Описываются высоко аффинные 2'фтор(2-F')-пиримидин-РНК-лиганды к сосудистому эндотелиальному фактору роста (VEGF). Использованный здесь способ идентификации таких Нуклеиновых Кислот-Лигандов называется SELEX, акроним для Systematic Evolution of Ligands by Exponential enrichment. Далее в данное изобретение включен способ получения терапевтического или диагностического Комплекса, состоящего из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, посредством идентификации Нуклеиновой Кислоты-Лиганда VEGF с помощью методологии SELEX и ковалентной сшивки Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением. Далее изобретение включает в себя Комплексы, состоящие из одной или более чем одной Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения. Кроме того, изобретение относится к усовершенствованию фармакокинетических свойств Нуклеиновой Кислоты-Лиганда VEGF посредством ковалентной сшивки Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением с образованием Комплекса. Далее изобретение относится к усовершенствованию фармакокинетических свойств Нуклеиновой Кислоты-Лиганда VEGF посредством использования Липидной Конструкции, содержащей Нуклеиновую Кислоту-Лиганд VEGF или Комплекс, содержащий Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение. Кроме того, данное изобретение относится к способу для направления терапевтического или диагностического агента к биологической мишени, которая экспрессирует VEGF, посредством соединения этого агента с Комплексом, состоящим из Нуклеиновой Кислоты-Лиганда VEGF и Липофильного Соединения или Неиммуногенного Соединения Высокой Молекулярной Массы, причем этот Комплекс дополнительно соединен с Липидной Конструкцией, а Нуклеиновая Кислота-Лиганд VEGF дополнительно соединена с наружной стороной Липидной Конструкции.
Предпосылки изобретения
A. SELEX
В течение многих лет догмой являлось то, что нуклеиновые кислоты играют прежде всего информационную роль. Благодаря способу, известному как систематическое выделение лигандов посредством экспоненциального обогащения (Systematic Evolution of Ligands by Exponential enrichment), названному SELEX, стало ясно, что нуклеиновые кислоты, подобно белкам, имеют трехмерное структурное многообразие. SELEX представляет собой способ выделения in vitro молекул Нуклеиновых Кислот с высокоспецифичным связыванием с молекулами-мишенями и описан в Заявке на патент США, порядковый N 07/536428, поданной 11 июня 1990, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment", в настоящее время отозвана, в Заявке на патент США, порядковый N 07/714131, поданной 10 июня 1991, озаглавленной "Nucleic Acid Ligands", в настоящее время патент США N 5475096, в Заявке на патент США, порядковый N 07/931473, поданной 17 августа 1992, озаглавленной "Method for Identifying Nucleic Acid Ligadns", в настоящее время патент США N 5270163 (см. также WO 91/19813), каждая из которых конкретно включена здесь путем ссылки. В каждой из этих заявок, на которые совокупно ссылаются здесь как на Заявки на патент SELEX, описывается фундаментально новый способ получения Нуклеиновой Кислоты-Лиганда к любой желаемой молекуле-мишени. Способ SELEX обеспечивает класс продуктов, на которые ссылаются как на Нуклеиновые Кислоты-Лиганды, причем каждый лиганд имеет уникальную последовательность, и которые обладают свойством специфического связывания с желаемым соединением-мишенью или с желаемой молекулой-мишенью. Каждая идентифицированная по SELEX Нуклеиновая Кислота-Лиганд представляет собой специфический лиганд данного соединения-мишени или данной молекулы-мишени. SELEX основан на уникальном представлении, что нуклеиновые кислоты имеют достаточную способность к образованию множества двух- и трехмерных структур и достаточную химическую многосторонность, возможную в пределах их мономеров, чтобы действовать в качестве лигандов (образовывать специфичные связывающиеся пары) фактически с любым химическим соединением, либо мономерным, либо полимерным. В качестве мишеней могут служить молекулы любого размера и состава.
Способ SELEX включает в себя отбор из смеси олигонуклеотидов-кандидатов и ступенчатые повторения связывания, разделения и амплификации с использованием одной и той же общей схемы отбора, чтобы достичь фактически любого желаемого критерия аффинности и избирательности связывания. Начиная со смеси Нуклеиновых Кислот, предпочтительно содержащей сегмент рандомизированной последовательности, способ SELEX включает в себя стадии приведения в контакт смеси с мишенью в условиях, благоприятных для связывания, отделения несвязанных Нуклеиновых Кислот от тех Нуклеиновых Кислот, которые специфически связались с молекулами-мишенями, диссоциации Комплексов Нуклеиновая Кислота-мишень, амплификации Нуклеиновых Кислот, диссоциированных из Комплексов Нуклеиновая Кислота-мишень, с получением смеси Нуклеиновых Кислот, обогащенной лигандами, затем повторения еще раз стадий связывания, разделения, диссоциации и амплификации в течение стольких циклов, сколько желательно для получения высокоспецифичных высоко аффинных Нуклеиновых Кислот-Лигандов к молекуле-мишени.
Авторами настоящего изобретения показано, что способ SELEX демонстрирует, что нуклеиновые кислоты как химические соединения могут образовывать большое множество форм, размеров и конфигураций и способны к значительно более широкому спектру связывания и к функциям, иным чем те, которые проявляют нуклеиновые кислоты в биологических системах.
Авторы настоящего изобретения показали, что SELEX или подобные SELEX способы могут применяться для того, чтобы идентифицировать нуклеиновые кислоты, которые могут способствовать любой выбранной реакции путем, подобным тому, при котором можно идентифицировать Нуклеиновые Кислоты-Лиганды для любой данной мишени. Авторы настоящего изобретения постулируют, что теоретически в пределах Смеси Кандидатов из примерно от 1013 до 1018 Нуклеиновых Кислот существует по меньшей мере одна Нуклеиновая Кислота подходящей формы, чтобы способствовать каждому из широкого разнообразия физических и химических взаимодействий.
Основной способ SELEX был модифицирован для достижения ряда конкретных целей. Например, в Заявке на патент США, порядковый N 07/960093, поданной 14 октября 1992, озаглавленной "Method for Selecting Nucleic Acids on the Basis of Structure", описывается применение SELEX вместе с гель-электрофорезом для отбора молекул Нуклеиновых Кислот со специфическими структурными характеристиками, таких как изогнутая ДНК. В Заявке на патент США, порядковый N 08/123935, поданной 17 сентября 1993, озаглавленной "Photoselection of Nucleic Acid Ligands", описывается способ на основе SELEX для отбора Нуклеиновых Кислот-Лигандов, содержащих фотореактивные группы, способные к связыванию и/или к поперечному фотосвязыванию и/или к фотоинактивации молекулы-мишени. В Заявке на патент США, порядковый N 08/134028, поданной 7 октября 1993, озаглавленной "High-Affinity Nucleic Acid Ligands That Discriminate Between Theophylline and Caffeine", в настоящее время патент США N 5580737, описывается способ идентификации высокоспецифичных Нуклеиновых Кислот-Лигандов, способных различать близкородственные молекулы, которые могут не являться пептидами, названный Counter-SELEX. В Заявке на патент США, порядковый N 08/143564, поданной 25 октября 1993, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Solution SELEX", в настоящее время патент США N 5567588, описывается способ на основе SELEX, с помощью которого достигается высокоэффективное разделение между олигонуклеотидами, имеющими высокую и низкую аффинность к молекуле-мишени.
Способ SELEX заключается в идентификации высоко аффинных Нуклеиновых Кислот-Лигандов, содержащих модифицированные нуклеотиды, придающие лиганду усовершенствованные характеристики, такие как улучшенная стабильность in vivo или улучшенные характеристики доставки. Примеры таких модификаций включают в себя химические замещения в положениях рибозы и/или фосфата и/или основания. Идентифицированные по SELEX Нуклеиновые Кислоты-Лиганды, содержащие модифицированные нуклеотиды, описываются в Заявке на патент США, порядковый N 08/177991, поданной 8 сентября 1993, озаглавленной "High Affinity Nucleic Acid Ligands Containing Modified Nucleotides", в настоящее время патент США N 5660985, где описываются олигонуклеотиды, содержащие производные нуклеотидов, химически модифицированных в 5- и 2'-положениях пиримидинов. В Заявке на патент США, порядковый N 08/134028, см. выше, описываются высокоспецифичные Нуклеиновые Кислоты-Лиганды, содержащие один или более чем один нуклеотид, модифицированный 2'-амино (2'-NH2), 2'-фторо(2'-F) и/или 2'-O-метил (2'-ОМе). В Заявке на патент США, порядковый N 08/264029, поданной 22 июня 1994, озаглавленной "Novel Method of Preparation of Known and Novel 2'-Modified Nucleosides by Intramolecular Nucleophilic Displacement", описываются олигонуклеотиды, содержащие различные 2'-модифицированные пиримидины.
Способ SELEX заключается в соединении выбранных олигонуклеотидов с другими выбранными олигонуклеотидами и не-олигонуклеотидными функциональными единицами, как описано в Заявке на патент США, порядковый N 08/284063, поданной 2 августа 1994, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Chimeric SELEX", в настоящее время патент США N 5637459, и в Заявке на патент США, порядковый N 08/234997, поданной 28 апреля 1994, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Blended SELEX", соответственно. Данные заявки дают возможность сочетания широкого множества форм и других свойств, а также свойств эффективной амплификации и репликации олигонуклеотидов с желаемыми свойствами других молекул.
Способ SELEX далее заключается в соединении отобранных Нуклеиновых Кислот-Лигандов с Липофильными Соединениями или Неиммуногенными Соединениями Высокой Молекулярной Массы в диагностический или терапевтический Комплекс, как описано в Заявке на патент США, порядковый N 08/434465, поданной 4 мая 1995, озаглавленной "Nucleic Acid Complexes". Нуклеиновые Кислоты-Лиганды VEGF, которые соединены с Липофильным Соединением, таким как диацилглицерин или диалкилглицерин, в диагностический или терапевтический Комплекс, описаны в Заявке на патент США, порядковый N 08/739109, поданной 25 октября 1996, озаглавленной "Vascular Endothelial Growth Factor (VEGF) Nucleic Acid Ligand Complexes". Нуклеиновые Кислоты-Лиганды VEGF, которые соединены с Неиммуногенным Соединением Высокой Молекулярной Массы, таким как Полиэтиленгликоль, или Липофильным Соединением, таким как Глицеролипид, фосфолипид или глицеринамидлипид, в диагностический или терапевтический Комплекс, описаны в Заявке на патент США, порядковый N 08/897351, поданной 21 июля 1997, озаглавленной "Vascular Endothelial Growth Factor (VEGF) Nucleic Acid Ligand Complexes". Каждая из вышеописанных Заявок на патент, в которых описываются модификации основной методики SELEX, конкретно включены здесь полностью путем ссылки.
Б. ЛИПИДНЫЕ КОНСТРУКЦИИ
Липидные бислойные везикулы представляют собой замкнутые, заполненные жидкостью микроскопические сферы, которые образуются главным образом из индивидуальных молекул, имеющих полярные (гидрофильные) и неполярные (липофильные) участки. Гидрофильные участки могут содержать фосфатную, глицерилфосфатную, карбокси, сульфатную, амино, гидрокси, холиновую или другие полярные группы. Примерами липофильных групп являются насыщенные или ненасыщенные углеводороды, такие как алкил, алкенил, или другие липидные группы. Стерины (например, холестерин) и другие фармацевтически приемлемые адъюванты (включая антиоксиданты, подобные альфа-токоферолу) также могут быть включены для улучшения стабильности везикул или придания других желаемых характеристик.
Липосомы представляют собой подгруппу этих бислойных везикул и содержат главным образом фосфолипидные молекулы, которые несут два гидрофобных хвоста, состоящих из цепей жирных кислот. При экспозиции с водой эти молекулы спонтанно выстраиваются с образованием сферических бислойных мембран с липофильными концами этих молекул в каждом слое, соединенными в центре мембраны, и противоположными полярными концами, образующими соответствующие внутреннюю и внешнюю поверхности этой двухслойной мембраны (мембран). Таким образом, каждая сторона мембраны представляет собой гидрофильную поверхность, тогда как внутри мембрана представляет собой липофильную среду. Эти мембраны могут быть организованы в ряд концентрических сферических мембран, разделенных тонким слоем воды, в некоторой степени подобно слоям луковицы, вокруг внутреннего водного пространства. Эти многослойные везикулы (MLV) можно превратить в маленькие или Однослойные Везикулы (UV) при приложении силы гидродинамического фрагментирования.
Терапевтическое применение липосом включает в себя доставку лекарственных средств, которые обычно являются токсичными в свободной форме. В форме липосом токсичное лекарственное средство является закрытым и может не затрагивать ткани, чувствительные к этому лекарственному средству, и доставлено к избранным областям. Липосомы также можно применять терапевтически для высвобождения лекарственных средств в течение длительного периода времени, уменьшая частоту введения. Кроме того, липосомы могут обеспечить способ получения водных дисперсий гидрофобных или амфифильных лекарственных средств, которые обычно являются неподходящими для внутривенного введения.
Для того чтобы многие лекарственные средства и визуализирующие агенты обладали терапевтическим или диагностическим потенциалом, необходимо, чтобы они были доставлены к правильному местоположению в организме, и, таким образом, липосома может быть легко инъецирована, что создает основу для длительного высвобождения и доставки лекарственного средства к клеткам специфических типов или частям организма. Можно использовать несколько методик для применения липосом для направления инкапсулированных лекарственных средств к избранным тканям хозяина и удаления от чувствительных тканей. Эти методики включают в себя манипулирование размером липосом, их поверхностным зарядом и путем их введения. MLV, прежде всего в связи с тем, что они являются относительно большими, обычно быстро поглощаются ретикулоэндотелиальной системой (в основном, печени и селезенки). С другой стороны, обнаружено, что UV демонстрируют повышенное время циркуляции, пониженные скорости клиренса и большее биологическое распространение относительно MLV.
Пассивная доставка липосом включает в себя применение различных путей введения, например внутривенного, подкожного, внутримышечного и местного. Каждый путь дает различия в локализации липосом. Два общих способа, используемых для активного направления липосом к избранным областям-мишеням, включают в себя присоединение к поверхности липосом либо антител, либо специфичных рецепторных лигандов. Известно, что антитела обладают высокой специфичностью к их соответствующему антигену, и их присоединяли к поверхности липосом, но результаты во многих случаях были менее чем успешными. Некоторые усилия, однако, были успешными при направлении липосом к опухолям без использования антител, см. , например, патент США N 5019369, патент США N 5441745 или патент США N 5435989.
Областью разработок, которыми энергично занимаются исследователи, является доставка агентов не только к специфическому типу клеток, но внутрь цитоплазмы клетки и, более того, в ядро. Это является особенно важным для доставки биологических агентов, таких как ДНК, РНК, рибозимы и белки. Многообещающее терапевтическое исследование в этой области включает в себя применение антисмысловых олигонуклеотидов ДНК и РНК для лечения заболевания. Однако, одна из главных проблем, встречающихся на пути эффективного применения антисмысловой технологии, состоит в том, что олигонуклеотиды в своей фосфодиэфирной форме быстро разрушаются в жидкостях организма, а также внутриклеточными и внеклеточными ферментами, такими как эндонуклеазы и экзонуклеазы, прежде чем достигают клетки-мишени. Внутривенное введение также приводит к быстрому почечному клиренсу из кровотока, и поглощение является недостаточным для получения эффективной внутриклеточной концентрации лекарственного средства. Липосомная инкапсуляция защищает олигонуклеотиды от расщепления ферментами, повышает период полураспада в кровообращении и повышает эффективность поглощения в результате фагоцитоза липосом. Таким образом, олигонуклеотиды способны достичь их желаемой мишени и доставляться к клеткам in vivo.
Сообщалось о нескольких случаях, когда исследователи присоединяли антисмысловые олигонуклеотиды к Липофильным Соединениям или Неиммуногенным Соединениям Высокой Молекулярной Массы. Антисмысловые олигонуклеотиды, однако, являются эффективными только как внутриклеточные агенты. Антисмысловые олигодезоксирибонуклеотиды, имеющие мишенью рецептор эпидермального фактора роста (EGF), инкапсулированные в липосомы, связанные с фолатом через полиэтиленгликолевый спейсер (фолат-ПЭГ-липосомы), доставляли к культивируемым клеткам KB через опосредованный рецептором фолата эндоцитоз (Wang et al. (1995) Proc. Natl. Acad. Sci. USA 92: 3318-3322). Кроме того, к олигонуклеотидам присоединяли алкилендиолы (Weiss et al. , патент США N 5245022). Кроме того, в литературе описано Липофильное Соединение, ковалентно присоединенное к антисмысловому олигонуклеотиду (ЕР 462145В1).
Загрузку биологических агентов в липосомы можно осуществлять путем включения в липидный препарат или загрузки в преформированные липосомы. Описано пассивное заякоривание олигопептидных и олигосахаридных лигандов во внешнюю поверхность липосом (Zalipsky et al. (1997) Bioconjug. Chem. , 8: 111: 118).
В. VEGF
Рост новых кровеносных сосудов из существующего эндотелия (ангиогенез) у здоровых взрослых людей жестко контролируется противоположными воздействиями позитивных и негативных регуляторов. При определенных патологических состояниях, включая пролиферативные ретинопатии, ревматоидный артрит, псориаз и рак, превалируют позитивные регуляторы, и ангиогенез вносит вклад в прогрессирование заболевания (обзор Folkman (1995) Nature Medicine 1: 27-31). Представление о том, что при раке ангиогенез представляет собой лимитирующую скорость стадию опухолевого роста и метастаза (Folkman (1971) New Engl. J. Med. 285: 1182-1186), в настоящее время подтверждается многочисленными экспериментальными данными (обзоры Aznavoorian et al. (1993) Cancer 71: 1368-1383; Fidler and Ellis (1994) Cell 79: 185-188; Folkman (1990) J. Natl. Cancer Inst. 82: 4-6).
Количество кровеносных сосудов в ткани опухоли является сильным отрицательным прогностическим признаком при раке молочной железы (Weidner et al. (1992) J. Natl. Cancer Inst. 84: 1875-1887), раке простаты (Weidner et al. (1993) Am. J. Pathol. 143: 401-409), опухолях головного мозга (Li et al. (1994) Lancet 344: 82-86) и меланоме (Foss et al. (1996) Cancer Res. 56: 2900-2903).
Оказывается, что ряд ангиогенных факторов роста, описанных к настоящему времени, среди которых находится сосудистый эндотелиальный фактор роста (VEGF), играют ключевую роль в качестве позитивного регулятора физиологического и патологического ангиогенеза (обзор в Brown et al. (1996) Control of Angiogenesis (ред. Goldberg and Rosen) Birkhauser, Basel, в печати: Thomas (1996) J. Biol. Chem. 271: 603-606). VEGF представляет собой секретируемый дисульфидно связанный гомодимер, который избирательно стимулирует эндотелиальные клетки к пролиферации, миграции и продуцированию ферментов, разрушающих матрикс (Conn et al. (1990) Рrос. Natl. Acad. Sci. USA 87: 1323-1327; Ferrara and Henzel (1989) Biochem. Biophys. Res. Commun. 161: 851-858; Gospodarowicz et al. (1989) Ргос. Natl. Acad. Sci. USA 7311-7315; Pepper et al. (1991) Biochem. Biophys. Res. Commun. 181: 902-906; Unemori et al. (1992) J. Cell. Physiol. 153: 557-562), причем все эти процессы требуются для образования новых сосудов. Кроме того, что VEGF является единственным известным митогеном, специфичным для эндотелиальных клеток, он является уникальным среди ангиогенных факторов роста по своей способности индуцировать в кровеносном сосуде временное повышение его проницаемости для макромолекул (отсюда его исходное и альтернативное название - фактор сосудистой проницаемости, VPF) (Dvorak et al. (1979) J. Immunol. 122: 166-174; Senger et al. (1983) Science 219: 983-985; Senger et al. (1986) Cancer Res. 46: 5629-5632). Повышенная сосудистая проницаемость и, в результате нее, отложение белков плазмы во внесосудистом пространстве содействуют образованию новых сосудов посредством создания временного матрикса для миграции эндотелиальных клеток (Dvorak et al. (1995) Am. J. Pathol. 146: 1029-1039). Сверхпроницаемость в действительности является характерной чертой новых сосудов, включая те, которые связаны с опухолями (Dvorak et al. (1995) Am. J. Pathol. 146: 1029-1039). Кроме того, в настоящее время известно, что компенсаторный ангиогенез, индуцированный гипоксией ткани, является опосредованным VEGF (Levy et al. (1996) J. Biol. Chem. 2746-2753); Shweiki et al. (1992) Nature 359: 843-845).
VEGF встречается в четырех формах (VEGF-121, VEGF-165, VEGF-189, VEGF-206) в результате альтернативного сплайсинга гена VEGF (Houck et al. (1991) Mol. Endocrin. 5: 1806-1814; Tischer et al. (1991) J. Biol. Chem. 266: 11947-11954). Две меньшие формы являются способными к диффузии, тогда как две большие формы остаются преимущественно локализованными на клеточной мембране как следствие их высокой аффинности к гепарину. VEGF-165 также связывается с гепарином и является наиболее часто встречающейся формой. VEGF-121, единственная форма, которая не связывается с гепарином, как оказывается, обладает более низкой аффинностью к рецепторам (Gitay-Goren et al. (1996) J. Biol. Chem. 271: 5519-5523), так же как и более низкой митогенной активностью (Keyt et al. (1996) J. Biol. Chem. 271: 7788-7795). Биологические воздействия VEGF опосредованы двумя тирозинкиназными рецепторами (Flt-1 и Flk-1/KDR), экспрессия которых в высокой степени ограничена клетками эндотелиального происхождения (de Vries et al. (1992) Science 255: 989-991; Millauer et al. (1993) Cell 72: 835-846; Terman et al. (1991) Oncogene 6: 519-524). Хотя для высоко аффинного связывания требуется экспрессия обоих функциональных рецепторов, хемотаксическая и митогенная передача сигнала в эндотелиальных клетках, как оказывается, осуществляется прежде всего через KDR рецептор (Park et al. (1994) J. Biol. Chem. 269: 25646-25654; Seetharam et al. (1995) Oncogene 10: 135-147; Waltenberger et al. (1994) J. Biol. Chem. 26988-26995). Значение VEGF и рецепторов VEGF для развития кровеносных сосудов недавно продемонстрировано на мышах, у которых отсутствует единственная аллель для гена VEGF (Carmeliet et al. (1996) Nature 380: 435-439; Ferrara et al. (1996) Nature 380: 439-442) или обе аллели Flt-1 (Fong et al. (1995) 376: 66-70) или Flk-1 генов (Shalaby et al. (1995) Nature 376: 62-66). В каждом случае наблюдались явные аномалии в образовании сосудов, приводящие к эмбриональной летальности.
VEGF продуцируется и секретируется в варьирующих количествах фактически всеми опухолевыми клетками (Brown et al. (1997) Regulation of Angiogenesis (ред. Goldberg and Rosen) Birkhauser, Basel, pp. 233-269). Прямое свидетельство того, что VEGF и его рецепторы вносят вклад в рост опухоли, было недавно получено путем демонстрации того, что рост ксенотрансплантатов опухоли человека у мышей nude может ингибироваться нейтрализующими антителами к VEGF (Kim et al. (1993) Nature 362: 841-844), экспрессией доминантно-негативного рецептора VEGF flk-1 (Millauer et al. (1996) Cancer Res. 56: 1615-1620; Millauer et al. (1994) Nature 367: 576-579), низкомолекулярными ингибиторами Flk-1 тирозинкиназной активности (Strawn et al. (1996) Cancer Res. 56: 3540-3545) или экспрессией антисмысловой последовательности к мРНК VEGF (Saleh et al. (1996) Cancer Res. 56: 393-401). Важно, что распространение метастазов опухоли, как было обнаружено, также резко снижается антагонистами VEGF (Claffey et al. (1996) Cancer Res. 56: 172-181).
Ингибиторы VEGF, помимо их использования в качестве противораковых агентов, могут быть полезными при широком разнообразии пролиферативных заболеваний, характеризующихся избыточным ангиогенезом, включая псориаз, глазные расстройства, коллагеново-сосудистые заболевания и ревматоидный артрит. Хотя известно, что большинство типов опухолей продуцируют VEGF, до недавнего времени не показано, что какой-либо из них экспрессирует функциональные рецепторы VEGF. Показано, что клетки саркомы Капоши (КS) не только продуцируют обильные количества VEGF, но также экспрессируют функциональные рецепторы VEGF и, следовательно, используют VEGF для аутокринного роста. Саркому Капоши обычно лечат с использованием общепринятых антиметаболических лекарственных средств. Однако, основной недостаток применения химиотерапии у пациентов с KS состоит в сопутствующей индукции иммуносупрессии, которая имеет серьезные последствия для пациентов, иммунная система которых уже подвержена риску. Потребность в альтернативных терапиях особенно велика на ранних стадиях этого заболевания, когда поражения KS начинают появляться, но в других отношениях пациенты чувствуют себя совершенно здоровыми. Недавно доказано, что в этом отношении инкапсуляция химиотерапевтических лекарственных средств, таких как даунорубицин (daunorubicin), в липосомы является многообещающим способом минимизации побочных эффектов химиотерапии при сохранении противоопухолевой эффективности. Лекарственные средства с низкой токсичностью, которые избирательно направляются к активированным клеткам эндотелиального происхождения, такие как описанные здесь Нуклеиновые Кислоты-Лиганды - антагонисты VEGF, должны представлять огромную ценность при лечении КS.
Другими областями возможного клинического применения Нуклеиновых Кислот-Лигандов VEGF являются глазные расстройства, характеризующиеся избыточным ангиогенезом. Примерами таких заболеваний являются дегенерация желтого пятна и диабетическая ретинопатия. При дегенерации желтого пятна прогрессирующий ангиогенез сосудистой оболочки под желтым пятном (частью сетчатки, ответственной за самую высокую остроту зрения) вредит зрению. При диабетической ретинопатии ангиогенез в сетчатке вредит зрению. Хотя начальные стимулы, которые инициируют рост кровеносных сосудов при дегенерации желтого пятна и диабетической ретинопатии, в настоящее время неизвестны, VEGF, как оказывается, является ключевым индуктором ангиогенеза (Lopez, P. F. et al. (1996) Invest. Ophthalmol. Visual Science 37, 855-868; Kliffen, M. et al. (1997) Br. J. Ophthalmol. 81, 154-162; Kvanta, A. et al. (1996) Invest. Ophthalmol. Visual Science 37, 1929-1934; Paques et al. (1997) Diabetes & Metabolism 23: 125-130). Ингибиторы VEGF, таким образом, могут быть полезными для ослабления ангиогенеза при дегенерации желтого пятна.
Сущность изобретения
Здесь описываются высоко аффинные 2'фтор(2'-F)-модифицированные-пиримидин-РНК-лиганды к сосудистому эндотелиальному фактору роста (VEGF). Способ, использованный здесь для идентификации таких Нуклеиновых Кислот-Лигандов, называется SELEX, акроним для Systematic Evolution of Ligands by Exponential enrichment. Лиганды, описанные здесь, были отобраны из исходного пула примерно 1014 молекул РНК, рандомизированных по 30 или 40 смежным положениям. В настоящее изобретение включены выделенные лиганды, которые представлены в таблицах 2-6. Далее, в настоящее изобретение включен способ получения Комплекса, состоящего из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, с помощью способа, при котором идентифицируют Нуклеиновую Кислоту-Лиганд из Смеси Кандидатов Нуклеиновых Кислот, где Нуклеиновая Кислота является лигандом VEGF, с помощью способа (а) приведения в контакт Смеси Кандидатов Нуклеиновых Кислот с VEGF, (б) отделения членов указанной Смеси Кандидатов на основе аффинности к VEGF и (в) амплификации отобранных молекул с получением смеси Нуклеиновых Кислот, обогащенной последовательностями Нуклеиновых Кислот с относительно высокой аффинностью для связывания с VEGF, и ковалентной сшивки указанной идентифицированной Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением. Далее изобретение включает в себя Комплекс, состоящий из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения.
