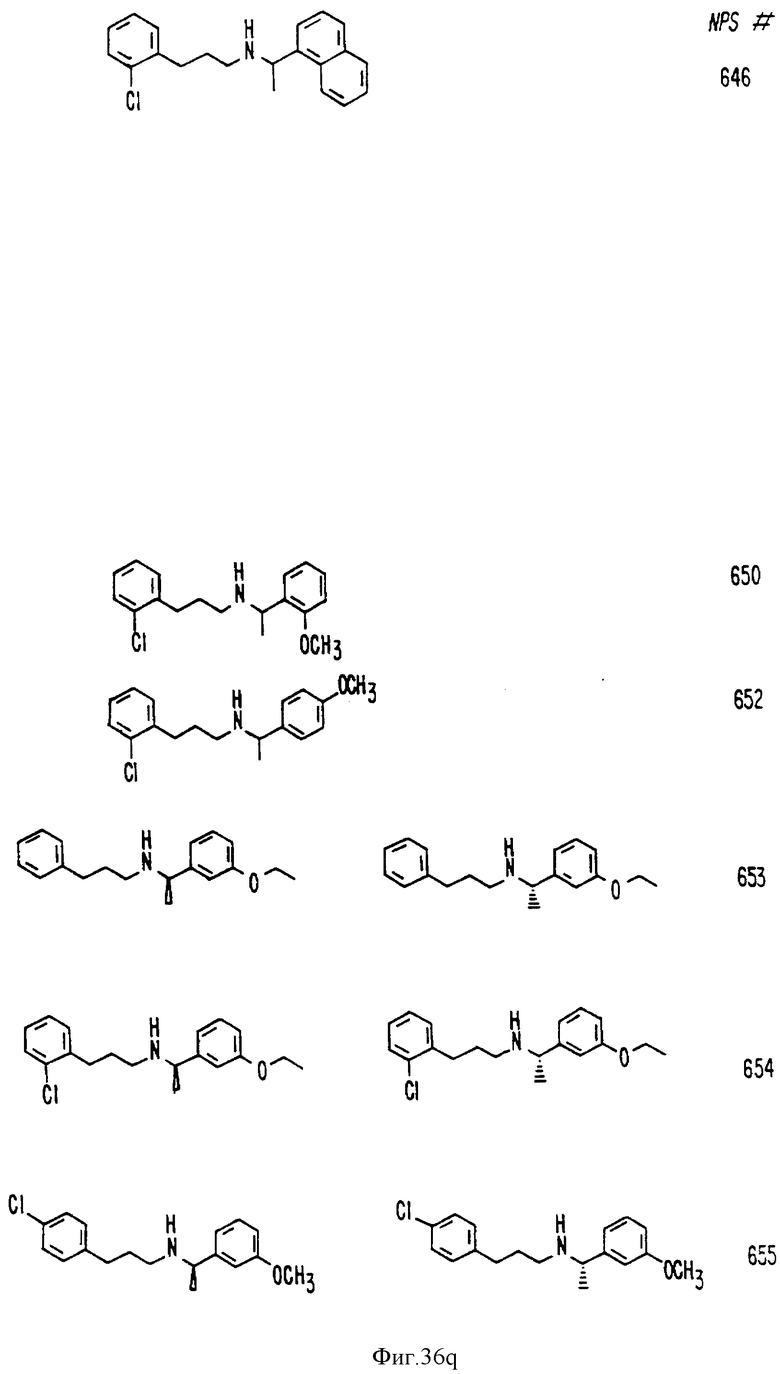
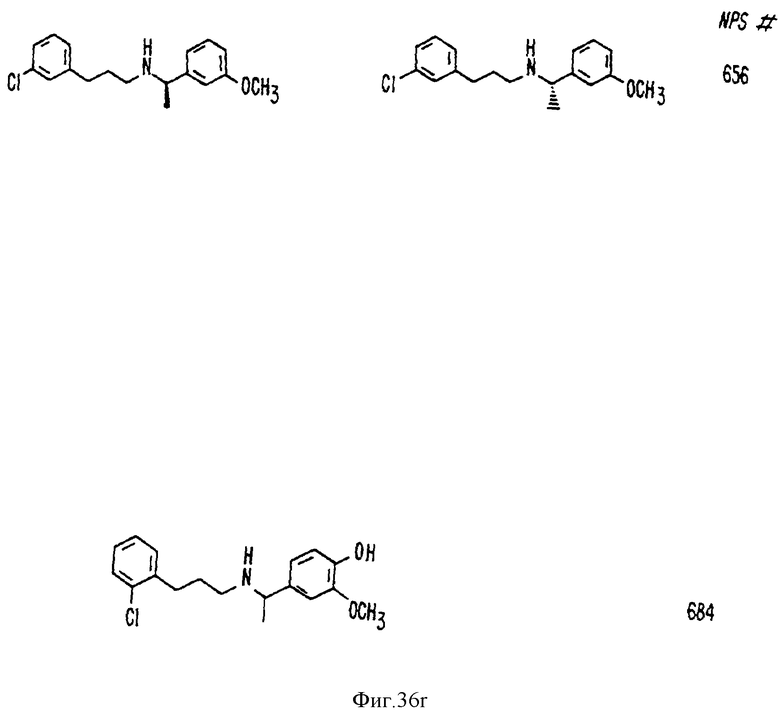
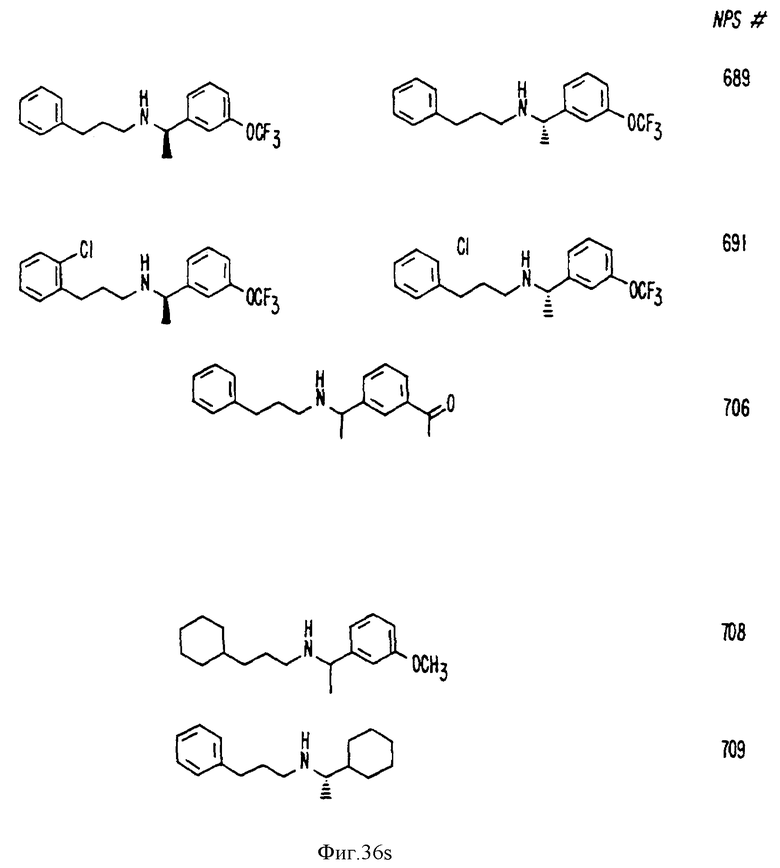



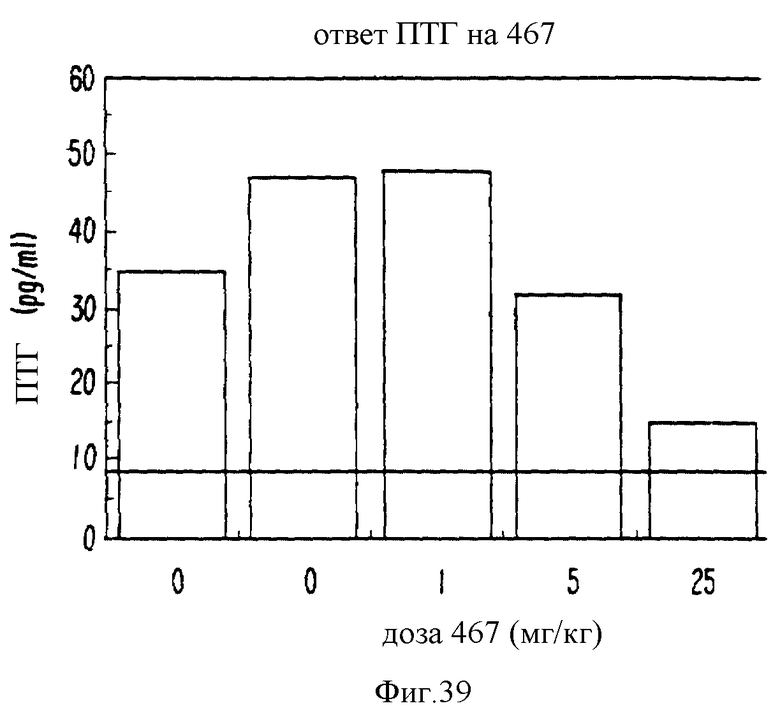




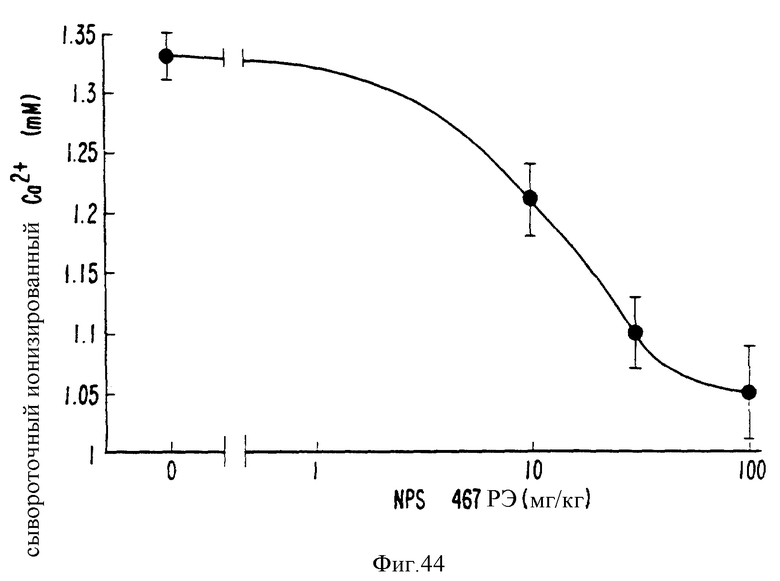
Соединение, способы и композиции для лечения пациентов, страдающих заболеваниями, которые характеризуются аномальным уровнем одного или более компонентов, активность которых регулируется или подвержена воздействию одного или более Са+2 рецепторов. Созданы также новые соединения для этих способов и композиций. Способ включает введение пациенту терапевтически эффективного количества молекул, действующих на один или более Са+2 рецепторов, и обладающих сродством к этим рецепторам или являющихся их антагонистами. Предпочтительно, молекула способна действовать или как вещество, обладающее