Изобретение относится к способам исследования микроорганизмов микроскопическими методами, в частности к способам определения общей концентрации (живых и мертвых) микробов подсчетом под микроскопом, и может быть использовано при производстве диагностических и лечебно-профилактических бактерийных препаратов, а также при стандартизации микробных культур в процессе проведения коллекционных работ.
Известен способ подготовки микробных взвесей для подсчета в гемоцитометрах Горяева, Тома и др., состоящий в разведении исследуемой суспензии физиологическим раствором или дистиллированной водой до конечного разведения, вводимого в камеру (Б. А. Фихман. Оптическая стандартизация бактерийных препаратов. - М., Бюро научной информации, 1960).
Общим с заявляемым способом является этап приготовления из исследуемой суспензии предварительного разведения необходимой кратности.
К недостаткам данного способа следует отнести низкую точность и плохую воспроизводимость результатов подсчета. Источником ошибок являются низкая осаждаемость на дно камеры клеток многих видов бактерий, связанная с высокой седиментационной устойчивостью их взвесей в дистиллированной воде или физиологическом растворе. Присущая таким микробам подвижность в среде суспендирования, обусловленная броуновским движением либо наличием у клеток жгутиков, приводит к их перемещению из одного поля зрения в другое, что препятствует подсчету и существенно снижает его точность.
Серьезным недостатком этого способа является опасность заражения в процессе подсчета, поскольку многие патогенные микробы способны длительно сохранять жизнеспособность в физиологическом растворе и дистиллированной воде. Подсчет таких микробов приходится проводить с использованием ватно-марлевых повязок либо респиратора "Лепесток-200". Дыхание оператора через эти средства защиты вызывает запотевание окуляров, для устранения которого требуется прерывать работу и протирать линзы. Все это не способствует экспрессности и точности анализа.
Известны также способы подготовки микробных взвесей для подсчета, основанные на использовании для диспергирования клеток коллоидных растворов, способных при застывании переходить в состояние геля, например 4 %-ного водного раствора поливинилового спирта (Б.А. Фихман. Новая среда для микроскопического счета бактерий в камере. - Журн. микробиол., эпидимиол. и иммунобиол. - 1962. N 2. -С. 55- 56).
Общим с заявляемым способом является этап предварительного разведения анализируемой суспензии до необходимой концентрации клеток во взвеси.
К недостаткам данного способа следует отнести трудоемкость и сложность подсчета, поскольку микробы фиксируются образовавшимся гелем по всей глубине гемоцитометра, равной 100 мкм. В результате лишь часть клеток попадает в поле зрения микроскопа. Для подсчета остальных клеток требуется многократно перефокусировать микроскоп и просматривать каждое поле зрения по всей глубине гемоцитометра с исключением подсчитанных микробов. Указанные сложности затрудняют подсчет и существенно снижают его точность.
Недостатком данного способа является сохранение жизнеспособности многими патогенными бактериями в 4 %-ном растворе поливинилового спирта, что приводит к описанным выше последствиям.
Известны также способы подготовки взвесей бактерий для подсчета, основанные на использовании растворов химических соединений, способных снижать седиментационную устойчивость микробных суспензий. Например, жидкости, состоящей из 10 см3 спирта, 30 см3 глицерина, 60 см3 дистиллированной воды и 10 см3 спиртового раствора генцианвиолета (D. Horvath. Ein eifaches Verfahres fur Bakterien - Zahlung. - Zbl. Bakteriol., parariterl., Jnfektions Krankh. 1. Abt. Jrig., 1930, Bd 1, H.3/4, S. 238-239).
Общим с заявляемым способом является этап приготовления из исследуемой суспензии предварительного разведения необходимой кратности.