Далее изобретение включает в себя Липидную Конструкцию, содержащую Нуклеиновую Кислоту-Лиганд VEGF или Комплекс. Кроме того, настоящее изобретение относится к способу получения Липидной Конструкции, содержащей Комплекс, где Комплекс состоит из Нуклеиновой Кислоты-Лиганда VEGF и Липофильного Соединения.
В другом воплощении данного изобретения предлагается способ усовершенствования фармакокинетических свойств Нуклеиновой Кислоты-Лиганда VEGF путем ковалентной сшивки Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением с образованием Комплекса и введения этого Комплекса пациенту. Далее изобретение относится к способу усовершенствования фармакокинетических свойств Нуклеиновой Кислоты-Лиганда VEGF путем дополнительного соединения этого Комплекса с Липидной Конструкцией.
Задачей настоящего изобретения является разработка Комплексов, содержащих одну или более чем одну Нуклеиновую Кислоту-Лиганд VEGF в соединении с одним или более чем одним Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением, и способов их получения. Дальнейшей задачей настоящего изобретения является разработка Липидных Конструкций, содержащих этот Комплекс. Следующей задачей настоящего изобретения является разработка одной или более чем одной Нуклеиновой Кислоты-Лиганда VEGF в соединении с одним или более чем одним Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением с усовершенствованными фармакокинетическими свойствами.
В воплощениях изобретения, направленных на Комплексы, состоящие из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы, предпочтительным является то, что Неиммуногенное Соединение Высокой Молекулярной Массы представляет собой полиалкиленгликоль, более предпочтительно полиэтиленгликоль (ПЭГ). Более предпочтительно этот ПЭГ имеет молекулярную массу примерно 10-80К. Наиболее предпочтительно ПЭГ имеет молекулярную массу примерно 20-45К. В воплощениях изобретения, направленных на Комплексы, состоящие из Нуклеиновой Кислоты-Лиганда VEGF и Липофильного Соединения, предпочтительным является то, что Липофильное Соединение представляет собой глицеролипид. В предпочтительных воплощениях изобретения Липидная Конструкция предпочтительно представляет собой липидную бислойную везикулу, и наиболее предпочтительно липосому. В предпочтительном воплощении Нуклеиновую Кислоту-Лиганд VEGF идентифицируют согласно способу SELEX.
В воплощениях изобретения, направленных на Комплексы, состоящие из Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, ковалентно сшитого с Нуклеиновой Кислотой-Лигандом или лигандами VEGF, Нуклеиновая Кислота-Лиганд или лиганды VEGF могут служить для направляющей способности.
Кроме того, Нуклеиновая Кислота-Лиганд VEGF может быть соединена через Ковалентные или Нековалентные Взаимодействия с Липидной Конструкцией, не являясь частью Комплекса.
Кроме того, в воплощениях изобретения, направленных на липидные конструкции, содержащие Нуклеиновую Кислоту-Лиганд VEGF или Комплекс Неиммуногенное Соединение Высокой Молекулярной Массы либо Липофильное Соединение/Нуклеиновая Кислота-Лиганд VEGF, где Липидная Конструкция представляет собой конструкцию такого типа, который имеет мембрану, определяющую внутренний компартмент, как например липидная бислойная везикула, Нуклеиновая Кислота-Лиганд VEGF или Комплекс в соединении с Липидной Конструкцией могут быть соединены с мембраной Липидной Конструкции или инкапсулированы внутри компартмента. В воплощениях, где Нуклеиновая Кислота-Лиганд VEGF находится в соединении с мембраной, Нуклеиновая Кислота-Лиганд VEGF может соединяться с ориентированной внутрь либо с ориентированной наружу частью мембраны, таким образом, что Нуклеиновая Кислота-Лиганд VEGF выступает внутри или снаружи везикулы. В некоторых воплощениях Комплекс Нуклеиновой Кислоты-Лиганда VEGF может быть пассивно загружен во внешнюю сторону преформированной Липидной Конструкции. В воплощениях, где Нуклеиновая Кислота-Лиганд VEGF выступает снаружи Липидной Конструкции, Нуклеиновая Кислота-Лиганд VEGF может служить для направляющей способности.
В воплощениях, где Нуклеиновая Кислота-Лиганд VEGF Липидной Конструкции служит для направляющей способности, Липидная Конструкция может быть соединена с дополнительными терапевтическими или диагностическими агентами. В одном воплощении этот терапевтический или диагностический агент соединен с внешней стороной Липидной Конструкции. В других воплощениях этот терапевтический или диагностический агент инкапсулирован в Липидную Конструкцию или соединен с внутренней стороной Липидной Конструкции. В еще одном дополнительном воплощении этот терапевтический или диагностический агент соединен с Комплексом. В одном воплощении этот терапевтический агент представляет собой лекарственное средство. В альтернативном воплощении этот терапевтический или диагностический агент представляет собой одну или более чем одну дополнительную Нуклеиновую Кислоту-Лиганд.
Дальнейшей задачей настоящего изобретения является разработка способа ингибирования ангиогенеза посредством введения Нуклеиновой Кислоты-Лиганда VEGF либо Комплекса, состоящего из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, либо Липидной Конструкции, содержащей Комплекс по настоящему изобретению. Еще одной дальнейшей задачей настоящего изобретения является разработка способа ингибирования роста опухолей посредством введения Нуклеиновой Кислоты-Лиганда VEGF либо Комплекса, состоящего из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, либо Липидной Конструкции, содержащей Комплекс по настоящему изобретению. Еще одной дальнейшей задачей настоящего изобретения является разработка способа ингибирования саркомы Капоши посредством введения Нуклеиновой Кислоты-Лиганда VEGF либо Комплекса, состоящего из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, либо Липидной Конструкции, содержащей Комплекс по настоящему изобретению. Еще одной дальнейшей задачей настоящего изобретения является разработка способа ингибирования дегенерации желтого пятна посредством введения Нуклеиновой Кислоты-Лиганда VEGF либо Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, либо Липидной Конструкции, содержащей Комплекс по настоящему изобретению. Еще одной дальнейшей задачей настоящего изобретения является разработка способа ингибирования диабетической ретинопатии посредством введения Нуклеиновой Кислоты-Лиганда VEGF либо Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, либо Липидной Конструкции, содержащей Комплекс по настоящему изобретению.
Следующей задачей настоящего изобретения является разработка способа направления терапевтического или диагностического агента к биологической мишени, которая экспрессирует VEGF, путем соединения агента с Комплексом, содержащим Нуклеиновую Кислоту-Лиганд VEGF и Липофильное Соединение или Неиммуногенное Соединение Высокой Молекулярной Массы, причем Комплекс дополнительно соединен с Липидной Конструкцией, а Нуклеиновая Кислота-Лиганд VEGF дополнительно соединена с внешней стороной Липидной Конструкции.
Эти и другие задачи, а также характер, объем и применение данного изобретения станут легко очевидными специалистам из дальнейшего описания и прилагаемой формулы изобретения.
Краткое описание графических материалов
На фиг. 1A-1Q показаны молекулярные изображения NX213 (фиг. 1А), NX278 (фиг. 1В), scNX278 (фиг. 1С), scNX213 (фиг. 1D), NX31838-PL (фиг. 1Е), NX31838 липидамид 1 (фиг. 1F), NX31838 липидамид 2 (фиг. 1G), NX31838-40K PEG (фиг. 1Н), NX31838-20K PEG (фиг. 1I), NX31838-40K PEG димер без линкера (NX31838d0) (фиг. 1J), NX31838-40K PEG димер с одним линкером С5 (NX31838d1) (фиг. 1К), NX31838-40K PEG димер с двумя линкерами С5 (NX31838d2) (фиг. 1L), С-5 аминолинкер (фиг. 1М), глицериндифосфатный линкер (фиг. 1N), 18-атомный спейсерный линкер (фиг. 1О), аминотетраэтиленгликолевый линкер (фиг. 1Р), 3'3'dT (фиг. 1Q) и NX31917 (фиг. 1R). 5'фосфатная группа лиганда изображена на фиг. mPEG означает метилполиэтиленгликоль. Буква в нижнем положении, предшествующая нуклеотиду, показывает следующее: m= 2'-O-метил, а= 2'-амино, r= рибо и f= 2'-фторо. Нуклеотид без предшествующей буквы показывает дезоксирибонуклеотид (2'Н). 3'3'-dT показывает 3'3' инвертированную фосфодиэфирную связь на 3'конце. S после нуклеотида означает модификацию остова, состоящую в фосфотиоатной межнуклеозидной связи.
На фиг. 2 показаны связывающие свойства различных Нуклеиновых Кислот-Лигандов по отношению к VEGF. Аффинности связывания немодифицированной Нуклеиновой Кислоты-Лиганда (NX213, незаштрихованная окружность), его диалкилглицерин-модифицированного аналога (NX278, незаштрихованный ромб) и липосомного NX278 (NX278-L, незаштрихованный квадрат) параллельно с последовательностью перемешанных (scrambled) контролей (sc) (scNX213 заштрихованная окружность; scNX278, заштрихованный ромб и scNX278-L, заштрихованный квадрат) определяли с помощью конкурентного анализа сдвига электрофоретической подвижности. NX213 представляет собой
5'-TsTsTsTsmAaCaCaCaUrGrAaUmGrGaUmAmGrAaCmGaCaCmGmGmGmGaUmGTsTsTsTsT-3' (SEQ ID NO: 1),
a scNX213 представляет собой
5'-TsTsTsTsmGaUaCmGmGaUmAaCrGmGrAmGaUmGrGrAaCnCmGaUaCmAaCmGTsTsTsTsT-3' (SEQ ID NO: 4)
32P 5= меченый по концу NX213 (1,5 нМ) инкубировали в связывающем буфере (физиологический раствор, забуференный фосфатом, с 0,01% сывороточного альбумина человека) при 37oС в течение 20 мин в присутствии VEGF (0,33 нМ) и конкурентного олигонуклеотида (5 пМ-0,33 мкМ). Комплекс 32P NX213/VEGF отделяли от свободного 32P NX213 с помощью электрофореза в 8% полиакриламидном геле (19: 1 акриламид: бис-акриламид, Трис-борат, 89 мМ, 1мМ ЭДТА в качестве буфера для пробега). Интенсивность полосы, соответствующей Комплексу 32P NX213/VEGF при варьирующих концентрациях конкурента, определяли количественно с помощью анализа на фосфорном формирователе изображения. Данные, нормализованные по количеству Комплекса, образованного в отсутствие конкурента, приводили к уравнению конкурентного связывания с помощью метода наименьших квадратов.
На фиг. 3 показано воздействие различных Нуклеиновых Кислот-Лигандов на индуцированные VEGF повышения сосудистой проницаемости. VEGF (20 нМ) с Нуклеиновыми Кислотами-Лигандами или без них инъецировали внутрикожно морским свинкам, которые до этого получили инъекцию синего красителя Эванса. Просачивание красителя определяли количественно путем измерения относительного количества света, поглощаемого кожей в сайте инъекции.
На фиг. 4 показано, что NX278-L ингибирует рост клеток КS. Рост клеток KSY-1 в присутствии различных концентраций NX213, NX278-L и scNX278-L. Клетки KSY-1 высевали на 24-луночные планшеты при плотности 1•104 клеток/лунку на 0 сутки. Свежую среду, обработанную идентично, заменяли на 1 и 3 сутки. Число клеток определяли с помощью трипсинизации клеток на 5 или 6 сутки культивирования с использованием счетчика частиц. Эксперименты ставили в трех повторностях несколько раз. Представленные результаты являются средним и SE (средней квадратичной ошибкой) репрезентативного эксперимента.
На фиг. 5А и 5В показано, что NX278 ингибирует рост клеток КS у бестимусных мышей. Бестимусным мышам имплантировали опухоль KS за передними лапами на сутки 1. Мышей обрабатывали NX278-L (50 мкг/сутки/мышь, фиг. 5А, и 150 мкг/сутки/мышь, фиг. 5В) путем внутрибрюшинной инъекции ежесуточно в течение пяти суток, начиная на сутки 2. Контрольных мышей обрабатывали пустыми липосомами с использованием того же количества липидов, что и в группе, обработанной Нуклеиновой Кислотой-Лигандом. Размеры опухоли измеряли через период в две недели. Опухоли удаляли на сутки 14 и измеряли.
На фиг. 6 суммируются данные по концентрации в плазме NX31838 20К PEG, 40К PEG NX31838 (минус PEG) как функция времени после болюсной инъекции.
На фиг. 7 суммируются данные по концентрации в плазме NX31838 PL как функция времени после болюсной инъекции.
На фиг. 8A-8D показаны изменения в сосудистой проницаемости, вызванные внутрикожной инъекцией белка VEGF (0,8 пмоль) ± Нуклеиновая Кислота-Лиганд/моноклональное антитело, как указано. Локальную транссудацию синего красителя Эванса определяли через 30 минут после инъекции с помощью трансиллюминации собранной кожи. На фиг. А, В, С и D показано воздействие смешанных вместе с белком NX31838-20K PEG, NX31838-40K PEG, NX31838-PL или NX31838d2-40K PEG за 30 минут до инъекции. Значения представляют собой среднее ± SEM (среднюю квадратичную ошибку). *Р<0,05 по сравнению с одним VEGF. См. молекулярные изображения на фиг. 1.
На фиг. 9А-9С показана оценка ослабления Нуклеиновой Кислотой-Лигандом VEGF-индуцированного роговичного ангиогенеза. Ноль или три пмоль белка VEGF включали в биополимер (Hydron) и имплантировали в собственное вещество роговицы. Животных обрабатывали внутривенно дважды в сутки либо ФСБ, либо Нуклеиновой Кислотой-Лигандом, как указано, в течение 5 суток. На фиг. А, В и С иллюстрируется воздействие системной обработки Нуклеиновой Кислотой-Лигандом NX31838-20K PEG, NX31838-40K PEG или NX31838-PL на образование новых сосудов. Значения представляют собой среднее ± SEM. *Р<0,05 по сравнению с 3 пмоль VEGF + ФСБ группой. Смотри молекулярные изображения на фиг. 1.
На фиг. 10 суммируются данные по концентрации в плазме ( ) или в стекловидном теле
) или в стекловидном теле  NX31838-40K PEG как функции времени после введения.
NX31838-40K PEG как функции времени после введения.
На фиг. 11 показаны кривые опухолевого роста опухолей человека А673, растущих подкожно (s. c. ) у мышей nude, обработанных 40 мг/кг или 10 мг/кг Нуклеиновой Кислоты-Лиганда VEGF NX31838-40K PEG (NX31838 NAL), вводимых дважды в сутки (BID). Отрицательный контроль заключался в перемешанной последовательности Нуклеиновой Кислоты-Лиганда VEGF NX31917 NAL (см. молекулярное изображение на фиг. 1R), дозированного по 40 мг/кг дважды в сутки, а положительный контроль заключался в моноклональном антителе анти-VEGF mAb 26503.11 (R& D Systems), дозированном по 100 мкг/мышь дважды в неделю. Поскольку не выявилось значительного различия между группой дозы 40 мг/кг и группой дозы 10 мг/кг, дальнейшее дозирование группы 40 мг/кг не повторяли после суток 14. Группам из 8 мышей имплантировали s. c. 1•107 опухолевых клеток А673 на 0 сутки, и обработку тестируемыми соединениями путем внутрибрюшинных инъекций начинали на 1 сутки эксперимента. Объем опухоли, выраженный в мм3, определяли с использованием формулы: Объем опухоли= L•W2/2.
На фиг. 12 показаны кривые опухолевого роста при различных схемах дозировки (сравнение дозирования дважды в сутки (BID) с дозированием один раз в сутки (QD)), партиях 40К PEG (сравнение партии NX31838.07 с новой партией NX31838.04) и различных лекарственных препаратах (сравнение липосомного VEGF NX31838PL NAL с VEGF NX31838 NAL 40K PEG) Нуклеиновой Кислоты-Лиганда (NAL) VEGF NX31838. Группам из 8 мышей имплантировали s. c. 1•107 опухолевых клеток А673 на сутки 0, и обработку тестируемыми соединениями путем внутрибрюшинных инъекций начинали на сутки 1 на время эксперимента. В нескольких группах были животные, у которых опухоли не росли, и, следовательно, для конечного анализа некоторые группы содержали только 7 (NX31838.04 10 мг/кг BID и NX31838.04 3 мг/кг BID) или 6 (NX31838.04 10 мг/кг QD и NX31838.07 10 мг/кг BID) животных. Объем опухоли, выраженный в мм3, определяли с использованием формулы: Объем опухоли= L•W2/2.
На фиг. 13 показано зависимое от дозы ингибирование опухолей А673, растущих подкожно (s. c. ), у мышей nude Нуклеиновой Кислотой-Лигандом VEGF NX31838 40K PEG (NX31838 NAL) при введении один раз в сутки. С помощью данного титрования не смогли достичь отсутствия эффекта дозы; ингибирование опухоли наблюдали еще при самой низкой дозе (0,03 мг/кг). Группам из 8 мышей имплантировали s. c. 1•107 опухолевых клеток А673 на 0 сутки, и обработку тестируемыми соединениями путем внутрибрюшинных инъекций начинали на 1 сутки эксперимента; в группе NX31838 NAL 3 мг/кг было 2 животных, у которых опухоли не росли, и, следовательно, она содержала только 6 животных. Объем опухоли, выраженный в мм3, определяли с использованием формулы: Объем опухоли= L•W2/2.
На фиг. 14 показаны кривые опухолевого роста, демонстрирующие ингибирование установившихся опухолей А673, растущих подкожно (s. c. ), у мышей nude Нуклеиновой Кислотой-Лигандом VEGF NX31838 40К PEG (NAL) при введении дважды в сутки. Положительный контроль заключался в моноклональном антителе анти-VEGF mAb 26503.11 (R& D Systems), дозированного по 100 мкг/мышь дважды в неделю. Мышам имплантировали 1•107 клеток А673 и давали опухолям возможность расти до объема 200±100 мм3, и в это время животных сортировали по массе, татуировали для постоянной идентификации, и обработку тестируемыми соединениями путем внутрибрюшинных инъекций начинали и продолжали на протяжении эксперимента. Каждая точка представляет среднее для 8 мышей. Объем опухоли, выраженный в мм3, определяли с использованием формулы: Объем опухоли= L•W2/2.
На фиг. 15 суммируются данные по концентрации в плазме NX213, NX278, NX-278-липосома после болюсной инъекции.
На фиг. 16 показаны кривые роста опухолей KSY-1, имплантированных подкожно, у мышей nude. Мышей обрабатывали путем внутрибрюшинных инъекций NX31917 40К PEG или NX31838 40К PEG (30 мг/кг) или ФСБ дважды в сутки на протяжении эксперимента. Обработку начинали через одни сутки после подкожной имплантации 2•107 клеток KSY-1 в заднюю боковую сторону мышей nude. В каждой группе использовали четырех мышей. Ошибки представляют собой SEM.
Определения:
"Ковалентная Связь" представляет собой химическую связь, образованную посредством обобществления электронов.
"Нековалентные Взаимодействия" представляют собой средства, с помощью которых молекулярные частицы держатся вместе путем взаимодействий, иных чем ковалентные связи, включая ионные взаимодействия и водородные связи.
"Липофильные Соединения" представляют собой соединения, которые имеют склонность к соединению или к распределению внутри липида и/или других материалов или фаз с низкими диэлектрическими константами, включая структуры, которые содержат по существу липофильные компоненты. Липофильные Соединения включают в себя как липиды, так и соединения, не содержащие липид, которые имеют склонность к соединению с липидом (и/или другими материалами или фазами с низкими диэлектрическими константами). Следующими примерами Липофильных Соединений являются холестерин, фосфолипид и глицеролипиды, такие как диалкилглицерин и диацилглицерин и глицеринамидлипиды. В одном предпочтительном воплощении изобретения Липофильное Соединение, ковалентно сшитое с Нуклеиновой Кислотой-Лигандом VEGF, представляет собой глицеринлипид, имеющий структуру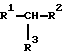
где R1 и R2 представляют собой СН3(СН2)n-O(РО3)-СН2- и СН3(СН2)n-CONH2-CH2-, СН3(СН2)nО-, СH3(CH2)nOCH2-, где n= 10-20,
R3 представляет собой -Х-, где -Х- независимо выбран из группы, состоящей из -(РО4)-, -О- и -СН2ОС(О)-. Когда по меньшей мере один из R1 и R2 представляет собой СН3(СН2)n-O(РО3)-СН2-, Липофильное Соединение представляет собой фосфолипид. Когда по меньшей мере один из R1 и R2 представляет собой CH3(CH2)n-CONH2-CH2-, Липофильное Соединение представляет собой гицеринамидлипид. Когда по меньшей мере один из R1 и R2 представляет собой СН3(СН2)nО- или СН3(СН2)nОСН2-, Липофильное Соединение представляет собой диалкилглицеринлипид.
"Комплекс", как использован здесь, описывает молекулярную частицу, образованную путем ковалентной сшивки Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением. В некоторых воплощениях настоящего изобретения этот Комплекс обозначается как A-B-Y, где А представляет собой Липофильное Соединение или Неиммуногенное Соединение Высокой Молекулярной Массы, как описано здесь; В является возможным и может представлять собой один или более чем один линкер Z; и Y представляет собой Нуклеиновую Кислоту-Лиганд VEGF.
"Липидные Конструкции" для целей настоящего изобретения представляют собой структуры, содержащие липиды, фосфолипиды или их производные, содержащие ряд различных структурных конфигураций, которые липиды, как известно, принимают в водной суспензии. Эти структуры включают в себя, не ограничиваясь ими, липидные бислойные везикулы, мицеллы, липосомы, эмульсии, липидные полоски или слои, и могут образовывать комплексы с рядом лекарственных средств и компонентов, которые известны как фармацевтически приемлемые. В предпочтительном воплощении Липидная Конструкция представляет собой Липосому. Предпочтительная Липосома является однослойной и имеет относительный размер менее чем 200 нм. Обычные дополнительные компоненты в Липидных Конструкциях включают в себя в числе прочих холестерин и альфа-токоферол. Липидные Конструкции можно применять отдельно или в любом сочетании, хорошо известном специалисту, чтобы обеспечить желаемые характеристики для конкретного применения. Кроме того, технические аспекты формирования Липидных Конструкций и Липосом хорошо известны специалистам, и любой из способов, обычно практикуемых в данной области, можно использовать для настоящего изобретения.
"Нуклеиновая Кислота-Лиганд", как использован здесь, представляет собой не встречающуюся в природе нуклеиновую кислоту, имеющую желаемое действие на мишень. Мишень по настоящему изобретению представляет собой VEGF, отсюда термин Нуклеиновая Кислота-Лиганд VEGF. Желаемое действие включает в себя, не ограничиваясь этим, связывание с мишенью, каталитическое изменение мишени, взаимодействие с мишенью путем, который модифицирует/изменяет мишень или функциональную активность мишени, ковалентное присоединение к мишени в качестве суицидного ингибитора, облегчение взаимодействия между мишенью и другой молекулой. В предпочтительном воплощении это действие представляет собой специфическую связывающую аффинность к VEGF, где Нуклеиновая Кислота-Лиганд не является нуклеиновой кислотой, имеющей известную физиологическую функцию, являясь связанной с VEGF.
В предпочтительных воплощениях изобретения Нуклеиновую Кислоту-Лиганд VEGF Комплексов и Липидных Конструкций по изобретению идентифицируют с помощью методологии SELEX. Нуклеиновые Кислоты-Лиганды VEGF идентифицируют из Смеси Кандидатов Нуклеиновых Кислот, где указанная Нуклеиновая Кислота является лигандом VEGF, с помощью способа, при котором а) приводят в контакт Смесь Кандидатов с VEGF, причем нуклеиновые кислоты, имеющие повышенную аффинность к VEGF относительно Смеси Кандидатов, можно отделить от остатка этой Смеси Кандидатов; б) отделяют нуклеиновые кислоты с повышенной аффинностью от остатка этой Смеси Кандидатов; и в) амплифицируют нуклеиновые кислоты с повышенной аффинностью с получением смеси Нуклеиновых Кислот, обогащенной лигандами (см. Заявку на патент США, порядковый N 08/233012, поданную 25 апреля 1994, озаглавленную "High Affinity Oligonucleotides to Vascular Endothelial Growth Factor (VEGF)", Заявку на патент США, порядковый N 08/447169, поданную 19 мая 1995, озаглавленную "High Affinity Oligonucleotide Ligands to Vascular Endothelial Growth Factor (VEGF)", которые настоящим включены здесь путем ссылки).
"Смесь Кандидатов" представляет собой смесь Нуклеиновых Кислот с различными последовательностями, из которой отбирают желаемый лиганд. Источником Смеси Кандидатов могут являться встречающиеся в природе нуклеиновые кислоты или их фрагменты, химически синтезированные нуклеиновые кислоты, ферментативно синтезированные нуклеиновые кислоты или нуклеиновые кислоты, полученные путем комбинирования вышеуказанных методик. В предпочтительном воплощении каждая Нуклеиновая Кислота имеет фиксированные последовательности, окружающие рандомизированный район, для облегчения процесса амплификации.
"Нуклеиновая Кислота" означает либо ДНК, либо РНК, однонитевую или двунитевую, и любые их химические модификации. Модификации включают в себя, не ограничиваясь ими, те, которые предусматривают другие химические группы, которые придают дополнительный заряд, поляризуемость, водородное связывание, электростатическое взаимодействие и плавкость основаниям Нуклеиновой Кислоты-Лиганда или Нуклеиновой Кислоте-Лиганду в целом. Такие модификации включают в себя, не ограничиваясь ими, модификации 2'-положения сахара, модификации 5-положения пиримидина, модификации 8-положения пурина, модификации по экзоциклическим аминам, замещение 4-тиоуридина, замещение 5-бромо или 5-иодоурацила, модификации остова, такие как межнуклеозидные фосфоротиоатные связи, метилирования, необычные комбинации спаривания оснований, такие как изооснования изоцитидин и изогуанидин, и тому подобные. Модификации также могут включать в себя 3' и 5' модификации, такие как кэппирование.
"Неиммуногенное Соединение Высокой Молекулярной Массы" представляет собой соединение между примерно 1000 Да и 1000000 Да, более предпочтительно примерно от 1000 Да до 500000 Да и наиболее предпочтительно примерно от 1000 Да до 200000 Да, которое обычно не вызывает иммуногенный ответ. Для целей данного изобретения иммуногенный ответ представляет собой такой ответ, который заставляет организм производить белки-антитела. Примеры Неиммуногенных Соединений Высокой Молекулярной Массы включают в себя полиалкиленгликоль и полиэтиленгликоль. В одном предпочтительном воплощении изобретения Неиммуногенное Соединение Высокой Молекулярной Массы, ковалентно сшитое с Нуклеиновой Кислотой-Лигандом VEGF, представляет собой полиалкиленгликоль и имеет структуру R(O(CH2)x)nO-, где R независимо выбран из группы, состоящей из Н и СН3, х= 2-5, a n≈MW (молекулярная масса) полиалкиленгликоля/16+14х. В предпочтительном воплощении настоящего изобретения молекулярная масса составляет примерно между 10-80 кДа. В наиболее предпочтительном воплощении молекулярная масса полиалкиленгликоля составляет примерно между 20-45 кДа. В наиболее предпочтительном воплощении х= 2, а n= 9•102. К одной и той же Нуклеиновой Кислоте-Лиганду VEGF может быть присоединен один или более чем один полиалкиленгликоль, причем сумма молекулярных масс предпочтительно составляет между 10-80 кДа, более предпочтительно 20-45 кДа. В некоторых воплощениях Неиммуногенное Соединение Высокой Молекулярной Массы может также представлять собой Нуклеиновую Кислоту-Лиганд.