сродством, или как антагонист на Са+2 рецепторы одного или более, но не всех типов клеток, выбранных из группы, состоящей из клеток паращитовидной железы, костных остеокластов, юкстагломерулярных почечных клеток, клеток проксимального поперечного канальца, кератиноцитов, парафолликулярных клеток щитовидной железы и трофобластов плаценты. В композициях настоящего изобретения присутствует также фармацевтически приемлемый носитель. Изобретение обеспечивает специфичность действия заявленных соединений, композиций, способов на их основе. 7 с. и 58 з.п. ф-лы, 3 табл., 44 фиг.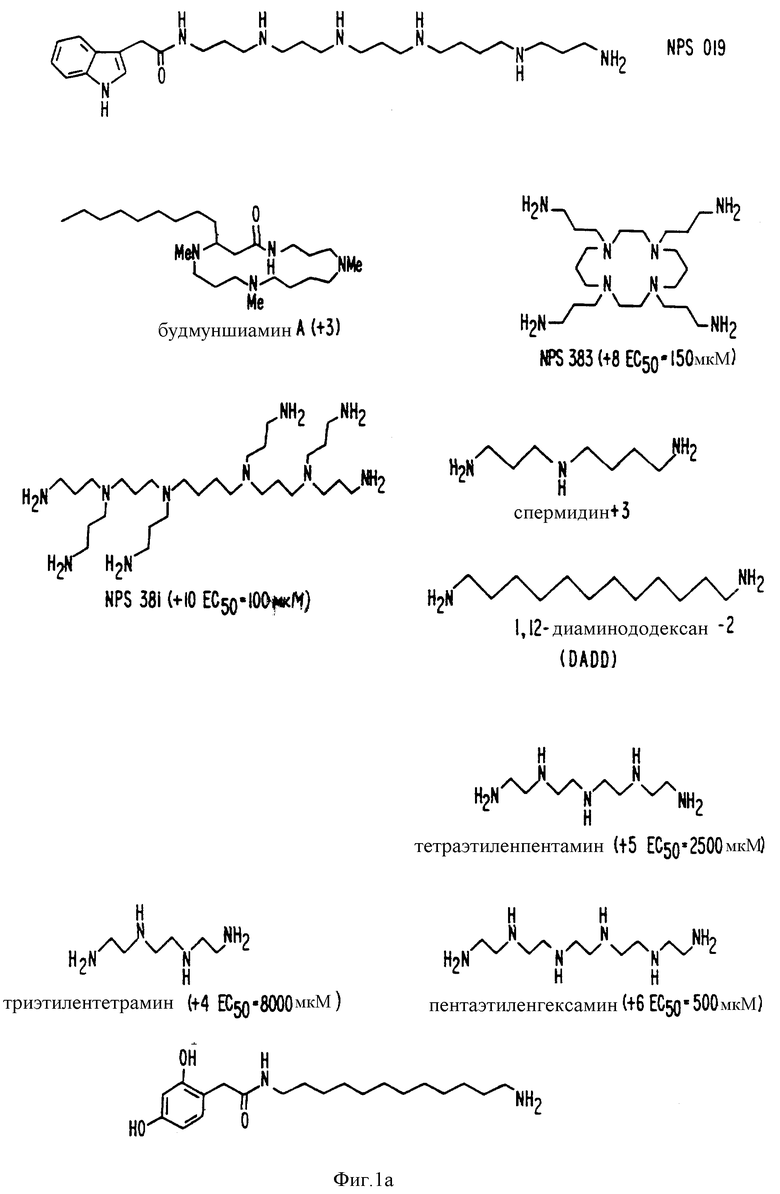
где alk - алкилен с прямой или разветвленной цепью из 1-6 атомов углерода;
R1 - низший алкил из 1-3 атомов углерода или низший галоалкил из 1-3 атомов углерода, замещенных 1-7 атомами галогена;