Недостатком данного метода является его низкая эффективность применительно к возбудителям сапа, туляремии и бруцеллеза. Рассматриваемая жидкость обеспечивает осаждение на дно гемоцитометров от 10 до 15% клеток указанных возбудителей. Почти половина микробов до завершения анализа (в течение 3 часов с момента приготовления взвеси) сохраняет жизнеспособность. Все это затрудняет подсчет и приводит к неприемлемой вариабельности результатов.
Наиболее близким к заявляемому решению является способ подготовки микробных взвесей для подсчета под микроскопом в гемоцитометре, основанный на использовании для суспендирования микробов жидкости Калиссона, состоящей из 2 см3 соляной кислоты, 100 см3 двухлористой ртути 1:500 с добавлением 1%-ного фуксина (J. Callison. A diluting fluid for standardization of vaccinnes with the hemacytometr. - J. Med. Res. 1912. V. 27. N 2. P. 225 - 227).
Общим с заявляемым способом является этап предварительного разведения анализируемой суспензии раствором, содержащим соляную кислоту.
К недостаткам рассматриваемого способа следует отнести плохую осаждаемость в гемоцитометрах клеток возбудителей сапа, туляремии и бруцеллеза, суспендированных в жидкости Каллисона. Так, через 24 часа после введения смеси в гемоцитометр подавляющее большинство 80...90% сапных, туляремийных и бруцеллезных микробов остается во взвешенном состоянии и подвержено броуновскому движению. Кроме того, не менее половины микробов указанных видов сохраняют свою жизнеспособность в течение 3 часов после суспендирования в жидкости Каллисона. Все это является источником значительных ошибок, неприемлемых при стандартизации бактерийных препаратов, и требует соблюдения строгих мер специальной техники безопасности.
Задачей изобретения является обеспечение достаточно быстрого (в течение 1 часа с момента введения в камеру) осаждения на дно гемоцитометра большинства клеток возбудителей сапа, туляремии и бруцеллеза с одновременной их инактивацией и сохранением четкости наблюдаемой микроскопической картины. Все это необходимо для точного и экспрессного подсчета.
Поставленная задача решается благодаря тому, что в способе подготовки взвесей клеток возбудителей сапа, туляремии и бруцеллеза для подсчета, включающем этап предварительного разведения анализируемых материалов, предусмотрены следующие отличия:
предварительная обработка микробов раствором едкого натра;
последующая обработка клеток раствором танина;
использование для приготовления окончательного разведения суспензии раствора, содержащего соляную и молочную кислоты;
выдерживание заполненного гемоцитометра в парах формалина.
Указанные отличия обусловлены следующими причинами:
седиментационная устойчивость в дисперсных средах сапных, туляремийных и бруцеллезных микробов определяется электрокинетическим потенциалом клеток, наличием у них мощных сольватных оболочек, обусловленной особой гидрофильностью биополимеров микрокапсул, и малыми их размерами (небольшой массой), что в совокупности препятствует действию гравитационных сил. Следовательно, для обеспечения осаждения микробов требуется устранить или уменьшить влияние указанных факторов. Среди этих факторов масса клеток наименее подвержена экспериментальным воздействиям, а более изменчивы электрокинетический потенциал и соль ватная оболочка;
для уменьшения гидрофильности поверхности клеток их вначале обрабатывают раствором едкого натра, обеспечивающим гидролиз биополимеров микрокапсул;
раствор танина способствует уменьшению размеров сольватной оболочки;
соляная и молочная кислоты обеспечивают pH среды, близкий к изоэлектрической точке микробов. Катионы водорода улучшают адсорбцию клеток на стекле;
соляная и молочная кислоты вызывают гибель клеток возбудителей сапа, туляремии и бруцеллеза;
выдерживание заполненного гемоцитометра в парах формалина в течение 1 часа обеспечивает обеззараживание его поверхностей, контаминирование которых могло произойти в процессе введения взвеси в камеру;
используемые растворы не вызывают лизиса клеток и их избыточной агрегации, характеризуются прозрачностью, низкими показателями светопреломления и обеспечивают высокий контраст изображения сапных, туляремийных и бруцеллезных микробов в фазово-контрастном микроскопе.