"Липидные Бислойные Везикулы" представляют собой замкнутые, заполненные жидкостью микроскопические сферы, которые образованы главным образом из индивидуальных молекул, имеющих полярные (гидрофильные) и неполярные (липофильные) участки. Гидрофильные участки могут содержать фосфатную, глицерилфосфатную, карбокси, сульфатную, амино, гидрокси, холиновую и другие полярные группы. Примерами неполярных групп являются насыщенные или ненасыщенные углеводороды, такие как алкил, алкенил, или другие липидные группы. Стерины (например, холестерин) и другие фармацевтически приемлемые компоненты (включая антиоксиданты, подобные альфа-токоферолу) также могут быть включены для улучшения стабильности везикул или придания других желаемых характеристик.
"Липосомы" представляют собой подгруппу Липидных Бислойных Везикул и содержат главным образом молекулы фосфолипидов, которые несут два гидрофобных хвоста, состоящих из длинных цепей жирных кислот. При экспозиции с водой эти молекулы спонтанно выстраиваются с образованием бислойной мембраны с липофильными концами этих молекул в каждом слое, соединенными в центре мембраны, и противоположными полярными концами, образующими соответствующие внутреннюю и внешнюю поверхности этой бислойной мембраны. Таким образом, каждая сторона мембраны представляет собой гидрофильную поверхность, тогда как внутри мембрана представляет собой липофильную среду. Эти мембраны при формировании, как правило, организованы в систему концентрических замкнутых мембран, разделенных межслойными водными фазами, до некоторой степени подобно слоям луковицы, вокруг внутреннего водного пространства. Эти многослойные везикулы (MLV) можно превратить в Однослойные Везикулы (UV) при приложении силы гидродинамического фрагментирования.
"Катионная Липосома" представляет собой липосому, которая содержит липидные компоненты, имеющие общий положительный заряд при физиологическом рН.
"SELEX" методология включает в себя сочетание отбора Нуклеиновых Кислот-Лигандов, которые взаимодействуют с мишенью желаемым образом, например путем связывания с белком, с амплификацией этих отобранных Нуклеиновых Кислот. Циклическое повторение этих стадий отбора/амплификации дает возможность отбора одной Нуклеиновой Кислоты или небольшого количества Нуклеиновых Кислот, которые более сильно взаимодействуют с мишенью, из пула, который содержит очень большое число Нуклеиновых Кислот. Проведение цикла процедуры отбора/амплификации продолжают, пока избранная цель не будет достигнута. Методология SELEX описана в Заявках на патент SELEX.
"Мишень" означает любое из представляющих интерес соединений или молекул, для которых лиганд является желаемым. Мишень может представлять собой белок (как например VEGF, тромбин и селектин), пептид, углевод, полисахарид, гликопротеин, гормон, рецептор, антиген, антитело, вирус, субстрат, метаболит, аналог в переходном состоянии, кофактор, ингибитор, лекарственное средство, краситель, питательное вещество, фактор роста и т. д. без ограничения. Главной мишенью по данному изобретению является VEGF.
"Усовершенствованные Фармакокинетические Свойства" означают, что Нуклеиновая Кислота-Лиганд VEGF, ковалентно сшитая с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением, либо в соединении с Липидной Конструкцией проявляет более длительный период полураспада в кровообращении in vivo относительно той же Нуклеиновой Кислоты-Лиганда VEGF, не соединенной с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением либо с Липидной Конструкцией.
"Линкер" представляет собой молекулярную частицу, которая соединяет две или более чем две молекулярные частицы через ковалентную связь или нековалентные взаимодействия и может дать возможность пространственного разделения этих молекулярных частиц таким способом, при котором сохраняются функциональные свойства одной или более чем одной молекулярной частицы. Линкер также может быть известен как спейсер. Примеры линкеров включают в себя, не ограничиваясь ими, структуры, показанные на фиг. 1М-1Р.
"Терапевтический", как использован здесь, включает в себя лечение и/или профилактику. Когда используется термин "терапевтический", он относится к людям и к другим животным.
Данное изобретение включает в себя РНК-лиганды к VEGF, которые содержат 2'F-модифицированные нуклеотиды. Данное изобретение, кроме того, включает в себя специфичные РНК-лиганды к VEGF, представленные в таблицах 2-6 (SEQ ID Nos: 15-132). Более конкретно данное изобретение включает в себя последовательности Нуклеиновых Кислот, которые являются по существу гомологичными специфичным Нуклеиновым Кислотам-Лигандам, представленным в таблицах 2-6, и которые обладают по существу такой же способностью связывать VEGF. Под существенной гомологией подразумевается степень гомологии первичной последовательности, превышающая 70%, более предпочтительно превышающая 80% и даже более предпочтительно превышающая 90, 95 или 99%. Процент гомологии, как описано здесь, вычисляют как процент нуклеотидов, находящихся в меньшей из двух последовательностей, которые выравниваются с идентичными нуклеотидными остатками в последовательности, с которой ее сравнивают, причем на длину в 10 нуклеотидов можно ввести 1 разрыв, чтобы способствовать этому выравниванию. По существу такая же способность к связыванию VEGF означает, что аффинность находится в пределах одного или двух порядков величины аффинности лигандов, описанных здесь. Определение того, обладает ли данная последовательность - по существу гомологичная конкретно описанным здесь - такой же способностью к связыванию VEGF, находится в пределах компетентности обычного специалиста в этой области.
Обзор гомологии последовательностей Нуклеиновых Кислот-Лигандов VEGF, представленных в таблицах 2-6 (SEO ID NOS: 15-132), показывает, что последовательности с небольшой первичной гомологией или с отсутствием первичной гомологии могут обладать по существу такой же способностью к связыванию VEGF. По этим причинам данное изобретение также включает в себя Нуклеиновые Кислоты-Лиганды, которые имеют по существу такую же постулированную структуру или структурные мотивы и способность связывать VEGF, как и Нуклеиновые Кислоты-Лиганды, представленные в таблицах 2-6. По существу такую же структуру или структурные мотивы можно постулировать по выравниванию последовательностей с использованием программы Zukerfold (см. Zuker (1989) Science 244: 48-52). Как должно быть известно специалистам, для предсказания вторичной структуры и структурных мотивов можно использовать другие компьютерные программы. По существу такую же структуру или структурный мотив Нуклеиновых Кислот-Лигандов в растворе или в виде связанной структуры также можно постулировать с использованием ЯМР или других методик, которые должны быть известны специалистам.
Далее в настоящее изобретение включен способ получения Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, с помощью способа, при котором идентифицируют Нуклеиновую Кислоту-Лиганд из Смеси Кандидатов Нуклеиновых Кислот, где Нуклеиновая Кислота является лигандом VEGF, с помощью способа (а) приведения в контакт Смеси Кандидатов Нуклеиновых Кислот с VEGF, (б) отделения членов указанной Смеси Кандидатов на основании аффинности к VEGF и (в) амплификации отобранных молекул с получением смеси Нуклеиновых Кислот, обогащенной последовательностями Нуклеиновых Кислот с относительно более высокой аффинностью для связывания с VEGF, а также ковалентного сшивания указанной идентифицированной Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением.
Следующей задачей настоящего изобретения является разработка Комплексов, содержащих одну или более чем одну Нуклеиновую Кислоту-Лиганд VEGF, ковалентно сшитую с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением. Такие Комплексы имеют одно или более чем одно из следующих преимуществ перед Нуклеиновой Кислотой-Лигандом VEGF, не соединенной с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением:
1) усовершенствованные фармакокинетические свойства и 2) усовершенствованная способность к внутриклеточной доставке или 3) усовершенствованная способность направления к мишени. Комплексы, дополнительно соединенные с Липидной Конструкцией, имеют такие же преимущества.
Эти Комплексы или Липидные Конструкции, содержащие Нуклеиновую Кислоту-Лиганд VEGF или Комплексы, могут быть выгодными из-за одного, двух или трех из этих преимуществ. Например, Липидная Конструкция по настоящему изобретению может состоять из а) липосомы, б) лекарственного средства, которое инкапсулировано во внутренней части этой липосомы, и в) Комплекса, в котором содержится Нуклеиновая Кислота-Лиганд VEGF и Липофильное Соединение, причем компонент Нуклеиновой Кислоты-Лиганда
VEGF соединен с внешней стороной Липидной Конструкции и выступает из нее. В этом случае Липидная Конструкция, содержащая Комплекс, будет 1) обладать усовершенствованными фармакокинетическими свойствами, 2) обладать усиленной способностью к внутриклеточной доставке инкапсулированного лекарственного средства и 3) специфически направляться в предварительно выбранное местоположение in vivo, где экспрессируется VEGF, с помощью присоединенной к внешней стороне Нуклеиновой Кислоты-Лиганда VEGF.
В предпочтительном воплощении данного изобретения предлагается способ усовершенствования фармакокинетических свойств Нуклеиновой Кислоты-Лиганда VEGF путем ковалентного сшивания Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением с образованием Комплекса и введения этого Комплекса пациенту. Далее изобретение относится к способу усовершенствования фармакокинетических свойств Нуклеиновой Кислоты-Лиганда VEGF посредством дополнительного соединения этого Комплекса с Липидной Конструкцией.
В другом воплощении Комплекс по настоящему изобретению содержит Нуклеиновую Кислоту-Лиганд VEGF, ковалентно присоединенную к Липофильному Соединению, такому как глицеролипид, или Неиммуногенному Соединению Высокой Молекулярной Массы, такому как полиалкиленгликоль или полиэтиленгликоль (ПЭГ). В этих случаях фармакокинетические свойства этого Комплекса будут усилены относительно одной Нуклеиновой Кислоты-Лиганда VEGF. В другом воплощении фармакокинетические свойства Нуклеиновой Кислоты-Лиганда VEGF усилены относительно одной Нуклеиновой Кислоты-Лиганда VEGF, когда Нуклеиновая Кислота-Лиганд VEGF ковалентно присоединена к Неиммуногенному Соединению Высокой Молекулярной Массы или Липофильному Соединению и дополнительно соединена с Липидной Конструкцией, либо Нуклеиновая Кислота-Лиганд VEGF инкапсулирована внутри Липидной Конструкции.
В воплощениях, где имеются множественные Нуклеиновые Кислоты-Лиганды VEGF, наблюдается повышение их авидности к VEGF вследствие множественных связывающих взаимодействий с VEGF. Кроме того, в воплощениях, где Комплекс содержит множественные Нуклеиновые Кислоты-Лиганды VEGF, фармакокинетические свойства этого Комплекса будут усовершенствованы относительно одной только Нуклеиновой Кислоты-Лиганда VEGF. В воплощениях, где Липидная Конструкция содержит множественные Нуклеиновые Кислоты-Лиганды или Комплексы, фармакокинетические свойства Нуклеиновой Кислоты-Лиганда VEGF можно усовершенствовать относительно Липидных Конструкций, в которых имеется только одна Нуклеиновая Кислота-Лиганд VEGF или только один Комплекс.
В некоторых воплощениях изобретения Комплекс по настоящему изобретению содержит Нуклеиновую Кислоту-Лиганд VEGF, присоединенную к одной (димерный) или более чем одной (мультимерный) другой Нуклеиновой Кислоте-Лиганду. Нуклеиновая Кислота может являться лигандом к VEGF или к другой мишени. В воплощениях, где имеются множественные Нуклеиновые Кислоты-Лиганды VEGF, наблюдается повышение их авидности вследствие множественных связывающих взаимодействий с VEGF. Кроме того, в воплощениях изобретения, где Комплекс содержит Нуклеиновую Кислоту-Лиганд VEGF, присоединенную к одной или более чем одной другой Нуклеиновой Кислоте-Лиганду VEGF, фармакокинетические свойства этого Комплекса будут усовершенствованы относительно одной Нуклеиновой Кислоты-Лиганда VEGF отдельно.
Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение может быть ковалентно сшитым с множеством положений на нуклеиновой кислоте-лиганде VEGF, как например с экзоциклической аминогруппой на основании, 5-положением пиримидинового нуклеотида, 8-положением пуринового нуклеотида, гидроксильной группой фосфата либо с гидроксильной группой или другой группой на 5' или 3'конце Нуклеиновой Кислоты-Лиганда VEGF. В воплощениях, где Липофильное Соединение представляет собой глицеролипид, либо Неиммуногенное Соединение Высокой Молекулярной Массы представляет собой полиалкиленгликоль или полиэтиленгликоль, предпочтительно оно связано с 5' или 3' гидроксилом его фосфатной группы. В наиболее предпочтительном воплощении Липофильное Соединение или Неиммуногенное Соединение Высокой Молекулярной Массы связано с 5' гидроксилом фосфатной группы Нуклеиновой Кислоты-Лиганда. Присоединение Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения к Нуклеиновой Кислоте-Лиганду VEGF можно осуществить непосредственно или с использованием линкеров или спейсеров. В воплощениях, где Липидная Конструкция содержит Комплекс или где Нуклеиновые Кислоты-Лиганды VEGF инкапсулированы внутри липосомы, предпочтительным является нековалентное взаимодействие между Нуклеиновой Кислотой-Лигандом VEGF или Комплексом и этой Липидной Конструкцией.
Одна из проблем, встречающихся на пути терапевтического применения Нуклеиновых Кислот, состоит в том, что олигонуклеотиды в их фосфодиэфирной форме могут легко расщепляться в жидкостях организма внутриклеточными и внеклеточными ферментами, такими как эндонуклеазы и экзонуклеазы, прежде чем проявляется желаемый эффект. Можно создать некоторые химические модификации Нуклеиновой Кислоты-Лиганда VEGF, чтобы повысить стабильность Нуклеиновой Кислоты-Лиганда VEGF in vivo либо чтобы усилить или опосредовать доставку Нуклеиновой Кислоты-Лиганда VEGF. Модификации Нуклеиновых Кислот-Лигандов VEGF, рассматриваемые в настоящем изобретении, включают в себя, не ограничиваясь ими, те, которые предусматривают другие химические группы, которые придают дополнительный заряд, поляризуемость, гидрофобность, водородное связывание, электростатическое взаимодействие и плавкость основаниям Нуклеиновой Кислоты-Лиганда VEGF или Нуклеиновой Кислоте-Лиганду VEGF в целом. Такие модификации включают в себя, не ограничиваясь ими, модификации 2'-положения сахара, модификации 5-положения пиримидина, модификации 8-положения пурина, модификации по экзоциклическим аминам, замещение 4-тиоуридина, замещение 5-бромо- или 5-иодоурацила, модификации остова, фосфоротиоатные или алкилфосфатные модификации, метилирования, необычные комбинации спаривания оснований, такие как изооснования изоцитидин и изогуанидин и тому подобные. Модификации также могут включать в себя 3' и 5' модификации, как например кэппирование.
Если Нуклеиновые Кислоты-Лиганды получены с помощью способа SELEX, эти модификации могут представлять собой пре- или пост-SELEX модификации. Пре-SELEX модификации дают Нуклеиновые Кислоты-Лиганды VEGF как со специфичностью к VEGF, так и с улучшенной стабильностью in vivo. Пост-SELEX модификации, сделанные в 2'-ОН Нуклеиновых Кислотах-Лигандах, могут привести к улучшенной стабильности in vivo без неблагоприятного влияния на связывающую способность Нуклеиновых Кислот-Лигандов. Предпочтительными модификациями Нуклеиновых Кислот-Лигандов VEGF по данному изобретению является 5' и 3'фосфоротиоатное кэппирование и/или 3'3'инвертированная фосфодиэфирная связь на 3'конце. В наиболее предпочтительном воплощении предпочтительной модификацией Нуклеиновой Кислоты-Лиганда VEGF является 3'3'инвертированная фосфодиэфирная связь на 3'конце. Дополнительная 2'-фтор (2'-F), 2'-амино (2'-NН2) и 2'-O-метил (2'-Оме) модификация нескольких или всех нуклеотидов является предпочтительной.
В другом аспекте настоящего изобретения ковалентная сшивка Нуклеиновой Кислоты-Лиганда VEGF с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением приводит к усовершенствованным фармакокинетическим свойствам (то есть, более медленная скорость клиренса) относительно Нуклеиновой Кислоты-Лиганда VEGF, не соединенной с Неиммуногенным Соединением Высокой Молекулярной Массы или Липофильным Соединением.
В другом аспекте настоящего изобретения Комплекс, содержащий Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, может быть дополнительно соединен с Липидной Конструкцией. Это соединение может привести к усовершенствованным фармакокинетическим свойствам относительно Нуклеиновой Кислоты-Лиганда VEGF или Комплекса, не соединенных с Липидной Конструкцией. Нуклеиновую Кислоту-Лиганд VEGF или Комплекс можно соединить с Липидной Конструкцией посредством ковалентных или нековалентных взаимодействий. В предпочтительном воплощении имеет место соединение через нековалентные взаимодействия. В предпочтительном воплощении Липидная Конструкция представляет собой Липидную Бислойную Везикулу. В наиболее предпочтительном воплощении Липидная Конструкция представляет собой липосому.
Липосомы для применения по настоящему изобретению можно получить с помощью любой из различных методик, известных в настоящее время специалистам или разработанных позже. Обычно их получают из фосфолипида, например дистеароилфосфатидилхолина, и они могут включать в себя другие материалы, такие как нейтральные липиды, например холестерин, а также поверхностные модификаторы, как например положительно заряженные (например, стериламин либо аминоманнозные или аминоманнитные производные холестерина) или отрицательно заряженные (например, диацетилфосфат, фосфатидилглицерин) соединения. Многослойные липосомы можно формировать с помощью общепринятых методик, которые представляют собой осаждение выбранного липида на внутренней стенке подходящего контейнера или сосуда путем растворения этого липида в подходящем растворителе, а затем выпаривания растворителя, с тем чтобы оставить тонкую пленку на внутренней стороне сосуда, либо путем аэрозольного высушивания. Водную фазу затем добавляют в сосуд при круговом или вихревом движении, которое приводит к образованию MLV. UV можно формировать с помощью гомогенизации, озвучивания или экструзии (через фильтры) MLV. Кроме того, UV можно формировать с помощью методик удаления детергентом.
В некоторых воплощениях данного изобретения Липидная Конструкция содержит направляющую Нуклеиновую Кислоту-Лиганд (Нуклеиновые Кислоты-Лиганды) к VEGF мишени, соединенные с поверхностью Липидной Конструкции, и инкапсулированный терапевтический или диагностический агент. Предпочтительно Липидная Конструкция представляет собой липосому. Преформированные липосомы могут быть модифицированы для соединения с Нуклеиновыми Кислотами-Лигандами VEGF. Например, Катионная Липосома соединяется через электростатические взаимодействия с Нуклеиновой Кислотой-Лигандом VEGF. Нуклеиновую Кислоту-Лиганд VEGF, ковалентно сшитую с Липофильным Соединением, таким как глицеролипид, можно добавить к преформированным липосомам, посредством чего глицеролипид, фосфолипид или глицеринамидлипид становится соединенным с липосомной мембраной. Альтернативно, Нуклеиновую Кислоту-Лиганд VEGF можно соединить с липосомой в процессе изготовления липосомы.
Специалистам хорошо известно, что липосомы являются полезными для инкапсулирования или включения широкого разнообразия терапевтических и диагностических агентов. Любое многообразие соединений можно заключить во внутренний водный компартмент липосом. Иллюстративные терапевтические агенты включают в себя антибиотики, противовирусные нуклеозиды, противогрибковые нуклеозиды, регуляторы метаболизма, иммуномодуляторы, химиотерапевтические лекарственные средства, антидоты токсинов, ДНК, РНК, антисмысловые олигонуклеотиды и т. д. Кроме того, Липидные Бислойные Везикулы можно загружать диагностическим радионуклидом (например, индий 111, иод 131, иттрий 90, фосфор 32 или гадолиний) и флюоресцентными материалами или другими материалами, которые являются обнаружимыми при применениях in vitro и in vivo. Следует понимать, что терапевтический или диагностический агент может быть инкапсулирован стенками липосомы в водном внутреннем пространстве. Альтернативно, переносимый агент может являться частью материалов, образующих стенку везикулы, то есть, диспергированным или растворенным в них.
В процессе формирования липосом водорастворимые агенты-носители могут быть инкапсулированы в водном внутреннем пространстве путем включения их в гидратирующий раствор, а липофильные молекулы могут быть включены в липидный бислой путем включения в липидный препарат. В случае некоторых молекул (например, катионных или анионных липофильных лекарственных средств) загрузку лекарственного средства в преформированные липосомы можно осуществить, например, с помощью способов, описанных в патенте США N 4946683, содержание которого включено здесь путем ссылки. После инкапсулирования лекарственного средства липосомы подвергают удалению неинкапсулированного лекарственного средства с помощью способов, таких как гель-хроматография или ультрафильтрация. Липосомы затем обычно стерильно фильтруют, чтобы удалить какие-либо микроорганизмы, которые могут присутствовать в суспензии. Микроорганизмы могут быть также удалены посредством асептической обработки.
Если желают инкапсулировать большие гидрофильные молекулы в липосомы, большие Однослойные Везикулы могут быть сформированы с помощью способов, таких как обращенно-фазное выпаривание (REV), или способы инфузии растворителя. Другие стандартные способы формирования липосом известны специалистам, например, способы коммерческого производства липосом включают в себя процедуру гомогенизации, описанную в патенте США N 4753788, и способ тонкопленочного выпаривания, описанный в патенте США N 4935171, которые включены здесь путем ссылки.
Следует понимать, что терапевтический или диагностический агент может быть также соединен с поверхностью Липидной Бислойной Везикулы. Например, лекарственное средство может быть присоединено к фосфолипиду или глицериду (пролекарство). Фосфолипидная или глицеридная часть этого пролекарства может быть включена в липидный бислой липосомы путем включения в липидный препарат или загрузки в преформированные липосомы (см. патенты США NN5194654 и 5223263, которые включены здесь путем ссылки).
Специалисту легко понятно, что конкретный способ получения липосомы будет зависеть от предназначенного применения и типа липидов, использованных для образования бислойной мембраны.
Lee and Low (1994, JBC, 269: 3198-3204) и DeFrees et al. (1996, JACS, 118: 6101-6104) впервые показали, что совместное изготовление лиганда-ПЭГ-липида с липидными компонентами давало липосомы как с внутренней, так и с внешней ориентацией ПЭГ-лиганда. Пассивное заякоривание показано Zalipsky et al. (1997, Bioconj. Chem. 8: 111-118) как способ для заякоривания олигопептидных и олигосахаридных лигандов исключительно на внешней поверхности липосом. Центральная концепция, присутствующая в их работе, состоит в том, что конъюгаты лиганд-ПЭГ-липид могут быть получены, а затем введены в состав преформированных липосом через спонтанное включение ("заякоривание") липидного хвоста в существующий липидный бислой. Липидная группа претерпевает это включение, чтобы достичь более низкого состояния свободной энергии посредством удаления ее гидрофобного липидного якоря из водного раствора и его последующей установки в гидрофобном липидном бислое. Ключевым преимуществом такой системы является то, что олиголипид заякоривается исключительно на внешней стороне липидного бислоя. Следовательно, никакие олиголипиды не теряются из-за недоступности для взаимодействий с их биологическими мишенями вследствие их нахождения в обращенной внутрь ориентации.
Эффективность доставки Нуклеиновой Кислоты-Лиганда VEGF к клеткам можно оптимизировать путем использования липидных препаратов и условий, известных для усиления слияния липосом с клеточными мембранами. Например, некоторые отрицательно заряженные липиды, такие как фосфатидилглицерин и фосфатидилсерин, способствуют слиянию, особенно в присутствии других веществ, способствующих слиянию (например, многовалентных катионов, подобных Са2+, свободных жирных кислот, вирусных белков слияния, ПЭГ с короткой цепью, лизолецитина, детергентов и сурфактантов). Фосфатидилэтаноламин также может быть включен в препарат липосомы для повышения слияния с мембраной и сопутствующего усиления клеточной доставки. Кроме того, свободные жирные кислоты и их производные, содержащие, например, карбоксилатные группировки, можно использовать для получения рН-чувствительных липосом, которые являются отрицательно заряженными при более высоком рН и нейтральными или протонированными при более низком рН. Известно, что такие рН-чувствительные липосомы обладают большей тенденцией к слиянию.
В предпочтительном воплощении Нуклеиновые Кислоты-Лиганды VEGF по настоящему изобретению получают по методологии SELEX. SELEX описан в Заявке на патент США, порядковый N 07/536428, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment", в настоящее время отозванной, в Заявке на патент США, порядковый N 07/714131, поданной 10 июня 1991, озаглавленной "Nucleic Acid Ligands", в настоящее время патент США N 5475096, в Заявке на патент США, порядковый N 07/931473, поданной 17 августа 1992, озаглавленной "Methods for Identifying Nucleic Acid Ligands", в настоящее время патент США N 5270163 (см. также WO 91/19813). Данные заявки, каждая из которых включена здесь путем ссылки, все вместе названы Заявки на патент SELEX.
Способ SELEX обеспечивает класс продуктов, которые представляют собой молекулы нуклеиновой кислоты, каждая из которых имеет уникальную последовательность, и каждая из которых обладает свойством специфичного связывания с желаемым соединением-мишенью или с желаемой молекулой-мишенью. Молекулы-мишени предпочтительно представляют собой белки, но могут также включать в себя в числе прочих другие углеводы, пептидогликаны и множество малых молекул. Методологию SELEX можно использовать также, чтобы делать мишенью биологические структуры, такие как клеточные поверхности или вирусы, посредством специфичного взаимодействия с молекулой, которая является интегральной частью этой биологической структуры.
В наиболее упрощенной форме способ SELEX можно определить как следующие серии стадий:
1) Получают Смесь Кандидатов Нуклеиновых Кислот с различной последовательностью. Смесь Кандидатов, как правило, включает в себя районы фиксированных последовательностей (то есть, каждый из членов Смеси Кандидатов содержит одинаковые последовательности в одной и той же локализации) и районы рандомизированных последовательностей. Районы фиксированных последовательностей выбирают, чтобы либо: (а) содействовать на стадиях амплификации, описанных ниже, (б) имитировать последовательность, известную как связывающуюся с мишенью, (в) повысить концентрацию данной структурной конфигурации Нуклеиновых Кислот в Смеси Кандидатов. Рандомизированные последовательности могут быть рандомизированными полностью (то есть, вероятность нахождения основания в любом положении составляет одну четвертую) или только частично рандомизированными (то есть, вероятность нахождения основания в любом положении может быть выбрана на любом уровне между 0 и 100 процентами).
2) Смесь Кандидатов приводят в контакт с выбранной мишенью в условиях, благоприятных для связывания между мишенью и членами Смеси Кандидатов. В этих условиях взаимодействие между мишенью и Нуклеиновыми Кислотами Смеси Кандидатов можно рассматривать как образование пар Нуклеиновая Кислота-мишень между мишенью и теми Нуклеиновыми Кислотами, которые обладают самой сильной аффинностью к мишени.
3) Нуклеиновые Кислоты с самой высокой аффинностью к мишени отделяют от Нуклеиновых Кислот с меньшей аффинностью к мишени. Поскольку только крайне малое число последовательностей (а возможно, только одна молекула Нуклеиновой Кислоты), соответствующих самым высоко аффинным Нуклеиновым Кислотам, существует в Смеси Кандидатов, как правило, является желательным установить критерий отделения таким образом, чтобы значительное количество Нуклеиновых Кислот в Смеси Кандидатов (примерно 5-50%) сохранялось в процессе разделения.