R2 и R3 - независимо моноциклический или бициклический карбоциклический арил или циклоалкильная группа, имеющая 5- или 6-членное кольцо, необязательно замещенное 1-5 заместителями, независимо выбранными из группы, содержащей OCF3, низший алкил из 1-3 атомов углерода, низший галоалкил из 1-3 атомов углерода, замещенный 1-7 атомами галогена, низший алкокси из 1-3 атомов углерода, галоген, нитро, амино, алкиламино, амидо, низший алкиламидо из 1-3 атомов углерода, циано, гидрокси, ацил из 2-4 атомов углерода, низший гидроксиалкил из 1-3 атомов углерода и низший тиоалкил из 1-3 атомов углерода;
причем, если R2 - фенил, R2 имеет 1-5 заместителей; если R3 - фенил, R3 имеет 1-5 заместителей, но не гидрокси, и указанные соединения не являются:
N-(метоксибензил)-1-(2-нитрофенил)этиламин,
N-альфа-(4-толил)этил-1-(4-толил)пропиламин),
N-альфа-(4-толил)этил-1-(4-толил)изопентиламин,
N-альфа-(4-толил)этил-1-(4-толил)изобутиламин,
бис[1-(4-толил)этиламин],
N-альфа-(2,6-дихлорфенил)этил-2-(3,4-диметоксифенил)этиламин,
N-альфа-(2-хлорфенил)этил-2-(3,4-диметоксифенил)этиламин,
бис-(1-циклогексилэтиламин),
N-бета-(3,4-диметоксифенил)этил-1-(1-нафтил)этиламин,
N-альфа-(1-нафтил)пропил-1-(3-этилфенил)пентиламин,
N-альфа-(3-хлорфенил)пропил-1-[1-(5-метилнафтил)]гексиламин,
N-альфа-[1-(6-метилнафтил)]этил-1-(4-хлорфенил)пропиламин,
N-альфа-[1-(7-метилнафтил)]этил-1-(4-метилфенил)бутиламин,
N-[1-(5-метилнафтил)]метил-1-(3-бромфенил)этиламин,
N-(1-нафтил)метил-1-(4-толил)этиламин,
N-(1-нафтил)метил-1-(4-пропилфенил)бутиламин,
N-циклогексилметил-1-(4-аминофенил)этиламин;
или его фармацевтически приемлемая соль.