Способ включает следующие этапы:
разведение исследуемой взвеси;
введение подготовленного конечного разведения в гемоцитометр;
выдерживание гемоцитометра в камере с парами формалина для обеспечения обеззараживания и осаждения клеток.
Способ выполняется следующим образом.
Исследуемый материал разбавляют в пробирке раствором едкого натра до концентрации, соответствующей по оптической плотности стандарту мутности ГИСК имени Л.А. Тарасевича на 10 ЕД. При этом учитывают разведение материала (величина Р). Микробы в растворе щелочи выдерживают 30 минут.
Затем готовят еще два десятикратных разведения.
Одно - на растворе танина, а из него - на растворе соляной и молочной кислот. В пробирках с танином и кислотами микробы выдерживают до 5 минут. Пробирки закрывают ватно-марлевыми пробками. Содержимое пробирок тщательно перемешивают путем периодического встряхивания.
Для обработки сапных микробов используют 0,05 М раствор едкого натра и 0,15%-ный раствор танина; для туляремийных микробов - 0,001 М раствор едкого натра и 0,2%-ный раствор танина; для бруцеллезных микробов - 0,01 М раствор едкого натра и 0,5%-ный раствор танина.
Заключительную обработку микробов всех указанных видов осуществляют 0,1 М раствором соляной кислоты с добавлением к нему 4 % молочной кислоты (по объему).
Растворы всех реактивов готовят на дистиллированной воде.
Взвесь клеток из пробирки с кислотами вводят при помощи пипеток в гемоцитометр с притертым покровным стеклом. Гемоцитометры и покровные стекла предварительно тщательно промывают и протирают с целью удаления посторонних частиц, имитирующих микробные клетки.
Заполненный гемоцитометр помещают в чашку Петри. Закрывают ее крышкой, на дно которой помещают диск фильтровальной бумаги, смоченной 2...3 см3 формалина. Чашки Петри с камерами выдерживают на рабочем столе в течение 1 часа.
Для контроля отсутствия загрязнения гемоцитометров и подсчета клеток используют оптическую систему фазово-контрастного микроскопа (микроскоп МБИ-3 либо МБР, МБ-1А, "Биолам Л-211", фазово-контрастное устройство КФ-4, объектив х 20Ф, бинокулярная насадка АУ-12, окуляры х 10). Настройку микроскопа осуществляют по описанию, прилагаемому к комплекту КФ-4.
Микроскоп фокусируют на сетку Горяева, добиваясь при помощи микровинта четкого изображения микробов. Они выглядят в виде черных палочек и кокков.
Подсчитывают клетки, расположенные в больших квадратах сетки Горяева по диагонали камеры. Вначале - клетки, находящиеся в квадрате сетки, а затем, медленно поднимая тубус микроскопа микровинтом, учитывают над тем же квадратом единичные микробы, которые могут прилипнуть к нижней поверхности покровного стекла.
При анализе не считают клетки, пересекаемые нижней и левой границами квадрата. Их принимают за клетки, лежащие в соседних квадратах.
Всего требуется учесть около 500 клеток (10...12 больших квадратов).
Искомую концентрацию микробов в исследуемом материале рассчитывают по формуле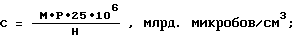
где C - концентрация клеток,
P - величина разведения материала раствором щелочи,
H - количество проанализированных квадратов,
M - количество учтенных клеток.