4) Нуклеиновые Кислоты, отобранные в процессе отделения как обладающие относительно более высокой аффинностью к мишени, затем амплифицируют с образованием новой Смеси Кандидатов, которая является обогащенной нуклеиновыми кислотами, обладающими относительно более высокой аффинностью к мишени.
5) В результате повторения вышеуказанных стадий разделения и амплификации вновь образованная Смесь Кандидатов содержит меньше и меньше уникальных последовательностей, и средняя степень аффинности Нуклеиновых Кислот к мишени, как правило, возрастает. Доведенный до конца, способ SELEX даст Смесь Кандидатов, содержащую одну или небольшое число уникальных Нуклеиновых Кислот, представляющих собой те нуклеиновые кислоты из исходной Смеси Кандидатов, которые обладают самой высокой аффинностью к молекуле-мишени.
Для решения ряда конкретных задач основной способ SELEX модифицирован. Например, в Заявке на патент США, порядковый N 07/960093, поданной 14 октября 1992, озаглавленной "Method for Selecting Nucleic Acids on the Basis of Structure", описывается применение SELEX в сочетании с гель-электрофорезом для отбора молекул нуклеиновой кислоты со специфическими структурными характеристиками, таких как изогнутая ДНК. В Заявке на патент США, порядковый N 08/123935, поданной 17 сентября 1993, озаглавленной "Photoselection of Nucleic Acid Ligands", описывается способ на основе SELEX для отбора Нуклеиновых Кислот-Лигандов, содержащих фотореактивные группы, способные к связыванию и/или к поперечному фотосвязыванию и/или к фотоинактивации молекулы-мишени. В Заявке на патент США, порядковый N 08/134028, поданной 7 октября 1993, озаглавленной "High-Affinity Nucleic Acid Ligands That discriminate Berween Theophylline and Caffeine", в настоящее время патент США N 5580737, описывается способ идентификации высокоспецифичных Нуклеиновых Кислот-Лигандов, способных различать между собой близкородственные молекулы, названный Counter-SELEX. В Заявке на патент США, порядковый N 08/143564, поданной 25 октября 1993, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Solution SELEX", в настоящее время патент США N 5567588, описывается способ на основе SELEX, с помощью которого достигается высокоэффективное отделение олигонуклеотидов, обладающих высокой и низкой аффинностью к молекуле-мишени. В Заявке на патент США, порядковый N 07/964624, поданной 21 октября 1992, озаглавленной "Nucleic Acid Ligands to HIV-RT and HINE-1 Rev", в настоящее время патент США N 5496938, описываются способы получения усовершенствованных Нуклеиновых Кислот-Лигандов после того, как проведен SELEX. В Заявке на патент США, порядковый N 08/400440, поданной 8 марта 1995, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Chemi-SELEX", описываются способы ковалентного сшивания лиганда с его мишенью.
Способ SELEX заключается в идентификации высоко аффинных Нуклеиновых Кислот-Лигандов, содержащих модифицированные нуклеотиды, придающие лиганду усовершенствованные характеристики, такие как улучшенная стабильность in vivo или улучшенные характеристики доставки. Примеры таких модификаций включают в себя химические замещения в положениях рибозы и/или фосфата и/или основания. Идентифицированные по SELEX Нуклеиновые Кислоты-Лиганды, содержащие модифицированные нуклеотиды, описаны в Заявке на патент США, порядковый N 08/117991, поданной 8 сентября 1993, озаглавленной "High Affinity Nucleic Acid Ligands Containing Modified Nucleotides", в настоящее время патент США N 5660985, в которой описываются олигонуклеотиды, содержащие производные нуклеотидов, химически модифицированные в 5- и 2'-положениях пиримидинов. В Заявке на патент США, порядковый N 08/134028, см. выше, описываются высоко специфичные Нуклеиновые Кислоты-Лиганды, содержащие один или более чем один нуклеотид, модифицированный 2'-амино (2'-NH2), 2'-фторо(2'-F) и/или 2'-O-метил (2'-Ome). В Заявке на патент США, порядковый N 08/264029, поданной 22 июня 1994, озаглавленной "Novel Method of Preparation of Known and в Novel 2'-Modified Nucleosides by Intramolecular Nucleophilic Displacement", описываются олигонуклеотиды, содержащие различные 2'-модифицированные пиримидины.
Способ SELEX заключается в сочетании выбранных олигонуклеотидов с другими выбранными олигонуклеотидами и не-олигонуклеотидными функциональными единицами, как описано в Заявке на патент США, порядковый N 08/284063, поданной 2 августа 1994, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Chimeric SELEX", в настоящее время патент США N 5637459, и в Заявке на патент США, порядковый N 08/234997, поданной 28 апреля 1994, озаглавленной "Systematic Evolution of Ligands by Exponential Enrichment: Blended SELEX", соответственно. В этих заявках предлагается сочетание широкого множества форм и других свойств, а также свойств эффективной амплификации и репликации олигонуклеотидов с желаемыми свойствами других молекул.
Способ SELEX далее заключается в сочетании выбранных Нуклеиновых Кислот-Лигандов с Липофильными Соединениями или Неиммуногенными Соединениями Высокой Молекулярной Массы в диагностическом или терапевтическом Комплексе, как описано в Заявке на патент США, порядковый N 08/434465, поданной 4 мая 1995, озаглавленной "Nucleic Acid Complexes". Способ SELEX далее заключается в сочетании отобранных Нуклеиновых Кислот-Лигандов VEGF с Липофильными Соединениями, такими как диацилглицерин или диалкилглицерин, как описано в Заявке на патент США, порядковый N 08/739109, поданной 25 октября 1996, озаглавленной "Vascular Endothelial Growth Factor (VEGF) Nucleic Acid Ligand Complexes". Нуклеиновые Кислоты-Лиганды VEGF, которые соединены с Неиммуногенным Соединением Высокой Молекулярной Массы, таким как полиэтиленгликоль, или с Липофильным Соединением, таким как глицеролипид, фосфолипид или глицеринамидлипид, в терапевтическом или диагностическом Комплексе описываются в Заявке на патент США, порядковый N 08/897351, поданной 21 июля 1997, озаглавленной "Vascular Endothelial Growth Factor (VEGF) Nucleic Acid Complexes". Каждая из вышеописанных Заявок на патент, в которых описываются модификации основной методики SELEX, конкретно включена здесь полностью путем ссылки.
SELEX идентифицирует Нуклеиновые Кислоты-Лиганды, которые способны связывать мишени с высокой аффинностью и с выдающейся специфичностью, которые представляют собой исключительное достижение, что является беспрецедентным в области исследования Нуклеиновых Кислот. Данные характеристики являются, несомненно, желаемыми свойствами, которые специалист будет искать в терапевтическом или диагностическом лиганде.
В целях производства Нуклеиновых Кислот-Лигандов, желаемых для применения в качестве фармацевтического средства, предпочтительным является, чтобы Нуклеиновая Кислота-Лиганд (1) связывалась с мишенью таким образом, чтобы достичь желаемого воздействия на мишень; (2) являлась настолько малой, насколько возможно, чтобы получить желаемое воздействие; (3) являлась настолько стабильной, насколько возможно; и (4) являлась специфичным лигандом к выбранной мишени. В большинстве ситуаций предпочтительным является, чтобы Нуклеиновая Кислота-Лиганд обладала самой высокой возможной аффинностью к мишени. Кроме того, Нуклеиновые Кислоты-Лиганды могут обладать вспомогательными свойствами.
В Заявке на патент США, порядковый N 07/964624, поданной 21 октября 1992 ('624), в настоящее время патент США N 5496938, описываются способы получения усовершенствованных Нуклеиновых Кислот-Лигандов после того, как проведен SELEX. Заявка '624, озаглавленная "Nucleic Acid Ligands to HIV-RT and HIV-1 Rev", конкретно включена здесь путем ссылки.
Способ SELEX применяли для идентификации группы высоко аффинных РНК-лигандов к VEGF из библиотек случайных 2'-аминопиримидиновых РНК и однонитевых ДНК-лигандов из библиотек случайных однонитевых ДНК (Заявка на патент США, порядковый N 08/447169, поданная 19 мая 1995, озаглавленная "High Affinity Oligonucleotide Ligands to Vascular Endothelial Growth Factor (VEGF)", которая является продолжением-частью патентной заявки США, порядковый N 08/233012, поданной 25 апреля 1994, озаглавленной "High Affinity Oligonucleotide Ligands to Vascular Endothelial Growth Factor (VEGF)", обе из которых включены здесь путем ссылки; см. также Green et al. (1995) Chemistry and Biology 2: 683-695).
В воплощениях, где Нуклеиновая Кислота-Лиганд (лиганды) VEGF может служить для направления к мишени, Нуклеиновые Кислоты-Лиганды VEGF принимают трехмерную структуру, которая должна сохраняться, для того чтобы Нуклеиновая Кислота-Лиганд VEGF была способна связывать свою мишень. В воплощениях, где Липидная Конструкция содержит Комплекс, а Нуклеиновая Кислота-Лиганд VEGF Комплекса выступает из поверхности этой Липидной Конструкции, Нуклеиновая Кислота-Лиганд VEGF должна быть надлежащим образом ориентирована по отношению к поверхности этой Липидной Конструкции таким образом, чтобы ее способность связывать мишень не подвергалась риску. Это можно осуществить путем присоединения Нуклеиновой Кислоты-Лиганда VEGF в положении, которое является удаленным от связывающего участка Нуклеиновой Кислоты-Лиганда VEGF. Эту трехмерную структуру и надлежащую ориентацию можно сохранить с помощью использования линкера или спейсера, как описано выше.
Любые разнообразные терапевтические или диагностические агенты можно присоединить к Комплексу для направленной доставки посредством этого Комплекса. Кроме того, любое разнообразие терапевтических или диагностических агентов можно присоединить, инкапсулировав либо включив их внутрь этой Липидной Конструкции, как обсуждалось выше, для направленной доставки посредством этой Липидной Конструкции.
В воплощениях, где Комплекс состоит из Липофильного Соединения и Нуклеиновой Кислоты-Лиганда VEGF в соединении с липосомой, например, Нуклеиновая Кислота-Лиганд VEGF может направляться к опухолевым клеткам, экспрессирующим VEGF (например, при саркоме Капоши) для доставки противоопухолевого лекарственного средства (например, даунорубицина) либо визуализирующего агента (например, радиоактивных меток). Следует отметить, что клетки и ткани, окружающие опухоль, также могут экспрессировать VEGF, и направленная доставка противоопухолевого лекарственного средства к этим клеткам также должна быть эффективной.
В альтернативном воплощении терапевтический или диагностический агент, который нужно доставить к клетке-мишени, может представлять собой другую Нуклеиновую Кислоту-Лиганд.
В данном изобретении также рассматривается, что агент, который нужно доставить, может быть включен в Комплекс таким образом, как например в соединении с наружной поверхностью липосомы (например, пролекарство, антагонист рецептора или радиоактивное вещество для лечения или визуализации). Как и Нуклеиновая Кислота-Лиганд VEGF, агент может быть присоединен посредством ковалентных или нековалентных взаимодействий. Липосома должна обеспечивать направленную доставку этого агента внеклеточно, причем липосома служит в качестве линкера.
В другом воплощении Неиммуногенное Соединение Высокой Молекулярной Массы (например, ПЭГ) может быть присоединено к липосоме, чтобы обеспечить усовершенствованные фармакокинетические свойства для Комплекса. Нуклеиновые Кислоты-Лиганды VEGF могут быть присоединены к мембране липосомы или могут быть присоединены к Неиммуногенному Соединению Высокой Молекулярной Массы, которое в свою очередь присоединено к мембране. Таким образом, этот Комплекс может быть экранирован от белков крови и, таким образом, это приводит к его циркуляции в течение продолжительных периодов времени, причем Нуклеиновая Кислота-Лиганд VEGF тем не менее является достаточно доступной, чтобы осуществить контакт и связаться со своей мишенью.
В другом воплощении настоящего изобретения более чем одна Нуклеиновая Кислота-Лиганд VEGF присоединена к поверхности одной и той же липосомы. Это обеспечивает возможность приведения одинаковых молекул VEGF в тесную близость друг к другу и может быть использовано для генерирования специфичных взаимодействий между молекулами VEGF.
В альтернативном воплощении настоящего изобретения Нуклеиновые Кислоты-Лиганды VEGF и Нуклеиновая Кислота-Лиганд к другой мишени могут быть присоединены к поверхности одной и той же липосомы. Это обеспечивает возможность приведения VEGF в тесную близость к другой мишени и может быть использовано для генерирования специфичных взаимодействий между VEGF и другой мишенью. Кроме использования липосом в качестве способа приведения мишеней в тесную близость, в липосому могут быть инкапсулированы агенты для повышения интенсивности взаимодействия.
Липидная Конструкция, содержащая Комплекс, дает возможность множественных связывающих взаимодействий с VEGF. Это, несомненно, зависит от числа Нуклеиновых Кислот-Лигандов VEGF на Комплекс, от числа Комплексов на Липидную Конструкцию и от подвижности Нуклеиновых Кислот-Лигандов VEGF и рецепторов в их соответствующих мембранах. Поскольку константа эффективного связывания может возрастать как произведение констант связывания для каждого сайта, имеется существенное преимущество в наличии множественных связывающих взаимодействий. Другими словами, в результате наличия многих Нуклеиновых Кислот-Лигандов VEGF, присоединенных к Липидной Конструкции, а следовательно, создания многовалентности, эффективная аффинность (то есть авидность) этого мультимерного Комплекса к его мишени может стать такой значительной, как произведение констант связывания для каждого сайта.
В некоторых воплощениях изобретения Комплекс по настоящему изобретению состоит из Нуклеиновой Кислоты-Лиганда VEGF, присоединенной к Липофильному Соединению, такому как глицеролипид. В этом случае фармакокинетические свойства этого Комплекса будут усовершенствованными относительно только Нуклеиновой Кислоты-Лиганда VEGF. Как обсуждалось выше, глицеролипид, фосфолипид или глицеринамидлипид может быть ковалентно сшит с Нуклеиновой Кислотой-Лигандом VEGF в различных положениях на нуклеиновой кислоте-лиганде VEGF. В воплощениях, где используется глицеролипид, предпочтительным является то, что Нуклеиновая Кислота-Лиганд VEGF связана с липидом через фосфодиэфирные связи.
В другом воплощении изобретения Липидная Конструкция содержит Нуклеиновую Кислоту-Лиганд VEGF или Комплекс. В данном воплощении глицеролипид может быть полезным при включении Нуклеиновой Кислоты-Лиганда VEGF в липосому вследствие склонности глицеролипида к соединению с другими Липофильными Соединениями. Глицеролипид в соединении с Нуклеиновой Кислотой-Лигандом VEGF может быть включен в липидный бислой липосомы путем включения в препарат или путем загрузки в преформированные липосомы. Глицеролипид может соединяться с мембраной липосомы таким образом, что Нуклеиновая Кислота-Лиганд VEGF выступает внутрь или наружу липосомы. В воплощениях, где Нуклеиновая Кислота-Лиганд VEGF выступает наружу этого Комплекса, Нуклеиновая Кислота-Лиганд VEGF может служить при направляющей способности. Следует понимать, что с Липидной Конструкцией могут быть соединены дополнительные соединения для дальнейшего усовершенствования фармакокинетических свойств этой Липидной Конструкции. Например, ПЭГ может быть присоединен к наружной стороне мембраны этой Липидной Конструкции.
В других воплощениях Комплекс по настоящему изобретению состоит из Нуклеиновой Кислоты-Лиганда VEGF, ковалентно сшитой с Неиммуногенным Соединением Высокой Молекулярной Массы, таким как полиалкиленгликоль или ПЭГ. В данном воплощении фармакокинетические свойства этого Комплекса являются усовершенствованными относительно только Нуклеиновой Кислоты-Лиганда VEGF. Полиалкиленгликоль или ПЭГ может быть ковалентно сшит с множеством положений на Нуклеиновой Кислоте-Лиганде VEGF. В воплощениях, где используются полиалкиленгликоль или ПЭГ, предпочтительным является то, что Нуклеиновая Кислота-Лиганд VEGF связана посредством 5'гидроксильной группы через фосфодиэфирную связь.
В некоторых воплощениях множество Нуклеиновых Кислот-Лигандов может быть соединено с одним Неиммуногенным Соединением Высокой Молекулярной Массы, таким как полиалкиленгликоль или ПЭГ, либо Липофильным Соединением, таким как глицеролипид. Нуклеиновые Кислоты-Лиганды могут быть все к VEGF или к VEGF и к другой мишени. В воплощениях, где имеются множественные Нуклеиновые Кислоты-Лиганды VEGF, наблюдается повышение авидности вследствие множественных связывающих взаимодействий с VEGF. В дополнительных воплощениях множество молекул полиалкиленгликоля, ПЭГ, глицеролипида может быть присоединено друг к другу. В этих воплощениях одна или более чем одна Нуклеиновая Кислота-Лиганд VEGF или Нуклеиновые Кислоты-Лиганды к VEGF и другим мишеням могут быть соединены с каждым из: полиалкиленгликоль, ПЭГ или глицеролипид. Это также приводит к повышению авидности каждой из Нуклеиновых Кислот-Лигандов к своей мишени. В воплощениях, где множественные Нуклеиновые Кислоты-Лиганды VEGF присоединены к полиалкиленгликолю, ПЭГ или глицеролипиду, существует возможность приведения молекул VEGF в тесную близость друг к другу для генерирования специфичных взаимодействий между VEGF. Если множественные Нуклеиновые Кислоты-Лиганды, специфичные к VEGF и другим мишеням, присоединены к полиалкиленгликолю, ПЭГ или глицеролипиду, существует возможность приведения VEGF и другой мишени в тесную близость друг к другу для генерирования специфичных взаимодействий между VEGF и другой мишенью. Кроме того, в воплощениях, где имеются Нуклеиновые Кислоты-Лиганды к VEGF или Нуклеиновые Кислоты-Лиганды к VEGF и другим мишеням, соединенные с полиалкиленгликолем, ПЭГ или глицеролипидом, лекарственное средство также может быть соединено с полиалкиленгликолем, ПЭГ или глицеролипидом. Таким образом, этот Комплекс должен обеспечивать направленную доставку этого лекарственного средства, причем полиалкиленгликоль, ПЭГ или глицеролипид служат в качестве линкера.
Нуклеиновые Кислоты-Лиганды VEGF избирательно связывают VEGF. Таким образом, Комплекс, содержащий Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, либо Липидная Конструкция, содержащая Нуклеиновую Кислоту-Лиганд VEGF или Комплекс, являются полезными в качестве фармацевтических средств или диагностических агентов. Настоящее изобретение, следовательно, включает в себя способы ингибирования ангиогенеза посредством введения Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, Липидной Конструкции, содержащей Нуклеиновую Кислоту-Лиганд VEGF или Комплекс, содержащий Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение. Комплексы и липидные конструкции, содержащие Нуклеиновую Кислоту-Лиганд VEGF, могут применяться для лечения, ингибирования, предупреждения или диагностики любого болезненного состояния, в которое вовлечено несоответствующее продуцирование VEGF, в частности, ангиогенеза. Ангиогенез редко встречается у здоровых взрослых людей, за исключением периода менструального цикла и заживления раны. Ангиогенез, однако, является главным признаком различных болезненных состояний, включая, но не ограничиваясь ими, рак, диабетическую ретинопатию, дегенерацию желтого пятна, псориаз и ревматоидный артрит. Настоящее изобретение, таким образом, также включает в себя, не ограничиваясь ими, способы лечения, ингибирования, предупреждения или диагностики диабетической ретинопатии, дегенерации желтого пятна, псориаза и ревматоидного артрита. Кроме того, VEGF продуцируется и секретируется в варьирующих количествах фактически всеми опухолевыми клетками. Следовательно, настоящее изобретение включает в себя способы лечения, ингибирования, предупреждения или диагностики рака посредством введения Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, Липидной Конструкции, содержащей Комплекс, или Нуклеиновой Кислоты-Лиганда VEGF, не являющейся частью Комплекса, в соединении с Липидной Конструкцией. Показано, что при таком типе рака, как саркома Капоши (KS), клетки не только продуцируют обильные количества VEGF, но также экспрессируют функциональные рецепторы VEGF и, следовательно, используют VEGF для аутокринного роста. Таким образом, настоящее изобретение включает в себя способ ингибирования саркомы Капоши посредством введения Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, Липидной Конструкции, содержащей Комплекс, или Нуклеиновой Кислоты-Лиганда VEGF, не являющейся частью Комплекса, в соединении с Липидной Конструкцией.
В одном воплощении настоящего изобретения Липидная Конструкция содержит Комплекс, состоящий из Нуклеиновой Кислоты-Лиганда VEGF и Липофильного Соединения с дополнительным диагностическим или терапевтическим агентом, инкапсулированным в эту Липидную Конструкцию или соединенным с внутренней стороной этой Липидной Конструкции. В предпочтительном воплощении Липидная Конструкция представляет собой Липидную Бислойную Везикулу, и наиболее предпочтительно липосому. Терапевтическое применение липосом включает в себя доставку лекарственных средств, которые обычно являются токсичными в свободной форме. В липосомной форме токсичное лекарственное средство является закрытым и может не затрагивать ткани, чувствительные к этому лекарственному средству, и направлено к выбранным областям. Липосомы могут также применяться терапевтически для высвобождения лекарственного средства в течение продолжительного периода времени, уменьшая частоту введения. Кроме того, липосомы могут обеспечить способ получения водных дисперсий гидрофобных или амфифильных лекарственных средств, которые обычно являются неподходящими для внутривенной доставки.
Чтобы многие лекарственные средства и визуализирующие агенты обладали терапевтическим или диагностическим потенциалом, необходимо, чтобы они доставлялись к надлежащему местоположению в организме, и липосома, таким образом, может быть легко инъецирована, что создает основу для длительного высвобождения и доставки лекарственного средства к клеткам специфических типов или частям организма. Чтобы применять липосомы для направления инкапсулированных лекарственных средств к выбранным тканям хозяина и удаления их от чувствительных тканей, можно использовать несколько методик. Эти методики включают в себя манипулирование размером липосом, их поверхностным зарядом и путем их введения. MLV, прежде всего поскольку они являются достаточно большими, обычно легко поглощаются ретикулоэндотелиальной системой (главным образом печени и селезенки). Обнаружено, что UV, с другой стороны, проявляют повышенное время циркуляции, сниженную скорость клиренса и большее биологическое распространение относительно MLV.
Пассивная доставка липосом включает в себя применение различных путей введения, например внутривенного, подкожного, внутримышечного и местного. Каждый путь дает различия в локализации липосом. Два распространенных способа, применяемых для активного направления липосом к выбранным областям-мишеням, включают в себя присоединение к поверхности липосом либо антител, либо специфических лигандов рецепторов и служит для направляющей способности. Дополнительные направляющие компоненты, такие как антитела или специфические лиганды рецепторов, могут находиться на поверхности липосом, что должно быть известно специалисту в данной области. В одном воплощении настоящего изобретения Нуклеиновая Кислота-Лиганд VEGF соединена с наружной поверхностью липосомы, как должно быть известно специалистам. Кроме того, некоторые усилия по направлению липосом к опухолям без использования антител были успешными, см. , например, патент США N 5019369, патент США N 5435989 и патент США N 4441775, и специалистам должно быть известно, что они включают в себя эти альтернативные способы направленной доставки.
Терапевтические или диагностические составы Комплекса, содержащего Нуклеиновую Кислоту-Лиганд VEGF и Неиммуногенное Соединение Высокой Молекулярной Массы или Липофильное Соединение, Липидной Конструкции, содержащей Комплекс, состоящий из Нуклеиновой Кислоты-Лиганда VEGF и Неиммуногенного Соединения Высокой Молекулярной Массы или Липофильного Соединения, и Нуклеиновой Кислоты-Лиганда VEGF, не являющейся частью Комплекса, в соединении с Липидной Конструкцией, можно вводить парентерально путем инъекции, хотя рассматриваются также другие эффективные формы введения, такие как внутрисуставная инъекция, ингаляционные аэрозоли, перорально активные препараты, трансдермальный лекарственный электрофорез или суппозитории. Одним предпочтительным носителем является физиологический раствор, но предполагается, что могут использоваться также другие фармацевтически приемлемые носители. В одном воплощении рассматривается, что носитель и Комплекс Нуклеиновой Кислоты-Лиганда VEGF составляют физиологически совместимый препарат медленного высвобождения. Первичный растворитель в таком носителе может быть по природе либо водным, либо неводным. Кроме того, носитель может содержать другие фармакологически приемлемые эксципиенты для модифицирования или поддержания рН, осмотического давления, вязкости, чистоты, цвета, стерильности, стабильности, скорости растворения или запаха препарата. Подобным образом носитель может содержать еще другие фармакологически приемлемые эксципиенты для модифицирования или поддержания стабильности, скорости растворения, высвобождения или поглощения Нуклеиновой Кислоты-Лиганда VEGF. Такие эксципиенты представляют собой те вещества, которые обычно и общепринято используются, чтобы изготавливать дозировки для парентерального введения либо в форме стандартной дозы, либо в форме множественных доз.
Как только терапевтический или диагностический состав изготовлен, его можно хранить в стерильных флаконах в виде раствора, суспензии, геля, эмульсии, твердого вещества либо дегидратированного или лиофилизированного порошка. Такие препараты можно хранить либо в готовой к употреблению форме, либо в форме, требующей восстановления немедленно перед введением. Путь введения препаратов, содержащих Нуклеиновую Кислоту-Лиганд VEGF для системной доставки, может быть подкожным, внутримышечным, внутривенным, интраназальным либо посредством вагинальных или ректальных суппозиториев.
Преимущества Комплексов и липидных конструкций по изобретению включают в себя: i) усовершенствование плазменной фармакокинетики Нуклеиновой Кислоты-Лиганда; ii) представление Нуклеиновых Кислот-Лигандов в виде многовалентного множества с целью повышения авидности к взаимодействию с их мишенями; iii) сочетание двух или более чем двух представляемых Нуклеиновых Кислот-Лигандов с различными специфичностями в одной и той же липосомной частице; iv) усиление доставки представляемых Нуклеиновых Кислот-Лигандов к опухолям, пользуясь преимуществом присущего липосомам свойства направления к опухоли, и v) использование высокой аффинности и специфичности представляемых Нуклеиновых Кислот-Лигандов, которая является сравнимой с таковой антител, для направления липосомного содержимого к специфическим мишеням. Представляемые Нуклеиновые Кислоты-Лиганды являются хорошо подходящими для описанных здесь видов препаратов, поскольку, в отличие от большинства белков, денатурация представляемых Нуклеиновых Кислот-Лигандов под действием нагревания, различных молекулярных денатурирующих агентов и органических растворителей является легко обратимой.