или его фармацевтически приемлемой солью.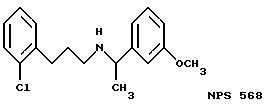

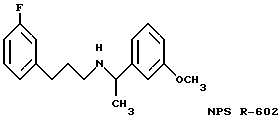
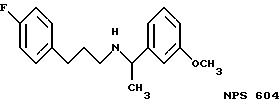


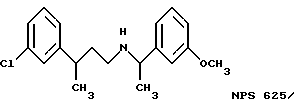
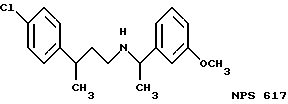
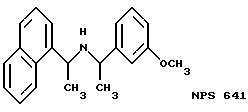

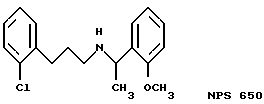


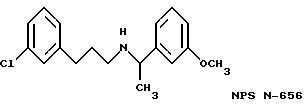
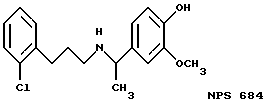
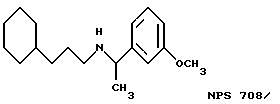
или его фармацевтически приемлемая соль.
или его фармацевтически приемлемой солью.
или его фармацевтически приемлемой солью.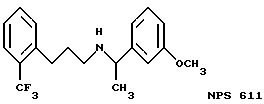
или его фармацевтически приемлемой солью.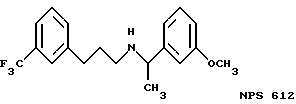
или его фармацевтически приемлемой солью.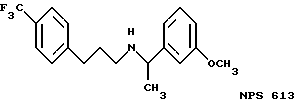
или его фармацевтически приемлемой солью.
или его фармацевтически приемлемой солью.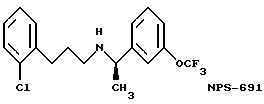
или его фармацевтически приемлемой солью.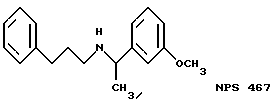

или его фармацевтически приемлемая соль.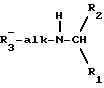
отличающаяся тем, что alk является алкиленом с простой или разветвленной цепью из 1-6 атомов углерода;
где R1 - низший алкил из 1-3 атомов углерода или низший галоалкил из 1-3 атомов углерода, замещенный 1-7 атомами галогена; и
R2 и R3 - независимо выбранные моноциклический или бициклический карбоциклический арил или циклоалкильная группы, имеющие 5- или 6-членное кольцо необязательно замещенные 1-5 заместителями независимо выбранными из группы, содержащей OCF3, низший алкил из 1-3 атомов углерода, низший галоалкил из 1-3 атомов углерода, замещенный 1-7 атомами галогена, низший алкокси из 1-3 атомов углерода, галоген, нитро, амино, алкиламино, амидо, низший алкиламидо из 1-3 атомов углерода, циано, гидрокси, ацил из 2-4 атомов углерода, низший гидроксиалкил из 1-3 атомов углерода и низший тиоалкил из 1-3 атомов углерода;
причем это указанное соединение не является N-(3,4,5-триметоксибензил)-1-фенэтиламином, N-бензил-1-фенилбутиламином, N-фенилпропил-1-фенилбутиламином или N-циклогексилметил-1-фенилпропиламином;
или его фармацевтически приемлемая соль.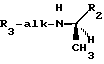
или его фармацевтически приемлемой солью.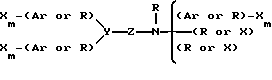
где каждый Х независимо либо H, CH3, CH3O, CH3CH2O, Br, Cl, F, CF3, CHF2, CH2F, CF3O, CH3S, OH, CH2OH, CONH2, CN, NO2, либо CH3CH2;
Ar каждый - гидрофобная структура;
R каждый независимо выбран из группы, содержащей: водород, метил, этил, пропил, изопропил, бутил, изобутил, циклопентил, циклогексил, циклогептил, циклооктил, инденил, инданил, дигидроиндолил, тиодигидроиндолил и 2-, 3- или 4-пиперид(ин)ил;
Y выбран из группы, содержащей СН, азот и ненасыщенный углерод;
Z выбран из группы, содержащей кислород, азот, серу,
где каждое n независимо между 1-4 включительно и каждое m независимо между 0 и 5 включительно;
или его фармацевтически приемлемая соль.
Приоритет по пунктам и признакам:
23.08.91 и 11.02.92 по пп.1 - 65 в соответствии с конкретными соединениями, указанными в них.
| Larsson, Biochim | |||
| et Biophys | |||
| Acta, 1985, 847, p.263-269 BROWN et al, Endocrinology, 1991, p.3047 | |||
| Способ получения вторичных аминов | 1973 |
|
SU455944A1 |
| US 4988730 A, 29.01.91 | |||
| US 4980354 A, 25.12.90. | |||