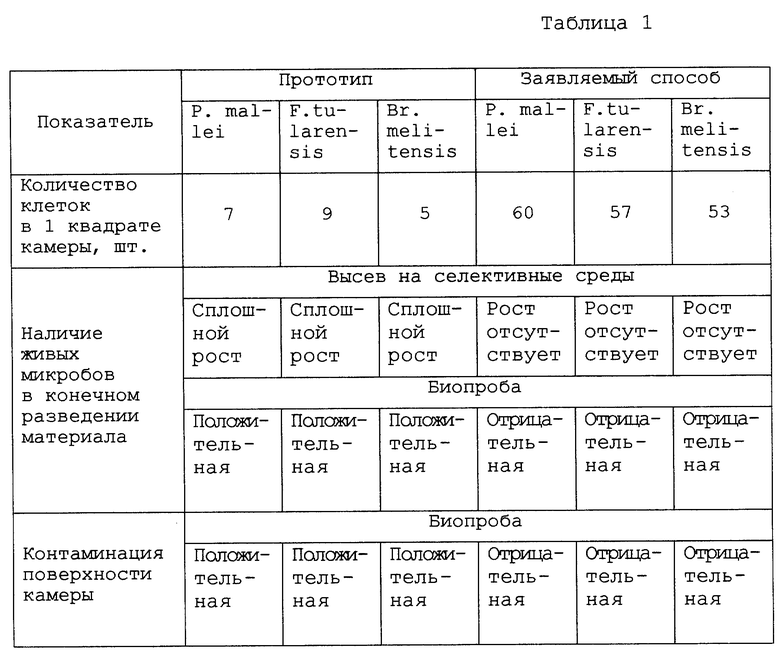
Для сравнения эффективности заявляемого способа и прототипа готовили разведения из одних и тех же материалов возбудителей сапа штамм Ц-5, туляремии штамм Schu и бруцеллеза штамм Br. melitensis M 16. С целью обеспечения равных концентраций клеток в конечных разведениях, вводимых в камеры, материалы в экспериментах "по прототипу" разводили таким же образом, как при заявленном способе, с той лишь разницей, что вместо предлагаемых растворов щелочи, танина, соляной и молочной кислот использовали жидкость Каллисона на всех этапах разведения. В каждом опыте анализировали по 10 полей зрения. Осаждаемость микробов характеризовали средним количеством клеток, приходящимся на один квадрат. Жизнеспособность микробов к началу подсчета оценивали методом высева на селективные питательные среды и биопробой на экспериментальных животных. Для контроля контаминации поверхности гемоцитометров делали смывы ватно-марлевыми тампонами. Полученные пробы высевали на плотные питательные среды и использовали для заражения. Микробы сапа высевали на агар, приготовленный на основе соляно-кислотного гидролизата казеина, туляремийные микробы - на основе эрититагара, бруцеллы - на кровяной агар. Для постановки биопроб в опытах с сапом, туляремией и бруцеллезом использовали соответственно золотистых хомячков, белых мышей и морских свинок. Сравнительные исследования показывают, что заявляемый способ по сравнению с прототипом, обеспечивает значительно большую осаждаемость микробов на дно гемоцитометра и их инактивацию в процессе подготовки к подсчету (табл. 1).