Следующие примеры предлагаются, чтобы объяснить и проиллюстрировать настоящее изобретение, и не должны рассматриваться как ограничивающие изобретение. Структуры Нуклеиновых Кислот-Лигандов, описываемые в приведенных ниже примерах, представлены на фиг. 1. В примере 1 описывается конъюгация Нуклеиновых Кислот-Лигандов с липидными реагентами. Способность диалкилглицеринового производного Нуклеиновой Кислоты-Лиганда VEGF (NX278) либо в виде свободного лиганда, либо включенного в бислой липосом (NX278-L), ингибировать активность VEGF in vitro и in vivo описывается в примере 2. В примере 3 описываются экспериментальные методики для получения 2'-F пиримидин-модифицированных РНК-лигандов к VEGF. В примере 4 описываются 2'-F пиримидин-модифицированные РНК-лиганды к VEGF. В примере 5 описывается синтез глицеролипид-, фосфолипид- и глицеринамидный липид-, а также ПЭГ-модифицированных Нуклеиновых Кислот-Лигандов VEGF. В примере 6 описываются фармакокинетические свойства фосфолипид-(РL) и ПЭГ-модифицированных Нуклеиновых Кислот-Лигандов. В примере 7 описываются препараты NX31838 PL-липосома Комплекса. В примерах 8-10 описывается эффективность in vivo Комплексов Нуклеиновых Кислот-Лигандов VEGF. В примере 11 описывается внутристекловидная фармакокинетика NX31838-40K PEG у кроликов.
Пример 1. Синтез диалкилглицерин (1,2-ди-O-октадецил-sn-глицерин) -модифицированной Нуклеиновой Кислоты-Лиганда VEGF.
В данном примере описывается конъюгация Нуклеиновых Кислот-Лигандов с липидными реагентами. Синтез (1,2-ди-О-октадецил-sn-глицерин) -модифицированной Нуклеиновой Кислоты-Лиганда VEGF (см. схему 1 в конце описания).
Тетраэтиленгликольмонотозилат (2а): Тетраэтиленгликоль (200 мл, 1,15 моль) растворяли в 500 мл пиридина, охлаждали до 0oC и обрабатывали 22,0 г (0,115 моль) п-толуолсульфонилхлорида. Когда растворение было полным, реакционную смесь хранили в холодильнике в течение ночи, а затем концентрировали в вакууме. Остаток растворяли в 800 мл ЕtOАс и экстрагировали 3•600 мл Н2О. Н2О фракции снова экстрагировали ЕtOАс, и объединенные ЕtOАс фракции экстрагировали насыщенным водным Na2HPO4. Органическую фазу высушивали над МgSО4 и концентрировали до бесцветного масла. Это масло очищали с помощью флэш-хроматографии с использованием 800 мл силикагеля и элюции гексаном, 25% ЕtOАс-50% ЕtOАс в гексане, затем ЕtOАс, затем 10% МеОН-20% МеОН в ЕtOАс с получением 23,7 г (60%) чистого продукта и 11% продукта, содержащего минорные примеси. 2а: 1H ЯМР (300 МГц, СDСl3) d 7.77 (d, J= 8.1 Гц, 2Н), 7.32 (d, J= 8.1 Гц, 2H), 4.13 (t, J= 4.8 Гц, 2Н), 3.68-3.53 (m, 14H), 2.58 (t, J= 5.6 Гц, 1Н), 2.42 (s, 3H); 13С ЯМР (75 МГц, CDCl3) d 168.2, 158.3, 144.8, 135.9, 133.8, 132.0, 129.9, 128.0, 127.7, 126.6, 123.1, 113.0, 85.9, 73.0, 70.6, 69.7, 67.8, 64.4, 55.1, 37.1; МС низкого разрешения, m/e, вычисленный для С15H24О8S (M+1): 349,1.
Тетраэтиленгликольмонофталимид (3а): К перемешанному раствору 31,96 г (0,092 моль) 2а в 400 мл безводного ДМФ добавляли 14,2 г (1,05 эквив. ) фталимида и 14,4 мл (1,05 эквив. ) 1,8-диазабицикло[5.4.0] ундец-7-ена (ДБУ). Раствор нагревали при 70oС в течение 18 ч, затем концентрировали в вакууме. Сырое желтое масло очищали с помощью флэш-хроматографии с использованием 1600 мл силикагеля и элюции 25% EtOAc-50% EtOAc-75% ЕtOАс в гексане, затем ЕtOАс, затем 10% МеОН-20% МеОН в ЕtOАс с получением 23,8 г (80%) 3а в виде масла. При стоянии 3а становился похожим на воск белым твердым веществом. 1H ЯМР (300 МГц, СDСl3) 7.84-7.78 (m, 2H), 7.70-7.66 (m, 2H), 3.86 (t, J= 5.6 Гц, 2H), 3.70 (t, J= 5.6 Гц, 2H), 3.64-3.51 (m, 12H), 2.67 (bs, 1H); 13С ЯМР (75 МГц, CDCl3) d 168.2, 133.8, 132.0, 123.1, 72.4, 70.5, 70.4, 70.2, 70.0, 67.8, 61.6, 37.2.
Синтез соединения 4а: Раствор 15 г (0,0464 моль) 3а в 150 мл ТГФ и 15 мл ДМФ охлаждали до 0oС в атмосфере Аr. К раствору добавляли аллилбромид (6,0 мл, 1,5 эквив. ), после чего добавляли 1,76 г (1,5 эквив. ) NaH в виде твердого вещества. Непрозрачную желтую суспензию перемешивали при 0oС в течение 30 минут, а затем при комнатной температуре в течение 18 ч. Добавляли МеОН (50-100 мл), и концентрированную после этого смесь концентрировали в вакууме. Сырой материал очищали с помощью флэш-хроматографии с использованием 1500 мл силикагеля и элюции 25% EtOAc-50% EtOAc-75% ЕtOАс в гексане, затем ЕtOАс, затем 10% МеОН в ЕtOАс с получением 11,05 г (65%) 4а в виде желтого масла. 1H ЯМР (300 МГц, СDСl3) 7.84-7.80 (m, 2H), 7.72-7.67 (m, 2H), 5.94-5.84 (m, 1H), 5.28-5.14 (m, 2H), 3.99 (d, J= 5.61 Гц, 2H), 3.88 (t, J= 5.85 Гц, 2H), 3.72 (t, J= 5.76 Гц, 2H), 3.64-3.54 (m, 13H); 13С ЯМР (75 МГц, CDCl3) d 168.0, 134.6, 133.7, 131.9, 123.0, 116.9, 72.0, 70.4, 69.9, 69.2, 67.7, 37.0.
1-Диметокситритил-3- (фталимидотетраэтиленгликолил)-sn- глицерин (9):
Согласно схеме 1 соединение 9 синтезировали следующим образом: К перемешанному раствору 4а (10,13 г, 0,0279 моль) в 100 мл ацетона и 1 мл Н2О добавляли 3,98 г (1,22 эквив. ) N-метилморфолин-N-оксида (NMO). К этой суспензии добавляли 1,75 мл (0,005 эквив. ) тетраоксида осмия в виде 2,5% раствора в изопропаноле (iPrOH). После добавления раствора OsO4 реакционная смесь становилась чисто желтой. После того как ТСХ-анализ показывал полное превращение 4а (примерно 16 ч), реакционную смесь обрабатывали 1,5 г гидросульфита натрия и 5,0 г флорисила и перемешивали в течение 30 минут. Суспензию фильтровали через флорисил, фильтрат концентрировали до масла. Этот сырой продукт объединяли с другой партией, полученной тем же способом из 1,0 г 4а. Две части по 100 мл пиридина совместно выпаривали с объединенными партиями, и остаток растворяли в 300 мл пиридина. Раствор охлаждали до 0oС и добавляли 10,89 г (1,05 эквив. ) 4,4'-диметокситритилхлорида. В колбу вставляли сушильную трубку, и реакционную смесь перемешивали при комнатной температуре в течение 16 ч. Раствор обрабатывали 20 мл МеОН и концентрировали в вакууме, держа температуру водяной бани ниже 40oС. Сырое масло очищали с помощью флэш-хроматографии с использованием 1100 мл силикагеля (упакованного в колонку влажным с использованием 3% триэтиламина в гексане) и элюции 10-100% ЕtOАс в гексане (все содержащие 3% триэтиламин) с получением 21,3 г (89% после двух стадий) 9 в виде желтого масла. 1H ЯМР (300 МГц, СDСl3) d 7.80-7.77 (m, 2H), 7.66-7.64 (m, 2H), 7.39-7.22 (m, 9H), 7.20-6.76 (m, 4H), 3.97 (bs, 1H), 3.84 (t, J= 5.97 Гц, 2H), 3.74 (s, 6H), 3.68 (t, J= 5.7 Гц, 2H), 3.60-3.49 (m, 14H), 3.13-2.76 (m, 2H), 2.00 (bs, 1H); 13C ЯМР (75 МГц, CDCl3) d 168.2, 158.3, 144.8, 135.9, 133.8, 132.0, 129.9, 128.0, 127.7, 126.6, 123.1, 113.0, 85.9, 73.0, 70.6, 70.4, 70.0, 69.7, 67.8, 64.4, 55.1, 37.1; МС низкого разрешения, m/e, вычисленный для C40H45O10N (M+NH4 +): 717.5.
1-Диметокситритил-3- (аминотетраэтиленгликолил)-sn- глицерин (10):
Согласно схеме 1 соединение 10 синтезировали следующим образом: Соединение 9 (5,2 г, 7,2 ммоль) растворяли в 50 мл 40% метиламина в Н2О и добавляли 10 мл метанола для солюбилизации исходного материала. Реакционную смесь нагревали при 50oС в течение 5 ч, а затем концентрировали в вакумме и выпаривали вместе с толуолом. Сырой материал очищали с помощью флэш-хроматографии на 200 мл силикагеля, элюируя 15% метаноламмиаком в дихлорметане. Собирали 3,94 г (96%) 10 в виде светло-желтого масла. 1H ЯМР (300 МГц, СDСl3) d 7.46-7.21 (m, 9H, DMT), 6.81 (d, 4H, DMT), 4.00 (m, 1H), 3.80 (s, 6H), 3.70-3.49 (перекрывающийся, 18Н), 3.20 (dd, J= 9.24, 5.49 Гц, 1H), 3.12 (dd, J= 9.21, 6.00 Гц, 1H), 2.84-2.80 (m, 3H); 13С ЯМР (75 МГц, CDCl3) d 158.30, 144.82, 136.01, 129.95, 128.04, 127.66, 126.61, 112.95, 85.85, 73.46, 72.85, 70.55, 70.45, 69.99, 69.51, 64.43, 55.10, 41.40; МС низкого разрешения, m/e, вычисленный для С32Р44O8N (М+1): 570.353, обнаружено 570,4.
Хлорформиат 19: К перемешанному раствору 3 г (5,03 ммоль) 1,2-ди-О-октадецил-sn-глицерина 18 в 60 мл толуола добавляли 20 мл 1,93 М раствора фосгена. Дополнительный раствор фосгена (2•10 мл: 14,5 эквив. фосгена всего) добавляли до тех пор, пока исходного спиртового раствора больше не оставалось (по 1H ЯМР анализу концентрированных аликвот). Избыток фосгена и HCl удаляли с помощью аспиратора, и реакционную смесь концентрировали в вакууме с получением 3,3 г (98%) желаемого хлорформиата 19 в виде белого порошка. 1H ЯМР (300 МГц, CDCl3) d 4.45 (dd, J= 11.22, 3.69 Гц, 1H), 4.34 (dd, J= 11.22, 6.15 Гц, 1Н), 3.65 (m, 1H), 3.56-3.40 (m, 6H), 1,53 (m, 4H), 1.24 (m, 62H), 0.87 (t, J= 6.36 Гц, 6H); 13С ЯМР (75 МГц, СDСl3) d 75.90, 71.91, 71.35, 70.93, 69.36, 31.99, 29.96-29.44 (перекрывающиеся сигналы от углеводородных цепей), 26.13, 26.04, 22.76, 14.18.
Конъюгат 20: К перемешанному раствору 2,25 г (3,95 ммоль) 10 в 60 мл пиридина добавляли 2,6 г дистеарилглицеринхлорформиата 18. 1H ЯМР анализ концентрированной аликвоты через 2 ч не выявлял оставшегося хлорформиата, и смесь концентрировали в вакууме. Сырой остаток объединяли с материалом, полученным подобным образом из 0,5 г (0,88 ммоль) 10 и 0,58 г хлорформиата, и объединенные партии очищали с помощью силикагелевой флэш-хроматографии на колонке со 100 мл силикагеля (упакованного в гексанах, содержащих 2% триэтиламина) с элюцией 200 мл гексанов, затем 250 мл каждого из 10-20 и 30% ЕtOАс в гексанах, 500 мл 40% ЕtOАс в гексанах, затем 250 мл каждого из 50-60-70 и 80% ЕtOАс в гексанах, и, наконец, 250 мл ЕtOАс. Фракции, содержащие продукт, концентрировали с получением 3,3 г (57%) конъюгата 20.
Фосфорамидит 21: К перемешанному раствору 3,8 г (3,26 ммоль) конъюгата в 25 мл СН2Сl2 добавляли 1,14 мл (6,52 ммоль) диизопропилэтиламинэтена (ДИПЭА), затем 1,09 мл (4,88 ммоль) 2-цианоэтил-N, N-диизопропилхлорфосфорамидита. Через 2 часа смесь разбавляли CH2Cl2 и промывали насыщенным раствором NаНСО3, высушивали над Na2SO4 и концентрировали. Сырой остаток очищали с помощью силикагелевой флэш-хроматографии на колонке со 125 мл силикагеля (упакованного в гексанах, содержащих 2% триэтиламина) с элюцией 100 мл гексанов, затем 250 мл каждого из 10 и 20% ЕtOАс в гексанах, 500 мл 30% ЕtOАс в гексанах, затем 250 мл 50% ЕtOАс в гексанах. Фракции, содержащие продукт, концентрировали с получением 4,2 г (95%) фосфорамидита 21. 31Р ЯМР (СDСl3) d 151.52, 151.08.
Нуклеиновая Кислота-Лиганд VЕGF-1,2-ди-O-октадецил-sn-глицерин конъюгат
1,2-ди-O-октадецил-sn-глицериновую группу конъюгировали с Нуклеиновой Кислотой-Лигандом VEGF NX213 (см. фиг. 1А) с использованием фосфорамидита 21 (схема 2). Полученный конъюгат был назван NX278 (см. фиг. 1В). NX278 (SEQ ID NO: 2) очищали с помощью обратнофазной ВЭЖХ, и его состав подтверждали с помощью электроспрей-масс-спектроскопии (наблюдаемое m/z= 11703±4, вычисленное m/z= 11720). Фосфоротиоатные межнуклеозидные связи использовались в положениях 8 в NX278 (на 3' и 5' концах), и различие в 0,16 массовых единиц между ожидаемой и наблюдаемой массами возможно связано с неполным окислением сульфурирующим агентом, в результате чего в среднем на одну фосфоротиоатную связь на молекулу было меньше, чем ожидалось.
Пример 2. Эффективность in vitro и in vivo Комплекса Нуклеиновая Кислота-Лиганд-липосома. Диалкилглицерин(DАG)-модифицированная Нуклеиновая Кислота-Лиганд VEGF (NX278), встроенная в бислой липосомы.
Комплекс NХ278-липосома получали путем инкубирования NX-278 (1 мг) (фиг. 1В; SEQ ID NO: 2) с высушенной распылением смесью DSРС: холестерин (50 мг/мл; 2: 1, моль: моль) в 25 мМ фосфатном буфере (рН 7,4), содержащем 9% сахарозы, и озвучивали в течение 15-30 мин при примерно 60oС с использованием источника ультразвука зондового типа, пока не был получен раствор опалового цвета. Контрольный Комплекс Нуклеиновая Кислота-Лиганд-липосома, содержащий аналог лиганда NX-278 с перемешанной последовательностью (scNX278) (фиг. 1С; SEQ ID NO: 3) получали подобным способом. В типичном препарате были получены липосомы со средним диаметром 50 нм и шириной распределения 20 нм при половине высоты. Размер липосомных частиц определяли в анализаторе частиц (Leeds & Northrup Model Microtrack UPA 150, Horsham, PA). Липосомы сравнимого гранулометрического состава получали с тем же липидным составом, но без конъюгированной с липидом Нуклеиновой Кислоты-Лиганда. Ожидается, что липосома 50 нм содержит в среднем 40 Нуклеиновых Кислот-Лигандов, расположенных по обеим сторонам бислоя. Это вычисление делали следующим образом. Принимая площадь поверхности 19oС для холестерина и 60oС для дистеарилфосфатидилхолина в липосоме, получили число липидных молекул на липосому 3,13•104, для сферической липосомы с внешним диаметром 50 нм и толщиной мембраны 20oС. Исходя из состава липосомы (2: 1 моль: моль дистеарилфосфатидилхолин (МW= 790,2): холестерин (MW= 386,7)), принимая распределение липидов за гомогенное, вычислили молекулярную массу для липосомы, равную 2,1•107.
Чтобы определить распределение Нуклеиновых Кислот-Лигандов между внутренней и наружной поверхностями липосом, исследовали доступность NX278 в липосомном препарате для Т1 рибонуклеазы. При двух рибогуанозинах в последовательности (Green et al. (1995) Chemistry and Biology 2: 683-695) NX278 эффективно расщепляется рибонуклеазой Т1. Простая инкубация NX278 с преформированными липосомами не защищает Нуклеиновую Кислоту-Лиганд от рибонуклеазы Т1. Однако, когда NX278 включен в липосомы в результате озвучивания (NX278-липосома), примерно 1/3 его защищена от нуклеазы. Добавление к NX278-липосома 0,1% Тритона Х-100, который разрушает липосомы, не влияя на активность нуклеазы, делает ранее защищенную Нуклеиновую Кислоту-Лиганд доступной для расщепления. Эти результаты согласуются с мнением, что Нуклеиновая Кислота-Лиганд распределена на обеих сторонах бислоя.
Связывающие аффинности к VEGF NX213, NX278 и NX278-липосома
Связывающие аффинности к VEGF NX213, NX278 и NX278-липосома изучали с использованием способа конкурентного сдвига электрофоретической подвижности (фиг. 2). Связывающая аффинность к VEGF у NX278 была сравнима с таковой у NX213. Видимая связывающая аффинность NX278-липосома была в 3 раза ниже по сравнению с NX278. Наблюдаемое снижение аффинности, отчасти, возможно, связано с ограничением фракции Нуклеиновой Кислоты-Лиганда внутренней стороной липосомы. Как ожидалось, аналоги с перемешанной последовательностью связывались с VEGF с существенно более низкими аффинностями (фиг. 2).
Фармакокинетические свойства NX213, NX278 и NХ278-липосома в плазме
Концентрации NX213, NX278 и NX278-липосома в плазме крыс Sprague-Dawley как функция времени показаны на фиг. 15, и параметры компартментного анализа суммированы в таблице 1. Большая часть NX213 быстро выводится в альфа-фазе с t1/2 7 минут и общей скоростью клиренса 6,8 мл/кг/мин. Конъюгация фосфолипидной группы с Нуклеиновой Кислотой-Лигандом приводит к высоко бифазному клиренсу из крови с повышенной β(t1/2) и несколько более низкой общей скоростью клиренса (4,95 мл/кг/мин) относительно NX213. При включении NX278 в липосому проявляется существенное дополнительное снижение клиренса Нуклеиновой Кислоты-Лиганда из плазмы (1,88 мл/кг/мин).
Воздействие NX278 на пролиферацию HUVEC и ангиогенез
Изучали воздействия NХ278-липосома, scNХ278-липосома и NX213 на пролиферацию эндотелиальных клеток пупочной вены человека (HUVEC). HUVEC выращивали в присутствии VEGF (10 н/мл) в среде IMDM (Среда Дульбекко, модифицированная по способу Исков): Наm's F12 (1: 1), содержащей 10% фетальной сыворотки теленка (FCS) и гепарин (45 мкг/мл). Клетки высевали в 24-луночные планшеты, покрытые желатином, при плотности 20000 клеток на лунку на сутки ноль и обрабатывали вышеуказанными лигандами в концентрациях между 0,1 нМ и 1 мкМ на сутки 1, 2 и 3 (заменяя среду параллельно с лигандами). NХ278-липосома ингибировал пролиферацию HUVEC при IС50≈300 нМ (эта концентрация относится к компоненту Нуклеиновой Кислоты-Лиганда); scNХ278-липосома и NX213 были значительно менее эффективными (IС50>1 мкМ).
VEGF индуцирует ангиогенез в тестах на аллантоисной мембране цыпленка (САМ), и данный тест можно использовать для исследования веществ, которые ингибируют ангиогенез. Этот тест проводят путем помещения дисков фильтров, пропитанных VEGF, на САМ, после чего можно количественно определить развитие новых кровеносных сосудов. NX278-липосома эффективно блокировал VEGF-индуцированный ангиогенез (данные не представлены), тогда как NX213, NX278 и scNX278-липосома не оказывали воздействия. Эти исследования вместе демонстрируют, что NX278 является специфичным ингибитором VEGF-индуцированной пролиферации клеток in vitro и образования новых сосудов in vivo.
Воздействие NX278 на VEGF-индуцированную капиллярную проницаемость
VEGF является единственным известным ангиогенным фактором, который временно усиливает капиллярную проницаемость. Изучали способность NX278-липосома ингибировать активность VEGF на сосудистую проницаемость in vivo. Тест на сосудистую проницаемость (так же известный как
тест Майлса (Miles, A. A. and Miles, E. M. (1952) J. Physiol. (London) 118: 228)) проводили на морских свинках по существу как описано (Senger, R. S. et al. (1983) Science 219: 983). NХ278-липосома, NX278 и NX213 в концентрации 1 мкМ инъецировали внутрикожно с VEGF (20 нМ) морским свинкам, которым предварительно был инъецирован синий краситель Эванса. В ответ на VEGF повышение сосудистой проницаемости вызывает транссудацию альбумин-связанного синего красителя Эванса, приводящую к синему пятну в сайте инъекции. Поскольку восстановление этого красителя с помощью экстракции органическим растворителем, как правило, является очень слабым, разработан количественный метод, при котором измеряют поглощение света через кожу. NX213, NX278, NX278-липосома и нейтрализующее моноклональное антитело к VEGF все значительно ингибировали VEGF-индуцированную сосудистую проницаемость, как показано на фиг. 3. Среди Нуклеиновых Кислот-Лигандов NХ278-липосома оказался наиболее сильным антагонистом. Последовательности перемешанных аналогов этих соединений не являлись ингибирующими. Различия являлись значительными и заметными невооруженным глазом.
NX278-L ингибирует клеточные линии саркомы Капоши in vitro
Ингибиторы VEGF имеют потенциальную пользу при ряде заболеваний, включая злокачественности, где прогрессирование опухоли и метастазы являются зависимыми от образования новых сосудов. Хотя известно, что большинство типов опухолей продуцируют VEGF, ранее не показано, что они экспрессируют функциональные рецепторы VEGF. Недавно показано, что клетки саркомы Капоши (KS) не только продуцируют обильные количества VEGF, но также экспрессируют функциональные рецепторы VEGF и, следовательно, используют VEGF для аутокринного роста. Клеточные линии KS, таким образом, обеспечивают уникальную возможность изучать способность NX278 прерывать аутокринную ростовую активность VEGF.
Изучали воздействия NX278-липосома, scNX278-липосома и NX213 на пролиферацию KS клеток. KS-клеточную линию KSY-1 высевали в 24-луночные платы, покрытые желатином, при плотности 7500-10000 клеток на лунку на сутки ноль в среде, содержащей RPMI 1640 с добавлением 2% FCS, L-глутамина, пенициллина и стрептомицина. Нуклеиновые Кислоты-Лиганды добавляли при концентрациях между 0,1 нМ и 1 мкМ в свежую среду на сутки 1, 2 и 3, и подсчет клеток проводили на сутки 4. NХ278-липосома ингибировала пролиферацию KS клеток при IC50 100 нМ; при 1 мкМ NХ278-липосома рост этих клеток полностью ингибировался. scNХ278-липосома и NX213 показывали значения IC50>1 мкМ (фиг. 4).
NX278-липосома ингибирует рост KS клеток in vivo
Поскольку VEGF является фактором роста для KS клеток, воздействие антагонистов VEGF на KS опухоли in vivo, вероятно, является двукратным: ингибирование паракринного ростового воздействия VEGF на связанные с опухолью эндотелиальные клетки и ингибирование аутокринного ростового воздействия VEGF на опухолевые клетки. KS опухоли могут быть, таким образом, особенно чувствительными к антагонистам VEGF. Для тестирования активности Нуклеиновых Кислот-Лигандов in vivo, троакары опухоли (3 мм3) имплантировали бестимусным мышам на первые сутки и обрабатывали в течение пяти последующих суток, начиная на сутки два, 50, 100 или 150 мкг/сутки/мышь. Скорость роста опухоли измеряли в течение периода в две недели. NХ278-липосома ингибировала рост опухоли в зависимости от дозы с очень малым ингибированием роста опухоли при самом низком уровне дозы 50 мкг/сутки/мышь (фиг. 5А) и значительным ингибированием роста опухоли при обоих уровнях дозы 100 и 150 мкг/сутки/мышь (фиг. 5В, показано 150 мкг/сутки/мышь). Пустые липосомы (фиг. 5А, В), scNХ278-липосома, так же как и NX213 и NX278, были неэффективны при всех изученных дозах. Кроме того, NX278-липосома блокировала VEGF-индуцированное протекание жидкости из кровеносных сосудов.
Пример 3. Экспериментальные методики для 2'-фтор-пиримидин-модифицированных РНК лигандов к VEGF.
В данном примере предлагаются общие методики, выполняемые и включенные в примере 4 для выделения 2'-фтор-модифицированных Нуклеиновых Кислот-Лигандов к VEGF.
Материалы
Рекомбинантный VEGF165 человека, очищенный из линии клеток насекомых Sf21, покупали у R& D Systems в виде свободного от носителя лиофилизированного порошка. Белок ресуспендировали в физиологическом растворе, забуференном фосфатом, до концентрации 10 мкМ и хранили при 20oС в набольших аликвотах до использования. Аликвоты хранили при 4oС до 4 недель после оттаивания. Sf21-экспрессируемый, VEGF164 мыши и E. coli-экспрессируемый VEGF121 человека, гетеродимер VEGF/PIGF и PIGF также покупали у R& D Systems в виде свободных от носителя лиофилизированных препаратов.
Олигонуклеотиды покупали у Operon Technologies, Inc. , или синтезировали с использованием синтезатора олигонуклеотидов Applied Biosystems Model 394 согласно оптимизированным протоколам. 2'-F- и 2'-ОМе-рибонуклеотидфосфорамидиты производились JBL Scientific, Inc. (San Luis Obispo, CA). 2'-F-пиримидиновые НТФ также покупали у JBL. Использовали 2'-ОН-пуриновые НТФ и дНТФ от Pharmacia Biotech, Piscataway, NJ.
Термостабильную ДНК полимеразу T. aquaticus (Taq-полимеразу) покупали у Perkin Elmer-Cetus (Foster City, CA): использовали обратную транскриптазу AMV (AMV RT) от Life Sciences, Inc. ; ДНК полимеразу Кленова от New England Biolabs, Beverly, MA. РНК полимеразу Т7 от Enzyco, Inc. (Denver, CO). Секвенирующая ДНК полимераза производилась United States Biochemical Corp. (Cleveland, OH).
α-[32Р] -АТФ и γ-[32P] -АТФ получали от New England Nuclear (Boston, MA).
Протокол SELEX
Методика SELEX описана в деталях в Заявках на патент SELEX. Получали библиотеки химически синтезированных ДНК олигонуклеотидов ("30N7" и "40N7") с рандомизированными районами 30 или 40 нуклеотидов, фланкированными общими 5' и 3' фиксированными последовательностями (5'-TAATACGACTCACTATAGGGAGGACGATGCGG(30 или 40 N) CAGACGACTCGCCCGA-3'; SEQ ID NO: 133 и 134.
Выделенные курсивом нуклеотиды на 5'конце каждой матрицы соответствуют промоторной последовательности Т7 РНК полимеразы. Олигонуклеотидные праймеры также синтезировали для использования при получении матрицы, амплификации и обратной транскрипции:
5'-TCGGGCGAGTCGTCTG-3' ("3N7"; SEQ ID NO: 135) и 5'-TAATACGACTCACTATAGGGAGGACGATGCGG-3'("5N7"; SEQ ID NO: 136).
Двунитевые ДНК матрицы получали путем отжига праймера 3N7 к библиотекам 30N7 или 40N7 и элонгации этого праймера с использованием ДНК полимеразы Кленова или AMV RT. Более высокая температура инкубации, использованная для AMV RT (45oС по сравнению с 37oС), может лучше стимулировать полную элонгацию через высоко структурированные матричные олигонуклеотиды. Библиотеки транскрибировали с использованием Т7 РНК полимеразы в присутствии 1 мМ каждого из 2'-ОН-АТФ и ГТФ, 3 мМ каждого из 2'-F-ЦТФ и УТФ и 50 микрокюри α-32Р-АТФ. РНК очищали из денатурирующих полиакриламидных гелей путем вырезания полоски геля, содержащей РНК, ее измельчения и вымачивания в течение длительного времени в 2 мМ ЭДТА (этилен диамин-тетра-уксусная кислота).
Способ SELEX отбора на аффинность с последующей амплификацией отобранного пула описан в деталях (см. Заявки на патент SELEX). Кратко, один раунд отбора и амплификации проводили следующим образом: VEGF смешивали с 5- или 10-кратным избытком РНК в физиологическом растворе, забуференном фосфатом, с 1 мМ MgCl2 (ФСБМ) (библиотеки 30N7 и 40N7 или в физиологическом растворе, забуференном Трис, с 1 мМ МgСl2, 1 мМ СаСl2 (ТСБМК) (только библиотека 30N7), и эту смесь последовательно разбавляли в три раза. После инкубации при 37oС в течение 15 минут смеси проводили через 0,45 мкм фильтры Type НА (Millipore), чтобы собрать Комплексы VEGF с РНК. РНК элюировали с отобранных фильтров путем инкубации в 2: 1 фенол, рН 7: 7М мочевина. После осаждения из водной фазы РНК подвергали отжигу с праймером 3N7 и обратной транскрипции с использованием AMV RT. Полученные в результате кДНК амплифицировали в 15 циклах полимеразной цепной реакции (ПЦР) с использованием праймеров 3N7 и 5N7 и Taq ДНК полимеразы. Транскрипцией продукта ПЦР получали новую библиотеку, обогащенную последовательностями с аффинностью к VEGF. При раунде 4 во всех трех отобранных пулах РНК в отсутствие VEGF появлялся существенный фоновый сигнал, связанный с фильтром. Чтобы истощить эти пулы связанных с фильтром РНК, раунды 5 и 6 проводили с использованием альтернативной схемы для отделения VEGF-связанных РНК от несвязанных молекул: после инкубации пула РНК с фактором роста каждую смесь наносили на 8% полиакриламидный не денатурирующий гель и проводили электрофорез при 10 Вт в течение 45-60 минут при 4oС. Комплексы VEGF/PHK мигрировали выше несвязанной РНК в данной системе и были визуализированы с помощью экспозиции рентгеновской пленки с гелем. Для этих раундов отобранные РНК очищали с помощью способа измельчения и вымачивания, как описано выше. После двенадцати раундов отбора и амплификации индивидуальные молекулы в этих отобранных пулах клонировали с использованием набора pCR-Script Cloning от Stratagene (La Jolla, CA). Плазмиды очищали с использованием способа щелочного лизиса (набор PERFECTprep Plasmid DNA, 5 Prime-->3 Prime, Boulder, CO), и последовательности клонированных районов получали с использованием набора Dye Terminator Cycle Sequencing, доступного от Perkin Elmer (foster City, CA). Флюоресцентные леддеры последовательности нуклеотидов прочитывали в National Jewish Center, laboratory of Brian Kotzin, Denver, CO. Последовательности группировали в семейства и выравнивали на глаз.
Измерение связывающих аффинностей
Нуклеиновые Кислоты-Лиганды радиоактивно метили в процессе транскрипции посредством включения α-[32Р] -меченых НТФ или после синтеза с использованием γ-[32Р] -АТФ и Т4 полинуклеотидкиназы, инкубировали при низкой концентрации (между 20 и 70 пМ) с варьирующими концентрациями VEGF или других факторов роста при 37oС в течение 15 минут. Инкубации проводили в ТСБ, ФСБ или физиологическом растворе, забуференном ГЭПЭС (N-2-гидроксиэтил-пиперазин-N-2-этансульфоновая кислота) (ГСБ), рН 7,4, с добавлением или без добавления дополнительных двухвалентных катионов. Образцы проводили через предварительно промытые 0,45 мкм фильтры Type НА (Millipore) с последующей промывкой 5-10 мл связывающего буфера. Фильтры погружали в сцинтиллятор и считали, чтобы количественно определить количество связанной с белком РНК, задержанной каждым фильтром. Константу равновесной диссоциации (КD) Нуклеиновой Кислоты-Лиганда, связывающую специфический белок, вычисляли исходя из данных, как описано в Green et al. (1996) Biochem. 35: 14413-14424.
Отбор фрагментов Нуклеиновых Кислот-Лигандов по аффинности
Десять пмоль радиоактивно меченных внутри транскриптов Нуклеиновых Кислот-Лигандов VEGF высокой аффинности частично расщепляли нуклеазой S7, чтобы получить смесь радиоактивно меченных фрагментов. Одну десятую фрагментированной РНК инкубировали с 10 пМ VEGF в 45 мл связывающего буфера, после чего фильтровали через нитроцеллюлозу. Отобранные фрагменты, отделенные от фильтра, разгоняли в денатурирующем полиакриламидном геле с высоким разрешением на следующей дорожке после дорожки, на которую был нанесен пул не отобранных фрагментов. Самые малые из отобранных полос индивидуально очищали из геля и дополнительно метили по их 5'концам с использованием полинуклеотидкиназы, чтобы повысить их специфическую активность. Половину образца подвергали отжигу с кДНК исходного транскрипта и элонгации до конца матрицы с использованием секвенирующей ДНК полимеразы. Сравнение миграции очищенного фрагмента и продукта его элонгации со стандартным леддером последовательности нуклеотидов использовали для определения вероятного размера и положения отобранного фрагмента в пределах исходного транскрипта. Синтетические олигонуклеотиды, соответствующие по последовательности отобранным по аффинности фрагментам, получали, чтобы подтвердить, что усеченная Нуклеиновая Кислота-Лиганд сохранила аффинность к VEGF.
2'-ОМе-замещение
Эксперименты по 2'-ОМе-замещению проводили, по существу, как описано в Green et al. (1995) Chem. Biol. 2: 683-695. Три или четыре библиотеки получали для каждого из трех усеченных лигандов (t22, t2, t44), в которых пять или шесть 2'-ОН пуриновых положений были частично замещены 2'-ОМе. Каждое пуриновое положение было частично 2'-ОМе-модифицировано только в одной из библиотек. Каждую 5'-радиоактивно меченную библиотеку инкубировали с VEGF, и замещенные олигонуклеотиды, связанные этим белком, собирали на нитроцеллюлозные фильтры. Отобранный пул и исходную не отобранную библиотеку частично гидролизовали щелочью, и продукты выявляли на полиакриламидном геле с высоким разрешением. Для каждого пуринового положения определяли "отношение интенсивности полос" путем деления сигнала, полученного на фосфорном формирователе изображения в результате гидролиза в этом положении для отобранного пула, на сигнал, полученный в том же положении для не отобранной библиотеки. Отношения интенсивности полос, которые лежат значительно выше уровня для конкретного положения, являются показателями смещения к 2'-ОН (против 2'-ОМе) в отобранном по аффинности пуле.
Константы скорости связывания
Малое количество (обычно менее чем 1 пмоль) 5'-радиоактивно меченных Нуклеиновых Кислот-Лигандов инкубировали с 1 нМ VEGF при 37oС в 1 мл забуференного физиологического раствора с добавлением двухвалентных катионов. В момент времени "ноль" 50 мкл фильтровали через нитроцеллюлозу, чтобы определить фракцию РНК, связанной с белком, затем добавляли избыток (100 или 500 нМ в различных экспериментах) немеченой Нуклеиновой Кислоты-Лиганда, и 50 мкл аликвоты фильтровали в последующие моменты времени. Фильтры считали в сцинтилляторе, чтобы определить количество радиоактивно меченой РНК, все еще связанной с VEGF в каждый момент времени. Данные, нанесенные на график как фракция связанной РНК (f) против времени, совпадали с уравнением для экспоненциального распада:
f(t)= f0 е-kt+b,
где f0 представляет собой фракцию РНК, связанной в момент времени ноль, k представляет собой константу скорости диссоциации (kd), a b представляет собой остаточное связывание радиоактивно меченной РНК с фильтром в конце эксперимента (эффективно, в отсутствие белка). Константы скорости ассоциации (kas) вычисляли исходя из измеренных значений kd и КD согласно уравнению:
ka= kd/kD.
Пример 4. 2'-фтор-модифицированные РНК лиганды к VEGF.
Отбор лигандов.
Лиганды к VEGF выделяли в трех отдельных SELEX экспериментах из библиотек 2'-F-пиримидин-модифицированных РНК, содержащих 30 или 40 случайных нуклеотидов. Отборы проводили в ФСБ с добавлением 1 мМ MgCl2 (30N и 40N библиотеки) или в физиологическом растворе, забуференном Трис, с 1 мМ MgCl2 и 1 мМ CaCl2 (только 30N библиотека). Примерно 1014 уникальных последовательностей были включены в первый цикл отбора каждого эксперимента. После десяти циклов аффинность между VEGF и каждым пулом РНК была улучшена примерно в 1000 раз относительно исходных пулов. Поскольку дальнейшего улучшения аффинности не наблюдалось после двух дополнительных циклов, индивидуальные члены пулов двенадцатого раунда клонировали и определяли последовательности примерно для 50 изолятов из каждого отбора.
Олигонуклеотидные лиганды к VEGF165 выделяли в трех отдельных SELEX экспериментах. Индивидуальные клоны выделяли и секвенировали, и последовательности группировали в семейства на основании общих мотивов первичной структуры (таблица 2). Название каждого лиганда указывает мишень (V= VEGF), буфер отбора (Р= ФСБ (фосфатно-солевой буфер), Т= ТСБ(Трис-солевой буфер)), длину рандомизированного района в библиотеке (30 или 40 нуклеотидов) и номер клона (после десятичной дроби). Частота, с которой последовательность встречалась среди проанализированных клонов, указана в круглых скобках; последовательности, которые отличаются только одним нуклеотидом, приписывали ПЦР мутагенезу общего предшественника и группировали вместе, причем вариабельное основание указывали в последовательности соответствующим символом (Y= U или С). Фиксированные последовательности, общие для всех лигандов, указаны строчными буквами сверху. Для индивидуальных клонов последовательность вариабельного района показана прописными буквами. Для некоторых лигандов последовательности фиксированного района в строчных буквах прилагаются к последовательности вариабельного района, где они вносят вклад в возможные вторичные структуры. Высокая аффинность Кd для связывания с VEGF показана для каждого лиганда. Один лиганд в каждом семействе был выбран для дальнейшего анализа (серый прямоугольник).
Из общего числа 143 проанализированных клонов получили 76 последовательностей, различающихся более чем одним нуклеотидом. 44 из этих последовательностей возможно было сгруппировать в три основных семейства на основании консервативных первичных структурных мотивов (таблица 2). Последовательности, которые можно сгруппировать в минорные семейства с пятью или менее чем с пятью членами, и последовательности-"сироты", которые являлись уникальными среди изолятов, показаны в таблице 6. Лиганды, содержащие первичный структурный мотив, определенный семействами 1 и 2, встречался во всех трех отборах по аффинности. Сходства между консервативными первичными структурами обоих семейств позволяет предположить, что они могут также иметь общие схожие вторичные структуры и/или они могут граничить с VEGF, используя схожие контактные районы. Члены семейства 2 имеют общую возможность образования короткого стержня из спаренных оснований, окружающего консервативный мотив последовательности в большой "петле" (подчеркнуто в таблице 2). За исключением замыкающей пары оснований A/U идентичность последовательности оснований в предполагаемых стержневых районах не является консервативной. Такая "ковариация" оснований, которая сохраняет вторичную структуру скорее чем первичную, подтверждает существование этого предполагаемого стержня и позволяет предположить, что данная структура может быть важна для высоко аффинной конформации данного семейства лигандов VEGF. Среди последовательностей семейства 1 не обнаружено подобных консервативных взаимодействий спаривания оснований. Третье семейство лигандов встретилось только при отборах, проводимых в ТСБМК (семейство 3, таблица 2). Кроме высоко консервативного первичного структурного мотива, у всех членов данного семейства последовательности 3' консервативного района имеют общую комплементарность спаривания оснований с нуклеотидами в 5' фиксированном районе (подчеркнуто в таблице 2). Поскольку для большинства лигандов нельзя сказать, что основания на 5'стороне предполагаемого стержня коварьируют с их партнерами по спариванию оснований, данное наблюдение является в меньшей степени предсказательным для общей вторичной структуры; тем не менее, в начальном предположении о минимальной высоко аффинной последовательности, происходящей из этого семейства (описано ниже), авторы изобретения руководствовались сильной консервативностью этого мотива. Аффинности индивидуальных РНК лигандов к VEGF оценивали на основании отдельного определения КD для их взаимодействия. За небольшими исключениями лиганды проявляли очень высокую аффинность к фактору роста с КD между 5 и 50 пМ.
Минимальные лиганды
Общие первичные и вторичные структурные мотивы, которые определяют каждое семейство последовательностей, указывают на минимальные элементы последовательности, требующиеся для высокой аффинности связывания с VEGF. Встроенные один в другой усеченные варианты репрезентативного лиганда из каждого семейства (указаны серыми прямоугольниками в таблице 2) получали путем химического синтеза и определяли их относительные аффинности к VEGF (таблица 3). Усеченные варианты лигандов VP30.22, VP30.2 и VT30.44 получали путем химического синтеза и определяли их аффинности к VEGF, как описано в примере 3. Исходные усеченные варианты (t22, t2, t44) далее усовершенствовали путем синтеза олигонуклеотидов с дополнительными основаниями, отсутствующими с 5' и/или с 3' конца. Чтобы инициировать химический синтез, крайний 3' нуклеотид нескольких из лигандов модифицировали либо путем замены 2'-ОН-цитидина на 2'-F-цитидин (подчеркнуто), либо путем добавления 3'-3'-сцепленного дезокситимидинового "кэпа"(звездочки). Показана длина каждого нуклеотида (минус "кэп") и его высокая аффинность КD для связывания с VEGF.
Начальное предсказание для минимальной последовательности из клона VP30.22 (семейство 1) было сделано путем картирования концов очищенного, отобранного по аффинности фрагмента полноразмерного лиганда (см. пример 3). Эта 29-нуклеотидная молекула ("t22") проявила приблизительно трехкратную потерю связывающей аффинности к VEGF относительно полноразмерного лиганда. Дальнейшее усечение на 3' конце данной молекулы вызвало стремительную потерю аффинности, но с 5' конца можно было удалить до 6 дополнительных нуклеотидов с незначительными последствиями или без последствий (таблица 3). Для клона VP30.2 из семейства 2 и клона VT30.44 из семейства 3 синтезировали усеченные лиганды "t2" и "t44", которые заключали в себе предполагаемый стержень из пяти пар оснований и весь консервативный мотив последовательности. Оба усеченных лиганда почти полностью сохранили связывающую активность полноразмерной последовательности. Дальнейшее усечение путем делетирования одной пары оснований сразу (один нуклеотид с каждого конца лиганда) вызывало последовательную потерю аффинности. Таким образом, для этих последовательностей усечения, основанные на возможных вторичных структурах, позволяют очень хорошо предсказать высоко аффинный минимальный лиганд и далее подтверждают гипотезу о том, что предполагаемые стержни вносят вклад в высоко аффинную конформацию этих лигандов.
2'-ОМе модификация
Наблюдали, что замещение в 2'-ОН положениях РНК олигонуклеотидов на 2'-ОМе улучшает их стабильность против нуклеаз, присутствующих в моче крыс, так же как и в других биологических жидкостях. Вероятно, что стабилизация олигонуклеотидов к нуклеазам является критической для их успешности в качестве терапевтических или диагностических агентов. К сожалению, 2'-ОМе-модифицированные нуклеозидтрифосфаты, как правило, не являются приемлемыми для РНК полимераз в качестве субстратов при стандартных условиях реакции. Однако, 2'-ОМе пурины могут быть введены в специфический олигонуклеотид с помощью химического синтеза. Замечено, что некоторые высоко аффинные 2'-ОН-пуриновые РНК-лиганды будут претерпевать необычайно высокий процент 2'-ОМе пуриновых замен с небольшой потерей аффинности к белку-мишени. Чтобы идентифицировать те пуриновые положения, для которых 2'-ОМе замещение является совместимым с высоко аффинным связыванием с VEGF, провели несколько синтезов лигандов t2, t22 и t44, в которых пять или шесть пуринов одновременно были частично замещены на модифицированный нуклеотид (описано в примере 3). Отбор по аффинности каждой частично замещенной библиотеки использовали, чтобы выделить те молекулы, которые сохранили значительную аффинность к VEGF. В таком отобранном по аффинности пуле положения, которые не претерпели замещение, имеют склонность к 2'-ОН, и, следовательно, они проявляют высокую чувствительность к гидролизу щелочью относительно такого же положения в не отобранной библиотеке. 5' радиоактивно меченные пулы, отобранные и не отобранные по аффинности, частично гидролизовали щелочью, и продукты выявляли на полиакриламидном геле с высоким разрешением. В лиганде t22 G10 и А12 показывали значительную склонность к 2'-ОН в отобранном по аффинности пуле, так же как А6 и G21 в лиганде t2 и А5 и А6 в лиганде t44. Хотя вышеописанный анализ идентифицирует те положения, которые, вероятно, не допускают замену на 2'-ОМе-нуклеотиды, исходя из этих данных нельзя предсказать, как подобные модификации всех других пуринов будут влиять на связывающую аффинность. В действительности, лиганд t22, синтезированный со всеми 2'-ОМе пуринами за исключением G10, А12 и G22 (которые проявляли крайнее предпочтение 2'-ОН), связывался с VEGF с аффинностью, равной, если не лучшей, чем вся 2'-ОН-пуриновая последовательность (таблица 4).
Усеченные олигонуклеотиды (t22, t2 и t44) синтезировали химически со всеми пуриновыми положениями, замененными на 2'-ОМе-пурины, кроме одного, двух или трех. Оставшиеся 2'-ОН-пурины указаны в названии каждого лиганда и показаны жирным шрифтом в последовательности лиганда. Показаны Kds для связывания каждого замещенного лиганда с VEGF. Дополнительное замещение в G22 имело небольшое воздействие на связывание с VEGF, но включение 2'-ОМе в G10 или А12, как предсказано, являлось пагубным для связывающей аффинности. Подобным образом лиганды t2 и t44 претерпевали 2'-ОМе-замещение во всех пуринах, кроме двух, с трех-четырехкратным воздействием на аффинность Нуклеиновой Кислоты-Лиганда к VEGF (таблица 4).
Связывающие аффинности и константы скорости для усеченных лигандов
В надежде на идентификацию высоко 2'-замещенных Нуклеиновых Кислот-Лигандов VEGF минимальной длины все 2'-ОМе замещения, которые резко не снижали связывание, были включены в усеченные лиганды t22c, t2a и t44a (см. таблица 3). 2'-ОН-нуклеотиды указаны жирным шрифтом, а 2'-ОМе-нуклеотиды указаны обычным текстом. Полученные в результате Нуклеиновые Кислоты-Лиганды, t22-OMe и t44-OMe, связывались с VEGF с KDs 67 пМ и 49 пМ соответственно, тогда как лиганд t2-OMe связывался с КD примерно 140 пМ (таблица 5). Эти КD выгодно сравниваются с таковой для NX-213 (КD= 140 пМ), 2'-NН2-пиримидин-, 2'-ОМе-пурин-замещенного олигонуклеотидного ингибитора VEGF, описанного ранее (см. Заявку на патент США No. 08/447169, которая включена здесь путем ссылки). Было обнаружено, что каждый из усеченных 2'-ОМе-замещенных олигонуклеотидов конкурирует с NX-213 и друг с другом за связывание с VEGF.
Константы скорости диссоциации (kd) определяли для каждого из трех 2'-ОМе-замещенных лигандов путем наблюдения за потерей преформированного Комплекса между радиоактивно меченным лигандом и VEGF при добавлении большого избытка немеченого лиганда. Лиганд t22-OMe проявлял самую быструю скорость диссоциации с периодом полураспада примерно 60 секунд. Лиганды t2-OMe и t44-OMe проявляли немного более медленные скорости диссоциации с периодами полураспада порядка 170 и 90 секунд соответственно. Константы скорости ассоциации (ka), вычисленные исходя из константы равновесия диссоциации и константы скорости диссоциации (KD= kd/ka), находились в пределах от 3•107 до 2•108 М-1сек-1 (таблица 5). Такие быстрые скорости ассоциации позволяют предположить связывающее взаимодействие, ограниченное близкой диффузией, между этими лигандами и VEGF, и находятся в соответствии с константами скорости ассоциации, наблюдаемыми для полученных по SELEX Нуклеиновых Кислот-Лигандов к другим мишеням.
Зависимость от двухвалентных катионов
Лиганды в семействах 1 и 2 были отобраны в присутствии катионов магния, тогда как лиганды семейства 3 были отобраны в буфере, содержащем и магний, и кальций. Поскольку двухвалентные катионы могут вносить вклад в РНК/белковые взаимодействия через неспецифическую или специфическую стабилизацию высоко аффинных РНК-структур, авторы изобретения поставили вопрос, магний и/или кальций требуется для высоко аффинного связывания репрезентативных лигандов с VEGF. Аффинности Нуклеиновых Кислот-Лигандов t22-OMe и t2-OMe (из семейств 1 и 2 соответственно) были неизменными в присутствии или в отсутствие дополнительных двухвалентных катионов или хелатирующего агента ЭДТА (данные не представлены). Однако, лиганды семейства 3, представленные лигандом t44-OMe, проявляли абсолютную зависимость от присутствия кальция для высоко аффинного связывания с VEGF. Связывание резко снижалось (КD>10-7), когда двухвалентные катионы в связывающем буфере были удалены ЭДТА. Добавление избытка MgCl2 к истощенному связывающему буферу с двухвалентными катионами не приводило к улучшению связывающей аффинности, но CaCl2 при двукратном молярном избытке над ЭДТА полностью восстанавливал связывающую активность. Идентичное поведение в связывании наблюдалось для немодифицированного лиганда t44 (данные не представлены).
Белковая специфичность
Описанные здесь олигонуклеотиды были отобраны на основании их аффинностей к VEGF165, самой большой из двух способных к диффузии изоформ фактора роста. У VEGF121, более малой изоформы, отсутствует один из экзонов VEGF165 и, в отличие от последней, она не связывается с гепарином. Ни один из трех усеченных 2'-ОМе-замещенных олигонуклеотидов не связывался с какой-либо измеримой аффинностью с VEGF121. Кроме того, нативная структура VEGF165 является существенной для связывания всех трех Нуклеиновых Кислот-Лигандов, поскольку связывания не наблюдается, если белок восстановлен ДТТ (дитиотрейтол) до инкубации с олигонуклеотидами.
VEGF является высоко консервативным белком на уровне видов, причем изоформы VEGF165 человека и VEGF164 мыши проявляют 88% идентичности последовательности. Усеченные 2'-ОМе-замещенные лиганды связываются одинаково хорошо с человеческим и мышиным VEGF. Однако, ни для одного из лигандов не наблюдалось связывания с гомодимерами PIGF, белка плацентарного происхождения, который имеет 53% идентичности последовательности с VEGF в пределах консервативного домена, подобного фактору роста, полученному из тромбоцитов. Гетеродимеры между VEGF и PIGF недавно выделены из супернатантов клеточных линий, как нормальных, так и опухолевого происхождения, и такие гетеродимеры проявляют активность при связывании с одним или двумя высоко аффинными рецепторами VEGF и при индуцировании ответов в культивируемых эндотелиальных клетках. Биологическое значение гетеродимеров VEGF/PIGF неизвестно. Существенное связывание, хотя со значительно сниженными аффинностями, наблюдалось с гетеродимерами VEGF/PIGF. Эти данные могут указывать на то, что Нуклеиновые Кислоты-Лиганды связываются на поверхности раздела между двумя субъединицами в димере либо около нее, и что в PIGF не присутствуют все сайты контакта, необходимые для высоко аффинного связывания. Альтернативно, структура субъединицы VEGF может быть изменена посредством участия в гетеродимере с PIGF с последующим нарушением поверхности связывания Нуклеиновой Кислоты-Лиганда.
Пример 5. Синтез фосфолипида, глицеринамид липида и ПЭГ-модифицированных Нуклеиновых Кислот-Лигандов VEGF.
Для синтеза различных Комплексов Липофильное Соединение/Нуклеиновая Кислота-Лиганд использовали три различных препарата, как следующие:


I. Фосфорамидит С-18 для синтеза препарата PL
Описание в общих чертах для получения фосфорамидита С-18 представлено на схеме 3. 1-Октадеканол фосфорилировали при стандартных условиях. После смешивания реакционной смеси остаток очищали на силикагелевой колонке с гексаном: этилацетатом: триэтиламином (90: 10: 5) с получением 21,5 г чистого продукта (выход 57%).
II. Синтез липидамида 1
Данный фосфорамидит, в отличие от вышеописанного PL, имеет амидные связи. Структура олиго, полученная в результате конъюгации данного липида, показана ниже.
Несколько экспериментов продемонстрировали, что высокая нерастворимость соединения 22 в органических растворителях сделала невозможной ЯМР и МС характеристику и дальнейшее фосфитилирование соединения 22 до DAG амидита 23, однако, исходя из результатов для получения липид-спейсер-амидита (схема 1), авторы изобретения ожидали, что фосфилирование соединения 22 с использованием хлор-(2-цианоэтокси)-N, N-диизопропиламино-фосфина могло бы идти, если бы смесь кипятили с обратным холодильником. Подход к получению DAG амидита показан на схеме 4.
N, N'-Бис(стеароил)-1,3-диамино-2- пропанол (22). Раствор стеароилхлорида (6,789 г, 22,41 ммоль) в СlCН2СН2Сl (50 мл) добавляли по каплям к раствору 1,3-диамино-2-гидроксипропана (1,0 г, 11,10 ммоль) в СlCН2СН2Сl (100 мл) и ТЭА (тетраэтиламмоний) (2,896 г, 22,41 ммоль) с перемешиванием при комнатной температуре. После того как добавление заканчивали, смесь нагревали до 70oС в течение ночи, и образовывался чистый раствор, этот раствор охлаждали до комнатной температуры, фильтровали и промывали твердые вещества СН2Сl2, СН3ОН, 5% NaHCО3 и этиловым эфиром и высушивали в вакууме с получением 22 (6,40 г, выход 93%) в виде белых твердых веществ. 1H ЯМР (пиридин-d5; 60oС, δ, млн-1): 3.82-3.78 (m, 1H), 2.37 (t, J= 7.5 Гц, 4Н), 1.81-1.76 (m, 4Н), 1.30-1.27 (m, 60H), 0.87 (t, J= 5.7 Гц, 6Н).