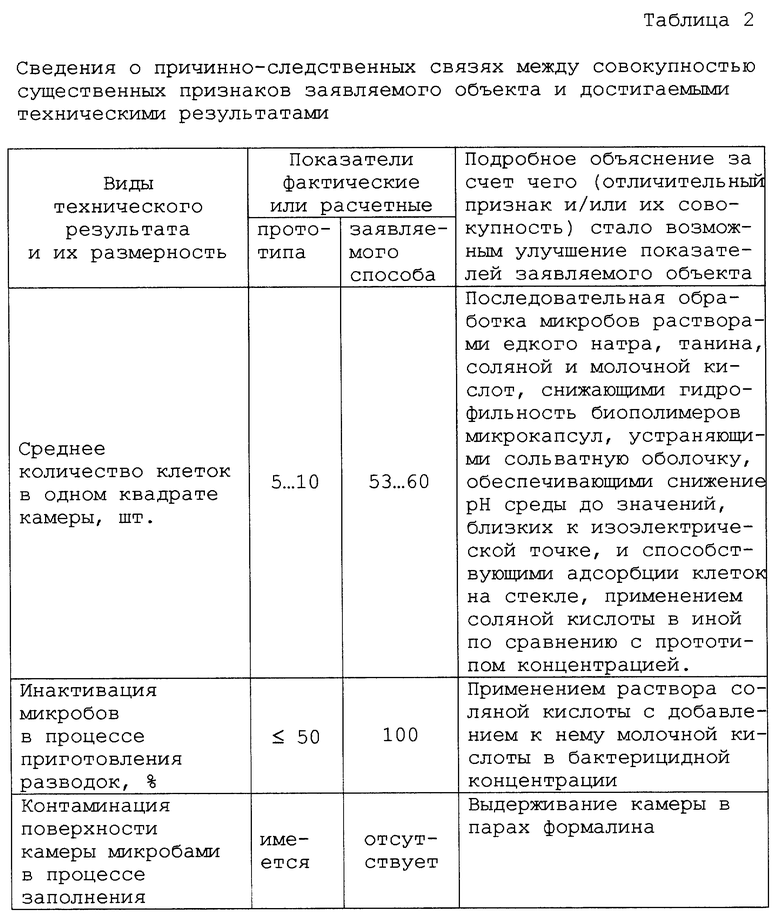
Наличие причинно-следственной связи между совокупностью существенных признаков заявляемого объекта и достигаемыми техническими результатами показано в табл. 2.
Предлагаемый способ подготовки взвесей клеток возбудителей сапа, туляремии и бруцеллеза для подсчета имеет следующие преимущества:
обеспечивает осаждение на дно камеры 80...90 % клеток через один час после введения подготовленного разведения в гемоцитометр. Остальные микробы адсорбируются нижней поверхностью покровного стекла; приводит к инактивации микробов указанных видов как в самой камере, так и на всей поверхности гемоцитометра, что позволяет проводить подсчет без соблюдения строгих мер защиты. Пробы анализируют вне вытяжного шкафа, без использования респираторов и ватно-марлевых повязок;
сохраняет внешнюю целостность микробов и четкость их микроскопической картины в используемой оптической системе, что в совокупности с иммобилизацией клеток на дне камеры и на покровном стекле обеспечивает условия, необходимые для осуществления подсчета под микроскопом. Выполнение анализа доступно лаборанту, имеющему навыки работы с фазово-контрастными микроскопами;
способ обеспечивает достаточную для практического применения точность подсчета, ошибка не превышает 10%, а расхождение результатов анализа между лабораториями - 12%;
затраты времени на определение общей концентрации микробов в пробе подсчетом под микроскопом составляют 2 часа с момента поступления ее в лабораторию, что позволяет охарактеризовать метод подсчета с предлагаемым способом подготовки взвесей как экспрессный.
Возможность осуществления предложенного способа показана на следующих примерах.
Пример 1. Подготовка взвесей сапных микробов для определения общей концентрации клеток в сапной вакцине.
В целях обеспечения удобного для подсчета количества клеток в одном квадрате гемоцитометра (примерно 30...80 особей) предварительно определяют оптическую концентрацию клеток в вакцине по стандарту мутности ГИСК им. Л.А. Тарасевича на 10 единиц мутности. В исследуемой серии она равна 5 млрд. микробов/см3. Затем вакцину разводят параллельно в двух пробирках 0,05 М раствором едкого натра в пять раз. Для этого в пробирки наливают по 4,0 см3 раствора щелочи и вносят по 1,0 см3 вакцины градуированной пипеткой на 1,0 см3. Величина разведения вакцины "P" составляет 5.
Пробирки закрывают ватно-марлевыми пробками. Содержимое периодически перемешивают путем встряхивания. Через 30 минут из пробирок со щелочью готовят два ряда двух последовательных десятикратных разведений. Первое из них готовят на 0,15%-ном растворе танина, а второе - на 0,1 М растворе соляной кислоты с добавлением к ней 4% молочной кислоты. Раствор танина и кислот разливают в пробирки по 4,5 см3. Градуированной пипеткой из средней части пробирки со щелочью отбирают по 0,5 см3 содержимого и переносят в пробирку с танином. Ее закрывают ватно-марлевой пробкой, содержимое перемешивают. Из этой пробирки аналогичным образом переносят 0,5 см3 взвеси клеток в пробирку с кислотами. Пробирки закрывают ватно-марлевыми пробками. Содержимое перемешивают встряхиванием.
Таким образом, для исследуемой вакцины готовят два ряда пробирок, содержащих по три параллельных пробирки.
После встряхивания из пробирок с кислотами переносят капли взвеси в гемоцитометр при помощи пипеток. В силу капиллярности жидкость с клетками затягивается в камеру. Гемоцитометр помещают в чашку Петри с фильтровальной бумагой, смоченной формалином. Через 1 час проводят подсчет.
Концентрацию клеток рассчитывают по формуле, представленной в описании, исходя из количества подсчитанных клеток, числа проанализированных квадратов и величины разведения вакцины. Из усреднения результатов двух параллельных определений устанавливают общую концентрацию клеток в вакцине.
Пример 2. Подготовка взвеси туляремийных микробов для определения их общей концентрации в культуре методом подсчета.
Предварительно определяют оптическую концентрацию клеток в культуре при помощи стандарта ГИСК им. Л.А. Тарасевича на 10 единиц мутности, чтобы обеспечить последующим разведением культуры удобное для подсчета количества клеток в одном квадрате камеры (30...80 особей). Оптическая концентрация оказалась равной 15 млрд. клеток/см3. Исходя из этих данных, культуру разводят параллельно в двух пробирках 0,001 М раствором едкого натра в 15 раз. Для этого в пробирки наливают по 7,0 см3 щелочи и вносят по 0,5 см3 исследуемой культуры. Величина разведения культуры "P" составляет 15. Последующие этапы выполнения способа подготовки взвесей являются аналогичными приведенным в примере 1, с той лишь разницей, что для разведения туляремийных микробов используется 0,2 %-ный раствор танина.
Пример 3. Подготовка взвесей бруцеллезных микробов для определения их общей концентрации в культуре методом подсчета.
Подобным образом, как в примерах 1 и 2, устанавливают оптическую концентрацию клеток в исследуемой культуре. Она оказалась равной 20 млрд. клеток/см3.
Исходя из этих данных, культуру разводят параллельно в двух пробирках 0,01 М раствором едкого натра в 20 раз. Для этого в пробирки разливают по 1,9 см3 щелочи и вносят по 0,1 см3 анализируемой культуры. Величина разведения культуры составляет 20.
Последующие десятикратные разведения готовят таким же образом, как в примере 1, с той лишь разницей, что для разведения бруцеллезных бактерий используют 0,5%-ный раствор танина.
В табл. 3 представлены результаты определения общей концентрации клеток методом подсчета и количества живых микробов в культурах P. mallei, F. tularensis и Br. melitensis высевом на чашки с плотными питательными средами. Взвеси микробов перед заполнением гемоцитометров готовили по заявляемому способу.