N, N'-Бис(стеароил)-O- (диизопропиламино-2- цианоэтоксифосфинил)- 1,3-диамино-2-пропанол (23). К соединению 22 (5,80 г, 9,31 ммоль), высушенному в течение ночи в вакууме, добавляли безводный СН2Сl2 (150,0 мл), и в него вбрызгивали N, N-диизопропилэтиламин (ДИПЭА) (4,2 мл, 18,62 ммоль). Смесь охлаждали на ледяной бане и вбрызгивали хлор-(2-цианоэтокси)-N, N-диизопропиламинофосфин (8,6 мл, 0,47 ммоль). После перемешивания в течение 30 мин смесь нагревали при 60oС в течение 90 мин. После охлаждения до комнатной температуры нерастворимые материалы отфильтровывали, и этот раствор промывали 5% NаНСО3 и рассолом, высушивали над Na2SO4 и концентрировали в вакууме. Сырой продукт очищали путем осаждения из CH3CN с получением чистого продукта (4,65 г, выход 61%) в виде белых твердых веществ. 31Р ЯМР (СDСl3, млн-1): 154.04.
I. Синтез DAG-спейсер-амидита, липидамида 2
Гекса(этиленгликоль) был включен в липидамидит, чтобы ослабить нерастворимость диамидного соединения 22, которое является непосредственным промежуточным соединением для липидамидита 23. Описание получения липид-спейсер-амидита 29 представлено на схеме 5. Стадия сочетания соединения 25 с 1,3-диамино-2-гидроксипропаном и трет-бутоксидом калия в ТГФ не идет хорошо, и выход составлял только примерно 20%. Одна попытка улучшить выход была сделана посредством взаимодействия 25 и диамида 22, однако, желаемый продукт не был обнаружен.
(4,4'-Диметокситритилокси)-гексаэтиленгликоль (24).
Гекса(этиленгликоль)(18,98 г, 67,05 ммоль) совместно выпаривали с безводным пиридином (3•50 мл), растворяли в безводном пиридине (400 мл) и после охлаждения до 0oС добавляли по каплям DMTrCI (23,85 г, 70,40 ммоль) в пиридине (50 мл) в течение 30 мин с перемешиванием в атмосфере аргона. Реакционную смесь выдерживали при комнатной температуре в течение ночи. Пиридин удаляли под высоким вакуумом, и остаток растворяли в CH2Cl2, затем его промывали 5% NaHCO3 и расссолом, высушивали над Na2SO4 и концентрировали в вакууме. Сырой продукт очищали влажной флэш-хроматографией на силикагелевой колонке в градиенте этилацетата, затем в CH2Cl2 и метаноле (95/5), содержащем 0,5% ТЭА. Подходящие фракции объединяли, выпаривали и высушивали в вакууме с получением 24 (26,1 г, выход 66,6%) в виде светло-желтого масла. 1H ЯМР (ДMCO-d6; δ, млн-1): 7.40 (d, J= 7.2 Гц, 2Н), 7.33-7.24 (m, 7Н), 6.89 (d, J= 8.9 Гц, 4H), 4.61 (t, J= 5.1 Гц, 1Н), 3.73 (s, 6H), 3.05 (m, 24H); 13С ЯМР (ДМСО-d6; δ, млн-1): 158.02, 145.02, 135.78, 129.67, 128.13, 127.71, 126.61, 113.14, 85.29, 72.33, 72.27, 70.06, 69.87, 69.80, 69.75, 69.70, 62.84, 60.25, 60.19, 55.01.
(4,4'-Диметокситритилокси)- гексаэтиленгликольтозилат (25). К охлажденному во льду (0oС) раствору 24 в безводном пиридине (50 мл) добавляли раствор толуолсульфонилхлорида в пиридине (30 мл). Через 2 ч при комнатной температуре раствор выпаривали до светло-желтого масла. Остаток растворяли в СН2Сl2 и промывали 5% NaHCO3 и рассолом, высушивали над Na2SO4, фильтровали и выпаривали в вакууме. Продукт очищали с помощью влажной флэш-хроматографии на силикагеле с элюцией этилацетатом с получением продукта (4,08 г, выход 93%) в виде светло-желтого масла. 1H ЯМР (ДМСО-d6; δ, млн-1): 7.78 (d, J= 8.3 Гц, 2Н), 7.46 (d, J= 8.1 Гц, 2Н), 7.40 (d, J= 7.4 Гц, 2Н), 7.32-7.23 (m, 7Н), 6.88 (d, J= 8.8 Гц, 4H), 4.09 (t, J= 4.3 Гц, 2Н), 3.72 (s, 6H), 3.06 (m, 22H), 2.40 (s, 3Н); 13С ЯМР (ДМСО-d6; δ, млн-1): 158.01, 145.01, 135.78, 132.38, 130.12, 129.67, 128.12, 128.02, 127.80, 127.70, 127.62, 113.13.
2-(4,4'-Диметокситритилокси)- гексаэтиленгликоль-1,3- диаминопропан (26). Смесь 1,3-диамино-2-гидроксипропана (74,7 мг, 8,28 ммоль) и трет-бутоксида калия (2,78 г, 24,84 ммоль) в безводном ТГФ нагревали до 70oС в течение 2 ч, а затем охлаждали до комнатной температуры. Соединение 25 (4,08 г, 5,25 ммоль) в ТГФ вбрызгивали, и смесь перемешивали при 70oС в течение ночи, пока ТСХ не показывала, что больше не осталось 25. После того как раствор охлаждали до комнатной температуры, ТГФ удаляли в вакууме и добавляли 25 мл CH2Cl2 и 25 мл воды. Слой CH2Cl2 отделяли, и водный слой экстрагировали СН2Сl2. Растворы CH2Cl2 объединяли, высушивали над Na2SO4 и выпаривали при пониженном давлении. Сырой продукт (2,43 г) непосредственно использовали для реакции без дальнейшей очистки. 1H ЯМР (ДMCO-d6; δ, млн-1): 7.41 (d, J= 7.7 Гц, 2Н), 7.32-7.21 (m, 7H), 6.87 (d, J= 8.8 Гц, 4H), 3.73 (s, 6H), 3.52-3.40 (m, 24H), 3.17 (s, 1H), 3.07-3.02 (m, 4H).
N, N'-Бис(стеароил)-2- (4,4'-диметокситритилокси)- гексаэтиленгликоль-1,3- диаминопропан (27). Раствор стеароилхлорида (3,363 г, 11,1 ммоль) в СlCН2СН2Сl вбрызгивали в раствор 26 в СlCН2СН2Сl и ТЭА (1,9 мл, 11,1 ммоль) с перемешиванием при комнатной температуре. Смесь выдерживали при комнатной температуре в течение 2 ч, затем нагревали до 70oС в течение ночи. После того как раствор охлаждали до комнатной температуры, этот раствор промывали 5% NаНСО3 и рассолом, высушивали над Na2SO4 и концентрировали в вакууме. Сырой продукт очищали с помощью влажной флэш-хроматографии на колонке с силикагелем в градиенте этилацетата и СН2Сl2 (50/50), а затем этилацетата и метанола (50/50). Вторую фракцию собирали, выпаривали и высушивали в вакууме с получением 27 (640 мг) в виде светло-желтого твердого вещества. 1H ЯМР (ДМСО-d6; δ, млн-1): 7.40 (d, J= 7.2 Гц, 2Н), 7.37-7.20 (m, 7H), 6.74 (d, J= 8.9 Гц, 4H), 3.71 (s, 6Н), 3.63-3.51 (m, 24 Н), 3.17 (s, 1Н), 3.16-3.13 (m, 4H), 2.12 (t, J= 7.3 Гц, 4H), 1.18 (m, 60H), 0.80 (t, J= 6.2 Гц, 6Н).
N, N'-Бис(стеароил)-2- гексаэтиленгликоль-1,3- диаминопропан (28). Смесь соединения 27 (640 мг), 2,5% раствора DCA в CH2Cl2 (5 мл) и тригексилсилана (2 мл) перемешивали при комнатной температуре, пока оранжевый цвет не превращался в бледный цвет. После удаления CH2Cl2 остаток повторно осаждали из гексана с получением светло-желтого твердого вещества (210 мг, выход 63%). 1H ЯМР (СDСl3, δ, млн-1): 3.69-3.59 (m, 24H), 3.17 (s, 1Н), 3.06-3.01 (m, 4H), 2.21 (t, J= 7.9 Гц, 4H), 1.18 (m, 60H), 0.81 (t, J= 6.3 Гц, 6Н).
N, N'-Бис(стеароил)-2- (диизопропиламино-2- цианоэтоксифосфинилгексаэтиленгликоль)-1,3- диаминопропан (29). Соединение 28 (210 мг, 0,237 ммоль) высушивали в течение ночи в вакууме, растворяли в безводном CH2Cl2 (5,0 мл) и добавляли N, N-диизопропилэтиламин (218 мкл, 1,25 ммоль). Раствор охлаждали на ледяной бане и вбрызгивали хлор-(2-цианоэтокси)-N, N-диизопропиламино-фосфин (106 мкл, 0,47 ммоль). После перемешивания в течение 30 мин реакционную смесь разбавляли СН2Сl2 и промывали 5% NаНСО3 и рассолом, высушивали над Na2SO4 и концентрировали в вакууме с получением соединения 29. 31Р ЯМР (СDСl3, млн-1): 154.04.
Конъюгация 20К или 40К ПЭГ NHS эфира с Нуклеиновыми Кислотами-Лигандами VEGF
Общая методика: VEGF олигонуклеотид подвергали обмену с получением триэтиламмониевой соли и лиофилизировали. Сырой олигонуклеотид растворяли в 100 мМ натрий-боратном буфере (рН 9) до концентрации 60 мг/мл. 2 эквивалента PEG NHS ester (Shearwater Polymers, Inc. ) растворяли в сухом ДМФ (отношение борат: ДМФ 1: 1), и смесь нагревали, чтобы растворить ПЭГ NHS эфир. Раствор олигонуклеотида быстро добавляли к раствору ПЭГ, и смесь энергично перемешивали при комнатной температуре в течение 10 мин. Примерно 90% олигонуклеотида становится конъюгированным с PEG NHS ester. См. Фиг. 1Н и 1I.
Синтез димерных Нуклеиновых Кислот-Лигандов VEGF
Димерные Нуклеиновые Кислоты-Лиганды VEGF, показанные на фиг. 1J, К и L, получали следующим образом (см. схему А)
Синтез 1,3-дипивалолил-2-O-диметокситритилглицерина 32
К перемешанному раствору соединения 31 в пиридине (62 г 70% чистого продукта, 200 ммоль, в 200 мл пиридина), приготовленному согласно McGee et al. (1988, Synthetic Communication, 1651), добавляли диметокситритилхлорид (84 г, 240 ммоль, 1,2-кратный избыток), и реакции давали перемешиваться при комнатной температуре в течение 16 часов. Реакционную смесь концентрировали при пониженном давлении, и остаток растворяли в СН2Сl2 (1 л), промывали водой, высушивали (MgSO4) и концентрировали. Сырую смесь (130 г) использовали как таковую в следующей реакции.
Синтез-2-О-диметокситритилглицерина 33
Смесь сырого соединения 32 (130 г), NaOMe (28 г) и метанола (900 мл) нагревали при 50oС в течение 16 часов. После того как реакция прошла полностью (ТСХ), смесь концентрировали до сухости, и остаток растворяли в воде и CH2Cl2 (1: 1). Органический слой отделяли, а водный слой промывали насыщенным NH4Cl, водой и рассолом и высушивали (MgSO4). Выпариванием растворителя получали смолистое соединение, которое очищали с помощью силикагелевой колонки с использованием 1: 1 гексана/этилацетата, содержащего 2% ТЭА, с получением соединения 33 при 75% отдельного выхода. 1H ЯМР (ДМСО-d6) 3.02-3.07 (m, 2Н), 3.17-3.23 (m, 2Н), 3.3-3.35 (m, 1H), 3.7 (s, 6H), 4.26 (t, J= 4.1 Гц, 2Н, D2O, взаимозаменяемы), 6.59-6.86 (m, 4H), 7.17-7.68 (m, 9H).
Синтез бисамидита 34
К охлажденному на льду перемешанному раствору спирта 33 (16,2 г, 41,1868 ммоль) в СH2Сl2 (125 мл) и диизопропилэтиламина (58 мл, 320 ммоль) добавляли фосфитилирующий реагент (20,5 мл, 90,62 ммоль), и раствор медленно нагревали до комнатной температуры и перемешивали в течение 2 ч при той же температуре. Реакционную смесь медленно наливали в колотый лед и экстрагировали СН2Сl2, промывали 5% NаНСО3, водой и рассолом и высушивали. Остаток, полученный после выпаривания растворителя, очищали с помощью хроматографии на силикагелевой колонке с использованием 1: 1 гексана/этилацетата, содержащего 2% ТЭА, с получением соединения 34 при 70% выходе. 1H ЯМР (ДМСО-d6) 1.03-1.12 (2d, 24H), 2.69-2.75 (2t, 4H), 3.1-3.33 (m, 4Н), 3.33-3.55 (m, 5H), 3.66-3.7 (m, 4Н), 3.72 (s, 6H), 6.83-6.89 (m, 4H), 7.19-7.48 (m, 9H). 31Р D3РO4 как внешний стандарт 153.64& 153.39.
Получение димеров VEGF
Синтез димеров VEGF проводили на 8800 автоматизированном ДНК/РНК синтезаторе. Получали NX31838, где rА означает аденозин, mG и mА означает 2'-O-метилгуанозин и аденозин соответственно, а fС и fU означает 2'-дезокси-2'-фторцитидин и 2'-фторуридин соответственно, и [3'-3'] означает межнуклеотидную связь. Синтез проводили при шкале 1 ммоль на автоматизированном синтезаторе Millipore 8800 с использованием 5'-DMT-2'-О-метил-N6-трет- бутилфеноксиацетиладенозин, 5'-DMT-2'-O-TBDMS-N2-трет- бутилфеноксиацетилгуанозин и 5'-DMT-2'-O-TBDMS-N6-тpeт- бутилфеноксиацетиладенозин 3'-N, N- диизопропил-(2-цианоэтил) -фосфорамидитов, а также 2'-дeзoкcи-2'-фтop-5'-DMT-N4- aцeтилцитидин и 2'-дезокси-2'-фтор-5'-DМТ- уридин 3'-N, N-диизопропил-(2-цианоэтил)- фосфорамидитов. Цикл синтеза представлял собой следующее. Активаторные препараты описаны в таблице 12. Синтез проводили с использованием носителя CPG с размером пор 600 Å, 80-120 меш и 60-70 мкмоль/г загруженного 5'-сукцинилтимидином. Объединенный цикл показан в таблице 12.
Пример 6. Фармакокинетические свойства фосфолипид (PL)- и ПЭГ-модифицированных Нуклеиновых Кислот-Лигандо в VEGF.
Из последовательностей, показанных в таблице 2, для дальнейшего исследования была выбрана последовательность VT 30.44 и переименована как NX31838. Фармакокинетические свойства Нуклеиновой Кислоты-Лиганда VEGF NX31838, конъюгированной с 20 и 40К ПЭГ, определяли у крыс Sprague-Dawley (см. молекулярные изображения на фиг. 1) (SEQ ID NOS: 8 и 9). Подобные исследования проводили также на NX31838, конъюгированном с PL липидом, в виде липосомного препарата и в виде свободного лекарственного средства (см. молекулярные изображения на фиг. 1Н и 1) (SEQ ID NOS: 8 и 9). В каждом исследовании олигонуклеотид разбавляли ФСБ до раствора концентрации 1,0 мг/мл на основании УФ поглощения при 260 нм и коэффициента экстинкции 0,037 мкг олиго/мл. Во всех исследованиях 9 крыс получали 1,0 мг олигонуклеотида/кг массы животного путем болюсной инъекции в хвостовую вену, и образцы плазмы брали в различные моменты времени от 2 минут до 24 часов. Образцы плазмы и образцы контроля качества анализировали с использованием гибридизационного анализа. В гибридизационном анализе использовали олигонуклеотид захвата, который содержит последовательность, комплементарную 5'-концу Нуклеиновой Кислоты-Лиганда VEGF, конъюгированный с каплей оксида железа (FeO) (FeO-спейсер-3'-d (GCC TTA GTC ACT T-5') (SEQ ID NO: 137), где спейсер = (dT)8), и детекторный олигонуклеотид, содержащий две молекулы биотина на 5'-конце (биотин-биотин-5'-d(спейсер-СGG ATG TAT AAG СА-3'), где спейсер = (dT)8 (SEQ ID NO: 138). После инкубации зонда захвата и детекторного зонда с образцом плазмы, содержащим Нуклеиновую Кислоту-Лиганд VEGF NX31838, количество биотинового олигонуклеотида, гибридизованного с каплей, количественно определяли стрептавидин-связанной щелочной фосфатазой с использованием CSPD-Sapphire в качестве люминесцентного субстрата.
Данные по концентрации в плазме свободной, ПЭГ 20К и ПЭГ 40К Нуклеиновых Кислот-Лигандов VEGF (NX31838) (SEQ ID NOS: 8 и 9) как функции времени после болюсной инъекции суммированы на фиг. 6. Конъюгат 40К ПЭГ выводился с моноэкспоненциальным t1/2 360 минут, тогда как вариант 20К ПЭГ выводился намного быстрее, причем 95% Нуклеиновой Кислоты-Лиганда выводилось с альфа t1/2 49 минут, и 5% выводилось с бета t1/2 192 минуты, что указывает на явное значение размера для клиренса. По сравнению с ПЭГ-конъюгированными Нуклеиновыми Кислотами-Лигандами свободный (неконъюгированный) NX31838 выводился из плазмы очень быстро с t1/2 в несколько минут. Концентрация олигонуклеотида в плазме как функция времени может быть значительно повышена посредством включения подходящих функциональных групп в олигонуклеотид.
Данные по концентрации в плазме PL липид-конъюгированной Нуклеиновой Кислоты-Лиганда VEGF (SEQ ID NOS: 5), изготовленной в виде препарата с липосомами или без липосом, как функции времени после болюсной инъекции суммированы на фиг. 7. Липосомы получали, как описано в примере 7А, с помощью обработки ультразвуком в присутствии Нуклеиновой Кислоты-Лиганда, и они содержат олигонуклеотид как внутри, так и снаружи. Липосомный препарат выводился намного медленнее, чем свободное лекарственное средство, бета t1/2 1161 минута и 131 минута, соответственно. Концентрация олигонуклеотида в плазме как функция времени может быть значительно повышена посредством липосомного препарата.
Пример 7. Получение Комплекса NX31838 PL-липосома
А. Получение липосом путем образования пленки
Липиды объединяют в соотношении 2 моля DSPC к 1 молю холестерина. NX31838 PL в воде добавляют к липидам в соотношении 1: 50 (масс. /масс. ). Материал объединяют посредством сольватирования раствором хлороформ: метанол: вода (1: 3: 1). Растворитель удаляют с помощью роторного испарения, оставляя гетерогенную пленку NX 31838 PL, перемешанного совместно с липидами. Пленку регидратируют до 50 мг/мл на основе липидов в растворе 9% сахарозы, забуференном 25 мМ фосфатом натрия при рН 7,4. Раствор энергично перемешивают, нагревают до 65oС, и полученный в результате белый, подобный молоку раствор обрабатывают ультразвуком в 75 мл аликвотах для сборки липидов в однослойные липосомы. За прогрессом в формировании липосом следят визуально, пока раствор не становится опалесцирующим, а затем с помощью измерения размера частиц посредством динамичного светорассеяния с использованием анализатора частиц (Leeds & Northrup Model Microtrack UPA 150, Horsham, PA). Размер липосом находится в диапазоне от 50 до 70 нм (по методу объемно-массового распределения).
Б. Получение липосом путем пассивного заякоривания
Тестировали scNX-278 (см. молекулярное изображение на фиг. 1С), чтобы увидеть, претерпевает ли он спонтанное включение в преформированные ("пустые") липосомы. Предварительные результаты с использованием ДЕАЕ анализа (для удаления Комплекса Нуклеиновая Кислота-Лиганд/глицеролипид) указывали на два важных открытия: 1) загрузка может быть достигнута и, что более важно, 2) по существу полная загрузка Комплекса Нуклеиновая Кислота-Лиганд/глицеролипид наблюдалась через 24 часа при комнатной температуре. Впоследствии было предпринято более детальное исследование для определения воздействий температуры на загрузку. Наблюдали, что температура явно воздействует на скорости включения. Хотя полная загрузка может быть достигнута через 24 часа при комнатной температуре, полное включение может быть достигнуто даже в течение минут при повышенных температурах (67oС). Это оказалось быстрым и эффективным способом включения Комплекса Нуклеиновая Кислота-Лиганд/Липофильное Соединение в преформированные липосомы.
Хроматографию по размеру затем использовали для отделения свободного scNX-278 от липосомо-ассоциированной формы. Предварительная работа была проведена с использованием загрузки scNX-278 в "пустые" 2: 1 DSРС: холестерин липосомы. Хроматограммы получали с использованием колонки Superdex S-200 при 22oС. За период в 22 часа наблюдалось постепенное включение scNX-278 в популяцию пустых липосом в виде сдвига в пиковых областях (данные не представлены). Эти результаты хорошо коррелируют с данными, полученными в ДЕАЕ анализе.
Предпринимали также исследования, чтобы определить, может ли дополнительный scNX-278 быть загружен в обработанные ультразвуком олиго-липосомы. Озвученный препарат scNX-278 получали путем совместного растворения олиго-липида с липидом и совместной обработки ультразвуком обоих. Полученные липосомы проявляли полное включение scNX-278. Этот обработанный ультразвуком препарат затем подвергали 2 отдельным раундам пассивного заякоривания с дополнительным свободным scNX-278, чтобы увидеть, может ли успешно включиться большее количество scNX-278. Во время первого раунда пассивного заякоривания весь свободный scNX-278 был пассивно заякорен в липосомах после инкубации в течение 1 часа при 65oС. Вторая попытка пассивного заякоривания дополнительного scNX-278 приводила к неполной загрузке.
Ключевым открытием из этих экспериментов является то, что Комплекс Нуклеиновая Кислота-Лиганд/Липофильное Соединение может быть пассивно заякорен в озвученные олиго-липосомы при высоких концентрациях, но что емкость липосом для поглощения дополнительных Комплексов Нуклеиновая Кислота-Лиганд/Липофильное Соединение может быть превышена. После 2 раундов пассивной загрузки (до примерно 3 мг липида-олиго/50 мг липида), липосомы очевидно достигают своей "емкости" поглощения дополнительного олиго-липида, поскольку некоторое количество свободного липида-олиго остается. Эти данные были подтверждены ДЕАЕ анализом на вращающейся колонке (данные не представлены). Выводами, которые можно сделать, являются: 1) озвученные липосомы обладают дополнительной емкостью для включения Комплексов Нуклеиновая Кислота-Лиганд/Липофильное Соединение; и 2) 100% включение Нуклеиновой Кислоты-Лиганда может быть достигнуто посредством озвучивания.
Последующие исследования проводились на NX31838 PL (см. молекулярное изображение на фиг. 1Е). NX31838 представляет значительный интерес, поскольку он обладает усовершенствованной фармакокинетикой (см. пример 6) и биологическим распространением против VEGF мишеней при включении в липосомы. Несколько исследований было проведено, чтобы лучше понять включение NX31838 в липосомы посредством пассивного заякоривания.
Исследования по кинетике NX31838 PL указывали на то, что пассивное заякоривание для данной молекулы было настолько быстрым, что считалось невозможным его измерение посредством какой-либо из хроматографических методик, известных в литературе (для всех из них требуется времени пробега минимум в несколько минут).
Чтобы определить ориентацию молекулы NX31838 PL (то есть, является ли компонент Нуклеиновой Кислоты-Лиганда выступающим снаружи липосомы или выступающим внутрь водного центра липосомы), использовали вводимую извне РНКазу, чтобы избирательно расщепить любой из компонентов Нуклеиновой Кислоты-Лиганда, который выступает снаружи из липосомы. В случае пассивно заякоренных липосом NX31838 PL вся Нуклеиновая Кислота-Лиганд является доступной для РНКазы I. Никакого дополнительного расщепления не наблюдалось после обработки Тритоном Х-100. Эти результаты указывают на то, что пассивно загруженный NX31838 PL ориентирован таким образом, что компонент Нуклеиновой Кислоты-Лиганда выступает снаружи из липосомы. Если пассивно заякоренные липосомы NX31838 PL предварительно переваривают РНКазой I, а затем пропускают через ДЕАЕ колонку, примерно 99% Нуклеиновой Кислоты-Лиганда удерживается колонкой, тогда как если тот же образец пропускают через ДЕАЕ колонку, но без преинкубации с РНКазой I, примерно 100% олиго способны пройти через колонку, не связавшись с ДЕАЕ. Липосома защищает олиго от ДЕАЕ. Липосома действует для защиты компонента Нуклеиновой Кислоты-Лиганда от ДЕАЕ, поскольку она соединяется с Нуклеиновой Кислотой-Лигандом с высокой аффинностью, значительно снижая ее доступность для ДЕАЕ групп.
Наконец, в качестве части разработки новых способов отделения свободного Комплекса Нуклеиновая Кислота-Лиганд/Липофильное Соединение от заякоренной в липосоме формы, авторы изобретения расщепляли NX31838.05 PL РНКазой I. Расщепленный олиго можно легко отделить с использованием хроматографии по размеру (смола S-1000) после удаления липидного хвоста, тогда как интактный Комплекс Нуклеиновая Кислота-Лиганд/Липофильное Соединение в идентичных условиях элюировался совместно с липосомой. Эти данные указывают на то, что Комплекс Нуклеиновая Кислота-Лиганд/Липофильное Соединение, вероятно, образует мицеллу при свободном нахождении в растворе. Это приводит к тому, что он совместно элюируется в свободном объеме колонки с липосомами. Удаление липидного хвоста позволяет ему войти в среду гель-фильтрации, разделить его соответственно размеру и сохранить.
Пример 8. Эффективность in vivo Комплексов Нуклеиновых Кислот-лигандов VEGF - анализ кожной сосудистой проницаемости
Способность нескольких различных препаратов Нуклеиновой Кислоты-Лиганда NX31838 к ослаблению VEGF-индуцированных изменений в проницаемости кожной сосудистой системы (анализ Майлса) проводили, как описано ранее (Senger et al. (1986) Cancer Research 46: 5629-5632), с незначительными модификациями. Кратко, взрослых самок морской свинки (3/исследование) анестезировали изофлюраном, и шерсть на спинной и боковой задних областях удаляли ножницами. Синий краситель Эванса (2,5 мг/морскую свинку) вводили внутривенно. Растворы для инъекций (препараты ФСБ, VEGF, NX31838 и моноклональное антитело анти-VEGF) готовили заранее за 30 мин, смешивали вместе, где указано, с конечными концентрациями, как показано. Каждый показанный раствор затем инъецировали внутрикожно (двойные инъекции/морскую свинку, 40 мкл/сайт) случайным образом по схеме решетки, нарисованной на выстриженной области. Морским свинкам давали восстановиться после анестезии и умерщвляли их посредством экспозиции с CO2 в течение 30 мин после завершения внутрикожных инъекций. Затем кожу собирали, очищали от подкожного слоя и подвергали трансиллюминации. Изображения снимали с использованием цветной камеры CCD (Hitachi Denshi KP-50U, Japan) и программного обеспечения Image-Pro Plus (версия 3.1, Media Cybernetics, Silver Springs, MD). Каждый образец кожи нормализовали по интенсивности, причем каждый сайт инъекции анализировали на оптическую плотность и вовлеченную область.