Из табл. 3 следует, что после обработки по заявляемому способу сапные, туляремийные и бруцеллезные микробы становятся доступными для подсчета при помощи микроскопа. Установленные значения общей концентрации (живых и мертвых) бактерий превышают данные чашечного метода о количестве живых микробов. Отмеченные расхождения могут быть объяснены частичной агрегацией клеток на поверхности агара, в результате которой образование некоторых колоний происходит не из одной клетки, а из их агрегатов, а также наличием в исследованных культурах не только живых, но и мертвых особей. Живые микробы в процессе приготовления взвесей по предлагаемому способу инактивируются, поэтому при подсчете они морфологически не могут быть отделены от мертвых особей, исходно имевшихся в исследованном материале.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ДИАГНОСТИКУМА ЭРИТРОЦИТАРНОГО АНТИГЕННОГО САПНОГО | 2001 |
|
RU2188036C1 |
| СПОСОБ ПРИГОТОВЛЕНИЯ ПРЕПАРАТОВ ДЛЯ ЦИТОРЕФРАКТОМЕТРИЧЕСКОГО ИССЛЕДОВАНИЯ САПНЫХ МИКРОБОВ | 1998 |
|
RU2149899C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ЖИВЫХ МИКРОБОВ В БИОПРЕПАРАТЕ | 1992 |
|
RU2037805C1 |
| СПОСОБ ОЦЕНКИ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ МИКРОБНОЙ ПОПУЛЯЦИИ | 2001 |
|
RU2195496C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОТИВОТУЛЯРЕМИЙНОЙ ГИПЕРИММУННОЙ СЫВОРОТКИ И СПОСОБ ПОЛУЧЕНИЯ ДИАГНОСТИКУМА ЭРИТРОЦИТАРНОГО ТУЛЯРЕМИЙНОГО ИММУНОГЛОБУЛИНОВОГО СУХОГО | 2002 |
|
RU2240822C2 |
| АНТИГЕННЫЙ ПРЕПАРАТ ДЛЯ ПРОФИЛАКТИКИ САПА У ЛЮДЕЙ И ЖИВОТНЫХ | 2004 |
|
RU2262949C1 |
| СПОСОБ ОЦЕНКИ КЛИНИЧЕСКОЙ ЭФФЕКТИВНОСТИ АНТИБАКТЕРИАЛЬНЫХ ПРЕПАРАТОВ ДЛЯ ВОЗБУДИТЕЛЕЙ ОСОБО ОПАСНЫХ ИНФЕКЦИЙ Francisella tularensis И Brucella spp | 2009 |
|
RU2417376C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛАТЕКСНОГО ДИАГНОСТИКУМА | 1991 |
|
RU2012888C1 |
| СПОСОБ ЭЛЮЦИИ ПАТОГЕНА С ИММОБИЛИЗОВАННОЙ МАГНИТНОЙ МАТРИЦЫ | 2013 |
|
RU2535070C1 |
| СПОСОБ ПОЛУЧЕНИЯ БИОМАГНОИММУНОСОРБЕНТА ДЛЯ ОБНАРУЖЕНИЯ БАКТЕРИАЛЬНЫХ АНТИГЕНОВ (ВАРИАНТЫ) | 2003 |
|
RU2271540C2 |
Изобретение предназначено для стандартизации микробных культур при производстве диагностических и лечебно-профилактических препаратов. Анализируемые микробные суспензии обрабатывают сначала растворами едкого натра. Затем готовят из них десятикратные разведения. Полученные разведения обрабатывают в течение 3-5 мин последовательно растворами танина и 0,1 М раствором соляной кислоты с добавлением 4% молочной кислоты. Обработанной микробной взвесью заполняют гемоцитометрическую камеру. Выдерживают ее в парах формалина не менее 1 ч. Для сапных, туляремийных и бруцеллезных микробов используют соответственно 0,05; 0,001; 0,01 М растворы едкого натра и 0,15; 0,2; 0,5%-ный растворы танина. Изобретение повышает точность и обеспечивает экспрессность подсчета микробных клеток. 3 з.п.ф-лы, 3 табл.
| RU 94004013 A, 10.12.1995 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ЖИВЫХ МИКРОБОВ В БИОПРЕПАРАТЕ | 1992 |
|
RU2037805C1 |
| Фихман Б.А | |||
| Новая среда для микроскопического счета бактерий в камере | |||
| ЖМЭИ | |||
| - М., 1962, N 2, с | |||
| Устройство двукратного усилителя с катодными лампами | 1920 |
|
SU55A1 |