На фиг. 8А-С показаны результаты ослабления Нуклеиновой Кислотой-Лигандом VEGF-индуцированного протекания из сосудов для NX31838-20K PEG, NX31838-40K PEG, NX31838 PL в липосомном препарате, как описано в примере 7А. Все препараты были способны значительно снижать протекание из кровеносных сосудов около или ниже контрольных уровней ФСБ при таких низких концентрациях, как 100 нМ. При 30 нМ блокирующее воздействие Нуклеиновой Кислоты-Лиганда исчезало. Липосомный препарат NX31838-PL не оценивали при этой концентрации, но оказалось, что он обладает сниженной блокирующей активностью при 100 нМ. Моноклональное антитело анти-VEGF также оценивали в этой модельной системе, и оно было также эффективно при 100 нМ с потерей активности при 30 нМ. Это позволяет предположить, что в этой модельной системе, в которой изучали NX31838 в различных препаратах, он является таким же эффективным, как антитело, при блокировании одного из функциональных воздействий белка VEGF.
Пример 9. Эффективность in vivo Комплексов Нуклеиновых Кислот-Лигандов VEGF - модель роговичного кармана
Препараты Нуклеиновой Кислоты-Лиганда VEGF (NX31838) тестировали на их способность снижать VEGF-индуцированный роговичный ангиогенез у крыс с природно бессосудистой роговицей. Кратко, биополимерные (Hydron) гранулы ± VEGF белок (3 пмоль) готовили примерно за 30 ч перед добавлением раствора белка или носителя к 12% биополимеру в 95% этаноле. Взрослых крыс Sprague-Dawley (200-240 г) анестезировали путем внутрибрюшинной инъекции кетамина HCl (50 мг/кг) и ксилазина (10 мг/кг). Затем готовили левый глаз путем местного введения тетракаина HCl для местной анестезии с последующим нанесением разбавленного раствора повидона-иода и последующей промывкой изотоническим физиологическим раствором. Делали вертикальный частичный надрез в толщину в середине роговицы. В среднем собственном веществе вырезали карман каудально по направлению к латеральному углу глазной щели, продолжающийся в пределах 1,5 мм от лимба. Затем гранулу вставляли и проталкивали внутрь к каудальной границе кармана. Оставшийся воздух мягко массировали из кармана. Затем в глаз наносили каплю глазного раствора хлорамфеникола. Животное переворачивали, и операцию повторяли на правом глазу со вставкой гранулы того же типа. После завершения вставки гранулы в каждый глаз каждому животному затем вводили либо ФСБ (объем, равный объему группы препаратов Нуклеиновой Кислоты-Лиганда), либо Нуклеиновую Кислоту-Лиганд (10 мг/кг) внутривенно дважды в сутки, как указано. Через 5 суток каждое животное анестезировали и получали фотографии с использованием 35 мм камеры (Minolta X9), смонтированной на рассекающем микроскопе (KAPS, Germany). Каждый глаз оценивали на ангиогенный ответ путем измерения максимальной длины сосудистого роста (0-5), плотности роста сосудов (1-4), прилежащих к имплантированной грануле, и окружность глаза с возникшим ангиогенезом (0-1). Затем определяли ангиогенный индекс как произведение длина•плотность•окружность.
Способность препаратов Нуклеиновой Кислоты-Лиганда блокировать VEGF-индуцированный ангиогенез видна на фиг. 9А-С. Несмотря на то, что NX31838-20K PEG являлся в равной степени эффективным, как другие препараты, при блокировании изменений сосудистой проницаемости, он являлся неэффективным при ослаблении ангиогенного ответа в природно бессосудистой роговице. Однако, как NX31838-40K PEG, так и липосомный NX31838-PL значительно снижали уровень ангиогенеза на 65-70%. Предполагают, что эти различия могут быть отнесены к соответствующим фармакокинетическим профилям Нуклеиновых Кислот-Лигандов.
Статистический анализ: Группы в анализе Майлса и моделях роговичного ангиогенеза сравнивали с использованием Rank ANOVA со сравнениями Dunnett.
Пример 10. Эффективность Нуклеиновой Кислоты-Лиганда in vivo в опухолевых моделях
Модель ксенотрансплантата опухоли человека: Способность Нуклеиновой Кислоты-Лиганда VEGF NX31838 40К PEG воздействовать на рост плотной опухоли определяли на подкожной опухолевой модели у мышей nude. Опухолевые клетки рабдомиосаркомы человека А673 выращивали в культуре ткани, собирали, и 1•107 жизнеспособных клеток имплантировали подкожно мышам nude проксимально к осевому району бока. Обработку тестируемыми соединениями начинали через 12 часов после этого и продолжали в течение всего эксперимента. Соединения дозировали внутрибрюшинно дважды в сутки по 10 и 40 мг/кг. Отрицательный контроль состоял из дозирования перемешанной последовательности соответствующего размера, NX31917-40K PEG (см. молекулярное изображение на фиг. 1R) по 40 мг/кг дважды в сутки, а положительный контроль состоял из антитела против VEGF Mab. 26503.11 (R& D Systems, Lot # LD03), дозированного по 100 мкг/мышь дважды в неделю. Обе группы, обработанные Нуклеиновой Кислотой-Лигандом, и группы, обработанные антителом, демонстрировали значительное замедление опухолевого роста относительно отрицательной контрольной группы перемешанной последовательности (фиг. 11). % Ингибирования опухолевого роста (ИОР) определили как 75% и 80% для BID (дважды в сутки) групп 40 мг/кг и 10 мг/кг и 83% для группы, обработанной моноклональным антителом (таблица 8). Поскольку между группой дозы 40 мг/кг и группой дозы 10 мг/кг не оказалось значительного различия, после суток 14 дальнейшее дозирование группы 40 мг/кг не повторяли. Как можно видеть на фиг. 11, несколько суток после прекращения дозировки опухоли росли быстро и имитировали скорость роста отрицательной контрольной группы, тогда как в группе 10 мг/кг Нуклеиновой Кислоты-Лиганда и в группе, обработанной антителом, они продолжали расти со сниженной скоростью.
Дополнительные исследования проводили с использованием той же опухолевой модели, где сравнивали новые партии Нуклеиновой Кислоты-Лиганда VEGF NX31838 40К PEG (обозначенные NX31838.04 и NX31838.07), а также дозы, титрованной с понижением от 10 мг/кг BID, 3 мг/кг BID и 1 мг/кг BID. Эксперимент также включал в себя дозу 10 мг/кг один раз в сутки, а также липосомальную форму Нуклеиновой Кислоты-Лиганда VEGF, NX31838 PL при дозе 10 мг/кг BID. Как можно видеть на фиг. 12 и в таблице 9, в обоих экспериментах достигалась одинаковая степень ингибирования опухолевого роста. Обе партии Нуклеиновой Кислоты-Лиганда VEGF являлись эквивалентными при сравнении в режиме дозирования дважды в сутки, со значениями ИОР 61% и 70% для старой и новой партии соответственно. Кроме того, определили, что дозирование один раз в сутки (SID) являлось таким же эффективным, как дозирование дважды в сутки.
Осуществили третий эксперимент, где дальнейшее понижающееся титрование Нуклеиновой Кислоты-Лиганда VEGF являлось способным продемонстрировать взаимоотношение доза/ответ в отношении опухолевого роста. В этом эксперименте Нуклеиновую Кислоту-Лиганд VEGF титровали с понижением, достигая отсутствия эффекта дозы при 0,03 мг/кг. Относительное ингибирование опухолевого роста может быть видно на фиг. 13 и суммировано в таблице 10.
Кроме трех исследований с неустановившейся опухолью, провели исследование с установившейся опухолью, где опухолям давали установиться и достичь 200+/-100 мм3 до начала обработки Нуклеиновой Кислотой-Лигандом VEGF. Группы дозы 10 мг/кг NX31838 40К PEG и 100 мкг дважды в неделю mAb 26503 (R& D Systems) достигали 59% и 69% ингибирования опухолевого роста соответственно (фиг. 14, таблица 11). Эти совокупные исследования демонстрируют, что Нуклеиновая Кислота-Лиганд VEGF является способной замедлять рост опухоли А673 с момента установления так же, как и ингибировать опухолевый рост, когда опухоли уже установились.
Модель саркомы Капоши: Исследовали также воздействие NX31838-40К PEG на подкожный рост клеточной линии саркомы Капоши KSY-1 у мышей nude. Клетки KSY-1 являются уникальными среди опухолевых клеточных линий в том, что их можно ингибировать в культуре антагонистами VEGF. Клетки KSY-1 выращивали в культуре, собирали и инъецировали подкожно (2•107 клеток/мышь) в заднюю боковую сторону мышей. Три группы мышей (4 мыши на группу) обрабатывали путем внутрибрюшинных инъекций каждые 12 часов либо 30 мг/кг NX31838-40K PEG, либо 30 мг/кг NX31917-40K PEG (см. молекулярное изображение на фиг. 1R), либо ФСБ в течение всего эксперимента. Обработку начинали через одни сутки после имплантации опухолевых клеток. При том, что опухолевый рост в обработанных ФСБ и в обработанных NX31917-40K PEG группах являлся сравнимым, значительное ингибирование опухолевого роста наблюдалось в обработанной NX31838-40K PEG группе (фиг. 16). NX31838-40K PEG ингибировал рост опухолей KSY-1 на 65% (по сравнению с обработанной ФСБ группой) или на 69% (по сравнению с обработанной NX31917-40K PEG группой) в то время, когда эксперимент был закончен (сутки 22).
Пример 11. Внутристекловидная фармакокинетика Нуклеиновой Кислоты-Лиганда VEGF NX31838+40K PEG у кроликов
Новозеландских белых кроликов обрабатывали Нуклеиновой Кислотой-Лигандом VEGF, конъюгированной с 40m ПЭГ, путем внутристекловидного введения дозы 0,5 мг/глаз. 40К ПЭГ конъюгировали с Нуклеиновой Кислотой-Лигандом VEGF, как описано в примере 5, и полученный в результате Комплекс являлся таким, как показан на фиг. 1Н (SEQ ID NO: 8). Кролики получали внутристекловидную инъекцию NX31838 40К PEG в каждый глаз. Время между дозами для данного животного не превышало 15 минут. Кровь и образцы стекловидного тела собирали, как описано в таблице 7.
Анализ образцов плазмы и стекловидного тела проводили с помощью использования двойного гибридизационного анализа. В этом анализе использовали два зонда гибридизации, зонд захвата, присоединенный к лункам 96-луночных плат, и биотинилированный детекторный зонд. Зонд захвата образует гибрид с 5' концом Нуклеиновой Кислоты-Лиганда. Данный анализ является высоко специфичным и чувствительным к полноразмерной Нуклеиновой Кислоте-Лиганду с получением положительного сигнала. Обычный предел количественного определения составляет примерно 2 фмоля в 5 мкл плазмы. Т










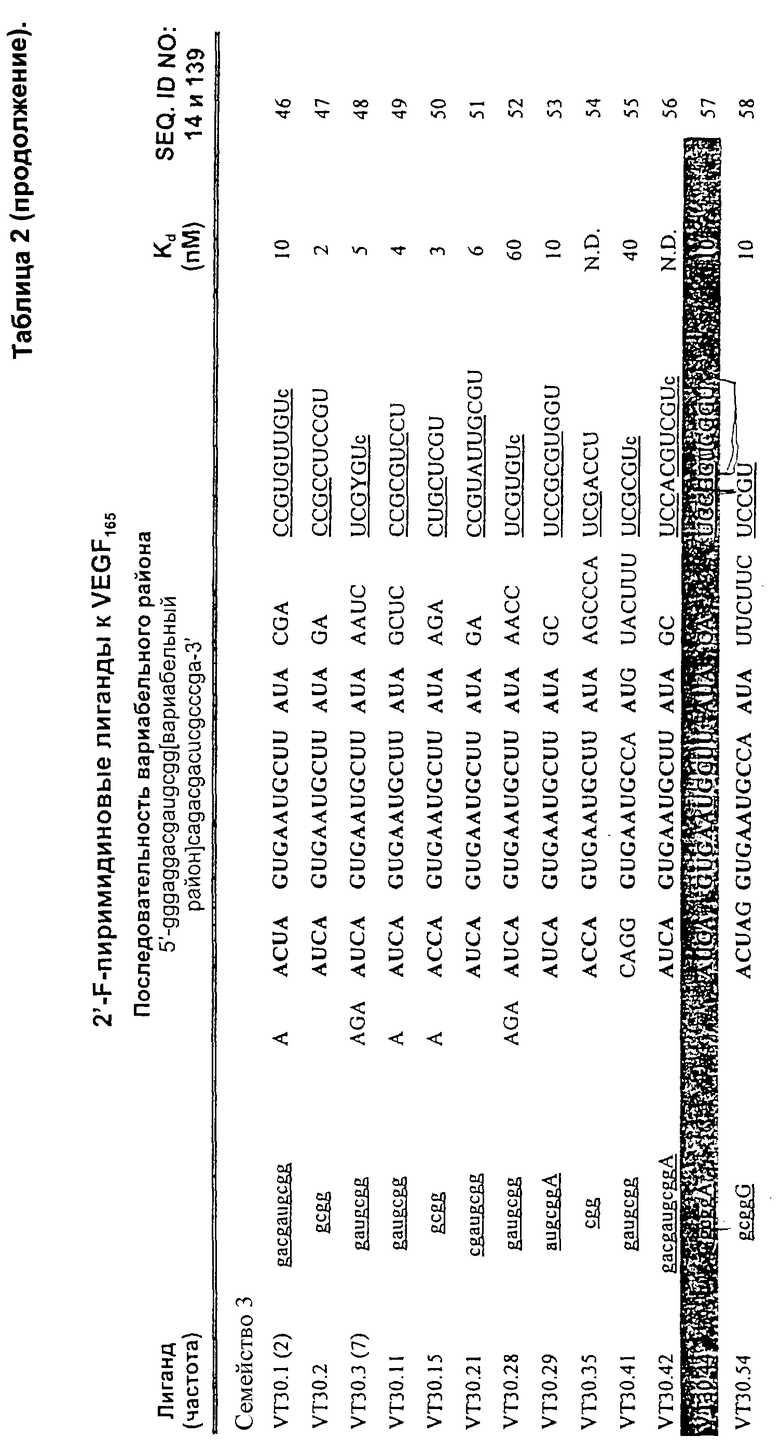













































































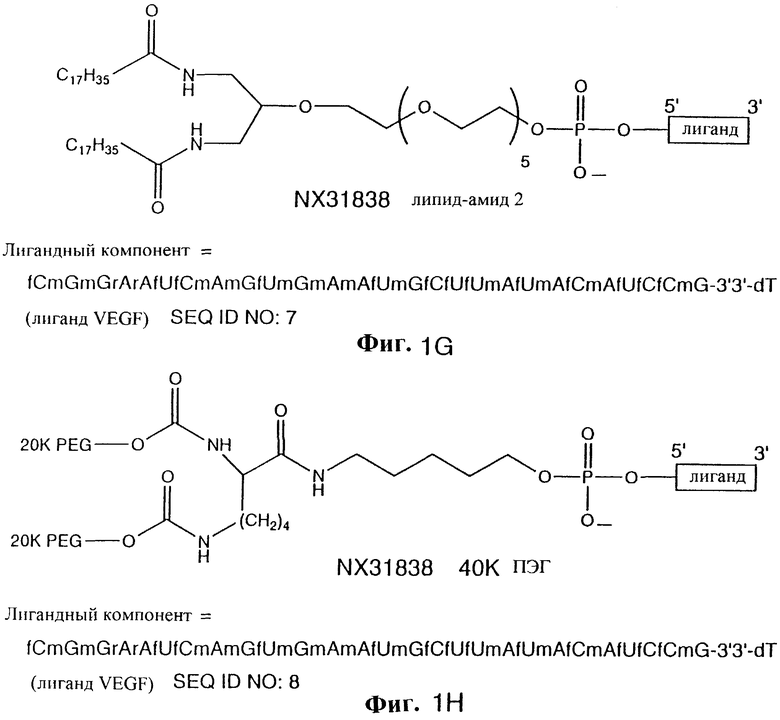



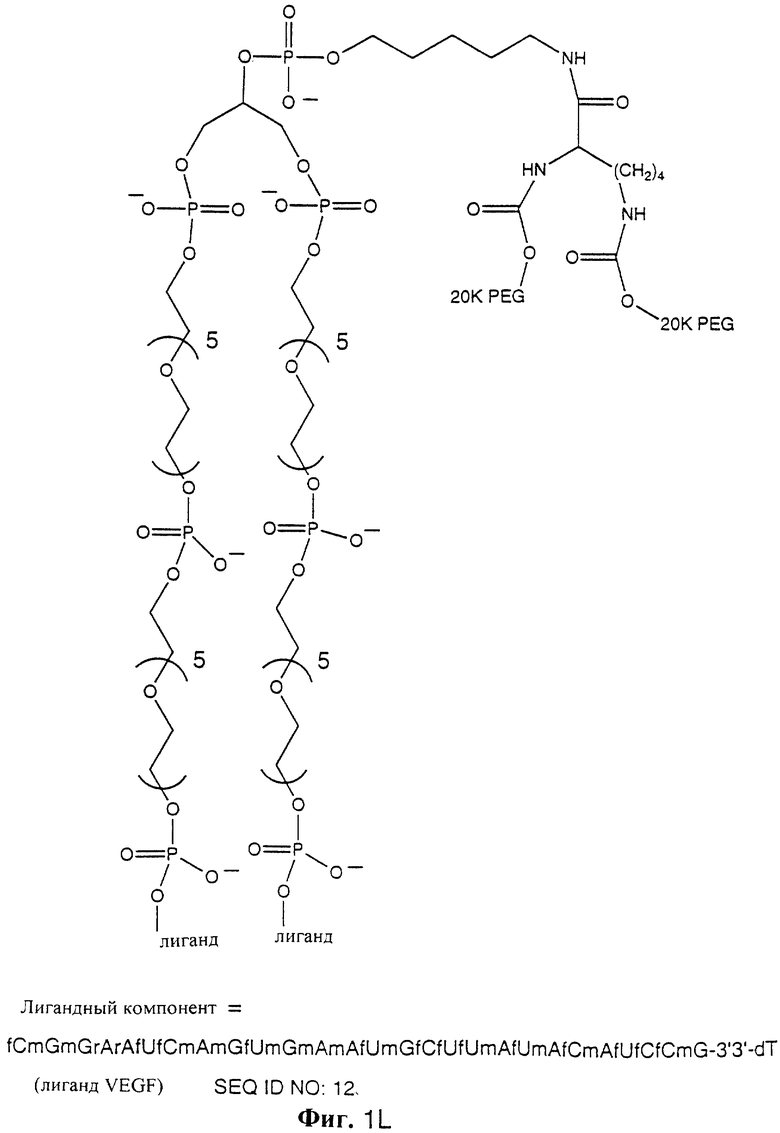












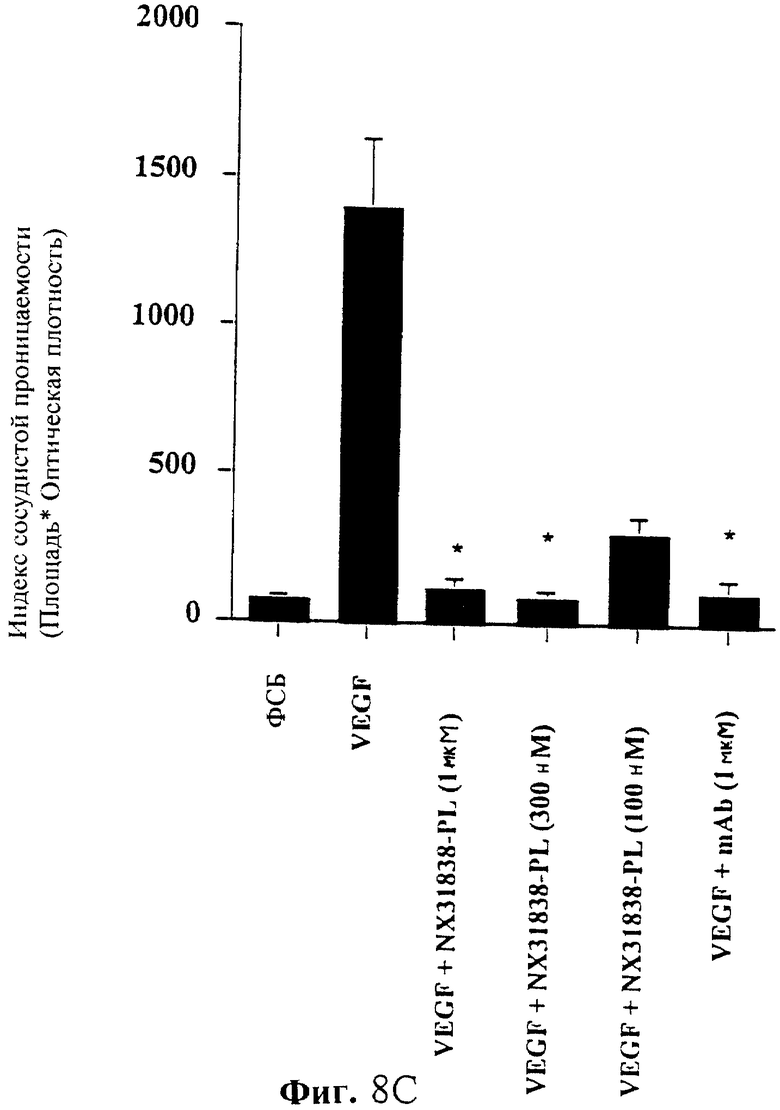


















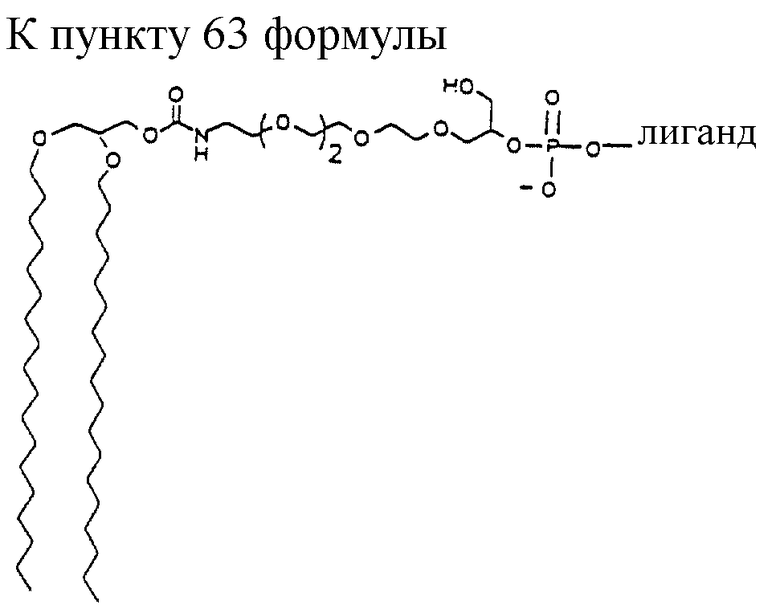
Изобретение относится к очищенным и выделенным не встречающимся в природе РНК-лигандам к сосудистому эндотелиальному фактору роста (VEGF) (указаны олигонуклеотидные последовательности). Описаны комплексы формулы А-В-Y, где А - полиалкиленгликоль или глицеринлипид, В - линкер(ы), Y - РНК-лиганд к VEGF. Описан способ получения этих комплексов, включающий идентификацию РНК-лиганда из смеси Кандидатов Нуклеиновых Кислот, обладающих повышенной аффинностью к VEGF, а также липидная конструкция, включающая этот комплекс. Комплексы по изобретению улучшают фармакокинетические свойства РНК-лигандов к VEGF и используются для лечения VEGF-опосредованных заболеваний, в том числе для ингибирования VEGF-опосредованного ангиогенеза, роста опухолей и дегенерации желтого пятна. Предлагаются также способ снижения клиренса из плазмы Нуклеиновой Кислоты Лиганда к VEGF, способ пролонгации его действия в глазе и способ направления терапевтического или диагностического агента к биологической мишени. 15 с. и 62 з. п. ф-лы, 39 ил. , 13 табл.
А-В-Y,
где А - полиалкиленгликоль, имеющий структуру R(O(CH2)x)nO-, где R независимо выбран из группы, состоящей из Н и СН3, х= 2-5, а n≈MW (молекулярная масса) полиалкиленгликоля/ 16+14х;
В - один или более линкеров,
Y - РНК-лиганд к VEGF по п. 1.
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF)
7. Комплекс по п. 3, где указанный Комплекс представляет собой
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF)
8. Комплекс по п. 3, где указанный Комплекс представляет собой (см. графическую часть).
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF)7
9. Комплекс по п. 3, где указанный Комплекс представляет собой (см. графическую часть).
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF)
10. Комплекс по п. 3, где указанный Комплекс представляет собой (см. графическую часть).
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF).

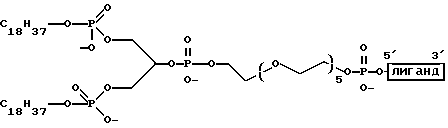


Лигандный компонент = 5'-TsTsTsTsmGaUaCmGmGaUmAaCrGmGrAmGaUmGrGrAaCaCmGaUaCmAaCmGTsTsTsTsT-3' (лиганд VEGF)
38. Способ лечения VEGF-опосредованного заболевания, при котором вводят фармацевтически эффективное количество Комплекса по п. 2 или 13.




Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF)
54. Способ по п. 50, где указанный Комплекс имеет структуру (см. графическую часть).
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF)
55. Способ по п. 50, где указанный Комплекс имеет структуру (см. графическую часть).
Лигандный компонент = fСmGmGrArAfUfCmAmGfUmGmAmAfUmGfCfUfUmAfUmAfCmAfUfCfCmG-3'3'-dT (лиганд VEGF).



Лигандный компонент = 5'-TsTsTsTsmAaCaCaUrGrAaUmGrGaUmAmGrAaCmGaCaCmGmGmGmGaUmGTsTsTsTsT-3' (лиганд VEGF).


Приоритет по признакам и пунктам:
25.10.1996 по п. 1: признаки, касающиеся последовательностей SEQ ID Nos 1-4;
по пп. 2-5, 11-20, 21 с признаками, касающимися последовательностей SEQ ID Nos 1-4;
по пп. 37 и 63;
06.06.1997 - признаки, касающиеся последовательностей SEQ ID Nos 5-12 и 15-90,
по пп. 6-10, 29, 33, 35, 38-42, 51-55, 64-66, 71-75, 77;
21.07.1997 - по пп. 2-5, 11-20, 21, 22-23, 45, 48-50 и 56 с признаками, касающимися последовательностей SEQ ID Nos 5-12 и 15-90;
по п. 76;
17.10.1997 - признаки, касающиеся последовательностей SEQ ID Nos 13-14 и 91-132;
с признаками, относящимися к последовательностям, кроме SEQ ID Nos 5-12 и 15-90;
по пп. 24-28, 30-32, 34, 36, 43, 44, 46-47, 57-59, 60-62, 67-70.
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| US 5459015 А, 17.10.1995 | |||
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| КОРШАК В.В | |||
| и др | |||
| Полимеры в процессах иммобилизации и модификации природных соединений | |||
| - М.: Наука, 1984, с.199 | |||
| 3-Фтор-2,3-дидезоксиаденозин,проявляющий цитостатическую активность | 1981 |
|
SU961354A1 |
| 3-Фтор-2,3-дидезоксигуанозин,проявляющий цитостатическую активность | 1982 |
|
SU1053474A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
